
プロテインA融合ポリペプチドの製造に利用するDNA配列、組換え発現ベクター、形質転換体、およびその融合ポリペプチドの製造方法
【課題】
本発明は、ブレビバチルス属細菌を用いた、効率の良いポリペプチドの分泌発現方法の提供を課題とする。
【解決手段】
プロテインAの免疫グロブリン結合ドメインまたはその部分的配列のC末端側に所望のポリペプチドを連結した融合生成物を、ブレビバチルス属細菌で分泌発現させる。本発明に従えば、所望のポリペプチドを含む融合生成物を効率良く生産でき、また、融合領域の有する親和性を利用して、当該融合生成物を培養液から簡便に精製できる。
本発明は、ブレビバチルス属細菌を用いた、効率の良いポリペプチドの分泌発現方法の提供を課題とする。
【解決手段】
プロテインAの免疫グロブリン結合ドメインまたはその部分的配列のC末端側に所望のポリペプチドを連結した融合生成物を、ブレビバチルス属細菌で分泌発現させる。本発明に従えば、所望のポリペプチドを含む融合生成物を効率良く生産でき、また、融合領域の有する親和性を利用して、当該融合生成物を培養液から簡便に精製できる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ブレビバチルス属細菌を使用して、プロテインAの免疫グロブリン結合ドメインもしくはその部分的配列と、所望するポリペプチドとの融合ポリペプチドを製造する際に利用するDNA配列、組換え発現ベクター、形質転換体、およびその融合ポリペプチドの製造方法に関する。本発明のDNA配列、組換え発現ベクター、または形質転換体を用いることにより、ブレビバチルス属細菌を使用して、産業上有用なポリペプチドを効率良く製造する事が可能である。
【背景技術】
【0002】
近年の遺伝子組換え技術の進歩により、各種の生物起源から得られたDNAを発現ベクターに連結し、適当な宿主細胞に導入することで、当該DNAにコードされるポリペプチドを生産出来るようになった。この遺伝子組換え技術の有効性から、現在までに様々な微生物を宿主とする数多くの宿主ベクター系が開発され、特に大腸菌および酵母を利用した系が確立されている(非特許文献1)。
【0003】
近年、鵜高らにより開発されたブレビバチルス・ブレビスを用いた遺伝子組換えポリペプチドの発現系は、ヒト上皮細胞増殖因子を、その生理活性を保持した状態で培養液中に大量に分泌発現できることが知られている(非特許文献1、2)。このブレビバチルス属細菌を用いたポリペプチド発現系は、宿主菌由来の細胞壁蛋白質であるMWPのプロモーター、シャイン・ダルガノ配列および、シグナル配列をコードするDNA配列を含む発現ベクターに、成熟型ポリペプチドをコードするDNA配列を連結し発現させることで、様々なポリペプチドの直接分泌発現を可能とする(非特許文献2)。
【0004】
しかし、ブレビバチルス属細菌のポリペプチド分泌発現系は、汎用なポリペプチド生産系として確立されてはいない。なぜならば、ブレビバチルス属細菌によるポリペプチドの分泌発現は、その所望するポリペプチドの性質や特性により発現量が大きく左右されるからである。すなわち原核生物由来で、ジスルフィド結合が存在しない、または少ないポリペプチドは比較的高い生産性が期待できるが、真核生物の分泌ポリペプチドのように複雑な高次構造を有するポリペプチドの分泌発現では、ヒト上皮細胞増殖因子やマグロ成長ホルモンなど一部の例外を除き、生産性が極めて低い(非特許文献2)。複雑な高次構造を有するポリペプチドに対する発現障害は、大腸菌や酵母などの他の微生物を宿主とした場合にも度々認められ、現在、発現障害を改善するために様々な方法が提案されている。
【0005】
効果的な方法として、所望するポリペプチドを融合ポリペプチドの形態で生産する方法(融合発現法)が提案されている。例えば、大腸菌の場合、カルモジュリン結合ペプチド(CBP)、グルタチオンS−トランスフェラーゼ(GST)、マルトース結合蛋白質(MBP)、セルロース結合ドメイン(CBD)、分子シャペロン(DNAJ,DNAK)、またはヒスチジンタグ(6から10残基程度のヒスチジン残基)等を、所望のポリペプチドのN末端、またはC末端へ結合させることにより、発現量を向上させる方法が開発されている(非特許文献1)。
【0006】
また、プロテインAが有する親和性を精製工程で利用し、所望のポリペプチドの分離、精製を容易にする方法も開発されている(特許文献1)。具体的には、免疫グロブリンに対する親和性を有するプロテインAとの融合生成物として、所望のポリペプチドを大腸菌またはスタフィロコッカス属細菌等の微生物宿主において発現させ、その融合生成物の精製工程において、プロテインAが有する親和性を利用して所望のポリペプチドを分離、精製する。
【0007】
ブレビバチルス属細菌のポリペプチド分泌発現系においても、所望のポリペプチドの生産性を向上させる方法として、(1)融合領域として数アミノ酸残基を用い、耐熱性エンドグルカナーゼを発現させる方法(特許文献2)、(2)ブレビバチルス・ブレビス47株由来の細胞壁蛋白質のN末端から数十アミノ酸残基を融合領域として用い、ヒト成長ホルモンまたはヒトプロインシュリンを融合発現させる方法(特許文献3、4)、(3)カビ由来のプロテインジスルフィドイソメラーゼ(PDI)を融合領域として用い、抗体L鎖を融合発現させる方法(特許文献5)が知られている。
【特許文献1】国際公開第WO8403103号パンフレット
【特許文献2】特開2004−129576号公報
【特許文献3】特開平11−341991号公報
【特許文献4】特開2003−79379号公報
【特許文献5】特開平11−75879号公報
【非特許文献1】塚越規弘編著,「組換えタンパク質生産法 生物化学実験法」,学会出版センター,2001年
【非特許文献2】Udakaら,Method.Enzymol.1993年,第217巻,23−33頁
【発明の開示】
【発明が解決しようとする課題】
【0008】
一般的に、融合ポリペプチドの発現量は、所望のポリペプチドの構造やアミノ酸配列と融合領域との相性に大きく左右され、選択した融合領域が適当でない場合、融合ポリペプチドの大量発現に結びつかないことが知られている。また、上記(1)から(3)で開示された融合領域では、それらが有する親和性を精製工程に利用することができず、所望の融合生成物の分離、精製を簡略化することができない。従って、ブレビバチルス属細菌のポリペプチド分泌発現系においても、多様なポリペプチドの融合発現に適用でき、かつ、融合生成物の分離、精製の簡略化が可能な、融合ペプチド領域の開発が望まれていた。
【0009】
本発明は、ブレビバチルス属細菌のポリペプチド分泌発現系において、多様なポリペプチドの融合発現に適用でき、かつ、融合生成物の分離、精製の簡略化が可能な、融合ペプチド領域の提供を一つの課題とする。さらに、当該融合ペプチド領域を利用した、ブレビバチルス属細菌による有用ポリペプチドの製造方法の提供を一つの課題とする。
【課題を解決するための手段】
【0010】
本発明者らは、プロテインAの免疫グロブリン結合ドメインまたはその部分的配列のC末端側に、所望のポリペプチドを連結した融合生成物を、ブレビバチルス属細菌で分泌発現させることにより、上記課題が解決されることを見いだした。本発明は、以下の内容を含む1または複数の特徴を有する。
【0011】
本発明は、ブレビバチルス属細菌で機能しうるシグナルペプチド、プロテインAの免疫グロブリン結合ドメインまたはその部分的配列、任意のアミノ酸配列からなるリンカー、および、所望のポリペプチドを、この順序で連結してなる融合ポリペプチドをコードするDNA配列を、さらに、ブレビバチルス属細菌で機能しうるプロモーターおよびシャイン・ダルガノ配列と作動可能に連結せしめたことを特徴とするDNA配列に関する。 また、本発明は、当該DNA配列を含むベクター、および、当該ベクターを用いてブレビバチルス属細菌を形質転換して得られる形質転換体に関する。さらに、本発明は、当該形質転換体を培養し、培地中に分泌される所望のポリペプチドの一部または全部を含むポリペプチドを回収することを特徴とする、ポリペプチドの製造方法に関する。
【発明の効果】
【0012】
本発明により、従来、ブレビバチルス属細菌のポリペプチド分泌発現系では著しく発現効率が低かったポリペプチドを、融合ポリペプチドとして大量に培養液中へ分泌発現させることが可能となった。さらに、抗体(免疫グロブリンG)などを固定化した担体を用いることにより、所望のポリペプチドを含む当該融合ポリペプチドを、培養液から簡便に分離精製することが可能となった。
【発明を実施するための最良の形態】
【0013】
以下に本発明の実施形態について詳しく説明する。
【0014】
1.DNA配列およびポリペプチド
本発明の第一の実施形態は、プロテインAの免疫グロブリン結合ドメインまたはその部分的配列のC末端側に、0または1以上のアミノ酸配列を介して所望のポリペプチドを連結した融合ポリペプチドを、ブレビバチルス属細菌で分泌発現させる機能を有するDNA配列である。詳細には、本発明の実施形態としてのDNA配列は、(a)ブレビバチルス属細菌で機能しうるプロモーター、(b)ブレビバチルス属細菌で機能しうるシャイン・ダルガノ配列、(c)ブレビバチルス属細菌で機能しうるシグナルペプチド、プロテインAの免疫グロブリン結合ドメインまたはその部分的配列、任意のアミノ酸配列からなるリンカー、および、所望のポリペプチドを、この順序で連結してなる融合ポリペプチドをコードするDNA配列、の3つの構成要素からなり、かつ、前記(a)から(c)の構成要素を互いに作動可能となるよう連結せしめたことを特徴とするDNA配列である。
【0015】
ここで、本発明における「作動可能となるよう連結せしめる」との記載は、前記(a)から(c)の構成要素がブレビバチルス属細菌内で機能することにより、所望のポリペプチドを含む融合ポリペプチドが分泌発現するよう、前記(a)から(c)の構成要素を必要に応じて適当なDNAを介して(またはDNAを介さず)連結せしめることを意味する。本発明における「作動可能となるよう連結」する概念には、前記(a)から(c)の構成要素に加えて、エンハンサー等を含む各種の調節エレメントまたは制御因子を連結する場合も含まれる。それら調節エレメントまたは制御因子の種類は、宿主によって変わり得ることは当業者に周知の事項である。
【0016】
本発明において用いられる「プロモーター」は、ブレビバチルス属細菌において作動可能であるプロモーターであればいかなる生物由来のプロモーターでもよい。好ましくはブレビバチルス属細菌由来のプロモーターが良く、より好ましくはブレビバチルス・ブレビス又はブレビバチルス・チョウシネンシス由来の細胞壁蛋白質のプロモーターが良く、更に好ましくはブレビバチルス・ブレビスの細胞壁蛋白質であるmiddle wall protein (MWP)、outer wall protein (OWP)、またはブレビバチルス・チョウシネンシスの細胞壁蛋白質HWP(J.Bacteriol.172:1312−1320(1990))のプロモーターが良い。
【0017】
本発明において用いられる「シャイン・ダルガノ配列」は、ブレビバチルス属細菌において機能する塩基配列であればいかなる原核生物種に由来するものでも良い。好ましくはブレビバチルス属細菌のシャイン・ダルガノ配列が良く、さらに好ましくはブレビバチルス・ブレビスまたはブレビバチルス・チョウシネンシスの細胞壁蛋白質をコードしているDNAの上流に存在するシャイン・ダルガノ配列が良い。更により好ましくはブレビバチルス・ブレビスの細胞壁蛋白質であるmiddle wall protein (MWP)、outer wall protein (OWP)、又はブレビバチルス・チョウシネンシス細胞壁蛋白質HWP(J.Bacteriol.172:1312−1320(1990))をコードする蛋白質のシャイン・ダルガノ配列が良い。ここでいうシャイン・ダルガノ配列とは、原核生物のmRNAにおいて、開始コドンの上流に見られる共通配列であり、プリン塩基に富んだ約3から9塩基の配列である。
【0018】
本発明において用いられる「シグナルペプチド」は、ブレビバチルス属細菌内で発現させた際、そのC末端側に連結されたポリペプチドを細胞外に分泌させる機能を有するペプチドであればいかなる生物種由来のものでも良い。好ましくはブレビバチルス属細菌のシグナルペプチドが良く、より好ましくはブレビバチルス・ブレビスの細胞壁蛋白質であるmiddle wall protein (MWP)またはouter wall protein (OWP)、さらにブレビバチルス・チョウシネンシスの細胞壁蛋白質(HWP)(J.Bacteriol.172:1312−1320(1990))のシグナルペプチドが良い。また天然の分泌シグナルペプチドのアミノ酸配列を改良した配列でも良い。具体的には、前記middle wall protein (MWP)のシグナルペプチドMet−Lys−Lys−Val−Val−Asn−Ser−Val−Leu−Ala−Ser−Ala−Leu−Ala−Leu−Thr−Val−Ala−Pro−Met−Ala−Phe−Alaに、Met−Lys−Lys−Arg(x)−Arg(x)−Val−Val−Asn−Asn−Ser−Val−Leu−Leu(x)−Leu(x)−Leu(x)−Leu(x)−Leu(x)−Ala(x)−Ser(x)−Ala−Leu−Ala−Leu−Thr−Val−Ala−Pro−Met−Ala−Phe−Ala、中の(x)部分で示すアミノ酸配列のように塩基性や疎水性アミノ酸残基を付加したシグナルペプチドを用いても良い。さらに、本発明において使用するプロテインAが本来有するシグナルペプチド配列を使用しても良い。
【0019】
本発明において用いられる「プロテインAの免疫グロブリン結合ドメイン」は、好ましくはスタフィロコッカス属細菌由来のプロテインAの免疫グロブリン結合ドメインであり、より好ましくは、図1(配列表の配列番号1、2)にて示されるスタフィロコッカス・アウレウス(Staphylococcus aureus) COWANI株
由来のプロテインAのE、D、A、B、またはCドメインである。しかし、本明細書において用いられる「プロテインAの免疫グロブリン結合ドメイン」なる用語は上記に限定されず、スタフィロコッカス・アウレウス COWANI株由来のプロテインAのE、D、A、B、またはCドメインと、それらドメインのいずれかと類似の構造および機能を有する分子すべてを包含する。具体的には、図1(配列番号1、2)にて示されるスタフィロコッカス・アウレウス COWAN1株由来のプロテインAのE、D、A、B、またはCドメインのアミノ酸配列に、1アミノ酸以上の置換、欠失、または挿入を施して得られ、かつ、本発明のDNA配列の構成要素の1つである融合領域として用いた場合に、ブレビバチルス属細菌内で所望のポリペプチドの分泌発現量を向上させる機能を有する変異体は、本明細書において用いられる用語「プロテインAの免疫グロブリン結合ドメイン」に包含される。このような変異体は、図1(配列番号1、2)にて示されるアミノ酸配列および塩基配列情報を基に、当業者であれば容易に作製、もしくは自然界より探索することができる。
【0020】
また、本発明において融合領域として使用される「プロテインAの免疫グロブリン結合ドメイン」は、ブレビバチルス属細菌内で所望のポリペプチドの分泌発現量を向上させる機能を有する限り、その個数および順列において何ら制限されるものでない。図1(配列番号1、2)にて示されるスタフィロコッカス・アウレウス COWANI株由来のプロテインAのE、D、A、B、またはCドメインを例として説明すると、E、D、A、B、またはCの任意の1ドメインを使用しても良いし、任意の2ドメインの順列として得られる25通りの配列のうちの1つを使用しても良いし、任意の3ドメインの順列として得られる125通りの配列のうちの1つを使用しても良いし、同様に4以上のドメインの順列として得られる配列のうちの1つを使用しても良い。好ましくは、Eドメイン(図2、3参照)、E、Dドメインをこの順で連結したもの(図4、5参照)、および、E、D、Aドメインをこの順で連結したもの(図6、7参照)が良い。
【0021】
本発明において用いられる「任意のアミノ酸配列からなるリンカー」とは、0または1以上のアミノ酸配列からなるペプチドを意味し、本発明のDNA配列がコードする融合ポリペプチド中に、当該リンカーが実質的に含まれないことをも意味する。当該リンカーを構成するアミノ酸配列に特に制限はないが、当該リンカー中に部位特異的切断に利用し得るアミノ酸配列を配置することにより、融合生成物から所望のポリペプチドを切り離す工程を容易にすることができる。
【0022】
本発明において用いられる「部位特異的切断に利用し得るアミノ酸配列」とは、化学的もしくは酵素的にポリペプチドを切断する際、切断箇所として特異的に認識されるアミノ酸配列を指す。化学的なポリペプチド切断法としては、臭化シアン、ヒドロキシルアミン、または蟻酸を使用する方法が挙げられ、これらの方法を用いる場合、特異的に認識されるアミノ酸配列として、それぞれ、[Met]、[Asn−Gly、−Leu、またはAla]、[Asp−Pro]を配置するのが好ましい。酵素的なポリペプチド切断方法としては、例えば、コラゲナーゼ、キモシン、カリクレインB、エンテロキナーゼ、トロンビン、ファクターXa、またはTEVペプチダーゼを使用する方法が挙げられ、これらの方法を用いる場合、特異的に認識されるアミノ酸配列として、それぞれ、[Pro−X−Gly−Pro(Xは任意のアミノ酸残基)]、[Met−Phe]、[X−Phe−Arg−Y(X、Yは任意のアミノ酸残基)]、[(Asp)n−Lys(n=2−4)]、[Leu−Val−Gln−Arg−Gly−Ser]、[Ile−Glu−Gly−Arg]、[Glu−Asn−Leu−Tyr−Phe−Gln−Gly、またはGlu−Asn−Leu−Tyr−Phe−Gln−Ser]を配置するのが好ましい。
【0023】
また、所望するポリペプチド自体の切断が問題とならなければ、より特異性の低いペプチダーゼ等を使用しても良い。具体的には、真核生物または原核生物由来のセリンペプチダーゼやシグナルペプチダーゼ、エンドペプチダーゼ、カルボキシルペプチダーゼ、アスパラギン酸ペプチダーゼ、システインペプチダーゼ、グルタミン酸ペプチダーゼ、金属ペプチダーゼ、スレオニンペプチダーゼのファミリーに属するペプチダーゼ群が利用できる。さらに、上記ペプチダーゼのアミノ酸配列を置換、欠失、付加、または変異導入することで基質特異性を改変させたペプチダーゼを利用することも可能である。
【0024】
本発明において用いられる「任意のアミノ酸配列からなるリンカー」中には、分離精製用のタグとして使用され得るアミノ酸配列を配置することもできる。「分離精製用のタグ」とは、ポリペプチドの精製を容易にするために付加される、特定物質に対する親和性を有するペプチドを意味し、その具体例としては、ニッケルメタルに対し親和性を有するヒスチジンが複数連結した構造のもの、またはアミロースに対し親和性を有するマルトース結合ポリペプチド等が挙げられる。本発明では前者が好適に用いられ、より好適には6個のヒスチジンの連続配列が用いられる。
【0025】
本発明において用いられる「任意のアミノ酸配列からなるリンカー」中には、前記「部位特異的切断に利用し得るアミノ酸配列」、または前記「分離精製用のタグ」のいずれか1つを配置してもよいし、両者を任意の順序で同時に配置してもよい。ただし、所望のポリペプチドから不要な融合領域を極力切除したい場合は、前記「任意のアミノ酸配列からなるリンカー」の最もC末端側に「部位特異的切断に利用し得るアミノ酸配列」を配置することが好ましい。
【0026】
本発明における「所望のポリペプチド」としては、例えば、各種の酵素類、ヒト成長ホルモン等のホルモン類、各種の血漿蛋白質類、各種のサイトカイン類、免疫グロブリンGのL鎖、Fd鎖等の部分抗体、一本鎖抗体等の修飾抗体などが挙げられる。
【0027】
2.ベクター
本発明の第二の実施形態は、本発明の第一の実施形態であるDNA配列を含むベクターである。本発明の第二の実施形態において用いられるベクターは、ブレビバチルス属細菌において自律複製可能であればいかなるものでも良いが、好ましくは、pHY500(特開平2‐31682号公報)、pNU200(日本農芸化学会誌61,669‐676(1987))、pNH326(図8参照)(Applied and Environmental Microbiology, 58:525−531.(1992))、pNH400(J. Bacteriol, 177:745−749(1995))、pNY700(特開平4−278091号公報)、pHT系プラスミド(特許第2727391号)、大腸菌とブレビバチルス属細菌とのシャトルベクターであるpNCO2(特開2002‐238569号公報)、またはこれらいずれかの誘導体が良い。ただし、本発明の第一の実施形態であるDNA配列を、ブレビバチルス属細菌の染色体へ組込むことを目的とするベクターの場合、ブレビバチルス属細菌において自律複製可能である必要はなく、このようなベクターも、本発明の第一の実施形態であるDNA配列を含む限り、本発明に包含される。
【0028】
3.形質転換体
本発明の第三の実施形態は、本発明の第二の実施形態であるベクターを用いてブレビバチルス属細菌を形質転換することにより得られる形質転換体である。本発明の第三の実施形態において用いられるブレビバチルス属細菌としては、ブレビバチルス・アグリ(Brevibacillus agri)、ブレビバチルス・ボルステレンシス(Brevibacillus borstelensis)、ブレビバチルス・ブレビス、ブレビバチルス・セントロスポルス(Brevibacillus centrosporus)、ブレビバチルス・チョウシネンシス、ブレビバチルス・フォーモサス(Brevibacillus formosus)、ブレビバチルス・インボカツス(Brevibacillus invocatus)、ブレビバチルス・ラテロスポルス(Brevibacillus laterosporus)、ブレビバチルス・リムノフィルス(Brevibacillus limnophilus)、ブレビバチルス・パラブレビス(Brevibacillus parabrevis)、ブレビバチルス・レウスゼリ(Brevibacillus reuszeri)、またはブレビバチルス・サーモルベル(Brevibacillus thermoruber)等が挙げられる。ブレビバチルス・ブレビス、またはブレビバチルス・チョウシネンシスが好ましく、ブレビバチルス・ブレビス47株、ブレビバチルス・ブレビス47K株、ブレビバチルス・ブレビス47−5Q株、ブレビバチルス・チョウシネンシスHPD31株、またはブレビバチルス・チョウシネンシスHPD31−OK株がより好ましい。また生産量の向上などの目的に応じて上記ブレビバチルス属細菌のプロテアーゼ欠損株や高分泌発現株のような変異株を使用しても良い。
【0029】
ブレビバチルス属細菌の形質転換は、例えば、Takahashiらの方法(J. Bacteriol. 156:1130−1134(1983))、Takagiらの方法(Agric. Biol. Chem, 53:3099−3100(1989))、またはOkamotoらの方法(Biosci. Biotechnol. Biochem. 61:202−203(1997))により実施することができる。
【0030】
4.ポリペプチド製造方法
本発明の第四の実施形態は、本発明の第三の実施形態である形質転換体を培養し、当該形質転換体から分泌される所望のポリペプチドの一部または全部を含むポリペプチドを回収することを特徴とする、ポリペプチドの製造方法である。
【0031】
当該形質転換体の培養に用いる培地は、当該形質転換体が所望のポリペプチドを効率良く分泌発現できる培地であれば特に制限は無い。具体的にはグルコース、蔗糖、グリセロールなどの炭素源や、ポリペプトン、肉エキス、酵母エキス、カザミノ酸などの窒素源を使用することができ、カリウム塩、ナトリウム塩、リン酸塩、マグネシウム塩、マンガン、亜鉛、鉄等の無機塩類が必要に応じて添加される。さらに、栄養要求性の宿主細胞を用いる場合は、生育に要求される栄養物質を添加すればよい。また、必要であればペニシリン、エリスロマイシン、クロラムフェニコール、ネオマイシンなどの抗生物質が添加されても良い。培養温度は15−42℃、好ましくは約28−37℃であり、通気および攪拌により好気的に培養を行うことが望ましいが、もし必要であれば通気を遮断し嫌気的に培養してもよい。
【0032】
当該形質転換体から分泌された、所望のポリペプチドの一部または全部を含むポリペプチドは、通常公知の蛋白質精製法を適当に組み合わせて用いることにより、当該形質転換体の培養液から回収、精製することができる。例えば、塩析、透析、限外濾過、ゲル濾過クロマトグラフィー、イオン交換クロマトグラフィー、逆相クロマトグラフィー、またはアフィニティークロマトグラフィー等の手法を利用できる。
【0033】
特に、本発明の第四の実施形態に従い分泌発現されたポリペプチドは、その分子内にプロテインAの免疫グロブリン結合ドメインまたはその部分的配列を有しているため、適当な担体に固定化された免疫グロブリンG、例えば免疫グロブリンGセファロース(アマシャムバイオサイエンス社)等を用いることで、効率良く精製され得る。
【0034】
また、本発明の第四の実施形態に従い分泌発現されたポリペプチドは、リンカー部分に精製用タグを含みうる。この場合、当該箇所に配置した精製用タグを利用して、当該ポリペプチドを精製することも可能である。例えば、精製用タグとしてヒスチジンの連続配列を配置した場合、ニッケルNTAカラム等を用いて精製することが可能である。
【0035】
さらに、本発明の第四の実施形態に従い分泌発現されたポリペプチドは、リンカー部分に部位特異的切断に利用し得るアミノ酸配列を含みうる。この場合、当該箇所に配置したアミノ酸配列に応じた方法により、当該箇所よりN末端側のポリペプチドを所望のポリペプチドから切除することが可能である。例えば、部位特異的切断に利用し得るアミノ酸配列として[Ile−Glu−Gly−Arg]を配置した場合、ファクターXaを用いて、当該箇所よりN末端側のポリペプチドを所望のポリペプチドから切除することが可能である。このような切断反応は、本発明の第四の実施形態に従い分泌発現されたポリペプチドの精製工程中のいかなる段階で実施しても良い。切断反応後の所望のポリペプチドは、通常公知の蛋白質精製法を適当に組み合わせて用いることにより、さらに精製することができる。
【0036】
上記のような本発明の各実施形態を利用することで、これまでブレビバチルス属細菌では効率的に生産できなかったポリペプチドを、極めて効率良く生産することが可能となった。従って、本発明の実施形態は、産業上有用なポリペプチドを製造する上で、非常に有用である。
【実施例】
【0037】
以下に参考例、実施例、図面に基づいて本発明の実施例を具体的に説明するが、これらが本発明の範囲を制限するものでないことは云うまでもない。本発明に関連する技術分野の当業者は、本発明の説明の範囲内で以下の実施例の変形を認め実行することができる。したがって、そのような変形は、本発明の範囲内に入るものとみなされる。
【0038】
本発明の実施例の実行にあたり、発現DNAの作製や操作などは特に断わらない限り下記の実験書に従って実施した。 (1)「Molecular Cloning/A Laboratory Manual」、第2版(1989)、Cold Spring Harbor Laboratory 刊(米国)。 (2) 村松正實 編著「ラボマニュアル遺伝子工学」、第3版(1996)、丸善株式会社刊。
(実施例1)プロテインAの免疫グロブリン結合ドメインをコードするDNA配列の単離
スタフィロコッカス・アウレウス Cowan I株を、T2液体培地(ポリペプトン 1%、イーストエキストラクト 0.2%、グルコース 1%、魚肉エキス 0.5%、pH7.0)を用いて、37℃で一晩振とう培養した。得られた培養液から遠心分離により菌体を回収し、トリス−塩酸緩衝液(pH8.0)で2度洗浄を行った。菌体を再度トリス−塩酸緩衝液にて懸濁後1%SDSで溶菌し、60℃にて30分間加熱後、フェノール抽出およびエタノール沈殿等の定法により全ゲノムDNAを抽出した。公知のプロテインAの塩基配列(配列番号1)に基づき、上記で調製したゲノムDNAを鋳型とし、フォワードプライマー(5’−TTGCTCCCATGGCTTTCGCTGCGCAACACGATGAAGCT−3’ (配列番号15))と、以下の3種のリバースプライマー(5’−TTTTCTGCAGTTTTGGAGCTTGAGAGTCATTAAGTTTTTGAGC −3’(配列番号16)、5’−TTTTCTGCAGTTTCGGTGCTTGAGATTCGTTTAATTTTTT−3’ (配列番号17)、または5’−TTTTCTGCAGTTTCGGTGCTTGAGATTCATTTAACTTTTT−3’ (配列番号18))を用い、PCR法により、Eドメイン、EDドメイン、またはEDAドメインをコードするDNAをそれぞれ増幅した。配列番号15は、プロテインAの免疫グロブリンG結合ドメインのPCR増幅用にデザインされたフォワードプライマーを示す。配列番号16は、プロテインAの免疫グロブリンG結合ドメインEのPCR増幅用にデザインされたリバースプライマーを示す。配列番号17は、プロテインAの免疫グロブリンG結合ドメインEDのPCR増幅用にデザインされたリバースプライマーを示す。配列番号18は、プロテインAの免疫グロブリンG結合ドメインEDAのPCR増幅用にデザインされたリバースプライマーを示す。
【0039】
また、同様に、Eドメイン、EDドメイン、またはEDAドメインをコードするDNAの3’側に6残基のヒスチジンを付加して増幅する場合は、上記ゲノムDNAを鋳型として、フォワードプライマー(5’−TTGCTCCCATGGCTTTCGCTGCGCAACACGATGAAGCT−3’ (配列番号15))と、以下の3種のリバースプライマー(5’−TTTTCTGCAGGTGATGATGATGATGATGTTTTGGAGCTTGAGAGTCATTAAGTTTTTGAGC−3’ (配列番号19)、 5’−TTTTCTGCAGGTGATGATGATGATGATGTTTCGGTGCTTGAGATTCGTTTAATTTTTT−3’ (配列番号20)、または5’−TTTTCTGCAGGTGATGATGATGATGATGTTTCGGTGCTTGAGATTCATTTAACTTTTT−3’ (配列番号21))を用いた。配列番号19は、プロテインAの免疫グロブリンG結合ドメインEのPCR増幅用にデザインされたリバースプライマーを示す。配列番号20は、プロテインAの免疫グロブリンG結合ドメインEDのPCR増幅用にデザインされたリバースプライマーを示す。配列番号21は、プロテインAの免疫グロブリンG結合ドメインEDAのPCR増幅用にデザインされたリバースプライマーを示す。
【0040】
得られたDNA断片は制限酵素NcoIおよびPstIにより消化した後、アガロースゲルより分離回収した。一方、発現ベクターpNH326(図8)(配列番号22、23)もまた同様に制限酵素NcoIおよびPstIにより消化後、精製回収してアルカリフォスファターゼ処理により脱リン酸化処理を行った。
(実施例2)プロテインA融合ポリペプチド発現ベクターの構築
実施例1にて、制限酵素処理したDNA断片および発現ベクターpNH326をT4DNAライゲースを用いて連結し、各免疫グロブリン結合ドメインを有するプロテインA融合ポリペプチド発現ベクターpProA3E(図2、配列番号3および4参照)、pProA4E(図3、配列番号5および6参照)、pProA3ED(図4、配列番号7および8参照)、pProA4ED(図5、配列番号9および10参照)、pProA3EDA(図6、配列番号11および12参照)、pProA4EDA(図7、配列番号13および14参照)を構築した。
(実施例3)プロテインA融合抗ヒトTNFα抗体L鎖発現ベクターの作成
プロテインA融合ポリペプチド発現ベクターの発現能を評価するために、ヒト・マウスキメラ抗ヒトTNFα抗体のL鎖(軽鎖)遺伝子(配列番号24、25)の発現を行った。ヒト・マウスキメラ抗ヒトTNFα抗体のL鎖遺伝子は、米国特許5698195号記載の抗ヒトTNFα抗体L鎖の遺伝子配列に従い作製されたpBluescript・抗ヒトTNFα抗体L鎖を鋳型にし、2つのオリゴヌクレオチドプライマー5’−AAAACTGCAGATCGAAGGTCGTGACATCTTGCTGACTCAGTCTCCAGCC−3’ (配列番号26)と5’−TTTGAATTCCTAACACTCTCCCCTGTTGAAGCTCTT−3’(配列番号27)とを用いて、PCR法により増幅した。配列番号26は、抗TNFα抗体のL鎖遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号27は、抗TNFα抗体のL鎖遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0041】
この増幅されたヒトTNFα抗体のL鎖をコードしているDNAの5’末端にはファクターXaの認識開裂サイトであるIle−Glu−Gly−ArgのペプチドをコードしているDNA配列(配列番号26における上記下線部ATCGAAGGTCGTの配列)を含むように設計されている。この抗ヒトTNFα抗体のL鎖遺伝子は制限酵素PstIとEcoRIにより切断した。また上記プロテインA融合ポリペプチド発現ベクターpProA4E(図3)のマルチクローニングサイトに存在する制限酵素PstIとEcoRIにより消化後、精製回収してアルカリフォスファターゼ処理により脱リン酸化処理を行った。制限酵素処理を行った上記のDNA断片とプロテインA融合ポリペプチド発現ベクターはT4DNAライゲースを用いて連結し、プロテインA融合抗ヒトTNFα抗体L鎖発現ベクターpProA4E−L(図9A)(配列番号28、29)を構築した。
(実施例4)プロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクターの作成
上記、抗ヒトTNFα抗体のFd遺伝子を有するプロテインA融合ポリペプチド発現ベクターの発現能を評価するため発現ベクターの構築を行った。抗ヒトTNFα抗体のFd鎖遺伝子は上記の抗TNFα抗体H鎖の遺伝子配列(配列番号30、31)に従い作製されたpBluescript・抗ヒトTNFα抗体H鎖を鋳型にし、2つのオリゴヌクレオチドプライマー5’−AAAACTGCAGATCGAAGGTCGTGAAGTGAAACTTGAGGAGTCTGAGGGA −3’ (配列番号32)と5’−CCGGAATTCTCACGGTGGGCATGTGTGAGTTTTGTCACA−3’(配列番号33)とを用いて、PCR法により増幅した。配列番号32は、抗TNFα抗体のFd鎖遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号33は、抗TNFα抗体のFd鎖遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0042】
この増幅された抗ヒトTNFα抗体Fd鎖をコードしているDNAの5’末端にはファクターXaの認識開裂サイトであるIle−Glu−Gly−ArgのペプチドをコードしているDNA配列(配列番号32の上記下線部ATCGAAGGTCGTの配列)を含むように設計されている。このヒトTNFα抗体のFd鎖遺伝子は制限酵素PstIとEcoRIにより切断した。また上記プロテインA融合ポリペプチド発現ベクターpProA3E(図2)、pProA3ED(図4)およびpProA3EDA(図6)のマルチクローニングサイトに存在する制限酵素PstIとEcoRIを用い消化後、精製回収してアルカリフォスファターゼ処理により脱リン酸化処理を行った。制限酵素処理を行った上記のDNA断片と各プロテインA融合ポリペプチド発現ベクターをT4DNAライゲースを用いて連結し、プロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクターpProA3E−Fd(図9B、配列番号34および35参照)、pProA3ED−Fd(図9C、配列番号36および37参照)、pProA3EDA−Fd(図9D、配列番号38および39参照)を構築した。
(実施例5)抗ヒトTNFα抗体鎖のL鎖またはFd鎖単独発現ベクターの作成
プロテインA融合型と従来からの目的とするポリペプチド単独で発現させる方法(融合型でない方法)とのポリペプチド発現能を比較検討するため、抗ヒトTNFα抗体L鎖鎖単独発現ベクター、および抗ヒトTNFα抗体Fd鎖単独発現ベクターを構築した。抗ヒトTNFα抗体のL鎖遺伝子は、先述のpBluescript・抗ヒトTNFα抗体L鎖を鋳型にし、2つのオリゴヌクレオチドプライマー5’− GCTCCCATGGCTTTCGCTGACATCTTGCTGACTCAGTCT −3’ (配列番号40)と5’−TTTCTGCAGCTAACACTCTCCCCTGTTGAAGCTCTT−3’(配列番号41)とを用いて、PCR法により増幅した。配列番号40は、抗TNFα抗体のL鎖遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号41は、抗TNFα抗体のL鎖遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0043】
また抗ヒトTNFα抗体のFd鎖遺伝子は、先述のpBluescript・抗ヒトTNFα抗体H鎖を鋳型にし、2つのオリゴヌクレオチドプライマー5’− GCTCCCATGGCTTTCGCTGAAGTGAAACTTGAGGAGTCT −3’ (配列番号42)と5’− AAACTGCAGTTATTCAGGTGCTGGGCACGG −3’(配列番号43)を用いて、PCR法により増幅した。配列番号42は、抗TNFα抗体のFd鎖遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号43は、抗TNFα抗体のFd鎖遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0044】
L鎖およびFd鎖をコードするDNAは制限酵素NcoIとPstIでそれぞれ消化した。発現ベクターとなるpNH326も同様に制限酵素NcoIとPstIでそれぞれ消化後、精製回収し、アルカリフォスファターゼ処理によりそれぞれ脱リン酸化処理を行った。制限酵素処理を行った上記のDNA断片と発現ベクターはT4DNAライゲースを用いて連結し、抗ヒトTNFα抗体L鎖発現ベクター、および、抗ヒトTNFα抗体Fd鎖発現ベクターpNH326−L(図10A、配列番号44および45)、pNH326−Fd(図10B、配列番号46および47)を構築した。
【0045】
(実施例6)プロテインA融合ヒト成長ホルモン発現ベクターの作成
上記抗体以外のポリペプチドに対する本発明の効果を示すために、ヒト成長ホルモンをコードするDNAを用いプロテインA融合発現試験を行った。ヒト成長ホルモンをコードするDNAを有するプラスミドpGH−L9(Proc.Natl.Acad.Sci.USA.,81,5956−5960 (1984))を鋳型とし、2つのオリゴヌクレオチドプライマー5’−AAAACTGCAGATCGAAGGTCGTTTCCCAACTATTCCACTGAGT−3’ (配列番号48)と5’−TCCCAAGCTTTTAGAAGCCACACGACCCTTCAACAGA−3’ (配列番号49)を用いて、PCR法によりヒト成長ホルモンをコードするDNA増幅した。配列番号48は、ヒト成長ホルモンをコードする遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号49は、ヒト成長ホルモンをコードする遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0046】
増幅されたヒト成長ホルモンをコードするDNAは制限酵素PstIとHindIIIで消化した。また発現ベクターとなるpProA3Eも同様に制限酵素PstIとHindIIIで消化後、精製回収し、アルカリフォスファターゼ処理により脱リン酸化処理を行った。制限酵素処理を行った上記のヒト成長ホルモンDNA断片とpProA3E発現ベクターはT4DNAライゲースを用いて連結し、プロテインA融合ヒト成長ホルモン発現ベクターpProA3E−hGH (図11A)(配列番号50、51)を構築した。
【0047】
(実施例7)ヒト成長ホルモン単独発現ベクター、およびMWP融合ヒト成長ホルモン発現ベクターの構築
本発明の有効性を示すため、ヒト成長ホルモン単独発現ベクター、および、特許文献5、6で開示されたブレビバチルス・ブレビス細胞壁蛋白質MWPを融合部分に用いたMWP融合ヒト成長ホルモン発現ベクターを構築した。ヒト成長ホルモン単独発現ベクターを構築するため、2つのオリゴヌクレオチドプライマー5’− TTGCTCCCATGGCTTTCGCTTTCCCAACTATTCCACTGAGT −3’ (配列番号52)と5’−TCCCAAGCTTTTAGAAGCCACACGACCCTTCAACAGA −3’ (配列番号53)を用い、先述のプラスミドpGH−L9を鋳型として、PCR法によりヒト成長ホルモンをコードするDNAを増幅した。配列番号52は、ヒト成長ホルモンをコードする遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号53は、ヒト成長ホルモンをコードする遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0048】
増幅されたヒト成長ホルモンをコードするDNAは制限酵素NcoIとHindIIIで消化した。同様に発現ベクターとなるpNH326も同様に制限酵素NcoIとHindIIIで消化後、精製回収し、アルカリフォスファターゼ処理により脱リン酸化処理を行った。制限酵素処理を行った上記のDNA断片と発現ベクターはT4DNAライゲースを用いて連結し、ヒト成長ホルモン単独発現ベクターpNH326−hGH(図11B)(配列番号54、55)を構築した。一方、MWP融合ヒト成長ホルモン発現ベクターとしては、特許文献5の実施例1および実施例4に記載の方法に従い、MWP20−His6−Linker−Clavage1−hGH/pNH326 (図11C)を構築した。
【0049】
(実施例8)各発現ベクターを有する形質転換体の作成
実施例2から7で構築した、プロテインA融合発現ベクター(pProA3E、pProA3ED、pProA3EDA、pProA4E、pNH326)、プロテインA融合抗ヒトTNFα抗体L鎖発現ベクター(pProA4E−L)、プロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクター(pProA3E−Fd、pProA3ED−Fd、pPro3EDA−Fd)、抗ヒトTNFα抗体L鎖単独発現ベクター(pNH326−L)、抗ヒトTNFα抗体Fd鎖単独発現ベクター(pNH326−Fd)、プロテインA融合ヒト成長ホルモン発現ベクター(pProA3E−hGH)、ヒト成長ホルモン単独発現ベクター(pNH326−hGH)、およびMWP融合ヒト成長ホルモン発現ベクター(MWP20−His6−Linker−Clavage1−hGH/pNH326)を、公知の方法に従い、それぞれブレビバチルス・チョウシネンシスHPD31−OK株へ導入し、形質転換体を得た。
【0050】
(実施例9)抗ヒトTNFα抗体のL鎖およびFd鎖の分泌発現
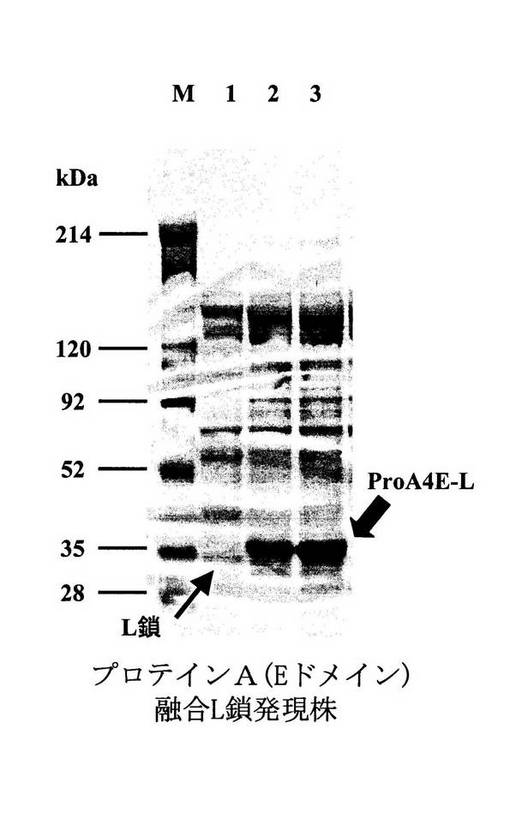
実施例8にて得られた、プロテインA融合抗ヒトTNFα抗体L鎖発現ベクターpProA4E−Lを有する形質転換体、プロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクターpProA3E−Fd、pProA3ED−Fd、pPro3EDA−Fdをそれぞれ有する形質転換体、抗ヒトTNFα抗体L鎖単独発現ベクターpNH326−Lを有する形質転換体、抗ヒトTNFα抗体Fd鎖単独発現ベクターpNH326−Fdを有する形質転換体、およびそれらの対照となるベクターpNH326を有する形質転換体の各々を、60mg/Lのネオマイシンを添加した3YC培地(ポリペプトンS 3%、酵母エキス0.5%、グルコース3%、MgSO4・7H2O 0.01%、CaCl2・7H2O 0.01%、MnSO4・4H2O 0.001%,FeSO4・7H2O 0.001%,ZnSO4・7H2O 0.0001% pH7.0)にて、30℃で2日間振とう培養した。当該培養液から遠心分離(10,000rpm,4℃,3分間)により菌体を除去し、得られた培養上清各5μlを定法に従い処理した後、10−20%グラジエントポリアクリルアミドゲルを用いたSDS−ポリアクリルアミドゲル電気泳動(SDS−PAGE)に供した。電気泳動後、当該ゲルをCBB(Coomassie Brilliant Blue)R−250で染色し、ポリペプチドのバンドを検出した(図12および、図13A、B、C)。
【0051】
図12は、CBB染色による、実施例8で得られた各プロテインA融合抗ヒトTNFα抗体L鎖発現株と抗ヒトTNFα抗体L鎖単独発現株との発現量の比較を示す。太い矢印は発現したプロテインA融合抗ヒトTNFα抗体L鎖を示す。レーンMは分子量マーカー(214,120,92,52,35,28kDa)を示す。レーン1は、抗ヒトTNFα抗体L鎖単独発現株からの培養液5μlを泳動したものであり、レーン2および3は、プロテインA(Eドメイン)融合抗ヒトTNFα抗体L鎖発現株からの培養液5μlを泳動したものである。
【0052】
図13は、CBB染色による、実施例8で得られた各プロテインA融合抗ヒトTNFα抗体Fd鎖発現株と抗ヒトTNFα抗体Fd鎖単独発現株との発現量の比較を示す。太い矢印は発現したプロテインA融合抗ヒトTNFα抗体Fd鎖[(図13A) プロテインA(Eドメイン)融合Fd鎖、(図13B) プロテインA(EDドメイン)融合Fd鎖、(図13C) プロテインA(EDAドメイン)融合Fd鎖]を示す。細い矢印は抗ヒトTNFα抗体Fd鎖単独発現株から、発現された抗ヒトTNFα抗体Fd鎖の移動度を示す。レーンMは分子量マーカー(214,120,92,52,35,28kDa)を示す。レーン1は、抗ヒトTNFα抗体Fd鎖単独発現株からの培養液5μlを泳動したもの、レーン2および3は、プロテインA(Eドメイン)融合抗ヒトTNFα抗体Fd鎖発現株からの培養液5μlを泳動したもの、レーン4は、抗ヒトTNFα抗体Fd鎖単独発現株からの培養液5μlを泳動したもの、レーン5および6は、プロテインA(EDドメイン)融合抗ヒトTNFα抗体Fd鎖発現株からの培養液5μlを泳動したもの、レーン7は、抗ヒトTNFα抗体Fd鎖単独発現株からの培養液5μlを泳動したもの、レーン8および9は、プロテインA(EDAドメイン)融合抗ヒトTNFα抗体Fd鎖発現株からの培養液5μlを泳動したものである。
【0053】
実験の結果、抗ヒトTNFα抗体L鎖単独発現ベクターpNH326−Lを有する形質転換体における抗ヒトTNFα抗体L鎖の発現量(図12参照)、および抗ヒトTNFα抗体Fd鎖単独発現ベクターpNH326−Fdを有する形質転換体における抗ヒトTNFα抗体Fd鎖の発現量は、そのバンドを確認することが困難なほど少なかった。一方、プロテインA融合抗ヒトTNFα抗体L鎖発現ベクターpProA4E−Lを有する形質転換体におけるプロテインA融合抗ヒトTNFα抗体L鎖の発現量(図12参照)、およびプロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクターpProA3E−Fd(図13A参照)、pProA3ED−Fd(図13B参照)、pPro3EDA−Fd(図13C参照)を有する形質転換体におけるプロテインA融合抗ヒトTNFα抗体Fd鎖の発現量は飛躍的に上昇した。また、3種類のプロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクターpProA3E−Fd、pProA3ED−Fd、およびpPro3EDA−Fdをそれぞれ有する形質転換体における、プロテインA融合抗ヒトTNFα抗体Fd鎖の発現量を比較すると、融合領域として使用するプロテインAの免疫グロブリン結合ドメインの数を増やす程、融合ポリペプチドの発現量が顕著に増加していた(図13A、B、C参照)。
【0054】
(実施例10)ヒト成長ホルモンの分泌発現
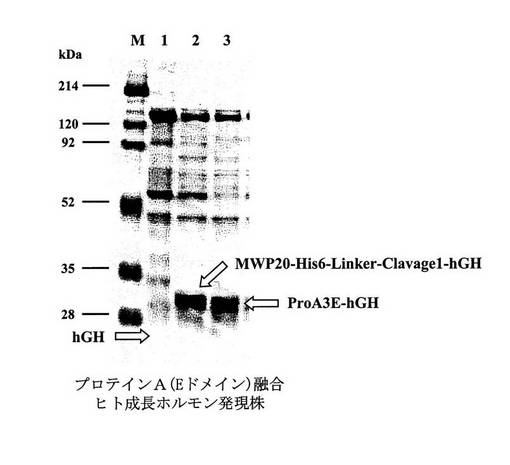
実施例8にて得られた、プロテインA融合ヒト成長ホルモン発現ベクターpProA3E−hGHを有する形質転換体、MWP融合ヒト成長ホルモン発現ベクターMWP20−His6−Linker−Clavage1−hGH/pNU211R2L5を有する形質転換体、およびヒト成長ホルモン単独発現ベクターpNH326−hGHを有する形質転換体の各々を、実施例9と同様に培養した。ただし、MWP融合ヒト成長ホルモン発現ベクターMWP20−His6−Linker−Clavage1−hGH/pNU211R2L5を有する形質転換体の培養のみ、培地に添加する薬剤を、ネオマイシン60mg/Lに替えて、エリスロマイシン10mg/Lとした。そして、それらの培養上清を実施例9と同様にSDS−PAGEにて解析した(図14)。
【0055】
図14は、CBB染色による、実施例8で得られたプロテインA融合ヒト成長ホルモン発現株、MWP融合ヒト成長ホルモン発現株、およびヒト成長ホルモン単独発現株の発現量の比較を示す図である。レーンMは分子量マーカー(214,120,92,52,35,28kDa)を示す。レーン1はヒト成長ホルモン単独発現株からの培養液5μl、レーン2はMWP融合ヒト成長ホルモン発現株からの培養液5μl、レーン3はプロテインA融合ヒト成長ホルモン発現株からの培養液5μlを泳動したものである。
【0056】
実験の結果、ヒト成長ホルモン単独発現ベクターpNH326−hGHを有する形質転換体におけるヒト成長ホルモンの発現は極めて少量であったが、MWP融合ヒト成長ホルモン発現ベクターMWP20−His6−Linker−Clavage1−hGH/pNU211R2L5を有する形質転換体、および、プロテインA融合ヒト成長ホルモン発現ベクターpProA3E−hGHを有する形質転換体の培養液上清には大量のヒト成長ホルモンが分泌生産されていた(図14参照)。
【0057】
(実施例11)プロテインA融合抗ヒトTNFα一本鎖抗体発現ベクターの作成
実施例3で作製したpBluescript・抗ヒトTNFα抗体L鎖を鋳型にし、2つのオリゴヌクレオチドプライマー5’−AAAACTGCAGATCGAAGGTCGTGACATCTTGCTGACTCAGTCTCCAGCC−3’ (配列番号26)と5‘−GGAACCACCGCCGCCGGAGCCGCCACCACCGGAGCCGCCACCGCCAGTGCGTTTTACTTC−3’(配列番号56)とを用いて、抗ヒトTNFα一本鎖抗体の前半部分をコードするDNAを、PCR法により増幅した。配列番号26は、抗TNFα抗体のL鎖遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号56は、scFv(single chain variable fragments)の前半部分の遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。scFvは、一般的に、抗体が抗原を認識するために必要な最小単位であるVHおよびVLで構成される可変領域(Fv)を、フレキシブルなペプチドリンカーで結合した単鎖可変領域フラグメントである。
【0058】
この時、抗ヒトTNFα一本鎖抗体のN末端に、ファクターXaの認識開裂サイトであるIle−Glu−Gly−Argからなる配列が付加されるよう、プライマーを設計した。同様に、実施例4で作製したpBluescript・抗ヒトTNFα抗体H鎖を鋳型にし、2つのオリゴヌクレオチドプライマー5’−GGTGGTGGCGGCTCCGGCGGCGGTGGTTCCGAAGTGAAACTGGAAGAATCTGGAGGAGGCTTG−3’(配列番号57)と5’−TTTCTGCAGTTAGTGATGATGATGATGATGGGTGGTGCCTTGGCCCCAGTAGTCGTA−3’(配列番号58)とを用いて、抗ヒトTNFα一本鎖抗体の後半部分をコードするDNAを、PCR法により増幅した。配列番号57は、scFvの後半部分の遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号58は、scFvの後半部分の遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0059】
この時、抗ヒトTNFα一本鎖抗体のC末端に、ヒスチジン6残基が付加されるよう、プライマーを設計した。そして、これら2つのDNA断片をPCR法により連結し、抗ヒトTNFα一本鎖抗をコードするDNAを作製した。
【0060】
作製したDNAを制限酵素PstIで消化した後、実施例2で作製したプロテインA融合発現ベクターpProA4E(図3)およびpProA4EDA(図7)のマルチクローニングサイトに存在する制限酵素PstI部位にそれぞれ挿入し、プロテインA融合抗ヒトTNFα一本鎖抗体発現ベクターpProA4E−scFv(図15A)(配列番号59、60)、およびpProA4EDA−scFv(図15B)(配列番号61、62)を構築した。
【0061】
(実施例12)抗ヒトTNFα一本鎖抗体単独発現ベクターの作成
オリゴヌクレオチドプライマー5’−AAAACTGCAGATCGAAGGTCGTGACATCTTGCTGACTCAGTCTCCAGCC−3’(配列番号26)に替えて、オリゴヌクレオチドプライマー5’− GCTCCCATGGCTTTCGCTGACATCTTGCTGACTCAGTCT −3’(配列番号41)を用いる以外は実施例11と同様の操作を行い、抗ヒトTNFα一本鎖抗をコードするDNAを作製した。配列番号41は、抗TNFα抗体のL鎖遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0062】
作製したDNAを制限酵素NcoIおよびPstIで消化した後、発現ベクターpNH326のマルチクローニングサイトに存在するNcoI−PstI部位に挿入し、抗ヒトTNFα一本鎖抗体単独発現ベクターpNH326−scFv(図15C)(配列番号63、64)を構築した。
【0063】
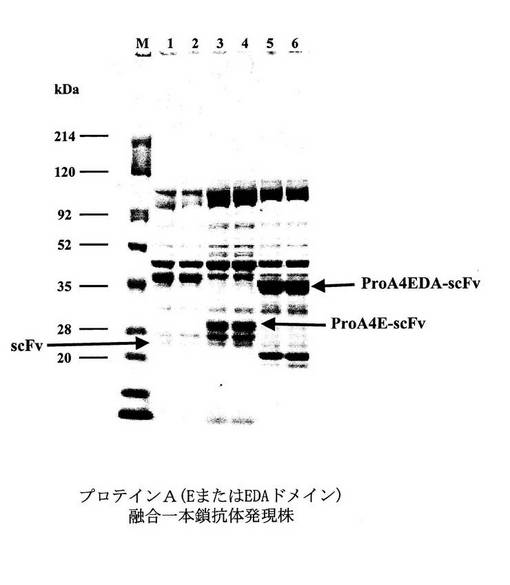
(実施例13)抗ヒトTNFα一本鎖抗体の分泌発現
実施例11にて得られたプロテインA融合抗ヒトTNFα一本鎖抗体発現ベクターpProA4E−scFvならびにpProA4EDA−scFv、および、実施例12にて得られた抗ヒトTNFα一本鎖抗体単独発現ベクターpNH326−scFvを、公知の方法に従い、それぞれブレビバチルス・チョウシネンシスHPD31−OK株へ導入し、形質転換体を得た。次に、これらの形質転換体を実施例9と同様に培養し、それらの培養上清を実施例9と同様にSDS−PAGEにて解析した(図16)。
【0064】
図16は、CBB染色による、実施例13で得られたプロテインA融合抗ヒトTNFα一本鎖抗体発現株、および抗ヒトTNFα一本鎖抗体単独発現株の発現量の比較を示す図である。レーンMは分子量マーカー(214,120,92,52,35,28,20kDa)を示す。レーン1および2は各々抗ヒトTNFα一本鎖抗体単独発現株の培養液5μl、レーン3および4は各々プロテインA(Eドメイン)融合抗ヒトTNFα一本鎖抗体発現株の培養液5μl、レーン5および6は各々プロテインA(EDAドメイン)融合抗ヒトTNFα一本鎖抗体発現株の培養液5μlを泳動したものである。
【0065】
実験の結果、抗ヒトTNFα一本鎖抗体単独発現ベクターpNH326−scFvを有する形質転換体における抗ヒトTNFα一本鎖抗体の発現量は、そのバンドを確認することが困難なほど少なかった。一方、プロテインA融合抗ヒトTNFα一本鎖抗体発現ベクターpProA4E−scFv、ならびにpProA4EDA−scFvを有する形質転換体におけるプロテインA融合抗ヒトTNFα一本鎖抗体の発現量は飛躍的に上昇した。
【図面の簡単な説明】
【0066】
【図1】スタフィロコッカス・アウレウス・コワン(Cowan)I株のプロテインAの塩基配列とアミノ酸配列図である。番号はアミノ酸残基数を示す。
【図2】プロテインA(Eドメイン)融合分泌発現ベクターpProA3Eの構造とそのマルチクローニングサイト付近のDNA配列を示す。分泌発現ベクターpProA3EはプロテインA免疫グロブリン結合Eドメインの後、ヒスチジンタグを有する。
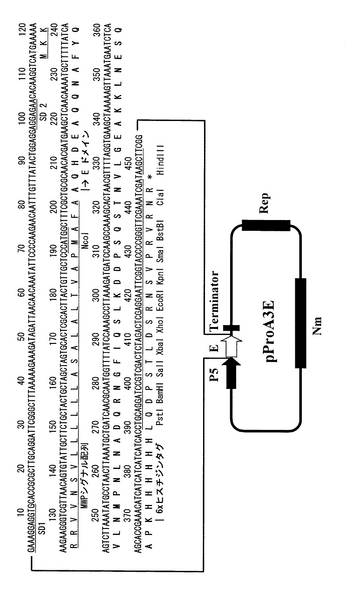
【図3】実施例2に記載のプロテインA(Eドメイン)融合分泌発現ベクターpProA4Eの構造とそのマルチクローニングサイト付近のDNA配列を示す。分泌発現ベクターpProA4EはプロテインA免疫グロブリン結合Eドメインの後、ヒスチジンタグを含まない。
【図4】実施例2に記載のプロテインA(EDドメイン)融合分泌発現ベクターpProA3EDの構造とそのマルチクローニングサイト付近のDNA配列を示す。分泌発現ベクターpProA3EDはプロテインA免疫グロブリン結合EDドメインの後、ヒスチジンタグを有する。
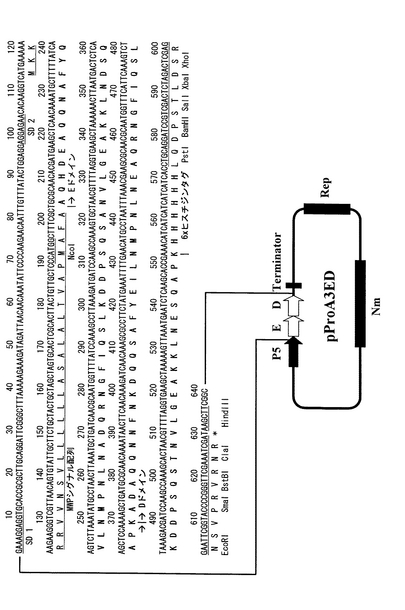
【図5】実施例2に記載のプロテインA(EDドメイン)融合分泌発現ベクターpProA4EDの構造とそのマルチクローニングサイト付近のDNA配列を示す。分泌発現ベクターpProA4EDはプロテインA免疫グロブリン結合EDドメインの後、ヒスチジンタグを含まない。
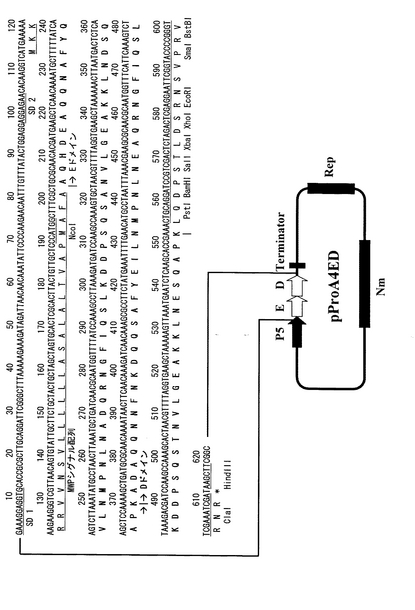
【図6】実施例2に記載のプロテインA(EDAドメイン)融合分泌発現ベクターpProA3EDAの構造とそのマルチクローニングサイト付近のDNA配列を示す。分泌発現ベクターpProA3EDAはプロテインA免疫グロブリン結合EDAドメインの後、ヒスチジンタグを有する。
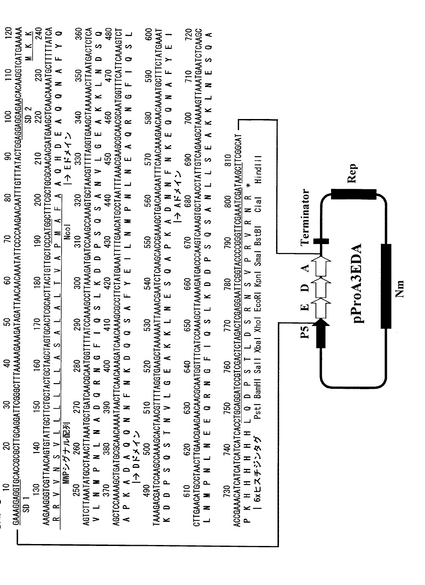
【図7】実施例2に記載のプロテインA(EDAドメイン)融合分泌発現ベクターpProA4EDAの構造とそのマルチクローニングサイト付近のDNA配列を示す。分泌発現ベクターpProA4EDAはプロテインA免疫グロブリン結合EDAドメインの後、ヒスチジンタグを含まない。
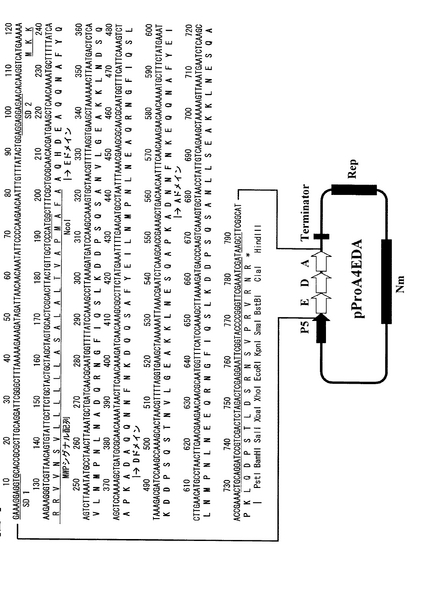
【図8】実施例2に記載のブレビバチルス・ブレビスポリペプチド分泌発現ベクターpNH326の構造とそのマルチクローニングサイト付近のDNA配列を示す。
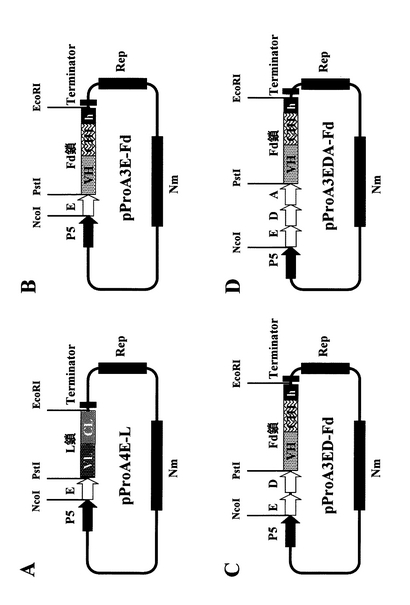
【図9】実施例3および実施例4に記載のプロテインA融合抗ヒトTNFα抗体L鎖発現ベクター(A) pProA4E−L、およびプロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクター(B) pProA3E−Fd、(C) pProA3ED−Fdおよび(D) pProA3EDA−Fdの構造を示す。
【図10】実施例5に記載の抗ヒトTNFα抗体鎖単独発現ベクター(A) pNH326−L、(B) pNH326‐Fdの構造を示す。
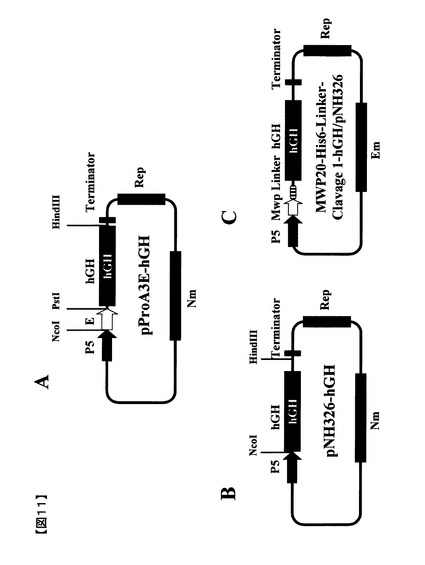
【図11】実施例6および7記載のプロテインA融合ヒト成長ホルモン発現ベクター(A) pProA3E−hGH、(B) ヒト成長ホルモン単独発現ベクターpNH326−hGH、および(C) MWP融合ヒト成長ホルモン発現ベクターMWP20−His6−Linker−Clavage1−hGH/pNU211R2L5、の構造を示す。
【図12】CBB染色による、実施例8で得られた各プロテインA融合抗ヒトTNFα抗体L鎖発現株と抗ヒトTNFα抗体L鎖単独発現株との発現量の比較を示す図である。
【0067】
【図13】CBB染色による、実施例8で得られた各プロテインA融合抗ヒトTNFα抗体Fd鎖発現株と抗ヒトTNFα抗体Fd鎖単独発現株との発現量の比較を示す図である。太い矢印は発現したプロテインA融合抗ヒトTNFα抗体Fd鎖[(図13A) プロテインA(Eドメイン)融合Fd鎖、(図13B) プロテインA(EDドメイン)融合Fd鎖、(図13C) プロテインA(EDAドメイン)融合Fd鎖]を示す。細い矢印は抗ヒトTNFα抗体鎖単独発現株から、発現された抗ヒトTNFα抗体Fd鎖の移動度を示す。
【図14】CBB染色による、実施例8で得られたプロテインA融合ヒト成長ホルモン発現株、MWP融合ヒト成長ホルモン発現株、およびヒト成長ホルモン単独発現株との発現量の比較を示す図である。
【図15】実施例11で作製したプロテインA融合抗ヒトTNFα一本鎖抗体発現ベクター(A)pProA4E−scFv、(B)pProA4EDA−scFv、および実施例12で作製した抗ヒトTNFα一本鎖抗体単独発現ベクター(C)pNH326−scFvの構造を示す。
【図16】CBB染色による、実施例13で得られたプロテインA融合抗ヒトTNFα一本鎖抗体発現株、および抗ヒトTNFα一本鎖抗体単独発現株の発現量の比較を示す図である。レーンMは分子量マーカー(214,120,92,52,35,28,20kDa)を示す。レーン1および2は各々抗ヒトTNFα一本鎖抗体単独発現株の培養液5μlを、レーン3および4は各々プロテインA(Eドメイン)融合抗ヒトTNFα一本鎖抗体発現株の培養液5μlを、レーン5および6は各々プロテインA(Eドメイン)融合抗ヒトTNFα一本鎖抗体発現株の培養液5μlを泳動した。
【符号の説明】
【0068】
NcoI、PstI、BamHI、SalI、XbaI、XhoI、EcoRI、KpnI:制限酵素。
【0069】
MWP:ブレビバチルス・ブレビス細菌の細胞壁蛋白質。
【0070】
P5:ブレビバチルス・ブレビス細菌の細胞壁蛋白質(MWP)のプロモーター5。
【0071】
Terminator:ヘヤピンループ翻訳終結領域。
【0072】
Rep:pUB110由来の複製起点。
【0073】
Nm:ネオマイシン耐性遺伝子。
【0074】
SD1およびSD2:シャイン・ダルガノ配列。
【0075】
VL:L鎖可変領域。
【0076】
CL:L鎖定常領域。
【0077】
VH:H鎖可変領域。
【0078】
CH1:H鎖定常領域。
【0079】
h:ヒンジ領域。
【技術分野】
【0001】
本発明は、ブレビバチルス属細菌を使用して、プロテインAの免疫グロブリン結合ドメインもしくはその部分的配列と、所望するポリペプチドとの融合ポリペプチドを製造する際に利用するDNA配列、組換え発現ベクター、形質転換体、およびその融合ポリペプチドの製造方法に関する。本発明のDNA配列、組換え発現ベクター、または形質転換体を用いることにより、ブレビバチルス属細菌を使用して、産業上有用なポリペプチドを効率良く製造する事が可能である。
【背景技術】
【0002】
近年の遺伝子組換え技術の進歩により、各種の生物起源から得られたDNAを発現ベクターに連結し、適当な宿主細胞に導入することで、当該DNAにコードされるポリペプチドを生産出来るようになった。この遺伝子組換え技術の有効性から、現在までに様々な微生物を宿主とする数多くの宿主ベクター系が開発され、特に大腸菌および酵母を利用した系が確立されている(非特許文献1)。
【0003】
近年、鵜高らにより開発されたブレビバチルス・ブレビスを用いた遺伝子組換えポリペプチドの発現系は、ヒト上皮細胞増殖因子を、その生理活性を保持した状態で培養液中に大量に分泌発現できることが知られている(非特許文献1、2)。このブレビバチルス属細菌を用いたポリペプチド発現系は、宿主菌由来の細胞壁蛋白質であるMWPのプロモーター、シャイン・ダルガノ配列および、シグナル配列をコードするDNA配列を含む発現ベクターに、成熟型ポリペプチドをコードするDNA配列を連結し発現させることで、様々なポリペプチドの直接分泌発現を可能とする(非特許文献2)。
【0004】
しかし、ブレビバチルス属細菌のポリペプチド分泌発現系は、汎用なポリペプチド生産系として確立されてはいない。なぜならば、ブレビバチルス属細菌によるポリペプチドの分泌発現は、その所望するポリペプチドの性質や特性により発現量が大きく左右されるからである。すなわち原核生物由来で、ジスルフィド結合が存在しない、または少ないポリペプチドは比較的高い生産性が期待できるが、真核生物の分泌ポリペプチドのように複雑な高次構造を有するポリペプチドの分泌発現では、ヒト上皮細胞増殖因子やマグロ成長ホルモンなど一部の例外を除き、生産性が極めて低い(非特許文献2)。複雑な高次構造を有するポリペプチドに対する発現障害は、大腸菌や酵母などの他の微生物を宿主とした場合にも度々認められ、現在、発現障害を改善するために様々な方法が提案されている。
【0005】
効果的な方法として、所望するポリペプチドを融合ポリペプチドの形態で生産する方法(融合発現法)が提案されている。例えば、大腸菌の場合、カルモジュリン結合ペプチド(CBP)、グルタチオンS−トランスフェラーゼ(GST)、マルトース結合蛋白質(MBP)、セルロース結合ドメイン(CBD)、分子シャペロン(DNAJ,DNAK)、またはヒスチジンタグ(6から10残基程度のヒスチジン残基)等を、所望のポリペプチドのN末端、またはC末端へ結合させることにより、発現量を向上させる方法が開発されている(非特許文献1)。
【0006】
また、プロテインAが有する親和性を精製工程で利用し、所望のポリペプチドの分離、精製を容易にする方法も開発されている(特許文献1)。具体的には、免疫グロブリンに対する親和性を有するプロテインAとの融合生成物として、所望のポリペプチドを大腸菌またはスタフィロコッカス属細菌等の微生物宿主において発現させ、その融合生成物の精製工程において、プロテインAが有する親和性を利用して所望のポリペプチドを分離、精製する。
【0007】
ブレビバチルス属細菌のポリペプチド分泌発現系においても、所望のポリペプチドの生産性を向上させる方法として、(1)融合領域として数アミノ酸残基を用い、耐熱性エンドグルカナーゼを発現させる方法(特許文献2)、(2)ブレビバチルス・ブレビス47株由来の細胞壁蛋白質のN末端から数十アミノ酸残基を融合領域として用い、ヒト成長ホルモンまたはヒトプロインシュリンを融合発現させる方法(特許文献3、4)、(3)カビ由来のプロテインジスルフィドイソメラーゼ(PDI)を融合領域として用い、抗体L鎖を融合発現させる方法(特許文献5)が知られている。
【特許文献1】国際公開第WO8403103号パンフレット
【特許文献2】特開2004−129576号公報
【特許文献3】特開平11−341991号公報
【特許文献4】特開2003−79379号公報
【特許文献5】特開平11−75879号公報
【非特許文献1】塚越規弘編著,「組換えタンパク質生産法 生物化学実験法」,学会出版センター,2001年
【非特許文献2】Udakaら,Method.Enzymol.1993年,第217巻,23−33頁
【発明の開示】
【発明が解決しようとする課題】
【0008】
一般的に、融合ポリペプチドの発現量は、所望のポリペプチドの構造やアミノ酸配列と融合領域との相性に大きく左右され、選択した融合領域が適当でない場合、融合ポリペプチドの大量発現に結びつかないことが知られている。また、上記(1)から(3)で開示された融合領域では、それらが有する親和性を精製工程に利用することができず、所望の融合生成物の分離、精製を簡略化することができない。従って、ブレビバチルス属細菌のポリペプチド分泌発現系においても、多様なポリペプチドの融合発現に適用でき、かつ、融合生成物の分離、精製の簡略化が可能な、融合ペプチド領域の開発が望まれていた。
【0009】
本発明は、ブレビバチルス属細菌のポリペプチド分泌発現系において、多様なポリペプチドの融合発現に適用でき、かつ、融合生成物の分離、精製の簡略化が可能な、融合ペプチド領域の提供を一つの課題とする。さらに、当該融合ペプチド領域を利用した、ブレビバチルス属細菌による有用ポリペプチドの製造方法の提供を一つの課題とする。
【課題を解決するための手段】
【0010】
本発明者らは、プロテインAの免疫グロブリン結合ドメインまたはその部分的配列のC末端側に、所望のポリペプチドを連結した融合生成物を、ブレビバチルス属細菌で分泌発現させることにより、上記課題が解決されることを見いだした。本発明は、以下の内容を含む1または複数の特徴を有する。
【0011】
本発明は、ブレビバチルス属細菌で機能しうるシグナルペプチド、プロテインAの免疫グロブリン結合ドメインまたはその部分的配列、任意のアミノ酸配列からなるリンカー、および、所望のポリペプチドを、この順序で連結してなる融合ポリペプチドをコードするDNA配列を、さらに、ブレビバチルス属細菌で機能しうるプロモーターおよびシャイン・ダルガノ配列と作動可能に連結せしめたことを特徴とするDNA配列に関する。 また、本発明は、当該DNA配列を含むベクター、および、当該ベクターを用いてブレビバチルス属細菌を形質転換して得られる形質転換体に関する。さらに、本発明は、当該形質転換体を培養し、培地中に分泌される所望のポリペプチドの一部または全部を含むポリペプチドを回収することを特徴とする、ポリペプチドの製造方法に関する。
【発明の効果】
【0012】
本発明により、従来、ブレビバチルス属細菌のポリペプチド分泌発現系では著しく発現効率が低かったポリペプチドを、融合ポリペプチドとして大量に培養液中へ分泌発現させることが可能となった。さらに、抗体(免疫グロブリンG)などを固定化した担体を用いることにより、所望のポリペプチドを含む当該融合ポリペプチドを、培養液から簡便に分離精製することが可能となった。
【発明を実施するための最良の形態】
【0013】
以下に本発明の実施形態について詳しく説明する。
【0014】
1.DNA配列およびポリペプチド
本発明の第一の実施形態は、プロテインAの免疫グロブリン結合ドメインまたはその部分的配列のC末端側に、0または1以上のアミノ酸配列を介して所望のポリペプチドを連結した融合ポリペプチドを、ブレビバチルス属細菌で分泌発現させる機能を有するDNA配列である。詳細には、本発明の実施形態としてのDNA配列は、(a)ブレビバチルス属細菌で機能しうるプロモーター、(b)ブレビバチルス属細菌で機能しうるシャイン・ダルガノ配列、(c)ブレビバチルス属細菌で機能しうるシグナルペプチド、プロテインAの免疫グロブリン結合ドメインまたはその部分的配列、任意のアミノ酸配列からなるリンカー、および、所望のポリペプチドを、この順序で連結してなる融合ポリペプチドをコードするDNA配列、の3つの構成要素からなり、かつ、前記(a)から(c)の構成要素を互いに作動可能となるよう連結せしめたことを特徴とするDNA配列である。
【0015】
ここで、本発明における「作動可能となるよう連結せしめる」との記載は、前記(a)から(c)の構成要素がブレビバチルス属細菌内で機能することにより、所望のポリペプチドを含む融合ポリペプチドが分泌発現するよう、前記(a)から(c)の構成要素を必要に応じて適当なDNAを介して(またはDNAを介さず)連結せしめることを意味する。本発明における「作動可能となるよう連結」する概念には、前記(a)から(c)の構成要素に加えて、エンハンサー等を含む各種の調節エレメントまたは制御因子を連結する場合も含まれる。それら調節エレメントまたは制御因子の種類は、宿主によって変わり得ることは当業者に周知の事項である。
【0016】
本発明において用いられる「プロモーター」は、ブレビバチルス属細菌において作動可能であるプロモーターであればいかなる生物由来のプロモーターでもよい。好ましくはブレビバチルス属細菌由来のプロモーターが良く、より好ましくはブレビバチルス・ブレビス又はブレビバチルス・チョウシネンシス由来の細胞壁蛋白質のプロモーターが良く、更に好ましくはブレビバチルス・ブレビスの細胞壁蛋白質であるmiddle wall protein (MWP)、outer wall protein (OWP)、またはブレビバチルス・チョウシネンシスの細胞壁蛋白質HWP(J.Bacteriol.172:1312−1320(1990))のプロモーターが良い。
【0017】
本発明において用いられる「シャイン・ダルガノ配列」は、ブレビバチルス属細菌において機能する塩基配列であればいかなる原核生物種に由来するものでも良い。好ましくはブレビバチルス属細菌のシャイン・ダルガノ配列が良く、さらに好ましくはブレビバチルス・ブレビスまたはブレビバチルス・チョウシネンシスの細胞壁蛋白質をコードしているDNAの上流に存在するシャイン・ダルガノ配列が良い。更により好ましくはブレビバチルス・ブレビスの細胞壁蛋白質であるmiddle wall protein (MWP)、outer wall protein (OWP)、又はブレビバチルス・チョウシネンシス細胞壁蛋白質HWP(J.Bacteriol.172:1312−1320(1990))をコードする蛋白質のシャイン・ダルガノ配列が良い。ここでいうシャイン・ダルガノ配列とは、原核生物のmRNAにおいて、開始コドンの上流に見られる共通配列であり、プリン塩基に富んだ約3から9塩基の配列である。
【0018】
本発明において用いられる「シグナルペプチド」は、ブレビバチルス属細菌内で発現させた際、そのC末端側に連結されたポリペプチドを細胞外に分泌させる機能を有するペプチドであればいかなる生物種由来のものでも良い。好ましくはブレビバチルス属細菌のシグナルペプチドが良く、より好ましくはブレビバチルス・ブレビスの細胞壁蛋白質であるmiddle wall protein (MWP)またはouter wall protein (OWP)、さらにブレビバチルス・チョウシネンシスの細胞壁蛋白質(HWP)(J.Bacteriol.172:1312−1320(1990))のシグナルペプチドが良い。また天然の分泌シグナルペプチドのアミノ酸配列を改良した配列でも良い。具体的には、前記middle wall protein (MWP)のシグナルペプチドMet−Lys−Lys−Val−Val−Asn−Ser−Val−Leu−Ala−Ser−Ala−Leu−Ala−Leu−Thr−Val−Ala−Pro−Met−Ala−Phe−Alaに、Met−Lys−Lys−Arg(x)−Arg(x)−Val−Val−Asn−Asn−Ser−Val−Leu−Leu(x)−Leu(x)−Leu(x)−Leu(x)−Leu(x)−Ala(x)−Ser(x)−Ala−Leu−Ala−Leu−Thr−Val−Ala−Pro−Met−Ala−Phe−Ala、中の(x)部分で示すアミノ酸配列のように塩基性や疎水性アミノ酸残基を付加したシグナルペプチドを用いても良い。さらに、本発明において使用するプロテインAが本来有するシグナルペプチド配列を使用しても良い。
【0019】
本発明において用いられる「プロテインAの免疫グロブリン結合ドメイン」は、好ましくはスタフィロコッカス属細菌由来のプロテインAの免疫グロブリン結合ドメインであり、より好ましくは、図1(配列表の配列番号1、2)にて示されるスタフィロコッカス・アウレウス(Staphylococcus aureus) COWANI株
由来のプロテインAのE、D、A、B、またはCドメインである。しかし、本明細書において用いられる「プロテインAの免疫グロブリン結合ドメイン」なる用語は上記に限定されず、スタフィロコッカス・アウレウス COWANI株由来のプロテインAのE、D、A、B、またはCドメインと、それらドメインのいずれかと類似の構造および機能を有する分子すべてを包含する。具体的には、図1(配列番号1、2)にて示されるスタフィロコッカス・アウレウス COWAN1株由来のプロテインAのE、D、A、B、またはCドメインのアミノ酸配列に、1アミノ酸以上の置換、欠失、または挿入を施して得られ、かつ、本発明のDNA配列の構成要素の1つである融合領域として用いた場合に、ブレビバチルス属細菌内で所望のポリペプチドの分泌発現量を向上させる機能を有する変異体は、本明細書において用いられる用語「プロテインAの免疫グロブリン結合ドメイン」に包含される。このような変異体は、図1(配列番号1、2)にて示されるアミノ酸配列および塩基配列情報を基に、当業者であれば容易に作製、もしくは自然界より探索することができる。
【0020】
また、本発明において融合領域として使用される「プロテインAの免疫グロブリン結合ドメイン」は、ブレビバチルス属細菌内で所望のポリペプチドの分泌発現量を向上させる機能を有する限り、その個数および順列において何ら制限されるものでない。図1(配列番号1、2)にて示されるスタフィロコッカス・アウレウス COWANI株由来のプロテインAのE、D、A、B、またはCドメインを例として説明すると、E、D、A、B、またはCの任意の1ドメインを使用しても良いし、任意の2ドメインの順列として得られる25通りの配列のうちの1つを使用しても良いし、任意の3ドメインの順列として得られる125通りの配列のうちの1つを使用しても良いし、同様に4以上のドメインの順列として得られる配列のうちの1つを使用しても良い。好ましくは、Eドメイン(図2、3参照)、E、Dドメインをこの順で連結したもの(図4、5参照)、および、E、D、Aドメインをこの順で連結したもの(図6、7参照)が良い。
【0021】
本発明において用いられる「任意のアミノ酸配列からなるリンカー」とは、0または1以上のアミノ酸配列からなるペプチドを意味し、本発明のDNA配列がコードする融合ポリペプチド中に、当該リンカーが実質的に含まれないことをも意味する。当該リンカーを構成するアミノ酸配列に特に制限はないが、当該リンカー中に部位特異的切断に利用し得るアミノ酸配列を配置することにより、融合生成物から所望のポリペプチドを切り離す工程を容易にすることができる。
【0022】
本発明において用いられる「部位特異的切断に利用し得るアミノ酸配列」とは、化学的もしくは酵素的にポリペプチドを切断する際、切断箇所として特異的に認識されるアミノ酸配列を指す。化学的なポリペプチド切断法としては、臭化シアン、ヒドロキシルアミン、または蟻酸を使用する方法が挙げられ、これらの方法を用いる場合、特異的に認識されるアミノ酸配列として、それぞれ、[Met]、[Asn−Gly、−Leu、またはAla]、[Asp−Pro]を配置するのが好ましい。酵素的なポリペプチド切断方法としては、例えば、コラゲナーゼ、キモシン、カリクレインB、エンテロキナーゼ、トロンビン、ファクターXa、またはTEVペプチダーゼを使用する方法が挙げられ、これらの方法を用いる場合、特異的に認識されるアミノ酸配列として、それぞれ、[Pro−X−Gly−Pro(Xは任意のアミノ酸残基)]、[Met−Phe]、[X−Phe−Arg−Y(X、Yは任意のアミノ酸残基)]、[(Asp)n−Lys(n=2−4)]、[Leu−Val−Gln−Arg−Gly−Ser]、[Ile−Glu−Gly−Arg]、[Glu−Asn−Leu−Tyr−Phe−Gln−Gly、またはGlu−Asn−Leu−Tyr−Phe−Gln−Ser]を配置するのが好ましい。
【0023】
また、所望するポリペプチド自体の切断が問題とならなければ、より特異性の低いペプチダーゼ等を使用しても良い。具体的には、真核生物または原核生物由来のセリンペプチダーゼやシグナルペプチダーゼ、エンドペプチダーゼ、カルボキシルペプチダーゼ、アスパラギン酸ペプチダーゼ、システインペプチダーゼ、グルタミン酸ペプチダーゼ、金属ペプチダーゼ、スレオニンペプチダーゼのファミリーに属するペプチダーゼ群が利用できる。さらに、上記ペプチダーゼのアミノ酸配列を置換、欠失、付加、または変異導入することで基質特異性を改変させたペプチダーゼを利用することも可能である。
【0024】
本発明において用いられる「任意のアミノ酸配列からなるリンカー」中には、分離精製用のタグとして使用され得るアミノ酸配列を配置することもできる。「分離精製用のタグ」とは、ポリペプチドの精製を容易にするために付加される、特定物質に対する親和性を有するペプチドを意味し、その具体例としては、ニッケルメタルに対し親和性を有するヒスチジンが複数連結した構造のもの、またはアミロースに対し親和性を有するマルトース結合ポリペプチド等が挙げられる。本発明では前者が好適に用いられ、より好適には6個のヒスチジンの連続配列が用いられる。
【0025】
本発明において用いられる「任意のアミノ酸配列からなるリンカー」中には、前記「部位特異的切断に利用し得るアミノ酸配列」、または前記「分離精製用のタグ」のいずれか1つを配置してもよいし、両者を任意の順序で同時に配置してもよい。ただし、所望のポリペプチドから不要な融合領域を極力切除したい場合は、前記「任意のアミノ酸配列からなるリンカー」の最もC末端側に「部位特異的切断に利用し得るアミノ酸配列」を配置することが好ましい。
【0026】
本発明における「所望のポリペプチド」としては、例えば、各種の酵素類、ヒト成長ホルモン等のホルモン類、各種の血漿蛋白質類、各種のサイトカイン類、免疫グロブリンGのL鎖、Fd鎖等の部分抗体、一本鎖抗体等の修飾抗体などが挙げられる。
【0027】
2.ベクター
本発明の第二の実施形態は、本発明の第一の実施形態であるDNA配列を含むベクターである。本発明の第二の実施形態において用いられるベクターは、ブレビバチルス属細菌において自律複製可能であればいかなるものでも良いが、好ましくは、pHY500(特開平2‐31682号公報)、pNU200(日本農芸化学会誌61,669‐676(1987))、pNH326(図8参照)(Applied and Environmental Microbiology, 58:525−531.(1992))、pNH400(J. Bacteriol, 177:745−749(1995))、pNY700(特開平4−278091号公報)、pHT系プラスミド(特許第2727391号)、大腸菌とブレビバチルス属細菌とのシャトルベクターであるpNCO2(特開2002‐238569号公報)、またはこれらいずれかの誘導体が良い。ただし、本発明の第一の実施形態であるDNA配列を、ブレビバチルス属細菌の染色体へ組込むことを目的とするベクターの場合、ブレビバチルス属細菌において自律複製可能である必要はなく、このようなベクターも、本発明の第一の実施形態であるDNA配列を含む限り、本発明に包含される。
【0028】
3.形質転換体
本発明の第三の実施形態は、本発明の第二の実施形態であるベクターを用いてブレビバチルス属細菌を形質転換することにより得られる形質転換体である。本発明の第三の実施形態において用いられるブレビバチルス属細菌としては、ブレビバチルス・アグリ(Brevibacillus agri)、ブレビバチルス・ボルステレンシス(Brevibacillus borstelensis)、ブレビバチルス・ブレビス、ブレビバチルス・セントロスポルス(Brevibacillus centrosporus)、ブレビバチルス・チョウシネンシス、ブレビバチルス・フォーモサス(Brevibacillus formosus)、ブレビバチルス・インボカツス(Brevibacillus invocatus)、ブレビバチルス・ラテロスポルス(Brevibacillus laterosporus)、ブレビバチルス・リムノフィルス(Brevibacillus limnophilus)、ブレビバチルス・パラブレビス(Brevibacillus parabrevis)、ブレビバチルス・レウスゼリ(Brevibacillus reuszeri)、またはブレビバチルス・サーモルベル(Brevibacillus thermoruber)等が挙げられる。ブレビバチルス・ブレビス、またはブレビバチルス・チョウシネンシスが好ましく、ブレビバチルス・ブレビス47株、ブレビバチルス・ブレビス47K株、ブレビバチルス・ブレビス47−5Q株、ブレビバチルス・チョウシネンシスHPD31株、またはブレビバチルス・チョウシネンシスHPD31−OK株がより好ましい。また生産量の向上などの目的に応じて上記ブレビバチルス属細菌のプロテアーゼ欠損株や高分泌発現株のような変異株を使用しても良い。
【0029】
ブレビバチルス属細菌の形質転換は、例えば、Takahashiらの方法(J. Bacteriol. 156:1130−1134(1983))、Takagiらの方法(Agric. Biol. Chem, 53:3099−3100(1989))、またはOkamotoらの方法(Biosci. Biotechnol. Biochem. 61:202−203(1997))により実施することができる。
【0030】
4.ポリペプチド製造方法
本発明の第四の実施形態は、本発明の第三の実施形態である形質転換体を培養し、当該形質転換体から分泌される所望のポリペプチドの一部または全部を含むポリペプチドを回収することを特徴とする、ポリペプチドの製造方法である。
【0031】
当該形質転換体の培養に用いる培地は、当該形質転換体が所望のポリペプチドを効率良く分泌発現できる培地であれば特に制限は無い。具体的にはグルコース、蔗糖、グリセロールなどの炭素源や、ポリペプトン、肉エキス、酵母エキス、カザミノ酸などの窒素源を使用することができ、カリウム塩、ナトリウム塩、リン酸塩、マグネシウム塩、マンガン、亜鉛、鉄等の無機塩類が必要に応じて添加される。さらに、栄養要求性の宿主細胞を用いる場合は、生育に要求される栄養物質を添加すればよい。また、必要であればペニシリン、エリスロマイシン、クロラムフェニコール、ネオマイシンなどの抗生物質が添加されても良い。培養温度は15−42℃、好ましくは約28−37℃であり、通気および攪拌により好気的に培養を行うことが望ましいが、もし必要であれば通気を遮断し嫌気的に培養してもよい。
【0032】
当該形質転換体から分泌された、所望のポリペプチドの一部または全部を含むポリペプチドは、通常公知の蛋白質精製法を適当に組み合わせて用いることにより、当該形質転換体の培養液から回収、精製することができる。例えば、塩析、透析、限外濾過、ゲル濾過クロマトグラフィー、イオン交換クロマトグラフィー、逆相クロマトグラフィー、またはアフィニティークロマトグラフィー等の手法を利用できる。
【0033】
特に、本発明の第四の実施形態に従い分泌発現されたポリペプチドは、その分子内にプロテインAの免疫グロブリン結合ドメインまたはその部分的配列を有しているため、適当な担体に固定化された免疫グロブリンG、例えば免疫グロブリンGセファロース(アマシャムバイオサイエンス社)等を用いることで、効率良く精製され得る。
【0034】
また、本発明の第四の実施形態に従い分泌発現されたポリペプチドは、リンカー部分に精製用タグを含みうる。この場合、当該箇所に配置した精製用タグを利用して、当該ポリペプチドを精製することも可能である。例えば、精製用タグとしてヒスチジンの連続配列を配置した場合、ニッケルNTAカラム等を用いて精製することが可能である。
【0035】
さらに、本発明の第四の実施形態に従い分泌発現されたポリペプチドは、リンカー部分に部位特異的切断に利用し得るアミノ酸配列を含みうる。この場合、当該箇所に配置したアミノ酸配列に応じた方法により、当該箇所よりN末端側のポリペプチドを所望のポリペプチドから切除することが可能である。例えば、部位特異的切断に利用し得るアミノ酸配列として[Ile−Glu−Gly−Arg]を配置した場合、ファクターXaを用いて、当該箇所よりN末端側のポリペプチドを所望のポリペプチドから切除することが可能である。このような切断反応は、本発明の第四の実施形態に従い分泌発現されたポリペプチドの精製工程中のいかなる段階で実施しても良い。切断反応後の所望のポリペプチドは、通常公知の蛋白質精製法を適当に組み合わせて用いることにより、さらに精製することができる。
【0036】
上記のような本発明の各実施形態を利用することで、これまでブレビバチルス属細菌では効率的に生産できなかったポリペプチドを、極めて効率良く生産することが可能となった。従って、本発明の実施形態は、産業上有用なポリペプチドを製造する上で、非常に有用である。
【実施例】
【0037】
以下に参考例、実施例、図面に基づいて本発明の実施例を具体的に説明するが、これらが本発明の範囲を制限するものでないことは云うまでもない。本発明に関連する技術分野の当業者は、本発明の説明の範囲内で以下の実施例の変形を認め実行することができる。したがって、そのような変形は、本発明の範囲内に入るものとみなされる。
【0038】
本発明の実施例の実行にあたり、発現DNAの作製や操作などは特に断わらない限り下記の実験書に従って実施した。 (1)「Molecular Cloning/A Laboratory Manual」、第2版(1989)、Cold Spring Harbor Laboratory 刊(米国)。 (2) 村松正實 編著「ラボマニュアル遺伝子工学」、第3版(1996)、丸善株式会社刊。
(実施例1)プロテインAの免疫グロブリン結合ドメインをコードするDNA配列の単離
スタフィロコッカス・アウレウス Cowan I株を、T2液体培地(ポリペプトン 1%、イーストエキストラクト 0.2%、グルコース 1%、魚肉エキス 0.5%、pH7.0)を用いて、37℃で一晩振とう培養した。得られた培養液から遠心分離により菌体を回収し、トリス−塩酸緩衝液(pH8.0)で2度洗浄を行った。菌体を再度トリス−塩酸緩衝液にて懸濁後1%SDSで溶菌し、60℃にて30分間加熱後、フェノール抽出およびエタノール沈殿等の定法により全ゲノムDNAを抽出した。公知のプロテインAの塩基配列(配列番号1)に基づき、上記で調製したゲノムDNAを鋳型とし、フォワードプライマー(5’−TTGCTCCCATGGCTTTCGCTGCGCAACACGATGAAGCT−3’ (配列番号15))と、以下の3種のリバースプライマー(5’−TTTTCTGCAGTTTTGGAGCTTGAGAGTCATTAAGTTTTTGAGC −3’(配列番号16)、5’−TTTTCTGCAGTTTCGGTGCTTGAGATTCGTTTAATTTTTT−3’ (配列番号17)、または5’−TTTTCTGCAGTTTCGGTGCTTGAGATTCATTTAACTTTTT−3’ (配列番号18))を用い、PCR法により、Eドメイン、EDドメイン、またはEDAドメインをコードするDNAをそれぞれ増幅した。配列番号15は、プロテインAの免疫グロブリンG結合ドメインのPCR増幅用にデザインされたフォワードプライマーを示す。配列番号16は、プロテインAの免疫グロブリンG結合ドメインEのPCR増幅用にデザインされたリバースプライマーを示す。配列番号17は、プロテインAの免疫グロブリンG結合ドメインEDのPCR増幅用にデザインされたリバースプライマーを示す。配列番号18は、プロテインAの免疫グロブリンG結合ドメインEDAのPCR増幅用にデザインされたリバースプライマーを示す。
【0039】
また、同様に、Eドメイン、EDドメイン、またはEDAドメインをコードするDNAの3’側に6残基のヒスチジンを付加して増幅する場合は、上記ゲノムDNAを鋳型として、フォワードプライマー(5’−TTGCTCCCATGGCTTTCGCTGCGCAACACGATGAAGCT−3’ (配列番号15))と、以下の3種のリバースプライマー(5’−TTTTCTGCAGGTGATGATGATGATGATGTTTTGGAGCTTGAGAGTCATTAAGTTTTTGAGC−3’ (配列番号19)、 5’−TTTTCTGCAGGTGATGATGATGATGATGTTTCGGTGCTTGAGATTCGTTTAATTTTTT−3’ (配列番号20)、または5’−TTTTCTGCAGGTGATGATGATGATGATGTTTCGGTGCTTGAGATTCATTTAACTTTTT−3’ (配列番号21))を用いた。配列番号19は、プロテインAの免疫グロブリンG結合ドメインEのPCR増幅用にデザインされたリバースプライマーを示す。配列番号20は、プロテインAの免疫グロブリンG結合ドメインEDのPCR増幅用にデザインされたリバースプライマーを示す。配列番号21は、プロテインAの免疫グロブリンG結合ドメインEDAのPCR増幅用にデザインされたリバースプライマーを示す。
【0040】
得られたDNA断片は制限酵素NcoIおよびPstIにより消化した後、アガロースゲルより分離回収した。一方、発現ベクターpNH326(図8)(配列番号22、23)もまた同様に制限酵素NcoIおよびPstIにより消化後、精製回収してアルカリフォスファターゼ処理により脱リン酸化処理を行った。
(実施例2)プロテインA融合ポリペプチド発現ベクターの構築
実施例1にて、制限酵素処理したDNA断片および発現ベクターpNH326をT4DNAライゲースを用いて連結し、各免疫グロブリン結合ドメインを有するプロテインA融合ポリペプチド発現ベクターpProA3E(図2、配列番号3および4参照)、pProA4E(図3、配列番号5および6参照)、pProA3ED(図4、配列番号7および8参照)、pProA4ED(図5、配列番号9および10参照)、pProA3EDA(図6、配列番号11および12参照)、pProA4EDA(図7、配列番号13および14参照)を構築した。
(実施例3)プロテインA融合抗ヒトTNFα抗体L鎖発現ベクターの作成
プロテインA融合ポリペプチド発現ベクターの発現能を評価するために、ヒト・マウスキメラ抗ヒトTNFα抗体のL鎖(軽鎖)遺伝子(配列番号24、25)の発現を行った。ヒト・マウスキメラ抗ヒトTNFα抗体のL鎖遺伝子は、米国特許5698195号記載の抗ヒトTNFα抗体L鎖の遺伝子配列に従い作製されたpBluescript・抗ヒトTNFα抗体L鎖を鋳型にし、2つのオリゴヌクレオチドプライマー5’−AAAACTGCAGATCGAAGGTCGTGACATCTTGCTGACTCAGTCTCCAGCC−3’ (配列番号26)と5’−TTTGAATTCCTAACACTCTCCCCTGTTGAAGCTCTT−3’(配列番号27)とを用いて、PCR法により増幅した。配列番号26は、抗TNFα抗体のL鎖遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号27は、抗TNFα抗体のL鎖遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0041】
この増幅されたヒトTNFα抗体のL鎖をコードしているDNAの5’末端にはファクターXaの認識開裂サイトであるIle−Glu−Gly−ArgのペプチドをコードしているDNA配列(配列番号26における上記下線部ATCGAAGGTCGTの配列)を含むように設計されている。この抗ヒトTNFα抗体のL鎖遺伝子は制限酵素PstIとEcoRIにより切断した。また上記プロテインA融合ポリペプチド発現ベクターpProA4E(図3)のマルチクローニングサイトに存在する制限酵素PstIとEcoRIにより消化後、精製回収してアルカリフォスファターゼ処理により脱リン酸化処理を行った。制限酵素処理を行った上記のDNA断片とプロテインA融合ポリペプチド発現ベクターはT4DNAライゲースを用いて連結し、プロテインA融合抗ヒトTNFα抗体L鎖発現ベクターpProA4E−L(図9A)(配列番号28、29)を構築した。
(実施例4)プロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクターの作成
上記、抗ヒトTNFα抗体のFd遺伝子を有するプロテインA融合ポリペプチド発現ベクターの発現能を評価するため発現ベクターの構築を行った。抗ヒトTNFα抗体のFd鎖遺伝子は上記の抗TNFα抗体H鎖の遺伝子配列(配列番号30、31)に従い作製されたpBluescript・抗ヒトTNFα抗体H鎖を鋳型にし、2つのオリゴヌクレオチドプライマー5’−AAAACTGCAGATCGAAGGTCGTGAAGTGAAACTTGAGGAGTCTGAGGGA −3’ (配列番号32)と5’−CCGGAATTCTCACGGTGGGCATGTGTGAGTTTTGTCACA−3’(配列番号33)とを用いて、PCR法により増幅した。配列番号32は、抗TNFα抗体のFd鎖遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号33は、抗TNFα抗体のFd鎖遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0042】
この増幅された抗ヒトTNFα抗体Fd鎖をコードしているDNAの5’末端にはファクターXaの認識開裂サイトであるIle−Glu−Gly−ArgのペプチドをコードしているDNA配列(配列番号32の上記下線部ATCGAAGGTCGTの配列)を含むように設計されている。このヒトTNFα抗体のFd鎖遺伝子は制限酵素PstIとEcoRIにより切断した。また上記プロテインA融合ポリペプチド発現ベクターpProA3E(図2)、pProA3ED(図4)およびpProA3EDA(図6)のマルチクローニングサイトに存在する制限酵素PstIとEcoRIを用い消化後、精製回収してアルカリフォスファターゼ処理により脱リン酸化処理を行った。制限酵素処理を行った上記のDNA断片と各プロテインA融合ポリペプチド発現ベクターをT4DNAライゲースを用いて連結し、プロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクターpProA3E−Fd(図9B、配列番号34および35参照)、pProA3ED−Fd(図9C、配列番号36および37参照)、pProA3EDA−Fd(図9D、配列番号38および39参照)を構築した。
(実施例5)抗ヒトTNFα抗体鎖のL鎖またはFd鎖単独発現ベクターの作成
プロテインA融合型と従来からの目的とするポリペプチド単独で発現させる方法(融合型でない方法)とのポリペプチド発現能を比較検討するため、抗ヒトTNFα抗体L鎖鎖単独発現ベクター、および抗ヒトTNFα抗体Fd鎖単独発現ベクターを構築した。抗ヒトTNFα抗体のL鎖遺伝子は、先述のpBluescript・抗ヒトTNFα抗体L鎖を鋳型にし、2つのオリゴヌクレオチドプライマー5’− GCTCCCATGGCTTTCGCTGACATCTTGCTGACTCAGTCT −3’ (配列番号40)と5’−TTTCTGCAGCTAACACTCTCCCCTGTTGAAGCTCTT−3’(配列番号41)とを用いて、PCR法により増幅した。配列番号40は、抗TNFα抗体のL鎖遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号41は、抗TNFα抗体のL鎖遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0043】
また抗ヒトTNFα抗体のFd鎖遺伝子は、先述のpBluescript・抗ヒトTNFα抗体H鎖を鋳型にし、2つのオリゴヌクレオチドプライマー5’− GCTCCCATGGCTTTCGCTGAAGTGAAACTTGAGGAGTCT −3’ (配列番号42)と5’− AAACTGCAGTTATTCAGGTGCTGGGCACGG −3’(配列番号43)を用いて、PCR法により増幅した。配列番号42は、抗TNFα抗体のFd鎖遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号43は、抗TNFα抗体のFd鎖遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0044】
L鎖およびFd鎖をコードするDNAは制限酵素NcoIとPstIでそれぞれ消化した。発現ベクターとなるpNH326も同様に制限酵素NcoIとPstIでそれぞれ消化後、精製回収し、アルカリフォスファターゼ処理によりそれぞれ脱リン酸化処理を行った。制限酵素処理を行った上記のDNA断片と発現ベクターはT4DNAライゲースを用いて連結し、抗ヒトTNFα抗体L鎖発現ベクター、および、抗ヒトTNFα抗体Fd鎖発現ベクターpNH326−L(図10A、配列番号44および45)、pNH326−Fd(図10B、配列番号46および47)を構築した。
【0045】
(実施例6)プロテインA融合ヒト成長ホルモン発現ベクターの作成
上記抗体以外のポリペプチドに対する本発明の効果を示すために、ヒト成長ホルモンをコードするDNAを用いプロテインA融合発現試験を行った。ヒト成長ホルモンをコードするDNAを有するプラスミドpGH−L9(Proc.Natl.Acad.Sci.USA.,81,5956−5960 (1984))を鋳型とし、2つのオリゴヌクレオチドプライマー5’−AAAACTGCAGATCGAAGGTCGTTTCCCAACTATTCCACTGAGT−3’ (配列番号48)と5’−TCCCAAGCTTTTAGAAGCCACACGACCCTTCAACAGA−3’ (配列番号49)を用いて、PCR法によりヒト成長ホルモンをコードするDNA増幅した。配列番号48は、ヒト成長ホルモンをコードする遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号49は、ヒト成長ホルモンをコードする遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0046】
増幅されたヒト成長ホルモンをコードするDNAは制限酵素PstIとHindIIIで消化した。また発現ベクターとなるpProA3Eも同様に制限酵素PstIとHindIIIで消化後、精製回収し、アルカリフォスファターゼ処理により脱リン酸化処理を行った。制限酵素処理を行った上記のヒト成長ホルモンDNA断片とpProA3E発現ベクターはT4DNAライゲースを用いて連結し、プロテインA融合ヒト成長ホルモン発現ベクターpProA3E−hGH (図11A)(配列番号50、51)を構築した。
【0047】
(実施例7)ヒト成長ホルモン単独発現ベクター、およびMWP融合ヒト成長ホルモン発現ベクターの構築
本発明の有効性を示すため、ヒト成長ホルモン単独発現ベクター、および、特許文献5、6で開示されたブレビバチルス・ブレビス細胞壁蛋白質MWPを融合部分に用いたMWP融合ヒト成長ホルモン発現ベクターを構築した。ヒト成長ホルモン単独発現ベクターを構築するため、2つのオリゴヌクレオチドプライマー5’− TTGCTCCCATGGCTTTCGCTTTCCCAACTATTCCACTGAGT −3’ (配列番号52)と5’−TCCCAAGCTTTTAGAAGCCACACGACCCTTCAACAGA −3’ (配列番号53)を用い、先述のプラスミドpGH−L9を鋳型として、PCR法によりヒト成長ホルモンをコードするDNAを増幅した。配列番号52は、ヒト成長ホルモンをコードする遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号53は、ヒト成長ホルモンをコードする遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0048】
増幅されたヒト成長ホルモンをコードするDNAは制限酵素NcoIとHindIIIで消化した。同様に発現ベクターとなるpNH326も同様に制限酵素NcoIとHindIIIで消化後、精製回収し、アルカリフォスファターゼ処理により脱リン酸化処理を行った。制限酵素処理を行った上記のDNA断片と発現ベクターはT4DNAライゲースを用いて連結し、ヒト成長ホルモン単独発現ベクターpNH326−hGH(図11B)(配列番号54、55)を構築した。一方、MWP融合ヒト成長ホルモン発現ベクターとしては、特許文献5の実施例1および実施例4に記載の方法に従い、MWP20−His6−Linker−Clavage1−hGH/pNH326 (図11C)を構築した。
【0049】
(実施例8)各発現ベクターを有する形質転換体の作成
実施例2から7で構築した、プロテインA融合発現ベクター(pProA3E、pProA3ED、pProA3EDA、pProA4E、pNH326)、プロテインA融合抗ヒトTNFα抗体L鎖発現ベクター(pProA4E−L)、プロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクター(pProA3E−Fd、pProA3ED−Fd、pPro3EDA−Fd)、抗ヒトTNFα抗体L鎖単独発現ベクター(pNH326−L)、抗ヒトTNFα抗体Fd鎖単独発現ベクター(pNH326−Fd)、プロテインA融合ヒト成長ホルモン発現ベクター(pProA3E−hGH)、ヒト成長ホルモン単独発現ベクター(pNH326−hGH)、およびMWP融合ヒト成長ホルモン発現ベクター(MWP20−His6−Linker−Clavage1−hGH/pNH326)を、公知の方法に従い、それぞれブレビバチルス・チョウシネンシスHPD31−OK株へ導入し、形質転換体を得た。
【0050】
(実施例9)抗ヒトTNFα抗体のL鎖およびFd鎖の分泌発現
実施例8にて得られた、プロテインA融合抗ヒトTNFα抗体L鎖発現ベクターpProA4E−Lを有する形質転換体、プロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクターpProA3E−Fd、pProA3ED−Fd、pPro3EDA−Fdをそれぞれ有する形質転換体、抗ヒトTNFα抗体L鎖単独発現ベクターpNH326−Lを有する形質転換体、抗ヒトTNFα抗体Fd鎖単独発現ベクターpNH326−Fdを有する形質転換体、およびそれらの対照となるベクターpNH326を有する形質転換体の各々を、60mg/Lのネオマイシンを添加した3YC培地(ポリペプトンS 3%、酵母エキス0.5%、グルコース3%、MgSO4・7H2O 0.01%、CaCl2・7H2O 0.01%、MnSO4・4H2O 0.001%,FeSO4・7H2O 0.001%,ZnSO4・7H2O 0.0001% pH7.0)にて、30℃で2日間振とう培養した。当該培養液から遠心分離(10,000rpm,4℃,3分間)により菌体を除去し、得られた培養上清各5μlを定法に従い処理した後、10−20%グラジエントポリアクリルアミドゲルを用いたSDS−ポリアクリルアミドゲル電気泳動(SDS−PAGE)に供した。電気泳動後、当該ゲルをCBB(Coomassie Brilliant Blue)R−250で染色し、ポリペプチドのバンドを検出した(図12および、図13A、B、C)。
【0051】
図12は、CBB染色による、実施例8で得られた各プロテインA融合抗ヒトTNFα抗体L鎖発現株と抗ヒトTNFα抗体L鎖単独発現株との発現量の比較を示す。太い矢印は発現したプロテインA融合抗ヒトTNFα抗体L鎖を示す。レーンMは分子量マーカー(214,120,92,52,35,28kDa)を示す。レーン1は、抗ヒトTNFα抗体L鎖単独発現株からの培養液5μlを泳動したものであり、レーン2および3は、プロテインA(Eドメイン)融合抗ヒトTNFα抗体L鎖発現株からの培養液5μlを泳動したものである。
【0052】
図13は、CBB染色による、実施例8で得られた各プロテインA融合抗ヒトTNFα抗体Fd鎖発現株と抗ヒトTNFα抗体Fd鎖単独発現株との発現量の比較を示す。太い矢印は発現したプロテインA融合抗ヒトTNFα抗体Fd鎖[(図13A) プロテインA(Eドメイン)融合Fd鎖、(図13B) プロテインA(EDドメイン)融合Fd鎖、(図13C) プロテインA(EDAドメイン)融合Fd鎖]を示す。細い矢印は抗ヒトTNFα抗体Fd鎖単独発現株から、発現された抗ヒトTNFα抗体Fd鎖の移動度を示す。レーンMは分子量マーカー(214,120,92,52,35,28kDa)を示す。レーン1は、抗ヒトTNFα抗体Fd鎖単独発現株からの培養液5μlを泳動したもの、レーン2および3は、プロテインA(Eドメイン)融合抗ヒトTNFα抗体Fd鎖発現株からの培養液5μlを泳動したもの、レーン4は、抗ヒトTNFα抗体Fd鎖単独発現株からの培養液5μlを泳動したもの、レーン5および6は、プロテインA(EDドメイン)融合抗ヒトTNFα抗体Fd鎖発現株からの培養液5μlを泳動したもの、レーン7は、抗ヒトTNFα抗体Fd鎖単独発現株からの培養液5μlを泳動したもの、レーン8および9は、プロテインA(EDAドメイン)融合抗ヒトTNFα抗体Fd鎖発現株からの培養液5μlを泳動したものである。
【0053】
実験の結果、抗ヒトTNFα抗体L鎖単独発現ベクターpNH326−Lを有する形質転換体における抗ヒトTNFα抗体L鎖の発現量(図12参照)、および抗ヒトTNFα抗体Fd鎖単独発現ベクターpNH326−Fdを有する形質転換体における抗ヒトTNFα抗体Fd鎖の発現量は、そのバンドを確認することが困難なほど少なかった。一方、プロテインA融合抗ヒトTNFα抗体L鎖発現ベクターpProA4E−Lを有する形質転換体におけるプロテインA融合抗ヒトTNFα抗体L鎖の発現量(図12参照)、およびプロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクターpProA3E−Fd(図13A参照)、pProA3ED−Fd(図13B参照)、pPro3EDA−Fd(図13C参照)を有する形質転換体におけるプロテインA融合抗ヒトTNFα抗体Fd鎖の発現量は飛躍的に上昇した。また、3種類のプロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクターpProA3E−Fd、pProA3ED−Fd、およびpPro3EDA−Fdをそれぞれ有する形質転換体における、プロテインA融合抗ヒトTNFα抗体Fd鎖の発現量を比較すると、融合領域として使用するプロテインAの免疫グロブリン結合ドメインの数を増やす程、融合ポリペプチドの発現量が顕著に増加していた(図13A、B、C参照)。
【0054】
(実施例10)ヒト成長ホルモンの分泌発現
実施例8にて得られた、プロテインA融合ヒト成長ホルモン発現ベクターpProA3E−hGHを有する形質転換体、MWP融合ヒト成長ホルモン発現ベクターMWP20−His6−Linker−Clavage1−hGH/pNU211R2L5を有する形質転換体、およびヒト成長ホルモン単独発現ベクターpNH326−hGHを有する形質転換体の各々を、実施例9と同様に培養した。ただし、MWP融合ヒト成長ホルモン発現ベクターMWP20−His6−Linker−Clavage1−hGH/pNU211R2L5を有する形質転換体の培養のみ、培地に添加する薬剤を、ネオマイシン60mg/Lに替えて、エリスロマイシン10mg/Lとした。そして、それらの培養上清を実施例9と同様にSDS−PAGEにて解析した(図14)。
【0055】
図14は、CBB染色による、実施例8で得られたプロテインA融合ヒト成長ホルモン発現株、MWP融合ヒト成長ホルモン発現株、およびヒト成長ホルモン単独発現株の発現量の比較を示す図である。レーンMは分子量マーカー(214,120,92,52,35,28kDa)を示す。レーン1はヒト成長ホルモン単独発現株からの培養液5μl、レーン2はMWP融合ヒト成長ホルモン発現株からの培養液5μl、レーン3はプロテインA融合ヒト成長ホルモン発現株からの培養液5μlを泳動したものである。
【0056】
実験の結果、ヒト成長ホルモン単独発現ベクターpNH326−hGHを有する形質転換体におけるヒト成長ホルモンの発現は極めて少量であったが、MWP融合ヒト成長ホルモン発現ベクターMWP20−His6−Linker−Clavage1−hGH/pNU211R2L5を有する形質転換体、および、プロテインA融合ヒト成長ホルモン発現ベクターpProA3E−hGHを有する形質転換体の培養液上清には大量のヒト成長ホルモンが分泌生産されていた(図14参照)。
【0057】
(実施例11)プロテインA融合抗ヒトTNFα一本鎖抗体発現ベクターの作成
実施例3で作製したpBluescript・抗ヒトTNFα抗体L鎖を鋳型にし、2つのオリゴヌクレオチドプライマー5’−AAAACTGCAGATCGAAGGTCGTGACATCTTGCTGACTCAGTCTCCAGCC−3’ (配列番号26)と5‘−GGAACCACCGCCGCCGGAGCCGCCACCACCGGAGCCGCCACCGCCAGTGCGTTTTACTTC−3’(配列番号56)とを用いて、抗ヒトTNFα一本鎖抗体の前半部分をコードするDNAを、PCR法により増幅した。配列番号26は、抗TNFα抗体のL鎖遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号56は、scFv(single chain variable fragments)の前半部分の遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。scFvは、一般的に、抗体が抗原を認識するために必要な最小単位であるVHおよびVLで構成される可変領域(Fv)を、フレキシブルなペプチドリンカーで結合した単鎖可変領域フラグメントである。
【0058】
この時、抗ヒトTNFα一本鎖抗体のN末端に、ファクターXaの認識開裂サイトであるIle−Glu−Gly−Argからなる配列が付加されるよう、プライマーを設計した。同様に、実施例4で作製したpBluescript・抗ヒトTNFα抗体H鎖を鋳型にし、2つのオリゴヌクレオチドプライマー5’−GGTGGTGGCGGCTCCGGCGGCGGTGGTTCCGAAGTGAAACTGGAAGAATCTGGAGGAGGCTTG−3’(配列番号57)と5’−TTTCTGCAGTTAGTGATGATGATGATGATGGGTGGTGCCTTGGCCCCAGTAGTCGTA−3’(配列番号58)とを用いて、抗ヒトTNFα一本鎖抗体の後半部分をコードするDNAを、PCR法により増幅した。配列番号57は、scFvの後半部分の遺伝子のPCR増幅用にデザインされたフォワードプライマーを示す。配列番号58は、scFvの後半部分の遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0059】
この時、抗ヒトTNFα一本鎖抗体のC末端に、ヒスチジン6残基が付加されるよう、プライマーを設計した。そして、これら2つのDNA断片をPCR法により連結し、抗ヒトTNFα一本鎖抗をコードするDNAを作製した。
【0060】
作製したDNAを制限酵素PstIで消化した後、実施例2で作製したプロテインA融合発現ベクターpProA4E(図3)およびpProA4EDA(図7)のマルチクローニングサイトに存在する制限酵素PstI部位にそれぞれ挿入し、プロテインA融合抗ヒトTNFα一本鎖抗体発現ベクターpProA4E−scFv(図15A)(配列番号59、60)、およびpProA4EDA−scFv(図15B)(配列番号61、62)を構築した。
【0061】
(実施例12)抗ヒトTNFα一本鎖抗体単独発現ベクターの作成
オリゴヌクレオチドプライマー5’−AAAACTGCAGATCGAAGGTCGTGACATCTTGCTGACTCAGTCTCCAGCC−3’(配列番号26)に替えて、オリゴヌクレオチドプライマー5’− GCTCCCATGGCTTTCGCTGACATCTTGCTGACTCAGTCT −3’(配列番号41)を用いる以外は実施例11と同様の操作を行い、抗ヒトTNFα一本鎖抗をコードするDNAを作製した。配列番号41は、抗TNFα抗体のL鎖遺伝子のPCR増幅用にデザインされたリバースプライマーを示す。
【0062】
作製したDNAを制限酵素NcoIおよびPstIで消化した後、発現ベクターpNH326のマルチクローニングサイトに存在するNcoI−PstI部位に挿入し、抗ヒトTNFα一本鎖抗体単独発現ベクターpNH326−scFv(図15C)(配列番号63、64)を構築した。
【0063】
(実施例13)抗ヒトTNFα一本鎖抗体の分泌発現
実施例11にて得られたプロテインA融合抗ヒトTNFα一本鎖抗体発現ベクターpProA4E−scFvならびにpProA4EDA−scFv、および、実施例12にて得られた抗ヒトTNFα一本鎖抗体単独発現ベクターpNH326−scFvを、公知の方法に従い、それぞれブレビバチルス・チョウシネンシスHPD31−OK株へ導入し、形質転換体を得た。次に、これらの形質転換体を実施例9と同様に培養し、それらの培養上清を実施例9と同様にSDS−PAGEにて解析した(図16)。
【0064】
図16は、CBB染色による、実施例13で得られたプロテインA融合抗ヒトTNFα一本鎖抗体発現株、および抗ヒトTNFα一本鎖抗体単独発現株の発現量の比較を示す図である。レーンMは分子量マーカー(214,120,92,52,35,28,20kDa)を示す。レーン1および2は各々抗ヒトTNFα一本鎖抗体単独発現株の培養液5μl、レーン3および4は各々プロテインA(Eドメイン)融合抗ヒトTNFα一本鎖抗体発現株の培養液5μl、レーン5および6は各々プロテインA(EDAドメイン)融合抗ヒトTNFα一本鎖抗体発現株の培養液5μlを泳動したものである。
【0065】
実験の結果、抗ヒトTNFα一本鎖抗体単独発現ベクターpNH326−scFvを有する形質転換体における抗ヒトTNFα一本鎖抗体の発現量は、そのバンドを確認することが困難なほど少なかった。一方、プロテインA融合抗ヒトTNFα一本鎖抗体発現ベクターpProA4E−scFv、ならびにpProA4EDA−scFvを有する形質転換体におけるプロテインA融合抗ヒトTNFα一本鎖抗体の発現量は飛躍的に上昇した。
【図面の簡単な説明】
【0066】
【図1】スタフィロコッカス・アウレウス・コワン(Cowan)I株のプロテインAの塩基配列とアミノ酸配列図である。番号はアミノ酸残基数を示す。
【図2】プロテインA(Eドメイン)融合分泌発現ベクターpProA3Eの構造とそのマルチクローニングサイト付近のDNA配列を示す。分泌発現ベクターpProA3EはプロテインA免疫グロブリン結合Eドメインの後、ヒスチジンタグを有する。
【図3】実施例2に記載のプロテインA(Eドメイン)融合分泌発現ベクターpProA4Eの構造とそのマルチクローニングサイト付近のDNA配列を示す。分泌発現ベクターpProA4EはプロテインA免疫グロブリン結合Eドメインの後、ヒスチジンタグを含まない。
【図4】実施例2に記載のプロテインA(EDドメイン)融合分泌発現ベクターpProA3EDの構造とそのマルチクローニングサイト付近のDNA配列を示す。分泌発現ベクターpProA3EDはプロテインA免疫グロブリン結合EDドメインの後、ヒスチジンタグを有する。
【図5】実施例2に記載のプロテインA(EDドメイン)融合分泌発現ベクターpProA4EDの構造とそのマルチクローニングサイト付近のDNA配列を示す。分泌発現ベクターpProA4EDはプロテインA免疫グロブリン結合EDドメインの後、ヒスチジンタグを含まない。
【図6】実施例2に記載のプロテインA(EDAドメイン)融合分泌発現ベクターpProA3EDAの構造とそのマルチクローニングサイト付近のDNA配列を示す。分泌発現ベクターpProA3EDAはプロテインA免疫グロブリン結合EDAドメインの後、ヒスチジンタグを有する。
【図7】実施例2に記載のプロテインA(EDAドメイン)融合分泌発現ベクターpProA4EDAの構造とそのマルチクローニングサイト付近のDNA配列を示す。分泌発現ベクターpProA4EDAはプロテインA免疫グロブリン結合EDAドメインの後、ヒスチジンタグを含まない。
【図8】実施例2に記載のブレビバチルス・ブレビスポリペプチド分泌発現ベクターpNH326の構造とそのマルチクローニングサイト付近のDNA配列を示す。
【図9】実施例3および実施例4に記載のプロテインA融合抗ヒトTNFα抗体L鎖発現ベクター(A) pProA4E−L、およびプロテインA融合抗ヒトTNFα抗体Fd鎖発現ベクター(B) pProA3E−Fd、(C) pProA3ED−Fdおよび(D) pProA3EDA−Fdの構造を示す。
【図10】実施例5に記載の抗ヒトTNFα抗体鎖単独発現ベクター(A) pNH326−L、(B) pNH326‐Fdの構造を示す。
【図11】実施例6および7記載のプロテインA融合ヒト成長ホルモン発現ベクター(A) pProA3E−hGH、(B) ヒト成長ホルモン単独発現ベクターpNH326−hGH、および(C) MWP融合ヒト成長ホルモン発現ベクターMWP20−His6−Linker−Clavage1−hGH/pNU211R2L5、の構造を示す。
【図12】CBB染色による、実施例8で得られた各プロテインA融合抗ヒトTNFα抗体L鎖発現株と抗ヒトTNFα抗体L鎖単独発現株との発現量の比較を示す図である。
【0067】
【図13】CBB染色による、実施例8で得られた各プロテインA融合抗ヒトTNFα抗体Fd鎖発現株と抗ヒトTNFα抗体Fd鎖単独発現株との発現量の比較を示す図である。太い矢印は発現したプロテインA融合抗ヒトTNFα抗体Fd鎖[(図13A) プロテインA(Eドメイン)融合Fd鎖、(図13B) プロテインA(EDドメイン)融合Fd鎖、(図13C) プロテインA(EDAドメイン)融合Fd鎖]を示す。細い矢印は抗ヒトTNFα抗体鎖単独発現株から、発現された抗ヒトTNFα抗体Fd鎖の移動度を示す。
【図14】CBB染色による、実施例8で得られたプロテインA融合ヒト成長ホルモン発現株、MWP融合ヒト成長ホルモン発現株、およびヒト成長ホルモン単独発現株との発現量の比較を示す図である。
【図15】実施例11で作製したプロテインA融合抗ヒトTNFα一本鎖抗体発現ベクター(A)pProA4E−scFv、(B)pProA4EDA−scFv、および実施例12で作製した抗ヒトTNFα一本鎖抗体単独発現ベクター(C)pNH326−scFvの構造を示す。
【図16】CBB染色による、実施例13で得られたプロテインA融合抗ヒトTNFα一本鎖抗体発現株、および抗ヒトTNFα一本鎖抗体単独発現株の発現量の比較を示す図である。レーンMは分子量マーカー(214,120,92,52,35,28,20kDa)を示す。レーン1および2は各々抗ヒトTNFα一本鎖抗体単独発現株の培養液5μlを、レーン3および4は各々プロテインA(Eドメイン)融合抗ヒトTNFα一本鎖抗体発現株の培養液5μlを、レーン5および6は各々プロテインA(Eドメイン)融合抗ヒトTNFα一本鎖抗体発現株の培養液5μlを泳動した。
【符号の説明】
【0068】
NcoI、PstI、BamHI、SalI、XbaI、XhoI、EcoRI、KpnI:制限酵素。
【0069】
MWP:ブレビバチルス・ブレビス細菌の細胞壁蛋白質。
【0070】
P5:ブレビバチルス・ブレビス細菌の細胞壁蛋白質(MWP)のプロモーター5。
【0071】
Terminator:ヘヤピンループ翻訳終結領域。
【0072】
Rep:pUB110由来の複製起点。
【0073】
Nm:ネオマイシン耐性遺伝子。
【0074】
SD1およびSD2:シャイン・ダルガノ配列。
【0075】
VL:L鎖可変領域。
【0076】
CL:L鎖定常領域。
【0077】
VH:H鎖可変領域。
【0078】
CH1:H鎖定常領域。
【0079】
h:ヒンジ領域。
【特許請求の範囲】
【請求項1】
ブレビバチルス(Brevibacillus)属細菌で機能しうるシグナルペプチド、プロテインAの免疫グロブリン結合ドメインまたはその部分的配列、任意のアミノ酸配列からなるリンカー、および、所望のポリペプチドを、この順序で連結してなる融合ポリペプチドをコードするDNA配列を、さらに、ブレビバチルス属細菌で機能しうるプロモーターおよびシャイン・ダルガノ配列と作動可能に連結せしめたことを特徴とするDNA配列。
【請求項2】
前記任意のアミノ酸配列からなるリンカー中に、部位特異的切断に利用し得るアミノ酸配列を含む、請求項1記載のDNA配列。
【請求項3】
前記部位特異的切断に利用し得るアミノ酸配列が、部位特異的プロテアーゼ、臭化シアン、ヒドロキシルアミン、または、蟻酸により切断され得るアミノ酸配列である、請求項2記載のDNA配列。
【請求項4】
前記任意のアミノ酸配列からなるリンカー中に、複数のヒスチジン残基からなる分離精製用タグをさらに含む、請求項1から3記載のDNA配列。
【請求項5】
前記ブレビバチルス属細菌で機能しうるプロモーターが、ブレビバチルス属細菌の細胞壁蛋白質のプロモーターである、請求項1から4のいずれかに記載のDNA配列。
【請求項6】
前記プロテインAがスタフィロコッカス(Staphylococcus)属細菌由来である、請求項1から5のいずれかに記載のDNA配列。
【請求項7】
請求項1から6のいずれかに記載のDNA配列を含むベクター。
【請求項8】
請求項7記載のベクターを用いてブレビバチルス属細菌を形質転換することにより得られる形質転換体。
【請求項9】
前記ブレビバチルス属細菌が、ブレビバチルス・ブレビス(Brevibacillus brevis)47株、ブレビバチルス・ブレビス47K株、ブレビバチルス・ブレビス47‐5Q株、ブレビバチルス・チョウシネンシス(Brevibacillus choshinensis)HPD31株またはブレビバチルス・チョウシネンシスHPD31‐OK株、並びにこれら由来の変異株からなる群より選択される、請求項8記載の形質転換体。
【請求項10】
請求項8または9記載の形質転換体を培養し、培地中に分泌される所望のポリペプチドの一部または全部を含む融合ポリペプチドを回収することを特徴とする、ポリペプチドの製造方法。
【請求項11】
前記所望のポリペプチドが、部分抗体あるいは一本鎖抗体である請求項10記載の製造方法。
【請求項12】
前記所望のポリペプチドが、哺乳類の成長ホルモンである請求項10記載の製造方法。
【請求項13】
請求項10から12のいずれかに記載の製造方法によって得られるポリペプチド。
【請求項1】
ブレビバチルス(Brevibacillus)属細菌で機能しうるシグナルペプチド、プロテインAの免疫グロブリン結合ドメインまたはその部分的配列、任意のアミノ酸配列からなるリンカー、および、所望のポリペプチドを、この順序で連結してなる融合ポリペプチドをコードするDNA配列を、さらに、ブレビバチルス属細菌で機能しうるプロモーターおよびシャイン・ダルガノ配列と作動可能に連結せしめたことを特徴とするDNA配列。
【請求項2】
前記任意のアミノ酸配列からなるリンカー中に、部位特異的切断に利用し得るアミノ酸配列を含む、請求項1記載のDNA配列。
【請求項3】
前記部位特異的切断に利用し得るアミノ酸配列が、部位特異的プロテアーゼ、臭化シアン、ヒドロキシルアミン、または、蟻酸により切断され得るアミノ酸配列である、請求項2記載のDNA配列。
【請求項4】
前記任意のアミノ酸配列からなるリンカー中に、複数のヒスチジン残基からなる分離精製用タグをさらに含む、請求項1から3記載のDNA配列。
【請求項5】
前記ブレビバチルス属細菌で機能しうるプロモーターが、ブレビバチルス属細菌の細胞壁蛋白質のプロモーターである、請求項1から4のいずれかに記載のDNA配列。
【請求項6】
前記プロテインAがスタフィロコッカス(Staphylococcus)属細菌由来である、請求項1から5のいずれかに記載のDNA配列。
【請求項7】
請求項1から6のいずれかに記載のDNA配列を含むベクター。
【請求項8】
請求項7記載のベクターを用いてブレビバチルス属細菌を形質転換することにより得られる形質転換体。
【請求項9】
前記ブレビバチルス属細菌が、ブレビバチルス・ブレビス(Brevibacillus brevis)47株、ブレビバチルス・ブレビス47K株、ブレビバチルス・ブレビス47‐5Q株、ブレビバチルス・チョウシネンシス(Brevibacillus choshinensis)HPD31株またはブレビバチルス・チョウシネンシスHPD31‐OK株、並びにこれら由来の変異株からなる群より選択される、請求項8記載の形質転換体。
【請求項10】
請求項8または9記載の形質転換体を培養し、培地中に分泌される所望のポリペプチドの一部または全部を含む融合ポリペプチドを回収することを特徴とする、ポリペプチドの製造方法。
【請求項11】
前記所望のポリペプチドが、部分抗体あるいは一本鎖抗体である請求項10記載の製造方法。
【請求項12】
前記所望のポリペプチドが、哺乳類の成長ホルモンである請求項10記載の製造方法。
【請求項13】
請求項10から12のいずれかに記載の製造方法によって得られるポリペプチド。
【図1】



【図2】
【図3】

【図4】
【図5】
【図6】
【図7】
【図8】

【図9】
【図10】

【図11】
【図15】

【図12】
【図13】

【図14】
【図16】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図15】
【図12】
【図13】
【図14】
【図16】
【公開番号】特開2006−320220(P2006−320220A)
【公開日】平成18年11月30日(2006.11.30)
【国際特許分類】
【出願番号】特願2005−144529(P2005−144529)
【出願日】平成17年5月17日(2005.5.17)
【出願人】(000000941)株式会社カネカ (3,932)
【出願人】(000120205)
【Fターム(参考)】
【公開日】平成18年11月30日(2006.11.30)
【国際特許分類】
【出願日】平成17年5月17日(2005.5.17)
【出願人】(000000941)株式会社カネカ (3,932)
【出願人】(000120205)
【Fターム(参考)】
[ Back to top ]