
植物ウイルスに基づいた誘導性発現システム
一つ又一つより多くの目的とするタンパク質を産生するプロセスであって:(a)RNAレプリコンをコードするヌクレオチド配列を含んでなる第一の異種ヌクレオチド配列、及び前記RNAレプリコンをコードする前記ヌクレオチド配列に機能可能なように連結された第一の誘導性プロモーターを含んでなる植物又は植物細胞を提供すること;前記RNAレプリコンは、前記植物中で前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない;前記RNAレプリコンはポリメラーゼ及び前記一つ又は一つより多くの目的とするタンパク質をコードし、前記ポリメラーゼは前記RNAレプリコンを複製するために適合されており;及び(b)工程(a)の前記植物又は植物細胞において前記誘導性プロモーターを誘導し、それにより前記植物又は植物細胞において前記一つ又は一つより多くの目的とするタンパク質を産生すること;を含んでなるプロセス。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ウイルス発現システムを使用し植物又は植物細胞中の一つ又は一つより多くの目的とするタンパク質を産生する又は発現するプロセスに関する。本発明は、さらにこのプロセスの為の植物又は植物細胞、特にトランスジェニック植物又は植物細胞に関する。本発明は、本発明の該植物又は植物細胞を産生するプロセスも提供する。
【背景技術】
【0002】
植物における異種タンパク質の高収率発現は、原則としてウイルスベクターを使用して達成し得る。しかしながら、異なった植物ウイルス発現システムの存在にもかかわらず、植物ウイルス発現システムは、主として感染に続いた植物中の目的とするタンパク質の一過性発現に(Donson et al.,1991, Proc Natl Acad Sci U S A, 88:7204-7208; Chapman, Kavanagh & Baulcombe, 1992, Plant J., 2:549-557)、又は組換えウイルスベクターを用いる宿主植物の形質移入(Marillonnet et al., 2005, Nat. Biotechnol., 23:718-723; Santi et al., 2006, Proc Natl Acad Sci U S A. 103:861-866; WO2005/049839) に使用されている。いくつかの科学出版物及び公開された特許出願にもかかわらず、主として二つ理由のため、容易にスケールアップでき、そして高収率を提供することに利用可能である商業的ウイルスに基づいた産生システムは未だ確立されていない:
第一に、一過性の植物ウイルスに基づいた発現システムは、一般的に特定の宿主に限定されており、それらの環境要因に対する感受性のため大規模培養には適していないであろう。さらに、それらは一般的に植物宿主の特定の部分に限定されており、それ故、産生プロセスからの植物バイオマスのほとんどを排除することになり、その結果、植物バイオマスの単位当たりの組換え産物の相対収率を最少化し、トランスジェニック植物において慣用的転写プロモーターを使用して達成されるレベルに匹敵するレベルまで下げている;
第二に、各細胞に安定に組み込まれたウイルスレプリコンを有するトランスジェニック宿主植物を発生させることによる、ウイルスに基づいた産生システムをスケールアップする試みは、特に、そのような位置の前記レプリコンの標準より低い働きのため、及びウイルスレプリコンの安定した形成が植物成長及び発達を損なうため、解決を提供していない。通常、一過性発現システムにおける浸透移行性ウイルスベクターは、異種核酸の比較的短い(1kbまで)挿入物に耐えることができ、それ故、比較的小さなタンパク質の発現に限定されている。トランスフェクションのために使用されるウイルスベクター(アグロバクテリウム仲介送達、WO2005/049839 )はより大きな挿入物を発現し得るが、全植物のアグロ浸潤を必要とする。明らかに、こうしたシステムは、使用する及びスケールアップするために短い時間しか必要としないので、抗原を含む多くの組換えタンパク質の産生に都合がよい、しかしウイルスベクターに基づいた発現システムのトランスジェニックバージョンが、多くの他の応用で有利であり得る。特に、大量に及び相対的に低コストで必要とされる組み換えタンパク質の産生には問題であり(例えば、異なったセルラーゼ及び他の技術的酵素)、アグロ浸潤 (WO2005/049839)に基づいた一過性発現システムは経済的に実行可能ではない。トランスジェニック宿主植物におけるウイルスベクターの発現は、通常植物生長及び発達のためには有害である。また、こうした発現は、最後には導入遺伝子サイレンシングを導くであろう。この問題の解決を見いだすために、転写後遺伝子サイレンシング(PTGS)サプレッサーの助けをかりて、植物染色体から発現停止したウイルスレプリコンを放出することが試みられた(US6395962; Mallory et al., 2002, Nat. Biotechnol., 20:622-625)。植物三要素RNAウイルス(Mori et al., 2001, Plant J., 27, 79-86)、ブロムモザイクウイルス(BMV)に基づいたグルココルチコイド誘導性発現システムは、おそらくPTGSのせいで、標準(非ウイルス性)転写プロモーターにより提供される収率に匹敵する、目的とするタンパク質の非常に低い収率(3〜4μg/g新鮮重量)しか与えなかった。
【0003】
現在のところ、細菌、真菌又は昆虫細胞発現システムのような他の大規模発現システムと市場で競争するために十分に高いであろう収率及び効率の大規模植物ウイルス発現システムはない。こうした植物発現は可能な限り良好に以下の基準を満たさなければならない:
(i) 高収率、可能な限り多くの植物組織中での、及び前記組織の多くの細胞中での、目的とするタンパク質の発現を含んで;
(ii)植物細胞の生存に対するタンパク質発現の悪影響を防止するため、目的とするタンパク質又は生成物の発現は、同時に処理された植物又は植物組織の全ての植物細胞中で開始されるべきである。
【0004】
典型的には、目的とするタンパク質又は生成物は、前記タンパク質又は生成物を産生している各細胞中で特定の点まで蓄積する。しかしながら、蓄積の間、分解プロセスがしばしば進み、それは目的とするタンパク質又は生成物の収量又は質を減少させる傾向がある。それ故、時間には至適時点があり、そこで目的とするタンパク質又は生成物を収穫すべきである。この時間の至適時点は、全プロセスを効率的に及び有益にするため、植物の全ての組織又は細胞で、及び選択されたロットの全ての植物で同時に到達可能であるべきである。
【発明の開示】
【0005】
本発明の一般的説明
それ故、大規模応用に容易に拡張可能で、発現されるべきタンパク質の高収率を与え、及び同時に、目的とする組み換えタンパク質の制御されない発現の確率は低いという点において生物学的に安全である植物システムにおいて、一つ又はそれより多くのタンパク質を発現するプロセスを提供することが一つの目的である。前記ウイルスレプリコンから、目的とするタンパク質を発現するために適したウイルスレプリコンをコードするトランスジェニック植物を産生する効率の良い方法を提供することが本発明のさらなる目的である。
【0006】
それ故、本発明は、一つの又は一つより多くの目的とするタンパク質を産生するプロセスを提供することであって:
(a)RNAレプリコンをコードするヌクレオチド配列を含んでなる第一の異種ヌクレオチド配列、及び前記RNAレプリコンをコードする前記ヌクレオチド配列に機能可能なように連結された第一の誘導性プロモーターを含んでなる植物又は植物細胞を提供すること;
前記RNAレプリコンは、前記植物における前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない;
前記RNAレプリコンはポリメラーゼ及び前記一つ又は一つより多くの目的とするタンパク質をコードし、前記ポリメラーゼは前記RNAレプリコンを複製するために適合されている;及び
(b)工程(a)の前記植物又は植物細胞において、前記誘導性プロモーターを誘導し、それにより、前記植物又は植物細胞において前記一つ又は一つより多くの目的とするタンパク質を産生すること;
を含んでなる。
【0007】
本発明はさらに、一つの又は一つより多くの目的とするタンパク質を産生するプロセスを提供することであって:
(a)植物を提供することであって、前記植物は、
(i)RNAレプリコンをコードするヌクレオチド配列、及び前記RNAレプリコンをコードする前記ヌクレオチド配列に機能可能なように連結された第一の誘導性プロモーターを含んでなる第一の異種ヌクレオチド配列;
前記RNAレプリコンは、前記植物における前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない;
前記RNAレプリコンはポリメラーゼ及び前記一つ又は一つより多くの目的とするタンパク質をコードし、前記ポリメラーゼは前記RNAレプリコンを複製するために適合されている;
(ii)前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードする配列を含んでなる第二の異種ヌクレオチド配列、ここで前記第二の異種ヌクレオチド配列は、前記RNAレプリコンの細胞間移行を可能にする前記タンパク質をコードする前記配列に機能可能なように連結された第二の誘導性プロモーターを含んでなる;
を含んでなり;及び
(b)工程(a)の前記植物において、前記第一及び第二の誘導性プロモーターを誘導し、それにより、前記植物において前記一つ又は一つより多くの目的とするタンパク質を産生すること;
を含んでなる。
【0008】
本発明はさらに、上記プロセスの工程(a)で提供された植物又は植物細胞を提供する。本発明はさらに、本発明の植物又は植物細胞を産生するプロセスを提供することであって、前記第一の異種ヌクレオチド配列及び任意選択で前記第二の異種ヌクレオチド配列を植物核染色体内に導入すること、続いて前記第一の及び任意選択で前記第二の異種ヌクレオチド配列を含有する形質転換植物を再生することを含んでなる。
【0009】
本発明の発明者は驚くことに、従来の技術では達成不可能であった発現レベルを達成する、植物又は植物細胞中で一つ又は一つより多くの目的とするタンパク質を産生するプロセスを発見した。同時に、本発明のプロセスは、生物学的に安全であり、及び工業的レベルへ拡張可能である。本発明の例外的な発現レベルは、今まである程度しか知られていない導入遺伝子サイレンシングのような、植物又は植物細胞による抗ウイルス応答を避けるウイルス発現システムにより達成可能である。本発明において、RNAレプリコンをコードするヌクレオチド配列からのRNAレプリコンの放出及び前記RNAレプリコンの細胞間移行を可能にする前記タンパク質の発現は、誘導性プロモーターにより制御されている。前記RNAレプリコンは、細胞間移行を提供する前記タンパク質が存在しない場合には前記植物中で又は前記植物細胞中で細胞間移行ができないので、前記RNAレプリコンをコードするヌクレオチド配列からのRNAレプリコンのリーキーな(leaky) 転写による、RNAレプリコンのいずれかの意図されない放出は、こうしたリーキーな転写が起こった細胞に限局されている。重要なことには、ウイルス配列は、誘導されない状態においては、導入遺伝子サイレンシングの引き金を引く事ことができるレベルでは発現されない。それ故、導入遺伝子サイレンシングは起こりそうもない。さらに、前記植物において前記RNAレプリコンの細胞間移行を提供するタンパク質を取り除くことによるRNAの発現収率に対する悪影響は、前記RNAレプリコンのトランスに前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードする配列を提供するすることにより補償されている。前記RNAレプリコンの細胞間移行を可能にする前記タンパク質は、誘導性プロモーターの制御下に発現される。このことは、前記ウイルスレプリコンのリーキーな発現が起こった細胞へ、こうしたリーキーな発現の限局を保存し、そして遺伝子サイレンシングを回避するが、誘導された状態においては前記目的とするタンパク質の高い発現レベルを可能にする。それ故、本発明において、目的とするタンパク質の収率に対する遺伝子サイレンシングの悪影響は、本質的に存在しない。
【0010】
さらに、本発明者は驚くべきことに、形質転換効率は、植物細胞が本発明の前記第一の異種ヌクレオチド配列で形質転換された場合、RNAレプリコンが細胞間移行を提供するタンパク質をコードする場合よりも高いことを発見した。いくつかの場合において、もしRNAレプリコンが細胞間移行を提供するタンパク質をコードするならば、前記第一の異種ヌクレオチド配列で一次形質転換体を全く得ることができなかった。形質転換効率を改良する本発明のこの効果は、細胞間移行についての本発明のRNAレプリコンの無能さによるものであろう。
【0011】
本発明の一つの態様において、前記植物又は植物細胞は、前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードするヌクレオチド配列を含んでなる第二の異種ヌクレオチド配列を含有し、前記第二の異種ヌクレオチド配列は、前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードする前記ヌクレオチド配列に機能可能なように連結された第二の誘導性プロモーターを含んでなる。本発明のこの態様は、前記一つ又は一つより多くの目的とするタンパク質の産生の収率をさらに増加させることを可能にする。
【0012】
一つ又は一つより多くの目的とするタンパク質を産生する本プロセスの工程(a)において、植物又は植物細胞に前記第一の異種ヌクレオチド配列が提供される。前記植物又は植物細胞には、トランスジェニック植物又はトランスジェニック植物細胞が得られるように、前記第一の異種ヌクレオチド配列が提供される。一つの態様において、前記トランスジェニック植物又は前記トランスジェニック植物細胞は、核染色体中に、前記第一の異種ヌクレオチド配列を含有する。
【0013】
前記植物又は植物細胞には、トランスジェニック植物又はトランスジェニック植物細胞が得られるように、前記第二の異種ヌクレオチド配列を提供することもできる。一つの態様において、前記トランスジェニック植物又は前記トランスジェニック植物細胞は、核染色体中に前記第二の異種ヌクレオチド配列を含有する。
【0014】
核染色体又はエピソーム内に安定に組み込まれた異種ヌクレオチド配列を含有しているトランスジェニック植物を発生させることは当該技術分野で公知である。典型的には、前記異種ヌクレオチド配列は、組み込まれた前記異種ヌクレオチド配列を有する植物細胞又は植物組織を選択するための選択可能マーカー遺伝子を含有するであろう。前記トランスジェニック植物の細胞中に前記異種ヌクレオチド配列を含有する全トランスジェニック植物は、次ぎに植物バイオテクノロジーの技術分野の標準法を使用して、形質転換細胞又は組織から再生することができる。本発明の前記異種ヌクレオチド配列は、典型的にはDNAである。
【0015】
もし植物が、前記第一の及び前記第二の異種ヌクレオチド配列を含有するように提供されるならば、前記第一の及び前記第二の異種ヌクレオチド配列は、植物細胞又は植物を形質転換するために使用された一つの大きな異種ヌクレオチド配列の一部であることができる。この態様において、前記大きな異種ヌクレオチド配列は、前記第一の及び前記第二の異種ヌクレオチド配列を含んでなる。もしくは、前記第一の異種ヌクレオチド配列を含有する第一の植物、及び前記第二の異種ヌクレオチド配列を含有する第二の植物を独立して発生させてもよい。前記第一及び前記第二植物は次ぎに交配することができて(例えば、性交配又は細胞融合により)、前記第一の及び前記第二の異種ヌクレオチド配列を含有する植物が得られる。さらなる代替法において、前記第一の及び前記第二の異種ヌクレオチド配列を含有する植物又は植物細胞を産生するため、前記第一の異種ヌクレオチド配列を含有するトランスジェニック植物又は植物細胞を前記第二の異種ヌクレオチド配列で形質転換することができ;又は前記第二の及び前記第一の異種ヌクレオチド配列を含有する植物又は植物細胞を産生するため、前記第二の異種ヌクレオチド配列を含有するトランスジェニック植物又は植物細胞を前記第一の異種ヌクレオチド配列で形質転換することができる。ここで、「異種」とは、前記植物について異種を意味する。
【0016】
前記第一の異種ヌクレオチド配列は、前記RNAレプリコンをコードするヌクレオチド配列セグメントを含んでなる。前記第一の異種ヌクレオチド配列はさらに、前記RNAレプリコンをコードする前記ヌクレオチド配列セグメントに機能可能なように連結されている第一の誘導性プロモーターを含んでなる。前記誘導性プロモーターは、本発明のプロセスの工程(b)において、前記RNAレプリコンをコードする前記ヌクレオチド配列セグメントの転写を誘導することを可能にする。転写は、前記RNAレプリコンをコードする前記ヌクレオチド配列セグメントから前記RNAレプリコンを放出する。前記RNAレプリコンはRNAレベル上のレプリコンである。植物細胞の核中で産生された前記RNAレプリコンは、次ぎに、前記RNAレプリコンによりコードされたタンパク質が産生され及びRNAレプリコンが複製されるサイトゾル内へ移行することができる。
【0017】
前記RNAレプリコンは、前記誘導性プロモーターを誘導することにより前記RNAレプリコンが放出された後に発現させることができる。本発明において、前記第一の異種ヌクレオチド配列によりコードされた前記第一のRNAレプリコンは、前記RNA配列を複製するためのポリメラーゼをコードするRNA配列であり、それにより、前記RNA配列はその上にコードされているポリメラーゼにより複製されるように適合される。それ故、前記ポリメラーゼは、本明細書において「レプリカーゼ」とも称されるRNA依存性RNAポリメラーゼである。前記RNAレプリコンは、好ましくは、前記ポリメラーゼを翻訳するための配列、及び前記植物の細胞又は前記植物細胞中での前記RNAレプリコンの複製を可能にするため、前記ポリメラーゼを結合するための配列を有する。前記RNAレプリコンはさらに、発現されるべき前記一つ又は一つより多くの目的とするタンパク質、ならびにサブゲノムプロモーター、転写エンハンサー又は翻訳エンハンサーのような、前記一つ又は一つより多くの目的とするタンパク質を発現するために必要とされる配列をコードする。
【0018】
好ましい態様において、前記RNAレプリコンは、一要素(monopartite)RNAウイルスのようなRNAウイルスに由来する。「一要素」とは、一要素ウイルスが一つの核酸分子から成るゲノムを有することを意味する。それ故、前記本発明のRNAレプリコンは、好ましくは、一要素RNAレプリコンであり、即ち、それは単一のタイプのRNA分子から成る。好ましい態様において、前記RNAレプリコンはプラスセンス一本鎖RNAウイルスに由来しており、こうしたウイルスは複製及び発現に必要とされる遺伝子要素を含有し、及び本発明の目的に進化により最適化されているからである。前記好ましい態様において、前記RNAレプリコンはプラスセンス一本鎖RNAレプリコンである。
【0019】
「由来する」とは、本発明に必要なRNAウイルスの遺伝子因子が使用され、一方、その他は欠失されるか又は機能障害性にされることを意味する。本発明において、前記ウイルスの細胞間移行を提供する前記タンパク質をコードする配列が、例えば、細胞間移行に必須な配列部分の部分的欠失又は突然変異により、欠失されるか又は機能障害性にされるであろう。もしくは、目的とするタンパク質をコードする配列を、完全に又は部分的に細胞間移行のための前記タンパク質をコードする配列で置き換えることができる。「由来する」は、植物ウイルスから取り出された前記RNAレプリコンの配列は、前記RNAウイルスの対応するRNA配列と同一である必要はなく、WO 2005/049839に記載されているように、前記レプリカーゼをコードする配列部分に挿入されたイントロンのような機能保存的相違を示すことができることを示唆する。前記相違は機能保存的であるので、前記配列は、好ましくは、前記RNAレプリコンが由来する前記RNAウイルス中で行うような、レプリコン機能を実施できるタンパク質をコードする。本発明の本発明の前記RNAレプリコン又はそれらのポリメラーゼを誘導することができる適したプラスセンス一本鎖RNAウイルスは、タバコモザイクウイルス(TMV)又は ジャガイモウイルスXである。前記RNAレプリコンを誘導することができるさらなる植物ウイルスは以下に与えられている。
【0020】
しかしながら、前記RNAレプリコン、又はそれらをコードするcDNAを人工的に合成することも可能であり、天然のウイルスの遺伝子因子を使用しても又はしなくてもよい。
【0021】
本発明の前記RNAレプリコンは、前記第一の異種ヌクレオチド配列の一つの連続したヌクレオチド配列セグメントによりコードされている必要はない。実際、前記第一の異種ヌクレオチド配列は、一緒になって前記RNAレプリコンをコードする二つ又はそれより多くの配列セグメントを有することができる。二つまたはそれより多くのこうした配列セグメントは、連続していてもよいし、又は別の配列部分により中断されていてもよい。前記植物の細胞中での前記RNAレプリコンの形成は、部位特異的DNA又はRNA組換えを含むことができる。DNA組換えの場合、前記RNAレプリコンは、前記RNAレプリコンが発現されることを阻止する配列部分の切除を介して形成させることができる。もしくは、不連続的に前記RNAレプリコンをコードする二つまたはそれより多くの配列セグメントの一つを組換えによりフリップさせることができ、それにより前記RNAレプリコンをコードする単一の連続配列セグメントを形成させる。前記RNAレプリコンは、そのどちらもがRNAレプリコンではない二つのレプリコン前駆体間の組換えにより形成することも可能である。こうした組換えは、WO02/097080 に記載されているリボザイム仲介トランススプライシングであってもよい。
【0022】
本発明の前記RNAレプリコンは、前記植物中での前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない。本発明のこの特色は、前記RNAレプリコンのいずれのリーキーな放出も、前記リーキーな放出の植物細胞に限局させる。前記植物において前記RNAレプリコンの細胞間移行を提供するタンパク質は、一般に「移行タンパク質」と称されている。植物ウイルスは通常、ウイルス又は前記ウイルスのゲノムRNAの細胞間拡散を許容する一つ又はそれより多くのタンパク質をコードする又は発現する。本発明のRNAレプリコンは、前記RNAレプリコンが前記植物において細胞間で有意に拡散するのを許容するであろう移行タンパク質を発現できるようにしてはならない。前記植物又は前記植物細胞は、誘導性プロモーターの制御下でない限り、前記植物において前記RNAレプリコンの細胞間移行提供するタンパク質を発現すべきではない。一つの態様において、前記RNAレプリコンは、天然のRNAウイルスの移行タンパク質の一部を含有することができるが、ただし、前記移行タンパク質の一部は、前記RNAレプリコンが前記植物において細胞間で有意に拡散するのを許容しない。
【0023】
本発明のプロセス及び植物の一つの態様において、前記植物は、前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードするヌクレオチド配列を含んでなる第二の異種ヌクレオチド配列を含んでなり、前記第二の異種ヌクレオチド配列は、前記RNAレプリコンの細胞間移行を可能にする前記タンパク質をコードする前記ヌクレオチド配列に機能可能なように連結された第二の誘導性プロモーターを含んでなる。前記タンパク質は、前記RNAレプリコンのポリメラーゼが誘導される同一の植物RNAウイルスに由来してもよいし、又は別のRNAウイルスに由来してもよい。
【0024】
RNAレプリコンをコードする前記ヌクレオチド配列及び細胞間移行を可能にするタンパク質をコードする前記ヌクレオチド配列は、一つの誘導性プロモーターからのリーキーな発現が移行タンパク質の発現及び前記RNAレプリコンの形成を導くことを避けるため、別の誘導性プロモーターの制御下にある。それ故、RNAレプリコンをコードする前記ヌクレオチド配列及び細胞間移行を可能にするタンパク質をコードする前記ヌクレオチド配列は、前記植物又は植物細胞において異なった発現カセット中に存在する。しかしながら、これら別々の誘導性プロモーターは、異なったタイプの誘導性プロモーターである必要はない。前記別々の誘導性プロモーターは、同一タイプであってもよいし、及び同一配列を有していてもよい。
【0025】
前記第一の、前記第二の前記誘導性プロモーター又はいずれかのさらなる誘導性プロモーターは、同一の又は異なった誘導シグナル又は剤により誘導性である。本発明に使用し得る誘導性プロモーターは以下に与えられている。一つの態様において、前記第一及び/又は前記第二の異種ヌクレオチド配列の誘導性プロモーターは化学的に誘導性である。別の態様において、前記第一及び/又は前記第二の異種ヌクレオチド配列の誘導性プロモーター(及び任意選択でさらなる異種ヌクレオチド配列)は、IPTG、エタノール、テトラサイクリン又はグルココルチコイドのような同一の誘導剤により誘導性である。
【0026】
本発明のプロセスの工程(b)において、工程(a)の前記植物又は植物細胞中で前記誘導性プロモーターが誘導され、それにより前記一つ又は一つより多くの目的とするタンパク質の発現が開始され、それにより前記植物又は植物細胞中で前記一つ又は一つより多くの目的とするタンパク質を産生する。誘導の様式は、誘導性プロモーターのタイプに依存する。もし誘導性プロモーターが化学的に誘導可能であれば、プロモーターを誘導することが可能な化学剤が前記植物又は前記植物細胞に提供される。もし前記第一の及び前記第二の異種ヌクレオチド配列について異なった誘導性プロモーターが使用されたならば、異なった化学剤を前記植物又は前記植物細胞に同時に適用することができる(例えば、異なった誘導剤の混合物として)。もし前記プロセスが液体培養液中の植物細胞で実施されるならば、該誘導剤を培養培地に加えることができる。もし前記プロセスが植物において実施されるならば、前記誘導剤の溶液又は懸濁液で前記植物をスプレーすることにより、前記植物に適用することができる。
【0027】
本発明のプロセスは、一つの目的とするタンパク質又は一つより多くの目的とするタンパク質を産生するために使用し得る。もし一つの目的とするタンパク質が産生されるべきならば、前記目的とするタンパク質をコードするヌクレオチド配列を、前記RNAレプリコンをコードする前記ヌクレオチド配列に含ませることができる。一つの態様において、前記目的とするタンパク質をコードするヌクレオチド配列は、前記RNAレプリコンが誘導される植物RNAウイルスの移行タンパク質遺伝子と置換することができる。もしくは、前記RNAレプリコンが誘導されるRNAウイルスのコートタンパク質遺伝子を、目的とするタンパク質をコードするヌクレオチド配列で置き換えることが可能である。もし二つの目的とするタンパク質が産生されるべきならば、移行タンパク質遺伝子及びコートタンパク質遺伝子の両方を目的とするタンパク質をコードするヌクレオチド配列で置き換える(完全に又は部分的に)ことが可能である。
【0028】
もし二つまたはそれより多くの目的とするタンパク質タンパク質が産生されるべきならば、前記植物又は植物細胞は、さらなるRNAレプリコン及び前記さらなるRNAレプリコンをコードする前記配列に機能可能なように連結された第三の誘導性プロモーターをコードするヌクレオチド配列を含んでなる第三の異種ヌクレオチド配列を含むことができる。前記さらなるRNAレプリコンは、好ましくは、前記植物中で前記RNAレプリコン又は前記さらなるRNAレプリコンの細胞間移行を提供するタンパク質をコードしていない。前記さらなるRNAレプリコンは次ぎに、一つ又はそれより多くのさらなる目的とするタンパク質をコードすることができる。もし前記植物が二つまたはそれより多くのRNAレプリコンをコードするならば、前記RNAレプリコンの細胞間移行を可能にするタンパク質は、前記さらなるRNAレプリコンの細胞間移行を可能にすることができる。もしくは、前記さらなるRNAレプリコンの細胞間移行は、その発現がさらなる誘導性プロモーターの制御下であることができる、前記さらなるRNAレプリコンの細胞間移行を可能にする別のタンパク質により可能にすることもできる。
【0029】
前記さらなるRNAレプリコンは、前記第一の異種ヌクレオチド配列によりコードされた前記RNAレプリコンによりコードされた前記ポリメラーゼにより複製可能であることができる。しかしながら一つの態様において、前記さらなるRNAレプリコンは、複製前記さらなるRNAレプリコンを複製するためのさらなるポリメラーゼをコードしており、前記さらなるポリメラーゼは、前記RNAレプリコンのものと異なっていることができる。
【0030】
一つの態様において、前記RNAレプリコン及び前記さらなるRNAレプリコンは、非競合RNAレプリコンである。非競合RNAレプリコン又は非競合ウイルスベクターからのタンパク質産生は、WO 2006/79546 (PCT/EP2006/000721) に記載されている。
【0031】
前記一つ又は一つより多くの目的とするタンパク質は、前記植物又は植物細胞中での産生後、望まれない細胞成分から精製することができる。植物又は植物細胞からタンパク質を精製する方法は当該技術分野で公知である。一つの方法において、目的とするタンパク質は、WO 03/020938 に記載されているように、植物アポプラストに方向付けることができる。
【0032】
本発明は原則として、感染性RNAウイルスが存在する及びウイルスベクターシステムが確立されたいずれの植物にも応用することができる。一つの態様において、双子葉植物が本発明を実施するために使用される。別の態様において、ナス科植物が使用される。好ましい植物は、ニコチニア・ベンサミアナ及びニコチニア・タバクムのようなニコチニア種であり;ニコチニア種以外の好ましい植物種は、ペチュニア・ヒブリダ、ブラシカ・カンペストリス、B.ユンセア、クレス、アルグラ、マスタード、イチゴほうれん草(Strawberry spinach)、ケノポディウム・カピターツム(Chenopodium capitatum)、アルファルファ、レタス、ヒマワリ、及びキュウリである。
【0033】
本発明の産生プロセスは、本明細書に記載した植物の細胞において実行することもできる。前記細胞は、葉のような植物組織の一部であってもよく、又は前記細胞は、懸濁培養のような細胞培養液中に存在していてもよい。
【0034】
適したRNAレプリコンは以下に与えたRNAウイルスのリストに由来することができる。本発明は、一要素植物RNAウイルスに適用し得る。本発明が基づくことができる最も好ましい植物RNAウイルスはトバモウイルス、特にタバコモザイクウイルス及びジャガイモウイルスXのようなポテクスウイルスである。タバコモザイクウイルスの場合、それは一般的には、発現されるべき前記目的とするタンパク質のORFにより置き換えられている移行タンパク質ORFであろう。コートタンパク質ORFも、目的とするタンパク質のORFにより除去される又は置き換えられることができる。
【0035】
本発明の主要な適用は、植物、植物の葉又は植物組織又は細胞培養における目的とするタンパク質の産生である。もし本発明のプロセスが植物において実施されるならば、ニコチニア種植物のような、ヒト又は動物の食物連鎖に入り込まない植物が好ましい。ヒト又は動物の食物連鎖に入り込まない植物は野外で耕作することが可能であり、及び前記RNAレプリコン放出の誘導後、特定の期間内で、植物組織中の一つ又は一つより多くの目的とするタンパク質の発現レベルがピークに達した時に採取される。好ましくは、全植物又は植物部分は閉ざされた環境に限局すべきである;例えば、発現の望まれるレベルを提供するために必要なインキュベーション期間のためのガラス温室又は設計されたチャンバー。
【0036】
本発明の生産プロセスの効率は、植物発現システムにおいて新たな局面が達成されるほどである。本発明で達成可能な発現レベルは、下流のプロセシング(目的とするタンパク質の分離及び精製を含んで)のための費用が、他の大規模発現システムとの競争力がある本発明のプロセスを作製するために十分低いほどである。ウイルスベクターで安定的に形質転換した植物を使用する従来技術の発現システムにおいては、これらのシステムの漏れやすさが、産生されるべきレプリコンの誘導されない状態をさえ許容するので、発現レベルは低く、それ故、収率を損なうPTGSの機構の引き金が引かれる。加えて、細胞間移行が可能であるRNAレプリコンは、植物染色体DNA内に安定的に取り込まれた前記レプリコンを運んでいる安定的に形質転換された植物細胞の産生を損なう。驚くべきことに、発明者は、細胞間移行が可能なRNAレプリコンを運んでいるトランスジェニック植物よりも、細胞間移行が不可能なRNAレプリコンをコードするベクターを含有するトランスジェニック植物を得ることがより容易であることを見いだした。この現象が、細胞間移行が不可能なウイルスベクターを含有するトランスジェニック植物中の高発現レベルに寄与することも非常にありそうである。本発明は、大規模で使用することが可能である、最初の高収率誘導性植物発現システムを提供する。
【0037】
発明の詳細な説明
この発明は、一要素RNAウイルスから誘導することができるRNAレプリコンを使用する、目的とするタンパク質の高収率、大規模生産のための誘導性発現システムを記述している。前記RNAレプリコンは、植物中で一つ又は一つより多くの目的とするタンパク質を発現できる。本発明のプロセスは、バイオセーフティー特色を有し、RNAレプリコン放出の厳格な制御を提供し、短距離(細胞間)移行及び、場合により、遠隔距離移行についてのその無能さのため、前記RNAレプリコンが他の植物に感染することを防止している。
【0038】
我々は驚くべきことに、前記RNAウイルスベクターからの細胞間移行機能の除去が、植物又は植物細胞の染色体DNA中に前記RNAレプリコンをコードする、安定に形質転換された植物の選択及び再生を容易にすることを発見した。本発明の基本原理は図1に示されている。細胞間移行が可能なウイルスベクターの場合、誘導性システムの漏れやすさは、サイトゾル内へのウイルスベクター放出を起こし、及びさらには隣接細胞に伝播する(図1A)。最終的に、このことは宿主植物内でのウイルスベクターの制御されない伝播を導き、前記植物の生長及び発達を損なう。こうした問題を克服している、及び転写後遺伝子サイレンシング(PTGS)のメカニズムによりウイルスベクター複製の制御を行っている宿主植物の場合、PTGSは、宿主植物中での目的とするタンパク質の発現レベルに対して負の効果を有しているであろう。本発明において、誘導性システムの漏れやすさは、本発明のRNAレプリコンは細胞間移行が欠損しており、それ故PTGSは実際に無視できるので、こうした劇的効果を有していない。前記RNAレプリコンは本質的にその細胞に限局されており、それはプロモーターの漏れやすさのために放出されており(図1B)、それ故、目的とするタンパク質の望まれない発現に対する制御を改良し、及び該システムの産生性に対するPTGSの潜在的負の効果を減少させる。
【0039】
本発明において、誘導性及び組織特異的プロモーターを、植物又は植物細胞において、目的とするタンパク質産生の引き金を引くために使用し得る。誘導性プロモーターは、それらの誘導条件に従って二つのカテゴリーに分割し得る;非生物的因子(温度、光、化学物質)により誘導可能なもの、及び生物的因子、例えば、病原体又は有害生物攻撃により誘導可能なもの。第一のカテゴリーの例には、限定されるわけではないが、熱誘導性(US 05187287) 及び低温誘導性(US05847102) プロモーター、銅誘導性システム(Mett et al., 1993, Proc. Natl. Acad. Sci., 90, 4567-4571) 、ステロイド誘導性システム(Aoyama & Chua, 1997, Plant J., 11, 605-612; McNellis et al., 1998, Plant J.,14, 247-257; US06063985) 、エタノール誘導性システム(Caddick et al., 1997, Nature Biotech., 16, 177-180; WO09321334; WO0109357; WO02064802) 、イソプロピル ベータ−D−チオガラクトピラノシド(IPTG)誘導性システム(Wilde et al., 1992, EMBO J., 11:1251-1259) 、及びテトラサイクリン誘導性システム(Weinmann et al., 1994, Plant J., 5, 559-569) が含まれる。植物についての化学的誘導性システムの領域における最近の開発の一つは、グルココルチコイド デキサメタゾンによりスイッチを入れることができ、及びテトラサイクリンによりスイッチを切ることができるキメラプロモーターである(Bohner et al., 1999, Plant J., 19, 87-95) 。化学的誘導性システムは本発明の実施に最も適している。化学的誘導性システムの総説は:Zuo & Chua, (2000, Current Opin. Biotechnol., 11, 146-151)、及びMoore et al., (2006, Plant J., 45: 651-683) を参照されたい。リプレッサー又はアクチベーターのような、選択された誘導性システムの機能性に必要とされるいずれのタンパク質も、誘導性システムを機能的にするためには前記植物又は前記植物細胞中で発現されなければならないことは、当業者には明白であろう(実施例参照)。
【0040】
本発明の一つの態様において、RNAレプリコンの放出及び目的とするタンパク質の産生を制御するためにIPTG誘導性システムを使用する。例示の構築物の設計は実施例1に記載されている。lacIリプレッサーをコードする細菌遺伝子を、強い構成的35Sプロモーターの制御下にクローン化した。二重のlacオペレーター配列を、本発明のRNAレプリコンの転写を駆動するシロイヌナズナアクチン2プロモーター内に挿入した。このシステムは、実施例2に記載されている一過性発現アッセイを使用して試験した。前記RNAレプリコンをコードする異種ヌクレオチド配列を、lacIリプレッサーを運んでいる構築物と一緒に植物内に浸潤させた場合、多分RNAレプリコンの形成がリプレッサーの合成に先行されるため、このシステムは何のRNAレプリコン放出の阻害も示さなかった(図3、右下)。実際、レプリコン形成を遅延させるために、RNAレプリコンのDNA前駆体への組立のために部位特異的リコンビナーゼを必要とするプロベクターシステム(Marillonnet et al., 2004, Proc. Natl. Acad. Sci. USA, 101:6852-6857) が使用された場合、レプリコン放出の抑制は明白であった(図3、右上)。lacIリプレッサーを提供するベクターを欠く場合(図3、左上)、プロベクターシステムのケースにおいてRNAレプリコン形成の抑制が全くないことが観察された。
【0041】
本発明の別の態様において(実施例3)、lacオペレーター配列を有するプロモーターの制御下、一過性に送達された第一の異種ヌクレオチド配列からのRNAレプリコンの放出を抑制するためのlacIリプレッサーを提供する構築物で安定的に形質転換されたトランスジェニック植物の能力を試験した。図4Aから、lacオペレーター配列含有構築物pICH17171のアグロ浸潤は、こうした配列を欠く構築物pICH16141での対照実験と比較して、効率的なRNAレプリコン放出を与えなかったことは明白である。同一構築物及び1mM IPTGでの前記トランスジェニック植物の同時浸潤は、pICH17171のケースにおいてはRNAレプリコン放出の誘導を導いたが(図4B、右パネル)、一方、IPTGなしの対照実験においては、RNAレプリコン放出は観察されなかった(図4B、左パネル)。細胞間移行が可能なRNAレプリコンを提供する構築物でのlacIリプレッサー含有植物の安定的再形質転換は、誘導されていない状態で重度のバックグラウンド発現を有する二重形質転換体を生み出した(図5)。LacI/lacOシステムについて得られたデータを要約すると、無視できるバックグラウンド発現でのRNAレプリコン放出に対する最良の制御が、細胞間移行が欠損したRNAレプリコンのケースで得られることが明白である(pICH17171、図4A;図4B、左パネル)。機能性MPを含有する構築物は、誘導されていない状態で重度のバックグラウンド発現を示した(pICH17424、図3、右上;図5、対照)。また、前記構築物を有する一次形質転換体は、最後には導入遺伝子発現を停止させ、それ故、目的とするタンパク質の高収率発現について役に立たなくなる。
【0042】
本発明の別の態様において、エタノール誘導性システムが、トランスジェニック植物におけるRNAレプリコン放出を制御するために使用された。構築物の設計は実施例5に記載されており、構築物の略図が図2Bに示されている。前記構築物の一過性発現実験の結果は、図6に示されている。alcRアクチベーターを有する構築物が同時浸潤されたところの実験にのみGFP発現を観察することができたので、エタノール誘導性システムはRNAレプリコン放出に対して厳格な制御を提供することが明白である。実際に、alcR構築物及び/又は化学的誘導剤エタノールがない場合には、バックグラウンド発現は観察されてなかった(無視できる発現が、alcR構築物の存在下で観察された)。4%エタノールの存在下、alcR構築物を有するRNAレプリコン(pICH18951)を提供する構築物が遺伝子導入されたN.ベンサミアナのアグロ浸潤は、強いGFP発現(図8)により報告されるRNAレプリコン形成を示した。驚くべきことに、エタノール誘導性システムにおいて、機能性MPを含有するウイルス構築物では一次形質転換体(RNAレプリコン放出に対する制御の漏れやすさと同等ではない)が得られなかった(実施例5)。このことは、RNAレプリコンの細胞間移行による植物カルス/細胞培養におけるシステムの漏れやすさにより説明し得る(Roberts et al., 2005, Plant Physiol., 138:1259-1267) 。この説明は、もしRNAレプリコンが使用された植物宿主中で細胞間移行のためのタンパク質をコードしていないならば、エタノール誘導性プロモーターの制御下でRNAレプリコンを提供する一次形質転換体が困難なく発生することができるという驚きの発見により支持される。我々は、RNAレプリコンを提供するベクターを有するトランスジェニック植物の交配子孫、及びalcR含有トランスジェニック体が、エタノール処理後、GFPの強いユビキタス発現を示すことを明らかにした(図9及び10)。この発明の別の態様において、我々は、我々のシステムが、GFPのようなレポーター遺伝子以外のタンパク質でも効率的に働くことを明らかにした。アプロチニンの発現のための構築物が図11に示されている。エタノール処理後、N.ベンサミアナ植物のF1子孫における組換えアプロチニン産生の分析は、クーマシー染色ゲル上の一つの主タンパク質バンドとして検出可能な高発現レベルを明らかにした。
【0043】
実施例において、我々はTMVに基づいたRNAレプリコンを使用した。しかしながら、異なった分類学的グループに属する多くの異なったウイルスを、本発明に従ったRNAウイルスに基づいたベクターの構築のために使用し得る。目、科及び属の名称は、ICTVにより承認されていればイタリック体である。引用符で囲まれている(及びイタリック体ではない)Taxa名は、この分類群がICTV国際承認名を有していないことを示している。種(その土地の言葉)名は通常書体で与えられている。属又は科への正式な帰属のないウイルスは示されている:
RNAウイルス:
ssRNAウイルス:科:ブロモウイルス科、属:アルファモウイルス属、基準種:アルファルファモザイクウイルス、属:イラルウイルス属、基準種:タバコ条斑病ウイルス、属:ブロモウイルス属、基準種:ブロムモザイクウイルス、属:ククモウイルス属、基準種:キュウリモザイクウイルス;
科:クロステロウイルス科、属:クロステロウイルス属、基準種:ビート萎黄ウイルス、属:クリニウイルス属、基準種:レタス伝染性黄斑ウイルス、科:コモウイルス科、属:コモウイルス属、基準種:ササゲモザイクウイルス、属:ファバウイルス属、基準種:ソラマメウイルトウイルス1、属:ネポウイルス属、基準種:タバコ輪点ウイルス;
科:ポティウイルス科、属:ポティウイルス属、基準種:ジャガイモYウイルス、属:ライモウイルス属、基準種:ドクムギモザイクウイルス、属:バイモウイルス属、基準種:大麦縞萎縮ウイルス;
科:セキウイルス科 、属:セキウイルス属、基準種:パースニップ黄色斑点ウイルス、属:ワイカウイルス属、基準種:イネ矮化ウイルス;科:トンブスウイルス科、属:カーモウイルス属、基準種:カーネーション斑紋ウイルス、属:ダイアントウイルス属、基準種:カーネーション輪点ウイルス、属:マクロモウイルス属、基準種:メイズクロロティック斑紋ウイルス、属:ネクロウイルス属、基準種:タバコ壊死ウイルス、属:トンブスウイルス属、基準種:トマトブッシースタントウイルス、ssRNAウイルスの割り当てられていない属、属:カピロウイルス、基準種:リンゴステムグルービング(stem grooving )ウイルス;
属:カーラウイルス属、基準種:カーネーション潜在ウイルス;属:エナモウイルス属、基準種:エンドウマメエネーション(enation )モザイクウイルス
属:フロウイルス属、基準種:ムギ類萎縮ウイルス、属:ホルデイウイルス属、基準種:オオムギ斑葉モザイクウイルス、属:イダエオウイルス属、基準種:ラズベリーブッシュドワーフウイルス(raspberry bushy dwarf virus);
属:ルテオウイルス属、基準種:オオムギ黄萎ウイルス;属:マラフィウイルス属、基準種:トウモロコシラヨドフィノ(rayodo fino )ウイルス;属:ポテクスウイルス属、基準種:ジャガイモウイルスX;属:ソベモウイルス属、基準種:インゲンマメ南部モザイクウイルス、属:テヌイウイルス属、基準種:イネ縞葉枯ウイルス、
属:トバモウイルス属、基準種:タバコモザイクウイルス、
属:トブラウイルス属、基準種:タバコ茎えそウイルス、
属:トリコウイルス属、基準種:リンゴクロロティックリーフスポット(chlorotic leaf spot )ウイルス;属:ティモウイルス属、基準種:ハクサイ黄化モザイクウイルス;属:アンブラウイルス属、基準種:ニンジン斑紋ウイルス;マイナスssRNAウイルス:目:モノネガウイルス目、科:ラブドウイルス科、属:サイトラブドウイルス属、基準種:レタス壊死性黄変病ウイルス、属:ヌクレオラブドウイルス属、基準種:ジャガイモ黄萎ウイルス;
マイナスssRNAウイルス:科:ブニヤウイルス科、属:トスポウイルス属、基準種:トマト黄化壊疽ウイルス;
dsRNAウイルス:科:パルティティウイルス科、属:アルファクリプトウイルス属、基準種:シロツメクサ潜在ウイルス1、属:ベータクリプトウイルス属、基準種:シロツメクサ潜在ウイルス2、科:レオウイルス科、属:フィジウイルス属、基準種:フィジー病ウイルス、属:ファイトレオウイルス属、基準種:創傷腫瘍ウイルス、属:イネウイルス属、基準種:イネラギッドスタント(ragged stunt )ウイルス;
割り当てられていないウイルス:ゲノムssDNA:種:バナナバンチートップ(bunchy top )ウイルス、種:ココナツ葉壊変ウイルス、種:サブテレーニアンクローバ スタント(subterranean clover stunt )ウイルス、
ゲノム:dsDNA、種:キュウリ葉脈イエローイング(yellowing )ウイルス;ゲノム:dsRNA、種:タバコ矮化ウイルス、
ゲノム:ssRNA、種:ニンニクウイルスA、B、C、D、種:ブドウつるフレックウイルス、種:トウモロコシホワイトライン(white line )モザイクウイルス、種:オリーブ潜在ウイルス2、種:ウルミアメロンウイルス、種:ペラルゴニウムゾナートスポット(zonate spot )ウイルス;
サテライト及びウイロイド:サテライト:ssRNAサテライトウイルス:サブグループ2サテライトウイルス、基準種:タバコネクローシスサテライト、
サテライトRNA、サブグループ2BタイプmRNAサテライト、サブグループ3Cタイプ直鎖RNAサテライト、サブグループ4Dタイプ環状RNAサテライト、
ウイロイド、基準種:ポテトスピンドルチュバーウィロイド(potato spindletuber viroid)。
【0044】
異なったRNAウイルスは、細胞間又は短距離移行のための一つ又はそれより多くの ウイルスタンパク質を有する。例えば、TMVの場合、一つのタンパク質(MP)が必要とされ;三要素ブロムモザイクウイルス(BMV)は、二つのタンパク質−3a及びCPを必要とする。一要素RNAウイルス、ジャガイモウイルスX(PVX)は、細胞間移行に関与する四つのタンパク質を有する:トリプルジーンブロック(triple gene block )(TGB)及びコートタンパク質(CP)によりコードされるタンパク質。しかしながら、細胞間移行に必要とされる二つまたはそれより多くのタンパク質の一つの欠損が、ウイルスベクターの効率的短距離移行を遮断するのに十分である。植物ウイルス移行タンパク質についてのさらなる詳細は、WJ Lucas (2006, Virology, 344:169-184) の最近の総説を参照されたい。
【0045】
本発明において、前記第一の異種ヌクレオチド配列は、前記RNAレプリコンをコードする配列セグメントを有する。もしくは、前記第一の異種ヌクレオチド配列は、一緒になって前記RNAレプリコンをコードする、一つより多くの配列セグメントを有することができ、即ち、前記RNAレプリコンは一つの連続したDNAによりコードされていない。代わりに、前記RNAレプリコンは二つまたはそれより多くの配列セグメントにより非連続的にコードされており、それにより前記セグメントはお互いに隣接して存在することができる。前記RNAレプリコンの形成は、例えば、組換えによる前記セグメントの再配列を必要とするであろう。前記組換えのためのリコンビナーゼは、工学処理された植物宿主により提供することができ、それ故、前記ウイルスレプリコンの誘導性発現を前記リコンビナーゼを発現することが可能な植物宿主に限局している。例としては、一つの配列セグメントは、前記RNAレプリコンの前記ポリメラーゼの一部をコードすることができ、及び前記ポリメラーゼの別の一部をコードする別の配列セグメントは、前記第一の異種ヌクレオチド配列中、第一の配列セグメントに関してフリップした(flipped)配向で存在することができる。フリップした部分に、組み換え部位が隣接することができる(WO2004/108934 参照)。この状況において、第一の異種ヌクレオチド配列又はそれらの配列セグメントの転写体は、機能性ポリメラーゼが転写体から翻訳できないので、RNAレプリコンではないであろう。組み換え部位を認識する部位特異的リコンビナーゼを提供することは、レプリコンが連続的にコードされるように前記セグメントの一つをフリップさせることを可能にする。この態様において、該リコンビナーゼは、RNAレプリコン形成及び誘導された状態の目的とする配列発現のスイッチを入れるためのスイッチとして機能することができ、高い生物学的安全性に寄与している。好ましくは、前記リコンビナーゼは誘導性プロモーターの制御下である。もしリコンビナーゼが本発明のプロセスのスイッチを入れるために使用されるならば、前記リコンビナーゼは前記植物又は植物の葉に一過性に提供することができ、それにより前記提供することが、前記一つ又は一つより多くの目的とするタンパク質を発現するためのスイッチとして働くことができる。好ましくは、こうしたリコンビナーゼは植物細胞中で安定にコードすることができ、構成的又は誘導性プロモーターの制御下でリコンビナーゼを発現する。前記プロモーターを誘導することによりリコンビナーゼ発現を誘導することは、次ぎに前記目的とする配列の発現を起こすことができる。一つの態様において、該リコンビナーゼは前記第一の異種ヌクレオチド配列によりコードされているであろうし、及び該リコンビナーゼの発現は前記第一の異種ヌクレオチド配列の誘導性プロモーターの制御下にあるであろう。
【0046】
もしくは、両方のセグメントは、異なった植物染色体に安定的に取り込まれた異なったT−DNA上に存在していてもよい。RNAレプリコンの形成は、両方のセグメントの転写及び両方の転写体のトランススプライシングを、前記RNAレプリコンを組み立てるために必要とするであろう。この態様は、WO02/097080 に記載されているように、子孫植物又は細胞中で一緒になって前記RNAレプリコンをコードするセグメントを迅速に分離するために使用することができ、それ故、システムの生物学的安全に寄与している。
【0047】
実施例において、植物ウイルスに由来する一つのタイプのRNAレプリコンをコードするトランスジェニック植物を記載する。しかしながら、本発明のプロセスにおいて、二つまたはそれより多くの異なったRNAレプリコン(特に二つまたはそれより多くの 異なった一要素RNAレプリコン)をトランスジェニック植物又は植物細胞において使用することができ、こうした異なったRNAレプリコンは、好ましくは異なった植物ウイルスから誘導される。前記異なったRNAレプリコンを誘導することができるこうした異なった植物ウイルスは、好ましくは相乗的又は非競合ウイルスである。本明細書において「相乗的」及び「非競合」は同意語として使用される。相乗的ウイルスは同一植物細胞中で同時に存在し、そして効率的に増幅することが可能である。同様に、相乗的RNAウイルスに由来するRNAレプリコンは同一植物細胞中で同時に存在し、そして効率的に増幅することが可能である。RNAレプリコンのこうした相乗的組み合わせの例は、一つのRNAレプリコンがTMVに由来し、及び他のRNAレプリコンがPVXに由来するRNAレプリコンの組み合わせである。こうした相乗的RNAレプリコンは、同一又は異なった誘導性プロモーターを利用する異なった発現カセットから放出し得る。相乗的RNAレプリコンは、同一植物細胞において、モノクローナル抗体の重鎖及び軽鎖のような、目的とする二つまたはそれより多くのタンパク質又はタンパク質サブユニットの発現のために使用することができる。異なった(非競合)ウイルスベクターを使用し、同一植物又は同一植物細胞中で二つまたはそれより多くの目的とするタンパク質を発現するプロセスは、その全体が本明細書において援用されるWO 2006/79546 (PCT/EP2006/000721)に記載されている。
【0048】
実施例において、植物細胞でのアグロバクテリウム仲介T−DNA送達を主に使用し、前記T−DNAは前記第一及び/又は前記第二の異種ヌクレオチド配列を含有する。微粒子銃、エレクトロポレーション又はプロトプラストのPEG仲介形質転換による細胞内への異種ヌクレオチド配列の直接導入のような多様な方法を、植物細胞内へのベクターの送達のために使用することができる。アグロバクテリウム仲介植物形質転換が好ましい。それ故、アグロバクテリウム(US 5,591,616; US 4,940,838; US 5,464,763)、粒子及び微粒子銃(US 05100792; EP 00444882B1; EP 00434616B1)により運ばれるTi−プラスミドベクターによるような多様な技術により、異種ヌクレオチド配列を植物細胞内に形質転換することができる。原則として、他の植物形質転換法、例えば、マイクロインジェクション(WO 09209696; WO 09400583A1; EP 175966B1)、エレクトロポレーション(EP00564595B1; EP00290395B1; WO 08706614A1)なども使用し得る。形質転換法の選択は、中でも、形質転換されるべき植物種に依存する。例えば、微粒子銃は単子葉植物形質転換のために好ましく、一方、双子葉植物にはアグロバクテリウム仲介形質転換が一般的により良好な結果を与える。
【0049】
本発明は好ましくは、より高等な多細胞植物で実施される。本発明での使用に好まし
い植物には、農学的及び園芸的に重要な種に与えられた優先度を有するいずれかの植物種が含まれる。本発明で用いる普通の作物には、アルファルファ、オオムギ、マメ、アブラナ、ササゲ、綿、トウモロコシ、クローバ、ハス、レンズマメ、ルピナス、キビ(millet )、カラスムギ、エンドウマメ、ピーナッツ、コメ、ライムギ、スイートクローバー、ヒマワリ、スイートピー、ダイズ、モロコシ、ライ小麦、クズイモ、ビロードマメ、カラスノエンドウ、コムギ、フジ及びナッツ植物が含まれる。本発明を実施するための好ましい植物種には、限定されるわけではないが:イネ科、キク科、ナス科及びバラ科の代表種が含まれる。加えて、本発明に使用するための好ましい種、ならびに上に特定された種には属:シロイヌナズナ属、コヌカグサ属、ネギ属、キンギョソウ属、セロリ属、ラッカセイ属、アスパラガス属、アトローパ属、アベナ属、バンブーサ属、アブラナ属、スズメノチャヒキ属、ブロワリア属、ツバキ属、カンナビス属、カプシクム属、ヒヨコマメ属、アカザ属、キクニガナ属、シトラス属、コーヒー属、ジュズダマ属、キュウリ属、カボチャ属、ギョウギシバ属、カモガヤ属、チョウセンアサガオ属、ニンジン属、ジギタリス属、ヤマノイモ属、アブラヤシ属、オヒシバ属、ウシノケグサ属、オランダイチゴ属、フウロソウ属、グリシン(Glycine)、ヒマワリ属、ワスレグサ属、ヘベア属、オオムギ属、ヒヨス属、サツマイモ属、アキノノゲシ属、レンズマメ属、ユリ属、アマ属、ドクムギ属、ハス属、リコペリシコン属、マジョラム(Majorana)、リンゴ属、マンゴー属、マニホット属、ウマゴヤシ属、ネメシア属、タバコ属、オノブリキス属 、イネ属、キビ属、ペラルゴニウム属、ペニセタム属、ペチュニア属 、エンドウ属、インゲンマメ属、アワガエリ属、イチゴツナギ属、サクラ属、キンポウゲ属、ダイコン属、スグリ属、トウゴマ属、キイチゴ属、サトウキビ属、サルメンバナ属、ライムギ属、セネキオ属、エノコログサ属、シロガラシ属、ナス属、モロコシ属、イヌシバ属、カカオ属、シャジクソウ属、レイリョウコウ属、コムギ属、ソラマメ属、ササゲ属、ブドウ属、トウモロコシ属及びオリレア属(Olyreae) 、ファルス亜科及び多くの他のものが含まれる。
【0050】
本発明の一つの態様において、TMVに由来するRNAレプリコンがタバコ属植物に使用される。別の態様において、PVXに由来するRNAレプリコンがタバコ属植物に使用される。
【0051】
本発明を使用し、センス又はアンチセンス配向で発現し得る、目的とするタンパク質又はそれらの断片には:デンプン修飾酵素(デンプンシンターゼ、デンプンリン酸化酵素、脱分枝酵素、デンプン分枝酵素、デンプン分枝酵素II、顆粒結合性デンプンシンターゼ)、スクロースリン酸シンターゼ、スクロースホスホリラーゼ、ポリガラクツロナーゼ、ポリフルクタンスクラーゼ、ADPグルコースピロホスホリラーゼ、シクロデキストリングリコシルトランスフェラーゼ、フルクトシルトランスフェラーゼ、グリコーゲンシンターゼ、ペクチンエステラーゼ、アプロチニン、アビジン、細菌レバンスクラーゼ、大腸菌glgAタンパク質、MAPK4及び相同分子種、窒素同化/代謝酵素、グルタミンシンターゼ、植物オスモチン、2Sアルブミン、タウマチン、部位特異的リコンビナーゼ/インテグラーゼ(FLP、Cre、Rリコンビナーゼ、Int、SSVIインテグラーゼR、インテグラーゼphiC31、又はそれらの活性フラグメント又は変異体)、イソペンテニルトランスフェラーゼ、ScaM5(ダイズカルモジュリン)、鞘翅目型毒素又は殺虫活性断片、ユビキチン結合酵素(E2)融合タンパク質、脂質を代謝する酵素、アミノ酸、糖、核酸及びポリサッカリド、スーパーオキシドジスムターゼ、プロテアーゼの不活性プロ酵素型、植物タンパク質毒素、繊維産生植物中の形質変形繊維、バチルス・チューリンゲンシスからの鞘翅目活性毒素(Bt2トキシン、殺虫性結晶タンパク(ICP)、CryICトキシン、デルタエンドトキシン、ポリペプチドトキシン、プロトキシンなど)、昆虫特異的トキシンAaIT、セルロース分解酵素、アシドサーマス・セルロリティカスからのE1セルラーゼ、リグニン修飾酵素、シンナモイルアルコールデヒドロゲナーゼ、トレハロース−6−リン酸シンターゼ、サイトカイニン代謝経路の酵素、HMG−CoAレダクターゼ、大腸菌無機ピロホスファターゼ、種子貯蔵タンパク質、エルウィニア・ヘルビコーラリコペンシンターゼ、ACCオキシダーゼ、pTOM36コードタンパク質、フィターゼ、ケトヒドロラーゼ、アセトアセチルCoAレダクターゼ、PHB(ポリヒドロキシブタノエート)シンターゼ、アシルキャリアータンパク質、ナピン、EA9、非高等植物フィトエンシンターゼ、pTOM5コードタンパク質、ETR(エチレンレセプター)、色素体ピルビン酸リン酸ジキナーゼ、線虫誘導性膜貫通ポアタンパク質、植物細胞の光合成又は色素体機能を増強する形質、スチルベンシンターゼ、フェノールのヒドロキシル化を可能にする酵素、カテコールジオキシゲナーゼ、カテコール 2,3−ジオキシゲナーゼ、クロロムコネートシクロイソメラーゼ、アントラニル酸シンターゼ、アブラナ属AGL15タンパク質、フルクトース 1,6−ジホスファターゼ(FBPase)、AMV RNA3、PVYレプリカーゼ、PLRVレプリカーゼ、ポティウイルスコートタンパク質、CMVコートタンパク質、TMVコートタンパク質、ルテオウイルスレプリカーゼ、MDMVメッセンジャーRNA、突然変異体ジェミニウイルスレプリカーゼ、ウンベルラリア・カルフォルニアC12:0選択性アシル−ACPチオエステラーゼ、植物C10又はC12:0選択性アシル−ACPチオエステラーゼ、C14:0選択性アシル−ACPチオエステラーゼ(luxD)、植物シンターゼ因子A、植物シンターゼ因子B、6−デサチュラーゼ、植物細胞中でペルオキシソーム−脂肪酸の酸化に酵素活性を有するタンパク質、アシル−CoAオキシダーゼ、3−ケトアシル−CoAチオラーゼ、リパーゼ、トウモロコシアセチル−CoA−カルボキシラーゼ、5−エノールピルビルシキミ酸−3−リン酸シンターゼ(EPSP)、ホスフィノチリシンアセチルトランスフェラーゼ(BAR、PAT)、CP4タンパク質、ACCデアミナーゼ、リボザイム、翻訳後切断部位を有するタンパク質、Gal4転写アクチベーターのDNA結合ドメイン及び転写活性化ドメインから成るタンパク質融合物、脂質相内へ融合タンパク質を標的化できるオレオシンタンパク質と目的とするタンパク質の翻訳融合物、スルホンアミド抵抗性を与えているDHPS遺伝子、細菌ニトリラーゼ、2,4−Dモノオキシゲナーゼ、アセト乳酸シンターゼ又はアセトヒドロキシ酸シンターゼ(ALS、AHAS)、ポリガラクツロナーゼ、細菌ニトリラーゼ、色素体の内部エンベロープ膜に存在する成熟リン酸輸送体タンパク質のアミノ末端疎水性領域と前記膜内へ標的化されるべき目的とするタンパク質との融合物が含まれる。
【0052】
いずれのヒト又は動物タンパク質も本発明のシステムを使用して発現し得る。例えば、こうした目的とするタンパク質の例には、中でも、以下のタンパク質(医薬タンパク質)が含まれる:免疫応答タンパク質(モノクローナル抗体、単鎖抗体、T細胞レセプターなど)、抗原、コロニー刺激因子、レラキシン、ポリペプチドホルモン、サイトカイン及びそれらのレセプター、インターフェロン、増殖因子及び凝固因子、酵素的に活性なリソソーム酵素、線維素溶解ポリペプチド、血液凝固因子、トリプシノーゲン、1−アンチトリプシン(AAT)、ならびに機能保存的タンパク質様融合物、上記タンパク質の突然変異体バージョン及び合成誘導体。
【0053】
本特許出願により優先権が主張される、2006年5月29日に出願された欧州特許出願番号06011002及び2006年6月2日に出願された米国仮特許出願60/810,398の開示は、それらの全体が本明細書において援用される。
【実施例】
【0054】
実施例1
IPTG誘導性lacシステム:構築物設計
lacリプレッサー (lacI, Acc. J01636)は、プライマーlacIpr1(配列番号:1)(5’-gat cca tgg aac cag taa cgt tat ac -3’)及びlacIpr2(配列番号:2)(5’-tc tgg atc ctc act gcc cgc ttt cca gtc g-3’)を使用するPCRにより増幅し、NcoI−BamHI断片として標準バイナリーベクターpICBV1内にクローン化すると、構築物pICH17155(図2A)を得た。核移行シグナル(NLS)は、lacIpr1の代わりにプライマーlacIpr5(配列番号:3)(5’-cgc cat ggg ccc taa gaa gaa gag gaa ggt tga acc agt aac gtt ata cga tgt c -3’)を使用することによりN末端に導入し、構築物pICH17401(図2A)を得た。この構築物は標準形質転換技術(Horsh et al., 1985, Science, 227, 1229-1231) を使用し、ニコチニア・タバクム及びN.ベンサミアナ植物内に安定的に形質転換した。
【0055】
合成lacオペレーター配列(配列番号:4)(aat tgt gag cgc tca caa tt)は、TATAボックス及びいくつかのウイルスベクターのアクチン2−プロモーター(An et al., 1996, Plant J., 10:107-121) の転写開始点間に導入した。このことは、プライマーA:brb4nosph(配列番号:5)(5’- gga acc ctg tgg ttg gca cat -3’)及びlacOact2pr2(配列番号:6)(5’- cga att gtg agc gct cac aat tta tat agg cgg gtt tat ctc -3’)、及びプライマーB:lacOactpr1(配列番号:7)(5’- taa att gtg agc gct cac aat tcg ctt tga agt ttt agt ttt att g -3’)及びrdrppr4(配列番号:8)(5’- ttt ctg cag gaa atg aaa ggc cgc gaa aca ag -3’)で作製した二つの重複PCR生成物を組み合わせることにより行われた。生じた生成物をKpn1−SphI断片としてウイルスベクターpICH16141内にクローン化して、ベクターpICH17171を得た。追加のウイルスベクターは、都合のよい制限酵素を使用し、プロモーターのサブクローニングによりpICH17171から誘導した。これらのベクターは発現のために最適化し(Marillonnet et al., 2005, Nat Biotechnol., 23:718-723)、そして完全MP(pICH18867)又はMPの欠失(pICH17171)を含有する。加えて、一過性試験に有用であることが証明されている5’−プロベクター(Marillonnet et al., 2004, Proc Natl Acad Sci USA, 101:6852-6857)を構築した(pICH17424)(実施例2参照)。
【0056】
実施例2
一過性システムにおける抑制効率の試験
一過性発現実験は、試験下で異なった構築物を宿しているアグロバクテリア株の混合物を使用して実施した。完全に組み立てられたウイルスベクターを使用して間は、一過性アッセイにおいて何の抑制も観察できなかった。多分、リプレッサーの翻訳に先立って、該構築物はウイルスRNAレプリコンにすでに転写されていた。それ故、部位特異的リコンビナーゼ(Marillonnet et al.,2004, Proc Natl Acad Sci USA, 101:6852-6857)の活性により、ウイルス性RNAレプリコンのDNA前駆体へ植物中で組み立てられる、ウイルスプロベクターpICH17424(図2A)、及びpICH6892(Marillonnet et al., 2004, Proc Natl Acad Sci U S A, 101:6852-6857)を使用した。この追加の工程はウイルスベクターの組立を遅延させるはずであり、及び翻訳される及びウイルス構築物中のオペレーター配列に結合される十分な時間をリプレッサーに提供する。実際、このアプローチを使用すると、ウイルス増幅の強い抑制が観察できた(図3)。
【0057】
実施例3
安定的に形質転換された植物におけるLacIリプレッサー活性及びIPTGによる誘導
リプレッサー構築物pICH17401(図2A)を、標準形質転換技術(Horsh et al., 1985, Science, 227, 1229-1231)を使用し、N.タバクム及びN.ベンサミアナ中に安定的に形質転換した。リプレッサー活性は、lacO含有ウイルス構築物での形質転換された植物のアグロ浸潤により示された(図4A)。誘導能は同一構築物でのアグロ浸潤により同様に試験され、浸潤緩衝液中にIPTGを含有していた(図4B)。
【0058】
実施例4
ウイルス構築物でのリプレッサー含有植物の再形質転換
ゲノム中に安定に組み込まれたlacIリプレッサー組換えDNA(pICH17401、図2A)を運んでいるN.タバクムを、lacオペレーター遺伝子(pICH18867、図2A)を含有する全ウイルスベクター構築物で2回目の形質転換を行った。いくつかの植物を再生できたが、それらのすべてが多かれ少なかれウイルスベクターの重度のバックグラウンド発現を示した。
【0059】
二重形質転換体の葉内へのIPTG(5mM)の浸潤は、トランスジェニック植物のいくつかにおいてRNAレプリコン増幅を実際に誘導できた(図5)。しかしながら、RNAレプリコンのリーキーな発現を有するすべての一次形質転換体は、最終的には導入遺伝子サイレンシングを受け、そして誘導条件下で何のRNAレプリコンの放出も示さない子孫を産生したことが観察された。
【0060】
実施例5
エタノール誘導性システムのための構築物の設計
エタノール誘導性システムの原理は、Caddick及び共同研究者(1998, Nat Biotechnol., 16:177-180)により記述されている。CaMV35Sプロモーターの制御下の転写アクチベーターalcR(構築物pICH18693、図2B)は、Caddick及び共同研究者(1998, Nat Biotechnol., 16:177-180)により記述されているように設計した。この構築物はN.タバクム及びN.ベンサミアナ植物内に安定的に形質転換した(Horsh et al.,1985, Science, 227, 1229-1231)。alcAプロモーターは、プライマーalcApr1(配列番号:9)(5’- cat gaa ttc tag gat tgg atg cat gcg g -3’)及びalcApr2(配列番号:10)(5’- cag ctc gag gtc gtc ctc tcc aaa tga aat g -3’)を使用するPCRにより増幅し、機能性MPを有する(pICH18969、図2B)又は有しない(pICH18951、図2B)TMVに基づいたウイルスベクターに、及び別に機能性ウイルスMP(pICH19940、図2B)にEcoRI−Xho1断片として融合した。加えて、構築物pICH18951及びpICH19940を一つのベクター(pICH20592、図2B)に結合させた。pICH18969を除いたこれらすべての構築物は、標準形質転換技術を使用してN.タバクム及びN.ベンサミアナ内の両方に形質転換した。機能性MPを伴ったRNAレプリコンをコードする構築物を有する一次形質転換体を得ることが可能であった。
【0061】
alcAプロモーターも、プライマーalcApr4(配列番号:11)(5’- cgc gca tgc tac tag gat tgg ata cat gcg gaa c -3’)及びalcApr5(配列番号:12)(5’- ttt ggt ctc atc aac tcc aaa tga aat gaa ctt cc -3’)を使用して増幅し、PVXに基づいたウイルスベクターpICH25233内にSphI−BsaI断片としてクローン化し、35Sプロモーターを置き換えて構築物pICH26022(図2C)を得た。alcAプロモーターの、細胞間移行に関与するPVXコートタンパク質への融合は(pICH26356、図2C)、pICH19940からのEcoRI−SacI断片のpICH22066内へのクローニングにより行われた。
【0062】
実施例6
エタノールにより誘導されるTMV構築物の一過性発現
上述の構築物はN.ベンサミアナ植物でアグロ浸潤により試験された(図6)。植物は2日後、4%エタノール溶液又は対照として水で処理した。ウイルスベクターの増幅及びGFPの発現はエタノール処理した植物において、及びアクチベーターalcRの存在下でのみ誘導された。非常に弱いバックグラウンド発現が、alcRの存在下、対照植物で観察された。
【0063】
実施例7
エタノールによって誘導されたPVX構築物の一過性発現
alcA−CP構築物(pICH26356)をalcR(pICH18693)及びCP欠損ウイルスベクター(pICH21692)と同時浸潤した。細胞間移行はエタノールで処理した植物でのみ検出でき、35Sプロモーター−CP構築物との相違は観察できなかった(図7)。
【0064】
実施例8
ウイルス構築物pICH18951及びpICH20592で安定的に形質転換した植物の分析
N.ベンサミアナ及びN.タバクム植物を、標準プロトコル(Horsh et al., 1985, Science, 227, 1229-1231) に従って形質転換した。再生した植物を、alcR構築物(pICH18693)でのアグロ浸潤、及びエタノール処理により導入遺伝子の存在を分析した。実際、ほとんどの植物は、葉の浸潤部分でGFP発現を示し、他の部分のバックグランドはなかった(図8)。
【0065】
実施例9
全トランスジェニック植物の誘導
pICH18951か又はpICH20592(実施例8に記載されている)を含有するトランスジェニック植物を、転写アクチベーターalcR(pICH18693)を含有する植物と交配した。これらの植物のF1子孫を、4%エタノールの噴霧により、又は根水浸(1%エタノールでの)及び噴霧(4%エタノール)の組み合わせによりエタノールで処理した。ウイルス増幅及びそれ故、GFP発現がこれらの植物のほとんど全ての部分で検出された(図9)。最も注目されたのは、強い発現が根水浸で処理した植物の茎及び葉柄でも検出された。植物のこれらの部分は通常、標準Magnifection法、即ち、全植物の減圧浸潤(Marillonnet et al., 2005, Nat. Biotechnol., 23:718-723) を使用すると、発現を示さないか又は弱い発現のみしか示さない。根水浸せずに4%エタノールで植物を噴霧すると、軟葉組織のみでのGFP発現を導き、茎及び葉柄では発現されない(図10)。
【0066】
実施例10
植物における組換えアプロチニンの発現のための誘導性ウイルスベクターシステムの使用
プラスミドpICH25408(図11)は、プラスミドpICH20592(実施例5参照)と同様の様式で設計した。N.ベンサミアナ植物を標準プロトコル(Horsh et al., 1985, Science, 227, 1229-1231)に従ってpICH25408で形質転換した。再生した植物を、alcR構築物(pICH18693)でのアグロ浸潤、及びエタノール処理により、続いてのポリアクリルアミドゲル(PAAG)電気泳動による組換えタンパク質発現の分析により導入遺伝子の存在を分析した。alcR構築物(pICH18693)でアグロ湿潤し、及びエタノールで処理した葉組織の一部を、2xLaemmli緩衝液(125mMトリス−HCl、pH7.8、10%メルカプトエタノール、20%グリセリン、0.001%ブロムフェノールブルー、10%SDS)による全可溶性タンパク質抽出、続いてのPAAGでの電気泳動分離に使用した。異なった一次形質転換体についてのこうした分析の結果は図12に示されている。
【0067】
次の工程において、pICH25408で形質転換した植物をalcR遺伝子(pICH18693)を運んでいる植物と交配した。エタノール処理後、F1子孫の組織葉から単離された全可溶性タンパク質をPAAGで分析した。この分析の結果は図13に示されている。アプロチニンが高レベルで発現されたことが明白である(約1mg/新鮮葉バイオマスのg)。
【図面の簡単な説明】
【0068】
【図1】本発明の一般原理。A−細胞間移行が可能なRNAレプリコンの伝搬;B−細胞への細胞間移行が欠損している本発明のRNAレプリコンの限局、ここでは前記RNAレプリコンの予定外形成が起こった。
【図2A】図2Aは、プラスミドpICH17155、pICH17401、pICH16141、pICH17171、pICH18867、pICH17424及びpICH17388のT−DNAドメインを描いている。
【図2B】図2Bは、プラスミドpICH18693、pICH18969、pICH18951、pICH19940及びpICH20592のT−DNAドメインを描いている。
【図2C】図2Cは、プラスミドpICH26022及びpICH26356のT−DNAドメインを描いている。TVCVポリメラーゼを示しているより大きな灰色のボックス中の白色ボックスは、核中で転写体を安定化し、そして結果として細胞核で形成されたRNAレプリコンをサイトゾルへより効率的に輸送させるイントロンを示す。3’Nos−ノパリンシンターゼ遺伝子の転写終結ドメイン;pNos−ノパリンシンターゼ遺伝子のプロモーター;p35S−CaMVの35Sプロモーター;pAct2−A.タリアナアクチン2遺伝子のプロモーター;NLS−核移行シグナル;lacO−大腸菌lacオペロンのオペレーター配列;lacI−大腸菌lacオペロンのレプレッサー遺伝子;BAR−除草剤ホスフィノトリシンに耐性を与えている遺伝子;int−イントロン配列の5’部分;AttP−インテグラーゼphC31により認識される組換え部位;NPTII−ネオマイシンホスホトランスフェラーゼII;sGFP−合成緑色蛍光タンパク質;NTR−トバモウイルス非翻訳領域;PalcA−アルコールデヒドロゲナーゼをコードする誘導性A.ニデュランスalcA遺伝子の誘導性プロモーター;alcR−アスペルギルス・ニデュランスのalcレギュロンの転写アクチベーター;TVCV MP−ハクサイ葉脈透化ウイルス移行タンパク質;PVX CP−ジャガイモウイルスXコートタンパク質;PVX Pol−ジャガイモウイルスX RNA依存性RNAポリメラーゼ;25K、12K、8K、トリプル遺伝子ブロック;sgp−サブゲノムプロモーター。
【図3】図3は、誘導性lacシステムでの一過性発現試験を示している。葉を異なった組み合わせ構築物で浸潤し、処理6日後、紫外光下でモニターした。以前暗いバックグラウンドであった光パッチはGFP蛍光を示している。葉の左側:lacリプレッサーなし;葉の右側:lacリプレッサーの存在下。pICH17424はlacOを有する5’−プロベクターであり、pICH17388はlacOがない対応する対照構築物である。pICH17401はlacリプレッサー構築物である。すべてのサンプルは、GFP3’−プロベクター及びインテグラーゼも含有する。
【図4A】図4Aは、lacリプレッサーで安定的に形質転換した植物の葉を示す。植物は、そのプロモーター中にlacO配列を含有するベクターpICH17171で、及びlacO配列を欠く、対応する対照構築物pICH16141でアグロ湿潤した。
【図4B】図4Bは、IPTG処理による抑制の解放を示す。lacリプレッサーで安定的に形質転換したN.ベンサミアナ植物を、そのプロモーター中にlacO配列を含有するベクターpICH17171で、及び対応する対照構築物pICH16141でアグロ湿潤した。誘導のため(右の写真)、5mM IPTGを浸潤緩衝液中に含ませた。左の写真は、誘導剤IPTGがないものである。
【図5】図5は、RNAレプリコンを提供する構築物での、lacIリプレッサー含有植物(pICH17155又はpICH17401)の再形質転換を示す。植物を5mM IPTGで浸潤した。系統N6(左の写真)は高い誘導能を示しているが、高いバックグラウンドも示し、一方、系統N8(右の写真)は低いバックグラウンド及び低い誘導能を示す。
【図6】図6は、TMVベクターに基づいたエタノール誘導性システムでの一過性発現試験を示す。対照:浸潤2日後に植物を水で処理をした;エタノール処理:浸潤2日後に植物を4%エタノールで処理をした。
【図7】図7は、PVXベクターに基づいたエタノール誘導性システムを使用する一過性発現試験を示す。対照:浸潤2日後に植物を水で処理をした;エタノール処理:浸潤2日後に植物を4%エタノールで処理をした。
【図8】図8は、紫外光下の、pICH18951のT−DNAを運んでいる安定な形質転換体の葉を示す。緑色(光)スポットは、pICH18693運搬アグロバクテリアで浸潤し及びアルコールを噴霧した領域に対応する。
【図9】図9は、水性アルコール溶液での処理後(灌水−1%アルコール、及び噴霧−4%アルコール)にGFPを発現している、トランスジェニックN.ベンサミアナ植物(F1子孫)を示す。
【図10】図10は、4%アルコール溶液で噴霧後、GFPを発現している、トランスジェニックN.タバクム植物(F1子孫)を示す。
【図11】図11は、プラスミドpICH25408のT−DNAドメインを描いている。TVCVポリメラーゼを示しているより大きな灰色のボックス中の白色ボックスは、核中で転写体を安定化し、そして結果として細胞核で形成されたRNAレプリコンをサイトゾルへ輸送させるイントロンを示す。TVCVポリメラーゼを示しているより大きな灰色のボックス中の白色ボックスは、核中で転写体を安定化し、そして結果として細胞核で形成されたRNAレプリコンをサイトゾルへ輸送させるイントロンを示す。3’Nos−ノパリンシンターゼ遺伝子の転写終結ドメイン;pNos−ノパリンシンターゼ遺伝子のプロモーター;3’Ocs−オクトピンシンターゼ遺伝子の転写終結ドメイン;NPTII−ネオマイシンホスホトランスフェラーゼII;NTR−トバモウイルス非翻訳領域;PalcA−アルコールデヒドロゲナーゼをコードする誘導性A.ニデュランスalcA遺伝子の誘導性プロモーター;alcR−アスペルギルス・ニデュランスのalcレギュロンの転写アクチベーター;TVCV MP−ハクサイ葉脈透化ウイルス移行タンパク質;sgp−サブゲノムプロモーター。
【図12】図12は、アプロチニン遺伝子を有するプラスミドpICH25408のT−DNAドメインを運んでいる異なったトランスジェニック植物(N18、N19及びN20)から抽出された全可溶性タンパク質の電気泳動分析の結果を伴ったクーマシー染色ポリアクリルアミドゲルを示す。レーン1−対照、未処理植物から単離された全可溶性タンパク質;レーン2−alcRアクチベーター(pICH18693)をコードするプラスミドを運んでいるアグロバクテリアで浸潤され、及び4%エタノールで処理された植物;レーン3−alcRアクチベーターをコードするプラスミド(pICH18693)、アプロチニン遺伝子を運んでいるウイルスベクターをコードするプラスミド(pICH25408)を運んでいるアグロバクテリアの混合物で浸潤され、及び4%エタノールで処理された植物。
【図13】図13は、アプロチニン遺伝子を有するウイルスベクターをコードするプラスミドpICH25408のT−DNA領域を運んでいるトランスジェニック植物、及びalcRアクチベーターをコードするT−DNA領域(pICH18693)を運んでいるトランスジェニック植物間の交配から得られたF1植物から抽出された全可溶性タンパク質の電気泳動分析の結果を伴ったクーマシー染色ポリアクリルアミドゲルを示す。
【技術分野】
【0001】
本発明は、ウイルス発現システムを使用し植物又は植物細胞中の一つ又は一つより多くの目的とするタンパク質を産生する又は発現するプロセスに関する。本発明は、さらにこのプロセスの為の植物又は植物細胞、特にトランスジェニック植物又は植物細胞に関する。本発明は、本発明の該植物又は植物細胞を産生するプロセスも提供する。
【背景技術】
【0002】
植物における異種タンパク質の高収率発現は、原則としてウイルスベクターを使用して達成し得る。しかしながら、異なった植物ウイルス発現システムの存在にもかかわらず、植物ウイルス発現システムは、主として感染に続いた植物中の目的とするタンパク質の一過性発現に(Donson et al.,1991, Proc Natl Acad Sci U S A, 88:7204-7208; Chapman, Kavanagh & Baulcombe, 1992, Plant J., 2:549-557)、又は組換えウイルスベクターを用いる宿主植物の形質移入(Marillonnet et al., 2005, Nat. Biotechnol., 23:718-723; Santi et al., 2006, Proc Natl Acad Sci U S A. 103:861-866; WO2005/049839) に使用されている。いくつかの科学出版物及び公開された特許出願にもかかわらず、主として二つ理由のため、容易にスケールアップでき、そして高収率を提供することに利用可能である商業的ウイルスに基づいた産生システムは未だ確立されていない:
第一に、一過性の植物ウイルスに基づいた発現システムは、一般的に特定の宿主に限定されており、それらの環境要因に対する感受性のため大規模培養には適していないであろう。さらに、それらは一般的に植物宿主の特定の部分に限定されており、それ故、産生プロセスからの植物バイオマスのほとんどを排除することになり、その結果、植物バイオマスの単位当たりの組換え産物の相対収率を最少化し、トランスジェニック植物において慣用的転写プロモーターを使用して達成されるレベルに匹敵するレベルまで下げている;
第二に、各細胞に安定に組み込まれたウイルスレプリコンを有するトランスジェニック宿主植物を発生させることによる、ウイルスに基づいた産生システムをスケールアップする試みは、特に、そのような位置の前記レプリコンの標準より低い働きのため、及びウイルスレプリコンの安定した形成が植物成長及び発達を損なうため、解決を提供していない。通常、一過性発現システムにおける浸透移行性ウイルスベクターは、異種核酸の比較的短い(1kbまで)挿入物に耐えることができ、それ故、比較的小さなタンパク質の発現に限定されている。トランスフェクションのために使用されるウイルスベクター(アグロバクテリウム仲介送達、WO2005/049839 )はより大きな挿入物を発現し得るが、全植物のアグロ浸潤を必要とする。明らかに、こうしたシステムは、使用する及びスケールアップするために短い時間しか必要としないので、抗原を含む多くの組換えタンパク質の産生に都合がよい、しかしウイルスベクターに基づいた発現システムのトランスジェニックバージョンが、多くの他の応用で有利であり得る。特に、大量に及び相対的に低コストで必要とされる組み換えタンパク質の産生には問題であり(例えば、異なったセルラーゼ及び他の技術的酵素)、アグロ浸潤 (WO2005/049839)に基づいた一過性発現システムは経済的に実行可能ではない。トランスジェニック宿主植物におけるウイルスベクターの発現は、通常植物生長及び発達のためには有害である。また、こうした発現は、最後には導入遺伝子サイレンシングを導くであろう。この問題の解決を見いだすために、転写後遺伝子サイレンシング(PTGS)サプレッサーの助けをかりて、植物染色体から発現停止したウイルスレプリコンを放出することが試みられた(US6395962; Mallory et al., 2002, Nat. Biotechnol., 20:622-625)。植物三要素RNAウイルス(Mori et al., 2001, Plant J., 27, 79-86)、ブロムモザイクウイルス(BMV)に基づいたグルココルチコイド誘導性発現システムは、おそらくPTGSのせいで、標準(非ウイルス性)転写プロモーターにより提供される収率に匹敵する、目的とするタンパク質の非常に低い収率(3〜4μg/g新鮮重量)しか与えなかった。
【0003】
現在のところ、細菌、真菌又は昆虫細胞発現システムのような他の大規模発現システムと市場で競争するために十分に高いであろう収率及び効率の大規模植物ウイルス発現システムはない。こうした植物発現は可能な限り良好に以下の基準を満たさなければならない:
(i) 高収率、可能な限り多くの植物組織中での、及び前記組織の多くの細胞中での、目的とするタンパク質の発現を含んで;
(ii)植物細胞の生存に対するタンパク質発現の悪影響を防止するため、目的とするタンパク質又は生成物の発現は、同時に処理された植物又は植物組織の全ての植物細胞中で開始されるべきである。
【0004】
典型的には、目的とするタンパク質又は生成物は、前記タンパク質又は生成物を産生している各細胞中で特定の点まで蓄積する。しかしながら、蓄積の間、分解プロセスがしばしば進み、それは目的とするタンパク質又は生成物の収量又は質を減少させる傾向がある。それ故、時間には至適時点があり、そこで目的とするタンパク質又は生成物を収穫すべきである。この時間の至適時点は、全プロセスを効率的に及び有益にするため、植物の全ての組織又は細胞で、及び選択されたロットの全ての植物で同時に到達可能であるべきである。
【発明の開示】
【0005】
本発明の一般的説明
それ故、大規模応用に容易に拡張可能で、発現されるべきタンパク質の高収率を与え、及び同時に、目的とする組み換えタンパク質の制御されない発現の確率は低いという点において生物学的に安全である植物システムにおいて、一つ又はそれより多くのタンパク質を発現するプロセスを提供することが一つの目的である。前記ウイルスレプリコンから、目的とするタンパク質を発現するために適したウイルスレプリコンをコードするトランスジェニック植物を産生する効率の良い方法を提供することが本発明のさらなる目的である。
【0006】
それ故、本発明は、一つの又は一つより多くの目的とするタンパク質を産生するプロセスを提供することであって:
(a)RNAレプリコンをコードするヌクレオチド配列を含んでなる第一の異種ヌクレオチド配列、及び前記RNAレプリコンをコードする前記ヌクレオチド配列に機能可能なように連結された第一の誘導性プロモーターを含んでなる植物又は植物細胞を提供すること;
前記RNAレプリコンは、前記植物における前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない;
前記RNAレプリコンはポリメラーゼ及び前記一つ又は一つより多くの目的とするタンパク質をコードし、前記ポリメラーゼは前記RNAレプリコンを複製するために適合されている;及び
(b)工程(a)の前記植物又は植物細胞において、前記誘導性プロモーターを誘導し、それにより、前記植物又は植物細胞において前記一つ又は一つより多くの目的とするタンパク質を産生すること;
を含んでなる。
【0007】
本発明はさらに、一つの又は一つより多くの目的とするタンパク質を産生するプロセスを提供することであって:
(a)植物を提供することであって、前記植物は、
(i)RNAレプリコンをコードするヌクレオチド配列、及び前記RNAレプリコンをコードする前記ヌクレオチド配列に機能可能なように連結された第一の誘導性プロモーターを含んでなる第一の異種ヌクレオチド配列;
前記RNAレプリコンは、前記植物における前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない;
前記RNAレプリコンはポリメラーゼ及び前記一つ又は一つより多くの目的とするタンパク質をコードし、前記ポリメラーゼは前記RNAレプリコンを複製するために適合されている;
(ii)前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードする配列を含んでなる第二の異種ヌクレオチド配列、ここで前記第二の異種ヌクレオチド配列は、前記RNAレプリコンの細胞間移行を可能にする前記タンパク質をコードする前記配列に機能可能なように連結された第二の誘導性プロモーターを含んでなる;
を含んでなり;及び
(b)工程(a)の前記植物において、前記第一及び第二の誘導性プロモーターを誘導し、それにより、前記植物において前記一つ又は一つより多くの目的とするタンパク質を産生すること;
を含んでなる。
【0008】
本発明はさらに、上記プロセスの工程(a)で提供された植物又は植物細胞を提供する。本発明はさらに、本発明の植物又は植物細胞を産生するプロセスを提供することであって、前記第一の異種ヌクレオチド配列及び任意選択で前記第二の異種ヌクレオチド配列を植物核染色体内に導入すること、続いて前記第一の及び任意選択で前記第二の異種ヌクレオチド配列を含有する形質転換植物を再生することを含んでなる。
【0009】
本発明の発明者は驚くことに、従来の技術では達成不可能であった発現レベルを達成する、植物又は植物細胞中で一つ又は一つより多くの目的とするタンパク質を産生するプロセスを発見した。同時に、本発明のプロセスは、生物学的に安全であり、及び工業的レベルへ拡張可能である。本発明の例外的な発現レベルは、今まである程度しか知られていない導入遺伝子サイレンシングのような、植物又は植物細胞による抗ウイルス応答を避けるウイルス発現システムにより達成可能である。本発明において、RNAレプリコンをコードするヌクレオチド配列からのRNAレプリコンの放出及び前記RNAレプリコンの細胞間移行を可能にする前記タンパク質の発現は、誘導性プロモーターにより制御されている。前記RNAレプリコンは、細胞間移行を提供する前記タンパク質が存在しない場合には前記植物中で又は前記植物細胞中で細胞間移行ができないので、前記RNAレプリコンをコードするヌクレオチド配列からのRNAレプリコンのリーキーな(leaky) 転写による、RNAレプリコンのいずれかの意図されない放出は、こうしたリーキーな転写が起こった細胞に限局されている。重要なことには、ウイルス配列は、誘導されない状態においては、導入遺伝子サイレンシングの引き金を引く事ことができるレベルでは発現されない。それ故、導入遺伝子サイレンシングは起こりそうもない。さらに、前記植物において前記RNAレプリコンの細胞間移行を提供するタンパク質を取り除くことによるRNAの発現収率に対する悪影響は、前記RNAレプリコンのトランスに前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードする配列を提供するすることにより補償されている。前記RNAレプリコンの細胞間移行を可能にする前記タンパク質は、誘導性プロモーターの制御下に発現される。このことは、前記ウイルスレプリコンのリーキーな発現が起こった細胞へ、こうしたリーキーな発現の限局を保存し、そして遺伝子サイレンシングを回避するが、誘導された状態においては前記目的とするタンパク質の高い発現レベルを可能にする。それ故、本発明において、目的とするタンパク質の収率に対する遺伝子サイレンシングの悪影響は、本質的に存在しない。
【0010】
さらに、本発明者は驚くべきことに、形質転換効率は、植物細胞が本発明の前記第一の異種ヌクレオチド配列で形質転換された場合、RNAレプリコンが細胞間移行を提供するタンパク質をコードする場合よりも高いことを発見した。いくつかの場合において、もしRNAレプリコンが細胞間移行を提供するタンパク質をコードするならば、前記第一の異種ヌクレオチド配列で一次形質転換体を全く得ることができなかった。形質転換効率を改良する本発明のこの効果は、細胞間移行についての本発明のRNAレプリコンの無能さによるものであろう。
【0011】
本発明の一つの態様において、前記植物又は植物細胞は、前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードするヌクレオチド配列を含んでなる第二の異種ヌクレオチド配列を含有し、前記第二の異種ヌクレオチド配列は、前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードする前記ヌクレオチド配列に機能可能なように連結された第二の誘導性プロモーターを含んでなる。本発明のこの態様は、前記一つ又は一つより多くの目的とするタンパク質の産生の収率をさらに増加させることを可能にする。
【0012】
一つ又は一つより多くの目的とするタンパク質を産生する本プロセスの工程(a)において、植物又は植物細胞に前記第一の異種ヌクレオチド配列が提供される。前記植物又は植物細胞には、トランスジェニック植物又はトランスジェニック植物細胞が得られるように、前記第一の異種ヌクレオチド配列が提供される。一つの態様において、前記トランスジェニック植物又は前記トランスジェニック植物細胞は、核染色体中に、前記第一の異種ヌクレオチド配列を含有する。
【0013】
前記植物又は植物細胞には、トランスジェニック植物又はトランスジェニック植物細胞が得られるように、前記第二の異種ヌクレオチド配列を提供することもできる。一つの態様において、前記トランスジェニック植物又は前記トランスジェニック植物細胞は、核染色体中に前記第二の異種ヌクレオチド配列を含有する。
【0014】
核染色体又はエピソーム内に安定に組み込まれた異種ヌクレオチド配列を含有しているトランスジェニック植物を発生させることは当該技術分野で公知である。典型的には、前記異種ヌクレオチド配列は、組み込まれた前記異種ヌクレオチド配列を有する植物細胞又は植物組織を選択するための選択可能マーカー遺伝子を含有するであろう。前記トランスジェニック植物の細胞中に前記異種ヌクレオチド配列を含有する全トランスジェニック植物は、次ぎに植物バイオテクノロジーの技術分野の標準法を使用して、形質転換細胞又は組織から再生することができる。本発明の前記異種ヌクレオチド配列は、典型的にはDNAである。
【0015】
もし植物が、前記第一の及び前記第二の異種ヌクレオチド配列を含有するように提供されるならば、前記第一の及び前記第二の異種ヌクレオチド配列は、植物細胞又は植物を形質転換するために使用された一つの大きな異種ヌクレオチド配列の一部であることができる。この態様において、前記大きな異種ヌクレオチド配列は、前記第一の及び前記第二の異種ヌクレオチド配列を含んでなる。もしくは、前記第一の異種ヌクレオチド配列を含有する第一の植物、及び前記第二の異種ヌクレオチド配列を含有する第二の植物を独立して発生させてもよい。前記第一及び前記第二植物は次ぎに交配することができて(例えば、性交配又は細胞融合により)、前記第一の及び前記第二の異種ヌクレオチド配列を含有する植物が得られる。さらなる代替法において、前記第一の及び前記第二の異種ヌクレオチド配列を含有する植物又は植物細胞を産生するため、前記第一の異種ヌクレオチド配列を含有するトランスジェニック植物又は植物細胞を前記第二の異種ヌクレオチド配列で形質転換することができ;又は前記第二の及び前記第一の異種ヌクレオチド配列を含有する植物又は植物細胞を産生するため、前記第二の異種ヌクレオチド配列を含有するトランスジェニック植物又は植物細胞を前記第一の異種ヌクレオチド配列で形質転換することができる。ここで、「異種」とは、前記植物について異種を意味する。
【0016】
前記第一の異種ヌクレオチド配列は、前記RNAレプリコンをコードするヌクレオチド配列セグメントを含んでなる。前記第一の異種ヌクレオチド配列はさらに、前記RNAレプリコンをコードする前記ヌクレオチド配列セグメントに機能可能なように連結されている第一の誘導性プロモーターを含んでなる。前記誘導性プロモーターは、本発明のプロセスの工程(b)において、前記RNAレプリコンをコードする前記ヌクレオチド配列セグメントの転写を誘導することを可能にする。転写は、前記RNAレプリコンをコードする前記ヌクレオチド配列セグメントから前記RNAレプリコンを放出する。前記RNAレプリコンはRNAレベル上のレプリコンである。植物細胞の核中で産生された前記RNAレプリコンは、次ぎに、前記RNAレプリコンによりコードされたタンパク質が産生され及びRNAレプリコンが複製されるサイトゾル内へ移行することができる。
【0017】
前記RNAレプリコンは、前記誘導性プロモーターを誘導することにより前記RNAレプリコンが放出された後に発現させることができる。本発明において、前記第一の異種ヌクレオチド配列によりコードされた前記第一のRNAレプリコンは、前記RNA配列を複製するためのポリメラーゼをコードするRNA配列であり、それにより、前記RNA配列はその上にコードされているポリメラーゼにより複製されるように適合される。それ故、前記ポリメラーゼは、本明細書において「レプリカーゼ」とも称されるRNA依存性RNAポリメラーゼである。前記RNAレプリコンは、好ましくは、前記ポリメラーゼを翻訳するための配列、及び前記植物の細胞又は前記植物細胞中での前記RNAレプリコンの複製を可能にするため、前記ポリメラーゼを結合するための配列を有する。前記RNAレプリコンはさらに、発現されるべき前記一つ又は一つより多くの目的とするタンパク質、ならびにサブゲノムプロモーター、転写エンハンサー又は翻訳エンハンサーのような、前記一つ又は一つより多くの目的とするタンパク質を発現するために必要とされる配列をコードする。
【0018】
好ましい態様において、前記RNAレプリコンは、一要素(monopartite)RNAウイルスのようなRNAウイルスに由来する。「一要素」とは、一要素ウイルスが一つの核酸分子から成るゲノムを有することを意味する。それ故、前記本発明のRNAレプリコンは、好ましくは、一要素RNAレプリコンであり、即ち、それは単一のタイプのRNA分子から成る。好ましい態様において、前記RNAレプリコンはプラスセンス一本鎖RNAウイルスに由来しており、こうしたウイルスは複製及び発現に必要とされる遺伝子要素を含有し、及び本発明の目的に進化により最適化されているからである。前記好ましい態様において、前記RNAレプリコンはプラスセンス一本鎖RNAレプリコンである。
【0019】
「由来する」とは、本発明に必要なRNAウイルスの遺伝子因子が使用され、一方、その他は欠失されるか又は機能障害性にされることを意味する。本発明において、前記ウイルスの細胞間移行を提供する前記タンパク質をコードする配列が、例えば、細胞間移行に必須な配列部分の部分的欠失又は突然変異により、欠失されるか又は機能障害性にされるであろう。もしくは、目的とするタンパク質をコードする配列を、完全に又は部分的に細胞間移行のための前記タンパク質をコードする配列で置き換えることができる。「由来する」は、植物ウイルスから取り出された前記RNAレプリコンの配列は、前記RNAウイルスの対応するRNA配列と同一である必要はなく、WO 2005/049839に記載されているように、前記レプリカーゼをコードする配列部分に挿入されたイントロンのような機能保存的相違を示すことができることを示唆する。前記相違は機能保存的であるので、前記配列は、好ましくは、前記RNAレプリコンが由来する前記RNAウイルス中で行うような、レプリコン機能を実施できるタンパク質をコードする。本発明の本発明の前記RNAレプリコン又はそれらのポリメラーゼを誘導することができる適したプラスセンス一本鎖RNAウイルスは、タバコモザイクウイルス(TMV)又は ジャガイモウイルスXである。前記RNAレプリコンを誘導することができるさらなる植物ウイルスは以下に与えられている。
【0020】
しかしながら、前記RNAレプリコン、又はそれらをコードするcDNAを人工的に合成することも可能であり、天然のウイルスの遺伝子因子を使用しても又はしなくてもよい。
【0021】
本発明の前記RNAレプリコンは、前記第一の異種ヌクレオチド配列の一つの連続したヌクレオチド配列セグメントによりコードされている必要はない。実際、前記第一の異種ヌクレオチド配列は、一緒になって前記RNAレプリコンをコードする二つ又はそれより多くの配列セグメントを有することができる。二つまたはそれより多くのこうした配列セグメントは、連続していてもよいし、又は別の配列部分により中断されていてもよい。前記植物の細胞中での前記RNAレプリコンの形成は、部位特異的DNA又はRNA組換えを含むことができる。DNA組換えの場合、前記RNAレプリコンは、前記RNAレプリコンが発現されることを阻止する配列部分の切除を介して形成させることができる。もしくは、不連続的に前記RNAレプリコンをコードする二つまたはそれより多くの配列セグメントの一つを組換えによりフリップさせることができ、それにより前記RNAレプリコンをコードする単一の連続配列セグメントを形成させる。前記RNAレプリコンは、そのどちらもがRNAレプリコンではない二つのレプリコン前駆体間の組換えにより形成することも可能である。こうした組換えは、WO02/097080 に記載されているリボザイム仲介トランススプライシングであってもよい。
【0022】
本発明の前記RNAレプリコンは、前記植物中での前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない。本発明のこの特色は、前記RNAレプリコンのいずれのリーキーな放出も、前記リーキーな放出の植物細胞に限局させる。前記植物において前記RNAレプリコンの細胞間移行を提供するタンパク質は、一般に「移行タンパク質」と称されている。植物ウイルスは通常、ウイルス又は前記ウイルスのゲノムRNAの細胞間拡散を許容する一つ又はそれより多くのタンパク質をコードする又は発現する。本発明のRNAレプリコンは、前記RNAレプリコンが前記植物において細胞間で有意に拡散するのを許容するであろう移行タンパク質を発現できるようにしてはならない。前記植物又は前記植物細胞は、誘導性プロモーターの制御下でない限り、前記植物において前記RNAレプリコンの細胞間移行提供するタンパク質を発現すべきではない。一つの態様において、前記RNAレプリコンは、天然のRNAウイルスの移行タンパク質の一部を含有することができるが、ただし、前記移行タンパク質の一部は、前記RNAレプリコンが前記植物において細胞間で有意に拡散するのを許容しない。
【0023】
本発明のプロセス及び植物の一つの態様において、前記植物は、前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードするヌクレオチド配列を含んでなる第二の異種ヌクレオチド配列を含んでなり、前記第二の異種ヌクレオチド配列は、前記RNAレプリコンの細胞間移行を可能にする前記タンパク質をコードする前記ヌクレオチド配列に機能可能なように連結された第二の誘導性プロモーターを含んでなる。前記タンパク質は、前記RNAレプリコンのポリメラーゼが誘導される同一の植物RNAウイルスに由来してもよいし、又は別のRNAウイルスに由来してもよい。
【0024】
RNAレプリコンをコードする前記ヌクレオチド配列及び細胞間移行を可能にするタンパク質をコードする前記ヌクレオチド配列は、一つの誘導性プロモーターからのリーキーな発現が移行タンパク質の発現及び前記RNAレプリコンの形成を導くことを避けるため、別の誘導性プロモーターの制御下にある。それ故、RNAレプリコンをコードする前記ヌクレオチド配列及び細胞間移行を可能にするタンパク質をコードする前記ヌクレオチド配列は、前記植物又は植物細胞において異なった発現カセット中に存在する。しかしながら、これら別々の誘導性プロモーターは、異なったタイプの誘導性プロモーターである必要はない。前記別々の誘導性プロモーターは、同一タイプであってもよいし、及び同一配列を有していてもよい。
【0025】
前記第一の、前記第二の前記誘導性プロモーター又はいずれかのさらなる誘導性プロモーターは、同一の又は異なった誘導シグナル又は剤により誘導性である。本発明に使用し得る誘導性プロモーターは以下に与えられている。一つの態様において、前記第一及び/又は前記第二の異種ヌクレオチド配列の誘導性プロモーターは化学的に誘導性である。別の態様において、前記第一及び/又は前記第二の異種ヌクレオチド配列の誘導性プロモーター(及び任意選択でさらなる異種ヌクレオチド配列)は、IPTG、エタノール、テトラサイクリン又はグルココルチコイドのような同一の誘導剤により誘導性である。
【0026】
本発明のプロセスの工程(b)において、工程(a)の前記植物又は植物細胞中で前記誘導性プロモーターが誘導され、それにより前記一つ又は一つより多くの目的とするタンパク質の発現が開始され、それにより前記植物又は植物細胞中で前記一つ又は一つより多くの目的とするタンパク質を産生する。誘導の様式は、誘導性プロモーターのタイプに依存する。もし誘導性プロモーターが化学的に誘導可能であれば、プロモーターを誘導することが可能な化学剤が前記植物又は前記植物細胞に提供される。もし前記第一の及び前記第二の異種ヌクレオチド配列について異なった誘導性プロモーターが使用されたならば、異なった化学剤を前記植物又は前記植物細胞に同時に適用することができる(例えば、異なった誘導剤の混合物として)。もし前記プロセスが液体培養液中の植物細胞で実施されるならば、該誘導剤を培養培地に加えることができる。もし前記プロセスが植物において実施されるならば、前記誘導剤の溶液又は懸濁液で前記植物をスプレーすることにより、前記植物に適用することができる。
【0027】
本発明のプロセスは、一つの目的とするタンパク質又は一つより多くの目的とするタンパク質を産生するために使用し得る。もし一つの目的とするタンパク質が産生されるべきならば、前記目的とするタンパク質をコードするヌクレオチド配列を、前記RNAレプリコンをコードする前記ヌクレオチド配列に含ませることができる。一つの態様において、前記目的とするタンパク質をコードするヌクレオチド配列は、前記RNAレプリコンが誘導される植物RNAウイルスの移行タンパク質遺伝子と置換することができる。もしくは、前記RNAレプリコンが誘導されるRNAウイルスのコートタンパク質遺伝子を、目的とするタンパク質をコードするヌクレオチド配列で置き換えることが可能である。もし二つの目的とするタンパク質が産生されるべきならば、移行タンパク質遺伝子及びコートタンパク質遺伝子の両方を目的とするタンパク質をコードするヌクレオチド配列で置き換える(完全に又は部分的に)ことが可能である。
【0028】
もし二つまたはそれより多くの目的とするタンパク質タンパク質が産生されるべきならば、前記植物又は植物細胞は、さらなるRNAレプリコン及び前記さらなるRNAレプリコンをコードする前記配列に機能可能なように連結された第三の誘導性プロモーターをコードするヌクレオチド配列を含んでなる第三の異種ヌクレオチド配列を含むことができる。前記さらなるRNAレプリコンは、好ましくは、前記植物中で前記RNAレプリコン又は前記さらなるRNAレプリコンの細胞間移行を提供するタンパク質をコードしていない。前記さらなるRNAレプリコンは次ぎに、一つ又はそれより多くのさらなる目的とするタンパク質をコードすることができる。もし前記植物が二つまたはそれより多くのRNAレプリコンをコードするならば、前記RNAレプリコンの細胞間移行を可能にするタンパク質は、前記さらなるRNAレプリコンの細胞間移行を可能にすることができる。もしくは、前記さらなるRNAレプリコンの細胞間移行は、その発現がさらなる誘導性プロモーターの制御下であることができる、前記さらなるRNAレプリコンの細胞間移行を可能にする別のタンパク質により可能にすることもできる。
【0029】
前記さらなるRNAレプリコンは、前記第一の異種ヌクレオチド配列によりコードされた前記RNAレプリコンによりコードされた前記ポリメラーゼにより複製可能であることができる。しかしながら一つの態様において、前記さらなるRNAレプリコンは、複製前記さらなるRNAレプリコンを複製するためのさらなるポリメラーゼをコードしており、前記さらなるポリメラーゼは、前記RNAレプリコンのものと異なっていることができる。
【0030】
一つの態様において、前記RNAレプリコン及び前記さらなるRNAレプリコンは、非競合RNAレプリコンである。非競合RNAレプリコン又は非競合ウイルスベクターからのタンパク質産生は、WO 2006/79546 (PCT/EP2006/000721) に記載されている。
【0031】
前記一つ又は一つより多くの目的とするタンパク質は、前記植物又は植物細胞中での産生後、望まれない細胞成分から精製することができる。植物又は植物細胞からタンパク質を精製する方法は当該技術分野で公知である。一つの方法において、目的とするタンパク質は、WO 03/020938 に記載されているように、植物アポプラストに方向付けることができる。
【0032】
本発明は原則として、感染性RNAウイルスが存在する及びウイルスベクターシステムが確立されたいずれの植物にも応用することができる。一つの態様において、双子葉植物が本発明を実施するために使用される。別の態様において、ナス科植物が使用される。好ましい植物は、ニコチニア・ベンサミアナ及びニコチニア・タバクムのようなニコチニア種であり;ニコチニア種以外の好ましい植物種は、ペチュニア・ヒブリダ、ブラシカ・カンペストリス、B.ユンセア、クレス、アルグラ、マスタード、イチゴほうれん草(Strawberry spinach)、ケノポディウム・カピターツム(Chenopodium capitatum)、アルファルファ、レタス、ヒマワリ、及びキュウリである。
【0033】
本発明の産生プロセスは、本明細書に記載した植物の細胞において実行することもできる。前記細胞は、葉のような植物組織の一部であってもよく、又は前記細胞は、懸濁培養のような細胞培養液中に存在していてもよい。
【0034】
適したRNAレプリコンは以下に与えたRNAウイルスのリストに由来することができる。本発明は、一要素植物RNAウイルスに適用し得る。本発明が基づくことができる最も好ましい植物RNAウイルスはトバモウイルス、特にタバコモザイクウイルス及びジャガイモウイルスXのようなポテクスウイルスである。タバコモザイクウイルスの場合、それは一般的には、発現されるべき前記目的とするタンパク質のORFにより置き換えられている移行タンパク質ORFであろう。コートタンパク質ORFも、目的とするタンパク質のORFにより除去される又は置き換えられることができる。
【0035】
本発明の主要な適用は、植物、植物の葉又は植物組織又は細胞培養における目的とするタンパク質の産生である。もし本発明のプロセスが植物において実施されるならば、ニコチニア種植物のような、ヒト又は動物の食物連鎖に入り込まない植物が好ましい。ヒト又は動物の食物連鎖に入り込まない植物は野外で耕作することが可能であり、及び前記RNAレプリコン放出の誘導後、特定の期間内で、植物組織中の一つ又は一つより多くの目的とするタンパク質の発現レベルがピークに達した時に採取される。好ましくは、全植物又は植物部分は閉ざされた環境に限局すべきである;例えば、発現の望まれるレベルを提供するために必要なインキュベーション期間のためのガラス温室又は設計されたチャンバー。
【0036】
本発明の生産プロセスの効率は、植物発現システムにおいて新たな局面が達成されるほどである。本発明で達成可能な発現レベルは、下流のプロセシング(目的とするタンパク質の分離及び精製を含んで)のための費用が、他の大規模発現システムとの競争力がある本発明のプロセスを作製するために十分低いほどである。ウイルスベクターで安定的に形質転換した植物を使用する従来技術の発現システムにおいては、これらのシステムの漏れやすさが、産生されるべきレプリコンの誘導されない状態をさえ許容するので、発現レベルは低く、それ故、収率を損なうPTGSの機構の引き金が引かれる。加えて、細胞間移行が可能であるRNAレプリコンは、植物染色体DNA内に安定的に取り込まれた前記レプリコンを運んでいる安定的に形質転換された植物細胞の産生を損なう。驚くべきことに、発明者は、細胞間移行が可能なRNAレプリコンを運んでいるトランスジェニック植物よりも、細胞間移行が不可能なRNAレプリコンをコードするベクターを含有するトランスジェニック植物を得ることがより容易であることを見いだした。この現象が、細胞間移行が不可能なウイルスベクターを含有するトランスジェニック植物中の高発現レベルに寄与することも非常にありそうである。本発明は、大規模で使用することが可能である、最初の高収率誘導性植物発現システムを提供する。
【0037】
発明の詳細な説明
この発明は、一要素RNAウイルスから誘導することができるRNAレプリコンを使用する、目的とするタンパク質の高収率、大規模生産のための誘導性発現システムを記述している。前記RNAレプリコンは、植物中で一つ又は一つより多くの目的とするタンパク質を発現できる。本発明のプロセスは、バイオセーフティー特色を有し、RNAレプリコン放出の厳格な制御を提供し、短距離(細胞間)移行及び、場合により、遠隔距離移行についてのその無能さのため、前記RNAレプリコンが他の植物に感染することを防止している。
【0038】
我々は驚くべきことに、前記RNAウイルスベクターからの細胞間移行機能の除去が、植物又は植物細胞の染色体DNA中に前記RNAレプリコンをコードする、安定に形質転換された植物の選択及び再生を容易にすることを発見した。本発明の基本原理は図1に示されている。細胞間移行が可能なウイルスベクターの場合、誘導性システムの漏れやすさは、サイトゾル内へのウイルスベクター放出を起こし、及びさらには隣接細胞に伝播する(図1A)。最終的に、このことは宿主植物内でのウイルスベクターの制御されない伝播を導き、前記植物の生長及び発達を損なう。こうした問題を克服している、及び転写後遺伝子サイレンシング(PTGS)のメカニズムによりウイルスベクター複製の制御を行っている宿主植物の場合、PTGSは、宿主植物中での目的とするタンパク質の発現レベルに対して負の効果を有しているであろう。本発明において、誘導性システムの漏れやすさは、本発明のRNAレプリコンは細胞間移行が欠損しており、それ故PTGSは実際に無視できるので、こうした劇的効果を有していない。前記RNAレプリコンは本質的にその細胞に限局されており、それはプロモーターの漏れやすさのために放出されており(図1B)、それ故、目的とするタンパク質の望まれない発現に対する制御を改良し、及び該システムの産生性に対するPTGSの潜在的負の効果を減少させる。
【0039】
本発明において、誘導性及び組織特異的プロモーターを、植物又は植物細胞において、目的とするタンパク質産生の引き金を引くために使用し得る。誘導性プロモーターは、それらの誘導条件に従って二つのカテゴリーに分割し得る;非生物的因子(温度、光、化学物質)により誘導可能なもの、及び生物的因子、例えば、病原体又は有害生物攻撃により誘導可能なもの。第一のカテゴリーの例には、限定されるわけではないが、熱誘導性(US 05187287) 及び低温誘導性(US05847102) プロモーター、銅誘導性システム(Mett et al., 1993, Proc. Natl. Acad. Sci., 90, 4567-4571) 、ステロイド誘導性システム(Aoyama & Chua, 1997, Plant J., 11, 605-612; McNellis et al., 1998, Plant J.,14, 247-257; US06063985) 、エタノール誘導性システム(Caddick et al., 1997, Nature Biotech., 16, 177-180; WO09321334; WO0109357; WO02064802) 、イソプロピル ベータ−D−チオガラクトピラノシド(IPTG)誘導性システム(Wilde et al., 1992, EMBO J., 11:1251-1259) 、及びテトラサイクリン誘導性システム(Weinmann et al., 1994, Plant J., 5, 559-569) が含まれる。植物についての化学的誘導性システムの領域における最近の開発の一つは、グルココルチコイド デキサメタゾンによりスイッチを入れることができ、及びテトラサイクリンによりスイッチを切ることができるキメラプロモーターである(Bohner et al., 1999, Plant J., 19, 87-95) 。化学的誘導性システムは本発明の実施に最も適している。化学的誘導性システムの総説は:Zuo & Chua, (2000, Current Opin. Biotechnol., 11, 146-151)、及びMoore et al., (2006, Plant J., 45: 651-683) を参照されたい。リプレッサー又はアクチベーターのような、選択された誘導性システムの機能性に必要とされるいずれのタンパク質も、誘導性システムを機能的にするためには前記植物又は前記植物細胞中で発現されなければならないことは、当業者には明白であろう(実施例参照)。
【0040】
本発明の一つの態様において、RNAレプリコンの放出及び目的とするタンパク質の産生を制御するためにIPTG誘導性システムを使用する。例示の構築物の設計は実施例1に記載されている。lacIリプレッサーをコードする細菌遺伝子を、強い構成的35Sプロモーターの制御下にクローン化した。二重のlacオペレーター配列を、本発明のRNAレプリコンの転写を駆動するシロイヌナズナアクチン2プロモーター内に挿入した。このシステムは、実施例2に記載されている一過性発現アッセイを使用して試験した。前記RNAレプリコンをコードする異種ヌクレオチド配列を、lacIリプレッサーを運んでいる構築物と一緒に植物内に浸潤させた場合、多分RNAレプリコンの形成がリプレッサーの合成に先行されるため、このシステムは何のRNAレプリコン放出の阻害も示さなかった(図3、右下)。実際、レプリコン形成を遅延させるために、RNAレプリコンのDNA前駆体への組立のために部位特異的リコンビナーゼを必要とするプロベクターシステム(Marillonnet et al., 2004, Proc. Natl. Acad. Sci. USA, 101:6852-6857) が使用された場合、レプリコン放出の抑制は明白であった(図3、右上)。lacIリプレッサーを提供するベクターを欠く場合(図3、左上)、プロベクターシステムのケースにおいてRNAレプリコン形成の抑制が全くないことが観察された。
【0041】
本発明の別の態様において(実施例3)、lacオペレーター配列を有するプロモーターの制御下、一過性に送達された第一の異種ヌクレオチド配列からのRNAレプリコンの放出を抑制するためのlacIリプレッサーを提供する構築物で安定的に形質転換されたトランスジェニック植物の能力を試験した。図4Aから、lacオペレーター配列含有構築物pICH17171のアグロ浸潤は、こうした配列を欠く構築物pICH16141での対照実験と比較して、効率的なRNAレプリコン放出を与えなかったことは明白である。同一構築物及び1mM IPTGでの前記トランスジェニック植物の同時浸潤は、pICH17171のケースにおいてはRNAレプリコン放出の誘導を導いたが(図4B、右パネル)、一方、IPTGなしの対照実験においては、RNAレプリコン放出は観察されなかった(図4B、左パネル)。細胞間移行が可能なRNAレプリコンを提供する構築物でのlacIリプレッサー含有植物の安定的再形質転換は、誘導されていない状態で重度のバックグラウンド発現を有する二重形質転換体を生み出した(図5)。LacI/lacOシステムについて得られたデータを要約すると、無視できるバックグラウンド発現でのRNAレプリコン放出に対する最良の制御が、細胞間移行が欠損したRNAレプリコンのケースで得られることが明白である(pICH17171、図4A;図4B、左パネル)。機能性MPを含有する構築物は、誘導されていない状態で重度のバックグラウンド発現を示した(pICH17424、図3、右上;図5、対照)。また、前記構築物を有する一次形質転換体は、最後には導入遺伝子発現を停止させ、それ故、目的とするタンパク質の高収率発現について役に立たなくなる。
【0042】
本発明の別の態様において、エタノール誘導性システムが、トランスジェニック植物におけるRNAレプリコン放出を制御するために使用された。構築物の設計は実施例5に記載されており、構築物の略図が図2Bに示されている。前記構築物の一過性発現実験の結果は、図6に示されている。alcRアクチベーターを有する構築物が同時浸潤されたところの実験にのみGFP発現を観察することができたので、エタノール誘導性システムはRNAレプリコン放出に対して厳格な制御を提供することが明白である。実際に、alcR構築物及び/又は化学的誘導剤エタノールがない場合には、バックグラウンド発現は観察されてなかった(無視できる発現が、alcR構築物の存在下で観察された)。4%エタノールの存在下、alcR構築物を有するRNAレプリコン(pICH18951)を提供する構築物が遺伝子導入されたN.ベンサミアナのアグロ浸潤は、強いGFP発現(図8)により報告されるRNAレプリコン形成を示した。驚くべきことに、エタノール誘導性システムにおいて、機能性MPを含有するウイルス構築物では一次形質転換体(RNAレプリコン放出に対する制御の漏れやすさと同等ではない)が得られなかった(実施例5)。このことは、RNAレプリコンの細胞間移行による植物カルス/細胞培養におけるシステムの漏れやすさにより説明し得る(Roberts et al., 2005, Plant Physiol., 138:1259-1267) 。この説明は、もしRNAレプリコンが使用された植物宿主中で細胞間移行のためのタンパク質をコードしていないならば、エタノール誘導性プロモーターの制御下でRNAレプリコンを提供する一次形質転換体が困難なく発生することができるという驚きの発見により支持される。我々は、RNAレプリコンを提供するベクターを有するトランスジェニック植物の交配子孫、及びalcR含有トランスジェニック体が、エタノール処理後、GFPの強いユビキタス発現を示すことを明らかにした(図9及び10)。この発明の別の態様において、我々は、我々のシステムが、GFPのようなレポーター遺伝子以外のタンパク質でも効率的に働くことを明らかにした。アプロチニンの発現のための構築物が図11に示されている。エタノール処理後、N.ベンサミアナ植物のF1子孫における組換えアプロチニン産生の分析は、クーマシー染色ゲル上の一つの主タンパク質バンドとして検出可能な高発現レベルを明らかにした。
【0043】
実施例において、我々はTMVに基づいたRNAレプリコンを使用した。しかしながら、異なった分類学的グループに属する多くの異なったウイルスを、本発明に従ったRNAウイルスに基づいたベクターの構築のために使用し得る。目、科及び属の名称は、ICTVにより承認されていればイタリック体である。引用符で囲まれている(及びイタリック体ではない)Taxa名は、この分類群がICTV国際承認名を有していないことを示している。種(その土地の言葉)名は通常書体で与えられている。属又は科への正式な帰属のないウイルスは示されている:
RNAウイルス:
ssRNAウイルス:科:ブロモウイルス科、属:アルファモウイルス属、基準種:アルファルファモザイクウイルス、属:イラルウイルス属、基準種:タバコ条斑病ウイルス、属:ブロモウイルス属、基準種:ブロムモザイクウイルス、属:ククモウイルス属、基準種:キュウリモザイクウイルス;
科:クロステロウイルス科、属:クロステロウイルス属、基準種:ビート萎黄ウイルス、属:クリニウイルス属、基準種:レタス伝染性黄斑ウイルス、科:コモウイルス科、属:コモウイルス属、基準種:ササゲモザイクウイルス、属:ファバウイルス属、基準種:ソラマメウイルトウイルス1、属:ネポウイルス属、基準種:タバコ輪点ウイルス;
科:ポティウイルス科、属:ポティウイルス属、基準種:ジャガイモYウイルス、属:ライモウイルス属、基準種:ドクムギモザイクウイルス、属:バイモウイルス属、基準種:大麦縞萎縮ウイルス;
科:セキウイルス科 、属:セキウイルス属、基準種:パースニップ黄色斑点ウイルス、属:ワイカウイルス属、基準種:イネ矮化ウイルス;科:トンブスウイルス科、属:カーモウイルス属、基準種:カーネーション斑紋ウイルス、属:ダイアントウイルス属、基準種:カーネーション輪点ウイルス、属:マクロモウイルス属、基準種:メイズクロロティック斑紋ウイルス、属:ネクロウイルス属、基準種:タバコ壊死ウイルス、属:トンブスウイルス属、基準種:トマトブッシースタントウイルス、ssRNAウイルスの割り当てられていない属、属:カピロウイルス、基準種:リンゴステムグルービング(stem grooving )ウイルス;
属:カーラウイルス属、基準種:カーネーション潜在ウイルス;属:エナモウイルス属、基準種:エンドウマメエネーション(enation )モザイクウイルス
属:フロウイルス属、基準種:ムギ類萎縮ウイルス、属:ホルデイウイルス属、基準種:オオムギ斑葉モザイクウイルス、属:イダエオウイルス属、基準種:ラズベリーブッシュドワーフウイルス(raspberry bushy dwarf virus);
属:ルテオウイルス属、基準種:オオムギ黄萎ウイルス;属:マラフィウイルス属、基準種:トウモロコシラヨドフィノ(rayodo fino )ウイルス;属:ポテクスウイルス属、基準種:ジャガイモウイルスX;属:ソベモウイルス属、基準種:インゲンマメ南部モザイクウイルス、属:テヌイウイルス属、基準種:イネ縞葉枯ウイルス、
属:トバモウイルス属、基準種:タバコモザイクウイルス、
属:トブラウイルス属、基準種:タバコ茎えそウイルス、
属:トリコウイルス属、基準種:リンゴクロロティックリーフスポット(chlorotic leaf spot )ウイルス;属:ティモウイルス属、基準種:ハクサイ黄化モザイクウイルス;属:アンブラウイルス属、基準種:ニンジン斑紋ウイルス;マイナスssRNAウイルス:目:モノネガウイルス目、科:ラブドウイルス科、属:サイトラブドウイルス属、基準種:レタス壊死性黄変病ウイルス、属:ヌクレオラブドウイルス属、基準種:ジャガイモ黄萎ウイルス;
マイナスssRNAウイルス:科:ブニヤウイルス科、属:トスポウイルス属、基準種:トマト黄化壊疽ウイルス;
dsRNAウイルス:科:パルティティウイルス科、属:アルファクリプトウイルス属、基準種:シロツメクサ潜在ウイルス1、属:ベータクリプトウイルス属、基準種:シロツメクサ潜在ウイルス2、科:レオウイルス科、属:フィジウイルス属、基準種:フィジー病ウイルス、属:ファイトレオウイルス属、基準種:創傷腫瘍ウイルス、属:イネウイルス属、基準種:イネラギッドスタント(ragged stunt )ウイルス;
割り当てられていないウイルス:ゲノムssDNA:種:バナナバンチートップ(bunchy top )ウイルス、種:ココナツ葉壊変ウイルス、種:サブテレーニアンクローバ スタント(subterranean clover stunt )ウイルス、
ゲノム:dsDNA、種:キュウリ葉脈イエローイング(yellowing )ウイルス;ゲノム:dsRNA、種:タバコ矮化ウイルス、
ゲノム:ssRNA、種:ニンニクウイルスA、B、C、D、種:ブドウつるフレックウイルス、種:トウモロコシホワイトライン(white line )モザイクウイルス、種:オリーブ潜在ウイルス2、種:ウルミアメロンウイルス、種:ペラルゴニウムゾナートスポット(zonate spot )ウイルス;
サテライト及びウイロイド:サテライト:ssRNAサテライトウイルス:サブグループ2サテライトウイルス、基準種:タバコネクローシスサテライト、
サテライトRNA、サブグループ2BタイプmRNAサテライト、サブグループ3Cタイプ直鎖RNAサテライト、サブグループ4Dタイプ環状RNAサテライト、
ウイロイド、基準種:ポテトスピンドルチュバーウィロイド(potato spindletuber viroid)。
【0044】
異なったRNAウイルスは、細胞間又は短距離移行のための一つ又はそれより多くの ウイルスタンパク質を有する。例えば、TMVの場合、一つのタンパク質(MP)が必要とされ;三要素ブロムモザイクウイルス(BMV)は、二つのタンパク質−3a及びCPを必要とする。一要素RNAウイルス、ジャガイモウイルスX(PVX)は、細胞間移行に関与する四つのタンパク質を有する:トリプルジーンブロック(triple gene block )(TGB)及びコートタンパク質(CP)によりコードされるタンパク質。しかしながら、細胞間移行に必要とされる二つまたはそれより多くのタンパク質の一つの欠損が、ウイルスベクターの効率的短距離移行を遮断するのに十分である。植物ウイルス移行タンパク質についてのさらなる詳細は、WJ Lucas (2006, Virology, 344:169-184) の最近の総説を参照されたい。
【0045】
本発明において、前記第一の異種ヌクレオチド配列は、前記RNAレプリコンをコードする配列セグメントを有する。もしくは、前記第一の異種ヌクレオチド配列は、一緒になって前記RNAレプリコンをコードする、一つより多くの配列セグメントを有することができ、即ち、前記RNAレプリコンは一つの連続したDNAによりコードされていない。代わりに、前記RNAレプリコンは二つまたはそれより多くの配列セグメントにより非連続的にコードされており、それにより前記セグメントはお互いに隣接して存在することができる。前記RNAレプリコンの形成は、例えば、組換えによる前記セグメントの再配列を必要とするであろう。前記組換えのためのリコンビナーゼは、工学処理された植物宿主により提供することができ、それ故、前記ウイルスレプリコンの誘導性発現を前記リコンビナーゼを発現することが可能な植物宿主に限局している。例としては、一つの配列セグメントは、前記RNAレプリコンの前記ポリメラーゼの一部をコードすることができ、及び前記ポリメラーゼの別の一部をコードする別の配列セグメントは、前記第一の異種ヌクレオチド配列中、第一の配列セグメントに関してフリップした(flipped)配向で存在することができる。フリップした部分に、組み換え部位が隣接することができる(WO2004/108934 参照)。この状況において、第一の異種ヌクレオチド配列又はそれらの配列セグメントの転写体は、機能性ポリメラーゼが転写体から翻訳できないので、RNAレプリコンではないであろう。組み換え部位を認識する部位特異的リコンビナーゼを提供することは、レプリコンが連続的にコードされるように前記セグメントの一つをフリップさせることを可能にする。この態様において、該リコンビナーゼは、RNAレプリコン形成及び誘導された状態の目的とする配列発現のスイッチを入れるためのスイッチとして機能することができ、高い生物学的安全性に寄与している。好ましくは、前記リコンビナーゼは誘導性プロモーターの制御下である。もしリコンビナーゼが本発明のプロセスのスイッチを入れるために使用されるならば、前記リコンビナーゼは前記植物又は植物の葉に一過性に提供することができ、それにより前記提供することが、前記一つ又は一つより多くの目的とするタンパク質を発現するためのスイッチとして働くことができる。好ましくは、こうしたリコンビナーゼは植物細胞中で安定にコードすることができ、構成的又は誘導性プロモーターの制御下でリコンビナーゼを発現する。前記プロモーターを誘導することによりリコンビナーゼ発現を誘導することは、次ぎに前記目的とする配列の発現を起こすことができる。一つの態様において、該リコンビナーゼは前記第一の異種ヌクレオチド配列によりコードされているであろうし、及び該リコンビナーゼの発現は前記第一の異種ヌクレオチド配列の誘導性プロモーターの制御下にあるであろう。
【0046】
もしくは、両方のセグメントは、異なった植物染色体に安定的に取り込まれた異なったT−DNA上に存在していてもよい。RNAレプリコンの形成は、両方のセグメントの転写及び両方の転写体のトランススプライシングを、前記RNAレプリコンを組み立てるために必要とするであろう。この態様は、WO02/097080 に記載されているように、子孫植物又は細胞中で一緒になって前記RNAレプリコンをコードするセグメントを迅速に分離するために使用することができ、それ故、システムの生物学的安全に寄与している。
【0047】
実施例において、植物ウイルスに由来する一つのタイプのRNAレプリコンをコードするトランスジェニック植物を記載する。しかしながら、本発明のプロセスにおいて、二つまたはそれより多くの異なったRNAレプリコン(特に二つまたはそれより多くの 異なった一要素RNAレプリコン)をトランスジェニック植物又は植物細胞において使用することができ、こうした異なったRNAレプリコンは、好ましくは異なった植物ウイルスから誘導される。前記異なったRNAレプリコンを誘導することができるこうした異なった植物ウイルスは、好ましくは相乗的又は非競合ウイルスである。本明細書において「相乗的」及び「非競合」は同意語として使用される。相乗的ウイルスは同一植物細胞中で同時に存在し、そして効率的に増幅することが可能である。同様に、相乗的RNAウイルスに由来するRNAレプリコンは同一植物細胞中で同時に存在し、そして効率的に増幅することが可能である。RNAレプリコンのこうした相乗的組み合わせの例は、一つのRNAレプリコンがTMVに由来し、及び他のRNAレプリコンがPVXに由来するRNAレプリコンの組み合わせである。こうした相乗的RNAレプリコンは、同一又は異なった誘導性プロモーターを利用する異なった発現カセットから放出し得る。相乗的RNAレプリコンは、同一植物細胞において、モノクローナル抗体の重鎖及び軽鎖のような、目的とする二つまたはそれより多くのタンパク質又はタンパク質サブユニットの発現のために使用することができる。異なった(非競合)ウイルスベクターを使用し、同一植物又は同一植物細胞中で二つまたはそれより多くの目的とするタンパク質を発現するプロセスは、その全体が本明細書において援用されるWO 2006/79546 (PCT/EP2006/000721)に記載されている。
【0048】
実施例において、植物細胞でのアグロバクテリウム仲介T−DNA送達を主に使用し、前記T−DNAは前記第一及び/又は前記第二の異種ヌクレオチド配列を含有する。微粒子銃、エレクトロポレーション又はプロトプラストのPEG仲介形質転換による細胞内への異種ヌクレオチド配列の直接導入のような多様な方法を、植物細胞内へのベクターの送達のために使用することができる。アグロバクテリウム仲介植物形質転換が好ましい。それ故、アグロバクテリウム(US 5,591,616; US 4,940,838; US 5,464,763)、粒子及び微粒子銃(US 05100792; EP 00444882B1; EP 00434616B1)により運ばれるTi−プラスミドベクターによるような多様な技術により、異種ヌクレオチド配列を植物細胞内に形質転換することができる。原則として、他の植物形質転換法、例えば、マイクロインジェクション(WO 09209696; WO 09400583A1; EP 175966B1)、エレクトロポレーション(EP00564595B1; EP00290395B1; WO 08706614A1)なども使用し得る。形質転換法の選択は、中でも、形質転換されるべき植物種に依存する。例えば、微粒子銃は単子葉植物形質転換のために好ましく、一方、双子葉植物にはアグロバクテリウム仲介形質転換が一般的により良好な結果を与える。
【0049】
本発明は好ましくは、より高等な多細胞植物で実施される。本発明での使用に好まし
い植物には、農学的及び園芸的に重要な種に与えられた優先度を有するいずれかの植物種が含まれる。本発明で用いる普通の作物には、アルファルファ、オオムギ、マメ、アブラナ、ササゲ、綿、トウモロコシ、クローバ、ハス、レンズマメ、ルピナス、キビ(millet )、カラスムギ、エンドウマメ、ピーナッツ、コメ、ライムギ、スイートクローバー、ヒマワリ、スイートピー、ダイズ、モロコシ、ライ小麦、クズイモ、ビロードマメ、カラスノエンドウ、コムギ、フジ及びナッツ植物が含まれる。本発明を実施するための好ましい植物種には、限定されるわけではないが:イネ科、キク科、ナス科及びバラ科の代表種が含まれる。加えて、本発明に使用するための好ましい種、ならびに上に特定された種には属:シロイヌナズナ属、コヌカグサ属、ネギ属、キンギョソウ属、セロリ属、ラッカセイ属、アスパラガス属、アトローパ属、アベナ属、バンブーサ属、アブラナ属、スズメノチャヒキ属、ブロワリア属、ツバキ属、カンナビス属、カプシクム属、ヒヨコマメ属、アカザ属、キクニガナ属、シトラス属、コーヒー属、ジュズダマ属、キュウリ属、カボチャ属、ギョウギシバ属、カモガヤ属、チョウセンアサガオ属、ニンジン属、ジギタリス属、ヤマノイモ属、アブラヤシ属、オヒシバ属、ウシノケグサ属、オランダイチゴ属、フウロソウ属、グリシン(Glycine)、ヒマワリ属、ワスレグサ属、ヘベア属、オオムギ属、ヒヨス属、サツマイモ属、アキノノゲシ属、レンズマメ属、ユリ属、アマ属、ドクムギ属、ハス属、リコペリシコン属、マジョラム(Majorana)、リンゴ属、マンゴー属、マニホット属、ウマゴヤシ属、ネメシア属、タバコ属、オノブリキス属 、イネ属、キビ属、ペラルゴニウム属、ペニセタム属、ペチュニア属 、エンドウ属、インゲンマメ属、アワガエリ属、イチゴツナギ属、サクラ属、キンポウゲ属、ダイコン属、スグリ属、トウゴマ属、キイチゴ属、サトウキビ属、サルメンバナ属、ライムギ属、セネキオ属、エノコログサ属、シロガラシ属、ナス属、モロコシ属、イヌシバ属、カカオ属、シャジクソウ属、レイリョウコウ属、コムギ属、ソラマメ属、ササゲ属、ブドウ属、トウモロコシ属及びオリレア属(Olyreae) 、ファルス亜科及び多くの他のものが含まれる。
【0050】
本発明の一つの態様において、TMVに由来するRNAレプリコンがタバコ属植物に使用される。別の態様において、PVXに由来するRNAレプリコンがタバコ属植物に使用される。
【0051】
本発明を使用し、センス又はアンチセンス配向で発現し得る、目的とするタンパク質又はそれらの断片には:デンプン修飾酵素(デンプンシンターゼ、デンプンリン酸化酵素、脱分枝酵素、デンプン分枝酵素、デンプン分枝酵素II、顆粒結合性デンプンシンターゼ)、スクロースリン酸シンターゼ、スクロースホスホリラーゼ、ポリガラクツロナーゼ、ポリフルクタンスクラーゼ、ADPグルコースピロホスホリラーゼ、シクロデキストリングリコシルトランスフェラーゼ、フルクトシルトランスフェラーゼ、グリコーゲンシンターゼ、ペクチンエステラーゼ、アプロチニン、アビジン、細菌レバンスクラーゼ、大腸菌glgAタンパク質、MAPK4及び相同分子種、窒素同化/代謝酵素、グルタミンシンターゼ、植物オスモチン、2Sアルブミン、タウマチン、部位特異的リコンビナーゼ/インテグラーゼ(FLP、Cre、Rリコンビナーゼ、Int、SSVIインテグラーゼR、インテグラーゼphiC31、又はそれらの活性フラグメント又は変異体)、イソペンテニルトランスフェラーゼ、ScaM5(ダイズカルモジュリン)、鞘翅目型毒素又は殺虫活性断片、ユビキチン結合酵素(E2)融合タンパク質、脂質を代謝する酵素、アミノ酸、糖、核酸及びポリサッカリド、スーパーオキシドジスムターゼ、プロテアーゼの不活性プロ酵素型、植物タンパク質毒素、繊維産生植物中の形質変形繊維、バチルス・チューリンゲンシスからの鞘翅目活性毒素(Bt2トキシン、殺虫性結晶タンパク(ICP)、CryICトキシン、デルタエンドトキシン、ポリペプチドトキシン、プロトキシンなど)、昆虫特異的トキシンAaIT、セルロース分解酵素、アシドサーマス・セルロリティカスからのE1セルラーゼ、リグニン修飾酵素、シンナモイルアルコールデヒドロゲナーゼ、トレハロース−6−リン酸シンターゼ、サイトカイニン代謝経路の酵素、HMG−CoAレダクターゼ、大腸菌無機ピロホスファターゼ、種子貯蔵タンパク質、エルウィニア・ヘルビコーラリコペンシンターゼ、ACCオキシダーゼ、pTOM36コードタンパク質、フィターゼ、ケトヒドロラーゼ、アセトアセチルCoAレダクターゼ、PHB(ポリヒドロキシブタノエート)シンターゼ、アシルキャリアータンパク質、ナピン、EA9、非高等植物フィトエンシンターゼ、pTOM5コードタンパク質、ETR(エチレンレセプター)、色素体ピルビン酸リン酸ジキナーゼ、線虫誘導性膜貫通ポアタンパク質、植物細胞の光合成又は色素体機能を増強する形質、スチルベンシンターゼ、フェノールのヒドロキシル化を可能にする酵素、カテコールジオキシゲナーゼ、カテコール 2,3−ジオキシゲナーゼ、クロロムコネートシクロイソメラーゼ、アントラニル酸シンターゼ、アブラナ属AGL15タンパク質、フルクトース 1,6−ジホスファターゼ(FBPase)、AMV RNA3、PVYレプリカーゼ、PLRVレプリカーゼ、ポティウイルスコートタンパク質、CMVコートタンパク質、TMVコートタンパク質、ルテオウイルスレプリカーゼ、MDMVメッセンジャーRNA、突然変異体ジェミニウイルスレプリカーゼ、ウンベルラリア・カルフォルニアC12:0選択性アシル−ACPチオエステラーゼ、植物C10又はC12:0選択性アシル−ACPチオエステラーゼ、C14:0選択性アシル−ACPチオエステラーゼ(luxD)、植物シンターゼ因子A、植物シンターゼ因子B、6−デサチュラーゼ、植物細胞中でペルオキシソーム−脂肪酸の酸化に酵素活性を有するタンパク質、アシル−CoAオキシダーゼ、3−ケトアシル−CoAチオラーゼ、リパーゼ、トウモロコシアセチル−CoA−カルボキシラーゼ、5−エノールピルビルシキミ酸−3−リン酸シンターゼ(EPSP)、ホスフィノチリシンアセチルトランスフェラーゼ(BAR、PAT)、CP4タンパク質、ACCデアミナーゼ、リボザイム、翻訳後切断部位を有するタンパク質、Gal4転写アクチベーターのDNA結合ドメイン及び転写活性化ドメインから成るタンパク質融合物、脂質相内へ融合タンパク質を標的化できるオレオシンタンパク質と目的とするタンパク質の翻訳融合物、スルホンアミド抵抗性を与えているDHPS遺伝子、細菌ニトリラーゼ、2,4−Dモノオキシゲナーゼ、アセト乳酸シンターゼ又はアセトヒドロキシ酸シンターゼ(ALS、AHAS)、ポリガラクツロナーゼ、細菌ニトリラーゼ、色素体の内部エンベロープ膜に存在する成熟リン酸輸送体タンパク質のアミノ末端疎水性領域と前記膜内へ標的化されるべき目的とするタンパク質との融合物が含まれる。
【0052】
いずれのヒト又は動物タンパク質も本発明のシステムを使用して発現し得る。例えば、こうした目的とするタンパク質の例には、中でも、以下のタンパク質(医薬タンパク質)が含まれる:免疫応答タンパク質(モノクローナル抗体、単鎖抗体、T細胞レセプターなど)、抗原、コロニー刺激因子、レラキシン、ポリペプチドホルモン、サイトカイン及びそれらのレセプター、インターフェロン、増殖因子及び凝固因子、酵素的に活性なリソソーム酵素、線維素溶解ポリペプチド、血液凝固因子、トリプシノーゲン、1−アンチトリプシン(AAT)、ならびに機能保存的タンパク質様融合物、上記タンパク質の突然変異体バージョン及び合成誘導体。
【0053】
本特許出願により優先権が主張される、2006年5月29日に出願された欧州特許出願番号06011002及び2006年6月2日に出願された米国仮特許出願60/810,398の開示は、それらの全体が本明細書において援用される。
【実施例】
【0054】
実施例1
IPTG誘導性lacシステム:構築物設計
lacリプレッサー (lacI, Acc. J01636)は、プライマーlacIpr1(配列番号:1)(5’-gat cca tgg aac cag taa cgt tat ac -3’)及びlacIpr2(配列番号:2)(5’-tc tgg atc ctc act gcc cgc ttt cca gtc g-3’)を使用するPCRにより増幅し、NcoI−BamHI断片として標準バイナリーベクターpICBV1内にクローン化すると、構築物pICH17155(図2A)を得た。核移行シグナル(NLS)は、lacIpr1の代わりにプライマーlacIpr5(配列番号:3)(5’-cgc cat ggg ccc taa gaa gaa gag gaa ggt tga acc agt aac gtt ata cga tgt c -3’)を使用することによりN末端に導入し、構築物pICH17401(図2A)を得た。この構築物は標準形質転換技術(Horsh et al., 1985, Science, 227, 1229-1231) を使用し、ニコチニア・タバクム及びN.ベンサミアナ植物内に安定的に形質転換した。
【0055】
合成lacオペレーター配列(配列番号:4)(aat tgt gag cgc tca caa tt)は、TATAボックス及びいくつかのウイルスベクターのアクチン2−プロモーター(An et al., 1996, Plant J., 10:107-121) の転写開始点間に導入した。このことは、プライマーA:brb4nosph(配列番号:5)(5’- gga acc ctg tgg ttg gca cat -3’)及びlacOact2pr2(配列番号:6)(5’- cga att gtg agc gct cac aat tta tat agg cgg gtt tat ctc -3’)、及びプライマーB:lacOactpr1(配列番号:7)(5’- taa att gtg agc gct cac aat tcg ctt tga agt ttt agt ttt att g -3’)及びrdrppr4(配列番号:8)(5’- ttt ctg cag gaa atg aaa ggc cgc gaa aca ag -3’)で作製した二つの重複PCR生成物を組み合わせることにより行われた。生じた生成物をKpn1−SphI断片としてウイルスベクターpICH16141内にクローン化して、ベクターpICH17171を得た。追加のウイルスベクターは、都合のよい制限酵素を使用し、プロモーターのサブクローニングによりpICH17171から誘導した。これらのベクターは発現のために最適化し(Marillonnet et al., 2005, Nat Biotechnol., 23:718-723)、そして完全MP(pICH18867)又はMPの欠失(pICH17171)を含有する。加えて、一過性試験に有用であることが証明されている5’−プロベクター(Marillonnet et al., 2004, Proc Natl Acad Sci USA, 101:6852-6857)を構築した(pICH17424)(実施例2参照)。
【0056】
実施例2
一過性システムにおける抑制効率の試験
一過性発現実験は、試験下で異なった構築物を宿しているアグロバクテリア株の混合物を使用して実施した。完全に組み立てられたウイルスベクターを使用して間は、一過性アッセイにおいて何の抑制も観察できなかった。多分、リプレッサーの翻訳に先立って、該構築物はウイルスRNAレプリコンにすでに転写されていた。それ故、部位特異的リコンビナーゼ(Marillonnet et al.,2004, Proc Natl Acad Sci USA, 101:6852-6857)の活性により、ウイルス性RNAレプリコンのDNA前駆体へ植物中で組み立てられる、ウイルスプロベクターpICH17424(図2A)、及びpICH6892(Marillonnet et al., 2004, Proc Natl Acad Sci U S A, 101:6852-6857)を使用した。この追加の工程はウイルスベクターの組立を遅延させるはずであり、及び翻訳される及びウイルス構築物中のオペレーター配列に結合される十分な時間をリプレッサーに提供する。実際、このアプローチを使用すると、ウイルス増幅の強い抑制が観察できた(図3)。
【0057】
実施例3
安定的に形質転換された植物におけるLacIリプレッサー活性及びIPTGによる誘導
リプレッサー構築物pICH17401(図2A)を、標準形質転換技術(Horsh et al., 1985, Science, 227, 1229-1231)を使用し、N.タバクム及びN.ベンサミアナ中に安定的に形質転換した。リプレッサー活性は、lacO含有ウイルス構築物での形質転換された植物のアグロ浸潤により示された(図4A)。誘導能は同一構築物でのアグロ浸潤により同様に試験され、浸潤緩衝液中にIPTGを含有していた(図4B)。
【0058】
実施例4
ウイルス構築物でのリプレッサー含有植物の再形質転換
ゲノム中に安定に組み込まれたlacIリプレッサー組換えDNA(pICH17401、図2A)を運んでいるN.タバクムを、lacオペレーター遺伝子(pICH18867、図2A)を含有する全ウイルスベクター構築物で2回目の形質転換を行った。いくつかの植物を再生できたが、それらのすべてが多かれ少なかれウイルスベクターの重度のバックグラウンド発現を示した。
【0059】
二重形質転換体の葉内へのIPTG(5mM)の浸潤は、トランスジェニック植物のいくつかにおいてRNAレプリコン増幅を実際に誘導できた(図5)。しかしながら、RNAレプリコンのリーキーな発現を有するすべての一次形質転換体は、最終的には導入遺伝子サイレンシングを受け、そして誘導条件下で何のRNAレプリコンの放出も示さない子孫を産生したことが観察された。
【0060】
実施例5
エタノール誘導性システムのための構築物の設計
エタノール誘導性システムの原理は、Caddick及び共同研究者(1998, Nat Biotechnol., 16:177-180)により記述されている。CaMV35Sプロモーターの制御下の転写アクチベーターalcR(構築物pICH18693、図2B)は、Caddick及び共同研究者(1998, Nat Biotechnol., 16:177-180)により記述されているように設計した。この構築物はN.タバクム及びN.ベンサミアナ植物内に安定的に形質転換した(Horsh et al.,1985, Science, 227, 1229-1231)。alcAプロモーターは、プライマーalcApr1(配列番号:9)(5’- cat gaa ttc tag gat tgg atg cat gcg g -3’)及びalcApr2(配列番号:10)(5’- cag ctc gag gtc gtc ctc tcc aaa tga aat g -3’)を使用するPCRにより増幅し、機能性MPを有する(pICH18969、図2B)又は有しない(pICH18951、図2B)TMVに基づいたウイルスベクターに、及び別に機能性ウイルスMP(pICH19940、図2B)にEcoRI−Xho1断片として融合した。加えて、構築物pICH18951及びpICH19940を一つのベクター(pICH20592、図2B)に結合させた。pICH18969を除いたこれらすべての構築物は、標準形質転換技術を使用してN.タバクム及びN.ベンサミアナ内の両方に形質転換した。機能性MPを伴ったRNAレプリコンをコードする構築物を有する一次形質転換体を得ることが可能であった。
【0061】
alcAプロモーターも、プライマーalcApr4(配列番号:11)(5’- cgc gca tgc tac tag gat tgg ata cat gcg gaa c -3’)及びalcApr5(配列番号:12)(5’- ttt ggt ctc atc aac tcc aaa tga aat gaa ctt cc -3’)を使用して増幅し、PVXに基づいたウイルスベクターpICH25233内にSphI−BsaI断片としてクローン化し、35Sプロモーターを置き換えて構築物pICH26022(図2C)を得た。alcAプロモーターの、細胞間移行に関与するPVXコートタンパク質への融合は(pICH26356、図2C)、pICH19940からのEcoRI−SacI断片のpICH22066内へのクローニングにより行われた。
【0062】
実施例6
エタノールにより誘導されるTMV構築物の一過性発現
上述の構築物はN.ベンサミアナ植物でアグロ浸潤により試験された(図6)。植物は2日後、4%エタノール溶液又は対照として水で処理した。ウイルスベクターの増幅及びGFPの発現はエタノール処理した植物において、及びアクチベーターalcRの存在下でのみ誘導された。非常に弱いバックグラウンド発現が、alcRの存在下、対照植物で観察された。
【0063】
実施例7
エタノールによって誘導されたPVX構築物の一過性発現
alcA−CP構築物(pICH26356)をalcR(pICH18693)及びCP欠損ウイルスベクター(pICH21692)と同時浸潤した。細胞間移行はエタノールで処理した植物でのみ検出でき、35Sプロモーター−CP構築物との相違は観察できなかった(図7)。
【0064】
実施例8
ウイルス構築物pICH18951及びpICH20592で安定的に形質転換した植物の分析
N.ベンサミアナ及びN.タバクム植物を、標準プロトコル(Horsh et al., 1985, Science, 227, 1229-1231) に従って形質転換した。再生した植物を、alcR構築物(pICH18693)でのアグロ浸潤、及びエタノール処理により導入遺伝子の存在を分析した。実際、ほとんどの植物は、葉の浸潤部分でGFP発現を示し、他の部分のバックグランドはなかった(図8)。
【0065】
実施例9
全トランスジェニック植物の誘導
pICH18951か又はpICH20592(実施例8に記載されている)を含有するトランスジェニック植物を、転写アクチベーターalcR(pICH18693)を含有する植物と交配した。これらの植物のF1子孫を、4%エタノールの噴霧により、又は根水浸(1%エタノールでの)及び噴霧(4%エタノール)の組み合わせによりエタノールで処理した。ウイルス増幅及びそれ故、GFP発現がこれらの植物のほとんど全ての部分で検出された(図9)。最も注目されたのは、強い発現が根水浸で処理した植物の茎及び葉柄でも検出された。植物のこれらの部分は通常、標準Magnifection法、即ち、全植物の減圧浸潤(Marillonnet et al., 2005, Nat. Biotechnol., 23:718-723) を使用すると、発現を示さないか又は弱い発現のみしか示さない。根水浸せずに4%エタノールで植物を噴霧すると、軟葉組織のみでのGFP発現を導き、茎及び葉柄では発現されない(図10)。
【0066】
実施例10
植物における組換えアプロチニンの発現のための誘導性ウイルスベクターシステムの使用
プラスミドpICH25408(図11)は、プラスミドpICH20592(実施例5参照)と同様の様式で設計した。N.ベンサミアナ植物を標準プロトコル(Horsh et al., 1985, Science, 227, 1229-1231)に従ってpICH25408で形質転換した。再生した植物を、alcR構築物(pICH18693)でのアグロ浸潤、及びエタノール処理により、続いてのポリアクリルアミドゲル(PAAG)電気泳動による組換えタンパク質発現の分析により導入遺伝子の存在を分析した。alcR構築物(pICH18693)でアグロ湿潤し、及びエタノールで処理した葉組織の一部を、2xLaemmli緩衝液(125mMトリス−HCl、pH7.8、10%メルカプトエタノール、20%グリセリン、0.001%ブロムフェノールブルー、10%SDS)による全可溶性タンパク質抽出、続いてのPAAGでの電気泳動分離に使用した。異なった一次形質転換体についてのこうした分析の結果は図12に示されている。
【0067】
次の工程において、pICH25408で形質転換した植物をalcR遺伝子(pICH18693)を運んでいる植物と交配した。エタノール処理後、F1子孫の組織葉から単離された全可溶性タンパク質をPAAGで分析した。この分析の結果は図13に示されている。アプロチニンが高レベルで発現されたことが明白である(約1mg/新鮮葉バイオマスのg)。
【図面の簡単な説明】
【0068】
【図1】本発明の一般原理。A−細胞間移行が可能なRNAレプリコンの伝搬;B−細胞への細胞間移行が欠損している本発明のRNAレプリコンの限局、ここでは前記RNAレプリコンの予定外形成が起こった。
【図2A】図2Aは、プラスミドpICH17155、pICH17401、pICH16141、pICH17171、pICH18867、pICH17424及びpICH17388のT−DNAドメインを描いている。
【図2B】図2Bは、プラスミドpICH18693、pICH18969、pICH18951、pICH19940及びpICH20592のT−DNAドメインを描いている。
【図2C】図2Cは、プラスミドpICH26022及びpICH26356のT−DNAドメインを描いている。TVCVポリメラーゼを示しているより大きな灰色のボックス中の白色ボックスは、核中で転写体を安定化し、そして結果として細胞核で形成されたRNAレプリコンをサイトゾルへより効率的に輸送させるイントロンを示す。3’Nos−ノパリンシンターゼ遺伝子の転写終結ドメイン;pNos−ノパリンシンターゼ遺伝子のプロモーター;p35S−CaMVの35Sプロモーター;pAct2−A.タリアナアクチン2遺伝子のプロモーター;NLS−核移行シグナル;lacO−大腸菌lacオペロンのオペレーター配列;lacI−大腸菌lacオペロンのレプレッサー遺伝子;BAR−除草剤ホスフィノトリシンに耐性を与えている遺伝子;int−イントロン配列の5’部分;AttP−インテグラーゼphC31により認識される組換え部位;NPTII−ネオマイシンホスホトランスフェラーゼII;sGFP−合成緑色蛍光タンパク質;NTR−トバモウイルス非翻訳領域;PalcA−アルコールデヒドロゲナーゼをコードする誘導性A.ニデュランスalcA遺伝子の誘導性プロモーター;alcR−アスペルギルス・ニデュランスのalcレギュロンの転写アクチベーター;TVCV MP−ハクサイ葉脈透化ウイルス移行タンパク質;PVX CP−ジャガイモウイルスXコートタンパク質;PVX Pol−ジャガイモウイルスX RNA依存性RNAポリメラーゼ;25K、12K、8K、トリプル遺伝子ブロック;sgp−サブゲノムプロモーター。
【図3】図3は、誘導性lacシステムでの一過性発現試験を示している。葉を異なった組み合わせ構築物で浸潤し、処理6日後、紫外光下でモニターした。以前暗いバックグラウンドであった光パッチはGFP蛍光を示している。葉の左側:lacリプレッサーなし;葉の右側:lacリプレッサーの存在下。pICH17424はlacOを有する5’−プロベクターであり、pICH17388はlacOがない対応する対照構築物である。pICH17401はlacリプレッサー構築物である。すべてのサンプルは、GFP3’−プロベクター及びインテグラーゼも含有する。
【図4A】図4Aは、lacリプレッサーで安定的に形質転換した植物の葉を示す。植物は、そのプロモーター中にlacO配列を含有するベクターpICH17171で、及びlacO配列を欠く、対応する対照構築物pICH16141でアグロ湿潤した。
【図4B】図4Bは、IPTG処理による抑制の解放を示す。lacリプレッサーで安定的に形質転換したN.ベンサミアナ植物を、そのプロモーター中にlacO配列を含有するベクターpICH17171で、及び対応する対照構築物pICH16141でアグロ湿潤した。誘導のため(右の写真)、5mM IPTGを浸潤緩衝液中に含ませた。左の写真は、誘導剤IPTGがないものである。
【図5】図5は、RNAレプリコンを提供する構築物での、lacIリプレッサー含有植物(pICH17155又はpICH17401)の再形質転換を示す。植物を5mM IPTGで浸潤した。系統N6(左の写真)は高い誘導能を示しているが、高いバックグラウンドも示し、一方、系統N8(右の写真)は低いバックグラウンド及び低い誘導能を示す。
【図6】図6は、TMVベクターに基づいたエタノール誘導性システムでの一過性発現試験を示す。対照:浸潤2日後に植物を水で処理をした;エタノール処理:浸潤2日後に植物を4%エタノールで処理をした。
【図7】図7は、PVXベクターに基づいたエタノール誘導性システムを使用する一過性発現試験を示す。対照:浸潤2日後に植物を水で処理をした;エタノール処理:浸潤2日後に植物を4%エタノールで処理をした。
【図8】図8は、紫外光下の、pICH18951のT−DNAを運んでいる安定な形質転換体の葉を示す。緑色(光)スポットは、pICH18693運搬アグロバクテリアで浸潤し及びアルコールを噴霧した領域に対応する。
【図9】図9は、水性アルコール溶液での処理後(灌水−1%アルコール、及び噴霧−4%アルコール)にGFPを発現している、トランスジェニックN.ベンサミアナ植物(F1子孫)を示す。
【図10】図10は、4%アルコール溶液で噴霧後、GFPを発現している、トランスジェニックN.タバクム植物(F1子孫)を示す。
【図11】図11は、プラスミドpICH25408のT−DNAドメインを描いている。TVCVポリメラーゼを示しているより大きな灰色のボックス中の白色ボックスは、核中で転写体を安定化し、そして結果として細胞核で形成されたRNAレプリコンをサイトゾルへ輸送させるイントロンを示す。TVCVポリメラーゼを示しているより大きな灰色のボックス中の白色ボックスは、核中で転写体を安定化し、そして結果として細胞核で形成されたRNAレプリコンをサイトゾルへ輸送させるイントロンを示す。3’Nos−ノパリンシンターゼ遺伝子の転写終結ドメイン;pNos−ノパリンシンターゼ遺伝子のプロモーター;3’Ocs−オクトピンシンターゼ遺伝子の転写終結ドメイン;NPTII−ネオマイシンホスホトランスフェラーゼII;NTR−トバモウイルス非翻訳領域;PalcA−アルコールデヒドロゲナーゼをコードする誘導性A.ニデュランスalcA遺伝子の誘導性プロモーター;alcR−アスペルギルス・ニデュランスのalcレギュロンの転写アクチベーター;TVCV MP−ハクサイ葉脈透化ウイルス移行タンパク質;sgp−サブゲノムプロモーター。
【図12】図12は、アプロチニン遺伝子を有するプラスミドpICH25408のT−DNAドメインを運んでいる異なったトランスジェニック植物(N18、N19及びN20)から抽出された全可溶性タンパク質の電気泳動分析の結果を伴ったクーマシー染色ポリアクリルアミドゲルを示す。レーン1−対照、未処理植物から単離された全可溶性タンパク質;レーン2−alcRアクチベーター(pICH18693)をコードするプラスミドを運んでいるアグロバクテリアで浸潤され、及び4%エタノールで処理された植物;レーン3−alcRアクチベーターをコードするプラスミド(pICH18693)、アプロチニン遺伝子を運んでいるウイルスベクターをコードするプラスミド(pICH25408)を運んでいるアグロバクテリアの混合物で浸潤され、及び4%エタノールで処理された植物。
【図13】図13は、アプロチニン遺伝子を有するウイルスベクターをコードするプラスミドpICH25408のT−DNA領域を運んでいるトランスジェニック植物、及びalcRアクチベーターをコードするT−DNA領域(pICH18693)を運んでいるトランスジェニック植物間の交配から得られたF1植物から抽出された全可溶性タンパク質の電気泳動分析の結果を伴ったクーマシー染色ポリアクリルアミドゲルを示す。
【特許請求の範囲】
【請求項1】
一つ又一つより多くの目的とするタンパク質を産生するプロセスであって:
(a)植物又は植物細胞を提供することであって、前記植物又は植物細胞が、
(i)RNAレプリコンをコードするヌクレオチド配列;及び
前記RNAレプリコンをコードする前記ヌクレオチド配列に機能可能なように連結された第一の誘導性プロモーター;
を含んでなる第一の異種ヌクレオチド配列;
前記RNAレプリコンは、前記植物中で前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない;
前記RNAレプリコンは、前記RNAレプリコン及び前記一つ又は一つより多くの目的とするタンパク質を複製するためのポリメラーゼをコードする;及び
(ii)前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードする配列を含んでなる第二の異種ヌクレオチド配列、ここで前記第二の異種ヌクレオチド配列は、前記RNAレプリコンの細胞間移行を可能にする前記タンパク質をコードする前記配列に機能可能なように連結された第二の誘導性プロモーターを含んでなる;
を含んでなり;及び
(b)工程(a)の前記植物又は植物細胞において、前記誘導性プロモーターを誘導し、それにより、前記植物又は前記植物細胞において前記一つ又は一つより多くの目的とするタンパク質を産生すること;
を含んでなる、前記プロセス。
【請求項2】
前記第一及び前記第二の誘導性プロモーターが同一の誘導シグナルにより、誘導可能である、請求項1に記載のプロセス。
【請求項3】
前記第一又は前記第二の誘導性プロモーターが化学的誘導性プロモーターである、請求項1又は2に記載のプロセス。
【請求項4】
前記化学的誘導性プロモーターが、エタノール誘導性プロモーター、IPTG誘導性プロモーター及びテトラサイクリン誘導性プロモーターから成る群より選択される、請求項3に記載のプロセス。
【請求項5】
前記誘導性プロモーターが熱ショック誘導性プロモーターである、請求項1から4のいずれか一項に記載のプロセス。
【請求項6】
前記RNAレプリコンがプラスセンス一本鎖RNAウイルスから誘導される、請求項1から5のいずれか一項に記載のプロセス。
【請求項7】
前記プラスセンス一本鎖RNAウイルスがタバコモザイクウイルス又はジャガイモウイルスXである、請求項6に記載のプロセス。
【請求項8】
前記植物が、さらなるRNAレプリコンをコードするヌクレオチド配列及び前記さらなるRNAレプリコンをコードする前記配列に機能可能なように連結された第三の誘導性プロモーター、を含んでなる第三の異種ヌクレオチド配列を含んでなり、前記さらなるRNAレプリコンが、前記植物中の前記RNAレプリコン又は前記さらなるRNAレプリコンの細胞間移行を提供するタンパク質をコードしておらず、前記さらなるRNAレプリコンが目的とするタンパク質をコードする、請求項1から7のいずれか一項に記載のプロセス。
【請求項9】
前記植物が、前記RNAレプリコンの細胞間移行を可能にする前記タンパク質をコードするヌクレオチド配列に機能可能なように連結された、前記さらなるRNAレプリコンの細胞間移行を可能にするタンパク質をコードするヌクレオチド配列を含んでなる異種ヌクレオチド配列を含んでなる、請求項8に記載のプロセス。
【請求項10】
前記RNAレプリコンの細胞間移行を可能にする前記タンパク質及び 前記さらなるRNAレプリコンの細胞間移行を可能にする前記タンパク質が同一タンパク質であるか又は異なったタンパク質である、請求項9に記載のプロセス。
【請求項11】
前記さらなるレプリコンが、前記さらなるRNAレプリコンを複製するためのポリメラーゼをコードする、請求項8に記載のプロセス。
【請求項12】
前記第三の誘導性プロモーターが、前記第一の誘導性プロモーターと同一の誘導剤により誘導可能である、請求項8から11のいずれか一項に記載のプロセス。
【請求項13】
前記RNAレプリコン及び前記さらなるRNAレプリコンが、非競合 レプリコンである、請求項8から12のいずれか一項に記載のプロセス。
【請求項14】
前記非競合レプリコンが、異なったウイルス属の植物ウイルスから誘導される植物ウイルスレプリコンである、請求項13に記載のプロセス。
【請求項15】
工程(a)が、前記第一の異種ヌクレオチド配列及び、任意選択で、前記第二の及び/又は第三の異種ヌクレオチド配列を含んでなる種子から植物を生長させることを含んでなる、請求項1から14のいずれか一項に記載のプロセス。
【請求項16】
植物又は植物細胞であって:
核染色体中に、RNAレプリコンをコードするヌクレオチド配列及び前記RNAレプリコンをコードする前記ヌクレオチド配列に機能可能なように連結された第一の誘導性プロモーターを含んでなる第一の異種ヌクレオチド配列;
前記RNAレプリコンは、前記植物中での前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない;
前記RNAレプリコンは、前記RNAレプリコンを複製するためのポリメラーゼをコードする;
を含んでなる、前記植物又は植物細胞。
【請求項17】
植物又は植物細胞であって;
(i)核染色体中に、RNAレプリコンをコードするヌクレオチド配列及び前記RNAレプリコンをコードする前記ヌクレオチド配列に機能可能なように連結された第一の誘導性プロモーターを含んでなる第一の異種ヌクレオチド配列;
前記RNAレプリコンは、前記植物中での前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない;
前記RNAレプリコンは、前記RNAレプリコンを複製するためのポリメラーゼをコードする;及び
(ii)前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードするヌクレオチド配列を含んでなる第二の異種ヌクレオチド配列、前記第二の異種ヌクレオチド配列は、前記RNAレプリコンの細胞間移行を可能にする前記タンパク質をコードする前記ヌクレオチド配列に機能可能なように連結された第二の誘導性プロモーターを含んでなる;
を含んでなる、前記植物又は植物細胞。
【請求項18】
タバコ属に属する、請求項16又は17に記載の植物又は植物細胞。
【請求項19】
請求項17で定義した植物又は植物細胞を産生するプロセスであって、植物核染色体内に請求項17で定義した第一の異種ヌクレオチド配列及び請求項17で定義した第二の異種ヌクレオチド配列を導入すること、及び前記第一の及び前記第二の異種ヌクレオチド配列を含有する形質転換された植物を再生することを含んでなる、前記プロセス。
【請求項20】
請求項8で定義した植物又は植物細胞産生するプロセスであって、前記第一の異種ヌクレオチド配列及び任意選択で、前記第二の異種ヌクレオチド配列を含んでなる第一の植物と、前記第三の異種ヌクレオチド配列を含んでなる第二の植物を交配することを含んでなる、前記プロセス。
【請求項1】
一つ又一つより多くの目的とするタンパク質を産生するプロセスであって:
(a)植物又は植物細胞を提供することであって、前記植物又は植物細胞が、
(i)RNAレプリコンをコードするヌクレオチド配列;及び
前記RNAレプリコンをコードする前記ヌクレオチド配列に機能可能なように連結された第一の誘導性プロモーター;
を含んでなる第一の異種ヌクレオチド配列;
前記RNAレプリコンは、前記植物中で前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない;
前記RNAレプリコンは、前記RNAレプリコン及び前記一つ又は一つより多くの目的とするタンパク質を複製するためのポリメラーゼをコードする;及び
(ii)前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードする配列を含んでなる第二の異種ヌクレオチド配列、ここで前記第二の異種ヌクレオチド配列は、前記RNAレプリコンの細胞間移行を可能にする前記タンパク質をコードする前記配列に機能可能なように連結された第二の誘導性プロモーターを含んでなる;
を含んでなり;及び
(b)工程(a)の前記植物又は植物細胞において、前記誘導性プロモーターを誘導し、それにより、前記植物又は前記植物細胞において前記一つ又は一つより多くの目的とするタンパク質を産生すること;
を含んでなる、前記プロセス。
【請求項2】
前記第一及び前記第二の誘導性プロモーターが同一の誘導シグナルにより、誘導可能である、請求項1に記載のプロセス。
【請求項3】
前記第一又は前記第二の誘導性プロモーターが化学的誘導性プロモーターである、請求項1又は2に記載のプロセス。
【請求項4】
前記化学的誘導性プロモーターが、エタノール誘導性プロモーター、IPTG誘導性プロモーター及びテトラサイクリン誘導性プロモーターから成る群より選択される、請求項3に記載のプロセス。
【請求項5】
前記誘導性プロモーターが熱ショック誘導性プロモーターである、請求項1から4のいずれか一項に記載のプロセス。
【請求項6】
前記RNAレプリコンがプラスセンス一本鎖RNAウイルスから誘導される、請求項1から5のいずれか一項に記載のプロセス。
【請求項7】
前記プラスセンス一本鎖RNAウイルスがタバコモザイクウイルス又はジャガイモウイルスXである、請求項6に記載のプロセス。
【請求項8】
前記植物が、さらなるRNAレプリコンをコードするヌクレオチド配列及び前記さらなるRNAレプリコンをコードする前記配列に機能可能なように連結された第三の誘導性プロモーター、を含んでなる第三の異種ヌクレオチド配列を含んでなり、前記さらなるRNAレプリコンが、前記植物中の前記RNAレプリコン又は前記さらなるRNAレプリコンの細胞間移行を提供するタンパク質をコードしておらず、前記さらなるRNAレプリコンが目的とするタンパク質をコードする、請求項1から7のいずれか一項に記載のプロセス。
【請求項9】
前記植物が、前記RNAレプリコンの細胞間移行を可能にする前記タンパク質をコードするヌクレオチド配列に機能可能なように連結された、前記さらなるRNAレプリコンの細胞間移行を可能にするタンパク質をコードするヌクレオチド配列を含んでなる異種ヌクレオチド配列を含んでなる、請求項8に記載のプロセス。
【請求項10】
前記RNAレプリコンの細胞間移行を可能にする前記タンパク質及び 前記さらなるRNAレプリコンの細胞間移行を可能にする前記タンパク質が同一タンパク質であるか又は異なったタンパク質である、請求項9に記載のプロセス。
【請求項11】
前記さらなるレプリコンが、前記さらなるRNAレプリコンを複製するためのポリメラーゼをコードする、請求項8に記載のプロセス。
【請求項12】
前記第三の誘導性プロモーターが、前記第一の誘導性プロモーターと同一の誘導剤により誘導可能である、請求項8から11のいずれか一項に記載のプロセス。
【請求項13】
前記RNAレプリコン及び前記さらなるRNAレプリコンが、非競合 レプリコンである、請求項8から12のいずれか一項に記載のプロセス。
【請求項14】
前記非競合レプリコンが、異なったウイルス属の植物ウイルスから誘導される植物ウイルスレプリコンである、請求項13に記載のプロセス。
【請求項15】
工程(a)が、前記第一の異種ヌクレオチド配列及び、任意選択で、前記第二の及び/又は第三の異種ヌクレオチド配列を含んでなる種子から植物を生長させることを含んでなる、請求項1から14のいずれか一項に記載のプロセス。
【請求項16】
植物又は植物細胞であって:
核染色体中に、RNAレプリコンをコードするヌクレオチド配列及び前記RNAレプリコンをコードする前記ヌクレオチド配列に機能可能なように連結された第一の誘導性プロモーターを含んでなる第一の異種ヌクレオチド配列;
前記RNAレプリコンは、前記植物中での前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない;
前記RNAレプリコンは、前記RNAレプリコンを複製するためのポリメラーゼをコードする;
を含んでなる、前記植物又は植物細胞。
【請求項17】
植物又は植物細胞であって;
(i)核染色体中に、RNAレプリコンをコードするヌクレオチド配列及び前記RNAレプリコンをコードする前記ヌクレオチド配列に機能可能なように連結された第一の誘導性プロモーターを含んでなる第一の異種ヌクレオチド配列;
前記RNAレプリコンは、前記植物中での前記RNAレプリコンの細胞間移行を提供するタンパク質をコードしていない;
前記RNAレプリコンは、前記RNAレプリコンを複製するためのポリメラーゼをコードする;及び
(ii)前記RNAレプリコンの細胞間移行を可能にするタンパク質をコードするヌクレオチド配列を含んでなる第二の異種ヌクレオチド配列、前記第二の異種ヌクレオチド配列は、前記RNAレプリコンの細胞間移行を可能にする前記タンパク質をコードする前記ヌクレオチド配列に機能可能なように連結された第二の誘導性プロモーターを含んでなる;
を含んでなる、前記植物又は植物細胞。
【請求項18】
タバコ属に属する、請求項16又は17に記載の植物又は植物細胞。
【請求項19】
請求項17で定義した植物又は植物細胞を産生するプロセスであって、植物核染色体内に請求項17で定義した第一の異種ヌクレオチド配列及び請求項17で定義した第二の異種ヌクレオチド配列を導入すること、及び前記第一の及び前記第二の異種ヌクレオチド配列を含有する形質転換された植物を再生することを含んでなる、前記プロセス。
【請求項20】
請求項8で定義した植物又は植物細胞産生するプロセスであって、前記第一の異種ヌクレオチド配列及び任意選択で、前記第二の異種ヌクレオチド配列を含んでなる第一の植物と、前記第三の異種ヌクレオチド配列を含んでなる第二の植物を交配することを含んでなる、前記プロセス。
【図2C】



【図9】

【図1】

【図2A】
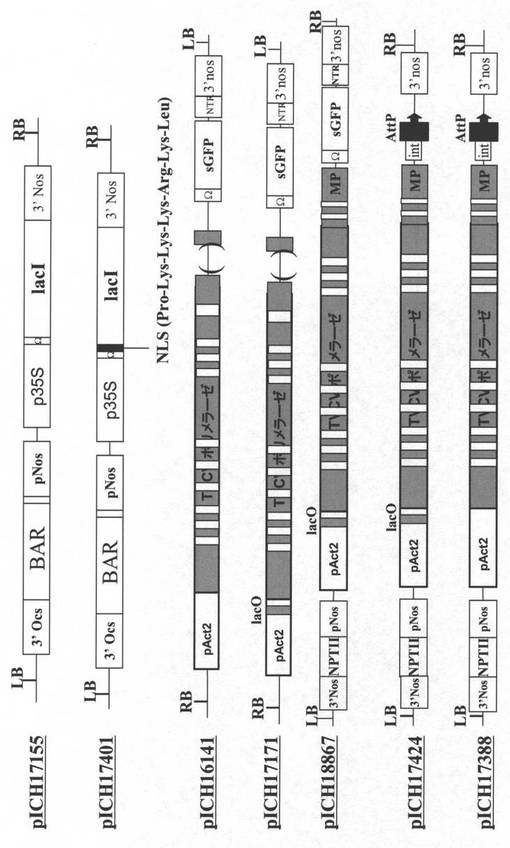
【図2B】
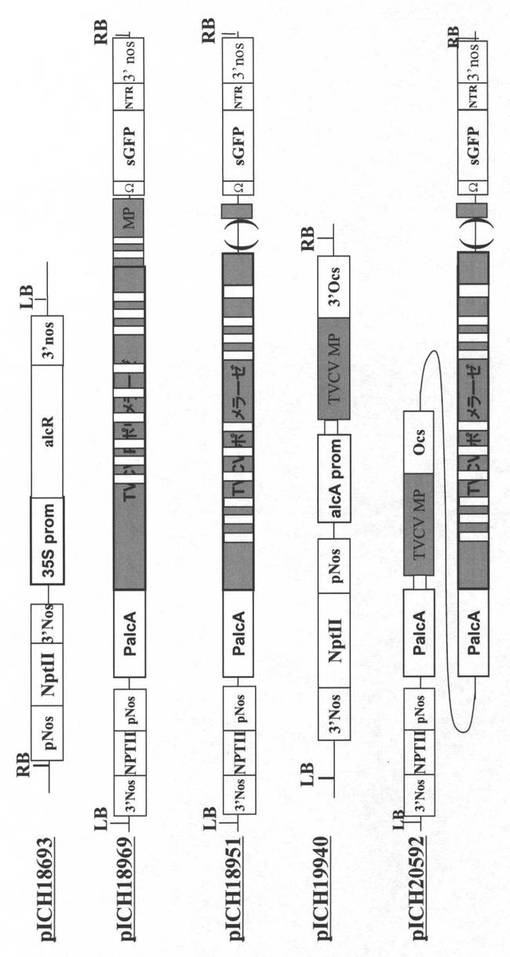
【図3】

【図4A】

【図4B】
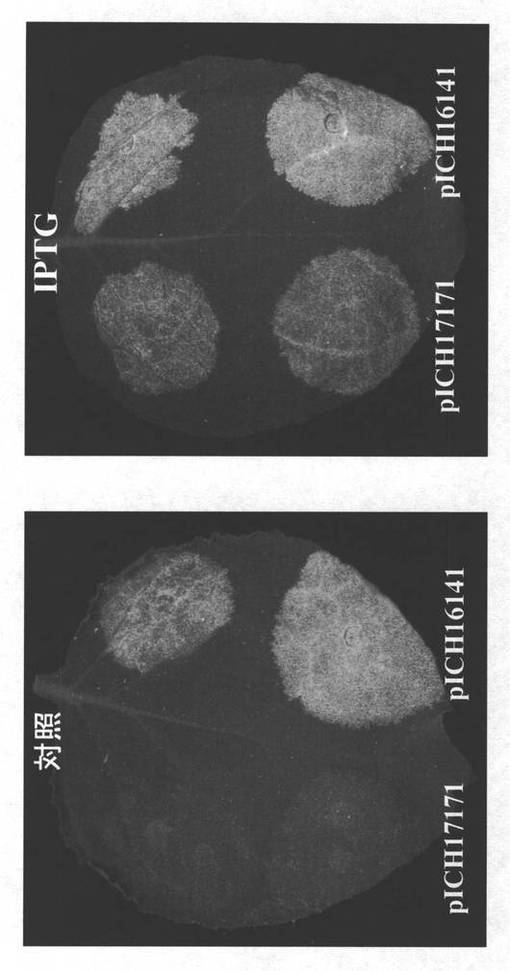
【図5】

【図6】

【図7】

【図8】

【図10】

【図11】

【図12】

【図13】

【図9】
【図1】
【図2A】
【図2B】
【図3】
【図4A】
【図4B】
【図5】
【図6】
【図7】
【図8】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2009−538606(P2009−538606A)
【公表日】平成21年11月12日(2009.11.12)
【国際特許分類】
【出願番号】特願2009−512470(P2009−512470)
【出願日】平成19年5月25日(2007.5.25)
【国際出願番号】PCT/EP2007/004688
【国際公開番号】WO2007/137788
【国際公開日】平成19年12月6日(2007.12.6)
【出願人】(507254997)アイコン・ジェネティクス・ゲーエムベーハー (5)
【Fターム(参考)】
【公表日】平成21年11月12日(2009.11.12)
【国際特許分類】
【出願日】平成19年5月25日(2007.5.25)
【国際出願番号】PCT/EP2007/004688
【国際公開番号】WO2007/137788
【国際公開日】平成19年12月6日(2007.12.6)
【出願人】(507254997)アイコン・ジェネティクス・ゲーエムベーハー (5)
【Fターム(参考)】
[ Back to top ]