
メチロトローフ酵母における蛋白質グリコシル化の修飾
本発明は、哺乳類様のグリコシル化を伴う糖蛋白質を生じさせるための方法及び遺伝子操作されたメチロトローフ酵母に関する。本発明は、哺乳類様のグリコシル化を伴う糖蛋白質を生じさせることができるメチロトローフ酵母株を生成するのに有用なベクターにも関する。遺伝子操作されたメチロトローフ酵母から生成した糖蛋白質も提供される。
【発明の詳細な説明】
【発明の開示】
【0001】
発明の分野
本発明は、哺乳類様グリコシル化により糖蛋白質を生産するための方法及び操作されたメチロトローフ酵母株に関する。本発明は、哺乳類様グリコシル化により糖蛋白質を生産することができるメチロトローフ酵母株を生成するのに有用なベクターにも関する。遺伝子工学によるメチロトローフ酵母株から生産された糖蛋白質も提供される。
発明の背景
ピキアパストリスを含むメチロトローフ酵母は、商業上又は医学上重要な組換え蛋白質の生産のために広く使用されてきた。しかしながら、幾つかの治療用糖蛋白質の生産及び医学上の応用は、これらの酵母と標的生物、例えば哺乳類又はヒト被験者の間の蛋白質結合糖質の生合成の違いにより妨げられ得る。
【0002】
蛋白質のN−グリコシル化は細胞質網状構造(ER)を起源とし、そこでは、ドリコール(脂質キャリアー中間体)上で集合したN結合オリゴサッカライド(Glc3Man9GlcNAc2)が隣の蛋白質の適当なAsnに転移される。これは、全ての真核生物のN−結合糖蛋白質に共通の事象である。3つのグルコース残基と一つの特定のα−1,2−結合マンノース残基がER中の特異的グルコシダーゼと一つの1,2−マンノシダーゼにより除去されて、コアのオリゴサッカライド構造Man8GlcNAc2をもたらす。このコアの糖構造を有する蛋白質はゴルジ体に運ばれて、そこで、糖部分が様々な修飾を受ける。酵母と高度真核生物の間ではゴルジ体内の糖鎖の修飾に顕著な違いがある。
【0003】
哺乳類細胞においては、糖鎖の修飾は3つの異なる経路により進行し、それが添加される蛋白質部分に依存する。即ち、(1)コアの糖鎖は変化しない;(2)UDP−N−アセチルグルコサミン(UDP−GlcNAc)からのN−アセチルグルコサミン−1−ホスフェート部分(GlcNAc−1−P)のコア糖鎖中のマンノースの6位への添加によりコアの糖鎖が変化し、続いてGlcNAc部分の除去による糖蛋白質中の酸性糖鎖の形成する;又は(3)マンノシダーゼIによる3マンノース残基の除去の結果としてコア糖鎖が最初にMan5GlcNAc2に変換されて;そしてGlcNAcの添加及び2つのさらなるマンノース残基の除去、続くGlcNAc、ガラクトース(Gal)、及びN−アセチルノイラミニ酸(シアル酸(NeuNAc)とも呼ばれる)の連続添加により様々なハイブリッド又は複合体糖鎖を形成することにより、Man5GlcNAc2がさらに修飾される(R.Kornfeld and S.Kornfeld,Ann.Rev.Biochem.54:631−664,1985;Chiba et al J.Biol.Chem.273:26298−26304,1998)。
【0004】
酵母においては、Man8GlcNAc2グリカンは削られない(not trimmed)。ゴルジ体内の糖鎖の修飾は、別々のマンノシルトランスフェラーゼによるマンノース残基の一連の添加(「外部鎖」グリコシル化)を含む。外部鎖グリコシル化の構造は生物に特異的であって、典型的にはサッカロミセスセレビシエにおいて50を超えるマンノース残基を伴い、そしてもっとも共通にはピキアパストリスにおいてMan14GlcNAc2より小さな構造を伴う。高いマンノース種のこの酵母特異的外部鎖グリコシル化は、高グリコシル化又は高マンノース化とも呼ばれる。
【0005】
グリコシル化は、蛋白質のフォールディング、安定性及び生物活性を修正するのに非常に重要である。ヒト体内において、グリコシル化は、蛋白質の薬力学特性、例えば組織分配及び血流からの浄化に、一部必須である。さらに、グリカン構造は、抗原応答に関与し得る。例えば、α−ガラクトースの糖蛋白質上の存在はブタからの異種移植に対する免疫反応に関する主要な理由である(Chen et al.,Curr Opin Chem Biol,3(6):650−658,1999)が、酵母に対する糖蛋白質に対する免疫反応は、主に、α−1,3−マンノース、β結合マンノース及び/又はホスホモノ−又はホスホジエステル結合の何れかの上のリン酸残基の存在による(Ballou,C.E.,Methods Enzymol,185:440−470,1990;Yip et l.,Proc Natl Acad Sci USA,91(7):2723−2727,1994)。
【0006】
高グリコシル化はしばしば望まれないが、何故ならば、糖質組成及び分子量の両方において組換え蛋白質生成物の不均質性を導き、蛋白質の精製を複雑にし得るからである。高グリコシル化された酵素の比活性(ユニット/質量)は、糖質の増加した部分により低下し得る。さらに、外部の鎖のグリコシル化はしばしば免疫原性が強く、治療への応用においては望まれないかもしれない。さらに、大きな外部の鎖状の糖(chain sugar)は治療用蛋白質の免疫原決定基を遮断し得る。例えば、ピキアパストリスにおいて発現されたインフルエンザノイラミニダーゼ(NA)は30−40までのマンノース残基を含むN−グリカンによりグリコシル化される。高グリコシル化されたNAは、NA分子の上部の可変性の免疫支配性表面ループがN−グリカンにより遮断されるために、マウスにおいて免疫原性が低下する(Martinet et al.Eur J.Biochem.247:332−338,1997)。
【0007】
よって、哺乳類(例えば、ヒト)の糖質構造に似せた糖質構造を有する組換え糖蛋白質を生じるメチロトローフ酵母株を遺伝子操作することが望まれる。
発明の概要
本発明は、哺乳類様のN−グリカンを伴う糖蛋白質を生じさせるための、遺伝子操作されたメチロトローフ酵母株及び方法に向けられる。本発明は、哺乳類様のN−グリカンを伴う糖蛋白質を生じさせることができる遺伝子操作されたメチロトローフ酵母株を生成するのに有用なベクター及びキットにも向けられる。
【0008】
本明細書の中で使用されている用語「メチロトローフ酵母」は、限定ではないが、メタノール上で生育可能な酵母株、例えば、属カンジダ、ハンセヌラ、トルロプシス、及びピキアを含む。
【0009】
一つの態様において、本発明は、コアのマンノース構造及び末端ガラクトース残基に結合した5又はそれより少ないマンノース残基及び少なくとも一つのN−アセチルグルコサミン残基(GlcNAc)を有することにより特徴付けされる、哺乳類様のN−グリカン構造を有する糖蛋白質を生じる、遺伝子操作されたメチロトローフ酵母を提供する。
【0010】
好ましい態様において、本発明は、哺乳類様のN−グリカン構造、GalGlcNAcMan5GlcNAc2を有する糖蛋白質を生じる遺伝子操作されたメチロトローフ酵母株を提供する。
【0011】
本発明によれば、GalGlcNAcMan5GlcNAc2を有する糖蛋白質を生じる上記メチロトローフ酵母株は、α−1,2−マンノシダーゼ又はその機能部分、N−アセチルグルコサミニルトランスフェラーゼI(又はGnTI)又はその機能部分、及びβ−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分を発現するように遺伝子操作される。好ましくは、当該メチロトローフ酵母株は、ゲノミックOCH1遺伝子が不活性化されるようにも遺伝子操作される。
【0012】
遺伝子操作されたメチロトローフ酵母株内での発現のためのα−1,2−マンノシダーゼ又はその機能部分は、あらゆる種の起源であることができ、マウス、ウサギ又はヒトのような哺乳類の種、及びアスペルギルス、又はトリコデルマリセイのような真菌を含む。本発明における使用のための好ましいα−1,2−マンノシダーゼは、トリコデルマリセイのα−1,2−マンノシダーゼである。好ましくは、α−1,2−マンノシダーゼ又はその機能部分は、その基質であるMan8GlcNAcが利用可能な分泌経路中の部位を標的化する。より好ましくは、α−1,2−マンノシダーゼ又はその機能部分は、ER滞留(retention)シグナルを含むように遺伝子操作されて、ERを標的化する。好ましいER滞留シグナルは、ペプチドHDEL(配列番号:1)である。
【0013】
遺伝子操作されたメチロトローフ酵母株における発現のためのGnTI又はその機能部分は、あらゆる種の起源であることができ、ウサギ、ラット、ヒト、植物、昆虫、線虫及び原生動物、例えば、リーシュマニアタレントレを含む。本発明における使用のための好ましいGnTIは、ヒトGnTIであり、配列番号:13に示される。好ましくは、GnTI又はその機能部分は、その基質であるMan5GlcNAc2が利用可能な分泌経路中の部位を標的化する。より好ましくは、GnTIまたはその機能部分は、ゴルジ滞留シグナルを含むように遺伝子操作されて、ゴルジ体を標的化する。好ましいゴルジ滞留シグナルは、配列番号:11に示されるペプチドであり、サッカロミセスセレビシエのKre2蛋白質の最初の100アミノ酸からなる。
【0014】
遺伝子操作されたメチロトローフ酵母株における発現のためのGalT又はその機能部分は、あらゆる種の起源であることができ、ヒト、植物(例えば、アラビドプシスサリアナ)、昆虫(例えば、ドロソフィラメラノガスター)を含む。本発明における使用のための好ましいGalTは、ヒトGalTであり、配列番号:21に示される。好ましくは、GalT又はその機能部分は、ゴルジ滞留シグナルを含むように遺伝子操作されて、ゴルジ体を標的化する。好ましいゴルジ滞留シグナルは、配列番号:11に示されるペプチドであり、サッカロミセスセレビシエのKre2蛋白質の最初の100アミノ酸からなる。
【0015】
メチロトローフ酵母株は、例えば、形質転換によりこれらの酵素をコードする株ヌクレオチド配列へ導入することにより上記の所望の酵素を発現するように遺伝子操作することができる。好ましくは、コーディング配列はベクター内に提供され、各配列は、酵母株中で機能するプロモーター配列及び3’ターミネーション配列に作動可能に連結されるように配置される。ベクター又はその直鎖状断片で、次に、株を形質転換する。
【0016】
本発明の好ましい態様によれば、上記メチロトローフ酵母株は、OCH1遺伝子が破壊されるようにも遺伝子操作される。遺伝子の破壊は、ゲノミックOCH1配列とノックアウトベクター中のOCH1配列の間の相同組換えにより達成することができる。
【0017】
さらなる側面において、本発明は、哺乳類様のN−グリカン構造を有する糖蛋白質生じるメチロトローフ酵母株を生成するのに有用なベクターを提供する。
一つの態様において、本発明は、発現される酵素、即ち、α−1,2−マンノシダーゼ、GnTI,GalT,又はこれらの蛋白質の何れかの機能部分をコードするヌクレオチド配列を含む「ノックイン」ベクターを提供する。コーディング配列は、コードされた蛋白質の発現のためにメチロトローフ酵母中で機能するプロモーター配列及び3’ターミネーション配列に作動可能に連結されるように配置される。2つ又はそれより多いコーディング配列をメチロトローフ酵母株へ同時形質転換するために同じベクター中に入れることができる。好ましくは、当該ベクターは、形質転換体の便利な選択のための選択可能なマーカー遺伝子も含む。
【0018】
別の態様において、本発明は、メチロトローフ酵母株に導入されたときにゲノミックOCH1遺伝子を不活性化するか又は破壊する不活性化ベクター(又は「ノックアウト」ベクター)を提供する。
【0019】
OCH1ノックアウトベクターは選択可能なマーカーを含むことができ、ゲノミックOCH1遺伝子との二重相同組換えを媒介するのに十分な長さのOCH1配列に対して両5’及び3’末端にて作動可能に連結する。或いは、OCH1不活性化ベクターは、破壊されるOCH1遺伝子の一部を含むことができ、その部分はOCH1蛋白質を一切コードしないか又はOCH1蛋白質の不活性断片をコードし、そして選択可能なマーカー遺伝子を含むことができる。OCH1部分は、何れかの公知のプロモーター配列に対して作動可能な連結はされず、そして上記ベクターの直鎖状断片の形質転換に際して、1回の相同組換えによりゲノミックOCH1座に組み込まれる。好ましくは、一つ又はそれより多い不活性化変異、例えば停止コドン又はフレームシフト変異も、ベクター中のOCH1配列内に導入されることにより、何れかの可能性のある活性OCH1ポリペプチドの生産を阻止する。
【0020】
さらに別の側面において、本発明は、哺乳類様のN−グリカン構造を有する糖蛋白質の生成方法を提供する。目的の糖蛋白質をコードするヌクレオチド配列を、哺乳類様N−グリカンを生じさせるように遺伝子操作されたメチロトローフ酵母株に導入することができる。或いは、目的の糖蛋白質を発現するメチロトローフ酵母株を修飾することにより、
所望の酵素(例えば、α−1,2−マンノシダーゼ、GnTI及びGalT)を発現させ、そしてゲノミックOCH1遺伝子を不活性化することができ、それにより、哺乳類様N−グリカンを伴う糖蛋白質を生成する。
【0021】
また別の側面において、本発明の方法を用いることにより生成された糖蛋白質、即ち、哺乳類様のN−グリカン、特にGalGlcNAcMan5GlcNAc2N−グリカンを有する糖蛋白質が、本発明により提供される。
【0022】
さらなる側面において、本発明は、本発明の一つ又はそれより多いベクター、又は本発明の遺伝子操作された一つ又はそれより多い株を含むキットを提供する。
発明の詳細な説明
本発明は、哺乳類様又はヒト様N−グリコシル化を伴う組換え糖蛋白質を作成するための、方法、ベクター及び遺伝子操作されたメチロトローフ酵母株に向けられる。
【0023】
「哺乳類」は、哺乳類のあらゆる種、例えば、ヒト、マウス、ラット、イヌ、ウサギ、ウシ、ヒツジ、ウマ等を含むことを意味する。
典型的には複雑な種類の哺乳類のグリカン、例えば、ヒトにおいて生じたグリカンは、Man3GlcNAc2の内部コア構造に結合したシアリル−N−アセチル−ラクトサミン配列を伴う2から6の外部ブランチを有する。哺乳類のN−グリカンは、ER中で形成されるコアオリゴサッカライド構造であるMan8GlcNAc2に由来する。このコアの糖構造を有する蛋白質はゴルジ体に輸送されて、そこで、ゴルジ体マンノシダーゼI(ゴルジ体α−1,2−マンノシダーゼ)により3つのマンノース残基を除去した結果として、Man8GlcNAc2がMan5GlcNAc2に変換される。蛋白質がゴルジ体を通して加工されるにつれ、Man5GlcNAc2は、さらに、GlcNAcの追加と2つのさらなるマンノース残基の除去、次に、GlcNAc,ガラクトース(Gal)及びシアル酸(SA)残基の追加により、修飾される。
【0024】
本明細書にて使用される用語「哺乳類様N−グリコシル化」は、遺伝子操作されたメチロトローフ酵母株内で生成された糖蛋白質のN−グリカンが、5つ又はそれ未満のマンノース残基を含み、そして哺乳類細胞、例えばヒト細胞において生じた蛋白質のN−グリカンの生合成におけるN−グリカン又は中間体糖質構造の特徴を有することを意味する。
【0025】
好ましい態様において、本発明の遺伝子操作されたメチロトローフ酵母株において生成された糖蛋白質は、5つ又はそれ未満のマンノース残基、及びマンノース残基を含むコア構造及びターミナルガラクトース残基に結合した少なくとも一つのN−アセチルグルコサミン残基(GlcNAc)を含む。例えば、遺伝子操作されたメチロトローフ酵母株内で生成された糖蛋白質は、GalGlcNAcMan5GlcNAc2を有し、図1に描写されるとおりである。この糖質(GalGlcNAcMan5GlcNAc2)のIUPACの命名は、Gal(β−1,4)GlcNac(β−1,2)Man(α−1,3){Man(α−1,3)[Man(α−1,6)]Man(α−1,6)}Man(β−1,4)GlcNac(β−1,4)GlcNacである。その拡大された命名は、β−D−Galp−(1→4)−β−D−GlcpNAc−(1→2)−α−D−Manp−(1→3)−{α−D−Manp−(1→3)−[α−D−Manp−(1→6)]−α−D−Manp−(1→6)}−β−D−Manp−(1→4)−β−D−GlcpNAc−(1→4)−D−GlcpNAcである。
【0026】
ピキア、特にピキアパストリスを含むメチロトローフ酵母の細胞質網状構造(ER)を通り過ぎる糖蛋白質上のN−グリカンの大多数は、Man8GlcNAc2オリゴサッカライド構造を有する。糖蛋白質がERからゴルジ体に輸送された後は、追加のマンノース残基が別のマンノシルトランスフェラーゼによりこのコア糖部分に追加されて、高マンノースコア又は延長されて分枝したマンナン外部鎖からなるオリゴサッカライド構造を有する糖蛋白質をもたらす。
【0027】
本発明によれば、哺乳類様グリコシル化を伴う組換え糖蛋白質を生成するためには、メチロトローフ酵母を修飾することにより、一連の工程において糖質構造Man8GlcNAc2を哺乳類様N−グリカンに変換する酵素を発現させる。好ましくは、メチロトローフ酵母は、高マンノース構造の生産に関与する一つ又はそれより多い酵素、例えばOCH1遺伝子によりコードされるα−1,6−マンノシルトランスフェラーゼの発現を不活性化するようにも修飾される。
【0028】
本明細書において使用される用語「メチロトローフ酵母」は、限定ではないが、メタノール上で生育できる酵母株、例えば、属カンジダ、ハンセヌラ、トルロプシス、及びピキアを含む。本発明の好ましいメチロトローフ酵母は、属ピキアの株である。特に好ましいのは、ピキアパストリス株GS115(NRRL Y−15851),GS190(NRRL Y−18014),PPF1(NRRL Y−18017),PPY120H,YGC4,及びそれらに由来する株である。
【0029】
一つの態様において、本発明は、コアのマンノース含有構造及びターミナルガラクトース残基に結合した5つ又はそれ未満のマンノース残基及び少なくとも一つのN−アセチルグルコサミン残基(GlcNAc)を有する特徴を有する、哺乳類様N−グリカン構造を有する糖蛋白質を生成する遺伝子操作されたメチロトローフ酵母株を提供する。
【0030】
好ましい態様において、本発明は、哺乳類様N−グリカン構造、GalGlcNAcMan5GlcNAc2を有する糖蛋白質を生じる遺伝子操作されたメチロトローフ酵母株を提供する。
【0031】
本発明によれば、GalGlcNAcMan5GlcNAc2を有する糖蛋白質を生じる上記のメチロトローフ酵母株は、α−1,2−マンノシダーゼ又はその機能部分、N−アセチルグルコサミニルトランスフェラーゼI(又はGnTI)又はその機能部分、及びβ−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分を発現するように遺伝子操作される。好ましくは、当該メチロトローフ酵母株は、ゲノミックOCH1遺伝子が不活性化されるようにも遺伝子操作される。
【0032】
α−1,2−マンノシダーゼは、Man8GlcNAc2の非還元末端においてα−1,2−結合したマンノース残基を分断し、そして糖蛋白質上のこのコアのオリゴサッカライドをMan5GlcNAc2に変換し、これは哺乳類N−アセチルグルコサミニルトランスフェラーゼIのアクセプター基質である。
【0033】
本発明によれば、メチロトローフ酵母株を操作して、当該株に、例えば形質転換により、α−1,2−マンノシダーゼ又はその機能部分をコードするヌクレオチド配列を導入することにより、α−1,2−マンノシダーゼ又はその機能部分をコードするヌクレオチド配列を発現することができる。α−1,2−マンノシダーゼ又はその機能部分をコードするヌクレオチド配列は、如何なる種に由来することもできる。多数のα−1,2−マンノシダーゼ遺伝子がクローン化されて、当業者には利用可能であり、例えば、マウスのα−1,2−マンノシダーゼ遺伝子(Herscovics et al.J.Biol.Chem.269:9864−9871,1994)、ウサギのα−1,2−マンノシダーゼ遺伝子(Lal et al.J.Biol.Chem.269:9−872−9881,1994)又はヒトα−1,2−マンノシダーゼ遺伝子(Tremblay et al.Glycobiology 8:585−595,1998)をコードする哺乳類遺伝子、並びに、例えば、アスペルギルスα−1,2−マンノシダーゼ遺伝子(msdS遺伝子)、又はトリコデルマリセイα−1,2−マンノシダーゼ遺伝子(Maras et al.J.Biotechnol.77:255−263,2000)を含む真菌遺伝子を含む。蛋白質の配列分析が、これまでに同定された真核生物α−1,2−マンノシダーゼ遺伝子の間の高度の保存を明らかにした。
【0034】
好ましくは、本ベクターにおける使用のためのヌクレオチド配列は、真菌のα−1,2−マンノシダーゼ、より好ましくは、トリコデルマリセイα−1,2−マンノシダーゼ、そしてより特定すれば、Maras et al.J.Biotechnol.77:255−63(2000)に記載されたトリコデルマリセイα−1,2−マンノシダーゼをコードする。
【0035】
「機能部分」は、完全長蛋白質の酵素活性を実質上保持したα−1,2−マンノシダーゼのポリペプチド断片を意味する。「実質上」は、完全長α−1,2−マンノシダーゼの酵素活性の少なくとも約40%、又は好ましくは少なくとも50%またはそれ以上が保持されることを意味する。多数のα−1,2−マンノシダーゼの、触媒ドメインを含む様々なドメインの特性決定は、記録されている。例えば、”Isolation of a mouse Golgi mannosidase cDNA, a member of a gene family conserved from yeast to mammalls”,Herscovics et al.,J Biol Chem 269:13 9864−71(1994);Isolation and expression of murine and rabbit cDNAs encoding an alpha 1,2−mannosidase involved in the processing of asparagine−linked oligosaccharides”,La et al.,J Biol Chem 269:13 9872−81(1994);”Molecula cloning and enzymatic characterization of a Trichoderma reeses 1,2−alpha−D−mannosidase”,Maras M et al.,J Biotechnol 77:255−63(2000);及び米国特許出願20020188109を参照されたく、引用により本明細書に編入される。当業者は、当業界で公知の技術の組み合わせを用いることによりα−1,2−マンノシダーゼの機能部分を容易に同定して作成することができる。適切な発現系から発現されて精製された目的のα−1,2−マンノシダーゼの一部の活性は、引用により本明細書に編入される米国特許出願20020188109に記載されたインビトロ又はインビボアッセイを用いることにより確認することができる。
【0036】
本発明によれば、メチロトローフ酵母内で発現されたα−1,2−マンノシダーゼ又はその機能部分は、好ましくは、Man8GlcNAc2(α−1,2−マンノシダーゼの基質)が糖蛋白質上で既に形成されているが、追加のマンノース残基により糖鎖を伸長するゴルジ体のグリコシルトランスフェラーゼには到達していない分泌経路中の部位を標的化する。本発明の好ましい態様において、α−1,2−マンノシダーゼ又はその機能部分は、α−1,2−マンノシダーゼがメチロトローフ酵母株内で発現されてERを標的化するように、ER滞留シグナルを含むように操作される。
【0037】
「ER滞留シグナル」は、ERに輸送されてER中で滞留するようにペプチド配列を有する蛋白質を指示するペプチド配列を意味する。そのようなER滞留配列は、しばしば、ER中に存在して機能する蛋白質の中に見いだされる。ER滞留シグナルの複数の選択が当業者に利用可能であり、例えば、サッカロミセスセレビシエのER蛋白質のMNS1の最初の21アミノ酸残基(Martinet et al.Biotechnology Letters 20:1171−1177,1998)及びペプチドHDEL(配列番号:1)である。
【0038】
本発明における使用のための好ましいER滞留シグナルは、ペプチドHDEL(配列番号:1)である。HDELペプチド配列は、多数の酵母蛋白質のC−末端に見いだされ、ERのための滞留/修正(retrieval)シグナルとして作用する(Pelham EMBO J.7:913−918,1988)。HDEL配列を有する蛋白質は、膜結合受容体(Erd2p)により結合して、次に、ゴルジ体からERに戻るための逆行(retrograde)輸送経路に入る。
【0039】
本発明における使用のためのα−1,2−マンノシダーゼは、さらに、例えば、抗体が利用可能なエピトープタグを含むように操作することができ、例えば、当業界でよく知られているMyc,HA,FLAG及びHis6タグである。エピトープタグを付加されたα−1,2−マンノシダーゼは、発現及び細胞外局在化の両方のために便利に精製、又は監視され得る。
【0040】
本発明によれば、ER滞留シグナルを、α−1,2−マンノシダーゼの蛋白質配列のどこか、しかし好ましくはα−1,2−マンノシダーゼのC末端において、遺伝子操作することにより配置することができる。
【0041】
ER滞留シグナル及びエピトープタグは、当業界公知の分子生物学の技術の何れかを用いて、α−1,2−マンノシダーゼ又はその機能部分をコードするヌクレオチド配列に、そのようなシグナル又はタグをコードするヌクレオチド配列を挿入することにより、α−1,2−マンノシダーゼ又はその機能部分に容易に挿入することができる。
【0042】
操作された酵母株内でのα−1,2−マンノシダーゼの発現は、mRNAレベルにおいては例えばノーザンブロット分析により、そして蛋白質レベルにおいてはウエスタンブロット分析により確認することができる。蛋白質の細胞内局在は様々な技術を用いることにより分析することができ、細胞成分分画(subcellular fractionation)及び免疫蛍光の実験を含む。α−1,2−マンノシダーゼもER中での局在は、細胞成分分画実験においてこの酵素と公知のER滞留蛋白質(例えば、プロテインジスルフィドイソメラーゼ)との共沈殿により決定することができる。ER中の局在は、ER滞留蛋白質の免疫蛍光染色パターン特性、典型的には核周囲染色パターンにより、決定することもできる。
【0043】
メチロトローフ酵母株中で発現されたα−1,2−マンノシダーゼ又はその機能部分が期待されたマンノーストリミング活性を有することを確認するために、インビトロ及びインビボ両方のアッセイを用いることができる。典型的には、インビトロアッセイは、インビトロで合成された基質、例えば、Man8GlcNAc2の免疫酵母株で発現されて精製された酵素による消化、及びそのような酵素がMan8GlcNAc2を例えばMan5GlcNAc2に削る能力を評価することを含む。インビボアッセイにおいては、α−1,2−マンノシダーゼ又はその一部をメチロトローフ酵母内で、そのような酵母内でN−グリカンを生じるターミナルα−1,2−結合したマンノース残基によりグリコシル化されることが知られている糖蛋白質と同時発現させる。そのようなα−1,2−マンノシダーゼ又はその一部の酵素活性は、糖蛋白質のN−グリカンの構造中のα−1,2−結合したマンノース残基の数の低下に基づいて測定することができる。インビトロ及びインビボの両アッセイにおいて、糖質基の組成は、当業界でよく知られており且つ本明細書の以下の実施例において例示される技術を用いて測定することができる。
【0044】
さらに、本発明によれば、GlcNAc−トランスフェラーゼI又はその機能部分をコードするヌクレオチド配列をマンノース酵母株に、例えば形質転換により導入することにより、GlcNAc−トランスフェラーゼI又はその機能部分を発現するようにマンノース酵母株を操作することができる。GlcNAc−トランスフェラーゼIはβ−1,2−GlcNAcのMan5GlcNAc2への付加に必須であり、糖蛋白質上のこのコアオリゴサッカライドをGlcNAcMan5GlcNAc2へ変換する。GlcNAcMan5GlcNAc2のマンノース残基は哺乳類ゴルジ体マンノシダーゼIIによりさらに削られて、追加の糖ユニット、例えばガラクトースが哺乳類糖蛋白質のハイブリッド−又はコンプレックス−タイプの糖ブランチ特性を形成するために(towards)追加される。
【0045】
メチロトローフ酵母株への導入のためのGlcNAc−トランスフェラーゼI(GnTI)又はその機能部分をコードするヌクレオチド配列は、あらゆる種、例えば、ウサギ、ラット、ヒト、植物、昆虫、線虫及び原生動物、例えばリーシュマニアタレントレを含む。好ましくは、本発明における使用のためのヌクレオチド配列は、ヒトGnTIであり、そしてより好ましくは配列番号:13に示されるヒトGnTIである。
【0046】
GnTIの「機能部分」は、完全長のGnTIの酵素活性を実質上保持するGnTIのポリペプチド断片を意味する。「実質上」は、少なくとも約40%、又は好ましくは少なくとも約50%またはそれ以上の完全長のGnTIの酵素活性が保持されることを意味する。GnTI又はその一部の酵素活性は、Reeves et al.(Proc.Natl.Acad.Sci.USA.99(21):13419−24,2002);Maras et al.(Eur J Biochem.249(3):701−7,1997),又は本明細書中の以下の実施例に記載されるアッセイにより測定することができる。当業者は、当業界で公知の技術の組み合わせを用いてGnTIの機能部分を容易に同定して作成することができる。例えば、本発明により例示されるとおり、ヒトGnTIの触媒ドメイン(最後の327残基を含む)はヒトGnTIの「機能部分」を構成する。
【0047】
本発明によれば、メチロトローフ酵母株において発現されたGnTI又はその機能部分は、好ましくは、Man5GlcNAc2(GnTIの基質)が糖蛋白質上に既に形成された分泌経路中の部位を標的化する。好ましくは、GnTI又はその機能部分はゴルジ体を標的化する。
【0048】
従って、本発明の好ましい態様においては、GnTI又はその機能部分がゴルジ体局在化シグナルを含むように操作される。
本明細書において使用される「ゴルジ体局在化シグナル」は、ペプチド配列であって、そのような配列を有する蛋白質をメチロトローフ酵母株のゴルジ体に振り向け(directs)、そして当該蛋白質をその中に保持するペプチド配列を意味する。そのようなゴルジ体局在化配列は、ゴルジ体に存在して機能する蛋白質中にしばしば見いだされる。
【0049】
ゴルジ体局在化シグナルの選択は、当業者に利用可能である。本発明における使用のための好ましいゴルジ体局在化シグナルは、サッカロミセスセレビシエのKre2蛋白質(ScKre2)のN末端部分に由来するペプチド;より好ましくは配列番号:10に示されるScKre2である。特に好ましいゴルジ体局在化シグナルは、配列番号:10に示されるScKre2蛋白質のアミノ酸1−100からなる、ペプチド(配列番号:11)である。
【0050】
本発明によれば、ゴルジ体局在化シグナルは、GnTI内の如何なる場所に配置されてもよいが、好ましくはGnTIの末端、そしてより好ましくはGnTIのN末端に配置され得る。
【0051】
本発明における使用のためのGnTIは、例えば、抗体が利用可能なエピトープタグ、例えば、当業界でよく知られているMyc,HA,FLAG及びHis6を含むようにさらに操作され得る。エピトープタグを付加されたGnTIは便利に精製されるか、又は発現及び細胞内局在に関して監視され得る。
【0052】
ゴルジ体局在化シグナル及びエピトープタグは、当業界公知の分子生物学の技術の何れかを用いて、GnTI又はその機能部分をコードするヌクレオチド配列に、そのようなシグナル又はタグをコードするヌクレオチド配列を挿入することにより、GnTI又はその機能部分に容易に挿入することができる。
【0053】
さらに、本発明によれば、メチロトローフ酵母株をβ−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分をコードするヌクレオチド配列をマンノース酵母株に、典型的には形質転換により導入することにより、β−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分を発現するようにマンノース酵母株を操作することができる。GalTはβ−1,4−ガラクトース残基をグリカン構造(GlcNAcMan5GlcNAc2)の左のアーム上のGlcNAcへの付加し、図1に描写されるとおりである。
【0054】
メチロトローフ酵母株への導入のためのGalT又はその機能部分をコードするヌクレオチド配列は、あらゆる種、例えば、哺乳類(例えば、ヒト、マウス)、植物(例えば、アラビドプシスサリアナ)、昆虫(例えば、ドロソフィラメラノガスター)、又は線虫(シーノラブディティスエレガンス)に由来することができる。好ましくは、本発明における使用のためのヌクレオチド配列は、ヒトGalT、そしてより好ましくは配列番号:21に示されたヒトGalTをコードする。
【0055】
GalTの「機能部分」は、完全長のGalTの酵素活性を実質上保持するGalTのポリペプチド断片を意味する。「実質上」は、少なくとも約40%、又は好ましくは少なくとも約50%またはそれ以上の完全長のGalTの酵素活性が保持されることを意味する。GalT又はその一部の酵素活性は、Maras et al.(Eur J Biochem.249(3):701−7,1997),又は本明細書中の以下の実施例に記載されるアッセイにより測定することができる。当業者は、当業界で公知の技術の組み合わせを用いてGalTの機能部分を容易に同定して作成することができる。例えば、本発明により例示されるとおり、ヒトGalTの触媒ドメインはヒトGalTの「機能部分」を構成する。
【0056】
本発明によれば、メチロトローフ酵母株において発現されたGalT又はその機能部分は、好ましくは、GlcNAcMan5GlcNAc2(GlcNAcMの基質)が糖蛋白質上に既に形成された分泌経路中の部位を標的化する。好ましくは、GalT又はその機能部分はゴルジ体を標的化する。
【0057】
従って、本発明の好ましい態様において、GalT又はその機能部分は、上で記載されたとおり、ゴルジ体局在化シグナルを含むように操作される。GalTをゴルジ体に標的化するための好ましいゴルジ体局在化シグナルは、配列番号:10に示されるScKre2蛋白質のアミノ酸1−100からなる、ペプチド(配列番号:11)である。
【0058】
本発明における使用のためのGalTは、例えば、抗体が利用可能なエピトープタグ、例えば、当業界でよく知られているMyc,HA,FLAG及びHis6を含むようにさらに操作され得る。エピトープタグを付加されたGalTは便利に精製されるか、又は発現及び細胞内局在に関して監視され得る。
【0059】
ゴルジ体局在化シグナル及びエピトープタグは、当業界公知の分子生物学の技術の何れかを用いて、GalT又はその機能部分をコードするヌクレオチド配列に、そのようなシグナル又はタグをコードするヌクレオチド配列を挿入することにより、GalT又はその機能部分に容易に挿入することができる。
【0060】
メチロトローフ酵母株内での所望の蛋白質(即ち、α−1,2−マンノシダーゼ、GnTI,GalT,又はこれらの酵素の何れかの機能部分)の発現を達成するためには、当該蛋白質をコードするヌクレオチド配列を、上記メチロトローフ酵母株内で機能するプロモーター及び3’終結配列に作動可能なような連結にてベクター内に配置させることができる。当該ベクターを、次に、メチロトローフ酵母株へ、例えば、形質転換により導入する。
【0061】
メチロトローフ酵母内で蛋白質の発現に適したプロモーターは、構成的プロモーター及び誘導可能なプロモーターの両方を含む。構成的プロモーターは、例えば、ピキアパストリスのグリセルアルデヒド−3−ホスフェートデヒドロゲナーゼプロモーター(「GAPプロモーター」)を含む。誘導可能なプロモーターの例は、例えば、ピキアパストリスの「アルコールオキシダーゼIプロモーター(「AOXプロモーター」)(米国特許第4,855,231号)、又はピキアパストリスのホルムアルデヒドデヒドロゲナーゼプロモーター(「FLDプロモーター」)(Shen et al.Gene 216:93−102,1998)を含む。
【0062】
3’終結配列は、当該配列が作動可能に連結した遺伝子のmRNA転写産物を安定化するために機能する構造遺伝子の停止コドンの3’の配列であり、例えば、ポリアデニル化を導く配列である。3’終結配列は、ピキア又はメチロトローフ酵母から得ることができる。本発明の実施に有用なピキアパストリスの3’終結配列の例は、AOX1遺伝子及びHIS4の遺伝子由来の終結配列を含む。
【0063】
ベクター又はその直鎖状断片の形質転換は、あらゆる公知の方法、例えば、Cregg ら(Mol.Cell.Biol.(12):3376−85,1985)により記載されたようなスフェロプラスト技術、又はEP312,934に記載されたピキアにおける使用のために修飾された、Itoら(Agric.Biol.Chem.48(2):341,(1984))により記載された全細胞塩化リチウム酵母形質転換系を用いて実施することができる。形質転換に有用な他の方法は、米国特許第4,929,555号;Hinnen et al.(Proc.Natl.Acad.Sci.USA 75:1929(1978));Ito et al.(J.Bacteriol.153:163(1983));米国特許第4,879,231号;及びSreekrishna et al.(Gene 59:115(1987))に記載された方法を含む。エレクトロポレーション及びPEG1000全細胞形質転換手法を使用することもできる。Cregg and Russel,Methods in Molecular Biology:Pichia Protocols,Chapter 3,Humana Press,Totowa,N.J.,pp.27−39(1998)を参照。
【0064】
形質転換された酵母細胞は、限定ではないが、(細胞の栄養要求性のために)必要とされる生化学生成物の不在下での形質転換後に栄養要求性細胞を培養すること、新規な表現型の選択及び検出、又は形質転換体内に含まれる耐性遺伝子の不在下で当該酵母に毒性の抗生物質の存在下で培養することを含む、適切な技術を用いることにより選択することができる。形質転換体はゲノム中の発現カセットの組み込みにより選択及び/又は確認することもでき、例えばサザンブロット又はPCR分析により評価することができる。
【0065】
上で記載されたとおり、α−1,2−マンノシダーゼ、及びN−アセチルグルコサミニルトランスフェラーゼI(又はGnTI)、β−1,4−ガラクトシルトランスフェラーゼ(GalT)、又はその機能部分の発現に加えて、上記メチロトローフ酵母株は、好ましくは、GlcNAcMan5GlcNAc2グリカンを有する糖蛋白質を有効に生成するためにゲノミックOCH1遺伝子を不活性化するように遺伝子操作もされる。
【0066】
OCH1遺伝子は、初期ゴルジ体複合体内に局在化してN−結合コアオリゴサッカライド(Man5GlcNAc2及びMan8GlcNAc2)へのα−1,6−ポリマンノース外部鎖付加を開始する膜結合性α−1,1,6−マンノシルトランスフェラーゼをコードする。サッカロミセスセレビシエのOCH1遺伝子及びピキアのOCH1遺伝子はクローン化されている(それぞれ、Nakayama et al.EMBO J.11:2511−19,1992、及び日本特許出願番号07145005)。当業者は、当業界で公知の技術を用いることにより、他のメチロトローフ酵母からOCH1遺伝子を単離することができる。
【0067】
本発明によれば、メチロトローフ酵母株のOCH1遺伝子の破壊は不活性蛋白質産物の生成か又は生成物なしの何れかをもたらし得る。当該破壊は、外来DNA配列の上記コーディング配列への挿入の形態及び/又は当該コーディング配列の幾つか又は全ての欠失の形態を採ってよい。遺伝子破壊は、相同組換えにより生じさせることができ、本質的にはRothstein(in Methods in Enzymology,Wu et al.,eds.,vol 101:202−211,1983)に記載される。
【0068】
二重相同組換えによりゲノミックOCH1遺伝子を破壊するためには、OCH1「ノックアウト」ベクターを構築することができ、それは相同組換えを媒介するのに十分な長さのOCH1遺伝子の一部に対してその5’及び3’末端の両方において作動可能に連結された選択可能なマーカー遺伝子を含む。当該選択可能マーカーは、宿主細胞栄養要求性を相補するか又は抗生物質耐性を提供するあらゆる数の遺伝子の一つであり得、URA3,ARG4,HIS4,ADE1,LEU2,HIS3,Sh ble(ストレプトアロテイカスヒンダスタヌスのブレオマイシン遺伝子)及びBSD(アスペルギルステレウスからのブラスチシジンSデアミナーゼ)遺伝子を含む。他の適切な選択可能マーカーは、メチロトローフ酵母の蔗糖上の生育を可能にさせるサッカロミセスセレビシエ由来のインベルターゼ遺伝子;活性βガラクトシダーゼの発現のために青いコロニーをもたらすlacZ遺伝子を含む。OCH1不活性化ベクターの直鎖状DNA断片は、その5’及び3’両末端においてOCH1配列と共に選択可能マーカーを含み、のちに、当業界で公知の形質転換方法の何れかを用いることにより宿主メチロトローフ酵母細胞内に導入される。直鎖状断片のゲノミックOCH1座への組み込み及びOCH1遺伝子の破壊は、選択可能マーカーに基づいて測定することができ、そして例えば、サザンブロット分析により確認することができる。
【0069】
或いは、OCH1遺伝子の一部を含み、但し当該一部はあらゆるOCH1プロモーター配列を欠き、そしてOCH1蛋白質を全くコードしないか又は不活性部分をコードする、OCH1遺伝子ノックアウトベクターを構築することができる。「不活性断片」は、完全長OCH1蛋白質の断片であって、当該断片が完全長のOCH1蛋白質の活性の、好ましくは約10%未満、そしてより好ましくは約0%を有するものを意味する。OCH1遺伝子のそのような部分は、メチロトローフ酵母において機能するあらゆるプロモーター配列に作動可能に連結されずにベクター内に挿入される。このベクターは、OCH1遺伝子内の部位においてのちに直鎖状化され、そして当業界で公知のあらゆる形質転換方法を用いてメチロトローフ酵母株を形質転換することができる。1回の相同組換えにより、この直鎖状化されたベクターは次にOCH1座に組み込まれて、染色体中に2つのoch1配列をもたらすが、何れもが活性なOch1p蛋白質を生成することができず、図3Aに描写されるとおりである。
【0070】
好ましくは、不活性化変異を直鎖状化された部位の5’(上流の)部位及びOCH1の翻訳開始コドンの3’(下流)の部位においてベクター中のoch1配列内にも挿入される。「不活性化変異(inactivating mutation)」は、停止コドンを導入する変異、フレームシフト変異又は読み枠の破壊を引き起こすあらゆる他の変異を意味する。そのような変異は、当業界で公知の部位特異的変異導入方法の何れかを用いてベクター内にoch1配列内に導入され得る。そのような不活性化変異は、たとえノックアウトベクター中のOch1配列に対して5’に幾つかのプロモーター配列が存在しようとも、機能しないOch1p蛋白質が相同組換え後に形成されることを保証する。
【0071】
本明細書において上で記載されたような遺伝子操作されたメチロトローフ酵母株は、所望であればさらに修飾することができる。例えば、あらゆるその他のピキアのマンノシルトランスフェラーゼをコードする追加の遺伝子の破壊を作成することができる。α−1,2−マンノシダーゼ、GnTI又はGalT以外の哺乳類グリコシル化経路中で機能する酵素をコードする遺伝子が、哺乳類様N−グリカンの比率を増加させるため、及び/又は、所望であれば哺乳類様N−グリカンをさらに修飾するために、導入することができる。例えば、上で記載された遺伝子操作されたメチロトローフ酵母株をさらに修飾することにより、UDP−グルコースをUDP−ガラクトースに、そして逆もまた同じく変換するサッカロミセスセレビシエのGAL−10にコードされた酵素を発現することができる。これは、サイトゾルのUDP−ガラクトースのレベルを増加させるかもしれず、次に、GalTの活性を刺激してGalGlcNacM5GlcNac2グリカンの比率を増加させる。さらに、上で記載された遺伝子操作されたメチロトローフ酵母は、ゴルジ体中でマンノシダーゼIIを発現するようにさらに修飾され得て、GalGlcNacM5GlcNac2から追加のマンノース残基を除去し、それにより他の糖残基の追加を許容する。
【0072】
遺伝子修飾の順序は本発明には必須ではない。α−1,2−マンノシダーゼ、GnTI及びGalTをコードするヌクレオチド配列の導入、及びゲノミックOCH1遺伝子の破壊は、連続してか、如何なる順序でも、又は同時に、2つ又はそれより多い別々のベクター又はコーディング配列により共形質転換(co-transformation)することによるか又は2つ又はそれより多い別々のコーディング配列を含む一つのベクターによる形質転換により行うことができる。
【0073】
さらなる側面において、本発明は、5又はそれ未満のマンノース残基及びコアマンノース含有構造並びにターミナルガラクトース残基に結合した少なくとも一つのN−アセチルグルコサミン残基(GlcNAc)を有するものとして特徴付けされる哺乳類様N−グリカン構造、例えば、GalGlcNAcMan5GlcNAc2を有する構造を生じるメチロトローフ酵母株を生じさせるのに有用なベクターを提供する。
【0074】
一つの態様において、本発明は、発現される酵素、即ち、α−1,2−マンノシダーゼ、GnTI,GalT,又はこれらの酵素の何れかの機能部分をコードするヌクレオチド配列を含むベクターを提供する。そのようなベクターは、「ノックイン」ベクターとも呼ばれる。コーディング配列は、コードされた蛋白質の発現のために宿主メチロトローフ酵母において機能するプロモーター及び3’終結配列に作動可能に連結して配置され得る。2つ又はそれより多いコーディング配列をメチロトローフ酵母株への同時形質転換のための同じベクター内に入れることができる。好ましくは、当該ベクターは、形質転換の便利な選択のための上で記載された選択可能なマーカー遺伝子の何れか一つも含む。
【0075】
本発明によれば、ノックインベクターは、メチロトローフ酵母株内で発現される所望の蛋白質をコードする配列を含み、組み込み型ベクター又は複製型ベクター(例えば、複製型環状プラスミド)の何れかであり得る。組み込み型ベクターは、例えば、米国特許第4,882,279号に開示されており、引用により本明細書に編入される。組み込み型ベクターは、通常、少なくとも第1の挿入可能DNA断片、選択可能なマーカー遺伝子、及び第2の挿入可能なDNA断片の連続して並んだ配置(serially arranged sequence)を含む。第1及び第2の挿入可能なDNA断片は、各々、約200ヌクレオチドの長さであり得て、形質転換される種のゲノミックDNAの一部に相補なヌクレオチド配列を有する。発現のための目的の構造遺伝子を含むヌクレオチド配列をこのベクター中で第1の挿入可能DNA断片と第2の挿入可能DNA断片の間に挿入するが、マーカー遺伝子の前又は後である。組み込み型ベクターは宿主細胞ゲノムへの目的のヌクレオチド配列の組み込みを促進するために酵母形質転換に先立って直鎖状にすることができる。
【0076】
別の態様において、本発明は、メチロトローフ酵母株に導入されたときにゲノムOCH1遺伝子を不活性化するか又は破壊する不活性化ベクター(又は「ノックアウト」ベクター)を提供する。
【0077】
ゲノミックOCH1遺伝子を不活性化するためのベクターは、上で記載されたとおり、相同組換えを媒介するのに十分な長さのOCH1遺伝子の一部に、その5’及び3’末端の両方において作動可能に連結した選択可能なマーカー遺伝子を含み得る。メチロトローフ酵母細胞の、その5’及び3’末端の両方においてOCH1配列を選択可能なマーカー遺伝子と共に含み得るそのようなOCH1不活性化ベクターの直鎖状DNA断片による形質転換は、ゲノミックOCH1座への直鎖状断片の組み込み及びゲノミックOCH1遺伝子の破壊を導く。
【0078】
或いは、OCH1不活性化ベクターは、破壊されるOCH1遺伝子の一部及び上で記載された選択可能なマーカー遺伝子の何れか一つを含むことができるが、上記一部はOCH1蛋白質を何もコードしないか又はOCH1蛋白質の不活性断片をコードする。そのようなOCH1遺伝子の一部は如何なるOCH1プロモーター配列をも欠き、そして如何なるプロモーター配列に対しても作動可能に連結していない。そのようなベクターはOCH1遺伝子内の部位において直鎖状化し、そして次にメチロトローフ酵母株を形質転換することができ、1回の相同組換え媒介性の組み込みにより、ゲノミックOCH1遺伝子の不活性化をもたらす。好ましくは、不活性化変異、例えば、停止コドン又はフレームシフト変異も、直鎖状化部位の5’(上流)の部位及びOCH1の翻訳開始コドンの3’(下流)の部位においてベクター中のOCH1配列内に導入される。
【0079】
所望であれば、メチロトローフ酵母株中で発現される酵素をコードするヌクレオチド配列は、「ノックインアンドノックアウト」ベクターを創製するために、同じベクター中でゲノミックOCH1遺伝子を不活性化することができるヌクレオチド配列と組み合わせることができる。
【0080】
ノックインベクター及びノックアウトベクターの両方を含む本発明のベクターは、細菌中で機能する選択可能なマーカー遺伝子、並びに細菌中で複製及び細胞外染色体の維持管理に必須の配列をも含むことができる。細菌の選択可能なマーカー遺伝子の例は、アンピシリン耐性(Ampr)、テトラサイクリン耐性(Tetr)、ハイグロマイシン耐性、ブラスチシジン耐性及びゼオシン耐性(Zeor)遺伝子を含む。
【0081】
さらに、上記のベクターの何れも、さらに、メチロトローフ酵母株中でのそのような糖蛋白質の発現のための、目的の糖蛋白質をコードするヌクレオチド配列を含むことができる。
【0082】
さらに別の側面において、本発明は、哺乳類様N−グリカン構造を有する糖蛋白質を生じさせる方法を提供する。
本明細書にて使用される「糖蛋白質」は、一つ又はそれより多いアスパラギン残基上又は一つ又はそれより多いセリン又はスレオニン残基、又はアスパラギン及びセリン又はスレオニン残基の両方の上の何れかで、メチロトローフ酵母株中において、グリコシル化される蛋白質を意味する。好ましくは、糖蛋白質は宿主メチロトローフ酵母株に対して異種である。
【0083】
本発明によれば、減少したグリコシル化を伴う目的の糖蛋白質の生産は、多数の様式において達成できる。例えば、目的の糖蛋白質をコードするヌクレオチド配列を、哺乳類様N−グリカンを応じるように予め操作されたメチロトローフ酵母株に導入することができる。
【0084】
糖蛋白質をコードするヌクレオチド配列は、宿主株中で機能するプロモーター配列及び3’終結配列に対して作動可能に配置することができる。当該ヌクレオチド配列は、追加の配列、例えば、蛋白質生成物の分泌が望まれる場合に運搬(transit)ペプチドをコードするシグナル配列を含むことができる。そのようなシグナル配列は、よく知られており、容易に利用可能であって、サッカロミセスセレビシエのアルファメイティング因子プレプロ(αmf)、ピキアパストリス酸ホスファターゼ(PHO1)シグナル配列等を含む。
【0085】
或いは、目的の糖蛋白質のコーディング配列を導入されたメイティング酵母株を修飾することにより、所望の酵素(即ち、a−1,2−マンノシダーゼ、GnTI及びGalT)を発現させ、そして上記のとおりにゲノミックOCH1遺伝子を不活性化し、それにより哺乳類様N−グリカンを有する糖蛋白質を生産することができる。
【0086】
メイティング酵母株中で生産された糖蛋白質は慣用の方法により精製することができる。精製プロトコルは、精製される特定の蛋白質の性質により決定することができる。そのような決定は当業者の範囲内である。例えば、細胞培養培地を細胞から分離し、そして当該細胞から分泌される蛋白質を日常の単離技術、例えば沈殿、免疫吸着、フラクショネーション又は様々なクロマトグラフ法により培地から単離することができる。
【0087】
本発明の方法により生産することができる糖蛋白質は、細菌、真菌又はウイルスの蛋白質又は抗原、例えば、バチルスアミロリクエファシエンスのα−アミラーゼ、サッカロミセスセレビシエのインベルターゼ、トリパノゾーマクルジのトランス−シアリダーゼ、HIVエンベロープ蛋白質、インフルエンザウイルスのAヘマグルチニン、インフルエンザのノイラミニダーゼ、ウシヘルペスウイルスのタイプ1糖蛋白質D;蛋白質、哺乳類起源の蛋白質、例えばヒト蛋白質、成長因子又は受容体、例えば、ヒトアンギオスタチン、ヒトB7−1,B7−2及びB−7受容体CTLA−4,ヒト組織因子、成長因子(例えば、血小板由来の成長因子)、組織プラスミノーゲン活性化因子、プラスミノーゲン活性化因子阻害剤−I、ウロキナーゼ、ヒトリソソーム蛋白質、例えば、α−ガラクトシダーゼ、プラスミノーゲン、トロンビン、因子XIII;及びイムノグロブリン又はイムノグロブリンの断片(例えば、Fab,Fab’,F(ab’)2)を含む。本発明の遺伝子操作されたピキア株において発現され得る追加の有用な糖蛋白質に関しては、Bretthauer and Castellino,Biotechnol.Appl.Biochem.30:193−200(1999),及びKukuruzinska et al.,Ann Rev.Biochem.56:915−944(1987)を参照されたい。
【0088】
本発明の方法を用いて生産される糖蛋白質、即ち、哺乳類様のN−グリカン、特にGalGlcNAcMan5GlcNAc2N−グリカンを有する糖蛋白質も、本発明の一部である。
【0089】
また別の側面において、本発明は、上で記載された本発明のノックインベクター、ノックアウトベクター、又はノックインアンドノックアウトベクターの一つ又はそれより多くを含むキットを提供する。
【0090】
より特定すれば、本発明のキットは、α−マンノシダーゼI又はその機能部分をコードする、好ましくはER滞留シグナルを含む、ヌクレオチド配列を有するベクター;GnTI又はその機能部分をコードする、好ましくはゴルジ体滞留シグナルを含む、ヌクレオチド配列を有するベクター;GalT又はその機能部分をコードする、好ましくはゴルジ体滞留シグナルを含む、ヌクレオチド配列を有するベクター;又はメチロトローフ酵母中でゲノミックOCH1遺伝子を破壊することができるベクター、又はその何れかの組み合わせを含む。
【0091】
上記キットは、目的の異種糖蛋白質をコードする配列を有する核酸分子を含むこともできる。どのような核酸分子は、本明細書において前で記載されたノッキングイン又はノッキングアウトのための配列を含む別のベクター又は同じベクター内に提供することができる。或いは、キット中のノックイン又はノックアウトベクターは、目的の異種蛋白質をコードするヌクレオチド配列の挿入のための便利なクローニング部位を有する。
【0092】
上記キットは、本明細書において前で記載された、ノックイン、ノックアウト又はノックインアンドノックアウトベクターの何れかにより形質転換され得るメチロトローフ酵母株を含むこともできる。或いは、上記キットは、哺乳類様N−グリカンを生じさせるために操作されたメチロトローフ酵母株を含むことができる。
【0093】
本発明は、さらに、以下の実施例により例示される。
実施例1
方法と材料
ベクターの構築と形質転換
ピキアパストリス配列はGenBankにて受け入れ番号E12456(配列番号:2)にて見いだされ、日本特許出願番号07145005に記載され、引用により本明細書に編入される。この配列は、α−1,6−マンノシルトランスフェラーゼの全ての典型的な特徴を示し、そしてサッカロミセスセレビシエのOCH1にほとんど相同であり、即ち、本明細書においてはピキアパストリスOCH1遺伝子と呼ぶ。
【0094】
ピキアパストリスOCH1遺伝子の完全ORFを、PCRにより、鋳型として株GS115から単離されたゲノミックDNA、及び以下のオリゴヌクレオチド5’GGAATTCAGCATGGAGTATGGATCATTGAGTCCGTTGGAAAGG(配列番号:4)及び5’GCCGCTCGAGCTAGCTTTCTTTAGTCC(配列番号:5)を用いて単離した。単離されたOCH1遺伝子をpUC18内にクローン化することにより、プラスミドpUC18pOCH1を得て、OCH1遺伝子配列の同定を配列決定により確認した。
【0095】
プラスミドpGlycoSwitchM8(2875bp,配列番号:6、図3Aに模式的に描写する)はAla25−Ala155をコードするピキアパストリスOCH1のORFの断片を含み、当該断片はpPICZB(インビトロジェン、カールスバッド、CA)のBglII部位とHindIII部位の間に挿入された。2つの停止コドンをインフレームにてコドンAla25の前に配置することにより、末端削除された蛋白質の起こり得る合成を阻止した。消化後のフィルイン及び再ライゲーションにより、pPICZBのポリリンカーのBstBI部位を予め排除した。クローン化されたORF1遺伝子の内部に位置する唯一のBstBI部位を上記プラスミドの直鎖状化に用いることができる(不活性化戦略の全体像に関しては図3Aを参照)。
【0096】
pGlycoSwitchM5(5485bp,配列番号:9、図3Bに模式的に描写する)を以下のとおりに構築した。ピキアパストリスHIS4転写終結配列を含むpPIC9(インビトロジェン、カールスバッド、CA)のXbaI/ClaI断片を、pGlycoSwitch M8のHindIII部位及びEcoRI部位の間に挿入した。のちに、GAPプロモーター及びpreMFマンノシダーゼHDELカセットを含むpGAPZMFManHDEL(Callewaert et al.,FEBS Lett,503(2−3):173−178,2001)の2.3kbのBglII/NotI断片をHindIII部位とNotI部位の間に挿入した。この構築物のために使用される全ての制限部位(NotI部位を除く)をクレノーDNAポリメラーゼによりフィルインした。pGAPZMFmanHDEL内の唯一のBstBI部位を、消化後のフィルイン及び再ライゲーションにより排除した。
【0097】
ヒトGlcNAc−トランスフェラーゼ(GnTI)をゴルジ体に標的化するため、GnTIのN−末端部分を、酵母のゴルジ体における直鎖状化に必須のサッカロミセスセレビシエのKre2のN−末端部分により置き換えた(Lussier et al.,J Cell Biol,131(4):913−927,1995)。プラスミドYEp352Kre2(Howard Bussey博士、McGill大学、モントリオール、カナダ、により提供)を生成したが、Kre2遺伝子のSacI/PvuII断片を、SalI(クレノーにより平滑化)及びSalIにより消化されていたYep352ベクターに挿入することにより生成した。YE352Kre2をSacI/PvuIにより消化してT4−ポリメラーゼにより平滑化した。Kre2遺伝子の5’末端を単離して、クレノー平滑化されたSgrAI/XbaIにより開いたpUChGnTI(Maras et al.,Eur J Biochem 249(3):701−707,1997)にクローン化した。2つのDNA断片の融合場所を標準手法を用いて配列決定した。結果のKre2−GnTIオープンリーディングフレームは、Kre2遺伝子のN−末端部分(配列番号:11に示されるとおり、Kre2蛋白質の最初の110アミノ酸をコードする)及びGnTIの触媒ドメイン(配列番号:13に示されるとおり、GnTI蛋白質の最後の327アミノ酸)を含み、EcoRV/HindIIIにより二重消化して、SalI/EcoRIで開いたpPIC6Aベクター(インビトロジェン)内へ、両断片をクレノーポリメラーゼにより平滑化した後に、連結した。結果のプラスミドはpPIC6AKrecoGnTIと命名した(配列番号:14、図3Cにおいて模式的に描写される)。それは、メタノールにより誘導可能なAOX1プロモーターの制御下にKre2GnTIのオープンリーディングフレームを、そして抗生物質ブラスチシジンに対して耐性のアスペルギルステレウスカラノBSD遺伝子を含む。
【0098】
GalTの局在化は、GnTIを標的化するために行われたのと同じ様式にて、Kre2pのN末端部分へのGalTの触媒ドメインの融合により達成された。β−1,4−ガラクトシルトランスフェラーゼをhepg2のcDNAライブラリーからオリゴヌクレオチド5’TTCGAAGCTTCGCTAGCTCGGTGTCCCGATGTC(配列番号:15)及び5’GAATTCGAAGGGAAGATGAGGCTTCGGGAGCC(配列番号:16)をスターター配列として用いて増幅した。増幅された断片をHindIII/EcoRIにてpUC18にクローン化された。GalT蛋白質のN末端の77アミノ酸を削除するため、以下のオリゴヌクレオチドをプライマーとして用いてPCRを実施した:5’TTCGAAGCTTCGCTAGCTCGGTGTCCCGATGTC(配列番号:15)及び5’CGTTCGCGACCGGAGGGGCCCGGCCGCC(配列番号:17)。増幅された断片をNruI/HindIIIに切断して、HindIII/SgrA Iクレノー平滑化pUCKreGnTIベクターに連結した。結果のKre2−GalT融合構築物を再びPCRによりプライマー:5’TCGATATCAAGCTTAGCTCGGTGTCCCGATGTC(配列番号:18)及び5’GAATTCGAACTTAAGATGGCCCTCTTTCTCAGTAAG(配列番号:19)を用いて増幅した。増幅された断片をEcoRV/BstB IにてpBLURA IX(Cereghino et al.,Gene,263:159−169,2001)(James Cregg(オレゴンインスティチュートオブサイエンスアンドテクノロジー、ビーバートン、USA)より提供)に挿入した。最後に、URA3遺伝子をカナマイシン耐性カセットにより置き換えたが、ベクターpFA6a−KanMX4からのSpeI/SmaI断片をSpeI/SspIにおいて開いたプラスミドに連結することによった。最後のプラスミドはpBIKanMX4KrehGalT(配列番号:7、図3Dに模式的に描写される)と呼ばれるが、AOX1プロモーターに作動可能に連結したKre2−GalT融合蛋白質をコードする配列を含んだ。当該融合蛋白質は、Kre2の最初の100アミノ酸及びGalTの最後の320アミノ酸から構成された。
【0099】
これらのプラスミドによる、様々な蛋白質を発現するGS115ピキア株に対する形質転換は、前に記載されたとおりに行った(Cregg et al.,Methods in Molecular Biology,103:27−39,1998)。PpOCH1座における正確なゲノミックインテグレーションは、ゲノミックDNA上のPCRにより確認した。
蛋白質の調製
分泌されたトリコデルマリセイのα−1,2−マンノシダーゼを、HIC、アニオン交換及びゲル濾過クロマトグラフの組み合わせを用いて、記載されたとおりに精製した(Maras et al.,J Biotechnol,77(2−3):255−263,2000;Van Petegem et al.,J Mol Biol 312(1):157−165,2001)。全てのSDS−PAGE実験は10%PAAゲル上において標準泳動条件下で行った。酵母の細胞壁のマンノ蛋白質をBallou(Methods Enzymol,185:440−470,1990)に記載されたとおりに放出させたが、0.9%のNaCl水溶液中の酵母細胞の大規模な洗浄、20mM クエン酸Na中の酵母細胞の延期されたオートクレーブ(90分)、続くメタノール沈殿(4容)を含んだ。
N−グリカン分析
ABI 377 DNA配列決定機(DSA−FACE)上でのレーザー誘導性DNA配列決定機補助によるフルオロフォア補助糖質電気泳動により、記載されたとおりに(Callewaert et al.,Glycobiology,11(4):275−281,2001)、Nグリカン分析を行った。簡単に言えば、糖蛋白質を、マルチスクリーンイムノビロン−P上に固定化し、そしてPNGase処理により脱グリコシル化した。N−グリカンを除去し、そしてAPTSにより誘導した。過剰の標識をセファデックスG10樹脂上のサイズ分画により除去した。APTS−標識されたオリゴサッカライドの蒸発後に、ROX−標識されたGENESCAN 500標準混合物(アプライドバイオシステムズ)を加えて、内部標準化を許容した。この混合物をABI 377A DNA配列決定機(アプライドバイオシステムズ)上で、12%ポリアクリルアミドゲルにより、89mM Tris,89mM 硼酸塩、2.2mM EDTAバッファー中で泳動した。各ゲル上で、ウシRNase BとマルトデキストロースラダーのN−グリカンをレファレンスとして泳動した。データ解析は、GENESCAN 3.1ソフトウエア(アプライドバイオシステムズ)を用いて実施した。β−N−アセチルヘキソサミニダーゼ(Glyko)及びβ−ガラクトシダーゼ(Prozyme)によるエキソグリコシダーゼ処理を、標識されたグリカン上で、一晩、37℃において20mM酢酸ナトリウムpH5.5中で実施した。慣用のFACE(N−グリカンのANTS標識と30%PAAミニゲル上の電気泳動)をJackson(Biochem J,270(3):705−713,1990)により記載されたとおりに実施した。DSA−FACE法は非常に高い解析度と感度を有するが、慣用のFACEは高分子量のN−グリカン(「高グリコシル化」)の複合混合物の検出によく適合されており、分解されていなかった(not resolved)ので、慣用のFACE内のゲル上では特徴的な「スメア」を形成した。即ち、DSA−FACEと慣用のFACE分析の組み合わせは、酵母により生じる糖蛋白質の特徴の、より完全な写真を与えた。
成長曲線の測定
新鮮な一晩酵母培養物を新鮮なYPD培地によりOD600 0.02まで希釈し、そして250rpmにて30℃において一晩生育させた(12時間、OD 600<3.0)。実験を開始するために、50mLのポリプロピレンチューブ中の10mLの新鮮なYPDに一晩酵母培養物を接種し、0.5のOD600値から始めた。アリコートを2時間ごとに採取し、そしてOD600値を測定した。全ての酵母株を同じ時間平行してランさせた。
【0100】
実施例2
OCH1の不活性化
ゲノミックピキアパストリスOCH1遺伝子の破壊は、以下のとおりにして1回の相同組換えにより達成した。プラスミド、pGlycoSwitchM8(図3A)を実施例1において記載されたとおりに生成したが、ピキアパストリスOCH1遺伝子の塩基対番号73−467を含み、その前に当該ベクター中の可能な初期翻訳開始部位からのリードスルーを回避するために2つのインフレームのナンセンスコドンを置いた。この断片は、形質転換前の上記ベクターの直鎖状化に有用な中心に位置するBstB I部位を含んだ。このベクターは、ピキアのゲノミックOCH1座への1回の相同組換えによるインテグレーションに際してベクター中に存在するOCH1遺伝子を複製するはずである。結果として、ピキア染色体内のOCH1遺伝子は2つのOch1配列に置き換わった。第1のOCH1配列は、最大で161のアミノ酸長の蛋白質生成物をコードし(そのうちの6アミノ酸はベクター中の配列からであった)、完全長OCH1遺伝子によりコードされたタイプIIの膜貫通蛋白質の触媒ドメインを含まなかった。第2のOCH1配列は、完全長の蛋白質の最初の25アミノ酸のコーディング配列を欠き、そして可能な上流の翻訳開始部位からの如何なるリードスルーをも阻止するはずの2つのインフレームの停止コドンを含んだ。
【0101】
株GS115をプラスミドpGlycoSwitchM8により形質転換した。形質転換体をGlycoSwitchM8と呼ぶか、又は短くM8株又はoch1株と呼んだ。図3Aにおいて特定されたプライマーの組み合わせによるゲノミックDNAのPCRは、3つの個別の実験により示されたところによれば、ゼオシン耐性形質転換体の約50%における、予測されたゲノミック座へのこの構築物の正確なインテグレーションを示した。
【0102】
細胞壁のマンノ蛋白質のN−グリカンの分析は、図4において演繹されるとおり、グリコシル化パターンの変化を明らかにした。優位なピークは、野生型GS115株からの細胞壁マンノ蛋白質に関してはMan9GlcNAc2であり、主要なピークはGlycoSwitchM8株に関してはMan8GlcNAc2である(図4のパネル2から3を比較)。N−グリカンにおけるこの変化は、M8株の完全長OCH1のORFによる形質転換の後に復帰した。
【0103】
M8株からの分泌された糖蛋白質の不均質性が低下したか否かを確認するために、野生型GS115株中の分泌された蛋白質である典型的には高グリコシル化された(Maras et al.,J Bacteriol,77(2−3):255−263,2000)トリコデルマリセイのα−1,2−マンノシダーゼをoch1のM8株を用いて分析した。トリコデルマリセイのα−1,2−マンノシダーゼをコードする配列により形質転換されたM8株の細胞の培養上清を、SDS−PAGEにより分離した(図5A)。ゲルは、高グリコシル化された蛋白質の特徴であるスメアがGlycoSwitchM8株において生成した蛋白質には不在であったことを明らかにした。平行して、分泌された糖蛋白質をPNGaseF処理により脱グリコシル化し、そしてグリカンをミニゲル上のFACE分析により分析した。典型的にはFACE分析においては、大きな高グリコシル構造が解析されず、一つのスメアバンドとして出現する(図5B)。スメアバンドはoch1株からの糖蛋白質においては不在であり、och1株からのN−グリカンの不均質性が低下したことが確認された。
【0104】
実施例3
ER滞留したマンノシダーゼ−HDELの発現
ピキアパストリスのN−グリカンをさらにヒト化するために、ER滞留されたトリコデルマリセイのα−1,2−マンノシダーゼ−HDELをoch1株中で発現させた。ピキアパストリス発現株の容易な変換のために、トリコデルマリセイのα−1,2−マンノシダーゼ−HDELをコードするヌクレオチド配列をoch1不活性化ベクターに挿入した。結果のコンビネーションベクターをpGlycoSwitchM5と呼び、その構成を実施例1に記載する。
【0105】
株GS115を直鎖状化されたpGlycoSwitchM5により形質転換した。当該ベクターの正確なインテグレーションはPCR分析により確認した。当該形質転換体からのマンノ蛋白質のN−グリカンをDSA−FACE法により分析した。グリカンのプロフィールは、均質なMan5GlcNAc2ピークを明らかにした(図4、パネル4)。Man5GlcNAc2ピークと、5から25までのグルコースユニットのサイズエリアのこの方法の検出限界を超える全ての小さなピークの積分(integration)は、この高等真核種の高マンノースグリカンがこの混合物中に存在する全N−グリカンプールの少なくとも90%を成したことを明らかにした。
【0106】
別のアプローチにおいては、マンノシダーゼ−HDELをメタノール誘導可能AOX1プロモーターの制御下で発現させた。2つのマンノシダーゼ発現株(即ち、構成的及び誘導可能)の間のN−グリカンプロフィールに明確な差異は検出できなかった。
【0107】
異種蛋白質のN−グリカン修飾を確認するため、Laroy et al(Protein Expr Purif,20(3):389−393,2000)に記載されたとおりに、pGlycoSwitchM5プラスミドで、トリパノゾーマクルジのトランス−シアリダーゼ発現ピキア株を形質転換した。この点に関して(here too)、Man5GlcNAc2が精製された蛋白質上に検出され、精製された蛋白質上の全N−グリカンの95%より多くを占める。
【0108】
振盪フラスコ培養におけるpGlycoSwitchM5形質転換株の成長曲線分析は、その倍加時間が野生型株のそれを密接に模倣したことを示した。しかしながら、操作された株は野生型株より約20%低い光学密度において定常期に到達したことから、高細胞密度のストレス条件に対していくらか感受性であり得たことを示す。にもかかわらず、そのストレス感受性の表現型はサッカロミセスセレビシエのoch1株よりもはるかに劣って表現された(much less pronounced)。
【0109】
実施例4
ゴルジ体局在N−アセチルグルコサミニルトランスフェラーゼ
(Kre2GnTI)の発現
GnTIをゴルジ体に標的化するために、サイトゾル部分、膜貫通領域及び管腔幹領域(luminal stem region)を含むGnTIのN−末端部分をコードするヌクレオチド配列を、サッカロミセスセレビシエのKre2シグナル配列をコードするヌクレオチド配列により置き換えた。これは、Kre2pからの最初の327アミノ酸及びGnTIの最後の327アミノ酸を有するキメラ蛋白質をコードするヌクレオチド配列をもたらした。
【0110】
ピキアパストリスにおける発現のために、Kre2−GnTIキメラ配列を、ブラスチシジン耐性マーカーを有するプラスミド中で強力なメタノール誘導可能性AOX1プロモーターの制御下に置いた。結果の構築物であるpPIC6KrecoGnTI(実施例1に記載されるとおり)をNsiI消化によるAOX1座内の直鎖状化の後にGS115 M5株を形質転換した。当該形質転換体内の上記構築物の存在は、AOX1の3’及び5’プライマーを用いたゲノミックDNA上でのPCRにより確認した。
【0111】
幾つかの形質転換体のマンノ蛋白質のN−グリカンはDSA−FACE法により分析した。優位なピークは、Man5GlcNAc2ピークより大きな約1つのグルコースユニットであった(図4、パネル5)。このピークが末端GlcNAcを有するか否かを決定するため、エキソグリコシダーゼ消化を、β−Glc−NAc結合を加水分解する酵素であるβ−N−アセチルヘキソサミニダーゼを用いて実施した。この酵素による消化に際して、上記ピークはMan5GlcNAc2にシフトして戻った(図4、パネル6)。これは、オリジナルピークがGlcNAcMan5GlcNAc2を表すことを示し、即ち、キメラGnTI酵素の正確なインビボ活性を証明する。
【0112】
Kre2GnTIキメラの過剰発現は、Man5GlcNAc2のGlcNAcMan5GlcNAc2へのほとんど完全な変換を導く。これは、十分なUDP−GlcNAcドナー基質がゴルジ体においてN−アセチルグルコサミニレート、ほとんど全てのグリカンに存在したことを示唆する。
【0113】
実施例5
ゴルジ体滞留β−1,4−ガラクトシルトランスフェラーゼの発現
ヒトβ−1,4−ガラクトシルトランスフェラーゼ1の、当該酵素の膜貫通ドメイン及びサイトゾル部分を含むN末端部分(最初の77アミノ酸)をコードするヌクレオチド配列を、サッカロミセスセレビシエのKre2シグナル配列をコードするヌクレオチド配列で置き換えた。このキメラ融合配列をAOX1プロモーター及びAOX1の3’末端をターミネーターとする制御下に置いた。最終的なプラスミドであるpB1KanMX4KrehGalT(実施例1に記載される)をPmeIによりM5−GnTI株の形質転換前に直鎖状化した。
【0114】
N−グリカン分析を幾つかの形質転換体からのマンノ蛋白質にて実施した。GlcNAcMan5GlcNAc2ピークより大きな約1ユニットのグルコースのピークが形質転換体において検出されたが、当該ピークは非形質転換株においては不在であった(図3、パネル7)。このピークが末端β−ガラクトースを含むグリカンであるか否かを決定するため、N−グリカンをβ−ガラクトシダーゼにより消化した。グリカンプロフィールの消化後に、このピークはGlcNAcMan5GlcNAc2位置にシフトして戻った(図4、パネル8、パネル7と比較)。β−ガラクトシダーゼ消化の前と後にGlcNAcMan5GlcNAc2ピークを積分することにより、GalGlcNAcMan5GlcNAc2の量を決定した。これらの2つのピークの差し引きは、約10%のGlcNAcMan5GlcNAc2がGalGlcNAcMan5GlcNAc2に変換されたことを明らかにした。0.2%ガラクトースを含む追加の培地はGal−含有オリゴサッカライドの量を増加させなかった。
【図面の簡単な説明】
【0115】
【図1】図1は、M8GlcNAc2,M5GlcNAc2,GlcNAcM5GlcNc2,及びGal GlcNAcM5GlcNAc2を描写する。
【図2】図2は、酵母とヒトのN−結合グリコシル化及びピキアパストリスのグリコシル化のヒト化のための戦略を模式的に描写する。糖操作工程は、α−1,6−マンノシルトランスフェラーゼOCH1の不活性化、HDEL標識されたα−1,2−マンノシダーゼの過剰発現及びゴルジ局在化GnTI及びGalTの過剰発現を含む。最後に部分的に得られたハイブリッド構造を組み立てる(framed)。
【図3】図3Aは、一回の相同組換えによりゲノミックOCH1遺伝子を不活性化するための戦略を模式的に描写する。図3Bは、ピキアパストリスのグリカン操作のために使用されたプラスミドpGlycoSwitchM5を模式的に描写する。pGlycoSwitchM5をBstBIにより直鎖状化し、次に形質転換し、そしてピキアパストリスのゲノムに正確に組み込むことにより、OCH1遺伝子が不活性化された。図3Cは、pPIC6AKrecoGnTIを模式的に描写する。 図3Dは、pB1KanMX4KrehGalTを模式的に描写する。
【図4】図4は、異なったグリカン操作をされたピキアパストリス株からのN−グリカンのDSA−FACE分析を模式的に描写する。パネル1:オリゴマルトースレファレンス。パネル2−9はN−グリカンを表し:2:−野生型株GS115からのN−グリカンを表し、主要ピークを表すMan9GlcNAc2を伴う;3:och1不活性化された株からのN−グリカンを表し、主要ピークを表すMan8GlcNAc2を伴う;4:och1不活性化されたManHDEL発現株からのN−グリカンを表し、主要ピークを表すMan5GlcNAc2を伴う;5:och1不活性化されたManHDEL,KreGnTI発現株からのN−グリカンを表し、主要ピークを表すGlcNAcMan5GlcNAc2を伴う;6:5と同じであるが、但し、グリカンがβ−N−アセチルヘキソサミニダーゼにより処理され、そしてGlcNAcMan5GlcNAc2ピークがMan5GlcNAc2位置にシフトし、ターミナルGlcNAcが存在したことを示すことを除く;7:och1不活性化されたManHDEL,KreGnTI,KreGalT発現株からのN−グリカンを表し、GalGlcNAcMan5GlcNAc2を表す追加のピークを伴うが、β−N−アセチルヘキソサミニダーゼによる処理により消失した;9:ウシRNaseBからのレファレンスグリカン(Man5−9GlcNAc2)。
【図5】図5Aは、ピキアパストリスOCH1の不活性化後のグリコシル化を示す。5A:トリコデルマリセイのマンノシダーゼを分泌するピキアパストリス株のCBB染色されたSDS−PAGEゲル。操作されていない株(WT)においては、明確なスメアが見えるが、このスメアはoch1不活性化された株(och1(M8))においては消失した。図5Bは、ピキアパストリスOCH1の不活性化後のグリコシル化を示す。5B:操作されていない株(WT)及びoch1株により分泌されたマンノシダーゼに由来するN−グリカンのFACE分析。高い電気泳動移動度のバンドはMan8及びMan9により示され、「コア」N−グリカン構造を表す。
【発明の開示】
【0001】
発明の分野
本発明は、哺乳類様グリコシル化により糖蛋白質を生産するための方法及び操作されたメチロトローフ酵母株に関する。本発明は、哺乳類様グリコシル化により糖蛋白質を生産することができるメチロトローフ酵母株を生成するのに有用なベクターにも関する。遺伝子工学によるメチロトローフ酵母株から生産された糖蛋白質も提供される。
発明の背景
ピキアパストリスを含むメチロトローフ酵母は、商業上又は医学上重要な組換え蛋白質の生産のために広く使用されてきた。しかしながら、幾つかの治療用糖蛋白質の生産及び医学上の応用は、これらの酵母と標的生物、例えば哺乳類又はヒト被験者の間の蛋白質結合糖質の生合成の違いにより妨げられ得る。
【0002】
蛋白質のN−グリコシル化は細胞質網状構造(ER)を起源とし、そこでは、ドリコール(脂質キャリアー中間体)上で集合したN結合オリゴサッカライド(Glc3Man9GlcNAc2)が隣の蛋白質の適当なAsnに転移される。これは、全ての真核生物のN−結合糖蛋白質に共通の事象である。3つのグルコース残基と一つの特定のα−1,2−結合マンノース残基がER中の特異的グルコシダーゼと一つの1,2−マンノシダーゼにより除去されて、コアのオリゴサッカライド構造Man8GlcNAc2をもたらす。このコアの糖構造を有する蛋白質はゴルジ体に運ばれて、そこで、糖部分が様々な修飾を受ける。酵母と高度真核生物の間ではゴルジ体内の糖鎖の修飾に顕著な違いがある。
【0003】
哺乳類細胞においては、糖鎖の修飾は3つの異なる経路により進行し、それが添加される蛋白質部分に依存する。即ち、(1)コアの糖鎖は変化しない;(2)UDP−N−アセチルグルコサミン(UDP−GlcNAc)からのN−アセチルグルコサミン−1−ホスフェート部分(GlcNAc−1−P)のコア糖鎖中のマンノースの6位への添加によりコアの糖鎖が変化し、続いてGlcNAc部分の除去による糖蛋白質中の酸性糖鎖の形成する;又は(3)マンノシダーゼIによる3マンノース残基の除去の結果としてコア糖鎖が最初にMan5GlcNAc2に変換されて;そしてGlcNAcの添加及び2つのさらなるマンノース残基の除去、続くGlcNAc、ガラクトース(Gal)、及びN−アセチルノイラミニ酸(シアル酸(NeuNAc)とも呼ばれる)の連続添加により様々なハイブリッド又は複合体糖鎖を形成することにより、Man5GlcNAc2がさらに修飾される(R.Kornfeld and S.Kornfeld,Ann.Rev.Biochem.54:631−664,1985;Chiba et al J.Biol.Chem.273:26298−26304,1998)。
【0004】
酵母においては、Man8GlcNAc2グリカンは削られない(not trimmed)。ゴルジ体内の糖鎖の修飾は、別々のマンノシルトランスフェラーゼによるマンノース残基の一連の添加(「外部鎖」グリコシル化)を含む。外部鎖グリコシル化の構造は生物に特異的であって、典型的にはサッカロミセスセレビシエにおいて50を超えるマンノース残基を伴い、そしてもっとも共通にはピキアパストリスにおいてMan14GlcNAc2より小さな構造を伴う。高いマンノース種のこの酵母特異的外部鎖グリコシル化は、高グリコシル化又は高マンノース化とも呼ばれる。
【0005】
グリコシル化は、蛋白質のフォールディング、安定性及び生物活性を修正するのに非常に重要である。ヒト体内において、グリコシル化は、蛋白質の薬力学特性、例えば組織分配及び血流からの浄化に、一部必須である。さらに、グリカン構造は、抗原応答に関与し得る。例えば、α−ガラクトースの糖蛋白質上の存在はブタからの異種移植に対する免疫反応に関する主要な理由である(Chen et al.,Curr Opin Chem Biol,3(6):650−658,1999)が、酵母に対する糖蛋白質に対する免疫反応は、主に、α−1,3−マンノース、β結合マンノース及び/又はホスホモノ−又はホスホジエステル結合の何れかの上のリン酸残基の存在による(Ballou,C.E.,Methods Enzymol,185:440−470,1990;Yip et l.,Proc Natl Acad Sci USA,91(7):2723−2727,1994)。
【0006】
高グリコシル化はしばしば望まれないが、何故ならば、糖質組成及び分子量の両方において組換え蛋白質生成物の不均質性を導き、蛋白質の精製を複雑にし得るからである。高グリコシル化された酵素の比活性(ユニット/質量)は、糖質の増加した部分により低下し得る。さらに、外部の鎖のグリコシル化はしばしば免疫原性が強く、治療への応用においては望まれないかもしれない。さらに、大きな外部の鎖状の糖(chain sugar)は治療用蛋白質の免疫原決定基を遮断し得る。例えば、ピキアパストリスにおいて発現されたインフルエンザノイラミニダーゼ(NA)は30−40までのマンノース残基を含むN−グリカンによりグリコシル化される。高グリコシル化されたNAは、NA分子の上部の可変性の免疫支配性表面ループがN−グリカンにより遮断されるために、マウスにおいて免疫原性が低下する(Martinet et al.Eur J.Biochem.247:332−338,1997)。
【0007】
よって、哺乳類(例えば、ヒト)の糖質構造に似せた糖質構造を有する組換え糖蛋白質を生じるメチロトローフ酵母株を遺伝子操作することが望まれる。
発明の概要
本発明は、哺乳類様のN−グリカンを伴う糖蛋白質を生じさせるための、遺伝子操作されたメチロトローフ酵母株及び方法に向けられる。本発明は、哺乳類様のN−グリカンを伴う糖蛋白質を生じさせることができる遺伝子操作されたメチロトローフ酵母株を生成するのに有用なベクター及びキットにも向けられる。
【0008】
本明細書の中で使用されている用語「メチロトローフ酵母」は、限定ではないが、メタノール上で生育可能な酵母株、例えば、属カンジダ、ハンセヌラ、トルロプシス、及びピキアを含む。
【0009】
一つの態様において、本発明は、コアのマンノース構造及び末端ガラクトース残基に結合した5又はそれより少ないマンノース残基及び少なくとも一つのN−アセチルグルコサミン残基(GlcNAc)を有することにより特徴付けされる、哺乳類様のN−グリカン構造を有する糖蛋白質を生じる、遺伝子操作されたメチロトローフ酵母を提供する。
【0010】
好ましい態様において、本発明は、哺乳類様のN−グリカン構造、GalGlcNAcMan5GlcNAc2を有する糖蛋白質を生じる遺伝子操作されたメチロトローフ酵母株を提供する。
【0011】
本発明によれば、GalGlcNAcMan5GlcNAc2を有する糖蛋白質を生じる上記メチロトローフ酵母株は、α−1,2−マンノシダーゼ又はその機能部分、N−アセチルグルコサミニルトランスフェラーゼI(又はGnTI)又はその機能部分、及びβ−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分を発現するように遺伝子操作される。好ましくは、当該メチロトローフ酵母株は、ゲノミックOCH1遺伝子が不活性化されるようにも遺伝子操作される。
【0012】
遺伝子操作されたメチロトローフ酵母株内での発現のためのα−1,2−マンノシダーゼ又はその機能部分は、あらゆる種の起源であることができ、マウス、ウサギ又はヒトのような哺乳類の種、及びアスペルギルス、又はトリコデルマリセイのような真菌を含む。本発明における使用のための好ましいα−1,2−マンノシダーゼは、トリコデルマリセイのα−1,2−マンノシダーゼである。好ましくは、α−1,2−マンノシダーゼ又はその機能部分は、その基質であるMan8GlcNAcが利用可能な分泌経路中の部位を標的化する。より好ましくは、α−1,2−マンノシダーゼ又はその機能部分は、ER滞留(retention)シグナルを含むように遺伝子操作されて、ERを標的化する。好ましいER滞留シグナルは、ペプチドHDEL(配列番号:1)である。
【0013】
遺伝子操作されたメチロトローフ酵母株における発現のためのGnTI又はその機能部分は、あらゆる種の起源であることができ、ウサギ、ラット、ヒト、植物、昆虫、線虫及び原生動物、例えば、リーシュマニアタレントレを含む。本発明における使用のための好ましいGnTIは、ヒトGnTIであり、配列番号:13に示される。好ましくは、GnTI又はその機能部分は、その基質であるMan5GlcNAc2が利用可能な分泌経路中の部位を標的化する。より好ましくは、GnTIまたはその機能部分は、ゴルジ滞留シグナルを含むように遺伝子操作されて、ゴルジ体を標的化する。好ましいゴルジ滞留シグナルは、配列番号:11に示されるペプチドであり、サッカロミセスセレビシエのKre2蛋白質の最初の100アミノ酸からなる。
【0014】
遺伝子操作されたメチロトローフ酵母株における発現のためのGalT又はその機能部分は、あらゆる種の起源であることができ、ヒト、植物(例えば、アラビドプシスサリアナ)、昆虫(例えば、ドロソフィラメラノガスター)を含む。本発明における使用のための好ましいGalTは、ヒトGalTであり、配列番号:21に示される。好ましくは、GalT又はその機能部分は、ゴルジ滞留シグナルを含むように遺伝子操作されて、ゴルジ体を標的化する。好ましいゴルジ滞留シグナルは、配列番号:11に示されるペプチドであり、サッカロミセスセレビシエのKre2蛋白質の最初の100アミノ酸からなる。
【0015】
メチロトローフ酵母株は、例えば、形質転換によりこれらの酵素をコードする株ヌクレオチド配列へ導入することにより上記の所望の酵素を発現するように遺伝子操作することができる。好ましくは、コーディング配列はベクター内に提供され、各配列は、酵母株中で機能するプロモーター配列及び3’ターミネーション配列に作動可能に連結されるように配置される。ベクター又はその直鎖状断片で、次に、株を形質転換する。
【0016】
本発明の好ましい態様によれば、上記メチロトローフ酵母株は、OCH1遺伝子が破壊されるようにも遺伝子操作される。遺伝子の破壊は、ゲノミックOCH1配列とノックアウトベクター中のOCH1配列の間の相同組換えにより達成することができる。
【0017】
さらなる側面において、本発明は、哺乳類様のN−グリカン構造を有する糖蛋白質生じるメチロトローフ酵母株を生成するのに有用なベクターを提供する。
一つの態様において、本発明は、発現される酵素、即ち、α−1,2−マンノシダーゼ、GnTI,GalT,又はこれらの蛋白質の何れかの機能部分をコードするヌクレオチド配列を含む「ノックイン」ベクターを提供する。コーディング配列は、コードされた蛋白質の発現のためにメチロトローフ酵母中で機能するプロモーター配列及び3’ターミネーション配列に作動可能に連結されるように配置される。2つ又はそれより多いコーディング配列をメチロトローフ酵母株へ同時形質転換するために同じベクター中に入れることができる。好ましくは、当該ベクターは、形質転換体の便利な選択のための選択可能なマーカー遺伝子も含む。
【0018】
別の態様において、本発明は、メチロトローフ酵母株に導入されたときにゲノミックOCH1遺伝子を不活性化するか又は破壊する不活性化ベクター(又は「ノックアウト」ベクター)を提供する。
【0019】
OCH1ノックアウトベクターは選択可能なマーカーを含むことができ、ゲノミックOCH1遺伝子との二重相同組換えを媒介するのに十分な長さのOCH1配列に対して両5’及び3’末端にて作動可能に連結する。或いは、OCH1不活性化ベクターは、破壊されるOCH1遺伝子の一部を含むことができ、その部分はOCH1蛋白質を一切コードしないか又はOCH1蛋白質の不活性断片をコードし、そして選択可能なマーカー遺伝子を含むことができる。OCH1部分は、何れかの公知のプロモーター配列に対して作動可能な連結はされず、そして上記ベクターの直鎖状断片の形質転換に際して、1回の相同組換えによりゲノミックOCH1座に組み込まれる。好ましくは、一つ又はそれより多い不活性化変異、例えば停止コドン又はフレームシフト変異も、ベクター中のOCH1配列内に導入されることにより、何れかの可能性のある活性OCH1ポリペプチドの生産を阻止する。
【0020】
さらに別の側面において、本発明は、哺乳類様のN−グリカン構造を有する糖蛋白質の生成方法を提供する。目的の糖蛋白質をコードするヌクレオチド配列を、哺乳類様N−グリカンを生じさせるように遺伝子操作されたメチロトローフ酵母株に導入することができる。或いは、目的の糖蛋白質を発現するメチロトローフ酵母株を修飾することにより、
所望の酵素(例えば、α−1,2−マンノシダーゼ、GnTI及びGalT)を発現させ、そしてゲノミックOCH1遺伝子を不活性化することができ、それにより、哺乳類様N−グリカンを伴う糖蛋白質を生成する。
【0021】
また別の側面において、本発明の方法を用いることにより生成された糖蛋白質、即ち、哺乳類様のN−グリカン、特にGalGlcNAcMan5GlcNAc2N−グリカンを有する糖蛋白質が、本発明により提供される。
【0022】
さらなる側面において、本発明は、本発明の一つ又はそれより多いベクター、又は本発明の遺伝子操作された一つ又はそれより多い株を含むキットを提供する。
発明の詳細な説明
本発明は、哺乳類様又はヒト様N−グリコシル化を伴う組換え糖蛋白質を作成するための、方法、ベクター及び遺伝子操作されたメチロトローフ酵母株に向けられる。
【0023】
「哺乳類」は、哺乳類のあらゆる種、例えば、ヒト、マウス、ラット、イヌ、ウサギ、ウシ、ヒツジ、ウマ等を含むことを意味する。
典型的には複雑な種類の哺乳類のグリカン、例えば、ヒトにおいて生じたグリカンは、Man3GlcNAc2の内部コア構造に結合したシアリル−N−アセチル−ラクトサミン配列を伴う2から6の外部ブランチを有する。哺乳類のN−グリカンは、ER中で形成されるコアオリゴサッカライド構造であるMan8GlcNAc2に由来する。このコアの糖構造を有する蛋白質はゴルジ体に輸送されて、そこで、ゴルジ体マンノシダーゼI(ゴルジ体α−1,2−マンノシダーゼ)により3つのマンノース残基を除去した結果として、Man8GlcNAc2がMan5GlcNAc2に変換される。蛋白質がゴルジ体を通して加工されるにつれ、Man5GlcNAc2は、さらに、GlcNAcの追加と2つのさらなるマンノース残基の除去、次に、GlcNAc,ガラクトース(Gal)及びシアル酸(SA)残基の追加により、修飾される。
【0024】
本明細書にて使用される用語「哺乳類様N−グリコシル化」は、遺伝子操作されたメチロトローフ酵母株内で生成された糖蛋白質のN−グリカンが、5つ又はそれ未満のマンノース残基を含み、そして哺乳類細胞、例えばヒト細胞において生じた蛋白質のN−グリカンの生合成におけるN−グリカン又は中間体糖質構造の特徴を有することを意味する。
【0025】
好ましい態様において、本発明の遺伝子操作されたメチロトローフ酵母株において生成された糖蛋白質は、5つ又はそれ未満のマンノース残基、及びマンノース残基を含むコア構造及びターミナルガラクトース残基に結合した少なくとも一つのN−アセチルグルコサミン残基(GlcNAc)を含む。例えば、遺伝子操作されたメチロトローフ酵母株内で生成された糖蛋白質は、GalGlcNAcMan5GlcNAc2を有し、図1に描写されるとおりである。この糖質(GalGlcNAcMan5GlcNAc2)のIUPACの命名は、Gal(β−1,4)GlcNac(β−1,2)Man(α−1,3){Man(α−1,3)[Man(α−1,6)]Man(α−1,6)}Man(β−1,4)GlcNac(β−1,4)GlcNacである。その拡大された命名は、β−D−Galp−(1→4)−β−D−GlcpNAc−(1→2)−α−D−Manp−(1→3)−{α−D−Manp−(1→3)−[α−D−Manp−(1→6)]−α−D−Manp−(1→6)}−β−D−Manp−(1→4)−β−D−GlcpNAc−(1→4)−D−GlcpNAcである。
【0026】
ピキア、特にピキアパストリスを含むメチロトローフ酵母の細胞質網状構造(ER)を通り過ぎる糖蛋白質上のN−グリカンの大多数は、Man8GlcNAc2オリゴサッカライド構造を有する。糖蛋白質がERからゴルジ体に輸送された後は、追加のマンノース残基が別のマンノシルトランスフェラーゼによりこのコア糖部分に追加されて、高マンノースコア又は延長されて分枝したマンナン外部鎖からなるオリゴサッカライド構造を有する糖蛋白質をもたらす。
【0027】
本発明によれば、哺乳類様グリコシル化を伴う組換え糖蛋白質を生成するためには、メチロトローフ酵母を修飾することにより、一連の工程において糖質構造Man8GlcNAc2を哺乳類様N−グリカンに変換する酵素を発現させる。好ましくは、メチロトローフ酵母は、高マンノース構造の生産に関与する一つ又はそれより多い酵素、例えばOCH1遺伝子によりコードされるα−1,6−マンノシルトランスフェラーゼの発現を不活性化するようにも修飾される。
【0028】
本明細書において使用される用語「メチロトローフ酵母」は、限定ではないが、メタノール上で生育できる酵母株、例えば、属カンジダ、ハンセヌラ、トルロプシス、及びピキアを含む。本発明の好ましいメチロトローフ酵母は、属ピキアの株である。特に好ましいのは、ピキアパストリス株GS115(NRRL Y−15851),GS190(NRRL Y−18014),PPF1(NRRL Y−18017),PPY120H,YGC4,及びそれらに由来する株である。
【0029】
一つの態様において、本発明は、コアのマンノース含有構造及びターミナルガラクトース残基に結合した5つ又はそれ未満のマンノース残基及び少なくとも一つのN−アセチルグルコサミン残基(GlcNAc)を有する特徴を有する、哺乳類様N−グリカン構造を有する糖蛋白質を生成する遺伝子操作されたメチロトローフ酵母株を提供する。
【0030】
好ましい態様において、本発明は、哺乳類様N−グリカン構造、GalGlcNAcMan5GlcNAc2を有する糖蛋白質を生じる遺伝子操作されたメチロトローフ酵母株を提供する。
【0031】
本発明によれば、GalGlcNAcMan5GlcNAc2を有する糖蛋白質を生じる上記のメチロトローフ酵母株は、α−1,2−マンノシダーゼ又はその機能部分、N−アセチルグルコサミニルトランスフェラーゼI(又はGnTI)又はその機能部分、及びβ−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分を発現するように遺伝子操作される。好ましくは、当該メチロトローフ酵母株は、ゲノミックOCH1遺伝子が不活性化されるようにも遺伝子操作される。
【0032】
α−1,2−マンノシダーゼは、Man8GlcNAc2の非還元末端においてα−1,2−結合したマンノース残基を分断し、そして糖蛋白質上のこのコアのオリゴサッカライドをMan5GlcNAc2に変換し、これは哺乳類N−アセチルグルコサミニルトランスフェラーゼIのアクセプター基質である。
【0033】
本発明によれば、メチロトローフ酵母株を操作して、当該株に、例えば形質転換により、α−1,2−マンノシダーゼ又はその機能部分をコードするヌクレオチド配列を導入することにより、α−1,2−マンノシダーゼ又はその機能部分をコードするヌクレオチド配列を発現することができる。α−1,2−マンノシダーゼ又はその機能部分をコードするヌクレオチド配列は、如何なる種に由来することもできる。多数のα−1,2−マンノシダーゼ遺伝子がクローン化されて、当業者には利用可能であり、例えば、マウスのα−1,2−マンノシダーゼ遺伝子(Herscovics et al.J.Biol.Chem.269:9864−9871,1994)、ウサギのα−1,2−マンノシダーゼ遺伝子(Lal et al.J.Biol.Chem.269:9−872−9881,1994)又はヒトα−1,2−マンノシダーゼ遺伝子(Tremblay et al.Glycobiology 8:585−595,1998)をコードする哺乳類遺伝子、並びに、例えば、アスペルギルスα−1,2−マンノシダーゼ遺伝子(msdS遺伝子)、又はトリコデルマリセイα−1,2−マンノシダーゼ遺伝子(Maras et al.J.Biotechnol.77:255−263,2000)を含む真菌遺伝子を含む。蛋白質の配列分析が、これまでに同定された真核生物α−1,2−マンノシダーゼ遺伝子の間の高度の保存を明らかにした。
【0034】
好ましくは、本ベクターにおける使用のためのヌクレオチド配列は、真菌のα−1,2−マンノシダーゼ、より好ましくは、トリコデルマリセイα−1,2−マンノシダーゼ、そしてより特定すれば、Maras et al.J.Biotechnol.77:255−63(2000)に記載されたトリコデルマリセイα−1,2−マンノシダーゼをコードする。
【0035】
「機能部分」は、完全長蛋白質の酵素活性を実質上保持したα−1,2−マンノシダーゼのポリペプチド断片を意味する。「実質上」は、完全長α−1,2−マンノシダーゼの酵素活性の少なくとも約40%、又は好ましくは少なくとも50%またはそれ以上が保持されることを意味する。多数のα−1,2−マンノシダーゼの、触媒ドメインを含む様々なドメインの特性決定は、記録されている。例えば、”Isolation of a mouse Golgi mannosidase cDNA, a member of a gene family conserved from yeast to mammalls”,Herscovics et al.,J Biol Chem 269:13 9864−71(1994);Isolation and expression of murine and rabbit cDNAs encoding an alpha 1,2−mannosidase involved in the processing of asparagine−linked oligosaccharides”,La et al.,J Biol Chem 269:13 9872−81(1994);”Molecula cloning and enzymatic characterization of a Trichoderma reeses 1,2−alpha−D−mannosidase”,Maras M et al.,J Biotechnol 77:255−63(2000);及び米国特許出願20020188109を参照されたく、引用により本明細書に編入される。当業者は、当業界で公知の技術の組み合わせを用いることによりα−1,2−マンノシダーゼの機能部分を容易に同定して作成することができる。適切な発現系から発現されて精製された目的のα−1,2−マンノシダーゼの一部の活性は、引用により本明細書に編入される米国特許出願20020188109に記載されたインビトロ又はインビボアッセイを用いることにより確認することができる。
【0036】
本発明によれば、メチロトローフ酵母内で発現されたα−1,2−マンノシダーゼ又はその機能部分は、好ましくは、Man8GlcNAc2(α−1,2−マンノシダーゼの基質)が糖蛋白質上で既に形成されているが、追加のマンノース残基により糖鎖を伸長するゴルジ体のグリコシルトランスフェラーゼには到達していない分泌経路中の部位を標的化する。本発明の好ましい態様において、α−1,2−マンノシダーゼ又はその機能部分は、α−1,2−マンノシダーゼがメチロトローフ酵母株内で発現されてERを標的化するように、ER滞留シグナルを含むように操作される。
【0037】
「ER滞留シグナル」は、ERに輸送されてER中で滞留するようにペプチド配列を有する蛋白質を指示するペプチド配列を意味する。そのようなER滞留配列は、しばしば、ER中に存在して機能する蛋白質の中に見いだされる。ER滞留シグナルの複数の選択が当業者に利用可能であり、例えば、サッカロミセスセレビシエのER蛋白質のMNS1の最初の21アミノ酸残基(Martinet et al.Biotechnology Letters 20:1171−1177,1998)及びペプチドHDEL(配列番号:1)である。
【0038】
本発明における使用のための好ましいER滞留シグナルは、ペプチドHDEL(配列番号:1)である。HDELペプチド配列は、多数の酵母蛋白質のC−末端に見いだされ、ERのための滞留/修正(retrieval)シグナルとして作用する(Pelham EMBO J.7:913−918,1988)。HDEL配列を有する蛋白質は、膜結合受容体(Erd2p)により結合して、次に、ゴルジ体からERに戻るための逆行(retrograde)輸送経路に入る。
【0039】
本発明における使用のためのα−1,2−マンノシダーゼは、さらに、例えば、抗体が利用可能なエピトープタグを含むように操作することができ、例えば、当業界でよく知られているMyc,HA,FLAG及びHis6タグである。エピトープタグを付加されたα−1,2−マンノシダーゼは、発現及び細胞外局在化の両方のために便利に精製、又は監視され得る。
【0040】
本発明によれば、ER滞留シグナルを、α−1,2−マンノシダーゼの蛋白質配列のどこか、しかし好ましくはα−1,2−マンノシダーゼのC末端において、遺伝子操作することにより配置することができる。
【0041】
ER滞留シグナル及びエピトープタグは、当業界公知の分子生物学の技術の何れかを用いて、α−1,2−マンノシダーゼ又はその機能部分をコードするヌクレオチド配列に、そのようなシグナル又はタグをコードするヌクレオチド配列を挿入することにより、α−1,2−マンノシダーゼ又はその機能部分に容易に挿入することができる。
【0042】
操作された酵母株内でのα−1,2−マンノシダーゼの発現は、mRNAレベルにおいては例えばノーザンブロット分析により、そして蛋白質レベルにおいてはウエスタンブロット分析により確認することができる。蛋白質の細胞内局在は様々な技術を用いることにより分析することができ、細胞成分分画(subcellular fractionation)及び免疫蛍光の実験を含む。α−1,2−マンノシダーゼもER中での局在は、細胞成分分画実験においてこの酵素と公知のER滞留蛋白質(例えば、プロテインジスルフィドイソメラーゼ)との共沈殿により決定することができる。ER中の局在は、ER滞留蛋白質の免疫蛍光染色パターン特性、典型的には核周囲染色パターンにより、決定することもできる。
【0043】
メチロトローフ酵母株中で発現されたα−1,2−マンノシダーゼ又はその機能部分が期待されたマンノーストリミング活性を有することを確認するために、インビトロ及びインビボ両方のアッセイを用いることができる。典型的には、インビトロアッセイは、インビトロで合成された基質、例えば、Man8GlcNAc2の免疫酵母株で発現されて精製された酵素による消化、及びそのような酵素がMan8GlcNAc2を例えばMan5GlcNAc2に削る能力を評価することを含む。インビボアッセイにおいては、α−1,2−マンノシダーゼ又はその一部をメチロトローフ酵母内で、そのような酵母内でN−グリカンを生じるターミナルα−1,2−結合したマンノース残基によりグリコシル化されることが知られている糖蛋白質と同時発現させる。そのようなα−1,2−マンノシダーゼ又はその一部の酵素活性は、糖蛋白質のN−グリカンの構造中のα−1,2−結合したマンノース残基の数の低下に基づいて測定することができる。インビトロ及びインビボの両アッセイにおいて、糖質基の組成は、当業界でよく知られており且つ本明細書の以下の実施例において例示される技術を用いて測定することができる。
【0044】
さらに、本発明によれば、GlcNAc−トランスフェラーゼI又はその機能部分をコードするヌクレオチド配列をマンノース酵母株に、例えば形質転換により導入することにより、GlcNAc−トランスフェラーゼI又はその機能部分を発現するようにマンノース酵母株を操作することができる。GlcNAc−トランスフェラーゼIはβ−1,2−GlcNAcのMan5GlcNAc2への付加に必須であり、糖蛋白質上のこのコアオリゴサッカライドをGlcNAcMan5GlcNAc2へ変換する。GlcNAcMan5GlcNAc2のマンノース残基は哺乳類ゴルジ体マンノシダーゼIIによりさらに削られて、追加の糖ユニット、例えばガラクトースが哺乳類糖蛋白質のハイブリッド−又はコンプレックス−タイプの糖ブランチ特性を形成するために(towards)追加される。
【0045】
メチロトローフ酵母株への導入のためのGlcNAc−トランスフェラーゼI(GnTI)又はその機能部分をコードするヌクレオチド配列は、あらゆる種、例えば、ウサギ、ラット、ヒト、植物、昆虫、線虫及び原生動物、例えばリーシュマニアタレントレを含む。好ましくは、本発明における使用のためのヌクレオチド配列は、ヒトGnTIであり、そしてより好ましくは配列番号:13に示されるヒトGnTIである。
【0046】
GnTIの「機能部分」は、完全長のGnTIの酵素活性を実質上保持するGnTIのポリペプチド断片を意味する。「実質上」は、少なくとも約40%、又は好ましくは少なくとも約50%またはそれ以上の完全長のGnTIの酵素活性が保持されることを意味する。GnTI又はその一部の酵素活性は、Reeves et al.(Proc.Natl.Acad.Sci.USA.99(21):13419−24,2002);Maras et al.(Eur J Biochem.249(3):701−7,1997),又は本明細書中の以下の実施例に記載されるアッセイにより測定することができる。当業者は、当業界で公知の技術の組み合わせを用いてGnTIの機能部分を容易に同定して作成することができる。例えば、本発明により例示されるとおり、ヒトGnTIの触媒ドメイン(最後の327残基を含む)はヒトGnTIの「機能部分」を構成する。
【0047】
本発明によれば、メチロトローフ酵母株において発現されたGnTI又はその機能部分は、好ましくは、Man5GlcNAc2(GnTIの基質)が糖蛋白質上に既に形成された分泌経路中の部位を標的化する。好ましくは、GnTI又はその機能部分はゴルジ体を標的化する。
【0048】
従って、本発明の好ましい態様においては、GnTI又はその機能部分がゴルジ体局在化シグナルを含むように操作される。
本明細書において使用される「ゴルジ体局在化シグナル」は、ペプチド配列であって、そのような配列を有する蛋白質をメチロトローフ酵母株のゴルジ体に振り向け(directs)、そして当該蛋白質をその中に保持するペプチド配列を意味する。そのようなゴルジ体局在化配列は、ゴルジ体に存在して機能する蛋白質中にしばしば見いだされる。
【0049】
ゴルジ体局在化シグナルの選択は、当業者に利用可能である。本発明における使用のための好ましいゴルジ体局在化シグナルは、サッカロミセスセレビシエのKre2蛋白質(ScKre2)のN末端部分に由来するペプチド;より好ましくは配列番号:10に示されるScKre2である。特に好ましいゴルジ体局在化シグナルは、配列番号:10に示されるScKre2蛋白質のアミノ酸1−100からなる、ペプチド(配列番号:11)である。
【0050】
本発明によれば、ゴルジ体局在化シグナルは、GnTI内の如何なる場所に配置されてもよいが、好ましくはGnTIの末端、そしてより好ましくはGnTIのN末端に配置され得る。
【0051】
本発明における使用のためのGnTIは、例えば、抗体が利用可能なエピトープタグ、例えば、当業界でよく知られているMyc,HA,FLAG及びHis6を含むようにさらに操作され得る。エピトープタグを付加されたGnTIは便利に精製されるか、又は発現及び細胞内局在に関して監視され得る。
【0052】
ゴルジ体局在化シグナル及びエピトープタグは、当業界公知の分子生物学の技術の何れかを用いて、GnTI又はその機能部分をコードするヌクレオチド配列に、そのようなシグナル又はタグをコードするヌクレオチド配列を挿入することにより、GnTI又はその機能部分に容易に挿入することができる。
【0053】
さらに、本発明によれば、メチロトローフ酵母株をβ−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分をコードするヌクレオチド配列をマンノース酵母株に、典型的には形質転換により導入することにより、β−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分を発現するようにマンノース酵母株を操作することができる。GalTはβ−1,4−ガラクトース残基をグリカン構造(GlcNAcMan5GlcNAc2)の左のアーム上のGlcNAcへの付加し、図1に描写されるとおりである。
【0054】
メチロトローフ酵母株への導入のためのGalT又はその機能部分をコードするヌクレオチド配列は、あらゆる種、例えば、哺乳類(例えば、ヒト、マウス)、植物(例えば、アラビドプシスサリアナ)、昆虫(例えば、ドロソフィラメラノガスター)、又は線虫(シーノラブディティスエレガンス)に由来することができる。好ましくは、本発明における使用のためのヌクレオチド配列は、ヒトGalT、そしてより好ましくは配列番号:21に示されたヒトGalTをコードする。
【0055】
GalTの「機能部分」は、完全長のGalTの酵素活性を実質上保持するGalTのポリペプチド断片を意味する。「実質上」は、少なくとも約40%、又は好ましくは少なくとも約50%またはそれ以上の完全長のGalTの酵素活性が保持されることを意味する。GalT又はその一部の酵素活性は、Maras et al.(Eur J Biochem.249(3):701−7,1997),又は本明細書中の以下の実施例に記載されるアッセイにより測定することができる。当業者は、当業界で公知の技術の組み合わせを用いてGalTの機能部分を容易に同定して作成することができる。例えば、本発明により例示されるとおり、ヒトGalTの触媒ドメインはヒトGalTの「機能部分」を構成する。
【0056】
本発明によれば、メチロトローフ酵母株において発現されたGalT又はその機能部分は、好ましくは、GlcNAcMan5GlcNAc2(GlcNAcMの基質)が糖蛋白質上に既に形成された分泌経路中の部位を標的化する。好ましくは、GalT又はその機能部分はゴルジ体を標的化する。
【0057】
従って、本発明の好ましい態様において、GalT又はその機能部分は、上で記載されたとおり、ゴルジ体局在化シグナルを含むように操作される。GalTをゴルジ体に標的化するための好ましいゴルジ体局在化シグナルは、配列番号:10に示されるScKre2蛋白質のアミノ酸1−100からなる、ペプチド(配列番号:11)である。
【0058】
本発明における使用のためのGalTは、例えば、抗体が利用可能なエピトープタグ、例えば、当業界でよく知られているMyc,HA,FLAG及びHis6を含むようにさらに操作され得る。エピトープタグを付加されたGalTは便利に精製されるか、又は発現及び細胞内局在に関して監視され得る。
【0059】
ゴルジ体局在化シグナル及びエピトープタグは、当業界公知の分子生物学の技術の何れかを用いて、GalT又はその機能部分をコードするヌクレオチド配列に、そのようなシグナル又はタグをコードするヌクレオチド配列を挿入することにより、GalT又はその機能部分に容易に挿入することができる。
【0060】
メチロトローフ酵母株内での所望の蛋白質(即ち、α−1,2−マンノシダーゼ、GnTI,GalT,又はこれらの酵素の何れかの機能部分)の発現を達成するためには、当該蛋白質をコードするヌクレオチド配列を、上記メチロトローフ酵母株内で機能するプロモーター及び3’終結配列に作動可能なような連結にてベクター内に配置させることができる。当該ベクターを、次に、メチロトローフ酵母株へ、例えば、形質転換により導入する。
【0061】
メチロトローフ酵母内で蛋白質の発現に適したプロモーターは、構成的プロモーター及び誘導可能なプロモーターの両方を含む。構成的プロモーターは、例えば、ピキアパストリスのグリセルアルデヒド−3−ホスフェートデヒドロゲナーゼプロモーター(「GAPプロモーター」)を含む。誘導可能なプロモーターの例は、例えば、ピキアパストリスの「アルコールオキシダーゼIプロモーター(「AOXプロモーター」)(米国特許第4,855,231号)、又はピキアパストリスのホルムアルデヒドデヒドロゲナーゼプロモーター(「FLDプロモーター」)(Shen et al.Gene 216:93−102,1998)を含む。
【0062】
3’終結配列は、当該配列が作動可能に連結した遺伝子のmRNA転写産物を安定化するために機能する構造遺伝子の停止コドンの3’の配列であり、例えば、ポリアデニル化を導く配列である。3’終結配列は、ピキア又はメチロトローフ酵母から得ることができる。本発明の実施に有用なピキアパストリスの3’終結配列の例は、AOX1遺伝子及びHIS4の遺伝子由来の終結配列を含む。
【0063】
ベクター又はその直鎖状断片の形質転換は、あらゆる公知の方法、例えば、Cregg ら(Mol.Cell.Biol.(12):3376−85,1985)により記載されたようなスフェロプラスト技術、又はEP312,934に記載されたピキアにおける使用のために修飾された、Itoら(Agric.Biol.Chem.48(2):341,(1984))により記載された全細胞塩化リチウム酵母形質転換系を用いて実施することができる。形質転換に有用な他の方法は、米国特許第4,929,555号;Hinnen et al.(Proc.Natl.Acad.Sci.USA 75:1929(1978));Ito et al.(J.Bacteriol.153:163(1983));米国特許第4,879,231号;及びSreekrishna et al.(Gene 59:115(1987))に記載された方法を含む。エレクトロポレーション及びPEG1000全細胞形質転換手法を使用することもできる。Cregg and Russel,Methods in Molecular Biology:Pichia Protocols,Chapter 3,Humana Press,Totowa,N.J.,pp.27−39(1998)を参照。
【0064】
形質転換された酵母細胞は、限定ではないが、(細胞の栄養要求性のために)必要とされる生化学生成物の不在下での形質転換後に栄養要求性細胞を培養すること、新規な表現型の選択及び検出、又は形質転換体内に含まれる耐性遺伝子の不在下で当該酵母に毒性の抗生物質の存在下で培養することを含む、適切な技術を用いることにより選択することができる。形質転換体はゲノム中の発現カセットの組み込みにより選択及び/又は確認することもでき、例えばサザンブロット又はPCR分析により評価することができる。
【0065】
上で記載されたとおり、α−1,2−マンノシダーゼ、及びN−アセチルグルコサミニルトランスフェラーゼI(又はGnTI)、β−1,4−ガラクトシルトランスフェラーゼ(GalT)、又はその機能部分の発現に加えて、上記メチロトローフ酵母株は、好ましくは、GlcNAcMan5GlcNAc2グリカンを有する糖蛋白質を有効に生成するためにゲノミックOCH1遺伝子を不活性化するように遺伝子操作もされる。
【0066】
OCH1遺伝子は、初期ゴルジ体複合体内に局在化してN−結合コアオリゴサッカライド(Man5GlcNAc2及びMan8GlcNAc2)へのα−1,6−ポリマンノース外部鎖付加を開始する膜結合性α−1,1,6−マンノシルトランスフェラーゼをコードする。サッカロミセスセレビシエのOCH1遺伝子及びピキアのOCH1遺伝子はクローン化されている(それぞれ、Nakayama et al.EMBO J.11:2511−19,1992、及び日本特許出願番号07145005)。当業者は、当業界で公知の技術を用いることにより、他のメチロトローフ酵母からOCH1遺伝子を単離することができる。
【0067】
本発明によれば、メチロトローフ酵母株のOCH1遺伝子の破壊は不活性蛋白質産物の生成か又は生成物なしの何れかをもたらし得る。当該破壊は、外来DNA配列の上記コーディング配列への挿入の形態及び/又は当該コーディング配列の幾つか又は全ての欠失の形態を採ってよい。遺伝子破壊は、相同組換えにより生じさせることができ、本質的にはRothstein(in Methods in Enzymology,Wu et al.,eds.,vol 101:202−211,1983)に記載される。
【0068】
二重相同組換えによりゲノミックOCH1遺伝子を破壊するためには、OCH1「ノックアウト」ベクターを構築することができ、それは相同組換えを媒介するのに十分な長さのOCH1遺伝子の一部に対してその5’及び3’末端の両方において作動可能に連結された選択可能なマーカー遺伝子を含む。当該選択可能マーカーは、宿主細胞栄養要求性を相補するか又は抗生物質耐性を提供するあらゆる数の遺伝子の一つであり得、URA3,ARG4,HIS4,ADE1,LEU2,HIS3,Sh ble(ストレプトアロテイカスヒンダスタヌスのブレオマイシン遺伝子)及びBSD(アスペルギルステレウスからのブラスチシジンSデアミナーゼ)遺伝子を含む。他の適切な選択可能マーカーは、メチロトローフ酵母の蔗糖上の生育を可能にさせるサッカロミセスセレビシエ由来のインベルターゼ遺伝子;活性βガラクトシダーゼの発現のために青いコロニーをもたらすlacZ遺伝子を含む。OCH1不活性化ベクターの直鎖状DNA断片は、その5’及び3’両末端においてOCH1配列と共に選択可能マーカーを含み、のちに、当業界で公知の形質転換方法の何れかを用いることにより宿主メチロトローフ酵母細胞内に導入される。直鎖状断片のゲノミックOCH1座への組み込み及びOCH1遺伝子の破壊は、選択可能マーカーに基づいて測定することができ、そして例えば、サザンブロット分析により確認することができる。
【0069】
或いは、OCH1遺伝子の一部を含み、但し当該一部はあらゆるOCH1プロモーター配列を欠き、そしてOCH1蛋白質を全くコードしないか又は不活性部分をコードする、OCH1遺伝子ノックアウトベクターを構築することができる。「不活性断片」は、完全長OCH1蛋白質の断片であって、当該断片が完全長のOCH1蛋白質の活性の、好ましくは約10%未満、そしてより好ましくは約0%を有するものを意味する。OCH1遺伝子のそのような部分は、メチロトローフ酵母において機能するあらゆるプロモーター配列に作動可能に連結されずにベクター内に挿入される。このベクターは、OCH1遺伝子内の部位においてのちに直鎖状化され、そして当業界で公知のあらゆる形質転換方法を用いてメチロトローフ酵母株を形質転換することができる。1回の相同組換えにより、この直鎖状化されたベクターは次にOCH1座に組み込まれて、染色体中に2つのoch1配列をもたらすが、何れもが活性なOch1p蛋白質を生成することができず、図3Aに描写されるとおりである。
【0070】
好ましくは、不活性化変異を直鎖状化された部位の5’(上流の)部位及びOCH1の翻訳開始コドンの3’(下流)の部位においてベクター中のoch1配列内にも挿入される。「不活性化変異(inactivating mutation)」は、停止コドンを導入する変異、フレームシフト変異又は読み枠の破壊を引き起こすあらゆる他の変異を意味する。そのような変異は、当業界で公知の部位特異的変異導入方法の何れかを用いてベクター内にoch1配列内に導入され得る。そのような不活性化変異は、たとえノックアウトベクター中のOch1配列に対して5’に幾つかのプロモーター配列が存在しようとも、機能しないOch1p蛋白質が相同組換え後に形成されることを保証する。
【0071】
本明細書において上で記載されたような遺伝子操作されたメチロトローフ酵母株は、所望であればさらに修飾することができる。例えば、あらゆるその他のピキアのマンノシルトランスフェラーゼをコードする追加の遺伝子の破壊を作成することができる。α−1,2−マンノシダーゼ、GnTI又はGalT以外の哺乳類グリコシル化経路中で機能する酵素をコードする遺伝子が、哺乳類様N−グリカンの比率を増加させるため、及び/又は、所望であれば哺乳類様N−グリカンをさらに修飾するために、導入することができる。例えば、上で記載された遺伝子操作されたメチロトローフ酵母株をさらに修飾することにより、UDP−グルコースをUDP−ガラクトースに、そして逆もまた同じく変換するサッカロミセスセレビシエのGAL−10にコードされた酵素を発現することができる。これは、サイトゾルのUDP−ガラクトースのレベルを増加させるかもしれず、次に、GalTの活性を刺激してGalGlcNacM5GlcNac2グリカンの比率を増加させる。さらに、上で記載された遺伝子操作されたメチロトローフ酵母は、ゴルジ体中でマンノシダーゼIIを発現するようにさらに修飾され得て、GalGlcNacM5GlcNac2から追加のマンノース残基を除去し、それにより他の糖残基の追加を許容する。
【0072】
遺伝子修飾の順序は本発明には必須ではない。α−1,2−マンノシダーゼ、GnTI及びGalTをコードするヌクレオチド配列の導入、及びゲノミックOCH1遺伝子の破壊は、連続してか、如何なる順序でも、又は同時に、2つ又はそれより多い別々のベクター又はコーディング配列により共形質転換(co-transformation)することによるか又は2つ又はそれより多い別々のコーディング配列を含む一つのベクターによる形質転換により行うことができる。
【0073】
さらなる側面において、本発明は、5又はそれ未満のマンノース残基及びコアマンノース含有構造並びにターミナルガラクトース残基に結合した少なくとも一つのN−アセチルグルコサミン残基(GlcNAc)を有するものとして特徴付けされる哺乳類様N−グリカン構造、例えば、GalGlcNAcMan5GlcNAc2を有する構造を生じるメチロトローフ酵母株を生じさせるのに有用なベクターを提供する。
【0074】
一つの態様において、本発明は、発現される酵素、即ち、α−1,2−マンノシダーゼ、GnTI,GalT,又はこれらの酵素の何れかの機能部分をコードするヌクレオチド配列を含むベクターを提供する。そのようなベクターは、「ノックイン」ベクターとも呼ばれる。コーディング配列は、コードされた蛋白質の発現のために宿主メチロトローフ酵母において機能するプロモーター及び3’終結配列に作動可能に連結して配置され得る。2つ又はそれより多いコーディング配列をメチロトローフ酵母株への同時形質転換のための同じベクター内に入れることができる。好ましくは、当該ベクターは、形質転換の便利な選択のための上で記載された選択可能なマーカー遺伝子の何れか一つも含む。
【0075】
本発明によれば、ノックインベクターは、メチロトローフ酵母株内で発現される所望の蛋白質をコードする配列を含み、組み込み型ベクター又は複製型ベクター(例えば、複製型環状プラスミド)の何れかであり得る。組み込み型ベクターは、例えば、米国特許第4,882,279号に開示されており、引用により本明細書に編入される。組み込み型ベクターは、通常、少なくとも第1の挿入可能DNA断片、選択可能なマーカー遺伝子、及び第2の挿入可能なDNA断片の連続して並んだ配置(serially arranged sequence)を含む。第1及び第2の挿入可能なDNA断片は、各々、約200ヌクレオチドの長さであり得て、形質転換される種のゲノミックDNAの一部に相補なヌクレオチド配列を有する。発現のための目的の構造遺伝子を含むヌクレオチド配列をこのベクター中で第1の挿入可能DNA断片と第2の挿入可能DNA断片の間に挿入するが、マーカー遺伝子の前又は後である。組み込み型ベクターは宿主細胞ゲノムへの目的のヌクレオチド配列の組み込みを促進するために酵母形質転換に先立って直鎖状にすることができる。
【0076】
別の態様において、本発明は、メチロトローフ酵母株に導入されたときにゲノムOCH1遺伝子を不活性化するか又は破壊する不活性化ベクター(又は「ノックアウト」ベクター)を提供する。
【0077】
ゲノミックOCH1遺伝子を不活性化するためのベクターは、上で記載されたとおり、相同組換えを媒介するのに十分な長さのOCH1遺伝子の一部に、その5’及び3’末端の両方において作動可能に連結した選択可能なマーカー遺伝子を含み得る。メチロトローフ酵母細胞の、その5’及び3’末端の両方においてOCH1配列を選択可能なマーカー遺伝子と共に含み得るそのようなOCH1不活性化ベクターの直鎖状DNA断片による形質転換は、ゲノミックOCH1座への直鎖状断片の組み込み及びゲノミックOCH1遺伝子の破壊を導く。
【0078】
或いは、OCH1不活性化ベクターは、破壊されるOCH1遺伝子の一部及び上で記載された選択可能なマーカー遺伝子の何れか一つを含むことができるが、上記一部はOCH1蛋白質を何もコードしないか又はOCH1蛋白質の不活性断片をコードする。そのようなOCH1遺伝子の一部は如何なるOCH1プロモーター配列をも欠き、そして如何なるプロモーター配列に対しても作動可能に連結していない。そのようなベクターはOCH1遺伝子内の部位において直鎖状化し、そして次にメチロトローフ酵母株を形質転換することができ、1回の相同組換え媒介性の組み込みにより、ゲノミックOCH1遺伝子の不活性化をもたらす。好ましくは、不活性化変異、例えば、停止コドン又はフレームシフト変異も、直鎖状化部位の5’(上流)の部位及びOCH1の翻訳開始コドンの3’(下流)の部位においてベクター中のOCH1配列内に導入される。
【0079】
所望であれば、メチロトローフ酵母株中で発現される酵素をコードするヌクレオチド配列は、「ノックインアンドノックアウト」ベクターを創製するために、同じベクター中でゲノミックOCH1遺伝子を不活性化することができるヌクレオチド配列と組み合わせることができる。
【0080】
ノックインベクター及びノックアウトベクターの両方を含む本発明のベクターは、細菌中で機能する選択可能なマーカー遺伝子、並びに細菌中で複製及び細胞外染色体の維持管理に必須の配列をも含むことができる。細菌の選択可能なマーカー遺伝子の例は、アンピシリン耐性(Ampr)、テトラサイクリン耐性(Tetr)、ハイグロマイシン耐性、ブラスチシジン耐性及びゼオシン耐性(Zeor)遺伝子を含む。
【0081】
さらに、上記のベクターの何れも、さらに、メチロトローフ酵母株中でのそのような糖蛋白質の発現のための、目的の糖蛋白質をコードするヌクレオチド配列を含むことができる。
【0082】
さらに別の側面において、本発明は、哺乳類様N−グリカン構造を有する糖蛋白質を生じさせる方法を提供する。
本明細書にて使用される「糖蛋白質」は、一つ又はそれより多いアスパラギン残基上又は一つ又はそれより多いセリン又はスレオニン残基、又はアスパラギン及びセリン又はスレオニン残基の両方の上の何れかで、メチロトローフ酵母株中において、グリコシル化される蛋白質を意味する。好ましくは、糖蛋白質は宿主メチロトローフ酵母株に対して異種である。
【0083】
本発明によれば、減少したグリコシル化を伴う目的の糖蛋白質の生産は、多数の様式において達成できる。例えば、目的の糖蛋白質をコードするヌクレオチド配列を、哺乳類様N−グリカンを応じるように予め操作されたメチロトローフ酵母株に導入することができる。
【0084】
糖蛋白質をコードするヌクレオチド配列は、宿主株中で機能するプロモーター配列及び3’終結配列に対して作動可能に配置することができる。当該ヌクレオチド配列は、追加の配列、例えば、蛋白質生成物の分泌が望まれる場合に運搬(transit)ペプチドをコードするシグナル配列を含むことができる。そのようなシグナル配列は、よく知られており、容易に利用可能であって、サッカロミセスセレビシエのアルファメイティング因子プレプロ(αmf)、ピキアパストリス酸ホスファターゼ(PHO1)シグナル配列等を含む。
【0085】
或いは、目的の糖蛋白質のコーディング配列を導入されたメイティング酵母株を修飾することにより、所望の酵素(即ち、a−1,2−マンノシダーゼ、GnTI及びGalT)を発現させ、そして上記のとおりにゲノミックOCH1遺伝子を不活性化し、それにより哺乳類様N−グリカンを有する糖蛋白質を生産することができる。
【0086】
メイティング酵母株中で生産された糖蛋白質は慣用の方法により精製することができる。精製プロトコルは、精製される特定の蛋白質の性質により決定することができる。そのような決定は当業者の範囲内である。例えば、細胞培養培地を細胞から分離し、そして当該細胞から分泌される蛋白質を日常の単離技術、例えば沈殿、免疫吸着、フラクショネーション又は様々なクロマトグラフ法により培地から単離することができる。
【0087】
本発明の方法により生産することができる糖蛋白質は、細菌、真菌又はウイルスの蛋白質又は抗原、例えば、バチルスアミロリクエファシエンスのα−アミラーゼ、サッカロミセスセレビシエのインベルターゼ、トリパノゾーマクルジのトランス−シアリダーゼ、HIVエンベロープ蛋白質、インフルエンザウイルスのAヘマグルチニン、インフルエンザのノイラミニダーゼ、ウシヘルペスウイルスのタイプ1糖蛋白質D;蛋白質、哺乳類起源の蛋白質、例えばヒト蛋白質、成長因子又は受容体、例えば、ヒトアンギオスタチン、ヒトB7−1,B7−2及びB−7受容体CTLA−4,ヒト組織因子、成長因子(例えば、血小板由来の成長因子)、組織プラスミノーゲン活性化因子、プラスミノーゲン活性化因子阻害剤−I、ウロキナーゼ、ヒトリソソーム蛋白質、例えば、α−ガラクトシダーゼ、プラスミノーゲン、トロンビン、因子XIII;及びイムノグロブリン又はイムノグロブリンの断片(例えば、Fab,Fab’,F(ab’)2)を含む。本発明の遺伝子操作されたピキア株において発現され得る追加の有用な糖蛋白質に関しては、Bretthauer and Castellino,Biotechnol.Appl.Biochem.30:193−200(1999),及びKukuruzinska et al.,Ann Rev.Biochem.56:915−944(1987)を参照されたい。
【0088】
本発明の方法を用いて生産される糖蛋白質、即ち、哺乳類様のN−グリカン、特にGalGlcNAcMan5GlcNAc2N−グリカンを有する糖蛋白質も、本発明の一部である。
【0089】
また別の側面において、本発明は、上で記載された本発明のノックインベクター、ノックアウトベクター、又はノックインアンドノックアウトベクターの一つ又はそれより多くを含むキットを提供する。
【0090】
より特定すれば、本発明のキットは、α−マンノシダーゼI又はその機能部分をコードする、好ましくはER滞留シグナルを含む、ヌクレオチド配列を有するベクター;GnTI又はその機能部分をコードする、好ましくはゴルジ体滞留シグナルを含む、ヌクレオチド配列を有するベクター;GalT又はその機能部分をコードする、好ましくはゴルジ体滞留シグナルを含む、ヌクレオチド配列を有するベクター;又はメチロトローフ酵母中でゲノミックOCH1遺伝子を破壊することができるベクター、又はその何れかの組み合わせを含む。
【0091】
上記キットは、目的の異種糖蛋白質をコードする配列を有する核酸分子を含むこともできる。どのような核酸分子は、本明細書において前で記載されたノッキングイン又はノッキングアウトのための配列を含む別のベクター又は同じベクター内に提供することができる。或いは、キット中のノックイン又はノックアウトベクターは、目的の異種蛋白質をコードするヌクレオチド配列の挿入のための便利なクローニング部位を有する。
【0092】
上記キットは、本明細書において前で記載された、ノックイン、ノックアウト又はノックインアンドノックアウトベクターの何れかにより形質転換され得るメチロトローフ酵母株を含むこともできる。或いは、上記キットは、哺乳類様N−グリカンを生じさせるために操作されたメチロトローフ酵母株を含むことができる。
【0093】
本発明は、さらに、以下の実施例により例示される。
実施例1
方法と材料
ベクターの構築と形質転換
ピキアパストリス配列はGenBankにて受け入れ番号E12456(配列番号:2)にて見いだされ、日本特許出願番号07145005に記載され、引用により本明細書に編入される。この配列は、α−1,6−マンノシルトランスフェラーゼの全ての典型的な特徴を示し、そしてサッカロミセスセレビシエのOCH1にほとんど相同であり、即ち、本明細書においてはピキアパストリスOCH1遺伝子と呼ぶ。
【0094】
ピキアパストリスOCH1遺伝子の完全ORFを、PCRにより、鋳型として株GS115から単離されたゲノミックDNA、及び以下のオリゴヌクレオチド5’GGAATTCAGCATGGAGTATGGATCATTGAGTCCGTTGGAAAGG(配列番号:4)及び5’GCCGCTCGAGCTAGCTTTCTTTAGTCC(配列番号:5)を用いて単離した。単離されたOCH1遺伝子をpUC18内にクローン化することにより、プラスミドpUC18pOCH1を得て、OCH1遺伝子配列の同定を配列決定により確認した。
【0095】
プラスミドpGlycoSwitchM8(2875bp,配列番号:6、図3Aに模式的に描写する)はAla25−Ala155をコードするピキアパストリスOCH1のORFの断片を含み、当該断片はpPICZB(インビトロジェン、カールスバッド、CA)のBglII部位とHindIII部位の間に挿入された。2つの停止コドンをインフレームにてコドンAla25の前に配置することにより、末端削除された蛋白質の起こり得る合成を阻止した。消化後のフィルイン及び再ライゲーションにより、pPICZBのポリリンカーのBstBI部位を予め排除した。クローン化されたORF1遺伝子の内部に位置する唯一のBstBI部位を上記プラスミドの直鎖状化に用いることができる(不活性化戦略の全体像に関しては図3Aを参照)。
【0096】
pGlycoSwitchM5(5485bp,配列番号:9、図3Bに模式的に描写する)を以下のとおりに構築した。ピキアパストリスHIS4転写終結配列を含むpPIC9(インビトロジェン、カールスバッド、CA)のXbaI/ClaI断片を、pGlycoSwitch M8のHindIII部位及びEcoRI部位の間に挿入した。のちに、GAPプロモーター及びpreMFマンノシダーゼHDELカセットを含むpGAPZMFManHDEL(Callewaert et al.,FEBS Lett,503(2−3):173−178,2001)の2.3kbのBglII/NotI断片をHindIII部位とNotI部位の間に挿入した。この構築物のために使用される全ての制限部位(NotI部位を除く)をクレノーDNAポリメラーゼによりフィルインした。pGAPZMFmanHDEL内の唯一のBstBI部位を、消化後のフィルイン及び再ライゲーションにより排除した。
【0097】
ヒトGlcNAc−トランスフェラーゼ(GnTI)をゴルジ体に標的化するため、GnTIのN−末端部分を、酵母のゴルジ体における直鎖状化に必須のサッカロミセスセレビシエのKre2のN−末端部分により置き換えた(Lussier et al.,J Cell Biol,131(4):913−927,1995)。プラスミドYEp352Kre2(Howard Bussey博士、McGill大学、モントリオール、カナダ、により提供)を生成したが、Kre2遺伝子のSacI/PvuII断片を、SalI(クレノーにより平滑化)及びSalIにより消化されていたYep352ベクターに挿入することにより生成した。YE352Kre2をSacI/PvuIにより消化してT4−ポリメラーゼにより平滑化した。Kre2遺伝子の5’末端を単離して、クレノー平滑化されたSgrAI/XbaIにより開いたpUChGnTI(Maras et al.,Eur J Biochem 249(3):701−707,1997)にクローン化した。2つのDNA断片の融合場所を標準手法を用いて配列決定した。結果のKre2−GnTIオープンリーディングフレームは、Kre2遺伝子のN−末端部分(配列番号:11に示されるとおり、Kre2蛋白質の最初の110アミノ酸をコードする)及びGnTIの触媒ドメイン(配列番号:13に示されるとおり、GnTI蛋白質の最後の327アミノ酸)を含み、EcoRV/HindIIIにより二重消化して、SalI/EcoRIで開いたpPIC6Aベクター(インビトロジェン)内へ、両断片をクレノーポリメラーゼにより平滑化した後に、連結した。結果のプラスミドはpPIC6AKrecoGnTIと命名した(配列番号:14、図3Cにおいて模式的に描写される)。それは、メタノールにより誘導可能なAOX1プロモーターの制御下にKre2GnTIのオープンリーディングフレームを、そして抗生物質ブラスチシジンに対して耐性のアスペルギルステレウスカラノBSD遺伝子を含む。
【0098】
GalTの局在化は、GnTIを標的化するために行われたのと同じ様式にて、Kre2pのN末端部分へのGalTの触媒ドメインの融合により達成された。β−1,4−ガラクトシルトランスフェラーゼをhepg2のcDNAライブラリーからオリゴヌクレオチド5’TTCGAAGCTTCGCTAGCTCGGTGTCCCGATGTC(配列番号:15)及び5’GAATTCGAAGGGAAGATGAGGCTTCGGGAGCC(配列番号:16)をスターター配列として用いて増幅した。増幅された断片をHindIII/EcoRIにてpUC18にクローン化された。GalT蛋白質のN末端の77アミノ酸を削除するため、以下のオリゴヌクレオチドをプライマーとして用いてPCRを実施した:5’TTCGAAGCTTCGCTAGCTCGGTGTCCCGATGTC(配列番号:15)及び5’CGTTCGCGACCGGAGGGGCCCGGCCGCC(配列番号:17)。増幅された断片をNruI/HindIIIに切断して、HindIII/SgrA Iクレノー平滑化pUCKreGnTIベクターに連結した。結果のKre2−GalT融合構築物を再びPCRによりプライマー:5’TCGATATCAAGCTTAGCTCGGTGTCCCGATGTC(配列番号:18)及び5’GAATTCGAACTTAAGATGGCCCTCTTTCTCAGTAAG(配列番号:19)を用いて増幅した。増幅された断片をEcoRV/BstB IにてpBLURA IX(Cereghino et al.,Gene,263:159−169,2001)(James Cregg(オレゴンインスティチュートオブサイエンスアンドテクノロジー、ビーバートン、USA)より提供)に挿入した。最後に、URA3遺伝子をカナマイシン耐性カセットにより置き換えたが、ベクターpFA6a−KanMX4からのSpeI/SmaI断片をSpeI/SspIにおいて開いたプラスミドに連結することによった。最後のプラスミドはpBIKanMX4KrehGalT(配列番号:7、図3Dに模式的に描写される)と呼ばれるが、AOX1プロモーターに作動可能に連結したKre2−GalT融合蛋白質をコードする配列を含んだ。当該融合蛋白質は、Kre2の最初の100アミノ酸及びGalTの最後の320アミノ酸から構成された。
【0099】
これらのプラスミドによる、様々な蛋白質を発現するGS115ピキア株に対する形質転換は、前に記載されたとおりに行った(Cregg et al.,Methods in Molecular Biology,103:27−39,1998)。PpOCH1座における正確なゲノミックインテグレーションは、ゲノミックDNA上のPCRにより確認した。
蛋白質の調製
分泌されたトリコデルマリセイのα−1,2−マンノシダーゼを、HIC、アニオン交換及びゲル濾過クロマトグラフの組み合わせを用いて、記載されたとおりに精製した(Maras et al.,J Biotechnol,77(2−3):255−263,2000;Van Petegem et al.,J Mol Biol 312(1):157−165,2001)。全てのSDS−PAGE実験は10%PAAゲル上において標準泳動条件下で行った。酵母の細胞壁のマンノ蛋白質をBallou(Methods Enzymol,185:440−470,1990)に記載されたとおりに放出させたが、0.9%のNaCl水溶液中の酵母細胞の大規模な洗浄、20mM クエン酸Na中の酵母細胞の延期されたオートクレーブ(90分)、続くメタノール沈殿(4容)を含んだ。
N−グリカン分析
ABI 377 DNA配列決定機(DSA−FACE)上でのレーザー誘導性DNA配列決定機補助によるフルオロフォア補助糖質電気泳動により、記載されたとおりに(Callewaert et al.,Glycobiology,11(4):275−281,2001)、Nグリカン分析を行った。簡単に言えば、糖蛋白質を、マルチスクリーンイムノビロン−P上に固定化し、そしてPNGase処理により脱グリコシル化した。N−グリカンを除去し、そしてAPTSにより誘導した。過剰の標識をセファデックスG10樹脂上のサイズ分画により除去した。APTS−標識されたオリゴサッカライドの蒸発後に、ROX−標識されたGENESCAN 500標準混合物(アプライドバイオシステムズ)を加えて、内部標準化を許容した。この混合物をABI 377A DNA配列決定機(アプライドバイオシステムズ)上で、12%ポリアクリルアミドゲルにより、89mM Tris,89mM 硼酸塩、2.2mM EDTAバッファー中で泳動した。各ゲル上で、ウシRNase BとマルトデキストロースラダーのN−グリカンをレファレンスとして泳動した。データ解析は、GENESCAN 3.1ソフトウエア(アプライドバイオシステムズ)を用いて実施した。β−N−アセチルヘキソサミニダーゼ(Glyko)及びβ−ガラクトシダーゼ(Prozyme)によるエキソグリコシダーゼ処理を、標識されたグリカン上で、一晩、37℃において20mM酢酸ナトリウムpH5.5中で実施した。慣用のFACE(N−グリカンのANTS標識と30%PAAミニゲル上の電気泳動)をJackson(Biochem J,270(3):705−713,1990)により記載されたとおりに実施した。DSA−FACE法は非常に高い解析度と感度を有するが、慣用のFACEは高分子量のN−グリカン(「高グリコシル化」)の複合混合物の検出によく適合されており、分解されていなかった(not resolved)ので、慣用のFACE内のゲル上では特徴的な「スメア」を形成した。即ち、DSA−FACEと慣用のFACE分析の組み合わせは、酵母により生じる糖蛋白質の特徴の、より完全な写真を与えた。
成長曲線の測定
新鮮な一晩酵母培養物を新鮮なYPD培地によりOD600 0.02まで希釈し、そして250rpmにて30℃において一晩生育させた(12時間、OD 600<3.0)。実験を開始するために、50mLのポリプロピレンチューブ中の10mLの新鮮なYPDに一晩酵母培養物を接種し、0.5のOD600値から始めた。アリコートを2時間ごとに採取し、そしてOD600値を測定した。全ての酵母株を同じ時間平行してランさせた。
【0100】
実施例2
OCH1の不活性化
ゲノミックピキアパストリスOCH1遺伝子の破壊は、以下のとおりにして1回の相同組換えにより達成した。プラスミド、pGlycoSwitchM8(図3A)を実施例1において記載されたとおりに生成したが、ピキアパストリスOCH1遺伝子の塩基対番号73−467を含み、その前に当該ベクター中の可能な初期翻訳開始部位からのリードスルーを回避するために2つのインフレームのナンセンスコドンを置いた。この断片は、形質転換前の上記ベクターの直鎖状化に有用な中心に位置するBstB I部位を含んだ。このベクターは、ピキアのゲノミックOCH1座への1回の相同組換えによるインテグレーションに際してベクター中に存在するOCH1遺伝子を複製するはずである。結果として、ピキア染色体内のOCH1遺伝子は2つのOch1配列に置き換わった。第1のOCH1配列は、最大で161のアミノ酸長の蛋白質生成物をコードし(そのうちの6アミノ酸はベクター中の配列からであった)、完全長OCH1遺伝子によりコードされたタイプIIの膜貫通蛋白質の触媒ドメインを含まなかった。第2のOCH1配列は、完全長の蛋白質の最初の25アミノ酸のコーディング配列を欠き、そして可能な上流の翻訳開始部位からの如何なるリードスルーをも阻止するはずの2つのインフレームの停止コドンを含んだ。
【0101】
株GS115をプラスミドpGlycoSwitchM8により形質転換した。形質転換体をGlycoSwitchM8と呼ぶか、又は短くM8株又はoch1株と呼んだ。図3Aにおいて特定されたプライマーの組み合わせによるゲノミックDNAのPCRは、3つの個別の実験により示されたところによれば、ゼオシン耐性形質転換体の約50%における、予測されたゲノミック座へのこの構築物の正確なインテグレーションを示した。
【0102】
細胞壁のマンノ蛋白質のN−グリカンの分析は、図4において演繹されるとおり、グリコシル化パターンの変化を明らかにした。優位なピークは、野生型GS115株からの細胞壁マンノ蛋白質に関してはMan9GlcNAc2であり、主要なピークはGlycoSwitchM8株に関してはMan8GlcNAc2である(図4のパネル2から3を比較)。N−グリカンにおけるこの変化は、M8株の完全長OCH1のORFによる形質転換の後に復帰した。
【0103】
M8株からの分泌された糖蛋白質の不均質性が低下したか否かを確認するために、野生型GS115株中の分泌された蛋白質である典型的には高グリコシル化された(Maras et al.,J Bacteriol,77(2−3):255−263,2000)トリコデルマリセイのα−1,2−マンノシダーゼをoch1のM8株を用いて分析した。トリコデルマリセイのα−1,2−マンノシダーゼをコードする配列により形質転換されたM8株の細胞の培養上清を、SDS−PAGEにより分離した(図5A)。ゲルは、高グリコシル化された蛋白質の特徴であるスメアがGlycoSwitchM8株において生成した蛋白質には不在であったことを明らかにした。平行して、分泌された糖蛋白質をPNGaseF処理により脱グリコシル化し、そしてグリカンをミニゲル上のFACE分析により分析した。典型的にはFACE分析においては、大きな高グリコシル構造が解析されず、一つのスメアバンドとして出現する(図5B)。スメアバンドはoch1株からの糖蛋白質においては不在であり、och1株からのN−グリカンの不均質性が低下したことが確認された。
【0104】
実施例3
ER滞留したマンノシダーゼ−HDELの発現
ピキアパストリスのN−グリカンをさらにヒト化するために、ER滞留されたトリコデルマリセイのα−1,2−マンノシダーゼ−HDELをoch1株中で発現させた。ピキアパストリス発現株の容易な変換のために、トリコデルマリセイのα−1,2−マンノシダーゼ−HDELをコードするヌクレオチド配列をoch1不活性化ベクターに挿入した。結果のコンビネーションベクターをpGlycoSwitchM5と呼び、その構成を実施例1に記載する。
【0105】
株GS115を直鎖状化されたpGlycoSwitchM5により形質転換した。当該ベクターの正確なインテグレーションはPCR分析により確認した。当該形質転換体からのマンノ蛋白質のN−グリカンをDSA−FACE法により分析した。グリカンのプロフィールは、均質なMan5GlcNAc2ピークを明らかにした(図4、パネル4)。Man5GlcNAc2ピークと、5から25までのグルコースユニットのサイズエリアのこの方法の検出限界を超える全ての小さなピークの積分(integration)は、この高等真核種の高マンノースグリカンがこの混合物中に存在する全N−グリカンプールの少なくとも90%を成したことを明らかにした。
【0106】
別のアプローチにおいては、マンノシダーゼ−HDELをメタノール誘導可能AOX1プロモーターの制御下で発現させた。2つのマンノシダーゼ発現株(即ち、構成的及び誘導可能)の間のN−グリカンプロフィールに明確な差異は検出できなかった。
【0107】
異種蛋白質のN−グリカン修飾を確認するため、Laroy et al(Protein Expr Purif,20(3):389−393,2000)に記載されたとおりに、pGlycoSwitchM5プラスミドで、トリパノゾーマクルジのトランス−シアリダーゼ発現ピキア株を形質転換した。この点に関して(here too)、Man5GlcNAc2が精製された蛋白質上に検出され、精製された蛋白質上の全N−グリカンの95%より多くを占める。
【0108】
振盪フラスコ培養におけるpGlycoSwitchM5形質転換株の成長曲線分析は、その倍加時間が野生型株のそれを密接に模倣したことを示した。しかしながら、操作された株は野生型株より約20%低い光学密度において定常期に到達したことから、高細胞密度のストレス条件に対していくらか感受性であり得たことを示す。にもかかわらず、そのストレス感受性の表現型はサッカロミセスセレビシエのoch1株よりもはるかに劣って表現された(much less pronounced)。
【0109】
実施例4
ゴルジ体局在N−アセチルグルコサミニルトランスフェラーゼ
(Kre2GnTI)の発現
GnTIをゴルジ体に標的化するために、サイトゾル部分、膜貫通領域及び管腔幹領域(luminal stem region)を含むGnTIのN−末端部分をコードするヌクレオチド配列を、サッカロミセスセレビシエのKre2シグナル配列をコードするヌクレオチド配列により置き換えた。これは、Kre2pからの最初の327アミノ酸及びGnTIの最後の327アミノ酸を有するキメラ蛋白質をコードするヌクレオチド配列をもたらした。
【0110】
ピキアパストリスにおける発現のために、Kre2−GnTIキメラ配列を、ブラスチシジン耐性マーカーを有するプラスミド中で強力なメタノール誘導可能性AOX1プロモーターの制御下に置いた。結果の構築物であるpPIC6KrecoGnTI(実施例1に記載されるとおり)をNsiI消化によるAOX1座内の直鎖状化の後にGS115 M5株を形質転換した。当該形質転換体内の上記構築物の存在は、AOX1の3’及び5’プライマーを用いたゲノミックDNA上でのPCRにより確認した。
【0111】
幾つかの形質転換体のマンノ蛋白質のN−グリカンはDSA−FACE法により分析した。優位なピークは、Man5GlcNAc2ピークより大きな約1つのグルコースユニットであった(図4、パネル5)。このピークが末端GlcNAcを有するか否かを決定するため、エキソグリコシダーゼ消化を、β−Glc−NAc結合を加水分解する酵素であるβ−N−アセチルヘキソサミニダーゼを用いて実施した。この酵素による消化に際して、上記ピークはMan5GlcNAc2にシフトして戻った(図4、パネル6)。これは、オリジナルピークがGlcNAcMan5GlcNAc2を表すことを示し、即ち、キメラGnTI酵素の正確なインビボ活性を証明する。
【0112】
Kre2GnTIキメラの過剰発現は、Man5GlcNAc2のGlcNAcMan5GlcNAc2へのほとんど完全な変換を導く。これは、十分なUDP−GlcNAcドナー基質がゴルジ体においてN−アセチルグルコサミニレート、ほとんど全てのグリカンに存在したことを示唆する。
【0113】
実施例5
ゴルジ体滞留β−1,4−ガラクトシルトランスフェラーゼの発現
ヒトβ−1,4−ガラクトシルトランスフェラーゼ1の、当該酵素の膜貫通ドメイン及びサイトゾル部分を含むN末端部分(最初の77アミノ酸)をコードするヌクレオチド配列を、サッカロミセスセレビシエのKre2シグナル配列をコードするヌクレオチド配列で置き換えた。このキメラ融合配列をAOX1プロモーター及びAOX1の3’末端をターミネーターとする制御下に置いた。最終的なプラスミドであるpB1KanMX4KrehGalT(実施例1に記載される)をPmeIによりM5−GnTI株の形質転換前に直鎖状化した。
【0114】
N−グリカン分析を幾つかの形質転換体からのマンノ蛋白質にて実施した。GlcNAcMan5GlcNAc2ピークより大きな約1ユニットのグルコースのピークが形質転換体において検出されたが、当該ピークは非形質転換株においては不在であった(図3、パネル7)。このピークが末端β−ガラクトースを含むグリカンであるか否かを決定するため、N−グリカンをβ−ガラクトシダーゼにより消化した。グリカンプロフィールの消化後に、このピークはGlcNAcMan5GlcNAc2位置にシフトして戻った(図4、パネル8、パネル7と比較)。β−ガラクトシダーゼ消化の前と後にGlcNAcMan5GlcNAc2ピークを積分することにより、GalGlcNAcMan5GlcNAc2の量を決定した。これらの2つのピークの差し引きは、約10%のGlcNAcMan5GlcNAc2がGalGlcNAcMan5GlcNAc2に変換されたことを明らかにした。0.2%ガラクトースを含む追加の培地はGal−含有オリゴサッカライドの量を増加させなかった。
【図面の簡単な説明】
【0115】
【図1】図1は、M8GlcNAc2,M5GlcNAc2,GlcNAcM5GlcNc2,及びGal GlcNAcM5GlcNAc2を描写する。
【図2】図2は、酵母とヒトのN−結合グリコシル化及びピキアパストリスのグリコシル化のヒト化のための戦略を模式的に描写する。糖操作工程は、α−1,6−マンノシルトランスフェラーゼOCH1の不活性化、HDEL標識されたα−1,2−マンノシダーゼの過剰発現及びゴルジ局在化GnTI及びGalTの過剰発現を含む。最後に部分的に得られたハイブリッド構造を組み立てる(framed)。
【図3】図3Aは、一回の相同組換えによりゲノミックOCH1遺伝子を不活性化するための戦略を模式的に描写する。図3Bは、ピキアパストリスのグリカン操作のために使用されたプラスミドpGlycoSwitchM5を模式的に描写する。pGlycoSwitchM5をBstBIにより直鎖状化し、次に形質転換し、そしてピキアパストリスのゲノムに正確に組み込むことにより、OCH1遺伝子が不活性化された。図3Cは、pPIC6AKrecoGnTIを模式的に描写する。 図3Dは、pB1KanMX4KrehGalTを模式的に描写する。
【図4】図4は、異なったグリカン操作をされたピキアパストリス株からのN−グリカンのDSA−FACE分析を模式的に描写する。パネル1:オリゴマルトースレファレンス。パネル2−9はN−グリカンを表し:2:−野生型株GS115からのN−グリカンを表し、主要ピークを表すMan9GlcNAc2を伴う;3:och1不活性化された株からのN−グリカンを表し、主要ピークを表すMan8GlcNAc2を伴う;4:och1不活性化されたManHDEL発現株からのN−グリカンを表し、主要ピークを表すMan5GlcNAc2を伴う;5:och1不活性化されたManHDEL,KreGnTI発現株からのN−グリカンを表し、主要ピークを表すGlcNAcMan5GlcNAc2を伴う;6:5と同じであるが、但し、グリカンがβ−N−アセチルヘキソサミニダーゼにより処理され、そしてGlcNAcMan5GlcNAc2ピークがMan5GlcNAc2位置にシフトし、ターミナルGlcNAcが存在したことを示すことを除く;7:och1不活性化されたManHDEL,KreGnTI,KreGalT発現株からのN−グリカンを表し、GalGlcNAcMan5GlcNAc2を表す追加のピークを伴うが、β−N−アセチルヘキソサミニダーゼによる処理により消失した;9:ウシRNaseBからのレファレンスグリカン(Man5−9GlcNAc2)。
【図5】図5Aは、ピキアパストリスOCH1の不活性化後のグリコシル化を示す。5A:トリコデルマリセイのマンノシダーゼを分泌するピキアパストリス株のCBB染色されたSDS−PAGEゲル。操作されていない株(WT)においては、明確なスメアが見えるが、このスメアはoch1不活性化された株(och1(M8))においては消失した。図5Bは、ピキアパストリスOCH1の不活性化後のグリコシル化を示す。5B:操作されていない株(WT)及びoch1株により分泌されたマンノシダーゼに由来するN−グリカンのFACE分析。高い電気泳動移動度のバンドはMan8及びMan9により示され、「コア」N−グリカン構造を表す。
【特許請求の範囲】
【請求項1】
コアのマンノース構造及び末端ガラクトース残基に結合した5又はそれより少ないマンノース残基及び少なくとも一つのN−アセチルグルコサミン残基(GlcNAc)を有することにより特徴付けされる、哺乳類様のN−グリカン構造を有する糖蛋白質を生じる、遺伝子操作されたメチロトローフ酵母株。
【請求項2】
哺乳類様のN−グリカン構造、GalGlcNAcMan5GlcNAc2を含む、請求項1記載の株。
【請求項3】
属カンジダ、ハンセヌラ、トルロプシス、及びピキアの株である、請求項1記載の株。
【請求項4】
ピキアパストリス株である、請求項3記載の株。
【請求項5】
(1)α−1,2−マンノシダーゼ又はその機能部分、(2)N−アセチルグルコサミニルトランスフェラーゼI(又はGnTI)又はその機能部分、及び(3)β−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分を発現する、請求項2記載の株。
【請求項6】
OCH1遺伝子が不活性化されている、請求項2記載の株。
【請求項7】
α−1,2−マンノシダーゼ又はその機能部分が哺乳類の種又は真菌の種の起源のものである、請求項5記載の株。
【請求項8】
哺乳類の種がマウス、ウサギ又はヒトから選択される、請求項7記載の株。
【請求項9】
真菌の種がアスペルギルス、又はトリコデルマリセイから選択される、請求項7記載の株。
【請求項10】
α−1,2−マンノシダーゼ又はその機能部分が上記株のER又はゴルジ体を標的化する、請求項5記載の株。
【請求項11】
α−1,2−マンノシダーゼ又はその機能部分がER滞留シグナルを含むように操作される、請求項10記載の株。
【請求項12】
ER滞留シグナルがHDEL(配列番号:1)を含む、請求項11記載の株。
【請求項13】
GnTI又はその機能部分が、ウサギ、ラット、ヒト、植物、昆虫、線虫及び原生動物からなる群から選択される種の起源のものである、請求項5記載の株。
【請求項14】
GnTI又はその機能部分がヒト起源のものである、請求項13記載の株。
【請求項15】
GnTI又はその機能部分が上記株のゴルジ体を標的化する、請求項5記載の株。
【請求項16】
GnTI又はその機能部分がゴルジ体滞留シグナルを含むように操作される、請求項15記載の株。
【請求項17】
ゴルジ体滞留シグナルが配列番号:11を含む、請求項16記載の株。
【請求項18】
GalT又はその機能部分が、ウサギ、ラット、ヒト、植物、昆虫及び線虫からなる群から選択される種の起源のものである、請求項5記載の株。
【請求項19】
GalT又はその機能部分がヒト起源のものである、請求項18記載の株。
【請求項20】
GalT又はその機能部分が上記株のゴルジ体を標的化する、請求項5記載の株。
【請求項21】
GalT又はその機能部分がゴルジ体滞留シグナルを含むように操作される、請求項20記載の株。
【請求項22】
ゴルジ体滞留シグナルが配列番号:11を含む、請求項21記載の株。
【請求項23】
α−1,2−マンノシダーゼ又はその機能部分がピキアパストリスのAOX1プロモーター、AOXIIプロモーター、GAPプロモーター、及びFLDプロモーターからなる群から選択されるプロモーターから発現される、請求項5記載の株。
【請求項24】
GnTI又はその機能部分がピキアパストリスのAOX1プロモーター、AOXIIプロモーター、GAPプロモーター、及びFLDプロモーターからなる群から選択されるプロモーターから発現される、請求項5記載の株。
【請求項25】
GalT又はその機能部分がピキアパストリスのAOX1プロモーター、AOXIIプロモーター、GAPプロモーター、及びFLDプロモーターからなる群から選択されるプロモーターから発現される、請求項5記載の株。
【請求項26】
α−1,2−マンノシダーゼ又はその機能部分がピキアパストリスのAOX1プロモーターから発現され、そして、GnTI又はその機能部分がピキアパストリスのGAPプロモーターから発現される、請求項5記載の株。
【請求項27】
哺乳類様グリカンがヒト様グリカンである、請求項1記載の株。
【請求項28】
哺乳類様のN−グリカン構造を有する糖蛋白質を組換えにより生成する方法であって、糖蛋白質をコードするヌクレオチド配列により請求項1−2又は5−6の何れか1項に記載の株を形質転換し、そして当該株中で糖蛋白質を発現させることを含む方法。
【請求項29】
糖蛋白質が細菌、真菌、ウイルス又は哺乳類起源のものである、請求項28記載の方法。
【請求項30】
糖蛋白質がイムノグロブリン又はその断片である、請求項28記載の方法。
【請求項31】
請求項28記載の方法により生産れた糖蛋白質。
【請求項32】
糖蛋白質がイムノグロブリン又はその断片である、請求項31記載の方法。
【請求項33】
哺乳類様のN−グリカン構造を有する糖蛋白質をメチロトローフ酵母株中で組換えにより生成する方法であって、糖蛋白質をコードするヌクレオチド配列により上記株を形質転換し;当該株を修飾するが、修飾された株が(1)α−1,2−マンノシダーゼ又はその機能部分、(2)N−アセチルグルコサミニルトランスフェラーゼI(又はGnTI)又はその機能部分、及び(3)β−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分を発現するようにし、そしてその際修飾された株中でのゲノミックOCH1遺伝子が不活性化されて;修飾された株中で糖蛋白質を生成する方法。
【請求項34】
糖蛋白質が細菌、真菌、ウイルス又は哺乳類起源のものである、請求項33記載の方法。
【請求項35】
糖蛋白質がイムノグロブリン又はその断片である、請求項33記載の方法。
【請求項36】
請求項33記載の方法により生産れた糖蛋白質。
【請求項37】
糖蛋白質がイムノグロブリン又はその断片である、請求項36記載の方法。
【請求項38】
プロモーター及び3’終結配列に対して作動可能なように連結したGalTをコードするヌクレオチド配列を含むベクターであって、当該プロモーター及び3’終結配列は当該株中で上記GalTの発現を達成するようにメチロトローフ酵母株中で機能する、上記ベクター。
【請求項39】
(1)α−1,2−マンノシダーゼ又はその機能部分をコードするヌクレオチド配列を含むベクター、(2)N−アセチルグルコサミニルトランスフェラーゼI(又はGnTI)又はその機能部分をコードするヌクレオチド配列を含むベクター、及び(3)β−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分をコードするヌクレオチド配列を含むベクターを含むキットであって、但し、ベクターの各々はコードされた蛋白質のメチロトローフ酵母株中での発現を指示することができる、キット。
【請求項40】
メチロトローフ酵母株のゲノミックOCH1遺伝子を破壊することができるベクターをさらに含む、請求項39記載のキット。
【請求項41】
メチロトローフ酵母株に対して異種の糖蛋白質をコードするベクターをさらに含む、請求項39記載のキット。
【請求項42】
メチロトローフ酵母株をさらに含む、請求項39記載のキット。
【請求項1】
コアのマンノース構造及び末端ガラクトース残基に結合した5又はそれより少ないマンノース残基及び少なくとも一つのN−アセチルグルコサミン残基(GlcNAc)を有することにより特徴付けされる、哺乳類様のN−グリカン構造を有する糖蛋白質を生じる、遺伝子操作されたメチロトローフ酵母株。
【請求項2】
哺乳類様のN−グリカン構造、GalGlcNAcMan5GlcNAc2を含む、請求項1記載の株。
【請求項3】
属カンジダ、ハンセヌラ、トルロプシス、及びピキアの株である、請求項1記載の株。
【請求項4】
ピキアパストリス株である、請求項3記載の株。
【請求項5】
(1)α−1,2−マンノシダーゼ又はその機能部分、(2)N−アセチルグルコサミニルトランスフェラーゼI(又はGnTI)又はその機能部分、及び(3)β−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分を発現する、請求項2記載の株。
【請求項6】
OCH1遺伝子が不活性化されている、請求項2記載の株。
【請求項7】
α−1,2−マンノシダーゼ又はその機能部分が哺乳類の種又は真菌の種の起源のものである、請求項5記載の株。
【請求項8】
哺乳類の種がマウス、ウサギ又はヒトから選択される、請求項7記載の株。
【請求項9】
真菌の種がアスペルギルス、又はトリコデルマリセイから選択される、請求項7記載の株。
【請求項10】
α−1,2−マンノシダーゼ又はその機能部分が上記株のER又はゴルジ体を標的化する、請求項5記載の株。
【請求項11】
α−1,2−マンノシダーゼ又はその機能部分がER滞留シグナルを含むように操作される、請求項10記載の株。
【請求項12】
ER滞留シグナルがHDEL(配列番号:1)を含む、請求項11記載の株。
【請求項13】
GnTI又はその機能部分が、ウサギ、ラット、ヒト、植物、昆虫、線虫及び原生動物からなる群から選択される種の起源のものである、請求項5記載の株。
【請求項14】
GnTI又はその機能部分がヒト起源のものである、請求項13記載の株。
【請求項15】
GnTI又はその機能部分が上記株のゴルジ体を標的化する、請求項5記載の株。
【請求項16】
GnTI又はその機能部分がゴルジ体滞留シグナルを含むように操作される、請求項15記載の株。
【請求項17】
ゴルジ体滞留シグナルが配列番号:11を含む、請求項16記載の株。
【請求項18】
GalT又はその機能部分が、ウサギ、ラット、ヒト、植物、昆虫及び線虫からなる群から選択される種の起源のものである、請求項5記載の株。
【請求項19】
GalT又はその機能部分がヒト起源のものである、請求項18記載の株。
【請求項20】
GalT又はその機能部分が上記株のゴルジ体を標的化する、請求項5記載の株。
【請求項21】
GalT又はその機能部分がゴルジ体滞留シグナルを含むように操作される、請求項20記載の株。
【請求項22】
ゴルジ体滞留シグナルが配列番号:11を含む、請求項21記載の株。
【請求項23】
α−1,2−マンノシダーゼ又はその機能部分がピキアパストリスのAOX1プロモーター、AOXIIプロモーター、GAPプロモーター、及びFLDプロモーターからなる群から選択されるプロモーターから発現される、請求項5記載の株。
【請求項24】
GnTI又はその機能部分がピキアパストリスのAOX1プロモーター、AOXIIプロモーター、GAPプロモーター、及びFLDプロモーターからなる群から選択されるプロモーターから発現される、請求項5記載の株。
【請求項25】
GalT又はその機能部分がピキアパストリスのAOX1プロモーター、AOXIIプロモーター、GAPプロモーター、及びFLDプロモーターからなる群から選択されるプロモーターから発現される、請求項5記載の株。
【請求項26】
α−1,2−マンノシダーゼ又はその機能部分がピキアパストリスのAOX1プロモーターから発現され、そして、GnTI又はその機能部分がピキアパストリスのGAPプロモーターから発現される、請求項5記載の株。
【請求項27】
哺乳類様グリカンがヒト様グリカンである、請求項1記載の株。
【請求項28】
哺乳類様のN−グリカン構造を有する糖蛋白質を組換えにより生成する方法であって、糖蛋白質をコードするヌクレオチド配列により請求項1−2又は5−6の何れか1項に記載の株を形質転換し、そして当該株中で糖蛋白質を発現させることを含む方法。
【請求項29】
糖蛋白質が細菌、真菌、ウイルス又は哺乳類起源のものである、請求項28記載の方法。
【請求項30】
糖蛋白質がイムノグロブリン又はその断片である、請求項28記載の方法。
【請求項31】
請求項28記載の方法により生産れた糖蛋白質。
【請求項32】
糖蛋白質がイムノグロブリン又はその断片である、請求項31記載の方法。
【請求項33】
哺乳類様のN−グリカン構造を有する糖蛋白質をメチロトローフ酵母株中で組換えにより生成する方法であって、糖蛋白質をコードするヌクレオチド配列により上記株を形質転換し;当該株を修飾するが、修飾された株が(1)α−1,2−マンノシダーゼ又はその機能部分、(2)N−アセチルグルコサミニルトランスフェラーゼI(又はGnTI)又はその機能部分、及び(3)β−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分を発現するようにし、そしてその際修飾された株中でのゲノミックOCH1遺伝子が不活性化されて;修飾された株中で糖蛋白質を生成する方法。
【請求項34】
糖蛋白質が細菌、真菌、ウイルス又は哺乳類起源のものである、請求項33記載の方法。
【請求項35】
糖蛋白質がイムノグロブリン又はその断片である、請求項33記載の方法。
【請求項36】
請求項33記載の方法により生産れた糖蛋白質。
【請求項37】
糖蛋白質がイムノグロブリン又はその断片である、請求項36記載の方法。
【請求項38】
プロモーター及び3’終結配列に対して作動可能なように連結したGalTをコードするヌクレオチド配列を含むベクターであって、当該プロモーター及び3’終結配列は当該株中で上記GalTの発現を達成するようにメチロトローフ酵母株中で機能する、上記ベクター。
【請求項39】
(1)α−1,2−マンノシダーゼ又はその機能部分をコードするヌクレオチド配列を含むベクター、(2)N−アセチルグルコサミニルトランスフェラーゼI(又はGnTI)又はその機能部分をコードするヌクレオチド配列を含むベクター、及び(3)β−1,4−ガラクトシルトランスフェラーゼ(GalT)又はその機能部分をコードするヌクレオチド配列を含むベクターを含むキットであって、但し、ベクターの各々はコードされた蛋白質のメチロトローフ酵母株中での発現を指示することができる、キット。
【請求項40】
メチロトローフ酵母株のゲノミックOCH1遺伝子を破壊することができるベクターをさらに含む、請求項39記載のキット。
【請求項41】
メチロトローフ酵母株に対して異種の糖蛋白質をコードするベクターをさらに含む、請求項39記載のキット。
【請求項42】
メチロトローフ酵母株をさらに含む、請求項39記載のキット。
【図1】



【図2】

【図3】
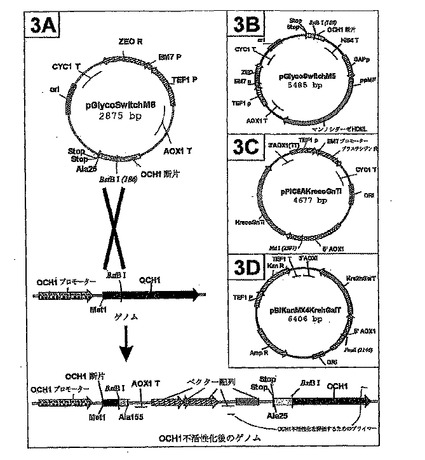
【図4】
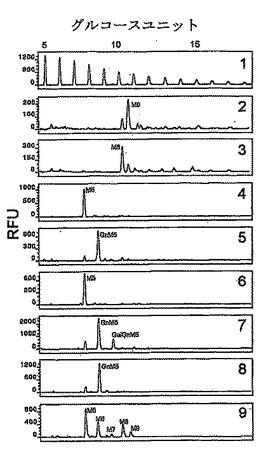
【図5】
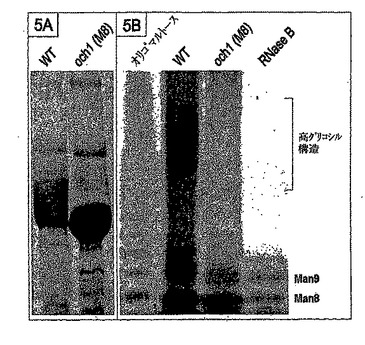
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2007−511223(P2007−511223A)
【公表日】平成19年5月10日(2007.5.10)
【国際特許分類】
【出願番号】特願2006−539939(P2006−539939)
【出願日】平成16年11月12日(2004.11.12)
【国際出願番号】PCT/US2004/037936
【国際公開番号】WO2005/049807
【国際公開日】平成17年6月2日(2005.6.2)
【出願人】(594063315)リサーチ・コーポレーション・テクノロジーズ・インコーポレーテッド (1)
【Fターム(参考)】
【公表日】平成19年5月10日(2007.5.10)
【国際特許分類】
【出願日】平成16年11月12日(2004.11.12)
【国際出願番号】PCT/US2004/037936
【国際公開番号】WO2005/049807
【国際公開日】平成17年6月2日(2005.6.2)
【出願人】(594063315)リサーチ・コーポレーション・テクノロジーズ・インコーポレーテッド (1)
【Fターム(参考)】
[ Back to top ]