
抗ルイスY抗イディオタイプ抗体およびその使用
本発明は、抗ルイスYモノクローナル抗体に特異的な抗イディオタイプ抗体を提供する。本発明は、hu3S193の可変領域に対して特異的に結合するハイブリドーマクローンによって生成されるmAbのELISAスクリーニング法、およびルイスY抗原に対するhu3S193結合を阻害する抗イディオタイプmAbの能力にも関する。また、本発明は、抗ルイスYモノクローナル抗体に特異的な抗イディオタイプ抗体を生成することが可能なハイブリドーマを提供する。本発明の別の態様は、LMH−1、LMH−2、LMH−3、またはLMH−4より成る群から選択される抗ルイスYモノクローナル抗体に特異的であるハイブリドーマを提供することである。本発明は、本発明の抗体を使用するHAMA、HACA、およびHAHA反応を検出する方法にも関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、分子生物学の分野、かつより具体的には抗体に関する。
【背景技術】
【0002】
25年前のモノクロナール抗体(mAb)技術の出現は、有用な研究用試薬の膨大なレパートリーを提供し、癌療法、自己免疫疾患、移植による拒絶反応、抗ウイルス予防における承認された医薬試薬として、および抗血栓剤として抗体を使用する機会を創造した(グレニー(Glennie)とジョンソン(Johnson)、2000年)。マウスmAbをキメラmAb(マウスV領域、ヒトC領域)およびmAb相補性決定領域(CDR)のみがマウス由来であるヒト化試薬へ変換する分子工学の応用は、mAb療法の臨床的成功に重要であった。工学的mAbは、免疫原性を大幅に削減または遠ざけ、血清中半減を増大させ、mAbのヒトFc部分は補体および細胞毒性細胞の免疫エフェクターを補充する可能性を増大させる(クラーク(Clark)、2000年)。生体内分布、薬物動態、および臨床的に投与されるmAbに対する免疫反応の誘導に対する調査には、医薬と内因性タンパク質とを区別する分析の開発が必要である。
【0003】
抗体は通常、それが認識する抗原に関して定義される。特定の抗原に対する抗体の特異性は、その抗原結合部位、抗原に接触する抗体分子の異なる領域よって判定される。この部位は、免疫グロブリンの重鎖および軽鎖の可変領域内に存在する。しかし、抗体は、抗体分子の単一特異的集団の独自の重鎖および軽鎖領域と関連した、そのイディオタイプ、イディオトープの集合体、または表面マーカーによっても定義される。抗体に特有の抗原決定基はイディオトープと呼ばれ、イディオトープを有する抗体との抗イディオタイプ抗体の反応によって定義される。イディオタイプは、研究者が免疫反応および遺伝性免疫グロブリン遺伝子における特定の抗体の外観および持続性を追跡できるという点で有用なマーカーである。イディオタイプは、抗イディオタイプ抗体の産生を促進しうる固有の決定基でもある。
【0004】
抗イディオタイプ抗体は、他の抗体の可変領域に結合し、免疫系において、内部(自己)抗原に対して生じる結合抗体における主要な役割を果たす(ジェルネ(Jerne)、1974年)。イディオタイプ抗体を誘発する可変領域によって誘発される抗イディオタイプ抗体は、3つのカテゴリー、すなわち、1)抗原結合部位(パラトープ)におけるエピトープの認識、2)パラトープ近く結合およびイディオタイプ抗体−抗原相互作用の立体的干渉、または3)イディオタイプ抗体(「内部イメージ」抗イディオタイプ抗体)によって認識される抗原の構造的模倣のうちの1つでありうる。後者のカテゴリーの抗イディオタイプは、細菌、ウイルス、および寄生虫感染に加えて腫瘍関連抗原に対する宿主の免疫反応の修飾におけるその可能性が与えられるワクチンとして生成および調査されている(バッタチャリヤ−チャタジー(Bhattacharya−Chatterjee)、チャタジー(Chatterjee)ら、2001年で概説)。齧歯類抗イディオタイプ抗体は、補助剤、および担体、通常、誘発される免疫または抗腫瘍反応に対するキーホールリンペットヘモシアニン(KLH)との抱合を必要とする。
【0005】
抗イディオタイプ抗体は、以下、ELISAと称される酵素免疫測定、注入イディオタイプ抗体の患者の血清レベルを検出するための免疫療法治験における試薬、または投与されるイディオタイプ抗体に対する免疫反応としても利用されうる。かかる免疫反応としては、ヒト抗マウス抗体、ヒト抗キメラ抗体、およびヒト抗ヒト化抗体(以下、それぞれ、HAMA、HACA、HAHAと呼ぶ)が挙げられる。抗イディオタイプ抗体NUH−82は、ヒト腎細胞癌におけるG250抗原を認識するマウスmAb G250の結合部位に対するものである(ウエムラ(Uemura)、オカジマ(Okajima)ら、1994年)。NUH−82は、cG250抗体放射免疫療法を受ける患者におけるHACA反応の測定に有用であることが証明されたが(ステフェンス(steffens)、ボーマン(Boerman)ら、1997年)、最近になって、NUH−82は薬物動態分析のために患者の血清におけるcG250の血清レベルを測定するサンドイッチELISAにおいて使用されている(リュー(Liu)、スミス(Smyth)ら、2002年)。
【0006】
hu3S193抗体は、乳癌、結腸癌、肺癌、前立腺癌、および卵巣癌を含む上皮腫瘍を発現するルイスY、ジフコシル化炭水化物抗原を標的するために開発された(キタムラ(Kitamura)、ストッカート(Stockert)ら、1994年、スコット(Scott)、ゲレイク(Geleick)ら、2000年)。ルイスYは、上皮由来の腫瘍細胞、例えば、肺癌、腸癌、乳癌、前立腺癌、または卵巣癌の表面上に発現される炭水化物分子である。ルイスY抗原は、正常組織において発現されるが、発現レベルは一部の腫瘍型においてより高く、抗原は一部の乳癌、結腸癌、胃癌、食道癌、膵癌、十二指腸癌、肺癌、膀胱癌、および腎癌のほか、胃および島細胞神経内分泌系腫瘍の細胞のマーカーとして使用されうる。ルイスY関連関連抗原は、その高い密度と一次および転移性病変における同質の発現により免疫療法の魅力的な標的である(キム(Kim)、ユアン(Yuan)ら、1986年、サカモト(Sakamoto)ら、1986年、チャン(Zhang)、コードン−カルド(Cordon−Cardo)ら、1997年)。
【0007】
hu3S193は、インビトロで強いADCCおよびCDCを誘発し、前臨床試験における選択充実腫瘍の免疫療法として有望であることが証明されおり(クラーク(Clarke)、リー(Lee)ら、2000年b、クラーク(Clarke)、リー(Lee)ら、2000年a)、現在、第I相用量増加臨床試験中である。前臨床生体内分布試験では放射性標識hu3S193がトレースされ、乳癌の動物モデルにおける安定化した放射性抱合体の腫瘍ターゲティング能を評価した(クラーク(Clarke)、リー(Lee)ら、2000年a)。病院では、hu3S193などmAbが放射性標識リングを使用してトレースされ、生体内分布および腫瘍イメージング、および血清からの放射性抱合体のクリアランスをmAb薬物動態の尺度として評価することができる(クラーク(Clarke)、リー(Lee)ら、2000年a、スコット(Scott)、ゲレイク(Geleick)ら、2000年、ホフマン(Hoffman)、スコット(Scott)ら、2001年)。しかし、臨床症状の非存在下には、HAHA免疫反応を検出することはできない。
【0008】
hu3S193の免疫エフェクター機能を考えると、初期治験における投与抗体の血清レベルの測定は将来の治療方式の計画を補助し、非標識hu3S193の薬物動態の測定を可能にすると同時に、HAHAを測定する能力はhu3S193の安全性評価を補助するであろう。
【0009】
具体的には、本発明は、それらが患者の循環において認識する投与イディオタイプの薬物動態をモニタリングするための診断用試薬として免疫療法治験における抗イディオタイプの使用にも関する。同様に、抗イディオタイプは、投与されたイディオタイプmAbに対するHAHA、HACA、またはHAMA免疫反応の陽性対照として使用されうる。かかる免疫反応は、患者集団の一部における臨床結果に対して全く影響を与えないか(グルーバー(Gruber)、ヴァンハーレム(van Haarlem)ら、2000年)、またはその後の治療を不可能にする免疫療法のmAbの薬物動態および生体内分布における過敏反応および劇的な変化と関連しうる(クラーク(Clark)、2000年)。
【0010】
(図面の簡単な記載)
図1A−B:抗−hu3S193モノクロナール抗体、LMH−1(●)、−2(▽)、−3(◇)、および抗−huIgG−4(◇)の結合特異性は、ELISAによって測定された。連続希釈ハイブリドーマ培養上清が、A)hu3S193(図1A)およびB)huIgG(図1B)でコーティングされたマイクロタイタープレートへの結合活性について検査された。二次抱合体(▲)および基質のみ(○)の対照も含めた。hu3S193に対する特異的結合が、LMH−1、−2、および−3で確認された。LMH−4は抗huIgG結合活性を示した。
【0011】
図2:クローン増殖後、ハイブリドーマ培養上清が、hu3S193抗原結合活性を中和する能力についてELISAによって3部で検査された。平均±SD結果は、合成Ley抗原でコーティングされたプレートへのhu3S193結合の遮断により抗hu3S193mAb、LMH−1、−2、および−3の拮抗薬活性を示した。
【0012】
図3A−D:3S193イディオトープに対するLMH−2およびLMH−3の特異性の確認。図3A。プロテインA精製LMH−3をELISAプレート上にコーティングし、連続希釈した10μg/mlのhu3S193、mu3S193、BR55.2の3部サンプル、および対照IgG1 mAb huA33およびmuA33をウェルに添加し、さらにLMH−3の結合特異性を特徴づけた。このクローンの追加の好ましい特徴は、ビオチニル化LMH−3がLMH−3で捕捉される結合3S193mAbの検出に利用されうることであった。図3B。hu3S193−Ley抗原結合に対する抗Leyおよび抗イディオタイプmAbによるLey四糖類コーティングプレートへの結合を阻止する拮抗薬結合。図3C。mu3S193−Ley抗原結合、および図3D。BR55.2−Ley抗原結合活性。
【0013】
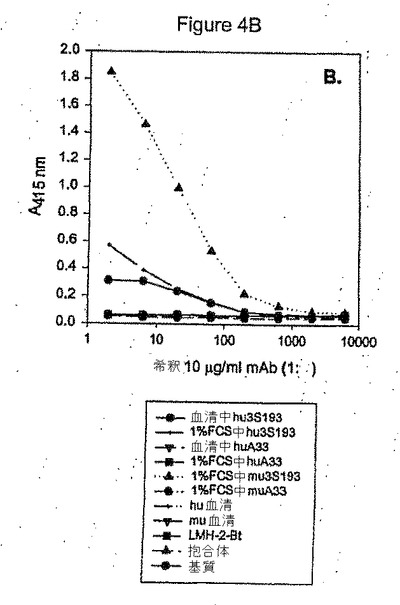
図4A−C:患者血清サンプルにおけるhu3S193を測定するためのELISAの開発。図4A。合成抗原Ley−BSA抱合体による捕捉、抗huIgGによる検出の抗イディオタイプ利用可能性より前に利用可能な方法は、患者の注入前血清サンプルにおけるhu3S193標準による低い感度および高いノイズを示す。図4B。抗hu3S193イディオタイプLMH−2による捕捉およびLMH−2−ビオチンによる検出。低い感度が確認される。図4C。抗hu3S193イディオタイプLMH−3による捕捉およびLMH−3−ビオチンによる検出は、最小のノイズおよびhu3S193血清中の3ng/mlに対する感度を示す。
【0014】
図5:hu3S193に対する血清HAHA反応のBiacore分析が最適化され、陽性対照としての抗イディオタイプLMH−3を使用して確認された。hu3S193をバイオセンサーチップ上に固定化し、ヒト血清μg/ml)中の抗イディオタイプmAb LMH−3(■)およびアイソタイプマッチド対照抗体(▲)の連続希釈を3部でチップ上を通した。
【発明の開示】
【発明が解決しようとする課題】
【0015】
本発明は、ヒト化抗ルイスYモノクローナル抗体、hu3S193に対する抗イディオタイプ抗体に関する。本発明は、hu3S193の可変領域に対して特異的に結合するハイブリドーマクローンによって生成されるmAbのELISAスクリーニング法、およびルイスY抗原に対するhu3S193結合を阻害する抗イディオタイプmABの能力にも関する。
【0016】
本発明は、本発明の抗体を使用するHAMA、HACA、およびHAHA反応物を検出する方法にも関する。
【課題を解決するための手段】
【0017】
本発明の1つの態様は、抗ルイスYモノクロナール抗体に特異的な抗イディオタイプ抗体を提供することである。本発明の別の態様は、抗ルイスYモノクロナール抗体の可変領域に結合する抗ルイスY抗体に特異的な抗イディオタイプ抗体を提供する。同様に、本発明の別の態様は、抗ルイスYモノクロナール抗体の結合を遮断する抗イディオタイプ抗体である。本発明の別の態様は、特異的にhu3S193に結合する抗イディオタイプ抗体を提供することである。具体的には、本発明において提供される抗イディオタイプ抗体は、モノクロナール抗体、キメラ抗体、ヒト抗体、ヒト化抗体、または一本鎖抗体より成る群から選択される。
【0018】
本発明の別の態様は、抗ルイスYモノクロナール抗体に特異的な抗イディオタイプ抗体を生成することが可能なハイブリドーマを提供することである。本発明の別の態様は、LMH−1、LMH−2、およびLMH−3より成る群から選択される抗ルイスYモノクロナール抗体に特異的であるハイブリドーマを提供することである。
【0019】
本発明の別の態様は、抗原への抗体の結合を阻害する抗イディオタイプ抗体の能力を検出する方法を提供する。本発明の別の態様は、結合イディオタイプ抗体を捕捉し、検出する抗イディオタイプの能力を検出するための方法を提供することである。本発明の別の態様は、イディオタイプ抗ルイスY抗体に結合する抗イディオタイプ抗体の能力を検出するための方法を提供することである。本発明の別の態様は、サンプル血清中の抗体の量を検出する方法である。
【0020】
本発明は、hu3S193を捕捉する抗原としてBSAに結合した合成ルイスY四糖類と、結合hu3S193の検出器としての市販の抗ヒト抗体製剤を使用するELISAによって、血清サンプル中のhu3S193を測定する方法を含む。アッセイの感度は、高いノイズをもたらす二次抗huIgG抗体によって検出される血清サンプルからのhuIgGの非特異的結合によって大きく制限されている。cG250に対するHACA反応および抗イディオタイプNUH−82を有するcG250の血清レベルの測定による経験に基づき、適切な抗イディオタイプhu3S193mAbの産生が不可欠となった。
【0021】
本発明は、本発明の抗体を使用するHAMA、HACA、およびHAHA反応物を検出する方法にも関する。
【発明を実施するための最良の形態】
【0022】
本発明は、抗ルイスYモノクロナール抗体に特異的な抗イディオタイプ抗体を提供する。これらの抗イディオタイプ抗体は、抗ルイスYモノクロナール抗体の可変領域に結合する抗ルイスY抗体に特異的である。同様に、本発明の別の態様は、抗ルイスYモノクロナール抗体の結合を遮断する抗イディオタイプ抗体である。本発明の別の態様は、特異的に抗ルイスYモノクロナール、hu3S193に結合する抗イディオタイプ抗体を提供することである。具体的には、本発明において提供される抗イディオタイプ抗体は、モノクロナール抗体、キメラ抗体、ヒト抗体、ヒト化抗体、または一本鎖抗体より成る群から選択されうる。
【0023】
また、本発明は、抗ルイスYモノクロナール抗体に特異的な抗イディオタイプ抗体を生成することが可能なハイブリドーマを提供する。本発明の別の態様は、LMH−1、LMH−2、およびLMH−3より成る群から選択される抗ルイスYモノクロナール抗体に特異的であるハイブリドーマを提供することである。
【0024】
本明細書で使用される「ヒト化」抗体という語は、非ヒト種免疫グロブリン由来のそのCDR(補体決定領域)を有する分子、および主にヒト免疫グロブリン由来の抗体分子の残りを指す。
【0025】
本明細書で使用される「抗体」という語は、別に指示がない限り、広く抗体分子およびさまざまな抗体由来分子の両方を指す。かかる抗体由来分子は、少なくとも1つの可変領域(重鎖または軽鎖可変領域のいずれか)およびFabフラグメント、Fab’フラグメント、F(ab’)2フラグメント、Fdフラグメント、Fabcフラグメント、Fdフラグメント、Fabcフラグメント、Sc抗体 (一本鎖抗体)、二重特異性抗体、個々の抗体軽鎖、個々の抗体重鎖、抗体鎖と他の分子とのキメラ融合などの分子を含む。
【0026】
免疫グロブリンに関連して本明細書で使用される「可変領域」という語は、免疫学の当業者によってこの語に与えられた通常の意味を有する。抗体重鎖および抗体軽鎖は「可変領域」および「定常領域」へ分割されうる。可変領域と重鎖領域との分割点は、抗体構造を記載した標準の教科書、例えば、カバト(Kabat)ら、「Sequence of Proteins of Immunological Interest:第5版」、米国保険福祉省、米国政府印刷局(1991年)を参照することにより当業者によって容易に決定されうる。
【0027】
本明細書で使用される「イディオタイプ」という語は、抗原に対するその特異性を決定する抗体分子の部分を指す。イディオタイプは、Fab領域に位置しており、その発現には通常、抗原結合部位を形成する重鎖および軽鎖の可変領域の関与が必要とされる。
【0028】
本明細書で使用される「アイソタイプ」という語は、免疫グロブリンの重鎖のクラスまたはサブクラス、または軽鎖の型および亜型を決定する抗原を指す。例えば、IgGの4つのアイソタイプは、IgG1、IgG2、IgG3、およびIgG4で指名される。
【0029】
ヒト化抗ルイスYモノクロナール抗体hu3S193は、ルイスY上皮抗原を特異的に標的する。hu3S193の薬物動態および病院において投与されるhu3S193に対する免疫反応の測定を補助するために、抗イディオタイプhu3S193抗体が生成され、HAHA分析における患者の血清サンプルおよび陽性対照におけるhu3S193を測定するためのELISA試薬としての適合性について特徴づけられた。hu3S193で免疫化したマウスからの脾細胞をSP2/0−AG14形質細胞腫細胞と融合し、結果として生じたハイブリドーマによって生成された抗イディオタイプ抗体をhu3S193への特異的結合およびルイスY抗原に対する競合結合ELISAにより選択した。9つのハイブリドーマを最初に選択し、LMH−1、−2、および−3と指名された3つは、hu3S193への特異的結合、およびルイスY抗原に結合するhu3S193との競合結合を示した。hu3S193イディオトープの認識は、抗ルイスYモノクロナール抗体、BR55.2、および他の無関係のヒトIgG、マウスIgG、ヒト化またはキメラモノクロナール抗体との交差反応の欠如によって特異的と証明された。
【0030】
抗ルイスYモノクロナール抗体、hu3S193に対するマウスモノクロナール抗イディオタイプが生成され、その特異性が規定され、薬物動態および病院で投与されたhu3S193に対する免疫反応をモニタリングするアッセイのための特定の抗イディオタイプの選択を可能にした。
【0031】
hu3S193に特異的に結合し、hu3S193およびmu3S193の結合を阻害するが、ルイスY抗原に対するBR55.2を阻害しない、LMH−1、−2、および−3で指名される3つの抗イディオタイプhu3S193抗体が生成された。抗huIgG mAb LMH−4も単離され、すべての試験キメラヒト化mAbまたは精製huIgGに対する強い結合活性が証明された。4つのAbはすべてマウスIgG1カッパアイソタイプとして同定された。ハイブリドーマLMH−3はローラー培養における1ml培養上清当たり13μgの精製LMH−3を産生し、LMH−4は15μg/mlを産生し、これらのクローンが大規模生産に適していることを示す。LMH−4は、huIgGの検出のための検査、またはウシIgGで汚染された組織培養上清からのモノクロナール抗体の親和性精製における試薬としての潜在能力を有する。BIAcore分析では、Ka=4.76×108M−1(Kd=2.10×10−9M)のhu3S193に対するLMH−3の高い明らかな親和性が測定された。この親和性は、捕捉のための精製LMH−3および検出のためのビオチニル化LMH−3を使用するhu3S193の血清濃度を測定するための高再現性、感受性、特異性ELISAアッセイの開発を可能にした。BIAcoreにより血清サンプルのHAHA分析の確立は、患者血清における免疫反応の定量のために陽性対照としてLMH−3を使用することにより可能であり、これらの抗イディオタイプを使用し、腫瘍生検の免疫組織化学的分析を通じて腫瘍へのhu3S193の浸透および結合を試験することもできた。各可変ドメインでの標的抗体を同時に結合することが可能な抗イディオタイプ抗体の生成は、臨床的に投与される標的抗体の安全性および薬物動態プロフィールの評価に高い感度を有する試薬を提供する。hu3S193免疫療法を受ける患者からの臨床サンプルを試験するための診断検査用試薬として重要な用途を有する抗イディオタイプhu3S193mAbが生成されている。
【0032】
本発明の別の態様は、抗原に対する抗体の結合を阻害する抗イディオタイプ抗体の能力を検出するための方法を提供する。本発明の別の態様は、結合イディオタイプ抗体を捕捉し、検出する抗イディオタイプの能力を検出するための方法を提供することである。本発明の別の態様は、イディオタイプ抗ルイスY抗体に結合する抗イディオタイプ抗体の能力を検出するための方法を提供することである。本発明の別の態様は、サンプル血清における抗体の量を検出する方法である。本発明は、本発明の抗体を使用するHAMA、HACA、およびHAHA反応を検出する方法にも関する。
【0033】
血清hu3S193レベルを測定するための2つのELISAアッセイを調査した。第1のアッセイには合成ルイスY抗原による捕捉が含まれ、第2のアッセイでは抗イディオタイプ抗体が使用された。アッセイの感度および特異性を比較し、LMH−3を使用する分析が、hu3S193に対する高い親和性により合成抗原よりも優れており、高い感度を可能にする非特異的結合の欠如からの低いノイズ、および3ng/mlの血清hu3S193の定量限界が見出された。BIAcore分析では、Ka=4.76×108M−1(Kd=2.10×10−9M)のhu3S193に対するLMH−3の明らかな親和性が測定された。ハイブリドーマは、抗体LMH−3の高いレベルを産生する。
【0034】
以下の実施例は本発明の例示のために示されており、本発明の限定と解釈されてはならない。
【実施例】
【0035】
実施例1
細胞培養
すべての分析用試薬を、別に指示がない限り、シグマ・ケミカル社(Sigma Chemical Co.)(St Louis、モンタナ州(MO)、米国)から購入した。
【0036】
SP2/0−Ag14形質細胞腫細胞系をアメリカン・タイプ・カルチャー・コレクション(ATCC、Manassas、バージニア州(VA)、米国)から購入した。SP2/0および選択ハイブリドーマクローンを正常RPMI−1640培地、すなわち、10%加熱不活性化ウシ胎仔血清(FCS、トレース・バイオサイエンス社(TRACE Biosciences Pty Ltd)、シドニー、オーストラリア)、100μg/mlストレプトマイシン、100U/mlペニシリン(ジブコ(Gibco)、および4mM L−グルタミンを含有するRPMI−1640培地(ジブコ・インビトロジェン社(Gibco Invitrogen Corp.)、Mt Waverley、ビクトリア州、オーストラリア)中で培養した。融合生成物の初期の数代継代を、上記のとおり20%FCS、ペニシリン、ストレプトマイシン、およびL−グルタミンを含有するRPMI−1640で培養した。HAT培地をハイブリドーマ選択のために使用した、20%FCS、HAT(ヒポキサンチン、アミノプテリン、チミジン)、OPI(オキサロ酢酸、ピルビン酸塩、インスリン培地補充)(シグマ(Sigma))、ペニシリン、ストレプトマイシン、L−グルタミンを含有するRPMI−1640培地。ハイブリドーマ増殖のために、細胞をHT培地、すなわち、20%FCS、HT(ヒポキサンチン、チミジン)、ペニシリン、ストレプトマイシン、L−グルタミンを含有するRPMI−1640培地で培養した。
【0037】
実施例2
免疫化およびスクリーニングのための抗体
ヒト化抗ルイスY IgG1抗体hu3S193(ロット:D01097、10mg/ml PBS)、マウス3S193(mu3S193)、対照アイソタイプマッチドヒト化mAb、huA33(ロット:5GMA33 10mg/ml)、および無関係のアイソタイプマッチドマウス:ヒトキメラIgG1抗体は、生物学的生産機関(Biological Production Facility)、ルートビッヒ癌研究協会(Ludwig Institute for Cancer Research)、メルボルン、オーストラリア)によって供給された。精製ヒトIgGはシグマ・ケミカル社(Sigma Chemical Co.)から購入された。F(ab)’2フラグメントを、メーカーの指示に従い免疫化ペプシンによる消化によって調製し(ピアス(Pierce)、Rockford、イリノイ州(IL)、米国)、非消化hu3S193および1mlプロテインAカラム(ファルマシア・バイオテック(Pharmacia Biotech)、Uppsala、スウェーデン)上の通過よってFc含有フラグメントから精製した。精製フラグメントをミリポア遠心フィルタ(バイオマックス(Biomax)10k膜、ミリポア(Millipore)、シドニー、オーストラリア)上で1mg/mlに濃縮した。
【0038】
実施例3
免疫化プロトコール
6〜8週齢の雌BALB/cマウスを繁殖用施設、ウォルター・アンド・エリザ・ホール研究所(Walter and Eliza Hall Institute)、メルボルン、オーストラリアから購入し、飼料および水を自由に与えて飼育した。本試験のために動物で実施されたすべての手順は、オースチン・アンド・レパトリエーション・メディカルセンター(Austin and Repatriation Medical Centre)の動物倫理委員会(Animal Ethics Committee)によって承認された。免疫化前静脈血液サンプル(200μl)を各動物から収集し、室温(RT)下に1時間、4℃下に一夜凝固させた。結果として生じる血清(約100μl/マウス)を収集し、−20℃下に保存した。
【0039】
免疫化手順をマウス6匹で実施した。0日目、40μg精製hu3S193:フロイント完全アジュバント(1:1、v:v)含有200μlの腹腔内(i.p.)注射によってBALB/cマウスを免疫化した。マウスに対し14日目および28日目にフロイント不完全アジュバントでhu3S193を40μg含有するブースター注射を投与した。35日目、免疫化後血液サンプルを尾静脈から収集し、マウス抗hu3S193抗体の血清力価をELISAによって測定した。免疫化前血清サンプルを対照として含めた。3回目、40μgのhu3S193のみを含有する最後の静脈内(i.v.)ブースター注射後4日にマウスを屠殺した。脾臓を無菌で5ml PBS/Gに収集し、融合用に脾細胞懸濁液を調製した。
【0040】
実施例4
融合およびハイブリドーマ生成
すべての手順を室温下に実施し、これらは以前に公表された方法に基づくものであった(12)。脾細胞を収集し、RPMI−1640培地で3回洗浄し、細胞濃度を測定した。同時に、融合パートナーSP2/0細胞を回収し、血清を含まない培地中で洗浄し、計算した。各細胞ペレットを血清を含まないRPMI 5ml中に再懸濁し、次いで50mlチューブに5:1比の脾細胞:SP2/0で混合した。細胞混合物を300×g、10分遠心分離し、結果として生じる細胞ペレットを軽くたたくことによって静かに破壊した。チューブを37℃の湯浴中にインキュベートし、培地(1ml)中50%PEGを1分間にわたって滴下した。次いで、予熱した血清を含まないRPMI(2ml)を2分にわたって添加した後、3分にわたって培地8mlを添加した。細胞を300×g、10分遠心分離し、HAT培地(約1×10 5脾細胞、2×10 4SP2/0骨髄腫細胞を含有する60ml HAT培地/マウス脾臓)中に静かに再懸濁した。混合細胞懸濁液を6個の96ウェル細胞培養プレート(ファルコン(Falcon)、ベクトン・ディッキンソン(Becton Dickinson)、Franklin Lakes、ニュージャージー州(NJ)、米国)へ等分し(100μl/ウェル)、加湿5%CO2/空気インキュベーター中37℃下に培養した。
【0041】
実施例5
モノクロナール抗体選択
ハイブリドーマ培養上清をhu3S193mAbおよび精製ヒトIgGとの陽性結合活性についてELISAによってスクリーニングした。選択クローンを24ウェル培養プレート(ファルコン(Falcon))中で増殖させ、その結合活性および競合結合活性をさらにELISAによって特徴づけた。培養上清中の選択抗イディオタイプhu3S193抗体のサブクラスをマウスモノクロナール抗体アイソタイピング用キット(アイソストリップ(IsoStrip)、ロシュ・ディアグノスティックス・ゲーエムベーハー(Roche Diagnostics GmbH)、Mannheim、ドイツ)で測定した。
【0042】
実施例6
限界希釈によるハイブリドーマのクローニング
一次希釈系列
選択ハイブリドーマクローンを96ウェルプレートの内側60ウェル中の200μl HAT培地中100細胞/ウェルの密度でプレーティングした。外側ウェルを抗生物質を含有するRPMI−1640培地で充填し、脱水およびウェルを含有するHAT培地/細胞の濃縮を阻止した。細胞成長がほぼ密集(約7日)であった場合、上清を収集し、ルイスY抗原結合のためのhu3S193への特異的結合およびhu3S193への拮抗薬結合について試験した。陽性クローンを継続的分析およびクローニングのために選択した。
【0043】
二次希釈系列
本ステップは、単一の細胞から成長したハイブリドーマを確立することを目的とし、培養では脾細胞支持細胞層が利用された。脾細胞を非免疫化BALB/cマウスから調製し、100mM 2−メルカプトエタノールを補充した10 6細胞/ml HAT培地に再懸濁し、滅菌環境下に室温で一夜インキュベートし、汚染感染がないことを確実にした。翌日、2つの96ウェルマイクロタイタープレートを、既述どおり、使用したプレートの内側60ウェルのみで各選択クローン用に調製した。HAT培地のアリコート(100μl)を列1に添加し、脾細胞懸濁液を列2−6(10 5細胞/ウェル)に添加し、プレートを37℃下に一夜インキュベートした。選択細胞をHAT培地中に希釈し、100、10、5、1、または0.5細胞を含有する100μlのアリコートを、それぞれ、支持細胞を含有する2から6までの列のウェルに添加した。プレートを37℃下に10−14日間インキュベートし、1または0.5細胞/ウェルで接種したウェルからクローンコロニーを選択した。
【0044】
三次希釈系列
選択クローンが単一の細胞から由来することを確実にするために、すべての選択細胞系は反復回の二次希釈手順を受けた。好ましい系を増殖させ、ストックを調製し、液体窒素下に保存した。
【0045】
実施例7
抗体産生
ハイブリドーマを産生する選択および再クローン化抗イディオタイプ抗体をローラーボトル(コーニング(Corning)、コーニング社(Corning Incorporated)、ニューヨーク州(NY)、米国)で培養し、培養上清を無菌で回収した。抗イディオタイプ抗体を、50mM Tris−HCl、pH8(バッファーA)で前平衡した5mlプロテインAセファロース迅速フロー(fast flow)カラム(アマシャム・ファルマシア・バイオテック(Amersham Pharmacia Biotech)、Uppsala、スウェーデン)における親和性クロマトグラフィーによって精製した。20カラム量バッファーAでの洗浄後、200mM NaCl含有100mMグリシン−HCL、pH2.7を使用して結合抗体を溶出し、1M Tris HCl、pH8を使用して直ちに中和した。PBSへの4℃での一夜透析後、抗体を遠心フィルタ(ミリポア(Millipore)、バイオマックス(Biomax)10k膜)で濃縮した。還元および非還元条件下にSDS−PAGEによって純度を検査した。クマシーブルー染色によってタンパク質を視覚化した。ビオチニル化抗イディオタイプ抗体をECLタンパク質ビオチニル化モジュールキットを使用し、メーカーの指示(アマシャム・ファルマシア・バイオテック(Amersham Pharmacia Biotech)に従い調製した。
【0046】
実施例8
結合特異性のELISA分析
ハイブリドーマ上清または精製抗イディオタイプ抗体の結合特異性をELISA分析によって特徴づけた。96ウェルプレート(ヌンク・マキシソープ(Nunc MaxiSorp)、Roskilde、デンマーク)をhu3S193(100μl、4.0μg/ml)、精製ヒトIgG、または炭酸バッファー(0.05M、pH9.6)中の他の対照モノクロナール抗体で4℃下に一夜コーティングした。非特異的結合を3%BSA/PBSで室温(23℃)下に2時間遮断した。上清または精製抗体の連続希釈(1:2−1:6300)を調製し、ウェル当たり100μlアリコートを添加した。陽性(huIgG)および陰性(抱合体および基質のみ)対照を各アッセイで含めた。プレートを室温下に1時間インキュベートし、0.05%Tween20/PBSで3回洗浄し、次いで1時間二次抗体(ヤギ抗マウスIgG抱合アルカリホスファターゼ(100μl/ウェル、1%BSA/PBS中1:3000希釈)と一緒にインキュベートした。結合抗体を発色団基質p−ニトロフェニルリン酸基質(ICNバイオメディカルズ社(ICN Biomedicals Inc.)、Aurora、オハイオ州(Ohaio)、米国)で検出した。光学密度をVmaxキネティックマイクロプレートリーダー(モレキュラー・デバイス社(Molecular Devices Corporation)、Sunyvale、カリフォルニア州)により450nmで測定した。
【0047】
実施例9
拮抗薬活性のELISA分析
選択抗イディオタイプ抗体をルイスY−抗原コートプレートへのhu3S193mAbの結合を遮断するその能力について試験した。合成ルイスY四糖類は、モル比約32:1でBSAに結合されて入手可能である(アルバータ・リサーチ・カウンシル(Alberta Research Council)、Edmonton、アルバータ州(Alberta)、カナダ)。ELISAプレート(ヌンク・マキシソープ(Nunc MaxiSorp))をルイスY−BSA 抗原(50μl 3.0μg/ml PBS)でコーティングし、4℃下に一夜インキュベートした。プレートを室温下に2時間、3%BSA/PBSで非特異的結合のために遮断した。ハーフログ連続希釈サンプル(上清または精製抗イディオタイプmAb)を各ウェルに添加した後、hu3S193mAb(50μl、1μg/ml最終濃度)を添加した。室温下に1時間のインキュベーション後、ヤギ抗−ヒトIgG−HRPを使用し、結合hu3S193mAbを検出し、広範な洗浄後、ABTS基質(2,2’−アジノ−ビス−(3−エチル−ベンズチアゾリン−6−スルホン酸、100μl 40μM/ウェル、ICN)で視覚化した。光学密度(OD)をVmaxキネティックマイクロプレートリーダーにより415nmで測定した。
【0048】
実施例10
Hu3S193の抗イディオタイプ特異性
抗ルイスY mAbへの結合の精製選択クローンの特異性をさらに、hu3S193、およびマウスmAb mu3S193およびBR55.2(ルートビッヒ癌研究協会(Ludwig Institute for Cancer Research)hu3S193を使用して検査した。特異性を2つのELISAで検査した。第1のELISAでは結合イディオタイプmAbの捕捉および検出のための抗イディオタイプが利用され、第2のELISAでは溶液中でイディオタイプ抗ルイスY mAbに結合し、ルイスY抗原結合を阻害する抗イディオタイプmAbの能力が検査された。
【0049】
第1のアッセイでは、ELISAマキシソープ(Maxisorp)プレートを選択抗イディオタイプmAbで一夜、4℃下にコーティングした(100μl 4μg/ml 0.05M炭酸バッファー、pH9.6)。3%FCS/PBSによる遮断後、連続希釈した10μg/mlのhu3S193、mu3S193、BR55.2の3部サンプル、および無関係の対照mAb huA33およびmuA33をウェルに添加し、次いで室温下に1時間インキュベートした。洗浄後、結合mAbをビオチニル化抗イディオタイプmAb(LMH−3−Bt、4℃下に一夜、1%FCS/PBS中100μl/ウェル 4μg/ml)で検出した。結合複合体ストレパビジン−HRPの(1%FCS/PBS中1:1000希釈、100μl/ウェル)検出の後、着色のためにABTSを添加した。OD415を既述どおりに測定した。
【0050】
特異性の第2の試験のために、3つのELISAプレートを合成ルイスY−BSA抗原でコーティングした(PBS中4.0μg/ml、4℃下に一夜)。次いで、連続希釈抗ルイスYmAbをプレートの各列上に等分した、プレート1、hu3S193、プレート2、mu3S193、プレート3 BR55.2最終濃度範囲(0.003−10μg/ml)。ルイスY抗原への結合の競合を試験するために、他の2つの抗ルイスY mAbおよび2つの抗イディオタイプmAbを2部で(最終濃度10μg/ml)添加し、次いで室温下に1時間インキュベートした。洗浄後、プレート1における結合hu3S193を抗huIgG−HRP(シグマ(Sigma)、1%FCS/PBS中100μl 1:1000)の後、着色のためのABTSで検出し、A415nmで測定した。プレート2における結合mu3S193およびプレート3におけるBR55.2をヤギ抗−mIgGアルカリホスファターゼで検出し、p−ニトロフェノール色原体をA405で測定した。
【0051】
実施例11
血清中のHu3S193のELISA測定
血清サンプル中のhu3S193mAbを試験するために2つのELISAアッセイが開発された。第1のアッセイでは、合成ルイスY−BSA抗原(PBS中3.0μg/ml、50μl/ウェル)をhu3S193捕捉のために利用した。健康ドナー血清中のhu3S193の連続希釈サンプルまたは3%BSA/PBSによるインキュベーション後、抗原に結合するhu3S193mAbの量をヤギ抗ヒトIgG−HRP二次抗体およびABTS基質で検出した。
【0052】
第2のアッセイには抗イディオタイプ抗体によるコーティングおよびビオチニル化抗イディオタイプ抗体による検出が含まれた。プレートを抗イディオタイプhu3S193抗体(0.05M炭酸バッファーpH9.6中3.0μg/ml、100μl/ウェル)で一夜、4℃下にコーティングし、次いで2時間、室温(23℃)下に3%BSA/PBSで遮断した。3%BSA/PBSまたは健康ドナー血清中のhu3S193(0.0315−10μg/ml)の連続希釈サンプルを3部でプレートに添加し、1時間、室温下にインキュベートした。戦場後、ビオチニル化抗イディオタイプhu3S193抗体を添加し(1%BSA/PBS中100μl 3μg/ml)、1時間、室温下にインキュベートした。結合複合体ストレパビジン−HRPの(1%BSA/PBS中1:1000希釈、100μl/ウェル)検出の後、着色のためにABTSを添加した。OD415を既述どおりに測定した。
【0053】
バイオセンサー分析:
カルボキシメチルデキストランコートセンサーチップ(CM5)上でBIAcore2000(BIAcore AB、Uppsala、スウェーデン)を使用してバイオセンサー分析を実施した。チップをチャンネル2でhu3S193、チャンネル3でLMH−3、およびチャンネル4でウシ血清アルブミン(アルバータ・リサーチ・カウンシル(Alberta Research Council)、Edmonton、アルバータ州(Alberta)、カナダ)に結合した合成ルイスy四糖類により、標準NHS/EDCアミン結合化学を使用して誘導体化した。hu3S193およびLMH−3のサンプルをHBSバッファー(10mM HEPES、pH7.4、150mM NaCl、3.4mMジ−Na−EDTA、0.005%Tween−20)、およびアリコート(30μl)を5μl/分の流量でセンサーチップ表面にわたって注入した。注入相後、300秒間チップ表面上にHBSバッファーを流すことによって解離をモニタリングした。結合抗体を溶出し、100mMグリシン/100mM NaCL、pH2.7、20μlの注入によってサンプル間でチップ表面を再生した。結合の動態分析にために、各種濃度のhu3S193およびLMH−3をセンサーチップ表面上に注入した。明らかな結合(ka)および解離(kd)速度定数を二価分析モデルを使用し、Rmaxの計算のための全体的、およびローカル適合により、BIA評価v3.0ソフトウェア(ファルマシア・バイオセンサー(Pharmacia Biosensor)、Uppsala、スウェーデン)を使用して計算した。
【0054】
投与されたモノクロナール抗体に対する患者の免疫反応は、患者血清のバイオコア(Biocore)(登録商標)分析によって評価されうる(リッター(Ritter)、コーエン(Cohen)ら、2001年)。抗イディオタイプ抗体は、これらの分析において陽性対照試薬として有用である。したがって、LMH−3を健康ドナー血清(3−50μg/ml)に添加し、免疫化hu3S193に対する結合反応を確認した。
【0055】
実施例12
マウス免疫化およびハイブリドーマクローン選択
3群のBALB/cマウスを繰り返しhu3S193mAbで免疫化し、その血清をマウス抗hu3S193mAb活性について評価した。血清サンプルの免疫化前後の免疫活性は、高力価のマウス抗無傷およびF(ab)’2hu3S193mAbの進展を示した。アイソタイプ対照ヒト化IgG(huA33)またはキメラIgGF(ab)’2でコーティングされたプレートも強い陽性結合を示した(結果は示さず)。マウスを屠殺し、その脾臓を除去した。脾細胞およびパートナー融合Sp2/0形質細胞腫細胞を融合し、これら3つの融合からのハイブリドーマをスクリーニングした。hu3S193mAbに対して強い結合活性を示すが、対照huA33または精製ヒトIgGに対しては弱い結合を示し、または結合を示さない3−4ウェルのみを選択し、増殖させた。増殖後、抗体特異性をさらにELISAアッセイで試験した(図1)。3つの対照ヒト化またはキメラ抗体を第1の融合からのクローンの選択中に使用した。交差結合活性が通常、精製ヒトIgGコーティングプレートで確認されたため、その後のスクリーニングにおいてhu3S193に加えて精製ヒトIgGを主に使用した。
【0056】
実施例13
選択抗イディオタイプ抗体の結合および遮断活性
すべての選択ハイブリドーマの結合および拮抗薬活性をさらに分析し、抗イディオタイプhu3S193抗体を産生するハイブリドーマを確認した。ハーフログ連続希釈を各ハイブリドーマ培養上清から調製し、ELISAによって評価した。最初に選択された12個のクローンのうち、hu3S193に結合するが、huIgGには結合せず、hu3S193のルイスY抗原への結合を阻害しうる3つを確認した(図1)。これらのハイブリドーマ、F1−1、F2−3、およびF3−27を限界希釈によって単一の細胞からクローン化し、それぞれ、ルートビッヒ癌研究協会(Ludwig Institute for Cancer Research)メルボルン(Melbourne)ハイブリドーマ(LMH)−1、LMH−2、およびLMH−3と指名された。クローンの1つ(FS2−1)は、すべての試験されたキメラヒト化mAbもしくは精製huIgGまたは(図1)に対する強い抗huIgG結合活性を示し、その後にhuIgG結合に対する陽性対照として使用された。このクローンは単一の細胞から調製され、LMH−4を命名された。LMH−1から−4までの抗体は、マウスモノクロナール抗体アイソタイピング用キットによってアイソタイプIgG1eとして確認され、プロテインA親和性クロマトグラフィーによって培養上清から精製された。4つのクローンの結合特異性がELISAによって確認された(図4A−C)。huIgG定常鎖との交差反応は、LMH−1、−2、またはー3では確認されず(図4B)、3つの抗体はすべて特異的にhu3S193mAbのみに結合された(図4A)。これらの抗体の拮抗薬活性は、競合ELISAアッセイによって分析された。LMH−1、−2、および−3抗hu3S193イディオタイプ抗体は、hu3S193mAbに結合し、hu3S193mAbのルイスY抗原への結合を用量依存的に阻害する(図2)。
【0057】
実施例14
抗体結合親和性の測定
溶液中でLMH−3と相互作用する固定化hu3S193のバイオセンサー結合実験の明らかな結合親和性では、4.76×108M−1(Kd=2.10×10−9M)のKaが測定された。チップ上に固定化されたLMH−3、および分析物として表面上を走るhu3S193との相互実験において、9.12×108M−1(Kd=1.10×10−9M)のKaが測定された。
【0058】
実施例15
Hu3S193に対する抗イディオタイプ特異性
3S193イディオタイプに対するクローンLMH−2およびLMH−3の特異性がこれらの実験で確認された(図5)。プロテインA精製LMH−3をELISAプレート上にコーティングし、連続希釈した10μg/mlのhu3S193、mu3S193、BR55.2の3部サンプル、および対照IgG1 mAb huA33およびmuA33をウェルに添加し、さらにLMH−3の結合特異性を特徴づけた。このクローンの追加の好ましい特徴は、ビオチニル化LMH−3がLMH−3で捕捉された結合3S193mAbの検出に利用されうることであった。ELISAの結果(図3A)は、LMH−3が特異的に抗ルイスY3S193イディオタイプに結合し、マウス抗ルイスYBR55.2との反応性は検出されず、対照muA33およびhuA33のマウスおよびヒトIgG1定常ドメインとの交差反応はなかったことを示した。hu3S193およぶmu3S193のルイスY抗原への結合は、10μg/ml LMH−2およびLMH−3によって溶液中で有効に競合したが、抗イディオタイプ結合部位の飽和時には、一部のhu3S193およびmu3S193の抗原への結合は確認されなかった(それぞれ、図3Bおよび3C)。マウスBR55.2mAbは一部の競合結合をもたらしたが、高い親和性のmu3S193は、最高濃度(10μg/ml)での場合を除き、hu3S193のルイスY抗原への結合を完全に遮断した(図3B)。プレート2相互実験においては、低い親和性のhu3S193は、mu3S193が最初にウェルに添加されると、ルイスY抗原に競合しえなかった(図3C)。プレート3からの結果(図3D)は、BR55.2のルイスY抗原への結合が、hu3S193または抗イディオタイプクローンLMH−2および−3と競合せず、またはこれらによって阻害されないことを示す。したがって、3S193イディオトープに対するこれらクローンの特異性が確認される。図3Bにおいて確認されるBR55.2による競合は、その異なるルイスYエピトープへの高い親和性結合に対する立体障害により達成されうる。
【0059】
実施例16
臨床サンプルにおけるHu3S193を測定するためのELISAアッセイの開発
第1のELISAアッセイでは、血清および検出用のペルオキシダーゼと抱合された二次ヤギ抗ヒトIgGにおけるhu3S193の捕捉用に使用された合成抗原(ルイスY四糖類−BSA抱合体)が調査された。合成ルイスY抗原は、記録された低い光学密度によって示されるアッセイ感度を削減した、hu3S193mAbに対する比較的低い親和性を有する。二次抱合抗体ではすべてのヒト免疫グロブリンが検出され、したがって高いノイズが分析したすべてサンプルで確認された(図4A)。高いノイズ干渉が、特に低い血清hu3S193濃度でアッセイの感度および精度を大幅に削減する。第2のELISAアッセイでは、コーティング用に精製抗イディオタイプhu3S193抗体LMH−1、2、および−3が使用され、二次抗体としてビオチニル化LMH−1、−2、または−3が使用され、かつ結合複合体の視覚化用にストレパビジン−HRPおよびABTS基質が使用された。LMH−1およびLMH−2抗イディオタイプ抗体は、血清中または3%BSA/PBS中でmu3S193mAbに対する強い結合示したが、hu3S193mAbに対しては弱い結合を示した。しかし、ヒト化mAb、キメラmAb、マウスmAb、ヒト血清由来のIgまたはマウス血清由来のIgを含む対照抗体では交差結合活性は確認されなかった(図4B)。コーティング抗体または二次抗体の濃度を変更し、かつ異なるコーティングバッファーを使用することによってLMH−1およびLMH−2によるELISAを最適化または改善する試みを調査した。一部の改善が達成されたが、反応パターンへの変化はもたらされなかった(データは示さず)。しかし、捕捉用のLMH−3抗イディオタイプmAbおよび二次抗体としてLMH−3−ビオチンを使用することにより、最小の非特異的ノイズ活性を有する血清中でhu3S193mAbおよびmu3S193mAbに対する特異的に強い結合が確認された(図4C)。高再現性のアッセイは優れた感度を示し、血清サンプルにおけるhu3S193mAbまたはmu3S193mAbに対する検出限界は3ng/mlであった。
【0060】
免疫反応のバイオコア分析
抗イディオタイプLMH−3を使用するhu3S193に対する血清中のHAHAの測定が開発された。高感度で再現性の方法が確立され、確認された(図5)。
【図面の簡単な説明】
【0061】
【図1】ELISAによって測定される抗−hu3S193モノクロナール抗体、LMH−1(●)、−2(▽)、−3(◇)、および抗−huIgG−4(◇)の結合特異性を示す。
【図2】ELISAによって検査されたハイブリドーマ培養上清のhu3S193抗原結合活性を中和する能力について示す。
【図3】3S193イディオトープに対するLMH−2およびLMH−3の特異性の確認を示す。
【図4A】患者血清サンプルにおけるhu3S193を測定するためのELISAの開発を示す。
【図4B】患者血清サンプルにおけるhu3S193を測定するためのELISAの開発を示す。
【図4C】患者血清サンプルにおけるhu3S193を測定するためのELISAの開発を示す。
【図5】hu3S193に対する血清HAHA反応のBiacore分析が最適化され、陽性対照としての抗イディオタイプLMH−3を使用して確認されたことを示す。
【技術分野】
【0001】
本発明は、分子生物学の分野、かつより具体的には抗体に関する。
【背景技術】
【0002】
25年前のモノクロナール抗体(mAb)技術の出現は、有用な研究用試薬の膨大なレパートリーを提供し、癌療法、自己免疫疾患、移植による拒絶反応、抗ウイルス予防における承認された医薬試薬として、および抗血栓剤として抗体を使用する機会を創造した(グレニー(Glennie)とジョンソン(Johnson)、2000年)。マウスmAbをキメラmAb(マウスV領域、ヒトC領域)およびmAb相補性決定領域(CDR)のみがマウス由来であるヒト化試薬へ変換する分子工学の応用は、mAb療法の臨床的成功に重要であった。工学的mAbは、免疫原性を大幅に削減または遠ざけ、血清中半減を増大させ、mAbのヒトFc部分は補体および細胞毒性細胞の免疫エフェクターを補充する可能性を増大させる(クラーク(Clark)、2000年)。生体内分布、薬物動態、および臨床的に投与されるmAbに対する免疫反応の誘導に対する調査には、医薬と内因性タンパク質とを区別する分析の開発が必要である。
【0003】
抗体は通常、それが認識する抗原に関して定義される。特定の抗原に対する抗体の特異性は、その抗原結合部位、抗原に接触する抗体分子の異なる領域よって判定される。この部位は、免疫グロブリンの重鎖および軽鎖の可変領域内に存在する。しかし、抗体は、抗体分子の単一特異的集団の独自の重鎖および軽鎖領域と関連した、そのイディオタイプ、イディオトープの集合体、または表面マーカーによっても定義される。抗体に特有の抗原決定基はイディオトープと呼ばれ、イディオトープを有する抗体との抗イディオタイプ抗体の反応によって定義される。イディオタイプは、研究者が免疫反応および遺伝性免疫グロブリン遺伝子における特定の抗体の外観および持続性を追跡できるという点で有用なマーカーである。イディオタイプは、抗イディオタイプ抗体の産生を促進しうる固有の決定基でもある。
【0004】
抗イディオタイプ抗体は、他の抗体の可変領域に結合し、免疫系において、内部(自己)抗原に対して生じる結合抗体における主要な役割を果たす(ジェルネ(Jerne)、1974年)。イディオタイプ抗体を誘発する可変領域によって誘発される抗イディオタイプ抗体は、3つのカテゴリー、すなわち、1)抗原結合部位(パラトープ)におけるエピトープの認識、2)パラトープ近く結合およびイディオタイプ抗体−抗原相互作用の立体的干渉、または3)イディオタイプ抗体(「内部イメージ」抗イディオタイプ抗体)によって認識される抗原の構造的模倣のうちの1つでありうる。後者のカテゴリーの抗イディオタイプは、細菌、ウイルス、および寄生虫感染に加えて腫瘍関連抗原に対する宿主の免疫反応の修飾におけるその可能性が与えられるワクチンとして生成および調査されている(バッタチャリヤ−チャタジー(Bhattacharya−Chatterjee)、チャタジー(Chatterjee)ら、2001年で概説)。齧歯類抗イディオタイプ抗体は、補助剤、および担体、通常、誘発される免疫または抗腫瘍反応に対するキーホールリンペットヘモシアニン(KLH)との抱合を必要とする。
【0005】
抗イディオタイプ抗体は、以下、ELISAと称される酵素免疫測定、注入イディオタイプ抗体の患者の血清レベルを検出するための免疫療法治験における試薬、または投与されるイディオタイプ抗体に対する免疫反応としても利用されうる。かかる免疫反応としては、ヒト抗マウス抗体、ヒト抗キメラ抗体、およびヒト抗ヒト化抗体(以下、それぞれ、HAMA、HACA、HAHAと呼ぶ)が挙げられる。抗イディオタイプ抗体NUH−82は、ヒト腎細胞癌におけるG250抗原を認識するマウスmAb G250の結合部位に対するものである(ウエムラ(Uemura)、オカジマ(Okajima)ら、1994年)。NUH−82は、cG250抗体放射免疫療法を受ける患者におけるHACA反応の測定に有用であることが証明されたが(ステフェンス(steffens)、ボーマン(Boerman)ら、1997年)、最近になって、NUH−82は薬物動態分析のために患者の血清におけるcG250の血清レベルを測定するサンドイッチELISAにおいて使用されている(リュー(Liu)、スミス(Smyth)ら、2002年)。
【0006】
hu3S193抗体は、乳癌、結腸癌、肺癌、前立腺癌、および卵巣癌を含む上皮腫瘍を発現するルイスY、ジフコシル化炭水化物抗原を標的するために開発された(キタムラ(Kitamura)、ストッカート(Stockert)ら、1994年、スコット(Scott)、ゲレイク(Geleick)ら、2000年)。ルイスYは、上皮由来の腫瘍細胞、例えば、肺癌、腸癌、乳癌、前立腺癌、または卵巣癌の表面上に発現される炭水化物分子である。ルイスY抗原は、正常組織において発現されるが、発現レベルは一部の腫瘍型においてより高く、抗原は一部の乳癌、結腸癌、胃癌、食道癌、膵癌、十二指腸癌、肺癌、膀胱癌、および腎癌のほか、胃および島細胞神経内分泌系腫瘍の細胞のマーカーとして使用されうる。ルイスY関連関連抗原は、その高い密度と一次および転移性病変における同質の発現により免疫療法の魅力的な標的である(キム(Kim)、ユアン(Yuan)ら、1986年、サカモト(Sakamoto)ら、1986年、チャン(Zhang)、コードン−カルド(Cordon−Cardo)ら、1997年)。
【0007】
hu3S193は、インビトロで強いADCCおよびCDCを誘発し、前臨床試験における選択充実腫瘍の免疫療法として有望であることが証明されおり(クラーク(Clarke)、リー(Lee)ら、2000年b、クラーク(Clarke)、リー(Lee)ら、2000年a)、現在、第I相用量増加臨床試験中である。前臨床生体内分布試験では放射性標識hu3S193がトレースされ、乳癌の動物モデルにおける安定化した放射性抱合体の腫瘍ターゲティング能を評価した(クラーク(Clarke)、リー(Lee)ら、2000年a)。病院では、hu3S193などmAbが放射性標識リングを使用してトレースされ、生体内分布および腫瘍イメージング、および血清からの放射性抱合体のクリアランスをmAb薬物動態の尺度として評価することができる(クラーク(Clarke)、リー(Lee)ら、2000年a、スコット(Scott)、ゲレイク(Geleick)ら、2000年、ホフマン(Hoffman)、スコット(Scott)ら、2001年)。しかし、臨床症状の非存在下には、HAHA免疫反応を検出することはできない。
【0008】
hu3S193の免疫エフェクター機能を考えると、初期治験における投与抗体の血清レベルの測定は将来の治療方式の計画を補助し、非標識hu3S193の薬物動態の測定を可能にすると同時に、HAHAを測定する能力はhu3S193の安全性評価を補助するであろう。
【0009】
具体的には、本発明は、それらが患者の循環において認識する投与イディオタイプの薬物動態をモニタリングするための診断用試薬として免疫療法治験における抗イディオタイプの使用にも関する。同様に、抗イディオタイプは、投与されたイディオタイプmAbに対するHAHA、HACA、またはHAMA免疫反応の陽性対照として使用されうる。かかる免疫反応は、患者集団の一部における臨床結果に対して全く影響を与えないか(グルーバー(Gruber)、ヴァンハーレム(van Haarlem)ら、2000年)、またはその後の治療を不可能にする免疫療法のmAbの薬物動態および生体内分布における過敏反応および劇的な変化と関連しうる(クラーク(Clark)、2000年)。
【0010】
(図面の簡単な記載)
図1A−B:抗−hu3S193モノクロナール抗体、LMH−1(●)、−2(▽)、−3(◇)、および抗−huIgG−4(◇)の結合特異性は、ELISAによって測定された。連続希釈ハイブリドーマ培養上清が、A)hu3S193(図1A)およびB)huIgG(図1B)でコーティングされたマイクロタイタープレートへの結合活性について検査された。二次抱合体(▲)および基質のみ(○)の対照も含めた。hu3S193に対する特異的結合が、LMH−1、−2、および−3で確認された。LMH−4は抗huIgG結合活性を示した。
【0011】
図2:クローン増殖後、ハイブリドーマ培養上清が、hu3S193抗原結合活性を中和する能力についてELISAによって3部で検査された。平均±SD結果は、合成Ley抗原でコーティングされたプレートへのhu3S193結合の遮断により抗hu3S193mAb、LMH−1、−2、および−3の拮抗薬活性を示した。
【0012】
図3A−D:3S193イディオトープに対するLMH−2およびLMH−3の特異性の確認。図3A。プロテインA精製LMH−3をELISAプレート上にコーティングし、連続希釈した10μg/mlのhu3S193、mu3S193、BR55.2の3部サンプル、および対照IgG1 mAb huA33およびmuA33をウェルに添加し、さらにLMH−3の結合特異性を特徴づけた。このクローンの追加の好ましい特徴は、ビオチニル化LMH−3がLMH−3で捕捉される結合3S193mAbの検出に利用されうることであった。図3B。hu3S193−Ley抗原結合に対する抗Leyおよび抗イディオタイプmAbによるLey四糖類コーティングプレートへの結合を阻止する拮抗薬結合。図3C。mu3S193−Ley抗原結合、および図3D。BR55.2−Ley抗原結合活性。
【0013】
図4A−C:患者血清サンプルにおけるhu3S193を測定するためのELISAの開発。図4A。合成抗原Ley−BSA抱合体による捕捉、抗huIgGによる検出の抗イディオタイプ利用可能性より前に利用可能な方法は、患者の注入前血清サンプルにおけるhu3S193標準による低い感度および高いノイズを示す。図4B。抗hu3S193イディオタイプLMH−2による捕捉およびLMH−2−ビオチンによる検出。低い感度が確認される。図4C。抗hu3S193イディオタイプLMH−3による捕捉およびLMH−3−ビオチンによる検出は、最小のノイズおよびhu3S193血清中の3ng/mlに対する感度を示す。
【0014】
図5:hu3S193に対する血清HAHA反応のBiacore分析が最適化され、陽性対照としての抗イディオタイプLMH−3を使用して確認された。hu3S193をバイオセンサーチップ上に固定化し、ヒト血清μg/ml)中の抗イディオタイプmAb LMH−3(■)およびアイソタイプマッチド対照抗体(▲)の連続希釈を3部でチップ上を通した。
【発明の開示】
【発明が解決しようとする課題】
【0015】
本発明は、ヒト化抗ルイスYモノクローナル抗体、hu3S193に対する抗イディオタイプ抗体に関する。本発明は、hu3S193の可変領域に対して特異的に結合するハイブリドーマクローンによって生成されるmAbのELISAスクリーニング法、およびルイスY抗原に対するhu3S193結合を阻害する抗イディオタイプmABの能力にも関する。
【0016】
本発明は、本発明の抗体を使用するHAMA、HACA、およびHAHA反応物を検出する方法にも関する。
【課題を解決するための手段】
【0017】
本発明の1つの態様は、抗ルイスYモノクロナール抗体に特異的な抗イディオタイプ抗体を提供することである。本発明の別の態様は、抗ルイスYモノクロナール抗体の可変領域に結合する抗ルイスY抗体に特異的な抗イディオタイプ抗体を提供する。同様に、本発明の別の態様は、抗ルイスYモノクロナール抗体の結合を遮断する抗イディオタイプ抗体である。本発明の別の態様は、特異的にhu3S193に結合する抗イディオタイプ抗体を提供することである。具体的には、本発明において提供される抗イディオタイプ抗体は、モノクロナール抗体、キメラ抗体、ヒト抗体、ヒト化抗体、または一本鎖抗体より成る群から選択される。
【0018】
本発明の別の態様は、抗ルイスYモノクロナール抗体に特異的な抗イディオタイプ抗体を生成することが可能なハイブリドーマを提供することである。本発明の別の態様は、LMH−1、LMH−2、およびLMH−3より成る群から選択される抗ルイスYモノクロナール抗体に特異的であるハイブリドーマを提供することである。
【0019】
本発明の別の態様は、抗原への抗体の結合を阻害する抗イディオタイプ抗体の能力を検出する方法を提供する。本発明の別の態様は、結合イディオタイプ抗体を捕捉し、検出する抗イディオタイプの能力を検出するための方法を提供することである。本発明の別の態様は、イディオタイプ抗ルイスY抗体に結合する抗イディオタイプ抗体の能力を検出するための方法を提供することである。本発明の別の態様は、サンプル血清中の抗体の量を検出する方法である。
【0020】
本発明は、hu3S193を捕捉する抗原としてBSAに結合した合成ルイスY四糖類と、結合hu3S193の検出器としての市販の抗ヒト抗体製剤を使用するELISAによって、血清サンプル中のhu3S193を測定する方法を含む。アッセイの感度は、高いノイズをもたらす二次抗huIgG抗体によって検出される血清サンプルからのhuIgGの非特異的結合によって大きく制限されている。cG250に対するHACA反応および抗イディオタイプNUH−82を有するcG250の血清レベルの測定による経験に基づき、適切な抗イディオタイプhu3S193mAbの産生が不可欠となった。
【0021】
本発明は、本発明の抗体を使用するHAMA、HACA、およびHAHA反応物を検出する方法にも関する。
【発明を実施するための最良の形態】
【0022】
本発明は、抗ルイスYモノクロナール抗体に特異的な抗イディオタイプ抗体を提供する。これらの抗イディオタイプ抗体は、抗ルイスYモノクロナール抗体の可変領域に結合する抗ルイスY抗体に特異的である。同様に、本発明の別の態様は、抗ルイスYモノクロナール抗体の結合を遮断する抗イディオタイプ抗体である。本発明の別の態様は、特異的に抗ルイスYモノクロナール、hu3S193に結合する抗イディオタイプ抗体を提供することである。具体的には、本発明において提供される抗イディオタイプ抗体は、モノクロナール抗体、キメラ抗体、ヒト抗体、ヒト化抗体、または一本鎖抗体より成る群から選択されうる。
【0023】
また、本発明は、抗ルイスYモノクロナール抗体に特異的な抗イディオタイプ抗体を生成することが可能なハイブリドーマを提供する。本発明の別の態様は、LMH−1、LMH−2、およびLMH−3より成る群から選択される抗ルイスYモノクロナール抗体に特異的であるハイブリドーマを提供することである。
【0024】
本明細書で使用される「ヒト化」抗体という語は、非ヒト種免疫グロブリン由来のそのCDR(補体決定領域)を有する分子、および主にヒト免疫グロブリン由来の抗体分子の残りを指す。
【0025】
本明細書で使用される「抗体」という語は、別に指示がない限り、広く抗体分子およびさまざまな抗体由来分子の両方を指す。かかる抗体由来分子は、少なくとも1つの可変領域(重鎖または軽鎖可変領域のいずれか)およびFabフラグメント、Fab’フラグメント、F(ab’)2フラグメント、Fdフラグメント、Fabcフラグメント、Fdフラグメント、Fabcフラグメント、Sc抗体 (一本鎖抗体)、二重特異性抗体、個々の抗体軽鎖、個々の抗体重鎖、抗体鎖と他の分子とのキメラ融合などの分子を含む。
【0026】
免疫グロブリンに関連して本明細書で使用される「可変領域」という語は、免疫学の当業者によってこの語に与えられた通常の意味を有する。抗体重鎖および抗体軽鎖は「可変領域」および「定常領域」へ分割されうる。可変領域と重鎖領域との分割点は、抗体構造を記載した標準の教科書、例えば、カバト(Kabat)ら、「Sequence of Proteins of Immunological Interest:第5版」、米国保険福祉省、米国政府印刷局(1991年)を参照することにより当業者によって容易に決定されうる。
【0027】
本明細書で使用される「イディオタイプ」という語は、抗原に対するその特異性を決定する抗体分子の部分を指す。イディオタイプは、Fab領域に位置しており、その発現には通常、抗原結合部位を形成する重鎖および軽鎖の可変領域の関与が必要とされる。
【0028】
本明細書で使用される「アイソタイプ」という語は、免疫グロブリンの重鎖のクラスまたはサブクラス、または軽鎖の型および亜型を決定する抗原を指す。例えば、IgGの4つのアイソタイプは、IgG1、IgG2、IgG3、およびIgG4で指名される。
【0029】
ヒト化抗ルイスYモノクロナール抗体hu3S193は、ルイスY上皮抗原を特異的に標的する。hu3S193の薬物動態および病院において投与されるhu3S193に対する免疫反応の測定を補助するために、抗イディオタイプhu3S193抗体が生成され、HAHA分析における患者の血清サンプルおよび陽性対照におけるhu3S193を測定するためのELISA試薬としての適合性について特徴づけられた。hu3S193で免疫化したマウスからの脾細胞をSP2/0−AG14形質細胞腫細胞と融合し、結果として生じたハイブリドーマによって生成された抗イディオタイプ抗体をhu3S193への特異的結合およびルイスY抗原に対する競合結合ELISAにより選択した。9つのハイブリドーマを最初に選択し、LMH−1、−2、および−3と指名された3つは、hu3S193への特異的結合、およびルイスY抗原に結合するhu3S193との競合結合を示した。hu3S193イディオトープの認識は、抗ルイスYモノクロナール抗体、BR55.2、および他の無関係のヒトIgG、マウスIgG、ヒト化またはキメラモノクロナール抗体との交差反応の欠如によって特異的と証明された。
【0030】
抗ルイスYモノクロナール抗体、hu3S193に対するマウスモノクロナール抗イディオタイプが生成され、その特異性が規定され、薬物動態および病院で投与されたhu3S193に対する免疫反応をモニタリングするアッセイのための特定の抗イディオタイプの選択を可能にした。
【0031】
hu3S193に特異的に結合し、hu3S193およびmu3S193の結合を阻害するが、ルイスY抗原に対するBR55.2を阻害しない、LMH−1、−2、および−3で指名される3つの抗イディオタイプhu3S193抗体が生成された。抗huIgG mAb LMH−4も単離され、すべての試験キメラヒト化mAbまたは精製huIgGに対する強い結合活性が証明された。4つのAbはすべてマウスIgG1カッパアイソタイプとして同定された。ハイブリドーマLMH−3はローラー培養における1ml培養上清当たり13μgの精製LMH−3を産生し、LMH−4は15μg/mlを産生し、これらのクローンが大規模生産に適していることを示す。LMH−4は、huIgGの検出のための検査、またはウシIgGで汚染された組織培養上清からのモノクロナール抗体の親和性精製における試薬としての潜在能力を有する。BIAcore分析では、Ka=4.76×108M−1(Kd=2.10×10−9M)のhu3S193に対するLMH−3の高い明らかな親和性が測定された。この親和性は、捕捉のための精製LMH−3および検出のためのビオチニル化LMH−3を使用するhu3S193の血清濃度を測定するための高再現性、感受性、特異性ELISAアッセイの開発を可能にした。BIAcoreにより血清サンプルのHAHA分析の確立は、患者血清における免疫反応の定量のために陽性対照としてLMH−3を使用することにより可能であり、これらの抗イディオタイプを使用し、腫瘍生検の免疫組織化学的分析を通じて腫瘍へのhu3S193の浸透および結合を試験することもできた。各可変ドメインでの標的抗体を同時に結合することが可能な抗イディオタイプ抗体の生成は、臨床的に投与される標的抗体の安全性および薬物動態プロフィールの評価に高い感度を有する試薬を提供する。hu3S193免疫療法を受ける患者からの臨床サンプルを試験するための診断検査用試薬として重要な用途を有する抗イディオタイプhu3S193mAbが生成されている。
【0032】
本発明の別の態様は、抗原に対する抗体の結合を阻害する抗イディオタイプ抗体の能力を検出するための方法を提供する。本発明の別の態様は、結合イディオタイプ抗体を捕捉し、検出する抗イディオタイプの能力を検出するための方法を提供することである。本発明の別の態様は、イディオタイプ抗ルイスY抗体に結合する抗イディオタイプ抗体の能力を検出するための方法を提供することである。本発明の別の態様は、サンプル血清における抗体の量を検出する方法である。本発明は、本発明の抗体を使用するHAMA、HACA、およびHAHA反応を検出する方法にも関する。
【0033】
血清hu3S193レベルを測定するための2つのELISAアッセイを調査した。第1のアッセイには合成ルイスY抗原による捕捉が含まれ、第2のアッセイでは抗イディオタイプ抗体が使用された。アッセイの感度および特異性を比較し、LMH−3を使用する分析が、hu3S193に対する高い親和性により合成抗原よりも優れており、高い感度を可能にする非特異的結合の欠如からの低いノイズ、および3ng/mlの血清hu3S193の定量限界が見出された。BIAcore分析では、Ka=4.76×108M−1(Kd=2.10×10−9M)のhu3S193に対するLMH−3の明らかな親和性が測定された。ハイブリドーマは、抗体LMH−3の高いレベルを産生する。
【0034】
以下の実施例は本発明の例示のために示されており、本発明の限定と解釈されてはならない。
【実施例】
【0035】
実施例1
細胞培養
すべての分析用試薬を、別に指示がない限り、シグマ・ケミカル社(Sigma Chemical Co.)(St Louis、モンタナ州(MO)、米国)から購入した。
【0036】
SP2/0−Ag14形質細胞腫細胞系をアメリカン・タイプ・カルチャー・コレクション(ATCC、Manassas、バージニア州(VA)、米国)から購入した。SP2/0および選択ハイブリドーマクローンを正常RPMI−1640培地、すなわち、10%加熱不活性化ウシ胎仔血清(FCS、トレース・バイオサイエンス社(TRACE Biosciences Pty Ltd)、シドニー、オーストラリア)、100μg/mlストレプトマイシン、100U/mlペニシリン(ジブコ(Gibco)、および4mM L−グルタミンを含有するRPMI−1640培地(ジブコ・インビトロジェン社(Gibco Invitrogen Corp.)、Mt Waverley、ビクトリア州、オーストラリア)中で培養した。融合生成物の初期の数代継代を、上記のとおり20%FCS、ペニシリン、ストレプトマイシン、およびL−グルタミンを含有するRPMI−1640で培養した。HAT培地をハイブリドーマ選択のために使用した、20%FCS、HAT(ヒポキサンチン、アミノプテリン、チミジン)、OPI(オキサロ酢酸、ピルビン酸塩、インスリン培地補充)(シグマ(Sigma))、ペニシリン、ストレプトマイシン、L−グルタミンを含有するRPMI−1640培地。ハイブリドーマ増殖のために、細胞をHT培地、すなわち、20%FCS、HT(ヒポキサンチン、チミジン)、ペニシリン、ストレプトマイシン、L−グルタミンを含有するRPMI−1640培地で培養した。
【0037】
実施例2
免疫化およびスクリーニングのための抗体
ヒト化抗ルイスY IgG1抗体hu3S193(ロット:D01097、10mg/ml PBS)、マウス3S193(mu3S193)、対照アイソタイプマッチドヒト化mAb、huA33(ロット:5GMA33 10mg/ml)、および無関係のアイソタイプマッチドマウス:ヒトキメラIgG1抗体は、生物学的生産機関(Biological Production Facility)、ルートビッヒ癌研究協会(Ludwig Institute for Cancer Research)、メルボルン、オーストラリア)によって供給された。精製ヒトIgGはシグマ・ケミカル社(Sigma Chemical Co.)から購入された。F(ab)’2フラグメントを、メーカーの指示に従い免疫化ペプシンによる消化によって調製し(ピアス(Pierce)、Rockford、イリノイ州(IL)、米国)、非消化hu3S193および1mlプロテインAカラム(ファルマシア・バイオテック(Pharmacia Biotech)、Uppsala、スウェーデン)上の通過よってFc含有フラグメントから精製した。精製フラグメントをミリポア遠心フィルタ(バイオマックス(Biomax)10k膜、ミリポア(Millipore)、シドニー、オーストラリア)上で1mg/mlに濃縮した。
【0038】
実施例3
免疫化プロトコール
6〜8週齢の雌BALB/cマウスを繁殖用施設、ウォルター・アンド・エリザ・ホール研究所(Walter and Eliza Hall Institute)、メルボルン、オーストラリアから購入し、飼料および水を自由に与えて飼育した。本試験のために動物で実施されたすべての手順は、オースチン・アンド・レパトリエーション・メディカルセンター(Austin and Repatriation Medical Centre)の動物倫理委員会(Animal Ethics Committee)によって承認された。免疫化前静脈血液サンプル(200μl)を各動物から収集し、室温(RT)下に1時間、4℃下に一夜凝固させた。結果として生じる血清(約100μl/マウス)を収集し、−20℃下に保存した。
【0039】
免疫化手順をマウス6匹で実施した。0日目、40μg精製hu3S193:フロイント完全アジュバント(1:1、v:v)含有200μlの腹腔内(i.p.)注射によってBALB/cマウスを免疫化した。マウスに対し14日目および28日目にフロイント不完全アジュバントでhu3S193を40μg含有するブースター注射を投与した。35日目、免疫化後血液サンプルを尾静脈から収集し、マウス抗hu3S193抗体の血清力価をELISAによって測定した。免疫化前血清サンプルを対照として含めた。3回目、40μgのhu3S193のみを含有する最後の静脈内(i.v.)ブースター注射後4日にマウスを屠殺した。脾臓を無菌で5ml PBS/Gに収集し、融合用に脾細胞懸濁液を調製した。
【0040】
実施例4
融合およびハイブリドーマ生成
すべての手順を室温下に実施し、これらは以前に公表された方法に基づくものであった(12)。脾細胞を収集し、RPMI−1640培地で3回洗浄し、細胞濃度を測定した。同時に、融合パートナーSP2/0細胞を回収し、血清を含まない培地中で洗浄し、計算した。各細胞ペレットを血清を含まないRPMI 5ml中に再懸濁し、次いで50mlチューブに5:1比の脾細胞:SP2/0で混合した。細胞混合物を300×g、10分遠心分離し、結果として生じる細胞ペレットを軽くたたくことによって静かに破壊した。チューブを37℃の湯浴中にインキュベートし、培地(1ml)中50%PEGを1分間にわたって滴下した。次いで、予熱した血清を含まないRPMI(2ml)を2分にわたって添加した後、3分にわたって培地8mlを添加した。細胞を300×g、10分遠心分離し、HAT培地(約1×10 5脾細胞、2×10 4SP2/0骨髄腫細胞を含有する60ml HAT培地/マウス脾臓)中に静かに再懸濁した。混合細胞懸濁液を6個の96ウェル細胞培養プレート(ファルコン(Falcon)、ベクトン・ディッキンソン(Becton Dickinson)、Franklin Lakes、ニュージャージー州(NJ)、米国)へ等分し(100μl/ウェル)、加湿5%CO2/空気インキュベーター中37℃下に培養した。
【0041】
実施例5
モノクロナール抗体選択
ハイブリドーマ培養上清をhu3S193mAbおよび精製ヒトIgGとの陽性結合活性についてELISAによってスクリーニングした。選択クローンを24ウェル培養プレート(ファルコン(Falcon))中で増殖させ、その結合活性および競合結合活性をさらにELISAによって特徴づけた。培養上清中の選択抗イディオタイプhu3S193抗体のサブクラスをマウスモノクロナール抗体アイソタイピング用キット(アイソストリップ(IsoStrip)、ロシュ・ディアグノスティックス・ゲーエムベーハー(Roche Diagnostics GmbH)、Mannheim、ドイツ)で測定した。
【0042】
実施例6
限界希釈によるハイブリドーマのクローニング
一次希釈系列
選択ハイブリドーマクローンを96ウェルプレートの内側60ウェル中の200μl HAT培地中100細胞/ウェルの密度でプレーティングした。外側ウェルを抗生物質を含有するRPMI−1640培地で充填し、脱水およびウェルを含有するHAT培地/細胞の濃縮を阻止した。細胞成長がほぼ密集(約7日)であった場合、上清を収集し、ルイスY抗原結合のためのhu3S193への特異的結合およびhu3S193への拮抗薬結合について試験した。陽性クローンを継続的分析およびクローニングのために選択した。
【0043】
二次希釈系列
本ステップは、単一の細胞から成長したハイブリドーマを確立することを目的とし、培養では脾細胞支持細胞層が利用された。脾細胞を非免疫化BALB/cマウスから調製し、100mM 2−メルカプトエタノールを補充した10 6細胞/ml HAT培地に再懸濁し、滅菌環境下に室温で一夜インキュベートし、汚染感染がないことを確実にした。翌日、2つの96ウェルマイクロタイタープレートを、既述どおり、使用したプレートの内側60ウェルのみで各選択クローン用に調製した。HAT培地のアリコート(100μl)を列1に添加し、脾細胞懸濁液を列2−6(10 5細胞/ウェル)に添加し、プレートを37℃下に一夜インキュベートした。選択細胞をHAT培地中に希釈し、100、10、5、1、または0.5細胞を含有する100μlのアリコートを、それぞれ、支持細胞を含有する2から6までの列のウェルに添加した。プレートを37℃下に10−14日間インキュベートし、1または0.5細胞/ウェルで接種したウェルからクローンコロニーを選択した。
【0044】
三次希釈系列
選択クローンが単一の細胞から由来することを確実にするために、すべての選択細胞系は反復回の二次希釈手順を受けた。好ましい系を増殖させ、ストックを調製し、液体窒素下に保存した。
【0045】
実施例7
抗体産生
ハイブリドーマを産生する選択および再クローン化抗イディオタイプ抗体をローラーボトル(コーニング(Corning)、コーニング社(Corning Incorporated)、ニューヨーク州(NY)、米国)で培養し、培養上清を無菌で回収した。抗イディオタイプ抗体を、50mM Tris−HCl、pH8(バッファーA)で前平衡した5mlプロテインAセファロース迅速フロー(fast flow)カラム(アマシャム・ファルマシア・バイオテック(Amersham Pharmacia Biotech)、Uppsala、スウェーデン)における親和性クロマトグラフィーによって精製した。20カラム量バッファーAでの洗浄後、200mM NaCl含有100mMグリシン−HCL、pH2.7を使用して結合抗体を溶出し、1M Tris HCl、pH8を使用して直ちに中和した。PBSへの4℃での一夜透析後、抗体を遠心フィルタ(ミリポア(Millipore)、バイオマックス(Biomax)10k膜)で濃縮した。還元および非還元条件下にSDS−PAGEによって純度を検査した。クマシーブルー染色によってタンパク質を視覚化した。ビオチニル化抗イディオタイプ抗体をECLタンパク質ビオチニル化モジュールキットを使用し、メーカーの指示(アマシャム・ファルマシア・バイオテック(Amersham Pharmacia Biotech)に従い調製した。
【0046】
実施例8
結合特異性のELISA分析
ハイブリドーマ上清または精製抗イディオタイプ抗体の結合特異性をELISA分析によって特徴づけた。96ウェルプレート(ヌンク・マキシソープ(Nunc MaxiSorp)、Roskilde、デンマーク)をhu3S193(100μl、4.0μg/ml)、精製ヒトIgG、または炭酸バッファー(0.05M、pH9.6)中の他の対照モノクロナール抗体で4℃下に一夜コーティングした。非特異的結合を3%BSA/PBSで室温(23℃)下に2時間遮断した。上清または精製抗体の連続希釈(1:2−1:6300)を調製し、ウェル当たり100μlアリコートを添加した。陽性(huIgG)および陰性(抱合体および基質のみ)対照を各アッセイで含めた。プレートを室温下に1時間インキュベートし、0.05%Tween20/PBSで3回洗浄し、次いで1時間二次抗体(ヤギ抗マウスIgG抱合アルカリホスファターゼ(100μl/ウェル、1%BSA/PBS中1:3000希釈)と一緒にインキュベートした。結合抗体を発色団基質p−ニトロフェニルリン酸基質(ICNバイオメディカルズ社(ICN Biomedicals Inc.)、Aurora、オハイオ州(Ohaio)、米国)で検出した。光学密度をVmaxキネティックマイクロプレートリーダー(モレキュラー・デバイス社(Molecular Devices Corporation)、Sunyvale、カリフォルニア州)により450nmで測定した。
【0047】
実施例9
拮抗薬活性のELISA分析
選択抗イディオタイプ抗体をルイスY−抗原コートプレートへのhu3S193mAbの結合を遮断するその能力について試験した。合成ルイスY四糖類は、モル比約32:1でBSAに結合されて入手可能である(アルバータ・リサーチ・カウンシル(Alberta Research Council)、Edmonton、アルバータ州(Alberta)、カナダ)。ELISAプレート(ヌンク・マキシソープ(Nunc MaxiSorp))をルイスY−BSA 抗原(50μl 3.0μg/ml PBS)でコーティングし、4℃下に一夜インキュベートした。プレートを室温下に2時間、3%BSA/PBSで非特異的結合のために遮断した。ハーフログ連続希釈サンプル(上清または精製抗イディオタイプmAb)を各ウェルに添加した後、hu3S193mAb(50μl、1μg/ml最終濃度)を添加した。室温下に1時間のインキュベーション後、ヤギ抗−ヒトIgG−HRPを使用し、結合hu3S193mAbを検出し、広範な洗浄後、ABTS基質(2,2’−アジノ−ビス−(3−エチル−ベンズチアゾリン−6−スルホン酸、100μl 40μM/ウェル、ICN)で視覚化した。光学密度(OD)をVmaxキネティックマイクロプレートリーダーにより415nmで測定した。
【0048】
実施例10
Hu3S193の抗イディオタイプ特異性
抗ルイスY mAbへの結合の精製選択クローンの特異性をさらに、hu3S193、およびマウスmAb mu3S193およびBR55.2(ルートビッヒ癌研究協会(Ludwig Institute for Cancer Research)hu3S193を使用して検査した。特異性を2つのELISAで検査した。第1のELISAでは結合イディオタイプmAbの捕捉および検出のための抗イディオタイプが利用され、第2のELISAでは溶液中でイディオタイプ抗ルイスY mAbに結合し、ルイスY抗原結合を阻害する抗イディオタイプmAbの能力が検査された。
【0049】
第1のアッセイでは、ELISAマキシソープ(Maxisorp)プレートを選択抗イディオタイプmAbで一夜、4℃下にコーティングした(100μl 4μg/ml 0.05M炭酸バッファー、pH9.6)。3%FCS/PBSによる遮断後、連続希釈した10μg/mlのhu3S193、mu3S193、BR55.2の3部サンプル、および無関係の対照mAb huA33およびmuA33をウェルに添加し、次いで室温下に1時間インキュベートした。洗浄後、結合mAbをビオチニル化抗イディオタイプmAb(LMH−3−Bt、4℃下に一夜、1%FCS/PBS中100μl/ウェル 4μg/ml)で検出した。結合複合体ストレパビジン−HRPの(1%FCS/PBS中1:1000希釈、100μl/ウェル)検出の後、着色のためにABTSを添加した。OD415を既述どおりに測定した。
【0050】
特異性の第2の試験のために、3つのELISAプレートを合成ルイスY−BSA抗原でコーティングした(PBS中4.0μg/ml、4℃下に一夜)。次いで、連続希釈抗ルイスYmAbをプレートの各列上に等分した、プレート1、hu3S193、プレート2、mu3S193、プレート3 BR55.2最終濃度範囲(0.003−10μg/ml)。ルイスY抗原への結合の競合を試験するために、他の2つの抗ルイスY mAbおよび2つの抗イディオタイプmAbを2部で(最終濃度10μg/ml)添加し、次いで室温下に1時間インキュベートした。洗浄後、プレート1における結合hu3S193を抗huIgG−HRP(シグマ(Sigma)、1%FCS/PBS中100μl 1:1000)の後、着色のためのABTSで検出し、A415nmで測定した。プレート2における結合mu3S193およびプレート3におけるBR55.2をヤギ抗−mIgGアルカリホスファターゼで検出し、p−ニトロフェノール色原体をA405で測定した。
【0051】
実施例11
血清中のHu3S193のELISA測定
血清サンプル中のhu3S193mAbを試験するために2つのELISAアッセイが開発された。第1のアッセイでは、合成ルイスY−BSA抗原(PBS中3.0μg/ml、50μl/ウェル)をhu3S193捕捉のために利用した。健康ドナー血清中のhu3S193の連続希釈サンプルまたは3%BSA/PBSによるインキュベーション後、抗原に結合するhu3S193mAbの量をヤギ抗ヒトIgG−HRP二次抗体およびABTS基質で検出した。
【0052】
第2のアッセイには抗イディオタイプ抗体によるコーティングおよびビオチニル化抗イディオタイプ抗体による検出が含まれた。プレートを抗イディオタイプhu3S193抗体(0.05M炭酸バッファーpH9.6中3.0μg/ml、100μl/ウェル)で一夜、4℃下にコーティングし、次いで2時間、室温(23℃)下に3%BSA/PBSで遮断した。3%BSA/PBSまたは健康ドナー血清中のhu3S193(0.0315−10μg/ml)の連続希釈サンプルを3部でプレートに添加し、1時間、室温下にインキュベートした。戦場後、ビオチニル化抗イディオタイプhu3S193抗体を添加し(1%BSA/PBS中100μl 3μg/ml)、1時間、室温下にインキュベートした。結合複合体ストレパビジン−HRPの(1%BSA/PBS中1:1000希釈、100μl/ウェル)検出の後、着色のためにABTSを添加した。OD415を既述どおりに測定した。
【0053】
バイオセンサー分析:
カルボキシメチルデキストランコートセンサーチップ(CM5)上でBIAcore2000(BIAcore AB、Uppsala、スウェーデン)を使用してバイオセンサー分析を実施した。チップをチャンネル2でhu3S193、チャンネル3でLMH−3、およびチャンネル4でウシ血清アルブミン(アルバータ・リサーチ・カウンシル(Alberta Research Council)、Edmonton、アルバータ州(Alberta)、カナダ)に結合した合成ルイスy四糖類により、標準NHS/EDCアミン結合化学を使用して誘導体化した。hu3S193およびLMH−3のサンプルをHBSバッファー(10mM HEPES、pH7.4、150mM NaCl、3.4mMジ−Na−EDTA、0.005%Tween−20)、およびアリコート(30μl)を5μl/分の流量でセンサーチップ表面にわたって注入した。注入相後、300秒間チップ表面上にHBSバッファーを流すことによって解離をモニタリングした。結合抗体を溶出し、100mMグリシン/100mM NaCL、pH2.7、20μlの注入によってサンプル間でチップ表面を再生した。結合の動態分析にために、各種濃度のhu3S193およびLMH−3をセンサーチップ表面上に注入した。明らかな結合(ka)および解離(kd)速度定数を二価分析モデルを使用し、Rmaxの計算のための全体的、およびローカル適合により、BIA評価v3.0ソフトウェア(ファルマシア・バイオセンサー(Pharmacia Biosensor)、Uppsala、スウェーデン)を使用して計算した。
【0054】
投与されたモノクロナール抗体に対する患者の免疫反応は、患者血清のバイオコア(Biocore)(登録商標)分析によって評価されうる(リッター(Ritter)、コーエン(Cohen)ら、2001年)。抗イディオタイプ抗体は、これらの分析において陽性対照試薬として有用である。したがって、LMH−3を健康ドナー血清(3−50μg/ml)に添加し、免疫化hu3S193に対する結合反応を確認した。
【0055】
実施例12
マウス免疫化およびハイブリドーマクローン選択
3群のBALB/cマウスを繰り返しhu3S193mAbで免疫化し、その血清をマウス抗hu3S193mAb活性について評価した。血清サンプルの免疫化前後の免疫活性は、高力価のマウス抗無傷およびF(ab)’2hu3S193mAbの進展を示した。アイソタイプ対照ヒト化IgG(huA33)またはキメラIgGF(ab)’2でコーティングされたプレートも強い陽性結合を示した(結果は示さず)。マウスを屠殺し、その脾臓を除去した。脾細胞およびパートナー融合Sp2/0形質細胞腫細胞を融合し、これら3つの融合からのハイブリドーマをスクリーニングした。hu3S193mAbに対して強い結合活性を示すが、対照huA33または精製ヒトIgGに対しては弱い結合を示し、または結合を示さない3−4ウェルのみを選択し、増殖させた。増殖後、抗体特異性をさらにELISAアッセイで試験した(図1)。3つの対照ヒト化またはキメラ抗体を第1の融合からのクローンの選択中に使用した。交差結合活性が通常、精製ヒトIgGコーティングプレートで確認されたため、その後のスクリーニングにおいてhu3S193に加えて精製ヒトIgGを主に使用した。
【0056】
実施例13
選択抗イディオタイプ抗体の結合および遮断活性
すべての選択ハイブリドーマの結合および拮抗薬活性をさらに分析し、抗イディオタイプhu3S193抗体を産生するハイブリドーマを確認した。ハーフログ連続希釈を各ハイブリドーマ培養上清から調製し、ELISAによって評価した。最初に選択された12個のクローンのうち、hu3S193に結合するが、huIgGには結合せず、hu3S193のルイスY抗原への結合を阻害しうる3つを確認した(図1)。これらのハイブリドーマ、F1−1、F2−3、およびF3−27を限界希釈によって単一の細胞からクローン化し、それぞれ、ルートビッヒ癌研究協会(Ludwig Institute for Cancer Research)メルボルン(Melbourne)ハイブリドーマ(LMH)−1、LMH−2、およびLMH−3と指名された。クローンの1つ(FS2−1)は、すべての試験されたキメラヒト化mAbもしくは精製huIgGまたは(図1)に対する強い抗huIgG結合活性を示し、その後にhuIgG結合に対する陽性対照として使用された。このクローンは単一の細胞から調製され、LMH−4を命名された。LMH−1から−4までの抗体は、マウスモノクロナール抗体アイソタイピング用キットによってアイソタイプIgG1eとして確認され、プロテインA親和性クロマトグラフィーによって培養上清から精製された。4つのクローンの結合特異性がELISAによって確認された(図4A−C)。huIgG定常鎖との交差反応は、LMH−1、−2、またはー3では確認されず(図4B)、3つの抗体はすべて特異的にhu3S193mAbのみに結合された(図4A)。これらの抗体の拮抗薬活性は、競合ELISAアッセイによって分析された。LMH−1、−2、および−3抗hu3S193イディオタイプ抗体は、hu3S193mAbに結合し、hu3S193mAbのルイスY抗原への結合を用量依存的に阻害する(図2)。
【0057】
実施例14
抗体結合親和性の測定
溶液中でLMH−3と相互作用する固定化hu3S193のバイオセンサー結合実験の明らかな結合親和性では、4.76×108M−1(Kd=2.10×10−9M)のKaが測定された。チップ上に固定化されたLMH−3、および分析物として表面上を走るhu3S193との相互実験において、9.12×108M−1(Kd=1.10×10−9M)のKaが測定された。
【0058】
実施例15
Hu3S193に対する抗イディオタイプ特異性
3S193イディオタイプに対するクローンLMH−2およびLMH−3の特異性がこれらの実験で確認された(図5)。プロテインA精製LMH−3をELISAプレート上にコーティングし、連続希釈した10μg/mlのhu3S193、mu3S193、BR55.2の3部サンプル、および対照IgG1 mAb huA33およびmuA33をウェルに添加し、さらにLMH−3の結合特異性を特徴づけた。このクローンの追加の好ましい特徴は、ビオチニル化LMH−3がLMH−3で捕捉された結合3S193mAbの検出に利用されうることであった。ELISAの結果(図3A)は、LMH−3が特異的に抗ルイスY3S193イディオタイプに結合し、マウス抗ルイスYBR55.2との反応性は検出されず、対照muA33およびhuA33のマウスおよびヒトIgG1定常ドメインとの交差反応はなかったことを示した。hu3S193およぶmu3S193のルイスY抗原への結合は、10μg/ml LMH−2およびLMH−3によって溶液中で有効に競合したが、抗イディオタイプ結合部位の飽和時には、一部のhu3S193およびmu3S193の抗原への結合は確認されなかった(それぞれ、図3Bおよび3C)。マウスBR55.2mAbは一部の競合結合をもたらしたが、高い親和性のmu3S193は、最高濃度(10μg/ml)での場合を除き、hu3S193のルイスY抗原への結合を完全に遮断した(図3B)。プレート2相互実験においては、低い親和性のhu3S193は、mu3S193が最初にウェルに添加されると、ルイスY抗原に競合しえなかった(図3C)。プレート3からの結果(図3D)は、BR55.2のルイスY抗原への結合が、hu3S193または抗イディオタイプクローンLMH−2および−3と競合せず、またはこれらによって阻害されないことを示す。したがって、3S193イディオトープに対するこれらクローンの特異性が確認される。図3Bにおいて確認されるBR55.2による競合は、その異なるルイスYエピトープへの高い親和性結合に対する立体障害により達成されうる。
【0059】
実施例16
臨床サンプルにおけるHu3S193を測定するためのELISAアッセイの開発
第1のELISAアッセイでは、血清および検出用のペルオキシダーゼと抱合された二次ヤギ抗ヒトIgGにおけるhu3S193の捕捉用に使用された合成抗原(ルイスY四糖類−BSA抱合体)が調査された。合成ルイスY抗原は、記録された低い光学密度によって示されるアッセイ感度を削減した、hu3S193mAbに対する比較的低い親和性を有する。二次抱合抗体ではすべてのヒト免疫グロブリンが検出され、したがって高いノイズが分析したすべてサンプルで確認された(図4A)。高いノイズ干渉が、特に低い血清hu3S193濃度でアッセイの感度および精度を大幅に削減する。第2のELISAアッセイでは、コーティング用に精製抗イディオタイプhu3S193抗体LMH−1、2、および−3が使用され、二次抗体としてビオチニル化LMH−1、−2、または−3が使用され、かつ結合複合体の視覚化用にストレパビジン−HRPおよびABTS基質が使用された。LMH−1およびLMH−2抗イディオタイプ抗体は、血清中または3%BSA/PBS中でmu3S193mAbに対する強い結合示したが、hu3S193mAbに対しては弱い結合を示した。しかし、ヒト化mAb、キメラmAb、マウスmAb、ヒト血清由来のIgまたはマウス血清由来のIgを含む対照抗体では交差結合活性は確認されなかった(図4B)。コーティング抗体または二次抗体の濃度を変更し、かつ異なるコーティングバッファーを使用することによってLMH−1およびLMH−2によるELISAを最適化または改善する試みを調査した。一部の改善が達成されたが、反応パターンへの変化はもたらされなかった(データは示さず)。しかし、捕捉用のLMH−3抗イディオタイプmAbおよび二次抗体としてLMH−3−ビオチンを使用することにより、最小の非特異的ノイズ活性を有する血清中でhu3S193mAbおよびmu3S193mAbに対する特異的に強い結合が確認された(図4C)。高再現性のアッセイは優れた感度を示し、血清サンプルにおけるhu3S193mAbまたはmu3S193mAbに対する検出限界は3ng/mlであった。
【0060】
免疫反応のバイオコア分析
抗イディオタイプLMH−3を使用するhu3S193に対する血清中のHAHAの測定が開発された。高感度で再現性の方法が確立され、確認された(図5)。
【図面の簡単な説明】
【0061】
【図1】ELISAによって測定される抗−hu3S193モノクロナール抗体、LMH−1(●)、−2(▽)、−3(◇)、および抗−huIgG−4(◇)の結合特異性を示す。
【図2】ELISAによって検査されたハイブリドーマ培養上清のhu3S193抗原結合活性を中和する能力について示す。
【図3】3S193イディオトープに対するLMH−2およびLMH−3の特異性の確認を示す。
【図4A】患者血清サンプルにおけるhu3S193を測定するためのELISAの開発を示す。
【図4B】患者血清サンプルにおけるhu3S193を測定するためのELISAの開発を示す。
【図4C】患者血清サンプルにおけるhu3S193を測定するためのELISAの開発を示す。
【図5】hu3S193に対する血清HAHA反応のBiacore分析が最適化され、陽性対照としての抗イディオタイプLMH−3を使用して確認されたことを示す。
【特許請求の範囲】
【請求項1】
抗ルイスYモノクロナール抗体に対する抗イディオタイプ抗体。
【請求項2】
抗ルイスYモノクロナール抗体の可変領域に結合する、請求項1記載の抗イディオタイプ抗体。
【請求項3】
抗ルイスYモノクロナール抗体の結合を遮断する、請求項1記載の抗イディオタイプ抗体。
【請求項4】
hu3S193と特異的に結合する、請求項1記載の抗イディオタイプ抗体。
【請求項5】
モノクロナール抗体、キメラ抗体、ヒト抗体、ヒト化抗体、または一本鎖抗体からなる群より選択される、請求項1記載の抗イディオタイプ抗体。
【請求項6】
モノクロナール抗体、キメラ抗体、ヒト抗体、ヒト化抗体、または一本鎖抗体からなる群より選択される、請求項2記載の抗イディオタイプ抗体。
【請求項7】
モノクロナール抗体、キメラ抗体、ヒト抗体、ヒト化抗体、または一本鎖抗体からなる群より選択される、請求項3記載の抗イディオタイプ抗体。
【請求項8】
モノクロナール抗体、キメラ抗体、ヒト抗体、ヒト化抗体、または一本鎖抗体からなる群より選択される、請求項4記載の抗イディオタイプ抗体。
【請求項9】
請求項1に記載の抗イディオタイプ抗体の産生能を有するハイブリドーマ。
【請求項10】
請求項2に記載の抗イディオタイプ抗体の産生能を有するハイブリドーマ。
【請求項11】
請求項3に記載の抗イディオタイプ抗体の産生能を有するハイブリドーマ。
【請求項12】
請求項4に記載の抗イディオタイプ抗体の産生能を有するハイブリドーマ。
【請求項13】
LMH−1、LMH−2、およびLMH−3からなる群から選択される、請求項12記載のハイブリドーマ。
【請求項14】
LMH−1、LMH−2、およびLMH−3からなる群より選択されるハイブリドーマによって産生される、請求項4記載の抗イディオタイプ抗体。
【請求項15】
抗イディオタイプ抗体の結合特異性を検出する方法であって、
a.Elisaプレートを抗ルイスY抗体、精製ヒトIgG、または他の対照Mabでコーティングし、
b.精製抗イディオタイプ抗体を添加し、
c.二次抗体と一緒にインキュベートし、および
d.抗イディオタイプ抗体の抗体との結合が結合特異性を証明するところの、結合した抗イディオタイプ抗体の量を検出する、
ことを含む、方法。
【請求項16】
前記mABがhu3S193である、請求項15記載の方法。
【請求項17】
抗イディオタイプ抗体が抗ルイスY抗原に対して方向付けられる、請求項15記載の方法。
【請求項18】
抗ルイスYモノクロナール抗体の結合の遮断能を有する抗イディオタイプ抗ルイスY抗体を検出する方法であって、
a.ElisaプレートをルイスY−BSA結合抗原でコーティングし、
b.抗イディオタイプ抗体を添加し、
c.モノクロナール抗ルイスYイディオタイプ抗体を添加し、および
d.抗イディオタイプ抗体の存在および非存在下にて抗原と結合した抗ルイスY抗体の量が、抗ルイスモノクロナール抗体の結合を遮断する抗イディオタイプ抗体の能力の証拠である、結合抗ルイスYモノクロナール抗体を検出する、
ことを含む、方法。
【請求項19】
抗−抗ルイスYモノクロナール抗体がhu3S193である、請求項18記載の方法。
【請求項20】
前記抗イディオタイプ抗体が抗ルイスY抗体に対して方向付けられる、請求項18記載の方法。
【請求項21】
血清サンプル中の抗ルイスY抗体を検出する方法であって、
a.ELISAプレートを合成ルイスY−BSA抗原でコーティングし、
b.血清サンプルを前記ELISAプレートに添加し、
c.ペルオキシダーゼ抱合抗ルイスY抗イディオタイプ抗体をELISAプレートに添加し、および
d.抗ルイスY抗体の存在を抗原をコーティングしたELISAプレートと結合したペルオキシダーゼ抱合抗ルイスY抗イディオタイプ抗体の量から測定する、
ことを含む、方法。
【請求項22】
前記モノクロナールイディオタイプ抗体がhu3S193である、請求項21記載の方法。
【請求項23】
ヒト化抗ルイスYモノクロナール抗体が投与されている対象における抗ルイスY HAHA反応を検出する方法であって、
a.血清サンプルを前記対象から収集し、
b.前記血清サンプルをペルオキシダーゼ抱合抗ルイスY抗体の存在または非存在下にてルイスY抗原コーティングのELISAプレートに反応させ、および
c.そこで抗イディオタイプ抗ルイス抗体の存在がHAHA反応を証明するところの、抗イディオタイプ抗ルイス抗体の存在または非存在を測定する
ことを含む、方法。
【請求項1】
抗ルイスYモノクロナール抗体に対する抗イディオタイプ抗体。
【請求項2】
抗ルイスYモノクロナール抗体の可変領域に結合する、請求項1記載の抗イディオタイプ抗体。
【請求項3】
抗ルイスYモノクロナール抗体の結合を遮断する、請求項1記載の抗イディオタイプ抗体。
【請求項4】
hu3S193と特異的に結合する、請求項1記載の抗イディオタイプ抗体。
【請求項5】
モノクロナール抗体、キメラ抗体、ヒト抗体、ヒト化抗体、または一本鎖抗体からなる群より選択される、請求項1記載の抗イディオタイプ抗体。
【請求項6】
モノクロナール抗体、キメラ抗体、ヒト抗体、ヒト化抗体、または一本鎖抗体からなる群より選択される、請求項2記載の抗イディオタイプ抗体。
【請求項7】
モノクロナール抗体、キメラ抗体、ヒト抗体、ヒト化抗体、または一本鎖抗体からなる群より選択される、請求項3記載の抗イディオタイプ抗体。
【請求項8】
モノクロナール抗体、キメラ抗体、ヒト抗体、ヒト化抗体、または一本鎖抗体からなる群より選択される、請求項4記載の抗イディオタイプ抗体。
【請求項9】
請求項1に記載の抗イディオタイプ抗体の産生能を有するハイブリドーマ。
【請求項10】
請求項2に記載の抗イディオタイプ抗体の産生能を有するハイブリドーマ。
【請求項11】
請求項3に記載の抗イディオタイプ抗体の産生能を有するハイブリドーマ。
【請求項12】
請求項4に記載の抗イディオタイプ抗体の産生能を有するハイブリドーマ。
【請求項13】
LMH−1、LMH−2、およびLMH−3からなる群から選択される、請求項12記載のハイブリドーマ。
【請求項14】
LMH−1、LMH−2、およびLMH−3からなる群より選択されるハイブリドーマによって産生される、請求項4記載の抗イディオタイプ抗体。
【請求項15】
抗イディオタイプ抗体の結合特異性を検出する方法であって、
a.Elisaプレートを抗ルイスY抗体、精製ヒトIgG、または他の対照Mabでコーティングし、
b.精製抗イディオタイプ抗体を添加し、
c.二次抗体と一緒にインキュベートし、および
d.抗イディオタイプ抗体の抗体との結合が結合特異性を証明するところの、結合した抗イディオタイプ抗体の量を検出する、
ことを含む、方法。
【請求項16】
前記mABがhu3S193である、請求項15記載の方法。
【請求項17】
抗イディオタイプ抗体が抗ルイスY抗原に対して方向付けられる、請求項15記載の方法。
【請求項18】
抗ルイスYモノクロナール抗体の結合の遮断能を有する抗イディオタイプ抗ルイスY抗体を検出する方法であって、
a.ElisaプレートをルイスY−BSA結合抗原でコーティングし、
b.抗イディオタイプ抗体を添加し、
c.モノクロナール抗ルイスYイディオタイプ抗体を添加し、および
d.抗イディオタイプ抗体の存在および非存在下にて抗原と結合した抗ルイスY抗体の量が、抗ルイスモノクロナール抗体の結合を遮断する抗イディオタイプ抗体の能力の証拠である、結合抗ルイスYモノクロナール抗体を検出する、
ことを含む、方法。
【請求項19】
抗−抗ルイスYモノクロナール抗体がhu3S193である、請求項18記載の方法。
【請求項20】
前記抗イディオタイプ抗体が抗ルイスY抗体に対して方向付けられる、請求項18記載の方法。
【請求項21】
血清サンプル中の抗ルイスY抗体を検出する方法であって、
a.ELISAプレートを合成ルイスY−BSA抗原でコーティングし、
b.血清サンプルを前記ELISAプレートに添加し、
c.ペルオキシダーゼ抱合抗ルイスY抗イディオタイプ抗体をELISAプレートに添加し、および
d.抗ルイスY抗体の存在を抗原をコーティングしたELISAプレートと結合したペルオキシダーゼ抱合抗ルイスY抗イディオタイプ抗体の量から測定する、
ことを含む、方法。
【請求項22】
前記モノクロナールイディオタイプ抗体がhu3S193である、請求項21記載の方法。
【請求項23】
ヒト化抗ルイスYモノクロナール抗体が投与されている対象における抗ルイスY HAHA反応を検出する方法であって、
a.血清サンプルを前記対象から収集し、
b.前記血清サンプルをペルオキシダーゼ抱合抗ルイスY抗体の存在または非存在下にてルイスY抗原コーティングのELISAプレートに反応させ、および
c.そこで抗イディオタイプ抗ルイス抗体の存在がHAHA反応を証明するところの、抗イディオタイプ抗ルイス抗体の存在または非存在を測定する
ことを含む、方法。
【図1】



【図2】

【図3】

【図4A】

【図4B】
【図4C】

【図5】

【図2】
【図3】
【図4A】
【図4B】
【図4C】
【図5】
【公表番号】特表2007−528868(P2007−528868A)
【公表日】平成19年10月18日(2007.10.18)
【国際特許分類】
【出願番号】特願2006−523289(P2006−523289)
【出願日】平成16年8月10日(2004.8.10)
【国際出願番号】PCT/US2004/025789
【国際公開番号】WO2005/019271
【国際公開日】平成17年3月3日(2005.3.3)
【出願人】(591011502)ワイス (573)
【氏名又は名称原語表記】Wyeth
【出願人】(505023777)ラドウィッグ・インスティテュート・フォー・キャンサー・リサーチ (2)
【氏名又は名称原語表記】Ludwig Institute for Cancer Research
【Fターム(参考)】
【公表日】平成19年10月18日(2007.10.18)
【国際特許分類】
【出願日】平成16年8月10日(2004.8.10)
【国際出願番号】PCT/US2004/025789
【国際公開番号】WO2005/019271
【国際公開日】平成17年3月3日(2005.3.3)
【出願人】(591011502)ワイス (573)
【氏名又は名称原語表記】Wyeth
【出願人】(505023777)ラドウィッグ・インスティテュート・フォー・キャンサー・リサーチ (2)
【氏名又は名称原語表記】Ludwig Institute for Cancer Research
【Fターム(参考)】
[ Back to top ]