
ベータ−ラクタマーゼの使用
クラスA ベータ-ラクタマーゼは、ベータ-ラクタム抗生物質およびベータ-ラクタマーゼ阻害剤の組み合わせでの抗生物質治療に付随する、腸における副作用を低減させるために使用することができる。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、腸管内の正常な微生物叢に対する抗生物質の有害作用を低減させることに関する。より正確には、本発明は、腸における副作用を低減させるための医薬を調製するための、クラスA ベータ-ラクタマーゼの使用に関する。腸における未吸収のベータ-ラクタム抗生物質の副作用を低減させる方法も開示される。
【背景技術】
【0002】
技術的背景
ベータ-ラクタム抗生物質は、細菌感染に対して最も広く用いられる抗生物質である。それらは全て、ベータ-ラクタム核を含有するという共通の構造的特徴を有する。ベータ-ラクタム抗生物質は細菌の細胞壁の生合成を阻害するが、一方で宿主に対しては非常に低い毒性しか有さない。しかし、ベータ-ラクタム治療に付随する一つの問題は、多くの細菌が、ベータ-ラクタム環のアミド結合を加水分解することによってベータ-ラクタム抗生物質を不活化することができるベータ-ラクタマーゼと称される酵素を産生することである。
【0003】
グラム陽性およびグラム陰性細菌のベータ-ラクタマーゼ産生株の有病率の増大が、ベータ-ラクタム抗生物質の有用性を制限してきた。それ故、ベータ-ラクタム抗生物質とベータ-ラクタマーゼ阻害剤の組み合わせを含有する医薬組成物が、ベータ-ラクタマーゼ産生細菌に依存しない効果的な治療を提供するために開発されてきた。既知の組み合わせは、例えばアモキシシリンとクラブラン酸、アンピシリンとスルバクタム、ピペラシリンとタゾバクタム、およびチカルシリンとクラブラン酸である(Higgins et al.、2004)。
【0004】
抗生物質処置に付随する別の問題は、抗生物質が腸管に到達すると、それらが腸の微生物叢に選択圧を及ぼすことによって抗生物質耐性を促進することである。経口的だけでなく非経口的に投与されたベータ-ラクタムも、恐らくはかなりの濃度で胆汁中に分泌されるため、腸管内の微生物叢の構成に対して有害作用を有し得る。それらは胆汁から腸内へ排出され、正常な腸管内ミクロフローラの崩壊を引き起こし得る。宿主と腸管内微生物叢の間の生態学的均衡の撹乱は、抗生物質に付随する下痢、病原性細菌、例えばバンコマイシン耐性腸球菌の異常増殖、ベータ-ラクタマーゼ産生グラム陰性桿菌の拡大、または正常な腸管内微生物叢もしくは病原体の間の抗生物質耐性の出現および蔓延につながり得る(Sullivan et al.、2001、Donskey、2006)。
【0005】
腸管内微生物叢の乱れ(disarrangements)を低減させるための一つの戦略は、非経口の抗生物質治療の間、最小の胆汁排泄を有する抗菌剤を選択することである(Rice et al.、2004)。別の戦略は、プロバイオティクスの使用を包含する。非病原性酵母サッカロマイセス・ボウラディ(Saccharomyces boulardii)および多数の乳酸発酵細菌、例えばラクトバチルス・ラムノサス(Lactobacillus rhamnosus) GG (LGG)を含む多くの異なるプロバイオティクスが、成人および小児における抗生物質に付随する下痢の予防および低減において評価されてきた。サッカロマイセス・ボウラディ(S. boulardii)処置は、成人において抗生物質に付随する下痢を繰り返す(diarrhea recurrent)クロストリジウム・ディフィシル(C. difficile)感染を防止すると考えられ、一方、LGG は小児における抗生物質に付随する下痢の処置において有用である(Katz、2006)。さらなる戦略にはウシの初乳に基づく免疫乳製品が包含され、それらは様々な抗生物質に付随する腸管内感染に対する予防法において効果的であることが証明されている(Korhonen et al.、2000)。
【0006】
腸におけるベータ-ラクタム抗生物質の有害作用を回避するためのよりさらなる戦略は、抗生物質とベータ-ラクタマーゼの同時投与である。ベータ-ラクタマーゼの経口投与は消化管における未吸収のベータ-ラクタムを不活化することを可能にし、それによって腸管内の正常な微生物叢の変化およびベータ-ラクタム耐性細菌の異常増殖を含む未吸収のベータ-ラクタムの副作用が低減される。ベータ-ラクタマーゼは、消化管の所望の部分において放出されるよう、使いやすく製剤化される(WO 93/13795)。
【0007】
イヌにおいて非経口アンピシリン治療と併用して経口投与されたベータ-ラクタマーゼは、血清中のアンピシリンレベルに影響を及ぼすことなく、胆汁排泄されたアンピシリンを用量依存的に分解することが示された(Harmoinen et al.、2003)。さらに、ベータ-ラクタマーゼ治療は、イヌモデルにおける非経口アンピシリンでの数日の処置の間、抗生物質に誘導される糞便の微生物叢の変化を妨げることも示された(Harmoinen et al.、2004)。ベータ-ラクタマーゼの、結腸に標的化された剤形を採用することによっても、同等の結果が得られている(US 2005/249716)。
【0008】
Harmoinen et al.、2003 and 2004 によって行われた研究において採用されたベータ-ラクタマーゼは、組換えのバチルス・リケニフォルミス(Bacillus licheniformis)のベータ-ラクタマーゼ(PenP)であり、Ambler のクラスA 酵素に属する (Ambler、1980)。それはペニシリン(penincillins)、アミノペニシリン、例えばアンピシリンおよびアモキシシリン、およびウレイドペニシリン、例えばピペラシリンに対する高い加水分解活性を有する。しかし、それは一般的なベータ-ラクタマーゼ阻害剤、例えばスルバクタム、クラブラン酸およびタゾバクタムによって容易に不活化される。
【0009】
ベータ-ラクタマーゼ阻害剤は、ベータ-ラクタマーゼ産生細菌によるベータ-ラクタムの不活化を防止するのに効果的である。そのため、ベータ-ラクタマーゼ阻害剤はベータ-ラクタムと併用され得る。一般的に、かかる組み合わせの構成成分はどちらも、体の様々な体液および組織についてのある程度類似した薬物動態パラメータおよびある程度類似した排出半減期を有し、この事は組み合わせ調製物の治療上有効性にとって必須の要件であると考えられる。しかし、胆汁排出に関しては、ベータ-ラクタムおよびベータ-ラクタマーゼ阻害剤の薬物動態特性は異なることが見出された。例えば、血清中におけるアンピシリンに対するスルバクタムの比はほぼ一定であることが見出されたが(およそ 1:2)、他方、胆汁中におけるスルバクタム/アンピシリンの比は、1:3 から 1:13 の範囲であった(Wildfeuer et al. 1988)。胆汁中における比の変動の大きさにもかかわらず、ベータ-ラクタムとベータ-ラクタマーゼ阻害剤との組み合わせは、胆道における感染に対する安全かつ効果的な治療であると考えられている(Morris et al.、1986.、Brogard et al.、1989、Westphal et al.、1997)。
【0010】
以上より、ベータ-ラクタム抗生物質の効果は、そうしなければ該抗生物質を不活化してしまうベータ-ラクタマーゼの効果を減少させるため、ベータ-ラクタム抗生物質をベータ-ラクタマーゼ阻害剤と併用することによって増強されたと結論づけられ得る。さらに、抗生物質処置の有害な副作用、例えば腸内の微生物叢の撹乱を減少させるための多くの方法が提案されている。有害な副作用を伴わない、より効果的な抗生物質処置に対する要求は未だ存在する。本発明はこれらのニーズに合致する。本発明は、ベータ-ラクタム抗生物質の使用に伴う重複感染のリスクおよび抗生物質耐性増大のリスクを低減させる。
【発明の概要】
【0011】
本発明の概要
本発明は、ベータ-ラクタマーゼ産生細菌による不活化に対し感受性でなく、かつ、腸内における正常な微生物学的フローラの平衡を乱さない、ベータ-ラクタム抗生物質治療に関する。ベータ-ラクタム抗生物質とベータ-ラクタマーゼ阻害剤の併用を伴う抗生物質処置に関連して、腸内に残存しているベータ-ラクタムを不活化するのにベータ-ラクタマーゼが効果的であることが現在判明した。これは驚くべき事であった; なぜなら、ベータ-ラクタムおよびその阻害剤が体から胆汁を介して小腸へと部分的に排出されること、および該阻害剤がインビトロでベータ-ラクタマーゼを不活化することが知られていたからである。
【0012】
本発明は、ベータ-ラクタム抗生物質とベータ-ラクタマーゼ阻害剤の併用を伴う処置に付随する、腸における副作用を低減させるための医薬を製造するためのクラスA ベータ-ラクタマーゼの使用を提供する。
【0013】
本発明は、ベータ-ラクタム抗生物質とベータ-ラクタマーゼ阻害剤の併用を伴う処置に付随する、腸における副作用を低減させる方法をさらに記載し、ここで、有効量のクラスA ベータ-ラクタマーゼがそれを必要とする対象に投与される。
【0014】
本発明の特定の態様は、従属クレームに記載される。本発明の他の目的、詳細および利点は、以下の図、詳細な説明および実施例から明らかになるであろう。
【図面の簡単な説明】
【0015】
【図1】図 1 は、分泌ベクター pKTH141 にクローン化されたバチルス・リケニフォルミス(Bacillus licheniformis)のベータ-ラクタマーゼ遺伝子のヌクレオチド配列および推定アミノ酸配列を示す。
【図2】図 2 は、ビーグル犬における、経口投与されたベータ-ラクタマーゼの非存在下または存在下でのアンピシリン/スルバクタムの組み合わせの非経口(parental)投与後の、空腸の消化粥内のアンピシリン濃度を示す。
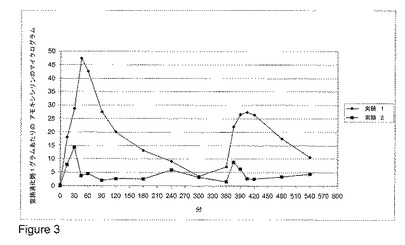
【図3】図 3 は、ビーグル犬における、経口投与されたベータ-ラクタマーゼの非存在下または存在下でのアモキシシリン/クラブラン酸の組み合わせの非経口(parental)投与後の、空腸の消化粥内のアモキシシリン濃度を示す。
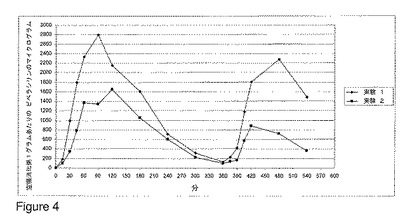
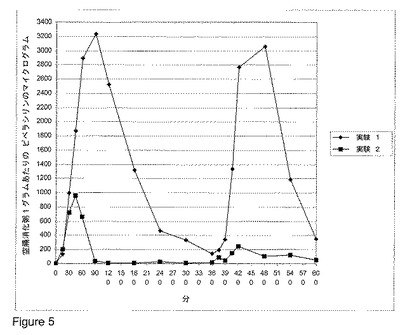
【図4】図 4 および 5 は、ビーグル犬における、様々な用量で経口投与されたベータ-ラクタマーゼの非存在下または存在下でのピペラシリン/タゾバクタムの組み合わせの非経口(parental)投与後の、空腸の消化粥内のピペラシリン濃度を示す。
【図5】図 4 および 5 は、ビーグル犬における、様々な用量で経口投与されたベータ-ラクタマーゼの非存在下または存在下でのピペラシリン/タゾバクタムの組み合わせの非経口(parental)投与後の、空腸の消化粥内のピペラシリン濃度を示す。
【発明を実施するための形態】
【0016】
本発明の詳細な説明
本発明は、ベータ-ラクタム抗生物質とベータ-ラクタマーゼ阻害剤の併用を伴う治療に由来する未吸収の残留ベータ-ラクタム抗生物質の腸管内微生物叢に対する有害作用を低減させるための医薬を調製するための、経口投与されるベータ-ラクタマーゼの使用に関する。経口投与されたベータ-ラクタマーゼの医薬組成物は、回腸の遠位部および結腸における主要な腸管内微生物叢に対するベータ-ラクタム/ベータ-ラクタマーゼ阻害剤の組み合わせの影響を減少させ、以下のように腸管内微生物叢の生態学的均衡を維持するよう意図される。したがって、ベータ-ラクタマーゼ治療を採用することにより、小腸および結腸内の未吸収の残留ベータ-ラクタム/ベータ-ラクタマーゼ阻害剤に付随する副作用が阻止される。
【0017】
ベータ-ラクタマーゼ
ベータ-ラクタマーゼは、EC 3.5.2.6 として分類されるベータ-ラクタム加水分解酵素である。ベータ-ラクタマーゼは、そのアミノ酸配列に基づいて A、B、C および D の4つのクラスにさらに分類される(Ambler、1980)。クラス A、C および D は、その活性部位中にセリン残基を有することから、セリン ベータ-ラクタマーゼとも称される。その一次構造とともに、基質の認識または触媒作用に重要な3つの保存されたペプチド配列が、3D 構造の比較によって同定されている(Colombo et al.、2004):
【表1】

【0018】
第1のエレメントは、全てのセリンベータ-ラクタマーゼの間で均一である。第1のエレメントは、その側鎖が活性部位の中へ向いている活性部位セリン(S)およびリジン(K)を含有している。第2のエレメントは、触媒的空洞(catalytic cavity)の1側面を形成する。それはクラスA ベータ-ラクタマーゼにおける SDN ループと称される。SDN ループは、少数の例外は別として、クラスA酵素の間でほぼ不変である。第3のエレメントは、ベータ-シートの最も内側の鎖上に存在し、触媒的空洞(catalytic cavity)の反対側の壁を形成する。それは一般的に KTG である。少数の例外的な場合において、リジン(K)はヒスチジン(H)またはアルギニン(R)によって置換されることがあり、いくつかのクラスA ベータ-ラクタマーゼにおいて、スレオニン(T)はセリン(S)によって置換されることがある(Matagne et al.、1998)。
【0019】
本発明のひとつの態様によれば、クラスA ベータ-ラクタマーゼはバチルス属の種に由来する。本発明の特定の態様によれば、クラスA ベータ-ラクタマーゼはバチルス・リケニフォルミス(Bacillus licheniformis)の PenP である。この酵素は、とりわけ Izui et al.、1980 によって記載されており、例えばバチルス・リケニフォルミス(B. licheniformis) 749/C (ATCC 25972)由来であり得る。749/C 株からの PenP のアミノ酸配列は、タンパク質配列データベース Swiss-Prot に配列番号 P00808 として記載されている。それはまた、本明細書において配列番号 1 として与えられる。対応する penP 遺伝子のヌクレオチド配列は、DDBJ/EMBL GenBank データベースにおいて配列 V00093 として与えられる。バチルス・リケニフォルミス(B. licheniformis)のベータ-ラクタマーゼはリポタンパクであり、タンパク質部分が膜の外側で折りたたまれるように脂肪酸尾部を介して該バチルスの細胞膜に固定されている。配列番号 1 は、26 アミノ酸の長いシグナル配列を含む該タンパク質の完全長アミノ酸配列を示す。この形は前駆体リポタンパクである。ジアシルグリセリドが位置 27 における NH2-末端のシステイン(C)に共有結合し、リポタンパク形態をもたらす。
【0020】
さらに、細胞外に分泌される該タンパク質のより短い形態が存在する。これらはエキソフォーム(exoforms)とも称される。該エキソフォーム(exoforms)は、細胞壁または培地中におけるプロテアーゼの加水分解活性の結果である。
【0021】
本明細書において用いる場合、“PenP”は、あらゆるベータ-ラクタマーゼの活性断片および/または明確に与えられるアミノ酸配列(配列番号 1)のバリアントを包含する。特にそれは、アミノ末端で切断されていることを意味する N-切断型の配列である。N-切断に加えて、それがベータ-ラクタマーゼ活性を有する限り、それは1以上のさらなるアミノ酸欠失、置換および/または挿入を含み得る。前記修飾は、自然に生じる変異または突然変異、または例えば遺伝子技術によって導入される人工的な修飾であり得る。異なったアミノ末端切断をされたエキソフォーム(exoforms)が、バチルス・リケニフォルミス(B. licheniformis)の増殖培地中において見出された。本明細書において、かかるエキソフォーム(exoforms)もまた PenP の用語によって包含される。Matagne et al.、1991 は、天然の宿主であるバチルス・リケニフォルミス(B. licheniformis) 749/C によって作り出される細胞外形態における、様々な程度の微小不均一性を記載している。異なる N-末端アミノ酸残基を有する、以下の5つの異なる分泌エキソフォーム(exoforms)が同定された:
SQPAEKNEKTEMKDD……KALNMNGK (アミノ酸 35-49...300-307)
EKTEMKDD……KALNMNGK (アミノ酸 42-49...300-307)
KTEMKDD……KALNMNGK (アミノ酸 43-49...300-307)
EMKDD……KALNMNGK (アミノ酸 45-49...300-307)
MKDD……KALNMNGK (アミノ酸 46-49...300-307)
【0022】
最初のアミノ酸残基は下線で表されている。C-末端のアミノ酸残基は右側に示される。アミノ酸の位置は配列番号 1 を参照する。位置 35 のセリン(S)から始まるエキソフォーム(exoform)は、バチルス・リケニフォルミス(B. licheniformis)のベータ-ラクタマーゼの“大分泌型”と称され、位置 43 のリジン(K)から始まるエキソフォームは“小分泌型”と称される。第1のアルファ へリックス(α1-へリックス)は位置 48 のアスパラギン酸(D)から始まり、最後のアルファ へリックス (α11-へリックス)の末端は位置 303 のアスパラギン(N)で終わる。本発明の一つの態様によれば、PenP は少なくとも、タンパク質の二次構造に関与する位置 48 から 303 のアミノ酸を含む(Knox et al.、1991)。本発明の別の態様によれば、1以上の前記位置 48 から 303 のアミノ酸は欠失しまたは置換されている。
【0023】
本発明のさらに別の態様によれば、PenP のアミノ末端は、NH2-KTEMKDD (配列番号 1 のアミノ酸 43-49)で始まる。このいわゆる ES-ベータ-L エキソフォーム(exoform)は、Gebhard et al.、2006 によって記載される通り、さらに 21 までの連続する残基を欠き得る。本発明の別の態様によれば、アミノ末端は配列番号 1 のグルタミン酸(E)で始まり、特にそれは NH2-EMKDD(配列番号 1 のアミノ酸 45-49)で始まるか、または NH2-MKDD (配列番号 1 のアミノ酸 46-49)で始まる。
【0024】
PenP タンパク質のカルボキシル末端における最後の4つのアミノ酸 MNGK-COOH は、二次構造の部分ではなく、そのため活性を失うことなく除去されることも可能である。別の態様において、9までの C-末端アミノ酸が除去され得る。C-切断型のタンパク質は Santos et al.、2004 によって記載されている。
【0025】
本明細書において用いる場合、ベータ-ラクタマーゼ活性を有するタンパク質の他の形態と共に、上で示されるベータ-ラクタマーゼの全ての異なる形態が PenP の用語によって包含される。本発明の1つの特定の態様によれば、ベータ-ラクタマーゼは、配列番号 1 またはそのベータ-ラクタマーゼ活性断片、特に位置 27 で始まるタンパク質の成熟した断片、および好ましくは配列番号 1 の位置 45 または 46 に対応する位置で始まるタンパク質の N-切断断片に対して少なくとも 40、50、60、70、80、90、95、97、98 または 99 % の配列同一性を有するアミノ酸配列を有する。配列同一性は、Altschul et al.、1997 に記載される通り、BLAST(Basic Local Alignment Search Tools)を用いて決定される。
【0026】
ベータ-ラクタマーゼ活性は、O'Callaghan et al.、1972 によって記載される通り、ニトロセフィンアッセイによって決定され得る。
【0027】
クラスA ベータ-ラクタマーゼは、組換えタンパク質として便利に生産される。好ましくは、それは医薬品生産に適したバチルス属の宿主株、例えばバチルス・アミロリクエファシエンス(B. amyloliquefaciens)、バチルス・プムリス(B. pumulis)、またはバチルス・スブティリス(B. subtilis)において生産される。非-胞子形成性のバチルス・スブティリス(B. subtilis)株においてベータ-ラクタマーゼを生産する一つの方法は、WO 03/040352 において記載されている。タンパク質は、過剰産生によってバチルス・リケニフォルミス(B. licheniformis)においても同様に(homologously)生産され得る。
【0028】
製剤
標的とするベータ-ラクタマーゼ製剤を得るため、ベータ-ラクタマーゼは腸溶性コーティングされた、経口投与される医薬組成物へと、例えば胃耐性のベータ-ラクタマーゼ ペレットとして都合良く製剤化される。本発明の一つの態様によれば、ベータ-ラクタマーゼは、例えば腸溶性コーティングされた硬いゼラチンカプセルに充填されたペレットとして都合良く投与される。腸溶性コーティングの剤形は、医薬産業における経口製品のうちでよく知られているものである。腸溶性コーティングを有する薬物製品は、インタクトな形で胃を通過(bypass)し、小腸、すなわち十二指腸、空腸および/または回腸において剤形の内容物を放出するよう設計される。腸溶性の固形剤を適用する理由は、薬剤を胃の酵素の破壊作用または胃の低 pH 環境から保護するため、または薬剤に誘導される胃粘膜の刺激、吐き気または出血を予防するため、または小腸内の標的部位に希釈されていない形で薬剤を送達するためである。これらの基準に基づけば、腸溶性コーティングされた薬物製品は遅延作用型の剤形と考えることができる。水に基づくコーティング形態は、親水性の PenP タンパク質のコーティング工程に最も好都合な材料であると思われる。腸溶性を獲得するために一般に用いられる水性ポリマーは、ポリメタクリレート、例えば Eudragit(登録商標)、セルロースに基づくポリマー、例えばセルロースエーテル、例えば Duodcell(登録商標)、またはセルロースエステル、例えば Aquateric(登録商標)、またはポリ酢酸ビニル コポリマー(copymers)例えば Opadry(登録商標) である。
【0029】
処置の方法
クラスA ベータ-ラクタマーゼは、ベータ-ラクタム抗生物質とベータ-ラクタマーゼ阻害剤の併用によって誘導される腸内における副作用を低減させるために用いられる。腸溶性コーティングされたベータ-ラクタマーゼは、腸内において、未吸収のベータ-ラクタム抗生物質を除去することが可能な量で放出され、それによって該抗生物質の有害作用が低減させられる。ベータ-ラクタマーゼは、例えば、下痢、病原性細菌、例えばバンコマイシン耐性腸球菌の異常増殖、ベータ-ラクタマーゼ産生グラム陰性桿菌の拡大、または正常な腸管内微生物叢または病原体の間での抗生物質耐性の出現および蔓延につながり得る、抗生物質に付随する宿主および腸管内微生物叢間の生態学的均衡の撹乱を減少させまたは阻止する。従って、ベータ-ラクタマーゼは、例えばクロストリジウム・ディフィシル(Clostridium difficile)および病原性酵母による重複感染を回避することを可能にし、このことは免疫抑制患者において特に重要である。標的とされ、腸溶性コーティングされたベータ-ラクタマーゼは、適切に非経口と併用して経口的に与えられるかあるいは経口投与される抗生物質およびベータ-ラクタマーゼ阻害剤である。ベータ-ラクタマーゼで処置される対象は、ヒトまたは動物、例えばベータ-ラクタム抗生物質とベータ-ラクタマーゼの阻害剤の併用で処置される家畜である。
【0030】
抗生物質および阻害剤
“ベータ-ラクタム抗生物質”は、4員のベータ-ラクタム(アゼチジン-2-オン)環を含有する抗菌性の化合物である。ベータ-ラクタム抗生物質は当該技術分野において周知であり、それらは天然、半合成または合成起源のものであり得る。ベータ-ラクタム抗生物質は一般に、そのさらなる構造的特徴に基づいて、ペニシリン、セファロスポリン、セファマイシン、オキサ-ベータ-ラクタム、カルバペネム、カルバセフェムおよびモノバクタムに分類され得る。好ましくは、抗生物質は非経口的に投与されるものである。ベータ-ラクタム抗生物質は、適切なベータ-ラクタマーゼ阻害剤と併用される。この目的に適した抗生物質は、例えばペニシリンであり、例えばペニシリン G、アモキシシリンおよびアンピシリンなどのアミノペニシリン、ピペラシリンなどのウレイドペニシリン、またはチカルシリンなどのアルファ-カルボキシペニシリンを含む。
【0031】
“ベータ-ラクタマーゼ阻害剤”は、ベータ-ラクタム抗生物質を加水分解することが可能なベータ-ラクタマーゼを阻害することができる化合物である。該阻害剤は一般的に、必ずしもそうではないが、ベータ-ラクタム抗生物質と構造的に関連しており、それ自体が弱い抗菌活性を有し得るが、組み合わせ治療におけるその機能は、存在している抗生物質を細菌のベータ-ラクタマーゼによる不活化から保護することである。本明細書において、阻害剤は特にクラスA ベータ-ラクタマーゼに対する阻害剤である。適切な阻害剤は、例えばスルバクタム、クラブラン酸およびタゾバクタムである。クラブラン酸は天然の類縁体であり、他方、スルバクタムおよびタゾバクタムは半合成のものである。ほとんどの阻害剤は非経口的に、すなわち静脈内にまたは筋肉内に投与される。クラブラン酸は、経口的にも投与され得る。いくつかのベータ-ラクタム抗生物質/ベータ-ラクタマーゼ阻害剤の組み合わせが当該技術分野において記載されており、臨床的に使用されている。
【0032】
抗生物質および前記阻害剤は、混合物として便利に投与される。市販のベータ-ラクタマーゼ阻害剤は、様々なベータ-ラクタムと組み合わせて臨床的に使用される。クラブラン酸はアモキシシリンまたはチカルシリンと組み合わせて使用され、同様に、スルバクタムはアンピシリンと、そしてタゾバクタムはピペラシリンと共に使用される。他の組み合わせもまた可能である。ベータ-ラクタマーゼは、抗生物質-阻害剤の組み合わせでの処置と同時に、またはその前に、経口投与され得る。好ましくは、それはベータ-ラクタム/阻害剤の組み合わせと同時に投与される。
【0033】
用量
腸管内微生物叢の撹乱の程度およびベータ-ラクタムとベータ-ラクタマーゼ阻害剤の組み合わせの投与による副作用の発生率は、薬剤の用量、投与の経路、およびベータ-ラクタムおよび前記阻害剤の薬物動態学的/動力学的特性を含む様々な要因に依存している。ベータ-ラクタマーゼは、小腸および結腸内において残存している未吸収のベータ-ラクタムに付随する副作用を低減させるのに有効な量で投与される。実施された実験において、約 0.1 mg のベータ-ラクタマーゼ/kg体重の用量が、空腸の消化粥中においてアンピシリンおよびアモキシシリンを検出限界を下回る濃度にまで除去するのに効果的であったが、他方、ピペラシリンを除去するのにはより高い用量が必要である。適切な用量は、0.1 - 1.0、特に 0.2 - 0.4 mg のベータ-ラクタマーゼ/kg体重であり得る。
【0034】
本発明は、以下の非限定的な実施例によってさらに説明される。しかし、上記の説明および実施例において提供される態様は説明を目的とするだけであること、および本発明の範囲内において様々な変更および修正が可能であることが理解されるべきである。試験結果は、ベータ-ラクタム/ベータ-ラクタマーゼ阻害剤治療に関して、未吸収のベータ-ラクタムに対するベータ-ラクタマーゼの予測できない効果を示す。結果は、ベータ-ラクタムとベータ-ラクタマーゼ阻害剤の組み合わせでの抗生物質治療に対するバチルス・リケニフォルミス(Bacillus licheniformis)のベータ-ラクタマーゼの使用の拡大を支持する。
【実施例】
【0035】
実施例 1
バチルス・リケニフォルミス(Bacillus licheniformis) 749/C 由来の組換えベータ-ラクタマーゼを、実験において用いた。タンパク質を、WO 03/040352 に記載される通りに、バチルス・スブティリス(Bacillus subtilis)の非-胞子形成性の株において生産した。
【0036】
バチルス・アミロリクエファシエンス(B. amyloliquefaciens) E18 のアミラーゼ遺伝子(amyQ)の強力な増殖期プロモーター(vegetative promoter)(amyQp)、リボソーム結合部位(RBS)、およびシグナル配列コード領域(amyQss)を保有する発現カセットを含む分泌ベクター pKTH141 を用いた。加えて、単一の HindIII 部位を有する合成オリゴヌクレオチドを、シグナル配列コード領域の 3'-末端に直接挿入した。このため、外来性タンパク質をコードする挿入断片を、アルファ-アミラーゼのシグナル配列と同じ読み枠で翻訳されるよう、HindIII 部位内にクローン化することができる。該 HindIII オリゴヌクレオチドは3つのアミノ酸残基 (NH2-QAS)をコードし、それは成熟タンパク質の NH2-末端の伸長を構成すると期待される。
【0037】
配列番号 1 の連続するアミノ酸残基 43-307 をコードする、バチルス・リケニフォルミス(Bacillus licheniformis)のベータ-ラクタマーゼ構造遺伝子(penP)を、鋳型としてバチルス・リケニフォルミス(B. licheniformis)の染色体 DNA を用いて、HindIII 制限部位を含有する適切なプライマーでの PCR によって増幅した。続いて、増幅された断片を HindIII で切断し、pKTH141 の対応する部位へ連結した結果、AmyQ シグナルペプチドをコードする配列と PenP タンパク質をコードする配列の間の枠融合がもたらされた。ベータ-ラクタマーゼ遺伝子のヌクレオチド配列を、自動 DNA シークエンサーでジデオキシ-鎖終結法によって決定した。組換えのバチルス・リケニフォルミス(B. licheniformis) 749/C のベータ-ラクタマーゼ遺伝子の完全ヌクレオチド配列および推定アミノ酸配列を配列番号 2 および 3 として表し、図 1 に示す。
【0038】
図 1 において、線の下のおよび括弧内に示される番号は、アミノ酸残基を参照する。NH2-QAS 伸長をコードする HindIII クローニング部位は、ヌクレオチド配列の上に示される。予測上のシグナルペプチダーゼ切断部位は、-31 の位置におけるアラニンの後ろである。
【0039】
オープンリーディングフレームは、amyQ 遺伝子の 31 アミノ酸残基の長いシグナル配列を有する 299 アミノ酸のポリペプチドをコードする。シグナルペプチダーゼの切断部位は、-1 の位置におけるアラニンの後ろに位置すると予測されている。成熟ベータ-ラクタマーゼは、位置 +1 におけるグルタミン(Q)から始まると期待された。従って、成熟ベータ-ラクタマーゼは、その NH2-QAS 伸長が HindIII クローニング部位によってコードされる 268 アミノ酸残基を含有すると期待された。
【0040】
精製された組換えベータ-ラクタマーゼの NH2-末端の配列を、タンパク質配列決定装置(sequenator)を用いて自動化エドマン分解によって決定した。解析によって、組換えベータ-ラクタマーゼはその推定アミノ末端における NH2-QASKT-ペンタペプチドを欠いていることが明らかになった。該結果は、組換えベータ-ラクタマーゼタンパク質の切断型が、細菌の細胞壁中および培地中の両方に存在するタンパク質分解酵素の翻訳後作用によって生成されることを示す。結論として、精製された組換えベータ-ラクタマーゼの大部分は 263 アミノ酸残基を構成し、29.3 kDa の分子質量を有する。決定されたアミノ末端の配列は、推定アミノ酸配列より5アミノ酸残基下流から始まる。精製された組換えベータ-ラクタマーゼの最初のアミノ酸残基は、図 1 における位置 +6 のグルタミン酸(E)である。
【0041】
該精製された酵素タンパク質を、P1A と命名する。それは本質的に(少なくとも約 95 重量-% は)配列番号 1 の連続するアミノ酸残基 45 から 307 より成る。残りは、本質的に配列番号 1 の連続するアミノ酸残基 46 から 307 より成る。該ベータ-ラクタマーゼを、Harmoinen et al.、2004 によって行われた研究において利用されたペレットと本質的に同様の、腸溶性コーティングされたペレットの形態で投与した。
【0042】
バチルス・リケニフォルミス(B. licheniformis)のベータ-ラクタマーゼが有する、アンピシリン-スルバクタムの組み合わせでの非経口治療の間に胆汁排泄されたアンピシリンを小腸中において除去する能力を、イヌモデルにおいて調査した。分析用サンプルの収集を可能にするため、実験用ビーグル犬の、幽門に対しおよそ 170 cm 遠位の空腸内に、ニップル弁(nipple valve)を外科的に挿入した。腸管手術は、腸管運動を変化させなかった。研究の間を通して、6頭のビーグル犬を利用した。研究を、2つの連続する処置として行った: 第1の実験において、2回の連続用量(consecutive doses)のアンピシリンとスルバクタムの組み合わせ(体重 kg あたり 40 mg のアンピシリンおよび 20 mg のスルバクタム)を、給餌後 6 時間 20 分の投与間隔で静脈内に投与した。7日後、同じ犬がアンピシリン/スルバクタムの注入の10分前にさらにベータ-ラクタマーゼを経口投与されたこと以外は第1の実験と同様である第2の実験を行った。体重kgあたり約 0.1 mg の活性なベータ-ラクタマーゼを含有する腸溶性コーティングされたペレットの単回投与量(single dose)を用いた。
【0043】
空腸の消化粥サンプルを様々な時点で収集した。消化粥サンプルを直ぐに凍結させ、分析を待つため -70℃で保存した。該消化粥サンプルを、固相抽出によって精製した。アンピシリンの定量化のため、UV 検出での逆相高速液体クロマトグラフィー(HPLC)法を行った。
【0044】
得られた結果は、ベータ-ラクタマーゼ治療を伴わずに実施された第1の実験において高レベルのアンピシリンが空腸のサンプル中で検出されることを示した一方、第2の実験は、経口投与されたベータ-ラクタマーゼが、空腸のアンピシリンレベルを定量限界(空腸の消化粥1グラムあたり 10 マイクログラムのアンピシリン)より下へ減少させることができることを示した。
【0045】
図 2 は、アンピシリン/スルバクタムの組み合わせ(体重1kgあたり 40 mg のアンピシリンおよび 20 mg のスルバクタム)の静脈内投与後におけるビーグル犬(n=6)の空腸消化粥中のアンピシリン濃度に対する、経口投与されたベータ-ラクタマーゼ ペレット(体重1kgあたり約 0.1 mg の活性なベータ-ラクタマーゼの用量)の効果を示す。両方の実験についての値は、様々な時点での平均の空腸アンピシリン濃度として表される。実験 1 におけるアンピシリンの値は、ベータ-ラクタマーゼ処置を伴わずに、6 時間の投与間隔でアンピシリン/スルバクタムを2回別々に投与した後に達成された、空腸のアンピシリン濃度を表す。実験 2 において、同時にベータ-ラクタマーゼ治療を伴うアンピシリン/スルバクタムの組み合わせでビーグル犬を処理した。採用された用量のベータ-ラクタマーゼは、第1のアンピシリン/スルバクタム処置の間におけるビーグル犬の空腸のアンピシリンの大部分を除去することができ、濃度は低下し、同時のベータ-ラクタマーゼ治療を伴う第2のアンピシリン/スルバクタム処置の間を通して定量レベルよりも低いままであった。
【0046】
該結果は、胆汁排泄された残余のベータ-ラクタマーゼ阻害剤が、ベータ-ラクタマーゼの活性に対して限られた影響しか有さないことを示す。
【0047】
実施例 2
アモキシシリンとクラブラン酸の組み合わせでの非経口治療の間に胆汁排泄されたアモキシシリンを不活化する、バチルス・リケニフォルミス(B. licheniformis)のベータ-ラクタマーゼ P1A の有効性を、アモキシシリン/クラブラン酸の組み合わせの単回投与量(single dose)が体重1kgあたり 25 mg のアモキシシリンおよび 5 mg のクラブラン酸を含有したこと以外は実施例 1 と本質的に同様にして調査し、アモキシシリンの分析に適したものとなるよう HPLC 分析法を作り上げた(was elaborated) (定量限界は空腸の消化粥1グラムあたり 2 マイクログラムであった)。
【0048】
得られた結果を図 3 に示し、それはアモキシシリン/クラブラン酸の組み合わせ(体重1kgあたり 25 mg のアモキシシリンおよび 5 mg のクラブラン酸)の静脈内投与後におけるビーグル犬(n=6)の空腸消化粥中のアモキシシリン濃度に対する、経口投与されたベータ-ラクタマーゼ ペレットの有効性を示す。両方の実験についての値を、様々な時点での平均の空腸アモキシシリン濃度として表す。実験 1 におけるアモキシシリンの値は、ベータ-ラクタマーゼ処置を伴わずに 6 時間の投与間隔でアモキシシリン/クラブラン酸を2回別々に投与した後に達成された空腸のアモキシシリン濃度を示す。実験 2 において、経口のベータ-ラクタマーゼ処置を、アモキシシリン/クラブラン酸の組み合わせの非経口治療と併用した。
【0049】
ベータ-ラクタマーゼ処置は、アモキシシリン/クラブラン酸の組み合わせでの非経口治療の間、胆汁排泄されたアモキシシリンの大部分を除去することができるという事が見出された。様々な時点でいくつかの空腸サンプルにおいて見出されるアモキシシリンの痕跡(traces)は、ベータ-ラクタマーゼの用量を増大させることによって除去することができる。結果は、バチルス・リケニフォルミス(B. licheniformis)のベータ-ラクタマーゼが、非経口のアモキシシリン/クラブラン酸の使用に関連する副作用を低減させるための原薬(drug substance)としての有力な候補であることを示唆する。
【0050】
実施例 3
同時のベータ-ラクタマーゼ治療を伴わずにおよび伴って、ピペラシリンおよびタゾバクタムの組み合わせでビーグル犬を処置した。該実験は、ピペラシリン/タゾバクタムの組み合わせの単回投与量(single dose)が体重1kgあたり 100 mg のピペラシリンおよび 12.5 mg のタゾバクタムを含有したこと以外は本質的に実施例 1 および 2 に記載された通りに行い、ピペラシリンの分析に適したものとなるよう HPLC 分析法を作り上げた(was elaborated)(定量限界は、空腸消化粥1グラムあたり 10 マイクログラムであった)。
【0051】
結果を図 4 に示し、それは、ピペラシリン/タゾバクタムの組み合わせ(体重1kgあたり 100 mg のピペラシリンおよび 12.5 mg のタゾバクタム)の静脈内投与後におけるビーグル犬(n=6)の空腸消化粥中のピペラシリン濃度に対する、経口投与されたベータ-ラクタマーゼ ペレットの有効性を示す。両方の実験についての値を、様々な時点での平均の空腸ピペラシリン濃度として表す。実験 1 におけるピペラシリンの値は、ベータ-ラクタマーゼ処置を伴わずに、6 時間の投与間隔でピペラシリン/タゾバクタムを2回別々に投与した後に達成された、空腸のピペラシリン濃度を示す。実験 2 において、同時のベータ-ラクタマーゼ治療と共に、ピペラシリン/タゾバクタムの組み合わせでビーグル犬を処置した。
【0052】
ベータ-ラクタマーゼを用いずに得られた結果(実験 1)は、ビーグル犬におけるピペラシリンの胆汁排出が、アンピシリンまたはアモキシシリンの胆汁排出よりもかなり高いことを示した。それにもかかわらず、ベータ-ラクタマーゼ処置は全ての時点において空腸のピペラシリン濃度を減少させた。しかし、ベータ-ラクタマーゼ処置の間中、ピペラシリン濃度は検出可能なままであった(実験 2)。従って、得られた結果により、ベータ-ラクタマーゼ治療はピペラシリン/タゾバクタムの組み合わせでの非経口治療の間に空腸のピペラシリンを除去することができるが、空腸のピペラシリン濃度を定量限界より下へ除去することができる処方(dosage formulation)を達成するためには、腸溶性コーティングされたペレットにおけるベータ-ラクタマーゼの量を増加させるべきであることが示された。
【0053】
ベータ-ラクタマーゼ ペレットの単回投与量(single dose)が体重1kgあたり約 0.3 mg の活性なベータ-ラクタマーゼを含有し、ピペラシリン/タゾバクタムの組み合わせの単回投与量(single dose)が体重1kgあたり 65.6 mg のピペラシリンおよび 9.4 mg のタゾバクタムを含有したことを除いて、前記実験を繰り返した。結果を図 5 に示し、それは、空腸のピペラシリンを除去するのにベータ-ラクタマーゼが非常に効果的であったことを示す。
【0054】
参考文献
Altschul S.F., Madden T.L., Schaffer A.A., Zhang J., Zhang Z., Miller W., Lipman D.J. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25: 3389-3402
Ambler, R.P. 1980. The structure of beta-lactamases. Philos. Trans. R. Soc. London B Biol. Sci 289:321-331
Brogard, J.M., Jehl, F., Blickle,J.F., Adloff M., Donner, M., and H. Montell. 1989. Biliary elimination of ticarcillin plus clavulanic acid (Claventin): experimental and clinical study. Int. J. Clin. Pharmacol. Ther. Toxicol. 27:135-144.
Colombo, M., L., Hanique, S., Baurin, S.L., Bauvois, C., De Vriendt, K., Van Beeumen, J.J., Frere, J.M., and B. Joris. 2004. The ybxI gene of Bacillus subtilis 168 encodes a class D beta-lactamase of low activity. Antimicrob. Agents Chemother.48:484-490
Donskey, C.J., 2006. Antibiotic regimens and intestinal colonization with antibiotic-resistamt Gram-negative bacilli. Clin. Infect. Dis. 43:62-69.
Harmoinen, J., Mentula, S., Heikkila, M., van der Rest, M., Rajala-Schultz, P.J., Donskey, C.J., Frias, R., Koski, P., Wickstrand, N., Jousimies-Somer, H., Westermarck, E., and K. Lindevall. 2004. Orally administered targeted recombinant beta-lactamase prevents ampicillin-induced selective pressure on the gut microbiota: a novel approach to reducing antimicrobial resistance. Antimicrob. Agents Chemother. 48:75-79
Harmoinen, J. Vaali, K., Koski, P., Syrjanen, K., Laitinen, O., Lindevall, K., and E. Westermarck. 2003. Enzymatic degradation of a β-lactam antibiotic, ampicillin, in the gut: a novel treatment modality. J.Antimicrob. Chemother. 51:287-292.
Izui, K., J.B.K. Nielsen, M.P. Gaulifield, and J.O. Lampen. 1980. Large exopenicillinase, initial extracellular form detected in growths of Bacillus licheniformis. Biochemistry 19:1882-1886
J.A. Katz. 2006. Probiotics for prevention of antibiotic-associated diarrhea and Clostridium difficile diarrhea. J. Clin. Gastroenterol. 40:249-255.
Gebhard L.G., Rrisso V.A. Sanots J., Ferreyra R.G., Noguera M.E. and Ermacora M.R., 2006. Mapping the distribution of conformational information throughout a protein sequence. J. Mol. Biol. 21:358(1)280-288
Higgins, P.G., Wisplinghoff, H., Stefanik, D., and H. Seifert. 2004. In Vitro activities of the β-lactamase inhibitors clavulanic acid, sulbactam, and tazobactam alone or in combination with β-lactams against epidemiologically characterized multidrug-resistant Acinetobacter baumannii strains. 48: 1586-1592
Knox, J.R., and P.C. Moews. 1991. Beta-lactamase of Bacillus licheniformis 749/C. Refinement at 2 Å resolution and analysis of hydration. J.Mol.Biol. 220:435-455
Korhonen, H., Marnila, P., and H.S. Gill. 2000. Bovine milk antibodies for health. Br J Nutr. 84: 135-146
Matagne, A., Joris, B., van Beeumen J., and J-M. Frere. 1991. Ragged N-termini and other variants of class A beta-lactamases analysed by chromatofocusing. Biochem. J. 273: 503-510
Matagne, A., Lamotte-Brasseur, J., and J.M. Frere. 1998. Catalytic properties of class A beta-lactamases: efficiency and diversity. Biochem. J. 330: 581-598
Morris, D.L., Ubhi, C.S., Robertson, C.S., and K.W. Brammer. 1986. Biliary pharmacokinetics of sulbactam in humans. Rev.Infect.Dis. 8:589-592.
O'Callaghan, C. H., Morris, A., Kirby, S.M., and A. H. Singler. 1972. Novel method for detection of beta-lactamases by using a chromogenic cephalosporin substrate. Antimicrob. Agents Chemother. 1:283-288
Rice, L.B., Hutton-Thomas,R., Lakticova, V., Helfand, M.S., and C.J. Donskey.2004. Beta-lactam antibiotics and gastrointestinal colonization of vancomycin-resistant enterococci. J.Infect.Dis. 189:1113-1118.
Santos, J, Gebhard L.G., Risso V.A., Fereyra R.G., Rossi J.P. and Ermacora M.R. 2004. Folding of an abridged beta-lactamase. Biochemistry 43(6):1715-123
Sullivan, Å., Edlund, C., and Nord, C.E. 2001. Effect of antimicrobial agents on the ecological balance of human microflora. Lancet 1:101-114.
Westphal, J.F., Brogard, J.M., Caro-Sampara, F., Adloff, M., Blickle, J.F., Monteil, H., and F. Jehl. 1997. Assessment of biliary excretion of piperacillin-tazobactam in humans. Antimicrob. Agents Chemother. 41:1636-1640.
Wildfeuer, A., Schwiersch, U., Engel, K., von Castell, E., Schilling, A., Potempa, J., and H. Lenders. 1988. Pharmacokinetics of sulbactam and ampicillin intravenously applied in combination to healthy volunteers and patients. Determination of the ratio of the two drugs in serum and in various tissues. Arzneimittelforschung. 38:1640-1643.
【技術分野】
【0001】
発明の分野
本発明は、腸管内の正常な微生物叢に対する抗生物質の有害作用を低減させることに関する。より正確には、本発明は、腸における副作用を低減させるための医薬を調製するための、クラスA ベータ-ラクタマーゼの使用に関する。腸における未吸収のベータ-ラクタム抗生物質の副作用を低減させる方法も開示される。
【背景技術】
【0002】
技術的背景
ベータ-ラクタム抗生物質は、細菌感染に対して最も広く用いられる抗生物質である。それらは全て、ベータ-ラクタム核を含有するという共通の構造的特徴を有する。ベータ-ラクタム抗生物質は細菌の細胞壁の生合成を阻害するが、一方で宿主に対しては非常に低い毒性しか有さない。しかし、ベータ-ラクタム治療に付随する一つの問題は、多くの細菌が、ベータ-ラクタム環のアミド結合を加水分解することによってベータ-ラクタム抗生物質を不活化することができるベータ-ラクタマーゼと称される酵素を産生することである。
【0003】
グラム陽性およびグラム陰性細菌のベータ-ラクタマーゼ産生株の有病率の増大が、ベータ-ラクタム抗生物質の有用性を制限してきた。それ故、ベータ-ラクタム抗生物質とベータ-ラクタマーゼ阻害剤の組み合わせを含有する医薬組成物が、ベータ-ラクタマーゼ産生細菌に依存しない効果的な治療を提供するために開発されてきた。既知の組み合わせは、例えばアモキシシリンとクラブラン酸、アンピシリンとスルバクタム、ピペラシリンとタゾバクタム、およびチカルシリンとクラブラン酸である(Higgins et al.、2004)。
【0004】
抗生物質処置に付随する別の問題は、抗生物質が腸管に到達すると、それらが腸の微生物叢に選択圧を及ぼすことによって抗生物質耐性を促進することである。経口的だけでなく非経口的に投与されたベータ-ラクタムも、恐らくはかなりの濃度で胆汁中に分泌されるため、腸管内の微生物叢の構成に対して有害作用を有し得る。それらは胆汁から腸内へ排出され、正常な腸管内ミクロフローラの崩壊を引き起こし得る。宿主と腸管内微生物叢の間の生態学的均衡の撹乱は、抗生物質に付随する下痢、病原性細菌、例えばバンコマイシン耐性腸球菌の異常増殖、ベータ-ラクタマーゼ産生グラム陰性桿菌の拡大、または正常な腸管内微生物叢もしくは病原体の間の抗生物質耐性の出現および蔓延につながり得る(Sullivan et al.、2001、Donskey、2006)。
【0005】
腸管内微生物叢の乱れ(disarrangements)を低減させるための一つの戦略は、非経口の抗生物質治療の間、最小の胆汁排泄を有する抗菌剤を選択することである(Rice et al.、2004)。別の戦略は、プロバイオティクスの使用を包含する。非病原性酵母サッカロマイセス・ボウラディ(Saccharomyces boulardii)および多数の乳酸発酵細菌、例えばラクトバチルス・ラムノサス(Lactobacillus rhamnosus) GG (LGG)を含む多くの異なるプロバイオティクスが、成人および小児における抗生物質に付随する下痢の予防および低減において評価されてきた。サッカロマイセス・ボウラディ(S. boulardii)処置は、成人において抗生物質に付随する下痢を繰り返す(diarrhea recurrent)クロストリジウム・ディフィシル(C. difficile)感染を防止すると考えられ、一方、LGG は小児における抗生物質に付随する下痢の処置において有用である(Katz、2006)。さらなる戦略にはウシの初乳に基づく免疫乳製品が包含され、それらは様々な抗生物質に付随する腸管内感染に対する予防法において効果的であることが証明されている(Korhonen et al.、2000)。
【0006】
腸におけるベータ-ラクタム抗生物質の有害作用を回避するためのよりさらなる戦略は、抗生物質とベータ-ラクタマーゼの同時投与である。ベータ-ラクタマーゼの経口投与は消化管における未吸収のベータ-ラクタムを不活化することを可能にし、それによって腸管内の正常な微生物叢の変化およびベータ-ラクタム耐性細菌の異常増殖を含む未吸収のベータ-ラクタムの副作用が低減される。ベータ-ラクタマーゼは、消化管の所望の部分において放出されるよう、使いやすく製剤化される(WO 93/13795)。
【0007】
イヌにおいて非経口アンピシリン治療と併用して経口投与されたベータ-ラクタマーゼは、血清中のアンピシリンレベルに影響を及ぼすことなく、胆汁排泄されたアンピシリンを用量依存的に分解することが示された(Harmoinen et al.、2003)。さらに、ベータ-ラクタマーゼ治療は、イヌモデルにおける非経口アンピシリンでの数日の処置の間、抗生物質に誘導される糞便の微生物叢の変化を妨げることも示された(Harmoinen et al.、2004)。ベータ-ラクタマーゼの、結腸に標的化された剤形を採用することによっても、同等の結果が得られている(US 2005/249716)。
【0008】
Harmoinen et al.、2003 and 2004 によって行われた研究において採用されたベータ-ラクタマーゼは、組換えのバチルス・リケニフォルミス(Bacillus licheniformis)のベータ-ラクタマーゼ(PenP)であり、Ambler のクラスA 酵素に属する (Ambler、1980)。それはペニシリン(penincillins)、アミノペニシリン、例えばアンピシリンおよびアモキシシリン、およびウレイドペニシリン、例えばピペラシリンに対する高い加水分解活性を有する。しかし、それは一般的なベータ-ラクタマーゼ阻害剤、例えばスルバクタム、クラブラン酸およびタゾバクタムによって容易に不活化される。
【0009】
ベータ-ラクタマーゼ阻害剤は、ベータ-ラクタマーゼ産生細菌によるベータ-ラクタムの不活化を防止するのに効果的である。そのため、ベータ-ラクタマーゼ阻害剤はベータ-ラクタムと併用され得る。一般的に、かかる組み合わせの構成成分はどちらも、体の様々な体液および組織についてのある程度類似した薬物動態パラメータおよびある程度類似した排出半減期を有し、この事は組み合わせ調製物の治療上有効性にとって必須の要件であると考えられる。しかし、胆汁排出に関しては、ベータ-ラクタムおよびベータ-ラクタマーゼ阻害剤の薬物動態特性は異なることが見出された。例えば、血清中におけるアンピシリンに対するスルバクタムの比はほぼ一定であることが見出されたが(およそ 1:2)、他方、胆汁中におけるスルバクタム/アンピシリンの比は、1:3 から 1:13 の範囲であった(Wildfeuer et al. 1988)。胆汁中における比の変動の大きさにもかかわらず、ベータ-ラクタムとベータ-ラクタマーゼ阻害剤との組み合わせは、胆道における感染に対する安全かつ効果的な治療であると考えられている(Morris et al.、1986.、Brogard et al.、1989、Westphal et al.、1997)。
【0010】
以上より、ベータ-ラクタム抗生物質の効果は、そうしなければ該抗生物質を不活化してしまうベータ-ラクタマーゼの効果を減少させるため、ベータ-ラクタム抗生物質をベータ-ラクタマーゼ阻害剤と併用することによって増強されたと結論づけられ得る。さらに、抗生物質処置の有害な副作用、例えば腸内の微生物叢の撹乱を減少させるための多くの方法が提案されている。有害な副作用を伴わない、より効果的な抗生物質処置に対する要求は未だ存在する。本発明はこれらのニーズに合致する。本発明は、ベータ-ラクタム抗生物質の使用に伴う重複感染のリスクおよび抗生物質耐性増大のリスクを低減させる。
【発明の概要】
【0011】
本発明の概要
本発明は、ベータ-ラクタマーゼ産生細菌による不活化に対し感受性でなく、かつ、腸内における正常な微生物学的フローラの平衡を乱さない、ベータ-ラクタム抗生物質治療に関する。ベータ-ラクタム抗生物質とベータ-ラクタマーゼ阻害剤の併用を伴う抗生物質処置に関連して、腸内に残存しているベータ-ラクタムを不活化するのにベータ-ラクタマーゼが効果的であることが現在判明した。これは驚くべき事であった; なぜなら、ベータ-ラクタムおよびその阻害剤が体から胆汁を介して小腸へと部分的に排出されること、および該阻害剤がインビトロでベータ-ラクタマーゼを不活化することが知られていたからである。
【0012】
本発明は、ベータ-ラクタム抗生物質とベータ-ラクタマーゼ阻害剤の併用を伴う処置に付随する、腸における副作用を低減させるための医薬を製造するためのクラスA ベータ-ラクタマーゼの使用を提供する。
【0013】
本発明は、ベータ-ラクタム抗生物質とベータ-ラクタマーゼ阻害剤の併用を伴う処置に付随する、腸における副作用を低減させる方法をさらに記載し、ここで、有効量のクラスA ベータ-ラクタマーゼがそれを必要とする対象に投与される。
【0014】
本発明の特定の態様は、従属クレームに記載される。本発明の他の目的、詳細および利点は、以下の図、詳細な説明および実施例から明らかになるであろう。
【図面の簡単な説明】
【0015】
【図1】図 1 は、分泌ベクター pKTH141 にクローン化されたバチルス・リケニフォルミス(Bacillus licheniformis)のベータ-ラクタマーゼ遺伝子のヌクレオチド配列および推定アミノ酸配列を示す。
【図2】図 2 は、ビーグル犬における、経口投与されたベータ-ラクタマーゼの非存在下または存在下でのアンピシリン/スルバクタムの組み合わせの非経口(parental)投与後の、空腸の消化粥内のアンピシリン濃度を示す。
【図3】図 3 は、ビーグル犬における、経口投与されたベータ-ラクタマーゼの非存在下または存在下でのアモキシシリン/クラブラン酸の組み合わせの非経口(parental)投与後の、空腸の消化粥内のアモキシシリン濃度を示す。
【図4】図 4 および 5 は、ビーグル犬における、様々な用量で経口投与されたベータ-ラクタマーゼの非存在下または存在下でのピペラシリン/タゾバクタムの組み合わせの非経口(parental)投与後の、空腸の消化粥内のピペラシリン濃度を示す。
【図5】図 4 および 5 は、ビーグル犬における、様々な用量で経口投与されたベータ-ラクタマーゼの非存在下または存在下でのピペラシリン/タゾバクタムの組み合わせの非経口(parental)投与後の、空腸の消化粥内のピペラシリン濃度を示す。
【発明を実施するための形態】
【0016】
本発明の詳細な説明
本発明は、ベータ-ラクタム抗生物質とベータ-ラクタマーゼ阻害剤の併用を伴う治療に由来する未吸収の残留ベータ-ラクタム抗生物質の腸管内微生物叢に対する有害作用を低減させるための医薬を調製するための、経口投与されるベータ-ラクタマーゼの使用に関する。経口投与されたベータ-ラクタマーゼの医薬組成物は、回腸の遠位部および結腸における主要な腸管内微生物叢に対するベータ-ラクタム/ベータ-ラクタマーゼ阻害剤の組み合わせの影響を減少させ、以下のように腸管内微生物叢の生態学的均衡を維持するよう意図される。したがって、ベータ-ラクタマーゼ治療を採用することにより、小腸および結腸内の未吸収の残留ベータ-ラクタム/ベータ-ラクタマーゼ阻害剤に付随する副作用が阻止される。
【0017】
ベータ-ラクタマーゼ
ベータ-ラクタマーゼは、EC 3.5.2.6 として分類されるベータ-ラクタム加水分解酵素である。ベータ-ラクタマーゼは、そのアミノ酸配列に基づいて A、B、C および D の4つのクラスにさらに分類される(Ambler、1980)。クラス A、C および D は、その活性部位中にセリン残基を有することから、セリン ベータ-ラクタマーゼとも称される。その一次構造とともに、基質の認識または触媒作用に重要な3つの保存されたペプチド配列が、3D 構造の比較によって同定されている(Colombo et al.、2004):
【表1】
【0018】
第1のエレメントは、全てのセリンベータ-ラクタマーゼの間で均一である。第1のエレメントは、その側鎖が活性部位の中へ向いている活性部位セリン(S)およびリジン(K)を含有している。第2のエレメントは、触媒的空洞(catalytic cavity)の1側面を形成する。それはクラスA ベータ-ラクタマーゼにおける SDN ループと称される。SDN ループは、少数の例外は別として、クラスA酵素の間でほぼ不変である。第3のエレメントは、ベータ-シートの最も内側の鎖上に存在し、触媒的空洞(catalytic cavity)の反対側の壁を形成する。それは一般的に KTG である。少数の例外的な場合において、リジン(K)はヒスチジン(H)またはアルギニン(R)によって置換されることがあり、いくつかのクラスA ベータ-ラクタマーゼにおいて、スレオニン(T)はセリン(S)によって置換されることがある(Matagne et al.、1998)。
【0019】
本発明のひとつの態様によれば、クラスA ベータ-ラクタマーゼはバチルス属の種に由来する。本発明の特定の態様によれば、クラスA ベータ-ラクタマーゼはバチルス・リケニフォルミス(Bacillus licheniformis)の PenP である。この酵素は、とりわけ Izui et al.、1980 によって記載されており、例えばバチルス・リケニフォルミス(B. licheniformis) 749/C (ATCC 25972)由来であり得る。749/C 株からの PenP のアミノ酸配列は、タンパク質配列データベース Swiss-Prot に配列番号 P00808 として記載されている。それはまた、本明細書において配列番号 1 として与えられる。対応する penP 遺伝子のヌクレオチド配列は、DDBJ/EMBL GenBank データベースにおいて配列 V00093 として与えられる。バチルス・リケニフォルミス(B. licheniformis)のベータ-ラクタマーゼはリポタンパクであり、タンパク質部分が膜の外側で折りたたまれるように脂肪酸尾部を介して該バチルスの細胞膜に固定されている。配列番号 1 は、26 アミノ酸の長いシグナル配列を含む該タンパク質の完全長アミノ酸配列を示す。この形は前駆体リポタンパクである。ジアシルグリセリドが位置 27 における NH2-末端のシステイン(C)に共有結合し、リポタンパク形態をもたらす。
【0020】
さらに、細胞外に分泌される該タンパク質のより短い形態が存在する。これらはエキソフォーム(exoforms)とも称される。該エキソフォーム(exoforms)は、細胞壁または培地中におけるプロテアーゼの加水分解活性の結果である。
【0021】
本明細書において用いる場合、“PenP”は、あらゆるベータ-ラクタマーゼの活性断片および/または明確に与えられるアミノ酸配列(配列番号 1)のバリアントを包含する。特にそれは、アミノ末端で切断されていることを意味する N-切断型の配列である。N-切断に加えて、それがベータ-ラクタマーゼ活性を有する限り、それは1以上のさらなるアミノ酸欠失、置換および/または挿入を含み得る。前記修飾は、自然に生じる変異または突然変異、または例えば遺伝子技術によって導入される人工的な修飾であり得る。異なったアミノ末端切断をされたエキソフォーム(exoforms)が、バチルス・リケニフォルミス(B. licheniformis)の増殖培地中において見出された。本明細書において、かかるエキソフォーム(exoforms)もまた PenP の用語によって包含される。Matagne et al.、1991 は、天然の宿主であるバチルス・リケニフォルミス(B. licheniformis) 749/C によって作り出される細胞外形態における、様々な程度の微小不均一性を記載している。異なる N-末端アミノ酸残基を有する、以下の5つの異なる分泌エキソフォーム(exoforms)が同定された:
SQPAEKNEKTEMKDD……KALNMNGK (アミノ酸 35-49...300-307)
EKTEMKDD……KALNMNGK (アミノ酸 42-49...300-307)
KTEMKDD……KALNMNGK (アミノ酸 43-49...300-307)
EMKDD……KALNMNGK (アミノ酸 45-49...300-307)
MKDD……KALNMNGK (アミノ酸 46-49...300-307)
【0022】
最初のアミノ酸残基は下線で表されている。C-末端のアミノ酸残基は右側に示される。アミノ酸の位置は配列番号 1 を参照する。位置 35 のセリン(S)から始まるエキソフォーム(exoform)は、バチルス・リケニフォルミス(B. licheniformis)のベータ-ラクタマーゼの“大分泌型”と称され、位置 43 のリジン(K)から始まるエキソフォームは“小分泌型”と称される。第1のアルファ へリックス(α1-へリックス)は位置 48 のアスパラギン酸(D)から始まり、最後のアルファ へリックス (α11-へリックス)の末端は位置 303 のアスパラギン(N)で終わる。本発明の一つの態様によれば、PenP は少なくとも、タンパク質の二次構造に関与する位置 48 から 303 のアミノ酸を含む(Knox et al.、1991)。本発明の別の態様によれば、1以上の前記位置 48 から 303 のアミノ酸は欠失しまたは置換されている。
【0023】
本発明のさらに別の態様によれば、PenP のアミノ末端は、NH2-KTEMKDD (配列番号 1 のアミノ酸 43-49)で始まる。このいわゆる ES-ベータ-L エキソフォーム(exoform)は、Gebhard et al.、2006 によって記載される通り、さらに 21 までの連続する残基を欠き得る。本発明の別の態様によれば、アミノ末端は配列番号 1 のグルタミン酸(E)で始まり、特にそれは NH2-EMKDD(配列番号 1 のアミノ酸 45-49)で始まるか、または NH2-MKDD (配列番号 1 のアミノ酸 46-49)で始まる。
【0024】
PenP タンパク質のカルボキシル末端における最後の4つのアミノ酸 MNGK-COOH は、二次構造の部分ではなく、そのため活性を失うことなく除去されることも可能である。別の態様において、9までの C-末端アミノ酸が除去され得る。C-切断型のタンパク質は Santos et al.、2004 によって記載されている。
【0025】
本明細書において用いる場合、ベータ-ラクタマーゼ活性を有するタンパク質の他の形態と共に、上で示されるベータ-ラクタマーゼの全ての異なる形態が PenP の用語によって包含される。本発明の1つの特定の態様によれば、ベータ-ラクタマーゼは、配列番号 1 またはそのベータ-ラクタマーゼ活性断片、特に位置 27 で始まるタンパク質の成熟した断片、および好ましくは配列番号 1 の位置 45 または 46 に対応する位置で始まるタンパク質の N-切断断片に対して少なくとも 40、50、60、70、80、90、95、97、98 または 99 % の配列同一性を有するアミノ酸配列を有する。配列同一性は、Altschul et al.、1997 に記載される通り、BLAST(Basic Local Alignment Search Tools)を用いて決定される。
【0026】
ベータ-ラクタマーゼ活性は、O'Callaghan et al.、1972 によって記載される通り、ニトロセフィンアッセイによって決定され得る。
【0027】
クラスA ベータ-ラクタマーゼは、組換えタンパク質として便利に生産される。好ましくは、それは医薬品生産に適したバチルス属の宿主株、例えばバチルス・アミロリクエファシエンス(B. amyloliquefaciens)、バチルス・プムリス(B. pumulis)、またはバチルス・スブティリス(B. subtilis)において生産される。非-胞子形成性のバチルス・スブティリス(B. subtilis)株においてベータ-ラクタマーゼを生産する一つの方法は、WO 03/040352 において記載されている。タンパク質は、過剰産生によってバチルス・リケニフォルミス(B. licheniformis)においても同様に(homologously)生産され得る。
【0028】
製剤
標的とするベータ-ラクタマーゼ製剤を得るため、ベータ-ラクタマーゼは腸溶性コーティングされた、経口投与される医薬組成物へと、例えば胃耐性のベータ-ラクタマーゼ ペレットとして都合良く製剤化される。本発明の一つの態様によれば、ベータ-ラクタマーゼは、例えば腸溶性コーティングされた硬いゼラチンカプセルに充填されたペレットとして都合良く投与される。腸溶性コーティングの剤形は、医薬産業における経口製品のうちでよく知られているものである。腸溶性コーティングを有する薬物製品は、インタクトな形で胃を通過(bypass)し、小腸、すなわち十二指腸、空腸および/または回腸において剤形の内容物を放出するよう設計される。腸溶性の固形剤を適用する理由は、薬剤を胃の酵素の破壊作用または胃の低 pH 環境から保護するため、または薬剤に誘導される胃粘膜の刺激、吐き気または出血を予防するため、または小腸内の標的部位に希釈されていない形で薬剤を送達するためである。これらの基準に基づけば、腸溶性コーティングされた薬物製品は遅延作用型の剤形と考えることができる。水に基づくコーティング形態は、親水性の PenP タンパク質のコーティング工程に最も好都合な材料であると思われる。腸溶性を獲得するために一般に用いられる水性ポリマーは、ポリメタクリレート、例えば Eudragit(登録商標)、セルロースに基づくポリマー、例えばセルロースエーテル、例えば Duodcell(登録商標)、またはセルロースエステル、例えば Aquateric(登録商標)、またはポリ酢酸ビニル コポリマー(copymers)例えば Opadry(登録商標) である。
【0029】
処置の方法
クラスA ベータ-ラクタマーゼは、ベータ-ラクタム抗生物質とベータ-ラクタマーゼ阻害剤の併用によって誘導される腸内における副作用を低減させるために用いられる。腸溶性コーティングされたベータ-ラクタマーゼは、腸内において、未吸収のベータ-ラクタム抗生物質を除去することが可能な量で放出され、それによって該抗生物質の有害作用が低減させられる。ベータ-ラクタマーゼは、例えば、下痢、病原性細菌、例えばバンコマイシン耐性腸球菌の異常増殖、ベータ-ラクタマーゼ産生グラム陰性桿菌の拡大、または正常な腸管内微生物叢または病原体の間での抗生物質耐性の出現および蔓延につながり得る、抗生物質に付随する宿主および腸管内微生物叢間の生態学的均衡の撹乱を減少させまたは阻止する。従って、ベータ-ラクタマーゼは、例えばクロストリジウム・ディフィシル(Clostridium difficile)および病原性酵母による重複感染を回避することを可能にし、このことは免疫抑制患者において特に重要である。標的とされ、腸溶性コーティングされたベータ-ラクタマーゼは、適切に非経口と併用して経口的に与えられるかあるいは経口投与される抗生物質およびベータ-ラクタマーゼ阻害剤である。ベータ-ラクタマーゼで処置される対象は、ヒトまたは動物、例えばベータ-ラクタム抗生物質とベータ-ラクタマーゼの阻害剤の併用で処置される家畜である。
【0030】
抗生物質および阻害剤
“ベータ-ラクタム抗生物質”は、4員のベータ-ラクタム(アゼチジン-2-オン)環を含有する抗菌性の化合物である。ベータ-ラクタム抗生物質は当該技術分野において周知であり、それらは天然、半合成または合成起源のものであり得る。ベータ-ラクタム抗生物質は一般に、そのさらなる構造的特徴に基づいて、ペニシリン、セファロスポリン、セファマイシン、オキサ-ベータ-ラクタム、カルバペネム、カルバセフェムおよびモノバクタムに分類され得る。好ましくは、抗生物質は非経口的に投与されるものである。ベータ-ラクタム抗生物質は、適切なベータ-ラクタマーゼ阻害剤と併用される。この目的に適した抗生物質は、例えばペニシリンであり、例えばペニシリン G、アモキシシリンおよびアンピシリンなどのアミノペニシリン、ピペラシリンなどのウレイドペニシリン、またはチカルシリンなどのアルファ-カルボキシペニシリンを含む。
【0031】
“ベータ-ラクタマーゼ阻害剤”は、ベータ-ラクタム抗生物質を加水分解することが可能なベータ-ラクタマーゼを阻害することができる化合物である。該阻害剤は一般的に、必ずしもそうではないが、ベータ-ラクタム抗生物質と構造的に関連しており、それ自体が弱い抗菌活性を有し得るが、組み合わせ治療におけるその機能は、存在している抗生物質を細菌のベータ-ラクタマーゼによる不活化から保護することである。本明細書において、阻害剤は特にクラスA ベータ-ラクタマーゼに対する阻害剤である。適切な阻害剤は、例えばスルバクタム、クラブラン酸およびタゾバクタムである。クラブラン酸は天然の類縁体であり、他方、スルバクタムおよびタゾバクタムは半合成のものである。ほとんどの阻害剤は非経口的に、すなわち静脈内にまたは筋肉内に投与される。クラブラン酸は、経口的にも投与され得る。いくつかのベータ-ラクタム抗生物質/ベータ-ラクタマーゼ阻害剤の組み合わせが当該技術分野において記載されており、臨床的に使用されている。
【0032】
抗生物質および前記阻害剤は、混合物として便利に投与される。市販のベータ-ラクタマーゼ阻害剤は、様々なベータ-ラクタムと組み合わせて臨床的に使用される。クラブラン酸はアモキシシリンまたはチカルシリンと組み合わせて使用され、同様に、スルバクタムはアンピシリンと、そしてタゾバクタムはピペラシリンと共に使用される。他の組み合わせもまた可能である。ベータ-ラクタマーゼは、抗生物質-阻害剤の組み合わせでの処置と同時に、またはその前に、経口投与され得る。好ましくは、それはベータ-ラクタム/阻害剤の組み合わせと同時に投与される。
【0033】
用量
腸管内微生物叢の撹乱の程度およびベータ-ラクタムとベータ-ラクタマーゼ阻害剤の組み合わせの投与による副作用の発生率は、薬剤の用量、投与の経路、およびベータ-ラクタムおよび前記阻害剤の薬物動態学的/動力学的特性を含む様々な要因に依存している。ベータ-ラクタマーゼは、小腸および結腸内において残存している未吸収のベータ-ラクタムに付随する副作用を低減させるのに有効な量で投与される。実施された実験において、約 0.1 mg のベータ-ラクタマーゼ/kg体重の用量が、空腸の消化粥中においてアンピシリンおよびアモキシシリンを検出限界を下回る濃度にまで除去するのに効果的であったが、他方、ピペラシリンを除去するのにはより高い用量が必要である。適切な用量は、0.1 - 1.0、特に 0.2 - 0.4 mg のベータ-ラクタマーゼ/kg体重であり得る。
【0034】
本発明は、以下の非限定的な実施例によってさらに説明される。しかし、上記の説明および実施例において提供される態様は説明を目的とするだけであること、および本発明の範囲内において様々な変更および修正が可能であることが理解されるべきである。試験結果は、ベータ-ラクタム/ベータ-ラクタマーゼ阻害剤治療に関して、未吸収のベータ-ラクタムに対するベータ-ラクタマーゼの予測できない効果を示す。結果は、ベータ-ラクタムとベータ-ラクタマーゼ阻害剤の組み合わせでの抗生物質治療に対するバチルス・リケニフォルミス(Bacillus licheniformis)のベータ-ラクタマーゼの使用の拡大を支持する。
【実施例】
【0035】
実施例 1
バチルス・リケニフォルミス(Bacillus licheniformis) 749/C 由来の組換えベータ-ラクタマーゼを、実験において用いた。タンパク質を、WO 03/040352 に記載される通りに、バチルス・スブティリス(Bacillus subtilis)の非-胞子形成性の株において生産した。
【0036】
バチルス・アミロリクエファシエンス(B. amyloliquefaciens) E18 のアミラーゼ遺伝子(amyQ)の強力な増殖期プロモーター(vegetative promoter)(amyQp)、リボソーム結合部位(RBS)、およびシグナル配列コード領域(amyQss)を保有する発現カセットを含む分泌ベクター pKTH141 を用いた。加えて、単一の HindIII 部位を有する合成オリゴヌクレオチドを、シグナル配列コード領域の 3'-末端に直接挿入した。このため、外来性タンパク質をコードする挿入断片を、アルファ-アミラーゼのシグナル配列と同じ読み枠で翻訳されるよう、HindIII 部位内にクローン化することができる。該 HindIII オリゴヌクレオチドは3つのアミノ酸残基 (NH2-QAS)をコードし、それは成熟タンパク質の NH2-末端の伸長を構成すると期待される。
【0037】
配列番号 1 の連続するアミノ酸残基 43-307 をコードする、バチルス・リケニフォルミス(Bacillus licheniformis)のベータ-ラクタマーゼ構造遺伝子(penP)を、鋳型としてバチルス・リケニフォルミス(B. licheniformis)の染色体 DNA を用いて、HindIII 制限部位を含有する適切なプライマーでの PCR によって増幅した。続いて、増幅された断片を HindIII で切断し、pKTH141 の対応する部位へ連結した結果、AmyQ シグナルペプチドをコードする配列と PenP タンパク質をコードする配列の間の枠融合がもたらされた。ベータ-ラクタマーゼ遺伝子のヌクレオチド配列を、自動 DNA シークエンサーでジデオキシ-鎖終結法によって決定した。組換えのバチルス・リケニフォルミス(B. licheniformis) 749/C のベータ-ラクタマーゼ遺伝子の完全ヌクレオチド配列および推定アミノ酸配列を配列番号 2 および 3 として表し、図 1 に示す。
【0038】
図 1 において、線の下のおよび括弧内に示される番号は、アミノ酸残基を参照する。NH2-QAS 伸長をコードする HindIII クローニング部位は、ヌクレオチド配列の上に示される。予測上のシグナルペプチダーゼ切断部位は、-31 の位置におけるアラニンの後ろである。
【0039】
オープンリーディングフレームは、amyQ 遺伝子の 31 アミノ酸残基の長いシグナル配列を有する 299 アミノ酸のポリペプチドをコードする。シグナルペプチダーゼの切断部位は、-1 の位置におけるアラニンの後ろに位置すると予測されている。成熟ベータ-ラクタマーゼは、位置 +1 におけるグルタミン(Q)から始まると期待された。従って、成熟ベータ-ラクタマーゼは、その NH2-QAS 伸長が HindIII クローニング部位によってコードされる 268 アミノ酸残基を含有すると期待された。
【0040】
精製された組換えベータ-ラクタマーゼの NH2-末端の配列を、タンパク質配列決定装置(sequenator)を用いて自動化エドマン分解によって決定した。解析によって、組換えベータ-ラクタマーゼはその推定アミノ末端における NH2-QASKT-ペンタペプチドを欠いていることが明らかになった。該結果は、組換えベータ-ラクタマーゼタンパク質の切断型が、細菌の細胞壁中および培地中の両方に存在するタンパク質分解酵素の翻訳後作用によって生成されることを示す。結論として、精製された組換えベータ-ラクタマーゼの大部分は 263 アミノ酸残基を構成し、29.3 kDa の分子質量を有する。決定されたアミノ末端の配列は、推定アミノ酸配列より5アミノ酸残基下流から始まる。精製された組換えベータ-ラクタマーゼの最初のアミノ酸残基は、図 1 における位置 +6 のグルタミン酸(E)である。
【0041】
該精製された酵素タンパク質を、P1A と命名する。それは本質的に(少なくとも約 95 重量-% は)配列番号 1 の連続するアミノ酸残基 45 から 307 より成る。残りは、本質的に配列番号 1 の連続するアミノ酸残基 46 から 307 より成る。該ベータ-ラクタマーゼを、Harmoinen et al.、2004 によって行われた研究において利用されたペレットと本質的に同様の、腸溶性コーティングされたペレットの形態で投与した。
【0042】
バチルス・リケニフォルミス(B. licheniformis)のベータ-ラクタマーゼが有する、アンピシリン-スルバクタムの組み合わせでの非経口治療の間に胆汁排泄されたアンピシリンを小腸中において除去する能力を、イヌモデルにおいて調査した。分析用サンプルの収集を可能にするため、実験用ビーグル犬の、幽門に対しおよそ 170 cm 遠位の空腸内に、ニップル弁(nipple valve)を外科的に挿入した。腸管手術は、腸管運動を変化させなかった。研究の間を通して、6頭のビーグル犬を利用した。研究を、2つの連続する処置として行った: 第1の実験において、2回の連続用量(consecutive doses)のアンピシリンとスルバクタムの組み合わせ(体重 kg あたり 40 mg のアンピシリンおよび 20 mg のスルバクタム)を、給餌後 6 時間 20 分の投与間隔で静脈内に投与した。7日後、同じ犬がアンピシリン/スルバクタムの注入の10分前にさらにベータ-ラクタマーゼを経口投与されたこと以外は第1の実験と同様である第2の実験を行った。体重kgあたり約 0.1 mg の活性なベータ-ラクタマーゼを含有する腸溶性コーティングされたペレットの単回投与量(single dose)を用いた。
【0043】
空腸の消化粥サンプルを様々な時点で収集した。消化粥サンプルを直ぐに凍結させ、分析を待つため -70℃で保存した。該消化粥サンプルを、固相抽出によって精製した。アンピシリンの定量化のため、UV 検出での逆相高速液体クロマトグラフィー(HPLC)法を行った。
【0044】
得られた結果は、ベータ-ラクタマーゼ治療を伴わずに実施された第1の実験において高レベルのアンピシリンが空腸のサンプル中で検出されることを示した一方、第2の実験は、経口投与されたベータ-ラクタマーゼが、空腸のアンピシリンレベルを定量限界(空腸の消化粥1グラムあたり 10 マイクログラムのアンピシリン)より下へ減少させることができることを示した。
【0045】
図 2 は、アンピシリン/スルバクタムの組み合わせ(体重1kgあたり 40 mg のアンピシリンおよび 20 mg のスルバクタム)の静脈内投与後におけるビーグル犬(n=6)の空腸消化粥中のアンピシリン濃度に対する、経口投与されたベータ-ラクタマーゼ ペレット(体重1kgあたり約 0.1 mg の活性なベータ-ラクタマーゼの用量)の効果を示す。両方の実験についての値は、様々な時点での平均の空腸アンピシリン濃度として表される。実験 1 におけるアンピシリンの値は、ベータ-ラクタマーゼ処置を伴わずに、6 時間の投与間隔でアンピシリン/スルバクタムを2回別々に投与した後に達成された、空腸のアンピシリン濃度を表す。実験 2 において、同時にベータ-ラクタマーゼ治療を伴うアンピシリン/スルバクタムの組み合わせでビーグル犬を処理した。採用された用量のベータ-ラクタマーゼは、第1のアンピシリン/スルバクタム処置の間におけるビーグル犬の空腸のアンピシリンの大部分を除去することができ、濃度は低下し、同時のベータ-ラクタマーゼ治療を伴う第2のアンピシリン/スルバクタム処置の間を通して定量レベルよりも低いままであった。
【0046】
該結果は、胆汁排泄された残余のベータ-ラクタマーゼ阻害剤が、ベータ-ラクタマーゼの活性に対して限られた影響しか有さないことを示す。
【0047】
実施例 2
アモキシシリンとクラブラン酸の組み合わせでの非経口治療の間に胆汁排泄されたアモキシシリンを不活化する、バチルス・リケニフォルミス(B. licheniformis)のベータ-ラクタマーゼ P1A の有効性を、アモキシシリン/クラブラン酸の組み合わせの単回投与量(single dose)が体重1kgあたり 25 mg のアモキシシリンおよび 5 mg のクラブラン酸を含有したこと以外は実施例 1 と本質的に同様にして調査し、アモキシシリンの分析に適したものとなるよう HPLC 分析法を作り上げた(was elaborated) (定量限界は空腸の消化粥1グラムあたり 2 マイクログラムであった)。
【0048】
得られた結果を図 3 に示し、それはアモキシシリン/クラブラン酸の組み合わせ(体重1kgあたり 25 mg のアモキシシリンおよび 5 mg のクラブラン酸)の静脈内投与後におけるビーグル犬(n=6)の空腸消化粥中のアモキシシリン濃度に対する、経口投与されたベータ-ラクタマーゼ ペレットの有効性を示す。両方の実験についての値を、様々な時点での平均の空腸アモキシシリン濃度として表す。実験 1 におけるアモキシシリンの値は、ベータ-ラクタマーゼ処置を伴わずに 6 時間の投与間隔でアモキシシリン/クラブラン酸を2回別々に投与した後に達成された空腸のアモキシシリン濃度を示す。実験 2 において、経口のベータ-ラクタマーゼ処置を、アモキシシリン/クラブラン酸の組み合わせの非経口治療と併用した。
【0049】
ベータ-ラクタマーゼ処置は、アモキシシリン/クラブラン酸の組み合わせでの非経口治療の間、胆汁排泄されたアモキシシリンの大部分を除去することができるという事が見出された。様々な時点でいくつかの空腸サンプルにおいて見出されるアモキシシリンの痕跡(traces)は、ベータ-ラクタマーゼの用量を増大させることによって除去することができる。結果は、バチルス・リケニフォルミス(B. licheniformis)のベータ-ラクタマーゼが、非経口のアモキシシリン/クラブラン酸の使用に関連する副作用を低減させるための原薬(drug substance)としての有力な候補であることを示唆する。
【0050】
実施例 3
同時のベータ-ラクタマーゼ治療を伴わずにおよび伴って、ピペラシリンおよびタゾバクタムの組み合わせでビーグル犬を処置した。該実験は、ピペラシリン/タゾバクタムの組み合わせの単回投与量(single dose)が体重1kgあたり 100 mg のピペラシリンおよび 12.5 mg のタゾバクタムを含有したこと以外は本質的に実施例 1 および 2 に記載された通りに行い、ピペラシリンの分析に適したものとなるよう HPLC 分析法を作り上げた(was elaborated)(定量限界は、空腸消化粥1グラムあたり 10 マイクログラムであった)。
【0051】
結果を図 4 に示し、それは、ピペラシリン/タゾバクタムの組み合わせ(体重1kgあたり 100 mg のピペラシリンおよび 12.5 mg のタゾバクタム)の静脈内投与後におけるビーグル犬(n=6)の空腸消化粥中のピペラシリン濃度に対する、経口投与されたベータ-ラクタマーゼ ペレットの有効性を示す。両方の実験についての値を、様々な時点での平均の空腸ピペラシリン濃度として表す。実験 1 におけるピペラシリンの値は、ベータ-ラクタマーゼ処置を伴わずに、6 時間の投与間隔でピペラシリン/タゾバクタムを2回別々に投与した後に達成された、空腸のピペラシリン濃度を示す。実験 2 において、同時のベータ-ラクタマーゼ治療と共に、ピペラシリン/タゾバクタムの組み合わせでビーグル犬を処置した。
【0052】
ベータ-ラクタマーゼを用いずに得られた結果(実験 1)は、ビーグル犬におけるピペラシリンの胆汁排出が、アンピシリンまたはアモキシシリンの胆汁排出よりもかなり高いことを示した。それにもかかわらず、ベータ-ラクタマーゼ処置は全ての時点において空腸のピペラシリン濃度を減少させた。しかし、ベータ-ラクタマーゼ処置の間中、ピペラシリン濃度は検出可能なままであった(実験 2)。従って、得られた結果により、ベータ-ラクタマーゼ治療はピペラシリン/タゾバクタムの組み合わせでの非経口治療の間に空腸のピペラシリンを除去することができるが、空腸のピペラシリン濃度を定量限界より下へ除去することができる処方(dosage formulation)を達成するためには、腸溶性コーティングされたペレットにおけるベータ-ラクタマーゼの量を増加させるべきであることが示された。
【0053】
ベータ-ラクタマーゼ ペレットの単回投与量(single dose)が体重1kgあたり約 0.3 mg の活性なベータ-ラクタマーゼを含有し、ピペラシリン/タゾバクタムの組み合わせの単回投与量(single dose)が体重1kgあたり 65.6 mg のピペラシリンおよび 9.4 mg のタゾバクタムを含有したことを除いて、前記実験を繰り返した。結果を図 5 に示し、それは、空腸のピペラシリンを除去するのにベータ-ラクタマーゼが非常に効果的であったことを示す。
【0054】
参考文献
Altschul S.F., Madden T.L., Schaffer A.A., Zhang J., Zhang Z., Miller W., Lipman D.J. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25: 3389-3402
Ambler, R.P. 1980. The structure of beta-lactamases. Philos. Trans. R. Soc. London B Biol. Sci 289:321-331
Brogard, J.M., Jehl, F., Blickle,J.F., Adloff M., Donner, M., and H. Montell. 1989. Biliary elimination of ticarcillin plus clavulanic acid (Claventin): experimental and clinical study. Int. J. Clin. Pharmacol. Ther. Toxicol. 27:135-144.
Colombo, M., L., Hanique, S., Baurin, S.L., Bauvois, C., De Vriendt, K., Van Beeumen, J.J., Frere, J.M., and B. Joris. 2004. The ybxI gene of Bacillus subtilis 168 encodes a class D beta-lactamase of low activity. Antimicrob. Agents Chemother.48:484-490
Donskey, C.J., 2006. Antibiotic regimens and intestinal colonization with antibiotic-resistamt Gram-negative bacilli. Clin. Infect. Dis. 43:62-69.
Harmoinen, J., Mentula, S., Heikkila, M., van der Rest, M., Rajala-Schultz, P.J., Donskey, C.J., Frias, R., Koski, P., Wickstrand, N., Jousimies-Somer, H., Westermarck, E., and K. Lindevall. 2004. Orally administered targeted recombinant beta-lactamase prevents ampicillin-induced selective pressure on the gut microbiota: a novel approach to reducing antimicrobial resistance. Antimicrob. Agents Chemother. 48:75-79
Harmoinen, J. Vaali, K., Koski, P., Syrjanen, K., Laitinen, O., Lindevall, K., and E. Westermarck. 2003. Enzymatic degradation of a β-lactam antibiotic, ampicillin, in the gut: a novel treatment modality. J.Antimicrob. Chemother. 51:287-292.
Izui, K., J.B.K. Nielsen, M.P. Gaulifield, and J.O. Lampen. 1980. Large exopenicillinase, initial extracellular form detected in growths of Bacillus licheniformis. Biochemistry 19:1882-1886
J.A. Katz. 2006. Probiotics for prevention of antibiotic-associated diarrhea and Clostridium difficile diarrhea. J. Clin. Gastroenterol. 40:249-255.
Gebhard L.G., Rrisso V.A. Sanots J., Ferreyra R.G., Noguera M.E. and Ermacora M.R., 2006. Mapping the distribution of conformational information throughout a protein sequence. J. Mol. Biol. 21:358(1)280-288
Higgins, P.G., Wisplinghoff, H., Stefanik, D., and H. Seifert. 2004. In Vitro activities of the β-lactamase inhibitors clavulanic acid, sulbactam, and tazobactam alone or in combination with β-lactams against epidemiologically characterized multidrug-resistant Acinetobacter baumannii strains. 48: 1586-1592
Knox, J.R., and P.C. Moews. 1991. Beta-lactamase of Bacillus licheniformis 749/C. Refinement at 2 Å resolution and analysis of hydration. J.Mol.Biol. 220:435-455
Korhonen, H., Marnila, P., and H.S. Gill. 2000. Bovine milk antibodies for health. Br J Nutr. 84: 135-146
Matagne, A., Joris, B., van Beeumen J., and J-M. Frere. 1991. Ragged N-termini and other variants of class A beta-lactamases analysed by chromatofocusing. Biochem. J. 273: 503-510
Matagne, A., Lamotte-Brasseur, J., and J.M. Frere. 1998. Catalytic properties of class A beta-lactamases: efficiency and diversity. Biochem. J. 330: 581-598
Morris, D.L., Ubhi, C.S., Robertson, C.S., and K.W. Brammer. 1986. Biliary pharmacokinetics of sulbactam in humans. Rev.Infect.Dis. 8:589-592.
O'Callaghan, C. H., Morris, A., Kirby, S.M., and A. H. Singler. 1972. Novel method for detection of beta-lactamases by using a chromogenic cephalosporin substrate. Antimicrob. Agents Chemother. 1:283-288
Rice, L.B., Hutton-Thomas,R., Lakticova, V., Helfand, M.S., and C.J. Donskey.2004. Beta-lactam antibiotics and gastrointestinal colonization of vancomycin-resistant enterococci. J.Infect.Dis. 189:1113-1118.
Santos, J, Gebhard L.G., Risso V.A., Fereyra R.G., Rossi J.P. and Ermacora M.R. 2004. Folding of an abridged beta-lactamase. Biochemistry 43(6):1715-123
Sullivan, Å., Edlund, C., and Nord, C.E. 2001. Effect of antimicrobial agents on the ecological balance of human microflora. Lancet 1:101-114.
Westphal, J.F., Brogard, J.M., Caro-Sampara, F., Adloff, M., Blickle, J.F., Monteil, H., and F. Jehl. 1997. Assessment of biliary excretion of piperacillin-tazobactam in humans. Antimicrob. Agents Chemother. 41:1636-1640.
Wildfeuer, A., Schwiersch, U., Engel, K., von Castell, E., Schilling, A., Potempa, J., and H. Lenders. 1988. Pharmacokinetics of sulbactam and ampicillin intravenously applied in combination to healthy volunteers and patients. Determination of the ratio of the two drugs in serum and in various tissues. Arzneimittelforschung. 38:1640-1643.
【特許請求の範囲】
【請求項1】
ベータ-ラクタム抗生物質およびベータ-ラクタマーゼ阻害剤の組み合わせでの処置に付随する腸における副作用を低減させるための医薬を製造するための、クラスA ベータ-ラクタマーゼの使用。
【請求項2】
前記クラスA ベータ-ラクタマーゼが、バチルス・リケニフォルミスの PenP である、請求項1に記載の使用。
【請求項3】
ベータ-ラクタム抗生物質が、ペニシリン、アミノペニシリン、ウレイドペニシリンおよびカルボキシペニシリンからなる群から選択される、請求項1に記載の使用。
【請求項4】
ベータ-ラクタム抗生物質が、ペニシリン G、アンピシリン、アモキシシリン、ピペラシリンおよびチカルシリンからなる群から選択される、請求項3に記載の使用。
【請求項5】
阻害剤が、クラスA ベータ-ラクタマーゼに対する阻害剤である、請求項1に記載の使用。
【請求項6】
阻害剤が、スルバクタム、クラブラン酸およびタゾバクタムからなる群から選択される、請求項5に記載の使用。
【請求項7】
ベータ-ラクタム抗生物質およびベータ-ラクタマーゼ阻害剤の組み合わせが、アンピシリンおよびスルバクタム; アモキシシリンおよびクラブラン酸; ピペラシリンおよびタゾバクタム; ならびにチカルシリンおよびクラブラン酸からなる群から選択される組み合わせである、請求項1に記載の使用。
【請求項8】
ベータ-ラクタマーゼが、バチルス・リケニフォルミス 749/C(ATCC 25972)由来である、請求項1−7のいずれか一項に記載の使用。
【請求項9】
ベータ-ラクタマーゼが、バチルス・スブティリス、バチルス・アミロリクエファシエンス、バチルス・プムリス、またはバチルス・リケニフォルミスにおいて生産された組換えベータ-ラクタマーゼである、請求項1−8のいずれか一項に記載の使用。
【請求項10】
ベータ-ラクタマーゼが、経口用医薬組成物として製造される、請求項1−9のいずれか一項に記載の使用。
【請求項11】
医薬組成物が、腸溶性コーティングされた組成物である、請求項10に記載の使用。
【請求項12】
ベータ-ラクタム抗生物質およびベータ-ラクタマーゼ阻害剤が非経口投与される、請求項1−11のいずれか一項に記載の使用。
【請求項13】
ベータ-ラクタム抗生物質およびベータ-ラクタマーゼ阻害剤の組み合わせでの処置に付随する腸における副作用を低減させる方法であって、必要としている対象に有効量のクラスA ベータ-ラクタマーゼが投与される、方法。
【請求項14】
前記クラスA ベータ-ラクタマーゼが、バチルス・リケニフォルミスの PenP である、請求項13に記載の方法。
【請求項1】
ベータ-ラクタム抗生物質およびベータ-ラクタマーゼ阻害剤の組み合わせでの処置に付随する腸における副作用を低減させるための医薬を製造するための、クラスA ベータ-ラクタマーゼの使用。
【請求項2】
前記クラスA ベータ-ラクタマーゼが、バチルス・リケニフォルミスの PenP である、請求項1に記載の使用。
【請求項3】
ベータ-ラクタム抗生物質が、ペニシリン、アミノペニシリン、ウレイドペニシリンおよびカルボキシペニシリンからなる群から選択される、請求項1に記載の使用。
【請求項4】
ベータ-ラクタム抗生物質が、ペニシリン G、アンピシリン、アモキシシリン、ピペラシリンおよびチカルシリンからなる群から選択される、請求項3に記載の使用。
【請求項5】
阻害剤が、クラスA ベータ-ラクタマーゼに対する阻害剤である、請求項1に記載の使用。
【請求項6】
阻害剤が、スルバクタム、クラブラン酸およびタゾバクタムからなる群から選択される、請求項5に記載の使用。
【請求項7】
ベータ-ラクタム抗生物質およびベータ-ラクタマーゼ阻害剤の組み合わせが、アンピシリンおよびスルバクタム; アモキシシリンおよびクラブラン酸; ピペラシリンおよびタゾバクタム; ならびにチカルシリンおよびクラブラン酸からなる群から選択される組み合わせである、請求項1に記載の使用。
【請求項8】
ベータ-ラクタマーゼが、バチルス・リケニフォルミス 749/C(ATCC 25972)由来である、請求項1−7のいずれか一項に記載の使用。
【請求項9】
ベータ-ラクタマーゼが、バチルス・スブティリス、バチルス・アミロリクエファシエンス、バチルス・プムリス、またはバチルス・リケニフォルミスにおいて生産された組換えベータ-ラクタマーゼである、請求項1−8のいずれか一項に記載の使用。
【請求項10】
ベータ-ラクタマーゼが、経口用医薬組成物として製造される、請求項1−9のいずれか一項に記載の使用。
【請求項11】
医薬組成物が、腸溶性コーティングされた組成物である、請求項10に記載の使用。
【請求項12】
ベータ-ラクタム抗生物質およびベータ-ラクタマーゼ阻害剤が非経口投与される、請求項1−11のいずれか一項に記載の使用。
【請求項13】
ベータ-ラクタム抗生物質およびベータ-ラクタマーゼ阻害剤の組み合わせでの処置に付随する腸における副作用を低減させる方法であって、必要としている対象に有効量のクラスA ベータ-ラクタマーゼが投与される、方法。
【請求項14】
前記クラスA ベータ-ラクタマーゼが、バチルス・リケニフォルミスの PenP である、請求項13に記載の方法。
【図1】


【図2】

【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2010−511020(P2010−511020A)
【公表日】平成22年4月8日(2010.4.8)
【国際特許分類】
【出願番号】特願2009−538736(P2009−538736)
【出願日】平成19年11月21日(2007.11.21)
【国際出願番号】PCT/FI2007/050627
【国際公開番号】WO2008/065247
【国際公開日】平成20年6月5日(2008.6.5)
【出願人】(504176324)
【氏名又は名称原語表記】IPSAT THERAPIES OY
【Fターム(参考)】
【公表日】平成22年4月8日(2010.4.8)
【国際特許分類】
【出願日】平成19年11月21日(2007.11.21)
【国際出願番号】PCT/FI2007/050627
【国際公開番号】WO2008/065247
【国際公開日】平成20年6月5日(2008.6.5)
【出願人】(504176324)
【氏名又は名称原語表記】IPSAT THERAPIES OY
【Fターム(参考)】
[ Back to top ]