
血友病Aモデルブタの作出
【課題】血友病の病態や効果的な治療方法を開発する上で有用な、自発性の出血症状を呈する血友病Aモデルブタを提供すること。
【解決手段】(a)第VIII因子遺伝子のエキソン16を標的としたターゲティングベクターを作製する工程;(b)胎児線維芽細胞に、工程(a)で作製したターゲティングベクターを導入し、相同組換えにより生じた遺伝子組換え細胞を選抜する工程;(c)採取した豚の体内成熟卵子から除核する工程;(d)工程(c)で除核された卵子に、工程(c)で選抜された遺伝子組換え細胞の細胞核を注入する工程;(e)工程(d)で細胞核が注入された卵子に活性化処理を施し、活性化処理後の核移植胚を雌豚の卵管又は子宮に移植する工程;を備えた血友病Aモデルブタの作出方法。
【解決手段】(a)第VIII因子遺伝子のエキソン16を標的としたターゲティングベクターを作製する工程;(b)胎児線維芽細胞に、工程(a)で作製したターゲティングベクターを導入し、相同組換えにより生じた遺伝子組換え細胞を選抜する工程;(c)採取した豚の体内成熟卵子から除核する工程;(d)工程(c)で除核された卵子に、工程(c)で選抜された遺伝子組換え細胞の細胞核を注入する工程;(e)工程(d)で細胞核が注入された卵子に活性化処理を施し、活性化処理後の核移植胚を雌豚の卵管又は子宮に移植する工程;を備えた血友病Aモデルブタの作出方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、X染色体上の第VIII因子遺伝子をノックアウトした血友病Aモデルブタ、特に自発性の出血症状(自然出血症状)を呈する血友病Aモデルブタに関する。
【背景技術】
【0002】
血友病は血液凝固因子欠乏症の1つで、難治性の先天性出血性疾患である。X染色体に座する第VIII因子遺伝子異常による血友病Aと第IX因子遺伝子異常による血友病Bに分けられる。19世紀中頃まで重症血友病男児は成人する前に死亡していたが、診断の進歩や有効な治療法の進歩により患者の生活は向上した。特に血液製剤の開発は血友病治療の転換点であり現在の治療の基礎となった。血液製剤の出現はまさに画期的なことであったが、一方で、多くの血友病患者が血液製剤輸注により肝炎ウイルスやHIVに感染し、患者は大きな犠牲を強いられることになった。その後、ウイルス不活化技術の向上と遺伝子組み換え凝固因子製剤の開発によりウイルス感染は激減し、既知のウイルスやプリオンへの暴露などの危険はほぼ無くなったと考えられる。現在は出血時に凝固因子製剤を投与するオンデマンド療法が中心であり、不測の致死的出血や傷害性出血を防ぐことは出来ない。また、補充する凝固因子の半減期が短いため(第VIII因子は8〜12時間、第IX因子は18〜24時間)、重症患者では凝固因子製剤の輸注回数が多くなることで、補充した凝固因子に対する同種抗体(インヒビター)が発生することがあり、治療が極めて困難になる。この他、製剤療法には、凝固因子製剤のコストの問題、製剤への未知の病原物質の混入の問題なども残っている。こうした問題を解決し、患者のQOL(Quality of Life)を向上させ治癒へとつながる次世代の治療法として遺伝子治療が期待されている。
【0003】
遺伝子治療の現在の技術レベルにおいて、治療レベルの遺伝子発現を得るためには、遺伝子導入にウイルスベクターを利用することが現実的である。ウイルスベクターの安全性については様々な改良がなされ、実用レベルに達している。しかし、各ベクターには搭載可能遺伝子サイズに制限があり、第VIII因子は遺伝子サイズが大きすぎる為、血友病A遺伝子治療研究は遅れていた。生体内で活性化される際に分解除去される部分の遺伝子を除く等の改変第VIII因子が考案され、海外では臨床治験も行われるようになったが、いずれのトライアルも一年以上に及ぶ長期間の第VIII因子活性の上昇は見られず、造血障害や肝障害が出現した症例もあった。血友病Aに対する遺伝子治療の臨床応用には、これらの課題をクリアすることが必須であり、更なる検討が必要であると考えられる。これまでの臨床治験は、第VIII因子遺伝子ノックアウトマウス(非特許文献1及び2参照)や自然発症の血友病イヌ(非特許文献3参照)を用いた実験結果を基に実施されている。
【0004】
他方、本発明者らは体細胞核直接導入法によるクローンブタの作出について報告(非特許文献4参照)し、また、EndoGalC遺伝子及びhDAF遺伝子からなる導入遺伝子を共発現する、超急性拒絶反応及び急性拒絶反応が抑制されたブタ臓器を提供しうるダブルトランスジェニックブタ(特許文献1参照)を報告している。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2008−220222号公報
【非特許文献】
【0006】
【非特許文献1】Nat Genet.10(1):119-21.1995
【非特許文献2】血栓止血誌8(6):517〜520,1997
【非特許文献3】Mol Ther. 5: S289. 2000
【非特許文献4】Science,289,1118-1119,2000
【発明の概要】
【発明が解決しようとする課題】
【0007】
凝固第VIII因子(FVIII)は種特異性が強いことから、よりヒトに近い実験動物を用いた前臨床試験で治療効果と安全性が評価された後にヒト臨床治験が行われるべきである。非ヒト霊長類であるサルには血友病が同定されておらず、導入したBDDFVIII抗原量の測定は可能になったが、FVIII活性の測定は不可能である。ヒト臨床試験開始前にはBDDFVIIIの止血能・治療効果の評価は必須であり、そのためにはヒトの類縁動物の欠損モデルが必要となる。しかし、霊長類で血友病モデルを作製するには、技術的にも倫理的にも大きな問題がある。
【0008】
本発明の課題は、血友病Aの病態や効果的な治療方法を開発する上で有用な、自然出血症状を呈する血友病Aモデルブタを提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、血友病Aの根治に向けた治療研究を進め、臨床への展開を加速させるために、ヒトに近い病態モデルとして近年本発明者らのグループで開発されたクローンブタ作製技術を用い、FVIII遺伝子を破壊した血友病Aブタの作製を検討した。種々の工夫をしてFVIIIノックアウトブタを作出した。その結果、自発的な出血症状の発症頻度が低いFVIII遺伝子欠損マウスに比べて、自発的な出血症状の発症頻度が極めて高いFVIII遺伝子欠損ブタが得られることを見いだし、本発明を完成した。これら血友病モデルブタは、出産後1日以内にFVIIIを投与し、その後2〜3日間隔でFVIIIを投与する必要があり、その措置を行わなければ、自然出血が多く観察されるため、死亡したり重篤な症状が観察される。また、飼養管理する柵内の床に通常とは異なる柔らかい素材のゴムマットを敷くなどの措置も講じる必要があることがわかった。
【0010】
すなわち、本発明は(1)X染色体上の第VIII因子遺伝子の一部もしくは全部が欠損し、野生型において発現される第VIII因子を発現する機能が失われたことを特徴とする血友病Aモデルブタや、(2)上記(1)記載の血友病Aモデルブタに由来する臓器、組織又は細胞に関する。
【0011】
また、本発明は、(3)以下の工程を備えた血友病Aモデルブタの作出方法:
(a)第VIII因子遺伝子のエキソン16を標的としたターゲティングベクターを作製する工程;
(b)胎児線維芽細胞に、工程(a)で作製したターゲティングベクターを導入し、相同組換えにより生じた遺伝子組換え細胞を選抜する工程;
(c)採取した豚の体内成熟卵子から除核する工程;
(d)工程(c)で除核された卵子に、工程(c)で選抜された遺伝子組換え細胞の細胞核を注入する工程;
(e)工程(d)で細胞核が注入された卵子に活性化処理を施し、活性化処理後の核移植胚を雌豚の卵管又は子宮に移植する工程;
や、(4)X染色体上の第VIII因子遺伝子の一部もしくは全部が欠損し、野生型において発現される第VIII因子を発現する機能が失われた第VIII因子ノックアウトブタに被検物質を投与し、該モデルブタにおける血液凝固機能の程度を評価することを特徴とする血友病の予防・治療剤のスクリーニング方法に関する。
【0012】
さらに、本発明は(5)ヒト第VIII因子とブタ第VIII因子とに交差反応性を有する抗第VIII因子抗体や、(6)ヒト第VIII因子により、X染色体上の第VIII因子遺伝子の一部もしくは全部が欠損し、野生型において発現される第VIII因子を発現する機能が失われた第VIII因子ノックアウトブタを免疫し、ヒト第VIII因子とブタ第VIII因子とに交差反応性を有する抗第VIII因子抗体を調製することを特徴とする抗第VIII因子抗体の製造方法に関する。
【発明の効果】
【0013】
ヒトに近い病態モデルが作製されれば、遺伝子治療研究だけでなく、新規治療法や新規製剤開発、インヒビター治療研究に絶大な貢献をもたらし、血友病A患者の望む治癒へ向けた治療研究が加速すると期待される。本発明によると、自然出血症状を呈する血友病Aモデルブタを提供することができる。また、本発明の血友病Aモデルブタに被検物質を投与し、該モデルブタにおける血液凝固機能の程度を比較・評価することにより、血友病の予防・治療剤をスクリーニングすることもできる。
【図面の簡単な説明】
【0014】
【図1】ブタFVIII遺伝子エキソン14-エキソン22制限酵素地図(F8)と遺伝子ターゲッティングベクター(F8−GTV)を示す図である。ブタFVIII遺伝子のエキソンは黒で示し、PGK(phosphoglycerate kinase)プロモーターで発現させるネオマイシン耐性遺伝子cDNAは斜線(右上から左下)で示している。HSV−TK(Herpes simplex virus thymidine kinase)自殺遺伝子cDNAは斜線(左上から右下)で示した。サザンブロットで用いたプローブの位置は二重下線で示した位置である。PCRスクリーニングで用いたプライマーの位置は鉤矢印で示した。また、制限酵素認識部位を矢印で示した。F8−GTVにおいてNeo下流のエキソン16は16塩基欠如させている。
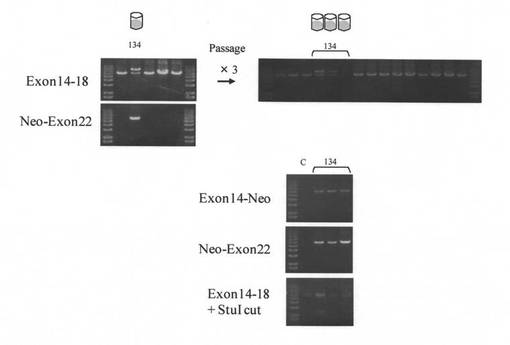
【図2】薬剤耐性コロニーのPCRスクリーニングの結果を示す図である。エキソン14−18で#134は野生型と同様のバンドと共に長いバンドが確認された。Neo−エキソン22により#134はNeoの挿入が示された。継代した3ウェルについてもエキソン14−18において2本のバンドが確認できる。Neoの挿入がエキソン14−Neo、Neo−エキソン22、エキソン14−18のStuIの切断によっても示された。C;野生型ブタゲノムDNA
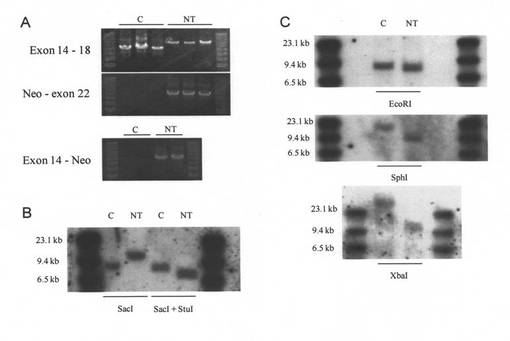
【図3】#134細胞核移植由来クローンブタ胎仔ゲノムDNAの解析結果を示す図である。A)PCRによる解析:エキソン14−18のPCRにおいてNTはCに比べてNeoの挿入とみられる長いバンドのみが認められた。Neo−エキソン22、エキソン14−NeoではNTにのみバンドが検出された。B)エキソン14プローブを用いたサザンブロット:SacIはエキソン14、イントロン18に存在するため、C(野生型ブタゲノムDNA)において7.7kbのバンドが検出され、NT(#134細胞核移植由来ブタ胎仔ゲノムDNA)ではNeoが挿入されたため、9.5kbにバンドがシフトしている。StuIはNeoにのみ存在するので、SacI+StuIではNTは7kbにバンドがシフトした。C)エキソン22プローブを用いたサザンブロット:EcoRIはNeoの3’に存在するため、NTでは僅かにバンドのシフトが認められる。SphIは5’−アームとNeoに、XbaIはNeoにのみ存在するのでNTにおいて減少するバンドのシフトが認められる。
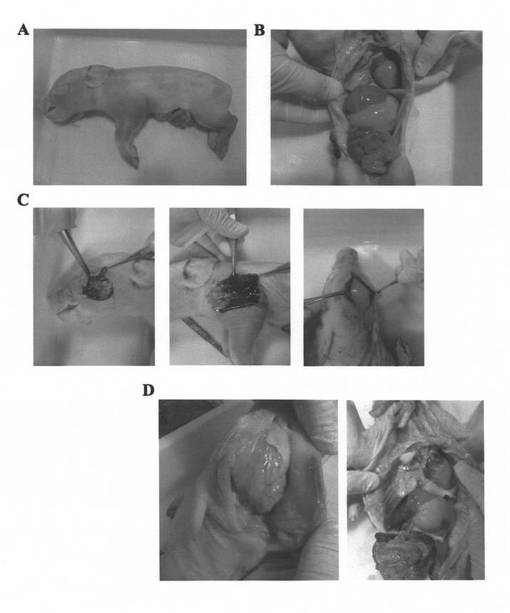
【図4】FVIII−KOブタ新生仔の解剖像の写真である。A)FVIII−KOブタ#1:全身に多くの内出血箇所が認められる。B)FVIII−KOブタ#1の臓器所見:臓器には異常は認められない。C)FVIII−KOブタ#1の内出血部位切開:内出血部位には血腫が認められた。D)FVIII−KOブタ#2の頭蓋内切開と臓器所見:頭蓋内には出血は認められず、皮下には出血が認められた。臓器に異常は認められない。
【図5】5ブタ血漿中FVIIIの解析結果を示す図である。A)ブタ血漿におけるFVIII抗原の検出:ブタ血漿ではFVIIIの検出はされなかった。ヒト血漿との混和(ブタ血漿/ヒト血漿:50/50、30/50、20/50、10/50、5/50、3/50、2/50、1/50、0/50)においてヒトFVIIIの検出はほぼ変化なく検出可能であった。B)ブタ血漿におけるAPPTとFVIII活性:ヒト血漿を100%とした場合、ブタ血漿は約3〜3.5倍のFVIII比活性を示した。
【発明を実施するための形態】
【0015】
本発明の血友病Aモデルブタとしては、X染色体上のFVIII遺伝子の一部もしくは全部が欠損し、野生型において発現されるFVIIIを発現する機能が失われたブタであれば特に制限されず、より具体的には雄ヘミ接合体ブタ(FVIII:X−/Y)や雌ホモ接合体(FVIII:X−/X−)を例示することができるが、これらの中でも自発性の出血症状を呈する血友病モデルブタを好適に例示することができる。上記雌ホモ接合体は、雌ヘテロ接合体ブタ(FVIII:X−/X+)と雄ヘミ接合体ブタ(FVIII:X−/Y)を掛け合わせると、25%の産生比率で出生する。
【0016】
上記血友病モデルブタは、(a)FVIII遺伝子のエキソン16を標的としたターゲティングベクターを作製する工程;(b)胎児線維芽細胞に、工程(a)で作製したターゲティングベクターを導入し、相同組換えにより生じた遺伝子組換え細胞を選抜する工程;(c)採取した豚の体内成熟卵子から除核する工程;(d)工程(c)で除核された卵子に、工程(c)で選抜された遺伝子組換え細胞の細胞核を注入する工程;(e)工程(d)で細胞核が注入された卵子に活性化処理を施し、活性化処理後の核移植胚を雌豚の卵管又は子宮に移植する工程;の各工程を備えたFVIIIノックアウトブタの作出方法により得ることができる。
【0017】
上記工程(a)における血友病モデルブタの作出に用いられるターゲティングベクターとしては、相同組換えによりFVIII遺伝子を破壊し、FVIII遺伝子機能を欠失させうるもので、FVIIIをコードする遺伝子中のエキソン16を含む領域が改変されたポリヌクレオチドを含有し、かつ該ポリヌクレオチドが、FVIIIの生理活性を欠損した産物を生じるものを挙げることができ、相同組換えにより生じた遺伝子組換え細胞をスクリーニングしやすくするために、エキソン16の中にネオマイシン耐性遺伝子(PGK−Neo)等の抗生物質耐性遺伝子やβガラクトシダーゼ遺伝子等の陽性選択マーカーを挿入させることができる他、HSVチミジンキナーゼ(HSV−TK)遺伝子、ジフテリア毒素(DT−A)遺伝子等の陰性選択マーカーを有するベクターが好ましい。
【0018】
上記工程(b)において、胎児線維芽細胞におけるターゲティングベクターによるFVIIIをコードする遺伝子の相同組換えは、ターゲティングベクターをブタ胎児線維芽細胞に導入することにより達成される。ターゲティングベクターを胎児線維芽細胞に導入する方法は、特に限定されるものではなく、例えば、エレクトロポレーション法、リポソーム法、リン酸カルシウム法、DEAE−デキストラン法等を挙げることができる。ターゲティングベクター導入後の胎児線維芽細胞を、細胞内での相同組換えが起こっているかを確認するためのサザンブロッティング法やPCR法等の常法によりスクリーニングすることにより、FVIIIをコードする遺伝子が欠損した胎児線維芽細胞(遺伝子組換え細胞)が得られる。上記胎児線維芽細胞はトリプシン処理で細胞を分散させたものが好ましく、また、培養細胞が線維芽細胞であることを、サイトケラチンとSSEA−1との陰性反応、ビメンチンでの陽性反応、線維芽細胞の特異的プライマーによるPCR分析等により確認することが好ましい。また、レシピエントや仮親と毛色の異なる品種のブタの線維芽細胞を核移植用ドナーとすると、毛色からノックアウトブタであるかどうかを簡便に判定することができる。
【0019】
また、核移植に用いるドナー線維芽細胞の細胞周期は特に制限されるものではないが、細胞周期G0/G01期に同調させた細胞が好ましい。細胞周期G0/G01期に同調させた細胞は、例えばコンフルエントな状態で培養液の交換なしに、線維芽細胞を16日間前後培養し続けることによって得ることができる。
【0020】
上記工程(c)におけるブタの卵子(レシピエント卵子)としては、ブタの成熟卵子であれば特に制限されるものではなく、PGF2α、PGF2α類縁体クロプロステロール、eCG等のホルモン投与による過排卵処理により得られる体内成熟卵子の他、屠場由来の卵巣から採取した卵子を体外成熟させたものも使用できるが、着床率の点からして体内成熟卵子、特に性成熟した雌ブタ、例えば生後160〜194日齢の春期発動前の雌ブタから採取した体内成熟卵子が好ましい。かかる体内成熟卵子は、過排卵処理により得られる雌ブタの子宮及び卵巣を、PBS溶液等を用いて卵管灌流を行うことにより採取することができるが、卵丘細胞が付着している卵子はヒアルロニダーゼ処理を行って、卵丘細胞を除去することが好ましい。
【0021】
上記ブタのレシピエント卵子からの除核は、細胞骨格形成阻害剤であるサイトカラシンB処理を施したブタの卵子から除核することが好ましく、より具体的にはサイトカラシンBを含有するPZM3液、NCSU23等の培地で体外成熟卵子等のレシピエント卵子を処理した後、除核操作用シャーレのサイトカラシン入りドロップに移してホールディングピペットで保定し、透明帯を迅速・的確に貫通することができる除核用ピペット(外径25〜30μm)を用いて、M(metaphase)II期の染色体を含む第一極体の付近を極体ごと吸引することにより行われる。なお、吸引した極体を調べることにより除核できていることを確認することが好ましく、また除核卵子からはサイトカラシンBを除去することが好ましい。
【0022】
上記工程(d)において、除核された卵子にブタ胎児遺伝子組換え線維芽細胞核を注入する方法としては、ブタ胎児遺伝子組換え線維芽細胞の核を除核されたレシピエント卵母細胞に直接注入(インジェクション)する方法を特に好適に例示することができる。かかる核の直接注入には、例えば、プライムテック株式会社製のPMM三次元マイクロマニピュレーションシステムを用いることができる。特に、インジェクションピペットとしては、透明帯の貫通が迅速・精確かつ簡単にでき、細胞質膜へのダメージを最小にすることができるものが好ましく、かかるインジェクションピペットとしてはピエゾマイクロマニピュレーター(プライムテック株式会社製)に取り付けた体細胞注入用ピペット(外径25〜30μm、先端を30〜45度の角度で研磨)を具体的に例示することができる。
【0023】
かかる工程(d)において体細胞核が注入された卵子には、上記工程(e)において活性化処理を施すことが好ましい。かかる活性化処理としては、従来公知の核移植胚の活性化処理方法であれば特に制限されるものではないが、電気パルス活性化処理を好適に例示することができる。電気パルス活性化処理としては、電荷の大きい1回の電気パルス、例えば1.5kV/cm、100μsec、1回を印加する方がそれより小さい電荷の電気パルスを2回印加するよりも胚活性の点で好ましく、また、電気パルス活性化処理における培地としてはNCSU23(J.Reprod.Fertil.Suppl.,48,61,1993)を用いることが高い胚盤胞形成率の点で好ましい。また、電気パルスによる活性化処理の場合、体内成熟卵子の方が体外成熟卵子に比べて胚盤胞の発生能の点で好ましい。さらに、レシピエント細胞として体内成熟卵子を用いる場合には、過排卵処理のために使用した最初のhCG投与後、50〜60時間後、好ましくは54〜55時間後に活性化処理をすることが望ましい。
【0024】
また、卵細胞からの除核時及び該除核細胞への体細胞核の直接注入時の2回にわたって損傷を受けた核移植胚の胚盤胞への発生率を高め、クローンブタを効率よく作出するために、活性化処理後の核移植胚は、卵管又は子宮への移植前に、包埋剤により包埋処理を施すこともできる。かかる包埋剤による包埋処理は、複数回の包埋処理を行い、複数被膜で包埋した核移植胚とすることが好ましく、特に3重包埋処理を行い、3重被膜核移植胚とすることが好ましい。また、かかる複数回の包埋処理を行うに際しては、包埋剤の濃度を順次高めていく包埋処理、すなわち外層膜ほど高濃度の包埋剤を用いて包埋し、その物理的強度を内層から外層へと順次高めていくことが好ましい。
【0025】
包埋処理に用いられる包埋剤としては、核移植胚を包埋することにより、2回にわたって損傷を受けた核移植胚の桑実胚や胚盤胞への発生率を高め、クローンブタを効率よく作出することができるものであれば特に制限されるものではないが、損傷を受けた透明帯の修復・保護機能を有するものや、包埋処理後の核移植胚を卵管及び子宮に移植した後、生体内での分解機能を有するものや、卵管及び子宮の膜運動による損耗からの保護機能を有するものや、白血球の攻撃からの防御機能を有するものや、包埋皮膜を通しての栄養分の透過・排泄物の排出機能を有するものの他、包埋処理時に核移植胚に影響を与えることなく包埋処理することができるものや、顕微鏡下で包埋皮膜が確認しやすいものや、顕微鏡下で包埋胚の操作がし易くなるものなどが好ましい。かかる包埋剤としては、アルギン酸、寒天等の天然多糖類の他、ムコタンパク質、ポリアミノ酸などの蛋白質、生分解性有機高分子等を例示することができるが、上記包埋剤としての好ましい機能を備えたアルギン酸が特に好ましい。その他、包埋剤の使用濃度についても特に制限されるものではないが、上記包埋剤としての好ましい機能を十分発揮しうる濃度が好ましい。なお、複数回の包埋処理を行う場合、例えばアルギン酸と寒天等、包埋剤の種類を変えて包埋処理をすることもできる。
【0026】
例えば、アルギン酸を包埋剤として3重包埋処理する方法としては、アルギン酸ナトリウムを所定濃度(例えば0.5%、1.5%、2.0%)となるようにリンゲル液等に溶解し、あらかじめ滅菌しておき、この滅菌済みのアルギン酸ナトリウム溶液(例えば0.5%)に核移植胚を浸漬して十分馴染ませた後、塩化カルシウム液等のカルシウムイオン含有液と接触させ、アルギン酸ゲル包埋胚とした後、この包埋胚を前記アルギン酸ナトリウム溶液よりも高濃度のアルギン酸ナトリウム溶液(例えば1.5%)に浸漬して十分馴染ませた後、塩化カルシウム液等のカルシウムイオン含有液と接触させ、アルギン酸ゲル2重包埋胚とし、次いでこの2重包埋胚を上記アルギン酸ナトリウム溶液よりも高濃度のアルギン酸ナトリウム溶液(例えば2.0%)に浸漬して十分馴染ませた後、塩化カルシウム液等のカルシウムイオン含有液と接触させ、アルギン酸ゲル3重包埋核移植胚とする方法を具体的に示すことができる。
【0027】
活性化処理後に、必要に応じて包埋剤による包埋処理をした核移植胚を、卵管又は子宮に移植する雌ブタとしては特に制限されるものではないが、人工授精させた後の妊娠21〜40日目にプロスタグランジンF2α等を用いて人工流産させ、同期化を行った雌ブタを用いることが好ましい。また、核移植胚を雌ブタの卵管又は子宮に移植するに際し、複数個の受精卵を核移植胚に混合して雌ブタの卵管又は子宮に移植する追い移植法を用いることが好ましい。
【0028】
また産仔したブタが、FVIIIノックアウトブタであることの確認は、産仔の毛色の他、FVIIIノックアウトクローンブタ、FVIIIノックアウトクローンブタの仮親の耳から採取したDNA並びにFVIIIノックアウトクローンブタを作出するために用いた線維芽細胞のDNAを採取し、PCRを行い、FVIIIノックアウトクローンブタが線維芽細胞と同一の遺伝子をもち、FVIII遺伝子が破壊され、自然出血症状を呈するかどうかを確認することにより同定することができる。
【0029】
核移植用ドナーとして雄の胎児線維芽細胞を用いたときには雄ヘミ接合体ブタ(FVIII:X−/Y)からなる血友病モデルブタを作出することができるが、雌の胎児線維芽細胞を用いたときには血友病症状を呈さない雌ヘテロ接合体ブタ(FVIII:X−/X+)が得られる。この場合、雄ヘミ接合体ブタと雌ヘテロ接合体ブタを掛け合わせると、血友病症状を呈する25%の雌ホモ接合体、25%の雌ヘテロ接合体、血友病症状を呈する25%の雄ヘミ接合体ブタ、25%の雄の正常ブタの出生割合で産仔が得られる。前記非特許文献2によると、雄ヘミ接合体マウスで約11%、雌ホモ接合体マウスで約7%が自発性の出血症状を呈するに過ぎず、8〜10週を経過したFVIIIKOマウスでは、尾を切断したり、尾を火傷させた場合でも全マウスが生存すると報告されているが、本発明によると、雄ヘミ接合体ブタで100%自発性の出血症状を呈し、5週を経過した後でも自然出血症状を維持していた。
【0030】
本発明はまた、本発明の血友病モデルブタに由来する臓器、組織又は細胞に関し、かかる臓器、組織又は細胞は、血友病モデルブタと同様に、血友病の予防・治療剤のスクリーニングに有利に用いることができる。また、臓器、組織または生殖細胞を含む細胞を用いることで、後代を含む個体作出に利用できる。
【0031】
本発明の血友病の予防・治療剤のスクリーニング方法としては、上記本発明の血友病モデルブタ(FVIIIノックアウトブタ)をに被検物質を投与し、該モデルブタにおける血液凝固機能の程度を、被検物質を投与しない場合と比較・評価する方法であれば特に制限されず、上記血液凝固機能の程度とは、自然出血や怪我等の外因性要因による出血の症状の止血の程度や止血の持続期間の程度をいい、血液凝固機能の程度を既存の血友病の予防・治療剤と比較・評価することができる。上記被検物質としては、FVIII遺伝子やその一部、又はFVIII遺伝子改変体が組み込まれたDNAベクター、例えば、組換えアデノ随伴ウイルス(adeno-associated virus:AAV)ベクター、組換えレンチウイルス(lentivirus)ベクター、組換えアデノウイルス(adeno virus)ベクター、ネイキッドDNAベクター等を好適に例示することができる。本発明は、上記本発明のスクリーニング方法により得られた血友病Aの予防・治療剤をも包含する。
【0032】
本発明の抗FVIII抗体としては、ヒトFVIIIとブタFVIIIとに交差反応性を有する抗体であれば特に制限されず、抗FVIIIモノクローナル抗体が好ましい。本発明の抗FVIII抗体は、ヒトFVIIIでFVIIIノックアウトブタを免疫することにより調製することができる。かかる本発明のヒトFVIIIとブタFVIIIとに交差反応性を有する抗体は、FVIIIインヒビターである抗FVIII抗体を無力化あるいは回避する血友病の予防・治療剤のスクリーニングに使用することができる。
【実施例1】
【0033】
[FVIII遺伝子KOクローンブタの作出]
1.FVIII遺伝子ターゲティングベクターの作製(ベクター名:F8−GTV)
胎齢65日目の雄ブタ胎仔線維芽細胞からDNAzol(Invitrogen社製)を用いてブタゲノムDNAを分離し、このゲノムDNAからFVIII遺伝子のエキソン16を標的とするベクターを構築した。Sus scrofa coagulation factor VIII cDNA sequence(Accession number;NM_214167)をもとにプライマーを作製し(表1)、エキソン14からエキソン21までと、エキソン16からエキソン22までをExpand Long Template PCR System(Roche Diagnostics社製)を用いたPCRにて増幅させ、pCR2.1-TOPO Cloning Kit(Invitrogen社製)またはpCRBluntII-TOPO Cloning Kit(Invitrogen社製)を用いてクローニング後、ABI PRISM Genetic Analyzer(Applied Biosystems社製)によるDNA塩基配列決定と、エキソン部位とcDNA配列とを照合した。さらにイントロン14部位におけるXhoI認識配列を付加したプライマー(表1)とエキソン21プライマーのPCRによりクローニングしたイントロン14−エキソン21をエキソン16内にあるXhoIで処理したイントロン14−エキソン16の5’−アームを、エキソン16プライマーとイントロン21部位におけるXbaI認識配列を付加したプライマー(表1)によりエキソン16−イントロン21の3’−アームをクローニングし、pBS-HSVTK-PGKNeo(京都大学ウイルス研究所 生田宏一教授からの供与)のXhoIとXbaI部位へ組み込み、エキソン16の中にPGK−Neoを挿入させ、5’−アーム上流に自殺遺伝子であるHSV−TKを配したFVIII遺伝子ターゲティングベクターを作製した。エレクトロポレーションに用いるため、NotI(TOYOBO社製)の消化により線状化し、フェノール/クロロフォルムによる精製後、HBS(HEPEs Buffered Saline)で25nMの濃度に希釈した。
【0034】
【表1】

【0035】
2.制限酵素地図の作成
5’−アームあるいは3’−アームをクローニングしたプラスミドを任意の制限酵素にて37℃、2時間処理後、アガロース電気泳動とエチジウムブロマイド染色にて消化バンドを確認し、DNA塩基配列決定した配列の制限酵素認識部位を照合させ、制限酵素地図を作成した。
【0036】
3.胎児線維芽細胞の採取
ランドレース♀×大ヨークシャー♂で交配後、妊娠65もしくは72日目の胎児を採取した。採取した胎児は、ハサミで筋肉組織を採取し、細切した。細切した胎児組織1g当たり10mlの0.25%トリプシン−0.04%EDTA−2Na液に懸濁し、一晩冷蔵した(4℃)。遠心処理(1500rpm、5min)により上清を除去した後、37℃に保温したウォーターバスにて20−30分間処理した。保温処理後、胎児組織1g当たり10mlのDMEM+10%FCSを加えてピペッティングし、セルストイレーナーにて濾過した。濾過液を遠心後、上清を除去し、沈殿物に10mlのDMEM+10%FCSを添加して再懸濁した。細胞数を計測し、適当量になるようDMEM+10%FCSにて調整した後、75cm2細胞培養用フラスコに播いた。その播いた細胞は、37℃で5%CO2でコンフルエントになるまで培養し、以下のエレクトロポレーションによる遺伝子導入に用いた。
【0037】
4.遺伝子導入とG418薬剤選択(FVIII遺伝子KO細胞の作出)
分離および増殖した胎児線維芽細胞は、0.25%トリプシン−0.04%EDTA−2Na液で剥離させ、DMEM+10%FCSにて懸濁した。細胞懸濁液を遠心処理(1500rpm、5min)した後、上清を除去し、冷HBSに再懸濁した。再懸濁液は、再度遠心処理を行って上清を除去した。細胞数が2.5×107細胞/mlになるようにHBSにて調整した。キュベットに細胞溶液0.4ml(1.0×107細胞/ml)と100μlの調整済みDNA液(25nMの直線化したターゲッティングベクターを添加、図1.)を加えた後、良く混和した。遺伝子導入には、電気穿孔法装置を300V、950μF、抵抗∞の条件のパルスにて行った。パルス付加後、10分間室温にて静置した。静置した細胞は、DMEM+10%FCSに懸濁後、3枚の10cmシャーレに分けて培養した。パルス後48時間目に800μg/mlのネオマイシン(G418)および2μMガンシクロビア(Denosine;Mitsubishi Tanabe Pharma社製)を培養液に添加し、耐性細胞株を7−12日間選択培養した。薬剤選択後、PBSで洗浄し、0.25%トリプシン−EDTA(Invitrogen社製)をしみ込ませたペーパーディスク(ADVANTEC社製)を細胞コロニーに乗せ、37℃で3分インキュベーションした後、培地を入れておいた24ウェル-プレート(Corning社製)へ1枚/ウェルとなるように継代した。培養5−6日目にフルコンフルエントになった各コロニーは1/6ずつ24ウェル−プレートの3ウェルに継代し、1/2の細胞についてゲノムDNAを抽出し、相同組換えにより生じた遺伝子組換え体についてスクリーニングした。同様に継代した3ウェルについてもフルコンフルエントにした後に細胞継代とゲノムDNAを抽出し、確認のPCRスクリーニングをした。PCRスクリーニングで陽性の結果が出たコロニーは、そのまま培養を継続し、核移植に供した。
【0038】
5.PCR解析によるターゲッティングの検出
PCRスクリーニングはエキソン14とエキソン18に設置したプライマー(表2)を用いたPCRによりFVIII遺伝子組み換え体コロニーを特定し、Neoに設置したプライマー(表2)とのPCRにより確認した。また、Neo遺伝子の挿入を確かめるため、エキソン14―18のPCRで得られたバンドをNeo遺伝子に存在するStuIの消化により確認した。FVIII遺伝子組換え体コロニーはG0期へ以降するまで増殖させ、核移植に使用した。核移植・胚移植由来の雄ブタ胎仔ゲノムについても同様のPCRにより解析した。細胞は、200g/mlのプロテナーゼKが含む50μlの超純水に再懸濁し、55℃で3時間保温した後、95℃10分間加熱してプロテナーゼを失活させた。PCR分析は、Roche社製Expand Long Template PCR Systemを用いた。25μl量の反応液中に消化した50μlのサンプルから鋳型として1μl(30〜500ng/μl)を用いた。PCRサイクルのパラメーターは、最初に94℃を2分間、次に94℃を10秒、68℃を6分の1サイクルを10サイクル、94℃を15秒、68℃を6分の1サイクルにおいて68℃については、1サイクル毎に20秒間増加させ25サイクル後、68℃を7分間処理した。プライマーはF8エキソン14F(5’−CATCAGAGGGACATAAGCCTTCCTAC−3’)およびF8エキソン18R(5’−GCCAGGGAGTGTATCCATCACATAG−3’)を用いた。このプライマーは、8kbの変異アレル特異的な産物を設定した。精製したDNAに対するPCRは、鋳型として100ngの精製DNAを使用した点を除いて同様に実施した。
【0039】
【表2】
【0040】
6.ホルモン処理による採卵方法
ランドレース(L)およびランドレース、大ヨークシャー(W)、デュロック(D)の三元交雑種(LWD)の生後160〜194日齢の春期発動前の雌ブタを用いた。供試ブタにeCG(ピーメックス、三共)を1500IU投与し、その72時間後にhCG(プペローゲン、三共)を500IU投与した。hCG投与48時間後に屠殺し、卵巣、卵管、子宮の結合組織を摘出した。また、性成熟に達した雌豚からの採卵は、予め発情時に雄希釈精液を用いて人工授精を行い、妊娠21−50日後に流産処置し行った。核移植予定5日前にPGF2α類縁体クロプロステロール約0.2mg(プラネート:2ml量、武田製薬社製)を臀部筋肉内に投与した。その24時間後に、同様にPGF2α類縁体クロプロステロールを注射し、同時にeCG(ピーメックス、三共製薬社製)を臀部筋肉内に1500IU投与した。PGF2α+eCG投与72時間後にhCG(プペローゲン、三共製薬社製)を500IU投与した。hCG投与46時間後に屠殺し、卵巣、卵管、子宮の結合組織を摘出した。
【0041】
7.体内成熟卵子の準備
摘出した卵巣、卵管、子宮の結合組織を直ちに実験室に持ち帰り、灌流液(PBS:Ca2+、Mg2+不含、(TAKARA社製)+0.1%ウシ血清アルブミン:BSA(Sigma社製、Cat.No.A−4503)+100mlあたり1ml添加抗生物質:Antibiotic,Antimycotic(Sigma社製、A−7292)で卵管を灌流し、卵子を採取した。得られた卵子は0.1%ヒアルロニダーゼを含む、灌流液を用いて卵丘細胞を裸化し、卵細胞質の均一な卵子を選抜した後、ヒアルロニダーゼを含まない灌流液中で洗浄した。その後、PZM−3液(Biol Reprod. 66:112-119. 2002)を用いてインキュベーター(38.5℃、5%CO2、5%O2、90%N2)内で供試するまで体外培養を行った。
【0042】
8.屠場卵巣由来卵胞卵子の採卵方法
屠場にて食肉用として屠殺された春機発動前の未経産雌ブタより卵巣を採取し、PBS(−)液にて35℃保温下で研究室に持ち帰った。持ち帰った卵巣は直ちに、37℃に保温したPBS(−)にて洗浄し、卵巣結合組織を除去した後、採卵作業を行った。直径3−6mmの卵胞を選び、16G注射針付き10mlシリンジで吸引した。吸引した卵胞液は、50mlディスポチューブに回収し、5分以上室温放置後、上清を除去した。卵子が含まれる沈殿物に対し、TALP−Hepes液を加えて懸濁後、沈殿物がチューブの底に溜まったのを確認し、沈殿物を吸引しないよう、パスツールピペットにて上清を除去した。この洗浄操作を2−3回繰り返した。卵丘細胞が3層以上密に付着した卵丘細胞卵子複合体(COC)とCOCに壁側顆粒層細胞が付着した壁側顆粒層細胞−卵丘細胞卵子複合体(GCOC)を選別・採卵をした。
【0043】
9.体外成熟培養
採取したCOCおよびGCOCは、約15〜20個/穴で改変NCSU37を100μl分注し、100μl流動パラフィンオイルカバーした6穴ディッシュを用いて成熟培養した。改変NCSU−37は、10%ブタ卵胞液、0.6mMシステイン、50μM βメルカプトエタノール、1mMジブチリルcAMP、10IU/ml PMSG(ピーメックス;1000IU三共製薬社製)、10IU/ml hCG(プペローゲン;1500IU、三共製薬社製)を添加して作製した。20〜22時間まで培養後、ホルモン及びジブチリルcAMP無添加改変NCSU37に移して供試時まで培養した。成熟培養は39℃、気相条件は、CO25%、O25%、N290%の条件で実施した。成熟培養時間は、42−44時間で行い、成熟卵子(第一極体放出卵子)のみを試験に使用した。
【0044】
10.卵子核の除去操作(除核操作)
除核操作前に、屠殺採卵した未受精卵子を、サイトカラシンB(Sigma社製:Cat.No.C−6762)が5μg/ml濃度含むPZM3液に移し、15分間以上処理した。除核操作も同濃度のサイトカラシンBが含まれるPZM3液で作製したドロップ内(10cmプラスチックシャーレの中心付近に50μlでドロップを作製し、ミネラルオイル20mlで被った)で行った。卵子は、第一極体が、12時から3時方向の位置になるようにホールディングピペットで保定した。ピエゾマイクロマニピュレーター(プライムテック株式会社製)に取り付けた除核用ピペット(外径25〜30μm、先端を30〜45度の角度で研磨)により、第一極体ごと細胞質を1/4〜1/3量程吸引した。除核用ピペットの操作性が悪くなった場合は、20%PVP液(Sigma社製:Cat.No.PVP−360)ドロップにて、吸引・排出を繰り返し、洗浄を行った。除核操作終了後、直ちにサイトカラシンBが含まれないPZM3液に移し、洗浄を行う。洗浄には、PZM3液が3ml入った35mmディッシュ(Falcon社製、1008)で2回以上、丁寧に洗浄し、サイトカラシンBを除去した。その後、注入操作まで、PZM3液ドロップ(35mmディッシュ:Falcon社製、3001に100μlのドロップを作製し、4mlミネラルオイルで被った)に移し、インキュベーター内で培養した。
【0045】
11.ドナー組換え体細胞の準備
PCR解析によりFVIII遺伝子がノックアウトされていると判定した雄胎児の線維芽細胞を核移植ドナー細胞として用いた。培養液にはDMEM+10% FCSを用い、植え継ぎ回数は0〜5回の細胞を用いた。核移植予定日に合わせて、植え継ぎを行った後、コンフルエント状態から培養液の交換を行わず、3〜14日間放置し、細胞周期をG1/G0期に同調した。核移植に用いる約1時間前にドナー体細胞の準備を行った。細胞処理は、培養容器の培養液を除去した後に、PBS(−)にて3回以上洗浄を行い、0.25%トリプシン+0.04%EDTA-2Na液にて細胞剥離処理を行った。剥離した細胞に10%FCS添加DMEMを加えて懸濁した後、遠心処理(1,500rpm、5min、室温)を行った。遠心処理後、上清を除去し、PZM3液を適量加えて懸濁し、核移植に用いるまで室温で放置した。体細胞核注入操作時に、適当な細胞数を注入操作用チャンバーのドロップに添加して使用した。
【0046】
12.体細胞核の注入操作
注入用チャンバー(10cmシャーレにPZM3液50μlでドロップを作製し、ミネラルオイル20mlで被った)のドロップに適量のドナー細胞を添加し、除核済み卵子50〜100個を入れて操作した。注入操作には、ピエゾマイクロマニュピレーターに体細胞核注入用ピペット(外径7−10μm、鈍端)を用いた。注入用ピペットをピエゾマイクロマニュピレーターに取り付ける前に、ピペット後端より5mmの位置にフロリナートを3−5mm幅になるように、充填して使用した。そのピペットをピエゾに取り付けて使用する時は、先端より培養液、フロリナート、空気、ミネラルオイルの順で満たされている状態にして操作した。注入操作を行う前に、必ず20%PVP液ドロップ内で注入用ピペットを洗浄(ピペッティングやPVPドロップへのピペット先端の出し入れ)した。浮遊している体細胞を吸引し、数回ピペッティングを行い、細胞膜を壊し、ホールディングピペットにて保定した除核済み卵子細胞質内へ注入した。体細胞核を注入した卵子は、活性化処理時間までインキュベーターで培養を行った(1〜3時間)。
【0047】
13.卵子の活性化処理
体細胞核を注入した卵子の活性化処理は直流パルス刺激(SSH−2、島津製作所製)を、1.5kV/cm 100μsec×1回の条件にて行った。チャンバーには2mm幅のステンレスワイヤー電極を用い、電気刺激用の電解質溶液には0.05mM CaCl2、0.1mM MgSO4ならびに0.02%BSAを含む0.28Mマンニトール溶液を用いた。
【0048】
14.活性化卵子の第2極体放出抑制処理
活性化処理卵子は5μg/mlサイトカラシンB(Sigma社製:Cat.No.C−6762)を含むPZM3液で2時間培養し、第2極体の放出を抑制させる処理を行った。
【0049】
15.活性化卵子の体外培養
活性化後2倍体処理をした卵子は、PZM3液にて体外培養を行い、活性化処理後2日目に分割胚のみ胚移植に用いた。
【0050】
16.ホルモン処理による仮腹の同期化と胚移植方法
仮腹の同期化は妊娠豚を人工流産処置することで行った。また、仮腹の発情は、採卵豚のものより、1日遅れるように発情同期化処置を行った。供試予定の雌ブタは、発情時に、雄豚希釈精液を用いて人工授精を行い妊娠させた。妊娠後21−50日目の妊娠豚を用いた。核移植予定6日前にPGF2α類縁体クロプロステロール約0.2mg(プラネート、武田製薬社製:2ml量)を臀部に注射した。その24時間後に、同様にPGF2α類縁体クロプロステロールを注射し、同時にeCG(ピーメックス、三共製薬社製)を1000IU投与した。PGF2α+eCG投与72時間後にhCG(プペローゲン、三共製薬社製)を500IU投与した。hCG投与、約68時間目に仮腹へケタラール投与後、ハロセン麻酔下で外科的に胚移植手術を行った。胚移植はPPカテーテル(フジヒラ社製)を卵管へ挿入して行った。
【0051】
17.組み換えクローン胎児の採取および細胞分離作業
同定したPCR陽性コロニーは、非相同組換え細胞が、様々な割合で含まれている可能性があり、体細胞核移植技術を用いた効率的な相同組換え動物の作出の妨げとなりうることが予想された。そこで、単一な相同組換え細胞のみの集団を得るため、妊娠した雌ブタの32日目もしくは39日目に胎児を採取した。採取した胎児は、3.と同様に細胞を分離すると同時にゲノムDNAを採取し、相同組み換え個体であるか5.と同様の条件でPCR解析した。PCR解析にて陽性であった胎児細胞を用いて再核移植を実施し、胚移植によりFVIIIノックアウトクローン個体の作出に供した。
【0052】
18.サザンブロット解析
核移植・胚移植により発生した雄ブタ胎仔線維芽細胞から抽出したゲノムDNAを用いてFVIII遺伝子の組換えをサザンブロットにて確認した。5mgのゲノムDNAを制限酵素SacI、StuI、EcoRI、SphI、あるいはXbaIで切断し、アガロース電気泳動後、アルカリトランスファー条件によるサザンブロッティングを実施した。バンドの検出はDIG PCR Labeling Kit(Roche社製)にてエキソン14またはエキソン22のDIGラベルプローブを、エキソンプライマー(表3)を用いたPCRにより作製し、DIG Easy Hyb(Roche社製)にてハイブリダイゼーション、DIG Detection Kit(Roche社製)により検出した。
【0053】
【表3】
【0054】
19.ブタ血漿の解析
血液と3.8%クエン酸ナトリウム溶液(Harasawa Pharmaceutical社製)を1/10となるようにブタから採血し、3000rpm、10分の遠心分離により血漿を分離した。活性化部分トロンボプラスチン時間(APTT)をCA500(Sysmex社製) にて測定し、ヒトFVIIIに対する抗体Sheep anti-human FVIII Antibody(Affinity Biologicals Inc.社製)とRabbit Anti-human FVIII(研究室で作製したポリクローナル抗体)、Goat anti-Rabbit IgG-HRP(BioSourse社製)を用いたELISAにより、FVIIIの検出を検討した。ヒト正常血漿(健常者ボランティアによる)においてAPTTのスタンダードを作製し、ヒト血漿FVIII活性を100%としてブタ血漿FVIII活性を算出した。ELISAでは、ブタ血漿とヒト血漿、それらの混和(ブタ/ヒト:1/1〜1/50)をサンプルとし、Benchmark plus(Bio-Rad Laboratories社製)にて吸光度415nmを測定し、ABTS(Kirkegaard&Perry Laboratories社製)の発色によりFVIIIを検出した。
【0055】
20.誕生したクローンブタのPCR判定
誕生したクローンブタから採取可能な組織(耳、臍帯組織)や血液等からDNAを抽出し、前記5.「PCR解析によるターゲッティングの検出」に準じてPCR解析を行った。
【0056】
(FVIII遺伝子ターゲッティングベクター)
ブタの全ゲノムDNA配列が解読・公開されていないため、現時点で公開されているブタ肝臓からクローニングされたFVIII cDNA配列をヒトFVIII cDNA配列との相同性によりエキソン部位を予測、PCRにより増幅・クローニングすることで遺伝子発現を欠損させるためのターゲッティングベクターを構築し、配列を解読した。cDNA配列とクローニングしたゲノムDNA配列の照合によってエキソン部位が特定された。
また、種々の制限酵素処理によってクローニングしたゲノムDNA領域の制限酵素地図を作製した(図1参照)。
【0057】
(FVIII遺伝子組換えブタ胎仔線維芽細胞の作出)
FVIII遺伝子ターゲティングベクターの導入と薬剤選択によって得られた936コロニーのうち増殖可能であった200コロニーをPCRスクリーニングした。遺伝子導入の回数と薬剤耐性コロニー数、増殖コロニー数、FVIII遺伝子組換え型コロニー数を表4に示す。エキソン14−18において、遺伝子組換え型ではエキソン16へのNeoの挿入により遺伝子増幅部位が長くなることを検証したところ、野生型と遺伝子組換え型の両方にバンドが出るコロニー#134が1つ得られた(図2)。コロニー#134におけるNeoの挿入はPCR(エキソン14−Neo、Neo-エキソン22)とエキソン14-18のPCR後のStuIの切断により陽性と確認された(図2)。さらに継代した3ウェルについても同様の結果が得られた(図2)。よって、得られたコロニー#134は野生型細胞とエキソン16にNeoが挿入されたFVIII遺伝子組換え細胞とを含む細胞集団であると考えられた。このコロニー#134を核移植に用いた。
【0058】
【表4】
【0059】
(FVIII遺伝子組換え細胞をもちいた核移植由来胎仔の解析)
FVIII遺伝子組換え細胞コロニー#134を用いた計5回の核移植・胚移植の結果、1匹のブタ胎仔が得られ、その線維芽細胞を分離・増殖・保存し、そのゲノムDNAの解析を検討した。PCRによる解析はエキソン14−18において遺伝子組換え型のみを示す結果が得られ、エキソン16へのNeoの挿入も陽性であった(図3A)。
また、サザンブロッティングによりFVIII遺伝子の組換えの確認したところ、エキソン14プローブまたはエキソン22プローブを用いた場合において、任意のバンドシフトが認められ(図3B、3C)、目的であるFVIII遺伝子のエキソン16にNeoが挿入されているという結果が得られた。よって、核移植に用いた細胞コロニーはFVIII遺伝子組換え細胞と野生型細胞が混在する細胞集団であったが、核移植後に発生したクローンブタ胎仔はFVIII遺伝子組換えブタ(FVIII−KOブタ)であることが示された。
【0060】
(FVIII遺伝子組換えブタ胎仔細胞の再核移植・胚移植)
作出した細胞由来のクローンブタ胎仔がFVIII遺伝子組換え(FVIII−KO)ブタであることがゲノムDNAの解析により確かめられたため、そのブタ胎仔線維芽細胞をもちいて計7回の核移植・胚移植を実施した。その結果、3頭の仮親が、妊娠し、分娩まで至った。4頭の生存クローンブタが誕生した。胚移植の実施日程と妊娠の有無、出産予定日、分娩頭数(死産)の結果を表5に示す。
【0061】
【表5】
【0062】
(FVIII−KOブタ新生仔の観察)
FVIII−KO胎仔線維芽細胞の核移植・胚移植により、4頭のFVIII−KOクローンブタが誕生した。しかし、出産直後あるいは2日目までに3頭が死亡した。
死亡したFVIII−KOブタ新生仔の剖検から、体の複数箇所に内出血が認められた(図4A)。母ブタとの接触による内出血と推察された。また、内出血部位を切開したところ、血腫が認められ(図4C)、FVIII−KOブタ自身には死亡前に出血傾向があるものと考えられた。
一方、頭蓋内には出血は認められず、臓器所見も異常がなかった(図4B、4D)。FVIII−KOブタ新生仔のDNAをエキソン14−18 PCRにより解析したところ、2頭ともFVIII−KOのクローンであることが確認された。
【0063】
(ブタ血漿の解析)
野生型ブタの血漿をヒトFVIIIを検出するELISAによって検出したところ、ブタFVIIIは検出されず、ヒト血漿との混和によるELISAにおいてもヒトFVIIIの検出を阻害するような結果は得られなかった(図5A)。また、野生型ブタ血漿のFVIII活性を測定したところ、ブタ血漿はヒト血漿の約3〜3.5倍のFVIII活性を有することが示された(図5B)。
生後2日に出血死したFVIII-KOクローンブタの血液の凝固因子活性(ヒト凝固因子活性換算)を測定したところ、第VIII因子活性は感度以下であり、正常対照となる野生型新生仔ブタの(生後1日)の凝固第VIII因子活性は200%以上であった。FVIIIKOクローンブタの血漿中で低下がみられた第VII因子活性と第XI因子活性は野生型新生仔ブタでも低値であった。FVIII-KOクローンブタでは第VIII因子活性のみが特異的に低下していたことは、得られたクローンブタがFVIII-KOクローンブタであることを確実に示し、出血症状は第VIII因子活性の低下に基づくものであるといえる。
【技術分野】
【0001】
本発明は、X染色体上の第VIII因子遺伝子をノックアウトした血友病Aモデルブタ、特に自発性の出血症状(自然出血症状)を呈する血友病Aモデルブタに関する。
【背景技術】
【0002】
血友病は血液凝固因子欠乏症の1つで、難治性の先天性出血性疾患である。X染色体に座する第VIII因子遺伝子異常による血友病Aと第IX因子遺伝子異常による血友病Bに分けられる。19世紀中頃まで重症血友病男児は成人する前に死亡していたが、診断の進歩や有効な治療法の進歩により患者の生活は向上した。特に血液製剤の開発は血友病治療の転換点であり現在の治療の基礎となった。血液製剤の出現はまさに画期的なことであったが、一方で、多くの血友病患者が血液製剤輸注により肝炎ウイルスやHIVに感染し、患者は大きな犠牲を強いられることになった。その後、ウイルス不活化技術の向上と遺伝子組み換え凝固因子製剤の開発によりウイルス感染は激減し、既知のウイルスやプリオンへの暴露などの危険はほぼ無くなったと考えられる。現在は出血時に凝固因子製剤を投与するオンデマンド療法が中心であり、不測の致死的出血や傷害性出血を防ぐことは出来ない。また、補充する凝固因子の半減期が短いため(第VIII因子は8〜12時間、第IX因子は18〜24時間)、重症患者では凝固因子製剤の輸注回数が多くなることで、補充した凝固因子に対する同種抗体(インヒビター)が発生することがあり、治療が極めて困難になる。この他、製剤療法には、凝固因子製剤のコストの問題、製剤への未知の病原物質の混入の問題なども残っている。こうした問題を解決し、患者のQOL(Quality of Life)を向上させ治癒へとつながる次世代の治療法として遺伝子治療が期待されている。
【0003】
遺伝子治療の現在の技術レベルにおいて、治療レベルの遺伝子発現を得るためには、遺伝子導入にウイルスベクターを利用することが現実的である。ウイルスベクターの安全性については様々な改良がなされ、実用レベルに達している。しかし、各ベクターには搭載可能遺伝子サイズに制限があり、第VIII因子は遺伝子サイズが大きすぎる為、血友病A遺伝子治療研究は遅れていた。生体内で活性化される際に分解除去される部分の遺伝子を除く等の改変第VIII因子が考案され、海外では臨床治験も行われるようになったが、いずれのトライアルも一年以上に及ぶ長期間の第VIII因子活性の上昇は見られず、造血障害や肝障害が出現した症例もあった。血友病Aに対する遺伝子治療の臨床応用には、これらの課題をクリアすることが必須であり、更なる検討が必要であると考えられる。これまでの臨床治験は、第VIII因子遺伝子ノックアウトマウス(非特許文献1及び2参照)や自然発症の血友病イヌ(非特許文献3参照)を用いた実験結果を基に実施されている。
【0004】
他方、本発明者らは体細胞核直接導入法によるクローンブタの作出について報告(非特許文献4参照)し、また、EndoGalC遺伝子及びhDAF遺伝子からなる導入遺伝子を共発現する、超急性拒絶反応及び急性拒絶反応が抑制されたブタ臓器を提供しうるダブルトランスジェニックブタ(特許文献1参照)を報告している。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2008−220222号公報
【非特許文献】
【0006】
【非特許文献1】Nat Genet.10(1):119-21.1995
【非特許文献2】血栓止血誌8(6):517〜520,1997
【非特許文献3】Mol Ther. 5: S289. 2000
【非特許文献4】Science,289,1118-1119,2000
【発明の概要】
【発明が解決しようとする課題】
【0007】
凝固第VIII因子(FVIII)は種特異性が強いことから、よりヒトに近い実験動物を用いた前臨床試験で治療効果と安全性が評価された後にヒト臨床治験が行われるべきである。非ヒト霊長類であるサルには血友病が同定されておらず、導入したBDDFVIII抗原量の測定は可能になったが、FVIII活性の測定は不可能である。ヒト臨床試験開始前にはBDDFVIIIの止血能・治療効果の評価は必須であり、そのためにはヒトの類縁動物の欠損モデルが必要となる。しかし、霊長類で血友病モデルを作製するには、技術的にも倫理的にも大きな問題がある。
【0008】
本発明の課題は、血友病Aの病態や効果的な治療方法を開発する上で有用な、自然出血症状を呈する血友病Aモデルブタを提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、血友病Aの根治に向けた治療研究を進め、臨床への展開を加速させるために、ヒトに近い病態モデルとして近年本発明者らのグループで開発されたクローンブタ作製技術を用い、FVIII遺伝子を破壊した血友病Aブタの作製を検討した。種々の工夫をしてFVIIIノックアウトブタを作出した。その結果、自発的な出血症状の発症頻度が低いFVIII遺伝子欠損マウスに比べて、自発的な出血症状の発症頻度が極めて高いFVIII遺伝子欠損ブタが得られることを見いだし、本発明を完成した。これら血友病モデルブタは、出産後1日以内にFVIIIを投与し、その後2〜3日間隔でFVIIIを投与する必要があり、その措置を行わなければ、自然出血が多く観察されるため、死亡したり重篤な症状が観察される。また、飼養管理する柵内の床に通常とは異なる柔らかい素材のゴムマットを敷くなどの措置も講じる必要があることがわかった。
【0010】
すなわち、本発明は(1)X染色体上の第VIII因子遺伝子の一部もしくは全部が欠損し、野生型において発現される第VIII因子を発現する機能が失われたことを特徴とする血友病Aモデルブタや、(2)上記(1)記載の血友病Aモデルブタに由来する臓器、組織又は細胞に関する。
【0011】
また、本発明は、(3)以下の工程を備えた血友病Aモデルブタの作出方法:
(a)第VIII因子遺伝子のエキソン16を標的としたターゲティングベクターを作製する工程;
(b)胎児線維芽細胞に、工程(a)で作製したターゲティングベクターを導入し、相同組換えにより生じた遺伝子組換え細胞を選抜する工程;
(c)採取した豚の体内成熟卵子から除核する工程;
(d)工程(c)で除核された卵子に、工程(c)で選抜された遺伝子組換え細胞の細胞核を注入する工程;
(e)工程(d)で細胞核が注入された卵子に活性化処理を施し、活性化処理後の核移植胚を雌豚の卵管又は子宮に移植する工程;
や、(4)X染色体上の第VIII因子遺伝子の一部もしくは全部が欠損し、野生型において発現される第VIII因子を発現する機能が失われた第VIII因子ノックアウトブタに被検物質を投与し、該モデルブタにおける血液凝固機能の程度を評価することを特徴とする血友病の予防・治療剤のスクリーニング方法に関する。
【0012】
さらに、本発明は(5)ヒト第VIII因子とブタ第VIII因子とに交差反応性を有する抗第VIII因子抗体や、(6)ヒト第VIII因子により、X染色体上の第VIII因子遺伝子の一部もしくは全部が欠損し、野生型において発現される第VIII因子を発現する機能が失われた第VIII因子ノックアウトブタを免疫し、ヒト第VIII因子とブタ第VIII因子とに交差反応性を有する抗第VIII因子抗体を調製することを特徴とする抗第VIII因子抗体の製造方法に関する。
【発明の効果】
【0013】
ヒトに近い病態モデルが作製されれば、遺伝子治療研究だけでなく、新規治療法や新規製剤開発、インヒビター治療研究に絶大な貢献をもたらし、血友病A患者の望む治癒へ向けた治療研究が加速すると期待される。本発明によると、自然出血症状を呈する血友病Aモデルブタを提供することができる。また、本発明の血友病Aモデルブタに被検物質を投与し、該モデルブタにおける血液凝固機能の程度を比較・評価することにより、血友病の予防・治療剤をスクリーニングすることもできる。
【図面の簡単な説明】
【0014】
【図1】ブタFVIII遺伝子エキソン14-エキソン22制限酵素地図(F8)と遺伝子ターゲッティングベクター(F8−GTV)を示す図である。ブタFVIII遺伝子のエキソンは黒で示し、PGK(phosphoglycerate kinase)プロモーターで発現させるネオマイシン耐性遺伝子cDNAは斜線(右上から左下)で示している。HSV−TK(Herpes simplex virus thymidine kinase)自殺遺伝子cDNAは斜線(左上から右下)で示した。サザンブロットで用いたプローブの位置は二重下線で示した位置である。PCRスクリーニングで用いたプライマーの位置は鉤矢印で示した。また、制限酵素認識部位を矢印で示した。F8−GTVにおいてNeo下流のエキソン16は16塩基欠如させている。
【図2】薬剤耐性コロニーのPCRスクリーニングの結果を示す図である。エキソン14−18で#134は野生型と同様のバンドと共に長いバンドが確認された。Neo−エキソン22により#134はNeoの挿入が示された。継代した3ウェルについてもエキソン14−18において2本のバンドが確認できる。Neoの挿入がエキソン14−Neo、Neo−エキソン22、エキソン14−18のStuIの切断によっても示された。C;野生型ブタゲノムDNA
【図3】#134細胞核移植由来クローンブタ胎仔ゲノムDNAの解析結果を示す図である。A)PCRによる解析:エキソン14−18のPCRにおいてNTはCに比べてNeoの挿入とみられる長いバンドのみが認められた。Neo−エキソン22、エキソン14−NeoではNTにのみバンドが検出された。B)エキソン14プローブを用いたサザンブロット:SacIはエキソン14、イントロン18に存在するため、C(野生型ブタゲノムDNA)において7.7kbのバンドが検出され、NT(#134細胞核移植由来ブタ胎仔ゲノムDNA)ではNeoが挿入されたため、9.5kbにバンドがシフトしている。StuIはNeoにのみ存在するので、SacI+StuIではNTは7kbにバンドがシフトした。C)エキソン22プローブを用いたサザンブロット:EcoRIはNeoの3’に存在するため、NTでは僅かにバンドのシフトが認められる。SphIは5’−アームとNeoに、XbaIはNeoにのみ存在するのでNTにおいて減少するバンドのシフトが認められる。
【図4】FVIII−KOブタ新生仔の解剖像の写真である。A)FVIII−KOブタ#1:全身に多くの内出血箇所が認められる。B)FVIII−KOブタ#1の臓器所見:臓器には異常は認められない。C)FVIII−KOブタ#1の内出血部位切開:内出血部位には血腫が認められた。D)FVIII−KOブタ#2の頭蓋内切開と臓器所見:頭蓋内には出血は認められず、皮下には出血が認められた。臓器に異常は認められない。
【図5】5ブタ血漿中FVIIIの解析結果を示す図である。A)ブタ血漿におけるFVIII抗原の検出:ブタ血漿ではFVIIIの検出はされなかった。ヒト血漿との混和(ブタ血漿/ヒト血漿:50/50、30/50、20/50、10/50、5/50、3/50、2/50、1/50、0/50)においてヒトFVIIIの検出はほぼ変化なく検出可能であった。B)ブタ血漿におけるAPPTとFVIII活性:ヒト血漿を100%とした場合、ブタ血漿は約3〜3.5倍のFVIII比活性を示した。
【発明を実施するための形態】
【0015】
本発明の血友病Aモデルブタとしては、X染色体上のFVIII遺伝子の一部もしくは全部が欠損し、野生型において発現されるFVIIIを発現する機能が失われたブタであれば特に制限されず、より具体的には雄ヘミ接合体ブタ(FVIII:X−/Y)や雌ホモ接合体(FVIII:X−/X−)を例示することができるが、これらの中でも自発性の出血症状を呈する血友病モデルブタを好適に例示することができる。上記雌ホモ接合体は、雌ヘテロ接合体ブタ(FVIII:X−/X+)と雄ヘミ接合体ブタ(FVIII:X−/Y)を掛け合わせると、25%の産生比率で出生する。
【0016】
上記血友病モデルブタは、(a)FVIII遺伝子のエキソン16を標的としたターゲティングベクターを作製する工程;(b)胎児線維芽細胞に、工程(a)で作製したターゲティングベクターを導入し、相同組換えにより生じた遺伝子組換え細胞を選抜する工程;(c)採取した豚の体内成熟卵子から除核する工程;(d)工程(c)で除核された卵子に、工程(c)で選抜された遺伝子組換え細胞の細胞核を注入する工程;(e)工程(d)で細胞核が注入された卵子に活性化処理を施し、活性化処理後の核移植胚を雌豚の卵管又は子宮に移植する工程;の各工程を備えたFVIIIノックアウトブタの作出方法により得ることができる。
【0017】
上記工程(a)における血友病モデルブタの作出に用いられるターゲティングベクターとしては、相同組換えによりFVIII遺伝子を破壊し、FVIII遺伝子機能を欠失させうるもので、FVIIIをコードする遺伝子中のエキソン16を含む領域が改変されたポリヌクレオチドを含有し、かつ該ポリヌクレオチドが、FVIIIの生理活性を欠損した産物を生じるものを挙げることができ、相同組換えにより生じた遺伝子組換え細胞をスクリーニングしやすくするために、エキソン16の中にネオマイシン耐性遺伝子(PGK−Neo)等の抗生物質耐性遺伝子やβガラクトシダーゼ遺伝子等の陽性選択マーカーを挿入させることができる他、HSVチミジンキナーゼ(HSV−TK)遺伝子、ジフテリア毒素(DT−A)遺伝子等の陰性選択マーカーを有するベクターが好ましい。
【0018】
上記工程(b)において、胎児線維芽細胞におけるターゲティングベクターによるFVIIIをコードする遺伝子の相同組換えは、ターゲティングベクターをブタ胎児線維芽細胞に導入することにより達成される。ターゲティングベクターを胎児線維芽細胞に導入する方法は、特に限定されるものではなく、例えば、エレクトロポレーション法、リポソーム法、リン酸カルシウム法、DEAE−デキストラン法等を挙げることができる。ターゲティングベクター導入後の胎児線維芽細胞を、細胞内での相同組換えが起こっているかを確認するためのサザンブロッティング法やPCR法等の常法によりスクリーニングすることにより、FVIIIをコードする遺伝子が欠損した胎児線維芽細胞(遺伝子組換え細胞)が得られる。上記胎児線維芽細胞はトリプシン処理で細胞を分散させたものが好ましく、また、培養細胞が線維芽細胞であることを、サイトケラチンとSSEA−1との陰性反応、ビメンチンでの陽性反応、線維芽細胞の特異的プライマーによるPCR分析等により確認することが好ましい。また、レシピエントや仮親と毛色の異なる品種のブタの線維芽細胞を核移植用ドナーとすると、毛色からノックアウトブタであるかどうかを簡便に判定することができる。
【0019】
また、核移植に用いるドナー線維芽細胞の細胞周期は特に制限されるものではないが、細胞周期G0/G01期に同調させた細胞が好ましい。細胞周期G0/G01期に同調させた細胞は、例えばコンフルエントな状態で培養液の交換なしに、線維芽細胞を16日間前後培養し続けることによって得ることができる。
【0020】
上記工程(c)におけるブタの卵子(レシピエント卵子)としては、ブタの成熟卵子であれば特に制限されるものではなく、PGF2α、PGF2α類縁体クロプロステロール、eCG等のホルモン投与による過排卵処理により得られる体内成熟卵子の他、屠場由来の卵巣から採取した卵子を体外成熟させたものも使用できるが、着床率の点からして体内成熟卵子、特に性成熟した雌ブタ、例えば生後160〜194日齢の春期発動前の雌ブタから採取した体内成熟卵子が好ましい。かかる体内成熟卵子は、過排卵処理により得られる雌ブタの子宮及び卵巣を、PBS溶液等を用いて卵管灌流を行うことにより採取することができるが、卵丘細胞が付着している卵子はヒアルロニダーゼ処理を行って、卵丘細胞を除去することが好ましい。
【0021】
上記ブタのレシピエント卵子からの除核は、細胞骨格形成阻害剤であるサイトカラシンB処理を施したブタの卵子から除核することが好ましく、より具体的にはサイトカラシンBを含有するPZM3液、NCSU23等の培地で体外成熟卵子等のレシピエント卵子を処理した後、除核操作用シャーレのサイトカラシン入りドロップに移してホールディングピペットで保定し、透明帯を迅速・的確に貫通することができる除核用ピペット(外径25〜30μm)を用いて、M(metaphase)II期の染色体を含む第一極体の付近を極体ごと吸引することにより行われる。なお、吸引した極体を調べることにより除核できていることを確認することが好ましく、また除核卵子からはサイトカラシンBを除去することが好ましい。
【0022】
上記工程(d)において、除核された卵子にブタ胎児遺伝子組換え線維芽細胞核を注入する方法としては、ブタ胎児遺伝子組換え線維芽細胞の核を除核されたレシピエント卵母細胞に直接注入(インジェクション)する方法を特に好適に例示することができる。かかる核の直接注入には、例えば、プライムテック株式会社製のPMM三次元マイクロマニピュレーションシステムを用いることができる。特に、インジェクションピペットとしては、透明帯の貫通が迅速・精確かつ簡単にでき、細胞質膜へのダメージを最小にすることができるものが好ましく、かかるインジェクションピペットとしてはピエゾマイクロマニピュレーター(プライムテック株式会社製)に取り付けた体細胞注入用ピペット(外径25〜30μm、先端を30〜45度の角度で研磨)を具体的に例示することができる。
【0023】
かかる工程(d)において体細胞核が注入された卵子には、上記工程(e)において活性化処理を施すことが好ましい。かかる活性化処理としては、従来公知の核移植胚の活性化処理方法であれば特に制限されるものではないが、電気パルス活性化処理を好適に例示することができる。電気パルス活性化処理としては、電荷の大きい1回の電気パルス、例えば1.5kV/cm、100μsec、1回を印加する方がそれより小さい電荷の電気パルスを2回印加するよりも胚活性の点で好ましく、また、電気パルス活性化処理における培地としてはNCSU23(J.Reprod.Fertil.Suppl.,48,61,1993)を用いることが高い胚盤胞形成率の点で好ましい。また、電気パルスによる活性化処理の場合、体内成熟卵子の方が体外成熟卵子に比べて胚盤胞の発生能の点で好ましい。さらに、レシピエント細胞として体内成熟卵子を用いる場合には、過排卵処理のために使用した最初のhCG投与後、50〜60時間後、好ましくは54〜55時間後に活性化処理をすることが望ましい。
【0024】
また、卵細胞からの除核時及び該除核細胞への体細胞核の直接注入時の2回にわたって損傷を受けた核移植胚の胚盤胞への発生率を高め、クローンブタを効率よく作出するために、活性化処理後の核移植胚は、卵管又は子宮への移植前に、包埋剤により包埋処理を施すこともできる。かかる包埋剤による包埋処理は、複数回の包埋処理を行い、複数被膜で包埋した核移植胚とすることが好ましく、特に3重包埋処理を行い、3重被膜核移植胚とすることが好ましい。また、かかる複数回の包埋処理を行うに際しては、包埋剤の濃度を順次高めていく包埋処理、すなわち外層膜ほど高濃度の包埋剤を用いて包埋し、その物理的強度を内層から外層へと順次高めていくことが好ましい。
【0025】
包埋処理に用いられる包埋剤としては、核移植胚を包埋することにより、2回にわたって損傷を受けた核移植胚の桑実胚や胚盤胞への発生率を高め、クローンブタを効率よく作出することができるものであれば特に制限されるものではないが、損傷を受けた透明帯の修復・保護機能を有するものや、包埋処理後の核移植胚を卵管及び子宮に移植した後、生体内での分解機能を有するものや、卵管及び子宮の膜運動による損耗からの保護機能を有するものや、白血球の攻撃からの防御機能を有するものや、包埋皮膜を通しての栄養分の透過・排泄物の排出機能を有するものの他、包埋処理時に核移植胚に影響を与えることなく包埋処理することができるものや、顕微鏡下で包埋皮膜が確認しやすいものや、顕微鏡下で包埋胚の操作がし易くなるものなどが好ましい。かかる包埋剤としては、アルギン酸、寒天等の天然多糖類の他、ムコタンパク質、ポリアミノ酸などの蛋白質、生分解性有機高分子等を例示することができるが、上記包埋剤としての好ましい機能を備えたアルギン酸が特に好ましい。その他、包埋剤の使用濃度についても特に制限されるものではないが、上記包埋剤としての好ましい機能を十分発揮しうる濃度が好ましい。なお、複数回の包埋処理を行う場合、例えばアルギン酸と寒天等、包埋剤の種類を変えて包埋処理をすることもできる。
【0026】
例えば、アルギン酸を包埋剤として3重包埋処理する方法としては、アルギン酸ナトリウムを所定濃度(例えば0.5%、1.5%、2.0%)となるようにリンゲル液等に溶解し、あらかじめ滅菌しておき、この滅菌済みのアルギン酸ナトリウム溶液(例えば0.5%)に核移植胚を浸漬して十分馴染ませた後、塩化カルシウム液等のカルシウムイオン含有液と接触させ、アルギン酸ゲル包埋胚とした後、この包埋胚を前記アルギン酸ナトリウム溶液よりも高濃度のアルギン酸ナトリウム溶液(例えば1.5%)に浸漬して十分馴染ませた後、塩化カルシウム液等のカルシウムイオン含有液と接触させ、アルギン酸ゲル2重包埋胚とし、次いでこの2重包埋胚を上記アルギン酸ナトリウム溶液よりも高濃度のアルギン酸ナトリウム溶液(例えば2.0%)に浸漬して十分馴染ませた後、塩化カルシウム液等のカルシウムイオン含有液と接触させ、アルギン酸ゲル3重包埋核移植胚とする方法を具体的に示すことができる。
【0027】
活性化処理後に、必要に応じて包埋剤による包埋処理をした核移植胚を、卵管又は子宮に移植する雌ブタとしては特に制限されるものではないが、人工授精させた後の妊娠21〜40日目にプロスタグランジンF2α等を用いて人工流産させ、同期化を行った雌ブタを用いることが好ましい。また、核移植胚を雌ブタの卵管又は子宮に移植するに際し、複数個の受精卵を核移植胚に混合して雌ブタの卵管又は子宮に移植する追い移植法を用いることが好ましい。
【0028】
また産仔したブタが、FVIIIノックアウトブタであることの確認は、産仔の毛色の他、FVIIIノックアウトクローンブタ、FVIIIノックアウトクローンブタの仮親の耳から採取したDNA並びにFVIIIノックアウトクローンブタを作出するために用いた線維芽細胞のDNAを採取し、PCRを行い、FVIIIノックアウトクローンブタが線維芽細胞と同一の遺伝子をもち、FVIII遺伝子が破壊され、自然出血症状を呈するかどうかを確認することにより同定することができる。
【0029】
核移植用ドナーとして雄の胎児線維芽細胞を用いたときには雄ヘミ接合体ブタ(FVIII:X−/Y)からなる血友病モデルブタを作出することができるが、雌の胎児線維芽細胞を用いたときには血友病症状を呈さない雌ヘテロ接合体ブタ(FVIII:X−/X+)が得られる。この場合、雄ヘミ接合体ブタと雌ヘテロ接合体ブタを掛け合わせると、血友病症状を呈する25%の雌ホモ接合体、25%の雌ヘテロ接合体、血友病症状を呈する25%の雄ヘミ接合体ブタ、25%の雄の正常ブタの出生割合で産仔が得られる。前記非特許文献2によると、雄ヘミ接合体マウスで約11%、雌ホモ接合体マウスで約7%が自発性の出血症状を呈するに過ぎず、8〜10週を経過したFVIIIKOマウスでは、尾を切断したり、尾を火傷させた場合でも全マウスが生存すると報告されているが、本発明によると、雄ヘミ接合体ブタで100%自発性の出血症状を呈し、5週を経過した後でも自然出血症状を維持していた。
【0030】
本発明はまた、本発明の血友病モデルブタに由来する臓器、組織又は細胞に関し、かかる臓器、組織又は細胞は、血友病モデルブタと同様に、血友病の予防・治療剤のスクリーニングに有利に用いることができる。また、臓器、組織または生殖細胞を含む細胞を用いることで、後代を含む個体作出に利用できる。
【0031】
本発明の血友病の予防・治療剤のスクリーニング方法としては、上記本発明の血友病モデルブタ(FVIIIノックアウトブタ)をに被検物質を投与し、該モデルブタにおける血液凝固機能の程度を、被検物質を投与しない場合と比較・評価する方法であれば特に制限されず、上記血液凝固機能の程度とは、自然出血や怪我等の外因性要因による出血の症状の止血の程度や止血の持続期間の程度をいい、血液凝固機能の程度を既存の血友病の予防・治療剤と比較・評価することができる。上記被検物質としては、FVIII遺伝子やその一部、又はFVIII遺伝子改変体が組み込まれたDNAベクター、例えば、組換えアデノ随伴ウイルス(adeno-associated virus:AAV)ベクター、組換えレンチウイルス(lentivirus)ベクター、組換えアデノウイルス(adeno virus)ベクター、ネイキッドDNAベクター等を好適に例示することができる。本発明は、上記本発明のスクリーニング方法により得られた血友病Aの予防・治療剤をも包含する。
【0032】
本発明の抗FVIII抗体としては、ヒトFVIIIとブタFVIIIとに交差反応性を有する抗体であれば特に制限されず、抗FVIIIモノクローナル抗体が好ましい。本発明の抗FVIII抗体は、ヒトFVIIIでFVIIIノックアウトブタを免疫することにより調製することができる。かかる本発明のヒトFVIIIとブタFVIIIとに交差反応性を有する抗体は、FVIIIインヒビターである抗FVIII抗体を無力化あるいは回避する血友病の予防・治療剤のスクリーニングに使用することができる。
【実施例1】
【0033】
[FVIII遺伝子KOクローンブタの作出]
1.FVIII遺伝子ターゲティングベクターの作製(ベクター名:F8−GTV)
胎齢65日目の雄ブタ胎仔線維芽細胞からDNAzol(Invitrogen社製)を用いてブタゲノムDNAを分離し、このゲノムDNAからFVIII遺伝子のエキソン16を標的とするベクターを構築した。Sus scrofa coagulation factor VIII cDNA sequence(Accession number;NM_214167)をもとにプライマーを作製し(表1)、エキソン14からエキソン21までと、エキソン16からエキソン22までをExpand Long Template PCR System(Roche Diagnostics社製)を用いたPCRにて増幅させ、pCR2.1-TOPO Cloning Kit(Invitrogen社製)またはpCRBluntII-TOPO Cloning Kit(Invitrogen社製)を用いてクローニング後、ABI PRISM Genetic Analyzer(Applied Biosystems社製)によるDNA塩基配列決定と、エキソン部位とcDNA配列とを照合した。さらにイントロン14部位におけるXhoI認識配列を付加したプライマー(表1)とエキソン21プライマーのPCRによりクローニングしたイントロン14−エキソン21をエキソン16内にあるXhoIで処理したイントロン14−エキソン16の5’−アームを、エキソン16プライマーとイントロン21部位におけるXbaI認識配列を付加したプライマー(表1)によりエキソン16−イントロン21の3’−アームをクローニングし、pBS-HSVTK-PGKNeo(京都大学ウイルス研究所 生田宏一教授からの供与)のXhoIとXbaI部位へ組み込み、エキソン16の中にPGK−Neoを挿入させ、5’−アーム上流に自殺遺伝子であるHSV−TKを配したFVIII遺伝子ターゲティングベクターを作製した。エレクトロポレーションに用いるため、NotI(TOYOBO社製)の消化により線状化し、フェノール/クロロフォルムによる精製後、HBS(HEPEs Buffered Saline)で25nMの濃度に希釈した。
【0034】
【表1】
【0035】
2.制限酵素地図の作成
5’−アームあるいは3’−アームをクローニングしたプラスミドを任意の制限酵素にて37℃、2時間処理後、アガロース電気泳動とエチジウムブロマイド染色にて消化バンドを確認し、DNA塩基配列決定した配列の制限酵素認識部位を照合させ、制限酵素地図を作成した。
【0036】
3.胎児線維芽細胞の採取
ランドレース♀×大ヨークシャー♂で交配後、妊娠65もしくは72日目の胎児を採取した。採取した胎児は、ハサミで筋肉組織を採取し、細切した。細切した胎児組織1g当たり10mlの0.25%トリプシン−0.04%EDTA−2Na液に懸濁し、一晩冷蔵した(4℃)。遠心処理(1500rpm、5min)により上清を除去した後、37℃に保温したウォーターバスにて20−30分間処理した。保温処理後、胎児組織1g当たり10mlのDMEM+10%FCSを加えてピペッティングし、セルストイレーナーにて濾過した。濾過液を遠心後、上清を除去し、沈殿物に10mlのDMEM+10%FCSを添加して再懸濁した。細胞数を計測し、適当量になるようDMEM+10%FCSにて調整した後、75cm2細胞培養用フラスコに播いた。その播いた細胞は、37℃で5%CO2でコンフルエントになるまで培養し、以下のエレクトロポレーションによる遺伝子導入に用いた。
【0037】
4.遺伝子導入とG418薬剤選択(FVIII遺伝子KO細胞の作出)
分離および増殖した胎児線維芽細胞は、0.25%トリプシン−0.04%EDTA−2Na液で剥離させ、DMEM+10%FCSにて懸濁した。細胞懸濁液を遠心処理(1500rpm、5min)した後、上清を除去し、冷HBSに再懸濁した。再懸濁液は、再度遠心処理を行って上清を除去した。細胞数が2.5×107細胞/mlになるようにHBSにて調整した。キュベットに細胞溶液0.4ml(1.0×107細胞/ml)と100μlの調整済みDNA液(25nMの直線化したターゲッティングベクターを添加、図1.)を加えた後、良く混和した。遺伝子導入には、電気穿孔法装置を300V、950μF、抵抗∞の条件のパルスにて行った。パルス付加後、10分間室温にて静置した。静置した細胞は、DMEM+10%FCSに懸濁後、3枚の10cmシャーレに分けて培養した。パルス後48時間目に800μg/mlのネオマイシン(G418)および2μMガンシクロビア(Denosine;Mitsubishi Tanabe Pharma社製)を培養液に添加し、耐性細胞株を7−12日間選択培養した。薬剤選択後、PBSで洗浄し、0.25%トリプシン−EDTA(Invitrogen社製)をしみ込ませたペーパーディスク(ADVANTEC社製)を細胞コロニーに乗せ、37℃で3分インキュベーションした後、培地を入れておいた24ウェル-プレート(Corning社製)へ1枚/ウェルとなるように継代した。培養5−6日目にフルコンフルエントになった各コロニーは1/6ずつ24ウェル−プレートの3ウェルに継代し、1/2の細胞についてゲノムDNAを抽出し、相同組換えにより生じた遺伝子組換え体についてスクリーニングした。同様に継代した3ウェルについてもフルコンフルエントにした後に細胞継代とゲノムDNAを抽出し、確認のPCRスクリーニングをした。PCRスクリーニングで陽性の結果が出たコロニーは、そのまま培養を継続し、核移植に供した。
【0038】
5.PCR解析によるターゲッティングの検出
PCRスクリーニングはエキソン14とエキソン18に設置したプライマー(表2)を用いたPCRによりFVIII遺伝子組み換え体コロニーを特定し、Neoに設置したプライマー(表2)とのPCRにより確認した。また、Neo遺伝子の挿入を確かめるため、エキソン14―18のPCRで得られたバンドをNeo遺伝子に存在するStuIの消化により確認した。FVIII遺伝子組換え体コロニーはG0期へ以降するまで増殖させ、核移植に使用した。核移植・胚移植由来の雄ブタ胎仔ゲノムについても同様のPCRにより解析した。細胞は、200g/mlのプロテナーゼKが含む50μlの超純水に再懸濁し、55℃で3時間保温した後、95℃10分間加熱してプロテナーゼを失活させた。PCR分析は、Roche社製Expand Long Template PCR Systemを用いた。25μl量の反応液中に消化した50μlのサンプルから鋳型として1μl(30〜500ng/μl)を用いた。PCRサイクルのパラメーターは、最初に94℃を2分間、次に94℃を10秒、68℃を6分の1サイクルを10サイクル、94℃を15秒、68℃を6分の1サイクルにおいて68℃については、1サイクル毎に20秒間増加させ25サイクル後、68℃を7分間処理した。プライマーはF8エキソン14F(5’−CATCAGAGGGACATAAGCCTTCCTAC−3’)およびF8エキソン18R(5’−GCCAGGGAGTGTATCCATCACATAG−3’)を用いた。このプライマーは、8kbの変異アレル特異的な産物を設定した。精製したDNAに対するPCRは、鋳型として100ngの精製DNAを使用した点を除いて同様に実施した。
【0039】
【表2】
【0040】
6.ホルモン処理による採卵方法
ランドレース(L)およびランドレース、大ヨークシャー(W)、デュロック(D)の三元交雑種(LWD)の生後160〜194日齢の春期発動前の雌ブタを用いた。供試ブタにeCG(ピーメックス、三共)を1500IU投与し、その72時間後にhCG(プペローゲン、三共)を500IU投与した。hCG投与48時間後に屠殺し、卵巣、卵管、子宮の結合組織を摘出した。また、性成熟に達した雌豚からの採卵は、予め発情時に雄希釈精液を用いて人工授精を行い、妊娠21−50日後に流産処置し行った。核移植予定5日前にPGF2α類縁体クロプロステロール約0.2mg(プラネート:2ml量、武田製薬社製)を臀部筋肉内に投与した。その24時間後に、同様にPGF2α類縁体クロプロステロールを注射し、同時にeCG(ピーメックス、三共製薬社製)を臀部筋肉内に1500IU投与した。PGF2α+eCG投与72時間後にhCG(プペローゲン、三共製薬社製)を500IU投与した。hCG投与46時間後に屠殺し、卵巣、卵管、子宮の結合組織を摘出した。
【0041】
7.体内成熟卵子の準備
摘出した卵巣、卵管、子宮の結合組織を直ちに実験室に持ち帰り、灌流液(PBS:Ca2+、Mg2+不含、(TAKARA社製)+0.1%ウシ血清アルブミン:BSA(Sigma社製、Cat.No.A−4503)+100mlあたり1ml添加抗生物質:Antibiotic,Antimycotic(Sigma社製、A−7292)で卵管を灌流し、卵子を採取した。得られた卵子は0.1%ヒアルロニダーゼを含む、灌流液を用いて卵丘細胞を裸化し、卵細胞質の均一な卵子を選抜した後、ヒアルロニダーゼを含まない灌流液中で洗浄した。その後、PZM−3液(Biol Reprod. 66:112-119. 2002)を用いてインキュベーター(38.5℃、5%CO2、5%O2、90%N2)内で供試するまで体外培養を行った。
【0042】
8.屠場卵巣由来卵胞卵子の採卵方法
屠場にて食肉用として屠殺された春機発動前の未経産雌ブタより卵巣を採取し、PBS(−)液にて35℃保温下で研究室に持ち帰った。持ち帰った卵巣は直ちに、37℃に保温したPBS(−)にて洗浄し、卵巣結合組織を除去した後、採卵作業を行った。直径3−6mmの卵胞を選び、16G注射針付き10mlシリンジで吸引した。吸引した卵胞液は、50mlディスポチューブに回収し、5分以上室温放置後、上清を除去した。卵子が含まれる沈殿物に対し、TALP−Hepes液を加えて懸濁後、沈殿物がチューブの底に溜まったのを確認し、沈殿物を吸引しないよう、パスツールピペットにて上清を除去した。この洗浄操作を2−3回繰り返した。卵丘細胞が3層以上密に付着した卵丘細胞卵子複合体(COC)とCOCに壁側顆粒層細胞が付着した壁側顆粒層細胞−卵丘細胞卵子複合体(GCOC)を選別・採卵をした。
【0043】
9.体外成熟培養
採取したCOCおよびGCOCは、約15〜20個/穴で改変NCSU37を100μl分注し、100μl流動パラフィンオイルカバーした6穴ディッシュを用いて成熟培養した。改変NCSU−37は、10%ブタ卵胞液、0.6mMシステイン、50μM βメルカプトエタノール、1mMジブチリルcAMP、10IU/ml PMSG(ピーメックス;1000IU三共製薬社製)、10IU/ml hCG(プペローゲン;1500IU、三共製薬社製)を添加して作製した。20〜22時間まで培養後、ホルモン及びジブチリルcAMP無添加改変NCSU37に移して供試時まで培養した。成熟培養は39℃、気相条件は、CO25%、O25%、N290%の条件で実施した。成熟培養時間は、42−44時間で行い、成熟卵子(第一極体放出卵子)のみを試験に使用した。
【0044】
10.卵子核の除去操作(除核操作)
除核操作前に、屠殺採卵した未受精卵子を、サイトカラシンB(Sigma社製:Cat.No.C−6762)が5μg/ml濃度含むPZM3液に移し、15分間以上処理した。除核操作も同濃度のサイトカラシンBが含まれるPZM3液で作製したドロップ内(10cmプラスチックシャーレの中心付近に50μlでドロップを作製し、ミネラルオイル20mlで被った)で行った。卵子は、第一極体が、12時から3時方向の位置になるようにホールディングピペットで保定した。ピエゾマイクロマニピュレーター(プライムテック株式会社製)に取り付けた除核用ピペット(外径25〜30μm、先端を30〜45度の角度で研磨)により、第一極体ごと細胞質を1/4〜1/3量程吸引した。除核用ピペットの操作性が悪くなった場合は、20%PVP液(Sigma社製:Cat.No.PVP−360)ドロップにて、吸引・排出を繰り返し、洗浄を行った。除核操作終了後、直ちにサイトカラシンBが含まれないPZM3液に移し、洗浄を行う。洗浄には、PZM3液が3ml入った35mmディッシュ(Falcon社製、1008)で2回以上、丁寧に洗浄し、サイトカラシンBを除去した。その後、注入操作まで、PZM3液ドロップ(35mmディッシュ:Falcon社製、3001に100μlのドロップを作製し、4mlミネラルオイルで被った)に移し、インキュベーター内で培養した。
【0045】
11.ドナー組換え体細胞の準備
PCR解析によりFVIII遺伝子がノックアウトされていると判定した雄胎児の線維芽細胞を核移植ドナー細胞として用いた。培養液にはDMEM+10% FCSを用い、植え継ぎ回数は0〜5回の細胞を用いた。核移植予定日に合わせて、植え継ぎを行った後、コンフルエント状態から培養液の交換を行わず、3〜14日間放置し、細胞周期をG1/G0期に同調した。核移植に用いる約1時間前にドナー体細胞の準備を行った。細胞処理は、培養容器の培養液を除去した後に、PBS(−)にて3回以上洗浄を行い、0.25%トリプシン+0.04%EDTA-2Na液にて細胞剥離処理を行った。剥離した細胞に10%FCS添加DMEMを加えて懸濁した後、遠心処理(1,500rpm、5min、室温)を行った。遠心処理後、上清を除去し、PZM3液を適量加えて懸濁し、核移植に用いるまで室温で放置した。体細胞核注入操作時に、適当な細胞数を注入操作用チャンバーのドロップに添加して使用した。
【0046】
12.体細胞核の注入操作
注入用チャンバー(10cmシャーレにPZM3液50μlでドロップを作製し、ミネラルオイル20mlで被った)のドロップに適量のドナー細胞を添加し、除核済み卵子50〜100個を入れて操作した。注入操作には、ピエゾマイクロマニュピレーターに体細胞核注入用ピペット(外径7−10μm、鈍端)を用いた。注入用ピペットをピエゾマイクロマニュピレーターに取り付ける前に、ピペット後端より5mmの位置にフロリナートを3−5mm幅になるように、充填して使用した。そのピペットをピエゾに取り付けて使用する時は、先端より培養液、フロリナート、空気、ミネラルオイルの順で満たされている状態にして操作した。注入操作を行う前に、必ず20%PVP液ドロップ内で注入用ピペットを洗浄(ピペッティングやPVPドロップへのピペット先端の出し入れ)した。浮遊している体細胞を吸引し、数回ピペッティングを行い、細胞膜を壊し、ホールディングピペットにて保定した除核済み卵子細胞質内へ注入した。体細胞核を注入した卵子は、活性化処理時間までインキュベーターで培養を行った(1〜3時間)。
【0047】
13.卵子の活性化処理
体細胞核を注入した卵子の活性化処理は直流パルス刺激(SSH−2、島津製作所製)を、1.5kV/cm 100μsec×1回の条件にて行った。チャンバーには2mm幅のステンレスワイヤー電極を用い、電気刺激用の電解質溶液には0.05mM CaCl2、0.1mM MgSO4ならびに0.02%BSAを含む0.28Mマンニトール溶液を用いた。
【0048】
14.活性化卵子の第2極体放出抑制処理
活性化処理卵子は5μg/mlサイトカラシンB(Sigma社製:Cat.No.C−6762)を含むPZM3液で2時間培養し、第2極体の放出を抑制させる処理を行った。
【0049】
15.活性化卵子の体外培養
活性化後2倍体処理をした卵子は、PZM3液にて体外培養を行い、活性化処理後2日目に分割胚のみ胚移植に用いた。
【0050】
16.ホルモン処理による仮腹の同期化と胚移植方法
仮腹の同期化は妊娠豚を人工流産処置することで行った。また、仮腹の発情は、採卵豚のものより、1日遅れるように発情同期化処置を行った。供試予定の雌ブタは、発情時に、雄豚希釈精液を用いて人工授精を行い妊娠させた。妊娠後21−50日目の妊娠豚を用いた。核移植予定6日前にPGF2α類縁体クロプロステロール約0.2mg(プラネート、武田製薬社製:2ml量)を臀部に注射した。その24時間後に、同様にPGF2α類縁体クロプロステロールを注射し、同時にeCG(ピーメックス、三共製薬社製)を1000IU投与した。PGF2α+eCG投与72時間後にhCG(プペローゲン、三共製薬社製)を500IU投与した。hCG投与、約68時間目に仮腹へケタラール投与後、ハロセン麻酔下で外科的に胚移植手術を行った。胚移植はPPカテーテル(フジヒラ社製)を卵管へ挿入して行った。
【0051】
17.組み換えクローン胎児の採取および細胞分離作業
同定したPCR陽性コロニーは、非相同組換え細胞が、様々な割合で含まれている可能性があり、体細胞核移植技術を用いた効率的な相同組換え動物の作出の妨げとなりうることが予想された。そこで、単一な相同組換え細胞のみの集団を得るため、妊娠した雌ブタの32日目もしくは39日目に胎児を採取した。採取した胎児は、3.と同様に細胞を分離すると同時にゲノムDNAを採取し、相同組み換え個体であるか5.と同様の条件でPCR解析した。PCR解析にて陽性であった胎児細胞を用いて再核移植を実施し、胚移植によりFVIIIノックアウトクローン個体の作出に供した。
【0052】
18.サザンブロット解析
核移植・胚移植により発生した雄ブタ胎仔線維芽細胞から抽出したゲノムDNAを用いてFVIII遺伝子の組換えをサザンブロットにて確認した。5mgのゲノムDNAを制限酵素SacI、StuI、EcoRI、SphI、あるいはXbaIで切断し、アガロース電気泳動後、アルカリトランスファー条件によるサザンブロッティングを実施した。バンドの検出はDIG PCR Labeling Kit(Roche社製)にてエキソン14またはエキソン22のDIGラベルプローブを、エキソンプライマー(表3)を用いたPCRにより作製し、DIG Easy Hyb(Roche社製)にてハイブリダイゼーション、DIG Detection Kit(Roche社製)により検出した。
【0053】
【表3】
【0054】
19.ブタ血漿の解析
血液と3.8%クエン酸ナトリウム溶液(Harasawa Pharmaceutical社製)を1/10となるようにブタから採血し、3000rpm、10分の遠心分離により血漿を分離した。活性化部分トロンボプラスチン時間(APTT)をCA500(Sysmex社製) にて測定し、ヒトFVIIIに対する抗体Sheep anti-human FVIII Antibody(Affinity Biologicals Inc.社製)とRabbit Anti-human FVIII(研究室で作製したポリクローナル抗体)、Goat anti-Rabbit IgG-HRP(BioSourse社製)を用いたELISAにより、FVIIIの検出を検討した。ヒト正常血漿(健常者ボランティアによる)においてAPTTのスタンダードを作製し、ヒト血漿FVIII活性を100%としてブタ血漿FVIII活性を算出した。ELISAでは、ブタ血漿とヒト血漿、それらの混和(ブタ/ヒト:1/1〜1/50)をサンプルとし、Benchmark plus(Bio-Rad Laboratories社製)にて吸光度415nmを測定し、ABTS(Kirkegaard&Perry Laboratories社製)の発色によりFVIIIを検出した。
【0055】
20.誕生したクローンブタのPCR判定
誕生したクローンブタから採取可能な組織(耳、臍帯組織)や血液等からDNAを抽出し、前記5.「PCR解析によるターゲッティングの検出」に準じてPCR解析を行った。
【0056】
(FVIII遺伝子ターゲッティングベクター)
ブタの全ゲノムDNA配列が解読・公開されていないため、現時点で公開されているブタ肝臓からクローニングされたFVIII cDNA配列をヒトFVIII cDNA配列との相同性によりエキソン部位を予測、PCRにより増幅・クローニングすることで遺伝子発現を欠損させるためのターゲッティングベクターを構築し、配列を解読した。cDNA配列とクローニングしたゲノムDNA配列の照合によってエキソン部位が特定された。
また、種々の制限酵素処理によってクローニングしたゲノムDNA領域の制限酵素地図を作製した(図1参照)。
【0057】
(FVIII遺伝子組換えブタ胎仔線維芽細胞の作出)
FVIII遺伝子ターゲティングベクターの導入と薬剤選択によって得られた936コロニーのうち増殖可能であった200コロニーをPCRスクリーニングした。遺伝子導入の回数と薬剤耐性コロニー数、増殖コロニー数、FVIII遺伝子組換え型コロニー数を表4に示す。エキソン14−18において、遺伝子組換え型ではエキソン16へのNeoの挿入により遺伝子増幅部位が長くなることを検証したところ、野生型と遺伝子組換え型の両方にバンドが出るコロニー#134が1つ得られた(図2)。コロニー#134におけるNeoの挿入はPCR(エキソン14−Neo、Neo-エキソン22)とエキソン14-18のPCR後のStuIの切断により陽性と確認された(図2)。さらに継代した3ウェルについても同様の結果が得られた(図2)。よって、得られたコロニー#134は野生型細胞とエキソン16にNeoが挿入されたFVIII遺伝子組換え細胞とを含む細胞集団であると考えられた。このコロニー#134を核移植に用いた。
【0058】
【表4】
【0059】
(FVIII遺伝子組換え細胞をもちいた核移植由来胎仔の解析)
FVIII遺伝子組換え細胞コロニー#134を用いた計5回の核移植・胚移植の結果、1匹のブタ胎仔が得られ、その線維芽細胞を分離・増殖・保存し、そのゲノムDNAの解析を検討した。PCRによる解析はエキソン14−18において遺伝子組換え型のみを示す結果が得られ、エキソン16へのNeoの挿入も陽性であった(図3A)。
また、サザンブロッティングによりFVIII遺伝子の組換えの確認したところ、エキソン14プローブまたはエキソン22プローブを用いた場合において、任意のバンドシフトが認められ(図3B、3C)、目的であるFVIII遺伝子のエキソン16にNeoが挿入されているという結果が得られた。よって、核移植に用いた細胞コロニーはFVIII遺伝子組換え細胞と野生型細胞が混在する細胞集団であったが、核移植後に発生したクローンブタ胎仔はFVIII遺伝子組換えブタ(FVIII−KOブタ)であることが示された。
【0060】
(FVIII遺伝子組換えブタ胎仔細胞の再核移植・胚移植)
作出した細胞由来のクローンブタ胎仔がFVIII遺伝子組換え(FVIII−KO)ブタであることがゲノムDNAの解析により確かめられたため、そのブタ胎仔線維芽細胞をもちいて計7回の核移植・胚移植を実施した。その結果、3頭の仮親が、妊娠し、分娩まで至った。4頭の生存クローンブタが誕生した。胚移植の実施日程と妊娠の有無、出産予定日、分娩頭数(死産)の結果を表5に示す。
【0061】
【表5】
【0062】
(FVIII−KOブタ新生仔の観察)
FVIII−KO胎仔線維芽細胞の核移植・胚移植により、4頭のFVIII−KOクローンブタが誕生した。しかし、出産直後あるいは2日目までに3頭が死亡した。
死亡したFVIII−KOブタ新生仔の剖検から、体の複数箇所に内出血が認められた(図4A)。母ブタとの接触による内出血と推察された。また、内出血部位を切開したところ、血腫が認められ(図4C)、FVIII−KOブタ自身には死亡前に出血傾向があるものと考えられた。
一方、頭蓋内には出血は認められず、臓器所見も異常がなかった(図4B、4D)。FVIII−KOブタ新生仔のDNAをエキソン14−18 PCRにより解析したところ、2頭ともFVIII−KOのクローンであることが確認された。
【0063】
(ブタ血漿の解析)
野生型ブタの血漿をヒトFVIIIを検出するELISAによって検出したところ、ブタFVIIIは検出されず、ヒト血漿との混和によるELISAにおいてもヒトFVIIIの検出を阻害するような結果は得られなかった(図5A)。また、野生型ブタ血漿のFVIII活性を測定したところ、ブタ血漿はヒト血漿の約3〜3.5倍のFVIII活性を有することが示された(図5B)。
生後2日に出血死したFVIII-KOクローンブタの血液の凝固因子活性(ヒト凝固因子活性換算)を測定したところ、第VIII因子活性は感度以下であり、正常対照となる野生型新生仔ブタの(生後1日)の凝固第VIII因子活性は200%以上であった。FVIIIKOクローンブタの血漿中で低下がみられた第VII因子活性と第XI因子活性は野生型新生仔ブタでも低値であった。FVIII-KOクローンブタでは第VIII因子活性のみが特異的に低下していたことは、得られたクローンブタがFVIII-KOクローンブタであることを確実に示し、出血症状は第VIII因子活性の低下に基づくものであるといえる。
【特許請求の範囲】
【請求項1】
X染色体上の第VIII因子遺伝子の一部もしくは全部が欠損し、野生型において発現される第VIII因子を発現する機能が失われたことを特徴とする血友病Aモデルブタ。
【請求項2】
請求項1記載の血友病Aモデルブタに由来する臓器、組織又は細胞。
【請求項3】
以下の工程を備えた血友病Aモデルブタの作出方法。
(a)第VIII因子遺伝子のエキソン16を標的としたターゲティングベクターを作製する工程;
(b)胎児線維芽細胞に、工程(a)で作製したターゲティングベクターを導入し、相同組換えにより生じた遺伝子組換え細胞を選抜する工程;
(c)採取した豚の体内成熟卵子から除核する工程;
(d)工程(c)で除核された卵子に、工程(c)で選抜された遺伝子組換え細胞の細胞核を注入する工程;
(e)工程(d)で細胞核が注入された卵子に活性化処理を施し、活性化処理後の核移植胚を雌豚の卵管又は子宮に移植する工程;
【請求項4】
X染色体上の第VIII因子遺伝子の一部もしくは全部が欠損し、野生型において発現される第VIII因子を発現する機能が失われた第VIII因子ノックアウトブタに被検物質を投与し、該モデルブタにおける血液凝固機能の程度を評価することを特徴とする血友病の予防・治療剤のスクリーニング方法。
【請求項5】
ヒト第VIII因子とブタ第VIII因子とに交差反応性を有する抗第VIII因子抗体。
【請求項6】
ヒト第VIII因子により、X染色体上の第VIII因子遺伝子の一部もしくは全部が欠損し、野生型において発現される第VIII因子を発現する機能が失われた第VIII因子ノックアウトブタを免疫し、ヒト第VIII因子とブタ第VIII因子とに交差反応性を有する抗第VIII因子抗体を調製することを特徴とする抗第VIII因子抗体の製造方法。
【請求項1】
X染色体上の第VIII因子遺伝子の一部もしくは全部が欠損し、野生型において発現される第VIII因子を発現する機能が失われたことを特徴とする血友病Aモデルブタ。
【請求項2】
請求項1記載の血友病Aモデルブタに由来する臓器、組織又は細胞。
【請求項3】
以下の工程を備えた血友病Aモデルブタの作出方法。
(a)第VIII因子遺伝子のエキソン16を標的としたターゲティングベクターを作製する工程;
(b)胎児線維芽細胞に、工程(a)で作製したターゲティングベクターを導入し、相同組換えにより生じた遺伝子組換え細胞を選抜する工程;
(c)採取した豚の体内成熟卵子から除核する工程;
(d)工程(c)で除核された卵子に、工程(c)で選抜された遺伝子組換え細胞の細胞核を注入する工程;
(e)工程(d)で細胞核が注入された卵子に活性化処理を施し、活性化処理後の核移植胚を雌豚の卵管又は子宮に移植する工程;
【請求項4】
X染色体上の第VIII因子遺伝子の一部もしくは全部が欠損し、野生型において発現される第VIII因子を発現する機能が失われた第VIII因子ノックアウトブタに被検物質を投与し、該モデルブタにおける血液凝固機能の程度を評価することを特徴とする血友病の予防・治療剤のスクリーニング方法。
【請求項5】
ヒト第VIII因子とブタ第VIII因子とに交差反応性を有する抗第VIII因子抗体。
【請求項6】
ヒト第VIII因子により、X染色体上の第VIII因子遺伝子の一部もしくは全部が欠損し、野生型において発現される第VIII因子を発現する機能が失われた第VIII因子ノックアウトブタを免疫し、ヒト第VIII因子とブタ第VIII因子とに交差反応性を有する抗第VIII因子抗体を調製することを特徴とする抗第VIII因子抗体の製造方法。
【図1】


【図2】
【図3】
【図4】
【図5】

【図2】
【図3】
【図4】
【図5】
【公開番号】特開2011−229456(P2011−229456A)
【公開日】平成23年11月17日(2011.11.17)
【国際特許分類】
【出願番号】特願2010−102569(P2010−102569)
【出願日】平成22年4月27日(2010.4.27)
【出願人】(500362442)プライムテック株式会社 (5)
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【出願人】(505246789)学校法人自治医科大学 (49)
【Fターム(参考)】
【公開日】平成23年11月17日(2011.11.17)
【国際特許分類】
【出願日】平成22年4月27日(2010.4.27)
【出願人】(500362442)プライムテック株式会社 (5)
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【出願人】(505246789)学校法人自治医科大学 (49)
【Fターム(参考)】
[ Back to top ]