
新規な調節タンパク質
【課題】R2R3型MYBファミリーに属し、かつベンゼノイドの生産に関するシキミ酸経路における調節タンパク質を提供する。
【解決手段】特定のポリペプチド配列またはその変異体もしくは誘導体を含む、R2R3型MYBファミリーに属し、かつシキミ酸経路を調節する、DNA結合活性を示すポリペプチド。前記シキミ酸経路を調節することを含む、必須アミノ酸、チロシン、フェニルアラニンおよびトリプトファンから誘導される芳香族および非芳香族化合物の生合成の調節のための方法、並びに、花の匂いを調節する方法または害虫もしくは病原性生物に対する耐性を調節する方法。
【解決手段】特定のポリペプチド配列またはその変異体もしくは誘導体を含む、R2R3型MYBファミリーに属し、かつシキミ酸経路を調節する、DNA結合活性を示すポリペプチド。前記シキミ酸経路を調節することを含む、必須アミノ酸、チロシン、フェニルアラニンおよびトリプトファンから誘導される芳香族および非芳香族化合物の生合成の調節のための方法、並びに、花の匂いを調節する方法または害虫もしくは病原性生物に対する耐性を調節する方法。
【発明の詳細な説明】
【発明の分野】
【0001】
本発明は、植物におけるmyb調節タンパク質に関する。より具体的には、本発明は、R2R3型のmybタンパク質、該タンパク質をコードする遺伝子ならびにこのタンパク質およびそれをコードする遺伝子の適用に関する。
【発明の背景】
【0002】
植物の匂いは、数多くの理由のために重要な性質である。例えば、花による揮発性化合物の産生は、繁殖のプロセスにおいて受粉する昆虫を誘引する重要な役割を担うことができる。同時にまた、良質および高収量の結実のための重要な役割を担うことができる。代わりに、植物はまた、害虫またはその捕食動物を誘引する生殖性および植物性部位において揮発性化合物を生産することもできる。このプロセスにおいて、揮発性の性質は、有害な生物体、例えば害虫、線虫または菌類に対する感受性または抵抗性を決定する。揮発性物質の合成および放出を干渉および修飾することは、関心の対象になり得る。例えば、植物/昆虫の関係について干渉し、生殖プロセスの向上や害虫に対する耐性を向上させる。
【0003】
匂いはまた、フレーバーおよびフレグランス、食品および化粧品産業にとって重要である。なぜなら、多くの場合において、これらの産業では、香水、食品、化粧品等々の使用に適したフレーバーおよびフレグランス成分の供給源として、花、ハーブ、果実および香辛料から天然の揮発性化合物を生産するからである。
【0004】
植物由来の揮発性化合物の重要なクラスは、いわゆるベンゼノイドである。これらのフェノール類化合物は、塩基性C6骨格を有し、特定の器官または特定の細胞種においてシキミ酸経路から生産されることが多い。ランおよびペチュニアのような多数の植物は、そのフレグランスの主要部としてベンゼノイドを生産する花を有する。多数のペチュニア雑種系統において、ベンゼノイド揮発性化合物は特異的に放出され、日/夜リズムにおいて午後の終わりおよび夜の間に花弁によって始まる。現在まで、唯一の制限された知見は、この経路に関与する分子および遺伝子プロセスについて存在する。数少ない構造的遺伝子のみがクローン化および特徴づけられ、現在まで調節遺伝子は同定されていない。従って、植物がベンゼノイドの生合成および放出をどのように調節するのかについては未だ大部分が知られていない。
【0005】
匂いの生合成を、再生的、効果的かつ費用効率的に遮断、増強または修飾する信頼ある方法が植物または他の(微小)生物体において達成され得るならば、これは非常に大きな利益になる。
【図面の簡単な説明】
【0006】
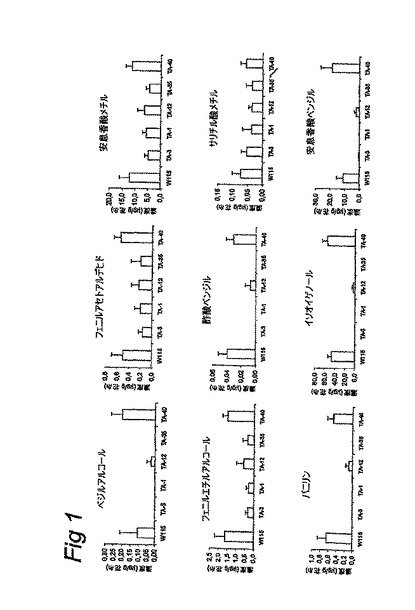
【図1】Mitchell (W115)による、揮発性ベンゼノイドの定量化された放出量。揮発性の減少した放出を示す4つのRNAi系統 (TA-1、TA-3、TA-12およびTA-35) およびベンゼノイドの放出において減少を示さない一つのRNAi系統(40)。
【図2】ODO1、ならびにシキミ酸経路、フェニルアラニンおよびt-ケイ皮酸の合成経路からの遺伝子、例えばDAHPシンターゼ(DAHPS)、EPSPシンターゼ(EPSPS)、コリスミ酸ムターゼ(CM) および二つのフェニルアラニンアンモニアリアーゼ遺伝子 (PAL1および2)についての;安息香酸ベンジルトランスフェラーゼ(BEBT)および安息香酸/サリチル酸メチルトランスフェラーゼ(BSMT)についての、Mitchell(M)およびRNAi系統1、3、12、35および40のRNAゲルブロット分析。花の結合タンパク質1(FBP1)の転写レベルが、RNAi系統におけるODO1抑制の特異性を証明するために示された。
【図3】機能的な保存の指標である保存された残基(暗色部)を明示したODO1相同体のR2R3ドメインのアライメント。
【詳細な説明】
【0007】
本発明は、配列番号1において示されたポリペプチド配列またはその変異体もしくは誘導体を含むDNA結合活性を示すポリペプチドに関する。本発明のこれらのポリペプチドは、R2R3型MYBファミリーに属し、かつシキミ酸経路を調節する。シキミ酸経路を通して、3つの芳香族アミノ酸であるチロシン、フェニルアラニンおよびトリプトファンが合成される。これらの化合物から、さらに他の芳香族化合物が合成される。ベンゼノイドに関するシキミ酸経路における調節タンパク質が同定されたのはこれが初めてである。従って、本発明の利点の1つは、シキミ酸経路における調節タンパク質を最初に提供し、かつ哺乳動物によって生成できない3つの必須アミノ酸の生合成を調節する手段を提供することである。同時に、これらの必須アミノ酸から誘導される芳香族および非芳香族化合物の生合成の調節のための道を開く。3つの芳香族アミノ酸から誘導される最も重要な化合物の例には、ケイ皮酸、クマル酸、カフェー酸、フェルラ酸のような化合物;それ自体が他の多くの産業的に興味ある化合物の中間体になり得るシキミ酸経路由来の化合物が含まれる。2〜3の例を挙げると、ベンゼノイドには、安息香酸メチル、サリチル酸メチル、ベンズアルデヒド、酢酸ベンジル、安息香酸ベンジル、バニリン、イソオイゲノール、ならびにフラボノールおよびアントシアニンを含むフェニルプロパノイドが含まれる。これらの化合物は一次および二次代謝の両方に関与するので、当業者は、本発明のタンパク質が多くの生合成のプロセスに影響を与える手段を提供することを理解するであろう。これには、フェノール酸起原であって、病原体に対する防御に関与する化学物質の調節が含まれる。また、揮発性のベンゼノイドの放出の調節に関与する化学物質の調節が含まれる。シキミ酸経路はまた、細菌および菌類の中に存在するので、本発明の教示はまたこのような系にまで及ぶ。
【0008】
[発明のポリペプチドの変異体および誘導体]
本明細書中で使用されるとき、「変異体」または「誘導体」には、列挙されたポリペプチドの性質が保持される限りにおいて、列挙されたポリペプチドとは置換、欠失、付加がある点で異なるかまたは一以上のアミノ酸との融合である点で異なるペプチドまたは非ペプチド化合物が含まれる。これらの用語にはまた、列挙されたポリペプチドの性質が保持される限りにおいて、幾つかのグリコシル化部位が導入または修飾される点で列挙されたポリペプチドとは異なるペプチドまたは非ペプチド化合物が含まれる。これらの用語にはまた、列挙されたポリペプチドの性質が保持される限りにおいて、修飾基がペプチド構造と共有結合または非共有結合している点で列挙されたポリペプチドとは異なるペプチドまたは非ペプチド化合物が含まれる。特に、変異体および誘導体は、ベンゼノイドに関するシキミ酸経路の遺伝子の転写レベルを操作する能力を保持している。上記転写レベルには、3-デオキシ-D-アラビノ-ヘプツロソナート-7-リン酸塩シンターゼ(DAHPS)、5-エノール-ピルビルシキマート-3-リン酸塩シンターゼ(EPSPS)、コリスミ酸ムターゼ(CM)およびL-フェニルアラニンアンモニアリアーゼ(PAL)の遺伝子の転写レベルが含まれる。
【0009】
一実施態様において、変異体または誘導体には、配列番号1のアミノ酸配列と少なくとも50%、55%、60%、65%、70%、75%、好ましくは80%、85%、90%、95%、97%、98%または99%の同一性を示すアミノ酸配列が含まれる。
【0010】
さらなる他の実施態様において、変異体または誘導体には、配列番号1のアミノ酸13-116と少なくとも90%、95%、97%、98%または99%の同一性を示すアミノ酸配列が含まれる。このアミノ酸の領域は、本発明のポリペプチドのDNA結合ドメイン(R2R3型のmyb DNA結合ドメイン)と一致する。このタイプのmybドメインについてのさらなる情報については、Stracke et al.(2001) Current Opinion in Plant Biology 4: 447-456を参照されたい。
【0011】
さらなる他の実施態様において、変異体または誘導体には、配列番号1のアミノ酸128〜アミノ酸294の領域と少なくとも70%、75%または80%の同一性を示すアミノ酸配列が含まれる。好ましくは、変異体または誘導体には、配列番号2のアミノ酸128〜アミノ酸294の領域と少なくとも85%、87%、89%または90%の同一性を示すアミノ酸配列が含まれる。最も好ましくは、変異体または誘導体には、配列番号1のアミノ酸128〜アミノ酸294の領域と少なくとも94%、97%、98%または99%の同一性を示すアミノ酸配列が含まれる。
【0012】
さらに他の実施態様において、変異体または誘導体は、保存的置換基においてのみ列挙されたポリペプチドとは異なるポリペプチドである。本明細書中において使用されたとき、「アミノ酸の保存的置換基」は、一つのアミノ酸の類似の性質をもつ他のアミノ酸との置換を意味する。例えば、疎水性性質を示すアミノ酸が疎水性性質を示す他のアミノによって置換される場合がある。
【0013】
用語「ペプチド」および「ポリペプチド」は、原則として本明細書中において交換可能に使用され得、アミノ酸の列を含む分子を意味する。
【0014】
アミノ酸の同一性は、これらに限定されないが、以下の既知の方法によって容易に計算し得る(Computational Molecular Biology, Lesk, A. M., ed., Oxford University Press, New York, 1988;Biocomputing: Infomatics and Genome Projects, Smith, D. W., ed., Academic Press, New York, 1993;Computer Analysis of Sequence Data, Part I, Griffin, A. M., and Griffin, H. G., eds., Humana Press, New Jersey, 1994;Sequence Analysis in Molecular Biology, von Heine, G., Academic Press, 1987;およびSequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991;およびCarillo, H., and Lipman, D., SIAM J. Applied Math., 48:1073 (1988))。同一性を決定するための好ましい方法は、試験対象の配列間の最も大きな一致を与えるよう設計される。同一性を決定するための方法は、公共的に利用可能なコンピュータープログラムにおいて体系化される。二配列間の同一性および類似性を決定する好ましいコンピュータープログラム方法には、これらに限定されないが、GCGプログラムパッケージ(Devereux, J., et al., Nucleic Acids Research 12 (1):387 (1984))、BestFit、BLASTP、BLASTN、およびFASTA (Altschul, S. F. et al., J. Mol. Biol. 215:403-410 (1990)) が含まれる。BLAST X プログラムは、NCBIおよび他のソース( BLAST Manual, Altschul, S., et al., NCBI NLM NIH Bethesda
, MD 20894;Altschul, S., et al., J. Mol. Biol. 215:403-410 (1990) )から公共的に利用可能である。既知のSmith Watermanアルゴリズムがまた、同一性を決定するために使用され得る。ポリペプチド配列の比較のための好ましいパラメーターには、以下のものが含まれる1) アルゴリズム: Needleman and Wunsch, J. Mol. Biol. 48:443-453 (1970) 比較マトリックス: BLOSSUM62 from Hentikoff and Hentikoff, Proc. Natl. Acad. Sci. USA. 89:10915-10919 (1992);ギャップペナルティー: 12;およびギャップ長ペナルティー: 4。これらのパラメーターで有用なプログラムは、ウィスコンシン州マディソン所在のGenetics Computer Group社製の「Ogap」プログラムとして公的に利用可能である。上述したパラメーターは、ペプチド比較のための初期設定パラメーター(エンドギャップについてペナルティーのない)である。
【0015】
例えば、トマトESTs SGNU217873およびLeHTM16 (図3を参照) は、ODO1の相同体である。両方のcDNAは、特徴的な一でR2R3ドメイン中のODO1ファミリータンパク質(PbMYB, AtMYB42およびAtMYB85)とホモロジーを共有する。このホモロジーは、機能的保存の指標である。トマトは、その花において大量のベンゼノイドを生成しないが、幾つかのベンゼノイドが成熟したトマト果肉中に蓄積される。SGNU217873のESTsは、花芽および子房においてのみ見出され(SGN転写データベース、http://www.sgn.cornell.edu/を参照)、その発現は、この遺伝子が果肉の成熟過程のベンゼノイド生産に関与することを示す成熟過程において増加する。LeHTM16は果肉成熟中に誘導されない。従って、このMYBは植物における他の箇所においてプロセスを制御するであろう。
【0016】
[本発明のポリヌクレオチド]
他の側面において、本発明は、配列番号2に示された配列をもつヌクレオチド配列または配列番号2と少なくとも50%、55%、60%、65%、70%、75%、好ましくは80%、85%、90%、95%、97%、98%もしくは99%の同一性を示す前記ヌクレオチド配列の変異体を含む、単離された、組み換えまたは合成ポリヌクレオチドであって、かつベンゼノイドに関するシキミ酸経路について制御活性を有するタンパク質をコードするポリヌクレオチドを提供する。
【0017】
一実施態様において、本発明のポリヌクレオチドには、配列番号1に示されたアミノ酸配列をもつポリペプチドをコードするポリヌクレオチド配列、またはベンゼノイドに関するシキミ酸経路について制御活性を有する前記ポリヌクレオチド配列の断片が含まれる。
【0018】
また、配列番号2のポリヌクレオチド配列と相補的である配列、例えば、アンチセンスRNAまたは他の阻害RNA(例えばRNAiに使用)を有するポリヌクレオチド配列は、本発明によって包含される。これらの相補的およびハイブリダイズする配列は任意の長さであってよく、当業者は、適切な長さがその配列が使用される目的に適うであろうことを理解する。例えば、任意の長さの転写後サイレンシング二本鎖RNAが植物において使用され得る。
【0019】
二つのヌクレオチド配列の同一性は、上述した方法を使用して決定される。ヌクレオチド配列、核酸およびポリヌクレオチドなる用語は、原則として本出願において交換可能に使用され得、ヌクレオチドの配列を意味する。本発明のポリヌクレオチドには、ゲノム配列、細胞外ゲノムおよびプラスミドコード配列ならびにタンパク質、ポリペプチド、ペプチド等々を発現するかまたは発現するのに適合されたより小さな加工された遺伝子セグメントが含まれうる。本発明のポリヌクレオチドは、一本鎖(コーディングまたはアンチセンス)または二本鎖であってもよく、DNA(ゲノム、cDNA)またはRNA分子であってもよい。これらは、単離された、組換え体または合成物であってもよい。RNA分子には、イントロンを含みかつ一対一のやり方においてDNA分子と一致する不均一な核RNA(hn RNA)分子、およびイントロンを含まないmRNA分子を含みうる。さらなるコーディングまたは非コーディング配列が、本発明のポリヌクレオチド内に存在していてもよい(但し、必ずしも存在する必要はない)。ポリヌクレオチドは、他の分子および/または支持材料と結合していてもよい(但し、必ずしも結合している必要はない)。
【0020】
「単離された」は、本明細書中において使用されるとき、ポリヌクレオチドが実質的に他の核酸分子から自由であることを意味する。また、ポリヌクレオチドが、関連のない配列の大きな部位、例えば、大きな染色体断片または他の機能的遺伝子またはポリペプチドコーディング領域を含まないことを意味する。当然ながら、これは、最初に単離されたポリヌクレオチド分子を意味し、ヒトの手によってセグメントに後に付加された遺伝子またはコーディング領域を排除するものではない。
【0021】
[本発明のベクター]
他の側面において、本発明は、本発明のポリヌクレオチドを含むベクターに関する。有効に使用され得るベクターには、周知の植物ベクター、例えばpK7GWIG2(I)およびpGreen、ならびに微生物においてタンパク質を形質転換および発現させるために使用されるベクターが含まれる。また、Arabidopsis, A laboratory manual Eds. Weigel & Glazebrook, Cold Spring Harbor Lab Press (2002) およびManiatis et al. Molecular Cloning, Cold Spring Harbor Lab (1982) を参照されたい。
【0022】
[本発明の宿主細胞]
さらに他の側面において、本発明は、本発明によるポリヌクレオチドまたはベクターを含む宿主細胞に関する。本発明による適切な宿主細胞には、植物細胞、酵母細胞、真菌細胞、藻類細胞、ヒト細胞および動物細胞が含まれる。適切な植物細胞の例には、トマトおよびシロイヌナズナが含まれる。適切な宿主細胞の例には、Saccharomyces cerevisiaeおよびPichia pastorisが含まれる。適切な真菌細胞の例には、Aspergillusが含まれる。適切な動物細胞の例には、昆虫細胞、例えば、Spodoptera frugiperda由来の細胞;哺乳類細胞、例えばチャイニーズハムスター卵巣細胞またはPERC6細胞が含まれる。当該技術分野の様々な細胞系統、例えばFlp-In細胞系統(Invitrogen)が使用され得る。上述したように、宿主細胞への本発明のポリヌクレオチドを導入するための様々なベクターが使用され得る。これらのベクターは、例えば、プラスミドDNAベクター、ウイルスDNAベクター(例えば、アデノウイルスまたはアデノ関連ベクター)、ウイルスRNAベクター(例えばレトロウイルス)またはウイルス植物ベクター、例えばタバコ(tobacco rattle)ウイルスおよびジャガイモウイルスXから選択され得る、クローニングベクター、発現ベクター、サイレンシングベクターにし得る。
【0023】
細胞遊離抽出物による本発明のポリペプチドの生産はまた、本発明によって包含される。細胞遊離抽出物における生産のための方法は、当該技術分野において知られている。例えばPelman and Jackson (1976) Eur.J.Biochem 67 : 247-56を参照されたい。
【0024】
本発明の宿主細胞は、本発明のポリペプチドを産生するために使用され得る。これは、ポリペプチドの産生を可能にし、かつ任意的にはポリペプチドを回収する条件下において本発明による宿主細胞を培養することを含む。好ましい実施態様において、DNA結合活性をもつ組換え体ポリペプチドが産生される。
【0025】
一実施態様において、宿主細胞は、本発明によるタンパク質をコードする遺伝子がサイレンシングされるトランスジェニック植物である。結果として、ベンゼノイドに関するシキミ酸経路における酵素が下方制御され、例えば、3-デオキシ-D-アラビノ-ヘプツロソナート-7-ホスファート シンターゼ(DAHPS)、5-エノール-ピルビルシキマート-3-ホスファート シンターゼ(EPSPS)、コリスマート ムターゼ(CM) およびL-フェニルアラニン アンモニア-リアーゼ(PAL) の遺伝子の転写レベルが減少し、植物の揮発性の性質が変化するであろう。
【0026】
他の実施態様において、宿主細胞は、本発明のタンパク質をコードする遺伝子が過剰発現し、かつシキミ酸経路の酵素が上方制御されるトランスジェニック植物である。例えば、3-デオキシ-D-アラビノ-ヘプツロソナート-7-ホスファート シンターゼ(DAHPS)、5-エノール-ピルビルシキマート-3-ホスファート シンターゼ(EPSPS)、コリスマート ムターゼ(CM) およびL-フェニルアラニン アンモニア-リアーゼ(PAL) の遺伝子の転写レベルが増加し、揮発性の性質がより強い匂いが産生されるように修飾される。EPSPS産生の増加したレベルをもつトランスジェニック植物は、化学的防御戦略において特別の関心を集める。とりわけこの戦略においては、グリフォセートを含む植物保護産物が使用される。増加したレベルのEPSPSをもつ植物はグリフォセートに対する増加した耐性を有するためである。
【0027】
さらに他の実施態様において、宿主細胞は、本発明のタンパク質をコードする遺伝子が過剰発現するトランスジェニック植物であり、揮発性の性質は、ベンゼノイド産物を増加させることによって病原生物体に対する植物の化学的防御系を強化するように修飾される。これには、殺虫剤、殺菌剤、殺線虫剤、軟体動物駆除剤または殺鼠剤として機能するベンゼノイドが含まれる。
【0028】
他の実施態様において、宿主細胞は、本発明のタンパク質をコードする遺伝子が過剰発現し、相互抑制効果が生じるトランスジェニック植物においてもたらされるトランスジェニック植物細胞である。結果として、遺伝子がサイレンスされ (Jorgensen et al. (1996) Plant Mol Biol 31: 957-973)、DAHPS、EPSPS、CMおよびPALについてのベンゼノイドに関するシキミ酸経路の遺伝子の転写レベルが減少する。
【0029】
他の実施態様において、宿主細胞は、本発明のタンパク質をコードする遺伝子が当該技術分野において知られた他の方法においてサイレンスされるトランスジェニック植物をもたらすトランスジェニック植物細胞である。
【0030】
さらに他の実施態様において、宿主細胞は、本発明のタンパク質をコードする遺伝子がサイレンスされ、かつ害虫が誘引されないように揮発性の性質が修飾される、トランスジェニック植物においてもたらされるトランスジェニック細胞である。さらに他の実施態様において、宿主細胞は、本発明のタンパク質をコードする遺伝子が、導入の前に該遺伝子を含まないかまたは活性形態において存在しない宿主において導入されるトランスジェニック植物をもたらすトランスジェニック植物細胞である。この方法において、シキミ酸経路および芳香族化合物の生合成経路を制御することが可能である。
【0031】
[抗体]
本発明のポリペプチドに対する抗体がまた、本発明において包含される。ポリクローナルおよびモノクローナル抗体を生成するための方法が当該技術において一般に知られている(例えばHarlow and Lane (1988) Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, New Yorkを参照)。抗体はそれ自体で使用され得るが、好ましくは抗体は検出可能なラベルでラベルされる。適切な抗体ラベルが当業者に知られ、これらに限定されないが、放射活性ラベル、電子密度ラベル、酵素ラベルおよび蛍光ラベルが含まれる。好ましい実施態様において、酵素または蛍光マーカー、例えばアルカリホスファターゼ、西洋ワサビペルオキシダーゼ、およびフルオレセインが使用される。
【0032】
また、細胞内で生産された抗体、いわゆる細胞内発現抗体(intrabody)が本発明によって包含される。細胞内発現抗体の構築は、当該技術、例えば米国特許6,004,940およびWO 01/48017において記載されている。
【0033】
本発明の抗体は、芳香花を同定または検出するための方法において使用され得る。当該方法では、植物材料を本発明の抗体と接触させ;本発明のポリペプチドと結合するか否かを検出する。このような方法はまた、本発明によって包含される。タンパク質は当該技術分野において既知の回収技術、例えばMethods Enzymol. vol. 182, Guide to protein purification. Eds. M.P. Deutscher (1990) Academic Press Inc.を使用して回収され得る。
【0034】
[発明の方法]
本発明によるポリペプチド、ポリヌクレオチド、ベクターもしくは抗体またはこれらの断片(集合的には本発明の化合物と呼ばれる)は、シキミ酸経路の遺伝子の転写レベルを操作する方法、例えば、3-デオキシ-D-アラビノ-ヘプツロソナート-7-ホスファート シンターゼ(DAHPS)、5-エノール-ピルビルシキマート-3-ホスファート シンターゼ(EPSPS)、コリスマート ムターゼ(CM) およびL-フェニルアラニン アンモニア-リアーゼ(PAL) についての遺伝子の転写レベルを操作する方法において使用され得る。これらの酵素を通して、下流生合成プロセスが影響を受ける。例えば、本発明の化合物は、花における匂いを制御する方法において使用され得る。あるいは本発明の化合物は、害虫または病原生物体に対する耐性を調節する方法において使用され得る。従って、植物中の揮発性匂い化合物のの性質を修飾するための;シキミ酸フェニルアラニン合成経路から遺伝子の転写レベルを調節するための;フェニルプロパノイド経路から遺伝子の転写レベルを調節するための;ベンゼノイド生合成に関与する遺伝子の転写レベルを調節するための;または芳香族アミノ酸の生合成、特にフェニルアラニン、チロシンおよびトリプトファンの生合成を調節するための、本発明によるポリペプチド、ポリヌクレオチド、ベクターもしくは抗体またはこれらの断片の使用はまた、本発明において包含される。
【0035】
一実施態様において、本発明の化合物は、揮発性匂い化合物の性質が修飾され得る植物を生産するための方法において使用される。該方法は、植物体に本発明のポリヌクレオチドを導入することを含む。一実施態様において、本発明のポリヌクレオチドは、植物体のゲノム内に導入される。
【0036】
さらに他の実施態様において、本発明の化合物は、匂いを発する植物と匂いを発しない植物とを識別するための方法において使用される。本発明には以下の工程が含まれる:
− 植物と本発明による化合物とを接触させる工程と、
− 上記化合物と結合することを検出するかまたは本発明のヌクレオチド内の多型を検出する工程。
【0037】
さらに他の実施態様において、本発明の化合物は、植物育種中における遺伝的分析またはマーカー補助的選別において使用される。特に、本発明の化合物は、PCRに基づいたマーカー補助的選別において好適に使用され得る。例えば、制限断片長多型(RFLP)、増幅断片長多型(AFLP)、ランダム増幅多型DNA(RAPD)、一塩基多型(SNP)およびマイクロサテライトがある。これらの技術のさらなる詳細については、例えば、Welsh & McClelland (1990) Nucleic Acids Research 18: 7213-7218;Vos et al. (1995) Nucleic Acids Research 23: 4407-4414 and Struss & Plieske (1998) Theoretical & Applied Genomics 97:308-315を参照されたい。二つの育種系統または品種の制限パターンまたはヌクレオチド配列の差異を比較することによって、本発明のポリペプチドをコードする遺伝子内の多型が同定され得る。これは、本発明のポリペプチドと関連した形質(例えば匂いまたは増加したベンゼノイドレベル)の非常に迅速な選択を可能にする。
【0038】
さらに他の実施態様において、本発明の化合物は、本発明によるポリペプチドの下方制御によって害虫に対する耐性を増加させる方法において使用される。下方制御は、揮発性ベンゼノイド(放出されるが、害虫をあまり誘引しないかまたは害虫の捕食者をあまり誘引しない)の性質を変化させるであろう。
【0039】
さらに他の実施態様において、本発明の化合物は、本発明によるポリペプチドの発現を上方制御することによって、病原性生物に対する耐性を増加させるための方法において使用される。上方制御は、病原性生物、例えば病原性細菌および真菌に対する植物の化学的防御メカニズムの一部である、生成されるベンゼノイドの性質における変化を導くであろう。
【0040】
植物および動物系統におけるポリペプチドの発現を上方および下方制御するための方法が、当該技術分野において知られている。上方制御は、植物全体または特定の植物部位(例えば、花弁および葉)における対象のポリペプチドの過剰発現に基づいている。
【0041】
下方制御は、DNAレベル、例えば転写を妨害することによって行われる。代わりに、下方制御は、RNAレベル、例えばタンパク質翻訳の部位へのRNAの転座を妨害することによって、あるいはRNAからのタンパク質の翻訳を妨害することによって、あるいはRNAのスプライシングを妨害し一以上のmRNA化学種を得ることによって行われる。このような発現に対する妨害の全体的効果は、遺伝子の発現における減少(抑制)である。RNAレベルでの妨害が好ましい。RNAレベルでの妨害を達成するための好適な方法は、二本鎖またはヘアピンRNAを使用するRNAiを通した方法;siRNAを使用するサイレンシングを通した方法;または相互抑制作用を通した方法である。例えば、Hammond & Hannon (2001) Nature Rev Gen 2: 110-119, Arabidopsis, A laboratory manual Eds. D. Weigel & J Glazebrook (2002), CSHL Press and Cogoni & Macino (2000) Genes Dev 10: 638-643を参照されたい。
【0042】
下方制御にはまた、翻訳阻害および翻訳後阻害が含まれる。翻訳阻害および翻訳後阻害についての方法は、当業者に周知である。好ましくは、翻訳のリプレッサーとして主に機能し、タンパク質の発現レベルのみに影響を与える内在性21−24 nt RNAであるmiRNAが使用され得;リン酸化、アセチル化、メチル化、グリコシル化、プロリル異性化、シアリル化、ヒドロキシル化、酸化、グルタチオニル化およびユビキチン化が使用され得;または抗体、抗体断片および化学的およびペプチド阻害剤がまた、この目的のために使用され得る。
【0043】
阻害物質を同定するための方法は当該技術分野において既知であり、ペプチド擬態、ペプチド、DNAのライブラリーまたはcDNA発現ライブラリー、コンビナトリアル・ケミストリーおよび、特に有用なファージディスプレーライブラリーをスクリーニングする方法が含まれる。これらのライブラリーは、ライブラリーを、実質的に精製されたポリペプチド、その断片またはその構造的類似体と接触させることによって結合分子についてスクリーニングされ得る。好ましい実施態様において、阻害物質は、本発明のポリペプチドのDNA結合ドメインを標的にする。本明細書中において使用されたとき、用語「阻害物質」には、本発明の化合物と結合するペプチド、ペプチド配列、ペプチド様分子および非ペプチド分子が含まれる。
【0044】
[例]
植物材料および形質転換
Petunia hybrida cv. Mitchell (W115系統ともいう;P. axillaris x (P. axillaris x P. hybrida Rose of Heaven)) およびW138系統植物は、Verdonk et al. Phytochemistry 62, 997-1008 (2003) において記載されたように生育された。少なくとも三つの成熟した花を有する植物が全ての実験において使用された。遺伝子組換えペチュニアは、Agrobacterium tumefaciens (株 GV3101::pMP90) を媒介した形質転換を介して得られた。具体的には、切断葉を細菌培養液中(約28℃、10倍稀釈) に浸漬する。遺伝子組換えカルスは、150 mg/ml カナマイシンを含むMS-培養液上で選択された。植物体は、Lucker et al.. Plant Journal 27, 315-324 (2001) に記載されたように遺伝子組換えカルスから再生された。発根植物は、PCRを使用してntpII 遺伝子およびRNAi構造物の存在について試験された。陽性の植物が温室に移された。
【0045】
ODO1の選別および同定
花弁特異的DNAマイクロアレイの構成、ラベリングおよび分析は、Verdonk et al. Phytochemistry 62, 997-1008 (2003) に記載されている。以下の三つの実験結果が比較されている: 9:00時のMitchell petalsと15:00時のMitchell petals;12:00時のpetalsと15:00時のpetalsおよび15:00時のMitchell petalsと15:00時のW138 (匂いを発しない栽培種) petals。匂いの放出で顕著に(Verdonk et al. Phytochemistry 62, 997-1008 (2003)を参照)および調整的に上方制御されたcDNAおよびW138において上方制御されていないcDNAがシークエンスされた。これらのうちの一つがMYB相同体として同定されたが、DAHPS、EPSPS、CM、PAL1 および2 ならびに BEBTは、これらのマイクロアレイ実験から選択された。RNAゲルブロット分析は、Verdonk et al. Phytochemistry 62, 997-1008 (2003) において記載されたように行われ;特定の3’ UTRプローブがPAL1および2について使用された。
【0046】
RNAiサイレンシング構築物の生成
Gateway(商標) (Invitrogen life technologies, Carlsbad, CA, USA) アダプターを含む二つのプライマーが、ODO1 オープンリーディングフレームのヌクレオチド573〜876の領域を増幅するように設計された。フォワードプライマー: 5’-aaa aag cag gct CAC CAC TGA TGA ATC CAA GC-3’;リバースプライマー: 5’-aga aag ctg ggt CCT GTT CTC TAC GTT ATC-3’ (小文字はプライマー中に構築されたGateway(商標)アダプターを表わす)。増幅したPCR産物は、pDONR207 ベクターにおいてクローニングされ、RNAiデスティネーションベクターpK7GWIWG2(I)に導入された(当該ベクターのnptII遺伝子は植物細胞に対してカナマイシン耐性を与える;VIB、Gent、Belgium)。上記操作は、製造業者用資料(Invitrogen life technologies) によって記載されたように大腸菌DH5α中において行われた。前記構築物がシークエンスされ、続いて標準的な分子生物学的技術を使用してA. tumefaciens GV3101::pMP90細胞に形質転換された。
【0047】
揮発性物質のサンプリング
揮発性物質は、水の入ったエルレンマイヤーグラス中において切り花を静置することによって収集され、続いて、ガラス製の空気の出入口を備えた蓋で閉じられた1リットルの容器内に静置された。炭素ろ過された空気が、容器の出口に真空を適用することによって容器内に導かれた。花の頭隙は、5mmのワイドガラス管中の150mg Tenax TA上で20時間にわたって出て行く空気を捕捉することによって収集された。Tenaxは、内部標準として8.37 ng/μl α-テルピネンを含んだ2mlペンタン:ジエチルエーテル(4:1)で稀釈された。溶出剤中の揮発物質は、キャピラリーガスクロマトグラフィー質量分光測定を通して分析された。1μlの稀釈剤が、Optic(ATAS, GL, International)注入ポートに250℃で注入された。分流は、2分間にわたって0 ml/分であり、流れの終わりまで25 ml/分であった。化合物は、40℃で3分間にわたってキャピラリーDB-5カラム(10×180μm、フィルム厚0.18μm;Hewlett Packard)上で分離され、その後キャリアガスとしてヘリウムを用いて30℃/分で250℃まで加熱された。カラム流は、2分間にわたって3ml/分であり、その後1.5ml/分とした。溶出化合物の質量スペクトルは、70eV(200℃のイオン源)で生成され、20スペクトル/秒の獲得速度で、-1597 eVで90秒の獲得遅延を示す飛行時間MS (Leco, Pegasus III, St.Joseph, MI, USA)上で収集された。化合物は、既知の濃度の合成外部標準および内部標準に基づいて、ならびにKant, et al. Plant Physiology 135, 483-495 (2004)に記載されているように同定および定量された。各系統は、少なくとも3回測定された。各実験について、花の新鮮重量が決定された。
【0048】
例1 ペチュニアの花の匂いの調節に関与した転写因子の同定および発現
ペチュニアの花の匂いの調節に関与した成分を同定するために、トランスクリプトミクス的手法が使用された。匂いを発した花のトランスクリプトームは、匂いを発する直前の花のトランスクリプトームおよび匂いを発しないペチュニア栽培種の花のトランスクリプトームと比較された。比較は専用の特異的なマイクロアレイを使用して行われた。匂いを発する直前に増加した転写レベルを示し、かつ匂いを発しないペチュニアにおいて非常に低い転写レベルを示す転写因子が選択された。一つの転写因子、ODO1 (ODORANT 1) が本明細書中において詳細に記載される。
【0049】
花の匂いを調節する役割と一致して、ODO1の転写レベルは、揮発性ベンゼノイドの放出の開始時点である正午〜14:00時にかけて増加した。ODO1の転写レベルは一時的に増加し、次の早朝に最も低いレベルに戻った。ODO1の発現は、花の管および花弁に制限された。花の発達中、ODO1の転写産物が、開花後〜老化(開花6日後)まで検出された。ODO1の転写レベルは、ペチュニアW138系統(匂いを発しない系統とみなすことができるペチュニア系統)では非常に低かった。
【0050】
例2 花の匂いの調節に関与する転写因子の特徴づけ
ODO1のシークエンスは、294アミノ酸(配列番号1)の推定上のタンパク質をコードすることを明らかにした。該タンパク質は、核局在化シグナルをもたないR2R3型MYBファミリーのメンバーと高い相同性を有する。N末端R2R3ドメイン(配列番号1のアミノ酸1〜128)には、高度に保存されたモチーフが含まれ、かつDNA結合作用を示し、可変コアモチーフおよびへリックス−ターン−へリックス構造の形成への関与が推定されるアミノ酸が含まれる。一方、C末端には、ジーンバンクのデータベースにおいて相同配列は存在しない。系統発生樹の分析によって、ODO1は、Pimpinella brachicarpa 由来のMYBならびにArabidopsis thaliana 由来のAtMYB42 およびAtMYB85 に最も近いことが解る。これらのタンパク質の機能は未知である。R2R3ドメインの17個の可変性に富むアミノ酸は、これらの三つのタンパク質において保存されている。
【0051】
例3 花の匂いの調節における役割を証明するための、ODO1遺伝子のサイレンシング
花の匂いの調節におけるODO1の役割を調べるために、遺伝子組換え手法が使用された。ODO1のMitchellにおける発現は、RNAiを通して抑制された。ODO1は管(tube)および花弁においてのみ発現され、かつ任意の他の組織中においては発現しないので、我々は、RNAi構造物に適した恒常的なプロモーターを使用した。このプロモーターは、データベースにおいて他の遺伝子と相同性を示さないC末端のODO1をコードする配列に作用し、ODO1転写産物の蓄積を抑制する。ネガティブコントロールとして、我々は、RNAi構造物に適したODO1のイントロンを使用してMitchell系統を形質転換した。各独立した形質転換体の花が、ODO1転写産物のレベルについての分析に供された。分析の時刻は、該転写産物が親のMitchell系統において大量に産生される時刻である17:00とした。各遺伝子組換え系統の個々の花による揮発性物質の生産量を調べるために、我々は、Verdonk et al. (2003) Phytochemistry 62, 997-1008. において記載されたように代謝学的手法を使用した。続いて、揮発性物質をあまり産生しない系統の花における揮発性物質の放出が定量化され、親系統の揮発性物質の放出と比較された。これらの転写物質および揮発性分析から、ODO1転写産物のレベルと揮発性ベンゼノイドの放出との間に明確な相関関係があることが明らかになった。
【0052】
例4 サイレンスされた遺伝子導入植物および対照植物による揮発性物質の放出の定量化
独立した遺伝子組換え系統についての放出された揮発性物質の定量的分析は、図1において示され、安息香酸メチル、安息香酸ベンジルおよびイソオイゲノールが、遺伝子組換え系統において最も強い影響を受けた揮発性物質であることを明らかにした。安息香酸メチルの放出は50%まで減少し、安息香酸ベンジルおよびイソオイゲノールの放出は95%まで減少した。バニリン(Mitchellによって低量において放出される物質)は、RNAi系統3では検出され得なかった。該系統におけるODO1の抑制によって、この放出の減少が最も強かった。ODO1を抑制しなかった系統では、揮発性物質の放出の減少は観察されなかった。Floral Binding Protein 1 (FBP1) (花の発達に関与するタンパク質)の転写レベルは、ODO1による明確な影響を受けなかった。この事実は、ODO1ターゲットが高い特異性を有することを示している。
【0053】
例5 シキミ酸経路における複数の酵素およびL-フェニルアラニンアンモニアリアーゼ(PAL)の転写レベルについてのODO1遺伝子のサイレンシングの効果
安息香酸メチル、安息香酸ベンジルおよびイソオイゲノールの合成を誘導する正確な経路は知られていないが、第一の前駆物質であるトランス−ケイ皮酸は、L-フェニルアラニンアンモニアリアーゼ(PAL)による、L-フェニルアラニンの転換によって生成される。シキミ酸経路は、L-フェニルアラニンの生合成を導く。ODO1がシキミ酸経路における複数の酵素の転写レベルおよびPALの転写レベルに影響を与えたか否かを調べるために、我々はRNA-ゲルブロット分析を行った。図2は、シキミ酸経路における第一の酵素である3-デオキシ-D-アラビノ-ヘプツロソナート-7-リン酸塩シンターゼ(DAHPS) の転写レベルが、Mitchell中よりもRNAi植物体において非常に低いことを示す。さらに、5-エノール-ピルビルシキマート-3-リン酸塩シンターゼ(EPSPS)およびPALの転写レベルがまた、RNAi植物において減少したことを示す(図2)。興味深いことに、これらの結果は、odo1が花の匂いを制御するだけでなく、シキミ酸経路の上流に位置する酵素のレベルをも制御することを明確に示す。
【発明の分野】
【0001】
本発明は、植物におけるmyb調節タンパク質に関する。より具体的には、本発明は、R2R3型のmybタンパク質、該タンパク質をコードする遺伝子ならびにこのタンパク質およびそれをコードする遺伝子の適用に関する。
【発明の背景】
【0002】
植物の匂いは、数多くの理由のために重要な性質である。例えば、花による揮発性化合物の産生は、繁殖のプロセスにおいて受粉する昆虫を誘引する重要な役割を担うことができる。同時にまた、良質および高収量の結実のための重要な役割を担うことができる。代わりに、植物はまた、害虫またはその捕食動物を誘引する生殖性および植物性部位において揮発性化合物を生産することもできる。このプロセスにおいて、揮発性の性質は、有害な生物体、例えば害虫、線虫または菌類に対する感受性または抵抗性を決定する。揮発性物質の合成および放出を干渉および修飾することは、関心の対象になり得る。例えば、植物/昆虫の関係について干渉し、生殖プロセスの向上や害虫に対する耐性を向上させる。
【0003】
匂いはまた、フレーバーおよびフレグランス、食品および化粧品産業にとって重要である。なぜなら、多くの場合において、これらの産業では、香水、食品、化粧品等々の使用に適したフレーバーおよびフレグランス成分の供給源として、花、ハーブ、果実および香辛料から天然の揮発性化合物を生産するからである。
【0004】
植物由来の揮発性化合物の重要なクラスは、いわゆるベンゼノイドである。これらのフェノール類化合物は、塩基性C6骨格を有し、特定の器官または特定の細胞種においてシキミ酸経路から生産されることが多い。ランおよびペチュニアのような多数の植物は、そのフレグランスの主要部としてベンゼノイドを生産する花を有する。多数のペチュニア雑種系統において、ベンゼノイド揮発性化合物は特異的に放出され、日/夜リズムにおいて午後の終わりおよび夜の間に花弁によって始まる。現在まで、唯一の制限された知見は、この経路に関与する分子および遺伝子プロセスについて存在する。数少ない構造的遺伝子のみがクローン化および特徴づけられ、現在まで調節遺伝子は同定されていない。従って、植物がベンゼノイドの生合成および放出をどのように調節するのかについては未だ大部分が知られていない。
【0005】
匂いの生合成を、再生的、効果的かつ費用効率的に遮断、増強または修飾する信頼ある方法が植物または他の(微小)生物体において達成され得るならば、これは非常に大きな利益になる。
【図面の簡単な説明】
【0006】
【図1】Mitchell (W115)による、揮発性ベンゼノイドの定量化された放出量。揮発性の減少した放出を示す4つのRNAi系統 (TA-1、TA-3、TA-12およびTA-35) およびベンゼノイドの放出において減少を示さない一つのRNAi系統(40)。
【図2】ODO1、ならびにシキミ酸経路、フェニルアラニンおよびt-ケイ皮酸の合成経路からの遺伝子、例えばDAHPシンターゼ(DAHPS)、EPSPシンターゼ(EPSPS)、コリスミ酸ムターゼ(CM) および二つのフェニルアラニンアンモニアリアーゼ遺伝子 (PAL1および2)についての;安息香酸ベンジルトランスフェラーゼ(BEBT)および安息香酸/サリチル酸メチルトランスフェラーゼ(BSMT)についての、Mitchell(M)およびRNAi系統1、3、12、35および40のRNAゲルブロット分析。花の結合タンパク質1(FBP1)の転写レベルが、RNAi系統におけるODO1抑制の特異性を証明するために示された。
【図3】機能的な保存の指標である保存された残基(暗色部)を明示したODO1相同体のR2R3ドメインのアライメント。
【詳細な説明】
【0007】
本発明は、配列番号1において示されたポリペプチド配列またはその変異体もしくは誘導体を含むDNA結合活性を示すポリペプチドに関する。本発明のこれらのポリペプチドは、R2R3型MYBファミリーに属し、かつシキミ酸経路を調節する。シキミ酸経路を通して、3つの芳香族アミノ酸であるチロシン、フェニルアラニンおよびトリプトファンが合成される。これらの化合物から、さらに他の芳香族化合物が合成される。ベンゼノイドに関するシキミ酸経路における調節タンパク質が同定されたのはこれが初めてである。従って、本発明の利点の1つは、シキミ酸経路における調節タンパク質を最初に提供し、かつ哺乳動物によって生成できない3つの必須アミノ酸の生合成を調節する手段を提供することである。同時に、これらの必須アミノ酸から誘導される芳香族および非芳香族化合物の生合成の調節のための道を開く。3つの芳香族アミノ酸から誘導される最も重要な化合物の例には、ケイ皮酸、クマル酸、カフェー酸、フェルラ酸のような化合物;それ自体が他の多くの産業的に興味ある化合物の中間体になり得るシキミ酸経路由来の化合物が含まれる。2〜3の例を挙げると、ベンゼノイドには、安息香酸メチル、サリチル酸メチル、ベンズアルデヒド、酢酸ベンジル、安息香酸ベンジル、バニリン、イソオイゲノール、ならびにフラボノールおよびアントシアニンを含むフェニルプロパノイドが含まれる。これらの化合物は一次および二次代謝の両方に関与するので、当業者は、本発明のタンパク質が多くの生合成のプロセスに影響を与える手段を提供することを理解するであろう。これには、フェノール酸起原であって、病原体に対する防御に関与する化学物質の調節が含まれる。また、揮発性のベンゼノイドの放出の調節に関与する化学物質の調節が含まれる。シキミ酸経路はまた、細菌および菌類の中に存在するので、本発明の教示はまたこのような系にまで及ぶ。
【0008】
[発明のポリペプチドの変異体および誘導体]
本明細書中で使用されるとき、「変異体」または「誘導体」には、列挙されたポリペプチドの性質が保持される限りにおいて、列挙されたポリペプチドとは置換、欠失、付加がある点で異なるかまたは一以上のアミノ酸との融合である点で異なるペプチドまたは非ペプチド化合物が含まれる。これらの用語にはまた、列挙されたポリペプチドの性質が保持される限りにおいて、幾つかのグリコシル化部位が導入または修飾される点で列挙されたポリペプチドとは異なるペプチドまたは非ペプチド化合物が含まれる。これらの用語にはまた、列挙されたポリペプチドの性質が保持される限りにおいて、修飾基がペプチド構造と共有結合または非共有結合している点で列挙されたポリペプチドとは異なるペプチドまたは非ペプチド化合物が含まれる。特に、変異体および誘導体は、ベンゼノイドに関するシキミ酸経路の遺伝子の転写レベルを操作する能力を保持している。上記転写レベルには、3-デオキシ-D-アラビノ-ヘプツロソナート-7-リン酸塩シンターゼ(DAHPS)、5-エノール-ピルビルシキマート-3-リン酸塩シンターゼ(EPSPS)、コリスミ酸ムターゼ(CM)およびL-フェニルアラニンアンモニアリアーゼ(PAL)の遺伝子の転写レベルが含まれる。
【0009】
一実施態様において、変異体または誘導体には、配列番号1のアミノ酸配列と少なくとも50%、55%、60%、65%、70%、75%、好ましくは80%、85%、90%、95%、97%、98%または99%の同一性を示すアミノ酸配列が含まれる。
【0010】
さらなる他の実施態様において、変異体または誘導体には、配列番号1のアミノ酸13-116と少なくとも90%、95%、97%、98%または99%の同一性を示すアミノ酸配列が含まれる。このアミノ酸の領域は、本発明のポリペプチドのDNA結合ドメイン(R2R3型のmyb DNA結合ドメイン)と一致する。このタイプのmybドメインについてのさらなる情報については、Stracke et al.(2001) Current Opinion in Plant Biology 4: 447-456を参照されたい。
【0011】
さらなる他の実施態様において、変異体または誘導体には、配列番号1のアミノ酸128〜アミノ酸294の領域と少なくとも70%、75%または80%の同一性を示すアミノ酸配列が含まれる。好ましくは、変異体または誘導体には、配列番号2のアミノ酸128〜アミノ酸294の領域と少なくとも85%、87%、89%または90%の同一性を示すアミノ酸配列が含まれる。最も好ましくは、変異体または誘導体には、配列番号1のアミノ酸128〜アミノ酸294の領域と少なくとも94%、97%、98%または99%の同一性を示すアミノ酸配列が含まれる。
【0012】
さらに他の実施態様において、変異体または誘導体は、保存的置換基においてのみ列挙されたポリペプチドとは異なるポリペプチドである。本明細書中において使用されたとき、「アミノ酸の保存的置換基」は、一つのアミノ酸の類似の性質をもつ他のアミノ酸との置換を意味する。例えば、疎水性性質を示すアミノ酸が疎水性性質を示す他のアミノによって置換される場合がある。
【0013】
用語「ペプチド」および「ポリペプチド」は、原則として本明細書中において交換可能に使用され得、アミノ酸の列を含む分子を意味する。
【0014】
アミノ酸の同一性は、これらに限定されないが、以下の既知の方法によって容易に計算し得る(Computational Molecular Biology, Lesk, A. M., ed., Oxford University Press, New York, 1988;Biocomputing: Infomatics and Genome Projects, Smith, D. W., ed., Academic Press, New York, 1993;Computer Analysis of Sequence Data, Part I, Griffin, A. M., and Griffin, H. G., eds., Humana Press, New Jersey, 1994;Sequence Analysis in Molecular Biology, von Heine, G., Academic Press, 1987;およびSequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991;およびCarillo, H., and Lipman, D., SIAM J. Applied Math., 48:1073 (1988))。同一性を決定するための好ましい方法は、試験対象の配列間の最も大きな一致を与えるよう設計される。同一性を決定するための方法は、公共的に利用可能なコンピュータープログラムにおいて体系化される。二配列間の同一性および類似性を決定する好ましいコンピュータープログラム方法には、これらに限定されないが、GCGプログラムパッケージ(Devereux, J., et al., Nucleic Acids Research 12 (1):387 (1984))、BestFit、BLASTP、BLASTN、およびFASTA (Altschul, S. F. et al., J. Mol. Biol. 215:403-410 (1990)) が含まれる。BLAST X プログラムは、NCBIおよび他のソース( BLAST Manual, Altschul, S., et al., NCBI NLM NIH Bethesda
, MD 20894;Altschul, S., et al., J. Mol. Biol. 215:403-410 (1990) )から公共的に利用可能である。既知のSmith Watermanアルゴリズムがまた、同一性を決定するために使用され得る。ポリペプチド配列の比較のための好ましいパラメーターには、以下のものが含まれる1) アルゴリズム: Needleman and Wunsch, J. Mol. Biol. 48:443-453 (1970) 比較マトリックス: BLOSSUM62 from Hentikoff and Hentikoff, Proc. Natl. Acad. Sci. USA. 89:10915-10919 (1992);ギャップペナルティー: 12;およびギャップ長ペナルティー: 4。これらのパラメーターで有用なプログラムは、ウィスコンシン州マディソン所在のGenetics Computer Group社製の「Ogap」プログラムとして公的に利用可能である。上述したパラメーターは、ペプチド比較のための初期設定パラメーター(エンドギャップについてペナルティーのない)である。
【0015】
例えば、トマトESTs SGNU217873およびLeHTM16 (図3を参照) は、ODO1の相同体である。両方のcDNAは、特徴的な一でR2R3ドメイン中のODO1ファミリータンパク質(PbMYB, AtMYB42およびAtMYB85)とホモロジーを共有する。このホモロジーは、機能的保存の指標である。トマトは、その花において大量のベンゼノイドを生成しないが、幾つかのベンゼノイドが成熟したトマト果肉中に蓄積される。SGNU217873のESTsは、花芽および子房においてのみ見出され(SGN転写データベース、http://www.sgn.cornell.edu/を参照)、その発現は、この遺伝子が果肉の成熟過程のベンゼノイド生産に関与することを示す成熟過程において増加する。LeHTM16は果肉成熟中に誘導されない。従って、このMYBは植物における他の箇所においてプロセスを制御するであろう。
【0016】
[本発明のポリヌクレオチド]
他の側面において、本発明は、配列番号2に示された配列をもつヌクレオチド配列または配列番号2と少なくとも50%、55%、60%、65%、70%、75%、好ましくは80%、85%、90%、95%、97%、98%もしくは99%の同一性を示す前記ヌクレオチド配列の変異体を含む、単離された、組み換えまたは合成ポリヌクレオチドであって、かつベンゼノイドに関するシキミ酸経路について制御活性を有するタンパク質をコードするポリヌクレオチドを提供する。
【0017】
一実施態様において、本発明のポリヌクレオチドには、配列番号1に示されたアミノ酸配列をもつポリペプチドをコードするポリヌクレオチド配列、またはベンゼノイドに関するシキミ酸経路について制御活性を有する前記ポリヌクレオチド配列の断片が含まれる。
【0018】
また、配列番号2のポリヌクレオチド配列と相補的である配列、例えば、アンチセンスRNAまたは他の阻害RNA(例えばRNAiに使用)を有するポリヌクレオチド配列は、本発明によって包含される。これらの相補的およびハイブリダイズする配列は任意の長さであってよく、当業者は、適切な長さがその配列が使用される目的に適うであろうことを理解する。例えば、任意の長さの転写後サイレンシング二本鎖RNAが植物において使用され得る。
【0019】
二つのヌクレオチド配列の同一性は、上述した方法を使用して決定される。ヌクレオチド配列、核酸およびポリヌクレオチドなる用語は、原則として本出願において交換可能に使用され得、ヌクレオチドの配列を意味する。本発明のポリヌクレオチドには、ゲノム配列、細胞外ゲノムおよびプラスミドコード配列ならびにタンパク質、ポリペプチド、ペプチド等々を発現するかまたは発現するのに適合されたより小さな加工された遺伝子セグメントが含まれうる。本発明のポリヌクレオチドは、一本鎖(コーディングまたはアンチセンス)または二本鎖であってもよく、DNA(ゲノム、cDNA)またはRNA分子であってもよい。これらは、単離された、組換え体または合成物であってもよい。RNA分子には、イントロンを含みかつ一対一のやり方においてDNA分子と一致する不均一な核RNA(hn RNA)分子、およびイントロンを含まないmRNA分子を含みうる。さらなるコーディングまたは非コーディング配列が、本発明のポリヌクレオチド内に存在していてもよい(但し、必ずしも存在する必要はない)。ポリヌクレオチドは、他の分子および/または支持材料と結合していてもよい(但し、必ずしも結合している必要はない)。
【0020】
「単離された」は、本明細書中において使用されるとき、ポリヌクレオチドが実質的に他の核酸分子から自由であることを意味する。また、ポリヌクレオチドが、関連のない配列の大きな部位、例えば、大きな染色体断片または他の機能的遺伝子またはポリペプチドコーディング領域を含まないことを意味する。当然ながら、これは、最初に単離されたポリヌクレオチド分子を意味し、ヒトの手によってセグメントに後に付加された遺伝子またはコーディング領域を排除するものではない。
【0021】
[本発明のベクター]
他の側面において、本発明は、本発明のポリヌクレオチドを含むベクターに関する。有効に使用され得るベクターには、周知の植物ベクター、例えばpK7GWIG2(I)およびpGreen、ならびに微生物においてタンパク質を形質転換および発現させるために使用されるベクターが含まれる。また、Arabidopsis, A laboratory manual Eds. Weigel & Glazebrook, Cold Spring Harbor Lab Press (2002) およびManiatis et al. Molecular Cloning, Cold Spring Harbor Lab (1982) を参照されたい。
【0022】
[本発明の宿主細胞]
さらに他の側面において、本発明は、本発明によるポリヌクレオチドまたはベクターを含む宿主細胞に関する。本発明による適切な宿主細胞には、植物細胞、酵母細胞、真菌細胞、藻類細胞、ヒト細胞および動物細胞が含まれる。適切な植物細胞の例には、トマトおよびシロイヌナズナが含まれる。適切な宿主細胞の例には、Saccharomyces cerevisiaeおよびPichia pastorisが含まれる。適切な真菌細胞の例には、Aspergillusが含まれる。適切な動物細胞の例には、昆虫細胞、例えば、Spodoptera frugiperda由来の細胞;哺乳類細胞、例えばチャイニーズハムスター卵巣細胞またはPERC6細胞が含まれる。当該技術分野の様々な細胞系統、例えばFlp-In細胞系統(Invitrogen)が使用され得る。上述したように、宿主細胞への本発明のポリヌクレオチドを導入するための様々なベクターが使用され得る。これらのベクターは、例えば、プラスミドDNAベクター、ウイルスDNAベクター(例えば、アデノウイルスまたはアデノ関連ベクター)、ウイルスRNAベクター(例えばレトロウイルス)またはウイルス植物ベクター、例えばタバコ(tobacco rattle)ウイルスおよびジャガイモウイルスXから選択され得る、クローニングベクター、発現ベクター、サイレンシングベクターにし得る。
【0023】
細胞遊離抽出物による本発明のポリペプチドの生産はまた、本発明によって包含される。細胞遊離抽出物における生産のための方法は、当該技術分野において知られている。例えばPelman and Jackson (1976) Eur.J.Biochem 67 : 247-56を参照されたい。
【0024】
本発明の宿主細胞は、本発明のポリペプチドを産生するために使用され得る。これは、ポリペプチドの産生を可能にし、かつ任意的にはポリペプチドを回収する条件下において本発明による宿主細胞を培養することを含む。好ましい実施態様において、DNA結合活性をもつ組換え体ポリペプチドが産生される。
【0025】
一実施態様において、宿主細胞は、本発明によるタンパク質をコードする遺伝子がサイレンシングされるトランスジェニック植物である。結果として、ベンゼノイドに関するシキミ酸経路における酵素が下方制御され、例えば、3-デオキシ-D-アラビノ-ヘプツロソナート-7-ホスファート シンターゼ(DAHPS)、5-エノール-ピルビルシキマート-3-ホスファート シンターゼ(EPSPS)、コリスマート ムターゼ(CM) およびL-フェニルアラニン アンモニア-リアーゼ(PAL) の遺伝子の転写レベルが減少し、植物の揮発性の性質が変化するであろう。
【0026】
他の実施態様において、宿主細胞は、本発明のタンパク質をコードする遺伝子が過剰発現し、かつシキミ酸経路の酵素が上方制御されるトランスジェニック植物である。例えば、3-デオキシ-D-アラビノ-ヘプツロソナート-7-ホスファート シンターゼ(DAHPS)、5-エノール-ピルビルシキマート-3-ホスファート シンターゼ(EPSPS)、コリスマート ムターゼ(CM) およびL-フェニルアラニン アンモニア-リアーゼ(PAL) の遺伝子の転写レベルが増加し、揮発性の性質がより強い匂いが産生されるように修飾される。EPSPS産生の増加したレベルをもつトランスジェニック植物は、化学的防御戦略において特別の関心を集める。とりわけこの戦略においては、グリフォセートを含む植物保護産物が使用される。増加したレベルのEPSPSをもつ植物はグリフォセートに対する増加した耐性を有するためである。
【0027】
さらに他の実施態様において、宿主細胞は、本発明のタンパク質をコードする遺伝子が過剰発現するトランスジェニック植物であり、揮発性の性質は、ベンゼノイド産物を増加させることによって病原生物体に対する植物の化学的防御系を強化するように修飾される。これには、殺虫剤、殺菌剤、殺線虫剤、軟体動物駆除剤または殺鼠剤として機能するベンゼノイドが含まれる。
【0028】
他の実施態様において、宿主細胞は、本発明のタンパク質をコードする遺伝子が過剰発現し、相互抑制効果が生じるトランスジェニック植物においてもたらされるトランスジェニック植物細胞である。結果として、遺伝子がサイレンスされ (Jorgensen et al. (1996) Plant Mol Biol 31: 957-973)、DAHPS、EPSPS、CMおよびPALについてのベンゼノイドに関するシキミ酸経路の遺伝子の転写レベルが減少する。
【0029】
他の実施態様において、宿主細胞は、本発明のタンパク質をコードする遺伝子が当該技術分野において知られた他の方法においてサイレンスされるトランスジェニック植物をもたらすトランスジェニック植物細胞である。
【0030】
さらに他の実施態様において、宿主細胞は、本発明のタンパク質をコードする遺伝子がサイレンスされ、かつ害虫が誘引されないように揮発性の性質が修飾される、トランスジェニック植物においてもたらされるトランスジェニック細胞である。さらに他の実施態様において、宿主細胞は、本発明のタンパク質をコードする遺伝子が、導入の前に該遺伝子を含まないかまたは活性形態において存在しない宿主において導入されるトランスジェニック植物をもたらすトランスジェニック植物細胞である。この方法において、シキミ酸経路および芳香族化合物の生合成経路を制御することが可能である。
【0031】
[抗体]
本発明のポリペプチドに対する抗体がまた、本発明において包含される。ポリクローナルおよびモノクローナル抗体を生成するための方法が当該技術において一般に知られている(例えばHarlow and Lane (1988) Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, New Yorkを参照)。抗体はそれ自体で使用され得るが、好ましくは抗体は検出可能なラベルでラベルされる。適切な抗体ラベルが当業者に知られ、これらに限定されないが、放射活性ラベル、電子密度ラベル、酵素ラベルおよび蛍光ラベルが含まれる。好ましい実施態様において、酵素または蛍光マーカー、例えばアルカリホスファターゼ、西洋ワサビペルオキシダーゼ、およびフルオレセインが使用される。
【0032】
また、細胞内で生産された抗体、いわゆる細胞内発現抗体(intrabody)が本発明によって包含される。細胞内発現抗体の構築は、当該技術、例えば米国特許6,004,940およびWO 01/48017において記載されている。
【0033】
本発明の抗体は、芳香花を同定または検出するための方法において使用され得る。当該方法では、植物材料を本発明の抗体と接触させ;本発明のポリペプチドと結合するか否かを検出する。このような方法はまた、本発明によって包含される。タンパク質は当該技術分野において既知の回収技術、例えばMethods Enzymol. vol. 182, Guide to protein purification. Eds. M.P. Deutscher (1990) Academic Press Inc.を使用して回収され得る。
【0034】
[発明の方法]
本発明によるポリペプチド、ポリヌクレオチド、ベクターもしくは抗体またはこれらの断片(集合的には本発明の化合物と呼ばれる)は、シキミ酸経路の遺伝子の転写レベルを操作する方法、例えば、3-デオキシ-D-アラビノ-ヘプツロソナート-7-ホスファート シンターゼ(DAHPS)、5-エノール-ピルビルシキマート-3-ホスファート シンターゼ(EPSPS)、コリスマート ムターゼ(CM) およびL-フェニルアラニン アンモニア-リアーゼ(PAL) についての遺伝子の転写レベルを操作する方法において使用され得る。これらの酵素を通して、下流生合成プロセスが影響を受ける。例えば、本発明の化合物は、花における匂いを制御する方法において使用され得る。あるいは本発明の化合物は、害虫または病原生物体に対する耐性を調節する方法において使用され得る。従って、植物中の揮発性匂い化合物のの性質を修飾するための;シキミ酸フェニルアラニン合成経路から遺伝子の転写レベルを調節するための;フェニルプロパノイド経路から遺伝子の転写レベルを調節するための;ベンゼノイド生合成に関与する遺伝子の転写レベルを調節するための;または芳香族アミノ酸の生合成、特にフェニルアラニン、チロシンおよびトリプトファンの生合成を調節するための、本発明によるポリペプチド、ポリヌクレオチド、ベクターもしくは抗体またはこれらの断片の使用はまた、本発明において包含される。
【0035】
一実施態様において、本発明の化合物は、揮発性匂い化合物の性質が修飾され得る植物を生産するための方法において使用される。該方法は、植物体に本発明のポリヌクレオチドを導入することを含む。一実施態様において、本発明のポリヌクレオチドは、植物体のゲノム内に導入される。
【0036】
さらに他の実施態様において、本発明の化合物は、匂いを発する植物と匂いを発しない植物とを識別するための方法において使用される。本発明には以下の工程が含まれる:
− 植物と本発明による化合物とを接触させる工程と、
− 上記化合物と結合することを検出するかまたは本発明のヌクレオチド内の多型を検出する工程。
【0037】
さらに他の実施態様において、本発明の化合物は、植物育種中における遺伝的分析またはマーカー補助的選別において使用される。特に、本発明の化合物は、PCRに基づいたマーカー補助的選別において好適に使用され得る。例えば、制限断片長多型(RFLP)、増幅断片長多型(AFLP)、ランダム増幅多型DNA(RAPD)、一塩基多型(SNP)およびマイクロサテライトがある。これらの技術のさらなる詳細については、例えば、Welsh & McClelland (1990) Nucleic Acids Research 18: 7213-7218;Vos et al. (1995) Nucleic Acids Research 23: 4407-4414 and Struss & Plieske (1998) Theoretical & Applied Genomics 97:308-315を参照されたい。二つの育種系統または品種の制限パターンまたはヌクレオチド配列の差異を比較することによって、本発明のポリペプチドをコードする遺伝子内の多型が同定され得る。これは、本発明のポリペプチドと関連した形質(例えば匂いまたは増加したベンゼノイドレベル)の非常に迅速な選択を可能にする。
【0038】
さらに他の実施態様において、本発明の化合物は、本発明によるポリペプチドの下方制御によって害虫に対する耐性を増加させる方法において使用される。下方制御は、揮発性ベンゼノイド(放出されるが、害虫をあまり誘引しないかまたは害虫の捕食者をあまり誘引しない)の性質を変化させるであろう。
【0039】
さらに他の実施態様において、本発明の化合物は、本発明によるポリペプチドの発現を上方制御することによって、病原性生物に対する耐性を増加させるための方法において使用される。上方制御は、病原性生物、例えば病原性細菌および真菌に対する植物の化学的防御メカニズムの一部である、生成されるベンゼノイドの性質における変化を導くであろう。
【0040】
植物および動物系統におけるポリペプチドの発現を上方および下方制御するための方法が、当該技術分野において知られている。上方制御は、植物全体または特定の植物部位(例えば、花弁および葉)における対象のポリペプチドの過剰発現に基づいている。
【0041】
下方制御は、DNAレベル、例えば転写を妨害することによって行われる。代わりに、下方制御は、RNAレベル、例えばタンパク質翻訳の部位へのRNAの転座を妨害することによって、あるいはRNAからのタンパク質の翻訳を妨害することによって、あるいはRNAのスプライシングを妨害し一以上のmRNA化学種を得ることによって行われる。このような発現に対する妨害の全体的効果は、遺伝子の発現における減少(抑制)である。RNAレベルでの妨害が好ましい。RNAレベルでの妨害を達成するための好適な方法は、二本鎖またはヘアピンRNAを使用するRNAiを通した方法;siRNAを使用するサイレンシングを通した方法;または相互抑制作用を通した方法である。例えば、Hammond & Hannon (2001) Nature Rev Gen 2: 110-119, Arabidopsis, A laboratory manual Eds. D. Weigel & J Glazebrook (2002), CSHL Press and Cogoni & Macino (2000) Genes Dev 10: 638-643を参照されたい。
【0042】
下方制御にはまた、翻訳阻害および翻訳後阻害が含まれる。翻訳阻害および翻訳後阻害についての方法は、当業者に周知である。好ましくは、翻訳のリプレッサーとして主に機能し、タンパク質の発現レベルのみに影響を与える内在性21−24 nt RNAであるmiRNAが使用され得;リン酸化、アセチル化、メチル化、グリコシル化、プロリル異性化、シアリル化、ヒドロキシル化、酸化、グルタチオニル化およびユビキチン化が使用され得;または抗体、抗体断片および化学的およびペプチド阻害剤がまた、この目的のために使用され得る。
【0043】
阻害物質を同定するための方法は当該技術分野において既知であり、ペプチド擬態、ペプチド、DNAのライブラリーまたはcDNA発現ライブラリー、コンビナトリアル・ケミストリーおよび、特に有用なファージディスプレーライブラリーをスクリーニングする方法が含まれる。これらのライブラリーは、ライブラリーを、実質的に精製されたポリペプチド、その断片またはその構造的類似体と接触させることによって結合分子についてスクリーニングされ得る。好ましい実施態様において、阻害物質は、本発明のポリペプチドのDNA結合ドメインを標的にする。本明細書中において使用されたとき、用語「阻害物質」には、本発明の化合物と結合するペプチド、ペプチド配列、ペプチド様分子および非ペプチド分子が含まれる。
【0044】
[例]
植物材料および形質転換
Petunia hybrida cv. Mitchell (W115系統ともいう;P. axillaris x (P. axillaris x P. hybrida Rose of Heaven)) およびW138系統植物は、Verdonk et al. Phytochemistry 62, 997-1008 (2003) において記載されたように生育された。少なくとも三つの成熟した花を有する植物が全ての実験において使用された。遺伝子組換えペチュニアは、Agrobacterium tumefaciens (株 GV3101::pMP90) を媒介した形質転換を介して得られた。具体的には、切断葉を細菌培養液中(約28℃、10倍稀釈) に浸漬する。遺伝子組換えカルスは、150 mg/ml カナマイシンを含むMS-培養液上で選択された。植物体は、Lucker et al.. Plant Journal 27, 315-324 (2001) に記載されたように遺伝子組換えカルスから再生された。発根植物は、PCRを使用してntpII 遺伝子およびRNAi構造物の存在について試験された。陽性の植物が温室に移された。
【0045】
ODO1の選別および同定
花弁特異的DNAマイクロアレイの構成、ラベリングおよび分析は、Verdonk et al. Phytochemistry 62, 997-1008 (2003) に記載されている。以下の三つの実験結果が比較されている: 9:00時のMitchell petalsと15:00時のMitchell petals;12:00時のpetalsと15:00時のpetalsおよび15:00時のMitchell petalsと15:00時のW138 (匂いを発しない栽培種) petals。匂いの放出で顕著に(Verdonk et al. Phytochemistry 62, 997-1008 (2003)を参照)および調整的に上方制御されたcDNAおよびW138において上方制御されていないcDNAがシークエンスされた。これらのうちの一つがMYB相同体として同定されたが、DAHPS、EPSPS、CM、PAL1 および2 ならびに BEBTは、これらのマイクロアレイ実験から選択された。RNAゲルブロット分析は、Verdonk et al. Phytochemistry 62, 997-1008 (2003) において記載されたように行われ;特定の3’ UTRプローブがPAL1および2について使用された。
【0046】
RNAiサイレンシング構築物の生成
Gateway(商標) (Invitrogen life technologies, Carlsbad, CA, USA) アダプターを含む二つのプライマーが、ODO1 オープンリーディングフレームのヌクレオチド573〜876の領域を増幅するように設計された。フォワードプライマー: 5’-aaa aag cag gct CAC CAC TGA TGA ATC CAA GC-3’;リバースプライマー: 5’-aga aag ctg ggt CCT GTT CTC TAC GTT ATC-3’ (小文字はプライマー中に構築されたGateway(商標)アダプターを表わす)。増幅したPCR産物は、pDONR207 ベクターにおいてクローニングされ、RNAiデスティネーションベクターpK7GWIWG2(I)に導入された(当該ベクターのnptII遺伝子は植物細胞に対してカナマイシン耐性を与える;VIB、Gent、Belgium)。上記操作は、製造業者用資料(Invitrogen life technologies) によって記載されたように大腸菌DH5α中において行われた。前記構築物がシークエンスされ、続いて標準的な分子生物学的技術を使用してA. tumefaciens GV3101::pMP90細胞に形質転換された。
【0047】
揮発性物質のサンプリング
揮発性物質は、水の入ったエルレンマイヤーグラス中において切り花を静置することによって収集され、続いて、ガラス製の空気の出入口を備えた蓋で閉じられた1リットルの容器内に静置された。炭素ろ過された空気が、容器の出口に真空を適用することによって容器内に導かれた。花の頭隙は、5mmのワイドガラス管中の150mg Tenax TA上で20時間にわたって出て行く空気を捕捉することによって収集された。Tenaxは、内部標準として8.37 ng/μl α-テルピネンを含んだ2mlペンタン:ジエチルエーテル(4:1)で稀釈された。溶出剤中の揮発物質は、キャピラリーガスクロマトグラフィー質量分光測定を通して分析された。1μlの稀釈剤が、Optic(ATAS, GL, International)注入ポートに250℃で注入された。分流は、2分間にわたって0 ml/分であり、流れの終わりまで25 ml/分であった。化合物は、40℃で3分間にわたってキャピラリーDB-5カラム(10×180μm、フィルム厚0.18μm;Hewlett Packard)上で分離され、その後キャリアガスとしてヘリウムを用いて30℃/分で250℃まで加熱された。カラム流は、2分間にわたって3ml/分であり、その後1.5ml/分とした。溶出化合物の質量スペクトルは、70eV(200℃のイオン源)で生成され、20スペクトル/秒の獲得速度で、-1597 eVで90秒の獲得遅延を示す飛行時間MS (Leco, Pegasus III, St.Joseph, MI, USA)上で収集された。化合物は、既知の濃度の合成外部標準および内部標準に基づいて、ならびにKant, et al. Plant Physiology 135, 483-495 (2004)に記載されているように同定および定量された。各系統は、少なくとも3回測定された。各実験について、花の新鮮重量が決定された。
【0048】
例1 ペチュニアの花の匂いの調節に関与した転写因子の同定および発現
ペチュニアの花の匂いの調節に関与した成分を同定するために、トランスクリプトミクス的手法が使用された。匂いを発した花のトランスクリプトームは、匂いを発する直前の花のトランスクリプトームおよび匂いを発しないペチュニア栽培種の花のトランスクリプトームと比較された。比較は専用の特異的なマイクロアレイを使用して行われた。匂いを発する直前に増加した転写レベルを示し、かつ匂いを発しないペチュニアにおいて非常に低い転写レベルを示す転写因子が選択された。一つの転写因子、ODO1 (ODORANT 1) が本明細書中において詳細に記載される。
【0049】
花の匂いを調節する役割と一致して、ODO1の転写レベルは、揮発性ベンゼノイドの放出の開始時点である正午〜14:00時にかけて増加した。ODO1の転写レベルは一時的に増加し、次の早朝に最も低いレベルに戻った。ODO1の発現は、花の管および花弁に制限された。花の発達中、ODO1の転写産物が、開花後〜老化(開花6日後)まで検出された。ODO1の転写レベルは、ペチュニアW138系統(匂いを発しない系統とみなすことができるペチュニア系統)では非常に低かった。
【0050】
例2 花の匂いの調節に関与する転写因子の特徴づけ
ODO1のシークエンスは、294アミノ酸(配列番号1)の推定上のタンパク質をコードすることを明らかにした。該タンパク質は、核局在化シグナルをもたないR2R3型MYBファミリーのメンバーと高い相同性を有する。N末端R2R3ドメイン(配列番号1のアミノ酸1〜128)には、高度に保存されたモチーフが含まれ、かつDNA結合作用を示し、可変コアモチーフおよびへリックス−ターン−へリックス構造の形成への関与が推定されるアミノ酸が含まれる。一方、C末端には、ジーンバンクのデータベースにおいて相同配列は存在しない。系統発生樹の分析によって、ODO1は、Pimpinella brachicarpa 由来のMYBならびにArabidopsis thaliana 由来のAtMYB42 およびAtMYB85 に最も近いことが解る。これらのタンパク質の機能は未知である。R2R3ドメインの17個の可変性に富むアミノ酸は、これらの三つのタンパク質において保存されている。
【0051】
例3 花の匂いの調節における役割を証明するための、ODO1遺伝子のサイレンシング
花の匂いの調節におけるODO1の役割を調べるために、遺伝子組換え手法が使用された。ODO1のMitchellにおける発現は、RNAiを通して抑制された。ODO1は管(tube)および花弁においてのみ発現され、かつ任意の他の組織中においては発現しないので、我々は、RNAi構造物に適した恒常的なプロモーターを使用した。このプロモーターは、データベースにおいて他の遺伝子と相同性を示さないC末端のODO1をコードする配列に作用し、ODO1転写産物の蓄積を抑制する。ネガティブコントロールとして、我々は、RNAi構造物に適したODO1のイントロンを使用してMitchell系統を形質転換した。各独立した形質転換体の花が、ODO1転写産物のレベルについての分析に供された。分析の時刻は、該転写産物が親のMitchell系統において大量に産生される時刻である17:00とした。各遺伝子組換え系統の個々の花による揮発性物質の生産量を調べるために、我々は、Verdonk et al. (2003) Phytochemistry 62, 997-1008. において記載されたように代謝学的手法を使用した。続いて、揮発性物質をあまり産生しない系統の花における揮発性物質の放出が定量化され、親系統の揮発性物質の放出と比較された。これらの転写物質および揮発性分析から、ODO1転写産物のレベルと揮発性ベンゼノイドの放出との間に明確な相関関係があることが明らかになった。
【0052】
例4 サイレンスされた遺伝子導入植物および対照植物による揮発性物質の放出の定量化
独立した遺伝子組換え系統についての放出された揮発性物質の定量的分析は、図1において示され、安息香酸メチル、安息香酸ベンジルおよびイソオイゲノールが、遺伝子組換え系統において最も強い影響を受けた揮発性物質であることを明らかにした。安息香酸メチルの放出は50%まで減少し、安息香酸ベンジルおよびイソオイゲノールの放出は95%まで減少した。バニリン(Mitchellによって低量において放出される物質)は、RNAi系統3では検出され得なかった。該系統におけるODO1の抑制によって、この放出の減少が最も強かった。ODO1を抑制しなかった系統では、揮発性物質の放出の減少は観察されなかった。Floral Binding Protein 1 (FBP1) (花の発達に関与するタンパク質)の転写レベルは、ODO1による明確な影響を受けなかった。この事実は、ODO1ターゲットが高い特異性を有することを示している。
【0053】
例5 シキミ酸経路における複数の酵素およびL-フェニルアラニンアンモニアリアーゼ(PAL)の転写レベルについてのODO1遺伝子のサイレンシングの効果
安息香酸メチル、安息香酸ベンジルおよびイソオイゲノールの合成を誘導する正確な経路は知られていないが、第一の前駆物質であるトランス−ケイ皮酸は、L-フェニルアラニンアンモニアリアーゼ(PAL)による、L-フェニルアラニンの転換によって生成される。シキミ酸経路は、L-フェニルアラニンの生合成を導く。ODO1がシキミ酸経路における複数の酵素の転写レベルおよびPALの転写レベルに影響を与えたか否かを調べるために、我々はRNA-ゲルブロット分析を行った。図2は、シキミ酸経路における第一の酵素である3-デオキシ-D-アラビノ-ヘプツロソナート-7-リン酸塩シンターゼ(DAHPS) の転写レベルが、Mitchell中よりもRNAi植物体において非常に低いことを示す。さらに、5-エノール-ピルビルシキマート-3-リン酸塩シンターゼ(EPSPS)およびPALの転写レベルがまた、RNAi植物において減少したことを示す(図2)。興味深いことに、これらの結果は、odo1が花の匂いを制御するだけでなく、シキミ酸経路の上流に位置する酵素のレベルをも制御することを明確に示す。
【特許請求の範囲】
【請求項1】
DNA結合活性を有するポリペプチドであって、以下の(a)〜(c)か選択されるポリペプチド:
(a)配列番号1のアミノ酸配列と少なくとも50%の同一性を示すポリペプチド;
(b)配列番号1のアミノ酸の領域13〜116と少なくとも90%の同一性を示すアミノ酸配列を含むポリペプチド;
(c)配列番号1のアミノ酸128からアミノ酸294の領域と少なくとも70%の同一性を示すアミノ酸配列を含むポリペプチド。
【請求項2】
以下の群(a)〜(d)から選択されるポリヌクレオチドを含む組換えまたは合成ポリヌクレオチド:
(a)配列番号2に示されたヌクレオチド配列と少なくとも50%の同一性を有するポリヌクレオチドまたはベンゼノイドに関するシキミ酸経路についての活性を調節するペプチドをコードする前記ポリヌクレオチドの断片;および
(b)請求項1に記載のポリペプチドをコードするポリヌクレオチド、またはベンゼノイドに関するシキミ酸経路についての活性を調節するポリペプチドの断片;
(c)配列番号2のポリヌクレオチド配列と相補的な配列を有するポリヌクレオチド;
(d)配列番号2の配列の部分とストリンジェントな条件下でハイブリダイズするポリヌクレオチド配列。
【請求項3】
請求項2に記載のポリヌクレオチドを含むベクター。
【請求項4】
請求項2に記載のポリヌクレオチドまたは請求項3に記載のベクターを含む宿主細胞。
【請求項5】
請求項4に記載の宿主細胞であって、前記宿主細胞が、植物細胞、細菌細胞、酵母細胞、真菌細胞または動物細胞である宿主細胞。
【請求項6】
請求項2に記載のポリヌクレオチドを含む遺伝子組換え植物。
【請求項7】
請求項2のポリヌクレオチドまたは請求項1のポリペプチドと結合する化合物であって、前記化合物は、好ましくは抗体、抗体の抗原結合性断片、またはこれらの誘導体;または請求項1に記載のポリヌクレオチドの配列の部分と相補的な配列を有するポリヌクレオチドである化合物。
【請求項8】
ポリペプチドの生産を可能にする条件下において請求項3または4に記載の宿主細胞を培養し、かつポリペプチドを回収することを含む、ベンゼノイドに関するシキミ酸経路についての活性を調節する組換え体ポリペプチドを生産する方法。
【請求項9】
請求項2に記載のポリヌクレオチド、請求項3に記載のベクター、請求項4または5に記載の宿主細胞、請求項1に記載のポリペプチド、または請求項8に記載の化合物の、植物中の揮発性匂い化合物の性質を修飾するための;シキミ酸フェニルアラニン合成経路からの遺伝子の転写レベルを調節するための;フェニルプロパノイド経路からの遺伝子の転写レベルを調節するための;ベンゼノイド生合成に関与する遺伝子の転写レベルを調節するための;または芳香族アミノ酸の生合成を調節するための、特にフェニルアラニン、チロシンおよびトリプトファンの生合成を調節するための、使用。
【請求項10】
前記揮発性匂い化合物の性質が修飾され得る植物を生産するための方法であって、前記方法が、請求項2に記載の(a)または(b)のもとで言及されたポリヌクレオチドを植物体のゲノムに導入することを含む方法。
【請求項11】
花の匂いを調節するための方法であって、前記方法が、請求項2に記載のポリヌクレオチドによってコードされたタンパク質の発現のレベルを操作することを含む方法。
【請求項12】
匂いを発する植物と匂いを発しない植物とを識別するための方法であって、
前記方法が、
− 植物と請求項8に記載の化合物とを接触することと、
− 請求項1に記載のポリペプチドと結合することを検出するかまたは請求項2に記載の(a)または(b)のもとで言及されたヌクレオチド内の多型を検出することと
を含む方法。
【請求項13】
害虫または病原性生物に対する植物の耐性を調節または修飾するための方法であって、前記方法が、請求項8に記載のポリペプチドの発現を修飾することを含む方法。
【請求項14】
植物中においてシキミ酸フェニルアラニン合成経路からの遺伝子の転写レベルを調節するための;フェニルプロパノイド経路からの遺伝子の転写レベルを調節するための;ベンゼノイド生合成に関与する遺伝子の転写レベルを調節するための;または芳香族アミノ酸の生合成を調節するための、特にフェニルアラニン、チロシンおよびトリプトファンの生合成を調節するための方法であって、
前記方法が、
(a)請求項2に記載の(a)または(b)のもとで言及されたポリヌクレオチドの転写レベルを修飾することと;
(b)請求項1に記載のポリペプチドの発現レベルを修飾することと;または
(c)請求項8に記載の化合物を植物に導入することと
を含む方法。
【請求項15】
遺伝的分析またはマーカー補助的選択のための方法における本発明のポリヌクレオチドの使用。
【請求項16】
植物育種における請求項1に記載のポリペプチドの使用。
【請求項1】
DNA結合活性を有するポリペプチドであって、以下の(a)〜(c)か選択されるポリペプチド:
(a)配列番号1のアミノ酸配列と少なくとも50%の同一性を示すポリペプチド;
(b)配列番号1のアミノ酸の領域13〜116と少なくとも90%の同一性を示すアミノ酸配列を含むポリペプチド;
(c)配列番号1のアミノ酸128からアミノ酸294の領域と少なくとも70%の同一性を示すアミノ酸配列を含むポリペプチド。
【請求項2】
以下の群(a)〜(d)から選択されるポリヌクレオチドを含む組換えまたは合成ポリヌクレオチド:
(a)配列番号2に示されたヌクレオチド配列と少なくとも50%の同一性を有するポリヌクレオチドまたはベンゼノイドに関するシキミ酸経路についての活性を調節するペプチドをコードする前記ポリヌクレオチドの断片;および
(b)請求項1に記載のポリペプチドをコードするポリヌクレオチド、またはベンゼノイドに関するシキミ酸経路についての活性を調節するポリペプチドの断片;
(c)配列番号2のポリヌクレオチド配列と相補的な配列を有するポリヌクレオチド;
(d)配列番号2の配列の部分とストリンジェントな条件下でハイブリダイズするポリヌクレオチド配列。
【請求項3】
請求項2に記載のポリヌクレオチドを含むベクター。
【請求項4】
請求項2に記載のポリヌクレオチドまたは請求項3に記載のベクターを含む宿主細胞。
【請求項5】
請求項4に記載の宿主細胞であって、前記宿主細胞が、植物細胞、細菌細胞、酵母細胞、真菌細胞または動物細胞である宿主細胞。
【請求項6】
請求項2に記載のポリヌクレオチドを含む遺伝子組換え植物。
【請求項7】
請求項2のポリヌクレオチドまたは請求項1のポリペプチドと結合する化合物であって、前記化合物は、好ましくは抗体、抗体の抗原結合性断片、またはこれらの誘導体;または請求項1に記載のポリヌクレオチドの配列の部分と相補的な配列を有するポリヌクレオチドである化合物。
【請求項8】
ポリペプチドの生産を可能にする条件下において請求項3または4に記載の宿主細胞を培養し、かつポリペプチドを回収することを含む、ベンゼノイドに関するシキミ酸経路についての活性を調節する組換え体ポリペプチドを生産する方法。
【請求項9】
請求項2に記載のポリヌクレオチド、請求項3に記載のベクター、請求項4または5に記載の宿主細胞、請求項1に記載のポリペプチド、または請求項8に記載の化合物の、植物中の揮発性匂い化合物の性質を修飾するための;シキミ酸フェニルアラニン合成経路からの遺伝子の転写レベルを調節するための;フェニルプロパノイド経路からの遺伝子の転写レベルを調節するための;ベンゼノイド生合成に関与する遺伝子の転写レベルを調節するための;または芳香族アミノ酸の生合成を調節するための、特にフェニルアラニン、チロシンおよびトリプトファンの生合成を調節するための、使用。
【請求項10】
前記揮発性匂い化合物の性質が修飾され得る植物を生産するための方法であって、前記方法が、請求項2に記載の(a)または(b)のもとで言及されたポリヌクレオチドを植物体のゲノムに導入することを含む方法。
【請求項11】
花の匂いを調節するための方法であって、前記方法が、請求項2に記載のポリヌクレオチドによってコードされたタンパク質の発現のレベルを操作することを含む方法。
【請求項12】
匂いを発する植物と匂いを発しない植物とを識別するための方法であって、
前記方法が、
− 植物と請求項8に記載の化合物とを接触することと、
− 請求項1に記載のポリペプチドと結合することを検出するかまたは請求項2に記載の(a)または(b)のもとで言及されたヌクレオチド内の多型を検出することと
を含む方法。
【請求項13】
害虫または病原性生物に対する植物の耐性を調節または修飾するための方法であって、前記方法が、請求項8に記載のポリペプチドの発現を修飾することを含む方法。
【請求項14】
植物中においてシキミ酸フェニルアラニン合成経路からの遺伝子の転写レベルを調節するための;フェニルプロパノイド経路からの遺伝子の転写レベルを調節するための;ベンゼノイド生合成に関与する遺伝子の転写レベルを調節するための;または芳香族アミノ酸の生合成を調節するための、特にフェニルアラニン、チロシンおよびトリプトファンの生合成を調節するための方法であって、
前記方法が、
(a)請求項2に記載の(a)または(b)のもとで言及されたポリヌクレオチドの転写レベルを修飾することと;
(b)請求項1に記載のポリペプチドの発現レベルを修飾することと;または
(c)請求項8に記載の化合物を植物に導入することと
を含む方法。
【請求項15】
遺伝的分析またはマーカー補助的選択のための方法における本発明のポリヌクレオチドの使用。
【請求項16】
植物育種における請求項1に記載のポリペプチドの使用。
【図1】


【図2】

【図3】

【図2】
【図3】
【公開番号】特開2012−115273(P2012−115273A)
【公開日】平成24年6月21日(2012.6.21)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−1354(P2012−1354)
【出願日】平成24年1月6日(2012.1.6)
【分割の表示】特願2007−529751(P2007−529751)の分割
【原出願日】平成17年9月1日(2005.9.1)
【出願人】(505477187)ケイヘーネ・エヌ・ブイ (10)
【Fターム(参考)】
【公開日】平成24年6月21日(2012.6.21)
【国際特許分類】
【出願番号】特願2012−1354(P2012−1354)
【出願日】平成24年1月6日(2012.1.6)
【分割の表示】特願2007−529751(P2007−529751)の分割
【原出願日】平成17年9月1日(2005.9.1)
【出願人】(505477187)ケイヘーネ・エヌ・ブイ (10)
【Fターム(参考)】
[ Back to top ]