
耐熱性チオレドキシン、耐熱性チオレドキシンレダクターゼ、耐熱性チオレドキシンペルオキシダーゼおよびこれらの酵素をコードするDNA

【課題】 耐熱性チオレドキシン、耐熱性チオレドキシンレダクターゼ及び耐熱性チオレドキシンペルオキシダーゼ、並びに、これらの酵素をコードするDNA等を提供する。
【解決手段】 90〜100℃という高温下で生育できる超好熱性古細菌Aeropyrum pernixか
らチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼを単離し、アミノ酸配列およびDNA配列を決定した。これらのタンパク質又は酵素は、非常
に耐熱性に優れ、また常温での安定性にも優れる。さらに有機溶媒に耐性である。本発明のチオレドキシン等は、耐熱性が高いことから、高温で効率的に反応を行うことができるとともに、医薬品、食品、化粧品などに添加した場合には加熱滅菌を行うことができる。
【解決手段】 90〜100℃という高温下で生育できる超好熱性古細菌Aeropyrum pernixか
らチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼを単離し、アミノ酸配列およびDNA配列を決定した。これらのタンパク質又は酵素は、非常
に耐熱性に優れ、また常温での安定性にも優れる。さらに有機溶媒に耐性である。本発明のチオレドキシン等は、耐熱性が高いことから、高温で効率的に反応を行うことができるとともに、医薬品、食品、化粧品などに添加した場合には加熱滅菌を行うことができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、高温下で機能できるチオレドキシン、チオレドキシンレダクターゼおよびチオレドキシンペルオキシダーゼ、これらの酵素をコードする各DNA、これらのDNAを組み込んだ各ベクター、これらのベクターで形質転換された各形質転換体、並びに、これらの各形質転換体を用いた耐熱性チオレドキシン、耐熱性チオレドキシンレダクターゼ及び耐熱性チオレドキシンペルオキシダーゼの生産方法に関する。
【背景技術】
【0002】
チオレドキシンは、大腸菌、酵母、高等植物及び高等動物等に普遍的に存在する分子量1万〜1.3万の電子伝達タンパク質である。チオレドキシンは、2個のアミノ酸を挟んで
2個のシステイン残基が存在する部位(―Cys―X―X―Cys―)を活性中心として有し、
ジチオールの可逆的な酸化還元反応を通して、生体内の酸化還元状態を制御していると考えられている(Laurent, T.C.,Moor,E.C.,&Reinchard,P.(1964)J.Biol.Chem.239,3436-3444;Holmgren,A.(1989)Ann.Rev.Biochem.54,237-271;Holmgren,A.(1989)J.Biol.Chem.264,13963-13966;Buchanan,B.B.,Schurmann,P.,Decottignies,P.&Lozano,R.M.(1994)Arch.Biochem.Biophys.314,257-260)。チオレドキシンは、活性中心の2個のシステインのチオール基がジスルフィドとなった酸化型またはチオール基となった還元型のどちらの状態でも存在している。
【0003】
チオレドキシンの生体内での機能の1例としてタンパク質の分子内または分子間S−S結合の開裂が挙げられる。この際、還元型チオレドキシンは、タンパク質のジスルフィドをジチオールに還元するのに伴い、酸化されて酸化型チオレドキシンとなる。生成した酸化型チオレドキシンは、チオレドキシンレダクターゼ及びNADPHにより還元されて還元型チ
オレドキシンに戻る。
【0004】
チオレドキシンの生体内での機能の他の例として過酸化水素や過酸化脂質などの過酸化物の消去が挙げられる。この際、還元型チオレドキシンは、過酸化物(ROOH)を水酸化物(ROH)に還元するとともに、酸化されて酸化型チオレドキシンとなる。この反応はチオ
レドキシンペルオキシダーゼにより行われる。酸化型チオレドキシンは、チオレドキシンレダクターゼ及びNADPHにより還元されて還元型チオレドキシンに戻る。
【0005】
また、チオレドキシンの生体内での機能の更に他の例として、紫外線照射による細胞障害防止機能、転写因子の制御機能等も挙げられる。
【0006】
このような機能を有するチオレドキシンは、例えば、酸化型システインの状態で活性を示す消化酵素インヒビタータンパク質の還元によるインヒビター活性の阻害、蛇毒タンパク質のシステイン残基間のS−S結合の除去による無毒化、紫外線照射による皮膚の炎症の予防等の目的で、医薬品として用いることが提案されている。また、食物アレルゲンの除去等の目的で食品に添加して用いたり、乾燥や紫外線等に起因する酸化ストレスによる肌荒れの改善剤等の目的で化粧品として用いることが提案されている(特開2001-288103号
公報、特表2001-520027号公報、特開2000-103743号公報)。
【0007】
しかし、固体状態または半固体状態の医薬品、食品及び化粧品等ではろ過滅菌が困難であるため、通常加熱滅菌が行われるところ、従来知られているチオレドキシンは、耐熱性が低い。従って、従来の易熱性のチオレドキシンを医薬品等として用いたり、食品等に添加して用いる場合には、高温での滅菌を行うことができないという難点がある。
【0008】
また、一般に溶質の水に対する溶解度は温度と共に上昇するため、高温下で機能できる耐熱性のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼを用いることができれば、タンパク質中のジスルフィドの還元による還元型タンパク質の合成反応、過酸化水素等の活性酸素の除去反応等を、高温下で調製した高濃度の基質溶液に該酵素を作用させることにより効率よく行うことができる。
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明は、耐熱性のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼ、これらの酵素をコードするDNA、これらのDNAを組み込んだベクター、これらのベクターで形質転換された形質転換体、並びに、これらの形質転換体を用いた耐熱性チオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの効率的な生産方法を提供することを主目的とする。
【課題を解決するための手段】
【0010】
前記目的を達成するために、本発明者は、90〜100℃という高温下で生育できる超好熱性古細菌に着目した。古細菌は、真核生物及び原核生物のいずれにも属さない第3の生物群に属する生物である。また、太古の地球の生物の末裔と考えられ、常温環境に対する進化・適用を免れた特殊な生物である。
【0011】
本発明者は、このような超好熱性古細菌由来のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼを初めて単離した。また、これらのタンパク質または酵素は、従来公知のチオレドキシン等に比べて非常に耐熱性が高く、また常温下での安定性にも優れること、有機溶媒の存在下でも活性を示すことを見出した。
【0012】
さらに、これらのタンパク質又は酵素をコードするDNA配列を決定し、ベクターに組み
込んで、これらを効率よく生産することに成功した。
【0013】
前記知見に基づき、本発明者は、以下の各項のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼ等を提供する。
【0014】
項1. 100℃で0.5時間熱処理した場合に50%以上活性が残存する耐熱性チオレドキシン。
【0015】
項2. 超好熱性古細菌Aeropyrum pernix由来の項1に記載の耐熱性チオレドキシン。
【0016】
項3. 以下の(1)または(2)のポリペプチド。
・ 配列表の配列番号2に示すアミノ酸配列を含むポリペプチド。
・ 配列表の配列番号2に示すアミノ酸配列において1若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含み、かつ、100℃で0.5時間熱処理した場合に50%以上チオレドキシン活性が残存するポリペプチド。
【0017】
項4. 以下の(3)または(4)のDNA。
・ 配列表の配列番号1に示す塩基配列を含むDNA。
・ 配列表の配列番号1に示す塩基配列を含むDNAとストリンジェントな条件下でハイ
ブリダイズし、かつ、100℃で0.5時間熱処理した場合に50%以上活性が残存する耐熱性チオレドキシンをコードするDNA。
【0018】
項5. 項3に記載のポリペプチドをコードするDNA。
【0019】
項6. 項3または5に記載のDNAを含有するベクター。
【0020】
項7. 項6に記載のベクターを含有する形質転換体。
【0021】
項8. 項7の形質転換体を培養し、該形質転換体から耐熱性チオレドキシンを分離および精製する耐熱性チオレドキシンの生産方法。
【0022】
項9. 70℃以上の温度下で、100 unit/mg以上の活性を有する耐熱性チオレドキシ
ンレダクターゼ。
【0023】
項10. 100℃で0.5時間熱処理した場合に50%以上活性が残存する項9に記載の耐熱性チオレドキシンレダクターゼ。
【0024】
項11. 至適温度が70℃以上である項9に記載の耐熱性チオレドキシンレダクターゼ。
【0025】
項12. 超好熱性古細菌Aeropyrum pernix由来の項9に記載の耐熱性チオレドキシンレダクターゼ。
【0026】
項13. 以下の(5)または(6)のポリペプチド。
・ 配列表の配列番号4に示すアミノ酸配列を含むポリペプチド。
・ 配列表の配列番号4に示すアミノ酸配列において1若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含み、かつ、70℃以上の温度下で、100 unit/mg以上のチオレドキシンレダクターゼ活性を有するポリペプチド。
【0027】
項14. 以下の(7)または(8)のDNA。
・ 配列表の配列番号3に示す塩基配列を含むDNA。
・ 配列表の配列番号3に示す塩基配列を含むDNAとストリンジェントな条件下でハイ
ブリダイズし、かつ、70℃以上の温度下で、100 unit/mg以上の活性を有する耐熱性チオレドキシンレダクターゼをコードするDNA。
【0028】
項15. 項13に記載のポリペプチドをコードするDNA。
【0029】
項16. 項13または15に記載のDNAを含有するベクター。
【0030】
項17. 項16に記載のベクターを含有する形質転換体。
【0031】
項18. 項17の形質転換体を培養し、該形質転換体から耐熱性チオレドキシンレダクターゼを分離および精製する耐熱性チオレドキシンレダクターゼの生産方法。
【0032】
項19. 70℃以上の温度下で、100unit/mg以上の活性を有する耐熱性チオレドキシ
ンペルオキシダーゼ。
【0033】
項20. 100 ℃で0.5時間熱処理した場合に50%以上活性が残存する項19に記載の耐熱性チオレドキシンペルオキシダーゼ。
【0034】
項21. 至適温度が70 ℃以上である項19に記載の耐熱性チオレドキシンペルオキシダーゼ。
【0035】
項22. 超好熱性古細菌Aeropyrum pernix由来の項19に記載の耐熱性チオレドキシンペルオキシダーゼ。
【0036】
項23. 以下の(9)または(10)のポリペプチド。
・ 配列表の配列番号6に示すアミノ酸配列を含むポリペプチド。
・ 配列表の配列番号6に示すアミノ酸配列において1若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含み、かつ、70℃以上の温度下で、100 unit/mg以上のチオレドキシンペルオキシダーゼ活性を有するポリペプチド。
【0037】
項24. 以下の(11)または(12)のDNA。
・ 配列表の配列番号5に示す塩基配列を含むDNA。
・ 配列表の配列番号5に示す塩基配列を含むDNAとストリンジェントな条件下でハイ
ブリダイズし、かつ、70 ℃以上の温度下で、100 unit/mg以上の活性を有する耐熱性チオレドキシンペルオキシダーゼをコードするDNA。
【0038】
項25. 項23に記載のポリペプチドをコードするDNA。
【0039】
項26. 項23または25に記載のDNAを含有するベクター。
【0040】
項27. 項26に記載のベクターを含有する形質転換体。
【0041】
項28. 項27の形質転換体を培養し、該形質転換体から耐熱性チオレドキシンペルオキシダーゼを分離および精製する耐熱性チオレドキシンペルオキシダーゼの生産方法。
【0042】
項29. 精製すべき耐熱性タンパク質を10〜30分間インキュベートした場合に60%以上活性が残存し、かつ、耐熱性タンパク質を生産する宿主の生育限界温度より10℃以上高い温度で、被精製蛋白溶液を10〜120分間インキュベートする加熱工程を含む耐熱性タン
パク質の精製方法。
【発明の効果】
【0043】
本発明によると優れた耐熱性を有するチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼを提供することができる。
従って、これらを医薬品や化粧品として用いたり、医薬品、食品及び化粧品に添加して用いる場合には、高温での殺菌を行うことができる。
詳述すれば、本発明の耐熱性チオレドキシンは、消化酵素インヒビターの阻害、蛇毒及びサソリ毒等の酸化による無毒化、紫外線照射による皮膚の炎症の治療及び予防等の目的で医薬品として用いたり、食物アレルゲンの除去等の目的で食品に添加して用いたり、乾燥や紫外線等に起因する酸化ストレスによる肌荒れの改善等の目的で化粧品に添加して用いたりすることができる。また、本発明のチオレドキシンレダクターゼは、チオレドキシン併用して医薬品等として用いることにより、チオレドキシンを活性な還元型チオレドキシンに保つことができる。また、本発明のチオレドキシンペルオキシダーゼは、医薬品、食品及び化粧品等の酸化防止の目的で、チオレドキシンとともにこれらに添加して用いることができる。これらの場合に、本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼは、耐熱性であることから加熱殺菌をすることができる。
【0044】
また、本発明のチオレドキシン及びチオレドキシンレダクターゼは、タンパク質中のシスチンの還元による還元型タンパク質の合成反応等を高温下で調製した高濃度の基質溶液に対して効率よく行うことができる。
【0045】
また、本発明のチオレドキシン及びチオレドキシンペルオキシダーゼは、過酸化水素等の活性酸素の除去反応等を、高温下で調製した高濃度の基質溶液に対して効率よく行うことができる。
【0046】
また、本発明のチオレドキシンペルオキシダーゼは、過酸化物と反応して発色するセンサーとして用いたり、ウェスタンブロット法において抗体に結合させるペルオキシダーゼとして用いることができる。この場合には、比較的高温下で酵素反応を行うことができるため、夾雑タンパク質の影響が少なくでき、検出感度が向上する。
【0047】
また、本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼは、常温での安定性にも優れるため、比較的長い期間保存することができ、また繰り返し使用にも耐えることができる。
【0048】
また、一般的にタンパク質または酵素は、有機溶媒の存在により失活し易いが、本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼは、有機溶媒に対しても安定である。従って、有機溶媒中あるいは有機溶媒を含む水溶液中で、反応を行うことができる。その結果、水溶液には溶解し難い物質に対しても作用させることができ、反応の適用対象が広がる。
【発明を実施するための最良の形態】
【0049】
(1)本発明のタンパク質・酵素
耐熱性
(i)チオレドキシン
本発明の耐熱性チオレドキシンは、非常に耐熱性が優れるタンパク質であり、100℃で0.5 時間熱処理した場合に50%以上活性が残存するタンパク質である。100℃で0.5 時間熱処理した場合に60%以上活性が残存するタンパク質であることがより好ましい。
【0050】
本明細書において、チオレドキシンの活性は、実施例に記載の方法で測定した値である。
(ii)チオレドキシンレダクターゼ
本発明の耐熱性チオレドキシンレダクターゼは、非常に耐熱性に優れる酵素であり、70
℃以上の温度下で、100 unit/mg以上の活性を有する酵素である。70℃以上の温度下で150 unit/mg以上の活性を有する酵素であることがより好ましい。活性を示す最高温度は、通常85 ℃程度である。また、酵素反応を行う緩衝液の種類によっても異なるが、酵素反応の初速度を測定した場合の至適温度が70℃以上の酵素であることが好ましい。
【0051】
また、本発明のチオレドキシンレダクターゼは、100℃で0.5 時間熱処理した場合に50
%以上活性が残存する酵素であることが好ましく、100℃で0.5時間熱処理した場合に60%以上活性が残存する酵素であることがより好ましい。
【0052】
本明細書において、チオレドキシンレダクターゼの活性は、実施例に記載の方法で測定した値である。
(iii)チオレドキシンペルオキシダーゼ
本発明の耐熱性チオレドキシンペルオキシダーゼは、非常に耐熱性に優れる酵素であり、70℃以上の温度下で、100 unit/mg以上の活性を有する酵素である。70℃以上の温度下で150 unit/mg以上の活性を有する酵素であることがより好ましい。活性を示す最高温度は、通常85℃程度である。また、酵素反応を行う緩衝液の種類によっても異なるが、酵素反応の初速度を測定した場合の至適温度が70℃以上の酵素であることが好ましい。
【0053】
本発明のチオレドキシンペルオキシダーゼは、100℃で0.5時間熱処理した場合に50 %
以上活性が残存する酵素であることが好ましく、100 ℃で0.5 時間熱処理した場合に60%以上活性が残存する酵素であることがより好ましい。
【0054】
本明細書において、チオレドキシンペルオキシダーゼの活性は、実施例に記載の方法で測定した値である。
常温での安定性
(i)チオレドキシン
本発明のチオレドキシンは、常温下での安定性にも優れる。例えば、本発明のチオレドキシンは、30℃の50 mMリン酸カリウム緩衝液(pH7.0)中で、12 時間インキュベート
した場合に、90%以上の活性が残存する酵素であることが好ましい。
(ii)チオレドキシンレダクターゼ
本発明のチオレドキシンレダクターゼは、常温下での安定性にも優れる。例えば、本発明のチオレドキシンレダクターゼは、30℃の50mMリン酸カリウム緩衝液(pH7.0)中で、12時間インキュベートした場合に、90%以上の活性が残存する酵素であることが好ましい
。
(iii)チオレドキシンペルオキシダーゼ
本発明のチオレドキシンペルオキシダーゼは、常温下での安定性にも優れる。例えば、本発明のチオレドキシンペルオキシダーゼは、30℃の50mMリン酸カリウム緩衝液(pH7.0
)中で、12時間インキュベートした場合に、90%以上の活性が残存する酵素であることが好ましい。
有機溶媒耐性
本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼは、有機溶媒に耐性である。例えば、エタノール、ブタノール、テトラヒドロフラン、酢酸エチル等の有機溶媒を30容量%以上含む緩衝液中で活性を示すタンパク質または酵素(以下、「タンパク質等」という。)であることが好ましい。本発明のこれらのタンパク質等が活性を示し得る、緩衝液中の有機溶媒の容量比率の上限は、当該タンパク質等が沈殿しない範囲である。なお、有機溶媒に対する耐性は、古細菌由来の酵素に特有の性質である。
活性
本発明のチオレドキシンは、還元型チオレドキシンの形で、種々のタンパク質のシステイン残基のジスルフィドをジチオールに還元できるタンパク質である。また本発明のチオレドキシンは、還元型チオレドキシンの形で、過酸化水素等を還元できるタンパク質であることが好ましい。
本発明のチオレドキシンレダクターゼは、酸化型チオレドキシンを、そのジスルフィドをジチオールに還元することにより還元型チオレドキシンに変換できる酵素である。また、本発明のチオレドキシンレダクターゼは、酸化型グルタチオン等を還元できる酵素であることが好ましい。これらの還元反応には、通常、NADPH、NADH、FADH及びFADH2等の補酵素が用いられる。補酵素は、チオレドキシンレダクターゼ1分子に対して、通常100〜100000モル程度の比率で用いることが好ましい。
本発明のチオレドキシンペルオキシダーゼは、還元型チオレドキシンの存在下に、過酸化水素を水に還元できる酵素である。また本発明のチオレドキシンペルオキシダーゼは、その他の活性酸素も還元できる酵素であることが好ましい。
生産細菌
また、本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼとしては、好熱性細菌、例えばAeropyrum属、Sufolobus属、Thermoplasma属、Thermoproteus属、Mastigocladus属、Bacillus属、Synechococcus属、Thermus属等の細菌により生産されたものが挙げられる。特に、超好熱性古細菌Aeropyrum pernixにより生産されたものが好ましい。
アミノ酸配列
(i)チオレドキシン
本発明のチオレドキシンとしては、以下の(1)又は(2)のアミノ酸配列を有するポリペプチドが挙げられる。
(1) 配列表の配列番号2に示すアミノ酸配列を含むポリペプチド。
・ 配列表の配列番号2に示すアミノ酸配列において1若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含み、かつ、100 ℃で0.5 時間熱処理した場合に50%以上チオレドキシン活性が残存するポリペプチド。
【0055】
(2)のポリペプチドのアミノ酸配列は、100 ℃で0.5時間熱処理した場合に50 %以上
チオレドキシン活性が残存するものであれば特に制限されないが、(1)のポリペプチドの
アミノ酸配列において、通常20%以下、特に10%以下のアミノ酸が欠失、置換若しくは付加されたものであることが好ましい。
【0056】
(1)のポリペプチドにおいて、配列番号2に示すアミノ酸配列を含むポリペプチドとし
ては、例えば、配列番号2に示すアミノ酸配列の3倍以下の長さのポリペプチドが挙げられる。同様に、(2)のポリペプチドにおいて、配列番号2に示すアミノ酸配列において1
若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含むポリペプチドとしては、例えば、当該アミノ酸配列の3倍以下の長さのポリペプチドが挙げられる。
(ii)チオレドキシンレダクターゼ
本発明のチオレドキシンレダクターゼとしては、以下の(3)又は(4)のアミノ酸配列を有するポリペプチドが挙げられる。
(3) 配列表の配列番号4に示すアミノ酸配列を含むポリペプチド。
(4) 配列表の配列番号4に示すアミノ酸配列において1若しくは2以上のアミノ酸が欠
失、置換若しくは付加されたアミノ酸配列を含み、かつ、70 ℃以上の温度下で、100 unit/mg以上のチオレドキシンレダクターゼ活性を有するポリペプチド
(4)のポリペプチドのアミノ酸配列は、チオレドキシンレダクターゼ活性を70℃以上の
温度下で100 unit/mg以上有するものであれば特に制限されないが、(3)のポリペプチドのアミノ酸配列において、通常20%以下、特に10%以下のアミノ酸が欠失、置換若しくは付加されたものであることが好ましい。
【0057】
(3)のポリペプチドにおいて、配列番号4に示すアミノ酸配列を含むポリペプチドとし
ては、例えば、配列番号4に示すアミノ酸配列の3倍以下の長さのポリペプチドが挙げられる。同様に、(4)のポリペプチドにおいて、配列表の配列番号4に示すアミノ酸配列に
おいて1若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含むポリペプチドとしては、例えば、当該アミノ酸配列の3倍以下の長さのポリペプチドが挙げられる。
(iii)チオレドキシンペルオキシダーゼ
本発明のチオレドキシンペルオキシダーゼとしては、以下の(5)又は(6)のアミノ酸配列を有するポリペプチドが挙げられる。
(5) 配列表の配列番号6に示すアミノ酸配列を含むポリペプチド。
(6) 配列表の配列番号6に示すアミノ酸配列において1若しくは2以上のアミノ酸が欠
失、置換若しくは付加されたアミノ酸配列を含み、かつ、70℃以上の温度下で、100 unit/mg以上のチオレドキシンペルオキシダーゼ活性を有するポリペプチド
(6)のポリペプチドのアミノ酸配列は、チオレドキシンペルオキシダーゼ活性を70 ℃
以上の温度下で100 unit/mg以上有するものであれば特に制限されないが、(5)のポリペ
プチドのアミノ酸配列において、通常20%以下、特に10%以下のアミノ酸が欠失、置換若しくは付加されたものであることが好ましい。
【0058】
(5)のポリペプチドにおいて、配列番号6に示すアミノ酸配列を含むポリペプチドとし
ては、例えば、配列番号6に示すアミノ酸配列の3倍以下の長さのポリペプチドが挙げら
れる。同様に、(6)のポリペプチドにおいて、配列表の配列番号6に示すアミノ酸配列に
おいて1若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含むポリペプチドとしては、例えば、当該アミノ酸配列の3倍以下の長さのポリペプチドが挙げられる。
【0059】
本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼは、これらのタンパク質等を生産する細菌を培養し、培養上清から精製することにより得られる。また、配列番号2、4または6のアミノ酸配列を基に、ペプチド化学合成により得られる。さらに、後述する本発明方法によっても得られる。
(2)本発明のDNA
(i)チオレドキシン
本発明のDNAは、前記説明した本発明の(1)又は(2)のポリペプチドをコードするDNAである。特に、 以下の(7)または(8)のDNAが好ましい。
(7) 配列表の配列番号1に示す塩基配列を含むDNA。
・ 配列表の配列番号1に示す塩基配列を含むDNAとストリンジェントな条件下でハイ
ブリダイズし、かつ、100℃で0.5 時間熱処理した場合に50%以上のチオレドキシン活性が残存するポリペプチドをコードするDNA。
【0060】
(8)のDNAは、100 ℃で0.5時間熱処理した場合に50%以上のチオレドキシン活性が残存するポリペプチドをコードするDNAであれば特に制限されないが、(7)のDNA配列において
、通常20%以下、特に10%以下のヌクレオチドが欠失、置換若しくは付加されたものであることが好ましい。
【0061】
(7)及び(8)のDNAにおいて、配列番号1に示す塩基配列を含むDNAとしては、例えば、配列番号1に示す塩基配列の3倍以下の長さのDNAが挙げられる。
(ii)チオレドキシンレダクターゼ
本発明のDNAは、前記説明した本発明の(3)又は(4)のポリペプチドをコードするDNAである。特に、 以下の(9)または(10)のDNAが好ましい。
(9) 配列表の配列番号3に示す塩基配列を含むDNA。
・ 配列表の配列番号3に示す塩基配列を含むDNAとストリンジェントな条件下でハイ
ブリダイズし、かつ、70℃以上の温度下で100unit/mg以上のチオレドキシンレダクター
ゼ活性を有するポリペプチドをコードするDNA。
【0062】
(10)のDNAは、70℃以上の温度下で100 unit/mg以上のチオレドキシンレダクターゼ活性を有するポリペプチドをコードするDNAであれば特に制限されないが、(9)のDNA配列に
おいて、通常20%以下、特に10%以下のヌクレオチドが欠失、置換若しくは付加されたものであることが好ましい。
【0063】
(9)及び(10)のDNAにおいて、配列番号3に示す塩基配列を含むDNAとしては、例えば、
配列番号3に示す塩基配列の3倍以下の長さのDNAが挙げられる。
(iii)チオレドキシンペルオキシダーゼ
本発明のDNAは、前記説明した本発明の(5)又は(6)のポリペプチドをコードするDNAである。特に、 以下の(11)または(12)のDNAが好ましい。
(11) 配列表の配列番号5に示す塩基配列を含むDNA。
(12) 配列表の配列番号5に示す塩基配列を含むDNAとストリンジェントな条件下でハイ
ブリダイズし、かつ、70 ℃以上の温度下で100unit/mg以上のチオレドキシンペルオキ
シダーゼ活性を有するポリペプチドをコードするDNA。
【0064】
(12)のDNAは、70℃以上の温度下で100 unit/mg以上のチオレドキシンペルオキシダーゼ比活性を有するポリペプチドをコードするDNAであれば特に制限されないが、(11)のDNA
配列において、通常20%以下、特に10%以下のヌクレオチドが欠失、置換若しくは付加されたものであることが好ましい。
【0065】
(11)及び(12)のDNAにおいて、配列番号5に示す塩基配列を含むDNAとしては、例えば、配列番号5に示す塩基配列の3倍以下の長さのDNAが挙げられる。
【0066】
本明細書において「ストリンジェントな条件」とは、例えば、通常のハイブリダイゼーション溶液中であれば68℃で行う条件が挙げられ、50%フォルムアミドを含むハイブリダイゼーション溶液中であれば42℃で行う条件が挙げられる。詳しくは、Molecular Cloning: A Laboratory Manual第2版第2巻に記載のサザンハイブリダイゼーションに用いられる条件が挙げられる。
【0067】
本発明のDNAは、例えばAeropyrum属、Sufolobus属、Thermoplasma属、Thermoproteus属、Mastigocladus属、Bacillus属、Synechococcus属、Thermus属等の好熱性細菌の染色体DNAのライブラリーからプローブを用いたハイブリダイゼーションにより単離することもできるが、配列表の配列番号1、3または5のDNA配列を基にPCR法により増幅することもできる。また、化学合成によっても取得できる。
【0068】
また、前記の(8)、(10)及び(12)の変異したDNAは、化学合成法、遺伝子工学的手法、突然変異誘発法などの公知の方法で作成することができる。遺伝子工学的手法としては、入手可能なチオレドキシン、チオレドキシンレダクターゼ又はチオレドキシンペルオキシダーゼに対して、エキソヌクレアーゼを用いたDNA欠失導入、リンカー導入、位置指定突然
変異導入、変異プライマーを用いたPCR法による塩基配列の改変などの公知の方法を挙げ
ることができる。
(3)本発明のベクター
本発明のベクターは、前記説明した本発明の各DNAを含有する組換えベクターである。
本発明のDNAが組み込まれるベクターは公知のものを広く利用でき、細菌用ベクターの他
、酵母用ベクター、動物細胞用ベクター等も利用できる。酵素の生産効率の点から、通常は細菌用ベクターを用いればよい。公知のベクターとしては、大腸菌ベクターのpBR322、pUC19、pKK233-2等、バチルス属細菌ベクターのpUB110、pC194、pE194、pTHT15、pBD16
等、酵母用ベクターYip5、Yrp17、Yep24等、動物細胞用ベクターpUC18、pUC19、M13mp18
等を例示できる。
(4)本発明の形質転換体
また、本発明の形質転換体は、前記説明した本発明の各組換えベクターを含有する形質転換体である。宿主としては、ベクターに応じて細菌、酵母、動物細胞等を利用できる。形質転換は、リン酸カルシウム法、プロトプラスト法、エレクトロポレーション法、スフェロプラスト法、酢酸リチウム法、リポフェクション法、マイクロインジェクション法等の公知の方法で行うことができる。これらの公知の方法の中から、宿主の種類に応じて選択すればよい。
(5)本発明のタンパク質または酵素の製造方法
本発明のチオレドキシン、チオレドキシンレダクターゼまたはチオレドキシンペルオキシダーゼの製造方法は、本発明の各形質転換体を培養し、この形質転換体から各タンパク質等を精製する方法である。
【0069】
これらの細菌が生産した酵素は、遠心分離、塩析、溶媒沈殿法、透析法、限外ろ過法、ゲルろ過法、イオン交換クロマトグラフィー法、アフィニティクロマトグラフィー法、逆相高速液体クロマトグラフィー法等の公知の蛋白精製方法を組み合わせて精製することができる。本発明の酵素が細胞内またはペリプラズム内に生産される場合には超音波処理、界面活性剤処理等の公知の方法で細胞を破壊した後に前記の精製を行えばよい。
【0070】
また、本発明の耐熱性タンパク質等を精製するにあたっては、目的耐熱性タンパク質等を10〜30分間程度(特に20分間程度)インキュベートした場合に通常60%以上、特に80%以上活性が残存し、かつ、当該耐熱性タンパク質等を生産する宿主の生育限界温度より通常10℃以上、特に15℃以上高い温度で、被精製タンパク質溶液を通常10〜120分間程度、
特に10〜30分間程度インキュベートすることが好ましい。これにより、目的タンパク質を殆ど失活させることなく、宿主が生産する夾雑タンパク質を変性、失活させることができる。この加熱処理工程後に、タンパク質溶液を、それには限定されないが、例えば15000rpm程度で20分間程度遠心することにより変性した夾雑タンパク質を沈殿させることができる。この加熱処理工程は、精製のいずれの段階で行ってもよい。
【0071】
このような加熱処理工程は、本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの精製に限らず、いずれの耐熱性タンパク質の精製においても行うことができ、これにより耐熱性タンパク質の精製度を飛躍的に向上させることができる。
実施例
(1)超好熱性古細菌Aeropyrum pernix K1株のチオレドキシン遺伝子、チオレドキシン
レダクターゼ遺伝子およびチオレドキシンペルオキシダーゼ遺伝子と他の対応酵素遺伝子との相同性
コンピューターによる相同性検索の結果、配列表の配列番号1に示す本発明のチオレドキシン遺伝子の塩基配列とSalmonella typhimurium LT2株のチオレドキシン遺伝子の塩基配列との間に31%の相同性が見出された。また、配列表の配列番号3に示す本発明のチオレドキシンレダクターゼ遺伝子の塩基配列とSulfolobus salfataricusのチオレドキシン
レダクターゼ遺伝子との間に48%の相同性が見出された。また、配列表の配列番号5に示す本発明のチオレドキシンペルオキシダーゼ遺伝子の塩基配列とSulfolobus tokodaiのチオレドキシンペルオキシダーゼ遺伝子との間に 62%の相同性が見出された。
(2)Aeropyrum pernix K1株のチオレドキシン遺伝子、チオレドキシンレダクターゼ遺
伝子及びチオレドキシンペルオキシダーゼ遺伝子のクローニング
<染色体DNAの調製>
バクトマリン培地(Difco)37.4g とNa2S2O3-5H2O 1.0g を水1リットルに溶かした後、pHを7.0-7.2に調整することにより培地を調製した。超好熱性古細菌Aeropyrum pernix K1株(理化学研究所においてJCM9820として登録)をこの培地に接種し、90℃で3日間振盪培養した。培養液を5,000rpmで10分間遠心分離することにより集菌した。
【0072】
菌体を10mM トリス(pH 7.5)−1mM EDTA 溶液で2回洗浄後インサートアガロース(InCert Agarose、FMC社製)ブロック中に封入した。このブロックを1%N−ラウロイルザルコシン−1mg/ml プロテアーゼK溶液中で処理することにより、染色体DNAがアガロースブロ
ック中に分離調製された。インサートアガロースブロックを用いた染色体DNAの分離条件
は、アガロースブロックの添付マニュアルに従った。
<遺伝子の増幅・組み換えベクターの構築>
(i)チオレドキシン
配列表の配列番号1の塩基配列を含むDNAをPCR法により増幅した。PCR条件は、PCRキットの添付マニュアルに従った。5’末端側に対応するプライマーとしては、配列表の配列
番号1に示すDNA配列において、塩基番号1から始まる(すなわち開始コドンから始まる
)オリゴヌクレオチドプライマーを合成した。また、3’末端側に対応するプライマーとしては、Aeropyrum pernix K1株の染色体DNAにおいて配列番号1の塩基配列の3’末端より下流域に対応するプライマーであって、増幅されたDNA中に制限酵素のBamHIサイトが生じるようなプライマーを合成した。PCR反応後、DNAを制限酵素のBamHIで37℃で3時間
処理することにより完全分解した。次いで、チオレドキシン遺伝子を精製カラムキットを用いて精製した。
【0073】
次いで、チオレドキシン遺伝子を組み込んだベクターを構築する目的で、ベクターのpET-8c(Novagen社製)を制限酵素NcoIで切断し、精製した後、T4 DNA ポリメラーゼを用いて平滑末端化した。精製したプラスミドを制限酵素BamHIで切断・精製した。次いで、BamHIで切断されたプラスミドpET-8cと BamHIで切断された上記のチオレドキシン遺伝子とをT4リガーゼを用いて16℃で、16時間反応させることにより連結した。連結したDNAを
用いて、大腸菌(E. coli) XL2-BlueMRF’ 株(Novagen社製)のコンピテントセルを形質
転換した。形質転換体は、0.05 mg/ml アンピシリンを含むLB寒天プレート上でのコロニ
ー形成を指標に選択した。形質転換体からシステイン合成酵素遺伝子含有プラスミドをアルカリ法で抽出した。
(ii)チオレドキシンレダクターゼ
前記説明したチオレドキシンのクローニング方法において、5’末端側に対応するPCRプライマーとして、配列番号3に示すDNA配列において塩基番号1から始まるオリゴヌクレ
オチドを使用し、3’末端側に対応するPCRプライマーとして、Aeropyrum pernix K1株
の染色体DNAにおいて配列番号3の塩基配列の3’末端より下流側に対応するプライマーであって、増幅されたDNA中にBamHIサイトが生じるようなプライマーを合成し、使用した。その他は、チオレドキシンの場合と同様にして、Aeropyrum pernix K1株由来のチオレ
ドキシンレダクターゼを組み込んだプラスミドpET-8cを保持する大腸菌L2-BlueMRF’株を得た。
(iii)チオレドキシンペルオキシダーゼ
前記説明したチオレドキシンのクローニング方法において、5’末端側に対応するPCRプライマーとして、配列番号5に示すDNA配列において塩基番号1から始まるオリゴヌクレ
オチドを使用し、3’末端側に対応するPCRプライマーとして、Aeropyrum pernix K1株
の染色体DNAにおいて配列番号5の塩基配列の3’末端より下流側に対応するプライマーであって、増幅されたDNA中にBamHIサイトが生じるようなプライマーを合成し、使用した。その他は、チオレドキシンの場合と同様にして、Aeropyrum pernix K1株由来のチオレ
ドキシンペルオキシダーゼを組み込んだプラスミドpET-8cを保持する大腸菌L2-BlueMRF’株を得た。
(3)チオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの生産
<形質転換体の作製>
1.5ml容チューブ内に、大腸菌(E. coli )Rosetta (DE3)株( Novagen社製)のコンピテントセル0.04ml(2,000,0000cfu/mg)と、上記調製したチオレドキシン遺伝子含有プ
ラスミド、チオレドキシンレダクターゼ遺伝子含有プラスミドまたはチオレドキシンペルオキシダーゼ遺伝子含有プラスミドのDNA溶液0.003ml(プラスミドDNA 8.4ng)を加え
氷中に30分間放置した後、42℃で、30秒間ヒートショックを与えた。次いで、チューブ内にSOCmedium を0.25ml加え、37℃で1時間震盪培養した。次いで、アンピシリン及
びクロラムフェニコールを含むLB寒天プレートに塗布し、37度で一晩培養することにより形質転換体を得た。
<タンパク質または酵素の精製>
(i)チオレドキシン
チオレドキシン遺伝子を含むプラスミドを保持する形質転換体をアンピシリン及びクロラムフェニコールを含むNZCYM培地に接種し、600nmにおける吸光度が0.5に達するまで、37℃で培養した後、プラスミドの発現量を高めるためにIPTG(Isopropyl-b-D-thiogalactopyranoside)を加えさらに4時間培養した。培養液を8,000rpmで10min遠心分
離することにより集菌した。
【0074】
集菌した菌体4 gに、1mM DTT、1mM EDTA を含む50mM Tris−HCl(pH8.0)を加え、
菌体を90Wの出力で5分間超音波破砕 した。破砕した菌液を 15,000rpmで30分間遠心分離し、上清を採取した。
【0075】
夾雑蛋白を沈殿させて除去する目的で、この上清を85℃で30分間加熱した後、15,000rpmで30分間遠心分離し、上清を採取した。上清を1 mM EDTAを含む50 mM(Tris - HCl
)緩衝液(pH8.0)で透析し、同緩衝液で平衡化した陰イオン交換樹脂のHitrapQ(ファルマシア社製)カラムにアプライしてイオン交換クロマトグラフィーを行った。さらに、活性画分を150 mM NaCl を含む50 mMリン酸ナトリウム緩衝液(pH7.0)中で透析し、同緩衝液で平衡化した。ゲル濾過材のSuperdex 200(ファルマシア社製)カラムにア
プライしてゲル濾過クロマトグラフィーを行った。活性画分には、SDS−ポリアクリルア
ミドゲル電気泳動により単一バンドを与える均一標品が含まれていた。
【0076】
また、ゲルろ過クロマトグラフィーの結果、本酵素の分子量は約37kDaであった。
(ii)チオレドキシンレダクターゼ
チオレドキシンレダクターゼ遺伝子を含むプラスミドを保持する形質転換体をチオレドキシン遺伝子の場合と同様にして培養、集菌した。
【0077】
集菌した菌体4 gに、1mM DTT、1mM EDTA を含む50mM Tris−HCl(pH8.5)を加え、
菌体を90Wの出力で5分間超音波破砕した。破砕した菌液を 15,000rpmで30分間遠心分離し、上清を採取した。
【0078】
夾雑蛋白を沈殿させて除去する目的で、この上清を85℃で30分間加熱した後、15,000rpmで20分間遠心分離し、上清を採取した。上清を1 mM EDTAを含む50 mM Tris - HCl緩衝液(pH8.0)で透析し、同緩衝液で平衡化した陰イオン交換樹脂のHitrapQ(ファルマシア社製)カラムにアプライしてイオン交換クロマトグラフィーを行った。さらに、活性画分を150mM NaClを含む50 mMリン酸ナトリウム緩衝液(pH7.0 )中で透析し、同緩
衝液で平衡化した。ゲル濾過材のSuperdex 200(ファルマシア社製)カラムにアプライ
してゲル濾過クロマトグラフィーを行った。活性画分には、SDS−ポリアクリルアミドゲ
ル電気泳動により単一バンドを与える均一標品が含まれていた。
【0079】
また、ゲルろ過クロマトグラフィーの結果、本酵素の分子量は約37kDaであった。
(iii)チオレドキシンペルオキシダーゼ
チオレドキシンペルオキシダーゼ遺伝子を含むプラスミドを保持する形質転換体をチオレドキシン遺伝子の場合と同様にして培養、集菌、超音波破砕、遠心による上清採取、加熱処理、遠心による上清採取を行った。
【0080】
得られた上清を1mM EDTAを含む50 mM(Tris - HCl )緩衝液(pH8.0)で透析し、同緩衝液で平衡化した陰イオン交換樹脂のHitrapQ(ファルマシア社製)カラムにアプライ
してイオン交換クロマトグラフィーを行った。さらに、活性画分を150mM NaCl を含む50mM リン酸ナトリウム緩衝液(pH7.0)中で透析し、同緩衝液で平衡化した。ゲル
濾過材のSephacryl S-100(ファルマシア社製)カラムにアプライしてゲル濾過クロマトグラフィーを行った。活性画分には、SDS−ポリアクリルアミドゲル電気泳動により単一
バンドを与える均一標品が含まれていた。
【0081】
また、ゲルろ過クロマトグラフィーの結果、本酵素の分子量は約29kDaであった。
<活性測定法>
本発明において、精製工程における目的タンパク質又は酵素の検出、至適温度の検討および安定性の検討における目的タンパク質又は酵素の活性検出は、以下の方法で行った。(i)チオレドキシン
チオレドキシンの活性測定は、還元型チオレドキシンによるインスリンサブユニット間のジスルフィド結合を還元する活性を測定するMolmgrenらの方法(Method in Enzymology 1993)に従った。
【0082】
すなわち、先ず0.4mM DTT及び0.4mg/mlウシ血清アルブミンを含む100mM Tris−塩酸バッファー(pH7.5)で37℃、15分間チオレドキシンサンプルを前処理して還元型チオレド
キシンとした。次いで、基質として1mg/mlのウシ膵臓インスリン(シグマ社製)を含む100mMTris−塩酸バッファー(pH7.5)に、チオレドキシンを200ng添加し、インスリンの還元分解による650nmにおける吸光度の上昇を室温で20分間測定した。
(ii)チオレドキシンレダクターゼ
基質としての1mMジチオビス2-ニトロベンゾイックアシッド(DTNB)0.2mM NADPH、1mM
EDTAを含む50mMリン酸カリウム緩衝液(pH7.0)中に、精製チオレドキシンレダクター
ゼ 2μgを添加し、NADPHの濃度の指標である340nmにおける吸光度の減少速度を5 分間測定することによりチオレドキシンレダクターゼの活性を測定した。
【0083】
精製工程では、70℃の緩衝液中で活性を測定した。
(iii)チオレドキシンペルオキシダーゼ
基質としての1mM過酸化水素、0.2mM NADPH、0.1μMの精製チオレドキシンレダクター
ゼ、5μMの精製チオレドキシンを含む50mMリン酸カリウム緩衝液(pH7.0)中に、精製チオレドキシンペルオキシダーゼ2 μgを添加し、NADPHの濃度の指標である340nmにおける
吸光度の減少速度を5 分間測定することによりチオレドキシンペルオキシダーゼの活性を測定した。
【0084】
精製工程では、70℃の緩衝液中で活性を測定した。
(4)タンパク質・酵素の性質
前記の方法により得られたAeropyrum pernix K1株由来のチオレドキシン、チオレドキ
シンレダクターゼ及びチオレドキシンペルオキシダーゼの諸特性を評価した。
<至適温度>
前述したチオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの活性測定方法において、酵素反応を行う緩衝液の温度を20〜90℃の範囲で変化させて、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの各活性を測定した。
【0085】
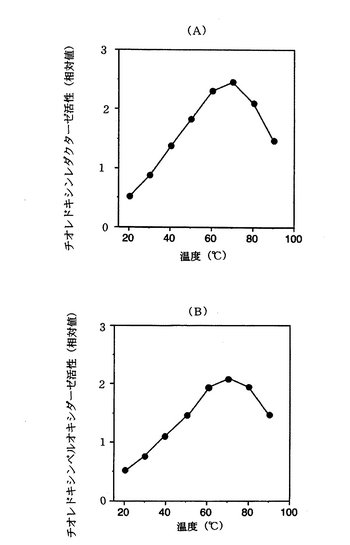
この結果、図1(A)及び(B)に示すように、チオレドキシンレダクターゼの至適温度は70℃(図1(A))、チオレドキシンペルオキシダーゼの至適温度は70℃(図1(B))であった。<耐熱性>
2 mM EDTAを含む50mMリン酸ナトリウム緩衝液(pH7.0)中に、チオレドキシンを0.1 mg/ml濃度になるように添加した酵素溶液サンプルを用意し、100℃でインキュベ
ートして、経時的に残存活性を測定した。チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼについても同様にして、インキュベートし、経時的に残存活性を測定した。
【0086】
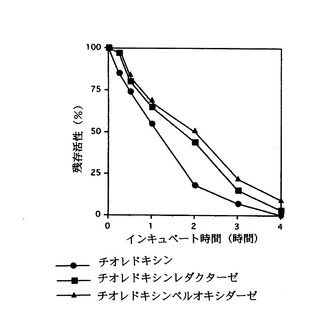
この結果、図2に示すように、チオレドキシンは、100℃でインキュベートした場合に1時間後に55%程度の活性が残り、チオレドキシンレダクターゼは、100℃でインキュベー
トした場合に1時間後に65%程度の活性が残り、チオレドキシンぺルオキシダーゼは、100℃でインキュベートした場合に1時間後に70%程度の活性が残った。
【図面の簡単な説明】
【0087】
【図1】(A)及び(B)は、それぞれ本発明の1実施例であるチオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの至適温度を示すグラフである。
【図2】本発明の1実施例であるチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの100℃における熱安定性を示すグラフである。
【技術分野】
【0001】
本発明は、高温下で機能できるチオレドキシン、チオレドキシンレダクターゼおよびチオレドキシンペルオキシダーゼ、これらの酵素をコードする各DNA、これらのDNAを組み込んだ各ベクター、これらのベクターで形質転換された各形質転換体、並びに、これらの各形質転換体を用いた耐熱性チオレドキシン、耐熱性チオレドキシンレダクターゼ及び耐熱性チオレドキシンペルオキシダーゼの生産方法に関する。
【背景技術】
【0002】
チオレドキシンは、大腸菌、酵母、高等植物及び高等動物等に普遍的に存在する分子量1万〜1.3万の電子伝達タンパク質である。チオレドキシンは、2個のアミノ酸を挟んで
2個のシステイン残基が存在する部位(―Cys―X―X―Cys―)を活性中心として有し、
ジチオールの可逆的な酸化還元反応を通して、生体内の酸化還元状態を制御していると考えられている(Laurent, T.C.,Moor,E.C.,&Reinchard,P.(1964)J.Biol.Chem.239,3436-3444;Holmgren,A.(1989)Ann.Rev.Biochem.54,237-271;Holmgren,A.(1989)J.Biol.Chem.264,13963-13966;Buchanan,B.B.,Schurmann,P.,Decottignies,P.&Lozano,R.M.(1994)Arch.Biochem.Biophys.314,257-260)。チオレドキシンは、活性中心の2個のシステインのチオール基がジスルフィドとなった酸化型またはチオール基となった還元型のどちらの状態でも存在している。
【0003】
チオレドキシンの生体内での機能の1例としてタンパク質の分子内または分子間S−S結合の開裂が挙げられる。この際、還元型チオレドキシンは、タンパク質のジスルフィドをジチオールに還元するのに伴い、酸化されて酸化型チオレドキシンとなる。生成した酸化型チオレドキシンは、チオレドキシンレダクターゼ及びNADPHにより還元されて還元型チ
オレドキシンに戻る。
【0004】
チオレドキシンの生体内での機能の他の例として過酸化水素や過酸化脂質などの過酸化物の消去が挙げられる。この際、還元型チオレドキシンは、過酸化物(ROOH)を水酸化物(ROH)に還元するとともに、酸化されて酸化型チオレドキシンとなる。この反応はチオ
レドキシンペルオキシダーゼにより行われる。酸化型チオレドキシンは、チオレドキシンレダクターゼ及びNADPHにより還元されて還元型チオレドキシンに戻る。
【0005】
また、チオレドキシンの生体内での機能の更に他の例として、紫外線照射による細胞障害防止機能、転写因子の制御機能等も挙げられる。
【0006】
このような機能を有するチオレドキシンは、例えば、酸化型システインの状態で活性を示す消化酵素インヒビタータンパク質の還元によるインヒビター活性の阻害、蛇毒タンパク質のシステイン残基間のS−S結合の除去による無毒化、紫外線照射による皮膚の炎症の予防等の目的で、医薬品として用いることが提案されている。また、食物アレルゲンの除去等の目的で食品に添加して用いたり、乾燥や紫外線等に起因する酸化ストレスによる肌荒れの改善剤等の目的で化粧品として用いることが提案されている(特開2001-288103号
公報、特表2001-520027号公報、特開2000-103743号公報)。
【0007】
しかし、固体状態または半固体状態の医薬品、食品及び化粧品等ではろ過滅菌が困難であるため、通常加熱滅菌が行われるところ、従来知られているチオレドキシンは、耐熱性が低い。従って、従来の易熱性のチオレドキシンを医薬品等として用いたり、食品等に添加して用いる場合には、高温での滅菌を行うことができないという難点がある。
【0008】
また、一般に溶質の水に対する溶解度は温度と共に上昇するため、高温下で機能できる耐熱性のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼを用いることができれば、タンパク質中のジスルフィドの還元による還元型タンパク質の合成反応、過酸化水素等の活性酸素の除去反応等を、高温下で調製した高濃度の基質溶液に該酵素を作用させることにより効率よく行うことができる。
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明は、耐熱性のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼ、これらの酵素をコードするDNA、これらのDNAを組み込んだベクター、これらのベクターで形質転換された形質転換体、並びに、これらの形質転換体を用いた耐熱性チオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの効率的な生産方法を提供することを主目的とする。
【課題を解決するための手段】
【0010】
前記目的を達成するために、本発明者は、90〜100℃という高温下で生育できる超好熱性古細菌に着目した。古細菌は、真核生物及び原核生物のいずれにも属さない第3の生物群に属する生物である。また、太古の地球の生物の末裔と考えられ、常温環境に対する進化・適用を免れた特殊な生物である。
【0011】
本発明者は、このような超好熱性古細菌由来のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼを初めて単離した。また、これらのタンパク質または酵素は、従来公知のチオレドキシン等に比べて非常に耐熱性が高く、また常温下での安定性にも優れること、有機溶媒の存在下でも活性を示すことを見出した。
【0012】
さらに、これらのタンパク質又は酵素をコードするDNA配列を決定し、ベクターに組み
込んで、これらを効率よく生産することに成功した。
【0013】
前記知見に基づき、本発明者は、以下の各項のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼ等を提供する。
【0014】
項1. 100℃で0.5時間熱処理した場合に50%以上活性が残存する耐熱性チオレドキシン。
【0015】
項2. 超好熱性古細菌Aeropyrum pernix由来の項1に記載の耐熱性チオレドキシン。
【0016】
項3. 以下の(1)または(2)のポリペプチド。
・ 配列表の配列番号2に示すアミノ酸配列を含むポリペプチド。
・ 配列表の配列番号2に示すアミノ酸配列において1若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含み、かつ、100℃で0.5時間熱処理した場合に50%以上チオレドキシン活性が残存するポリペプチド。
【0017】
項4. 以下の(3)または(4)のDNA。
・ 配列表の配列番号1に示す塩基配列を含むDNA。
・ 配列表の配列番号1に示す塩基配列を含むDNAとストリンジェントな条件下でハイ
ブリダイズし、かつ、100℃で0.5時間熱処理した場合に50%以上活性が残存する耐熱性チオレドキシンをコードするDNA。
【0018】
項5. 項3に記載のポリペプチドをコードするDNA。
【0019】
項6. 項3または5に記載のDNAを含有するベクター。
【0020】
項7. 項6に記載のベクターを含有する形質転換体。
【0021】
項8. 項7の形質転換体を培養し、該形質転換体から耐熱性チオレドキシンを分離および精製する耐熱性チオレドキシンの生産方法。
【0022】
項9. 70℃以上の温度下で、100 unit/mg以上の活性を有する耐熱性チオレドキシ
ンレダクターゼ。
【0023】
項10. 100℃で0.5時間熱処理した場合に50%以上活性が残存する項9に記載の耐熱性チオレドキシンレダクターゼ。
【0024】
項11. 至適温度が70℃以上である項9に記載の耐熱性チオレドキシンレダクターゼ。
【0025】
項12. 超好熱性古細菌Aeropyrum pernix由来の項9に記載の耐熱性チオレドキシンレダクターゼ。
【0026】
項13. 以下の(5)または(6)のポリペプチド。
・ 配列表の配列番号4に示すアミノ酸配列を含むポリペプチド。
・ 配列表の配列番号4に示すアミノ酸配列において1若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含み、かつ、70℃以上の温度下で、100 unit/mg以上のチオレドキシンレダクターゼ活性を有するポリペプチド。
【0027】
項14. 以下の(7)または(8)のDNA。
・ 配列表の配列番号3に示す塩基配列を含むDNA。
・ 配列表の配列番号3に示す塩基配列を含むDNAとストリンジェントな条件下でハイ
ブリダイズし、かつ、70℃以上の温度下で、100 unit/mg以上の活性を有する耐熱性チオレドキシンレダクターゼをコードするDNA。
【0028】
項15. 項13に記載のポリペプチドをコードするDNA。
【0029】
項16. 項13または15に記載のDNAを含有するベクター。
【0030】
項17. 項16に記載のベクターを含有する形質転換体。
【0031】
項18. 項17の形質転換体を培養し、該形質転換体から耐熱性チオレドキシンレダクターゼを分離および精製する耐熱性チオレドキシンレダクターゼの生産方法。
【0032】
項19. 70℃以上の温度下で、100unit/mg以上の活性を有する耐熱性チオレドキシ
ンペルオキシダーゼ。
【0033】
項20. 100 ℃で0.5時間熱処理した場合に50%以上活性が残存する項19に記載の耐熱性チオレドキシンペルオキシダーゼ。
【0034】
項21. 至適温度が70 ℃以上である項19に記載の耐熱性チオレドキシンペルオキシダーゼ。
【0035】
項22. 超好熱性古細菌Aeropyrum pernix由来の項19に記載の耐熱性チオレドキシンペルオキシダーゼ。
【0036】
項23. 以下の(9)または(10)のポリペプチド。
・ 配列表の配列番号6に示すアミノ酸配列を含むポリペプチド。
・ 配列表の配列番号6に示すアミノ酸配列において1若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含み、かつ、70℃以上の温度下で、100 unit/mg以上のチオレドキシンペルオキシダーゼ活性を有するポリペプチド。
【0037】
項24. 以下の(11)または(12)のDNA。
・ 配列表の配列番号5に示す塩基配列を含むDNA。
・ 配列表の配列番号5に示す塩基配列を含むDNAとストリンジェントな条件下でハイ
ブリダイズし、かつ、70 ℃以上の温度下で、100 unit/mg以上の活性を有する耐熱性チオレドキシンペルオキシダーゼをコードするDNA。
【0038】
項25. 項23に記載のポリペプチドをコードするDNA。
【0039】
項26. 項23または25に記載のDNAを含有するベクター。
【0040】
項27. 項26に記載のベクターを含有する形質転換体。
【0041】
項28. 項27の形質転換体を培養し、該形質転換体から耐熱性チオレドキシンペルオキシダーゼを分離および精製する耐熱性チオレドキシンペルオキシダーゼの生産方法。
【0042】
項29. 精製すべき耐熱性タンパク質を10〜30分間インキュベートした場合に60%以上活性が残存し、かつ、耐熱性タンパク質を生産する宿主の生育限界温度より10℃以上高い温度で、被精製蛋白溶液を10〜120分間インキュベートする加熱工程を含む耐熱性タン
パク質の精製方法。
【発明の効果】
【0043】
本発明によると優れた耐熱性を有するチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼを提供することができる。
従って、これらを医薬品や化粧品として用いたり、医薬品、食品及び化粧品に添加して用いる場合には、高温での殺菌を行うことができる。
詳述すれば、本発明の耐熱性チオレドキシンは、消化酵素インヒビターの阻害、蛇毒及びサソリ毒等の酸化による無毒化、紫外線照射による皮膚の炎症の治療及び予防等の目的で医薬品として用いたり、食物アレルゲンの除去等の目的で食品に添加して用いたり、乾燥や紫外線等に起因する酸化ストレスによる肌荒れの改善等の目的で化粧品に添加して用いたりすることができる。また、本発明のチオレドキシンレダクターゼは、チオレドキシン併用して医薬品等として用いることにより、チオレドキシンを活性な還元型チオレドキシンに保つことができる。また、本発明のチオレドキシンペルオキシダーゼは、医薬品、食品及び化粧品等の酸化防止の目的で、チオレドキシンとともにこれらに添加して用いることができる。これらの場合に、本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼは、耐熱性であることから加熱殺菌をすることができる。
【0044】
また、本発明のチオレドキシン及びチオレドキシンレダクターゼは、タンパク質中のシスチンの還元による還元型タンパク質の合成反応等を高温下で調製した高濃度の基質溶液に対して効率よく行うことができる。
【0045】
また、本発明のチオレドキシン及びチオレドキシンペルオキシダーゼは、過酸化水素等の活性酸素の除去反応等を、高温下で調製した高濃度の基質溶液に対して効率よく行うことができる。
【0046】
また、本発明のチオレドキシンペルオキシダーゼは、過酸化物と反応して発色するセンサーとして用いたり、ウェスタンブロット法において抗体に結合させるペルオキシダーゼとして用いることができる。この場合には、比較的高温下で酵素反応を行うことができるため、夾雑タンパク質の影響が少なくでき、検出感度が向上する。
【0047】
また、本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼは、常温での安定性にも優れるため、比較的長い期間保存することができ、また繰り返し使用にも耐えることができる。
【0048】
また、一般的にタンパク質または酵素は、有機溶媒の存在により失活し易いが、本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼは、有機溶媒に対しても安定である。従って、有機溶媒中あるいは有機溶媒を含む水溶液中で、反応を行うことができる。その結果、水溶液には溶解し難い物質に対しても作用させることができ、反応の適用対象が広がる。
【発明を実施するための最良の形態】
【0049】
(1)本発明のタンパク質・酵素
耐熱性
(i)チオレドキシン
本発明の耐熱性チオレドキシンは、非常に耐熱性が優れるタンパク質であり、100℃で0.5 時間熱処理した場合に50%以上活性が残存するタンパク質である。100℃で0.5 時間熱処理した場合に60%以上活性が残存するタンパク質であることがより好ましい。
【0050】
本明細書において、チオレドキシンの活性は、実施例に記載の方法で測定した値である。
(ii)チオレドキシンレダクターゼ
本発明の耐熱性チオレドキシンレダクターゼは、非常に耐熱性に優れる酵素であり、70
℃以上の温度下で、100 unit/mg以上の活性を有する酵素である。70℃以上の温度下で150 unit/mg以上の活性を有する酵素であることがより好ましい。活性を示す最高温度は、通常85 ℃程度である。また、酵素反応を行う緩衝液の種類によっても異なるが、酵素反応の初速度を測定した場合の至適温度が70℃以上の酵素であることが好ましい。
【0051】
また、本発明のチオレドキシンレダクターゼは、100℃で0.5 時間熱処理した場合に50
%以上活性が残存する酵素であることが好ましく、100℃で0.5時間熱処理した場合に60%以上活性が残存する酵素であることがより好ましい。
【0052】
本明細書において、チオレドキシンレダクターゼの活性は、実施例に記載の方法で測定した値である。
(iii)チオレドキシンペルオキシダーゼ
本発明の耐熱性チオレドキシンペルオキシダーゼは、非常に耐熱性に優れる酵素であり、70℃以上の温度下で、100 unit/mg以上の活性を有する酵素である。70℃以上の温度下で150 unit/mg以上の活性を有する酵素であることがより好ましい。活性を示す最高温度は、通常85℃程度である。また、酵素反応を行う緩衝液の種類によっても異なるが、酵素反応の初速度を測定した場合の至適温度が70℃以上の酵素であることが好ましい。
【0053】
本発明のチオレドキシンペルオキシダーゼは、100℃で0.5時間熱処理した場合に50 %
以上活性が残存する酵素であることが好ましく、100 ℃で0.5 時間熱処理した場合に60%以上活性が残存する酵素であることがより好ましい。
【0054】
本明細書において、チオレドキシンペルオキシダーゼの活性は、実施例に記載の方法で測定した値である。
常温での安定性
(i)チオレドキシン
本発明のチオレドキシンは、常温下での安定性にも優れる。例えば、本発明のチオレドキシンは、30℃の50 mMリン酸カリウム緩衝液(pH7.0)中で、12 時間インキュベート
した場合に、90%以上の活性が残存する酵素であることが好ましい。
(ii)チオレドキシンレダクターゼ
本発明のチオレドキシンレダクターゼは、常温下での安定性にも優れる。例えば、本発明のチオレドキシンレダクターゼは、30℃の50mMリン酸カリウム緩衝液(pH7.0)中で、12時間インキュベートした場合に、90%以上の活性が残存する酵素であることが好ましい
。
(iii)チオレドキシンペルオキシダーゼ
本発明のチオレドキシンペルオキシダーゼは、常温下での安定性にも優れる。例えば、本発明のチオレドキシンペルオキシダーゼは、30℃の50mMリン酸カリウム緩衝液(pH7.0
)中で、12時間インキュベートした場合に、90%以上の活性が残存する酵素であることが好ましい。
有機溶媒耐性
本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼは、有機溶媒に耐性である。例えば、エタノール、ブタノール、テトラヒドロフラン、酢酸エチル等の有機溶媒を30容量%以上含む緩衝液中で活性を示すタンパク質または酵素(以下、「タンパク質等」という。)であることが好ましい。本発明のこれらのタンパク質等が活性を示し得る、緩衝液中の有機溶媒の容量比率の上限は、当該タンパク質等が沈殿しない範囲である。なお、有機溶媒に対する耐性は、古細菌由来の酵素に特有の性質である。
活性
本発明のチオレドキシンは、還元型チオレドキシンの形で、種々のタンパク質のシステイン残基のジスルフィドをジチオールに還元できるタンパク質である。また本発明のチオレドキシンは、還元型チオレドキシンの形で、過酸化水素等を還元できるタンパク質であることが好ましい。
本発明のチオレドキシンレダクターゼは、酸化型チオレドキシンを、そのジスルフィドをジチオールに還元することにより還元型チオレドキシンに変換できる酵素である。また、本発明のチオレドキシンレダクターゼは、酸化型グルタチオン等を還元できる酵素であることが好ましい。これらの還元反応には、通常、NADPH、NADH、FADH及びFADH2等の補酵素が用いられる。補酵素は、チオレドキシンレダクターゼ1分子に対して、通常100〜100000モル程度の比率で用いることが好ましい。
本発明のチオレドキシンペルオキシダーゼは、還元型チオレドキシンの存在下に、過酸化水素を水に還元できる酵素である。また本発明のチオレドキシンペルオキシダーゼは、その他の活性酸素も還元できる酵素であることが好ましい。
生産細菌
また、本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼとしては、好熱性細菌、例えばAeropyrum属、Sufolobus属、Thermoplasma属、Thermoproteus属、Mastigocladus属、Bacillus属、Synechococcus属、Thermus属等の細菌により生産されたものが挙げられる。特に、超好熱性古細菌Aeropyrum pernixにより生産されたものが好ましい。
アミノ酸配列
(i)チオレドキシン
本発明のチオレドキシンとしては、以下の(1)又は(2)のアミノ酸配列を有するポリペプチドが挙げられる。
(1) 配列表の配列番号2に示すアミノ酸配列を含むポリペプチド。
・ 配列表の配列番号2に示すアミノ酸配列において1若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含み、かつ、100 ℃で0.5 時間熱処理した場合に50%以上チオレドキシン活性が残存するポリペプチド。
【0055】
(2)のポリペプチドのアミノ酸配列は、100 ℃で0.5時間熱処理した場合に50 %以上
チオレドキシン活性が残存するものであれば特に制限されないが、(1)のポリペプチドの
アミノ酸配列において、通常20%以下、特に10%以下のアミノ酸が欠失、置換若しくは付加されたものであることが好ましい。
【0056】
(1)のポリペプチドにおいて、配列番号2に示すアミノ酸配列を含むポリペプチドとし
ては、例えば、配列番号2に示すアミノ酸配列の3倍以下の長さのポリペプチドが挙げられる。同様に、(2)のポリペプチドにおいて、配列番号2に示すアミノ酸配列において1
若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含むポリペプチドとしては、例えば、当該アミノ酸配列の3倍以下の長さのポリペプチドが挙げられる。
(ii)チオレドキシンレダクターゼ
本発明のチオレドキシンレダクターゼとしては、以下の(3)又は(4)のアミノ酸配列を有するポリペプチドが挙げられる。
(3) 配列表の配列番号4に示すアミノ酸配列を含むポリペプチド。
(4) 配列表の配列番号4に示すアミノ酸配列において1若しくは2以上のアミノ酸が欠
失、置換若しくは付加されたアミノ酸配列を含み、かつ、70 ℃以上の温度下で、100 unit/mg以上のチオレドキシンレダクターゼ活性を有するポリペプチド
(4)のポリペプチドのアミノ酸配列は、チオレドキシンレダクターゼ活性を70℃以上の
温度下で100 unit/mg以上有するものであれば特に制限されないが、(3)のポリペプチドのアミノ酸配列において、通常20%以下、特に10%以下のアミノ酸が欠失、置換若しくは付加されたものであることが好ましい。
【0057】
(3)のポリペプチドにおいて、配列番号4に示すアミノ酸配列を含むポリペプチドとし
ては、例えば、配列番号4に示すアミノ酸配列の3倍以下の長さのポリペプチドが挙げられる。同様に、(4)のポリペプチドにおいて、配列表の配列番号4に示すアミノ酸配列に
おいて1若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含むポリペプチドとしては、例えば、当該アミノ酸配列の3倍以下の長さのポリペプチドが挙げられる。
(iii)チオレドキシンペルオキシダーゼ
本発明のチオレドキシンペルオキシダーゼとしては、以下の(5)又は(6)のアミノ酸配列を有するポリペプチドが挙げられる。
(5) 配列表の配列番号6に示すアミノ酸配列を含むポリペプチド。
(6) 配列表の配列番号6に示すアミノ酸配列において1若しくは2以上のアミノ酸が欠
失、置換若しくは付加されたアミノ酸配列を含み、かつ、70℃以上の温度下で、100 unit/mg以上のチオレドキシンペルオキシダーゼ活性を有するポリペプチド
(6)のポリペプチドのアミノ酸配列は、チオレドキシンペルオキシダーゼ活性を70 ℃
以上の温度下で100 unit/mg以上有するものであれば特に制限されないが、(5)のポリペ
プチドのアミノ酸配列において、通常20%以下、特に10%以下のアミノ酸が欠失、置換若しくは付加されたものであることが好ましい。
【0058】
(5)のポリペプチドにおいて、配列番号6に示すアミノ酸配列を含むポリペプチドとし
ては、例えば、配列番号6に示すアミノ酸配列の3倍以下の長さのポリペプチドが挙げら
れる。同様に、(6)のポリペプチドにおいて、配列表の配列番号6に示すアミノ酸配列に
おいて1若しくは2以上のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を含むポリペプチドとしては、例えば、当該アミノ酸配列の3倍以下の長さのポリペプチドが挙げられる。
【0059】
本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼは、これらのタンパク質等を生産する細菌を培養し、培養上清から精製することにより得られる。また、配列番号2、4または6のアミノ酸配列を基に、ペプチド化学合成により得られる。さらに、後述する本発明方法によっても得られる。
(2)本発明のDNA
(i)チオレドキシン
本発明のDNAは、前記説明した本発明の(1)又は(2)のポリペプチドをコードするDNAである。特に、 以下の(7)または(8)のDNAが好ましい。
(7) 配列表の配列番号1に示す塩基配列を含むDNA。
・ 配列表の配列番号1に示す塩基配列を含むDNAとストリンジェントな条件下でハイ
ブリダイズし、かつ、100℃で0.5 時間熱処理した場合に50%以上のチオレドキシン活性が残存するポリペプチドをコードするDNA。
【0060】
(8)のDNAは、100 ℃で0.5時間熱処理した場合に50%以上のチオレドキシン活性が残存するポリペプチドをコードするDNAであれば特に制限されないが、(7)のDNA配列において
、通常20%以下、特に10%以下のヌクレオチドが欠失、置換若しくは付加されたものであることが好ましい。
【0061】
(7)及び(8)のDNAにおいて、配列番号1に示す塩基配列を含むDNAとしては、例えば、配列番号1に示す塩基配列の3倍以下の長さのDNAが挙げられる。
(ii)チオレドキシンレダクターゼ
本発明のDNAは、前記説明した本発明の(3)又は(4)のポリペプチドをコードするDNAである。特に、 以下の(9)または(10)のDNAが好ましい。
(9) 配列表の配列番号3に示す塩基配列を含むDNA。
・ 配列表の配列番号3に示す塩基配列を含むDNAとストリンジェントな条件下でハイ
ブリダイズし、かつ、70℃以上の温度下で100unit/mg以上のチオレドキシンレダクター
ゼ活性を有するポリペプチドをコードするDNA。
【0062】
(10)のDNAは、70℃以上の温度下で100 unit/mg以上のチオレドキシンレダクターゼ活性を有するポリペプチドをコードするDNAであれば特に制限されないが、(9)のDNA配列に
おいて、通常20%以下、特に10%以下のヌクレオチドが欠失、置換若しくは付加されたものであることが好ましい。
【0063】
(9)及び(10)のDNAにおいて、配列番号3に示す塩基配列を含むDNAとしては、例えば、
配列番号3に示す塩基配列の3倍以下の長さのDNAが挙げられる。
(iii)チオレドキシンペルオキシダーゼ
本発明のDNAは、前記説明した本発明の(5)又は(6)のポリペプチドをコードするDNAである。特に、 以下の(11)または(12)のDNAが好ましい。
(11) 配列表の配列番号5に示す塩基配列を含むDNA。
(12) 配列表の配列番号5に示す塩基配列を含むDNAとストリンジェントな条件下でハイ
ブリダイズし、かつ、70 ℃以上の温度下で100unit/mg以上のチオレドキシンペルオキ
シダーゼ活性を有するポリペプチドをコードするDNA。
【0064】
(12)のDNAは、70℃以上の温度下で100 unit/mg以上のチオレドキシンペルオキシダーゼ比活性を有するポリペプチドをコードするDNAであれば特に制限されないが、(11)のDNA
配列において、通常20%以下、特に10%以下のヌクレオチドが欠失、置換若しくは付加されたものであることが好ましい。
【0065】
(11)及び(12)のDNAにおいて、配列番号5に示す塩基配列を含むDNAとしては、例えば、配列番号5に示す塩基配列の3倍以下の長さのDNAが挙げられる。
【0066】
本明細書において「ストリンジェントな条件」とは、例えば、通常のハイブリダイゼーション溶液中であれば68℃で行う条件が挙げられ、50%フォルムアミドを含むハイブリダイゼーション溶液中であれば42℃で行う条件が挙げられる。詳しくは、Molecular Cloning: A Laboratory Manual第2版第2巻に記載のサザンハイブリダイゼーションに用いられる条件が挙げられる。
【0067】
本発明のDNAは、例えばAeropyrum属、Sufolobus属、Thermoplasma属、Thermoproteus属、Mastigocladus属、Bacillus属、Synechococcus属、Thermus属等の好熱性細菌の染色体DNAのライブラリーからプローブを用いたハイブリダイゼーションにより単離することもできるが、配列表の配列番号1、3または5のDNA配列を基にPCR法により増幅することもできる。また、化学合成によっても取得できる。
【0068】
また、前記の(8)、(10)及び(12)の変異したDNAは、化学合成法、遺伝子工学的手法、突然変異誘発法などの公知の方法で作成することができる。遺伝子工学的手法としては、入手可能なチオレドキシン、チオレドキシンレダクターゼ又はチオレドキシンペルオキシダーゼに対して、エキソヌクレアーゼを用いたDNA欠失導入、リンカー導入、位置指定突然
変異導入、変異プライマーを用いたPCR法による塩基配列の改変などの公知の方法を挙げ
ることができる。
(3)本発明のベクター
本発明のベクターは、前記説明した本発明の各DNAを含有する組換えベクターである。
本発明のDNAが組み込まれるベクターは公知のものを広く利用でき、細菌用ベクターの他
、酵母用ベクター、動物細胞用ベクター等も利用できる。酵素の生産効率の点から、通常は細菌用ベクターを用いればよい。公知のベクターとしては、大腸菌ベクターのpBR322、pUC19、pKK233-2等、バチルス属細菌ベクターのpUB110、pC194、pE194、pTHT15、pBD16
等、酵母用ベクターYip5、Yrp17、Yep24等、動物細胞用ベクターpUC18、pUC19、M13mp18
等を例示できる。
(4)本発明の形質転換体
また、本発明の形質転換体は、前記説明した本発明の各組換えベクターを含有する形質転換体である。宿主としては、ベクターに応じて細菌、酵母、動物細胞等を利用できる。形質転換は、リン酸カルシウム法、プロトプラスト法、エレクトロポレーション法、スフェロプラスト法、酢酸リチウム法、リポフェクション法、マイクロインジェクション法等の公知の方法で行うことができる。これらの公知の方法の中から、宿主の種類に応じて選択すればよい。
(5)本発明のタンパク質または酵素の製造方法
本発明のチオレドキシン、チオレドキシンレダクターゼまたはチオレドキシンペルオキシダーゼの製造方法は、本発明の各形質転換体を培養し、この形質転換体から各タンパク質等を精製する方法である。
【0069】
これらの細菌が生産した酵素は、遠心分離、塩析、溶媒沈殿法、透析法、限外ろ過法、ゲルろ過法、イオン交換クロマトグラフィー法、アフィニティクロマトグラフィー法、逆相高速液体クロマトグラフィー法等の公知の蛋白精製方法を組み合わせて精製することができる。本発明の酵素が細胞内またはペリプラズム内に生産される場合には超音波処理、界面活性剤処理等の公知の方法で細胞を破壊した後に前記の精製を行えばよい。
【0070】
また、本発明の耐熱性タンパク質等を精製するにあたっては、目的耐熱性タンパク質等を10〜30分間程度(特に20分間程度)インキュベートした場合に通常60%以上、特に80%以上活性が残存し、かつ、当該耐熱性タンパク質等を生産する宿主の生育限界温度より通常10℃以上、特に15℃以上高い温度で、被精製タンパク質溶液を通常10〜120分間程度、
特に10〜30分間程度インキュベートすることが好ましい。これにより、目的タンパク質を殆ど失活させることなく、宿主が生産する夾雑タンパク質を変性、失活させることができる。この加熱処理工程後に、タンパク質溶液を、それには限定されないが、例えば15000rpm程度で20分間程度遠心することにより変性した夾雑タンパク質を沈殿させることができる。この加熱処理工程は、精製のいずれの段階で行ってもよい。
【0071】
このような加熱処理工程は、本発明のチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの精製に限らず、いずれの耐熱性タンパク質の精製においても行うことができ、これにより耐熱性タンパク質の精製度を飛躍的に向上させることができる。
実施例
(1)超好熱性古細菌Aeropyrum pernix K1株のチオレドキシン遺伝子、チオレドキシン
レダクターゼ遺伝子およびチオレドキシンペルオキシダーゼ遺伝子と他の対応酵素遺伝子との相同性
コンピューターによる相同性検索の結果、配列表の配列番号1に示す本発明のチオレドキシン遺伝子の塩基配列とSalmonella typhimurium LT2株のチオレドキシン遺伝子の塩基配列との間に31%の相同性が見出された。また、配列表の配列番号3に示す本発明のチオレドキシンレダクターゼ遺伝子の塩基配列とSulfolobus salfataricusのチオレドキシン
レダクターゼ遺伝子との間に48%の相同性が見出された。また、配列表の配列番号5に示す本発明のチオレドキシンペルオキシダーゼ遺伝子の塩基配列とSulfolobus tokodaiのチオレドキシンペルオキシダーゼ遺伝子との間に 62%の相同性が見出された。
(2)Aeropyrum pernix K1株のチオレドキシン遺伝子、チオレドキシンレダクターゼ遺
伝子及びチオレドキシンペルオキシダーゼ遺伝子のクローニング
<染色体DNAの調製>
バクトマリン培地(Difco)37.4g とNa2S2O3-5H2O 1.0g を水1リットルに溶かした後、pHを7.0-7.2に調整することにより培地を調製した。超好熱性古細菌Aeropyrum pernix K1株(理化学研究所においてJCM9820として登録)をこの培地に接種し、90℃で3日間振盪培養した。培養液を5,000rpmで10分間遠心分離することにより集菌した。
【0072】
菌体を10mM トリス(pH 7.5)−1mM EDTA 溶液で2回洗浄後インサートアガロース(InCert Agarose、FMC社製)ブロック中に封入した。このブロックを1%N−ラウロイルザルコシン−1mg/ml プロテアーゼK溶液中で処理することにより、染色体DNAがアガロースブロ
ック中に分離調製された。インサートアガロースブロックを用いた染色体DNAの分離条件
は、アガロースブロックの添付マニュアルに従った。
<遺伝子の増幅・組み換えベクターの構築>
(i)チオレドキシン
配列表の配列番号1の塩基配列を含むDNAをPCR法により増幅した。PCR条件は、PCRキットの添付マニュアルに従った。5’末端側に対応するプライマーとしては、配列表の配列
番号1に示すDNA配列において、塩基番号1から始まる(すなわち開始コドンから始まる
)オリゴヌクレオチドプライマーを合成した。また、3’末端側に対応するプライマーとしては、Aeropyrum pernix K1株の染色体DNAにおいて配列番号1の塩基配列の3’末端より下流域に対応するプライマーであって、増幅されたDNA中に制限酵素のBamHIサイトが生じるようなプライマーを合成した。PCR反応後、DNAを制限酵素のBamHIで37℃で3時間
処理することにより完全分解した。次いで、チオレドキシン遺伝子を精製カラムキットを用いて精製した。
【0073】
次いで、チオレドキシン遺伝子を組み込んだベクターを構築する目的で、ベクターのpET-8c(Novagen社製)を制限酵素NcoIで切断し、精製した後、T4 DNA ポリメラーゼを用いて平滑末端化した。精製したプラスミドを制限酵素BamHIで切断・精製した。次いで、BamHIで切断されたプラスミドpET-8cと BamHIで切断された上記のチオレドキシン遺伝子とをT4リガーゼを用いて16℃で、16時間反応させることにより連結した。連結したDNAを
用いて、大腸菌(E. coli) XL2-BlueMRF’ 株(Novagen社製)のコンピテントセルを形質
転換した。形質転換体は、0.05 mg/ml アンピシリンを含むLB寒天プレート上でのコロニ
ー形成を指標に選択した。形質転換体からシステイン合成酵素遺伝子含有プラスミドをアルカリ法で抽出した。
(ii)チオレドキシンレダクターゼ
前記説明したチオレドキシンのクローニング方法において、5’末端側に対応するPCRプライマーとして、配列番号3に示すDNA配列において塩基番号1から始まるオリゴヌクレ
オチドを使用し、3’末端側に対応するPCRプライマーとして、Aeropyrum pernix K1株
の染色体DNAにおいて配列番号3の塩基配列の3’末端より下流側に対応するプライマーであって、増幅されたDNA中にBamHIサイトが生じるようなプライマーを合成し、使用した。その他は、チオレドキシンの場合と同様にして、Aeropyrum pernix K1株由来のチオレ
ドキシンレダクターゼを組み込んだプラスミドpET-8cを保持する大腸菌L2-BlueMRF’株を得た。
(iii)チオレドキシンペルオキシダーゼ
前記説明したチオレドキシンのクローニング方法において、5’末端側に対応するPCRプライマーとして、配列番号5に示すDNA配列において塩基番号1から始まるオリゴヌクレ
オチドを使用し、3’末端側に対応するPCRプライマーとして、Aeropyrum pernix K1株
の染色体DNAにおいて配列番号5の塩基配列の3’末端より下流側に対応するプライマーであって、増幅されたDNA中にBamHIサイトが生じるようなプライマーを合成し、使用した。その他は、チオレドキシンの場合と同様にして、Aeropyrum pernix K1株由来のチオレ
ドキシンペルオキシダーゼを組み込んだプラスミドpET-8cを保持する大腸菌L2-BlueMRF’株を得た。
(3)チオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの生産
<形質転換体の作製>
1.5ml容チューブ内に、大腸菌(E. coli )Rosetta (DE3)株( Novagen社製)のコンピテントセル0.04ml(2,000,0000cfu/mg)と、上記調製したチオレドキシン遺伝子含有プ
ラスミド、チオレドキシンレダクターゼ遺伝子含有プラスミドまたはチオレドキシンペルオキシダーゼ遺伝子含有プラスミドのDNA溶液0.003ml(プラスミドDNA 8.4ng)を加え
氷中に30分間放置した後、42℃で、30秒間ヒートショックを与えた。次いで、チューブ内にSOCmedium を0.25ml加え、37℃で1時間震盪培養した。次いで、アンピシリン及
びクロラムフェニコールを含むLB寒天プレートに塗布し、37度で一晩培養することにより形質転換体を得た。
<タンパク質または酵素の精製>
(i)チオレドキシン
チオレドキシン遺伝子を含むプラスミドを保持する形質転換体をアンピシリン及びクロラムフェニコールを含むNZCYM培地に接種し、600nmにおける吸光度が0.5に達するまで、37℃で培養した後、プラスミドの発現量を高めるためにIPTG(Isopropyl-b-D-thiogalactopyranoside)を加えさらに4時間培養した。培養液を8,000rpmで10min遠心分
離することにより集菌した。
【0074】
集菌した菌体4 gに、1mM DTT、1mM EDTA を含む50mM Tris−HCl(pH8.0)を加え、
菌体を90Wの出力で5分間超音波破砕 した。破砕した菌液を 15,000rpmで30分間遠心分離し、上清を採取した。
【0075】
夾雑蛋白を沈殿させて除去する目的で、この上清を85℃で30分間加熱した後、15,000rpmで30分間遠心分離し、上清を採取した。上清を1 mM EDTAを含む50 mM(Tris - HCl
)緩衝液(pH8.0)で透析し、同緩衝液で平衡化した陰イオン交換樹脂のHitrapQ(ファルマシア社製)カラムにアプライしてイオン交換クロマトグラフィーを行った。さらに、活性画分を150 mM NaCl を含む50 mMリン酸ナトリウム緩衝液(pH7.0)中で透析し、同緩衝液で平衡化した。ゲル濾過材のSuperdex 200(ファルマシア社製)カラムにア
プライしてゲル濾過クロマトグラフィーを行った。活性画分には、SDS−ポリアクリルア
ミドゲル電気泳動により単一バンドを与える均一標品が含まれていた。
【0076】
また、ゲルろ過クロマトグラフィーの結果、本酵素の分子量は約37kDaであった。
(ii)チオレドキシンレダクターゼ
チオレドキシンレダクターゼ遺伝子を含むプラスミドを保持する形質転換体をチオレドキシン遺伝子の場合と同様にして培養、集菌した。
【0077】
集菌した菌体4 gに、1mM DTT、1mM EDTA を含む50mM Tris−HCl(pH8.5)を加え、
菌体を90Wの出力で5分間超音波破砕した。破砕した菌液を 15,000rpmで30分間遠心分離し、上清を採取した。
【0078】
夾雑蛋白を沈殿させて除去する目的で、この上清を85℃で30分間加熱した後、15,000rpmで20分間遠心分離し、上清を採取した。上清を1 mM EDTAを含む50 mM Tris - HCl緩衝液(pH8.0)で透析し、同緩衝液で平衡化した陰イオン交換樹脂のHitrapQ(ファルマシア社製)カラムにアプライしてイオン交換クロマトグラフィーを行った。さらに、活性画分を150mM NaClを含む50 mMリン酸ナトリウム緩衝液(pH7.0 )中で透析し、同緩
衝液で平衡化した。ゲル濾過材のSuperdex 200(ファルマシア社製)カラムにアプライ
してゲル濾過クロマトグラフィーを行った。活性画分には、SDS−ポリアクリルアミドゲ
ル電気泳動により単一バンドを与える均一標品が含まれていた。
【0079】
また、ゲルろ過クロマトグラフィーの結果、本酵素の分子量は約37kDaであった。
(iii)チオレドキシンペルオキシダーゼ
チオレドキシンペルオキシダーゼ遺伝子を含むプラスミドを保持する形質転換体をチオレドキシン遺伝子の場合と同様にして培養、集菌、超音波破砕、遠心による上清採取、加熱処理、遠心による上清採取を行った。
【0080】
得られた上清を1mM EDTAを含む50 mM(Tris - HCl )緩衝液(pH8.0)で透析し、同緩衝液で平衡化した陰イオン交換樹脂のHitrapQ(ファルマシア社製)カラムにアプライ
してイオン交換クロマトグラフィーを行った。さらに、活性画分を150mM NaCl を含む50mM リン酸ナトリウム緩衝液(pH7.0)中で透析し、同緩衝液で平衡化した。ゲル
濾過材のSephacryl S-100(ファルマシア社製)カラムにアプライしてゲル濾過クロマトグラフィーを行った。活性画分には、SDS−ポリアクリルアミドゲル電気泳動により単一
バンドを与える均一標品が含まれていた。
【0081】
また、ゲルろ過クロマトグラフィーの結果、本酵素の分子量は約29kDaであった。
<活性測定法>
本発明において、精製工程における目的タンパク質又は酵素の検出、至適温度の検討および安定性の検討における目的タンパク質又は酵素の活性検出は、以下の方法で行った。(i)チオレドキシン
チオレドキシンの活性測定は、還元型チオレドキシンによるインスリンサブユニット間のジスルフィド結合を還元する活性を測定するMolmgrenらの方法(Method in Enzymology 1993)に従った。
【0082】
すなわち、先ず0.4mM DTT及び0.4mg/mlウシ血清アルブミンを含む100mM Tris−塩酸バッファー(pH7.5)で37℃、15分間チオレドキシンサンプルを前処理して還元型チオレド
キシンとした。次いで、基質として1mg/mlのウシ膵臓インスリン(シグマ社製)を含む100mMTris−塩酸バッファー(pH7.5)に、チオレドキシンを200ng添加し、インスリンの還元分解による650nmにおける吸光度の上昇を室温で20分間測定した。
(ii)チオレドキシンレダクターゼ
基質としての1mMジチオビス2-ニトロベンゾイックアシッド(DTNB)0.2mM NADPH、1mM
EDTAを含む50mMリン酸カリウム緩衝液(pH7.0)中に、精製チオレドキシンレダクター
ゼ 2μgを添加し、NADPHの濃度の指標である340nmにおける吸光度の減少速度を5 分間測定することによりチオレドキシンレダクターゼの活性を測定した。
【0083】
精製工程では、70℃の緩衝液中で活性を測定した。
(iii)チオレドキシンペルオキシダーゼ
基質としての1mM過酸化水素、0.2mM NADPH、0.1μMの精製チオレドキシンレダクター
ゼ、5μMの精製チオレドキシンを含む50mMリン酸カリウム緩衝液(pH7.0)中に、精製チオレドキシンペルオキシダーゼ2 μgを添加し、NADPHの濃度の指標である340nmにおける
吸光度の減少速度を5 分間測定することによりチオレドキシンペルオキシダーゼの活性を測定した。
【0084】
精製工程では、70℃の緩衝液中で活性を測定した。
(4)タンパク質・酵素の性質
前記の方法により得られたAeropyrum pernix K1株由来のチオレドキシン、チオレドキ
シンレダクターゼ及びチオレドキシンペルオキシダーゼの諸特性を評価した。
<至適温度>
前述したチオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの活性測定方法において、酵素反応を行う緩衝液の温度を20〜90℃の範囲で変化させて、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの各活性を測定した。
【0085】
この結果、図1(A)及び(B)に示すように、チオレドキシンレダクターゼの至適温度は70℃(図1(A))、チオレドキシンペルオキシダーゼの至適温度は70℃(図1(B))であった。<耐熱性>
2 mM EDTAを含む50mMリン酸ナトリウム緩衝液(pH7.0)中に、チオレドキシンを0.1 mg/ml濃度になるように添加した酵素溶液サンプルを用意し、100℃でインキュベ
ートして、経時的に残存活性を測定した。チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼについても同様にして、インキュベートし、経時的に残存活性を測定した。
【0086】
この結果、図2に示すように、チオレドキシンは、100℃でインキュベートした場合に1時間後に55%程度の活性が残り、チオレドキシンレダクターゼは、100℃でインキュベー
トした場合に1時間後に65%程度の活性が残り、チオレドキシンぺルオキシダーゼは、100℃でインキュベートした場合に1時間後に70%程度の活性が残った。
【図面の簡単な説明】
【0087】
【図1】(A)及び(B)は、それぞれ本発明の1実施例であるチオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの至適温度を示すグラフである。
【図2】本発明の1実施例であるチオレドキシン、チオレドキシンレダクターゼ及びチオレドキシンペルオキシダーゼの100℃における熱安定性を示すグラフである。
【特許請求の範囲】
【請求項1】
100℃で0.5時間熱処理した場合に50%以上チオレドキシンペルオキシダーゼ活性が残存する耐熱性チオレドキシンペルオキシダーゼ。
【請求項2】
至適温度が70℃以上である請求項1に記載の耐熱性チオレドキシンペルオキシダーゼ。
【請求項3】
超好熱性古細菌Aeropyrum pernix由来の請求項1又は2に記載の耐熱性チオレドキシンペルオキシダーゼ。
【請求項4】
配列番号6に示すアミノ酸配列からなる耐熱性チオレドキシンペルオキシダーゼ。
【請求項5】
配列番号6に示すアミノ酸配列において、1個若しくは2個以上であって、10%以下のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、100℃で0.5時間熱処理した場合に50%以上チオレドキシンペルオキシダーゼ活性が残存する耐熱性チオレドキシンペルオキシダーゼ。
【請求項6】
配列番号5に示す塩基配列のDNAからなる耐熱性チオレドキシンペルオキシダーゼ遺伝子。
【請求項7】
配列番号5に示す塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし
、かつ、100℃で0.5時間熱処理した場合に50%以上チオレドキシンペルオキシダーゼ活性が残存する耐熱性チオレドキシンペルオキシダーゼをコードするDNAからなる耐熱性チオ
レドキシンペルオキシダーゼ遺伝子。
【請求項8】
請求項6又は7に耐熱性チオレドキシンペルオキシダーゼ遺伝子を含有するベクター。
【請求項9】
請求項8に記載のベクターを含有する形質転換体。
【請求項10】
請求項9の形質転換体を培養し、該形質転換体から耐熱性チオレドキシンペルオキシダーゼを分離する耐熱性チオレドキシンペルオキシダーゼの生産方法。
【請求項1】
100℃で0.5時間熱処理した場合に50%以上チオレドキシンペルオキシダーゼ活性が残存する耐熱性チオレドキシンペルオキシダーゼ。
【請求項2】
至適温度が70℃以上である請求項1に記載の耐熱性チオレドキシンペルオキシダーゼ。
【請求項3】
超好熱性古細菌Aeropyrum pernix由来の請求項1又は2に記載の耐熱性チオレドキシンペルオキシダーゼ。
【請求項4】
配列番号6に示すアミノ酸配列からなる耐熱性チオレドキシンペルオキシダーゼ。
【請求項5】
配列番号6に示すアミノ酸配列において、1個若しくは2個以上であって、10%以下のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ、100℃で0.5時間熱処理した場合に50%以上チオレドキシンペルオキシダーゼ活性が残存する耐熱性チオレドキシンペルオキシダーゼ。
【請求項6】
配列番号5に示す塩基配列のDNAからなる耐熱性チオレドキシンペルオキシダーゼ遺伝子。
【請求項7】
配列番号5に示す塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし
、かつ、100℃で0.5時間熱処理した場合に50%以上チオレドキシンペルオキシダーゼ活性が残存する耐熱性チオレドキシンペルオキシダーゼをコードするDNAからなる耐熱性チオ
レドキシンペルオキシダーゼ遺伝子。
【請求項8】
請求項6又は7に耐熱性チオレドキシンペルオキシダーゼ遺伝子を含有するベクター。
【請求項9】
請求項8に記載のベクターを含有する形質転換体。
【請求項10】
請求項9の形質転換体を培養し、該形質転換体から耐熱性チオレドキシンペルオキシダーゼを分離する耐熱性チオレドキシンペルオキシダーゼの生産方法。
【図1】


【図2】
【図2】
【公開番号】特開2006−20650(P2006−20650A)
【公開日】平成18年1月26日(2006.1.26)
【国際特許分類】
【出願番号】特願2005−289541(P2005−289541)
【出願日】平成17年10月3日(2005.10.3)
【分割の表示】特願2001−379462(P2001−379462)の分割
【原出願日】平成13年12月13日(2001.12.13)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成18年1月26日(2006.1.26)
【国際特許分類】
【出願日】平成17年10月3日(2005.10.3)
【分割の表示】特願2001−379462(P2001−379462)の分割
【原出願日】平成13年12月13日(2001.12.13)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]