
キメラ第VII因子分子
本発明は、キメラ第VII因子ポリペプチドおよびそれを使用する方法に関する。

【発明の詳細な説明】
【技術分野】
【0001】
[優先権の主張]
本出願は、35 U.S.C.§119(e)の下に、その内容全体を引用することにより本明細書の一部をなすものとする、2009年6月25日出願の米国特許仮出願第61/220,278号の優先権を主張する。
【0002】
[政府支援の言明]
本発明は、米国国立衛生研究所によって授与された認可番号5−P01−HL06350の下での政府支援を得て為された。米国政府は本発明に一定の権利を有する。
【0003】
本発明は、現在使用可能な第VII因子ポリペプチドよりも高い凝固活性を有し、血栓合併症がより少ないキメラヒト凝固第VII因子(FVII)ポリペプチド、ならびにかかるポリペプチドをコードするポリヌクレオチド構築物、該ポリヌクレオチドを含み、これを発現するベクターおよび宿主細胞、医薬組成物、使用および治療の方法に関する。
【背景技術】
【0004】
血液凝固は、最終的にフィブリン血餅を生じさせる、様々な血液成分(または因子)の複雑な相互作用からなるプロセスである。一般に、凝固「カスケード」と称されてきたものに関与する血液成分は、アクチベータ(これ自体は活性化された凝固因子である)の作用によってタンパク質分解酵素に変換される、酵素的に不活性なタンパク質(プロ酵素またはチモーゲン)である。かかる変換を受けた凝固因子は、一般に「活性因子」と称され、凝固因子の名称に「a」の文字を付加することによって表される(例えば活性化第VII因子は第VIIa因子またはFVIIaと表される)。
【0005】
通常、止血プロセスの開始は、組織因子と第VIIa因子との間の複合体の形成によって媒介される。この複合体は、次に、第IX因子(FIX)および第X因子(FX)をそれらの活性型に変換する。第Xa因子(FXa)は、組織因子担持細胞上で限られた量のプロトロンビンをトロンビンに変換する。トロンビンは血小板を活性化し、第V因子(FV)および第VIII因子(FVIII)を第Va因子(FVa)および第VIIIa因子(FVIIIa)へと活性化して、これらの両補因子がさらなるプロセスにおいて完全なトロンビンバーストを導く。このプロセスは、第IXa因子(FIXa)(第VIIIa因子との複合体として)による第Xa因子の産生を含み、活性化された血小板の表面上で起こる。トロンビンは、最終的にフィブリノーゲンをフィブリンへと変換し、フィブリン血餅の形成をもたらす。近年、第VII因子および組織因子が血液凝固の主たるイニシエータであることが見出された。
【0006】
第VII因子は、一本鎖チモーゲンとして血液中を循環する血漿糖タンパク質である。チモーゲンは最小限の触媒活性を有する。一本鎖第VII因子は、インビトロで第Xa因子、第XIIa因子、第IXa因子、第VIIa因子またはトロンビンによって二本鎖第VIIa因子に変換され得る。第Xa因子は第VII因子の主要な生理的アクチベータであると考えられている。止血に関与するいくつかの他の血漿タンパク質と同様に、第VII因子はその活性に関してビタミンKに依存し、該活性は該タンパク質のアミノ末端に近接してクラスタ形成している複数のグルタミン酸残基のγ−カルボキシル化のために必要である。これらのγ−カルボキシル化されたグルタミン酸は、リン脂質と第VII因子との金属イオン誘導性相互作用に必要とされる。チモーゲン第VII因子の活性化二本鎖分子への変換は、内部のArg152−Ile153ペプチド結合の切断によって生じる。加えて、高濃度の第VII因子がインビトロで自己活性化を導くことは公知である。組織因子、リン脂質およびカルシウムイオンの存在下で、二本鎖第VIIa因子は、限定的なタンパク質分解によって第X因子または第IX因子を速やかに活性化する。
【0007】
ヒトFVII(hFVII)をコードする遺伝子は、第13番染色体のq34−qter9にマッピングされている(de Grouchy et al.,Hum Genet 1984;66:230−233)。この遺伝子は9個のエクソンを含み、12.8Kbにわたる(O’Hara et al.,Proc Natl Acad Sci USA 1987;84:5158−5162)。FVIIの遺伝子構成およびタンパク質構造は他のビタミンK依存性プロ凝固タンパク質のものに類似し、エクソン1aおよび1bはシグナル配列をコードし、エクソン2はプロペプチドおよびGLAドメイン、エクソン3は短い疎水性領域、エクソン4および5は上皮増殖因子様ドメイン、ならびにエクソン6から8はセリンプロテアーゼ触媒ドメインをコードする(Yoshitake et al.,Biochemistry 1985;24:3736−3750)。
【0008】
第IX因子(クリスマス因子)は、正常な止血において活性なセリンプロテアーゼのチモーゲンであり、その酵素活性は特定のグルタミン酸残基のカルボキシル化を必要とする。第IX、X、VII因子およびプロテインCは、同じファミリーのセリンプロテアーゼの密接に関連するパラログであり、これらのタンパク質をコードする遺伝子の高い度合のアミノ酸配列同一性およびイントロン−エクソン配置を有する。これらの密接に関連するタンパク質は、アミノ末端からカルボキシル末端までの機能性ドメインの類似構造を有し、γ−カルボキシグルタミン酸(GLA)ドメイン、2つの上皮増殖因子様(EGF)ドメイン、活性化ペプチドおよび触媒ドメインを含む。プロテインSは、666アミノ酸の、GLAドメイン、4つのEGF様ドメイン、トロンビン感受性領域および2つのラミニンドメインを有するビタミンK依存性タンパク質である。
【0009】
ビタミンK依存性凝固血漿タンパク質は、膜へのタンパク質結合の部位として機能するGLAドメインを含み、GLAドメインは様々な凝固タンパク質の間で高度に保存されている。それらの類似性にもかかわらず、GLAドメインはリン脂質に対して広い範囲の親和性を示し、プロテインSのGLAドメインがリン脂質に対して最も高い親和性を有する(Ellison et al.,Biochemistry,1998;37:7997−8003)、(McDonald et al.,Biochemistry 1997;36:5120−27)。
【0010】
被験体において凝固カスケードを刺激するまたは改善することがしばしば望ましい。第VIIa因子は、凝固因子欠損症(例えば血友病AおよびBまたは凝固第XI因子もしくは第VII因子の欠損)または凝固因子インヒビターなどのいくつかの原因を有する出血性疾患を管理するために使用されてきた。第VIIa因子はまた、正常に機能する血液凝固カスケードを有する(凝固因子欠損、または凝固因子のいずれかに対するインヒビターを有さない)被験体において生じる過剰の出血を制御するためにも使用されてきた。かかる出血は、例えば、血小板機能欠損、血小板減少症またはフォン・ヴィレブランド病によって引き起こされ得る。
【0011】
出血はまた、手術および他の形態の外傷に関連する重大な問題である。例えば、第VII因子はイラクおよびアフガニスタンで負傷した兵士を治療するために広く使用された(Perkins JG,et al.The Journal of Trauma.2007;62:1095−9;discussion 9−101)。その使用は多くの命を救ったことで高い評価を得ているが、大部分の医学的治療に関しては、治療後の脳卒中または他の血栓事象などの副作用が存在する。しかし、FVIIaを使用する医師の全体的な印象は、その使用はそのために失われた命よりも多くの命を救ってきたというものである。おそらく、このことを最もよく表しているのは、過去の戦争において負傷者の約30%が損傷のために死亡しているのに対し、現在の湾岸戦争ではその数が約10%に減少したことである(Gawande A,et al.,N Engl J Med.2004;351:2471−5)。
【0012】
第VIIa因子を発現するトランスジェニック血友病Bマウスの試験は、低レベル(1.5μg/ml未満)の持続的な第VIIa因子発現が血友病Bマウスにおいて凝固活性を回復させることを明らかにした。しかし、野生型または血友病Bマウスでの2μg/mlを上回る第VIIa因子レベルは、心臓および肺における血栓症を導いた。心臓および肺はどちらも、組織因子の高発現部位である。これは、循環中の高レベルの第VIIa因子は、心臓および肺の血管損傷後に暴露される組織因子に接触した場合血栓症を誘発することを示唆する(Margaritas et al.,J.Clin.Invest.2004;113:1025−31)。さらに、血友病のイヌにおけるベクターを介したイヌ第VIIa因子の遺伝子導入が、短期および中期的に安全かつ有効であることが試験で示された(Margaritas et al.,Gene Therapy 2009;113:3682−3689)。
【0013】
第VII因子による治療に関する警告が、現在、規制認可を求める製品について提案されている。例えば、欧州医薬品庁、人体用医薬品評価ユニット(European Medicines Agency,Human Medicines Evaluation Unit)は、現在の第VII因子治療には、特に第VII因子が冠状動脈性心疾患または肝疾患の病歴を有する患者、術後患者、新生児ならびに血栓症および播種性血管内凝固症候群の危険性がある患者に投与される状況では、血栓症および播種性血管内凝固症候群の危険性の警告を付すことを推奨している。例えば、2004年7月のCore SPC for Human Plasma Derived Coagulation Factor VII Products(CPMP/BPWG/2048/01)参照。
【0014】
これまでに、第VIIa因子のEGF−1ドメインが組織因子に対する第VIIa因子の親和性に重要な役割を果たすことが示されている。組織因子の存在下および不存在下の両方で、合成基質と第X因子の両方を使用して、第IX因子のEGF−1ドメインを有する第VIIa因子ポリペプチドは野生型第VIIa因子よりも低い触媒活性を有していた(Jin et al.,Biochemistry,1999,28:1185−92)。一見したところ、これは出血を治療するためのキメラ構築物の使用を排除すると思われる。しかし、Monroe(British Journal of Haematology 1997;99:542−549)は、血友病および出血を治療するうえでのFVIIaの機構は組織因子非依存性であると提案した。しかし、第VIIa因子が組織因子とは独立して活性でないかどうかに関しては当技術分野で見解が分かれる。
【0015】
ヒト組換えFVIIaの市販製剤は、NovoSeven(登録商標)およびNovoSeven(登録商標)RTとして販売されている。NovoSeven(登録商標)およびNovoSeven(登録商標)RTは、血友病AまたはB患者における出血エピソードの治療に適用され、出血エピソードの治療のための市場で入手可能な唯一のrFVIIaである。近年、NovoSeven(登録商標)が、第VII因子を損傷部位に局在化させるために組み合わせて投与できる、再水和された凍結乾燥血小板に結合し得ることが明らかにされた(Fischer et al.,Platelets,2008;19:182−91)。加えて、第VII因子の選択的なPEG化は血漿半減期を上昇させ得ること(Stennicke et al.,Thromb.Haemost,2008;100:920−28)および3個のアミノ酸置換を有する組換えヒト第VII因子は血小板の表面上で高い活性を有すること(Moss et al.,J.Thromb.Haemost.,2009;7:299−305)が示された。本発明のキメラ第VIIa因子分子のPEG化は、同様に血漿半減期を上昇させるように働くと期待される。同じように、コンジュゲートを形成する非ポリペプチド部分の共有結合などの、当技術分野で知られているタンパク質の他の修飾、例えばグリコシル化は、本発明のキメラ第VIIa因子分子において同様に機能すると期待され、すなわち非ポリペプチド部分の共有結合によってタンパク質に与えられる特性がキメラ第VIIa因子分子に付与されると予想される。
【発明の概要】
【発明が解決しようとする課題】
【0016】
利用可能な治療に結びつく、比較的低い用量で投与できる高い凝固活性を有する第VIIa因子の変異体、および血栓合併症などの望ましくない副作用を生じさせることが少ない変異体が必要とされている。
【課題を解決するための手段】
【0017】
本出願は、1以上の望ましい恩恵を提供する、キメラFVIIa分子、特にhFVIIaドメインと凝固系の1以上のタンパク質からのドメインを含むキメラhFVIIa分子を開示する。従って、本発明のキメラFVIIa分子は、市販のrFVIIaと比較して1以上の改善された特性を有し、かかる特性は、より高い凝固活性を有することおよび/または比較的低い用量で投与できることおよび/または血栓合併症を生じさせることがより少ないことを含む。その結果として、本発明のキメラによる医学的治療は、潜在的により低い用量および/またはより少ない望ましくない副作用などの、現在使用可能なrFVIIa化合物を上回る利点を提供する。
【0018】
本発明の代表的なキメラFVIIaポリペプチドは、FVIIのEGF−2および触媒ドメインと、ビタミンK依存性凝固タンパク質のGLAドメインと、ビタミンK依存性凝固タンパク質のEGF−1ドメインとを含むキメラFVIIaを包含する。本発明の特定の実施形態では、本発明のキメラFVIIaポリペプチドは、1)FIXのGLAおよびEGF−1ドメインとFVIIのEGF−2および触媒ドメインとを含むキメラFVIIa、2)FIXのEGF−1ドメインとFVIIのGLA、EGF−2および触媒ドメインとを含むキメラFVIIa、3)プロテインSのGLAドメインと、FIXのEGF−1ドメインと、FVIIのEGF−2および触媒ドメインとを含むキメラFVIIa、4)プロテインSのGLAおよびEGF−1ドメインとFVIIのEGF−2および触媒ドメインとを含むキメラFVIIa、5)プロテインSのEGF−1ドメインとFVIIのGLA、EGF−2および触媒ドメインとを含むキメラFVIIa、ならびに6)プロテインSのEGF−1ドメインと、FIXのGLAドメインと、FVIIのEGF−2および触媒ドメインとを含むキメラFVIIaを包含する。本発明の代表的なキメラFVIIaポリペプチドはまた、EGF−1ドメインまたはGLAドメイン内にアミノ酸置換を有する、野生型FVIIaまたは上述したキメラFVIIaポリペプチドのいずれかを包含し得る。置換は、保存的置換または非保存的置換であり得る。かかる置換は、EGF−1ドメインの残基69のイソロイシンのアラニンによる置換および/または残基79のアルギニンのアラニンによる置換を含み得る。さらなる置換は、GLAドメインにIV型コラーゲンに対するより高い結合親和性をもたらすが、血小板結合には影響を及ぼさないと思われる、残基5のリシンのアルギニンによる置換(Gui et al.,J.Thromb Haemost.2009;7:1843−1851)、残基306のメチオニンの別のアミノ酸による置換、好ましくは保存的アミノ酸置換、より好ましくは、組織因子に対する親和性をさらに低下させるアラニンによる置換、ならびにより高い比活性を有する第VIIa因子を生じさせる、残基158のバリンのアスパラギン酸による置換、残基296のグルタミン酸のバリンによる置換および/または残基298のメチオニンのグルタミンによる置換を含み得る。
【0019】
また、本明細書で述べるキメラFVIIaポリペプチドの1以上を投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法も提供される。出血性疾患を治療する方法は、本発明のキメラ第VIIa因子ポリペプチドをコードするヌクレオチド配列を含む核酸分子を被験体に投与する方法を含み得る。
【0020】
加えて、本発明は、GLAドメインを含むタンパク質を投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法を提供し、該タンパク質は、血餅の部位に標的されるおよび/または被験体の血漿中の循環タンパク質の濃度に比べて血餅の部位付近で濃縮されている。これは、血餅形成部位でまたはその付近で認められる、現在使用可能な第VII因子ポリペプチドよりも高い親和性で血小板に結合するドメインを使用することを含み得る。かかるドメインの例は、血小板の表面に位置する負に荷電したリン脂質層に結合する様々な凝固タンパク質のGLAドメインである。かかるGLAドメインの非限定的な例は、FVIIaのGLAドメインよりもはるかに密接に血小板およびリン脂質に結合する、FIXのGLAドメイン(Melton et al.「Location of the platelet binding site in zymogen coagulation factor IX」Blood Coagul Fibrinolysis 12:237−243(2001))ならびに任意の他の公知のGLAドメインよりも密接にリン脂質に結合する、プロテインSのGLAドメイン(McDonald et al.「Comparison of naturally occurring vitamin K−dependent proteins:correlation of amino acid sequences and membrane binding properties suggests a membrane contact site」Biochemistry:36:5120−5127(1997))を含む。GLAドメインを含むタンパク質の非限定的な例は、FIXもしくはプロテインSのGLAドメインを含む組換えタンパク質、FIXもしくはプロテインSのGLAドメインを含むキメラタンパク質、および/またはFIXもしくはプロテインSのGLAドメインを含む本発明のキメラFVIIaポリペプチドを包含する。
【0021】
もう1つの実施形態では、本発明は、第VII因子ポリペプチドに由来する触媒ドメインおよび1以上の血小板標的ドメインを含む本発明のキメラ第VIIa因子ポリペプチドを投与することにより、出血性疾患を有する被験体を治療する方法を提供する。かかる血小板標的ドメインは、血小板の表面と相互作用するまたは結合するタンパク質からのドメインを含む。血小板の表面とのかかる相互作用または表面への結合は、血小板の膜リン脂質を通してまたは血小板細胞表面タンパク質および/もしくは受容体を通して媒介され得る。かかるドメインの非限定的な例は、血小板糖タンパク質Ibについての主要結合部位である、フォン・ヴィレブランド因子のA1ドメイン(Emsley et al.,JBC,273:10396−10401(1998))ならびに血小板膜タンパク質および/または受容体に結合する抗体、例えば血小板膜リン脂質に結合する抗体のFabフラグメント(ドメイン)(Out et al.,Blood,77:2655−2659(1991))を含む。
【0022】
血小板に結合し、組織因子に対して低い親和性を有するドメインを使用した、血餅部位への凝固タンパク質のかかる標的化は、血栓形成性などの、現在の第VII因子療法に関連する合併症の危険度を低下させるという付加的な、しかし予想外の利益を有する。このアプローチはまた、他の治療上有用なポリペプチドまたは他の分子を血小板または血餅部位に標的するためにも使用できる。かかる治療上有用な分子は、抗凝固剤等を含み得る。
【0023】
本発明はさらに、第VII因子の血栓形成性に比べて低い血栓形成性(例えば、天然第VII因子の血栓形成性と比較して少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、90%低い等)を有するポリペプチドを被験体に投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法を提供する。「低い血栓形成性」は、当技術分野で知られている方法に従って様々なやり方で、例えば、対照と比較して、血餅の測定数がより少ない、血餅がより小さい、血餅が生じるのにより長い時間を要する(インビボまたはインビトロアッセイにおいて)、血栓形成による被験体の死亡がより少ないおよび/または生存期間が長いことを含むが、これらに限定されない方法で測定できる。第VII因子と比較して低い血栓形成性を有するポリペプチドは、例えば、本発明のキメラ第VII因子ポリペプチドまたはその活性フラグメントであり得る。かかるポリペプチドの非限定的な例は、FIXのGLAおよびEGF−1ドメインとFVIIのEGF−2および触媒ドメインとを含むキメラFVIIa、ならびにプロテインSのGLAドメインと、FIXのEGF−1ドメインと、FVIIのEGF−2および触媒ドメインとを含むキメラFVIIaを包含する。
【図面の簡単な説明】
【0024】
【図1】図1は、本発明において有用なキメラFVIIa分子の図式的表示を示す。FVII:第VII因子、FIX:第IX因子。
【図2】図2は、様々な濃度の野生型第VIIa因子についての血友病の細胞モデル系における正常条件および血友病条件(第IX因子および第VIII因子の除去)でのトロンビン産生(nM)を示す。
【図3】図3は、50nMの野生型第VIIa因子または10nMのキメラ第VIIa因子を投与した血友病の細胞モデル系における正常条件および血友病条件(第IX因子および第VIII因子の除去)でのトロンビン産生(nM)を示す。
【図4】図4は、50nMの野生型第VIIa因子または10nMのキメラ第VIIa因子の投与後の血友病の細胞モデル系において産生される、対照値と比較した、ピークトロンビンを示す。
【図5】図5は、処置していない血友病Bマウス、2mg/kgのNovoSeven(登録商標)を投与した血友病Bマウス、2mg/kgのキメラFVIIa分子を投与した血友病Bマウスおよび処置していない野生型マウスについての凝固アッセイにおける崩壊止血時間(disruption hemostasis time)を示す。
【図6】図6は、シグナル、プロペプチド、第IX因子のGLAおよびEGF1ドメイン(下線)を含む、本発明の例示的なキメラ第VII因子の配列(配列番号:1)を示す。
【図7】図7は、シグナル、プロペプチドおよび第IX因子のEGF1ドメイン(下線)およびプロテインSのGLAドメイン(太字)を含む、本発明の例示的なキメラ第VII因子の配列(配列番号:2)を示す。
【発明を実施するための形態】
【0025】
本発明を以下でより詳細に説明する。しかし本発明は、種々の形態で具体化でき、本明細書で述べる実施形態に限定されると解釈されるべきではない。むしろ、これらの実施形態は、この開示が詳細かつ完全となり、当業者に本発明の範囲を十分に伝えるために提供されるものである。
【0026】
本明細書で本発明の説明において使用される用語は、特定の実施形態を説明することだけを目的とし、本発明の限定を意図されない。本発明の説明および付随の特許請求の範囲において使用される、「a」、「an」および「the」という単数形態は、文脈によって明確に指示されない限り、複数形態も包含することが意図されている。
【0027】
特に定義されない限り、本明細書で使用されるすべての用語(技術および学術用語を含む)は、本発明が属する技術分野の当業者によって一般的に理解されるのと同じ意味を有する。一般的に使用される辞書において定義されるもののような用語は、本出願および関連技術分野におけるそれらの意味と一致する意味を有すると解釈されるべきであり、本明細書において明白に定義されない限り理想的なまたは過度に正式な意味に解釈されるべきではないことがさらに了解される。本明細書で本発明の説明において使用される用語は、特定の実施形態を説明することを目的とし、本発明の限定を意図されない。本明細書で言及するすべての出版物、特許出願、特許および他の参考文献は、それらの全体を引用することにより本明細書の一部をなすものとする。
【0028】
また本明細書で使用される、「および/または」は、関連して列挙される項目の1以上のありとあらゆる可能な組合せ、ならびに選択的に解釈される場合(「または」)は組合せの欠如を指し、これらを包含する。
【0029】
文脈によって特に指示されない限り、本明細書で述べる本発明の様々な特徴は任意の組合せで使用できることが明確に意図されている。
【0030】
さらに、本発明はまた、本発明の一部の実施形態では、本明細書で述べる任意の特徴または特徴の組合せが排除または除外され得ることを企図する。例示すると、明細書において複合体が成分A、BおよびCを含むと記述される場合、A、BまたはCのいずれかまたはそれらの組合せが、単独でまたは任意の組合せで除外され、排除され得ることが明確に意図されている。
【0031】
本明細書で使用される、「〜から実質的になる」という移行句(および文法的変形)は、列挙される物質または段階ならびに特許請求される本発明の「基本的なおよび新規の特徴に実質的に影響を及ぼさないもの」を包含すると解釈されるべきである。In re Herz,537 F.2d 549,551−52,190 U.S.P.Q.461,463(CCPA 1976)(原文での強調)参照、MPEP §2111.03も参照のこと。従って、本明細書で使用される「〜から実質的になる」という用語は、「含む」と等価であると解釈されるべきではない。
【0032】
量または濃度等のような測定可能な値に言及する場合に本明細書で使用される、「約」という用語は、特定される量の20%、10%、5%、1%、0.5%またはさらに0.1%の変動を包含することが意図されている。
【0033】
本明細書で使用される「活性」という用語は、第VII因子ポリペプチドがその基質である第X因子を活性な第Xa因子に変換する能力を意味する。
【0034】
「固有活性」という用語はまた、組織因子の不存在下に活性化血小板の表面でトロンビンを産生する能力も包含する。
【0035】
「N末端GLAドメイン」という用語は、配列番号:18のアミノ酸配列の約アミノ酸残基61から約アミノ酸残基105までのアミノ酸配列、配列番号:19のアミノ酸配列の約アミノ酸残基47から約アミノ酸残基92までのアミノ酸配列、同定されたアミノ酸配列への任意の翻訳後修飾、同定されたアミノ酸配列における任意の保存的アミノ酸置換、同定されたアミノ酸配列へのアミノ酸残基の付加、同定されたアミノ酸配列からのアミノ酸残基の欠失、またはリン脂質膜に結合する凝固カスケードタンパク質からの任意の他のアミノ酸配列を包含する。
【0036】
「EGF−1」という用語は、ジスルフィド結合1〜3、2〜4および5〜6を含むほとんどすべての公知のEGF様ドメインを含む、EGF(上皮増殖因子)中ならびにまた細胞シグナル伝達および凝固タンパク質に関与する一連のタンパク質中でもともと認められる6個のシステインを含む30〜40アミノ酸の領域を表す。第VII因子のEGF−1ドメインは、配列番号:18のアミノ酸配列の約アミノ酸残基106から約アミノ酸残基142までである。第IX因子のEGF−1ドメインは、配列番号:19のアミノ酸配列の約アミノ酸残基93から約アミノ酸残基129までである。プロテインSのEGF−1ドメインは、配列番号:20のアミノ酸配列の約アミノ酸残基117から約アミノ酸残基155までである。これらのアミノ酸配列は、同定されたアミノ酸配列への任意の翻訳後修飾、同定されたアミノ酸配列における任意の保存的アミノ酸置換、同定されたアミノ酸配列へのアミノ酸残基の任意の付加および/または同定されたアミノ酸配列からのアミノ酸残基の任意の欠失を含み得る。
【0037】
「EGF−2」という用語は、(2またはそれ以上のEGF様ドメインの)一続きにおける2番目のEGF様ドメインを意味する。第VII因子のEGF−2ドメインは、配列番号:18のアミノ酸配列の約アミノ酸残基147から約アミノ酸残基188までである。第IX因子のEGF−2ドメインは、配列番号:19のアミノ酸配列の約アミノ酸残基130から約アミノ酸残基171までである。プロテインSのEGF−2ドメインは、配列番号:20のアミノ酸配列の約アミノ酸残基157から約アミノ酸残基200までである。これらのアミノ酸配列は、同定されたアミノ酸配列への任意の翻訳後修飾、同定されたアミノ酸配列における任意の保存的アミノ酸置換、同定されたアミノ酸配列へのアミノ酸残基の付加および/または同定されたアミノ酸配列からのアミノ酸残基の任意の欠失を含み得る。
【0038】
本明細書で使用される「触媒ドメイン」という用語は、ペプチド結合の切断を媒介するタンパク質中のドメインを意味する。第VII因子の触媒ドメインは、配列番号:18のアミノ酸配列の約アミノ酸残基213から約アミノ酸残基452までである。第IX因子の触媒ドメインは、配列番号:19のアミノ酸配列の約アミノ酸残基227から約アミノ酸残基459までである。これらのアミノ酸配列は、同定されたアミノ酸配列への任意の翻訳後修飾、同定されたアミノ酸配列における任意の保存的アミノ酸置換、同定されたアミノ酸配列へのアミノ酸残基の任意の付加および/または同定されたアミノ酸配列からのアミノ酸残基の任意の欠失を含み得る。
【0039】
「GLA」という3文字表記は4−カルボキシグルタミン酸(γ−カルボキシグルタミン酸)を意味する。
【0040】
「プロテアーゼドメイン」という用語は、一般に配列番号:18の約アミノ酸残基213からカルボキシ末端アミノ酸残基までとみなされる、ペプチド結合の切断を媒介するタンパク質中のドメインを意味する(第VIIa因子の重鎖)。本明細書で使用される「第VII因子ポリペプチド」という用語は、天然ヒト第VII因子のアミノ酸配列61〜466(配列番号:18)を含む任意のタンパク質またはその変異体もしくはフラグメントを意味する。これは、ヒト第VII因子、ヒト第VIIa因子およびそれらの変異体を含むが、これらに限定されない。本明細書で使用される「第VII因子」という用語は、不活性な一本鎖チモーゲン第VII因子分子ならびに活性化二本鎖第VII因子分子(第VIIa因子)を含むことが意図されている。これは、天然ヒト第VII因子または第VIIa因子のアミノ酸配列61〜466(配列番号:18)を有するタンパク質を包含する。当業者は、小さな配列変化は同様に機能すると予想されること、ならびに関与するドメイン、ポリペプチドおよび第VII因子は、本発明から逸脱することなくわずかに短縮もしくは伸長され得るまたは置換を含み得ることを認識する。従って、この定義はまた、わずかに修飾されたアミノ酸配列、例えばN末端アミノ酸欠失または付加を含む修飾されたN末端を有するタンパク質およびペプチドも包含し、一部の実施形態ではそれらのタンパク質は第VIIa因子の活性を実質的に保持する(例えば天然第VIIa因子の活性の約50%、60%、70%、80%、90%、95%等を保持する)。本明細書で使用される「第VIIa因子」または「FVIIa」という用語は、活性化形態(第VIIa因子)からなる生成物を意味する。上記定義内の「第VII因子」または「第VIIa因子」はまた、個体によって存在し、生じ得る天然の対立遺伝子変異も包含する。また、グリコシル化または他の翻訳後修飾の程度と位置は、発現の選択される宿主細胞、組織または動物種および宿主の細胞または組織環境の性質に依存して異なり得る。
【0041】
本明細書で使用される「ドメイン」という用語は、タンパク質鎖の残りの部分とは独立して進化し、機能し、存在し得るタンパク質配列および構造の一部を包含することが意図されている。ドメインはコンパクトな三次元構造を形成することができ、しばしば独立して安定化され、折りたたまれ得る。1つのドメインが様々な進化的に関連するタンパク質中に出現し得る。ドメインの長さは、約25アミノ酸から約500アミノ酸長までにわたる。「ドメイン」はまた、単数または複数のアミノ酸残基が保存的置換によって置き換えられた野生型タンパク質からのドメインも包含し得る。それらはタンパク質媒質中で自己安定性であるので、ドメインは、キメラタンパク質を作製するためにあるタンパク質と別のタンパク質との間で遺伝子操作によって「交換」され得る。
【0042】
本明細書で使用される「変異体(単数または複数)」という用語は、天然ヒト第VII因子または第VIIa因子のアミノ酸配列61〜466(配列番号:18)を有する第VII因子を表すことが意図され、ここで、親タンパク質の1もしくはそれ以上のアミノ酸が別のアミノ酸によって置換されている、および/または、親タンパク質の1もしくはそれ以上のアミノ酸が欠失している、および/または、1もしくはそれ以上のアミノ酸がタンパク質に挿入されている、および/または、1もしくはそれ以上のアミノ酸が親タンパク質に付加されている、および/または、GLAドメインが異なるタンパク質からのGLAドメイン(例えば血小板膜もしくはリン脂質膜に結合するGLAドメイン)で置換されている、および/または、GLAドメインが異なるタンパク質からの血小板結合ドメイン(例えば血小板糖タンパク質Ibについての主要結合部位である、フォン・ヴィレブランド因子のA1ドメイン(Emsley et al.,JBC,273:10396−10401(1998))ならびに血小板膜タンパク質および/もしくは受容体に結合する抗体、例えば血小板膜リン脂質に結合する抗体のFabフラグメント(ドメイン)(Out et al.,Blood,77:2655−2659(1991))で置換されている、および/または、第VII因子のEGF−1ドメインが異なるタンパク質からのEGF−1ドメイン(例えば第IX因子のEGF−1ドメインなどの、組織因子により低い親和性で結合するEGF−1ドメイン)で置換されている。かかる付加は、親タンパク質のN末端またはC末端またはその両方で、ならびに内部で生じ得る。従って、一部の実施形態では、本発明の「変異体(単数または複数)」は、活性化形態のFVIIの凝固活性をまだ有し得る。一部の実施形態では、本発明の変異体(単数または複数)は凝固活性を有さなくてもよい。
【0043】
従って、一部の実施形態では、変異体は、天然ヒト第VII因子または第VIIa因子のアミノ酸配列61〜466(配列番号:18)と少なくとも40%、50%、60%または70%同一である。例えば、FVIIのEGF−1ドメインはFIXのEGF−1ドメインと65.7%の同一性を有し、FVIIのGLAドメインはFIXのGLAドメインと58.6%の同一性およびプロテインSのGLAドメインと51%の同一性を有する。1つの実施形態では、変異体は、天然ヒト第VII因子または第VIIa因子のアミノ酸配列61〜466(配列番号:18)と少なくとも80%同一である。もう1つの実施形態では、変異体は、天然ヒト第VII因子または第VIIa因子のアミノ酸配列61〜466(配列番号:18)と少なくとも90%同一である。さらなる実施形態では、変異体は、天然ヒト第VII因子または第VIIa因子のアミノ酸配列61〜466(配列番号:18)と少なくとも95%同一である。
【0044】
本明細書で使用される「任意の他のアミノ酸」という用語は、その位置に天然に存在するアミノ酸とは異なる1つのアミノ酸を意味する。これは、ポリヌクレオチドによってコードされ得るアミノ酸を含むが、これに限定されない。好ましくは、異なるアミノ酸は天然L型であり、ポリヌクレオチドによってコードされ得る。具体的な例はL−システイン(Cys)である。
【0045】
本明細書で使用される、「作動可能に連結された」という用語は、配列の正常な機能を果たすことができるような相互の立体配置での、酵素的連結または別の方法による2またはそれ以上のヌクレオチド配列の共有結合を指す。例えば、プレ配列または分泌リーダーをコードするヌクレオチド配列は、それがポリペプチドの分泌に関与するプレタンパク質として発現される場合は、ポリペプチドをコードするヌクレオチド配列に作動可能に連結されている。プロモータまたはエンハンサは、それが対象とする配列の転写に影響を及ぼす場合は、コード配列に作動可能に連結されている。リボソーム結合部位は、それが対象とするタンパク質またはペプチドへの翻訳を促進するように位置づけられている場合は、コード配列に作動可能に連結されている。一般に、「作動可能に連結されている」とは、連結されているヌクレオチド配列が連続していることを意味し、分泌リーダーの場合は、連続しかつ読み枠内にあることを意味する。連結は、好都合な制限部位での結合によって最も容易に達成される。かかる部位が存在しない場合は、合成オリゴヌクレオチドアダプターまたはリンカーを標準的な組換えDNA法と共に使用する。
【0046】
本明細書で使用される「ベクター」という用語は、宿主細胞において増幅することができる任意の核酸実体を意味する。従って、ベクターは、自律複製ベクター、すなわち、その複製が染色体の複製から独立している、染色体外実体として存在するベクター、例えばプラスミドであり得る。あるいは、ベクターは、宿主細胞に導入された場合、宿主細胞のゲノムに組み込まれ、組み込まれた染色体と共に複製されるものであってもよい。ベクターの選択は、それが導入される宿主細胞に依存することが多い。ベクターは、プラスミドベクター、ファージベクター、ウイルスまたはコスミドベクターを含むが、これらに限定されない。ベクターは通常、複製起点および少なくとも1つの選択可能な遺伝子、すなわち容易に検出可能であるかまたはその存在が細胞増殖に必須である生成物をコードする遺伝子を含む。
【0047】
本明細書で使用される「宿主細胞」という用語は、雑種細胞を含む、異種DNAがその中で発現され得る任意の細胞を意味する。典型的な宿主細胞は、昆虫細胞、酵母細胞、BHK細胞、CHO細胞、HEK細胞およびCOS細胞などのヒト細胞を含む哺乳動物細胞を含むが、これらに限定されない。本発明を実施する場合、培養される宿主細胞は、好ましくは哺乳動物細胞、より好ましくは、限定されることなく、CHO細胞株(例えばATCC CCL 61)、COS−1細胞株(例えばATCC CRL 1650)、ベビーハムスター腎(BHK)細胞株およびHEK293細胞株(例えばATCC CRL 1573;Graham et al.,J.Gen.Virol.36:59−72,1977)を含む、樹立哺乳動物細胞株である。適切なBHK細胞株は、本明細書中ではBHK 570細胞と称する、tk−ts13BHK細胞株である(Waechter and Baserga,Proc.Natl.Acad.Sci.USA 79:1106−1110,1982)。BHK 570細胞株は、ATCCアクセッション番号CRL 10314の下に、American Type Culture Collection,10801 University,Boulevard,Manassas,Va.20110から入手可能である。tk−ts13BHK細胞株も、アクセッション番号CRL 1632の下にATCCから入手可能である。他の適切な細胞株は、限定されることなく、Rat Hep I細胞(ラット肝細胞癌;ATCC CRL 1600)、Rat Hep II細胞(ラット肝細胞癌;ATCC CRL 1548)、TCMK細胞(ATCC CCL 139)、ヒト肺細胞(ATCC HB 8065)、NCTC 1469細胞(ATCC CCL 9.1)およびDUKX細胞(Urlaub and Chasin,Proc.Natl.Acad.Sci.USA 77:4216−4220,1980)を含む。また、3T3細胞、ナマルバ細胞、骨髄腫細胞および骨髄腫細胞と他の細胞の融合物も有用である。
【0048】
本明細書で使用される場合、「適切な増殖培地」という用語は、細胞の増殖および本発明の第VII因子ポリペプチドをコードする核酸配列の発現のために必要な栄養素および他の成分を含む培地を意味する。
【0049】
「コンジュゲート」(または交換可能に「コンジュゲート化ポリペプチド」)という用語は、1以上のポリペプチドの、ポリマー分子、親油性化合物、糖成分または有機誘導体化剤などの1以上の非ポリペプチド部分への共有結合によって形成される異種(複合体またはキメラという意味で)分子を示すことが意図されている。好ましくは、コンジュゲートは適切な濃度および条件で可溶性である、すなわち血液などの生理的液体に可溶性である。本発明のコンジュゲート化ポリペプチドの例は、グリコシル化および/またはPEG化ポリペプチドを含む。
【0050】
「共有結合」という用語は、ポリペプチドと非ポリペプチド部分が互いに直接共有結合で連結されているか、または架橋、スペーサーもしくは単数もしくは複数の連結部分などの、単数もしくは複数の介在部分を介して互いに間接的に共有結合で連結されていることを意味する。「非コンジュゲート化ポリペプチド」という用語は、コンジュゲートのポリペプチド部分を指すために使用できる。
【0051】
本明細書で使用される場合、「非ポリペプチド部分」という用語は、本発明のポリペプチドの結合基にコンジュゲート化することができる分子を意味する。かかる分子の適切な例は、ポリマー分子、糖成分、親油性化合物または有機誘導体化剤を含む。本発明のコンジュゲートに関連して使用される場合、非ポリペプチド部分はポリペプチドの結合基を介してコンジュゲートのポリペプチド部分に連結されることが了解される。上記で説明したように、非ポリペプチド部分は、結合基に直接共有結合で連結され得るかまたは架橋、スペーサーもしくは単数もしくは複数の連結部分などの、単数もしくは複数の介在部分を介して間接的に結合基に共有結合で連結され得る。
【0052】
「ポリマー分子」は、2またはそれ以上のモノマーの共有結合によって形成される分子であり、ポリマーがヒトアルブミンまたは別の豊富な血漿タンパク質である場合を除き、いずれのモノマーもアミノ酸残基ではない。「ポリマー」という用語は、「ポリマー分子」という用語と交換可能に使用できる。この用語は、インビトログリコシル化、すなわち、任意選択的に架橋剤を使用して、通常はポリペプチドの結合基に炭水化物分子を共有結合連結することを含む、インビトロで実施される合成的グリコシル化によって結合された炭水化物分子を包含することが意図されている。
【0053】
N−グリコシル化またはO−グリコシル化(以下でさらに説明する)などのインビボでのグリコシル化によって結合された炭水化物分子を本明細書では「糖成分」と称する。コンジュゲート中のポリマー分子または糖成分などの非ポリペプチド部分の数が明白に示されている場合を除き、本発明で使用されるコンジュゲートまたは別のものに含まれる「非ポリペプチド部分」へのあらゆる言及は、ポリマー分子または糖成分などの1以上の非ポリペプチド部分への言及であるものとする。
【0054】
「結合基」という用語は、ポリマー分子、親油性分子、糖成分または有機誘導体化剤などの非ポリペプチド部分に結合することができる、ポリペプチドの官能基、特にそのアミノ酸残基または炭水化物部分の官能基を示すことが意図されている。
【0055】
インビボでのN−グリコシル化に関しては、「結合基」という用語は通常と異なるように使用され、N−グリコシル化部位(配列N−X−S/T/C[式中、Xはプロリンを除く任意のアミノ酸残基であり、Nはアスパラギンであり、およびS/T/Cはセリン、トレオニンまたはシステインのいずれか、好ましくはセリンまたはトレオニン、最も好ましくはトレオニンである]を有する)を構成するアミノ酸残基を示す。N−グリコシル化部位のアスパラギン残基は、グリコシル化の間に糖成分が結合される残基であるが、かかる結合は、N−グリコシル化部位のその他のアミノ酸残基が存在しない限り達成され得ない。
【0056】
従って、非ポリペプチド部分が糖成分であり、コンジュゲート化がN−グリコシル化によって行われる場合、対象とするポリペプチドのアミノ酸配列の変化に関連して使用される「非ポリペプチド部分のための結合基を含むアミノ酸残基」という用語は、N−グリコシル化部位を構成する1以上のアミノ酸残基が、機能的N−グリコシル化部位がアミノ酸配列内に導入されるかまたは該配列から除去されるように変化していることを意味すると理解される。
【0057】
本明細書において、「治療する」または「治療」という用語は、出血を阻止するまたは最小限に抑えることを目的とした、手術におけるような予想される出血の管理および/または予防、ならびに外傷または血友病患者のためのオンデマンド治療におけるような既に発生している出血の制御の両方を含むことが意図されている。従って本発明による第VIIa因子ポリペプチドの予防的投与は、「治療」の用語に包含される。
【0058】
「出血エピソード」という用語は、制御されない、かつ過剰の出血を含むことが意図されている。出血エピソードは、手術および他の形態の組織損傷の両方に関して重大な問題であり得る。制御されない、かつ過剰の出血は、正常な凝固系を有する被験体および凝固または出血性疾患を有する被験体において起こり得る。
【0059】
本明細書で使用される「出血性疾患」という用語は、出血として現れる、細胞、生理または分子起源の、先天性、後天性または誘導された何らかの欠損を反映する。例としては、凝固因子欠損症(例えば血友病AおよびBまたは凝固第XIもしくは第VII因子の欠損症)、凝固因子インヒビター、血小板機能欠損、血小板減少症、フォン・ヴィレブランド病、または手術もしくは外傷によって誘発される出血である。
【0060】
過剰の出血はまた、正常に機能する血液凝固カスケードを有する(凝固因子欠損または凝固因子のいずれかに対するインヒビターを有さない)被験体においても発生し、血小板機能欠損、血小板減少症またはフォン・ヴィレブランド病によって引き起こされ得る。かかる場合、出血は血友病によって引き起こされる出血にたとえられることがあり、というのは、止血系が、血友病におけるように、欠如しているかまたは、重大な出血を引き起こす、必須の凝固「化合物」(血小板またはフォン・ヴィレブランド因子タンパク質など)の異常を有しているからである。手術または外傷に関連する広範囲の組織損傷を経験した被験体では、正常な止血機構が即時止血の必要性によって圧倒されることがあり、該被験体は、正常な止血機構にも関わらず出血を発現し得る。十分な止血を達成することは、外科的止血の可能性が限られる、脳、内耳領域および眼などの器官で出血が起こる場合にも問題である。同じ問題が、様々な器官(肝臓、肺、腫瘍組織、胃腸管)から生検を採取する過程ならびに腹腔鏡手術において生じ得る。これらすべての状況に共通するのは、外科的手術手技(縫合、クリップ等)によって止血を行うことの困難さであり、これはまた、出血が広汎性である場合(出血性胃炎および大量の子宮出血)にもあてはまる。急性かつ大量の出血は、抗凝固療法時に、与えられた治療によって不完全な止血が誘発された被験体においても起こり得る。かかる被験体は、抗凝固作用を速やかに中和しなければならない場合は外科的処置を必要とし得る。根治的恥骨後前立腺切除術は、限局性前立腺癌を有する被験体に対して一般的に実施される手技である。この手術は、重大な、時として大量の失血によってしばしば困難になる。前立腺切除術の間の多量の失血は、主として、外科的止血が容易にアクセスできない様々な密に血管形成された部位を伴い、大きな領域から広範な出血を生じさせ得る、複雑な解剖学的状況に関連する。また、脳内出血は脳卒中の最も治療が難しい形態であり、脳内出血後数時間以内に高い死亡率と血腫成長に結びつく。rFVIIaによる治療は、血腫の成長を制限し、死亡率を低下させ、90日目の機能転帰を改善することができる。しかしながら、現在使用可能なrFVIIaに関しては、血栓塞栓性有害事象の危険性が存在する。血栓形成性がより低い本発明のキメラ第VII因子分子は、脳卒中治療におけるこの問題を克服し得る。不十分な止血の場合に問題を引き起こし得るもう1つの状況は、正常な止血機構を有する被験体が血栓塞栓症を予防するために抗凝固療法を受ける場合である。かかる療法は、ヘパリン、他の形態のプロテオグリカン、ワルファリンまたは他の形態のビタミンKアンタゴニストならびにアスピリンおよび他の血小板凝固阻害剤を含み得る。
【0061】
本発明の1つの実施形態では、出血は血友病に関連する。もう1つの実施形態では、出血は、後天性インヒビターを有する血友病に関連する。もう1つの実施形態では、出血は血小板減少症に関連する。もう1つの実施形態では、出血はフォン・ヴィレブランド病に関連する。もう1つの実施形態では、出血は重症組織損傷に関連する。もう1つの実施形態では、出血は重度の外傷に関連する。もう1つの実施形態では、出血は手術に関連する。もう1つの実施形態では、出血は腹腔鏡手術に関連する。もう1つの実施形態では、出血は出血性胃炎に関連する。もう1つの実施形態では、出血は大量の子宮出血に関連する。もう1つの実施形態では、出血は、機械的止血の可能性が限られている器官で起こる。もう1つの実施形態では、出血は脳、内耳領域または眼において生じる。もう1つの実施形態では、出血は生検を採取する過程に関連する。もう1つの実施形態では、出血は抗凝固療法に関連する。
【0062】
本明細書で使用される「被験体」という用語は、任意の動物、特にヒトなどの哺乳動物を意味することが意図され、適切な場合は、「患者」という用語と交換可能に使用され得る。
【0063】
「正常な止血系の増強」という用語は、トロンビンを産生するまたは機能性血餅を生成する能力の改善を意味する。
【0064】
「遺伝子治療」という用語は、遺伝子の外因性投与によって内因性遺伝子の発現を変化させる方法を指す。本明細書で使用される、「遺伝子治療」はまた、その必要のある個体の体細胞または幹細胞に、欠陥があるまたは欠如している遺伝子に対応する機能性遺伝子を導入することによる、欠陥タンパク質をコードする欠陥遺伝子の置換または欠如遺伝子の補充を指す。遺伝子治療は、分化した細胞または体性幹細胞を個体の体内から取り出し、続いて、ウイルスベクターを遺伝子送達ビヒクルとして用いて欠陥遺伝子の正常なコピーを外植された細胞に導入する、エクスビボ法によって実施され得る。加えて、インビボでの直接遺伝子導入技術は、治療効果を達成するために広範囲のウイルスベクター、リポソーム、タンパク質DNA複合体または裸のDNAを用いてインサイチューで個体の細胞に遺伝子導入することを可能にする。「遺伝子治療」という用語はまた、欠陥遺伝子またはタンパク質が欠陥でなかった場合に機能するはずであるのと実質的に同じように機能するポリヌクレオチドを、その必要のある個体の体細胞または幹細胞に導入することによる、欠陥タンパク質をコードする欠陥遺伝子の置換を指す。
【0065】
本発明において、遺伝子治療は、本発明のキメラFVIIをコードするヌクレオチド配列を含む核酸分子を被験体に投与することに関連して用いられる。従って、本発明は、本発明のキメラFVIIタンパク質をコードするヌクレオチド配列を含む単離された核酸分子を提供する。かかる核酸分子は核酸構築物(例えばベクターまたはプラスミド)中に存在し得る。かかる核酸構築物は細胞中に存在し得る。さらに、本発明のキメラFVIIタンパク質を細胞に送達する方法が本明細書で提供され、該方法は、キメラFVIIタンパク質をコードするヌクレオチド配列を含む核酸分子を、該ヌクレオチド配列が細胞において発現されてキメラFVIIタンパク質を生成する条件下で細胞に導入することを含む。細胞は、被験体に導入される細胞であり得るおよび/または被験体において既に存在する細胞であり得る。
【0066】
[キメラ第VII因子の作製]
本明細書で述べるキメラ第VII因子ポリペプチドは、組換え核酸技術によって作製され得る。一般に、クローニングされた野生型第VII因子核酸配列を、所望タンパク質をコードするように修飾する。次に、この修飾された配列を発現ベクターに挿入し、今度はこの発現ベクターを宿主細胞に形質転換またはトランスフェクトする。高等真核細胞、特に培養された哺乳動物細胞が宿主細胞として適切である。ヒト第VII因子についての完全なヌクレオチド配列およびアミノ酸配列が知られており(組換えヒト第VII因子のクローニングおよび発現が記載されている米国特許第4,784,950号、ならびにGenBank(登録商標)アクセッション番号J02933およびAAA51983ならびにSwissProtアクセッション番号P08709−1参照)、該アミノ酸配列を本明細書で配列番号:18として示す。ウシ第VII因子配列は、Takeya et al.,J.Biol.Chem.263:14868−14872(1988))に記載されている。第IX因子についての完全なヌクレオチド配列およびアミノ酸配列が知られており(Davie et al.,Proc.Natl.Acad.Sci.U.S.A 1982;79:6461−6464;Jaye et al.,Nucleic Acids Res.1983;11:2325−2335;およびMcGraw et al.,Proc.Natl.Acad.Sci.U.S.A.1985;82:2847−2851ならびにGenBank(登録商標)アクセッション番号J00136およびAAA98726ならびにSwissProtアクセッション番号P00740参照)、該アミノ酸配列を本明細書で配列番号:19として提供する。プロテインSについても完全なヌクレオチド配列およびアミノ酸配列が知られており(Hoskins et al.,Proc.Natl.Acad.Sci.U.S.A.1987;84:349−353ならびにGenBank(登録商標)アクセッション番号M15036およびAAA36479ならびにSwissProtアクセッション番号P07225参照)、該アミノ酸配列を本明細書で配列番号:20として提供する。他の凝固タンパク質についての完全なヌクレオチド配列およびアミノ酸配列も知られており、SwissProtを用いてまたはNCBIウエブページで見ることができる。
【0067】
アミノ酸配列の変化は様々な技術によって達成され得る。核酸配列の修飾は、部位特異的突然変異誘発によって実施し得る。部位特異的突然変異誘発のための技術は当技術分野において公知であり、例えば、Zoller and Smith(DNA 3:479−488,1984)または「Splicing by extension overlap」,Horton et al.,Gene 77,1989,pp.61−68に記載されている。従って、第VII因子のヌクレオチド配列およびアミノ酸配列を使用して、選択した変化を導入し得る。同様に、特異的なプライマーを使用したポリメラーゼ連鎖反応を用いてDNA構築物を作製するための手順も当業者に公知である(PCR Protocols,1990,Academic Press,San Diego,Calif.,USA参照)。加えて、本発明のキメラ第VII因子ポリペプチドは、様々なポリペプチドをコードするヌクレオチド配列にユニーク制限部位を導入することによって作製でき、これを使用して、様々なポリペプチドのドメイン全体をコードするヌクレオチド配列のフラグメントを単離することができる。その後、これらの様々なヌクレオチド配列を組み換えて、本発明の様々なキメラ第VII因子ポリペプチドを作製することができる。
【0068】
例えば、GLA、EGF−1、EGF−2および触媒ドメインを様々な凝固タンパク質の間で自由に交換し得るように、様々な凝固タンパク質をコードするヌクレオチド配列にユニーク制限部位を導入し得る。EGF−1ドメインはEGF−2ドメインの周囲を自由に回転することができるが(Pike et al.Proc Natl.Acad.Sci.1999;96:8925−8930)、GLAドメインおよびEGF−1ドメインは、カルシウムが存在する場合は互いの周囲を自由に回転できない(Sunnerhagen et al.Nature Structural Biology 1995;2:504−509)。この実施形態では、GLAおよびEGF−1ドメインは同じ凝固タンパク質に由来し得るが、EGF−2および触媒ドメインとは異なるタンパク質に由来し得る。従って、本発明のキメラ第VIIa因子ポリペプチドは、1つのドメインの少なくとも50%が第VII因子ドメインであることを条件として、凝固タンパク質の任意のものに由来するドメインを含み得る。さらに、本発明のキメラ第VIIa因子ポリペプチドは、同じタンパク質または異なるタンパク質に由来するGLAおよびEGF−1ドメインを含み得る。
【0069】
本発明のキメラ第VII因子ポリペプチドをコードする核酸構築物は、適切には、例えば標準的な技術に従ってゲノムライブラリーまたはcDNAライブラリーを作製し、合成オリゴヌクレオチドプローブを用いたハイブリダイゼーションによって様々なポリペプチドの全部または一部をコードするDNA配列をスクリーニングすることによって得られる、ゲノムまたはcDNA起源のものであり得る(Sambrook et al.,Molecular Cloning:A Laboratory Manual,2nd.Ed.Cold Spring Harbor Laboratory,Cold Spring Harbor,N.Y.,1989参照)。
【0070】
キメラ第VII因子ポリペプチドをコードする核酸構築物はまた、確立された標準的な方法、例えばBeaucage and Caruthers,Tetrahedron Letters 22(1981),1859−1869によって述べられているホスホルアミダイト法またはMatthes et al.,EMBO Journal 3(1984),801−805によって述べられている方法によって合成的に作製し得る。ホスホルアミダイト法によれば、例えば自動DNA合成装置において、オリゴヌクレオチドを合成し、精製して、アニーリングし、連結して、適切なベクターにクローニングする。
【0071】
さらに、核酸構築物は、標準的な技術に従って、核酸構築物全体の様々な部分に対応するフラグメントである、合成、ゲノムまたはcDNA起源(適宜に)のフラグメントを連結することによって作製される、合成とゲノムの混合型、合成とcDNA起源の混合型またはゲノムとcDNA起源の混合型であり得る。
【0072】
本発明によるキメラ第VII因子ポリペプチドを作製する際に使用するためのDNA配列は、典型的には、適切な翻訳後プロセシング(例えばグルタミン酸残基のγ−カルボキシル化)および宿主細胞からの分泌を得るために第VII因子のアミノ末端でプレプロポリペプチドをコードする。該プレプロポリペプチドは、第VII因子または別のビタミンK依存性血漿タンパク質、例えば第IX因子、第X因子、プロトロンビン、プロテインCもしくはプロテインSなどのものであり得る。当業者に認識されるように、付加的な修飾は、それらの修飾が凝固因子として働くタンパク質の能力を有意に損なわない場合は、キメラ第VII因子ポリペプチドのアミノ酸配列において為され得る。
【0073】
第VIIa因子変異体を発現する際に使用するための発現ベクターは、クローニングされた遺伝子またはcDNAの転写を指令することができるプロモータを含む。培養された哺乳動物細胞における使用のための適切なプロモータは、ウイルスプロモータおよび細胞プロモータを含む。ウイルスプロモータは、SV40プロモータ(Subramani et al.,Mol.Cell.Biol.1:854−864,1981)およびCMVプロモータ(Boshart et al.,Cell 41:521−530,1985)を含む。特に適切なウイルスプロモータは、アデノウイルス2由来の主要後期プロモータである(Kaufman and Sharp,Mol.Cell.Biol.2:1304−1319,1982)。もう1つの特に好ましいプロモータは、チャイニーズハムスター伸長因子1−α(CHEF1)プロモータである。細胞プロモータは、マウスκ遺伝子プロモータ(Bergman et al.,Proc.Natl.Acad.Sci.USA 81:7041−7045,1983)およびマウスVHプロモータ(Loh et al.,Cell 33:85−93,1983)を含む。特に適切な細胞プロモータは、マウスメタロチオネイン−Iプロモータである(Palmiter et al.,Science 222:809−814,1983)。発現ベクターはまた、プロモータから下流でかつキメラ第VII因子配列自体の挿入部位から上流に位置する一組のRNAスプライス部位を含み得る。適切なRNAスプライス部位は、アデノウイルスおよび/または免疫グロブリン遺伝子から得られ得る。また、挿入部位の下流に位置するポリアデニル化シグナルも発現ベクター中に含まれる。特に適切なポリアデニル化シグナルは、SV40由来の早期または後期ポリアデニル化シグナル(Kaufman and Sharp)、アデノウイルス5 Elb領域由来のポリアデニル化シグナル、ヒト成長ホルモン遺伝子ターミネーター(DeNoto et al.,Nucl.Acids Res.9:3719−3730,1981)またはヒト第VII因子遺伝子もしくはウシ第VII因子遺伝子由来のポリアデニル化シグナルを含む。発現ベクターはまた、プロモータとRNAスプライス部位の間に位置するアデノウイルス2の3分節リーダーなどの非コードウイルスリーダー配列、およびSV40エンハンサなどのエンハンサ配列も含み得る。
【0074】
クローニングされたDNA配列を、例えばリン酸カルシウムを介したトランスフェクション(Wigler et al.,Cell 14:725−732,1978;Corsaro and Pearson,Somatic Cell Genetics 7:603−616,1981;Graham and Van der Eb,Virology 52d:456−467,1973)または電気穿孔法(Neumann et al.,EMBO J.1:841−845,1982)によって、培養された哺乳動物細胞に導入する。外因性DNAを発現する細胞を同定し、選択するために、選択可能な表現型を付与する遺伝子(選択マーカー)を、一般に、対象とする遺伝子またはcDNAと共に細胞に導入する。適切な選択マーカーは、ネオマイシン、ハイグロマイシンおよびメトトレキサートなどの薬剤に対する耐性を付与する遺伝子を含む。選択マーカーは、増幅可能な選択マーカーであり得る。適切な増幅可能選択マーカーは、ジヒドロ葉酸レダクターゼ(DHFR)配列である。もう1つの適切な選択マーカーはヒスチジノールである。選択マーカーは、Thilly(引用することにより本明細書の一部をなすものとする、Mammalian Cell Technology,Butterworth Publishers,Stoneham,MA)によって総説されている。当業者は、適切な選択マーカーを容易に選択することができる。
【0075】
選択マーカーは、対象とする遺伝子と同時に別々のプラスミド上で細胞に導入され得るか、または同じプラスミド上で導入され得る。同じプラスミド上の場合、選択マーカーと対象遺伝子は異なるプロモータまたは同じプロモータの制御下にあってよく、後者の配置はジシストロニックメッセージを生成する。このタイプの構築物は当技術分野において知られている(例えば、Levinson and Simonsen,米国特許第4,713,339号)。また、「担体DNA」として知られている付加的なDNAを、細胞に導入される混合物に添加することも好都合であり得る。
【0076】
細胞はDNAを取り込んだ後、適切な増殖培地において、典型的には1〜2日間増殖され、対象とする遺伝子の発現を開始する。細胞を培養するために使用する培地は、適切な添加物を含有する最小または複合培地などの、宿主細胞を増殖させるのに適した任意の従来培地であり得る。適切な培地は、商業的供給者から入手可能であるか、または公表されている配合(例えばAmerican Type Culture Collectionのカタログにおいて)に従って調製され得る。培地は当技術分野において知られている手順を用いて調製される(例えば、細菌および酵母に関する参考文献;Bennett,J.W.and LaSure,L.,editors,More Gene Manipulations in Fungi,Academic Press,CA, 1991参照)。増殖培地は一般に、炭素源、窒素源、必須アミノ酸、必須糖、ビタミン、塩、リン脂質、タンパク質および増殖因子を含有する。γ−カルボキシル化されたキメラ第VII因子ポリペプチドの生成のためには、培地はビタミンKを、好ましくは約0.1ng/mlから約20μg/mlの濃度で含有する。次に薬剤選択を適用して、選択マーカーを安定に発現している細胞の増殖物を選択する。増幅可能な選択マーカーでトランスフェクトした細胞に関しては、増大したコピー数のクローン化配列を選択するために、薬剤濃度を高めてもよく、それによって発現レベルを上昇させ得る。次に、安定にトランスフェクトされた細胞のクローンを、所望のキメラ第VII因子ポリペプチドの発現に関してスクリーニングする。
【0077】
適切な哺乳動物細胞株は、CHO細胞株(ATCC CCL 61)、COS−1細胞株(ATCC CRL 1650)、ベビーハムスター腎(BHK)細胞株および293細胞株(ATCC CRL 1573;Graham et al.,J.Gen.Virol.36:59−72,1977)を含む。適切なBHK細胞株は、本明細書中以下ではBHK570細胞と称する、tk.sup.−ts13 BHK細胞株である(Waechter and Baserga,Proc.Natl.Acad.Sci.USA 79:1106−1110,1982)。BHK 570細胞株は、ATCCアクセッション番号CRL 10314の下にAmerican Type Culture Collection,12301 Parklawn Dr.,Manassas,Va.20852から入手可能である。tk−ts13BHK細胞株も、アクセッション番号CRL 1632の下にATCCから入手可能である。加えて、Rat Hep I細胞(ラット肝細胞癌;ATCC CRL 1600)、Rat Hep II細胞(ラット肝細胞癌;ATCC CRL 1548)、TCMK細胞(ATCC CCL 139)、ヒト肺細胞(ATCC HB 8065)、NCTC 1469(ATCC CCL 9.1)およびDUKX細胞(Urlaub and Chasin,Proc.Natl.Acad.Sci.USA 77:4216−4220,1980)を含む、多くの他の細胞株を使用し得る。
【0078】
本発明のキメラ第VII因子ポリペプチドは、インビボでのポリペプチドの半減期を延長させるためにコンジュゲート化し得る。かかるコンジュゲート化は、キメラ第VII因子ポリペプチドの投与頻度を低減する一方で、腎クリアランスを低下させ、インビボで存在する第VII因子の量を増加させ得る。適切なコンジュゲートおよびコンジュゲート化第VII因子ポリペプチドを作製する方法は、その全体を引用することにより本明細書の一部をなすものとする、米国特許第7,442,524号に記載されている。
【0079】
[トランスジェニック動物]
本発明のキメラ第VII因子ポリペプチドを作製するためにトランスジェニック動物技術を使用し得る。宿主雌性哺乳動物の乳腺中でタンパク質を生成することが好ましい。乳腺中での発現およびその後の乳中への対象タンパクの分泌は、他の供給源からタンパク質を単離する際に遭遇する多くの困難を克服する。乳は容易に収集され、大量に入手可能であり、生化学的に広く特徴付けられている。さらに、主要な乳タンパク質は乳中に高濃度で存在する(典型的には約1〜15g/l)。本発明のキメラ第VII因子ポリペプチドはまた、宿主動物の尿中でも生成でき、これは、雄性と雌性の両方の動物が尿を生成することおよび単離のためには乳中よりも尿中の方が失われるタンパク質が少ないという点で乳中での生成よりも有利である。
【0080】
本発明のキメラ第VII因子ポリペプチドの供給源として乳を使用することを考慮する場合、商業的見地から、大量の乳算出量を有する種を宿主として用いることが明らかに好ましい。マウスおよびラットなどのより小さな動物も使用できる(および原理段階の証明においては好ましい)が、ブタ、ヤギ、ヒツジおよびウシを含むがこれらに限定されない、家畜哺乳動物を用いることが好ましい。ヒツジは、この種における遺伝子組換えの前歴、乳算出量、コストおよびヒツジの乳を収集するための装置が容易に利用可能であることなどの因子のために、特に好ましい(例えば、宿主の種の選択に影響を及ぼす因子の比較に関しては国際公開公報第WO88/00239号参照)。イーストフリースランドシープ(East Friesland sheep)のような、酪農用に育種された宿主動物の品種を選択すること、または後日にトランスジェニック系統の交配によって搾乳用家畜を導入することが一般に望ましい。いずれの場合も、既知の、健康状態の良好な動物を使用すべきである。
【0081】
乳腺における発現を得るために、乳タンパク質遺伝子由来の転写プロモータを使用する。乳タンパク質遺伝子は、カゼイン(米国特許第5,304,489号参照)、β−ラクトグロブリン、α−ラクトアルブミンおよび乳清酸性タンパク質をコードする遺伝子を含む。β−ラクトグロブリン(BLG)プロモータが適切である。ヒツジβ−ラクトグロブリン遺伝子の場合は、遺伝子の5’フランキング配列の少なくとも近位の406bpの領域が一般的に使用されるが、β−ラクトグロブリン遺伝子の5’フランキングプロモータおよび非コード部分を包含する約4.25kbpのDNAセグメントなどの、約5kbpまでの5’フランキング配列のより大きな部分が適切である(Whitelaw et al.,Biochem.J.286:31−39(1992)参照)。他の種に由来するプロモータDNAの同様のフラグメントも適切である。
【0082】
尿における発現を得るために、尿路上皮特異的プロモータを使用する。例えば、ウロプラキン関連遺伝子の発現を駆動するプロモータが使用できる(例えば、その全体を引用することにより本明細書の一部をなすものとする、米国特許第6,001,646号参照)。
【0083】
β−ラクトグロブリン遺伝子の他の領域も、発現される遺伝子のゲノム領域と同様に、構築物に組み込み得る。イントロンを欠く構築物は、例えば、かかるDNA配列を含むものと比較して十分に発現されないことが当技術分野で一般的に認められている(各々を引用することにより本明細書の一部をなすものとする、Brinster et al.,Proc.Natl.Acad.Sci.USA 85:836−840(1988);Palmiter et al.,Proc.Natl.Acad.Sci.USA 88:478−482(1991);Whitelaw et al.,Transgenic Res.1:3−13(1991);国際公開公報第WO89/01343号;および国際公開公報第WO91/02318号参照)。これに関して、可能な場合は、対象とするタンパク質またはポリペプチドをコードする遺伝子の天然イントロンの全部または一部を含むゲノム配列を使用することが一般に好ましく、従って、例えばβ−ラクトグロブリン遺伝子由来の、少なくともいくつかのイントロンをさらに含むことが好ましい。かかる領域の1つは、ヒツジβ−ラクトグロブリン遺伝子の3’非コード領域由来の、イントロンのスプライシングおよびRNAのポリアデニル化を提供するDNAセグメントである。遺伝子の天然3’非コード配列を置換した場合、このヒツジβ−ラクトグロブリンセグメントは、対象タンパク質またはポリペプチドの発現レベルを高めかつ安定化することができる。他の実施形態では、キメラ第VII因子配列の開始ATGの周囲の領域を、乳特異的タンパク質遺伝子由来の対応する配列で置換する。かかる置換は、発現を増強する推定上の組織特異的開始環境を提供する。キメラ第VII因子プレプロ配列全体および5’非コード配列を、例えばBLG遺伝子のもので置換することは好都合であるが、より小さな領域を置換してもよい。
【0084】
トランスジェニック動物におけるキメラFVIIaポリペプチドの発現のために、キメラ第VIIa因子をコードするDNAセグメントを、発現ユニットを生成するためにその発現に必要とされる付加的なDNAセグメントに作動可能に連結する。かかる付加的なセグメントは、上記で挙げたプロモータ、ならびにmRNAの転写およびポリアデニル化の終結をもたらす配列を含む。発現ユニットは、キメラ第VIIa因子をコードするセグメントに作動可能に連結された分泌シグナル配列をコードするDNAセグメントをさらに含む。この分泌シグナル配列は、天然第VII因子分泌シグナル配列であり得るかまたは乳タンパク質などの別のタンパク質のものであり得る(例えば、引用することにより本明細書の一部をなすものとする、von Heijne,Nucl.Acids Res.14:4683−4690(1986);およびMeade et al.,米国特許第4,873,316号参照)。
【0085】
トランスジェニック動物に使用するための発現ユニットの構築は、キメラ第VII因子配列を、付加的なDNAセグメントを含むプラスミドまたはファージベクターに挿入することによって好都合に実施されるが、発現ユニットは、基本的には任意の一連の連結によって構築され得る。乳タンパク質をコードするDNAセグメントを含むベクターを提供し、乳タンパク質についてのコード配列をキメラ第VII因子ポリペプチドのもので置換して、それにより乳タンパク質遺伝子の発現制御配列を含む遺伝子融合物を生成することは特に好都合である。いずれの場合も、プラスミドまたは他のベクターへの発現ユニットのクローニングは、キメラ第VII因子配列の増幅を促進する。増幅は、好都合には細菌(例えば大腸菌(E.coli))宿主細胞において実施され、従ってベクターは、典型的には細菌宿主細胞において機能性の複製起点および選択マーカーを含む。発現ユニットを、次に、選択した宿主種の受精卵(初期胚を含む)に導入する。異種DNAの導入は、マイクロインジェクション(例えば米国特許第4,873,191号)、レトロウイルス感染(Jaenisch,Science 240:1468−1474(1988))または胚性幹(ES)細胞を用いた部位指定組込み(Bradley et al.,Bio/Technology 10:534−539(1992)によって総説されている)を含む、いくつかの経路の1つによって実施できる。該卵をその後、偽妊娠雌性動物の卵管または子宮に移植し、出産まで発育させる。生殖細胞系に導入されたDNAを担持する子は、そのDNAを正常なメンデルの様式で子孫に伝えることができ、トランスジェニック集団の発生を可能にする。トランスジェニック動物を作製するための一般的な手順は当技術分野において知られている(例えば、Hogan et al.,Manipulating the Mouse Embryo:A Laboratory Manual,Cold Spring Harbor Laboratory,1986;Simons et al.,Bio/Technology 6:179−183(1988);Wall et al.,Biol.Reprod.32:645−651(1985);Buhler et al.,Bio/Technology 8:140−143(1990);Ebert et al.,Bio/Technology 9:835−838(1991);Krimpenfort et al.,Bio/Technology 9:844−847(1991);Wall et al.,J.Cell.Biochem.49:113−120(1992);米国特許第4,873,191号;米国特許第4,873,316号;国際公開公報第WO88/00239号、国際公開公報第WO90/05188号、国際公開公報第WO92/11757号;およびイギリス特許公開公報第GB87/00458号参照)。外来性DNA配列を哺乳動物およびそれらの生殖細胞に導入するための技術はマウスにおいて最初に開発された(例えば、Gordon et al.,Proc.Natl.Acad.Sci.USA 77:7380−7384(1980);Gordon and Ruddle,Science 214:1244−1246(1981);Palmiter and Brinster,Cell 41:343−345(1985);Brinster et al.,Proc.Natl.Acad.Sci.USA 82:4438−4442(1985);およびHogan et al.参照)。これらの技術は、その後、家畜種を含むより大きな動物に関する使用のために適合された(例えば国際公開公報第WO88/00239号、国際公開公報第WO90/05188号および国際公開公報第WO92/11757号;ならびにSimons et al.,Bio/Technology 6:179−183(1988)参照)。要約すると、トランスジェニックマウスまたは家畜の作製においてこれまでに使用された最も効率的な経路では、対象とするDNAの数百の直鎖状分子を、確立された技術に従って受精卵の前核の1つに注入する。接合子の細胞質へのDNAの注入も使用し得る。
【0086】
トランスジェニック植物における生産も使用し得る。発現は、植物全体または塊茎などの特定の器官を対象とし得る(Hiatt,Nature 344:469−479(1990);Edelbaum et al.,J.Interferon Res.12:449−453(1992);Sijmons et al.,Bio/Technology 8:217−221(1990);および欧州特許第EP 0 255 378号参照)。
【0087】
本発明のキメラ第VII因子ポリペプチドの作製はまた、例えばウサギ網状赤血球溶解物、コムギ胚芽抽出物および大腸菌無細胞系等による、インビトロでの翻訳を介して達成され得る。インビトロ翻訳はまた、連鎖または共役させ得る。連鎖インビトロ翻訳は、バクテリオファージポリメラーゼでの転写、それに続くウサギ網状赤血球溶解物またはコムギ胚芽溶解物系での翻訳に基づく。共役インビトロ翻訳は大腸菌無細胞系に基づく。
【0088】
[回収および活性化]
本発明のキメラ第VII因子ポリペプチドは、細胞培養培地または乳から回収される。本発明のキメラ第VII因子ポリペプチドは、当技術分野において知られている様々な方法によって精製でき、かかる方法は、クロマトグラフィ(例えばイオン交換、アフィニティ、疎水性、クロマトフォーカシングおよびサイズ排除)、電気泳動法(例えば分取等電点電気泳動(IEF)、示差溶解度(例えば硫安沈殿)または抽出(例えば、Protein Purification,J.−C.Janson and Lars Ryden,editors,VCH Publishers,New York,1989参照)を含むが、これらに限定されない。さらなる精製は、高速液体クロマトグラフィなどの従来の化学的精製手段によって達成され得る。クエン酸バリウム沈殿を含む他の精製方法が当技術分野において知られており、本明細書で述べる新規キメラ第VII因子ポリペプチドの精製に適用し得る(例えば、Scopes,R.,Protein Purification,Springer−Verlag,N.Y.,1982参照)。
【0089】
治療目的には、本発明のキメラ第VII因子は実質的に純粋であることが好ましい。従って、本発明の1つの実施形態では、本発明の第VII因子変異体は、少なくとも約90〜95%の純度、好ましくは少なくとも約98%の純度に精製される。純度は、例えばゲル電気泳動、アミノ末端のアミノ酸配列決定および逆相HPLCによって評価し得る。
【0090】
キメラ第VII因子ポリペプチドは、二本鎖の形態に変換するためにその活性化部位で切断される。活性化は、Osterud,et al.,Biochemistry 11:2853−2857(1972);Thomas,米国特許第4,456,591号;Hedner and Kisiel,J.Clin.Invest.71:1836−1841(1983);またはKisiel and Fujikawa,Behring Inst.Mitt.73:29−42(1983)によって開示されているような、当技術分野で知られている手順に従って実施し得る。あるいは、キメラ第VII因子ポリペプチドを濃縮し、正に荷電した表面または樹脂と接触させることによってキメラ第VII因子ポリペプチドを活性化することができ、例えばBjoern et al.(Research Disclosure,269 September 1986,pp.564−565)によって述べられているように、第VII因子をMono Q(登録商標)(Pharmacia fine Chemicals)等のようなイオン交換クロマトグラフィカラムに通すことによって活性化し得る。キメラ第VII因子ポリペプチドはまた、キメラ第VII因子ポリペプチドの実質的に精製された製剤を含有する溶液を得、その溶液にアミン化合物、Ca2+を約5mM〜約50mM(約10mM〜約30mMなど)の最終濃度になるように添加して、溶液の最終pHを約7.2〜8.6(約7.6〜約8.2など)に調整し、生じた活性化混合物を約2℃〜約25℃で、少なくとも90%のキメラFVIIポリペプチドをキメラFVIIaポリペプチドに変換するのに十分な時間インキュベートして、任意選択的に、活性化混合物からFVIIaを単離することによって溶液中で活性化し得る。生じた活性化キメラ第VII因子ポリペプチドを、その後、以下のように製剤し、投与し得る。
【0091】
[遺伝子治療]
本発明のキメラ第VII因子ポリペプチドはまた、遺伝子治療によって出血性疾患を治療する方法においても使用し得る。本発明のこの実施形態では、本発明のキメラ第VII因子ポリペプチドは、エクスビボでの導入またはインビボでの導入によって被験体の細胞に導入され得る核酸分子によってコードされる。
【0092】
当技術分野で利用可能な遺伝子治療のための方法のいずれもが本発明に従って使用できる。例示的な方法を以下で述べる。遺伝子治療の方法の一般的な総説については、Goldspiel et al.,Clinical Pharmacy 1993,12:488−505;Wu and Wu,Biotherapy 1991,3:87−95;Tolstoshev,Ann.Rev.Pharmacol.Toxicol.1993,32:573−596;Mulligan,Science 1993,260:926−932;およびMorgan and Anderson,Ann.Rev.Biochem.1993,62:191−217;May,TIBTECH 1993,11:155−215参照。使用できる組換えDNA技術の当技術分野で一般的に知られている方法は、Ausubel et al.,(eds.),1993,Current Protocols in Molecular Biology,John Wiley & Sons,NY;Kriegler,1990,Gene Transfer and Expression,A Laboratory Manual,Stockton Press,NY;およびin Chapters 12 and 13,Dracopoli et al.,(eds.),1994,Current Protocols in Human Genetics,John Wiley & Sons,NY;Colosimo et al.,Biotechniques 2000;29(2):314−8,320−2,324に記載されている。
【0093】
キメラ第VII因子ポリペプチドをコードするポリヌクレオチドは、適切なクローニングベクターに挿入し得る。遺伝子治療に適するベクターは、アデノウイルス、アデノ随伴ウイルス(AAV)、ワクシニアウイルス、ヘルペスウイルス、バキュロウイルスおよびレトロウイルス、パルボウイルス、レンチウイルスなどのウイルス、バクテリオファージ、コスミド、プラスミド、真菌ベクターならびに様々な真核および原核生物宿主における発現に関して記述されている、遺伝子治療ならびに単純なタンパク質発現のために使用し得る当技術分野で典型的に使用される他の組換えビヒクルを含む。
【0094】
1つの実施形態では、該ベクターはウイルスベクターである。ウイルスベクター、特にアデノウイルスベクターは、標的細胞のウイルス感染の効率を上昇させる、カチオン性脂質、ポリL−リシン(PLL)およびジエチルアミノエチルデキストラン(DELAE−デキストラン)などのカチオン性両親媒物質と複合体形成することができる(例えば、引用することにより本明細書の一部をなすものとする、1997年11月20日出願の国際特許出願第PCT/US97/21496号参照)。本発明において使用するための適切なウイルスベクターは、ワクシニアウイルス、ヘルペスウイルス、AAVおよびレトロウイルス由来のベクターを含む。遺伝子治療におけるウイルスベクターの総説については、Mah et al.,Clin.Pharmacokinet.2002;41(12):901−11;Scott et al.,Neuromuscul.Disord.2002;12 Suppl 1:S23−9参照。
【0095】
1つの実施形態では、コード配列および任意の他の所望配列がゲノム内の所望部位での相同的組換えを促進する領域によって隣接され、それによりゲノムに組み込まれた核酸分子からの構築物の発現を提供するベクターが使用される(各々その全体を引用することにより本明細書の一部をなすものとする、Koller and Smithies,Proc.Natl.Acad.Sci.USA 1989,86:8932−8935;Zijlstra et al.,Nature 1989,342:435−438;Zarling et al.への米国特許第6,244,113号;およびPati et al.への米国特許第6,200,812号)。
【0096】
患者へのベクターの送達は、患者をベクターもしくは送達複合体に直接暴露する直接的送達、または細胞を最初にインビトロにてベクターで形質転換し、その後患者に移植する間接的送達のいずれかであり得る。これら2つのアプローチは、それぞれインビボおよびエクスビボ遺伝子治療として知られている。
【0097】
1つの実施形態では、ベクターをインビボで直接投与し、そこでベクターは被験体の細胞に進入して、遺伝子の発現を媒介する。これは、当技術分野で知られている数多くの方法および上記で論じた方法のいずれかによって、例えばそれを適切な発現ベクターの一部として構築し、例えば欠陥もしくは弱毒化レトロウイルスまたは他のウイルスベクターを用いた感染によって(例えば米国特許第4,980,286号参照)、または裸のDNAの直接注入によって、または微粒子ボンバードメント(例えば遺伝子銃;Biolistic,Dupont)の使用によって、または脂質もしくは細胞表面受容体もしくはトランスフェクション剤でのコーティング、バイオポリマー(例えばポリ−β−1−64−N−アセチルグルコサミン多糖;米国特許第5,635,493号参照)への封入、リポソーム、微粒子もしくはマイクロカプセルへの封入の使用によって、ペプチドもしくは核内に進入することが公知の他のリガンドに連結して投与することによって、または受容体媒介性エンドサイトーシスを受けるリガンドに連結して投与すること(例えば、Wu and Wu,J.Biol.Chem.1987,62:4429−4432)等によって、細胞内に組み込まれるように投与することによって実施できる。もう1つの実施形態では、リガンドが、エンドソームを崩壊させ、核酸がリソソーム分解を回避することを可能にする膜融合性ウイルスペプチド、または治療用DNAを細胞に導入するために使用できる、例えばアンテナペディア由来の、カチオン性12量体ペプチドを含む、核酸−リガンド複合体を形成することができる(Mi et al.,Mol.Therapy 2000,2:339−47参照)。さらにもう1つの実施形態では、特定の受容体を標的することにより、細胞特異的な取込みおよび発現のために核酸をインビボで標的することができる(例えば、国際公開公報第WO92/06180号、国際公開公報第WO92/22635号、国際公開公報第WO92/20316号および国際公開公報第WO93/14188号参照)。加えて、マグネトフェクションと称される技術を、ベクターを哺乳動物に送達するために使用し得る。この技術は、磁場の影響下で送達するためにベクターと超常磁性ナノ粒子を結合させる。この適用は、送達時間を短縮し、ベクターの有効性を高める(Scherer et al.,Gene Therapy 2002;9:102−9)。
【0098】
1つの実施形態では、核酸は脂質担体を用いて投与し得る。細胞膜の通過を促進するために脂質担体を裸の核酸(例えばプラスミドDNA)と結合することができる。カチオン性、アニオン性または中性脂質がこのために使用できる。しかし、カチオン性脂質は、一般に負の電荷を有するDNAとより良好に結合することが示されているため、適切である。カチオン性脂質はまた、プラスミドDNAの細胞内送達を媒介することが示されている(Felgner and Ringold,Nature 1989;337:387)。マウスへのカチオン性脂質−プラスミド複合体の静脈内注射は、肺におけるDNAの発現を生じさせることが示された(Brigham et al.,Am.J.Med.Sci.1989;298:278)。また、Osaka et al.,J.Pharm.Sci.1996;85(6):612−618;San et al.,Human Gene Therapy 1993;4:781−788;Senior et al.,Biochemica et Biophysica Acta 1991;1070:173−179);Kabanov and Kabanov,Bioconjugate Chem.1995;6:7−20;Liu et al.,Pharmaceut.Res.1996;13;Remy et al.,Bioconjugate Chem.1994;5:647−654;Behr,J−P.,Bioconjugate Chem 1994;5:382−389;Wyman et al.,Biochem.1997;36:3008−3017;Marshall et al.への米国特許第5,939,401号;Scheule et al.への米国特許第6,331,524号も参照のこと。
【0099】
代表的なカチオン性脂質は、例えば、その開示を引用することにより本明細書の一部をなすものとする、米国特許第5,283,185号および米国特許第5,767,099号に開示されているものを含む。1つの実施形態では、カチオン性脂質は、米国特許第5,767,099号に開示されているN4−スペルミンコレステリルカルバメート(GL−67)である。さらなる適切な脂質は、N4−スペルミジンコレステリルカルバメート(GL−53)および1−(N4−スペルミン)−2,3−ジラウリルグリセロールカルバメート(GL−89)を含む。
【0100】
ウイルスベクターのインビボでの投与に関して、ウイルスベクターおよびトランスフェクトされた細胞の免疫脱活性化を回避するために、適切な免疫抑制治療をウイルスベクター、例えばアデノウイルスベクターと組み合わせて使用し得る。例えば、インターロイキン12(IL−12)、インターフェロンγ(IFN−γ)または抗CD4抗体などの免疫抑制性サイトカインが、ウイルスベクターに対する体液性または細胞性免疫応答をブロックするために投与し得る。それに関して、ごく少数の抗原を発現するように遺伝子操作されたウイルスベクターを用いることが好都合である。
【0101】
体細胞は、上述した方法のいずれかを用いて本発明のキメラ第VII因子ポリペプチドをコードする構築物でエクスビボにて遺伝子操作し、個体に再移植し得る。この方法は、Selden et al.への国際公開公報第WO93/09222号において一般的に説明されている。加えて、この技術は、Payumo et al.,Clin.Orthopaed.and Related Res.2002;403S:S228−S242に記載されている、細胞ベースの送達についての特許ImPACT技術で使用される。かかる遺伝子治療系では、体細胞(例えば線維芽細胞、肝細胞または内皮細胞)を患者から取り出し、インビトロで培養して、治療対象の遺伝子でトランスフェクトし、特徴付けて、患者に再び導入する。一次細胞(個体または組織に由来し、継代の前に遺伝子操作された)および二次細胞(インビボでの導入の前にインビトロで継代した)の両方、ならびに当技術分野で知られている不死化細胞株を使用し得る。本発明の方法のために有用な体細胞は、線維芽細胞、ケラチノサイト、上皮細胞、内皮細胞、肝細胞、血液の有形成分、筋肉細胞、培養できる他の体細胞および体細胞前駆体などの体細胞を含むが、これらに限定されない。1つの実施形態では、細胞は肝細胞である。
【0102】
本発明のキメラFVIIaポリペプチドをコードするポリヌクレオチド、および、任意選択的に、選択マーカーをコードする核酸を、受容者の一次または二次細胞におけるキメラFVIIaの発現のために必要な付加的配列と共に含む構築物を使用して、コードされた産物が生成される一次または二次細胞をトランスフェクトする。かかる構築物は、レトロウイルス、ヘルペスウイルス、アデノウイルス、アデノ随伴ウイルス、耳下腺炎ウイルスおよびポリオウイルスベクターなどの感染性ベクターを含むがこれらに限定されず、この目的に使用し得る。
【0103】
経皮送達は、ケラチノサイト、メラノサイトおよび樹状細胞を含む表皮の細胞型を用いた間接的導入に特に適する(Pfutzner et al.,Expert Opin.Investig.Drugs 2000;9:2069−83)。
【0104】
1つの実施形態では、遺伝子治療のためのベクターは、一本鎖自己相補性アデノ随伴ウイルスである。加えて、この自己相補性アデノ随伴ウイルスは、小さなトランスサイレチンプロモータ−エンハンサ、マウス微小ウイルスからのイントロンおよびウシ成長ホルモンからのポリアデニル化シグナルを含み得る。ベクターはまた、第VII因子を活性化形態、すなわち第VIIa因子として分泌させる、Margaritis et al.によって記述されたRKRRKR配列も含み得る(Margaritis et al.,J.Clin.Invest.,2004;113:1025−31)。
【0105】
[投与および医薬組成物]
本発明によるキメラ第VII因子ポリペプチドは、凝固因子欠損症(例えば血友病AおよびBまたは凝固第XI因子もしくは第VII因子の欠損)または凝固因子インヒビターなどのいくつかの原因を有する出血性疾患を管理するために使用され得、あるいはそれらは、正常に機能する血液凝固カスケードを有する(凝固因子欠損または凝固因子のいずれかに対するインヒビターを有さない)被験体において生じる過剰の出血を制御するために使用され得る。出血は、血小板機能欠損、血小板減少症またはフォン・ヴィレブランド病によって引き起こされ得る。出血はまた、様々な刺激によって線維素溶解活性の上昇が誘発された被験体でも見られ得る。
【0106】
手術または外傷に関連する広範囲の組織損傷を経験した被験体では、正常な止血機構が即時止血の必要性によって圧倒されることがあり、該被験体は、正常な止血機構にも関わらず出血を発現し得る。十分な止血を達成することは、脳、内耳領域および眼などの器官で出血が起こる場合にも問題であり、また、発生源を同定することが困難である広汎性出血(出血性胃炎および大量の子宮出血)の場合にも問題であり得る。同じ問題が、様々な器官(肝臓、肺、腫瘍組織、胃腸管)から生検を採取する過程ならびに腹腔鏡手術において生じ得る。これらの状況は、外科的手術手技(縫合、クリップ等)によって止血を行うことが困難であるという共通点を有する。急性かつ大量の出血はまた、抗凝固療法時に、与えられた治療によって不完全な止血が誘発された被験体においても起こり得る。かかる被験体は、抗凝固作用を速やかに中和しなければならない場合は外科的処置を必要とし得る。不十分な止血の場合に問題を引き起こし得るもう1つの状況は、正常な止血機構を有する被験体が血栓塞栓症を予防するために抗凝固療法を受ける場合である。かかる療法は、ヘパリン、他の形態のプロテオグリカン、ワルファリンまたは他の形態のビタミンKアンタゴニストならびにアスピリンおよび他の血小板凝固阻害剤を含み得る。
【0107】
計画的処置に関連する治療に関して、本発明のキメラ第VII因子ポリペプチドは、典型的には処置を実施する前の約24時間以内、およびその後7日間またはそれ以上投与される。凝固薬としての投与は、本明細書で述べる様々な経路によって実施できる。
【0108】
キメラ第VII因子ポリペプチドの用量は、被験体の体重および状態の重症度に依存して、負荷用量および維持用量として70kgの被験体について約0.05mg/日〜約500mg/日、好ましくは約1mg/日〜約200mg/日、より好ましくは約10mg/日〜約175mg/日の範囲である。当業者は、所与の被験体および所与の状態についての最適用量を決定することができる。
【0109】
医薬組成物は、主として予防的および/または治療的処置のための非経口投与を意図される。好ましくは、医薬組成物は非経口的に、すなわち静脈内、皮下もしくは筋肉内経路で投与されるか、または持続的もしくはパルス注入によって投与され得る。あるいは、医薬組成物は、経口的、皮下的、静脈内、脳内、鼻内、経皮的、腹腔内、筋肉内、肺内、経膣的、経直腸的、眼内経路または任意の他の許容される方法を含むがこれらに限定されない、様々な方法での投与用に製剤され得る。
【0110】
非経口投与用の組成物は、医薬的に許容される担体、好ましくは水性担体と組み合わせた、好ましくはその中に溶解された、本発明のキメラ第VII因子ポリペプチドを含有する。水、緩衝水、0.4%食塩水、0.3%グリシン等のような、様々な水性担体を使用し得る。本発明のキメラ第VII因子ポリペプチドはまた、メチオニンおよびスクロースなどの、安定性および保存を延長させる組成物と共に製剤化され得る。本発明のキメラ第VII因子ポリペプチドはまた、損傷の部位への送達または標的化のためにリポソーム製剤に製剤することができる。リポソーム製剤は、例えば米国特許第4,837,028号、米国特許第4,501,728号および米国特許第4,975,282号において一般的に説明されている。組成物は、従来の公知の滅菌技術によって滅菌され得る。生じた水溶液を、使用のために包装し得るか、または無菌条件下でろ過し、凍結乾燥して、凍結乾燥製剤を投与の前に滅菌水溶液と組み合わせ得る。組成物は、pH調整剤および緩衝剤、張度調整剤等のような、生理的条件に近付けるために必要な医薬的に許容される補助物質、例えば酢酸ナトリウム、乳酸ナトリウム、塩化ナトリウム、塩化カリウム、塩化カルシウム等を含有し得る。組成物はまた、防腐剤、等張化剤、非イオン性界面活性剤または洗浄剤、抗酸化剤および/または他の種々の添加物も含有し得る。
【0111】
これらの製剤中のキメラ第VII因子ポリペプチドの濃度は広く異なり、すなわち約0.5重量%未満、通常は約1重量%またはそれ以下から約15または20重量%までにわたり得、選択される特定の投与方法に従い、主として液体の容量、粘度等によって選択される。従って、静脈内注入用の典型的な医薬組成物は、滅菌リンガー液250mlおよびキメラ第VII因子ポリペプチド10mgを含有するように作製され得る。非経口的に投与可能な組成物を調製するための実際の方法は、公知であるかまたは当業者に明らかであり、例えばRemington’s Pharmaceutical Sciences,18th ed.,Mack Publishing Company,Easton,Pa.(1990)の中でより詳細に説明されている。
【0112】
本発明のキメラ第VII因子ポリペプチドを含有する組成物は、予防的および/または治療的処置のために投与できる。治療適用では、組成物を、上述したように既に疾患に罹患している被験体に、疾患およびその合併症を治療する、緩和するまたは部分的に停止させるのに十分な量で投与する。これを達成するのに適切な量は、「治療有効量」と定義される。当業者に理解されるように、このために有効な量は、疾患または損傷の重症度ならびに被験体の体重および全身状態に依存する。しかし一般に、有効量は、70kgの被験体についてキメラ第VII因子ポリペプチド約0.05mg/日〜約500mg/日の範囲であり、キメラ第VII因子ポリペプチド約1.0mg/日〜約200mg/日の用量がより一般的に使用される。
【0113】
本発明の物質は、一般に、重篤な疾患または損傷状態、すなわち生命を脅かすまたは潜在的に生命を脅かす状況において使用され得ることに留意しなければならない。かかる場合、外来性物質が最小限であることおよび一般にヒトにおいてヒト第VII因子の免疫原性がないことを考慮すると、実質的に過剰なこれらのキメラ第VII因子ポリペプチド組成物を投与することが可能であり、治療医師はこれを望ましいと考えるはずである。
【0114】
予防適用では、本発明のキメラ第VII因子ポリペプチドを含有する組成物を、被験体自身の凝固能力を増強するために疾患状態または損傷を生じやすいまたはその危険性がある被験体に投与する。かかる量は、「予防有効量」と定義される。予防適用では、正確な量は、やはり被験体の健康状態と体重に依存するが、用量は一般に70kgの被験体について約0.05mg/日〜約500mg/日、より一般的には70kgの被験体について約1.0mg/日〜約200mg/日の範囲である。
【0115】
組成物の単回または多回投与を、治療医師によって選択される用量レベルおよびパターンで実施し得る。毎日の維持レベルを必要とする通院被験体については、キメラ第VII因子ポリペプチドを、例えば携帯ポンプシステムを用いた持続注入によって投与し得る。
【0116】
本発明のキメラ第VII因子ポリペプチドはまた、徐放性または持続放出性製剤に製剤化し得る。徐放性または持続放出性製剤を製剤化する方法は当技術分野において知られており、ポリペプチドを含有する固体疎水性粒子の半透性マトリックスを含むが、これに限定されない。
【0117】
例えば局所適用などの、本発明のキメラ第VIIa因子ポリペプチドの局所送達は、例えば、スプレー、灌流、二重バルーンカテーテル、ステントによって、人工血管またはステントに組み込んで、バルーンカテーテルを被覆するのに使用されるヒドロゲルによって、または他の広く確立された方法によって実施し得る。いずれの場合も、医薬組成物は、被験体を有効に治療するのに十分な量の第VII因子変異体を提供すべきである。
【0118】
以下の実施例は本発明を説明するために含まれるものである。以下の実施例の特定の態様を、本発明の実施において良好に機能するように発明者が見出したまたは企図した技術および手順に関して述べる。本開示および当技術分野の一般的な技術レベルに照らして、当業者は、以下の実施例が例示だけを意図し、本発明の範囲から逸脱することなく数多くの変更、修正および改変を用い得ることを認識する。
【0119】
[実施例1:キメラ第VIIa因子ポリペプチドの構築および発現]
ヒト第VII因子およびヒト第IX因子の両方のcDNAを入手し、ドメイン交換を容易にするために以下のユニーク制限部位を導入した。
BstEII部位をEGF−1ドメインの5’末端に(第VII因子の残基47〜49、第IX因子の残基48〜50)、
SacI部位をEGF−1とEGF−2ドメインの接合部に(第VII因子の残基82〜83、第IX因子の残基83〜84)、
NotI部位をEGF−2ドメインの3’末端に(第VII因子の残基135〜137、第IX因子の残基132〜134)導入した。
【0120】
第VII因子と第IX因子の間でドメインを交換するためのこれらの制限部位を用いて様々なキメラを構築し、図1に示している。同じかまたは同様の制限部位を使用して第VII因子とプロテインSとの間でドメインを交換し得る。
【0121】
生じたキメラのアミノ酸配列を配列番号:1(第IX因子のシグナル、プロペプチド、GLAおよびEGF−1ドメインと、第VII因子のEGF−2および触媒ドメイン)および配列番号:2(第IX因子のシグナルおよびプロペプチドドメインと、プロテインSのGLAドメインと、第IX因子のEGF−1ドメインと、第VII因子のEGF−2および触媒ドメイン)に示す。これらの組換えDNAから選択したものを、哺乳動物細胞のトランスフェクションのために発現ベクターpCMV5にサブクローニングした。
【0122】
生じた組換え構築物を、次に、pSV2neoおよびpCMVhGCを用いて293ヒト腎細胞株(ATCC CRL 1573)に同時トランスフェクトした。高レベルの各構築物を発現するクローンを選択し、先に述べられているようにスクリーニングした(Toomey et al.,1991,J.Biol.Chem.,266:19198−19202)。各々のスクリーニングしたクローンを、大規模生産のために900cm2のローラーボトルに広げ、組換えタンパク質を、Fast Flow Q−Sepharoseを用いた偽アフィニティクロマトグラフィ法、および、カルシウム勾配、次いでNaCl勾配での溶出によって精製した。
【0123】
[実施例2:第VIIa因子ポリペプチドのトロンビン産生]
第VIIa因子のトロンビン産生を、血友病のインビトロモデルを用いて測定した。単球を組織因子の供給源として使用し、不活性化血小板と、血漿濃度の第V、第VII、第IX、第VIIIおよび第XI因子ならびに血漿濃度の抗トロンビンおよびTFPIを含有する合成血漿とを組み合わせた。第VIIIおよび第IX因子を除外することによって血友病状態を作り出した。
【0124】
健常個体から血液4mlを採取し、クエン酸ナトリウム管に入れて単球を調製した。血液を15ml円錐管中のAccu−Prep(商標)リンパ球分離培地(Accurate Chemicals,NY,USA)3mlの上に注意深く層状に重ね、1500rpmで30分間遠心分離した。次に単核層を取り出し、4℃で等容量のVersene(Lineberger Tissue Culture)に添加して、800rpmで10分間遠心分離した。生じたペレットを4℃でVersene 5mlに再懸濁し、800rpmで10分間遠心分離した。生じたペレットを、500ng/mlのリポ多糖(LPS)(Sigma−Aldrich,MO,USA)を添加したマクロファージSFM培地(Life Technologies,CA,USA)4mlに再懸濁し、細胞を96ウェルプレートに200μl/ウェルで播種した。プレートを37℃、5%CO2で2時間インキュベートし、次にマクロファージSFM培地で3回洗浄して、その後一晩インキュベートした。
【0125】
健常個体から血液4mlを採取し、クエン酸ナトリウム管に入れて血小板を調製した。血液を、15ml円錐管中のAccu−Prep(商標)リンパ球分離培地(Accurate Chemicals,NY,USA)5mlの上に、注意深く層状に重ね、1500rpmで30分間遠心分離した。次に血小板層を取り出し、等容量のクエン酸−グルコース−食塩水および10μg/mlのプロスタグランジンE1(PGE1)(Sigma−Aldrich,MO,USA)に添加して、800rpmで10分間遠心分離した。生じたペレットを廃棄し、1mg/mlオボアルブミンを添加したカルシウム不含タイロード緩衝液中でセファロースゲルろ過によって血漿タンパク質から血小板を単離した。回収した血小板を37℃で保存した。
【0126】
血友病を模倣するインビトロアッセイのための合成血漿を、5μg/mlの濃度の第XI因子C−1エステラーゼインヒビターを使用して、あらかじめ調製しておいた単球と共に1時間インキュベートした後、200μg/mlの抗トロンビン、0.07μg/mlのTFPI、100mg/mlのプロトロンビン、8μg/mlの第X因子および0.5μg/mlの第VII因子を添加して作製した。一晩インキュベートした後、第V因子を7μg/mlの濃度で添加した。正常条件については、1時間インキュベートした後、第IX因子を4μg/mlの濃度で添加し、一晩インキュベートした後、第VIII因子を1U/mlで添加した。
【0127】
血小板260μlを上記合成血漿溶液に添加し、生じた懸濁液を単球に添加して反応を開始させた。一定の時間間隔で試料10μlを取り出し、1mM EDTAおよび0.5mM Gly−Pro−Arg−pNA(Centerchem Inc.,CT,USA)を含有するトロンビンアッセイ緩衝液90μlに添加した。50%酢酸100μlを添加して合成基質切断を停止させ、405nmでODを測定した。以下の式を用いてトロンビン濃度を決定した。
[トロンビン]=希釈×((A405−バックグラウンド)/(停止−開始))/(変換係数)
式中、変換係数が1nMである場合、トロンビンは405nmで0.0117OD/分の変化量を与える。
【0128】
正常条件(第VIIIおよび第IX因子を含む)でのトロンビン産生は、短い遅滞期でピークトロンビン産生を示す(図2)。野生型第VIIa因子の濃度上昇は、血友病条件でのトロンビン産生を改善した(図2)。
【0129】
[実施例3:キメラ第VIIa因子ポリペプチドのトロンビン産生]
実施例1で作製したキメラタンパク質のトロンビン産生を、実施例2で述べた血友病のインビトロモデルを用いて測定した。10nMの、第VII因子のEGF−2および触媒ドメインと第IX因子のGLAおよびEGF−1ドメインとを含むキメラ第VIIa因子、または第VII因子のGLA、EGF−2および触媒ドメインと第IX因子のEGF−1ドメインとを含むキメラ第VIIa因子の添加は、50nMの野生型第VIIa因子と同様の活性を有していた(図3)。
【0130】
対照と比較してピークトロンビン産生を検討すると、野生型第VIIa因子または試験したキメラ第VIIa因子ポリペプチドのいずれもが、対照レベルまでのピークトロンビン産生を回復していなかった。しかし、10nMの、第VII因子のEGF−2および触媒ドメインと第IX因子のGLAおよびEGF−1ドメインとを含むキメラ第VIIa因子、または第VII因子のGLA、EGF−2および触媒ドメインと第IX因子のEGF−1ドメインとを含むキメラ第VIIa因子は、50nMの野生型第VIIa因子によって産生されるのと同様のレベルまでピークトロンビン産生を回復していた(図4)。
【0131】
[実施例4:キメラ第VIIa因子ポリペプチドの凝固アッセイ]
キメラ第VIIa因子ポリペプチドの凝固活性を、Buyue,Y.,et al.,2008,Blood,112:3234−3241によって述べられているように、凝固アッセイにおいてインビボで評価した。簡単に述べると、第VII因子を、脚の静脈に挿入したカテーテルを介して、血友病Bマウスに投与した(2mg/kgのNovoSeven(登録商標)、2mg/kgの、第IX因子のGLAおよびEGF−1ドメインと第VIIa因子のEGF−2および触媒ドメインとを含むキメラ第VIIa因子、または1.4mg/kgの、プロテインSのGLAドメインと、第IX因子のEGF−1ドメインと、第VIIa因子のEGF−2および触媒ドメインとを含むキメラ第VIIa因子)。野生型マウスを対照として使用した。マウスを麻酔し、他方の脚の伏在静脈を、小さなゲージの針を押し通すことによって離断した。次に鋏の先端を静脈に挿入し、小さな杯を作製するように鋏で切って遠位部分を切断した。血餅を形成させ、出血が停止するまでの時間を記録した。次に30ゲージの針を用いて血餅を除去し、出血が停止するまでの時間を再び記録した。この工程を30分間反復した。血餅の数を崩壊の数として記録した。記録した時間を平均し、平均時間として記録した。実験を、野生型マウス、ならびに、プロテインSのGLAドメインと第IX因子のEGF−1ドメインと第VIIa因子のEGF−2および触媒ドメインとを含むキメラ第VIIa因子について1匹のマウスで実施した。第IX因子のGLAおよびEGF−1ドメインと第VIIa因子のEGF−2および触媒ドメインとを含むキメラ第VIIa因子については、2匹の血友病Bマウスで実験を実施し、同じ結果を得た。第VII因子を投与しなかった血友病Bマウスに関して、以下の表1において記録した0.25の崩壊は、4匹のマウスのうち1匹だけが初期損傷で出血しなかったことを示す。
【0132】
【表1】
【0133】
処置を行わなかった1匹の血友病Bマウス、2mg/kgのNovoSeven(登録商標)を投与した1匹の血友病Bマウス、2mg/kgの、第IX因子のGLAおよびEGF−1ドメインと第VIIa因子のEGF−2および触媒ドメインとを含むキメラ第VIIa因子を投与した1匹の血友病Bマウスならびに処置を行わなかった1匹の野生型マウスに関して実験を反復した。2mg/kgの、第IX因子のGLAおよびEGF−1ドメインと第VIIa因子のEGF−2および触媒ドメインとを含むキメラ第VIIa因子を投与した血友病Bマウスは、2mg/kgのNovoSeven(登録商標)を投与した血友病Bマウスおよび野生型マウスよりも出血の停止までの平均時間がより短かった(図5)。
【0134】
[実施例5:キメラ第VIIa因子ポリペプチドの血栓形成性]
本発明のキメラ第VIIa因子ポリペプチドの血栓形成性を、高レベルのこれらのタンパク質を発現するマウスモデルにおいて試験することができる。近年、2μg/ml以上の組換え野生型マウス第VIIa因子を発現するマウスの50%が16か月以内に血栓症で死亡することが示された。同様のレベルの第VIIa因子を発現する血友病Bマウスは同じ死亡率を有し、マウスをC57BL/6Jマウス系統に戻し交配した場合、はるかに早期に(4か月未満)血栓症が起こった(Aljamali MN,et al.J Clin Invest.2008;118:1825−1834)。
【0135】
2〜10μgまたはそれ以上のキメラポリペプチドを発現する血友病Bマウス(C57BL/6J系統)を作製することができ、キメラ第VII因子ポリペプチドの発現が血栓症を引き起こす場合、これらのマウスは早期に(4か月未満)血栓症を発現するはずである。
【0136】
UNCにおけるthe Gene Therapy CenterのDr.Paul Monahanによって開発された新しい自己相補性アデノ随伴ベクター(AAV)を使用することによってマウス系統を開発し得る(Wu Z,et al.Mol Ther.2008;16:280−289)。このベクターは、小さなトランスサイレチンプロモータ−エンハンサ(TTRプロモータ)、マウスの微小ウイルスからのイントロン、MVMおよびウシ成長ホルモンpAシグナルを含む。このベクターに基づく、1×1011のベクターゲノムの門脈への注入は、感染から8週間以内に7μg/mlの第IX因子の持続的発現を生じさせた(Wu Z,et al.Mol Ther.2008;16:280−289)。
【0137】
マウスおよびヒトの両方の第VIIa因子ならびに本発明のキメラ第VII因子ポリペプチドのcDNAを、血栓形成性を評価するために使用し得る。第VII因子を活性化形態、すなわち第VIIa因子として分泌させる、Margaritis et al.によって記述されたRKRRKR(配列番号:21)配列も使用し得る(Margaritis et al.,J.Clin.Invest.,2004;113:1025−31)。
【0138】
この仮説を検証するため、野生型およびキメラ第VII因子マウスポリペプチドを作製することができる。マウス組織因子はヒト第VII因子と反応しないことが報告されているので、組織因子は外傷において投与される第VIIa因子の作用とは無関係であるが、血栓症に関係することを明らかにするためにマウスポリペプチドを使用する。この仮説を検証するために使用できるマウスポリペプチドは、野生型マウス第VII因子(配列番号:3)、ポリペプチドを二本鎖の活性形態(FVIIa)で分泌させるRKRRKR配列を有する野生型マウス第VII因子(配列番号:4)、マウス第IX因子のシグナル、プロペプチド、GLAおよびEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列(配列番号:5)、ならびにマウス第IX因子のシグナルおよびプロペプチドドメイン、プロテインSのGLAドメイン、第IX因子のEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列(配列番号:6)を包含する。キメラマウス第VII因子ポリペプチド(配列番号:5および配列番号:6)は、組織因子に対して低い親和性を有し(おそらく野生型FVII分子と比較して100倍低い)、これは望ましくない血栓症を予防するのに十分なはずである。
【0139】
キメラ第VII因子ポリペプチドはまた、組織因子に対する親和性をさらに低減するように付加的に修飾し得る。かかる突然変異の一例は、残基306のメチオニンをアラニンに変更することである。第VIIa因子のより高い比活性を生じさせるキメラ第VII因子ポリペプチドの他の修飾も含み得る。かかる修飾の例は、ヒト配列の残基158、296および298に対する以下の等価物、V158D、E296VおよびM298Qを含む。
【0140】
上述した野生型およびキメラマウス第VII因子ポリペプチドは、マウス第VII因子cDNAを鋳型として使用し、様々なドメインを増幅するためのオリゴヌクレオチドプライマーを用い、次いで、この構築物をpSC−TTR−mvmベクターに挿入することによって構築でき、該構築は、Wu et al.,Molecular Therapy,2008;16:280−289に記載されている。具体的には、野生型マウス第VII因子は、以下のプライマー、5’−TGAGGATCCCCACCATGGTTCCACAGGCGCATGGGCT−3’(配列番号:7)および5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)を使用してマウス第VII因子cDNAから増幅できる。次に、増幅したフラグメントをBamHI/SphIで消化し、BglII/SphI部位の間でpSC−TTR−mvmベクターに挿入し得る。その後、BGHポリアデニル化配列を、生じたベクターにSphI部位で挿入し得る。
【0141】
RKRRKR配列を有する野生型マウス第VII因子構築物は、同様の方法で構築し得る。具体的には、RKRRKR配列を有する野生型マウス第VII因子は、以下のプライマー、
1番目のフラグメントを作製するための、5’−TGAGGATCCCCACCATGGTTCCACAGGCGCATGGGCT−3’(配列番号:7)および5’−ACAATGCGTTTTCGCCGCTTACGGCGGCCTTGGCGGCTGCTGGAGT−3’(配列番号:9)、ならびに
2番目のフラグメントを作製するための、5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)および5’−GCCGCCGTAAGCGGCGAAAACGCATTGTGGGAGGCAACGTGTGCCC−3’(配列番号:10)
を使用してマウス第VII因子cDNAから増幅できる。次に、これらの2つのフラグメントを鋳型として使用し、以下のプライマー、
5’−TGAGGATCCCCACCATGGTTCCACAGGCGCATGGGCT−3’(配列番号:7)および5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)
を用いてオーバーラップPCRを実施する。次に、生じた増幅フラグメントをBamHI/SphIで消化し、BglII/SphI部位の間でpSC−TTR−mvmベクターに挿入し得る。その後、BGHポリアデニル化配列を、生じたベクターにSphI部位で挿入し得る。
【0142】
マウス第IX因子のシグナル、プロペプチド、GLAおよびEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列を含む構築物(配列番号:5)を同様に構築する。具体的には、1番目のフラグメントを、マウス第IX因子cDNAを鋳型として使用し、以下のプライマー、
5’−AGGCCTGAAGATCTCCACCATGAAGCACCTGAACACCGTC−3’(配列番号:11)および
5’−GATCAGCTGCTCATTCTTGCTTTTTTCACAGTTCCTTCCTTCAAATC−3’(配列番号:12)
を用いて作製する。2番目のフラグメントは、マウス第VII因子を鋳型として使用し、以下のプライマー、
5’−GATTTGAAGGAAGGAACTGTGAAAAAAGCAAGAATGAGCAGCTGATC−3’(配列番号:13)および
5’−ACAATGCGTTTTCGCCGCTTACGGCGGCCTTGGCGGCTGCTGGAGT−3’(配列番号:9)
を用いて作製する。
【0143】
3番目のフラグメントは、マウス第VII因子を鋳型として使用し、以下のプライマー、
5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)および
5’−GCCGCCGTAAGCGGCGAAAACGCATTGTGGGAGGCAACGTGTGCCC−3’(配列番号:10)
を用いて作製できる。次に、これらの3つフラグメントを鋳型として使用し、以下のプライマー、
5’−AGGCCTGAAGATCTCCACCATGAAGCACCTGAACACCGTC−3’(配列番号:11)および
5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)
を用いてオーバーラップPCRを実施し得る。次に、生じた増幅フラグメントをBamHI/SphIで消化し、BglII/SphI部位の間でpSC−TTR−mvmベクターに挿入する。その後、BGHポリアデニル化配列を、生じたベクターにSphI部位で挿入する。
【0144】
同様の工程を使用して、マウス第IX因子のシグナルおよびプロペプチドドメイン、プロテインSのGLAドメイン、第IX因子のEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列を含む構築物(配列番号:6)を作製し得る。具体的には、1番目のフラグメントを、マウス第IX因子cDNAおよび以下のプライマー、
5’−AGGCCTGAAGATCTCCACCATGAAGCACCTGAACACCGTC−3’(配列番号:11)および
5’−GTTTCTTCGAACAAGGTATTTGCTCTCTTTGGACGGGTAAGAATTTTG−3’(配列番号:14)
を使用して作製し得る。2番目のフラグメントは、マウスプロテインSを鋳型として使用し、以下のプライマー、
5’−CAAAATTCTTACCCGTCCAAAGAGAGCAAATACCTTGTTCGAAGAAAC−3’(配列番号:15)および
5’−GATCTCCATCAACATACTGCTTATAAAAATAATCCGTCTCGGGATT−3’(配列番号:16)
を用いて作製し得る。3番目のフラグメントは、マウス第IX因子のシグナル、プロペプチド、GLAおよびEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列を有するベクターを鋳型として使用し、以下のプライマー、
5’−AATCCCGAGACGGATTATTTTTATAAGCAGTATGTTGATGGAGATC−3’(配列番号:17)および
5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)
を用いて作製し得る。次に、これらの3つフラグメントを鋳型として使用し、以下のプライマー、
5’−AGGCCTGAAGATCTCCACCATGAAGCACCTGAACACCGTC−3’(配列番号:11)および
5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)
を用いてオーバーラップPCRを実施する。次に、生じた増幅フラグメントをBamHI/SphIで消化し、BglII/SphI部位の間でpSC−TTR−mvmベクターに挿入する。その後、BGHポリアデニル化配列を、生じたベクターにSphI部位で挿入し得る。
【0145】
野生型マウス第VII因子(配列番号:3)およびRKRRKR配列を有する野生型マウス第VII因子(配列番号:4)のベクター構築物を、各々、年齢が適合する2匹の雄性血友病Bマウスに注射した(マウスにつき5×1011ベクターゲノム)。マウス第IX因子のシグナル、プロペプチド、GLAおよびEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列(配列番号:5)、ならびにマウス第IX因子のシグナルおよびプロペプチドドメイン、プロテインSのGLAドメイン、第IX因子のEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列(配列番号:6)のベクター構築物を、各々、年齢が適合する3匹の雄性血友病Bマウスに注射した(マウスにつき5×1011ベクターゲノム)。注射の約3週間後、野生型マウス第VII因子(配列番号:3)またはRKRRKR配列を有する野生型マウス第VII因子(配列番号:4)のいずれかを注射したすべてのマウスが死亡した。マウス第IX因子のシグナル、プロペプチド、GLAおよびEGF−1ドメインと、第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列(配列番号:5)、またはマウス第IX因子のシグナルおよびプロペプチドドメイン、プロテインSのGLAドメイン、第IX因子のEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列(配列番号:6)の、いずれかを注射したマウスは、注射の約5週間後に生存し、有害症状を示さなかった。マウスから週に1回血液を採取し、注射の6〜8週間後にマウス第VII因子ポリペプチドのレベルを測定し得る。また、マウス第VII因子ポリペプチドに対する抗原レベルは、注射したマウスの血液試料からインビトロで凝固活性を測定することによって間接的に測定し得る(Margaritas et al.,Gene Therapy 2009;113:3682−3689)。
【0146】
上記は本発明の説明であり、本発明を限定すると解釈されるべきではない。本発明は、以下の特許請求の範囲およびその中に含まれる特許請求の範囲の等価物によって定義される。本明細書で引用されるすべての出版物、特許出願、特許、特許公開、GenBank(登録商標)データベースおよび/またはSNPアクセッション番号によって同定される配列、ならびに他の参考文献は、参考文献が提示される文および/または段落に関連する教示に関してそれらの全体を引用することにより本明細書の一部をなすものとする。
【技術分野】
【0001】
[優先権の主張]
本出願は、35 U.S.C.§119(e)の下に、その内容全体を引用することにより本明細書の一部をなすものとする、2009年6月25日出願の米国特許仮出願第61/220,278号の優先権を主張する。
【0002】
[政府支援の言明]
本発明は、米国国立衛生研究所によって授与された認可番号5−P01−HL06350の下での政府支援を得て為された。米国政府は本発明に一定の権利を有する。
【0003】
本発明は、現在使用可能な第VII因子ポリペプチドよりも高い凝固活性を有し、血栓合併症がより少ないキメラヒト凝固第VII因子(FVII)ポリペプチド、ならびにかかるポリペプチドをコードするポリヌクレオチド構築物、該ポリヌクレオチドを含み、これを発現するベクターおよび宿主細胞、医薬組成物、使用および治療の方法に関する。
【背景技術】
【0004】
血液凝固は、最終的にフィブリン血餅を生じさせる、様々な血液成分(または因子)の複雑な相互作用からなるプロセスである。一般に、凝固「カスケード」と称されてきたものに関与する血液成分は、アクチベータ(これ自体は活性化された凝固因子である)の作用によってタンパク質分解酵素に変換される、酵素的に不活性なタンパク質(プロ酵素またはチモーゲン)である。かかる変換を受けた凝固因子は、一般に「活性因子」と称され、凝固因子の名称に「a」の文字を付加することによって表される(例えば活性化第VII因子は第VIIa因子またはFVIIaと表される)。
【0005】
通常、止血プロセスの開始は、組織因子と第VIIa因子との間の複合体の形成によって媒介される。この複合体は、次に、第IX因子(FIX)および第X因子(FX)をそれらの活性型に変換する。第Xa因子(FXa)は、組織因子担持細胞上で限られた量のプロトロンビンをトロンビンに変換する。トロンビンは血小板を活性化し、第V因子(FV)および第VIII因子(FVIII)を第Va因子(FVa)および第VIIIa因子(FVIIIa)へと活性化して、これらの両補因子がさらなるプロセスにおいて完全なトロンビンバーストを導く。このプロセスは、第IXa因子(FIXa)(第VIIIa因子との複合体として)による第Xa因子の産生を含み、活性化された血小板の表面上で起こる。トロンビンは、最終的にフィブリノーゲンをフィブリンへと変換し、フィブリン血餅の形成をもたらす。近年、第VII因子および組織因子が血液凝固の主たるイニシエータであることが見出された。
【0006】
第VII因子は、一本鎖チモーゲンとして血液中を循環する血漿糖タンパク質である。チモーゲンは最小限の触媒活性を有する。一本鎖第VII因子は、インビトロで第Xa因子、第XIIa因子、第IXa因子、第VIIa因子またはトロンビンによって二本鎖第VIIa因子に変換され得る。第Xa因子は第VII因子の主要な生理的アクチベータであると考えられている。止血に関与するいくつかの他の血漿タンパク質と同様に、第VII因子はその活性に関してビタミンKに依存し、該活性は該タンパク質のアミノ末端に近接してクラスタ形成している複数のグルタミン酸残基のγ−カルボキシル化のために必要である。これらのγ−カルボキシル化されたグルタミン酸は、リン脂質と第VII因子との金属イオン誘導性相互作用に必要とされる。チモーゲン第VII因子の活性化二本鎖分子への変換は、内部のArg152−Ile153ペプチド結合の切断によって生じる。加えて、高濃度の第VII因子がインビトロで自己活性化を導くことは公知である。組織因子、リン脂質およびカルシウムイオンの存在下で、二本鎖第VIIa因子は、限定的なタンパク質分解によって第X因子または第IX因子を速やかに活性化する。
【0007】
ヒトFVII(hFVII)をコードする遺伝子は、第13番染色体のq34−qter9にマッピングされている(de Grouchy et al.,Hum Genet 1984;66:230−233)。この遺伝子は9個のエクソンを含み、12.8Kbにわたる(O’Hara et al.,Proc Natl Acad Sci USA 1987;84:5158−5162)。FVIIの遺伝子構成およびタンパク質構造は他のビタミンK依存性プロ凝固タンパク質のものに類似し、エクソン1aおよび1bはシグナル配列をコードし、エクソン2はプロペプチドおよびGLAドメイン、エクソン3は短い疎水性領域、エクソン4および5は上皮増殖因子様ドメイン、ならびにエクソン6から8はセリンプロテアーゼ触媒ドメインをコードする(Yoshitake et al.,Biochemistry 1985;24:3736−3750)。
【0008】
第IX因子(クリスマス因子)は、正常な止血において活性なセリンプロテアーゼのチモーゲンであり、その酵素活性は特定のグルタミン酸残基のカルボキシル化を必要とする。第IX、X、VII因子およびプロテインCは、同じファミリーのセリンプロテアーゼの密接に関連するパラログであり、これらのタンパク質をコードする遺伝子の高い度合のアミノ酸配列同一性およびイントロン−エクソン配置を有する。これらの密接に関連するタンパク質は、アミノ末端からカルボキシル末端までの機能性ドメインの類似構造を有し、γ−カルボキシグルタミン酸(GLA)ドメイン、2つの上皮増殖因子様(EGF)ドメイン、活性化ペプチドおよび触媒ドメインを含む。プロテインSは、666アミノ酸の、GLAドメイン、4つのEGF様ドメイン、トロンビン感受性領域および2つのラミニンドメインを有するビタミンK依存性タンパク質である。
【0009】
ビタミンK依存性凝固血漿タンパク質は、膜へのタンパク質結合の部位として機能するGLAドメインを含み、GLAドメインは様々な凝固タンパク質の間で高度に保存されている。それらの類似性にもかかわらず、GLAドメインはリン脂質に対して広い範囲の親和性を示し、プロテインSのGLAドメインがリン脂質に対して最も高い親和性を有する(Ellison et al.,Biochemistry,1998;37:7997−8003)、(McDonald et al.,Biochemistry 1997;36:5120−27)。
【0010】
被験体において凝固カスケードを刺激するまたは改善することがしばしば望ましい。第VIIa因子は、凝固因子欠損症(例えば血友病AおよびBまたは凝固第XI因子もしくは第VII因子の欠損)または凝固因子インヒビターなどのいくつかの原因を有する出血性疾患を管理するために使用されてきた。第VIIa因子はまた、正常に機能する血液凝固カスケードを有する(凝固因子欠損、または凝固因子のいずれかに対するインヒビターを有さない)被験体において生じる過剰の出血を制御するためにも使用されてきた。かかる出血は、例えば、血小板機能欠損、血小板減少症またはフォン・ヴィレブランド病によって引き起こされ得る。
【0011】
出血はまた、手術および他の形態の外傷に関連する重大な問題である。例えば、第VII因子はイラクおよびアフガニスタンで負傷した兵士を治療するために広く使用された(Perkins JG,et al.The Journal of Trauma.2007;62:1095−9;discussion 9−101)。その使用は多くの命を救ったことで高い評価を得ているが、大部分の医学的治療に関しては、治療後の脳卒中または他の血栓事象などの副作用が存在する。しかし、FVIIaを使用する医師の全体的な印象は、その使用はそのために失われた命よりも多くの命を救ってきたというものである。おそらく、このことを最もよく表しているのは、過去の戦争において負傷者の約30%が損傷のために死亡しているのに対し、現在の湾岸戦争ではその数が約10%に減少したことである(Gawande A,et al.,N Engl J Med.2004;351:2471−5)。
【0012】
第VIIa因子を発現するトランスジェニック血友病Bマウスの試験は、低レベル(1.5μg/ml未満)の持続的な第VIIa因子発現が血友病Bマウスにおいて凝固活性を回復させることを明らかにした。しかし、野生型または血友病Bマウスでの2μg/mlを上回る第VIIa因子レベルは、心臓および肺における血栓症を導いた。心臓および肺はどちらも、組織因子の高発現部位である。これは、循環中の高レベルの第VIIa因子は、心臓および肺の血管損傷後に暴露される組織因子に接触した場合血栓症を誘発することを示唆する(Margaritas et al.,J.Clin.Invest.2004;113:1025−31)。さらに、血友病のイヌにおけるベクターを介したイヌ第VIIa因子の遺伝子導入が、短期および中期的に安全かつ有効であることが試験で示された(Margaritas et al.,Gene Therapy 2009;113:3682−3689)。
【0013】
第VII因子による治療に関する警告が、現在、規制認可を求める製品について提案されている。例えば、欧州医薬品庁、人体用医薬品評価ユニット(European Medicines Agency,Human Medicines Evaluation Unit)は、現在の第VII因子治療には、特に第VII因子が冠状動脈性心疾患または肝疾患の病歴を有する患者、術後患者、新生児ならびに血栓症および播種性血管内凝固症候群の危険性がある患者に投与される状況では、血栓症および播種性血管内凝固症候群の危険性の警告を付すことを推奨している。例えば、2004年7月のCore SPC for Human Plasma Derived Coagulation Factor VII Products(CPMP/BPWG/2048/01)参照。
【0014】
これまでに、第VIIa因子のEGF−1ドメインが組織因子に対する第VIIa因子の親和性に重要な役割を果たすことが示されている。組織因子の存在下および不存在下の両方で、合成基質と第X因子の両方を使用して、第IX因子のEGF−1ドメインを有する第VIIa因子ポリペプチドは野生型第VIIa因子よりも低い触媒活性を有していた(Jin et al.,Biochemistry,1999,28:1185−92)。一見したところ、これは出血を治療するためのキメラ構築物の使用を排除すると思われる。しかし、Monroe(British Journal of Haematology 1997;99:542−549)は、血友病および出血を治療するうえでのFVIIaの機構は組織因子非依存性であると提案した。しかし、第VIIa因子が組織因子とは独立して活性でないかどうかに関しては当技術分野で見解が分かれる。
【0015】
ヒト組換えFVIIaの市販製剤は、NovoSeven(登録商標)およびNovoSeven(登録商標)RTとして販売されている。NovoSeven(登録商標)およびNovoSeven(登録商標)RTは、血友病AまたはB患者における出血エピソードの治療に適用され、出血エピソードの治療のための市場で入手可能な唯一のrFVIIaである。近年、NovoSeven(登録商標)が、第VII因子を損傷部位に局在化させるために組み合わせて投与できる、再水和された凍結乾燥血小板に結合し得ることが明らかにされた(Fischer et al.,Platelets,2008;19:182−91)。加えて、第VII因子の選択的なPEG化は血漿半減期を上昇させ得ること(Stennicke et al.,Thromb.Haemost,2008;100:920−28)および3個のアミノ酸置換を有する組換えヒト第VII因子は血小板の表面上で高い活性を有すること(Moss et al.,J.Thromb.Haemost.,2009;7:299−305)が示された。本発明のキメラ第VIIa因子分子のPEG化は、同様に血漿半減期を上昇させるように働くと期待される。同じように、コンジュゲートを形成する非ポリペプチド部分の共有結合などの、当技術分野で知られているタンパク質の他の修飾、例えばグリコシル化は、本発明のキメラ第VIIa因子分子において同様に機能すると期待され、すなわち非ポリペプチド部分の共有結合によってタンパク質に与えられる特性がキメラ第VIIa因子分子に付与されると予想される。
【発明の概要】
【発明が解決しようとする課題】
【0016】
利用可能な治療に結びつく、比較的低い用量で投与できる高い凝固活性を有する第VIIa因子の変異体、および血栓合併症などの望ましくない副作用を生じさせることが少ない変異体が必要とされている。
【課題を解決するための手段】
【0017】
本出願は、1以上の望ましい恩恵を提供する、キメラFVIIa分子、特にhFVIIaドメインと凝固系の1以上のタンパク質からのドメインを含むキメラhFVIIa分子を開示する。従って、本発明のキメラFVIIa分子は、市販のrFVIIaと比較して1以上の改善された特性を有し、かかる特性は、より高い凝固活性を有することおよび/または比較的低い用量で投与できることおよび/または血栓合併症を生じさせることがより少ないことを含む。その結果として、本発明のキメラによる医学的治療は、潜在的により低い用量および/またはより少ない望ましくない副作用などの、現在使用可能なrFVIIa化合物を上回る利点を提供する。
【0018】
本発明の代表的なキメラFVIIaポリペプチドは、FVIIのEGF−2および触媒ドメインと、ビタミンK依存性凝固タンパク質のGLAドメインと、ビタミンK依存性凝固タンパク質のEGF−1ドメインとを含むキメラFVIIaを包含する。本発明の特定の実施形態では、本発明のキメラFVIIaポリペプチドは、1)FIXのGLAおよびEGF−1ドメインとFVIIのEGF−2および触媒ドメインとを含むキメラFVIIa、2)FIXのEGF−1ドメインとFVIIのGLA、EGF−2および触媒ドメインとを含むキメラFVIIa、3)プロテインSのGLAドメインと、FIXのEGF−1ドメインと、FVIIのEGF−2および触媒ドメインとを含むキメラFVIIa、4)プロテインSのGLAおよびEGF−1ドメインとFVIIのEGF−2および触媒ドメインとを含むキメラFVIIa、5)プロテインSのEGF−1ドメインとFVIIのGLA、EGF−2および触媒ドメインとを含むキメラFVIIa、ならびに6)プロテインSのEGF−1ドメインと、FIXのGLAドメインと、FVIIのEGF−2および触媒ドメインとを含むキメラFVIIaを包含する。本発明の代表的なキメラFVIIaポリペプチドはまた、EGF−1ドメインまたはGLAドメイン内にアミノ酸置換を有する、野生型FVIIaまたは上述したキメラFVIIaポリペプチドのいずれかを包含し得る。置換は、保存的置換または非保存的置換であり得る。かかる置換は、EGF−1ドメインの残基69のイソロイシンのアラニンによる置換および/または残基79のアルギニンのアラニンによる置換を含み得る。さらなる置換は、GLAドメインにIV型コラーゲンに対するより高い結合親和性をもたらすが、血小板結合には影響を及ぼさないと思われる、残基5のリシンのアルギニンによる置換(Gui et al.,J.Thromb Haemost.2009;7:1843−1851)、残基306のメチオニンの別のアミノ酸による置換、好ましくは保存的アミノ酸置換、より好ましくは、組織因子に対する親和性をさらに低下させるアラニンによる置換、ならびにより高い比活性を有する第VIIa因子を生じさせる、残基158のバリンのアスパラギン酸による置換、残基296のグルタミン酸のバリンによる置換および/または残基298のメチオニンのグルタミンによる置換を含み得る。
【0019】
また、本明細書で述べるキメラFVIIaポリペプチドの1以上を投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法も提供される。出血性疾患を治療する方法は、本発明のキメラ第VIIa因子ポリペプチドをコードするヌクレオチド配列を含む核酸分子を被験体に投与する方法を含み得る。
【0020】
加えて、本発明は、GLAドメインを含むタンパク質を投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法を提供し、該タンパク質は、血餅の部位に標的されるおよび/または被験体の血漿中の循環タンパク質の濃度に比べて血餅の部位付近で濃縮されている。これは、血餅形成部位でまたはその付近で認められる、現在使用可能な第VII因子ポリペプチドよりも高い親和性で血小板に結合するドメインを使用することを含み得る。かかるドメインの例は、血小板の表面に位置する負に荷電したリン脂質層に結合する様々な凝固タンパク質のGLAドメインである。かかるGLAドメインの非限定的な例は、FVIIaのGLAドメインよりもはるかに密接に血小板およびリン脂質に結合する、FIXのGLAドメイン(Melton et al.「Location of the platelet binding site in zymogen coagulation factor IX」Blood Coagul Fibrinolysis 12:237−243(2001))ならびに任意の他の公知のGLAドメインよりも密接にリン脂質に結合する、プロテインSのGLAドメイン(McDonald et al.「Comparison of naturally occurring vitamin K−dependent proteins:correlation of amino acid sequences and membrane binding properties suggests a membrane contact site」Biochemistry:36:5120−5127(1997))を含む。GLAドメインを含むタンパク質の非限定的な例は、FIXもしくはプロテインSのGLAドメインを含む組換えタンパク質、FIXもしくはプロテインSのGLAドメインを含むキメラタンパク質、および/またはFIXもしくはプロテインSのGLAドメインを含む本発明のキメラFVIIaポリペプチドを包含する。
【0021】
もう1つの実施形態では、本発明は、第VII因子ポリペプチドに由来する触媒ドメインおよび1以上の血小板標的ドメインを含む本発明のキメラ第VIIa因子ポリペプチドを投与することにより、出血性疾患を有する被験体を治療する方法を提供する。かかる血小板標的ドメインは、血小板の表面と相互作用するまたは結合するタンパク質からのドメインを含む。血小板の表面とのかかる相互作用または表面への結合は、血小板の膜リン脂質を通してまたは血小板細胞表面タンパク質および/もしくは受容体を通して媒介され得る。かかるドメインの非限定的な例は、血小板糖タンパク質Ibについての主要結合部位である、フォン・ヴィレブランド因子のA1ドメイン(Emsley et al.,JBC,273:10396−10401(1998))ならびに血小板膜タンパク質および/または受容体に結合する抗体、例えば血小板膜リン脂質に結合する抗体のFabフラグメント(ドメイン)(Out et al.,Blood,77:2655−2659(1991))を含む。
【0022】
血小板に結合し、組織因子に対して低い親和性を有するドメインを使用した、血餅部位への凝固タンパク質のかかる標的化は、血栓形成性などの、現在の第VII因子療法に関連する合併症の危険度を低下させるという付加的な、しかし予想外の利益を有する。このアプローチはまた、他の治療上有用なポリペプチドまたは他の分子を血小板または血餅部位に標的するためにも使用できる。かかる治療上有用な分子は、抗凝固剤等を含み得る。
【0023】
本発明はさらに、第VII因子の血栓形成性に比べて低い血栓形成性(例えば、天然第VII因子の血栓形成性と比較して少なくとも約1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、90%低い等)を有するポリペプチドを被験体に投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法を提供する。「低い血栓形成性」は、当技術分野で知られている方法に従って様々なやり方で、例えば、対照と比較して、血餅の測定数がより少ない、血餅がより小さい、血餅が生じるのにより長い時間を要する(インビボまたはインビトロアッセイにおいて)、血栓形成による被験体の死亡がより少ないおよび/または生存期間が長いことを含むが、これらに限定されない方法で測定できる。第VII因子と比較して低い血栓形成性を有するポリペプチドは、例えば、本発明のキメラ第VII因子ポリペプチドまたはその活性フラグメントであり得る。かかるポリペプチドの非限定的な例は、FIXのGLAおよびEGF−1ドメインとFVIIのEGF−2および触媒ドメインとを含むキメラFVIIa、ならびにプロテインSのGLAドメインと、FIXのEGF−1ドメインと、FVIIのEGF−2および触媒ドメインとを含むキメラFVIIaを包含する。
【図面の簡単な説明】
【0024】
【図1】図1は、本発明において有用なキメラFVIIa分子の図式的表示を示す。FVII:第VII因子、FIX:第IX因子。
【図2】図2は、様々な濃度の野生型第VIIa因子についての血友病の細胞モデル系における正常条件および血友病条件(第IX因子および第VIII因子の除去)でのトロンビン産生(nM)を示す。
【図3】図3は、50nMの野生型第VIIa因子または10nMのキメラ第VIIa因子を投与した血友病の細胞モデル系における正常条件および血友病条件(第IX因子および第VIII因子の除去)でのトロンビン産生(nM)を示す。
【図4】図4は、50nMの野生型第VIIa因子または10nMのキメラ第VIIa因子の投与後の血友病の細胞モデル系において産生される、対照値と比較した、ピークトロンビンを示す。
【図5】図5は、処置していない血友病Bマウス、2mg/kgのNovoSeven(登録商標)を投与した血友病Bマウス、2mg/kgのキメラFVIIa分子を投与した血友病Bマウスおよび処置していない野生型マウスについての凝固アッセイにおける崩壊止血時間(disruption hemostasis time)を示す。
【図6】図6は、シグナル、プロペプチド、第IX因子のGLAおよびEGF1ドメイン(下線)を含む、本発明の例示的なキメラ第VII因子の配列(配列番号:1)を示す。
【図7】図7は、シグナル、プロペプチドおよび第IX因子のEGF1ドメイン(下線)およびプロテインSのGLAドメイン(太字)を含む、本発明の例示的なキメラ第VII因子の配列(配列番号:2)を示す。
【発明を実施するための形態】
【0025】
本発明を以下でより詳細に説明する。しかし本発明は、種々の形態で具体化でき、本明細書で述べる実施形態に限定されると解釈されるべきではない。むしろ、これらの実施形態は、この開示が詳細かつ完全となり、当業者に本発明の範囲を十分に伝えるために提供されるものである。
【0026】
本明細書で本発明の説明において使用される用語は、特定の実施形態を説明することだけを目的とし、本発明の限定を意図されない。本発明の説明および付随の特許請求の範囲において使用される、「a」、「an」および「the」という単数形態は、文脈によって明確に指示されない限り、複数形態も包含することが意図されている。
【0027】
特に定義されない限り、本明細書で使用されるすべての用語(技術および学術用語を含む)は、本発明が属する技術分野の当業者によって一般的に理解されるのと同じ意味を有する。一般的に使用される辞書において定義されるもののような用語は、本出願および関連技術分野におけるそれらの意味と一致する意味を有すると解釈されるべきであり、本明細書において明白に定義されない限り理想的なまたは過度に正式な意味に解釈されるべきではないことがさらに了解される。本明細書で本発明の説明において使用される用語は、特定の実施形態を説明することを目的とし、本発明の限定を意図されない。本明細書で言及するすべての出版物、特許出願、特許および他の参考文献は、それらの全体を引用することにより本明細書の一部をなすものとする。
【0028】
また本明細書で使用される、「および/または」は、関連して列挙される項目の1以上のありとあらゆる可能な組合せ、ならびに選択的に解釈される場合(「または」)は組合せの欠如を指し、これらを包含する。
【0029】
文脈によって特に指示されない限り、本明細書で述べる本発明の様々な特徴は任意の組合せで使用できることが明確に意図されている。
【0030】
さらに、本発明はまた、本発明の一部の実施形態では、本明細書で述べる任意の特徴または特徴の組合せが排除または除外され得ることを企図する。例示すると、明細書において複合体が成分A、BおよびCを含むと記述される場合、A、BまたはCのいずれかまたはそれらの組合せが、単独でまたは任意の組合せで除外され、排除され得ることが明確に意図されている。
【0031】
本明細書で使用される、「〜から実質的になる」という移行句(および文法的変形)は、列挙される物質または段階ならびに特許請求される本発明の「基本的なおよび新規の特徴に実質的に影響を及ぼさないもの」を包含すると解釈されるべきである。In re Herz,537 F.2d 549,551−52,190 U.S.P.Q.461,463(CCPA 1976)(原文での強調)参照、MPEP §2111.03も参照のこと。従って、本明細書で使用される「〜から実質的になる」という用語は、「含む」と等価であると解釈されるべきではない。
【0032】
量または濃度等のような測定可能な値に言及する場合に本明細書で使用される、「約」という用語は、特定される量の20%、10%、5%、1%、0.5%またはさらに0.1%の変動を包含することが意図されている。
【0033】
本明細書で使用される「活性」という用語は、第VII因子ポリペプチドがその基質である第X因子を活性な第Xa因子に変換する能力を意味する。
【0034】
「固有活性」という用語はまた、組織因子の不存在下に活性化血小板の表面でトロンビンを産生する能力も包含する。
【0035】
「N末端GLAドメイン」という用語は、配列番号:18のアミノ酸配列の約アミノ酸残基61から約アミノ酸残基105までのアミノ酸配列、配列番号:19のアミノ酸配列の約アミノ酸残基47から約アミノ酸残基92までのアミノ酸配列、同定されたアミノ酸配列への任意の翻訳後修飾、同定されたアミノ酸配列における任意の保存的アミノ酸置換、同定されたアミノ酸配列へのアミノ酸残基の付加、同定されたアミノ酸配列からのアミノ酸残基の欠失、またはリン脂質膜に結合する凝固カスケードタンパク質からの任意の他のアミノ酸配列を包含する。
【0036】
「EGF−1」という用語は、ジスルフィド結合1〜3、2〜4および5〜6を含むほとんどすべての公知のEGF様ドメインを含む、EGF(上皮増殖因子)中ならびにまた細胞シグナル伝達および凝固タンパク質に関与する一連のタンパク質中でもともと認められる6個のシステインを含む30〜40アミノ酸の領域を表す。第VII因子のEGF−1ドメインは、配列番号:18のアミノ酸配列の約アミノ酸残基106から約アミノ酸残基142までである。第IX因子のEGF−1ドメインは、配列番号:19のアミノ酸配列の約アミノ酸残基93から約アミノ酸残基129までである。プロテインSのEGF−1ドメインは、配列番号:20のアミノ酸配列の約アミノ酸残基117から約アミノ酸残基155までである。これらのアミノ酸配列は、同定されたアミノ酸配列への任意の翻訳後修飾、同定されたアミノ酸配列における任意の保存的アミノ酸置換、同定されたアミノ酸配列へのアミノ酸残基の任意の付加および/または同定されたアミノ酸配列からのアミノ酸残基の任意の欠失を含み得る。
【0037】
「EGF−2」という用語は、(2またはそれ以上のEGF様ドメインの)一続きにおける2番目のEGF様ドメインを意味する。第VII因子のEGF−2ドメインは、配列番号:18のアミノ酸配列の約アミノ酸残基147から約アミノ酸残基188までである。第IX因子のEGF−2ドメインは、配列番号:19のアミノ酸配列の約アミノ酸残基130から約アミノ酸残基171までである。プロテインSのEGF−2ドメインは、配列番号:20のアミノ酸配列の約アミノ酸残基157から約アミノ酸残基200までである。これらのアミノ酸配列は、同定されたアミノ酸配列への任意の翻訳後修飾、同定されたアミノ酸配列における任意の保存的アミノ酸置換、同定されたアミノ酸配列へのアミノ酸残基の付加および/または同定されたアミノ酸配列からのアミノ酸残基の任意の欠失を含み得る。
【0038】
本明細書で使用される「触媒ドメイン」という用語は、ペプチド結合の切断を媒介するタンパク質中のドメインを意味する。第VII因子の触媒ドメインは、配列番号:18のアミノ酸配列の約アミノ酸残基213から約アミノ酸残基452までである。第IX因子の触媒ドメインは、配列番号:19のアミノ酸配列の約アミノ酸残基227から約アミノ酸残基459までである。これらのアミノ酸配列は、同定されたアミノ酸配列への任意の翻訳後修飾、同定されたアミノ酸配列における任意の保存的アミノ酸置換、同定されたアミノ酸配列へのアミノ酸残基の任意の付加および/または同定されたアミノ酸配列からのアミノ酸残基の任意の欠失を含み得る。
【0039】
「GLA」という3文字表記は4−カルボキシグルタミン酸(γ−カルボキシグルタミン酸)を意味する。
【0040】
「プロテアーゼドメイン」という用語は、一般に配列番号:18の約アミノ酸残基213からカルボキシ末端アミノ酸残基までとみなされる、ペプチド結合の切断を媒介するタンパク質中のドメインを意味する(第VIIa因子の重鎖)。本明細書で使用される「第VII因子ポリペプチド」という用語は、天然ヒト第VII因子のアミノ酸配列61〜466(配列番号:18)を含む任意のタンパク質またはその変異体もしくはフラグメントを意味する。これは、ヒト第VII因子、ヒト第VIIa因子およびそれらの変異体を含むが、これらに限定されない。本明細書で使用される「第VII因子」という用語は、不活性な一本鎖チモーゲン第VII因子分子ならびに活性化二本鎖第VII因子分子(第VIIa因子)を含むことが意図されている。これは、天然ヒト第VII因子または第VIIa因子のアミノ酸配列61〜466(配列番号:18)を有するタンパク質を包含する。当業者は、小さな配列変化は同様に機能すると予想されること、ならびに関与するドメイン、ポリペプチドおよび第VII因子は、本発明から逸脱することなくわずかに短縮もしくは伸長され得るまたは置換を含み得ることを認識する。従って、この定義はまた、わずかに修飾されたアミノ酸配列、例えばN末端アミノ酸欠失または付加を含む修飾されたN末端を有するタンパク質およびペプチドも包含し、一部の実施形態ではそれらのタンパク質は第VIIa因子の活性を実質的に保持する(例えば天然第VIIa因子の活性の約50%、60%、70%、80%、90%、95%等を保持する)。本明細書で使用される「第VIIa因子」または「FVIIa」という用語は、活性化形態(第VIIa因子)からなる生成物を意味する。上記定義内の「第VII因子」または「第VIIa因子」はまた、個体によって存在し、生じ得る天然の対立遺伝子変異も包含する。また、グリコシル化または他の翻訳後修飾の程度と位置は、発現の選択される宿主細胞、組織または動物種および宿主の細胞または組織環境の性質に依存して異なり得る。
【0041】
本明細書で使用される「ドメイン」という用語は、タンパク質鎖の残りの部分とは独立して進化し、機能し、存在し得るタンパク質配列および構造の一部を包含することが意図されている。ドメインはコンパクトな三次元構造を形成することができ、しばしば独立して安定化され、折りたたまれ得る。1つのドメインが様々な進化的に関連するタンパク質中に出現し得る。ドメインの長さは、約25アミノ酸から約500アミノ酸長までにわたる。「ドメイン」はまた、単数または複数のアミノ酸残基が保存的置換によって置き換えられた野生型タンパク質からのドメインも包含し得る。それらはタンパク質媒質中で自己安定性であるので、ドメインは、キメラタンパク質を作製するためにあるタンパク質と別のタンパク質との間で遺伝子操作によって「交換」され得る。
【0042】
本明細書で使用される「変異体(単数または複数)」という用語は、天然ヒト第VII因子または第VIIa因子のアミノ酸配列61〜466(配列番号:18)を有する第VII因子を表すことが意図され、ここで、親タンパク質の1もしくはそれ以上のアミノ酸が別のアミノ酸によって置換されている、および/または、親タンパク質の1もしくはそれ以上のアミノ酸が欠失している、および/または、1もしくはそれ以上のアミノ酸がタンパク質に挿入されている、および/または、1もしくはそれ以上のアミノ酸が親タンパク質に付加されている、および/または、GLAドメインが異なるタンパク質からのGLAドメイン(例えば血小板膜もしくはリン脂質膜に結合するGLAドメイン)で置換されている、および/または、GLAドメインが異なるタンパク質からの血小板結合ドメイン(例えば血小板糖タンパク質Ibについての主要結合部位である、フォン・ヴィレブランド因子のA1ドメイン(Emsley et al.,JBC,273:10396−10401(1998))ならびに血小板膜タンパク質および/もしくは受容体に結合する抗体、例えば血小板膜リン脂質に結合する抗体のFabフラグメント(ドメイン)(Out et al.,Blood,77:2655−2659(1991))で置換されている、および/または、第VII因子のEGF−1ドメインが異なるタンパク質からのEGF−1ドメイン(例えば第IX因子のEGF−1ドメインなどの、組織因子により低い親和性で結合するEGF−1ドメイン)で置換されている。かかる付加は、親タンパク質のN末端またはC末端またはその両方で、ならびに内部で生じ得る。従って、一部の実施形態では、本発明の「変異体(単数または複数)」は、活性化形態のFVIIの凝固活性をまだ有し得る。一部の実施形態では、本発明の変異体(単数または複数)は凝固活性を有さなくてもよい。
【0043】
従って、一部の実施形態では、変異体は、天然ヒト第VII因子または第VIIa因子のアミノ酸配列61〜466(配列番号:18)と少なくとも40%、50%、60%または70%同一である。例えば、FVIIのEGF−1ドメインはFIXのEGF−1ドメインと65.7%の同一性を有し、FVIIのGLAドメインはFIXのGLAドメインと58.6%の同一性およびプロテインSのGLAドメインと51%の同一性を有する。1つの実施形態では、変異体は、天然ヒト第VII因子または第VIIa因子のアミノ酸配列61〜466(配列番号:18)と少なくとも80%同一である。もう1つの実施形態では、変異体は、天然ヒト第VII因子または第VIIa因子のアミノ酸配列61〜466(配列番号:18)と少なくとも90%同一である。さらなる実施形態では、変異体は、天然ヒト第VII因子または第VIIa因子のアミノ酸配列61〜466(配列番号:18)と少なくとも95%同一である。
【0044】
本明細書で使用される「任意の他のアミノ酸」という用語は、その位置に天然に存在するアミノ酸とは異なる1つのアミノ酸を意味する。これは、ポリヌクレオチドによってコードされ得るアミノ酸を含むが、これに限定されない。好ましくは、異なるアミノ酸は天然L型であり、ポリヌクレオチドによってコードされ得る。具体的な例はL−システイン(Cys)である。
【0045】
本明細書で使用される、「作動可能に連結された」という用語は、配列の正常な機能を果たすことができるような相互の立体配置での、酵素的連結または別の方法による2またはそれ以上のヌクレオチド配列の共有結合を指す。例えば、プレ配列または分泌リーダーをコードするヌクレオチド配列は、それがポリペプチドの分泌に関与するプレタンパク質として発現される場合は、ポリペプチドをコードするヌクレオチド配列に作動可能に連結されている。プロモータまたはエンハンサは、それが対象とする配列の転写に影響を及ぼす場合は、コード配列に作動可能に連結されている。リボソーム結合部位は、それが対象とするタンパク質またはペプチドへの翻訳を促進するように位置づけられている場合は、コード配列に作動可能に連結されている。一般に、「作動可能に連結されている」とは、連結されているヌクレオチド配列が連続していることを意味し、分泌リーダーの場合は、連続しかつ読み枠内にあることを意味する。連結は、好都合な制限部位での結合によって最も容易に達成される。かかる部位が存在しない場合は、合成オリゴヌクレオチドアダプターまたはリンカーを標準的な組換えDNA法と共に使用する。
【0046】
本明細書で使用される「ベクター」という用語は、宿主細胞において増幅することができる任意の核酸実体を意味する。従って、ベクターは、自律複製ベクター、すなわち、その複製が染色体の複製から独立している、染色体外実体として存在するベクター、例えばプラスミドであり得る。あるいは、ベクターは、宿主細胞に導入された場合、宿主細胞のゲノムに組み込まれ、組み込まれた染色体と共に複製されるものであってもよい。ベクターの選択は、それが導入される宿主細胞に依存することが多い。ベクターは、プラスミドベクター、ファージベクター、ウイルスまたはコスミドベクターを含むが、これらに限定されない。ベクターは通常、複製起点および少なくとも1つの選択可能な遺伝子、すなわち容易に検出可能であるかまたはその存在が細胞増殖に必須である生成物をコードする遺伝子を含む。
【0047】
本明細書で使用される「宿主細胞」という用語は、雑種細胞を含む、異種DNAがその中で発現され得る任意の細胞を意味する。典型的な宿主細胞は、昆虫細胞、酵母細胞、BHK細胞、CHO細胞、HEK細胞およびCOS細胞などのヒト細胞を含む哺乳動物細胞を含むが、これらに限定されない。本発明を実施する場合、培養される宿主細胞は、好ましくは哺乳動物細胞、より好ましくは、限定されることなく、CHO細胞株(例えばATCC CCL 61)、COS−1細胞株(例えばATCC CRL 1650)、ベビーハムスター腎(BHK)細胞株およびHEK293細胞株(例えばATCC CRL 1573;Graham et al.,J.Gen.Virol.36:59−72,1977)を含む、樹立哺乳動物細胞株である。適切なBHK細胞株は、本明細書中ではBHK 570細胞と称する、tk−ts13BHK細胞株である(Waechter and Baserga,Proc.Natl.Acad.Sci.USA 79:1106−1110,1982)。BHK 570細胞株は、ATCCアクセッション番号CRL 10314の下に、American Type Culture Collection,10801 University,Boulevard,Manassas,Va.20110から入手可能である。tk−ts13BHK細胞株も、アクセッション番号CRL 1632の下にATCCから入手可能である。他の適切な細胞株は、限定されることなく、Rat Hep I細胞(ラット肝細胞癌;ATCC CRL 1600)、Rat Hep II細胞(ラット肝細胞癌;ATCC CRL 1548)、TCMK細胞(ATCC CCL 139)、ヒト肺細胞(ATCC HB 8065)、NCTC 1469細胞(ATCC CCL 9.1)およびDUKX細胞(Urlaub and Chasin,Proc.Natl.Acad.Sci.USA 77:4216−4220,1980)を含む。また、3T3細胞、ナマルバ細胞、骨髄腫細胞および骨髄腫細胞と他の細胞の融合物も有用である。
【0048】
本明細書で使用される場合、「適切な増殖培地」という用語は、細胞の増殖および本発明の第VII因子ポリペプチドをコードする核酸配列の発現のために必要な栄養素および他の成分を含む培地を意味する。
【0049】
「コンジュゲート」(または交換可能に「コンジュゲート化ポリペプチド」)という用語は、1以上のポリペプチドの、ポリマー分子、親油性化合物、糖成分または有機誘導体化剤などの1以上の非ポリペプチド部分への共有結合によって形成される異種(複合体またはキメラという意味で)分子を示すことが意図されている。好ましくは、コンジュゲートは適切な濃度および条件で可溶性である、すなわち血液などの生理的液体に可溶性である。本発明のコンジュゲート化ポリペプチドの例は、グリコシル化および/またはPEG化ポリペプチドを含む。
【0050】
「共有結合」という用語は、ポリペプチドと非ポリペプチド部分が互いに直接共有結合で連結されているか、または架橋、スペーサーもしくは単数もしくは複数の連結部分などの、単数もしくは複数の介在部分を介して互いに間接的に共有結合で連結されていることを意味する。「非コンジュゲート化ポリペプチド」という用語は、コンジュゲートのポリペプチド部分を指すために使用できる。
【0051】
本明細書で使用される場合、「非ポリペプチド部分」という用語は、本発明のポリペプチドの結合基にコンジュゲート化することができる分子を意味する。かかる分子の適切な例は、ポリマー分子、糖成分、親油性化合物または有機誘導体化剤を含む。本発明のコンジュゲートに関連して使用される場合、非ポリペプチド部分はポリペプチドの結合基を介してコンジュゲートのポリペプチド部分に連結されることが了解される。上記で説明したように、非ポリペプチド部分は、結合基に直接共有結合で連結され得るかまたは架橋、スペーサーもしくは単数もしくは複数の連結部分などの、単数もしくは複数の介在部分を介して間接的に結合基に共有結合で連結され得る。
【0052】
「ポリマー分子」は、2またはそれ以上のモノマーの共有結合によって形成される分子であり、ポリマーがヒトアルブミンまたは別の豊富な血漿タンパク質である場合を除き、いずれのモノマーもアミノ酸残基ではない。「ポリマー」という用語は、「ポリマー分子」という用語と交換可能に使用できる。この用語は、インビトログリコシル化、すなわち、任意選択的に架橋剤を使用して、通常はポリペプチドの結合基に炭水化物分子を共有結合連結することを含む、インビトロで実施される合成的グリコシル化によって結合された炭水化物分子を包含することが意図されている。
【0053】
N−グリコシル化またはO−グリコシル化(以下でさらに説明する)などのインビボでのグリコシル化によって結合された炭水化物分子を本明細書では「糖成分」と称する。コンジュゲート中のポリマー分子または糖成分などの非ポリペプチド部分の数が明白に示されている場合を除き、本発明で使用されるコンジュゲートまたは別のものに含まれる「非ポリペプチド部分」へのあらゆる言及は、ポリマー分子または糖成分などの1以上の非ポリペプチド部分への言及であるものとする。
【0054】
「結合基」という用語は、ポリマー分子、親油性分子、糖成分または有機誘導体化剤などの非ポリペプチド部分に結合することができる、ポリペプチドの官能基、特にそのアミノ酸残基または炭水化物部分の官能基を示すことが意図されている。
【0055】
インビボでのN−グリコシル化に関しては、「結合基」という用語は通常と異なるように使用され、N−グリコシル化部位(配列N−X−S/T/C[式中、Xはプロリンを除く任意のアミノ酸残基であり、Nはアスパラギンであり、およびS/T/Cはセリン、トレオニンまたはシステインのいずれか、好ましくはセリンまたはトレオニン、最も好ましくはトレオニンである]を有する)を構成するアミノ酸残基を示す。N−グリコシル化部位のアスパラギン残基は、グリコシル化の間に糖成分が結合される残基であるが、かかる結合は、N−グリコシル化部位のその他のアミノ酸残基が存在しない限り達成され得ない。
【0056】
従って、非ポリペプチド部分が糖成分であり、コンジュゲート化がN−グリコシル化によって行われる場合、対象とするポリペプチドのアミノ酸配列の変化に関連して使用される「非ポリペプチド部分のための結合基を含むアミノ酸残基」という用語は、N−グリコシル化部位を構成する1以上のアミノ酸残基が、機能的N−グリコシル化部位がアミノ酸配列内に導入されるかまたは該配列から除去されるように変化していることを意味すると理解される。
【0057】
本明細書において、「治療する」または「治療」という用語は、出血を阻止するまたは最小限に抑えることを目的とした、手術におけるような予想される出血の管理および/または予防、ならびに外傷または血友病患者のためのオンデマンド治療におけるような既に発生している出血の制御の両方を含むことが意図されている。従って本発明による第VIIa因子ポリペプチドの予防的投与は、「治療」の用語に包含される。
【0058】
「出血エピソード」という用語は、制御されない、かつ過剰の出血を含むことが意図されている。出血エピソードは、手術および他の形態の組織損傷の両方に関して重大な問題であり得る。制御されない、かつ過剰の出血は、正常な凝固系を有する被験体および凝固または出血性疾患を有する被験体において起こり得る。
【0059】
本明細書で使用される「出血性疾患」という用語は、出血として現れる、細胞、生理または分子起源の、先天性、後天性または誘導された何らかの欠損を反映する。例としては、凝固因子欠損症(例えば血友病AおよびBまたは凝固第XIもしくは第VII因子の欠損症)、凝固因子インヒビター、血小板機能欠損、血小板減少症、フォン・ヴィレブランド病、または手術もしくは外傷によって誘発される出血である。
【0060】
過剰の出血はまた、正常に機能する血液凝固カスケードを有する(凝固因子欠損または凝固因子のいずれかに対するインヒビターを有さない)被験体においても発生し、血小板機能欠損、血小板減少症またはフォン・ヴィレブランド病によって引き起こされ得る。かかる場合、出血は血友病によって引き起こされる出血にたとえられることがあり、というのは、止血系が、血友病におけるように、欠如しているかまたは、重大な出血を引き起こす、必須の凝固「化合物」(血小板またはフォン・ヴィレブランド因子タンパク質など)の異常を有しているからである。手術または外傷に関連する広範囲の組織損傷を経験した被験体では、正常な止血機構が即時止血の必要性によって圧倒されることがあり、該被験体は、正常な止血機構にも関わらず出血を発現し得る。十分な止血を達成することは、外科的止血の可能性が限られる、脳、内耳領域および眼などの器官で出血が起こる場合にも問題である。同じ問題が、様々な器官(肝臓、肺、腫瘍組織、胃腸管)から生検を採取する過程ならびに腹腔鏡手術において生じ得る。これらすべての状況に共通するのは、外科的手術手技(縫合、クリップ等)によって止血を行うことの困難さであり、これはまた、出血が広汎性である場合(出血性胃炎および大量の子宮出血)にもあてはまる。急性かつ大量の出血は、抗凝固療法時に、与えられた治療によって不完全な止血が誘発された被験体においても起こり得る。かかる被験体は、抗凝固作用を速やかに中和しなければならない場合は外科的処置を必要とし得る。根治的恥骨後前立腺切除術は、限局性前立腺癌を有する被験体に対して一般的に実施される手技である。この手術は、重大な、時として大量の失血によってしばしば困難になる。前立腺切除術の間の多量の失血は、主として、外科的止血が容易にアクセスできない様々な密に血管形成された部位を伴い、大きな領域から広範な出血を生じさせ得る、複雑な解剖学的状況に関連する。また、脳内出血は脳卒中の最も治療が難しい形態であり、脳内出血後数時間以内に高い死亡率と血腫成長に結びつく。rFVIIaによる治療は、血腫の成長を制限し、死亡率を低下させ、90日目の機能転帰を改善することができる。しかしながら、現在使用可能なrFVIIaに関しては、血栓塞栓性有害事象の危険性が存在する。血栓形成性がより低い本発明のキメラ第VII因子分子は、脳卒中治療におけるこの問題を克服し得る。不十分な止血の場合に問題を引き起こし得るもう1つの状況は、正常な止血機構を有する被験体が血栓塞栓症を予防するために抗凝固療法を受ける場合である。かかる療法は、ヘパリン、他の形態のプロテオグリカン、ワルファリンまたは他の形態のビタミンKアンタゴニストならびにアスピリンおよび他の血小板凝固阻害剤を含み得る。
【0061】
本発明の1つの実施形態では、出血は血友病に関連する。もう1つの実施形態では、出血は、後天性インヒビターを有する血友病に関連する。もう1つの実施形態では、出血は血小板減少症に関連する。もう1つの実施形態では、出血はフォン・ヴィレブランド病に関連する。もう1つの実施形態では、出血は重症組織損傷に関連する。もう1つの実施形態では、出血は重度の外傷に関連する。もう1つの実施形態では、出血は手術に関連する。もう1つの実施形態では、出血は腹腔鏡手術に関連する。もう1つの実施形態では、出血は出血性胃炎に関連する。もう1つの実施形態では、出血は大量の子宮出血に関連する。もう1つの実施形態では、出血は、機械的止血の可能性が限られている器官で起こる。もう1つの実施形態では、出血は脳、内耳領域または眼において生じる。もう1つの実施形態では、出血は生検を採取する過程に関連する。もう1つの実施形態では、出血は抗凝固療法に関連する。
【0062】
本明細書で使用される「被験体」という用語は、任意の動物、特にヒトなどの哺乳動物を意味することが意図され、適切な場合は、「患者」という用語と交換可能に使用され得る。
【0063】
「正常な止血系の増強」という用語は、トロンビンを産生するまたは機能性血餅を生成する能力の改善を意味する。
【0064】
「遺伝子治療」という用語は、遺伝子の外因性投与によって内因性遺伝子の発現を変化させる方法を指す。本明細書で使用される、「遺伝子治療」はまた、その必要のある個体の体細胞または幹細胞に、欠陥があるまたは欠如している遺伝子に対応する機能性遺伝子を導入することによる、欠陥タンパク質をコードする欠陥遺伝子の置換または欠如遺伝子の補充を指す。遺伝子治療は、分化した細胞または体性幹細胞を個体の体内から取り出し、続いて、ウイルスベクターを遺伝子送達ビヒクルとして用いて欠陥遺伝子の正常なコピーを外植された細胞に導入する、エクスビボ法によって実施され得る。加えて、インビボでの直接遺伝子導入技術は、治療効果を達成するために広範囲のウイルスベクター、リポソーム、タンパク質DNA複合体または裸のDNAを用いてインサイチューで個体の細胞に遺伝子導入することを可能にする。「遺伝子治療」という用語はまた、欠陥遺伝子またはタンパク質が欠陥でなかった場合に機能するはずであるのと実質的に同じように機能するポリヌクレオチドを、その必要のある個体の体細胞または幹細胞に導入することによる、欠陥タンパク質をコードする欠陥遺伝子の置換を指す。
【0065】
本発明において、遺伝子治療は、本発明のキメラFVIIをコードするヌクレオチド配列を含む核酸分子を被験体に投与することに関連して用いられる。従って、本発明は、本発明のキメラFVIIタンパク質をコードするヌクレオチド配列を含む単離された核酸分子を提供する。かかる核酸分子は核酸構築物(例えばベクターまたはプラスミド)中に存在し得る。かかる核酸構築物は細胞中に存在し得る。さらに、本発明のキメラFVIIタンパク質を細胞に送達する方法が本明細書で提供され、該方法は、キメラFVIIタンパク質をコードするヌクレオチド配列を含む核酸分子を、該ヌクレオチド配列が細胞において発現されてキメラFVIIタンパク質を生成する条件下で細胞に導入することを含む。細胞は、被験体に導入される細胞であり得るおよび/または被験体において既に存在する細胞であり得る。
【0066】
[キメラ第VII因子の作製]
本明細書で述べるキメラ第VII因子ポリペプチドは、組換え核酸技術によって作製され得る。一般に、クローニングされた野生型第VII因子核酸配列を、所望タンパク質をコードするように修飾する。次に、この修飾された配列を発現ベクターに挿入し、今度はこの発現ベクターを宿主細胞に形質転換またはトランスフェクトする。高等真核細胞、特に培養された哺乳動物細胞が宿主細胞として適切である。ヒト第VII因子についての完全なヌクレオチド配列およびアミノ酸配列が知られており(組換えヒト第VII因子のクローニングおよび発現が記載されている米国特許第4,784,950号、ならびにGenBank(登録商標)アクセッション番号J02933およびAAA51983ならびにSwissProtアクセッション番号P08709−1参照)、該アミノ酸配列を本明細書で配列番号:18として示す。ウシ第VII因子配列は、Takeya et al.,J.Biol.Chem.263:14868−14872(1988))に記載されている。第IX因子についての完全なヌクレオチド配列およびアミノ酸配列が知られており(Davie et al.,Proc.Natl.Acad.Sci.U.S.A 1982;79:6461−6464;Jaye et al.,Nucleic Acids Res.1983;11:2325−2335;およびMcGraw et al.,Proc.Natl.Acad.Sci.U.S.A.1985;82:2847−2851ならびにGenBank(登録商標)アクセッション番号J00136およびAAA98726ならびにSwissProtアクセッション番号P00740参照)、該アミノ酸配列を本明細書で配列番号:19として提供する。プロテインSについても完全なヌクレオチド配列およびアミノ酸配列が知られており(Hoskins et al.,Proc.Natl.Acad.Sci.U.S.A.1987;84:349−353ならびにGenBank(登録商標)アクセッション番号M15036およびAAA36479ならびにSwissProtアクセッション番号P07225参照)、該アミノ酸配列を本明細書で配列番号:20として提供する。他の凝固タンパク質についての完全なヌクレオチド配列およびアミノ酸配列も知られており、SwissProtを用いてまたはNCBIウエブページで見ることができる。
【0067】
アミノ酸配列の変化は様々な技術によって達成され得る。核酸配列の修飾は、部位特異的突然変異誘発によって実施し得る。部位特異的突然変異誘発のための技術は当技術分野において公知であり、例えば、Zoller and Smith(DNA 3:479−488,1984)または「Splicing by extension overlap」,Horton et al.,Gene 77,1989,pp.61−68に記載されている。従って、第VII因子のヌクレオチド配列およびアミノ酸配列を使用して、選択した変化を導入し得る。同様に、特異的なプライマーを使用したポリメラーゼ連鎖反応を用いてDNA構築物を作製するための手順も当業者に公知である(PCR Protocols,1990,Academic Press,San Diego,Calif.,USA参照)。加えて、本発明のキメラ第VII因子ポリペプチドは、様々なポリペプチドをコードするヌクレオチド配列にユニーク制限部位を導入することによって作製でき、これを使用して、様々なポリペプチドのドメイン全体をコードするヌクレオチド配列のフラグメントを単離することができる。その後、これらの様々なヌクレオチド配列を組み換えて、本発明の様々なキメラ第VII因子ポリペプチドを作製することができる。
【0068】
例えば、GLA、EGF−1、EGF−2および触媒ドメインを様々な凝固タンパク質の間で自由に交換し得るように、様々な凝固タンパク質をコードするヌクレオチド配列にユニーク制限部位を導入し得る。EGF−1ドメインはEGF−2ドメインの周囲を自由に回転することができるが(Pike et al.Proc Natl.Acad.Sci.1999;96:8925−8930)、GLAドメインおよびEGF−1ドメインは、カルシウムが存在する場合は互いの周囲を自由に回転できない(Sunnerhagen et al.Nature Structural Biology 1995;2:504−509)。この実施形態では、GLAおよびEGF−1ドメインは同じ凝固タンパク質に由来し得るが、EGF−2および触媒ドメインとは異なるタンパク質に由来し得る。従って、本発明のキメラ第VIIa因子ポリペプチドは、1つのドメインの少なくとも50%が第VII因子ドメインであることを条件として、凝固タンパク質の任意のものに由来するドメインを含み得る。さらに、本発明のキメラ第VIIa因子ポリペプチドは、同じタンパク質または異なるタンパク質に由来するGLAおよびEGF−1ドメインを含み得る。
【0069】
本発明のキメラ第VII因子ポリペプチドをコードする核酸構築物は、適切には、例えば標準的な技術に従ってゲノムライブラリーまたはcDNAライブラリーを作製し、合成オリゴヌクレオチドプローブを用いたハイブリダイゼーションによって様々なポリペプチドの全部または一部をコードするDNA配列をスクリーニングすることによって得られる、ゲノムまたはcDNA起源のものであり得る(Sambrook et al.,Molecular Cloning:A Laboratory Manual,2nd.Ed.Cold Spring Harbor Laboratory,Cold Spring Harbor,N.Y.,1989参照)。
【0070】
キメラ第VII因子ポリペプチドをコードする核酸構築物はまた、確立された標準的な方法、例えばBeaucage and Caruthers,Tetrahedron Letters 22(1981),1859−1869によって述べられているホスホルアミダイト法またはMatthes et al.,EMBO Journal 3(1984),801−805によって述べられている方法によって合成的に作製し得る。ホスホルアミダイト法によれば、例えば自動DNA合成装置において、オリゴヌクレオチドを合成し、精製して、アニーリングし、連結して、適切なベクターにクローニングする。
【0071】
さらに、核酸構築物は、標準的な技術に従って、核酸構築物全体の様々な部分に対応するフラグメントである、合成、ゲノムまたはcDNA起源(適宜に)のフラグメントを連結することによって作製される、合成とゲノムの混合型、合成とcDNA起源の混合型またはゲノムとcDNA起源の混合型であり得る。
【0072】
本発明によるキメラ第VII因子ポリペプチドを作製する際に使用するためのDNA配列は、典型的には、適切な翻訳後プロセシング(例えばグルタミン酸残基のγ−カルボキシル化)および宿主細胞からの分泌を得るために第VII因子のアミノ末端でプレプロポリペプチドをコードする。該プレプロポリペプチドは、第VII因子または別のビタミンK依存性血漿タンパク質、例えば第IX因子、第X因子、プロトロンビン、プロテインCもしくはプロテインSなどのものであり得る。当業者に認識されるように、付加的な修飾は、それらの修飾が凝固因子として働くタンパク質の能力を有意に損なわない場合は、キメラ第VII因子ポリペプチドのアミノ酸配列において為され得る。
【0073】
第VIIa因子変異体を発現する際に使用するための発現ベクターは、クローニングされた遺伝子またはcDNAの転写を指令することができるプロモータを含む。培養された哺乳動物細胞における使用のための適切なプロモータは、ウイルスプロモータおよび細胞プロモータを含む。ウイルスプロモータは、SV40プロモータ(Subramani et al.,Mol.Cell.Biol.1:854−864,1981)およびCMVプロモータ(Boshart et al.,Cell 41:521−530,1985)を含む。特に適切なウイルスプロモータは、アデノウイルス2由来の主要後期プロモータである(Kaufman and Sharp,Mol.Cell.Biol.2:1304−1319,1982)。もう1つの特に好ましいプロモータは、チャイニーズハムスター伸長因子1−α(CHEF1)プロモータである。細胞プロモータは、マウスκ遺伝子プロモータ(Bergman et al.,Proc.Natl.Acad.Sci.USA 81:7041−7045,1983)およびマウスVHプロモータ(Loh et al.,Cell 33:85−93,1983)を含む。特に適切な細胞プロモータは、マウスメタロチオネイン−Iプロモータである(Palmiter et al.,Science 222:809−814,1983)。発現ベクターはまた、プロモータから下流でかつキメラ第VII因子配列自体の挿入部位から上流に位置する一組のRNAスプライス部位を含み得る。適切なRNAスプライス部位は、アデノウイルスおよび/または免疫グロブリン遺伝子から得られ得る。また、挿入部位の下流に位置するポリアデニル化シグナルも発現ベクター中に含まれる。特に適切なポリアデニル化シグナルは、SV40由来の早期または後期ポリアデニル化シグナル(Kaufman and Sharp)、アデノウイルス5 Elb領域由来のポリアデニル化シグナル、ヒト成長ホルモン遺伝子ターミネーター(DeNoto et al.,Nucl.Acids Res.9:3719−3730,1981)またはヒト第VII因子遺伝子もしくはウシ第VII因子遺伝子由来のポリアデニル化シグナルを含む。発現ベクターはまた、プロモータとRNAスプライス部位の間に位置するアデノウイルス2の3分節リーダーなどの非コードウイルスリーダー配列、およびSV40エンハンサなどのエンハンサ配列も含み得る。
【0074】
クローニングされたDNA配列を、例えばリン酸カルシウムを介したトランスフェクション(Wigler et al.,Cell 14:725−732,1978;Corsaro and Pearson,Somatic Cell Genetics 7:603−616,1981;Graham and Van der Eb,Virology 52d:456−467,1973)または電気穿孔法(Neumann et al.,EMBO J.1:841−845,1982)によって、培養された哺乳動物細胞に導入する。外因性DNAを発現する細胞を同定し、選択するために、選択可能な表現型を付与する遺伝子(選択マーカー)を、一般に、対象とする遺伝子またはcDNAと共に細胞に導入する。適切な選択マーカーは、ネオマイシン、ハイグロマイシンおよびメトトレキサートなどの薬剤に対する耐性を付与する遺伝子を含む。選択マーカーは、増幅可能な選択マーカーであり得る。適切な増幅可能選択マーカーは、ジヒドロ葉酸レダクターゼ(DHFR)配列である。もう1つの適切な選択マーカーはヒスチジノールである。選択マーカーは、Thilly(引用することにより本明細書の一部をなすものとする、Mammalian Cell Technology,Butterworth Publishers,Stoneham,MA)によって総説されている。当業者は、適切な選択マーカーを容易に選択することができる。
【0075】
選択マーカーは、対象とする遺伝子と同時に別々のプラスミド上で細胞に導入され得るか、または同じプラスミド上で導入され得る。同じプラスミド上の場合、選択マーカーと対象遺伝子は異なるプロモータまたは同じプロモータの制御下にあってよく、後者の配置はジシストロニックメッセージを生成する。このタイプの構築物は当技術分野において知られている(例えば、Levinson and Simonsen,米国特許第4,713,339号)。また、「担体DNA」として知られている付加的なDNAを、細胞に導入される混合物に添加することも好都合であり得る。
【0076】
細胞はDNAを取り込んだ後、適切な増殖培地において、典型的には1〜2日間増殖され、対象とする遺伝子の発現を開始する。細胞を培養するために使用する培地は、適切な添加物を含有する最小または複合培地などの、宿主細胞を増殖させるのに適した任意の従来培地であり得る。適切な培地は、商業的供給者から入手可能であるか、または公表されている配合(例えばAmerican Type Culture Collectionのカタログにおいて)に従って調製され得る。培地は当技術分野において知られている手順を用いて調製される(例えば、細菌および酵母に関する参考文献;Bennett,J.W.and LaSure,L.,editors,More Gene Manipulations in Fungi,Academic Press,CA, 1991参照)。増殖培地は一般に、炭素源、窒素源、必須アミノ酸、必須糖、ビタミン、塩、リン脂質、タンパク質および増殖因子を含有する。γ−カルボキシル化されたキメラ第VII因子ポリペプチドの生成のためには、培地はビタミンKを、好ましくは約0.1ng/mlから約20μg/mlの濃度で含有する。次に薬剤選択を適用して、選択マーカーを安定に発現している細胞の増殖物を選択する。増幅可能な選択マーカーでトランスフェクトした細胞に関しては、増大したコピー数のクローン化配列を選択するために、薬剤濃度を高めてもよく、それによって発現レベルを上昇させ得る。次に、安定にトランスフェクトされた細胞のクローンを、所望のキメラ第VII因子ポリペプチドの発現に関してスクリーニングする。
【0077】
適切な哺乳動物細胞株は、CHO細胞株(ATCC CCL 61)、COS−1細胞株(ATCC CRL 1650)、ベビーハムスター腎(BHK)細胞株および293細胞株(ATCC CRL 1573;Graham et al.,J.Gen.Virol.36:59−72,1977)を含む。適切なBHK細胞株は、本明細書中以下ではBHK570細胞と称する、tk.sup.−ts13 BHK細胞株である(Waechter and Baserga,Proc.Natl.Acad.Sci.USA 79:1106−1110,1982)。BHK 570細胞株は、ATCCアクセッション番号CRL 10314の下にAmerican Type Culture Collection,12301 Parklawn Dr.,Manassas,Va.20852から入手可能である。tk−ts13BHK細胞株も、アクセッション番号CRL 1632の下にATCCから入手可能である。加えて、Rat Hep I細胞(ラット肝細胞癌;ATCC CRL 1600)、Rat Hep II細胞(ラット肝細胞癌;ATCC CRL 1548)、TCMK細胞(ATCC CCL 139)、ヒト肺細胞(ATCC HB 8065)、NCTC 1469(ATCC CCL 9.1)およびDUKX細胞(Urlaub and Chasin,Proc.Natl.Acad.Sci.USA 77:4216−4220,1980)を含む、多くの他の細胞株を使用し得る。
【0078】
本発明のキメラ第VII因子ポリペプチドは、インビボでのポリペプチドの半減期を延長させるためにコンジュゲート化し得る。かかるコンジュゲート化は、キメラ第VII因子ポリペプチドの投与頻度を低減する一方で、腎クリアランスを低下させ、インビボで存在する第VII因子の量を増加させ得る。適切なコンジュゲートおよびコンジュゲート化第VII因子ポリペプチドを作製する方法は、その全体を引用することにより本明細書の一部をなすものとする、米国特許第7,442,524号に記載されている。
【0079】
[トランスジェニック動物]
本発明のキメラ第VII因子ポリペプチドを作製するためにトランスジェニック動物技術を使用し得る。宿主雌性哺乳動物の乳腺中でタンパク質を生成することが好ましい。乳腺中での発現およびその後の乳中への対象タンパクの分泌は、他の供給源からタンパク質を単離する際に遭遇する多くの困難を克服する。乳は容易に収集され、大量に入手可能であり、生化学的に広く特徴付けられている。さらに、主要な乳タンパク質は乳中に高濃度で存在する(典型的には約1〜15g/l)。本発明のキメラ第VII因子ポリペプチドはまた、宿主動物の尿中でも生成でき、これは、雄性と雌性の両方の動物が尿を生成することおよび単離のためには乳中よりも尿中の方が失われるタンパク質が少ないという点で乳中での生成よりも有利である。
【0080】
本発明のキメラ第VII因子ポリペプチドの供給源として乳を使用することを考慮する場合、商業的見地から、大量の乳算出量を有する種を宿主として用いることが明らかに好ましい。マウスおよびラットなどのより小さな動物も使用できる(および原理段階の証明においては好ましい)が、ブタ、ヤギ、ヒツジおよびウシを含むがこれらに限定されない、家畜哺乳動物を用いることが好ましい。ヒツジは、この種における遺伝子組換えの前歴、乳算出量、コストおよびヒツジの乳を収集するための装置が容易に利用可能であることなどの因子のために、特に好ましい(例えば、宿主の種の選択に影響を及ぼす因子の比較に関しては国際公開公報第WO88/00239号参照)。イーストフリースランドシープ(East Friesland sheep)のような、酪農用に育種された宿主動物の品種を選択すること、または後日にトランスジェニック系統の交配によって搾乳用家畜を導入することが一般に望ましい。いずれの場合も、既知の、健康状態の良好な動物を使用すべきである。
【0081】
乳腺における発現を得るために、乳タンパク質遺伝子由来の転写プロモータを使用する。乳タンパク質遺伝子は、カゼイン(米国特許第5,304,489号参照)、β−ラクトグロブリン、α−ラクトアルブミンおよび乳清酸性タンパク質をコードする遺伝子を含む。β−ラクトグロブリン(BLG)プロモータが適切である。ヒツジβ−ラクトグロブリン遺伝子の場合は、遺伝子の5’フランキング配列の少なくとも近位の406bpの領域が一般的に使用されるが、β−ラクトグロブリン遺伝子の5’フランキングプロモータおよび非コード部分を包含する約4.25kbpのDNAセグメントなどの、約5kbpまでの5’フランキング配列のより大きな部分が適切である(Whitelaw et al.,Biochem.J.286:31−39(1992)参照)。他の種に由来するプロモータDNAの同様のフラグメントも適切である。
【0082】
尿における発現を得るために、尿路上皮特異的プロモータを使用する。例えば、ウロプラキン関連遺伝子の発現を駆動するプロモータが使用できる(例えば、その全体を引用することにより本明細書の一部をなすものとする、米国特許第6,001,646号参照)。
【0083】
β−ラクトグロブリン遺伝子の他の領域も、発現される遺伝子のゲノム領域と同様に、構築物に組み込み得る。イントロンを欠く構築物は、例えば、かかるDNA配列を含むものと比較して十分に発現されないことが当技術分野で一般的に認められている(各々を引用することにより本明細書の一部をなすものとする、Brinster et al.,Proc.Natl.Acad.Sci.USA 85:836−840(1988);Palmiter et al.,Proc.Natl.Acad.Sci.USA 88:478−482(1991);Whitelaw et al.,Transgenic Res.1:3−13(1991);国際公開公報第WO89/01343号;および国際公開公報第WO91/02318号参照)。これに関して、可能な場合は、対象とするタンパク質またはポリペプチドをコードする遺伝子の天然イントロンの全部または一部を含むゲノム配列を使用することが一般に好ましく、従って、例えばβ−ラクトグロブリン遺伝子由来の、少なくともいくつかのイントロンをさらに含むことが好ましい。かかる領域の1つは、ヒツジβ−ラクトグロブリン遺伝子の3’非コード領域由来の、イントロンのスプライシングおよびRNAのポリアデニル化を提供するDNAセグメントである。遺伝子の天然3’非コード配列を置換した場合、このヒツジβ−ラクトグロブリンセグメントは、対象タンパク質またはポリペプチドの発現レベルを高めかつ安定化することができる。他の実施形態では、キメラ第VII因子配列の開始ATGの周囲の領域を、乳特異的タンパク質遺伝子由来の対応する配列で置換する。かかる置換は、発現を増強する推定上の組織特異的開始環境を提供する。キメラ第VII因子プレプロ配列全体および5’非コード配列を、例えばBLG遺伝子のもので置換することは好都合であるが、より小さな領域を置換してもよい。
【0084】
トランスジェニック動物におけるキメラFVIIaポリペプチドの発現のために、キメラ第VIIa因子をコードするDNAセグメントを、発現ユニットを生成するためにその発現に必要とされる付加的なDNAセグメントに作動可能に連結する。かかる付加的なセグメントは、上記で挙げたプロモータ、ならびにmRNAの転写およびポリアデニル化の終結をもたらす配列を含む。発現ユニットは、キメラ第VIIa因子をコードするセグメントに作動可能に連結された分泌シグナル配列をコードするDNAセグメントをさらに含む。この分泌シグナル配列は、天然第VII因子分泌シグナル配列であり得るかまたは乳タンパク質などの別のタンパク質のものであり得る(例えば、引用することにより本明細書の一部をなすものとする、von Heijne,Nucl.Acids Res.14:4683−4690(1986);およびMeade et al.,米国特許第4,873,316号参照)。
【0085】
トランスジェニック動物に使用するための発現ユニットの構築は、キメラ第VII因子配列を、付加的なDNAセグメントを含むプラスミドまたはファージベクターに挿入することによって好都合に実施されるが、発現ユニットは、基本的には任意の一連の連結によって構築され得る。乳タンパク質をコードするDNAセグメントを含むベクターを提供し、乳タンパク質についてのコード配列をキメラ第VII因子ポリペプチドのもので置換して、それにより乳タンパク質遺伝子の発現制御配列を含む遺伝子融合物を生成することは特に好都合である。いずれの場合も、プラスミドまたは他のベクターへの発現ユニットのクローニングは、キメラ第VII因子配列の増幅を促進する。増幅は、好都合には細菌(例えば大腸菌(E.coli))宿主細胞において実施され、従ってベクターは、典型的には細菌宿主細胞において機能性の複製起点および選択マーカーを含む。発現ユニットを、次に、選択した宿主種の受精卵(初期胚を含む)に導入する。異種DNAの導入は、マイクロインジェクション(例えば米国特許第4,873,191号)、レトロウイルス感染(Jaenisch,Science 240:1468−1474(1988))または胚性幹(ES)細胞を用いた部位指定組込み(Bradley et al.,Bio/Technology 10:534−539(1992)によって総説されている)を含む、いくつかの経路の1つによって実施できる。該卵をその後、偽妊娠雌性動物の卵管または子宮に移植し、出産まで発育させる。生殖細胞系に導入されたDNAを担持する子は、そのDNAを正常なメンデルの様式で子孫に伝えることができ、トランスジェニック集団の発生を可能にする。トランスジェニック動物を作製するための一般的な手順は当技術分野において知られている(例えば、Hogan et al.,Manipulating the Mouse Embryo:A Laboratory Manual,Cold Spring Harbor Laboratory,1986;Simons et al.,Bio/Technology 6:179−183(1988);Wall et al.,Biol.Reprod.32:645−651(1985);Buhler et al.,Bio/Technology 8:140−143(1990);Ebert et al.,Bio/Technology 9:835−838(1991);Krimpenfort et al.,Bio/Technology 9:844−847(1991);Wall et al.,J.Cell.Biochem.49:113−120(1992);米国特許第4,873,191号;米国特許第4,873,316号;国際公開公報第WO88/00239号、国際公開公報第WO90/05188号、国際公開公報第WO92/11757号;およびイギリス特許公開公報第GB87/00458号参照)。外来性DNA配列を哺乳動物およびそれらの生殖細胞に導入するための技術はマウスにおいて最初に開発された(例えば、Gordon et al.,Proc.Natl.Acad.Sci.USA 77:7380−7384(1980);Gordon and Ruddle,Science 214:1244−1246(1981);Palmiter and Brinster,Cell 41:343−345(1985);Brinster et al.,Proc.Natl.Acad.Sci.USA 82:4438−4442(1985);およびHogan et al.参照)。これらの技術は、その後、家畜種を含むより大きな動物に関する使用のために適合された(例えば国際公開公報第WO88/00239号、国際公開公報第WO90/05188号および国際公開公報第WO92/11757号;ならびにSimons et al.,Bio/Technology 6:179−183(1988)参照)。要約すると、トランスジェニックマウスまたは家畜の作製においてこれまでに使用された最も効率的な経路では、対象とするDNAの数百の直鎖状分子を、確立された技術に従って受精卵の前核の1つに注入する。接合子の細胞質へのDNAの注入も使用し得る。
【0086】
トランスジェニック植物における生産も使用し得る。発現は、植物全体または塊茎などの特定の器官を対象とし得る(Hiatt,Nature 344:469−479(1990);Edelbaum et al.,J.Interferon Res.12:449−453(1992);Sijmons et al.,Bio/Technology 8:217−221(1990);および欧州特許第EP 0 255 378号参照)。
【0087】
本発明のキメラ第VII因子ポリペプチドの作製はまた、例えばウサギ網状赤血球溶解物、コムギ胚芽抽出物および大腸菌無細胞系等による、インビトロでの翻訳を介して達成され得る。インビトロ翻訳はまた、連鎖または共役させ得る。連鎖インビトロ翻訳は、バクテリオファージポリメラーゼでの転写、それに続くウサギ網状赤血球溶解物またはコムギ胚芽溶解物系での翻訳に基づく。共役インビトロ翻訳は大腸菌無細胞系に基づく。
【0088】
[回収および活性化]
本発明のキメラ第VII因子ポリペプチドは、細胞培養培地または乳から回収される。本発明のキメラ第VII因子ポリペプチドは、当技術分野において知られている様々な方法によって精製でき、かかる方法は、クロマトグラフィ(例えばイオン交換、アフィニティ、疎水性、クロマトフォーカシングおよびサイズ排除)、電気泳動法(例えば分取等電点電気泳動(IEF)、示差溶解度(例えば硫安沈殿)または抽出(例えば、Protein Purification,J.−C.Janson and Lars Ryden,editors,VCH Publishers,New York,1989参照)を含むが、これらに限定されない。さらなる精製は、高速液体クロマトグラフィなどの従来の化学的精製手段によって達成され得る。クエン酸バリウム沈殿を含む他の精製方法が当技術分野において知られており、本明細書で述べる新規キメラ第VII因子ポリペプチドの精製に適用し得る(例えば、Scopes,R.,Protein Purification,Springer−Verlag,N.Y.,1982参照)。
【0089】
治療目的には、本発明のキメラ第VII因子は実質的に純粋であることが好ましい。従って、本発明の1つの実施形態では、本発明の第VII因子変異体は、少なくとも約90〜95%の純度、好ましくは少なくとも約98%の純度に精製される。純度は、例えばゲル電気泳動、アミノ末端のアミノ酸配列決定および逆相HPLCによって評価し得る。
【0090】
キメラ第VII因子ポリペプチドは、二本鎖の形態に変換するためにその活性化部位で切断される。活性化は、Osterud,et al.,Biochemistry 11:2853−2857(1972);Thomas,米国特許第4,456,591号;Hedner and Kisiel,J.Clin.Invest.71:1836−1841(1983);またはKisiel and Fujikawa,Behring Inst.Mitt.73:29−42(1983)によって開示されているような、当技術分野で知られている手順に従って実施し得る。あるいは、キメラ第VII因子ポリペプチドを濃縮し、正に荷電した表面または樹脂と接触させることによってキメラ第VII因子ポリペプチドを活性化することができ、例えばBjoern et al.(Research Disclosure,269 September 1986,pp.564−565)によって述べられているように、第VII因子をMono Q(登録商標)(Pharmacia fine Chemicals)等のようなイオン交換クロマトグラフィカラムに通すことによって活性化し得る。キメラ第VII因子ポリペプチドはまた、キメラ第VII因子ポリペプチドの実質的に精製された製剤を含有する溶液を得、その溶液にアミン化合物、Ca2+を約5mM〜約50mM(約10mM〜約30mMなど)の最終濃度になるように添加して、溶液の最終pHを約7.2〜8.6(約7.6〜約8.2など)に調整し、生じた活性化混合物を約2℃〜約25℃で、少なくとも90%のキメラFVIIポリペプチドをキメラFVIIaポリペプチドに変換するのに十分な時間インキュベートして、任意選択的に、活性化混合物からFVIIaを単離することによって溶液中で活性化し得る。生じた活性化キメラ第VII因子ポリペプチドを、その後、以下のように製剤し、投与し得る。
【0091】
[遺伝子治療]
本発明のキメラ第VII因子ポリペプチドはまた、遺伝子治療によって出血性疾患を治療する方法においても使用し得る。本発明のこの実施形態では、本発明のキメラ第VII因子ポリペプチドは、エクスビボでの導入またはインビボでの導入によって被験体の細胞に導入され得る核酸分子によってコードされる。
【0092】
当技術分野で利用可能な遺伝子治療のための方法のいずれもが本発明に従って使用できる。例示的な方法を以下で述べる。遺伝子治療の方法の一般的な総説については、Goldspiel et al.,Clinical Pharmacy 1993,12:488−505;Wu and Wu,Biotherapy 1991,3:87−95;Tolstoshev,Ann.Rev.Pharmacol.Toxicol.1993,32:573−596;Mulligan,Science 1993,260:926−932;およびMorgan and Anderson,Ann.Rev.Biochem.1993,62:191−217;May,TIBTECH 1993,11:155−215参照。使用できる組換えDNA技術の当技術分野で一般的に知られている方法は、Ausubel et al.,(eds.),1993,Current Protocols in Molecular Biology,John Wiley & Sons,NY;Kriegler,1990,Gene Transfer and Expression,A Laboratory Manual,Stockton Press,NY;およびin Chapters 12 and 13,Dracopoli et al.,(eds.),1994,Current Protocols in Human Genetics,John Wiley & Sons,NY;Colosimo et al.,Biotechniques 2000;29(2):314−8,320−2,324に記載されている。
【0093】
キメラ第VII因子ポリペプチドをコードするポリヌクレオチドは、適切なクローニングベクターに挿入し得る。遺伝子治療に適するベクターは、アデノウイルス、アデノ随伴ウイルス(AAV)、ワクシニアウイルス、ヘルペスウイルス、バキュロウイルスおよびレトロウイルス、パルボウイルス、レンチウイルスなどのウイルス、バクテリオファージ、コスミド、プラスミド、真菌ベクターならびに様々な真核および原核生物宿主における発現に関して記述されている、遺伝子治療ならびに単純なタンパク質発現のために使用し得る当技術分野で典型的に使用される他の組換えビヒクルを含む。
【0094】
1つの実施形態では、該ベクターはウイルスベクターである。ウイルスベクター、特にアデノウイルスベクターは、標的細胞のウイルス感染の効率を上昇させる、カチオン性脂質、ポリL−リシン(PLL)およびジエチルアミノエチルデキストラン(DELAE−デキストラン)などのカチオン性両親媒物質と複合体形成することができる(例えば、引用することにより本明細書の一部をなすものとする、1997年11月20日出願の国際特許出願第PCT/US97/21496号参照)。本発明において使用するための適切なウイルスベクターは、ワクシニアウイルス、ヘルペスウイルス、AAVおよびレトロウイルス由来のベクターを含む。遺伝子治療におけるウイルスベクターの総説については、Mah et al.,Clin.Pharmacokinet.2002;41(12):901−11;Scott et al.,Neuromuscul.Disord.2002;12 Suppl 1:S23−9参照。
【0095】
1つの実施形態では、コード配列および任意の他の所望配列がゲノム内の所望部位での相同的組換えを促進する領域によって隣接され、それによりゲノムに組み込まれた核酸分子からの構築物の発現を提供するベクターが使用される(各々その全体を引用することにより本明細書の一部をなすものとする、Koller and Smithies,Proc.Natl.Acad.Sci.USA 1989,86:8932−8935;Zijlstra et al.,Nature 1989,342:435−438;Zarling et al.への米国特許第6,244,113号;およびPati et al.への米国特許第6,200,812号)。
【0096】
患者へのベクターの送達は、患者をベクターもしくは送達複合体に直接暴露する直接的送達、または細胞を最初にインビトロにてベクターで形質転換し、その後患者に移植する間接的送達のいずれかであり得る。これら2つのアプローチは、それぞれインビボおよびエクスビボ遺伝子治療として知られている。
【0097】
1つの実施形態では、ベクターをインビボで直接投与し、そこでベクターは被験体の細胞に進入して、遺伝子の発現を媒介する。これは、当技術分野で知られている数多くの方法および上記で論じた方法のいずれかによって、例えばそれを適切な発現ベクターの一部として構築し、例えば欠陥もしくは弱毒化レトロウイルスまたは他のウイルスベクターを用いた感染によって(例えば米国特許第4,980,286号参照)、または裸のDNAの直接注入によって、または微粒子ボンバードメント(例えば遺伝子銃;Biolistic,Dupont)の使用によって、または脂質もしくは細胞表面受容体もしくはトランスフェクション剤でのコーティング、バイオポリマー(例えばポリ−β−1−64−N−アセチルグルコサミン多糖;米国特許第5,635,493号参照)への封入、リポソーム、微粒子もしくはマイクロカプセルへの封入の使用によって、ペプチドもしくは核内に進入することが公知の他のリガンドに連結して投与することによって、または受容体媒介性エンドサイトーシスを受けるリガンドに連結して投与すること(例えば、Wu and Wu,J.Biol.Chem.1987,62:4429−4432)等によって、細胞内に組み込まれるように投与することによって実施できる。もう1つの実施形態では、リガンドが、エンドソームを崩壊させ、核酸がリソソーム分解を回避することを可能にする膜融合性ウイルスペプチド、または治療用DNAを細胞に導入するために使用できる、例えばアンテナペディア由来の、カチオン性12量体ペプチドを含む、核酸−リガンド複合体を形成することができる(Mi et al.,Mol.Therapy 2000,2:339−47参照)。さらにもう1つの実施形態では、特定の受容体を標的することにより、細胞特異的な取込みおよび発現のために核酸をインビボで標的することができる(例えば、国際公開公報第WO92/06180号、国際公開公報第WO92/22635号、国際公開公報第WO92/20316号および国際公開公報第WO93/14188号参照)。加えて、マグネトフェクションと称される技術を、ベクターを哺乳動物に送達するために使用し得る。この技術は、磁場の影響下で送達するためにベクターと超常磁性ナノ粒子を結合させる。この適用は、送達時間を短縮し、ベクターの有効性を高める(Scherer et al.,Gene Therapy 2002;9:102−9)。
【0098】
1つの実施形態では、核酸は脂質担体を用いて投与し得る。細胞膜の通過を促進するために脂質担体を裸の核酸(例えばプラスミドDNA)と結合することができる。カチオン性、アニオン性または中性脂質がこのために使用できる。しかし、カチオン性脂質は、一般に負の電荷を有するDNAとより良好に結合することが示されているため、適切である。カチオン性脂質はまた、プラスミドDNAの細胞内送達を媒介することが示されている(Felgner and Ringold,Nature 1989;337:387)。マウスへのカチオン性脂質−プラスミド複合体の静脈内注射は、肺におけるDNAの発現を生じさせることが示された(Brigham et al.,Am.J.Med.Sci.1989;298:278)。また、Osaka et al.,J.Pharm.Sci.1996;85(6):612−618;San et al.,Human Gene Therapy 1993;4:781−788;Senior et al.,Biochemica et Biophysica Acta 1991;1070:173−179);Kabanov and Kabanov,Bioconjugate Chem.1995;6:7−20;Liu et al.,Pharmaceut.Res.1996;13;Remy et al.,Bioconjugate Chem.1994;5:647−654;Behr,J−P.,Bioconjugate Chem 1994;5:382−389;Wyman et al.,Biochem.1997;36:3008−3017;Marshall et al.への米国特許第5,939,401号;Scheule et al.への米国特許第6,331,524号も参照のこと。
【0099】
代表的なカチオン性脂質は、例えば、その開示を引用することにより本明細書の一部をなすものとする、米国特許第5,283,185号および米国特許第5,767,099号に開示されているものを含む。1つの実施形態では、カチオン性脂質は、米国特許第5,767,099号に開示されているN4−スペルミンコレステリルカルバメート(GL−67)である。さらなる適切な脂質は、N4−スペルミジンコレステリルカルバメート(GL−53)および1−(N4−スペルミン)−2,3−ジラウリルグリセロールカルバメート(GL−89)を含む。
【0100】
ウイルスベクターのインビボでの投与に関して、ウイルスベクターおよびトランスフェクトされた細胞の免疫脱活性化を回避するために、適切な免疫抑制治療をウイルスベクター、例えばアデノウイルスベクターと組み合わせて使用し得る。例えば、インターロイキン12(IL−12)、インターフェロンγ(IFN−γ)または抗CD4抗体などの免疫抑制性サイトカインが、ウイルスベクターに対する体液性または細胞性免疫応答をブロックするために投与し得る。それに関して、ごく少数の抗原を発現するように遺伝子操作されたウイルスベクターを用いることが好都合である。
【0101】
体細胞は、上述した方法のいずれかを用いて本発明のキメラ第VII因子ポリペプチドをコードする構築物でエクスビボにて遺伝子操作し、個体に再移植し得る。この方法は、Selden et al.への国際公開公報第WO93/09222号において一般的に説明されている。加えて、この技術は、Payumo et al.,Clin.Orthopaed.and Related Res.2002;403S:S228−S242に記載されている、細胞ベースの送達についての特許ImPACT技術で使用される。かかる遺伝子治療系では、体細胞(例えば線維芽細胞、肝細胞または内皮細胞)を患者から取り出し、インビトロで培養して、治療対象の遺伝子でトランスフェクトし、特徴付けて、患者に再び導入する。一次細胞(個体または組織に由来し、継代の前に遺伝子操作された)および二次細胞(インビボでの導入の前にインビトロで継代した)の両方、ならびに当技術分野で知られている不死化細胞株を使用し得る。本発明の方法のために有用な体細胞は、線維芽細胞、ケラチノサイト、上皮細胞、内皮細胞、肝細胞、血液の有形成分、筋肉細胞、培養できる他の体細胞および体細胞前駆体などの体細胞を含むが、これらに限定されない。1つの実施形態では、細胞は肝細胞である。
【0102】
本発明のキメラFVIIaポリペプチドをコードするポリヌクレオチド、および、任意選択的に、選択マーカーをコードする核酸を、受容者の一次または二次細胞におけるキメラFVIIaの発現のために必要な付加的配列と共に含む構築物を使用して、コードされた産物が生成される一次または二次細胞をトランスフェクトする。かかる構築物は、レトロウイルス、ヘルペスウイルス、アデノウイルス、アデノ随伴ウイルス、耳下腺炎ウイルスおよびポリオウイルスベクターなどの感染性ベクターを含むがこれらに限定されず、この目的に使用し得る。
【0103】
経皮送達は、ケラチノサイト、メラノサイトおよび樹状細胞を含む表皮の細胞型を用いた間接的導入に特に適する(Pfutzner et al.,Expert Opin.Investig.Drugs 2000;9:2069−83)。
【0104】
1つの実施形態では、遺伝子治療のためのベクターは、一本鎖自己相補性アデノ随伴ウイルスである。加えて、この自己相補性アデノ随伴ウイルスは、小さなトランスサイレチンプロモータ−エンハンサ、マウス微小ウイルスからのイントロンおよびウシ成長ホルモンからのポリアデニル化シグナルを含み得る。ベクターはまた、第VII因子を活性化形態、すなわち第VIIa因子として分泌させる、Margaritis et al.によって記述されたRKRRKR配列も含み得る(Margaritis et al.,J.Clin.Invest.,2004;113:1025−31)。
【0105】
[投与および医薬組成物]
本発明によるキメラ第VII因子ポリペプチドは、凝固因子欠損症(例えば血友病AおよびBまたは凝固第XI因子もしくは第VII因子の欠損)または凝固因子インヒビターなどのいくつかの原因を有する出血性疾患を管理するために使用され得、あるいはそれらは、正常に機能する血液凝固カスケードを有する(凝固因子欠損または凝固因子のいずれかに対するインヒビターを有さない)被験体において生じる過剰の出血を制御するために使用され得る。出血は、血小板機能欠損、血小板減少症またはフォン・ヴィレブランド病によって引き起こされ得る。出血はまた、様々な刺激によって線維素溶解活性の上昇が誘発された被験体でも見られ得る。
【0106】
手術または外傷に関連する広範囲の組織損傷を経験した被験体では、正常な止血機構が即時止血の必要性によって圧倒されることがあり、該被験体は、正常な止血機構にも関わらず出血を発現し得る。十分な止血を達成することは、脳、内耳領域および眼などの器官で出血が起こる場合にも問題であり、また、発生源を同定することが困難である広汎性出血(出血性胃炎および大量の子宮出血)の場合にも問題であり得る。同じ問題が、様々な器官(肝臓、肺、腫瘍組織、胃腸管)から生検を採取する過程ならびに腹腔鏡手術において生じ得る。これらの状況は、外科的手術手技(縫合、クリップ等)によって止血を行うことが困難であるという共通点を有する。急性かつ大量の出血はまた、抗凝固療法時に、与えられた治療によって不完全な止血が誘発された被験体においても起こり得る。かかる被験体は、抗凝固作用を速やかに中和しなければならない場合は外科的処置を必要とし得る。不十分な止血の場合に問題を引き起こし得るもう1つの状況は、正常な止血機構を有する被験体が血栓塞栓症を予防するために抗凝固療法を受ける場合である。かかる療法は、ヘパリン、他の形態のプロテオグリカン、ワルファリンまたは他の形態のビタミンKアンタゴニストならびにアスピリンおよび他の血小板凝固阻害剤を含み得る。
【0107】
計画的処置に関連する治療に関して、本発明のキメラ第VII因子ポリペプチドは、典型的には処置を実施する前の約24時間以内、およびその後7日間またはそれ以上投与される。凝固薬としての投与は、本明細書で述べる様々な経路によって実施できる。
【0108】
キメラ第VII因子ポリペプチドの用量は、被験体の体重および状態の重症度に依存して、負荷用量および維持用量として70kgの被験体について約0.05mg/日〜約500mg/日、好ましくは約1mg/日〜約200mg/日、より好ましくは約10mg/日〜約175mg/日の範囲である。当業者は、所与の被験体および所与の状態についての最適用量を決定することができる。
【0109】
医薬組成物は、主として予防的および/または治療的処置のための非経口投与を意図される。好ましくは、医薬組成物は非経口的に、すなわち静脈内、皮下もしくは筋肉内経路で投与されるか、または持続的もしくはパルス注入によって投与され得る。あるいは、医薬組成物は、経口的、皮下的、静脈内、脳内、鼻内、経皮的、腹腔内、筋肉内、肺内、経膣的、経直腸的、眼内経路または任意の他の許容される方法を含むがこれらに限定されない、様々な方法での投与用に製剤され得る。
【0110】
非経口投与用の組成物は、医薬的に許容される担体、好ましくは水性担体と組み合わせた、好ましくはその中に溶解された、本発明のキメラ第VII因子ポリペプチドを含有する。水、緩衝水、0.4%食塩水、0.3%グリシン等のような、様々な水性担体を使用し得る。本発明のキメラ第VII因子ポリペプチドはまた、メチオニンおよびスクロースなどの、安定性および保存を延長させる組成物と共に製剤化され得る。本発明のキメラ第VII因子ポリペプチドはまた、損傷の部位への送達または標的化のためにリポソーム製剤に製剤することができる。リポソーム製剤は、例えば米国特許第4,837,028号、米国特許第4,501,728号および米国特許第4,975,282号において一般的に説明されている。組成物は、従来の公知の滅菌技術によって滅菌され得る。生じた水溶液を、使用のために包装し得るか、または無菌条件下でろ過し、凍結乾燥して、凍結乾燥製剤を投与の前に滅菌水溶液と組み合わせ得る。組成物は、pH調整剤および緩衝剤、張度調整剤等のような、生理的条件に近付けるために必要な医薬的に許容される補助物質、例えば酢酸ナトリウム、乳酸ナトリウム、塩化ナトリウム、塩化カリウム、塩化カルシウム等を含有し得る。組成物はまた、防腐剤、等張化剤、非イオン性界面活性剤または洗浄剤、抗酸化剤および/または他の種々の添加物も含有し得る。
【0111】
これらの製剤中のキメラ第VII因子ポリペプチドの濃度は広く異なり、すなわち約0.5重量%未満、通常は約1重量%またはそれ以下から約15または20重量%までにわたり得、選択される特定の投与方法に従い、主として液体の容量、粘度等によって選択される。従って、静脈内注入用の典型的な医薬組成物は、滅菌リンガー液250mlおよびキメラ第VII因子ポリペプチド10mgを含有するように作製され得る。非経口的に投与可能な組成物を調製するための実際の方法は、公知であるかまたは当業者に明らかであり、例えばRemington’s Pharmaceutical Sciences,18th ed.,Mack Publishing Company,Easton,Pa.(1990)の中でより詳細に説明されている。
【0112】
本発明のキメラ第VII因子ポリペプチドを含有する組成物は、予防的および/または治療的処置のために投与できる。治療適用では、組成物を、上述したように既に疾患に罹患している被験体に、疾患およびその合併症を治療する、緩和するまたは部分的に停止させるのに十分な量で投与する。これを達成するのに適切な量は、「治療有効量」と定義される。当業者に理解されるように、このために有効な量は、疾患または損傷の重症度ならびに被験体の体重および全身状態に依存する。しかし一般に、有効量は、70kgの被験体についてキメラ第VII因子ポリペプチド約0.05mg/日〜約500mg/日の範囲であり、キメラ第VII因子ポリペプチド約1.0mg/日〜約200mg/日の用量がより一般的に使用される。
【0113】
本発明の物質は、一般に、重篤な疾患または損傷状態、すなわち生命を脅かすまたは潜在的に生命を脅かす状況において使用され得ることに留意しなければならない。かかる場合、外来性物質が最小限であることおよび一般にヒトにおいてヒト第VII因子の免疫原性がないことを考慮すると、実質的に過剰なこれらのキメラ第VII因子ポリペプチド組成物を投与することが可能であり、治療医師はこれを望ましいと考えるはずである。
【0114】
予防適用では、本発明のキメラ第VII因子ポリペプチドを含有する組成物を、被験体自身の凝固能力を増強するために疾患状態または損傷を生じやすいまたはその危険性がある被験体に投与する。かかる量は、「予防有効量」と定義される。予防適用では、正確な量は、やはり被験体の健康状態と体重に依存するが、用量は一般に70kgの被験体について約0.05mg/日〜約500mg/日、より一般的には70kgの被験体について約1.0mg/日〜約200mg/日の範囲である。
【0115】
組成物の単回または多回投与を、治療医師によって選択される用量レベルおよびパターンで実施し得る。毎日の維持レベルを必要とする通院被験体については、キメラ第VII因子ポリペプチドを、例えば携帯ポンプシステムを用いた持続注入によって投与し得る。
【0116】
本発明のキメラ第VII因子ポリペプチドはまた、徐放性または持続放出性製剤に製剤化し得る。徐放性または持続放出性製剤を製剤化する方法は当技術分野において知られており、ポリペプチドを含有する固体疎水性粒子の半透性マトリックスを含むが、これに限定されない。
【0117】
例えば局所適用などの、本発明のキメラ第VIIa因子ポリペプチドの局所送達は、例えば、スプレー、灌流、二重バルーンカテーテル、ステントによって、人工血管またはステントに組み込んで、バルーンカテーテルを被覆するのに使用されるヒドロゲルによって、または他の広く確立された方法によって実施し得る。いずれの場合も、医薬組成物は、被験体を有効に治療するのに十分な量の第VII因子変異体を提供すべきである。
【0118】
以下の実施例は本発明を説明するために含まれるものである。以下の実施例の特定の態様を、本発明の実施において良好に機能するように発明者が見出したまたは企図した技術および手順に関して述べる。本開示および当技術分野の一般的な技術レベルに照らして、当業者は、以下の実施例が例示だけを意図し、本発明の範囲から逸脱することなく数多くの変更、修正および改変を用い得ることを認識する。
【0119】
[実施例1:キメラ第VIIa因子ポリペプチドの構築および発現]
ヒト第VII因子およびヒト第IX因子の両方のcDNAを入手し、ドメイン交換を容易にするために以下のユニーク制限部位を導入した。
BstEII部位をEGF−1ドメインの5’末端に(第VII因子の残基47〜49、第IX因子の残基48〜50)、
SacI部位をEGF−1とEGF−2ドメインの接合部に(第VII因子の残基82〜83、第IX因子の残基83〜84)、
NotI部位をEGF−2ドメインの3’末端に(第VII因子の残基135〜137、第IX因子の残基132〜134)導入した。
【0120】
第VII因子と第IX因子の間でドメインを交換するためのこれらの制限部位を用いて様々なキメラを構築し、図1に示している。同じかまたは同様の制限部位を使用して第VII因子とプロテインSとの間でドメインを交換し得る。
【0121】
生じたキメラのアミノ酸配列を配列番号:1(第IX因子のシグナル、プロペプチド、GLAおよびEGF−1ドメインと、第VII因子のEGF−2および触媒ドメイン)および配列番号:2(第IX因子のシグナルおよびプロペプチドドメインと、プロテインSのGLAドメインと、第IX因子のEGF−1ドメインと、第VII因子のEGF−2および触媒ドメイン)に示す。これらの組換えDNAから選択したものを、哺乳動物細胞のトランスフェクションのために発現ベクターpCMV5にサブクローニングした。
【0122】
生じた組換え構築物を、次に、pSV2neoおよびpCMVhGCを用いて293ヒト腎細胞株(ATCC CRL 1573)に同時トランスフェクトした。高レベルの各構築物を発現するクローンを選択し、先に述べられているようにスクリーニングした(Toomey et al.,1991,J.Biol.Chem.,266:19198−19202)。各々のスクリーニングしたクローンを、大規模生産のために900cm2のローラーボトルに広げ、組換えタンパク質を、Fast Flow Q−Sepharoseを用いた偽アフィニティクロマトグラフィ法、および、カルシウム勾配、次いでNaCl勾配での溶出によって精製した。
【0123】
[実施例2:第VIIa因子ポリペプチドのトロンビン産生]
第VIIa因子のトロンビン産生を、血友病のインビトロモデルを用いて測定した。単球を組織因子の供給源として使用し、不活性化血小板と、血漿濃度の第V、第VII、第IX、第VIIIおよび第XI因子ならびに血漿濃度の抗トロンビンおよびTFPIを含有する合成血漿とを組み合わせた。第VIIIおよび第IX因子を除外することによって血友病状態を作り出した。
【0124】
健常個体から血液4mlを採取し、クエン酸ナトリウム管に入れて単球を調製した。血液を15ml円錐管中のAccu−Prep(商標)リンパ球分離培地(Accurate Chemicals,NY,USA)3mlの上に注意深く層状に重ね、1500rpmで30分間遠心分離した。次に単核層を取り出し、4℃で等容量のVersene(Lineberger Tissue Culture)に添加して、800rpmで10分間遠心分離した。生じたペレットを4℃でVersene 5mlに再懸濁し、800rpmで10分間遠心分離した。生じたペレットを、500ng/mlのリポ多糖(LPS)(Sigma−Aldrich,MO,USA)を添加したマクロファージSFM培地(Life Technologies,CA,USA)4mlに再懸濁し、細胞を96ウェルプレートに200μl/ウェルで播種した。プレートを37℃、5%CO2で2時間インキュベートし、次にマクロファージSFM培地で3回洗浄して、その後一晩インキュベートした。
【0125】
健常個体から血液4mlを採取し、クエン酸ナトリウム管に入れて血小板を調製した。血液を、15ml円錐管中のAccu−Prep(商標)リンパ球分離培地(Accurate Chemicals,NY,USA)5mlの上に、注意深く層状に重ね、1500rpmで30分間遠心分離した。次に血小板層を取り出し、等容量のクエン酸−グルコース−食塩水および10μg/mlのプロスタグランジンE1(PGE1)(Sigma−Aldrich,MO,USA)に添加して、800rpmで10分間遠心分離した。生じたペレットを廃棄し、1mg/mlオボアルブミンを添加したカルシウム不含タイロード緩衝液中でセファロースゲルろ過によって血漿タンパク質から血小板を単離した。回収した血小板を37℃で保存した。
【0126】
血友病を模倣するインビトロアッセイのための合成血漿を、5μg/mlの濃度の第XI因子C−1エステラーゼインヒビターを使用して、あらかじめ調製しておいた単球と共に1時間インキュベートした後、200μg/mlの抗トロンビン、0.07μg/mlのTFPI、100mg/mlのプロトロンビン、8μg/mlの第X因子および0.5μg/mlの第VII因子を添加して作製した。一晩インキュベートした後、第V因子を7μg/mlの濃度で添加した。正常条件については、1時間インキュベートした後、第IX因子を4μg/mlの濃度で添加し、一晩インキュベートした後、第VIII因子を1U/mlで添加した。
【0127】
血小板260μlを上記合成血漿溶液に添加し、生じた懸濁液を単球に添加して反応を開始させた。一定の時間間隔で試料10μlを取り出し、1mM EDTAおよび0.5mM Gly−Pro−Arg−pNA(Centerchem Inc.,CT,USA)を含有するトロンビンアッセイ緩衝液90μlに添加した。50%酢酸100μlを添加して合成基質切断を停止させ、405nmでODを測定した。以下の式を用いてトロンビン濃度を決定した。
[トロンビン]=希釈×((A405−バックグラウンド)/(停止−開始))/(変換係数)
式中、変換係数が1nMである場合、トロンビンは405nmで0.0117OD/分の変化量を与える。
【0128】
正常条件(第VIIIおよび第IX因子を含む)でのトロンビン産生は、短い遅滞期でピークトロンビン産生を示す(図2)。野生型第VIIa因子の濃度上昇は、血友病条件でのトロンビン産生を改善した(図2)。
【0129】
[実施例3:キメラ第VIIa因子ポリペプチドのトロンビン産生]
実施例1で作製したキメラタンパク質のトロンビン産生を、実施例2で述べた血友病のインビトロモデルを用いて測定した。10nMの、第VII因子のEGF−2および触媒ドメインと第IX因子のGLAおよびEGF−1ドメインとを含むキメラ第VIIa因子、または第VII因子のGLA、EGF−2および触媒ドメインと第IX因子のEGF−1ドメインとを含むキメラ第VIIa因子の添加は、50nMの野生型第VIIa因子と同様の活性を有していた(図3)。
【0130】
対照と比較してピークトロンビン産生を検討すると、野生型第VIIa因子または試験したキメラ第VIIa因子ポリペプチドのいずれもが、対照レベルまでのピークトロンビン産生を回復していなかった。しかし、10nMの、第VII因子のEGF−2および触媒ドメインと第IX因子のGLAおよびEGF−1ドメインとを含むキメラ第VIIa因子、または第VII因子のGLA、EGF−2および触媒ドメインと第IX因子のEGF−1ドメインとを含むキメラ第VIIa因子は、50nMの野生型第VIIa因子によって産生されるのと同様のレベルまでピークトロンビン産生を回復していた(図4)。
【0131】
[実施例4:キメラ第VIIa因子ポリペプチドの凝固アッセイ]
キメラ第VIIa因子ポリペプチドの凝固活性を、Buyue,Y.,et al.,2008,Blood,112:3234−3241によって述べられているように、凝固アッセイにおいてインビボで評価した。簡単に述べると、第VII因子を、脚の静脈に挿入したカテーテルを介して、血友病Bマウスに投与した(2mg/kgのNovoSeven(登録商標)、2mg/kgの、第IX因子のGLAおよびEGF−1ドメインと第VIIa因子のEGF−2および触媒ドメインとを含むキメラ第VIIa因子、または1.4mg/kgの、プロテインSのGLAドメインと、第IX因子のEGF−1ドメインと、第VIIa因子のEGF−2および触媒ドメインとを含むキメラ第VIIa因子)。野生型マウスを対照として使用した。マウスを麻酔し、他方の脚の伏在静脈を、小さなゲージの針を押し通すことによって離断した。次に鋏の先端を静脈に挿入し、小さな杯を作製するように鋏で切って遠位部分を切断した。血餅を形成させ、出血が停止するまでの時間を記録した。次に30ゲージの針を用いて血餅を除去し、出血が停止するまでの時間を再び記録した。この工程を30分間反復した。血餅の数を崩壊の数として記録した。記録した時間を平均し、平均時間として記録した。実験を、野生型マウス、ならびに、プロテインSのGLAドメインと第IX因子のEGF−1ドメインと第VIIa因子のEGF−2および触媒ドメインとを含むキメラ第VIIa因子について1匹のマウスで実施した。第IX因子のGLAおよびEGF−1ドメインと第VIIa因子のEGF−2および触媒ドメインとを含むキメラ第VIIa因子については、2匹の血友病Bマウスで実験を実施し、同じ結果を得た。第VII因子を投与しなかった血友病Bマウスに関して、以下の表1において記録した0.25の崩壊は、4匹のマウスのうち1匹だけが初期損傷で出血しなかったことを示す。
【0132】
【表1】
【0133】
処置を行わなかった1匹の血友病Bマウス、2mg/kgのNovoSeven(登録商標)を投与した1匹の血友病Bマウス、2mg/kgの、第IX因子のGLAおよびEGF−1ドメインと第VIIa因子のEGF−2および触媒ドメインとを含むキメラ第VIIa因子を投与した1匹の血友病Bマウスならびに処置を行わなかった1匹の野生型マウスに関して実験を反復した。2mg/kgの、第IX因子のGLAおよびEGF−1ドメインと第VIIa因子のEGF−2および触媒ドメインとを含むキメラ第VIIa因子を投与した血友病Bマウスは、2mg/kgのNovoSeven(登録商標)を投与した血友病Bマウスおよび野生型マウスよりも出血の停止までの平均時間がより短かった(図5)。
【0134】
[実施例5:キメラ第VIIa因子ポリペプチドの血栓形成性]
本発明のキメラ第VIIa因子ポリペプチドの血栓形成性を、高レベルのこれらのタンパク質を発現するマウスモデルにおいて試験することができる。近年、2μg/ml以上の組換え野生型マウス第VIIa因子を発現するマウスの50%が16か月以内に血栓症で死亡することが示された。同様のレベルの第VIIa因子を発現する血友病Bマウスは同じ死亡率を有し、マウスをC57BL/6Jマウス系統に戻し交配した場合、はるかに早期に(4か月未満)血栓症が起こった(Aljamali MN,et al.J Clin Invest.2008;118:1825−1834)。
【0135】
2〜10μgまたはそれ以上のキメラポリペプチドを発現する血友病Bマウス(C57BL/6J系統)を作製することができ、キメラ第VII因子ポリペプチドの発現が血栓症を引き起こす場合、これらのマウスは早期に(4か月未満)血栓症を発現するはずである。
【0136】
UNCにおけるthe Gene Therapy CenterのDr.Paul Monahanによって開発された新しい自己相補性アデノ随伴ベクター(AAV)を使用することによってマウス系統を開発し得る(Wu Z,et al.Mol Ther.2008;16:280−289)。このベクターは、小さなトランスサイレチンプロモータ−エンハンサ(TTRプロモータ)、マウスの微小ウイルスからのイントロン、MVMおよびウシ成長ホルモンpAシグナルを含む。このベクターに基づく、1×1011のベクターゲノムの門脈への注入は、感染から8週間以内に7μg/mlの第IX因子の持続的発現を生じさせた(Wu Z,et al.Mol Ther.2008;16:280−289)。
【0137】
マウスおよびヒトの両方の第VIIa因子ならびに本発明のキメラ第VII因子ポリペプチドのcDNAを、血栓形成性を評価するために使用し得る。第VII因子を活性化形態、すなわち第VIIa因子として分泌させる、Margaritis et al.によって記述されたRKRRKR(配列番号:21)配列も使用し得る(Margaritis et al.,J.Clin.Invest.,2004;113:1025−31)。
【0138】
この仮説を検証するため、野生型およびキメラ第VII因子マウスポリペプチドを作製することができる。マウス組織因子はヒト第VII因子と反応しないことが報告されているので、組織因子は外傷において投与される第VIIa因子の作用とは無関係であるが、血栓症に関係することを明らかにするためにマウスポリペプチドを使用する。この仮説を検証するために使用できるマウスポリペプチドは、野生型マウス第VII因子(配列番号:3)、ポリペプチドを二本鎖の活性形態(FVIIa)で分泌させるRKRRKR配列を有する野生型マウス第VII因子(配列番号:4)、マウス第IX因子のシグナル、プロペプチド、GLAおよびEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列(配列番号:5)、ならびにマウス第IX因子のシグナルおよびプロペプチドドメイン、プロテインSのGLAドメイン、第IX因子のEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列(配列番号:6)を包含する。キメラマウス第VII因子ポリペプチド(配列番号:5および配列番号:6)は、組織因子に対して低い親和性を有し(おそらく野生型FVII分子と比較して100倍低い)、これは望ましくない血栓症を予防するのに十分なはずである。
【0139】
キメラ第VII因子ポリペプチドはまた、組織因子に対する親和性をさらに低減するように付加的に修飾し得る。かかる突然変異の一例は、残基306のメチオニンをアラニンに変更することである。第VIIa因子のより高い比活性を生じさせるキメラ第VII因子ポリペプチドの他の修飾も含み得る。かかる修飾の例は、ヒト配列の残基158、296および298に対する以下の等価物、V158D、E296VおよびM298Qを含む。
【0140】
上述した野生型およびキメラマウス第VII因子ポリペプチドは、マウス第VII因子cDNAを鋳型として使用し、様々なドメインを増幅するためのオリゴヌクレオチドプライマーを用い、次いで、この構築物をpSC−TTR−mvmベクターに挿入することによって構築でき、該構築は、Wu et al.,Molecular Therapy,2008;16:280−289に記載されている。具体的には、野生型マウス第VII因子は、以下のプライマー、5’−TGAGGATCCCCACCATGGTTCCACAGGCGCATGGGCT−3’(配列番号:7)および5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)を使用してマウス第VII因子cDNAから増幅できる。次に、増幅したフラグメントをBamHI/SphIで消化し、BglII/SphI部位の間でpSC−TTR−mvmベクターに挿入し得る。その後、BGHポリアデニル化配列を、生じたベクターにSphI部位で挿入し得る。
【0141】
RKRRKR配列を有する野生型マウス第VII因子構築物は、同様の方法で構築し得る。具体的には、RKRRKR配列を有する野生型マウス第VII因子は、以下のプライマー、
1番目のフラグメントを作製するための、5’−TGAGGATCCCCACCATGGTTCCACAGGCGCATGGGCT−3’(配列番号:7)および5’−ACAATGCGTTTTCGCCGCTTACGGCGGCCTTGGCGGCTGCTGGAGT−3’(配列番号:9)、ならびに
2番目のフラグメントを作製するための、5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)および5’−GCCGCCGTAAGCGGCGAAAACGCATTGTGGGAGGCAACGTGTGCCC−3’(配列番号:10)
を使用してマウス第VII因子cDNAから増幅できる。次に、これらの2つのフラグメントを鋳型として使用し、以下のプライマー、
5’−TGAGGATCCCCACCATGGTTCCACAGGCGCATGGGCT−3’(配列番号:7)および5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)
を用いてオーバーラップPCRを実施する。次に、生じた増幅フラグメントをBamHI/SphIで消化し、BglII/SphI部位の間でpSC−TTR−mvmベクターに挿入し得る。その後、BGHポリアデニル化配列を、生じたベクターにSphI部位で挿入し得る。
【0142】
マウス第IX因子のシグナル、プロペプチド、GLAおよびEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列を含む構築物(配列番号:5)を同様に構築する。具体的には、1番目のフラグメントを、マウス第IX因子cDNAを鋳型として使用し、以下のプライマー、
5’−AGGCCTGAAGATCTCCACCATGAAGCACCTGAACACCGTC−3’(配列番号:11)および
5’−GATCAGCTGCTCATTCTTGCTTTTTTCACAGTTCCTTCCTTCAAATC−3’(配列番号:12)
を用いて作製する。2番目のフラグメントは、マウス第VII因子を鋳型として使用し、以下のプライマー、
5’−GATTTGAAGGAAGGAACTGTGAAAAAAGCAAGAATGAGCAGCTGATC−3’(配列番号:13)および
5’−ACAATGCGTTTTCGCCGCTTACGGCGGCCTTGGCGGCTGCTGGAGT−3’(配列番号:9)
を用いて作製する。
【0143】
3番目のフラグメントは、マウス第VII因子を鋳型として使用し、以下のプライマー、
5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)および
5’−GCCGCCGTAAGCGGCGAAAACGCATTGTGGGAGGCAACGTGTGCCC−3’(配列番号:10)
を用いて作製できる。次に、これらの3つフラグメントを鋳型として使用し、以下のプライマー、
5’−AGGCCTGAAGATCTCCACCATGAAGCACCTGAACACCGTC−3’(配列番号:11)および
5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)
を用いてオーバーラップPCRを実施し得る。次に、生じた増幅フラグメントをBamHI/SphIで消化し、BglII/SphI部位の間でpSC−TTR−mvmベクターに挿入する。その後、BGHポリアデニル化配列を、生じたベクターにSphI部位で挿入する。
【0144】
同様の工程を使用して、マウス第IX因子のシグナルおよびプロペプチドドメイン、プロテインSのGLAドメイン、第IX因子のEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列を含む構築物(配列番号:6)を作製し得る。具体的には、1番目のフラグメントを、マウス第IX因子cDNAおよび以下のプライマー、
5’−AGGCCTGAAGATCTCCACCATGAAGCACCTGAACACCGTC−3’(配列番号:11)および
5’−GTTTCTTCGAACAAGGTATTTGCTCTCTTTGGACGGGTAAGAATTTTG−3’(配列番号:14)
を使用して作製し得る。2番目のフラグメントは、マウスプロテインSを鋳型として使用し、以下のプライマー、
5’−CAAAATTCTTACCCGTCCAAAGAGAGCAAATACCTTGTTCGAAGAAAC−3’(配列番号:15)および
5’−GATCTCCATCAACATACTGCTTATAAAAATAATCCGTCTCGGGATT−3’(配列番号:16)
を用いて作製し得る。3番目のフラグメントは、マウス第IX因子のシグナル、プロペプチド、GLAおよびEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列を有するベクターを鋳型として使用し、以下のプライマー、
5’−AATCCCGAGACGGATTATTTTTATAAGCAGTATGTTGATGGAGATC−3’(配列番号:17)および
5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)
を用いて作製し得る。次に、これらの3つフラグメントを鋳型として使用し、以下のプライマー、
5’−AGGCCTGAAGATCTCCACCATGAAGCACCTGAACACCGTC−3’(配列番号:11)および
5’−TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC−3’(配列番号:8)
を用いてオーバーラップPCRを実施する。次に、生じた増幅フラグメントをBamHI/SphIで消化し、BglII/SphI部位の間でpSC−TTR−mvmベクターに挿入する。その後、BGHポリアデニル化配列を、生じたベクターにSphI部位で挿入し得る。
【0145】
野生型マウス第VII因子(配列番号:3)およびRKRRKR配列を有する野生型マウス第VII因子(配列番号:4)のベクター構築物を、各々、年齢が適合する2匹の雄性血友病Bマウスに注射した(マウスにつき5×1011ベクターゲノム)。マウス第IX因子のシグナル、プロペプチド、GLAおよびEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列(配列番号:5)、ならびにマウス第IX因子のシグナルおよびプロペプチドドメイン、プロテインSのGLAドメイン、第IX因子のEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列(配列番号:6)のベクター構築物を、各々、年齢が適合する3匹の雄性血友病Bマウスに注射した(マウスにつき5×1011ベクターゲノム)。注射の約3週間後、野生型マウス第VII因子(配列番号:3)またはRKRRKR配列を有する野生型マウス第VII因子(配列番号:4)のいずれかを注射したすべてのマウスが死亡した。マウス第IX因子のシグナル、プロペプチド、GLAおよびEGF−1ドメインと、第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列(配列番号:5)、またはマウス第IX因子のシグナルおよびプロペプチドドメイン、プロテインSのGLAドメイン、第IX因子のEGF−1ドメインと第VII因子のEGF−2および触媒ドメインならびにRKRRKR配列(配列番号:6)の、いずれかを注射したマウスは、注射の約5週間後に生存し、有害症状を示さなかった。マウスから週に1回血液を採取し、注射の6〜8週間後にマウス第VII因子ポリペプチドのレベルを測定し得る。また、マウス第VII因子ポリペプチドに対する抗原レベルは、注射したマウスの血液試料からインビトロで凝固活性を測定することによって間接的に測定し得る(Margaritas et al.,Gene Therapy 2009;113:3682−3689)。
【0146】
上記は本発明の説明であり、本発明を限定すると解釈されるべきではない。本発明は、以下の特許請求の範囲およびその中に含まれる特許請求の範囲の等価物によって定義される。本明細書で引用されるすべての出版物、特許出願、特許、特許公開、GenBank(登録商標)データベースおよび/またはSNPアクセッション番号によって同定される配列、ならびに他の参考文献は、参考文献が提示される文および/または段落に関連する教示に関してそれらの全体を引用することにより本明細書の一部をなすものとする。
【特許請求の範囲】
【請求項1】
キメラ第VIIa因子ポリペプチドを投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法。
【請求項2】
前記キメラ第VIIa因子ポリペプチドが第VII因子の触媒ドメインを含む、請求項1に記載の方法。
【請求項3】
前記キメラ第VIIa因子ポリペプチドが第VII因子のEGF−2ドメインをさらに含む、請求項2に記載の方法。
【請求項4】
前記キメラ第VIIa因子ポリペプチドが、ビタミンK依存性凝固タンパク質のGLAドメインをさらに含む、請求項3に記載の方法。
【請求項5】
前記キメラ第VIIa因子ポリペプチドが、ビタミンK依存性凝固タンパク質のEGF−1ドメインをさらに含む、請求項3に記載の方法。
【請求項6】
前記GLAドメインが、第IX因子のGLAドメインおよびプロテインSのGLAドメインからなる群より選択される、請求項4に記載の方法。
【請求項7】
前記EGF−1ドメインが、第IX因子のEGF−1ドメインおよびプロテインSのEGF−1ドメインからなる群より選択される、請求項5に記載の方法。
【請求項8】
キメラ第VIIa因子ポリペプチドを投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法であって、該キメラ第VIIa因子ポリペプチドが、第VII因子のEGF−2ドメインおよび触媒ドメインと、第IX因子のGLAドメインおよびプロテインSのGLAドメインからなる群より選択されるGLAドメインと、第IX因子のEGF−1ドメインおよびプロテインSのEGF−1ドメインからなる群より選択されるEGF−1ドメインとを含む、方法。
【請求項9】
前記出血性疾患が、凝固因子欠損症、血小板機能欠損、血小板減少症、フォン・ヴィレブランド病、凝固因子の阻害、手術によって誘発される出血、および外傷によって誘発される出血からなる群より選択される、請求項1に記載の方法。
【請求項10】
前記出血性疾患が凝固因子欠損症である、請求項9に記載の方法。
【請求項11】
前記凝固因子欠損症が血友病である、請求項10に記載の方法。
【請求項12】
前記出血性疾患の治療が、キメラ第VIIa因子ポリペプチドをコードするヌクレオチド配列を含む核酸分子を前記被験体に投与することを含む、請求項1に記載の方法。
【請求項13】
GLAドメインを含むタンパク質を投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法であって、該タンパク質が、循環中の該タンパク質の濃度に比べて血餅の部位付近で濃縮されている、方法。
【請求項14】
プロテインSのGLAドメインと、
プロテインSのEGF−1ドメインおよび第IX因子のEGF−1ドメインから選択されるEGF−1ドメインと、
第VII因子のEGF−2ドメインおよび触媒ドメインと
を含む、キメラ凝固タンパク質。
【請求項15】
請求項14に記載のキメラ凝固タンパク質をコードする核酸分子。
【請求項16】
請求項15に記載の核酸分子を含むベクター。
【請求項17】
請求項14に記載のキメラ凝固タンパク質を含む細胞。
【請求項18】
請求項16に記載のベクターを含む細胞。
【請求項19】
第VII因子の血栓形成性に比べて低い血栓形成性を有するポリペプチドを投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法。
【請求項1】
キメラ第VIIa因子ポリペプチドを投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法。
【請求項2】
前記キメラ第VIIa因子ポリペプチドが第VII因子の触媒ドメインを含む、請求項1に記載の方法。
【請求項3】
前記キメラ第VIIa因子ポリペプチドが第VII因子のEGF−2ドメインをさらに含む、請求項2に記載の方法。
【請求項4】
前記キメラ第VIIa因子ポリペプチドが、ビタミンK依存性凝固タンパク質のGLAドメインをさらに含む、請求項3に記載の方法。
【請求項5】
前記キメラ第VIIa因子ポリペプチドが、ビタミンK依存性凝固タンパク質のEGF−1ドメインをさらに含む、請求項3に記載の方法。
【請求項6】
前記GLAドメインが、第IX因子のGLAドメインおよびプロテインSのGLAドメインからなる群より選択される、請求項4に記載の方法。
【請求項7】
前記EGF−1ドメインが、第IX因子のEGF−1ドメインおよびプロテインSのEGF−1ドメインからなる群より選択される、請求項5に記載の方法。
【請求項8】
キメラ第VIIa因子ポリペプチドを投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法であって、該キメラ第VIIa因子ポリペプチドが、第VII因子のEGF−2ドメインおよび触媒ドメインと、第IX因子のGLAドメインおよびプロテインSのGLAドメインからなる群より選択されるGLAドメインと、第IX因子のEGF−1ドメインおよびプロテインSのEGF−1ドメインからなる群より選択されるEGF−1ドメインとを含む、方法。
【請求項9】
前記出血性疾患が、凝固因子欠損症、血小板機能欠損、血小板減少症、フォン・ヴィレブランド病、凝固因子の阻害、手術によって誘発される出血、および外傷によって誘発される出血からなる群より選択される、請求項1に記載の方法。
【請求項10】
前記出血性疾患が凝固因子欠損症である、請求項9に記載の方法。
【請求項11】
前記凝固因子欠損症が血友病である、請求項10に記載の方法。
【請求項12】
前記出血性疾患の治療が、キメラ第VIIa因子ポリペプチドをコードするヌクレオチド配列を含む核酸分子を前記被験体に投与することを含む、請求項1に記載の方法。
【請求項13】
GLAドメインを含むタンパク質を投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法であって、該タンパク質が、循環中の該タンパク質の濃度に比べて血餅の部位付近で濃縮されている、方法。
【請求項14】
プロテインSのGLAドメインと、
プロテインSのEGF−1ドメインおよび第IX因子のEGF−1ドメインから選択されるEGF−1ドメインと、
第VII因子のEGF−2ドメインおよび触媒ドメインと
を含む、キメラ凝固タンパク質。
【請求項15】
請求項14に記載のキメラ凝固タンパク質をコードする核酸分子。
【請求項16】
請求項15に記載の核酸分子を含むベクター。
【請求項17】
請求項14に記載のキメラ凝固タンパク質を含む細胞。
【請求項18】
請求項16に記載のベクターを含む細胞。
【請求項19】
第VII因子の血栓形成性に比べて低い血栓形成性を有するポリペプチドを投与することにより、出血性疾患を有する被験体において該出血性疾患を治療する方法。
【図1】

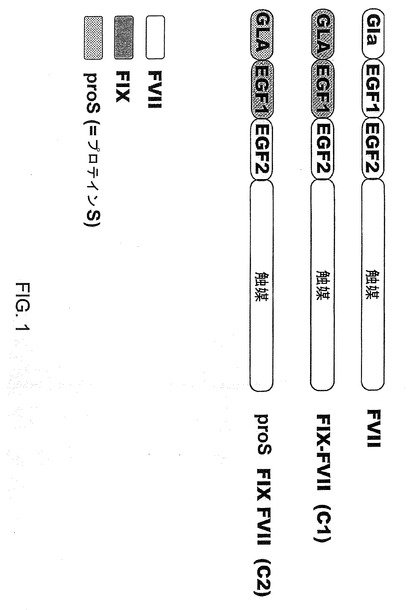
【図2】
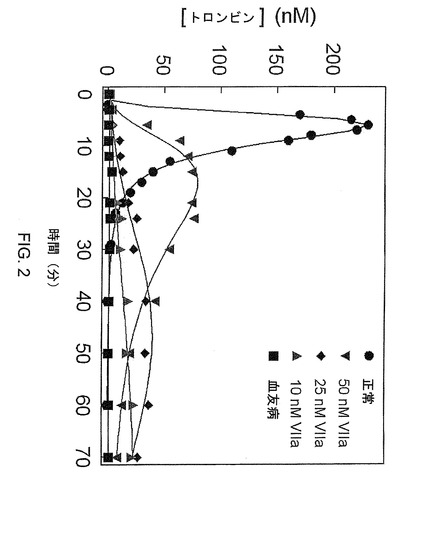
【図3】
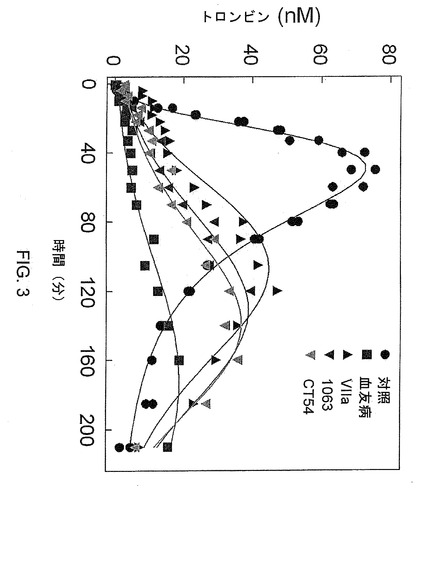
【図4】
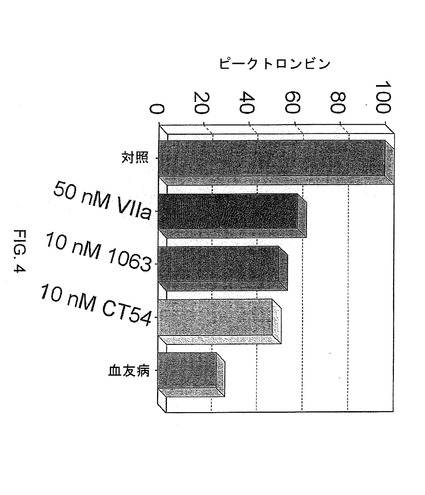
【図5】

【図6】

【図7】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2012−531206(P2012−531206A)
【公表日】平成24年12月10日(2012.12.10)
【国際特許分類】
【出願番号】特願2012−517759(P2012−517759)
【出願日】平成22年6月25日(2010.6.25)
【国際出願番号】PCT/US2010/039934
【国際公開番号】WO2010/151736
【国際公開日】平成22年12月29日(2010.12.29)
【出願人】(500093834)ザ・ユニヴァーシティ・オヴ・ノース・キャロライナ・アト・チャペル・ヒル (14)
【Fターム(参考)】
【公表日】平成24年12月10日(2012.12.10)
【国際特許分類】
【出願日】平成22年6月25日(2010.6.25)
【国際出願番号】PCT/US2010/039934
【国際公開番号】WO2010/151736
【国際公開日】平成22年12月29日(2010.12.29)
【出願人】(500093834)ザ・ユニヴァーシティ・オヴ・ノース・キャロライナ・アト・チャペル・ヒル (14)
【Fターム(参考)】
[ Back to top ]