
耐熱性脱硫酵素とそれをコードする遺伝子

【解決手段】 チオフェン系化合物を分解する機能を有する酵素及びそれをコードする遺伝子、並びに前記遺伝子を含むベクター及び形質転換体を提供する。
【効果】 化石燃料中の硫黄を容易に遊離させることができるようになる。
【効果】 化石燃料中の硫黄を容易に遊離させることができるようになる。
【発明の詳細な説明】
【0001】
【発明の属する技術分野】本発明は、微生物を利用するチオフェン系化合物、すなわちベンゾチオフェン、ジベンゾチオフェン(以下「DBT 」という)およびこれらの置換体、又はそれらの誘導体を分解する機能を有する酵素及びそれをコードする遺伝子に関するものである。本発明の酵素及び遺伝子を利用することにより、石油等の化石燃料中に含まれるベンゾチオフェンやDBT およびこれらの置換体、又はそれらの誘導体中の硫黄を遊離させることができるので、通常石油・石炭等の化石燃料の燃焼により空気中に拡散すると言われる硫黄を、化石燃料中から容易に除去することができるようになる。
【0002】
【従来の技術】石油のような炭化水素燃料から硫黄を除去する脱硫のための方法としては、アルカリ洗浄や溶剤脱硫などの方法も知られているが、現在では水素化脱硫が主流となっている。水素化脱硫は、石油留分中の硫黄化合物を触媒の存在下で水素と反応させ、硫化水素として除去して製品の低硫黄化を図る方法である。触媒としては、アルミナを担体としてコバルト、モリブデン、ニッケル、タングステン、などの金属触媒が使用される。モリブデン担持アルミナ触媒の場合には、触媒性能を向上させるために、通常コバルトやニッケルが助触媒として加えられる。金属触媒を用いた水素化脱硫は、現在世界中で広く使用されているきわめて完成度の高いプロセスであることは疑いのないことである。しかし、より厳しい環境規制に対応した石油製品を作るためのプロセスという観点からは、いくつかの問題点がある。以下にその例を簡単に記載する。
【0003】金属触媒は、一般にその基質特異性が低く、このため多様な種類の硫黄化合物を分解し、化石燃料全体の硫黄含量を低下させる目的には適しているが、特定のグループの硫黄化合物、すなわちベンゾチオフェンやDBT のような複素環式硫黄化合物類およびそれらのアルキル誘導体類に対してはその脱硫効果が不十分となることがあると考えられる。たとえば、脱硫後の軽油中にはなおも種々の複素環式有機硫黄化合物が残存している。このように金属触媒による脱硫効果が不十分となる原因の一つは、これらの有機硫黄化合物中の硫黄原子の周囲に存在する置換基による立体障害が考えられる。これらの置換基のうち、メチル置換基の存在が水素化脱硫における金属触媒の反応性に及ぼす影響は、チオフェン、ベンゾチオフェン、DBT などについて検討されている。それらの結果によると、一般的には置換基の数が増すほど脱硫反応は減少するが、置換基の位置が反応性に及ぼす影響もきわめて大きいことが明らかである。メチルDBT 類の脱硫反応性を比較し、置換基による立体障害が金属触媒の反応性に及ぼす影響が非常に大きいことを示した報告は、たとえば、Houalla, M., Broderick, D.H., Sapre, A.V., Nag,N.K., de Beer, V.H., Gates, B.C., Kwart, H.J., Catalt., 61, 523-527(1980) に見られる。実際、これらのDBT の種々のアルキル化誘導体が軽油中にかなりの量存在することが知られている(たとえば、Kabe, T., Ishihara, A. and Tajima, H. lnd. Eng. Chem. Res., 31, 1577-1580(1992))。
【0004】上記のように水素化脱硫に抵抗性を示す有機硫黄化合物を脱硫するためには、現在用いられているよりも高い反応温度や圧力が必要とされ、また、添加する水素の量も非常に増大すると考えられている。このような水素化脱硫プロセスの改良は、ばく大な設備投資と運転コストを必要とすることが予想される。このような水素化脱硫に抵抗性を示す有機硫黄化合物を主たる硫黄化合物種として含むものとしては、たとえば、軽油があり、軽油のより高度な脱硫(超深度脱硫)を行う場合には上記のような水素化脱硫プロセスの大幅な改良が要求される。
【0005】一方、生物が行う酵素反応は比較的穏和な条件下で進行し、しかも酵素反応の速度自体は、化学触媒を用いた反応の速度と遜色のないという特徴を有している。さらに、生体内で起こる多種多様の生物反応に適切に対応する必要があるため、非常に多くの種類の酵素が存在し、それらの酵素は一般的に非常に高い基質特異性を示すことが知られている。このような特徴は、微生物を用いて化石燃料中に含まれる硫黄化合物中の硫黄の除去を行ういわゆるバイオ脱硫反応においても活かされるものと期待されている(Monticello, D.J., Hydrocarbon Processing39-45(1994))。
【0006】一方、細菌を用いて石油の成分である複素環式硫黄化合物から硫黄を除去する方法については、多数の報告があるが、それらは環分解(C-C 結合切断)型反応とC-S 結合切断型反応とに大別される。C-C 結合攻撃型脱硫活性を有する細菌としては、例えば、Pseudomonas sp., Pseudomonas aeruginosa, Beijerinckia sp., Pseudomonas alcaligenes, Pseudomonas stutzeri, Pseudomonas putida, Brevibacterium sp.などが知られている。これらの細菌は、DBT で代表される複素環式硫黄化合物中のC-C 結合の切断を行い、ベンゼン環を分解し、その後の酸化反応カスケードにより、硫黄塩を放出するというタイプの代謝を行うものである。これらの炭素骨格攻撃型経路の反応機構は芳香環の水酸化(DBT →1,2-ジヒドロキシジベンゾチオフェン)、環の開裂、水溶性産物への酸化(1,2-ジヒドロキシジベンゾチオフェン →トランス-4[2-(3-ヒドロキシ)チアンナフテニル]-2- オキソ- ブテノイン酸、3-ヒドロキシ-2- ホルミルベンゾチオフェン)といったものであり、Kodama経路と呼ばれている。このタイプの反応により、DBT のベンゼン環中のC-C 結合が攻撃を受け、油から抽出可能な種々の水溶性物質を生じる。しかし、この反応により、油中の他の芳香族分子が攻撃を受け、その結果かなりの量の炭化水素が液相に移動することになる(Hartdegen, F.J., Coburn,J.M. and Roberts, R.L. Chem. Eng. Progress, 80, 63-67(1984)) 。このようなことは石油の総熱量単位の低下を招くことになり、工業的には非効率的な反応である。また、このタイプのDBT 酸化分解菌は、児玉らが報告しているように酸化産物として水溶性のチオフェン化合物(主として3-ヒドロキシ-2- ホルミルベンゾチオフェン)を与えるが、これは液相から除去するのが困難な物質でもある。更に、DBT の炭素環の攻撃は、しばしばアルキル置換基やアリル置換基を持つDBT の 2位及び 3位の位置で起こるため、これらの位置で置換されたDBT はKodama経路の基質とはならない。
【0007】原油や石炭のみならず硫黄を含んだモデル化合物を分解し、ヘテロ原子である硫黄を選択的に除去して、硫酸塩や水酸化化合物を産生する微生物類が報告されている。このタイプの反応は、その代謝産物の構造から考えて、硫黄化合物中のC-S 結合を特異的に切断して、その結果硫黄を硫酸塩の形で遊離する反応であると考えられる。現在までに、表1 に示すような硫黄攻撃型のバイオ脱硫反応系の報告がある。
【0008】
【表1】

【0009】以上記載したバイオ脱硫はすべて、30℃近辺の温度条件下で進行する微生物代謝反応を利用するものである。一方、化学反応の速度は一般に温度に依存して増大することが知られている。また、石油精製プロセス中の脱硫工程では、高温・高圧条件下で分別蒸留や脱硫反応が行われる。従って、石油精製プロセス中にバイオ脱硫工程を組み込むとすると、常温近くにまで石油留分を冷却することなしに、冷却途中のより高い温度でバイオ脱硫反応ができる方が望ましいと考えられる。高温バイオ脱硫に関する報告には以下のようなものがある。
【0010】微生物を用いて高温で脱硫反応を行わせる試みのほとんどは、石炭脱硫において見ることができる。石炭中には種々の硫黄化合物が含まれている。主要な無機硫黄化合物は黄鉄鉱であるが、有機硫黄化合物に関しては多種多様のものが混在しており、多くがチオール、スルフィド、ジスルフィド、チオフェン基を含んでいることが知られている。用いられた微生物は、Sulfolobus属の細菌で、これらはすべて好熱性細菌である。鉱物スルフィドからの金属のリーチング(BrierleyC.L. & Murr, L.E., Science 179, 448-490(1973)) や石炭からの黄鉄鉱の硫黄除去などに種々の異なったSulfolobus株を用いた例が報告されている(Kargi, F. & Robinson, J.M., Biotechnol. Bioeng, 24, 2115-2121(1982); Kargi, F. &Robinson, J.M., Appl. Environ. Microbiol., 44, 878-883(1982); Kargi, F.& Cervoni, T.D., Biotechnol. Letters 5, 33-38(1983); Kargi, F.およびRobinson, J.M., Biotechnol. Bioeng., 26, 687-690(1984); Kargi, F. & Robinson, J.M., Biotechnol. Bioeng. 27, 41-49(1985); Kargi, F., Biotechnol. Lett., 9, 478-482(1987))。Kargi とRobinson (Kargi, F.およびRobinson, J.M.,Appl. Environ. Microbiol., 44, 878-883(1982))によれば、米国のイエローストーン国立公園の酸性温泉から分離されたSulfolobus acidocaldarius (以下、「S.acidocaldarius」という)のある株は、45〜70℃で生育するが、至適pH2 で元素状硫黄を酸化する。また、別の 2種のS. acidocaldarius 株による黄鉄鉱の酸化も報告されている(Tobita, M., Yokozeki, M., Nishikawa, N. & Kawakami, Y., Biosci. Biotech. Biochem. 58, 771-772(1994))。
【0011】化石燃料中に含まれる有機硫黄化合物のうち、DBT およびその置換体又はそれらの誘導体は通常の石油精製プロセスにおいて水素化脱硫を受けにくいことが知られている。そのDBT のS. acidocaldariusによる高温分解も報告されている(Kargi, & Robinson, J.M., Biotechnol. Bioeng, 26, 687-690(1984); Kargi, F., Biotechnol. Letters 9, 478-482(1987))。
【0012】これらの報告によれば、チアントレン、チオキサンテン、DBT などのモデル芳香族複素環式硫黄化合物を高温でこの微生物と反応させると、これらの硫黄化合物は酸化されて、分解する。S. acidocaldarius によるこれらの芳香族複素環式硫黄化合物の酸化は、70℃で観察されており、反応産物として硫酸イオンを生じる。しかし、この反応は硫黄化合物の他には炭素源を含まない培地中での反応であり、硫黄化合物を炭素源としても利用している。すなわち硫黄化合物中のC-C結合を分解していることは明瞭である。さらに、このS. acidocaldarius は酸性の培地でのみ増殖でき、DBT の酸化分解反応は、きびしい酸性条件下(pH2.5)での進行を要求する。このようなきびしい条件は石油製品の劣化を引き起こすと同時に脱硫に関わる工程に耐酸性材料を必要とするためプロセス上望ましくないと考えられる。S. acidocaldarius を、独立栄養条件下で増殖させると、必要なエネルギーを還元された鉄・硫黄化合物から獲得し、炭素源として二酸化炭素を利用する。しかし、S. acidocaldarius は、従属栄養条件下に増殖させると、炭素源およびエネルギー源として種々の有機化合物を利用することができる。すなわち、化石燃料が存在すると炭素源として資化されるものと考えられる。
【0013】Finnertyらは、Pseudomonas stutzeri、Pseudomonas alcaligenes 、Pseudomonas putidaに属する株がDBT 、ベンゾチオフェン、チオキサンテン、チアントレンを分解して、水溶性の物質に変換することを報告している(Finnerty, W.R.,Shockiey, K., Attaway, H. in Microbial Enhanced Oil Recovery, Zajic, J.E. ら(編)、Penwell. Tuisa, Okia, 83-91(1983))。この場合、酸化反応は55℃でも進むとしている。しかし、これらのPseudomonas 菌株によるDBT の分解産物は、Kodamaらが報告している3-ヒドロキシ-2- ホルミルベンゾチオフェンである(Monticello, D.J., Bakker, D., Finnerty, W.R. Appl. Environ. Microbiol.,49, 756-760(1985))。これらのPseudomonas 菌株によるDBT の酸化活性は、硫黄を含まない芳香族炭化水素であるナフタレンやサリチル酸により誘導を受け、クロラムフェニコールにより阻止される。このことから、これらのPseudomonas 菌株によるDBT の分解反応は、芳香環中のC-C 結合を切断することによる分解を基礎としていることが分かる。また、硫黄化合物以外にも石油留分中に含まれる貴重な芳香族炭化水素を同時に分解するおそれもあり、これは、燃料の価値や石油留分の品質を低下させることになる。
【0014】このように、今までに発見されている高温でDBT を分解できる菌は、DBT 分子中のC-C 結合を切断し、炭素源として利用する反応を触媒するものである。C-S結合を特異的に切断するが、C-C 結合は切断しないでそのまま残すタイプの有機硫黄化合物の分解反応が実際の石油の脱硫方法として望ましいことは上述の通りである。すなわち、高温でDBT およびそのアルキル置換体、又はそれらの誘導体分子中のC-S 結合を切断する活性を示し、水溶性の物質の形で、脱硫産物を生じる微生物を利用するのがバイオ脱硫プロセスとして最も望ましい。
【0015】前述のように、C-S結合切断型のDBT分解反応を行う微生物は、いくつかの属の細菌で知られている。たとえば、Rhodococcus sp. のATCC53968 はよく調べられたDBT分解菌株であり、DBTの硫黄原子に酸素原子を付加し、ジベンゾチオフェンスルホキシド(以下、「DBTO」という)からジベンゾチオフェンスルホン(以下、「DBTO2」という)を生成し、ついで2‐(2'-ヒドロキシフェニル)ベンゼンスルフィン酸塩を経て2-ヒドロキシビフェニル(以下、「2-HBP」という)を生成する反応を行う。しかし、この菌も通常の培養温度である30℃よりも少し高い37℃および43℃でさえ、48時間培養すると非常に生育が遅れるか、生育しなくなることが報告されている(特開平6-54695号公報)。このことから、高温脱硫反応を行わせるためには、高温で生育でき、しかも高温で有機硫黄化合物、特にDBTおよびその置換体、またはそれらの誘導体化合物を含む複素環式硫黄化合物類をC-S結合特異的に切断できる微生物を用いるのが最適であると考えられる。
【0016】Suzukiらは、Paenibacillus sp. A11-2株が、高温条件下で生育でき、かつ、有機硫黄化合物のC-S結合を特異的に切断できることを報告している(Suzuki,M., Konishi, J., Ishii, Y., Okumura, K., Appl. Environ. Microbiol., 63,3164-3169(1997))。この報告によると、この菌株は30〜60℃の温度条件下で生育でき、DBTまたは、4-メチルジベンゾチオフェンおよび4,6-ジメチルジベンゾチオフェンなどのDBT誘導体のC-S結合を特異的に切断できる。よって、この菌株は高温脱硫反応を行うのに適している。しかし、この菌株は、比較的低い温度、とりわけ30℃付近において、脱硫活性が著しく低下する。
【0017】バイオ脱硫プロセスにおいて、高温域(以降、「高温域」とは、50〜60℃付近の温度のことを指す)を含む広範な温度条件下(30〜60℃)で高い脱硫活性を維持していることは、石油留分の温度調整を簡略化できるという面で有効である。Furuyaらは、Bacillus subtilis WU-S2B株(受託番号FERM P-17041)が、高温域を含む広範な温度条件下で比較的高い脱硫活性を維持し、かつ、有機硫黄化合物のC-S結合を特異的に切断できることを報告している(古屋, 西井, 中川, 桐村,宇佐美, 日本農芸化学会大会講演要旨集, 384(1999))。この報告によると、この菌株は30〜60℃の広範な温度条件下で比較的高い脱硫活性を維持し、DBTまたは、2,8-ジメチルDBTおよび4,6-ジメチルDBTなどのDBT誘導体のC-S結合を特異的に切断できる。
【0018】C-S 結合切断型の脱硫反応を起こすことが知られている細菌で、そのDBT 分解反応に関与する酵素活性をコードする遺伝子が同定され、その塩基配列が決定されているものとしては、例えば、Rhodococcus sp. IGTS8 株のdsz 遺伝子(Denome, S., Oldfleld., C., Nash, L.J.およびYoung, K.D. J.Bacteriol., 176:6707-6716, 1994; Piddington, C.S., Kovacevich, B.R.およびRambosek, J. Appl.Environ. Microbiol., 61:468-475, 1995)、Sphingomonas sp. AD109株のdsz遺伝子(米国特許第6,133,016号)およびPaenibacillus sp. A11-2株のtds遺伝子(Y. Ishii, J. Konishi, H. Okada, K. Hirasawa, T. Onaka, M. Suzuki., Biochem. Biophy. Res. Comm., 270 (1), 81-88, 2000)が挙げられる。
【0019】IGTS8 株によるDBT 分解反応は、DBT からDBTOを経てDBTO2への変換を触媒するDszC、DBTO2から2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸への変換を触媒するDszAおよび2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸から2-HBP への変換を触媒するDszBの3つの酵素により触媒される(Denome, S., Oldfield., C., Nash, L.J.およびYoung, K.D. J.Bacteriol., 176:6707-6716, 1994; Gray, K.A., Pogrebinshy, O.S., Mrachko, G.T., Xi, L. Monticello, D.J.and Squires, C.H. Nat Biotechnol., 14:1705-1709, 1996; Oldfield, C., Pogrebinsky, O., Simmonds, J., Olson, E.S.および Kulpa, C.F., Microbiology,143:2961-2973, 1997)。それぞれ対応する遺伝子はdszC, dszA, dszBと呼ばれている。DszCとDszAはモノオキシゲナーゼで、両者ともその酸素添加反応にはNADH-FMNオキシドレダクターゼ活性の共存を必要とすることが知られている(Gray, K.A., Pogrebinsky, O.S., Mrachko, G.T., Xi, L. Monticello, D.J. およびSquires, C.H. Nat Biotechnol., 14:1705-1709, 1996; Xi, L. Squires, C.H., Monticello, D.J. and Chids, J.D. Biochem. Biophys. Res Commun., 230:73-76, 1997) 。これらのdsz 遺伝子を大腸菌で温度シフトにより誘導発現させた場合、菌体培養によるDszA活性は39℃で最大となり、42℃では顕著に低下することが報告されている(Denome, S., Oldfield., D., Nash, L.J. および Young,K.D. J. Bacteriol., 176:6707-6716, 1994) 。この結果は、IGTS8 株の有する脱硫酵素活性は常温付近で最大になり、より高温では活性は低下し、50℃以上ではまったく脱硫活性は見られなくなるという休止菌体反応系の実験結果(Konishi, J., Ishii, Y., Onaka, T., Okumura, K. および Suzuki, M. Appl. Environ. Microbiol., 63:3164-3169, 1997)と一致する。
【0020】A11-2株のDBT分解酵素をコードする遺伝子tdsA,tdsB,tdsCもIGTS8株のDBT分解経路と同様の経路によりDBTを分解脱硫する酵素を産生する。tdsA,tdsB,tdsCによりコードされるDBT分解酵素TdsA,TdsB,TdsCは、それぞれIGTS8株のDszA,DszB,DszCと同様の変換反応を触媒する。(Y. Ishii, J. Konishi, H. Okada, K. Hirasawa, T. Onaka, M. Suzuki., Biochem. Biophy. Res. Comm., 270 (1), 81-88, 2000)これらtds遺伝子を大腸菌内で発現させる場合、50℃以上で活性を有し、55℃で最適温度が見られるものの、45℃以下では顕著に活性が低下する。従ってdsz遺伝子、tds遺伝子の両者の温度特異性から示されるように、40〜50℃の温度条件下で効率的にC-S 結合特異的なDBT 分解活性を指令する遺伝子は従来報告されていないものである。
【0021】
【発明が解決しようとする課題】本発明の課題は、ベンゾチオフェン、DBT 系化合物に作用し、それらを高温で分解する能力を有する微生物から高温脱硫反応に関与する遺伝子を単離し、その構造(特に塩基配列)を特定し、また、これらの遺伝子をそれが単離されたのとは異なる微生物に導入し、脱硫能を賦与することにより、新規な脱硫微生物を創製することである。また、このような微生物を実際にベンゾチオフェン、DBT およびそれらのアルキル誘導体に作用させて、これらの化合物のC-S 結合を切断することにより、硫黄を遊離させる方法を確立することである。
【0022】
【課題を解決するための手段】本発明者らは、上記課題を解決するために鋭意検討を重ねた結果、高温脱硫細菌 Bacillus subtilis WU-S2B株(受託番号:FERM P-17041)から脱硫反応に関与する遺伝子群を単離することに成功し、本発明を完成するに至った。
【0023】即ち、本発明は、以下の1〜10を提供する。
1.以下の(a) 又は(b) のタンパク質をコードする遺伝子。
(a) 配列番号2記載のアミノ酸配列により表されるタンパク質(b) 配列番号2記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBTO2を2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸に変換する機能を有するタンパク質
【0024】2.以下の(a) 又は(b) のタンパク質をコードする遺伝子。
(a) 配列番号4記載のアミノ酸配列により表されるタンパク質(b) 配列番号4記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を2-HBPに変換する機能を有するタンパク質
【0025】3.以下の(a) 又は(b) のタンパク質をコードする遺伝子。
(a) 配列番号6記載のアミノ酸配列により表されるタンパク質(b) 配列番号6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBTをDBTO2に変換する機能を有するタンパク質
【0026】4.上記の1、2又は3に記載の遺伝子を含むベクター。
5.上記の4に記載のベクターを含有する形質転換体。
6.以下の(a) 又は(b) に示すタンパク質。
(a) 配列番号2記載のアミノ酸配列により表されるタンパク質(b) 配列番号2記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBTO2を2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸に変換する機能を有するタンパク質
【0027】7.以下の(a) 又は(b) に示すタンパク質。
(a) 配列番号4記載のアミノ酸配列により表されるタンパク質(b) 配列番号4記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を2-HBPに変換する機能を有するタンパク質
【0028】8.以下の(a) 又は(b) に示すタンパク質。
(a) 配列番号6記載のアミノ酸配列により表されるタンパク質(b) 配列番号6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBTをDBTO2に変換する機能を有するタンパク質
【0029】9.以下の性質を有するタンパク質。
(1) 作用:DBTをDBTO2にする(2) 至適pH:8.0 、安定pH:8〜11(3) 至適温度:45〜50℃、安定温度:53℃以下(4) 分子量:200,000(ゲル濾過法による)
(5) 活性阻害:キレート剤、SH阻害剤によって阻害される(6) 補酵素の要求性:フラビンレダクターゼによって供給される FMNH2 を要求する
【0030】10.以下の性質を有するタンパク質。
(1) 作用:DBTO2を2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸にする(2) 至適pH:7.5、安定pH:6〜10(3) 至適温度:50℃、安定温度:45℃以下(4) 分子量:174,000(ゲル濾過法による)
(5) 活性阻害:キレート剤、SH阻害剤によって阻害される(6) 補酵素の要求性:フラビンレダクターゼによって供給される FMNH2 を要求する
【0031】
【発明の実施の形態】以下、本発明を詳細に説明する。
(1)耐熱性脱硫酵素をコードする遺伝子本発明の遺伝子には、以下の3種類の遺伝子が含まれる。第一の遺伝子(以下、「bdsA」という)は、(a) 配列番号2記載のアミノ酸配列により表されるタンパク質、又は(b) 配列番号2記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBTO2 を2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸に変換する機能を有するタンパク質をコードするものである。
【0032】第二の遺伝子(以下、「bdsB」という)は、(a) 配列番号4記載のアミノ酸配列により表されるタンパク質、又は配列番号4記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を2-HBP に変換する機能を有するタンパク質をコードするものである。
【0033】第三の遺伝子(以下、「bdsC」という)は、(a) 配列番号6記載のアミノ酸配列により表されるタンパク質、又は(b) 配列番号6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBT をDBTOを経てDBTO2に変換する機能を有するタンパク質をコードするものである。
【0034】上記bdsA、bdsB、及びbdsC遺伝子は、Rhodococcus sp.IGTS8株由来のdszA、dszB、dszCおよびSphingomonas sp. AD109株由来のdszA、dszB、dszCおよびPaenibacillus sp. A11-2株由来のtdsA、tdsB、tdsCと一定の相同性を示すが、後述するようにこれらの遺伝子がコードするタンパク質は、2種のdszA、dszB、dszC、およびtdsA、tdsB、tdsCがコードするタンパク質とはその性質において異なる。
【0035】本発明の遺伝子のうち、配列番号2、4及び6記載のアミノ酸配列をコードする遺伝子については、本明細書の実施例に記載された方法により得ることができる。また、これらの遺伝子の塩基配列は既に決定されているので、これらの配列を基に適当なプライマーを合成し、Bacillus subtilis WU-S2B 株(受託番号:FERM P-17041)から調製されたDNA を鋳型としてPCR を行うことによっても得ることができる。
【0036】配列番号2、4及び6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列をコードする遺伝子は、本願の出願時において常用される技術、例えば、部位特異的変異誘発法(Zoller et al., Nucleic Acids Res. 10 6487-6500, 1982)により配列番号2、4及び6記載のアミノ酸配列をコードする遺伝子を改変することにより得ることができる。本発明の遺伝子は、DBT の分解に関与する酵素をコードするので、石油の脱硫に利用することができる。
【0037】(2)脱硫酵素をコードする遺伝子を含むベクター本発明のベクターは、上記のbdsA、bdsB又はbdsC遺伝子を含む。このようなベクターは、本発明のbdsA、bdsB又はbdsC遺伝子を含むDNA 断片を、公知のベクターに挿入することにより作製することができる。DNA 断片を挿入するベクターは、形質転換する宿主に応じて決めればよい。例えば、本発明に係るベクターとしては、プラスミドベクター、ファージミドベクター、コスミドベクター及び酵母ベクターなどが挙げられる。宿主として大腸菌を使用するのであれば、以下のようなベクターを使用するのが好ましい。強力なプロモーターとして、例えば、lac 、lacUV5、trp 、tac 、trc 、λpL、T7、rrnB、などを含むpUR 系、pGEX系、pUC 系、pET 系、pT7 系、pBluescript 系、pKK 系、pBS 系、pBC 系、pCAL系などのベクターを使用するのが好ましい。
【0038】(3)脱硫酵素をコードする遺伝子を含むベクターを含有する形質転換体本発明の形質転換体は、上記ベクターを含有する。形質転換体の宿主とする細胞は、植物細胞や動物細胞などであってもよいが、大腸菌などの微生物が好ましい。代表的な菌株としては、Sambrook等の成書Molecular Cloning Laboratory Mannual 2nd ed.に記載されている、71/18、BB4、BHB2668、BHB2690、BL21(DE3)、BNNl02(C600hflA)、C-1a、C600(BNN93)、CES200、CES201、CJ236、CSH18、DH1、DH5、DH5 α、DP50supF、ED8654、ED8767、HB101、HMS174、JM101、JM105、JM107、JM109、JM110、K802、KK2186、LE392、LG90、M5219、MBM7014.5、MC1061、MM294、MV1184、MV1193、MZ-1、NM531、NM538、NM539、Q358、Q359、R594、RB791、RR1、SMR10、TAP90、TG1、TG2、XL1-Blue、XS101、XS127、Y1089、Y1090hsdR、YK537 などが挙げられる。
【0039】本発明の形質転換体は、電気パルス法、コンピテントセル法、接合伝達法、プロトプラスト法など当技術分野において公知の技術を用いて宿主細胞を形質転換又は形質導入することにより取得することができる。これらの方法は、例えば、Sambrookらの成書(前掲)に記載されている。
【0040】(4)脱硫酵素本発明の脱硫酵素には、以下の3種類のタンパク質が含まれる。第一のタンパク質(以下、「BdsA」という)は、配列番号2記載のアミノ酸配列により表されるタンパク質と、配列番号2記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBTO2を2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸に変換する機能を有するタンパク質とを包含する。
【0041】第二のタンパク質(以下、「BdsB」という)は、配列番号4記載のアミノ酸配列により表されるタンパク質と、配列番号4記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を2-HBP に変換する機能を有するタンパク質とを包含する。
【0042】第三のタンパク質(以下、「BdsC」という)は、配列番号6記載のアミノ酸配列により表されるタンパク質と、配列番号6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBT をDBTO2に変換する機能を有するタンパク質とを包含する。
【0043】上記BdsA、BdsB、及びBdsCは、Rhodococcus sp.IGTS8株由来の脱硫酵素DszA、DszB、DszCと一定の相同性を示し、また、酵素としての作用も同一であるが、以下の点で明確に相違する。
(イ)DszA、DszB、DszCでは、難脱硫物質である4,6-ジブチルジベンゾチオフェンを脱硫できないが、本発明のBdsA、BdsB、及びBdsCでは脱硫可能である。
(ロ)DszA、DszB、DszCは、常温領域で脱硫活性を示すが、本発明のBdsA、BdsB、及びBdsCは常温−高温の中間領域で脱硫活性を示す。
【0044】本発明の脱硫酵素は、上述の本発明の脱硫酵素をコードする遺伝子を利用して製造することができる。また、配列番号2、4、及び6に記載のアミノ酸配列により表される脱硫酵素は、Bacillus subtilis WU-S2B 株(受託番号:FERM P-17041)から常法に従って調製することも可能である。
【0045】本発明のBdsAに包含される一タンパク質の性質を以下に示す。
(1) 作用:DBTO2を2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸にする(2) pH特性:図8及び9に示す通り、至適pHは7.5、安定pHは6〜10である(3) 温度特性:図6及び7に示す通り、至適温度は50℃、安定温度は45℃以下である(4) 分子量:174,000(ゲル濾過法による)
(5) 活性阻害:キレート剤、SH阻害剤によって阻害される(6) 補酵素の要求性:フラビンレダクターゼによって供給される FMNH2 を要求する
【0046】本発明のBdsCに包含される一タンパク質の性質を以下に示す。
(1) 作用:DBTをDBTO2にする(2) pH特性:図4及び5に示す通り、至適pHは8、安定pHは8〜11である(3) 温度特性:図2及び3に示す通り、至適温度は45〜50℃、安定温度は53℃以下である(4) 分子量:200,000(ゲル濾過法による)
(5) 活性阻害:キレート剤、SH阻害剤によって阻害される(6) 補酵素の要求性:フラビンレダクターゼによって供給される FMNH2 を要求する
【0047】本明細書において、脱硫酵素の「至適pH」とは、特定の温度(例えば、50℃)において最も好適な活性を有するpHを指す。同様に、脱硫酵素の「至適温度」とは、特定のpH(例えば、pH 7.0)において最も好適な活性を有する温度を指す。
【0048】また、本明細書において、脱硫酵素の「安定pH」とは、該酵素溶液を種々のpHに一定時間(例えば、30分間)放置した後、特定のpH(例えば、pH 7.0)に再調整し、特定の温度(例えば、50℃)において残存する酵素活性を測定したときに、最も高い活性レベルを100%として、好ましくは60%以上、より好ましくは70%以上、最も好ましくは80%以上の活性レベルを示すpHの範囲を指す。同様に、脱硫酵素の「安定温度」とは、該酵素溶液を種々の温度に一定時間(例えば、30分間)放置した後、特定の温度(例えば、50℃)に再調整し、特定のpH(例えば、pH 7.0)において残存する酵素活性を測定したときに、最も高い活性レベルを100%として、好ましくは60%以上、より好ましくは70%以上、最も好ましくは80%以上の活性レベルを示す温度範囲を指す。
【0049】
【実施例】以下、本発明を実施例により具体的に説明する。実施例中の遺伝子操作に関連した実験は、主にSambrookらの成書(前掲)に詳述されている方法に従って行った。
〔実施例1〕耐熱性脱硫酵素をコードする遺伝子断片のクローニング常温脱硫菌であるRhodococcus sp. IGTS8株や高温脱硫菌であるPaenibacillussp. A11-2株は、中間体としてDBTO2および2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を経由して、DBTを2-HBPへと変換する活性を有することが知られている。
【0050】一方、本発明者らは、本発明で用いられるBacillus subtilis WU-S2B株もRhodococcus sp. IGTS8株やPaenibacillus sp. A11-2株と同様の経路によりDBTを2-HBPに変化する能力を有することを明らかにしている(特開2000-139450)。ここで、上に挙げたDBTを2-HBPに変換する活性に関与する酵素およびその遺伝子に着目すると、Rhodococcus sp. IGTS8株やPaenibacillus sp. A11-2株では、既に各酵素のアミノ酸配列および遺伝子の塩基配列が判明しており、それらは50〜65%の相同性を有することが知られている。
【0051】そこで、Rhodococcus sp. IGTS8株やPaenibacillus sp. A11-2株と同様の活性を有するBacillus subtilis WU-S2B株においても、ある程度の相同性を有することが考えられたため、Rhodococcus sp. IGTS8株やPaenibacillus sp. A11-2株の酵素のアミノ酸配列で相同性の高い部分を検索したところ、以下に示すアミノ酸配列が見つかった。
【0052】
Paenibacillus sp. A11-2 株 TdsA WNVVTSLNNAEARNFG(配列番号7)
Rhodococcus sp. IGTS8 株 DszA WNVVTSLNDAEARNFG(配列番号8)
Paenibacillus sp. A11-2 株 TdsC GFDRFWRDARTHTLHDPV(配列番号9)
Rhodococcus sp. IGTS8 株 DszC GFDRFWRNVRTHSLHDPV(配列番号10)
【0053】そこで、上記のアミノ酸配列をもとに以下の塩基配列を有するPCR 用センスプライマーおよびアンチセンスプライマーをそれぞれ設計・合成した。
センスプライマー:WUS2-SI1 5'-GCI GAR GCI MGI AAY TTY GG-3'(配列番号11)(Iはデオキシイノシンを表す)アンチセンスプライマー:WUS2-C2 5'-CGT IGC GCC AIA AGC GGT C-3'(配列番号12)(Iはデオキシイノシンを表す)
【0054】このセンスプライマーとアンチセンスプライマーを使用して、Bacillus subtilis WU-S2B 株から抽出したDNA を鋳型としてPCRを行った。Bacillus subtilisWU-S2B 株からのDNA の調製は以下のように行った。Bacillus subtilis WU-S2B株を、DBT を含むAII培地(組成は表2に示す)で50℃で18時間培養して、菌体を回収した。
【0055】
【表2】
【0056】得られた菌体を3.7mlのSET緩衝液(6.7% スクロース, 1mM EDTA, 50mM Tris-HCl, pH 8.0)に懸濁させた。この懸濁液に、6mg/mlのリゾチームと4mg/mlのアクロモペプチダーゼを溶解させた1mlのTE緩衝液(1mM EDTA, 10mM Tris-HCl, pH8.0)を添加して、37℃で5分間反応させた。反応液に0.5mlのTE緩衝液(25mM EDTA, 50mM Tris-HCl, pH 8.0)と0.3mlのSDS溶液(25mM EDTA, 20%(w/v)SDS,50mM Tris-HCl, pH 8.0)を添加、攪拌混合して、37℃で10分間反応させ、30秒間ミキサーで攪拌した後、0.3mlの3N NaOHを添加し、10分間断続的に転倒混和した後、0.5mlのTris緩衝液(2M Tris-HCl, pH 7.0)を添加、3分間断続的に転倒混和した後、0.7mlの5M NaClを添加、撹拌混合して菌体反応液を調製した。菌体反応液に7mlのフェノール試薬を添加、撹拌混合して、12,000rpm で5分間遠心を行い、得られた上清に対して、7mlのフェノールクロロホルム試薬を添加、撹拌混合して、12,000rpm で5分間遠心を行った。得られた上清に対して、7mlのクロロホルム試薬を添加、撹拌混合して、12,000rpm で5分間遠心を行い、DNA溶液を調製した。DNA溶液に7mlのイソプロパノールを添加して、0℃で30分間放置した後、15,000rpm で30分間遠心を行い、DNA を沈澱させた。沈殿したDNA を冷70%(w/v) エタノールで洗浄後、減圧乾燥した後、100μlのTE緩衝液に溶解して、DNA溶液を調製した。調製したBacillus subtilis WU-S2B 株DNA を鋳型として用いて行ったPCR の条件は以下の通りである。
【0057】
反応液組成:10mM Tris-HCl 1.5mM MgCl2 各0.2mM dNTP Mixture 1 μM センスプライマー 1 μM アンチセンスプライマー 1μg 鋳型DNA 2.5U Taq DNA polymeraseアニーリング温度:52℃に固定してPCR を行った。
PCR サイクル:95℃ 3min 1回 95℃ 1min ↓ 52℃ 1.5min この間を25回繰り返し 72℃ 3min ↑ 72℃ 10min 1回DNA 増幅機:DNA Thermal Cycler 温度サイクラー(PERKIN ELMER CETUS社製)
【0058】上記の条件でPCR を行った結果、約3.2kb の増幅断片を与えることが確認された。この3.2kb のPCR 産物を、pGEM-T ベクターを用いて大腸菌JM109 株にクローニングした。この取得したDNA 断片の塩基配列を決定した結果、この塩基配列は、Rhodococcus sp. IGTS8株のdsz遺伝子と69.2%、Paenibacillus sp. A11-2株のtds遺伝子と56.9%の相同性を有することが明らかとなった。
【0059】決定されたDNA 塩基配列から、この断片にコードされるアミノ酸配列を推定し、これとRhodococcus sp. IGTS8 株からクローニングされているdsz 遺伝子によりコードされる蛋白質であるDszA、DszB およびDszC のアミノ酸配列とを比較した。その結果、DszA、DszBおよびDszCにそれぞれ76.3%、67.9% および71.3%の相同性を示す推定アミノ酸配列が存在することが確認された。さらに、この推定アミノ酸配列は、Paenibacillus sp. A11-2株のtds 遺伝子によりコードされる蛋白質であるTdsA、TdsB およびTdsC のアミノ酸配列と比較した結果、TdsA、TdsB およびTdsC とそれぞれ60.0%、52.7%および48.9%の相同性を示した。Rhodococcus sp. IGTS8 株およびPaenibacillus sp. A11-2株の脱硫遺伝子の塩基配列および脱硫酵素のアミノ酸配列との相同性が見つかったことから、Bacillussubtilis WU-S2B 株からクローニングされたこのDNA 断片に脱硫酵素がコードされている可能性が高く、これをプローブとして用いて隣接するDNA 領域つまり脱硫遺伝子全域をクローニングすることにした。
【0060】〔実施例2〕全DNA ライブラリーの作製Bacillus subtilis WU-S2B 株からのDNA の調製は以下のように行った。DBTおよびツニカマイシンを含むAII培地(組成は表2に示す)で50℃で18時間培養して菌体を回収した。得られた菌体を1mlのB1緩衝液(50mM EDTA, 50mM Tris-HCl, 0.5% Triton X-100, 0.2mg/ml RNaseA, pH 8.0)に懸濁させた。この懸濁液に、100mg/mlのリゾチーム溶液を20μl と20mg/ml のProteinase K溶液を45μl添加して、37℃で10分間反応させた。反応液に0.35mlのB2緩衝液(800mM GuHCl,20% Tween-20, pH 5.5 )を添加、攪拌混合して、50℃で30分間反応させ、5秒間ミキサーで攪拌して、菌体反応液を調製した。陰イオン交換樹脂が充填されたQIAGEN GENOMIC-TIP20/G(QIAGEN社製)カラムを2mlのQBT 緩衝液(750mM NaCl, 50mM MOPS, 15% ethanol, 0.15% Triton X-100, pH7.0 )で平衡化して、菌体反応液をカラムに注入した。カラムを3mlのQC緩衝液(1.0M NaCl, 50mM MOPS,15% ethanol, pH7.0)で洗浄したのち、2mlのQF緩衝液(1.25M NaCl, 50mM Tris-HCl, 15% ethanol, pH 8.5)でゲノムDNA 溶液を溶出した。ゲノムDNA 溶液に1.4ml のイソプロパノールを添加してDNA を沈殿させたのち、ガラス棒で巻きとり回収した。回収したDNA を50μl のTE緩衝液(10mM Tris-HCl, 1mM EDTA, pH8.0)に溶解してゲノムDNA 溶液を調製した。
【0061】Bacillus subtilis WU-S2B 株の全DNA ライブラリーは以下のようにして作製した。Bacillus subtilis WU-S2B 株の全DNA 標品約 50μg を5 ユニットのSau3AIで各々6分、7分、8分消化した後、消化物をフェノール-クロロホルムで抽出しエタノール沈殿を行って遠心分離して回収されたDNA 断片を3.5ユニットの子ウシ小腸由来のアルカリ性ホスファターゼで、37℃18時間処理することにより脱リン酸化を行った。アルカリ性ホスファターゼ処理後フェノール-クロロホルム処理によりDNA を抽出し、エタノール沈殿によりこれを回収した。得られたDNA 断片約 0.2μg をλDASHI/BamHI アーム約2μg と2ユニットのT4DNA リガーゼ存在下に4℃18時間反応させた。反応混合物をGigapack III Gold packaging Extractと反応させることによりin vitroパッケージングを行い、ファージライブラリーを作製した。得られたファージライブラリーの力価は、105 pfu/mlであった。
【0062】〔実施例3〕全DNA ライブラリーのスクリーニングファージライブラリーのスクリーニングを行うためのDNA プローブは以下のようにして作製した。実施例1で記載した方法により取得したDNA断片とRhodococcus sp. IGTS8株のdsz遺伝子塩基配列の間で特に相同性の高かった領域を選択して以下に示すセンスプライマー(Rhodococcus sp. IGTS8株のdszAの5'端側より427 番目から447 番目の塩基配列に相当)とアンチセンスプライマー(Rhodococcus sp. IGTS8株のdszCの5'端側より1135番目から1153番目の塩基配列に相当)を作製した:センスプライマー:WUS2-NSI1 5'-GCA TGA CAT CCG ATA CG-3'(配列番号13)
アンチセンスプライマーWUS2-NC2 5'-TAG TTT GGG TGG GTT CC-3'(配列番号14)。
【0063】このプライマーを用いてBacillus subtilis WU-S2B 株から調製したDNA を鋳型としたPCR を行うことにより、実施例1で取得したDNA断片の内側領域のDNA断片を増幅させた。得られたPCR 産物を鋳型としてランダムプライム法(マルチプライム法)によりジオキシゲニン(DIG)で標識されたDSZACプローブを調製した。DIG 標識プローブの調製法は、Boehringer Mannheim 社のプロトコールに従った。DIG 標識プローブの調製方法を以下に示す。
【0064】得られたPCR 産物5μl を沸騰した熱湯中で10分間熱変性させ、塩を含んだ氷上で冷却した。得られた変性DNA 溶液に、2μl のヘキサヌクレオチド混合液(0.5M Tris-HCl, 0.1M MgCl2, 1mM Dithioerythritol, 2mg/ml BSA, 3.143mg/mlRandom Primer, pH7.2 )、2μl のdNTP標識混合液(1mM dATP, 1mM dCTP, 1mMdGTP, 0.65mM dTTP, 0.35mM DIG-dUTP, pH7.5 )、10μl の滅菌蒸留水及び1μl のKlenow酵素(2units )を添加して、37℃で18時間反応させた。反応液に、2μl の0.2M EDTA 溶液を添加して反応を停止させた。次に2μl の4M LiClと60 μl 冷エタノール(-20℃)を添加して、-70℃で30分間放置したのち、15,000rpm で30分間遠心を行い、DNA を沈殿させた。沈殿したDNA を冷70%(w/v)エタノールで洗浄後、吸引乾燥したのち、50μl のTE緩衝液に溶解して、DIG 標識プローブを調製した。
【0065】Bacillus subtilis WU-S2B株の脱硫遺伝子のスクリーニングは上述の方法で調製したDIG 標識DSZACプローブを用い、Hybond N+ メンブレンに転写されたプラークに対するプラークハイブリダイゼーションにより行った。ハイブリダイズするクローンの検出にはDIG-ELISA (Boehringer Mannheim)を用いた。ゲノムライブラリーより約2000個のファージプラークをDSZACプローブを用いてスクリーニングしたところ、 3個の陽性プラークが検出された。この 3個のプラークについて単プラーク分離を行い、再度プラークハイブリダイゼーションを行った結果、3個とも全て陽性プラークであることが確認された。検出されたDSZACプローブ陽性プラークを用いてファージクローンを調製し、それらのクローンからQIAGENLambda キットを用いてファージDNA を抽出した。 3個の陽性プラークを用いて調製したファージDNA をEcoRI、NotIで切断し、図1に示す制限酵素地図を作成した。さらに、これら 3種のファージDNA をNotIおよびNheIなどの様々な制限酵素を用いて消化して得られたDNA 断片についてDSZACプローブを用いたサザンブロット分析を行ったところ、約7.1kb のNheI 断片にハイブリダイズすることが確認された。これらの制限酵素地図およびサザンブロット分析の結果から、No.1のファージDNA の約7.1kb のNheI 断片にdsz 遺伝子がコードされているものと考えられた。
【0066】〔実施例4〕脱硫活性の確認次にNheI断片に脱硫遺伝子が存在するかを確認するため組換え大腸菌によるDBT分解能を検定した。プラスミドベクターpUC19のXbaI部位に約7.1kbのNheI断片を挿入してpU1Nhを作製した(図1参照)。このpU1Nhを保有する組換え大腸菌JM109株を、0.1mg/ml のアンピシリンを含むLB培地(Sambrook等の成書Molecularcloning Laboratory Manual 2nd に記載)(LB-Amp培地)に基質としてそれぞれDBTを添加した培地で、37℃で 1晩培養した。また、対照株としてベクターpUC19のみを保有するJM109株を同様の条件で培養した。LB-Amp培地(上述)で37℃、1晩前培養を行い、前培養液を得た。次に、1/100 容量の前培養液を検定用の培地(LB-Amp培地に基質としてDBT を添加した培地)に添加し、37℃で18時間培養後、酢酸エチルを用いて分解産物の抽出を行い、高速液体クロマトグラフィー分析を行った。その結果、pU1Nhを保有する組換え大腸菌では、基質としてDBTを含む培地で培養した場合2-HBP が生成することが確認された。また、ベクターのみの組換え大腸菌ではそのような変換活性はまったく示さなかった。このことから、pU1Nhに挿入されている約7.1kbのNheI断片はDBT から2-HBP への一連の変換反応を触媒する活性をすべてコードできる配列を持っていることが証明された。
【0067】〔実施例5〕脱硫遺伝子の塩基配列の決定陽性プラークから調製したファージDNAおよびpU1Nhを用いて、プライマー歩行法により、約7.1kbのNheI断片のうち約4.1kbの塩基配列の決定を行った。クローンのシークエンシング反応はBigDye Terminator Cycle Sequencing Kit(AppliedBiosystems)を用いて行い、ABI PRISM 310 Genetic Analyzer(Applied Biosystems) により塩基配列を決定した(配列番号1、3及び5)。得られた塩基配列データは、GENETYX-MAC/ATSQ v3.0 およびGENETYX-MAC v10.0を用いて解析した。
【0068】決定した約4.1kbの配列中のORF を探索した結果、1kb以上の長さのORF が同一方向に 3個見つかった。これらのORF を上流側からORF 1, 2, 3 と命名した。ORF 1, 2, 3 は、各々 453個、 356個、 415個のアミノ酸をコードする。ORF 1の翻訳終始コドンTGA とORF 2 の翻訳開始コドンATG は、部分的に重なっており、5'-ATGA-3'という配列になっており、Rhodococcus sp. IGTS8株のdsz オペロン中およびPaenibacillus sp. A11-2株のtdsオペロン中の塩基配列と同様の構成を有していることが確認された。これらのORF についてRhodococcus sp. IGTS8株のdsz 遺伝子との塩基配列相同性の解析を行ったところ、ORF 1, 2, 3 は、それぞれIGTS8株のdsz A, B, C と72.8%、67.4%、71.3%の相同性を示した。また、Paenibacillus sp. A11-2 株のtds 遺伝子との塩基配列相同性の解析を行ったところ、ORF 1, 2, 3 は、それぞれPaenibacillus sp. A11-2株のtds A, B, Cと61.3%、56.8%、56.0%の相同性を示した。さらに、Bacillus subtilis WU-S2B 株遺伝子の塩基配列を基礎としてそれらにコードされる蛋白質のアミノ酸配列を推定したところ、ORF 1, 2, 3 によりコードされるポリペプチドはそれぞれRhodococcus sp. IGTS8 株のDsz A, B, Cと78.6%、67.9%、72.0%の相同性を示した。また、Paenibacillus sp. A11-2 株のTds A, B, Cと63.7%、52.7%、50.6%の相同性を示した。
【0069】Bacillus subtilis WU-S2B 株のORF でコードされる蛋白質のアミノ酸配列とRhodococcus sp. IGTS8 のdsz 配列でコードされる蛋白質のアミノ酸配列とを比較すると、いくつかの点で特徴的な差異が見いだされる。まず、ORF 1 によりコードされるBdsA とDszAとを比較すると、カルボキシル末端側において、アミノ酸の異なる部分が連続して見られる。また、BdsAとTdsAとを比較すると、BdsAのアミノ末端の配列の方がTdsAのアミノ末端の配列より長くなっており、さらに、特にカルボキシル末端側において、アミノ酸の異なる部分が連続して見られる。DszBとORF 2 によりコードされるBdsB のアミノ酸配列については、DszBのアミノ末端の配列の方がBdsBのアミノ末端の配列より延びて長くなっている。また、TdsBとBdsBとを比較すると、BdsBのカルボキシル末端の配列の方がTdsBのカルボキシル末端の配列より長くなっている。また、特にカルボキシル末端側のアミノ酸配列の相同性が低くなっている。DszCとORF 3 によりコードされるBdsC のアミノ酸配列とを比較すると、全長のサイズはほとんど同じであるが、アミノ末端側のアミノ酸配列の相同性が低くなっている。また、TdsCとBdsCとを比較すると、BdsCのアミノ末端の配列の方がTdsCのアミノ末端の配列より延びて長くなっている。
【0070】〔実施例7〕
(1)酵素活性測定に必要なフラビンレダクターゼの精製酵素活性を 50°C で測定するためには、この温度でも活性を有するフラビンレダクターゼが必要である。そのために、Paenibacillus sp. A11-2 株由来のフラビンレダクターゼ(TdsD)高発現株から同酵素を精製した。高発現株を 50 μg/ml のアンピシリンを含む LB 培地中、37°C で一晩振盪培養後、0.2 mM IPTGを添加してさらに 4 時間培養後、菌体を取得した。湿重量 11 g の菌体を緩衝液 A(50 mM トリス塩酸, pH 7.0、10% グリセロール、1 mM ジチオスレイトール)に懸濁し、超音波破砕機(ブランソン、モデル 450)で 4°C、15 分間破砕を行った。10,000 x g で 30 分間遠心分離を行った後の上清を無細胞抽出液とした。この無細胞抽出液を緩衝液 A に対して充分透析をした後、同じ緩衝液で平衡化した Q-Sepharose カラムにアプライした。同緩衝液および、0.05 M、0.1M の塩化カリウムを含む緩衝液 A でそれぞれ洗浄後、0.15 M 塩化カリウムを含む緩衝液 Aで溶出を行った。活性画分を集めて、限外ろ過により濃縮し、引き続き限外ろ過を用いて酵素が溶解している緩衝液を 1.25 M 硫酸アンモニウムを含む緩衝液 A にした。この溶液を同緩衝液で平衡化した Phenyl-Toyopearl カラムにアプライした。同緩衝液で洗浄後、硫酸アンモニウムを含まない緩衝液 Aまでの硫酸アンモニウムによるリニアグラジエント溶出を行い、活性画分を集めて、限外ろ過により濃縮した。引き続き限外ろ過を用いて酵素が溶解している緩衝液を 0.15 M 塩化ナトリウムを含む緩衝液 A にした。この酵素溶液を同緩衝液で平衡化した Superdex200 HR10/30 カラムにアプライし、同緩衝液で溶出し、活性画分を集めて、限外ろ過により濃縮した。この結果、活性画分は電気泳動的に均一であることが確認された。
【0071】酵素活性の測定は20 mM リン酸バッファー、pH 7.0、0.5 mM NADH、0.02 mM FMN を含む反応液中で行い、70°C における NADH の減少速度を 340 nm の吸光度の減少速度を求めることにより算出した。1分間に 1 μmol の NADH を減少させる酵素量を 1 unitと定義した。
【0072】(2)BdsCの精製B. subtilis WU-S2B 株を AII 培地(表2参照)に 0.54 mM になるように DBT/エタノール溶液を添加した培地中で 40°C、2 日間、ジャーファーメンターを用いて通気攪拌培養を行い、菌体 334 g(湿菌体重量)を得た。得られた菌体を緩衝液 A(50 mM トリス塩酸, pH 7.0、10% グリセロール、1 mM ジチオスレイトール)に懸濁し、超音波破砕機(ブランソン、モデル 450)で 4°C、75 分間破砕を行った。10,000 x g で 60 分間遠心分離を行った後の上清を無細胞抽出液とした。この無細胞抽出液を 0.1 M 塩化カリウムを含む緩衝液 A に対して充分透析をした後、同じ緩衝液で平衡化したDEAE-Sepharose カラムにアプライした。同緩衝液で洗浄後、0.25 M 塩化カリウムを含む緩衝液 A までの塩化カリウムによるリニアグラジエント溶出を行った。活性画分を集めて、限外ろ過により濃縮した。この溶液を0.25 M 塩化カリウムを含む緩衝液 A に対して充分透析をした後、同緩衝液で平衡化したQ-Sepharose カラムにアプライした。同緩衝液で溶出し、下記に示す蛋白質Aを含む画分と蛋白質Cを含む画分とに分けて回収して、限外ろ過により濃縮し、引き続き限外ろ過を用いて酵素が溶解している緩衝液を 1 M 硫酸アンモニウムを含む緩衝液 A にした。この溶液を同緩衝液で平衡化した Phenyl-Toyopearl カラムにアプライした。同緩衝液で洗浄後、硫酸アンモニウムを含まない緩衝液 A までの硫酸アンモニウムによるリニアグラジエント溶出を行い、活性画分を集めて、限外ろ過により濃縮した。この結果、活性画分は電気泳動的に均一であることが確認された。
【0073】酵素活性の測定は、10μg精製酵素と、100 mM リン酸バッファー pH 7.0、0.01 mM FMN、6 mM NADH、0.45 units/ml TdsD、0.3 mM DBT とを含む酵素反応液で、50°C で振盪して行った。10 分の 1 量の 12 M HCl を添加して反応を停止後、10 分の 8 量の酢酸エチルを加え、よく混合した後、12,000 回転で 3 分間遠心し、上層を HPLC による分析に供した。1分間に 1 nmol の DBTO2を生成させる酵素量を 1 unit と定義した。精製段階における酵素活性の測定においては、上記の反応液を用いて活性を確認するのに十分な適当量の酵素を使用した。各精製段階における酵素活性を表3に示す。また、上記の精製酵素反応液を、種々の温度で活性の測定を行うことにより、該酵素の温度依存性について検討した。BdsC酵素活性についての温度依存性を図2に示す。上記の精製酵素溶液を種々の温度で30分間放置した後の残存活性を、上記のように測定し、該酵素の温度安定性について検討した。該酵素についての温度安定性を図3に示す。さらに、種々のpHを有する100 mMの各バッファー、0.01 mM FMN、6 mM FADH、0.45 units/ml TdsD、0.3 mM DBTを含む反応液を、50℃で振盪して、酵素活性を測定することにより、該酵素のpH依存性について検討した。pH依存性を図4に示す。また、種々のpHを有する100 mMの各バッファーと精製酵素を混合し、30分間放置した後、100mM リン酸バッファー pH 7.0、0.01 mM FMN、6 mM NADH、0.45 units/ml TdsD、0.3 mM DBT を含む反応液で、50°C で振盪して、該酵素のpH安定性について検討した。該酵素のpH安定性を図5に示す。
【0074】
【表3】
【0075】〔実施例8〕
(1)BdsAの精製BdsCの精製途中における Q-Sepharose の段階で回収した活性画分を 0.1 M 塩化カリウムを含む緩衝液 A に対して透析を行った後、同緩衝液で平衡化した Q-Sepharose カラムにアプライした。同緩衝液で洗浄後、0.3 M 塩化カリウムを含む緩衝液 A までの塩化カリウムによるリニアグラジエント溶出を行った。活性画分を集めて、限外ろ過により濃縮した。引き続き限外ろ過を用いて酵素が溶解している緩衝液を 0.7 M 硫酸アンモニウムを含む緩衝液 A にした。この溶液を同緩衝液で平衡化した Phenyl-Toyopearl カラムにアプライした。同緩衝液および、0.5 M、0.4 M、0.3 M の硫酸アンモニウムを含む緩衝液 A でそれぞれ洗浄後、0.2 M 硫酸アンモニウムを含む緩衝液 A で溶出し、画分を集めて、限外ろ過により濃縮した。引き続き限外ろ過を用いて酵素が溶解している緩衝液を 0.15 M 塩化ナトリウムを含む緩衝液 A にした。この酵素溶液を同緩衝液で平衡化した Superdex200 HR10/30 カラムにアプライし、同緩衝液で溶出し、活性画分を集めて、限外ろ過により濃縮した。さらに、限外ろ過を用いて酵素が溶解している緩衝液を 0.25 M 塩化ナトリウムを含む緩衝液 A にした。この酵素溶液を同緩衝液で平衡化した MonoQ HR10/10 カラムにアプライし、同緩衝液で洗浄後、0.35 M 塩化ナトリウムを含む緩衝液 A までの塩化ナトリウムによるリニアグラジエント溶出を行った。活性画分を集めて、限外ろ過により濃縮した。この結果、活性画分は電気泳動的に均一であることが確認された。
【0076】酵素活性の測定は基質として DBT の代わりにDBTO2を添加する以外は、上述したBdsCの活性測定法と同様に行った。各段階における酵素活性を表4に示す。BdsA酵素活性についての温度依存性を図6に、温度安定性を図7に、pH依存性を図8に、pH安定性を図9に、それぞれ示す。
【0077】
【表4】
【0078】
【発明の効果】本発明は、脱硫に関与する新規な遺伝子及び酵素を提供する。これらの遺伝子及び酵素を利用することにより、化石燃料中の硫黄を容易に遊離させることができるようになる。
【0079】
【配列表】
SEQUENCE LISTING <110> Petroleum Energy Center<120> Thermostable desulfurising enzyme and the gene encoding thereof<130> P01-0068<160> 14<170> PatentIn Ver. 2.0<210> 1<211> 4031<212> DNA<213> Bacillus subtilis<220> <221> CDS<222> (312)..(1670)<400> 1ctagcgaggt gcgagttccg cgcggtagcc atccgcttca ttgattttta catcgatacc 60 gccgcggtgg ccgtcagggt gtcgaagccg ggcggaggtg cacaccccgc cggttggtga 120 tcttgctcac aggtggcgct ttgacggcac tcgtccgggg gcgagaatcg gcgaaggtcg 180 ggtgggtcgt cgggaggaag tggcgtgttg ggtgcccctt tgaatcatcg ggatcctaac 240 tctgttgggg tcggttgcag tagccgagca ttggtctccg gtaactgatc cccacggagg 300 cactgctgat t atg gct gaa caa cgc caa cta cat tta gcc ggg ttt ttc 350 Met Ala Glu Gln Arg Gln Leu His Leu Ala Gly Phe Phe 1 5 10tcg gcc ggc aac gtc act cat gcg cat ggc gcg tgg cgc cat gtc ggt 398 Ser Ala Gly Asn Val Thr His Ala His Gly Ala Trp Arg His Val Gly 15 20 25 gcg acg aac ggt ttt ctg act ggc gag ttc tac aag cag att gcg cgg 446 Ala Thr Asn Gly Phe Leu Thr Gly Glu Phe Tyr Lys Gln Ile Ala Arg 30 35 40 45acc ctg gag cgg ggc aag ttt gat ctg ttg ttt ctt ccc gat ggc ctg 494 Thr Leu Glu Arg Gly Lys Phe Asp Leu Leu Phe Leu Pro Asp Gly Leu 50 55 60 gcc atc gaa gat agc tat ggc gat aat ttg gag acg ggg gtc ggt ctg 542 Ala Ile Glu Asp Ser Tyr Gly Asp Asn Leu Glu Thr Gly Val Gly Leu 65 70 75 ggt ggc cag ggc gcg gtg gcg ctg gag ccg acc agc gtg ata gcg acg 590 Gly Gly Gln Gly Ala Val Ala Leu Glu Pro Thr Ser Val Ile Ala Thr 80 85 90 atg gct gcg gtg acc cag cga tta ggt ctg ggc gct acg gtt tcg acg 638 Met Ala Ala Val Thr Gln Arg Leu Gly Leu Gly Ala Thr Val Ser Thr 95 100 105 acc tac tac ccg ccc tat cat gtg gct cgg gtg ttt gcc acg ctc gac 686 Thr Tyr Tyr Pro Pro Tyr His Val Ala Arg Val Phe Ala Thr Leu Asp110 115 120 125aat ctg tct gat ggc cgg atc tcg tgg aat gtg gtc acg tcg ctg aac 734 Asn Leu Ser Asp Gly Arg Ile Ser Trp Asn Val Val Thr Ser Leu Asn 130 135 140 gac tcc gag gca cgc aac ttc ggt gtg gat gag cac ctt gag cat gac 782 Asp Ser Glu Ala Arg Asn Phe Gly Val Asp Glu His Leu Glu His Asp 145 150 155 atc cga tac gac cgg gct gac gaa ttc ttg gag gcc gtc aag aag cta 830 Ile Arg Tyr Asp Arg Ala Asp Glu Phe Leu Glu Ala Val Lys Lys Leu 160 165 170 tgg agt tct tgg tcc gag gat gcc ttg ttg ttg gac aag gtt ggc ggt 878 Trp Ser Ser Trp Ser Glu Asp Ala Leu Leu Leu Asp Lys Val Gly Gly 175 180 185 cgg ttc gct gat ccc aag aag gtt caa tac gtc aat cat cgc ggc cgc 926 Arg Phe Ala Asp Pro Lys Lys Val Gln Tyr Val Asn His Arg Gly Arg190 195 200 205tgg ttg tcg gtc cgt ggc ccg ttg cag gtt cca cgg tcc cgt cag ggt 974 Trp Leu Ser Val Arg Gly Pro Leu Gln Val Pro Arg Ser Arg Gln Gly 210 215 220 gaa ccg gtc atc ttg cag gcc ggc ttg tcg ccg cgg ggg cgt cgc ttc 1022 Glu Pro Val Ile Leu Gln Ala Gly Leu Ser Pro Arg Gly Arg Arg Phe 225 230 235 gcg ggg cgc tgg gct gaa gcg gta ttc agc gtc tcg ccc aac ctc gac 1070 Ala Gly Arg Trp Ala Glu Ala Val Phe Ser Val Ser Pro Asn Leu Asp 240 245 250 atc atg cgc gcg gtc tat cag gac atc aaa gct cac gtt gcg gcc gcg 1118 Ile Met Arg Ala Val Tyr Gln Asp Ile Lys Ala His Val Ala Ala Ala 255 260 265 ggg cgt gac ccg gag caa aca aag gta ttc acc gcg gtg atg ccg gtg 1166 Gly Arg Asp Pro Glu Gln Thr Lys Val Phe Thr Ala Val Met Pro Val270 275 280 285ctc ggt gag aca gag cag gtg gcc cgg gaa cgt ctg gaa tat ctc aat 1214 Leu Gly Glu Thr Glu Gln Val Ala Arg Glu Arg Leu Glu Tyr Leu Asn 290 295 300 tcg ctg gtg cat ccc gaa gtg ggt ttg tcg acg cta tcc agc cac agc 1262 Ser Leu Val His Pro Glu Val Gly Leu Ser Thr Leu Ser Ser His Ser 305 310 315 ggc ttg aat ctc tcc aag tat ccg ctg gat acg aag ttt tcc gac atc 1310 Gly Leu Asn Leu Ser Lys Tyr Pro Leu Asp Thr Lys Phe Ser Asp Ile 320 325 330 gtc gcc gat ctc ggg gat cgt cac gtg ccg acc atg ttg cag atg ttt 1358 Val Ala Asp Leu Gly Asp Arg His Val Pro Thr Met Leu Gln Met Phe 335 340 345 tcc gcc gtg gcc ggt ggc ggt gcg gac ctg acg ctg gcc gaa ctg ggc 1406 Ser Ala Val Ala Gly Gly Gly Ala Asp Leu Thr Leu Ala Glu Leu Gly350 355 360 365cgc cga tac ggc acc aac gtc ggg ttt gtg ccg cag tgg gcc gga act 1454 Arg Arg Tyr Gly Thr Asn Val Gly Phe Val Pro Gln Trp Ala Gly Thr 370 375 380 gcc gaa cag atc gct gat caa ttg att agt cac ttt gag gcc ggg gcc 1502 Ala Glu Gln Ile Ala Asp Gln Leu Ile Ser His Phe Glu Ala Gly Ala 385 390 395 gct gac gga ttc atc att tcg ccg gcg tat ctg ccg ggt att tac gag 1550 Ala Asp Gly Phe Ile Ile Ser Pro Ala Tyr Leu Pro Gly Ile Tyr Glu 400 405 410 gag ttc gtc gac cag gtg gtt ccg ctg ctg cag caa cgt ggt gtg ttc 1598 Glu Phe Val Asp Gln Val Val Pro Leu Leu Gln Gln Arg Gly Val Phe 415 420 425 cgt acc gag tac gaa ggg aca acg ctg cgt gag cac ctt ggg ttg gct 1646 Arg Thr Glu Tyr Glu Gly Thr Thr Leu Arg Glu His Leu Gly Leu Ala430 435 440 445cac cct gaa gtc ggg agc atc cga tgaccaccac cgatgttgat catgacgttc 1700 His Pro Glu Val Gly Ser Ile Arg 450 ttgcctacag caactgcccg gttcccaatg cactactgac agcgctggag tcaaacctgt 1760 tggccggcaa cgggatttcg ctgaatgtgc tcagcggcgc gcaagcaggg ttgcatttca 1820 cctacgatca tcccgcttac acccgctttg gtggcgaaat tccacctctt atcagtgagg 1880 gcctgcgggc gccgggacgc acccgcctgc taggcatcac ccctctggcc ggtcggcaag 1940 ggatctatgt ccgtgttgac agcccggtga catcaccgga gcagctgcgg ggccggcggg 2000 tgggtgtgtc gggtgcagcg atccgcatcc tcaccggcga gttgggcgat tatcggcact 2060 tggacccgtg gcggcaaaca ctgatcgcgc tgggtacctg ggaggcgcgg ggacttttgc 2120 agaccctgca catcgggggg ataggggtcg acgacgtcga gttggtgcgt atcgaaagcc 2180 ctggcgtcga cgtgcccgaa gagcggctgg aagcagcggc cagtgtcaaa ggcgctgacc 2240 tgtttcccga cgtggccggc caccagaggg acatcctgga tagcggcaac gttgatgcgc 2300 tgttcacctg gttgccctgg gcggcggagt tagaagatct cagcggagcg cgggtgctcg 2360 ccgatctcgg cgatgaccaa cgaagccggt atgccagtgt ctggacggtc agcgctcagc 2420 tggttgacga gcgacccgac ctggtgcaac gactcgtcga cgcggcggtc caagccagcc 2480 gttgggcaca ggcccatccc gaggaaaccg ttggcatcca tgccgccaac ctcggggtcg 2540 cgccctcggc gatcggacgc ggtttcggcg ctgatttcgc ccaacatctg atcccacggc 2600 ttgacgactc ggcactggcc gttgtcgacc aaactcaaca gttcctcctc gaccacaatc 2660 tcctcgacca tccggtggac ctcacgcagt gggcggcacc gcaattcctc actcaatcca 2720 cgactggaga ccagcaatga ccctgaccga tgacaccgcc acggcacaga acaataggaa 2780 tggcgacccc atcgaggtgg cgcgcgagct gacgcgaaag tggcaagcta ccgtcgtcga 2840 gcgcgacaag gctggtggtt cggcgacaga agagcgtgag gatctacgcg ccagtgggct 2900 gctctcggtg accgttcccc ggcacctggg cggttgggga gccgactggc ctaccgctct 2960 ggaagtggtg cgtgagatcg ccaaggtcga tggatcgctg ggccacttgt tcggatacca 3020 cctcagcacc ccagccgtta ttgatctgtg gggttcaccc gagcaaagtg aacgcctact 3080 tcgtcaactg gccgaaaaca actggtggac cgggaacgcc tccagcgaga acaacagcca 3140 catcctcgaa tggaaggtaa ctgccacccc gaccgacgac ggcgggtacc ttctcaacgg 3200 tatcaagcac ttcagcagtg gagccaaggg ctcagatctg ctcttggtgt tcggcgtgat 3260 accggagggt tccccacagc aaggcgccat cgttgccgcg gccattccca ccaccagaga 3320 aggcgttcaa acaaacgacg actggcaggc gctagggatg cggcgcaccg acagcggcac 3380 caccgaattc cacaacgtcg tagtccgtcc cgacgaggtg ctgggcaaac ccaatgccgt 3440 tgtcgaggcg ttcctggcgt ccgggcgcgg cagcctattc ggcccgatcg tgcagttggt 3500 gttcgcaacg gtgtatctgg ggatcgcgcg tggtgcgctc gaaacagcgc gcgagtacac 3560 ccgaacgcag gcacgcccct ggacgccggc tggggttact caagccgtcg aggatccgta 3620 caccattcgc agctacggcg aattcggtat cgaactacag gctggcgatg ctgcagcacg 3680 tgaggctgcg cagctgctac agaccgcgtg ggacaaaggt gacgcattaa ctccacaaga 3740 acgcggcgaa ctcgcggtac aggtagccgg agtcaaagcc atagccacca aagcgggcct 3800 ggacgtgacc agccgaattt tcgaggtcat cggtgcccgg ggaacccacc caaactatgg 3860 attcgaccgg ttctggcgca acatccgcac ccacacgctg cacgatcccg tctcttacaa 3920 gatcgctgaa gtcggaaact acgtccttaa ccagcgctac ccgatacccg gtttcacttc 3980 ctgactccag gaccgcgctg ccgggcacgc gttaatgcct gagaacggtg g 4031 <210> 2<211> 453<212> PRT<213> Bacillus subtilis<400> 2Met Ala Glu Gln Arg Gln Leu His Leu Ala Gly Phe Phe Ser Ala Gly 1 5 10 15 Asn Val Thr His Ala His Gly Ala Trp Arg His Val Gly Ala Thr Asn 20 25 30 Gly Phe Leu Thr Gly Glu Phe Tyr Lys Gln Ile Ala Arg Thr Leu Glu 35 40 45 Arg Gly Lys Phe Asp Leu Leu Phe Leu Pro Asp Gly Leu Ala Ile Glu 50 55 60 Asp Ser Tyr Gly Asp Asn Leu Glu Thr Gly Val Gly Leu Gly Gly Gln 65 70 75 80 Gly Ala Val Ala Leu Glu Pro Thr Ser Val Ile Ala Thr Met Ala Ala 85 90 95 Val Thr Gln Arg Leu Gly Leu Gly Ala Thr Val Ser Thr Thr Tyr Tyr 100 105 110 Pro Pro Tyr His Val Ala Arg Val Phe Ala Thr Leu Asp Asn Leu Ser 115 120 125 Asp Gly Arg Ile Ser Trp Asn Val Val Thr Ser Leu Asn Asp Ser Glu 130 135 140 Ala Arg Asn Phe Gly Val Asp Glu His Leu Glu His Asp Ile Arg Tyr 145 150 155 160 Asp Arg Ala Asp Glu Phe Leu Glu Ala Val Lys Lys Leu Trp Ser Ser 165 170 175 Trp Ser Glu Asp Ala Leu Leu Leu Asp Lys Val Gly Gly Arg Phe Ala 180 185 190 Asp Pro Lys Lys Val Gln Tyr Val Asn His Arg Gly Arg Trp Leu Ser 195 200 205 Val Arg Gly Pro Leu Gln Val Pro Arg Ser Arg Gln Gly Glu Pro Val 210 215 220 Ile Leu Gln Ala Gly Leu Ser Pro Arg Gly Arg Arg Phe Ala Gly Arg 225 230 235 240 Trp Ala Glu Ala Val Phe Ser Val Ser Pro Asn Leu Asp Ile Met Arg 245 250 255 Ala Val Tyr Gln Asp Ile Lys Ala His Val Ala Ala Ala Gly Arg Asp 260 265 270 Pro Glu Gln Thr Lys Val Phe Thr Ala Val Met Pro Val Leu Gly Glu 275 280 285 Thr Glu Gln Val Ala Arg Glu Arg Leu Glu Tyr Leu Asn Ser Leu Val 290 295 300 His Pro Glu Val Gly Leu Ser Thr Leu Ser Ser His Ser Gly Leu Asn 305 310 315 320 Leu Ser Lys Tyr Pro Leu Asp Thr Lys Phe Ser Asp Ile Val Ala Asp 325 330 335 Leu Gly Asp Arg His Val Pro Thr Met Leu Gln Met Phe Ser Ala Val 340 345 350 Ala Gly Gly Gly Ala Asp Leu Thr Leu Ala Glu Leu Gly Arg Arg Tyr 355 360 365 Gly Thr Asn Val Gly Phe Val Pro Gln Trp Ala Gly Thr Ala Glu Gln 370 375 380 Ile Ala Asp Gln Leu Ile Ser His Phe Glu Ala Gly Ala Ala Asp Gly 385 390 395 400 Phe Ile Ile Ser Pro Ala Tyr Leu Pro Gly Ile Tyr Glu Glu Phe Val 405 410 415 Asp Gln Val Val Pro Leu Leu Gln Gln Arg Gly Val Phe Arg Thr Glu 420 425 430 Tyr Glu Gly Thr Thr Leu Arg Glu His Leu Gly Leu Ala His Pro Glu 435 440 445 Val Gly Ser Ile Arg 450 <210> 3<211> 4031<212> DNA<213> Bacillus subtilis<220> <221> CDS<222> (1670)..(2737)<400> 3ctagcgaggt gcgagttccg cgcggtagcc atccgcttca ttgattttta catcgatacc 60 gccgcggtgg ccgtcagggt gtcgaagccg ggcggaggtg cacaccccgc cggttggtga 120 tcttgctcac aggtggcgct ttgacggcac tcgtccgggg gcgagaatcg gcgaaggtcg 180 ggtgggtcgt cgggaggaag tggcgtgttg ggtgcccctt tgaatcatcg ggatcctaac 240 tctgttgggg tcggttgcag tagccgagca ttggtctccg gtaactgatc cccacggagg 300 cactgctgat tatggctgaa caacgccaac tacatttagc cgggtttttc tcggccggca 360 acgtcactca tgcgcatggc gcgtggcgcc atgtcggtgc gacgaacggt tttctgactg 420 gcgagttcta caagcagatt gcgcggaccc tggagcgggg caagtttgat ctgttgtttc 480 ttcccgatgg cctggccatc gaagatagct atggcgataa tttggagacg ggggtcggtc 540 tgggtggcca gggcgcggtg gcgctggagc cgaccagcgt gatagcgacg atggctgcgg 600 tgacccagcg attaggtctg ggcgctacgg tttcgacgac ctactacccg ccctatcatg 660 tggctcgggt gtttgccacg ctcgacaatc tgtctgatgg ccggatctcg tggaatgtgg 720 tcacgtcgct gaacgactcc gaggcacgca acttcggtgt ggatgagcac cttgagcatg 780 acatccgata cgaccgggct gacgaattct tggaggccgt caagaagcta tggagttctt 840 ggtccgagga tgccttgttg ttggacaagg ttggcggtcg gttcgctgat cccaagaagg 900 ttcaatacgt caatcatcgc ggccgctggt tgtcggtccg tggcccgttg caggttccac 960 ggtcccgtca gggtgaaccg gtcatcttgc aggccggctt gtcgccgcgg gggcgtcgct 1020 tcgcggggcg ctgggctgaa gcggtattca gcgtctcgcc caacctcgac atcatgcgcg 1080 cggtctatca ggacatcaaa gctcacgttg cggccgcggg gcgtgacccg gagcaaacaa 1140 aggtattcac cgcggtgatg ccggtgctcg gtgagacaga gcaggtggcc cgggaacgtc 1200 tggaatatct caattcgctg gtgcatcccg aagtgggttt gtcgacgcta tccagccaca 1260 gcggcttgaa tctctccaag tatccgctgg atacgaagtt ttccgacatc gtcgccgatc 1320 tcggggatcg tcacgtgccg accatgttgc agatgttttc cgccgtggcc ggtggcggtg 1380 cggacctgac gctggccgaa ctgggccgcc gatacggcac caacgtcggg tttgtgccgc 1440 agtgggccgg aactgccgaa cagatcgctg atcaattgat tagtcacttt gaggccgggg 1500 ccgctgacgg attcatcatt tcgccggcgt atctgccggg tatttacgag gagttcgtcg 1560 accaggtggt tccgctgctg cagcaacgtg gtgtgttccg taccgagtac gaagggacaa 1620 cgctgcgtga gcaccttggg ttggctcacc ctgaagtcgg gagcatccg atg acc acc 1678 Met Thr Thr 1 acc gat gtt gat cat gac gtt ctt gcc tac agc aac tgc ccg gtt ccc 1726 Thr Asp Val Asp His Asp Val Leu Ala Tyr Ser Asn Cys Pro Val Pro 5 10 15 aat gca cta ctg aca gcg ctg gag tca aac ctg ttg gcc ggc aac ggg 1774 Asn Ala Leu Leu Thr Ala Leu Glu Ser Asn Leu Leu Ala Gly Asn Gly 20 25 30 35att tcg ctg aat gtg ctc agc ggc gcg caa gca ggg ttg cat ttc acc 1822 Ile Ser Leu Asn Val Leu Ser Gly Ala Gln Ala Gly Leu His Phe Thr 40 45 50 tac gat cat ccc gct tac acc cgc ttt ggt ggc gaa att cca cct ctt 1870 Tyr Asp His Pro Ala Tyr Thr Arg Phe Gly Gly Glu Ile Pro Pro Leu 55 60 65 atc agt gag ggc ctg cgg gcg ccg gga cgc acc cgc ctg cta ggc atc 1918 Ile Ser Glu Gly Leu Arg Ala Pro Gly Arg Thr Arg Leu Leu Gly Ile 70 75 80 acc cct ctg gcc ggt cgg caa ggg atc tat gtc cgt gtt gac agc ccg 1966 Thr Pro Leu Ala Gly Arg Gln Gly Ile Tyr Val Arg Val Asp Ser Pro 85 90 95 gtg aca tca ccg gag cag ctg cgg ggc cgg cgg gtg ggt gtg tcg ggt 2014 Val Thr Ser Pro Glu Gln Leu Arg Gly Arg Arg Val Gly Val Ser Gly100 105 110 115gca gcg atc cgc atc ctc acc ggc gag ttg ggc gat tat cgg cac ttg 2062 Ala Ala Ile Arg Ile Leu Thr Gly Glu Leu Gly Asp Tyr Arg His Leu 120 125 130 gac ccg tgg cgg caa aca ctg atc gcg ctg ggt acc tgg gag gcg cgg 2110 Asp Pro Trp Arg Gln Thr Leu Ile Ala Leu Gly Thr Trp Glu Ala Arg 135 140 145 gga ctt ttg cag acc ctg cac atc ggg ggg ata ggg gtc gac gac gtc 2158 Gly Leu Leu Gln Thr Leu His Ile Gly Gly Ile Gly Val Asp Asp Val 150 155 160 gag ttg gtg cgt atc gaa agc cct ggc gtc gac gtg ccc gaa gag cgg 2206 Glu Leu Val Arg Ile Glu Ser Pro Gly Val Asp Val Pro Glu Glu Arg 165 170 175 ctg gaa gca gcg gcc agt gtc aaa ggc gct gac ctg ttt ccc gac gtg 2254 Leu Glu Ala Ala Ala Ser Val Lys Gly Ala Asp Leu Phe Pro Asp Val180 185 190 195gcc ggc cac cag agg gac atc ctg gat agc ggc aac gtt gat gcg ctg 2302 Ala Gly His Gln Arg Asp Ile Leu Asp Ser Gly Asn Val Asp Ala Leu 200 205 210 ttc acc tgg ttg ccc tgg gcg gcg gag tta gaa gat ctc agc gga gcg 2350 Phe Thr Trp Leu Pro Trp Ala Ala Glu Leu Glu Asp Leu Ser Gly Ala 215 220 225 cgg gtg ctc gcc gat ctc ggc gat gac caa cga agc cgg tat gcc agt 2398 Arg Val Leu Ala Asp Leu Gly Asp Asp Gln Arg Ser Arg Tyr Ala Ser 230 235 240 gtc tgg acg gtc agc gct cag ctg gtt gac gag cga ccc gac ctg gtg 2446 Val Trp Thr Val Ser Ala Gln Leu Val Asp Glu Arg Pro Asp Leu Val 245 250 255 caa cga ctc gtc gac gcg gcg gtc caa gcc agc cgt tgg gca cag gcc 2494 Gln Arg Leu Val Asp Ala Ala Val Gln Ala Ser Arg Trp Ala Gln Ala260 265 270 275cat ccc gag gaa acc gtt ggc atc cat gcc gcc aac ctc ggg gtc gcg 2542 His Pro Glu Glu Thr Val Gly Ile His Ala Ala Asn Leu Gly Val Ala 280 285 290 ccc tcg gcg atc gga cgc ggt ttc ggc gct gat ttc gcc caa cat ctg 2590 Pro Ser Ala Ile Gly Arg Gly Phe Gly Ala Asp Phe Ala Gln His Leu 295 300 305 atc cca cgg ctt gac gac tcg gca ctg gcc gtt gtc gac caa act caa 2638 Ile Pro Arg Leu Asp Asp Ser Ala Leu Ala Val Val Asp Gln Thr Gln 310 315 320 cag ttc ctc ctc gac cac aat ctc ctc gac cat ccg gtg gac ctc acg 2686 Gln Phe Leu Leu Asp His Asn Leu Leu Asp His Pro Val Asp Leu Thr 325 330 335 cag tgg gcg gca ccg caa ttc ctc act caa tcc acg act gga gac cag 2734 Gln Trp Ala Ala Pro Gln Phe Leu Thr Gln Ser Thr Thr Gly Asp Gln340 345 350 355caa tgaccctgac cgatgacacc gccacggcac agaacaatag gaatggcgac 2787 Gln cccatcgagg tggcgcgcga gctgacgcga aagtggcaag ctaccgtcgt cgagcgcgac 2847 aaggctggtg gttcggcgac agaagagcgt gaggatctac gcgccagtgg gctgctctcg 2907 gtgaccgttc cccggcacct gggcggttgg ggagccgact ggcctaccgc tctggaagtg 2967 gtgcgtgaga tcgccaaggt cgatggatcg ctgggccact tgttcggata ccacctcagc 3027 accccagccg ttattgatct gtggggttca cccgagcaaa gtgaacgcct acttcgtcaa 3087 ctggccgaaa acaactggtg gaccgggaac gcctccagcg agaacaacag ccacatcctc 3147 gaatggaagg taactgccac cccgaccgac gacggcgggt accttctcaa cggtatcaag 3207 cacttcagca gtggagccaa gggctcagat ctgctcttgg tgttcggcgt gataccggag 3267 ggttccccac agcaaggcgc catcgttgcc gcggccattc ccaccaccag agaaggcgtt 3327 caaacaaacg acgactggca ggcgctaggg atgcggcgca ccgacagcgg caccaccgaa 3387 ttccacaacg tcgtagtccg tcccgacgag gtgctgggca aacccaatgc cgttgtcgag 3447 gcgttcctgg cgtccgggcg cggcagccta ttcggcccga tcgtgcagtt ggtgttcgca 3507 acggtgtatc tggggatcgc gcgtggtgcg ctcgaaacag cgcgcgagta cacccgaacg 3567 caggcacgcc cctggacgcc ggctggggtt actcaagccg tcgaggatcc gtacaccatt 3627 cgcagctacg gcgaattcgg tatcgaacta caggctggcg atgctgcagc acgtgaggct 3687 gcgcagctgc tacagaccgc gtgggacaaa ggtgacgcat taactccaca agaacgcggc 3747 gaactcgcgg tacaggtagc cggagtcaaa gccatagcca ccaaagcggg cctggacgtg 3807 accagccgaa ttttcgaggt catcggtgcc cggggaaccc acccaaacta tggattcgac 3867 cggttctggc gcaacatccg cacccacacg ctgcacgatc ccgtctctta caagatcgct 3927 gaagtcggaa actacgtcct taaccagcgc tacccgatac ccggtttcac ttcctgactc 3987 caggaccgcg ctgccgggca cgcgttaatg cctgagaacg gtgg 4031 <210> 4<211> 356<212> PRT<213> Bacillus subtilis<400> 4Met Thr Thr Thr Asp Val Asp His Asp Val Leu Ala Tyr Ser Asn Cys 1 5 10 15 Pro Val Pro Asn Ala Leu Leu Thr Ala Leu Glu Ser Asn Leu Leu Ala 20 25 30 Gly Asn Gly Ile Ser Leu Asn Val Leu Ser Gly Ala Gln Ala Gly Leu 35 40 45 His Phe Thr Tyr Asp His Pro Ala Tyr Thr Arg Phe Gly Gly Glu Ile 50 55 60 Pro Pro Leu Ile Ser Glu Gly Leu Arg Ala Pro Gly Arg Thr Arg Leu 65 70 75 80 Leu Gly Ile Thr Pro Leu Ala Gly Arg Gln Gly Ile Tyr Val Arg Val 85 90 95 Asp Ser Pro Val Thr Ser Pro Glu Gln Leu Arg Gly Arg Arg Val Gly 100 105 110 Val Ser Gly Ala Ala Ile Arg Ile Leu Thr Gly Glu Leu Gly Asp Tyr 115 120 125 Arg His Leu Asp Pro Trp Arg Gln Thr Leu Ile Ala Leu Gly Thr Trp 130 135 140 Glu Ala Arg Gly Leu Leu Gln Thr Leu His Ile Gly Gly Ile Gly Val 145 150 155 160 Asp Asp Val Glu Leu Val Arg Ile Glu Ser Pro Gly Val Asp Val Pro 165 170 175 Glu Glu Arg Leu Glu Ala Ala Ala Ser Val Lys Gly Ala Asp Leu Phe 180 185 190 Pro Asp Val Ala Gly His Gln Arg Asp Ile Leu Asp Ser Gly Asn Val 195 200 205 Asp Ala Leu Phe Thr Trp Leu Pro Trp Ala Ala Glu Leu Glu Asp Leu 210 215 220 Ser Gly Ala Arg Val Leu Ala Asp Leu Gly Asp Asp Gln Arg Ser Arg 225 230 235 240 Tyr Ala Ser Val Trp Thr Val Ser Ala Gln Leu Val Asp Glu Arg Pro 245 250 255 Asp Leu Val Gln Arg Leu Val Asp Ala Ala Val Gln Ala Ser Arg Trp 260 265 270 Ala Gln Ala His Pro Glu Glu Thr Val Gly Ile His Ala Ala Asn Leu 275 280 285 Gly Val Ala Pro Ser Ala Ile Gly Arg Gly Phe Gly Ala Asp Phe Ala 290 295 300 Gln His Leu Ile Pro Arg Leu Asp Asp Ser Ala Leu Ala Val Val Asp 305 310 315 320 Gln Thr Gln Gln Phe Leu Leu Asp His Asn Leu Leu Asp His Pro Val 325 330 335 Asp Leu Thr Gln Trp Ala Ala Pro Gln Phe Leu Thr Gln Ser Thr Thr 340 345 350 Gly Asp Gln Gln 355 <210> 5<211> 4031<212> DNA<213> Bacillus subtilis<220> <221> CDS<222> (2737)..(3981)<400> 5ctagcgaggt gcgagttccg cgcggtagcc atccgcttca ttgattttta catcgatacc 60 gccgcggtgg ccgtcagggt gtcgaagccg ggcggaggtg cacaccccgc cggttggtga 120 tcttgctcac aggtggcgct ttgacggcac tcgtccgggg gcgagaatcg gcgaaggtcg 180 ggtgggtcgt cgggaggaag tggcgtgttg ggtgcccctt tgaatcatcg ggatcctaac 240 tctgttgggg tcggttgcag tagccgagca ttggtctccg gtaactgatc cccacggagg 300 cactgctgat tatggctgaa caacgccaac tacatttagc cgggtttttc tcggccggca 360 acgtcactca tgcgcatggc gcgtggcgcc atgtcggtgc gacgaacggt tttctgactg 420 gcgagttcta caagcagatt gcgcggaccc tggagcgggg caagtttgat ctgttgtttc 480 ttcccgatgg cctggccatc gaagatagct atggcgataa tttggagacg ggggtcggtc 540 tgggtggcca gggcgcggtg gcgctggagc cgaccagcgt gatagcgacg atggctgcgg 600 tgacccagcg attaggtctg ggcgctacgg tttcgacgac ctactacccg ccctatcatg 660 tggctcgggt gtttgccacg ctcgacaatc tgtctgatgg ccggatctcg tggaatgtgg 720 tcacgtcgct gaacgactcc gaggcacgca acttcggtgt ggatgagcac cttgagcatg 780 acatccgata cgaccgggct gacgaattct tggaggccgt caagaagcta tggagttctt 840 ggtccgagga tgccttgttg ttggacaagg ttggcggtcg gttcgctgat cccaagaagg 900 ttcaatacgt caatcatcgc ggccgctggt tgtcggtccg tggcccgttg caggttccac 960 ggtcccgtca gggtgaaccg gtcatcttgc aggccggctt gtcgccgcgg gggcgtcgct 1020 tcgcggggcg ctgggctgaa gcggtattca gcgtctcgcc caacctcgac atcatgcgcg 1080 cggtctatca ggacatcaaa gctcacgttg cggccgcggg gcgtgacccg gagcaaacaa 1140 aggtattcac cgcggtgatg ccggtgctcg gtgagacaga gcaggtggcc cgggaacgtc 1200 tggaatatct caattcgctg gtgcatcccg aagtgggttt gtcgacgcta tccagccaca 1260 gcggcttgaa tctctccaag tatccgctgg atacgaagtt ttccgacatc gtcgccgatc 1320 tcggggatcg tcacgtgccg accatgttgc agatgttttc cgccgtggcc ggtggcggtg 1380 cggacctgac gctggccgaa ctgggccgcc gatacggcac caacgtcggg tttgtgccgc 1440 agtgggccgg aactgccgaa cagatcgctg atcaattgat tagtcacttt gaggccgggg 1500 ccgctgacgg attcatcatt tcgccggcgt atctgccggg tatttacgag gagttcgtcg 1560 accaggtggt tccgctgctg cagcaacgtg gtgtgttccg taccgagtac gaagggacaa 1620 cgctgcgtga gcaccttggg ttggctcacc ctgaagtcgg gagcatccga tgaccaccac 1680 cgatgttgat catgacgttc ttgcctacag caactgcccg gttcccaatg cactactgac 1740 agcgctggag tcaaacctgt tggccggcaa cgggatttcg ctgaatgtgc tcagcggcgc 1800 gcaagcaggg ttgcatttca cctacgatca tcccgcttac acccgctttg gtggcgaaat 1860 tccacctctt atcagtgagg gcctgcgggc gccgggacgc acccgcctgc taggcatcac 1920 ccctctggcc ggtcggcaag ggatctatgt ccgtgttgac agcccggtga catcaccgga 1980 gcagctgcgg ggccggcggg tgggtgtgtc gggtgcagcg atccgcatcc tcaccggcga 2040 gttgggcgat tatcggcact tggacccgtg gcggcaaaca ctgatcgcgc tgggtacctg 2100 ggaggcgcgg ggacttttgc agaccctgca catcgggggg ataggggtcg acgacgtcga 2160 gttggtgcgt atcgaaagcc ctggcgtcga cgtgcccgaa gagcggctgg aagcagcggc 2220 cagtgtcaaa ggcgctgacc tgtttcccga cgtggccggc caccagaggg acatcctgga 2280 tagcggcaac gttgatgcgc tgttcacctg gttgccctgg gcggcggagt tagaagatct 2340 cagcggagcg cgggtgctcg ccgatctcgg cgatgaccaa cgaagccggt atgccagtgt 2400 ctggacggtc agcgctcagc tggttgacga gcgacccgac ctggtgcaac gactcgtcga 2460 cgcggcggtc caagccagcc gttgggcaca ggcccatccc gaggaaaccg ttggcatcca 2520 tgccgccaac ctcggggtcg cgccctcggc gatcggacgc ggtttcggcg ctgatttcgc 2580 ccaacatctg atcccacggc ttgacgactc ggcactggcc gttgtcgacc aaactcaaca 2640 gttcctcctc gaccacaatc tcctcgacca tccggtggac ctcacgcagt gggcggcacc 2700 gcaattcctc actcaatcca cgactggaga ccagca atg acc ctg acc gat gac 2754 Met Thr Leu Thr Asp Asp 1 5 acc gcc acg gca cag aac aat agg aat ggc gac ccc atc gag gtg gcg 2802 Thr Ala Thr Ala Gln Asn Asn Arg Asn Gly Asp Pro Ile Glu Val Ala 10 15 20 cgc gag ctg acg cga aag tgg caa gct acc gtc gtc gag cgc gac aag 2850 Arg Glu Leu Thr Arg Lys Trp Gln Ala Thr Val Val Glu Arg Asp Lys 25 30 35 gct ggt ggt tcg gcg aca gaa gag cgt gag gat cta cgc gcc agt ggg 2898 Ala Gly Gly Ser Ala Thr Glu Glu Arg Glu Asp Leu Arg Ala Ser Gly 40 45 50 ctg ctc tcg gtg acc gtt ccc cgg cac ctg ggc ggt tgg gga gcc gac 2946 Leu Leu Ser Val Thr Val Pro Arg His Leu Gly Gly Trp Gly Ala Asp 55 60 65 70tgg cct acc gct ctg gaa gtg gtg cgt gag atc gcc aag gtc gat gga 2994 Trp Pro Thr Ala Leu Glu Val Val Arg Glu Ile Ala Lys Val Asp Gly 75 80 85 tcg ctg ggc cac ttg ttc gga tac cac ctc agc acc cca gcc gtt att 3042 Ser Leu Gly His Leu Phe Gly Tyr His Leu Ser Thr Pro Ala Val Ile 90 95 100 gat ctg tgg ggt tca ccc gag caa agt gaa cgc cta ctt cgt caa ctg 3090 Asp Leu Trp Gly Ser Pro Glu Gln Ser Glu Arg Leu Leu Arg Gln Leu 105 110 115 gcc gaa aac aac tgg tgg acc ggg aac gcc tcc agc gag aac aac agc 3138 Ala Glu Asn Asn Trp Trp Thr Gly Asn Ala Ser Ser Glu Asn Asn Ser 120 125 130 cac atc ctc gaa tgg aag gta act gcc acc ccg acc gac gac ggc ggg 3186 His Ile Leu Glu Trp Lys Val Thr Ala Thr Pro Thr Asp Asp Gly Gly135 140 145 150tac ctt ctc aac ggt atc aag cac ttc agc agt gga gcc aag ggc tca 3234 Tyr Leu Leu Asn Gly Ile Lys His Phe Ser Ser Gly Ala Lys Gly Ser 155 160 165 gat ctg ctc ttg gtg ttc ggc gtg ata ccg gag ggt tcc cca cag caa 3282 Asp Leu Leu Leu Val Phe Gly Val Ile Pro Glu Gly Ser Pro Gln Gln 170 175 180 ggc gcc atc gtt gcc gcg gcc att ccc acc acc aga gaa ggc gtt caa 3330 Gly Ala Ile Val Ala Ala Ala Ile Pro Thr Thr Arg Glu Gly Val Gln 185 190 195 aca aac gac gac tgg cag gcg cta ggg atg cgg cgc acc gac agc ggc 3378 Thr Asn Asp Asp Trp Gln Ala Leu Gly Met Arg Arg Thr Asp Ser Gly 200 205 210 acc acc gaa ttc cac aac gtc gta gtc cgt ccc gac gag gtg ctg ggc 3426 Thr Thr Glu Phe His Asn Val Val Val Arg Pro Asp Glu Val Leu Gly215 220 225 230aaa ccc aat gcc gtt gtc gag gcg ttc ctg gcg tcc ggg cgc ggc agc 3474 Lys Pro Asn Ala Val Val Glu Ala Phe Leu Ala Ser Gly Arg Gly Ser 235 240 245 cta ttc ggc ccg atc gtg cag ttg gtg ttc gca acg gtg tat ctg ggg 3522 Leu Phe Gly Pro Ile Val Gln Leu Val Phe Ala Thr Val Tyr Leu Gly 250 255 260 atc gcg cgt ggt gcg ctc gaa aca gcg cgc gag tac acc cga acg cag 3570 Ile Ala Arg Gly Ala Leu Glu Thr Ala Arg Glu Tyr Thr Arg Thr Gln 265 270 275 gca cgc ccc tgg acg ccg gct ggg gtt act caa gcc gtc gag gat ccg 3618 Ala Arg Pro Trp Thr Pro Ala Gly Val Thr Gln Ala Val Glu Asp Pro 280 285 290 tac acc att cgc agc tac ggc gaa ttc ggt atc gaa cta cag gct ggc 3666 Tyr Thr Ile Arg Ser Tyr Gly Glu Phe Gly Ile Glu Leu Gln Ala Gly295 300 305 310gat gct gca gca cgt gag gct gcg cag ctg cta cag acc gcg tgg gac 3714 Asp Ala Ala Ala Arg Glu Ala Ala Gln Leu Leu Gln Thr Ala Trp Asp 315 320 325 aaa ggt gac gca tta act cca caa gaa cgc ggc gaa ctc gcg gta cag 3762 Lys Gly Asp Ala Leu Thr Pro Gln Glu Arg Gly Glu Leu Ala Val Gln 330 335 340 gta gcc gga gtc aaa gcc ata gcc acc aaa gcg ggc ctg gac gtg acc 3810 Val Ala Gly Val Lys Ala Ile Ala Thr Lys Ala Gly Leu Asp Val Thr 345 350 355 agc cga att ttc gag gtc atc ggt gcc cgg gga acc cac cca aac tat 3858 Ser Arg Ile Phe Glu Val Ile Gly Ala Arg Gly Thr His Pro Asn Tyr 360 365 370 gga ttc gac cgg ttc tgg cgc aac atc cgc acc cac acg ctg cac gat 3906 Gly Phe Asp Arg Phe Trp Arg Asn Ile Arg Thr His Thr Leu His Asp375 380 385 390ccc gtc tct tac aag atc gct gaa gtc gga aac tac gtc ctt aac cag 3954 Pro Val Ser Tyr Lys Ile Ala Glu Val Gly Asn Tyr Val Leu Asn Gln 395 400 405 cgc tac ccg ata ccc ggt ttc act tcc tgactccagg accgcgctgc 4001 Arg Tyr Pro Ile Pro Gly Phe Thr Ser 410 415 cgggcacgcg ttaatgcctg agaacggtgg 4031 <210> 6<211> 415<212> PRT<213> Bacillus subtilis<400> 6Met Thr Leu Thr Asp Asp Thr Ala Thr Ala Gln Asn Asn Arg Asn Gly 1 5 10 15 Asp Pro Ile Glu Val Ala Arg Glu Leu Thr Arg Lys Trp Gln Ala Thr 20 25 30 Val Val Glu Arg Asp Lys Ala Gly Gly Ser Ala Thr Glu Glu Arg Glu 35 40 45 Asp Leu Arg Ala Ser Gly Leu Leu Ser Val Thr Val Pro Arg His Leu 50 55 60 Gly Gly Trp Gly Ala Asp Trp Pro Thr Ala Leu Glu Val Val Arg Glu 65 70 75 80 Ile Ala Lys Val Asp Gly Ser Leu Gly His Leu Phe Gly Tyr His Leu 85 90 95 Ser Thr Pro Ala Val Ile Asp Leu Trp Gly Ser Pro Glu Gln Ser Glu 100 105 110 Arg Leu Leu Arg Gln Leu Ala Glu Asn Asn Trp Trp Thr Gly Asn Ala 115 120 125 Ser Ser Glu Asn Asn Ser His Ile Leu Glu Trp Lys Val Thr Ala Thr 130 135 140 Pro Thr Asp Asp Gly Gly Tyr Leu Leu Asn Gly Ile Lys His Phe Ser 145 150 155 160 Ser Gly Ala Lys Gly Ser Asp Leu Leu Leu Val Phe Gly Val Ile Pro 165 170 175 Glu Gly Ser Pro Gln Gln Gly Ala Ile Val Ala Ala Ala Ile Pro Thr 180 185 190 Thr Arg Glu Gly Val Gln Thr Asn Asp Asp Trp Gln Ala Leu Gly Met 195 200 205 Arg Arg Thr Asp Ser Gly Thr Thr Glu Phe His Asn Val Val Val Arg 210 215 220 Pro Asp Glu Val Leu Gly Lys Pro Asn Ala Val Val Glu Ala Phe Leu 225 230 235 240 Ala Ser Gly Arg Gly Ser Leu Phe Gly Pro Ile Val Gln Leu Val Phe 245 250 255 Ala Thr Val Tyr Leu Gly Ile Ala Arg Gly Ala Leu Glu Thr Ala Arg 260 265 270 Glu Tyr Thr Arg Thr Gln Ala Arg Pro Trp Thr Pro Ala Gly Val Thr 275 280 285 Gln Ala Val Glu Asp Pro Tyr Thr Ile Arg Ser Tyr Gly Glu Phe Gly 290 295 300 Ile Glu Leu Gln Ala Gly Asp Ala Ala Ala Arg Glu Ala Ala Gln Leu 305 310 315 320 Leu Gln Thr Ala Trp Asp Lys Gly Asp Ala Leu Thr Pro Gln Glu Arg 325 330 335 Gly Glu Leu Ala Val Gln Val Ala Gly Val Lys Ala Ile Ala Thr Lys 340 345 350 Ala Gly Leu Asp Val Thr Ser Arg Ile Phe Glu Val Ile Gly Ala Arg 355 360 365 Gly Thr His Pro Asn Tyr Gly Phe Asp Arg Phe Trp Arg Asn Ile Arg 370 375 380 Thr His Thr Leu His Asp Pro Val Ser Tyr Lys Ile Ala Glu Val Gly 385 390 395 400 Asn Tyr Val Leu Asn Gln Arg Tyr Pro Ile Pro Gly Phe Thr Ser 405 410 415 <210> 7<211> 16<212> PRT<213> Paenibacillus sp.<400> 7Trp Asn Val Val Thr Ser Leu Asn Asn Ala Glu Ala Arg Asn Phe Gly 1 5 10 15 <210> 8<211> 16<212> PRT<213> Rhodococcus sp. IGTS8<400> 8Trp Asn Val Val Thr Ser Leu Asn Asp Ala Glu Ala Arg Asn Phe Gly 1 5 10 15 <210> 9<211> 18<212> PRT<213> Paenibacillus sp.<400> 9Gly Phe Asp Arg Phe Trp Arg Asp Ala Arg Thr His Thr Leu His Asp 1 5 10 15 Pro Val <210> 10<211> 18<212> PRT<213> Rhodococcus sp. IGTS8<400> 10Gly Phe Asp Arg Phe Trp Arg Asn Val Arg Thr His Ser Leu His Asp 1 5 10 15 Pro Val <210> 11<211> 20<212> DNA<213> Artificial Sequence<220> <223> Description of Artificial Sequence: sense primer for PCR<220><221> modified base<222> n = inosine<223> (3),(9) and (12)<400> 11gcngargcnm gnaayttygg 20 <210> 12<211> 19<212> DNA<213> Artificial Sequence<220> <223> Description of Artificial Sequence: antisense primer for PCR<220><221> modified base<222> n = inosine<223> (4) and (11)<400> 12cgtngcgcca naagcggtc 19 <210> 13<211> 17<212> DNA<213> Artificial Sequence<220> <223> Description of Artificial Sequence: sense primer for PCR<400> 13gcatgacatc cgatacg 17 <210> 14<211> 17<212> DNA<213> Artificial Sequence<220> <223> Description of Artificial Sequence: antisense primer for PCR<400> 14tagtttgggt gggttcc 17
【0080】
【配列表フリーテキスト】配列番号1−bdsABC遺伝子を含むヌクレオチド配列配列番号2−bdsA遺伝子によってコードされるタンパク質BdsAのアミノ酸配列配列番号3−bdsABC遺伝子を含むヌクレオチド配列配列番号4−bdsB遺伝子によってコードされるタンパク質BdsBのアミノ酸配列配列番号5−bdsABC遺伝子を含むヌクレオチド配列配列番号6−bdsC遺伝子によってコードされるタンパク質BdsCのアミノ酸配列配列番号7−TdsAの部分アミノ酸配列配列番号8−DszAの部分アミノ酸配列配列番号9−TdsCの部分アミノ酸配列配列番号10−DszCの部分アミノ酸配列配列番号11−人工配列の説明:センスプライマー(3、9及び12番目のNはデオキシイノシンを表す)配列番号12−人工配列の説明:アンチセンスプライマー(4及び11番目のNはデオキシイノシンを表す)配列番号13−人工配列の説明:センスプライマー配列番号14−人工配列の説明:アンチセンスプライマー
【図面の簡単な説明】
【図1】脱硫遺伝子bdsABCを含む発現プラスミドpU1Nhの制限酵素地図を示す。
【図2】BdsC酵素活性の温度依存性を示す。
【図3】BdsC酵素活性の温度安定性を示す。
【図4】BdsC酵素活性のpH依存性を示す。
【図5】BdsC酵素活性のpH安定性を示す。
【図6】BdsA酵素活性の温度依存性を示す。
【図7】BdsA酵素活性の温度安定性を示す。
【図8】BdsA酵素活性のpH依存性を示す。
【図9】BdsA酵素活性のpH安定性を示す。
【0001】
【発明の属する技術分野】本発明は、微生物を利用するチオフェン系化合物、すなわちベンゾチオフェン、ジベンゾチオフェン(以下「DBT 」という)およびこれらの置換体、又はそれらの誘導体を分解する機能を有する酵素及びそれをコードする遺伝子に関するものである。本発明の酵素及び遺伝子を利用することにより、石油等の化石燃料中に含まれるベンゾチオフェンやDBT およびこれらの置換体、又はそれらの誘導体中の硫黄を遊離させることができるので、通常石油・石炭等の化石燃料の燃焼により空気中に拡散すると言われる硫黄を、化石燃料中から容易に除去することができるようになる。
【0002】
【従来の技術】石油のような炭化水素燃料から硫黄を除去する脱硫のための方法としては、アルカリ洗浄や溶剤脱硫などの方法も知られているが、現在では水素化脱硫が主流となっている。水素化脱硫は、石油留分中の硫黄化合物を触媒の存在下で水素と反応させ、硫化水素として除去して製品の低硫黄化を図る方法である。触媒としては、アルミナを担体としてコバルト、モリブデン、ニッケル、タングステン、などの金属触媒が使用される。モリブデン担持アルミナ触媒の場合には、触媒性能を向上させるために、通常コバルトやニッケルが助触媒として加えられる。金属触媒を用いた水素化脱硫は、現在世界中で広く使用されているきわめて完成度の高いプロセスであることは疑いのないことである。しかし、より厳しい環境規制に対応した石油製品を作るためのプロセスという観点からは、いくつかの問題点がある。以下にその例を簡単に記載する。
【0003】金属触媒は、一般にその基質特異性が低く、このため多様な種類の硫黄化合物を分解し、化石燃料全体の硫黄含量を低下させる目的には適しているが、特定のグループの硫黄化合物、すなわちベンゾチオフェンやDBT のような複素環式硫黄化合物類およびそれらのアルキル誘導体類に対してはその脱硫効果が不十分となることがあると考えられる。たとえば、脱硫後の軽油中にはなおも種々の複素環式有機硫黄化合物が残存している。このように金属触媒による脱硫効果が不十分となる原因の一つは、これらの有機硫黄化合物中の硫黄原子の周囲に存在する置換基による立体障害が考えられる。これらの置換基のうち、メチル置換基の存在が水素化脱硫における金属触媒の反応性に及ぼす影響は、チオフェン、ベンゾチオフェン、DBT などについて検討されている。それらの結果によると、一般的には置換基の数が増すほど脱硫反応は減少するが、置換基の位置が反応性に及ぼす影響もきわめて大きいことが明らかである。メチルDBT 類の脱硫反応性を比較し、置換基による立体障害が金属触媒の反応性に及ぼす影響が非常に大きいことを示した報告は、たとえば、Houalla, M., Broderick, D.H., Sapre, A.V., Nag,N.K., de Beer, V.H., Gates, B.C., Kwart, H.J., Catalt., 61, 523-527(1980) に見られる。実際、これらのDBT の種々のアルキル化誘導体が軽油中にかなりの量存在することが知られている(たとえば、Kabe, T., Ishihara, A. and Tajima, H. lnd. Eng. Chem. Res., 31, 1577-1580(1992))。
【0004】上記のように水素化脱硫に抵抗性を示す有機硫黄化合物を脱硫するためには、現在用いられているよりも高い反応温度や圧力が必要とされ、また、添加する水素の量も非常に増大すると考えられている。このような水素化脱硫プロセスの改良は、ばく大な設備投資と運転コストを必要とすることが予想される。このような水素化脱硫に抵抗性を示す有機硫黄化合物を主たる硫黄化合物種として含むものとしては、たとえば、軽油があり、軽油のより高度な脱硫(超深度脱硫)を行う場合には上記のような水素化脱硫プロセスの大幅な改良が要求される。
【0005】一方、生物が行う酵素反応は比較的穏和な条件下で進行し、しかも酵素反応の速度自体は、化学触媒を用いた反応の速度と遜色のないという特徴を有している。さらに、生体内で起こる多種多様の生物反応に適切に対応する必要があるため、非常に多くの種類の酵素が存在し、それらの酵素は一般的に非常に高い基質特異性を示すことが知られている。このような特徴は、微生物を用いて化石燃料中に含まれる硫黄化合物中の硫黄の除去を行ういわゆるバイオ脱硫反応においても活かされるものと期待されている(Monticello, D.J., Hydrocarbon Processing39-45(1994))。
【0006】一方、細菌を用いて石油の成分である複素環式硫黄化合物から硫黄を除去する方法については、多数の報告があるが、それらは環分解(C-C 結合切断)型反応とC-S 結合切断型反応とに大別される。C-C 結合攻撃型脱硫活性を有する細菌としては、例えば、Pseudomonas sp., Pseudomonas aeruginosa, Beijerinckia sp., Pseudomonas alcaligenes, Pseudomonas stutzeri, Pseudomonas putida, Brevibacterium sp.などが知られている。これらの細菌は、DBT で代表される複素環式硫黄化合物中のC-C 結合の切断を行い、ベンゼン環を分解し、その後の酸化反応カスケードにより、硫黄塩を放出するというタイプの代謝を行うものである。これらの炭素骨格攻撃型経路の反応機構は芳香環の水酸化(DBT →1,2-ジヒドロキシジベンゾチオフェン)、環の開裂、水溶性産物への酸化(1,2-ジヒドロキシジベンゾチオフェン →トランス-4[2-(3-ヒドロキシ)チアンナフテニル]-2- オキソ- ブテノイン酸、3-ヒドロキシ-2- ホルミルベンゾチオフェン)といったものであり、Kodama経路と呼ばれている。このタイプの反応により、DBT のベンゼン環中のC-C 結合が攻撃を受け、油から抽出可能な種々の水溶性物質を生じる。しかし、この反応により、油中の他の芳香族分子が攻撃を受け、その結果かなりの量の炭化水素が液相に移動することになる(Hartdegen, F.J., Coburn,J.M. and Roberts, R.L. Chem. Eng. Progress, 80, 63-67(1984)) 。このようなことは石油の総熱量単位の低下を招くことになり、工業的には非効率的な反応である。また、このタイプのDBT 酸化分解菌は、児玉らが報告しているように酸化産物として水溶性のチオフェン化合物(主として3-ヒドロキシ-2- ホルミルベンゾチオフェン)を与えるが、これは液相から除去するのが困難な物質でもある。更に、DBT の炭素環の攻撃は、しばしばアルキル置換基やアリル置換基を持つDBT の 2位及び 3位の位置で起こるため、これらの位置で置換されたDBT はKodama経路の基質とはならない。
【0007】原油や石炭のみならず硫黄を含んだモデル化合物を分解し、ヘテロ原子である硫黄を選択的に除去して、硫酸塩や水酸化化合物を産生する微生物類が報告されている。このタイプの反応は、その代謝産物の構造から考えて、硫黄化合物中のC-S 結合を特異的に切断して、その結果硫黄を硫酸塩の形で遊離する反応であると考えられる。現在までに、表1 に示すような硫黄攻撃型のバイオ脱硫反応系の報告がある。
【0008】
【表1】
【0009】以上記載したバイオ脱硫はすべて、30℃近辺の温度条件下で進行する微生物代謝反応を利用するものである。一方、化学反応の速度は一般に温度に依存して増大することが知られている。また、石油精製プロセス中の脱硫工程では、高温・高圧条件下で分別蒸留や脱硫反応が行われる。従って、石油精製プロセス中にバイオ脱硫工程を組み込むとすると、常温近くにまで石油留分を冷却することなしに、冷却途中のより高い温度でバイオ脱硫反応ができる方が望ましいと考えられる。高温バイオ脱硫に関する報告には以下のようなものがある。
【0010】微生物を用いて高温で脱硫反応を行わせる試みのほとんどは、石炭脱硫において見ることができる。石炭中には種々の硫黄化合物が含まれている。主要な無機硫黄化合物は黄鉄鉱であるが、有機硫黄化合物に関しては多種多様のものが混在しており、多くがチオール、スルフィド、ジスルフィド、チオフェン基を含んでいることが知られている。用いられた微生物は、Sulfolobus属の細菌で、これらはすべて好熱性細菌である。鉱物スルフィドからの金属のリーチング(BrierleyC.L. & Murr, L.E., Science 179, 448-490(1973)) や石炭からの黄鉄鉱の硫黄除去などに種々の異なったSulfolobus株を用いた例が報告されている(Kargi, F. & Robinson, J.M., Biotechnol. Bioeng, 24, 2115-2121(1982); Kargi, F. &Robinson, J.M., Appl. Environ. Microbiol., 44, 878-883(1982); Kargi, F.& Cervoni, T.D., Biotechnol. Letters 5, 33-38(1983); Kargi, F.およびRobinson, J.M., Biotechnol. Bioeng., 26, 687-690(1984); Kargi, F. & Robinson, J.M., Biotechnol. Bioeng. 27, 41-49(1985); Kargi, F., Biotechnol. Lett., 9, 478-482(1987))。Kargi とRobinson (Kargi, F.およびRobinson, J.M.,Appl. Environ. Microbiol., 44, 878-883(1982))によれば、米国のイエローストーン国立公園の酸性温泉から分離されたSulfolobus acidocaldarius (以下、「S.acidocaldarius」という)のある株は、45〜70℃で生育するが、至適pH2 で元素状硫黄を酸化する。また、別の 2種のS. acidocaldarius 株による黄鉄鉱の酸化も報告されている(Tobita, M., Yokozeki, M., Nishikawa, N. & Kawakami, Y., Biosci. Biotech. Biochem. 58, 771-772(1994))。
【0011】化石燃料中に含まれる有機硫黄化合物のうち、DBT およびその置換体又はそれらの誘導体は通常の石油精製プロセスにおいて水素化脱硫を受けにくいことが知られている。そのDBT のS. acidocaldariusによる高温分解も報告されている(Kargi, & Robinson, J.M., Biotechnol. Bioeng, 26, 687-690(1984); Kargi, F., Biotechnol. Letters 9, 478-482(1987))。
【0012】これらの報告によれば、チアントレン、チオキサンテン、DBT などのモデル芳香族複素環式硫黄化合物を高温でこの微生物と反応させると、これらの硫黄化合物は酸化されて、分解する。S. acidocaldarius によるこれらの芳香族複素環式硫黄化合物の酸化は、70℃で観察されており、反応産物として硫酸イオンを生じる。しかし、この反応は硫黄化合物の他には炭素源を含まない培地中での反応であり、硫黄化合物を炭素源としても利用している。すなわち硫黄化合物中のC-C結合を分解していることは明瞭である。さらに、このS. acidocaldarius は酸性の培地でのみ増殖でき、DBT の酸化分解反応は、きびしい酸性条件下(pH2.5)での進行を要求する。このようなきびしい条件は石油製品の劣化を引き起こすと同時に脱硫に関わる工程に耐酸性材料を必要とするためプロセス上望ましくないと考えられる。S. acidocaldarius を、独立栄養条件下で増殖させると、必要なエネルギーを還元された鉄・硫黄化合物から獲得し、炭素源として二酸化炭素を利用する。しかし、S. acidocaldarius は、従属栄養条件下に増殖させると、炭素源およびエネルギー源として種々の有機化合物を利用することができる。すなわち、化石燃料が存在すると炭素源として資化されるものと考えられる。
【0013】Finnertyらは、Pseudomonas stutzeri、Pseudomonas alcaligenes 、Pseudomonas putidaに属する株がDBT 、ベンゾチオフェン、チオキサンテン、チアントレンを分解して、水溶性の物質に変換することを報告している(Finnerty, W.R.,Shockiey, K., Attaway, H. in Microbial Enhanced Oil Recovery, Zajic, J.E. ら(編)、Penwell. Tuisa, Okia, 83-91(1983))。この場合、酸化反応は55℃でも進むとしている。しかし、これらのPseudomonas 菌株によるDBT の分解産物は、Kodamaらが報告している3-ヒドロキシ-2- ホルミルベンゾチオフェンである(Monticello, D.J., Bakker, D., Finnerty, W.R. Appl. Environ. Microbiol.,49, 756-760(1985))。これらのPseudomonas 菌株によるDBT の酸化活性は、硫黄を含まない芳香族炭化水素であるナフタレンやサリチル酸により誘導を受け、クロラムフェニコールにより阻止される。このことから、これらのPseudomonas 菌株によるDBT の分解反応は、芳香環中のC-C 結合を切断することによる分解を基礎としていることが分かる。また、硫黄化合物以外にも石油留分中に含まれる貴重な芳香族炭化水素を同時に分解するおそれもあり、これは、燃料の価値や石油留分の品質を低下させることになる。
【0014】このように、今までに発見されている高温でDBT を分解できる菌は、DBT 分子中のC-C 結合を切断し、炭素源として利用する反応を触媒するものである。C-S結合を特異的に切断するが、C-C 結合は切断しないでそのまま残すタイプの有機硫黄化合物の分解反応が実際の石油の脱硫方法として望ましいことは上述の通りである。すなわち、高温でDBT およびそのアルキル置換体、又はそれらの誘導体分子中のC-S 結合を切断する活性を示し、水溶性の物質の形で、脱硫産物を生じる微生物を利用するのがバイオ脱硫プロセスとして最も望ましい。
【0015】前述のように、C-S結合切断型のDBT分解反応を行う微生物は、いくつかの属の細菌で知られている。たとえば、Rhodococcus sp. のATCC53968 はよく調べられたDBT分解菌株であり、DBTの硫黄原子に酸素原子を付加し、ジベンゾチオフェンスルホキシド(以下、「DBTO」という)からジベンゾチオフェンスルホン(以下、「DBTO2」という)を生成し、ついで2‐(2'-ヒドロキシフェニル)ベンゼンスルフィン酸塩を経て2-ヒドロキシビフェニル(以下、「2-HBP」という)を生成する反応を行う。しかし、この菌も通常の培養温度である30℃よりも少し高い37℃および43℃でさえ、48時間培養すると非常に生育が遅れるか、生育しなくなることが報告されている(特開平6-54695号公報)。このことから、高温脱硫反応を行わせるためには、高温で生育でき、しかも高温で有機硫黄化合物、特にDBTおよびその置換体、またはそれらの誘導体化合物を含む複素環式硫黄化合物類をC-S結合特異的に切断できる微生物を用いるのが最適であると考えられる。
【0016】Suzukiらは、Paenibacillus sp. A11-2株が、高温条件下で生育でき、かつ、有機硫黄化合物のC-S結合を特異的に切断できることを報告している(Suzuki,M., Konishi, J., Ishii, Y., Okumura, K., Appl. Environ. Microbiol., 63,3164-3169(1997))。この報告によると、この菌株は30〜60℃の温度条件下で生育でき、DBTまたは、4-メチルジベンゾチオフェンおよび4,6-ジメチルジベンゾチオフェンなどのDBT誘導体のC-S結合を特異的に切断できる。よって、この菌株は高温脱硫反応を行うのに適している。しかし、この菌株は、比較的低い温度、とりわけ30℃付近において、脱硫活性が著しく低下する。
【0017】バイオ脱硫プロセスにおいて、高温域(以降、「高温域」とは、50〜60℃付近の温度のことを指す)を含む広範な温度条件下(30〜60℃)で高い脱硫活性を維持していることは、石油留分の温度調整を簡略化できるという面で有効である。Furuyaらは、Bacillus subtilis WU-S2B株(受託番号FERM P-17041)が、高温域を含む広範な温度条件下で比較的高い脱硫活性を維持し、かつ、有機硫黄化合物のC-S結合を特異的に切断できることを報告している(古屋, 西井, 中川, 桐村,宇佐美, 日本農芸化学会大会講演要旨集, 384(1999))。この報告によると、この菌株は30〜60℃の広範な温度条件下で比較的高い脱硫活性を維持し、DBTまたは、2,8-ジメチルDBTおよび4,6-ジメチルDBTなどのDBT誘導体のC-S結合を特異的に切断できる。
【0018】C-S 結合切断型の脱硫反応を起こすことが知られている細菌で、そのDBT 分解反応に関与する酵素活性をコードする遺伝子が同定され、その塩基配列が決定されているものとしては、例えば、Rhodococcus sp. IGTS8 株のdsz 遺伝子(Denome, S., Oldfleld., C., Nash, L.J.およびYoung, K.D. J.Bacteriol., 176:6707-6716, 1994; Piddington, C.S., Kovacevich, B.R.およびRambosek, J. Appl.Environ. Microbiol., 61:468-475, 1995)、Sphingomonas sp. AD109株のdsz遺伝子(米国特許第6,133,016号)およびPaenibacillus sp. A11-2株のtds遺伝子(Y. Ishii, J. Konishi, H. Okada, K. Hirasawa, T. Onaka, M. Suzuki., Biochem. Biophy. Res. Comm., 270 (1), 81-88, 2000)が挙げられる。
【0019】IGTS8 株によるDBT 分解反応は、DBT からDBTOを経てDBTO2への変換を触媒するDszC、DBTO2から2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸への変換を触媒するDszAおよび2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸から2-HBP への変換を触媒するDszBの3つの酵素により触媒される(Denome, S., Oldfield., C., Nash, L.J.およびYoung, K.D. J.Bacteriol., 176:6707-6716, 1994; Gray, K.A., Pogrebinshy, O.S., Mrachko, G.T., Xi, L. Monticello, D.J.and Squires, C.H. Nat Biotechnol., 14:1705-1709, 1996; Oldfield, C., Pogrebinsky, O., Simmonds, J., Olson, E.S.および Kulpa, C.F., Microbiology,143:2961-2973, 1997)。それぞれ対応する遺伝子はdszC, dszA, dszBと呼ばれている。DszCとDszAはモノオキシゲナーゼで、両者ともその酸素添加反応にはNADH-FMNオキシドレダクターゼ活性の共存を必要とすることが知られている(Gray, K.A., Pogrebinsky, O.S., Mrachko, G.T., Xi, L. Monticello, D.J. およびSquires, C.H. Nat Biotechnol., 14:1705-1709, 1996; Xi, L. Squires, C.H., Monticello, D.J. and Chids, J.D. Biochem. Biophys. Res Commun., 230:73-76, 1997) 。これらのdsz 遺伝子を大腸菌で温度シフトにより誘導発現させた場合、菌体培養によるDszA活性は39℃で最大となり、42℃では顕著に低下することが報告されている(Denome, S., Oldfield., D., Nash, L.J. および Young,K.D. J. Bacteriol., 176:6707-6716, 1994) 。この結果は、IGTS8 株の有する脱硫酵素活性は常温付近で最大になり、より高温では活性は低下し、50℃以上ではまったく脱硫活性は見られなくなるという休止菌体反応系の実験結果(Konishi, J., Ishii, Y., Onaka, T., Okumura, K. および Suzuki, M. Appl. Environ. Microbiol., 63:3164-3169, 1997)と一致する。
【0020】A11-2株のDBT分解酵素をコードする遺伝子tdsA,tdsB,tdsCもIGTS8株のDBT分解経路と同様の経路によりDBTを分解脱硫する酵素を産生する。tdsA,tdsB,tdsCによりコードされるDBT分解酵素TdsA,TdsB,TdsCは、それぞれIGTS8株のDszA,DszB,DszCと同様の変換反応を触媒する。(Y. Ishii, J. Konishi, H. Okada, K. Hirasawa, T. Onaka, M. Suzuki., Biochem. Biophy. Res. Comm., 270 (1), 81-88, 2000)これらtds遺伝子を大腸菌内で発現させる場合、50℃以上で活性を有し、55℃で最適温度が見られるものの、45℃以下では顕著に活性が低下する。従ってdsz遺伝子、tds遺伝子の両者の温度特異性から示されるように、40〜50℃の温度条件下で効率的にC-S 結合特異的なDBT 分解活性を指令する遺伝子は従来報告されていないものである。
【0021】
【発明が解決しようとする課題】本発明の課題は、ベンゾチオフェン、DBT 系化合物に作用し、それらを高温で分解する能力を有する微生物から高温脱硫反応に関与する遺伝子を単離し、その構造(特に塩基配列)を特定し、また、これらの遺伝子をそれが単離されたのとは異なる微生物に導入し、脱硫能を賦与することにより、新規な脱硫微生物を創製することである。また、このような微生物を実際にベンゾチオフェン、DBT およびそれらのアルキル誘導体に作用させて、これらの化合物のC-S 結合を切断することにより、硫黄を遊離させる方法を確立することである。
【0022】
【課題を解決するための手段】本発明者らは、上記課題を解決するために鋭意検討を重ねた結果、高温脱硫細菌 Bacillus subtilis WU-S2B株(受託番号:FERM P-17041)から脱硫反応に関与する遺伝子群を単離することに成功し、本発明を完成するに至った。
【0023】即ち、本発明は、以下の1〜10を提供する。
1.以下の(a) 又は(b) のタンパク質をコードする遺伝子。
(a) 配列番号2記載のアミノ酸配列により表されるタンパク質(b) 配列番号2記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBTO2を2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸に変換する機能を有するタンパク質
【0024】2.以下の(a) 又は(b) のタンパク質をコードする遺伝子。
(a) 配列番号4記載のアミノ酸配列により表されるタンパク質(b) 配列番号4記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を2-HBPに変換する機能を有するタンパク質
【0025】3.以下の(a) 又は(b) のタンパク質をコードする遺伝子。
(a) 配列番号6記載のアミノ酸配列により表されるタンパク質(b) 配列番号6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBTをDBTO2に変換する機能を有するタンパク質
【0026】4.上記の1、2又は3に記載の遺伝子を含むベクター。
5.上記の4に記載のベクターを含有する形質転換体。
6.以下の(a) 又は(b) に示すタンパク質。
(a) 配列番号2記載のアミノ酸配列により表されるタンパク質(b) 配列番号2記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBTO2を2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸に変換する機能を有するタンパク質
【0027】7.以下の(a) 又は(b) に示すタンパク質。
(a) 配列番号4記載のアミノ酸配列により表されるタンパク質(b) 配列番号4記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を2-HBPに変換する機能を有するタンパク質
【0028】8.以下の(a) 又は(b) に示すタンパク質。
(a) 配列番号6記載のアミノ酸配列により表されるタンパク質(b) 配列番号6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBTをDBTO2に変換する機能を有するタンパク質
【0029】9.以下の性質を有するタンパク質。
(1) 作用:DBTをDBTO2にする(2) 至適pH:8.0 、安定pH:8〜11(3) 至適温度:45〜50℃、安定温度:53℃以下(4) 分子量:200,000(ゲル濾過法による)
(5) 活性阻害:キレート剤、SH阻害剤によって阻害される(6) 補酵素の要求性:フラビンレダクターゼによって供給される FMNH2 を要求する
【0030】10.以下の性質を有するタンパク質。
(1) 作用:DBTO2を2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸にする(2) 至適pH:7.5、安定pH:6〜10(3) 至適温度:50℃、安定温度:45℃以下(4) 分子量:174,000(ゲル濾過法による)
(5) 活性阻害:キレート剤、SH阻害剤によって阻害される(6) 補酵素の要求性:フラビンレダクターゼによって供給される FMNH2 を要求する
【0031】
【発明の実施の形態】以下、本発明を詳細に説明する。
(1)耐熱性脱硫酵素をコードする遺伝子本発明の遺伝子には、以下の3種類の遺伝子が含まれる。第一の遺伝子(以下、「bdsA」という)は、(a) 配列番号2記載のアミノ酸配列により表されるタンパク質、又は(b) 配列番号2記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBTO2 を2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸に変換する機能を有するタンパク質をコードするものである。
【0032】第二の遺伝子(以下、「bdsB」という)は、(a) 配列番号4記載のアミノ酸配列により表されるタンパク質、又は配列番号4記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を2-HBP に変換する機能を有するタンパク質をコードするものである。
【0033】第三の遺伝子(以下、「bdsC」という)は、(a) 配列番号6記載のアミノ酸配列により表されるタンパク質、又は(b) 配列番号6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBT をDBTOを経てDBTO2に変換する機能を有するタンパク質をコードするものである。
【0034】上記bdsA、bdsB、及びbdsC遺伝子は、Rhodococcus sp.IGTS8株由来のdszA、dszB、dszCおよびSphingomonas sp. AD109株由来のdszA、dszB、dszCおよびPaenibacillus sp. A11-2株由来のtdsA、tdsB、tdsCと一定の相同性を示すが、後述するようにこれらの遺伝子がコードするタンパク質は、2種のdszA、dszB、dszC、およびtdsA、tdsB、tdsCがコードするタンパク質とはその性質において異なる。
【0035】本発明の遺伝子のうち、配列番号2、4及び6記載のアミノ酸配列をコードする遺伝子については、本明細書の実施例に記載された方法により得ることができる。また、これらの遺伝子の塩基配列は既に決定されているので、これらの配列を基に適当なプライマーを合成し、Bacillus subtilis WU-S2B 株(受託番号:FERM P-17041)から調製されたDNA を鋳型としてPCR を行うことによっても得ることができる。
【0036】配列番号2、4及び6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列をコードする遺伝子は、本願の出願時において常用される技術、例えば、部位特異的変異誘発法(Zoller et al., Nucleic Acids Res. 10 6487-6500, 1982)により配列番号2、4及び6記載のアミノ酸配列をコードする遺伝子を改変することにより得ることができる。本発明の遺伝子は、DBT の分解に関与する酵素をコードするので、石油の脱硫に利用することができる。
【0037】(2)脱硫酵素をコードする遺伝子を含むベクター本発明のベクターは、上記のbdsA、bdsB又はbdsC遺伝子を含む。このようなベクターは、本発明のbdsA、bdsB又はbdsC遺伝子を含むDNA 断片を、公知のベクターに挿入することにより作製することができる。DNA 断片を挿入するベクターは、形質転換する宿主に応じて決めればよい。例えば、本発明に係るベクターとしては、プラスミドベクター、ファージミドベクター、コスミドベクター及び酵母ベクターなどが挙げられる。宿主として大腸菌を使用するのであれば、以下のようなベクターを使用するのが好ましい。強力なプロモーターとして、例えば、lac 、lacUV5、trp 、tac 、trc 、λpL、T7、rrnB、などを含むpUR 系、pGEX系、pUC 系、pET 系、pT7 系、pBluescript 系、pKK 系、pBS 系、pBC 系、pCAL系などのベクターを使用するのが好ましい。
【0038】(3)脱硫酵素をコードする遺伝子を含むベクターを含有する形質転換体本発明の形質転換体は、上記ベクターを含有する。形質転換体の宿主とする細胞は、植物細胞や動物細胞などであってもよいが、大腸菌などの微生物が好ましい。代表的な菌株としては、Sambrook等の成書Molecular Cloning Laboratory Mannual 2nd ed.に記載されている、71/18、BB4、BHB2668、BHB2690、BL21(DE3)、BNNl02(C600hflA)、C-1a、C600(BNN93)、CES200、CES201、CJ236、CSH18、DH1、DH5、DH5 α、DP50supF、ED8654、ED8767、HB101、HMS174、JM101、JM105、JM107、JM109、JM110、K802、KK2186、LE392、LG90、M5219、MBM7014.5、MC1061、MM294、MV1184、MV1193、MZ-1、NM531、NM538、NM539、Q358、Q359、R594、RB791、RR1、SMR10、TAP90、TG1、TG2、XL1-Blue、XS101、XS127、Y1089、Y1090hsdR、YK537 などが挙げられる。
【0039】本発明の形質転換体は、電気パルス法、コンピテントセル法、接合伝達法、プロトプラスト法など当技術分野において公知の技術を用いて宿主細胞を形質転換又は形質導入することにより取得することができる。これらの方法は、例えば、Sambrookらの成書(前掲)に記載されている。
【0040】(4)脱硫酵素本発明の脱硫酵素には、以下の3種類のタンパク質が含まれる。第一のタンパク質(以下、「BdsA」という)は、配列番号2記載のアミノ酸配列により表されるタンパク質と、配列番号2記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBTO2を2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸に変換する機能を有するタンパク質とを包含する。
【0041】第二のタンパク質(以下、「BdsB」という)は、配列番号4記載のアミノ酸配列により表されるタンパク質と、配列番号4記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を2-HBP に変換する機能を有するタンパク質とを包含する。
【0042】第三のタンパク質(以下、「BdsC」という)は、配列番号6記載のアミノ酸配列により表されるタンパク質と、配列番号6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつDBT をDBTO2に変換する機能を有するタンパク質とを包含する。
【0043】上記BdsA、BdsB、及びBdsCは、Rhodococcus sp.IGTS8株由来の脱硫酵素DszA、DszB、DszCと一定の相同性を示し、また、酵素としての作用も同一であるが、以下の点で明確に相違する。
(イ)DszA、DszB、DszCでは、難脱硫物質である4,6-ジブチルジベンゾチオフェンを脱硫できないが、本発明のBdsA、BdsB、及びBdsCでは脱硫可能である。
(ロ)DszA、DszB、DszCは、常温領域で脱硫活性を示すが、本発明のBdsA、BdsB、及びBdsCは常温−高温の中間領域で脱硫活性を示す。
【0044】本発明の脱硫酵素は、上述の本発明の脱硫酵素をコードする遺伝子を利用して製造することができる。また、配列番号2、4、及び6に記載のアミノ酸配列により表される脱硫酵素は、Bacillus subtilis WU-S2B 株(受託番号:FERM P-17041)から常法に従って調製することも可能である。
【0045】本発明のBdsAに包含される一タンパク質の性質を以下に示す。
(1) 作用:DBTO2を2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸にする(2) pH特性:図8及び9に示す通り、至適pHは7.5、安定pHは6〜10である(3) 温度特性:図6及び7に示す通り、至適温度は50℃、安定温度は45℃以下である(4) 分子量:174,000(ゲル濾過法による)
(5) 活性阻害:キレート剤、SH阻害剤によって阻害される(6) 補酵素の要求性:フラビンレダクターゼによって供給される FMNH2 を要求する
【0046】本発明のBdsCに包含される一タンパク質の性質を以下に示す。
(1) 作用:DBTをDBTO2にする(2) pH特性:図4及び5に示す通り、至適pHは8、安定pHは8〜11である(3) 温度特性:図2及び3に示す通り、至適温度は45〜50℃、安定温度は53℃以下である(4) 分子量:200,000(ゲル濾過法による)
(5) 活性阻害:キレート剤、SH阻害剤によって阻害される(6) 補酵素の要求性:フラビンレダクターゼによって供給される FMNH2 を要求する
【0047】本明細書において、脱硫酵素の「至適pH」とは、特定の温度(例えば、50℃)において最も好適な活性を有するpHを指す。同様に、脱硫酵素の「至適温度」とは、特定のpH(例えば、pH 7.0)において最も好適な活性を有する温度を指す。
【0048】また、本明細書において、脱硫酵素の「安定pH」とは、該酵素溶液を種々のpHに一定時間(例えば、30分間)放置した後、特定のpH(例えば、pH 7.0)に再調整し、特定の温度(例えば、50℃)において残存する酵素活性を測定したときに、最も高い活性レベルを100%として、好ましくは60%以上、より好ましくは70%以上、最も好ましくは80%以上の活性レベルを示すpHの範囲を指す。同様に、脱硫酵素の「安定温度」とは、該酵素溶液を種々の温度に一定時間(例えば、30分間)放置した後、特定の温度(例えば、50℃)に再調整し、特定のpH(例えば、pH 7.0)において残存する酵素活性を測定したときに、最も高い活性レベルを100%として、好ましくは60%以上、より好ましくは70%以上、最も好ましくは80%以上の活性レベルを示す温度範囲を指す。
【0049】
【実施例】以下、本発明を実施例により具体的に説明する。実施例中の遺伝子操作に関連した実験は、主にSambrookらの成書(前掲)に詳述されている方法に従って行った。
〔実施例1〕耐熱性脱硫酵素をコードする遺伝子断片のクローニング常温脱硫菌であるRhodococcus sp. IGTS8株や高温脱硫菌であるPaenibacillussp. A11-2株は、中間体としてDBTO2および2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を経由して、DBTを2-HBPへと変換する活性を有することが知られている。
【0050】一方、本発明者らは、本発明で用いられるBacillus subtilis WU-S2B株もRhodococcus sp. IGTS8株やPaenibacillus sp. A11-2株と同様の経路によりDBTを2-HBPに変化する能力を有することを明らかにしている(特開2000-139450)。ここで、上に挙げたDBTを2-HBPに変換する活性に関与する酵素およびその遺伝子に着目すると、Rhodococcus sp. IGTS8株やPaenibacillus sp. A11-2株では、既に各酵素のアミノ酸配列および遺伝子の塩基配列が判明しており、それらは50〜65%の相同性を有することが知られている。
【0051】そこで、Rhodococcus sp. IGTS8株やPaenibacillus sp. A11-2株と同様の活性を有するBacillus subtilis WU-S2B株においても、ある程度の相同性を有することが考えられたため、Rhodococcus sp. IGTS8株やPaenibacillus sp. A11-2株の酵素のアミノ酸配列で相同性の高い部分を検索したところ、以下に示すアミノ酸配列が見つかった。
【0052】
Paenibacillus sp. A11-2 株 TdsA WNVVTSLNNAEARNFG(配列番号7)
Rhodococcus sp. IGTS8 株 DszA WNVVTSLNDAEARNFG(配列番号8)
Paenibacillus sp. A11-2 株 TdsC GFDRFWRDARTHTLHDPV(配列番号9)
Rhodococcus sp. IGTS8 株 DszC GFDRFWRNVRTHSLHDPV(配列番号10)
【0053】そこで、上記のアミノ酸配列をもとに以下の塩基配列を有するPCR 用センスプライマーおよびアンチセンスプライマーをそれぞれ設計・合成した。
センスプライマー:WUS2-SI1 5'-GCI GAR GCI MGI AAY TTY GG-3'(配列番号11)(Iはデオキシイノシンを表す)アンチセンスプライマー:WUS2-C2 5'-CGT IGC GCC AIA AGC GGT C-3'(配列番号12)(Iはデオキシイノシンを表す)
【0054】このセンスプライマーとアンチセンスプライマーを使用して、Bacillus subtilis WU-S2B 株から抽出したDNA を鋳型としてPCRを行った。Bacillus subtilisWU-S2B 株からのDNA の調製は以下のように行った。Bacillus subtilis WU-S2B株を、DBT を含むAII培地(組成は表2に示す)で50℃で18時間培養して、菌体を回収した。
【0055】
【表2】
【0056】得られた菌体を3.7mlのSET緩衝液(6.7% スクロース, 1mM EDTA, 50mM Tris-HCl, pH 8.0)に懸濁させた。この懸濁液に、6mg/mlのリゾチームと4mg/mlのアクロモペプチダーゼを溶解させた1mlのTE緩衝液(1mM EDTA, 10mM Tris-HCl, pH8.0)を添加して、37℃で5分間反応させた。反応液に0.5mlのTE緩衝液(25mM EDTA, 50mM Tris-HCl, pH 8.0)と0.3mlのSDS溶液(25mM EDTA, 20%(w/v)SDS,50mM Tris-HCl, pH 8.0)を添加、攪拌混合して、37℃で10分間反応させ、30秒間ミキサーで攪拌した後、0.3mlの3N NaOHを添加し、10分間断続的に転倒混和した後、0.5mlのTris緩衝液(2M Tris-HCl, pH 7.0)を添加、3分間断続的に転倒混和した後、0.7mlの5M NaClを添加、撹拌混合して菌体反応液を調製した。菌体反応液に7mlのフェノール試薬を添加、撹拌混合して、12,000rpm で5分間遠心を行い、得られた上清に対して、7mlのフェノールクロロホルム試薬を添加、撹拌混合して、12,000rpm で5分間遠心を行った。得られた上清に対して、7mlのクロロホルム試薬を添加、撹拌混合して、12,000rpm で5分間遠心を行い、DNA溶液を調製した。DNA溶液に7mlのイソプロパノールを添加して、0℃で30分間放置した後、15,000rpm で30分間遠心を行い、DNA を沈澱させた。沈殿したDNA を冷70%(w/v) エタノールで洗浄後、減圧乾燥した後、100μlのTE緩衝液に溶解して、DNA溶液を調製した。調製したBacillus subtilis WU-S2B 株DNA を鋳型として用いて行ったPCR の条件は以下の通りである。
【0057】
反応液組成:10mM Tris-HCl 1.5mM MgCl2 各0.2mM dNTP Mixture 1 μM センスプライマー 1 μM アンチセンスプライマー 1μg 鋳型DNA 2.5U Taq DNA polymeraseアニーリング温度:52℃に固定してPCR を行った。
PCR サイクル:95℃ 3min 1回 95℃ 1min ↓ 52℃ 1.5min この間を25回繰り返し 72℃ 3min ↑ 72℃ 10min 1回DNA 増幅機:DNA Thermal Cycler 温度サイクラー(PERKIN ELMER CETUS社製)
【0058】上記の条件でPCR を行った結果、約3.2kb の増幅断片を与えることが確認された。この3.2kb のPCR 産物を、pGEM-T ベクターを用いて大腸菌JM109 株にクローニングした。この取得したDNA 断片の塩基配列を決定した結果、この塩基配列は、Rhodococcus sp. IGTS8株のdsz遺伝子と69.2%、Paenibacillus sp. A11-2株のtds遺伝子と56.9%の相同性を有することが明らかとなった。
【0059】決定されたDNA 塩基配列から、この断片にコードされるアミノ酸配列を推定し、これとRhodococcus sp. IGTS8 株からクローニングされているdsz 遺伝子によりコードされる蛋白質であるDszA、DszB およびDszC のアミノ酸配列とを比較した。その結果、DszA、DszBおよびDszCにそれぞれ76.3%、67.9% および71.3%の相同性を示す推定アミノ酸配列が存在することが確認された。さらに、この推定アミノ酸配列は、Paenibacillus sp. A11-2株のtds 遺伝子によりコードされる蛋白質であるTdsA、TdsB およびTdsC のアミノ酸配列と比較した結果、TdsA、TdsB およびTdsC とそれぞれ60.0%、52.7%および48.9%の相同性を示した。Rhodococcus sp. IGTS8 株およびPaenibacillus sp. A11-2株の脱硫遺伝子の塩基配列および脱硫酵素のアミノ酸配列との相同性が見つかったことから、Bacillussubtilis WU-S2B 株からクローニングされたこのDNA 断片に脱硫酵素がコードされている可能性が高く、これをプローブとして用いて隣接するDNA 領域つまり脱硫遺伝子全域をクローニングすることにした。
【0060】〔実施例2〕全DNA ライブラリーの作製Bacillus subtilis WU-S2B 株からのDNA の調製は以下のように行った。DBTおよびツニカマイシンを含むAII培地(組成は表2に示す)で50℃で18時間培養して菌体を回収した。得られた菌体を1mlのB1緩衝液(50mM EDTA, 50mM Tris-HCl, 0.5% Triton X-100, 0.2mg/ml RNaseA, pH 8.0)に懸濁させた。この懸濁液に、100mg/mlのリゾチーム溶液を20μl と20mg/ml のProteinase K溶液を45μl添加して、37℃で10分間反応させた。反応液に0.35mlのB2緩衝液(800mM GuHCl,20% Tween-20, pH 5.5 )を添加、攪拌混合して、50℃で30分間反応させ、5秒間ミキサーで攪拌して、菌体反応液を調製した。陰イオン交換樹脂が充填されたQIAGEN GENOMIC-TIP20/G(QIAGEN社製)カラムを2mlのQBT 緩衝液(750mM NaCl, 50mM MOPS, 15% ethanol, 0.15% Triton X-100, pH7.0 )で平衡化して、菌体反応液をカラムに注入した。カラムを3mlのQC緩衝液(1.0M NaCl, 50mM MOPS,15% ethanol, pH7.0)で洗浄したのち、2mlのQF緩衝液(1.25M NaCl, 50mM Tris-HCl, 15% ethanol, pH 8.5)でゲノムDNA 溶液を溶出した。ゲノムDNA 溶液に1.4ml のイソプロパノールを添加してDNA を沈殿させたのち、ガラス棒で巻きとり回収した。回収したDNA を50μl のTE緩衝液(10mM Tris-HCl, 1mM EDTA, pH8.0)に溶解してゲノムDNA 溶液を調製した。
【0061】Bacillus subtilis WU-S2B 株の全DNA ライブラリーは以下のようにして作製した。Bacillus subtilis WU-S2B 株の全DNA 標品約 50μg を5 ユニットのSau3AIで各々6分、7分、8分消化した後、消化物をフェノール-クロロホルムで抽出しエタノール沈殿を行って遠心分離して回収されたDNA 断片を3.5ユニットの子ウシ小腸由来のアルカリ性ホスファターゼで、37℃18時間処理することにより脱リン酸化を行った。アルカリ性ホスファターゼ処理後フェノール-クロロホルム処理によりDNA を抽出し、エタノール沈殿によりこれを回収した。得られたDNA 断片約 0.2μg をλDASHI/BamHI アーム約2μg と2ユニットのT4DNA リガーゼ存在下に4℃18時間反応させた。反応混合物をGigapack III Gold packaging Extractと反応させることによりin vitroパッケージングを行い、ファージライブラリーを作製した。得られたファージライブラリーの力価は、105 pfu/mlであった。
【0062】〔実施例3〕全DNA ライブラリーのスクリーニングファージライブラリーのスクリーニングを行うためのDNA プローブは以下のようにして作製した。実施例1で記載した方法により取得したDNA断片とRhodococcus sp. IGTS8株のdsz遺伝子塩基配列の間で特に相同性の高かった領域を選択して以下に示すセンスプライマー(Rhodococcus sp. IGTS8株のdszAの5'端側より427 番目から447 番目の塩基配列に相当)とアンチセンスプライマー(Rhodococcus sp. IGTS8株のdszCの5'端側より1135番目から1153番目の塩基配列に相当)を作製した:センスプライマー:WUS2-NSI1 5'-GCA TGA CAT CCG ATA CG-3'(配列番号13)
アンチセンスプライマーWUS2-NC2 5'-TAG TTT GGG TGG GTT CC-3'(配列番号14)。
【0063】このプライマーを用いてBacillus subtilis WU-S2B 株から調製したDNA を鋳型としたPCR を行うことにより、実施例1で取得したDNA断片の内側領域のDNA断片を増幅させた。得られたPCR 産物を鋳型としてランダムプライム法(マルチプライム法)によりジオキシゲニン(DIG)で標識されたDSZACプローブを調製した。DIG 標識プローブの調製法は、Boehringer Mannheim 社のプロトコールに従った。DIG 標識プローブの調製方法を以下に示す。
【0064】得られたPCR 産物5μl を沸騰した熱湯中で10分間熱変性させ、塩を含んだ氷上で冷却した。得られた変性DNA 溶液に、2μl のヘキサヌクレオチド混合液(0.5M Tris-HCl, 0.1M MgCl2, 1mM Dithioerythritol, 2mg/ml BSA, 3.143mg/mlRandom Primer, pH7.2 )、2μl のdNTP標識混合液(1mM dATP, 1mM dCTP, 1mMdGTP, 0.65mM dTTP, 0.35mM DIG-dUTP, pH7.5 )、10μl の滅菌蒸留水及び1μl のKlenow酵素(2units )を添加して、37℃で18時間反応させた。反応液に、2μl の0.2M EDTA 溶液を添加して反応を停止させた。次に2μl の4M LiClと60 μl 冷エタノール(-20℃)を添加して、-70℃で30分間放置したのち、15,000rpm で30分間遠心を行い、DNA を沈殿させた。沈殿したDNA を冷70%(w/v)エタノールで洗浄後、吸引乾燥したのち、50μl のTE緩衝液に溶解して、DIG 標識プローブを調製した。
【0065】Bacillus subtilis WU-S2B株の脱硫遺伝子のスクリーニングは上述の方法で調製したDIG 標識DSZACプローブを用い、Hybond N+ メンブレンに転写されたプラークに対するプラークハイブリダイゼーションにより行った。ハイブリダイズするクローンの検出にはDIG-ELISA (Boehringer Mannheim)を用いた。ゲノムライブラリーより約2000個のファージプラークをDSZACプローブを用いてスクリーニングしたところ、 3個の陽性プラークが検出された。この 3個のプラークについて単プラーク分離を行い、再度プラークハイブリダイゼーションを行った結果、3個とも全て陽性プラークであることが確認された。検出されたDSZACプローブ陽性プラークを用いてファージクローンを調製し、それらのクローンからQIAGENLambda キットを用いてファージDNA を抽出した。 3個の陽性プラークを用いて調製したファージDNA をEcoRI、NotIで切断し、図1に示す制限酵素地図を作成した。さらに、これら 3種のファージDNA をNotIおよびNheIなどの様々な制限酵素を用いて消化して得られたDNA 断片についてDSZACプローブを用いたサザンブロット分析を行ったところ、約7.1kb のNheI 断片にハイブリダイズすることが確認された。これらの制限酵素地図およびサザンブロット分析の結果から、No.1のファージDNA の約7.1kb のNheI 断片にdsz 遺伝子がコードされているものと考えられた。
【0066】〔実施例4〕脱硫活性の確認次にNheI断片に脱硫遺伝子が存在するかを確認するため組換え大腸菌によるDBT分解能を検定した。プラスミドベクターpUC19のXbaI部位に約7.1kbのNheI断片を挿入してpU1Nhを作製した(図1参照)。このpU1Nhを保有する組換え大腸菌JM109株を、0.1mg/ml のアンピシリンを含むLB培地(Sambrook等の成書Molecularcloning Laboratory Manual 2nd に記載)(LB-Amp培地)に基質としてそれぞれDBTを添加した培地で、37℃で 1晩培養した。また、対照株としてベクターpUC19のみを保有するJM109株を同様の条件で培養した。LB-Amp培地(上述)で37℃、1晩前培養を行い、前培養液を得た。次に、1/100 容量の前培養液を検定用の培地(LB-Amp培地に基質としてDBT を添加した培地)に添加し、37℃で18時間培養後、酢酸エチルを用いて分解産物の抽出を行い、高速液体クロマトグラフィー分析を行った。その結果、pU1Nhを保有する組換え大腸菌では、基質としてDBTを含む培地で培養した場合2-HBP が生成することが確認された。また、ベクターのみの組換え大腸菌ではそのような変換活性はまったく示さなかった。このことから、pU1Nhに挿入されている約7.1kbのNheI断片はDBT から2-HBP への一連の変換反応を触媒する活性をすべてコードできる配列を持っていることが証明された。
【0067】〔実施例5〕脱硫遺伝子の塩基配列の決定陽性プラークから調製したファージDNAおよびpU1Nhを用いて、プライマー歩行法により、約7.1kbのNheI断片のうち約4.1kbの塩基配列の決定を行った。クローンのシークエンシング反応はBigDye Terminator Cycle Sequencing Kit(AppliedBiosystems)を用いて行い、ABI PRISM 310 Genetic Analyzer(Applied Biosystems) により塩基配列を決定した(配列番号1、3及び5)。得られた塩基配列データは、GENETYX-MAC/ATSQ v3.0 およびGENETYX-MAC v10.0を用いて解析した。
【0068】決定した約4.1kbの配列中のORF を探索した結果、1kb以上の長さのORF が同一方向に 3個見つかった。これらのORF を上流側からORF 1, 2, 3 と命名した。ORF 1, 2, 3 は、各々 453個、 356個、 415個のアミノ酸をコードする。ORF 1の翻訳終始コドンTGA とORF 2 の翻訳開始コドンATG は、部分的に重なっており、5'-ATGA-3'という配列になっており、Rhodococcus sp. IGTS8株のdsz オペロン中およびPaenibacillus sp. A11-2株のtdsオペロン中の塩基配列と同様の構成を有していることが確認された。これらのORF についてRhodococcus sp. IGTS8株のdsz 遺伝子との塩基配列相同性の解析を行ったところ、ORF 1, 2, 3 は、それぞれIGTS8株のdsz A, B, C と72.8%、67.4%、71.3%の相同性を示した。また、Paenibacillus sp. A11-2 株のtds 遺伝子との塩基配列相同性の解析を行ったところ、ORF 1, 2, 3 は、それぞれPaenibacillus sp. A11-2株のtds A, B, Cと61.3%、56.8%、56.0%の相同性を示した。さらに、Bacillus subtilis WU-S2B 株遺伝子の塩基配列を基礎としてそれらにコードされる蛋白質のアミノ酸配列を推定したところ、ORF 1, 2, 3 によりコードされるポリペプチドはそれぞれRhodococcus sp. IGTS8 株のDsz A, B, Cと78.6%、67.9%、72.0%の相同性を示した。また、Paenibacillus sp. A11-2 株のTds A, B, Cと63.7%、52.7%、50.6%の相同性を示した。
【0069】Bacillus subtilis WU-S2B 株のORF でコードされる蛋白質のアミノ酸配列とRhodococcus sp. IGTS8 のdsz 配列でコードされる蛋白質のアミノ酸配列とを比較すると、いくつかの点で特徴的な差異が見いだされる。まず、ORF 1 によりコードされるBdsA とDszAとを比較すると、カルボキシル末端側において、アミノ酸の異なる部分が連続して見られる。また、BdsAとTdsAとを比較すると、BdsAのアミノ末端の配列の方がTdsAのアミノ末端の配列より長くなっており、さらに、特にカルボキシル末端側において、アミノ酸の異なる部分が連続して見られる。DszBとORF 2 によりコードされるBdsB のアミノ酸配列については、DszBのアミノ末端の配列の方がBdsBのアミノ末端の配列より延びて長くなっている。また、TdsBとBdsBとを比較すると、BdsBのカルボキシル末端の配列の方がTdsBのカルボキシル末端の配列より長くなっている。また、特にカルボキシル末端側のアミノ酸配列の相同性が低くなっている。DszCとORF 3 によりコードされるBdsC のアミノ酸配列とを比較すると、全長のサイズはほとんど同じであるが、アミノ末端側のアミノ酸配列の相同性が低くなっている。また、TdsCとBdsCとを比較すると、BdsCのアミノ末端の配列の方がTdsCのアミノ末端の配列より延びて長くなっている。
【0070】〔実施例7〕
(1)酵素活性測定に必要なフラビンレダクターゼの精製酵素活性を 50°C で測定するためには、この温度でも活性を有するフラビンレダクターゼが必要である。そのために、Paenibacillus sp. A11-2 株由来のフラビンレダクターゼ(TdsD)高発現株から同酵素を精製した。高発現株を 50 μg/ml のアンピシリンを含む LB 培地中、37°C で一晩振盪培養後、0.2 mM IPTGを添加してさらに 4 時間培養後、菌体を取得した。湿重量 11 g の菌体を緩衝液 A(50 mM トリス塩酸, pH 7.0、10% グリセロール、1 mM ジチオスレイトール)に懸濁し、超音波破砕機(ブランソン、モデル 450)で 4°C、15 分間破砕を行った。10,000 x g で 30 分間遠心分離を行った後の上清を無細胞抽出液とした。この無細胞抽出液を緩衝液 A に対して充分透析をした後、同じ緩衝液で平衡化した Q-Sepharose カラムにアプライした。同緩衝液および、0.05 M、0.1M の塩化カリウムを含む緩衝液 A でそれぞれ洗浄後、0.15 M 塩化カリウムを含む緩衝液 Aで溶出を行った。活性画分を集めて、限外ろ過により濃縮し、引き続き限外ろ過を用いて酵素が溶解している緩衝液を 1.25 M 硫酸アンモニウムを含む緩衝液 A にした。この溶液を同緩衝液で平衡化した Phenyl-Toyopearl カラムにアプライした。同緩衝液で洗浄後、硫酸アンモニウムを含まない緩衝液 Aまでの硫酸アンモニウムによるリニアグラジエント溶出を行い、活性画分を集めて、限外ろ過により濃縮した。引き続き限外ろ過を用いて酵素が溶解している緩衝液を 0.15 M 塩化ナトリウムを含む緩衝液 A にした。この酵素溶液を同緩衝液で平衡化した Superdex200 HR10/30 カラムにアプライし、同緩衝液で溶出し、活性画分を集めて、限外ろ過により濃縮した。この結果、活性画分は電気泳動的に均一であることが確認された。
【0071】酵素活性の測定は20 mM リン酸バッファー、pH 7.0、0.5 mM NADH、0.02 mM FMN を含む反応液中で行い、70°C における NADH の減少速度を 340 nm の吸光度の減少速度を求めることにより算出した。1分間に 1 μmol の NADH を減少させる酵素量を 1 unitと定義した。
【0072】(2)BdsCの精製B. subtilis WU-S2B 株を AII 培地(表2参照)に 0.54 mM になるように DBT/エタノール溶液を添加した培地中で 40°C、2 日間、ジャーファーメンターを用いて通気攪拌培養を行い、菌体 334 g(湿菌体重量)を得た。得られた菌体を緩衝液 A(50 mM トリス塩酸, pH 7.0、10% グリセロール、1 mM ジチオスレイトール)に懸濁し、超音波破砕機(ブランソン、モデル 450)で 4°C、75 分間破砕を行った。10,000 x g で 60 分間遠心分離を行った後の上清を無細胞抽出液とした。この無細胞抽出液を 0.1 M 塩化カリウムを含む緩衝液 A に対して充分透析をした後、同じ緩衝液で平衡化したDEAE-Sepharose カラムにアプライした。同緩衝液で洗浄後、0.25 M 塩化カリウムを含む緩衝液 A までの塩化カリウムによるリニアグラジエント溶出を行った。活性画分を集めて、限外ろ過により濃縮した。この溶液を0.25 M 塩化カリウムを含む緩衝液 A に対して充分透析をした後、同緩衝液で平衡化したQ-Sepharose カラムにアプライした。同緩衝液で溶出し、下記に示す蛋白質Aを含む画分と蛋白質Cを含む画分とに分けて回収して、限外ろ過により濃縮し、引き続き限外ろ過を用いて酵素が溶解している緩衝液を 1 M 硫酸アンモニウムを含む緩衝液 A にした。この溶液を同緩衝液で平衡化した Phenyl-Toyopearl カラムにアプライした。同緩衝液で洗浄後、硫酸アンモニウムを含まない緩衝液 A までの硫酸アンモニウムによるリニアグラジエント溶出を行い、活性画分を集めて、限外ろ過により濃縮した。この結果、活性画分は電気泳動的に均一であることが確認された。
【0073】酵素活性の測定は、10μg精製酵素と、100 mM リン酸バッファー pH 7.0、0.01 mM FMN、6 mM NADH、0.45 units/ml TdsD、0.3 mM DBT とを含む酵素反応液で、50°C で振盪して行った。10 分の 1 量の 12 M HCl を添加して反応を停止後、10 分の 8 量の酢酸エチルを加え、よく混合した後、12,000 回転で 3 分間遠心し、上層を HPLC による分析に供した。1分間に 1 nmol の DBTO2を生成させる酵素量を 1 unit と定義した。精製段階における酵素活性の測定においては、上記の反応液を用いて活性を確認するのに十分な適当量の酵素を使用した。各精製段階における酵素活性を表3に示す。また、上記の精製酵素反応液を、種々の温度で活性の測定を行うことにより、該酵素の温度依存性について検討した。BdsC酵素活性についての温度依存性を図2に示す。上記の精製酵素溶液を種々の温度で30分間放置した後の残存活性を、上記のように測定し、該酵素の温度安定性について検討した。該酵素についての温度安定性を図3に示す。さらに、種々のpHを有する100 mMの各バッファー、0.01 mM FMN、6 mM FADH、0.45 units/ml TdsD、0.3 mM DBTを含む反応液を、50℃で振盪して、酵素活性を測定することにより、該酵素のpH依存性について検討した。pH依存性を図4に示す。また、種々のpHを有する100 mMの各バッファーと精製酵素を混合し、30分間放置した後、100mM リン酸バッファー pH 7.0、0.01 mM FMN、6 mM NADH、0.45 units/ml TdsD、0.3 mM DBT を含む反応液で、50°C で振盪して、該酵素のpH安定性について検討した。該酵素のpH安定性を図5に示す。
【0074】
【表3】
【0075】〔実施例8〕
(1)BdsAの精製BdsCの精製途中における Q-Sepharose の段階で回収した活性画分を 0.1 M 塩化カリウムを含む緩衝液 A に対して透析を行った後、同緩衝液で平衡化した Q-Sepharose カラムにアプライした。同緩衝液で洗浄後、0.3 M 塩化カリウムを含む緩衝液 A までの塩化カリウムによるリニアグラジエント溶出を行った。活性画分を集めて、限外ろ過により濃縮した。引き続き限外ろ過を用いて酵素が溶解している緩衝液を 0.7 M 硫酸アンモニウムを含む緩衝液 A にした。この溶液を同緩衝液で平衡化した Phenyl-Toyopearl カラムにアプライした。同緩衝液および、0.5 M、0.4 M、0.3 M の硫酸アンモニウムを含む緩衝液 A でそれぞれ洗浄後、0.2 M 硫酸アンモニウムを含む緩衝液 A で溶出し、画分を集めて、限外ろ過により濃縮した。引き続き限外ろ過を用いて酵素が溶解している緩衝液を 0.15 M 塩化ナトリウムを含む緩衝液 A にした。この酵素溶液を同緩衝液で平衡化した Superdex200 HR10/30 カラムにアプライし、同緩衝液で溶出し、活性画分を集めて、限外ろ過により濃縮した。さらに、限外ろ過を用いて酵素が溶解している緩衝液を 0.25 M 塩化ナトリウムを含む緩衝液 A にした。この酵素溶液を同緩衝液で平衡化した MonoQ HR10/10 カラムにアプライし、同緩衝液で洗浄後、0.35 M 塩化ナトリウムを含む緩衝液 A までの塩化ナトリウムによるリニアグラジエント溶出を行った。活性画分を集めて、限外ろ過により濃縮した。この結果、活性画分は電気泳動的に均一であることが確認された。
【0076】酵素活性の測定は基質として DBT の代わりにDBTO2を添加する以外は、上述したBdsCの活性測定法と同様に行った。各段階における酵素活性を表4に示す。BdsA酵素活性についての温度依存性を図6に、温度安定性を図7に、pH依存性を図8に、pH安定性を図9に、それぞれ示す。
【0077】
【表4】
【0078】
【発明の効果】本発明は、脱硫に関与する新規な遺伝子及び酵素を提供する。これらの遺伝子及び酵素を利用することにより、化石燃料中の硫黄を容易に遊離させることができるようになる。
【0079】
【配列表】
SEQUENCE LISTING <110> Petroleum Energy Center<120> Thermostable desulfurising enzyme and the gene encoding thereof<130> P01-0068<160> 14<170> PatentIn Ver. 2.0<210> 1<211> 4031<212> DNA<213> Bacillus subtilis<220> <221> CDS<222> (312)..(1670)<400> 1ctagcgaggt gcgagttccg cgcggtagcc atccgcttca ttgattttta catcgatacc 60 gccgcggtgg ccgtcagggt gtcgaagccg ggcggaggtg cacaccccgc cggttggtga 120 tcttgctcac aggtggcgct ttgacggcac tcgtccgggg gcgagaatcg gcgaaggtcg 180 ggtgggtcgt cgggaggaag tggcgtgttg ggtgcccctt tgaatcatcg ggatcctaac 240 tctgttgggg tcggttgcag tagccgagca ttggtctccg gtaactgatc cccacggagg 300 cactgctgat t atg gct gaa caa cgc caa cta cat tta gcc ggg ttt ttc 350 Met Ala Glu Gln Arg Gln Leu His Leu Ala Gly Phe Phe 1 5 10tcg gcc ggc aac gtc act cat gcg cat ggc gcg tgg cgc cat gtc ggt 398 Ser Ala Gly Asn Val Thr His Ala His Gly Ala Trp Arg His Val Gly 15 20 25 gcg acg aac ggt ttt ctg act ggc gag ttc tac aag cag att gcg cgg 446 Ala Thr Asn Gly Phe Leu Thr Gly Glu Phe Tyr Lys Gln Ile Ala Arg 30 35 40 45acc ctg gag cgg ggc aag ttt gat ctg ttg ttt ctt ccc gat ggc ctg 494 Thr Leu Glu Arg Gly Lys Phe Asp Leu Leu Phe Leu Pro Asp Gly Leu 50 55 60 gcc atc gaa gat agc tat ggc gat aat ttg gag acg ggg gtc ggt ctg 542 Ala Ile Glu Asp Ser Tyr Gly Asp Asn Leu Glu Thr Gly Val Gly Leu 65 70 75 ggt ggc cag ggc gcg gtg gcg ctg gag ccg acc agc gtg ata gcg acg 590 Gly Gly Gln Gly Ala Val Ala Leu Glu Pro Thr Ser Val Ile Ala Thr 80 85 90 atg gct gcg gtg acc cag cga tta ggt ctg ggc gct acg gtt tcg acg 638 Met Ala Ala Val Thr Gln Arg Leu Gly Leu Gly Ala Thr Val Ser Thr 95 100 105 acc tac tac ccg ccc tat cat gtg gct cgg gtg ttt gcc acg ctc gac 686 Thr Tyr Tyr Pro Pro Tyr His Val Ala Arg Val Phe Ala Thr Leu Asp110 115 120 125aat ctg tct gat ggc cgg atc tcg tgg aat gtg gtc acg tcg ctg aac 734 Asn Leu Ser Asp Gly Arg Ile Ser Trp Asn Val Val Thr Ser Leu Asn 130 135 140 gac tcc gag gca cgc aac ttc ggt gtg gat gag cac ctt gag cat gac 782 Asp Ser Glu Ala Arg Asn Phe Gly Val Asp Glu His Leu Glu His Asp 145 150 155 atc cga tac gac cgg gct gac gaa ttc ttg gag gcc gtc aag aag cta 830 Ile Arg Tyr Asp Arg Ala Asp Glu Phe Leu Glu Ala Val Lys Lys Leu 160 165 170 tgg agt tct tgg tcc gag gat gcc ttg ttg ttg gac aag gtt ggc ggt 878 Trp Ser Ser Trp Ser Glu Asp Ala Leu Leu Leu Asp Lys Val Gly Gly 175 180 185 cgg ttc gct gat ccc aag aag gtt caa tac gtc aat cat cgc ggc cgc 926 Arg Phe Ala Asp Pro Lys Lys Val Gln Tyr Val Asn His Arg Gly Arg190 195 200 205tgg ttg tcg gtc cgt ggc ccg ttg cag gtt cca cgg tcc cgt cag ggt 974 Trp Leu Ser Val Arg Gly Pro Leu Gln Val Pro Arg Ser Arg Gln Gly 210 215 220 gaa ccg gtc atc ttg cag gcc ggc ttg tcg ccg cgg ggg cgt cgc ttc 1022 Glu Pro Val Ile Leu Gln Ala Gly Leu Ser Pro Arg Gly Arg Arg Phe 225 230 235 gcg ggg cgc tgg gct gaa gcg gta ttc agc gtc tcg ccc aac ctc gac 1070 Ala Gly Arg Trp Ala Glu Ala Val Phe Ser Val Ser Pro Asn Leu Asp 240 245 250 atc atg cgc gcg gtc tat cag gac atc aaa gct cac gtt gcg gcc gcg 1118 Ile Met Arg Ala Val Tyr Gln Asp Ile Lys Ala His Val Ala Ala Ala 255 260 265 ggg cgt gac ccg gag caa aca aag gta ttc acc gcg gtg atg ccg gtg 1166 Gly Arg Asp Pro Glu Gln Thr Lys Val Phe Thr Ala Val Met Pro Val270 275 280 285ctc ggt gag aca gag cag gtg gcc cgg gaa cgt ctg gaa tat ctc aat 1214 Leu Gly Glu Thr Glu Gln Val Ala Arg Glu Arg Leu Glu Tyr Leu Asn 290 295 300 tcg ctg gtg cat ccc gaa gtg ggt ttg tcg acg cta tcc agc cac agc 1262 Ser Leu Val His Pro Glu Val Gly Leu Ser Thr Leu Ser Ser His Ser 305 310 315 ggc ttg aat ctc tcc aag tat ccg ctg gat acg aag ttt tcc gac atc 1310 Gly Leu Asn Leu Ser Lys Tyr Pro Leu Asp Thr Lys Phe Ser Asp Ile 320 325 330 gtc gcc gat ctc ggg gat cgt cac gtg ccg acc atg ttg cag atg ttt 1358 Val Ala Asp Leu Gly Asp Arg His Val Pro Thr Met Leu Gln Met Phe 335 340 345 tcc gcc gtg gcc ggt ggc ggt gcg gac ctg acg ctg gcc gaa ctg ggc 1406 Ser Ala Val Ala Gly Gly Gly Ala Asp Leu Thr Leu Ala Glu Leu Gly350 355 360 365cgc cga tac ggc acc aac gtc ggg ttt gtg ccg cag tgg gcc gga act 1454 Arg Arg Tyr Gly Thr Asn Val Gly Phe Val Pro Gln Trp Ala Gly Thr 370 375 380 gcc gaa cag atc gct gat caa ttg att agt cac ttt gag gcc ggg gcc 1502 Ala Glu Gln Ile Ala Asp Gln Leu Ile Ser His Phe Glu Ala Gly Ala 385 390 395 gct gac gga ttc atc att tcg ccg gcg tat ctg ccg ggt att tac gag 1550 Ala Asp Gly Phe Ile Ile Ser Pro Ala Tyr Leu Pro Gly Ile Tyr Glu 400 405 410 gag ttc gtc gac cag gtg gtt ccg ctg ctg cag caa cgt ggt gtg ttc 1598 Glu Phe Val Asp Gln Val Val Pro Leu Leu Gln Gln Arg Gly Val Phe 415 420 425 cgt acc gag tac gaa ggg aca acg ctg cgt gag cac ctt ggg ttg gct 1646 Arg Thr Glu Tyr Glu Gly Thr Thr Leu Arg Glu His Leu Gly Leu Ala430 435 440 445cac cct gaa gtc ggg agc atc cga tgaccaccac cgatgttgat catgacgttc 1700 His Pro Glu Val Gly Ser Ile Arg 450 ttgcctacag caactgcccg gttcccaatg cactactgac agcgctggag tcaaacctgt 1760 tggccggcaa cgggatttcg ctgaatgtgc tcagcggcgc gcaagcaggg ttgcatttca 1820 cctacgatca tcccgcttac acccgctttg gtggcgaaat tccacctctt atcagtgagg 1880 gcctgcgggc gccgggacgc acccgcctgc taggcatcac ccctctggcc ggtcggcaag 1940 ggatctatgt ccgtgttgac agcccggtga catcaccgga gcagctgcgg ggccggcggg 2000 tgggtgtgtc gggtgcagcg atccgcatcc tcaccggcga gttgggcgat tatcggcact 2060 tggacccgtg gcggcaaaca ctgatcgcgc tgggtacctg ggaggcgcgg ggacttttgc 2120 agaccctgca catcgggggg ataggggtcg acgacgtcga gttggtgcgt atcgaaagcc 2180 ctggcgtcga cgtgcccgaa gagcggctgg aagcagcggc cagtgtcaaa ggcgctgacc 2240 tgtttcccga cgtggccggc caccagaggg acatcctgga tagcggcaac gttgatgcgc 2300 tgttcacctg gttgccctgg gcggcggagt tagaagatct cagcggagcg cgggtgctcg 2360 ccgatctcgg cgatgaccaa cgaagccggt atgccagtgt ctggacggtc agcgctcagc 2420 tggttgacga gcgacccgac ctggtgcaac gactcgtcga cgcggcggtc caagccagcc 2480 gttgggcaca ggcccatccc gaggaaaccg ttggcatcca tgccgccaac ctcggggtcg 2540 cgccctcggc gatcggacgc ggtttcggcg ctgatttcgc ccaacatctg atcccacggc 2600 ttgacgactc ggcactggcc gttgtcgacc aaactcaaca gttcctcctc gaccacaatc 2660 tcctcgacca tccggtggac ctcacgcagt gggcggcacc gcaattcctc actcaatcca 2720 cgactggaga ccagcaatga ccctgaccga tgacaccgcc acggcacaga acaataggaa 2780 tggcgacccc atcgaggtgg cgcgcgagct gacgcgaaag tggcaagcta ccgtcgtcga 2840 gcgcgacaag gctggtggtt cggcgacaga agagcgtgag gatctacgcg ccagtgggct 2900 gctctcggtg accgttcccc ggcacctggg cggttgggga gccgactggc ctaccgctct 2960 ggaagtggtg cgtgagatcg ccaaggtcga tggatcgctg ggccacttgt tcggatacca 3020 cctcagcacc ccagccgtta ttgatctgtg gggttcaccc gagcaaagtg aacgcctact 3080 tcgtcaactg gccgaaaaca actggtggac cgggaacgcc tccagcgaga acaacagcca 3140 catcctcgaa tggaaggtaa ctgccacccc gaccgacgac ggcgggtacc ttctcaacgg 3200 tatcaagcac ttcagcagtg gagccaaggg ctcagatctg ctcttggtgt tcggcgtgat 3260 accggagggt tccccacagc aaggcgccat cgttgccgcg gccattccca ccaccagaga 3320 aggcgttcaa acaaacgacg actggcaggc gctagggatg cggcgcaccg acagcggcac 3380 caccgaattc cacaacgtcg tagtccgtcc cgacgaggtg ctgggcaaac ccaatgccgt 3440 tgtcgaggcg ttcctggcgt ccgggcgcgg cagcctattc ggcccgatcg tgcagttggt 3500 gttcgcaacg gtgtatctgg ggatcgcgcg tggtgcgctc gaaacagcgc gcgagtacac 3560 ccgaacgcag gcacgcccct ggacgccggc tggggttact caagccgtcg aggatccgta 3620 caccattcgc agctacggcg aattcggtat cgaactacag gctggcgatg ctgcagcacg 3680 tgaggctgcg cagctgctac agaccgcgtg ggacaaaggt gacgcattaa ctccacaaga 3740 acgcggcgaa ctcgcggtac aggtagccgg agtcaaagcc atagccacca aagcgggcct 3800 ggacgtgacc agccgaattt tcgaggtcat cggtgcccgg ggaacccacc caaactatgg 3860 attcgaccgg ttctggcgca acatccgcac ccacacgctg cacgatcccg tctcttacaa 3920 gatcgctgaa gtcggaaact acgtccttaa ccagcgctac ccgatacccg gtttcacttc 3980 ctgactccag gaccgcgctg ccgggcacgc gttaatgcct gagaacggtg g 4031 <210> 2<211> 453<212> PRT<213> Bacillus subtilis<400> 2Met Ala Glu Gln Arg Gln Leu His Leu Ala Gly Phe Phe Ser Ala Gly 1 5 10 15 Asn Val Thr His Ala His Gly Ala Trp Arg His Val Gly Ala Thr Asn 20 25 30 Gly Phe Leu Thr Gly Glu Phe Tyr Lys Gln Ile Ala Arg Thr Leu Glu 35 40 45 Arg Gly Lys Phe Asp Leu Leu Phe Leu Pro Asp Gly Leu Ala Ile Glu 50 55 60 Asp Ser Tyr Gly Asp Asn Leu Glu Thr Gly Val Gly Leu Gly Gly Gln 65 70 75 80 Gly Ala Val Ala Leu Glu Pro Thr Ser Val Ile Ala Thr Met Ala Ala 85 90 95 Val Thr Gln Arg Leu Gly Leu Gly Ala Thr Val Ser Thr Thr Tyr Tyr 100 105 110 Pro Pro Tyr His Val Ala Arg Val Phe Ala Thr Leu Asp Asn Leu Ser 115 120 125 Asp Gly Arg Ile Ser Trp Asn Val Val Thr Ser Leu Asn Asp Ser Glu 130 135 140 Ala Arg Asn Phe Gly Val Asp Glu His Leu Glu His Asp Ile Arg Tyr 145 150 155 160 Asp Arg Ala Asp Glu Phe Leu Glu Ala Val Lys Lys Leu Trp Ser Ser 165 170 175 Trp Ser Glu Asp Ala Leu Leu Leu Asp Lys Val Gly Gly Arg Phe Ala 180 185 190 Asp Pro Lys Lys Val Gln Tyr Val Asn His Arg Gly Arg Trp Leu Ser 195 200 205 Val Arg Gly Pro Leu Gln Val Pro Arg Ser Arg Gln Gly Glu Pro Val 210 215 220 Ile Leu Gln Ala Gly Leu Ser Pro Arg Gly Arg Arg Phe Ala Gly Arg 225 230 235 240 Trp Ala Glu Ala Val Phe Ser Val Ser Pro Asn Leu Asp Ile Met Arg 245 250 255 Ala Val Tyr Gln Asp Ile Lys Ala His Val Ala Ala Ala Gly Arg Asp 260 265 270 Pro Glu Gln Thr Lys Val Phe Thr Ala Val Met Pro Val Leu Gly Glu 275 280 285 Thr Glu Gln Val Ala Arg Glu Arg Leu Glu Tyr Leu Asn Ser Leu Val 290 295 300 His Pro Glu Val Gly Leu Ser Thr Leu Ser Ser His Ser Gly Leu Asn 305 310 315 320 Leu Ser Lys Tyr Pro Leu Asp Thr Lys Phe Ser Asp Ile Val Ala Asp 325 330 335 Leu Gly Asp Arg His Val Pro Thr Met Leu Gln Met Phe Ser Ala Val 340 345 350 Ala Gly Gly Gly Ala Asp Leu Thr Leu Ala Glu Leu Gly Arg Arg Tyr 355 360 365 Gly Thr Asn Val Gly Phe Val Pro Gln Trp Ala Gly Thr Ala Glu Gln 370 375 380 Ile Ala Asp Gln Leu Ile Ser His Phe Glu Ala Gly Ala Ala Asp Gly 385 390 395 400 Phe Ile Ile Ser Pro Ala Tyr Leu Pro Gly Ile Tyr Glu Glu Phe Val 405 410 415 Asp Gln Val Val Pro Leu Leu Gln Gln Arg Gly Val Phe Arg Thr Glu 420 425 430 Tyr Glu Gly Thr Thr Leu Arg Glu His Leu Gly Leu Ala His Pro Glu 435 440 445 Val Gly Ser Ile Arg 450 <210> 3<211> 4031<212> DNA<213> Bacillus subtilis<220> <221> CDS<222> (1670)..(2737)<400> 3ctagcgaggt gcgagttccg cgcggtagcc atccgcttca ttgattttta catcgatacc 60 gccgcggtgg ccgtcagggt gtcgaagccg ggcggaggtg cacaccccgc cggttggtga 120 tcttgctcac aggtggcgct ttgacggcac tcgtccgggg gcgagaatcg gcgaaggtcg 180 ggtgggtcgt cgggaggaag tggcgtgttg ggtgcccctt tgaatcatcg ggatcctaac 240 tctgttgggg tcggttgcag tagccgagca ttggtctccg gtaactgatc cccacggagg 300 cactgctgat tatggctgaa caacgccaac tacatttagc cgggtttttc tcggccggca 360 acgtcactca tgcgcatggc gcgtggcgcc atgtcggtgc gacgaacggt tttctgactg 420 gcgagttcta caagcagatt gcgcggaccc tggagcgggg caagtttgat ctgttgtttc 480 ttcccgatgg cctggccatc gaagatagct atggcgataa tttggagacg ggggtcggtc 540 tgggtggcca gggcgcggtg gcgctggagc cgaccagcgt gatagcgacg atggctgcgg 600 tgacccagcg attaggtctg ggcgctacgg tttcgacgac ctactacccg ccctatcatg 660 tggctcgggt gtttgccacg ctcgacaatc tgtctgatgg ccggatctcg tggaatgtgg 720 tcacgtcgct gaacgactcc gaggcacgca acttcggtgt ggatgagcac cttgagcatg 780 acatccgata cgaccgggct gacgaattct tggaggccgt caagaagcta tggagttctt 840 ggtccgagga tgccttgttg ttggacaagg ttggcggtcg gttcgctgat cccaagaagg 900 ttcaatacgt caatcatcgc ggccgctggt tgtcggtccg tggcccgttg caggttccac 960 ggtcccgtca gggtgaaccg gtcatcttgc aggccggctt gtcgccgcgg gggcgtcgct 1020 tcgcggggcg ctgggctgaa gcggtattca gcgtctcgcc caacctcgac atcatgcgcg 1080 cggtctatca ggacatcaaa gctcacgttg cggccgcggg gcgtgacccg gagcaaacaa 1140 aggtattcac cgcggtgatg ccggtgctcg gtgagacaga gcaggtggcc cgggaacgtc 1200 tggaatatct caattcgctg gtgcatcccg aagtgggttt gtcgacgcta tccagccaca 1260 gcggcttgaa tctctccaag tatccgctgg atacgaagtt ttccgacatc gtcgccgatc 1320 tcggggatcg tcacgtgccg accatgttgc agatgttttc cgccgtggcc ggtggcggtg 1380 cggacctgac gctggccgaa ctgggccgcc gatacggcac caacgtcggg tttgtgccgc 1440 agtgggccgg aactgccgaa cagatcgctg atcaattgat tagtcacttt gaggccgggg 1500 ccgctgacgg attcatcatt tcgccggcgt atctgccggg tatttacgag gagttcgtcg 1560 accaggtggt tccgctgctg cagcaacgtg gtgtgttccg taccgagtac gaagggacaa 1620 cgctgcgtga gcaccttggg ttggctcacc ctgaagtcgg gagcatccg atg acc acc 1678 Met Thr Thr 1 acc gat gtt gat cat gac gtt ctt gcc tac agc aac tgc ccg gtt ccc 1726 Thr Asp Val Asp His Asp Val Leu Ala Tyr Ser Asn Cys Pro Val Pro 5 10 15 aat gca cta ctg aca gcg ctg gag tca aac ctg ttg gcc ggc aac ggg 1774 Asn Ala Leu Leu Thr Ala Leu Glu Ser Asn Leu Leu Ala Gly Asn Gly 20 25 30 35att tcg ctg aat gtg ctc agc ggc gcg caa gca ggg ttg cat ttc acc 1822 Ile Ser Leu Asn Val Leu Ser Gly Ala Gln Ala Gly Leu His Phe Thr 40 45 50 tac gat cat ccc gct tac acc cgc ttt ggt ggc gaa att cca cct ctt 1870 Tyr Asp His Pro Ala Tyr Thr Arg Phe Gly Gly Glu Ile Pro Pro Leu 55 60 65 atc agt gag ggc ctg cgg gcg ccg gga cgc acc cgc ctg cta ggc atc 1918 Ile Ser Glu Gly Leu Arg Ala Pro Gly Arg Thr Arg Leu Leu Gly Ile 70 75 80 acc cct ctg gcc ggt cgg caa ggg atc tat gtc cgt gtt gac agc ccg 1966 Thr Pro Leu Ala Gly Arg Gln Gly Ile Tyr Val Arg Val Asp Ser Pro 85 90 95 gtg aca tca ccg gag cag ctg cgg ggc cgg cgg gtg ggt gtg tcg ggt 2014 Val Thr Ser Pro Glu Gln Leu Arg Gly Arg Arg Val Gly Val Ser Gly100 105 110 115gca gcg atc cgc atc ctc acc ggc gag ttg ggc gat tat cgg cac ttg 2062 Ala Ala Ile Arg Ile Leu Thr Gly Glu Leu Gly Asp Tyr Arg His Leu 120 125 130 gac ccg tgg cgg caa aca ctg atc gcg ctg ggt acc tgg gag gcg cgg 2110 Asp Pro Trp Arg Gln Thr Leu Ile Ala Leu Gly Thr Trp Glu Ala Arg 135 140 145 gga ctt ttg cag acc ctg cac atc ggg ggg ata ggg gtc gac gac gtc 2158 Gly Leu Leu Gln Thr Leu His Ile Gly Gly Ile Gly Val Asp Asp Val 150 155 160 gag ttg gtg cgt atc gaa agc cct ggc gtc gac gtg ccc gaa gag cgg 2206 Glu Leu Val Arg Ile Glu Ser Pro Gly Val Asp Val Pro Glu Glu Arg 165 170 175 ctg gaa gca gcg gcc agt gtc aaa ggc gct gac ctg ttt ccc gac gtg 2254 Leu Glu Ala Ala Ala Ser Val Lys Gly Ala Asp Leu Phe Pro Asp Val180 185 190 195gcc ggc cac cag agg gac atc ctg gat agc ggc aac gtt gat gcg ctg 2302 Ala Gly His Gln Arg Asp Ile Leu Asp Ser Gly Asn Val Asp Ala Leu 200 205 210 ttc acc tgg ttg ccc tgg gcg gcg gag tta gaa gat ctc agc gga gcg 2350 Phe Thr Trp Leu Pro Trp Ala Ala Glu Leu Glu Asp Leu Ser Gly Ala 215 220 225 cgg gtg ctc gcc gat ctc ggc gat gac caa cga agc cgg tat gcc agt 2398 Arg Val Leu Ala Asp Leu Gly Asp Asp Gln Arg Ser Arg Tyr Ala Ser 230 235 240 gtc tgg acg gtc agc gct cag ctg gtt gac gag cga ccc gac ctg gtg 2446 Val Trp Thr Val Ser Ala Gln Leu Val Asp Glu Arg Pro Asp Leu Val 245 250 255 caa cga ctc gtc gac gcg gcg gtc caa gcc agc cgt tgg gca cag gcc 2494 Gln Arg Leu Val Asp Ala Ala Val Gln Ala Ser Arg Trp Ala Gln Ala260 265 270 275cat ccc gag gaa acc gtt ggc atc cat gcc gcc aac ctc ggg gtc gcg 2542 His Pro Glu Glu Thr Val Gly Ile His Ala Ala Asn Leu Gly Val Ala 280 285 290 ccc tcg gcg atc gga cgc ggt ttc ggc gct gat ttc gcc caa cat ctg 2590 Pro Ser Ala Ile Gly Arg Gly Phe Gly Ala Asp Phe Ala Gln His Leu 295 300 305 atc cca cgg ctt gac gac tcg gca ctg gcc gtt gtc gac caa act caa 2638 Ile Pro Arg Leu Asp Asp Ser Ala Leu Ala Val Val Asp Gln Thr Gln 310 315 320 cag ttc ctc ctc gac cac aat ctc ctc gac cat ccg gtg gac ctc acg 2686 Gln Phe Leu Leu Asp His Asn Leu Leu Asp His Pro Val Asp Leu Thr 325 330 335 cag tgg gcg gca ccg caa ttc ctc act caa tcc acg act gga gac cag 2734 Gln Trp Ala Ala Pro Gln Phe Leu Thr Gln Ser Thr Thr Gly Asp Gln340 345 350 355caa tgaccctgac cgatgacacc gccacggcac agaacaatag gaatggcgac 2787 Gln cccatcgagg tggcgcgcga gctgacgcga aagtggcaag ctaccgtcgt cgagcgcgac 2847 aaggctggtg gttcggcgac agaagagcgt gaggatctac gcgccagtgg gctgctctcg 2907 gtgaccgttc cccggcacct gggcggttgg ggagccgact ggcctaccgc tctggaagtg 2967 gtgcgtgaga tcgccaaggt cgatggatcg ctgggccact tgttcggata ccacctcagc 3027 accccagccg ttattgatct gtggggttca cccgagcaaa gtgaacgcct acttcgtcaa 3087 ctggccgaaa acaactggtg gaccgggaac gcctccagcg agaacaacag ccacatcctc 3147 gaatggaagg taactgccac cccgaccgac gacggcgggt accttctcaa cggtatcaag 3207 cacttcagca gtggagccaa gggctcagat ctgctcttgg tgttcggcgt gataccggag 3267 ggttccccac agcaaggcgc catcgttgcc gcggccattc ccaccaccag agaaggcgtt 3327 caaacaaacg acgactggca ggcgctaggg atgcggcgca ccgacagcgg caccaccgaa 3387 ttccacaacg tcgtagtccg tcccgacgag gtgctgggca aacccaatgc cgttgtcgag 3447 gcgttcctgg cgtccgggcg cggcagccta ttcggcccga tcgtgcagtt ggtgttcgca 3507 acggtgtatc tggggatcgc gcgtggtgcg ctcgaaacag cgcgcgagta cacccgaacg 3567 caggcacgcc cctggacgcc ggctggggtt actcaagccg tcgaggatcc gtacaccatt 3627 cgcagctacg gcgaattcgg tatcgaacta caggctggcg atgctgcagc acgtgaggct 3687 gcgcagctgc tacagaccgc gtgggacaaa ggtgacgcat taactccaca agaacgcggc 3747 gaactcgcgg tacaggtagc cggagtcaaa gccatagcca ccaaagcggg cctggacgtg 3807 accagccgaa ttttcgaggt catcggtgcc cggggaaccc acccaaacta tggattcgac 3867 cggttctggc gcaacatccg cacccacacg ctgcacgatc ccgtctctta caagatcgct 3927 gaagtcggaa actacgtcct taaccagcgc tacccgatac ccggtttcac ttcctgactc 3987 caggaccgcg ctgccgggca cgcgttaatg cctgagaacg gtgg 4031 <210> 4<211> 356<212> PRT<213> Bacillus subtilis<400> 4Met Thr Thr Thr Asp Val Asp His Asp Val Leu Ala Tyr Ser Asn Cys 1 5 10 15 Pro Val Pro Asn Ala Leu Leu Thr Ala Leu Glu Ser Asn Leu Leu Ala 20 25 30 Gly Asn Gly Ile Ser Leu Asn Val Leu Ser Gly Ala Gln Ala Gly Leu 35 40 45 His Phe Thr Tyr Asp His Pro Ala Tyr Thr Arg Phe Gly Gly Glu Ile 50 55 60 Pro Pro Leu Ile Ser Glu Gly Leu Arg Ala Pro Gly Arg Thr Arg Leu 65 70 75 80 Leu Gly Ile Thr Pro Leu Ala Gly Arg Gln Gly Ile Tyr Val Arg Val 85 90 95 Asp Ser Pro Val Thr Ser Pro Glu Gln Leu Arg Gly Arg Arg Val Gly 100 105 110 Val Ser Gly Ala Ala Ile Arg Ile Leu Thr Gly Glu Leu Gly Asp Tyr 115 120 125 Arg His Leu Asp Pro Trp Arg Gln Thr Leu Ile Ala Leu Gly Thr Trp 130 135 140 Glu Ala Arg Gly Leu Leu Gln Thr Leu His Ile Gly Gly Ile Gly Val 145 150 155 160 Asp Asp Val Glu Leu Val Arg Ile Glu Ser Pro Gly Val Asp Val Pro 165 170 175 Glu Glu Arg Leu Glu Ala Ala Ala Ser Val Lys Gly Ala Asp Leu Phe 180 185 190 Pro Asp Val Ala Gly His Gln Arg Asp Ile Leu Asp Ser Gly Asn Val 195 200 205 Asp Ala Leu Phe Thr Trp Leu Pro Trp Ala Ala Glu Leu Glu Asp Leu 210 215 220 Ser Gly Ala Arg Val Leu Ala Asp Leu Gly Asp Asp Gln Arg Ser Arg 225 230 235 240 Tyr Ala Ser Val Trp Thr Val Ser Ala Gln Leu Val Asp Glu Arg Pro 245 250 255 Asp Leu Val Gln Arg Leu Val Asp Ala Ala Val Gln Ala Ser Arg Trp 260 265 270 Ala Gln Ala His Pro Glu Glu Thr Val Gly Ile His Ala Ala Asn Leu 275 280 285 Gly Val Ala Pro Ser Ala Ile Gly Arg Gly Phe Gly Ala Asp Phe Ala 290 295 300 Gln His Leu Ile Pro Arg Leu Asp Asp Ser Ala Leu Ala Val Val Asp 305 310 315 320 Gln Thr Gln Gln Phe Leu Leu Asp His Asn Leu Leu Asp His Pro Val 325 330 335 Asp Leu Thr Gln Trp Ala Ala Pro Gln Phe Leu Thr Gln Ser Thr Thr 340 345 350 Gly Asp Gln Gln 355 <210> 5<211> 4031<212> DNA<213> Bacillus subtilis<220> <221> CDS<222> (2737)..(3981)<400> 5ctagcgaggt gcgagttccg cgcggtagcc atccgcttca ttgattttta catcgatacc 60 gccgcggtgg ccgtcagggt gtcgaagccg ggcggaggtg cacaccccgc cggttggtga 120 tcttgctcac aggtggcgct ttgacggcac tcgtccgggg gcgagaatcg gcgaaggtcg 180 ggtgggtcgt cgggaggaag tggcgtgttg ggtgcccctt tgaatcatcg ggatcctaac 240 tctgttgggg tcggttgcag tagccgagca ttggtctccg gtaactgatc cccacggagg 300 cactgctgat tatggctgaa caacgccaac tacatttagc cgggtttttc tcggccggca 360 acgtcactca tgcgcatggc gcgtggcgcc atgtcggtgc gacgaacggt tttctgactg 420 gcgagttcta caagcagatt gcgcggaccc tggagcgggg caagtttgat ctgttgtttc 480 ttcccgatgg cctggccatc gaagatagct atggcgataa tttggagacg ggggtcggtc 540 tgggtggcca gggcgcggtg gcgctggagc cgaccagcgt gatagcgacg atggctgcgg 600 tgacccagcg attaggtctg ggcgctacgg tttcgacgac ctactacccg ccctatcatg 660 tggctcgggt gtttgccacg ctcgacaatc tgtctgatgg ccggatctcg tggaatgtgg 720 tcacgtcgct gaacgactcc gaggcacgca acttcggtgt ggatgagcac cttgagcatg 780 acatccgata cgaccgggct gacgaattct tggaggccgt caagaagcta tggagttctt 840 ggtccgagga tgccttgttg ttggacaagg ttggcggtcg gttcgctgat cccaagaagg 900 ttcaatacgt caatcatcgc ggccgctggt tgtcggtccg tggcccgttg caggttccac 960 ggtcccgtca gggtgaaccg gtcatcttgc aggccggctt gtcgccgcgg gggcgtcgct 1020 tcgcggggcg ctgggctgaa gcggtattca gcgtctcgcc caacctcgac atcatgcgcg 1080 cggtctatca ggacatcaaa gctcacgttg cggccgcggg gcgtgacccg gagcaaacaa 1140 aggtattcac cgcggtgatg ccggtgctcg gtgagacaga gcaggtggcc cgggaacgtc 1200 tggaatatct caattcgctg gtgcatcccg aagtgggttt gtcgacgcta tccagccaca 1260 gcggcttgaa tctctccaag tatccgctgg atacgaagtt ttccgacatc gtcgccgatc 1320 tcggggatcg tcacgtgccg accatgttgc agatgttttc cgccgtggcc ggtggcggtg 1380 cggacctgac gctggccgaa ctgggccgcc gatacggcac caacgtcggg tttgtgccgc 1440 agtgggccgg aactgccgaa cagatcgctg atcaattgat tagtcacttt gaggccgggg 1500 ccgctgacgg attcatcatt tcgccggcgt atctgccggg tatttacgag gagttcgtcg 1560 accaggtggt tccgctgctg cagcaacgtg gtgtgttccg taccgagtac gaagggacaa 1620 cgctgcgtga gcaccttggg ttggctcacc ctgaagtcgg gagcatccga tgaccaccac 1680 cgatgttgat catgacgttc ttgcctacag caactgcccg gttcccaatg cactactgac 1740 agcgctggag tcaaacctgt tggccggcaa cgggatttcg ctgaatgtgc tcagcggcgc 1800 gcaagcaggg ttgcatttca cctacgatca tcccgcttac acccgctttg gtggcgaaat 1860 tccacctctt atcagtgagg gcctgcgggc gccgggacgc acccgcctgc taggcatcac 1920 ccctctggcc ggtcggcaag ggatctatgt ccgtgttgac agcccggtga catcaccgga 1980 gcagctgcgg ggccggcggg tgggtgtgtc gggtgcagcg atccgcatcc tcaccggcga 2040 gttgggcgat tatcggcact tggacccgtg gcggcaaaca ctgatcgcgc tgggtacctg 2100 ggaggcgcgg ggacttttgc agaccctgca catcgggggg ataggggtcg acgacgtcga 2160 gttggtgcgt atcgaaagcc ctggcgtcga cgtgcccgaa gagcggctgg aagcagcggc 2220 cagtgtcaaa ggcgctgacc tgtttcccga cgtggccggc caccagaggg acatcctgga 2280 tagcggcaac gttgatgcgc tgttcacctg gttgccctgg gcggcggagt tagaagatct 2340 cagcggagcg cgggtgctcg ccgatctcgg cgatgaccaa cgaagccggt atgccagtgt 2400 ctggacggtc agcgctcagc tggttgacga gcgacccgac ctggtgcaac gactcgtcga 2460 cgcggcggtc caagccagcc gttgggcaca ggcccatccc gaggaaaccg ttggcatcca 2520 tgccgccaac ctcggggtcg cgccctcggc gatcggacgc ggtttcggcg ctgatttcgc 2580 ccaacatctg atcccacggc ttgacgactc ggcactggcc gttgtcgacc aaactcaaca 2640 gttcctcctc gaccacaatc tcctcgacca tccggtggac ctcacgcagt gggcggcacc 2700 gcaattcctc actcaatcca cgactggaga ccagca atg acc ctg acc gat gac 2754 Met Thr Leu Thr Asp Asp 1 5 acc gcc acg gca cag aac aat agg aat ggc gac ccc atc gag gtg gcg 2802 Thr Ala Thr Ala Gln Asn Asn Arg Asn Gly Asp Pro Ile Glu Val Ala 10 15 20 cgc gag ctg acg cga aag tgg caa gct acc gtc gtc gag cgc gac aag 2850 Arg Glu Leu Thr Arg Lys Trp Gln Ala Thr Val Val Glu Arg Asp Lys 25 30 35 gct ggt ggt tcg gcg aca gaa gag cgt gag gat cta cgc gcc agt ggg 2898 Ala Gly Gly Ser Ala Thr Glu Glu Arg Glu Asp Leu Arg Ala Ser Gly 40 45 50 ctg ctc tcg gtg acc gtt ccc cgg cac ctg ggc ggt tgg gga gcc gac 2946 Leu Leu Ser Val Thr Val Pro Arg His Leu Gly Gly Trp Gly Ala Asp 55 60 65 70tgg cct acc gct ctg gaa gtg gtg cgt gag atc gcc aag gtc gat gga 2994 Trp Pro Thr Ala Leu Glu Val Val Arg Glu Ile Ala Lys Val Asp Gly 75 80 85 tcg ctg ggc cac ttg ttc gga tac cac ctc agc acc cca gcc gtt att 3042 Ser Leu Gly His Leu Phe Gly Tyr His Leu Ser Thr Pro Ala Val Ile 90 95 100 gat ctg tgg ggt tca ccc gag caa agt gaa cgc cta ctt cgt caa ctg 3090 Asp Leu Trp Gly Ser Pro Glu Gln Ser Glu Arg Leu Leu Arg Gln Leu 105 110 115 gcc gaa aac aac tgg tgg acc ggg aac gcc tcc agc gag aac aac agc 3138 Ala Glu Asn Asn Trp Trp Thr Gly Asn Ala Ser Ser Glu Asn Asn Ser 120 125 130 cac atc ctc gaa tgg aag gta act gcc acc ccg acc gac gac ggc ggg 3186 His Ile Leu Glu Trp Lys Val Thr Ala Thr Pro Thr Asp Asp Gly Gly135 140 145 150tac ctt ctc aac ggt atc aag cac ttc agc agt gga gcc aag ggc tca 3234 Tyr Leu Leu Asn Gly Ile Lys His Phe Ser Ser Gly Ala Lys Gly Ser 155 160 165 gat ctg ctc ttg gtg ttc ggc gtg ata ccg gag ggt tcc cca cag caa 3282 Asp Leu Leu Leu Val Phe Gly Val Ile Pro Glu Gly Ser Pro Gln Gln 170 175 180 ggc gcc atc gtt gcc gcg gcc att ccc acc acc aga gaa ggc gtt caa 3330 Gly Ala Ile Val Ala Ala Ala Ile Pro Thr Thr Arg Glu Gly Val Gln 185 190 195 aca aac gac gac tgg cag gcg cta ggg atg cgg cgc acc gac agc ggc 3378 Thr Asn Asp Asp Trp Gln Ala Leu Gly Met Arg Arg Thr Asp Ser Gly 200 205 210 acc acc gaa ttc cac aac gtc gta gtc cgt ccc gac gag gtg ctg ggc 3426 Thr Thr Glu Phe His Asn Val Val Val Arg Pro Asp Glu Val Leu Gly215 220 225 230aaa ccc aat gcc gtt gtc gag gcg ttc ctg gcg tcc ggg cgc ggc agc 3474 Lys Pro Asn Ala Val Val Glu Ala Phe Leu Ala Ser Gly Arg Gly Ser 235 240 245 cta ttc ggc ccg atc gtg cag ttg gtg ttc gca acg gtg tat ctg ggg 3522 Leu Phe Gly Pro Ile Val Gln Leu Val Phe Ala Thr Val Tyr Leu Gly 250 255 260 atc gcg cgt ggt gcg ctc gaa aca gcg cgc gag tac acc cga acg cag 3570 Ile Ala Arg Gly Ala Leu Glu Thr Ala Arg Glu Tyr Thr Arg Thr Gln 265 270 275 gca cgc ccc tgg acg ccg gct ggg gtt act caa gcc gtc gag gat ccg 3618 Ala Arg Pro Trp Thr Pro Ala Gly Val Thr Gln Ala Val Glu Asp Pro 280 285 290 tac acc att cgc agc tac ggc gaa ttc ggt atc gaa cta cag gct ggc 3666 Tyr Thr Ile Arg Ser Tyr Gly Glu Phe Gly Ile Glu Leu Gln Ala Gly295 300 305 310gat gct gca gca cgt gag gct gcg cag ctg cta cag acc gcg tgg gac 3714 Asp Ala Ala Ala Arg Glu Ala Ala Gln Leu Leu Gln Thr Ala Trp Asp 315 320 325 aaa ggt gac gca tta act cca caa gaa cgc ggc gaa ctc gcg gta cag 3762 Lys Gly Asp Ala Leu Thr Pro Gln Glu Arg Gly Glu Leu Ala Val Gln 330 335 340 gta gcc gga gtc aaa gcc ata gcc acc aaa gcg ggc ctg gac gtg acc 3810 Val Ala Gly Val Lys Ala Ile Ala Thr Lys Ala Gly Leu Asp Val Thr 345 350 355 agc cga att ttc gag gtc atc ggt gcc cgg gga acc cac cca aac tat 3858 Ser Arg Ile Phe Glu Val Ile Gly Ala Arg Gly Thr His Pro Asn Tyr 360 365 370 gga ttc gac cgg ttc tgg cgc aac atc cgc acc cac acg ctg cac gat 3906 Gly Phe Asp Arg Phe Trp Arg Asn Ile Arg Thr His Thr Leu His Asp375 380 385 390ccc gtc tct tac aag atc gct gaa gtc gga aac tac gtc ctt aac cag 3954 Pro Val Ser Tyr Lys Ile Ala Glu Val Gly Asn Tyr Val Leu Asn Gln 395 400 405 cgc tac ccg ata ccc ggt ttc act tcc tgactccagg accgcgctgc 4001 Arg Tyr Pro Ile Pro Gly Phe Thr Ser 410 415 cgggcacgcg ttaatgcctg agaacggtgg 4031 <210> 6<211> 415<212> PRT<213> Bacillus subtilis<400> 6Met Thr Leu Thr Asp Asp Thr Ala Thr Ala Gln Asn Asn Arg Asn Gly 1 5 10 15 Asp Pro Ile Glu Val Ala Arg Glu Leu Thr Arg Lys Trp Gln Ala Thr 20 25 30 Val Val Glu Arg Asp Lys Ala Gly Gly Ser Ala Thr Glu Glu Arg Glu 35 40 45 Asp Leu Arg Ala Ser Gly Leu Leu Ser Val Thr Val Pro Arg His Leu 50 55 60 Gly Gly Trp Gly Ala Asp Trp Pro Thr Ala Leu Glu Val Val Arg Glu 65 70 75 80 Ile Ala Lys Val Asp Gly Ser Leu Gly His Leu Phe Gly Tyr His Leu 85 90 95 Ser Thr Pro Ala Val Ile Asp Leu Trp Gly Ser Pro Glu Gln Ser Glu 100 105 110 Arg Leu Leu Arg Gln Leu Ala Glu Asn Asn Trp Trp Thr Gly Asn Ala 115 120 125 Ser Ser Glu Asn Asn Ser His Ile Leu Glu Trp Lys Val Thr Ala Thr 130 135 140 Pro Thr Asp Asp Gly Gly Tyr Leu Leu Asn Gly Ile Lys His Phe Ser 145 150 155 160 Ser Gly Ala Lys Gly Ser Asp Leu Leu Leu Val Phe Gly Val Ile Pro 165 170 175 Glu Gly Ser Pro Gln Gln Gly Ala Ile Val Ala Ala Ala Ile Pro Thr 180 185 190 Thr Arg Glu Gly Val Gln Thr Asn Asp Asp Trp Gln Ala Leu Gly Met 195 200 205 Arg Arg Thr Asp Ser Gly Thr Thr Glu Phe His Asn Val Val Val Arg 210 215 220 Pro Asp Glu Val Leu Gly Lys Pro Asn Ala Val Val Glu Ala Phe Leu 225 230 235 240 Ala Ser Gly Arg Gly Ser Leu Phe Gly Pro Ile Val Gln Leu Val Phe 245 250 255 Ala Thr Val Tyr Leu Gly Ile Ala Arg Gly Ala Leu Glu Thr Ala Arg 260 265 270 Glu Tyr Thr Arg Thr Gln Ala Arg Pro Trp Thr Pro Ala Gly Val Thr 275 280 285 Gln Ala Val Glu Asp Pro Tyr Thr Ile Arg Ser Tyr Gly Glu Phe Gly 290 295 300 Ile Glu Leu Gln Ala Gly Asp Ala Ala Ala Arg Glu Ala Ala Gln Leu 305 310 315 320 Leu Gln Thr Ala Trp Asp Lys Gly Asp Ala Leu Thr Pro Gln Glu Arg 325 330 335 Gly Glu Leu Ala Val Gln Val Ala Gly Val Lys Ala Ile Ala Thr Lys 340 345 350 Ala Gly Leu Asp Val Thr Ser Arg Ile Phe Glu Val Ile Gly Ala Arg 355 360 365 Gly Thr His Pro Asn Tyr Gly Phe Asp Arg Phe Trp Arg Asn Ile Arg 370 375 380 Thr His Thr Leu His Asp Pro Val Ser Tyr Lys Ile Ala Glu Val Gly 385 390 395 400 Asn Tyr Val Leu Asn Gln Arg Tyr Pro Ile Pro Gly Phe Thr Ser 405 410 415 <210> 7<211> 16<212> PRT<213> Paenibacillus sp.<400> 7Trp Asn Val Val Thr Ser Leu Asn Asn Ala Glu Ala Arg Asn Phe Gly 1 5 10 15 <210> 8<211> 16<212> PRT<213> Rhodococcus sp. IGTS8<400> 8Trp Asn Val Val Thr Ser Leu Asn Asp Ala Glu Ala Arg Asn Phe Gly 1 5 10 15 <210> 9<211> 18<212> PRT<213> Paenibacillus sp.<400> 9Gly Phe Asp Arg Phe Trp Arg Asp Ala Arg Thr His Thr Leu His Asp 1 5 10 15 Pro Val <210> 10<211> 18<212> PRT<213> Rhodococcus sp. IGTS8<400> 10Gly Phe Asp Arg Phe Trp Arg Asn Val Arg Thr His Ser Leu His Asp 1 5 10 15 Pro Val <210> 11<211> 20<212> DNA<213> Artificial Sequence<220> <223> Description of Artificial Sequence: sense primer for PCR<220><221> modified base<222> n = inosine<223> (3),(9) and (12)<400> 11gcngargcnm gnaayttygg 20 <210> 12<211> 19<212> DNA<213> Artificial Sequence<220> <223> Description of Artificial Sequence: antisense primer for PCR<220><221> modified base<222> n = inosine<223> (4) and (11)<400> 12cgtngcgcca naagcggtc 19 <210> 13<211> 17<212> DNA<213> Artificial Sequence<220> <223> Description of Artificial Sequence: sense primer for PCR<400> 13gcatgacatc cgatacg 17 <210> 14<211> 17<212> DNA<213> Artificial Sequence<220> <223> Description of Artificial Sequence: antisense primer for PCR<400> 14tagtttgggt gggttcc 17
【0080】
【配列表フリーテキスト】配列番号1−bdsABC遺伝子を含むヌクレオチド配列配列番号2−bdsA遺伝子によってコードされるタンパク質BdsAのアミノ酸配列配列番号3−bdsABC遺伝子を含むヌクレオチド配列配列番号4−bdsB遺伝子によってコードされるタンパク質BdsBのアミノ酸配列配列番号5−bdsABC遺伝子を含むヌクレオチド配列配列番号6−bdsC遺伝子によってコードされるタンパク質BdsCのアミノ酸配列配列番号7−TdsAの部分アミノ酸配列配列番号8−DszAの部分アミノ酸配列配列番号9−TdsCの部分アミノ酸配列配列番号10−DszCの部分アミノ酸配列配列番号11−人工配列の説明:センスプライマー(3、9及び12番目のNはデオキシイノシンを表す)配列番号12−人工配列の説明:アンチセンスプライマー(4及び11番目のNはデオキシイノシンを表す)配列番号13−人工配列の説明:センスプライマー配列番号14−人工配列の説明:アンチセンスプライマー
【図面の簡単な説明】
【図1】脱硫遺伝子bdsABCを含む発現プラスミドpU1Nhの制限酵素地図を示す。
【図2】BdsC酵素活性の温度依存性を示す。
【図3】BdsC酵素活性の温度安定性を示す。
【図4】BdsC酵素活性のpH依存性を示す。
【図5】BdsC酵素活性のpH安定性を示す。
【図6】BdsA酵素活性の温度依存性を示す。
【図7】BdsA酵素活性の温度安定性を示す。
【図8】BdsA酵素活性のpH依存性を示す。
【図9】BdsA酵素活性のpH安定性を示す。
【特許請求の範囲】
【請求項1】 以下の(a) 又は(b) のタンパク質をコードする遺伝子。
(a) 配列番号2記載のアミノ酸配列により表されるタンパク質(b) 配列番号2記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつジベンゾチオフェンスルホンを2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸に変換する機能を有するタンパク質
【請求項2】 以下の(a) 又は(b) のタンパク質をコードする遺伝子。
(a) 配列番号4記載のアミノ酸配列により表されるタンパク質(b) 配列番号4記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を2-ヒドロキシビフェニルに変換する機能を有するタンパク質
【請求項3】 以下の(a) 又は(b) のタンパク質をコードする遺伝子。
(a) 配列番号6記載のアミノ酸配列により表されるタンパク質(b) 配列番号6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつジベンゾチオフェンをジベンゾチオフェンスルホンに変換する機能を有するタンパク質
【請求項4】 請求項1、2又は3に記載の遺伝子を含むベクター。
【請求項5】 請求項4記載のベクターを含有する形質転換体。
【請求項6】 以下の(a) 又は(b) に示すタンパク質。
(a) 配列番号2記載のアミノ酸配列により表されるタンパク質(b) 配列番号2記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつジベンゾチオフェンスルホンを2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸に変換する機能を有するタンパク質
【請求項7】 以下の(a) 又は(b) に示すタンパク質。
(a) 配列番号4記載のアミノ酸配列により表されるタンパク質(b) 配列番号4記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を2-ヒドロキシビフェニルに変換する機能を有するタンパク質
【請求項8】 以下の(a) 又は(b) に示すタンパク質。
(a) 配列番号6記載のアミノ酸配列により表されるタンパク質(b) 配列番号6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつジベンゾチオフェンをジベンゾチオフェンスルホンに変換する機能を有するタンパク質
【請求項9】 以下の性質を有するタンパク質。
(1) 作用:ジベンゾチオフェンをジベンゾチオフェンスルホンにする(2) 至適pH:8.0 、安定pH:8〜11(3) 至適温度:45〜50℃、安定温度:53℃以下(4) 分子量:200,000(ゲル濾過法による)
(5) 活性阻害:キレート剤、SH阻害剤によって阻害される(6) 補酵素の要求性:フラビンレダクターゼによって供給される FMNH2 を要求する
【請求項10】 以下の性質を有するタンパク質。
(1) 作用:ジベンゾチオフェンスルホンを2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸にする(2) 至適pH:7.5、安定pH:6〜10(3) 至適温度:50℃、安定温度:45℃以下(4) 分子量:174,000(ゲル濾過法による)
(5) 活性阻害:キレート剤、SH阻害剤によって阻害される(6) 補酵素の要求性:フラビンレダクターゼによって供給される FMNH2 を要求する
【請求項1】 以下の(a) 又は(b) のタンパク質をコードする遺伝子。
(a) 配列番号2記載のアミノ酸配列により表されるタンパク質(b) 配列番号2記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつジベンゾチオフェンスルホンを2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸に変換する機能を有するタンパク質
【請求項2】 以下の(a) 又は(b) のタンパク質をコードする遺伝子。
(a) 配列番号4記載のアミノ酸配列により表されるタンパク質(b) 配列番号4記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を2-ヒドロキシビフェニルに変換する機能を有するタンパク質
【請求項3】 以下の(a) 又は(b) のタンパク質をコードする遺伝子。
(a) 配列番号6記載のアミノ酸配列により表されるタンパク質(b) 配列番号6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつジベンゾチオフェンをジベンゾチオフェンスルホンに変換する機能を有するタンパク質
【請求項4】 請求項1、2又は3に記載の遺伝子を含むベクター。
【請求項5】 請求項4記載のベクターを含有する形質転換体。
【請求項6】 以下の(a) 又は(b) に示すタンパク質。
(a) 配列番号2記載のアミノ酸配列により表されるタンパク質(b) 配列番号2記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつジベンゾチオフェンスルホンを2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸に変換する機能を有するタンパク質
【請求項7】 以下の(a) 又は(b) に示すタンパク質。
(a) 配列番号4記載のアミノ酸配列により表されるタンパク質(b) 配列番号4記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつ2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸を2-ヒドロキシビフェニルに変換する機能を有するタンパク質
【請求項8】 以下の(a) 又は(b) に示すタンパク質。
(a) 配列番号6記載のアミノ酸配列により表されるタンパク質(b) 配列番号6記載のアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつジベンゾチオフェンをジベンゾチオフェンスルホンに変換する機能を有するタンパク質
【請求項9】 以下の性質を有するタンパク質。
(1) 作用:ジベンゾチオフェンをジベンゾチオフェンスルホンにする(2) 至適pH:8.0 、安定pH:8〜11(3) 至適温度:45〜50℃、安定温度:53℃以下(4) 分子量:200,000(ゲル濾過法による)
(5) 活性阻害:キレート剤、SH阻害剤によって阻害される(6) 補酵素の要求性:フラビンレダクターゼによって供給される FMNH2 を要求する
【請求項10】 以下の性質を有するタンパク質。
(1) 作用:ジベンゾチオフェンスルホンを2-(2'-ヒドロキシフェニル)ベンゼンスルフィン酸にする(2) 至適pH:7.5、安定pH:6〜10(3) 至適温度:50℃、安定温度:45℃以下(4) 分子量:174,000(ゲル濾過法による)
(5) 活性阻害:キレート剤、SH阻害剤によって阻害される(6) 補酵素の要求性:フラビンレダクターゼによって供給される FMNH2 を要求する
【図1】

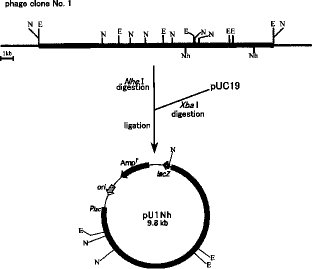
【図2】

【図3】

【図4】
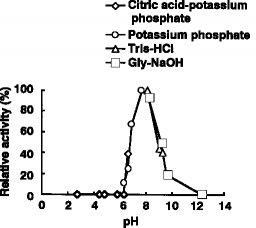
【図6】

【図7】

【図5】
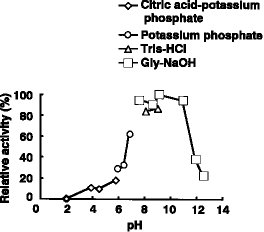
【図8】
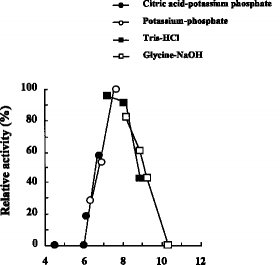
【図9】
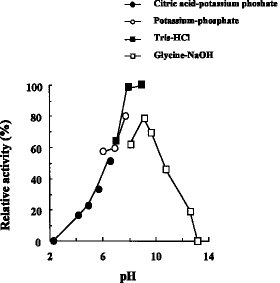
【図2】
【図3】
【図4】
【図6】
【図7】
【図5】
【図8】
【図9】
【公開番号】特開2002−253247(P2002−253247A)
【公開日】平成14年9月10日(2002.9.10)
【国際特許分類】
【出願番号】特願2001−55379(P2001−55379)
【出願日】平成13年2月28日(2001.2.28)
【出願人】(590000455)財団法人石油産業活性化センター (249)
【Fターム(参考)】
【公開日】平成14年9月10日(2002.9.10)
【国際特許分類】
【出願日】平成13年2月28日(2001.2.28)
【出願人】(590000455)財団法人石油産業活性化センター (249)
【Fターム(参考)】
[ Back to top ]