
新規なプロモーター、これを有する組み換え七面鳥ヘルペスウイルス、及びこれを利用した家禽用ワクチン
【課題】 複数の外来遺伝子が挿入されても安定な組み換えHVTを得るための外来遺伝子発現用プロモーター及びそれを利用した安定な組み換え七面鳥ヘルペスウイルスを得る。
【解決手段】 2つの異なる外来遺伝子と、当該2つの外来遺伝子を両方とも支配するように、2つの外来遺伝子の間に配置された、マレック病ウイルス由来の1つのプロモーターとを有する発現遺伝子カセットが、ゲノム中に1つ以上挿入された組み換え七面鳥ヘルペスウイルス。
【解決手段】 2つの異なる外来遺伝子と、当該2つの外来遺伝子を両方とも支配するように、2つの外来遺伝子の間に配置された、マレック病ウイルス由来の1つのプロモーターとを有する発現遺伝子カセットが、ゲノム中に1つ以上挿入された組み換え七面鳥ヘルペスウイルス。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、七面鳥ヘルペスウイルス(以下、HVTということがある)ゲノムに2つ以上の外来遺伝子を組み込んだ組み換えHVTと、外来遺伝子を発現させるために用いるマレック病ウイルス由来のプロモーター、及びそれを利用したワクチンに関する。
【背景技術】
【0002】
米国等の養鶏業においては、既に、ウイルスベクターワクチンである組み換え鶏痘ウイルス(FPV)や組み換え七面鳥ヘルペスウイルスが実用化されている。中でもHVTは家禽での安全性が高く、高マレック病ウイルスワクチンとしても利用されるほど、ワクチン特性も良好であること、鶏へのワクチン接種法の一つである発育鶏卵内にワクチンを接種する卵内接種が可能であることから、鳥類に対するワクチン用のウイルスベクターとしても注目されている。
【0003】
また、HVTの感染様式として、感染細胞から他の細胞にウイルスが感染する際、隣り合う細胞へ細胞−細胞間で感染が成立するため、血流中に存在するHVT特異的抗体の影響を受けにくい。従って、これまでのウイルス生ワクチンの効果を減弱させる大きな要因であった親鳥からの移行抗体の存在下でも十分な効果を発揮するという有利な点もある。
【0004】
更に、HVTを含むヘルペスウイルス属は、潜伏感染(latent infection)や永続感染(permanent infection) の状態で感染動物の体内に永続的に生存し続けることが知られていて、このことも、HVTのワクチンとしての有効性、つまり、感染期間が長いため長期にわたってワクチン効果を付与できる特徴も有している。従って本発明者らもこの組み換えHVTベクターワクチンの研究を行い、報告してきた(特許第3587065号、米国特許第7153511号)。この内、特許第3587065号では、複数の外来抗原遺伝子(例えばニューカッスル病ウイルス(NDV)のHN蛋白質やF蛋白質、ガンボロ病ウイルス(IBDV)のVP2蛋白質の遺伝子)を挿入した組み換えHVT(HF006、HF007)を純化し、これら外来抗原遺伝子の発現やワクチン効果を報告している。
【0005】
しかしながら、複数の外来遺伝子を発現させるような組み換えHVTは、不安定である場合が多く、更に、純化した組み換えHVTを培養細胞で繰り返し継代するうちに、挿入させた外来遺伝子の一部を喪失したウイルスに変遷してしまうことが度々あった。
【0006】
【特許文献1】特許第3587065号(米国特許第6632664号)公報
【特許文献2】米国特許第7153511号公報
【特許文献3】国際公開公報WO94/23019号(米国特許第5871742号)公報
【特許文献4】特開2004−111号(米国特許第6764684号、米国特許第7153511号)公報
【特許文献5】国際公開公報WO2005/093070号(欧州公開第1731612号)公報
【0007】
【非特許文献1】Avian Dis.36:858−870(1992)
【非特許文献2】J.Virology 74:7980−7988(2000)
【非特許文献3】J.Gen.Virology 79:1997−2001(1998)
【非特許文献4】J.Gen.Virology 80:2417−2422(1999)
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明者は、上記の課題を解決すべく鋭意研究を重ねた結果、プロモーターとして、双方向の転写活性を有するものを採用し、このプロモーターを挟むように2つの外来遺伝子を連結させて、1つのプロモーターによって2つの外来遺伝子を支配する発現遺伝子カセットを組み込んだ組み換え七面鳥ヘルペスウイルスが、挿入した遺伝子の脱落を起こさない極めて安定な組み換えウイルスであり、これがワクチンとして使用できることを見出し、本発明を完成したものである。
【課題を解決するための手段】
【0009】
かくして本発明によれば、2つの異なる外来遺伝子と、当該2つの外来遺伝子を両方とも支配するように、2つの外来遺伝子の間に配置された、マレック病ウイルス由来の1つのプロモーターとを有する発現遺伝子カセットが、ゲノム中に1つ以上挿入された組み換え七面鳥ヘルペスウイルスが提供される。
前記プロモーターとしては、プロモーターが配列番号1又は配列番号2が好ましい。これら二つのプロモーターは、いずれも新規なプロモーターである。
また、前記発現遺伝子カセットの挿入部位は、七面鳥ヘルペスウイルスのUL45とUL46のオープン・リーディング・フレームの間であるのが好ましい。
挿入される発現遺伝子カセット中の2つの異なる外来遺伝子のうち、少なくとも1つが鳥類感染性の病原体に由来する抗原遺伝子であるものは、家禽用ワクチンとして用いることができる。
【発明を実施するための最良の形態】
【0010】
以下に本発明を詳述する。
(1)七面鳥ヘルペスウイルス(HVT)
本発明の組み換え七面鳥ヘルペスウイルスの親株として使用する七面鳥ヘルペスウイルスは、天然に得られるものの他、細胞・微生物・遺伝子バンクであるAmerican Type Culture Collection(ATCC)などから有償または無償で入手できるものなどであればよく、特に限定されるものではない。
HVTの好ましい例としては、ガンマヘルペスウイルス亜科に属し、生来非病原性であり、かつ非腫瘍性のウイルスで家禽用のワクチンとして用いられているものが挙げられる。具体的には、FC126(ATCCVR−584B)、PB−THV1、H−2、YT−7、WTHV−1、HPRS−26などの株が挙げられ、例えば、FC126株を好適に使用することができる。
【0011】
(2)マレック病ウイルス(MDV)
本発明において、二つの外来遺伝子を発現させるプロモーターの由来となるマレック病ウイルスもまた、天然に得られるものの他、ATCCなどから有償または無償で入手できるものなどであればよく、特に限定されるものではない。好ましい例としては、家禽用ワクチンとして用いられているものが挙げられ、具体的には、CVI988株やSB1株などを挙げることができる。
【0012】
(3)プロモーター
本発明の組み換えHVTにおいて、2つの異なる外来遺伝子を発現させるために、その間に置かれるMDV由来プロモーターは、マレック病ウイルスゲノムから得られたプロモーターであって、双方向の転写活性を有するものである。マレック病ウイルスゲノムから得られたプロモーターを、プロモーターとしての活性が維持できる範囲で、一部を切り縮めるなどの改変をしてもよい。
双方向の転写活性を有するプロモーターは、マレック病ウイルスゲノムの中から、読み枠が反対方向を向いている、二つのオープン・リーディング・フレームを探し、その間に位置するプロモーター領域をプロモーター候補として絞り込み、後述する実施例中の、プロモーターのクローニング方法に従ってクローニングすることにより得られる。
また、本発明に用いるプロモーターは、組み換え体の安定性の観点から、遺伝子長が1kbpより短いのが好ましく、800bpより短いのが更に好ましい。
このようなプロモーターの具体例として、MDVのゲノム配列(ジーンバンクのアクセッション番号:AF243438)の91827番目から92196番目までの塩基領域(配列番号1)や、同じく98757番目から99412番目までの塩基領域(配列番号2)が挙げられる。
【0013】
更に、ゲノムから得られたプロモーターの両末端に異なる制限酵素認識サイトを導入すると、このプロモーターの前後に異なる外来遺伝子を容易に配置できるようになり実用性が向上するので、極めて好ましい。そのように改変されたプロモーターの具体例としては、配列番号19の60番目から482番目までの配列、又は配列番号22の60番目から768番目までの配列が挙げられる。
【0014】
(4)発現遺伝子カセット
本発明の組み換えHVTに挿入される遺伝子は、異なる2つの外来遺伝子とその間にMDV由来の1つのプロモーターからなる発現遺伝子カセットとして挿入されたものである。挿入されるカセットの数に格別な制限はないが、組み換えウイルスの安定性の観点から、挿入するカセットの全長(複数個ある時は、その合計の長さ)が10Kbpを超えないことが好ましい。
【0015】
発現遺伝子カセット中の外来遺伝子は、本来、HVT由来でない遺伝子であり、発現調節遺伝子以外の遺伝子であれば特に限定されないが、家禽用ワクチンを得る場合、鳥類の感染症の病原体に由来する抗原遺伝子が好ましい。鳥類の感染症を引き起こす病原体としては、ウイルス、細菌、真菌、原虫などを挙げることができる。
【0016】
このような病原体としては、具体的には、ニューキャッスル病ウイルス(NDV)、ガンボロ病ウイルス(IBDV)、伝染性喉頭気管炎ウイルス(ILTV)、伝染性気管支炎ウイルス(IBV)、マイコプラズマ(MG)、およびコクシジウムなどを挙げることができる。
【0017】
特に、中和抗原遺伝子または感染防御抗原の遺伝子を挿入した組み換えHVTはワクチンとして使用することが可能になるので最適である。
具体的な例を挙げれば、NDVでは、HN蛋白質、F蛋白質、NP蛋白質が、また、IBVでは、N蛋白質、S蛋白質が挙げられる。IBDVでは、VP2タンパク質、ILTVではgB蛋白質、マイコプラズマでは40K蛋白質、66K蛋白質、67K蛋白質など(国際公開公報WO94/23019号に配列が記載)などが挙げられる。
【0018】
抗原遺伝子以外に好適に用いられる外来遺伝子としては、組み換えHVTであることの指標となるようなマーカー蛋白質の遺伝子も好適である。マーカー蛋白質の具体例としては、緑色蛍光を発するオワンクラゲ由来のGFP蛋白質や、βーガラクトシダーゼなどの酵素が挙げられる。
【0019】
本発明の発現遺伝子カセットは、前述したとおり、異なる2つの外来遺伝子とその間にMDV由来の1つのプロモーターから構成されるが、それ以外にカセットの両末端にポリAシグナルと制限酵素サイトからなる短い端末配列(組み換え体の安定性の観点から、通常100bp以下、好ましくは5〜50bp)を有することが、より望ましい(図1)。その端末配列の一例は配列番号3と配列番号4である。
【0020】
(5)組み換えHVTの作製法
組み換えHVTの作製法に格別な制限はなく、その具体例としては、Avian Dis.36:858−870(1992)や特許第3587065号公報に記載された、親ウイルスゲノムの非必須領域の一部の中に、目的の抗原遺伝子等が挿入されたプラスミドを構築し、ウイルスに相同組み換えによって、抗原遺伝子を挿入する一般的な組み換えウイルスの作製方法が挙げられる。
【0021】
組み換えHVTを作製するためのプラスミド(本発明では、ホモロジーベクター又はカセット含有プラスミドと呼ぶことがある)は、挿入部位を含むHVTゲノムDNAの一部をクローニングしたプラスミドの挿入部位に、発現遺伝子カセットを1つ以上組み込んだプラスミドである。ここで用いられるプラスミドは、特に限定されるものではなく、例えば、pBR322、pBR325、pBR327、pBR328、pUC18、pUC19、pUC7、pUC8、およびpUC9などのプラスミド、pHC79などのコスミド等を例示することができる。
【0022】
発現遺伝子カセットを挿入するHVTゲノム中の挿入部位としては、特許第3587065号公報に記載された挿入部位が好適であり、特に、発現遺伝子カセットの脱落が極めて起こりにくい、安定した組み換え体が得られる観点から、HVTのUL45とUL46のオープン・リーディング・フレームの間がより好ましい。
【0023】
カセット含有プラスミドを用いて組み換えHVTを作製及び純化する方法は、前述の通り、Avian Dis.36:858−870(1992)や特許第3587065号公報に記載されている。
【0024】
(6)組み換え七面鳥ヘルペスウイルスを有効成分とする家禽用ワクチン
本発明の組み換え七面鳥ヘルペスウイルスを有効成分とする家禽用ワクチンの調製方法は、特に限定されないが、例えば次の方法によって調製することができる。
【0025】
本発明の組み換えHVTの感染細胞を当該ウイルスが生育できる細胞(以下、宿主細胞という)に感染させ、増殖させた後、細胞をスクレーパーまたはトリプシンではがし、遠心分離によって感染細胞と上清とに分離する。
【0026】
宿主細胞としては、トリ由来の細胞が好ましく、CEF(鶏胚繊維芽細胞)、鶏腎細胞などを好適に使用することができる。使用する培地は、Eagle’s MEM、ライボビッツL−15/マッコイ5A(1:1混合)培地(以下LM培地という場合がある)などが好適である。得られた感染細胞は、10%のジメチルスルフォキシド(DMSO)を含む培地に、懸濁し、液体窒素存在下で凍結保存する。ワクチンとして使用するときは100倍量のリン酸緩衝液(PBS)にこの凍結保存品を溶かして使用する。液体窒素下で上記感染細胞を保存するための安定剤やその他の成分は、ウイルス感染細胞が安定的に生存でき、かつ接種を受ける家禽にとって薬理学的に問題のない成分であれば特に限定されない。
【0027】
本発明のワクチンの家禽への投与方法もまた特に限定されないが、皮下に注射により接種する方法や、発育鶏卵に接種する方法が一般的に用いられており、マレック病に対するワクチンとして用いられている従来ワクチンと同じである。接種量も従来ワクチンと同様でよい。即ち、10〜105p.f.u./dose、好ましくは102〜104p.f.u./doseになるように適量のリン酸緩衝液などに溶かして、初日齢の鶏の頸部皮下に、あるいは孵化前の卵に注射や接種用機器を使って投与する。
【0028】
本発明のワクチンは、親ウイルスであるHVTによるマレック病に対するワクチン効果だけでなく、挿入された発現遺伝子カセット中の抗原遺伝子が発現することによって、それらの遺伝子の由来病原体による疾病に対するワクチン効果も発現するため、多価ワクチンとして使用することができる。
【実施例】
【0029】
以下に実施例を挙げて、本発明を説明するが、本発明はこれに限定されるものではない。
本発明のプラスミド構築は特に記載がない限り、Molecular Cloning:A Laboratory Manual.2nd Edition.(Cold Spring Harbor Laboratory.Cold Spring Harbor.N.Y.1989)記載の標準的な分子生物学手法を用いて行った。尚、制限酵素断片はアガロースゲルからQIAquick Gel Extraction Kit(QIAGEN社製、カタログ番号#28704)を使って精製した。
七面鳥ヘルペスウイルス(HVT)は、アメリカ農務省Avian Disease and Oncology LaboratoryのDr.Witterが分離した七面鳥ヘルペスウイルスFC126株を使用した(Witter R.L.et al.Am.J.Vet.Res.1970.31.525−538)。
【0030】
1.MDV由来プロモーターのクローニング
マレック病ウイルスのゲノム構造は、Tulmanらによって報告されている(J.Virology 74:7980−7988(2000))が、MDVのゲノム上にあるプロモーターの明確な位置や活性についてはあまり解析がなされていない。数少ない例として、IzumiyaらによるUL41からUL51遺伝子間のmRNA解析論文(J.Gen.Virology 79:1997−2001(1998))や、UL30からUL40遺伝子間のmRNA解析論文(J.Gen.Virology 80:2417−2422(1999))が報告されている程度である。上記3つの論文から、MDV由来プロモーターとして、MDVのゲノム配列(ジーンバンクのアクセッション番号:AF243438)の91827番目から92196番目までの塩基領域、及び98757番目から99412番目までの塩基配列領域が候補として挙げられた。
【0031】
マレック病ワクチンとして市販されているマレック病ウイルスCVI988株を直径15cmの培養皿のCEF細胞に感染させウイルスを増やした後、PBSで洗浄後、リシスバッファー(0.5%SDS10mMトリス(pH8)、100mMNaCl、1mMEDTA、200μg/mlプロテナーゼ)を4ml加えて、CO2インキュベーター(37℃)で4時間インキュベートし、細胞溶解物を50mlファルコンチューブに移し、等量のフェノール(TE飽和)を加え、15分間ゆっくり回転し、混合した。次に、1500×Gで5分間遠心後、水層をピペットでゆっくり吸い取り、新しいチューブに移した。クロロフォルム:イソアミルアルコール=24:1を等量加え、15分間ゆっくり混合した。その後、1500×Gで5分間遠心し、水層をピペットでゆっくり吸い取り、新しいチューブに移し、100%エタノールを水層の倍量加え、良く混ぜてから1500×Gで5分間遠心した。その沈殿を70%エタノールでリンスした後、軽く風乾してMDVのゲノムDNAを含む核酸画分を調製した。
【0032】
上記で調製したMDVのゲノムDNA画分をテンプレートにして、MD50F(配列番号5)とMD51R(配列番号6)のプライマーセットでPCRを行い、MDVのゲノム配列の91709番目から92261番目までのDNA領域を増幅させ、それをpPCR−Script Ampベクター(ストラタジーン社製)にライゲーションし、JM109を形質転換した。そうして、得られた形質転換株3株からプラスミドを調製して、挿入DNAの塩基配列分析を行った。3株のうち、2株の挿入DNAの塩基配列が完全に一致した(配列番号7)。それらのプラスミドをpPCR−Pmd50と命名した。
【0033】
同様にして、MD54F(配列番号8)とMD55R(配列番号9)のプライマーセットでPCRを行い、MDVのゲノム配列の98715番目から99464番目までのDNA領域を増幅させ、それをpPCR−Script Ampベクターにクローニングした。挿入されたDNA配列は、ジーンバンクのAF243438の配列とは1塩基異なるが、3クローン間で完全に一致した。その配列を配列番号10に示す。こうして得られたプラスミドをpPCR−Pmd54と命名した。
【0034】
2.MDV由来プロモーターの改変
pPCR−Pmd50FとpPCR−Pmd54Fをそれぞれ制限酵素KpnIで切断後、更にSalIで切断して回収したプラスミドに、Ad_Kp−Bg−SaU(配列番号11)と、Ad_Kp−Bg−SaL(配列番号12)の2つの合成ヌクレオチドをアニールさせた合成DNAアダプターを挿入した。このようにして得られたプラスミドをそれぞれpPmd50M1、pPmd54M1と命名した。
【0035】
pPmd50M1とpPmd54M1をそれぞれ制限酵素SacIで切断後、更にNotIで切断して回収したプラスミドに、Ad_No−Sf−SaU(配列番号13)と、Ad_No−Sf−SaL(配列番号14)の2つの合成ヌクレオチドをアニールさせた合成DNAアダプターを挿入した。このようにして得られたプラスミドをそれぞれpPmd50M2、pPmd54M2と命名した。
【0036】
次いで、pPmd50M2とpPmd54M2をそれぞれ制限酵素NcoI/NotIで二重切断して回収したプラスミドに、Ad_Nc−Bg−Xh−NoU(配列番号15)と、Ad_Nc−Bg−Xh−NoL(配列番号16)の2つの合成ヌクレオチドをアニールさせた合成DNAアダプターを挿入した。このようにして得られたプラスミドをそれぞれpPmd50M3、pPmd54M3と命名した
【0037】
pPCR−Pmd50をテンプレートにして、Pmd50F(配列番号17)とPmd51R(配列番号18)のPCRプライマーでPCRを実施した。増幅されたPCR産物をBamHIとNcoIで二重切断して得られた約380bpのDNA断片を、BamHIとNcoIで二重切断したpPmd50M3とライゲーションして、pPmd50Fを構築した。このpPmd50FのKpnIサイトからSacIサイトまでの塩基配列が、配列番号19である。
【0038】
同様に、pPCR−Pmd54をテンプレートにして、Pmd54F(配列番号20)とPmd55R(配列番号21)のPCRプライマーでPCRを実施した。増幅されたPCR産物をBamHIとNcoIで二重切断してうられた約670bpのDNA断片を、BamHIとNcoIで二重切断したpPmd54M3とライゲーションして、pPmd54Fを構築した。このpPmd54FのKpnIサイトからSacIサイトまでの塩基配列が、配列番号22である。
【0039】
3.2つの外来遺伝子を発現する発現遺伝子カセットプラスミドの構築
オワンクラゲ由来の緑色蛍光蛋白質GFP遺伝子を持った市販プラスミドベクターpGFP(クロンテック社製、カタログ番号:6097−1)をテンプレートにして、GFPBspF(配列番号23)とGFPSalR(配列番号24)のプライマーセットでPCRを実施した。増幅されたPCR産物を制限酵素BspHIとSalIで二重切断し、NcoIとXhoIで二重切断したpPmd50FとライゲーションしてpPmd51GFPを作製した。
【0040】
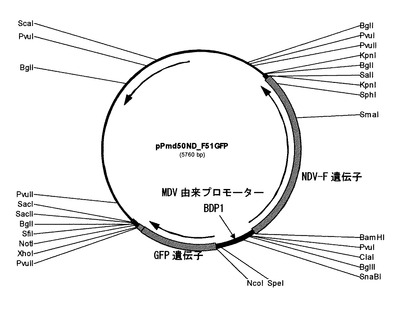
特許第3587065号の実施例7に記載されているpRSV−Fのプラスミドに含まれているニューカッスル病ウイルス(NDV)のF遺伝子を制限酵素BamHIとSalIで二重切断してアガロースゲルから回収し、BamHIとSalIで二重切断したpPmd51GFPとライゲーションして、pPmd50ND_F51GFP(図2)を作製した。
このプラスミドpPmd50ND_F51GFPは、オワンクラゲ由来のGFP遺伝子とNDV由来のF遺伝子という2つの外来遺伝子とその間にMDV由来の1つのプロモーター(BDP1:配列番号1)から構成される発現遺伝子カセットを有するプラスミドである。
【0041】
特開2004−111号公報記載のプラスミドp45/46bacVP2−Eをテンプレートにして、BspVP2−F(配列番号25)とVP2Xho−R(配列番号26)のプライマーセットでPCRを実施した。増幅されたPCR産物(伝染性ファブリキウス嚢病ウイルス(IBDV)のVP2蛋白質遺伝子;約1.4kbp)を制限酵素BspHIとXhoIで二重切断し、NcoIとXhoIで二重切断したpPmd54FとライゲーションしてpPmd55IBD_VP2を作製した。
【0042】
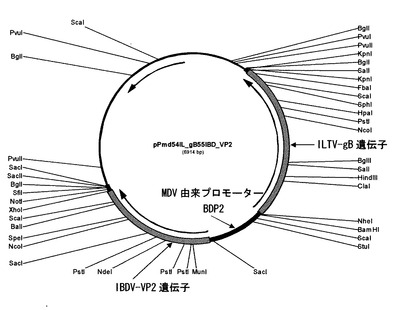
国際公開WO2005/093070号公報記載のホモロジーベクターp45/46PecILgBdellac+STPから制限酵素BamHIとSalIで伝染性喉頭気管炎ウイルス(ILTV)のgB遺伝子(約1.9kbp)を切り出してアガロースゲルから回収し、BamHIとSalIで二重切断したpPmd55IBD_VP2とライゲーションして、pPmd54IL_gB55IBD_VP2(図3)を作製した。
このプラスミドpPmd54IL_gB55IBD_VP2は、ILTV由来のgB遺伝子とIBDV由来のVP2遺伝子という2つの外来遺伝子とその間にMDV由来の1つのプロモーター(BDP2:配列番号2)から構成される発現遺伝子カセットを有するプラスミドである。
【0043】
4.組み換えHVTを作製するためのホモロジーベクター(カセット含有プラスミド)の構築
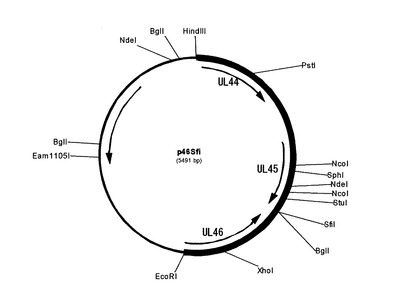
特許第3587065号公報の実施例1の記載に従って、HVT−DNAを調整し、それをテンプレートにして、HVT45Sph(配列番号27)とHVT46Xho(配列番号28)のプライマーセットでPCRを実施した。増幅したPCR産物(約1.0Kbp)をSphIとXhoIで二重切断して、同じくSphIとXhoIで二重切断したpNZ45/46BacpA(特開2004−111の実施例3.2)とライゲーションしてプラスミドp46を得た。これをテンプレートにして、HVT45Sph(配列番号27)と45SfiR(配列番号29)のプライマーセットでPCRを実施した。同様にして、46SfiF(配列番号30)とHVT46Xho(配列番号28)のプライマーセットで、PCRを実施した。それぞれ0.4kbpと0.6kbpのDNA断片が増幅したので、更にその2断片をテンプレートにして、HVT45Sph(配列番号27)とHVT46Xho(配列番号28)のプライマーセットで乗り換えPCRを実施してSfiIサイトを導入した。その結果生じた1.0kbpのPCR産物を制限酵素SphIとXhoIで二重切断して、p46のSphIとXhoIの間のDNAと置き換えたプラスミドp46Sfi(図4)を作製した。
【0044】
プラスミドpPmd50NDV_F51GFPを制限酵素BglIで切断して、約3.1kbpの発現カセットをアガロースゲルから回収した。そのカセットを制限酵素SfiIで切断したp46Sfiにライゲーションしてp46md50ND_F51GFPを作製した。
【0045】
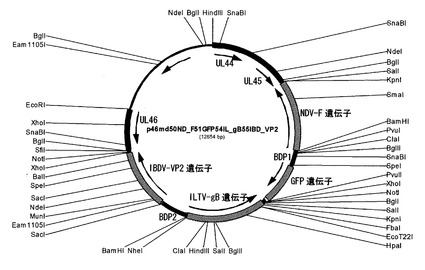
次にプラスミドpPmd54IL_gB55IBD_VP2を制限酵素BglIで切断して、約4.0kbpの発現カセットをアガロースゲルから回収し、制限酵素SfiIで切断したp46md50ND_F51GFPにライゲーションしてp46md50ND_F51GFP54IL_gB55IBD_VP2を作製した。このプラスミドp46md50ND_F51GFP54IL_gB55IBD_VP2(図5)は、2つの発現遺伝子カセットを含有する組み換えHVTを作製するためのホモロジーベクターである。
【0046】
5.組み換えHVT(FW101)の作製・純化
トリプシンで剥がした単層のCEF細胞を、SalineG(0.14M塩化ナトリウム、0.5mM塩化カリウム、1.1mMリン酸一水素二ナトリウム、15mMリン酸二水素一ナトリウム、0.5mM塩化マグネシウム・6水和物、0.011%グルコース)に懸濁し、細胞懸濁液を調製した。この細胞懸濁液(2×107個)に、実施例4で作製したカセット含有プラスミドp46md50ND_F51GFP54IL_gB55IBD_VP2を25μgと、特許第3587065号公報の実施例1記載の方法で調製したHVTのDNA15μgを混合して、ジーンパルサー(BioRad社製)を用いて、0.5kVcm−1、0.5msecの条件下でエレクトロポレーションした。
【0047】
プラスミド及びHVTのDNAを導入した細胞を、107個のCEF細胞と混合して、培養用96ウェル平底マルチプレート(ファルコン社製)5枚にまいて37℃にてHVT特有のプラークが形成されるまで約4日〜5日間培養した。
【0048】
倒立型落射蛍光観察装置(オリンパス社製「IX−FLA」)付の倒立顕微鏡(オリンパス社製「IX70」)を使って、出現したHVTのプラークを観察し、蛍光蛋白を発現しているプラークを選択した。選択したプラークの入っているウェルに1%トリプシンを加えて組み換えHVT感染細胞を含むCEF細胞をそれぞれ回収し、2×106個の細胞と混合し、それぞれのプラークごとに、1枚の培養用96ウェル平底マルチプレートにまいて、37℃にて約3日〜5日間培養した。
【0049】
再び出現したHVTのプラークを蛍光顕微鏡にて、各プラークが蛍光蛋白を発現しているか否かを観察した。各ウェルに存在するHVTプラークが全て蛍光蛋白を発現するプラークとなっていなければ、蛍光蛋白を発現するプラークの割合が最も多いウェルを選択して、再び、1%トリプシンを加えて組み換えHVT感染細胞を含むCEF細胞をそれぞれ回収し、2×106個の細胞と混合し、それぞれのプラークごとに、1枚の培養用96ウェル平底マルチプレートにまいて、37℃にて約3日〜5日間培養した。
【0050】
このようにして、各ウェルに存在するHVTプラークが全て蛍光蛋白を発現するプラークとなるまで、スクリーニング操作を繰り返した。
【0051】
ウェル中の全HVTプラークが蛍光蛋白を発現するようになったら、そのウェルのCEF細胞をトリプシンで回収し、培養用6ウェルプレート(ファルコン社製)の1ウェルを使って感染細胞を増殖させた後、SPGA−SBTバッファー(100ml当たり、7.46gシュークロース、5.2mg KH2PO4、125.4mg K2HPO4、91.7mg L−グルタミン・Na、1g BSA、10gソルビトール)2mlに回収し、1mlをエッペンドルフチューブに移した。
【0052】
そのチューブを超音波破砕器(三光純薬社製「BIOSONIC」)にセットし、最大出力で3分間超音波破砕した。その後、5000rpm5分間遠心分離し、上清を回収した。これにより、セルフリーウイルスが回収された。回収した上清をSPGA−SBTバッファーで段階希釈して、別途用意したCEF細胞を単層培養した96ウェル平底マルチプレートに各ウェルあたり40μlずつ重層して、2時間インキュベーターで感染させた。液をすて、LM培地を200μlずつ加えてプラークが出現するまで培養した。
【0053】
出現したHVTのプラークを蛍光顕微鏡にて観察し、全ウェル中に1つのプラークで、且つ、そのプラークが蛍光蛋白を発現しているウェルを選択した。これで、このウェル中のHVTは100%純度になったと判断され、このようにして純化した組み換えHVTをFW101と命名した。
【0054】
6.組み換えHVTのゲノム構造の確認とその安定性試験
6.1 サザンハイブリダイゼーション
実施例5で純化が終了したFW101をCEF細胞とともに2枚の150mm径培養皿で全面にプラークが出現するまで培養し、その感染細胞をスクレーパーで培養皿から剥がして遠心分離(500×G、5分)して感染細胞を集め、PBSで洗浄後、1.2mlのPBSに再懸濁し、溶解バッファー(1.25%TritonX−100、250mM 2−メルカプトエタノール、50mM EDTAがPBSに入ったもの)0.8mlを加えて攪拌して細胞を可溶化した。3000rpm、5分間で細胞残さを除去し、上澄をエッペンドルフチューブに移して15000rpm、20分間、22℃で遠心してウイルスを落とした。12.5mM トリス(pH7.5)を1ml加えて、ヌクレアーゼ液(0.25mg/ml DNaseI、0.25mg/ml RNaseA、150mM 塩化ナトリウム)を4μl加えて37℃、30分間静置した。その後、0.5M EDTAを25μl、10%SDSを125μl、水87μl、10mg/mlの10mg/mlのプロテアーゼKを12.5μl、2−メルカプトエタノール0.5μl加えて、55℃、30分間静置した。その液をフェノール:クロロホルム(=1:1混合)で2回抽出した。
【0055】
その水層に5M塩化ナトリウム16μlを加え、水層の2.5倍量の(−20℃で冷やした)100%エタノールを加えて沈殿させ、遠心分離で沈殿させた沈殿物(DNA)を70%エタノールで洗浄後、再度遠心分離して沈殿させた後乾燥させ、TE緩衝液(10mMトリス(pH8.0)、1mM EDTA)50μlに溶かした。
こうして回収されたウイルスDNAを制限酵素XhoI/StuIで切断し、0.8%アガロースゲル電気泳動を行い、制限酵素で切断されたDNA断片をナイロンメンブレンにブロットし、サザンハイブリダイゼーションを行った。プローブは“PCR DIG Probe Synthesis Kit”(ROCHE DIAGNOSTICS社製、カタログ番号#1636090)の試薬を用いて2種類のDIG標識されたDNAプローブを調製した。そのDNAプローブは、VP2 cDNA領域内のプローブ(以下VP2プローブ)と外来遺伝子の挿入部位のHVTゲノム領域(以下45/46プローブ)である。前者のプローブはVP2−F(配列番号31)とVP2−R(配列番号32)の2本のプライマー及びプラスミドp46md50ND_F51GFP54IL_gB55IBD_VP2をテンプレートとして、後者は、45/46−F(配列番号33)と45/46−R(配列番号34)の2本のプライマー及びプラスミドp46SfiをテンプレートとしてPCR反応によって調製した。
【0056】
この2種類のプローブを使ったサザンハイブリダイゼーションの結果、VP2プローブでは2.0kbpのバンドがハイブリダイズし、45/46プローブでは0.6kbと1.7kbの2本のバンドがハイブリダイズした。この結果は、どちらのプローブも予期した通りのバンドであったので、組み換えHVTは設計どおり正しい組み換え体であることが確認された。
【0057】
6.2 ゲノム安定性試験
組み換えHVTをCEF細胞で20代継代した組み換えウイルスについても前記6.1と同様にサザンブロッティングを実施し、20代継代後の組み換えウイルスも正しい遺伝子構造を持っていることが確認された。一方、特許第3587065号公報記載の組み換えHVTのHF004〜HF007の組み換え体について、同様に20代継代したところ、これら全ての組み換えHVTのゲノム構造は、継代前の組み換えHVTの構造と異なっていた。これらの組み換えHVTについては、継代途中に遡って安定性を調べたところ、HF004とHF005は10代、HF006とHF007については、5代継代した時点で、継代前の構造と異なっており、不安定であることがわかった。従って、20代継代してもゲノム構造が変化しないFW101は極めて安定な組み換えHVTであることがわかった。
【0058】
7.組み換えHVTの蛋白発現確認
7.1 FITC標識抗体を使った蛍光検出法
特許第3587065号公報の実施例16に記載した方法に従って、組み換えウイルスFW101が外来抗原遺伝子であるNDVのF蛋白質、IBDVのVP2蛋白質を発現していることを確認した。
【0059】
7.2 ウエスタンブロット法
9cm径ディッシュに新鮮な107個のCEF細胞と1×105p.f.u.のFW101をまき、約72時間後に、感染細胞をスクレーパーにて回収した。遠心分離(2000rpmで5分)で感染細胞を回収し、10mlのPBSで細胞を洗浄し、再び遠心分離で細胞を集めた。その細胞ペレットに2×サンプルバッファー(0.112Mトリス−アセテート(pH7)、2%SDS、5%メルカプトエタノール、5%シュークロース、0.005%ブロモフェノールブルー )を0.1ml添加し、100℃で5分煮沸した。
【0060】
煮沸したサンプル5μlを10%SDS−ポリアクリルアミドゲルにアプライした。電気泳動終了後、PVDFメンブレン(ミリポア)にエレクトロブロッティングし、ブロットしたPVDFメンブレンをIBDVのVP2に対するモノクローナル抗体R−63(ハイブリドーマはATCCから入手、ATCC:HB−9490)を用いてウエスタンブロットを行った。その結果、約40kDaの大きさの蛋白質が検出された。この大きさは、文献で知られているIBDVのVP2の大きさであり、FW101は期待通り目的とする大きさのVP2が発現していた。
【0061】
また、GFP蛋白に対する市販モノクローナル抗体JL−8(クロンテック社製)を使って同じくブロットしたPVDFメンブレンと反応させたところ、約26kDaの大きさの蛋白質が検出された。この大きさはGFP蛋白の分子量と一致し、FW101が期待通りの大きさのGFP蛋白質も発現していることを確認した。
【0062】
8.FW101のワクチン効果動物試験
FW101を鶏に免疫してワクチン効果を調べた。3000p.f.u./doseのFW101(対照は親株HVT)をSPF鶏の発育鶏卵(日生研)の孵化後の鶏の背部皮下に接種した。接種後、1週間隔で採血をし、その血中の抗体価を市販ELISAキット(ゲン・コーポレーション社製、IBDエリーザキット)で測定した。そのデータ(S/P value)を表1に示す。
【0063】
【表1】

【0064】
表1に示したように、挿入した抗原遺伝子の蛋白質に対する抗体が上昇し、ワクチン活性が認められた。
【図面の簡単な説明】
【0065】
【図1】図1は、発現遺伝子カセットの概念図である。
【図2】図2は、発現遺伝子カセットプラスミドpPmd50ND_F51GFPの模式図。
【図3】図3は、発現遺伝子カセットプラスミドpPmd54IL_gB55IBD_VP2の模式図である。
【図4】図4は、挿入部位を含むHVT−DNA含有プラスミドp46Sfiの模式図である。
【図5】図5は、FW101作製用ホモロジーベクター p46md50ND_F51GFP54IL_gB55IBD_VP2の模式図である。
【技術分野】
【0001】
本発明は、七面鳥ヘルペスウイルス(以下、HVTということがある)ゲノムに2つ以上の外来遺伝子を組み込んだ組み換えHVTと、外来遺伝子を発現させるために用いるマレック病ウイルス由来のプロモーター、及びそれを利用したワクチンに関する。
【背景技術】
【0002】
米国等の養鶏業においては、既に、ウイルスベクターワクチンである組み換え鶏痘ウイルス(FPV)や組み換え七面鳥ヘルペスウイルスが実用化されている。中でもHVTは家禽での安全性が高く、高マレック病ウイルスワクチンとしても利用されるほど、ワクチン特性も良好であること、鶏へのワクチン接種法の一つである発育鶏卵内にワクチンを接種する卵内接種が可能であることから、鳥類に対するワクチン用のウイルスベクターとしても注目されている。
【0003】
また、HVTの感染様式として、感染細胞から他の細胞にウイルスが感染する際、隣り合う細胞へ細胞−細胞間で感染が成立するため、血流中に存在するHVT特異的抗体の影響を受けにくい。従って、これまでのウイルス生ワクチンの効果を減弱させる大きな要因であった親鳥からの移行抗体の存在下でも十分な効果を発揮するという有利な点もある。
【0004】
更に、HVTを含むヘルペスウイルス属は、潜伏感染(latent infection)や永続感染(permanent infection) の状態で感染動物の体内に永続的に生存し続けることが知られていて、このことも、HVTのワクチンとしての有効性、つまり、感染期間が長いため長期にわたってワクチン効果を付与できる特徴も有している。従って本発明者らもこの組み換えHVTベクターワクチンの研究を行い、報告してきた(特許第3587065号、米国特許第7153511号)。この内、特許第3587065号では、複数の外来抗原遺伝子(例えばニューカッスル病ウイルス(NDV)のHN蛋白質やF蛋白質、ガンボロ病ウイルス(IBDV)のVP2蛋白質の遺伝子)を挿入した組み換えHVT(HF006、HF007)を純化し、これら外来抗原遺伝子の発現やワクチン効果を報告している。
【0005】
しかしながら、複数の外来遺伝子を発現させるような組み換えHVTは、不安定である場合が多く、更に、純化した組み換えHVTを培養細胞で繰り返し継代するうちに、挿入させた外来遺伝子の一部を喪失したウイルスに変遷してしまうことが度々あった。
【0006】
【特許文献1】特許第3587065号(米国特許第6632664号)公報
【特許文献2】米国特許第7153511号公報
【特許文献3】国際公開公報WO94/23019号(米国特許第5871742号)公報
【特許文献4】特開2004−111号(米国特許第6764684号、米国特許第7153511号)公報
【特許文献5】国際公開公報WO2005/093070号(欧州公開第1731612号)公報
【0007】
【非特許文献1】Avian Dis.36:858−870(1992)
【非特許文献2】J.Virology 74:7980−7988(2000)
【非特許文献3】J.Gen.Virology 79:1997−2001(1998)
【非特許文献4】J.Gen.Virology 80:2417−2422(1999)
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明者は、上記の課題を解決すべく鋭意研究を重ねた結果、プロモーターとして、双方向の転写活性を有するものを採用し、このプロモーターを挟むように2つの外来遺伝子を連結させて、1つのプロモーターによって2つの外来遺伝子を支配する発現遺伝子カセットを組み込んだ組み換え七面鳥ヘルペスウイルスが、挿入した遺伝子の脱落を起こさない極めて安定な組み換えウイルスであり、これがワクチンとして使用できることを見出し、本発明を完成したものである。
【課題を解決するための手段】
【0009】
かくして本発明によれば、2つの異なる外来遺伝子と、当該2つの外来遺伝子を両方とも支配するように、2つの外来遺伝子の間に配置された、マレック病ウイルス由来の1つのプロモーターとを有する発現遺伝子カセットが、ゲノム中に1つ以上挿入された組み換え七面鳥ヘルペスウイルスが提供される。
前記プロモーターとしては、プロモーターが配列番号1又は配列番号2が好ましい。これら二つのプロモーターは、いずれも新規なプロモーターである。
また、前記発現遺伝子カセットの挿入部位は、七面鳥ヘルペスウイルスのUL45とUL46のオープン・リーディング・フレームの間であるのが好ましい。
挿入される発現遺伝子カセット中の2つの異なる外来遺伝子のうち、少なくとも1つが鳥類感染性の病原体に由来する抗原遺伝子であるものは、家禽用ワクチンとして用いることができる。
【発明を実施するための最良の形態】
【0010】
以下に本発明を詳述する。
(1)七面鳥ヘルペスウイルス(HVT)
本発明の組み換え七面鳥ヘルペスウイルスの親株として使用する七面鳥ヘルペスウイルスは、天然に得られるものの他、細胞・微生物・遺伝子バンクであるAmerican Type Culture Collection(ATCC)などから有償または無償で入手できるものなどであればよく、特に限定されるものではない。
HVTの好ましい例としては、ガンマヘルペスウイルス亜科に属し、生来非病原性であり、かつ非腫瘍性のウイルスで家禽用のワクチンとして用いられているものが挙げられる。具体的には、FC126(ATCCVR−584B)、PB−THV1、H−2、YT−7、WTHV−1、HPRS−26などの株が挙げられ、例えば、FC126株を好適に使用することができる。
【0011】
(2)マレック病ウイルス(MDV)
本発明において、二つの外来遺伝子を発現させるプロモーターの由来となるマレック病ウイルスもまた、天然に得られるものの他、ATCCなどから有償または無償で入手できるものなどであればよく、特に限定されるものではない。好ましい例としては、家禽用ワクチンとして用いられているものが挙げられ、具体的には、CVI988株やSB1株などを挙げることができる。
【0012】
(3)プロモーター
本発明の組み換えHVTにおいて、2つの異なる外来遺伝子を発現させるために、その間に置かれるMDV由来プロモーターは、マレック病ウイルスゲノムから得られたプロモーターであって、双方向の転写活性を有するものである。マレック病ウイルスゲノムから得られたプロモーターを、プロモーターとしての活性が維持できる範囲で、一部を切り縮めるなどの改変をしてもよい。
双方向の転写活性を有するプロモーターは、マレック病ウイルスゲノムの中から、読み枠が反対方向を向いている、二つのオープン・リーディング・フレームを探し、その間に位置するプロモーター領域をプロモーター候補として絞り込み、後述する実施例中の、プロモーターのクローニング方法に従ってクローニングすることにより得られる。
また、本発明に用いるプロモーターは、組み換え体の安定性の観点から、遺伝子長が1kbpより短いのが好ましく、800bpより短いのが更に好ましい。
このようなプロモーターの具体例として、MDVのゲノム配列(ジーンバンクのアクセッション番号:AF243438)の91827番目から92196番目までの塩基領域(配列番号1)や、同じく98757番目から99412番目までの塩基領域(配列番号2)が挙げられる。
【0013】
更に、ゲノムから得られたプロモーターの両末端に異なる制限酵素認識サイトを導入すると、このプロモーターの前後に異なる外来遺伝子を容易に配置できるようになり実用性が向上するので、極めて好ましい。そのように改変されたプロモーターの具体例としては、配列番号19の60番目から482番目までの配列、又は配列番号22の60番目から768番目までの配列が挙げられる。
【0014】
(4)発現遺伝子カセット
本発明の組み換えHVTに挿入される遺伝子は、異なる2つの外来遺伝子とその間にMDV由来の1つのプロモーターからなる発現遺伝子カセットとして挿入されたものである。挿入されるカセットの数に格別な制限はないが、組み換えウイルスの安定性の観点から、挿入するカセットの全長(複数個ある時は、その合計の長さ)が10Kbpを超えないことが好ましい。
【0015】
発現遺伝子カセット中の外来遺伝子は、本来、HVT由来でない遺伝子であり、発現調節遺伝子以外の遺伝子であれば特に限定されないが、家禽用ワクチンを得る場合、鳥類の感染症の病原体に由来する抗原遺伝子が好ましい。鳥類の感染症を引き起こす病原体としては、ウイルス、細菌、真菌、原虫などを挙げることができる。
【0016】
このような病原体としては、具体的には、ニューキャッスル病ウイルス(NDV)、ガンボロ病ウイルス(IBDV)、伝染性喉頭気管炎ウイルス(ILTV)、伝染性気管支炎ウイルス(IBV)、マイコプラズマ(MG)、およびコクシジウムなどを挙げることができる。
【0017】
特に、中和抗原遺伝子または感染防御抗原の遺伝子を挿入した組み換えHVTはワクチンとして使用することが可能になるので最適である。
具体的な例を挙げれば、NDVでは、HN蛋白質、F蛋白質、NP蛋白質が、また、IBVでは、N蛋白質、S蛋白質が挙げられる。IBDVでは、VP2タンパク質、ILTVではgB蛋白質、マイコプラズマでは40K蛋白質、66K蛋白質、67K蛋白質など(国際公開公報WO94/23019号に配列が記載)などが挙げられる。
【0018】
抗原遺伝子以外に好適に用いられる外来遺伝子としては、組み換えHVTであることの指標となるようなマーカー蛋白質の遺伝子も好適である。マーカー蛋白質の具体例としては、緑色蛍光を発するオワンクラゲ由来のGFP蛋白質や、βーガラクトシダーゼなどの酵素が挙げられる。
【0019】
本発明の発現遺伝子カセットは、前述したとおり、異なる2つの外来遺伝子とその間にMDV由来の1つのプロモーターから構成されるが、それ以外にカセットの両末端にポリAシグナルと制限酵素サイトからなる短い端末配列(組み換え体の安定性の観点から、通常100bp以下、好ましくは5〜50bp)を有することが、より望ましい(図1)。その端末配列の一例は配列番号3と配列番号4である。
【0020】
(5)組み換えHVTの作製法
組み換えHVTの作製法に格別な制限はなく、その具体例としては、Avian Dis.36:858−870(1992)や特許第3587065号公報に記載された、親ウイルスゲノムの非必須領域の一部の中に、目的の抗原遺伝子等が挿入されたプラスミドを構築し、ウイルスに相同組み換えによって、抗原遺伝子を挿入する一般的な組み換えウイルスの作製方法が挙げられる。
【0021】
組み換えHVTを作製するためのプラスミド(本発明では、ホモロジーベクター又はカセット含有プラスミドと呼ぶことがある)は、挿入部位を含むHVTゲノムDNAの一部をクローニングしたプラスミドの挿入部位に、発現遺伝子カセットを1つ以上組み込んだプラスミドである。ここで用いられるプラスミドは、特に限定されるものではなく、例えば、pBR322、pBR325、pBR327、pBR328、pUC18、pUC19、pUC7、pUC8、およびpUC9などのプラスミド、pHC79などのコスミド等を例示することができる。
【0022】
発現遺伝子カセットを挿入するHVTゲノム中の挿入部位としては、特許第3587065号公報に記載された挿入部位が好適であり、特に、発現遺伝子カセットの脱落が極めて起こりにくい、安定した組み換え体が得られる観点から、HVTのUL45とUL46のオープン・リーディング・フレームの間がより好ましい。
【0023】
カセット含有プラスミドを用いて組み換えHVTを作製及び純化する方法は、前述の通り、Avian Dis.36:858−870(1992)や特許第3587065号公報に記載されている。
【0024】
(6)組み換え七面鳥ヘルペスウイルスを有効成分とする家禽用ワクチン
本発明の組み換え七面鳥ヘルペスウイルスを有効成分とする家禽用ワクチンの調製方法は、特に限定されないが、例えば次の方法によって調製することができる。
【0025】
本発明の組み換えHVTの感染細胞を当該ウイルスが生育できる細胞(以下、宿主細胞という)に感染させ、増殖させた後、細胞をスクレーパーまたはトリプシンではがし、遠心分離によって感染細胞と上清とに分離する。
【0026】
宿主細胞としては、トリ由来の細胞が好ましく、CEF(鶏胚繊維芽細胞)、鶏腎細胞などを好適に使用することができる。使用する培地は、Eagle’s MEM、ライボビッツL−15/マッコイ5A(1:1混合)培地(以下LM培地という場合がある)などが好適である。得られた感染細胞は、10%のジメチルスルフォキシド(DMSO)を含む培地に、懸濁し、液体窒素存在下で凍結保存する。ワクチンとして使用するときは100倍量のリン酸緩衝液(PBS)にこの凍結保存品を溶かして使用する。液体窒素下で上記感染細胞を保存するための安定剤やその他の成分は、ウイルス感染細胞が安定的に生存でき、かつ接種を受ける家禽にとって薬理学的に問題のない成分であれば特に限定されない。
【0027】
本発明のワクチンの家禽への投与方法もまた特に限定されないが、皮下に注射により接種する方法や、発育鶏卵に接種する方法が一般的に用いられており、マレック病に対するワクチンとして用いられている従来ワクチンと同じである。接種量も従来ワクチンと同様でよい。即ち、10〜105p.f.u./dose、好ましくは102〜104p.f.u./doseになるように適量のリン酸緩衝液などに溶かして、初日齢の鶏の頸部皮下に、あるいは孵化前の卵に注射や接種用機器を使って投与する。
【0028】
本発明のワクチンは、親ウイルスであるHVTによるマレック病に対するワクチン効果だけでなく、挿入された発現遺伝子カセット中の抗原遺伝子が発現することによって、それらの遺伝子の由来病原体による疾病に対するワクチン効果も発現するため、多価ワクチンとして使用することができる。
【実施例】
【0029】
以下に実施例を挙げて、本発明を説明するが、本発明はこれに限定されるものではない。
本発明のプラスミド構築は特に記載がない限り、Molecular Cloning:A Laboratory Manual.2nd Edition.(Cold Spring Harbor Laboratory.Cold Spring Harbor.N.Y.1989)記載の標準的な分子生物学手法を用いて行った。尚、制限酵素断片はアガロースゲルからQIAquick Gel Extraction Kit(QIAGEN社製、カタログ番号#28704)を使って精製した。
七面鳥ヘルペスウイルス(HVT)は、アメリカ農務省Avian Disease and Oncology LaboratoryのDr.Witterが分離した七面鳥ヘルペスウイルスFC126株を使用した(Witter R.L.et al.Am.J.Vet.Res.1970.31.525−538)。
【0030】
1.MDV由来プロモーターのクローニング
マレック病ウイルスのゲノム構造は、Tulmanらによって報告されている(J.Virology 74:7980−7988(2000))が、MDVのゲノム上にあるプロモーターの明確な位置や活性についてはあまり解析がなされていない。数少ない例として、IzumiyaらによるUL41からUL51遺伝子間のmRNA解析論文(J.Gen.Virology 79:1997−2001(1998))や、UL30からUL40遺伝子間のmRNA解析論文(J.Gen.Virology 80:2417−2422(1999))が報告されている程度である。上記3つの論文から、MDV由来プロモーターとして、MDVのゲノム配列(ジーンバンクのアクセッション番号:AF243438)の91827番目から92196番目までの塩基領域、及び98757番目から99412番目までの塩基配列領域が候補として挙げられた。
【0031】
マレック病ワクチンとして市販されているマレック病ウイルスCVI988株を直径15cmの培養皿のCEF細胞に感染させウイルスを増やした後、PBSで洗浄後、リシスバッファー(0.5%SDS10mMトリス(pH8)、100mMNaCl、1mMEDTA、200μg/mlプロテナーゼ)を4ml加えて、CO2インキュベーター(37℃)で4時間インキュベートし、細胞溶解物を50mlファルコンチューブに移し、等量のフェノール(TE飽和)を加え、15分間ゆっくり回転し、混合した。次に、1500×Gで5分間遠心後、水層をピペットでゆっくり吸い取り、新しいチューブに移した。クロロフォルム:イソアミルアルコール=24:1を等量加え、15分間ゆっくり混合した。その後、1500×Gで5分間遠心し、水層をピペットでゆっくり吸い取り、新しいチューブに移し、100%エタノールを水層の倍量加え、良く混ぜてから1500×Gで5分間遠心した。その沈殿を70%エタノールでリンスした後、軽く風乾してMDVのゲノムDNAを含む核酸画分を調製した。
【0032】
上記で調製したMDVのゲノムDNA画分をテンプレートにして、MD50F(配列番号5)とMD51R(配列番号6)のプライマーセットでPCRを行い、MDVのゲノム配列の91709番目から92261番目までのDNA領域を増幅させ、それをpPCR−Script Ampベクター(ストラタジーン社製)にライゲーションし、JM109を形質転換した。そうして、得られた形質転換株3株からプラスミドを調製して、挿入DNAの塩基配列分析を行った。3株のうち、2株の挿入DNAの塩基配列が完全に一致した(配列番号7)。それらのプラスミドをpPCR−Pmd50と命名した。
【0033】
同様にして、MD54F(配列番号8)とMD55R(配列番号9)のプライマーセットでPCRを行い、MDVのゲノム配列の98715番目から99464番目までのDNA領域を増幅させ、それをpPCR−Script Ampベクターにクローニングした。挿入されたDNA配列は、ジーンバンクのAF243438の配列とは1塩基異なるが、3クローン間で完全に一致した。その配列を配列番号10に示す。こうして得られたプラスミドをpPCR−Pmd54と命名した。
【0034】
2.MDV由来プロモーターの改変
pPCR−Pmd50FとpPCR−Pmd54Fをそれぞれ制限酵素KpnIで切断後、更にSalIで切断して回収したプラスミドに、Ad_Kp−Bg−SaU(配列番号11)と、Ad_Kp−Bg−SaL(配列番号12)の2つの合成ヌクレオチドをアニールさせた合成DNAアダプターを挿入した。このようにして得られたプラスミドをそれぞれpPmd50M1、pPmd54M1と命名した。
【0035】
pPmd50M1とpPmd54M1をそれぞれ制限酵素SacIで切断後、更にNotIで切断して回収したプラスミドに、Ad_No−Sf−SaU(配列番号13)と、Ad_No−Sf−SaL(配列番号14)の2つの合成ヌクレオチドをアニールさせた合成DNAアダプターを挿入した。このようにして得られたプラスミドをそれぞれpPmd50M2、pPmd54M2と命名した。
【0036】
次いで、pPmd50M2とpPmd54M2をそれぞれ制限酵素NcoI/NotIで二重切断して回収したプラスミドに、Ad_Nc−Bg−Xh−NoU(配列番号15)と、Ad_Nc−Bg−Xh−NoL(配列番号16)の2つの合成ヌクレオチドをアニールさせた合成DNAアダプターを挿入した。このようにして得られたプラスミドをそれぞれpPmd50M3、pPmd54M3と命名した
【0037】
pPCR−Pmd50をテンプレートにして、Pmd50F(配列番号17)とPmd51R(配列番号18)のPCRプライマーでPCRを実施した。増幅されたPCR産物をBamHIとNcoIで二重切断して得られた約380bpのDNA断片を、BamHIとNcoIで二重切断したpPmd50M3とライゲーションして、pPmd50Fを構築した。このpPmd50FのKpnIサイトからSacIサイトまでの塩基配列が、配列番号19である。
【0038】
同様に、pPCR−Pmd54をテンプレートにして、Pmd54F(配列番号20)とPmd55R(配列番号21)のPCRプライマーでPCRを実施した。増幅されたPCR産物をBamHIとNcoIで二重切断してうられた約670bpのDNA断片を、BamHIとNcoIで二重切断したpPmd54M3とライゲーションして、pPmd54Fを構築した。このpPmd54FのKpnIサイトからSacIサイトまでの塩基配列が、配列番号22である。
【0039】
3.2つの外来遺伝子を発現する発現遺伝子カセットプラスミドの構築
オワンクラゲ由来の緑色蛍光蛋白質GFP遺伝子を持った市販プラスミドベクターpGFP(クロンテック社製、カタログ番号:6097−1)をテンプレートにして、GFPBspF(配列番号23)とGFPSalR(配列番号24)のプライマーセットでPCRを実施した。増幅されたPCR産物を制限酵素BspHIとSalIで二重切断し、NcoIとXhoIで二重切断したpPmd50FとライゲーションしてpPmd51GFPを作製した。
【0040】
特許第3587065号の実施例7に記載されているpRSV−Fのプラスミドに含まれているニューカッスル病ウイルス(NDV)のF遺伝子を制限酵素BamHIとSalIで二重切断してアガロースゲルから回収し、BamHIとSalIで二重切断したpPmd51GFPとライゲーションして、pPmd50ND_F51GFP(図2)を作製した。
このプラスミドpPmd50ND_F51GFPは、オワンクラゲ由来のGFP遺伝子とNDV由来のF遺伝子という2つの外来遺伝子とその間にMDV由来の1つのプロモーター(BDP1:配列番号1)から構成される発現遺伝子カセットを有するプラスミドである。
【0041】
特開2004−111号公報記載のプラスミドp45/46bacVP2−Eをテンプレートにして、BspVP2−F(配列番号25)とVP2Xho−R(配列番号26)のプライマーセットでPCRを実施した。増幅されたPCR産物(伝染性ファブリキウス嚢病ウイルス(IBDV)のVP2蛋白質遺伝子;約1.4kbp)を制限酵素BspHIとXhoIで二重切断し、NcoIとXhoIで二重切断したpPmd54FとライゲーションしてpPmd55IBD_VP2を作製した。
【0042】
国際公開WO2005/093070号公報記載のホモロジーベクターp45/46PecILgBdellac+STPから制限酵素BamHIとSalIで伝染性喉頭気管炎ウイルス(ILTV)のgB遺伝子(約1.9kbp)を切り出してアガロースゲルから回収し、BamHIとSalIで二重切断したpPmd55IBD_VP2とライゲーションして、pPmd54IL_gB55IBD_VP2(図3)を作製した。
このプラスミドpPmd54IL_gB55IBD_VP2は、ILTV由来のgB遺伝子とIBDV由来のVP2遺伝子という2つの外来遺伝子とその間にMDV由来の1つのプロモーター(BDP2:配列番号2)から構成される発現遺伝子カセットを有するプラスミドである。
【0043】
4.組み換えHVTを作製するためのホモロジーベクター(カセット含有プラスミド)の構築
特許第3587065号公報の実施例1の記載に従って、HVT−DNAを調整し、それをテンプレートにして、HVT45Sph(配列番号27)とHVT46Xho(配列番号28)のプライマーセットでPCRを実施した。増幅したPCR産物(約1.0Kbp)をSphIとXhoIで二重切断して、同じくSphIとXhoIで二重切断したpNZ45/46BacpA(特開2004−111の実施例3.2)とライゲーションしてプラスミドp46を得た。これをテンプレートにして、HVT45Sph(配列番号27)と45SfiR(配列番号29)のプライマーセットでPCRを実施した。同様にして、46SfiF(配列番号30)とHVT46Xho(配列番号28)のプライマーセットで、PCRを実施した。それぞれ0.4kbpと0.6kbpのDNA断片が増幅したので、更にその2断片をテンプレートにして、HVT45Sph(配列番号27)とHVT46Xho(配列番号28)のプライマーセットで乗り換えPCRを実施してSfiIサイトを導入した。その結果生じた1.0kbpのPCR産物を制限酵素SphIとXhoIで二重切断して、p46のSphIとXhoIの間のDNAと置き換えたプラスミドp46Sfi(図4)を作製した。
【0044】
プラスミドpPmd50NDV_F51GFPを制限酵素BglIで切断して、約3.1kbpの発現カセットをアガロースゲルから回収した。そのカセットを制限酵素SfiIで切断したp46Sfiにライゲーションしてp46md50ND_F51GFPを作製した。
【0045】
次にプラスミドpPmd54IL_gB55IBD_VP2を制限酵素BglIで切断して、約4.0kbpの発現カセットをアガロースゲルから回収し、制限酵素SfiIで切断したp46md50ND_F51GFPにライゲーションしてp46md50ND_F51GFP54IL_gB55IBD_VP2を作製した。このプラスミドp46md50ND_F51GFP54IL_gB55IBD_VP2(図5)は、2つの発現遺伝子カセットを含有する組み換えHVTを作製するためのホモロジーベクターである。
【0046】
5.組み換えHVT(FW101)の作製・純化
トリプシンで剥がした単層のCEF細胞を、SalineG(0.14M塩化ナトリウム、0.5mM塩化カリウム、1.1mMリン酸一水素二ナトリウム、15mMリン酸二水素一ナトリウム、0.5mM塩化マグネシウム・6水和物、0.011%グルコース)に懸濁し、細胞懸濁液を調製した。この細胞懸濁液(2×107個)に、実施例4で作製したカセット含有プラスミドp46md50ND_F51GFP54IL_gB55IBD_VP2を25μgと、特許第3587065号公報の実施例1記載の方法で調製したHVTのDNA15μgを混合して、ジーンパルサー(BioRad社製)を用いて、0.5kVcm−1、0.5msecの条件下でエレクトロポレーションした。
【0047】
プラスミド及びHVTのDNAを導入した細胞を、107個のCEF細胞と混合して、培養用96ウェル平底マルチプレート(ファルコン社製)5枚にまいて37℃にてHVT特有のプラークが形成されるまで約4日〜5日間培養した。
【0048】
倒立型落射蛍光観察装置(オリンパス社製「IX−FLA」)付の倒立顕微鏡(オリンパス社製「IX70」)を使って、出現したHVTのプラークを観察し、蛍光蛋白を発現しているプラークを選択した。選択したプラークの入っているウェルに1%トリプシンを加えて組み換えHVT感染細胞を含むCEF細胞をそれぞれ回収し、2×106個の細胞と混合し、それぞれのプラークごとに、1枚の培養用96ウェル平底マルチプレートにまいて、37℃にて約3日〜5日間培養した。
【0049】
再び出現したHVTのプラークを蛍光顕微鏡にて、各プラークが蛍光蛋白を発現しているか否かを観察した。各ウェルに存在するHVTプラークが全て蛍光蛋白を発現するプラークとなっていなければ、蛍光蛋白を発現するプラークの割合が最も多いウェルを選択して、再び、1%トリプシンを加えて組み換えHVT感染細胞を含むCEF細胞をそれぞれ回収し、2×106個の細胞と混合し、それぞれのプラークごとに、1枚の培養用96ウェル平底マルチプレートにまいて、37℃にて約3日〜5日間培養した。
【0050】
このようにして、各ウェルに存在するHVTプラークが全て蛍光蛋白を発現するプラークとなるまで、スクリーニング操作を繰り返した。
【0051】
ウェル中の全HVTプラークが蛍光蛋白を発現するようになったら、そのウェルのCEF細胞をトリプシンで回収し、培養用6ウェルプレート(ファルコン社製)の1ウェルを使って感染細胞を増殖させた後、SPGA−SBTバッファー(100ml当たり、7.46gシュークロース、5.2mg KH2PO4、125.4mg K2HPO4、91.7mg L−グルタミン・Na、1g BSA、10gソルビトール)2mlに回収し、1mlをエッペンドルフチューブに移した。
【0052】
そのチューブを超音波破砕器(三光純薬社製「BIOSONIC」)にセットし、最大出力で3分間超音波破砕した。その後、5000rpm5分間遠心分離し、上清を回収した。これにより、セルフリーウイルスが回収された。回収した上清をSPGA−SBTバッファーで段階希釈して、別途用意したCEF細胞を単層培養した96ウェル平底マルチプレートに各ウェルあたり40μlずつ重層して、2時間インキュベーターで感染させた。液をすて、LM培地を200μlずつ加えてプラークが出現するまで培養した。
【0053】
出現したHVTのプラークを蛍光顕微鏡にて観察し、全ウェル中に1つのプラークで、且つ、そのプラークが蛍光蛋白を発現しているウェルを選択した。これで、このウェル中のHVTは100%純度になったと判断され、このようにして純化した組み換えHVTをFW101と命名した。
【0054】
6.組み換えHVTのゲノム構造の確認とその安定性試験
6.1 サザンハイブリダイゼーション
実施例5で純化が終了したFW101をCEF細胞とともに2枚の150mm径培養皿で全面にプラークが出現するまで培養し、その感染細胞をスクレーパーで培養皿から剥がして遠心分離(500×G、5分)して感染細胞を集め、PBSで洗浄後、1.2mlのPBSに再懸濁し、溶解バッファー(1.25%TritonX−100、250mM 2−メルカプトエタノール、50mM EDTAがPBSに入ったもの)0.8mlを加えて攪拌して細胞を可溶化した。3000rpm、5分間で細胞残さを除去し、上澄をエッペンドルフチューブに移して15000rpm、20分間、22℃で遠心してウイルスを落とした。12.5mM トリス(pH7.5)を1ml加えて、ヌクレアーゼ液(0.25mg/ml DNaseI、0.25mg/ml RNaseA、150mM 塩化ナトリウム)を4μl加えて37℃、30分間静置した。その後、0.5M EDTAを25μl、10%SDSを125μl、水87μl、10mg/mlの10mg/mlのプロテアーゼKを12.5μl、2−メルカプトエタノール0.5μl加えて、55℃、30分間静置した。その液をフェノール:クロロホルム(=1:1混合)で2回抽出した。
【0055】
その水層に5M塩化ナトリウム16μlを加え、水層の2.5倍量の(−20℃で冷やした)100%エタノールを加えて沈殿させ、遠心分離で沈殿させた沈殿物(DNA)を70%エタノールで洗浄後、再度遠心分離して沈殿させた後乾燥させ、TE緩衝液(10mMトリス(pH8.0)、1mM EDTA)50μlに溶かした。
こうして回収されたウイルスDNAを制限酵素XhoI/StuIで切断し、0.8%アガロースゲル電気泳動を行い、制限酵素で切断されたDNA断片をナイロンメンブレンにブロットし、サザンハイブリダイゼーションを行った。プローブは“PCR DIG Probe Synthesis Kit”(ROCHE DIAGNOSTICS社製、カタログ番号#1636090)の試薬を用いて2種類のDIG標識されたDNAプローブを調製した。そのDNAプローブは、VP2 cDNA領域内のプローブ(以下VP2プローブ)と外来遺伝子の挿入部位のHVTゲノム領域(以下45/46プローブ)である。前者のプローブはVP2−F(配列番号31)とVP2−R(配列番号32)の2本のプライマー及びプラスミドp46md50ND_F51GFP54IL_gB55IBD_VP2をテンプレートとして、後者は、45/46−F(配列番号33)と45/46−R(配列番号34)の2本のプライマー及びプラスミドp46SfiをテンプレートとしてPCR反応によって調製した。
【0056】
この2種類のプローブを使ったサザンハイブリダイゼーションの結果、VP2プローブでは2.0kbpのバンドがハイブリダイズし、45/46プローブでは0.6kbと1.7kbの2本のバンドがハイブリダイズした。この結果は、どちらのプローブも予期した通りのバンドであったので、組み換えHVTは設計どおり正しい組み換え体であることが確認された。
【0057】
6.2 ゲノム安定性試験
組み換えHVTをCEF細胞で20代継代した組み換えウイルスについても前記6.1と同様にサザンブロッティングを実施し、20代継代後の組み換えウイルスも正しい遺伝子構造を持っていることが確認された。一方、特許第3587065号公報記載の組み換えHVTのHF004〜HF007の組み換え体について、同様に20代継代したところ、これら全ての組み換えHVTのゲノム構造は、継代前の組み換えHVTの構造と異なっていた。これらの組み換えHVTについては、継代途中に遡って安定性を調べたところ、HF004とHF005は10代、HF006とHF007については、5代継代した時点で、継代前の構造と異なっており、不安定であることがわかった。従って、20代継代してもゲノム構造が変化しないFW101は極めて安定な組み換えHVTであることがわかった。
【0058】
7.組み換えHVTの蛋白発現確認
7.1 FITC標識抗体を使った蛍光検出法
特許第3587065号公報の実施例16に記載した方法に従って、組み換えウイルスFW101が外来抗原遺伝子であるNDVのF蛋白質、IBDVのVP2蛋白質を発現していることを確認した。
【0059】
7.2 ウエスタンブロット法
9cm径ディッシュに新鮮な107個のCEF細胞と1×105p.f.u.のFW101をまき、約72時間後に、感染細胞をスクレーパーにて回収した。遠心分離(2000rpmで5分)で感染細胞を回収し、10mlのPBSで細胞を洗浄し、再び遠心分離で細胞を集めた。その細胞ペレットに2×サンプルバッファー(0.112Mトリス−アセテート(pH7)、2%SDS、5%メルカプトエタノール、5%シュークロース、0.005%ブロモフェノールブルー )を0.1ml添加し、100℃で5分煮沸した。
【0060】
煮沸したサンプル5μlを10%SDS−ポリアクリルアミドゲルにアプライした。電気泳動終了後、PVDFメンブレン(ミリポア)にエレクトロブロッティングし、ブロットしたPVDFメンブレンをIBDVのVP2に対するモノクローナル抗体R−63(ハイブリドーマはATCCから入手、ATCC:HB−9490)を用いてウエスタンブロットを行った。その結果、約40kDaの大きさの蛋白質が検出された。この大きさは、文献で知られているIBDVのVP2の大きさであり、FW101は期待通り目的とする大きさのVP2が発現していた。
【0061】
また、GFP蛋白に対する市販モノクローナル抗体JL−8(クロンテック社製)を使って同じくブロットしたPVDFメンブレンと反応させたところ、約26kDaの大きさの蛋白質が検出された。この大きさはGFP蛋白の分子量と一致し、FW101が期待通りの大きさのGFP蛋白質も発現していることを確認した。
【0062】
8.FW101のワクチン効果動物試験
FW101を鶏に免疫してワクチン効果を調べた。3000p.f.u./doseのFW101(対照は親株HVT)をSPF鶏の発育鶏卵(日生研)の孵化後の鶏の背部皮下に接種した。接種後、1週間隔で採血をし、その血中の抗体価を市販ELISAキット(ゲン・コーポレーション社製、IBDエリーザキット)で測定した。そのデータ(S/P value)を表1に示す。
【0063】
【表1】
【0064】
表1に示したように、挿入した抗原遺伝子の蛋白質に対する抗体が上昇し、ワクチン活性が認められた。
【図面の簡単な説明】
【0065】
【図1】図1は、発現遺伝子カセットの概念図である。
【図2】図2は、発現遺伝子カセットプラスミドpPmd50ND_F51GFPの模式図。
【図3】図3は、発現遺伝子カセットプラスミドpPmd54IL_gB55IBD_VP2の模式図である。
【図4】図4は、挿入部位を含むHVT−DNA含有プラスミドp46Sfiの模式図である。
【図5】図5は、FW101作製用ホモロジーベクター p46md50ND_F51GFP54IL_gB55IBD_VP2の模式図である。
【特許請求の範囲】
【請求項1】
2つの異なる外来遺伝子と、当該2つの外来遺伝子を両方とも支配するように、2つの外来遺伝子の間に配置された、マレック病ウイルス由来の1つのプロモーターとを有する発現遺伝子カセットが、ゲノム中に1つ以上挿入された組み換え七面鳥ヘルペスウイルス。
【請求項2】
プロモーターが配列番号1又は配列番号2に記載された塩基配列からなるものである請求項1記載の組み換え七面鳥ヘルペスウイルス。
【請求項3】
発現遺伝子カセットの挿入部位が、七面鳥ヘルペスウイルスのUL45とUL46のオープン・リーディング・フレームの間である請求項1又は2記載の組み換え七面鳥ヘルペスウイルス。
【請求項4】
挿入される発現遺伝子カセット中の2つの異なる外来遺伝子のうち、少なくとも1つが鳥類感染性の病原体に由来する抗原遺伝子である請求項1から3記載の組み換え七面鳥ヘルペスウイルス。
【請求項5】
請求項4に記載された組み換え七面鳥ヘルペスウイルスを有効成分とする家禽用ワクチン
【請求項6】
配列番号1記載の塩基配列からなるプロモーター。
【請求項7】
配列番号2記載の塩基配列からなるプロモーター。
【請求項1】
2つの異なる外来遺伝子と、当該2つの外来遺伝子を両方とも支配するように、2つの外来遺伝子の間に配置された、マレック病ウイルス由来の1つのプロモーターとを有する発現遺伝子カセットが、ゲノム中に1つ以上挿入された組み換え七面鳥ヘルペスウイルス。
【請求項2】
プロモーターが配列番号1又は配列番号2に記載された塩基配列からなるものである請求項1記載の組み換え七面鳥ヘルペスウイルス。
【請求項3】
発現遺伝子カセットの挿入部位が、七面鳥ヘルペスウイルスのUL45とUL46のオープン・リーディング・フレームの間である請求項1又は2記載の組み換え七面鳥ヘルペスウイルス。
【請求項4】
挿入される発現遺伝子カセット中の2つの異なる外来遺伝子のうち、少なくとも1つが鳥類感染性の病原体に由来する抗原遺伝子である請求項1から3記載の組み換え七面鳥ヘルペスウイルス。
【請求項5】
請求項4に記載された組み換え七面鳥ヘルペスウイルスを有効成分とする家禽用ワクチン
【請求項6】
配列番号1記載の塩基配列からなるプロモーター。
【請求項7】
配列番号2記載の塩基配列からなるプロモーター。
【図1】


【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2008−228658(P2008−228658A)
【公開日】平成20年10月2日(2008.10.2)
【国際特許分類】
【出願番号】特願2007−73429(P2007−73429)
【出願日】平成19年3月20日(2007.3.20)
【出願人】(000229117)日本ゼオン株式会社 (1,870)
【Fターム(参考)】
【公開日】平成20年10月2日(2008.10.2)
【国際特許分類】
【出願日】平成19年3月20日(2007.3.20)
【出願人】(000229117)日本ゼオン株式会社 (1,870)
【Fターム(参考)】
[ Back to top ]