
タンパク質発現系
本発明は、2分節のRNAウイルスのRNA−2ゲノムセグメント由来の発現エンハンサー配列に基づき、そのRNA−2ゲノムセグメントにおける標的開始部位は変異される。主要なRNA2翻訳開始の上流の適切な開始コドンの欠失が、ウイルス複製を必要とせずに、外来性タンパク質の蓄積を非常に増加させ得る。「高翻訳可能(hyper−translatable)」ササゲモザイクウイルス(CPMV−HT)ベースのタンパク質発現系を含む、方法、ベクターおよび系も提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、概して、植物および他の真核細胞において遺伝子発現を増加させるための方法および物質、特に、ウイルス由来の配列、例えば、目的のタンパク質をコードする異種遺伝子の配列に関する。
【背景技術】
【0002】
コモウイルス(CPMV)
コモウイルスは、2分節ゲノムを有するRNAウイルスである。コモウイルスRNAゲノムのセグメントは、RNA−1およびRNA−2と呼ばれている。RNA−1は、VPg、レプリカーゼおよびプロテアーゼタンパク質をコードする(非特許文献1)。レプリカーゼは、ウイルスゲノムの複製のためにウイルスによって必要とされる。コモウイルスのササゲモザイクウイルス(cowpea mosaic virus)(CPMV)のRNA−2は、58Kおよび48Kのタンパク質、ならびに2つのウイルスコートタンパク質LおよびSをコードする。
【0003】
全てのコモウイルスのRNA−2の翻訳の開始は、2つのカルボキシ共末端(coterminal)タンパク質の合成を生じる、同じトリプレットリーディングフレームに位置する2つの異なる開始部位で起こる。この二重の開始現象は、翻訳の間のリボソームによる「漏出スキャニング(leaky scanning)」の結果として生じる。
【0004】
CPMVのRNA−2における5’末端開始コドン(AUG)は、115位、161位、512位および524位で生じる。161位および512位における開始コドンは、同じトリプレットリーディングフレーム内にある。161位での開始コドンにおける開始は、105Kのポリタンパク質の合成を生じるが、一方、512位での開始コドンにおける開始は、95Kのポリタンパク質の合成に関する。両方のポリタンパク質の合成は3299位での同じ終止コドンで停止されるため、105Kおよび95Kのタンパク質は、カルボキシ共末端である。524位におけるAUGコドンは、512位におけるAUGが欠失される場合、イニシエーターとして役立ち得る。しかしながら、AUG512の存在下において、これは機能せず、単にアミノ酸メチオニンをコードするだけである(非特許文献2;特許文献3)。115位での開始コドンは、ウイルス複製に必須ではない(非特許文献3)。
【0005】
CPMVのRNA−2ゲノムセグメントによってコードされる105Kおよび95Kのタンパク質は、後にRNA−1にコードされるタンパク質分解活性によって分解されて、プロセシングされる105Kまたは95Kのポリタンパク質であるかどうかに依存する、58Kまたは48Kのタンパク質のいずれか、および2つのウイルスコートタンパク質LおよびSを生じる一次翻訳産物である。CPMVにおける512位での開始コドンにおける翻訳の開始は、161位での開始より効果的であり、105Kのポリタンパク質より多い95Kのポリタンパク質の産生を生じる。
【0006】
CPMVのRNA−2における115位の開始コドンは、161位および512位における開始部位の上流に存在し、異なるリーディングフレーム内にある。この開始コドンは、175位における停止コドンと同相であるため、この部位での開始は、20アミノ酸のペプチドの産生を生じ得る。しかしながら、このようなペプチドの産生は、今までに検出されていなかった。
【0007】
AUG間のフレームを維持する必要性
突然変異誘発実験により、CPMVのRNA−2における161位および512位での開始部位の間のフレームの維持が、RNA−1にコードされるレプリカーゼによって、RNA−2の効果的な複製に必須であることが示された(非特許文献2;非特許文献4;非特許文献5;非特許文献3)。この条件は、CPMVのRNA−2(以下に示す)に基づく発現ベクターにおける512位の開始コドンの上流に挿入され得る配列の長さを制限し、そのようなベクターに外因性遺伝子をクローニングすることを、理想的なものよりも困難にさせる。例えば、それらの使用がしばしば、それらの開始部位の間のオープンリーディングフレーム(ORF)を変化させるため、ポリリンカーの使用を妨げる。
【0008】
CPMVベクター
CPMVは、植物における異種ポリペプチドの産生に適切なベクター系の開発のための基礎として役立っている(非特許文献6;非特許文献7)。これらの系は、RNA−2の修飾に基づくが、完全長か欠失した型を使用するか否かにより異なる。しかしながら、両方の場合において、修飾したRNA−2の複製は、RNA−1との共接種によりなされる。RNA−2の完全長型に基づいた発現系は、RNA2由来のポリペプチドのC末端に対する外来性タンパク質の融合に関与する。N末端ポリペプチドの解放は、口蹄疫ウイルス由来の2A触媒ペプチド配列の作用により媒介される(非特許文献8)。得られるRNA−2分子は、植物内および植物間の両方で分離できる。この戦略は、cowpea植物において、Hepatitis Bコア抗原(HBcAg)および小さな免疫タンパク質(SIP)などの多くの組み換えタンパク質を発現するために使用されている(非特許文献9;非特許文献10;非特許文献11)。成功的ではあるが、完全長のウイルスベクターの使用は、挿入される配列のサイズ制限の点で不都合であり、生物学的封じこめについての問題がある。
【0009】
これらに対処するために、CPMV RNA−2の欠失した型に基づく系が最近開発されている(非特許文献12)。この系において、移動タンパク質およびコートタンパク質の両方をコードするRNA−2の領域は除去される。しかしながら、欠失した分子はさらに、RNA−1にコードされるレプリカーゼによる複製に必要なcis作動性の配列を有し、それにより、高レベルの遺伝子増幅が、環境を汚染する修飾されたウイルスの可能性を付随することなく、維持される。RNA−1に加えて接種材料において、PVY由来のHcPro(非特許文献13)などの遺伝子サイレンシングの抑制因子の封入とともに、欠失したCPMVベクターが、一過性の発現系(特許文献1、Bipartite System,Method And Composition For The Constitutive And Inducible Expression Of High Levels Of Foreign Proteins In Plants;非特許文献14)として使用され得る。しかしながら、完全長のRNA−2に基づいたベクターを用いる状況と対照的に、複製は接種レベルに制限される。これらのCPMVベクターは、単一のタイプのポリペプチドからなる複数の鎖の複合体を発現するのに使用されている。
【0010】
CPMV RNA−2の完全長または欠失した型のいずれかに基づいたベクターの複数のコピーはまた、植物における異種タンパク質の産生に適切であることが示されている(非特許文献15)。RNA−1の存在下においてNicotiana benthamiana内への異なるマーカー遺伝子を含む2つの完全長のRNA−2構築物の同時浸潤は、2つの外来性タンパク質が、接種組織において同じ細胞内で効果的に発現され得ることを示すために使用される。さらに、タンパク質は、ヘテロマー形成に必須条件である、同じサブ細胞区画に同時に局在化され得る。
【0011】
植物におけるヘテロマータンパク質の発現のための異なるCPMV RNA−2ベクターの適合性も研究されている。RNA−2の完全長および欠失した型内へのIgGの重鎖および軽鎖の挿入により、接種した組織において完全なサイズのIgG分子を蓄積するための両方のアプローチが示されたが、欠失したRNA−2ベクターが使用された場合、そのレベルは顕著に高かった。従って、浸透して広がる完全長のRNA−2構築物の能力は、ヘテロマータンパク質の産生に重要ではないように見られ、RNA−2の欠失した型の使用が、明らかに利点がある。特に、それらはまた、生物学的封じ込めの利点を与える。
【0012】
従って、ベクター系に基づく公知のCPMVは、植物において目的のタンパク質をコードする異種遺伝子の発現に有用な手段であることを表す。しかしながら、改良を必要とする最適化されるベクター系のための当該分野における必要性、例えば発現される異種タンパク質の収率およびベクターの使用の容易さが依然として存在する。
【先行技術文献】
【特許文献】
【0013】
【特許文献1】国際公開第2007/135480号パンフレット
【非特許文献】
【0014】
【非特許文献1】Lomonossoff,G.P.& Shanks,M.(1983),The nucleotide sequence of cowpea mosaic virus B RNA.EMBO Journal 2,2253−2258
【非特許文献2】Holness,C.L.,Lomonossoff,G.P.,Evans,D.およびMaule,A.J.(1989).Identification of the initiation codons for translation cowpea mosaic virus middle component RNA using site directed mutagenesis of an infectious cDNA clone.Virology 172,311 320
【非特許文献3】Wellink J,Verver J,van Kammen A.(1993).Mutational analysis of AUG codons of cowpea mosaic virus M RNA.Biochimie.75(8):741−7
【非特許文献4】van Bokhoven,H.,Le Gall,O,Kasteel,D.,Verver,J.,Wellink,J.およびvan Kammen,A.(1993).Cis−and Trans−acting Elements in Cowpea Mosaic Virus RNA Replication.Virology 195,377−386
【非特許文献5】Rohll,J.B.,Holness,C.L.,Lomonossoff,G.P.およびMaule,A.J.(1993).3’terminal nucleotide sequences important for the accumulation of cowpea mosaic virus M−RNA.Virology 193,672−679
【非特許文献6】Liu,L.,Canizares,M.C.,Monger,W.,Perrin,Y.,Tsakiris,E.,Porta,C.,Shariat,N.,Nicholson,L.およびLomonossoff,G.P.(2005).Cowpea mosaic virus−based systems for the production of antigens and antibodies in plants.Vaccine 23,1788−1792
【非特許文献7】Sainsbury,F.,Canizares,M.C.およびLomonossoff,G.P.(2007)Cowpea mosaic virus−based expression vectors.In:Virus Expression Vectors(Hefferon,K.ed),pp.339−555.Kerala,India:Transworld Research Network.
【非特許文献8】Gopinath,K.,Wellink,J.,Porta,C.,Taylor,K.M.,Lomonossoff,G.P.およびvan Kammen,A.(2000)Engineering cowpea mosaic virus RNA−2 into a vector to express heterologous proteins in plants.Virology 267:159−173
【非特許文献9】Mechtcheriakova,I.A.,Eldarov,M.A.,Nicholson,L.,Shanks,M.,Skryabin,K.G.およびLomonossoff,G.P.(2006)The use of viral vectors to produce hepatitis B virus core particles in plants.J.Virol.Methods 131,10−15
【非特許文献10】Monger,W.,Alamillo,J.M.,Sola,I.,Perrin,Y.,Bestagno,M.,Burrone,O.R.,Sabella,P.,Plana−Duran,J.,Enjuanes,L.,Garcia,J.A.およびLomonossoff,G.P.(2006)An antibody derivative expressed from viral vectors passively immunizes pigs against transmissible gastroenteritis virus infection when supplied orally in crude plant extrancts.Plant Biotechnol.J.4,623−631
【非特許文献11】Alamillo,J.M.,Monger,W.,Sola,I.,Garcia,B.,Perrin,Y.,Bestagno,M.,Burrone,O.R.,Plana−Duran,J.,Enjuanes,L.,Lomonossoff,P.G.およびGarcia,J.A.(2006)Use of virus vectors for the expression in plants of active full−length and single chain anticoronavirus antibodies.Biotechnol.J.1,1103−1111
【非特許文献12】Canizares,M.C.,Liu,L.,Perrin,Y.,Tsakiris,E.およびLomonossoff,G.P.(2006).A bipartite system for the constitutive and inducible expression of high levels of foreign proteins in plants.Plant Biotechnol.J.4,183−193
【非特許文献13】Brigneti,G.,Voinnet,O.,Li,W.X.,Ji,L.H.,Ding,S.WおよびBaulcombe,D.C.(1998)Viral pathogenicity determinants are suppressors of transgene silencing in Nicotiana benthamiana.EMBO Journal 17:6739−6746
【非特許文献14】Sainsbury,F.,Liu,L.およびLomonossoff G.P.(2009)Cowpea mosaic virus−based expression of antigens and antibodies in plants.In:Methods in Molecular Biology Vol.483:Recombinant Pharmaceutical Proteins from Plants(Faye,L.およびGomord,V.eds),pp25−39,NY:Humana Press
【非特許文献15】Sainsbury,F.およびLomonossoff,G.P.(2008).Extremely high−level and rapid transient protein production in plants without the use of viral replication.Plant Physiology 148,1212−1218
【発明の概要】
【課題を解決するための手段】
【0015】
本発明者らは、驚くべきことに、CPMV RNA−2ベクターにおける161位での開始コドンの変異が、512位での開始コドンの後に挿入される遺伝子によってコードされるタンパク質の発現レベルを著しく増加させることを見出した。タンパク質発現のレベルは、161位での開始コドンがインタクトである(SainsburyおよびLomonossoff,2008)ことのみが異なる、CPMV RNA−2ベクター由来の同じタンパク質の発現と比較して、約20〜30倍増加した。本発明は、ウイルス複製を必要とせず、外来性タンパク質の高レベルの産生を可能にする。
【0016】
本発明者らはまた、161位での開始コドンの変異が、161での変異された開始コドンと、512での開始コドンの位置の間のフレームを維持するための必要性を無効にし、これにより、161位での変異された開始コドンの後の任意の長さの配列の挿入を可能にすることを見出した。このことは特に、任意の長さのポリリンカーを、変異された開始コドンの後にRNA−2ベクター内に挿入することを可能にし、次いで、ベクター内に目的の遺伝子のクローニングを容易にするのに使用できるという利点がある。
【0017】
さらに、本発明者らは、タンパク質発現の増加にもかかわらず、161位での変異した開始コドンを含むCPMV RNA−2ベクターで形質転換した植物が健康的であるように見えることを見出した。すなわち、公知のCPMV RNA−2ベクターで形質転換した植物より壊死をほとんど示さなかった。植物の健康は、長期間生存する健康的な植物として植物由来のタンパク質の発現に重要な因子である。さらに、植物の健康はまた、タンパク質精製を妨げ得る壊死の結果として放出されるタンニンなどの植物由来のタンパク質の精製に重要である(SainsburyおよびLomonossoff,2008)。
【0018】
従って、本発明は、修飾された2分節のウイルス配列に基づいた改良されたタンパク質産生系および方法に関連する。
【0019】
従って、本発明の種々の態様において、発現エンハンサー配列が与えられるか、または利用され、その配列は、コモウイルスなどの2分節RNAウイルスのRNA−2ゲノムセグメント由来(またはそれらと相同性を共有する)であり、その配列において、標的開始部位は変異されている。
【0020】
本発明はさらに、2分節ウイルスのRNA−2ゲノムセグメント由来の配列の発現または翻訳を増強する活性を増加させるためのプロセスを与え、そのプロセスは、配列内の標的開始部位を変異させることを含む。
【0021】
本発明の一部の特定の定義および実施形態は、ここでより詳細に記載される。
【図面の簡単な説明】
【0022】
【図1】図1は、CPMV発現ベクター00、10、01および11の概略図を示す。00発現ベクターにおいて、115位および161位の開始部位はインタクトである。10発現ベクターにおいて、115位の開始部位は変異されているが、161位の開始部位はインタクトである。01発現ベクターにおいて、161位の開始部位は変異されているが、115位の開始部位はインタクトである。11発現ベクターにおいて、115位および161位の開始部位は両方変異されている。CPMV発現ベクター00、10、01および11はまた、512(FSC2−152)、513(FSC2−513)または514(FSC2−514)のいずれかの位置に開始部位を含む。バーは、タンパク質の発現が起こる開始部位を示すために使用される。
【図2】図2は、図1に概略的に示されるCPMV発現ベクターでトランスフェクトされた植物で発現される可溶性の緑色蛍光タンパク質(GFP)のレベルを示す。発現ベクターFSC2−512、FSC2−513、およびFSC2−514において、GFPをコードする遺伝子を、それぞれ、512位、513位および514位の開始コドンの後に挿入した。SDS−PAGEゲルのレーンに、00、10、01および11をマークし、115位および161位でのCPMVベクターにおける開始部位のものに応じて、インタクトまたは変異される。「500ng」とマークしたレーンは、500ngのGFPタンパク質に対応するバンドの位置を示し、それによって、CPMV発現ベクターから発現されるGFPタンパク質の予想される位置を示す。各SDS−PAGEゲルの左側のレーンは、タンパク質のサイズマーカーの位置を示す。
【図3】図3は、図2に示される実験で使用される同じCPMV発現ベクターでトランスフェクトされたNicotiana benthamianaの葉におけるGFP発現のレベルを示す。葉の先端の薄い領域はGFP発現の領域に対応した。発現ベクター10、01および11における115位および/または161位の開始部位を不活性化するために変異が行われ、それも示す。
【図4】図4は、緑色蛍光タンパク質(GFP)、Discosoma赤色蛍光タンパク質(DsRed)、およびB型肝炎コア抗原(HBcAg)を一過性発現するためのDel−RNA−2(図1における発現ベクター00[FSC2−512])およびHT(図1における発現ベクター01[FSC2−512])の比較を示す。各タンパク質についてのdelRNA−2またはHTクローンに、サイレンシング抑制因子P19を浸潤させた。(A)DsRedまたはHBcAgが発現された場合、delRNA−2構築物での浸潤後7日の組織は壊死したが、これはHTにより駆動された発現についての場合ではない。実際に、HTによってDsRedを発現する組織は、昼間の明かりの条件下で明白に赤く見える。(B)タンパク質発現のCoomassie染色したSDS−PAGE分析。示したように組み換えタンパク質に対応する突出したバンドが、ウェスタンブロッティングによって確認された。1−マーカー、2−浸潤されていない組織、3−delRNA−2構築物、4−HT構築物、5−利用可能な場合、市販の標準物質。約5mgの浸潤した組織のからの粗抽出物を、レーンごとに負荷し、同様に2μgのGFP標準物質および2μgのHBcAg標準物質を負荷した。DsRedについての標準物質は今回、利用可能ではなかった。
【図5】図5は、ヒトの抗ヒト免疫不全ウイルス抗体2G12を一過性発現するためのDel−RNA−2(図1における発現ベクター00[FSC2−512])およびHT(図1における発現ベクター01[FSC2−512])の初期の比較を示す。IgG重鎖は、天然形態(HL)またはER保有(HEL)のいずれかであり、軽鎖およびP19を浸潤させた。(A)del−RNA−2による2G12−HELの発現により、浸潤された組織の壊死が導かれるが、これはHT発現については生じない。(B)抗体重鎖(delRNA−2またはHT)とP19が浸潤した組織の粗抽出物のSDS−PAGE分析。各試料について、5mgの浸潤した組織からの粗抽出物を1μgの標準ヒトIgGとして負荷した。2G12に対応するバンドは、coomassie染色後、容易に目に見える。(C)5日後に抗体2G12の蓄積を、プロテインAに対する捕捉および表面プラズモン共鳴分光法により測定し、2容量の緩衝液(PBS、5mM EDTA)での抽出後の濃度を表す。従って、本発明者らは、植物インキュベーションまたは抽出を1つも最適化せずに、150mg/kg(HT HELについて)に近づく初期の重量濃度を導くことができる。3つの試料を各処置について測定した。
【図6】図6は、集合したHBcAg粒子の電子顕微鏡写真を示し、これは、本明細書に記載されるHT(図1における発現ベクター01[FSC2−512])発現系を用いて発現された。集合したHBcAg粒子は、直径約30nmの中空球のように見える。HBcAg粒子を含む樹液(sap)は、電子顕微鏡写真を撮る前に濃縮されなかったが、望ましくない塩は除去した。従って、電子顕微鏡写真は樹液(sap)中のHBcAg粒子の濃度を表す。
【図7】図7は、ベクターpM81−FSC1を示す。
【図8】図8は、ベクターpM81−FSC2を示す。
【図9】図9は、pEAQの構築物の概略図を示す。(A)灰色で示された外来配列を有する開始pBINPLUSベースのプラスミド。(B)バイナリーベクターの必須エレメントを含むPCR産物。(C)末端を合わせた(end−tailored)PCR産物の3部のライゲーション後の中間体プラスミド。(D)増幅およびその後の中間体からの2つのフラグメントのライゲーション後の最終プラスミド。
【図10】図10は、主なpEAQ誘導体のT−DNAの概略図を示す。T−DNAは、示した制限部位内に起こり得るクローニングを残す、示したP19およびNPTIIカセットのいずれかまたは両方を含む。
【図11】図11は、その親プラスミドPBB−FSC2−512−HTと比較したpEAQベクターによって産生したGFPの発現レベルを示す。組織を、pEAQexpressを除いて、P19および示したベクターでの浸潤後、6日に分析し、それを、標準的なODまたは2倍の希釈で単独で浸潤させた。(A)UV照射下で視覚化した葉、(B)coomassie染色した12%SDS−PAGE、および(C)蛍光分光分析。
【図12】図12は、野生型P19と比較したGFP発現を向上させるP19(R43W)の能力を示す。組織は、2倍希釈でのpEAQselectK(selK−P19)、pEAQselectKおよびP19(selK)、pEAQspecialK(spK)、2倍希釈でのpEAQspecialK(spK1:2)、pEAQspecialKm(spKm)、2倍希釈でのpEAQspecialKm(spKm1:2)、pEAQexpress(ex)、および2倍希釈でのpEAQexpress(ex1:2)での浸潤後6日に分析した。(A)UV照射下で視覚化した葉および(B)蛍光分光分析。
【図13】図13は、単一のpEAQプラスミドを有する完全なサイズのIgG、2G12の発現を示す。(A)2G12を発現するために構築された2つのpEAQexpress由来プラスミドの概略図。(B)希釈物を示す浸潤スキームおよび各プラスミドの組み合わせについてのそれらの代表的なOD、ならびに各ODでの侵潤後に作製されたタンパク質抽出物の濃度(±SD)。(C)Coomassie染色した12〜4%のSDS−PAGE分析および(D)2G12重鎖(γ)の免疫学的検出および(E)浸潤後8日の2G12Fab領域(Fab)鎖の免疫学的検出。M、示したサイズを有するマーカー;C、コントロール抽出物;Std、CHO産生2G12。coomassie染色に関して、3mgの浸潤組織の等価物からのタンパク質は、1μgのCHO2G12を有する各レーンに位置し、250ngのCHO2G12の各レーンにおけるウェスタンブロッティングについて0.75mgの組織の等価物。推測した集合/分解産物を示す。
【図14】図14は、ネイティブおよびhis−タグ化変異体におけるpEAQ−HT由来のGFPのクローニングおよび発現を示す。(A)ポリリンカーの詳細を有するpEAQ−HT T−DNAの概略図。(B)GFP発現の蛍光分光分析。spK=pEAQspecialK−GFP−HT、GFP、HisGFP、およびGFPHis=pEAQ−HTクローン。(C)12%SDS−PAGEおよびGFP発現のウェスタン分析。C=コントロール抽出物。
【図15】図15は、バキュロウイルスベクターの一部としての昆虫細胞における使用に適切な本発明の核酸構築物を示す。
【発明を実施するための形態】
【0023】
本明細書中で言及する場合、「エンハンサー」配列(またはエンハンサー要素)とは、コモウイルスなどの2分節のRNAウイルスのRNA−2ゲノムセグメント由来の(またはそれらと相同性を共有する)配列であり、その配列において、標的開始部位は変異される。そのような配列は、それらが結合される異種ORFの下流の発現を向上できる。限定せずに、転写RNAに存在する場合、そのような配列は、それらが結合される異種ORFの翻訳を向上できると考えられる。
【0024】
本明細書中で言及する場合、「標的開始部位」とは、野生型RNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端タンパク質のより長い方の産生(翻訳)のための開始部位として役立つ、問題のあるエンハンサー配列が由来する2分節のウイルス(例えばコモウイルス)の野生型RNA−2ゲノムセグメントにおける開始部位(開始コドン)である。
【0025】
上記のように、CPMV RNA−2によってコードされる2つのカルボキシ共末端タンパク質のより長い方である、105Kのタンパク質の産生は、野生型CPMV RNA−2ゲノムセグメント内の161位での開始部位で開始される。従って、CPMV RNA−2ゲノムセグメント由来のエンハンサー配列における標的開始部位は、野生型CPMV RNA−2における161位での開始部位である。
【0026】
161位の開始コドン付近の変異は、161位の開始コドン自体を変異することと同じ(または同様の)効果を有することができる。例えば、この開始コドン付近の状態を破壊することは、開始コドンがより頻繁にバイパスされることを意味し得る。
【0027】
本発明の一態様において、従って、標的開始部位は、標的開始部位の上流および/または下流の1つ以上のヌクレオチドを変異することによって、間接的に「変異され」得るが、野生型標的開始部位は残ったままであり、これらのヌクレオチドを変異させる効果は、標的開始部位自体が変異される場合に観測される効果と同じまたは類似である。
【0028】
標的開始部位は、野生型RNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端タンパク質のより長い方の産生のための開始部位として役立つため、標的開始部位が、野生型RNA−2によってコードされる2つのカルボキシ共末端タンパク質のより短い方の産生のための開始部位として役立つ、同じ野生型RNA−2ゲノムセグメント上の第2の開始部位を有するフレーム内(同相)にあるという結果となる。2つの開始部位は、それらが同じトリプレットリーディングフレーム内にある場合、フレーム内にある。
【0029】
野生型CPMV RNA−2ゲノムセグメント由来のエンハンサー配列における標的開始部位、すなわち161位の開始部位は、野生型CPMV RNA−2ゲノムセグメントにおけるCPMV RNA−2によってコードされる2つのカルボキシ共末端タンパク質のより短い方(95Kのタンパク質)の産生のための開始部位として役立つ、512位に開始部位を有するフレーム内にある。
【0030】
従って、標的開始部位は、野生型RNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端ポリタンパク質のより短い方の産生のための開始部位として役立つ、エンハンサー配列が由来する野生型RNA−2ゲノムセグメントにおける第2の開始部位の上流(5’)に位置する。さらに、標的開始部位はまた、エンハンサー配列が由来する野生型RNA−2ゲノムの第3の開始部位の下流(3’)に位置し得る。CPMVにおいて、標的開始部位、すなわち、161位の開始部位は、95Kのタンパク質の産生のための開始部位として役立つ512位の第2の開始部位の上流、および115位の第3の開始部位の下流に位置する。
【0031】
従って、2分節のウイルスのRNA−2ゲノムセグメント由来のエンハンサー配列における標的開始部位は、野生型RNA−2によってコードされる2つのカルボキシ共末端タンパク質の産生のための2つの開始部位のうちの第1である。この文脈において「第1の」とは、野生型RNA−2ゲノムセグメントの5’末端近くに位置する開始部位を指す。
【0032】
所望の場合、配列中の1つより多い開始部位が変異されてもよい。例えば、115位(またはそれに相当する)「第3の」開始部位が、欠失または変更されてもよい。AUG161の除去に加えてAUG115の除去が、発現をさらに向上させることが示されている(SainsburyおよびLomonossoff,2008)。
【0033】
本発明のエンハンサー配列は、2分節のRNAウイルスのRNA−2ゲノムセグメント由来の修飾された配列に基づく。
【0034】
本明細書中で言及される場合、2分節のウイルス、または2分節のゲノムを有するウイルスは、Comoviridaeファミリーのメンバーであってもよい。Comoviridaeファミリーの全ての属は、2つのカルボキシ共末端タンパク質をコードするようである。Comoviridaeファミリーの属としては、コモウイルス、ネポウイルス、ファバウイルス、ケラウイルス(Cheravirus)、およびサドワウイルスが挙げられる。コモウイルスとしては、Cowpea mosaic virus(CPMV)、Cowpea severe mosaic virus(CPSMV)、Squash mosaic virus(SqMV)、Red clover mottle virus(RCMV)、Bean pod mottle virus(BPMV)が挙げられる。好ましくは、2分節のウイルス(またはコモウイルス)はCPMVである。
【0035】
これらのコモウイルスおよびいくつかの特定の株のRNA−2ゲノムセグメントの配列は、括弧に記載される受託番号の下でNCBIデータベースから利用可能である:cowpea mosaic virus RNA−2(NC_003550)、cowpea severe mosaic virus RNA−2(NC_003544)、squash mosaic virus RNA−2(NC_003800)、squash mosaic virus strain Kimble RNA−2(AF059533)、squash mosaic virus strain Arizona RNA−2(AF059532)、red clover mottle virus RNA−2(NC_003738)、bean pod mottle virus RNA−2(NC_003495)、bean pod mottle virus strain K−Hopkins1 RNA−2(AF394609)、bean pod mottle virus strain K−Hancock1 RNA−2(AF394607)、Andean potato mottle virus(APMoV:L16239)およびRadish mosaic virus(RaMV;AB295644)。同様に、bean rugose mosaic virus(BRMV;AF263548)およびコモウイルス属、turnip ringspot virus(EF191015)の暫定的なメンバーから利用可能な部分的なRNA−2配列も存在する。Comoviridaeファミリーにおける他の属由来の多数の配列も利用可能である。
【0036】
今まで、研究されてきた全てのコモウイルスは、それらのRNA−2ゲノムセグメント由来の2つのカルボキシ共末端ポリタンパク質を発現するための2つの別の開始コドンを有することが示されている。特に、CPMV、CPSMV、BPMV、SqMVおよびRCMVのRNA−2ゲノムセグメントは、2つのカルボキシ共末端ポリタンパク質を発現するための2つの別の開始コドンを含むことが知られている。
【0037】
従って、CPMV(すなわち、それに対応する)の野生型RNA−2セグメントにおける161位の開始部位と同等である、他のコモウイルスにおける標的開始部位は、当該分野において公知の方法によって同定され得る。例えば、標的開始部位は、CPMVの野生型RNA−2ゲノムセグメント配列と、別のコモウイルスのRNA−2ゲノムセグメント配列との間の配列アラインメントによって同定され得る。次いで、このような配列アラインメントは、少なくともそのアラインメントにおいて、野生型CPMV RNA−2における161位の標的開始部位と近いか、または同じ位置である、開始部位を同定することによって、コモウイルスRNA−2ゲノムセグメント配列における標的開始部位を同定するために使用され得る。
【0038】
他のコモウイルスにおける標的開始部位もまた、野生型コモウイルスRNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端タンパク質のより長い方の合成のための開始部位として役立つ開始コドンを決定することによって同定され得る。このアプローチはまた、上記のアラインメントと合わせて使用され得る。すなわち、このアプローチは、CPMV RNA−2を有するアラインメントによって同定されるコモウイルス開始部位が標的開始部位であることを確認するために使用され得る。
【0039】
もちろん、上記の方法はまた、野生型CPMV RNA−2ゲノムセグメントにおける512位の開始部位と同等である、他のコモウイルスRNA−2ゲノムセグメントにおける開始部位を同定するために使用され得る。しかしながら、野生型コモウイルスRNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端タンパク質のより長い方を合成するための開始部位として役立つ開始コドンを同定する代わりに、野生型コモウイルスRNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端タンパク質のより短い方を合成するための開始部位として役立つ開始コドンが同定される。
【0040】
一旦、CPMV RNA−2における161位および512位の開始部位と同等である可能性がある2つのコモウイルスRNA−2開始部位が同定されると、その標的開始部位の同定は、2つの開始部位が同じフレーム内、すなわち、同じトリプレットリーディングフレーム内にあることを調べることによって確認され得る。なぜなら、この場合、それらは、2つのカルボキシ共末端タンパク質を産生するための開始部位としてのみ機能するからである。
【0041】
本発明の一実施形態において、エンハンサー配列は、CPMV RNA−2ゲノムセグメントのヌクレオチド1〜512を含み(表1を参照のこと)、161位の標的開始部位は変異される。本発明の別の実施形態において、エンハンサー配列は、別のコモウイルス由来の等価配列を含み、CPMVの161位の開始コドンと同等の標的開始部位は変異される。その標的開始部位は、置換、欠失または挿入によって変異されてもよい。好ましくは、その標的開始部位は、点突然変異によって変異される。
【0042】
本発明の代替の実施形態において、エンハンサー配列は、変異標的開始部位を有するコモウイルスRNA−2ゲノムセグメント配列のヌクレオチド10〜512、20〜512、30〜512、40〜512、50〜512、100〜512、150〜512、1〜514、10〜514、20〜514、30〜514、40〜514、50〜514、100〜514、150〜514、1〜511、10〜511、20〜511、30〜511、40〜511、50〜511、100〜511、150〜511、1〜509、10〜509、20〜509、30〜509、40〜509、50〜509、100〜509、150〜509、1〜507、10〜507、20〜507、30〜507、40〜507、50〜507、100〜507、または150〜507を含む。本発明の他の実施形態において、エンハンサー配列は、表1に示すCPMV RNA−2ゲノムセグメント配列のヌクレオチド10〜512、20〜512、30〜512、40〜512、50〜512、100〜512、150〜512、1〜514、10〜514、20〜514、30〜514、40〜514、50〜514、100〜514、150〜514、1〜511、10〜511、20〜511、30〜511、40〜511、50〜511、100〜511、150〜511、1〜509、10〜509、20〜509、30〜509、40〜509、50〜509、100〜509、150〜509、1〜507、10〜507、20〜507、30〜507、40〜507、50〜507、100〜507、または150〜507を含み、野生型CPMV RNA−2ゲノムセグメントにおける161位の標的開始部位は変異される。
【0043】
本発明のさらなる実施形態において、エンハンサー配列は、変異標的開始部位を有するコモウイルスRNA−2ゲノムセグメント配列のヌクレオチド1〜500、1〜490、1〜480、1〜470、1〜460、1〜450、1〜400、1〜350、1〜300、1〜250、1〜200、または1〜100を含む。
【0044】
本発明の代替の実施形態において、エンハンサー配列は、表1に示すCPMV RNA−2ゲノムセグメント配列のヌクレオチド1〜500、1〜490、1〜480、1〜470、1〜460、1〜450、1〜400、1〜350、1〜300、1〜250、1〜200、または1〜100を含み、野生型CPMV RNA−2ゲノムセグメントにおける161位の標的開始部位は変異される。
【0045】
変異標的開始部位を有するコモウイルスRNA−2ゲノムセグメントの少なくとも100または200、少なくとも300、少なくとも350、少なくとも400、少なくとも450、少なくとも460、少なくとも470、少なくとも480、少なくとも490、あるいは少なくとも500のヌクレオチドを含むエンハンサー配列もまた、本発明の実施形態である。
【0046】
さらに、野生型CPMV RNA−2ゲノムセグメントにおける161位の標的開始部位が変異される、表1に示すCPMV RNA−2ゲノムセグメント配列の少なくとも100または200、少なくとも300、少なくとも350、少なくとも400、少なくとも450、少なくとも460、少なくとも470、少なくとも480、少なくとも490あるいは少なくとも500のヌクレオチドを含むエンハンサー配列もまた、本発明の実施形態である。
【0047】
本発明の代替の実施形態は、野生型CPMV RNA−2ゲノムセグメントにおける161位の標的開始部位が変異される、表1に示すCPMV RNA−2ゲノムセグメント配列と少なくとも99%、98%、97%、96%、95%、90%、85%、80%、75%、70%、65%、60%、55%または50%の同一性を有するエンハンサー配列である。
【0048】
特定の配列に対して言及する場合、用語「%の類似性」、「%の同一性」および「%の相同性」は、University of Wisconsin GCGソフトウェアプログラムに記載されるように使用される。従って、エンハンサー配列は、表1に示すCPMV RNA−2ゲノムセグメント配列の相補的配列と特異的にハイブリダイズする。ただし、野生型CPMV RNA−2ゲノムセグメントにおける161位に対応する標的開始部位は変異される。
【0049】
用語「特異的にハイブリダイズする」とは、当該分野において一般に使用される予め決定された条件下で、そのようなハイブリダイゼーションを可能にするのに十分な相補的配列(時々、「実質的に相補的な」といわれる)の2つの一本鎖核酸分子の間の会合を指す。特に、この用語は、本発明の1本鎖DNAまたはRNA分子内に含まれる実質的に相補的な配列を有するオリゴヌクレオチドのハイブリダイゼーションを指し、非相補的配列の一本鎖核酸を有するオリゴヌクレオチドのハイブリダイゼーションの実質的な排除を指す。「相補的な」とは、塩基対合(相補的配列T−C−AとのA−G−T対)による核酸配列の天然の会合を指す。2つの一本鎖分子間の相補性は、核酸対の一部のみが相補的である場合、部分的であってもよいか、または全ての塩基対が相補的である場合、完全であってもよい。相補性の程度は、ハイブリダイゼーションおよび増幅反応の効果および強度に影響を与える。
【0050】
本発明のエンハンサー配列における標的開部位は、翻訳開始部位としてもはや機能しなくなるように、欠失、挿入または置換によって変異されてもよい。例えば、点突然変異が、エンハンサー配列における標的開始部位の位置でなされてもよい。あるいは、エンハンサー配列における標的開始部位は、部分的または完全のいずれかで欠失されてもよい。例えば、エンハンサー配列における標的開始部位にわたる欠失がなされてもよい。開始部位にわたる欠失は、エンハンサー配列が由来する野生型RNA−2ゲノムセグメントの配列と比較した場合、5まで、10まで、15まで、20まで、25まで、30まで、35まで、40まで、45まで、または50までのヌクレオチド長でなされてもよい。
【0051】
理論に束縛されることを望まないなら、CPMVにおける161位の開始コドンの変異は、不活化翻訳抑制因子の下流に位置する開始コドンからの翻訳の向上した開始を生じる、翻訳抑制因子の不活化を導くと考えられる。
【0052】
従って、本発明はさらに、2分節ウイルスのRNA−2ゲノムセグメント由来のエンハンサー配列を提供し、そのエンハンサー配列は、不活化翻訳抑制因子配列を含む。
【0053】
本発明はさらに、2分節ウイルスのRNA−2ゲノムセグメント由来の配列の発現または翻訳を増強する活性を増加させるためのプロセスを提供し、そのプロセスは、配列内の翻訳抑制因子配列を不活化する工程を含む。
【0054】
既に上で述べたように、CPMV RNA−2ゲノムセグメントにおける161位の開始部位の変異は、CPMV RNA−2に通常存在する翻訳抑制因子の不活化を導くと考えられる。
【0055】
本明細書中で言及する場合、翻訳抑制因子配列とは、問題のあるエンハンサー配列が由来する2分節のウイルス(例えばコモウイルス)の野生型RNA−2ゲノムセグメントにおける配列であり、野生型RNA−2ゲノムセグメントによってコードされる2つのカボキシ共末端タンパク質のより長い方を産生(翻訳)するための制限部位を含むか、それから構成される。
【0056】
CPMV RNA−2ゲノムセグメント由来のエンハンサー配列における翻訳抑制因子配列は、上記の標的開始部位を含むか、またはそれから構成される配列である。従って、翻訳抑制因子配列は、上記の標的開始部位を含むか、それから構成され、上記の変異誘発によって不活性化され得る。
【0057】
上記のエンハンサー配列は、以下のような本発明の種々の態様および実施形態において使用されてもよい。
【0058】
従って、本発明の一態様において、上記の発現エンハンサー配列から構成されるか、またはそれから実質的に構成される単離された核酸が提供または利用される。
【0059】
本明細書中で使用される場合、「核酸」または「核酸分子」とは、一本鎖または二本鎖のいずれかである任意のDNAまたはRNA分子を指し、一本鎖の場合、その相補的配列の分子は、直線状または環状の形態のいずれかである。核酸分子を考慮する際に、特定の核酸分子の配列または構造が、通常の従来の5’から3’方向における配列を与えることによって本明細書中に記載され得る。本発明の核酸に関して、用語「単離された核酸」が時々使用される。DNAに当てはめる場合、この用語は、起源となる生物の天然のゲノムにすぐに隣接する配列から分離されるDNA分子を指す。
【0060】
例えば、「単離された核酸」は、プラスミドまたはウイルスベクターなどのベクターに挿入されるか、あるいは原核細胞もしくは真核細胞または宿主生物のゲノムDNAに組み込まれるDNA分子を含んでもよい。
【0061】
RNAに当てはめる場合、この用語「単離された核酸」とは主に、上記の単離されたDNA分子によってコードされるRNA分子を指す。あるいは、この用語は、天然の状態(すなわち、細胞または組織中)で会合する他の核酸から十分に分離されているRNA分子を指す場合もある。「単離された核酸」(DNAまたはRNAのいずれか)はさらに、生物学的または合成的手段によって直接産生され、その産生の間に存在する他の化合物から分離される分子を表す場合もある。
【0062】
従って、核酸は、エンハンサーが由来する2分節のRNAウイルスのRNA−2ゲノムセグメントの一部またはそのフラグメントを含むか、または実質的に含んでもよい。例えば、一実施形態において、核酸は、由来するRNA−2ゲノムセグメントのコード領域の少なくとも一部を含まない。コード領域は、2つのカルボキシ共末端タンパク質のより短い方をコードするRNA−2ゲノムセグメントの領域であり得る。核酸は、野生型RNA−2ゲノムセグメントの5’末端から、野生型RNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端タンパク質のより短い方の産生(翻訳)が開始される開始部位まで伸長する2分節のウイルスのRNA−2ゲノムセグメントの一部から構成されてもよいか、またはそれから実質的に構成されてもよい。
【0063】
特定のヌクレオチドまたはアミノ酸を言及する場合、用語「から実質的に構成される」とは、所定の配列番号の特性を有する配列を意味する。例えば、アミノ酸配列に関して使用される場合、この用語は、配列の基本的かつ新規の特徴に影響を及ぼさない配列それ自体および分子修飾を含む。例えば、核酸に対して使用される場合、この用語は、配列のエンハンサー機能に影響を及ぼさないか、またはさらなる(追加的な)機能を与えない配列それ自体ならびに少しの変化および/または伸長物を含む。
【0064】
本発明はさらに、本発明のエンハンサー配列を含む遺伝子発現系に関する。
【0065】
従って、別の態様において、本発明は、上記のエンハンサー配列を含む遺伝子発現系を提供する。
【0066】
遺伝子発現系はまた、エンハンサー配列の下流に挿入される目的のタンパク質をコードする遺伝子を含んでもよい。目的のタンパク質をコードする挿入される配列は、任意のサイズであってもよい。
【0067】
従って、さらなる態様において、本発明は、
(a)上記のエンハンサー配列、および(b)目的のタンパク質をコードする遺伝子を含む遺伝子発現系を提供し、ここで、その遺伝子はエンハンサー配列の下流に位置する。
【0068】
目的の遺伝子およびタンパク質は異種であってもよい。すなわち、エンハンサー配列が由来する野生型の2分節のRNAウイルスによってコードされない。
【0069】
遺伝子発現系は、宿主生物において目的のタンパク質を発現するために使用され得る。この場合、目的のタンパク質はまた、問題の宿主細胞に対して異種であってもよい。すなわち遺伝子操作を用いて、すなわちヒトの介入によって、問題の細胞(例えば、植物またはその原種)に組み込まれる。生物内の異種遺伝子は、内因性の等価遺伝子(すなわち、同じまたは類似の機能を正常に果たすもの)と置き換えられてもよいか、または挿入される配列は内因性遺伝子もしくは他の配列に対して付加的であってもよい。
【0070】
当業者は、目的の遺伝子の発現が、発現される遺伝子の上流に位置する開始部位(AUG)の存在を必要とすることを理解するだろう。そのような開始部位は、エンハンサー配列の一部または目的のタンパク質をコードする遺伝子の一部のいずれかとして与えられてもよい。
【0071】
宿主生物は植物であってもよい。しかしながら、翻訳機構が真核生物にわたって十分に保存されているため、遺伝子発現系はまた、植物以外の真核宿主生物、例えば修飾されたバキュロウイルスベクターなどの昆虫細胞、または酵母もしくは哺乳動物細胞において目的のタンパク質を発現するために使用されてもよい。
【0072】
遺伝子発現系は、プロモーターおよび終結配列に作動可能に連結されてもよい。
【0073】
従って、遺伝子発現系はさらに、終止配列を含んでもよく、目的のタンパク質をコードする遺伝子は、エンハンサー配列と終止配列との間、すなわちエンハンサー配列の下流(3’)および終止配列の上流(5’)に位置してもよい。
【0074】
従って、本発明はさらに、
(i)作動可能に連結されるプロモーター
(ii)上記のエンハンサー配列
(iii)発現することが所望される目的の遺伝子
(iv)終結配列
を含む、発現カセットを提供する。
【0075】
好ましくは、目的の遺伝子を駆動するのに使用されるプロモーターは、強力なプロモーターである。植物に使用するための強力なプロモーターの例としては、
(1)p35S:Odellら、1985
(2)Cassava Vein Mosaic Virusプロモーター、pCAS、Verdaguerら、1996
(3)リブロース二リン酸カルボキシラーザの小さいサブユニットのプロモーター、pRbcS:Outchkourovら、2003
が挙げられる。
【0076】
他の強力なプロモーターとしては、pUbi(単子葉植物および双子葉植物について)およびpActinが挙げられる。
【0077】
好ましい実施形態において、プロモーターは、誘導プロモーターである。
【0078】
プロモーターに適用される場合、用語「誘導」は当業者に十分に理解されている。本質的に、誘導プロモーターの制御下での発現は、与えられた刺激に応答して「スイッチオン」または増加される。刺激の性質はプロモーター間で変化する。一部の誘導プロモーターは、適切な刺激の非存在下において、少し、または検出できないレベルの発現(または発現なし)を生じる。他の誘導プロモーターは、刺激の非存在下において、検出可能な構成的発現を生じる。発現レベルは刺激の非存在下にかかわらず、任意の誘導プロモーター由来の発現は、適切な刺激の存在下において増加する。
【0079】
終止(終結)配列は、2分節RNAウイルス、例えばコモウイルスのRNA−2ゲノムセグメント由来の終止配列であり得る。一実施形態において、終止配列は、エンハンサー配列が由来する同じ2分節RNAウイルス由来であってもよい。終止配列は、停止コドンを含んでもよい。終止配列はまた、ポリアデニル化信号を伴ってもよい。
【0080】
本発明の遺伝子発現カセット、遺伝子発現構築物および遺伝子発現系はまた、非翻訳領域(UTR)を含んでもよい。UTRは、遺伝子発現カセット、遺伝子発現構築物または遺伝子発現系に存在する終結配列の上流に位置してもよい。遺伝子発現カセット、遺伝子発現構築物または遺伝子発現系が、目的のタンパク質をコードする遺伝子を含む場合、UTRは前記遺伝子の下流に位置してもよい。
【0081】
従って、UTRは、目的のタンパク質をコードする遺伝子と終結配列との間に位置してもよい。UTRは、2分節RNAウイルス由来、例えば2分節RNAウイルスのRNA−2ゲノムセグメント由来であってもよい。UTRは、遺伝子発現カセット、遺伝子発現構築物または遺伝子発現系に存在するエンハンサー配列が由来する同じRNA−2ゲノムセグメントの3’UTRであってもよい。好ましくは、UTRは、コモウイルスRNA−2ゲノムセグメントの3’UTR、例えばCPMV RNA−2ゲノムセグメントの3’UTRである。
【0082】
上記のように、RNA−2における161位および512位の開始部位の間のフレームが維持されること、すなわち、2つの開始部位が同じトリプルリーディングフレームに残っていることは、CPMV RNA−1にコードされるレプリカーゼによるCPMV RNA−2の効率的な複製のために必須であることが以前に示されている(Holnessら,1989;van Bokhovenら,1993;Rohllら,1993)。この条件は、CPMVに基づいた発現ベクターにおける512位の開始部位の上流に挿入できる配列の長さを制限した。特に、ポリリンカーの使用がしばしば、2つの開始部位の間のオープンリーディングフレーム(ORF)を変化させるため、それらの使用が妨げられる。
【0083】
本発明は、CPMV RNA−2における161位および512位の開始部位の間のリーディングフレームの維持がまた、512位の開始部位における翻訳の効率的な開始に必要とされること、すなわち、CPMVによってコードされる2つのカルボキシ共末端タンパク質より短い方(95Kタンパク質)の効率的な発現に必要とされることも示す。
【0084】
しかしながら、本発明はまた、CPMV RNA−2における161位の開始部位の変異が、95Kタンパク質の発現レベルに対していかなる負の影響も与えずに、変異された開始コドンと512位の開始部位との間のフレームを変化させる、512位の開始部位の上流の配列の挿入を可能にすることも示す。従って、161位の開始部位の変異は、512位の開始部位の上流に挿入できる配列の長さをもはや制限しないことを意味する。
【0085】
2つのカルボキシ共末端タンパク質をコードする開始部位の間のリーディングフレームの維持もまた、他の2分節ウイルスに必要とされる場合、この条件はまた、ウイルスRNA−2ゲノムセグメントによってコードされるより2つのカルボキシ共末端タンパク質のより長い方を産生するための開始部位として役立つAUGを変異させることによって克服され得る。従って、別の態様において、本発明は、
(a)上記のエンハンサー配列、および
(b)遺伝子発現系に目的のタンパク質をコードする遺伝子の挿入を促進するための異種配列を含む遺伝子発現構築物を提供し、ここで、その異種配列は、エンハンサー配列における変異された標的開始部位の下流に位置する。
【0086】
異種配列は開始コドンの上流に位置してもよく、その開始コドンから2つのカルボキシ共末端タンパク質のうちのより短い方の産生が、遺伝子発現系のエンハンサー配列が由来する野生型RNA−2ゲノムにおいて開始される。あるいは、異種配列は、開始コドンの部位付近に与えられてもよいか、または開始コドンと置換されてもよく、その開始コドンから2つのカルボキシ共末端タンパク質のより短い方が、遺伝子発現系のエンハンサー配列が由来する野生型RNA−2ゲノムセグメントにおいて開始される。CPMVのRNA−2由来のエンハンサー配列を有する遺伝子発現系において、異種配列は、開始コドンの部位の上流、その付近に与えられてもよいか、または開始コドンと置換されてもよく、その開始コドンは、野生型RNA−2 CPMVゲノムセグメントにおける512位にある。
【0087】
異種配列は、ポリリンカーまたは複数のクローニング部位、すなわち、発現系に目的のタンパク質をコードする遺伝子のクローニングを促進する配列であってもよい。
【0088】
例えば、以下に記載するように、本発明は、CPMVに基づいた発現カセットの5’リーダーと3’UTRとの間にポリリンカーを含む構築物を与える。以下に記載するように、任意のポリリンカーは、必要に応じて、タンパク質の精製を促進するためにNまたはC末端His−タグの融合を可能にするように複数のxヒスチジン残基の1つ以上のセットをコードしてもよい。
【0089】
好ましくは、上記の発現構築物はベクターに存在し、好ましくは、それは、生物ゲノムへの発現カセットの転写および組み込みを可能にする境界配列を含む。
【0090】
好ましくは、構築物は植物のバイナリーベクターである。好ましくは、バイナリー形質転換ベクターはpPZPに基づく(Hajdukiewiczら,1994)。他の例示的な構築物としては、pBin19が挙げられる(Frisch,D.A.,L.W.Harris−Hallerら,(1995).「Complete Sequence of the binary vector Bin19.」Plant Molecular Biology 27:405−409を参照のこと)。
【0091】
本明細書に記載するように、本発明は、存在するpBin発現カセットに必須の要素を有する発現カセットを移動させることによって実施されてもよいか、または他の実施形態において、pBin発現ベクターの直接クローニングが利用されてもよい。
【0092】
例えば、以下に記載するように、本発明は、本明細書に記載されるエンハンサー配列とともに使用する(それらに制限されないが)ために設計されたモジュラーバイナリーベクターを有する。それらは、pBINPLUSベクターに対する改良に基づき、従って、複製およびTDNA転写についての機能を妥協することなしに、ベクターのサイズを劇的に減少させることが可能であることが示されている。さらに、(いわゆる「CPMV−HT」系によって例示されるように)エンハンサー系の要素が、複数のタンパク質が単一のT−DNAから発現され得るように、モジュラー方式で、得られるベクター内に組み込まれてもよい。それらの改良は、外来性遺伝子の効率的な直接クローニングを可能にする多用途の高レベル発現ベクターの産生を導く。
【0093】
これらの例は好ましいバイナリー植物ベクターを表す。好ましくは、それらには、ColEI複製起点が含まれるが、高コピー数も生じる他の複製起点を含むプラスミド(例えば、pRiに基づくプラスミド、LeeおよびGelvin,2008)、特に一過性発現系も好ましい。
【0094】
所望の場合、選択可能な遺伝子マーカーは、抗生物質または除草剤(例えば、カナマイシン、ハイグロマイシン、ホスフィノトリシン、クロルスルフロン、メトトレキサート、ゲンタマイシン、スペクチノマイシン、イミダゾリノンおよびグリホサート)に対する耐性などの選択可能な表現型を与えるものなどの構築物に含まれてもよい。
【0095】
最も好ましいベクターは以下に記載するpEAQベクターであり、遺伝子サイレンシングの抑制因子およびNPTIIカセットも含むT−DNAに配置される、本発明の翻訳エンハンサーを含む発現カセットの5’リーダーと3’UTRとの間のポリリンカーの使用による直接クローニング型を可能にする。ポリリンカーはまた、NまたはC末端His−タグの融合により、タンパク質精製を促進できる1つまたは2つのセットの6xヒスチジン残基をコードする。
【0096】
pEAQ由来ベクターの利点は、IgGなどの複数鎖タンパク質の各要素が、各感染細胞に自動的に送達され得ることである。
【0097】
本発明はまた、植物、酵母、昆虫または哺乳動物細胞などの宿主生物において、本発明の遺伝子発現系を用いて、タンパク質、例えば異種タンパク質を発現する方法を提供する。
【0098】
本発明はさらに、上記のRNA2由来の配列に作動可能に連結される、目的の異種タンパク質をコードする遺伝子またはORFからその目的の異種タンパク質の翻訳を向上させる方法を提供し、その方法は、RNA2由来の配列における標的開始部位を変異させることを含む。
【0099】
本明細書に記載されるエンハンサー配列はまた、WO/2007/135480に記載される2分節の発現系に使用され得る。従って、本発明は、切断されたRNA2−遺伝子セグメントに基づいた遺伝子発現系にも関連し、必要に応じてさらに、プロモーターおよび終結配列に作動可能に連結された遺伝子サイレンシングの抑制因子をコードする第2の遺伝子構築物を含む。
【0100】
従って、さらなる態様において、本発明は、
(a)プロモーターおよび終結配列に作動可能に連結される目的の異種タンパク質をコードする少なくとも1つの外来性遺伝子を保有する2分節のウイルスゲノムの切断されたRNA−2を含む第1の遺伝子構築物であって、その第1の遺伝子構築物は、外来性遺伝子の上流に変異された標的開始部位を含む、第1の遺伝子構築物と、必要に応じて、
(b)プロモーターおよび終結配列に作動可能に連結される、前記2分節のウイルスゲノムのRNA−1を含む第2の遺伝子構築物と、必要に応じて、
(c)前記第1の遺伝子構築物、前記第2の遺伝子構築物またはその両方内に必要に応じて組み込まれる第3の遺伝子構築物であって、プロモーターおよび終結配列に作動可能に連結される遺伝子サイレンシングの抑制因子を含む、第3の遺伝子構築物と、
を含む遺伝子発現系に関連する。
【0101】
本発明の遺伝子発現系(上記のもののいずれかを含む)における遺伝子サイレンシングの抑制因子の存在は好ましいが、必須ではない。従って、上記の遺伝子発現系は、好ましくは、前記第1の遺伝子構築物、前記第2の遺伝子構築物、またはその両方内に必要に応じて組み込まれる第3の遺伝子構築物であって、プロモーターおよび終結配列に作動可能に連結される遺伝子サイレンシングの抑制因子を含む、第3の遺伝子構築物を含む。
【0102】
従って、別の態様において、本発明は、以下の工程
(a)本発明の遺伝子発現構築物を植物細胞に導入する工程と、必要に応じて、
(b)プロモーターおよび終結配列に作動可能に連結される、前記2分節のウイルスゲノムのRNA−1を含む第2の遺伝子構築物を植物細胞に導入する工程と、必要に応じて、
(c)前記第1の遺伝子構築物、前記第2の遺伝子構築物、またはその両方内に必要に応じて組み込まれる第3の遺伝子構築物であって、プロモーターおよび終結配列に作動可能に連結される遺伝子サイレンシングの抑制因子を含む第3の遺伝子構築物を植物細胞に組み込む工程と、
を含む植物においてタンパク質を発現する方法を提供する。
【0103】
好ましくは、上記の植物においてタンパク質を発現する方法は、
前記第1の遺伝子構築物、前記第2の遺伝子構築物、またはその両方内に必要に応じて組み込まれる第3の遺伝子構築物であって、プロモーターおよび終結配列に作動可能に連結される遺伝子サイレンシングの抑制因子を含む第3の遺伝子構築物を植物細胞に導入する工程を含む。
【0104】
本発明はまた、このような構築物を植物細胞に導入することを含む方法を提供する。
【0105】
本発明は、翻訳エンハンサーとして同じT−DNAに目的の遺伝子およびサイレンシングの抑制因子を組み込むことによって非常に高い発現レベルを示す。従って、好ましい実施形態は、同じT−DNAに存在する全てのこれらの要素を利用することであり得る。
【0106】
さらに、RNA−1が、本明細書に記載される系において、高レベルの発現を必要とされず、実際には、本明細書に記載される「CPMV−HT」系はRNA−1の作用によってではないことは理解されるべきである。
【0107】
従って、さらなる態様において、本発明は、
(a)プロモーターおよび終結配列に作動可能に連結される目的の異種タンパク質をコードする少なくとも1つの外来性遺伝子を保有する2分節のウイルスゲノムの切断されたRNA−2を含む第1の遺伝子構築物であって、その遺伝子構築物は、外因性遺伝子の上流に変異された標的開始部位を含む、第1の遺伝子構築物と、必要に応じて、
(b)前記第1の遺伝子構築物内に必要に応じて組み込まれる第2の遺伝子構築物、プロモーターおよび終結配列に作動可能に連結される遺伝子サイレンシングの抑制因子と、
を含む遺伝子発現系に関する。
【0108】
従って、別の態様において、本発明は、以下の工程
(a)本発明の遺伝子発現構築物を植物細胞に導入する工程と、必要に応じて、
(b)前記第1の遺伝子構築物内に必要に応じて第2の遺伝子構築物を導入する工程であって、前記第2の遺伝子構築物は、植物細胞内のプロモーターおよび終結配列に作動可能に連結される遺伝子サイレンシングの抑制因子を含む、工程と、
を含む植物においてタンパク質を発現する方法を提供する。
【0109】
これらの態様において有用な遺伝子サイレンシングの抑制因子は、当該分野において公知であり、WO/2007/135480に記載される。それらとしては、ジャガイモウイルス(Potato virus)Y由来のHcPro、TEV由来のHe−Pro、TBSV由来のP19、rgsCam、FHV由来のB2タンパク質、CPMVの小コートタンパク質、およびTCV由来のコートタンパク質が挙げられる。最も好ましくは、この系のRNA−2は、感染ウイルスが産生されないように切断されている。
【0110】
安定なトランスジェニック植物を産生する場合、好ましい抑制因子は、R43W変異を組み込んでいるP19抑制因子である。
【0111】
本発明のさらなる態様において、本発明による異種構築物を含む宿主細胞が開示される。
【0112】
本発明の遺伝子発現ベクターは、植物細胞内に一過性または安定に組み込まれてもよい。
【0113】
スモールスケールの産生に関しては、本発明の構築物を有する葉の機械的アグロ浸潤法(agroinfiltration)である。スケールアップは、例えば減圧浸潤の使用により達成される。
【0114】
他の実施形態において、本発明の発現ベクターは、トランスジェニック植物または植物細胞のゲノム内に安定に組み込まれてもよい。
【0115】
一態様において、本発明は、形質転換した植物細胞から植物を再生する工程をさらに含んでもよい。
【0116】
植物で広範に成功している以前に使用されている特定の手順およびベクターは、GuerineauおよびMullineaux(1993)(Plant transformation and expression vectors.In:Plant Molecular Biology Labfax(Croy RRD ed)Oxford,BIOS Scientific Publishers,pp121−148)に記載されている。適切なベクターは、植物ウイルス由来のベクターを含んでもよい(例えば欧州特許EP−A−194809を参照のこと)。所望の場合、選択可能な遺伝子マーカーは、抗生物質または除草剤(例えば、カナマイシン、ハイグロマイシン、ホスフィノトリシン、クロルスルフロン、メトトレキサート、ゲンタマイシン、スペクチノマイシン、イミダゾリノンおよびグリホサート)に対する耐性などの選択可能な表現型を与えるものなどの構築物に含まれてもよい。
【0117】
核酸は、その天然の遺伝子導入の能力を利用するアグロバクテリウムによって運搬される安全化されたTi−プラスミドベクター(欧州特許EP−A−270355、欧州特許EP−A−0116718、NAR12(22)8711−87215 1984;the floral dip method of Clough and Bent,1998)、粒子衝突または微粒子銃(米国特許第5100792号、EP−A−444882、EP−A−434616)、微量注入法(WO92/09696、WO94/00583、欧州特許EP331083、欧州特許EP175966、Greenら(1987)Plant Tissue and Cell Culture,Academic Press)、エレクトロポレーション(欧州特許EP290395、WO8706614 Gelvin Debeyser)、直接的なDNA取り込みの他の形態(独国特許第4005152号、WO9012096、米国特許第4684611号)、リポソームを介したDNA取り込み(例えば、Freemanら、Plant Cell Physiol.29:1353(1984))、または渦法(vortexing method)(例えば、Kindle,PNAS U.S.A.87:1228(1990d)Physical methods for the transformation of plant cells are reviewed in Oard,1991,Biotech.Adv.9:1−11)などの任意の適切な技術を用いて植物細胞内に組み込むことができる。Ti−プラスミド、特に2分節のベクターは、以下により詳細に記載される。
【0118】
アグロバクテリウム形質転換は、双子葉植物の種を形質転換するために、当業者によって広範に使用されている。しかしながら、ほとんど全ての経済的に関係のある単子葉植物における安定で、繁殖性のトランスジェニック植物の所定の産生にもかなり成功している(例えば、Hieiら(1994)The Plant Journal 6,271−282を参照のこと)。アグロバクテリウム単独が、非効率かまたは効果がない場合、微粒子銃、エレクトロポレーション、および直接的なDNA取り込みが好ましい。あるいは、異なる技術の組み合わせ、例えば、アグロバクテリウムでコートされた微小粒子での照射(欧州特許EP−A−486234)または損傷後のアグロバクテリウムとの共培養を誘発する微粒子銃(欧州特許EP−A−486233)が、形質転換プロセスの効率を向上させるために利用されてもよい。
【0119】
形質転換技術の特定の選択は、特定の植物種を形質転換するその効率性ならびに特定の方法を選択する本発明を実施する人の経験および好みによって決定される。
【0120】
核酸を植物細胞に導入する形質転換系の特定の選択は、本発明に必須ではなく、または本発明の限定ではなく、植物再生についての技術の選択も本発明に必須ではなく、または本発明の限定でもないことは当業者には明らかであろう。本発明者らによって実施される実験において、向上した発現の効果は、T−DNAの種々の組み込みパターンにおいて見られる。
【0121】
従って、本発明の種々の態様は、植物組織(例えば植物細胞)への本発明の構築物の導入、ならびに本発明による核酸をゲノムへ導入するためにベクターと植物細胞ゲノムとの間の組み換えを生じることまたは可能にすること、を含む植物細胞を形質転換する方法を提供する。これは、一過性発現をもたらすように行われ得る。
【0122】
あるいは、植物組織の形質転換後、植物は、当該分野において標準的な例えば、単一の細胞、カルス組織または葉片から再生されてもよい。ほとんどのあらゆる植物は、植物の細胞、組織および器官から完全に再生され得る。利用可能な技術は、Vasilら,Cell Culture and Somatic Cell Genetics of Plants,Vol I,IIおよびIII,Laboratory Procedures and Their Applications,Academic Press,1984,ならびにWeissbachおよびWeissbach,Methods for Plant Molecular Biology,Academic Press,1989に概説されている。
【0123】
繁殖性のトランスジェニック植物の産生は、コメ、トウモロコシ、コムギ、カラスムギ、およびオオムギなどの穀物、さらに多くの他の植物種においてなされている(Shimamoto,K.(1994)Current Opinion in Biotechnology 5,158−162;Vasilら(1992)Bio/Technology 10,667−674;Vainら,1995,Biotehnology Advances 13(4):653−671;Vasil,1996,Nature Biotechnology 14,702ページに概説されている)。
【0124】
再生された植物またはその一部は、クローン、種子、自己またはハイブリッド子孫(progenyおよびdescendant)(例えば、F1およびF2子孫)、挿し木(例えば、食用部分)、栄養繁殖体などを提供するために使用されてもよい。
【0125】
本発明はさらに、トランスジェニック生物(例えば、本明細書に記載される方法によって得たか、または得られる)を提供し、このトランスジェニック生物に発現ベクターまたはカセットが導入され、そのカセットにおける異種遺伝子が向上したレベルで発現される。
【0126】
本発明はさらに、目的のタンパク質を産生する方法を含み、その方法は、上記の方法を実施する(または生物を使用する)工程と、必要に応じて少なくとも、目的のタンパク質が発現される組織を収集する工程と、その組織から目的のタンパク質を単離する工程を含む。
【0127】
従って、具体的には、本発明は、トランスジェニック植物または本発明の発現ベクターで一過性にトランスフェクトされた植物細胞を提供する。
【0128】
さらなる態様において、本発明はまた、トランスジェニック植物または本発明の発現ベクターで安定に形質転換された植物細胞を提供する。
【0129】
本発明はまた、挿し木、種子などを含む、複製または増殖、有性または無性に使用され得る任意の部分である、そのような植物由来の植物栄養繁殖体を提供する。本発明はまた、上記の植物細胞または異種DNAを含むそれらの植物の任意の部分を提供する。
【0130】
従って、種々の態様において、(限定されないが)本発明は以下を提供する。
・本発明のエンハンサー配列から構成されるか、またはエンハンサー配列から実質的に構成される核酸(そのエンハンサー配列は、(例えば)CPMV RNA−2ゲノムセグメントのヌクレオチド1〜512から構成されるか、あるいはそれら由来であるか、または2分節のRNAウイルスの別のRNA−2ゲノムセグメント由来であり、各々の場合において、CPMV RNA−2の161位に対応する標的開始部位が変位される)。
・例えば、目的のタンパク質、またはポリリンカー、および必要に応じてターミネーターをコードするORFの上流にそのようなエンハンサー配列を含む遺伝子発現系。
・本明細書に記載されるエンハンサー配列を使用するために本発明に従って修飾されたWO/2007/135480に記載される2分節の発現系。
・(i)作動可能に連結されるプロモーター、(ii)上記のエンハンサー配列、(iii)発現することが望まれる目的のポリリンカーまたは遺伝子、(iv)同系の3’UTR(すなわち、CPMV RNA−2ゲノムセグメントの3’UTR由来)、(v)終結配列を含む、発現カセット。
・本発明の遺伝子発現系またはベクターを用いる植物などの宿主生物におけるタンパク質、例えば異種タンパク質を発現する方法。
・本発明の遺伝子発現系またはベクター由来のタンパク質を発現する宿主細胞および生物(例えば、植物または酵母)ならびにそれらを産生する方法。
【0131】
文脈が他に要求しない限り、「遺伝子」とは、ペプチド、ポリペプチドまたはタンパク質への翻訳のための遺伝情報をコードする任意の核酸を指す。従って、文脈が他に要求しない限り、それは「ORF」と交換可能に用いられる。
【0132】
発現することが所望され得る遺伝子は、導入遺伝子または内因性であってもよい。
【0133】
目的の遺伝子には、農業形質、虫害抵抗性、病害抵抗性、除草剤抵抗性、無菌性、粒子特性などをコードする遺伝子が含まれる。この遺伝子は、油、デンプン、炭水化物、栄養素などの代謝に関与してもよい。従って、目的の遺伝子または特徴としては、限定されないが、環境またはストレス関連の形質、疾患関連の形質、および農業生産力に影響を与える形質が挙げられる。標的配列にはまた、タンパク質、ペプチド、脂肪酸、脂質、ワックス、油、デンプン、糖、炭水化物、香味料、香気、毒素、カロテノイド、ホルモン、ポリマー、フラボノイド、貯蔵タンパク質、フェノール酸、アルカロイド、リグニン、タンニン、セルロース、糖タンパク質、糖脂質などの合成に関与する遺伝子が含まれる。
【0134】
最も好ましくは、単子葉植物および/または双子葉植物における標的遺伝子としては、セイヨウアブラナ、ヒマワリ、ダイズおよびトウモロコシなどの植物中の油産生に関与する酵素;ジャガイモ、トウモロコシ、穀物などの植物中のデンプン合成に関与する酵素;医薬製剤または獣医用製剤などの天然の薬剤を合成する酵素、またはそれらのタンパク質自体をコードする遺伝子が挙げられる。
【0135】
異種核酸は、とりわけ、細菌、真菌、植物または動物起源の遺伝子をコードすることができる。ポリペプチドは、(例えば、害虫感受性、活力、組織分化、繁殖性、栄養価などに関連する植物の特徴を修飾するために)植物体で利用されてもよいか、または植物は、他に使用するためにその植物から精製できるポリペプチドを産生するための中間体であってもよい。そのようなタンパク質としては、限定されないが、網膜芽細胞腫タンパク質、p53、アンギオスタチン、およびレプチンが挙げられる。同様に、本発明の方法は、哺乳動物の調節タンパク質を産生するために使用されてもよい。目的の他の配列としては、タンパク質、ホルモン、成長因子、サイトカイン、血清アルブミン、ヘモグロビン、コラーゲンなどが挙げられる。
【0136】
従って、標的遺伝子またはヌクレオチド配列は、好ましくは、目的のタンパク質:虫害抵抗性タンパク質;病害抵抗性タンパク質;除草剤抵抗性タンパク質;哺乳動物タンパク質をコードする。
【0137】
「ベクター」は、とりわけ、二本鎖または一本鎖の直線状または環状形態で任意のプラスミド、コスミド、ファージ、ウイルスまたはアグロバクテリウムバイナリーベクターを含むと定義され、それは、自己感染性または可動性であってもなくてもよく、細胞ゲノムまたは存在する染色体外への組み込みによって原核生物または真核生物宿主のいずれかを形質転換できる(例えば、複製起点を有するプラスミドの自立的複製)。使用される構築物は、完全または部分的に合成される。特にそれらは、天然に一緒に見出されない(隣接しない)核酸配列が連結されるか、またはそうでなければ人工的に合成される組み換え体である。他に指定されない限り、本発明によるベクターは、特に、ベクターが、ゲノムへの組み込みのために核酸を細胞に組み込むために使用される場合、必ずしも、プロモーターまたは他の調節配列を含まなくてもよい。
【0138】
「バイナリーベクター」は当業者に周知であり、バイナリーベクター系には、(a)所望のヌクレオチド配列の植物細胞ゲノムへの移入を可能にする境界配列;(b)(i)作動可能に連結される植物活性プロモーターと、(ii)適切な場合、標的配列および/またはエンハンサーの発現カセットと、を一般的に含む所望のヌクレオチド配列自体、が含まれる。所望のヌクレオチド配列は境界配列の間に位置し、適切な条件下で植物ゲノム内に挿入できる。バイナリーベクター系は、通常、組み込みをもたらすために他の配列(A.tumefaciens由来)を必要とする。通常、このことは、アグロバクテリウムによって媒介される一過性形質転換を使用する、いわゆる「アグロ浸潤法」の使用によりなされ得る。つまり、この技術は、核DNAに組み込む場合、そのDNA(「T−DNA」)の一部を宿主細胞へ移入するAgrobacterium tumefaciensの性質に基づく。T−DNAは、21〜23ヌクレオチド長付近の左および右の境界配列によって定義される。浸潤は、例えば、シリンジ(葉)または真空(完全植物)によってなされ得る。本発明において、境界配列は、通常、アグロ浸潤法によって植物物質へ組み込まれる1つ以上のベクターを有する所望のヌクレオチド配列(T−DNA)に含まれる。
【0139】
「発現カセット」とは、核酸が、微生物または植物細胞などの宿主細胞内の転写のための適切なプロモーターまたは他の調節エレメントの制御下であり、作動可能に連結される状態を指す。
【0140】
「プロモーター」は、下流に(すなわち、二本鎖DNAのセンス鎖の3’方向において)作動可能に連結されるDNAの転写が開始され得るヌクレオチド配列である。
【0141】
「作動可能に連結される」とは、同じ核酸分子の一部として、プロモーターから開始される転写のための安定な位置および方向に結合されることを意味する。
【0142】
目的の「植物」種としては、限定されないが、トウモロコシ(Zea mays)、Brassica sp.(例えば、B.napus、B.rapa、B.juncea)特に種油の源として有用なそれらのBrassica種、ムラサキウマゴヤシ(Medicago sativa)、コメ(Oryza sativa)、ライムギ(Secale cereale)、モロコシ(Sorghum bicolor、Sorghum vulgare)、ヒエ(例えば、トウジンビエ(Pennisetum glaucum))、キビ(Panicum miliaceum)、アワ(Setaria italica)、シコクビエ(Eleusine coracana))、ヒマワリ(Helianthus annuus)、ベニバナ(Carthamus tinctorius)、コムギ(Triticum aestivum)、ダイズ(Glycine max)、タバコ(Nicotiana tabacum)、Nicotiana benthamiana、ジャガイモ(Solanum tuberosum)、ピーナッツ(Arachis hypogaea)、綿(Gossypium barbadense、Gossypium hirsutum)、サツマイモ(Ipomoea batatus)、キャッサバ(Manihot esculenta)、コーヒー(Coffea spp.)、ココナツ(Cocos nucifera)、パイナップル(Ananas comosus)、シトラスの木(Citrus spp.)、ココア(Theobroma cacao)、茶(Camellia sinensis)、バナナ(Musa spp.)、アボガド(Persea americana)、イチジク(Ficus casica)、グアバ(Psidium guajava)、マンゴー(Mangifera indica)、オリーブ(Olea europaea)、パパイヤ(Carica papaya)、カシュー(Anacardium occidentale)、マカダミア(Macadamia integrifolia)、アーモンド(Prunus amygdalus)、サトウダイコン(Beta vulgaris)、サトウキビ(Saccharum spp.)、オートムギ、オオムギ、野菜、観賞植物、および針葉樹が挙げられる。当業者は、本明細書に記載されるウイルスベクターの向性が変化することは理解するだろう。しかしながら、そのようなウイルスに対する感受性を決定することは、当業者に周知である。さらに、植物細胞内へのウイルス浸入を促進するレセプターを組み換え発現することによってそのような特異性を変更することは可能であり得る。
【0143】
本発明は、ここで、以下の非限定的な図および実施例を参照して詳細に記載される。本発明の他の実施形態は、これらの観点から当業者が想到するだろう。
【0144】
本発明を実施する当業者に使用され得る限り、本明細書に引用される全ての参考文献の開示は、相互参照として本明細書に具体的に援用される。
【実施例】
【0145】
(実施例1)
1.1方法
発現ベクターFSC2およびその誘導体の作製
pBinP−1−GFPベースのプラスミド(Canizaresら,2006)由来の外来性タンパク質の発現のために有用なクローニングベクターを、カリフラワーモザイクウイルス(CaMV)35SプロモーターおよびpBinP−S2NT(LiuおよびLomonossoff,2002)由来のノパリン合成(nos)ターミネーターに隣接するRNA−2の完全な配列を切断し、それを、Ascl/Pacl断片として変異誘発プラスミドpM81W(LiuおよびLomonossoff,2006)に挿入することによって作製した。得られたプラスミド、pM81W−S2NTに、4つの変更を同時に導入する単一ラウンドの変異誘発(LiuおよびLomonossoff,2006における方法を参照のこと)を行って、pM81B−S2NT−1を得た。変異誘発により、ベクターバックボーンから2つのBspHI部位を除去し、AUG512付近にBspHI部位(T/CATGA)およびUAA3299後にStuI部位(AGG/CCT)、RNA−2によりコードされるポリタンパク質についての終止コドンを導入した。続いて、BamHI/Ascl断片を、pBinP−NS−1(Liuら,2005)から切断し、同様に消化したpM81B−S2NT−1にライゲーションして、pM81−FSC−1を得た。このベクターにより、AUG512の下流のRNA−2 ORFの全てを、BspHIおよびStuIでの消化により切断でき、BspHIおよびStuI(平滑)適合性末端((blunt)−compatible end)を有する任意の配列と置換できる。BspHI部位の使用は重要である。なぜなら、それは、512位のAUGを保存し、このイニシエーターは、挿入した遺伝子の翻訳を駆動するために使用されるからである。植物において外来性遺伝子を発現するために、pM81−FSC−1由来プラスミドはAsclおよびPaclで消化され、同様に消化されたpBINPLUSに移入された外来性配列を含む発現カセットを有する断片および得られるプラスミドは、最終的に、A.tumefaciensに形質転換される。
【0146】
クローニングの容易さを向上させ、適切な制限酵素の選択を広げ、外来性遺伝子発現に対するリーディングフレームの効果を調べるために、全てのRNA−2 ORFを短いポリリンカーと置換した。オリゴヌクレオチド挿入およびpM81−FSC−2を生じる部位特異的突然変異誘発法の組み合わせにより、NruI(TCG/CGA)およびXhoI(C/TCGAG)またはStuIのいずれかでのクローニングが可能となる。NruI部位の末端のアデニンは512位に存在するので、ここに見られるAUGを保存する。この修飾により、5’から512位のAUGまで即座にヌクレオチドを変更させるが、十分な状況が維持される。翻訳が、512位、513位、514位または515位のAUGから開始されるようにpM81−FSC−2内にGFPをクローニングすることにより、pM81−FSC−1由来の構築物であるpM81−FSC2−512、pM81−FSC2−513、pM81−FSC2−514、およびpM81−FSC2−515を得た。これらのpM81ベースのプラスミドは、発現カセットを含むクローニングベクターであり、次いで、これをバイナリーベクターに移入して、図2および3に示す実験に使用する発現ベクターFSC2−512、FSC2−513およびFSC2−514を産生した。CPMVの野生型RNA−2ゲノムセグメントならびにpM81−FSC1およびpM81−FSC2ベクターの配列間の差異は表3に示す。野生型CPMV配列と比較したベクター内でのヌクレオチドの変更は大文字として示す。
【0147】
(pM81−FSC−1について上で概説したように)pBINPLUS内の動員後のアグロバクテリウムによって媒介される一過性形質転換により、AUG161と下流のAUGとの間のフレーム連続性が維持されない場合、より低いタンパク質レベルが得られることが示された。AUG161および512に対して+1位および+2位から翻訳したGFPの量は著しく減少したが、+3位からの翻訳(つまり、515からでフレーム内の後方)は、512位のAUGからの翻訳と同じ効率であった。このことが、513位および(より少ない程度の)514位の弱まった状況に起因していないことを示すために、FSC2−513のような同じ不十分な状況ではなく、FSC2−515+を+3位から開始するために作製した。FSC2−515+からの発現は、FSC2−512または515から達成したものと同じ高さであり、これは、低い状況がFSC2−513および514からの発現の減少ではないことを示す。
【0148】
公知の機構によって、翻訳が第1のAUGルールを逃れることができることを考慮すると、フレーム連続性を必要とすることが知られておらず、欠失したRNA−2ベースのベクターからの効果的な翻訳が、AUG161と下流のAUGとの間のフレーム連続性に依存することは興味をそそられる。この減少を理解して、望ましくは克服するために、一連の変異体を、RNA−2の5’配列に対する修飾を用いて作製した。オリゴヌクレオチドの相補対(表2を参照のこと)を、pM81−FSC2−512、513および514の部位特異的突然変異誘発法に使用した。それらの変異体のAUG115(uORFについての開始コドン)、AUG161(uORFのアミノ酸配列の変更なし)のいずれか、またはそれらの上流の開始部位の両方を除去した。2つの変異を、U162Cオリゴを用いてA115G変異体を変異誘発することによって作製した(表2)。
【0149】
これらの変異体転写産物からの一過性発現を、以前のpM81−FSC−2構築物について記載したように行った。coomassie染色したSDS−PAGE(図2)または葉の全てを視覚化するためにUV照射(図3)を用いるこれらの変異体からのGFPの発現の分析により、161位のAUGの非存在下において発現の強力な増加が示される。さらに、AUG161単独またはAUG115および161の両方の除去は、AUG161と下流のAUGとの間のフレーム連続性に対する依存性を軽減する。対照的に、単なるAUG115の除去はこの依存性を高め、そして全体的に翻訳を抑制するように見える。結論として、uORFは、AUG161からの翻訳を下方制御するために機能するように見え、それは全体的に抑制性があり、フレーム連続性に対する依存性を与える。
【0150】
HBcAG粒子を含む樹液(sap)の電子顕微鏡法
図6に示した集合したHBcAg粒子の電子顕微鏡写真に使用した樹液(sap)は以下のように調製した。葉組織を、2容量のTris/NaCl緩衝液中に抽出し、100kDa MWCOカラムで濃縮せずにTEと交換した。HBcAgの最終濃度は、coomassie染色したSDS−PAGEゲルでの標準物との比較により判断した場合、約0.2mg/mlであった。
【0151】
1.2結果
(1)161位(AUG161)および512位(AUG512)における開始部位の相対位相を変化させる効果
これを達成するために、余分なヌクレオチドを、AUG512(FSC2−512)のすぐ上流に挿入して、AUGを513位(FSC2−513)、514位(FSC2−514)および515位(FSC2−515)に移動させた(図1)。AUG161と位相をずらしてAUG512(FSC2−513およびFSC2−514)を配置すると、蛍光(図3)およびCoomassie染色したゲル(図2)によって判断した場合、GFP発現は低かった。位相(FSC2−515)を元に戻すと、天然の状態(FSC2−512)で見られたレベルの発現に戻った。結論としては、AUG161が存在し、161位のAUGと位相が一致する場合、下流のAUGでの開始が最も効率的である。
【0152】
(2)161位(AUG161)および512位(AUG512)の開始部位の相対位相を変化させることに加えて115位(AUG115)の開始部位の除去
GFP発現がAUG512から駆動される場合、すなわち、この第2のAUGがAUG161と位相が一致する場合、AUG115の除去は、ほとんどまたは全く影響を与えない(図2および図3の標識した10のレーンを参照のこと)。しかしながら、第2のAUGがAUG161(513、514)と位相がずれる場合、AUG115の欠失は、実質的にGFP発現を生じない(図2および図3の標識した10のレーンを参照のこと)。結論:AUG115は、何らかの形で、AUG161を回避し、AUG512に到達することによってリボソームの能力に関与する。しかしながら、このことは、正確な位相に存在する下流のAUGを必要とする。
【0153】
(3)161位(AUG161)における開始部位の除去の効果
この変異の効果は信じられないほど劇的であり、GFP発現レベルが、AUG161が存在する場合に見られる量の20〜30倍に到達する(図2および図3における標識した01のレーンを参照のこと)。さらに、AUG161の非存在にもかかわらず、位相AUG512はもはや問題ではないように見え、位相の考えは大して重要ではない。さらに、AUG115の存在または非存在に差異はない(図2および図3における標識した11のレーンを参照のこと)。DsRedおよびHBcAg発現についてのdelRNA−2(図1における発現ベクター00[FSC2−512])構築物を用いる場合、5日以内に、浸潤したパッチは膨圧を失い、クロロティック(chlorotic)(薄い)になった。7日までに、組織は灰色のように見え、完全に死んだ。HT(図1における発現ベクター01[FSC2−512])を用いる場合、組織は7日後に膨張し、ストレスのサインのみが、HBcAg発現組織をわずかに白化する。2G12 IgGの重鎖が、delRNA−2によって発現され、ERに残存する場合、白化は7日後に明らかであるが、HTについては、これは観測されない。従って、異種タンパク質を発現するためにHTを用いる場合、植物に見られる壊死のレベルは、より高いレベルの異種タンパク質発現が達成されるにもかかわらず、異種タンパク質を発現するためにdelRNA−2を用いる場合より非常に低い。
【0154】
考察
非常に高レベルの外来性遺伝子発現が、AUG161を欠失することによってdelRNA−2構築物から発現され得る。現在のところ、GFPを用いて、本発明者らは、全可溶性タンパク質(TSP)のうちの25〜30%または1Kgの葉あたり約1gの発現されたタンパク質のレベルと測定する。これは非常に高いレベルであり、かつ、本発明者らが用いるアプローチは非常に簡単である。本発明者らがもはやリーディングフレームを保存する必要がないという事実は、ポリリンカーを有する、使用者によって使いやすいベクターが産生され得るということを意味する。
【0155】
(実施例2)
2.1背景
実施例1に記載したように、効果的な発現に必要なCPMC RNA−2の5’非翻訳領域(UTR)に必要な特徴を調べるために、本発明者らは、主要な開始部位の上流の5’リーダー配列内に見られる2つのAUGコドンの役割について扱った。本発明者らは、フレーム内の主要な翻訳開始部位(512)の上流の開始コドン(161)の欠失が、外来性タンパク質蓄積の大幅な増加をもたらすことを実証した。
【0156】
この系を用いて、本発明者らは、浸潤後6日によって、完全なサイズのIgGおよび自己集合性ウイルス様粒子を含む、多くの関係のないタンパク質が、それぞれ、全抽出タンパク質のうちの10%より多く、および20%発現されたことを示した。従って、この系は、転写産物のウイルス複製に頼らない高レベルの発現のための理想的な媒体を提供する。
【0157】
この新規の系(図1における発現ベクター01[FSC−512]によって例示される)を、高翻訳可能(hyper−translatable)なササゲモザイクウイルス(Cow Pea Mosaic Virus)タンパク質発現系のための「CPMV−HT」と呼ぶ。
【0158】
HT−CPMV系は、タンパク質レベルの劇的な増加を示すので、植物における外来性タンパク質の迅速で、高レベル発現のための優れた方法である。
【0159】
バイナリーベクターの増殖アレイは、過去25年にわたって植物形質転換のために開発されている(Hellensら,2000b;Veluthambiら,2003;LeeおよびGelvin,2008)。従って、これらの開発の主な目的は、例えば、アグロバクテリウムのための宿主範囲を拡大すること(Hieiら、1994)、選択マーカー、発現カセットおよび融合タンパク質の選択を可能にする一連のベクターを作製すること(オープンソースバイナリーベクターのpCAMBIA範囲;http://www.cambia.org/daisy/bioforge_legacy/3725.htmlによって例示される)、または外来性DNA組み込みおよびマーカーフリーの形質転換を最小化するための系を開発すること(例えばpCLEAN;Tholeら、2007)によって、安定な組み込みを向上させることにかなり焦点をあてられている。
【0160】
バイナリーベクターはまた、同じ導入遺伝子の複数の組み込み事象の頻度を低減させるために、低コピー数で複製するために操作される。なぜならこれにより、遺伝子サイレンシングが導かれ得るからである(JohansenおよびCarrington、2001)。
【0161】
しかしながら、一過性発現に関して、宿主核への効果的な組み込みを確実にすることおよび植物体選択におけるマーカーの存在は厳格に必要とされていない。さらに、アグロ浸潤法(agro−infiltration)の際に、各細胞は、ゲノムを組み込まなくても核において転写に適合すると考えられるT−DNA分子で満たされる(JanssenおよびGardner,1989;Narasimhuluら,1996)。このことは、一過性発現が、より高いコピー数のバイナリープラスミドから利益を受けることができることを示唆する。
【0162】
バイナリーベクターの改良の別の領域は、ベクターバックボーンのサイズの減少である。より小さなプラスミドの利点を実証し続けている2つの優れた例は、pPZP(Hajdukiewiczら、1994)およびpGREEN(Helensら、2000a)である。クローニング手順および細菌形質転換の効率を向上させることに加えて、これらのベクターは、単一のT−DANに存在する複数のカセットに頼る発現系のための鋳型を提供している(Tzfiraら,2005;Tholeら,2007)。
【0163】
本実施例は、クローニングベクター(例えばpM81−FSC2)と、発現系(例えばPBINPLUS)との間の発現カセットのサブクローニングを必要とするよりむしろ、単一の工程でなされるクローニングを可能にすることによって、このベクターの実用的用途を容易にするそのベクターの進歩性のある改良を開示する。より具体的には、本明細書の結果は、複製およびTDNA移入に関する実施を妥協することなく、pBINPLUSのサイズを劇的に減少させることが可能であったことを示す。さらに、CPMV−HT系の要素は、モジュール方式で、得られるベクターに組み込まれるので、複数のタンパク質が単一のT−DNAから発現され得る。これらの改良により、外来性遺伝子の効率的な直接クローニングを可能にする多用で、高レベルの発現ベクターの作製がもたらされる。
【0164】
2.2材料および方法
pBD−FSC2−512−U162C(HT)は、pBINPLUSのPacI/AscI部位に挿入されるFSC2−512−U162Cカセット(実施例1を参照のこと)を含む(van Engelenら、1995)。このプラスミドの本質的なセグメント(以下を参照のこと)を、再ライゲーションのための特有の制限酵素部位をコードするオリゴヌクレオチドを用いて、高忠実度のポリメラーゼ、PHUSION(New England Biolabs)で増幅させた(表4.1)。T−DNA領域を、特有のAhdI部位(pBD−LB−F)の上流の配列と相同のセンスプライマーおよびApaI部位(pBD−RB−ApaI−R)を含むアンチセンスプライマーで増幅させた。ColEI複製起点、NPTIII遺伝子、およびTrfA遺伝子座を含む領域を、ApaI部位(pBD−ColEI−ApaI−F)を含むセンスプライマー、およびSpeI部位(pBD−TrfA−SpeI−R)を含むアンチセンスプライマーで増幅させた。RK2複製起点(OriV)を、SpeI部位(pBD−oriVSpeI−F)を含むセンスプライマーおよびAhdI部位(pBD−oriV−AhdI−R)を含むアンチセンスプライマーで増幅させた。精製後、それらの産物を、それらの末端でコードされ、3点のライゲーションのために混合した特有の制限部位に従って消化した。これにより、プラスミドpEAQbetaが得られ、そのためのライゲーション結合を、シークエンシングにより確認した。CPMV−GFP−HTカセットのnosターミネーターの一部を除去したT−DNAからの約1.2kbの欠失を検出した。従って、プライマーpBD−ColEI−ApaI−FおよびpMini>pMicroBIN−Rを用いて、右側の境界を含むpEAQbetaバックボーンのように、pBD−FSC2−GFP−HTからの右側の境界を含むターミネーターの一部を、プライマーpMini>pMicroBIN−F2およびpBD−RB−ApaI−Rで再び増幅させた(表4.1)。精製した産物を、ApaIおよびFseIで消化して、pEAQを得るためにライゲーションした(図9)。
【0165】
35Sプロモーターおよび35Sターミネーターに隣接するP19遺伝子を、プライマーとして35SP19−PacI−Fおよび35SP19−AsclR、または35SP19−FseI−Fおよび35S−P19−FseI−Rのいずれかを用いてpBIN61−P19(Voinnetら、2003)から増幅させた(表4.1)。nosプロモーターおよびターミネーターに隣接するNPTII遺伝子を、プライマーpBD−NPTII−FseI−FおよびpBD−NPTII−FseI−Rを用いてpBD−FSC2−GFPHTから増幅させた(表4.1)。A−テイリング(tailing)後、増幅させたカセットを、pGEM−T easy(Promega)にライゲーションした。FseIを用いてpGEM−T easyから切断したP19カセットを、FseIで消化したpEAQ−GFP−HTにライゲーションして、pEAQexpress−GFP−HTを得た。FseIで切断したNPTIIカセットを、両方向においてFseIで消化したpEAQ−GFP−HTにライゲーションして、pEAQselectK−GFP−HTおよびpEAQselectK(rev)−GFP−HTを得た。NPTIIカセットをまたPacl/Asclで切断し、pEAQselectK−GFP−HTのAsiSI/Mlul部位にライゲーションして、pEAQspecialK−GFP−HTを得た。pGEM−T中のP19を、QUICKCHANGE法(Stratagene)により部位特異的突然変異誘発し、プライマーP19−R43W−FおよびP19−R43W−Rを用いてアルギニン43をトリプトファン残基に変換した。変異P19カセットをPacl/Ascl消化で解放し、pEAQselectK−GFP−HTのAsiSI/Mlul部位に挿入して、pEAQspecialKm−GFP−HTを得た。
【0166】
短いポリリンカーのセンスおよびアンチセンス鎖をコードするオリゴヌクレオチド(表4.1)をアニールし、5’末端におけるNruI部位の下流の半分および3’末端におけるXhoIのものと適合するオーバーハングを残した。アニールしたオリゴを、NruI/XhoIで消化したpM81−FSC2−A115G−U162C(上記を参照)とライゲーションして、pM81−FSC2−POWを得た。NruI部位を、プライマーP19−ΔNruI−FおよびP19−ΔNruI−Rを用いて部位特異的突然変異誘発(QUICKCHANGE;Stratagene)によりpGEM−T中のP19カセットから除去し、pEAQselectK−GFP−HTのAsiSI/Mlul部位に再挿入して、pEAQspecialK−GFP−HTと比べて発現の減少を示さないpEAQspecialKΔNruI−GFP−HTを得た(データは示さず)。次いで、pM81−FSC2−POW由来のPacl/Ascl断片を解放し、同様に消化したpEAQspecialKΔNruI−GFP−HTに挿入し、それにより、GFP HT発現カセットを置換し、pEAQ−HTを得た。GFPを、pEAQ−HT:GFP−AgeI−FおよびGFP−XhoI−R;GFP−AgeI−FおよびGFP−XmaI−R;ならびにGFP−XmaI−FおよびGFP−XhoI−Rに挿入するために3つの組み合わせで4つのプライマーセット(表4.1)を用いてpBD−FSC2−GFP−HTから増幅させた。精製したPCR産物を、それらのプライマーに特有の酵素で消化し、適切に消化したpEAQ−HTに挿入して、pEAQ−HT−GFP、pEAQ−HT−GFPHisおよびpEAQ−HT−HisGFPを得た。
【0167】
表4.1.増幅および変異誘発に使用したオリゴヌクレオチド。制限酵素部位またはその部分は、下の例で示し、変異はイタリックで下線をひいた。
【表1−1】

【0168】
【表1−2】
【0169】
【表1−3】
【0170】
2.3 結果
2.3.1 pBINPLUSは、少なくとも7.4kbの外来性配列を含む
CPMV−HTからの発現は、非常に高レベルの組み換えタンパク質の産生を可能にする。それにも関わらず、一過性形質転換のための系およびその使用をさらに改良することが望まれた。
【0171】
改良の第1の領域は、小さいプラスミドが、ライゲーション反応および細菌形質転換手順において大きいものより効果的であるという事実に関する。小さいバイナリーベクターの構造との比較により、pBINPLUSが、外来性配列の有意な量を含む可能性があることが示された。pBINPLUSの4つの要素は、バイナリーベクターとしての適切な機能:T−DNA、RK2(OriV)広範な宿主範囲の複製起点、カナマイシン耐性を与えるNPTIII遺伝子(Trieu−CuotおよびCourvalin,1983)、および複製を促進するRK2由来のTrfAに必須であると決定した(図9)。残存しているバックボーン部分のバイオインフォマティクス分析により、それらが、親プラスミド内の適切な制限部位の存在で信頼される、pBIN19の構築物の人工物であることが示される(Bevan,1984)。これらの観測は、pBIN19の完全なシークエンシングでの報告により確認される(Frischら、1995)。pBINPLUSは、E.coliにおけるより高いコピー数のための必須でないColEI複製起点を含む。約2.6kbの余分なDNAを、T−DNA内に見出すことができる。これは、一過性発現に必要ではない植物形質転換のためのNPTII選択マーカーを含む。全体として、pBINPLUS内の外因性配列の全量は、7.2kbを超えているようである。
【0172】
2.3.2 pEAQシリーズの構築物
ベクターに対する修飾を生じる発現の効果をモニターするために、本発明者らは、pBINPLUS由来プラスミド、pBD−FSC2−512−U162C(HT)で開始することを選択した。T−DNA、RK2(OriV)複製起点、ならびにColEI起点、NPTIIIおよびTrfAを含むセグメントからなる3つの領域を、pBD−FSC2−GFP−HTからPCRによって増幅させた。これらの3つのフラグメントのライゲーションにより、プラスミドpEAQbeta(図9)が得られ、それは、その親プラスミドより4584bp小さい。pEAQbetaのPCR増幅のさらなるラウンドにより、T−DNA領域由来の2639bpの必須でない配列を除去し、3つの特有の制限部位、AsiSI、MlulおよびFseIを挿入した。AsiSI/Mlul消化は、Pacl/Ascl断片の挿入と適合するので、全ての以前のCPMVクローニングベクターから複数のカセットをクローニングするのに非常に有用である。FseIは、異なる選択マーカーまたはサイレンシング抑制因子カセットを置き換えるのに有用な特有の8塩基の認識部位を提供する。得られるpEAQGFP−HTプラスミドは、pBINPLUSのサイズの半分より小さく、CPMVHT発現カセットを有さず、5137bpのみで、最も小さい公知のバイナリーベクターの1つである(図9)。pEAQプラスミド全体の配列を決定し、RK2複製起点が以前に報告されたもの(Frischら、1995)と逆方向であることを発見したので、pEAQ−GFP−HTの正確な方向を示す。
【0173】
pEAQ−GFP−HTを、T−DNA内に種々のさらなる特徴を包含するための開始点として用いた(図10)。pBINPLUS由来のNPTIIカセットを、GFP−HTカセットに対して順方向および逆方向の両方においてpEAQのFseI部位に再挿入して、pEAQselectK−GFP−HTおよびpEAQselectK(rev)−GFP−HTを得た。
【0174】
35S−P19カセットをFseI部位に挿入して、pEAQexpress−GFP−HTを得た。最終的に、35S−P19カセットを、pEAQselectK−GFP−HTのMlu/AsiSI部位に挿入して、pEAQspecialK−GFP−HTを得た。このように、一過性発現を容易かつ迅速にするための一連の小さなバイナリーベクターを構築した。
【0175】
2.3.3 サイズの減少は、pEAQからの一過性発現を妥協しない
pEAQシリーズのベクターのアグロ浸潤法(agro−infiltration)により、サイズの大幅な減少が一過性アッセイにおいて発現レベルをほとんど妥協しないことが示される。pEAQ−GFP−HT、およびpBIN61−P19によって提供されるP19を有するpEAQselectK(rev)−GFP−HTの同時浸潤は、pBD−FSC2−512−HTおよびP19の同時浸潤とほとんど変わらない発現レベルを生じる。これは、UV照射下(図11A)、SDS−PAGE(図11B)、およびタンパク質抽出物におけるGFPのスペクトル蛍光測定(図11C)で見られ得る。興味深いことに、T−DNA内のNPTIIカセットの方向は、発現レベルに影響を与えるようである。pEAQselectKは、GFP蓄積の減少を生じる他の同等のpEAQselectK(rev)と比べて著しい向上を示す。
【0176】
理論的には、pEAQへのサイレンシングカセットの抑制因子の組み込みは、同じT−DNAから発現される外因性遺伝子からの一過性発現レベルを向上させるその能力に影響を与えるはずはない。実際には、pEAQexpress−GFP−HT単独の浸潤はまた、pBD−FSC2−GFP−HTと同様、またはそれより高い発現レベルを生じる(図7.3)。さらに、pEAQexpressの効果を試験するために、アグロバクテリウム培養物を2倍希釈し、それにより、最終的な吸光度(OD)は、同時浸潤の各々の個々の培養物のものであった。
【0177】
予想されるように、これは、同様の高い発現レベルを生じ、同じT−DNA上の目的の遺伝子およびサイレンシングの抑制因子の両方の組み込みが、アグロバクテリウムの半分の量の使用を可能にすることを示す(図11)。従って、全ての要素が同じT−DNAに存在する場合、CPMV−HTは、高レベルの外来性タンパク質を発現させるために使用できる。
【0178】
2.3.4 変異P19は一過性アッセイにおける導入遺伝子のサイレンシングを抑制できる
T−DNA内のNPTIIカセットの順方向によって得られる発現の増加を利用するために、P19カセットを、pEAQselectK−GFP−HTにおけるAsiSIとMlul部位との間に挿入して、pEAQspecialK−GFP−HTを得た(図10)。GFPの配列として同じT−DNA上のP19の存在により、P19で同時浸潤したpEAQselectK−GFPHTと同様のレベルの発現が生じる(図12)。これは、pEAQexpress−GFP−HTによって産生される発現より多く、NPTIIカセットの存在に起因するように見える(図12)。対照的に、pEAQexpressからの低い発現は、T−DNA内のP19カセットの異なる位置および方向に起因し得る。それにも関わらず、pEAQexpressと同様に、pEAQspecialKベクターは、2つの培養物が同時浸潤されなければならない場合に使用されるものの最終的なODの半分のアグロバクテリウムの懸濁液で高レベルの発現を与える。
【0179】
外来性遺伝子発現カセットと、P19カセットおよび選択マーカーを組み合わせることにより、トランスジェニック植物におけるCPMV−HTの機能を試験することが可能となる。しかしながら、P19などのサイレンシングの抑制因子の構成的発現は、発生プロセスに関連する内因性遺伝子サイレンシングとのそれらの干渉に起因する厳しい表現型を生じ得る(SilhavyおよびBurgyan,2004)。最近、特徴付けられたP19の変異(R43W)は、内因性遺伝子サイレンシングに対して減少した活性を有することが提案されているので、安定な形質転換における導入遺伝子サイレンシングの抑制のためのより良い候補となり得る(Scholthof、2007)。CPMV−HT系を用いる安定な形質転換の実行可能性を調べるために、野生型および変異型の両方のP19を、pEAQselectK−GFP−HTのT−DNAに挿入して、一過性に変異体をアッセイした。示すように、浸潤した葉のUV照射、タンパク質抽出物のSDS−PAGE、およびGFPレベルの蛍光分光測定によって、pEAQspecialKmの変異体P19は、pEAQspecialKの野生型P19の外来性遺伝子の発現の向上の効率の約半分である(図12)。これは、導入遺伝子のサイレンシングを抑制する能力に対するP19におけるR43W変異の効果の第1の研究を表す。
【0180】
(実施例3)
単一のプラスミドからの高レベルIgG発現
pEAQシリーズ、ヒト抗HIV IgGのERを保有する重鎖(HE)および軽鎖(L)を含むCPMV−HT発現カセットのモジュールの性質を利用するために、2G12を、pEAQexpressのPacl/AsclおよびAsiSI/Mlul部位に挿入した。挿入部位が発現レベルに影響を与えるか否かを決定するために、LおよびHE鎖を、pEAQex−2G12HELおよびpEAQex−2G12LHEを生じる両方の位置に挿入した(図13A)。上記のプラスミドを含む単一のアグロバクテリウム培養物でのN.benthamianaの浸潤により、各々が個々の要素、L、HEおよびP19を発現する3つのアグロバクテリウム培養物を混合することにより産生された2G12とサイズが同一の完全に集合した2G12抗体の形成が生じた(図13C)。各レーンに負荷したタンパク質は、90mgの浸潤した組織から得た抽出物の1/30または1gの組織から潜在的に得られるタンパク質の1/333を示す。3つの株の混合物から産生された集合したIgGの最大量は、coomassie染色した減少していないSDS−PAGEゲルでの1μgのCHOにより産生された2G12に相当する。このことは、325mg/kgの新鮮な重量の組織を超える2G12の発現レベルを示唆し、SPRにより測定された濃度と一致する。pEAQex−2G12HELの使用は、このすでに高レベルの抗体の蓄積を抑制するように見える。
【0181】
IgGなどの複数鎖タンパク質の各要素であるpEAQ由来のベクターの利点は、自動的に各感染細胞に送達され得ることである。従って、複数の培養物が使用されなければならない場合より、高発現レベルが、より高い希釈のアグロバクテリウム懸濁液で維持されなければならない。実際にこれが起こる場合を試験するために、最初にOD1.2に再懸濁し、必要な場合、混合した培養物を、2回連続して3倍希釈した(図13B)。これにより、3つの培養混合物において、各々最終ODが0.4、0.13および0.04である個々の培養物が得られた。pEAQexpress構築物を有する単一の培養物を、1.2、0.4および0.13のODで浸潤した。3つの別の培養物を使用した場合、集合した2G12のレベルは連続希釈で顕著に減少する。対照的に、pEAQex−2G12HELおよびpEAQex−2G12LHEからの2G12発現を、非常に少しの希釈でいくらか減少させて、常に高レベルに維持した(図13C〜E)。希釈に対する感受性の欠如により、同じT−DNAに全ての3つの発現カセットを配置することにより得られる向上した効果を確認する。興味深いことに、浸潤組織から抽出した全タンパク質の量は、浸潤物のODが1.2〜0.4まで減少した場合、ほとんど半分であった。これは、より高いODの懸濁液が浸潤される組織由来の抽出物中のタンパク質の有意な画分が、より高い濃度のアグロバクテリウムに対して産生されるアグロバクテリウム由来のタンパク質または植物タンパク質から構成され得ることを示唆する。
【0182】
図13Cの検査は、T−DNA内のカセットの相対位置が発現レベルに影響を与え得ることを示唆する。pEAQex−2G12LHEからの全発現は、pEAQex−2G12HELからのものよりわずかに低い。このことは、減少していない試料のウェスタンブロッティングにより確認し、また、分解産物および組み込まれていない免疫グロブリン鎖の量においていくらかの差異が示された(図13C〜E)。pEAQex−2G12LHEで浸潤した組織は、約70〜80kDaの重鎖に特異的な分解産物を欠いているように見える(図13D)。また、HL2集合中間体および十分に遊離している軽鎖がほとんどないように見える(図13E)。重鎖は、植物中の2G12集合において制限されることが知られており(Markus Sack,pers.comm.,RWTH,Aachen,Germany)、これは、全ての試料における認識可能な遊離重鎖の欠如によって確認されているため、これらの結果は、pEAQex−2G12LHEが、pEAQex−2G12HELより少ない重鎖を産生することを示す。これは、T−DNAの左側の境界に近いCPMV−HTカセットからの減少した発現に起因し得る。
【0183】
他の実施形態において(データは示さず)、CPMV−HT系はまた、以下を発現するために、N.benthamianaの一過性形態において首尾よく使用される。
・ブルータングウイルス(Bluetongue Virus)(血清型10)VP2、VP3、VP5、VP7およびNS1。
・ロタウイルスNSP5
・Medicago truncatula由来のカルモジュリン(これは後で精製された)
・発現するのが困難なヒトFcγ受容体1(CD64)の細胞外ドメイン−これは精製され、抗体結合研究において機能的であることが示されている。
・CPMV小(S)および大(L)コートタンパク質が同時発現され、ウイルス様粒子内で集合することが示されている(データは示さず)。
【0184】
(実施例4)
CPMV−HT発現ベクターへの直接クローニング
単一のプラスミド上に系の要素を組み合わせるが、上記のベクターはなお、バイナリープラスミドに発現される配列を組み込むために2段階のクローニング手順を必要とした。本実施例は、目的の遺伝子が直接挿入され得るバイナリープラスミドを提供する。そのプラスミドはpEAQベースのプラスミドへ直接挿入できないポリリンカーを組み込むが、所望の場合、CまたはN末端ヒスチジンタグの融合も可能にする(pEAQ−HT;図14A)。ポリリンカーをまず、アニールしたオリゴヌクレオチドとしてpM81−FSC2−512(A115G)(U162C)に挿入し、pM81−FSC−POWを得た。この構築物はなお、複数のポリペプチドを発現させるためのpEAQベースの構築物を発生させるための標準的な2段階クローニング手順に使用できる。さらに、二重変異した5’リーダーの使用は、単一の変異で得られるよりも、さらに高発現レベルを得ることができる。次いで、CPMV−HTカセットを、Pacl/Ascl部位を介してpEAQspecialKに移して、pEAQ−HTを得た。pEAQ−HTのポリリンカー内の全ての3つの位置へのGFPの挿入により、非標識GFP、および5’(HisGFP)および3’(GFPHis)His−タグ融合を得た。
【0185】
予想されるように、非標識GFPは、pEAQspecialK−GFP−HTで得られるものより高いレベルまで発現し、1.6g/kg FW組織を超えた(図14B)。これは、pEAQ−HTのCPMV5’リーダーが、AUG161に加えて除去される場合、AUG115を除去する余分な変異を含み、さらに発現を向上するという事実に起因するようである。
【0186】
ウェスタンブロッティングによって検出される場合、His−タグの存在により、ポリリンカーによってコードされるアミノ酸残基のNおよびC末端の両方において正確な融合を確認した。全ての3つのGFP変異体は抗GFP抗体で検出可能であるが、HisGFPおよびGFPHisのみが抗His抗体で検出可能であり(図14C)、His−タグの存在により、予想された量でSDS−PAGEにおいてGFPバンドの流動性が減少した。この標識はまた、蛍光の分析によって検出されるGFPの量を減少させた(図14B)。この効果はN末端Hisタグについてより顕著であった。coomassie染色したバンドの強度により、これは、GFPの蛍光特性を干渉するというよりむしろ、標識されたGFP蓄積(図14C)の減少を表すことが示唆される。それにも関わらず、Hisタグしたタンパク質のレベルは依然として非常に高く、1kgのFW組織あたり0.6および1.0のGFPを超えた。
【0187】
実施例2〜4の考察
CPMV−HT発現系の使用の容易さおよび機能を改良するために、ベクターのモジュラーセットが、容易かつ迅速な植物発現のために作製される。
【0188】
バイナリーベクター、pBINPLUS由来のプラスミドバックボーンの半分より多く、および一過性発現に必須ではないT−DNA領域の一部を除去することにより、発現レベルで妥協しない公知の最も小さいバイナリーTiプラスミドが得られる。
【0189】
バックボーンの同様の割合は、機能を損失せずにpBIN19から以前に除去されている(Xiangら、1999)。しかしながら、pBINPLUSは、pBIN19に対して2つの有意な改良点を有し(van Engelenら、1995);E.coliにおける増加したコピー数はColEI複製起点の添加のおかげであり、目的の遺伝子を確保する再構築したT−DNAはさらに、植物体における広範な欠失を受け得る左側の境界に由来する(Rossiら、1996)。同様のサイズのpEAQプラスミドは、それらのコピー数に対する顕著な効果を有さないが、それらは、10kb未満のプラスミドに最も効果的であるため(データは示さず)、市販のプラスミド抽出キットを用いてクローニング手順の間に非常に向上した収率を与える。
【0190】
pEAQバイナリーベクターのモジュールの性質は、必要な場合、任意のサイレンシング抑制因子および/またはマーカー遺伝子を、1つまたは2つのCPMV−HTカセットで同時発現させることによって、CPMV−HT発現に対する機能性を加える。例えば、pEAQexpress−GFP−HTのAsiSI/Mlul部位への異種配列を含む第2のHTカセットの挿入は、GFP蛍光での発現の追跡を可能にする。
【0191】
さらに、ベクターの柔軟性は、単一のアグロバクテリウム構築物の浸潤を必要とすることのみによって、一過性発現のための系を単純化し、T−DNA内に存在する発現カセットの数に比例して必要とされる浸潤の量を減少させることによって、効果を向上させる。FseI部位を占めるP19とともに、クローニングベクター(例えばpM81−FSC2−U162C)からHTカセットを受容するための2つのクローニング部位の存在はまた、完全なサイズの抗体などの複数のサブユニットタンパク質のさらにより効果的な発現を可能にする。
【0192】
導入遺伝子の発現レベルを向上させることに対するP19の効果は十分に特徴付けられている(Voinnetら、2003)。しかしながら、この研究は、同じTDNA上の各細胞に同時送達される場合、その有効性の第1の実証を示す。以前の研究により、導入遺伝子を含むT−DNAとして同じアグロバクテリウム内の別のTDNAからのP19の同時送達が報告されている(Hellensら、2005)。しかしながら、浸潤後6日までP19の効果はなく、T−DNAの非効率の移入が示唆される。本研究はまた、導入遺伝子の発現を向上させるためのR43W変異体19の第1の使用を実証する。その変異体が、野生型P19のGFPの発現を増加させる効果の約半分であるという発見は、活性のその公知の減少を満たし、TBSVの感染性(Chuら、2000)、および短い干渉RNAのより小さいクラス(21〜22nts)に結合するタンパク質の能力(Omarovら、2006)の両方を妥協する。しかしながら、この特徴は潜在的に、R43W変異体を、安定な形質転換に関する適用に、より適切にすることを可能にする。発達に関連するマイクロRNAはまた、より小さいサイズのクラスであり(Vaucheret,2006;Zhangら、2006)、従って、発達プロセスは、野生型によってほど、変異体P19の存在によってそんなに影響を受けない(Scholthof,2007)。さらに、この変異体は、潜在的に細胞毒性の外来性タンパク質の一過性発現を制御する方法を提供することができる。
【0193】
単一のプラスミドからの2G12の発現は、単一のアグロバクテリウム培養物で浸潤された植物組織からの抗体の最も高い報告された収率を表す。CPMV−HT発現のための3つのアグロバクテリウム培養物を用いることとは別に、別の系を用いて同様のレベルを達成する唯一の方法は、6つの別の培養物の浸潤およびウイルスベクターのアプローチに関連した(Giritchら、2006)。さらに、単一のプラスミドの使用により、複数の発現カセットの同時送達を確実にするのに必要とされる細菌の量の減少が得られ、これは、工業生産レベルで著しいコスト節約を与える。この浸潤プロセスはまた、葉組織の細胞間隙の少ない詰まりに起因して、より希釈された培養物を用いて実施するのに物理的に容易である。さらに、全ODの0.4までの希釈により、浸潤により誘導されるタンパク質汚染の量が減少した。各ODでの9つの別の浸潤の分析により、培養物のODが1.2〜0.4まで減少した場合、2.7±0.2〜1.5±0.1mg/mlからの抽出物のタンパク質濃度の減少が示された。pEAQexpressの使用は、OD0.4において、1.2の浸潤ODでの3つの培養物系と同じ量の2G12を産生するため、組み換え標的タンパク質は、pEAQexpressを用いる汚染タンパク質の量の半分のみから精製されなければならない。このことは、下流のプロセシングについての非常に有用かつ予想されない利点を与える。pEAQexpressからの2G12の発現はまた、得られる発現レベルでpEAQベクターのT−DNA内の発現カセットの位置の効果を示す。pEAQex−2G12LHEからの遊離軽鎖蓄積の増加により、少ない重鎖がこの構築物を用いて発現されることが示唆され、少ない集合した抗体を生じるように見える。これは、T−DNA上の発現カセットの配置に起因し得る。代替的に、一定の割合のT−DNAは、左側の境界で核酸分解しやすい(Rossiら、1996)。T−DNA内のNPTIIカセットの再挿入は、その配向に依存する発現に対する注目すべき効果を有するように見えた。クローニング操作の間、pEAQselectK−GFP−HTは、pEAQselectK(rev)−GFP−HTの約1.5倍のE.coliのプラスミドコピー数に到達したようであった(QIAprepキット、QIAGENを用いて実施した3つの別のプラスミド調製物の収率測定から決定した)。これは大まかに、2つのベクター間で観測された発現レベルの差異と相互関連する。プラスミドがアグロバクテリウムに移入される場合、増加したコピー数に起因するか、または実際に差異も存在するか否かは知られていない。しかしながら、これらの観測は、プラスミドコピー数が、効果的なアグロバクテリウム媒介性一過性発現に重要であり得ることを示唆する。これに関して、pBIN19およびその誘導体によるRK2オリジン(図9におけるoriV)の使用により、一過性発現のための良い選択が行われる。なぜなら、RK2プラスミドは、アグロバクテリウムにおいて7〜10コピーまで蓄積することが知られているからである(Veluthambiら、1987)。これは、pPZPによって利用されるpVS1オリジンと同様であり、pSaオリジン(LeeおよびGelvin、2008)によって生成されるより約2〜5倍高く、広範に使用されるpGREENバイナリーベクターに存在する(Helensら、2000)。pRiベースのプラスミドなどのより高いコピー数を生じる複製起点を含むプラスミド(LeeおよびGelvin、2008)は、一過性発現にさらにより適切であり得る。
【0194】
CPMVベースの発現、または実際に一般の植物ベースの発現を以前に経験していない実験室用に容易に入手可能なpEAQベクターを用いる高レベルの発現を行うために、pEAQの直接クローニング型を作製した。これは、P19およびNPTIIカセットを同様に含むT−DNA上に位置するCPMVHT発現カセットの5’リーダーと3’UTRとの間にポリリンカーを挿入することにより達成された。NPTIIカセットの存在が感知できるほどに発現を向上させるように見えるため(上記を参照のこと)、そのNPTIIカセットが含まれた。ポリリンカーはまた、タンパク質精製を促進するために、NまたはC末端Hisタグの融合を可能にするために、2セットの6×ヒスチジン残基をコードする。得られた構築物はまた、HTに対して発現を向上させる5’リーダーにおいて第2の変異体からの利点を有する。
【0195】
これらの向上した発現カセットはまた、任意のpEAQプラスミド内にクローニングベクター、pM81−FSC−POWからサブクローニングされてもよい。pEAQHTの使用により、ただ単一の変異体(U162C)を含む、pEAQspecialKと比較して増加したGFP発現を生じた。さらに、ポリリンカー設計はまた、1段階のクローニング手順を用いるHisタグした変異体の発現を可能にした。本明細書に示したモジュールバイナリーベクターは、制限されないが、CPMV−HT発現での使用のために特別に設計される。非常に高レベルの発現は、向上したクローニング効果および使用の容易さと関連する。この系は、ウイルス増幅の複雑化がされることなく、植物における付加価値タンパク質の一過性発現のための最も効果的かつ容易な方法を提供する。これにより、植物成長施設で利用可能な任意の分子生物学実験と同一の配列の2週間以内のミリグラム量の組み換えタンパク質が可能となる。従って、大学および工業的環境の両方において非常に有益な手段を与えることが予想される。
【0196】
(実施例5)
pEAQプラスミドおよびトランスジェニック植物での安定な組み込み
pEAQベクターシリーズを、一過性発現を考慮して設計したが、T−DNA内にNPTIIカセットの再挿入をして、ゲノム組み込みのための選択マーカーを与えた。これは、それらのより小さく、安定な植物および植物細胞培養形質転換に使用するための、より有用なバイナリーベクターを潜在的に可能にする。N.benthamiana葉片を形質転換するために使用する場合、T−DNA内にNPTIIカセットを含むpEAQベクターは、pBINPLUSベースの構築物と同じ効率を有する選択下でカルス形成を誘導できた。さらに、GFP発現は、UV光下でこれらの組織において検出可能であった(データは示さず)。これは、pEAQベクター由来のマルチカセットT−DNA分子が、植物ゲノム内に安定して組み込み、外来性遺伝子の発現を駆動することができることを実証する。
【0197】
蛍光植物もまた再生される。一次形質転換体(T0)の葉はUV光下で蛍光を発し、高レベルのGFP発現を示した。自家受粉したT0植物からの種子は成長でき、導入遺伝子を保有する得られたT1苗もまた、蛍光を発する(結果は示さず)。
【0198】
(実施例6)
バキュロウイルスベクターを用いるCPMVベースのHT系の使用
図15は、昆虫細胞におけるバキュロウイルスベクターを用いるCPMVベースのHT系を利用するのに適切な構築物を示す。p10プロモーターの制御下で、HyperTrans CPMV RNA−2 UTRもまた、バキュロウイルス発現系を用いる昆虫細胞におけるGFPの発現を向上させる。フローサイトメトリーによって測定される場合、バキュロウイルス感染sf21細胞の蛍光レベルの約5倍の向上を、CPMV−HTカセットを有さない構築物と比較して得た。
【0199】
(参考文献)
Alamillo,J.M.,Monger,W.,Sola,I.,Garcia,B.,Perrin,Y.,Bestagno,M.,Burrone,O.R.,Plana−Duran,J.,Enjuanes,L.,Lomonossoff,P.G. and Garcia,J.A.(2006)Use of virus vectors for the expression in plants of active full−length and single chain anticoronavirus antibodies. Biotechnol.J.1,1103−1111.
Brigneti,G.,Voinnet,O.,Li,W.X.,Ji,LH.,Ding,S.W and Baulcombe,D.C.(1998)Viral pathogenicity determinants are suppressors of transgene silencing in Nicotiana benthamiana. EMBO Journal 17:6739−6746.
Canizares,M.C,Liu,L.,Perrin,Y.,Tsakiris,E. and Lomonossoff,GP.(2006).A bipartite system for the constitutive and inducible expression of high levels of foreign proteins in plants.Plant Biotechnol.J.4,183−193.
Gopinath,K.,Wellink,J.,Porta,C,Taylor,K.M.,Lomonossoff,G.P. and van Kammen,A.(2000)Engineering cowpea mosaic virus RNA−2 into a vector to express heterologous proteins in plants.Virology 267:159−173.
Holness,C.L.,Lomonossoff,G.P.,Evans,D. and Maule,A.J.(1989).Identification of the initiation codons for translation of cowpea mosaic virus middle component RNA using site directed mutagenesis of an infectious cDNA clone.Virology 172,311 320.
Liu, L. and Lomonossoff,G.P.(2002)Agroinfection as a rapid method for propagating Cowpea mosaic virus−based constructs.J.Virol.Methods 105,343−348.
Liu,L. and Lomonossoff,G.P.(2006) A site−directed mutagenesis method utilising large double−stranded DNA templates for the simultaneous introduction of multiple changes and sequential multiple rounds of mutation:Application to the study of whole viral genomes.J.Virol.Methods 137,63−71.
Liu,L.,Canizares,M.C.,Monger,W.,Perrin,Y.,Tsakiris,E.,Porta,C,Shariat,N.,Nicholson,L. and Lomonossoff,G.P.(2005).Cowpea mosaic virus−based systems for the production of antigens and antibodies in plants.Vaccine 23,1788−1792.
Lomonossoff,G.P. & Shanks,M.(1983).The nucleotide sequence of cowpea mosaic virus B RNA.EMBO Journal 2,2253−2258.
Mechtcheriakova,I.A.,Eldarov,M.A.,Nicholson,L.,Shanks,M.,Skryabin,K.G. and Lomonossoff,G.P.(2006) The use of viral vectors to produce hepatitis B virus core particles in plants.J.Virol.Methods 131,10−15.
Monger,W.,Alamillo,J.M.,Sola,I.,Perrin,Y.,Bestagno,M.,Burrone,O.R.,Sabella,P.,Plana−Duran,J.,Enjuanes,L.,Garcia,J.A. and Lomonossoff,G.P.(2006) An antibody derivative expressed from viral vectors passively immunizes pigs against transmissible gastroenteritis virus infection when supplied orally in crude plant extracts. Plant Biotechnol.J.4, 623−631.
Rohll,J.B.,Holness,C.L.,Lomonossoff,G.P. and Maule,A.J.(1993).3’ terminal nucleotide sequences important for the accumulation of cowpea mosaic virus M−RNA.Virology 193, 672−679.
Sainsbury,F.,Lavoie,P−O.,D’Aoust,M−A.,Vezina,L−P. and Lomonossoff,G.P.(2008).Expression of Multiple Proteins Using Full−Length and Deleted Versions of Cowpea Mosaic Virus RNA−2.Plant Biotechnology Journal,6:82−92.
Sainsbury,F.,Canizares,M.C. and Lomonossoff,G.P.(2007) Cowpea mosaic virus−based expression vectors. In:Virus Expression Vectors(Hefferon, K. ed),pp.339−555. Kerala,India:Transworld Research Network.
Sainsbury,F. and Lomonossoff,G.P.(2008). Extremely high−level and rapid transient protein production in plants without the use of viral replication. Plant Physiology 148,1212−1218.
Sainsbury,F.,Liu,L. and Lomonossoff G.P.(2009) Cowpea mosaic virus−based expression of antigens and antibodies in plants. In:Methods in Molecular Biology Vol.483:Recombinant Pharmaceutical Proteins from Plants(Faye,L. and Gomord,V. eds),pp25−39,NY:Humana Press.
van Bokhoven,H.,Le Gall,O,Kasteel,D.,Verver,J.,Wellink,J. and van Kammen,A.(1993).Cis− and Trans−acting Elements in Cowpea Mosaic Virus RNA Replication. Virology 195,377−386.
Wellink J,Verver J, van Kammen A.(1993).Mutational analysis of AUG codons of cowpea mosaic virus M RNA. Biochimie.75(8):741−7.
さらなる参考文献:
Lee LY,Gelvin SB(2008)T−DNA binary vectors and systems. Plant Physiology 146:325−332
Hajdukiewicz P,Svab Z,Maliga P(1994)The small,versatile pPZP family of Agrobacterium binary vectors for plant transformation. Plant Molecular Biology 25:989−994
Hellens R,Mullineaux P,Klee H(2000b)A guide to Agrobacterium binary Ti vectors. Trends in Plant Science 5:446−451
Hiei Y,Ohta S,Komari T,Kumashiro T(1994)Efficient transformation of rice(Oryza−sativa L)mediated by Agrobacterium and sequence−analysis of the boundaries of the T−DNA. Plant Journal 6:271−282
Veluthambi K,Jayaswal RK,Gelvin SB(1987)Virulence genes−a,gene−G, and gene−D mediate the double−stranded border cleavage of T−DNA from the Agrobacterium Ti plasmid. Proceedings of the National Academy of Sciences of the United States of America 84: 1881−1885
Thole V,Worland B,Snape JW,Vain P(2007) The pCLEAN dual binary vector system for Agrobacterium−Mediated plant transformation. Plant Physiology 145:1211−1219
Johansen LK,Carrington JC(2001)Silencing on the spot. Induction and suppression of RNA silencing in the Agrobacterium−mediated transient expression system. Plant Physiology 126:930−938
Janssen BJ,Gardner RC(1989)Localized transient expression of Gus in leaf−disks following cocultivation with Agrobacterium. Plant Molecular Biology 14:61−72
Narasimhulu SB,Deng X,Sarria R,Gelvin SB(1996)Early transcription of Agrobacterium T−DNA genes in tobacco and maize. Plant Cell 8:873−886
Hellens RP,Edwards EA,Leyland NR,Bean S,Mullineaux PM(2000a)pGreen:a versatile and flexible binary Ti vector for Agrobacterium−mediated plant transformation. Plant Molecular Biology 42:819−832
Tzfira T,Tian GW,Lacroix B,Vyas S,Li JX,Leitner−Dagan Y, Krichevsky A,Taylor T,Vainstein A,Citovsky V(2005)pSAT vectors:a modular series of plasmids for autofluorescent protein tagging and expression of multiple genes in plants. Plant Molecular Biology 57: 503−516
Voinnet O,Rivas S,Mestre P,Baulcombe D(2003) An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant Journal 33:949−956
Frisch DA,Harrishaller LW,Yokubaitis NT,Thomas TL,Hardin SH,Hall TC(1995)Complete sequence of the binary vector BIN−19. Plant Molecular Biology 27:405−409
Trieu−cuot P,Courvalin P(1983)Nucleotide−sequence of the Streptococcus−faecalis plasmid gene encoding the 3’5’’−Aminoglycoside phosphotransferase type−Ill.Gene 23:331−341
Bevan M(1984)Binary Agrobacterium vectors for plant transformation. Nucleic Acids Research 12:8711−8721
Silhavy D,Burgyan J(2004)Effects and side−effects of viral RNA silencing suppressors on short RNAs. Trends in Plant Science 9:76−83
Scholthof HB(2007)Heterologous expression of viral RNA interference suppressors:RISC management. Plant Physiology 145:1110−1117
van Engelen FA,Molthoff JW,Conner AJ,Nap JP,Pereira A,Stiekema WJ(1995)pBINPLUS−an improved plant transformation vector based on pBIN19. Transgenic Research 4:288−290
Rossi L,Hohn B,Tinland B(1996)Integration of complete transferred DNA units is dependent on the activity of virulence E2 protein of Agrobacterium tumefaciens.
Proceedings of the National Academy of Sciences of the United States of America 93:126−130
Xiang CB,Han P,Lutziger I,Wang K,Oliver DJ(1999)A mini binary vector series for plant transformation. Plant Molecular Biology 40:711−717
Hellens RP,Allan AC,Friel EN,Bolitho K,Grafton K,Templeton MD,Karunairetnam S,Gleave AP,Laing WA(2005)Transient expression vectors for functional genomics,quantification of promoter activity and RNA silencing in plants. Plant Methods 1:13
Chu M,Desvoyes B,Turina M,Noad R,Scholthof HB(2000)Genetic dissection of tomato bushy stunt virus p19−protein−mediated host−dependent symptom induction and systemic invasion. Virology 266:79−87
Omarov R,Sparks K,Smith L,Zindovic J,Scholthof HB(2006)Biological relevance of a stable biochemical interaction between the tombusvirus−encoded P19 and short interfering RNAs. Journal of Virology 80:3000−3008
Vaucheret H(2006)Post−transcriptional small RNA pathways in plants:mechanisms and regulations. Genes & Development 20:759−771
Zhang BH,Pan XP,Cobb GP,Anderson TA(2006)Plant microRNA:A small regulatory molecule with big impact. Developmental Biology 289:3−16
Giritch A,Marillonnet S,Engler C, van Eldik G,Botterman J,Klimyuk V,Gleba Y(2006)Rapid high−yield expression of full−size IgG antibodies in plants coinfected with noncompeting viral vectors. Proceedings of the National Academy of Sciences of the United States of America 103:14701−14706
【0200】
【表2−1】
【0201】
【表2−2】
【0202】
CPMV RNA−2ゲノムセグメントの115位、161位、512位および524位の開始コドンを太字および下線で示す。
【0203】
【表3】
【0204】
pM81−FSC−2クローンの5’領域の変異誘発に使用されるオリゴヌクレオチドの変異ヌクレオチドを太字で示す。
【0205】
【表4】
【0206】
pM81−FSC−1およびpM81−FSC−2ベクターの配列と、表1からのCPMV野生型配列との間のヌクレオチドの相違を大文字として示す。
【0207】
【表5】
【0208】
【表6】
【0209】
【表7】
【0210】
【表8】
【0211】
【表9】
【0212】
【表10】
【0213】
【表11】
【0214】
【表12】
【技術分野】
【0001】
本発明は、概して、植物および他の真核細胞において遺伝子発現を増加させるための方法および物質、特に、ウイルス由来の配列、例えば、目的のタンパク質をコードする異種遺伝子の配列に関する。
【背景技術】
【0002】
コモウイルス(CPMV)
コモウイルスは、2分節ゲノムを有するRNAウイルスである。コモウイルスRNAゲノムのセグメントは、RNA−1およびRNA−2と呼ばれている。RNA−1は、VPg、レプリカーゼおよびプロテアーゼタンパク質をコードする(非特許文献1)。レプリカーゼは、ウイルスゲノムの複製のためにウイルスによって必要とされる。コモウイルスのササゲモザイクウイルス(cowpea mosaic virus)(CPMV)のRNA−2は、58Kおよび48Kのタンパク質、ならびに2つのウイルスコートタンパク質LおよびSをコードする。
【0003】
全てのコモウイルスのRNA−2の翻訳の開始は、2つのカルボキシ共末端(coterminal)タンパク質の合成を生じる、同じトリプレットリーディングフレームに位置する2つの異なる開始部位で起こる。この二重の開始現象は、翻訳の間のリボソームによる「漏出スキャニング(leaky scanning)」の結果として生じる。
【0004】
CPMVのRNA−2における5’末端開始コドン(AUG)は、115位、161位、512位および524位で生じる。161位および512位における開始コドンは、同じトリプレットリーディングフレーム内にある。161位での開始コドンにおける開始は、105Kのポリタンパク質の合成を生じるが、一方、512位での開始コドンにおける開始は、95Kのポリタンパク質の合成に関する。両方のポリタンパク質の合成は3299位での同じ終止コドンで停止されるため、105Kおよび95Kのタンパク質は、カルボキシ共末端である。524位におけるAUGコドンは、512位におけるAUGが欠失される場合、イニシエーターとして役立ち得る。しかしながら、AUG512の存在下において、これは機能せず、単にアミノ酸メチオニンをコードするだけである(非特許文献2;特許文献3)。115位での開始コドンは、ウイルス複製に必須ではない(非特許文献3)。
【0005】
CPMVのRNA−2ゲノムセグメントによってコードされる105Kおよび95Kのタンパク質は、後にRNA−1にコードされるタンパク質分解活性によって分解されて、プロセシングされる105Kまたは95Kのポリタンパク質であるかどうかに依存する、58Kまたは48Kのタンパク質のいずれか、および2つのウイルスコートタンパク質LおよびSを生じる一次翻訳産物である。CPMVにおける512位での開始コドンにおける翻訳の開始は、161位での開始より効果的であり、105Kのポリタンパク質より多い95Kのポリタンパク質の産生を生じる。
【0006】
CPMVのRNA−2における115位の開始コドンは、161位および512位における開始部位の上流に存在し、異なるリーディングフレーム内にある。この開始コドンは、175位における停止コドンと同相であるため、この部位での開始は、20アミノ酸のペプチドの産生を生じ得る。しかしながら、このようなペプチドの産生は、今までに検出されていなかった。
【0007】
AUG間のフレームを維持する必要性
突然変異誘発実験により、CPMVのRNA−2における161位および512位での開始部位の間のフレームの維持が、RNA−1にコードされるレプリカーゼによって、RNA−2の効果的な複製に必須であることが示された(非特許文献2;非特許文献4;非特許文献5;非特許文献3)。この条件は、CPMVのRNA−2(以下に示す)に基づく発現ベクターにおける512位の開始コドンの上流に挿入され得る配列の長さを制限し、そのようなベクターに外因性遺伝子をクローニングすることを、理想的なものよりも困難にさせる。例えば、それらの使用がしばしば、それらの開始部位の間のオープンリーディングフレーム(ORF)を変化させるため、ポリリンカーの使用を妨げる。
【0008】
CPMVベクター
CPMVは、植物における異種ポリペプチドの産生に適切なベクター系の開発のための基礎として役立っている(非特許文献6;非特許文献7)。これらの系は、RNA−2の修飾に基づくが、完全長か欠失した型を使用するか否かにより異なる。しかしながら、両方の場合において、修飾したRNA−2の複製は、RNA−1との共接種によりなされる。RNA−2の完全長型に基づいた発現系は、RNA2由来のポリペプチドのC末端に対する外来性タンパク質の融合に関与する。N末端ポリペプチドの解放は、口蹄疫ウイルス由来の2A触媒ペプチド配列の作用により媒介される(非特許文献8)。得られるRNA−2分子は、植物内および植物間の両方で分離できる。この戦略は、cowpea植物において、Hepatitis Bコア抗原(HBcAg)および小さな免疫タンパク質(SIP)などの多くの組み換えタンパク質を発現するために使用されている(非特許文献9;非特許文献10;非特許文献11)。成功的ではあるが、完全長のウイルスベクターの使用は、挿入される配列のサイズ制限の点で不都合であり、生物学的封じこめについての問題がある。
【0009】
これらに対処するために、CPMV RNA−2の欠失した型に基づく系が最近開発されている(非特許文献12)。この系において、移動タンパク質およびコートタンパク質の両方をコードするRNA−2の領域は除去される。しかしながら、欠失した分子はさらに、RNA−1にコードされるレプリカーゼによる複製に必要なcis作動性の配列を有し、それにより、高レベルの遺伝子増幅が、環境を汚染する修飾されたウイルスの可能性を付随することなく、維持される。RNA−1に加えて接種材料において、PVY由来のHcPro(非特許文献13)などの遺伝子サイレンシングの抑制因子の封入とともに、欠失したCPMVベクターが、一過性の発現系(特許文献1、Bipartite System,Method And Composition For The Constitutive And Inducible Expression Of High Levels Of Foreign Proteins In Plants;非特許文献14)として使用され得る。しかしながら、完全長のRNA−2に基づいたベクターを用いる状況と対照的に、複製は接種レベルに制限される。これらのCPMVベクターは、単一のタイプのポリペプチドからなる複数の鎖の複合体を発現するのに使用されている。
【0010】
CPMV RNA−2の完全長または欠失した型のいずれかに基づいたベクターの複数のコピーはまた、植物における異種タンパク質の産生に適切であることが示されている(非特許文献15)。RNA−1の存在下においてNicotiana benthamiana内への異なるマーカー遺伝子を含む2つの完全長のRNA−2構築物の同時浸潤は、2つの外来性タンパク質が、接種組織において同じ細胞内で効果的に発現され得ることを示すために使用される。さらに、タンパク質は、ヘテロマー形成に必須条件である、同じサブ細胞区画に同時に局在化され得る。
【0011】
植物におけるヘテロマータンパク質の発現のための異なるCPMV RNA−2ベクターの適合性も研究されている。RNA−2の完全長および欠失した型内へのIgGの重鎖および軽鎖の挿入により、接種した組織において完全なサイズのIgG分子を蓄積するための両方のアプローチが示されたが、欠失したRNA−2ベクターが使用された場合、そのレベルは顕著に高かった。従って、浸透して広がる完全長のRNA−2構築物の能力は、ヘテロマータンパク質の産生に重要ではないように見られ、RNA−2の欠失した型の使用が、明らかに利点がある。特に、それらはまた、生物学的封じ込めの利点を与える。
【0012】
従って、ベクター系に基づく公知のCPMVは、植物において目的のタンパク質をコードする異種遺伝子の発現に有用な手段であることを表す。しかしながら、改良を必要とする最適化されるベクター系のための当該分野における必要性、例えば発現される異種タンパク質の収率およびベクターの使用の容易さが依然として存在する。
【先行技術文献】
【特許文献】
【0013】
【特許文献1】国際公開第2007/135480号パンフレット
【非特許文献】
【0014】
【非特許文献1】Lomonossoff,G.P.& Shanks,M.(1983),The nucleotide sequence of cowpea mosaic virus B RNA.EMBO Journal 2,2253−2258
【非特許文献2】Holness,C.L.,Lomonossoff,G.P.,Evans,D.およびMaule,A.J.(1989).Identification of the initiation codons for translation cowpea mosaic virus middle component RNA using site directed mutagenesis of an infectious cDNA clone.Virology 172,311 320
【非特許文献3】Wellink J,Verver J,van Kammen A.(1993).Mutational analysis of AUG codons of cowpea mosaic virus M RNA.Biochimie.75(8):741−7
【非特許文献4】van Bokhoven,H.,Le Gall,O,Kasteel,D.,Verver,J.,Wellink,J.およびvan Kammen,A.(1993).Cis−and Trans−acting Elements in Cowpea Mosaic Virus RNA Replication.Virology 195,377−386
【非特許文献5】Rohll,J.B.,Holness,C.L.,Lomonossoff,G.P.およびMaule,A.J.(1993).3’terminal nucleotide sequences important for the accumulation of cowpea mosaic virus M−RNA.Virology 193,672−679
【非特許文献6】Liu,L.,Canizares,M.C.,Monger,W.,Perrin,Y.,Tsakiris,E.,Porta,C.,Shariat,N.,Nicholson,L.およびLomonossoff,G.P.(2005).Cowpea mosaic virus−based systems for the production of antigens and antibodies in plants.Vaccine 23,1788−1792
【非特許文献7】Sainsbury,F.,Canizares,M.C.およびLomonossoff,G.P.(2007)Cowpea mosaic virus−based expression vectors.In:Virus Expression Vectors(Hefferon,K.ed),pp.339−555.Kerala,India:Transworld Research Network.
【非特許文献8】Gopinath,K.,Wellink,J.,Porta,C.,Taylor,K.M.,Lomonossoff,G.P.およびvan Kammen,A.(2000)Engineering cowpea mosaic virus RNA−2 into a vector to express heterologous proteins in plants.Virology 267:159−173
【非特許文献9】Mechtcheriakova,I.A.,Eldarov,M.A.,Nicholson,L.,Shanks,M.,Skryabin,K.G.およびLomonossoff,G.P.(2006)The use of viral vectors to produce hepatitis B virus core particles in plants.J.Virol.Methods 131,10−15
【非特許文献10】Monger,W.,Alamillo,J.M.,Sola,I.,Perrin,Y.,Bestagno,M.,Burrone,O.R.,Sabella,P.,Plana−Duran,J.,Enjuanes,L.,Garcia,J.A.およびLomonossoff,G.P.(2006)An antibody derivative expressed from viral vectors passively immunizes pigs against transmissible gastroenteritis virus infection when supplied orally in crude plant extrancts.Plant Biotechnol.J.4,623−631
【非特許文献11】Alamillo,J.M.,Monger,W.,Sola,I.,Garcia,B.,Perrin,Y.,Bestagno,M.,Burrone,O.R.,Plana−Duran,J.,Enjuanes,L.,Lomonossoff,P.G.およびGarcia,J.A.(2006)Use of virus vectors for the expression in plants of active full−length and single chain anticoronavirus antibodies.Biotechnol.J.1,1103−1111
【非特許文献12】Canizares,M.C.,Liu,L.,Perrin,Y.,Tsakiris,E.およびLomonossoff,G.P.(2006).A bipartite system for the constitutive and inducible expression of high levels of foreign proteins in plants.Plant Biotechnol.J.4,183−193
【非特許文献13】Brigneti,G.,Voinnet,O.,Li,W.X.,Ji,L.H.,Ding,S.WおよびBaulcombe,D.C.(1998)Viral pathogenicity determinants are suppressors of transgene silencing in Nicotiana benthamiana.EMBO Journal 17:6739−6746
【非特許文献14】Sainsbury,F.,Liu,L.およびLomonossoff G.P.(2009)Cowpea mosaic virus−based expression of antigens and antibodies in plants.In:Methods in Molecular Biology Vol.483:Recombinant Pharmaceutical Proteins from Plants(Faye,L.およびGomord,V.eds),pp25−39,NY:Humana Press
【非特許文献15】Sainsbury,F.およびLomonossoff,G.P.(2008).Extremely high−level and rapid transient protein production in plants without the use of viral replication.Plant Physiology 148,1212−1218
【発明の概要】
【課題を解決するための手段】
【0015】
本発明者らは、驚くべきことに、CPMV RNA−2ベクターにおける161位での開始コドンの変異が、512位での開始コドンの後に挿入される遺伝子によってコードされるタンパク質の発現レベルを著しく増加させることを見出した。タンパク質発現のレベルは、161位での開始コドンがインタクトである(SainsburyおよびLomonossoff,2008)ことのみが異なる、CPMV RNA−2ベクター由来の同じタンパク質の発現と比較して、約20〜30倍増加した。本発明は、ウイルス複製を必要とせず、外来性タンパク質の高レベルの産生を可能にする。
【0016】
本発明者らはまた、161位での開始コドンの変異が、161での変異された開始コドンと、512での開始コドンの位置の間のフレームを維持するための必要性を無効にし、これにより、161位での変異された開始コドンの後の任意の長さの配列の挿入を可能にすることを見出した。このことは特に、任意の長さのポリリンカーを、変異された開始コドンの後にRNA−2ベクター内に挿入することを可能にし、次いで、ベクター内に目的の遺伝子のクローニングを容易にするのに使用できるという利点がある。
【0017】
さらに、本発明者らは、タンパク質発現の増加にもかかわらず、161位での変異した開始コドンを含むCPMV RNA−2ベクターで形質転換した植物が健康的であるように見えることを見出した。すなわち、公知のCPMV RNA−2ベクターで形質転換した植物より壊死をほとんど示さなかった。植物の健康は、長期間生存する健康的な植物として植物由来のタンパク質の発現に重要な因子である。さらに、植物の健康はまた、タンパク質精製を妨げ得る壊死の結果として放出されるタンニンなどの植物由来のタンパク質の精製に重要である(SainsburyおよびLomonossoff,2008)。
【0018】
従って、本発明は、修飾された2分節のウイルス配列に基づいた改良されたタンパク質産生系および方法に関連する。
【0019】
従って、本発明の種々の態様において、発現エンハンサー配列が与えられるか、または利用され、その配列は、コモウイルスなどの2分節RNAウイルスのRNA−2ゲノムセグメント由来(またはそれらと相同性を共有する)であり、その配列において、標的開始部位は変異されている。
【0020】
本発明はさらに、2分節ウイルスのRNA−2ゲノムセグメント由来の配列の発現または翻訳を増強する活性を増加させるためのプロセスを与え、そのプロセスは、配列内の標的開始部位を変異させることを含む。
【0021】
本発明の一部の特定の定義および実施形態は、ここでより詳細に記載される。
【図面の簡単な説明】
【0022】
【図1】図1は、CPMV発現ベクター00、10、01および11の概略図を示す。00発現ベクターにおいて、115位および161位の開始部位はインタクトである。10発現ベクターにおいて、115位の開始部位は変異されているが、161位の開始部位はインタクトである。01発現ベクターにおいて、161位の開始部位は変異されているが、115位の開始部位はインタクトである。11発現ベクターにおいて、115位および161位の開始部位は両方変異されている。CPMV発現ベクター00、10、01および11はまた、512(FSC2−152)、513(FSC2−513)または514(FSC2−514)のいずれかの位置に開始部位を含む。バーは、タンパク質の発現が起こる開始部位を示すために使用される。
【図2】図2は、図1に概略的に示されるCPMV発現ベクターでトランスフェクトされた植物で発現される可溶性の緑色蛍光タンパク質(GFP)のレベルを示す。発現ベクターFSC2−512、FSC2−513、およびFSC2−514において、GFPをコードする遺伝子を、それぞれ、512位、513位および514位の開始コドンの後に挿入した。SDS−PAGEゲルのレーンに、00、10、01および11をマークし、115位および161位でのCPMVベクターにおける開始部位のものに応じて、インタクトまたは変異される。「500ng」とマークしたレーンは、500ngのGFPタンパク質に対応するバンドの位置を示し、それによって、CPMV発現ベクターから発現されるGFPタンパク質の予想される位置を示す。各SDS−PAGEゲルの左側のレーンは、タンパク質のサイズマーカーの位置を示す。
【図3】図3は、図2に示される実験で使用される同じCPMV発現ベクターでトランスフェクトされたNicotiana benthamianaの葉におけるGFP発現のレベルを示す。葉の先端の薄い領域はGFP発現の領域に対応した。発現ベクター10、01および11における115位および/または161位の開始部位を不活性化するために変異が行われ、それも示す。
【図4】図4は、緑色蛍光タンパク質(GFP)、Discosoma赤色蛍光タンパク質(DsRed)、およびB型肝炎コア抗原(HBcAg)を一過性発現するためのDel−RNA−2(図1における発現ベクター00[FSC2−512])およびHT(図1における発現ベクター01[FSC2−512])の比較を示す。各タンパク質についてのdelRNA−2またはHTクローンに、サイレンシング抑制因子P19を浸潤させた。(A)DsRedまたはHBcAgが発現された場合、delRNA−2構築物での浸潤後7日の組織は壊死したが、これはHTにより駆動された発現についての場合ではない。実際に、HTによってDsRedを発現する組織は、昼間の明かりの条件下で明白に赤く見える。(B)タンパク質発現のCoomassie染色したSDS−PAGE分析。示したように組み換えタンパク質に対応する突出したバンドが、ウェスタンブロッティングによって確認された。1−マーカー、2−浸潤されていない組織、3−delRNA−2構築物、4−HT構築物、5−利用可能な場合、市販の標準物質。約5mgの浸潤した組織のからの粗抽出物を、レーンごとに負荷し、同様に2μgのGFP標準物質および2μgのHBcAg標準物質を負荷した。DsRedについての標準物質は今回、利用可能ではなかった。
【図5】図5は、ヒトの抗ヒト免疫不全ウイルス抗体2G12を一過性発現するためのDel−RNA−2(図1における発現ベクター00[FSC2−512])およびHT(図1における発現ベクター01[FSC2−512])の初期の比較を示す。IgG重鎖は、天然形態(HL)またはER保有(HEL)のいずれかであり、軽鎖およびP19を浸潤させた。(A)del−RNA−2による2G12−HELの発現により、浸潤された組織の壊死が導かれるが、これはHT発現については生じない。(B)抗体重鎖(delRNA−2またはHT)とP19が浸潤した組織の粗抽出物のSDS−PAGE分析。各試料について、5mgの浸潤した組織からの粗抽出物を1μgの標準ヒトIgGとして負荷した。2G12に対応するバンドは、coomassie染色後、容易に目に見える。(C)5日後に抗体2G12の蓄積を、プロテインAに対する捕捉および表面プラズモン共鳴分光法により測定し、2容量の緩衝液(PBS、5mM EDTA)での抽出後の濃度を表す。従って、本発明者らは、植物インキュベーションまたは抽出を1つも最適化せずに、150mg/kg(HT HELについて)に近づく初期の重量濃度を導くことができる。3つの試料を各処置について測定した。
【図6】図6は、集合したHBcAg粒子の電子顕微鏡写真を示し、これは、本明細書に記載されるHT(図1における発現ベクター01[FSC2−512])発現系を用いて発現された。集合したHBcAg粒子は、直径約30nmの中空球のように見える。HBcAg粒子を含む樹液(sap)は、電子顕微鏡写真を撮る前に濃縮されなかったが、望ましくない塩は除去した。従って、電子顕微鏡写真は樹液(sap)中のHBcAg粒子の濃度を表す。
【図7】図7は、ベクターpM81−FSC1を示す。
【図8】図8は、ベクターpM81−FSC2を示す。
【図9】図9は、pEAQの構築物の概略図を示す。(A)灰色で示された外来配列を有する開始pBINPLUSベースのプラスミド。(B)バイナリーベクターの必須エレメントを含むPCR産物。(C)末端を合わせた(end−tailored)PCR産物の3部のライゲーション後の中間体プラスミド。(D)増幅およびその後の中間体からの2つのフラグメントのライゲーション後の最終プラスミド。
【図10】図10は、主なpEAQ誘導体のT−DNAの概略図を示す。T−DNAは、示した制限部位内に起こり得るクローニングを残す、示したP19およびNPTIIカセットのいずれかまたは両方を含む。
【図11】図11は、その親プラスミドPBB−FSC2−512−HTと比較したpEAQベクターによって産生したGFPの発現レベルを示す。組織を、pEAQexpressを除いて、P19および示したベクターでの浸潤後、6日に分析し、それを、標準的なODまたは2倍の希釈で単独で浸潤させた。(A)UV照射下で視覚化した葉、(B)coomassie染色した12%SDS−PAGE、および(C)蛍光分光分析。
【図12】図12は、野生型P19と比較したGFP発現を向上させるP19(R43W)の能力を示す。組織は、2倍希釈でのpEAQselectK(selK−P19)、pEAQselectKおよびP19(selK)、pEAQspecialK(spK)、2倍希釈でのpEAQspecialK(spK1:2)、pEAQspecialKm(spKm)、2倍希釈でのpEAQspecialKm(spKm1:2)、pEAQexpress(ex)、および2倍希釈でのpEAQexpress(ex1:2)での浸潤後6日に分析した。(A)UV照射下で視覚化した葉および(B)蛍光分光分析。
【図13】図13は、単一のpEAQプラスミドを有する完全なサイズのIgG、2G12の発現を示す。(A)2G12を発現するために構築された2つのpEAQexpress由来プラスミドの概略図。(B)希釈物を示す浸潤スキームおよび各プラスミドの組み合わせについてのそれらの代表的なOD、ならびに各ODでの侵潤後に作製されたタンパク質抽出物の濃度(±SD)。(C)Coomassie染色した12〜4%のSDS−PAGE分析および(D)2G12重鎖(γ)の免疫学的検出および(E)浸潤後8日の2G12Fab領域(Fab)鎖の免疫学的検出。M、示したサイズを有するマーカー;C、コントロール抽出物;Std、CHO産生2G12。coomassie染色に関して、3mgの浸潤組織の等価物からのタンパク質は、1μgのCHO2G12を有する各レーンに位置し、250ngのCHO2G12の各レーンにおけるウェスタンブロッティングについて0.75mgの組織の等価物。推測した集合/分解産物を示す。
【図14】図14は、ネイティブおよびhis−タグ化変異体におけるpEAQ−HT由来のGFPのクローニングおよび発現を示す。(A)ポリリンカーの詳細を有するpEAQ−HT T−DNAの概略図。(B)GFP発現の蛍光分光分析。spK=pEAQspecialK−GFP−HT、GFP、HisGFP、およびGFPHis=pEAQ−HTクローン。(C)12%SDS−PAGEおよびGFP発現のウェスタン分析。C=コントロール抽出物。
【図15】図15は、バキュロウイルスベクターの一部としての昆虫細胞における使用に適切な本発明の核酸構築物を示す。
【発明を実施するための形態】
【0023】
本明細書中で言及する場合、「エンハンサー」配列(またはエンハンサー要素)とは、コモウイルスなどの2分節のRNAウイルスのRNA−2ゲノムセグメント由来の(またはそれらと相同性を共有する)配列であり、その配列において、標的開始部位は変異される。そのような配列は、それらが結合される異種ORFの下流の発現を向上できる。限定せずに、転写RNAに存在する場合、そのような配列は、それらが結合される異種ORFの翻訳を向上できると考えられる。
【0024】
本明細書中で言及する場合、「標的開始部位」とは、野生型RNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端タンパク質のより長い方の産生(翻訳)のための開始部位として役立つ、問題のあるエンハンサー配列が由来する2分節のウイルス(例えばコモウイルス)の野生型RNA−2ゲノムセグメントにおける開始部位(開始コドン)である。
【0025】
上記のように、CPMV RNA−2によってコードされる2つのカルボキシ共末端タンパク質のより長い方である、105Kのタンパク質の産生は、野生型CPMV RNA−2ゲノムセグメント内の161位での開始部位で開始される。従って、CPMV RNA−2ゲノムセグメント由来のエンハンサー配列における標的開始部位は、野生型CPMV RNA−2における161位での開始部位である。
【0026】
161位の開始コドン付近の変異は、161位の開始コドン自体を変異することと同じ(または同様の)効果を有することができる。例えば、この開始コドン付近の状態を破壊することは、開始コドンがより頻繁にバイパスされることを意味し得る。
【0027】
本発明の一態様において、従って、標的開始部位は、標的開始部位の上流および/または下流の1つ以上のヌクレオチドを変異することによって、間接的に「変異され」得るが、野生型標的開始部位は残ったままであり、これらのヌクレオチドを変異させる効果は、標的開始部位自体が変異される場合に観測される効果と同じまたは類似である。
【0028】
標的開始部位は、野生型RNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端タンパク質のより長い方の産生のための開始部位として役立つため、標的開始部位が、野生型RNA−2によってコードされる2つのカルボキシ共末端タンパク質のより短い方の産生のための開始部位として役立つ、同じ野生型RNA−2ゲノムセグメント上の第2の開始部位を有するフレーム内(同相)にあるという結果となる。2つの開始部位は、それらが同じトリプレットリーディングフレーム内にある場合、フレーム内にある。
【0029】
野生型CPMV RNA−2ゲノムセグメント由来のエンハンサー配列における標的開始部位、すなわち161位の開始部位は、野生型CPMV RNA−2ゲノムセグメントにおけるCPMV RNA−2によってコードされる2つのカルボキシ共末端タンパク質のより短い方(95Kのタンパク質)の産生のための開始部位として役立つ、512位に開始部位を有するフレーム内にある。
【0030】
従って、標的開始部位は、野生型RNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端ポリタンパク質のより短い方の産生のための開始部位として役立つ、エンハンサー配列が由来する野生型RNA−2ゲノムセグメントにおける第2の開始部位の上流(5’)に位置する。さらに、標的開始部位はまた、エンハンサー配列が由来する野生型RNA−2ゲノムの第3の開始部位の下流(3’)に位置し得る。CPMVにおいて、標的開始部位、すなわち、161位の開始部位は、95Kのタンパク質の産生のための開始部位として役立つ512位の第2の開始部位の上流、および115位の第3の開始部位の下流に位置する。
【0031】
従って、2分節のウイルスのRNA−2ゲノムセグメント由来のエンハンサー配列における標的開始部位は、野生型RNA−2によってコードされる2つのカルボキシ共末端タンパク質の産生のための2つの開始部位のうちの第1である。この文脈において「第1の」とは、野生型RNA−2ゲノムセグメントの5’末端近くに位置する開始部位を指す。
【0032】
所望の場合、配列中の1つより多い開始部位が変異されてもよい。例えば、115位(またはそれに相当する)「第3の」開始部位が、欠失または変更されてもよい。AUG161の除去に加えてAUG115の除去が、発現をさらに向上させることが示されている(SainsburyおよびLomonossoff,2008)。
【0033】
本発明のエンハンサー配列は、2分節のRNAウイルスのRNA−2ゲノムセグメント由来の修飾された配列に基づく。
【0034】
本明細書中で言及される場合、2分節のウイルス、または2分節のゲノムを有するウイルスは、Comoviridaeファミリーのメンバーであってもよい。Comoviridaeファミリーの全ての属は、2つのカルボキシ共末端タンパク質をコードするようである。Comoviridaeファミリーの属としては、コモウイルス、ネポウイルス、ファバウイルス、ケラウイルス(Cheravirus)、およびサドワウイルスが挙げられる。コモウイルスとしては、Cowpea mosaic virus(CPMV)、Cowpea severe mosaic virus(CPSMV)、Squash mosaic virus(SqMV)、Red clover mottle virus(RCMV)、Bean pod mottle virus(BPMV)が挙げられる。好ましくは、2分節のウイルス(またはコモウイルス)はCPMVである。
【0035】
これらのコモウイルスおよびいくつかの特定の株のRNA−2ゲノムセグメントの配列は、括弧に記載される受託番号の下でNCBIデータベースから利用可能である:cowpea mosaic virus RNA−2(NC_003550)、cowpea severe mosaic virus RNA−2(NC_003544)、squash mosaic virus RNA−2(NC_003800)、squash mosaic virus strain Kimble RNA−2(AF059533)、squash mosaic virus strain Arizona RNA−2(AF059532)、red clover mottle virus RNA−2(NC_003738)、bean pod mottle virus RNA−2(NC_003495)、bean pod mottle virus strain K−Hopkins1 RNA−2(AF394609)、bean pod mottle virus strain K−Hancock1 RNA−2(AF394607)、Andean potato mottle virus(APMoV:L16239)およびRadish mosaic virus(RaMV;AB295644)。同様に、bean rugose mosaic virus(BRMV;AF263548)およびコモウイルス属、turnip ringspot virus(EF191015)の暫定的なメンバーから利用可能な部分的なRNA−2配列も存在する。Comoviridaeファミリーにおける他の属由来の多数の配列も利用可能である。
【0036】
今まで、研究されてきた全てのコモウイルスは、それらのRNA−2ゲノムセグメント由来の2つのカルボキシ共末端ポリタンパク質を発現するための2つの別の開始コドンを有することが示されている。特に、CPMV、CPSMV、BPMV、SqMVおよびRCMVのRNA−2ゲノムセグメントは、2つのカルボキシ共末端ポリタンパク質を発現するための2つの別の開始コドンを含むことが知られている。
【0037】
従って、CPMV(すなわち、それに対応する)の野生型RNA−2セグメントにおける161位の開始部位と同等である、他のコモウイルスにおける標的開始部位は、当該分野において公知の方法によって同定され得る。例えば、標的開始部位は、CPMVの野生型RNA−2ゲノムセグメント配列と、別のコモウイルスのRNA−2ゲノムセグメント配列との間の配列アラインメントによって同定され得る。次いで、このような配列アラインメントは、少なくともそのアラインメントにおいて、野生型CPMV RNA−2における161位の標的開始部位と近いか、または同じ位置である、開始部位を同定することによって、コモウイルスRNA−2ゲノムセグメント配列における標的開始部位を同定するために使用され得る。
【0038】
他のコモウイルスにおける標的開始部位もまた、野生型コモウイルスRNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端タンパク質のより長い方の合成のための開始部位として役立つ開始コドンを決定することによって同定され得る。このアプローチはまた、上記のアラインメントと合わせて使用され得る。すなわち、このアプローチは、CPMV RNA−2を有するアラインメントによって同定されるコモウイルス開始部位が標的開始部位であることを確認するために使用され得る。
【0039】
もちろん、上記の方法はまた、野生型CPMV RNA−2ゲノムセグメントにおける512位の開始部位と同等である、他のコモウイルスRNA−2ゲノムセグメントにおける開始部位を同定するために使用され得る。しかしながら、野生型コモウイルスRNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端タンパク質のより長い方を合成するための開始部位として役立つ開始コドンを同定する代わりに、野生型コモウイルスRNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端タンパク質のより短い方を合成するための開始部位として役立つ開始コドンが同定される。
【0040】
一旦、CPMV RNA−2における161位および512位の開始部位と同等である可能性がある2つのコモウイルスRNA−2開始部位が同定されると、その標的開始部位の同定は、2つの開始部位が同じフレーム内、すなわち、同じトリプレットリーディングフレーム内にあることを調べることによって確認され得る。なぜなら、この場合、それらは、2つのカルボキシ共末端タンパク質を産生するための開始部位としてのみ機能するからである。
【0041】
本発明の一実施形態において、エンハンサー配列は、CPMV RNA−2ゲノムセグメントのヌクレオチド1〜512を含み(表1を参照のこと)、161位の標的開始部位は変異される。本発明の別の実施形態において、エンハンサー配列は、別のコモウイルス由来の等価配列を含み、CPMVの161位の開始コドンと同等の標的開始部位は変異される。その標的開始部位は、置換、欠失または挿入によって変異されてもよい。好ましくは、その標的開始部位は、点突然変異によって変異される。
【0042】
本発明の代替の実施形態において、エンハンサー配列は、変異標的開始部位を有するコモウイルスRNA−2ゲノムセグメント配列のヌクレオチド10〜512、20〜512、30〜512、40〜512、50〜512、100〜512、150〜512、1〜514、10〜514、20〜514、30〜514、40〜514、50〜514、100〜514、150〜514、1〜511、10〜511、20〜511、30〜511、40〜511、50〜511、100〜511、150〜511、1〜509、10〜509、20〜509、30〜509、40〜509、50〜509、100〜509、150〜509、1〜507、10〜507、20〜507、30〜507、40〜507、50〜507、100〜507、または150〜507を含む。本発明の他の実施形態において、エンハンサー配列は、表1に示すCPMV RNA−2ゲノムセグメント配列のヌクレオチド10〜512、20〜512、30〜512、40〜512、50〜512、100〜512、150〜512、1〜514、10〜514、20〜514、30〜514、40〜514、50〜514、100〜514、150〜514、1〜511、10〜511、20〜511、30〜511、40〜511、50〜511、100〜511、150〜511、1〜509、10〜509、20〜509、30〜509、40〜509、50〜509、100〜509、150〜509、1〜507、10〜507、20〜507、30〜507、40〜507、50〜507、100〜507、または150〜507を含み、野生型CPMV RNA−2ゲノムセグメントにおける161位の標的開始部位は変異される。
【0043】
本発明のさらなる実施形態において、エンハンサー配列は、変異標的開始部位を有するコモウイルスRNA−2ゲノムセグメント配列のヌクレオチド1〜500、1〜490、1〜480、1〜470、1〜460、1〜450、1〜400、1〜350、1〜300、1〜250、1〜200、または1〜100を含む。
【0044】
本発明の代替の実施形態において、エンハンサー配列は、表1に示すCPMV RNA−2ゲノムセグメント配列のヌクレオチド1〜500、1〜490、1〜480、1〜470、1〜460、1〜450、1〜400、1〜350、1〜300、1〜250、1〜200、または1〜100を含み、野生型CPMV RNA−2ゲノムセグメントにおける161位の標的開始部位は変異される。
【0045】
変異標的開始部位を有するコモウイルスRNA−2ゲノムセグメントの少なくとも100または200、少なくとも300、少なくとも350、少なくとも400、少なくとも450、少なくとも460、少なくとも470、少なくとも480、少なくとも490、あるいは少なくとも500のヌクレオチドを含むエンハンサー配列もまた、本発明の実施形態である。
【0046】
さらに、野生型CPMV RNA−2ゲノムセグメントにおける161位の標的開始部位が変異される、表1に示すCPMV RNA−2ゲノムセグメント配列の少なくとも100または200、少なくとも300、少なくとも350、少なくとも400、少なくとも450、少なくとも460、少なくとも470、少なくとも480、少なくとも490あるいは少なくとも500のヌクレオチドを含むエンハンサー配列もまた、本発明の実施形態である。
【0047】
本発明の代替の実施形態は、野生型CPMV RNA−2ゲノムセグメントにおける161位の標的開始部位が変異される、表1に示すCPMV RNA−2ゲノムセグメント配列と少なくとも99%、98%、97%、96%、95%、90%、85%、80%、75%、70%、65%、60%、55%または50%の同一性を有するエンハンサー配列である。
【0048】
特定の配列に対して言及する場合、用語「%の類似性」、「%の同一性」および「%の相同性」は、University of Wisconsin GCGソフトウェアプログラムに記載されるように使用される。従って、エンハンサー配列は、表1に示すCPMV RNA−2ゲノムセグメント配列の相補的配列と特異的にハイブリダイズする。ただし、野生型CPMV RNA−2ゲノムセグメントにおける161位に対応する標的開始部位は変異される。
【0049】
用語「特異的にハイブリダイズする」とは、当該分野において一般に使用される予め決定された条件下で、そのようなハイブリダイゼーションを可能にするのに十分な相補的配列(時々、「実質的に相補的な」といわれる)の2つの一本鎖核酸分子の間の会合を指す。特に、この用語は、本発明の1本鎖DNAまたはRNA分子内に含まれる実質的に相補的な配列を有するオリゴヌクレオチドのハイブリダイゼーションを指し、非相補的配列の一本鎖核酸を有するオリゴヌクレオチドのハイブリダイゼーションの実質的な排除を指す。「相補的な」とは、塩基対合(相補的配列T−C−AとのA−G−T対)による核酸配列の天然の会合を指す。2つの一本鎖分子間の相補性は、核酸対の一部のみが相補的である場合、部分的であってもよいか、または全ての塩基対が相補的である場合、完全であってもよい。相補性の程度は、ハイブリダイゼーションおよび増幅反応の効果および強度に影響を与える。
【0050】
本発明のエンハンサー配列における標的開部位は、翻訳開始部位としてもはや機能しなくなるように、欠失、挿入または置換によって変異されてもよい。例えば、点突然変異が、エンハンサー配列における標的開始部位の位置でなされてもよい。あるいは、エンハンサー配列における標的開始部位は、部分的または完全のいずれかで欠失されてもよい。例えば、エンハンサー配列における標的開始部位にわたる欠失がなされてもよい。開始部位にわたる欠失は、エンハンサー配列が由来する野生型RNA−2ゲノムセグメントの配列と比較した場合、5まで、10まで、15まで、20まで、25まで、30まで、35まで、40まで、45まで、または50までのヌクレオチド長でなされてもよい。
【0051】
理論に束縛されることを望まないなら、CPMVにおける161位の開始コドンの変異は、不活化翻訳抑制因子の下流に位置する開始コドンからの翻訳の向上した開始を生じる、翻訳抑制因子の不活化を導くと考えられる。
【0052】
従って、本発明はさらに、2分節ウイルスのRNA−2ゲノムセグメント由来のエンハンサー配列を提供し、そのエンハンサー配列は、不活化翻訳抑制因子配列を含む。
【0053】
本発明はさらに、2分節ウイルスのRNA−2ゲノムセグメント由来の配列の発現または翻訳を増強する活性を増加させるためのプロセスを提供し、そのプロセスは、配列内の翻訳抑制因子配列を不活化する工程を含む。
【0054】
既に上で述べたように、CPMV RNA−2ゲノムセグメントにおける161位の開始部位の変異は、CPMV RNA−2に通常存在する翻訳抑制因子の不活化を導くと考えられる。
【0055】
本明細書中で言及する場合、翻訳抑制因子配列とは、問題のあるエンハンサー配列が由来する2分節のウイルス(例えばコモウイルス)の野生型RNA−2ゲノムセグメントにおける配列であり、野生型RNA−2ゲノムセグメントによってコードされる2つのカボキシ共末端タンパク質のより長い方を産生(翻訳)するための制限部位を含むか、それから構成される。
【0056】
CPMV RNA−2ゲノムセグメント由来のエンハンサー配列における翻訳抑制因子配列は、上記の標的開始部位を含むか、またはそれから構成される配列である。従って、翻訳抑制因子配列は、上記の標的開始部位を含むか、それから構成され、上記の変異誘発によって不活性化され得る。
【0057】
上記のエンハンサー配列は、以下のような本発明の種々の態様および実施形態において使用されてもよい。
【0058】
従って、本発明の一態様において、上記の発現エンハンサー配列から構成されるか、またはそれから実質的に構成される単離された核酸が提供または利用される。
【0059】
本明細書中で使用される場合、「核酸」または「核酸分子」とは、一本鎖または二本鎖のいずれかである任意のDNAまたはRNA分子を指し、一本鎖の場合、その相補的配列の分子は、直線状または環状の形態のいずれかである。核酸分子を考慮する際に、特定の核酸分子の配列または構造が、通常の従来の5’から3’方向における配列を与えることによって本明細書中に記載され得る。本発明の核酸に関して、用語「単離された核酸」が時々使用される。DNAに当てはめる場合、この用語は、起源となる生物の天然のゲノムにすぐに隣接する配列から分離されるDNA分子を指す。
【0060】
例えば、「単離された核酸」は、プラスミドまたはウイルスベクターなどのベクターに挿入されるか、あるいは原核細胞もしくは真核細胞または宿主生物のゲノムDNAに組み込まれるDNA分子を含んでもよい。
【0061】
RNAに当てはめる場合、この用語「単離された核酸」とは主に、上記の単離されたDNA分子によってコードされるRNA分子を指す。あるいは、この用語は、天然の状態(すなわち、細胞または組織中)で会合する他の核酸から十分に分離されているRNA分子を指す場合もある。「単離された核酸」(DNAまたはRNAのいずれか)はさらに、生物学的または合成的手段によって直接産生され、その産生の間に存在する他の化合物から分離される分子を表す場合もある。
【0062】
従って、核酸は、エンハンサーが由来する2分節のRNAウイルスのRNA−2ゲノムセグメントの一部またはそのフラグメントを含むか、または実質的に含んでもよい。例えば、一実施形態において、核酸は、由来するRNA−2ゲノムセグメントのコード領域の少なくとも一部を含まない。コード領域は、2つのカルボキシ共末端タンパク質のより短い方をコードするRNA−2ゲノムセグメントの領域であり得る。核酸は、野生型RNA−2ゲノムセグメントの5’末端から、野生型RNA−2ゲノムセグメントによってコードされる2つのカルボキシ共末端タンパク質のより短い方の産生(翻訳)が開始される開始部位まで伸長する2分節のウイルスのRNA−2ゲノムセグメントの一部から構成されてもよいか、またはそれから実質的に構成されてもよい。
【0063】
特定のヌクレオチドまたはアミノ酸を言及する場合、用語「から実質的に構成される」とは、所定の配列番号の特性を有する配列を意味する。例えば、アミノ酸配列に関して使用される場合、この用語は、配列の基本的かつ新規の特徴に影響を及ぼさない配列それ自体および分子修飾を含む。例えば、核酸に対して使用される場合、この用語は、配列のエンハンサー機能に影響を及ぼさないか、またはさらなる(追加的な)機能を与えない配列それ自体ならびに少しの変化および/または伸長物を含む。
【0064】
本発明はさらに、本発明のエンハンサー配列を含む遺伝子発現系に関する。
【0065】
従って、別の態様において、本発明は、上記のエンハンサー配列を含む遺伝子発現系を提供する。
【0066】
遺伝子発現系はまた、エンハンサー配列の下流に挿入される目的のタンパク質をコードする遺伝子を含んでもよい。目的のタンパク質をコードする挿入される配列は、任意のサイズであってもよい。
【0067】
従って、さらなる態様において、本発明は、
(a)上記のエンハンサー配列、および(b)目的のタンパク質をコードする遺伝子を含む遺伝子発現系を提供し、ここで、その遺伝子はエンハンサー配列の下流に位置する。
【0068】
目的の遺伝子およびタンパク質は異種であってもよい。すなわち、エンハンサー配列が由来する野生型の2分節のRNAウイルスによってコードされない。
【0069】
遺伝子発現系は、宿主生物において目的のタンパク質を発現するために使用され得る。この場合、目的のタンパク質はまた、問題の宿主細胞に対して異種であってもよい。すなわち遺伝子操作を用いて、すなわちヒトの介入によって、問題の細胞(例えば、植物またはその原種)に組み込まれる。生物内の異種遺伝子は、内因性の等価遺伝子(すなわち、同じまたは類似の機能を正常に果たすもの)と置き換えられてもよいか、または挿入される配列は内因性遺伝子もしくは他の配列に対して付加的であってもよい。
【0070】
当業者は、目的の遺伝子の発現が、発現される遺伝子の上流に位置する開始部位(AUG)の存在を必要とすることを理解するだろう。そのような開始部位は、エンハンサー配列の一部または目的のタンパク質をコードする遺伝子の一部のいずれかとして与えられてもよい。
【0071】
宿主生物は植物であってもよい。しかしながら、翻訳機構が真核生物にわたって十分に保存されているため、遺伝子発現系はまた、植物以外の真核宿主生物、例えば修飾されたバキュロウイルスベクターなどの昆虫細胞、または酵母もしくは哺乳動物細胞において目的のタンパク質を発現するために使用されてもよい。
【0072】
遺伝子発現系は、プロモーターおよび終結配列に作動可能に連結されてもよい。
【0073】
従って、遺伝子発現系はさらに、終止配列を含んでもよく、目的のタンパク質をコードする遺伝子は、エンハンサー配列と終止配列との間、すなわちエンハンサー配列の下流(3’)および終止配列の上流(5’)に位置してもよい。
【0074】
従って、本発明はさらに、
(i)作動可能に連結されるプロモーター
(ii)上記のエンハンサー配列
(iii)発現することが所望される目的の遺伝子
(iv)終結配列
を含む、発現カセットを提供する。
【0075】
好ましくは、目的の遺伝子を駆動するのに使用されるプロモーターは、強力なプロモーターである。植物に使用するための強力なプロモーターの例としては、
(1)p35S:Odellら、1985
(2)Cassava Vein Mosaic Virusプロモーター、pCAS、Verdaguerら、1996
(3)リブロース二リン酸カルボキシラーザの小さいサブユニットのプロモーター、pRbcS:Outchkourovら、2003
が挙げられる。
【0076】
他の強力なプロモーターとしては、pUbi(単子葉植物および双子葉植物について)およびpActinが挙げられる。
【0077】
好ましい実施形態において、プロモーターは、誘導プロモーターである。
【0078】
プロモーターに適用される場合、用語「誘導」は当業者に十分に理解されている。本質的に、誘導プロモーターの制御下での発現は、与えられた刺激に応答して「スイッチオン」または増加される。刺激の性質はプロモーター間で変化する。一部の誘導プロモーターは、適切な刺激の非存在下において、少し、または検出できないレベルの発現(または発現なし)を生じる。他の誘導プロモーターは、刺激の非存在下において、検出可能な構成的発現を生じる。発現レベルは刺激の非存在下にかかわらず、任意の誘導プロモーター由来の発現は、適切な刺激の存在下において増加する。
【0079】
終止(終結)配列は、2分節RNAウイルス、例えばコモウイルスのRNA−2ゲノムセグメント由来の終止配列であり得る。一実施形態において、終止配列は、エンハンサー配列が由来する同じ2分節RNAウイルス由来であってもよい。終止配列は、停止コドンを含んでもよい。終止配列はまた、ポリアデニル化信号を伴ってもよい。
【0080】
本発明の遺伝子発現カセット、遺伝子発現構築物および遺伝子発現系はまた、非翻訳領域(UTR)を含んでもよい。UTRは、遺伝子発現カセット、遺伝子発現構築物または遺伝子発現系に存在する終結配列の上流に位置してもよい。遺伝子発現カセット、遺伝子発現構築物または遺伝子発現系が、目的のタンパク質をコードする遺伝子を含む場合、UTRは前記遺伝子の下流に位置してもよい。
【0081】
従って、UTRは、目的のタンパク質をコードする遺伝子と終結配列との間に位置してもよい。UTRは、2分節RNAウイルス由来、例えば2分節RNAウイルスのRNA−2ゲノムセグメント由来であってもよい。UTRは、遺伝子発現カセット、遺伝子発現構築物または遺伝子発現系に存在するエンハンサー配列が由来する同じRNA−2ゲノムセグメントの3’UTRであってもよい。好ましくは、UTRは、コモウイルスRNA−2ゲノムセグメントの3’UTR、例えばCPMV RNA−2ゲノムセグメントの3’UTRである。
【0082】
上記のように、RNA−2における161位および512位の開始部位の間のフレームが維持されること、すなわち、2つの開始部位が同じトリプルリーディングフレームに残っていることは、CPMV RNA−1にコードされるレプリカーゼによるCPMV RNA−2の効率的な複製のために必須であることが以前に示されている(Holnessら,1989;van Bokhovenら,1993;Rohllら,1993)。この条件は、CPMVに基づいた発現ベクターにおける512位の開始部位の上流に挿入できる配列の長さを制限した。特に、ポリリンカーの使用がしばしば、2つの開始部位の間のオープンリーディングフレーム(ORF)を変化させるため、それらの使用が妨げられる。
【0083】
本発明は、CPMV RNA−2における161位および512位の開始部位の間のリーディングフレームの維持がまた、512位の開始部位における翻訳の効率的な開始に必要とされること、すなわち、CPMVによってコードされる2つのカルボキシ共末端タンパク質より短い方(95Kタンパク質)の効率的な発現に必要とされることも示す。
【0084】
しかしながら、本発明はまた、CPMV RNA−2における161位の開始部位の変異が、95Kタンパク質の発現レベルに対していかなる負の影響も与えずに、変異された開始コドンと512位の開始部位との間のフレームを変化させる、512位の開始部位の上流の配列の挿入を可能にすることも示す。従って、161位の開始部位の変異は、512位の開始部位の上流に挿入できる配列の長さをもはや制限しないことを意味する。
【0085】
2つのカルボキシ共末端タンパク質をコードする開始部位の間のリーディングフレームの維持もまた、他の2分節ウイルスに必要とされる場合、この条件はまた、ウイルスRNA−2ゲノムセグメントによってコードされるより2つのカルボキシ共末端タンパク質のより長い方を産生するための開始部位として役立つAUGを変異させることによって克服され得る。従って、別の態様において、本発明は、
(a)上記のエンハンサー配列、および
(b)遺伝子発現系に目的のタンパク質をコードする遺伝子の挿入を促進するための異種配列を含む遺伝子発現構築物を提供し、ここで、その異種配列は、エンハンサー配列における変異された標的開始部位の下流に位置する。
【0086】
異種配列は開始コドンの上流に位置してもよく、その開始コドンから2つのカルボキシ共末端タンパク質のうちのより短い方の産生が、遺伝子発現系のエンハンサー配列が由来する野生型RNA−2ゲノムにおいて開始される。あるいは、異種配列は、開始コドンの部位付近に与えられてもよいか、または開始コドンと置換されてもよく、その開始コドンから2つのカルボキシ共末端タンパク質のより短い方が、遺伝子発現系のエンハンサー配列が由来する野生型RNA−2ゲノムセグメントにおいて開始される。CPMVのRNA−2由来のエンハンサー配列を有する遺伝子発現系において、異種配列は、開始コドンの部位の上流、その付近に与えられてもよいか、または開始コドンと置換されてもよく、その開始コドンは、野生型RNA−2 CPMVゲノムセグメントにおける512位にある。
【0087】
異種配列は、ポリリンカーまたは複数のクローニング部位、すなわち、発現系に目的のタンパク質をコードする遺伝子のクローニングを促進する配列であってもよい。
【0088】
例えば、以下に記載するように、本発明は、CPMVに基づいた発現カセットの5’リーダーと3’UTRとの間にポリリンカーを含む構築物を与える。以下に記載するように、任意のポリリンカーは、必要に応じて、タンパク質の精製を促進するためにNまたはC末端His−タグの融合を可能にするように複数のxヒスチジン残基の1つ以上のセットをコードしてもよい。
【0089】
好ましくは、上記の発現構築物はベクターに存在し、好ましくは、それは、生物ゲノムへの発現カセットの転写および組み込みを可能にする境界配列を含む。
【0090】
好ましくは、構築物は植物のバイナリーベクターである。好ましくは、バイナリー形質転換ベクターはpPZPに基づく(Hajdukiewiczら,1994)。他の例示的な構築物としては、pBin19が挙げられる(Frisch,D.A.,L.W.Harris−Hallerら,(1995).「Complete Sequence of the binary vector Bin19.」Plant Molecular Biology 27:405−409を参照のこと)。
【0091】
本明細書に記載するように、本発明は、存在するpBin発現カセットに必須の要素を有する発現カセットを移動させることによって実施されてもよいか、または他の実施形態において、pBin発現ベクターの直接クローニングが利用されてもよい。
【0092】
例えば、以下に記載するように、本発明は、本明細書に記載されるエンハンサー配列とともに使用する(それらに制限されないが)ために設計されたモジュラーバイナリーベクターを有する。それらは、pBINPLUSベクターに対する改良に基づき、従って、複製およびTDNA転写についての機能を妥協することなしに、ベクターのサイズを劇的に減少させることが可能であることが示されている。さらに、(いわゆる「CPMV−HT」系によって例示されるように)エンハンサー系の要素が、複数のタンパク質が単一のT−DNAから発現され得るように、モジュラー方式で、得られるベクター内に組み込まれてもよい。それらの改良は、外来性遺伝子の効率的な直接クローニングを可能にする多用途の高レベル発現ベクターの産生を導く。
【0093】
これらの例は好ましいバイナリー植物ベクターを表す。好ましくは、それらには、ColEI複製起点が含まれるが、高コピー数も生じる他の複製起点を含むプラスミド(例えば、pRiに基づくプラスミド、LeeおよびGelvin,2008)、特に一過性発現系も好ましい。
【0094】
所望の場合、選択可能な遺伝子マーカーは、抗生物質または除草剤(例えば、カナマイシン、ハイグロマイシン、ホスフィノトリシン、クロルスルフロン、メトトレキサート、ゲンタマイシン、スペクチノマイシン、イミダゾリノンおよびグリホサート)に対する耐性などの選択可能な表現型を与えるものなどの構築物に含まれてもよい。
【0095】
最も好ましいベクターは以下に記載するpEAQベクターであり、遺伝子サイレンシングの抑制因子およびNPTIIカセットも含むT−DNAに配置される、本発明の翻訳エンハンサーを含む発現カセットの5’リーダーと3’UTRとの間のポリリンカーの使用による直接クローニング型を可能にする。ポリリンカーはまた、NまたはC末端His−タグの融合により、タンパク質精製を促進できる1つまたは2つのセットの6xヒスチジン残基をコードする。
【0096】
pEAQ由来ベクターの利点は、IgGなどの複数鎖タンパク質の各要素が、各感染細胞に自動的に送達され得ることである。
【0097】
本発明はまた、植物、酵母、昆虫または哺乳動物細胞などの宿主生物において、本発明の遺伝子発現系を用いて、タンパク質、例えば異種タンパク質を発現する方法を提供する。
【0098】
本発明はさらに、上記のRNA2由来の配列に作動可能に連結される、目的の異種タンパク質をコードする遺伝子またはORFからその目的の異種タンパク質の翻訳を向上させる方法を提供し、その方法は、RNA2由来の配列における標的開始部位を変異させることを含む。
【0099】
本明細書に記載されるエンハンサー配列はまた、WO/2007/135480に記載される2分節の発現系に使用され得る。従って、本発明は、切断されたRNA2−遺伝子セグメントに基づいた遺伝子発現系にも関連し、必要に応じてさらに、プロモーターおよび終結配列に作動可能に連結された遺伝子サイレンシングの抑制因子をコードする第2の遺伝子構築物を含む。
【0100】
従って、さらなる態様において、本発明は、
(a)プロモーターおよび終結配列に作動可能に連結される目的の異種タンパク質をコードする少なくとも1つの外来性遺伝子を保有する2分節のウイルスゲノムの切断されたRNA−2を含む第1の遺伝子構築物であって、その第1の遺伝子構築物は、外来性遺伝子の上流に変異された標的開始部位を含む、第1の遺伝子構築物と、必要に応じて、
(b)プロモーターおよび終結配列に作動可能に連結される、前記2分節のウイルスゲノムのRNA−1を含む第2の遺伝子構築物と、必要に応じて、
(c)前記第1の遺伝子構築物、前記第2の遺伝子構築物またはその両方内に必要に応じて組み込まれる第3の遺伝子構築物であって、プロモーターおよび終結配列に作動可能に連結される遺伝子サイレンシングの抑制因子を含む、第3の遺伝子構築物と、
を含む遺伝子発現系に関連する。
【0101】
本発明の遺伝子発現系(上記のもののいずれかを含む)における遺伝子サイレンシングの抑制因子の存在は好ましいが、必須ではない。従って、上記の遺伝子発現系は、好ましくは、前記第1の遺伝子構築物、前記第2の遺伝子構築物、またはその両方内に必要に応じて組み込まれる第3の遺伝子構築物であって、プロモーターおよび終結配列に作動可能に連結される遺伝子サイレンシングの抑制因子を含む、第3の遺伝子構築物を含む。
【0102】
従って、別の態様において、本発明は、以下の工程
(a)本発明の遺伝子発現構築物を植物細胞に導入する工程と、必要に応じて、
(b)プロモーターおよび終結配列に作動可能に連結される、前記2分節のウイルスゲノムのRNA−1を含む第2の遺伝子構築物を植物細胞に導入する工程と、必要に応じて、
(c)前記第1の遺伝子構築物、前記第2の遺伝子構築物、またはその両方内に必要に応じて組み込まれる第3の遺伝子構築物であって、プロモーターおよび終結配列に作動可能に連結される遺伝子サイレンシングの抑制因子を含む第3の遺伝子構築物を植物細胞に組み込む工程と、
を含む植物においてタンパク質を発現する方法を提供する。
【0103】
好ましくは、上記の植物においてタンパク質を発現する方法は、
前記第1の遺伝子構築物、前記第2の遺伝子構築物、またはその両方内に必要に応じて組み込まれる第3の遺伝子構築物であって、プロモーターおよび終結配列に作動可能に連結される遺伝子サイレンシングの抑制因子を含む第3の遺伝子構築物を植物細胞に導入する工程を含む。
【0104】
本発明はまた、このような構築物を植物細胞に導入することを含む方法を提供する。
【0105】
本発明は、翻訳エンハンサーとして同じT−DNAに目的の遺伝子およびサイレンシングの抑制因子を組み込むことによって非常に高い発現レベルを示す。従って、好ましい実施形態は、同じT−DNAに存在する全てのこれらの要素を利用することであり得る。
【0106】
さらに、RNA−1が、本明細書に記載される系において、高レベルの発現を必要とされず、実際には、本明細書に記載される「CPMV−HT」系はRNA−1の作用によってではないことは理解されるべきである。
【0107】
従って、さらなる態様において、本発明は、
(a)プロモーターおよび終結配列に作動可能に連結される目的の異種タンパク質をコードする少なくとも1つの外来性遺伝子を保有する2分節のウイルスゲノムの切断されたRNA−2を含む第1の遺伝子構築物であって、その遺伝子構築物は、外因性遺伝子の上流に変異された標的開始部位を含む、第1の遺伝子構築物と、必要に応じて、
(b)前記第1の遺伝子構築物内に必要に応じて組み込まれる第2の遺伝子構築物、プロモーターおよび終結配列に作動可能に連結される遺伝子サイレンシングの抑制因子と、
を含む遺伝子発現系に関する。
【0108】
従って、別の態様において、本発明は、以下の工程
(a)本発明の遺伝子発現構築物を植物細胞に導入する工程と、必要に応じて、
(b)前記第1の遺伝子構築物内に必要に応じて第2の遺伝子構築物を導入する工程であって、前記第2の遺伝子構築物は、植物細胞内のプロモーターおよび終結配列に作動可能に連結される遺伝子サイレンシングの抑制因子を含む、工程と、
を含む植物においてタンパク質を発現する方法を提供する。
【0109】
これらの態様において有用な遺伝子サイレンシングの抑制因子は、当該分野において公知であり、WO/2007/135480に記載される。それらとしては、ジャガイモウイルス(Potato virus)Y由来のHcPro、TEV由来のHe−Pro、TBSV由来のP19、rgsCam、FHV由来のB2タンパク質、CPMVの小コートタンパク質、およびTCV由来のコートタンパク質が挙げられる。最も好ましくは、この系のRNA−2は、感染ウイルスが産生されないように切断されている。
【0110】
安定なトランスジェニック植物を産生する場合、好ましい抑制因子は、R43W変異を組み込んでいるP19抑制因子である。
【0111】
本発明のさらなる態様において、本発明による異種構築物を含む宿主細胞が開示される。
【0112】
本発明の遺伝子発現ベクターは、植物細胞内に一過性または安定に組み込まれてもよい。
【0113】
スモールスケールの産生に関しては、本発明の構築物を有する葉の機械的アグロ浸潤法(agroinfiltration)である。スケールアップは、例えば減圧浸潤の使用により達成される。
【0114】
他の実施形態において、本発明の発現ベクターは、トランスジェニック植物または植物細胞のゲノム内に安定に組み込まれてもよい。
【0115】
一態様において、本発明は、形質転換した植物細胞から植物を再生する工程をさらに含んでもよい。
【0116】
植物で広範に成功している以前に使用されている特定の手順およびベクターは、GuerineauおよびMullineaux(1993)(Plant transformation and expression vectors.In:Plant Molecular Biology Labfax(Croy RRD ed)Oxford,BIOS Scientific Publishers,pp121−148)に記載されている。適切なベクターは、植物ウイルス由来のベクターを含んでもよい(例えば欧州特許EP−A−194809を参照のこと)。所望の場合、選択可能な遺伝子マーカーは、抗生物質または除草剤(例えば、カナマイシン、ハイグロマイシン、ホスフィノトリシン、クロルスルフロン、メトトレキサート、ゲンタマイシン、スペクチノマイシン、イミダゾリノンおよびグリホサート)に対する耐性などの選択可能な表現型を与えるものなどの構築物に含まれてもよい。
【0117】
核酸は、その天然の遺伝子導入の能力を利用するアグロバクテリウムによって運搬される安全化されたTi−プラスミドベクター(欧州特許EP−A−270355、欧州特許EP−A−0116718、NAR12(22)8711−87215 1984;the floral dip method of Clough and Bent,1998)、粒子衝突または微粒子銃(米国特許第5100792号、EP−A−444882、EP−A−434616)、微量注入法(WO92/09696、WO94/00583、欧州特許EP331083、欧州特許EP175966、Greenら(1987)Plant Tissue and Cell Culture,Academic Press)、エレクトロポレーション(欧州特許EP290395、WO8706614 Gelvin Debeyser)、直接的なDNA取り込みの他の形態(独国特許第4005152号、WO9012096、米国特許第4684611号)、リポソームを介したDNA取り込み(例えば、Freemanら、Plant Cell Physiol.29:1353(1984))、または渦法(vortexing method)(例えば、Kindle,PNAS U.S.A.87:1228(1990d)Physical methods for the transformation of plant cells are reviewed in Oard,1991,Biotech.Adv.9:1−11)などの任意の適切な技術を用いて植物細胞内に組み込むことができる。Ti−プラスミド、特に2分節のベクターは、以下により詳細に記載される。
【0118】
アグロバクテリウム形質転換は、双子葉植物の種を形質転換するために、当業者によって広範に使用されている。しかしながら、ほとんど全ての経済的に関係のある単子葉植物における安定で、繁殖性のトランスジェニック植物の所定の産生にもかなり成功している(例えば、Hieiら(1994)The Plant Journal 6,271−282を参照のこと)。アグロバクテリウム単独が、非効率かまたは効果がない場合、微粒子銃、エレクトロポレーション、および直接的なDNA取り込みが好ましい。あるいは、異なる技術の組み合わせ、例えば、アグロバクテリウムでコートされた微小粒子での照射(欧州特許EP−A−486234)または損傷後のアグロバクテリウムとの共培養を誘発する微粒子銃(欧州特許EP−A−486233)が、形質転換プロセスの効率を向上させるために利用されてもよい。
【0119】
形質転換技術の特定の選択は、特定の植物種を形質転換するその効率性ならびに特定の方法を選択する本発明を実施する人の経験および好みによって決定される。
【0120】
核酸を植物細胞に導入する形質転換系の特定の選択は、本発明に必須ではなく、または本発明の限定ではなく、植物再生についての技術の選択も本発明に必須ではなく、または本発明の限定でもないことは当業者には明らかであろう。本発明者らによって実施される実験において、向上した発現の効果は、T−DNAの種々の組み込みパターンにおいて見られる。
【0121】
従って、本発明の種々の態様は、植物組織(例えば植物細胞)への本発明の構築物の導入、ならびに本発明による核酸をゲノムへ導入するためにベクターと植物細胞ゲノムとの間の組み換えを生じることまたは可能にすること、を含む植物細胞を形質転換する方法を提供する。これは、一過性発現をもたらすように行われ得る。
【0122】
あるいは、植物組織の形質転換後、植物は、当該分野において標準的な例えば、単一の細胞、カルス組織または葉片から再生されてもよい。ほとんどのあらゆる植物は、植物の細胞、組織および器官から完全に再生され得る。利用可能な技術は、Vasilら,Cell Culture and Somatic Cell Genetics of Plants,Vol I,IIおよびIII,Laboratory Procedures and Their Applications,Academic Press,1984,ならびにWeissbachおよびWeissbach,Methods for Plant Molecular Biology,Academic Press,1989に概説されている。
【0123】
繁殖性のトランスジェニック植物の産生は、コメ、トウモロコシ、コムギ、カラスムギ、およびオオムギなどの穀物、さらに多くの他の植物種においてなされている(Shimamoto,K.(1994)Current Opinion in Biotechnology 5,158−162;Vasilら(1992)Bio/Technology 10,667−674;Vainら,1995,Biotehnology Advances 13(4):653−671;Vasil,1996,Nature Biotechnology 14,702ページに概説されている)。
【0124】
再生された植物またはその一部は、クローン、種子、自己またはハイブリッド子孫(progenyおよびdescendant)(例えば、F1およびF2子孫)、挿し木(例えば、食用部分)、栄養繁殖体などを提供するために使用されてもよい。
【0125】
本発明はさらに、トランスジェニック生物(例えば、本明細書に記載される方法によって得たか、または得られる)を提供し、このトランスジェニック生物に発現ベクターまたはカセットが導入され、そのカセットにおける異種遺伝子が向上したレベルで発現される。
【0126】
本発明はさらに、目的のタンパク質を産生する方法を含み、その方法は、上記の方法を実施する(または生物を使用する)工程と、必要に応じて少なくとも、目的のタンパク質が発現される組織を収集する工程と、その組織から目的のタンパク質を単離する工程を含む。
【0127】
従って、具体的には、本発明は、トランスジェニック植物または本発明の発現ベクターで一過性にトランスフェクトされた植物細胞を提供する。
【0128】
さらなる態様において、本発明はまた、トランスジェニック植物または本発明の発現ベクターで安定に形質転換された植物細胞を提供する。
【0129】
本発明はまた、挿し木、種子などを含む、複製または増殖、有性または無性に使用され得る任意の部分である、そのような植物由来の植物栄養繁殖体を提供する。本発明はまた、上記の植物細胞または異種DNAを含むそれらの植物の任意の部分を提供する。
【0130】
従って、種々の態様において、(限定されないが)本発明は以下を提供する。
・本発明のエンハンサー配列から構成されるか、またはエンハンサー配列から実質的に構成される核酸(そのエンハンサー配列は、(例えば)CPMV RNA−2ゲノムセグメントのヌクレオチド1〜512から構成されるか、あるいはそれら由来であるか、または2分節のRNAウイルスの別のRNA−2ゲノムセグメント由来であり、各々の場合において、CPMV RNA−2の161位に対応する標的開始部位が変位される)。
・例えば、目的のタンパク質、またはポリリンカー、および必要に応じてターミネーターをコードするORFの上流にそのようなエンハンサー配列を含む遺伝子発現系。
・本明細書に記載されるエンハンサー配列を使用するために本発明に従って修飾されたWO/2007/135480に記載される2分節の発現系。
・(i)作動可能に連結されるプロモーター、(ii)上記のエンハンサー配列、(iii)発現することが望まれる目的のポリリンカーまたは遺伝子、(iv)同系の3’UTR(すなわち、CPMV RNA−2ゲノムセグメントの3’UTR由来)、(v)終結配列を含む、発現カセット。
・本発明の遺伝子発現系またはベクターを用いる植物などの宿主生物におけるタンパク質、例えば異種タンパク質を発現する方法。
・本発明の遺伝子発現系またはベクター由来のタンパク質を発現する宿主細胞および生物(例えば、植物または酵母)ならびにそれらを産生する方法。
【0131】
文脈が他に要求しない限り、「遺伝子」とは、ペプチド、ポリペプチドまたはタンパク質への翻訳のための遺伝情報をコードする任意の核酸を指す。従って、文脈が他に要求しない限り、それは「ORF」と交換可能に用いられる。
【0132】
発現することが所望され得る遺伝子は、導入遺伝子または内因性であってもよい。
【0133】
目的の遺伝子には、農業形質、虫害抵抗性、病害抵抗性、除草剤抵抗性、無菌性、粒子特性などをコードする遺伝子が含まれる。この遺伝子は、油、デンプン、炭水化物、栄養素などの代謝に関与してもよい。従って、目的の遺伝子または特徴としては、限定されないが、環境またはストレス関連の形質、疾患関連の形質、および農業生産力に影響を与える形質が挙げられる。標的配列にはまた、タンパク質、ペプチド、脂肪酸、脂質、ワックス、油、デンプン、糖、炭水化物、香味料、香気、毒素、カロテノイド、ホルモン、ポリマー、フラボノイド、貯蔵タンパク質、フェノール酸、アルカロイド、リグニン、タンニン、セルロース、糖タンパク質、糖脂質などの合成に関与する遺伝子が含まれる。
【0134】
最も好ましくは、単子葉植物および/または双子葉植物における標的遺伝子としては、セイヨウアブラナ、ヒマワリ、ダイズおよびトウモロコシなどの植物中の油産生に関与する酵素;ジャガイモ、トウモロコシ、穀物などの植物中のデンプン合成に関与する酵素;医薬製剤または獣医用製剤などの天然の薬剤を合成する酵素、またはそれらのタンパク質自体をコードする遺伝子が挙げられる。
【0135】
異種核酸は、とりわけ、細菌、真菌、植物または動物起源の遺伝子をコードすることができる。ポリペプチドは、(例えば、害虫感受性、活力、組織分化、繁殖性、栄養価などに関連する植物の特徴を修飾するために)植物体で利用されてもよいか、または植物は、他に使用するためにその植物から精製できるポリペプチドを産生するための中間体であってもよい。そのようなタンパク質としては、限定されないが、網膜芽細胞腫タンパク質、p53、アンギオスタチン、およびレプチンが挙げられる。同様に、本発明の方法は、哺乳動物の調節タンパク質を産生するために使用されてもよい。目的の他の配列としては、タンパク質、ホルモン、成長因子、サイトカイン、血清アルブミン、ヘモグロビン、コラーゲンなどが挙げられる。
【0136】
従って、標的遺伝子またはヌクレオチド配列は、好ましくは、目的のタンパク質:虫害抵抗性タンパク質;病害抵抗性タンパク質;除草剤抵抗性タンパク質;哺乳動物タンパク質をコードする。
【0137】
「ベクター」は、とりわけ、二本鎖または一本鎖の直線状または環状形態で任意のプラスミド、コスミド、ファージ、ウイルスまたはアグロバクテリウムバイナリーベクターを含むと定義され、それは、自己感染性または可動性であってもなくてもよく、細胞ゲノムまたは存在する染色体外への組み込みによって原核生物または真核生物宿主のいずれかを形質転換できる(例えば、複製起点を有するプラスミドの自立的複製)。使用される構築物は、完全または部分的に合成される。特にそれらは、天然に一緒に見出されない(隣接しない)核酸配列が連結されるか、またはそうでなければ人工的に合成される組み換え体である。他に指定されない限り、本発明によるベクターは、特に、ベクターが、ゲノムへの組み込みのために核酸を細胞に組み込むために使用される場合、必ずしも、プロモーターまたは他の調節配列を含まなくてもよい。
【0138】
「バイナリーベクター」は当業者に周知であり、バイナリーベクター系には、(a)所望のヌクレオチド配列の植物細胞ゲノムへの移入を可能にする境界配列;(b)(i)作動可能に連結される植物活性プロモーターと、(ii)適切な場合、標的配列および/またはエンハンサーの発現カセットと、を一般的に含む所望のヌクレオチド配列自体、が含まれる。所望のヌクレオチド配列は境界配列の間に位置し、適切な条件下で植物ゲノム内に挿入できる。バイナリーベクター系は、通常、組み込みをもたらすために他の配列(A.tumefaciens由来)を必要とする。通常、このことは、アグロバクテリウムによって媒介される一過性形質転換を使用する、いわゆる「アグロ浸潤法」の使用によりなされ得る。つまり、この技術は、核DNAに組み込む場合、そのDNA(「T−DNA」)の一部を宿主細胞へ移入するAgrobacterium tumefaciensの性質に基づく。T−DNAは、21〜23ヌクレオチド長付近の左および右の境界配列によって定義される。浸潤は、例えば、シリンジ(葉)または真空(完全植物)によってなされ得る。本発明において、境界配列は、通常、アグロ浸潤法によって植物物質へ組み込まれる1つ以上のベクターを有する所望のヌクレオチド配列(T−DNA)に含まれる。
【0139】
「発現カセット」とは、核酸が、微生物または植物細胞などの宿主細胞内の転写のための適切なプロモーターまたは他の調節エレメントの制御下であり、作動可能に連結される状態を指す。
【0140】
「プロモーター」は、下流に(すなわち、二本鎖DNAのセンス鎖の3’方向において)作動可能に連結されるDNAの転写が開始され得るヌクレオチド配列である。
【0141】
「作動可能に連結される」とは、同じ核酸分子の一部として、プロモーターから開始される転写のための安定な位置および方向に結合されることを意味する。
【0142】
目的の「植物」種としては、限定されないが、トウモロコシ(Zea mays)、Brassica sp.(例えば、B.napus、B.rapa、B.juncea)特に種油の源として有用なそれらのBrassica種、ムラサキウマゴヤシ(Medicago sativa)、コメ(Oryza sativa)、ライムギ(Secale cereale)、モロコシ(Sorghum bicolor、Sorghum vulgare)、ヒエ(例えば、トウジンビエ(Pennisetum glaucum))、キビ(Panicum miliaceum)、アワ(Setaria italica)、シコクビエ(Eleusine coracana))、ヒマワリ(Helianthus annuus)、ベニバナ(Carthamus tinctorius)、コムギ(Triticum aestivum)、ダイズ(Glycine max)、タバコ(Nicotiana tabacum)、Nicotiana benthamiana、ジャガイモ(Solanum tuberosum)、ピーナッツ(Arachis hypogaea)、綿(Gossypium barbadense、Gossypium hirsutum)、サツマイモ(Ipomoea batatus)、キャッサバ(Manihot esculenta)、コーヒー(Coffea spp.)、ココナツ(Cocos nucifera)、パイナップル(Ananas comosus)、シトラスの木(Citrus spp.)、ココア(Theobroma cacao)、茶(Camellia sinensis)、バナナ(Musa spp.)、アボガド(Persea americana)、イチジク(Ficus casica)、グアバ(Psidium guajava)、マンゴー(Mangifera indica)、オリーブ(Olea europaea)、パパイヤ(Carica papaya)、カシュー(Anacardium occidentale)、マカダミア(Macadamia integrifolia)、アーモンド(Prunus amygdalus)、サトウダイコン(Beta vulgaris)、サトウキビ(Saccharum spp.)、オートムギ、オオムギ、野菜、観賞植物、および針葉樹が挙げられる。当業者は、本明細書に記載されるウイルスベクターの向性が変化することは理解するだろう。しかしながら、そのようなウイルスに対する感受性を決定することは、当業者に周知である。さらに、植物細胞内へのウイルス浸入を促進するレセプターを組み換え発現することによってそのような特異性を変更することは可能であり得る。
【0143】
本発明は、ここで、以下の非限定的な図および実施例を参照して詳細に記載される。本発明の他の実施形態は、これらの観点から当業者が想到するだろう。
【0144】
本発明を実施する当業者に使用され得る限り、本明細書に引用される全ての参考文献の開示は、相互参照として本明細書に具体的に援用される。
【実施例】
【0145】
(実施例1)
1.1方法
発現ベクターFSC2およびその誘導体の作製
pBinP−1−GFPベースのプラスミド(Canizaresら,2006)由来の外来性タンパク質の発現のために有用なクローニングベクターを、カリフラワーモザイクウイルス(CaMV)35SプロモーターおよびpBinP−S2NT(LiuおよびLomonossoff,2002)由来のノパリン合成(nos)ターミネーターに隣接するRNA−2の完全な配列を切断し、それを、Ascl/Pacl断片として変異誘発プラスミドpM81W(LiuおよびLomonossoff,2006)に挿入することによって作製した。得られたプラスミド、pM81W−S2NTに、4つの変更を同時に導入する単一ラウンドの変異誘発(LiuおよびLomonossoff,2006における方法を参照のこと)を行って、pM81B−S2NT−1を得た。変異誘発により、ベクターバックボーンから2つのBspHI部位を除去し、AUG512付近にBspHI部位(T/CATGA)およびUAA3299後にStuI部位(AGG/CCT)、RNA−2によりコードされるポリタンパク質についての終止コドンを導入した。続いて、BamHI/Ascl断片を、pBinP−NS−1(Liuら,2005)から切断し、同様に消化したpM81B−S2NT−1にライゲーションして、pM81−FSC−1を得た。このベクターにより、AUG512の下流のRNA−2 ORFの全てを、BspHIおよびStuIでの消化により切断でき、BspHIおよびStuI(平滑)適合性末端((blunt)−compatible end)を有する任意の配列と置換できる。BspHI部位の使用は重要である。なぜなら、それは、512位のAUGを保存し、このイニシエーターは、挿入した遺伝子の翻訳を駆動するために使用されるからである。植物において外来性遺伝子を発現するために、pM81−FSC−1由来プラスミドはAsclおよびPaclで消化され、同様に消化されたpBINPLUSに移入された外来性配列を含む発現カセットを有する断片および得られるプラスミドは、最終的に、A.tumefaciensに形質転換される。
【0146】
クローニングの容易さを向上させ、適切な制限酵素の選択を広げ、外来性遺伝子発現に対するリーディングフレームの効果を調べるために、全てのRNA−2 ORFを短いポリリンカーと置換した。オリゴヌクレオチド挿入およびpM81−FSC−2を生じる部位特異的突然変異誘発法の組み合わせにより、NruI(TCG/CGA)およびXhoI(C/TCGAG)またはStuIのいずれかでのクローニングが可能となる。NruI部位の末端のアデニンは512位に存在するので、ここに見られるAUGを保存する。この修飾により、5’から512位のAUGまで即座にヌクレオチドを変更させるが、十分な状況が維持される。翻訳が、512位、513位、514位または515位のAUGから開始されるようにpM81−FSC−2内にGFPをクローニングすることにより、pM81−FSC−1由来の構築物であるpM81−FSC2−512、pM81−FSC2−513、pM81−FSC2−514、およびpM81−FSC2−515を得た。これらのpM81ベースのプラスミドは、発現カセットを含むクローニングベクターであり、次いで、これをバイナリーベクターに移入して、図2および3に示す実験に使用する発現ベクターFSC2−512、FSC2−513およびFSC2−514を産生した。CPMVの野生型RNA−2ゲノムセグメントならびにpM81−FSC1およびpM81−FSC2ベクターの配列間の差異は表3に示す。野生型CPMV配列と比較したベクター内でのヌクレオチドの変更は大文字として示す。
【0147】
(pM81−FSC−1について上で概説したように)pBINPLUS内の動員後のアグロバクテリウムによって媒介される一過性形質転換により、AUG161と下流のAUGとの間のフレーム連続性が維持されない場合、より低いタンパク質レベルが得られることが示された。AUG161および512に対して+1位および+2位から翻訳したGFPの量は著しく減少したが、+3位からの翻訳(つまり、515からでフレーム内の後方)は、512位のAUGからの翻訳と同じ効率であった。このことが、513位および(より少ない程度の)514位の弱まった状況に起因していないことを示すために、FSC2−513のような同じ不十分な状況ではなく、FSC2−515+を+3位から開始するために作製した。FSC2−515+からの発現は、FSC2−512または515から達成したものと同じ高さであり、これは、低い状況がFSC2−513および514からの発現の減少ではないことを示す。
【0148】
公知の機構によって、翻訳が第1のAUGルールを逃れることができることを考慮すると、フレーム連続性を必要とすることが知られておらず、欠失したRNA−2ベースのベクターからの効果的な翻訳が、AUG161と下流のAUGとの間のフレーム連続性に依存することは興味をそそられる。この減少を理解して、望ましくは克服するために、一連の変異体を、RNA−2の5’配列に対する修飾を用いて作製した。オリゴヌクレオチドの相補対(表2を参照のこと)を、pM81−FSC2−512、513および514の部位特異的突然変異誘発法に使用した。それらの変異体のAUG115(uORFについての開始コドン)、AUG161(uORFのアミノ酸配列の変更なし)のいずれか、またはそれらの上流の開始部位の両方を除去した。2つの変異を、U162Cオリゴを用いてA115G変異体を変異誘発することによって作製した(表2)。
【0149】
これらの変異体転写産物からの一過性発現を、以前のpM81−FSC−2構築物について記載したように行った。coomassie染色したSDS−PAGE(図2)または葉の全てを視覚化するためにUV照射(図3)を用いるこれらの変異体からのGFPの発現の分析により、161位のAUGの非存在下において発現の強力な増加が示される。さらに、AUG161単独またはAUG115および161の両方の除去は、AUG161と下流のAUGとの間のフレーム連続性に対する依存性を軽減する。対照的に、単なるAUG115の除去はこの依存性を高め、そして全体的に翻訳を抑制するように見える。結論として、uORFは、AUG161からの翻訳を下方制御するために機能するように見え、それは全体的に抑制性があり、フレーム連続性に対する依存性を与える。
【0150】
HBcAG粒子を含む樹液(sap)の電子顕微鏡法
図6に示した集合したHBcAg粒子の電子顕微鏡写真に使用した樹液(sap)は以下のように調製した。葉組織を、2容量のTris/NaCl緩衝液中に抽出し、100kDa MWCOカラムで濃縮せずにTEと交換した。HBcAgの最終濃度は、coomassie染色したSDS−PAGEゲルでの標準物との比較により判断した場合、約0.2mg/mlであった。
【0151】
1.2結果
(1)161位(AUG161)および512位(AUG512)における開始部位の相対位相を変化させる効果
これを達成するために、余分なヌクレオチドを、AUG512(FSC2−512)のすぐ上流に挿入して、AUGを513位(FSC2−513)、514位(FSC2−514)および515位(FSC2−515)に移動させた(図1)。AUG161と位相をずらしてAUG512(FSC2−513およびFSC2−514)を配置すると、蛍光(図3)およびCoomassie染色したゲル(図2)によって判断した場合、GFP発現は低かった。位相(FSC2−515)を元に戻すと、天然の状態(FSC2−512)で見られたレベルの発現に戻った。結論としては、AUG161が存在し、161位のAUGと位相が一致する場合、下流のAUGでの開始が最も効率的である。
【0152】
(2)161位(AUG161)および512位(AUG512)の開始部位の相対位相を変化させることに加えて115位(AUG115)の開始部位の除去
GFP発現がAUG512から駆動される場合、すなわち、この第2のAUGがAUG161と位相が一致する場合、AUG115の除去は、ほとんどまたは全く影響を与えない(図2および図3の標識した10のレーンを参照のこと)。しかしながら、第2のAUGがAUG161(513、514)と位相がずれる場合、AUG115の欠失は、実質的にGFP発現を生じない(図2および図3の標識した10のレーンを参照のこと)。結論:AUG115は、何らかの形で、AUG161を回避し、AUG512に到達することによってリボソームの能力に関与する。しかしながら、このことは、正確な位相に存在する下流のAUGを必要とする。
【0153】
(3)161位(AUG161)における開始部位の除去の効果
この変異の効果は信じられないほど劇的であり、GFP発現レベルが、AUG161が存在する場合に見られる量の20〜30倍に到達する(図2および図3における標識した01のレーンを参照のこと)。さらに、AUG161の非存在にもかかわらず、位相AUG512はもはや問題ではないように見え、位相の考えは大して重要ではない。さらに、AUG115の存在または非存在に差異はない(図2および図3における標識した11のレーンを参照のこと)。DsRedおよびHBcAg発現についてのdelRNA−2(図1における発現ベクター00[FSC2−512])構築物を用いる場合、5日以内に、浸潤したパッチは膨圧を失い、クロロティック(chlorotic)(薄い)になった。7日までに、組織は灰色のように見え、完全に死んだ。HT(図1における発現ベクター01[FSC2−512])を用いる場合、組織は7日後に膨張し、ストレスのサインのみが、HBcAg発現組織をわずかに白化する。2G12 IgGの重鎖が、delRNA−2によって発現され、ERに残存する場合、白化は7日後に明らかであるが、HTについては、これは観測されない。従って、異種タンパク質を発現するためにHTを用いる場合、植物に見られる壊死のレベルは、より高いレベルの異種タンパク質発現が達成されるにもかかわらず、異種タンパク質を発現するためにdelRNA−2を用いる場合より非常に低い。
【0154】
考察
非常に高レベルの外来性遺伝子発現が、AUG161を欠失することによってdelRNA−2構築物から発現され得る。現在のところ、GFPを用いて、本発明者らは、全可溶性タンパク質(TSP)のうちの25〜30%または1Kgの葉あたり約1gの発現されたタンパク質のレベルと測定する。これは非常に高いレベルであり、かつ、本発明者らが用いるアプローチは非常に簡単である。本発明者らがもはやリーディングフレームを保存する必要がないという事実は、ポリリンカーを有する、使用者によって使いやすいベクターが産生され得るということを意味する。
【0155】
(実施例2)
2.1背景
実施例1に記載したように、効果的な発現に必要なCPMC RNA−2の5’非翻訳領域(UTR)に必要な特徴を調べるために、本発明者らは、主要な開始部位の上流の5’リーダー配列内に見られる2つのAUGコドンの役割について扱った。本発明者らは、フレーム内の主要な翻訳開始部位(512)の上流の開始コドン(161)の欠失が、外来性タンパク質蓄積の大幅な増加をもたらすことを実証した。
【0156】
この系を用いて、本発明者らは、浸潤後6日によって、完全なサイズのIgGおよび自己集合性ウイルス様粒子を含む、多くの関係のないタンパク質が、それぞれ、全抽出タンパク質のうちの10%より多く、および20%発現されたことを示した。従って、この系は、転写産物のウイルス複製に頼らない高レベルの発現のための理想的な媒体を提供する。
【0157】
この新規の系(図1における発現ベクター01[FSC−512]によって例示される)を、高翻訳可能(hyper−translatable)なササゲモザイクウイルス(Cow Pea Mosaic Virus)タンパク質発現系のための「CPMV−HT」と呼ぶ。
【0158】
HT−CPMV系は、タンパク質レベルの劇的な増加を示すので、植物における外来性タンパク質の迅速で、高レベル発現のための優れた方法である。
【0159】
バイナリーベクターの増殖アレイは、過去25年にわたって植物形質転換のために開発されている(Hellensら,2000b;Veluthambiら,2003;LeeおよびGelvin,2008)。従って、これらの開発の主な目的は、例えば、アグロバクテリウムのための宿主範囲を拡大すること(Hieiら、1994)、選択マーカー、発現カセットおよび融合タンパク質の選択を可能にする一連のベクターを作製すること(オープンソースバイナリーベクターのpCAMBIA範囲;http://www.cambia.org/daisy/bioforge_legacy/3725.htmlによって例示される)、または外来性DNA組み込みおよびマーカーフリーの形質転換を最小化するための系を開発すること(例えばpCLEAN;Tholeら、2007)によって、安定な組み込みを向上させることにかなり焦点をあてられている。
【0160】
バイナリーベクターはまた、同じ導入遺伝子の複数の組み込み事象の頻度を低減させるために、低コピー数で複製するために操作される。なぜならこれにより、遺伝子サイレンシングが導かれ得るからである(JohansenおよびCarrington、2001)。
【0161】
しかしながら、一過性発現に関して、宿主核への効果的な組み込みを確実にすることおよび植物体選択におけるマーカーの存在は厳格に必要とされていない。さらに、アグロ浸潤法(agro−infiltration)の際に、各細胞は、ゲノムを組み込まなくても核において転写に適合すると考えられるT−DNA分子で満たされる(JanssenおよびGardner,1989;Narasimhuluら,1996)。このことは、一過性発現が、より高いコピー数のバイナリープラスミドから利益を受けることができることを示唆する。
【0162】
バイナリーベクターの改良の別の領域は、ベクターバックボーンのサイズの減少である。より小さなプラスミドの利点を実証し続けている2つの優れた例は、pPZP(Hajdukiewiczら、1994)およびpGREEN(Helensら、2000a)である。クローニング手順および細菌形質転換の効率を向上させることに加えて、これらのベクターは、単一のT−DANに存在する複数のカセットに頼る発現系のための鋳型を提供している(Tzfiraら,2005;Tholeら,2007)。
【0163】
本実施例は、クローニングベクター(例えばpM81−FSC2)と、発現系(例えばPBINPLUS)との間の発現カセットのサブクローニングを必要とするよりむしろ、単一の工程でなされるクローニングを可能にすることによって、このベクターの実用的用途を容易にするそのベクターの進歩性のある改良を開示する。より具体的には、本明細書の結果は、複製およびTDNA移入に関する実施を妥協することなく、pBINPLUSのサイズを劇的に減少させることが可能であったことを示す。さらに、CPMV−HT系の要素は、モジュール方式で、得られるベクターに組み込まれるので、複数のタンパク質が単一のT−DNAから発現され得る。これらの改良により、外来性遺伝子の効率的な直接クローニングを可能にする多用で、高レベルの発現ベクターの作製がもたらされる。
【0164】
2.2材料および方法
pBD−FSC2−512−U162C(HT)は、pBINPLUSのPacI/AscI部位に挿入されるFSC2−512−U162Cカセット(実施例1を参照のこと)を含む(van Engelenら、1995)。このプラスミドの本質的なセグメント(以下を参照のこと)を、再ライゲーションのための特有の制限酵素部位をコードするオリゴヌクレオチドを用いて、高忠実度のポリメラーゼ、PHUSION(New England Biolabs)で増幅させた(表4.1)。T−DNA領域を、特有のAhdI部位(pBD−LB−F)の上流の配列と相同のセンスプライマーおよびApaI部位(pBD−RB−ApaI−R)を含むアンチセンスプライマーで増幅させた。ColEI複製起点、NPTIII遺伝子、およびTrfA遺伝子座を含む領域を、ApaI部位(pBD−ColEI−ApaI−F)を含むセンスプライマー、およびSpeI部位(pBD−TrfA−SpeI−R)を含むアンチセンスプライマーで増幅させた。RK2複製起点(OriV)を、SpeI部位(pBD−oriVSpeI−F)を含むセンスプライマーおよびAhdI部位(pBD−oriV−AhdI−R)を含むアンチセンスプライマーで増幅させた。精製後、それらの産物を、それらの末端でコードされ、3点のライゲーションのために混合した特有の制限部位に従って消化した。これにより、プラスミドpEAQbetaが得られ、そのためのライゲーション結合を、シークエンシングにより確認した。CPMV−GFP−HTカセットのnosターミネーターの一部を除去したT−DNAからの約1.2kbの欠失を検出した。従って、プライマーpBD−ColEI−ApaI−FおよびpMini>pMicroBIN−Rを用いて、右側の境界を含むpEAQbetaバックボーンのように、pBD−FSC2−GFP−HTからの右側の境界を含むターミネーターの一部を、プライマーpMini>pMicroBIN−F2およびpBD−RB−ApaI−Rで再び増幅させた(表4.1)。精製した産物を、ApaIおよびFseIで消化して、pEAQを得るためにライゲーションした(図9)。
【0165】
35Sプロモーターおよび35Sターミネーターに隣接するP19遺伝子を、プライマーとして35SP19−PacI−Fおよび35SP19−AsclR、または35SP19−FseI−Fおよび35S−P19−FseI−Rのいずれかを用いてpBIN61−P19(Voinnetら、2003)から増幅させた(表4.1)。nosプロモーターおよびターミネーターに隣接するNPTII遺伝子を、プライマーpBD−NPTII−FseI−FおよびpBD−NPTII−FseI−Rを用いてpBD−FSC2−GFPHTから増幅させた(表4.1)。A−テイリング(tailing)後、増幅させたカセットを、pGEM−T easy(Promega)にライゲーションした。FseIを用いてpGEM−T easyから切断したP19カセットを、FseIで消化したpEAQ−GFP−HTにライゲーションして、pEAQexpress−GFP−HTを得た。FseIで切断したNPTIIカセットを、両方向においてFseIで消化したpEAQ−GFP−HTにライゲーションして、pEAQselectK−GFP−HTおよびpEAQselectK(rev)−GFP−HTを得た。NPTIIカセットをまたPacl/Asclで切断し、pEAQselectK−GFP−HTのAsiSI/Mlul部位にライゲーションして、pEAQspecialK−GFP−HTを得た。pGEM−T中のP19を、QUICKCHANGE法(Stratagene)により部位特異的突然変異誘発し、プライマーP19−R43W−FおよびP19−R43W−Rを用いてアルギニン43をトリプトファン残基に変換した。変異P19カセットをPacl/Ascl消化で解放し、pEAQselectK−GFP−HTのAsiSI/Mlul部位に挿入して、pEAQspecialKm−GFP−HTを得た。
【0166】
短いポリリンカーのセンスおよびアンチセンス鎖をコードするオリゴヌクレオチド(表4.1)をアニールし、5’末端におけるNruI部位の下流の半分および3’末端におけるXhoIのものと適合するオーバーハングを残した。アニールしたオリゴを、NruI/XhoIで消化したpM81−FSC2−A115G−U162C(上記を参照)とライゲーションして、pM81−FSC2−POWを得た。NruI部位を、プライマーP19−ΔNruI−FおよびP19−ΔNruI−Rを用いて部位特異的突然変異誘発(QUICKCHANGE;Stratagene)によりpGEM−T中のP19カセットから除去し、pEAQselectK−GFP−HTのAsiSI/Mlul部位に再挿入して、pEAQspecialK−GFP−HTと比べて発現の減少を示さないpEAQspecialKΔNruI−GFP−HTを得た(データは示さず)。次いで、pM81−FSC2−POW由来のPacl/Ascl断片を解放し、同様に消化したpEAQspecialKΔNruI−GFP−HTに挿入し、それにより、GFP HT発現カセットを置換し、pEAQ−HTを得た。GFPを、pEAQ−HT:GFP−AgeI−FおよびGFP−XhoI−R;GFP−AgeI−FおよびGFP−XmaI−R;ならびにGFP−XmaI−FおよびGFP−XhoI−Rに挿入するために3つの組み合わせで4つのプライマーセット(表4.1)を用いてpBD−FSC2−GFP−HTから増幅させた。精製したPCR産物を、それらのプライマーに特有の酵素で消化し、適切に消化したpEAQ−HTに挿入して、pEAQ−HT−GFP、pEAQ−HT−GFPHisおよびpEAQ−HT−HisGFPを得た。
【0167】
表4.1.増幅および変異誘発に使用したオリゴヌクレオチド。制限酵素部位またはその部分は、下の例で示し、変異はイタリックで下線をひいた。
【表1−1】
【0168】
【表1−2】
【0169】
【表1−3】
【0170】
2.3 結果
2.3.1 pBINPLUSは、少なくとも7.4kbの外来性配列を含む
CPMV−HTからの発現は、非常に高レベルの組み換えタンパク質の産生を可能にする。それにも関わらず、一過性形質転換のための系およびその使用をさらに改良することが望まれた。
【0171】
改良の第1の領域は、小さいプラスミドが、ライゲーション反応および細菌形質転換手順において大きいものより効果的であるという事実に関する。小さいバイナリーベクターの構造との比較により、pBINPLUSが、外来性配列の有意な量を含む可能性があることが示された。pBINPLUSの4つの要素は、バイナリーベクターとしての適切な機能:T−DNA、RK2(OriV)広範な宿主範囲の複製起点、カナマイシン耐性を与えるNPTIII遺伝子(Trieu−CuotおよびCourvalin,1983)、および複製を促進するRK2由来のTrfAに必須であると決定した(図9)。残存しているバックボーン部分のバイオインフォマティクス分析により、それらが、親プラスミド内の適切な制限部位の存在で信頼される、pBIN19の構築物の人工物であることが示される(Bevan,1984)。これらの観測は、pBIN19の完全なシークエンシングでの報告により確認される(Frischら、1995)。pBINPLUSは、E.coliにおけるより高いコピー数のための必須でないColEI複製起点を含む。約2.6kbの余分なDNAを、T−DNA内に見出すことができる。これは、一過性発現に必要ではない植物形質転換のためのNPTII選択マーカーを含む。全体として、pBINPLUS内の外因性配列の全量は、7.2kbを超えているようである。
【0172】
2.3.2 pEAQシリーズの構築物
ベクターに対する修飾を生じる発現の効果をモニターするために、本発明者らは、pBINPLUS由来プラスミド、pBD−FSC2−512−U162C(HT)で開始することを選択した。T−DNA、RK2(OriV)複製起点、ならびにColEI起点、NPTIIIおよびTrfAを含むセグメントからなる3つの領域を、pBD−FSC2−GFP−HTからPCRによって増幅させた。これらの3つのフラグメントのライゲーションにより、プラスミドpEAQbeta(図9)が得られ、それは、その親プラスミドより4584bp小さい。pEAQbetaのPCR増幅のさらなるラウンドにより、T−DNA領域由来の2639bpの必須でない配列を除去し、3つの特有の制限部位、AsiSI、MlulおよびFseIを挿入した。AsiSI/Mlul消化は、Pacl/Ascl断片の挿入と適合するので、全ての以前のCPMVクローニングベクターから複数のカセットをクローニングするのに非常に有用である。FseIは、異なる選択マーカーまたはサイレンシング抑制因子カセットを置き換えるのに有用な特有の8塩基の認識部位を提供する。得られるpEAQGFP−HTプラスミドは、pBINPLUSのサイズの半分より小さく、CPMVHT発現カセットを有さず、5137bpのみで、最も小さい公知のバイナリーベクターの1つである(図9)。pEAQプラスミド全体の配列を決定し、RK2複製起点が以前に報告されたもの(Frischら、1995)と逆方向であることを発見したので、pEAQ−GFP−HTの正確な方向を示す。
【0173】
pEAQ−GFP−HTを、T−DNA内に種々のさらなる特徴を包含するための開始点として用いた(図10)。pBINPLUS由来のNPTIIカセットを、GFP−HTカセットに対して順方向および逆方向の両方においてpEAQのFseI部位に再挿入して、pEAQselectK−GFP−HTおよびpEAQselectK(rev)−GFP−HTを得た。
【0174】
35S−P19カセットをFseI部位に挿入して、pEAQexpress−GFP−HTを得た。最終的に、35S−P19カセットを、pEAQselectK−GFP−HTのMlu/AsiSI部位に挿入して、pEAQspecialK−GFP−HTを得た。このように、一過性発現を容易かつ迅速にするための一連の小さなバイナリーベクターを構築した。
【0175】
2.3.3 サイズの減少は、pEAQからの一過性発現を妥協しない
pEAQシリーズのベクターのアグロ浸潤法(agro−infiltration)により、サイズの大幅な減少が一過性アッセイにおいて発現レベルをほとんど妥協しないことが示される。pEAQ−GFP−HT、およびpBIN61−P19によって提供されるP19を有するpEAQselectK(rev)−GFP−HTの同時浸潤は、pBD−FSC2−512−HTおよびP19の同時浸潤とほとんど変わらない発現レベルを生じる。これは、UV照射下(図11A)、SDS−PAGE(図11B)、およびタンパク質抽出物におけるGFPのスペクトル蛍光測定(図11C)で見られ得る。興味深いことに、T−DNA内のNPTIIカセットの方向は、発現レベルに影響を与えるようである。pEAQselectKは、GFP蓄積の減少を生じる他の同等のpEAQselectK(rev)と比べて著しい向上を示す。
【0176】
理論的には、pEAQへのサイレンシングカセットの抑制因子の組み込みは、同じT−DNAから発現される外因性遺伝子からの一過性発現レベルを向上させるその能力に影響を与えるはずはない。実際には、pEAQexpress−GFP−HT単独の浸潤はまた、pBD−FSC2−GFP−HTと同様、またはそれより高い発現レベルを生じる(図7.3)。さらに、pEAQexpressの効果を試験するために、アグロバクテリウム培養物を2倍希釈し、それにより、最終的な吸光度(OD)は、同時浸潤の各々の個々の培養物のものであった。
【0177】
予想されるように、これは、同様の高い発現レベルを生じ、同じT−DNA上の目的の遺伝子およびサイレンシングの抑制因子の両方の組み込みが、アグロバクテリウムの半分の量の使用を可能にすることを示す(図11)。従って、全ての要素が同じT−DNAに存在する場合、CPMV−HTは、高レベルの外来性タンパク質を発現させるために使用できる。
【0178】
2.3.4 変異P19は一過性アッセイにおける導入遺伝子のサイレンシングを抑制できる
T−DNA内のNPTIIカセットの順方向によって得られる発現の増加を利用するために、P19カセットを、pEAQselectK−GFP−HTにおけるAsiSIとMlul部位との間に挿入して、pEAQspecialK−GFP−HTを得た(図10)。GFPの配列として同じT−DNA上のP19の存在により、P19で同時浸潤したpEAQselectK−GFPHTと同様のレベルの発現が生じる(図12)。これは、pEAQexpress−GFP−HTによって産生される発現より多く、NPTIIカセットの存在に起因するように見える(図12)。対照的に、pEAQexpressからの低い発現は、T−DNA内のP19カセットの異なる位置および方向に起因し得る。それにも関わらず、pEAQexpressと同様に、pEAQspecialKベクターは、2つの培養物が同時浸潤されなければならない場合に使用されるものの最終的なODの半分のアグロバクテリウムの懸濁液で高レベルの発現を与える。
【0179】
外来性遺伝子発現カセットと、P19カセットおよび選択マーカーを組み合わせることにより、トランスジェニック植物におけるCPMV−HTの機能を試験することが可能となる。しかしながら、P19などのサイレンシングの抑制因子の構成的発現は、発生プロセスに関連する内因性遺伝子サイレンシングとのそれらの干渉に起因する厳しい表現型を生じ得る(SilhavyおよびBurgyan,2004)。最近、特徴付けられたP19の変異(R43W)は、内因性遺伝子サイレンシングに対して減少した活性を有することが提案されているので、安定な形質転換における導入遺伝子サイレンシングの抑制のためのより良い候補となり得る(Scholthof、2007)。CPMV−HT系を用いる安定な形質転換の実行可能性を調べるために、野生型および変異型の両方のP19を、pEAQselectK−GFP−HTのT−DNAに挿入して、一過性に変異体をアッセイした。示すように、浸潤した葉のUV照射、タンパク質抽出物のSDS−PAGE、およびGFPレベルの蛍光分光測定によって、pEAQspecialKmの変異体P19は、pEAQspecialKの野生型P19の外来性遺伝子の発現の向上の効率の約半分である(図12)。これは、導入遺伝子のサイレンシングを抑制する能力に対するP19におけるR43W変異の効果の第1の研究を表す。
【0180】
(実施例3)
単一のプラスミドからの高レベルIgG発現
pEAQシリーズ、ヒト抗HIV IgGのERを保有する重鎖(HE)および軽鎖(L)を含むCPMV−HT発現カセットのモジュールの性質を利用するために、2G12を、pEAQexpressのPacl/AsclおよびAsiSI/Mlul部位に挿入した。挿入部位が発現レベルに影響を与えるか否かを決定するために、LおよびHE鎖を、pEAQex−2G12HELおよびpEAQex−2G12LHEを生じる両方の位置に挿入した(図13A)。上記のプラスミドを含む単一のアグロバクテリウム培養物でのN.benthamianaの浸潤により、各々が個々の要素、L、HEおよびP19を発現する3つのアグロバクテリウム培養物を混合することにより産生された2G12とサイズが同一の完全に集合した2G12抗体の形成が生じた(図13C)。各レーンに負荷したタンパク質は、90mgの浸潤した組織から得た抽出物の1/30または1gの組織から潜在的に得られるタンパク質の1/333を示す。3つの株の混合物から産生された集合したIgGの最大量は、coomassie染色した減少していないSDS−PAGEゲルでの1μgのCHOにより産生された2G12に相当する。このことは、325mg/kgの新鮮な重量の組織を超える2G12の発現レベルを示唆し、SPRにより測定された濃度と一致する。pEAQex−2G12HELの使用は、このすでに高レベルの抗体の蓄積を抑制するように見える。
【0181】
IgGなどの複数鎖タンパク質の各要素であるpEAQ由来のベクターの利点は、自動的に各感染細胞に送達され得ることである。従って、複数の培養物が使用されなければならない場合より、高発現レベルが、より高い希釈のアグロバクテリウム懸濁液で維持されなければならない。実際にこれが起こる場合を試験するために、最初にOD1.2に再懸濁し、必要な場合、混合した培養物を、2回連続して3倍希釈した(図13B)。これにより、3つの培養混合物において、各々最終ODが0.4、0.13および0.04である個々の培養物が得られた。pEAQexpress構築物を有する単一の培養物を、1.2、0.4および0.13のODで浸潤した。3つの別の培養物を使用した場合、集合した2G12のレベルは連続希釈で顕著に減少する。対照的に、pEAQex−2G12HELおよびpEAQex−2G12LHEからの2G12発現を、非常に少しの希釈でいくらか減少させて、常に高レベルに維持した(図13C〜E)。希釈に対する感受性の欠如により、同じT−DNAに全ての3つの発現カセットを配置することにより得られる向上した効果を確認する。興味深いことに、浸潤組織から抽出した全タンパク質の量は、浸潤物のODが1.2〜0.4まで減少した場合、ほとんど半分であった。これは、より高いODの懸濁液が浸潤される組織由来の抽出物中のタンパク質の有意な画分が、より高い濃度のアグロバクテリウムに対して産生されるアグロバクテリウム由来のタンパク質または植物タンパク質から構成され得ることを示唆する。
【0182】
図13Cの検査は、T−DNA内のカセットの相対位置が発現レベルに影響を与え得ることを示唆する。pEAQex−2G12LHEからの全発現は、pEAQex−2G12HELからのものよりわずかに低い。このことは、減少していない試料のウェスタンブロッティングにより確認し、また、分解産物および組み込まれていない免疫グロブリン鎖の量においていくらかの差異が示された(図13C〜E)。pEAQex−2G12LHEで浸潤した組織は、約70〜80kDaの重鎖に特異的な分解産物を欠いているように見える(図13D)。また、HL2集合中間体および十分に遊離している軽鎖がほとんどないように見える(図13E)。重鎖は、植物中の2G12集合において制限されることが知られており(Markus Sack,pers.comm.,RWTH,Aachen,Germany)、これは、全ての試料における認識可能な遊離重鎖の欠如によって確認されているため、これらの結果は、pEAQex−2G12LHEが、pEAQex−2G12HELより少ない重鎖を産生することを示す。これは、T−DNAの左側の境界に近いCPMV−HTカセットからの減少した発現に起因し得る。
【0183】
他の実施形態において(データは示さず)、CPMV−HT系はまた、以下を発現するために、N.benthamianaの一過性形態において首尾よく使用される。
・ブルータングウイルス(Bluetongue Virus)(血清型10)VP2、VP3、VP5、VP7およびNS1。
・ロタウイルスNSP5
・Medicago truncatula由来のカルモジュリン(これは後で精製された)
・発現するのが困難なヒトFcγ受容体1(CD64)の細胞外ドメイン−これは精製され、抗体結合研究において機能的であることが示されている。
・CPMV小(S)および大(L)コートタンパク質が同時発現され、ウイルス様粒子内で集合することが示されている(データは示さず)。
【0184】
(実施例4)
CPMV−HT発現ベクターへの直接クローニング
単一のプラスミド上に系の要素を組み合わせるが、上記のベクターはなお、バイナリープラスミドに発現される配列を組み込むために2段階のクローニング手順を必要とした。本実施例は、目的の遺伝子が直接挿入され得るバイナリープラスミドを提供する。そのプラスミドはpEAQベースのプラスミドへ直接挿入できないポリリンカーを組み込むが、所望の場合、CまたはN末端ヒスチジンタグの融合も可能にする(pEAQ−HT;図14A)。ポリリンカーをまず、アニールしたオリゴヌクレオチドとしてpM81−FSC2−512(A115G)(U162C)に挿入し、pM81−FSC−POWを得た。この構築物はなお、複数のポリペプチドを発現させるためのpEAQベースの構築物を発生させるための標準的な2段階クローニング手順に使用できる。さらに、二重変異した5’リーダーの使用は、単一の変異で得られるよりも、さらに高発現レベルを得ることができる。次いで、CPMV−HTカセットを、Pacl/Ascl部位を介してpEAQspecialKに移して、pEAQ−HTを得た。pEAQ−HTのポリリンカー内の全ての3つの位置へのGFPの挿入により、非標識GFP、および5’(HisGFP)および3’(GFPHis)His−タグ融合を得た。
【0185】
予想されるように、非標識GFPは、pEAQspecialK−GFP−HTで得られるものより高いレベルまで発現し、1.6g/kg FW組織を超えた(図14B)。これは、pEAQ−HTのCPMV5’リーダーが、AUG161に加えて除去される場合、AUG115を除去する余分な変異を含み、さらに発現を向上するという事実に起因するようである。
【0186】
ウェスタンブロッティングによって検出される場合、His−タグの存在により、ポリリンカーによってコードされるアミノ酸残基のNおよびC末端の両方において正確な融合を確認した。全ての3つのGFP変異体は抗GFP抗体で検出可能であるが、HisGFPおよびGFPHisのみが抗His抗体で検出可能であり(図14C)、His−タグの存在により、予想された量でSDS−PAGEにおいてGFPバンドの流動性が減少した。この標識はまた、蛍光の分析によって検出されるGFPの量を減少させた(図14B)。この効果はN末端Hisタグについてより顕著であった。coomassie染色したバンドの強度により、これは、GFPの蛍光特性を干渉するというよりむしろ、標識されたGFP蓄積(図14C)の減少を表すことが示唆される。それにも関わらず、Hisタグしたタンパク質のレベルは依然として非常に高く、1kgのFW組織あたり0.6および1.0のGFPを超えた。
【0187】
実施例2〜4の考察
CPMV−HT発現系の使用の容易さおよび機能を改良するために、ベクターのモジュラーセットが、容易かつ迅速な植物発現のために作製される。
【0188】
バイナリーベクター、pBINPLUS由来のプラスミドバックボーンの半分より多く、および一過性発現に必須ではないT−DNA領域の一部を除去することにより、発現レベルで妥協しない公知の最も小さいバイナリーTiプラスミドが得られる。
【0189】
バックボーンの同様の割合は、機能を損失せずにpBIN19から以前に除去されている(Xiangら、1999)。しかしながら、pBINPLUSは、pBIN19に対して2つの有意な改良点を有し(van Engelenら、1995);E.coliにおける増加したコピー数はColEI複製起点の添加のおかげであり、目的の遺伝子を確保する再構築したT−DNAはさらに、植物体における広範な欠失を受け得る左側の境界に由来する(Rossiら、1996)。同様のサイズのpEAQプラスミドは、それらのコピー数に対する顕著な効果を有さないが、それらは、10kb未満のプラスミドに最も効果的であるため(データは示さず)、市販のプラスミド抽出キットを用いてクローニング手順の間に非常に向上した収率を与える。
【0190】
pEAQバイナリーベクターのモジュールの性質は、必要な場合、任意のサイレンシング抑制因子および/またはマーカー遺伝子を、1つまたは2つのCPMV−HTカセットで同時発現させることによって、CPMV−HT発現に対する機能性を加える。例えば、pEAQexpress−GFP−HTのAsiSI/Mlul部位への異種配列を含む第2のHTカセットの挿入は、GFP蛍光での発現の追跡を可能にする。
【0191】
さらに、ベクターの柔軟性は、単一のアグロバクテリウム構築物の浸潤を必要とすることのみによって、一過性発現のための系を単純化し、T−DNA内に存在する発現カセットの数に比例して必要とされる浸潤の量を減少させることによって、効果を向上させる。FseI部位を占めるP19とともに、クローニングベクター(例えばpM81−FSC2−U162C)からHTカセットを受容するための2つのクローニング部位の存在はまた、完全なサイズの抗体などの複数のサブユニットタンパク質のさらにより効果的な発現を可能にする。
【0192】
導入遺伝子の発現レベルを向上させることに対するP19の効果は十分に特徴付けられている(Voinnetら、2003)。しかしながら、この研究は、同じTDNA上の各細胞に同時送達される場合、その有効性の第1の実証を示す。以前の研究により、導入遺伝子を含むT−DNAとして同じアグロバクテリウム内の別のTDNAからのP19の同時送達が報告されている(Hellensら、2005)。しかしながら、浸潤後6日までP19の効果はなく、T−DNAの非効率の移入が示唆される。本研究はまた、導入遺伝子の発現を向上させるためのR43W変異体19の第1の使用を実証する。その変異体が、野生型P19のGFPの発現を増加させる効果の約半分であるという発見は、活性のその公知の減少を満たし、TBSVの感染性(Chuら、2000)、および短い干渉RNAのより小さいクラス(21〜22nts)に結合するタンパク質の能力(Omarovら、2006)の両方を妥協する。しかしながら、この特徴は潜在的に、R43W変異体を、安定な形質転換に関する適用に、より適切にすることを可能にする。発達に関連するマイクロRNAはまた、より小さいサイズのクラスであり(Vaucheret,2006;Zhangら、2006)、従って、発達プロセスは、野生型によってほど、変異体P19の存在によってそんなに影響を受けない(Scholthof,2007)。さらに、この変異体は、潜在的に細胞毒性の外来性タンパク質の一過性発現を制御する方法を提供することができる。
【0193】
単一のプラスミドからの2G12の発現は、単一のアグロバクテリウム培養物で浸潤された植物組織からの抗体の最も高い報告された収率を表す。CPMV−HT発現のための3つのアグロバクテリウム培養物を用いることとは別に、別の系を用いて同様のレベルを達成する唯一の方法は、6つの別の培養物の浸潤およびウイルスベクターのアプローチに関連した(Giritchら、2006)。さらに、単一のプラスミドの使用により、複数の発現カセットの同時送達を確実にするのに必要とされる細菌の量の減少が得られ、これは、工業生産レベルで著しいコスト節約を与える。この浸潤プロセスはまた、葉組織の細胞間隙の少ない詰まりに起因して、より希釈された培養物を用いて実施するのに物理的に容易である。さらに、全ODの0.4までの希釈により、浸潤により誘導されるタンパク質汚染の量が減少した。各ODでの9つの別の浸潤の分析により、培養物のODが1.2〜0.4まで減少した場合、2.7±0.2〜1.5±0.1mg/mlからの抽出物のタンパク質濃度の減少が示された。pEAQexpressの使用は、OD0.4において、1.2の浸潤ODでの3つの培養物系と同じ量の2G12を産生するため、組み換え標的タンパク質は、pEAQexpressを用いる汚染タンパク質の量の半分のみから精製されなければならない。このことは、下流のプロセシングについての非常に有用かつ予想されない利点を与える。pEAQexpressからの2G12の発現はまた、得られる発現レベルでpEAQベクターのT−DNA内の発現カセットの位置の効果を示す。pEAQex−2G12LHEからの遊離軽鎖蓄積の増加により、少ない重鎖がこの構築物を用いて発現されることが示唆され、少ない集合した抗体を生じるように見える。これは、T−DNA上の発現カセットの配置に起因し得る。代替的に、一定の割合のT−DNAは、左側の境界で核酸分解しやすい(Rossiら、1996)。T−DNA内のNPTIIカセットの再挿入は、その配向に依存する発現に対する注目すべき効果を有するように見えた。クローニング操作の間、pEAQselectK−GFP−HTは、pEAQselectK(rev)−GFP−HTの約1.5倍のE.coliのプラスミドコピー数に到達したようであった(QIAprepキット、QIAGENを用いて実施した3つの別のプラスミド調製物の収率測定から決定した)。これは大まかに、2つのベクター間で観測された発現レベルの差異と相互関連する。プラスミドがアグロバクテリウムに移入される場合、増加したコピー数に起因するか、または実際に差異も存在するか否かは知られていない。しかしながら、これらの観測は、プラスミドコピー数が、効果的なアグロバクテリウム媒介性一過性発現に重要であり得ることを示唆する。これに関して、pBIN19およびその誘導体によるRK2オリジン(図9におけるoriV)の使用により、一過性発現のための良い選択が行われる。なぜなら、RK2プラスミドは、アグロバクテリウムにおいて7〜10コピーまで蓄積することが知られているからである(Veluthambiら、1987)。これは、pPZPによって利用されるpVS1オリジンと同様であり、pSaオリジン(LeeおよびGelvin、2008)によって生成されるより約2〜5倍高く、広範に使用されるpGREENバイナリーベクターに存在する(Helensら、2000)。pRiベースのプラスミドなどのより高いコピー数を生じる複製起点を含むプラスミド(LeeおよびGelvin、2008)は、一過性発現にさらにより適切であり得る。
【0194】
CPMVベースの発現、または実際に一般の植物ベースの発現を以前に経験していない実験室用に容易に入手可能なpEAQベクターを用いる高レベルの発現を行うために、pEAQの直接クローニング型を作製した。これは、P19およびNPTIIカセットを同様に含むT−DNA上に位置するCPMVHT発現カセットの5’リーダーと3’UTRとの間にポリリンカーを挿入することにより達成された。NPTIIカセットの存在が感知できるほどに発現を向上させるように見えるため(上記を参照のこと)、そのNPTIIカセットが含まれた。ポリリンカーはまた、タンパク質精製を促進するために、NまたはC末端Hisタグの融合を可能にするために、2セットの6×ヒスチジン残基をコードする。得られた構築物はまた、HTに対して発現を向上させる5’リーダーにおいて第2の変異体からの利点を有する。
【0195】
これらの向上した発現カセットはまた、任意のpEAQプラスミド内にクローニングベクター、pM81−FSC−POWからサブクローニングされてもよい。pEAQHTの使用により、ただ単一の変異体(U162C)を含む、pEAQspecialKと比較して増加したGFP発現を生じた。さらに、ポリリンカー設計はまた、1段階のクローニング手順を用いるHisタグした変異体の発現を可能にした。本明細書に示したモジュールバイナリーベクターは、制限されないが、CPMV−HT発現での使用のために特別に設計される。非常に高レベルの発現は、向上したクローニング効果および使用の容易さと関連する。この系は、ウイルス増幅の複雑化がされることなく、植物における付加価値タンパク質の一過性発現のための最も効果的かつ容易な方法を提供する。これにより、植物成長施設で利用可能な任意の分子生物学実験と同一の配列の2週間以内のミリグラム量の組み換えタンパク質が可能となる。従って、大学および工業的環境の両方において非常に有益な手段を与えることが予想される。
【0196】
(実施例5)
pEAQプラスミドおよびトランスジェニック植物での安定な組み込み
pEAQベクターシリーズを、一過性発現を考慮して設計したが、T−DNA内にNPTIIカセットの再挿入をして、ゲノム組み込みのための選択マーカーを与えた。これは、それらのより小さく、安定な植物および植物細胞培養形質転換に使用するための、より有用なバイナリーベクターを潜在的に可能にする。N.benthamiana葉片を形質転換するために使用する場合、T−DNA内にNPTIIカセットを含むpEAQベクターは、pBINPLUSベースの構築物と同じ効率を有する選択下でカルス形成を誘導できた。さらに、GFP発現は、UV光下でこれらの組織において検出可能であった(データは示さず)。これは、pEAQベクター由来のマルチカセットT−DNA分子が、植物ゲノム内に安定して組み込み、外来性遺伝子の発現を駆動することができることを実証する。
【0197】
蛍光植物もまた再生される。一次形質転換体(T0)の葉はUV光下で蛍光を発し、高レベルのGFP発現を示した。自家受粉したT0植物からの種子は成長でき、導入遺伝子を保有する得られたT1苗もまた、蛍光を発する(結果は示さず)。
【0198】
(実施例6)
バキュロウイルスベクターを用いるCPMVベースのHT系の使用
図15は、昆虫細胞におけるバキュロウイルスベクターを用いるCPMVベースのHT系を利用するのに適切な構築物を示す。p10プロモーターの制御下で、HyperTrans CPMV RNA−2 UTRもまた、バキュロウイルス発現系を用いる昆虫細胞におけるGFPの発現を向上させる。フローサイトメトリーによって測定される場合、バキュロウイルス感染sf21細胞の蛍光レベルの約5倍の向上を、CPMV−HTカセットを有さない構築物と比較して得た。
【0199】
(参考文献)
Alamillo,J.M.,Monger,W.,Sola,I.,Garcia,B.,Perrin,Y.,Bestagno,M.,Burrone,O.R.,Plana−Duran,J.,Enjuanes,L.,Lomonossoff,P.G. and Garcia,J.A.(2006)Use of virus vectors for the expression in plants of active full−length and single chain anticoronavirus antibodies. Biotechnol.J.1,1103−1111.
Brigneti,G.,Voinnet,O.,Li,W.X.,Ji,LH.,Ding,S.W and Baulcombe,D.C.(1998)Viral pathogenicity determinants are suppressors of transgene silencing in Nicotiana benthamiana. EMBO Journal 17:6739−6746.
Canizares,M.C,Liu,L.,Perrin,Y.,Tsakiris,E. and Lomonossoff,GP.(2006).A bipartite system for the constitutive and inducible expression of high levels of foreign proteins in plants.Plant Biotechnol.J.4,183−193.
Gopinath,K.,Wellink,J.,Porta,C,Taylor,K.M.,Lomonossoff,G.P. and van Kammen,A.(2000)Engineering cowpea mosaic virus RNA−2 into a vector to express heterologous proteins in plants.Virology 267:159−173.
Holness,C.L.,Lomonossoff,G.P.,Evans,D. and Maule,A.J.(1989).Identification of the initiation codons for translation of cowpea mosaic virus middle component RNA using site directed mutagenesis of an infectious cDNA clone.Virology 172,311 320.
Liu, L. and Lomonossoff,G.P.(2002)Agroinfection as a rapid method for propagating Cowpea mosaic virus−based constructs.J.Virol.Methods 105,343−348.
Liu,L. and Lomonossoff,G.P.(2006) A site−directed mutagenesis method utilising large double−stranded DNA templates for the simultaneous introduction of multiple changes and sequential multiple rounds of mutation:Application to the study of whole viral genomes.J.Virol.Methods 137,63−71.
Liu,L.,Canizares,M.C.,Monger,W.,Perrin,Y.,Tsakiris,E.,Porta,C,Shariat,N.,Nicholson,L. and Lomonossoff,G.P.(2005).Cowpea mosaic virus−based systems for the production of antigens and antibodies in plants.Vaccine 23,1788−1792.
Lomonossoff,G.P. & Shanks,M.(1983).The nucleotide sequence of cowpea mosaic virus B RNA.EMBO Journal 2,2253−2258.
Mechtcheriakova,I.A.,Eldarov,M.A.,Nicholson,L.,Shanks,M.,Skryabin,K.G. and Lomonossoff,G.P.(2006) The use of viral vectors to produce hepatitis B virus core particles in plants.J.Virol.Methods 131,10−15.
Monger,W.,Alamillo,J.M.,Sola,I.,Perrin,Y.,Bestagno,M.,Burrone,O.R.,Sabella,P.,Plana−Duran,J.,Enjuanes,L.,Garcia,J.A. and Lomonossoff,G.P.(2006) An antibody derivative expressed from viral vectors passively immunizes pigs against transmissible gastroenteritis virus infection when supplied orally in crude plant extracts. Plant Biotechnol.J.4, 623−631.
Rohll,J.B.,Holness,C.L.,Lomonossoff,G.P. and Maule,A.J.(1993).3’ terminal nucleotide sequences important for the accumulation of cowpea mosaic virus M−RNA.Virology 193, 672−679.
Sainsbury,F.,Lavoie,P−O.,D’Aoust,M−A.,Vezina,L−P. and Lomonossoff,G.P.(2008).Expression of Multiple Proteins Using Full−Length and Deleted Versions of Cowpea Mosaic Virus RNA−2.Plant Biotechnology Journal,6:82−92.
Sainsbury,F.,Canizares,M.C. and Lomonossoff,G.P.(2007) Cowpea mosaic virus−based expression vectors. In:Virus Expression Vectors(Hefferon, K. ed),pp.339−555. Kerala,India:Transworld Research Network.
Sainsbury,F. and Lomonossoff,G.P.(2008). Extremely high−level and rapid transient protein production in plants without the use of viral replication. Plant Physiology 148,1212−1218.
Sainsbury,F.,Liu,L. and Lomonossoff G.P.(2009) Cowpea mosaic virus−based expression of antigens and antibodies in plants. In:Methods in Molecular Biology Vol.483:Recombinant Pharmaceutical Proteins from Plants(Faye,L. and Gomord,V. eds),pp25−39,NY:Humana Press.
van Bokhoven,H.,Le Gall,O,Kasteel,D.,Verver,J.,Wellink,J. and van Kammen,A.(1993).Cis− and Trans−acting Elements in Cowpea Mosaic Virus RNA Replication. Virology 195,377−386.
Wellink J,Verver J, van Kammen A.(1993).Mutational analysis of AUG codons of cowpea mosaic virus M RNA. Biochimie.75(8):741−7.
さらなる参考文献:
Lee LY,Gelvin SB(2008)T−DNA binary vectors and systems. Plant Physiology 146:325−332
Hajdukiewicz P,Svab Z,Maliga P(1994)The small,versatile pPZP family of Agrobacterium binary vectors for plant transformation. Plant Molecular Biology 25:989−994
Hellens R,Mullineaux P,Klee H(2000b)A guide to Agrobacterium binary Ti vectors. Trends in Plant Science 5:446−451
Hiei Y,Ohta S,Komari T,Kumashiro T(1994)Efficient transformation of rice(Oryza−sativa L)mediated by Agrobacterium and sequence−analysis of the boundaries of the T−DNA. Plant Journal 6:271−282
Veluthambi K,Jayaswal RK,Gelvin SB(1987)Virulence genes−a,gene−G, and gene−D mediate the double−stranded border cleavage of T−DNA from the Agrobacterium Ti plasmid. Proceedings of the National Academy of Sciences of the United States of America 84: 1881−1885
Thole V,Worland B,Snape JW,Vain P(2007) The pCLEAN dual binary vector system for Agrobacterium−Mediated plant transformation. Plant Physiology 145:1211−1219
Johansen LK,Carrington JC(2001)Silencing on the spot. Induction and suppression of RNA silencing in the Agrobacterium−mediated transient expression system. Plant Physiology 126:930−938
Janssen BJ,Gardner RC(1989)Localized transient expression of Gus in leaf−disks following cocultivation with Agrobacterium. Plant Molecular Biology 14:61−72
Narasimhulu SB,Deng X,Sarria R,Gelvin SB(1996)Early transcription of Agrobacterium T−DNA genes in tobacco and maize. Plant Cell 8:873−886
Hellens RP,Edwards EA,Leyland NR,Bean S,Mullineaux PM(2000a)pGreen:a versatile and flexible binary Ti vector for Agrobacterium−mediated plant transformation. Plant Molecular Biology 42:819−832
Tzfira T,Tian GW,Lacroix B,Vyas S,Li JX,Leitner−Dagan Y, Krichevsky A,Taylor T,Vainstein A,Citovsky V(2005)pSAT vectors:a modular series of plasmids for autofluorescent protein tagging and expression of multiple genes in plants. Plant Molecular Biology 57: 503−516
Voinnet O,Rivas S,Mestre P,Baulcombe D(2003) An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant Journal 33:949−956
Frisch DA,Harrishaller LW,Yokubaitis NT,Thomas TL,Hardin SH,Hall TC(1995)Complete sequence of the binary vector BIN−19. Plant Molecular Biology 27:405−409
Trieu−cuot P,Courvalin P(1983)Nucleotide−sequence of the Streptococcus−faecalis plasmid gene encoding the 3’5’’−Aminoglycoside phosphotransferase type−Ill.Gene 23:331−341
Bevan M(1984)Binary Agrobacterium vectors for plant transformation. Nucleic Acids Research 12:8711−8721
Silhavy D,Burgyan J(2004)Effects and side−effects of viral RNA silencing suppressors on short RNAs. Trends in Plant Science 9:76−83
Scholthof HB(2007)Heterologous expression of viral RNA interference suppressors:RISC management. Plant Physiology 145:1110−1117
van Engelen FA,Molthoff JW,Conner AJ,Nap JP,Pereira A,Stiekema WJ(1995)pBINPLUS−an improved plant transformation vector based on pBIN19. Transgenic Research 4:288−290
Rossi L,Hohn B,Tinland B(1996)Integration of complete transferred DNA units is dependent on the activity of virulence E2 protein of Agrobacterium tumefaciens.
Proceedings of the National Academy of Sciences of the United States of America 93:126−130
Xiang CB,Han P,Lutziger I,Wang K,Oliver DJ(1999)A mini binary vector series for plant transformation. Plant Molecular Biology 40:711−717
Hellens RP,Allan AC,Friel EN,Bolitho K,Grafton K,Templeton MD,Karunairetnam S,Gleave AP,Laing WA(2005)Transient expression vectors for functional genomics,quantification of promoter activity and RNA silencing in plants. Plant Methods 1:13
Chu M,Desvoyes B,Turina M,Noad R,Scholthof HB(2000)Genetic dissection of tomato bushy stunt virus p19−protein−mediated host−dependent symptom induction and systemic invasion. Virology 266:79−87
Omarov R,Sparks K,Smith L,Zindovic J,Scholthof HB(2006)Biological relevance of a stable biochemical interaction between the tombusvirus−encoded P19 and short interfering RNAs. Journal of Virology 80:3000−3008
Vaucheret H(2006)Post−transcriptional small RNA pathways in plants:mechanisms and regulations. Genes & Development 20:759−771
Zhang BH,Pan XP,Cobb GP,Anderson TA(2006)Plant microRNA:A small regulatory molecule with big impact. Developmental Biology 289:3−16
Giritch A,Marillonnet S,Engler C, van Eldik G,Botterman J,Klimyuk V,Gleba Y(2006)Rapid high−yield expression of full−size IgG antibodies in plants coinfected with noncompeting viral vectors. Proceedings of the National Academy of Sciences of the United States of America 103:14701−14706
【0200】
【表2−1】
【0201】
【表2−2】
【0202】
CPMV RNA−2ゲノムセグメントの115位、161位、512位および524位の開始コドンを太字および下線で示す。
【0203】
【表3】
【0204】
pM81−FSC−2クローンの5’領域の変異誘発に使用されるオリゴヌクレオチドの変異ヌクレオチドを太字で示す。
【0205】
【表4】
【0206】
pM81−FSC−1およびpM81−FSC−2ベクターの配列と、表1からのCPMV野生型配列との間のヌクレオチドの相違を大文字として示す。
【0207】
【表5】
【0208】
【表6】
【0209】
【表7】
【0210】
【表8】
【0211】
【表9】
【0212】
【表10】
【0213】
【表11】
【0214】
【表12】
【特許請求の範囲】
【請求項1】
2分節RNAウイルスのRNA−2ゲノムセグメント由来の発現エンハンサー配列であって、前記RNA−2ゲノムセグメントの標的開始部位が変異される、発現エンハンサー配列。
【請求項2】
前記2分節RNAウイルスがコモウイルスである、請求項1に記載の発現エンハンサー配列。
【請求項3】
前記コモウイルスがササゲモザイクウイルスである、請求項2に記載の発現エンハンサー配列。
【請求項4】
請求項1〜3のいずれか1項に記載の発現エンハンサー配列から構成されるか、またはそれらから実質的に構成される、単離された核酸。
【請求項5】
2分節ウイルスのRNA−2ゲノムセグメント由来の配列の発現または翻訳を増強する活性を増加させるためのプロセスであって、前記配列内の標的開始部位を変異させることを含む、プロセス。
【請求項6】
前記2分節RNAウイルスがコモウイルスである、請求項5に記載のプロセス。
【請求項7】
前記コモウイルスがササゲモザイクウイルスである、請求項6に記載のプロセス。
【請求項8】
(a)請求項1〜3のいずれか1項に記載のエンハンサー配列と、
(b)目的のタンパク質をコードする遺伝子の、遺伝子発現系への挿入を促進するための異種配列であって、前記エンハンサー配列内の変異された標的開始部位の下流に位置する異種配列と、必要に応じて、
(c)3’UTRと、
を含む、遺伝子発現構築物。
【請求項9】
(i)作動可能に連結されるプロモーターと、
(ii)請求項1〜3のいずれか1項に記載のエンハンサー配列と、
(iii)発現することが所望される目的の遺伝子と、
(iv)終結配列と、必要に応じて、
(v)前記終結配列の上流に位置する3’UTRと、
を含む、発現カセット。
【請求項10】
請求項1〜3のいずれか1項に記載のエンハンサー配列を含む、遺伝子発現系。
【請求項11】
目的のタンパク質をコードする遺伝子をさらに含み、前記目的のタンパク質をコードする遺伝子は、前記エンハンサー配列の下流に位置する、請求項10に記載の遺伝子発現系。
【請求項12】
前記目的のタンパク質をコードする遺伝子は、プロモーターおよび終結配列に作動可能に連結される、請求項11に記載の遺伝子発現系。
【請求項13】
前記目的のタンパク質をコードする遺伝子は、前記エンハンサー配列の下流で、かつ、前記終結配列の上流に位置する、請求項12に記載の遺伝子発現系。
【請求項14】
3’UTRを含むか、またはさらに含み、前記3’UTRは必要に応じて、同じ2分節RNAウイルス由来である、請求項8〜13のいずれか1項に記載の遺伝子発現系。
【請求項15】
請求項8または9に記載の発現構築物または発現カセットを含む、遺伝子発現系。
【請求項16】
請求項10〜15のいずれか1項に記載の遺伝子発現系を用いて、宿主生物内で目的のタンパク質を発現させるための方法。
【請求項17】
前記宿主生物は真核性宿主であり、前記真核性宿主は必要に応じて植物または昆虫である、請求項16に記載の方法。
【請求項18】
RNA2由来の配列に作動可能に連結される、目的の異種タンパク質をコードする遺伝子またはオープンリーディングフレーム(ORF)から前記目的の異種タンパク質の翻訳を増強する方法であって、前記RNA2由来の配列内の標的開始部位を変異させることを含む、方法。
【請求項19】
(a)プロモーターおよび終結配列に作動可能に連結された目的の異種タンパク質をコードする少なくとも1つの外来性遺伝子を保有する2分節ウイルスゲノムの切断されたRNA−2を含む第1の遺伝子構築物であって、前記遺伝子構築物は前記外来性遺伝子の上流に変異された標的開始部位を含む、第1の遺伝子構築物と、必要に応じて、
(b)プロモーターおよび終結配列に作動可能に連結された遺伝子サイレンシングの抑制因子を含む、前記第1の遺伝子構築物内に必要に応じて組み込まれる第2の遺伝子構築物と、
を含む、遺伝子発現系。
【請求項20】
以下の工程:
(a)プロモーターおよび終結配列に作動可能に連結された目的の異種タンパク質をコードする少なくとも1つの外来性遺伝子を保有する2分節ウイルスゲノムの切断されたRNA−2を含む第1の遺伝子構築物を含む遺伝子発現構築物を導入する工程であって、前記遺伝子構築物は、植物細胞内の前記外来性遺伝子の上流に変異された標的開始部位を含む工程と、必要に応じて、
(b)前記植物細胞内のプロモーターおよび終結配列に作動可能に連結された遺伝子サイレンシングの抑制因子を含む、前記第1の遺伝子構築物内に必要に応じて組み込まれる第2の遺伝子構築物を導入する工程と、
を含む、植物においてタンパク質を発現させる方法。
【請求項21】
前記遺伝子発現系を前記宿主生物内に導入する工程を含む、請求項16または17に記載の方法。
【請求項22】
前記目的のタンパク質をコードする遺伝子が、向上したレベルで発現される、請求項20または21に記載の方法によって得たか、または得られる宿主生物。
【請求項23】
請求項10〜15および19のいずれか1項に記載の遺伝子発現系で一過性にトランスフェクトされる宿主生物。
【請求項24】
前記宿主生物は植物または植物細胞である、請求項22または23に記載の宿主生物。
【請求項25】
請求項10〜15および19のいずれか1項に記載の遺伝子発現系で安定に形質転換されたトランスジェニック宿主生物。
【請求項26】
請求項22〜24のいずれか1項に記載の宿主生物を使用することと、必要に応じて、前記目的のタンパク質が発現される組織を収集することと、前記組織から前記目的のタンパク質を単離することと、を含む、目的のタンパク質を産生するための方法。
【請求項27】
(a)プロモーターと、
(b)表1に示されるササゲモザイクウイルスRNA−2ゲノムセグメント配列のヌクレオチド1〜507であって、161位のAUGは表2に示されるように変異され、前記プロモーターの下流に位置する、ヌクレオチド1〜507と、
(c)(b)に規定される配列の下流に位置する目的のタンパク質をコードする遺伝子と、
(d)前記目的のタンパク質をコードする遺伝子の下流に位置する、表1に示されるササゲモザイクウイルスRNA−2ゲノムセグメント配列のヌクレオチド3302〜3481と、
(e)(d)に規定される配列の下流に位置するノパリン合成ターミネーターと、
を含む遺伝子発現系。
【請求項28】
前記遺伝子発現系は、
(a)プロモーターと、
(b)表1に示されるササゲモザイクウイルスRNA−2ゲノムセグメント配列のヌクレオチド1〜507と少なくとも70%の同一性を有する発現エンハンサー配列であって、161位のAUGは変異され、前記プロモーターの下流に位置する、発現エンハンサー配列と、
(c)(b)に規定された配列の下流に位置する目的のタンパク質をコードする遺伝子と、
(d)表1に示され、前記目的のタンパク質をコードする遺伝子の下流に位置する、ササゲモザイクウイルスRNA−2ゲノムセグメント配列のヌクレオチド3302〜3481と、
(e)(d)に規定される配列の下流に位置するノパリン合成ターミネーターと、
を含む、請求項27に記載の遺伝子発現系の変異体。
【請求項29】
請求項27または28に記載の遺伝子発現系を用いて、植物において目的のタンパク質を発現させる方法。
【請求項1】
2分節RNAウイルスのRNA−2ゲノムセグメント由来の発現エンハンサー配列であって、前記RNA−2ゲノムセグメントの標的開始部位が変異される、発現エンハンサー配列。
【請求項2】
前記2分節RNAウイルスがコモウイルスである、請求項1に記載の発現エンハンサー配列。
【請求項3】
前記コモウイルスがササゲモザイクウイルスである、請求項2に記載の発現エンハンサー配列。
【請求項4】
請求項1〜3のいずれか1項に記載の発現エンハンサー配列から構成されるか、またはそれらから実質的に構成される、単離された核酸。
【請求項5】
2分節ウイルスのRNA−2ゲノムセグメント由来の配列の発現または翻訳を増強する活性を増加させるためのプロセスであって、前記配列内の標的開始部位を変異させることを含む、プロセス。
【請求項6】
前記2分節RNAウイルスがコモウイルスである、請求項5に記載のプロセス。
【請求項7】
前記コモウイルスがササゲモザイクウイルスである、請求項6に記載のプロセス。
【請求項8】
(a)請求項1〜3のいずれか1項に記載のエンハンサー配列と、
(b)目的のタンパク質をコードする遺伝子の、遺伝子発現系への挿入を促進するための異種配列であって、前記エンハンサー配列内の変異された標的開始部位の下流に位置する異種配列と、必要に応じて、
(c)3’UTRと、
を含む、遺伝子発現構築物。
【請求項9】
(i)作動可能に連結されるプロモーターと、
(ii)請求項1〜3のいずれか1項に記載のエンハンサー配列と、
(iii)発現することが所望される目的の遺伝子と、
(iv)終結配列と、必要に応じて、
(v)前記終結配列の上流に位置する3’UTRと、
を含む、発現カセット。
【請求項10】
請求項1〜3のいずれか1項に記載のエンハンサー配列を含む、遺伝子発現系。
【請求項11】
目的のタンパク質をコードする遺伝子をさらに含み、前記目的のタンパク質をコードする遺伝子は、前記エンハンサー配列の下流に位置する、請求項10に記載の遺伝子発現系。
【請求項12】
前記目的のタンパク質をコードする遺伝子は、プロモーターおよび終結配列に作動可能に連結される、請求項11に記載の遺伝子発現系。
【請求項13】
前記目的のタンパク質をコードする遺伝子は、前記エンハンサー配列の下流で、かつ、前記終結配列の上流に位置する、請求項12に記載の遺伝子発現系。
【請求項14】
3’UTRを含むか、またはさらに含み、前記3’UTRは必要に応じて、同じ2分節RNAウイルス由来である、請求項8〜13のいずれか1項に記載の遺伝子発現系。
【請求項15】
請求項8または9に記載の発現構築物または発現カセットを含む、遺伝子発現系。
【請求項16】
請求項10〜15のいずれか1項に記載の遺伝子発現系を用いて、宿主生物内で目的のタンパク質を発現させるための方法。
【請求項17】
前記宿主生物は真核性宿主であり、前記真核性宿主は必要に応じて植物または昆虫である、請求項16に記載の方法。
【請求項18】
RNA2由来の配列に作動可能に連結される、目的の異種タンパク質をコードする遺伝子またはオープンリーディングフレーム(ORF)から前記目的の異種タンパク質の翻訳を増強する方法であって、前記RNA2由来の配列内の標的開始部位を変異させることを含む、方法。
【請求項19】
(a)プロモーターおよび終結配列に作動可能に連結された目的の異種タンパク質をコードする少なくとも1つの外来性遺伝子を保有する2分節ウイルスゲノムの切断されたRNA−2を含む第1の遺伝子構築物であって、前記遺伝子構築物は前記外来性遺伝子の上流に変異された標的開始部位を含む、第1の遺伝子構築物と、必要に応じて、
(b)プロモーターおよび終結配列に作動可能に連結された遺伝子サイレンシングの抑制因子を含む、前記第1の遺伝子構築物内に必要に応じて組み込まれる第2の遺伝子構築物と、
を含む、遺伝子発現系。
【請求項20】
以下の工程:
(a)プロモーターおよび終結配列に作動可能に連結された目的の異種タンパク質をコードする少なくとも1つの外来性遺伝子を保有する2分節ウイルスゲノムの切断されたRNA−2を含む第1の遺伝子構築物を含む遺伝子発現構築物を導入する工程であって、前記遺伝子構築物は、植物細胞内の前記外来性遺伝子の上流に変異された標的開始部位を含む工程と、必要に応じて、
(b)前記植物細胞内のプロモーターおよび終結配列に作動可能に連結された遺伝子サイレンシングの抑制因子を含む、前記第1の遺伝子構築物内に必要に応じて組み込まれる第2の遺伝子構築物を導入する工程と、
を含む、植物においてタンパク質を発現させる方法。
【請求項21】
前記遺伝子発現系を前記宿主生物内に導入する工程を含む、請求項16または17に記載の方法。
【請求項22】
前記目的のタンパク質をコードする遺伝子が、向上したレベルで発現される、請求項20または21に記載の方法によって得たか、または得られる宿主生物。
【請求項23】
請求項10〜15および19のいずれか1項に記載の遺伝子発現系で一過性にトランスフェクトされる宿主生物。
【請求項24】
前記宿主生物は植物または植物細胞である、請求項22または23に記載の宿主生物。
【請求項25】
請求項10〜15および19のいずれか1項に記載の遺伝子発現系で安定に形質転換されたトランスジェニック宿主生物。
【請求項26】
請求項22〜24のいずれか1項に記載の宿主生物を使用することと、必要に応じて、前記目的のタンパク質が発現される組織を収集することと、前記組織から前記目的のタンパク質を単離することと、を含む、目的のタンパク質を産生するための方法。
【請求項27】
(a)プロモーターと、
(b)表1に示されるササゲモザイクウイルスRNA−2ゲノムセグメント配列のヌクレオチド1〜507であって、161位のAUGは表2に示されるように変異され、前記プロモーターの下流に位置する、ヌクレオチド1〜507と、
(c)(b)に規定される配列の下流に位置する目的のタンパク質をコードする遺伝子と、
(d)前記目的のタンパク質をコードする遺伝子の下流に位置する、表1に示されるササゲモザイクウイルスRNA−2ゲノムセグメント配列のヌクレオチド3302〜3481と、
(e)(d)に規定される配列の下流に位置するノパリン合成ターミネーターと、
を含む遺伝子発現系。
【請求項28】
前記遺伝子発現系は、
(a)プロモーターと、
(b)表1に示されるササゲモザイクウイルスRNA−2ゲノムセグメント配列のヌクレオチド1〜507と少なくとも70%の同一性を有する発現エンハンサー配列であって、161位のAUGは変異され、前記プロモーターの下流に位置する、発現エンハンサー配列と、
(c)(b)に規定された配列の下流に位置する目的のタンパク質をコードする遺伝子と、
(d)表1に示され、前記目的のタンパク質をコードする遺伝子の下流に位置する、ササゲモザイクウイルスRNA−2ゲノムセグメント配列のヌクレオチド3302〜3481と、
(e)(d)に規定される配列の下流に位置するノパリン合成ターミネーターと、
を含む、請求項27に記載の遺伝子発現系の変異体。
【請求項29】
請求項27または28に記載の遺伝子発現系を用いて、植物において目的のタンパク質を発現させる方法。
【図1】


【図2】
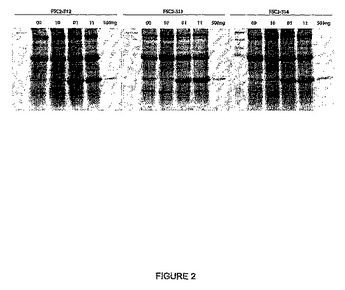
【図3】
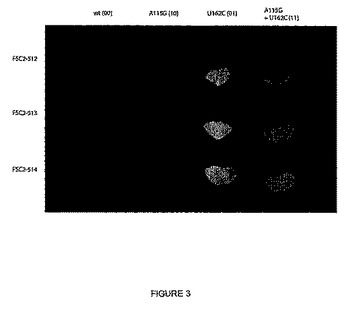
【図4A】

【図4B】
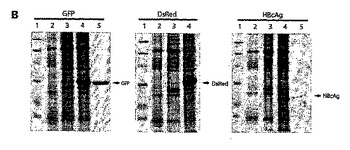
【図5】
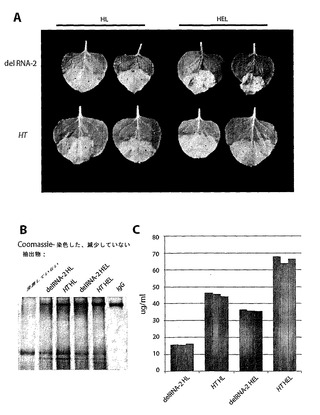
【図6】

【図7】
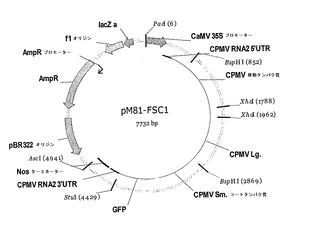
【図8】
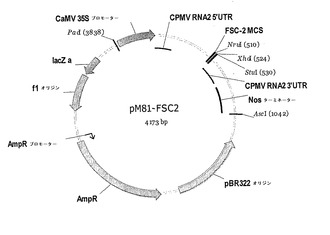
【図9】
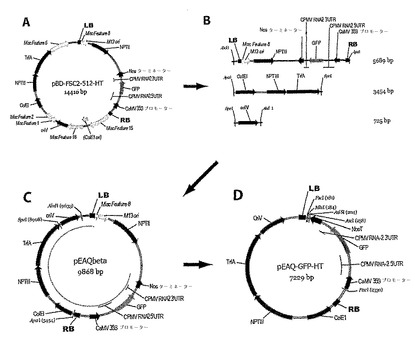
【図10】
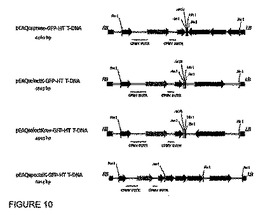
【図11】
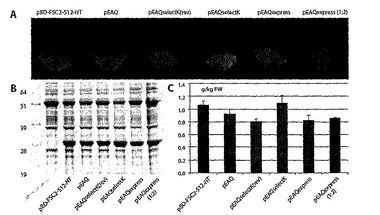
【図12】

【図13】

【図14】
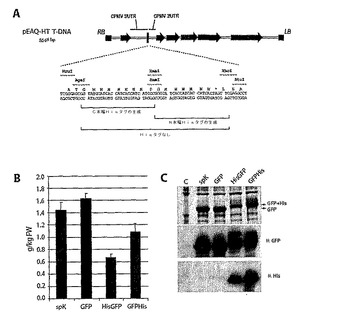
【図15】

【図2】
【図3】
【図4A】
【図4B】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公表番号】特表2011−509081(P2011−509081A)
【公表日】平成23年3月24日(2011.3.24)
【国際特許分類】
【出願番号】特願2010−541103(P2010−541103)
【出願日】平成21年1月8日(2009.1.8)
【国際出願番号】PCT/GB2009/000060
【国際公開番号】WO2009/087391
【国際公開日】平成21年7月16日(2009.7.16)
【出願人】(500123706)プラント・バイオサイエンス・リミテッド (5)
【Fターム(参考)】
【公表日】平成23年3月24日(2011.3.24)
【国際特許分類】
【出願日】平成21年1月8日(2009.1.8)
【国際出願番号】PCT/GB2009/000060
【国際公開番号】WO2009/087391
【国際公開日】平成21年7月16日(2009.7.16)
【出願人】(500123706)プラント・バイオサイエンス・リミテッド (5)
【Fターム(参考)】
[ Back to top ]