
ヒト化抗NGF抗体
【課題】結晶学的研究に由来し実験的に得られる構造データに一貫して基づく手法による、ヒトに対して実質的に免疫原性でない最適化されたヒト化形態の免疫グロブリンを得るための方法の提案。
【解決手段】NGFに対する結合活性を維持するヒト化抗NGF抗体またはその断片であって、
(c) アミノ酸配列GFSLTNNNVNWを含む第一のCDR、アミノ酸配列GVWAGGATDYNSALKSを含む第二のCDR、アミノ酸配列DGGYSSSTLYAMDAを含む第三のCDRを有するVH領域;及び
(d) アミノ酸配列RASEDIYNALAを含む第一のCDR、アミノ酸配列HTを含む第二のCDR、アミノ酸配列QHYFHYPRTを含む第三のCDRを有するVL領域
を含むヒト化抗NGF抗体の提供。
【解決手段】NGFに対する結合活性を維持するヒト化抗NGF抗体またはその断片であって、
(c) アミノ酸配列GFSLTNNNVNWを含む第一のCDR、アミノ酸配列GVWAGGATDYNSALKSを含む第二のCDR、アミノ酸配列DGGYSSSTLYAMDAを含む第三のCDRを有するVH領域;及び
(d) アミノ酸配列RASEDIYNALAを含む第一のCDR、アミノ酸配列HTを含む第二のCDR、アミノ酸配列QHYFHYPRTを含む第三のCDRを有するVL領域
を含むヒト化抗NGF抗体の提供。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、三次元構造を決定し、比較することによる抗体のヒト化方法、それによって得たヒト化抗体ならびに治療および診断におけるそれらのin vivoでの使用に関する。
【背景技術】
【0002】
動物起源のモノクローナル抗体のヒトにおける治療的および診断的応用は、特に反復投与を必要とする治療レジメンでは基本的に禁忌である。特に、マウスモノクローナル抗体は半減期が比較的短く、ヒトで使用される場合は免疫グロブリンの基本的な機能特性、例えば補体依存性細胞傷害作用および細胞媒介性細胞傷害作用の一部が失われる。
【0003】
さらに、ヒト以外の起源のモノクローナル抗体は、患者に注入された場合に免疫原性のアミノ酸配列を含む。多数の研究により、外来性抗体の注射の後、対象は抗体自体に対してかなり強い免疫反応(HAMA(ヒト抗マウス抗体)反応として知られる)を起こし、免疫複合体の形成、薬物動態の変更、アレルギー反応の生成などによりその治療的な有用性が完全に失われることが明らかにされている。さらに、異なる病理の治療のために益々多くの異なるモノクローナル抗体がマウスまたは他の哺乳類(したがってヒトに抗原性の)で開発されていることを考慮すると、相互に関係のない療法についても、交差反応性のために治療は無効になるか危険にさえなる可能性がある。いわゆるキメラ抗体(ヒト起源の定常部に連結したマウスの可変部)の生成は多少のプラスの結果をもたらしたが、かなりの免疫原性の問題がまだ残されている。
【0004】
ヒト化抗体は、ヒトの治療的使用の分野で、動物起源の抗体に関して少なくとも3つの考えられる利点を有する。最初に、ヒトのエフェクター領域はヒト免疫系の他の部分とよりよく相互作用することができ、補体依存性細胞傷害作用または細胞媒介性抗体依存性細胞傷害性によって標的細胞をより効率的に破壊する。さらに、ヒト免疫系はヒト化抗体のフレームワーク部または定常部(C)を外来性であると認識せず、それ故に、マウス抗体(まったく外来性)に対する反応およびキメラ抗体(部分的に外来性)によって誘導される反応と比較して、ヒト化抗体に対する抗体反応は最小化されている。
【0005】
ヒトに注入されるマウス抗体は正常抗体よりも非常に短い半減期を有することが報告されている(Shawら、1987)。ヒト化抗体は天然ヒト抗体に非常に類似した半減期を有し、より低い頻度での投与およびより低い用量を可能にする。
【0006】
ヒト化の基本原理は、ヒト免疫グロブリンとの関連で抗原認識の特異性、すなわちCDRドメインを移動させる構成である(「CDRグラフト」、WinterおよびMilstein、1991)。免疫原性の問題を解決するために試作されたいくつかのヒト化抗体の例が報告されている(Maedaら、1991;Singerら、1993;Tempestら、1994;Kettleboroughら、1996;Hsiaoら、1994;Bacaら、1997;Legerら、1997;Ellisら、1995;Satoら、1994;Jonesら、1986;Benharら、1994;ShaおよびXiang、1994;Shearmanら、1991;Rosokら、1996;Gussow & Seemann、1991;Coutoら、1994;Kashmiriら、1995;Bakerら、1994;Riechmannら、1988;Gormanら、1991;Verhoeyenら、1988;Foote & Winter、1992;Lewis & Crowe、1991;Coら、1991;Coら、1991;Verhoeyenら、1991;Eigenbrotら、1994;Hamiltonら、1997;Tempestら、1995;Verhoeyenら、1993;Cookら、1996;Poulら、1995;Coら、1992;Grazianoら、1995;Prestaら、1993;Hakimiら、1993;Roguskaら、1996;Adairら、1994;Satoら、1993;Tempestら、1991;Satoら、1996;Kolbingerら、1993;ZhuおよびCarter、1995;Simsら、1993;Routledgeら、1991;Roguskaら、1994;Queenら、1989;Carterら、1992)。
【0007】
動物(一般にマウス)からヒト化への抗体の変換は対立する要求事項間の妥協であり、その解決方法は個々に異なる。免疫原性を最小にするために、免疫グロブリンは受容側のヒト配列をできるだけ多く維持するものとする。いずれにせよ、元の結合持性を保持するために、免疫グロブリンフレームワークは、CDR部の高次構造が供与体マウスの免疫グロブリンのそれとできるだけ類似するように、受容側のヒト配列に十分な数の変異を含むべきである。これらの対立する考慮事項の結果として、多くのヒト化抗体では、対応するマウス抗体に関して結合親和性のかなりの減少が報告されている(Jonesら、1986;Shearmanら、1991;Kettleborough、1991;Gormanら、1991;Riechmannら、1988)。
現在、ヒト化免疫グロブリンの最も一般的な生成方法は、適当なゲノム配列、合成配列ならびにcDNAの使用に基づく(Reichmannら、1988)。
特許出願EP592106は、齧歯動物からの抗体のヒト化方法を開示している。この方法はヒト化する抗体の三次元構造の表面に露出しているアミノ酸残基の同定、対応するヒト抗体上の同じ位置のアミノ酸残基の同定、および齧歯動物抗体の配列で同定された残基のヒト抗体で同定されたそれらによる置換に基づく。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】EP592106
【特許文献2】EP1181318
【特許文献3】国際公開91/08216
【特許文献4】U.S.Ser.No.07/290,968
【非特許文献】
【0009】
【非特許文献1】Shawら、1987
【非特許文献2】WinterおよびMilstein、1991
【非特許文献3】Maedaら、1991
【非特許文献4】Singerら、1993
【非特許文献5】Tempestら、1994
【非特許文献6】Kettleboroughら、1996
【非特許文献7】Hsiaoら、1994
【非特許文献8】Bacaら、1997
【非特許文献9】Legerら、1997
【非特許文献10】Ellisら、1995
【非特許文献11】Satoら、1994
【非特許文献12】Jonesら、1986
【非特許文献13】Benharら、1994
【非特許文献14】ShaおよびXiang、1994
【非特許文献15】Shearmanら、1991
【非特許文献16】Rosokら、1996
【非特許文献17】Gussow & Seemann、1991
【非特許文献18】Coutoら、1994
【非特許文献19】Kashmiriら、1995
【非特許文献20】Bakerら、1994
【非特許文献21】Riechmannら、1988
【非特許文献22】Gormanら、1991
【非特許文献23】Verhoeyenら、1988
【非特許文献24】Foote & Winter、1992
【非特許文献25】Lewis & Crowe、1991
【非特許文献26】Coら、1991
【非特許文献27】Coら、1991
【非特許文献28】Verhoeyenら、1991
【非特許文献29】Eigenbrotら、1994
【非特許文献30】Hamiltonら、1997
【非特許文献31】Tempestら、1995
【非特許文献32】Verhoeyenら、1993
【非特許文献33】Cookら、1996
【非特許文献34】Poulら、1995
【非特許文献35】Coら、1992
【非特許文献36】Grazianoら、1995
【非特許文献37】Prestaら、1993
【非特許文献38】Hakimiら、1993
【非特許文献39】Roguskaら、1996
【非特許文献40】Adairら、1994
【非特許文献41】Satoら、1993
【非特許文献42】Tempestら、1991
【非特許文献43】Satoら、1996
【非特許文献44】Kolbingerら、1993
【非特許文献45】ZhuおよびCarter、1995
【非特許文献46】Simsら、1993
【非特許文献47】Routledgeら、1991
【非特許文献48】Roguskaら、1994
【非特許文献49】Queenら、1989
【非特許文献50】Carterら、1992
【非特許文献51】Sequences of Proteins of Immunological Interest(National Institutes of Health、Bethesda MD、1987、1991)
【非特許文献52】Paul、1993
【非特許文献53】Kabatら、1987
【非特許文献54】ChothiaおよびLesk、1987
【非特許文献55】Kabat(1987)
【非特許文献56】Kabat(1991)
【非特許文献57】Chothiaら(1989)
【非特許文献58】Lanzavecchiaら、1987
【非特許文献59】Hoodら、1984
【非特許文献60】HarlowおよびLane、1988
【非特許文献61】HunkapillerおよびHood、1986
【非特許文献62】American Type Culture Collection(Catalogue of Cell Lines and Hybridomas、第5版(1985)Rockville、Maryland、USA
【非特許文献63】Satoら、1994
【非特許文献64】CarugoおよびPongor、2001
【非特許文献65】CarugoおよびPongor、2003
【非特許文献66】Daughertyら、1991
【非特許文献67】De Martinoら、1991
【非特許文献68】Gillman & Smith、1979
【非特許文献69】Robertsら、1987
【非特許文献70】Hustonら、1988
【非特許文献71】Birdら、1988
【非特許文献72】BuchnerおよびRudolph、1991
【非特許文献73】Skerraら、1991
【非特許文献74】Cesareni、1992
【非特許文献75】Swimmerら、1992
【非特許文献76】Gramら、1992
【非特許文献77】Clacksonら、1991
【非特許文献78】Scott & Smith、1990
【非特許文献79】Garrardら、1991
【非特許文献80】Putlitzら、1990
【非特許文献81】Larrick & Fry、1991
【非特許文献82】Benvenutoら、1991
【非特許文献83】Durinら、1990
【非特許文献84】Hiattら、1989
【非特許文献85】Winnacker、1987
【非特許文献86】Queenら、1986
【非特許文献87】Buhlerら、1990
【非特許文献88】Meadeら、1990
【非特許文献89】Scopes、1982
【非特許文献90】LefkovitsおよびPernis、1979
【非特許文献91】LefkovitsおよびPernis、1981
【非特許文献92】Chaudharyら、1989
【非特許文献93】Thorpeら(1982)
【非特許文献94】BaldwinおよびByers、1985;;
【非特許文献95】OlsnesおよびPhil、1982
【非特許文献96】Chenら、1995
【非特許文献97】Angelesら、1996
【非特許文献98】Goding(1986)
【非特許文献99】Paikら(1982)
【非特許文献100】Jancarik & Kim、1991
【非特許文献101】Otwinowski & Minor、1997
【非特許文献102】Bermanら、2000
【非特許文献103】Kodandapaniら、1999
【非特許文献104】Bentley、1990
【非特許文献105】Navaza、1994
【非特許文献106】KleywegtおよびJones、1994
【非特許文献107】Brungerら、1998
【非特許文献108】Laskowskiら、1993
【非特許文献109】Diederichs、1995
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明者らは、結晶学的研究に由来し実験的に得られる構造データに一貫して基づく手法による、ヒトに対して実質的に免疫原性でない最適化されたヒト化形態の免疫グロブリンを得るための方法を提案した。本発明の方法は、治療製剤ならびに他の医学的および診断の用途に適応した形態の抗体の入手を可能にする。
【課題を解決するための手段】
【0011】
本発明は、ヒト化の最初の設計段階(一般によりエラーが起こりやすい)を行うための、完全に構造データに基づく方法に関するものである。ヒト化免疫グロブリンは、軽鎖および重鎖の間に2対のヘテロダイマーを有し、少なくとも1つの鎖は機能的にヒト起源のフレームワーク部の部分と結合した1つまたは複数の動物起源のCDRを有す。例えば、動物起源の元の免疫グロブリンの親和性と同等の親和性でそれぞれの抗原と結合することができるヒト化免疫グロブリンを生成するために、動物起源のCDRを、同じく動物起源で自然に結合しているアミノ酸残基と共に、ヒト起源のフレームワーク部に導入する。
【0012】
本発明の方法は、治療および診断の用途のために適当なヒト化抗体の入手を導いた。特に、それぞれTrkAおよびNGFと高い特異性で結合してリガンドと受容体の間の相互作用を中和することができる、抗TrkA抗体(特許EP1181318)および抗NGF抗体に由来するヒト化免疫グロブリンが得られた。そのような分子は、NGF/TrkAに依存する腫瘍、慢性疼痛、炎症性形態の治療のために、および診断目的のために、例えばTrkA陽性腫瘍または、アルツハイマー病の早発マーカーとしての前脳基底核のin vivo画像化のために有用である。詳細には、ヒト化抗TrkA抗体は、尿路および骨盤領域の炎症性形態で特異的な治療および診断の用途を有す。詳細には、ヒト化抗NGF抗体は、HIVウイルスによって誘導される病理状態で、HIVに感染したNGF依存性マクロファージなどの免疫細胞のアポトーシスを誘導するための、特異的な治療および診断の用途を有す。
【0013】
したがって、本発明の目的は、既知の配列の動物抗体のVHおよびVL可変部のヒト化方法を提供することであり、これは、
a)入手不能であるならば、動物抗体のVHおよびVL部の結晶構造を得る段階と、
b)その構造が3Å以上の分解能で実験的に測定されたヒト起源またはヒト化抗体の0からn個の一連の可能なフレームワーク受容体を、動物抗体のフレームワークの一次配列との最高水準の相同性および同一性に基づいて予備選択する段階と、
c)動物抗体のVHおよびVL可変部とb)で得られるVHおよびVL部との間の構造比較をそれぞれ行い、各比較についてRMSを計算してより小さなRMSを有するヒト起源のVHおよびVL部を同定する段階と、
d)c)で同定したヒト配列内の適当な位置に前記動物抗体のCDR部の配列を挿入する段階と、
e)必要に応じて、c)で同定したヒトVHおよびVL部の1つまたは複数のアミノ酸残基を復帰変異させる段階とを含む。
【0014】
好ましくは、抗体の修飾は、組換えDNA技術によって起こる。
【0015】
好ましい一実施形態では、動物抗体は抗NGF抗体であり、好ましくは、それはαD11抗体であり、ヒト化配列は本質的に以下のVH配列;HumαD11VH、
EVQLVESGGGLVQPGGSLRLSCAASGFSLTNNNVNWVRQAPGKGLEWVGGVWAGGATDYNSALKSRFTISRDNSKNTAYLQMNSLRAEDTAVYYCARDGGYSSSTLYAMDAWGQGTLVTVSS、(配列番号17)
およびVL;HumαD11 Vk
DIQMTQSPSSLSASVGDRVTITCRASEDIYNALAWYQQKPGKAPKLLIYNTDTLHTGVPSRFSGSGSGTDYTLTISSLQPEDFATYFCQHYFHYPRTFGQGTKVEIK(配列番号18)。
を有す。
【0016】
他の実施形態では、動物抗体は抗TrkA抗体であり、好ましくは、それはαMNAC13抗体であり、ヒト化配列は本質的に以下のVH配列;HumMNAC13VH、
EVQLLESGGGLVQPGGSLRLSCAASGFTFSTYTMSWARQAPGKGLEWVAYISKGGGSTYYPDTVKGRFTISRDNSKNTLYLQMNSLRAEDSAVYYCARGAMFGNDFFFPMDRWGQGTLVTVSSA、(配列番号37)
およびVL;HumMNAC13Vk
DIVLTQSPSSLSASVGDRVTITCSASSSVSYMHWYQQKPGQAPKLLIYTTSNLASGVPSRFSGSGSGTDYTLTISSLQPEDVATYYCHQWSSYPWTFGGGTKVEIK(配列番号38)。
を有する。
【0017】
本発明のヒト化免疫グロブリン(または結合活性を維持している誘導断片および他の誘導可能な化合物)は、公知の組換えDNA技術によって生成することができる。その後のヒト化免疫グロブリンの使用の機能としては、トランスジェニック動物またはトランスフェクション細胞をそれらの発現のために使用することができ、好ましくは不死化された真核細胞(例えば骨髄腫またはハイブリドーマ細胞)であるが、原核生物の宿主、昆虫または植物細胞でもよい。得られたヒト化免疫グロブリン配列をコードするポリヌクレオチドは、合成によっても得ることができる。
【0018】
本発明のヒト化免疫グロブリンは、単独でも、または他の治療薬と併用しても使用することができる。抗腫瘍剤として使用する場合は化学療法剤が好ましく、それは薬理学的用途(例えばアントラサイクリン、パクリタキセル、シスプラチン、ゲムシタビン、非ステロイド性コルチコステロイド抗炎症薬または免疫抑制薬)ならびに各特定の病理状態の療法で現在適用されているすべての薬剤に従い異なる。ヒト化免疫グロブリンまたはそれらの複合体は、薬理学的に許容できる投薬形態で調製することができ、それは投与の型に従い異なる。
【0019】
定義
2つのポリヌクレオチドまたはポリペプチド(それぞれヒト化免疫グロブリンをコードするDNAの配列またはヒト化免疫グロブリンのアミノ酸配列またはそれらの一部)に関連する用語「実質的に同一の」は、最大の対応で比較および整列させたときに、ヌクレオチドまたはアミノ酸残基が最低80%(好ましくは90〜95%以上)の同一性を有する2個以上の配列を指す。通常、「実質的な同一性」は少なくとも50残基の長さの領域で、より好ましくは少なくとも100残基の領域で、または最適条件では150以上の残基で、または完全な配列で検証される。下記のように、抗体のいずれか2つの配列は、カバットのナンバリング方式を使用して1つの方法だけで整列させることができる。したがって、抗体については、同一性割合は特有の明確な意味を有する。成熟免疫グロブリンの重鎖および軽鎖の可変部のアミノ酸はHxおよびLxと称され、xは、カバットのナンバリング方式、Sequences of Proteins of Immunological Interest(National Institutes of Health、Bethesda MD、1987、1991)によるアミノ酸位置を指定する数である。カバットは、コンセンサス配列を作成するために、各サブグループの抗体のアミノ酸配列のリスト、ならびに各サブグループの各位置で最も頻出するアミノ酸のリストを決定した。カバットは数字をリスト内の各配列の各アミノ酸に割り当てる方法を使用するが、この各残基の数字を割り当てる方法はこの分野で標準の方法になっている。カバットの方式は彼の研究には存在しない他の抗体にまで拡張することができ、保存されたアミノ酸に基づいてカバットによって特定されるコンセンサス配列の1つと問題の抗体を整列させることができる。カバットのナンバリング方式を使用すると、異なる抗体中の同等の位置のアミノ酸の特定が容易になる。例えば、ヒト起源の抗体のL10位置のアミノ酸は、マウス起源の抗体のL10位置の同等のアミノ酸位置を占める。
【0020】
抗体の基本的な構造単位が四量体を含むことは公知である。各四量体は2つの同一の対のポリペプチド鎖によって構成され、それぞれは軽鎖(25kDa)および重鎖(50〜75kDa)によって構成される。各鎖のアミノ末端領域は約100〜110以上のアミノ酸の可変部を含み、それは抗原認識に関係している。各鎖のカルボキシ末端領域は、エフェクター機能を媒介する定常部を含む。軽鎖および重鎖の各対の可変部は、抗体の結合部位を形成する。したがって、手つかずの抗体は2つの結合部位を有する。
【0021】
軽鎖は、κまたはλに分類されている。重鎖はγ、μ、α、εに分類され、それらは抗体のイソタイプをそれぞれIgG、IgM、IgA、IgDおよびIgEと定義する。軽鎖および重鎖の中では、可変部および定常部は少なくとも約12のアミノ酸の「J」領域で接続され、重鎖だけは約10アミノ酸の「D」領域を含む(Paul、1993)。
【0022】
軽鎖および重鎖の各対の可変部は、抗体の結合部位を形成する。それらは、相補性決定部(CDR)と呼ばれている3つの超可変部が接続しているフレームワーク(FR)と呼ばれている比較的保存された領域で構成される同じ一般構造を特徴とする(Kabatら、1987;ChothiaおよびLesk、1987)。各対の2つの鎖のCDRはフレームワーク部によって整列させられ、特定のエピトープと結合する機能を取得している。アミノ末端領域からカルボキシ末端領域の方へかけて、軽鎖および重鎖の両方の可変ドメインは、交互に出現するFRおよびCDR領域、すなわちFR、CDR、FR、CDR、FR、CDR、FRを含む。したがって、重鎖および軽鎖の両方は3つのCDR、すなわちそれぞれCDRH1、CDRH2、CDRH3およびCDRL1、CDRL2、CDRL3を特徴とする。各領域へのアミノ酸の帰属は、Kabat(1987および1991)および/またはChothia & Lesk(1987)、Chothiaら(1989)による定義に従って行われた。
【0023】
好ましくは、例示されたヒト化免疫グロブリンの類似体は、保存的なアミノ酸の置換のために元の免疫グロブリンとは異なる。アミノ酸置換を保存的または非保存的に分類するために、アミノ酸は以下の通りに分類することができる。
群I(疎水性側鎖):M、A、V、L、I;
群II(中性の親水性側鎖):C、S、T、N、Q;
群III(酸性側鎖):D、E;
群IV(塩基性側鎖):K、R;
群V(主鎖の配向性に影響する残基):G、P;
群VI(芳香族側鎖):F、Y、W。
【0024】
保存的なアミノ酸置換は同じクラスのアミノ酸の間の置換に関係があるが、非保存的なアミノ酸置換は異なるクラスのメンバーの間の交換を伴う。
【0025】
用語「エピトープ」は、免疫グロブリンと特異的に結合することができるいかなるタンパク質決定要素を含む。通常、エピトープは高分子の化学的に活性な表面の組、例えばアミノ酸または糖の側鎖によって形成され、それらは通常、特異的な化学物理特性および配座特性を有す。
【0026】
用語「免疫グロブリン」は、免疫グロブリン遺伝子によってコードされた1つまたは複数のポリペプチドから成るタンパク質を指す。四量体抗体の形態に加えて、免疫グロブリンは様々な形態で存在することができる。例えば、それらは断片Fv、FabおよびF(ab')、ならびに二機能性ハイブリッド抗体(Lanzavecchiaら、1987)および単鎖Fv断片(Hoodら、1984;HarlowおよびLane、1988;HunkapillerおよびHood、1986)を含む。
【0027】
キメラ抗体は、その軽鎖および重鎖の遺伝子が異なる種に属している免疫グロブリンの遺伝子領域から始まるように操作された抗体である。例えば、マウスのモノクローナル抗体の遺伝子の可変部分(V)は、ヒト起源の抗体の定常部(C)に連結させることができる。治療用のキメラ抗体は、したがって、マウス抗体に由来する抗原を認識するドメインVおよびヒト抗体に由来するエフェクタードメインCから成るハイブリッドタンパク質である(他の哺乳類の組合せも使用することができる)。
【0028】
カバットの定義によれば、用語「フレームワーク」は、種内の異なる免疫グロブリンの間で比較的保存されている、免疫グロブリンの軽鎖および重鎖の可変部の部分(CDRに属さない)を指す。それ故に、ヒトフレームワークは、ヒト抗体で天然に見られるフレームワークと実質的に同一(少なくとも85%以上)なフレームワーク部である。
用語「ヒト化免疫グロブリン」は、ヒトフレームワークおよび非ヒト抗体に由来している少なくとも1つのCDRを含み、存在する各定常部がヒト免疫グロブリンの領域と実質的に同一(少なくとも85%、好ましくは少なくとも90〜95%同一)である免疫グロブリンを指す。それ故に、CDR以外のヒト化免疫グロブリンのすべての部分は、天然のヒト免疫グロブリンの1つまたは複数の配列の対応する領域と、実質的に同一である。例えば、マウス可変部およびヒト起源の定常部によって構成されるキメラ抗体は、ヒト化免疫グロブリンには含まれない。
【図面の簡単な説明】
【0029】
【図1A】変性条件下でのポリアクリルアミド(12% SDS-PAGE)およびクーマシーブルーでの染色による、MNAC13抗体Fab断片の精製結果の分析を示す図である(ウェル1:パパインでタンパク分解的に消化されたMNAC13抗体の試料;ウェル2:DEAE Sephacellイオン交換樹脂に結合し、250mMのNaClで溶出した分画;ウェル3:分子量;ウェル4:精製され濃縮されたMNAC13抗体のFab断片)。
【図1B】MNAC13抗体のFab断片の典型的結晶を示す図である。
【図1C】MNAC13抗体のFab断片の結晶で得られる高分解能回折スペクトラムを示す図である。
【図1D】MNAC13抗体のFab断片の重鎖および軽鎖ドメインの主鎖のねじれ角のラマチャンドランチャートである。
【図2A】変性条件下でのポリアクリルアミド(12% SDS-PAGE)およびクーマシーブルーでの染色による、αD11抗体Fab断片の精製結果の分析を示す図である(ウェル1:パパインでタンパク分解的に消化されたαD11抗体の試料;ウェル2:精製され濃縮されたαD11抗体のFab断片;ウェル3:分子量)。
【図2B】αD11抗体のFab断片の典型的結晶を示す図である。
【図2C】αD11抗体のFab断片の結晶で得られる高分解能回折スペクトラムを示す図である。
【図2D】αD11抗体のFab断片の重鎖および軽鎖ドメインの主鎖のねじれ角のラマチャンドランチャートである。
【図3−1】図3Aは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図3Bは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図3Cは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図3Dは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図3Eは、MNAC13の仮定的最適値からのヒト化またはヒト起源の抗体の逸脱を示す図である(全体の同一性および相同性の程度-青-およびフレームワークレベル-マゼンタ-を考慮して計算)。図3Fは、MNAC13の仮定的最適値からのヒト化またはヒト起源の抗体の逸脱を示す図である(全体の同一性および相同性の程度-青-およびフレームワークレベル-マゼンタ-を考慮して計算)。
【図3−2】図3Gは、マウス抗体との同一性および相同性の程度および利用できる構造データの分解能の程度に従って選択された、ヒト化またはヒト起源の抗体のそれぞれの領域のMNAC13のFv断片との構造的整列を示す図である。図3Hは、H)は選択されたヒト化抗体1AD0(赤で示す)のそれぞれの領域のMNAC13のFv断片(シアンで示す)との構造的整列を示す図である。図3Iは、I)はCDR移植の後に生じた抗体モデル(フレームワークレベルでは黄色で、CDRレベルでは白で示されている)のMNAC13のFv断片との構造的整列を示す図である。図3Lは、選択されたフレームワーク内の推定される復帰変異の同定の結果として得られたMNAC13ヒト化抗体のFv断片のモデルを示す図である(ヒト起源残基は緑で示し、マウス起源残基はマゼンタで示す)。
【図4−1】図4Aは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図4Bは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図4Cは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図4Dは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図4Eは、αD11の仮定的最適値からのヒト化またはヒト起源の抗体の逸脱を示す図である(全体の同一性および相同性の程度-青-およびフレームワークレベル-マゼンタ-を考慮して計算)。図4Fは、αD11の仮定的最適値からのヒト化またはヒト起源の抗体の逸脱を示す図である(全体の同一性および相同性の程度-青-およびフレームワークレベル-マゼンタ-を考慮して計算)。
【図4−2】図4Gは、マウス抗体との同一性および相同性の程度および利用できる構造データの分解能の程度に従って選択された、ヒト化またはヒト起源の抗体のそれぞれの領域のαD11のFv断片との構造的整列を示す図である。図4Hは、H)は選択されたヒト化抗体1JPS(赤で示す)のそれぞれの領域のαD11のFv断片(シアンで示す)との構造的整列を示す図である。図4Iは、I)はCDR移植の後に生じた抗体のモデル(フレームワークレベルでは黄色で、CDRレベルでは白で示されている)のαD11のFv断片との構造的整列を示す図である。図4Lは、選択されたフレームワーク内の推定される復帰変異の同定の結果として得られたαD11ヒト化抗体のFv断片のモデルを示す図である(ヒト起源残基はシアンで示し、マウス起源残基は紫で示す)。
【図5】MNAC13(配列番号22、配列番号24)、ヒト化のために選択されたヒト化抗体(1AD0;配列番号39、配列番号40)、1AD0のフレームワーク上へのCDR移植および記載されている復帰変異および変異の後のMNAC13のヒト化形態(Hum MNAC13:配列番号37、配列番号38)の重鎖(A)および軽鎖(B)それぞれの可変部の一次構造の整列を示す図である。CDRは、MNAC13の2つの鎖のヒト化形態の配列内に下線付き文字で強調されている。
【図6】αD11(配列番号2、配列番号4)、ヒト化のために選択されたヒト化抗体(1JPS;配列番号19、配列番号20)、1AD0のフレームワーク上へのCDR移植および記載されている復帰変異および変異の後のαD11のヒト化形態(Hum αD11:配列番号17、配列番号18)の重鎖(A)および軽鎖(B)それぞれの可変部の一次構造の整列を示す図である。CDRは、αD11の2つの鎖のヒト化形態の配列内に下線付き文字で強調されている。
【図7A】MNAC13のマウス形態の軽鎖可変部のcDNAのヌクレオチド配列を示す図である(配列番号23)。
【図7B】MNAC13のマウス形態の重鎖可変部のcDNAのヌクレオチド配列を示す図である(配列番号21)。
【図7C】MNAC13の軽鎖可変部(配列番号38)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:L1S:配列番号31;L2AS:配列番号32;L3S:配列番号33;L4AS:配列番号34;L5S:配列番号35;L6AS:配列番号36を、対応するアミノ酸配列への翻訳と共に示す図である。
【図7D】MNAC13の重鎖可変部(配列番号37)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:H1S:配列番号25;H2AS:配列番号26;H3S:配列番号27;H4AS:配列番号28;H5S:配列番号29;H6AS:配列番号30を、対応するアミノ酸配列への翻訳と共に示す図である。
【図7E】MNAC13の軽鎖可変部(配列番号38)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:L1S:配列番号31;L2AS:配列番号32;L3S:配列番号33;L4AS:配列番号34;L5S:配列番号35;L6AS:配列番号36を、対応するアミノ酸配列への翻訳と共に示す図である。
【図7F】MNAC13の重鎖可変部(配列番号37)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:H1S:配列番号25;H2AS:配列番号26;H3S:配列番号27;H4AS:配列番号28;H5S:配列番号29;H6AS:配列番号30を、対応するアミノ酸配列への翻訳と共に示す図である。
【図8A】αD11のラット形態の軽鎖可変部のcDNAのヌクレオチド配列を示す図である(配列番号3)。
【図8B】αD11のマウス形態の重鎖可変部のcDNAのヌクレオチド配列を示す図である(配列番号1)。
【図8C】αD11の軽鎖可変部(配列番号18)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:L1S:配列番号11;L2AS:配列番号12;L3S:配列番号13;L4AS:配列番号14;L5S:配列番号15;L6AS:配列番号16を、対応するアミノ酸配列への翻訳と共に示す図である。
【図8D】αD11の重鎖可変部(配列番号17)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:H1S:配列番号5;H2AS:配列番号6;H3S:配列番号7;H4AS:配列番号8;H5S:配列番号9;H6AS:配列番号10を、対応するアミノ酸配列への翻訳と共に示す図である。
【図8E】αD11の軽鎖可変部(配列番号18)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:L1S:配列番号11;L2AS:配列番号12;L3S:配列番号13;L4AS:配列番号14;L5S:配列番号15;L6AS:配列番号16を、対応するアミノ酸配列への翻訳と共に示す図である。
【図8F】αD11の重鎖可変部(配列番号17)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:H1S:配列番号5;H2AS:配列番号6;H3S:配列番号7;H4AS:配列番号8;H5S:配列番号9;H6AS:配列番号10を、対応するアミノ酸配列への翻訳と共に示す図である。
【図9A】オーバーラップ-アセンブリーPCRによって得られた両抗体のヒト化可変部の配列をクローニングするために使用したプラスミドの地図である。A)軽鎖可変ドメインのためのpVLexpress。
【図9B】オーバーラップ-アセンブリーPCRによって得られた両抗体のヒト化可変部の配列をクローニングするために使用したプラスミドの地図である。B)重鎖可変ドメインのためのpVHexpress。
【図9C】オーバーラップ-アセンブリーPCRによって得られた両抗体のヒト化可変部の配列をクローニングするために使用したプラスミドの地図である。C)pVLexpressでの各ヒト化抗体の軽鎖可変部のクローニングから生じたプラスミド。
【図9D】オーバーラップ-アセンブリーPCRによって得られた両抗体のヒト化可変部の配列をクローニングするために使用したプラスミドの地図である。D)pVHexpressでの各ヒト化抗体の重鎖可変部のクローニングの結果得られた、1)無傷の免疫グロブリンの形態IgG1での発現のための、2)Fab断片の形態での発現のための、3)免疫毒素の形態での発現のための代替構築物。
【図10】プラスチック上に免疫付着因子の形態のTrkAを固定化して行ったELISAアッセイによる、キメラ態およびヒト化形態のMNAC13抗体の結合活性の比較を示す図である。A)トランスフェクションされたコス細胞の上清の連続希釈間の濃縮後の比較;B)Gセファロースタンパク質で精製された、トランスフェクションされたコス細胞の上清の連続希釈間の比較。
【図11】プラスチック上にNGFを固定化して行ったELISAアッセイによる、ヒト化形態のαD11抗体の結合活性の検定を示す図である。
【発明を実施するための形態】
【0030】
この方法は、in vivo治療および診断上の関心のある抗体のヒト化のための、高分解能構造比較に基づく。さらに、それぞれの抗原(すなわちNGFニューロトロフィンおよびそのTrkA受容体)に対して特異的に反応することができる、ヒト化免疫グロブリンが提供される。ヒト化免疫グロブリンはヒト起源のフレームワークを有し、またそれらは、元の免疫グロブリン(すなわちNGFに対して特異的に反応するラットの免疫グロブリンαD11、およびTrkAを特異的に認識するマウスの免疫グロブリンMNAC13)のそれぞれに由来する1つまたは複数の相補性決定部(CDR)を有す。したがって、大規模に容易に生産することができる本発明の免疫グロブリンは、NGF/TrkA依存性腫瘍形態の治療だけでなく、慢性の痛みおよび炎症形態の治療においても治療用途を有す。さらに、受容体に特異的なヒト化免疫グロブリンは、TrkA陽性の腫瘍および前脳基底核細胞(アルツハイマー病の早発マーカーとして)のin vivo画像化のために、さらなる診断用途を有す。
【0031】
本発明は、モノクローナル抗体αD11およびMNAC13(それぞれラットおよびマウス)の場合と同様に、NGF上およびTrkA上両方の関心のエピトープと結合することができる軽鎖および/または重鎖のCDR領域をコードしているDNAの組換え部分を使用する。これらの領域をコードしているDNA断片は、ヒト起源の適当なフレームワーク部をコードしているDNA断片と連結させる。モノクローナル抗体MNAC13およびαD11の軽鎖および重鎖のCDRを含んでいるポリペプチド鎖をコードするDNA配列は、それぞれ図7A、7Bおよび8A、8Bに含まれる。遺伝子コードの変性および重要ではないアミノ酸の置換により、DNA配列は容易に修飾することができる。
【0032】
さらに、一般的にDNA断片は、ヒト化免疫グロブリンのコード配列に作動可能に結合し、異種プロモーターまたは天然の関連プロモーターの領域を含んでいる発現のためのさらなる制御配列を含む。好ましくは、発現制御配列は真核生物の宿主細胞を形質転換またはトランスフェクションすることができるベクター内に真核生物プロモーターを有する系であるが、原核生物の制御配列も同様に使用することができる。一旦ベクターが適当な宿主に組み込まれると、宿主は高レベルな発現を確実にするために適当な条件で維持される。二量体、手つかずの抗体の形態または他の免疫グロブリンの形態で、軽鎖および重鎖個々のさらなる精製が続く。
【0033】
ヒト定常部をコードするDNAの配列は、様々なヒト細胞から公知の手法で単離することができるが、不死化B細胞から開始することが好ましい。本発明の免疫グロブリンのCDRは、それぞれNGFおよびTrkAと結合することができ、またそれぞれラットおよびマウスの生成物である、モノクローナル抗体αD11およびMNAC13から同様に誘導される。免疫グロブリンの発現および分泌に適当な宿主細胞は、American Type Culture Collection(Catalogue of Cell Lines and Hybridomas、第5版(1985)Rockville、Maryland、USA)など多くのソースから得ることができる。好ましくは、ヒト化抗体に組み込まれるCDRは、αD11およびMNAC13のCDR配列に対応している配列を有し、抗体自体の対応するアミノ酸配列をコードしている変性ヌクレオチド配列を含むことができる。
【0034】
通常、ヒト化の設計手順は周期的かつ反復的であり、以下を含む。
マウス抗体のアミノ酸配列の分析。
対応するFv領域のモデリング。
ヒト抗体の受容体フレームワークのアミノ酸配列の分析および選択。
選択されたフレームワーク内の推定される復帰変異の同定。
ヒト化抗体の設計および実際の構築。
in vitroおよび/またはin vivoアッセイによる維持された結合親和性および特異性の検証。
【0035】
これらの活性がヒトフレームワークによって負の影響を受けるならば、受容体ヒト抗体のフレームワークの選択を変更するか、補償変異を導入する必要がある。
【0036】
ヒトフレームワークの選択がサイクルの中で最も重要な相として構築されているとしても、一般規則は現在まで確立されていない。このことは、様々な選択(患者における免疫原性に関して)の利点が臨床視点から正確に研究されていない事実に依存する。したがって、フレームワークの正しい選択を実行するために、一連の手法だけが利用でき、これらはこれまでに得られた結果と組み合わせなければならない。
【0037】
詳細には、固定されたフレームワーク(それらの構造は長い間利用されてきているので、通常重鎖のためにはNEW、軽鎖のためにはREI)を使用することが可能である。
【0038】
他の手法は、ヒト化する抗体と配列が最も相同的であることが判明したフレームワークの使用を可能にする。相同性のヒト抗体を検索するための多くのデータベースが存在する。選択では、供与体および受容体の配列間でのより高い同一性割合に加えて、一般にCDRの長さ、正規残基のレベルでのおよび境界面レベルの残基の同一性を考慮する。これらの2つの方法の間の比較については、Grazianoら(1995)を参照。
【0039】
さらに、第2の手法の変異形によれば、軽鎖および重鎖は、高い配列相同性を特徴とする2つの異なるヒト抗体から選択することができる。この手法は、Riechmannら(1988)およびShearmanら(1991)によって提案された。この点に関しては、一般に、2つの鎖の間の境界面がよく保存されているという事実は正しい相互作用を等しく保証することができるが、同じ抗体に由来する軽鎖および重鎖は異なる抗体に由来する軽鎖および重鎖に比して正しく結合して機能的結合部位を形成する可能性がより高い。これらの2つの方法の間の比較については、Roguskaら(1996および1996)を参照。
【0040】
手法を特定のヒト抗体に由来しているフレームワークに制限することは、たとえフレームワークがヒト起源であるとしても、免疫原性エピトープを生成する体細胞変異を引き起こすリスクを伴う恐れがある。代替手法はヒトコンセンサス配列に基づくフレームワークを使用することであり、そこでは、特有の体細胞変異は除去されている。2つの手法が比較された。1つのケースでは結合親和力の差を認めなかった(Kolbingerら、1993)が、他のケースでは、代わりに結合は個々のフレームワークの場合優れていることが判明した(Satoら、1994)。
【0041】
いずれにせよ、コンセンサス配列自体は人工的であり、したがって、たとえそれらが特有の残基を有しないとしても、それらは免疫原性である非天然のモチーフを形成することができる。代替手段(Rosokら、1996)は、V-BASEデータベースで収集された生殖細胞系ヒト配列を使用することである。
【0042】
ヒト起源のフレームワーク可変部とマウスCDR部との不自然な並置は、自然界では現れない配座制限をもたらす可能性があり、それらは特定のアミノ酸残基の置換によって修正されない限り、結合親和性の損失を決定付ける。置換するアミノ酸残基の選択は、一部コンピュータモデリングによって決定される。免疫グロブリン分子の三次元画像を作製するための、ハードおよびソフトが利用できる。一般に、分子模型は、免疫グロブリンドメインまたは鎖の解決済みの結晶構造から始まって作製される。モデル化する鎖は解決済みの三次元構造の鎖またはドメインとのアミノ酸類似性に基づいて比較され、配列に関して最も高い類似を示す鎖またはドメインが分子模型の構築の出発点として選択される。しかし、抗体構造の予測は常に正確とは限らない。特に、第3のCDR部はモデル化が困難であり、それは抗体の構造予測において常に不確実な点になっている(Chothiaら、1987)。この理由から、概して、ヒト化抗体は第一次近似としては、抗原に対する結合親和性および/または特異性は出発モノクローナル抗体よりもかなり低い。このことは、完全には合理化することができない試行錯誤手法により出発抗体の特性を再構成する試みのために、多くの連続した点突然変異サイクルを必要とする。
【0043】
利用できるヒトおよびヒト化抗体の増大する高分解能X線構造を考慮すると、その意図はコンピュータモデリングの使用に由来する不確実性および曖昧さを避けて、X線結晶学手段により本発明の両抗体のFabフラグメントの高分解能構造データを得ることであった。この目的のために、両抗体をハイブリドーマから精製し、パパイン(重鎖のCH1およびCH2のドメイン間の接合部で切断するプロテアーゼ)でタンパク分解的に処理し、Fabフラグメントを生成した。さらなる精製の結果として両Fabフラグメントが結晶化され、2つのデータベース(低および高分解能)から、Molecular Substitution法で構造を決定してその後それらを精製することができた。
【0044】
実験的に得られた構造データに基づく本発明で提案されている手法は、受容体ヒト抗体フレームワークを選択する重要な相において、および両中和抗体のヒト化過程で選択されたフレームワーク内の推定される復帰変異の同定のために、非常により堅実で合理的な出発点を提供する。
【0045】
ヒト抗体フレームワークの選択の指針となるための既報告の様々な基準の中で、構造的整列に基づく分析によりその結果を拡張して完成するために使用されたものは、マウスおよびヒト起源の抗体間の一次配列における同一性の程度であった。元の基準に関連して対応する構造の比較分析は、非常により正確な比較を、したがって得られたヒト化抗体が元のマウスの抗体の親和性および特異性の特性を保存するより大きな可能性を保証するものである。したがって、使用された手法は、アミノ酸配列の分析および比較に由来する同一性の程度および相同性レベルに関する情報を、それぞれの三次元構造の比較と組み合わせる。
【0046】
詳細には、一次構造の最適アラインメントに由来する情報は、二重の役割を有する。第一に、この分析は、比較する可能な三次構造の数を減らし、高い相同性および同一性を特徴とするものに分析を限定することが可能になる。一次構造レベルでの最適アラインメントを特徴とし構造データが入手可能なこれらの配列の中で、高分解能により、さもなければ筆者らが得た構造のそれに相当する分解能(すなわち2.5Å以下)により解決された構造のみを重視して、さらなる選択を実施した。この手法は三次構造のずっと正確な整列およびRMS(根平均二乗偏差:平均二乗偏差の平方根;CarugoおよびPongor、2001および2003)で表される構造差のずっと有意な推定値を保証するものである。低分解能データは、空間内の個々の原子の実際の相対位置に関して、むしろ暗示的で明らかにより不正確な情報を提供する。
【0047】
ヒト起源または操作した個々の構造の重なり程度を評価するために、それぞれのアミノ酸骨格を構成しているα炭素原子の間でRMSを計算したが、2Åを上回っているRMSの原子対は無視した。この分析から、したがって構造間の多様性(RMSの値によって表される)だけでなく、各RMSを計算する際に実際に使用されたα炭素原子のパーセンテージも考慮しなければならない情報が得られる。
【0048】
これらの三次構造レベルの類似性データは、同一性および相同性に関する一次配列の比較分析と関連付けされた。
【0049】
ヒト化のための最適フレームワークの選択が3変数問題として構成され、それはしたがって相同性レベルおよび同一性の程度を構造的整列と関連させる場合に空間で表すことができると推測される。次に、2種類の整列における問題の領域をそれぞれのフレームワークの領域に減らして、この種の分析を実行した。
【0050】
分析した3つの変数(それぞれ、RMS値、RMSが計算された原子の割合、および一次構造間の類似指数、すなわち全体の同一性、全体の相同性、フレームワークレベルの同一性、フレームワークレベルの相同性の割合)の空間における検討された抗体の分布を、それがヒト起源だとするならば各抗体が占めるであろう3変数の空間における最適な位置と比較すると、一次および三次構造レベルでこの理想位置に最も近づくヒト起源抗体を同定することはおそらく明らかである。この結果を合理的に説明するために、4つの分析のそれぞれで、検討されるヒト化またはヒト起源の抗体の各位置について、仮定上の最適位置からの逸脱が計算される。
【0051】
この選択法に基づいて、与えられた抗体のヒト化のための以降のCDR移植過程で受容体フレームワークを選択することが可能である。
【0052】
一般に、マウス起源の残基によるヒト起源アミノ酸残基の置換を最小化する必要があるが、その理由は、マウス残基の導入は抗体がヒト患者でHAMA反応を誘発するリスクを増加させるからである。他方、相補性決定部(CDR)は抗原と相互作用するより大きな可能性を有する残基を含み、この理由のために、それらはヒト化抗体で維持されなければならない。それらは、カバットに従う配列によって、またはChothiaによる構造によって確定される。それらを確定するために第2の系を使用する利点は、一般にCDRがより短く、それ故にヒト化抗体は異種断片のより小さい部分によって構成されることである。いずれにせよ、一般にカバットの定義に従うと、ヒト化のために必要なサイクル数を大幅に減らすことが可能なことが証明された。一旦CDRが確定されると、それらが属している正規クラス(ChothiaおよびLeskによって規定されている)を同定し、その後ヒト化抗体で正規残基を維持することが必要である。
【0053】
可変ドメインの軽鎖および重鎖の間の相互作用を媒介する残基を分析し(表1)、ヒト化抗体内のいかなる特異な残基をも維持することが必須である(Singerら、1993;Daughertyら、1991;De Martinoら、1991)。
【0054】
さらに、維持するさらなるアミノ酸は、CDRの高次構造に及ぼすそれらの可能な影響および/または抗原との相互作用に基づいて選択される。アミノ酸が動物起源のフレームワークおよびヒト起源の同等の受容体フレームワークの間で異なる場合は、そのアミノ酸が抗原と非共有結合的に直接接触しているか、またはCDR部に隣接しているか、またはいずれにせよCDR部と相互作用している(CDR部から4〜6Åの範囲内にある)と予想することが合理的ならば、受容体フレームワークのアミノ酸は同等のマウス残基によって置換するべきである。
【0055】
【表1】

【0056】
詳細には、さらなる分析は、CDRの構造を安定させるゾーンであるいわゆるバーニヤ(Vernier)ゾーンを定義する他の残基を含む。この領域の特性を維持することが重要である。
【0057】
変異のための他の残基候補は、受容体フレームワークのその位置ではヒト免疫グロブリンにとって普通ではないアミノ酸である。これらの残基はより典型的なヒト免疫グロブリンの同等の位置に由来するアミノ酸で置換することができ、あるいは供与体フレームワークの同等の位置に由来する残基を受容体フレームワークに導入することができるが、それはそれらのアミノ酸がそれらの特定の位置でヒト免疫グロブリンにとって典型的な場合である。
【0058】
さらに、ヒト免疫グロブリンのコンセンサス配列に基づき、供与体および受容体フレームワークに存在する普通でない残基の代わりに、ヒトで保存されている残基を挿入する変異をヒト化形態に導入する。
【0059】
次に結晶構造のそれぞれの対を修飾するが、先ず、動物起源のCDRをヒトフレームワークに移植する。その後、上述のすべての変異および復帰変異を導入する。修飾された構造体を次に複合免疫グロブリンに組み立てる。得られたモデルは、力場を使用して機械的エネルギー(ねじれ角および結合角度および距離に関して)を最小にすることにより洗練する。
【0060】
上で議論した特定のアミノ酸置換と異なる他のすべての領域については、ヒト化免疫グロブリンのフレームワーク部は、通常、それらが由来するヒト抗体のフレームワーク部と実質的に同一である。いずれにしても、移植によって得られるこれらの改変タンパク質では、末端または中間の多くのアミノ酸置換、欠失または挿入、および他の変化のために、フレームワーク部は未変性の配列と比較して一次構造レベルで変動する可能性がある。当然、抗体の特異性または親和性に対するフレームワーク部の大部分の残基の貢献は、非常に小さいか皆無である。したがって、フレームワークの残基内の多くの個々の保存的置換は、生じるヒト化免疫グロブリンにおいて特異性または親和性をかなり変更することなく、耐容される。しかしながら、そのような置換は一般に望ましくない。広く使用されている様々な技術、例えば部位特異的突然変異(Gillman & Smith、1979;Robertsら、1987)を使用して、ヌクレオチド配列を修飾することが可能である。
【0061】
抗体の一次構造の一部だけを含んでいる代わりのポリペプチド断片を生成することができ、それら断片は免疫グロブリンの1つまたは複数の特有の活性(例えば結合活性)を保持する。これらのポリペプチド断片はタンパク分解的消化によって生成することができ、これは未変性の抗体、または部位特異的突然変異(詳細には、Fab断片を生成するためにはCH1領域の後、または(Fab')2断片を生成するためにはヒンジ部の後)による重鎖および軽鎖の可変部のコードDNA配列を含む担体内の所望の位置への終止コドンの挿入から始まる。scFvの形の抗体は、重鎖および軽鎖の可変部をリンカーにより連結することによって得ることができる(Hustonら、1988;Birdら、1988)。FvまたはFab断片は、大腸菌(BuchnerおよびRudolph、1991;Skerraら、1991)、または真核細胞、好ましくは哺乳類由来のものにおいても発現させることができる。他の多くの遺伝子と同様に、免疫グロブリンスーパーファミリーの遺伝子がそれぞれ1つまたは複数の特異的生物活性を特徴とする他とは異なる機能的領域を含むことを考えると、遺伝子は、新しい特性を備えている融合タンパク質(例えば免疫毒素)を生成するために、他の遺伝子(例えば酵素)に由来する機能的領域に融合させることができる。
【0062】
細菌内でのヒト化免疫グロブリン配列の発現は、CDR部に突然変異を起こさせる高親和性を特徴としファージディスプレイのためにファージライブラリーを生成する、ヒト化免疫グロブリン配列を選択するために使用することができる。これらのライブラリーを使用して、抗原に対して高い親和性および/または結合特異性を有するヒト化免疫グロブリンのCDRレベルでの、変異体を探索するスクリーニングを実行することが可能である。免疫グロブリンの可変部の配列を含むファージディスプレイライブラリーを得る方法は、豊富に報告されている(Cesareni、1992;Swimmerら、1992;Gramら、1992;Clacksonら、1991;Scott & Smith、1990;Garrardら、1991)。それらのCDRがこのように改造されたヒト化免疫グロブリンの変異体から生じる配列は、その後その高い発現を保障するために適当な宿主で発現される。
【0063】
上述の通り、DNA配列は発現制御配列に動作可能に結合された後(すなわち、それらの機能が保障されるような方法で置かれた後)に、宿主細胞で発現される。一般的にこれらの担体はエピソームとして、または染色体DNAの不可欠の部分として、宿主生物内で複製される。通常、発現担体は、関心のDNA配列で形質転換された細胞の特定を可能にする選択可能なマーカーを含む。
【0064】
本発明のヒト化免疫グロブリンのscFvの組換え態またはFab形態での生成のためには、原核生物の系が好ましい。大腸菌は、本発明のDNA配列をクローニングするために特に有用である原核生物宿主の1つである。さらに、lacもしくはtrpオペロンまたはβラクタマーゼまたはλファージなど、多数の特徴が明らかなプロモーターが利用できる。一般的に、これらのプロモーターは転写および翻訳の正しい開始および終了のために、リボソームのために発現を制御して結合部位を運ぶ。ポリエチレングリコール(PEG)とのコンジュゲーションによって、原核生物の系で産生される本発明のヒト化免疫グロブリンの半減期を増加させることが可能である。
【0065】
酵母など他の単細胞生物も発現のために使用することができる。最適な宿主は、発現調節、複製終止および開始の配列を備えている適当な担体を使用するサッカロミセス(Saccharomyces)である。
【0066】
昆虫細胞培養も本発明のヒト化免疫グロブリンを生成するために使用することができ、一般には、安定的にトランスフェクションされたS2ショウジョウバエ(Drosophila)の細胞、またはバキュロウイルス(Baculovirus)に基づく発現系を有するSpodoptera frugiperdaの細胞を使用する(Putlitzら、1990)。
【0067】
植物および植物細胞培養は、本発明のヒト化免疫グロブリンの発現のために使用することができる。(Larrick & Fry、1991;Benvenutoら、1991;Durinら、1990;Hiattら、1989)。
【0068】
しかし、これらすべてのケースで、ヒト免疫系の活性化におけるエフェクター機能を保障するのに必要な、正しい種類のグリコシル化を得ることは不可能である。このために、免疫反応の誘導における免疫グロブリンの中で最も有効なイソタイプであると判明したIgG1の完全な形態で本発明のポリペプチドを発現するために、哺乳類細胞の組織培養を使用することが可能である(Winnacker、1987)。イソタイプが抗体の溶解能を決定することを考えると、通常、IgG1イソタイプは治療目的のために使用され(細胞および補体系によって媒介される免疫反応を誘導するので)、一方、IgG4は診断用途のために使用されることを強調すべきである(Riechmannら、1988)。詳細には、手つかずの免疫グロブリンの分泌のために開発された多数の宿主細胞系を考えると、哺乳類細胞、中でもCHO細胞系、COSのいくつかの系、HeLa細胞、骨髄腫細胞系(NS0、SP/2、YB/0、P3X63.Ag8.653)、形質転換B細胞またはハイブリドーマが好ましい。これらの細胞のための発現担体としては、複製開始点、プロモーター、エンハンサーなどの発現制御配列(Queenら、1986)、およびリボソーム結合、RNAスプライシングおよびポリアデニル化のために必要な配列、および転写終止のための配列が含まれ得る。最適な発現制御配列は、免疫グロブリン遺伝子およびSV40、アデノウイルス、ウシ乳頭腫ウイルス、サイトメガロウイルスなどのウイルスに由来するプロモーターである。通常、発現ベクターは選択可能なマーカー、例えばネオマイシン耐性を含む。
【0069】
ヒト化抗体の発現のためには、無血清培地で哺乳類細胞系を培養することが好ましい。例えば、HUDREG-55細胞系は、Sigma(St.Louis、Mo)製の無血清および無タンパク質ハイブリドーマ培地Cat.No.S-2897で容易に増殖させることができる。
【0070】
本発明のヒト化免疫グロブリンをコードしている遺伝子は、関心のヒト化免疫グロブリンを、一般的にミルクまたは血清などの回収可能な体液内に発現する非ヒトトランスジェニック動物を作製するために使用することができる。そのような導入遺伝子は、通常エンハンサー配列と共にプロモーター、例えば齧歯動物の免疫グロブリンまたはカゼイン遺伝子のプロモーター/エンハンサーと動作可能に結合したヒト化免疫グロブリンをコードしているポリヌクレオチド配列を含む(Buhlerら、1990;Meadeら、1990)。導入遺伝子は、相同組換え構築物によって細胞または胚へ移すことができる。使用される非ヒト動物としては、マウス、ラット、羊、ウシおよびヤギがある(国際公開91/08216)。
【0071】
それらが一旦手つかずの抗体、それらの二量体、個々の軽鎖および重鎖、または他の形態で発現されると、本発明の免疫グロブリンは硫酸アンモニウムによる沈殿、アフィニティーカラム、カラムクロマトグラフィーなどの標準方法で精製することができる(Scopes、1982)。薬剤用途のためには、最低90から95%、好ましくは98から99%、またはさらに高い均一性を有する、実質的に純粋な免疫グロブリンが必要である。部分的に、または所望の均一性が得られるまで一旦精製されると、タンパク質は治療用途(また体外で)、診断用途(腫瘍またはアルツハイマー病の診断のための画像化)または生化学アッセイ、免疫蛍光染色などの開発および実施のために使用することができる(一般には、LefkovitsおよびPernis、1979および1981を参照)。
【0072】
本発明の医薬用途は、TrkA発現細胞(膵臓および前立腺腫瘍の場合)を除去するための、免疫毒素の形のヒト化免疫グロブリンMNAC13の使用に関係する。免疫毒素は2つの成分を特徴とし、in vitroおよびin vivoで特定の細胞を殺すために特に好適である。細胞に対して通常致死性の細胞傷害剤の1成分は、吸収されるかまたは細胞表面と相互に作用する。第2の成分は、毒剤を特異的な標的細胞型、例えばTrkA受容体のエピトープを発現する細胞に向かわせるための手段を提供する。2つの成分は、利用できる多種多様な化学手法のいずれか1つによって、互いに化学的に結合させる。例えば、細胞傷害剤がタンパク質であり第2の成分が無傷の免疫グロブリンである場合、結合は交差結合およびヘテロ二機能性剤(SPDP、カルボジイミド、グルタルアルデヒド)に媒介させることができる。あるいは、2つの成分を遺伝的に結合させることができる(Chaudharyら、1989)。様々な免疫毒素の生成がThorpeら(1982)によって報告されている。
【0073】
多数の細胞傷害剤が、免疫毒素としての用途に適当である。細胞傷害剤としては、ヨウ素131または他のヨウ素同位体、イットリウム90、レニウム188およびビスマス212または他の同位体などのα粒子を放出する放射性核種、多数の化学療法薬、例えばビンデシン(vindesin)、メトトレキセート、アドリアマイシンおよびシスプラチン;ならびに細胞傷害性タンパク質、例えばリボソームを阻害するタンパク質(例えばヤマゴボウ抗ウイルスタンパク質、シュードモナス外毒素Aおよびジフテリア毒素、リシンAおよび植物起源のクラビン(clavin))、または細胞表面レベルで活性を示す剤(例えばホスホリパーゼCなどのホスホリパーゼ酵素)がある。BaldwinおよびByers、1985;U.S.Ser.No.07/290,968;OlsnesおよびPhil、1982。免疫毒素の細胞傷害性領域はそれ自体免疫原性であるので、慢性または長期療法の場合には融合タンパク質の臨床有用性を制限する可能性があることを強調するべきである。毒素の免疫原性の問題を避ける代替手段は、DNAと相互作用することができるタンパク質を抗体の結合ドメインと融合させて発現させ、毒素発現カセットを含む発現担体をその融合タンパク質に結合させることである。DNAと結合するヒトタンパク質であるプロタミンの多数の正電荷はDNAの陰電荷と安定的に相互作用することができ、毒素自体よりずっと安定でしかも免疫原性の低い中性抗体の融合パートナーを生成する。受容体媒介エンドサイトーシスによる抗体-プラスミド複合体の内部移行の後、毒素の発現は細胞の死を引き起こす。さらに、除去すべき標的細胞に対する選択性は、誘導可能なまたは細胞特異性のプロモーターを毒素発現カセットに挿入することによって、さらに強化することができる。この手法は毒性副作用を最小にしつつ、腫瘍細胞の選択的な除去を最大化することを目的とする(Chenら、1995)。
【0074】
免疫毒素を正しい目標に向かわせる成分には、手つかずの免疫グロブリンまたは結合断片の形の、あるいは、FabまたはFv断片としての本発明のMNAC13ヒト化免疫グロブリンが含まれる。一般的に、免疫毒素内の抗体はヒトイソタイプIgMまたはIgGであるが、他の定常部も同様に使用することができる。
【0075】
本発明の抗体および医薬組成物は、病理に関係する組織のレベルで抗体を送達するための、いかなる有効な方法に従う投与に特に有用である。これには、それには限定されないが、腹腔内、筋肉内、静脈内、皮下、気管内、経口、経腸、腸管外、鼻腔内または経皮投与が含まれる。本発明の抗体は、液状製剤の注射による局所投与(腹腔内または脳内-一般的に脳室-または心膜内または嚢内)のために、あるいは、固形剤(ピル、錠剤、カプセルの剤形)または液状製剤(乳濁液および溶液の剤形)の摂取により投与することができる。非経口投与のための組成物は、通常、好ましくは水性の適合する溶液に溶かした免疫グロブリンの溶液を含む。これらの製剤内の抗体濃度は0.005%未満から15〜20%の範囲であり、主に液体容積、その粘度、その他に従って、また選択される特定の投与様式に従って選択される。
【0076】
あるいは、抗体は固体剤形での投与のために調製することができる。抗体は異なる不活性物質または添加物質と組み合わせることができ、その例としてはリガンド、例えば微結晶性セルロース、ゼラチンまたはアラビアゴム;受容体、例えばラクトースまたは澱粉;アルギン酸、Primogelまたはコーンスターチなどの剤;滑剤、例えばステアリン酸マグネシウム、コロイド状二酸化ケイ素;甘味料、例えばサッカロースまたはサッカリン;または着香剤、例えばミントおよびサリチル酸メチルがある。他の薬剤投与系としては、ヒドロゲル、ヒドロキシメチルセルロース、リポソーム、マイクロカプセル、マイクロエマルジョン、マイクロスフェアー、その他がある。腫瘍などの罹患組織への直接的局所注射は、本発明の抗体の投与で優先される方法である。
【0077】
本発明の抗体は、凍結または凍結乾燥させ、適当な緩衝液で使用直前に再構成することができる。凍結乾燥および再構成が抗体の活性低下の変動量を決定する(従来の免疫グロブリンでは、クラスIgM抗体は、クラスIgG抗体よりも活性の損失は大きい傾向がある)ことを考えれば、投与レベルはこの事実を補償するために調整されなければならない。
【0078】
それらの高い遮断能力のおかげで、本発明の抗体を含んでいる組成物は、病理的状態と関連する炎症要素または慢性疼痛、特に慢性内臓痛(月経困難症、消化不良、胃腸逆流、膵臓炎、内臓痛または過敏性腸症候群などの生理障害に関連する)を予防するか減らすかするための予防法および/または治療法のために、投与することができる。
【0079】
予防用途では、本発明の抗体を含んでいる組成物は、特定の病理状態がまだ起きていない患者に対して抵抗性を強化するために投与される。
【0080】
本発明の抗体は、前立腺または膵臓の腫瘍の体積を減らすための方法、およびさらなる腫瘍増殖を予防するかまたは腫瘍増殖率を低下させる方法も提供する。この効果は、本発明の両方のヒト化抗体によって媒介されるが、その理由はそれらが、自己分泌またはパラクリンの様式で腫瘍の増殖および進行を持続させるのに必要なNGFおよびTrkAの間の相互作用の中和に、極めて効果が高いからである。さらに、MNAC13のヒト化形態は膜受容体と相互に作用するが故に腫瘍細胞の直接的除去のためにも使用することができるが、その理由は、それらが宿主の免疫反応(IgG1の形で投与された場合)を活性化するか、または細胞傷害剤を運んでそれを癌性塊のレベルで局在化する(免疫毒素の形で投与された場合)ことができるからである。
【0081】
腫瘍部位へのそれらの投与は、好ましくは組織または腫瘍部位の近くへの直接的局所注射を通して起こる。全身投与のためには日用量は0.05mg/kgから500mg/kgの範囲であるが、投与するのがより簡単であるのでその範囲の低い方の領域の投薬量が好ましい。例えば抗体の特定の血漿レベル(約5〜30mg/ml、好ましくは10〜15mg/mlの範囲)を保証し、臨床成績が達成されるまで与えられた期間このレベルを維持するために、投薬量を調整することができる。ヒト化抗体はずっとゆっくり除去されるべきで、血漿中有効レベルを維持するのにより低い投薬量を必要とするべきである。さらに、高親和性を考慮すると、より低い親和性を有する抗体よりも投与の頻度は低く量も少ない。各抗体の治療的有効投薬量は、腫瘍の体積の減少または腫瘍増殖率に基づいて、または、理想的には癌性病理状態の完全消失に基づいて治療期間中に決定することができる。膵臓または前立腺の腫瘍の段階を測定または評価するための効果的な方法は、血液中の前立腺特異抗原(PSA)の測定、膵臓腫瘍の場合は生存期間の測定、両腫瘍の転移の拡散の遅延化または阻害の測定に基づく。
【0082】
腫瘍部位レベルでの直接注射については、投薬量は腫瘍の型、段階および体積を含む種々の因子、ならびに他の多くの変数によって決まる。腫瘍体積に従い、典型的治療量は0.01mg/mmから10mg/mmの範囲の注射であり、必要な頻度で投与することができる。特定の治療法の効果を評価する別の方法は、例えばELISAアッセイによりその活性を測定することによってTrkA受容体の阻害を評価することである(Angelesら、1996)。
【0083】
重要なことは、TrkAは治療上の標的としてだけでなく、in vivo画像化、例えばTrkA陽性腫瘍(腫瘍型および起源に従い陽性または陰性マーカーとして)の画像化、および前脳基底核細胞の画像化(アルツハイマー病の襲撃の早発マーカーとして)のための診断上の標的としても構成されていることである。本発明のMNAC13ヒト化抗体は、多種多様なin vitro用途も有す(ELISA、IRMA、RIA、免疫組織化学)。
【0084】
診断目的のために、抗体に印を付けても付けなくてもよい。印がない抗体は、ヒト化抗体またはヒト抗体(例えばヒト免疫グロブリンの定常部に対する特異抗体)に対して反応性である他の印付きの抗体(二次抗体)と組み合わせて使用することができる。あるいは、抗体は直接印を付けることができる。多種多様な標識、例えば放射性核種、蛍光団、色素、酵素、酵素基質、酵素因子、酵素阻害剤、リガンド(特にアプテニック(aptenic)な)、その他を使用することができる。多種類の免疫アッセイがこの分野で利用できる。
【0085】
詳細には、画像診断用途については、Goding(1986)およびPaikら(1982)によって記載されているように、検出可能であるかまたは同位体(ヨウ素、インジウム、テクネチウムの放射性同位体)もしくは常磁体(常磁性の原子またはイオン、例えば遷移元素、アクチニドおよび希土類;特に、マンガン(II)、銅(II)およびコバルト(II))で標識された剤を抗体とコンジュゲートする。画像化手法は静脈内、腹腔内または皮下の注射(リンパ液のドレナージ領域ではリンパ節転移を同定するため)を伴い、それらは、免疫シンチグラフィーの場合、放射性核種放出の検出器(例えばシンチレーションβカウンター)を使う。常磁性の標識を代わりに使用するならば、NMR(核磁気共鳴)スペクトロメータを使用する。
【0086】
以下の図面を参照しながら、本発明はその非限定的な実施形態で説明されるものとする。
【0087】
結果
MNAC13およびαD11モノクローナル抗体のFab断片のX線構造
標準手順により両方のモノクローナル抗体を得て精製した。MNAC13 IgG1およびαD11 IgG2a免疫グロブリンを、ハイブリドーマ細胞の培養により上清で発現させ、29%硫酸アンモニウムによる沈殿で濃縮し、その後PBSで透析した。両免疫グロブリンは、Protein G Sepharose(Pharmacia)のカラムを使用してアフィニティークロマトグラフィーにより精製した。
【0088】
Spectra-Por 12/14K膜(Spectrum)を使用した4℃における10mMリン酸緩衝液pH7、20mM EDTA中での透析の後、各試料はCentricon 50kDa限外濾過ユニット(Amicon)によって濃縮し、13mMのシステインと一緒にインキュベートし、37℃で5h、固定化パパイン(Pierce)(1:15の酵素:基質比率)で処理した。それぞれのFab断片を精製する手法は多様であるが、イオン交換クロマトグラフィーに常に基づいている。
【0089】
MNAC13の場合、100mMトリス塩酸pH8.0に対する透析の後、同じ緩衝液で平衡化したDEAE-Sephacelカラム(Pharmacia)を通してFc断片を除去することが可能であった。FabMNAC13は排除液中で収集し、Fc断片および未消化IgGl部分は250mMのNaClで溶出した。Fab断片は、100mMトリス塩酸pH8.0、150mM NaClで平衡化したSuperdex G75(Pharmacia)カラム上でのゲル濾過によって、未消化のIgGlから分離した。これら分画の均一性および純度は、12%ポリアクリルアミドゲル上での電気泳動分離とその後のクーマシー染色によって制御された(図1A)。精製されたタンパク質の濃度は、Lowryアッセイ(Bio-Rad)で測定した。11のハイブリドーマ上清から、最高3mgのMNAC13 Fabを得ることができた(99%を超える純度)。
【0090】
αD11抗体のFab断片の精製に関して、パパインで処理した試料は、10mMリン酸緩衝液pH7.8に対して透析した。Fc断片は、この同じ緩衝液で平衡化したDEAE-Sephacelカラム(Pharmacia)を通して除去した。αD11のFab断片は排除液中で収集し、Fc断片および未消化IgG2a部分は250mMのリン酸緩衝液pH6.8で溶出した。Fab断片は、10mMリン酸緩衝液pH7.8、150mM NaClで平衡化したSuperdex G75カラム(Pharmacia)上でのゲル濾過によって、未消化のIgG2aから分離した。これら分画の均一性および純度は、12%ポリアクリルアミドゲル上での電気泳動分離とその後のクーマシー染色によって制御された(図2A)。精製されたタンパク質の濃度は、Lowryアッセイ(Bio-Rad)で測定した。11のハイブリドーマ上清から、最高6mgのαD11 Fabを得ることができた(99%を超える純度)。
【0091】
10mMのトリスpH8.0、50mM NaCl中で精製したMNAC13抗体のFab断片、および10mMのリン酸ナトリウムpH7.8および50mMのNaCl中で精製したαD11抗体のFab断片を、Centricon 30kDa限外濾過ユニット(Amicon)により5〜10mg/mlまで濃縮した。Crystal Screen IおよびII(Hampton Research - Laguna Niguel、CA、USA-)ならびにスクリーニングキット(Jena BioSciences)を使い、要因組合せ(factorial combination)手法(Jancarik & Kim、1991)による16℃での懸滴(hanging-drop)法に従って、結晶化実験を実施した。
【0092】
濃縮したタンパク試料2μlを、沈殿剤を含んでいる等量の溶液に滴下し、24穴Linbroプレートの貯水槽(0.7ml)内の溶液との拡散によって平衡化した。
【0093】
MNAC13抗体Fab断片に関して、等量のタンパク質と2M硫酸アンモニウム、5%v/vイソプロパノールを含んでいる沈殿剤(Crystal Screen II、Reactant #5)とで得られた最も有望な初期の成果を、図1Bで示されているものと同様に約1週で成長する結晶が得られるまで最適化した。
【0094】
αD11抗体Fab断片に関して、等量のタンパク質と、20%のPEG4000、0.6MのNaCl、100mMのMES pH6.5を含んでいる沈殿剤(キット番号4、溶液C2)とで得られた最も有望な初期の成果は、図2Bで示されているものと同様に約1週間で成長する結晶が得られるまで、沈殿剤の組成をPEG4000、0.6MのNaCl、100mMのBTP pH5.5およびタンパク質および沈殿剤溶液比(1.5:1)に修飾する長い最適化過程を必要とした。
【0095】
両方の場合で、初期の低分解データセットがELETTRAシンクロトロン(Trieste、Italy)のXRD1回折線上で収集され、その後第2のより高い分解能のより完全なデータセットがESRFシンクロトロン(Grenoble、France)のID14-EH1回折線上で収集された。結晶は、MNAC13抗体のFab断片の場合は低温保護体として2.2Mの硫酸アンモニウム、6%v/vイソプロパノールおよび20%v/vグリセリンを含んでいる溶液を使用して、Oxford Crysosystems(Oxford、UK)製の冷却システムにより液体窒素下で凍結した。各タンパク質の代表的な高分解能回折スペクトルを図1Bおよび2Bに示す。
【0096】
4つのX線回折データセットすべてを処理、指数化、積算し、その後それぞれDENZOおよびSCALEPACKプログラム(Otwinowski & Minor、1997)を使用して評価し、一方、データ整理のためにはCCP4(Collaborative Computational Project、Number 4、1994)パッケージを使用した。MNAC13抗体のFab断片の結晶の高分解能および低分解能データの収集および処理の統計値を、以下の表で示す。
X線源 ELETTRA ESRF
波長(Å) 1.000 0.934
検出器 mar345 marCCD
空間群 P212121 P212121
単一細胞のパラメータ
a(Å) 52.78 52.73
b(Å) 67.53 67.55
c(Å) 111.51 111.43
Mosaicity(°) 0.40 0.47
分解能間隔(Å) 12.0〜2.50 17.0〜1.80
(2.59〜2.50) (1.83〜1.80)
測定数 98688 414115
I≧0で観察される反射の数 56918 227914
I≧0での特有な反射の数 14203(1371) 38392(1893)
完全性(%) 99.5(99.3) 99.5(99.6)
リダンダンシー 4.0(4.0) 5.9(4.9)
測定データの<I/σ(I)> 9.4(4.7) 8.2(1.1)
Rsym(%) 5.7(15.2) 6.3(39.8)
【0097】
同様に、αD11抗体のFab断片の結晶の高分解能および低分解能データの収集および処理の統計値を、以下の表で示す。
X線源 ELETTRA ESRF
波長(Å) 1.000 0.934
検出器 marCCD marCCD
空間群 P1 C2
単一細胞のパラメータ
a(Å) 42.685 114.801
b(Å) 50.626 69.354
c(Å) 102.697 64.104
α(°) 81.977 90
β(°) 89.116 117.02
γ(°) 85.957 90
Mosaicity(°) 0.44 0.40
分解能間隔(Å) 47.6〜2.57 17.0〜1.70
(2.8〜2.7) (1.75〜1.70)
測定数 124456 492594
I≧0で観察される反射の数 74241 399184
I≧0での特有な反射の数 23413(2162) 47951(3198)
完全性(%) 98.2(92.4) 97.2(78.4)
リダンダンシー 5.7(5.2) 6.7(7.5)
測定データの<I/σ(I)> 29.6(6.7) 9.5(2.1)
Rsym(%) 11.0(33.5) 5.8(27.8)
括弧内の値は最高分解能のシェルを指す。
【0098】
Fab断片の利用できる構造の数が多いことを考慮すると、両タンパク質の構造を決定する最も便利な方法は、Molecular Substitutionであった。相同構造に関するプロテインデータバンク(Bermanら、2000)での研究において、選択基準は同等の分解能および最高レベルの配列同一性の間の組合せを優先した。それぞれこれらに基づいて下記が選択された。
MNAC13のためには:1BM3:Opg2 Fab免疫グロブリンのFab断片およびそれによって認識されるペプチド間の複合体の構造(Kodandapaniら、1999)、分解能2.0Åで分解され、重鎖および軽鎖についてそれぞれ70および88%の配列同一性を有する。
αD11のためには:1CIC:イディオタイプ-抗イディオタイプFab断片FabD1.3-FabE225の複合体の構造(Bentley、1990)、分解能2.5Åで分解され、重鎖および軽鎖についてそれぞれ82および82.65%の配列同一性を有する。
【0099】
両方の構造の決定は、AMoReプログラム(Navaza、1994)を使用してMolecular Substitution法によって得られたが、可変部および定常部の間のバイナリー擬似対称軸によって形成される角度の極端な変動性を考慮して、それぞれのモデルでは定常ドメインおよび可変ドメインを別々に使用した。剛体による洗練後の、MNAC13のFab断片の構造決定において得られたソリューションを以下の表で示す。
【0100】
【表2】
【0101】
同様に、空間群C2のための、剛体による洗練後の、αD11のFab断片の構造決定において得られたソリューションを以下の表で示す。
【0102】
【表3】
【0103】
2つの構造の以降の洗練は循環手法によって得られ、下記2つの交互相を含む。コンピュータグラフィクスのためのインタラクティブなソフトウェア「O」(KleywegtおよびJones、1994)を使用したモデルの手動構築。CNSセット、Crystallography and NMR System(Brungerら、1998)の自動プロトコルを使った位置的洗練およびB等方性温度因子の洗練。剛体による洗練のいくつかの相の後の手法は、異なる洗練サイクルを企図したものである。一旦すべての変異の挿入および欠失が終了してモデルが完成すると、水分子、あらゆるイオンおよびリガンドのローカライゼーションを行った。終わりに、モデルを立体化学的に理想値のできるだけ近くに保って、位置的重量waおよび熱因子B r-重量を最適化した。MNAC13抗体のFab断片のために得られたモデルの品質を記載する統計量および最終パラメータは、以下の表に要約する。
タンパク原子数 3244
溶媒原子数 351
硫酸イオン数 4
トリス分子数 1
イソプロパノール分子数 1
分解能間隔(Å) 39〜1.778
最終R因子 19.35%
最終Rfree因子(データの10%で計算) 23.22%
Rms偏差
結合距離(Å) 0.008
結合角度(°) 1.456
二面角(°) 27.29
非固有角度(°) 0.928
平均等方性温度因子(A2)
完全タンパク質 23.55
軽鎖 24.14
重鎖 22.99
水分子 31.95
イオン(硫酸塩) 55.94
トリス 46.06
イソプロパノール 32.60
【0104】
同様に、αD11抗体のFab断片のために得られたモデルの品質を記載する統計量および最終パラメータを以下の表に要約する。
タンパク原子数 3229
溶媒原子数 403
塩素イオン数 1
分解能間隔(Å) 30〜1.70
最終R因子 19.54%
最終Rfree因子(データの10%で計算) 24.22%
Rms偏差
結合距離(Å) 0.0096
Angoli di legame(°) 1.6571
結合角度(°) 1.6571
Angoli die二面角(°) 27.40
非固有角度(°) 1.048
平均等方性温度因子(A2)
完全タンパク質 25.58
軽鎖 24.14
重鎖 22.99
水分子 38.80
イオン(塩化物) 20.58
【0105】
さらに、両モデルは、それぞれの表およびそれぞれのラマチャンドランチャート(図1Dおよび2D)で示すように、PROCHECKセット(Laskowskiら、1993)による最終幾何分析によって検討した。
【0106】
ヒト起源のフレームワークの選択におけるMNAC13およびαD11モノクローナル抗体のFab断片のX線構造の使用
ヒト抗体フレームワークの選択において、マウスおよびヒト起源の抗体間での一次配列レベルでの同一性の程度をポリペプチド骨格の構造類似性の程度と組み合わせる、上述の手法に従った。
【0107】
詳細には、ヒト起源のまたはヒト化された抗体の一連の可能な受容体フレームワークを、BLASTデータベースの検索により一次構造の最高レベルの相同性および同一性に基づいて選択した。この選択は、抗体の可変部全体を考慮し、また検索をフレームワーク部に狭めることにより、両遮断抗体について行った。
【0108】
選択された抗体の各群の中で、高分解能による、さもなければ筆者らが得た構造のそれに相当する分解能(すなわち2.5Å以下)による構造データが入手可能なものだけを考慮し、PDB(タンパクデータバンク)を検索した。次に、「スーパーインポーズ」ソフトウェア(Diederichs、1995)を使用してそれぞれのアミノ酸骨格をスーパーインポーズした。
【0109】
図3Gおよび4Gは、一次構造のヒト化する抗体との最適な整列および利用できる構造データの高分解能に基づいて選択された、Fv部(それぞれMNAC13およびαD11の)およびヒト化またはヒト起源の抗体のα炭素原子骨格の三次構造の間の整列の結果を示す。
【0110】
ヒト起源または操作した個々の構造のMNAC13およびαD11との重なり程度を評価するために、それぞれのアミノ酸骨格を構成しているα炭素原子の間でRMSを計算したが、2Åを上回っているRMSの原子対は無視した。
【0111】
ヒト化のための最適フレームワークの選択が3変数問題として構成され、それはしたがって相同性レベルおよび同一性の程度を構造的整列と関連させる場合に空間で表すことができる。次に、2種類の整列における問題の領域をそれぞれのフレームワークの領域に減らして、この種の分析を実行した。
【0112】
図3および4で示すように、分析した3つの変数(それぞれ、RMS値、RMSが計算された原子の割合および一次構造間の類似指数、すなわち全体の同一性-A-、全体の相同性-C-、フレームワークレベルの同一性-B-、フレームワークレベルの相同性-D-の割合)の空間で検討された抗体の分布は、検討した両ケースで相互に首尾一貫して整合的である。
【0113】
さらに、これらの分布を、それがヒト起源であるならば各抗体が占めるであろう3変数の空間における最適な位置と比較すると、一次および三次構造レベルでこの理想位置に最も近づくヒト起源抗体を明らかに同定することは可能である。両抗体の場合についてこの結果を合理的に説明するために、4つの分析のそれぞれで、検討したヒト化またはヒト起源の抗体の各位置について、仮定上の最適位置からの逸脱を計算した(MNAC13については図3Eおよび3F、αD11については図4Eおよび4F)。この場合もまた、結果は首尾一貫しており、以前の指摘を確認するものである。
【0114】
この選択法に基づいて、NGF/TrkA相互作用を中和する2つの抗体のヒト化のための以降のCDR移植過程で、2つの異なるヒト化抗体が受容体フレームワークとして選択された。詳細には、図3Hおよび4Hは、Fv部のレベルにおける2つの遮断抗体と選択されたヒト化抗体それぞれとの構造的整列を示し、すなわち、αD11のためにはPDB 1JPSコードが、MNAC13のためには1AD0コードが使用された。図3Iおよび4Iでは、マウス抗体の同じ領域とCDR移植後の同じ抗体のモデルとが比較される。
【0115】
一旦CDRが確定されると、それらが属している正規クラス(ChothiaおよびLeskによって規定されている)を同定し、その後ヒト化抗体内の正規残基を維持した。図5および図6では、各抗体についてそれらは下線付き文字で強調されている。
【0116】
導入する復帰変異の以降の分析に関して、可変ドメインの軽鎖および重鎖の間の相互作用を媒介する残基を維持するために、2つのドメイン間の境界面を維持するために以下の復帰変異を挿入した。
MNAC13のためにL46およびH35、H37
αD11のためにL34、L46、L92およびH35。
【0117】
さらに、バーニヤゾーンの特性を維持するために、以下の復帰変異を起こした。
MNAC13のためにL98
αD11のためにH71(これはいずれにせよ、ヒトコンセンサス配列で表されるアミノ酸残基の置換に関係がある)。
【0118】
その後、ヒト免疫グロブリンの主なコンセンサス配列との比較の後、以下の復帰変異を起こした。
MNAC13のためにL1、L2、L13、L50、L73、L104およびH24、H48、H49、H73、H76、H82B、H87、H90、H93
αD11のためにL56
【0119】
さらに、同じくヒト免疫グロブリンのコンセンサス配列に基づき、供与体および受容体フレームワークに存在する普通でない残基の代わりに、ヒトで保存されている残基を挿入する以下の変異をMNAC13のヒト化形態に導入した。
L42(L→Q)、L83(I→V)およびH83(Q→R)、H89(I→V)。
【0120】
同じ理由から、以下の変異をαD11のヒト化形態に導入した。H67(V→F)。
【0121】
結晶構造のそれぞれの対を修飾したが、先ず、動物起源のCDRをヒトフレームワークにグラフトした。その後、上述のすべての変異および復帰変異を導入した。修飾された構造体を次に複合免疫グロブリンに組み立てた。得られたモデルは、力場を使用して機械的エネルギー(ねじれ角および結合角度および距離に関して)を最小にすることにより洗練した。
【0122】
MNAC13およびαD11モノクローナル抗体のヒト化
MNAC13のCDR移植を達成するために供与体フレームワークのヒト化抗体を選択した後に、マウスMNAC13のCDRを上で述べた変異により修飾したヒト化抗体のフレームワークと組み合わせた可変部をそれぞれ設計する。αD11についても類似した手法に従った。本質的に、約80塩基のオリゴヌクレオチドを用いたオーバーラップ-アセンブリー(overlap assembly PCR)法に基づく手法によって2つのヒト化可変部を得ることができるが、これらの塩基ではセンスおよびアンチセンスのフィラメントが交互し、ハイブリッド動作の結果として部分的に二重のフィラメント分子の形成を可能にするような方法で20塩基の長さで連続的に重複する(図7Bおよび8B)。Ventポリメラーゼによって不連続部を埋めた後に、軽鎖の可変ドメインのためにはpVLexpressプラスミド(図9A)、重鎖の可変ドメインのためにはpVHexpressプラスミド(図9B)における、制限酵素による消化の後の方向性クローニングのために適当な制限部位(それぞれ軽鎖の可変ドメインのクローニングのためにはApaLI/BgIII、重鎖の可変ドメインのクローニングのためにはBssHII/BstEII)と共にプライマーとして二重フィラメント自体の5'の配列を含む2つの短いオリゴヌクレオチドを使用して、PCRのために二重フィラメントを増幅する。これらの担体は、それぞれヒト起源の定常ドメインCκおよびCH1、CH2およびCH3それぞれのクローン配列との融合による発現を可能にする。したがって、これらのベクターを使って、上で述べたようなヒト細胞系でIgG1分子(図9Cおよび9D1)の形で両抗体を発現することが可能である。
【0123】
Fab断片の形で両方のヒト化抗体を得るために、重鎖がクローニングされる担体だけに作用することで十分である。詳細には、SacII-XbaI方向性クローニングのための制限部位を両端に備えている特定のプライマーを使用して、定常部全体をPCRのために増幅した唯一のCH1ドメインで置換することが可能である(図9D2で示す)。
【0124】
最後に、免疫毒素の形でMNAC13ヒト化抗体を得るために、非方向性XbaIクローニングのために端に制限部位を備えた特定のプライマーを使用して、PCRのために増幅した塩基性プロタミンタンパク質を、定常ドメインCH1のC末端で発現することが可能である(図9D3で示す)。
【0125】
MNACおよびαD11ヒト化抗体の発現および結合アッセイ
推奨されたプロトコル(Roche)に従ってFuGENEにより、250,000のコス細胞を、各ヒト化抗体のVKでVHのために1μgのコードプラスミドDNAでコトランスフェクションした(合計2μg)。構築物は、IgG1の形のヒト化抗体を得るために使用した。
【0126】
上述の構築物のコトランスフェクションと並行して、対応するキメラ形態を各抗体についてコトランスフェクションした。すなわち、
マウスMNAC13のVHはCMV pVHexpressに、マウスMNAC13のVkはCMV pVkexpressにクローニングする。
αD11に関しては、ラットVHはヒト起源のCγとの融合によりpcDNA1に、ラットVkはヒト起源のCγとの融合によりpcDNA1にクローニングする。
【0127】
トランスフェクションから72時間後に、宿主細胞によって発現された免疫グロブリンを含んでいる上清を収集し、Centriprep 50(Amicon)を使用して濃縮した。
【0128】
2つのヒト化抗体のそれぞれのリガンドを認識する能力を、ELISAアッセイによって検証し、それぞれのキメラ形態と比較した。結果を図10および11に示す。
【0129】
プラスチック上への固定化のために、96穴Maxisorbプレートを、0.1M炭酸ナトリウム緩衝液pH 9.6で10μg/mlの濃度にした2つの抗体(精製された組換え免疫付着因子TrkA Camelおよび顎下腺から精製されたマウスNGF)のそれぞれのリガンドを含んでいる溶液と、4℃で一晩インキュベートした。
【0130】
3%のミルクを含んでいるPBS(MPBS)により周囲温度で1時間ブロックした後に、濃縮した上清を連続希釈(1:2、1.20;1:200)と、また並行してトランスフェクションしていないコス細胞(陰性対照)の上清とインキュベートした。
【0131】
一次抗体(ヒト起源の定常Cγ領域を認識する)および二次抗体(ペルオキシダーゼとコンジュゲートした抗ウサギ抗体)とのインキュベーションの後、結合活性をTMB基質(TECNA)とのインキュベーションによる450nm(OD450)における光学密度として検出することが可能である。500ng/mlの濃度のそれぞれのモノクローナル抗体を、陽性対照として含めた。
【0132】
図10Bは、発現した免疫グロブリンのアフィニティークロマトグラフィーによる精製の後に行われた、類似したELISAアッセイの結果を示す。
【0133】
詳細には、トランスフェクション細胞の上清を収集し、遠心処理によって細胞破片を除去した後に、それらを100μlのProtein Gセファロースとインキュベートし、PBSで十分に水洗の後、各タンパク質を1mM HClで溶出し、pHは1MのトリスpH 8.8で溶出直後に中和した。それぞれの濃度は、Lowryアッセイで推定した。
(参考文献)
【技術分野】
【0001】
本発明は、三次元構造を決定し、比較することによる抗体のヒト化方法、それによって得たヒト化抗体ならびに治療および診断におけるそれらのin vivoでの使用に関する。
【背景技術】
【0002】
動物起源のモノクローナル抗体のヒトにおける治療的および診断的応用は、特に反復投与を必要とする治療レジメンでは基本的に禁忌である。特に、マウスモノクローナル抗体は半減期が比較的短く、ヒトで使用される場合は免疫グロブリンの基本的な機能特性、例えば補体依存性細胞傷害作用および細胞媒介性細胞傷害作用の一部が失われる。
【0003】
さらに、ヒト以外の起源のモノクローナル抗体は、患者に注入された場合に免疫原性のアミノ酸配列を含む。多数の研究により、外来性抗体の注射の後、対象は抗体自体に対してかなり強い免疫反応(HAMA(ヒト抗マウス抗体)反応として知られる)を起こし、免疫複合体の形成、薬物動態の変更、アレルギー反応の生成などによりその治療的な有用性が完全に失われることが明らかにされている。さらに、異なる病理の治療のために益々多くの異なるモノクローナル抗体がマウスまたは他の哺乳類(したがってヒトに抗原性の)で開発されていることを考慮すると、相互に関係のない療法についても、交差反応性のために治療は無効になるか危険にさえなる可能性がある。いわゆるキメラ抗体(ヒト起源の定常部に連結したマウスの可変部)の生成は多少のプラスの結果をもたらしたが、かなりの免疫原性の問題がまだ残されている。
【0004】
ヒト化抗体は、ヒトの治療的使用の分野で、動物起源の抗体に関して少なくとも3つの考えられる利点を有する。最初に、ヒトのエフェクター領域はヒト免疫系の他の部分とよりよく相互作用することができ、補体依存性細胞傷害作用または細胞媒介性抗体依存性細胞傷害性によって標的細胞をより効率的に破壊する。さらに、ヒト免疫系はヒト化抗体のフレームワーク部または定常部(C)を外来性であると認識せず、それ故に、マウス抗体(まったく外来性)に対する反応およびキメラ抗体(部分的に外来性)によって誘導される反応と比較して、ヒト化抗体に対する抗体反応は最小化されている。
【0005】
ヒトに注入されるマウス抗体は正常抗体よりも非常に短い半減期を有することが報告されている(Shawら、1987)。ヒト化抗体は天然ヒト抗体に非常に類似した半減期を有し、より低い頻度での投与およびより低い用量を可能にする。
【0006】
ヒト化の基本原理は、ヒト免疫グロブリンとの関連で抗原認識の特異性、すなわちCDRドメインを移動させる構成である(「CDRグラフト」、WinterおよびMilstein、1991)。免疫原性の問題を解決するために試作されたいくつかのヒト化抗体の例が報告されている(Maedaら、1991;Singerら、1993;Tempestら、1994;Kettleboroughら、1996;Hsiaoら、1994;Bacaら、1997;Legerら、1997;Ellisら、1995;Satoら、1994;Jonesら、1986;Benharら、1994;ShaおよびXiang、1994;Shearmanら、1991;Rosokら、1996;Gussow & Seemann、1991;Coutoら、1994;Kashmiriら、1995;Bakerら、1994;Riechmannら、1988;Gormanら、1991;Verhoeyenら、1988;Foote & Winter、1992;Lewis & Crowe、1991;Coら、1991;Coら、1991;Verhoeyenら、1991;Eigenbrotら、1994;Hamiltonら、1997;Tempestら、1995;Verhoeyenら、1993;Cookら、1996;Poulら、1995;Coら、1992;Grazianoら、1995;Prestaら、1993;Hakimiら、1993;Roguskaら、1996;Adairら、1994;Satoら、1993;Tempestら、1991;Satoら、1996;Kolbingerら、1993;ZhuおよびCarter、1995;Simsら、1993;Routledgeら、1991;Roguskaら、1994;Queenら、1989;Carterら、1992)。
【0007】
動物(一般にマウス)からヒト化への抗体の変換は対立する要求事項間の妥協であり、その解決方法は個々に異なる。免疫原性を最小にするために、免疫グロブリンは受容側のヒト配列をできるだけ多く維持するものとする。いずれにせよ、元の結合持性を保持するために、免疫グロブリンフレームワークは、CDR部の高次構造が供与体マウスの免疫グロブリンのそれとできるだけ類似するように、受容側のヒト配列に十分な数の変異を含むべきである。これらの対立する考慮事項の結果として、多くのヒト化抗体では、対応するマウス抗体に関して結合親和性のかなりの減少が報告されている(Jonesら、1986;Shearmanら、1991;Kettleborough、1991;Gormanら、1991;Riechmannら、1988)。
現在、ヒト化免疫グロブリンの最も一般的な生成方法は、適当なゲノム配列、合成配列ならびにcDNAの使用に基づく(Reichmannら、1988)。
特許出願EP592106は、齧歯動物からの抗体のヒト化方法を開示している。この方法はヒト化する抗体の三次元構造の表面に露出しているアミノ酸残基の同定、対応するヒト抗体上の同じ位置のアミノ酸残基の同定、および齧歯動物抗体の配列で同定された残基のヒト抗体で同定されたそれらによる置換に基づく。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】EP592106
【特許文献2】EP1181318
【特許文献3】国際公開91/08216
【特許文献4】U.S.Ser.No.07/290,968
【非特許文献】
【0009】
【非特許文献1】Shawら、1987
【非特許文献2】WinterおよびMilstein、1991
【非特許文献3】Maedaら、1991
【非特許文献4】Singerら、1993
【非特許文献5】Tempestら、1994
【非特許文献6】Kettleboroughら、1996
【非特許文献7】Hsiaoら、1994
【非特許文献8】Bacaら、1997
【非特許文献9】Legerら、1997
【非特許文献10】Ellisら、1995
【非特許文献11】Satoら、1994
【非特許文献12】Jonesら、1986
【非特許文献13】Benharら、1994
【非特許文献14】ShaおよびXiang、1994
【非特許文献15】Shearmanら、1991
【非特許文献16】Rosokら、1996
【非特許文献17】Gussow & Seemann、1991
【非特許文献18】Coutoら、1994
【非特許文献19】Kashmiriら、1995
【非特許文献20】Bakerら、1994
【非特許文献21】Riechmannら、1988
【非特許文献22】Gormanら、1991
【非特許文献23】Verhoeyenら、1988
【非特許文献24】Foote & Winter、1992
【非特許文献25】Lewis & Crowe、1991
【非特許文献26】Coら、1991
【非特許文献27】Coら、1991
【非特許文献28】Verhoeyenら、1991
【非特許文献29】Eigenbrotら、1994
【非特許文献30】Hamiltonら、1997
【非特許文献31】Tempestら、1995
【非特許文献32】Verhoeyenら、1993
【非特許文献33】Cookら、1996
【非特許文献34】Poulら、1995
【非特許文献35】Coら、1992
【非特許文献36】Grazianoら、1995
【非特許文献37】Prestaら、1993
【非特許文献38】Hakimiら、1993
【非特許文献39】Roguskaら、1996
【非特許文献40】Adairら、1994
【非特許文献41】Satoら、1993
【非特許文献42】Tempestら、1991
【非特許文献43】Satoら、1996
【非特許文献44】Kolbingerら、1993
【非特許文献45】ZhuおよびCarter、1995
【非特許文献46】Simsら、1993
【非特許文献47】Routledgeら、1991
【非特許文献48】Roguskaら、1994
【非特許文献49】Queenら、1989
【非特許文献50】Carterら、1992
【非特許文献51】Sequences of Proteins of Immunological Interest(National Institutes of Health、Bethesda MD、1987、1991)
【非特許文献52】Paul、1993
【非特許文献53】Kabatら、1987
【非特許文献54】ChothiaおよびLesk、1987
【非特許文献55】Kabat(1987)
【非特許文献56】Kabat(1991)
【非特許文献57】Chothiaら(1989)
【非特許文献58】Lanzavecchiaら、1987
【非特許文献59】Hoodら、1984
【非特許文献60】HarlowおよびLane、1988
【非特許文献61】HunkapillerおよびHood、1986
【非特許文献62】American Type Culture Collection(Catalogue of Cell Lines and Hybridomas、第5版(1985)Rockville、Maryland、USA
【非特許文献63】Satoら、1994
【非特許文献64】CarugoおよびPongor、2001
【非特許文献65】CarugoおよびPongor、2003
【非特許文献66】Daughertyら、1991
【非特許文献67】De Martinoら、1991
【非特許文献68】Gillman & Smith、1979
【非特許文献69】Robertsら、1987
【非特許文献70】Hustonら、1988
【非特許文献71】Birdら、1988
【非特許文献72】BuchnerおよびRudolph、1991
【非特許文献73】Skerraら、1991
【非特許文献74】Cesareni、1992
【非特許文献75】Swimmerら、1992
【非特許文献76】Gramら、1992
【非特許文献77】Clacksonら、1991
【非特許文献78】Scott & Smith、1990
【非特許文献79】Garrardら、1991
【非特許文献80】Putlitzら、1990
【非特許文献81】Larrick & Fry、1991
【非特許文献82】Benvenutoら、1991
【非特許文献83】Durinら、1990
【非特許文献84】Hiattら、1989
【非特許文献85】Winnacker、1987
【非特許文献86】Queenら、1986
【非特許文献87】Buhlerら、1990
【非特許文献88】Meadeら、1990
【非特許文献89】Scopes、1982
【非特許文献90】LefkovitsおよびPernis、1979
【非特許文献91】LefkovitsおよびPernis、1981
【非特許文献92】Chaudharyら、1989
【非特許文献93】Thorpeら(1982)
【非特許文献94】BaldwinおよびByers、1985;;
【非特許文献95】OlsnesおよびPhil、1982
【非特許文献96】Chenら、1995
【非特許文献97】Angelesら、1996
【非特許文献98】Goding(1986)
【非特許文献99】Paikら(1982)
【非特許文献100】Jancarik & Kim、1991
【非特許文献101】Otwinowski & Minor、1997
【非特許文献102】Bermanら、2000
【非特許文献103】Kodandapaniら、1999
【非特許文献104】Bentley、1990
【非特許文献105】Navaza、1994
【非特許文献106】KleywegtおよびJones、1994
【非特許文献107】Brungerら、1998
【非特許文献108】Laskowskiら、1993
【非特許文献109】Diederichs、1995
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明者らは、結晶学的研究に由来し実験的に得られる構造データに一貫して基づく手法による、ヒトに対して実質的に免疫原性でない最適化されたヒト化形態の免疫グロブリンを得るための方法を提案した。本発明の方法は、治療製剤ならびに他の医学的および診断の用途に適応した形態の抗体の入手を可能にする。
【課題を解決するための手段】
【0011】
本発明は、ヒト化の最初の設計段階(一般によりエラーが起こりやすい)を行うための、完全に構造データに基づく方法に関するものである。ヒト化免疫グロブリンは、軽鎖および重鎖の間に2対のヘテロダイマーを有し、少なくとも1つの鎖は機能的にヒト起源のフレームワーク部の部分と結合した1つまたは複数の動物起源のCDRを有す。例えば、動物起源の元の免疫グロブリンの親和性と同等の親和性でそれぞれの抗原と結合することができるヒト化免疫グロブリンを生成するために、動物起源のCDRを、同じく動物起源で自然に結合しているアミノ酸残基と共に、ヒト起源のフレームワーク部に導入する。
【0012】
本発明の方法は、治療および診断の用途のために適当なヒト化抗体の入手を導いた。特に、それぞれTrkAおよびNGFと高い特異性で結合してリガンドと受容体の間の相互作用を中和することができる、抗TrkA抗体(特許EP1181318)および抗NGF抗体に由来するヒト化免疫グロブリンが得られた。そのような分子は、NGF/TrkAに依存する腫瘍、慢性疼痛、炎症性形態の治療のために、および診断目的のために、例えばTrkA陽性腫瘍または、アルツハイマー病の早発マーカーとしての前脳基底核のin vivo画像化のために有用である。詳細には、ヒト化抗TrkA抗体は、尿路および骨盤領域の炎症性形態で特異的な治療および診断の用途を有す。詳細には、ヒト化抗NGF抗体は、HIVウイルスによって誘導される病理状態で、HIVに感染したNGF依存性マクロファージなどの免疫細胞のアポトーシスを誘導するための、特異的な治療および診断の用途を有す。
【0013】
したがって、本発明の目的は、既知の配列の動物抗体のVHおよびVL可変部のヒト化方法を提供することであり、これは、
a)入手不能であるならば、動物抗体のVHおよびVL部の結晶構造を得る段階と、
b)その構造が3Å以上の分解能で実験的に測定されたヒト起源またはヒト化抗体の0からn個の一連の可能なフレームワーク受容体を、動物抗体のフレームワークの一次配列との最高水準の相同性および同一性に基づいて予備選択する段階と、
c)動物抗体のVHおよびVL可変部とb)で得られるVHおよびVL部との間の構造比較をそれぞれ行い、各比較についてRMSを計算してより小さなRMSを有するヒト起源のVHおよびVL部を同定する段階と、
d)c)で同定したヒト配列内の適当な位置に前記動物抗体のCDR部の配列を挿入する段階と、
e)必要に応じて、c)で同定したヒトVHおよびVL部の1つまたは複数のアミノ酸残基を復帰変異させる段階とを含む。
【0014】
好ましくは、抗体の修飾は、組換えDNA技術によって起こる。
【0015】
好ましい一実施形態では、動物抗体は抗NGF抗体であり、好ましくは、それはαD11抗体であり、ヒト化配列は本質的に以下のVH配列;HumαD11VH、
EVQLVESGGGLVQPGGSLRLSCAASGFSLTNNNVNWVRQAPGKGLEWVGGVWAGGATDYNSALKSRFTISRDNSKNTAYLQMNSLRAEDTAVYYCARDGGYSSSTLYAMDAWGQGTLVTVSS、(配列番号17)
およびVL;HumαD11 Vk
DIQMTQSPSSLSASVGDRVTITCRASEDIYNALAWYQQKPGKAPKLLIYNTDTLHTGVPSRFSGSGSGTDYTLTISSLQPEDFATYFCQHYFHYPRTFGQGTKVEIK(配列番号18)。
を有す。
【0016】
他の実施形態では、動物抗体は抗TrkA抗体であり、好ましくは、それはαMNAC13抗体であり、ヒト化配列は本質的に以下のVH配列;HumMNAC13VH、
EVQLLESGGGLVQPGGSLRLSCAASGFTFSTYTMSWARQAPGKGLEWVAYISKGGGSTYYPDTVKGRFTISRDNSKNTLYLQMNSLRAEDSAVYYCARGAMFGNDFFFPMDRWGQGTLVTVSSA、(配列番号37)
およびVL;HumMNAC13Vk
DIVLTQSPSSLSASVGDRVTITCSASSSVSYMHWYQQKPGQAPKLLIYTTSNLASGVPSRFSGSGSGTDYTLTISSLQPEDVATYYCHQWSSYPWTFGGGTKVEIK(配列番号38)。
を有する。
【0017】
本発明のヒト化免疫グロブリン(または結合活性を維持している誘導断片および他の誘導可能な化合物)は、公知の組換えDNA技術によって生成することができる。その後のヒト化免疫グロブリンの使用の機能としては、トランスジェニック動物またはトランスフェクション細胞をそれらの発現のために使用することができ、好ましくは不死化された真核細胞(例えば骨髄腫またはハイブリドーマ細胞)であるが、原核生物の宿主、昆虫または植物細胞でもよい。得られたヒト化免疫グロブリン配列をコードするポリヌクレオチドは、合成によっても得ることができる。
【0018】
本発明のヒト化免疫グロブリンは、単独でも、または他の治療薬と併用しても使用することができる。抗腫瘍剤として使用する場合は化学療法剤が好ましく、それは薬理学的用途(例えばアントラサイクリン、パクリタキセル、シスプラチン、ゲムシタビン、非ステロイド性コルチコステロイド抗炎症薬または免疫抑制薬)ならびに各特定の病理状態の療法で現在適用されているすべての薬剤に従い異なる。ヒト化免疫グロブリンまたはそれらの複合体は、薬理学的に許容できる投薬形態で調製することができ、それは投与の型に従い異なる。
【0019】
定義
2つのポリヌクレオチドまたはポリペプチド(それぞれヒト化免疫グロブリンをコードするDNAの配列またはヒト化免疫グロブリンのアミノ酸配列またはそれらの一部)に関連する用語「実質的に同一の」は、最大の対応で比較および整列させたときに、ヌクレオチドまたはアミノ酸残基が最低80%(好ましくは90〜95%以上)の同一性を有する2個以上の配列を指す。通常、「実質的な同一性」は少なくとも50残基の長さの領域で、より好ましくは少なくとも100残基の領域で、または最適条件では150以上の残基で、または完全な配列で検証される。下記のように、抗体のいずれか2つの配列は、カバットのナンバリング方式を使用して1つの方法だけで整列させることができる。したがって、抗体については、同一性割合は特有の明確な意味を有する。成熟免疫グロブリンの重鎖および軽鎖の可変部のアミノ酸はHxおよびLxと称され、xは、カバットのナンバリング方式、Sequences of Proteins of Immunological Interest(National Institutes of Health、Bethesda MD、1987、1991)によるアミノ酸位置を指定する数である。カバットは、コンセンサス配列を作成するために、各サブグループの抗体のアミノ酸配列のリスト、ならびに各サブグループの各位置で最も頻出するアミノ酸のリストを決定した。カバットは数字をリスト内の各配列の各アミノ酸に割り当てる方法を使用するが、この各残基の数字を割り当てる方法はこの分野で標準の方法になっている。カバットの方式は彼の研究には存在しない他の抗体にまで拡張することができ、保存されたアミノ酸に基づいてカバットによって特定されるコンセンサス配列の1つと問題の抗体を整列させることができる。カバットのナンバリング方式を使用すると、異なる抗体中の同等の位置のアミノ酸の特定が容易になる。例えば、ヒト起源の抗体のL10位置のアミノ酸は、マウス起源の抗体のL10位置の同等のアミノ酸位置を占める。
【0020】
抗体の基本的な構造単位が四量体を含むことは公知である。各四量体は2つの同一の対のポリペプチド鎖によって構成され、それぞれは軽鎖(25kDa)および重鎖(50〜75kDa)によって構成される。各鎖のアミノ末端領域は約100〜110以上のアミノ酸の可変部を含み、それは抗原認識に関係している。各鎖のカルボキシ末端領域は、エフェクター機能を媒介する定常部を含む。軽鎖および重鎖の各対の可変部は、抗体の結合部位を形成する。したがって、手つかずの抗体は2つの結合部位を有する。
【0021】
軽鎖は、κまたはλに分類されている。重鎖はγ、μ、α、εに分類され、それらは抗体のイソタイプをそれぞれIgG、IgM、IgA、IgDおよびIgEと定義する。軽鎖および重鎖の中では、可変部および定常部は少なくとも約12のアミノ酸の「J」領域で接続され、重鎖だけは約10アミノ酸の「D」領域を含む(Paul、1993)。
【0022】
軽鎖および重鎖の各対の可変部は、抗体の結合部位を形成する。それらは、相補性決定部(CDR)と呼ばれている3つの超可変部が接続しているフレームワーク(FR)と呼ばれている比較的保存された領域で構成される同じ一般構造を特徴とする(Kabatら、1987;ChothiaおよびLesk、1987)。各対の2つの鎖のCDRはフレームワーク部によって整列させられ、特定のエピトープと結合する機能を取得している。アミノ末端領域からカルボキシ末端領域の方へかけて、軽鎖および重鎖の両方の可変ドメインは、交互に出現するFRおよびCDR領域、すなわちFR、CDR、FR、CDR、FR、CDR、FRを含む。したがって、重鎖および軽鎖の両方は3つのCDR、すなわちそれぞれCDRH1、CDRH2、CDRH3およびCDRL1、CDRL2、CDRL3を特徴とする。各領域へのアミノ酸の帰属は、Kabat(1987および1991)および/またはChothia & Lesk(1987)、Chothiaら(1989)による定義に従って行われた。
【0023】
好ましくは、例示されたヒト化免疫グロブリンの類似体は、保存的なアミノ酸の置換のために元の免疫グロブリンとは異なる。アミノ酸置換を保存的または非保存的に分類するために、アミノ酸は以下の通りに分類することができる。
群I(疎水性側鎖):M、A、V、L、I;
群II(中性の親水性側鎖):C、S、T、N、Q;
群III(酸性側鎖):D、E;
群IV(塩基性側鎖):K、R;
群V(主鎖の配向性に影響する残基):G、P;
群VI(芳香族側鎖):F、Y、W。
【0024】
保存的なアミノ酸置換は同じクラスのアミノ酸の間の置換に関係があるが、非保存的なアミノ酸置換は異なるクラスのメンバーの間の交換を伴う。
【0025】
用語「エピトープ」は、免疫グロブリンと特異的に結合することができるいかなるタンパク質決定要素を含む。通常、エピトープは高分子の化学的に活性な表面の組、例えばアミノ酸または糖の側鎖によって形成され、それらは通常、特異的な化学物理特性および配座特性を有す。
【0026】
用語「免疫グロブリン」は、免疫グロブリン遺伝子によってコードされた1つまたは複数のポリペプチドから成るタンパク質を指す。四量体抗体の形態に加えて、免疫グロブリンは様々な形態で存在することができる。例えば、それらは断片Fv、FabおよびF(ab')、ならびに二機能性ハイブリッド抗体(Lanzavecchiaら、1987)および単鎖Fv断片(Hoodら、1984;HarlowおよびLane、1988;HunkapillerおよびHood、1986)を含む。
【0027】
キメラ抗体は、その軽鎖および重鎖の遺伝子が異なる種に属している免疫グロブリンの遺伝子領域から始まるように操作された抗体である。例えば、マウスのモノクローナル抗体の遺伝子の可変部分(V)は、ヒト起源の抗体の定常部(C)に連結させることができる。治療用のキメラ抗体は、したがって、マウス抗体に由来する抗原を認識するドメインVおよびヒト抗体に由来するエフェクタードメインCから成るハイブリッドタンパク質である(他の哺乳類の組合せも使用することができる)。
【0028】
カバットの定義によれば、用語「フレームワーク」は、種内の異なる免疫グロブリンの間で比較的保存されている、免疫グロブリンの軽鎖および重鎖の可変部の部分(CDRに属さない)を指す。それ故に、ヒトフレームワークは、ヒト抗体で天然に見られるフレームワークと実質的に同一(少なくとも85%以上)なフレームワーク部である。
用語「ヒト化免疫グロブリン」は、ヒトフレームワークおよび非ヒト抗体に由来している少なくとも1つのCDRを含み、存在する各定常部がヒト免疫グロブリンの領域と実質的に同一(少なくとも85%、好ましくは少なくとも90〜95%同一)である免疫グロブリンを指す。それ故に、CDR以外のヒト化免疫グロブリンのすべての部分は、天然のヒト免疫グロブリンの1つまたは複数の配列の対応する領域と、実質的に同一である。例えば、マウス可変部およびヒト起源の定常部によって構成されるキメラ抗体は、ヒト化免疫グロブリンには含まれない。
【図面の簡単な説明】
【0029】
【図1A】変性条件下でのポリアクリルアミド(12% SDS-PAGE)およびクーマシーブルーでの染色による、MNAC13抗体Fab断片の精製結果の分析を示す図である(ウェル1:パパインでタンパク分解的に消化されたMNAC13抗体の試料;ウェル2:DEAE Sephacellイオン交換樹脂に結合し、250mMのNaClで溶出した分画;ウェル3:分子量;ウェル4:精製され濃縮されたMNAC13抗体のFab断片)。
【図1B】MNAC13抗体のFab断片の典型的結晶を示す図である。
【図1C】MNAC13抗体のFab断片の結晶で得られる高分解能回折スペクトラムを示す図である。
【図1D】MNAC13抗体のFab断片の重鎖および軽鎖ドメインの主鎖のねじれ角のラマチャンドランチャートである。
【図2A】変性条件下でのポリアクリルアミド(12% SDS-PAGE)およびクーマシーブルーでの染色による、αD11抗体Fab断片の精製結果の分析を示す図である(ウェル1:パパインでタンパク分解的に消化されたαD11抗体の試料;ウェル2:精製され濃縮されたαD11抗体のFab断片;ウェル3:分子量)。
【図2B】αD11抗体のFab断片の典型的結晶を示す図である。
【図2C】αD11抗体のFab断片の結晶で得られる高分解能回折スペクトラムを示す図である。
【図2D】αD11抗体のFab断片の重鎖および軽鎖ドメインの主鎖のねじれ角のラマチャンドランチャートである。
【図3−1】図3Aは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図3Bは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図3Cは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図3Dは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図3Eは、MNAC13の仮定的最適値からのヒト化またはヒト起源の抗体の逸脱を示す図である(全体の同一性および相同性の程度-青-およびフレームワークレベル-マゼンタ-を考慮して計算)。図3Fは、MNAC13の仮定的最適値からのヒト化またはヒト起源の抗体の逸脱を示す図である(全体の同一性および相同性の程度-青-およびフレームワークレベル-マゼンタ-を考慮して計算)。
【図3−2】図3Gは、マウス抗体との同一性および相同性の程度および利用できる構造データの分解能の程度に従って選択された、ヒト化またはヒト起源の抗体のそれぞれの領域のMNAC13のFv断片との構造的整列を示す図である。図3Hは、H)は選択されたヒト化抗体1AD0(赤で示す)のそれぞれの領域のMNAC13のFv断片(シアンで示す)との構造的整列を示す図である。図3Iは、I)はCDR移植の後に生じた抗体モデル(フレームワークレベルでは黄色で、CDRレベルでは白で示されている)のMNAC13のFv断片との構造的整列を示す図である。図3Lは、選択されたフレームワーク内の推定される復帰変異の同定の結果として得られたMNAC13ヒト化抗体のFv断片のモデルを示す図である(ヒト起源残基は緑で示し、マウス起源残基はマゼンタで示す)。
【図4−1】図4Aは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図4Bは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図4Cは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図4Dは、分析された3つの変数に従うヒト化またはヒト起源の抗体(それらの結晶構造のPDBコードを使用して命名)の分布を示す図である。図4Eは、αD11の仮定的最適値からのヒト化またはヒト起源の抗体の逸脱を示す図である(全体の同一性および相同性の程度-青-およびフレームワークレベル-マゼンタ-を考慮して計算)。図4Fは、αD11の仮定的最適値からのヒト化またはヒト起源の抗体の逸脱を示す図である(全体の同一性および相同性の程度-青-およびフレームワークレベル-マゼンタ-を考慮して計算)。
【図4−2】図4Gは、マウス抗体との同一性および相同性の程度および利用できる構造データの分解能の程度に従って選択された、ヒト化またはヒト起源の抗体のそれぞれの領域のαD11のFv断片との構造的整列を示す図である。図4Hは、H)は選択されたヒト化抗体1JPS(赤で示す)のそれぞれの領域のαD11のFv断片(シアンで示す)との構造的整列を示す図である。図4Iは、I)はCDR移植の後に生じた抗体のモデル(フレームワークレベルでは黄色で、CDRレベルでは白で示されている)のαD11のFv断片との構造的整列を示す図である。図4Lは、選択されたフレームワーク内の推定される復帰変異の同定の結果として得られたαD11ヒト化抗体のFv断片のモデルを示す図である(ヒト起源残基はシアンで示し、マウス起源残基は紫で示す)。
【図5】MNAC13(配列番号22、配列番号24)、ヒト化のために選択されたヒト化抗体(1AD0;配列番号39、配列番号40)、1AD0のフレームワーク上へのCDR移植および記載されている復帰変異および変異の後のMNAC13のヒト化形態(Hum MNAC13:配列番号37、配列番号38)の重鎖(A)および軽鎖(B)それぞれの可変部の一次構造の整列を示す図である。CDRは、MNAC13の2つの鎖のヒト化形態の配列内に下線付き文字で強調されている。
【図6】αD11(配列番号2、配列番号4)、ヒト化のために選択されたヒト化抗体(1JPS;配列番号19、配列番号20)、1AD0のフレームワーク上へのCDR移植および記載されている復帰変異および変異の後のαD11のヒト化形態(Hum αD11:配列番号17、配列番号18)の重鎖(A)および軽鎖(B)それぞれの可変部の一次構造の整列を示す図である。CDRは、αD11の2つの鎖のヒト化形態の配列内に下線付き文字で強調されている。
【図7A】MNAC13のマウス形態の軽鎖可変部のcDNAのヌクレオチド配列を示す図である(配列番号23)。
【図7B】MNAC13のマウス形態の重鎖可変部のcDNAのヌクレオチド配列を示す図である(配列番号21)。
【図7C】MNAC13の軽鎖可変部(配列番号38)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:L1S:配列番号31;L2AS:配列番号32;L3S:配列番号33;L4AS:配列番号34;L5S:配列番号35;L6AS:配列番号36を、対応するアミノ酸配列への翻訳と共に示す図である。
【図7D】MNAC13の重鎖可変部(配列番号37)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:H1S:配列番号25;H2AS:配列番号26;H3S:配列番号27;H4AS:配列番号28;H5S:配列番号29;H6AS:配列番号30を、対応するアミノ酸配列への翻訳と共に示す図である。
【図7E】MNAC13の軽鎖可変部(配列番号38)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:L1S:配列番号31;L2AS:配列番号32;L3S:配列番号33;L4AS:配列番号34;L5S:配列番号35;L6AS:配列番号36を、対応するアミノ酸配列への翻訳と共に示す図である。
【図7F】MNAC13の重鎖可変部(配列番号37)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:H1S:配列番号25;H2AS:配列番号26;H3S:配列番号27;H4AS:配列番号28;H5S:配列番号29;H6AS:配列番号30を、対応するアミノ酸配列への翻訳と共に示す図である。
【図8A】αD11のラット形態の軽鎖可変部のcDNAのヌクレオチド配列を示す図である(配列番号3)。
【図8B】αD11のマウス形態の重鎖可変部のcDNAのヌクレオチド配列を示す図である(配列番号1)。
【図8C】αD11の軽鎖可変部(配列番号18)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:L1S:配列番号11;L2AS:配列番号12;L3S:配列番号13;L4AS:配列番号14;L5S:配列番号15;L6AS:配列番号16を、対応するアミノ酸配列への翻訳と共に示す図である。
【図8D】αD11の重鎖可変部(配列番号17)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:H1S:配列番号5;H2AS:配列番号6;H3S:配列番号7;H4AS:配列番号8;H5S:配列番号9;H6AS:配列番号10を、対応するアミノ酸配列への翻訳と共に示す図である。
【図8E】αD11の軽鎖可変部(配列番号18)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:L1S:配列番号11;L2AS:配列番号12;L3S:配列番号13;L4AS:配列番号14;L5S:配列番号15;L6AS:配列番号16を、対応するアミノ酸配列への翻訳と共に示す図である。
【図8F】αD11の重鎖可変部(配列番号17)のヒト化形態を得るためにオーバーラップ-アセンブリーPCR法により引き抜いたオリゴヌクレオチド配列:H1S:配列番号5;H2AS:配列番号6;H3S:配列番号7;H4AS:配列番号8;H5S:配列番号9;H6AS:配列番号10を、対応するアミノ酸配列への翻訳と共に示す図である。
【図9A】オーバーラップ-アセンブリーPCRによって得られた両抗体のヒト化可変部の配列をクローニングするために使用したプラスミドの地図である。A)軽鎖可変ドメインのためのpVLexpress。
【図9B】オーバーラップ-アセンブリーPCRによって得られた両抗体のヒト化可変部の配列をクローニングするために使用したプラスミドの地図である。B)重鎖可変ドメインのためのpVHexpress。
【図9C】オーバーラップ-アセンブリーPCRによって得られた両抗体のヒト化可変部の配列をクローニングするために使用したプラスミドの地図である。C)pVLexpressでの各ヒト化抗体の軽鎖可変部のクローニングから生じたプラスミド。
【図9D】オーバーラップ-アセンブリーPCRによって得られた両抗体のヒト化可変部の配列をクローニングするために使用したプラスミドの地図である。D)pVHexpressでの各ヒト化抗体の重鎖可変部のクローニングの結果得られた、1)無傷の免疫グロブリンの形態IgG1での発現のための、2)Fab断片の形態での発現のための、3)免疫毒素の形態での発現のための代替構築物。
【図10】プラスチック上に免疫付着因子の形態のTrkAを固定化して行ったELISAアッセイによる、キメラ態およびヒト化形態のMNAC13抗体の結合活性の比較を示す図である。A)トランスフェクションされたコス細胞の上清の連続希釈間の濃縮後の比較;B)Gセファロースタンパク質で精製された、トランスフェクションされたコス細胞の上清の連続希釈間の比較。
【図11】プラスチック上にNGFを固定化して行ったELISAアッセイによる、ヒト化形態のαD11抗体の結合活性の検定を示す図である。
【発明を実施するための形態】
【0030】
この方法は、in vivo治療および診断上の関心のある抗体のヒト化のための、高分解能構造比較に基づく。さらに、それぞれの抗原(すなわちNGFニューロトロフィンおよびそのTrkA受容体)に対して特異的に反応することができる、ヒト化免疫グロブリンが提供される。ヒト化免疫グロブリンはヒト起源のフレームワークを有し、またそれらは、元の免疫グロブリン(すなわちNGFに対して特異的に反応するラットの免疫グロブリンαD11、およびTrkAを特異的に認識するマウスの免疫グロブリンMNAC13)のそれぞれに由来する1つまたは複数の相補性決定部(CDR)を有す。したがって、大規模に容易に生産することができる本発明の免疫グロブリンは、NGF/TrkA依存性腫瘍形態の治療だけでなく、慢性の痛みおよび炎症形態の治療においても治療用途を有す。さらに、受容体に特異的なヒト化免疫グロブリンは、TrkA陽性の腫瘍および前脳基底核細胞(アルツハイマー病の早発マーカーとして)のin vivo画像化のために、さらなる診断用途を有す。
【0031】
本発明は、モノクローナル抗体αD11およびMNAC13(それぞれラットおよびマウス)の場合と同様に、NGF上およびTrkA上両方の関心のエピトープと結合することができる軽鎖および/または重鎖のCDR領域をコードしているDNAの組換え部分を使用する。これらの領域をコードしているDNA断片は、ヒト起源の適当なフレームワーク部をコードしているDNA断片と連結させる。モノクローナル抗体MNAC13およびαD11の軽鎖および重鎖のCDRを含んでいるポリペプチド鎖をコードするDNA配列は、それぞれ図7A、7Bおよび8A、8Bに含まれる。遺伝子コードの変性および重要ではないアミノ酸の置換により、DNA配列は容易に修飾することができる。
【0032】
さらに、一般的にDNA断片は、ヒト化免疫グロブリンのコード配列に作動可能に結合し、異種プロモーターまたは天然の関連プロモーターの領域を含んでいる発現のためのさらなる制御配列を含む。好ましくは、発現制御配列は真核生物の宿主細胞を形質転換またはトランスフェクションすることができるベクター内に真核生物プロモーターを有する系であるが、原核生物の制御配列も同様に使用することができる。一旦ベクターが適当な宿主に組み込まれると、宿主は高レベルな発現を確実にするために適当な条件で維持される。二量体、手つかずの抗体の形態または他の免疫グロブリンの形態で、軽鎖および重鎖個々のさらなる精製が続く。
【0033】
ヒト定常部をコードするDNAの配列は、様々なヒト細胞から公知の手法で単離することができるが、不死化B細胞から開始することが好ましい。本発明の免疫グロブリンのCDRは、それぞれNGFおよびTrkAと結合することができ、またそれぞれラットおよびマウスの生成物である、モノクローナル抗体αD11およびMNAC13から同様に誘導される。免疫グロブリンの発現および分泌に適当な宿主細胞は、American Type Culture Collection(Catalogue of Cell Lines and Hybridomas、第5版(1985)Rockville、Maryland、USA)など多くのソースから得ることができる。好ましくは、ヒト化抗体に組み込まれるCDRは、αD11およびMNAC13のCDR配列に対応している配列を有し、抗体自体の対応するアミノ酸配列をコードしている変性ヌクレオチド配列を含むことができる。
【0034】
通常、ヒト化の設計手順は周期的かつ反復的であり、以下を含む。
マウス抗体のアミノ酸配列の分析。
対応するFv領域のモデリング。
ヒト抗体の受容体フレームワークのアミノ酸配列の分析および選択。
選択されたフレームワーク内の推定される復帰変異の同定。
ヒト化抗体の設計および実際の構築。
in vitroおよび/またはin vivoアッセイによる維持された結合親和性および特異性の検証。
【0035】
これらの活性がヒトフレームワークによって負の影響を受けるならば、受容体ヒト抗体のフレームワークの選択を変更するか、補償変異を導入する必要がある。
【0036】
ヒトフレームワークの選択がサイクルの中で最も重要な相として構築されているとしても、一般規則は現在まで確立されていない。このことは、様々な選択(患者における免疫原性に関して)の利点が臨床視点から正確に研究されていない事実に依存する。したがって、フレームワークの正しい選択を実行するために、一連の手法だけが利用でき、これらはこれまでに得られた結果と組み合わせなければならない。
【0037】
詳細には、固定されたフレームワーク(それらの構造は長い間利用されてきているので、通常重鎖のためにはNEW、軽鎖のためにはREI)を使用することが可能である。
【0038】
他の手法は、ヒト化する抗体と配列が最も相同的であることが判明したフレームワークの使用を可能にする。相同性のヒト抗体を検索するための多くのデータベースが存在する。選択では、供与体および受容体の配列間でのより高い同一性割合に加えて、一般にCDRの長さ、正規残基のレベルでのおよび境界面レベルの残基の同一性を考慮する。これらの2つの方法の間の比較については、Grazianoら(1995)を参照。
【0039】
さらに、第2の手法の変異形によれば、軽鎖および重鎖は、高い配列相同性を特徴とする2つの異なるヒト抗体から選択することができる。この手法は、Riechmannら(1988)およびShearmanら(1991)によって提案された。この点に関しては、一般に、2つの鎖の間の境界面がよく保存されているという事実は正しい相互作用を等しく保証することができるが、同じ抗体に由来する軽鎖および重鎖は異なる抗体に由来する軽鎖および重鎖に比して正しく結合して機能的結合部位を形成する可能性がより高い。これらの2つの方法の間の比較については、Roguskaら(1996および1996)を参照。
【0040】
手法を特定のヒト抗体に由来しているフレームワークに制限することは、たとえフレームワークがヒト起源であるとしても、免疫原性エピトープを生成する体細胞変異を引き起こすリスクを伴う恐れがある。代替手法はヒトコンセンサス配列に基づくフレームワークを使用することであり、そこでは、特有の体細胞変異は除去されている。2つの手法が比較された。1つのケースでは結合親和力の差を認めなかった(Kolbingerら、1993)が、他のケースでは、代わりに結合は個々のフレームワークの場合優れていることが判明した(Satoら、1994)。
【0041】
いずれにせよ、コンセンサス配列自体は人工的であり、したがって、たとえそれらが特有の残基を有しないとしても、それらは免疫原性である非天然のモチーフを形成することができる。代替手段(Rosokら、1996)は、V-BASEデータベースで収集された生殖細胞系ヒト配列を使用することである。
【0042】
ヒト起源のフレームワーク可変部とマウスCDR部との不自然な並置は、自然界では現れない配座制限をもたらす可能性があり、それらは特定のアミノ酸残基の置換によって修正されない限り、結合親和性の損失を決定付ける。置換するアミノ酸残基の選択は、一部コンピュータモデリングによって決定される。免疫グロブリン分子の三次元画像を作製するための、ハードおよびソフトが利用できる。一般に、分子模型は、免疫グロブリンドメインまたは鎖の解決済みの結晶構造から始まって作製される。モデル化する鎖は解決済みの三次元構造の鎖またはドメインとのアミノ酸類似性に基づいて比較され、配列に関して最も高い類似を示す鎖またはドメインが分子模型の構築の出発点として選択される。しかし、抗体構造の予測は常に正確とは限らない。特に、第3のCDR部はモデル化が困難であり、それは抗体の構造予測において常に不確実な点になっている(Chothiaら、1987)。この理由から、概して、ヒト化抗体は第一次近似としては、抗原に対する結合親和性および/または特異性は出発モノクローナル抗体よりもかなり低い。このことは、完全には合理化することができない試行錯誤手法により出発抗体の特性を再構成する試みのために、多くの連続した点突然変異サイクルを必要とする。
【0043】
利用できるヒトおよびヒト化抗体の増大する高分解能X線構造を考慮すると、その意図はコンピュータモデリングの使用に由来する不確実性および曖昧さを避けて、X線結晶学手段により本発明の両抗体のFabフラグメントの高分解能構造データを得ることであった。この目的のために、両抗体をハイブリドーマから精製し、パパイン(重鎖のCH1およびCH2のドメイン間の接合部で切断するプロテアーゼ)でタンパク分解的に処理し、Fabフラグメントを生成した。さらなる精製の結果として両Fabフラグメントが結晶化され、2つのデータベース(低および高分解能)から、Molecular Substitution法で構造を決定してその後それらを精製することができた。
【0044】
実験的に得られた構造データに基づく本発明で提案されている手法は、受容体ヒト抗体フレームワークを選択する重要な相において、および両中和抗体のヒト化過程で選択されたフレームワーク内の推定される復帰変異の同定のために、非常により堅実で合理的な出発点を提供する。
【0045】
ヒト抗体フレームワークの選択の指針となるための既報告の様々な基準の中で、構造的整列に基づく分析によりその結果を拡張して完成するために使用されたものは、マウスおよびヒト起源の抗体間の一次配列における同一性の程度であった。元の基準に関連して対応する構造の比較分析は、非常により正確な比較を、したがって得られたヒト化抗体が元のマウスの抗体の親和性および特異性の特性を保存するより大きな可能性を保証するものである。したがって、使用された手法は、アミノ酸配列の分析および比較に由来する同一性の程度および相同性レベルに関する情報を、それぞれの三次元構造の比較と組み合わせる。
【0046】
詳細には、一次構造の最適アラインメントに由来する情報は、二重の役割を有する。第一に、この分析は、比較する可能な三次構造の数を減らし、高い相同性および同一性を特徴とするものに分析を限定することが可能になる。一次構造レベルでの最適アラインメントを特徴とし構造データが入手可能なこれらの配列の中で、高分解能により、さもなければ筆者らが得た構造のそれに相当する分解能(すなわち2.5Å以下)により解決された構造のみを重視して、さらなる選択を実施した。この手法は三次構造のずっと正確な整列およびRMS(根平均二乗偏差:平均二乗偏差の平方根;CarugoおよびPongor、2001および2003)で表される構造差のずっと有意な推定値を保証するものである。低分解能データは、空間内の個々の原子の実際の相対位置に関して、むしろ暗示的で明らかにより不正確な情報を提供する。
【0047】
ヒト起源または操作した個々の構造の重なり程度を評価するために、それぞれのアミノ酸骨格を構成しているα炭素原子の間でRMSを計算したが、2Åを上回っているRMSの原子対は無視した。この分析から、したがって構造間の多様性(RMSの値によって表される)だけでなく、各RMSを計算する際に実際に使用されたα炭素原子のパーセンテージも考慮しなければならない情報が得られる。
【0048】
これらの三次構造レベルの類似性データは、同一性および相同性に関する一次配列の比較分析と関連付けされた。
【0049】
ヒト化のための最適フレームワークの選択が3変数問題として構成され、それはしたがって相同性レベルおよび同一性の程度を構造的整列と関連させる場合に空間で表すことができると推測される。次に、2種類の整列における問題の領域をそれぞれのフレームワークの領域に減らして、この種の分析を実行した。
【0050】
分析した3つの変数(それぞれ、RMS値、RMSが計算された原子の割合、および一次構造間の類似指数、すなわち全体の同一性、全体の相同性、フレームワークレベルの同一性、フレームワークレベルの相同性の割合)の空間における検討された抗体の分布を、それがヒト起源だとするならば各抗体が占めるであろう3変数の空間における最適な位置と比較すると、一次および三次構造レベルでこの理想位置に最も近づくヒト起源抗体を同定することはおそらく明らかである。この結果を合理的に説明するために、4つの分析のそれぞれで、検討されるヒト化またはヒト起源の抗体の各位置について、仮定上の最適位置からの逸脱が計算される。
【0051】
この選択法に基づいて、与えられた抗体のヒト化のための以降のCDR移植過程で受容体フレームワークを選択することが可能である。
【0052】
一般に、マウス起源の残基によるヒト起源アミノ酸残基の置換を最小化する必要があるが、その理由は、マウス残基の導入は抗体がヒト患者でHAMA反応を誘発するリスクを増加させるからである。他方、相補性決定部(CDR)は抗原と相互作用するより大きな可能性を有する残基を含み、この理由のために、それらはヒト化抗体で維持されなければならない。それらは、カバットに従う配列によって、またはChothiaによる構造によって確定される。それらを確定するために第2の系を使用する利点は、一般にCDRがより短く、それ故にヒト化抗体は異種断片のより小さい部分によって構成されることである。いずれにせよ、一般にカバットの定義に従うと、ヒト化のために必要なサイクル数を大幅に減らすことが可能なことが証明された。一旦CDRが確定されると、それらが属している正規クラス(ChothiaおよびLeskによって規定されている)を同定し、その後ヒト化抗体で正規残基を維持することが必要である。
【0053】
可変ドメインの軽鎖および重鎖の間の相互作用を媒介する残基を分析し(表1)、ヒト化抗体内のいかなる特異な残基をも維持することが必須である(Singerら、1993;Daughertyら、1991;De Martinoら、1991)。
【0054】
さらに、維持するさらなるアミノ酸は、CDRの高次構造に及ぼすそれらの可能な影響および/または抗原との相互作用に基づいて選択される。アミノ酸が動物起源のフレームワークおよびヒト起源の同等の受容体フレームワークの間で異なる場合は、そのアミノ酸が抗原と非共有結合的に直接接触しているか、またはCDR部に隣接しているか、またはいずれにせよCDR部と相互作用している(CDR部から4〜6Åの範囲内にある)と予想することが合理的ならば、受容体フレームワークのアミノ酸は同等のマウス残基によって置換するべきである。
【0055】
【表1】
【0056】
詳細には、さらなる分析は、CDRの構造を安定させるゾーンであるいわゆるバーニヤ(Vernier)ゾーンを定義する他の残基を含む。この領域の特性を維持することが重要である。
【0057】
変異のための他の残基候補は、受容体フレームワークのその位置ではヒト免疫グロブリンにとって普通ではないアミノ酸である。これらの残基はより典型的なヒト免疫グロブリンの同等の位置に由来するアミノ酸で置換することができ、あるいは供与体フレームワークの同等の位置に由来する残基を受容体フレームワークに導入することができるが、それはそれらのアミノ酸がそれらの特定の位置でヒト免疫グロブリンにとって典型的な場合である。
【0058】
さらに、ヒト免疫グロブリンのコンセンサス配列に基づき、供与体および受容体フレームワークに存在する普通でない残基の代わりに、ヒトで保存されている残基を挿入する変異をヒト化形態に導入する。
【0059】
次に結晶構造のそれぞれの対を修飾するが、先ず、動物起源のCDRをヒトフレームワークに移植する。その後、上述のすべての変異および復帰変異を導入する。修飾された構造体を次に複合免疫グロブリンに組み立てる。得られたモデルは、力場を使用して機械的エネルギー(ねじれ角および結合角度および距離に関して)を最小にすることにより洗練する。
【0060】
上で議論した特定のアミノ酸置換と異なる他のすべての領域については、ヒト化免疫グロブリンのフレームワーク部は、通常、それらが由来するヒト抗体のフレームワーク部と実質的に同一である。いずれにしても、移植によって得られるこれらの改変タンパク質では、末端または中間の多くのアミノ酸置換、欠失または挿入、および他の変化のために、フレームワーク部は未変性の配列と比較して一次構造レベルで変動する可能性がある。当然、抗体の特異性または親和性に対するフレームワーク部の大部分の残基の貢献は、非常に小さいか皆無である。したがって、フレームワークの残基内の多くの個々の保存的置換は、生じるヒト化免疫グロブリンにおいて特異性または親和性をかなり変更することなく、耐容される。しかしながら、そのような置換は一般に望ましくない。広く使用されている様々な技術、例えば部位特異的突然変異(Gillman & Smith、1979;Robertsら、1987)を使用して、ヌクレオチド配列を修飾することが可能である。
【0061】
抗体の一次構造の一部だけを含んでいる代わりのポリペプチド断片を生成することができ、それら断片は免疫グロブリンの1つまたは複数の特有の活性(例えば結合活性)を保持する。これらのポリペプチド断片はタンパク分解的消化によって生成することができ、これは未変性の抗体、または部位特異的突然変異(詳細には、Fab断片を生成するためにはCH1領域の後、または(Fab')2断片を生成するためにはヒンジ部の後)による重鎖および軽鎖の可変部のコードDNA配列を含む担体内の所望の位置への終止コドンの挿入から始まる。scFvの形の抗体は、重鎖および軽鎖の可変部をリンカーにより連結することによって得ることができる(Hustonら、1988;Birdら、1988)。FvまたはFab断片は、大腸菌(BuchnerおよびRudolph、1991;Skerraら、1991)、または真核細胞、好ましくは哺乳類由来のものにおいても発現させることができる。他の多くの遺伝子と同様に、免疫グロブリンスーパーファミリーの遺伝子がそれぞれ1つまたは複数の特異的生物活性を特徴とする他とは異なる機能的領域を含むことを考えると、遺伝子は、新しい特性を備えている融合タンパク質(例えば免疫毒素)を生成するために、他の遺伝子(例えば酵素)に由来する機能的領域に融合させることができる。
【0062】
細菌内でのヒト化免疫グロブリン配列の発現は、CDR部に突然変異を起こさせる高親和性を特徴としファージディスプレイのためにファージライブラリーを生成する、ヒト化免疫グロブリン配列を選択するために使用することができる。これらのライブラリーを使用して、抗原に対して高い親和性および/または結合特異性を有するヒト化免疫グロブリンのCDRレベルでの、変異体を探索するスクリーニングを実行することが可能である。免疫グロブリンの可変部の配列を含むファージディスプレイライブラリーを得る方法は、豊富に報告されている(Cesareni、1992;Swimmerら、1992;Gramら、1992;Clacksonら、1991;Scott & Smith、1990;Garrardら、1991)。それらのCDRがこのように改造されたヒト化免疫グロブリンの変異体から生じる配列は、その後その高い発現を保障するために適当な宿主で発現される。
【0063】
上述の通り、DNA配列は発現制御配列に動作可能に結合された後(すなわち、それらの機能が保障されるような方法で置かれた後)に、宿主細胞で発現される。一般的にこれらの担体はエピソームとして、または染色体DNAの不可欠の部分として、宿主生物内で複製される。通常、発現担体は、関心のDNA配列で形質転換された細胞の特定を可能にする選択可能なマーカーを含む。
【0064】
本発明のヒト化免疫グロブリンのscFvの組換え態またはFab形態での生成のためには、原核生物の系が好ましい。大腸菌は、本発明のDNA配列をクローニングするために特に有用である原核生物宿主の1つである。さらに、lacもしくはtrpオペロンまたはβラクタマーゼまたはλファージなど、多数の特徴が明らかなプロモーターが利用できる。一般的に、これらのプロモーターは転写および翻訳の正しい開始および終了のために、リボソームのために発現を制御して結合部位を運ぶ。ポリエチレングリコール(PEG)とのコンジュゲーションによって、原核生物の系で産生される本発明のヒト化免疫グロブリンの半減期を増加させることが可能である。
【0065】
酵母など他の単細胞生物も発現のために使用することができる。最適な宿主は、発現調節、複製終止および開始の配列を備えている適当な担体を使用するサッカロミセス(Saccharomyces)である。
【0066】
昆虫細胞培養も本発明のヒト化免疫グロブリンを生成するために使用することができ、一般には、安定的にトランスフェクションされたS2ショウジョウバエ(Drosophila)の細胞、またはバキュロウイルス(Baculovirus)に基づく発現系を有するSpodoptera frugiperdaの細胞を使用する(Putlitzら、1990)。
【0067】
植物および植物細胞培養は、本発明のヒト化免疫グロブリンの発現のために使用することができる。(Larrick & Fry、1991;Benvenutoら、1991;Durinら、1990;Hiattら、1989)。
【0068】
しかし、これらすべてのケースで、ヒト免疫系の活性化におけるエフェクター機能を保障するのに必要な、正しい種類のグリコシル化を得ることは不可能である。このために、免疫反応の誘導における免疫グロブリンの中で最も有効なイソタイプであると判明したIgG1の完全な形態で本発明のポリペプチドを発現するために、哺乳類細胞の組織培養を使用することが可能である(Winnacker、1987)。イソタイプが抗体の溶解能を決定することを考えると、通常、IgG1イソタイプは治療目的のために使用され(細胞および補体系によって媒介される免疫反応を誘導するので)、一方、IgG4は診断用途のために使用されることを強調すべきである(Riechmannら、1988)。詳細には、手つかずの免疫グロブリンの分泌のために開発された多数の宿主細胞系を考えると、哺乳類細胞、中でもCHO細胞系、COSのいくつかの系、HeLa細胞、骨髄腫細胞系(NS0、SP/2、YB/0、P3X63.Ag8.653)、形質転換B細胞またはハイブリドーマが好ましい。これらの細胞のための発現担体としては、複製開始点、プロモーター、エンハンサーなどの発現制御配列(Queenら、1986)、およびリボソーム結合、RNAスプライシングおよびポリアデニル化のために必要な配列、および転写終止のための配列が含まれ得る。最適な発現制御配列は、免疫グロブリン遺伝子およびSV40、アデノウイルス、ウシ乳頭腫ウイルス、サイトメガロウイルスなどのウイルスに由来するプロモーターである。通常、発現ベクターは選択可能なマーカー、例えばネオマイシン耐性を含む。
【0069】
ヒト化抗体の発現のためには、無血清培地で哺乳類細胞系を培養することが好ましい。例えば、HUDREG-55細胞系は、Sigma(St.Louis、Mo)製の無血清および無タンパク質ハイブリドーマ培地Cat.No.S-2897で容易に増殖させることができる。
【0070】
本発明のヒト化免疫グロブリンをコードしている遺伝子は、関心のヒト化免疫グロブリンを、一般的にミルクまたは血清などの回収可能な体液内に発現する非ヒトトランスジェニック動物を作製するために使用することができる。そのような導入遺伝子は、通常エンハンサー配列と共にプロモーター、例えば齧歯動物の免疫グロブリンまたはカゼイン遺伝子のプロモーター/エンハンサーと動作可能に結合したヒト化免疫グロブリンをコードしているポリヌクレオチド配列を含む(Buhlerら、1990;Meadeら、1990)。導入遺伝子は、相同組換え構築物によって細胞または胚へ移すことができる。使用される非ヒト動物としては、マウス、ラット、羊、ウシおよびヤギがある(国際公開91/08216)。
【0071】
それらが一旦手つかずの抗体、それらの二量体、個々の軽鎖および重鎖、または他の形態で発現されると、本発明の免疫グロブリンは硫酸アンモニウムによる沈殿、アフィニティーカラム、カラムクロマトグラフィーなどの標準方法で精製することができる(Scopes、1982)。薬剤用途のためには、最低90から95%、好ましくは98から99%、またはさらに高い均一性を有する、実質的に純粋な免疫グロブリンが必要である。部分的に、または所望の均一性が得られるまで一旦精製されると、タンパク質は治療用途(また体外で)、診断用途(腫瘍またはアルツハイマー病の診断のための画像化)または生化学アッセイ、免疫蛍光染色などの開発および実施のために使用することができる(一般には、LefkovitsおよびPernis、1979および1981を参照)。
【0072】
本発明の医薬用途は、TrkA発現細胞(膵臓および前立腺腫瘍の場合)を除去するための、免疫毒素の形のヒト化免疫グロブリンMNAC13の使用に関係する。免疫毒素は2つの成分を特徴とし、in vitroおよびin vivoで特定の細胞を殺すために特に好適である。細胞に対して通常致死性の細胞傷害剤の1成分は、吸収されるかまたは細胞表面と相互に作用する。第2の成分は、毒剤を特異的な標的細胞型、例えばTrkA受容体のエピトープを発現する細胞に向かわせるための手段を提供する。2つの成分は、利用できる多種多様な化学手法のいずれか1つによって、互いに化学的に結合させる。例えば、細胞傷害剤がタンパク質であり第2の成分が無傷の免疫グロブリンである場合、結合は交差結合およびヘテロ二機能性剤(SPDP、カルボジイミド、グルタルアルデヒド)に媒介させることができる。あるいは、2つの成分を遺伝的に結合させることができる(Chaudharyら、1989)。様々な免疫毒素の生成がThorpeら(1982)によって報告されている。
【0073】
多数の細胞傷害剤が、免疫毒素としての用途に適当である。細胞傷害剤としては、ヨウ素131または他のヨウ素同位体、イットリウム90、レニウム188およびビスマス212または他の同位体などのα粒子を放出する放射性核種、多数の化学療法薬、例えばビンデシン(vindesin)、メトトレキセート、アドリアマイシンおよびシスプラチン;ならびに細胞傷害性タンパク質、例えばリボソームを阻害するタンパク質(例えばヤマゴボウ抗ウイルスタンパク質、シュードモナス外毒素Aおよびジフテリア毒素、リシンAおよび植物起源のクラビン(clavin))、または細胞表面レベルで活性を示す剤(例えばホスホリパーゼCなどのホスホリパーゼ酵素)がある。BaldwinおよびByers、1985;U.S.Ser.No.07/290,968;OlsnesおよびPhil、1982。免疫毒素の細胞傷害性領域はそれ自体免疫原性であるので、慢性または長期療法の場合には融合タンパク質の臨床有用性を制限する可能性があることを強調するべきである。毒素の免疫原性の問題を避ける代替手段は、DNAと相互作用することができるタンパク質を抗体の結合ドメインと融合させて発現させ、毒素発現カセットを含む発現担体をその融合タンパク質に結合させることである。DNAと結合するヒトタンパク質であるプロタミンの多数の正電荷はDNAの陰電荷と安定的に相互作用することができ、毒素自体よりずっと安定でしかも免疫原性の低い中性抗体の融合パートナーを生成する。受容体媒介エンドサイトーシスによる抗体-プラスミド複合体の内部移行の後、毒素の発現は細胞の死を引き起こす。さらに、除去すべき標的細胞に対する選択性は、誘導可能なまたは細胞特異性のプロモーターを毒素発現カセットに挿入することによって、さらに強化することができる。この手法は毒性副作用を最小にしつつ、腫瘍細胞の選択的な除去を最大化することを目的とする(Chenら、1995)。
【0074】
免疫毒素を正しい目標に向かわせる成分には、手つかずの免疫グロブリンまたは結合断片の形の、あるいは、FabまたはFv断片としての本発明のMNAC13ヒト化免疫グロブリンが含まれる。一般的に、免疫毒素内の抗体はヒトイソタイプIgMまたはIgGであるが、他の定常部も同様に使用することができる。
【0075】
本発明の抗体および医薬組成物は、病理に関係する組織のレベルで抗体を送達するための、いかなる有効な方法に従う投与に特に有用である。これには、それには限定されないが、腹腔内、筋肉内、静脈内、皮下、気管内、経口、経腸、腸管外、鼻腔内または経皮投与が含まれる。本発明の抗体は、液状製剤の注射による局所投与(腹腔内または脳内-一般的に脳室-または心膜内または嚢内)のために、あるいは、固形剤(ピル、錠剤、カプセルの剤形)または液状製剤(乳濁液および溶液の剤形)の摂取により投与することができる。非経口投与のための組成物は、通常、好ましくは水性の適合する溶液に溶かした免疫グロブリンの溶液を含む。これらの製剤内の抗体濃度は0.005%未満から15〜20%の範囲であり、主に液体容積、その粘度、その他に従って、また選択される特定の投与様式に従って選択される。
【0076】
あるいは、抗体は固体剤形での投与のために調製することができる。抗体は異なる不活性物質または添加物質と組み合わせることができ、その例としてはリガンド、例えば微結晶性セルロース、ゼラチンまたはアラビアゴム;受容体、例えばラクトースまたは澱粉;アルギン酸、Primogelまたはコーンスターチなどの剤;滑剤、例えばステアリン酸マグネシウム、コロイド状二酸化ケイ素;甘味料、例えばサッカロースまたはサッカリン;または着香剤、例えばミントおよびサリチル酸メチルがある。他の薬剤投与系としては、ヒドロゲル、ヒドロキシメチルセルロース、リポソーム、マイクロカプセル、マイクロエマルジョン、マイクロスフェアー、その他がある。腫瘍などの罹患組織への直接的局所注射は、本発明の抗体の投与で優先される方法である。
【0077】
本発明の抗体は、凍結または凍結乾燥させ、適当な緩衝液で使用直前に再構成することができる。凍結乾燥および再構成が抗体の活性低下の変動量を決定する(従来の免疫グロブリンでは、クラスIgM抗体は、クラスIgG抗体よりも活性の損失は大きい傾向がある)ことを考えれば、投与レベルはこの事実を補償するために調整されなければならない。
【0078】
それらの高い遮断能力のおかげで、本発明の抗体を含んでいる組成物は、病理的状態と関連する炎症要素または慢性疼痛、特に慢性内臓痛(月経困難症、消化不良、胃腸逆流、膵臓炎、内臓痛または過敏性腸症候群などの生理障害に関連する)を予防するか減らすかするための予防法および/または治療法のために、投与することができる。
【0079】
予防用途では、本発明の抗体を含んでいる組成物は、特定の病理状態がまだ起きていない患者に対して抵抗性を強化するために投与される。
【0080】
本発明の抗体は、前立腺または膵臓の腫瘍の体積を減らすための方法、およびさらなる腫瘍増殖を予防するかまたは腫瘍増殖率を低下させる方法も提供する。この効果は、本発明の両方のヒト化抗体によって媒介されるが、その理由はそれらが、自己分泌またはパラクリンの様式で腫瘍の増殖および進行を持続させるのに必要なNGFおよびTrkAの間の相互作用の中和に、極めて効果が高いからである。さらに、MNAC13のヒト化形態は膜受容体と相互に作用するが故に腫瘍細胞の直接的除去のためにも使用することができるが、その理由は、それらが宿主の免疫反応(IgG1の形で投与された場合)を活性化するか、または細胞傷害剤を運んでそれを癌性塊のレベルで局在化する(免疫毒素の形で投与された場合)ことができるからである。
【0081】
腫瘍部位へのそれらの投与は、好ましくは組織または腫瘍部位の近くへの直接的局所注射を通して起こる。全身投与のためには日用量は0.05mg/kgから500mg/kgの範囲であるが、投与するのがより簡単であるのでその範囲の低い方の領域の投薬量が好ましい。例えば抗体の特定の血漿レベル(約5〜30mg/ml、好ましくは10〜15mg/mlの範囲)を保証し、臨床成績が達成されるまで与えられた期間このレベルを維持するために、投薬量を調整することができる。ヒト化抗体はずっとゆっくり除去されるべきで、血漿中有効レベルを維持するのにより低い投薬量を必要とするべきである。さらに、高親和性を考慮すると、より低い親和性を有する抗体よりも投与の頻度は低く量も少ない。各抗体の治療的有効投薬量は、腫瘍の体積の減少または腫瘍増殖率に基づいて、または、理想的には癌性病理状態の完全消失に基づいて治療期間中に決定することができる。膵臓または前立腺の腫瘍の段階を測定または評価するための効果的な方法は、血液中の前立腺特異抗原(PSA)の測定、膵臓腫瘍の場合は生存期間の測定、両腫瘍の転移の拡散の遅延化または阻害の測定に基づく。
【0082】
腫瘍部位レベルでの直接注射については、投薬量は腫瘍の型、段階および体積を含む種々の因子、ならびに他の多くの変数によって決まる。腫瘍体積に従い、典型的治療量は0.01mg/mmから10mg/mmの範囲の注射であり、必要な頻度で投与することができる。特定の治療法の効果を評価する別の方法は、例えばELISAアッセイによりその活性を測定することによってTrkA受容体の阻害を評価することである(Angelesら、1996)。
【0083】
重要なことは、TrkAは治療上の標的としてだけでなく、in vivo画像化、例えばTrkA陽性腫瘍(腫瘍型および起源に従い陽性または陰性マーカーとして)の画像化、および前脳基底核細胞の画像化(アルツハイマー病の襲撃の早発マーカーとして)のための診断上の標的としても構成されていることである。本発明のMNAC13ヒト化抗体は、多種多様なin vitro用途も有す(ELISA、IRMA、RIA、免疫組織化学)。
【0084】
診断目的のために、抗体に印を付けても付けなくてもよい。印がない抗体は、ヒト化抗体またはヒト抗体(例えばヒト免疫グロブリンの定常部に対する特異抗体)に対して反応性である他の印付きの抗体(二次抗体)と組み合わせて使用することができる。あるいは、抗体は直接印を付けることができる。多種多様な標識、例えば放射性核種、蛍光団、色素、酵素、酵素基質、酵素因子、酵素阻害剤、リガンド(特にアプテニック(aptenic)な)、その他を使用することができる。多種類の免疫アッセイがこの分野で利用できる。
【0085】
詳細には、画像診断用途については、Goding(1986)およびPaikら(1982)によって記載されているように、検出可能であるかまたは同位体(ヨウ素、インジウム、テクネチウムの放射性同位体)もしくは常磁体(常磁性の原子またはイオン、例えば遷移元素、アクチニドおよび希土類;特に、マンガン(II)、銅(II)およびコバルト(II))で標識された剤を抗体とコンジュゲートする。画像化手法は静脈内、腹腔内または皮下の注射(リンパ液のドレナージ領域ではリンパ節転移を同定するため)を伴い、それらは、免疫シンチグラフィーの場合、放射性核種放出の検出器(例えばシンチレーションβカウンター)を使う。常磁性の標識を代わりに使用するならば、NMR(核磁気共鳴)スペクトロメータを使用する。
【0086】
以下の図面を参照しながら、本発明はその非限定的な実施形態で説明されるものとする。
【0087】
結果
MNAC13およびαD11モノクローナル抗体のFab断片のX線構造
標準手順により両方のモノクローナル抗体を得て精製した。MNAC13 IgG1およびαD11 IgG2a免疫グロブリンを、ハイブリドーマ細胞の培養により上清で発現させ、29%硫酸アンモニウムによる沈殿で濃縮し、その後PBSで透析した。両免疫グロブリンは、Protein G Sepharose(Pharmacia)のカラムを使用してアフィニティークロマトグラフィーにより精製した。
【0088】
Spectra-Por 12/14K膜(Spectrum)を使用した4℃における10mMリン酸緩衝液pH7、20mM EDTA中での透析の後、各試料はCentricon 50kDa限外濾過ユニット(Amicon)によって濃縮し、13mMのシステインと一緒にインキュベートし、37℃で5h、固定化パパイン(Pierce)(1:15の酵素:基質比率)で処理した。それぞれのFab断片を精製する手法は多様であるが、イオン交換クロマトグラフィーに常に基づいている。
【0089】
MNAC13の場合、100mMトリス塩酸pH8.0に対する透析の後、同じ緩衝液で平衡化したDEAE-Sephacelカラム(Pharmacia)を通してFc断片を除去することが可能であった。FabMNAC13は排除液中で収集し、Fc断片および未消化IgGl部分は250mMのNaClで溶出した。Fab断片は、100mMトリス塩酸pH8.0、150mM NaClで平衡化したSuperdex G75(Pharmacia)カラム上でのゲル濾過によって、未消化のIgGlから分離した。これら分画の均一性および純度は、12%ポリアクリルアミドゲル上での電気泳動分離とその後のクーマシー染色によって制御された(図1A)。精製されたタンパク質の濃度は、Lowryアッセイ(Bio-Rad)で測定した。11のハイブリドーマ上清から、最高3mgのMNAC13 Fabを得ることができた(99%を超える純度)。
【0090】
αD11抗体のFab断片の精製に関して、パパインで処理した試料は、10mMリン酸緩衝液pH7.8に対して透析した。Fc断片は、この同じ緩衝液で平衡化したDEAE-Sephacelカラム(Pharmacia)を通して除去した。αD11のFab断片は排除液中で収集し、Fc断片および未消化IgG2a部分は250mMのリン酸緩衝液pH6.8で溶出した。Fab断片は、10mMリン酸緩衝液pH7.8、150mM NaClで平衡化したSuperdex G75カラム(Pharmacia)上でのゲル濾過によって、未消化のIgG2aから分離した。これら分画の均一性および純度は、12%ポリアクリルアミドゲル上での電気泳動分離とその後のクーマシー染色によって制御された(図2A)。精製されたタンパク質の濃度は、Lowryアッセイ(Bio-Rad)で測定した。11のハイブリドーマ上清から、最高6mgのαD11 Fabを得ることができた(99%を超える純度)。
【0091】
10mMのトリスpH8.0、50mM NaCl中で精製したMNAC13抗体のFab断片、および10mMのリン酸ナトリウムpH7.8および50mMのNaCl中で精製したαD11抗体のFab断片を、Centricon 30kDa限外濾過ユニット(Amicon)により5〜10mg/mlまで濃縮した。Crystal Screen IおよびII(Hampton Research - Laguna Niguel、CA、USA-)ならびにスクリーニングキット(Jena BioSciences)を使い、要因組合せ(factorial combination)手法(Jancarik & Kim、1991)による16℃での懸滴(hanging-drop)法に従って、結晶化実験を実施した。
【0092】
濃縮したタンパク試料2μlを、沈殿剤を含んでいる等量の溶液に滴下し、24穴Linbroプレートの貯水槽(0.7ml)内の溶液との拡散によって平衡化した。
【0093】
MNAC13抗体Fab断片に関して、等量のタンパク質と2M硫酸アンモニウム、5%v/vイソプロパノールを含んでいる沈殿剤(Crystal Screen II、Reactant #5)とで得られた最も有望な初期の成果を、図1Bで示されているものと同様に約1週で成長する結晶が得られるまで最適化した。
【0094】
αD11抗体Fab断片に関して、等量のタンパク質と、20%のPEG4000、0.6MのNaCl、100mMのMES pH6.5を含んでいる沈殿剤(キット番号4、溶液C2)とで得られた最も有望な初期の成果は、図2Bで示されているものと同様に約1週間で成長する結晶が得られるまで、沈殿剤の組成をPEG4000、0.6MのNaCl、100mMのBTP pH5.5およびタンパク質および沈殿剤溶液比(1.5:1)に修飾する長い最適化過程を必要とした。
【0095】
両方の場合で、初期の低分解データセットがELETTRAシンクロトロン(Trieste、Italy)のXRD1回折線上で収集され、その後第2のより高い分解能のより完全なデータセットがESRFシンクロトロン(Grenoble、France)のID14-EH1回折線上で収集された。結晶は、MNAC13抗体のFab断片の場合は低温保護体として2.2Mの硫酸アンモニウム、6%v/vイソプロパノールおよび20%v/vグリセリンを含んでいる溶液を使用して、Oxford Crysosystems(Oxford、UK)製の冷却システムにより液体窒素下で凍結した。各タンパク質の代表的な高分解能回折スペクトルを図1Bおよび2Bに示す。
【0096】
4つのX線回折データセットすべてを処理、指数化、積算し、その後それぞれDENZOおよびSCALEPACKプログラム(Otwinowski & Minor、1997)を使用して評価し、一方、データ整理のためにはCCP4(Collaborative Computational Project、Number 4、1994)パッケージを使用した。MNAC13抗体のFab断片の結晶の高分解能および低分解能データの収集および処理の統計値を、以下の表で示す。
X線源 ELETTRA ESRF
波長(Å) 1.000 0.934
検出器 mar345 marCCD
空間群 P212121 P212121
単一細胞のパラメータ
a(Å) 52.78 52.73
b(Å) 67.53 67.55
c(Å) 111.51 111.43
Mosaicity(°) 0.40 0.47
分解能間隔(Å) 12.0〜2.50 17.0〜1.80
(2.59〜2.50) (1.83〜1.80)
測定数 98688 414115
I≧0で観察される反射の数 56918 227914
I≧0での特有な反射の数 14203(1371) 38392(1893)
完全性(%) 99.5(99.3) 99.5(99.6)
リダンダンシー 4.0(4.0) 5.9(4.9)
測定データの<I/σ(I)> 9.4(4.7) 8.2(1.1)
Rsym(%) 5.7(15.2) 6.3(39.8)
【0097】
同様に、αD11抗体のFab断片の結晶の高分解能および低分解能データの収集および処理の統計値を、以下の表で示す。
X線源 ELETTRA ESRF
波長(Å) 1.000 0.934
検出器 marCCD marCCD
空間群 P1 C2
単一細胞のパラメータ
a(Å) 42.685 114.801
b(Å) 50.626 69.354
c(Å) 102.697 64.104
α(°) 81.977 90
β(°) 89.116 117.02
γ(°) 85.957 90
Mosaicity(°) 0.44 0.40
分解能間隔(Å) 47.6〜2.57 17.0〜1.70
(2.8〜2.7) (1.75〜1.70)
測定数 124456 492594
I≧0で観察される反射の数 74241 399184
I≧0での特有な反射の数 23413(2162) 47951(3198)
完全性(%) 98.2(92.4) 97.2(78.4)
リダンダンシー 5.7(5.2) 6.7(7.5)
測定データの<I/σ(I)> 29.6(6.7) 9.5(2.1)
Rsym(%) 11.0(33.5) 5.8(27.8)
括弧内の値は最高分解能のシェルを指す。
【0098】
Fab断片の利用できる構造の数が多いことを考慮すると、両タンパク質の構造を決定する最も便利な方法は、Molecular Substitutionであった。相同構造に関するプロテインデータバンク(Bermanら、2000)での研究において、選択基準は同等の分解能および最高レベルの配列同一性の間の組合せを優先した。それぞれこれらに基づいて下記が選択された。
MNAC13のためには:1BM3:Opg2 Fab免疫グロブリンのFab断片およびそれによって認識されるペプチド間の複合体の構造(Kodandapaniら、1999)、分解能2.0Åで分解され、重鎖および軽鎖についてそれぞれ70および88%の配列同一性を有する。
αD11のためには:1CIC:イディオタイプ-抗イディオタイプFab断片FabD1.3-FabE225の複合体の構造(Bentley、1990)、分解能2.5Åで分解され、重鎖および軽鎖についてそれぞれ82および82.65%の配列同一性を有する。
【0099】
両方の構造の決定は、AMoReプログラム(Navaza、1994)を使用してMolecular Substitution法によって得られたが、可変部および定常部の間のバイナリー擬似対称軸によって形成される角度の極端な変動性を考慮して、それぞれのモデルでは定常ドメインおよび可変ドメインを別々に使用した。剛体による洗練後の、MNAC13のFab断片の構造決定において得られたソリューションを以下の表で示す。
【0100】
【表2】
【0101】
同様に、空間群C2のための、剛体による洗練後の、αD11のFab断片の構造決定において得られたソリューションを以下の表で示す。
【0102】
【表3】
【0103】
2つの構造の以降の洗練は循環手法によって得られ、下記2つの交互相を含む。コンピュータグラフィクスのためのインタラクティブなソフトウェア「O」(KleywegtおよびJones、1994)を使用したモデルの手動構築。CNSセット、Crystallography and NMR System(Brungerら、1998)の自動プロトコルを使った位置的洗練およびB等方性温度因子の洗練。剛体による洗練のいくつかの相の後の手法は、異なる洗練サイクルを企図したものである。一旦すべての変異の挿入および欠失が終了してモデルが完成すると、水分子、あらゆるイオンおよびリガンドのローカライゼーションを行った。終わりに、モデルを立体化学的に理想値のできるだけ近くに保って、位置的重量waおよび熱因子B r-重量を最適化した。MNAC13抗体のFab断片のために得られたモデルの品質を記載する統計量および最終パラメータは、以下の表に要約する。
タンパク原子数 3244
溶媒原子数 351
硫酸イオン数 4
トリス分子数 1
イソプロパノール分子数 1
分解能間隔(Å) 39〜1.778
最終R因子 19.35%
最終Rfree因子(データの10%で計算) 23.22%
Rms偏差
結合距離(Å) 0.008
結合角度(°) 1.456
二面角(°) 27.29
非固有角度(°) 0.928
平均等方性温度因子(A2)
完全タンパク質 23.55
軽鎖 24.14
重鎖 22.99
水分子 31.95
イオン(硫酸塩) 55.94
トリス 46.06
イソプロパノール 32.60
【0104】
同様に、αD11抗体のFab断片のために得られたモデルの品質を記載する統計量および最終パラメータを以下の表に要約する。
タンパク原子数 3229
溶媒原子数 403
塩素イオン数 1
分解能間隔(Å) 30〜1.70
最終R因子 19.54%
最終Rfree因子(データの10%で計算) 24.22%
Rms偏差
結合距離(Å) 0.0096
Angoli di legame(°) 1.6571
結合角度(°) 1.6571
Angoli die二面角(°) 27.40
非固有角度(°) 1.048
平均等方性温度因子(A2)
完全タンパク質 25.58
軽鎖 24.14
重鎖 22.99
水分子 38.80
イオン(塩化物) 20.58
【0105】
さらに、両モデルは、それぞれの表およびそれぞれのラマチャンドランチャート(図1Dおよび2D)で示すように、PROCHECKセット(Laskowskiら、1993)による最終幾何分析によって検討した。
【0106】
ヒト起源のフレームワークの選択におけるMNAC13およびαD11モノクローナル抗体のFab断片のX線構造の使用
ヒト抗体フレームワークの選択において、マウスおよびヒト起源の抗体間での一次配列レベルでの同一性の程度をポリペプチド骨格の構造類似性の程度と組み合わせる、上述の手法に従った。
【0107】
詳細には、ヒト起源のまたはヒト化された抗体の一連の可能な受容体フレームワークを、BLASTデータベースの検索により一次構造の最高レベルの相同性および同一性に基づいて選択した。この選択は、抗体の可変部全体を考慮し、また検索をフレームワーク部に狭めることにより、両遮断抗体について行った。
【0108】
選択された抗体の各群の中で、高分解能による、さもなければ筆者らが得た構造のそれに相当する分解能(すなわち2.5Å以下)による構造データが入手可能なものだけを考慮し、PDB(タンパクデータバンク)を検索した。次に、「スーパーインポーズ」ソフトウェア(Diederichs、1995)を使用してそれぞれのアミノ酸骨格をスーパーインポーズした。
【0109】
図3Gおよび4Gは、一次構造のヒト化する抗体との最適な整列および利用できる構造データの高分解能に基づいて選択された、Fv部(それぞれMNAC13およびαD11の)およびヒト化またはヒト起源の抗体のα炭素原子骨格の三次構造の間の整列の結果を示す。
【0110】
ヒト起源または操作した個々の構造のMNAC13およびαD11との重なり程度を評価するために、それぞれのアミノ酸骨格を構成しているα炭素原子の間でRMSを計算したが、2Åを上回っているRMSの原子対は無視した。
【0111】
ヒト化のための最適フレームワークの選択が3変数問題として構成され、それはしたがって相同性レベルおよび同一性の程度を構造的整列と関連させる場合に空間で表すことができる。次に、2種類の整列における問題の領域をそれぞれのフレームワークの領域に減らして、この種の分析を実行した。
【0112】
図3および4で示すように、分析した3つの変数(それぞれ、RMS値、RMSが計算された原子の割合および一次構造間の類似指数、すなわち全体の同一性-A-、全体の相同性-C-、フレームワークレベルの同一性-B-、フレームワークレベルの相同性-D-の割合)の空間で検討された抗体の分布は、検討した両ケースで相互に首尾一貫して整合的である。
【0113】
さらに、これらの分布を、それがヒト起源であるならば各抗体が占めるであろう3変数の空間における最適な位置と比較すると、一次および三次構造レベルでこの理想位置に最も近づくヒト起源抗体を明らかに同定することは可能である。両抗体の場合についてこの結果を合理的に説明するために、4つの分析のそれぞれで、検討したヒト化またはヒト起源の抗体の各位置について、仮定上の最適位置からの逸脱を計算した(MNAC13については図3Eおよび3F、αD11については図4Eおよび4F)。この場合もまた、結果は首尾一貫しており、以前の指摘を確認するものである。
【0114】
この選択法に基づいて、NGF/TrkA相互作用を中和する2つの抗体のヒト化のための以降のCDR移植過程で、2つの異なるヒト化抗体が受容体フレームワークとして選択された。詳細には、図3Hおよび4Hは、Fv部のレベルにおける2つの遮断抗体と選択されたヒト化抗体それぞれとの構造的整列を示し、すなわち、αD11のためにはPDB 1JPSコードが、MNAC13のためには1AD0コードが使用された。図3Iおよび4Iでは、マウス抗体の同じ領域とCDR移植後の同じ抗体のモデルとが比較される。
【0115】
一旦CDRが確定されると、それらが属している正規クラス(ChothiaおよびLeskによって規定されている)を同定し、その後ヒト化抗体内の正規残基を維持した。図5および図6では、各抗体についてそれらは下線付き文字で強調されている。
【0116】
導入する復帰変異の以降の分析に関して、可変ドメインの軽鎖および重鎖の間の相互作用を媒介する残基を維持するために、2つのドメイン間の境界面を維持するために以下の復帰変異を挿入した。
MNAC13のためにL46およびH35、H37
αD11のためにL34、L46、L92およびH35。
【0117】
さらに、バーニヤゾーンの特性を維持するために、以下の復帰変異を起こした。
MNAC13のためにL98
αD11のためにH71(これはいずれにせよ、ヒトコンセンサス配列で表されるアミノ酸残基の置換に関係がある)。
【0118】
その後、ヒト免疫グロブリンの主なコンセンサス配列との比較の後、以下の復帰変異を起こした。
MNAC13のためにL1、L2、L13、L50、L73、L104およびH24、H48、H49、H73、H76、H82B、H87、H90、H93
αD11のためにL56
【0119】
さらに、同じくヒト免疫グロブリンのコンセンサス配列に基づき、供与体および受容体フレームワークに存在する普通でない残基の代わりに、ヒトで保存されている残基を挿入する以下の変異をMNAC13のヒト化形態に導入した。
L42(L→Q)、L83(I→V)およびH83(Q→R)、H89(I→V)。
【0120】
同じ理由から、以下の変異をαD11のヒト化形態に導入した。H67(V→F)。
【0121】
結晶構造のそれぞれの対を修飾したが、先ず、動物起源のCDRをヒトフレームワークにグラフトした。その後、上述のすべての変異および復帰変異を導入した。修飾された構造体を次に複合免疫グロブリンに組み立てた。得られたモデルは、力場を使用して機械的エネルギー(ねじれ角および結合角度および距離に関して)を最小にすることにより洗練した。
【0122】
MNAC13およびαD11モノクローナル抗体のヒト化
MNAC13のCDR移植を達成するために供与体フレームワークのヒト化抗体を選択した後に、マウスMNAC13のCDRを上で述べた変異により修飾したヒト化抗体のフレームワークと組み合わせた可変部をそれぞれ設計する。αD11についても類似した手法に従った。本質的に、約80塩基のオリゴヌクレオチドを用いたオーバーラップ-アセンブリー(overlap assembly PCR)法に基づく手法によって2つのヒト化可変部を得ることができるが、これらの塩基ではセンスおよびアンチセンスのフィラメントが交互し、ハイブリッド動作の結果として部分的に二重のフィラメント分子の形成を可能にするような方法で20塩基の長さで連続的に重複する(図7Bおよび8B)。Ventポリメラーゼによって不連続部を埋めた後に、軽鎖の可変ドメインのためにはpVLexpressプラスミド(図9A)、重鎖の可変ドメインのためにはpVHexpressプラスミド(図9B)における、制限酵素による消化の後の方向性クローニングのために適当な制限部位(それぞれ軽鎖の可変ドメインのクローニングのためにはApaLI/BgIII、重鎖の可変ドメインのクローニングのためにはBssHII/BstEII)と共にプライマーとして二重フィラメント自体の5'の配列を含む2つの短いオリゴヌクレオチドを使用して、PCRのために二重フィラメントを増幅する。これらの担体は、それぞれヒト起源の定常ドメインCκおよびCH1、CH2およびCH3それぞれのクローン配列との融合による発現を可能にする。したがって、これらのベクターを使って、上で述べたようなヒト細胞系でIgG1分子(図9Cおよび9D1)の形で両抗体を発現することが可能である。
【0123】
Fab断片の形で両方のヒト化抗体を得るために、重鎖がクローニングされる担体だけに作用することで十分である。詳細には、SacII-XbaI方向性クローニングのための制限部位を両端に備えている特定のプライマーを使用して、定常部全体をPCRのために増幅した唯一のCH1ドメインで置換することが可能である(図9D2で示す)。
【0124】
最後に、免疫毒素の形でMNAC13ヒト化抗体を得るために、非方向性XbaIクローニングのために端に制限部位を備えた特定のプライマーを使用して、PCRのために増幅した塩基性プロタミンタンパク質を、定常ドメインCH1のC末端で発現することが可能である(図9D3で示す)。
【0125】
MNACおよびαD11ヒト化抗体の発現および結合アッセイ
推奨されたプロトコル(Roche)に従ってFuGENEにより、250,000のコス細胞を、各ヒト化抗体のVKでVHのために1μgのコードプラスミドDNAでコトランスフェクションした(合計2μg)。構築物は、IgG1の形のヒト化抗体を得るために使用した。
【0126】
上述の構築物のコトランスフェクションと並行して、対応するキメラ形態を各抗体についてコトランスフェクションした。すなわち、
マウスMNAC13のVHはCMV pVHexpressに、マウスMNAC13のVkはCMV pVkexpressにクローニングする。
αD11に関しては、ラットVHはヒト起源のCγとの融合によりpcDNA1に、ラットVkはヒト起源のCγとの融合によりpcDNA1にクローニングする。
【0127】
トランスフェクションから72時間後に、宿主細胞によって発現された免疫グロブリンを含んでいる上清を収集し、Centriprep 50(Amicon)を使用して濃縮した。
【0128】
2つのヒト化抗体のそれぞれのリガンドを認識する能力を、ELISAアッセイによって検証し、それぞれのキメラ形態と比較した。結果を図10および11に示す。
【0129】
プラスチック上への固定化のために、96穴Maxisorbプレートを、0.1M炭酸ナトリウム緩衝液pH 9.6で10μg/mlの濃度にした2つの抗体(精製された組換え免疫付着因子TrkA Camelおよび顎下腺から精製されたマウスNGF)のそれぞれのリガンドを含んでいる溶液と、4℃で一晩インキュベートした。
【0130】
3%のミルクを含んでいるPBS(MPBS)により周囲温度で1時間ブロックした後に、濃縮した上清を連続希釈(1:2、1.20;1:200)と、また並行してトランスフェクションしていないコス細胞(陰性対照)の上清とインキュベートした。
【0131】
一次抗体(ヒト起源の定常Cγ領域を認識する)および二次抗体(ペルオキシダーゼとコンジュゲートした抗ウサギ抗体)とのインキュベーションの後、結合活性をTMB基質(TECNA)とのインキュベーションによる450nm(OD450)における光学密度として検出することが可能である。500ng/mlの濃度のそれぞれのモノクローナル抗体を、陽性対照として含めた。
【0132】
図10Bは、発現した免疫グロブリンのアフィニティークロマトグラフィーによる精製の後に行われた、類似したELISAアッセイの結果を示す。
【0133】
詳細には、トランスフェクション細胞の上清を収集し、遠心処理によって細胞破片を除去した後に、それらを100μlのProtein Gセファロースとインキュベートし、PBSで十分に水洗の後、各タンパク質を1mM HClで溶出し、pHは1MのトリスpH 8.8で溶出直後に中和した。それぞれの濃度は、Lowryアッセイで推定した。
(参考文献)
【特許請求の範囲】
【請求項1】
NGFに対する結合活性を維持するヒト化抗NGF抗体またはその断片であって、
(a) アミノ酸配列GFSLTNNNVNWを含む第一のCDR、アミノ酸配列GVWAGGATDYNSALKSを含む第二のCDR、アミノ酸配列DGGYSSSTLYAMDAを含む第三のCDRを有するVH領域;及び
(b) アミノ酸配列RASEDIYNALAを含む第一のCDR、アミノ酸配列HTを含む第二のCDR、アミノ酸配列QHYFHYPRTを含む第三のCDRを有するVL領域
を含むヒト化抗NGF抗体。
【請求項1】
NGFに対する結合活性を維持するヒト化抗NGF抗体またはその断片であって、
(a) アミノ酸配列GFSLTNNNVNWを含む第一のCDR、アミノ酸配列GVWAGGATDYNSALKSを含む第二のCDR、アミノ酸配列DGGYSSSTLYAMDAを含む第三のCDRを有するVH領域;及び
(b) アミノ酸配列RASEDIYNALAを含む第一のCDR、アミノ酸配列HTを含む第二のCDR、アミノ酸配列QHYFHYPRTを含む第三のCDRを有するVL領域
を含むヒト化抗NGF抗体。
【図1A】

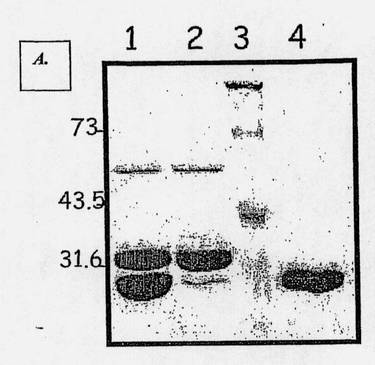
【図1B】

【図1C】
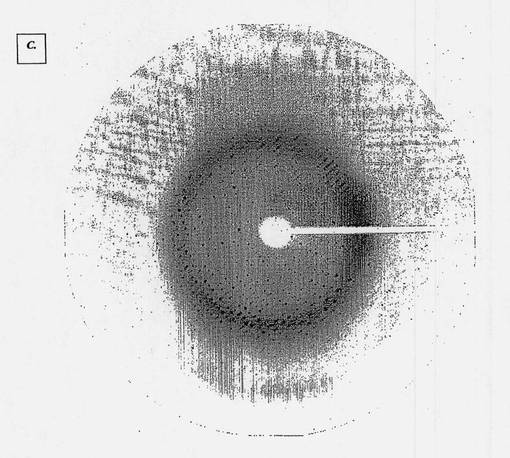
【図1D】
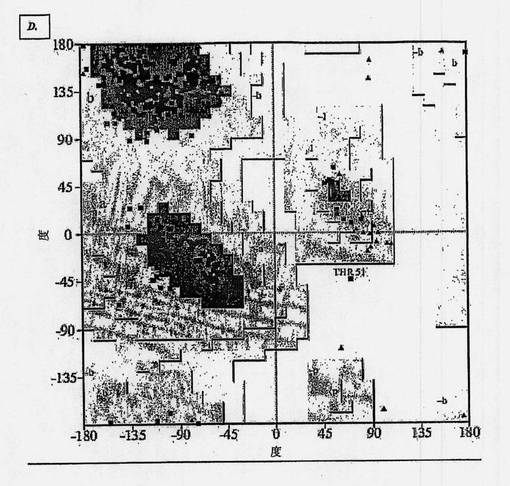
【図2A】
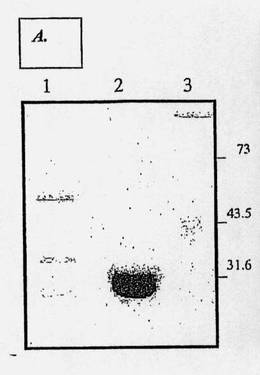
【図2B】
【図2C】

【図2D】
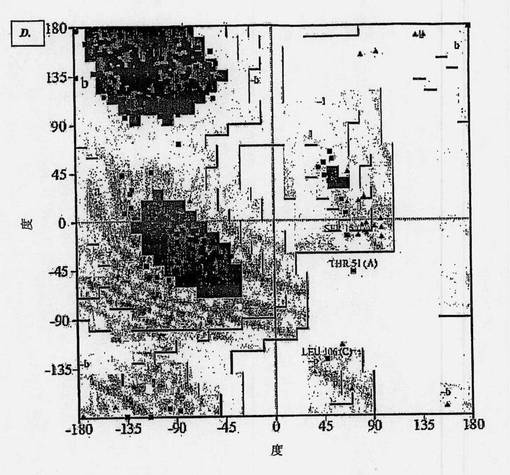
【図3−1】
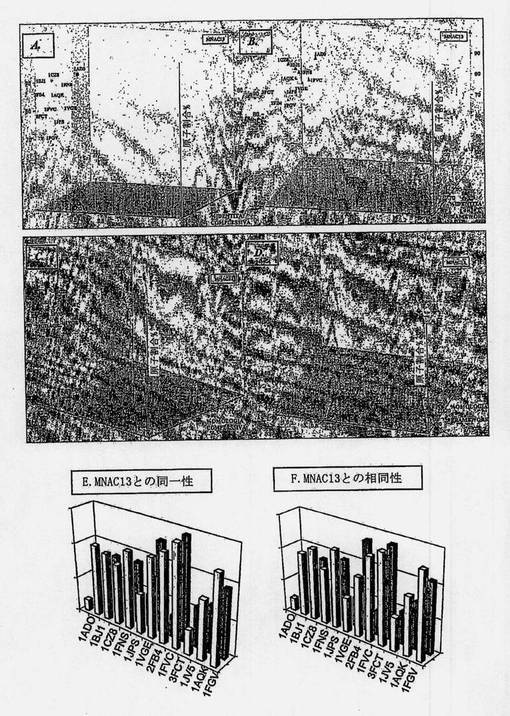
【図3−2】
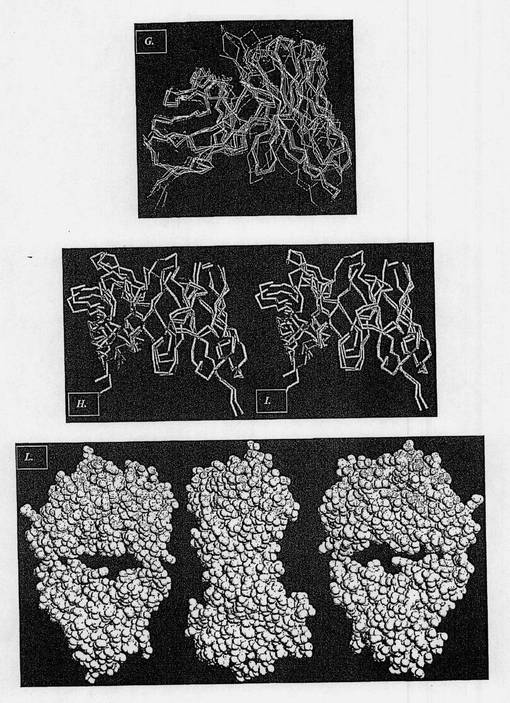
【図4−1】

【図4−2】
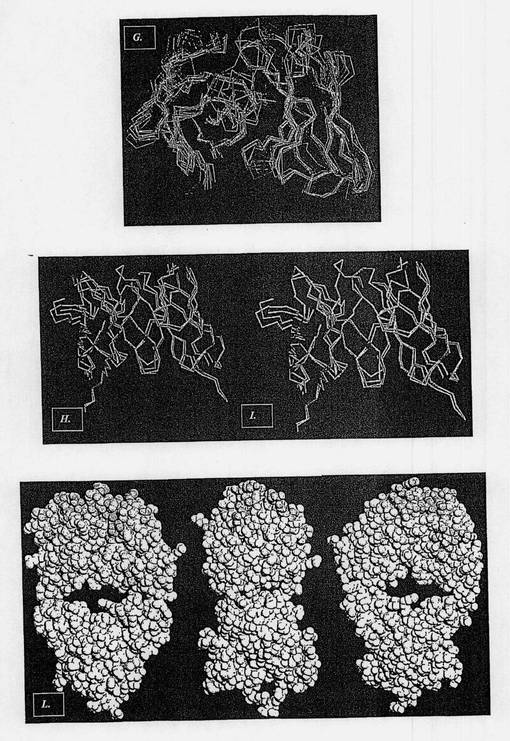
【図5】

【図6】
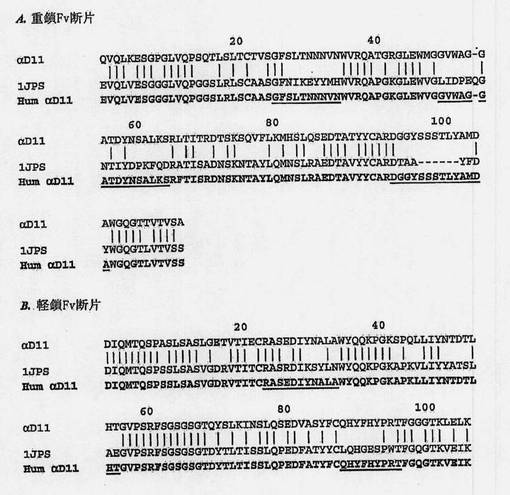
【図7A】

【図7B】

【図7C】

【図7D】

【図7E】

【図7F】

【図8A】

【図8B】

【図8C】

【図8D】

【図8E】

【図8F】

【図9A】
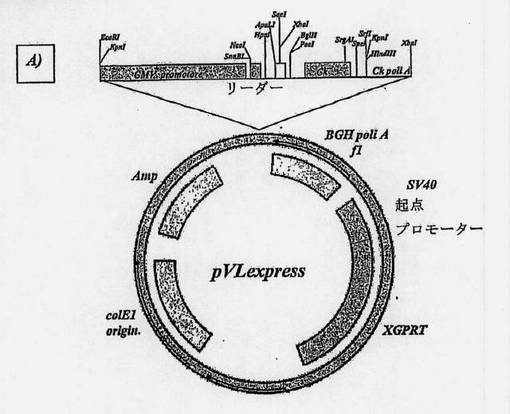
【図9B】
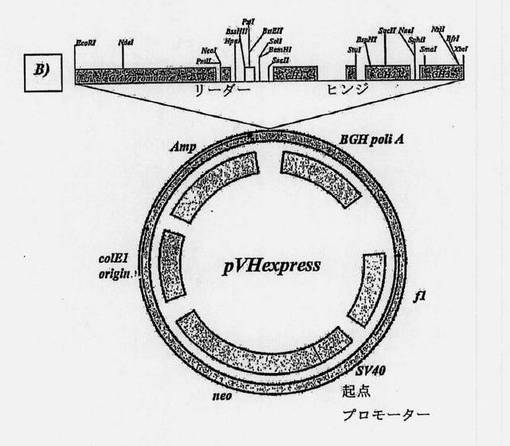
【図9C】
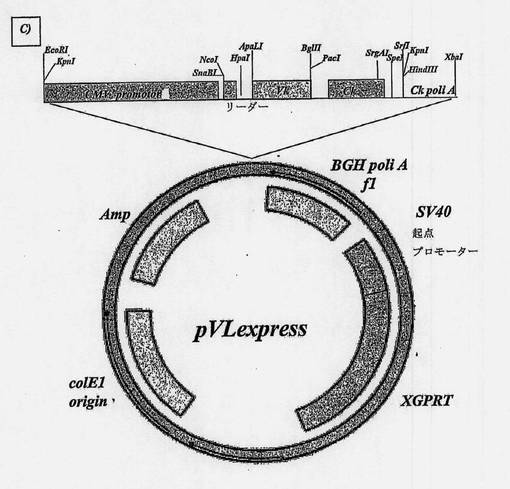
【図9D】
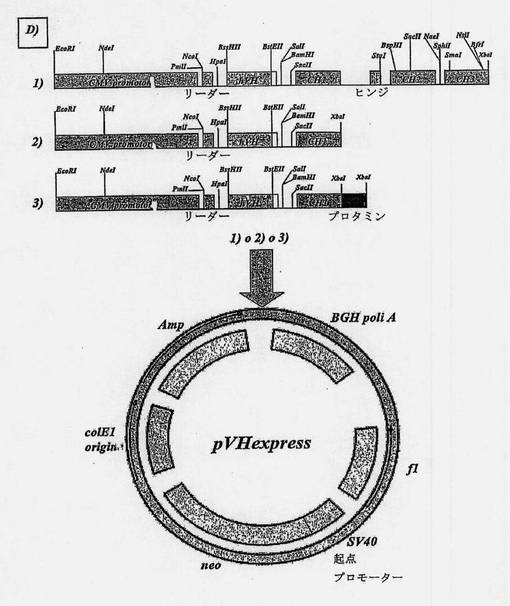
【図10】
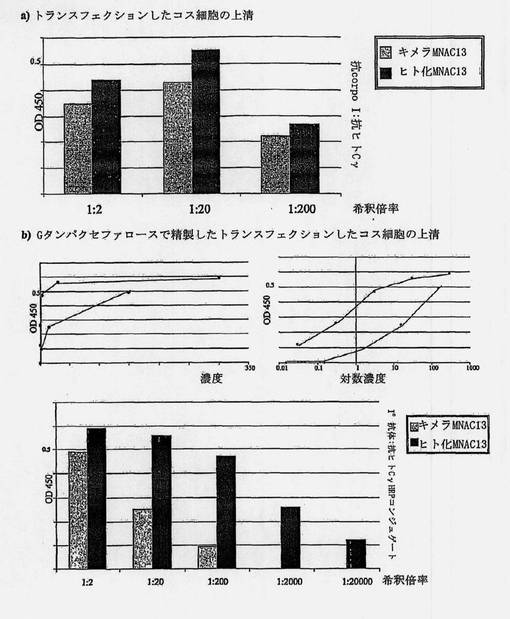
【図11】

【図1B】
【図1C】
【図1D】
【図2A】
【図2B】
【図2C】
【図2D】
【図3−1】
【図3−2】
【図4−1】
【図4−2】
【図5】
【図6】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図7F】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図8F】
【図9A】
【図9B】
【図9C】
【図9D】
【図10】
【図11】
【公開番号】特開2012−126726(P2012−126726A)
【公開日】平成24年7月5日(2012.7.5)
【国際特許分類】
【出願番号】特願2012−15219(P2012−15219)
【出願日】平成24年1月27日(2012.1.27)
【分割の表示】特願2009−213454(P2009−213454)の分割
【原出願日】平成16年12月23日(2004.12.23)
【出願人】(512022893)
【Fターム(参考)】
【公開日】平成24年7月5日(2012.7.5)
【国際特許分類】
【出願日】平成24年1月27日(2012.1.27)
【分割の表示】特願2009−213454(P2009−213454)の分割
【原出願日】平成16年12月23日(2004.12.23)
【出願人】(512022893)
【Fターム(参考)】
[ Back to top ]