
モノミコリルグリセロール(MMG)のアジュバントとしての使用
本発明において、本発明者らは、MMG並びにそのα及びケトミコール酸誘導体を、極めて低用量でヒトDCを刺激及び活性化し得るウシ型結核菌(コペンハーゲン株)に由来する高度に生物活性な脂質として同定する。ヒトDCの免疫刺激剤としてのそれらの直接的役割に加えて、本発明者らは、ヒトへの投与に適した新世代のアジュバントの開発におけるそれらの使用を示す。本発明者らは、癌治療において、並びに感染性疾患及びアルツハイマー病のような疾患の両方に対するワクチンアジュバントとして、大なる可能性を有する多数の高度に活性な合成MMG同族体をさらに同定する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、免疫調節剤、アジュバント及びワクチン又はこのアジュバントを含む送達系を調製するための、モノミコリルグリセロール(MMG)又はその合成同族体、類似体及び改変体の使用を開示する。
【背景技術】
【0002】
最初のワクチンは、生きた弱毒化された病原体からなるものであった。弱毒化形態は、天然に生ずる密接に関係する生物であるか又は培養で連続継代により得られるかのいずれかであった。例えば、ヒトにおける結核(TB)は、多年にわたり、80年を超える以前に開発されたウシ型結核菌(M.bovis BCG)の弱毒株BCGワクチンによるワクチン接種により対処されてきた。しかしながら、30億を超える用量のBCGが投与されたにも拘わらず(他のいかなるワクチンも超える)、それは、どの集団においてもヒトTBに対する満足な抵抗性を必ずしも提供しない。
【0003】
現在、最新の手法は、高度に精製された物質、例えば精製された組み替えタンパク質又はペプチドを使用することである。これらのワクチンは、十分解明されており、副作用は最小化されている。残念ながら、多くの高度に精製された物質は、免疫原性が強くなくて、防御を与えるのに十分な免疫応答を誘発しない。これを行うために、抗原は、アジュバントと呼ばれる免疫応答強化剤から幾らかの助力を必要とする。病原体に応じて、防御は、液性の又は細胞媒介の応答のいずれが支配的であるかを要求することができる。免疫血清により移入され得る免疫応答は液性免疫と称され、感染性因子に関連する抗原性物質と結合して、それにより抗原性物質に対する免疫応答を始動させる抗体により媒介される抵抗性を意味する。細胞媒介免疫(CMI)は、免疫応答を担う免疫系細胞に依存する。CMI、又はTヘルパー(Th)1の免疫応答は、一般的には、リーシュマニア及び結核を含む細胞内病原体との闘いに関連しているが、他のタイプの感染例えば酵母感染カンジダとの闘いにも役割を有する。液性、又はTh2免疫応答は、細胞外病原体、例えば蠕虫感染に対する防御のために必要とされる。
【0004】
多くの場合、例えばインフルエンザ、肝炎C(HCV)、ヒト免疫不全ウイルス(HIV)、クラミジア及びマラリアの場合に、感染のステージに依存して、混合したTh1/Th2応答が必要になり得る(Mosmann and Sad 1996)。これら場合、それらの生活環の一部は細胞内であるが、それらは細胞外の相を通っても移動、例えば細胞間伝播するので、Th1及びTh2の両方が必要になる。
【0005】
特定の種類の免疫応答(液性又は細胞媒介)の開発は、アジュバントの選択により決定され得る。例えば、結核菌(M.tuberculosis)のような細胞内病原体に対する防御免疫は、細胞媒介免疫応答を必要とし、TBを目的とするサブユニットワクチン用の適当なアジュバントは、Th1応答を増強すべきである(Lindblad et al,1997)。
【0006】
多数のアジュバントが存在するが、それらの大部分は、それらのヒトにおける使用を妨げる数多くの問題に悩まされている。少数のアジュバントだけ、例えばアルミニウム系アジュバント(AlOH塩)及びMF−59が、ヒトへの使用を許容されているが、しかし、それらは両方ともTh2に偏った応答を誘発し、そのことがそれらを、Th1応答を必要とするTBワクチンおよび他のワクチンとして不適当にしている(Lindblad et al,1997)。
【0007】
過去20〜30年の間、多数の新しいアジュバント系が確認されて、それらの幾つかは現在開発中である。それにも拘わらず、新アジュバント系に対する必要性は、臨床用途に利用可能な選択が不足している中で、未だなお認識されており(Moingeon et al.,2001)、明らかである。
【0008】
アジュバント(adjuvant:ラテン語のadjuvareに由来し、助けるという意味)は、ワクチン中で投与されたときに、特異的な免疫応答を指示、促進、延長、及び/又は増強するのに役立つ任意の物質と定義することができる。アジュバントは、送達系又は免疫調節剤/免疫刺激剤のいずれか二つの主なカテゴリーに分けられている。送達系は、例えば、エマルション、ポリスチレン粒子、ニオソーム(niosome)、ISCOM(免疫刺激複合体)、ビロソーム(virosome)、ミクロスフェア、又は脂質二重層で構成される小胞である界面活性剤様リポソームにすることがでる。リポソームは、抗原(小胞内か又は表面に付着しているかのいずれか)の担体として作用し、接種部位でデポを形成することができて、抗原の緩やかな連続的放出を可能にする。注射及び食作用後の短時間中に、リポソームの提示により、特定量の抗原が単一抗原提示細胞に行き渡ることが確実になる(Gluck,1995)。免疫調節剤は、独特の細胞又は受容体、例えばAPCの表面上のToll様受容体を標的とする。送達系と免疫調節剤とは、一緒に、例えばGlaxoの一連のアジュバントにおけるように使用することができる。したがって、送達系は、ワクチン抗原の送達に加えて、免疫調節剤を送達するために使用することもできる。
【0009】
ワクチン中の成分であることに加えて、免疫調節剤は、抗原(単数又は複数)なしで投与することもできる。この手法により、免疫系を局所的に活性化することが可能であり、例えば、抗腫瘍及び抗ウイルス活性にとって重要な、抗原提示細胞の成熟、サイトカイン産生として見られる。したがって、免疫調節剤の投与は、例えば癌及び皮膚疾患の根絶における支援になり得る。局所投与できる免疫調節剤の例は、タキサン類、例えばタキソール、Toll様受容体7/8リガンドのレシキモッド(Resiquimod)、イミキモッド(Imiquimod)、ガルジキモッド(Gardiquimod)である。
【0010】
ジメチルジオクタデシルアンモニウム−ブロミド、−クロリド、−ホスフェート、−アセテート又は他の有機若しくは無機塩(DDA)は、親油性第四級アンモニウム化合物であり、水溶液中において40℃を超える温度でカチオン性リポソームを形成する。DDAは、APC中へのワクチン抗原の取り込みを増大させる非常に効率的な送達系である。DDAと免疫調節剤との組合せは記載がある。DDAを含むArquad 2HTのヒトにおける投与は有望であり、明確な副作用を誘発しなかった(Stanfield,1973)。DDAとTDBとの又はDDAとMPLとの組合せは、送達ビヒクル(DDA)と免疫調節剤(TDB又はMPL)との間の非常に明瞭な相乗効果を示し、どちらかの成分単独で得られる応答に比較して、高度に上昇したレベルのCMI応答があった。したがって、DDAは、例えば、TB及び他の細胞内病原体に対するワクチンのためのアジュバント系の開発において、ワクチン抗原及び免疫調節剤のための有望な送達ビヒクルである。
【0011】
マイコバクテリア由来の種々の化合物が、免疫調節性であると報告されている。ウシ型結核菌BCGから抽出された脂質がアジュバントとして使用されたとき、卵白アルブミンに対する皮膚試験応答が、モルモットで得られた(Hiu 1975)。ウシ型結核菌BCGの全極性脂質から温度を上げて形成されたリポソームは、卵白アルブミンに対する液性応答を生じさせることができて、これらの極性脂質から調製されたワクチンは、マウスにおいて腫瘍細胞でチャレンジしたとき、防御を与えた(WO03/011336)。モルモットに対する実験的TBワクチンにおける結核菌H37Rv由来の全脂質の抗原としての効果が、Dascher et al(2003)により研究された。この研究においては、コレステロールと1,2−ジステアロイル−sn−グリセロ−3−ホスドコリン(DSPC)とに基づくリポソームが、結核菌H37R.Vの全脂質抽出物と混合された。溶媒を除去した後、脂質は、ジメチルジオクタデシルアンモニウム(DDA)をアジュバントとして、PBS緩衝液中で再構成された。このワクチンで免疫されたモルモットは、細菌の有意の減少を示さず、DDAと混合されたリポソームのこの製剤は、強力な抗原を欠くこと、又はマイコバクテリアの脂質とコレステロール/DSPCとの製剤は、DDAのアジュバント効果を妨げることを示唆した。また、種々の脂質の混合物が投与されたとき、有効な脂質が、全脂質の構成要素となり得る比率はあまりに限られている。
【0012】
マイコバクテリア由来の種々の精製された成分は、それらのアジュバント活性についても研究されている。精製されたタンパク質誘導体(PPD)は、それ自体では遅延型過敏性反応を誘発しなかったが、ワックスD(マイコバクテリアの細胞壁のペプチドグリカンフラグメント−アラビノガラクタン−ミコール酸複合体)がアジュバントとして添加されたとき、強い反応が観察された(Yamazaki S,1969)。免疫調節剤SSM又はZ−100、結核菌から抽出された脂質アラビノマンナンは、抗腫瘍活性を有する(Suzuki F,1986)。他のマイコバクテリア細胞由来の化合物は、トレハロース6,6’−ジミコレート(TDM)(コード因子;ミコール酸含有糖脂質)である(Saito et al,1976)。TDM(又は合成類似体)も免疫刺激効果を有し、種々のアジュバント製剤中に含まれてきた(McBride et al,1998)(Koike et al,1998)。
【0013】
Silva et al(1985)による論文においては、ウシ型結核菌BCGから精製された5つの成分が、脂質で被覆されたチャーコール粒子として静脈内に注射されて、マウスの肺内に炎症性反応を生じさせた。5つの成分には、TDM、トレハロースモノミコレート(MMT)、グルコースモノミコレート(MMGlc)、アラビノースモノミコレート(MMAr)、及びグリセロールモノミコレート(MMGlyc)が含まれた。この論文は、モノグリコリルグリセロール頭部基を記載しているが、ミコール酸の組成物の定義が不十分で、構造データも提供されていない。それに加えて、脂質を投与したときの反応だけは、肺における炎症性活性として記載されているが、アジュバント効果として知られている特異的免疫応答を増強する性能は記載されていない。
【0014】
マイコバクテリア由来の脂質の免疫刺激性及び炎症性活性は、動物(マウス)モデルにおいて免疫応答を刺激し得る脂質についての増加し続ける文献で何十年間も認められてきたが、今日まで、ヒト樹状細胞(DC)を刺激する能力を有する個々の脂質(単数又は複数)は同定されていない。例として、TDMは、炎症誘発性応答に関して最も活性なマイコバクテリア脂質であることが示されているにも拘わらず、TDMで刺激したとき樹状細胞の活性化は観察されていない(Uehori et al,2003)。このように、TDMは、数編の論文において炎症性活性を示しているが、この脂質は、免疫応答を開始するために不可欠な樹状細胞を活性化する能力を明らかに欠いている。ヒト樹状細胞を活性化する能力を有するような脂質の同定は、それがヒトにおける使用に適した新規アジュバント系の一部として使用することができることを示唆するであろう。さらに、ヒト使用に適したTh1誘発アジュバントの欠如により、Th1助長能力を有する単独のマイコバクテリア由来の脂質の同定は大いなる発見になる。
【0015】
DCは、結核菌などの病原体に感染したときに免疫応答を指示する必須の役割を演ずる専門の抗原提示細胞(APC)である。したがって、活性化されたDCによるIL−12の産生は、それが、マクロファージの活性化を促進する、Th−1細胞によるIFN−γの産生の推進にこの上なく重要なサイトカインなので、結核菌感染の制御に不可欠なステップである(Nathan et al,1983)。さらに、近年、免疫応答を調節する試みの中で、マイコバクテリアはDCも標的とすることを示す証拠が見出されて、この過程におけるマイコバクテリアの脂質にとって極めて重要な役割が確立された。
【0016】
マイコバクテリアの細胞エンベロープの乾燥重量の40%までが、脂質からなる(Minnikin 1982)。これらの脂質は、この科の生物の独特の病原性と長らく関連づけられており、宿主のマイコバクテリア感染に対する応答において実質的役割を演ずることが知られている(Brennan and Goren 1979)。これらの脂質の中で重要なのは、フチオセロールジミコセロセート(PDIM:phthiocerol dimycocerosate)ワックスであり(Minnikin et al.2002)、その存在は病原性と相関することが示されている。PDIM欠損結核菌変異体は、マウス中で減弱した増殖を示す(Sirakova et al.2003)。PDIMに密接に関係するのは、いわゆる「フェノール性糖脂質」(PGL)であり、適当な例は、ウシ型結核菌中で見出された2−メチルラムノシルフェノール−フチオセロールジミコセロセート(「マイコサイド(mycoside)B」)である。このモノグリコシルPGLと結核菌のある種の分離菌の高毒性との間の関連が、最近、明らかにされた(Reed et al.2004)。
【0017】
特に興味ある他の種類の脂質は、トレハロース−6,6−ジミコレート(TDM)、いわゆる「コード因子」である。TDMは、TNF−α、IL−6及びIL−12などの炎症誘発性サイトカイン、及びTh−1促進サイトカインであるIFN−γ(Lima et al.2001)の放出を刺激することにより肉芽腫性病変の維持を助長し、ファゴソーム−リソソーム融合を阻害することにより、マクロファージ内の結核菌の生存を長引かせる役割を有する(Indrigo et al.2002)。TDMのミコレート成分の微細構造は、初期感染期におけるマクロファージの炎症誘発性活性化において重要である(Rao et al.2005)。
【0018】
マイコバクテリアの生存の増加におけるそれらの役割にも拘わらず、マイコバクテリア脂質の免疫調節力は、新世代のTh1誘発アジュバントを創出することに利用することもできる。強力な免疫刺激活性を有する個々の脂質の同定において、強力なアジュバント活性をなお維持しながら、油分と混合された加熱殺菌結核菌の全細胞(フロイント完全アジュバント(CFA))の使用に伴う毒性ゆえの問題を回避することは可能であるかもしれない。実際、ウシ型結核菌バチルス・カルメット・ゲラン(bacillus Calmette−Guerin)(BCG)の極性脂質から形成されたリポソームは、マウスの骨髄由来の樹状細胞(BM−DC)を活性化することが最近示された。この活性の大部分は、糖リン脂質であるホスファチジルイノシトールジマンノシドという脂質に起因し得ることが見出された(Sprott et al.2004)。
【0019】
本発明者らの研究室における最近の研究は、強力な液性及び細胞媒介成分で複雑な遅延免疫応答を促進することができる、ウシ型結核菌BCGからのマイコバクテリア脂質抽出物とジメチルジオクタデシルアンモニウムブロミド(DDA)との新規なアジュバントの組合せの特性を明らかにした(Rosenkrands et al.2005)。全BCG脂質のアジュバント活性の大部分は、無極性脂質に起因することが見出された。
【0020】
これらの研究はマイコバクテリア脂質がアジュバントとして作用する可能性をさらに確かなものにするが、最善の解決は、単独で強力な活性を有する、単独の最も免疫刺激性の脂質を同定することであろう。このことは、さらに単純で廉価なアジュバントを提示し、世界中で投与されるワクチンで使用するためのアジュバントに要求される大量に産生され得る脂質の合成類似体を作製して、より無害な系を可能にする可能性も浮揚させるであろう。
【発明の概要】
【0021】
本発明は、ヒトDCを活性化し得る免疫刺激性脂質、モノミコリルグリセロール(MMG)及びそれらの合成同族体、類似体及び改変体を開示する。MMGは全BCG脂質の無極性分画に由来し、これらの脂質に関連する、アジュバントの及び防御の効果を誘発する原因である。小さめの炭素主鎖を有する合成MMGは、in vitroにおいて天然MMGのヒトDCに対する刺激性を増強することができ、マウスモデルにおいて、in vivoで、TBに対する長く継続する防御免疫応答につながる強いTh1応答も誘発する。
【図面の簡単な説明】
【0022】
【図1】ウシ型結核菌BCGの無極性及び極性脂質の単離及び免疫刺激活性のアセスメント。ウシ型結核菌BCGコペンハーゲン株から抽出された極性及び無極性脂質抽出物を2次元TLCにより分析した。極性脂質分画中において、1〜4はホスファチジルイノシトールマンノシドであり、PIはホスファチジルイノシトールであり、PEはホスファチジルエタノールアミンであり、及びDPGはジホスファチジルグリセロール(カルジオリピド(cardiolipid))である。PGは、L−α−ホスファチジル−DL−グリセロールであるが、5及び6は、未知のリン脂質である。無極性分画において、TAGはトリアシルグリセロールであり、PDIMはフチオセロールジミコセロセートA、B及びCであり、MMGはモノミコリルグリセロールであり、PGLはフェノール性糖脂質であり、FFAは遊離脂肪酸である(パネルA)。iDCは、培地単独、LPS(0.1μg/ml)、MPL(100μg/ml)、コード因子(CF)(100μg/ml)、無極性脂質(0.1−100μg/ml)又は極性脂質(0.1−100μg/ml)の存在下に、24時間インキュベートした。処理後のDC上の表面マーカーの幾何平均蛍光強度(MFI)のレベルを示す。3名の異なる提供者を使用した3つの実験のうち1つの代表的実験から得られたデータ(パネルB)。100μg/mlの無極性又は極性脂質による処理後に得られた培養上清を、サイトカインIL−6、TNF−α及びIL−12の存在についてELISAにより分析した。3名の異なる提供者を使用して3重で実施された3つのうち1つの代表的実験から得られたデータ(±s.e.m)を示す(パネルC)。PPD陰性の提供者からの、100μg/mlの無極性又は極性脂質で24時間処理されたDCとインキュベーション後の、MLRアッセイにおける、T細胞の増殖(パネルD)及びIFN−γ放出(パネルE)。3名の異なる提供者を使用して3重で実施された3つのうち1つの代表的実験から得られたデータ(±s.e.m)を示す。
【図2】MMG、TAG、PDIM及びPGLの構造。個々の脂質PDIM、TAG、PGL及びMMGの2次元TLC分析(パネルA)並びにMMG、PGL、PDIM及びTAGの代表的構造(パネルB)。
【図3】MMGによるヒト樹状細胞の活性化。iDCを、培地単独で(点線)又はMMG、PDIM、PGL若しくはTAG(10μg/ml)の存在下に24時間インキュベートした。脂質の調製物は、エンドドキシン汚染のないことが認められた(<0.001ngLPS/μg脂質)。6名中1名の代表的提供者について、処理後のDC上の表面マーカーのレベルのMFIを示す(パネルA)。MMG、PDIMA、PGL又はTAG(10μg/ml)で処理後に得られた培養上清を、サイトカインIL−6、TNF−α及びIL−12の存在について、ELISAにより分析した(パネルB)。異なる提供者を使用して3重で実施した3つ又は4つの実験から得られたデータの平均(±s.e.m.)を示す。データはチューキー検定を使用して分析した。
【図4】MMGのα及びケトミコレートは免疫刺激性である。iDCは、培地単独及びMMGのα−又はケトミコレート(10μg/ml)の存在下に24時間インキュベートした。処理後のDC上の表面マーカーのMFIを示す。
【図5】ウシ型結核菌BCGコペンハーゲン株から単離されたMMGにより誘発されたIFN−γ放出。C57BL/6マウスを、BCGコペンハーゲン株から単離された脂質に基づくアジュバントとの組合せでDDAリポソームに組み込まれたAg85B−ESAT−6で免疫した。ワクチン接種7日後に排出するリンパ節から単離されたPBMCによるIFN−γ放出を測定した。
【図6】有毒TB感染に対するMMGアジュバントを用いる防御。C57BL/6マウスを、DDAと10μg又は50μgのMMGとに基づくアジュバントとを組み合せて、Ag85B−ESAT−6で3回免疫した。最終ワクチン接種の6週間後に、マウスは結核菌を用いるエアロゾルのチャレンジを受けた。6週間後に細菌の数を肺及び脾臓で測定した。標準的BCGワクチン接種を受けるマウスは陽性対照として、及び抗原なしでDDA/MMG(10μg)により免疫されるマウスは陰性対照として含まれた。実験的ワクチンの防御の有効性は、免疫されないマウスと比較して、肺における細菌負荷の減少のLog10として表す。結果は、各群中の6匹のマウスについての平均値±SEMである。免疫されていない対照と有意に異なる値には、*印を付けてある。P<0.05。
【図7】IFN−γ放出は、MMGとDDAとを組み合わせることにより増大する。2通りの実験において、C57BL/6マウスは、DDA若しくはDDA/MMG中(パネルA)又はMMG若しくはDDA/MMG中(パネルB)のAg85B−ESAT−6で免疫した。最終ワクチン接種の3週間後に、血液から単離された(パネルA)又は脾臓中の(パネルB)PBMCによるIFN−γ放出を測定した。
【図8】IFN−γ放出は、MMG/DDAの組合せにTDBを添加することにより増大する。C57BL/6マウスは、MMGが組み込まれたDDAリポソーム中、又はTDBを含有し、MMGが組み込まれたDDAリポソーム中で投与されたAg85B−ESAT−6により免疫した。ワクチン接種の5ヵ月後に血液から単離されたPBMCによるIFN−γ放出を測定した。
【図9】合成MMG類似体の構造の例。
【図10】免疫応答は、天然及び合成MMG類似体で同等である。C57BL/6マウスは、DDA、DDA/MMG中(10μg)、DDA/MMG C36中(10μg)又はDDA/MMG C16中(10μg)中のAg85B−ESAT−6を用いて免疫した。最終免疫の1週間後に血液から単離されたPBMCによるIFN−γ放出を測定した。
【図11】鎖長が短いほど免疫応答は高い。C57BL/6マウスを、DDA中、又は8から36の範囲の鎖長の種々のMMG類似体を含むDDA中のAg85B−ESAT−6で免疫した(1μg/用量)。最終免疫の3週間後に血液から単離されたPBMCによるIFN−γ放出を測定した。
【発明を実施するための形態】
【0023】
本発明は、免疫調節剤、アジュバント及びワクチン、又はヒト樹状細胞を刺激する特有の能力を有するこのアジュバントを含む送達系を調製するための、モノミコリルグリセロール(MMG)又はそれらの合成同族体、類似体及び改変体の使用を開示する。
【0024】
免疫調節剤として、MMG又はそれらの合成同族体,類似体及び改変体は、抗原(単数又は複数)なしでも投与される。この手法により、免疫系を局所的に活性化することが可能であり、例えば、抗腫瘍及び抗ウイルス活性にとって重要な、抗原提示細胞の成熟、サイトカイン産生として見られる。
【0025】
アジュバント(adjuvant:ラテン語のadjuvareに由来し、助けるという意味)は、ワクチン中で投与されたときに、特定の免疫応答を指示、促進、延長、及び/又は増強するのに役立つ任意の物質と定義することができる。アジュバントの性質に依存して、それは、細胞媒介免疫応答、液性免疫応答又はその2つの混合した応答を促進することができる。ワクチンアジュバントとして使用されるとき、抗原性成分がアジュバントに添加される。アジュバントにより媒介される免疫応答の増強は非特異的であるから、同じアジュバントが異なる抗原とともに使用されて、異なる標的に対する応答を助長すること、例えば、結核菌由来の抗原とともに結核菌に対する免疫を助長すること、又は腫瘍由来の抗原とともにその特定の種類の腫瘍に対する免疫を助長することができることは、当分野においてよく理解されている。
【0026】
本発明により開示された好ましいアジュバントは、MMG又はそれらの合成同族体、類似体又は改変体を含むアジュバントであり、それらには、送達ビヒクル、例えばエマルション、ポリスチレン粒子、ニオソーム、IS−COMS、ビロソーム、ミクロスフェア、又は界面活性剤様リポソームがさらに含まれる。好ましい界面活性剤は、ジメチルジオクタデシルアンモニウムブロミド又はこのクロリド(DDA−B又はDDA−C)又はスルフェート、ホスフェート又はアセテート塩(DDA−X)、又はジメチルジオクタデセニルアンモニウムブロミド又はこのクロリド(DODA−B又はDODA−C)又はスルフェート、ホスフェート又はアセテート化合物(DODA−X)に基づくカチオン性脂質であることが最も好ましい。この発明において使用される他のタイプの好ましいカチオン性脂質には、1,2−ジオレイル−3−トリメチルアンモニウムプロパン(DOTAP)、1,2−ジミリストイル−3−トリメチルアンモニウム−プロパン,1,2−ジパルミトイル−3−トリメチルアンモニウム−プロパン、1,2−ジステアロイル−3−トリメチルアンモニウム−プロパン及びジオレイル−3−ジメチルアンモニウムプロパン(DODAP)及びN−[1−(2,3−ジオレイルオキシ)プロピル]−N,N,N−トリメチルアンモニウム(DOTMA)が含まれるが、これらに限定はされない。他の界面活性剤は、DXPC、DXPE、DXPG又はこれらの組合せの中から選択され、ここで、Xは、鎖長記載についての置き換え、例えばP=パルミトイル(16C)、S=ステアロイル(18C)、A=アラキドイル(20C)である。
【0027】
送達ビヒクルは、TLR及び非TLRリガンド様MPL(モノホスホリル脂質A)、ポリイノシンポリシチジン酸(ポリ−IC)、ムラミルジペプチド(MDP)、ザイモサン、二重螺旋RNA(dsRNA)、DC−Chol、CpGオリゴデオキシヌクレオチド、カチオン性ペプチド、TDM、TDB、タモキシフェン又はこれらの分子の任意の類似体など他の免疫調節剤のためにも使用することができる。したがって、好ましいアジュバントには、送達ビヒクル中のMMG又はそれらの同族体、類似体、又は改変体が含まれ、さらにTLR−又は非TLRリガンドが含まれる。
【0028】
MMG又はそれらの合成同族体、類似体及び改変体を含む送達系は、癌、自己免疫障害、神経障害例えばアルツハイマー病、気道炎、炎症性疾患、感染性疾患、皮膚疾患、アレルギー、喘息、又は病原体により惹起される疾患を治療するために使用することができる。MMG又はそれらの合成同族体、類似体及び改変体は、1つまたは複数ワクチン、抗原、抗体、細胞傷害性因子、アレルゲン、抗生物質、アンチセンスオリゴヌクレオチド、TLR−及び非TLRアゴニスト、TLR−及び非TLRアンタゴニスト、ペプチド、タンパク質、遺伝子療法ベクター、DNAワクチン、又は共刺激分子との組合せで投与される。
【0029】
抗原性成分又は抗原性物質は、予め形成された抗体及び/又はT細胞及びB細胞上の特異的受容体と反応する分子である。ワクチン接種との関係においては、特定のT細胞又はB細胞の発生を刺激する分子が、抗原が免疫細胞と2度目に遭遇した場合に、より速い「記憶」応答を促進する免疫細胞の記憶集団の形成をもたらすことができる。記憶集団がクローン性であることは稀なので、実際には、このことは、抗原は、以前それに曝されたことがある個体の免疫細胞に再度遭遇したときに、免疫応答における増強を刺激することができる任意の分子又は分子の集合であることを意味する。
【0030】
本発明は、非経口、経口若しくは粘膜投与又は送達系のための、アジュバントを含むワクチンをさらに開示する。好ましいワクチンには、細胞内病原体に由来する抗原性エピトープ、例えば有毒マイコバクテリウム(例えば、融合生成物Ag85b_TB10.4、Ag85b_ESAT−6_Rv2660、Ag85b_TB10.4_Rv2660及びAg85a_TB10.4_Rv2660)、熱帯熱マラリア原虫(Plasmodium falciparum)(Msp1、Msp2、Msp3、Ama1、GLURP、LSA1、LSA3又はCSP)、トラコーマクラミジア(Chlamydia trachomatis)(例えばCT184、CT521、CT443、CT520、CT521、CT375、CT583、CT603、CT610又はCT681)、HIV、インフルエンザ又はB型若しくはC型肝炎ウイルスに由来する抗原性エピトープが含まれる。アジュバント又は送達系は、癌、アレルギー又は自己免疫疾患を治療するためのワクチンにおいて使用することもできる。
【0031】
全マイコバクテリア脂質抽出物は、マイコバクテリア、例えばBCG、M.ミクロティ(M.microti)、結核菌及びM.バッカエ(M.vaccae)から、化学的又は物理的方法により得られる脂質の混合物である。本研究において、抽出に使用した方法は、有機溶媒の作用であるが(以下に記載する)、当業者に知られた他の可能性も可能である。
【0032】
無極性脂質の分画は、極性のない脂質と定義される。無極性脂質の分画は、マイコバクテリアをメタノール/食塩水と石油エーテルとの二相混合物で処理することにより得られる。石油エーテル抽出物は、無極性(非極性)脂質を含む。その後、マイコバクテリア及び残留水相にクロロホルムを添加することにより、極性脂質の分画が得られる。クロロホルム抽出物は、残留する極性脂質を含有する。無極性脂質の分画中の主成分は、フチオセロールジミコセロセート、トリアシルグリセロール、トレハロースミコリペネート(trehalose mycolipenate)及びメナキノンである。極性脂質分画の主成分は、ホスファチジルエタノールアミン、ホスファチジルグリセロール、及びホスファチジルイノシトールなどのリン脂質である。中間的極性の脂質は、スルホリピド、トレハロースミコレート、グリコシル化フェノールフチオセロール(フェノール性糖脂質、PGLを含む)及びアシル化トレハロースである(Dobson et al,1985)。
【0033】
MMGは、無極性脂質の分画から得られる脂質モノミコリルグリセロール、及びそれらの誘導体、例えばα−MMG及びケトMMG並びに天然及び合成類似体を指す。MMGは、トルエン/アセトン(95:5)でTLC処理により単離することができる。PGL及びMMGは、この方法により一緒に抽出されるが、クロロホルム:メタノール:0.880アンモニア(97:3:0.5)での1次元TLCにより分離することができる。MMGの誘導体、α−MMG及びケト−MMGは、16×100mmの試験管中で5%TBAH水溶液(2.5ml)とともに、100℃で終夜加熱することにより得ることができる(Minnikin 1988)。
【0034】
MMGの合成同族体、類似体又は改変体は、化学合成の任意の従来法により製造することができる。類似体とは、構造は類似しているが、基本的組成に関して異なる一群の化合物の一つを指し、同族体とは、同族列の化合物の任意の構成要素を指す。これらの化合物は、炭素鎖の長さを変えることができる。特にサイズを小さくすることは毒性の減少を伴い、したがって、類似体のいかなる顕性毒性も減少させることに役立ち得る。したがって、合成体は、例えば、8〜36個の炭素を有し、且つ各脂質尾部に0〜3個の二重結合を有するアルキル鎖に基づくことができる。あるいは、脂質尾部の一つを除去することにより、単純化された形態を得ることができる。合成MMGの炭素主鎖サイズは、好ましくはC8〜C36、例えば3−ヒドロキシ−2−エチル−ヘキサン酸−2,3−ジヒドロキシプロピルエステル(C8)、3−ヒドロキシ−2−ブチル−オクタン酸−2,3−ジヒドロキシプロピルエステル(C12)、3−ヒドロキシ−2−ヘキシル−デカン酸−2,3−ジヒドロキシプロピルエステル(C16)、3−ヒドロキシ−2−ヘプチル−ウンデカン酸−2,3−ジヒドロキシプロピルエステル(C18)、3−ヒドロキシ−2−テトラデシル−オクタデカン酸−2,3−ジヒドロキシプロピルエステル(C32)又は3−ヒドロキシ−2−ヘキサデシル−アイコサン酸−2,3−ジヒドロキシプロピルエステル(C36)、及び最も好ましくはC8又はC16である。改変体は、グリセロール部分を他のポリオール頭部群、例えば、ポリプロピレングリコール及びポリエチレングリセロールで置き換えることにより調製することができる。合成モノミコレートのC2及びC3の周囲の立体化学は、グリセロールにおけると同様に変更することができる。以下において、単にMMGと書かれたものは、MMGの上記の合成同族体、類似体又は改変体も意味する。
【0035】
抗原性成分又は物質は、ポリペプチド又はポリペプチドの部分であってよく、それは、動物又は人類において、及び/又は生物学的試料において、本明細書中に記載する生物学的アッセイのいずれかにより測定される免疫応答を生じさせる。ポリペプチドの免疫原性部分は、T細胞エピトープ又はB細胞エピトープであってよい。免疫応答の間に認識される関連のあるT細胞エピトープを同定するために、「力づく」の方法を使用することが可能である。即ち、T細胞エピトープは線状であるから、ポリペプチドの欠失変異体が体系的に構成されれば、例えば、これらの欠失変異体を、例えば本明細書中に記載したIFN−γアッセイにかけることにより、ポリペプチドのどの領域が免疫認識において必須であるかが明らかになるであろう。他の方法は、ポリペプチド由来の重複オリゴペプチド(好ましくは、例えば20アミノ酸の長さの残基を有する合成体)を利用する。これらのペプチドは、生物学的アッセイ(例えば本明細書中に記載したIFN−γアッセイ)で試験することができ、これらの一部はペプチド中のT細胞エピトープの存在に対する確証として、陽性の応答(したがって免疫原性である)を与えるであろう。線状のB細胞エピトープは、例えばHarboe et al,1998に記載されたように、関心のあるポリペプチドを包含する重複ペプチドに対するB細胞の認識を分析することにより、決定することができる。
【0036】
T細胞エピトープの最小長さは、少なくとも6アミノ酸であることが示されているが、そのようなエピトープがそれより長いアミノ酸のつながりで構成されていることは普通である。したがって、本発明のポリペプチドフラグメントは、少なくとも7アミノ酸残基、例えば、少なくとも8、少なくとも9、少なくとも10、少なくとも12、少なくとも14、少なくとも16、少なくとも18、少なくとも20、少なくとも22、少なくとも24、及び少なくとも30アミノ酸残基などの長さを有することが好ましい。したがって、本発明の方法の重要な実施形態において、ポリペプチドフラグメントは、最大で50アミノ酸残基、例えば、最大で、40、35、30、25、及び20アミノ酸残基などの長さを有することが好ましい。10と20との間のアミノ酸残基の長さを有するペプチドは、診断の手段として最も有効であると証明されることが予想され、したがって、本発明の方法で使用されるポリペプチドフラグメントの特に好ましい長さは、18、例えば、15、14、13、12さらに11アミノ酸などである。
【0037】
具体的に、抗原性物質は、代謝する結核菌、ウシ型結核菌及び他の環境マイコバクテリア、例えばトリ型結核菌(Mycobacterium avium)などの培養液から取り出すことができる。そのようなマイコバクテリアの濾液からの特に興味ある物質は、ESAT−6遺伝子ファミリーのタンパク質(ESAT6及びTB10.4など)、並びにTB患者及び種々の動物モデルにおける結核の初期相において、細胞媒介免疫にとって主要な標的であるAg85A、Ag85B、ORF2c、Rv1036及びRv0285などの他の初期抗原である。TB感染の後期段階の期間中、主要な標的であるRv2653、Rv2655、Rv2656、Rv2657、Rv2658、Rv2659、Rv2660など他の抗原も関連する。それらの免疫原性は、それ自体では低いが、この明細書の以下の詳細な部分で示すように、本発明のアジュバントの組合せとの組合せで、結核に対する高く且つ持続する免疫を誘発するための有力な候補であることが判明した。ESAT−6遺伝子ファミリーのタンパク質並びに本発明のアジュバントの組合せとの組合せに適用可能な他の抗原は、現在、人工的に、例えば、合成的に又は遺伝子組み替え技法により製造することができる。
【0038】
融合タンパク質は、ワクチンにおける抗原性物質として特に十分に適していることが証明されており、例えば、融合生成物Ag85b_TB10.4、Ag85b_ESAT−6_Rv2660、Ag85b_TB10.4_Rv2660及びAg85a_TB10.4_Rv2660は、TBに対して非常に有効であることが証明されている。
【0039】
ワクチンは、疾患に対する免疫を生じさせる接種のための、死滅したか弱毒化されたか若しくは他の方法で改変された微生物(細菌、ウイルス、又はリケッチャ)又はそれらの部分の懸濁液と定義される。ワクチンは、疾患を防止するための予防用ワクチン又は癌若しくは潜在性の感染性疾患など既存の疾患に対処するための治療用ワクチンのいずれかとしてだけでなく、アレルギー及び自己免疫疾患と関連して投与することができる。ワクチンは、免疫応答を強化するために、適当なアジュバント中に乳化することができる。
【0040】
ワクチンは、投与製剤と適合した様式で、且つ治療的に有効で免疫原性であるような量で投与される。投与されるべき量は、例えば、個体の免疫系の免疫応答を開始する能力、及び所望の防御の程度を含めて治療される対象に依存する。適当な用量範囲は、ワクチン接種当たり数百マイクログラムの桁の有効成分であり、好ましい範囲は約0.1μgから1000μg、例えば約1μgから300μgの範囲など、特に約1μgから50μgの範囲内である。初期投与及びブースター投与に適当な投与計画も調節可能であるが、初期投与に類型化され、その後の接種又は他の投与が続く。
【0041】
適用の様式は広範に変えることができる。ワクチン投与の任意の従来法が適用可能である。これらの方法は、生理学的に許容される固体基剤による経口若しくは粘膜適用、又は生理学的に許容される分散剤での、非経口的な、注射による等の適用を含むと考えられる。ワクチンの用量は、投与経路に依存し、また、ワクチン接種されるヒトの年齢、及び程度は低いが、ワクチン接種されるヒトの体格によって変わる。
【0042】
ワクチンは、非経口的に、注射により、例えば皮下又は筋肉内のいずれかに投与するのが便利である。他の様式の投与に適したさらなる製剤には、坐剤及び、ある場合には、経口又は粘膜用製剤が含まれる。坐剤に対して、在来の結合剤及び担体は、例えば、ポリアルキレングリコール又はトリグリセリドが含むことができ、そのような坐剤は、0.5%から10%、好ましくは1〜2%の範囲内の有効成分を含有する混合物から形成することができる。経口製剤には、例えば、医薬品グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウム等のような普通に使用される賦形剤が含まれる。これらの組成物は、溶液剤、懸濁液剤、錠剤、丸薬、カプセル剤、徐放性製剤又は散剤の形態をとり、10〜95%、好ましくは25〜70%の有効成分を含有することが有利である。
【0043】
選択のワクチンには、例えば、次のものがなり得る。
タンパク質ワクチン:ポリペプチド(又はそれらの少なくとも1つ免疫原性部分)、ペプチド混合物又は融合ポリペプチドを含むワクチン組成物;
組み替え生ワクチン:非病原性微生物又はウイルス中におけるワクチンに関連ある抗原の発現。そのような微生物の周知の例は、ウシ型結核菌BCG、サルモネラ及びシュードモナス(Pseudomonas)であり、ウイルスの例は、ワクシニアウイルス及びアデノウイルスである。
【0044】
これらのワクチン構築物の全てに対して、適当なアジュバントの添加は、結果として、ワクチンの有効性を増強した(Brandt et al.2000)、(van Rooij et al.2002)、(Bennekov et al.2006)。
【0045】
リポソーム(即ち、脂質小胞)は、脂質二重層により封入された水性区画である。脂質成分は、コレステロール及び他の荷電脂質でしばしば補完された、通常リン脂質又は界面活性剤など他の両親媒性物質である。リポソームは、水に及び脂質に可溶性の化合物を捕捉することができ、その結果、リポソームは担体として作用することが可能になる。リポソームは、免疫アジュバント、感染性疾患及び炎症の治療、癌療法、及び遺伝子療法など薬理学及び医学における送達系として使用されてきた(Gregoriadis et al,1995)。リポソームのアジュバント効果に対して影響を有し得る要因は、リポソームのサイズ、脂質組成物、及び表面電荷である。さらに、抗原の所在も(例えば、それが、リポソーム表面に吸着されているか若しくは共有結合で結合しているか、又はリポソームの水性区画中に封入されているかどうか)重要であり得る。樹状細胞は、抗原送達ビヒクルとして使用することができる。抗原を樹状細胞などの抗原提示細胞に負荷することは、抗腫瘍免疫において役割を有する活性T細胞を発生させるのに有効な方法であることを示した。
【0046】
第四級アンモニウム化合物、例えばジメチルジオクタデシルアンモニウムブロミド、クロリド若しくはこれらの他の有機若しくは無機塩(DDA−B、DDA−C又はDDA−X)、ジメチルオクタデセニルアンモニウムクロリド、ブロミド若しくはこれらの他の有機若しくは無機塩(DODA−C、DODA−B又はDODA−X)、又は1,2−ジオレイル−3−トリメチルアンモニウムプロパン(DOTAP)、1,2−ジミリストイル−3−トリメチルアンモニウム−プロパン、1,2−ジパルミトイル−3−トリメチルアンモニウム−プロパン、1,2−ジステアロイル−3−トリメチルアンモニウム−プロパン及びジオレイル−3−ジメチルアンモニウムプロパン(DODAP)及びN−[1−(2,3−ジオレイルオキシ)プロピル]−N,N,N−トリメチルアンモニウム(DOTMA)は、水性媒体に分散されたときに、脂質二重層、単層及び多層両方の全てのタイプのリポソーム、ミセルなどの脂質集合体を形成することができる。これらの構造の脂質膜は、糖脂質、例えば、小胞の分散を安定化することが示されているMMG又はα,α’−トレハロース6,6’−ジベヘネート(TDB)(Davidsen et al,PCT/DK2005/000467)など他の両親媒性化合物の封入体のための優れた基質を提供する。
【0047】
MMGと送達系との組合せは相乗的に作用して、例えば、DDAが単独で投与されたときの免疫応答を増強することができる。その結果、DDAは、低レベルのIFN−γ産生を促進するが、しかしながらMMGとの組合せで、IFN−γ産生は劇的に増大する。
【0048】
本発明のリポソームは、当技術分野において周知の種々の方法(Davidsen et al,PCT/DK2005/000467)により作製することができる。MMGのリポソーム/送達系中への組み込みは、リポソームとMMGとの単なる混合を含む、当技術分野において周知の種々の方法により行うことができる。特に、MMGのリポソーム中への組み込みは、Davidsen et al,PCT/DK2005/000467に記載されたようにして行うことができる。
【0049】
疾患に対する免疫を提供するのに加えて、本発明のアジュバントの組合せは、それ自体では免疫原性の不十分な物質である化合物に対する抗体を産生するために使用することもでき、そのような抗体は、例えば医学及び分析化学において問題の化合物の検出及び定量のために使用することができる。
【実施例】
【0050】
材料及び方法:
ウシ型結核菌BCGからの無極性及び極性の脂質の抽出
ウシ型結核菌BCG(コペンハーゲン株)をSauton変法培地中で培養した。マイコバクテリアを2〜3週間後に収集してPBS中に懸濁させ、60℃で1.5時間インキュベートすることにより殺菌した。無極性及び極性の脂質を、標準的手順(Dobson et al,1985)(Rosenkrands et al.2005)に従って抽出した。
【0051】
無極性脂質を抽出するために、0.3%NaCl:メタノール(440ml)及び440mlの石油エーテルを20gのマイコバクテリア(湿重量)に加えて、混合物を2時間攪拌した。遠心分離後、上層を除去して下層を440mlの石油エーテルで再抽出した。両方の抽出で得た上清相を合わせて蒸発させ、無極性脂質を得た。
【0052】
極性脂質を抽出するために、生物由来資源を含むメタノール性食塩水溶液を、沸騰水浴中100℃で10分間加熱し、続いて37℃で10分間冷却した。体積520mlのクロロホルム:0.3%NaCl:メタノール(9:10:3)を加えて、混合物を終夜攪拌した。全混合物を焼結ガラス漏斗を通して濾過し、濾過ケーキを捕集して170mlのクロロホルム:メタノール:0.3%NaCl(5:10:4)で2回再抽出した。3つの水性メタノール性クロロホルム相全てを合わせて、580mlのクロロホルム:0.3%NaCl水溶液(1:1)を加え、10分間攪拌した。放置して相を分離させた後、上の水層を除去して廃棄した。下の有機層を蒸発乾固すると極性脂質が得られた。
【0053】
個々の無極性脂質の精製
PDIM及びTAGを石油エーテル/アセトン(98:2)におけるTLC処理を使用して単離した。フチオセロールAに基づくPDIMの主成分のみ回収された。PGL及びMMGは、トルエン/アセトン(95:5)におけるTLC処理により一緒に分離された。PGLとMMGとは、クロロホルム:メタノール:0.880アンモニア(97:3:0.5)で1次元TLCにより分離された。PDIM、TAG、PGL及びMMGを、500MHz 1H及び13C核磁気共鳴(NMR)(Bruker drx500)及びMALDI−TOF質量分析(MS)(Bruker Biflex IV)にかけた。再水和した脂質抽出物(1mg/ml)の試料(10μl)を、SDS−PAGE(Laemmli et al,1970)及び銀染色(Blum et al,1987)により分析し、残留タンパク質含有量を求めた。脂質調製物は、エンドドキシン汚染のないことが認められた(<0.001ngLPS/μg脂質)。
【0054】
モノミコリルグリセロールの加水分解
モノミコリルグリセロールを16×100mmの試験管中で5%TBAH水溶液(2.5ml)とともに100℃で終夜加熱した(Minnikin et al,1988)。冷却後、混合物を水(2ml)で希釈し、ジクロロメタン中ヨードメタンの10%溶液(3ml)を加えて、試験管を回転装置上に1時間置いた。上の水層を廃棄して、下層を等量の1MのHCl及び水で洗浄し、蒸発乾固すると残渣が生じて、TLC(石油エーテル:アセトン95:5)により、α−及びケトミコレートメチルエステルを含有することが示された。分取TLC(上記の)により、αメチルミコレート及びケトメチルミコレートが得られた。
【0055】
α−MMG及びケト−MMGを分離するために、トリメチルシリル誘導体化を使用した。MMG混合物と600mlのTRI−SIL試薬(Pierce)とを75℃で20分間加熱した。次に、冷却した溶液を窒素気流下で乾燥し、分取TLCプレート上に負荷して石油エーテル:トルエン(50:50)で展開し、0.01%ローダミンを使用してα−MMG及びケト−MMGのバンドを可視化し、長波長の蛍光で検査した。対応するバンドをTLCプレートから掻き落とし、ジエチルエーテルを使用してα−MMG及びケト−MMGを、シリカゲルから3回抽出した(3×5mL)。合わせた抽出物を乾燥し、ヘプタン:メタノール(1:1)及びパラ−トルエンスルホン酸の少量の結晶を添加して、1時間攪拌することによりトリメチルシリル基を除去した。ヘプタン層を回収して蒸発乾固すると、精製されたα−MMG及びケト−MMGが得られた。
【0056】
C36の生成を示すMMGの合成
3−ヒドロキシ−2−テトラデシル−オクタデカン酸(Datta et al,1991に従って合成された合成C32コリノミコール酸)(100mg、0.20mmol、1eq)及び4−ピロリジノピリジン(100mg、3eq)を50mlの丸底フラスコ中に入れて、モレキュラーシーブ4Aと一緒にジクロロメタン(500μl)中50μlの2,2−ジメチル−4−ヒドロキシメチル−1,3−ジオキソラン(sn−イソプロピリデングリセロール)溶液を加えた。その混合物を、高真空下に室温で完全に乾固させ、N’,N−ジシクロヘキシルカルボジイミダゾール(DCC)(15ml、DCM中0.1MのDCC、5eq)を加えて、反応混合物を室温で終夜攪拌するままにした。モレキュラーシーブを濾過により除去し、反応混合物を真空で濃縮して乾固し、残渣を、フラッシュカラムクロマトグラフィ(Fluka 60741シリカゲル60)を使用して、ヘキサンからヘキサン:酢酸エチル(8:2)を使用して5%増分で溶出させて精製し、純粋なイソプロピリデン保護化合物(3−ヒドロキシ−2−テトラデシル−オクタデカン酸−2,2−ジメチル−[1,3]−ジオキソラン−4−イルメチルエステル)を収率56%で得た(68mg)。1H NMR(CDCl3、300MHz)δH0.90(t、6H、CH3)、1.20(s、54H、CH2)、1.40(s、3H、CH3)、1.45(s、3H、CH3)、2.50(m、1H、CH)、4.05〜4.40(m、5H、CH2、CH);13C NMR(CDCl3、75MHz)δC15.0(CH3)、22.1、28.8、28.9、29.0、31.4(CH2)、29.1(CH3)、52.1(CH(CH2)13CH3)、63.2(CH2OCO)、69.3(CH2O)、73.4(CH(CH2)14CH3)、174.3(C=O);m/z(EI)633.55[M+Na+](100%);HRMS C38H74O5Na[M+Na+]に対する計算値633.5536、実測値633.5527。
【0057】
3−ヒドロキシ−2−テトラデシル−オクタデカン酸−2,2−ジメチル−[1,3]−ジオキソラン−4−イルメチルエステル(68mg、1eq)を6mlのトリフルオロ酢酸:テトラヒドロフラン:水(体積で8:17:3)溶液中に溶解して、室温で終夜攪拌した。その溶液を飽和重炭酸ナトリウム水溶液で中和して、混合物をクロロホルムで2回抽出した。有機抽出物を水及び塩水で洗浄して乾燥し、真空中で濃縮して粗生成物を白色固体として得、それを、10gのシリカゲルVarian Bond Elut 12256026カートリッジでフラッシュカラムクロマトグラフィにより、ヘキサンからヘキサン:酢酸エチル(7:3)を使用して、5%増分で溶出させることにより精製し、表題化合物を白色固体として収率49%で得た(32mg)。融点72〜74℃。1H NMR(CDCl3、300MHz)δH0.90(t、6H、CH3)、1.25(s、54H、CH2)、2.50(m、1H、CH)、3.45〜3.85(m、3H、CH、CH2)、4.25(m、2H、CH2);13C NMR(CDCl3、75MHz)δC15.0(CH3)、26.3、30.9、31.3、33.5(CH2)、47.5(CH(CH2)13CH3)、68.4(CH2)、69.5(CH(CH2)14CH3)、72.5(CH2O)、76.4(CH)、175.4(C−1);m/z(EI)593.50[M+Na+](100%);HRMS C35H70O5Na[M+Na+]に対する計算値593.5121、実測値593.5143。
【0058】
樹状細胞アッセイ
ヒトPBMC由来のDCを、Romani et al.,1994から改良した方法に従って得た。末梢血はバフィーコートから得た。簡単に述べると、単球は、Ficoll−Hypaque遠心分離により単離し(Lymphoprep 1077密度媒質、Nycomed、オスロー、ノルウエイ)、続いて抗CD14標識磁性ビーズ(MACS;Miltenyi Biotech、Bergesh Gladbach、ドイツ)を使用してCD14陽性細胞を分離した。単球は、10%FCS、50μMの2−メルカプトエタノール、100U/mlのペニシリン、100μg/mlのストレプトマイシン、2mMのL−グルタミン(全てGibco)(CM)で補完した完全RPMI 1640中で、100ng/mlのヒト組み替えGM−CSF(Prepotech、Rocky Hill、NJ、USA)及び50ng/mlのヒト組み替えIL−4(Becton Dickinson(BD))を存在させて、37℃、5%CO2で7日間培養した。
【0059】
7日目に、iDC(1×105細胞/ml)を、リポ多糖(LPS)(大腸菌O127:B8)(Sigma−Aldrich、Brondby、デンマーク)又はウシ型結核菌BCG由来の脂質とともに、さらに24時間培養した。脂質抽出物は、乾燥ウシ型結核菌脂質材料をクロロホルム:メタノール(2:1)で再溶解し、続いて溶媒を蒸発させ、CM中にプローブ超音波処理することにより調製した。脂質を、未成熟DCに0.1ないし100μg/mlで加えた。
【0060】
フローサイトメトリー分析
DCを先ず関連するmAb(BD Pharmingen)と(30分、4℃)、続いて1/20希釈FITC結合ヤギ抗マウスIg(Jackson ImmunoResearch Laboratories、West Grove、PA)と(30分、4℃)インキュベートすることにより、表面マーカーを染色した。非特異的Abの結合は、関連する1次ヒトmAbの添加前に、10%ウシ胎児血清溶液で阻止した(15分、4℃)。染色された細胞は、直ちにFACScanフローサイトメトーター(BD)を使用して、フローサイトメトリーにより検査し、CellQuestソフトウェアで分析した。
【0061】
サイトカイン測定
DC培養上清を収集して、−20℃で貯蔵した。分泌されたIL−12p70、IL−6及びTNF−αを、ELISA(BD)によりメーカーの説明書に従って測定した。
【0062】
混合白血球反応(MLR)アッセイ
混合リンパ球反応(MLR)アッセイのためのiDCは、上で概説したようにして、単球から発生させた。生成した細胞(iDC)は、同じ培地中で、又は脂質(10μg又は100μg/ml)を含有する培地中で24時間培養した。0.125×105から2×105まで漸増させたDCを、37℃/5%CO2でPPD陰性の提供者からの同種異系のT細胞(105細胞/ウェル)と平底96ウェルのマイクロタイタープレート中でインキュベートした。T細胞は、Pan−T細胞単離キット(Miltenyi)を使用し、メーカーの説明書に従って単離した。DCと同種異系T細胞との共培養は、6日間インキュベートした。上清を収集して、分泌されたIFN−γがELISA(BD)によりメーカーの説明書に従って測定されるまで、−20℃で貯蔵した。次に、両方のアッセイには、18時間の最終培養の間、1μCi/ウェルの[3H]チミジンを含む培地をパルスで適用した。細胞を収集して、T細胞の増殖を液体シンチレーション計数(Microbeta Systems)により測定した。全てのアッセイは、少なくとも3名の異なる提供者を使用して3重で実施した。
【0063】
抗原
Ag85BとESAT−6との融合タンパク質(以下、Ag85B−ESAT−6と称する)は、前に記載したように(Olsen et al,2001)組み替えタンパク質として産生された。
【0064】
動物
8ないし12週齢の雌BALB/c又はC57BL/6マウスを、Bomholtgaard(Ry、デンマーク)又はHarlan Scandinavia(デンマーク)から入手した。感染させたマウスは、BL−3層流安全密閉箱内のケージ中で飼育した。
【0065】
免疫
マウスは、尾部の底部で皮下に(s.c.)、各免疫化間2週間の間隔で最高3回免疫した。ワクチン(0.2ml/マウス)は、特に断らない限り、250μgのDDAと10μgの再水和脂質抽出物との中で乳化された2μgの融合タンパク質Ag85B−ESAT−6からなるものであった。幾つかの場合には、11mol%のTDBがDDAリポソーム中に組み込まれた(Davidsen et al、PCT/DK2005/000467)。結核菌感染を含む実験における陽性対照として、1群のマウスが、尾部の底部皮下に、1用量のBCGデンマーク株1331の注射を受けた。全ての又は個々の脂質抽出物は、乾燥ウシ型結核菌脂質材料を1mg/ml又は5mg/mlのMilli Q水で再水和し、続いてプローブ超音波処理することにより調製した。標準的脂質ワクチンは、抗原を食塩水と混合し、続いて再水和した脂質抽出物及びDDAを添加して、ボルテックスで混合することにより調製した。ワクチンを終夜放置して、抗原を吸着させた。
【0066】
リンパ球培養
血液試料又は鼠径部リンパ節を、最終免疫の7〜150日後のマウスから採取し、前に記載したようにして(Rosenkrands et al,2005)調製した。細胞培養は、2−メルカプトエタノール、グルタミン、ペニシリン−ストレプトマイシン、Hepes、及び10%ウシ胎児血清で補完された、体積200μlのRPMI中に2×105の細胞を入れた丸底マイクロタイターウェル中で、3重で実施した。再刺激のための抗原は、5μg/mlの濃度で使用した。培地のみ又は5μg/mlのConAを入れたウェルが、全ての実験でそれぞれ陰性及び陽性対照として含まれた。培養上清は、並行する培養液から、抗原を存在させて72時間インキュベートした後収集して、IFN−γの量を酵素結合免疫吸着測定法(Brandt et al,2000)により測定した。
【0067】
実験的感染
ワクチンの有効性を評価するために、マウスに、最初の免疫の2.5ヵ月後に、約25CFUの有毒結核菌Erdman株を投入するように較正されたGlas−Colの吸入曝露系で、エアロゾル経路により肺にチャレンジした。脾臓及び肺における細菌負荷は、BCGの増殖を選択的に阻害するために1ml当たり2μlの2−チオフェン−カルボン酸ヒドラジドで補完したMiddlebrook 7H11アガー上に、段階希釈液を蒔くことにより、6週間後に測定した。37℃で2〜3週間インキュベーション後に、コロニーを計数した。
【0068】
統計分析
感染マウスと対照マウスとの間のコロニー数の差を、変動分析により検定した。有意の効果が示されたときは、平均間の差をダネット検定により評価した。ヒトDCアッセイについては、異なる脂質に対する応答におけるサイトカイン放出の差を、変動分析により検定し、有意の効果が示されたときは、平均間の差をチューキー検定により評価した。
【実施例1】
【0069】
ウシ型結核菌BCG由来の無極性脂質の単離及び免疫刺激活性
ウシ型結核菌BCGの全脂質を、極性及び無極性分画に分離した。極性分画中で同定することができた脂質は、ホスファチジルイノシトールマンノシド(1〜4)、ホスファチジルイノシトール(PI)、ホスファチジルエタノールアミン(PE)、ジホスファチジルジグリセロール(DPG)及びL−α−ホスファチジル−DL−グリセロール(PG)であった。多数の未知のリン脂質も同定された(7及び8)(図1A)。無極性分画中で同定された主な脂質は、フチオセロールジミコセロセート(PDIM)、トリアシルグリセロール(TAG)、フェノール性糖脂質(PGL)及びモノミコリルグリセロール(MMG)(図1)であった。FFAは遊離脂肪酸である。
【0070】
無極性及び極性脂質の比較のための免疫刺激性活性は、ヒト末梢血単球由来の未成熟DC(iDC)を使用して検査した(図1)。無極性脂質で処理した結果、活性化マーカーCD86、CD40及びHLA−DRのレベルは、処理しなかった対照と比較して、用量に依存して上昇した(図1B)。100μg/mlの無極性脂質の用量は、強力な免疫刺激性分子LPS(0.1μg/ml)で観察されるのと同等の、並びにマイコバクテリアのコード因子(TDM)及びMPLと比較して優れたDC活性化を生じた。これらの分子の上方制御は、炎症誘発性メディエーターの腫瘍壊死因子−α(TNF−α)、インターロイキン(IL)−6及びIL−12の分泌を伴った(図1C)。極性脂質で処理されたiDCの上清中のこれらの炎症誘発性サイトカインのレベルは、このアッセイを使用する検出の限界未満であった。最後に、本発明者らは、DC活性化の他の表示として、PPD陰性の提供者からの同族異系T細胞を使用する混合白血球反応(MLR)を使用した(図1DおよびE)。無極性脂質で処理したDCの高い活性化状態をさらに支持することとして、本発明者らは、高レベルの増殖及びIFN−γ放出を認めたが、MLRは極性分画によっては誘発されなかった。
【実施例2】
【0071】
ウシ型結核菌BCGの無極性脂質抽出物からの個々の脂質の単離及びMMGの特徴づけ
免疫刺激性の無極性分画の脂質は、分取TLCを使用して単離され、フチオセロールジミコセロセートA(PDIMA)、TAG、PGL及びMMGの純粋な試料を与えた。少量のフチオセロールB及びフチオジオロンAも検出されたが、利用した分取方法では回収されなかった(図2A)。脂質の構造及び同一性は、1H核磁気共鳴(NMR)及び質量分析法(MS)により確認された(図2B及び不掲載データ)。MMG成分は、1−モノアシルグリセロールに特徴的な1H及び13C NMRスペクトルを示した(Gunstone et al,1991)。全MMG分画のNMRデータと結びつけたMALDI−TOFMS(表1)は、α−ミコレート並びにcis及びtrans型のケト−ミコレートの存在を明らかにした。主成分のおよその比は、それぞれ1.00:0.29:0.24であった。
【0072】
表1:精製されたMMGのMALDI−TOF質量分析。シグナルはM+Na+イオンに対するm/zである。系列の多量成分は太字で示し、その主成分には下線を引いた。

【実施例3】
【0073】
MMGによるヒト樹状細胞の活性化
精製されたMMG、PDIMA、PGL及びTAGを、ヒトiDCを活性化するそれらの能力について評価した。これらのアッセイにおいて、MMGは、CD86、CD40及びHLA−DRの顕著な上方制御に通ずる、DC活性化の最も強力な誘発因子であることが一貫して認められた(図3A)。MMGは、マイコバクテリアの病原性と長らく関連づけられてきた脂質であるPDIMAよりさらに大きくDCを活性化し(Cox et al,1999)、PDIMAは2番目に活性な脂質となったが、一方、PGL及びTAGが誘発する活性化はそれより小さかった。6名の個々の提供者における活性化の順は、MMG>PDIMA>PGL>TAGと認められ、CD86レベルにおいて、未処理iDCに関するレベルを超える平均の上昇倍率は、それぞれ1.91±0.29、1.82±0.43、1.52±0.26及び1.32±0.14であった。サイトカイン誘発も全体的に同じ傾向をたどり(図3B)、MMGは最も強力な免疫刺激剤脂質として卓越している。IL−6は、MMGに曝されたDCにより、PGL又はTAGにより誘発されるレベルを有意に超えるレベルで(P<0.05)放出された。他の脂質によるサイトカイン誘発の間で、有意の差は記録されなかった。したがって、MMGは、ウシ型結核菌BCGの無極性脂質分画中で最も強力な免疫刺激性脂質として分類することができる。
【実施例4】
【0074】
α−及びケト−MMGの免疫刺激活性
この実施例において、本発明者らは、MMGの免疫刺激特性を精査し、その強力な免疫刺激性能の原因となる有効成分を同定することをさらに求めた。α−MMGとケト−MMGとを、MMGのトリメチルシリルエーテルの調製、分取TLC処理及びそれに続くトリメチルシリルエーテル保護基の加水分解によって分離し、α−MMG及びケト−MMGを得た。α及びケト−ミコレートの構造を図2Bに示す。ヒトiDCを活性化するそれらの能力を評価したとき、α−及びケト−MMGは、活性化マーカーのレベルにおいて2〜3倍の程度で刺激した(図4)。したがって、MMGの2つの下位成分も、ヒトDCを刺激する顕著な能力を示す。
【実施例5】
【0075】
ウシ型結核菌BCGから単離されたMMGによるTh1免疫応答の誘発
MMGのアジュバント活性を研究するために、単離された脂質を、マウスにおいてIFN−γ産生を誘発するそれらの能力について試験した。C57BL/6マウスに、10μgの全脂質又は個々の脂質を投与した。In vivoにおいて、DDAは、脂質を入れて送達するビヒクルとして役立つ。それゆえ、250μgのDDA中で乳化された2μgの融合タンパク質Ag85B−ESAT−6及び10μgの再水和脂質抽出物を、皮下経路により投与した。DDAリポソーム中に組み込まれた10μgのMMGの用量は、排出するリンパ節から単離されたPBMCの再刺激で、10ng/mlのIFN−γレベルを生じた。このレベルは、DDA中の全脂質に対して記録されたレベルと、等用量で同等である(図5)。PDIM−Aも、より低いレベルとはいえ、IFN−γ産生を誘発したが、TAG又はPGLの組み込まれたDDAリポソームは、IFN−γ放出を殆ど助長しなかった(図5及び不記載データ)。これらの個々の脂質は、全脂質抽出物又は個々の脂質のいずれで再刺激しても記憶応答は観察されなかったので、アジュバントとして作用しているように思われることは注目すべきである(不記載データ)。したがって、MMGは、in vivoにおいて最も活性な無極性脂質でもあるとして確認されて、BCG由来の全脂質のアジュバント活性の大部分を単独で説明することができた。
【実施例6】
【0076】
MMGに基づくアジュバントの防御有効性
MMGに基づくアジュバントのTB感染に対する防御提供能力を評価するために、C57BL/6マウスを、MMG(2つの異なる用量)及びDDAに送達されたAg85B−ESAT−6により免疫した。BCGワクチン接種及びアジュバント単独を受けるマウスの群は、それぞれ陽性及び陰性対照として含まれた。最後のワクチン接種の6週間後に、マウスに、生結核菌をエアロゾル経路によりチャレンジした。ワクチンが細菌負荷を減少させる能力を、その6週間後に肺及び脾臓で測定した。これらのデータは、MMG/DDAをアジュバントとして有意の防御レベル、及びBCGのレベルと同等の防御レベルを示した(図6)。予想されたように、抗原なしでアジュバントを接種されたマウスは、細菌の増殖を阻止することができなかったので、この効果は特異的である。
【実施例7】
【0077】
MMGとDDAとを組み合わせることによるさらに優れた効果
免疫調節剤(MMG)と送達系(DDA)とを組み合わせる効果を評価するために、DDA単独若しくはDDA/MMGの組合せで(実験1、図7A)、又は実験2(図7B)においてはMMG単独若しくはDDA/MMGの組合せで、C57BL/6マウスにワクチンを接種した。これらの実験から、免疫応答は、DDAとMMGとを組み合わせることにより、劇的に増強されることが明らかである。
【実施例8】
【0078】
TDB/第3成分のMMGとDDAとのリポソーム中への組み込みによる免疫応答の増強
他の免疫刺激性成分と組み合わせたときの、MMGのアジュバント活性に対する効果を研究するために、C57BL/6マウスにおける免疫応答を、DDAリポソーム中に組み込まれたAg85B−ESAT−6及び10μgのMMG、又は免疫調節剤TDBの組み込まれたDDAリポソーム(Davidsen et al,PCT/DK2005/000467)を使用して皮下で免疫した後、最初のワクチン接種の5ヵ月後に評価した。DDAリポソーム中に組み込まれたMMGという組合せは、血液から単離されたPBMCの再刺激で、約25ng/mlのIFN−γレベルを生じたが、IFN−γの放出は、TDBの組み込まれたDDAリポソームが使用されたとき、劇的に上昇した(図8)。したがって、MMG、DDA及びTDBの間で相乗的効果が観察され、MMGとDDAとの組合せに対する第3成分の添加は、アジュバント活性をさらに増強させ得ることを示した。
【実施例9】
【0079】
MMG類似体のアジュバント活性は、天然MMGの活性と同等である
合成MMG類似体の免疫学的効果を評価するために、C57BL/6マウスを、天然MMG、16個の炭素を有する合成MMG類似体(図9に示した)及び36個の炭素を有する合成MMG類似体(図9に示した)(全て10μg/DDA/MMG)を加えたDDA中のAg85B−ESAT−6で免疫した。免疫応答は、最後のワクチン接種の1週間後に血液で測定され、3種のMMGに基づくアジュバントで同等のレベルの応答を示したが、DDAは、それだけではより低い効果を再び示した。
【実施例10】
【0080】
鎖長が短いほど免疫応答は高い
鎖長がより短い合成MMG類似体の免疫学的効果を評価するために、単独のDDA中、天然MMGを加えたDDA中、C8ないしC36の範囲の異なる合成類似体を加えたDDA中のAg85B−ESAT−6で(全て1μg/用量)、C57BL/6マウスを免疫した。免疫応答は、最後のワクチン接種の3週間後に血液で測定され、合成MMG類似体は1μgの用量レベルでも活性であることを示した。さらに、これらの結果は、より短い合成MMG類似体(16C以下)は、天然MMGと比較してより有効であることも示す。***P<0.001(図11)
【0081】
[参考文献]
Bennekov, T., J. Dietrich, I. Rosenkrands, A. Stryhn, T.M. Doherty, and P. Andersen. 2006. Alteration of epitope recognition pattern in Ag85B and ESAT−6 has a profound influence on vaccine−induced protection against Mycobacterium tuberculosis. Eur. J. Immunol. 36( 12): 3346−55.
Gregoriadis, G. 1995. Engineering liposomes for drug delivery: progress and problems. Trends Biotechnol. 13(12): 527−37.
Blum, H., H. Beier, and H. J. Gross. 1987. Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis 8: 93−99.
Brandt. L., M. Elhay, I. Rosenkrands, E. B. Lindblad, and P. Andersen. 2000. ESAT−6 subunit vaccination against Mycobacterium tuberculosis. Infect. Immun. 68: 791−795.
Brennan, P. J. and M. B. Goren. 1979. Structural studies on the type−specific antigens and lipids of the mycobacterium avium.Mycobacterium intracellulare.Mycobacterium scrofulaceum serocomplex. Mycobacterium intracellulare serotvpe 9. J. Biol. Chem. 254(10): 4205−4211.
Cox, J. S., B. Chen, M. McNeil, and W.R. Jacobs, Jr. 1999. Complex lipid determines tissue−specific replication of Mycobacterium tuberculosis in mice. Nature 402: 79−83.
Dascher, C. C, K. Hiromatsu, et al. 2003. Immunization with a mycobacterial lipid vaccine improves pulmonary pathology in the guinea pig model of tuberculosis. Int. Immunol. 15(8): 915−925.
Davidsen, J., Rosenkrands, L, and P. Andersen. (PCT/DK2005/000467)
Dobson, G., D. E. Minnikin, S. M. Minnikin, J. H. Parlett, M. Goodfellow, and M. Ridell, a. M. M. Systematic analysis of complex mycobacterial lipids (ed. Minnikin, M. G. a. D. E.) 237−265 (Academic Press, London, 1985).
Gluck, R. 1995. Liposomal presentation of antigens for human vaccines. Pharm Biotechnol. 6: 325−345.
Gunstone, F. D. 1991. Chemistry and Physics of Lipids 58: 219−224.
Harboe, M., A.S. Malin, H.S. Dockrell, H.G. Wiker, G. Ulvsund, A. Holm, M.C. Jβrgensen, Andersen P. B−cell epitope and quantification of the ESAT−6 protein of Mycobacterium tuberculosis. Inft. Immun. 66(2):717−23.
Hiu, I. J. 1975. Mycobacterial adjuvant and its carrier. Experientia. 31(8): 983−5.
Indrigo, J., R. L. Hunter, Jr, et al. 2002. Influence of trehalose 6,6’−dimycolate (TDM) during mycobacterial infection of bone marrow macrophages. Microbiology 148(7): 1991−1998.
Koike, Y., Y. C. Yoo, et al. 1998. Enhancing activity of mycobacterial cell−derived adjuvants on immunogenicity of recombinant human hepatitis B virus vaccine. Vaccine 16(20): 1982−1989.
Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680−685.
Lima, V. M. F., V. L. D. Bonato, et al. 2001. Role of Trehalose Dimycolate in Recruitment of Cells and Modulation of Production of Cytokines and NO in Tuberculosis. Infect. Immun. 69(9): 5305−5312.
Lindblad, E., M. Elhay, et al. 1997. Adjuvant modulation of immune responses to tuberculosis subunit vaccines. Infect. Immun. 65(2): 623−629.
McBride, B. W., A. Mogg, et al. 1998. Protective efficacy of a recombinant protective antigen against Bacillus anthracis challenge and assessment of immunological markers. Vaccine 16(8): 810−817.
Minnikin, D. E. 1982. Lipids: complex lipids, their chemistry, biosynthesis and roles. The Biology of the Mycobacteria. C. R. a. J. L. Stanford. London, Academic Press.
Minnikin, D. E. in Bacterial Cell Surface Techniques (ed. Hancock, I. C, Poxton, I. R.) 125−135 (Wiley, Chichester, 1988).
Minnikin, D. E. 1988. Isolation and purification of mycobacterial wall lipids. Bacterial Cell Surface Techniques. I. C. Hancock, Poxton, I. R. Chichester, Wiley: 125−135.
Minnikin, D. E., L. Kremer, et al. 2002. ”The Methyl−Branched Fortifications of Mycobacterium tuberculosis. Chemistry & Biology 9(5): 545−553.
Moingeon, P., J. Haensler, et al. 2001. Towards the rational design of ThI adjuvants. Vaccine 19(31): 4363−4372.
Mosmann, T. R. and S. Sad 1996. The expanding universe of T−cell subsets: ThI, Th2 and more. Immunology Today 17(3): 138−146.
Nathan, C, H. Murray, et al. 1983. Identification of interferon−gamma as the lymphokine that activates human macrophage oxidative metabolism and antimicrobial activity. J. Exp. Med. 158(3): 670−689.
Olsen, A. W., L. A. H. van Pinxteren, L. M. Okkels, P. B. Rasmussen, and P. Andersen. 2001. Protection of Mice with a Tuberculosis Subunit Vaccine Based on a Fusion Protein of Antigen 85B and ESAT−6. Infect. Immun. 69:2773−2778.
Rao V, F. N., Porcelli SA, Glickman MS 2005. Mycobacterium tuberculosis controls host innate immune activation through cyclopropane modification of a glycolipid effector molecule. J Exp Med 201: 535−543.
Reed, M. B., P. Domenech, et al. 2004. A glycolipid of hypervirulent tuberculosis strains that inhibits the innate immune response. Nature 431(7004): 84−87.
Romani, N., S. Gruner, D. Brang, E. Kampgen, A. Lenz, B. Trockenbacher, G. Konwalinker, P.O. Fritsch, R.M. Steinman, and G. Schuler. 1994. Proliferating dendritic cell progenitors in human blood. J.Exp. Med. 180: 83−93.
Rosenkrands, L, E.M. Agger, A.W. Olsen, K.S. Korsholm, CS. Andersen, K.T. Jensen, and P. Andersen. 2005. Cationic Liposomes Containing Mycobac− terial Lipids: a New Powerful ThI Adjuvant System. Infect. Immun. 73: 5817− 5826.
Saito, R., Tanaka, A., Sugiyama, K., Azuma, I. and Yamamura, Y. 1976. Adjuvant effect of cord factor, a mycobacterial lipid. Infect Immun. 13(3): 776−781.
Silva, CL. 1985. Inflamation induced by mycolic acid−containing glycolipids of Mycobacterium bovis (BCG). Brazilian J.Med.Biol.Res. 18: 327−335.
Sirakova, T. D., V. S. Dubey, et al. 2003. The Largest Open Reading Frame (pksl2) in the Mycobacterium tuberculosis Genome Is Involved in Pathogenesis and Dimycocerosyl Phthiocerol Synthesis. Infect. Immun. 71(7): 3794−3801.
Sprott, G. D., C J. Dicaire, et al. 2004. Activation of Dendritic Cells by Liposomes Prepared from Phosphatidylinositol Mannosides from Mycobacterium bo− vis Bacillus Calmette−Guerin and Adjuvant Activity In Vivo. Infect. Immun. 72(9): 5235−5246.
Stanfield, J.P., D. Gall, Bracken P.M. 1973. Single−dose antenatal tetanus immunisation. Lancet l(7797):215−9.
Suzuki F, B. R., Pollard RB. 1986. Importance of Lyt 1+ T−cells in the antitumor activity of an immunomodulator, SSM, extracted from human−type Tubercle bacilli.1986 77(2): 441−7.
Uehori, J., M. Matsumoto, S. Tsuji, T. Akazawa, O. Takeuchi, S. Akira, T. Kawata, I. Azuma, K. Toyoshima, and T. Seya. 2003. Simultaneous blocking of human Toll−like receptors 2 and 4 suppresses myeloid dendritic cell activation induced by Mycobacterium bovis Bacillus Calmette−Guerin peptidoglycan. Infect. Immun. 71:4238−4249.
van Rooij, E. M. A., H. L. Glansbeek, et al. 2002. Protective Antiviral Immune Responses to Pseudorabies Virus Induced by DNA Vaccination Using Di− methyldioctadecylammonium Bromide as an Adjuvant.” J. Virol. 76(20): 10540− 10545.
Yamazaki S, K. K., Someya S, Azuma I, Yamamura Y. 1969. Studies on the allergenicity of various tuberculoprotein derivatives and the adjuvanticity of wax D fractions of Mycobacterium tuberculosis. Am Rev Respir Pis. 100(5): 691−8.
【技術分野】
【0001】
本発明は、免疫調節剤、アジュバント及びワクチン又はこのアジュバントを含む送達系を調製するための、モノミコリルグリセロール(MMG)又はその合成同族体、類似体及び改変体の使用を開示する。
【背景技術】
【0002】
最初のワクチンは、生きた弱毒化された病原体からなるものであった。弱毒化形態は、天然に生ずる密接に関係する生物であるか又は培養で連続継代により得られるかのいずれかであった。例えば、ヒトにおける結核(TB)は、多年にわたり、80年を超える以前に開発されたウシ型結核菌(M.bovis BCG)の弱毒株BCGワクチンによるワクチン接種により対処されてきた。しかしながら、30億を超える用量のBCGが投与されたにも拘わらず(他のいかなるワクチンも超える)、それは、どの集団においてもヒトTBに対する満足な抵抗性を必ずしも提供しない。
【0003】
現在、最新の手法は、高度に精製された物質、例えば精製された組み替えタンパク質又はペプチドを使用することである。これらのワクチンは、十分解明されており、副作用は最小化されている。残念ながら、多くの高度に精製された物質は、免疫原性が強くなくて、防御を与えるのに十分な免疫応答を誘発しない。これを行うために、抗原は、アジュバントと呼ばれる免疫応答強化剤から幾らかの助力を必要とする。病原体に応じて、防御は、液性の又は細胞媒介の応答のいずれが支配的であるかを要求することができる。免疫血清により移入され得る免疫応答は液性免疫と称され、感染性因子に関連する抗原性物質と結合して、それにより抗原性物質に対する免疫応答を始動させる抗体により媒介される抵抗性を意味する。細胞媒介免疫(CMI)は、免疫応答を担う免疫系細胞に依存する。CMI、又はTヘルパー(Th)1の免疫応答は、一般的には、リーシュマニア及び結核を含む細胞内病原体との闘いに関連しているが、他のタイプの感染例えば酵母感染カンジダとの闘いにも役割を有する。液性、又はTh2免疫応答は、細胞外病原体、例えば蠕虫感染に対する防御のために必要とされる。
【0004】
多くの場合、例えばインフルエンザ、肝炎C(HCV)、ヒト免疫不全ウイルス(HIV)、クラミジア及びマラリアの場合に、感染のステージに依存して、混合したTh1/Th2応答が必要になり得る(Mosmann and Sad 1996)。これら場合、それらの生活環の一部は細胞内であるが、それらは細胞外の相を通っても移動、例えば細胞間伝播するので、Th1及びTh2の両方が必要になる。
【0005】
特定の種類の免疫応答(液性又は細胞媒介)の開発は、アジュバントの選択により決定され得る。例えば、結核菌(M.tuberculosis)のような細胞内病原体に対する防御免疫は、細胞媒介免疫応答を必要とし、TBを目的とするサブユニットワクチン用の適当なアジュバントは、Th1応答を増強すべきである(Lindblad et al,1997)。
【0006】
多数のアジュバントが存在するが、それらの大部分は、それらのヒトにおける使用を妨げる数多くの問題に悩まされている。少数のアジュバントだけ、例えばアルミニウム系アジュバント(AlOH塩)及びMF−59が、ヒトへの使用を許容されているが、しかし、それらは両方ともTh2に偏った応答を誘発し、そのことがそれらを、Th1応答を必要とするTBワクチンおよび他のワクチンとして不適当にしている(Lindblad et al,1997)。
【0007】
過去20〜30年の間、多数の新しいアジュバント系が確認されて、それらの幾つかは現在開発中である。それにも拘わらず、新アジュバント系に対する必要性は、臨床用途に利用可能な選択が不足している中で、未だなお認識されており(Moingeon et al.,2001)、明らかである。
【0008】
アジュバント(adjuvant:ラテン語のadjuvareに由来し、助けるという意味)は、ワクチン中で投与されたときに、特異的な免疫応答を指示、促進、延長、及び/又は増強するのに役立つ任意の物質と定義することができる。アジュバントは、送達系又は免疫調節剤/免疫刺激剤のいずれか二つの主なカテゴリーに分けられている。送達系は、例えば、エマルション、ポリスチレン粒子、ニオソーム(niosome)、ISCOM(免疫刺激複合体)、ビロソーム(virosome)、ミクロスフェア、又は脂質二重層で構成される小胞である界面活性剤様リポソームにすることがでる。リポソームは、抗原(小胞内か又は表面に付着しているかのいずれか)の担体として作用し、接種部位でデポを形成することができて、抗原の緩やかな連続的放出を可能にする。注射及び食作用後の短時間中に、リポソームの提示により、特定量の抗原が単一抗原提示細胞に行き渡ることが確実になる(Gluck,1995)。免疫調節剤は、独特の細胞又は受容体、例えばAPCの表面上のToll様受容体を標的とする。送達系と免疫調節剤とは、一緒に、例えばGlaxoの一連のアジュバントにおけるように使用することができる。したがって、送達系は、ワクチン抗原の送達に加えて、免疫調節剤を送達するために使用することもできる。
【0009】
ワクチン中の成分であることに加えて、免疫調節剤は、抗原(単数又は複数)なしで投与することもできる。この手法により、免疫系を局所的に活性化することが可能であり、例えば、抗腫瘍及び抗ウイルス活性にとって重要な、抗原提示細胞の成熟、サイトカイン産生として見られる。したがって、免疫調節剤の投与は、例えば癌及び皮膚疾患の根絶における支援になり得る。局所投与できる免疫調節剤の例は、タキサン類、例えばタキソール、Toll様受容体7/8リガンドのレシキモッド(Resiquimod)、イミキモッド(Imiquimod)、ガルジキモッド(Gardiquimod)である。
【0010】
ジメチルジオクタデシルアンモニウム−ブロミド、−クロリド、−ホスフェート、−アセテート又は他の有機若しくは無機塩(DDA)は、親油性第四級アンモニウム化合物であり、水溶液中において40℃を超える温度でカチオン性リポソームを形成する。DDAは、APC中へのワクチン抗原の取り込みを増大させる非常に効率的な送達系である。DDAと免疫調節剤との組合せは記載がある。DDAを含むArquad 2HTのヒトにおける投与は有望であり、明確な副作用を誘発しなかった(Stanfield,1973)。DDAとTDBとの又はDDAとMPLとの組合せは、送達ビヒクル(DDA)と免疫調節剤(TDB又はMPL)との間の非常に明瞭な相乗効果を示し、どちらかの成分単独で得られる応答に比較して、高度に上昇したレベルのCMI応答があった。したがって、DDAは、例えば、TB及び他の細胞内病原体に対するワクチンのためのアジュバント系の開発において、ワクチン抗原及び免疫調節剤のための有望な送達ビヒクルである。
【0011】
マイコバクテリア由来の種々の化合物が、免疫調節性であると報告されている。ウシ型結核菌BCGから抽出された脂質がアジュバントとして使用されたとき、卵白アルブミンに対する皮膚試験応答が、モルモットで得られた(Hiu 1975)。ウシ型結核菌BCGの全極性脂質から温度を上げて形成されたリポソームは、卵白アルブミンに対する液性応答を生じさせることができて、これらの極性脂質から調製されたワクチンは、マウスにおいて腫瘍細胞でチャレンジしたとき、防御を与えた(WO03/011336)。モルモットに対する実験的TBワクチンにおける結核菌H37Rv由来の全脂質の抗原としての効果が、Dascher et al(2003)により研究された。この研究においては、コレステロールと1,2−ジステアロイル−sn−グリセロ−3−ホスドコリン(DSPC)とに基づくリポソームが、結核菌H37R.Vの全脂質抽出物と混合された。溶媒を除去した後、脂質は、ジメチルジオクタデシルアンモニウム(DDA)をアジュバントとして、PBS緩衝液中で再構成された。このワクチンで免疫されたモルモットは、細菌の有意の減少を示さず、DDAと混合されたリポソームのこの製剤は、強力な抗原を欠くこと、又はマイコバクテリアの脂質とコレステロール/DSPCとの製剤は、DDAのアジュバント効果を妨げることを示唆した。また、種々の脂質の混合物が投与されたとき、有効な脂質が、全脂質の構成要素となり得る比率はあまりに限られている。
【0012】
マイコバクテリア由来の種々の精製された成分は、それらのアジュバント活性についても研究されている。精製されたタンパク質誘導体(PPD)は、それ自体では遅延型過敏性反応を誘発しなかったが、ワックスD(マイコバクテリアの細胞壁のペプチドグリカンフラグメント−アラビノガラクタン−ミコール酸複合体)がアジュバントとして添加されたとき、強い反応が観察された(Yamazaki S,1969)。免疫調節剤SSM又はZ−100、結核菌から抽出された脂質アラビノマンナンは、抗腫瘍活性を有する(Suzuki F,1986)。他のマイコバクテリア細胞由来の化合物は、トレハロース6,6’−ジミコレート(TDM)(コード因子;ミコール酸含有糖脂質)である(Saito et al,1976)。TDM(又は合成類似体)も免疫刺激効果を有し、種々のアジュバント製剤中に含まれてきた(McBride et al,1998)(Koike et al,1998)。
【0013】
Silva et al(1985)による論文においては、ウシ型結核菌BCGから精製された5つの成分が、脂質で被覆されたチャーコール粒子として静脈内に注射されて、マウスの肺内に炎症性反応を生じさせた。5つの成分には、TDM、トレハロースモノミコレート(MMT)、グルコースモノミコレート(MMGlc)、アラビノースモノミコレート(MMAr)、及びグリセロールモノミコレート(MMGlyc)が含まれた。この論文は、モノグリコリルグリセロール頭部基を記載しているが、ミコール酸の組成物の定義が不十分で、構造データも提供されていない。それに加えて、脂質を投与したときの反応だけは、肺における炎症性活性として記載されているが、アジュバント効果として知られている特異的免疫応答を増強する性能は記載されていない。
【0014】
マイコバクテリア由来の脂質の免疫刺激性及び炎症性活性は、動物(マウス)モデルにおいて免疫応答を刺激し得る脂質についての増加し続ける文献で何十年間も認められてきたが、今日まで、ヒト樹状細胞(DC)を刺激する能力を有する個々の脂質(単数又は複数)は同定されていない。例として、TDMは、炎症誘発性応答に関して最も活性なマイコバクテリア脂質であることが示されているにも拘わらず、TDMで刺激したとき樹状細胞の活性化は観察されていない(Uehori et al,2003)。このように、TDMは、数編の論文において炎症性活性を示しているが、この脂質は、免疫応答を開始するために不可欠な樹状細胞を活性化する能力を明らかに欠いている。ヒト樹状細胞を活性化する能力を有するような脂質の同定は、それがヒトにおける使用に適した新規アジュバント系の一部として使用することができることを示唆するであろう。さらに、ヒト使用に適したTh1誘発アジュバントの欠如により、Th1助長能力を有する単独のマイコバクテリア由来の脂質の同定は大いなる発見になる。
【0015】
DCは、結核菌などの病原体に感染したときに免疫応答を指示する必須の役割を演ずる専門の抗原提示細胞(APC)である。したがって、活性化されたDCによるIL−12の産生は、それが、マクロファージの活性化を促進する、Th−1細胞によるIFN−γの産生の推進にこの上なく重要なサイトカインなので、結核菌感染の制御に不可欠なステップである(Nathan et al,1983)。さらに、近年、免疫応答を調節する試みの中で、マイコバクテリアはDCも標的とすることを示す証拠が見出されて、この過程におけるマイコバクテリアの脂質にとって極めて重要な役割が確立された。
【0016】
マイコバクテリアの細胞エンベロープの乾燥重量の40%までが、脂質からなる(Minnikin 1982)。これらの脂質は、この科の生物の独特の病原性と長らく関連づけられており、宿主のマイコバクテリア感染に対する応答において実質的役割を演ずることが知られている(Brennan and Goren 1979)。これらの脂質の中で重要なのは、フチオセロールジミコセロセート(PDIM:phthiocerol dimycocerosate)ワックスであり(Minnikin et al.2002)、その存在は病原性と相関することが示されている。PDIM欠損結核菌変異体は、マウス中で減弱した増殖を示す(Sirakova et al.2003)。PDIMに密接に関係するのは、いわゆる「フェノール性糖脂質」(PGL)であり、適当な例は、ウシ型結核菌中で見出された2−メチルラムノシルフェノール−フチオセロールジミコセロセート(「マイコサイド(mycoside)B」)である。このモノグリコシルPGLと結核菌のある種の分離菌の高毒性との間の関連が、最近、明らかにされた(Reed et al.2004)。
【0017】
特に興味ある他の種類の脂質は、トレハロース−6,6−ジミコレート(TDM)、いわゆる「コード因子」である。TDMは、TNF−α、IL−6及びIL−12などの炎症誘発性サイトカイン、及びTh−1促進サイトカインであるIFN−γ(Lima et al.2001)の放出を刺激することにより肉芽腫性病変の維持を助長し、ファゴソーム−リソソーム融合を阻害することにより、マクロファージ内の結核菌の生存を長引かせる役割を有する(Indrigo et al.2002)。TDMのミコレート成分の微細構造は、初期感染期におけるマクロファージの炎症誘発性活性化において重要である(Rao et al.2005)。
【0018】
マイコバクテリアの生存の増加におけるそれらの役割にも拘わらず、マイコバクテリア脂質の免疫調節力は、新世代のTh1誘発アジュバントを創出することに利用することもできる。強力な免疫刺激活性を有する個々の脂質の同定において、強力なアジュバント活性をなお維持しながら、油分と混合された加熱殺菌結核菌の全細胞(フロイント完全アジュバント(CFA))の使用に伴う毒性ゆえの問題を回避することは可能であるかもしれない。実際、ウシ型結核菌バチルス・カルメット・ゲラン(bacillus Calmette−Guerin)(BCG)の極性脂質から形成されたリポソームは、マウスの骨髄由来の樹状細胞(BM−DC)を活性化することが最近示された。この活性の大部分は、糖リン脂質であるホスファチジルイノシトールジマンノシドという脂質に起因し得ることが見出された(Sprott et al.2004)。
【0019】
本発明者らの研究室における最近の研究は、強力な液性及び細胞媒介成分で複雑な遅延免疫応答を促進することができる、ウシ型結核菌BCGからのマイコバクテリア脂質抽出物とジメチルジオクタデシルアンモニウムブロミド(DDA)との新規なアジュバントの組合せの特性を明らかにした(Rosenkrands et al.2005)。全BCG脂質のアジュバント活性の大部分は、無極性脂質に起因することが見出された。
【0020】
これらの研究はマイコバクテリア脂質がアジュバントとして作用する可能性をさらに確かなものにするが、最善の解決は、単独で強力な活性を有する、単独の最も免疫刺激性の脂質を同定することであろう。このことは、さらに単純で廉価なアジュバントを提示し、世界中で投与されるワクチンで使用するためのアジュバントに要求される大量に産生され得る脂質の合成類似体を作製して、より無害な系を可能にする可能性も浮揚させるであろう。
【発明の概要】
【0021】
本発明は、ヒトDCを活性化し得る免疫刺激性脂質、モノミコリルグリセロール(MMG)及びそれらの合成同族体、類似体及び改変体を開示する。MMGは全BCG脂質の無極性分画に由来し、これらの脂質に関連する、アジュバントの及び防御の効果を誘発する原因である。小さめの炭素主鎖を有する合成MMGは、in vitroにおいて天然MMGのヒトDCに対する刺激性を増強することができ、マウスモデルにおいて、in vivoで、TBに対する長く継続する防御免疫応答につながる強いTh1応答も誘発する。
【図面の簡単な説明】
【0022】
【図1】ウシ型結核菌BCGの無極性及び極性脂質の単離及び免疫刺激活性のアセスメント。ウシ型結核菌BCGコペンハーゲン株から抽出された極性及び無極性脂質抽出物を2次元TLCにより分析した。極性脂質分画中において、1〜4はホスファチジルイノシトールマンノシドであり、PIはホスファチジルイノシトールであり、PEはホスファチジルエタノールアミンであり、及びDPGはジホスファチジルグリセロール(カルジオリピド(cardiolipid))である。PGは、L−α−ホスファチジル−DL−グリセロールであるが、5及び6は、未知のリン脂質である。無極性分画において、TAGはトリアシルグリセロールであり、PDIMはフチオセロールジミコセロセートA、B及びCであり、MMGはモノミコリルグリセロールであり、PGLはフェノール性糖脂質であり、FFAは遊離脂肪酸である(パネルA)。iDCは、培地単独、LPS(0.1μg/ml)、MPL(100μg/ml)、コード因子(CF)(100μg/ml)、無極性脂質(0.1−100μg/ml)又は極性脂質(0.1−100μg/ml)の存在下に、24時間インキュベートした。処理後のDC上の表面マーカーの幾何平均蛍光強度(MFI)のレベルを示す。3名の異なる提供者を使用した3つの実験のうち1つの代表的実験から得られたデータ(パネルB)。100μg/mlの無極性又は極性脂質による処理後に得られた培養上清を、サイトカインIL−6、TNF−α及びIL−12の存在についてELISAにより分析した。3名の異なる提供者を使用して3重で実施された3つのうち1つの代表的実験から得られたデータ(±s.e.m)を示す(パネルC)。PPD陰性の提供者からの、100μg/mlの無極性又は極性脂質で24時間処理されたDCとインキュベーション後の、MLRアッセイにおける、T細胞の増殖(パネルD)及びIFN−γ放出(パネルE)。3名の異なる提供者を使用して3重で実施された3つのうち1つの代表的実験から得られたデータ(±s.e.m)を示す。
【図2】MMG、TAG、PDIM及びPGLの構造。個々の脂質PDIM、TAG、PGL及びMMGの2次元TLC分析(パネルA)並びにMMG、PGL、PDIM及びTAGの代表的構造(パネルB)。
【図3】MMGによるヒト樹状細胞の活性化。iDCを、培地単独で(点線)又はMMG、PDIM、PGL若しくはTAG(10μg/ml)の存在下に24時間インキュベートした。脂質の調製物は、エンドドキシン汚染のないことが認められた(<0.001ngLPS/μg脂質)。6名中1名の代表的提供者について、処理後のDC上の表面マーカーのレベルのMFIを示す(パネルA)。MMG、PDIMA、PGL又はTAG(10μg/ml)で処理後に得られた培養上清を、サイトカインIL−6、TNF−α及びIL−12の存在について、ELISAにより分析した(パネルB)。異なる提供者を使用して3重で実施した3つ又は4つの実験から得られたデータの平均(±s.e.m.)を示す。データはチューキー検定を使用して分析した。
【図4】MMGのα及びケトミコレートは免疫刺激性である。iDCは、培地単独及びMMGのα−又はケトミコレート(10μg/ml)の存在下に24時間インキュベートした。処理後のDC上の表面マーカーのMFIを示す。
【図5】ウシ型結核菌BCGコペンハーゲン株から単離されたMMGにより誘発されたIFN−γ放出。C57BL/6マウスを、BCGコペンハーゲン株から単離された脂質に基づくアジュバントとの組合せでDDAリポソームに組み込まれたAg85B−ESAT−6で免疫した。ワクチン接種7日後に排出するリンパ節から単離されたPBMCによるIFN−γ放出を測定した。
【図6】有毒TB感染に対するMMGアジュバントを用いる防御。C57BL/6マウスを、DDAと10μg又は50μgのMMGとに基づくアジュバントとを組み合せて、Ag85B−ESAT−6で3回免疫した。最終ワクチン接種の6週間後に、マウスは結核菌を用いるエアロゾルのチャレンジを受けた。6週間後に細菌の数を肺及び脾臓で測定した。標準的BCGワクチン接種を受けるマウスは陽性対照として、及び抗原なしでDDA/MMG(10μg)により免疫されるマウスは陰性対照として含まれた。実験的ワクチンの防御の有効性は、免疫されないマウスと比較して、肺における細菌負荷の減少のLog10として表す。結果は、各群中の6匹のマウスについての平均値±SEMである。免疫されていない対照と有意に異なる値には、*印を付けてある。P<0.05。
【図7】IFN−γ放出は、MMGとDDAとを組み合わせることにより増大する。2通りの実験において、C57BL/6マウスは、DDA若しくはDDA/MMG中(パネルA)又はMMG若しくはDDA/MMG中(パネルB)のAg85B−ESAT−6で免疫した。最終ワクチン接種の3週間後に、血液から単離された(パネルA)又は脾臓中の(パネルB)PBMCによるIFN−γ放出を測定した。
【図8】IFN−γ放出は、MMG/DDAの組合せにTDBを添加することにより増大する。C57BL/6マウスは、MMGが組み込まれたDDAリポソーム中、又はTDBを含有し、MMGが組み込まれたDDAリポソーム中で投与されたAg85B−ESAT−6により免疫した。ワクチン接種の5ヵ月後に血液から単離されたPBMCによるIFN−γ放出を測定した。
【図9】合成MMG類似体の構造の例。
【図10】免疫応答は、天然及び合成MMG類似体で同等である。C57BL/6マウスは、DDA、DDA/MMG中(10μg)、DDA/MMG C36中(10μg)又はDDA/MMG C16中(10μg)中のAg85B−ESAT−6を用いて免疫した。最終免疫の1週間後に血液から単離されたPBMCによるIFN−γ放出を測定した。
【図11】鎖長が短いほど免疫応答は高い。C57BL/6マウスを、DDA中、又は8から36の範囲の鎖長の種々のMMG類似体を含むDDA中のAg85B−ESAT−6で免疫した(1μg/用量)。最終免疫の3週間後に血液から単離されたPBMCによるIFN−γ放出を測定した。
【発明を実施するための形態】
【0023】
本発明は、免疫調節剤、アジュバント及びワクチン、又はヒト樹状細胞を刺激する特有の能力を有するこのアジュバントを含む送達系を調製するための、モノミコリルグリセロール(MMG)又はそれらの合成同族体、類似体及び改変体の使用を開示する。
【0024】
免疫調節剤として、MMG又はそれらの合成同族体,類似体及び改変体は、抗原(単数又は複数)なしでも投与される。この手法により、免疫系を局所的に活性化することが可能であり、例えば、抗腫瘍及び抗ウイルス活性にとって重要な、抗原提示細胞の成熟、サイトカイン産生として見られる。
【0025】
アジュバント(adjuvant:ラテン語のadjuvareに由来し、助けるという意味)は、ワクチン中で投与されたときに、特定の免疫応答を指示、促進、延長、及び/又は増強するのに役立つ任意の物質と定義することができる。アジュバントの性質に依存して、それは、細胞媒介免疫応答、液性免疫応答又はその2つの混合した応答を促進することができる。ワクチンアジュバントとして使用されるとき、抗原性成分がアジュバントに添加される。アジュバントにより媒介される免疫応答の増強は非特異的であるから、同じアジュバントが異なる抗原とともに使用されて、異なる標的に対する応答を助長すること、例えば、結核菌由来の抗原とともに結核菌に対する免疫を助長すること、又は腫瘍由来の抗原とともにその特定の種類の腫瘍に対する免疫を助長することができることは、当分野においてよく理解されている。
【0026】
本発明により開示された好ましいアジュバントは、MMG又はそれらの合成同族体、類似体又は改変体を含むアジュバントであり、それらには、送達ビヒクル、例えばエマルション、ポリスチレン粒子、ニオソーム、IS−COMS、ビロソーム、ミクロスフェア、又は界面活性剤様リポソームがさらに含まれる。好ましい界面活性剤は、ジメチルジオクタデシルアンモニウムブロミド又はこのクロリド(DDA−B又はDDA−C)又はスルフェート、ホスフェート又はアセテート塩(DDA−X)、又はジメチルジオクタデセニルアンモニウムブロミド又はこのクロリド(DODA−B又はDODA−C)又はスルフェート、ホスフェート又はアセテート化合物(DODA−X)に基づくカチオン性脂質であることが最も好ましい。この発明において使用される他のタイプの好ましいカチオン性脂質には、1,2−ジオレイル−3−トリメチルアンモニウムプロパン(DOTAP)、1,2−ジミリストイル−3−トリメチルアンモニウム−プロパン,1,2−ジパルミトイル−3−トリメチルアンモニウム−プロパン、1,2−ジステアロイル−3−トリメチルアンモニウム−プロパン及びジオレイル−3−ジメチルアンモニウムプロパン(DODAP)及びN−[1−(2,3−ジオレイルオキシ)プロピル]−N,N,N−トリメチルアンモニウム(DOTMA)が含まれるが、これらに限定はされない。他の界面活性剤は、DXPC、DXPE、DXPG又はこれらの組合せの中から選択され、ここで、Xは、鎖長記載についての置き換え、例えばP=パルミトイル(16C)、S=ステアロイル(18C)、A=アラキドイル(20C)である。
【0027】
送達ビヒクルは、TLR及び非TLRリガンド様MPL(モノホスホリル脂質A)、ポリイノシンポリシチジン酸(ポリ−IC)、ムラミルジペプチド(MDP)、ザイモサン、二重螺旋RNA(dsRNA)、DC−Chol、CpGオリゴデオキシヌクレオチド、カチオン性ペプチド、TDM、TDB、タモキシフェン又はこれらの分子の任意の類似体など他の免疫調節剤のためにも使用することができる。したがって、好ましいアジュバントには、送達ビヒクル中のMMG又はそれらの同族体、類似体、又は改変体が含まれ、さらにTLR−又は非TLRリガンドが含まれる。
【0028】
MMG又はそれらの合成同族体、類似体及び改変体を含む送達系は、癌、自己免疫障害、神経障害例えばアルツハイマー病、気道炎、炎症性疾患、感染性疾患、皮膚疾患、アレルギー、喘息、又は病原体により惹起される疾患を治療するために使用することができる。MMG又はそれらの合成同族体、類似体及び改変体は、1つまたは複数ワクチン、抗原、抗体、細胞傷害性因子、アレルゲン、抗生物質、アンチセンスオリゴヌクレオチド、TLR−及び非TLRアゴニスト、TLR−及び非TLRアンタゴニスト、ペプチド、タンパク質、遺伝子療法ベクター、DNAワクチン、又は共刺激分子との組合せで投与される。
【0029】
抗原性成分又は抗原性物質は、予め形成された抗体及び/又はT細胞及びB細胞上の特異的受容体と反応する分子である。ワクチン接種との関係においては、特定のT細胞又はB細胞の発生を刺激する分子が、抗原が免疫細胞と2度目に遭遇した場合に、より速い「記憶」応答を促進する免疫細胞の記憶集団の形成をもたらすことができる。記憶集団がクローン性であることは稀なので、実際には、このことは、抗原は、以前それに曝されたことがある個体の免疫細胞に再度遭遇したときに、免疫応答における増強を刺激することができる任意の分子又は分子の集合であることを意味する。
【0030】
本発明は、非経口、経口若しくは粘膜投与又は送達系のための、アジュバントを含むワクチンをさらに開示する。好ましいワクチンには、細胞内病原体に由来する抗原性エピトープ、例えば有毒マイコバクテリウム(例えば、融合生成物Ag85b_TB10.4、Ag85b_ESAT−6_Rv2660、Ag85b_TB10.4_Rv2660及びAg85a_TB10.4_Rv2660)、熱帯熱マラリア原虫(Plasmodium falciparum)(Msp1、Msp2、Msp3、Ama1、GLURP、LSA1、LSA3又はCSP)、トラコーマクラミジア(Chlamydia trachomatis)(例えばCT184、CT521、CT443、CT520、CT521、CT375、CT583、CT603、CT610又はCT681)、HIV、インフルエンザ又はB型若しくはC型肝炎ウイルスに由来する抗原性エピトープが含まれる。アジュバント又は送達系は、癌、アレルギー又は自己免疫疾患を治療するためのワクチンにおいて使用することもできる。
【0031】
全マイコバクテリア脂質抽出物は、マイコバクテリア、例えばBCG、M.ミクロティ(M.microti)、結核菌及びM.バッカエ(M.vaccae)から、化学的又は物理的方法により得られる脂質の混合物である。本研究において、抽出に使用した方法は、有機溶媒の作用であるが(以下に記載する)、当業者に知られた他の可能性も可能である。
【0032】
無極性脂質の分画は、極性のない脂質と定義される。無極性脂質の分画は、マイコバクテリアをメタノール/食塩水と石油エーテルとの二相混合物で処理することにより得られる。石油エーテル抽出物は、無極性(非極性)脂質を含む。その後、マイコバクテリア及び残留水相にクロロホルムを添加することにより、極性脂質の分画が得られる。クロロホルム抽出物は、残留する極性脂質を含有する。無極性脂質の分画中の主成分は、フチオセロールジミコセロセート、トリアシルグリセロール、トレハロースミコリペネート(trehalose mycolipenate)及びメナキノンである。極性脂質分画の主成分は、ホスファチジルエタノールアミン、ホスファチジルグリセロール、及びホスファチジルイノシトールなどのリン脂質である。中間的極性の脂質は、スルホリピド、トレハロースミコレート、グリコシル化フェノールフチオセロール(フェノール性糖脂質、PGLを含む)及びアシル化トレハロースである(Dobson et al,1985)。
【0033】
MMGは、無極性脂質の分画から得られる脂質モノミコリルグリセロール、及びそれらの誘導体、例えばα−MMG及びケトMMG並びに天然及び合成類似体を指す。MMGは、トルエン/アセトン(95:5)でTLC処理により単離することができる。PGL及びMMGは、この方法により一緒に抽出されるが、クロロホルム:メタノール:0.880アンモニア(97:3:0.5)での1次元TLCにより分離することができる。MMGの誘導体、α−MMG及びケト−MMGは、16×100mmの試験管中で5%TBAH水溶液(2.5ml)とともに、100℃で終夜加熱することにより得ることができる(Minnikin 1988)。
【0034】
MMGの合成同族体、類似体又は改変体は、化学合成の任意の従来法により製造することができる。類似体とは、構造は類似しているが、基本的組成に関して異なる一群の化合物の一つを指し、同族体とは、同族列の化合物の任意の構成要素を指す。これらの化合物は、炭素鎖の長さを変えることができる。特にサイズを小さくすることは毒性の減少を伴い、したがって、類似体のいかなる顕性毒性も減少させることに役立ち得る。したがって、合成体は、例えば、8〜36個の炭素を有し、且つ各脂質尾部に0〜3個の二重結合を有するアルキル鎖に基づくことができる。あるいは、脂質尾部の一つを除去することにより、単純化された形態を得ることができる。合成MMGの炭素主鎖サイズは、好ましくはC8〜C36、例えば3−ヒドロキシ−2−エチル−ヘキサン酸−2,3−ジヒドロキシプロピルエステル(C8)、3−ヒドロキシ−2−ブチル−オクタン酸−2,3−ジヒドロキシプロピルエステル(C12)、3−ヒドロキシ−2−ヘキシル−デカン酸−2,3−ジヒドロキシプロピルエステル(C16)、3−ヒドロキシ−2−ヘプチル−ウンデカン酸−2,3−ジヒドロキシプロピルエステル(C18)、3−ヒドロキシ−2−テトラデシル−オクタデカン酸−2,3−ジヒドロキシプロピルエステル(C32)又は3−ヒドロキシ−2−ヘキサデシル−アイコサン酸−2,3−ジヒドロキシプロピルエステル(C36)、及び最も好ましくはC8又はC16である。改変体は、グリセロール部分を他のポリオール頭部群、例えば、ポリプロピレングリコール及びポリエチレングリセロールで置き換えることにより調製することができる。合成モノミコレートのC2及びC3の周囲の立体化学は、グリセロールにおけると同様に変更することができる。以下において、単にMMGと書かれたものは、MMGの上記の合成同族体、類似体又は改変体も意味する。
【0035】
抗原性成分又は物質は、ポリペプチド又はポリペプチドの部分であってよく、それは、動物又は人類において、及び/又は生物学的試料において、本明細書中に記載する生物学的アッセイのいずれかにより測定される免疫応答を生じさせる。ポリペプチドの免疫原性部分は、T細胞エピトープ又はB細胞エピトープであってよい。免疫応答の間に認識される関連のあるT細胞エピトープを同定するために、「力づく」の方法を使用することが可能である。即ち、T細胞エピトープは線状であるから、ポリペプチドの欠失変異体が体系的に構成されれば、例えば、これらの欠失変異体を、例えば本明細書中に記載したIFN−γアッセイにかけることにより、ポリペプチドのどの領域が免疫認識において必須であるかが明らかになるであろう。他の方法は、ポリペプチド由来の重複オリゴペプチド(好ましくは、例えば20アミノ酸の長さの残基を有する合成体)を利用する。これらのペプチドは、生物学的アッセイ(例えば本明細書中に記載したIFN−γアッセイ)で試験することができ、これらの一部はペプチド中のT細胞エピトープの存在に対する確証として、陽性の応答(したがって免疫原性である)を与えるであろう。線状のB細胞エピトープは、例えばHarboe et al,1998に記載されたように、関心のあるポリペプチドを包含する重複ペプチドに対するB細胞の認識を分析することにより、決定することができる。
【0036】
T細胞エピトープの最小長さは、少なくとも6アミノ酸であることが示されているが、そのようなエピトープがそれより長いアミノ酸のつながりで構成されていることは普通である。したがって、本発明のポリペプチドフラグメントは、少なくとも7アミノ酸残基、例えば、少なくとも8、少なくとも9、少なくとも10、少なくとも12、少なくとも14、少なくとも16、少なくとも18、少なくとも20、少なくとも22、少なくとも24、及び少なくとも30アミノ酸残基などの長さを有することが好ましい。したがって、本発明の方法の重要な実施形態において、ポリペプチドフラグメントは、最大で50アミノ酸残基、例えば、最大で、40、35、30、25、及び20アミノ酸残基などの長さを有することが好ましい。10と20との間のアミノ酸残基の長さを有するペプチドは、診断の手段として最も有効であると証明されることが予想され、したがって、本発明の方法で使用されるポリペプチドフラグメントの特に好ましい長さは、18、例えば、15、14、13、12さらに11アミノ酸などである。
【0037】
具体的に、抗原性物質は、代謝する結核菌、ウシ型結核菌及び他の環境マイコバクテリア、例えばトリ型結核菌(Mycobacterium avium)などの培養液から取り出すことができる。そのようなマイコバクテリアの濾液からの特に興味ある物質は、ESAT−6遺伝子ファミリーのタンパク質(ESAT6及びTB10.4など)、並びにTB患者及び種々の動物モデルにおける結核の初期相において、細胞媒介免疫にとって主要な標的であるAg85A、Ag85B、ORF2c、Rv1036及びRv0285などの他の初期抗原である。TB感染の後期段階の期間中、主要な標的であるRv2653、Rv2655、Rv2656、Rv2657、Rv2658、Rv2659、Rv2660など他の抗原も関連する。それらの免疫原性は、それ自体では低いが、この明細書の以下の詳細な部分で示すように、本発明のアジュバントの組合せとの組合せで、結核に対する高く且つ持続する免疫を誘発するための有力な候補であることが判明した。ESAT−6遺伝子ファミリーのタンパク質並びに本発明のアジュバントの組合せとの組合せに適用可能な他の抗原は、現在、人工的に、例えば、合成的に又は遺伝子組み替え技法により製造することができる。
【0038】
融合タンパク質は、ワクチンにおける抗原性物質として特に十分に適していることが証明されており、例えば、融合生成物Ag85b_TB10.4、Ag85b_ESAT−6_Rv2660、Ag85b_TB10.4_Rv2660及びAg85a_TB10.4_Rv2660は、TBに対して非常に有効であることが証明されている。
【0039】
ワクチンは、疾患に対する免疫を生じさせる接種のための、死滅したか弱毒化されたか若しくは他の方法で改変された微生物(細菌、ウイルス、又はリケッチャ)又はそれらの部分の懸濁液と定義される。ワクチンは、疾患を防止するための予防用ワクチン又は癌若しくは潜在性の感染性疾患など既存の疾患に対処するための治療用ワクチンのいずれかとしてだけでなく、アレルギー及び自己免疫疾患と関連して投与することができる。ワクチンは、免疫応答を強化するために、適当なアジュバント中に乳化することができる。
【0040】
ワクチンは、投与製剤と適合した様式で、且つ治療的に有効で免疫原性であるような量で投与される。投与されるべき量は、例えば、個体の免疫系の免疫応答を開始する能力、及び所望の防御の程度を含めて治療される対象に依存する。適当な用量範囲は、ワクチン接種当たり数百マイクログラムの桁の有効成分であり、好ましい範囲は約0.1μgから1000μg、例えば約1μgから300μgの範囲など、特に約1μgから50μgの範囲内である。初期投与及びブースター投与に適当な投与計画も調節可能であるが、初期投与に類型化され、その後の接種又は他の投与が続く。
【0041】
適用の様式は広範に変えることができる。ワクチン投与の任意の従来法が適用可能である。これらの方法は、生理学的に許容される固体基剤による経口若しくは粘膜適用、又は生理学的に許容される分散剤での、非経口的な、注射による等の適用を含むと考えられる。ワクチンの用量は、投与経路に依存し、また、ワクチン接種されるヒトの年齢、及び程度は低いが、ワクチン接種されるヒトの体格によって変わる。
【0042】
ワクチンは、非経口的に、注射により、例えば皮下又は筋肉内のいずれかに投与するのが便利である。他の様式の投与に適したさらなる製剤には、坐剤及び、ある場合には、経口又は粘膜用製剤が含まれる。坐剤に対して、在来の結合剤及び担体は、例えば、ポリアルキレングリコール又はトリグリセリドが含むことができ、そのような坐剤は、0.5%から10%、好ましくは1〜2%の範囲内の有効成分を含有する混合物から形成することができる。経口製剤には、例えば、医薬品グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウム等のような普通に使用される賦形剤が含まれる。これらの組成物は、溶液剤、懸濁液剤、錠剤、丸薬、カプセル剤、徐放性製剤又は散剤の形態をとり、10〜95%、好ましくは25〜70%の有効成分を含有することが有利である。
【0043】
選択のワクチンには、例えば、次のものがなり得る。
タンパク質ワクチン:ポリペプチド(又はそれらの少なくとも1つ免疫原性部分)、ペプチド混合物又は融合ポリペプチドを含むワクチン組成物;
組み替え生ワクチン:非病原性微生物又はウイルス中におけるワクチンに関連ある抗原の発現。そのような微生物の周知の例は、ウシ型結核菌BCG、サルモネラ及びシュードモナス(Pseudomonas)であり、ウイルスの例は、ワクシニアウイルス及びアデノウイルスである。
【0044】
これらのワクチン構築物の全てに対して、適当なアジュバントの添加は、結果として、ワクチンの有効性を増強した(Brandt et al.2000)、(van Rooij et al.2002)、(Bennekov et al.2006)。
【0045】
リポソーム(即ち、脂質小胞)は、脂質二重層により封入された水性区画である。脂質成分は、コレステロール及び他の荷電脂質でしばしば補完された、通常リン脂質又は界面活性剤など他の両親媒性物質である。リポソームは、水に及び脂質に可溶性の化合物を捕捉することができ、その結果、リポソームは担体として作用することが可能になる。リポソームは、免疫アジュバント、感染性疾患及び炎症の治療、癌療法、及び遺伝子療法など薬理学及び医学における送達系として使用されてきた(Gregoriadis et al,1995)。リポソームのアジュバント効果に対して影響を有し得る要因は、リポソームのサイズ、脂質組成物、及び表面電荷である。さらに、抗原の所在も(例えば、それが、リポソーム表面に吸着されているか若しくは共有結合で結合しているか、又はリポソームの水性区画中に封入されているかどうか)重要であり得る。樹状細胞は、抗原送達ビヒクルとして使用することができる。抗原を樹状細胞などの抗原提示細胞に負荷することは、抗腫瘍免疫において役割を有する活性T細胞を発生させるのに有効な方法であることを示した。
【0046】
第四級アンモニウム化合物、例えばジメチルジオクタデシルアンモニウムブロミド、クロリド若しくはこれらの他の有機若しくは無機塩(DDA−B、DDA−C又はDDA−X)、ジメチルオクタデセニルアンモニウムクロリド、ブロミド若しくはこれらの他の有機若しくは無機塩(DODA−C、DODA−B又はDODA−X)、又は1,2−ジオレイル−3−トリメチルアンモニウムプロパン(DOTAP)、1,2−ジミリストイル−3−トリメチルアンモニウム−プロパン、1,2−ジパルミトイル−3−トリメチルアンモニウム−プロパン、1,2−ジステアロイル−3−トリメチルアンモニウム−プロパン及びジオレイル−3−ジメチルアンモニウムプロパン(DODAP)及びN−[1−(2,3−ジオレイルオキシ)プロピル]−N,N,N−トリメチルアンモニウム(DOTMA)は、水性媒体に分散されたときに、脂質二重層、単層及び多層両方の全てのタイプのリポソーム、ミセルなどの脂質集合体を形成することができる。これらの構造の脂質膜は、糖脂質、例えば、小胞の分散を安定化することが示されているMMG又はα,α’−トレハロース6,6’−ジベヘネート(TDB)(Davidsen et al,PCT/DK2005/000467)など他の両親媒性化合物の封入体のための優れた基質を提供する。
【0047】
MMGと送達系との組合せは相乗的に作用して、例えば、DDAが単独で投与されたときの免疫応答を増強することができる。その結果、DDAは、低レベルのIFN−γ産生を促進するが、しかしながらMMGとの組合せで、IFN−γ産生は劇的に増大する。
【0048】
本発明のリポソームは、当技術分野において周知の種々の方法(Davidsen et al,PCT/DK2005/000467)により作製することができる。MMGのリポソーム/送達系中への組み込みは、リポソームとMMGとの単なる混合を含む、当技術分野において周知の種々の方法により行うことができる。特に、MMGのリポソーム中への組み込みは、Davidsen et al,PCT/DK2005/000467に記載されたようにして行うことができる。
【0049】
疾患に対する免疫を提供するのに加えて、本発明のアジュバントの組合せは、それ自体では免疫原性の不十分な物質である化合物に対する抗体を産生するために使用することもでき、そのような抗体は、例えば医学及び分析化学において問題の化合物の検出及び定量のために使用することができる。
【実施例】
【0050】
材料及び方法:
ウシ型結核菌BCGからの無極性及び極性の脂質の抽出
ウシ型結核菌BCG(コペンハーゲン株)をSauton変法培地中で培養した。マイコバクテリアを2〜3週間後に収集してPBS中に懸濁させ、60℃で1.5時間インキュベートすることにより殺菌した。無極性及び極性の脂質を、標準的手順(Dobson et al,1985)(Rosenkrands et al.2005)に従って抽出した。
【0051】
無極性脂質を抽出するために、0.3%NaCl:メタノール(440ml)及び440mlの石油エーテルを20gのマイコバクテリア(湿重量)に加えて、混合物を2時間攪拌した。遠心分離後、上層を除去して下層を440mlの石油エーテルで再抽出した。両方の抽出で得た上清相を合わせて蒸発させ、無極性脂質を得た。
【0052】
極性脂質を抽出するために、生物由来資源を含むメタノール性食塩水溶液を、沸騰水浴中100℃で10分間加熱し、続いて37℃で10分間冷却した。体積520mlのクロロホルム:0.3%NaCl:メタノール(9:10:3)を加えて、混合物を終夜攪拌した。全混合物を焼結ガラス漏斗を通して濾過し、濾過ケーキを捕集して170mlのクロロホルム:メタノール:0.3%NaCl(5:10:4)で2回再抽出した。3つの水性メタノール性クロロホルム相全てを合わせて、580mlのクロロホルム:0.3%NaCl水溶液(1:1)を加え、10分間攪拌した。放置して相を分離させた後、上の水層を除去して廃棄した。下の有機層を蒸発乾固すると極性脂質が得られた。
【0053】
個々の無極性脂質の精製
PDIM及びTAGを石油エーテル/アセトン(98:2)におけるTLC処理を使用して単離した。フチオセロールAに基づくPDIMの主成分のみ回収された。PGL及びMMGは、トルエン/アセトン(95:5)におけるTLC処理により一緒に分離された。PGLとMMGとは、クロロホルム:メタノール:0.880アンモニア(97:3:0.5)で1次元TLCにより分離された。PDIM、TAG、PGL及びMMGを、500MHz 1H及び13C核磁気共鳴(NMR)(Bruker drx500)及びMALDI−TOF質量分析(MS)(Bruker Biflex IV)にかけた。再水和した脂質抽出物(1mg/ml)の試料(10μl)を、SDS−PAGE(Laemmli et al,1970)及び銀染色(Blum et al,1987)により分析し、残留タンパク質含有量を求めた。脂質調製物は、エンドドキシン汚染のないことが認められた(<0.001ngLPS/μg脂質)。
【0054】
モノミコリルグリセロールの加水分解
モノミコリルグリセロールを16×100mmの試験管中で5%TBAH水溶液(2.5ml)とともに100℃で終夜加熱した(Minnikin et al,1988)。冷却後、混合物を水(2ml)で希釈し、ジクロロメタン中ヨードメタンの10%溶液(3ml)を加えて、試験管を回転装置上に1時間置いた。上の水層を廃棄して、下層を等量の1MのHCl及び水で洗浄し、蒸発乾固すると残渣が生じて、TLC(石油エーテル:アセトン95:5)により、α−及びケトミコレートメチルエステルを含有することが示された。分取TLC(上記の)により、αメチルミコレート及びケトメチルミコレートが得られた。
【0055】
α−MMG及びケト−MMGを分離するために、トリメチルシリル誘導体化を使用した。MMG混合物と600mlのTRI−SIL試薬(Pierce)とを75℃で20分間加熱した。次に、冷却した溶液を窒素気流下で乾燥し、分取TLCプレート上に負荷して石油エーテル:トルエン(50:50)で展開し、0.01%ローダミンを使用してα−MMG及びケト−MMGのバンドを可視化し、長波長の蛍光で検査した。対応するバンドをTLCプレートから掻き落とし、ジエチルエーテルを使用してα−MMG及びケト−MMGを、シリカゲルから3回抽出した(3×5mL)。合わせた抽出物を乾燥し、ヘプタン:メタノール(1:1)及びパラ−トルエンスルホン酸の少量の結晶を添加して、1時間攪拌することによりトリメチルシリル基を除去した。ヘプタン層を回収して蒸発乾固すると、精製されたα−MMG及びケト−MMGが得られた。
【0056】
C36の生成を示すMMGの合成
3−ヒドロキシ−2−テトラデシル−オクタデカン酸(Datta et al,1991に従って合成された合成C32コリノミコール酸)(100mg、0.20mmol、1eq)及び4−ピロリジノピリジン(100mg、3eq)を50mlの丸底フラスコ中に入れて、モレキュラーシーブ4Aと一緒にジクロロメタン(500μl)中50μlの2,2−ジメチル−4−ヒドロキシメチル−1,3−ジオキソラン(sn−イソプロピリデングリセロール)溶液を加えた。その混合物を、高真空下に室温で完全に乾固させ、N’,N−ジシクロヘキシルカルボジイミダゾール(DCC)(15ml、DCM中0.1MのDCC、5eq)を加えて、反応混合物を室温で終夜攪拌するままにした。モレキュラーシーブを濾過により除去し、反応混合物を真空で濃縮して乾固し、残渣を、フラッシュカラムクロマトグラフィ(Fluka 60741シリカゲル60)を使用して、ヘキサンからヘキサン:酢酸エチル(8:2)を使用して5%増分で溶出させて精製し、純粋なイソプロピリデン保護化合物(3−ヒドロキシ−2−テトラデシル−オクタデカン酸−2,2−ジメチル−[1,3]−ジオキソラン−4−イルメチルエステル)を収率56%で得た(68mg)。1H NMR(CDCl3、300MHz)δH0.90(t、6H、CH3)、1.20(s、54H、CH2)、1.40(s、3H、CH3)、1.45(s、3H、CH3)、2.50(m、1H、CH)、4.05〜4.40(m、5H、CH2、CH);13C NMR(CDCl3、75MHz)δC15.0(CH3)、22.1、28.8、28.9、29.0、31.4(CH2)、29.1(CH3)、52.1(CH(CH2)13CH3)、63.2(CH2OCO)、69.3(CH2O)、73.4(CH(CH2)14CH3)、174.3(C=O);m/z(EI)633.55[M+Na+](100%);HRMS C38H74O5Na[M+Na+]に対する計算値633.5536、実測値633.5527。
【0057】
3−ヒドロキシ−2−テトラデシル−オクタデカン酸−2,2−ジメチル−[1,3]−ジオキソラン−4−イルメチルエステル(68mg、1eq)を6mlのトリフルオロ酢酸:テトラヒドロフラン:水(体積で8:17:3)溶液中に溶解して、室温で終夜攪拌した。その溶液を飽和重炭酸ナトリウム水溶液で中和して、混合物をクロロホルムで2回抽出した。有機抽出物を水及び塩水で洗浄して乾燥し、真空中で濃縮して粗生成物を白色固体として得、それを、10gのシリカゲルVarian Bond Elut 12256026カートリッジでフラッシュカラムクロマトグラフィにより、ヘキサンからヘキサン:酢酸エチル(7:3)を使用して、5%増分で溶出させることにより精製し、表題化合物を白色固体として収率49%で得た(32mg)。融点72〜74℃。1H NMR(CDCl3、300MHz)δH0.90(t、6H、CH3)、1.25(s、54H、CH2)、2.50(m、1H、CH)、3.45〜3.85(m、3H、CH、CH2)、4.25(m、2H、CH2);13C NMR(CDCl3、75MHz)δC15.0(CH3)、26.3、30.9、31.3、33.5(CH2)、47.5(CH(CH2)13CH3)、68.4(CH2)、69.5(CH(CH2)14CH3)、72.5(CH2O)、76.4(CH)、175.4(C−1);m/z(EI)593.50[M+Na+](100%);HRMS C35H70O5Na[M+Na+]に対する計算値593.5121、実測値593.5143。
【0058】
樹状細胞アッセイ
ヒトPBMC由来のDCを、Romani et al.,1994から改良した方法に従って得た。末梢血はバフィーコートから得た。簡単に述べると、単球は、Ficoll−Hypaque遠心分離により単離し(Lymphoprep 1077密度媒質、Nycomed、オスロー、ノルウエイ)、続いて抗CD14標識磁性ビーズ(MACS;Miltenyi Biotech、Bergesh Gladbach、ドイツ)を使用してCD14陽性細胞を分離した。単球は、10%FCS、50μMの2−メルカプトエタノール、100U/mlのペニシリン、100μg/mlのストレプトマイシン、2mMのL−グルタミン(全てGibco)(CM)で補完した完全RPMI 1640中で、100ng/mlのヒト組み替えGM−CSF(Prepotech、Rocky Hill、NJ、USA)及び50ng/mlのヒト組み替えIL−4(Becton Dickinson(BD))を存在させて、37℃、5%CO2で7日間培養した。
【0059】
7日目に、iDC(1×105細胞/ml)を、リポ多糖(LPS)(大腸菌O127:B8)(Sigma−Aldrich、Brondby、デンマーク)又はウシ型結核菌BCG由来の脂質とともに、さらに24時間培養した。脂質抽出物は、乾燥ウシ型結核菌脂質材料をクロロホルム:メタノール(2:1)で再溶解し、続いて溶媒を蒸発させ、CM中にプローブ超音波処理することにより調製した。脂質を、未成熟DCに0.1ないし100μg/mlで加えた。
【0060】
フローサイトメトリー分析
DCを先ず関連するmAb(BD Pharmingen)と(30分、4℃)、続いて1/20希釈FITC結合ヤギ抗マウスIg(Jackson ImmunoResearch Laboratories、West Grove、PA)と(30分、4℃)インキュベートすることにより、表面マーカーを染色した。非特異的Abの結合は、関連する1次ヒトmAbの添加前に、10%ウシ胎児血清溶液で阻止した(15分、4℃)。染色された細胞は、直ちにFACScanフローサイトメトーター(BD)を使用して、フローサイトメトリーにより検査し、CellQuestソフトウェアで分析した。
【0061】
サイトカイン測定
DC培養上清を収集して、−20℃で貯蔵した。分泌されたIL−12p70、IL−6及びTNF−αを、ELISA(BD)によりメーカーの説明書に従って測定した。
【0062】
混合白血球反応(MLR)アッセイ
混合リンパ球反応(MLR)アッセイのためのiDCは、上で概説したようにして、単球から発生させた。生成した細胞(iDC)は、同じ培地中で、又は脂質(10μg又は100μg/ml)を含有する培地中で24時間培養した。0.125×105から2×105まで漸増させたDCを、37℃/5%CO2でPPD陰性の提供者からの同種異系のT細胞(105細胞/ウェル)と平底96ウェルのマイクロタイタープレート中でインキュベートした。T細胞は、Pan−T細胞単離キット(Miltenyi)を使用し、メーカーの説明書に従って単離した。DCと同種異系T細胞との共培養は、6日間インキュベートした。上清を収集して、分泌されたIFN−γがELISA(BD)によりメーカーの説明書に従って測定されるまで、−20℃で貯蔵した。次に、両方のアッセイには、18時間の最終培養の間、1μCi/ウェルの[3H]チミジンを含む培地をパルスで適用した。細胞を収集して、T細胞の増殖を液体シンチレーション計数(Microbeta Systems)により測定した。全てのアッセイは、少なくとも3名の異なる提供者を使用して3重で実施した。
【0063】
抗原
Ag85BとESAT−6との融合タンパク質(以下、Ag85B−ESAT−6と称する)は、前に記載したように(Olsen et al,2001)組み替えタンパク質として産生された。
【0064】
動物
8ないし12週齢の雌BALB/c又はC57BL/6マウスを、Bomholtgaard(Ry、デンマーク)又はHarlan Scandinavia(デンマーク)から入手した。感染させたマウスは、BL−3層流安全密閉箱内のケージ中で飼育した。
【0065】
免疫
マウスは、尾部の底部で皮下に(s.c.)、各免疫化間2週間の間隔で最高3回免疫した。ワクチン(0.2ml/マウス)は、特に断らない限り、250μgのDDAと10μgの再水和脂質抽出物との中で乳化された2μgの融合タンパク質Ag85B−ESAT−6からなるものであった。幾つかの場合には、11mol%のTDBがDDAリポソーム中に組み込まれた(Davidsen et al、PCT/DK2005/000467)。結核菌感染を含む実験における陽性対照として、1群のマウスが、尾部の底部皮下に、1用量のBCGデンマーク株1331の注射を受けた。全ての又は個々の脂質抽出物は、乾燥ウシ型結核菌脂質材料を1mg/ml又は5mg/mlのMilli Q水で再水和し、続いてプローブ超音波処理することにより調製した。標準的脂質ワクチンは、抗原を食塩水と混合し、続いて再水和した脂質抽出物及びDDAを添加して、ボルテックスで混合することにより調製した。ワクチンを終夜放置して、抗原を吸着させた。
【0066】
リンパ球培養
血液試料又は鼠径部リンパ節を、最終免疫の7〜150日後のマウスから採取し、前に記載したようにして(Rosenkrands et al,2005)調製した。細胞培養は、2−メルカプトエタノール、グルタミン、ペニシリン−ストレプトマイシン、Hepes、及び10%ウシ胎児血清で補完された、体積200μlのRPMI中に2×105の細胞を入れた丸底マイクロタイターウェル中で、3重で実施した。再刺激のための抗原は、5μg/mlの濃度で使用した。培地のみ又は5μg/mlのConAを入れたウェルが、全ての実験でそれぞれ陰性及び陽性対照として含まれた。培養上清は、並行する培養液から、抗原を存在させて72時間インキュベートした後収集して、IFN−γの量を酵素結合免疫吸着測定法(Brandt et al,2000)により測定した。
【0067】
実験的感染
ワクチンの有効性を評価するために、マウスに、最初の免疫の2.5ヵ月後に、約25CFUの有毒結核菌Erdman株を投入するように較正されたGlas−Colの吸入曝露系で、エアロゾル経路により肺にチャレンジした。脾臓及び肺における細菌負荷は、BCGの増殖を選択的に阻害するために1ml当たり2μlの2−チオフェン−カルボン酸ヒドラジドで補完したMiddlebrook 7H11アガー上に、段階希釈液を蒔くことにより、6週間後に測定した。37℃で2〜3週間インキュベーション後に、コロニーを計数した。
【0068】
統計分析
感染マウスと対照マウスとの間のコロニー数の差を、変動分析により検定した。有意の効果が示されたときは、平均間の差をダネット検定により評価した。ヒトDCアッセイについては、異なる脂質に対する応答におけるサイトカイン放出の差を、変動分析により検定し、有意の効果が示されたときは、平均間の差をチューキー検定により評価した。
【実施例1】
【0069】
ウシ型結核菌BCG由来の無極性脂質の単離及び免疫刺激活性
ウシ型結核菌BCGの全脂質を、極性及び無極性分画に分離した。極性分画中で同定することができた脂質は、ホスファチジルイノシトールマンノシド(1〜4)、ホスファチジルイノシトール(PI)、ホスファチジルエタノールアミン(PE)、ジホスファチジルジグリセロール(DPG)及びL−α−ホスファチジル−DL−グリセロール(PG)であった。多数の未知のリン脂質も同定された(7及び8)(図1A)。無極性分画中で同定された主な脂質は、フチオセロールジミコセロセート(PDIM)、トリアシルグリセロール(TAG)、フェノール性糖脂質(PGL)及びモノミコリルグリセロール(MMG)(図1)であった。FFAは遊離脂肪酸である。
【0070】
無極性及び極性脂質の比較のための免疫刺激性活性は、ヒト末梢血単球由来の未成熟DC(iDC)を使用して検査した(図1)。無極性脂質で処理した結果、活性化マーカーCD86、CD40及びHLA−DRのレベルは、処理しなかった対照と比較して、用量に依存して上昇した(図1B)。100μg/mlの無極性脂質の用量は、強力な免疫刺激性分子LPS(0.1μg/ml)で観察されるのと同等の、並びにマイコバクテリアのコード因子(TDM)及びMPLと比較して優れたDC活性化を生じた。これらの分子の上方制御は、炎症誘発性メディエーターの腫瘍壊死因子−α(TNF−α)、インターロイキン(IL)−6及びIL−12の分泌を伴った(図1C)。極性脂質で処理されたiDCの上清中のこれらの炎症誘発性サイトカインのレベルは、このアッセイを使用する検出の限界未満であった。最後に、本発明者らは、DC活性化の他の表示として、PPD陰性の提供者からの同族異系T細胞を使用する混合白血球反応(MLR)を使用した(図1DおよびE)。無極性脂質で処理したDCの高い活性化状態をさらに支持することとして、本発明者らは、高レベルの増殖及びIFN−γ放出を認めたが、MLRは極性分画によっては誘発されなかった。
【実施例2】
【0071】
ウシ型結核菌BCGの無極性脂質抽出物からの個々の脂質の単離及びMMGの特徴づけ
免疫刺激性の無極性分画の脂質は、分取TLCを使用して単離され、フチオセロールジミコセロセートA(PDIMA)、TAG、PGL及びMMGの純粋な試料を与えた。少量のフチオセロールB及びフチオジオロンAも検出されたが、利用した分取方法では回収されなかった(図2A)。脂質の構造及び同一性は、1H核磁気共鳴(NMR)及び質量分析法(MS)により確認された(図2B及び不掲載データ)。MMG成分は、1−モノアシルグリセロールに特徴的な1H及び13C NMRスペクトルを示した(Gunstone et al,1991)。全MMG分画のNMRデータと結びつけたMALDI−TOFMS(表1)は、α−ミコレート並びにcis及びtrans型のケト−ミコレートの存在を明らかにした。主成分のおよその比は、それぞれ1.00:0.29:0.24であった。
【0072】
表1:精製されたMMGのMALDI−TOF質量分析。シグナルはM+Na+イオンに対するm/zである。系列の多量成分は太字で示し、その主成分には下線を引いた。
【実施例3】
【0073】
MMGによるヒト樹状細胞の活性化
精製されたMMG、PDIMA、PGL及びTAGを、ヒトiDCを活性化するそれらの能力について評価した。これらのアッセイにおいて、MMGは、CD86、CD40及びHLA−DRの顕著な上方制御に通ずる、DC活性化の最も強力な誘発因子であることが一貫して認められた(図3A)。MMGは、マイコバクテリアの病原性と長らく関連づけられてきた脂質であるPDIMAよりさらに大きくDCを活性化し(Cox et al,1999)、PDIMAは2番目に活性な脂質となったが、一方、PGL及びTAGが誘発する活性化はそれより小さかった。6名の個々の提供者における活性化の順は、MMG>PDIMA>PGL>TAGと認められ、CD86レベルにおいて、未処理iDCに関するレベルを超える平均の上昇倍率は、それぞれ1.91±0.29、1.82±0.43、1.52±0.26及び1.32±0.14であった。サイトカイン誘発も全体的に同じ傾向をたどり(図3B)、MMGは最も強力な免疫刺激剤脂質として卓越している。IL−6は、MMGに曝されたDCにより、PGL又はTAGにより誘発されるレベルを有意に超えるレベルで(P<0.05)放出された。他の脂質によるサイトカイン誘発の間で、有意の差は記録されなかった。したがって、MMGは、ウシ型結核菌BCGの無極性脂質分画中で最も強力な免疫刺激性脂質として分類することができる。
【実施例4】
【0074】
α−及びケト−MMGの免疫刺激活性
この実施例において、本発明者らは、MMGの免疫刺激特性を精査し、その強力な免疫刺激性能の原因となる有効成分を同定することをさらに求めた。α−MMGとケト−MMGとを、MMGのトリメチルシリルエーテルの調製、分取TLC処理及びそれに続くトリメチルシリルエーテル保護基の加水分解によって分離し、α−MMG及びケト−MMGを得た。α及びケト−ミコレートの構造を図2Bに示す。ヒトiDCを活性化するそれらの能力を評価したとき、α−及びケト−MMGは、活性化マーカーのレベルにおいて2〜3倍の程度で刺激した(図4)。したがって、MMGの2つの下位成分も、ヒトDCを刺激する顕著な能力を示す。
【実施例5】
【0075】
ウシ型結核菌BCGから単離されたMMGによるTh1免疫応答の誘発
MMGのアジュバント活性を研究するために、単離された脂質を、マウスにおいてIFN−γ産生を誘発するそれらの能力について試験した。C57BL/6マウスに、10μgの全脂質又は個々の脂質を投与した。In vivoにおいて、DDAは、脂質を入れて送達するビヒクルとして役立つ。それゆえ、250μgのDDA中で乳化された2μgの融合タンパク質Ag85B−ESAT−6及び10μgの再水和脂質抽出物を、皮下経路により投与した。DDAリポソーム中に組み込まれた10μgのMMGの用量は、排出するリンパ節から単離されたPBMCの再刺激で、10ng/mlのIFN−γレベルを生じた。このレベルは、DDA中の全脂質に対して記録されたレベルと、等用量で同等である(図5)。PDIM−Aも、より低いレベルとはいえ、IFN−γ産生を誘発したが、TAG又はPGLの組み込まれたDDAリポソームは、IFN−γ放出を殆ど助長しなかった(図5及び不記載データ)。これらの個々の脂質は、全脂質抽出物又は個々の脂質のいずれで再刺激しても記憶応答は観察されなかったので、アジュバントとして作用しているように思われることは注目すべきである(不記載データ)。したがって、MMGは、in vivoにおいて最も活性な無極性脂質でもあるとして確認されて、BCG由来の全脂質のアジュバント活性の大部分を単独で説明することができた。
【実施例6】
【0076】
MMGに基づくアジュバントの防御有効性
MMGに基づくアジュバントのTB感染に対する防御提供能力を評価するために、C57BL/6マウスを、MMG(2つの異なる用量)及びDDAに送達されたAg85B−ESAT−6により免疫した。BCGワクチン接種及びアジュバント単独を受けるマウスの群は、それぞれ陽性及び陰性対照として含まれた。最後のワクチン接種の6週間後に、マウスに、生結核菌をエアロゾル経路によりチャレンジした。ワクチンが細菌負荷を減少させる能力を、その6週間後に肺及び脾臓で測定した。これらのデータは、MMG/DDAをアジュバントとして有意の防御レベル、及びBCGのレベルと同等の防御レベルを示した(図6)。予想されたように、抗原なしでアジュバントを接種されたマウスは、細菌の増殖を阻止することができなかったので、この効果は特異的である。
【実施例7】
【0077】
MMGとDDAとを組み合わせることによるさらに優れた効果
免疫調節剤(MMG)と送達系(DDA)とを組み合わせる効果を評価するために、DDA単独若しくはDDA/MMGの組合せで(実験1、図7A)、又は実験2(図7B)においてはMMG単独若しくはDDA/MMGの組合せで、C57BL/6マウスにワクチンを接種した。これらの実験から、免疫応答は、DDAとMMGとを組み合わせることにより、劇的に増強されることが明らかである。
【実施例8】
【0078】
TDB/第3成分のMMGとDDAとのリポソーム中への組み込みによる免疫応答の増強
他の免疫刺激性成分と組み合わせたときの、MMGのアジュバント活性に対する効果を研究するために、C57BL/6マウスにおける免疫応答を、DDAリポソーム中に組み込まれたAg85B−ESAT−6及び10μgのMMG、又は免疫調節剤TDBの組み込まれたDDAリポソーム(Davidsen et al,PCT/DK2005/000467)を使用して皮下で免疫した後、最初のワクチン接種の5ヵ月後に評価した。DDAリポソーム中に組み込まれたMMGという組合せは、血液から単離されたPBMCの再刺激で、約25ng/mlのIFN−γレベルを生じたが、IFN−γの放出は、TDBの組み込まれたDDAリポソームが使用されたとき、劇的に上昇した(図8)。したがって、MMG、DDA及びTDBの間で相乗的効果が観察され、MMGとDDAとの組合せに対する第3成分の添加は、アジュバント活性をさらに増強させ得ることを示した。
【実施例9】
【0079】
MMG類似体のアジュバント活性は、天然MMGの活性と同等である
合成MMG類似体の免疫学的効果を評価するために、C57BL/6マウスを、天然MMG、16個の炭素を有する合成MMG類似体(図9に示した)及び36個の炭素を有する合成MMG類似体(図9に示した)(全て10μg/DDA/MMG)を加えたDDA中のAg85B−ESAT−6で免疫した。免疫応答は、最後のワクチン接種の1週間後に血液で測定され、3種のMMGに基づくアジュバントで同等のレベルの応答を示したが、DDAは、それだけではより低い効果を再び示した。
【実施例10】
【0080】
鎖長が短いほど免疫応答は高い
鎖長がより短い合成MMG類似体の免疫学的効果を評価するために、単独のDDA中、天然MMGを加えたDDA中、C8ないしC36の範囲の異なる合成類似体を加えたDDA中のAg85B−ESAT−6で(全て1μg/用量)、C57BL/6マウスを免疫した。免疫応答は、最後のワクチン接種の3週間後に血液で測定され、合成MMG類似体は1μgの用量レベルでも活性であることを示した。さらに、これらの結果は、より短い合成MMG類似体(16C以下)は、天然MMGと比較してより有効であることも示す。***P<0.001(図11)
【0081】
[参考文献]
Bennekov, T., J. Dietrich, I. Rosenkrands, A. Stryhn, T.M. Doherty, and P. Andersen. 2006. Alteration of epitope recognition pattern in Ag85B and ESAT−6 has a profound influence on vaccine−induced protection against Mycobacterium tuberculosis. Eur. J. Immunol. 36( 12): 3346−55.
Gregoriadis, G. 1995. Engineering liposomes for drug delivery: progress and problems. Trends Biotechnol. 13(12): 527−37.
Blum, H., H. Beier, and H. J. Gross. 1987. Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis 8: 93−99.
Brandt. L., M. Elhay, I. Rosenkrands, E. B. Lindblad, and P. Andersen. 2000. ESAT−6 subunit vaccination against Mycobacterium tuberculosis. Infect. Immun. 68: 791−795.
Brennan, P. J. and M. B. Goren. 1979. Structural studies on the type−specific antigens and lipids of the mycobacterium avium.Mycobacterium intracellulare.Mycobacterium scrofulaceum serocomplex. Mycobacterium intracellulare serotvpe 9. J. Biol. Chem. 254(10): 4205−4211.
Cox, J. S., B. Chen, M. McNeil, and W.R. Jacobs, Jr. 1999. Complex lipid determines tissue−specific replication of Mycobacterium tuberculosis in mice. Nature 402: 79−83.
Dascher, C. C, K. Hiromatsu, et al. 2003. Immunization with a mycobacterial lipid vaccine improves pulmonary pathology in the guinea pig model of tuberculosis. Int. Immunol. 15(8): 915−925.
Davidsen, J., Rosenkrands, L, and P. Andersen. (PCT/DK2005/000467)
Dobson, G., D. E. Minnikin, S. M. Minnikin, J. H. Parlett, M. Goodfellow, and M. Ridell, a. M. M. Systematic analysis of complex mycobacterial lipids (ed. Minnikin, M. G. a. D. E.) 237−265 (Academic Press, London, 1985).
Gluck, R. 1995. Liposomal presentation of antigens for human vaccines. Pharm Biotechnol. 6: 325−345.
Gunstone, F. D. 1991. Chemistry and Physics of Lipids 58: 219−224.
Harboe, M., A.S. Malin, H.S. Dockrell, H.G. Wiker, G. Ulvsund, A. Holm, M.C. Jβrgensen, Andersen P. B−cell epitope and quantification of the ESAT−6 protein of Mycobacterium tuberculosis. Inft. Immun. 66(2):717−23.
Hiu, I. J. 1975. Mycobacterial adjuvant and its carrier. Experientia. 31(8): 983−5.
Indrigo, J., R. L. Hunter, Jr, et al. 2002. Influence of trehalose 6,6’−dimycolate (TDM) during mycobacterial infection of bone marrow macrophages. Microbiology 148(7): 1991−1998.
Koike, Y., Y. C. Yoo, et al. 1998. Enhancing activity of mycobacterial cell−derived adjuvants on immunogenicity of recombinant human hepatitis B virus vaccine. Vaccine 16(20): 1982−1989.
Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680−685.
Lima, V. M. F., V. L. D. Bonato, et al. 2001. Role of Trehalose Dimycolate in Recruitment of Cells and Modulation of Production of Cytokines and NO in Tuberculosis. Infect. Immun. 69(9): 5305−5312.
Lindblad, E., M. Elhay, et al. 1997. Adjuvant modulation of immune responses to tuberculosis subunit vaccines. Infect. Immun. 65(2): 623−629.
McBride, B. W., A. Mogg, et al. 1998. Protective efficacy of a recombinant protective antigen against Bacillus anthracis challenge and assessment of immunological markers. Vaccine 16(8): 810−817.
Minnikin, D. E. 1982. Lipids: complex lipids, their chemistry, biosynthesis and roles. The Biology of the Mycobacteria. C. R. a. J. L. Stanford. London, Academic Press.
Minnikin, D. E. in Bacterial Cell Surface Techniques (ed. Hancock, I. C, Poxton, I. R.) 125−135 (Wiley, Chichester, 1988).
Minnikin, D. E. 1988. Isolation and purification of mycobacterial wall lipids. Bacterial Cell Surface Techniques. I. C. Hancock, Poxton, I. R. Chichester, Wiley: 125−135.
Minnikin, D. E., L. Kremer, et al. 2002. ”The Methyl−Branched Fortifications of Mycobacterium tuberculosis. Chemistry & Biology 9(5): 545−553.
Moingeon, P., J. Haensler, et al. 2001. Towards the rational design of ThI adjuvants. Vaccine 19(31): 4363−4372.
Mosmann, T. R. and S. Sad 1996. The expanding universe of T−cell subsets: ThI, Th2 and more. Immunology Today 17(3): 138−146.
Nathan, C, H. Murray, et al. 1983. Identification of interferon−gamma as the lymphokine that activates human macrophage oxidative metabolism and antimicrobial activity. J. Exp. Med. 158(3): 670−689.
Olsen, A. W., L. A. H. van Pinxteren, L. M. Okkels, P. B. Rasmussen, and P. Andersen. 2001. Protection of Mice with a Tuberculosis Subunit Vaccine Based on a Fusion Protein of Antigen 85B and ESAT−6. Infect. Immun. 69:2773−2778.
Rao V, F. N., Porcelli SA, Glickman MS 2005. Mycobacterium tuberculosis controls host innate immune activation through cyclopropane modification of a glycolipid effector molecule. J Exp Med 201: 535−543.
Reed, M. B., P. Domenech, et al. 2004. A glycolipid of hypervirulent tuberculosis strains that inhibits the innate immune response. Nature 431(7004): 84−87.
Romani, N., S. Gruner, D. Brang, E. Kampgen, A. Lenz, B. Trockenbacher, G. Konwalinker, P.O. Fritsch, R.M. Steinman, and G. Schuler. 1994. Proliferating dendritic cell progenitors in human blood. J.Exp. Med. 180: 83−93.
Rosenkrands, L, E.M. Agger, A.W. Olsen, K.S. Korsholm, CS. Andersen, K.T. Jensen, and P. Andersen. 2005. Cationic Liposomes Containing Mycobac− terial Lipids: a New Powerful ThI Adjuvant System. Infect. Immun. 73: 5817− 5826.
Saito, R., Tanaka, A., Sugiyama, K., Azuma, I. and Yamamura, Y. 1976. Adjuvant effect of cord factor, a mycobacterial lipid. Infect Immun. 13(3): 776−781.
Silva, CL. 1985. Inflamation induced by mycolic acid−containing glycolipids of Mycobacterium bovis (BCG). Brazilian J.Med.Biol.Res. 18: 327−335.
Sirakova, T. D., V. S. Dubey, et al. 2003. The Largest Open Reading Frame (pksl2) in the Mycobacterium tuberculosis Genome Is Involved in Pathogenesis and Dimycocerosyl Phthiocerol Synthesis. Infect. Immun. 71(7): 3794−3801.
Sprott, G. D., C J. Dicaire, et al. 2004. Activation of Dendritic Cells by Liposomes Prepared from Phosphatidylinositol Mannosides from Mycobacterium bo− vis Bacillus Calmette−Guerin and Adjuvant Activity In Vivo. Infect. Immun. 72(9): 5235−5246.
Stanfield, J.P., D. Gall, Bracken P.M. 1973. Single−dose antenatal tetanus immunisation. Lancet l(7797):215−9.
Suzuki F, B. R., Pollard RB. 1986. Importance of Lyt 1+ T−cells in the antitumor activity of an immunomodulator, SSM, extracted from human−type Tubercle bacilli.1986 77(2): 441−7.
Uehori, J., M. Matsumoto, S. Tsuji, T. Akazawa, O. Takeuchi, S. Akira, T. Kawata, I. Azuma, K. Toyoshima, and T. Seya. 2003. Simultaneous blocking of human Toll−like receptors 2 and 4 suppresses myeloid dendritic cell activation induced by Mycobacterium bovis Bacillus Calmette−Guerin peptidoglycan. Infect. Immun. 71:4238−4249.
van Rooij, E. M. A., H. L. Glansbeek, et al. 2002. Protective Antiviral Immune Responses to Pseudorabies Virus Induced by DNA Vaccination Using Di− methyldioctadecylammonium Bromide as an Adjuvant.” J. Virol. 76(20): 10540− 10545.
Yamazaki S, K. K., Someya S, Azuma I, Yamamura Y. 1969. Studies on the allergenicity of various tuberculoprotein derivatives and the adjuvanticity of wax D fractions of Mycobacterium tuberculosis. Am Rev Respir Pis. 100(5): 691−8.
【特許請求の範囲】
【請求項1】
アジュバント又は免疫調節剤を調製するための、モノミコリルグリセロール(MMG)又はそれらの合成同族体、類似体及び改変体の使用。
【請求項2】
脂質尾部を場合により有する、8〜36個の炭素を有するアルキル鎖に基づく合成MMG同族体、類似体又は改変体であって、最も好ましくは8〜16個の炭素を有するアルキル鎖を有し、各脂質尾部に0〜3個の二重結合を場合により有する、請求項1に記載の合成MMG同族体、類似体又は改変体の使用。
【請求項3】
図9に示されたような、8ないし36個の炭素を有するアルキル鎖に基づく合成MMG同族体、類似体又は改変体の、請求項2に記載の使用。
【請求項4】
請求項1〜3のいずれか一項に記載に従って調製されたアジュバント又は免疫調節剤。
【請求項5】
界面活性剤をさらに含む、請求項4に記載のアジュバント又は免疫調節剤。
【請求項6】
界面活性剤が、DDA−B、DDA−C、DDA−X、DODA−B、DODA−C、DODA−X、DOTAP、DODAP、DOTMA、DXPC、DXPE、DXPG又はそれらの組合せである、請求項5に記載のアジュバント又は免疫調節剤。
【請求項7】
非TLRリガンド例えばTDB及びMDP、又はTLRリガンド例えばPoly I:C、並びに/又はこれらの任意の類似体である追加の免疫調節剤を含む、請求項4〜6のいずれか一項に記載の、アジュバント又は免疫調節剤。
【請求項8】
各脂質尾部に0〜3個の二重結合を有する脂質尾部を場合により有する、8〜36個の炭素を有するアルキル鎖に基づく合成MMG同族体、類似体又は改変体。
【請求項9】
図9に示したような、8〜16個の炭素又は最大で36個の炭素を有する、請求項8に記載の合成MMG類似体。
【請求項10】
非経口、経口、粘膜投与、舌下、経皮、局所、吸入、鼻腔内、エアロゾル、眼内、気管内、直腸内、膣内のための、又は遺伝子銃、皮膚パッチによる、又は点眼剤若しくはうがい薬の形態における、請求項4〜7のいずれか一項に記載のアジュバント又は免疫調節剤を含むワクチン。
【請求項11】
抗原性成分が細胞内病原体由来の抗原性エピトープを含む、請求項10に記載のワクチン。
【請求項12】
抗原性成分が、有毒のマイコバクテリウム、例えば結核菌、ウシ型結核菌又はマイコバクテリウム・アフリカヌム(M.africanum)由来の抗原性エピトープを含む、請求項11に記載のワクチン。
【請求項13】
抗原性成分が、融合生成物Ag85b_TB10.4、Ag85b_ESAT−6_Rv2660、Ag85b_TB10.4_Rv2660及びAg85a_TB10.4_Rv2660の中から選択される、請求項12に記載のワクチン。
【請求項14】
抗原性成分が、抗原性エピトープ、即ち、熱帯熱マラリア原虫(Plasmodium falciparum)(例えばMsp1、Msp2、Msp3、Ama1、GLURP、LSA1、LSA3又はCSP)、トラコーマクラミジア(Chlamydia trachomatis)(例えばCT184、CT521、CT443、CT520、CT521、CT375、CT583、CT603、CT610又はCT681)、HIV、インフルエンザ又は肝炎B若しくはC由来の抗原性エピトープを含む、請求項11に記載のワクチン。
【請求項15】
癌、自己免疫障害、神経障害例えばアルツハイマー病、気道炎症、炎症性障害、感染性疾患、皮膚障害、アレルギー、喘息又は病原体により惹起される疾患を治療するための、請求項10に記載のワクチン。
【請求項16】
癌、自己免疫障害、神経疾患例えばアルツハイマー病、気道炎症、炎症性障害、感染性疾患、皮膚障害、アレルギー、喘息又は病原体により惹起される疾患を治療するための、請求項4〜7のいずれか一項に記載のアジュバント又は免疫調節剤を含む送達系。
【請求項17】
MMG又はそれらの合成同族体、類似体及び改変体が、1つまたは複数のワクチン、抗原、抗体、細胞傷害性因子、アレルゲン、抗生物質、アンチセンスオリゴヌクレオチド、TLR及び非TLRアゴニスト、TLR及び非TLRアンタゴニスト、ペプチド、タンパク質、遺伝子療法ベクター、DNAワクチン、免疫調節剤又は共刺激性分子との組合せで投与される、請求項14に記載の送達系。
【請求項1】
アジュバント又は免疫調節剤を調製するための、モノミコリルグリセロール(MMG)又はそれらの合成同族体、類似体及び改変体の使用。
【請求項2】
脂質尾部を場合により有する、8〜36個の炭素を有するアルキル鎖に基づく合成MMG同族体、類似体又は改変体であって、最も好ましくは8〜16個の炭素を有するアルキル鎖を有し、各脂質尾部に0〜3個の二重結合を場合により有する、請求項1に記載の合成MMG同族体、類似体又は改変体の使用。
【請求項3】
図9に示されたような、8ないし36個の炭素を有するアルキル鎖に基づく合成MMG同族体、類似体又は改変体の、請求項2に記載の使用。
【請求項4】
請求項1〜3のいずれか一項に記載に従って調製されたアジュバント又は免疫調節剤。
【請求項5】
界面活性剤をさらに含む、請求項4に記載のアジュバント又は免疫調節剤。
【請求項6】
界面活性剤が、DDA−B、DDA−C、DDA−X、DODA−B、DODA−C、DODA−X、DOTAP、DODAP、DOTMA、DXPC、DXPE、DXPG又はそれらの組合せである、請求項5に記載のアジュバント又は免疫調節剤。
【請求項7】
非TLRリガンド例えばTDB及びMDP、又はTLRリガンド例えばPoly I:C、並びに/又はこれらの任意の類似体である追加の免疫調節剤を含む、請求項4〜6のいずれか一項に記載の、アジュバント又は免疫調節剤。
【請求項8】
各脂質尾部に0〜3個の二重結合を有する脂質尾部を場合により有する、8〜36個の炭素を有するアルキル鎖に基づく合成MMG同族体、類似体又は改変体。
【請求項9】
図9に示したような、8〜16個の炭素又は最大で36個の炭素を有する、請求項8に記載の合成MMG類似体。
【請求項10】
非経口、経口、粘膜投与、舌下、経皮、局所、吸入、鼻腔内、エアロゾル、眼内、気管内、直腸内、膣内のための、又は遺伝子銃、皮膚パッチによる、又は点眼剤若しくはうがい薬の形態における、請求項4〜7のいずれか一項に記載のアジュバント又は免疫調節剤を含むワクチン。
【請求項11】
抗原性成分が細胞内病原体由来の抗原性エピトープを含む、請求項10に記載のワクチン。
【請求項12】
抗原性成分が、有毒のマイコバクテリウム、例えば結核菌、ウシ型結核菌又はマイコバクテリウム・アフリカヌム(M.africanum)由来の抗原性エピトープを含む、請求項11に記載のワクチン。
【請求項13】
抗原性成分が、融合生成物Ag85b_TB10.4、Ag85b_ESAT−6_Rv2660、Ag85b_TB10.4_Rv2660及びAg85a_TB10.4_Rv2660の中から選択される、請求項12に記載のワクチン。
【請求項14】
抗原性成分が、抗原性エピトープ、即ち、熱帯熱マラリア原虫(Plasmodium falciparum)(例えばMsp1、Msp2、Msp3、Ama1、GLURP、LSA1、LSA3又はCSP)、トラコーマクラミジア(Chlamydia trachomatis)(例えばCT184、CT521、CT443、CT520、CT521、CT375、CT583、CT603、CT610又はCT681)、HIV、インフルエンザ又は肝炎B若しくはC由来の抗原性エピトープを含む、請求項11に記載のワクチン。
【請求項15】
癌、自己免疫障害、神経障害例えばアルツハイマー病、気道炎症、炎症性障害、感染性疾患、皮膚障害、アレルギー、喘息又は病原体により惹起される疾患を治療するための、請求項10に記載のワクチン。
【請求項16】
癌、自己免疫障害、神経疾患例えばアルツハイマー病、気道炎症、炎症性障害、感染性疾患、皮膚障害、アレルギー、喘息又は病原体により惹起される疾患を治療するための、請求項4〜7のいずれか一項に記載のアジュバント又は免疫調節剤を含む送達系。
【請求項17】
MMG又はそれらの合成同族体、類似体及び改変体が、1つまたは複数のワクチン、抗原、抗体、細胞傷害性因子、アレルゲン、抗生物質、アンチセンスオリゴヌクレオチド、TLR及び非TLRアゴニスト、TLR及び非TLRアンタゴニスト、ペプチド、タンパク質、遺伝子療法ベクター、DNAワクチン、免疫調節剤又は共刺激性分子との組合せで投与される、請求項14に記載の送達系。
【図1】


【図2】
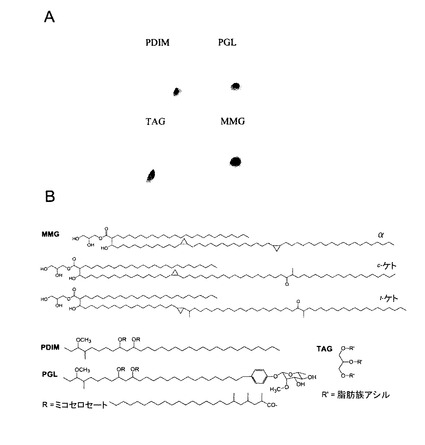
【図3】

【図4】

【図5】

【図6】
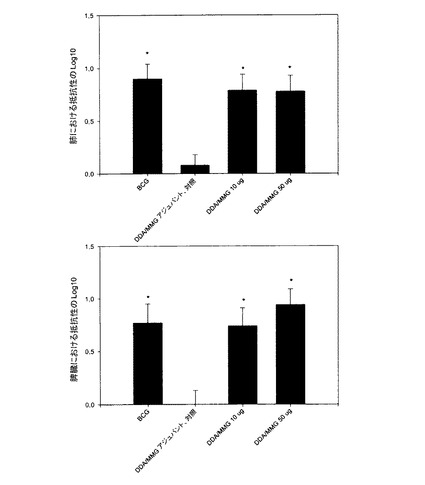
【図7】

【図8】

【図9】

【図10】

【図11】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2010−531820(P2010−531820A)
【公表日】平成22年9月30日(2010.9.30)
【国際特許分類】
【出願番号】特願2010−513651(P2010−513651)
【出願日】平成20年6月26日(2008.6.26)
【国際出願番号】PCT/DK2008/000239
【国際公開番号】WO2009/003474
【国際公開日】平成21年1月8日(2009.1.8)
【出願人】(507006422)
【Fターム(参考)】
【公表日】平成22年9月30日(2010.9.30)
【国際特許分類】
【出願日】平成20年6月26日(2008.6.26)
【国際出願番号】PCT/DK2008/000239
【国際公開番号】WO2009/003474
【国際公開日】平成21年1月8日(2009.1.8)
【出願人】(507006422)
【Fターム(参考)】
[ Back to top ]