
植物レクチンタンパク質の生産方法

【課題】植物レクチンタンパク質を効率的かつ安定的に生産するための手段を提供する。
【解決手段】ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換工程と、前記第一形質転換工程によって得られた形質転換体の中から前記転写因子を発現する形質転換体を選抜する選抜工程と、RNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる植物レクチンタンパク質発現用DNA断片を、前記選抜工程によって選抜された形質転換体に導入する第二形質転換工程とからなることを特徴とする植物レクチンタンパク質生産用形質転換体の生産方法に関する。
【解決手段】ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換工程と、前記第一形質転換工程によって得られた形質転換体の中から前記転写因子を発現する形質転換体を選抜する選抜工程と、RNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる植物レクチンタンパク質発現用DNA断片を、前記選抜工程によって選抜された形質転換体に導入する第二形質転換工程とからなることを特徴とする植物レクチンタンパク質生産用形質転換体の生産方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は植物レクチンタンパク質を効率的に大量生産するための技術を提供する。
本発明また、1kbを越える鎖長のDNAからなる遺伝子によりコードされるタンパク質を効率的に大量生産するための技術を提供する。
【背景技術】
【0002】
レクチンは糖鎖と特異的に結合するタンパク質であり、細胞の凝集や糖複合体を沈降させる性質を有する。レクチンは動植物界に広く存在している。レクチンは、近年の糖鎖研究の展開に伴い重要度が増しており、医学や生物学におけるツールとして様々な場面で利用されている。
【0003】
本発明者らはゴーヤ(「ツルレイシ」とも呼ばれる; Momordica charantia)由来のレクチンを単離し、アミノ酸配列及び該アミノ酸配列をコードする遺伝子の塩基配列を特定している(特許文献1)。
【0004】
現在市販されているレクチンの大部分は植物等から抽出し精製されたものである。しかしながら、このような分離法による製造方法では、原料が天然物であるためにロット間の品質の差異や供給量に対する懸念がある。
【0005】
一方、天然物由来のタンパク質を効率的に生産するための技術として本発明者らはこれまでに、転写因子をコードする遺伝子が、当該転写因子の発現に最も適した染色体上の位置に組み込まれた形質転換体(細胞)をはじめに作出し、得られた形質転換体に、緑色蛍光タンパク質をコードする遺伝子が挿入されたウイルスベクターを導入すれば、高効率でウイルスベクターを誘導発現し、緑色蛍光タンパク質を高発現する形質転換体(細胞)を高確率で作出できることを見出し、特許出願している(特許文献2)。
【0006】
しかしながら、植物レクチンタンパク質を遺伝子組み換え技術を用いて効率的に生産する技術は未だに確立されていない。従来の微生物を用いた発現系は、植物レクチンタンパク質の機能を保持した状態で発現させることが困難であった。また、植物による従来の発現系では、宿主植物にとって導入する植物レクチンタンパク質が生育に好ましくないものであるため、生産可能な植物レクチンの種類が限定されるという問題があった。
【0007】
一方、1kbを越える鎖長のDNAからなる遺伝子によりコードされるタンパク質は、ウイルスベクターを用いて細胞に生産させることは従来困難であった。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特開2009−39006号公報
【特許文献2】特開2005−245228号公報
【特許文献3】特許第3752223号公報
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明は、植物レクチンタンパク質を効率的かつ安定的に生産するための手段を提供することを目的とする。
【0010】
本発明はまた、1kbを越える鎖長のDNAからなる遺伝子によりコードされるタンパク質を効率的かつ安定的に生産するための手段を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明は以下の発明を包含する。
(1)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換工程と、
前記第一形質転換工程によって得られた形質転換体の中から前記転写因子を発現する形質転換体を選抜する選抜工程と、
RNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる植物レクチンタンパク質発現用DNA断片を、前記選抜工程によって選抜された形質転換体に導入する第二形質転換工程とからなることを特徴とする植物レクチンタンパク質生産用形質転換体の生産方法。
【0012】
(2)上記ホルモンが、エストロジェンまたはステロイドホルモンであることを特徴とする(1)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0013】
(3)上記エストロジェンで活性化される性質を有する転写因子としてLexA−VP16−hERを用い、上記転写誘導型プロモーターとしてOLexA−46を用いることを特徴とする(2)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0014】
(4)上記ウイルスベクターは、一本鎖プラス鎖RNAを遺伝子とするウイルス由来であることを特徴とする(1)〜(3)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
【0015】
(5)上記ウイルスベクターが、植物ウイルス由来であることを特徴とする(4)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0016】
(6)上記ウイルスベクターが、植物のサイレンシングを抑制する因子を持つ植物ウイルス由来であることを特徴とする(5)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0017】
(7)上記ウイルスベクターが、トバモウイルス属に属するウイルス由来であることを特徴とする(6)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0018】
(8)上記ウイルスベクターが、トマトモザイクウイルスベクターまたはタバコモザイクウイルスベクターであることを特徴とする(7)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0019】
(9)上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、植物レクチンタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターである、(1)〜(8)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
【0020】
(10)上記ウイルスベクターのcDNAの3’末端に、リボザイム配列が結合していることを特徴とする(1)〜(9)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
【0021】
(11)上記リボザイム配列が、肝炎デルタウイルスのリボザイム配列、またはサテライトタバコリングスポットウイルスのリボザイム配列であることを特徴とする(10)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0022】
(12)上記転写因子発現用DNA断片、および上記植物レクチンタンパク質発現用DNA断片が、アグロバクテリウム法により導入されていることを特徴とする(1)〜(11)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
【0023】
(13)上記宿主細胞および形質転換体が、植物体または植物由来培養細胞であることを特徴とする(1)〜(12)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
【0024】
(14)上記植物由来培養細胞がタバコ由来細胞であることを特徴とする(13)の植物レクチンタンパク質生産用形質転換体の生産方法。
(15)上記タバコ由来細胞がタバコBY2細胞であることを特徴とする(14)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0025】
(16)上記宿主細胞がタバコBY2細胞であり、
上記転写因子がLexA−VP16−hERであり、
上記転写誘導型プロモーターがOLexA−46であり、
上記ウイルスベクターが、トバモウイルス属に属するウイルス由来であり、
上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、植物レクチンタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターであり、
上記ウイルスベクターのcDNAの3’末端に、リボザイム配列が結合しており、かつ
上記転写因子発現用DNA断片、および上記植物レクチンタンパク質発現用DNA断片が、アグロバクテリウム法により導入される、
ことを特徴とする、(1)〜(15)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
【0026】
(17)上記植物レクチンタンパク質がゴーヤ由来レクチンタンパク質である、(1)〜(16)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
(18)(1)〜(17)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法によって生産された植物レクチンタンパク質生産用形質転換体。
【0027】
(19)(18)の植物レクチンタンパク質生産用形質転換体を用いることを特徴とする植物レクチンタンパク質の生産方法。
(20)(1)〜(17)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法を行うためのキット。
【0028】
(21)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換工程と、
前記第一形質転換工程によって得られた形質転換体の中から前記転写因子を発現する形質転換体を選抜する選抜工程と、
RNAを遺伝子とするウイルスに、生産しようとするタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなるタンパク質発現用DNA断片を、前記選抜工程によって選抜された形質転換体に導入する第二形質転換工程とからなり、
上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、生産しようとするタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターであることを特徴とする、タンパク質生産用形質転換体の生産方法。
【0029】
(22)生産しようとするタンパク質をコードする遺伝子が、1kbを越える鎖長のDNAからなる遺伝子である、(21)のタンパク質生産用形質転換体の生産方法
【0030】
(23)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片と、
RNAを遺伝子とするウイルスに、生産しようとするタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなるタンパク質発現用DNA断片であって、上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、生産しようとするタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターである、タンパク質発現用DNA断片と
が遺伝子DNAに導入されていることを特徴とする、タンパク質生産用形質転換体。
【発明の効果】
【0031】
本発明の方法により提供される植物レクチンタンパク質生産用形質転換体により、植物レクチンタンパク質を効率的かつ安定的に生産することが可能である。本発明の方法により生産される植物レクチンタンパク質は、植物から分離された植物レクチンタンパク質と実質的に同一の活性を維持している。
【0032】
本発明により提供されるウイルスベクターを用いて形質転換された細胞は、1kbを越える鎖長のDNAからなる遺伝子によりコードされるタンパク質を効率的かつ安定的に生産することができる。
【図面の簡単な説明】
【0033】
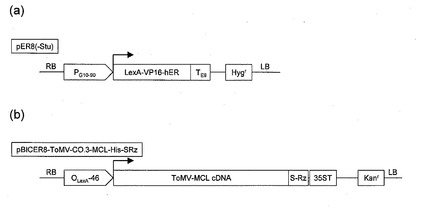
【図1】図1(a)はpER8(−Stu)の一部分の構造を示す。図1(b)はpBICER8−ToMV−CO.3−MCL−His−SRZの一部分の構造を示す。
【図2−1】図2−1はToMV変異体のcDNAの構造を示す。
【図2−2】図2−2はToMV変異体のcDNAの構造を示す(図2−1のつづき)。
【図3】図3は本発明の発現系由来レクチンの電気泳動の結果を示す。
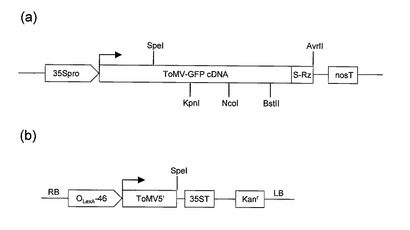
【図4】図4(a)はpiL.erG3(SF3)SRz(Avr)の一部分の構造を示す。図4(b)はpBICER8−ToMV5’−Speの一部分の構造を示す。
【発明を実施するための形態】
【0034】
本発明の実施の形態について具体的に説明する。
(A)本発明にかかる植物レクチンタンパク質生産用形質転換体の生産方法
本発明にかかる植物レクチンタンパク質生産用形質転換体の生産方法(以下、本形質転換体の生産方法と称する)は、(1)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を細胞に導入する第一形質転換工程と、(2)前記第一形質転換工程によって得られた形質転換体の中から前記転写因子を発現する形質転換体を選抜する選抜工程と、(3)RNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる植物レクチンタンパク質発現用DNA断片を、前記選抜工程によって選抜された形質転換体に導入する第二形質転換工程とからなる。以下、工程ごとに説明する。
【0035】
(A−1)第一形質転換工程
本第一形質転換工程では、ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、該転写因子の発現を担うプロモーター(転写因子発現用プロモーター)とを連結して構築したDNA断片(転写因子発現用DNA断片)を、適当な宿主細胞に導入する工程である。当該工程によって、後述するウイルスベクター発現用プロモーターの転写誘導を行なう転写因子を安定的に高発現する形質転換体(細胞)の生産(作出)を行なう。すなわち宿主細胞の転写因子の発現に最適な染色体上の位置に、該転写因子をコードする遺伝子が組み込まれた形質転換体(細胞)候補を生産(作出)する工程である。
【0036】
<転写因子発現用DNA断片>
次に本第一形質転換工程で使用する転写因子発現用DNA断片について説明する。転写因子発現用DNA断片は、転写因子発現用プロモーターの下流に転写因子をコードする遺伝子が連結されている。またこの他、ベクター配列、ターミネーター、薬剤耐性マーカー等のDNAセグメントが含まれていてもよい。かかる転写因子発現用DNA断片の構築方法は、通常の遺伝子工学的手法を用いて行なえばよい。
【0037】
ここで転写因子発現用プロモーターは、転写因子を発現することが可能なものであれば特に限定されるものではなく、また恒常的にプロモーター活性を有するもの(以下、恒常的プロモーターと称する)であってもよいし、さらに転写因子によってプロモーター活性が誘導されるものであってもよい。ただし、転写因子の発現をさらに別の転写因子で制御することは、植物レクチンタンパク質の発現系自体が複雑化すること、コスト面で不利等の理由により、上記転写発現用プロモーターとしては、恒常的プロモーターの方がより好ましいといえる。恒常的プロモーターの例としては、カリフラワーモザイクウイルス35Sプロモーター、PG10−90(Ishige, F., Takaichi, M., Foster, R.,Chua, N. H. and Oeda, K.(1999) A G-box motif(GCCACGTGCC) tetramer confers high-level constitutive expression in dicot and monocot plants. Plant J. 20, 127-133. 参照)、ユビキチンプロモーター、アクチンプロモーター等が挙げられる。
【0038】
一方、使用する転写因子は、ホルモンによって活性化される性質を有する転写因子であって、後述する第二形質転換工程において導入する植物レクチンタンパク質発現用DNA断片中に含まれるプロモーターを転写誘導するものであれば特に限定されない。転写因子を活性化するホルモンとしては、エストロジェン、ステロイドホルモン、エクジソン等が挙げられる。かかる転写因子は、ホルモンが存在しない状態においては不活性型でありプロモーターの転写誘導を行なうことができないが、ホルモンが存在することによって活性型に変化しプロモーターの転写誘導を行なうことができる。この性質を利用すれば植物レクチンタンパク質の生産をより綿密に制御できるため、より安全に植物レクチンタンパク質の生産を行なうことができるといえる。すなわち、植物レクチンタンパク質生産を行なう必要がないとき、あるいは植物レクチンタンパク質生産を行なってはいけないときには、植物レクチンタンパク質生産系にホルモンを添加しなければよい。
【0039】
かかるホルモンによって活性化される性質を有する転写因子、および該転写因子によって転写誘導されるプロモーターの組み合わせとしては、例えば、ステロイドホルモンで活性化される転写因子であるGVGと該転写因子によって転写誘導されるプロモーターである6xUASgal4の組み合わせ、エストロジェンで活性化される転写因子LexA−VP16−hERと該転写因子によって転写誘導可能なプロモーターであるOLexA−46(Zuo J, Niu QW, Chua NH. “An estrogen receptor-based transactivator XVE mediates highly inducible gene exprssion in transgenic plants.” Plant J.2000, 24: 265-273 参照)の組み合わせ、エクジソンで活性化される転写因子であるエクジソンレセプターGR Act and DBDとヘルペスウイルストランスアクチベーションドメインHecR LBDのキメラタンパク質と該転写因子によって転写誘導可能なプロモーターGREの組み合わせ等が挙げられる。ただし、植物レクチンタンパク質生産用の宿主として植物体および植物細胞を利用する場合においては、活性化に用いるホルモンによる宿主への悪影響が少ないという理由からエストロジェンで活性化される転写因子LexA−VP16−hERと該転写因子によって転写誘導可能なプロモーターであるOLexA−46の組み合わせが好ましいといえる。
【0040】
図1(a)に転写因子発現用DNAの一例を示す。図1(a)は、形質転換用ベクターであるTiプラスミドpER8(−Stu)の一部分を示している。同図中左から転写因子発現用の恒常的プロモーターPG10−90、その下流にエストロジェンレセプターを含む融合転写因子LexA−VP16−hER、その3’末端にターミネーター配列TE9、またその下流に薬剤耐性マーカーとしてハイグロマイシン耐性遺伝子Hygrがある。
【0041】
<転写因子発現用DNA断片を導入する宿主細胞およびDNA断片導入方法>
上記転写因子発現用DNA断片が導入される宿主細胞は特に限定されるものではなく、動物由来細胞であっても植物由来細胞であってもよい。ただし、植物由来細胞は、動物由来細胞に比して、増殖速度が速くコンタミネーションのリスクが少ない点、培地作成費用が非常に安価であるという点において、植物由来細胞がより好ましい。なお、動物由来細胞および植物由来細胞とは、細胞、組織、並びに器官も含む意味である。特に液体培地等で培養可能な細胞(培養細胞)が好ましい。
【0042】
また由来となる動物としては、特に限定されるものでないが、ヒト、サル、イヌ、ヒツジ、ヤギ、ウサギ、マウス、ラット、モルモット、チャイニーズハムスター、ウシ、ウマ、ブタ、メダカやゼブラフィッシュ等の魚類、カイコ、夜蛾(Spodoptera frugiperda)等が挙げられる。一方由来となる植物としては、特に限定されるものではないが、イネ、シロイナズナ、オオムギ、コムギ、タバコ、トマト、キュウリ、ダイズ、ジャガイモ、トウモロコシ、ニチニチソウ、シロイヌナズナ、アルファルファが挙げられる。その他、枯草菌や乳酸菌などの菌類、酵母など単細胞生物が宿主細胞として利用できる。
【0043】
また動物由来細胞の例としては特に限定されるものではないが、HeLa細胞、CHO細胞、メラノーマ細胞、マウス3T3細胞が挙げられる。また植物由来細胞の例としては、タバコBY2細胞、ジャガイモ由来、イネ由来、サツマイモ由来、ダイズ由来、パセリ由来、シロイヌナズナ由来、コムギ由来、トウモロコシ由来細胞、ニチニチソウ由来細胞が挙げられる。
【0044】
後述する実施例において、タバコBY2細胞を宿主として用いている。タバコBY2細胞(Toshiyuki nagata, Yasuyuki Nemoto, and Seiichiro Hasezawa “Tobacco BY-2 Cell Line as the “Hela” Cell in the Cell Biology of Higher Plants” International Review of cytology, vol.132, p.p. 1-30 (1992) 、および http://www.riken.go.jp/r-world/info/release/press/2003/030620/ 参照)は、植物培養細胞株としては、世界中で最も広く用いられているものであり、最も増殖速度が速いこと、遺伝子操作が容易なこと、大量培養を容易に行うことができるという理由から採用した。
【0045】
また、転写因子発現用DNA断片を宿主細胞へ導入する形質転換方法は特に限定されるものではなく、宿主細胞の種類に応じた適切な形質転換方法を用いればよい。例えば、特別なベクター配列を有しないDNA断片を導入する場合には、エレクトロポレーション法、パーティクルガン法、リン酸カルシウム法等を用いることができる。また植物由来細胞への一般的な形質転換法としては、アグロバクテリウムを用いた形質転換法(アグロバクテリウム法)を挙げることができ、本発明でもアグロバクテリウム法を好適に用いることができる。ただしアグロバクテリウム法を用いて形質転換する場合には、本発明にかかるDNA断片を含むTiプラスミドを構築する必要がある。またその他プロトプラスト/スフェロプラスト法、リン酸カルシウム法、リポソーム法、DEAEデキストラン法等の従来公知の方法を好適に用いることができる。
【0046】
ただし転写因子発現用DNA断片は、宿主細胞のゲノムに組み込まれることが好ましい。転写因子発現用DNA断片がゲノムに組み込まれることにより、細胞分裂後の娘細胞にもベクターの構成に含まれる遺伝子を確実に伝達することが可能となり、転写因子(タンパク質)の生産効率を維持することが可能となるからである。ゲノムは染色体(核ゲノム)に限定されるものではなく、ミトコンドリアゲノムや葉緑体ゲノムも含まれる。
【0047】
また転写因子発現用DNA断片が宿主細胞に導入されたか否かを確認する方法は、特に限定されるものではなく、公知の各種の方法を用いることができる。具体的には、各種マーカーを用いればよい。例えば、宿主細胞中で欠失している遺伝子をマーカーとして用い、このマーカーと組み換え植物ウイルス遺伝子とを含むプラスミド等を発現ベクターとして宿主細胞に導入する。これによってマーカー遺伝子の発現から本発明の遺伝子の導入を確認することができる。例えば後述する実施例においては、薬剤耐性マーカー(ハイグロマイシン耐性遺伝子、Hygr)を用いており、ハイグロマイシンを含有する培地中で、形質転換候補株を培養することにより、生育してきた細胞株を形質転換体として選抜することが可能となる。その他のマーカーとしては、ビアラホス耐性マーカー、カナマイシン耐性マーカー等が植物細胞の選抜に有効であり、ピューロマイシン耐性マーカー、ブレオマイシン耐性マーカー、XGPRT遺伝子、DHFR遺伝子、チミジンキナーゼ遺伝子等が動物細胞の選抜には有効である。さらに酵母などでは、ウラシル要求性マーカーをはじめとする栄養要求性マーカーを選抜に用いることができる。但しこれら形質転換体の選抜方法は、限定されるものではなく、発現ベクターを導入する宿主等に応じて適宜選択して用いればよい。その他、宿主細胞から調製したゲノムDNAを鋳型とし、導入したタンパク質(転写因子)の遺伝子全長を特異的に増幅するいわゆるジェノミックPCR法を挙げることができる。この方法によって、目的タンパク質(転写因子)をコードする遺伝子が増幅されてくることを電気泳動法等によって確認できれば、該遺伝子の導入を確認することができる。
【0048】
(A−2)選抜工程
本形質転換体の生産方法における第2工程として、当該選抜工程がある。当該選抜工程では、上記第一形質転換工程によって得られた形質転換体(細胞)の中から前記転写因子を安定的に高発現する形質転換体(細胞)の選抜を行なう。すなわち転写因子の発現に最適な宿主細胞の染色体上の位置に、該転写因子をコードする遺伝子が組み込まれた形質転換体(細胞)を選抜(スクリーニング)する工程である。
【0049】
当該選抜工程における具体的選抜方法は上記第一形質転換工程で得られた形質転換体(細胞)の中から目的とする転写因子を安定的に高発現する形質転換体を選抜する方法であれば特に限定されるものではない。
【0050】
例えば、形質転換体(細胞)から発現タンパク質を抽出し、該転写因子特異抗体を用いてその発現量を検出する方法として、ウエスタンブロット法、エライザ法、ドットブロット法等が適用可能である。また、形質転換体(細胞)から発現RNAを抽出し、該転写因子の遺伝子配列に相補的なプローブを用いて転写因子のmRNAの発現量を検出する方法として、例えばノーザンブロット法、ドットブロット法等が適用可能である。さらに、例えばRT−PCR法、リアルタイムPCR法、マイクロアレイ法等によって該転写因子mRNAの発現量を検出することも可能である。
【0051】
(A−3)第二形質転換工程
当該第二形質転換工程は、上記選抜工程により選抜された転写因子を高発現する形質転換体(細胞)(以下、転写因子高発現形質転換体(細胞)と称する)に、RNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子高発現形質転換体が発現する転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる植物レクチンタンパク質発現用DNA断片を導入する工程である。あらかじめ選抜された、転写因子を高発現する形質転換体に、目的とする植物レクチンタンパク質をコードする遺伝子が挿入されたウイルスベクターのcDNAを導入することにより、高確率に植物レクチンタンパク質を高生産する形質転換体(細胞)を生産することができる。
【0052】
<植物レクチンタンパク質発現用DNA断片>
本植物レクチンタンパク質発現用DNA断片は、形質転換体(細胞)に植物レクチンタンパク質を生産させるために使用するDNA断片であり、少なくともRNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子高発現形質転換体(細胞)が発現する転写因子で転写誘導される転写誘導型プロモーターとからなる。この他、植物レクチンタンパク質をコードする遺伝子挿入用のクローニングサイト(好ましくはマルチクローニングサイト)・ベクター配列・ターミネーター・薬剤耐性マーカー等のDNAセグメントが含まれていてもよい。かかる転写因子発現用DNA断片の調製方法は、通常の遺伝子工学的手法を用いて行なえばよい。
【0053】
転写誘導型プロモーターは、転写因子高発現形質転換体が生産する転写因子に応じて選択すればよく、その組み合わせの一例については、(A−1)第一形質転換工程の項で述べたとおりである。
【0054】
細胞に生産させる植物レクチンタンパク質は特に限定されない。例えば、ゴーヤ由来抗Hレクチン(配列番号1及び2)、コムギ由来WGA、トウゴマ由来RCA、ピーナツ由来PNA、タチナタマメ由来ConA、ダイズ由来SBA、ハリエニシダ由来UEA、ジャガイモ由来STL等の種々の植物レクチンタンパク質が例示できる。
【0055】
ホルモンによって活性化される性質を有する転写因子を用いて植物レクチンタンパク質の転写誘導を行う本発明の方法は、各種植物に由来するレクチンタンパク質を生産するための形質転換体の生産に用いることができる。植物レクチンには細胞毒性を示すものがあり、異種発現系を構築することが困難であるが、この問題は、恒常的発現プロモーターではなく誘導型の発現プロモーターを用いることにより解消される。プロモーターとして植物において良く用いられている恒常的発現プロモーター(カリフラワーモザイクウイルス由来35Sリボソーム発現プロモーター;PCaMV35S)を用いて植物レクチンの発現を試みたところ、レクチンの蓄積量は極めて低い結果となった。また、作製した細胞を長期間継代培養すると、細胞が暗黒色に変色し、増殖性が著しく低下した。これは、継代培養している間もレクチンを生産し続ける為であると推察される。ホルモンにて発現調整が可能な誘導型プロモーターを使用することにより、細胞を十分に増殖させた後にレクチンを発現させることが可能となり、生産能力が著しく向上する。また、継代培養を行っている間は、発現を抑制させることができるため、植物レクチンが細胞に悪影響を与えることなく、継代培養を行うことが可能である。なかでも、宿主細胞として植物由来培養細胞(特に好ましくは、BY2細胞等のタバコ由来細胞)を用いる実施形態は、各種植物に由来するレクチンタンパク質を生産するための形質転換体の生産に特に適している。植物レクチンの発現は、異種発現系において一般的に用いられている微生物宿主(大腸菌や酵母)にて構築している例は少ない。これは、植物レクチンを微生物細胞で発現させることがタンパク質フォールディングや翻訳後修飾の違いにより困難であるからと推察される。植物細胞を宿主として用いることにより、この問題は解消される。植物細胞として好適なものは、細胞増殖性が極めて高いタバコBY-2細胞である。植物細胞を宿主として用いることにより、フォールディングや翻訳後修飾などが正常に行われ、種子由来のものと同様の糖鎖特異性を示す植物レクチンが生産可能となる。したがって、本発明の発現系、特に、宿主細胞として植物由来培養細胞(特に好ましくは、BY2細胞等のタバコ由来細胞)を用いる発現系は、実施例において用いられたゴーヤ由来抗Hレクチンタンパク質の発現だけでなく、他の植物に由来するレクチンタンパク質の発現にも適した発現系であるといえる。
【0056】
ウイルスベクターとしては、RNAを遺伝子とするウイルス由来のウイルスベクターであれば特に限定されるものではなく、二本鎖RNAウイルス、一本鎖マイナス鎖RNAウイルス、一本鎖プラス鎖RNAウイルス由来のウイルスベクターを用いることができる。中でも細胞内でcDNAから転写されたRNA自体がmRNAとして機能するという理由から、一本鎖プラス鎖RNAを遺伝子とするウイルス由来のウイルスベクターであることが特に好ましい。
【0057】
また、ウイルスベクターは植物ウイルス由来のウイルスベクターに限定されるものではなく、動物ウイルス、ファージを含むあらゆるRNAウイルス由来のウイルスベクターを用いることが可能である。しかしながら、植物細胞に植物レクチンタンパク質を生産させる目的で使用する場合には、植物ウイルス由来のウイルスベクターであることが好ましく、中でも植物のサイレンシングを抑制する因子(サプレッサー)を持つウイルス由来のウイルスベクターであることが特に好ましい。サイレンシングのサプレッサーを持つウイルスベクターを用いることにより、増幅の後半においてもmRNAが分解される現象が生じない。サイレンシングのサプレッサーを持つ植物ウイルスとしては、例えば、ポティ属(Potyvirus属)ウイルス、ククモウイルス属(Cucumovirus属)ウイルス(例えばキュウリモザイクウイルス(CMV))、ポテックスウイルス属(Potexvirus属)ウイルス(例えばジャガイモXウイルス(PVX))、トンブスウイルス属(Tombusvirus属)ウイルス(例えばトマトブッシースタントウイルス(TBSV)、Cymbidiumu ringspot virus (CymRSV))、カルモウイルス属(Carmovirus属)ウイルス(例えば、Turnip crinkle virus (TCV))、トバモウイルス属(Tobamovirus属)ウイルス(例えば、タバコモザイクウイルス(TMV)、トマトモザイクウイルス(ToMV))が挙げられる。
【0058】
また上記ウイルスベクターのcDNAの3’末端には、リボザイム配列が結合していることが好ましい。形質転換体の細胞内でcDNAから転写されるウイルスRNAの3’末端に付加されウイルスの複製能力低下の原因となるターミネーター由来の配列やポリA配列を切断することができるからである。その結果ウイルスの複製能力の低下を防止することができ、植物レクチンタンパク質の高生産が可能となる。結合されるリボザイム配列は、上記3’末端に付加された余分な配列を切断できるものであればよく、特に限定されるものではない。例えば、肝炎デルタウイルスのリボザイム配列(GenBank accession No. X77627他)またはサテライトタバコリングスポットウイルスのリボザイム配列(GenBank accession No. M17439)を用いることができる。
【0059】
また植物レクチンタンパク質をコードする遺伝子は、ウイルスの外被タンパク質をコードする遺伝子と置換されていることが特に好ましい。この部位に植物レクチンタンパク質遺伝子を挿入することによりウイルスの外被タンパク質が産生されなくなり、増幅されるウイルス遺伝子は粒子化されず他の植物に感染することがなくなるため、ウイルスの外界への飛散という問題を解消することが可能となる。
【0060】
更に、ウイルスベクターにおいて、ウイルスの移行タンパク質(MP)をコードする遺伝子が削除されており、MPサブゲノムプロモーター領域を構成する塩基の少なくとも一部が、複製酵素である180Kタンパク質のアミノ酸配列は変えないように置換されており、ウイルスの外被タンパク質の開始コドンが他の塩基配列に置換されて無効化されており、かつ、ウイルスの外被タンパク質をコードする遺伝子が植物レクチンタンパク質をコードする遺伝子に置換されていることが好ましい。本発明者らは、この特徴を有するウイルスベクターをMPDベクターと称する(MPが削除(Delete)されているため)。MPDベクターから発現されるウイルスRNAは従来の旧型ベクターと同様のウイルス複製能力を持つ。なおMP遺伝子は誘導ウイルスベクターシステムでは不必要な遺伝子である。MPDベクターは、MP遺伝子が削除されているためにcDNAの鎖長は短い。このため、MPDベクターは宿主細胞において複製され易くかつ植物におけるcDNAの転写の効率が高いものと考えられる。
【0061】
図1(b)に植物レクチンタンパク質発現用DNA断片の一例を示す。図1(b)は、形質転換用ベクターであるTiプラスミドpBICER8−ToMV−CO.3−MCL−His−SRZの一部分を示している。同図中左からエストロジェンによって活性化される融合転写因子LexA−VP16−hERによって転写誘導されるプロモーターOLexA−46、その下流にゴーヤ由来抗Hレクチン遺伝子(以下、MCL遺伝子と称する)を組み込んだトマトモザイクウイルスベクターToMV−MCL cDNA、その3’末端にサテライトタバコリングスポットウイルスリボザイム配列S−Rz、さらに35Sターミネーター配列35ST、またその下流に薬剤耐性マーカーとしてカナマイシン耐性遺伝子Kanrがある。なお、上記植物レクチンタンパク質発現用DNA断片の転写因子高発現形質転換体への導入方法は、(A−1)第一形質転換工程の項で述べた方法と同様に行なえばよい。
【0062】
上記本形質転換体(細胞)の生産方法は種々の植物レクチンタンパク質の発現系に適用可能である。当該生産方法により、人工的に遺伝子に変異を加えて調製される新規糖鎖認識特異性をもつレクチンの効率的な生産が可能となる。
【0063】
本形質転換体(細胞)の生産方法の特に好ましい実施形態としては
宿主細胞がタバコBY2細胞であり、
転写因子がLexA−VP16−hERであり、
転写誘導型プロモーターがOLexA−46であり、
ウイルスベクターが、トバモウイルス属に属するウイルス由来であり、
ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、植物レクチンタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターであり、
ウイルスベクターのcDNAの3’末端に、リボザイム配列が結合しており、かつ
転写因子発現用DNA断片、および植物レクチンタンパク質発現用DNA断片が、アグロバクテリウム法により導入される
ことを特徴とする実施形態が挙げられる。この実施形態によれば、植物レクチンタンパク質を、天然物中と同様の活性を維持しつつ、特に効率的に生産することが可能である。
【0064】
通常、ウイルスベクターは導入する遺伝子のサイズが増加するにしたがい複製能力が低下することが知られている。しかしながら、植物レクチンは遺伝子DNAのサイズが比較的大きい(1.5k)にもかかわらず、本発明の方法によれば、予想に反して植物レクチンタンパク質を高発現可能な形質転換体を生産することができた。この理由は明らかではないが、本発明の発現系は、ウイルスベクターを用いるにも関わらず、予想に反して植物レクチンの発現に適していることが確認された。In vitro転写したウイルスベクターをBYIIプロトプラストに接種した場合にはMPを欠失させる効果はほとんど認められないが、形質転換体においてはレクチンをはじめ1kbを越えるタンパク質はMPDベクターでのみ効率よく生産することができた。本ベクターは従来ベクターでは効率よく合成することができなかった遺伝子DNAの塩基配列が1kbを越えるレクチン以外の2種のタンパク質(アラビドプシスのクチナーゼ、ヒトアクチン)を合成することに成功している。すなわち、本ベクターを用いた形質転換細胞は、植物レクチンタンパク質に限らず、アラビドプシスのクチナーゼ、ヒトアクチン等の、遺伝子DNAの塩基配列が1kbを越える(好ましくは、2kb以下である)タンパク質を効率的に生産することが可能である。
【0065】
また、下記に示す実施例では、通常の2倍の数のセルラインを選抜し、その結果、特に高い発現を示すラインを一つえることに成功した。本結果より、比較的サイズの大きなレクチンに対しても、同様に、多くのラインから選抜することにより、特に優れた形質転換体を選抜できる可能性が高まることが考えられ、このことは、植物レクチン全般に応用可能であると思われる。したがって、第二形質転換工程において得られた形質転換体から、所望の植物レクチン発現能力を有する形質転換体を選抜するためにはセルライン200以上であることが好ましいといえる。
【0066】
(B)本発明にかかる植物レクチンタンパク質生産用形質転換体およびその利用
本発明にかかる植物レクチンタンパク質生産用形質転換体(以下、本形質転換体)は、上記本形質転換体の生産方法によって生産された植物レクチンタンパク質生産用形質転換体(細胞)である。本形質転換体は、転写因子をコードする遺伝子およびウイルスベクターをコードする遺伝子の両者が、それぞれ発現に最適な染色体上の位置に組み込まれた形質転換体であるため、本形質転換体を用いることによって、植物レクチンタンパク質を高生産することができる。
【0067】
また本発明にかかる植物レクチンタンパク質の生産方法(以下、本植物レクチンタンパク質の生産方法)は、本形質転換体を用いることを特徴としている。すなわち、本植物レクチンタンパク質の生産方法は、植物レクチンタンパク質をコードする遺伝子が導入された本形質転換体から植物レクチンタンパク質を回収することにより行なう。さらに植物レクチンタンパク質をより大量に取得するためには、本形質転換体の数を培養・栽培・育成等によって増加させ、増加させた本形質転換体から植物レクチンタンパク質を回収することにより行なうことが好ましい。本形質転換体の培養・栽培・育成条件等は特に限定されるものではなく、本形質転換体に好適な条件を適宜選択して用いればよい。植物由来培養細胞を培養する際の培地としては特に限定されるものではないが、無機塩類、炭素源、ビタミン類、アミノ酸が加えられている場合がある。さらに、ココナツミルクや酵母エキスを加えて成長を促進させる場合がある。その他、オーキシンとサイトカイニン、ジベレリン、アブシジン酸、エチレン等の植物ホルモンを添加する場合がある。また培養条件であるが、光、温度、通気の有無等を培養する細胞に応じて最適なものを採用すればよい。例えば、タバコBY2細胞を培養条件の一例としては、370mg/lリン酸二水素カリウム、1mg/lチアミン塩酸、3%スクロース、0.2mg/l 2,4−Dを含むMS培地を用い、暗所、26℃、135回転/分で旋回振盪培養後、1/100量を一週間ごとに継代することが挙げられる。
【0068】
また、本形質転換体の種類としては特に限定されるものではなく、植物・動物の個体であっても、植物・動物由来培養細胞であってもよい。ただし、迅速に大量に植物レクチンタンパク質を生産するためには、形質転換体は培養可能な細胞(培養細胞)であることが好ましい。さらには、細胞の取り扱いのしやすさ・培地が安価である等の理由から、植物由来培養細胞であることが好ましい。上記植物由来培養細胞としては、例えばタバコ由来BY2細胞が挙げられる。
【0069】
(C)本発明にかかる植物レクチンタンパク質生産用形質転換体の生産キット
本発明にかかる植物レクチンタンパク質生産用形質転換体の生産キット(以下、本キットと称する)は、上記本形質転換体の生産方法を行なうためのキットである。本キットを構成するものについては特に限定されるものではないが、転写因子をコードする遺伝子と転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片、および/またはRNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる植物レクチンタンパク質発現用DNA断片により構成されていることが好ましい。具体的には本キットが、転写因子発現用DNA断片の一例であるpER8(−Stu)、および/または植物レクチンタンパク質発現用DNA断片の一例であるpBICER8−ToMV−CO.3−MCL−His−SRZにより構成されている例が挙げられる。
【0070】
本キットの利用者は、(a)転写因子発現用DNA断片を適当な宿主細胞に導入する工程、(b)その形質転換体の中から転写因子を高発現する形質転換体を選抜する工程、(c)該形質転換体に植物レクチンタンパク質をコードする遺伝子が挿入されている植物レクチンタンパク質発現用DNA断片を導入する工程を経て植物レクチンタンパク質生産用形質転換体が得られることとなる。なお本キットに含まれる転写因子発現用DNA断片に植物レクチンタンパク質をコードする遺伝子を挿入する方法は、遺伝子工学的手法により行なえばよい。
【0071】
さらに本キットは、上記転写因子高発現形質転換体および/または植物レクチンタンパク質発現用DNA断片により構成されていることが好ましい。より具体的には後述する実施例において取得した転写因子高発現形質転換体の一例である転写因子高発現タバコBY2細胞、および/または植物レクチンタンパク質発現用DNA断片の一例であるpBICER8−ToMV−CO.3−MCL−His−SRZにより構成されている例が挙げられる。
【0072】
転写因子高発現形質転換体が既に取得されているため、本キットの利用者は上記(c)のみを行なえばよく、より迅速かつ簡便に植物レクチンタンパク質生産用形質転換体を得ることができる。
【0073】
この他本キットには、例えば、細胞、培地、制限酵素、修飾酵素類、転写誘導用化学物質(ステロイドホルモン、エストロジェン等)、培養フラスコ、アグロバクテリウム(植物細胞の場合)等が含まれていてもよい。
【0074】
本発明は上述した各実施形態に限定されるものではなく、請求項に示した範囲で種々の変更が可能であり、異なる実施形態にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態についても本発明の技術的範囲に含まれる。
【実施例】
【0075】
1. ゴーヤレクチン遺伝子(MCL)発現系の作製方法
(1)転写因子発現用DNA断片導入用ベクターの構築
転写因子発現用DNA断片を宿主細胞(タバコBY2細胞)に導入するためのベクター(以下転写因子発現用DNA断片導入用ベクターと称する)として、TiプラスミドpER8(−Stu)を用いた。pER8(−Stu)は、恒常的プロモーターPG10-90の下流にエストロジェンレセプターを含む融合転写因子LexA−VP16−hERをコードする遺伝子と、ターミネーターTE9とを連結し、さらに薬剤耐性マーカーとしてハイグロマイシン耐性遺伝子(Hygr)を組み込んで構築した。図1(a)にpER8(−Stu)の一部分の構造を模式図として示した。
【0076】
なお、TiプラスミドpER8(−Stu)、恒常的プロモーターPG10-90、LexA−VP16−hER、ターミネーターTE9、Hygrはいずれも公知の方法により調製可能である。TiプラスミドpER8(−Stu)は、Dohi, K., Nishikiori, M., Tamai, A., Ishikawa, M., Meshi, T., Mori, T. (2006) Inducible virus-mediated expression of a foreign protein in suspension-cultured cells. Archives of Virology 151, 1075-1084に記載されている。恒常的プロモーターPG10-90はIshige, F., Takaichi, M., Foster, R.,Chua, N. H. and Oeda, K.(1999) A G-box motif(GCCACGTGCC) tetramer confers high-level constitutive expression in dicot and monocot plants. Plant J. 20, 127-133に記載されている。LexA−VP16−hER及びターミネーターTE9はZuo J, Niu QW, Chua NH. “An estrogen receptor-based transactivator XVE mediates highly inducible gene exprssion in transgenic plants.” Plant J.2000, 24: 265-273 に記載されている。
【0077】
(2)植物レクチンタンパク質発現用DNA断片導入用ベクターの構築
トマトモザイクウイルス(ToMV)の外被タンパク質をコードする遺伝子を、MCLをコードする遺伝子(配列表の配列番号1、以下、MCL遺伝子と称する)で置換したToMV変異体を用いた。配列番号1の塩基配列のうち23番〜1664番の塩基配列をMCL遺伝子として用いた。
【0078】
ToMV変異体のcDNAの塩基配列を図2及び配列表の配列番号3に示す。図2において、下線を引いた箇所はToMVのcDNAの塩基配列から変更した箇所である。下線部(b)は移行タンパク質(MP)遺伝子を削除し、MPサブゲノムプロモーター領域の塩基配列を変更した(ただし複製酵素である180Kタンパク質のアミノ酸配列は変えないように変更した)箇所である。下線部(d)は外被タンパク質(CP)の開始コドンをATCに置換して無効化した箇所である。(e)で示す箇所は外来遺伝子クローニング用に導入されたPmeIサイトである。下線部(f)はMCLをコードするMCL遺伝子(配列番号1の第23番〜1664番塩基)を示す。下線部(h)はヒスチジンタグ(His−tag)をコードする塩基配列である。枠により強調した箇所(j)、(a)、(c)、(e)、(g)、(i)はいずれも構築に関与する制限酵素サイトであって、(j)はSpeIのサイトであり、(a)はKpnIのサイトであり、(c)はNcoIのサイトであり、(e)はPmeIのサイトであり、(g)および(i)はBstEIIのサイトである。
【0079】
図2に示すToMV変異体のcDNAを含有するプラスミドは次の手順で作成した。
(1)KpnI(a)からNcoI(c)までの領域を人工合成した。
(2)NcoI(c)からPmeI(e)/BstEII(i)までの領域のPCR断片を、外被タンパク質(CP)の開始コドン(ATG)をATCに置換した変異を導入して作成した。具体的には、Dohi, K., Nishikiori, M., Tamai, A., Ishikawa, M., Meshi, T., Mori, T. (2006) Inducible virus-mediated expression of a foreign protein in suspension-cultured cells. Archives of Virology 151, 1075-1084に記載されているプラスミドpiL.erG3(SF3)SRz(Avr)を鋳型とし、以下のプライマーセットを用いてNcoI(c)からPmeI(e)/BstEII(i)までの領域のPCR断片を作成した。
フォワードプライマー: 5'-GACCCATGGGAAGTTGTTGATGAGTTCA-3’
リバースプライマー: 5'-TATGGTTACCGTTTAAACAAAAACACAAATTGCGATGGAGAAGTGATTGAGTAAGAGATATTTAATACGAATCA-3’
フォワードプライマー中、細下線部がNcoIサイト(c)に対応する配列である。
【0080】
リバースプライマー中、細下線部がクローニングサイトPmeI(e)/BstEII(i)に対応する配列であり、太下線部がATGからATCへの変異を導入するための配列である。
【0081】
プラスミドpiL.erG3(SF3)SRz(Avr)のうち、本発明に関連する部分の構造を図4(a)に示す。piL.erG3(SF3)SRz(Avr)に含まれるToMV−GFP cDNAは、トマトモザイクウイルス(ToMV)の外被タンパク質をコードする遺伝子を、緑色蛍光タンパク質をコードする遺伝子で置換したToMV変異体のcDNAである。
【0082】
(3)プラスミドpiL.erG3(SF3)SRz(Avr)において、KpnIからBstEIIまでの部分を、上記(1)で得られたKpnI(a)−NcoI(c)DNA断片と、上記(2)で得られたNcoI(c)−PmeI(e)/BstEII(i)DNA断片とに入れ替えた(3点ligation)。
【0083】
(4)上記(3)で得られたプラスミドのPmeI(e)/BstEII(i)部位に、ゴーヤ由来抗Hレクチン(MCL)のDNA断片(5’端PmeI、3’端BstEII)とHis−tagのDNA断片(5’−GTCACCTCACCATCACCATCACCATTG−3’と5’−GTTACCAATGGTGATGGTGATGGTGAG−3’をアニールしたもの)を挿入した(3点ligation)。
【0084】
5’端にPmeIサイト、3’端にBstEIIサイトがそれぞれ導入されたMCLのDNA断片は、MCLをコードするDNA(特許文献1)を鋳型とし、以下のプライマーセットを用いたPCRにより作製した。
フォワードプライマー: 5'-ATAGTTTAAACATGAGAATGAGAGTGTTAG-3’
リバースプライマー: 5'-TATGGTGACCAATAGAACACCGTCCATT-3’
【0085】
(5)上記(4)で得られたプラスミドのSpeI−AvrIIの領域を切り出し、pBICER8−ToMV5’−SpeのSpeI部位に挿入した。ここで用いたプラスミドpBICER8−ToMV5’−Speは、上記(2)で言及した文献に記載されており、エストロジェンで転写誘導可能なプロモーターOLexA−46(Zuo J, Niu QW, Chua NH. “An estrogen receptor-based transactivator XVE mediates highly inducible gene exprssion in transgenic plants.” Plant J.2000, 24: 265-273 参照)を有するTiプラスミドにおいて、OLexA−46の下流に、ToMVの5’末端からSpeIサイトまでの領域(図2の5’端から(j)までの領域に相当する)が組み込まれたものである。pBICER8−ToMV5’−Speのうち、本発明に関連する部分の構造を図4(b)に示す。
【0086】
こうして、植物レクチンタンパク質発現用DNA断片を宿主細胞(タバコBY2細胞)に導入するためのベクター(以下、植物レクチンタンパク質発現用DNA断片導入用ベクターと称する)pBICER8−ToMV−CO.3−MCL−His−SRZを構築した。図1(b)にpBICER8−ToMV−CO.3−MCL−His−SRZの一部分の構造を模式図として示した。
【0087】
このプラスミドpBICER8−ToMV−CO.3−MCL−His−SRZは、図1(b)に示すように、エストロジェンで転写誘導可能なプロモーターOLexA−46を有するTiプラスミドにおいて、OLexA−46の下流に、ToMV変異体(図2)のcDNAが連結され、さらにその3’末端にサテライトタバコリングスポットウイルスのリボザイム配列S−Rz(GenBank accession No. M17439)、35Sターミネーター(35ST)、カナマイシン耐性遺伝子(Kanr)が組み込まれた構造を有する。
【0088】
(3)第一形質転換工程:転写因子発現用DNA断片の宿主細胞への導入
転写因子発現用DNA断片導入用ベクターpER8(−Stu)を、タバコBY2細胞にアグロバクテリウム法により導入した。まずpER8(−Stu)をエレクトロポレーション法によってAgrobacterium tumefacience LBA4404系統に導入した。これをカナマイシン(50mg/l)を含むAB sucrose培地で前培養した。次にタバコBY2細胞と混合してシャーレに移し、26℃、暗所で42〜48時間静置してタバコBY2細胞を形質転換した。タバコBY2細胞用培地にて洗浄した後、カルベニシリン(100mg/l)およびハイグロマイシン(20mg/l)を含むタバコBY2細胞用固形培地に広げ、形質転換タバコBY2細胞を増殖させた。
【0089】
(4)選抜工程:転写因子高発現形質転換体の選抜
形質転換タバコBY2細胞のうち、ノーザンブロット法の結果、転写因子の発現量の高い細胞ライン(ER43)を選抜した。ここで「細胞ライン」とは、形質転換細胞を増殖させることにより形成された個々のコロニーのことを意味する。
【0090】
(5)第二形質転換工程:植物レクチンタンパク質発現用DNA断片の導入
上記で得られた転写因子高発現タバコBY2細胞ライン(ER43)に、ウイルスベクター(pBICER8−ToMV−CO.3−MCL−His−SRZ)をアグロバクテリウム法により導入し形質転換細胞(カルス)を192ライン得た。
【0091】
(6)組換えウイルス高発現形質転換カルスの選抜
得られた192ラインの形質転換カルスは、エストロジェン(0.01mM)を添加したBY2個体培地に移植し、移植2日後にカルスから全RNAを抽出した。抽出したRNAをToMVの3’非翻訳領域約200塩基に相補的なDIGRNAプローブを用いてノーザン分析を行った。その結果、ウイルスRNAの蓄積量の多い4ライン(10−1−3−11−13、10−1−3−12−9、10−1−3−5−17、10−1−3−14−20)を選抜し、選抜したカルスを15mlの液体培地に移し、細胞が飽和した後、1/100量になるように継代培養した。継代7日後1/100量になるようにさらに継代培養し、同様の継代培養により細胞を維持した。
【0092】
(7)MCLの誘導発現
上記で得られた4ラインの各形質転換細胞を1/100量になるよう継代し、継代培養後7日間前培養した細胞を、新しい培地に1/30量になるように移した。3日間培養した後、エストロジェンを終濃度が0.01mMとなるように加え、さらに4日間培養した後MCLを抽出し、ヒト赤血球凝集(HA)活性を検出した。その結果、10−1−3−11−13株において最も高いHA活性が認められた。
【0093】
上記において、転写因子高発現タバコBY2細胞ラインER43を形質転換して取得したMCL高発現細胞(10−1−3−11−13株)を用いて、MCLのエストロジェンによる誘導発現を試みた。10−1−3−11−13株を1/100量になるよう継代し、前培養を3日間行なった培養液に対して、エストロジェンを終濃度0.01mM添加しMCLの誘導発現を行なった。エストロジェン添加後48時間の10−1−3−11−13株から全RNAを抽出し、ノーザンブロット法によりToMVに特異的なRNAを検出した。解析には、ToMVの3’非翻訳領域約200塩基に相補的なRNAプローブを用いた。プローブのラベリングは、ロシュ・ディアグノスティック社のDIG RNA Labeling Kitを用いて行なった。検出には、同社のDIG Luminescent Ditection KitおよびCDP−Starを用い、それぞれキットのマニュアルに従って行なった。また、エストロジェン添加後48時間の10−1−3−11−13株から全タンパク質を抽出し、HA活性の検出を行なった。その結果、エストロジェンによるToMVの明らかな転写誘導およびHA活性の検出が確認された。10−1−3−11−13株は、増殖と発現誘導を行い、以降のレクチン抽出および評価に用いた。
【0094】
2.発現系由来レクチンの評価
(1)発現系由来レクチンの抽出
10−1−3−11−13株の培養液から細胞破砕装置またはフレンチプレスを用いてレクチンを抽出し、10,000×gで30分間、遠心分離後の上清を粗抽出液とした。
【0095】
(2)発現系由来粗抽出液のヒト赤血球凝集(HA)活性
この粗抽出液についてレクチン活性を評価した。レクチン活性は、連続希釈法によるヒトHA活性により測定した。すなわち、マイクロタイタープレートを用いて被検体のPBSによる2倍希釈列を作製し、これに等量の3%ヒト赤血球浮遊液(オーソ社製)を加えて、室温で30分間反応させ、肉眼で凝集の有無を判定した。その結果、発現系由来粗抽出液のヒトO型HA活性は256 titerを示した。これは、恒常的発現プロモーターpH35CGによる発現系を用いた下記比較例(粗抽出液の力価8)に比べて大幅に改善したものであった。本培養液1 Lから得られるレクチン活性量は、乾燥種子200−300 gから得られる抽出液の活性量に相当するものと推定された。適度に希釈した本粗抽出液のA1、A2、BおよびOの各血液型に対して、それぞれ2、8、8、16の力価を示し、種子由来レクチンの場合と特性が類似していたことから、種子由来レクチン遺伝子が機能を保持したまま発現していることが推定された。
【0096】
(3)発現系由来レクチンの精製
上記(1)で得られた粗抽出液は、PBSに対して透析した後、0.5mL/minの流速にてガラクトースセファロースカラム(30mL)に流し、ガラクトースに親和性のある成分を吸着させた。このカラムをPBSにて十分に洗浄後、0.2Mガラクトースを含むPBSを流すことにより吸着したガラクトース結合画分を溶出させた。この溶出画分は蒸留水に対して透析し凍結乾燥した。凍結乾燥により得られた粉末をPBSに溶解したものは、ヒトO型HA活性を有していたことから、レクチンであると認められた。約90mLの培養液からは、約1mg(PBSに1 mg/mLの濃度で溶解した際の力価は2,048)の発現系由来レクチンが得られた。
【0097】
(4)発現系由来レクチンの電気泳動特性
還元条件化のSDS−PAGE分析の結果、発現系由来レクチンは、種子由来レクチンの4本のバンドのうちの2本が強く観察された(図3)。
【0098】
(5)発現系由来レクチンの血球特異性
発現系由来レクチンおよび比較のために用いた種子由来レクチンはPBSにて希釈し、O型赤血球に対する力価を32 titerに調整した。発現系由来レクチンのA1、A2、BおよびOの各血液型に対するHA活性は、それぞれ4、16、8、32の力価を示し、種子由来レクチンの場合と同等であった(表1)。
【0099】
(6)発現系由来レクチンの糖鎖特異性
発現系由来レクチンの糖鎖特異性は、各種糖類によるHA阻害試験により評価した。(5)で調整した各レクチン溶液の12.5μlは、終濃度が100mMから0.01mMになるように希釈した糖類溶液の12.5μlと混和後、室温で1時間攪拌した。その後、3%ヒトO型赤血球浮遊液を添加し、30分後に凝集の有無を肉眼判定し、血球の凝集を阻害した最低濃度を最低HA阻害濃度として求めた。なお、用いた糖類は、D−ガラクトース、D−フコース、N−アセチル−D−ガラクトサミン、N−アセチルノイラミン酸、メチル−α−D−ガラクトピラノシド、L−アラビノース、D−ラクトース、ラクト−N−ビオース、N−アセチル−D−ラクトサミン、ヒトB抗原三糖、ヒトA抗原三糖、ヒトH抗原二糖、ヒトH抗原三糖、D−グルコース、D−マンノース、L−フコース、N−アセチル−D−グルコサミン、D−キシロース、L−ラフィノース、D−フルクトース、スクロース、マルトース、N,N−ジアセチルキトビオースである。その結果、両者の糖鎖特異性は極めて類似し(表2)、H抗原三糖に対して高い特異性を示した。以上のことから、本発現系に導入されたレクチン遺伝子は、その機能を保持したまま発現していることが確認された。
【0100】
【表1】

【0101】
【表2】
【0102】
比較例
恒常発現プロモーターを用いたゴーヤ由来レクチンタンパク質の発現
MCLのcDNAを、バイナリーベクターpH35CG(インプランタイノベーションズ)を用いて、微粒子発射装置により、タバコBY−2(Nicotiana tabacum L. cv. Bright Yellow−2)細胞の懸濁液中で発現させた。当該ベクターは、カリフラワーモザイクウイルス35Sプロモーターに連結されたMCLのcDNAと、ノパリン合成酵素(NOS)ターミネーターとを含む。ハイグロマイシンに対する耐性を付与するハイグロマイシンホスホトランスフェラーゼ(HPT)遺伝子を、NOSプロモーターに連結する。当該ベクターにより金粒子(直径1.5〜3μm、アルドリッチケミカル)を被覆した。減圧空気:−0.1MPa、標的距離:9.6cm、ヘリウム圧:5Kg/cm2の条件で、1試験区当り1回の発射を実施した。発射を受けた細胞は、50mg/mlハイグロマイシンを含有する固化LS培地上に載せ、続いて、14日間、同一の培地上において数回継代培養を行った。選択的条件での継代培養を50日間行った後、ロータリーシェーカーにより、100mlのLS培地を加えた500ml容三角フラスコ中で27℃にて10日間、ハイグロマイシン耐性細胞を培養した。懸濁した細胞を集め、Tissue ruptor(Quiagen)を用いて破砕した。破砕された細胞を、20,000×gにて10分間遠心沈降させた。上澄み液の赤血球凝集活性を、上記の手順で測定した。
【技術分野】
【0001】
本発明は植物レクチンタンパク質を効率的に大量生産するための技術を提供する。
本発明また、1kbを越える鎖長のDNAからなる遺伝子によりコードされるタンパク質を効率的に大量生産するための技術を提供する。
【背景技術】
【0002】
レクチンは糖鎖と特異的に結合するタンパク質であり、細胞の凝集や糖複合体を沈降させる性質を有する。レクチンは動植物界に広く存在している。レクチンは、近年の糖鎖研究の展開に伴い重要度が増しており、医学や生物学におけるツールとして様々な場面で利用されている。
【0003】
本発明者らはゴーヤ(「ツルレイシ」とも呼ばれる; Momordica charantia)由来のレクチンを単離し、アミノ酸配列及び該アミノ酸配列をコードする遺伝子の塩基配列を特定している(特許文献1)。
【0004】
現在市販されているレクチンの大部分は植物等から抽出し精製されたものである。しかしながら、このような分離法による製造方法では、原料が天然物であるためにロット間の品質の差異や供給量に対する懸念がある。
【0005】
一方、天然物由来のタンパク質を効率的に生産するための技術として本発明者らはこれまでに、転写因子をコードする遺伝子が、当該転写因子の発現に最も適した染色体上の位置に組み込まれた形質転換体(細胞)をはじめに作出し、得られた形質転換体に、緑色蛍光タンパク質をコードする遺伝子が挿入されたウイルスベクターを導入すれば、高効率でウイルスベクターを誘導発現し、緑色蛍光タンパク質を高発現する形質転換体(細胞)を高確率で作出できることを見出し、特許出願している(特許文献2)。
【0006】
しかしながら、植物レクチンタンパク質を遺伝子組み換え技術を用いて効率的に生産する技術は未だに確立されていない。従来の微生物を用いた発現系は、植物レクチンタンパク質の機能を保持した状態で発現させることが困難であった。また、植物による従来の発現系では、宿主植物にとって導入する植物レクチンタンパク質が生育に好ましくないものであるため、生産可能な植物レクチンの種類が限定されるという問題があった。
【0007】
一方、1kbを越える鎖長のDNAからなる遺伝子によりコードされるタンパク質は、ウイルスベクターを用いて細胞に生産させることは従来困難であった。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特開2009−39006号公報
【特許文献2】特開2005−245228号公報
【特許文献3】特許第3752223号公報
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明は、植物レクチンタンパク質を効率的かつ安定的に生産するための手段を提供することを目的とする。
【0010】
本発明はまた、1kbを越える鎖長のDNAからなる遺伝子によりコードされるタンパク質を効率的かつ安定的に生産するための手段を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明は以下の発明を包含する。
(1)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換工程と、
前記第一形質転換工程によって得られた形質転換体の中から前記転写因子を発現する形質転換体を選抜する選抜工程と、
RNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる植物レクチンタンパク質発現用DNA断片を、前記選抜工程によって選抜された形質転換体に導入する第二形質転換工程とからなることを特徴とする植物レクチンタンパク質生産用形質転換体の生産方法。
【0012】
(2)上記ホルモンが、エストロジェンまたはステロイドホルモンであることを特徴とする(1)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0013】
(3)上記エストロジェンで活性化される性質を有する転写因子としてLexA−VP16−hERを用い、上記転写誘導型プロモーターとしてOLexA−46を用いることを特徴とする(2)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0014】
(4)上記ウイルスベクターは、一本鎖プラス鎖RNAを遺伝子とするウイルス由来であることを特徴とする(1)〜(3)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
【0015】
(5)上記ウイルスベクターが、植物ウイルス由来であることを特徴とする(4)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0016】
(6)上記ウイルスベクターが、植物のサイレンシングを抑制する因子を持つ植物ウイルス由来であることを特徴とする(5)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0017】
(7)上記ウイルスベクターが、トバモウイルス属に属するウイルス由来であることを特徴とする(6)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0018】
(8)上記ウイルスベクターが、トマトモザイクウイルスベクターまたはタバコモザイクウイルスベクターであることを特徴とする(7)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0019】
(9)上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、植物レクチンタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターである、(1)〜(8)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
【0020】
(10)上記ウイルスベクターのcDNAの3’末端に、リボザイム配列が結合していることを特徴とする(1)〜(9)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
【0021】
(11)上記リボザイム配列が、肝炎デルタウイルスのリボザイム配列、またはサテライトタバコリングスポットウイルスのリボザイム配列であることを特徴とする(10)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0022】
(12)上記転写因子発現用DNA断片、および上記植物レクチンタンパク質発現用DNA断片が、アグロバクテリウム法により導入されていることを特徴とする(1)〜(11)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
【0023】
(13)上記宿主細胞および形質転換体が、植物体または植物由来培養細胞であることを特徴とする(1)〜(12)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
【0024】
(14)上記植物由来培養細胞がタバコ由来細胞であることを特徴とする(13)の植物レクチンタンパク質生産用形質転換体の生産方法。
(15)上記タバコ由来細胞がタバコBY2細胞であることを特徴とする(14)の植物レクチンタンパク質生産用形質転換体の生産方法。
【0025】
(16)上記宿主細胞がタバコBY2細胞であり、
上記転写因子がLexA−VP16−hERであり、
上記転写誘導型プロモーターがOLexA−46であり、
上記ウイルスベクターが、トバモウイルス属に属するウイルス由来であり、
上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、植物レクチンタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターであり、
上記ウイルスベクターのcDNAの3’末端に、リボザイム配列が結合しており、かつ
上記転写因子発現用DNA断片、および上記植物レクチンタンパク質発現用DNA断片が、アグロバクテリウム法により導入される、
ことを特徴とする、(1)〜(15)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
【0026】
(17)上記植物レクチンタンパク質がゴーヤ由来レクチンタンパク質である、(1)〜(16)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法。
(18)(1)〜(17)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法によって生産された植物レクチンタンパク質生産用形質転換体。
【0027】
(19)(18)の植物レクチンタンパク質生産用形質転換体を用いることを特徴とする植物レクチンタンパク質の生産方法。
(20)(1)〜(17)のいずれかの植物レクチンタンパク質生産用形質転換体の生産方法を行うためのキット。
【0028】
(21)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換工程と、
前記第一形質転換工程によって得られた形質転換体の中から前記転写因子を発現する形質転換体を選抜する選抜工程と、
RNAを遺伝子とするウイルスに、生産しようとするタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなるタンパク質発現用DNA断片を、前記選抜工程によって選抜された形質転換体に導入する第二形質転換工程とからなり、
上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、生産しようとするタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターであることを特徴とする、タンパク質生産用形質転換体の生産方法。
【0029】
(22)生産しようとするタンパク質をコードする遺伝子が、1kbを越える鎖長のDNAからなる遺伝子である、(21)のタンパク質生産用形質転換体の生産方法
【0030】
(23)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片と、
RNAを遺伝子とするウイルスに、生産しようとするタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなるタンパク質発現用DNA断片であって、上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、生産しようとするタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターである、タンパク質発現用DNA断片と
が遺伝子DNAに導入されていることを特徴とする、タンパク質生産用形質転換体。
【発明の効果】
【0031】
本発明の方法により提供される植物レクチンタンパク質生産用形質転換体により、植物レクチンタンパク質を効率的かつ安定的に生産することが可能である。本発明の方法により生産される植物レクチンタンパク質は、植物から分離された植物レクチンタンパク質と実質的に同一の活性を維持している。
【0032】
本発明により提供されるウイルスベクターを用いて形質転換された細胞は、1kbを越える鎖長のDNAからなる遺伝子によりコードされるタンパク質を効率的かつ安定的に生産することができる。
【図面の簡単な説明】
【0033】
【図1】図1(a)はpER8(−Stu)の一部分の構造を示す。図1(b)はpBICER8−ToMV−CO.3−MCL−His−SRZの一部分の構造を示す。
【図2−1】図2−1はToMV変異体のcDNAの構造を示す。
【図2−2】図2−2はToMV変異体のcDNAの構造を示す(図2−1のつづき)。
【図3】図3は本発明の発現系由来レクチンの電気泳動の結果を示す。
【図4】図4(a)はpiL.erG3(SF3)SRz(Avr)の一部分の構造を示す。図4(b)はpBICER8−ToMV5’−Speの一部分の構造を示す。
【発明を実施するための形態】
【0034】
本発明の実施の形態について具体的に説明する。
(A)本発明にかかる植物レクチンタンパク質生産用形質転換体の生産方法
本発明にかかる植物レクチンタンパク質生産用形質転換体の生産方法(以下、本形質転換体の生産方法と称する)は、(1)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を細胞に導入する第一形質転換工程と、(2)前記第一形質転換工程によって得られた形質転換体の中から前記転写因子を発現する形質転換体を選抜する選抜工程と、(3)RNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる植物レクチンタンパク質発現用DNA断片を、前記選抜工程によって選抜された形質転換体に導入する第二形質転換工程とからなる。以下、工程ごとに説明する。
【0035】
(A−1)第一形質転換工程
本第一形質転換工程では、ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、該転写因子の発現を担うプロモーター(転写因子発現用プロモーター)とを連結して構築したDNA断片(転写因子発現用DNA断片)を、適当な宿主細胞に導入する工程である。当該工程によって、後述するウイルスベクター発現用プロモーターの転写誘導を行なう転写因子を安定的に高発現する形質転換体(細胞)の生産(作出)を行なう。すなわち宿主細胞の転写因子の発現に最適な染色体上の位置に、該転写因子をコードする遺伝子が組み込まれた形質転換体(細胞)候補を生産(作出)する工程である。
【0036】
<転写因子発現用DNA断片>
次に本第一形質転換工程で使用する転写因子発現用DNA断片について説明する。転写因子発現用DNA断片は、転写因子発現用プロモーターの下流に転写因子をコードする遺伝子が連結されている。またこの他、ベクター配列、ターミネーター、薬剤耐性マーカー等のDNAセグメントが含まれていてもよい。かかる転写因子発現用DNA断片の構築方法は、通常の遺伝子工学的手法を用いて行なえばよい。
【0037】
ここで転写因子発現用プロモーターは、転写因子を発現することが可能なものであれば特に限定されるものではなく、また恒常的にプロモーター活性を有するもの(以下、恒常的プロモーターと称する)であってもよいし、さらに転写因子によってプロモーター活性が誘導されるものであってもよい。ただし、転写因子の発現をさらに別の転写因子で制御することは、植物レクチンタンパク質の発現系自体が複雑化すること、コスト面で不利等の理由により、上記転写発現用プロモーターとしては、恒常的プロモーターの方がより好ましいといえる。恒常的プロモーターの例としては、カリフラワーモザイクウイルス35Sプロモーター、PG10−90(Ishige, F., Takaichi, M., Foster, R.,Chua, N. H. and Oeda, K.(1999) A G-box motif(GCCACGTGCC) tetramer confers high-level constitutive expression in dicot and monocot plants. Plant J. 20, 127-133. 参照)、ユビキチンプロモーター、アクチンプロモーター等が挙げられる。
【0038】
一方、使用する転写因子は、ホルモンによって活性化される性質を有する転写因子であって、後述する第二形質転換工程において導入する植物レクチンタンパク質発現用DNA断片中に含まれるプロモーターを転写誘導するものであれば特に限定されない。転写因子を活性化するホルモンとしては、エストロジェン、ステロイドホルモン、エクジソン等が挙げられる。かかる転写因子は、ホルモンが存在しない状態においては不活性型でありプロモーターの転写誘導を行なうことができないが、ホルモンが存在することによって活性型に変化しプロモーターの転写誘導を行なうことができる。この性質を利用すれば植物レクチンタンパク質の生産をより綿密に制御できるため、より安全に植物レクチンタンパク質の生産を行なうことができるといえる。すなわち、植物レクチンタンパク質生産を行なう必要がないとき、あるいは植物レクチンタンパク質生産を行なってはいけないときには、植物レクチンタンパク質生産系にホルモンを添加しなければよい。
【0039】
かかるホルモンによって活性化される性質を有する転写因子、および該転写因子によって転写誘導されるプロモーターの組み合わせとしては、例えば、ステロイドホルモンで活性化される転写因子であるGVGと該転写因子によって転写誘導されるプロモーターである6xUASgal4の組み合わせ、エストロジェンで活性化される転写因子LexA−VP16−hERと該転写因子によって転写誘導可能なプロモーターであるOLexA−46(Zuo J, Niu QW, Chua NH. “An estrogen receptor-based transactivator XVE mediates highly inducible gene exprssion in transgenic plants.” Plant J.2000, 24: 265-273 参照)の組み合わせ、エクジソンで活性化される転写因子であるエクジソンレセプターGR Act and DBDとヘルペスウイルストランスアクチベーションドメインHecR LBDのキメラタンパク質と該転写因子によって転写誘導可能なプロモーターGREの組み合わせ等が挙げられる。ただし、植物レクチンタンパク質生産用の宿主として植物体および植物細胞を利用する場合においては、活性化に用いるホルモンによる宿主への悪影響が少ないという理由からエストロジェンで活性化される転写因子LexA−VP16−hERと該転写因子によって転写誘導可能なプロモーターであるOLexA−46の組み合わせが好ましいといえる。
【0040】
図1(a)に転写因子発現用DNAの一例を示す。図1(a)は、形質転換用ベクターであるTiプラスミドpER8(−Stu)の一部分を示している。同図中左から転写因子発現用の恒常的プロモーターPG10−90、その下流にエストロジェンレセプターを含む融合転写因子LexA−VP16−hER、その3’末端にターミネーター配列TE9、またその下流に薬剤耐性マーカーとしてハイグロマイシン耐性遺伝子Hygrがある。
【0041】
<転写因子発現用DNA断片を導入する宿主細胞およびDNA断片導入方法>
上記転写因子発現用DNA断片が導入される宿主細胞は特に限定されるものではなく、動物由来細胞であっても植物由来細胞であってもよい。ただし、植物由来細胞は、動物由来細胞に比して、増殖速度が速くコンタミネーションのリスクが少ない点、培地作成費用が非常に安価であるという点において、植物由来細胞がより好ましい。なお、動物由来細胞および植物由来細胞とは、細胞、組織、並びに器官も含む意味である。特に液体培地等で培養可能な細胞(培養細胞)が好ましい。
【0042】
また由来となる動物としては、特に限定されるものでないが、ヒト、サル、イヌ、ヒツジ、ヤギ、ウサギ、マウス、ラット、モルモット、チャイニーズハムスター、ウシ、ウマ、ブタ、メダカやゼブラフィッシュ等の魚類、カイコ、夜蛾(Spodoptera frugiperda)等が挙げられる。一方由来となる植物としては、特に限定されるものではないが、イネ、シロイナズナ、オオムギ、コムギ、タバコ、トマト、キュウリ、ダイズ、ジャガイモ、トウモロコシ、ニチニチソウ、シロイヌナズナ、アルファルファが挙げられる。その他、枯草菌や乳酸菌などの菌類、酵母など単細胞生物が宿主細胞として利用できる。
【0043】
また動物由来細胞の例としては特に限定されるものではないが、HeLa細胞、CHO細胞、メラノーマ細胞、マウス3T3細胞が挙げられる。また植物由来細胞の例としては、タバコBY2細胞、ジャガイモ由来、イネ由来、サツマイモ由来、ダイズ由来、パセリ由来、シロイヌナズナ由来、コムギ由来、トウモロコシ由来細胞、ニチニチソウ由来細胞が挙げられる。
【0044】
後述する実施例において、タバコBY2細胞を宿主として用いている。タバコBY2細胞(Toshiyuki nagata, Yasuyuki Nemoto, and Seiichiro Hasezawa “Tobacco BY-2 Cell Line as the “Hela” Cell in the Cell Biology of Higher Plants” International Review of cytology, vol.132, p.p. 1-30 (1992) 、および http://www.riken.go.jp/r-world/info/release/press/2003/030620/ 参照)は、植物培養細胞株としては、世界中で最も広く用いられているものであり、最も増殖速度が速いこと、遺伝子操作が容易なこと、大量培養を容易に行うことができるという理由から採用した。
【0045】
また、転写因子発現用DNA断片を宿主細胞へ導入する形質転換方法は特に限定されるものではなく、宿主細胞の種類に応じた適切な形質転換方法を用いればよい。例えば、特別なベクター配列を有しないDNA断片を導入する場合には、エレクトロポレーション法、パーティクルガン法、リン酸カルシウム法等を用いることができる。また植物由来細胞への一般的な形質転換法としては、アグロバクテリウムを用いた形質転換法(アグロバクテリウム法)を挙げることができ、本発明でもアグロバクテリウム法を好適に用いることができる。ただしアグロバクテリウム法を用いて形質転換する場合には、本発明にかかるDNA断片を含むTiプラスミドを構築する必要がある。またその他プロトプラスト/スフェロプラスト法、リン酸カルシウム法、リポソーム法、DEAEデキストラン法等の従来公知の方法を好適に用いることができる。
【0046】
ただし転写因子発現用DNA断片は、宿主細胞のゲノムに組み込まれることが好ましい。転写因子発現用DNA断片がゲノムに組み込まれることにより、細胞分裂後の娘細胞にもベクターの構成に含まれる遺伝子を確実に伝達することが可能となり、転写因子(タンパク質)の生産効率を維持することが可能となるからである。ゲノムは染色体(核ゲノム)に限定されるものではなく、ミトコンドリアゲノムや葉緑体ゲノムも含まれる。
【0047】
また転写因子発現用DNA断片が宿主細胞に導入されたか否かを確認する方法は、特に限定されるものではなく、公知の各種の方法を用いることができる。具体的には、各種マーカーを用いればよい。例えば、宿主細胞中で欠失している遺伝子をマーカーとして用い、このマーカーと組み換え植物ウイルス遺伝子とを含むプラスミド等を発現ベクターとして宿主細胞に導入する。これによってマーカー遺伝子の発現から本発明の遺伝子の導入を確認することができる。例えば後述する実施例においては、薬剤耐性マーカー(ハイグロマイシン耐性遺伝子、Hygr)を用いており、ハイグロマイシンを含有する培地中で、形質転換候補株を培養することにより、生育してきた細胞株を形質転換体として選抜することが可能となる。その他のマーカーとしては、ビアラホス耐性マーカー、カナマイシン耐性マーカー等が植物細胞の選抜に有効であり、ピューロマイシン耐性マーカー、ブレオマイシン耐性マーカー、XGPRT遺伝子、DHFR遺伝子、チミジンキナーゼ遺伝子等が動物細胞の選抜には有効である。さらに酵母などでは、ウラシル要求性マーカーをはじめとする栄養要求性マーカーを選抜に用いることができる。但しこれら形質転換体の選抜方法は、限定されるものではなく、発現ベクターを導入する宿主等に応じて適宜選択して用いればよい。その他、宿主細胞から調製したゲノムDNAを鋳型とし、導入したタンパク質(転写因子)の遺伝子全長を特異的に増幅するいわゆるジェノミックPCR法を挙げることができる。この方法によって、目的タンパク質(転写因子)をコードする遺伝子が増幅されてくることを電気泳動法等によって確認できれば、該遺伝子の導入を確認することができる。
【0048】
(A−2)選抜工程
本形質転換体の生産方法における第2工程として、当該選抜工程がある。当該選抜工程では、上記第一形質転換工程によって得られた形質転換体(細胞)の中から前記転写因子を安定的に高発現する形質転換体(細胞)の選抜を行なう。すなわち転写因子の発現に最適な宿主細胞の染色体上の位置に、該転写因子をコードする遺伝子が組み込まれた形質転換体(細胞)を選抜(スクリーニング)する工程である。
【0049】
当該選抜工程における具体的選抜方法は上記第一形質転換工程で得られた形質転換体(細胞)の中から目的とする転写因子を安定的に高発現する形質転換体を選抜する方法であれば特に限定されるものではない。
【0050】
例えば、形質転換体(細胞)から発現タンパク質を抽出し、該転写因子特異抗体を用いてその発現量を検出する方法として、ウエスタンブロット法、エライザ法、ドットブロット法等が適用可能である。また、形質転換体(細胞)から発現RNAを抽出し、該転写因子の遺伝子配列に相補的なプローブを用いて転写因子のmRNAの発現量を検出する方法として、例えばノーザンブロット法、ドットブロット法等が適用可能である。さらに、例えばRT−PCR法、リアルタイムPCR法、マイクロアレイ法等によって該転写因子mRNAの発現量を検出することも可能である。
【0051】
(A−3)第二形質転換工程
当該第二形質転換工程は、上記選抜工程により選抜された転写因子を高発現する形質転換体(細胞)(以下、転写因子高発現形質転換体(細胞)と称する)に、RNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子高発現形質転換体が発現する転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる植物レクチンタンパク質発現用DNA断片を導入する工程である。あらかじめ選抜された、転写因子を高発現する形質転換体に、目的とする植物レクチンタンパク質をコードする遺伝子が挿入されたウイルスベクターのcDNAを導入することにより、高確率に植物レクチンタンパク質を高生産する形質転換体(細胞)を生産することができる。
【0052】
<植物レクチンタンパク質発現用DNA断片>
本植物レクチンタンパク質発現用DNA断片は、形質転換体(細胞)に植物レクチンタンパク質を生産させるために使用するDNA断片であり、少なくともRNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子高発現形質転換体(細胞)が発現する転写因子で転写誘導される転写誘導型プロモーターとからなる。この他、植物レクチンタンパク質をコードする遺伝子挿入用のクローニングサイト(好ましくはマルチクローニングサイト)・ベクター配列・ターミネーター・薬剤耐性マーカー等のDNAセグメントが含まれていてもよい。かかる転写因子発現用DNA断片の調製方法は、通常の遺伝子工学的手法を用いて行なえばよい。
【0053】
転写誘導型プロモーターは、転写因子高発現形質転換体が生産する転写因子に応じて選択すればよく、その組み合わせの一例については、(A−1)第一形質転換工程の項で述べたとおりである。
【0054】
細胞に生産させる植物レクチンタンパク質は特に限定されない。例えば、ゴーヤ由来抗Hレクチン(配列番号1及び2)、コムギ由来WGA、トウゴマ由来RCA、ピーナツ由来PNA、タチナタマメ由来ConA、ダイズ由来SBA、ハリエニシダ由来UEA、ジャガイモ由来STL等の種々の植物レクチンタンパク質が例示できる。
【0055】
ホルモンによって活性化される性質を有する転写因子を用いて植物レクチンタンパク質の転写誘導を行う本発明の方法は、各種植物に由来するレクチンタンパク質を生産するための形質転換体の生産に用いることができる。植物レクチンには細胞毒性を示すものがあり、異種発現系を構築することが困難であるが、この問題は、恒常的発現プロモーターではなく誘導型の発現プロモーターを用いることにより解消される。プロモーターとして植物において良く用いられている恒常的発現プロモーター(カリフラワーモザイクウイルス由来35Sリボソーム発現プロモーター;PCaMV35S)を用いて植物レクチンの発現を試みたところ、レクチンの蓄積量は極めて低い結果となった。また、作製した細胞を長期間継代培養すると、細胞が暗黒色に変色し、増殖性が著しく低下した。これは、継代培養している間もレクチンを生産し続ける為であると推察される。ホルモンにて発現調整が可能な誘導型プロモーターを使用することにより、細胞を十分に増殖させた後にレクチンを発現させることが可能となり、生産能力が著しく向上する。また、継代培養を行っている間は、発現を抑制させることができるため、植物レクチンが細胞に悪影響を与えることなく、継代培養を行うことが可能である。なかでも、宿主細胞として植物由来培養細胞(特に好ましくは、BY2細胞等のタバコ由来細胞)を用いる実施形態は、各種植物に由来するレクチンタンパク質を生産するための形質転換体の生産に特に適している。植物レクチンの発現は、異種発現系において一般的に用いられている微生物宿主(大腸菌や酵母)にて構築している例は少ない。これは、植物レクチンを微生物細胞で発現させることがタンパク質フォールディングや翻訳後修飾の違いにより困難であるからと推察される。植物細胞を宿主として用いることにより、この問題は解消される。植物細胞として好適なものは、細胞増殖性が極めて高いタバコBY-2細胞である。植物細胞を宿主として用いることにより、フォールディングや翻訳後修飾などが正常に行われ、種子由来のものと同様の糖鎖特異性を示す植物レクチンが生産可能となる。したがって、本発明の発現系、特に、宿主細胞として植物由来培養細胞(特に好ましくは、BY2細胞等のタバコ由来細胞)を用いる発現系は、実施例において用いられたゴーヤ由来抗Hレクチンタンパク質の発現だけでなく、他の植物に由来するレクチンタンパク質の発現にも適した発現系であるといえる。
【0056】
ウイルスベクターとしては、RNAを遺伝子とするウイルス由来のウイルスベクターであれば特に限定されるものではなく、二本鎖RNAウイルス、一本鎖マイナス鎖RNAウイルス、一本鎖プラス鎖RNAウイルス由来のウイルスベクターを用いることができる。中でも細胞内でcDNAから転写されたRNA自体がmRNAとして機能するという理由から、一本鎖プラス鎖RNAを遺伝子とするウイルス由来のウイルスベクターであることが特に好ましい。
【0057】
また、ウイルスベクターは植物ウイルス由来のウイルスベクターに限定されるものではなく、動物ウイルス、ファージを含むあらゆるRNAウイルス由来のウイルスベクターを用いることが可能である。しかしながら、植物細胞に植物レクチンタンパク質を生産させる目的で使用する場合には、植物ウイルス由来のウイルスベクターであることが好ましく、中でも植物のサイレンシングを抑制する因子(サプレッサー)を持つウイルス由来のウイルスベクターであることが特に好ましい。サイレンシングのサプレッサーを持つウイルスベクターを用いることにより、増幅の後半においてもmRNAが分解される現象が生じない。サイレンシングのサプレッサーを持つ植物ウイルスとしては、例えば、ポティ属(Potyvirus属)ウイルス、ククモウイルス属(Cucumovirus属)ウイルス(例えばキュウリモザイクウイルス(CMV))、ポテックスウイルス属(Potexvirus属)ウイルス(例えばジャガイモXウイルス(PVX))、トンブスウイルス属(Tombusvirus属)ウイルス(例えばトマトブッシースタントウイルス(TBSV)、Cymbidiumu ringspot virus (CymRSV))、カルモウイルス属(Carmovirus属)ウイルス(例えば、Turnip crinkle virus (TCV))、トバモウイルス属(Tobamovirus属)ウイルス(例えば、タバコモザイクウイルス(TMV)、トマトモザイクウイルス(ToMV))が挙げられる。
【0058】
また上記ウイルスベクターのcDNAの3’末端には、リボザイム配列が結合していることが好ましい。形質転換体の細胞内でcDNAから転写されるウイルスRNAの3’末端に付加されウイルスの複製能力低下の原因となるターミネーター由来の配列やポリA配列を切断することができるからである。その結果ウイルスの複製能力の低下を防止することができ、植物レクチンタンパク質の高生産が可能となる。結合されるリボザイム配列は、上記3’末端に付加された余分な配列を切断できるものであればよく、特に限定されるものではない。例えば、肝炎デルタウイルスのリボザイム配列(GenBank accession No. X77627他)またはサテライトタバコリングスポットウイルスのリボザイム配列(GenBank accession No. M17439)を用いることができる。
【0059】
また植物レクチンタンパク質をコードする遺伝子は、ウイルスの外被タンパク質をコードする遺伝子と置換されていることが特に好ましい。この部位に植物レクチンタンパク質遺伝子を挿入することによりウイルスの外被タンパク質が産生されなくなり、増幅されるウイルス遺伝子は粒子化されず他の植物に感染することがなくなるため、ウイルスの外界への飛散という問題を解消することが可能となる。
【0060】
更に、ウイルスベクターにおいて、ウイルスの移行タンパク質(MP)をコードする遺伝子が削除されており、MPサブゲノムプロモーター領域を構成する塩基の少なくとも一部が、複製酵素である180Kタンパク質のアミノ酸配列は変えないように置換されており、ウイルスの外被タンパク質の開始コドンが他の塩基配列に置換されて無効化されており、かつ、ウイルスの外被タンパク質をコードする遺伝子が植物レクチンタンパク質をコードする遺伝子に置換されていることが好ましい。本発明者らは、この特徴を有するウイルスベクターをMPDベクターと称する(MPが削除(Delete)されているため)。MPDベクターから発現されるウイルスRNAは従来の旧型ベクターと同様のウイルス複製能力を持つ。なおMP遺伝子は誘導ウイルスベクターシステムでは不必要な遺伝子である。MPDベクターは、MP遺伝子が削除されているためにcDNAの鎖長は短い。このため、MPDベクターは宿主細胞において複製され易くかつ植物におけるcDNAの転写の効率が高いものと考えられる。
【0061】
図1(b)に植物レクチンタンパク質発現用DNA断片の一例を示す。図1(b)は、形質転換用ベクターであるTiプラスミドpBICER8−ToMV−CO.3−MCL−His−SRZの一部分を示している。同図中左からエストロジェンによって活性化される融合転写因子LexA−VP16−hERによって転写誘導されるプロモーターOLexA−46、その下流にゴーヤ由来抗Hレクチン遺伝子(以下、MCL遺伝子と称する)を組み込んだトマトモザイクウイルスベクターToMV−MCL cDNA、その3’末端にサテライトタバコリングスポットウイルスリボザイム配列S−Rz、さらに35Sターミネーター配列35ST、またその下流に薬剤耐性マーカーとしてカナマイシン耐性遺伝子Kanrがある。なお、上記植物レクチンタンパク質発現用DNA断片の転写因子高発現形質転換体への導入方法は、(A−1)第一形質転換工程の項で述べた方法と同様に行なえばよい。
【0062】
上記本形質転換体(細胞)の生産方法は種々の植物レクチンタンパク質の発現系に適用可能である。当該生産方法により、人工的に遺伝子に変異を加えて調製される新規糖鎖認識特異性をもつレクチンの効率的な生産が可能となる。
【0063】
本形質転換体(細胞)の生産方法の特に好ましい実施形態としては
宿主細胞がタバコBY2細胞であり、
転写因子がLexA−VP16−hERであり、
転写誘導型プロモーターがOLexA−46であり、
ウイルスベクターが、トバモウイルス属に属するウイルス由来であり、
ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、植物レクチンタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターであり、
ウイルスベクターのcDNAの3’末端に、リボザイム配列が結合しており、かつ
転写因子発現用DNA断片、および植物レクチンタンパク質発現用DNA断片が、アグロバクテリウム法により導入される
ことを特徴とする実施形態が挙げられる。この実施形態によれば、植物レクチンタンパク質を、天然物中と同様の活性を維持しつつ、特に効率的に生産することが可能である。
【0064】
通常、ウイルスベクターは導入する遺伝子のサイズが増加するにしたがい複製能力が低下することが知られている。しかしながら、植物レクチンは遺伝子DNAのサイズが比較的大きい(1.5k)にもかかわらず、本発明の方法によれば、予想に反して植物レクチンタンパク質を高発現可能な形質転換体を生産することができた。この理由は明らかではないが、本発明の発現系は、ウイルスベクターを用いるにも関わらず、予想に反して植物レクチンの発現に適していることが確認された。In vitro転写したウイルスベクターをBYIIプロトプラストに接種した場合にはMPを欠失させる効果はほとんど認められないが、形質転換体においてはレクチンをはじめ1kbを越えるタンパク質はMPDベクターでのみ効率よく生産することができた。本ベクターは従来ベクターでは効率よく合成することができなかった遺伝子DNAの塩基配列が1kbを越えるレクチン以外の2種のタンパク質(アラビドプシスのクチナーゼ、ヒトアクチン)を合成することに成功している。すなわち、本ベクターを用いた形質転換細胞は、植物レクチンタンパク質に限らず、アラビドプシスのクチナーゼ、ヒトアクチン等の、遺伝子DNAの塩基配列が1kbを越える(好ましくは、2kb以下である)タンパク質を効率的に生産することが可能である。
【0065】
また、下記に示す実施例では、通常の2倍の数のセルラインを選抜し、その結果、特に高い発現を示すラインを一つえることに成功した。本結果より、比較的サイズの大きなレクチンに対しても、同様に、多くのラインから選抜することにより、特に優れた形質転換体を選抜できる可能性が高まることが考えられ、このことは、植物レクチン全般に応用可能であると思われる。したがって、第二形質転換工程において得られた形質転換体から、所望の植物レクチン発現能力を有する形質転換体を選抜するためにはセルライン200以上であることが好ましいといえる。
【0066】
(B)本発明にかかる植物レクチンタンパク質生産用形質転換体およびその利用
本発明にかかる植物レクチンタンパク質生産用形質転換体(以下、本形質転換体)は、上記本形質転換体の生産方法によって生産された植物レクチンタンパク質生産用形質転換体(細胞)である。本形質転換体は、転写因子をコードする遺伝子およびウイルスベクターをコードする遺伝子の両者が、それぞれ発現に最適な染色体上の位置に組み込まれた形質転換体であるため、本形質転換体を用いることによって、植物レクチンタンパク質を高生産することができる。
【0067】
また本発明にかかる植物レクチンタンパク質の生産方法(以下、本植物レクチンタンパク質の生産方法)は、本形質転換体を用いることを特徴としている。すなわち、本植物レクチンタンパク質の生産方法は、植物レクチンタンパク質をコードする遺伝子が導入された本形質転換体から植物レクチンタンパク質を回収することにより行なう。さらに植物レクチンタンパク質をより大量に取得するためには、本形質転換体の数を培養・栽培・育成等によって増加させ、増加させた本形質転換体から植物レクチンタンパク質を回収することにより行なうことが好ましい。本形質転換体の培養・栽培・育成条件等は特に限定されるものではなく、本形質転換体に好適な条件を適宜選択して用いればよい。植物由来培養細胞を培養する際の培地としては特に限定されるものではないが、無機塩類、炭素源、ビタミン類、アミノ酸が加えられている場合がある。さらに、ココナツミルクや酵母エキスを加えて成長を促進させる場合がある。その他、オーキシンとサイトカイニン、ジベレリン、アブシジン酸、エチレン等の植物ホルモンを添加する場合がある。また培養条件であるが、光、温度、通気の有無等を培養する細胞に応じて最適なものを採用すればよい。例えば、タバコBY2細胞を培養条件の一例としては、370mg/lリン酸二水素カリウム、1mg/lチアミン塩酸、3%スクロース、0.2mg/l 2,4−Dを含むMS培地を用い、暗所、26℃、135回転/分で旋回振盪培養後、1/100量を一週間ごとに継代することが挙げられる。
【0068】
また、本形質転換体の種類としては特に限定されるものではなく、植物・動物の個体であっても、植物・動物由来培養細胞であってもよい。ただし、迅速に大量に植物レクチンタンパク質を生産するためには、形質転換体は培養可能な細胞(培養細胞)であることが好ましい。さらには、細胞の取り扱いのしやすさ・培地が安価である等の理由から、植物由来培養細胞であることが好ましい。上記植物由来培養細胞としては、例えばタバコ由来BY2細胞が挙げられる。
【0069】
(C)本発明にかかる植物レクチンタンパク質生産用形質転換体の生産キット
本発明にかかる植物レクチンタンパク質生産用形質転換体の生産キット(以下、本キットと称する)は、上記本形質転換体の生産方法を行なうためのキットである。本キットを構成するものについては特に限定されるものではないが、転写因子をコードする遺伝子と転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片、および/またはRNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる植物レクチンタンパク質発現用DNA断片により構成されていることが好ましい。具体的には本キットが、転写因子発現用DNA断片の一例であるpER8(−Stu)、および/または植物レクチンタンパク質発現用DNA断片の一例であるpBICER8−ToMV−CO.3−MCL−His−SRZにより構成されている例が挙げられる。
【0070】
本キットの利用者は、(a)転写因子発現用DNA断片を適当な宿主細胞に導入する工程、(b)その形質転換体の中から転写因子を高発現する形質転換体を選抜する工程、(c)該形質転換体に植物レクチンタンパク質をコードする遺伝子が挿入されている植物レクチンタンパク質発現用DNA断片を導入する工程を経て植物レクチンタンパク質生産用形質転換体が得られることとなる。なお本キットに含まれる転写因子発現用DNA断片に植物レクチンタンパク質をコードする遺伝子を挿入する方法は、遺伝子工学的手法により行なえばよい。
【0071】
さらに本キットは、上記転写因子高発現形質転換体および/または植物レクチンタンパク質発現用DNA断片により構成されていることが好ましい。より具体的には後述する実施例において取得した転写因子高発現形質転換体の一例である転写因子高発現タバコBY2細胞、および/または植物レクチンタンパク質発現用DNA断片の一例であるpBICER8−ToMV−CO.3−MCL−His−SRZにより構成されている例が挙げられる。
【0072】
転写因子高発現形質転換体が既に取得されているため、本キットの利用者は上記(c)のみを行なえばよく、より迅速かつ簡便に植物レクチンタンパク質生産用形質転換体を得ることができる。
【0073】
この他本キットには、例えば、細胞、培地、制限酵素、修飾酵素類、転写誘導用化学物質(ステロイドホルモン、エストロジェン等)、培養フラスコ、アグロバクテリウム(植物細胞の場合)等が含まれていてもよい。
【0074】
本発明は上述した各実施形態に限定されるものではなく、請求項に示した範囲で種々の変更が可能であり、異なる実施形態にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態についても本発明の技術的範囲に含まれる。
【実施例】
【0075】
1. ゴーヤレクチン遺伝子(MCL)発現系の作製方法
(1)転写因子発現用DNA断片導入用ベクターの構築
転写因子発現用DNA断片を宿主細胞(タバコBY2細胞)に導入するためのベクター(以下転写因子発現用DNA断片導入用ベクターと称する)として、TiプラスミドpER8(−Stu)を用いた。pER8(−Stu)は、恒常的プロモーターPG10-90の下流にエストロジェンレセプターを含む融合転写因子LexA−VP16−hERをコードする遺伝子と、ターミネーターTE9とを連結し、さらに薬剤耐性マーカーとしてハイグロマイシン耐性遺伝子(Hygr)を組み込んで構築した。図1(a)にpER8(−Stu)の一部分の構造を模式図として示した。
【0076】
なお、TiプラスミドpER8(−Stu)、恒常的プロモーターPG10-90、LexA−VP16−hER、ターミネーターTE9、Hygrはいずれも公知の方法により調製可能である。TiプラスミドpER8(−Stu)は、Dohi, K., Nishikiori, M., Tamai, A., Ishikawa, M., Meshi, T., Mori, T. (2006) Inducible virus-mediated expression of a foreign protein in suspension-cultured cells. Archives of Virology 151, 1075-1084に記載されている。恒常的プロモーターPG10-90はIshige, F., Takaichi, M., Foster, R.,Chua, N. H. and Oeda, K.(1999) A G-box motif(GCCACGTGCC) tetramer confers high-level constitutive expression in dicot and monocot plants. Plant J. 20, 127-133に記載されている。LexA−VP16−hER及びターミネーターTE9はZuo J, Niu QW, Chua NH. “An estrogen receptor-based transactivator XVE mediates highly inducible gene exprssion in transgenic plants.” Plant J.2000, 24: 265-273 に記載されている。
【0077】
(2)植物レクチンタンパク質発現用DNA断片導入用ベクターの構築
トマトモザイクウイルス(ToMV)の外被タンパク質をコードする遺伝子を、MCLをコードする遺伝子(配列表の配列番号1、以下、MCL遺伝子と称する)で置換したToMV変異体を用いた。配列番号1の塩基配列のうち23番〜1664番の塩基配列をMCL遺伝子として用いた。
【0078】
ToMV変異体のcDNAの塩基配列を図2及び配列表の配列番号3に示す。図2において、下線を引いた箇所はToMVのcDNAの塩基配列から変更した箇所である。下線部(b)は移行タンパク質(MP)遺伝子を削除し、MPサブゲノムプロモーター領域の塩基配列を変更した(ただし複製酵素である180Kタンパク質のアミノ酸配列は変えないように変更した)箇所である。下線部(d)は外被タンパク質(CP)の開始コドンをATCに置換して無効化した箇所である。(e)で示す箇所は外来遺伝子クローニング用に導入されたPmeIサイトである。下線部(f)はMCLをコードするMCL遺伝子(配列番号1の第23番〜1664番塩基)を示す。下線部(h)はヒスチジンタグ(His−tag)をコードする塩基配列である。枠により強調した箇所(j)、(a)、(c)、(e)、(g)、(i)はいずれも構築に関与する制限酵素サイトであって、(j)はSpeIのサイトであり、(a)はKpnIのサイトであり、(c)はNcoIのサイトであり、(e)はPmeIのサイトであり、(g)および(i)はBstEIIのサイトである。
【0079】
図2に示すToMV変異体のcDNAを含有するプラスミドは次の手順で作成した。
(1)KpnI(a)からNcoI(c)までの領域を人工合成した。
(2)NcoI(c)からPmeI(e)/BstEII(i)までの領域のPCR断片を、外被タンパク質(CP)の開始コドン(ATG)をATCに置換した変異を導入して作成した。具体的には、Dohi, K., Nishikiori, M., Tamai, A., Ishikawa, M., Meshi, T., Mori, T. (2006) Inducible virus-mediated expression of a foreign protein in suspension-cultured cells. Archives of Virology 151, 1075-1084に記載されているプラスミドpiL.erG3(SF3)SRz(Avr)を鋳型とし、以下のプライマーセットを用いてNcoI(c)からPmeI(e)/BstEII(i)までの領域のPCR断片を作成した。
フォワードプライマー: 5'-GACCCATGGGAAGTTGTTGATGAGTTCA-3’
リバースプライマー: 5'-TATGGTTACCGTTTAAACAAAAACACAAATTGCGATGGAGAAGTGATTGAGTAAGAGATATTTAATACGAATCA-3’
フォワードプライマー中、細下線部がNcoIサイト(c)に対応する配列である。
【0080】
リバースプライマー中、細下線部がクローニングサイトPmeI(e)/BstEII(i)に対応する配列であり、太下線部がATGからATCへの変異を導入するための配列である。
【0081】
プラスミドpiL.erG3(SF3)SRz(Avr)のうち、本発明に関連する部分の構造を図4(a)に示す。piL.erG3(SF3)SRz(Avr)に含まれるToMV−GFP cDNAは、トマトモザイクウイルス(ToMV)の外被タンパク質をコードする遺伝子を、緑色蛍光タンパク質をコードする遺伝子で置換したToMV変異体のcDNAである。
【0082】
(3)プラスミドpiL.erG3(SF3)SRz(Avr)において、KpnIからBstEIIまでの部分を、上記(1)で得られたKpnI(a)−NcoI(c)DNA断片と、上記(2)で得られたNcoI(c)−PmeI(e)/BstEII(i)DNA断片とに入れ替えた(3点ligation)。
【0083】
(4)上記(3)で得られたプラスミドのPmeI(e)/BstEII(i)部位に、ゴーヤ由来抗Hレクチン(MCL)のDNA断片(5’端PmeI、3’端BstEII)とHis−tagのDNA断片(5’−GTCACCTCACCATCACCATCACCATTG−3’と5’−GTTACCAATGGTGATGGTGATGGTGAG−3’をアニールしたもの)を挿入した(3点ligation)。
【0084】
5’端にPmeIサイト、3’端にBstEIIサイトがそれぞれ導入されたMCLのDNA断片は、MCLをコードするDNA(特許文献1)を鋳型とし、以下のプライマーセットを用いたPCRにより作製した。
フォワードプライマー: 5'-ATAGTTTAAACATGAGAATGAGAGTGTTAG-3’
リバースプライマー: 5'-TATGGTGACCAATAGAACACCGTCCATT-3’
【0085】
(5)上記(4)で得られたプラスミドのSpeI−AvrIIの領域を切り出し、pBICER8−ToMV5’−SpeのSpeI部位に挿入した。ここで用いたプラスミドpBICER8−ToMV5’−Speは、上記(2)で言及した文献に記載されており、エストロジェンで転写誘導可能なプロモーターOLexA−46(Zuo J, Niu QW, Chua NH. “An estrogen receptor-based transactivator XVE mediates highly inducible gene exprssion in transgenic plants.” Plant J.2000, 24: 265-273 参照)を有するTiプラスミドにおいて、OLexA−46の下流に、ToMVの5’末端からSpeIサイトまでの領域(図2の5’端から(j)までの領域に相当する)が組み込まれたものである。pBICER8−ToMV5’−Speのうち、本発明に関連する部分の構造を図4(b)に示す。
【0086】
こうして、植物レクチンタンパク質発現用DNA断片を宿主細胞(タバコBY2細胞)に導入するためのベクター(以下、植物レクチンタンパク質発現用DNA断片導入用ベクターと称する)pBICER8−ToMV−CO.3−MCL−His−SRZを構築した。図1(b)にpBICER8−ToMV−CO.3−MCL−His−SRZの一部分の構造を模式図として示した。
【0087】
このプラスミドpBICER8−ToMV−CO.3−MCL−His−SRZは、図1(b)に示すように、エストロジェンで転写誘導可能なプロモーターOLexA−46を有するTiプラスミドにおいて、OLexA−46の下流に、ToMV変異体(図2)のcDNAが連結され、さらにその3’末端にサテライトタバコリングスポットウイルスのリボザイム配列S−Rz(GenBank accession No. M17439)、35Sターミネーター(35ST)、カナマイシン耐性遺伝子(Kanr)が組み込まれた構造を有する。
【0088】
(3)第一形質転換工程:転写因子発現用DNA断片の宿主細胞への導入
転写因子発現用DNA断片導入用ベクターpER8(−Stu)を、タバコBY2細胞にアグロバクテリウム法により導入した。まずpER8(−Stu)をエレクトロポレーション法によってAgrobacterium tumefacience LBA4404系統に導入した。これをカナマイシン(50mg/l)を含むAB sucrose培地で前培養した。次にタバコBY2細胞と混合してシャーレに移し、26℃、暗所で42〜48時間静置してタバコBY2細胞を形質転換した。タバコBY2細胞用培地にて洗浄した後、カルベニシリン(100mg/l)およびハイグロマイシン(20mg/l)を含むタバコBY2細胞用固形培地に広げ、形質転換タバコBY2細胞を増殖させた。
【0089】
(4)選抜工程:転写因子高発現形質転換体の選抜
形質転換タバコBY2細胞のうち、ノーザンブロット法の結果、転写因子の発現量の高い細胞ライン(ER43)を選抜した。ここで「細胞ライン」とは、形質転換細胞を増殖させることにより形成された個々のコロニーのことを意味する。
【0090】
(5)第二形質転換工程:植物レクチンタンパク質発現用DNA断片の導入
上記で得られた転写因子高発現タバコBY2細胞ライン(ER43)に、ウイルスベクター(pBICER8−ToMV−CO.3−MCL−His−SRZ)をアグロバクテリウム法により導入し形質転換細胞(カルス)を192ライン得た。
【0091】
(6)組換えウイルス高発現形質転換カルスの選抜
得られた192ラインの形質転換カルスは、エストロジェン(0.01mM)を添加したBY2個体培地に移植し、移植2日後にカルスから全RNAを抽出した。抽出したRNAをToMVの3’非翻訳領域約200塩基に相補的なDIGRNAプローブを用いてノーザン分析を行った。その結果、ウイルスRNAの蓄積量の多い4ライン(10−1−3−11−13、10−1−3−12−9、10−1−3−5−17、10−1−3−14−20)を選抜し、選抜したカルスを15mlの液体培地に移し、細胞が飽和した後、1/100量になるように継代培養した。継代7日後1/100量になるようにさらに継代培養し、同様の継代培養により細胞を維持した。
【0092】
(7)MCLの誘導発現
上記で得られた4ラインの各形質転換細胞を1/100量になるよう継代し、継代培養後7日間前培養した細胞を、新しい培地に1/30量になるように移した。3日間培養した後、エストロジェンを終濃度が0.01mMとなるように加え、さらに4日間培養した後MCLを抽出し、ヒト赤血球凝集(HA)活性を検出した。その結果、10−1−3−11−13株において最も高いHA活性が認められた。
【0093】
上記において、転写因子高発現タバコBY2細胞ラインER43を形質転換して取得したMCL高発現細胞(10−1−3−11−13株)を用いて、MCLのエストロジェンによる誘導発現を試みた。10−1−3−11−13株を1/100量になるよう継代し、前培養を3日間行なった培養液に対して、エストロジェンを終濃度0.01mM添加しMCLの誘導発現を行なった。エストロジェン添加後48時間の10−1−3−11−13株から全RNAを抽出し、ノーザンブロット法によりToMVに特異的なRNAを検出した。解析には、ToMVの3’非翻訳領域約200塩基に相補的なRNAプローブを用いた。プローブのラベリングは、ロシュ・ディアグノスティック社のDIG RNA Labeling Kitを用いて行なった。検出には、同社のDIG Luminescent Ditection KitおよびCDP−Starを用い、それぞれキットのマニュアルに従って行なった。また、エストロジェン添加後48時間の10−1−3−11−13株から全タンパク質を抽出し、HA活性の検出を行なった。その結果、エストロジェンによるToMVの明らかな転写誘導およびHA活性の検出が確認された。10−1−3−11−13株は、増殖と発現誘導を行い、以降のレクチン抽出および評価に用いた。
【0094】
2.発現系由来レクチンの評価
(1)発現系由来レクチンの抽出
10−1−3−11−13株の培養液から細胞破砕装置またはフレンチプレスを用いてレクチンを抽出し、10,000×gで30分間、遠心分離後の上清を粗抽出液とした。
【0095】
(2)発現系由来粗抽出液のヒト赤血球凝集(HA)活性
この粗抽出液についてレクチン活性を評価した。レクチン活性は、連続希釈法によるヒトHA活性により測定した。すなわち、マイクロタイタープレートを用いて被検体のPBSによる2倍希釈列を作製し、これに等量の3%ヒト赤血球浮遊液(オーソ社製)を加えて、室温で30分間反応させ、肉眼で凝集の有無を判定した。その結果、発現系由来粗抽出液のヒトO型HA活性は256 titerを示した。これは、恒常的発現プロモーターpH35CGによる発現系を用いた下記比較例(粗抽出液の力価8)に比べて大幅に改善したものであった。本培養液1 Lから得られるレクチン活性量は、乾燥種子200−300 gから得られる抽出液の活性量に相当するものと推定された。適度に希釈した本粗抽出液のA1、A2、BおよびOの各血液型に対して、それぞれ2、8、8、16の力価を示し、種子由来レクチンの場合と特性が類似していたことから、種子由来レクチン遺伝子が機能を保持したまま発現していることが推定された。
【0096】
(3)発現系由来レクチンの精製
上記(1)で得られた粗抽出液は、PBSに対して透析した後、0.5mL/minの流速にてガラクトースセファロースカラム(30mL)に流し、ガラクトースに親和性のある成分を吸着させた。このカラムをPBSにて十分に洗浄後、0.2Mガラクトースを含むPBSを流すことにより吸着したガラクトース結合画分を溶出させた。この溶出画分は蒸留水に対して透析し凍結乾燥した。凍結乾燥により得られた粉末をPBSに溶解したものは、ヒトO型HA活性を有していたことから、レクチンであると認められた。約90mLの培養液からは、約1mg(PBSに1 mg/mLの濃度で溶解した際の力価は2,048)の発現系由来レクチンが得られた。
【0097】
(4)発現系由来レクチンの電気泳動特性
還元条件化のSDS−PAGE分析の結果、発現系由来レクチンは、種子由来レクチンの4本のバンドのうちの2本が強く観察された(図3)。
【0098】
(5)発現系由来レクチンの血球特異性
発現系由来レクチンおよび比較のために用いた種子由来レクチンはPBSにて希釈し、O型赤血球に対する力価を32 titerに調整した。発現系由来レクチンのA1、A2、BおよびOの各血液型に対するHA活性は、それぞれ4、16、8、32の力価を示し、種子由来レクチンの場合と同等であった(表1)。
【0099】
(6)発現系由来レクチンの糖鎖特異性
発現系由来レクチンの糖鎖特異性は、各種糖類によるHA阻害試験により評価した。(5)で調整した各レクチン溶液の12.5μlは、終濃度が100mMから0.01mMになるように希釈した糖類溶液の12.5μlと混和後、室温で1時間攪拌した。その後、3%ヒトO型赤血球浮遊液を添加し、30分後に凝集の有無を肉眼判定し、血球の凝集を阻害した最低濃度を最低HA阻害濃度として求めた。なお、用いた糖類は、D−ガラクトース、D−フコース、N−アセチル−D−ガラクトサミン、N−アセチルノイラミン酸、メチル−α−D−ガラクトピラノシド、L−アラビノース、D−ラクトース、ラクト−N−ビオース、N−アセチル−D−ラクトサミン、ヒトB抗原三糖、ヒトA抗原三糖、ヒトH抗原二糖、ヒトH抗原三糖、D−グルコース、D−マンノース、L−フコース、N−アセチル−D−グルコサミン、D−キシロース、L−ラフィノース、D−フルクトース、スクロース、マルトース、N,N−ジアセチルキトビオースである。その結果、両者の糖鎖特異性は極めて類似し(表2)、H抗原三糖に対して高い特異性を示した。以上のことから、本発現系に導入されたレクチン遺伝子は、その機能を保持したまま発現していることが確認された。
【0100】
【表1】
【0101】
【表2】
【0102】
比較例
恒常発現プロモーターを用いたゴーヤ由来レクチンタンパク質の発現
MCLのcDNAを、バイナリーベクターpH35CG(インプランタイノベーションズ)を用いて、微粒子発射装置により、タバコBY−2(Nicotiana tabacum L. cv. Bright Yellow−2)細胞の懸濁液中で発現させた。当該ベクターは、カリフラワーモザイクウイルス35Sプロモーターに連結されたMCLのcDNAと、ノパリン合成酵素(NOS)ターミネーターとを含む。ハイグロマイシンに対する耐性を付与するハイグロマイシンホスホトランスフェラーゼ(HPT)遺伝子を、NOSプロモーターに連結する。当該ベクターにより金粒子(直径1.5〜3μm、アルドリッチケミカル)を被覆した。減圧空気:−0.1MPa、標的距離:9.6cm、ヘリウム圧:5Kg/cm2の条件で、1試験区当り1回の発射を実施した。発射を受けた細胞は、50mg/mlハイグロマイシンを含有する固化LS培地上に載せ、続いて、14日間、同一の培地上において数回継代培養を行った。選択的条件での継代培養を50日間行った後、ロータリーシェーカーにより、100mlのLS培地を加えた500ml容三角フラスコ中で27℃にて10日間、ハイグロマイシン耐性細胞を培養した。懸濁した細胞を集め、Tissue ruptor(Quiagen)を用いて破砕した。破砕された細胞を、20,000×gにて10分間遠心沈降させた。上澄み液の赤血球凝集活性を、上記の手順で測定した。
【特許請求の範囲】
【請求項1】
ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換工程と、
前記第一形質転換工程によって得られた形質転換体の中から前記転写因子を発現する形質転換体を選抜する選抜工程と、
RNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる植物レクチンタンパク質発現用DNA断片を、前記選抜工程によって選抜された形質転換体に導入する第二形質転換工程とからなることを特徴とする植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項2】
上記ホルモンが、エストロジェンまたはステロイドホルモンであることを特徴とする請求項1の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項3】
上記エストロジェンで活性化される性質を有する転写因子としてLexA−VP16−hERを用い、上記転写誘導型プロモーターとしてOLexA−46を用いることを特徴とする請求項2の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項4】
上記ウイルスベクターは、一本鎖プラス鎖RNAを遺伝子とするウイルス由来であることを特徴とする請求項1〜3のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項5】
上記ウイルスベクターが、植物ウイルス由来であることを特徴とする請求項4の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項6】
上記ウイルスベクターが、植物のサイレンシングを抑制する因子を持つ植物ウイルス由来であることを特徴とする請求項5の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項7】
上記ウイルスベクターが、トバモウイルス属に属するウイルス由来であることを特徴とする請求項6の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項8】
上記ウイルスベクターが、トマトモザイクウイルスベクターまたはタバコモザイクウイルスベクターであることを特徴とする請求項7の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項9】
上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、植物レクチンタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターである、請求項1〜8のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項10】
上記ウイルスベクターのcDNAの3’末端に、リボザイム配列が結合していることを特徴とする請求項1〜9のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項11】
上記リボザイム配列が、肝炎デルタウイルスのリボザイム配列、またはサテライトタバコリングスポットウイルスのリボザイム配列であることを特徴とする請求項10の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項12】
上記転写因子発現用DNA断片、および上記植物レクチンタンパク質発現用DNA断片が、アグロバクテリウム法により導入されていることを特徴とする請求項1〜11のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項13】
上記宿主細胞および形質転換体が、植物体または植物由来培養細胞であることを特徴とする請求項1〜12のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項14】
上記植物由来培養細胞がタバコ由来細胞であることを特徴とする請求項13の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項15】
上記タバコ由来細胞がタバコBY2細胞であることを特徴とする請求項14の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項16】
上記宿主細胞がタバコBY2細胞であり、
上記転写因子がLexA−VP16−hERであり、
上記転写誘導型プロモーターがOLexA−46であり、
上記ウイルスベクターが、トバモウイルス属に属するウイルス由来であり、
上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、植物レクチンタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターであり、
上記ウイルスベクターのcDNAの3’末端に、リボザイム配列が結合しており、かつ
上記転写因子発現用DNA断片、および上記植物レクチンタンパク質発現用DNA断片が、アグロバクテリウム法により導入される、
ことを特徴とする、請求項1〜15のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項17】
上記植物レクチンタンパク質がゴーヤ由来レクチンタンパク質である、請求項1〜16のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項18】
請求項1〜17のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法によって生産された植物レクチンタンパク質生産用形質転換体。
【請求項19】
請求項18の植物レクチンタンパク質生産用形質転換体を用いることを特徴とする植物レクチンタンパク質の生産方法。
【請求項20】
請求項1〜17のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法を行うためのキット。
【請求項21】
ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換工程と、
前記第一形質転換工程によって得られた形質転換体の中から前記転写因子を発現する形質転換体を選抜する選抜工程と、
RNAを遺伝子とするウイルスに、生産しようとするタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなるタンパク質発現用DNA断片を、前記選抜工程によって選抜された形質転換体に導入する第二形質転換工程とからなり、
上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、生産しようとするタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターであることを特徴とする、タンパク質生産用形質転換体の生産方法。
【請求項22】
生産しようとするタンパク質をコードする遺伝子が、1kbを越える鎖長のDNAからなる遺伝子である、請求項21のタンパク質生産用形質転換体の生産方法。
【請求項23】
ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片と、
RNAを遺伝子とするウイルスに、生産しようとするタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなるタンパク質発現用DNA断片であって、上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、生産しようとするタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターである、タンパク質発現用DNA断片と
が遺伝子DNAに導入されていることを特徴とする、タンパク質生産用形質転換体。
【請求項1】
ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換工程と、
前記第一形質転換工程によって得られた形質転換体の中から前記転写因子を発現する形質転換体を選抜する選抜工程と、
RNAを遺伝子とするウイルスに植物レクチンタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる植物レクチンタンパク質発現用DNA断片を、前記選抜工程によって選抜された形質転換体に導入する第二形質転換工程とからなることを特徴とする植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項2】
上記ホルモンが、エストロジェンまたはステロイドホルモンであることを特徴とする請求項1の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項3】
上記エストロジェンで活性化される性質を有する転写因子としてLexA−VP16−hERを用い、上記転写誘導型プロモーターとしてOLexA−46を用いることを特徴とする請求項2の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項4】
上記ウイルスベクターは、一本鎖プラス鎖RNAを遺伝子とするウイルス由来であることを特徴とする請求項1〜3のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項5】
上記ウイルスベクターが、植物ウイルス由来であることを特徴とする請求項4の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項6】
上記ウイルスベクターが、植物のサイレンシングを抑制する因子を持つ植物ウイルス由来であることを特徴とする請求項5の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項7】
上記ウイルスベクターが、トバモウイルス属に属するウイルス由来であることを特徴とする請求項6の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項8】
上記ウイルスベクターが、トマトモザイクウイルスベクターまたはタバコモザイクウイルスベクターであることを特徴とする請求項7の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項9】
上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、植物レクチンタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターである、請求項1〜8のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項10】
上記ウイルスベクターのcDNAの3’末端に、リボザイム配列が結合していることを特徴とする請求項1〜9のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項11】
上記リボザイム配列が、肝炎デルタウイルスのリボザイム配列、またはサテライトタバコリングスポットウイルスのリボザイム配列であることを特徴とする請求項10の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項12】
上記転写因子発現用DNA断片、および上記植物レクチンタンパク質発現用DNA断片が、アグロバクテリウム法により導入されていることを特徴とする請求項1〜11のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項13】
上記宿主細胞および形質転換体が、植物体または植物由来培養細胞であることを特徴とする請求項1〜12のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項14】
上記植物由来培養細胞がタバコ由来細胞であることを特徴とする請求項13の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項15】
上記タバコ由来細胞がタバコBY2細胞であることを特徴とする請求項14の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項16】
上記宿主細胞がタバコBY2細胞であり、
上記転写因子がLexA−VP16−hERであり、
上記転写誘導型プロモーターがOLexA−46であり、
上記ウイルスベクターが、トバモウイルス属に属するウイルス由来であり、
上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、植物レクチンタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターであり、
上記ウイルスベクターのcDNAの3’末端に、リボザイム配列が結合しており、かつ
上記転写因子発現用DNA断片、および上記植物レクチンタンパク質発現用DNA断片が、アグロバクテリウム法により導入される、
ことを特徴とする、請求項1〜15のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項17】
上記植物レクチンタンパク質がゴーヤ由来レクチンタンパク質である、請求項1〜16のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法。
【請求項18】
請求項1〜17のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法によって生産された植物レクチンタンパク質生産用形質転換体。
【請求項19】
請求項18の植物レクチンタンパク質生産用形質転換体を用いることを特徴とする植物レクチンタンパク質の生産方法。
【請求項20】
請求項1〜17のいずれか1項の植物レクチンタンパク質生産用形質転換体の生産方法を行うためのキット。
【請求項21】
ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換工程と、
前記第一形質転換工程によって得られた形質転換体の中から前記転写因子を発現する形質転換体を選抜する選抜工程と、
RNAを遺伝子とするウイルスに、生産しようとするタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなるタンパク質発現用DNA断片を、前記選抜工程によって選抜された形質転換体に導入する第二形質転換工程とからなり、
上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、生産しようとするタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターであることを特徴とする、タンパク質生産用形質転換体の生産方法。
【請求項22】
生産しようとするタンパク質をコードする遺伝子が、1kbを越える鎖長のDNAからなる遺伝子である、請求項21のタンパク質生産用形質転換体の生産方法。
【請求項23】
ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片と、
RNAを遺伝子とするウイルスに、生産しようとするタンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子で転写誘導される転写誘導型プロモーターとが連結されてなるタンパク質発現用DNA断片であって、上記ウイルスベクターが、ウイルスの移行タンパク質をコードする遺伝子が削除されており、ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、生産しようとするタンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターである、タンパク質発現用DNA断片と
が遺伝子DNAに導入されていることを特徴とする、タンパク質生産用形質転換体。
【図1】

【図2−1】

【図2−2】

【図3】

【図4】
【図2−1】
【図2−2】
【図3】
【図4】
【公開番号】特開2011−15640(P2011−15640A)
【公開日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願番号】特願2009−162672(P2009−162672)
【出願日】平成21年7月9日(2009.7.9)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成18年度、独立行政法人科学技術振興機構、「地域イノベーション創出総合支援事業 重点地域研究開発推進プロジェクト」委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(504224153)国立大学法人 宮崎大学 (239)
【出願人】(591040236)石川県 (70)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
【公開日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願日】平成21年7月9日(2009.7.9)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成18年度、独立行政法人科学技術振興機構、「地域イノベーション創出総合支援事業 重点地域研究開発推進プロジェクト」委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(504224153)国立大学法人 宮崎大学 (239)
【出願人】(591040236)石川県 (70)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
[ Back to top ]