
真菌細胞において遺伝子を発現させるための真菌プロモーター
本発明は、単離された真菌プロモーターDNA配列、DNA構築物、ベクター、およびポリペプチドをコードするコード配列と作動可能に会合されるこれらのプロモーターを含む真菌宿主細胞に関する。本発明はまた、単離された新規のプロモーターを使用して遺伝子を発現させるおよび/またはポリペプチドを産生させるための方法に関する。本発明はまた、本発明の新規のプロモーターを使用して、転写レベルおよび/または内因性遺伝子の調節を改変するための方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、DNA配列、特に、単離された真菌プロモーター、およびDNA構築物、ベクター、ならびにポリペプチドをコードするコード配列と作動可能に会合されるこれらのプロモーターを含む真菌宿主細胞に関する。本発明はまた、遺伝子を発現させるおよび/またはポリペプチドを産生させるための方法に関する。
【背景技術】
【0002】
真菌宿主細胞における組換えポリペプチドの産生は、ポリペプチドをコードするDNAが、宿主細胞に適切なプロモーターの発現制御下に配置される発現カセットを構築することによって、通常達成される。発現カセットは、プラスミドまたはベクターによって仲介される形質転換により、宿主細胞に導入することができる。次いで、ポリペプチドの産生は、発現カセットに含有されるプロモーターの適切な機能化に必要な誘導条件下で形質転換された宿主細胞を培養することによって、達成することができる。
【0003】
各真菌宿主細胞について、形質転換によって真菌宿主に導入されているコード配列の発現およびこのコード配列によってコードされる組換えポリペプチドの産生には、機能的プロモーターが利用可能であることが必要である。多数のプロモーターが、真菌宿主細胞において機能的であることが既に公知である。プロモーターの異種間使用の例が存在する。すなわち、アスペルギルス・ニダランス(Aspergillus nidulans)(A.ニダランス(A.nidulans)gpdA遺伝子のプロモーターは、アスペルギルス・ニガー(Aspergillus niger)(A.ニガー(A.niger))において機能的であることが公知である(J Biotechnol.1991年1月;17(1):19−33.Intracellular and extracellular production of proteins in Aspergillus under the control of expression signals of the highly expressed A.nidulans gpdA gene.プント PJ(Punt PJ)、ゼガー ND(Zegers ND)、ブッチャー M(Busscher M)、ポウウェルズ PH(Pouwels PH)、バンデンホンデル CA(van den Hondel CA))。もう1つの例は、A.ニガー(A.niger)およびA.ニダランス(A.nidulans)において使用されるA.ニガー(A.niger)β−キシロシダーゼxlnDプロモーター「Transcriptional regulation of the xylanolytic enzyme system of Aspergillus」、ヴァンペイジ,NNME(van Peij,NNME)博士−thesis Landbouwuniversiteit Wageningen、蘭国、ISBN90−5808−154−0ならびにA.ニガー(A.niger)、A.ニダランス(A.nidulans)およびクラドスポリウム・フルブム(Cladosporium fulvum)における大腸菌(Escherichia coli)β−グルクロニダーゼ遺伝子の発現(Curr Genet、1989年3月;15(3):177−80:ロバーツ IN(Roberts IN)、オリバー RP(Oliver RP)、プント PJ(Punt PJ)、バンデンホンデル CA(van den Hondel CA)「Expression of the Escherichia coli beta−glucuronidase gene in industrial and phytopathogenic filamentous fungi」において記載される)である。
【0004】
しかし、誘導される遺伝子の発現制御するため、内因性遺伝子の発現のレベルを制御するため、内因性遺伝子の発現の調節を制御するためかもしくは内因性遺伝子の不活化を仲介するため、またはポリペプチドを産生させるため、あるいは先のアプリケーションの組み合わせのための改善されたプロモーターに必要性がなお存在する。これらの改善されたプロモーターは、例えば、これまでの公知のプロモーターよりも強力であり得る。それらはまた、特異的な簡便な基質または化合物によって誘導可能であり得る。単一の真菌宿主において多用な遺伝子を同時に過剰発現させようと考える場合、いくらかの機能的プロモーターについて知ることもまた有利である。スケルチング(squelching)(特定の転写因子のタイトレーション)を防止するためには、複数の個別のプロモーター(発現させようとする1つの遺伝子に対してそれぞれ1つの特異的プロモーター)を使用することが好適である。
【発明の詳細な説明】
【0005】
本発明は、以下からなる群から選択されるプロモーターDNA配列に関する。すなわち、
(a)配列番号1、配列番号2、配列番号3、配列番号4、配列番号5のヌクレオチド配列を含むDNA配列、
(b)(a)のDNA配列にハイブリダイズすることが可能なDNA配列、
(c)(a)のDNA配列に少なくとも50%相同であるDNA配列、
(d)(a)〜(c)のDNA配列のいずれかの変異体、および
(e)(a)〜(d)のDNA配列のいずれかのサブ配列。
【0006】
本発明に関して、プロモーターDNA配列がコード配列と作動可能に会合される場合、このプロモーターDNA配列は、このコード配列の発現を制御することが可能なDNA配列である。用語「作動可能に会合」は、本明細書において、プロモーターDNA配列がコード配列によってコードされるポリペプチドの産生を指令するようなコード配列に対する位置に、プロモーターDNA配列が適切に配置される配置形態として規定される。
【0007】
用語「コード配列」は、本明細書において、適切な制御配列の制御下に配置される場合、ポリペプチドに翻訳されるmRNAに転写される核酸配列として規定される。コード配列の境界は、一般に、通常、mRNAの5’末端におけるオープンリーディングフレームの開始部であるATG開始コドン、およびmRNAの3’末端におけるオープンリーディングフレームの直ぐ下流に位置する転写ターミネーター配列によって決定される。コード配列としては、ゲノムDNA、cDNA、半合成、合成、および組換え核酸配列を挙げることができるが、これらに限定されない。
【0008】
より具体的には、用語「プロモーター」は、本明細書において、RNAポリメラーゼに結合し、転写を開始するポリペプチドをコードするコード配列の正確な下流の転写開始部位にポリメラーゼを指向するDNA配列として規定される。RNAポリメラーゼは、コード領域の適切なDNA鎖に相補的なメッセンジャーRNAの集成を効果的に触媒する。用語「プロモーター」はまた、mRNAへの転写後の翻訳のための(プロモーターと転写開始部との間の)5’非コード領域、エンハンサーなどのシス作用性転写制御エレメント、および転写因子と相互作用することが可能な他のヌクレオチド配列を含むことが理解されよう。
【0009】
好適な実施態様では、本発明のプロモーターDNA配列は、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5に記載の配列である。
【0010】
別の好適な実施態様に従えば、本発明のプロモーターDNA配列は、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5にハイブリダイズすることが可能であり、なおプロモーター活性を保持するDNA配列である。
【0011】
プロモーター活性は、好ましくは、プロモーターと作動可能に会合されるコード配列によってコードされるタンパク質の濃度を測定することによって、決定される。あるいは、プロモーター活性は、プロモーターと作動可能に会合されるコード配列によってコードされるタンパク質の酵素活性を測定することによって、決定される。好適な実施態様に従えば、プロモーター活性(およびその強度)は、lacZレポーター遺伝子のコード配列の発現を測定することによって決定される(ルオ(Luo)(Gene 163(1995年)127−131)。別の好適な実施態様に従えば、プロモーター活性は、コード配列として緑色蛍光タンパク質を使用することによって決定される(Microbiology.1999年3月;145(Pt3):729−34、サンテリレ アンリクセンAL(Santerre Henriksen AL)、エヴェン S(Even S)、マラー C(Muller C)、プント PJ(Punt PJ)、バンデンホンデル CA(van den Hondel CA)、ニールセン J.スタディ(Nielsen J.Study))。
【0012】
さらに、プロモーター活性は、プロモーターの制御下で作製される転写物のmRNAレベルを測定することによって、決定することができる。mRNAレベルは、例えば、ノーザンブロットを介して測定することができる(J.サンブルック(J.Sambrook),E.F.フリッチュ(E.F.Fritsch)、およびT.マニアツス(T.Maniatus)、1989年、Molecular Cloning,A Laboratory Manual、第2版、Cold Spring Harbor、ニューヨーク州)。
【0013】
本発明は、極めて低いストリンジェンシー条件、好ましくは低いストリンジェンシー条件、より好ましくは中等度のストリンジェンシー条件、より好ましくは中等度〜高いストリンジェンシー条件、さらにより好ましくは高いストリンジェンシー条件、および最も好ましくは極めて高いストリンジェンシー条件下で、(i)配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5のヌクレオチド1〜2000、好ましくはヌクレオチド100〜1990、より好ましくは200〜1980、さらにより好ましくは300〜1970、さらにより好ましくは350〜1950および最も好ましくは360〜1900に対応するか、(ii)(i)のサブ配列であるか、または(iii)(i)、(ii)の相補鎖である核酸プローブにハイブリダイズする(単離された)プロモーターDNA配列を包含する(J.サンブルック(J.Sambrook)、E.F.フリッチュ(E.F.Fritsch)、およびT.マニアティス(T. Maniatis)、1989年、Molecular Cloning,A Laboratory Manual、第2版、Cold Spring Harbor、ニューヨーク州)。配列番号1、配列番号2、配列番号3、配列番号4または配列番号5のサブ配列は、少なくとも100ヌクレオチド、好ましくは少なくとも200ヌクレオチド、より好ましくは少なくとも300ヌクレオチド、さらにより好ましくは少なくとも400ヌクレオチド、および最も好ましくは少なくとも500ヌクレオチドであり得る。
【0014】
配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5の核酸配列またはそのサブ配列を使用して、核酸プローブを設計し、当該分野において周知の方法に従って、異なる属あるいは種の株からDNAプロモーターを同定およびクローニングしてもよい。特に、そのようなプローブは、本明細書に記載の対応する遺伝子を同定および単離するために、標準的なサザンブロッティング手順に従って、目的の属または種のゲノムもしくはcDNAとのハイブリダイゼーションのために使用することができる。そのようなプローブは、全配列よりもかなり短くあり得るが、少なくとも15、好ましくは少なくとも25、およびより好ましくは少なくとも35ヌクレオチド長であるべきである。さらに、そのようなプローブを使用して、PCRを介してDNAプロモーターを増幅することができる。PCRを介してプロモーターをクローニングする例については、本明細書に記載されている(実施例1.3を参照のこと)。また、より長いプローブを使用することもできる。DNA、RNAおよびペプチド核酸(PNA)プローブを使用することもできる。プローブは、典型的に、対応する遺伝子を、(例えば、32P、33P、3H、35S、ビオチン、もしくはアビジンまたは蛍光マーカーで)検出するために標識される。かかるプローブも本発明に包含される。
【0015】
従って、そのような他の生物体から調製されるゲノムDNAまたはcDNAライブラリーを、上記のプローブにハイブリダイズし、ポリペプチドをコードするDNAを対象にスクリーニングしてもよい。そのような他の生物体由来のゲノムまたは他のDNAは、アガロースもしくはポリアクリルアミドゲル電気泳動、または他の分離技術によって分離することができる。ライブラリー由来のDNAまたは分離されたDNAを、ニトロセルロースまたは他の適切なキャリア材料に移して固定化してもよい。配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5、またはそのサブ配列に相同であるクローンあるいはDNAを同定するために、キャリア材料を、サザンブロットにおいて使用してもよい。
【0016】
本発明の目的のために、ハイブリダイゼーションは、核酸配列が、配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5に示される核酸配列、その相補鎖、またはそのサブ配列に対応する標識核酸プローブに、極めて低い〜極めて高いストリンジェンシー条件下で、ハイブリダイズすることを示す。核酸プローブがこれらの条件下でハイブリダイズする分子は、例えば、X線フィルムを使用して、検出される。また、検出のために蛍光ならびに支持体としてガラススライド(glass sides)および/またはDNAマイクロアレイを使用する技術などの他のハイブリダイゼーション技術を使用することもできる。DNAマイクロアレイハイブリダイゼーション検出の例は、FEMS Yeast Res.2003年12月;4(3):259−69(ダラン−ラプジャデ P(Daran−Lapujade P)、ダラン JM(Daran JM)、コッテル P(Kotter P)、ペティット T(Petit T)、パイパー MD(Piper MD)、プランク JT(Pronk JT).「Comparative genotyping of the Saccharomyces cerevisiae laboratory strains S288C and CEN.PK113−7D using oligonucleotide microarrays」)に認められる。さらに、ハイブリダイゼーションのためのPNAマイクロアレイの使用については、Nucleic Acids Res.2003年10月1日;31(19):e119(ブラント O(Brandt O)、フェルトナー J(Feldner J)、シュテファン A(Stephan A)、シュレーダー M(Schroder M)、シュノルザー M(Schnolzer M)、アーリンハウス HF(Arlinghaus HF)、ホヘイセル JD(Hoheisel JD)、ヤコブ A(Jacob A).PNA microarrays for hybridisation of unlabelled DNA samples.)に記載されている。
【0017】
好適な実施態様では、核酸プローブは、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5の核酸配列である。好適な別の実施態様では、核酸プローブは、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5のヌクレオチド20〜1980、より好ましくは配列番号1、配列番号2、配列番号3、配列番号4または配列番号5のヌクレオチド500〜1950、さらにより好ましくは配列番号1、配列番号2、配列番号3、配列番号4または配列番号5のヌクレオチド800〜1920、および最も好ましくは配列番号1、配列番号2、配列番号3、配列番号4または配列番号5のヌクレオチド900〜1900を有する配列である。別の好適なプローブは、転写部位の前のDNA配列の部分である。
【0018】
少なくとも100ヌクレオチド長の長いプローブでは、極めて低い〜極めて高いストリンジェンシー条件は、標準的なサザンブロッティング手順に従う5×SSPE、0.3%SDS、200.mu.g/ml剪断および変性サケ精子DNA、ならびに極めて低いおよび低いストリンジェンシーでは25%ホルムアミド、中等度および中等度〜高いストリンジェンシーでは35%ホルムアミド、または高いおよび極めて高いストリンジェンシーでは50%ホルムアミドのいずれかにおける42℃でのプレハイブリダイゼーションおよびハイブリダイゼーションとして規定される。
【0019】
少なくとも100ヌクレオチド長の長いプローブでは、キャリア材料は、それぞれ、15分間、2×SSC、0.2%SDSを使用して、好ましくは少なくとも45℃(極めて低いストリンジェンシー)、より好ましくは少なくとも50℃(低いストリンジェンシー)、より好ましくは少なくとも55℃(中等度のストリンジェンシー)、より好ましくは少なくとも60℃(中等度〜高いストリンジェンシー)、さらにより好ましくは少なくとも65℃(高いストリンジェンシー)、および最も好ましくは少なくとも70℃(極めて高いストリンジェンシー)で、最終的に3回洗浄する。
【0020】
約15ヌクレオチド〜約70ヌクレオチド長である短いプローブでは、ストリンジェンシー条件は、標準的なサザンブロッティング手順に従う、0.9M NaCl、0.09M Tris−HCl pH7.6、6mM EDTA、0.5%NP−40、1×デンハルト溶液(Denhardt’s solution)、1mMピロリン酸ナトリウム、1mMリン酸二水素ナトリウム、0.1mM ATP、および1mlあたり0.2mgの酵母RNAにおいて、ボールトン(Bolton)およびマッカーシー(McCarthy)(1962年、Proceedings of the National Academy of Sciences USA 48:1390)に従う計算を使用して算出されるTmより5℃〜10℃低い温度でのプレハイブリダイゼーション、ハイブリダイゼーション、およびハイブリダイゼーション後洗浄として規定される。
【0021】
約15ヌクレオチド〜約70ヌクレオチド長である短いプローブでは、キャリア材料は、6×SCC+0.1%SDS中、15分間で1回、算出されたTmより5℃〜10℃低い温度で6×SSCを使用して、それぞれ15分間、2回洗浄される。
【0022】
別の好適な実施態様に従えば、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5を最初に使用して、それと作動可能に会合される生来の遺伝子、コード配列もしくはその一部をクローニングする。これは、先に規定されるような配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5、またはそのサブ配列のいずれかから開始し、プローブとしてこの配列を使用して、行うことができる。プローブは、cDNAまたは所定の宿主、アスペルギルス・ニガー(Aspergillus niger)かもしくは本出願において規定される他の任意の真菌宿主のいずれかのゲノムライブラリーにハイブリダイズされる。一旦、生来の遺伝子またはその一部がクローニングされれば、続いて、それ自体をプローブとして使用して、本明細書に記載のハイブリダイゼーション実験によって、他の真菌から誘導されるその相同遺伝子をクローニングすることができる。
【0023】
本発明に関して、相同遺伝子とは、生来の遺伝子に対して少なくとも50%相同(同一)である遺伝子を意味する。好ましくは、相同遺伝子は、生来の遺伝子に対して、少なくとも55%相同、より好ましくは少なくとも60%、より好ましくは少なくとも65%、より好ましくは少なくとも70%、さらにより好ましくは少なくとも75%、好ましくは約80%、より好ましくは約90%、さらにより好ましくは約95%、および最も好ましくは約97%相同である。
【0024】
相同遺伝子のコード配列の上流の配列は、本発明に包含されるプロモーターである。あるいは、本発明のプロモーターに作動可能に会合される生来の遺伝子の配列、コード配列またはその一部を、先に規定されるような配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5またはそのサブ配列を使用して同定し、例えば、本明細書に記載のアラインメントまたはBLASTアルゴリズムを使用して、ゲノムデータベースを検索することができる。続いて、この同定される配列を使用して、本出願において規定される他の任意の真菌宿主において、オーソログまたは相同遺伝子を同定することができる。同定されるオーソログまたは相同遺伝子のコード配列の上流の配列は、本発明に包含されるプロモーターである。
【0025】
別の好適な実施態様に従えば、本発明のプロモーターDNA配列は、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5に少なくとも50%相同(同一)である単離されたDNA配列である。好ましくは、DNA配列は、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5に対して、少なくとも55%相同、より好ましくは少なくとも60%、より好ましくは少なくとも65%、より好ましくは少なくとも70%、さらにより好ましくは少なくとも75%、好ましくは約80%、より好ましくは約90%、さらにより好ましくは約95%、および最も好ましくは約97%相同である。
【0026】
本発明の目的のために、2つの核酸配列間の相同性(同一性)の程度は、好ましくは、BLASTプログラムによって決定される。BLAST解析を実施するためのソフトウェアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)を介して公的に入手することができる。BLASTアルゴリズムパラメータW、TおよびXは、アラインメントの感度および速度を決定する。BLASTプログラムは、デフォルトとして、11のワード長(W)、50のBLOSUM62スコアリングマトリックス(ヘニコフ(Henikoff)およびヘニコフ(Henikoff)、Proc.Natl.Acad.Sci.USA89:10915(1989年)を参照のこと)アラインメント(B)、10の期待値(E)、M=5、N=−4、および両鎖の比較を使用する。
【0027】
好適な別の実施態様では、プロモーターは、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5のサブ配列であり、該サブ配列はなお、プロモーター活性を有する。サブ配列は、好ましくは、少なくとも約100ヌクレオチド、より好ましくは少なくとも約200ヌクレオチド、および最も好ましくは少なくとも約300ヌクレオチドを含有する。
【0028】
好適な別の実施態様では、サブ配列は、5’および/または3’末端からの1もしくはそれ以上のヌクレオチドが欠失されていることを除いて、配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5に包含される核酸配列であり、前記DNA配列はなおプロモーター活性を有する。
【0029】
好適な別の実施態様では、プロモーターサブ配列は、「トリミングした」のサブ配列、即ち、翻訳開始部および/または転写開始部より上流にある配列フラグメントである。プロモーターをトリミングし、それについて機能的に分析する例は、Gene.1994年8月5日;145(2):179−87:the effect of multiple copies of the upstream region on expression of the Aspergillus niger glucoamylase−encoding gene.ベルダズ JC(Verdoes JC)、プント PJ(Punt PJ)、ストウサメル AH(Stouthamer AH)、バンデンホンデル CA(van den Hondel CA))に記載されている。
【0030】
本発明の別の実施態様では、プロモーターDNA配列は、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5の変異体である。
【0031】
用語「変異体」または「変異体プロモーター」は、本明細書において、親プロモーターの1もしくはそれ以上のヌクレオチドの置換、欠失および/または挿入を含むヌクレオチド配列を有するプロモーターとして規定され、ここで、変異体プロモーターは、対応する親プロモーターよりも高いまたは低いプロモーター活性を有する。用語「変異体プロモーター」は、天然の変異体ならびに古典的な変異誘発、部位特異的変異誘発、およびDNAシャフリングなどの当該分野において周知の方法を使用して入手されるインビトロで作製される変異体を包含する。変異体プロモーターは、1つもしくはそれ以上の変種を有してもよい。各変種は、ヌクレオチドの独立した置換、欠失、および/または挿入である。
【0032】
好適な実施態様に従えば、変異体プロモーターは、最初に同定されたプロモーター配列(配列番号1、配列番号2、配列番号3、配列番号4または配列番号5)と比較して少なくとも修飾された調節部位を有するプロモーターである。そのような調節部位は、その全体を取り出すかまたは上記で説明されるように特異的に変異することができる。従って、かかるプロモーター変異体の調節は、例えば、それがもはやグルコースによって誘導されないように修飾される。かかるプロモーター変異体およびそれらを入手する仕方に関する技術の例は、EP673429号明細書または国際公開第94/04673号パンフレットに記載されている。これらの特許は、参照により本明細書に援用される。
【0033】
プロモーター変異体は対立遺伝子変異体であり得る。対立遺伝子変異体は、同じ染色体の遺伝子座を占有する遺伝子の2つもしくはそれ以上の代替的形態のいずれかを示す。対立遺伝子の変異体は変異を介して自然に生じ、集団内において多型を生じ得る。変異体プロモーターは、(a)DNAを、極めて低い、低い、中等度、中等度〜高い、高い、もしくは極めて高いストリンジェンシー条件下で、(i)配列番号1、配列番号2、配列番号3、配列番号4または配列番号5、(ii)(i)のサブ配列あるいは(iii)(i)、(ii)の相補鎖とハイブリダイズさせること、および(b)DNAから変異体プロモーターを単離することによって、入手することができる。ストリンジェンシーおよび洗浄条件は、本明細書において規定されるとおりである。
【0034】
本発明のプロモーターは、その配列が、プロモーター配列と、ポリペプチドをコードする核酸配列のコード領域との連結を容易にする特定の制限部位を誘導する目的のためのリンカーを伴って提供され得るプロモーターであり得る。
【0035】
本明細書において提供される配列情報は、誤って同定される塩基の夾在を要するようにし、あまりに限られた範囲で解釈すべきではない。本発明において開示される特定の配列は、糸状菌、特に、アスペルギルス・ニガー(Aspergillus niger)由来の本来のDNA配列を単離するために容易に使用することができ、さらなる配列解析に供し、それによって、シークエンシングエラーを同定することができる。
【0036】
他で示さない限り、本明細書においてDNA分子を配列決定することによって決定されるすべてのヌクレオチド配列は、自動化DNAシークエンサーを使用して決定した。従って、この自動化アプローチによって決定される任意のDNA配列について当該分野において公知であるように、本明細書において決定される任意のヌクレオチド配列は、いくつかのエラーを含有してもよい。自動化によって決定されるヌクレオチド配列は、配列決定されたDNA分子の実際のヌクレオチド配列に対して、典型的には、少なくとも約90%同一、より典型的には、少なくとも約95%〜少なくとも約99.9%同一である。実際の配列は、当該分野において周知の手動のDNA配列決定方法を含む他のアプローチによって、より正確に決定することができる。
【0037】
当業者は、そのように誤って同定される塩基を同定することが可能であり、そのようなエラーを補正する仕方を知っている。
【0038】
本発明は、本発明が係わるプロモーターの生物学的機能を改変しない変異を典型的に含有する機能的プロモーター等価物を包含する。用語「機能的等価物」はまた、A.ニガー(A.niger)DNA配列のオーソログを包含する。A.ニガー(A.niger)DNA配列のオーソログは、他の株または種から単離することができ、類似または同一の生物学的活性を所有するDNA配列である。
【0039】
本発明のプロモーター配列は、任意の属の微生物から入手してもよい。本発明の目的のために、所定の供給源に関して本明細書において使用する用語「から入手する」は、ポリペプチドが、供給源または供給源由来の遺伝子が挿入されている細胞によって産生されることを意味すべきである。
【0040】
プロモーター配列は、真菌供給源、好ましくは、カンジダ(Candida)、ハンセヌラ(Hansenula)、クリヴェロミセス(Kluyveromyces)、ピキア(Pichia)、サッカロミセス(Saccharomyces)、シゾサッカロマイセス(Schizosaccharomyces)、またはヤロウイア(Yarrowia)株などの酵母株、より好ましくは、サッカロマイセス・カールスベルゲンシス(Saccharomyces carlsbergensis)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、サッカロミセス・ディアスタティカス(Saccharomyces diastaticus)、サッカロミセス・ダグラシー(Saccharomyces douglasii)、サッカロミセス・クリヴェリ(Saccharomyces kluyveri)、サッカロミセス・ノルベンシス(Saccharomyces norbensis)またはサッカロミセス・オヴィフォルミス(Saccharomyces oviformis)株から入手することができる。
【0041】
好適な別の実施態様では、プロモーター配列は、アクレモニウム(Acremonium)、アスペルギルス(Aspergillus)、オーレオバシディウム(Aureobasidium)、クリプトコッカス(Cryptococcus)、フィリバシディウム(Filibasidium)、フザリウム(Fusarium)、フミコーラ(Humicola)、マグナポルテ(Magnaporthe)、ケカビ(Mucor)、ミセリオフトラ(Myceliophthora)、ネオカリマスティクス(Neocallimastix)、ニューロスポラ(Neurospora)、ペシロマイセス(Paecilomyces)、ペニシリウム(Penicillium)、ピロミセス(Piromyces)、スエヒロタケ(Schizophyllum)、タラロミセス(Talaromyces)、サーモアスカス(Thermoascus)、シーラビア(Thielavia)、トイポクラディウム(Tolypocladium)、またはトリコデルマ(Trichoderma)株などの糸状菌株、より好ましくは、アスペルギルス・アクレアータス(Aspergillus aculeatus)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・フォエティダス(Aspergillus foetidus)、アスペルギルス・ジャポニクス(Aspergillus japonicus)、A.ニダランス(A.nidulans)、A.ニガー(A.niger)、麹菌(Aspergillus oryzae)(麹菌(A.oryzae))、フミコーラ・インソレンス(Humicola insolens)、フミコーラ・ラヌギノサ(Humicola lanuginosa)、ムコール・ミエヘイ(Mucor miehei)、ミセリオフトラ・サーモフィラ(Myceliophthora thermophila)、アカパンカビ(Neurospora crassa)、ペニシリウム・パルロゼラム(Penicillium purpurogenum)、トリコデルマ・ハルジアナム(Trichoderma harzianum)、トリコデルマ・コニンギ(Trichoderma koningii)、トリコデルマ・ロンギプラチアタム(Trichoderma Iongibrachiatum)、トリコデルマ・リーセイ(Trichoderma reesei)、またはトリコデルマ・ビリデ(Trichoderma viride)株から入手される。
【0042】
好適な別の実施態様では、プロモーター配列は、フザリウム・バクトリディオデス(Fusarium bactridioides)、フザリウム・セレアリス(Fusarium cerealis)、フザリウム・クルックウェルエンス(Fusarium crookwellense)、フザリウム・クルモラム(Fusarium culmorum)、フザリウム・グラミネアラム(Fusarium graminearum)、フザリウム・グラミヌム(Fusarium graminum)、フザリウム・ヘテロスポルム(Fusarium heterosporum)フザリウム・ネグンディ(Fusarium negundi)、フザリウム・オキシスポラム(Fusarium oxysporum)、フザリウム・レティクラツム(Fusarium reticulatum)、フザリウム・ロゼウム(Fusarium roseum)、フザリウム・サンブシヌム(Fusarium sambucinum)、フザリウム・サルコチロウム(Fusarium sarcochroum)、フザリウム・スポロトリキオイデス(Fusarium sporotrichioides)、フザリウム・スルプレウム(Fusarium sulphureum)、フザリウム・トルローサム(Fusarium torulosum)、フザリウム・トリコテシオイデス(Fusarium trichothecioides)、フザリウム・ヴェネナツム(Fusarium venenatum)株から入手される。
【0043】
上記の種について、本発明は、完全および不完全な状態、ならびに名称が公知である種であるかどうかに係わらず、他の分類学的等価物、例えば、アナモルフを包含することが理解される。当業者であれば、適切な等価物の同一性を容易に認識する。これらの種の株は、米国菌培養収集所(the American Type Culture Collection)(ATCC)、Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(DSM)、Centraalbureau Voor Schimmelcultures(CBS)、および農業研究サービス機関(Agricultural Research Service Patent Culture Collection,Northern Regional Research Center)(NRRL)などの多くの培養コレクションにおいて、公的に容易に利用可能である。
【0044】
さらに、本発明に従うプロモーター配列は、上記のプローブを使用して、自然(例えば、土壌、堆肥、水など)から単離される微生物を含む他の供給源から同定および入手することができる。天然の生育地から微生物を単離するための技術は、当該分野において周知である。次いで、核酸配列は、別の微生物のゲノムDNAライブラリーを同様にスクリーニングすることによって、誘導することができる。一旦、プロモーターをコードする核酸配列がプローブで検出されたら、当業者に公知である技術を利用することによって、配列を単離またはクローニングすることができる(例えば、サンブルック(Sambrook)ら、1989年、上掲を参照のこと)。
【0045】
本発明では、プロモーターDNA配列はまた、本発明の1つもしくはそれ以上のプロモーターの一部;本発明のプロモーターの一部および別の既知のプロモーターの一部、例えば、一方のプロモーターのリーダー配列および他方のプロモーターの転写開始部位;または本発明の1つもしくはそれ以上のプロモーターの一部および1つもしくはそれ以上の他のプロモーターの一部を含むハイブリッドプロモーターであってもよい。他のプロモーターは、変異体、短縮型、ハイブリッドプロモーターを含む、好適な真菌宿主細胞において転写活性を示す任意のプロモーター配列であってもよく、宿主細胞に対して同種または異種のいずれかの細胞外もしくは細胞内ポリペプチドをコードする遺伝子から入手してもよい。他のプロモーター配列は、ポリペプチドをコードする核酸配列に生来または外来性であってもよく、細胞に対して生来または外来性であってもよい。
【0046】
好適な実施態様として、同定されるプロモーターの重要な調節サブ配列を、他の「基本」プロモーターに融合して、それらのプロモーター活性を増強することができる(例えば、Mol Microbiol.1994年5月;12(3):479−90.Regulation of the xylanase−encoding xlnA gene of Aspergillus tubigensis.デグラフ LH(de Graaff LH)、ファンデンブルック HC(van den Broeck HC)、ファンオオイジェン AJ(van Ooijen AJ)、ヴィセル J(Visser J)に記載される)。
【0047】
本発明のプロモーターとのハイブリッドプロモーターの構築において有用な他方のプロモーターの他の例として、麹菌(A.oryzae)TAKAアミラーゼ、リゾムコール・ミエヘイ(Rhizomucor miehei)アスパラギン酸プロテイナーゼ、A.ニガー(A.niger)中性α−アミラーゼ、A.ニガー(A.niger)酸性α−アミラーゼ、A.ニガー(A.niger)またはアスペルギルス・アワモリ(Aspergillus awamori)グルコアミラーゼ(glaA)、A.ニガー(A.niger)gpdA、A.ニガー(A.niger)グルコースオキシダーゼgoxC、リゾムコール・ミエヘイ(Rhizomucor miehei)リパーゼ、麹菌(A.oryzae)アルカリプロテアーゼ、麹菌(A.oryzae)トリオースリン酸イソメラーゼ、A.ニダランス(A.nidulans)アセトアミダーゼ、およびフザリウム・オキシスポラム(Fusarium oxysporum)トリプシン様プロテアーゼ(国際公開第96/00787号パンフレット)の遺伝子から入手されるプロモーター、ならびにNA2−tpiプロモーター(A.ニガー(A.niger)中性α−アミラーゼおよび麹菌(A.oryzae)トリオースリン酸イソメラーゼの遺伝子由来のプロモーターのハイブリッド)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)エノラーゼ(ENO−1)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)ガラクトキナーゼ(GAL1)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)アルコールデヒドロゲナーゼ/グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(ADH2/GAP)、およびサッカロミセス・セレビシエ(Saccharomyces cerevisiae)3−ホスホグリセリン酸キナーゼ、ならびにその変種、短縮型、およびハイブリッドプロモーターが挙げられる。酵母宿主細胞のための他の有用なプロモーターについては、ローマノス(Romanoset)ら、1992年、Yeast8:423−488に記載されている。
【0048】
本発明では、プロモーターDNA配列はまた、「タンデムプロモーター」であってもよい。「タンデムプロモーター」は、本明細書において、2つもしくはそれ以上のプロモーター配列として規定され、それらのそれぞれが、コード配列と作動可能に会合され、コード配列からmRNAへの転写を仲介する。
【0049】
タンデムプロモーターは、本発明の2つもしくはそれ以上のプロモーター、あるいは本発明の1つもしくはそれ以上のプロモーターおよびハイブリッドプロモーターの構築に有用な上記で例示されるような1つもしくはそれ以上の他の既知のプロモーターを含む。タンデムプロモーターの2つもしくはそれ以上のプロモーター配列は、核酸配列の転写を同時に促進することができる。あるいは、タンデムプロモーターのプロモーター配列のうちの1つもしくはそれ以上は、細胞の成長の異なる段階または菌糸体の形態学的に異なる部分での核酸配列の転写を促進することができる。
【0050】
本発明では、プロモーターは、目的のポリペプチドをコードするコード配列および/または真菌宿主細胞に対して外来性であってもよい。野生型プロモーターがコード配列または真菌宿主細胞に対して生来である場合であっても、本発明の変異体、ハイブリッド、またはタンデムプロモーターは、ポリペプチドをコードするコード配列に対して外来性であることが理解される。
【0051】
本発明の変異体、ハイブリッド、またはタンデムプロモーターは、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5を有するプロモーターのプロモーター活性の少なくとも約20%、好ましくは少なくとも約40%、より好ましくは少なくとも約60%、より好ましくは少なくとも約80%、より好ましくは少なくとも約90%、より好ましくは少なくとも約100%、さらにより好ましくは少なくとも約200%、最も好ましくは少なくとも約300%、およびさらに最も好ましくは少なくとも約400%を有する。
【0052】
本発明は、コード配列が、所定の真菌宿主細胞においてプロモーターDNA配列の制御下で発現され得るように、上記のように規定される少なくとも1つのプロモーターDNA配列、および前記プロモーターDNA配列と作動可能に会合されるコード配列を含むDNA構築物にさらに関する。
【0053】
コード配列は、目的の真菌宿主細胞に対して生来または異種であり得るポリペプチドをコードする。
【0054】
用語「ポリペプチド」は、本明細書においては、特定の長さのコードされた産物を指すことを意図するのではなく、従って、ペプチド、オリゴペプチド、およびタンパク質を包含する。用語「異種ポリペプチド」は、本明細書において、真菌細胞に対して生来ではないポリペプチド、生来の配列を改変するために修飾が施されている生来のポリペプチド、または発現が、組換えDNA技術による真菌細胞の操作の結果として量的に改変される生来のポリペプチドとして規定される。例えば、生来のポリペプチドは、例えば、ポリペプチドをコードする配列を本発明のプロモーターの制御下に配置して、ポリペプチドの発現を増強し、シグナル配列の使用によって、目的の生来のポリペプチドの細胞外への輸送を促進し、細胞によって通常産生されるポリペプチドをコードする遺伝子のコピー数を増加することによって、組換え的に産生され得る。真菌細胞は、ポリペプチドをコードする1コピーもしくはそれ以上のコード配列を含有してもよい。
【0055】
好ましくは、コード配列は、ペプチドホルモンもしくはその変異体、酵素、細胞内タンパク質、分泌プロセスに関与するタンパク質、ホールディングプロセスに関与するタンパク質、チャペロン、ペプチドアミノ酸トランスポーター、グリコシル化因子、転写因子、受容体もしくはその一部、抗体もしくはその一部、またはレポータータンパク質をコードする。
【0056】
好適な実施態様では、ポリペプチドは細胞外に分泌される。
【0057】
より好適な実施態様では、ポリペプチドは酵素である。酵素の例には、セルラーゼ、例えば、エンドグルカナーゼ、β−グルカナーゼ、セロビオヒドロラーゼまたはβ−グルコシダーゼ;ヘミセルラーゼまたはペクチン分解酵素、例えば、キシラナーゼ、キシロシダーゼ、マンナナーゼ、ガラクタナーゼ、ガラクトシダーゼ、ペクチンメチルエステラーゼ、ペクチンリアーゼ、ペクチン酸リアーゼ、エンドポリガラクツロナーゼ、エキソポリガラクツロナーゼ、ラムノガラクツロナーゼ、アラバナーゼ、アラビノフラノシダーゼ、アラビノキシランヒドロラーゼ、ガラクツロナーゼ、リアーゼ;デンプン分解酵素;ホスファターゼ、例えば、フィターゼ、エステラーゼ、例えば、リパーゼ、タンパク質分解酵素、例えば、プロテアーゼ、ペプチダーゼ、酸化還元酵素、例えば、オキシダーゼ、トランスフェラーゼ、またはイソメラーゼがある。
【0058】
あるいは、コード配列は、例えば、チャペロンまたは転写因子などの細胞内タンパク質をコードしてもよい。この例は、Appl Microbiol Biotechnol.1998年10月;50(4):447−54(「Analysis of the role of the gene bipA,encoding the major endoplasmic reticulum chaperone protein in the secretion of homologous and heterologous proteins in black Aspergilli.」プント PJ(Punt PJ)、バンゲメレン IA(van Gemeren IA)、ドリント−クイジュヴェンホヴェン J(Drint−Kuijvenhoven J)、ヘッシング JG(Hessing JG)、バンムイジュゥ−ハートベルト GM(van Muijlwijk−Harteveld GM)、ベイジェルスベルゲン A(Beijersbergen A)、ヴェリップス CT(Verrips CT)、バンデンホンデル CA(van den Hondel CA))に記載されている。チャペロンまたは転写因子などのこのコード配列が、タンパク質産生の制限要因であることが公知であった場合、これを使用して、例えば、タンパク質生産者としての宿主細胞の効率を改善することができる。
【0059】
コード配列はまた、有機酸、カロテノイド、(β−ラクタム)抗生物質、ビタミンなどの一次または二次代謝物の合成に関与する酵素をコードしてもよい。
【0060】
目的のポリペプチドをコードするコード配列は、任意の原核生物、真核生物、または他の供給源から入手することができる。
【0061】
あるいは、コード配列は、アンチセンスRNAおよび/またはRNAi(RNA干渉)構築物の発現をコードし得る。アンチセンス−RNAを発現させる例については、Appl Environ Microbiol.2000年2月;66(2):775−82.(Characterization of a foldase,protein disulfide isomerase A,in the protein secretory pathway of Aspergillus niger.ンギアム C(Ngiam C)、ジーネス DJ(Jeenes DJ)、プント PJ(Punt PJ)、バンデンホンデル CA(Van Den Hondel CA)、アーチャー DB(Archer DB))または(ズレンナー R(Zrenner R)、ウィルミツァー L(Willmitzer L)、ゾンネバルト U(Sonnewald U).Analysis of the expression of potato uridinediphosphate−glucose pyrophosphorylase and its inhibition by antisense RNA.Planta.(1993年);190(2):247−52.)に認められる。遺伝子の発現の完全な不活化は、例えば、(β−ラクタム)抗生物質またはカロテノイドなどの特定の二次代謝物の産生を増加するための、例えば、代謝経路の所望されない側枝を制御する遺伝子の不活化に有用である。完全な不活化はまた、毒性または所望されない化合物の産生を減少するのに有用である(ペニシリウム(Penicillium)におけるクリソゲニン(chrysogenin);アスペルギルス(Aspergillus)におけるアフラトキシン(Aflatoxin):マクドナルド KD(MacDonald KD)ら:heterokaryon studies and the genetic control of penicillin and chrysogenin production in Penicillium chrysogenum.J Gen Microbiol.(1963年)33:375−83)。完全な不活化はまた、発酵プロセスおよび下流プロセッシングが改善されるような方法で生物体の形態学を改変するのに有用である。
【0062】
本発明の別の実施態様は、真菌細胞の広範な代謝再プログラミングまたは操作に関する。完全な新規の経路の導入および/または所望されない経路の修飾は、タンパク質もしくは代謝物などの特定の化合物の産生に特異的に適応された細胞を提供する。
【0063】
本発明の方法では、コード配列がポリペプチドをコードする場合、前記ポリペプチドもまた、別のポリペプチドがポリペプチドまたはそのフラグメントのN末端もしくはC末端で融合される融合されたあるいはハイブリッドポリペプチドを含み得る。融合されたポリペプチドは、1つのポリペプチドをコードする核酸配列(またはその一部)を別のポリペプチドをコードする核酸配列(またはその一部)に融合させることによって、産生される。融合ポリペプチドを産生させるための技術は当該分野において公知であり、ポリペプチドをコードするコード配列を、それらがフレーム内にあって、融合されたポリペプチドの発現が同じプロモーターおよびターミネーターの制御下にあるように連結することを含む。ハイブリッドポリペプチドは、少なくとも2つの異なるポリペプチドから入手される部分的または完全なポリペプチド配列の組み合わせを含み得、ここで、1つもしくはそれ以上は、真菌細胞に対して異種であってもよい。
【0064】
DNA構築物は、プロモーターDNA配列に加えて、制御配列に適合する条件下で、適切な宿主細胞においてコード配列の発現を指令する1つもしくはそれ以上の制御配列を含み得る。発現は、転写、転写後修飾、翻訳、翻訳後修飾、および分泌を含むがこれらに限定されないポリペプチドの産生に関与する任意の工程を含むことが理解される。1つもしくはそれ以上の制御配列は、コード配列または宿主に対して生来であってもよい。あるいは、1つもしくはそれ以上の制御配列は、宿主細胞におけるコード配列の発現を改善するために、核酸配列に対して外来性の1つもしくはそれ以上の制御配列で置換されていてもよい。
【0065】
「DNA構築物」は、本明細書において、天然に存在する遺伝子から単離されるかまたはそうでなければ天然には存在しない様式で組み合わされ、並置される核酸のセグメントを含有するように修飾されている一本鎖もしくは二本鎖のいずれかの核酸分子として規定される。DNA構築物という用語は、DNA構築物がコード配列およびコード配列の発現に要求されるすべての制御配列を含有する場合、発現カセットという用語と同義である。
【0066】
用語「制御配列」は、本明細書において、コード配列の発現に必要または有利であるすべての成分を含むように規定され、本発明のプロモーターを含む。各制御配列は、ポリペプチドをコードする核酸配列に生来または外来性であってもよい。かかる制御配列として、リーダー、(コザク(Kozak)、1991年、J.Biol.Chem.266:19867−19870に記載のような)至適翻訳開始配列、ポリアデニル化配列、プロペプチド配列、シグナルペプチド配列、上流活性化配列、変異体、フラグメント、ならびにそれから誘導されるハイブリッドおよびタンデムプロモーターを含む本発明のプロモーターならびに転写ターミネーターが挙げられるが、それらに限定されない。少なくとも、制御配列は、転写および翻訳停止シグナルならびに本発明のプロモーター(の一部)を含む。制御配列は、制御配列とポリペプチドをコードする核酸配列のコード領域との連結を容易にする特定の制限部位を誘導する目的のためのリンカーを伴って提供され得る。
【0067】
制御配列は、適切な転写ターミネーター配列、即ち、転写を終結するために宿主細胞によって認識される配列であってもよい。ターミネーター配列は、ポリペプチドをコードするコード配列の3’末端と作動可能に会合される。好適な宿主細胞において機能的であるいずれのターミネーターも本発明において使用することができる。
【0068】
糸状真菌宿主細胞のための好適なターミネーターは、麹菌(A.oryzae)TAKAアミラーゼ、A.ニガー(A.niger)グルコアミラーゼ、A.ニダランス(A.nidulans)アントラニル酸シンターゼ、A.ニガー(A.niger)α−グルコシダーゼ、trpC遺伝子、およびフザリウム・オキシスポラム(Fusarium oxysporum)トリプシン様プロテアーゼの遺伝子から入手される。
【0069】
酵母宿主細胞のための好適なターミネーターは、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)エノラーゼ、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)シトクロムC(CYC1)、およびサッカロミセス・セレビシエ(Saccharomyces cerevisiae)グリセルアルデヒド(glyceraidehyde)−3−リン酸デヒドロゲナーゼの遺伝子から入手される。酵母宿主細胞のための他の有用なターミネーターについては、ローマノス(Romanos)ら、1992年、上掲に記載されている。
【0070】
制御配列はまた、適切なリーダー配列、即ち、宿主細胞による翻訳に重要であるmRNAの5’非翻訳領域であってもよい。リーダー配列は、ポリペプチドをコードする核酸配列の5’末端と作動可能に会合される。好適な宿主細胞において機能的であるいずれのリーダー配列も本発明において使用することができる。
【0071】
糸状真菌宿主細胞のための好適なリーダーは、麹菌(A.oryzae)TAKAアミラーゼ、A.ニダランス(A.nidulans)トリオースリン酸イソメラーゼおよびA.ニガー(A.niger)glaAの遺伝子から入手される。
【0072】
酵母宿主細胞のための適切なリーダーは、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)エノラーゼ(ENO−1)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)3−ホスホグリセリン酸キナーゼ、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)α−因子、およびサッカロミセス・セレビシエ(Saccharomyces cerevisiae)アルコールデヒドロゲナーゼ/グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(ADH2/GAP)の遺伝子から入手される。
【0073】
制御配列はまた、ポリアデニル化配列、核酸配列の3’末端と作動可能に会合され、転写される場合、転写されるmRNAにポリアデノシン残基を添加するためのシグナルとして宿主細胞に認識される配列であってもよい。好適な宿主細胞において機能的であるいずれのポリアデニル化配列も本発明において使用することができる。
【0074】
糸状真菌宿主細胞のための好適なポリアデニル化配列は、麹菌(A.oryzae)TAKAアミラーゼ、A.ニガー(A.niger)グルコアミラーゼ、A.ニダランス(A.nidulans)アントラニル酸シンターゼ、フザリウム・オキシスポラム(Fusarium oxysporum)トリプシン様プロテアーゼ、およびA.ニガー(A.niger)α−グルコシダーゼの遺伝子から入手される。
【0075】
酵母宿主細胞のための有用なポリアデニル化配列については、クオ(Guo)およびシャーマン(Sherman)、1995年、Molecular Cellular Biology15:5983−5990に記載されている。
【0076】
制御配列はまた、ポリペプチドのアミノ末端に連結されたアミノ酸配列をコードし、コードされるポリペプチドを細胞の分泌経路に指向するシグナルペプチドコード領域であってもよい。核酸配列のコード配列の5’末端は、分泌されるポリペプチドをコードするコード領域のセグメントを伴う翻訳リーディングフレームにおいて天然に連結されるシグナルペプチドコード領域を本来的に含有し得る。あるいは、コード配列の5’末端は、コード配列に対して外来性であるシグナルペプチドコード領域を含有してもよい。外来性のシグナルペプチドコード領域は、コード配列がシグナルペプチドコード領域を天然には含有しない場合に要求され得る。あるいは、外来性シグナルペプチドコード領域は、ポリペプチドの分泌を増強するために、天然のシグナルペプチドコード領域と単純に置き換わってもよい。しかし、発現されるポリペプチドを好適な宿主細胞の分泌経路に指向するいずれのシグナルペプチドコード領域も本発明において使用することができる。
【0077】
糸状真菌宿主細胞のための効果的なシグナルペプチドコード領域は、麹菌(A.oryzae)TAKAアミラーゼ、A.ニガー(A.niger)中性アミラーゼ、A.フィクウム(A.ficuum)フィターゼ、A.ニガー(A.niger)グルコアミラーゼ、A.ニガー(A.niger)エンドキシラナーゼ、リゾムコール・ミエヘイ(Rhizomucor miehei)アスパラギン酸プロテイナーゼ、フミコーラ・インソレンス(Humicola insolens)セルラーゼ、およびフミコーラ・ラヌギノサ(Humicola lanuginosa)リパーゼの遺伝子から入手されるシグナルペプチドコード領域である。
【0078】
酵母宿主細胞のための有用なシグナルペプチドは、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)α−因子およびサッカロミセス・セレビシエ(Saccharomyces cerevisiae)インベルターゼの遺伝子から入手される。他の有用なシグナルペプチドコード領域については、ローマノス(Romanos)ら、1992年、上掲により記載されている。
【0079】
制御配列はまた、ポリペプチドのアミノ末端に位置するアミノ酸配列をコードするプロペプチドコード領域であってもよい。得られるポリペプチドは、プロ酵素もしくはプロポリペプチド(または場合によりチモーゲン)として公知である。プロポリペプチドは一般に不活性であり、プロポリペプチドからのプロペプチドの触媒的または自己触媒的切断によって、成熟した活性なポリペプチドに変換することができる。プロペプチドコード領域は、枯草菌(Bacillus subtilis)アルカリプロテアーゼ(aprE)、枯草菌(Bacillus subtilis)中性プロテアーゼ(nprT)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)α−因子、リゾムコール・ミエヘイ(Rhizomucor miehei)アスパラギン酸プロテイナーゼ、ミセリオフトラ・サーモフィラ(Myceliophthora thermophila)ラッカーゼ(国際公開第95/33836号パンフレット)およびA.ニガー(A.niger)エンドキシラナーゼ(endo1)の遺伝子から入手することができる。
【0080】
シグナルペプチドおよびプロペプチド領域の両方がポリペプチドのアミノ末端に存在する場合、プロペプチド領域はポリペプチドのアミノ末端の隣に位置し、シグナルペプチド領域はプロペプチド領域のアミノ末端の隣に位置する。
【0081】
宿主細胞の増殖に関するポリペプチドの発現の調節を可能にする調節配列を添加することもまた所望され得る。調節系の例は、遺伝子の発現を、調節化合物の存在を含む化学的または物理的刺激に応答してオンもしくはオフに転向させる調節システムである。原核生物系における調節系としては、lac、およびtrpオペレーター系が挙げられる。酵母では、ADH2系またはGAL1系を使用することができる。糸状菌では、TAKAα−アミラーゼプロモーター、A.ニガー(A.niger)グルコアミラーゼプロモーター、麹菌(A.oryzae)グルコアミラーゼプロモーター、A.ツビンゲンシス(A.tubingensis)エンドキシラナーゼ(x/nA)プロモーター、A.ニガー(A.niger)硝酸レダクターゼ(niaD)プロモーター、トリコデルマ・リーセイ(Trichoderma reesei)セロビオヒドロラーゼプロモーターおよびA.ニダランス(A.nidulans)アルコールおよびアルデヒドデヒドロゲナーゼ(それぞれalcAおよびaldA)プロモーター(米国特許第5,503,991号明細書に記載される)を調節配列として使用することができる。調節配列の他の例は、遺伝子増幅を可能にする調節配列である。真核生物系では、これらは、メトトレキサートの存在下で増幅されるジヒドロ葉酸レダクターゼ遺伝子、および重金属で増幅されるメタロチオネイン遺伝子を含む。これらの場合、ポリペプチドをコードする核酸配列は調節配列と作動可能に会合される。
【0082】
creA結合部位の取り出し(EP673429号明細書において先に記載されているような炭素カタボライト抑制)、pacCおよびareAの変更(pHおよび窒素調節のため)は重要であり得る。
【0083】
本発明はまた、本発明のプロモーター、ポリペプチドをコードするコード配列、ならびに転写および翻訳停止シグナルを含む組換え発現ベクターに関する。上記の多様なコードおよび制御配列を共に接続して、1つもしくはそれ以上の簡便な制限部位を含んで、そのような部位における、プロモーターおよび/またはポリペプチドをコードするコード配列の挿入もしくは置換を可能にし得る組換え発現ベクターを産生させてもよい。あるいは、コード配列とプロモーターとの融合は、例えば、PCR(SOE−PCR)を使用する配列、重複伸長(Gene.1989年4月15日;77(1):51−9.ホウ SN(Ho SN)、ハント HD(Hunt HD)、ホートン RM(Horton RM)、プーレン JK(Pullen JK)、ピーズ LR(Pease LR)「Site−directed mutagenesis by overlap extension using the polymerase chain reaction」に記載される)によってかまたはGatewayTMクローニングシステム(Invitrogen)を使用するクローニングによって、行うことができる。あるいは、コード配列は、コード配列あるいはプロモーターおよび/またはコード配列を含むDNA構築物を、発現のための適切なベクターに挿入することによって、発現させてもよい。発現ベクターを作製する際に、コード配列が本発明のプロモーターおよび発現のための1つもしくはそれ以上の適切な制御配列と作動可能に会合されるように、コード配列がベクターに位置される。
【0084】
組換え発現ベクターは、組換えDNA手順に簡便に供され得、コード配列の発現をもたらし得る任意のベクター(例えば、プラスミドまたはウイルス)であってもよい。ベクターの選択は、典型的に、ベクターと、ベクターを導入しようとする宿主細胞との適合可能性に依存する。ベクターは、線状であってもまたは閉環状プラスミドであってもよい。
【0085】
ベクターは、自律的複製型ベクター、即ち、その複製が染色体複製とは独立する細胞外染色体実体として存在するベクター、例えば、プラスミド、染色体外エレメント、ミニ染色体、または人工染色体であってもよい。自律複製のために、ベクターは、問題の宿主細胞において、ベクターが自立的に複製することを可能にする複製開始点を含み得る。酵母宿主細胞において使用される複製開始点の例として、2マイクロン複製開始点、ARS1、ARS4、ARS1およびCEN3の組み合わせ、ならびにARS4およびCEN6の組み合わせがある。複製開始点は、その機能性を宿主細胞において温度感受性にする変異を有するものであってもよい(例えば、エールリヒ(Ehrlich)、1978年、Proceedings of the National Academy of Sciences USA75:1433を参照のこと)。糸状真菌において自律的に維持されるクローニングベクターの例は、AMA1配列を含むクローニングベクターである。AMA1は、アスペルギルス(Aspergillus)における自律的維持が可能であるA.ニダランス(A.nidulans)から単離される6.0−kbゲノムDNAフラグメントである(例えば、アレクセンコ(Aleksenko)およびクラッターバック(Clutterbuck)(1997年)、Fungal Genet.Biol.21:373−397を参照のこと)。
【0086】
あるいは、ベクターは、宿主細胞に導入される場合、ゲノムに組込まれ、それが組込まれている染色体と共に複製するものであってもよい。さらに、宿主細胞のゲノムに導入しようとする全DNAを共に含有する単一のベクターまたはプラスミドあるいは2つもしくはそれ以上のベクターまたはプラスミド、あるいはトランスポゾンを使用してもよい。
【0087】
本発明のベクターは、好ましくは、形質転換された細胞の容易な選択を可能にする1つもしくはそれ以上の選択マーカーを含有する。宿主は、少なくとも2つのベクター(一方は選択マーカーを含む)で同時形質転換してもよい。選択マーカーは、その産物が殺生物剤耐性またはウイルス耐性、重金属に対する耐性、栄養要求体に栄養非要求性、などを提供する遺伝子である。酵母宿主細胞に適切なマーカーは、ADE2、HIS3、LEU2、LYS2、MET3、TRP1、およびURA3である。糸状真菌宿主細胞において使用される選択マーカーとして、amdS(アセトアミダーゼ)、argB(オルニチンカルバモイルトランスフェラーゼ)、bar(ホスフィノトリシンアセチルトランスフェラーゼ)、hygB(ハイグロマイシンホスホトランスフェラーゼ)、niaD(硝酸レダクターゼ)、pyrG(オロチジン−5’−リン酸デカルボキシラーゼ)、sC(硫酸アデニルトランスフェラーゼ)、trpC(アントラニル酸シンターゼ)、ならびにそれらの等価物が挙げられるが、それらに限定されない。また、例えば、フレオマイシン、ハイグロマイシンBまたはG418に対する耐性を付与するマーカーを使用することもできる。アスペルギルス(Aspergillus)細胞における使用には、A.ニダランス(A.nidulans)または麹菌(A.oryzae)のamdSおよびpyrG遺伝子ならびにストレプトマイセス・ヒグロスコピカス(Streptomyces hygroscopicus)の遺伝子が好適である。amdSマーカー遺伝子は、好ましくは、EP635574号明細書または国際公開第97/0626号パンフレットに記載の技術を適用して、使用される。好適な選択マーカー遺伝子は、A.ニダランス(A.nidulans)gpdAプロモーターに融合されたA.ニダランス(A.nidulans)amdSコード配列である(EP635574号明細書)。他の糸状真菌由来のAmdS遺伝子を使用してもよい(国際公開第97/06261号パンフレット)。
【0088】
宿主細胞ゲノムへの組込みのために、ベクターは、プロモーター配列および/またはポリペプチドをコードするコード配列あるいは相同もしくは非相同組換えによるゲノムへのベクターの安定な組込みのためのベクターの他の任意のエレメントに依存し得る。あるいは、ベクターは、宿主細胞のゲノムへの相同組換えによる組込みを指令するためのさらなる核酸配列を含有してもよい。さらなる核酸配列は、染色体上の正確な位置で宿主細胞ゲノムにベクターを組込むことを可能にする。正確な位置での組込みの可能性を高めるために、組込みエレメントは、好ましくは、充分な数の核酸、例えば、100〜1,500塩基対、好ましくは400〜1,500塩基対、より好ましくは800〜1,500塩基対、および最も好ましくは少なくとも2kbを含有すべきであり、これは対応する標的配列と高度に相同であり、相同組換えの確率を増強する。組込みエレメントは、宿主細胞のゲノムにおける標的配列と相同である任意の配列であってもよい。さらに、組込みエレメントは、非コードまたはコード核酸配列であり得る。標的化組込みを促進するために、クローニングベクターは、好ましくは、宿主細胞の形質転換の前に線状化される。線状化は、好ましくは、少なくとも1つ、但し、好ましくは、クローニングベクターのいずれか一方の末端が標的遺伝子座に相同な配列によって隣接されるように、実施される。
【0089】
好ましくは、標的遺伝子座に相同であるクローニングベクターにおける組込みエレメントが、高度に発現される遺伝子座から誘導され、これは、それらが、真菌宿主細胞において高い発現レベルが可能である遺伝子から誘導されることを意味する。高い発現レベルが可能な遺伝子、即ち、高度に発現される遺伝子は、本明細書において、例えば、誘導条件下で、そのmRNAが少なくとも0.5%(w/w)の全細胞mRNAを構成し得る遺伝子、あるいは、その遺伝子産物が少なくとも1%(w/w)の全細胞性タンパク質を構成し得るか、または、分泌される遺伝子産物の場合、少なくとも0.1g/lのレベルまで分泌され得る遺伝子として規定される(欧州特許第357127B1号明細書に記載される)。多くの好適な高度に発現される真菌遺伝子は、次の例によって示される。すなわち、アスペルギルス(Aspergilli)またはトリコデルマ(Trichoderma)由来のアミラーゼ、グルコアミラーゼ、アルコールデヒドロゲナーゼ、キシラナーゼ、グリセルアルデヒド−リン酸デヒドロゲナーゼもしくはセロビオヒドロラーゼ遺伝子。これらの目的のための最も好適な高度に発現される遺伝子は、グルコアミラーゼ遺伝子、好ましくはA.ニガー(A.niger)グルコアミラーゼ遺伝子、麹菌(A.oryzae)TAKA−アミラーゼ遺伝子、A.ニダランス(A.nidulans)gpdA遺伝子、配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5の遺伝子座、配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5のA.ニガー(A.niger)遺伝子座、またはトリコデルマ・リーセイ(Trichoderma reesei)セロビオヒドロラーゼ遺伝子である。
【0090】
一方、ベクターは、非相同組換えによって、宿主細胞のゲノムに組込むことができる。
【0091】
ポリペプチドをコードする1コピーを超える核酸配列を宿主細胞に挿入して、遺伝子産物の産生を増加させてもよい。これは、好ましくは、DNA配列のそのゲノムコピーに組込むことによって、より好ましくは、高度に発現される遺伝子座、好ましくはグルコアミラーゼ遺伝子座または配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5の遺伝子座でDNA配列の組込みを標的化することによって、行うことができる。あるいは、これは、核酸配列を伴う増幅可能な選択マーカー遺伝子を含むことによって行うことができ、ここで、選択マーカー遺伝子の増幅されたコピー、およびそれによる核酸配列のさらなるコピーを含有する細胞を、適切な選択可能因子の存在下で細胞を培養することによって、選択することができる。過剰発現させようとするDNA配列のコピー数をさらにより増加するために、国際公開第98/46772号パンフレットに記載の遺伝子変換技術を使用してもよい。
【0092】
上記のエレメントを連結して、本発明の組換え発現ベクターを構築するために使用される手順は、当業者に周知である(例えば、サンブルック(Sambrook)ら、1989年、上掲を参照のこと)。
【0093】
本発明はまた、ポリペプチドをコードするコード配列と作動可能に会合される本発明のプロモーターDNA配列を含む組換え宿主細胞に関し、これは、ポリペプチドの産生において有利に使用される。ポリペプチドをコードするコード配列と作動可能に会合される本発明のプロモーターを含むベクターは、ベクターが染色体組込み体としてかまたは先に記載のような自己複製染色体外ベクターとして維持されるように、宿主細胞に導入される。用語「宿主細胞」は、複製中に生じる変異のため親細胞とは同一ではない親細胞の任意の子孫を包含する。宿主細胞の選択は、ポリペプチドをコードする遺伝子およびその供給源に大きな程度で依存する。
【0094】
本発明はまた、2つ以上の本発明のプロモーターDNA配列を含む組換え宿主細胞に関し、それぞれのプロモーターは、ポリペプチドをコードするコード配列と作動可能に会合される。かかる宿主細胞は、少なくとも1つのポリペプチドの組換え産生において有利に使用することができる。好ましくは少なくとも1つのプロモーターおよびその会合されるコード配列は、ベクター上に存在する。ベクターは、それが、染色体組込み体としておよび/または先に記載のような自己複製染色体外ベクターとして維持されるように、宿主細胞に導入される。
【0095】
別の好適な実施態様に従えば、宿主細胞は、(β−ラクタム)抗生物質、ビタミンもしくはカロテノイドなどの特定の一次または二次代謝物の産生のために使用される。
【0096】
宿主細胞は、本発明の方法において有用ないずれの真菌細胞であってもよい。本明細書において使用する「真菌」としては、子嚢菌門(Ascomycota)、担子菌門(Basidiomycota)、ツボカビ門(Chytridiomycota)、および接合菌門(Zygomycota)(ホークスワース(Hawksworth)ら、Ainsworth and Bisby’s Dictionary of The Fungi、第8版、1995年、CAB International,University Press,Cambridge、英国により記載される)ならびに卵菌門(Oomycota)(ホークスワース(Hawksworth)ら、1995年、上掲、171頁において引用される)およびすべての栄養胞子形成菌(mitosporic fungi)(ホークスワース(Hawksworth)ら、1995年、上掲)が挙げられる。
【0097】
好適な実施態様では、真菌宿主細胞は酵母細胞である。本明細書において使用する「酵母」としては、有子嚢胞子(ascosporogenous)酵母(エンドミセタレス目(Endomycetales))、担子胞子形成(basidiosporogenous)酵母、および不完全菌類(Fungi lmperfecti)(不完全酵母菌綱(Blastomycetes))に属する酵母が挙げられる。酵母の分類は将来変化し得るため、本発明の目的のために、酵母は、Biology and Activities of Yeast(スキナー,F.A.(Skinner,F.A.)、パスモア,S.M.(Passmore,S.M.)、およびダベンポート,R.R.(Davenport,R.R.)編、Soc.App.Bacteriol.第9回シンポジウム、1980年)において記載のように規定されるべきである。
【0098】
より好適な実施態様では、酵母宿主細胞は、カンジダ(Candida)、ハンセヌラ(Hansenula)、クリヴェロミセス(Kluyveromyces)、ピキア(Pichia)、サッカロミセス(Saccharomyces)、シゾサッカロマイセス(Schizosaccharomyces)、またはヤロウイア(Yarrowia)細胞である。
【0099】
最も好適な実施態様では、酵母宿主細胞は、サッカロマイセス・カールスベルゲンシス(Saccharomyces carlsbergensis)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、サッカロミセス・ディアスタティカス(Saccharomyces diastaticus)、サッカロミセス・ダグラシー(Saccharomyces douglasii)、サッカロミセス・クリヴェリ(Saccharomyces kluyveri)、サッカロミセス・ノルベンシス(Saccharomyces norbensis)またはサッカロミセス・オヴィフォルミス(Saccharomyces oviformis)細胞である。別の最も好適な実施態様では、酵母宿主細胞は、クリュイベロミセス・ラクティス(Kluyveromyces lactis)細胞である。別の最も好適な実施態様では、酵母宿主細胞は、ヤロウイア・リポリティカ(Yarrowia lipolytica)細胞である。
【0100】
別の好適な実施態様では、真菌宿主細胞は糸状真菌細胞である。「糸状菌」は、亜門の真菌門(Eumycota)および卵菌門(Oomycota)のすべての糸状形態を含む(ホークスワース(Hawksworth)ら、1995年、上掲により規定される)。糸状菌は、キチン、セルロース、グルカン、キトサン、マンナン、および他の複雑な多糖からなる菌糸体の壁によって特徴付けられる。栄養生長は菌糸伸長により、炭素の異化は偏性好気性である。対照的に、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)などの酵母による栄養生長は単細胞葉状体の出芽により、炭素の異化は発酵性であり得る。
【0101】
より好適な実施態様では、糸状真菌宿主細胞は、アクレモニウム(Acremonium)、アスペルギルス(Aspergillus)、フザリウム(Fusarium)、フミコーラ(Humicola)、ケカビ(Mucor)、ミセリオフトラ(Myceliophthora)、ニューロスポラ(Neurospora)、ペニシリウム(Penicillium)、シーラビア(Thielavia)、トイポクラディウム(Tolypocladium)、またはトリコデルマ(Trichoderma)の種の細胞であるが、これらに限定されない。
【0102】
最も好適な実施態様では、糸状真菌宿主細胞は、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・フォエティダス(Aspergillus foetidus)、アスペルギルス・ジャポニクス(Aspergillus japonicus)、A.ニダランス(A.nidulans)、A.ニガー(A.niger)または麹菌(A.oryzae)細胞である。最も好適な別の実施態様では、糸状真菌宿主細胞は、フザリウム・バクトリディオデス(Fusarium bactridioides)、フザリウム・セレアリス(Fusarium cerealis)、フザリウム・クルックウェルエンス(Fusarium crookwellense)、フザリウム・クルモラム(Fusarium culmorum)、フザリウム・グラミネアラム(Fusarium graminearum)、フザリウム・グラミヌム(Fusarium graminum)、フザリウム・ヘテロスポルム(Fusarium heterosporum)、フザリウム・ネグンディ(Fusarium negundi)、フザリウム・オキシスポラム(Fusarium oxysporum)、フザリウム・レティクラツム(Fusarium reticulatun)、フザリウム・ロゼウム(Fusarium roseum)、フザリウム・サンブシヌム(Fusarium sambucinum)、フザリウム・サルコチロウム(Fusarium sarcochroum)、フザリウム・スポロトリキオイデス(Fusarium sporotrichioides)、フザリウム・スルプレウム(Fusarium sulphureum)、フザリウム・トルローサム(Fusarium torulosum)、フザリウム・トリコテシオイデス(Fusarium trichothecioides)、またはフザリウム・ヴェネナツム(Fusarium venenatum)細胞である。最も好適な別の実施態様では、糸状真菌宿主細胞は、フミコーラ・インソレンス(Humicola insolens)、フミコーラ・ラヌギノサ(Humicola lanuginosa)、ムコール・ミエヘイ(Mucor miehei)、ミセリオフトラ・サーモフィラ(Myceliophthora thermophila)、アカパンカビ(Neurospora crassa)、ペニシリウム・パルロゼラム(Penicillium purpurogenum)、シーラビア・テレストリス(Thielavia terrestris)、トリコデルマ・ハルジアナム(Trichoderma harzianum)、トリコデルマ・コニンギ(Trichoderma koningii)、トリコデルマ・ロンギプラチアタム(Trichoderma longibrachiatum)、トリコデルマ・リーセイ(Trichoderma reesei)、またはトリコデルマ・ビリデ(Trichoderma viride)細胞である。
【0103】
真菌細胞は、プロトプラスト形成、プロトプラストの形質転換、およびそれ自体公知の様式での細胞壁の再生を包含するプロセスによって形質転換することができる。アスペルギルス(Aspergillus)宿主細胞の形質転換に適切な手順については、EP238023号明細書およびエルトン(Yelton)ら、1984年、Proceedings of the National Academy of Sciences USA81:1470−1474に記載されている。アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)を使用するアスペルギルス(Aspergillus)および他の糸状真菌宿主細胞の形質転換に適切な手順については、例えば、Nat Biotechnol.1998年9月;16(9):839−42.Nat Biotechnol 1998年11月;16(11):1074に掲載の訂正表.Agrobacterium tumefaciens−mediated transformation of filamentous fungi、デグルート MJ(de Groot MJ)、バンドドック P(Bundock P)、フーカス PJ(Hooykaas PJ)、ベイエルスベルゲン AG(Beijersbergen AG).Unilever Research Laboratory Vlaardingen,オランダに記載されている。フザリウム(Fusarium)種を形質転換するための適切な方法については、メーラディール(Malardier)ら、1989年、Gene78:147−156および国際公開第96/00787号パンフレットにより記載されている。酵母は、ベッカー(Becker)およびグアレンテ(Guarente)、編者:エイブルソン,J.N.(Abelson,J.N.)およびシモン,M.I.(Simon,M.I.)、Guide to Yeast Genetics and Molecular Biology,Methods in Enzymology、第194巻、182−187頁、Academic Press,Inc.、ニューヨーク;イトウら、1983年、Journal of Bacteriology153:163;ならびにヒネン(Hinnen)ら、1978年、Proceedings of the National Academy of Sciences USA75:1920に記載の手順を使用して、形質転換してもよい。
【0104】
本発明は、宿主細胞におけるコード配列の発現のための方法にさらに関する。方法は、以下の工程を含む。すなわち、
(a)本発明のプロモーターDNA配列および上記のコード配列を含むDNA構築物を提供すること、
(b)前記DNA構築物で適切な宿主細胞を形質転換すること、および
(c)前記プロモーターDNA配列の制御下でコード配列を発現させること。
【0105】
本発明はまた、適切な真菌宿主において本発明のプロモーターの制御下にあるコード配列によってコードされるポリペプチドの産生のための方法に関する。方法は、以下の工程を含む。すなわち、
(a)本発明のプロモーターDNA配列および上記のポリペプチドをコードするコード配列を含むDNA構築物を提供すること、
(b)前記DNA構築物で適切な真菌宿主細胞を形質転換すること、
(c)ポリペプチドの発現をもたらす適切な培養条件下で適切な真菌宿主を培養すること、
(d)培養ブロスからポリペプチドを回収すること。
【0106】
本発明はまた、以下を含む、適切な宿主における二次代謝物の産生のための方法に関する。すなわち、
(a)本発明のプロモーターDNA配列および上記の二次代謝物の産生に関与する酵素をコードするコード配列を含むDNA構築物を提供すること、
(b)前記DNA構築物で適切な宿主細胞を形質転換すること、
(c)二次代謝物の産生をもたらす適切な培養条件下で適切な真菌宿主を培養すること、および
(d)培養ブロスから二次代謝物を回収すること。
【0107】
本発明の産生方法では、当該分野において既知の方法を使用して、細胞は、ポリペプチドまたは代謝物の産生に適切な栄養培地中で培養される。例えば、細胞は、適切な培地において、ならびにコード配列を発現させるおよび/またはポリペプチドを単離することが可能な条件下で、実験室あるいは産業発酵槽における振盪フラスコ培養、小規模または大規模発酵(連続、バッチ、流加、もしくは固体発酵を含む)によって、培養することができる。培養は、当該分野において既知の手順を使用して、炭素および窒素源ならびに無機塩を含む適切な栄養培地において行われる。適切な培地は販売者から入手可能であり、または(例えば、米国菌培養収集所(the American Type Culture Collection)のカタログにおいて)公開された組成に従って、調製してもよい。ポリペプチドまたは代謝物が栄養培地に分泌される場合、ポリペプチドまたは代謝物は、培地から直接回収することができる。ポリペプチドまたは代謝物が分泌されない場合、それは、細胞溶解物から回収することができる。
【0108】
ポリペプチドは、ポリペプチドに特異的である当該分野において既知の方法を使用して、検出することができる。これらの検出方法は、特異的抗体の使用、酵素生成物の形成、または酵素基質の消失を含み得る。
【0109】
得られるポリペプチドまたは代謝物は、当該分野において既知の方法によって回収することができる。例えば、ポリペプチドまたは代謝物は、遠心分離、ろ過、抽出、噴霧乾燥、エバポレーション、または沈殿を含むがこれらに限定されない従来の手順によって、栄養培地から回収することができる。
【0110】
ポリペプチドは、クロマトグラフィー(例えば、イオン交換、アフィニティー、疎水性、クロマトフォーカシング、およびサイズ排除)、電気泳動手順(例えば、分取等電点電気泳動)、差分溶解性(differential solubility)(例えば、硫酸アンモニウム沈殿)、SDS−PAGE、または抽出(例えば、Protein Purification、編者:J.−C.ヤンソン(J.−C.Janson)およびラーズ・ライデン(Lars Ryden)、VCH Publishers、ニューヨーク、1989年を参照のこと)を含むが、これらに限定されない当該技術分野で既知の様々な手順によって、精製することができる。
【0111】
本発明はまた、真菌宿主細胞に対して内因性であるポリペプチドをコードするコード配列の発現を改変するためのDNA構築物に関する。構築物は、内因性遺伝子の発現を改変するのに必要な最小数の成分を含有し得る。
【0112】
1つの実施態様では、核酸構築物は、好ましくは、(a)ターゲティング配列、(b)本発明のプロモーターDNA配列、(c)エキソン、および(d)スプライスドナー部位を含有する。核酸構築物の細胞への導入時に、構築物は、相同組換えによって、内因性遺伝子部位で細胞ゲノムに組込む。ターゲティング配列は、エレメント(b)〜(d)が内因性遺伝子と作動可能に会合されるように、エレメント(a)〜(d)の内因性遺伝子への組込みを指令する。
【0113】
別の実施態様では、核酸構築物は、(a)ターゲティング配列、(b)本発明のプロモーターDNA配列、(c)エキソン、(d)スプライスドナー部位、(e)イントロン、および(f)スプライスアクセプター部位を含有し、ここで、ターゲティング配列は、エレメント(b)〜(f)が内因性遺伝子と作動可能に会合されるように、エレメント(a)〜(f)の組込みを指令する。しかし、構築物は、選択マーカーなどのさらなる成分を含有してもよい。使用することができる選択マーカーについては、先に記載された。
【0114】
両方の実施態様において、これらの成分の導入は、内因性遺伝子の発現が改変される新たな転写単位の生成を生じる。本質的に、新たな転写単位は、ターゲティング構築物によって誘導される配列と内因性遺伝子との融合産物である。内因性遺伝子が改変される1つの実施態様では、遺伝子が活性化される。本実施態様では、相同組換えを使用して、対応する親細胞において認められるレベルよりも高いレベルで遺伝子を発現させる調節配列の挿入を介して、親細胞の内因性遺伝子に、通常会合される調節領域を置換、破壊、または不能にする。
【0115】
ターゲティング配列は、内因性遺伝子内にあるか、遺伝子の直ぐ隣にあるか、上流の遺伝子内にあるか、または内因性遺伝子の上流かつ離れた場所にあり得る。1つもしくはそれ以上のターゲティング配列を使用することもできる。例えば、環状プラスミドまたはDNAフラグメントは、好ましくは、単一のターゲティング配列を用いる一方、線状プラスミドまたはDNAフラグメントは、好ましくは、2つのターゲティング配列を用いる。
【0116】
構築物は、内因性遺伝子の1つもしくはそれ以上のエキソンをさらに含有する。エキソンは、エキソン配列が内因性遺伝子のコード領域を伴うフレーム内に存在するように、RNAにコピーされ、成熟mRNA分子に存在するDNA配列として規定される。エキソンは、場合により、1つもしくはそれ以上のアミノ酸をコードするおよび/またはアミノ酸を部分的にコードするDNAを含有し得る。あるいは、エキソンは、5’非コード領域に対応するDNAを含有する。外因性エキソンが1つもしくはそれ以上のアミノ酸および/またはアミノ酸の部分をコードする場合、第2のエキソンから誘導されるmRNAの部分の適切なリーディングフレームが変更されないように、転写およびスプライシング時に、リーディングフレームが、内因性遺伝子のコード領域を伴うフレーム内にあるように、核酸構築物が設計される。構築物のスプライスドナー部位は、1つのエキソンの別のエキソンへのスプライシングを指令する。典型的に、第1のエキソンは第2のエキソンの5’にあり、第1のエキソンの3’側に重複および隣接するスプライスドナー部位は、第2のエキソンの5’側で第2のエキソンに隣接するスプライスアクセプター部位を認識する。スプライスアクセプター部位は、スプライスドナー部位と同様に、1つのエキソンの別のエキソンへのスプライシングを指令する配列である。スプライシング装置は、スプライスドナー部位と協働して作用し、スプライスアクセプター部位を使用して、イントロンの取り出しを生じさせる。
【0117】
所定のDNA配列の発現を改変するための好適なストラテジーは、所定のDNA配列の欠失および/または本発明のプロモーターなどの修飾されたプロモーターDNA配列による所定のDNA配列の内因性プロモーター配列の置換を含む。欠失および置換は、好ましくは、EP0357127号明細書に記載の遺伝子置換技術によって実施される。遺伝子および/またはプロモーター配列の特異的欠失は、好ましくは、EP635574号明細書に記載のように、amdS遺伝子を選択マーカー遺伝子として使用して実施される。EP635574号明細書に記載のようなフルオロアセトアミド(fluoracetamide)培地に対する対抗選択によって、得られる株は、選択マーカーを含まず、さらなる遺伝子修飾に使用することができる。
【0118】
あるいは、他の言及した技術との組み合わせで、大腸菌(E.coli)におけるコスミドのインビボ組換えに基づく技術を使用することができ、これについては:A rapid method for efficient gene replacement in the filamentous fungus A.nidulans(2000年)チャベロチェ,M−K(Chaveroche,M−K)、チコ,J−M.(Ghico,J−M.)およびダンフェール C(d’Enfert C);Nucleic acids Research、第28巻、22号に記載されている。この技術は、例えば、A.ニガー(A.niger)のような他の糸状菌にも適用可能である。本明細書において開示される特定の実施態様は、本発明のいくつかの態様の例示として意図されるため、本明細書において説明および特許請求される本発明は、これらの実施態様によってその範囲が限定されるべきではない。任意の等価な実施態様は、本発明の範囲内にあることが意図される。実際、本明細書において示しかつ説明される改変の他にも、本発明の多様な改変が存在することは、先の記載から当業者には明らかであろう。かかる改変もまた、添付の特許請求の範囲内に含まれることが意図される。抵触する場合については、定義を含む本発明の開示内容が優先される。
【0119】
本明細書において言及した特許および刊行物において開示されるすべての配列および方法を含むすべての特許および刊行物は、参照により明確に援用される。これらの特許および刊行物には次のものが含まれる。すなわち、EP357127号明細書、EP673429号明細書、EP635574号明細書、国際公開第97/06261号パンフレット、国際公開第98/46772号パンフレット、国際公開第94/04673号パンフレット。
【実施例】
【0120】
実験の情報
株
WT1:このA.ニガー(A.niger)株を野生型株として使用する。この株は、寄託番号CBS513.88でCBS研究所(CBS Institute)に寄託される。
【0121】
WT2:このA.ニガー(A.niger)株は、グルコアミラーゼをコードする遺伝子(glaA)の欠失を含むWT1株である。WT2は、EP0635574号明細書に記載のように、「マーカー遺伝子を含まない(MARKER−GENE FREE)」アプローチを使用して、構築した。本特許では、CBS513.88のゲノムにおけるglaA特異的DNA配列を欠失する仕方について、広範に説明する。該手順により、最終的に外来性のDNA配列を全く所有しない、マーカーを含まない(MARKER−GENE FREE)(?)glaA組換えA.ニガー(A.niger)CBS513.88株が得られた。
【0122】
グルコアミラーゼ活性アッセイ
国際公開第98/46772号パンフレットに記載のように、p−ニトロフェニルa−D−グルコピラノシド(シグマ(Sigma))を使用して、グルコアミラーゼ活性を決定した。
【0123】
実施例1
コード配列と作動可能に会合される本発明のプロモーターを含むDNA構築物の構築
本実施例では、本発明のプロモーターの制御下にある発現構築物の構築について説明する。ここで使用するコード配列またはレポーター構築物は、A.ニガー(A.niger)グルコアミラーゼ酵素をコードするglaA遺伝子である。グルコアミラーゼは、本発明のプロモーターの活性を測定することが可能であるレポーター酵素として使用する。
【0124】
1.1 集成的グルコアミラーゼ発現ベクター(pGBTOPGLA)の説明
グルコアミラーゼプロモーターおよびA.ニガー(A.niger)由来の遺伝子glaAをコードするグルコアミラーゼを、国際公開第99/32617号パンフレットに記載の発現ベクターpGBTOP−8にクローニングした。既知の原理および日常的なクローニング技術に従って、クローニングを実施し、プラスミドpGBTOPGLA(図1を参照のこと)を得た。本質的に、この発現ベクターは、大腸菌(E.coli)ベクターにおける3’および3’’glaAターゲティング部位に隣接されるグルコアミラーゼプロモーター、コード配列およびターミネーター領域を含む。
【0125】
1.2 複数のクローニング部位MCSを伴う集成的グルコアミラーゼ発現ベクター(pGBTOPGLA−2)の構築
テンプレートとして、オリゴヌクレオチド5’−ATgCggCCgCCTCgAgTTAATTAAggCCAggCCggCCggCgCgCCTCAgCAATgTCgTTCCgA−3’(配列番号6として同定される)および5’−AGCCATTGACTTCTTCCCAG−3’(配列番号7として同定される)および1ngのベクターpGBTOPGLAを使用して、glaAコード配列の部分を含有するPCRフラグメントを作製した。このフラグメントを、XhoIおよびBglIIで消化し、XhoIおよびBglII消化したベクターpGBTOPGLAに導入し、ベクターpGBTOPGLA−2(図2を参照のこと)を得た。MCSおよびglaAコード配列の一部を含む導入されたPCRフラグメントの配列を、配列解析によって確認した。
【0126】
1.3 グルコアミラーゼコード配列と作動可能に会合される本発明のプロモーターを伴う集成的発現ベクター(pGBTOPGLA−3)の構築
株CBS513.88のゲノムDNAを配列決定し、解析した。表において(オリゴ配列番号)以下のように同定されるオリゴヌクレオチドの組み合わせおよび株CBS513.88のゲノムDNAをテンプレートとして使用し、PCR増幅により、適切な制限部位を本発明のプロモーターに付着させた。配列番号1、配列番号2、配列番号3、配列番号4または配列番号5において同定される配列は、以下の表において同定されるように、約2kbの得られるフラグメントの配列を含む。すべての5つの得られるフラグメントを、AscIおよびXhoIで消化し、AscIおよびXhoI消化したベクターpGBTOPGLA−2に導入し、表(ベクターの名称)において同定されるように、ベクターpGBTOPGLA−4、pGBTOPGLA−5、pGBTOPGLA−7、GBTOPGLA−9またはpGBTOPGLA−10を得た。図3では、5つのすべてのベクターのレイアウトを例示する図を見出すことができる。本発明のプロモーターを含む多様な導入されたPCRフラグメントの配列を、配列解析によって確認した。
【0127】
【表1】

【0128】
実施例2.DNA構築物で形質転換された真菌宿主細胞
pGBTOPGLA、pGBTOPGLA−4、pGBTOPGLA−5、pGBTOPGLA−7、pGBTOPGLA−9またはpGBTOPGLA−10ベクターをWT2に導入するために、形質転換およびその後の形質転換体選択を、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットに記載のとおりに行った。原則として、NotIによる消化後、すべてのベクターの線状DNAを単離し、pGBAAS−1で示されるamdS選択マーカー遺伝子含有ベクターで同時形質転換した(EP635574号明細書に記載のとおりに構築した)。両方のベクターは、WT2における短縮型glaA遺伝子座への標的化を指令するためのA.ニガー(A.niger)宿主株のglaA遺伝子座に相同な2つのDNAドメインを含む。形質転換体をアセトアミド培地上で選択し、標準的手順に従ってコロニーを精製した。胞子をフルオロアセトアミド培地上に配置し、amdSマーカーを消失した株を選択した。増殖中のコロニーを、glaA遺伝子座での組込みおよびコピー数について診断した。類似の予想されるコピー数を伴うpGBTOPGLA、pGBTOPGLA−4、pGBTOPGLA−5、pGBTOPGLA−7、pGBTOPGLA−9またはpGBTOPGLA−10の形質転換体を選択した。好ましくは、単一コピーを伴う若干の形質転換体(1A、1B、1C)およびおそらく、複数コピーを伴う1つの形質転換体(2A)を選択した。
【0129】
さらに、選択マーカー遺伝子および本発明のプロモーターによって制御される目的の遺伝子は1つの構築物上に存在している。この例を図4に示す。
【0130】
実施例3.真菌宿主細胞における本発明のプロモーターの制御下でglaAコード配列によってコードされるグルコアミラーゼポリペプチドの産生
多くの選択された上記のようなWT2の形質転換体、および両方の5つの株WT1およびWT2を使用し、500mlバッフル付き振盪フラスコを使用する恒温振盪器において、EP635574号明細書に記載のような100mlの培地中、34℃および170rpmで、振盪フラスコ実験を実施した。発酵の4および5日後、サンプルを採取し、上記のようにグルコアミラーゼ活性を決定した。グルコアミラーゼ活性を、4日目のWT1の活性に対して正規化した。WT1、WT2、ならびにpGBTOPGLA、pGBTOPGLA−5、pGBTOPGLA−7、およびpGBTOPGLA−10について多くの選択された形質転換体の正規化された活性を図5に示す。
【0131】
グルコアミラーゼレポーターの測定される活性から結論し得るように、本発明は、真菌細胞において、目的の遺伝子の高い発現のための強力なプロモーターを提供する。従って、本発明のプロモーターは、真菌細胞において、目的の遺伝子の高い発現のための代替的およびさらなるプロモーターを提供する。
【0132】
実施例4.本発明のプロモーターを含むプロモーター置換構築物pGBDEL−PGLAAの構築
宿主細胞における所定の遺伝子の発現レベルを改変するために、本発明のプロモーターは、前記所定の遺伝子の内因性プロモーターと置き換わることができる。本実験では、本発明のプロモーターは、真菌宿主細胞においてglaA遺伝子をコードするグルコアミラーゼのプロモーターと置き換わる。実施例4、5および6では、このプロセスにおける多くの異なる工程について説明している。
【0133】
グルコアミラーゼプロモーターのための置換ベクターを、既知の原理に従って設計し、日常的なクローニング手順に従って構築した(図6を参照のこと)。本質的に、glaAプロモーター置換ベクターpGBDEL−PGLAAは、予め定められた遺伝子座での相同組換えを介して本発明のプロモーター(配列番号4に含まれる)によって置換されるべきglaAプロモーター配列の約1000bpフランキング領域を含む。ここで使用されるフランキング領域(図6を参照のこと)は、glaAプロモーターの5’上流領域であり、glaAコード配列の部分である。さらに、置換ベクターは、A.ニダランス(A.nidulans)両方向性amdS選択マーカーを中間的なダイレクトリピートで含有する。本実施例で使用されるダイレクトリピートは、glaAコード配列の部分である。これらの欠失ベクターの一般的な設計については、EP635574号明細書および国際公開第98/46772号パンフレットにおいて先に記載された。
【0134】
実施例5.真菌宿主細胞における本発明のプロモーターによるglaAプロモーターの置換
NotI消化した欠失ベクターpGBDEL−PGLAAの線状DNAを単離し、WT1(CBS513.88)を形質転換するために使用した。この線状DNAは、glaA遺伝子座でゲノムに組込むことができ、従って、glaAプロモーター領域をamdSおよび本発明のプロモーターを含有する構築物で置換する(図7を参照のこと)。形質転換体をアセトアミド培地上で選択し、標準的手順に従ってコロニーを精製した。増殖中のコロニーを、glaA遺伝子座での組込みについて、PCRによって診断した。glaAプロモーターの欠失は、本発明のプロモーターに特異的なサイズを伴うバンドの増幅、およびglaAプロモーターに特異的なバンドの消失によって、検出した。胞子をフルオロアセトアミド培地上に配置し、amdSマーカーを消失した株を選択した。候補株を、グルコアミラーゼプロモーターの適切な欠失および配列番号4に含まれる本発明のプロモーターによる置換について、サザン分析を使用して試験した。株dPGLAAを、glaAプロモーターが本発明のプロモーターによって置換され、回復された機能的glaAコード配列を有する代表的株として選択した(図7を参照のこと)。
【0135】
実施例6.真菌宿主細胞における本発明の置換されたプロモーターの制御下でglaAコード配列によってコードされるグルコアミラーゼポリペプチドの産生
選択されたdPGLAA株(実施例5において単離されたWT1の適切なpGBDEL−PGLAA形質転換体)および株WT1を使用して、500mlバッフル付き振盪フラスコを使用する恒温振盪器において、欧州特許第635574B1号明細書に記載のような100mlの培地中、34℃および170rpmで、振盪フラスコ実験を実施した。さらなる条件および活性測定は、実施例3に記載のとおりであった。WT1の選択されたpGBDEL−PGLAA形質転換体におけるグルコアミラーゼ活性は、発酵の両方の日にWT1について測定した活性と比較して増加していた(データ示さず)。
【0136】
実施例7.真菌宿主細胞における本発明のプロモーターの制御下におけるさらなるglaA遺伝子の付加
宿主細胞において所定の遺伝子の発現レベルを改変するために、本発明のプロモーターに作動可能に連結された複数のさらなるコピーの前記遺伝子を、内因的に付与された遺伝子に付加することができる。本実施例では、配列番号4に含まれ、glaAコード配列に作動可能に連結される本発明のプロモーターを、真菌宿主細胞において内因的に存在するglaA遺伝子をコードするグルコアミラーゼの隣に導入する。実施例7、および8では、このプロセスにおける多くの異なる工程について説明している。
【0137】
図8に示される環状構築物を単離し、WT1(CBS513.88)を形質転換するために使用した。この線状DNAは、glaAコード配列においてゲノムに組込むことができ、従って、本発明のプロモーターの制御下の第2のglaA遺伝子を、選択マーカーamdSの隣に付加することができる(図8を参照のこと)。形質転換体をアセトアミド培地上で選択し、標準的手順に従ってコロニーを精製した。増殖中のコロニーを、glaA遺伝子座での組込みについて、PCRによって診断した。glaA遺伝子座の組込みを、PCRおよびサザン分析によって検出した。株P2GLAAを、glaA遺伝子座で組込まれる本発明のプロモーターの制御下にある少なくとも第2のglaA遺伝子を伴う代表的株として選択した。
【0138】
実施例8.真菌宿主細胞における本発明のプロモーターおよび内因性glaAプロモーターの制御下でglaAコード配列によってコードされるグルコアミラーゼポリペプチドの産生
実施例7において単離された選択されたP2GLAA株、および株WT1を使用して、実施例3に記載のように、100mlの培地において、振盪フラスコ実験を実施した。発酵の4および6日後、サンプルを採取し、グルコアミラーゼ活性を決定した。WT1の選択されたP2GLAA形質転換体におけるグルコアミラーゼ活性は、発酵の4または5日後のいずれかで、WT1について測定した活性と比較して増加した。グルコアミラーゼレポーターの活性の増加は、本発明のプロモーターが、真菌細胞において強力なプロモーター下で既に発現されている目的の遺伝子の発現をさらに増加する手段を提供することを示す。
【図面の簡単な説明】
【0139】
【図1】集成的グルコアミラーゼ発現ベクターであるpGBTOPGLAのプラスミドマップを表す。
【図2】複数のクローニング部位を伴う集成的グルコアミラーゼ発現ベクターであるpGBTOPGLA−2のプラスミドマップを表す。
【図3】グルコアミラーゼコード配列と作動可能に会合される本発明のプロモーターを含有する集成的発現ベクターであるpGBTOPGLA−3のプラスミドマップを表す。
【図4】単一の相同組換えを介する組み込みの略図を表す。
【図5】WT1、WT2および多様なpGBTOPGLAベクターの形質転換体のグルコアミラーゼ活性。4日目のWT1の活性を100%と設定した場合の正規化された活性を示す。
【図6】置換ベクターであるpGBDEL−PGGLAAのプラスミドマップを表す。
【図7】プロモーター置換の略図を表す。
【図8】相同組換えを介する組み込みの略図を表す。
【技術分野】
【0001】
本発明は、DNA配列、特に、単離された真菌プロモーター、およびDNA構築物、ベクター、ならびにポリペプチドをコードするコード配列と作動可能に会合されるこれらのプロモーターを含む真菌宿主細胞に関する。本発明はまた、遺伝子を発現させるおよび/またはポリペプチドを産生させるための方法に関する。
【背景技術】
【0002】
真菌宿主細胞における組換えポリペプチドの産生は、ポリペプチドをコードするDNAが、宿主細胞に適切なプロモーターの発現制御下に配置される発現カセットを構築することによって、通常達成される。発現カセットは、プラスミドまたはベクターによって仲介される形質転換により、宿主細胞に導入することができる。次いで、ポリペプチドの産生は、発現カセットに含有されるプロモーターの適切な機能化に必要な誘導条件下で形質転換された宿主細胞を培養することによって、達成することができる。
【0003】
各真菌宿主細胞について、形質転換によって真菌宿主に導入されているコード配列の発現およびこのコード配列によってコードされる組換えポリペプチドの産生には、機能的プロモーターが利用可能であることが必要である。多数のプロモーターが、真菌宿主細胞において機能的であることが既に公知である。プロモーターの異種間使用の例が存在する。すなわち、アスペルギルス・ニダランス(Aspergillus nidulans)(A.ニダランス(A.nidulans)gpdA遺伝子のプロモーターは、アスペルギルス・ニガー(Aspergillus niger)(A.ニガー(A.niger))において機能的であることが公知である(J Biotechnol.1991年1月;17(1):19−33.Intracellular and extracellular production of proteins in Aspergillus under the control of expression signals of the highly expressed A.nidulans gpdA gene.プント PJ(Punt PJ)、ゼガー ND(Zegers ND)、ブッチャー M(Busscher M)、ポウウェルズ PH(Pouwels PH)、バンデンホンデル CA(van den Hondel CA))。もう1つの例は、A.ニガー(A.niger)およびA.ニダランス(A.nidulans)において使用されるA.ニガー(A.niger)β−キシロシダーゼxlnDプロモーター「Transcriptional regulation of the xylanolytic enzyme system of Aspergillus」、ヴァンペイジ,NNME(van Peij,NNME)博士−thesis Landbouwuniversiteit Wageningen、蘭国、ISBN90−5808−154−0ならびにA.ニガー(A.niger)、A.ニダランス(A.nidulans)およびクラドスポリウム・フルブム(Cladosporium fulvum)における大腸菌(Escherichia coli)β−グルクロニダーゼ遺伝子の発現(Curr Genet、1989年3月;15(3):177−80:ロバーツ IN(Roberts IN)、オリバー RP(Oliver RP)、プント PJ(Punt PJ)、バンデンホンデル CA(van den Hondel CA)「Expression of the Escherichia coli beta−glucuronidase gene in industrial and phytopathogenic filamentous fungi」において記載される)である。
【0004】
しかし、誘導される遺伝子の発現制御するため、内因性遺伝子の発現のレベルを制御するため、内因性遺伝子の発現の調節を制御するためかもしくは内因性遺伝子の不活化を仲介するため、またはポリペプチドを産生させるため、あるいは先のアプリケーションの組み合わせのための改善されたプロモーターに必要性がなお存在する。これらの改善されたプロモーターは、例えば、これまでの公知のプロモーターよりも強力であり得る。それらはまた、特異的な簡便な基質または化合物によって誘導可能であり得る。単一の真菌宿主において多用な遺伝子を同時に過剰発現させようと考える場合、いくらかの機能的プロモーターについて知ることもまた有利である。スケルチング(squelching)(特定の転写因子のタイトレーション)を防止するためには、複数の個別のプロモーター(発現させようとする1つの遺伝子に対してそれぞれ1つの特異的プロモーター)を使用することが好適である。
【発明の詳細な説明】
【0005】
本発明は、以下からなる群から選択されるプロモーターDNA配列に関する。すなわち、
(a)配列番号1、配列番号2、配列番号3、配列番号4、配列番号5のヌクレオチド配列を含むDNA配列、
(b)(a)のDNA配列にハイブリダイズすることが可能なDNA配列、
(c)(a)のDNA配列に少なくとも50%相同であるDNA配列、
(d)(a)〜(c)のDNA配列のいずれかの変異体、および
(e)(a)〜(d)のDNA配列のいずれかのサブ配列。
【0006】
本発明に関して、プロモーターDNA配列がコード配列と作動可能に会合される場合、このプロモーターDNA配列は、このコード配列の発現を制御することが可能なDNA配列である。用語「作動可能に会合」は、本明細書において、プロモーターDNA配列がコード配列によってコードされるポリペプチドの産生を指令するようなコード配列に対する位置に、プロモーターDNA配列が適切に配置される配置形態として規定される。
【0007】
用語「コード配列」は、本明細書において、適切な制御配列の制御下に配置される場合、ポリペプチドに翻訳されるmRNAに転写される核酸配列として規定される。コード配列の境界は、一般に、通常、mRNAの5’末端におけるオープンリーディングフレームの開始部であるATG開始コドン、およびmRNAの3’末端におけるオープンリーディングフレームの直ぐ下流に位置する転写ターミネーター配列によって決定される。コード配列としては、ゲノムDNA、cDNA、半合成、合成、および組換え核酸配列を挙げることができるが、これらに限定されない。
【0008】
より具体的には、用語「プロモーター」は、本明細書において、RNAポリメラーゼに結合し、転写を開始するポリペプチドをコードするコード配列の正確な下流の転写開始部位にポリメラーゼを指向するDNA配列として規定される。RNAポリメラーゼは、コード領域の適切なDNA鎖に相補的なメッセンジャーRNAの集成を効果的に触媒する。用語「プロモーター」はまた、mRNAへの転写後の翻訳のための(プロモーターと転写開始部との間の)5’非コード領域、エンハンサーなどのシス作用性転写制御エレメント、および転写因子と相互作用することが可能な他のヌクレオチド配列を含むことが理解されよう。
【0009】
好適な実施態様では、本発明のプロモーターDNA配列は、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5に記載の配列である。
【0010】
別の好適な実施態様に従えば、本発明のプロモーターDNA配列は、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5にハイブリダイズすることが可能であり、なおプロモーター活性を保持するDNA配列である。
【0011】
プロモーター活性は、好ましくは、プロモーターと作動可能に会合されるコード配列によってコードされるタンパク質の濃度を測定することによって、決定される。あるいは、プロモーター活性は、プロモーターと作動可能に会合されるコード配列によってコードされるタンパク質の酵素活性を測定することによって、決定される。好適な実施態様に従えば、プロモーター活性(およびその強度)は、lacZレポーター遺伝子のコード配列の発現を測定することによって決定される(ルオ(Luo)(Gene 163(1995年)127−131)。別の好適な実施態様に従えば、プロモーター活性は、コード配列として緑色蛍光タンパク質を使用することによって決定される(Microbiology.1999年3月;145(Pt3):729−34、サンテリレ アンリクセンAL(Santerre Henriksen AL)、エヴェン S(Even S)、マラー C(Muller C)、プント PJ(Punt PJ)、バンデンホンデル CA(van den Hondel CA)、ニールセン J.スタディ(Nielsen J.Study))。
【0012】
さらに、プロモーター活性は、プロモーターの制御下で作製される転写物のmRNAレベルを測定することによって、決定することができる。mRNAレベルは、例えば、ノーザンブロットを介して測定することができる(J.サンブルック(J.Sambrook),E.F.フリッチュ(E.F.Fritsch)、およびT.マニアツス(T.Maniatus)、1989年、Molecular Cloning,A Laboratory Manual、第2版、Cold Spring Harbor、ニューヨーク州)。
【0013】
本発明は、極めて低いストリンジェンシー条件、好ましくは低いストリンジェンシー条件、より好ましくは中等度のストリンジェンシー条件、より好ましくは中等度〜高いストリンジェンシー条件、さらにより好ましくは高いストリンジェンシー条件、および最も好ましくは極めて高いストリンジェンシー条件下で、(i)配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5のヌクレオチド1〜2000、好ましくはヌクレオチド100〜1990、より好ましくは200〜1980、さらにより好ましくは300〜1970、さらにより好ましくは350〜1950および最も好ましくは360〜1900に対応するか、(ii)(i)のサブ配列であるか、または(iii)(i)、(ii)の相補鎖である核酸プローブにハイブリダイズする(単離された)プロモーターDNA配列を包含する(J.サンブルック(J.Sambrook)、E.F.フリッチュ(E.F.Fritsch)、およびT.マニアティス(T. Maniatis)、1989年、Molecular Cloning,A Laboratory Manual、第2版、Cold Spring Harbor、ニューヨーク州)。配列番号1、配列番号2、配列番号3、配列番号4または配列番号5のサブ配列は、少なくとも100ヌクレオチド、好ましくは少なくとも200ヌクレオチド、より好ましくは少なくとも300ヌクレオチド、さらにより好ましくは少なくとも400ヌクレオチド、および最も好ましくは少なくとも500ヌクレオチドであり得る。
【0014】
配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5の核酸配列またはそのサブ配列を使用して、核酸プローブを設計し、当該分野において周知の方法に従って、異なる属あるいは種の株からDNAプロモーターを同定およびクローニングしてもよい。特に、そのようなプローブは、本明細書に記載の対応する遺伝子を同定および単離するために、標準的なサザンブロッティング手順に従って、目的の属または種のゲノムもしくはcDNAとのハイブリダイゼーションのために使用することができる。そのようなプローブは、全配列よりもかなり短くあり得るが、少なくとも15、好ましくは少なくとも25、およびより好ましくは少なくとも35ヌクレオチド長であるべきである。さらに、そのようなプローブを使用して、PCRを介してDNAプロモーターを増幅することができる。PCRを介してプロモーターをクローニングする例については、本明細書に記載されている(実施例1.3を参照のこと)。また、より長いプローブを使用することもできる。DNA、RNAおよびペプチド核酸(PNA)プローブを使用することもできる。プローブは、典型的に、対応する遺伝子を、(例えば、32P、33P、3H、35S、ビオチン、もしくはアビジンまたは蛍光マーカーで)検出するために標識される。かかるプローブも本発明に包含される。
【0015】
従って、そのような他の生物体から調製されるゲノムDNAまたはcDNAライブラリーを、上記のプローブにハイブリダイズし、ポリペプチドをコードするDNAを対象にスクリーニングしてもよい。そのような他の生物体由来のゲノムまたは他のDNAは、アガロースもしくはポリアクリルアミドゲル電気泳動、または他の分離技術によって分離することができる。ライブラリー由来のDNAまたは分離されたDNAを、ニトロセルロースまたは他の適切なキャリア材料に移して固定化してもよい。配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5、またはそのサブ配列に相同であるクローンあるいはDNAを同定するために、キャリア材料を、サザンブロットにおいて使用してもよい。
【0016】
本発明の目的のために、ハイブリダイゼーションは、核酸配列が、配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5に示される核酸配列、その相補鎖、またはそのサブ配列に対応する標識核酸プローブに、極めて低い〜極めて高いストリンジェンシー条件下で、ハイブリダイズすることを示す。核酸プローブがこれらの条件下でハイブリダイズする分子は、例えば、X線フィルムを使用して、検出される。また、検出のために蛍光ならびに支持体としてガラススライド(glass sides)および/またはDNAマイクロアレイを使用する技術などの他のハイブリダイゼーション技術を使用することもできる。DNAマイクロアレイハイブリダイゼーション検出の例は、FEMS Yeast Res.2003年12月;4(3):259−69(ダラン−ラプジャデ P(Daran−Lapujade P)、ダラン JM(Daran JM)、コッテル P(Kotter P)、ペティット T(Petit T)、パイパー MD(Piper MD)、プランク JT(Pronk JT).「Comparative genotyping of the Saccharomyces cerevisiae laboratory strains S288C and CEN.PK113−7D using oligonucleotide microarrays」)に認められる。さらに、ハイブリダイゼーションのためのPNAマイクロアレイの使用については、Nucleic Acids Res.2003年10月1日;31(19):e119(ブラント O(Brandt O)、フェルトナー J(Feldner J)、シュテファン A(Stephan A)、シュレーダー M(Schroder M)、シュノルザー M(Schnolzer M)、アーリンハウス HF(Arlinghaus HF)、ホヘイセル JD(Hoheisel JD)、ヤコブ A(Jacob A).PNA microarrays for hybridisation of unlabelled DNA samples.)に記載されている。
【0017】
好適な実施態様では、核酸プローブは、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5の核酸配列である。好適な別の実施態様では、核酸プローブは、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5のヌクレオチド20〜1980、より好ましくは配列番号1、配列番号2、配列番号3、配列番号4または配列番号5のヌクレオチド500〜1950、さらにより好ましくは配列番号1、配列番号2、配列番号3、配列番号4または配列番号5のヌクレオチド800〜1920、および最も好ましくは配列番号1、配列番号2、配列番号3、配列番号4または配列番号5のヌクレオチド900〜1900を有する配列である。別の好適なプローブは、転写部位の前のDNA配列の部分である。
【0018】
少なくとも100ヌクレオチド長の長いプローブでは、極めて低い〜極めて高いストリンジェンシー条件は、標準的なサザンブロッティング手順に従う5×SSPE、0.3%SDS、200.mu.g/ml剪断および変性サケ精子DNA、ならびに極めて低いおよび低いストリンジェンシーでは25%ホルムアミド、中等度および中等度〜高いストリンジェンシーでは35%ホルムアミド、または高いおよび極めて高いストリンジェンシーでは50%ホルムアミドのいずれかにおける42℃でのプレハイブリダイゼーションおよびハイブリダイゼーションとして規定される。
【0019】
少なくとも100ヌクレオチド長の長いプローブでは、キャリア材料は、それぞれ、15分間、2×SSC、0.2%SDSを使用して、好ましくは少なくとも45℃(極めて低いストリンジェンシー)、より好ましくは少なくとも50℃(低いストリンジェンシー)、より好ましくは少なくとも55℃(中等度のストリンジェンシー)、より好ましくは少なくとも60℃(中等度〜高いストリンジェンシー)、さらにより好ましくは少なくとも65℃(高いストリンジェンシー)、および最も好ましくは少なくとも70℃(極めて高いストリンジェンシー)で、最終的に3回洗浄する。
【0020】
約15ヌクレオチド〜約70ヌクレオチド長である短いプローブでは、ストリンジェンシー条件は、標準的なサザンブロッティング手順に従う、0.9M NaCl、0.09M Tris−HCl pH7.6、6mM EDTA、0.5%NP−40、1×デンハルト溶液(Denhardt’s solution)、1mMピロリン酸ナトリウム、1mMリン酸二水素ナトリウム、0.1mM ATP、および1mlあたり0.2mgの酵母RNAにおいて、ボールトン(Bolton)およびマッカーシー(McCarthy)(1962年、Proceedings of the National Academy of Sciences USA 48:1390)に従う計算を使用して算出されるTmより5℃〜10℃低い温度でのプレハイブリダイゼーション、ハイブリダイゼーション、およびハイブリダイゼーション後洗浄として規定される。
【0021】
約15ヌクレオチド〜約70ヌクレオチド長である短いプローブでは、キャリア材料は、6×SCC+0.1%SDS中、15分間で1回、算出されたTmより5℃〜10℃低い温度で6×SSCを使用して、それぞれ15分間、2回洗浄される。
【0022】
別の好適な実施態様に従えば、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5を最初に使用して、それと作動可能に会合される生来の遺伝子、コード配列もしくはその一部をクローニングする。これは、先に規定されるような配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5、またはそのサブ配列のいずれかから開始し、プローブとしてこの配列を使用して、行うことができる。プローブは、cDNAまたは所定の宿主、アスペルギルス・ニガー(Aspergillus niger)かもしくは本出願において規定される他の任意の真菌宿主のいずれかのゲノムライブラリーにハイブリダイズされる。一旦、生来の遺伝子またはその一部がクローニングされれば、続いて、それ自体をプローブとして使用して、本明細書に記載のハイブリダイゼーション実験によって、他の真菌から誘導されるその相同遺伝子をクローニングすることができる。
【0023】
本発明に関して、相同遺伝子とは、生来の遺伝子に対して少なくとも50%相同(同一)である遺伝子を意味する。好ましくは、相同遺伝子は、生来の遺伝子に対して、少なくとも55%相同、より好ましくは少なくとも60%、より好ましくは少なくとも65%、より好ましくは少なくとも70%、さらにより好ましくは少なくとも75%、好ましくは約80%、より好ましくは約90%、さらにより好ましくは約95%、および最も好ましくは約97%相同である。
【0024】
相同遺伝子のコード配列の上流の配列は、本発明に包含されるプロモーターである。あるいは、本発明のプロモーターに作動可能に会合される生来の遺伝子の配列、コード配列またはその一部を、先に規定されるような配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5またはそのサブ配列を使用して同定し、例えば、本明細書に記載のアラインメントまたはBLASTアルゴリズムを使用して、ゲノムデータベースを検索することができる。続いて、この同定される配列を使用して、本出願において規定される他の任意の真菌宿主において、オーソログまたは相同遺伝子を同定することができる。同定されるオーソログまたは相同遺伝子のコード配列の上流の配列は、本発明に包含されるプロモーターである。
【0025】
別の好適な実施態様に従えば、本発明のプロモーターDNA配列は、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5に少なくとも50%相同(同一)である単離されたDNA配列である。好ましくは、DNA配列は、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5に対して、少なくとも55%相同、より好ましくは少なくとも60%、より好ましくは少なくとも65%、より好ましくは少なくとも70%、さらにより好ましくは少なくとも75%、好ましくは約80%、より好ましくは約90%、さらにより好ましくは約95%、および最も好ましくは約97%相同である。
【0026】
本発明の目的のために、2つの核酸配列間の相同性(同一性)の程度は、好ましくは、BLASTプログラムによって決定される。BLAST解析を実施するためのソフトウェアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)を介して公的に入手することができる。BLASTアルゴリズムパラメータW、TおよびXは、アラインメントの感度および速度を決定する。BLASTプログラムは、デフォルトとして、11のワード長(W)、50のBLOSUM62スコアリングマトリックス(ヘニコフ(Henikoff)およびヘニコフ(Henikoff)、Proc.Natl.Acad.Sci.USA89:10915(1989年)を参照のこと)アラインメント(B)、10の期待値(E)、M=5、N=−4、および両鎖の比較を使用する。
【0027】
好適な別の実施態様では、プロモーターは、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5のサブ配列であり、該サブ配列はなお、プロモーター活性を有する。サブ配列は、好ましくは、少なくとも約100ヌクレオチド、より好ましくは少なくとも約200ヌクレオチド、および最も好ましくは少なくとも約300ヌクレオチドを含有する。
【0028】
好適な別の実施態様では、サブ配列は、5’および/または3’末端からの1もしくはそれ以上のヌクレオチドが欠失されていることを除いて、配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5に包含される核酸配列であり、前記DNA配列はなおプロモーター活性を有する。
【0029】
好適な別の実施態様では、プロモーターサブ配列は、「トリミングした」のサブ配列、即ち、翻訳開始部および/または転写開始部より上流にある配列フラグメントである。プロモーターをトリミングし、それについて機能的に分析する例は、Gene.1994年8月5日;145(2):179−87:the effect of multiple copies of the upstream region on expression of the Aspergillus niger glucoamylase−encoding gene.ベルダズ JC(Verdoes JC)、プント PJ(Punt PJ)、ストウサメル AH(Stouthamer AH)、バンデンホンデル CA(van den Hondel CA))に記載されている。
【0030】
本発明の別の実施態様では、プロモーターDNA配列は、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5の変異体である。
【0031】
用語「変異体」または「変異体プロモーター」は、本明細書において、親プロモーターの1もしくはそれ以上のヌクレオチドの置換、欠失および/または挿入を含むヌクレオチド配列を有するプロモーターとして規定され、ここで、変異体プロモーターは、対応する親プロモーターよりも高いまたは低いプロモーター活性を有する。用語「変異体プロモーター」は、天然の変異体ならびに古典的な変異誘発、部位特異的変異誘発、およびDNAシャフリングなどの当該分野において周知の方法を使用して入手されるインビトロで作製される変異体を包含する。変異体プロモーターは、1つもしくはそれ以上の変種を有してもよい。各変種は、ヌクレオチドの独立した置換、欠失、および/または挿入である。
【0032】
好適な実施態様に従えば、変異体プロモーターは、最初に同定されたプロモーター配列(配列番号1、配列番号2、配列番号3、配列番号4または配列番号5)と比較して少なくとも修飾された調節部位を有するプロモーターである。そのような調節部位は、その全体を取り出すかまたは上記で説明されるように特異的に変異することができる。従って、かかるプロモーター変異体の調節は、例えば、それがもはやグルコースによって誘導されないように修飾される。かかるプロモーター変異体およびそれらを入手する仕方に関する技術の例は、EP673429号明細書または国際公開第94/04673号パンフレットに記載されている。これらの特許は、参照により本明細書に援用される。
【0033】
プロモーター変異体は対立遺伝子変異体であり得る。対立遺伝子変異体は、同じ染色体の遺伝子座を占有する遺伝子の2つもしくはそれ以上の代替的形態のいずれかを示す。対立遺伝子の変異体は変異を介して自然に生じ、集団内において多型を生じ得る。変異体プロモーターは、(a)DNAを、極めて低い、低い、中等度、中等度〜高い、高い、もしくは極めて高いストリンジェンシー条件下で、(i)配列番号1、配列番号2、配列番号3、配列番号4または配列番号5、(ii)(i)のサブ配列あるいは(iii)(i)、(ii)の相補鎖とハイブリダイズさせること、および(b)DNAから変異体プロモーターを単離することによって、入手することができる。ストリンジェンシーおよび洗浄条件は、本明細書において規定されるとおりである。
【0034】
本発明のプロモーターは、その配列が、プロモーター配列と、ポリペプチドをコードする核酸配列のコード領域との連結を容易にする特定の制限部位を誘導する目的のためのリンカーを伴って提供され得るプロモーターであり得る。
【0035】
本明細書において提供される配列情報は、誤って同定される塩基の夾在を要するようにし、あまりに限られた範囲で解釈すべきではない。本発明において開示される特定の配列は、糸状菌、特に、アスペルギルス・ニガー(Aspergillus niger)由来の本来のDNA配列を単離するために容易に使用することができ、さらなる配列解析に供し、それによって、シークエンシングエラーを同定することができる。
【0036】
他で示さない限り、本明細書においてDNA分子を配列決定することによって決定されるすべてのヌクレオチド配列は、自動化DNAシークエンサーを使用して決定した。従って、この自動化アプローチによって決定される任意のDNA配列について当該分野において公知であるように、本明細書において決定される任意のヌクレオチド配列は、いくつかのエラーを含有してもよい。自動化によって決定されるヌクレオチド配列は、配列決定されたDNA分子の実際のヌクレオチド配列に対して、典型的には、少なくとも約90%同一、より典型的には、少なくとも約95%〜少なくとも約99.9%同一である。実際の配列は、当該分野において周知の手動のDNA配列決定方法を含む他のアプローチによって、より正確に決定することができる。
【0037】
当業者は、そのように誤って同定される塩基を同定することが可能であり、そのようなエラーを補正する仕方を知っている。
【0038】
本発明は、本発明が係わるプロモーターの生物学的機能を改変しない変異を典型的に含有する機能的プロモーター等価物を包含する。用語「機能的等価物」はまた、A.ニガー(A.niger)DNA配列のオーソログを包含する。A.ニガー(A.niger)DNA配列のオーソログは、他の株または種から単離することができ、類似または同一の生物学的活性を所有するDNA配列である。
【0039】
本発明のプロモーター配列は、任意の属の微生物から入手してもよい。本発明の目的のために、所定の供給源に関して本明細書において使用する用語「から入手する」は、ポリペプチドが、供給源または供給源由来の遺伝子が挿入されている細胞によって産生されることを意味すべきである。
【0040】
プロモーター配列は、真菌供給源、好ましくは、カンジダ(Candida)、ハンセヌラ(Hansenula)、クリヴェロミセス(Kluyveromyces)、ピキア(Pichia)、サッカロミセス(Saccharomyces)、シゾサッカロマイセス(Schizosaccharomyces)、またはヤロウイア(Yarrowia)株などの酵母株、より好ましくは、サッカロマイセス・カールスベルゲンシス(Saccharomyces carlsbergensis)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、サッカロミセス・ディアスタティカス(Saccharomyces diastaticus)、サッカロミセス・ダグラシー(Saccharomyces douglasii)、サッカロミセス・クリヴェリ(Saccharomyces kluyveri)、サッカロミセス・ノルベンシス(Saccharomyces norbensis)またはサッカロミセス・オヴィフォルミス(Saccharomyces oviformis)株から入手することができる。
【0041】
好適な別の実施態様では、プロモーター配列は、アクレモニウム(Acremonium)、アスペルギルス(Aspergillus)、オーレオバシディウム(Aureobasidium)、クリプトコッカス(Cryptococcus)、フィリバシディウム(Filibasidium)、フザリウム(Fusarium)、フミコーラ(Humicola)、マグナポルテ(Magnaporthe)、ケカビ(Mucor)、ミセリオフトラ(Myceliophthora)、ネオカリマスティクス(Neocallimastix)、ニューロスポラ(Neurospora)、ペシロマイセス(Paecilomyces)、ペニシリウム(Penicillium)、ピロミセス(Piromyces)、スエヒロタケ(Schizophyllum)、タラロミセス(Talaromyces)、サーモアスカス(Thermoascus)、シーラビア(Thielavia)、トイポクラディウム(Tolypocladium)、またはトリコデルマ(Trichoderma)株などの糸状菌株、より好ましくは、アスペルギルス・アクレアータス(Aspergillus aculeatus)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・フォエティダス(Aspergillus foetidus)、アスペルギルス・ジャポニクス(Aspergillus japonicus)、A.ニダランス(A.nidulans)、A.ニガー(A.niger)、麹菌(Aspergillus oryzae)(麹菌(A.oryzae))、フミコーラ・インソレンス(Humicola insolens)、フミコーラ・ラヌギノサ(Humicola lanuginosa)、ムコール・ミエヘイ(Mucor miehei)、ミセリオフトラ・サーモフィラ(Myceliophthora thermophila)、アカパンカビ(Neurospora crassa)、ペニシリウム・パルロゼラム(Penicillium purpurogenum)、トリコデルマ・ハルジアナム(Trichoderma harzianum)、トリコデルマ・コニンギ(Trichoderma koningii)、トリコデルマ・ロンギプラチアタム(Trichoderma Iongibrachiatum)、トリコデルマ・リーセイ(Trichoderma reesei)、またはトリコデルマ・ビリデ(Trichoderma viride)株から入手される。
【0042】
好適な別の実施態様では、プロモーター配列は、フザリウム・バクトリディオデス(Fusarium bactridioides)、フザリウム・セレアリス(Fusarium cerealis)、フザリウム・クルックウェルエンス(Fusarium crookwellense)、フザリウム・クルモラム(Fusarium culmorum)、フザリウム・グラミネアラム(Fusarium graminearum)、フザリウム・グラミヌム(Fusarium graminum)、フザリウム・ヘテロスポルム(Fusarium heterosporum)フザリウム・ネグンディ(Fusarium negundi)、フザリウム・オキシスポラム(Fusarium oxysporum)、フザリウム・レティクラツム(Fusarium reticulatum)、フザリウム・ロゼウム(Fusarium roseum)、フザリウム・サンブシヌム(Fusarium sambucinum)、フザリウム・サルコチロウム(Fusarium sarcochroum)、フザリウム・スポロトリキオイデス(Fusarium sporotrichioides)、フザリウム・スルプレウム(Fusarium sulphureum)、フザリウム・トルローサム(Fusarium torulosum)、フザリウム・トリコテシオイデス(Fusarium trichothecioides)、フザリウム・ヴェネナツム(Fusarium venenatum)株から入手される。
【0043】
上記の種について、本発明は、完全および不完全な状態、ならびに名称が公知である種であるかどうかに係わらず、他の分類学的等価物、例えば、アナモルフを包含することが理解される。当業者であれば、適切な等価物の同一性を容易に認識する。これらの種の株は、米国菌培養収集所(the American Type Culture Collection)(ATCC)、Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(DSM)、Centraalbureau Voor Schimmelcultures(CBS)、および農業研究サービス機関(Agricultural Research Service Patent Culture Collection,Northern Regional Research Center)(NRRL)などの多くの培養コレクションにおいて、公的に容易に利用可能である。
【0044】
さらに、本発明に従うプロモーター配列は、上記のプローブを使用して、自然(例えば、土壌、堆肥、水など)から単離される微生物を含む他の供給源から同定および入手することができる。天然の生育地から微生物を単離するための技術は、当該分野において周知である。次いで、核酸配列は、別の微生物のゲノムDNAライブラリーを同様にスクリーニングすることによって、誘導することができる。一旦、プロモーターをコードする核酸配列がプローブで検出されたら、当業者に公知である技術を利用することによって、配列を単離またはクローニングすることができる(例えば、サンブルック(Sambrook)ら、1989年、上掲を参照のこと)。
【0045】
本発明では、プロモーターDNA配列はまた、本発明の1つもしくはそれ以上のプロモーターの一部;本発明のプロモーターの一部および別の既知のプロモーターの一部、例えば、一方のプロモーターのリーダー配列および他方のプロモーターの転写開始部位;または本発明の1つもしくはそれ以上のプロモーターの一部および1つもしくはそれ以上の他のプロモーターの一部を含むハイブリッドプロモーターであってもよい。他のプロモーターは、変異体、短縮型、ハイブリッドプロモーターを含む、好適な真菌宿主細胞において転写活性を示す任意のプロモーター配列であってもよく、宿主細胞に対して同種または異種のいずれかの細胞外もしくは細胞内ポリペプチドをコードする遺伝子から入手してもよい。他のプロモーター配列は、ポリペプチドをコードする核酸配列に生来または外来性であってもよく、細胞に対して生来または外来性であってもよい。
【0046】
好適な実施態様として、同定されるプロモーターの重要な調節サブ配列を、他の「基本」プロモーターに融合して、それらのプロモーター活性を増強することができる(例えば、Mol Microbiol.1994年5月;12(3):479−90.Regulation of the xylanase−encoding xlnA gene of Aspergillus tubigensis.デグラフ LH(de Graaff LH)、ファンデンブルック HC(van den Broeck HC)、ファンオオイジェン AJ(van Ooijen AJ)、ヴィセル J(Visser J)に記載される)。
【0047】
本発明のプロモーターとのハイブリッドプロモーターの構築において有用な他方のプロモーターの他の例として、麹菌(A.oryzae)TAKAアミラーゼ、リゾムコール・ミエヘイ(Rhizomucor miehei)アスパラギン酸プロテイナーゼ、A.ニガー(A.niger)中性α−アミラーゼ、A.ニガー(A.niger)酸性α−アミラーゼ、A.ニガー(A.niger)またはアスペルギルス・アワモリ(Aspergillus awamori)グルコアミラーゼ(glaA)、A.ニガー(A.niger)gpdA、A.ニガー(A.niger)グルコースオキシダーゼgoxC、リゾムコール・ミエヘイ(Rhizomucor miehei)リパーゼ、麹菌(A.oryzae)アルカリプロテアーゼ、麹菌(A.oryzae)トリオースリン酸イソメラーゼ、A.ニダランス(A.nidulans)アセトアミダーゼ、およびフザリウム・オキシスポラム(Fusarium oxysporum)トリプシン様プロテアーゼ(国際公開第96/00787号パンフレット)の遺伝子から入手されるプロモーター、ならびにNA2−tpiプロモーター(A.ニガー(A.niger)中性α−アミラーゼおよび麹菌(A.oryzae)トリオースリン酸イソメラーゼの遺伝子由来のプロモーターのハイブリッド)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)エノラーゼ(ENO−1)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)ガラクトキナーゼ(GAL1)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)アルコールデヒドロゲナーゼ/グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(ADH2/GAP)、およびサッカロミセス・セレビシエ(Saccharomyces cerevisiae)3−ホスホグリセリン酸キナーゼ、ならびにその変種、短縮型、およびハイブリッドプロモーターが挙げられる。酵母宿主細胞のための他の有用なプロモーターについては、ローマノス(Romanoset)ら、1992年、Yeast8:423−488に記載されている。
【0048】
本発明では、プロモーターDNA配列はまた、「タンデムプロモーター」であってもよい。「タンデムプロモーター」は、本明細書において、2つもしくはそれ以上のプロモーター配列として規定され、それらのそれぞれが、コード配列と作動可能に会合され、コード配列からmRNAへの転写を仲介する。
【0049】
タンデムプロモーターは、本発明の2つもしくはそれ以上のプロモーター、あるいは本発明の1つもしくはそれ以上のプロモーターおよびハイブリッドプロモーターの構築に有用な上記で例示されるような1つもしくはそれ以上の他の既知のプロモーターを含む。タンデムプロモーターの2つもしくはそれ以上のプロモーター配列は、核酸配列の転写を同時に促進することができる。あるいは、タンデムプロモーターのプロモーター配列のうちの1つもしくはそれ以上は、細胞の成長の異なる段階または菌糸体の形態学的に異なる部分での核酸配列の転写を促進することができる。
【0050】
本発明では、プロモーターは、目的のポリペプチドをコードするコード配列および/または真菌宿主細胞に対して外来性であってもよい。野生型プロモーターがコード配列または真菌宿主細胞に対して生来である場合であっても、本発明の変異体、ハイブリッド、またはタンデムプロモーターは、ポリペプチドをコードするコード配列に対して外来性であることが理解される。
【0051】
本発明の変異体、ハイブリッド、またはタンデムプロモーターは、配列番号1、配列番号2、配列番号3、配列番号4または配列番号5を有するプロモーターのプロモーター活性の少なくとも約20%、好ましくは少なくとも約40%、より好ましくは少なくとも約60%、より好ましくは少なくとも約80%、より好ましくは少なくとも約90%、より好ましくは少なくとも約100%、さらにより好ましくは少なくとも約200%、最も好ましくは少なくとも約300%、およびさらに最も好ましくは少なくとも約400%を有する。
【0052】
本発明は、コード配列が、所定の真菌宿主細胞においてプロモーターDNA配列の制御下で発現され得るように、上記のように規定される少なくとも1つのプロモーターDNA配列、および前記プロモーターDNA配列と作動可能に会合されるコード配列を含むDNA構築物にさらに関する。
【0053】
コード配列は、目的の真菌宿主細胞に対して生来または異種であり得るポリペプチドをコードする。
【0054】
用語「ポリペプチド」は、本明細書においては、特定の長さのコードされた産物を指すことを意図するのではなく、従って、ペプチド、オリゴペプチド、およびタンパク質を包含する。用語「異種ポリペプチド」は、本明細書において、真菌細胞に対して生来ではないポリペプチド、生来の配列を改変するために修飾が施されている生来のポリペプチド、または発現が、組換えDNA技術による真菌細胞の操作の結果として量的に改変される生来のポリペプチドとして規定される。例えば、生来のポリペプチドは、例えば、ポリペプチドをコードする配列を本発明のプロモーターの制御下に配置して、ポリペプチドの発現を増強し、シグナル配列の使用によって、目的の生来のポリペプチドの細胞外への輸送を促進し、細胞によって通常産生されるポリペプチドをコードする遺伝子のコピー数を増加することによって、組換え的に産生され得る。真菌細胞は、ポリペプチドをコードする1コピーもしくはそれ以上のコード配列を含有してもよい。
【0055】
好ましくは、コード配列は、ペプチドホルモンもしくはその変異体、酵素、細胞内タンパク質、分泌プロセスに関与するタンパク質、ホールディングプロセスに関与するタンパク質、チャペロン、ペプチドアミノ酸トランスポーター、グリコシル化因子、転写因子、受容体もしくはその一部、抗体もしくはその一部、またはレポータータンパク質をコードする。
【0056】
好適な実施態様では、ポリペプチドは細胞外に分泌される。
【0057】
より好適な実施態様では、ポリペプチドは酵素である。酵素の例には、セルラーゼ、例えば、エンドグルカナーゼ、β−グルカナーゼ、セロビオヒドロラーゼまたはβ−グルコシダーゼ;ヘミセルラーゼまたはペクチン分解酵素、例えば、キシラナーゼ、キシロシダーゼ、マンナナーゼ、ガラクタナーゼ、ガラクトシダーゼ、ペクチンメチルエステラーゼ、ペクチンリアーゼ、ペクチン酸リアーゼ、エンドポリガラクツロナーゼ、エキソポリガラクツロナーゼ、ラムノガラクツロナーゼ、アラバナーゼ、アラビノフラノシダーゼ、アラビノキシランヒドロラーゼ、ガラクツロナーゼ、リアーゼ;デンプン分解酵素;ホスファターゼ、例えば、フィターゼ、エステラーゼ、例えば、リパーゼ、タンパク質分解酵素、例えば、プロテアーゼ、ペプチダーゼ、酸化還元酵素、例えば、オキシダーゼ、トランスフェラーゼ、またはイソメラーゼがある。
【0058】
あるいは、コード配列は、例えば、チャペロンまたは転写因子などの細胞内タンパク質をコードしてもよい。この例は、Appl Microbiol Biotechnol.1998年10月;50(4):447−54(「Analysis of the role of the gene bipA,encoding the major endoplasmic reticulum chaperone protein in the secretion of homologous and heterologous proteins in black Aspergilli.」プント PJ(Punt PJ)、バンゲメレン IA(van Gemeren IA)、ドリント−クイジュヴェンホヴェン J(Drint−Kuijvenhoven J)、ヘッシング JG(Hessing JG)、バンムイジュゥ−ハートベルト GM(van Muijlwijk−Harteveld GM)、ベイジェルスベルゲン A(Beijersbergen A)、ヴェリップス CT(Verrips CT)、バンデンホンデル CA(van den Hondel CA))に記載されている。チャペロンまたは転写因子などのこのコード配列が、タンパク質産生の制限要因であることが公知であった場合、これを使用して、例えば、タンパク質生産者としての宿主細胞の効率を改善することができる。
【0059】
コード配列はまた、有機酸、カロテノイド、(β−ラクタム)抗生物質、ビタミンなどの一次または二次代謝物の合成に関与する酵素をコードしてもよい。
【0060】
目的のポリペプチドをコードするコード配列は、任意の原核生物、真核生物、または他の供給源から入手することができる。
【0061】
あるいは、コード配列は、アンチセンスRNAおよび/またはRNAi(RNA干渉)構築物の発現をコードし得る。アンチセンス−RNAを発現させる例については、Appl Environ Microbiol.2000年2月;66(2):775−82.(Characterization of a foldase,protein disulfide isomerase A,in the protein secretory pathway of Aspergillus niger.ンギアム C(Ngiam C)、ジーネス DJ(Jeenes DJ)、プント PJ(Punt PJ)、バンデンホンデル CA(Van Den Hondel CA)、アーチャー DB(Archer DB))または(ズレンナー R(Zrenner R)、ウィルミツァー L(Willmitzer L)、ゾンネバルト U(Sonnewald U).Analysis of the expression of potato uridinediphosphate−glucose pyrophosphorylase and its inhibition by antisense RNA.Planta.(1993年);190(2):247−52.)に認められる。遺伝子の発現の完全な不活化は、例えば、(β−ラクタム)抗生物質またはカロテノイドなどの特定の二次代謝物の産生を増加するための、例えば、代謝経路の所望されない側枝を制御する遺伝子の不活化に有用である。完全な不活化はまた、毒性または所望されない化合物の産生を減少するのに有用である(ペニシリウム(Penicillium)におけるクリソゲニン(chrysogenin);アスペルギルス(Aspergillus)におけるアフラトキシン(Aflatoxin):マクドナルド KD(MacDonald KD)ら:heterokaryon studies and the genetic control of penicillin and chrysogenin production in Penicillium chrysogenum.J Gen Microbiol.(1963年)33:375−83)。完全な不活化はまた、発酵プロセスおよび下流プロセッシングが改善されるような方法で生物体の形態学を改変するのに有用である。
【0062】
本発明の別の実施態様は、真菌細胞の広範な代謝再プログラミングまたは操作に関する。完全な新規の経路の導入および/または所望されない経路の修飾は、タンパク質もしくは代謝物などの特定の化合物の産生に特異的に適応された細胞を提供する。
【0063】
本発明の方法では、コード配列がポリペプチドをコードする場合、前記ポリペプチドもまた、別のポリペプチドがポリペプチドまたはそのフラグメントのN末端もしくはC末端で融合される融合されたあるいはハイブリッドポリペプチドを含み得る。融合されたポリペプチドは、1つのポリペプチドをコードする核酸配列(またはその一部)を別のポリペプチドをコードする核酸配列(またはその一部)に融合させることによって、産生される。融合ポリペプチドを産生させるための技術は当該分野において公知であり、ポリペプチドをコードするコード配列を、それらがフレーム内にあって、融合されたポリペプチドの発現が同じプロモーターおよびターミネーターの制御下にあるように連結することを含む。ハイブリッドポリペプチドは、少なくとも2つの異なるポリペプチドから入手される部分的または完全なポリペプチド配列の組み合わせを含み得、ここで、1つもしくはそれ以上は、真菌細胞に対して異種であってもよい。
【0064】
DNA構築物は、プロモーターDNA配列に加えて、制御配列に適合する条件下で、適切な宿主細胞においてコード配列の発現を指令する1つもしくはそれ以上の制御配列を含み得る。発現は、転写、転写後修飾、翻訳、翻訳後修飾、および分泌を含むがこれらに限定されないポリペプチドの産生に関与する任意の工程を含むことが理解される。1つもしくはそれ以上の制御配列は、コード配列または宿主に対して生来であってもよい。あるいは、1つもしくはそれ以上の制御配列は、宿主細胞におけるコード配列の発現を改善するために、核酸配列に対して外来性の1つもしくはそれ以上の制御配列で置換されていてもよい。
【0065】
「DNA構築物」は、本明細書において、天然に存在する遺伝子から単離されるかまたはそうでなければ天然には存在しない様式で組み合わされ、並置される核酸のセグメントを含有するように修飾されている一本鎖もしくは二本鎖のいずれかの核酸分子として規定される。DNA構築物という用語は、DNA構築物がコード配列およびコード配列の発現に要求されるすべての制御配列を含有する場合、発現カセットという用語と同義である。
【0066】
用語「制御配列」は、本明細書において、コード配列の発現に必要または有利であるすべての成分を含むように規定され、本発明のプロモーターを含む。各制御配列は、ポリペプチドをコードする核酸配列に生来または外来性であってもよい。かかる制御配列として、リーダー、(コザク(Kozak)、1991年、J.Biol.Chem.266:19867−19870に記載のような)至適翻訳開始配列、ポリアデニル化配列、プロペプチド配列、シグナルペプチド配列、上流活性化配列、変異体、フラグメント、ならびにそれから誘導されるハイブリッドおよびタンデムプロモーターを含む本発明のプロモーターならびに転写ターミネーターが挙げられるが、それらに限定されない。少なくとも、制御配列は、転写および翻訳停止シグナルならびに本発明のプロモーター(の一部)を含む。制御配列は、制御配列とポリペプチドをコードする核酸配列のコード領域との連結を容易にする特定の制限部位を誘導する目的のためのリンカーを伴って提供され得る。
【0067】
制御配列は、適切な転写ターミネーター配列、即ち、転写を終結するために宿主細胞によって認識される配列であってもよい。ターミネーター配列は、ポリペプチドをコードするコード配列の3’末端と作動可能に会合される。好適な宿主細胞において機能的であるいずれのターミネーターも本発明において使用することができる。
【0068】
糸状真菌宿主細胞のための好適なターミネーターは、麹菌(A.oryzae)TAKAアミラーゼ、A.ニガー(A.niger)グルコアミラーゼ、A.ニダランス(A.nidulans)アントラニル酸シンターゼ、A.ニガー(A.niger)α−グルコシダーゼ、trpC遺伝子、およびフザリウム・オキシスポラム(Fusarium oxysporum)トリプシン様プロテアーゼの遺伝子から入手される。
【0069】
酵母宿主細胞のための好適なターミネーターは、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)エノラーゼ、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)シトクロムC(CYC1)、およびサッカロミセス・セレビシエ(Saccharomyces cerevisiae)グリセルアルデヒド(glyceraidehyde)−3−リン酸デヒドロゲナーゼの遺伝子から入手される。酵母宿主細胞のための他の有用なターミネーターについては、ローマノス(Romanos)ら、1992年、上掲に記載されている。
【0070】
制御配列はまた、適切なリーダー配列、即ち、宿主細胞による翻訳に重要であるmRNAの5’非翻訳領域であってもよい。リーダー配列は、ポリペプチドをコードする核酸配列の5’末端と作動可能に会合される。好適な宿主細胞において機能的であるいずれのリーダー配列も本発明において使用することができる。
【0071】
糸状真菌宿主細胞のための好適なリーダーは、麹菌(A.oryzae)TAKAアミラーゼ、A.ニダランス(A.nidulans)トリオースリン酸イソメラーゼおよびA.ニガー(A.niger)glaAの遺伝子から入手される。
【0072】
酵母宿主細胞のための適切なリーダーは、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)エノラーゼ(ENO−1)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)3−ホスホグリセリン酸キナーゼ、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)α−因子、およびサッカロミセス・セレビシエ(Saccharomyces cerevisiae)アルコールデヒドロゲナーゼ/グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(ADH2/GAP)の遺伝子から入手される。
【0073】
制御配列はまた、ポリアデニル化配列、核酸配列の3’末端と作動可能に会合され、転写される場合、転写されるmRNAにポリアデノシン残基を添加するためのシグナルとして宿主細胞に認識される配列であってもよい。好適な宿主細胞において機能的であるいずれのポリアデニル化配列も本発明において使用することができる。
【0074】
糸状真菌宿主細胞のための好適なポリアデニル化配列は、麹菌(A.oryzae)TAKAアミラーゼ、A.ニガー(A.niger)グルコアミラーゼ、A.ニダランス(A.nidulans)アントラニル酸シンターゼ、フザリウム・オキシスポラム(Fusarium oxysporum)トリプシン様プロテアーゼ、およびA.ニガー(A.niger)α−グルコシダーゼの遺伝子から入手される。
【0075】
酵母宿主細胞のための有用なポリアデニル化配列については、クオ(Guo)およびシャーマン(Sherman)、1995年、Molecular Cellular Biology15:5983−5990に記載されている。
【0076】
制御配列はまた、ポリペプチドのアミノ末端に連結されたアミノ酸配列をコードし、コードされるポリペプチドを細胞の分泌経路に指向するシグナルペプチドコード領域であってもよい。核酸配列のコード配列の5’末端は、分泌されるポリペプチドをコードするコード領域のセグメントを伴う翻訳リーディングフレームにおいて天然に連結されるシグナルペプチドコード領域を本来的に含有し得る。あるいは、コード配列の5’末端は、コード配列に対して外来性であるシグナルペプチドコード領域を含有してもよい。外来性のシグナルペプチドコード領域は、コード配列がシグナルペプチドコード領域を天然には含有しない場合に要求され得る。あるいは、外来性シグナルペプチドコード領域は、ポリペプチドの分泌を増強するために、天然のシグナルペプチドコード領域と単純に置き換わってもよい。しかし、発現されるポリペプチドを好適な宿主細胞の分泌経路に指向するいずれのシグナルペプチドコード領域も本発明において使用することができる。
【0077】
糸状真菌宿主細胞のための効果的なシグナルペプチドコード領域は、麹菌(A.oryzae)TAKAアミラーゼ、A.ニガー(A.niger)中性アミラーゼ、A.フィクウム(A.ficuum)フィターゼ、A.ニガー(A.niger)グルコアミラーゼ、A.ニガー(A.niger)エンドキシラナーゼ、リゾムコール・ミエヘイ(Rhizomucor miehei)アスパラギン酸プロテイナーゼ、フミコーラ・インソレンス(Humicola insolens)セルラーゼ、およびフミコーラ・ラヌギノサ(Humicola lanuginosa)リパーゼの遺伝子から入手されるシグナルペプチドコード領域である。
【0078】
酵母宿主細胞のための有用なシグナルペプチドは、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)α−因子およびサッカロミセス・セレビシエ(Saccharomyces cerevisiae)インベルターゼの遺伝子から入手される。他の有用なシグナルペプチドコード領域については、ローマノス(Romanos)ら、1992年、上掲により記載されている。
【0079】
制御配列はまた、ポリペプチドのアミノ末端に位置するアミノ酸配列をコードするプロペプチドコード領域であってもよい。得られるポリペプチドは、プロ酵素もしくはプロポリペプチド(または場合によりチモーゲン)として公知である。プロポリペプチドは一般に不活性であり、プロポリペプチドからのプロペプチドの触媒的または自己触媒的切断によって、成熟した活性なポリペプチドに変換することができる。プロペプチドコード領域は、枯草菌(Bacillus subtilis)アルカリプロテアーゼ(aprE)、枯草菌(Bacillus subtilis)中性プロテアーゼ(nprT)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)α−因子、リゾムコール・ミエヘイ(Rhizomucor miehei)アスパラギン酸プロテイナーゼ、ミセリオフトラ・サーモフィラ(Myceliophthora thermophila)ラッカーゼ(国際公開第95/33836号パンフレット)およびA.ニガー(A.niger)エンドキシラナーゼ(endo1)の遺伝子から入手することができる。
【0080】
シグナルペプチドおよびプロペプチド領域の両方がポリペプチドのアミノ末端に存在する場合、プロペプチド領域はポリペプチドのアミノ末端の隣に位置し、シグナルペプチド領域はプロペプチド領域のアミノ末端の隣に位置する。
【0081】
宿主細胞の増殖に関するポリペプチドの発現の調節を可能にする調節配列を添加することもまた所望され得る。調節系の例は、遺伝子の発現を、調節化合物の存在を含む化学的または物理的刺激に応答してオンもしくはオフに転向させる調節システムである。原核生物系における調節系としては、lac、およびtrpオペレーター系が挙げられる。酵母では、ADH2系またはGAL1系を使用することができる。糸状菌では、TAKAα−アミラーゼプロモーター、A.ニガー(A.niger)グルコアミラーゼプロモーター、麹菌(A.oryzae)グルコアミラーゼプロモーター、A.ツビンゲンシス(A.tubingensis)エンドキシラナーゼ(x/nA)プロモーター、A.ニガー(A.niger)硝酸レダクターゼ(niaD)プロモーター、トリコデルマ・リーセイ(Trichoderma reesei)セロビオヒドロラーゼプロモーターおよびA.ニダランス(A.nidulans)アルコールおよびアルデヒドデヒドロゲナーゼ(それぞれalcAおよびaldA)プロモーター(米国特許第5,503,991号明細書に記載される)を調節配列として使用することができる。調節配列の他の例は、遺伝子増幅を可能にする調節配列である。真核生物系では、これらは、メトトレキサートの存在下で増幅されるジヒドロ葉酸レダクターゼ遺伝子、および重金属で増幅されるメタロチオネイン遺伝子を含む。これらの場合、ポリペプチドをコードする核酸配列は調節配列と作動可能に会合される。
【0082】
creA結合部位の取り出し(EP673429号明細書において先に記載されているような炭素カタボライト抑制)、pacCおよびareAの変更(pHおよび窒素調節のため)は重要であり得る。
【0083】
本発明はまた、本発明のプロモーター、ポリペプチドをコードするコード配列、ならびに転写および翻訳停止シグナルを含む組換え発現ベクターに関する。上記の多様なコードおよび制御配列を共に接続して、1つもしくはそれ以上の簡便な制限部位を含んで、そのような部位における、プロモーターおよび/またはポリペプチドをコードするコード配列の挿入もしくは置換を可能にし得る組換え発現ベクターを産生させてもよい。あるいは、コード配列とプロモーターとの融合は、例えば、PCR(SOE−PCR)を使用する配列、重複伸長(Gene.1989年4月15日;77(1):51−9.ホウ SN(Ho SN)、ハント HD(Hunt HD)、ホートン RM(Horton RM)、プーレン JK(Pullen JK)、ピーズ LR(Pease LR)「Site−directed mutagenesis by overlap extension using the polymerase chain reaction」に記載される)によってかまたはGatewayTMクローニングシステム(Invitrogen)を使用するクローニングによって、行うことができる。あるいは、コード配列は、コード配列あるいはプロモーターおよび/またはコード配列を含むDNA構築物を、発現のための適切なベクターに挿入することによって、発現させてもよい。発現ベクターを作製する際に、コード配列が本発明のプロモーターおよび発現のための1つもしくはそれ以上の適切な制御配列と作動可能に会合されるように、コード配列がベクターに位置される。
【0084】
組換え発現ベクターは、組換えDNA手順に簡便に供され得、コード配列の発現をもたらし得る任意のベクター(例えば、プラスミドまたはウイルス)であってもよい。ベクターの選択は、典型的に、ベクターと、ベクターを導入しようとする宿主細胞との適合可能性に依存する。ベクターは、線状であってもまたは閉環状プラスミドであってもよい。
【0085】
ベクターは、自律的複製型ベクター、即ち、その複製が染色体複製とは独立する細胞外染色体実体として存在するベクター、例えば、プラスミド、染色体外エレメント、ミニ染色体、または人工染色体であってもよい。自律複製のために、ベクターは、問題の宿主細胞において、ベクターが自立的に複製することを可能にする複製開始点を含み得る。酵母宿主細胞において使用される複製開始点の例として、2マイクロン複製開始点、ARS1、ARS4、ARS1およびCEN3の組み合わせ、ならびにARS4およびCEN6の組み合わせがある。複製開始点は、その機能性を宿主細胞において温度感受性にする変異を有するものであってもよい(例えば、エールリヒ(Ehrlich)、1978年、Proceedings of the National Academy of Sciences USA75:1433を参照のこと)。糸状真菌において自律的に維持されるクローニングベクターの例は、AMA1配列を含むクローニングベクターである。AMA1は、アスペルギルス(Aspergillus)における自律的維持が可能であるA.ニダランス(A.nidulans)から単離される6.0−kbゲノムDNAフラグメントである(例えば、アレクセンコ(Aleksenko)およびクラッターバック(Clutterbuck)(1997年)、Fungal Genet.Biol.21:373−397を参照のこと)。
【0086】
あるいは、ベクターは、宿主細胞に導入される場合、ゲノムに組込まれ、それが組込まれている染色体と共に複製するものであってもよい。さらに、宿主細胞のゲノムに導入しようとする全DNAを共に含有する単一のベクターまたはプラスミドあるいは2つもしくはそれ以上のベクターまたはプラスミド、あるいはトランスポゾンを使用してもよい。
【0087】
本発明のベクターは、好ましくは、形質転換された細胞の容易な選択を可能にする1つもしくはそれ以上の選択マーカーを含有する。宿主は、少なくとも2つのベクター(一方は選択マーカーを含む)で同時形質転換してもよい。選択マーカーは、その産物が殺生物剤耐性またはウイルス耐性、重金属に対する耐性、栄養要求体に栄養非要求性、などを提供する遺伝子である。酵母宿主細胞に適切なマーカーは、ADE2、HIS3、LEU2、LYS2、MET3、TRP1、およびURA3である。糸状真菌宿主細胞において使用される選択マーカーとして、amdS(アセトアミダーゼ)、argB(オルニチンカルバモイルトランスフェラーゼ)、bar(ホスフィノトリシンアセチルトランスフェラーゼ)、hygB(ハイグロマイシンホスホトランスフェラーゼ)、niaD(硝酸レダクターゼ)、pyrG(オロチジン−5’−リン酸デカルボキシラーゼ)、sC(硫酸アデニルトランスフェラーゼ)、trpC(アントラニル酸シンターゼ)、ならびにそれらの等価物が挙げられるが、それらに限定されない。また、例えば、フレオマイシン、ハイグロマイシンBまたはG418に対する耐性を付与するマーカーを使用することもできる。アスペルギルス(Aspergillus)細胞における使用には、A.ニダランス(A.nidulans)または麹菌(A.oryzae)のamdSおよびpyrG遺伝子ならびにストレプトマイセス・ヒグロスコピカス(Streptomyces hygroscopicus)の遺伝子が好適である。amdSマーカー遺伝子は、好ましくは、EP635574号明細書または国際公開第97/0626号パンフレットに記載の技術を適用して、使用される。好適な選択マーカー遺伝子は、A.ニダランス(A.nidulans)gpdAプロモーターに融合されたA.ニダランス(A.nidulans)amdSコード配列である(EP635574号明細書)。他の糸状真菌由来のAmdS遺伝子を使用してもよい(国際公開第97/06261号パンフレット)。
【0088】
宿主細胞ゲノムへの組込みのために、ベクターは、プロモーター配列および/またはポリペプチドをコードするコード配列あるいは相同もしくは非相同組換えによるゲノムへのベクターの安定な組込みのためのベクターの他の任意のエレメントに依存し得る。あるいは、ベクターは、宿主細胞のゲノムへの相同組換えによる組込みを指令するためのさらなる核酸配列を含有してもよい。さらなる核酸配列は、染色体上の正確な位置で宿主細胞ゲノムにベクターを組込むことを可能にする。正確な位置での組込みの可能性を高めるために、組込みエレメントは、好ましくは、充分な数の核酸、例えば、100〜1,500塩基対、好ましくは400〜1,500塩基対、より好ましくは800〜1,500塩基対、および最も好ましくは少なくとも2kbを含有すべきであり、これは対応する標的配列と高度に相同であり、相同組換えの確率を増強する。組込みエレメントは、宿主細胞のゲノムにおける標的配列と相同である任意の配列であってもよい。さらに、組込みエレメントは、非コードまたはコード核酸配列であり得る。標的化組込みを促進するために、クローニングベクターは、好ましくは、宿主細胞の形質転換の前に線状化される。線状化は、好ましくは、少なくとも1つ、但し、好ましくは、クローニングベクターのいずれか一方の末端が標的遺伝子座に相同な配列によって隣接されるように、実施される。
【0089】
好ましくは、標的遺伝子座に相同であるクローニングベクターにおける組込みエレメントが、高度に発現される遺伝子座から誘導され、これは、それらが、真菌宿主細胞において高い発現レベルが可能である遺伝子から誘導されることを意味する。高い発現レベルが可能な遺伝子、即ち、高度に発現される遺伝子は、本明細書において、例えば、誘導条件下で、そのmRNAが少なくとも0.5%(w/w)の全細胞mRNAを構成し得る遺伝子、あるいは、その遺伝子産物が少なくとも1%(w/w)の全細胞性タンパク質を構成し得るか、または、分泌される遺伝子産物の場合、少なくとも0.1g/lのレベルまで分泌され得る遺伝子として規定される(欧州特許第357127B1号明細書に記載される)。多くの好適な高度に発現される真菌遺伝子は、次の例によって示される。すなわち、アスペルギルス(Aspergilli)またはトリコデルマ(Trichoderma)由来のアミラーゼ、グルコアミラーゼ、アルコールデヒドロゲナーゼ、キシラナーゼ、グリセルアルデヒド−リン酸デヒドロゲナーゼもしくはセロビオヒドロラーゼ遺伝子。これらの目的のための最も好適な高度に発現される遺伝子は、グルコアミラーゼ遺伝子、好ましくはA.ニガー(A.niger)グルコアミラーゼ遺伝子、麹菌(A.oryzae)TAKA−アミラーゼ遺伝子、A.ニダランス(A.nidulans)gpdA遺伝子、配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5の遺伝子座、配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5のA.ニガー(A.niger)遺伝子座、またはトリコデルマ・リーセイ(Trichoderma reesei)セロビオヒドロラーゼ遺伝子である。
【0090】
一方、ベクターは、非相同組換えによって、宿主細胞のゲノムに組込むことができる。
【0091】
ポリペプチドをコードする1コピーを超える核酸配列を宿主細胞に挿入して、遺伝子産物の産生を増加させてもよい。これは、好ましくは、DNA配列のそのゲノムコピーに組込むことによって、より好ましくは、高度に発現される遺伝子座、好ましくはグルコアミラーゼ遺伝子座または配列番号1、配列番号2、配列番号3、配列番号4もしくは配列番号5の遺伝子座でDNA配列の組込みを標的化することによって、行うことができる。あるいは、これは、核酸配列を伴う増幅可能な選択マーカー遺伝子を含むことによって行うことができ、ここで、選択マーカー遺伝子の増幅されたコピー、およびそれによる核酸配列のさらなるコピーを含有する細胞を、適切な選択可能因子の存在下で細胞を培養することによって、選択することができる。過剰発現させようとするDNA配列のコピー数をさらにより増加するために、国際公開第98/46772号パンフレットに記載の遺伝子変換技術を使用してもよい。
【0092】
上記のエレメントを連結して、本発明の組換え発現ベクターを構築するために使用される手順は、当業者に周知である(例えば、サンブルック(Sambrook)ら、1989年、上掲を参照のこと)。
【0093】
本発明はまた、ポリペプチドをコードするコード配列と作動可能に会合される本発明のプロモーターDNA配列を含む組換え宿主細胞に関し、これは、ポリペプチドの産生において有利に使用される。ポリペプチドをコードするコード配列と作動可能に会合される本発明のプロモーターを含むベクターは、ベクターが染色体組込み体としてかまたは先に記載のような自己複製染色体外ベクターとして維持されるように、宿主細胞に導入される。用語「宿主細胞」は、複製中に生じる変異のため親細胞とは同一ではない親細胞の任意の子孫を包含する。宿主細胞の選択は、ポリペプチドをコードする遺伝子およびその供給源に大きな程度で依存する。
【0094】
本発明はまた、2つ以上の本発明のプロモーターDNA配列を含む組換え宿主細胞に関し、それぞれのプロモーターは、ポリペプチドをコードするコード配列と作動可能に会合される。かかる宿主細胞は、少なくとも1つのポリペプチドの組換え産生において有利に使用することができる。好ましくは少なくとも1つのプロモーターおよびその会合されるコード配列は、ベクター上に存在する。ベクターは、それが、染色体組込み体としておよび/または先に記載のような自己複製染色体外ベクターとして維持されるように、宿主細胞に導入される。
【0095】
別の好適な実施態様に従えば、宿主細胞は、(β−ラクタム)抗生物質、ビタミンもしくはカロテノイドなどの特定の一次または二次代謝物の産生のために使用される。
【0096】
宿主細胞は、本発明の方法において有用ないずれの真菌細胞であってもよい。本明細書において使用する「真菌」としては、子嚢菌門(Ascomycota)、担子菌門(Basidiomycota)、ツボカビ門(Chytridiomycota)、および接合菌門(Zygomycota)(ホークスワース(Hawksworth)ら、Ainsworth and Bisby’s Dictionary of The Fungi、第8版、1995年、CAB International,University Press,Cambridge、英国により記載される)ならびに卵菌門(Oomycota)(ホークスワース(Hawksworth)ら、1995年、上掲、171頁において引用される)およびすべての栄養胞子形成菌(mitosporic fungi)(ホークスワース(Hawksworth)ら、1995年、上掲)が挙げられる。
【0097】
好適な実施態様では、真菌宿主細胞は酵母細胞である。本明細書において使用する「酵母」としては、有子嚢胞子(ascosporogenous)酵母(エンドミセタレス目(Endomycetales))、担子胞子形成(basidiosporogenous)酵母、および不完全菌類(Fungi lmperfecti)(不完全酵母菌綱(Blastomycetes))に属する酵母が挙げられる。酵母の分類は将来変化し得るため、本発明の目的のために、酵母は、Biology and Activities of Yeast(スキナー,F.A.(Skinner,F.A.)、パスモア,S.M.(Passmore,S.M.)、およびダベンポート,R.R.(Davenport,R.R.)編、Soc.App.Bacteriol.第9回シンポジウム、1980年)において記載のように規定されるべきである。
【0098】
より好適な実施態様では、酵母宿主細胞は、カンジダ(Candida)、ハンセヌラ(Hansenula)、クリヴェロミセス(Kluyveromyces)、ピキア(Pichia)、サッカロミセス(Saccharomyces)、シゾサッカロマイセス(Schizosaccharomyces)、またはヤロウイア(Yarrowia)細胞である。
【0099】
最も好適な実施態様では、酵母宿主細胞は、サッカロマイセス・カールスベルゲンシス(Saccharomyces carlsbergensis)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、サッカロミセス・ディアスタティカス(Saccharomyces diastaticus)、サッカロミセス・ダグラシー(Saccharomyces douglasii)、サッカロミセス・クリヴェリ(Saccharomyces kluyveri)、サッカロミセス・ノルベンシス(Saccharomyces norbensis)またはサッカロミセス・オヴィフォルミス(Saccharomyces oviformis)細胞である。別の最も好適な実施態様では、酵母宿主細胞は、クリュイベロミセス・ラクティス(Kluyveromyces lactis)細胞である。別の最も好適な実施態様では、酵母宿主細胞は、ヤロウイア・リポリティカ(Yarrowia lipolytica)細胞である。
【0100】
別の好適な実施態様では、真菌宿主細胞は糸状真菌細胞である。「糸状菌」は、亜門の真菌門(Eumycota)および卵菌門(Oomycota)のすべての糸状形態を含む(ホークスワース(Hawksworth)ら、1995年、上掲により規定される)。糸状菌は、キチン、セルロース、グルカン、キトサン、マンナン、および他の複雑な多糖からなる菌糸体の壁によって特徴付けられる。栄養生長は菌糸伸長により、炭素の異化は偏性好気性である。対照的に、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)などの酵母による栄養生長は単細胞葉状体の出芽により、炭素の異化は発酵性であり得る。
【0101】
より好適な実施態様では、糸状真菌宿主細胞は、アクレモニウム(Acremonium)、アスペルギルス(Aspergillus)、フザリウム(Fusarium)、フミコーラ(Humicola)、ケカビ(Mucor)、ミセリオフトラ(Myceliophthora)、ニューロスポラ(Neurospora)、ペニシリウム(Penicillium)、シーラビア(Thielavia)、トイポクラディウム(Tolypocladium)、またはトリコデルマ(Trichoderma)の種の細胞であるが、これらに限定されない。
【0102】
最も好適な実施態様では、糸状真菌宿主細胞は、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・フォエティダス(Aspergillus foetidus)、アスペルギルス・ジャポニクス(Aspergillus japonicus)、A.ニダランス(A.nidulans)、A.ニガー(A.niger)または麹菌(A.oryzae)細胞である。最も好適な別の実施態様では、糸状真菌宿主細胞は、フザリウム・バクトリディオデス(Fusarium bactridioides)、フザリウム・セレアリス(Fusarium cerealis)、フザリウム・クルックウェルエンス(Fusarium crookwellense)、フザリウム・クルモラム(Fusarium culmorum)、フザリウム・グラミネアラム(Fusarium graminearum)、フザリウム・グラミヌム(Fusarium graminum)、フザリウム・ヘテロスポルム(Fusarium heterosporum)、フザリウム・ネグンディ(Fusarium negundi)、フザリウム・オキシスポラム(Fusarium oxysporum)、フザリウム・レティクラツム(Fusarium reticulatun)、フザリウム・ロゼウム(Fusarium roseum)、フザリウム・サンブシヌム(Fusarium sambucinum)、フザリウム・サルコチロウム(Fusarium sarcochroum)、フザリウム・スポロトリキオイデス(Fusarium sporotrichioides)、フザリウム・スルプレウム(Fusarium sulphureum)、フザリウム・トルローサム(Fusarium torulosum)、フザリウム・トリコテシオイデス(Fusarium trichothecioides)、またはフザリウム・ヴェネナツム(Fusarium venenatum)細胞である。最も好適な別の実施態様では、糸状真菌宿主細胞は、フミコーラ・インソレンス(Humicola insolens)、フミコーラ・ラヌギノサ(Humicola lanuginosa)、ムコール・ミエヘイ(Mucor miehei)、ミセリオフトラ・サーモフィラ(Myceliophthora thermophila)、アカパンカビ(Neurospora crassa)、ペニシリウム・パルロゼラム(Penicillium purpurogenum)、シーラビア・テレストリス(Thielavia terrestris)、トリコデルマ・ハルジアナム(Trichoderma harzianum)、トリコデルマ・コニンギ(Trichoderma koningii)、トリコデルマ・ロンギプラチアタム(Trichoderma longibrachiatum)、トリコデルマ・リーセイ(Trichoderma reesei)、またはトリコデルマ・ビリデ(Trichoderma viride)細胞である。
【0103】
真菌細胞は、プロトプラスト形成、プロトプラストの形質転換、およびそれ自体公知の様式での細胞壁の再生を包含するプロセスによって形質転換することができる。アスペルギルス(Aspergillus)宿主細胞の形質転換に適切な手順については、EP238023号明細書およびエルトン(Yelton)ら、1984年、Proceedings of the National Academy of Sciences USA81:1470−1474に記載されている。アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)を使用するアスペルギルス(Aspergillus)および他の糸状真菌宿主細胞の形質転換に適切な手順については、例えば、Nat Biotechnol.1998年9月;16(9):839−42.Nat Biotechnol 1998年11月;16(11):1074に掲載の訂正表.Agrobacterium tumefaciens−mediated transformation of filamentous fungi、デグルート MJ(de Groot MJ)、バンドドック P(Bundock P)、フーカス PJ(Hooykaas PJ)、ベイエルスベルゲン AG(Beijersbergen AG).Unilever Research Laboratory Vlaardingen,オランダに記載されている。フザリウム(Fusarium)種を形質転換するための適切な方法については、メーラディール(Malardier)ら、1989年、Gene78:147−156および国際公開第96/00787号パンフレットにより記載されている。酵母は、ベッカー(Becker)およびグアレンテ(Guarente)、編者:エイブルソン,J.N.(Abelson,J.N.)およびシモン,M.I.(Simon,M.I.)、Guide to Yeast Genetics and Molecular Biology,Methods in Enzymology、第194巻、182−187頁、Academic Press,Inc.、ニューヨーク;イトウら、1983年、Journal of Bacteriology153:163;ならびにヒネン(Hinnen)ら、1978年、Proceedings of the National Academy of Sciences USA75:1920に記載の手順を使用して、形質転換してもよい。
【0104】
本発明は、宿主細胞におけるコード配列の発現のための方法にさらに関する。方法は、以下の工程を含む。すなわち、
(a)本発明のプロモーターDNA配列および上記のコード配列を含むDNA構築物を提供すること、
(b)前記DNA構築物で適切な宿主細胞を形質転換すること、および
(c)前記プロモーターDNA配列の制御下でコード配列を発現させること。
【0105】
本発明はまた、適切な真菌宿主において本発明のプロモーターの制御下にあるコード配列によってコードされるポリペプチドの産生のための方法に関する。方法は、以下の工程を含む。すなわち、
(a)本発明のプロモーターDNA配列および上記のポリペプチドをコードするコード配列を含むDNA構築物を提供すること、
(b)前記DNA構築物で適切な真菌宿主細胞を形質転換すること、
(c)ポリペプチドの発現をもたらす適切な培養条件下で適切な真菌宿主を培養すること、
(d)培養ブロスからポリペプチドを回収すること。
【0106】
本発明はまた、以下を含む、適切な宿主における二次代謝物の産生のための方法に関する。すなわち、
(a)本発明のプロモーターDNA配列および上記の二次代謝物の産生に関与する酵素をコードするコード配列を含むDNA構築物を提供すること、
(b)前記DNA構築物で適切な宿主細胞を形質転換すること、
(c)二次代謝物の産生をもたらす適切な培養条件下で適切な真菌宿主を培養すること、および
(d)培養ブロスから二次代謝物を回収すること。
【0107】
本発明の産生方法では、当該分野において既知の方法を使用して、細胞は、ポリペプチドまたは代謝物の産生に適切な栄養培地中で培養される。例えば、細胞は、適切な培地において、ならびにコード配列を発現させるおよび/またはポリペプチドを単離することが可能な条件下で、実験室あるいは産業発酵槽における振盪フラスコ培養、小規模または大規模発酵(連続、バッチ、流加、もしくは固体発酵を含む)によって、培養することができる。培養は、当該分野において既知の手順を使用して、炭素および窒素源ならびに無機塩を含む適切な栄養培地において行われる。適切な培地は販売者から入手可能であり、または(例えば、米国菌培養収集所(the American Type Culture Collection)のカタログにおいて)公開された組成に従って、調製してもよい。ポリペプチドまたは代謝物が栄養培地に分泌される場合、ポリペプチドまたは代謝物は、培地から直接回収することができる。ポリペプチドまたは代謝物が分泌されない場合、それは、細胞溶解物から回収することができる。
【0108】
ポリペプチドは、ポリペプチドに特異的である当該分野において既知の方法を使用して、検出することができる。これらの検出方法は、特異的抗体の使用、酵素生成物の形成、または酵素基質の消失を含み得る。
【0109】
得られるポリペプチドまたは代謝物は、当該分野において既知の方法によって回収することができる。例えば、ポリペプチドまたは代謝物は、遠心分離、ろ過、抽出、噴霧乾燥、エバポレーション、または沈殿を含むがこれらに限定されない従来の手順によって、栄養培地から回収することができる。
【0110】
ポリペプチドは、クロマトグラフィー(例えば、イオン交換、アフィニティー、疎水性、クロマトフォーカシング、およびサイズ排除)、電気泳動手順(例えば、分取等電点電気泳動)、差分溶解性(differential solubility)(例えば、硫酸アンモニウム沈殿)、SDS−PAGE、または抽出(例えば、Protein Purification、編者:J.−C.ヤンソン(J.−C.Janson)およびラーズ・ライデン(Lars Ryden)、VCH Publishers、ニューヨーク、1989年を参照のこと)を含むが、これらに限定されない当該技術分野で既知の様々な手順によって、精製することができる。
【0111】
本発明はまた、真菌宿主細胞に対して内因性であるポリペプチドをコードするコード配列の発現を改変するためのDNA構築物に関する。構築物は、内因性遺伝子の発現を改変するのに必要な最小数の成分を含有し得る。
【0112】
1つの実施態様では、核酸構築物は、好ましくは、(a)ターゲティング配列、(b)本発明のプロモーターDNA配列、(c)エキソン、および(d)スプライスドナー部位を含有する。核酸構築物の細胞への導入時に、構築物は、相同組換えによって、内因性遺伝子部位で細胞ゲノムに組込む。ターゲティング配列は、エレメント(b)〜(d)が内因性遺伝子と作動可能に会合されるように、エレメント(a)〜(d)の内因性遺伝子への組込みを指令する。
【0113】
別の実施態様では、核酸構築物は、(a)ターゲティング配列、(b)本発明のプロモーターDNA配列、(c)エキソン、(d)スプライスドナー部位、(e)イントロン、および(f)スプライスアクセプター部位を含有し、ここで、ターゲティング配列は、エレメント(b)〜(f)が内因性遺伝子と作動可能に会合されるように、エレメント(a)〜(f)の組込みを指令する。しかし、構築物は、選択マーカーなどのさらなる成分を含有してもよい。使用することができる選択マーカーについては、先に記載された。
【0114】
両方の実施態様において、これらの成分の導入は、内因性遺伝子の発現が改変される新たな転写単位の生成を生じる。本質的に、新たな転写単位は、ターゲティング構築物によって誘導される配列と内因性遺伝子との融合産物である。内因性遺伝子が改変される1つの実施態様では、遺伝子が活性化される。本実施態様では、相同組換えを使用して、対応する親細胞において認められるレベルよりも高いレベルで遺伝子を発現させる調節配列の挿入を介して、親細胞の内因性遺伝子に、通常会合される調節領域を置換、破壊、または不能にする。
【0115】
ターゲティング配列は、内因性遺伝子内にあるか、遺伝子の直ぐ隣にあるか、上流の遺伝子内にあるか、または内因性遺伝子の上流かつ離れた場所にあり得る。1つもしくはそれ以上のターゲティング配列を使用することもできる。例えば、環状プラスミドまたはDNAフラグメントは、好ましくは、単一のターゲティング配列を用いる一方、線状プラスミドまたはDNAフラグメントは、好ましくは、2つのターゲティング配列を用いる。
【0116】
構築物は、内因性遺伝子の1つもしくはそれ以上のエキソンをさらに含有する。エキソンは、エキソン配列が内因性遺伝子のコード領域を伴うフレーム内に存在するように、RNAにコピーされ、成熟mRNA分子に存在するDNA配列として規定される。エキソンは、場合により、1つもしくはそれ以上のアミノ酸をコードするおよび/またはアミノ酸を部分的にコードするDNAを含有し得る。あるいは、エキソンは、5’非コード領域に対応するDNAを含有する。外因性エキソンが1つもしくはそれ以上のアミノ酸および/またはアミノ酸の部分をコードする場合、第2のエキソンから誘導されるmRNAの部分の適切なリーディングフレームが変更されないように、転写およびスプライシング時に、リーディングフレームが、内因性遺伝子のコード領域を伴うフレーム内にあるように、核酸構築物が設計される。構築物のスプライスドナー部位は、1つのエキソンの別のエキソンへのスプライシングを指令する。典型的に、第1のエキソンは第2のエキソンの5’にあり、第1のエキソンの3’側に重複および隣接するスプライスドナー部位は、第2のエキソンの5’側で第2のエキソンに隣接するスプライスアクセプター部位を認識する。スプライスアクセプター部位は、スプライスドナー部位と同様に、1つのエキソンの別のエキソンへのスプライシングを指令する配列である。スプライシング装置は、スプライスドナー部位と協働して作用し、スプライスアクセプター部位を使用して、イントロンの取り出しを生じさせる。
【0117】
所定のDNA配列の発現を改変するための好適なストラテジーは、所定のDNA配列の欠失および/または本発明のプロモーターなどの修飾されたプロモーターDNA配列による所定のDNA配列の内因性プロモーター配列の置換を含む。欠失および置換は、好ましくは、EP0357127号明細書に記載の遺伝子置換技術によって実施される。遺伝子および/またはプロモーター配列の特異的欠失は、好ましくは、EP635574号明細書に記載のように、amdS遺伝子を選択マーカー遺伝子として使用して実施される。EP635574号明細書に記載のようなフルオロアセトアミド(fluoracetamide)培地に対する対抗選択によって、得られる株は、選択マーカーを含まず、さらなる遺伝子修飾に使用することができる。
【0118】
あるいは、他の言及した技術との組み合わせで、大腸菌(E.coli)におけるコスミドのインビボ組換えに基づく技術を使用することができ、これについては:A rapid method for efficient gene replacement in the filamentous fungus A.nidulans(2000年)チャベロチェ,M−K(Chaveroche,M−K)、チコ,J−M.(Ghico,J−M.)およびダンフェール C(d’Enfert C);Nucleic acids Research、第28巻、22号に記載されている。この技術は、例えば、A.ニガー(A.niger)のような他の糸状菌にも適用可能である。本明細書において開示される特定の実施態様は、本発明のいくつかの態様の例示として意図されるため、本明細書において説明および特許請求される本発明は、これらの実施態様によってその範囲が限定されるべきではない。任意の等価な実施態様は、本発明の範囲内にあることが意図される。実際、本明細書において示しかつ説明される改変の他にも、本発明の多様な改変が存在することは、先の記載から当業者には明らかであろう。かかる改変もまた、添付の特許請求の範囲内に含まれることが意図される。抵触する場合については、定義を含む本発明の開示内容が優先される。
【0119】
本明細書において言及した特許および刊行物において開示されるすべての配列および方法を含むすべての特許および刊行物は、参照により明確に援用される。これらの特許および刊行物には次のものが含まれる。すなわち、EP357127号明細書、EP673429号明細書、EP635574号明細書、国際公開第97/06261号パンフレット、国際公開第98/46772号パンフレット、国際公開第94/04673号パンフレット。
【実施例】
【0120】
実験の情報
株
WT1:このA.ニガー(A.niger)株を野生型株として使用する。この株は、寄託番号CBS513.88でCBS研究所(CBS Institute)に寄託される。
【0121】
WT2:このA.ニガー(A.niger)株は、グルコアミラーゼをコードする遺伝子(glaA)の欠失を含むWT1株である。WT2は、EP0635574号明細書に記載のように、「マーカー遺伝子を含まない(MARKER−GENE FREE)」アプローチを使用して、構築した。本特許では、CBS513.88のゲノムにおけるglaA特異的DNA配列を欠失する仕方について、広範に説明する。該手順により、最終的に外来性のDNA配列を全く所有しない、マーカーを含まない(MARKER−GENE FREE)(?)glaA組換えA.ニガー(A.niger)CBS513.88株が得られた。
【0122】
グルコアミラーゼ活性アッセイ
国際公開第98/46772号パンフレットに記載のように、p−ニトロフェニルa−D−グルコピラノシド(シグマ(Sigma))を使用して、グルコアミラーゼ活性を決定した。
【0123】
実施例1
コード配列と作動可能に会合される本発明のプロモーターを含むDNA構築物の構築
本実施例では、本発明のプロモーターの制御下にある発現構築物の構築について説明する。ここで使用するコード配列またはレポーター構築物は、A.ニガー(A.niger)グルコアミラーゼ酵素をコードするglaA遺伝子である。グルコアミラーゼは、本発明のプロモーターの活性を測定することが可能であるレポーター酵素として使用する。
【0124】
1.1 集成的グルコアミラーゼ発現ベクター(pGBTOPGLA)の説明
グルコアミラーゼプロモーターおよびA.ニガー(A.niger)由来の遺伝子glaAをコードするグルコアミラーゼを、国際公開第99/32617号パンフレットに記載の発現ベクターpGBTOP−8にクローニングした。既知の原理および日常的なクローニング技術に従って、クローニングを実施し、プラスミドpGBTOPGLA(図1を参照のこと)を得た。本質的に、この発現ベクターは、大腸菌(E.coli)ベクターにおける3’および3’’glaAターゲティング部位に隣接されるグルコアミラーゼプロモーター、コード配列およびターミネーター領域を含む。
【0125】
1.2 複数のクローニング部位MCSを伴う集成的グルコアミラーゼ発現ベクター(pGBTOPGLA−2)の構築
テンプレートとして、オリゴヌクレオチド5’−ATgCggCCgCCTCgAgTTAATTAAggCCAggCCggCCggCgCgCCTCAgCAATgTCgTTCCgA−3’(配列番号6として同定される)および5’−AGCCATTGACTTCTTCCCAG−3’(配列番号7として同定される)および1ngのベクターpGBTOPGLAを使用して、glaAコード配列の部分を含有するPCRフラグメントを作製した。このフラグメントを、XhoIおよびBglIIで消化し、XhoIおよびBglII消化したベクターpGBTOPGLAに導入し、ベクターpGBTOPGLA−2(図2を参照のこと)を得た。MCSおよびglaAコード配列の一部を含む導入されたPCRフラグメントの配列を、配列解析によって確認した。
【0126】
1.3 グルコアミラーゼコード配列と作動可能に会合される本発明のプロモーターを伴う集成的発現ベクター(pGBTOPGLA−3)の構築
株CBS513.88のゲノムDNAを配列決定し、解析した。表において(オリゴ配列番号)以下のように同定されるオリゴヌクレオチドの組み合わせおよび株CBS513.88のゲノムDNAをテンプレートとして使用し、PCR増幅により、適切な制限部位を本発明のプロモーターに付着させた。配列番号1、配列番号2、配列番号3、配列番号4または配列番号5において同定される配列は、以下の表において同定されるように、約2kbの得られるフラグメントの配列を含む。すべての5つの得られるフラグメントを、AscIおよびXhoIで消化し、AscIおよびXhoI消化したベクターpGBTOPGLA−2に導入し、表(ベクターの名称)において同定されるように、ベクターpGBTOPGLA−4、pGBTOPGLA−5、pGBTOPGLA−7、GBTOPGLA−9またはpGBTOPGLA−10を得た。図3では、5つのすべてのベクターのレイアウトを例示する図を見出すことができる。本発明のプロモーターを含む多様な導入されたPCRフラグメントの配列を、配列解析によって確認した。
【0127】
【表1】
【0128】
実施例2.DNA構築物で形質転換された真菌宿主細胞
pGBTOPGLA、pGBTOPGLA−4、pGBTOPGLA−5、pGBTOPGLA−7、pGBTOPGLA−9またはpGBTOPGLA−10ベクターをWT2に導入するために、形質転換およびその後の形質転換体選択を、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットに記載のとおりに行った。原則として、NotIによる消化後、すべてのベクターの線状DNAを単離し、pGBAAS−1で示されるamdS選択マーカー遺伝子含有ベクターで同時形質転換した(EP635574号明細書に記載のとおりに構築した)。両方のベクターは、WT2における短縮型glaA遺伝子座への標的化を指令するためのA.ニガー(A.niger)宿主株のglaA遺伝子座に相同な2つのDNAドメインを含む。形質転換体をアセトアミド培地上で選択し、標準的手順に従ってコロニーを精製した。胞子をフルオロアセトアミド培地上に配置し、amdSマーカーを消失した株を選択した。増殖中のコロニーを、glaA遺伝子座での組込みおよびコピー数について診断した。類似の予想されるコピー数を伴うpGBTOPGLA、pGBTOPGLA−4、pGBTOPGLA−5、pGBTOPGLA−7、pGBTOPGLA−9またはpGBTOPGLA−10の形質転換体を選択した。好ましくは、単一コピーを伴う若干の形質転換体(1A、1B、1C)およびおそらく、複数コピーを伴う1つの形質転換体(2A)を選択した。
【0129】
さらに、選択マーカー遺伝子および本発明のプロモーターによって制御される目的の遺伝子は1つの構築物上に存在している。この例を図4に示す。
【0130】
実施例3.真菌宿主細胞における本発明のプロモーターの制御下でglaAコード配列によってコードされるグルコアミラーゼポリペプチドの産生
多くの選択された上記のようなWT2の形質転換体、および両方の5つの株WT1およびWT2を使用し、500mlバッフル付き振盪フラスコを使用する恒温振盪器において、EP635574号明細書に記載のような100mlの培地中、34℃および170rpmで、振盪フラスコ実験を実施した。発酵の4および5日後、サンプルを採取し、上記のようにグルコアミラーゼ活性を決定した。グルコアミラーゼ活性を、4日目のWT1の活性に対して正規化した。WT1、WT2、ならびにpGBTOPGLA、pGBTOPGLA−5、pGBTOPGLA−7、およびpGBTOPGLA−10について多くの選択された形質転換体の正規化された活性を図5に示す。
【0131】
グルコアミラーゼレポーターの測定される活性から結論し得るように、本発明は、真菌細胞において、目的の遺伝子の高い発現のための強力なプロモーターを提供する。従って、本発明のプロモーターは、真菌細胞において、目的の遺伝子の高い発現のための代替的およびさらなるプロモーターを提供する。
【0132】
実施例4.本発明のプロモーターを含むプロモーター置換構築物pGBDEL−PGLAAの構築
宿主細胞における所定の遺伝子の発現レベルを改変するために、本発明のプロモーターは、前記所定の遺伝子の内因性プロモーターと置き換わることができる。本実験では、本発明のプロモーターは、真菌宿主細胞においてglaA遺伝子をコードするグルコアミラーゼのプロモーターと置き換わる。実施例4、5および6では、このプロセスにおける多くの異なる工程について説明している。
【0133】
グルコアミラーゼプロモーターのための置換ベクターを、既知の原理に従って設計し、日常的なクローニング手順に従って構築した(図6を参照のこと)。本質的に、glaAプロモーター置換ベクターpGBDEL−PGLAAは、予め定められた遺伝子座での相同組換えを介して本発明のプロモーター(配列番号4に含まれる)によって置換されるべきglaAプロモーター配列の約1000bpフランキング領域を含む。ここで使用されるフランキング領域(図6を参照のこと)は、glaAプロモーターの5’上流領域であり、glaAコード配列の部分である。さらに、置換ベクターは、A.ニダランス(A.nidulans)両方向性amdS選択マーカーを中間的なダイレクトリピートで含有する。本実施例で使用されるダイレクトリピートは、glaAコード配列の部分である。これらの欠失ベクターの一般的な設計については、EP635574号明細書および国際公開第98/46772号パンフレットにおいて先に記載された。
【0134】
実施例5.真菌宿主細胞における本発明のプロモーターによるglaAプロモーターの置換
NotI消化した欠失ベクターpGBDEL−PGLAAの線状DNAを単離し、WT1(CBS513.88)を形質転換するために使用した。この線状DNAは、glaA遺伝子座でゲノムに組込むことができ、従って、glaAプロモーター領域をamdSおよび本発明のプロモーターを含有する構築物で置換する(図7を参照のこと)。形質転換体をアセトアミド培地上で選択し、標準的手順に従ってコロニーを精製した。増殖中のコロニーを、glaA遺伝子座での組込みについて、PCRによって診断した。glaAプロモーターの欠失は、本発明のプロモーターに特異的なサイズを伴うバンドの増幅、およびglaAプロモーターに特異的なバンドの消失によって、検出した。胞子をフルオロアセトアミド培地上に配置し、amdSマーカーを消失した株を選択した。候補株を、グルコアミラーゼプロモーターの適切な欠失および配列番号4に含まれる本発明のプロモーターによる置換について、サザン分析を使用して試験した。株dPGLAAを、glaAプロモーターが本発明のプロモーターによって置換され、回復された機能的glaAコード配列を有する代表的株として選択した(図7を参照のこと)。
【0135】
実施例6.真菌宿主細胞における本発明の置換されたプロモーターの制御下でglaAコード配列によってコードされるグルコアミラーゼポリペプチドの産生
選択されたdPGLAA株(実施例5において単離されたWT1の適切なpGBDEL−PGLAA形質転換体)および株WT1を使用して、500mlバッフル付き振盪フラスコを使用する恒温振盪器において、欧州特許第635574B1号明細書に記載のような100mlの培地中、34℃および170rpmで、振盪フラスコ実験を実施した。さらなる条件および活性測定は、実施例3に記載のとおりであった。WT1の選択されたpGBDEL−PGLAA形質転換体におけるグルコアミラーゼ活性は、発酵の両方の日にWT1について測定した活性と比較して増加していた(データ示さず)。
【0136】
実施例7.真菌宿主細胞における本発明のプロモーターの制御下におけるさらなるglaA遺伝子の付加
宿主細胞において所定の遺伝子の発現レベルを改変するために、本発明のプロモーターに作動可能に連結された複数のさらなるコピーの前記遺伝子を、内因的に付与された遺伝子に付加することができる。本実施例では、配列番号4に含まれ、glaAコード配列に作動可能に連結される本発明のプロモーターを、真菌宿主細胞において内因的に存在するglaA遺伝子をコードするグルコアミラーゼの隣に導入する。実施例7、および8では、このプロセスにおける多くの異なる工程について説明している。
【0137】
図8に示される環状構築物を単離し、WT1(CBS513.88)を形質転換するために使用した。この線状DNAは、glaAコード配列においてゲノムに組込むことができ、従って、本発明のプロモーターの制御下の第2のglaA遺伝子を、選択マーカーamdSの隣に付加することができる(図8を参照のこと)。形質転換体をアセトアミド培地上で選択し、標準的手順に従ってコロニーを精製した。増殖中のコロニーを、glaA遺伝子座での組込みについて、PCRによって診断した。glaA遺伝子座の組込みを、PCRおよびサザン分析によって検出した。株P2GLAAを、glaA遺伝子座で組込まれる本発明のプロモーターの制御下にある少なくとも第2のglaA遺伝子を伴う代表的株として選択した。
【0138】
実施例8.真菌宿主細胞における本発明のプロモーターおよび内因性glaAプロモーターの制御下でglaAコード配列によってコードされるグルコアミラーゼポリペプチドの産生
実施例7において単離された選択されたP2GLAA株、および株WT1を使用して、実施例3に記載のように、100mlの培地において、振盪フラスコ実験を実施した。発酵の4および6日後、サンプルを採取し、グルコアミラーゼ活性を決定した。WT1の選択されたP2GLAA形質転換体におけるグルコアミラーゼ活性は、発酵の4または5日後のいずれかで、WT1について測定した活性と比較して増加した。グルコアミラーゼレポーターの活性の増加は、本発明のプロモーターが、真菌細胞において強力なプロモーター下で既に発現されている目的の遺伝子の発現をさらに増加する手段を提供することを示す。
【図面の簡単な説明】
【0139】
【図1】集成的グルコアミラーゼ発現ベクターであるpGBTOPGLAのプラスミドマップを表す。
【図2】複数のクローニング部位を伴う集成的グルコアミラーゼ発現ベクターであるpGBTOPGLA−2のプラスミドマップを表す。
【図3】グルコアミラーゼコード配列と作動可能に会合される本発明のプロモーターを含有する集成的発現ベクターであるpGBTOPGLA−3のプラスミドマップを表す。
【図4】単一の相同組換えを介する組み込みの略図を表す。
【図5】WT1、WT2および多様なpGBTOPGLAベクターの形質転換体のグルコアミラーゼ活性。4日目のWT1の活性を100%と設定した場合の正規化された活性を示す。
【図6】置換ベクターであるpGBDEL−PGGLAAのプラスミドマップを表す。
【図7】プロモーター置換の略図を表す。
【図8】相同組換えを介する組み込みの略図を表す。
【特許請求の範囲】
【請求項1】
(a)配列番号1、配列番号2、配列番号3、配列番号4、配列番号5のヌクレオチド配列を含むDNA配列、
(b)(a)のDNA配列にハイブリダイズすることが可能なDNA配列、
(c)(a)のDNA配列に少なくとも50%相同であるDNA配列、
(d)(a)〜(c)のDNA配列のいずれかの変異体、および
(e)(a)〜(d)のDNA配列のいずれかのサブ配列、
からなる群から選択されるプロモーターDNA配列。
【請求項2】
コード配列がプロモーターDNA配列の制御下で発現され得るように、請求項1に記載のプロモーターDNA配列および前記プロモーターDNA配列と作動可能に会合されるコード配列を含む、DNA構築物。
【請求項3】
請求項2に記載のDNA構築物を含む、宿主細胞。
【請求項4】
宿主細胞はアスペルギルス(Aspergillus)、ペニシリウム(Penicillium)またはトリコデルマ(Trichoderma)種である、請求項3に記載の宿主細胞。
【請求項5】
アスペルギルス(Aspergillus)はアスペルギルス・ニガー(Aspergillus niger)または麹菌(Aspergillus oryzae)種である、請求項4に記載の宿主細胞。
【請求項6】
(a)請求項2に記載のDNA構築物を提供すること、
(b)前記DNA構築物で適切な宿主細胞を形質転換すること、および
(c)前記プロモーターDNA配列の制御下で遺伝子を発現させること、
を含む、宿主細胞における遺伝子の発現のための方法。
【請求項7】
(a)ポリペプチドをコードするコード配列を含む請求項2に記載のDNA構築物を提供すること、
(b)前記DNA構築物で適切な宿主細胞を形質転換すること、
(e)ポリペプチドの発現をもたらす培養条件下で適切な宿主を培養すること、および
(f)培養ブロスからポリペプチドを回収すること、
を含む、適切な宿主におけるポリペプチドの産生のための方法。
【請求項8】
(a)二次代謝物の産生に関与する酵素をコードするコード配列を含む、請求項2に記載のDNA構築物を提供すること、
(b)前記DNA構築物で適切な宿主細胞を形質転換すること、
(c)二次代謝物の産生をもたらす培養条件下で適切な宿主を培養すること、および
(d)培養ブロスから二次代謝物を回収すること、
を含む、適切な宿主における二次代謝物の産生のための方法。
【請求項1】
(a)配列番号1、配列番号2、配列番号3、配列番号4、配列番号5のヌクレオチド配列を含むDNA配列、
(b)(a)のDNA配列にハイブリダイズすることが可能なDNA配列、
(c)(a)のDNA配列に少なくとも50%相同であるDNA配列、
(d)(a)〜(c)のDNA配列のいずれかの変異体、および
(e)(a)〜(d)のDNA配列のいずれかのサブ配列、
からなる群から選択されるプロモーターDNA配列。
【請求項2】
コード配列がプロモーターDNA配列の制御下で発現され得るように、請求項1に記載のプロモーターDNA配列および前記プロモーターDNA配列と作動可能に会合されるコード配列を含む、DNA構築物。
【請求項3】
請求項2に記載のDNA構築物を含む、宿主細胞。
【請求項4】
宿主細胞はアスペルギルス(Aspergillus)、ペニシリウム(Penicillium)またはトリコデルマ(Trichoderma)種である、請求項3に記載の宿主細胞。
【請求項5】
アスペルギルス(Aspergillus)はアスペルギルス・ニガー(Aspergillus niger)または麹菌(Aspergillus oryzae)種である、請求項4に記載の宿主細胞。
【請求項6】
(a)請求項2に記載のDNA構築物を提供すること、
(b)前記DNA構築物で適切な宿主細胞を形質転換すること、および
(c)前記プロモーターDNA配列の制御下で遺伝子を発現させること、
を含む、宿主細胞における遺伝子の発現のための方法。
【請求項7】
(a)ポリペプチドをコードするコード配列を含む請求項2に記載のDNA構築物を提供すること、
(b)前記DNA構築物で適切な宿主細胞を形質転換すること、
(e)ポリペプチドの発現をもたらす培養条件下で適切な宿主を培養すること、および
(f)培養ブロスからポリペプチドを回収すること、
を含む、適切な宿主におけるポリペプチドの産生のための方法。
【請求項8】
(a)二次代謝物の産生に関与する酵素をコードするコード配列を含む、請求項2に記載のDNA構築物を提供すること、
(b)前記DNA構築物で適切な宿主細胞を形質転換すること、
(c)二次代謝物の産生をもたらす培養条件下で適切な宿主を培養すること、および
(d)培養ブロスから二次代謝物を回収すること、
を含む、適切な宿主における二次代謝物の産生のための方法。
【図1】


【図2】
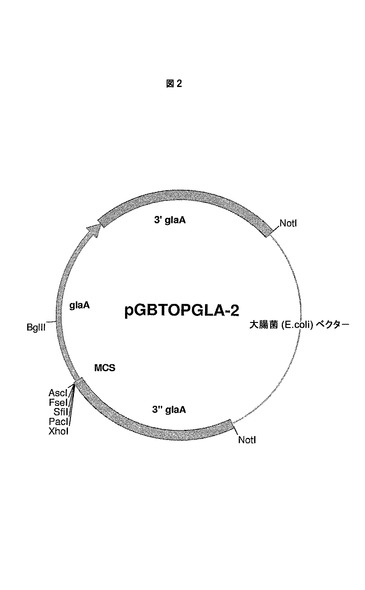
【図3】

【図4】

【図5】

【図6】
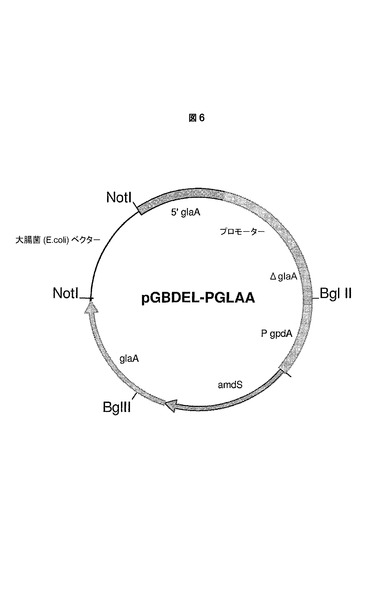
【図7】
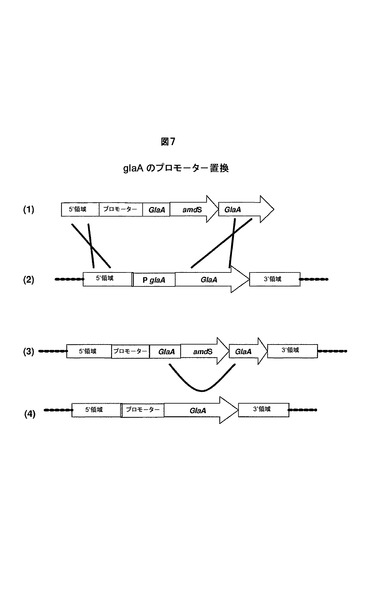
【図8】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公表番号】特表2007−532124(P2007−532124A)
【公表日】平成19年11月15日(2007.11.15)
【国際特許分類】
【出願番号】特願2007−507807(P2007−507807)
【出願日】平成17年4月18日(2005.4.18)
【国際出願番号】PCT/EP2005/051696
【国際公開番号】WO2005/100573
【国際公開日】平成17年10月27日(2005.10.27)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
【公表日】平成19年11月15日(2007.11.15)
【国際特許分類】
【出願日】平成17年4月18日(2005.4.18)
【国際出願番号】PCT/EP2005/051696
【国際公開番号】WO2005/100573
【国際公開日】平成17年10月27日(2005.10.27)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
[ Back to top ]