
腸管毒素原性大腸菌の定着因子(CF)抗原を発現するためのハイブリッドオペロン
遺伝子集合体を含む組み換えオペロンを開示し、ここで、腸管毒素原性大腸菌(ETEC)に関連した定着因子抗原(CFs)の少なくとも2つの主要サブユニットをコードする少なくとも2つの構造遺伝子が存在する。更に、かかる組み換えオペロンを含むように遺伝子操作された、大腸菌細胞等の宿主細胞を開示し、ここで、前記オペロンが、プラスミド等のエピソームエレメント上に位置するか又は前記宿主細胞の染色体に組み込まれている。また、腸管毒素原性大腸菌(ETEC)に関連した定着因子抗原(CFs)の少なくとも2つの主要サブユニットを前記オペロンから発現することが可能な宿主細胞の製造方法を開示する。加えて、薬学的に容認される賦形剤、緩衝剤及び/又は希釈剤と共に、かかる宿主細胞を少なくとも一つ含む抗下痢症ワクチン組成物を開示する。最後に、かかるワクチンの製造における前記オペロンの使用を開示する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、腸管毒素原性大腸菌の定着因子(CF)抗原を発現するためのハイブリッドオペロンに関し、特には、異なる2つのCFsを発現するための少なくとも2つの構造遺伝子を含む遺伝子を備える組み換えオペロンに関するものである。更に、本発明は、かかるオペロンを宿主細胞のゲノム上に又は宿主細胞に組み込まれたプラスミド中に備えた、大腸菌等の宿主細胞に関するものである。
【背景技術】
【0002】
腸管毒素原性大腸菌(ETEC)は、旅行者下痢症、並びに世界各地の流行地域における小児の下痢罹患率及び下痢死亡率の主たる原因となる。該細菌の病原性は、腸管への細菌接着を媒介する線毛定着因子(CFs)の発現と関連しており(Gaastra and Svennerholm,1996)、また、腸内の電解質及び体液輸送過程に影響を与えることによって疾病の下痢症特性の原因となる熱不安定性(LT)及び/又は熱安定性(ST)毒素の分泌と関連している(Qadri et al,2005a;Sanchez and Holmgren,2005)。
【0003】
ETECの疾病からの保護は、CFsに対するLT及び免疫応答の抗体媒介性の中和と関係している(Levine et al,1994,Svennerholm and Holmgren,1995;Svennerholm and Savarino,2004)。一般に、ワクチンの目的は、受容体の免疫応答を誘発することであり、実際の病原菌のその後の投与に対する保護を提供する。これは、弱毒化された生きている病原菌株、即ち、効果的な免疫応答を刺激しつつ疾病を引き起こさないように病原性が低減された株の接種によって、又は感染毒性株に対して効果的な防御免疫応答を誘発できる一つ若しくはそれ以上の死滅した病原性株の投与によって達成され得る。腸内感染症に対する免疫化のため、好ましくは腸粘膜において局所的に効果的な免疫応答を効率よく刺激する経口経路によってワクチンを与えるべきであるが、それのみならず、他の粘膜経路や非経口経路、更には経皮経路でさえ防御免疫を誘発するのに使用できる。
【0004】
ETECによって引き起こされる疾病から保護する効果的なワクチンの開発は難しい。100を超えた異なる血清型は、病原菌株に関連してきた。更に、それらの株は、腸内の感染の確立を促進する多数のCFs(各々は抗原性が異なる)のうち1つ又はそれ以上を保有することができる(Qadri et al,2005a)。
【0005】
CFsに対して向けられる免疫応答が保護的である、及び腸内の粘膜免疫応答が保護のために特に重要であるというかなりの証拠がある(Svennerholm et al, 1988,1990;Levine et al,1994;Svennerholm and Savarino,2004)。そのような応答を誘発するため、好ましくはETECワクチンを経口で投与すべきである。我々は、細胞全体が死滅した経口ETECワクチンを以前に開発しており、一般的なETEC血清型を表す五つの株を含有し、組み換えコレラ毒素Bサブユニット(CTB,ETEC LTのBサブユニットと非常に相同である)と共に最も一般的に遭遇するCFs(幾つかの場合においては、通常、コリ表面[CS]タンパク質と呼ばれる)、即ちCFA/I、CS1、CS2、CS3、CS4及びCS5のうち幾つかを発現する(Svennerholm and Holmgren,1995;Svennerholm and Savarino,2004)。このワクチンの初期臨床試験は、CTB及びワクチン内に存在する特異的CFsの両方に対して有意な免疫応答を起こし、これはスウェーデン人のボランティアにおいて見られたほか、続いてエジプト、バングラデシュの成人及び小児においても見られた(Jertborn et al,1993;Ahren et al,1998;Savarino et al,1998,1999,Qadri et al,2005a,2005b)。また、該ワクチンは、メキシコ及びグアテマラへのアメリカ人旅行者の日常の活動を妨害する十分に深刻な下痢症に対して、有意な保護を提供した(Sack et al,2002;Svennerholm and Savarino 2004)。しかしながら、エジプト人の生後6〜18か月の乳児において、該ワクチンの保護効果が低いことが分かった(Savarino et al,によって発表される)。このことは、該ワクチンが旅行者のより深刻な疾病に対して効果的である一方で、流行地域に住む乳児を保護するのには十分に強力でないことを示唆した(Svennerholm and Steele,2004)。
【0006】
前述のETECワクチンの有効性が乳児において低いことについては、この年齢層においてCF抗原に対して見られる抗体応答が比較的弱いことが理由の一つとして考えられている(Savarino and Svennerholm,2004)。この応答の低さは、異なるCFsの用量を増やすことで改善されるであろうことから、ワクチン投与量中の該抗体の量を増加させることが、細胞全体が死滅したETECワクチンの今後の開発における優先事項である。不活性化大腸菌(E.coli K12プラシーボ調製物でさえ)を生後6〜18か月の幼児に多量に与えると、おそらくエンドトキシン(LPS)の量が多いことにより、嘔吐という形での拒絶反応が起きることが分かったことから、各ワクチン用量で投与されるETEC細菌の数をただ単に増やすことは適切ではない。それら有害作用は、細菌の用量を少なくした(4倍少なくした)場合、観察されなかった(Qadri et al 2005b,Savarino et al,によって発表される)。
【0007】
当該技術分野において周知のように、ETECのヒト病原性株に関連したCFsには幾つかのタイプがあるが、CFA/I、CFA/II及びCFA/IVが主なタイプであり、現在のところ、およそ40〜80%の臨床分離株と関連している。CFA/Iは、単一の線毛抗原であり、一方、CFA/II及びCFA/IVは、CF/CSタンパク質の二つ以上のタイプから構成される場合がある。
【0008】
野生型ETECにおけるCFの発現は、天然株のみが最大で2種又は3種のCF抗原を特定の組み合わせで発現するように制限されていると考えられる。従って、天然のCFA/II ETEC株は、一般にCS1とCS3、CS2とCS3又はCS3単独で発現する。同様に、天然のCFA/IV ETEC株は、一般にCS4とCS6、CS5とCS6又はCS6単独で発現する。しかしながら、例えば、CS1とCS2は、同一の野生型株中で見つかったことがなく、同様に、CS4とCS5は、自然発生の株において同時に発現されない。更に、CS4、CS5又はCS6がCS1、CS2又はCS3と共に発現することは、野生型株について説明されていない。
【0009】
ETECに対するワクチンに提案された最低限の必要条件は、CFA/IとCFA/II及びCFA/IVの異なるサブ成分、即ちCS1〜CS6とを発現するETEC株に対する保護を誘発する可能性を有することである。従って、野生型株に基づいたETECワクチンは、少なくとも5つの細菌株が最低限必要となり得、CFA/I、CS1+CS3、CS2+CS3、CS4+CS6及びCS5+CS6を発現する。
【発明の概要】
【課題を解決するための手段】
【0010】
この問題を解決するための一つの方策は、我々の係属中の国際特許出願PCT/SE2007/050051において述べられており、そこでは、高濃度のETEC CFsを発現する大腸菌株が使用された。従って、ワクチン中の大腸菌の総数を増やすことなく、それら抗原の量をワクチン中で増加させることができる。この方策を、ETEC CFを発現する少なくとも一つの組み換えプラスミドの他のETEC CFを発現する細菌細胞中への挿入と組み合わせて、1つの細菌株から発現CFsの不自然な組み合わせを提供する。
【0011】
本願は、抗下痢症ワクチンに用いるETEC線毛を多数発現する大腸菌株の構築上の問題に対する他の解決策を与える。
【0012】
本発明は、一の態様において、遺伝子集合体を含む組み換えオペロンを提供するものであり、ここで、腸管毒素原性大腸菌(ETEC)に関連した定着因子抗原(CFs)の少なくとも2つの主要サブユニットをコードする少なくとも2つの構造遺伝子が存在する。このため、このオペロンからCFsの少なくとも2つの主要サブユニットをコードする少なくとも2つの構造遺伝子を発現することができ、ワクチン組成物中に必要とされる細菌細胞の数の低減を可能にする。
【0013】
本発明の他の態様においては、宿主細胞が、上述の組み換えオペロンを含むように遺伝子操作されており、ここで、前記オペロンは、エピソームエレメント上に位置するか又は前記宿主細胞の染色体に組み込まれている。
【0014】
本発明の第1の実施態様においては、上記宿主細胞中のエピソームエレメントがプラスミドである。
【0015】
本発明の第2の実施態様においては、上記定着因子抗原(CFs)の少なくとも2つの主要サブユニットが、同じものであるか又は異なるものである。例えば、あるCFのオペロン1がA、B、C及びDを備え、ここで、Bが主要サブユニットであり、他のCFのオペロン2がA'、B'、C'及びD'を備え、ここで、B'が主要サブユニットである場合、そのときのオペロン1及び2をA、B、B'、C及びD又はA'、B'、B、C'及びD'に組み合わせることができる。いずれの場合においても、2つの異なる主要サブユニットが、同一のオペロンから発現されることになる。定着因子の2つの主要サブユニットが同一である場合、該オペロンは、例えば、Bが主要サブユニットである場合、A、B、B、C、Dとなる。これは、より少量の細胞から主要サブユニットBに対するより強い免疫応答を誘発することになり得る。遺伝子断片の順序の他の例は、B、A、C、Dがあり得、主要サブユニットの2つの構造遺伝子を持つオペロンが、それぞれB、B、A、C、D又はB'、B、A、C、Dとなる。CFsの主要サブユニットをコードする幾つかの構造遺伝子が同一のオペロン中に含まれる場合、それらはB''、B'''等と名付けられる。この方法の一の例を後述すると、CS2の主要サブユニットをコードする構造遺伝子がCFA/Iオペロンに含まれる場合、ハイブリット線毛CFA/I+CS2の発現をもたらす。
【0016】
本発明の第3の実施態様においては、上記CFsが、CFA/I、CS1、CS2、CS3、CS4、CS5、CS6、CS14、CS17、CS19、及び推定定着因子O71(PCFO71)からなる群から選択される。これらのうちCFA/1、CS1、CS2、CS4、CS14、CS17、CS19及び推定定着因子O71(PCDO71)は、同様に構築されるオペロンを有する。従って、それらを相互に組み合わせることが好ましい。
【0017】
本発明の第4の実施態様においては、上記宿主細胞が、オペロン中でCFsの少なくとも2つの主要サブユニットを発現する。
【0018】
本発明の第5の実施態様においては、上記宿主細胞が、細菌及び単細胞真核生物からなる群から選択される生存可能な微生物である。該細菌は、コレラ菌、大腸菌等の種でよく、また、単細胞真核生物は、酵母等の種、特には出芽酵母(Saccharomyces cerevisiae)、分裂酵母(Schizosaccharomyces pombe)、ピキア・パストリス(Pichia pastoris)等の酵母種でよい。
【0019】
本発明の第6の実施態様においては、上記宿主細胞が、大腸菌細胞である。
【0020】
本発明の第7の実施態様においては、上記宿主細胞が、毒素非産生性の大腸菌細胞である。
【0021】
本発明の第8の実施態様においては、上記宿主細胞が、抗生物質耐性遺伝子を発現しない。
【0022】
本発明の第9の実施態様においては、上記宿主細胞が、一つ又はそれ以上のプラスミドによって補完される一つ又はそれ以上の補完可能な染色体欠失又は染色体変異を保有する。該染色体欠失は、例えば、必須アミノ酸の産生に必要な遺伝子が位置している場所に位置し得る。
【0023】
本発明の他の態様は、腸管毒素原性大腸菌(ETEC)に関連した定着因子抗原(CFs)の少なくとも2つの主要サブユニットを発現することが可能な組み換えプラスミドを少なくとも一つ保有する宿主細胞の製造方法に関するものであり、オペロン内にハイブリッドETEC CFの発現に必要な遺伝子又は遺伝子断片を集合させる工程と、上記サブユニットの発現を制御するプロモーターを集合させる工程と、上記宿主細胞のゲノムに上記オペロンを組み込むか又は上記オペロン、プラスミドを維持するための選択マーカー及び複製開始点を備えるプラスミドで上記宿主細胞を形質転換させる工程とを含む。
【0024】
本発明に従って産生される宿主細胞は、ETEC下痢症に対するワクチンの製造に使用されることができる。
【0025】
従って、本発明の更に他の態様は、ワクチンの経口送達用に選択されるような、薬学的に容認される賦形剤、緩衝剤及び/又は希釈剤と共に、本発明に従う宿主細胞を少なくとも一つ含む抗下痢症ワクチンを対象とする。ワクチンに適した賦形剤、緩衝剤及び/又は希釈剤は、ヨーロッパ薬局方又はアメリカ薬局方で見つけることができる。
【0026】
本発明の最後の態様においては、ワクチンの製造における本発明に従うオペロンの使用を提供するものである。
【0027】
上記の方法は、自然に発生する特定の組み合わせにおいてCF抗原の発現についての先の制限を回避するため、本発明は、わずか1〜2つの宿主株を備える抗下痢症ワクチンを提供する場合があり、CFA/I、CS1、CS2、CS4の主要サブユニット、即ちCFsの少なくとも2つの主要サブユニットをそれぞれの株で一緒に発現する。従って、上記ワクチンは、合計で、より少ない株を含むことになり、おそらく、以前試験された死滅したETECワクチンの場合と比べてそれぞれの株をより少ない用量で使用することができるという追加の利点がある。
【0028】
本発明に従うワクチンの製造のために意図され発現CFsは、哺乳類、特にはヒトの腸内感染症及び疾病を引き起こすETECと関連している。
【0029】
好ましくは、本発明に従う細胞は、宿主細胞の表面上に、前記CFsを発現する。
【0030】
本発明で得られる宿主細胞の表面上におけるCFsの発現レベルは、免疫学的な方法により、例えば阻害ELISAアッセイを適用することで検出できる。
【0031】
本発明の一の実施態様においては、本発明の細胞により発現されるCFsの主要サブユニットが、腸内ETEC感染症の哺乳類の排泄物から元来単離されるETEC株からのCFsの対応する主要サブユニットに対して産生される特異的抗体と反応することのできる形で発現される。
【0032】
本発明の他の実施形態において、本発明の細胞により発現されるCFは、該細胞が哺乳類の免疫化に効果的な量で使用される場合に、腸内ETEC感染症の哺乳類の排泄物から元来単離されるETEC株からのCFsの対応する主要サブユニットと反応できるCFsの発現主要サブユニットに対する抗体の形成を導く形で発現される。
【0033】
本発明の更に他の実施態様において、本発明の細胞により発現されるハイブリッドCFsは、ホルマリン処理又は他の手段による細胞の不活性化後に、腸内ETEC感染症の哺乳類の排泄物から元来単離されるETEC株からのCFsの対応する主要サブユニットに対して産生される特異的抗体と反応することのできる形で発現される。
【0034】
本発明の更に他の実施態様において、本発明の細胞により発現されるCFsは、ホルマリン処理又は他の手段による細胞の不活性化後、該細胞が哺乳類の免疫化に効果的な量で使用される場合に、腸内ETEC感染症の哺乳類の排泄物から元来単離されるETEC株からの対応するCFsと反応できる発現CFsに対する抗体の形成を導く形で発現される。
【0035】
本発明に従う宿主細胞は、前記ハイブリッドCFsの発現を提供する液体培地において該細胞を生体外で培養するための方法によって培養される。
【0036】
本発明の培養細胞は、ホルマリン若しくはフェノール又は他の手段による穏やかな処理を用いて該細胞の複製を防ぎ、不活性化前の細胞と実質的に同一の量(元の量の少なくとも50%)で発現ハイブリッドCFsを保有し、実質的に同一の抗体との反応性と、ほぼ同一の免疫原性とを持つ細胞をもたらすことによって不活性化できる。
【0037】
本発明の宿主細胞の一つ又は幾つかは、哺乳類に下痢症のワクチン接種をする方法での使用に特に適しており、それは、本発明に従う細胞の株又は組み合わせを哺乳類に投与することを含む。
【0038】
本発明の一の実施態様において、本発明の宿主細胞の一つ又は幾つかは、子豚、子牛、子羊又は馬、特にはヒト等、哺乳類のワクチン接種用のワクチンとして、単独で又は組み合わせて使用される。かかるワクチンは、経口経路で投与されるのが好ましい。
【0039】
以下、図面の説明、図面、配列表、材料及び方法、実施例、並びに表によって、本発明を説明することになるが、該発明が開示されるいかなる詳細事項によっても限定されるものでないことを理解すべきである。
【図面の簡単な説明】
【0040】
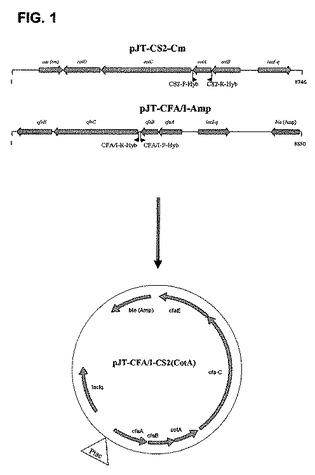
【図1】ハイブリッド発現ベクターpJT−CFA/I−CS2(CotA)の構築を示す。CotA及びpJT−CFA/I−Amp全体を増幅するための特異的プライマーを用いることで、2つの断片を増幅させて連結した。
【発明を実施するための形態】
【0041】
材料及び方法
菌種及び培養条件
本願に記載の株を表1に収載する。線毛ファミリーのメンバーのうち2つの主要サブユニットを含むハイブリッドタンパク質を発現する組み換え細胞の構築は、CFA/I及びCS2双方の主要サブユニットを発現する株の構築によって例証される。
【0042】
株は、使用するまでグリセロール含有凍結培地中−70℃にて冷凍保存された。増殖及び純度を確認するために37℃にて一晩寒天板を接種した後、細菌を37℃で16〜18時間振とうしながらCFA培養液(リットル当たり、カザミノ酸10g、酵母抽出物1.5g、MgSO47H2O 102mg、MnCl24H2O 8mg)中で増殖させた。必要な場合には、該培地をクロラムフェニコール(12.5μg/ml)又はアンピシリン(100μg/ml)で補った。
【0043】
発現ベクターにおけるETEC CFsオペロンのクローン化
ハイブリッド線毛としてのCFA/I+CS2の産生
CS2の主要サブユニット、並びにアシャー、シャペロン、CFA/Iのマイナーサブユニット及び主要サブユニットからなるハイブリッド線毛の産生は、幾つかの工程で行われた。Expand High Fidelity PCRシステム(ロシュ・ダイアグノスティックス・GmbH)、プライマーCS2−F−Hyb及びCS2−R−Hyb(表1)を用い、またテンプレートとしてプラスミドpJT−CS2−Cm(係属中の特許出願PCT/SE2007/050051参照)を用いたPCRによって、CS2の主要サブユニットを含有する断片CotAを増幅した。加えて、プラスミドpJT−CFA/I−Amp(係属中の特許出願PCT/SE2007/050051参照)に対して、プライマーCFA/I−F−Hyb及びCFA/I−R−Hyb(表1)を用いたリバースPCRを行い、最初のプラスミドを含有する断片が得られた。Eco31Iを持つ両断片の制限を受けて、両断片を連結し、CfaBの下流にCFA/I及びCotAのオペロン全体を含有するプラスミドが得られた。
【0044】
CFsの発現
各組み換えTOP10株の一晩培養物を、アンピシリン又はクロラムフェニコールがそれぞれ100又は12.5μg/mlで補われたCFA培養液(表1)中で1/100に希釈し、37℃及び150rev/minで2時間インキュベートし、次いで1mMの最終濃度までIPTGを加えて、同一の条件で更に4時間インキュベートした。その後、細菌を収集し、PBS中で再懸濁した。
【0045】
ドットブロットテスト
先に説明されるように、特異的モノクローナル抗CFAs抗体を用いて、クローン化株における各CFAsの発現を評価した(Binsztein et al 1991)。簡潔に述べると、PBSで洗浄し、CFA/I発現のためにIPTGで誘発した細菌培養物(PBS中109細菌/ml)2μlをニトロセルロース濾紙に塗布し、MAbsと、次いでHRPに抱合されたヤギの抗マウスIgGと共にそれぞれ1.5時間インキュベートした。最後の展開をTBS中の4−クロロ−1−ナフトール−H2O2によって最大で15分間行った。
【0046】
血球凝集
ヒト又はウシの新鮮な赤血球を0.85%のNaCl中で2度洗浄し、1%マンノースの生理食塩水中で3%に懸濁させた。この混合液10μlと、CFAsの発現のために導入されPBSで洗浄した、同体積の被験細菌のPBS懸濁液(109細菌/ml)とを混合し、2分後、室温にて血球凝集を観察した。
【0047】
電子顕微鏡検査
PBSで一度洗浄した各細菌の懸濁液(PBS中1010細菌/ml)10μlをパラフィルム上に塗布した。フォルムバール被覆グリッドを2分間懸濁液に置いた。次いで、該グリッドを、パラフィルム上のPBS−1%BSA 25μlに適用することで、10秒ずつ2回洗浄し、次いでPBS−Tween0.05%−BSA0.1%中に希釈された特異的モノクローナル抗体25μlと共に15分間該グリッドをインキュベートした。該グリッドを上記のPBS−1%BSAで6回洗浄し、次いでPBS−0.1%BSA−0.05%Tween中の抗マウスIgG−金抱合体(Amersham International,Amersham,UK)と共に15分間インキュベートした。その後、グリッドをPBS−0.1%BSAで3回、蒸留水で3回洗浄した。該グリッドを1%モリブデン酸アンモニウム(pH7.0)25μlに50〜60秒間適用して、ネガティブ染色を行い、次いでグリッドを濾紙上で5分間空気乾燥させた。電子顕微鏡検査を行うまで、グリッドを4℃で貯蔵した。
【0048】
ELISA
先に説明されるように、細菌表面上のCFA/I又はCS2の量を阻害ELISAアッセイで定量化し(Lopez−Vidal et al 1998)、血清中のIgA又はIgG+M抗体の力価をELISAアッセイで決定した(Rudin et al,1994)。
【0049】
ホルマリンによる細菌の不活性化
細菌を死滅させるため、各株の培養物をPBSで洗浄して、PBS中1010細菌/mlの密度まで再懸濁させた。ホルマリンを0.1Mの最終濃度まで加え、該懸濁液を37℃で2時間インキュベートして60rpmで撹拌し、次いで該懸濁液を4℃にて3日間インキュベートした。その後、細菌を洗浄し、同体積のPBSで再懸濁させ、その細菌懸濁液100μlを血液寒天培地上に広げて、37℃にて最大で1週間までインキュベートし、増殖を調べた。
【0050】
マウスの免疫化
メスのBalb/cマウス(生後6〜8週)を経口経路の免疫化に用いた。誘発してホルマリンで死滅した参照株325542−3及び58R957と組み換え株TOP10−CFA/I−CS2の培養物を洗浄し、所望の細菌密度までPBS中に再懸濁させた。先に説明されるように、経口経路による免疫化のため、109の細菌を10μgのCTと一緒に用いた(Rhagavan et al,200)。全てのマウスに2つの同一の免疫化を2週間おいて行い、第一の投与の直前と、第二の投与の2週間後に出血を収集した。
【0051】
統計的分析
全てのELISA実験及び阻害ELISA実験を二つ組で行い、異なる日に少なくとも3回繰り返した。特定のテストそれぞれについてのドットブロット実験を少なくとも2回繰り返した。学生のt−テストによって統計的分析を行い、P<0.05を有意性ありとみなした。
【実施例】
【0052】
実施例1
ハイブリッド線毛の産生:我々は、CFA/I及びCS2双方の主要サブユニットからなるハイブリッド線毛を発現させる可能性について調べた。かかるハイブリッド線毛を発現する発現ベクターの構築は、材料及び方法において説明されており、図1において表現される。次いで、該ベクターをTop10株中で増殖し、CFA/I及びCS2双方の主要サブユニットからなる線毛を発現する組み換え株が得られた。透過電子顕微鏡法(TEM)と、各主要サブユニットに対する特異的MAbs、即ち、α-CFA/I MAb(1:6)〜ヤギのα−マウスIgG 20mn 金及びビオチン化α-CS2 MAb(10:3)〜ストレプトアビジン 10nm 金を用いて、発現を検証した。
【0053】
ここで引用される参考文献の教示は、参照することによりこの明細書に組み込まれる。
【0054】
【表1】
【先行技術文献】
【非特許文献】
【0055】
【非特許文献1】Ahren, C. M. Jertborn, and A.M. Svennerholm. 1998. Intestinal immune responses to an inactivated oral enterotoxigenic Escherichia coli vaccine and associated immunoglobulin A responses in blood. Infect. Immun. 66:3311-3316.
【非特許文献2】Binsztein, N., M.J. Jouve, G.I. Viboud, L.L. Moral, M. Rivas, I. Orskov, C. Ahren, and A-M. Svennerholm. 1991. Colonization factors of enterotoxigenic Escherichia coli isolated from children with diarrhea in Argentina. J. Clin. Microbiol. 29:1893-1898.
【非特許文献3】Gaastra, W., and A.M. Svennerholm. 1996. Colonization factors of human enterotoxigenic Escherichia coli (ETEC). Trends Microbiol. 4:444-452.
【非特許文献4】Jertborn, M., C. Ahren, J. Holmgren, A.M. Svennerholm. 1993. Safety and immunogenicity of an oral inactivated enterotoxigenic Escherichia coli vaccine. Vaccine 16: 255-60.
【非特許文献5】Levine, M.M., J.A. Giron, and F. Noriega. 1994. Fimbrial vaccines,. In Fimbriae: adhesion, biogenics, genetics and vaccines, P. Klemm (ed.), pp.255-270. CRC Press, Boca Raton, Fla.
【非特許文献6】Qadri F., A.M. Svennerholm, A. S. G. Faruque, and Sack R.B. 2005a. Enterotoxigenic Escherichia coli in developing countries: epidemiology, microbiology, clinical features, treatment, and prevention. Clin. Microbiol. Rev. 18:465-483.
【非特許文献7】Qadri, F., T. Ahmed, F. Ahmed, Y.A. Begum, D.A. Sack, A.M. Svennerholm and the PTE Study Group. 2005b. Reduced doses of oral killed enterotoxigenic Escherichia coli plus cholera toxin B subunit vaccine is safe and immunogenic in Bangladeshi infants 6-17 months of age: Dosing studies in different age groups. Vaccine 24:1726-1733.
【非特許文献8】Rhagavan S, M. Hjulstroem, J. Holmgren and A.M. Svennerholm. 2002. Protection against experimental Helicobacter pylori infection after immunization with inactivated H. pylori whole cell vaccines. Infect. Immun. 70:6383-6388.
【非特許文献9】Rudin, A., M.M. McConnell, A.M. Svennerholm. 1994. Monoclonal antibodies against enterotoxigenic Escherichia coli colonization factor antigen I (CFA/I) that cross-react immunologically with heterologous CFAs. Infect. Immun. 62:4339-4346.
【非特許文献10】Sack, D.A., J. Shimko, O. Torres et al. 2002. Safety and efficacy of a killed oral vaccine for enterotoxigenic E. coli diarrhea in adult travelers to Guatemala and Mexico. 42nd Interscience Conference on Antimcrobial Agents and Chemotherapy. San Diego, CA. Abstract.
【非特許文献11】Sanchez J., and Holmgren. 2005. Virulence factors, pathogenesis and vaccine protection in cholera and ETEC diarrhea. Curr. Opin. Immunol. 17:388-98.
【非特許文献12】Savarino, S.J., F.M. Brown, E. Hall, S. Bassily, F. Youssef, T. Wierzba, L. Peruski, N.A. El-Masry, M. Safwat, M. Rao, M. Jertborn, A.M. Svennerholm, Y.J. Lee, and J.D. Clemens. 1998. Safety and immunogenicity of an oral, killed enterotoxigenic Escherichia coli-cholera toxin B subunit vaccine in Egyptian adults. J. Infect. Dis. 177:796-799.
【非特許文献13】Savarino, S.J., E. Hall,. S. Bassily,. F.M. Brown, F. Youssef, T.F. Wierzba,. L. Peruski, N.A. El-Masry, M. Safwat, M. Rao, H. El Mohamady, R. Abu-Elyazeed, A. Naficy, A.M. Svennerholm, M. Jertborn, Y.J. Lee, and J.D. Clemens. 1999. Oral, inactivated, whole cell enterotoxigenic Escherichia coli plus cholera toxin B subunit vaccine: results of the initial evaluation in children. J. Infect. Dis. 179:107-14.
【非特許文献14】Svennerholm, A.-M., Y.L. Vidal, J. Holmgren, M.M. McConnell, and B. Rowe. 1988. Role of PCF8775 antigen and its coli surface subcomponents for colonization, disease, and protective immunogenicity of enterotoxigenic Escherichia coli in rabbits. Infect. Immun. 56:523-528.
【非特許文献15】Svennerholm, A.-M., C. Wenneras, J. Holmgren, M.M. McConnell, and B. Rowe. 1990. Roles of different coli surface antigens of colonization factor antigen II in colonization by and protective immunogenicity of enterotoxigenic Escherichia coli in rabbits. Infect. Immun. 58:341-346.
【非特許文献16】Svennerholm, A.-M., and J. Holmgren. 1995. Oral vaccines against cholera and enterotoxigenic Escherichia coli diarrhea. Adv. Exp. Med. Biol. 371B:1623-1628.
【非特許文献17】Svennerholm, A.-M., and S. J. Savarino. 2004. Oral inactivated whole cell B subunit combination vaccine against enterotoxigenic Escherichia coli. In New Generation Vaccines: 3rd ed. Eds: Levine M.M. et al., pp. 737-750, Marcel Decker, New York, NY.
【非特許文献18】Svennerholm, A.-M., and D. Steele. 2004. Progress in enteric vaccine development. In: Microbial-gut interactions in health and disease, Ed. M. Farthing. Best Pract. Res Clin. Gastroenterol. 18: 421-445.
【技術分野】
【0001】
本発明は、腸管毒素原性大腸菌の定着因子(CF)抗原を発現するためのハイブリッドオペロンに関し、特には、異なる2つのCFsを発現するための少なくとも2つの構造遺伝子を含む遺伝子を備える組み換えオペロンに関するものである。更に、本発明は、かかるオペロンを宿主細胞のゲノム上に又は宿主細胞に組み込まれたプラスミド中に備えた、大腸菌等の宿主細胞に関するものである。
【背景技術】
【0002】
腸管毒素原性大腸菌(ETEC)は、旅行者下痢症、並びに世界各地の流行地域における小児の下痢罹患率及び下痢死亡率の主たる原因となる。該細菌の病原性は、腸管への細菌接着を媒介する線毛定着因子(CFs)の発現と関連しており(Gaastra and Svennerholm,1996)、また、腸内の電解質及び体液輸送過程に影響を与えることによって疾病の下痢症特性の原因となる熱不安定性(LT)及び/又は熱安定性(ST)毒素の分泌と関連している(Qadri et al,2005a;Sanchez and Holmgren,2005)。
【0003】
ETECの疾病からの保護は、CFsに対するLT及び免疫応答の抗体媒介性の中和と関係している(Levine et al,1994,Svennerholm and Holmgren,1995;Svennerholm and Savarino,2004)。一般に、ワクチンの目的は、受容体の免疫応答を誘発することであり、実際の病原菌のその後の投与に対する保護を提供する。これは、弱毒化された生きている病原菌株、即ち、効果的な免疫応答を刺激しつつ疾病を引き起こさないように病原性が低減された株の接種によって、又は感染毒性株に対して効果的な防御免疫応答を誘発できる一つ若しくはそれ以上の死滅した病原性株の投与によって達成され得る。腸内感染症に対する免疫化のため、好ましくは腸粘膜において局所的に効果的な免疫応答を効率よく刺激する経口経路によってワクチンを与えるべきであるが、それのみならず、他の粘膜経路や非経口経路、更には経皮経路でさえ防御免疫を誘発するのに使用できる。
【0004】
ETECによって引き起こされる疾病から保護する効果的なワクチンの開発は難しい。100を超えた異なる血清型は、病原菌株に関連してきた。更に、それらの株は、腸内の感染の確立を促進する多数のCFs(各々は抗原性が異なる)のうち1つ又はそれ以上を保有することができる(Qadri et al,2005a)。
【0005】
CFsに対して向けられる免疫応答が保護的である、及び腸内の粘膜免疫応答が保護のために特に重要であるというかなりの証拠がある(Svennerholm et al, 1988,1990;Levine et al,1994;Svennerholm and Savarino,2004)。そのような応答を誘発するため、好ましくはETECワクチンを経口で投与すべきである。我々は、細胞全体が死滅した経口ETECワクチンを以前に開発しており、一般的なETEC血清型を表す五つの株を含有し、組み換えコレラ毒素Bサブユニット(CTB,ETEC LTのBサブユニットと非常に相同である)と共に最も一般的に遭遇するCFs(幾つかの場合においては、通常、コリ表面[CS]タンパク質と呼ばれる)、即ちCFA/I、CS1、CS2、CS3、CS4及びCS5のうち幾つかを発現する(Svennerholm and Holmgren,1995;Svennerholm and Savarino,2004)。このワクチンの初期臨床試験は、CTB及びワクチン内に存在する特異的CFsの両方に対して有意な免疫応答を起こし、これはスウェーデン人のボランティアにおいて見られたほか、続いてエジプト、バングラデシュの成人及び小児においても見られた(Jertborn et al,1993;Ahren et al,1998;Savarino et al,1998,1999,Qadri et al,2005a,2005b)。また、該ワクチンは、メキシコ及びグアテマラへのアメリカ人旅行者の日常の活動を妨害する十分に深刻な下痢症に対して、有意な保護を提供した(Sack et al,2002;Svennerholm and Savarino 2004)。しかしながら、エジプト人の生後6〜18か月の乳児において、該ワクチンの保護効果が低いことが分かった(Savarino et al,によって発表される)。このことは、該ワクチンが旅行者のより深刻な疾病に対して効果的である一方で、流行地域に住む乳児を保護するのには十分に強力でないことを示唆した(Svennerholm and Steele,2004)。
【0006】
前述のETECワクチンの有効性が乳児において低いことについては、この年齢層においてCF抗原に対して見られる抗体応答が比較的弱いことが理由の一つとして考えられている(Savarino and Svennerholm,2004)。この応答の低さは、異なるCFsの用量を増やすことで改善されるであろうことから、ワクチン投与量中の該抗体の量を増加させることが、細胞全体が死滅したETECワクチンの今後の開発における優先事項である。不活性化大腸菌(E.coli K12プラシーボ調製物でさえ)を生後6〜18か月の幼児に多量に与えると、おそらくエンドトキシン(LPS)の量が多いことにより、嘔吐という形での拒絶反応が起きることが分かったことから、各ワクチン用量で投与されるETEC細菌の数をただ単に増やすことは適切ではない。それら有害作用は、細菌の用量を少なくした(4倍少なくした)場合、観察されなかった(Qadri et al 2005b,Savarino et al,によって発表される)。
【0007】
当該技術分野において周知のように、ETECのヒト病原性株に関連したCFsには幾つかのタイプがあるが、CFA/I、CFA/II及びCFA/IVが主なタイプであり、現在のところ、およそ40〜80%の臨床分離株と関連している。CFA/Iは、単一の線毛抗原であり、一方、CFA/II及びCFA/IVは、CF/CSタンパク質の二つ以上のタイプから構成される場合がある。
【0008】
野生型ETECにおけるCFの発現は、天然株のみが最大で2種又は3種のCF抗原を特定の組み合わせで発現するように制限されていると考えられる。従って、天然のCFA/II ETEC株は、一般にCS1とCS3、CS2とCS3又はCS3単独で発現する。同様に、天然のCFA/IV ETEC株は、一般にCS4とCS6、CS5とCS6又はCS6単独で発現する。しかしながら、例えば、CS1とCS2は、同一の野生型株中で見つかったことがなく、同様に、CS4とCS5は、自然発生の株において同時に発現されない。更に、CS4、CS5又はCS6がCS1、CS2又はCS3と共に発現することは、野生型株について説明されていない。
【0009】
ETECに対するワクチンに提案された最低限の必要条件は、CFA/IとCFA/II及びCFA/IVの異なるサブ成分、即ちCS1〜CS6とを発現するETEC株に対する保護を誘発する可能性を有することである。従って、野生型株に基づいたETECワクチンは、少なくとも5つの細菌株が最低限必要となり得、CFA/I、CS1+CS3、CS2+CS3、CS4+CS6及びCS5+CS6を発現する。
【発明の概要】
【課題を解決するための手段】
【0010】
この問題を解決するための一つの方策は、我々の係属中の国際特許出願PCT/SE2007/050051において述べられており、そこでは、高濃度のETEC CFsを発現する大腸菌株が使用された。従って、ワクチン中の大腸菌の総数を増やすことなく、それら抗原の量をワクチン中で増加させることができる。この方策を、ETEC CFを発現する少なくとも一つの組み換えプラスミドの他のETEC CFを発現する細菌細胞中への挿入と組み合わせて、1つの細菌株から発現CFsの不自然な組み合わせを提供する。
【0011】
本願は、抗下痢症ワクチンに用いるETEC線毛を多数発現する大腸菌株の構築上の問題に対する他の解決策を与える。
【0012】
本発明は、一の態様において、遺伝子集合体を含む組み換えオペロンを提供するものであり、ここで、腸管毒素原性大腸菌(ETEC)に関連した定着因子抗原(CFs)の少なくとも2つの主要サブユニットをコードする少なくとも2つの構造遺伝子が存在する。このため、このオペロンからCFsの少なくとも2つの主要サブユニットをコードする少なくとも2つの構造遺伝子を発現することができ、ワクチン組成物中に必要とされる細菌細胞の数の低減を可能にする。
【0013】
本発明の他の態様においては、宿主細胞が、上述の組み換えオペロンを含むように遺伝子操作されており、ここで、前記オペロンは、エピソームエレメント上に位置するか又は前記宿主細胞の染色体に組み込まれている。
【0014】
本発明の第1の実施態様においては、上記宿主細胞中のエピソームエレメントがプラスミドである。
【0015】
本発明の第2の実施態様においては、上記定着因子抗原(CFs)の少なくとも2つの主要サブユニットが、同じものであるか又は異なるものである。例えば、あるCFのオペロン1がA、B、C及びDを備え、ここで、Bが主要サブユニットであり、他のCFのオペロン2がA'、B'、C'及びD'を備え、ここで、B'が主要サブユニットである場合、そのときのオペロン1及び2をA、B、B'、C及びD又はA'、B'、B、C'及びD'に組み合わせることができる。いずれの場合においても、2つの異なる主要サブユニットが、同一のオペロンから発現されることになる。定着因子の2つの主要サブユニットが同一である場合、該オペロンは、例えば、Bが主要サブユニットである場合、A、B、B、C、Dとなる。これは、より少量の細胞から主要サブユニットBに対するより強い免疫応答を誘発することになり得る。遺伝子断片の順序の他の例は、B、A、C、Dがあり得、主要サブユニットの2つの構造遺伝子を持つオペロンが、それぞれB、B、A、C、D又はB'、B、A、C、Dとなる。CFsの主要サブユニットをコードする幾つかの構造遺伝子が同一のオペロン中に含まれる場合、それらはB''、B'''等と名付けられる。この方法の一の例を後述すると、CS2の主要サブユニットをコードする構造遺伝子がCFA/Iオペロンに含まれる場合、ハイブリット線毛CFA/I+CS2の発現をもたらす。
【0016】
本発明の第3の実施態様においては、上記CFsが、CFA/I、CS1、CS2、CS3、CS4、CS5、CS6、CS14、CS17、CS19、及び推定定着因子O71(PCFO71)からなる群から選択される。これらのうちCFA/1、CS1、CS2、CS4、CS14、CS17、CS19及び推定定着因子O71(PCDO71)は、同様に構築されるオペロンを有する。従って、それらを相互に組み合わせることが好ましい。
【0017】
本発明の第4の実施態様においては、上記宿主細胞が、オペロン中でCFsの少なくとも2つの主要サブユニットを発現する。
【0018】
本発明の第5の実施態様においては、上記宿主細胞が、細菌及び単細胞真核生物からなる群から選択される生存可能な微生物である。該細菌は、コレラ菌、大腸菌等の種でよく、また、単細胞真核生物は、酵母等の種、特には出芽酵母(Saccharomyces cerevisiae)、分裂酵母(Schizosaccharomyces pombe)、ピキア・パストリス(Pichia pastoris)等の酵母種でよい。
【0019】
本発明の第6の実施態様においては、上記宿主細胞が、大腸菌細胞である。
【0020】
本発明の第7の実施態様においては、上記宿主細胞が、毒素非産生性の大腸菌細胞である。
【0021】
本発明の第8の実施態様においては、上記宿主細胞が、抗生物質耐性遺伝子を発現しない。
【0022】
本発明の第9の実施態様においては、上記宿主細胞が、一つ又はそれ以上のプラスミドによって補完される一つ又はそれ以上の補完可能な染色体欠失又は染色体変異を保有する。該染色体欠失は、例えば、必須アミノ酸の産生に必要な遺伝子が位置している場所に位置し得る。
【0023】
本発明の他の態様は、腸管毒素原性大腸菌(ETEC)に関連した定着因子抗原(CFs)の少なくとも2つの主要サブユニットを発現することが可能な組み換えプラスミドを少なくとも一つ保有する宿主細胞の製造方法に関するものであり、オペロン内にハイブリッドETEC CFの発現に必要な遺伝子又は遺伝子断片を集合させる工程と、上記サブユニットの発現を制御するプロモーターを集合させる工程と、上記宿主細胞のゲノムに上記オペロンを組み込むか又は上記オペロン、プラスミドを維持するための選択マーカー及び複製開始点を備えるプラスミドで上記宿主細胞を形質転換させる工程とを含む。
【0024】
本発明に従って産生される宿主細胞は、ETEC下痢症に対するワクチンの製造に使用されることができる。
【0025】
従って、本発明の更に他の態様は、ワクチンの経口送達用に選択されるような、薬学的に容認される賦形剤、緩衝剤及び/又は希釈剤と共に、本発明に従う宿主細胞を少なくとも一つ含む抗下痢症ワクチンを対象とする。ワクチンに適した賦形剤、緩衝剤及び/又は希釈剤は、ヨーロッパ薬局方又はアメリカ薬局方で見つけることができる。
【0026】
本発明の最後の態様においては、ワクチンの製造における本発明に従うオペロンの使用を提供するものである。
【0027】
上記の方法は、自然に発生する特定の組み合わせにおいてCF抗原の発現についての先の制限を回避するため、本発明は、わずか1〜2つの宿主株を備える抗下痢症ワクチンを提供する場合があり、CFA/I、CS1、CS2、CS4の主要サブユニット、即ちCFsの少なくとも2つの主要サブユニットをそれぞれの株で一緒に発現する。従って、上記ワクチンは、合計で、より少ない株を含むことになり、おそらく、以前試験された死滅したETECワクチンの場合と比べてそれぞれの株をより少ない用量で使用することができるという追加の利点がある。
【0028】
本発明に従うワクチンの製造のために意図され発現CFsは、哺乳類、特にはヒトの腸内感染症及び疾病を引き起こすETECと関連している。
【0029】
好ましくは、本発明に従う細胞は、宿主細胞の表面上に、前記CFsを発現する。
【0030】
本発明で得られる宿主細胞の表面上におけるCFsの発現レベルは、免疫学的な方法により、例えば阻害ELISAアッセイを適用することで検出できる。
【0031】
本発明の一の実施態様においては、本発明の細胞により発現されるCFsの主要サブユニットが、腸内ETEC感染症の哺乳類の排泄物から元来単離されるETEC株からのCFsの対応する主要サブユニットに対して産生される特異的抗体と反応することのできる形で発現される。
【0032】
本発明の他の実施形態において、本発明の細胞により発現されるCFは、該細胞が哺乳類の免疫化に効果的な量で使用される場合に、腸内ETEC感染症の哺乳類の排泄物から元来単離されるETEC株からのCFsの対応する主要サブユニットと反応できるCFsの発現主要サブユニットに対する抗体の形成を導く形で発現される。
【0033】
本発明の更に他の実施態様において、本発明の細胞により発現されるハイブリッドCFsは、ホルマリン処理又は他の手段による細胞の不活性化後に、腸内ETEC感染症の哺乳類の排泄物から元来単離されるETEC株からのCFsの対応する主要サブユニットに対して産生される特異的抗体と反応することのできる形で発現される。
【0034】
本発明の更に他の実施態様において、本発明の細胞により発現されるCFsは、ホルマリン処理又は他の手段による細胞の不活性化後、該細胞が哺乳類の免疫化に効果的な量で使用される場合に、腸内ETEC感染症の哺乳類の排泄物から元来単離されるETEC株からの対応するCFsと反応できる発現CFsに対する抗体の形成を導く形で発現される。
【0035】
本発明に従う宿主細胞は、前記ハイブリッドCFsの発現を提供する液体培地において該細胞を生体外で培養するための方法によって培養される。
【0036】
本発明の培養細胞は、ホルマリン若しくはフェノール又は他の手段による穏やかな処理を用いて該細胞の複製を防ぎ、不活性化前の細胞と実質的に同一の量(元の量の少なくとも50%)で発現ハイブリッドCFsを保有し、実質的に同一の抗体との反応性と、ほぼ同一の免疫原性とを持つ細胞をもたらすことによって不活性化できる。
【0037】
本発明の宿主細胞の一つ又は幾つかは、哺乳類に下痢症のワクチン接種をする方法での使用に特に適しており、それは、本発明に従う細胞の株又は組み合わせを哺乳類に投与することを含む。
【0038】
本発明の一の実施態様において、本発明の宿主細胞の一つ又は幾つかは、子豚、子牛、子羊又は馬、特にはヒト等、哺乳類のワクチン接種用のワクチンとして、単独で又は組み合わせて使用される。かかるワクチンは、経口経路で投与されるのが好ましい。
【0039】
以下、図面の説明、図面、配列表、材料及び方法、実施例、並びに表によって、本発明を説明することになるが、該発明が開示されるいかなる詳細事項によっても限定されるものでないことを理解すべきである。
【図面の簡単な説明】
【0040】
【図1】ハイブリッド発現ベクターpJT−CFA/I−CS2(CotA)の構築を示す。CotA及びpJT−CFA/I−Amp全体を増幅するための特異的プライマーを用いることで、2つの断片を増幅させて連結した。
【発明を実施するための形態】
【0041】
材料及び方法
菌種及び培養条件
本願に記載の株を表1に収載する。線毛ファミリーのメンバーのうち2つの主要サブユニットを含むハイブリッドタンパク質を発現する組み換え細胞の構築は、CFA/I及びCS2双方の主要サブユニットを発現する株の構築によって例証される。
【0042】
株は、使用するまでグリセロール含有凍結培地中−70℃にて冷凍保存された。増殖及び純度を確認するために37℃にて一晩寒天板を接種した後、細菌を37℃で16〜18時間振とうしながらCFA培養液(リットル当たり、カザミノ酸10g、酵母抽出物1.5g、MgSO47H2O 102mg、MnCl24H2O 8mg)中で増殖させた。必要な場合には、該培地をクロラムフェニコール(12.5μg/ml)又はアンピシリン(100μg/ml)で補った。
【0043】
発現ベクターにおけるETEC CFsオペロンのクローン化
ハイブリッド線毛としてのCFA/I+CS2の産生
CS2の主要サブユニット、並びにアシャー、シャペロン、CFA/Iのマイナーサブユニット及び主要サブユニットからなるハイブリッド線毛の産生は、幾つかの工程で行われた。Expand High Fidelity PCRシステム(ロシュ・ダイアグノスティックス・GmbH)、プライマーCS2−F−Hyb及びCS2−R−Hyb(表1)を用い、またテンプレートとしてプラスミドpJT−CS2−Cm(係属中の特許出願PCT/SE2007/050051参照)を用いたPCRによって、CS2の主要サブユニットを含有する断片CotAを増幅した。加えて、プラスミドpJT−CFA/I−Amp(係属中の特許出願PCT/SE2007/050051参照)に対して、プライマーCFA/I−F−Hyb及びCFA/I−R−Hyb(表1)を用いたリバースPCRを行い、最初のプラスミドを含有する断片が得られた。Eco31Iを持つ両断片の制限を受けて、両断片を連結し、CfaBの下流にCFA/I及びCotAのオペロン全体を含有するプラスミドが得られた。
【0044】
CFsの発現
各組み換えTOP10株の一晩培養物を、アンピシリン又はクロラムフェニコールがそれぞれ100又は12.5μg/mlで補われたCFA培養液(表1)中で1/100に希釈し、37℃及び150rev/minで2時間インキュベートし、次いで1mMの最終濃度までIPTGを加えて、同一の条件で更に4時間インキュベートした。その後、細菌を収集し、PBS中で再懸濁した。
【0045】
ドットブロットテスト
先に説明されるように、特異的モノクローナル抗CFAs抗体を用いて、クローン化株における各CFAsの発現を評価した(Binsztein et al 1991)。簡潔に述べると、PBSで洗浄し、CFA/I発現のためにIPTGで誘発した細菌培養物(PBS中109細菌/ml)2μlをニトロセルロース濾紙に塗布し、MAbsと、次いでHRPに抱合されたヤギの抗マウスIgGと共にそれぞれ1.5時間インキュベートした。最後の展開をTBS中の4−クロロ−1−ナフトール−H2O2によって最大で15分間行った。
【0046】
血球凝集
ヒト又はウシの新鮮な赤血球を0.85%のNaCl中で2度洗浄し、1%マンノースの生理食塩水中で3%に懸濁させた。この混合液10μlと、CFAsの発現のために導入されPBSで洗浄した、同体積の被験細菌のPBS懸濁液(109細菌/ml)とを混合し、2分後、室温にて血球凝集を観察した。
【0047】
電子顕微鏡検査
PBSで一度洗浄した各細菌の懸濁液(PBS中1010細菌/ml)10μlをパラフィルム上に塗布した。フォルムバール被覆グリッドを2分間懸濁液に置いた。次いで、該グリッドを、パラフィルム上のPBS−1%BSA 25μlに適用することで、10秒ずつ2回洗浄し、次いでPBS−Tween0.05%−BSA0.1%中に希釈された特異的モノクローナル抗体25μlと共に15分間該グリッドをインキュベートした。該グリッドを上記のPBS−1%BSAで6回洗浄し、次いでPBS−0.1%BSA−0.05%Tween中の抗マウスIgG−金抱合体(Amersham International,Amersham,UK)と共に15分間インキュベートした。その後、グリッドをPBS−0.1%BSAで3回、蒸留水で3回洗浄した。該グリッドを1%モリブデン酸アンモニウム(pH7.0)25μlに50〜60秒間適用して、ネガティブ染色を行い、次いでグリッドを濾紙上で5分間空気乾燥させた。電子顕微鏡検査を行うまで、グリッドを4℃で貯蔵した。
【0048】
ELISA
先に説明されるように、細菌表面上のCFA/I又はCS2の量を阻害ELISAアッセイで定量化し(Lopez−Vidal et al 1998)、血清中のIgA又はIgG+M抗体の力価をELISAアッセイで決定した(Rudin et al,1994)。
【0049】
ホルマリンによる細菌の不活性化
細菌を死滅させるため、各株の培養物をPBSで洗浄して、PBS中1010細菌/mlの密度まで再懸濁させた。ホルマリンを0.1Mの最終濃度まで加え、該懸濁液を37℃で2時間インキュベートして60rpmで撹拌し、次いで該懸濁液を4℃にて3日間インキュベートした。その後、細菌を洗浄し、同体積のPBSで再懸濁させ、その細菌懸濁液100μlを血液寒天培地上に広げて、37℃にて最大で1週間までインキュベートし、増殖を調べた。
【0050】
マウスの免疫化
メスのBalb/cマウス(生後6〜8週)を経口経路の免疫化に用いた。誘発してホルマリンで死滅した参照株325542−3及び58R957と組み換え株TOP10−CFA/I−CS2の培養物を洗浄し、所望の細菌密度までPBS中に再懸濁させた。先に説明されるように、経口経路による免疫化のため、109の細菌を10μgのCTと一緒に用いた(Rhagavan et al,200)。全てのマウスに2つの同一の免疫化を2週間おいて行い、第一の投与の直前と、第二の投与の2週間後に出血を収集した。
【0051】
統計的分析
全てのELISA実験及び阻害ELISA実験を二つ組で行い、異なる日に少なくとも3回繰り返した。特定のテストそれぞれについてのドットブロット実験を少なくとも2回繰り返した。学生のt−テストによって統計的分析を行い、P<0.05を有意性ありとみなした。
【実施例】
【0052】
実施例1
ハイブリッド線毛の産生:我々は、CFA/I及びCS2双方の主要サブユニットからなるハイブリッド線毛を発現させる可能性について調べた。かかるハイブリッド線毛を発現する発現ベクターの構築は、材料及び方法において説明されており、図1において表現される。次いで、該ベクターをTop10株中で増殖し、CFA/I及びCS2双方の主要サブユニットからなる線毛を発現する組み換え株が得られた。透過電子顕微鏡法(TEM)と、各主要サブユニットに対する特異的MAbs、即ち、α-CFA/I MAb(1:6)〜ヤギのα−マウスIgG 20mn 金及びビオチン化α-CS2 MAb(10:3)〜ストレプトアビジン 10nm 金を用いて、発現を検証した。
【0053】
ここで引用される参考文献の教示は、参照することによりこの明細書に組み込まれる。
【0054】
【表1】
【先行技術文献】
【非特許文献】
【0055】
【非特許文献1】Ahren, C. M. Jertborn, and A.M. Svennerholm. 1998. Intestinal immune responses to an inactivated oral enterotoxigenic Escherichia coli vaccine and associated immunoglobulin A responses in blood. Infect. Immun. 66:3311-3316.
【非特許文献2】Binsztein, N., M.J. Jouve, G.I. Viboud, L.L. Moral, M. Rivas, I. Orskov, C. Ahren, and A-M. Svennerholm. 1991. Colonization factors of enterotoxigenic Escherichia coli isolated from children with diarrhea in Argentina. J. Clin. Microbiol. 29:1893-1898.
【非特許文献3】Gaastra, W., and A.M. Svennerholm. 1996. Colonization factors of human enterotoxigenic Escherichia coli (ETEC). Trends Microbiol. 4:444-452.
【非特許文献4】Jertborn, M., C. Ahren, J. Holmgren, A.M. Svennerholm. 1993. Safety and immunogenicity of an oral inactivated enterotoxigenic Escherichia coli vaccine. Vaccine 16: 255-60.
【非特許文献5】Levine, M.M., J.A. Giron, and F. Noriega. 1994. Fimbrial vaccines,. In Fimbriae: adhesion, biogenics, genetics and vaccines, P. Klemm (ed.), pp.255-270. CRC Press, Boca Raton, Fla.
【非特許文献6】Qadri F., A.M. Svennerholm, A. S. G. Faruque, and Sack R.B. 2005a. Enterotoxigenic Escherichia coli in developing countries: epidemiology, microbiology, clinical features, treatment, and prevention. Clin. Microbiol. Rev. 18:465-483.
【非特許文献7】Qadri, F., T. Ahmed, F. Ahmed, Y.A. Begum, D.A. Sack, A.M. Svennerholm and the PTE Study Group. 2005b. Reduced doses of oral killed enterotoxigenic Escherichia coli plus cholera toxin B subunit vaccine is safe and immunogenic in Bangladeshi infants 6-17 months of age: Dosing studies in different age groups. Vaccine 24:1726-1733.
【非特許文献8】Rhagavan S, M. Hjulstroem, J. Holmgren and A.M. Svennerholm. 2002. Protection against experimental Helicobacter pylori infection after immunization with inactivated H. pylori whole cell vaccines. Infect. Immun. 70:6383-6388.
【非特許文献9】Rudin, A., M.M. McConnell, A.M. Svennerholm. 1994. Monoclonal antibodies against enterotoxigenic Escherichia coli colonization factor antigen I (CFA/I) that cross-react immunologically with heterologous CFAs. Infect. Immun. 62:4339-4346.
【非特許文献10】Sack, D.A., J. Shimko, O. Torres et al. 2002. Safety and efficacy of a killed oral vaccine for enterotoxigenic E. coli diarrhea in adult travelers to Guatemala and Mexico. 42nd Interscience Conference on Antimcrobial Agents and Chemotherapy. San Diego, CA. Abstract.
【非特許文献11】Sanchez J., and Holmgren. 2005. Virulence factors, pathogenesis and vaccine protection in cholera and ETEC diarrhea. Curr. Opin. Immunol. 17:388-98.
【非特許文献12】Savarino, S.J., F.M. Brown, E. Hall, S. Bassily, F. Youssef, T. Wierzba, L. Peruski, N.A. El-Masry, M. Safwat, M. Rao, M. Jertborn, A.M. Svennerholm, Y.J. Lee, and J.D. Clemens. 1998. Safety and immunogenicity of an oral, killed enterotoxigenic Escherichia coli-cholera toxin B subunit vaccine in Egyptian adults. J. Infect. Dis. 177:796-799.
【非特許文献13】Savarino, S.J., E. Hall,. S. Bassily,. F.M. Brown, F. Youssef, T.F. Wierzba,. L. Peruski, N.A. El-Masry, M. Safwat, M. Rao, H. El Mohamady, R. Abu-Elyazeed, A. Naficy, A.M. Svennerholm, M. Jertborn, Y.J. Lee, and J.D. Clemens. 1999. Oral, inactivated, whole cell enterotoxigenic Escherichia coli plus cholera toxin B subunit vaccine: results of the initial evaluation in children. J. Infect. Dis. 179:107-14.
【非特許文献14】Svennerholm, A.-M., Y.L. Vidal, J. Holmgren, M.M. McConnell, and B. Rowe. 1988. Role of PCF8775 antigen and its coli surface subcomponents for colonization, disease, and protective immunogenicity of enterotoxigenic Escherichia coli in rabbits. Infect. Immun. 56:523-528.
【非特許文献15】Svennerholm, A.-M., C. Wenneras, J. Holmgren, M.M. McConnell, and B. Rowe. 1990. Roles of different coli surface antigens of colonization factor antigen II in colonization by and protective immunogenicity of enterotoxigenic Escherichia coli in rabbits. Infect. Immun. 58:341-346.
【非特許文献16】Svennerholm, A.-M., and J. Holmgren. 1995. Oral vaccines against cholera and enterotoxigenic Escherichia coli diarrhea. Adv. Exp. Med. Biol. 371B:1623-1628.
【非特許文献17】Svennerholm, A.-M., and S. J. Savarino. 2004. Oral inactivated whole cell B subunit combination vaccine against enterotoxigenic Escherichia coli. In New Generation Vaccines: 3rd ed. Eds: Levine M.M. et al., pp. 737-750, Marcel Decker, New York, NY.
【非特許文献18】Svennerholm, A.-M., and D. Steele. 2004. Progress in enteric vaccine development. In: Microbial-gut interactions in health and disease, Ed. M. Farthing. Best Pract. Res Clin. Gastroenterol. 18: 421-445.
【特許請求の範囲】
【請求項1】
腸管毒素原性大腸菌(ETEC)に関連した定着因子抗原(CFs)の少なくとも2つの主要サブユニットをコードする少なくとも2つの構造遺伝子が存在することを特徴とする遺伝子集合体を含む組み換えオペロン。
【請求項2】
請求項1に記載の組み換えオペロンを含むように遺伝子操作された宿主細胞であって、前記オペロンがエピソームエレメント上に位置するか又は前記宿主細胞の染色体に組み込まれていることを特徴とする宿主細胞。
【請求項3】
前記エピソームエレメントがプラスミドであることを特徴とする請求項1又は2に記載の宿主細胞。
【請求項4】
前記定着因子抗原(CFs)の少なくとも2つの主要サブユニットが同じものであるか又は異なるものであることを特徴とする請求項2又は3に記載の宿主細胞。
【請求項5】
前記CFsが、CFA/I、CS1、CS2、CS4、CS14、CS17、CS19及び推定定着因子O71(PCFO71)からなる群から選択されることを特徴とする請求項2〜4のいずれかに記載の宿主細胞。
【請求項6】
前記細胞が、前記CFsの少なくとも2つの主要サブユニットを発現することを特徴とする請求項2〜5のいずれかに記載の宿主細胞。
【請求項7】
前記宿主細胞が、細菌及び単細胞真核生物からなる群から選択される生存可能な微生物であることを特徴とする請求項2〜6のいずれかに記載の宿主細胞。
【請求項8】
前記宿主細胞が、大腸菌細胞であることを特徴とする請求項7に記載の宿主細胞。
【請求項9】
前記大腸菌細胞が、毒素非産生性の大腸菌細胞であることを特徴とする請求項8に記載の宿主細胞。
【請求項10】
前記細胞が、抗生物質耐性遺伝子を発現しないことを特徴とする請求項2〜9のいずれかに記載の宿主細胞。
【請求項11】
前記宿主細胞が、一つ又はそれ以上のプラスミドによって補完される一つ又はそれ以上の補完可能な染色体欠失又は染色体変異を保有することを特徴とする請求項2〜10のいずれかに記載の宿主細胞。
【請求項12】
腸管毒素原性大腸菌(ETEC)に関連した定着因子抗原(CFs)の少なくとも2つの主要サブユニットを発現することが可能な宿主細胞の製造方法であって、
オペロン内にハイブリッドETEC CFの発現に必要な遺伝子又は遺伝子断片を集合させる工程と、前記サブユニットの発現を制御するETECプロモーターを集合させる工程と、前記宿主細胞のゲノムに前記オペロンを組み込むか又は前記オペロン、プラスミドを維持するための選択マーカー及び複製開始点を備えるプラスミドで前記宿主細胞を形質転換させる工程とを含むことを特徴とする方法。
【請求項13】
薬学的に容認される賦形剤、緩衝剤及び/又は希釈剤と共に、請求項2〜11のいずれかに記載の宿主細胞を少なくとも一つ含むことを特徴とする抗下痢症ワクチン組成物。
【請求項14】
薬学的に容認される賦形剤、緩衝剤及び/又は希釈剤が、ワクチンの経口送達用に選択されることを特徴とする請求項13に記載のワクチン。
【請求項15】
ワクチンの製造における請求項1に記載のオペロンの使用。
【請求項1】
腸管毒素原性大腸菌(ETEC)に関連した定着因子抗原(CFs)の少なくとも2つの主要サブユニットをコードする少なくとも2つの構造遺伝子が存在することを特徴とする遺伝子集合体を含む組み換えオペロン。
【請求項2】
請求項1に記載の組み換えオペロンを含むように遺伝子操作された宿主細胞であって、前記オペロンがエピソームエレメント上に位置するか又は前記宿主細胞の染色体に組み込まれていることを特徴とする宿主細胞。
【請求項3】
前記エピソームエレメントがプラスミドであることを特徴とする請求項1又は2に記載の宿主細胞。
【請求項4】
前記定着因子抗原(CFs)の少なくとも2つの主要サブユニットが同じものであるか又は異なるものであることを特徴とする請求項2又は3に記載の宿主細胞。
【請求項5】
前記CFsが、CFA/I、CS1、CS2、CS4、CS14、CS17、CS19及び推定定着因子O71(PCFO71)からなる群から選択されることを特徴とする請求項2〜4のいずれかに記載の宿主細胞。
【請求項6】
前記細胞が、前記CFsの少なくとも2つの主要サブユニットを発現することを特徴とする請求項2〜5のいずれかに記載の宿主細胞。
【請求項7】
前記宿主細胞が、細菌及び単細胞真核生物からなる群から選択される生存可能な微生物であることを特徴とする請求項2〜6のいずれかに記載の宿主細胞。
【請求項8】
前記宿主細胞が、大腸菌細胞であることを特徴とする請求項7に記載の宿主細胞。
【請求項9】
前記大腸菌細胞が、毒素非産生性の大腸菌細胞であることを特徴とする請求項8に記載の宿主細胞。
【請求項10】
前記細胞が、抗生物質耐性遺伝子を発現しないことを特徴とする請求項2〜9のいずれかに記載の宿主細胞。
【請求項11】
前記宿主細胞が、一つ又はそれ以上のプラスミドによって補完される一つ又はそれ以上の補完可能な染色体欠失又は染色体変異を保有することを特徴とする請求項2〜10のいずれかに記載の宿主細胞。
【請求項12】
腸管毒素原性大腸菌(ETEC)に関連した定着因子抗原(CFs)の少なくとも2つの主要サブユニットを発現することが可能な宿主細胞の製造方法であって、
オペロン内にハイブリッドETEC CFの発現に必要な遺伝子又は遺伝子断片を集合させる工程と、前記サブユニットの発現を制御するETECプロモーターを集合させる工程と、前記宿主細胞のゲノムに前記オペロンを組み込むか又は前記オペロン、プラスミドを維持するための選択マーカー及び複製開始点を備えるプラスミドで前記宿主細胞を形質転換させる工程とを含むことを特徴とする方法。
【請求項13】
薬学的に容認される賦形剤、緩衝剤及び/又は希釈剤と共に、請求項2〜11のいずれかに記載の宿主細胞を少なくとも一つ含むことを特徴とする抗下痢症ワクチン組成物。
【請求項14】
薬学的に容認される賦形剤、緩衝剤及び/又は希釈剤が、ワクチンの経口送達用に選択されることを特徴とする請求項13に記載のワクチン。
【請求項15】
ワクチンの製造における請求項1に記載のオペロンの使用。
【図1】

【公表番号】特表2010−531653(P2010−531653A)
【公表日】平成22年9月30日(2010.9.30)
【国際特許分類】
【出願番号】特願2010−513967(P2010−513967)
【出願日】平成20年7月1日(2008.7.1)
【国際出願番号】PCT/EP2008/058438
【国際公開番号】WO2009/004002
【国際公開日】平成21年1月8日(2009.1.8)
【出願人】(510003667)クルセル スウェーデン アーベー (1)
【Fターム(参考)】
【公表日】平成22年9月30日(2010.9.30)
【国際特許分類】
【出願日】平成20年7月1日(2008.7.1)
【国際出願番号】PCT/EP2008/058438
【国際公開番号】WO2009/004002
【国際公開日】平成21年1月8日(2009.1.8)
【出願人】(510003667)クルセル スウェーデン アーベー (1)
【Fターム(参考)】
[ Back to top ]