
セルラーゼ活性を有するポリペプチド
本発明は、CBH IIキメラ融合ポリペプチド、該ポリペプチドをコードする核酸、およびポリペプチドを生成するための宿主細胞に関する。

【発明の詳細な説明】
【技術分野】
【0001】
<関連出願の優先権主張>
本願は35U.S.C.§119に基づいて、2009年4月6日に出願された米国仮出願第61/166,993号、および2009年5月13日に出願された米国仮出願第61/177,882号を参照して取り込むものとする。
【0002】
<連邦支援の研究に関する記載>
アメリカ政府は、国立衛生研究所によって与えられる研究費番号GM068664およびARO−US Army Robert Morris Acquisition Centerによって与えられる研究費番号DDAD19−03−0D−0004に従って、本発明において特定の権利を有する。
【0003】
本発明は、分子生物工学および分子生物デザイン、並びに改変タンパク質および改変核酸に関する。
【背景技術】
【0004】
バイオマス変換工程におけるセルラーゼ混合物の性能は、セルロース基質の物理的な状態および組成に加えて、安定性、生成阻害、多様なセルラーゼ成分間の相乗効果、非生産的な吸着に対する生産的な結合、およびpH依存性を含む多くの酵素特性に依存する。多様な適用および原料に対して酵素の形態を最適化するため、セルロース加水分解に多変量的な特性を付与し、多様なセルラーゼから選択することが望まれる。
【発明の概要】
【課題を解決するための手段】
【0005】
本発明は、セルラーゼ活性があり、野生型タンパク質と比較して、高い熱安定性および活性がある組み換え型ポリペプチドを提供する。本発明は、長時間の加水分解試験において、CBH IIのC末端(例えば天然タンパク質配列に応じてアミノ酸約310−315であり、例えば配列番号2および4を参照とする)で、CysからSerへの突然変異があるキメラおよび天然酵素が、天然酵素と比較してより確実なセルロース加水分解を提供することを示す。
【0006】
本発明は、配列GECDG(配列番号2の312−316)を含有するモチーフ中のC末端領域にC→S置換を含む組み換えポリペプチドであって、該変異体が、野生型セロビオヒドラーゼと比較して高い熱安定性およびセルラーゼ活性があるものを提供する。例えば、本発明は高い熱安定性およびセルラーゼ活性があるポリペプチドであって、C314Sを含有する配列番号2と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C311Sを含有する配列番号4と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C310Sを含有する配列番号12と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C312Sを含有する配列番号13と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C314Sを含有する配列番号14と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C315Sを含有する配列番号15と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C313Sを含有する配列番号16と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C311Sを含有する配列番号17と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C313Sを含有する配列番号19と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C311Sを含有する配列番号22と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号64と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C407Sを含有する配列番号65と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C394Sを含有する配列番号66と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号67と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号68と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号69と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号70と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号71と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号72と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号73と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号74と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号75と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C407Sを含有する配列番号76と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C394Sを含有する配列番号77と少なくとも85%、90%、95%、98%、99%の一致があるものを含むか;C412Sを含有する配列番号78と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;上記ポリペプチドは、セルラーゼ活性があり、それに相当するCys→Serの突然変異が欠如している親(野生型)タンパク質と比較して熱安定性が改良されている。
【0007】
本発明は、組み換えにより生成されたか、合成されたか、他の非天然的な製造方法により生成された、いずれかの実質的に精製されたポリペプチドも提供する。該ポリペプチドは、C→S置換の存在位置を除く、以下に示す1−10、10−20または20−30の保存的な配列のアミノ酸置換基を含む:C314Sを含む配列番号2;C311Sを含む配列番号4;C310Sを含む配列番号12;C312Sを含む配列番号13;C314Sを含む配列番号14;C315Sを含む配列番号15;C313Sを含む配列番号16;C311Sを含む配列番号17;C313Sを含む配列番号19;C312Sを含む配列番号21;C311Sを含む配列番号22;C400Sを含む配列番号64;C407Sを含む配列番号65;C394Sを含む配列番号66;C400Sを含む配列番号67;C400Sを含む配列番号68;C400Sを含む配列番号69;C400Sを含む配列番号70;C400Sを含む配列番号71;C400Sを含む配列番号72;C400Sを含む配列番号73;C400Sを含む配列番号74;C400Sを含む配列番号75;C407Sを含む配列番号76;C394Sを含む配列番号77;またはC412Sを含む配列番号78。
【0008】
本発明は、(a)位置314にSerがある配列番号2と少なくとも85%以上の一致があるポリペプチドであり、前記ポリペプチドはセルラーゼ活性がある;(b)位置311にSerがある配列番号4と少なくとも70%以上の一致があるポリペプチドであり、前記ポリペプチドはセルラーゼ活性がある;(c)(i)位置310にSerがある配列番号12、(ii)位置312にSerがある配列番号13、(iii)位置314にSerがある配列番号14、(iv)位置315にSerがある配列番号15、(v)位置313にSerがある配列番号16、(vi)位置311にSerがある配列番号17、(vii)位置313にSerがある配列番号19、(viii)位置312にSerがある配列番号21、(ix)位置311にSerがある配列番号22からなる群から選択される配列と70%以上の一致があるポリペプチドであり、それぞれ上記ポリペプチドは、セルラーゼ活性がある;(d)2つの異なるセロビオヒドラーゼポリペプチドからの少なくとも2つのドメインを含むキメラポリペプチドであり、ドメインはNからC末端までに:(断片1)−(断片2)−(断片3)−(断片4)−(断片5)−(断片6)−(断片7)−(断片8)を含み;断片1は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)のアミノ酸残基約1から約x1までと少なくとも50−100%の一致がある配列を含み;断片2は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)のアミノ酸残基約x1から約x2までと少なくとも50−100%の一致がある配列を含み;断片3は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)のアミノ酸残基約x2から約x3までと少なくとも50−100%の一致がある配列を含み;断片4は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)のアミノ酸残基約x3から約x4までと少なくとも50−100%の一致がある配列を含み;断片5は、配列番号2(“1”)、配列番号4(「2」)または配列番号(「3」)のアミノ酸残基約x4から約x5までと少なくとも50−100%の一致がある配列を含み;断片6は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)のアミノ酸残基約x5から約x6までと少なくとも50−100%の一致がある配列を含み;断片7は、配列番号2(「1」)または、配列番号4(「2」)のアミノ酸残基約x6から約x7までと少なくとも50−100%の一致がある配列を含み;断片8は、配列番号2(「1」)または配列番号4(「2」)のアミノ酸残基約x7から約x8までと少なくとも50−100%の一致がある配列を含み;x1は配列番号2の残基43,44,45,46,47であるか、配列番号4または配列番号6の残基42,43,44,45,46である;x2は配列番号2の残基70,71,72,73,74であるか、配列番号4もしくは配列番号6の残基68,69,70,71,72,73,74;x3は配列番号2の残基113,114,115,116,117,118であるか、配列番号4もしくは配列番号6の残基110,111,112,113,114,115,116;x4は配列番号2の残基153,154,155,156,157であるか、配列番号4もしくは配列番号6の残基149,150,151,152,153,154,155,156;x5は配列番号2の残基220,221,222,223または224であるか、配列番号4もしくは配列番号6の残基216,217,218,219,220,221,222,223;x6は配列番号2の残基256,257,258,259,260であるか、配列番号4もしくは配列番号6の残基253,254,255,256,257,258,259,260;x7は配列番号2の残基312,313,314,315または316であるか、配列番号4の残基309,310,311,312,313,314,315,318;およびx8はポリペプチドのC末端に相当するアミノ酸残基が配列番号2、配列番号4もしくは配列番号6の配列であり、キメラポリペプチドは、配列番号2の位置314または配列番号4の位置311でSerであり、キメラポリペプチドは配列番号2,4もしくは6を含むCBH IIポリペプチドと比較してセルラーゼ活性、改良した熱安定性および/またはpHの安定性があるものからなる群から選択される配列を含む組換え型ポリペプチドを提供する。
【0009】
本発明は、配列番号12−62または63の配列からなる組換え型ポリペプチドも提供する。
【0010】
本発明は、また上記いずれかのポリペプチドをコード化するポリヌクレオチド、該ポリヌクレオチドを含むベクター、および該ポリヌクレオチドまたは該ベクターを含む宿主細胞を提供する。
【0011】
本発明は、本発明の実質的に精製された形態、または細胞溶解物の一部としてポリペプチドを含む酵素製剤も提供する。
【0012】
本発明は、またセルロースを含むバイオマスの処理方法を提供し、該方法は本開示のポリペプチドまたは酵素液とバイオマスの接触工程を備える。
【図面の簡単な説明】
【0013】
【図1】候補遺伝子たるCBH IIの親遺伝子の酵母発現培養上清のSDS−PAGEゲルを示す図である。(A)ゲルレーン(左から右):1−H. jecorina、2−空ベクター、3−H. insolens、4−C. thermophilum、5−H. jecorina(二重)、6−P. chrysosporium、7−T. emersonil、8−空ベクター(二重)、9−H. jecorina(三重)。ゲルの下部の数字は、2時間、50℃でのPASC加水分解試験後に、反応物中に存在する還元糖の濃度(μg/mL)を示す。その後、BSAを標準としたSDS−PAGEによる比較は、H. insolens発現レベル5−10mg/Lの評価を可能とした。(B)S. cerevisiae CBH II発現培養上清のSDS−PAGEゲル分析を示す図である。CBH IIバンドが、ちょうど60kDa分子量基準値未満で現れた。レーンは、左から右に向かって、1−野生型H. jeco、2−H. jeco B7P3、3−H. jeco C311S、4−野生型C. ther、5−野生型H. inso、6−H. inso B7P3、7−H. inso C314Sである。数値は、50℃、50mM酢酸ナトリウム、pH4.8の条件下で、PASC(1mg/mL)を用いて100分間インキュベーションした際に生じるμgグルコース等量/SDCAAを発現する培養上清等量の1mL当たりのmL反応容量を示す。レーン1−4に関する値は、加えられた濃縮培養上清の容積の2倍に修正するため、2で除算する。かかる修正を省略すると、H. insolens酵素に関する特異的活性値が低下するように思われる。
【図2】CBH IIキメラライブラリーブロックの限度を示す図である。(A)色分けされたブロックを用いたH. insolens CBH II触媒ドメインのリボンダイアグラムを示す。CBH II酵素は、セロビオ由来のイソファゴミングリコシダーゼ阻害剤と複合体を形成する。(B)2次構造要素、ジスルフィド結合および黒矢印によって示されるブロックの区別を示す、H. insolens触媒ドメインの直線図である。(C)組み換えに応じて分解することができる接続部分(4.5A範囲内の側鎖重原子)を示す側鎖接続マップである。分解された接触部の大半は、連続したブロックの間に生じる。
【図3】23個の分泌/活性型および15個の非分泌/不活性型のサンプルセットのキメラに関して分割した接続部(E)の数および最も近い親(m)からの突然変異の数を示す図である。
【図4】親CBH II酵素および3つの熱安定性のあるキメラに関するpHの関数として、pH5.0で基準化した特異的活性を示す図である。提示するデータは、2つの測定の平均であり、HJPlusおよびH. jecoに関するエラーバーは、2つの独立した試験の値を示す。16時間反応物において、300μg酵素/g PASC、50℃での12.5mMクエン酸ナトリウム/12.5リン酸ナトリウム緩衝剤中において示すようなpHである。
【図5】親および広範囲にわたる温度での熱安定性のあるキメラに関する長期セルロース加水分解分析の結果を示す図である。エラーバーは、HJPlusおよびH. insolens CBH II酵素の3つの測定に関する標準誤差を示す。pH4.8における50mM酢酸ナトリウム中の100μg酵素/g PASCの40時間反応物である。
【図6】63℃で12時間インキュベーションした後、検証するためのキメラセットに関する標準化した残留活性を示す図である。50℃で25mM酢酸ナトリウム緩衝剤、pH4.8中、2時間の分析においてPASCを基質として用いて測定された濃縮培養上清におけるCBH II酵素に関する残留活性を示す。
【図7】親およびキメラCBH II酵素発現ベクターYep352/PGK91−1―rssに関するマップを示す図である。図示のベクターは、野生型H. jecorina ce16a(CBH II酵素)遺伝子を含む。キメラおよび親CBH II酵素の双方に関して、ss Lys−Arg Kex2部位に続くCBD/リンカーアミノ酸の配列は、以下の通りである:ASCSSVWGQCGGQNWSGPTCCASGSTCVYSNDYYSQCLPGAASSSSSTRAASTTSRVSPTTSRSSSATPPPGSTTTRVPPVGSGTATYS (配列番号8)。
【図8】CBH II親および51個のCBH IIキメラに関する観測されたか、予測されるT50値である。線は線形回帰モデル式を示す(Table7におけるパラメーター)。親CBH IIのT50値は、正方形で示す。
【図9】アビセルに対する、温度によるCBH II特異的活性を示す図であり、(a)CBH II親およびキメラの特異的活性である。(b)CBH II親、C311S突然変異体およびB7P3単一ブロック置換キメラの特異的活性である。アビセル濃度15mg/mLを用いた50mM酢酸ナトリウム、pH4.8中で16時間、反応を行った。(c)温度の関数としてアビセルに対するCBH II親、単一点突然変異体および単一阻害置換キメラ活性(μg/グルコース/mL反応)である。150分間、50mM酢酸ナトリウム、pH4.8条件下、アビセル濃度15mg/mLを用いて、反応を行った。CBH II酵母培養上清を、55℃でほぼ等量の還元糖産物の濃度を達成するため投与した。提示されるデータは、それぞれの温度点に関して二重の活性値を示すエラーバーと2つの独立した測定の平均である。
【図10】親1であるH.insolensおよび親3であるC. thermophilumからのブロック7ClustalW複数配列の配置である。矢印は、復帰突然変異における残基変異を示す。
【図11】21111331キメラ点突然変異に関するT50値を示す図である。値は、2つの独立した二重試験の平均値として示され、エラーバーは点突然変異に関する二重試験のそれぞれのT50値を示す。試験温度の50mM酢酸ナトリウム中で、pH4.8条件下、10分間実施された。残留活性は、上記緩衝剤中、1g/Lリン酸膨潤セルロース(PASC)と50℃で100分間のインキュベーションによって測定された。
【図12】H. insolensおよびH. jecorinaの親CBH II、Ser単一点突然変異およびB7P3ブロック置換キメラに関するT50値を示す図である。3つの独立した測定の平均、エラーバーとして示した値は、それぞれのCBH IIに関する1つの標準偏差を示す。不活性化は、試験される温度50mM酢酸ナトリウム中で、pH4.8条件下で、10分間実施された。残留活性は、上記緩衝剤中、1g/Lのリン酸、膨潤セルロース(PASC)と50℃で、100分間のインキュベーションによって測定された。
【図13】P. chrysosporium(H. jecorina CBMと融合した)およびS. cerevisiaeから非相同的に分泌されたCBH IIキメラ31311112,13231111および野生型CBH II触媒ドメインに関するT50値を示す。2つの独立した試験の測定値をエラーバーを用いて示す。不活性化は、試験される温度50mM酢酸ナトリウム中で、pH4.8条件下で10分間実施された。残留活性は、上記緩衝剤中、1g/Lリン酸、膨潤セルロース(PASC)を用いて50℃で100分インキュベーションすることによって測定された。
【図14】CBH II組み換え型ブロックの接合部位を示す図である。(a)新規非親残基の組み合わせが可能な場合(結合した球体)、ブロックの間の部位は、通常、その表面が曝露されており、潜在的に溶媒が接合部位をスクリーンすることを可能とする。(b)接合部位(B5−B6)の例は、バックボーン(位H. jecorinaおよびH. insolensに配置図)、の保存、表面上の可変的な残留物ならびに残基173および253(矢印)における新規な埋没した疎水性の対の比較的稀有な可能性を示す。(c)H. jecorina(図中、黒色で示す)からのブロック1−4は、基質結合と関連する移動が、B3部位(黄色)において観測される(矢印)にも関わらず、同属のH. insolensブロック(色分けされた図)と大きな偏差なく適合する。(d)B6、B7接点(矢印)での挿入欠失が構造変化を要するが、同属のブロック5−8も同様である。
【図15】C314S突然変異体およびその安定性への効果の構造分析を示す図である。(a)微細構造のH. insolens構造(1ocn)の水素の位置を、図1に示した。(b)類似したセリン構造の再構成された結合構造を、PyMOL(http: (//) www .pymol . org)でモデル化した。SHARPEN2モデリング・プラットフォーム(全ての原子ロゼッタエネルギー機能を有する)において、側鎖を最適化することで、Cys314およびSer314の双方が、Pro339のカルボニルの水素結合に寄与し、Gly316のアミドから水素結合を受容することを示唆した。セリンの優れた水素結合性能は、セリンを含む変異体のより高い安定性に影響し得る。他の説明は、補助的な位置を示すものである。特に、1ocnからのCys位置は、側鎖が顕著に曲がり(即ち、N−C−Cα−Cβからの不適切な上反角が、標準位置から6°である)、Proカルボニルからの距離が増加した、立体構造種を示す。図内の文字の後でない数値は、水素結合距離(オングストローム)を意味する。
【発明を実施するための形態】
【0014】
本明細書および特許請求の範囲で使用する、単数形“ある(a,an)”、および“この(the)”の語は、明確に記載しない限り、複数の指示物を含む。従って、例えば「ドメイン」の用語は複数のドメイン、「タンパク質」の用語は1つ以上のタンパク質を含む。
【0015】
また「または」の用語は、他に記載されない限り「および/または」を意味する。同様に「含む(comprise,comprises, comprising,include,includesおよびincluding)」は、同じ意味で用いられ、制限することは意図しない。
【0016】
さらに様々な実施形態の記載においては、「含む」の用語を使用し、当業者は、いくつかの特定の場合、実施形態が或いは「実質的になる」または「なる」の言語を使用して記載されていると理解するであろう。
【0017】
ここに記載されたものと同様または対応する方法および材料を、開示の方法および組成物の実施に使用することができ、ここでは、例示的な方法、装置および材料を開示するものである。
【0018】
他に規定がない限り、ここに使用される全ての専門的且つ化学的な用語は、一般的にこの開示が属する分野の当業者に理解されるものと同様の意味を有する。したがって、本出願にわたって使用される以下の用語は、以下の意味を有するものとする。
【0019】
最近の研究は、研究室レベルのバイオマス変換過程において、中等温度好性の対照物と比較して、好熱性菌からのセルラーゼが熱安定性に優れ、その高い安定性により、適温または高温状態における長期の活性保持を有することを示してきた。菌のセルラーゼについては、40g/Lまでの上清濃度で、高度に活性があり、菌宿主、例えばHypocrea jecorina(アナモルフ・トリコデルマ・リーゼイ)に発現することができることため、高い関心が寄せられている。不都合にも、熱安定性のある菌セルラーゼを発現する試料は、その数が少ない。。加工性のセロビオヒドラーゼクラスII(CBH II)酵素の場合、10個未満の天然熱安定遺伝子配列が、CAZyデータベースに掲載されている。
【0020】
ここに十分に記載するように、再帰的キメラポリペプチドの生成ならびに特定の安定したドメインおよび最終的な特定のアミノ酸の分析により、熱安定性に付与、活性化の改善がみられた。
【0021】
以下により詳細に記載するように、本発明は、少なくとも一部において、セルロースの加水分解に触媒作用を生じる新規酵素の生成および発現に基づくものである。一つの実施形態において、高温条件下でセルロースを加水分解するように改変された新規ポリペプチドを提供する。該ポリペプチドは、アミノ酸置換基を特定の残基で含むように変えられたセロビオヒドラーゼの変異体を含む。これらの変異体については以下でより詳細に記載されているが、本発明のポリペプチドは1つ以上の修飾されたアミノ酸を含んでもよいことが理解される。修飾されたアミノ酸の存在は、例えば(a)ポリペプチド半減期の長期化、(b)熱安定性および(c)基質の補充の増加において有利である。アミノ酸は、例えば、共翻訳中または翻訳後において、組み換え生産(例えば、哺乳類細胞での発現中の、N――X――S/TモチーフのN−結合型グリコシル化)の際に修飾されるか、合成手段によってによって修飾される。従って、「突然変異」、「変異体」または「修飾された」タンパク質、酵素、ポリヌクレオチド、遺伝子または細胞は、親のタンパク質、酵素、ポリヌクレオチド、遺伝子または細胞から選択されるか、由来するか、ある場合においては異なるか、変化した、タンパク質、酵素、ポリヌクレオチド、遺伝子または細胞を意味する。突然変異体もしくは修飾されたタンパク質または酵素は、必然的でないが、通常、突然変異体のポリヌクレオチドまたは遺伝子から発現する。
【0022】
「突然変異」は、突然変異タンパク質、酵素、ポリヌクレオチド、遺伝子または細胞を生じる、あらゆる工程または機構を意味する。これは、タンパク質、酵素、ポリヌクレオチドまたは遺伝子配列が変化するあらゆる突然変異、および、かかる突然変異から起因する細胞におけるあらゆる検出可能な変化を含む。通常、突然変異は、ポリヌクレオチドまたは遺伝子配列での、単一または複数のヌクレオチド残基の点変異、欠失または挿入によって生じる。突然変異は、タンパク質をコードする遺伝子の領域内で生じるポリヌクレオチドの変性、ならびにタンパク質コード配列外、例えば、調節またはプロモーター配列(これらに限定しない)領域の変性を含む。遺伝子内における突然変異は、「サイレント」であってもよく、即ち発現に応じてアミノ酸変性に反映されず、遺伝子の「配列保存的な」変異体をなってもよい。これは、通常、1個のアミノ酸が1種を超えるコドンに対応する場合において生じる。
【0023】
修飾されたアミノ酸の非制限的な例としては、グリコシル化アミノ酸、硫酸化アミノ酸、プレニル化(例えばファルネシル化、ゲラニルゲラニル化)アミノ酸、ビオチン化アミノ酸、カルボキシル化アミノ酸、リン酸化アミノ酸等が挙げられる。アミノ酸の修飾について、当業者を導くのに参照し得る文献が存在する。プロトコールの例は、WalkerのProtein Protocols(1998)(Humana Press, Towata、ニュージャージー州)のCD−ROMから取得可能である。
【0024】
本発明には、修飾したセロビオヒドラーゼポリペプチドを生成・分離する組み換え法を記載する。組み換え生成に加えて、固相技術を用いた直接的なペプチド合成(例えば、Stewart et al. (1969) Solid-Phase Peptide Synthesis (WH Freeman Co, San Francisco);およびMerrifield (1963) J. Am. Chem. Soc. 85: 2149-2154)によって、ポリペプチドを生成することができる。ペプチド合成は、手動または自動の方法を用いて実施することができる。自動合成は、例えばアプライドバイオシステムズ431Aペプチド合成装置(パーキンエルマー、カリフォルニア州フォスターシティ)を用いて、製造業者の提供する説明書に従って実施することができる。
【0025】
「セロビオヒドラーゼII」または「CBH II酵素」は、細菌類および菌類において多岐にわたるセルラーゼファミリー6タンパク質における酵素を意味する。酵素は、セルロースの加水分解に用いられる。
【0026】
「セルラーゼ活性」は、セルロースを加水分解できる酵素を意味する。セルラーゼは、セルロースの加水分解を触媒する菌類、細菌類および原生動物によって生成される酵素ファミリーを指す。しかしながら、他の種類の生物、例えば植物および動物によって生成されるセルラーゼもある。この群のEC番号は、EC3.2.1.4である。触媒作用に基づく、5つの一般的な種類のセルラーゼが存在する:エンドセルラーゼ;エキソセルラーゼであり、この部類内には2つの主な種類のエキソセルラーゼ(またはセロビオヒドラーゼ、省略してCBH)があり、1つの種類は還元端から進むように働き、もう1つの種類はセルロースの非還元末端から進むように働く;セロビアーゼまたはβグルコシダーゼ加水分解;酸化セルラーゼ;および、水の代わりにリン酸を用いてセルロース単量体へと分解するセルロースホスホリラーゼ。多くの菌のセルラーゼは、2つのドメイン;フレキシブルなリンカーによって結合している触媒ドメインおよびセルロース結合ドメインを有する。本発明のある実施形態において、セルラーゼ活性は、CBH活性である。ここに記載の配列は、いくつかの場合において、セルロース結合ドメインおよび触媒ドメインの双方、または触媒ドメインのみを含む。いくつかの場合において、触媒ドメイン配列のみを提供する場合、配列番号8で示されるようなセルロース結合ドメイン(CBD)が、直接又はリンカーを通して、触媒ドメインに機能的に連結し得ること(コード配列の一部とするか、後に融合するか)が認識されている。
【0027】
ここに用いられる「タンパク質」または「ポリペプチド」は、同じ意味で用いられており、1つ以上の化学基礎単位の鎖であり、化学結合であり、ペプチド結合と称するものによって共に連結されている複数アミノ酸を含む。「酵素」は、高度にまたは低度に特異的に1つ以上の化学または生化学反応を触媒または促進するあらゆる物質であり、好ましくは完全にまたは大半がタンパク質から構成されるものを意味する。「天然」または「野生型」タンパク質、酵素、ポリヌクレオチド、遺伝子または細胞は、自然に産出されるタンパク質、酵素、ポリヌクレオチド、遺伝子または細胞を意味する。
【0028】
「アミノ酸配列」は、アミノ酸のポリマー(タンパク質、ポリペプチド等)またはアミノ酸ポリマーを示す特徴となる文字列(character string)である。「タンパク質」、「ポリペプチド」および「ペプチド」の用語は、ここでは交換可能に同じ意味で使用される。「アミノ酸」は、中心の炭素原子が水素原子、カルボン酸基(ここに「カルボキシル炭素原子」として指す炭素原子)、アミノ基(ここに「網の窒素原子」として指す窒素原子)および側鎖基であるRの基に結合する構造を有する」分子である。アミノ酸をペプチド、ポリペプチドまたはタンパク質に取り入れる場合、1つのアミノ酸をもう1つのアミノ酸に結合させる脱水反応において、1つ以上のそのカルボン酸基の原子が欠失する。そのため、タンパク質に取り入れられたアミノ酸は「アミノ酸残基」と表される。
【0029】
一定のタンパク質の特定のアミノ酸配列(即ち、アミノ基末端からカルボキシ基末端まで表示される場合のポリペプチドの「一次構造」)は、コーディング部位のmRNAのヌクレオチド配列によって決定され、同様に遺伝情報、特にゲノムDNA(細胞小器官DNA、例えばミトコンドリアまたは葉緑体DNAを含む)によって特定される。従って、遺伝子の配列を決定することは、対応するポリペプチドの一次配列、より詳細には、遺伝子またはポリヌクレオチド配列によってコード化されるポリペプチドまたはタンパク質の役割または活性の予見を可能とする。
【0030】
特定の配列の「保存的アミノ酸置換」または単なる「保存的な変異体」は、アミノ酸配列と実質的に同一である1つ以上のアミノ酸または一連のアミノ酸の置換を示す。当業者は、アミノ酸の欠失、アミノ酸の付加または化学的に類似のアミノ酸との置換が起こる「保存的な変異体」を生じる、コード化配列における単一のアミノ酸または一定の割合のアミノ酸の変化、付加または欠失保存的を認識するであろう。
【0031】
機能的に同類のアミノ酸の保存的置換を示す表は、従来技術において既知である。例えば、ある保存的置換基は、アラニン(A)、セリン(S)およびトレオニン(T)を含む。他の保存的置換基は、アスパラギン酸(D)およびグルタミン酸(E)を含む。他の保存的置換基は、アスパラギン(N)およびグルタミン(Q)を含む。また他の保存的置換基は、アルギニン(R)およびリジン(K)を含む。他の保存的置換基はイソロイシン(I)、ロイシン(L)、メチオニン(M)およびバリン(V)を含む。他の保存的置換基は、フェニルアラニン(F)、チロシン(Y)およびトリプトファン(W)を含む。
【0032】
従って、列挙されたポリペプチド配列(例えば配列番号2,4,6および12−78)の「保存的アミノ酸置換基」は、同じ保存的置換基のアミノ酸から保存的に選択され、ポリペプチド配列のアミノ酸の割合としては、通常10%未満で含む。従って、本発明のポリペプチドの多様な保存的置換変異体は、同じ保存的置換基の多様な保存的置換変異体において、100,75,50,25または10個の置換を含み得る。
【0033】
核酸分子のコード化された活性を変えない配列の付加、例えば非機能的付加または非コード配列は、塩基性の核酸の保存的変異体であると理解されている。酵素の「活性」は、その反応を触媒する能力、即ち「機能」の測定値であり、反応生成物が生成される速度として表すことができる。例えば、酵素活性は、時間当たり、または酵素の単位(例えば濃度または重量)当たりについて、生成される生成物の量、または親和性、または解離定数に関して表すことができる。ここに同様の意味で用いられる「セロビオヒドラーゼ活性またはセルラーゼ活性」、「セロビオヒドラーゼまたはセルラーゼの生物学的活性」または「セロビオヒドラーゼまたはセルラーゼの機能的活性」は、特定の実施形態においては、タンパク質、セルラーゼ活性を有するポリペプチドであり、標準の技術によって生体内または生体外で決定される、セロセルロース基質へのビオヒドラーゼ活性によって及ぼす活性を指す。
【0034】
当業者は、開示される、機能的に同一の構造を有する核酸構造の多くの保存的変異体を理解するであろう。例えば、上記のように、遺伝子のコーディングの縮退のため、「サイレント置換」(即ち、核酸配列の置換であり、コードされたポリペプチドには変異を生じない)は、アミノ酸をコードする各々の核酸配列の特徴を包含する。同様に、アミノ酸配列における1つまたは数個のアミノ酸の「保存的アミノ酸置換」は、高い相同性の特徴を有する異なるアミノ酸と置換され、また開示の構造と高い相同性を有することが容易に同定される。それぞれ開示の配列の、このような保存的変異体は、ここに提供するポリペプチドの特性である。
【0035】
「保存的変異体」は、タンパク質または酵素の全体の構造および機能を変えることなく、アミノ酸残基を変化させた、タンパク質または酵素であり、同様の特性、極性または非極性、サイズ、形状、電荷等(これらに限定されない)を有するアミノ酸への置換を包含する。保存的なものとして示されるアミノ酸以外は、タンパク質または酵素において異なっていてもよく、あらゆる2つの同様の機能のタンパク質間のタンパク質またはアミノ酸配列の類似性は、異なっていてもよく、配列図によって決定されるように、例えば少なくとも30%、少なくとも50%、少なくとも70%、少なくとも80%または少なくとも90%であってもよい。本明細書において、「配列類似性」は、ヌクレオチドまたはタンパク質配列が連関する範囲を意味する。2つの配列の類似範囲は、配列類似性のパーセントおよび/または保守性に基づくことができる。ここに「配列相同性」は、2つのヌクレオチドまたはアミノ酸配列の範囲に差異がないことを意味する。「配列のアラインメント」は、最大限の一致を達成するため(アミノ酸配列の場合保存性)、類似性の程度を評価することを目的とする2つ以上の配列を揃える工程を意味する。配列を揃えて、類似性/相同性を評価するための数々の方法は、従来技術、例えばCluster法において既知であり、類似性はMEGALIGNアルゴリズムならびにBLASTN、BLASTPおよびFASTA(Lipman and Pearson, 1985; Pearson and Lipman, 1988)に基づくものが知られる。このプログラム全てを使用して、最も高い配列相同性を生じるものが好ましい設定といえる。
【0036】
特定のポリペプチドの非保存的な修飾としては、保存的置換を特徴としないあらゆるアミノ酸の置換が挙げられる。例えば、上記の6つの群の範囲を超えるあらゆる置換が挙げられる。これらは、塩基性または酸性のアミノ酸から中性のアミノ酸(例えば、Asp、Glu、AsnまたはGlnからVal、Ile、LeuまたはMet)、芳香族アミノ酸から塩基性または酸性アミノ酸(例えば、Phe、TyrまたはTrpからAsp、Asn、GluまたはGln)への置換、またはアミノ酸とその類似するアミノ酸との置換ではない、あらゆる他の置換を含有する。塩基性側鎖としては、リジン(K)、アルギニン(R)、ヒスチジン(H);酸性側鎖としては、アスパラギンサン(D)、グルタミン酸(E);電荷を有さない極性側鎖としては、グリシン(G)、アスパラギン(N)、グルタミン(Q)、セリン(S)、トレオニン(T)、チロシン(Y)、システイン(C);非極性側鎖としては、アラニン(A)、バリン(V)、ロイシン(L)、イソロイシン(I)、プロリン(P)、フェニルアラニン(F)、メチオニン(M)、トリプトファン(W);β岐側鎖としては、トレオニン(T)、バリン(V)、イソロイシン(I);芳香性の側鎖としては、チロシン(Y)、フェニルアラニン(F)、トリプトファン(W)、ヒスチジン(H)が挙げられる。
【0037】
「親」タンパク質、酵素、ポリヌクレオチド、遺伝子または細胞は、あらゆる他のタンパク質、酵素、ポリヌクレオチド、遺伝子または細胞に由来するか、あらゆる方法、装置または技術を用いて生成され、親そのものが天然か、突然変異体である任意のタンパク質、酵素、ポリヌクレオチド、遺伝子または細胞である。親ポリヌクレオチドまたは遺伝子は、親タンパク質または酵素をコードする。
【0038】
CBH IIポリペプチドの変異体を提供することに加えて、1)第1の親鎖から分離され、アミノ酸置換基を含むように修飾された変異体ドメイン;および2)第2の親鎖から分離され、新規活性またはドメインを補足する活性を含有するために非修飾または修飾のいずれかから分離されたドメインを含むキメラポリペプチド;が提供される。本発明のキメラポリペプチドの技術にかかる方法をここに開示する。
【0039】
本発明は、野生型、または親タンパク質と比較して高い熱安定性を有するセルラーゼおよびセロビオヒドラーゼ(CBH)II変異体、突然変異体およびキメラを開示し、野生型タンパク質は配列番号2,4または6からなる。変異体は、配列GEXDGのモチーフを有するC末端領域であって、セリンを含有し、前記Xは、C、AまたはG(配列番号107)であり、Xは、セリン、セルラーゼ活性を含む変異体と置換され、ポリペプチドは配列GEXDG(配列番号107)においてセリンを欠如している野生型セルラーゼと比較して高い熱安定性を有する。一つの実施形態において、変異体は多くのセルラーゼおよびセロビオヒドラーゼIIタンパク質において見出されるモチーフGECDG(以下により完全に記載する)において少なくともCys→Serの突然変異(例えば、配列番号2のアミノ酸312−316を参照のこと)を含み、熱安定性または活性を向上させた付加突然変異を含んでいてもよい。セルラーゼ間の相同性は、非常に低くてもよい。上記セリン置換基は、配列番号107のモチーフ(例えば、ポリペプチドが配列番号2または4と少なくとも60%以上の相同性がある)を有するあらゆるセルラーゼに適用できる。
【0040】
例えば、本発明は高い熱安定性およびセルラーゼ活性を有し、C314Sを含む配列番号2と少なくとも85%、90%、95%、98%、99%の相同性があり;C311Sを含む配列番号4と少なくとも85%、90%、95%、98%、99%の相同性があり;C310Sを含む配列番号12と少なくとも85%、90%、95%、98%、99%の相同性があり;C312Sを含む配列番号13と少なくとも85%、90%、95%、98%、99%の相同性があり;C314Sを含む配列番号14と少なくとも85%、90%、95%、98%、99%の相同性があり;C315Sを含む配列番号15と少なくとも85%、90%、95%、98%、99%の相同性があり;C313Sを含む配列番号16と少なくとも85%、90%、95%、98%、99%の相同性があり;C311Sを含む配列番号17と少なくとも85%、90%、95%、98%、99%の相同性があり;C313Sを含む配列番号19と少なくとも85%、90%、95%、98%、99%の相同性があり;C314Sを含む配列番号2と少なくとも85%、90%、95%、98%、99%の相同性があり;C312Sを含む配列番号21と少なくとも85%、90%、95%、98%、99%の相同性があり;C311Sを含む配列番号22と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号64と少なくとも85%、90%、95%、98%、99%の相同性があり;C407Sを含む配列番号65と少なくとも85%、90%、95%、98%、99%の相同性があり;C394Sを含む配列番号66と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号67と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号68と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号69と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号70と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号71と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号72と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号73と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号74と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号75と少なくとも85%、90%、95%、98%、99%の相同性があり;C407Sを含む配列番号76と少なくとも85%、90%、95%、98%、99%の相同性があり;C394Sを含む配列番号77と少なくとも85%、90%、95%、98%、99%の相同性があり;またはC412Sを含む配列番号78と少なくとも85%、90%、95%、98%、99%の相同性がある配列を提供し、前記ポリペプチドは、セルラーゼ活性およびその相当するCys→Ser突然変異を欠如している親(野生型)タンパク質と比較して高い熱安定性を有する。
【0041】
さらに別の実施形態において、本発明は上記ポリペプチドを提供するが、それらはさらに配列番号64の配置として同定される少なくとも1つの付加的な突然変異を含み、配列番号64は位置413でPro、位置231でSerまたはThr、位置305でSerまたはThr、位置410でGlnまたはAsn、位置82でGlu、または上記のあらゆる組み合わせ置換を包含する。同様の置換は、アミノ酸配列の配列番号2,4,6,12−63および65−78と、配列番号64の配列の配置として同定する。
【0042】
本発明は、組み換えにより生成されたか、合成されたか、非天然的に生成されたか、のいずれの実質的に生成されたポリペプチドをも提供するものであり、該ポリペプチドは、C→S置換位置を除き、下記の1−10、10−20または20−30の位置の同定された配列の保存アミノ酸置換を有する:
C314Sを含む配列番号2;C311Sを含む配列番号4;C310Sを含む配列番号12;C312Sを含む配列番号13;C314Sを含む配列番号14;C315Sを含む配列番号15;C313Sを含む配列番号16;C311Sを含む配列番号17;C313Sを含む配列番号19;C312Sを含む配列番号21;C310Sを含む配列番号22;C400Sを含む配列番号64;C407Sを含む配列番号65;C394Sを含む配列番号66;C400Sを含む配列番号67;C400Sを含む配列番号68;C400Sを含む配列番号69;C400Sを含む配列番号70;C400Sを含む配列番号71;C400Sを含む配列番号72;C400Sを含む配列番号73;C400Sを含む配列番号74;C400Sを含む配列番号75;C407Sを含む配列番号76;C394Sを含む配列番号77;またはC412Sを含む配列番号78。
【0043】
「分離されたポリペプチド」は、自然に伴う他の混入物、例えばたんぱく質、脂質およびポリヌクレオチドから抽出されたポリペプチドを指す。用語は、それら天然に分泌された環境または発現系(例えば宿主細胞または生体外合成)から除去されたか、精製されたポリペプチドを包含する。
【0044】
「実質的に純粋なポリペプチド」は、ポリペプチドの種類が主成分として存在する組成物(即ち、モルまたは重量に基づいて、ポリペプチドがあらゆる他の各々の高分子成分より組成物において多量に存在する)を指し、対象となる成分がモルまたは%重量によって示される高分子成分の少なくとも約50パーセントを含む場合、一般的に実質的に精製された組成物とする。概して、実質的に純粋なポリペプチド組成物は、モルまたは重量%で組成物中に存在する全ての高分子成分の約60%以上、約70%以上、約80%以上、約90%以上、約95%以上、約98%以上含まれる。いくつかの実施形態において、目的の成分は、実質的に均質となるまで精製され(即ち、混入物成分は従来の検出法によって組成物中に検出されない)、組成物は実質的に単一の高分子成分からなる。溶媒の種類、低分子成分(<500ダルトン)および基本的なイオン成分は、高分子成分としては考慮されない。
【0045】
「参照配列」は、配列比較の基準として使用される、規定された配列を指す。参照配列は、例えば、遺伝子全長またはポリペプチド配列全長の断片のような、より長鎖の配列のサブセットとしてもよい。概して、参照配列の長さは、少なくとも20ヌクレオチドまたはアミノ酸残基、少なくとも25ヌクレオチドまたはアミノ酸残基、少なくとも50ヌクレオチドまたはアミノ酸残基、または核酸またはポリペプチドの全長とすることができる。2つのポリヌクレオチドまたはポリペプチドは、それぞれ(1)2つの配列の間で類似の配列(即ち、完全な配列の一部)を含み、(2)2つの配列の間で異なる配列をさらに含むため、2つ(またはそれ以上)のポリヌクレオチドまたはポリペプチドの間の配列の比較は、配列類似性の局部領域を同定し、比較するため、典型的に2つのポリヌクレオチドまたはポリペプチドの配列を「比較領域」にわたって比較することによって実施される。
【0046】
「配列相同性」は、2つのアミノ酸配列が、比較領域にわたって実質的に相同性(即ち、アミノ酸−アミノ酸比較に基づく相同性)があることを意味する。「配列類似性」の用語は、同じ生物物理学的な特徴を共有する同様のアミノ酸を指す。「配列相同性のパーセンテージ」または「配列類似性のパーセンテージ」の用語は、2つの最適に配置された配列を比較領域にわたって比較し、適合する位置を計数するため、相同性残基(または同一残基)が双方のポリペプチド配列存在する位置を計数し、比較領域(即ち、領域サイズ)における適合する位置の数を総数で除算し、配列の一致のパーセンテージ(または配列類似性のパーセンテージ)を算出し、結果を100で乗算して、得ることができる。ポリヌクレオチド配列に関して、配列の相同性および類似性の用語は、タンパク質配列に関して記載されるそれと同様に、比較領域にわたって、2つのポリヌクレオチド配列が同一(ヌクレオチド−ヌクレオチド比較に基づく)である「配列相同性のパーセンテージ」の意味を有する。このように、ポリヌクレオチド配列の相同性のパーセンテージ(またはポリヌクレオチド配列の類似性のパーセンテージ、例えば、分析アルゴリズムに基づいたサイレント置換または他の置換)も算出することができる。最大の相関性は、ここに記載の1つ以上の配列アルゴリズム(または当業者が利用できる他のアルゴリズム)の使用、または目視によって決定することができる。
【0047】
ポリペプチドに適用する場合、実質的な相同性または実質的な類似性の用語は、最適な配列を有する場合の2つのペプチド配列が、初期条件のギャップ重量(gap weight)を用いたBLAST、GAPまたはBESTFITのプログラムや、目視等において、配列の相同性または類似性を共有することを意味する。同様に、二つの核酸において、実質的相同性または実質的類似性の用語は、最適な配列を有する場合の2つの核酸配列が、初期状態のギャップ重量を用いたBLAST、GAPまたはBESTFITのプログラムやまたは目視等において、配列相同性または類似性を共有することを意味する。
【0048】
配列相同性または配列類似性のパーセンテージを決定するために適切なアルゴリズムの1つの例は、Pearson, W. R. & Lipman, D. J., (1988) Proc. Natl. Acad. Sci. USA 85:2444に記載されたFASTアルゴリズムである。またW. R. Pearson, (1996) Methods Enzymology 266:227-258を参照してもよい。相同性または類似性のパーセンテージを算出するため、DNA配列のFASTAアライメントにおいて使用するのに好ましいパラメーターは、以下の様に最適化される:BL50 Matrix 15: -5, k-tuple=2; joining penalty=40, optimization=28 ; gap penalty -12, gap length penalty=-2;およびwidth=16。
【0049】
実用的なアルゴリズムの他の例として、PILEUPが挙げられる。PILEUPは、進行する対となる配置(アライメント)を使用して、連関する配列群からの多重配列アライメントを作成することにより、配列群の相関性および配列相同性と配列類似性のパーセンテージを示す。それは、進行するアライメントを形成するためのクラスター関係を示す、樹状図または系統樹もプロットする。PILEUPは、Feng & Doolittle, (1987) J. Mol. Evol. 35:351-360の進行アライメント法を簡略化したもの使用する。使用される方法は、Higgins & Sharp, CABIOS 5:151- 153, 1989に記載された方法に類似の方法である。プログラムは、最高300の配列を配置(アライン)することができ、それぞれ最長5000のヌクレオチドまたはアミノ酸を配置することができる。多重アラインメント法は、2つの最も相同性のある配列対のアライメントから始まり、2つのアラインされた配列のクラスターを生成する。その後、該クラスターは、次に最も関連した配列またはクラスター配列のクラスターにアラインする。2つのクラスターの配列は、2つの個々の配列の対アライメントの単純な延長によって配置される。最終的なアライメントは、一連の前進的な対アライメントによって達成される。プログラムは、配列比較領域における、特定の配列およびそのアミノ酸もしくはヌクレオチドの連携を設計すること、およびプログラムパラメーターを設計することによって実行される。PILEUPを使用することで、参照配列を以下のパラメーターを使用した関連の他の試験配列と比較し、パーセント配列相同性のパーセンテージ(または配列類似性のパーセンテージ)を決定した。:初期状態のギャップ重量(3.00)、初期状態のギャップ長さ(0.10)および重量化した末端のギャップ。PILEUPは、GCG配列分析ソフトウェアパッケージ、例えばバージョン7.0から得られる。
【0050】
多数のDNAおよびアミノ酸配列に適したアルゴリズムの他の例としては、CLUSTALWプログラムが挙げられる。CLUSTALWは、配列群間の複数の対比較を実行し、それらを多重のアライメント配列の相同性に基づいて組み立てる。ギャップオープンおよびギャップエクステンションのペナルティーは、それぞれ10および0.05であった。アミノ酸アライメントに関して、BLOSUMアルゴリズムをタンパク質重量マトリックスとして使用することができる。
【0051】
上記のように、セロビオヒドラーゼおよびセルラーゼファミリーのメンバーは、配列アライメントおよびGECDGモチーフへの置換(例えば、配列番号2のアミノ酸312−316を参照のこと)によって同定することができる。修飾されたポリペプチドは、有利な活性を導入するように修飾されたものを同定するため、その後以下に記載されるような様々な温度および条件で、その活性を分析される。典型的な配列は、以下のGenBank登録番号であり、参照によりここに援用される配列から見出すことができる。
【0052】
【表1】
【0053】
さらに他の実施形態において、改良した熱安定性のある多様なセルラーゼポリペプチドファミリーは、C→S、G→SまたはA→S置換を有する以下の表に示すものを含む。さらに、以下の表に一致する置換、セルラーゼ活性および熱安定性のあるものを有する、以下いずれかの配列と85%、90%、95%、98%または99%配列の一致を有するポリペプチドも本発明に包含される。
下表は、H. jecorina CBH IIと高い相同性を有するタンパク質配列に関するアミノ酸の枠内(frame bracketing)H. jecorina CBH II Cys311のアライメントをしめす。残基311に想到する位置を下線太文字で示す。この試験において検討された組み換え型H. insolensおよびP. chrys CBHIIの配列は、H. insoおよびP. chrysとして示す。250個のうち、最も相同性のある配列の54個は、重複しているため省略した(即ち、構造研究のための点突然変異体または>95%一致するイソタイプ)。セルラーゼに関する受入番号が同定され、相当する配列が、ここに参考文献によって受入番号から直接複製されるように、本発明に組み込む。受入番号と関連する配列が、配列番号79−106として参照される。太字下線部の残基(例えばC、AまたはG)は、Sに置換される。配列番号は、下記の配列中の数値による:
【0054】
【表2】
【0055】
開示の目的のため、親セルラーゼより少なくとも約4℃または少なくとも約9℃高いT50、例えば約4℃から約30℃高い、もしくはその間のあらゆる値のT50を有するセロビオヒドラーゼ、または約9℃から約30℃高い、もしくはその間のあらゆる値のT50を有するセロビオヒドラーゼの場合、親セロビオヒドラーゼと比較したとき、開示のポリペプチドは、相当する親ポリペプチドに関して改良された熱安定性を示す。T50は、15分間のプレインキュベーション後、修飾された酵素または天然の酵素が、その残留活性の50%を保持する温度であり、以下の実施例または従来技術における詳細な分析によって測定される。
【0056】
開示の修飾されたセロビオヒドラーゼまたはセルラーゼは、相当する親セロビオヒドラーゼ(例えば配列番号2,4または6)より約4℃から約30℃高いか、もしくはその間の範囲にあり、約5℃から約20℃高いか、もしくはその間の範囲にあり、約8℃から約15℃高いか、もしくはその間の範囲にあり、約9℃から約15℃高いか、もしくはその間の範囲にあるT50を有していてもよい。例えば、修飾されたセルラーゼは、相当する親セロビオヒドラーゼのものより少なくとも約4,5,6,7,8,9,10,12,14,16,18,20,22,24,26,28または30℃高いT50を有することがあり得る。
【0057】
ここに同定される変異体も、セロビオヒドラーゼを生成するために使用することができる。例えば、SCHEMAは、何百もの活性β−ラクタマーゼおよび活性シトクロームP450の酵素キメラのファミリーを形成するため、前もって使用されてきた。SCHEMAは、ブロックが異なる親同士で交換されたときに分解されるアミノ酸側鎖接触のライブラリー平均数である<E>を最小化する、接触したアミノ酸の「ブロック」の境を規定するため、タンパク質構造データを使用する。β−ラクタマーゼキメラが折り畳まれ、活性を有する可能性が、その配列のEの値に反比例することが示された。突然変異のライブラリー平均数<m>に対する<E>を最小化するようなブロック境界を同定するため、RASPP(最短距離問題としての組み換え)アルゴリズムが使用された。3つの親からの8つのブロック(38=6,561通りの配列)を含む、β−ラクタマーゼの収集物を特徴とする500個までの20%を超える固有のキメラは、触媒活性を有する。同様の方法により、2,300の新規の、触媒活性がある酵素を含む3つの親からの8ブロックのシトクロームP450キメラファミリーを生成した。これら2つの収集物からのキメラは、最も近い親から、それぞれ平均して66および72の多数のアミノ酸配列の突然変異を有することを特徴とする。したがって、SCHEMA/RASPPは、著しい配列多様性、およびかなりの部分の機能メンバーを有するキメラファミリーの設計を可能とした。
【0058】
SCHEMAキメラの熱安定性が、配列の小さいサンプルからの配列安定性データに基づいて予測されることが、示されてきた。184のシトクロームP450キメラの熱安定性に関する直線回帰モデルは、SCHEMAブロックが、熱安定性に付加的な寄与をすることを示した。300を超えるキメラが、このモデルによって熱安定性であると予測され、試験された全44が最も安定した親と比較して、より高い安定性を有した。わずか35の熱安定性試験の使用によって、最も熱安定性のあるキメラを予測することができた。さらに、熱安定性のあるP450キメラは、キメラ形成がさらなる実用的な酵素特性を導くことができることを証明する、固有の活性および特性プロフィールを示した。ここでSCHEMA組換え型のCBH II酵素は、高温で、長期間にわたって、幅広い範囲のpH条件下のリン酸膨潤セルロース(PASC)において、活性を有するキメラセルラーゼを生じることができる。
【0059】
ここに記載の方法を使用して、セロビオヒドラーゼ活性を有する多くのキメラポリペプチドを、野生型親CBH IIタンパク質と比較して、改良した特性を有するものとして産生した。
【0060】
新規CBH II酵素の多様なファミリーが、3つの菌のCBH II酵素からの配列のブロックを交換することによって作成された。このセットから採取された48のキメラ配列のうちの23個は、S. cerevisiaeによって活性形状で分泌され、5つは63℃で最も安定な親より長い半減期を有する。この48のメンバーのサンプルセットが、存在し得る全6,561配列の1%未満であるので、このキメラの収集物は、100もの活性型、熱安定性のあるCBH II酵素、CAZyデータベースにおけるより少数化された約20の菌CBH II酵素を有することが予測される。
【0061】
熱安定性を促進するブロックを同定するためにサンプルセット配列安定性データを使用する方法は、第2のCBH II検証セットにおける全10個の触媒活性キメラが、最も安定な親である好熱性菌H. insolensからの天然の熱安定性のあるCBH IIより熱安定性であることにより検証された。したがって、本発明は、S. cerevisiaeから触媒活性を有する形態で発現する全33の新規CBH II酵素を生じ、そのうち15は、それらが生成される最も安定な親より熱安定性である。これら15の熱安定性のある酵素は、互いにおよび、それらに最も近い天然相同物から、それぞれおよそ94および58アミノ酸の位置で異なる。
【0062】
組み合わせたサンプルおよび検証セットにおけるCBH IIキメラの熱安定性の分析は、同じキメラに存在する場合B1P1、B6P3、B7P3およびB8P2として同定された4つの熱安定のあるブロックが、熱安定性に累積的に寄与することを示す。H. insolens CBH IIと比較してより熱安定性のある5つのサンプルセットキメラのうちの4つは、2つまたは3つの安定性のあるブロックを含む(Table1)。H. insolens酵素より比較的安定な全ての検証セットの10個の活性型メンバーは、すべてH. insolens酵素より安定しているが、3つまたは4つの安定性のあるブロックを含有するこの群において、最も熱安定性のある6つのキメラのうちの5つで、少なくとも2つの安定性のあるブロックを含有する。
【0063】
本発明は、安定性ブロックが組みかえられ、新規の、高い安定性を有する活性型セルラーゼが生成されることを示す。安定性回帰モデルは、CBH II SCHEMAライブラリーが親酵素の最も安定なものと比較しても、より安定が2.026のキメラを有することを予測する。これらのキメラは多様であり、生来のセルラーゼと明確に区別できる:それらは親から8から72箇所の突然変異(平均50箇所)、キメラ間では平均63箇所の突然変異を有して多様化される。このセットから計33個の遺伝子がS. cerevisiaeにおいて合成され、発現される:これらキメラCBH IIの全てについて、63℃またはT50での不活性化の半減期が測定され、いずれも好熱性菌H. insolens由来の最も安定な親セルラーゼと比較しても、より安定であることが見出された。8個のみのブロックのキメラを作成して配列の複雑化を減少させることで、配列安定性モデルの生成および単一の高い安定性のある配列ブロックの同定が可能となる。このブロックにおける10個のみのアミノ酸置換の試験をすることによって、単一の高い安定化効果のある置換を同定した。キメラおよび生来のP. chrysosporium、H. insolensおよびH. jecorina CBH II酵素にわたってC313S(配列番号6;C314S、配列番号2;C311S、配列番号4)の置換による非常に大きな安定性効果が観測され、この位置のあらゆる残基のSerへの突然変異が、それが導入されたあらゆるセルラーゼ6ファミリーを安定化させることを示す。
【0064】
組み換えにおける接触分離の数を最小化することは(図2C)、ブロックがそれぞれ独立してタンパク質全体の安定性を高める、分離されたユニットに近い状態となることを可能としこれにより、累積的またはさらに付加的なキメラの熱安定性向上効果につながる。このCBH II酵素の組み換えに関して、SCHEMAは前記の接触分離を最小化するのに効果的であった:H. insolens親CBH II結晶構造において規定されるブロックの間の接触は計303ヶ所あり、CBH II SCHEMAライブラリーのデザインによると、潜在的な接所分離は33ヶ所のみであった。CBH II酵素の親が、そのサブドメインに明らかな構造上の特徴を有さず、且つ、8つのブロックのうちの4つのみ(1,5,7および8)が小型の構造ユニットまたはモジュール様の構造を示す場合、接触分離が低値であることはブロックの数が下位構造区分の数を超える場合においてSCHEMA/RASPPアルゴリズムが効果的であることを示した。β−ラクタマーゼおよびシトクロームP450キメラに関して前もって観測されるように、低いE値により、キメラの折り畳みおよび活性が予測可能である。ここでは使用しないが、この関係は、高い割合の活性型メンバーを含むキメラサンプルセットを設計するのに有用である。
【0065】
本発明において、キメラを用いて、pHの安定性をCBH II酵素において改良できるかを検討した。H. jecorina CBH IIの特異的活性は、pHが最適値5を超えて上昇するにつれて急激に減少し、一方、安定性のあるブロックを最も産業的に関連のあるH. jecorina CBH II酵素上に置換することによって作成されるHJプラスは、高いpHで顕著にその活性を保持する(図4)。熱安定性のある11113132および13311332キメラ、またH. insolensおよびC. thermophilum CBH IIセルラーゼ親は、HJプラスと比較してより幅広いpH/活性特性を有する。H. jecorina CBH IIの制限されたpH/活性特性は、いくつかのカルボキシルカルボン酸対の脱プロトン化に起因し、pH6を超えるタンパク質を不安定化する。HJプラスのブロック7における親3の置換は、カルボキシルカルボン酸対である(ブロック8の)D277およびD316の間のカルボキシルカルボン酸の対を解除し、アスパラギンサン277をヒスチジンに変更する。正荷電のヒスチジンとD277を置換することは、H. jecorina CBH IIと比較してより高いpHで、HJプラスが活性を保持することを可能とし、非酸性pHにおける電荷斥力の不安定化を予防することができる。残存する2つの熱安定性のあるキメラならびにH. insolensおよびC. thermophilum親CBH II酵素のより幅広いpH/活性特性は、HJプラスおよびH. jecorina CBH IIのE57−E119カルボキシルカルボン酸の対に相応する位置での酸残基の欠如によるものであろう。
【0066】
HJプラスは、比較的高い特異的活性および高い熱安定性の双方を示す。図5は、これらの特性の長期にわたる加水分解実験における優れた機能を示す:HJプラスは、親CBH II酵素と比較して7−15℃高い温度でセルロースを加水分解し、最適温度において、全ての親と比較して著しく向上した長期活性を示した(H. jecorina CBH IIの1.7倍)。HJプラスキメラの特異的活性がH. jecorina CBH II親のものより低い場合、この向上した長期活性は、HJプラスの熱安定性に由来し、最適な加水分解温度での比較的長い反応時間にわたる活性を保持できる。
【0067】
他の2つの熱安定性のあるキメラは、HJプラスの幅広い温度範囲を共有した。この観測は、上昇した温度t1/2と最高動作温度の間の正の相関を支持し、6,561のCBH IIキメラ配列中の多くの熱安定性キメラもセルロースを高い温度で分解できることを示唆する。高い温度での非結晶性PASC基質を加水分解するこの特性が、熱安定性のある菌CBH IIキメラの潜在的有用性をよく予見させる。一方、結晶基質およびリグニン含有基質の比較的困難な研究は、この新規CBH II酵素グループのバイオマス分解への応用に関して、より完全な評価を可能とする。
【0068】
バイオマス変換工程の大半は、高いレベルのセルロース加水分解を達成するため、菌セルラーゼの混合物(主にCBH II、セロビオヒドラーゼクラスI(CBH I)、エンドグルカナーゼおよびβ−グルコシダーゼ)を使用する。熱安定性のあるCBH II酵素キメラの多様な群を生成することは、酵素混合物を特定の応用および貯蔵のために調製し、最適化することができる、安定で、且つ活性の高いセルラーゼのストックを作成する第1工程である。
【0069】
「ペプチド断片」は、比較的大きなポリペプチドまたはタンパク質の一部分もしくは断片を指す。ペプチド断片は、それ自身、機能活性を有する必要性がないが、いくつかの場合において、ペプチド断片は、生物学的活性を有するポリペプチドドメインに相当する。安定性に関するペプチド断片は、ペプチド断片を有しないポリペプチドと比較して、安定性、機能または折り畳み度合いを促進するポリペプチドにおいて見出されるペプチド断片である。不安定性にペプチド断片は、ポリペプチド内に存在する場合、安定性、機能または折り畳み度合いを欠く原因として同定されるペプチド断片である。
【0070】
「融合」、「作動可能に結合された(operably linked)」および「作動可能に連結された(operably associated)」は、2つの明白なドメインまたはペプチド断片の化学的または物理学的な結合を幅広く指すため、ここに同じ意味で用いられて使用することができ、作動可能に結合している場合、それぞれのドメインまたはペプチドの断片は、望ましい活性を有する機能的なポリペプチドを提供する。ドメインまたはペプチド断片は、それらが機能的であるようにリンカーペプチドによって結合していてもよく、他の中間体または化学結合によって融合することができる。例えば、2つのドメインは、同じコード配列の一部であってもよく、ポリヌクレオチドは、単一のmRNAに転写されるようにコード化され、双方のドメインを含む単一のポリペプチドに翻訳されるように、同じ「フレーム」内に存在する。或いは、両方のドメインを各々のポリペプチドとして別々に発現させ、化学的手法により互いに融合させてもよい。具体的には、コーディングドメインは、「フレーム」内で直接的に結合するか、またはペプチドリンカーを間に挟んで隔離されており、単一のポリヌクレオチドをコーディングする形態とする。ペプチドリンカーおよびペプチドのコード配列は、従来技術において既知である。
【0071】
「ポリヌクレオチド」または「核酸配列」は、ヌクレオチドの多量体型を指す。ある場合において、ポリヌクレオチドは、由来生物の天然に生じるゲノムにおいて、それが隣接する2つのコード配列(1つは5’末端側、もう1つは3’末端側で隣接する)のいずれかには直接隣接しないものを指す。従って、前記用語は、例えばベクター;独立して複製するプラスミドまたはウイルス;原核生物もしくは真核生物のゲノムDNAに組み込まれる組み換え型DNA、または他の配列から独立して別の分子(例えばcDNA)として存在するものを含む。本発明のヌクレオチドは、リボヌクレオチド、デオキシリボヌクレオチド、または修飾されたいずれかのヌクレオチドであってもよい。ここに使用されるポリヌクレオチドは、とりわけ、一本鎖および二本鎖のDNA、一本鎖領域および二本鎖領域が混合しているDNA、一本鎖および二本鎖のRNA、および一本鎖領域および二本鎖領域が混合しているRNA、一本鎖、二本鎖、または一本鎖領域および二本鎖領域が混同したDNAおよびRNAのハイブリッドを指す。ポリヌクレオチドの用語は、ゲノムDNAまたはRNA(生物、即ちウイルスのRNAゲノム)並びにゲノムDNAおよびcDNAにコード化されたmRNAを包含する。
【0072】
「核酸断片」、「オリゴヌクレオチド断片」または「ポリヌクレオチド断片」は、比較的大きなポリヌクレオチド分子の一部を指す。ポリヌクレオチド断片は、タンパク質の機能ドメインをコードする必要はないが、タンパク質の機能ドメインをコードすることもあろう。ポリヌクレオチド断片は、約6ヌクレオチド以上の長さ(例えば、6−20,20−50,50−100,100−200,200−300,300−400ヌクレオチド以上の長さ)としてもよい。安定性に関するペプチド断片は、安定性に関するポリヌクレオチド断片によってコードされてもよく、ペプチド断片は、ペプチド断片を欠如するポリペプチドと比較して、ポリペプチドの安定性、機能または折り畳み度合いを促進する。
【0073】
「キメラ」は、少なくとも2つの異なる親タンパク質の少なくとも2つの断片の組み合わせを指す。当業者に理解されるように、それが関連のある特定の配列であって、そのものが物理的に核酸でないものであれば、実際、断片はそれぞれの親から由来する必要はない。例えば、キメラ菌クラスIIセロビオヒドラーゼ(CBH IIセルラーゼ)は、2つの異なる親CBH IIポリペプチドからの少なくとも2つの断片を有する。2つの断片は、セルラーゼ活性を有する新規ポリペプチドを生じるように結合している。即ち、タンパク質は、いずれかの親タンパク質の全長に一致する配列を有する場合、キメラとはいない。キメラポリペプチドは、2つ以上の異なる親タンパク質からの2つ以上の断片を含む。例えば、それぞれ最終的なキメラまたはキメラのライブラリーにつき、2,3,4,5−10,10−20以上の親がある。それぞれの親ポリペプチドの断片は、非常に短いか、非常に長いこともあり、断片はタンパク質の全長の1〜90%、95%、98%または99%範囲の連続した長さのアミノ酸配列を有し得る。ある実施形態において、最短の長さは10アミノ酸である。ある実施形態においては、2つの親タンパクについて単一のクロスオーバー点が定義される。クロスオーバーの位置は、一方の親のアミノ酸断片が終結し、他方の親のアミノ酸断片が始まるところに規定される。従って、単純なキメラは、クロスオーバー位置前の断片が第1の親、クロスオーバー点後の断片が第2の親に属する場所にクロスオーバー位置がある。ある実施形態において、キメラは複数の交差位置を有する。例えば、2,3,4,5,6,7,8,9,10,11−30個、またはこれを超えるクロスオーバー位置を有する。該クロスオーバーの位置および命名は、いずれも以下に記載するように規定する。2つのクロスオーバー位置および2つの親がある実施形態において、第1の親からの第1の連続断片に続いて、第2の親からの第2の連続断片が続き、これに第1の親由来の第3の連続断片が続く。「連続」とは、顕著な中断断片が存在しないことを示す。これら連続断片は、連続したアミノ酸配列を形成するために結合する。例えば、2つのクロスオーバー100および150を有するHumicola insolens(以下「1」とする)およびH. jecori(以下「2」とする)由来のCBH IIキメラは、1からの最初の100個のアミノ酸に、2から次の50個のアミノ酸が続き、さらにこれに1から残りのアミノ酸が続く形態をとり得て、1つの連続したアミノ酸の鎖として全てが結合している。或いは、CBH IIキメラは、2から最初の100アミノ酸、1から次の50アミノ酸、次いで2由来の残りに順に続く形態をとってもよい。当業者に理解されるように、キメラの変異体は、本来の配列として同程度に存在し得る。従って、それが変異体キメラの場合、100%でないそれぞれの断片が、最終キメラ内に存在すべきである。残基の付加、あるいは残基の除去または変性によって変更できる量は、変異体の用語が定義されるのと同様に定義される。当然、当業者に理解されるように、上記のものは、アミノ酸のみでなく、アミノ酸をコードする核酸にも応用することができる。
【0074】
本発明は、特定の変異体に加えて、CBH IIキメラの作成に使用可能な変異体を示す。誘導されたSCHEMA組み換え型ライブラリーは、セロビオヒドラーゼ酵素の生成に使用され、特に良く研究されたこの多様な酵素ファミリーのメンバー、具体的には下記のセロビオヒドラーゼII酵素に基づく。H. insolensは親「1」(配列番号2)、H. jecorinaは親「2」(配列番号4)およびC. thermophilumは親「3」(配列番号6)。SCHEMAは、タンパク質の構造一体性に影響を及ぼさずに組みかえることができる、相応するタンパク質の断片の位置を予測する計算方法である(Meyre et al. (2003) Protein Sci., 12: 1686-1693)。この計算は、CBH II親タンパク質の同定された7つの組み換え点に関してなされ、これにより各ポリペプチドが8つの断片を含むCBH IIキメラポリペプチドのライブラリーの作成を可能とした。高い安定性を有するキメラは、配列安定性データの線形回帰、または折り畳まれたタンパク質対折り畳まれていないタンパク質のMSAコンセンサス依存分析によって、全体的な安定性への各断片の付加的寄与を決定することで同定できる。SCHEMA組換えは、耐性配列の交換が行われても、重要な機能的残基の保存により、キメラが生物学的機能を保持し、高い配列の多様性を示す。
【0075】
従って、ここに様々な実施形態によって示すように、本発明は、親ドメインのキメラを含むCBH IIポリペプチドであり、上記のように、親鎖または結果として生じるキメラのコード配列が、C→S置換を含むように修飾することができるものを提供する。いくつかの実施形態において、ポリペプチドは、異なる親CBH IIタンパク質からのN末端からC末端までの複数のドメインを有するキメラを含む:(断片1)−(断片2)−(断片3)−(断片4)−(断片5)−(断片6)−(断片7)−(断片8);
断片1は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約1から約x1アミノ酸残基;断片2は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x1から約x2アミノ酸残基;断片3は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x2から約x3アミノ酸残基;断片4は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x3から約x4アミノ酸残基;断片5は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x4から約x5アミノ酸残基;断片6は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x5から約x6アミノ酸残基;断片7は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x6から約x7アミノ酸残基;および断片8は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x7から約x8アミノ酸残基を含み;
x1は配列番号2の残基43,44,45,46,47または配列番号6の残基42,43,44,45,46;x2は配列番号2の残基70,71,72,73,74または配列番号4もしくは配列番号6の残基68,69,70,71,72,73,74;x3は配列番号2の残基113,114,115,116,117,118または配列番号4もしくは配列番号6の残基110,111,112,113,114,115,116;x4は配列番号2の残基153,154,155,156,157または配列番号4もしくは配列番号6の残基149,150,151,152,153,154,155,156;x5は配列番号2の残基220,221,222,223,224または配列番号4もしくは配列番号6の残基216,217,218,219,220,221,222,223;x6は配列番号2の残基256,257,258,259,260,261または配列番号4もしくは配列番号6の残基253,254,255,256,257,258,259,260;x7は配列番号2の残基312,313,314,315,316または配列番号4もしくは配列番号6の残基309,310,311,312,313,314,315,316を含み;x8はポリペプチドのC末端に相当するアミノ酸残基が配列番号2、配列番号4または配列番号6を有する。
【0076】
以下のドメインの参照を用いて、Table1に示すように、多くのキメラ構造が作成された。
H. insolens親CBH IIに関して測定された64.8CのT50値を予測するT50値を有する1,588CBH IIキメラ配列。
【0077】
【表3】
【0078】
【表4】
【0079】
【表5】
【0080】
【表6】
【0081】
【表7】
【0082】
上記表において、それぞれの桁は、キメラCBH IIポリペプチドのドメインを指す。数字は、ドメインが由来する親鎖を示す。例えば、配列12111131を有するキメラCBH IIキメラポリペプチドは、ポリペプチドがN末端からC末端に向かって:配列番号2(「1」)のアミノ酸配列約1からx1に結合した配列番号4(「2」)のアミノ酸配列約x1からX2、これに結合した配列番号6(「3」)のアミノ酸配列約x2からx3、これに結合した配列番号2のアミノ酸配列約x3からx4、これに結合した配列番号2のアミノ酸配列約x4からx5、これに結合した配列番号2のアミノ酸配列約x5からx6、これに結合した配列番号6(「3」)のアミノ酸配列約x6からx7、これに結合した配列番号2のアミノ酸配列約x7からx8(例えばC末端)を含む。
【0083】
いくつかの実施形態において、CBH IIポリペプチドは、以下からなる群から選択されるキメラ断片構造を有する。
【0084】
【表8】
【0085】
いくつかの実施形態において、ポリペプチドは、配列番号2,4または6の野生型ポリペプチドと比較して改良した熱安定性を有する。ポリペプチドの活性は、実施例に記載されているように、1つの、または組み合わされた任意の基質を用いて測定することができる。当業者にとって明白であるように、実施例に記載されたものに例示される化合物のクラスの他の化合物を試験することができる。
【0086】
いくつかの実施形態において、ポリペプチドは、参照配列のアミノ酸配列に様々な変更を加えた形態であってもよい。この変更は、1つ以上のアミノ酸の置換、欠如または挿入であってもよい。この変更が置換である場合、変更は保存、非保存の置換、または保存、非保存の組み合された置換の組み合わせである。例えば、キメラは、配列番号2のC314または配列番号4のC311の位置でC→S置換を含むことができる。
【0087】
従って、いくつかの実施形態において、ポリペプチドはN末端からC末端において一般的な構造を含むことができる:(断片1)−(断片2)−(断片3)−(断片4)−(断片5)−(断片6)−(断片7)−(断片8);
断片1は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約1から約x1アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;断片2は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x1から約x2アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;断片3は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x2から約x3アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;断片4は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x3から約x4アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;断片5は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x4から約x5アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;断片6は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x5から約x6アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;断片7は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x6から約x7アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;ならびに断片8は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x7から約x8アミノ酸残基を含み、1−10個の保存されたアミノ酸の置換を有する;
【0088】
従って、いくつかの実施形態において、ポリペプチドは、N末端からC末端までの一般的な構造を含むことができ、x1は配列番号2の残基43,44,45,46,47または配列番号6の残基42,43,44,45,46;x2は配列番号2の残基70,71,72,73,74または配列番号4もしくは配列番号6の残基68,69,70,71,72,73,74;x3は配列番号2の残基113,114,115,116,117,118または配列番号4もしくは配列番号6の残基110,111,112,113,114,115,116;x4は配列番号2の残基153,154,155,156,157または配列番号4もしくは配列番号6の残基149,150,151,152,153,154,155,156;x5は配列番号2の残基220,221,222,223,224または配列番号4もしくは配列番号6の残基216,217,218,219,220,221,222,223;x6は配列番号2の残基256,257,258,259,260,261または配列番号4もしくは配列番号6の残基253,254,255,256,257,258,259,260;x7は配列番号2の残基312,313,314,315,316または配列番号4もしくは配列番号6の残基309,310,311,312,313,314,315,316を含み;x8はポリペプチドのC末端に相当するアミノ酸残基が配列番号2、配列番号4または配列番号6を有し、キメラは以下のTable1のようなアルゴリズムを有し、キメラは配列番号2のC314または配列番号4のC311に相当するC→S置換を含む。
【0089】
いくつかの実施形態において、アミノ酸置換される置換基の数は2,3,4,5,6,8,9,10個またはこれを超える数となり得る(例えば、10−20,21−30,31−40および同類のアミノ酸置換)。
【0090】
いくつかの実施形態において、機能的なCBH IIポリペプチドは、例えば実施例に記載の基質に対して、高い熱安定性を有すると共に、セルラーゼ活性を有することができ、また参照するセロビオヒドラーゼまたはその断片に一致する一定のレベルのアミノ酸を有することもできる。参照とする酵素またはその断片は、野生型(例えば天然発生)または改変された酵素由来のもとを使用できる。従って、いくつかの実施形態において、本発明のポリペプチドは、N末端からC末端への一般的な構造を含むことができる:
断片1は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約1から約x1のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片2は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約x1から約x2のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片3は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約x2から約x3のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片4は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約x3から約x4のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片5は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約x4から約x5のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片6は、配列番号2(「1」)、配列番号(「2」)、配列番号6(「3」)の約x5から約x6のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片7は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約x6から約x7のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片8は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約x7から約x8のアミノ酸残基で少なくとも50−100%一致がある配列を含み;
x1は、配列番号2の残基43,44,45,46,47、配列番号4もしくは配列番号6の残基42,43,44,45,46;x2は、配列番号2の残基70,71,72,73,74、配列番号4もしくは配列番号6の残基68,69,70,71,72,73,74;x3は、配列番号2の残基113,114,115,116,117,118、配列番号4もしくは配列番号6の残基119,111,112,113,114,115,116;x3は、配列番号2の残基113,114,115,116,117,118、配列番号4もしくは配列番号6の残基119,111,112,113,114,115,116;x4は、配列番号2の残基153,154,155,156,157、配列番号4もしくは配列番号6の残基149,150,151,152,153,154,155,156;x5は、配列番号2の残基220,221,222,223,224、配列番号4もしくは配列番号6の残基216,217,218,219,220,221,222,223;x6は、配列番号2の残基256,257,258,259,260,261、配列番号4もしくは配列番号6の残基253,254,255,256,257,258,259,260;x7は、配列番号2の残基312,313,314,315,316、配列番号4もしくは配列番号6の残基309,310,311,312,313,314,315,318;x8は、ポリペプチドのC末端に相当するアミノ酸残基が、配列番号2、配列番号4または配列番号6を有するものであり、キメラは以下に記載された表1のようなアルゴリズムを有し、キメラは配列番号2のC314または配列番号4のC311の位置にC→S置換を含む。
【0091】
いくつかの実施形態において、キメラポリペプチドのそれぞれの断片は、それぞれ配列番号2、配列番号4または配列番号6の(断片1)、(断片2)、(断片3)、(断片4)、(断片5)、(断片6)、(断片7)および(断片8)を参照とする断片と比較して、少なくとも60%、70%、80%、90%、95%、96%、97%、98%、99%、またはこれを超える配列の一致を有することができる。
【0092】
いくつかの実施形態において、ポリペプチド変異体は、配列番号2,4または6の野生型ポリペプチドと比較して改良した熱安定性を有するものであり;キメラは配列番号2のC314または配列番号4のC311の位置のC→S置換を含む。
【0093】
ここに記載のキメラ酵素は、様々な形態、例えば溶解物、粗抽出液または精製物として調製することができる。ポリペプチドは、適切な溶液に溶解することができて;粉末、例えばアセトン粉末(安定剤存在下、または不存在下で)として調製することができるか、凍結乾燥物として調製することができる。いくつかの実施形態において、ポリペプチドは単離されたポリペプチドであることができる。
【0094】
いくつかの実施形態においてポリペプチドは、アレイの形態とすることができる。酵素は、溶解した状態で、例えばミクロタイタープレートのウェル内の溶液としたり、または基質上に固相化したりすることができる。基質としては、有機ポリマー、例えばポリスチレン、ポリエチレン、ポリプロピレン、ポリフルオロエチレン、ポリエチレンオキシおよびポリアクリルアミドならびにその共重合体、およびそのグラフトから構成することができる、固体基質または多孔性基質(例えば膜)が挙げられる。固体サポートは、無機、例えばガラス、二酸化ケイ素、制御多孔質ガラス(CPG)、逆相シリカまたは金もしくはプラチナ等の金属としてもよい。基質の構造は、ビーズ、球体、粒子、顆粒剤、ジェル、膜または面の形態としてもよい。面は平ら、実質的に平らまたは非平面とすることができる。固体サポートは、多孔性であっても無孔であってもよく、膨張性であっても非膨張性であってもよい。固体サポートは、ウェル、くぼみ、または他の容器、入れ物の構成とでき、その特性および場所を変えることもできる。複数のサポートは、試薬のロボットデリバリー、または検出法および/または計測器がアドレス可能な、多様な箇所のアレイ上に構成することができる。
【0095】
本発明は、ここに開示した改変CBH IIポリペプチドをコード化するポリヌクレオチドも提供する。ポリヌクレオチドは、ポリペプチドを発現することができる組み換え型ポリヌクレオチドのため、遺伝子発現を制御する1つ以上の異種の調節または制御配列に作動可能なように結合することができる。CBH IIキメラをコード化する異種ポリヌクレオチドを含む発現構造は、ポリペプチドを発現するため、適切な宿主細胞内に導入することができる。
【0096】
CBH IIキメラ酵素の特定の配列の知識を有する場合(例えばキメラCBH IIの断片構造)、ポリヌクレオチド配列は、
形成されるCBH IIキメラ酵素のアミノ酸配列は、当業者にとって自明の形状となるであろう。様々なアミノ酸に相当するコドンの知識とポリペプチドのアミノ酸配列の知識とを組み合わせれば、当業者は、本発明のポリヌクペプチドをコード化する異なるポリヌクレオチドを作成することが可能である。従って、本発明は、コドンの組み合わせを選択することによって、いくつかの可能性のある構成のポリヌクレオチドの変異体を想起させるものであり、そのような変異体はすべて、ここに記載のポリペプチドとして、ここに明確に開示されているものと理解されるべきである。
【0097】
いくつかの実施形態において、ポリヌクレオチドは、ここに記載のポリペプチドをコード化するが、参照するキメラポリペプチドのCBH II変異体をコード化し、上記のようにC→S置換(例えば、ポリペプチドまたはキメラは、配列番号2のC314または配列番号4のC311の位置のC→S置換を含む)を有するポリペプチドと、ヌクレオチドレベルで、約80%以上、約85%以上、約90%以上、約91%以上、約92%以上、約93%以上、94%以上、約95%以上、約96%以上、約97%以上、約98%以上、約99%以上の配列の一致を有する。
【0098】
いくつかの実施形態において、ポリペプチドをコード化する分離されたポリヌクレオチドは、ポリペプチドの発現を提供するため、様々な方法にて処理することができる。ベクターへの挿入前に単離されたポリヌクレオチドを処理する工程は、発現ベクターによっては、必要とされ、、または必須とされる。組み換えDNA法を用いて、ポリヌクレオチドおよび核酸配列を修飾する技術は、従来技術において周知である。この技術に対する教示は、Sambrook et al . , 2001, Molecular Cloning: A Laboratory Manual, 3rd Ed., Cold Spring Harbor Laboratory Press; and Current Protocols in Molecular Biology, Ausubel. F. ed., Greene Pub. Associates, 1998, updates to 2007に提供されている。
【0099】
いくつかの実施形態において、ポリヌクレオチドは、ポリヌクレオチドおよび/またはポリペプチドの発現に関する制御配列に機能的に関連している。いくつかの実施形態において、制御配列は、細胞外または細胞内のポリペプチドをコード化し、宿主細胞に相同性または異種性のいずれかの遺伝子から得られる適当なプロモーター配列とすることができる。細菌宿主細胞に対して、本発明の核酸構造の転写を誘導するのに適切なプロモーターとしては、大腸菌ラクトースオペロン、枯草菌xylAおよびxylB遺伝子、バチルスメガテイルムキシロースを用いる遺伝子(例えばRygus et al., (1991) Appl . Microbiol. Biotechnol. 35:594-599; Meinhardt et al., (1989) Appl. Microbiol. Biotechnol . 30:343-350)、原核生物βラクタマーゼ遺伝子(Villa- Kamaroff et al., (1978) Proc . Natl Acad. Sci . USA 75: 3727-3731)ならびにtacプロモーター(DeBoerその他, (1983) Proc. Natl Acad. Sci. USA 80: 21-25)が挙げられる。様々な適切なプロモーターが、Scientific American, 1980, 242:74-94の"Useful proteins from recombinant bacteria";およびSambrookその他の文献に記載されている。
【0100】
いくつかの実施形態において、制御配列はまた適切な転写停止(ターミネーター)配列であり、宿主細胞によって転写を停止することが認識される配列としてもよい。停止配列は、ポリペプチドをコード化する核酸配列の3’末端に作動可能に結合している。選択した宿主細胞において機能的なあらゆるターミネーターが使用可能である。
【0101】
いくつかの実施形態において、制御配列は、適切なリーダー配列、宿主細胞による翻訳に重要なmRNAの非翻訳領域でもあり得る。リーダー配列は、ポリペプチドをコード化する核酸配列の5’末端に作動可能に結合される。選択した宿主細胞において機能的なあらゆるリーダー配列が使用可能である。
【0102】
いくつかの実施形態において、制御配列は、ポリペプチドのアミノ末端に結合したアミノ酸配列をコード化するシグナルペプチドコード領域としてもよく、コードされたポリペプチドを細胞の分泌経路へ誘導する。核酸配列のコード配列の5’末端は、本質的に分泌されたポリペプチドをコード化するコード領域の断片と翻訳リーディングフレーム内の、天然に結合したシグナルペプチドコード領域を含むことができる。或いは、コード配列の5’末端は、コード配列とは異質のコード領域のシグナルペプチドを含むことができる。コード配列がシグナルペプチドコード領域を天然に含まない部分において、異質のシグナルペプチド領域が必要とされる。細菌の宿主細胞に関する効果的なシグナルペプチドのコード領域は、バチルス属NCIB11837マルトース生成アミラーゼ、バチルス・ステアロサーモファイルスαアミラーゼ、バチルス・リケニフォルミス・スブチリシン、バチルス・リケニフォルミス・βラクタマーゼ、バシラス・ステアロサーモフィラス中性プロテアーゼ(nprT、nprS、nprM)および枯草菌prsAの遺伝子から得られるシグナルペプチドコードとしてもよい。さらなるシグナルペプチドは、Simonen and Palva, (1993) Microbiol Rev 57: 109-137に記載されている。
【0103】
さらに、本発明は、導入される宿主の種類に応じて、形成されたCBH II変異体またはキメラポリペプチドをコード化するポリヌクレオチドを含む組み換え型発現ベクター、および、例えばプロモーターおよびターミネーター、複写原点等のうちの1つ以上の、発現制御領域について示す。発現ベクター作成において、コード配列は、コード配列が発現のための適切な制御配列と作動可能に結合するようにベクター内に局在する。
【0104】
組み換え型発現ベクターは、有効に組み換え型DNA方法の対象として使用可能であり、ポリヌクレオチド配列の発現をもたらすあらゆるベクター(例えばプラスミドまたはウイルス)を使用できる。通常、ベクターの選択肢は、ベクターが導入される宿主細胞とベクターの適合性に依存する。ベクターとしては、線状または閉鎖環状プラスミドを使用してもよい。
【0105】
発現ベクターとしては、独立した複製ベクター、即ち染色体複製から独立して複製される染色体外のものとして存在するベクター、例えばプラスミド、染色体外の要素、ミニ染色体または人口染色体が使用可能である。ベクターは、自己再生を確実にするためのあらゆる手段を含むことができる。或いは、ベクターは宿主細胞内に導入されたとき、ゲノム内に組み込まれ、それが組み込まれた染色体と共に複製されるものを含むことができる。さらに宿主細胞のゲノム内に導入するため、全DNAを共に含む単一のベクター、プラスミドまたは2つ以上のベクターまたはプラスミドまたはトランスポゾンが使用可能である。
【0106】
いくつかの実施形態において、本発明の発現ベクターは、形質転換細胞の選択を容易とする、1つ以上の選択可能なマーカーを含有する。選択可能なマーカーは、殺生物性またはウイルス抵抗性、重金属抵抗性、独立栄養体への原栄養性、等を与える遺伝子産物である。細菌性の選択可能なマーカーの例は、枯草菌もしくはバチルス・リケニフォルミスからのdal遺伝子、抗生物質、例えばアンピシリン、カナマイシン、クロラムフェニコール(実施例1)への抵抗性またはテトラサイクリンへの抵抗性を与えるマーカーである。他の実用的なマーカーは、当業者にとって明白であろう。
【0107】
他の実施形態において、本発明はCBH II変異体またはキメラポリペプチドをコード化するポリヌクレオチドを含む宿主細胞を提供し、前記ポリヌクレオチドは、宿主細胞中におけるポリペプチドの発現に関する1つ以上の制御配列に作動可能に結合されるものを含む。本発明の発現ベクターによってコード化されるポリペプチドを発現するために使用される宿主細胞は、従来技術において周知であり、大腸菌および巨大菌等の細菌細胞;酵母細胞、CHO細胞等の真核生物細胞;ショウジョウバエS2およびスポドプテラSf9細胞等の昆虫細胞;CHO、COS、BHK、293およびボーズ・メラノーマ細胞等の動物細胞;および植物細胞が挙げられるが、これらに限定されるものではない。他の適切な宿主細胞は、当業者にとって明白であろう。上記宿主細胞への適切な培養培地および成長条件は、従来技術において周知である。
【0108】
本発明のCBH II変異体またはキメラポリペプチドは、従来技術において周知の方法を用いて作成することができる。ポリヌクレオチドは、例えばSambrook et al., 2001, Molecular Cloning: A Laboratory Manual, 3rd Ed., Cold Spring Harbor Laboratory Press; and Current Protocols in Molecular Biology, Ausubel. F. ed., Greene Pub. Associates, 1998, updates to 2007に提供されるような組み換え技術によって合成することができる。酵素をコード化するポリヌクレオチドまたは増幅のためのプライマーについても、既知の合成方法、例えばBeaucage et al., (1981) Tet Lett 22:1859-69に記載のホスホラミダイト法、またはMatthes et al., (1984) EMBO J. 3:801-05に記載の方法のような、標準的な固相法によって調製可能であり、通常、これらの方法は、自動化合成法として実施される。さらに実質的なあらゆる核酸は、様々な商業的供給源、例えばThe Midland Certified Reagent Company(テキサス州ミッドランド)、The Great American Gene Company(カリフォルニア州ラモナ)、ExpressGen Inc.(イリノイ州シカゴ)、Operon Technologies Inc.(カリフォルニア州アラメダ)等の多様な供給原から入手可能である。
【0109】
宿主細胞内に発現する改変酵素は、とりわけリゾチーム処理、超音波処理、濾過、塩析、超遠心分離、クロマトグラフィーおよび親和性分離(例えば、基質に結合した抗体)等の既知の技術のタンパク質精製手段を用いて、細胞および/または培養培地から回収することができる。細菌、例えば大腸菌からのタンパク質を溶解するために適切な溶液および高効率抽出液が、ミズーリ州セントルイスのシグマアルドリッチ社から商品名CelLytic BTMとして市販されている。
【0110】
ポリペプチドの分離に関するクロマトグラフィー技術は、とりわけ逆相クロマトグラフィー、高速液体クロマトグラフィー、イオン交換クロマトグラフィー、ゲル電気泳動および親和性クロマトグラフィーを含む。特定の酵素を精製するための条件が、実際の荷電、疎水性、親水性、分子量、分子形状などの要因に部分的に依存することは、当業者にとって明白である。
【0111】
キメラポリペプチドの組み換えおよび合成に係るSCHEMAの開示は、本明細書およびOteyその他, (2006), PLoS Biol. 4(5): ell2; Meyer et al., (2003) Protein Sci., 12:1686-1693;2008年2月1日に出願された米国特許出願番号第12/024,515号;2008年2月7日に出願された米国特許出願番号第12/027,885号の実施例に記載されており、その内容の全てをここに参照して取り込む。
【0112】
上記のように、ポリペプチドは、様々な応用、例えばとりわけ生物燃料生成、セルロースの分解等に使用することができる。
【0113】
以下の実施例は、本発明についての付加的な説明をするものであり、前記開示または添付の特許請求の範囲に制限するものではない。
【実施例】
【0114】
<CBH II発現プラスミドの作成>
酵母発現ベクターYEp352/PGK91−1−αss内で、CBH II酵素をコード化する親およびキメラ遺伝子をクローン化した(図6)。親およびキメラCBH II触媒ドメインをコード化するDNA配列を、S. cerevisiaeコドンバイアスで、GeneDesignerソフトウェア(DNA2.0)およびDNA2.0を用いた合成により設計した。CBH II触媒ドメイン遺伝子を、XhoIおよびKpnIで切断して、ベクター中のXhoIおよびKpnI部位の間にライゲーションし、大腸菌XL−1Blue(Stratagene)内に形質転換した。CBH II遺伝子は、プライマーCBH2L(5'- GCTGAACGTGTCATCGGTTAC-3' (配列番号9)およびRSQ3080(5'- GCAACACCTGGCAATTCCTTACC-3' (配列番号10))を用いて配列決定した。C末端His6親およびキメラCBH II構造は、CBH II遺伝子を前方プライマーCBH2LPCR (5'' -GCTGAACGTGTCATCGTTACTTAG-3' (配列番号11))と、His6オーバーハングと終止コドンを伴う適切なCBH II遺伝子に相補的な逆方向プライマーを増幅することによって作成された。PCR産物は、上記のようにライゲーションされ、形質変換され、配列決定された。
【0115】
<S. cerevisiaeにおけるCBH II酵素の発現>
S. cerevisiae菌YDR483W BY4742 (Matα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0 ΔKRE2、ATCC No.4014317)は、EZ Yeast II Transformation Kit(Zymo Research)を使用してコンピテント化され、プラスミドDNAを用いて形質転換され、合成ウラシルを欠く寒天培地上に播種された。コロニーは、20μg/mLトリプトファンを添加した5mL合成デキストロースカザミノ酸(SDCAA)培地(20g/Lデキストロース、6.7g/Lデフコ酵母窒素塩基、5g/Lバクトカザミノ酸、5.4g/L Na2HPO4、8.56g/L NaH2PO4−H2O)に入れ、一晩、30℃、250rpmで培養した。5mL培養物は、250mLチューンエアーフラスコ(Shelton Scientific)中の40mL SDCAAで希釈し、30℃、250rpmで48時間振盪した。培養物を遠心分離し、清をt1/2分析のため、30kDa PES膜を取り付けたAmicon限外ろ過膜を使用して、500μLにまで濃縮した。濃縮した上清は、1mMフェニルメチルスルホニルおよび0.02%NaN3に供した。His6タグを付けたCBH IIタンパク質を、Ni−NTAスピンカラム(Qiagen)を用いて製造元のプロトコールに従って精製し、次いで、タンパク質をZeba−Spin脱塩カラム(Pierce)を使用して50mM酢酸ナトリウム、pH4.8中に交換した。精製されたタンパク質濃度を、基準としてBSAとPierce Coomassie Plusタンパク質試薬を使用して測定した。SDS−PAGE分析は、濃縮された培養上清20μLまたは精製されたCBH II酵素約5μgのいずれかを、7.5%Tris−HClゲル(Biorad)上に加え、シンプリーブルーセーフステイン(Invitrogen)で染色することによって実行した。CBH II上清または精製されたタンパク質は、EndoH(New England Biolabs)を用いて、1時間、37℃、製造元のプロトコールに従って処理した。濃縮された酵母培養上清中のCBH II酵素活性は、37.5μLの濃縮された培養上清を37.5μL PASCに添加し、2時間50℃でインキュベーションして測定した。形成された還元糖等価物は、以下のようなソモギ・ネルソン分析によって測定した。
【0116】
<半減期、特異的活性、pH活性および長期セルロース加水分解の測定値>
リン酸膨潤セルロース(PASC)を調製した。基質へのCBH II酵素活性を向上させるため、PASCを10g/Lの濃度で50mM酢酸ナトリウム、pH4.8中の10mg/mLA. nigerエンドヌークレアーゼ(Sigma)を用いて、37℃で1時間プレインキュベーションした。エンドグルカナーゼを、95℃まで加熱し、15分間処理することで不活性化し、PASCを2回、50mM酢酸緩衝剤を用いて洗浄し、脱イオン水中に10g/Lとなるように再懸濁した。
【0117】
CBH II酵素t1/2は、以下に記載するように、処理されたPASCとインキュベーションした後、濃縮したCBH II発現培養上清を50mM酢酸ナトリウム(ソモギ・ネルソン還元糖分析で測定されたA520が0.5となる濃度でpH4.8)に添加することによって測定した。37.5μLのCBH II酵素/緩衝剤の混合物を、63℃の水浴で不活性化した。不活性化後、37.5μLのエンドグルカナーゼ処理したPASCを添加し、2時間、50℃の条件下で加水分解をで実施した。反応上清は、Multiscreen HTSプレート(ミリポア)を通してろ過した。SpectraMaxマイクロプレートリーダー(Molecular Devices)を用いて得られたソモギ・ネルソン分析log(A520)値を、バックグラウンドの吸光度について修正処理を施し、マイクロソフト・エクセルを使用して、時間軸に対してプロットすることで、線形回帰から得られるCBH II酵素の半減期のデータを得た。
【0118】
特異的活性測定のため、精製CBH II酵素をPASCに添加し、PASCの最終反応量が5g/Lで、CBH II 酵素濃度が3mg/gであり、酢酸ナトリウム濃度25mM、pH4.8のPASCを得た。50℃水浴中でのインキュベーションを2時間行い、還元糖濃度を測定した。pH/活性特性の測定のため、精製CBH II酵素を、75μL反応容積中の濃度300μg/gのPASCに添加した。反応は、12.5mMクエン酸ナトリウム/12.5mMリン酸ナトリウムの緩衝液中で、50℃で16時間実施し、還元糖濃度を測定した。長時間のセルロース加水分解測定は、100mM酢酸ナトリウム、pH4.8、20mM NaClの、1g/L濃度の処理済みPASC300μLを用いて行った。精製CBH II酵素を、100μg/gの濃度でPASCに添加し、還元糖測定の40時間前に水浴中にて反応させた。
【0119】
CBH II酵素をコードする5つの親遺伝子候補を、S. cerevisiaeコドンバイアスを用いて合成した。5つ全ては、1−89残基がセルロース結合モジュール(CBM)、弾性リンカー領域およびH. jecorina触媒ドメインの5つのN末端残基に相当する、N末端コード配列を含む。Humicol insolensおよびChaetomiun thermophilumからの2つのCBH II酵素候補は、Hypocrea jecorina、Phanerochaete chrysosporiumおよびTalaromyces emersonilによる3つの他のものより比較的高いレベルで、S. cerevisiaeから分泌された(図1)。3つのわずかに発現されている候補の親に関するSDS−PAGEゲルのバンドは、認識困難なため、濃縮培養上清における活性分析は、活性型セルラーゼの存在を確認するため、リン酸膨潤セルロース(PASC)とインキュベートされたものを用いて実施された。図1に示す形成された還元糖からの値は、T. emersonii CBH II以外の全ての酵素に関して、濃縮S. cerevisiae培養上清における活性型CBH IIの存在が確認された。H. insolensおよびC. thermophilum配列は、最も産業的に関連したH. jecorinaからの菌CBH II酵素と組み合わせて選択した。触媒ドメインのそれぞれの配列の一致は、64%(1:2)、66%(2:3)および82%(1:3)であり、H. insolensは親1、H. jecorinaは親2、C. thermophilumは親3とする。これらそれぞれの触媒ドメインは、360,358,359アミノ酸残基を含む。
【0120】
産業使用のためにセルラーゼを生成するのに最も頻繁に使用される生命体である糸状菌H. jecorinaにおける異種タンパク質発現は、酵母と比較して非常に困難である。組換え型セルラーゼ活性を減少することが報告されている、高度なグリコシル化を最小限にするため、組換え型CBH II遺伝子をグリコシル化機能を欠損させたdKRE2 S. cerevisiae株で発現させた。該株は、野生株と比較して、より短いマンノースオリゴマーがN−結合型およびO−結合型の両方のグリコシル化サイトに結合すると考えられ、これは野生型H. jecorinaのCBH II 酵素のグリコシル化に極めて類似している、高マンノース構造を取り除くためのEndoH処理を伴う条件、および伴わない条件において、CBH IIタンパク質のSDS−PAGEゲル分析を行ったところ、EndoH処理によっては、前記菌から分泌された酵素の電気泳動的な可動性が上昇しないことが判明し、野生型S. cerevisiae菌が組換え型タンパク質のグリコシル化部位に結合する分岐型マンノース部分を欠くことを確認した。
【0121】
H. insolensの高分解性の構造(pdb)SCHEMAのテンプレートとして使用して、組み換えに応じて引離される接触を同定した。RASPPは、それぞれ15未満の<E>を有する4つの候補ライブラリーを返した。全ての候補ライブラリーは、以前作成されたキメラライブラリーより低い<E>を示し、折り畳まれた活性型キメラの許容可能な部分は、比較的高い<m>を有して得られることが示された。キメラ配列の多様性は、最も大きな<m>=50を導くようなブロックの境界を選択すると、最大となった。この設計のためのブロックは、図2Bに例示され、Table2に詳細に示される。
Table2:親CBH II酵素触媒ドメインのためのClustalW複数配列アライメント。ブロック2,4,6および8は、ボックスおよびグレーの斜線で示す。ブロック1,3,5および7は、されていない(H. inso:配列番号2、H. Jeco:配列番号4およびC. Ther.:配列番号6)。
【0122】
【表9】
【0123】
H. insolens CBH II触媒ドメインは、8つの螺旋がバレル周縁を規定し、7つの平行のβシートが活性部位を形成するα/βバレル構造を有する(図2A)。2つの延長したループは、活性部位上にルーフを形成し、加水分解中に基質セルロース鎖が通過する通路を形成する。7つのブロック境界のうちの5つは、2次構造の要素の間に収まり、ブロック4は連続的なα螺旋の中央部分から開始し、終結する(図2A、2B)。ブロック間の側鎖接触の大多数は、初期構造において調節可能なブロック間に生じる(図2C)。
【0124】
48個のキメラ遺伝子のサンプルセットは、1つの親由来の5つのブロックと、残り2つのうちの1つまたは両方の親由来の3つのブロックを有する、16個のキメラの3セットとして設計された(Table3);それぞれの親の表記がそれぞれのブロックの位置で等しくなるように、配列を選択した。相当する遺伝子を合成し、発現させた。
Table3:サンプルセットCBH II酵素キメラの配列
【0125】
【表10】
【0126】
48個のサンプルセットのS. cerevisiaeのうち23個の濃縮培養上清は、PASCに対して加水分解活性を示した。これらの結果は、6,561個のCBH IIキメラ配列のうち数千(例えば、Table1を参照のこと)が活性型酵素をコード化することを示す。23の活性型CBH IIサンプルセットのキメラは、顕著な配列多様性を示し、最も近い親の配列と、少なくとも23および36、54および123相当のアミノ酸置換を有する。それらの平均突然変異レベル<m>は36である。
【0127】
Meyerらが、E、mおよびキメラが折り畳まれて活性を有する確率の間の相関を見出したように、同類の相互関係が、サンプルセットCBH IIキメラに関して存在するかの分析を実施した。PASC活性の分析によって測定されるように、濃縮発現培養上清におけるCBH II酵素活性の値は、SDS−PAGEゲルにおけるCBH IIバンドの強度に相関した(図1)。H. jecorina CBH II親と同様に、活性はいくつかのCBH IIキメラに関して検出できないゲルバンドで検出できる。ゲルバンドは示したが、活性を欠失するCBH IIキメラの存在が観察された。CBH IIキメラが活性型の形状で分泌される確率は、Eおよびmの双方に逆相関した。
【0128】
熱不活性の半減期(t1/2)は、63℃における親および活性型キメラのCBH II酵素の濃縮培養上清について測定された。H. insolens、H. jecorinaおよびC. thermophilumのCBH II親の半減期は、それぞれ95,2,25分であった。活性型サンプルセットのキメラは、1分未満から3,000超の幅広い半減期を示した。23個の活性キメラのうちの5つは、最も熱安定のある親であるH. insolens CBH IIを超える半減期を有した。
【0129】
CBH IIキメラの半減期のための予測定量的モデルの作成を試みる際、5つの異なる線形回帰データのモデリングアルゴリズムを使用した(表4)。それぞれのアルゴリズムを使用して、それぞれのサンプルセットCBH IIキメラおよび親のlog(t1/2)のブロック組成物に関するモデルを作成した。これらのモデルは、ブロックのlog(t1/2)への寄与を定量化した、熱安定性重量値を構築した。全て5つのモデリングアルゴリズムについて、この工程が、2つのランダムに選択された配列を省略して1,000回繰り返され、それぞれのアルゴリズムは、24個のブロックそれぞれに1,000重量値を示した。それぞれのブロックの熱安定性重量値について、平均値および標準偏差(SD)を算出した。それぞれのモデルアルゴリズムの予測精度は、それぞれのモデルが省略された配列のt1/2をどれだけ正確に予測できるかで評価した。1,000アルゴリズムの繰り返しについての測定値および予測値の間の相関を、モデルアルゴリズムの相互検証スコアとする。5つのモデル全てについての相互検証スコア(X−val)は、0.057以下であり(Table4)、線形回帰モデリングが、この小さな23個のキメラt1/2データセットの定量的CBH IIキメラの半減期予測については適応できないことを示した。
Table4:5つの線形回帰アルゴリズムのCBH II酵素キメラのブロック安定性スコアへの使用のための相互検証値。アルゴリズムの省略:リッジ回帰(RR)、部分最小二乗回帰(PLSR)、サポートベクターマシン回帰(SVMR)、リニアプログラミング・サポートベクターマシン回帰(LPSVMR)およびリニアプログラミングブースティング回帰(LPBoostR)。
【0130】
【表11】
【0131】
線形回帰モデリングを使用して、ブロックを質的に安定、不安定または中立に分類した。それぞれのブロックのキメラの熱安定性への影響は、それぞれの回帰アルゴリズムによって測定される熱安定性への寄与を提示するスコアリングを使用して評価した。それぞれのアルゴリズムに関して、中立から1SDを超える熱安定性重量値を有するブロックは、「+1」として記録し、中立から1SD以内の熱安定性重量値を有するブロックは0とし、中立から1SDを超えて低い熱安定性重量値を有するブロックは、「−1」として記録した。それぞれのブロックに関する「安定性スコア」は、それぞれ5つのモデルからの1,0、−1の安定性スコアを合計することによって得た。Table5は、それぞれのブロックに関するスコアを概説する。ブロックI/親1(B1P1)、B6P3、B7P3およびB8P2は、比較的高い安定性効果を有するものとして同定された、一方、B1P3、B2P1、B3P2、B6P2、B7P1、B7P2およびB8P3は、最も強くブロックを不安定化することを見出した。
Table5:5つの線形回帰アリゴリズム1によって生じたサンプルセットCBH II酵素キメラに関する質的なブロック分類。スコア+1は、熱安定性重量を有するブロック(キメラの熱安定性へのブロック寄与に関する無次元測定)であり、中立から1SDを超えるもの(安定性)を示し、スコア0は、中立から1SD内の重量値であるブロックを示し、−1は中性から1SD超低い重量値を有するブロックを示す(不安定性)。
【0132】
【表12】
【0133】
CBH II酵素キメラをコードする第2セットの遺伝子を合成し、予測された安定ブロックを評価して、最も安定な親より熱安定のあるセルラーゼを同定した。この検証セットに含まれる24個のキメラ(Table6)は、最も不安定であると予測される7つのブロックが全くなく、4つの最も安定なブロックに富んでいた。後者のブロックの発現は、安定なスコアに偏っていた。さらに「HJPlus」12222332キメラは、予測された最も安定なブロックをH. jecorina CBH II酵素内(親2)に置換することによって作成された。
Table6:24個の検証セットCBH II酵素キメラの配列中の9つは、活性型で発現した。
【0134】
【表13】
【0135】
24個の検証のセットのキメラの9つのS. cerevisiae発現培地の濃縮上清は、HJPlusキメラと同様に、PASCに対して活性を示した(Table6)。活性が検出されていない15個のキメラのうちの9つは、ブロックB4P2を有した。初期のサンプルセットのB4P2を含有する16個のキメラのうち1つのみが、PASCに対して活性を示した。キメラセットおよびHJPlusの両方を合計して、26個のキメラのうち2つのみが活性化していて、この特定のブロックがS. cerevisiaeにおける活性化セルラーゼの発現を阻害することを示した。
【0136】
検証セットからの10個の機能的キメラCBH II酵素の安定性を評価した。安定酵素は既に50時間を越える半減期を有したため、63℃、12時間の熱不活性化後のPASCに対する残留加水分解活性を、測定の予備的評価として使用した。この12時間のインキュベーションによって、サンプルセットの最も熱安定性のあるキメラである11113132の活性において、測定可能な減少が生じ、不活性型の熱安定性のあるH. insolens親CBH IIを完全に不活性化した。全10個の機能的な検証セットのキメラは、それらの活性として最も安定な親であるH. insolens CBH IIより多い活性を保持していた。
【0137】
精製された酵素を使用する選択された熱安定性のあるキメラの活性を分析した。親CBH II酵素および3つの熱安定性のあるキメラ(最も熱安定性のあるサンプルセットのキメラ11113132、最も熱安定性のある検証セットのキメラ13311332)およびHJプラスキメラ12222332を、C末端His6精製タグとともに発現させ、精製した。活性試験中のCBH II酵素の熱不活性化を最小限にするため、2時間(比較的短い時間)、50℃、pH4.8でのPASC基質とのインキュベーションを実施した。Table3に示すように、親およびキメラCBH IIの特異的活性は、最も活性のあるH. jecorinaからの親CBH II酵素のうちの4つの要素内にあった。HJプラスの特異的活性は、H. jecorina CBH II以外の全ての被験のCBH II酵素を超えていた。
【0138】
幅広い範囲のpH/活性特性により、CBH IIキメラの広範囲にわたる潜在的なセルロース加水分解条件下での使用を可能とするためには、セルラーゼ活性のpH依存性が重要である。H. jecorina CBH IIは、最適化された活性をpHの範囲4から6で有することが観測され、一方この範囲外ではその活性が著しく現象することが観察されてきた(文献16)。図4は、H. insolensおよびC. thermophilum CBH II酵素ならびに全3つの精製された熱安定性のあるCBH IIキメラが、H. jecorina CBH IIのものより相当広いpH/活性特性を有することを示した。Liuらは、C. thermophilum CBH IIに対してpH4を最適条件として報告したが、ここでの組み換え型酵素の最適なpHは、7付近であった。天然H. insolens CBH IIは、約pH9で最大活性、pH4で最大活性の約60%を示す幅広いpH/活性特性を有する。同様の幅広い特性が、組み換え型酵素について観測された。HJプラスキメラは、H. jecorina CBH IIと比較してより幅広いpH/活性特性を有し、他の2つの親CBH II酵素と同様のpH依存性を示した。
【0139】
高温での活性の達成と、長いタイムインターバルを超えた活性の維持は、改変した高安定性CBH II酵素を実現するための2つの初期目標である。セルロース加水分解における熱安定性のあるCBH IIキメラの性能を、温度範囲全体で、40時間のインターバルで試験した。図5に示すように、3つ全ての熱安定性キメラは、親CBH II酵素より比較的高い温度でPASCに活性を有した。H. jecorina CBH IIは、57℃長ではPASCを加水分解せず、安定なH. insolens酵素は63℃超ではPASCを全く加水分解しなかったが、キメラは70℃で活性を保持した。。長時間のセルロース加水分解分析におけるHJプラスの活性は、各々の最適な温度における全ての親の活性を上回った。
【0140】
CBH IIライブラリーは、いくつかの理由により、崩壊の可能性が比較的低い。CBH II親配列の高い相同性に加えて、CBH II折り畳みバレルの立体的構造は、組み換えによって分離することができる長範囲の接触の数を制限する。ブロック間の接触(4.5オングストローム範囲内の重原子)は、H. insolens構造1ocnに由来する接触マップ全体の27%(503/1831)のみを含む。残基の組み合わせに係る接触は、キメラにおいてのみ計数可能であり、ブロック内の総計は23%まで減少した(68/294)。さらに、これら相互作用のほとんどは、タンパク質の表面上の残基間に生じ、溶媒スクリーニングの可能性は、さらに残基間の相互作用の飛躍的な分離の可能性を減少させる(図14a)。1つの例外として、位置176−位置256間の埋没した相互作用が、図14bに示す。この部位で、B6P2と、B5P1またはB5P3のいずれかとを有するキメラは、Met173:Trp253の対を有する(親のMet176:Phe256の対、またはLeu173:Trp253の対と比較して大きなアミノ酸)。しかしながら、親の結晶学的なモデルの精査により、Trp253側に位置するタンパク質バックボーン部の可動性や、Met側鎖の内因的な可撓性のため、この部位における立体衝突は生じにくいと考えられる。特に、1つの特徴的なキメラは、このパターン(13333232)に適合し、回帰モデル適合性(68℃)よれば、親(67℃)よりも安定である。
【0141】
カップリングを生じる他のメカニズムは、ブロック構造の相違であるが、これはブロック接点での新規の残基対の存在に依存するものではない。そのかわり、親配列の相違により、内因性ブロック構造は、モジュール・ブロックの移動を妨げて、多様化することができる。CBH IIライブラリーの場合、親の対の高い配列相同性の値(82%、66%および64%)において、構造の偏差は軽度とすることができる(<1オングストロームRMSD)。この可能性は、H. insolensおよびH. jecorina CBH IIの結晶学的な構造を比較することによって、評価することができる(C. thermophilum CBH IIは結晶構造を欠如しているが、H. insolensと82%の相同性がある)。それぞれの親からの構造による配列ブロック(1ocnおよび1cb2)において、αカーボンRMSD値は低くなる(0.5,0.5,0.6,0.5,0.3,0.7,0.3および0.4A RMSD)。H. insolens上に重ねられたH. jecorinaブロックを、補足図5cに示す。背景に依存した効果を確認するため、対立する宿主構造上にそれぞれアラインされたブロックをスプライシングして、シリコ構造(silico structure)の組み換えを実行した。ブロック6および7の間にAsnを挿入して(図14D)B7P2(11111121)を使用した場合、軽度の衝突(2.65オングストローム)を除き、全ての単一ブロック置換キメラ(例えば11112111または22122222)に関する衝突しない構造モデルを作成することが可能である(αカーボン>3オングストローム間隔)。
【0142】
様々なブロック/断片のキメラの安定性への寄与を決定するためにさらなる実験を実施し、熱安定性および/またはpH安定性を改良した。CBH II酵素をコードする親およびキメラ遺伝子を、酵母発現ベクターYEp352/PGK91−1−αss内にクローン化し、合成デキストロースカザミノ酸(SDCAA)培地中で発現させた。アビセル活性分析のため、酵母ペプトンデキストロース(YPD)培養上清は、1mMフェニルメチルスルホニルおよび0.02%NaN3溶液にして、濃縮せずに使用した。濃縮SDCAA酵母培養上清中のCBH II酵素活性を、37.5μLのPASCおよび225μLの50mM酢酸ナトリウム、pH4.8に添加することで濃縮培養上清を希釈し、2時間、50℃でインキュベーションして測定した。形成した還元糖の等量を、ソモギ・ネルソン分析によって測定した。
【0143】
CBH II酵素のT50値を、エンドヌークレアーゼ処理したPASCとインキュベーションした後、濃縮したCBH IIのSDCAA発現培養上清を50mM酢酸ナトリウムに添加し、ソモギ・ネルソン還元糖分析の測定値A520が0.5となる濃度でpH4.8となるように調整して測定した。200μLのCBH II酵素/緩衝剤混合物を、適温で10分間、水浴中にてインキュベートした。インキュベーション後、37.5μLのエンドグルカナーゼ処理されたPASCおよび62.5μLの50mM酢酸ナトリウム溶液を添加し、50℃で2時間加水分解を実施した。酵素の半分の活性が欠如したときのインキュベーション温度を、ソモギ・ネルソン分析のA520値の対温度プロットの線形補間より算出した。
【0144】
長時間のアビセルPH101(Fluka)加水分解の測定のため、0.3μgの精製されたCBH IIを、水浴に配置されたPCRチューブ内で、270μLの50mM酢酸ナトリウム(pH4.8)において3mgのアビセルとともに16時間インキュベートした。チューブを室温の水浴中で10分間冷却し、1000gで10分間遠心分離し、収集した上清を還元糖分析に供した。
【0145】
YPD発現培養上清中のCBH II活性の評価のため、2mL〜40mLの上清を、コニカルチューブにて、50mM酢酸ナトリウム(pH4.8)に懸濁された33mg/mLのアビセル800μLに添加された。CBH IIは、アビセルと4℃、1時間で結合し、2000g、2分間遠心分離し、50mM酢酸ナトリウム(pH4.8)で2回洗浄された。2回洗浄した後、CBH IIと結合したアビセルを、2.75mL酢酸ナトリウム緩衝剤中に再懸濁し、270μLの分量で分注して、50℃で2.5時間インキュベートした。遠心分離および上清還元糖の分析は、上記のように実施した。
【0146】
Mathematicaにおける線形回帰パッケージを使用して、CBH IIキメラT50データを17パラメーターに適合し、相互検証分析を行った。ブロック効果は、親1(H. insolens CBH II)を基準として、親2および3からのそれぞれ8つのブロックの置換を示す16パラメーターを用いて、報告された。
【0147】
10分間のインキュベーションにより酵素が50%の活性を欠如する温度として規定されるT50値は、3つの親セロビオヒドラーゼについて決定した。前の実験からの33個の活性型CBH IIキメラおよび質的に安定なモデルが最も熱安定性があると予測したうちの18個のさらなるキメラ(即ち不安定と予測されたブロックは一切含まれず、安定性ブロックと予測された4つのうち3つまたは4つが含まれる)についてT50値を測定した。全51個のキメラ配列を表8に列挙した。以前に分泌されないと分類された全ての熱安定性と予測されるキメラの再培養および再濃縮により、T50測定のため、十分量の12112132,13111132および13322332CBH IIを得ることを可能とした。キメラおよび親のCBH IIに関するT50値の完全なセットをが、Table8に示す。全ての前記CBH IIのアミノ酸配列を、Table7に記載する。試験された全31個の熱安定性と予測されたキメラは、最も熱安定性のある親酵素(64.8℃)より2度高いT50値を有する。Tableは、熱安定を向上させるため、Serに変異することができるブロック/ドメイン7におけるCys残基も同定する。従って、本発明は、以下の配列のあらゆるポリペプチドを提供し、下線/イタリック体/太字のCysはSer残基と置換され、結果として生じたポリペプチドは、野生型酵素と比較して、高い熱安定性を有する。
Table7:CBH II親およびキメラ触媒ドメインのためのアミノ酸配列が、Table8に示される。
Tableは、P. chrysosporium CBH IIに関する触媒ドメインも含む。全ての組み換え型CBH IIは、天然H. jecorina CBH IIからのN末端CBMおよびリンカーCSSVWGQCGGQNWSGPTCCASGSTCVYSNDYYSQCLPGAASSSSSTRAASTTSRVSPTTSRSSSATPP PGSTTTRVPPVGSGTATYS (配列番号8)を共有する。
【0148】
【表14】
【0149】
【表15】
【0150】
【表16】
【0151】
【表17】
【0152】
【表18】
【0153】
【表19】
【0154】
【表20】
【0155】
【表21】
【0156】
【表22】
【0157】
Table8:
親CBH IIに関する2つの独立した二重のT50値(℃)、23個の初めのサンプルセットCBH IIキメラおよび予測される熱安定性のあるCBH IIキメラ。この研究のために合成された18個のキメラは、星印の後に記載される。
【0158】
【表23】
【0159】
線形回帰の配列安定性データへの適応は、R2=0.88で観測されたT50値に適合する10−パラメーターモデルを生じた。トレーニングセット外の回帰モデルの予測性能をよりよく推定するため、11の折り畳み構造の相互検証をR2が0.57となるように実施し、2つの外れ値を取り除き(11313121および22222222)、相互検証R2を0.76へと増加させた。回帰モデルは、最も安定な親1(H. insolens)を基準状態T50として使用して、p値≦0.1を有する9つのさらなる項を含む。モデルパラメーター(Table9)は、親3からのブロック7の単一のブロックが、H. insolens CBH IIと比較してキメラの熱安定性に最も強く寄与することを示す。C. thermophilum CBH IIからのこのブロックは、それを含むキメラの熱安定性を約8.5℃上昇させる。p値≦0.1の8つの残りのブロックのうち2つによる熱安定性の上昇は、1.2℃および2.7℃と小さかったが、他の6つはその安定性を減少させた。
Table9:T50線状回帰モデルパラメーターおよびp値。図1の回帰適合線を算出するために使用されたp<0.1のパラメーター値を太字体で示す。ブロック効果は、親1(H. insolens CBH II)を基準状態として、親2および3からのそれぞれ8つのブロックの置換を示す16パラメーターを用いて報告された。
【0160】
【表24】
【0161】
B7P1およびB7P3配列のアライメント(図10)は、H. insolensおよびC. thermophilum酵素において、ブロック7の56個のアミノ酸のうち、10箇所で相違する。最も高いT50値を有する21311131のキメラのバックボーンにおいて、B7P3のそれぞれの残基(親の断片7(配列番号6))は、B7P1において相当する残基に個々に変異され、それぞれの点突然変異体についてT50値が得られた。突然変異体S313Cは、キメラの熱安定性を著しく変化させた:この単一の突然変異体は21311131のT50を約10℃減少させた(図11)。
【0162】
復帰突然変異の効果を異なる背景において研究するため、C313S置換をコードするH. insolensおよびH. jecorinaの親CBH II遺伝子(H. insolensにおいてC314S、およびH. jecorinaにおいてC311S)を作成し、発現させ、酵素のT50値を測定した。安定性のB7P3を野生型H. insolensおよびH. jecorinaの酵素内に置換したキメラ、11111131および22222232の安定性についても定量化した。B7P3ブロック置換およびCys−Ser点突然変異体は、親CBH IIを著しく安定化させた;最も大きな効果は、C311S置換を含むH. jecorina CBH IIへのT50における8℃までの上昇であった。またCys−Ser突然変異は、B7P3を含まない2つのキメラである、31311112および13231111ならびに組み換え型の親のセットでない相同するCBH II(Phaneroachete chrysosporiumから)において試験した。P. chyrsosporium CBH IIの触媒ドメインは、親CBH II触媒ドメインに55−56%のみの相同性がある。これら全ての酵素は、Cys−Ser置換によって安定化された;P. chrysosporium CBH IIは10℃と高い安定化を見せた(図13)。
【0163】
同等のC313S突然変異を含む8つの熱安定性のあるCBH IIキメラおよび親の酵素は、His6のタグを付け、精製することで、特異的活性を測定できた。Table10Aに示すように、特異的活性は、50℃で非結晶性セルロース(PASC)について観測されるように、Cys−Ser突然変異体を含むこれらキメラおよび天然酵素について、野生型親のものと同等であった。従って、高い熱安定性は、特異的活性の代償として現れるものではない。
Table10A:天然、点突然変異体および選択される熱安定性のあるキメラCBH IIに関する特異的活性値(μgグルコース還元糖等価/(μg CBH II酵素×min×10))。エラーバーは、標準誤差を示し、標準誤差は3つの複製に関する標準偏差/sqrt(n)として定義される。2時間の反応、3mg酵素/g PASC、50℃、25mM酢酸ナトリウム、pH4.8。
【0164】
【表25】
【0165】
Table10B:H. jecroinaおよびH. insolens野生型、C313S点突然変異体およびB7P3ブロック置換CBH IIに関する合成(SDCAA)および豊富な(YPD)発現培養培地上清の双方における全活性。示す値は、セルラーゼ活性分析に添加される発現培養上清CBH II等価のmL当たりのμgグルコース/mLセルラーゼ活性分析値である。SDCAA培養については、濃縮されたSDCAA培養上清が使用され、50mM酢酸ナトリウム(pH4.8)において50℃で100分間のリン酸膨潤セルロース(1mg/mL)に対する活性が測定された。YPD上清CBH IIは、アビセルに結合させることによって濃縮し、50mM酢酸ナトリウム(pH4.8)において55℃で150分間、アビセル(15mg/mL)に対する活性を測定した。
【0166】
【表26】
【0167】
これらの8つの熱安定性のある同一のキメラ(最も安定な親よりT50が2−10℃高い)は、その後結晶性セルロースへの活性に関して、16時間のインキュベーションの間、親酵素がほとんど活性を示さない温度を含む幅広い温度範囲で試験した。図9aは、8つの試験された熱安定性のあるキメラのうちの7つが、60−65℃でアビセルに対して最大の活性を示す、8つのキメラ全てについて、70℃(試験中の最高温度)で、活性を保持していることを示した。対照的に、3つの親CBH IIは、50℃で最大の活性を示したが、70℃で完全にまたはほぼ完全に不活性となった。さらに高い最適活性温度を有する7つのキメラは、あらゆる親CBH II酵素と比較して非常の多量のアビセルを加水分解した。図9bに示すように、同様の挙動が、Cys−Ser点突然変異を含むH. insolensおよびH. jecorinaの親においても観測された。Cys−Ser点突然変異は、アビセル加水分解およびP. chrysoporium CBH IIにおける最高反応温度を上昇させた。H. insolensおよびH. jecorina親におけるP3B7ブロック置換により、反応温度およびH. insolens CBH IIの加水分解は上昇したが、上昇した最高反応温度は、H. jecorina酵素による全体的なセルロース加水分解を向上させるものではなかった。
【0168】
cerevisae発現宿主からの野生型H. jecorina CBH IIの分泌が低い(<1mg/L)ことが観測された。野生型H. jecorina CBH II酵素におけるC311S突然変異は、CBH IIの全分泌量の活性を著しく上昇させた(表11)。豊富な培地(YPD)における活性の上昇は10倍であるが、合成(SDCAA)培地において、C311SおよびB7P3置換は、H. jecorina CBH II全分泌活性を2倍に上昇させる。他の2つの親CBH IIと比較して高いレベルで発現しているH. insolens CBH II親について、B7P3ブロック置換は活性を減少させたが、C314S突然変異は、活性を1.5倍まで上昇させた。H. insolensおよびH. jecorina野生型およびCys−Ser突然変異の全てが同様の特異的活性を有するため(図10)、全分泌セルラーゼ活性の上昇は、機能的酵素のより優れた分泌の結果である。S. cerevisiae異種タンパク質分布とタンパク質の安定性との間の相互関係が観測され、Cys−Ser突然変異CBH IIの上昇した分泌が、それらの最も高い安定性に反映し得ることを示唆した。
Table11:天然、点突然変異体および選択される熱安定性のあるキメラCBH IIに関する特異的活性値(μgグルコース還元糖等価/(μg CBH II酵素×min×102))。エラーバーは、3つの複製に関する標準誤差を示し、標準誤差は標準偏差/sqrt(n)を規定する。2時間の反応、3μg酵素/g PASC、50℃、25mM酢酸ナトリウム、pH4.8。
【0169】
【表27】
【0170】
Cys−Ser突然変異をモデル化するため、微細構造のH. insolens CBH II 1ocn結晶構造を使用した。第1に、水素結合ネットワークをREDUCEで最適化した。C314はPro339のカルボニル基と水素結合を形成することを予測した。この予測を検証するため、側鎖パッケージングを、モデリング・プラットフォームSHARPENを用いて最適化した。Ser314は、より強い水素結合、およびより有利な結合構造が生じるように、Cys314と同様の相互作用を奏することが予測された。
【0171】
数々の効果は、Cys−Ser突然変異が、天然CBH IIおよびキメラを含む幅広いCBH IIを安定化することができる理由を説明することができる。CysおよびSerは同様であり(等比体積でない)、他の代価物と比較して、これら2つのアミノ酸は配列アライメントをこの位置で抑制する。この残基への水素結合パートナーは、バックボーン要素(Gly316のアミドおよびPro339のカルボニル)であり、従ってあまり第3者のアミノ酸変異体に依存しそうにない。さらにこのポケットに直接隣接した側鎖(Asn283、Pro339、Phe345)が、研究された全4つの天然CBH IIセルラーゼにおいて保存されている。
【0172】
微細構造(1.3オングストローム)のH. insolens結晶構造(pdb挿入1ocn6)は、Cys314が水素結合ネットワークの一部であることを示した(図15)。Cysと比較して高いSerの水素結合性能は、安定化における比較的強い水素結合相互作用を奏することを示す。結晶構造においても、立体構造の観点からSerが好適に使用し得ることを示す。特に、Cys側鎖が元来の結合角度を有するとき、CysがPro339のカルボニルに押しつけられるように挿入されることで、6°の角度はなくなるが、望まれざる立体構造の相互作用を生じた。
【0173】
H. jecorina CBH IIと最も多くの一致を共有する196個のタンパク質配列のアライメント。最も一致のある配列の250のうちの54については、重複しているためアライメントから排除した(即ち、構造研究のための点突然変異体または95%超で一致するアイソフォーム)。Ser311に対するバイアスがある:158個の配列はSer、20個はAla、10個はCys、5つは欠失、3つはGlyを有する。しかしながら、42個の他の配置があり、最も多くの選択肢は、H. jecorinaのアミノ酸の選択肢の2倍を超えて生じる。
【0174】
Cys−Ser突然変異の高い安定化効果は、この位置でのSerが、天然セルラーゼの熱安定性の全体的な指標となる可能性を向上させる。しかしながら、この位置にCysを有することを特徴とするH. insolens CBH IIのT50値が64.8℃であるが、これはC. thermophilum CBH II(64.0℃)より高く、Serが安定化決定の唯一の要因ではないことを示している。
【0175】
熱安定性だけが、産業用セルラーゼの重要な特徴ではない。特異的活性、セルロース結合に与える変化、発現に対する効果、および生成阻害は、全て同様に重要である。キメラおよび本明細書のデータは、組換えにより、短時間(例えば2時間)で測定されたセルロース加水分解分析において、特異的活性を損なうことなく向上した熱安定性を奏する、CBH IIキメラを得ることを示す。同様の現象が、熱安定性のあるCys−Ser突然変異を含むCBH IIに関してもみられた。長時間の加水分解分析において、いくつかのCBH IIキメラ、および3つの被験Cys−Ser突然変異CBH IIについて、天然CBH IIと比較してより多くセルロースを加水分解することを発見した。この優れた性能は、酵素が高温でより長時間で機能し続けることを可能とする高い熱安定性とともに、親CBH IIに匹敵する特異的活性を有することに起因すると考えられる。これらの分析は、等量の精製された親、キメラおよびCys−Ser突然変異体酵素で実施されたため、観測された高温での加水分解の向上は、S. cerevisiae発現宿主からの分泌量の増加によるものではない。熱安定性のあるキメラおよびCys−Ser突然変異体は、従って、セルロース分解のための酵素製剤に有用な要素であることが証明し得る。
【0176】
様々な特定の実施形態が示され、記載される一方で、この発明の思考と範囲から逸脱することなく、様々な変化がされることが理解される。
【図1A】
【図1B】
【図2A】
【図2B】
【図2C】
【技術分野】
【0001】
<関連出願の優先権主張>
本願は35U.S.C.§119に基づいて、2009年4月6日に出願された米国仮出願第61/166,993号、および2009年5月13日に出願された米国仮出願第61/177,882号を参照して取り込むものとする。
【0002】
<連邦支援の研究に関する記載>
アメリカ政府は、国立衛生研究所によって与えられる研究費番号GM068664およびARO−US Army Robert Morris Acquisition Centerによって与えられる研究費番号DDAD19−03−0D−0004に従って、本発明において特定の権利を有する。
【0003】
本発明は、分子生物工学および分子生物デザイン、並びに改変タンパク質および改変核酸に関する。
【背景技術】
【0004】
バイオマス変換工程におけるセルラーゼ混合物の性能は、セルロース基質の物理的な状態および組成に加えて、安定性、生成阻害、多様なセルラーゼ成分間の相乗効果、非生産的な吸着に対する生産的な結合、およびpH依存性を含む多くの酵素特性に依存する。多様な適用および原料に対して酵素の形態を最適化するため、セルロース加水分解に多変量的な特性を付与し、多様なセルラーゼから選択することが望まれる。
【発明の概要】
【課題を解決するための手段】
【0005】
本発明は、セルラーゼ活性があり、野生型タンパク質と比較して、高い熱安定性および活性がある組み換え型ポリペプチドを提供する。本発明は、長時間の加水分解試験において、CBH IIのC末端(例えば天然タンパク質配列に応じてアミノ酸約310−315であり、例えば配列番号2および4を参照とする)で、CysからSerへの突然変異があるキメラおよび天然酵素が、天然酵素と比較してより確実なセルロース加水分解を提供することを示す。
【0006】
本発明は、配列GECDG(配列番号2の312−316)を含有するモチーフ中のC末端領域にC→S置換を含む組み換えポリペプチドであって、該変異体が、野生型セロビオヒドラーゼと比較して高い熱安定性およびセルラーゼ活性があるものを提供する。例えば、本発明は高い熱安定性およびセルラーゼ活性があるポリペプチドであって、C314Sを含有する配列番号2と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C311Sを含有する配列番号4と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C310Sを含有する配列番号12と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C312Sを含有する配列番号13と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C314Sを含有する配列番号14と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C315Sを含有する配列番号15と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C313Sを含有する配列番号16と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C311Sを含有する配列番号17と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C313Sを含有する配列番号19と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C311Sを含有する配列番号22と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号64と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C407Sを含有する配列番号65と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C394Sを含有する配列番号66と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号67と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号68と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号69と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号70と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号71と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号72と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号73と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号74と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C400Sを含有する配列番号75と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C407Sを含有する配列番号76と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;C394Sを含有する配列番号77と少なくとも85%、90%、95%、98%、99%の一致があるものを含むか;C412Sを含有する配列番号78と少なくとも85%、90%、95%、98%、99%の一致があるものを含み;上記ポリペプチドは、セルラーゼ活性があり、それに相当するCys→Serの突然変異が欠如している親(野生型)タンパク質と比較して熱安定性が改良されている。
【0007】
本発明は、組み換えにより生成されたか、合成されたか、他の非天然的な製造方法により生成された、いずれかの実質的に精製されたポリペプチドも提供する。該ポリペプチドは、C→S置換の存在位置を除く、以下に示す1−10、10−20または20−30の保存的な配列のアミノ酸置換基を含む:C314Sを含む配列番号2;C311Sを含む配列番号4;C310Sを含む配列番号12;C312Sを含む配列番号13;C314Sを含む配列番号14;C315Sを含む配列番号15;C313Sを含む配列番号16;C311Sを含む配列番号17;C313Sを含む配列番号19;C312Sを含む配列番号21;C311Sを含む配列番号22;C400Sを含む配列番号64;C407Sを含む配列番号65;C394Sを含む配列番号66;C400Sを含む配列番号67;C400Sを含む配列番号68;C400Sを含む配列番号69;C400Sを含む配列番号70;C400Sを含む配列番号71;C400Sを含む配列番号72;C400Sを含む配列番号73;C400Sを含む配列番号74;C400Sを含む配列番号75;C407Sを含む配列番号76;C394Sを含む配列番号77;またはC412Sを含む配列番号78。
【0008】
本発明は、(a)位置314にSerがある配列番号2と少なくとも85%以上の一致があるポリペプチドであり、前記ポリペプチドはセルラーゼ活性がある;(b)位置311にSerがある配列番号4と少なくとも70%以上の一致があるポリペプチドであり、前記ポリペプチドはセルラーゼ活性がある;(c)(i)位置310にSerがある配列番号12、(ii)位置312にSerがある配列番号13、(iii)位置314にSerがある配列番号14、(iv)位置315にSerがある配列番号15、(v)位置313にSerがある配列番号16、(vi)位置311にSerがある配列番号17、(vii)位置313にSerがある配列番号19、(viii)位置312にSerがある配列番号21、(ix)位置311にSerがある配列番号22からなる群から選択される配列と70%以上の一致があるポリペプチドであり、それぞれ上記ポリペプチドは、セルラーゼ活性がある;(d)2つの異なるセロビオヒドラーゼポリペプチドからの少なくとも2つのドメインを含むキメラポリペプチドであり、ドメインはNからC末端までに:(断片1)−(断片2)−(断片3)−(断片4)−(断片5)−(断片6)−(断片7)−(断片8)を含み;断片1は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)のアミノ酸残基約1から約x1までと少なくとも50−100%の一致がある配列を含み;断片2は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)のアミノ酸残基約x1から約x2までと少なくとも50−100%の一致がある配列を含み;断片3は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)のアミノ酸残基約x2から約x3までと少なくとも50−100%の一致がある配列を含み;断片4は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)のアミノ酸残基約x3から約x4までと少なくとも50−100%の一致がある配列を含み;断片5は、配列番号2(“1”)、配列番号4(「2」)または配列番号(「3」)のアミノ酸残基約x4から約x5までと少なくとも50−100%の一致がある配列を含み;断片6は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)のアミノ酸残基約x5から約x6までと少なくとも50−100%の一致がある配列を含み;断片7は、配列番号2(「1」)または、配列番号4(「2」)のアミノ酸残基約x6から約x7までと少なくとも50−100%の一致がある配列を含み;断片8は、配列番号2(「1」)または配列番号4(「2」)のアミノ酸残基約x7から約x8までと少なくとも50−100%の一致がある配列を含み;x1は配列番号2の残基43,44,45,46,47であるか、配列番号4または配列番号6の残基42,43,44,45,46である;x2は配列番号2の残基70,71,72,73,74であるか、配列番号4もしくは配列番号6の残基68,69,70,71,72,73,74;x3は配列番号2の残基113,114,115,116,117,118であるか、配列番号4もしくは配列番号6の残基110,111,112,113,114,115,116;x4は配列番号2の残基153,154,155,156,157であるか、配列番号4もしくは配列番号6の残基149,150,151,152,153,154,155,156;x5は配列番号2の残基220,221,222,223または224であるか、配列番号4もしくは配列番号6の残基216,217,218,219,220,221,222,223;x6は配列番号2の残基256,257,258,259,260であるか、配列番号4もしくは配列番号6の残基253,254,255,256,257,258,259,260;x7は配列番号2の残基312,313,314,315または316であるか、配列番号4の残基309,310,311,312,313,314,315,318;およびx8はポリペプチドのC末端に相当するアミノ酸残基が配列番号2、配列番号4もしくは配列番号6の配列であり、キメラポリペプチドは、配列番号2の位置314または配列番号4の位置311でSerであり、キメラポリペプチドは配列番号2,4もしくは6を含むCBH IIポリペプチドと比較してセルラーゼ活性、改良した熱安定性および/またはpHの安定性があるものからなる群から選択される配列を含む組換え型ポリペプチドを提供する。
【0009】
本発明は、配列番号12−62または63の配列からなる組換え型ポリペプチドも提供する。
【0010】
本発明は、また上記いずれかのポリペプチドをコード化するポリヌクレオチド、該ポリヌクレオチドを含むベクター、および該ポリヌクレオチドまたは該ベクターを含む宿主細胞を提供する。
【0011】
本発明は、本発明の実質的に精製された形態、または細胞溶解物の一部としてポリペプチドを含む酵素製剤も提供する。
【0012】
本発明は、またセルロースを含むバイオマスの処理方法を提供し、該方法は本開示のポリペプチドまたは酵素液とバイオマスの接触工程を備える。
【図面の簡単な説明】
【0013】
【図1】候補遺伝子たるCBH IIの親遺伝子の酵母発現培養上清のSDS−PAGEゲルを示す図である。(A)ゲルレーン(左から右):1−H. jecorina、2−空ベクター、3−H. insolens、4−C. thermophilum、5−H. jecorina(二重)、6−P. chrysosporium、7−T. emersonil、8−空ベクター(二重)、9−H. jecorina(三重)。ゲルの下部の数字は、2時間、50℃でのPASC加水分解試験後に、反応物中に存在する還元糖の濃度(μg/mL)を示す。その後、BSAを標準としたSDS−PAGEによる比較は、H. insolens発現レベル5−10mg/Lの評価を可能とした。(B)S. cerevisiae CBH II発現培養上清のSDS−PAGEゲル分析を示す図である。CBH IIバンドが、ちょうど60kDa分子量基準値未満で現れた。レーンは、左から右に向かって、1−野生型H. jeco、2−H. jeco B7P3、3−H. jeco C311S、4−野生型C. ther、5−野生型H. inso、6−H. inso B7P3、7−H. inso C314Sである。数値は、50℃、50mM酢酸ナトリウム、pH4.8の条件下で、PASC(1mg/mL)を用いて100分間インキュベーションした際に生じるμgグルコース等量/SDCAAを発現する培養上清等量の1mL当たりのmL反応容量を示す。レーン1−4に関する値は、加えられた濃縮培養上清の容積の2倍に修正するため、2で除算する。かかる修正を省略すると、H. insolens酵素に関する特異的活性値が低下するように思われる。
【図2】CBH IIキメラライブラリーブロックの限度を示す図である。(A)色分けされたブロックを用いたH. insolens CBH II触媒ドメインのリボンダイアグラムを示す。CBH II酵素は、セロビオ由来のイソファゴミングリコシダーゼ阻害剤と複合体を形成する。(B)2次構造要素、ジスルフィド結合および黒矢印によって示されるブロックの区別を示す、H. insolens触媒ドメインの直線図である。(C)組み換えに応じて分解することができる接続部分(4.5A範囲内の側鎖重原子)を示す側鎖接続マップである。分解された接触部の大半は、連続したブロックの間に生じる。
【図3】23個の分泌/活性型および15個の非分泌/不活性型のサンプルセットのキメラに関して分割した接続部(E)の数および最も近い親(m)からの突然変異の数を示す図である。
【図4】親CBH II酵素および3つの熱安定性のあるキメラに関するpHの関数として、pH5.0で基準化した特異的活性を示す図である。提示するデータは、2つの測定の平均であり、HJPlusおよびH. jecoに関するエラーバーは、2つの独立した試験の値を示す。16時間反応物において、300μg酵素/g PASC、50℃での12.5mMクエン酸ナトリウム/12.5リン酸ナトリウム緩衝剤中において示すようなpHである。
【図5】親および広範囲にわたる温度での熱安定性のあるキメラに関する長期セルロース加水分解分析の結果を示す図である。エラーバーは、HJPlusおよびH. insolens CBH II酵素の3つの測定に関する標準誤差を示す。pH4.8における50mM酢酸ナトリウム中の100μg酵素/g PASCの40時間反応物である。
【図6】63℃で12時間インキュベーションした後、検証するためのキメラセットに関する標準化した残留活性を示す図である。50℃で25mM酢酸ナトリウム緩衝剤、pH4.8中、2時間の分析においてPASCを基質として用いて測定された濃縮培養上清におけるCBH II酵素に関する残留活性を示す。
【図7】親およびキメラCBH II酵素発現ベクターYep352/PGK91−1―rssに関するマップを示す図である。図示のベクターは、野生型H. jecorina ce16a(CBH II酵素)遺伝子を含む。キメラおよび親CBH II酵素の双方に関して、ss Lys−Arg Kex2部位に続くCBD/リンカーアミノ酸の配列は、以下の通りである:ASCSSVWGQCGGQNWSGPTCCASGSTCVYSNDYYSQCLPGAASSSSSTRAASTTSRVSPTTSRSSSATPPPGSTTTRVPPVGSGTATYS (配列番号8)。
【図8】CBH II親および51個のCBH IIキメラに関する観測されたか、予測されるT50値である。線は線形回帰モデル式を示す(Table7におけるパラメーター)。親CBH IIのT50値は、正方形で示す。
【図9】アビセルに対する、温度によるCBH II特異的活性を示す図であり、(a)CBH II親およびキメラの特異的活性である。(b)CBH II親、C311S突然変異体およびB7P3単一ブロック置換キメラの特異的活性である。アビセル濃度15mg/mLを用いた50mM酢酸ナトリウム、pH4.8中で16時間、反応を行った。(c)温度の関数としてアビセルに対するCBH II親、単一点突然変異体および単一阻害置換キメラ活性(μg/グルコース/mL反応)である。150分間、50mM酢酸ナトリウム、pH4.8条件下、アビセル濃度15mg/mLを用いて、反応を行った。CBH II酵母培養上清を、55℃でほぼ等量の還元糖産物の濃度を達成するため投与した。提示されるデータは、それぞれの温度点に関して二重の活性値を示すエラーバーと2つの独立した測定の平均である。
【図10】親1であるH.insolensおよび親3であるC. thermophilumからのブロック7ClustalW複数配列の配置である。矢印は、復帰突然変異における残基変異を示す。
【図11】21111331キメラ点突然変異に関するT50値を示す図である。値は、2つの独立した二重試験の平均値として示され、エラーバーは点突然変異に関する二重試験のそれぞれのT50値を示す。試験温度の50mM酢酸ナトリウム中で、pH4.8条件下、10分間実施された。残留活性は、上記緩衝剤中、1g/Lリン酸膨潤セルロース(PASC)と50℃で100分間のインキュベーションによって測定された。
【図12】H. insolensおよびH. jecorinaの親CBH II、Ser単一点突然変異およびB7P3ブロック置換キメラに関するT50値を示す図である。3つの独立した測定の平均、エラーバーとして示した値は、それぞれのCBH IIに関する1つの標準偏差を示す。不活性化は、試験される温度50mM酢酸ナトリウム中で、pH4.8条件下で、10分間実施された。残留活性は、上記緩衝剤中、1g/Lのリン酸、膨潤セルロース(PASC)と50℃で、100分間のインキュベーションによって測定された。
【図13】P. chrysosporium(H. jecorina CBMと融合した)およびS. cerevisiaeから非相同的に分泌されたCBH IIキメラ31311112,13231111および野生型CBH II触媒ドメインに関するT50値を示す。2つの独立した試験の測定値をエラーバーを用いて示す。不活性化は、試験される温度50mM酢酸ナトリウム中で、pH4.8条件下で10分間実施された。残留活性は、上記緩衝剤中、1g/Lリン酸、膨潤セルロース(PASC)を用いて50℃で100分インキュベーションすることによって測定された。
【図14】CBH II組み換え型ブロックの接合部位を示す図である。(a)新規非親残基の組み合わせが可能な場合(結合した球体)、ブロックの間の部位は、通常、その表面が曝露されており、潜在的に溶媒が接合部位をスクリーンすることを可能とする。(b)接合部位(B5−B6)の例は、バックボーン(位H. jecorinaおよびH. insolensに配置図)、の保存、表面上の可変的な残留物ならびに残基173および253(矢印)における新規な埋没した疎水性の対の比較的稀有な可能性を示す。(c)H. jecorina(図中、黒色で示す)からのブロック1−4は、基質結合と関連する移動が、B3部位(黄色)において観測される(矢印)にも関わらず、同属のH. insolensブロック(色分けされた図)と大きな偏差なく適合する。(d)B6、B7接点(矢印)での挿入欠失が構造変化を要するが、同属のブロック5−8も同様である。
【図15】C314S突然変異体およびその安定性への効果の構造分析を示す図である。(a)微細構造のH. insolens構造(1ocn)の水素の位置を、図1に示した。(b)類似したセリン構造の再構成された結合構造を、PyMOL(http: (//) www .pymol . org)でモデル化した。SHARPEN2モデリング・プラットフォーム(全ての原子ロゼッタエネルギー機能を有する)において、側鎖を最適化することで、Cys314およびSer314の双方が、Pro339のカルボニルの水素結合に寄与し、Gly316のアミドから水素結合を受容することを示唆した。セリンの優れた水素結合性能は、セリンを含む変異体のより高い安定性に影響し得る。他の説明は、補助的な位置を示すものである。特に、1ocnからのCys位置は、側鎖が顕著に曲がり(即ち、N−C−Cα−Cβからの不適切な上反角が、標準位置から6°である)、Proカルボニルからの距離が増加した、立体構造種を示す。図内の文字の後でない数値は、水素結合距離(オングストローム)を意味する。
【発明を実施するための形態】
【0014】
本明細書および特許請求の範囲で使用する、単数形“ある(a,an)”、および“この(the)”の語は、明確に記載しない限り、複数の指示物を含む。従って、例えば「ドメイン」の用語は複数のドメイン、「タンパク質」の用語は1つ以上のタンパク質を含む。
【0015】
また「または」の用語は、他に記載されない限り「および/または」を意味する。同様に「含む(comprise,comprises, comprising,include,includesおよびincluding)」は、同じ意味で用いられ、制限することは意図しない。
【0016】
さらに様々な実施形態の記載においては、「含む」の用語を使用し、当業者は、いくつかの特定の場合、実施形態が或いは「実質的になる」または「なる」の言語を使用して記載されていると理解するであろう。
【0017】
ここに記載されたものと同様または対応する方法および材料を、開示の方法および組成物の実施に使用することができ、ここでは、例示的な方法、装置および材料を開示するものである。
【0018】
他に規定がない限り、ここに使用される全ての専門的且つ化学的な用語は、一般的にこの開示が属する分野の当業者に理解されるものと同様の意味を有する。したがって、本出願にわたって使用される以下の用語は、以下の意味を有するものとする。
【0019】
最近の研究は、研究室レベルのバイオマス変換過程において、中等温度好性の対照物と比較して、好熱性菌からのセルラーゼが熱安定性に優れ、その高い安定性により、適温または高温状態における長期の活性保持を有することを示してきた。菌のセルラーゼについては、40g/Lまでの上清濃度で、高度に活性があり、菌宿主、例えばHypocrea jecorina(アナモルフ・トリコデルマ・リーゼイ)に発現することができることため、高い関心が寄せられている。不都合にも、熱安定性のある菌セルラーゼを発現する試料は、その数が少ない。。加工性のセロビオヒドラーゼクラスII(CBH II)酵素の場合、10個未満の天然熱安定遺伝子配列が、CAZyデータベースに掲載されている。
【0020】
ここに十分に記載するように、再帰的キメラポリペプチドの生成ならびに特定の安定したドメインおよび最終的な特定のアミノ酸の分析により、熱安定性に付与、活性化の改善がみられた。
【0021】
以下により詳細に記載するように、本発明は、少なくとも一部において、セルロースの加水分解に触媒作用を生じる新規酵素の生成および発現に基づくものである。一つの実施形態において、高温条件下でセルロースを加水分解するように改変された新規ポリペプチドを提供する。該ポリペプチドは、アミノ酸置換基を特定の残基で含むように変えられたセロビオヒドラーゼの変異体を含む。これらの変異体については以下でより詳細に記載されているが、本発明のポリペプチドは1つ以上の修飾されたアミノ酸を含んでもよいことが理解される。修飾されたアミノ酸の存在は、例えば(a)ポリペプチド半減期の長期化、(b)熱安定性および(c)基質の補充の増加において有利である。アミノ酸は、例えば、共翻訳中または翻訳後において、組み換え生産(例えば、哺乳類細胞での発現中の、N――X――S/TモチーフのN−結合型グリコシル化)の際に修飾されるか、合成手段によってによって修飾される。従って、「突然変異」、「変異体」または「修飾された」タンパク質、酵素、ポリヌクレオチド、遺伝子または細胞は、親のタンパク質、酵素、ポリヌクレオチド、遺伝子または細胞から選択されるか、由来するか、ある場合においては異なるか、変化した、タンパク質、酵素、ポリヌクレオチド、遺伝子または細胞を意味する。突然変異体もしくは修飾されたタンパク質または酵素は、必然的でないが、通常、突然変異体のポリヌクレオチドまたは遺伝子から発現する。
【0022】
「突然変異」は、突然変異タンパク質、酵素、ポリヌクレオチド、遺伝子または細胞を生じる、あらゆる工程または機構を意味する。これは、タンパク質、酵素、ポリヌクレオチドまたは遺伝子配列が変化するあらゆる突然変異、および、かかる突然変異から起因する細胞におけるあらゆる検出可能な変化を含む。通常、突然変異は、ポリヌクレオチドまたは遺伝子配列での、単一または複数のヌクレオチド残基の点変異、欠失または挿入によって生じる。突然変異は、タンパク質をコードする遺伝子の領域内で生じるポリヌクレオチドの変性、ならびにタンパク質コード配列外、例えば、調節またはプロモーター配列(これらに限定しない)領域の変性を含む。遺伝子内における突然変異は、「サイレント」であってもよく、即ち発現に応じてアミノ酸変性に反映されず、遺伝子の「配列保存的な」変異体をなってもよい。これは、通常、1個のアミノ酸が1種を超えるコドンに対応する場合において生じる。
【0023】
修飾されたアミノ酸の非制限的な例としては、グリコシル化アミノ酸、硫酸化アミノ酸、プレニル化(例えばファルネシル化、ゲラニルゲラニル化)アミノ酸、ビオチン化アミノ酸、カルボキシル化アミノ酸、リン酸化アミノ酸等が挙げられる。アミノ酸の修飾について、当業者を導くのに参照し得る文献が存在する。プロトコールの例は、WalkerのProtein Protocols(1998)(Humana Press, Towata、ニュージャージー州)のCD−ROMから取得可能である。
【0024】
本発明には、修飾したセロビオヒドラーゼポリペプチドを生成・分離する組み換え法を記載する。組み換え生成に加えて、固相技術を用いた直接的なペプチド合成(例えば、Stewart et al. (1969) Solid-Phase Peptide Synthesis (WH Freeman Co, San Francisco);およびMerrifield (1963) J. Am. Chem. Soc. 85: 2149-2154)によって、ポリペプチドを生成することができる。ペプチド合成は、手動または自動の方法を用いて実施することができる。自動合成は、例えばアプライドバイオシステムズ431Aペプチド合成装置(パーキンエルマー、カリフォルニア州フォスターシティ)を用いて、製造業者の提供する説明書に従って実施することができる。
【0025】
「セロビオヒドラーゼII」または「CBH II酵素」は、細菌類および菌類において多岐にわたるセルラーゼファミリー6タンパク質における酵素を意味する。酵素は、セルロースの加水分解に用いられる。
【0026】
「セルラーゼ活性」は、セルロースを加水分解できる酵素を意味する。セルラーゼは、セルロースの加水分解を触媒する菌類、細菌類および原生動物によって生成される酵素ファミリーを指す。しかしながら、他の種類の生物、例えば植物および動物によって生成されるセルラーゼもある。この群のEC番号は、EC3.2.1.4である。触媒作用に基づく、5つの一般的な種類のセルラーゼが存在する:エンドセルラーゼ;エキソセルラーゼであり、この部類内には2つの主な種類のエキソセルラーゼ(またはセロビオヒドラーゼ、省略してCBH)があり、1つの種類は還元端から進むように働き、もう1つの種類はセルロースの非還元末端から進むように働く;セロビアーゼまたはβグルコシダーゼ加水分解;酸化セルラーゼ;および、水の代わりにリン酸を用いてセルロース単量体へと分解するセルロースホスホリラーゼ。多くの菌のセルラーゼは、2つのドメイン;フレキシブルなリンカーによって結合している触媒ドメインおよびセルロース結合ドメインを有する。本発明のある実施形態において、セルラーゼ活性は、CBH活性である。ここに記載の配列は、いくつかの場合において、セルロース結合ドメインおよび触媒ドメインの双方、または触媒ドメインのみを含む。いくつかの場合において、触媒ドメイン配列のみを提供する場合、配列番号8で示されるようなセルロース結合ドメイン(CBD)が、直接又はリンカーを通して、触媒ドメインに機能的に連結し得ること(コード配列の一部とするか、後に融合するか)が認識されている。
【0027】
ここに用いられる「タンパク質」または「ポリペプチド」は、同じ意味で用いられており、1つ以上の化学基礎単位の鎖であり、化学結合であり、ペプチド結合と称するものによって共に連結されている複数アミノ酸を含む。「酵素」は、高度にまたは低度に特異的に1つ以上の化学または生化学反応を触媒または促進するあらゆる物質であり、好ましくは完全にまたは大半がタンパク質から構成されるものを意味する。「天然」または「野生型」タンパク質、酵素、ポリヌクレオチド、遺伝子または細胞は、自然に産出されるタンパク質、酵素、ポリヌクレオチド、遺伝子または細胞を意味する。
【0028】
「アミノ酸配列」は、アミノ酸のポリマー(タンパク質、ポリペプチド等)またはアミノ酸ポリマーを示す特徴となる文字列(character string)である。「タンパク質」、「ポリペプチド」および「ペプチド」の用語は、ここでは交換可能に同じ意味で使用される。「アミノ酸」は、中心の炭素原子が水素原子、カルボン酸基(ここに「カルボキシル炭素原子」として指す炭素原子)、アミノ基(ここに「網の窒素原子」として指す窒素原子)および側鎖基であるRの基に結合する構造を有する」分子である。アミノ酸をペプチド、ポリペプチドまたはタンパク質に取り入れる場合、1つのアミノ酸をもう1つのアミノ酸に結合させる脱水反応において、1つ以上のそのカルボン酸基の原子が欠失する。そのため、タンパク質に取り入れられたアミノ酸は「アミノ酸残基」と表される。
【0029】
一定のタンパク質の特定のアミノ酸配列(即ち、アミノ基末端からカルボキシ基末端まで表示される場合のポリペプチドの「一次構造」)は、コーディング部位のmRNAのヌクレオチド配列によって決定され、同様に遺伝情報、特にゲノムDNA(細胞小器官DNA、例えばミトコンドリアまたは葉緑体DNAを含む)によって特定される。従って、遺伝子の配列を決定することは、対応するポリペプチドの一次配列、より詳細には、遺伝子またはポリヌクレオチド配列によってコード化されるポリペプチドまたはタンパク質の役割または活性の予見を可能とする。
【0030】
特定の配列の「保存的アミノ酸置換」または単なる「保存的な変異体」は、アミノ酸配列と実質的に同一である1つ以上のアミノ酸または一連のアミノ酸の置換を示す。当業者は、アミノ酸の欠失、アミノ酸の付加または化学的に類似のアミノ酸との置換が起こる「保存的な変異体」を生じる、コード化配列における単一のアミノ酸または一定の割合のアミノ酸の変化、付加または欠失保存的を認識するであろう。
【0031】
機能的に同類のアミノ酸の保存的置換を示す表は、従来技術において既知である。例えば、ある保存的置換基は、アラニン(A)、セリン(S)およびトレオニン(T)を含む。他の保存的置換基は、アスパラギン酸(D)およびグルタミン酸(E)を含む。他の保存的置換基は、アスパラギン(N)およびグルタミン(Q)を含む。また他の保存的置換基は、アルギニン(R)およびリジン(K)を含む。他の保存的置換基はイソロイシン(I)、ロイシン(L)、メチオニン(M)およびバリン(V)を含む。他の保存的置換基は、フェニルアラニン(F)、チロシン(Y)およびトリプトファン(W)を含む。
【0032】
従って、列挙されたポリペプチド配列(例えば配列番号2,4,6および12−78)の「保存的アミノ酸置換基」は、同じ保存的置換基のアミノ酸から保存的に選択され、ポリペプチド配列のアミノ酸の割合としては、通常10%未満で含む。従って、本発明のポリペプチドの多様な保存的置換変異体は、同じ保存的置換基の多様な保存的置換変異体において、100,75,50,25または10個の置換を含み得る。
【0033】
核酸分子のコード化された活性を変えない配列の付加、例えば非機能的付加または非コード配列は、塩基性の核酸の保存的変異体であると理解されている。酵素の「活性」は、その反応を触媒する能力、即ち「機能」の測定値であり、反応生成物が生成される速度として表すことができる。例えば、酵素活性は、時間当たり、または酵素の単位(例えば濃度または重量)当たりについて、生成される生成物の量、または親和性、または解離定数に関して表すことができる。ここに同様の意味で用いられる「セロビオヒドラーゼ活性またはセルラーゼ活性」、「セロビオヒドラーゼまたはセルラーゼの生物学的活性」または「セロビオヒドラーゼまたはセルラーゼの機能的活性」は、特定の実施形態においては、タンパク質、セルラーゼ活性を有するポリペプチドであり、標準の技術によって生体内または生体外で決定される、セロセルロース基質へのビオヒドラーゼ活性によって及ぼす活性を指す。
【0034】
当業者は、開示される、機能的に同一の構造を有する核酸構造の多くの保存的変異体を理解するであろう。例えば、上記のように、遺伝子のコーディングの縮退のため、「サイレント置換」(即ち、核酸配列の置換であり、コードされたポリペプチドには変異を生じない)は、アミノ酸をコードする各々の核酸配列の特徴を包含する。同様に、アミノ酸配列における1つまたは数個のアミノ酸の「保存的アミノ酸置換」は、高い相同性の特徴を有する異なるアミノ酸と置換され、また開示の構造と高い相同性を有することが容易に同定される。それぞれ開示の配列の、このような保存的変異体は、ここに提供するポリペプチドの特性である。
【0035】
「保存的変異体」は、タンパク質または酵素の全体の構造および機能を変えることなく、アミノ酸残基を変化させた、タンパク質または酵素であり、同様の特性、極性または非極性、サイズ、形状、電荷等(これらに限定されない)を有するアミノ酸への置換を包含する。保存的なものとして示されるアミノ酸以外は、タンパク質または酵素において異なっていてもよく、あらゆる2つの同様の機能のタンパク質間のタンパク質またはアミノ酸配列の類似性は、異なっていてもよく、配列図によって決定されるように、例えば少なくとも30%、少なくとも50%、少なくとも70%、少なくとも80%または少なくとも90%であってもよい。本明細書において、「配列類似性」は、ヌクレオチドまたはタンパク質配列が連関する範囲を意味する。2つの配列の類似範囲は、配列類似性のパーセントおよび/または保守性に基づくことができる。ここに「配列相同性」は、2つのヌクレオチドまたはアミノ酸配列の範囲に差異がないことを意味する。「配列のアラインメント」は、最大限の一致を達成するため(アミノ酸配列の場合保存性)、類似性の程度を評価することを目的とする2つ以上の配列を揃える工程を意味する。配列を揃えて、類似性/相同性を評価するための数々の方法は、従来技術、例えばCluster法において既知であり、類似性はMEGALIGNアルゴリズムならびにBLASTN、BLASTPおよびFASTA(Lipman and Pearson, 1985; Pearson and Lipman, 1988)に基づくものが知られる。このプログラム全てを使用して、最も高い配列相同性を生じるものが好ましい設定といえる。
【0036】
特定のポリペプチドの非保存的な修飾としては、保存的置換を特徴としないあらゆるアミノ酸の置換が挙げられる。例えば、上記の6つの群の範囲を超えるあらゆる置換が挙げられる。これらは、塩基性または酸性のアミノ酸から中性のアミノ酸(例えば、Asp、Glu、AsnまたはGlnからVal、Ile、LeuまたはMet)、芳香族アミノ酸から塩基性または酸性アミノ酸(例えば、Phe、TyrまたはTrpからAsp、Asn、GluまたはGln)への置換、またはアミノ酸とその類似するアミノ酸との置換ではない、あらゆる他の置換を含有する。塩基性側鎖としては、リジン(K)、アルギニン(R)、ヒスチジン(H);酸性側鎖としては、アスパラギンサン(D)、グルタミン酸(E);電荷を有さない極性側鎖としては、グリシン(G)、アスパラギン(N)、グルタミン(Q)、セリン(S)、トレオニン(T)、チロシン(Y)、システイン(C);非極性側鎖としては、アラニン(A)、バリン(V)、ロイシン(L)、イソロイシン(I)、プロリン(P)、フェニルアラニン(F)、メチオニン(M)、トリプトファン(W);β岐側鎖としては、トレオニン(T)、バリン(V)、イソロイシン(I);芳香性の側鎖としては、チロシン(Y)、フェニルアラニン(F)、トリプトファン(W)、ヒスチジン(H)が挙げられる。
【0037】
「親」タンパク質、酵素、ポリヌクレオチド、遺伝子または細胞は、あらゆる他のタンパク質、酵素、ポリヌクレオチド、遺伝子または細胞に由来するか、あらゆる方法、装置または技術を用いて生成され、親そのものが天然か、突然変異体である任意のタンパク質、酵素、ポリヌクレオチド、遺伝子または細胞である。親ポリヌクレオチドまたは遺伝子は、親タンパク質または酵素をコードする。
【0038】
CBH IIポリペプチドの変異体を提供することに加えて、1)第1の親鎖から分離され、アミノ酸置換基を含むように修飾された変異体ドメイン;および2)第2の親鎖から分離され、新規活性またはドメインを補足する活性を含有するために非修飾または修飾のいずれかから分離されたドメインを含むキメラポリペプチド;が提供される。本発明のキメラポリペプチドの技術にかかる方法をここに開示する。
【0039】
本発明は、野生型、または親タンパク質と比較して高い熱安定性を有するセルラーゼおよびセロビオヒドラーゼ(CBH)II変異体、突然変異体およびキメラを開示し、野生型タンパク質は配列番号2,4または6からなる。変異体は、配列GEXDGのモチーフを有するC末端領域であって、セリンを含有し、前記Xは、C、AまたはG(配列番号107)であり、Xは、セリン、セルラーゼ活性を含む変異体と置換され、ポリペプチドは配列GEXDG(配列番号107)においてセリンを欠如している野生型セルラーゼと比較して高い熱安定性を有する。一つの実施形態において、変異体は多くのセルラーゼおよびセロビオヒドラーゼIIタンパク質において見出されるモチーフGECDG(以下により完全に記載する)において少なくともCys→Serの突然変異(例えば、配列番号2のアミノ酸312−316を参照のこと)を含み、熱安定性または活性を向上させた付加突然変異を含んでいてもよい。セルラーゼ間の相同性は、非常に低くてもよい。上記セリン置換基は、配列番号107のモチーフ(例えば、ポリペプチドが配列番号2または4と少なくとも60%以上の相同性がある)を有するあらゆるセルラーゼに適用できる。
【0040】
例えば、本発明は高い熱安定性およびセルラーゼ活性を有し、C314Sを含む配列番号2と少なくとも85%、90%、95%、98%、99%の相同性があり;C311Sを含む配列番号4と少なくとも85%、90%、95%、98%、99%の相同性があり;C310Sを含む配列番号12と少なくとも85%、90%、95%、98%、99%の相同性があり;C312Sを含む配列番号13と少なくとも85%、90%、95%、98%、99%の相同性があり;C314Sを含む配列番号14と少なくとも85%、90%、95%、98%、99%の相同性があり;C315Sを含む配列番号15と少なくとも85%、90%、95%、98%、99%の相同性があり;C313Sを含む配列番号16と少なくとも85%、90%、95%、98%、99%の相同性があり;C311Sを含む配列番号17と少なくとも85%、90%、95%、98%、99%の相同性があり;C313Sを含む配列番号19と少なくとも85%、90%、95%、98%、99%の相同性があり;C314Sを含む配列番号2と少なくとも85%、90%、95%、98%、99%の相同性があり;C312Sを含む配列番号21と少なくとも85%、90%、95%、98%、99%の相同性があり;C311Sを含む配列番号22と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号64と少なくとも85%、90%、95%、98%、99%の相同性があり;C407Sを含む配列番号65と少なくとも85%、90%、95%、98%、99%の相同性があり;C394Sを含む配列番号66と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号67と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号68と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号69と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号70と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号71と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号72と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号73と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号74と少なくとも85%、90%、95%、98%、99%の相同性があり;C400Sを含む配列番号75と少なくとも85%、90%、95%、98%、99%の相同性があり;C407Sを含む配列番号76と少なくとも85%、90%、95%、98%、99%の相同性があり;C394Sを含む配列番号77と少なくとも85%、90%、95%、98%、99%の相同性があり;またはC412Sを含む配列番号78と少なくとも85%、90%、95%、98%、99%の相同性がある配列を提供し、前記ポリペプチドは、セルラーゼ活性およびその相当するCys→Ser突然変異を欠如している親(野生型)タンパク質と比較して高い熱安定性を有する。
【0041】
さらに別の実施形態において、本発明は上記ポリペプチドを提供するが、それらはさらに配列番号64の配置として同定される少なくとも1つの付加的な突然変異を含み、配列番号64は位置413でPro、位置231でSerまたはThr、位置305でSerまたはThr、位置410でGlnまたはAsn、位置82でGlu、または上記のあらゆる組み合わせ置換を包含する。同様の置換は、アミノ酸配列の配列番号2,4,6,12−63および65−78と、配列番号64の配列の配置として同定する。
【0042】
本発明は、組み換えにより生成されたか、合成されたか、非天然的に生成されたか、のいずれの実質的に生成されたポリペプチドをも提供するものであり、該ポリペプチドは、C→S置換位置を除き、下記の1−10、10−20または20−30の位置の同定された配列の保存アミノ酸置換を有する:
C314Sを含む配列番号2;C311Sを含む配列番号4;C310Sを含む配列番号12;C312Sを含む配列番号13;C314Sを含む配列番号14;C315Sを含む配列番号15;C313Sを含む配列番号16;C311Sを含む配列番号17;C313Sを含む配列番号19;C312Sを含む配列番号21;C310Sを含む配列番号22;C400Sを含む配列番号64;C407Sを含む配列番号65;C394Sを含む配列番号66;C400Sを含む配列番号67;C400Sを含む配列番号68;C400Sを含む配列番号69;C400Sを含む配列番号70;C400Sを含む配列番号71;C400Sを含む配列番号72;C400Sを含む配列番号73;C400Sを含む配列番号74;C400Sを含む配列番号75;C407Sを含む配列番号76;C394Sを含む配列番号77;またはC412Sを含む配列番号78。
【0043】
「分離されたポリペプチド」は、自然に伴う他の混入物、例えばたんぱく質、脂質およびポリヌクレオチドから抽出されたポリペプチドを指す。用語は、それら天然に分泌された環境または発現系(例えば宿主細胞または生体外合成)から除去されたか、精製されたポリペプチドを包含する。
【0044】
「実質的に純粋なポリペプチド」は、ポリペプチドの種類が主成分として存在する組成物(即ち、モルまたは重量に基づいて、ポリペプチドがあらゆる他の各々の高分子成分より組成物において多量に存在する)を指し、対象となる成分がモルまたは%重量によって示される高分子成分の少なくとも約50パーセントを含む場合、一般的に実質的に精製された組成物とする。概して、実質的に純粋なポリペプチド組成物は、モルまたは重量%で組成物中に存在する全ての高分子成分の約60%以上、約70%以上、約80%以上、約90%以上、約95%以上、約98%以上含まれる。いくつかの実施形態において、目的の成分は、実質的に均質となるまで精製され(即ち、混入物成分は従来の検出法によって組成物中に検出されない)、組成物は実質的に単一の高分子成分からなる。溶媒の種類、低分子成分(<500ダルトン)および基本的なイオン成分は、高分子成分としては考慮されない。
【0045】
「参照配列」は、配列比較の基準として使用される、規定された配列を指す。参照配列は、例えば、遺伝子全長またはポリペプチド配列全長の断片のような、より長鎖の配列のサブセットとしてもよい。概して、参照配列の長さは、少なくとも20ヌクレオチドまたはアミノ酸残基、少なくとも25ヌクレオチドまたはアミノ酸残基、少なくとも50ヌクレオチドまたはアミノ酸残基、または核酸またはポリペプチドの全長とすることができる。2つのポリヌクレオチドまたはポリペプチドは、それぞれ(1)2つの配列の間で類似の配列(即ち、完全な配列の一部)を含み、(2)2つの配列の間で異なる配列をさらに含むため、2つ(またはそれ以上)のポリヌクレオチドまたはポリペプチドの間の配列の比較は、配列類似性の局部領域を同定し、比較するため、典型的に2つのポリヌクレオチドまたはポリペプチドの配列を「比較領域」にわたって比較することによって実施される。
【0046】
「配列相同性」は、2つのアミノ酸配列が、比較領域にわたって実質的に相同性(即ち、アミノ酸−アミノ酸比較に基づく相同性)があることを意味する。「配列類似性」の用語は、同じ生物物理学的な特徴を共有する同様のアミノ酸を指す。「配列相同性のパーセンテージ」または「配列類似性のパーセンテージ」の用語は、2つの最適に配置された配列を比較領域にわたって比較し、適合する位置を計数するため、相同性残基(または同一残基)が双方のポリペプチド配列存在する位置を計数し、比較領域(即ち、領域サイズ)における適合する位置の数を総数で除算し、配列の一致のパーセンテージ(または配列類似性のパーセンテージ)を算出し、結果を100で乗算して、得ることができる。ポリヌクレオチド配列に関して、配列の相同性および類似性の用語は、タンパク質配列に関して記載されるそれと同様に、比較領域にわたって、2つのポリヌクレオチド配列が同一(ヌクレオチド−ヌクレオチド比較に基づく)である「配列相同性のパーセンテージ」の意味を有する。このように、ポリヌクレオチド配列の相同性のパーセンテージ(またはポリヌクレオチド配列の類似性のパーセンテージ、例えば、分析アルゴリズムに基づいたサイレント置換または他の置換)も算出することができる。最大の相関性は、ここに記載の1つ以上の配列アルゴリズム(または当業者が利用できる他のアルゴリズム)の使用、または目視によって決定することができる。
【0047】
ポリペプチドに適用する場合、実質的な相同性または実質的な類似性の用語は、最適な配列を有する場合の2つのペプチド配列が、初期条件のギャップ重量(gap weight)を用いたBLAST、GAPまたはBESTFITのプログラムや、目視等において、配列の相同性または類似性を共有することを意味する。同様に、二つの核酸において、実質的相同性または実質的類似性の用語は、最適な配列を有する場合の2つの核酸配列が、初期状態のギャップ重量を用いたBLAST、GAPまたはBESTFITのプログラムやまたは目視等において、配列相同性または類似性を共有することを意味する。
【0048】
配列相同性または配列類似性のパーセンテージを決定するために適切なアルゴリズムの1つの例は、Pearson, W. R. & Lipman, D. J., (1988) Proc. Natl. Acad. Sci. USA 85:2444に記載されたFASTアルゴリズムである。またW. R. Pearson, (1996) Methods Enzymology 266:227-258を参照してもよい。相同性または類似性のパーセンテージを算出するため、DNA配列のFASTAアライメントにおいて使用するのに好ましいパラメーターは、以下の様に最適化される:BL50 Matrix 15: -5, k-tuple=2; joining penalty=40, optimization=28 ; gap penalty -12, gap length penalty=-2;およびwidth=16。
【0049】
実用的なアルゴリズムの他の例として、PILEUPが挙げられる。PILEUPは、進行する対となる配置(アライメント)を使用して、連関する配列群からの多重配列アライメントを作成することにより、配列群の相関性および配列相同性と配列類似性のパーセンテージを示す。それは、進行するアライメントを形成するためのクラスター関係を示す、樹状図または系統樹もプロットする。PILEUPは、Feng & Doolittle, (1987) J. Mol. Evol. 35:351-360の進行アライメント法を簡略化したもの使用する。使用される方法は、Higgins & Sharp, CABIOS 5:151- 153, 1989に記載された方法に類似の方法である。プログラムは、最高300の配列を配置(アライン)することができ、それぞれ最長5000のヌクレオチドまたはアミノ酸を配置することができる。多重アラインメント法は、2つの最も相同性のある配列対のアライメントから始まり、2つのアラインされた配列のクラスターを生成する。その後、該クラスターは、次に最も関連した配列またはクラスター配列のクラスターにアラインする。2つのクラスターの配列は、2つの個々の配列の対アライメントの単純な延長によって配置される。最終的なアライメントは、一連の前進的な対アライメントによって達成される。プログラムは、配列比較領域における、特定の配列およびそのアミノ酸もしくはヌクレオチドの連携を設計すること、およびプログラムパラメーターを設計することによって実行される。PILEUPを使用することで、参照配列を以下のパラメーターを使用した関連の他の試験配列と比較し、パーセント配列相同性のパーセンテージ(または配列類似性のパーセンテージ)を決定した。:初期状態のギャップ重量(3.00)、初期状態のギャップ長さ(0.10)および重量化した末端のギャップ。PILEUPは、GCG配列分析ソフトウェアパッケージ、例えばバージョン7.0から得られる。
【0050】
多数のDNAおよびアミノ酸配列に適したアルゴリズムの他の例としては、CLUSTALWプログラムが挙げられる。CLUSTALWは、配列群間の複数の対比較を実行し、それらを多重のアライメント配列の相同性に基づいて組み立てる。ギャップオープンおよびギャップエクステンションのペナルティーは、それぞれ10および0.05であった。アミノ酸アライメントに関して、BLOSUMアルゴリズムをタンパク質重量マトリックスとして使用することができる。
【0051】
上記のように、セロビオヒドラーゼおよびセルラーゼファミリーのメンバーは、配列アライメントおよびGECDGモチーフへの置換(例えば、配列番号2のアミノ酸312−316を参照のこと)によって同定することができる。修飾されたポリペプチドは、有利な活性を導入するように修飾されたものを同定するため、その後以下に記載されるような様々な温度および条件で、その活性を分析される。典型的な配列は、以下のGenBank登録番号であり、参照によりここに援用される配列から見出すことができる。
【0052】
【表1】
【0053】
さらに他の実施形態において、改良した熱安定性のある多様なセルラーゼポリペプチドファミリーは、C→S、G→SまたはA→S置換を有する以下の表に示すものを含む。さらに、以下の表に一致する置換、セルラーゼ活性および熱安定性のあるものを有する、以下いずれかの配列と85%、90%、95%、98%または99%配列の一致を有するポリペプチドも本発明に包含される。
下表は、H. jecorina CBH IIと高い相同性を有するタンパク質配列に関するアミノ酸の枠内(frame bracketing)H. jecorina CBH II Cys311のアライメントをしめす。残基311に想到する位置を下線太文字で示す。この試験において検討された組み換え型H. insolensおよびP. chrys CBHIIの配列は、H. insoおよびP. chrysとして示す。250個のうち、最も相同性のある配列の54個は、重複しているため省略した(即ち、構造研究のための点突然変異体または>95%一致するイソタイプ)。セルラーゼに関する受入番号が同定され、相当する配列が、ここに参考文献によって受入番号から直接複製されるように、本発明に組み込む。受入番号と関連する配列が、配列番号79−106として参照される。太字下線部の残基(例えばC、AまたはG)は、Sに置換される。配列番号は、下記の配列中の数値による:
【0054】
【表2】
【0055】
開示の目的のため、親セルラーゼより少なくとも約4℃または少なくとも約9℃高いT50、例えば約4℃から約30℃高い、もしくはその間のあらゆる値のT50を有するセロビオヒドラーゼ、または約9℃から約30℃高い、もしくはその間のあらゆる値のT50を有するセロビオヒドラーゼの場合、親セロビオヒドラーゼと比較したとき、開示のポリペプチドは、相当する親ポリペプチドに関して改良された熱安定性を示す。T50は、15分間のプレインキュベーション後、修飾された酵素または天然の酵素が、その残留活性の50%を保持する温度であり、以下の実施例または従来技術における詳細な分析によって測定される。
【0056】
開示の修飾されたセロビオヒドラーゼまたはセルラーゼは、相当する親セロビオヒドラーゼ(例えば配列番号2,4または6)より約4℃から約30℃高いか、もしくはその間の範囲にあり、約5℃から約20℃高いか、もしくはその間の範囲にあり、約8℃から約15℃高いか、もしくはその間の範囲にあり、約9℃から約15℃高いか、もしくはその間の範囲にあるT50を有していてもよい。例えば、修飾されたセルラーゼは、相当する親セロビオヒドラーゼのものより少なくとも約4,5,6,7,8,9,10,12,14,16,18,20,22,24,26,28または30℃高いT50を有することがあり得る。
【0057】
ここに同定される変異体も、セロビオヒドラーゼを生成するために使用することができる。例えば、SCHEMAは、何百もの活性β−ラクタマーゼおよび活性シトクロームP450の酵素キメラのファミリーを形成するため、前もって使用されてきた。SCHEMAは、ブロックが異なる親同士で交換されたときに分解されるアミノ酸側鎖接触のライブラリー平均数である<E>を最小化する、接触したアミノ酸の「ブロック」の境を規定するため、タンパク質構造データを使用する。β−ラクタマーゼキメラが折り畳まれ、活性を有する可能性が、その配列のEの値に反比例することが示された。突然変異のライブラリー平均数<m>に対する<E>を最小化するようなブロック境界を同定するため、RASPP(最短距離問題としての組み換え)アルゴリズムが使用された。3つの親からの8つのブロック(38=6,561通りの配列)を含む、β−ラクタマーゼの収集物を特徴とする500個までの20%を超える固有のキメラは、触媒活性を有する。同様の方法により、2,300の新規の、触媒活性がある酵素を含む3つの親からの8ブロックのシトクロームP450キメラファミリーを生成した。これら2つの収集物からのキメラは、最も近い親から、それぞれ平均して66および72の多数のアミノ酸配列の突然変異を有することを特徴とする。したがって、SCHEMA/RASPPは、著しい配列多様性、およびかなりの部分の機能メンバーを有するキメラファミリーの設計を可能とした。
【0058】
SCHEMAキメラの熱安定性が、配列の小さいサンプルからの配列安定性データに基づいて予測されることが、示されてきた。184のシトクロームP450キメラの熱安定性に関する直線回帰モデルは、SCHEMAブロックが、熱安定性に付加的な寄与をすることを示した。300を超えるキメラが、このモデルによって熱安定性であると予測され、試験された全44が最も安定した親と比較して、より高い安定性を有した。わずか35の熱安定性試験の使用によって、最も熱安定性のあるキメラを予測することができた。さらに、熱安定性のあるP450キメラは、キメラ形成がさらなる実用的な酵素特性を導くことができることを証明する、固有の活性および特性プロフィールを示した。ここでSCHEMA組換え型のCBH II酵素は、高温で、長期間にわたって、幅広い範囲のpH条件下のリン酸膨潤セルロース(PASC)において、活性を有するキメラセルラーゼを生じることができる。
【0059】
ここに記載の方法を使用して、セロビオヒドラーゼ活性を有する多くのキメラポリペプチドを、野生型親CBH IIタンパク質と比較して、改良した特性を有するものとして産生した。
【0060】
新規CBH II酵素の多様なファミリーが、3つの菌のCBH II酵素からの配列のブロックを交換することによって作成された。このセットから採取された48のキメラ配列のうちの23個は、S. cerevisiaeによって活性形状で分泌され、5つは63℃で最も安定な親より長い半減期を有する。この48のメンバーのサンプルセットが、存在し得る全6,561配列の1%未満であるので、このキメラの収集物は、100もの活性型、熱安定性のあるCBH II酵素、CAZyデータベースにおけるより少数化された約20の菌CBH II酵素を有することが予測される。
【0061】
熱安定性を促進するブロックを同定するためにサンプルセット配列安定性データを使用する方法は、第2のCBH II検証セットにおける全10個の触媒活性キメラが、最も安定な親である好熱性菌H. insolensからの天然の熱安定性のあるCBH IIより熱安定性であることにより検証された。したがって、本発明は、S. cerevisiaeから触媒活性を有する形態で発現する全33の新規CBH II酵素を生じ、そのうち15は、それらが生成される最も安定な親より熱安定性である。これら15の熱安定性のある酵素は、互いにおよび、それらに最も近い天然相同物から、それぞれおよそ94および58アミノ酸の位置で異なる。
【0062】
組み合わせたサンプルおよび検証セットにおけるCBH IIキメラの熱安定性の分析は、同じキメラに存在する場合B1P1、B6P3、B7P3およびB8P2として同定された4つの熱安定のあるブロックが、熱安定性に累積的に寄与することを示す。H. insolens CBH IIと比較してより熱安定性のある5つのサンプルセットキメラのうちの4つは、2つまたは3つの安定性のあるブロックを含む(Table1)。H. insolens酵素より比較的安定な全ての検証セットの10個の活性型メンバーは、すべてH. insolens酵素より安定しているが、3つまたは4つの安定性のあるブロックを含有するこの群において、最も熱安定性のある6つのキメラのうちの5つで、少なくとも2つの安定性のあるブロックを含有する。
【0063】
本発明は、安定性ブロックが組みかえられ、新規の、高い安定性を有する活性型セルラーゼが生成されることを示す。安定性回帰モデルは、CBH II SCHEMAライブラリーが親酵素の最も安定なものと比較しても、より安定が2.026のキメラを有することを予測する。これらのキメラは多様であり、生来のセルラーゼと明確に区別できる:それらは親から8から72箇所の突然変異(平均50箇所)、キメラ間では平均63箇所の突然変異を有して多様化される。このセットから計33個の遺伝子がS. cerevisiaeにおいて合成され、発現される:これらキメラCBH IIの全てについて、63℃またはT50での不活性化の半減期が測定され、いずれも好熱性菌H. insolens由来の最も安定な親セルラーゼと比較しても、より安定であることが見出された。8個のみのブロックのキメラを作成して配列の複雑化を減少させることで、配列安定性モデルの生成および単一の高い安定性のある配列ブロックの同定が可能となる。このブロックにおける10個のみのアミノ酸置換の試験をすることによって、単一の高い安定化効果のある置換を同定した。キメラおよび生来のP. chrysosporium、H. insolensおよびH. jecorina CBH II酵素にわたってC313S(配列番号6;C314S、配列番号2;C311S、配列番号4)の置換による非常に大きな安定性効果が観測され、この位置のあらゆる残基のSerへの突然変異が、それが導入されたあらゆるセルラーゼ6ファミリーを安定化させることを示す。
【0064】
組み換えにおける接触分離の数を最小化することは(図2C)、ブロックがそれぞれ独立してタンパク質全体の安定性を高める、分離されたユニットに近い状態となることを可能としこれにより、累積的またはさらに付加的なキメラの熱安定性向上効果につながる。このCBH II酵素の組み換えに関して、SCHEMAは前記の接触分離を最小化するのに効果的であった:H. insolens親CBH II結晶構造において規定されるブロックの間の接触は計303ヶ所あり、CBH II SCHEMAライブラリーのデザインによると、潜在的な接所分離は33ヶ所のみであった。CBH II酵素の親が、そのサブドメインに明らかな構造上の特徴を有さず、且つ、8つのブロックのうちの4つのみ(1,5,7および8)が小型の構造ユニットまたはモジュール様の構造を示す場合、接触分離が低値であることはブロックの数が下位構造区分の数を超える場合においてSCHEMA/RASPPアルゴリズムが効果的であることを示した。β−ラクタマーゼおよびシトクロームP450キメラに関して前もって観測されるように、低いE値により、キメラの折り畳みおよび活性が予測可能である。ここでは使用しないが、この関係は、高い割合の活性型メンバーを含むキメラサンプルセットを設計するのに有用である。
【0065】
本発明において、キメラを用いて、pHの安定性をCBH II酵素において改良できるかを検討した。H. jecorina CBH IIの特異的活性は、pHが最適値5を超えて上昇するにつれて急激に減少し、一方、安定性のあるブロックを最も産業的に関連のあるH. jecorina CBH II酵素上に置換することによって作成されるHJプラスは、高いpHで顕著にその活性を保持する(図4)。熱安定性のある11113132および13311332キメラ、またH. insolensおよびC. thermophilum CBH IIセルラーゼ親は、HJプラスと比較してより幅広いpH/活性特性を有する。H. jecorina CBH IIの制限されたpH/活性特性は、いくつかのカルボキシルカルボン酸対の脱プロトン化に起因し、pH6を超えるタンパク質を不安定化する。HJプラスのブロック7における親3の置換は、カルボキシルカルボン酸対である(ブロック8の)D277およびD316の間のカルボキシルカルボン酸の対を解除し、アスパラギンサン277をヒスチジンに変更する。正荷電のヒスチジンとD277を置換することは、H. jecorina CBH IIと比較してより高いpHで、HJプラスが活性を保持することを可能とし、非酸性pHにおける電荷斥力の不安定化を予防することができる。残存する2つの熱安定性のあるキメラならびにH. insolensおよびC. thermophilum親CBH II酵素のより幅広いpH/活性特性は、HJプラスおよびH. jecorina CBH IIのE57−E119カルボキシルカルボン酸の対に相応する位置での酸残基の欠如によるものであろう。
【0066】
HJプラスは、比較的高い特異的活性および高い熱安定性の双方を示す。図5は、これらの特性の長期にわたる加水分解実験における優れた機能を示す:HJプラスは、親CBH II酵素と比較して7−15℃高い温度でセルロースを加水分解し、最適温度において、全ての親と比較して著しく向上した長期活性を示した(H. jecorina CBH IIの1.7倍)。HJプラスキメラの特異的活性がH. jecorina CBH II親のものより低い場合、この向上した長期活性は、HJプラスの熱安定性に由来し、最適な加水分解温度での比較的長い反応時間にわたる活性を保持できる。
【0067】
他の2つの熱安定性のあるキメラは、HJプラスの幅広い温度範囲を共有した。この観測は、上昇した温度t1/2と最高動作温度の間の正の相関を支持し、6,561のCBH IIキメラ配列中の多くの熱安定性キメラもセルロースを高い温度で分解できることを示唆する。高い温度での非結晶性PASC基質を加水分解するこの特性が、熱安定性のある菌CBH IIキメラの潜在的有用性をよく予見させる。一方、結晶基質およびリグニン含有基質の比較的困難な研究は、この新規CBH II酵素グループのバイオマス分解への応用に関して、より完全な評価を可能とする。
【0068】
バイオマス変換工程の大半は、高いレベルのセルロース加水分解を達成するため、菌セルラーゼの混合物(主にCBH II、セロビオヒドラーゼクラスI(CBH I)、エンドグルカナーゼおよびβ−グルコシダーゼ)を使用する。熱安定性のあるCBH II酵素キメラの多様な群を生成することは、酵素混合物を特定の応用および貯蔵のために調製し、最適化することができる、安定で、且つ活性の高いセルラーゼのストックを作成する第1工程である。
【0069】
「ペプチド断片」は、比較的大きなポリペプチドまたはタンパク質の一部分もしくは断片を指す。ペプチド断片は、それ自身、機能活性を有する必要性がないが、いくつかの場合において、ペプチド断片は、生物学的活性を有するポリペプチドドメインに相当する。安定性に関するペプチド断片は、ペプチド断片を有しないポリペプチドと比較して、安定性、機能または折り畳み度合いを促進するポリペプチドにおいて見出されるペプチド断片である。不安定性にペプチド断片は、ポリペプチド内に存在する場合、安定性、機能または折り畳み度合いを欠く原因として同定されるペプチド断片である。
【0070】
「融合」、「作動可能に結合された(operably linked)」および「作動可能に連結された(operably associated)」は、2つの明白なドメインまたはペプチド断片の化学的または物理学的な結合を幅広く指すため、ここに同じ意味で用いられて使用することができ、作動可能に結合している場合、それぞれのドメインまたはペプチドの断片は、望ましい活性を有する機能的なポリペプチドを提供する。ドメインまたはペプチド断片は、それらが機能的であるようにリンカーペプチドによって結合していてもよく、他の中間体または化学結合によって融合することができる。例えば、2つのドメインは、同じコード配列の一部であってもよく、ポリヌクレオチドは、単一のmRNAに転写されるようにコード化され、双方のドメインを含む単一のポリペプチドに翻訳されるように、同じ「フレーム」内に存在する。或いは、両方のドメインを各々のポリペプチドとして別々に発現させ、化学的手法により互いに融合させてもよい。具体的には、コーディングドメインは、「フレーム」内で直接的に結合するか、またはペプチドリンカーを間に挟んで隔離されており、単一のポリヌクレオチドをコーディングする形態とする。ペプチドリンカーおよびペプチドのコード配列は、従来技術において既知である。
【0071】
「ポリヌクレオチド」または「核酸配列」は、ヌクレオチドの多量体型を指す。ある場合において、ポリヌクレオチドは、由来生物の天然に生じるゲノムにおいて、それが隣接する2つのコード配列(1つは5’末端側、もう1つは3’末端側で隣接する)のいずれかには直接隣接しないものを指す。従って、前記用語は、例えばベクター;独立して複製するプラスミドまたはウイルス;原核生物もしくは真核生物のゲノムDNAに組み込まれる組み換え型DNA、または他の配列から独立して別の分子(例えばcDNA)として存在するものを含む。本発明のヌクレオチドは、リボヌクレオチド、デオキシリボヌクレオチド、または修飾されたいずれかのヌクレオチドであってもよい。ここに使用されるポリヌクレオチドは、とりわけ、一本鎖および二本鎖のDNA、一本鎖領域および二本鎖領域が混合しているDNA、一本鎖および二本鎖のRNA、および一本鎖領域および二本鎖領域が混合しているRNA、一本鎖、二本鎖、または一本鎖領域および二本鎖領域が混同したDNAおよびRNAのハイブリッドを指す。ポリヌクレオチドの用語は、ゲノムDNAまたはRNA(生物、即ちウイルスのRNAゲノム)並びにゲノムDNAおよびcDNAにコード化されたmRNAを包含する。
【0072】
「核酸断片」、「オリゴヌクレオチド断片」または「ポリヌクレオチド断片」は、比較的大きなポリヌクレオチド分子の一部を指す。ポリヌクレオチド断片は、タンパク質の機能ドメインをコードする必要はないが、タンパク質の機能ドメインをコードすることもあろう。ポリヌクレオチド断片は、約6ヌクレオチド以上の長さ(例えば、6−20,20−50,50−100,100−200,200−300,300−400ヌクレオチド以上の長さ)としてもよい。安定性に関するペプチド断片は、安定性に関するポリヌクレオチド断片によってコードされてもよく、ペプチド断片は、ペプチド断片を欠如するポリペプチドと比較して、ポリペプチドの安定性、機能または折り畳み度合いを促進する。
【0073】
「キメラ」は、少なくとも2つの異なる親タンパク質の少なくとも2つの断片の組み合わせを指す。当業者に理解されるように、それが関連のある特定の配列であって、そのものが物理的に核酸でないものであれば、実際、断片はそれぞれの親から由来する必要はない。例えば、キメラ菌クラスIIセロビオヒドラーゼ(CBH IIセルラーゼ)は、2つの異なる親CBH IIポリペプチドからの少なくとも2つの断片を有する。2つの断片は、セルラーゼ活性を有する新規ポリペプチドを生じるように結合している。即ち、タンパク質は、いずれかの親タンパク質の全長に一致する配列を有する場合、キメラとはいない。キメラポリペプチドは、2つ以上の異なる親タンパク質からの2つ以上の断片を含む。例えば、それぞれ最終的なキメラまたはキメラのライブラリーにつき、2,3,4,5−10,10−20以上の親がある。それぞれの親ポリペプチドの断片は、非常に短いか、非常に長いこともあり、断片はタンパク質の全長の1〜90%、95%、98%または99%範囲の連続した長さのアミノ酸配列を有し得る。ある実施形態において、最短の長さは10アミノ酸である。ある実施形態においては、2つの親タンパクについて単一のクロスオーバー点が定義される。クロスオーバーの位置は、一方の親のアミノ酸断片が終結し、他方の親のアミノ酸断片が始まるところに規定される。従って、単純なキメラは、クロスオーバー位置前の断片が第1の親、クロスオーバー点後の断片が第2の親に属する場所にクロスオーバー位置がある。ある実施形態において、キメラは複数の交差位置を有する。例えば、2,3,4,5,6,7,8,9,10,11−30個、またはこれを超えるクロスオーバー位置を有する。該クロスオーバーの位置および命名は、いずれも以下に記載するように規定する。2つのクロスオーバー位置および2つの親がある実施形態において、第1の親からの第1の連続断片に続いて、第2の親からの第2の連続断片が続き、これに第1の親由来の第3の連続断片が続く。「連続」とは、顕著な中断断片が存在しないことを示す。これら連続断片は、連続したアミノ酸配列を形成するために結合する。例えば、2つのクロスオーバー100および150を有するHumicola insolens(以下「1」とする)およびH. jecori(以下「2」とする)由来のCBH IIキメラは、1からの最初の100個のアミノ酸に、2から次の50個のアミノ酸が続き、さらにこれに1から残りのアミノ酸が続く形態をとり得て、1つの連続したアミノ酸の鎖として全てが結合している。或いは、CBH IIキメラは、2から最初の100アミノ酸、1から次の50アミノ酸、次いで2由来の残りに順に続く形態をとってもよい。当業者に理解されるように、キメラの変異体は、本来の配列として同程度に存在し得る。従って、それが変異体キメラの場合、100%でないそれぞれの断片が、最終キメラ内に存在すべきである。残基の付加、あるいは残基の除去または変性によって変更できる量は、変異体の用語が定義されるのと同様に定義される。当然、当業者に理解されるように、上記のものは、アミノ酸のみでなく、アミノ酸をコードする核酸にも応用することができる。
【0074】
本発明は、特定の変異体に加えて、CBH IIキメラの作成に使用可能な変異体を示す。誘導されたSCHEMA組み換え型ライブラリーは、セロビオヒドラーゼ酵素の生成に使用され、特に良く研究されたこの多様な酵素ファミリーのメンバー、具体的には下記のセロビオヒドラーゼII酵素に基づく。H. insolensは親「1」(配列番号2)、H. jecorinaは親「2」(配列番号4)およびC. thermophilumは親「3」(配列番号6)。SCHEMAは、タンパク質の構造一体性に影響を及ぼさずに組みかえることができる、相応するタンパク質の断片の位置を予測する計算方法である(Meyre et al. (2003) Protein Sci., 12: 1686-1693)。この計算は、CBH II親タンパク質の同定された7つの組み換え点に関してなされ、これにより各ポリペプチドが8つの断片を含むCBH IIキメラポリペプチドのライブラリーの作成を可能とした。高い安定性を有するキメラは、配列安定性データの線形回帰、または折り畳まれたタンパク質対折り畳まれていないタンパク質のMSAコンセンサス依存分析によって、全体的な安定性への各断片の付加的寄与を決定することで同定できる。SCHEMA組換えは、耐性配列の交換が行われても、重要な機能的残基の保存により、キメラが生物学的機能を保持し、高い配列の多様性を示す。
【0075】
従って、ここに様々な実施形態によって示すように、本発明は、親ドメインのキメラを含むCBH IIポリペプチドであり、上記のように、親鎖または結果として生じるキメラのコード配列が、C→S置換を含むように修飾することができるものを提供する。いくつかの実施形態において、ポリペプチドは、異なる親CBH IIタンパク質からのN末端からC末端までの複数のドメインを有するキメラを含む:(断片1)−(断片2)−(断片3)−(断片4)−(断片5)−(断片6)−(断片7)−(断片8);
断片1は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約1から約x1アミノ酸残基;断片2は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x1から約x2アミノ酸残基;断片3は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x2から約x3アミノ酸残基;断片4は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x3から約x4アミノ酸残基;断片5は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x4から約x5アミノ酸残基;断片6は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x5から約x6アミノ酸残基;断片7は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x6から約x7アミノ酸残基;および断片8は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x7から約x8アミノ酸残基を含み;
x1は配列番号2の残基43,44,45,46,47または配列番号6の残基42,43,44,45,46;x2は配列番号2の残基70,71,72,73,74または配列番号4もしくは配列番号6の残基68,69,70,71,72,73,74;x3は配列番号2の残基113,114,115,116,117,118または配列番号4もしくは配列番号6の残基110,111,112,113,114,115,116;x4は配列番号2の残基153,154,155,156,157または配列番号4もしくは配列番号6の残基149,150,151,152,153,154,155,156;x5は配列番号2の残基220,221,222,223,224または配列番号4もしくは配列番号6の残基216,217,218,219,220,221,222,223;x6は配列番号2の残基256,257,258,259,260,261または配列番号4もしくは配列番号6の残基253,254,255,256,257,258,259,260;x7は配列番号2の残基312,313,314,315,316または配列番号4もしくは配列番号6の残基309,310,311,312,313,314,315,316を含み;x8はポリペプチドのC末端に相当するアミノ酸残基が配列番号2、配列番号4または配列番号6を有する。
【0076】
以下のドメインの参照を用いて、Table1に示すように、多くのキメラ構造が作成された。
H. insolens親CBH IIに関して測定された64.8CのT50値を予測するT50値を有する1,588CBH IIキメラ配列。
【0077】
【表3】
【0078】
【表4】
【0079】
【表5】
【0080】
【表6】
【0081】
【表7】
【0082】
上記表において、それぞれの桁は、キメラCBH IIポリペプチドのドメインを指す。数字は、ドメインが由来する親鎖を示す。例えば、配列12111131を有するキメラCBH IIキメラポリペプチドは、ポリペプチドがN末端からC末端に向かって:配列番号2(「1」)のアミノ酸配列約1からx1に結合した配列番号4(「2」)のアミノ酸配列約x1からX2、これに結合した配列番号6(「3」)のアミノ酸配列約x2からx3、これに結合した配列番号2のアミノ酸配列約x3からx4、これに結合した配列番号2のアミノ酸配列約x4からx5、これに結合した配列番号2のアミノ酸配列約x5からx6、これに結合した配列番号6(「3」)のアミノ酸配列約x6からx7、これに結合した配列番号2のアミノ酸配列約x7からx8(例えばC末端)を含む。
【0083】
いくつかの実施形態において、CBH IIポリペプチドは、以下からなる群から選択されるキメラ断片構造を有する。
【0084】
【表8】
【0085】
いくつかの実施形態において、ポリペプチドは、配列番号2,4または6の野生型ポリペプチドと比較して改良した熱安定性を有する。ポリペプチドの活性は、実施例に記載されているように、1つの、または組み合わされた任意の基質を用いて測定することができる。当業者にとって明白であるように、実施例に記載されたものに例示される化合物のクラスの他の化合物を試験することができる。
【0086】
いくつかの実施形態において、ポリペプチドは、参照配列のアミノ酸配列に様々な変更を加えた形態であってもよい。この変更は、1つ以上のアミノ酸の置換、欠如または挿入であってもよい。この変更が置換である場合、変更は保存、非保存の置換、または保存、非保存の組み合された置換の組み合わせである。例えば、キメラは、配列番号2のC314または配列番号4のC311の位置でC→S置換を含むことができる。
【0087】
従って、いくつかの実施形態において、ポリペプチドはN末端からC末端において一般的な構造を含むことができる:(断片1)−(断片2)−(断片3)−(断片4)−(断片5)−(断片6)−(断片7)−(断片8);
断片1は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約1から約x1アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;断片2は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x1から約x2アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;断片3は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x2から約x3アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;断片4は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x3から約x4アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;断片5は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x4から約x5アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;断片6は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x5から約x6アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;断片7は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x6から約x7アミノ酸残基および1−10個の保存されたアミノ酸の置換を有し;ならびに断片8は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x7から約x8アミノ酸残基を含み、1−10個の保存されたアミノ酸の置換を有する;
【0088】
従って、いくつかの実施形態において、ポリペプチドは、N末端からC末端までの一般的な構造を含むことができ、x1は配列番号2の残基43,44,45,46,47または配列番号6の残基42,43,44,45,46;x2は配列番号2の残基70,71,72,73,74または配列番号4もしくは配列番号6の残基68,69,70,71,72,73,74;x3は配列番号2の残基113,114,115,116,117,118または配列番号4もしくは配列番号6の残基110,111,112,113,114,115,116;x4は配列番号2の残基153,154,155,156,157または配列番号4もしくは配列番号6の残基149,150,151,152,153,154,155,156;x5は配列番号2の残基220,221,222,223,224または配列番号4もしくは配列番号6の残基216,217,218,219,220,221,222,223;x6は配列番号2の残基256,257,258,259,260,261または配列番号4もしくは配列番号6の残基253,254,255,256,257,258,259,260;x7は配列番号2の残基312,313,314,315,316または配列番号4もしくは配列番号6の残基309,310,311,312,313,314,315,316を含み;x8はポリペプチドのC末端に相当するアミノ酸残基が配列番号2、配列番号4または配列番号6を有し、キメラは以下のTable1のようなアルゴリズムを有し、キメラは配列番号2のC314または配列番号4のC311に相当するC→S置換を含む。
【0089】
いくつかの実施形態において、アミノ酸置換される置換基の数は2,3,4,5,6,8,9,10個またはこれを超える数となり得る(例えば、10−20,21−30,31−40および同類のアミノ酸置換)。
【0090】
いくつかの実施形態において、機能的なCBH IIポリペプチドは、例えば実施例に記載の基質に対して、高い熱安定性を有すると共に、セルラーゼ活性を有することができ、また参照するセロビオヒドラーゼまたはその断片に一致する一定のレベルのアミノ酸を有することもできる。参照とする酵素またはその断片は、野生型(例えば天然発生)または改変された酵素由来のもとを使用できる。従って、いくつかの実施形態において、本発明のポリペプチドは、N末端からC末端への一般的な構造を含むことができる:
断片1は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約1から約x1のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片2は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約x1から約x2のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片3は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約x2から約x3のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片4は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約x3から約x4のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片5は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約x4から約x5のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片6は、配列番号2(「1」)、配列番号(「2」)、配列番号6(「3」)の約x5から約x6のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片7は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約x6から約x7のアミノ酸残基少なくとも50−100%の一致がある配列を含み、断片8は、配列番号2(「1」)、配列番号4(「2」)、配列番号6(「3」)の約x7から約x8のアミノ酸残基で少なくとも50−100%一致がある配列を含み;
x1は、配列番号2の残基43,44,45,46,47、配列番号4もしくは配列番号6の残基42,43,44,45,46;x2は、配列番号2の残基70,71,72,73,74、配列番号4もしくは配列番号6の残基68,69,70,71,72,73,74;x3は、配列番号2の残基113,114,115,116,117,118、配列番号4もしくは配列番号6の残基119,111,112,113,114,115,116;x3は、配列番号2の残基113,114,115,116,117,118、配列番号4もしくは配列番号6の残基119,111,112,113,114,115,116;x4は、配列番号2の残基153,154,155,156,157、配列番号4もしくは配列番号6の残基149,150,151,152,153,154,155,156;x5は、配列番号2の残基220,221,222,223,224、配列番号4もしくは配列番号6の残基216,217,218,219,220,221,222,223;x6は、配列番号2の残基256,257,258,259,260,261、配列番号4もしくは配列番号6の残基253,254,255,256,257,258,259,260;x7は、配列番号2の残基312,313,314,315,316、配列番号4もしくは配列番号6の残基309,310,311,312,313,314,315,318;x8は、ポリペプチドのC末端に相当するアミノ酸残基が、配列番号2、配列番号4または配列番号6を有するものであり、キメラは以下に記載された表1のようなアルゴリズムを有し、キメラは配列番号2のC314または配列番号4のC311の位置にC→S置換を含む。
【0091】
いくつかの実施形態において、キメラポリペプチドのそれぞれの断片は、それぞれ配列番号2、配列番号4または配列番号6の(断片1)、(断片2)、(断片3)、(断片4)、(断片5)、(断片6)、(断片7)および(断片8)を参照とする断片と比較して、少なくとも60%、70%、80%、90%、95%、96%、97%、98%、99%、またはこれを超える配列の一致を有することができる。
【0092】
いくつかの実施形態において、ポリペプチド変異体は、配列番号2,4または6の野生型ポリペプチドと比較して改良した熱安定性を有するものであり;キメラは配列番号2のC314または配列番号4のC311の位置のC→S置換を含む。
【0093】
ここに記載のキメラ酵素は、様々な形態、例えば溶解物、粗抽出液または精製物として調製することができる。ポリペプチドは、適切な溶液に溶解することができて;粉末、例えばアセトン粉末(安定剤存在下、または不存在下で)として調製することができるか、凍結乾燥物として調製することができる。いくつかの実施形態において、ポリペプチドは単離されたポリペプチドであることができる。
【0094】
いくつかの実施形態においてポリペプチドは、アレイの形態とすることができる。酵素は、溶解した状態で、例えばミクロタイタープレートのウェル内の溶液としたり、または基質上に固相化したりすることができる。基質としては、有機ポリマー、例えばポリスチレン、ポリエチレン、ポリプロピレン、ポリフルオロエチレン、ポリエチレンオキシおよびポリアクリルアミドならびにその共重合体、およびそのグラフトから構成することができる、固体基質または多孔性基質(例えば膜)が挙げられる。固体サポートは、無機、例えばガラス、二酸化ケイ素、制御多孔質ガラス(CPG)、逆相シリカまたは金もしくはプラチナ等の金属としてもよい。基質の構造は、ビーズ、球体、粒子、顆粒剤、ジェル、膜または面の形態としてもよい。面は平ら、実質的に平らまたは非平面とすることができる。固体サポートは、多孔性であっても無孔であってもよく、膨張性であっても非膨張性であってもよい。固体サポートは、ウェル、くぼみ、または他の容器、入れ物の構成とでき、その特性および場所を変えることもできる。複数のサポートは、試薬のロボットデリバリー、または検出法および/または計測器がアドレス可能な、多様な箇所のアレイ上に構成することができる。
【0095】
本発明は、ここに開示した改変CBH IIポリペプチドをコード化するポリヌクレオチドも提供する。ポリヌクレオチドは、ポリペプチドを発現することができる組み換え型ポリヌクレオチドのため、遺伝子発現を制御する1つ以上の異種の調節または制御配列に作動可能なように結合することができる。CBH IIキメラをコード化する異種ポリヌクレオチドを含む発現構造は、ポリペプチドを発現するため、適切な宿主細胞内に導入することができる。
【0096】
CBH IIキメラ酵素の特定の配列の知識を有する場合(例えばキメラCBH IIの断片構造)、ポリヌクレオチド配列は、
形成されるCBH IIキメラ酵素のアミノ酸配列は、当業者にとって自明の形状となるであろう。様々なアミノ酸に相当するコドンの知識とポリペプチドのアミノ酸配列の知識とを組み合わせれば、当業者は、本発明のポリヌクペプチドをコード化する異なるポリヌクレオチドを作成することが可能である。従って、本発明は、コドンの組み合わせを選択することによって、いくつかの可能性のある構成のポリヌクレオチドの変異体を想起させるものであり、そのような変異体はすべて、ここに記載のポリペプチドとして、ここに明確に開示されているものと理解されるべきである。
【0097】
いくつかの実施形態において、ポリヌクレオチドは、ここに記載のポリペプチドをコード化するが、参照するキメラポリペプチドのCBH II変異体をコード化し、上記のようにC→S置換(例えば、ポリペプチドまたはキメラは、配列番号2のC314または配列番号4のC311の位置のC→S置換を含む)を有するポリペプチドと、ヌクレオチドレベルで、約80%以上、約85%以上、約90%以上、約91%以上、約92%以上、約93%以上、94%以上、約95%以上、約96%以上、約97%以上、約98%以上、約99%以上の配列の一致を有する。
【0098】
いくつかの実施形態において、ポリペプチドをコード化する分離されたポリヌクレオチドは、ポリペプチドの発現を提供するため、様々な方法にて処理することができる。ベクターへの挿入前に単離されたポリヌクレオチドを処理する工程は、発現ベクターによっては、必要とされ、、または必須とされる。組み換えDNA法を用いて、ポリヌクレオチドおよび核酸配列を修飾する技術は、従来技術において周知である。この技術に対する教示は、Sambrook et al . , 2001, Molecular Cloning: A Laboratory Manual, 3rd Ed., Cold Spring Harbor Laboratory Press; and Current Protocols in Molecular Biology, Ausubel. F. ed., Greene Pub. Associates, 1998, updates to 2007に提供されている。
【0099】
いくつかの実施形態において、ポリヌクレオチドは、ポリヌクレオチドおよび/またはポリペプチドの発現に関する制御配列に機能的に関連している。いくつかの実施形態において、制御配列は、細胞外または細胞内のポリペプチドをコード化し、宿主細胞に相同性または異種性のいずれかの遺伝子から得られる適当なプロモーター配列とすることができる。細菌宿主細胞に対して、本発明の核酸構造の転写を誘導するのに適切なプロモーターとしては、大腸菌ラクトースオペロン、枯草菌xylAおよびxylB遺伝子、バチルスメガテイルムキシロースを用いる遺伝子(例えばRygus et al., (1991) Appl . Microbiol. Biotechnol. 35:594-599; Meinhardt et al., (1989) Appl. Microbiol. Biotechnol . 30:343-350)、原核生物βラクタマーゼ遺伝子(Villa- Kamaroff et al., (1978) Proc . Natl Acad. Sci . USA 75: 3727-3731)ならびにtacプロモーター(DeBoerその他, (1983) Proc. Natl Acad. Sci. USA 80: 21-25)が挙げられる。様々な適切なプロモーターが、Scientific American, 1980, 242:74-94の"Useful proteins from recombinant bacteria";およびSambrookその他の文献に記載されている。
【0100】
いくつかの実施形態において、制御配列はまた適切な転写停止(ターミネーター)配列であり、宿主細胞によって転写を停止することが認識される配列としてもよい。停止配列は、ポリペプチドをコード化する核酸配列の3’末端に作動可能に結合している。選択した宿主細胞において機能的なあらゆるターミネーターが使用可能である。
【0101】
いくつかの実施形態において、制御配列は、適切なリーダー配列、宿主細胞による翻訳に重要なmRNAの非翻訳領域でもあり得る。リーダー配列は、ポリペプチドをコード化する核酸配列の5’末端に作動可能に結合される。選択した宿主細胞において機能的なあらゆるリーダー配列が使用可能である。
【0102】
いくつかの実施形態において、制御配列は、ポリペプチドのアミノ末端に結合したアミノ酸配列をコード化するシグナルペプチドコード領域としてもよく、コードされたポリペプチドを細胞の分泌経路へ誘導する。核酸配列のコード配列の5’末端は、本質的に分泌されたポリペプチドをコード化するコード領域の断片と翻訳リーディングフレーム内の、天然に結合したシグナルペプチドコード領域を含むことができる。或いは、コード配列の5’末端は、コード配列とは異質のコード領域のシグナルペプチドを含むことができる。コード配列がシグナルペプチドコード領域を天然に含まない部分において、異質のシグナルペプチド領域が必要とされる。細菌の宿主細胞に関する効果的なシグナルペプチドのコード領域は、バチルス属NCIB11837マルトース生成アミラーゼ、バチルス・ステアロサーモファイルスαアミラーゼ、バチルス・リケニフォルミス・スブチリシン、バチルス・リケニフォルミス・βラクタマーゼ、バシラス・ステアロサーモフィラス中性プロテアーゼ(nprT、nprS、nprM)および枯草菌prsAの遺伝子から得られるシグナルペプチドコードとしてもよい。さらなるシグナルペプチドは、Simonen and Palva, (1993) Microbiol Rev 57: 109-137に記載されている。
【0103】
さらに、本発明は、導入される宿主の種類に応じて、形成されたCBH II変異体またはキメラポリペプチドをコード化するポリヌクレオチドを含む組み換え型発現ベクター、および、例えばプロモーターおよびターミネーター、複写原点等のうちの1つ以上の、発現制御領域について示す。発現ベクター作成において、コード配列は、コード配列が発現のための適切な制御配列と作動可能に結合するようにベクター内に局在する。
【0104】
組み換え型発現ベクターは、有効に組み換え型DNA方法の対象として使用可能であり、ポリヌクレオチド配列の発現をもたらすあらゆるベクター(例えばプラスミドまたはウイルス)を使用できる。通常、ベクターの選択肢は、ベクターが導入される宿主細胞とベクターの適合性に依存する。ベクターとしては、線状または閉鎖環状プラスミドを使用してもよい。
【0105】
発現ベクターとしては、独立した複製ベクター、即ち染色体複製から独立して複製される染色体外のものとして存在するベクター、例えばプラスミド、染色体外の要素、ミニ染色体または人口染色体が使用可能である。ベクターは、自己再生を確実にするためのあらゆる手段を含むことができる。或いは、ベクターは宿主細胞内に導入されたとき、ゲノム内に組み込まれ、それが組み込まれた染色体と共に複製されるものを含むことができる。さらに宿主細胞のゲノム内に導入するため、全DNAを共に含む単一のベクター、プラスミドまたは2つ以上のベクターまたはプラスミドまたはトランスポゾンが使用可能である。
【0106】
いくつかの実施形態において、本発明の発現ベクターは、形質転換細胞の選択を容易とする、1つ以上の選択可能なマーカーを含有する。選択可能なマーカーは、殺生物性またはウイルス抵抗性、重金属抵抗性、独立栄養体への原栄養性、等を与える遺伝子産物である。細菌性の選択可能なマーカーの例は、枯草菌もしくはバチルス・リケニフォルミスからのdal遺伝子、抗生物質、例えばアンピシリン、カナマイシン、クロラムフェニコール(実施例1)への抵抗性またはテトラサイクリンへの抵抗性を与えるマーカーである。他の実用的なマーカーは、当業者にとって明白であろう。
【0107】
他の実施形態において、本発明はCBH II変異体またはキメラポリペプチドをコード化するポリヌクレオチドを含む宿主細胞を提供し、前記ポリヌクレオチドは、宿主細胞中におけるポリペプチドの発現に関する1つ以上の制御配列に作動可能に結合されるものを含む。本発明の発現ベクターによってコード化されるポリペプチドを発現するために使用される宿主細胞は、従来技術において周知であり、大腸菌および巨大菌等の細菌細胞;酵母細胞、CHO細胞等の真核生物細胞;ショウジョウバエS2およびスポドプテラSf9細胞等の昆虫細胞;CHO、COS、BHK、293およびボーズ・メラノーマ細胞等の動物細胞;および植物細胞が挙げられるが、これらに限定されるものではない。他の適切な宿主細胞は、当業者にとって明白であろう。上記宿主細胞への適切な培養培地および成長条件は、従来技術において周知である。
【0108】
本発明のCBH II変異体またはキメラポリペプチドは、従来技術において周知の方法を用いて作成することができる。ポリヌクレオチドは、例えばSambrook et al., 2001, Molecular Cloning: A Laboratory Manual, 3rd Ed., Cold Spring Harbor Laboratory Press; and Current Protocols in Molecular Biology, Ausubel. F. ed., Greene Pub. Associates, 1998, updates to 2007に提供されるような組み換え技術によって合成することができる。酵素をコード化するポリヌクレオチドまたは増幅のためのプライマーについても、既知の合成方法、例えばBeaucage et al., (1981) Tet Lett 22:1859-69に記載のホスホラミダイト法、またはMatthes et al., (1984) EMBO J. 3:801-05に記載の方法のような、標準的な固相法によって調製可能であり、通常、これらの方法は、自動化合成法として実施される。さらに実質的なあらゆる核酸は、様々な商業的供給源、例えばThe Midland Certified Reagent Company(テキサス州ミッドランド)、The Great American Gene Company(カリフォルニア州ラモナ)、ExpressGen Inc.(イリノイ州シカゴ)、Operon Technologies Inc.(カリフォルニア州アラメダ)等の多様な供給原から入手可能である。
【0109】
宿主細胞内に発現する改変酵素は、とりわけリゾチーム処理、超音波処理、濾過、塩析、超遠心分離、クロマトグラフィーおよび親和性分離(例えば、基質に結合した抗体)等の既知の技術のタンパク質精製手段を用いて、細胞および/または培養培地から回収することができる。細菌、例えば大腸菌からのタンパク質を溶解するために適切な溶液および高効率抽出液が、ミズーリ州セントルイスのシグマアルドリッチ社から商品名CelLytic BTMとして市販されている。
【0110】
ポリペプチドの分離に関するクロマトグラフィー技術は、とりわけ逆相クロマトグラフィー、高速液体クロマトグラフィー、イオン交換クロマトグラフィー、ゲル電気泳動および親和性クロマトグラフィーを含む。特定の酵素を精製するための条件が、実際の荷電、疎水性、親水性、分子量、分子形状などの要因に部分的に依存することは、当業者にとって明白である。
【0111】
キメラポリペプチドの組み換えおよび合成に係るSCHEMAの開示は、本明細書およびOteyその他, (2006), PLoS Biol. 4(5): ell2; Meyer et al., (2003) Protein Sci., 12:1686-1693;2008年2月1日に出願された米国特許出願番号第12/024,515号;2008年2月7日に出願された米国特許出願番号第12/027,885号の実施例に記載されており、その内容の全てをここに参照して取り込む。
【0112】
上記のように、ポリペプチドは、様々な応用、例えばとりわけ生物燃料生成、セルロースの分解等に使用することができる。
【0113】
以下の実施例は、本発明についての付加的な説明をするものであり、前記開示または添付の特許請求の範囲に制限するものではない。
【実施例】
【0114】
<CBH II発現プラスミドの作成>
酵母発現ベクターYEp352/PGK91−1−αss内で、CBH II酵素をコード化する親およびキメラ遺伝子をクローン化した(図6)。親およびキメラCBH II触媒ドメインをコード化するDNA配列を、S. cerevisiaeコドンバイアスで、GeneDesignerソフトウェア(DNA2.0)およびDNA2.0を用いた合成により設計した。CBH II触媒ドメイン遺伝子を、XhoIおよびKpnIで切断して、ベクター中のXhoIおよびKpnI部位の間にライゲーションし、大腸菌XL−1Blue(Stratagene)内に形質転換した。CBH II遺伝子は、プライマーCBH2L(5'- GCTGAACGTGTCATCGGTTAC-3' (配列番号9)およびRSQ3080(5'- GCAACACCTGGCAATTCCTTACC-3' (配列番号10))を用いて配列決定した。C末端His6親およびキメラCBH II構造は、CBH II遺伝子を前方プライマーCBH2LPCR (5'' -GCTGAACGTGTCATCGTTACTTAG-3' (配列番号11))と、His6オーバーハングと終止コドンを伴う適切なCBH II遺伝子に相補的な逆方向プライマーを増幅することによって作成された。PCR産物は、上記のようにライゲーションされ、形質変換され、配列決定された。
【0115】
<S. cerevisiaeにおけるCBH II酵素の発現>
S. cerevisiae菌YDR483W BY4742 (Matα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0 ΔKRE2、ATCC No.4014317)は、EZ Yeast II Transformation Kit(Zymo Research)を使用してコンピテント化され、プラスミドDNAを用いて形質転換され、合成ウラシルを欠く寒天培地上に播種された。コロニーは、20μg/mLトリプトファンを添加した5mL合成デキストロースカザミノ酸(SDCAA)培地(20g/Lデキストロース、6.7g/Lデフコ酵母窒素塩基、5g/Lバクトカザミノ酸、5.4g/L Na2HPO4、8.56g/L NaH2PO4−H2O)に入れ、一晩、30℃、250rpmで培養した。5mL培養物は、250mLチューンエアーフラスコ(Shelton Scientific)中の40mL SDCAAで希釈し、30℃、250rpmで48時間振盪した。培養物を遠心分離し、清をt1/2分析のため、30kDa PES膜を取り付けたAmicon限外ろ過膜を使用して、500μLにまで濃縮した。濃縮した上清は、1mMフェニルメチルスルホニルおよび0.02%NaN3に供した。His6タグを付けたCBH IIタンパク質を、Ni−NTAスピンカラム(Qiagen)を用いて製造元のプロトコールに従って精製し、次いで、タンパク質をZeba−Spin脱塩カラム(Pierce)を使用して50mM酢酸ナトリウム、pH4.8中に交換した。精製されたタンパク質濃度を、基準としてBSAとPierce Coomassie Plusタンパク質試薬を使用して測定した。SDS−PAGE分析は、濃縮された培養上清20μLまたは精製されたCBH II酵素約5μgのいずれかを、7.5%Tris−HClゲル(Biorad)上に加え、シンプリーブルーセーフステイン(Invitrogen)で染色することによって実行した。CBH II上清または精製されたタンパク質は、EndoH(New England Biolabs)を用いて、1時間、37℃、製造元のプロトコールに従って処理した。濃縮された酵母培養上清中のCBH II酵素活性は、37.5μLの濃縮された培養上清を37.5μL PASCに添加し、2時間50℃でインキュベーションして測定した。形成された還元糖等価物は、以下のようなソモギ・ネルソン分析によって測定した。
【0116】
<半減期、特異的活性、pH活性および長期セルロース加水分解の測定値>
リン酸膨潤セルロース(PASC)を調製した。基質へのCBH II酵素活性を向上させるため、PASCを10g/Lの濃度で50mM酢酸ナトリウム、pH4.8中の10mg/mLA. nigerエンドヌークレアーゼ(Sigma)を用いて、37℃で1時間プレインキュベーションした。エンドグルカナーゼを、95℃まで加熱し、15分間処理することで不活性化し、PASCを2回、50mM酢酸緩衝剤を用いて洗浄し、脱イオン水中に10g/Lとなるように再懸濁した。
【0117】
CBH II酵素t1/2は、以下に記載するように、処理されたPASCとインキュベーションした後、濃縮したCBH II発現培養上清を50mM酢酸ナトリウム(ソモギ・ネルソン還元糖分析で測定されたA520が0.5となる濃度でpH4.8)に添加することによって測定した。37.5μLのCBH II酵素/緩衝剤の混合物を、63℃の水浴で不活性化した。不活性化後、37.5μLのエンドグルカナーゼ処理したPASCを添加し、2時間、50℃の条件下で加水分解をで実施した。反応上清は、Multiscreen HTSプレート(ミリポア)を通してろ過した。SpectraMaxマイクロプレートリーダー(Molecular Devices)を用いて得られたソモギ・ネルソン分析log(A520)値を、バックグラウンドの吸光度について修正処理を施し、マイクロソフト・エクセルを使用して、時間軸に対してプロットすることで、線形回帰から得られるCBH II酵素の半減期のデータを得た。
【0118】
特異的活性測定のため、精製CBH II酵素をPASCに添加し、PASCの最終反応量が5g/Lで、CBH II 酵素濃度が3mg/gであり、酢酸ナトリウム濃度25mM、pH4.8のPASCを得た。50℃水浴中でのインキュベーションを2時間行い、還元糖濃度を測定した。pH/活性特性の測定のため、精製CBH II酵素を、75μL反応容積中の濃度300μg/gのPASCに添加した。反応は、12.5mMクエン酸ナトリウム/12.5mMリン酸ナトリウムの緩衝液中で、50℃で16時間実施し、還元糖濃度を測定した。長時間のセルロース加水分解測定は、100mM酢酸ナトリウム、pH4.8、20mM NaClの、1g/L濃度の処理済みPASC300μLを用いて行った。精製CBH II酵素を、100μg/gの濃度でPASCに添加し、還元糖測定の40時間前に水浴中にて反応させた。
【0119】
CBH II酵素をコードする5つの親遺伝子候補を、S. cerevisiaeコドンバイアスを用いて合成した。5つ全ては、1−89残基がセルロース結合モジュール(CBM)、弾性リンカー領域およびH. jecorina触媒ドメインの5つのN末端残基に相当する、N末端コード配列を含む。Humicol insolensおよびChaetomiun thermophilumからの2つのCBH II酵素候補は、Hypocrea jecorina、Phanerochaete chrysosporiumおよびTalaromyces emersonilによる3つの他のものより比較的高いレベルで、S. cerevisiaeから分泌された(図1)。3つのわずかに発現されている候補の親に関するSDS−PAGEゲルのバンドは、認識困難なため、濃縮培養上清における活性分析は、活性型セルラーゼの存在を確認するため、リン酸膨潤セルロース(PASC)とインキュベートされたものを用いて実施された。図1に示す形成された還元糖からの値は、T. emersonii CBH II以外の全ての酵素に関して、濃縮S. cerevisiae培養上清における活性型CBH IIの存在が確認された。H. insolensおよびC. thermophilum配列は、最も産業的に関連したH. jecorinaからの菌CBH II酵素と組み合わせて選択した。触媒ドメインのそれぞれの配列の一致は、64%(1:2)、66%(2:3)および82%(1:3)であり、H. insolensは親1、H. jecorinaは親2、C. thermophilumは親3とする。これらそれぞれの触媒ドメインは、360,358,359アミノ酸残基を含む。
【0120】
産業使用のためにセルラーゼを生成するのに最も頻繁に使用される生命体である糸状菌H. jecorinaにおける異種タンパク質発現は、酵母と比較して非常に困難である。組換え型セルラーゼ活性を減少することが報告されている、高度なグリコシル化を最小限にするため、組換え型CBH II遺伝子をグリコシル化機能を欠損させたdKRE2 S. cerevisiae株で発現させた。該株は、野生株と比較して、より短いマンノースオリゴマーがN−結合型およびO−結合型の両方のグリコシル化サイトに結合すると考えられ、これは野生型H. jecorinaのCBH II 酵素のグリコシル化に極めて類似している、高マンノース構造を取り除くためのEndoH処理を伴う条件、および伴わない条件において、CBH IIタンパク質のSDS−PAGEゲル分析を行ったところ、EndoH処理によっては、前記菌から分泌された酵素の電気泳動的な可動性が上昇しないことが判明し、野生型S. cerevisiae菌が組換え型タンパク質のグリコシル化部位に結合する分岐型マンノース部分を欠くことを確認した。
【0121】
H. insolensの高分解性の構造(pdb)SCHEMAのテンプレートとして使用して、組み換えに応じて引離される接触を同定した。RASPPは、それぞれ15未満の<E>を有する4つの候補ライブラリーを返した。全ての候補ライブラリーは、以前作成されたキメラライブラリーより低い<E>を示し、折り畳まれた活性型キメラの許容可能な部分は、比較的高い<m>を有して得られることが示された。キメラ配列の多様性は、最も大きな<m>=50を導くようなブロックの境界を選択すると、最大となった。この設計のためのブロックは、図2Bに例示され、Table2に詳細に示される。
Table2:親CBH II酵素触媒ドメインのためのClustalW複数配列アライメント。ブロック2,4,6および8は、ボックスおよびグレーの斜線で示す。ブロック1,3,5および7は、されていない(H. inso:配列番号2、H. Jeco:配列番号4およびC. Ther.:配列番号6)。
【0122】
【表9】
【0123】
H. insolens CBH II触媒ドメインは、8つの螺旋がバレル周縁を規定し、7つの平行のβシートが活性部位を形成するα/βバレル構造を有する(図2A)。2つの延長したループは、活性部位上にルーフを形成し、加水分解中に基質セルロース鎖が通過する通路を形成する。7つのブロック境界のうちの5つは、2次構造の要素の間に収まり、ブロック4は連続的なα螺旋の中央部分から開始し、終結する(図2A、2B)。ブロック間の側鎖接触の大多数は、初期構造において調節可能なブロック間に生じる(図2C)。
【0124】
48個のキメラ遺伝子のサンプルセットは、1つの親由来の5つのブロックと、残り2つのうちの1つまたは両方の親由来の3つのブロックを有する、16個のキメラの3セットとして設計された(Table3);それぞれの親の表記がそれぞれのブロックの位置で等しくなるように、配列を選択した。相当する遺伝子を合成し、発現させた。
Table3:サンプルセットCBH II酵素キメラの配列
【0125】
【表10】
【0126】
48個のサンプルセットのS. cerevisiaeのうち23個の濃縮培養上清は、PASCに対して加水分解活性を示した。これらの結果は、6,561個のCBH IIキメラ配列のうち数千(例えば、Table1を参照のこと)が活性型酵素をコード化することを示す。23の活性型CBH IIサンプルセットのキメラは、顕著な配列多様性を示し、最も近い親の配列と、少なくとも23および36、54および123相当のアミノ酸置換を有する。それらの平均突然変異レベル<m>は36である。
【0127】
Meyerらが、E、mおよびキメラが折り畳まれて活性を有する確率の間の相関を見出したように、同類の相互関係が、サンプルセットCBH IIキメラに関して存在するかの分析を実施した。PASC活性の分析によって測定されるように、濃縮発現培養上清におけるCBH II酵素活性の値は、SDS−PAGEゲルにおけるCBH IIバンドの強度に相関した(図1)。H. jecorina CBH II親と同様に、活性はいくつかのCBH IIキメラに関して検出できないゲルバンドで検出できる。ゲルバンドは示したが、活性を欠失するCBH IIキメラの存在が観察された。CBH IIキメラが活性型の形状で分泌される確率は、Eおよびmの双方に逆相関した。
【0128】
熱不活性の半減期(t1/2)は、63℃における親および活性型キメラのCBH II酵素の濃縮培養上清について測定された。H. insolens、H. jecorinaおよびC. thermophilumのCBH II親の半減期は、それぞれ95,2,25分であった。活性型サンプルセットのキメラは、1分未満から3,000超の幅広い半減期を示した。23個の活性キメラのうちの5つは、最も熱安定のある親であるH. insolens CBH IIを超える半減期を有した。
【0129】
CBH IIキメラの半減期のための予測定量的モデルの作成を試みる際、5つの異なる線形回帰データのモデリングアルゴリズムを使用した(表4)。それぞれのアルゴリズムを使用して、それぞれのサンプルセットCBH IIキメラおよび親のlog(t1/2)のブロック組成物に関するモデルを作成した。これらのモデルは、ブロックのlog(t1/2)への寄与を定量化した、熱安定性重量値を構築した。全て5つのモデリングアルゴリズムについて、この工程が、2つのランダムに選択された配列を省略して1,000回繰り返され、それぞれのアルゴリズムは、24個のブロックそれぞれに1,000重量値を示した。それぞれのブロックの熱安定性重量値について、平均値および標準偏差(SD)を算出した。それぞれのモデルアルゴリズムの予測精度は、それぞれのモデルが省略された配列のt1/2をどれだけ正確に予測できるかで評価した。1,000アルゴリズムの繰り返しについての測定値および予測値の間の相関を、モデルアルゴリズムの相互検証スコアとする。5つのモデル全てについての相互検証スコア(X−val)は、0.057以下であり(Table4)、線形回帰モデリングが、この小さな23個のキメラt1/2データセットの定量的CBH IIキメラの半減期予測については適応できないことを示した。
Table4:5つの線形回帰アルゴリズムのCBH II酵素キメラのブロック安定性スコアへの使用のための相互検証値。アルゴリズムの省略:リッジ回帰(RR)、部分最小二乗回帰(PLSR)、サポートベクターマシン回帰(SVMR)、リニアプログラミング・サポートベクターマシン回帰(LPSVMR)およびリニアプログラミングブースティング回帰(LPBoostR)。
【0130】
【表11】
【0131】
線形回帰モデリングを使用して、ブロックを質的に安定、不安定または中立に分類した。それぞれのブロックのキメラの熱安定性への影響は、それぞれの回帰アルゴリズムによって測定される熱安定性への寄与を提示するスコアリングを使用して評価した。それぞれのアルゴリズムに関して、中立から1SDを超える熱安定性重量値を有するブロックは、「+1」として記録し、中立から1SD以内の熱安定性重量値を有するブロックは0とし、中立から1SDを超えて低い熱安定性重量値を有するブロックは、「−1」として記録した。それぞれのブロックに関する「安定性スコア」は、それぞれ5つのモデルからの1,0、−1の安定性スコアを合計することによって得た。Table5は、それぞれのブロックに関するスコアを概説する。ブロックI/親1(B1P1)、B6P3、B7P3およびB8P2は、比較的高い安定性効果を有するものとして同定された、一方、B1P3、B2P1、B3P2、B6P2、B7P1、B7P2およびB8P3は、最も強くブロックを不安定化することを見出した。
Table5:5つの線形回帰アリゴリズム1によって生じたサンプルセットCBH II酵素キメラに関する質的なブロック分類。スコア+1は、熱安定性重量を有するブロック(キメラの熱安定性へのブロック寄与に関する無次元測定)であり、中立から1SDを超えるもの(安定性)を示し、スコア0は、中立から1SD内の重量値であるブロックを示し、−1は中性から1SD超低い重量値を有するブロックを示す(不安定性)。
【0132】
【表12】
【0133】
CBH II酵素キメラをコードする第2セットの遺伝子を合成し、予測された安定ブロックを評価して、最も安定な親より熱安定のあるセルラーゼを同定した。この検証セットに含まれる24個のキメラ(Table6)は、最も不安定であると予測される7つのブロックが全くなく、4つの最も安定なブロックに富んでいた。後者のブロックの発現は、安定なスコアに偏っていた。さらに「HJPlus」12222332キメラは、予測された最も安定なブロックをH. jecorina CBH II酵素内(親2)に置換することによって作成された。
Table6:24個の検証セットCBH II酵素キメラの配列中の9つは、活性型で発現した。
【0134】
【表13】
【0135】
24個の検証のセットのキメラの9つのS. cerevisiae発現培地の濃縮上清は、HJPlusキメラと同様に、PASCに対して活性を示した(Table6)。活性が検出されていない15個のキメラのうちの9つは、ブロックB4P2を有した。初期のサンプルセットのB4P2を含有する16個のキメラのうち1つのみが、PASCに対して活性を示した。キメラセットおよびHJPlusの両方を合計して、26個のキメラのうち2つのみが活性化していて、この特定のブロックがS. cerevisiaeにおける活性化セルラーゼの発現を阻害することを示した。
【0136】
検証セットからの10個の機能的キメラCBH II酵素の安定性を評価した。安定酵素は既に50時間を越える半減期を有したため、63℃、12時間の熱不活性化後のPASCに対する残留加水分解活性を、測定の予備的評価として使用した。この12時間のインキュベーションによって、サンプルセットの最も熱安定性のあるキメラである11113132の活性において、測定可能な減少が生じ、不活性型の熱安定性のあるH. insolens親CBH IIを完全に不活性化した。全10個の機能的な検証セットのキメラは、それらの活性として最も安定な親であるH. insolens CBH IIより多い活性を保持していた。
【0137】
精製された酵素を使用する選択された熱安定性のあるキメラの活性を分析した。親CBH II酵素および3つの熱安定性のあるキメラ(最も熱安定性のあるサンプルセットのキメラ11113132、最も熱安定性のある検証セットのキメラ13311332)およびHJプラスキメラ12222332を、C末端His6精製タグとともに発現させ、精製した。活性試験中のCBH II酵素の熱不活性化を最小限にするため、2時間(比較的短い時間)、50℃、pH4.8でのPASC基質とのインキュベーションを実施した。Table3に示すように、親およびキメラCBH IIの特異的活性は、最も活性のあるH. jecorinaからの親CBH II酵素のうちの4つの要素内にあった。HJプラスの特異的活性は、H. jecorina CBH II以外の全ての被験のCBH II酵素を超えていた。
【0138】
幅広い範囲のpH/活性特性により、CBH IIキメラの広範囲にわたる潜在的なセルロース加水分解条件下での使用を可能とするためには、セルラーゼ活性のpH依存性が重要である。H. jecorina CBH IIは、最適化された活性をpHの範囲4から6で有することが観測され、一方この範囲外ではその活性が著しく現象することが観察されてきた(文献16)。図4は、H. insolensおよびC. thermophilum CBH II酵素ならびに全3つの精製された熱安定性のあるCBH IIキメラが、H. jecorina CBH IIのものより相当広いpH/活性特性を有することを示した。Liuらは、C. thermophilum CBH IIに対してpH4を最適条件として報告したが、ここでの組み換え型酵素の最適なpHは、7付近であった。天然H. insolens CBH IIは、約pH9で最大活性、pH4で最大活性の約60%を示す幅広いpH/活性特性を有する。同様の幅広い特性が、組み換え型酵素について観測された。HJプラスキメラは、H. jecorina CBH IIと比較してより幅広いpH/活性特性を有し、他の2つの親CBH II酵素と同様のpH依存性を示した。
【0139】
高温での活性の達成と、長いタイムインターバルを超えた活性の維持は、改変した高安定性CBH II酵素を実現するための2つの初期目標である。セルロース加水分解における熱安定性のあるCBH IIキメラの性能を、温度範囲全体で、40時間のインターバルで試験した。図5に示すように、3つ全ての熱安定性キメラは、親CBH II酵素より比較的高い温度でPASCに活性を有した。H. jecorina CBH IIは、57℃長ではPASCを加水分解せず、安定なH. insolens酵素は63℃超ではPASCを全く加水分解しなかったが、キメラは70℃で活性を保持した。。長時間のセルロース加水分解分析におけるHJプラスの活性は、各々の最適な温度における全ての親の活性を上回った。
【0140】
CBH IIライブラリーは、いくつかの理由により、崩壊の可能性が比較的低い。CBH II親配列の高い相同性に加えて、CBH II折り畳みバレルの立体的構造は、組み換えによって分離することができる長範囲の接触の数を制限する。ブロック間の接触(4.5オングストローム範囲内の重原子)は、H. insolens構造1ocnに由来する接触マップ全体の27%(503/1831)のみを含む。残基の組み合わせに係る接触は、キメラにおいてのみ計数可能であり、ブロック内の総計は23%まで減少した(68/294)。さらに、これら相互作用のほとんどは、タンパク質の表面上の残基間に生じ、溶媒スクリーニングの可能性は、さらに残基間の相互作用の飛躍的な分離の可能性を減少させる(図14a)。1つの例外として、位置176−位置256間の埋没した相互作用が、図14bに示す。この部位で、B6P2と、B5P1またはB5P3のいずれかとを有するキメラは、Met173:Trp253の対を有する(親のMet176:Phe256の対、またはLeu173:Trp253の対と比較して大きなアミノ酸)。しかしながら、親の結晶学的なモデルの精査により、Trp253側に位置するタンパク質バックボーン部の可動性や、Met側鎖の内因的な可撓性のため、この部位における立体衝突は生じにくいと考えられる。特に、1つの特徴的なキメラは、このパターン(13333232)に適合し、回帰モデル適合性(68℃)よれば、親(67℃)よりも安定である。
【0141】
カップリングを生じる他のメカニズムは、ブロック構造の相違であるが、これはブロック接点での新規の残基対の存在に依存するものではない。そのかわり、親配列の相違により、内因性ブロック構造は、モジュール・ブロックの移動を妨げて、多様化することができる。CBH IIライブラリーの場合、親の対の高い配列相同性の値(82%、66%および64%)において、構造の偏差は軽度とすることができる(<1オングストロームRMSD)。この可能性は、H. insolensおよびH. jecorina CBH IIの結晶学的な構造を比較することによって、評価することができる(C. thermophilum CBH IIは結晶構造を欠如しているが、H. insolensと82%の相同性がある)。それぞれの親からの構造による配列ブロック(1ocnおよび1cb2)において、αカーボンRMSD値は低くなる(0.5,0.5,0.6,0.5,0.3,0.7,0.3および0.4A RMSD)。H. insolens上に重ねられたH. jecorinaブロックを、補足図5cに示す。背景に依存した効果を確認するため、対立する宿主構造上にそれぞれアラインされたブロックをスプライシングして、シリコ構造(silico structure)の組み換えを実行した。ブロック6および7の間にAsnを挿入して(図14D)B7P2(11111121)を使用した場合、軽度の衝突(2.65オングストローム)を除き、全ての単一ブロック置換キメラ(例えば11112111または22122222)に関する衝突しない構造モデルを作成することが可能である(αカーボン>3オングストローム間隔)。
【0142】
様々なブロック/断片のキメラの安定性への寄与を決定するためにさらなる実験を実施し、熱安定性および/またはpH安定性を改良した。CBH II酵素をコードする親およびキメラ遺伝子を、酵母発現ベクターYEp352/PGK91−1−αss内にクローン化し、合成デキストロースカザミノ酸(SDCAA)培地中で発現させた。アビセル活性分析のため、酵母ペプトンデキストロース(YPD)培養上清は、1mMフェニルメチルスルホニルおよび0.02%NaN3溶液にして、濃縮せずに使用した。濃縮SDCAA酵母培養上清中のCBH II酵素活性を、37.5μLのPASCおよび225μLの50mM酢酸ナトリウム、pH4.8に添加することで濃縮培養上清を希釈し、2時間、50℃でインキュベーションして測定した。形成した還元糖の等量を、ソモギ・ネルソン分析によって測定した。
【0143】
CBH II酵素のT50値を、エンドヌークレアーゼ処理したPASCとインキュベーションした後、濃縮したCBH IIのSDCAA発現培養上清を50mM酢酸ナトリウムに添加し、ソモギ・ネルソン還元糖分析の測定値A520が0.5となる濃度でpH4.8となるように調整して測定した。200μLのCBH II酵素/緩衝剤混合物を、適温で10分間、水浴中にてインキュベートした。インキュベーション後、37.5μLのエンドグルカナーゼ処理されたPASCおよび62.5μLの50mM酢酸ナトリウム溶液を添加し、50℃で2時間加水分解を実施した。酵素の半分の活性が欠如したときのインキュベーション温度を、ソモギ・ネルソン分析のA520値の対温度プロットの線形補間より算出した。
【0144】
長時間のアビセルPH101(Fluka)加水分解の測定のため、0.3μgの精製されたCBH IIを、水浴に配置されたPCRチューブ内で、270μLの50mM酢酸ナトリウム(pH4.8)において3mgのアビセルとともに16時間インキュベートした。チューブを室温の水浴中で10分間冷却し、1000gで10分間遠心分離し、収集した上清を還元糖分析に供した。
【0145】
YPD発現培養上清中のCBH II活性の評価のため、2mL〜40mLの上清を、コニカルチューブにて、50mM酢酸ナトリウム(pH4.8)に懸濁された33mg/mLのアビセル800μLに添加された。CBH IIは、アビセルと4℃、1時間で結合し、2000g、2分間遠心分離し、50mM酢酸ナトリウム(pH4.8)で2回洗浄された。2回洗浄した後、CBH IIと結合したアビセルを、2.75mL酢酸ナトリウム緩衝剤中に再懸濁し、270μLの分量で分注して、50℃で2.5時間インキュベートした。遠心分離および上清還元糖の分析は、上記のように実施した。
【0146】
Mathematicaにおける線形回帰パッケージを使用して、CBH IIキメラT50データを17パラメーターに適合し、相互検証分析を行った。ブロック効果は、親1(H. insolens CBH II)を基準として、親2および3からのそれぞれ8つのブロックの置換を示す16パラメーターを用いて、報告された。
【0147】
10分間のインキュベーションにより酵素が50%の活性を欠如する温度として規定されるT50値は、3つの親セロビオヒドラーゼについて決定した。前の実験からの33個の活性型CBH IIキメラおよび質的に安定なモデルが最も熱安定性があると予測したうちの18個のさらなるキメラ(即ち不安定と予測されたブロックは一切含まれず、安定性ブロックと予測された4つのうち3つまたは4つが含まれる)についてT50値を測定した。全51個のキメラ配列を表8に列挙した。以前に分泌されないと分類された全ての熱安定性と予測されるキメラの再培養および再濃縮により、T50測定のため、十分量の12112132,13111132および13322332CBH IIを得ることを可能とした。キメラおよび親のCBH IIに関するT50値の完全なセットをが、Table8に示す。全ての前記CBH IIのアミノ酸配列を、Table7に記載する。試験された全31個の熱安定性と予測されたキメラは、最も熱安定性のある親酵素(64.8℃)より2度高いT50値を有する。Tableは、熱安定を向上させるため、Serに変異することができるブロック/ドメイン7におけるCys残基も同定する。従って、本発明は、以下の配列のあらゆるポリペプチドを提供し、下線/イタリック体/太字のCysはSer残基と置換され、結果として生じたポリペプチドは、野生型酵素と比較して、高い熱安定性を有する。
Table7:CBH II親およびキメラ触媒ドメインのためのアミノ酸配列が、Table8に示される。
Tableは、P. chrysosporium CBH IIに関する触媒ドメインも含む。全ての組み換え型CBH IIは、天然H. jecorina CBH IIからのN末端CBMおよびリンカーCSSVWGQCGGQNWSGPTCCASGSTCVYSNDYYSQCLPGAASSSSSTRAASTTSRVSPTTSRSSSATPP PGSTTTRVPPVGSGTATYS (配列番号8)を共有する。
【0148】
【表14】
【0149】
【表15】
【0150】
【表16】
【0151】
【表17】
【0152】
【表18】
【0153】
【表19】
【0154】
【表20】
【0155】
【表21】
【0156】
【表22】
【0157】
Table8:
親CBH IIに関する2つの独立した二重のT50値(℃)、23個の初めのサンプルセットCBH IIキメラおよび予測される熱安定性のあるCBH IIキメラ。この研究のために合成された18個のキメラは、星印の後に記載される。
【0158】
【表23】
【0159】
線形回帰の配列安定性データへの適応は、R2=0.88で観測されたT50値に適合する10−パラメーターモデルを生じた。トレーニングセット外の回帰モデルの予測性能をよりよく推定するため、11の折り畳み構造の相互検証をR2が0.57となるように実施し、2つの外れ値を取り除き(11313121および22222222)、相互検証R2を0.76へと増加させた。回帰モデルは、最も安定な親1(H. insolens)を基準状態T50として使用して、p値≦0.1を有する9つのさらなる項を含む。モデルパラメーター(Table9)は、親3からのブロック7の単一のブロックが、H. insolens CBH IIと比較してキメラの熱安定性に最も強く寄与することを示す。C. thermophilum CBH IIからのこのブロックは、それを含むキメラの熱安定性を約8.5℃上昇させる。p値≦0.1の8つの残りのブロックのうち2つによる熱安定性の上昇は、1.2℃および2.7℃と小さかったが、他の6つはその安定性を減少させた。
Table9:T50線状回帰モデルパラメーターおよびp値。図1の回帰適合線を算出するために使用されたp<0.1のパラメーター値を太字体で示す。ブロック効果は、親1(H. insolens CBH II)を基準状態として、親2および3からのそれぞれ8つのブロックの置換を示す16パラメーターを用いて報告された。
【0160】
【表24】
【0161】
B7P1およびB7P3配列のアライメント(図10)は、H. insolensおよびC. thermophilum酵素において、ブロック7の56個のアミノ酸のうち、10箇所で相違する。最も高いT50値を有する21311131のキメラのバックボーンにおいて、B7P3のそれぞれの残基(親の断片7(配列番号6))は、B7P1において相当する残基に個々に変異され、それぞれの点突然変異体についてT50値が得られた。突然変異体S313Cは、キメラの熱安定性を著しく変化させた:この単一の突然変異体は21311131のT50を約10℃減少させた(図11)。
【0162】
復帰突然変異の効果を異なる背景において研究するため、C313S置換をコードするH. insolensおよびH. jecorinaの親CBH II遺伝子(H. insolensにおいてC314S、およびH. jecorinaにおいてC311S)を作成し、発現させ、酵素のT50値を測定した。安定性のB7P3を野生型H. insolensおよびH. jecorinaの酵素内に置換したキメラ、11111131および22222232の安定性についても定量化した。B7P3ブロック置換およびCys−Ser点突然変異体は、親CBH IIを著しく安定化させた;最も大きな効果は、C311S置換を含むH. jecorina CBH IIへのT50における8℃までの上昇であった。またCys−Ser突然変異は、B7P3を含まない2つのキメラである、31311112および13231111ならびに組み換え型の親のセットでない相同するCBH II(Phaneroachete chrysosporiumから)において試験した。P. chyrsosporium CBH IIの触媒ドメインは、親CBH II触媒ドメインに55−56%のみの相同性がある。これら全ての酵素は、Cys−Ser置換によって安定化された;P. chrysosporium CBH IIは10℃と高い安定化を見せた(図13)。
【0163】
同等のC313S突然変異を含む8つの熱安定性のあるCBH IIキメラおよび親の酵素は、His6のタグを付け、精製することで、特異的活性を測定できた。Table10Aに示すように、特異的活性は、50℃で非結晶性セルロース(PASC)について観測されるように、Cys−Ser突然変異体を含むこれらキメラおよび天然酵素について、野生型親のものと同等であった。従って、高い熱安定性は、特異的活性の代償として現れるものではない。
Table10A:天然、点突然変異体および選択される熱安定性のあるキメラCBH IIに関する特異的活性値(μgグルコース還元糖等価/(μg CBH II酵素×min×10))。エラーバーは、標準誤差を示し、標準誤差は3つの複製に関する標準偏差/sqrt(n)として定義される。2時間の反応、3mg酵素/g PASC、50℃、25mM酢酸ナトリウム、pH4.8。
【0164】
【表25】
【0165】
Table10B:H. jecroinaおよびH. insolens野生型、C313S点突然変異体およびB7P3ブロック置換CBH IIに関する合成(SDCAA)および豊富な(YPD)発現培養培地上清の双方における全活性。示す値は、セルラーゼ活性分析に添加される発現培養上清CBH II等価のmL当たりのμgグルコース/mLセルラーゼ活性分析値である。SDCAA培養については、濃縮されたSDCAA培養上清が使用され、50mM酢酸ナトリウム(pH4.8)において50℃で100分間のリン酸膨潤セルロース(1mg/mL)に対する活性が測定された。YPD上清CBH IIは、アビセルに結合させることによって濃縮し、50mM酢酸ナトリウム(pH4.8)において55℃で150分間、アビセル(15mg/mL)に対する活性を測定した。
【0166】
【表26】
【0167】
これらの8つの熱安定性のある同一のキメラ(最も安定な親よりT50が2−10℃高い)は、その後結晶性セルロースへの活性に関して、16時間のインキュベーションの間、親酵素がほとんど活性を示さない温度を含む幅広い温度範囲で試験した。図9aは、8つの試験された熱安定性のあるキメラのうちの7つが、60−65℃でアビセルに対して最大の活性を示す、8つのキメラ全てについて、70℃(試験中の最高温度)で、活性を保持していることを示した。対照的に、3つの親CBH IIは、50℃で最大の活性を示したが、70℃で完全にまたはほぼ完全に不活性となった。さらに高い最適活性温度を有する7つのキメラは、あらゆる親CBH II酵素と比較して非常の多量のアビセルを加水分解した。図9bに示すように、同様の挙動が、Cys−Ser点突然変異を含むH. insolensおよびH. jecorinaの親においても観測された。Cys−Ser点突然変異は、アビセル加水分解およびP. chrysoporium CBH IIにおける最高反応温度を上昇させた。H. insolensおよびH. jecorina親におけるP3B7ブロック置換により、反応温度およびH. insolens CBH IIの加水分解は上昇したが、上昇した最高反応温度は、H. jecorina酵素による全体的なセルロース加水分解を向上させるものではなかった。
【0168】
cerevisae発現宿主からの野生型H. jecorina CBH IIの分泌が低い(<1mg/L)ことが観測された。野生型H. jecorina CBH II酵素におけるC311S突然変異は、CBH IIの全分泌量の活性を著しく上昇させた(表11)。豊富な培地(YPD)における活性の上昇は10倍であるが、合成(SDCAA)培地において、C311SおよびB7P3置換は、H. jecorina CBH II全分泌活性を2倍に上昇させる。他の2つの親CBH IIと比較して高いレベルで発現しているH. insolens CBH II親について、B7P3ブロック置換は活性を減少させたが、C314S突然変異は、活性を1.5倍まで上昇させた。H. insolensおよびH. jecorina野生型およびCys−Ser突然変異の全てが同様の特異的活性を有するため(図10)、全分泌セルラーゼ活性の上昇は、機能的酵素のより優れた分泌の結果である。S. cerevisiae異種タンパク質分布とタンパク質の安定性との間の相互関係が観測され、Cys−Ser突然変異CBH IIの上昇した分泌が、それらの最も高い安定性に反映し得ることを示唆した。
Table11:天然、点突然変異体および選択される熱安定性のあるキメラCBH IIに関する特異的活性値(μgグルコース還元糖等価/(μg CBH II酵素×min×102))。エラーバーは、3つの複製に関する標準誤差を示し、標準誤差は標準偏差/sqrt(n)を規定する。2時間の反応、3μg酵素/g PASC、50℃、25mM酢酸ナトリウム、pH4.8。
【0169】
【表27】
【0170】
Cys−Ser突然変異をモデル化するため、微細構造のH. insolens CBH II 1ocn結晶構造を使用した。第1に、水素結合ネットワークをREDUCEで最適化した。C314はPro339のカルボニル基と水素結合を形成することを予測した。この予測を検証するため、側鎖パッケージングを、モデリング・プラットフォームSHARPENを用いて最適化した。Ser314は、より強い水素結合、およびより有利な結合構造が生じるように、Cys314と同様の相互作用を奏することが予測された。
【0171】
数々の効果は、Cys−Ser突然変異が、天然CBH IIおよびキメラを含む幅広いCBH IIを安定化することができる理由を説明することができる。CysおよびSerは同様であり(等比体積でない)、他の代価物と比較して、これら2つのアミノ酸は配列アライメントをこの位置で抑制する。この残基への水素結合パートナーは、バックボーン要素(Gly316のアミドおよびPro339のカルボニル)であり、従ってあまり第3者のアミノ酸変異体に依存しそうにない。さらにこのポケットに直接隣接した側鎖(Asn283、Pro339、Phe345)が、研究された全4つの天然CBH IIセルラーゼにおいて保存されている。
【0172】
微細構造(1.3オングストローム)のH. insolens結晶構造(pdb挿入1ocn6)は、Cys314が水素結合ネットワークの一部であることを示した(図15)。Cysと比較して高いSerの水素結合性能は、安定化における比較的強い水素結合相互作用を奏することを示す。結晶構造においても、立体構造の観点からSerが好適に使用し得ることを示す。特に、Cys側鎖が元来の結合角度を有するとき、CysがPro339のカルボニルに押しつけられるように挿入されることで、6°の角度はなくなるが、望まれざる立体構造の相互作用を生じた。
【0173】
H. jecorina CBH IIと最も多くの一致を共有する196個のタンパク質配列のアライメント。最も一致のある配列の250のうちの54については、重複しているためアライメントから排除した(即ち、構造研究のための点突然変異体または95%超で一致するアイソフォーム)。Ser311に対するバイアスがある:158個の配列はSer、20個はAla、10個はCys、5つは欠失、3つはGlyを有する。しかしながら、42個の他の配置があり、最も多くの選択肢は、H. jecorinaのアミノ酸の選択肢の2倍を超えて生じる。
【0174】
Cys−Ser突然変異の高い安定化効果は、この位置でのSerが、天然セルラーゼの熱安定性の全体的な指標となる可能性を向上させる。しかしながら、この位置にCysを有することを特徴とするH. insolens CBH IIのT50値が64.8℃であるが、これはC. thermophilum CBH II(64.0℃)より高く、Serが安定化決定の唯一の要因ではないことを示している。
【0175】
熱安定性だけが、産業用セルラーゼの重要な特徴ではない。特異的活性、セルロース結合に与える変化、発現に対する効果、および生成阻害は、全て同様に重要である。キメラおよび本明細書のデータは、組換えにより、短時間(例えば2時間)で測定されたセルロース加水分解分析において、特異的活性を損なうことなく向上した熱安定性を奏する、CBH IIキメラを得ることを示す。同様の現象が、熱安定性のあるCys−Ser突然変異を含むCBH IIに関してもみられた。長時間の加水分解分析において、いくつかのCBH IIキメラ、および3つの被験Cys−Ser突然変異CBH IIについて、天然CBH IIと比較してより多くセルロースを加水分解することを発見した。この優れた性能は、酵素が高温でより長時間で機能し続けることを可能とする高い熱安定性とともに、親CBH IIに匹敵する特異的活性を有することに起因すると考えられる。これらの分析は、等量の精製された親、キメラおよびCys−Ser突然変異体酵素で実施されたため、観測された高温での加水分解の向上は、S. cerevisiae発現宿主からの分泌量の増加によるものではない。熱安定性のあるキメラおよびCys−Ser突然変異体は、従って、セルロース分解のための酵素製剤に有用な要素であることが証明し得る。
【0176】
様々な特定の実施形態が示され、記載される一方で、この発明の思考と範囲から逸脱することなく、様々な変化がされることが理解される。
【図1A】
【図1B】
【図2A】
【図2B】
【図2C】
【特許請求の範囲】
【請求項1】
配列GEXDGを有するモチーフのC末端にセリンを含み、XはC、AまたはGであり(配列番号107)、Xはセリンと置換され、変異体は、セルラーゼ活性を有し、ポリペプチドは、配列GEXDG(配列番号107)においてセリンを欠如する野生型セルラーゼと比して高い熱安定性を有する、組み換え型ポリペプチド。
【請求項2】
前記ポリペプチドは、C314Sを含む配列番号2と少なくとも85%、90%、95%、98%、99%の相同性があり、C311Sを含む配列番号4と少なくとも85%、90%、95%、98%、99%の相同性があり、C310Sを含む配列番号12と少なくとも85%、90%、95%、98%、99%の相同性があり、C312Sを含む配列番号13と少なくとも85%、90%、95%、98%、99%の相同性があり、C314Sを含む配列番号14と少なくとも85%、90%、95%、98%、99%の相同性があり、C315Sを含む配列番号15と少なくとも85%、90%、95%、98%、99%の相同性があり、C313Sを含む配列番号16と少なくとも85%、90%、95%、98%、99%の相同性があり、C311Sを含む配列番号17と少なくとも85%、90%、95%、98%、99%の相同性があり、C313Sを含む配列番号19と少なくとも85%、90%、95%、98%、99%の相同性があり、C312Sを含む配列番号21と少なくとも85%、90%、95%、98%、99%の相同性があり、C311Sを含む配列番号22と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号64と少なくとも85%、90%、95%、98%、99%の相同性があり、C407Sを含む配列番号65と少なくとも85%、90%、95%、98%、99%の相同性があり、C394Sを含む配列番号66と少なくとも85%、90%、95%、98%、99%の相同性一致があり、C400Sを含む配列番号67と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号68と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号69と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号70と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号71と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号72と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号73と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号74と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号75と少なくとも85%、90%、95%、98%、99%の相同性があり、C407Sを含む配列番号76と少なくとも85%、90%、95%、98%、99%の相同性があり、C394Sを含む配列番号77と少なくとも85%、90%、95%、98%、99%の相同性があり、C412Sを含む配列番号78と少なくとも85%、90%、95%、98%、99%の相同性があり、前記ポリペプチドは、それらに相当するCys→Ser突然変異を欠如する親(野生型)タンパク質と比して、同等のセルラーゼ活性および高い熱安定性を有する、請求項1に記載の組み換え型ポリペプチド。
【請求項3】
以下に示すC→S置換位置以外で、1−10,10−20または20−30の保存的なアミノ酸の置換を有する、以下に示される配列を含む、組換え型ポリペプチド:C314Sを含む配列番号2;C311Sを含む配列番号4;C310Sを含む配列番号12;C312Sを含む配列番号13;C314Sを含む配列番号14;C315Sを含む配列番号15;C313Sを含む配列番号16;C311Sを含む配列番号17;C313Sを含む配列番号19;C312Sを含む配列番号21;C312Sを含む配列番号13;C311Sを含む配列番号22;C400Sを含む配列番号64;C407Sを含む配列番号65;C394Sを含む配列番号66;C400Sを含む配列番号67;C400Sを含む配列番号68;C400Sを含む配列番号69;C400Sを含む配列番号70;C400Sを含む配列番号71;C400Sを含む配列番号72;C400Sを含む配列番号73;C400Sを含む配列番号74;C400Sを含む配列番号75;C407Sを含む配列番号76;C394Sを含む配列番号77;またはC412Sを含む配列番号78。
【請求項4】
(a)配列番号2と少なくとも85%以上の相同性があり、位置314にSerを有し、セルラーゼ活性を有するポリペプチド;
(b)配列番号4と少なくとも70%以上の相同性があり、位置311にSerを有し、セルラーゼ活性を有するポリペプチド;
(c)(i)配列番号12で位置310にSerがある配列;(ii)配列番号13で位置312にSerがある配列;(iii)配列番号14で位置314にSerがある配列;(iv)配列番号15で位置315にSerがある配列;(v)配列番号16で位置313にSerがある配列;(vi)配列番号17で位置311にSerがある配列;(vii)配列番号19で位置313にSerがある配列;(viii)配列番号21で位置312にSerがある配列;(ix)配列番号22で位置311にSerがある配列からなる群から選択される配列と、少なくとも70%以上の相同性があり、位置314でSerを有するポリペプチド;
(d)2つの異なる親のセロビオヒドラーゼポリペプチド由来の少なくとも2つのドメインを含むキメラポリペプチドであり、ドメインはN末端側からC末端にかけて(断片1)−(断片2)−(断片3)−(断片4)−(断片5)−(断片6)−(断片7)−(断片8)を含み;
断片1は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約1から約x1までと少なくとも50−100%の相同性がある配列を含み;断片2は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x1から約x2までと少なくとも50−100%の相同性がある配列を含み;断片3は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x2から約x3までと少なくとも50−100%の相同性がある配列を含み;断片4は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x3から約x4までと少なくとも50−100%の相同性がある配列を含み;断片5は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x4から約x5までと少なくとも50−100%の相同性がある配列を含み;断片6は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x5から約x6までと少なくとも50−100%の相同性がある配列を含み;断片7は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x6から約x7までと少なくとも50−100%の相同性がある配列を含み;断片8は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x7から約x8までと少なくとも50−100%の相同性がある配列を含み;
x1は、配列番号2の残基43,44,45,46若しくは47、または配列番号4若しくは配列番号6の残基42,43,44,45若しくは46であり、x2は、配列番号2の残基70,71,72,73若しくは74、または配列番号4若しくは配列番号6の残基68,69,70,71,72,73若しくは74であり、x3は、配列番号2の残基113,114,115,116,117若しくは118、または配列番号4若しくは配列番号6の残基110,111,112,113,114,115若しくは116であり、x4は、配列番号2の残基153,154,155,156若しくは157、または配列番号4若しくは配列番号6の残基149,150,151,153,153,154若しくは155であり、x5は、配列番号2の残基220,221,222,223若しくは224、または配列番号4若しくは配列番号6の残基216,217,218,219,220,221,222若しくは223であり、x6は、配列番号2の残基256,257,258,259,260若しくは261、または配列番号4若しくは配列番号6の残基253,254,255,256,257,258,259若しくは260であり、x7は、配列番号2の残基312,313,314,315若しくは316、または配列番号4若しくは配列番号6の残基309,310,311,312,313,314,315または318であり;x8は、配列番号2、配列番号4または配列番号6を有するポリペプチドのC末端に相当するアミノ酸残基であり、
前記キメラポリペプチドが、配列番号2の位置314または配列番号4の位置311にSerを含み、配列番号2,4または6を含むCBH IIポリペプチドと比して、セルラーゼ活性および高い熱安定性および/またはpH安定性を有する、ポリペプチド;
からなる群から選択される配列を有する、
請求項1に記載の組み換え型ポリペプチド。
【請求項5】
前記断片1は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約1から約x1までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片2は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x1から約x2までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片3は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x2から約x3までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片4は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x3から約x4までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片5は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x4から約x5までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片6は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x5から約x6までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片7は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x6から約x7までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片8は、配列番号2の位置314、配列番号4の位置311または配列番号6の位置313を除く、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x7から約x8までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含む、請求項4に記載の組み換え型ポリペプチド。
【請求項6】
前記キメラポリペプチドは、配列番号12−62および63からなる群から選択される配列と、少なくとも80%、90%、95%、98%または99%の相同性がある、請求項4に記載の組み換え型ポリペプチド。
【請求項7】
前記ポリペプチドは、配列番号79−105または106と少なくとも85%の相同性を有し、配列番号107の配列のXの位置にセリンを有する配列を含む、請求項1に記載の組み換え型ポリペプチド。
【請求項8】
請求項1,2,3,4または7に記載のポリペプチドをコードするポリヌクレオチド。
【請求項9】
請求項8に記載のポリヌクレオチドを含むベクター。
【請求項10】
請求項8に記載のポリヌクレオチドを含む宿主細胞。
【請求項11】
請求項9に記載のベクターを含む宿主細胞。
【請求項12】
請求項1,2,3,4または7に記載のポリペプチドを含む酵素液。
【請求項13】
請求項10に記載の宿主細胞によって生成されるポリペプチドを含む酵素液。
【請求項14】
請求項1,2,3,4または7に記載のポリペプチドとバイオマスとの接触を備える、セルロース含有バイオマスの処理方法。
【請求項15】
請求項12に記載の酵素液とバイオマスとの接触を備える、セルロース含有バイオマスの処理方法。
【請求項1】
配列GEXDGを有するモチーフのC末端にセリンを含み、XはC、AまたはGであり(配列番号107)、Xはセリンと置換され、変異体は、セルラーゼ活性を有し、ポリペプチドは、配列GEXDG(配列番号107)においてセリンを欠如する野生型セルラーゼと比して高い熱安定性を有する、組み換え型ポリペプチド。
【請求項2】
前記ポリペプチドは、C314Sを含む配列番号2と少なくとも85%、90%、95%、98%、99%の相同性があり、C311Sを含む配列番号4と少なくとも85%、90%、95%、98%、99%の相同性があり、C310Sを含む配列番号12と少なくとも85%、90%、95%、98%、99%の相同性があり、C312Sを含む配列番号13と少なくとも85%、90%、95%、98%、99%の相同性があり、C314Sを含む配列番号14と少なくとも85%、90%、95%、98%、99%の相同性があり、C315Sを含む配列番号15と少なくとも85%、90%、95%、98%、99%の相同性があり、C313Sを含む配列番号16と少なくとも85%、90%、95%、98%、99%の相同性があり、C311Sを含む配列番号17と少なくとも85%、90%、95%、98%、99%の相同性があり、C313Sを含む配列番号19と少なくとも85%、90%、95%、98%、99%の相同性があり、C312Sを含む配列番号21と少なくとも85%、90%、95%、98%、99%の相同性があり、C311Sを含む配列番号22と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号64と少なくとも85%、90%、95%、98%、99%の相同性があり、C407Sを含む配列番号65と少なくとも85%、90%、95%、98%、99%の相同性があり、C394Sを含む配列番号66と少なくとも85%、90%、95%、98%、99%の相同性一致があり、C400Sを含む配列番号67と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号68と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号69と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号70と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号71と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号72と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号73と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号74と少なくとも85%、90%、95%、98%、99%の相同性があり、C400Sを含む配列番号75と少なくとも85%、90%、95%、98%、99%の相同性があり、C407Sを含む配列番号76と少なくとも85%、90%、95%、98%、99%の相同性があり、C394Sを含む配列番号77と少なくとも85%、90%、95%、98%、99%の相同性があり、C412Sを含む配列番号78と少なくとも85%、90%、95%、98%、99%の相同性があり、前記ポリペプチドは、それらに相当するCys→Ser突然変異を欠如する親(野生型)タンパク質と比して、同等のセルラーゼ活性および高い熱安定性を有する、請求項1に記載の組み換え型ポリペプチド。
【請求項3】
以下に示すC→S置換位置以外で、1−10,10−20または20−30の保存的なアミノ酸の置換を有する、以下に示される配列を含む、組換え型ポリペプチド:C314Sを含む配列番号2;C311Sを含む配列番号4;C310Sを含む配列番号12;C312Sを含む配列番号13;C314Sを含む配列番号14;C315Sを含む配列番号15;C313Sを含む配列番号16;C311Sを含む配列番号17;C313Sを含む配列番号19;C312Sを含む配列番号21;C312Sを含む配列番号13;C311Sを含む配列番号22;C400Sを含む配列番号64;C407Sを含む配列番号65;C394Sを含む配列番号66;C400Sを含む配列番号67;C400Sを含む配列番号68;C400Sを含む配列番号69;C400Sを含む配列番号70;C400Sを含む配列番号71;C400Sを含む配列番号72;C400Sを含む配列番号73;C400Sを含む配列番号74;C400Sを含む配列番号75;C407Sを含む配列番号76;C394Sを含む配列番号77;またはC412Sを含む配列番号78。
【請求項4】
(a)配列番号2と少なくとも85%以上の相同性があり、位置314にSerを有し、セルラーゼ活性を有するポリペプチド;
(b)配列番号4と少なくとも70%以上の相同性があり、位置311にSerを有し、セルラーゼ活性を有するポリペプチド;
(c)(i)配列番号12で位置310にSerがある配列;(ii)配列番号13で位置312にSerがある配列;(iii)配列番号14で位置314にSerがある配列;(iv)配列番号15で位置315にSerがある配列;(v)配列番号16で位置313にSerがある配列;(vi)配列番号17で位置311にSerがある配列;(vii)配列番号19で位置313にSerがある配列;(viii)配列番号21で位置312にSerがある配列;(ix)配列番号22で位置311にSerがある配列からなる群から選択される配列と、少なくとも70%以上の相同性があり、位置314でSerを有するポリペプチド;
(d)2つの異なる親のセロビオヒドラーゼポリペプチド由来の少なくとも2つのドメインを含むキメラポリペプチドであり、ドメインはN末端側からC末端にかけて(断片1)−(断片2)−(断片3)−(断片4)−(断片5)−(断片6)−(断片7)−(断片8)を含み;
断片1は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約1から約x1までと少なくとも50−100%の相同性がある配列を含み;断片2は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x1から約x2までと少なくとも50−100%の相同性がある配列を含み;断片3は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x2から約x3までと少なくとも50−100%の相同性がある配列を含み;断片4は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x3から約x4までと少なくとも50−100%の相同性がある配列を含み;断片5は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x4から約x5までと少なくとも50−100%の相同性がある配列を含み;断片6は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x5から約x6までと少なくとも50−100%の相同性がある配列を含み;断片7は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x6から約x7までと少なくとも50−100%の相同性がある配列を含み;断片8は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x7から約x8までと少なくとも50−100%の相同性がある配列を含み;
x1は、配列番号2の残基43,44,45,46若しくは47、または配列番号4若しくは配列番号6の残基42,43,44,45若しくは46であり、x2は、配列番号2の残基70,71,72,73若しくは74、または配列番号4若しくは配列番号6の残基68,69,70,71,72,73若しくは74であり、x3は、配列番号2の残基113,114,115,116,117若しくは118、または配列番号4若しくは配列番号6の残基110,111,112,113,114,115若しくは116であり、x4は、配列番号2の残基153,154,155,156若しくは157、または配列番号4若しくは配列番号6の残基149,150,151,153,153,154若しくは155であり、x5は、配列番号2の残基220,221,222,223若しくは224、または配列番号4若しくは配列番号6の残基216,217,218,219,220,221,222若しくは223であり、x6は、配列番号2の残基256,257,258,259,260若しくは261、または配列番号4若しくは配列番号6の残基253,254,255,256,257,258,259若しくは260であり、x7は、配列番号2の残基312,313,314,315若しくは316、または配列番号4若しくは配列番号6の残基309,310,311,312,313,314,315または318であり;x8は、配列番号2、配列番号4または配列番号6を有するポリペプチドのC末端に相当するアミノ酸残基であり、
前記キメラポリペプチドが、配列番号2の位置314または配列番号4の位置311にSerを含み、配列番号2,4または6を含むCBH IIポリペプチドと比して、セルラーゼ活性および高い熱安定性および/またはpH安定性を有する、ポリペプチド;
からなる群から選択される配列を有する、
請求項1に記載の組み換え型ポリペプチド。
【請求項5】
前記断片1は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約1から約x1までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片2は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x1から約x2までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片3は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x2から約x3までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片4は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x3から約x4までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片5は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x4から約x5までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片6は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x5から約x6までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片7は、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x6から約x7までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含み、前記断片8は、配列番号2の位置314、配列番号4の位置311または配列番号6の位置313を除く、配列番号2(「1」)、配列番号4(「2」)または配列番号6(「3」)の約x7から約x8までのアミノ酸残基および1−10の保存的なアミノ酸の置換を含む、請求項4に記載の組み換え型ポリペプチド。
【請求項6】
前記キメラポリペプチドは、配列番号12−62および63からなる群から選択される配列と、少なくとも80%、90%、95%、98%または99%の相同性がある、請求項4に記載の組み換え型ポリペプチド。
【請求項7】
前記ポリペプチドは、配列番号79−105または106と少なくとも85%の相同性を有し、配列番号107の配列のXの位置にセリンを有する配列を含む、請求項1に記載の組み換え型ポリペプチド。
【請求項8】
請求項1,2,3,4または7に記載のポリペプチドをコードするポリヌクレオチド。
【請求項9】
請求項8に記載のポリヌクレオチドを含むベクター。
【請求項10】
請求項8に記載のポリヌクレオチドを含む宿主細胞。
【請求項11】
請求項9に記載のベクターを含む宿主細胞。
【請求項12】
請求項1,2,3,4または7に記載のポリペプチドを含む酵素液。
【請求項13】
請求項10に記載の宿主細胞によって生成されるポリペプチドを含む酵素液。
【請求項14】
請求項1,2,3,4または7に記載のポリペプチドとバイオマスとの接触を備える、セルロース含有バイオマスの処理方法。
【請求項15】
請求項12に記載の酵素液とバイオマスとの接触を備える、セルロース含有バイオマスの処理方法。
【図3】

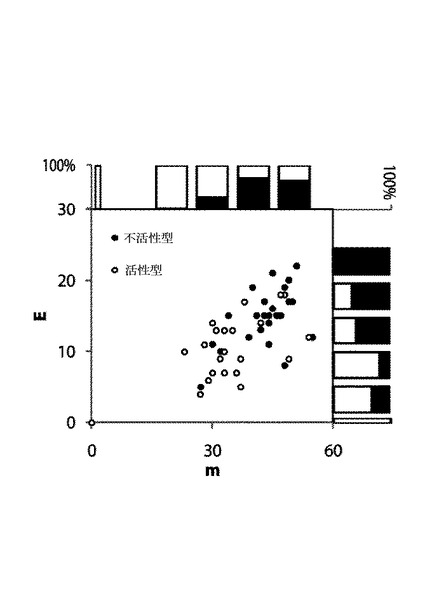
【図4】

【図5】
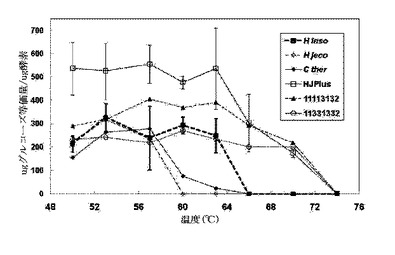
【図6】

【図7】
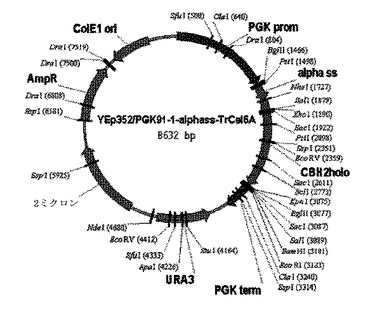
【図8】

【図9A】

【図9B】
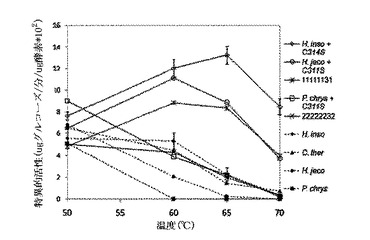
【図9C】

【図10】

【図11】
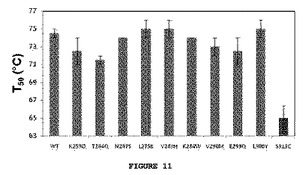
【図12】
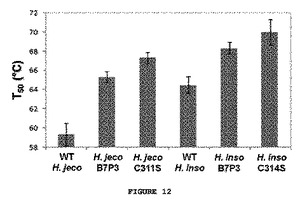
【図13】
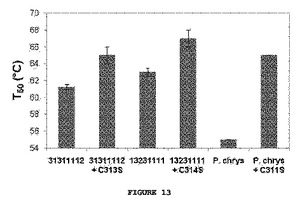
【図14A−D】

【図15】

【図4】
【図5】
【図6】
【図7】
【図8】
【図9A】
【図9B】
【図9C】
【図10】
【図11】
【図12】
【図13】
【図14A−D】
【図15】
【公表番号】特表2012−522521(P2012−522521A)
【公表日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願番号】特願2012−503780(P2012−503780)
【出願日】平成22年4月6日(2010.4.6)
【国際出願番号】PCT/US2010/030133
【国際公開番号】WO2010/118058
【国際公開日】平成22年10月14日(2010.10.14)
【出願人】(301059570)カリフォルニア インスティチュート オブ テクノロジー (14)
【Fターム(参考)】
【公表日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願日】平成22年4月6日(2010.4.6)
【国際出願番号】PCT/US2010/030133
【国際公開番号】WO2010/118058
【国際公開日】平成22年10月14日(2010.10.14)
【出願人】(301059570)カリフォルニア インスティチュート オブ テクノロジー (14)
【Fターム(参考)】
[ Back to top ]