
造血幹細胞増殖調節因子及びそれをコードするポリヌクレオチド
【課題】ストローマ細胞の産生する造血幹細胞増殖調節因子を解析し、cDNAライブラリーを作成し、新規な造血関連遺伝子を単離し、提供する。
【解決手段】以下の(a)又は(b)の蛋白質をコードする遺伝子: (a)特定な配列から成る群から選択される一つのアミノ酸配列、 (b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【解決手段】以下の(a)又は(b)の蛋白質をコードする遺伝子: (a)特定な配列から成る群から選択される一つのアミノ酸配列、 (b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、造血微小環境(niche)を形成し造血因子・接着因子を介して造血幹細胞又は造血前駆細胞の増殖・分化を制御していると考えられている骨髄ストローマ細胞が産生する、新規な造血幹細胞増殖調節因子及びそれをコードする遺伝子(ポリヌクレオチド)等に関する。
【0002】
骨髄内に存在する造血幹細胞及び間葉系幹細胞は終生に渡り、それぞれ血球系細胞、骨・軟骨、脂肪細胞等に分化し、それらを供給し続けることで成体の生命を維持している。しかし近年、これら臓器特異的幹細胞がある条件下で多臓器の細胞(神経、肝臓、肺、腸管、心筋、筋肉等)にも分化し得ることが示され、その増殖機構の解明が、移植治療及び再生医療への応用という見地からも大変に重要であると考えられている。
【0003】
これまで生体外にて造血幹細胞を増幅する試みが数多くなされ、一定の成果を挙げつつある。骨髄造血組織においては、骨髄ストローマ細胞が造血微小環境(niche)を形成し造血因子・接着因子を介して造血前駆細胞の増殖・分化を制御していると考えられている。ストローマ細胞の産生する造血因子として、IL−3,IL−6などのサイトカイン、SCF、Flt−3 ligandなどの受容体型チロシンキナーゼリガンド(非特許文献1)、分化抑制因子であるNotchリガンドのjagged−1(非特許文献2)が報告されている。
【0004】
しかしながら、上述の様に造血に重要な膜結合型因子・接着分子に関してはまだ多くが未同定であると考えられている。また、現時点では既知の因子の組み合わせだけで、造血幹細胞を長期に培養することは不可能といえ、一方、造血支持細胞であるストローマ細胞と共培養させることにより、それが一定期間可能となることから、ストローマ細胞には未だ同定されていない造血関連遺伝子を発現していることが期待される。
【非特許文献1】Gibson, F. M. et al., Br J Haematol 1995
【非特許文献2】Varnum-Finney, B. et.al., Blood 1998
【発明の開示】
【発明が解決しようとする課題】
【0005】
従って、本発明は、上記課題を解決する為に、ストローマ細胞の産生する造血幹細胞増殖調節因子を解析し、cDNAライブラリーを作成し、新規な造血関連遺伝子の単離を行うことによって、将来的には造血幹細胞の生体外増幅、造血器悪性腫瘍の移植治療、幹細胞による再生医学、遺伝子治療への応用を目的としたものである。
【課題を解決するための手段】
【0006】
本発明者は、上記の目的を達成する為に、鋭意研究の結果、造血幹細胞増殖調節活性を有する蛋白質をコードする新規な遺伝子を骨髄ストローマ細胞で見出し、本発明を完成した。
【0007】
即ち、本発明は、以下の(a)又は(b)の蛋白質をコードする遺伝子に係る:
(a)配列番号2、4、6、8、10及び12から成る群から選択される一つの配列番号に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【0008】
本発明は更に、以下の(a)又は(b)のDNAを含む遺伝子に係る:
(a)配列番号1、3、5、7、9及び11から成る群から選択される一つの配列番号に示される塩基配列からなるDNA
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNA。
【0009】
本明細書において、「造血幹細胞増殖調節活性」とは、例えば、造血幹細胞の増殖を増幅又は増加する能力、及び、逆に造血幹細胞の増殖を抑制する能力を含み、造血幹細胞の増殖に対して何らかの影響を与える機能を意味する。更に、「造血幹細胞」とは、狭義の造血幹細胞のみならず、例えば、後述のCobblestone area forming cell (CAFC) で検出できるような、1段階分化した骨髄球系、赤芽球系、巨核球系、リンパ球系前駆細胞を含むものである。
【0010】
本発明は更に、これら遺伝子又はDNAにコードされる蛋白質に係る。本発明で初めて見出されたこれら6種類の蛋白質は、ストローマ細胞に由来し、後述する実施例で示されるように造血幹細胞増殖調節活性を有するものであることから、夫々、SDHF(Stromal cell Derived Hematopoietic Factor)−1、SDHF−2、SDHF−3、SDHF−4、SDHF−5及びSDHF−6と命名される。これら各蛋白質をコードする塩基配列及び対応するアミノ酸配列は、それぞれ、配列番号1及び2、配列番号3及び4、配列番号5及び6、配列番号7及び8、配列番号9及び10、並びに、配列番号11及び12で示されている。
【0011】
本発明は更に、少なくとも一種の上記のいずれかの遺伝子を含有する組換え発現ビークル、該発現ビークルによって形質転換された形質転換体、及び、該形質転換体を培養することを含む、上記のいずれかの蛋白質の製造方法に係る。
【0012】
本発明は更に、少なくとも一種の上記のいずれかの遺伝子によりストローマ細胞を形質転換することから成る、該ストローマ細胞の造血幹細胞増殖活性の調節方法に係る。形質転換した該ストローマ細胞において、本発明の遺伝子が発現し、その造血幹細胞増殖調節活性によって、該ストローマ細胞の造血幹細胞増殖活性が増幅又は増加したり、抑制されたりするのである。尚、形質転換は、例えば、上記の組換え発現ビークルを使用して、当業者に公知の方法で行なうことが出来る。
【0013】
又、配列番号13に示されるアミノ酸配列をコードする塩基配列は既に報告されており(1999 Genomics 61 (1), pp37-43)、ISLRと呼ばれているものであるが、その機能がこれまで全くわかっておらず、今回、本発明者によって、以下の実施例に示されるように、造血幹細胞増殖調節活性を有していることが初めて実証されたものである。
【0014】
従って、本発明は、又、以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号13に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質、
でストローマ細胞を形質転換することから成る、該ストローマ細胞の造血幹細胞増殖活性の調節方法に係る。尚、形質転換は、例えば、上記の組換え発現ビークルを使用して、当業者に公知の方法で行なうことが出来る。
【0015】
本発明は更に、上記SDHF−1ないしSDHF−6及びISLRから選択される少なくとも一種の蛋白質を有効成分として含有する造血幹細胞増殖調節用組成物に係る。
【0016】
本明細書中で、「造血幹細胞増殖活性」とは、造血幹細胞の増殖に対して何らかの影響を与える機能を意味し、「調節」とは、該活性を増幅又は増加すること、及び、逆に抑制すること等の何らかの変化を意味する。
【0017】
本発明は、更に、上記SDHF−1、−2、−3、−4、−5及び−6等の本発明の各蛋白質に対するポリクローナル抗体及びモノクローナル抗体等の各種抗体に係る。
尚、図23に、本発明のSDHF−1からSDHF−6の分子構造を模式的に示す。
【発明を実施するための最良の形態】
【0018】
本発明遺伝子又はDNAは、以下の実施例に示すように、例えば、培養中に白血病阻害因子(LIF)で刺激された、マウス・ストローマ細胞株OP9細胞のようなストローマ細胞由来のmRNAから、cDNAライブラリーを作成して、これに基づき、例えば、以下の実施例に示された方法で調製することが出来る。
【0019】
或いは、本明細書で開示された配列情報に基づいて、当該技術分野における周知手段を用いた化学合成等によっても調製することが可能である。当業者であれば、特定のアミノ酸配列における1若しくは数個のアミノ酸を欠失、置換若しくは付加することも、当該技術分野における周知手段を用いて容易に実施することができる。
【0020】
本発明において、特定の遺伝子又はDNAは、当業者に周知の緩衝液中で、適当な温度及び塩濃度等の諸条件下のストリンジェントな条件でハイブリダイズさせることができる。
【0021】
このようなストリンジェントな条件下で本発明の遺伝子又はDNAとハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNAの例として、例えば、かかる遺伝子と相同性が70%以上、好ましくは90%以上であるようなDNAを挙げることができる。
【0022】
本発明の遺伝子又はDNAに、遺伝子組換え操作において当業者に公知である様々な配列、例えば、プロモーター及びエンハンサーなどの調節因子、制限酵素部位、並びに選択マーカー(マーカー酵素等)遺伝子等が適宜結合されたDNA分子も本発明の範囲である。
【0023】
更に、本発明の遺伝子、DNA又は上記DNA分子を含有する組換えプラスミドベクター又はレトロウイルス等を使用する組換えウイルスベクター等の発現ビークル、及び該発現ビークルを含みそれによって形質転換された大腸菌等の細菌、マウスCOS−7細胞又はマウス・ストローマ細胞OP9等の真核細胞等の形質転換体も本発明に含まれる。これらのDNA分子、発現ビークル及び形質転換体は、当該技術分野における公知の手段によって、容易に調製することができる。更に、本発明の蛋白質は、当業者であれば、上記の形質転換体を当該技術分野における公知手段によって培養し、得られた培養物から精製することにより、容易に製造することができる。
【0024】
上記蛋白質を有効成分として含有する、本発明の造血幹細胞増殖調節活性化組成物には、造血因子として公知のその他のサイトカイン、並びに当該技術分野で公知の適当なキャリア及び助剤等を目的に応じて適宜含有させることが出来る。蛋白質を有効成分としての本発明蛋白質及びその他の成分の含有量及び含有割合等は、使用目的などに応じて当業者が適宜選択することが出来る。
【0025】
上記SDHF−1、−2、−3、−4、−5及び−6等の本発明の各蛋白質に対する、本発明のポリクローナル抗体及びモノクローナル抗体等の各種抗体は、当業者に公知の任意の方法で製造することができる。例えば、ポリクローナル抗体の場合には、上記各蛋白質又はその部分ペプチドを免疫源としてウサギ等の各種実験動物に接種することによって容易に製造することができる。又、モノクローナル抗体の場合には、このような免疫源を使用して同様に調製した抗体産生細胞を適当な親細胞(不死化細胞株)と当業者に周知の各種方法によって細胞融合させてハイブリドーマを作成し、この中から各蛋白質と特異的に反応するクローンを選択することによって容易に得ることができる。尚、部分ペプチドは各蛋白質の任意の部分のアミノ酸配列に相当するものであり、そのアミノ酸数に特に制限はないが、少なくとも10個以上のアミノ酸を有するものが好ましく、通常、10〜20個の範囲のものが適当である。又、各蛋白質との反応性を考慮すると、免疫源として使用する部分ペプチドは各蛋白質の細胞外領域から選択することが好ましい。
【0026】
このような抗体は、骨髄幹細胞、間葉系幹細胞、及び神経幹細胞等の各種成体細胞、並びに骨髄造血支持細胞等の幹細胞支持細胞(ストローマ細胞)を単離又は純化する方法に使用することができる。これらの単離又は純化方法は、具体的には、当業者に公知の任意の各種手段、例えば、アフィニティカラムによる精製、及びFACSによるソーティング等により実施することができる。
【0027】
実施例
以下に実施例に即して本発明を詳述するが、これら実施例は本発明の技術的範囲を如何なる意味でも何ら限定するものではない。
【0028】
cDNAライブラリーの構築とSignal Sequence Trap (SST) 法
マウス・ストローマ細胞株OP9細胞はAlpha−MEM培地に20%の胎児牛血清を添加して培養する。PLAT−E細胞(2000 森田らGene Therapy vol7 1063-6)はDMEMに10%の胎児牛血清を添加した培地にて培養した。OP9細胞よりのpolyA RNAの抽出はInvitrogen社のFAST TRACK2.0mRNA Isolation Kitを用い、その添付プロトコールに従い行った。cDNAライブラリーの材料としmouse Leukemia Inhibitory Factor (LIF) 10ng/ml37℃60分にて刺激したOP9細胞1x108個から抽出したpolyARNA5μgを用いた。cDNAライブラリーの構築はSUPERSCRIPT II (Invitrogen社)を用い、ランダムヘキサマーをプライマーとして行った。得られたcDNAライブラリーにBstXIアダプタープライマー(Invitrogen社)を付加し、レトロウィルスベクターであるpMX−SSTベクター(1999 小嶋らNat Biotech vol 17 p487)のBstXI部位に組み込みプラスミド・ライブラリーを構築した。
【0029】
作製したcDNAライブラリーをPLAT−E細胞にFuGene6(Roche社)を用いて遺伝子導入して上清にレトロウィルスの形で回収した。得られたレトロウィルスcDNAライブラリーをインターロイキン3(IL-3)依存性のマウスプロB細胞であるBa/F3細胞に感染させた。pMX−SSTベクターはシグナルペプチドを欠損した恒常的活性化型c−Mpl遺伝子を有しており、ライブラリー由来の遺伝子がシグナルペプチドを有している場合にcMpl遺伝子が活性化し、Ba/F3細胞がIL−3非存在下において不死化する(1999 小嶋らNat Biotech vol 17 p487-90)。レトロウィルス・ライブラリーを感染させたBa/F3細胞からIL−3を除去し96ウェルプレートに1ウェルあたり10000個ずつ分けて培養、細胞が増殖してきたものを単離し、ゲノムDNAを抽出した。シグナルペプチドを有した遺伝子はレトロウィルスにてBa/F3細胞のゲノムDNAに挿入されているのでpMX−SSTベクターの配列をもとにして設計したプライマー(GGGGGTGGACCATCCTCTA, CGCGCAGCTGTAAACGGTAG)にてゲノムDNA PCR反応(LA-Taq, TAKARA 98℃ 20 sec 68℃ 300 sec; 35 cycles)を行い回収した。
【0030】
遺伝子配列の決定
シグナルペプチドを有したクローンは上記PCR反応にてDNA断片として得られるのでこの塩基配列をABI社373A シーケンサーにて決定した。得られたPCR断片をPCR purification kit (Quiagen社)にて精製しプライマーを除去した後、ABI社のプロトコールに従いdye terminator法にて配列を決定した。得られた配列情報はNCBIのBLASTプログラム(http://www.ncbi.nlm.nih.gov/BLAST/)にて解析し、既知配列、未知配列の判定を行った。未知配列の一部は得られた配列を元に設計したプライマーを用いて3’RACE法にて決定した。3’RACE法は、上記PCRクローンの配列をもとにして設計したプライマーとオリゴdTプライマーを用いたPCR法にて未知遺伝子のpolyA領域までを同定する方法である。得られたPCR断片は上記と同様にABI社373Aシーケンサーにて塩基配列を決定した。
【0031】
その結果、上記のSST法により既知・未知合わせて234クローンの遺伝子を単離した。その内訳は、既知遺伝子181、及び未知遺伝子42、解析できなかったもの11であった。既知遺伝子の内訳としては、増殖因子49、受容体23、接着因子71、及びその他38であった。
【0032】
未知遺伝子の中に、本発明の遺伝子である、6種類のSDHF(Stromal cell derived hematopoietic factor)遺伝子、及びISLRが含まれていた。尚、第1図〜第6図及び配列13に示されるように、6種類のSDHF及びISLR遺伝子がコードするアミノ酸配列のN末端には、シグナルペプチド(四角の枠部で示された部分)が含まれていることが判明した。以上の結果を表1及び表2に示す。
【0033】
【表1】

【0034】
【表2】
【0035】
マイクロアレイ法
Signal Sequence Trap法にて得られたPCR断片は全てスライドグラス上にスポットし(以上の操作は北海道システムサイエンス社に委託した。)OP9細胞をLIF10ng/ml 37℃60分による刺激前後の条件にて調整した細胞より作製した蛍光標識cDNAプローブにて発現の変化を検出した。プローブは上記と同様に抽出したpolyA RNAよりオリゴdTプライマーにてSuperScript IIにて逆転写反応を行い、Fluorolink Cy-dUTPを取り込ませることにより標識した後Microcon30(Millipore社)にて精製し用いた。得られた結果を第7図に示した。
【0036】
ノーザンブロッティング
OP9細胞をLIF 10ng/ml 37℃60分にて刺激後、0分、30分、1時間、3時間、8時間、24時間後にそれぞれtotal RNAをTrizol液(Invitrogen社)を用い、そのプロトコールに従い抽出した。抽出したtotal RNAは濃度測定後10μgずつ1%アガロースゲル上に電気泳動後、Hybond N(Amersham社)上に転写し、それぞれの遺伝子のDNA断片をプローブにして標準的な方法にてノーザンブロッティングを行った。その結果、第8図に示されるように、SDHF−1、SDHF−2、及びSDHF−5遺伝子について発現の増加が観察された。一方、対照的に、SDHF−6遺伝子について発現の減少が観察された。
【0037】
ウェスタン解析法
得られた遺伝子の全長配列よりアミノ酸をコードしている領域(Open Reading Frame: ORF)を決定し、そのカルボキシル末端にFLAGタグ配列(DYKDDDDK)をPCR法にて付加した。これらのcDNAを発現ベクターpUC−CAGGSに挿入して作製したプラスミドをFuGene6(ロシュ社)にてCOS−7細胞に一過性に発現した。ウェスタン解析は過去に報告した方法(1995 Ueno, JBC vol 270pp20135-42)にて行った。細胞はTriton lysis buffer (0.5% (v/v) Triton X-100, 50 mM Tris-HCl pH 7.4, 2 mM PMSF, 10 U/ml aprotinin 1mM EDTA) にて可溶化した後、50μgをSDS-polyacrylamide gel elestrophoresis (PAGE)にて展開し、Immobilon−Pメンブレインに転写後抗FLAG抗体、M2(SIGMA社)にて検出した。その結果、第9図に示されるように、本発明のいずれの遺伝子もCOS7細胞にて発現されウェスタン解析にてバンドが検出された。
【0038】
免疫染色法
上記と同条件にてFLAGタグ配列を付加した遺伝子をCOS−7細胞にFuGene試薬を用いて一過性に発現させ(上記参照)、3.7%ホルムアルデヒドにて固定後1%NP40にてpermeabilizationを行い、抗FLAG抗体を用いて染色した(二次抗体はTexas Red標識抗マウス抗体、Jackson社)。核はDAPI(フナコシ社)染色を行った。その結果を第10図に示す。
【0039】
Cobblestone Area Forming Cell Assay : CAFC(敷石状領域形成細胞アッセイ)
OP9細胞に上記にて得られた遺伝子をレトロウィルスベクター(pMX-puro) にて遺伝子導入し6wellディッシュに2.5x105ずつ蒔き、一晩培養した後、5−8週齢のC57BL/6マウス大腿骨より採取した骨髄細胞1x105まき、6日後に形成されたCAFC数を測定した。
【0040】
CAFC解析の結果
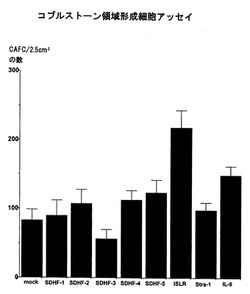
ストローマ細胞と骨髄細胞を共培養した際、骨髄細胞がストローマ細胞の下部に潜り込み増殖する現象をCobblestone area forming cell (CAFC) という。CAFC形成数が多い場合、ストローマ細胞が未熟な造血支持細胞を増幅していることを意味している。ただしCAFCを形成している細胞は造血幹細胞だけでなく、1段階分化した骨髄球系、赤芽球系、巨核球系、リンパ球系前駆細胞を含んでいるため、厳密にはCAFCの増加=造血幹細胞の増幅ではない。造血支持能を有するマウス骨髄ストローマ細胞株のOP9細胞に上記遺伝子をレトロウィルスpMX−puroにて導入し、ピューロマイシン5μg/mlにて薬剤耐性となった細胞についてCAFC形成能を解析した。その結果、第11図に示されるように、ISLRが最も高値を示しSDHF−1、SDHF−2、SDHF−4、及びSDHF−5についても軽度の高値を示した。これらの遺伝子の造血前駆細胞増幅能力が示唆された。逆にSDHF−3ではCAFC形成が抑制され、造血幹細胞増殖抑制機能を有すると考えられた。
【0041】
細胞培養上清の分泌蛋白質のメチオニン標識による検出
COS−7細胞を6cmディッシュに1x105個まき、一晩培養した。FuGene法(Roche社)を用いて遺伝子導入した。これらの遺伝子のカルボキシル末端にはFLAGタグが付加されており、前述したように、抗FLAG抗体によるウェスタン解析にて発現した蛋白質を検出することが出来る。36時間の培養後、上清を除去し、PBSにて2回洗浄後、0.5 ml DMEM(Methionine−, SIGMA社)中の0.5mCi[35S]methionine(ICN社TRAN-S)を加え、4時間培養した。培養上清から遠心にて細胞成分を除去し、Micron 10(ミリポア社)にて蛋白質を濃縮した。15%SDS−PAGEにて電気泳動後、標識された蛋白質をBAS2000(富士フィルム社)にて検出した。
【0042】
その結果、第12図及び第13図に示されるように、SDHF−1及びSDHF−5にて、培養上清への蛋白質分泌が検出された。これらの遺伝子は構造的に膜型蛋白質であること、分泌蛋白質の分子量がウェスタン解析で検出された成熟型蛋白質よりも約20kDa小さくなっていることから、これらの分泌蛋白質はカルボキシル末端の膜貫通領域が切断されたものと考えられる。カルボキシル末端の膜貫通領域が切断されて分泌されるタイプの増殖因子としてSCFが知られており、一方、接着蛋白質ではこうした例は認められないことから、本発明の遺伝子がコードする蛋白質は、液性因子として他の細胞に情報を伝達するタイプの蛋白質であることを強く示唆するものと考えられる。
【0043】
RT−PCR法による発現部位の検定
各SDHF遺伝子の塩基配列に基づきプライマーを設定、Multiple Tissue cDNA (MTC) Panels (クロンテック社)をtemplateとしてPCR反応(LA-Taq, TAKARA)を行った。また、骨髄内の細胞は蛍光標識したモノクローナル抗体(抗B220, 抗CD3, 抗Gr1, 抗MAC1, 抗Sca1抗体、Phamingen社)を用い、FACS Vantage(BectonDickinson社)にてソーティングしたTrizol試薬(Invitrogen社)にてRNAを抽出し、ランダムヘキサマーを用いてRT反応を行った上で、同様にPCR反応を行った。
【0044】
その結果、第14図に示される様に全てのSDHF遺伝子はOP9細胞での発現が確認された。特にSDHF−4は発現が脳と骨髄に限局して発現しており、また骨髄内においてはストローマ細胞に限局して発現しているなど興味深い。
【0045】
Long Term Culture-Initiating Cells Assay(LTC−IC法)による幹細胞支持機能の検定
上記のCAFC法では比較的短期間の造血支持機能を反映しているにすぎないため、より長期の幹細胞支持機能評価法としてLTC−IC法を行った。上記と同様に単離した遺伝子をレトロウィルス発現ベクターpMX−puroを用いてOP9細胞に高発現させLTC−IC法を行った。
【0046】
まず造血幹細胞を純化する。6週齢から8週齢のC57BL/6マウスの大腿骨より骨髄細胞を採取し、Ficoll法(比重1.100) にて単核細胞を遠心分離した。PBSにて洗浄後、ビオチン標識した1次抗体(抗Lineage抗体カクテル:抗CD3, 抗CD4, 抗CD8, 抗B220, 抗Ter119, 抗Gr1,抗 MAC1抗体、以上Phamingen社)およびストレプトアビジン標識磁気ビーズにそれぞれ4℃15分間反応させた。MACS法にてカラムを通過する分画(LIN(-)細胞)を回収し、遠心洗浄後、さらにPE標識抗Sca1抗体、FITC標識抗c−kit抗体、Per−CP−Cy5.5標識ストレプトアビジン(以上Phamingen社)にて4℃15分間反応させ、FACS Vantage(Becton Dickinson社)にてLIN(−),Sca1(+),c−kit(+)分画(以上にて得られた細胞をKSL細胞と定義する。)SDHF−1−6のカルボキシル末端にFLAGペプチドを付加したcDNAをレトロウィルスベクター(pMX-puro) にてOP9細胞に遺伝子導入して樹立した安定発現株を6穴ディッシュに2.5x105ずつ蒔き、一晩培養した後、KSL細胞を100個ずつまき、3週間培養した。その後細胞を回収し、γ線を20Gy照射したOP9細胞を1穴に3000個ずつまいておいた96穴ディッシュに、8穴ずつペアで2分の1ずつ薄めて蒔いた。5週間後、各穴から細胞を回収しそれぞれメチルセルロース培地(IMDM, メチルセルロース1%、牛胎児血清15%、牛血清アルブミン 1%、ヒトエリスロポイエチン3U/ml、ヒトIL-6 10ng/ml、マウスIL-3 10ng/ml、マウスSCF 100ng/ml牛インスリン10μg/ml)に蒔いて12日後に血球コロニー(CFU-C)の有無で判定した。幹細胞の頻度はL−Calcソフトウェア(Stemcell社)にて行った。
【0047】
その結果、第15図に示されるようにSDHF−4遺伝子に著明な造血支持能が観察された。この遺伝子の造血支持機能が示された。
【0048】
SDHF−4細胞外領域・リコンビナントタンパク質によるマウス造血幹細胞の増幅効果
SDHF−4の細胞膜貫通領域をPSORT IIプログラム(http://psort.ims.u-tokyo.ac.jp/form2.html)で予測し(アミノ酸524-540)、細胞外領域(アミノ酸1-523)にヒト免疫グロブリンFc領域と融合タンパク質(SDHF-4-ΔTM-Fc)を作製、発現ベクターpSSRaに組み込み、CHO−k1細胞に安定的に遺伝子導入した。無血清培地CD−CHO(Invitrogen社)にて培養することで、上清から融合リコンビナントタンパク質をProteinAカラムにて精製した(第16図)。マウス幹細胞はLTC−IC法の項に記載した様にマウス骨髄LIN(−),Sca1(+),c−kit(+)分画(KSL細胞)100個ずつ、SCF(50ng/ml),IL−3(10ng/ml),IL−6(10ng/ml)及びSDHF−4−ΔTM−Fc(100ng/ml)を添加したStem Span培地(StemCell社)にて培養した。7日後、細胞を回収し、FACS CaliburにてLIN,Sca1,c−kitの発現を解析した。
【0049】
その結果、第17図に示されるように、SCF,IL−3,IL−6を添加したStemSpan培地にて培養したLSK細胞はSDHF−4−ΔTM−Fc100ng/mlの存在下において非存在下と比較してLIN(−)細胞の割合が増加するだけでなく、LIN(−),Sca1(+),c−kit(+)分画の割合も増加することが判明した。この結果は、SDHF−4細胞外領域・リコンビナントタンパク質にマウス造血幹細胞の未分化状態を維持する作用があることを示している。
【0050】
脂肪細胞分化の検定とオイルレッドO染色法
SDHF−6遺伝子をレトロウィルスベクター(pMX-puro)導入した細胞とベクターのみを導入した細胞をそれぞれAlpha−MEM+20%牛胎児血清の条件にて4週間以上継代せずにConfluentの状態で培養し続けた。細胞は10%ホルマリンにて固定後オイルレッドOにて染色した。
その結果、第18図に示されるように、SDHF−6をレトロウィルス発現ベクターpMX−puroを用いてOP9細胞に高発現させると約4週間の培養の後、効率に脂肪細胞に分化させる効果があることが判明した。この遺伝子が、造血幹細胞よりもむしろ間葉系の細胞に作用して脂肪細胞への分化に関与していると考えられた。
【0051】
SDHF−4に対するポリクローナル抗体の作製及びそれによる骨髄内造血支持細胞の単離
SDHF−4のアミノ酸配列をもとに、合成ペプチド(GYMAKDKFRRMNEGQVY(第32-48番目のアミノ酸に相当する))を設計しラビットに免疫してポリクローナル抗体を作製した。
このペプチドを Epoxy-activated Sepharose 6B(ファルマシア社)に結合させ、抗原カラムを作製、これにより、得られたポリクローナル抗体をアフィニティー精製した。この抗体を用い、CHO−k1細胞にpSSRα−bsrベクターを用いてSDHF−4を発現させた細胞(SDHF4-cl12)とベクターだけ導入した細胞(mock)を比較してウェスタン解析を施行したところ、非常に特異的にSDHF−4遺伝子産物を認識できることがわかった(第19図)。また、得られた精製抗体1mgを LinKit Fluoro-Link (ISL社) によりFITC標識し、同じ細胞をFACS calibur(Becton Dickinson社)にて解析したところ、この抗体はFlowcytemetryにも使用可能であることがわかった(第20図; ○線がmock、■線がSDHF4-cl12)。この抗体を用いてC57BL/6Jマウスの骨髄細胞を染色し、FACS Vantage(Becton Dickinson社)によりSDHF4を発現している細胞をソーティングして回収したところ(第21図)、これらの細胞は培養ディッシュに付着し、形態学的にも骨髄造血支持細胞に相当する間葉系の細胞であった(第22図)。
【産業上の利用可能性】
【0052】
本発明によって、ストローマ細胞に由来する、新規な造血関連遺伝子の単離を行うことに成功し、それらが造血幹細胞増殖調節活性を有することが実証された。これによって、造血幹細胞の生体外増幅、造血器悪性腫瘍の移植治療、幹細胞による再生医学、遺伝子治療、免疫療法、細胞移植、遺伝子治療、神経疾患(アルツハイマー病、脳梗塞、変性神経疾患等)、肝疾患(肝硬変等)、肺疾患、心筋疾患、糖尿病、骨疾患、及び慢性腎疾患等の広範囲な応用を可能としたものである。
【図面の簡単な説明】
【0053】
【図1】第1図は、SDHF−1のアミノ酸配列を示す。
【図2】第2図は、SDHF−2のアミノ酸配列を示す。
【図3】第3図は、SDHF−3のアミノ酸配列を示す。
【図4】第4図は、SDHF−4のアミノ酸配列を示す。
【図5】第5図は、SDHF−5のアミノ酸配列を示す。
【図6】第6図は、SDHF−6のアミノ酸配列を示す。
【図7】第7図は、マイクロアレイ法により得られた結果を示す。
【図8−1】第8図は、ノーザンブロッティングにより得られた結果を示す。
【図8−2】第8図は、ノーザンブロッティングにより得られた結果を示す。
【図9】第9図は、ウェスタン解析により得られた結果を示す。
【図10】第10図は、免疫染色法により得られた結果を示す。
【図11】第11図は、CAFC解析の結果を示す。
【図12】第12図は、ウェスタン解析により検出された本発明遺伝子SDHF−1、SDHF−5の蛋白質翻訳後修飾の結果を示す。★は糖鎖が付加されて分子量が大きくなった成熟型の蛋白質を示し、▲は糖鎖の付加される以前の生成過程の蛋白質を示し、▼は切断後の残存蛋白質を示し、●は分泌型蛋白質を示す。
【図13】第13図は、細胞培養上清の分泌蛋白質のメチオニン標識による検出の結果を示す。●は分泌型蛋白質を示し、→は切断後培養上清に分泌された蛋白質を示す。
【図14−1】第14図は、RT−PCR法による本発明遺伝子の発現部位の検定の結果を示す、電気泳動の写真である。
【図14−2】第14図は、RT−PCR法による本発明遺伝子の発現部位の検定の結果を示す、電気泳動の写真である。
【図15】第15図は、LTC−IC法による本発明遺伝子の幹細胞支持機能の検定の結果を示す。
【図16】第16図は、SDHF−4細胞外領域・リコンビナントタンパク質をProteinAカラムにて精製した様子を示す、電気泳動の写真である。
【図17】第17図は、SDHF−4細胞外領域・リコンビナントタンパク質がマウス造血幹細胞の未分化状態を維持する作用を有することを示すグラフである。
【図18−1】第18図は、SDHF−6をレトロウィルス発現ベクターpMX−puroを用いてOP9細胞に高発現させると約4週間の培養の後、効率に脂肪細胞に分化させる効果があることを示す、顕微鏡写真(倍率:×40)である。
【図18−2】第18図は、SDHF−6をレトロウィルス発現ベクターpMX−puroを用いてOP9細胞に高発現させると約4週間の培養の後、効率に脂肪細胞に分化させる効果があることを示す、顕微鏡写真(倍率:×40)である。
【図19】第19図は、SDHF−4の部分ペプチドに対する抗体を用い、CHO−k1細胞にpSSRα−bsrベクターを用いてSDHF−4を発現させた結果を示す、ウェスタン解析による写真である。
【図20】第20図は、上記の精製抗体をFITC標識し、SDHF−4を発現させたCHO−k1細胞をFACS calibur (BectonDickinson社)にて解析した結果を示す。
【図21】第21図は、同抗体を用いてC57BL/6Jマウスの骨髄細胞を染色し、FACS Vantage (BectonDickinson社) によりSDHF4を発現している細胞をソーティングした結果を示す。
【図22】第22図は、ソーティングにより選択された上記細胞の顕微鏡写真(倍率:×200倍)である。
【図23】第23図は、SDHF−1からSDHF−6の分子構造を模式的に表したものである。
【技術分野】
【0001】
本発明は、造血微小環境(niche)を形成し造血因子・接着因子を介して造血幹細胞又は造血前駆細胞の増殖・分化を制御していると考えられている骨髄ストローマ細胞が産生する、新規な造血幹細胞増殖調節因子及びそれをコードする遺伝子(ポリヌクレオチド)等に関する。
【0002】
骨髄内に存在する造血幹細胞及び間葉系幹細胞は終生に渡り、それぞれ血球系細胞、骨・軟骨、脂肪細胞等に分化し、それらを供給し続けることで成体の生命を維持している。しかし近年、これら臓器特異的幹細胞がある条件下で多臓器の細胞(神経、肝臓、肺、腸管、心筋、筋肉等)にも分化し得ることが示され、その増殖機構の解明が、移植治療及び再生医療への応用という見地からも大変に重要であると考えられている。
【0003】
これまで生体外にて造血幹細胞を増幅する試みが数多くなされ、一定の成果を挙げつつある。骨髄造血組織においては、骨髄ストローマ細胞が造血微小環境(niche)を形成し造血因子・接着因子を介して造血前駆細胞の増殖・分化を制御していると考えられている。ストローマ細胞の産生する造血因子として、IL−3,IL−6などのサイトカイン、SCF、Flt−3 ligandなどの受容体型チロシンキナーゼリガンド(非特許文献1)、分化抑制因子であるNotchリガンドのjagged−1(非特許文献2)が報告されている。
【0004】
しかしながら、上述の様に造血に重要な膜結合型因子・接着分子に関してはまだ多くが未同定であると考えられている。また、現時点では既知の因子の組み合わせだけで、造血幹細胞を長期に培養することは不可能といえ、一方、造血支持細胞であるストローマ細胞と共培養させることにより、それが一定期間可能となることから、ストローマ細胞には未だ同定されていない造血関連遺伝子を発現していることが期待される。
【非特許文献1】Gibson, F. M. et al., Br J Haematol 1995
【非特許文献2】Varnum-Finney, B. et.al., Blood 1998
【発明の開示】
【発明が解決しようとする課題】
【0005】
従って、本発明は、上記課題を解決する為に、ストローマ細胞の産生する造血幹細胞増殖調節因子を解析し、cDNAライブラリーを作成し、新規な造血関連遺伝子の単離を行うことによって、将来的には造血幹細胞の生体外増幅、造血器悪性腫瘍の移植治療、幹細胞による再生医学、遺伝子治療への応用を目的としたものである。
【課題を解決するための手段】
【0006】
本発明者は、上記の目的を達成する為に、鋭意研究の結果、造血幹細胞増殖調節活性を有する蛋白質をコードする新規な遺伝子を骨髄ストローマ細胞で見出し、本発明を完成した。
【0007】
即ち、本発明は、以下の(a)又は(b)の蛋白質をコードする遺伝子に係る:
(a)配列番号2、4、6、8、10及び12から成る群から選択される一つの配列番号に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【0008】
本発明は更に、以下の(a)又は(b)のDNAを含む遺伝子に係る:
(a)配列番号1、3、5、7、9及び11から成る群から選択される一つの配列番号に示される塩基配列からなるDNA
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNA。
【0009】
本明細書において、「造血幹細胞増殖調節活性」とは、例えば、造血幹細胞の増殖を増幅又は増加する能力、及び、逆に造血幹細胞の増殖を抑制する能力を含み、造血幹細胞の増殖に対して何らかの影響を与える機能を意味する。更に、「造血幹細胞」とは、狭義の造血幹細胞のみならず、例えば、後述のCobblestone area forming cell (CAFC) で検出できるような、1段階分化した骨髄球系、赤芽球系、巨核球系、リンパ球系前駆細胞を含むものである。
【0010】
本発明は更に、これら遺伝子又はDNAにコードされる蛋白質に係る。本発明で初めて見出されたこれら6種類の蛋白質は、ストローマ細胞に由来し、後述する実施例で示されるように造血幹細胞増殖調節活性を有するものであることから、夫々、SDHF(Stromal cell Derived Hematopoietic Factor)−1、SDHF−2、SDHF−3、SDHF−4、SDHF−5及びSDHF−6と命名される。これら各蛋白質をコードする塩基配列及び対応するアミノ酸配列は、それぞれ、配列番号1及び2、配列番号3及び4、配列番号5及び6、配列番号7及び8、配列番号9及び10、並びに、配列番号11及び12で示されている。
【0011】
本発明は更に、少なくとも一種の上記のいずれかの遺伝子を含有する組換え発現ビークル、該発現ビークルによって形質転換された形質転換体、及び、該形質転換体を培養することを含む、上記のいずれかの蛋白質の製造方法に係る。
【0012】
本発明は更に、少なくとも一種の上記のいずれかの遺伝子によりストローマ細胞を形質転換することから成る、該ストローマ細胞の造血幹細胞増殖活性の調節方法に係る。形質転換した該ストローマ細胞において、本発明の遺伝子が発現し、その造血幹細胞増殖調節活性によって、該ストローマ細胞の造血幹細胞増殖活性が増幅又は増加したり、抑制されたりするのである。尚、形質転換は、例えば、上記の組換え発現ビークルを使用して、当業者に公知の方法で行なうことが出来る。
【0013】
又、配列番号13に示されるアミノ酸配列をコードする塩基配列は既に報告されており(1999 Genomics 61 (1), pp37-43)、ISLRと呼ばれているものであるが、その機能がこれまで全くわかっておらず、今回、本発明者によって、以下の実施例に示されるように、造血幹細胞増殖調節活性を有していることが初めて実証されたものである。
【0014】
従って、本発明は、又、以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号13に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質、
でストローマ細胞を形質転換することから成る、該ストローマ細胞の造血幹細胞増殖活性の調節方法に係る。尚、形質転換は、例えば、上記の組換え発現ビークルを使用して、当業者に公知の方法で行なうことが出来る。
【0015】
本発明は更に、上記SDHF−1ないしSDHF−6及びISLRから選択される少なくとも一種の蛋白質を有効成分として含有する造血幹細胞増殖調節用組成物に係る。
【0016】
本明細書中で、「造血幹細胞増殖活性」とは、造血幹細胞の増殖に対して何らかの影響を与える機能を意味し、「調節」とは、該活性を増幅又は増加すること、及び、逆に抑制すること等の何らかの変化を意味する。
【0017】
本発明は、更に、上記SDHF−1、−2、−3、−4、−5及び−6等の本発明の各蛋白質に対するポリクローナル抗体及びモノクローナル抗体等の各種抗体に係る。
尚、図23に、本発明のSDHF−1からSDHF−6の分子構造を模式的に示す。
【発明を実施するための最良の形態】
【0018】
本発明遺伝子又はDNAは、以下の実施例に示すように、例えば、培養中に白血病阻害因子(LIF)で刺激された、マウス・ストローマ細胞株OP9細胞のようなストローマ細胞由来のmRNAから、cDNAライブラリーを作成して、これに基づき、例えば、以下の実施例に示された方法で調製することが出来る。
【0019】
或いは、本明細書で開示された配列情報に基づいて、当該技術分野における周知手段を用いた化学合成等によっても調製することが可能である。当業者であれば、特定のアミノ酸配列における1若しくは数個のアミノ酸を欠失、置換若しくは付加することも、当該技術分野における周知手段を用いて容易に実施することができる。
【0020】
本発明において、特定の遺伝子又はDNAは、当業者に周知の緩衝液中で、適当な温度及び塩濃度等の諸条件下のストリンジェントな条件でハイブリダイズさせることができる。
【0021】
このようなストリンジェントな条件下で本発明の遺伝子又はDNAとハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNAの例として、例えば、かかる遺伝子と相同性が70%以上、好ましくは90%以上であるようなDNAを挙げることができる。
【0022】
本発明の遺伝子又はDNAに、遺伝子組換え操作において当業者に公知である様々な配列、例えば、プロモーター及びエンハンサーなどの調節因子、制限酵素部位、並びに選択マーカー(マーカー酵素等)遺伝子等が適宜結合されたDNA分子も本発明の範囲である。
【0023】
更に、本発明の遺伝子、DNA又は上記DNA分子を含有する組換えプラスミドベクター又はレトロウイルス等を使用する組換えウイルスベクター等の発現ビークル、及び該発現ビークルを含みそれによって形質転換された大腸菌等の細菌、マウスCOS−7細胞又はマウス・ストローマ細胞OP9等の真核細胞等の形質転換体も本発明に含まれる。これらのDNA分子、発現ビークル及び形質転換体は、当該技術分野における公知の手段によって、容易に調製することができる。更に、本発明の蛋白質は、当業者であれば、上記の形質転換体を当該技術分野における公知手段によって培養し、得られた培養物から精製することにより、容易に製造することができる。
【0024】
上記蛋白質を有効成分として含有する、本発明の造血幹細胞増殖調節活性化組成物には、造血因子として公知のその他のサイトカイン、並びに当該技術分野で公知の適当なキャリア及び助剤等を目的に応じて適宜含有させることが出来る。蛋白質を有効成分としての本発明蛋白質及びその他の成分の含有量及び含有割合等は、使用目的などに応じて当業者が適宜選択することが出来る。
【0025】
上記SDHF−1、−2、−3、−4、−5及び−6等の本発明の各蛋白質に対する、本発明のポリクローナル抗体及びモノクローナル抗体等の各種抗体は、当業者に公知の任意の方法で製造することができる。例えば、ポリクローナル抗体の場合には、上記各蛋白質又はその部分ペプチドを免疫源としてウサギ等の各種実験動物に接種することによって容易に製造することができる。又、モノクローナル抗体の場合には、このような免疫源を使用して同様に調製した抗体産生細胞を適当な親細胞(不死化細胞株)と当業者に周知の各種方法によって細胞融合させてハイブリドーマを作成し、この中から各蛋白質と特異的に反応するクローンを選択することによって容易に得ることができる。尚、部分ペプチドは各蛋白質の任意の部分のアミノ酸配列に相当するものであり、そのアミノ酸数に特に制限はないが、少なくとも10個以上のアミノ酸を有するものが好ましく、通常、10〜20個の範囲のものが適当である。又、各蛋白質との反応性を考慮すると、免疫源として使用する部分ペプチドは各蛋白質の細胞外領域から選択することが好ましい。
【0026】
このような抗体は、骨髄幹細胞、間葉系幹細胞、及び神経幹細胞等の各種成体細胞、並びに骨髄造血支持細胞等の幹細胞支持細胞(ストローマ細胞)を単離又は純化する方法に使用することができる。これらの単離又は純化方法は、具体的には、当業者に公知の任意の各種手段、例えば、アフィニティカラムによる精製、及びFACSによるソーティング等により実施することができる。
【0027】
実施例
以下に実施例に即して本発明を詳述するが、これら実施例は本発明の技術的範囲を如何なる意味でも何ら限定するものではない。
【0028】
cDNAライブラリーの構築とSignal Sequence Trap (SST) 法
マウス・ストローマ細胞株OP9細胞はAlpha−MEM培地に20%の胎児牛血清を添加して培養する。PLAT−E細胞(2000 森田らGene Therapy vol7 1063-6)はDMEMに10%の胎児牛血清を添加した培地にて培養した。OP9細胞よりのpolyA RNAの抽出はInvitrogen社のFAST TRACK2.0mRNA Isolation Kitを用い、その添付プロトコールに従い行った。cDNAライブラリーの材料としmouse Leukemia Inhibitory Factor (LIF) 10ng/ml37℃60分にて刺激したOP9細胞1x108個から抽出したpolyARNA5μgを用いた。cDNAライブラリーの構築はSUPERSCRIPT II (Invitrogen社)を用い、ランダムヘキサマーをプライマーとして行った。得られたcDNAライブラリーにBstXIアダプタープライマー(Invitrogen社)を付加し、レトロウィルスベクターであるpMX−SSTベクター(1999 小嶋らNat Biotech vol 17 p487)のBstXI部位に組み込みプラスミド・ライブラリーを構築した。
【0029】
作製したcDNAライブラリーをPLAT−E細胞にFuGene6(Roche社)を用いて遺伝子導入して上清にレトロウィルスの形で回収した。得られたレトロウィルスcDNAライブラリーをインターロイキン3(IL-3)依存性のマウスプロB細胞であるBa/F3細胞に感染させた。pMX−SSTベクターはシグナルペプチドを欠損した恒常的活性化型c−Mpl遺伝子を有しており、ライブラリー由来の遺伝子がシグナルペプチドを有している場合にcMpl遺伝子が活性化し、Ba/F3細胞がIL−3非存在下において不死化する(1999 小嶋らNat Biotech vol 17 p487-90)。レトロウィルス・ライブラリーを感染させたBa/F3細胞からIL−3を除去し96ウェルプレートに1ウェルあたり10000個ずつ分けて培養、細胞が増殖してきたものを単離し、ゲノムDNAを抽出した。シグナルペプチドを有した遺伝子はレトロウィルスにてBa/F3細胞のゲノムDNAに挿入されているのでpMX−SSTベクターの配列をもとにして設計したプライマー(GGGGGTGGACCATCCTCTA, CGCGCAGCTGTAAACGGTAG)にてゲノムDNA PCR反応(LA-Taq, TAKARA 98℃ 20 sec 68℃ 300 sec; 35 cycles)を行い回収した。
【0030】
遺伝子配列の決定
シグナルペプチドを有したクローンは上記PCR反応にてDNA断片として得られるのでこの塩基配列をABI社373A シーケンサーにて決定した。得られたPCR断片をPCR purification kit (Quiagen社)にて精製しプライマーを除去した後、ABI社のプロトコールに従いdye terminator法にて配列を決定した。得られた配列情報はNCBIのBLASTプログラム(http://www.ncbi.nlm.nih.gov/BLAST/)にて解析し、既知配列、未知配列の判定を行った。未知配列の一部は得られた配列を元に設計したプライマーを用いて3’RACE法にて決定した。3’RACE法は、上記PCRクローンの配列をもとにして設計したプライマーとオリゴdTプライマーを用いたPCR法にて未知遺伝子のpolyA領域までを同定する方法である。得られたPCR断片は上記と同様にABI社373Aシーケンサーにて塩基配列を決定した。
【0031】
その結果、上記のSST法により既知・未知合わせて234クローンの遺伝子を単離した。その内訳は、既知遺伝子181、及び未知遺伝子42、解析できなかったもの11であった。既知遺伝子の内訳としては、増殖因子49、受容体23、接着因子71、及びその他38であった。
【0032】
未知遺伝子の中に、本発明の遺伝子である、6種類のSDHF(Stromal cell derived hematopoietic factor)遺伝子、及びISLRが含まれていた。尚、第1図〜第6図及び配列13に示されるように、6種類のSDHF及びISLR遺伝子がコードするアミノ酸配列のN末端には、シグナルペプチド(四角の枠部で示された部分)が含まれていることが判明した。以上の結果を表1及び表2に示す。
【0033】
【表1】
【0034】
【表2】
【0035】
マイクロアレイ法
Signal Sequence Trap法にて得られたPCR断片は全てスライドグラス上にスポットし(以上の操作は北海道システムサイエンス社に委託した。)OP9細胞をLIF10ng/ml 37℃60分による刺激前後の条件にて調整した細胞より作製した蛍光標識cDNAプローブにて発現の変化を検出した。プローブは上記と同様に抽出したpolyA RNAよりオリゴdTプライマーにてSuperScript IIにて逆転写反応を行い、Fluorolink Cy-dUTPを取り込ませることにより標識した後Microcon30(Millipore社)にて精製し用いた。得られた結果を第7図に示した。
【0036】
ノーザンブロッティング
OP9細胞をLIF 10ng/ml 37℃60分にて刺激後、0分、30分、1時間、3時間、8時間、24時間後にそれぞれtotal RNAをTrizol液(Invitrogen社)を用い、そのプロトコールに従い抽出した。抽出したtotal RNAは濃度測定後10μgずつ1%アガロースゲル上に電気泳動後、Hybond N(Amersham社)上に転写し、それぞれの遺伝子のDNA断片をプローブにして標準的な方法にてノーザンブロッティングを行った。その結果、第8図に示されるように、SDHF−1、SDHF−2、及びSDHF−5遺伝子について発現の増加が観察された。一方、対照的に、SDHF−6遺伝子について発現の減少が観察された。
【0037】
ウェスタン解析法
得られた遺伝子の全長配列よりアミノ酸をコードしている領域(Open Reading Frame: ORF)を決定し、そのカルボキシル末端にFLAGタグ配列(DYKDDDDK)をPCR法にて付加した。これらのcDNAを発現ベクターpUC−CAGGSに挿入して作製したプラスミドをFuGene6(ロシュ社)にてCOS−7細胞に一過性に発現した。ウェスタン解析は過去に報告した方法(1995 Ueno, JBC vol 270pp20135-42)にて行った。細胞はTriton lysis buffer (0.5% (v/v) Triton X-100, 50 mM Tris-HCl pH 7.4, 2 mM PMSF, 10 U/ml aprotinin 1mM EDTA) にて可溶化した後、50μgをSDS-polyacrylamide gel elestrophoresis (PAGE)にて展開し、Immobilon−Pメンブレインに転写後抗FLAG抗体、M2(SIGMA社)にて検出した。その結果、第9図に示されるように、本発明のいずれの遺伝子もCOS7細胞にて発現されウェスタン解析にてバンドが検出された。
【0038】
免疫染色法
上記と同条件にてFLAGタグ配列を付加した遺伝子をCOS−7細胞にFuGene試薬を用いて一過性に発現させ(上記参照)、3.7%ホルムアルデヒドにて固定後1%NP40にてpermeabilizationを行い、抗FLAG抗体を用いて染色した(二次抗体はTexas Red標識抗マウス抗体、Jackson社)。核はDAPI(フナコシ社)染色を行った。その結果を第10図に示す。
【0039】
Cobblestone Area Forming Cell Assay : CAFC(敷石状領域形成細胞アッセイ)
OP9細胞に上記にて得られた遺伝子をレトロウィルスベクター(pMX-puro) にて遺伝子導入し6wellディッシュに2.5x105ずつ蒔き、一晩培養した後、5−8週齢のC57BL/6マウス大腿骨より採取した骨髄細胞1x105まき、6日後に形成されたCAFC数を測定した。
【0040】
CAFC解析の結果
ストローマ細胞と骨髄細胞を共培養した際、骨髄細胞がストローマ細胞の下部に潜り込み増殖する現象をCobblestone area forming cell (CAFC) という。CAFC形成数が多い場合、ストローマ細胞が未熟な造血支持細胞を増幅していることを意味している。ただしCAFCを形成している細胞は造血幹細胞だけでなく、1段階分化した骨髄球系、赤芽球系、巨核球系、リンパ球系前駆細胞を含んでいるため、厳密にはCAFCの増加=造血幹細胞の増幅ではない。造血支持能を有するマウス骨髄ストローマ細胞株のOP9細胞に上記遺伝子をレトロウィルスpMX−puroにて導入し、ピューロマイシン5μg/mlにて薬剤耐性となった細胞についてCAFC形成能を解析した。その結果、第11図に示されるように、ISLRが最も高値を示しSDHF−1、SDHF−2、SDHF−4、及びSDHF−5についても軽度の高値を示した。これらの遺伝子の造血前駆細胞増幅能力が示唆された。逆にSDHF−3ではCAFC形成が抑制され、造血幹細胞増殖抑制機能を有すると考えられた。
【0041】
細胞培養上清の分泌蛋白質のメチオニン標識による検出
COS−7細胞を6cmディッシュに1x105個まき、一晩培養した。FuGene法(Roche社)を用いて遺伝子導入した。これらの遺伝子のカルボキシル末端にはFLAGタグが付加されており、前述したように、抗FLAG抗体によるウェスタン解析にて発現した蛋白質を検出することが出来る。36時間の培養後、上清を除去し、PBSにて2回洗浄後、0.5 ml DMEM(Methionine−, SIGMA社)中の0.5mCi[35S]methionine(ICN社TRAN-S)を加え、4時間培養した。培養上清から遠心にて細胞成分を除去し、Micron 10(ミリポア社)にて蛋白質を濃縮した。15%SDS−PAGEにて電気泳動後、標識された蛋白質をBAS2000(富士フィルム社)にて検出した。
【0042】
その結果、第12図及び第13図に示されるように、SDHF−1及びSDHF−5にて、培養上清への蛋白質分泌が検出された。これらの遺伝子は構造的に膜型蛋白質であること、分泌蛋白質の分子量がウェスタン解析で検出された成熟型蛋白質よりも約20kDa小さくなっていることから、これらの分泌蛋白質はカルボキシル末端の膜貫通領域が切断されたものと考えられる。カルボキシル末端の膜貫通領域が切断されて分泌されるタイプの増殖因子としてSCFが知られており、一方、接着蛋白質ではこうした例は認められないことから、本発明の遺伝子がコードする蛋白質は、液性因子として他の細胞に情報を伝達するタイプの蛋白質であることを強く示唆するものと考えられる。
【0043】
RT−PCR法による発現部位の検定
各SDHF遺伝子の塩基配列に基づきプライマーを設定、Multiple Tissue cDNA (MTC) Panels (クロンテック社)をtemplateとしてPCR反応(LA-Taq, TAKARA)を行った。また、骨髄内の細胞は蛍光標識したモノクローナル抗体(抗B220, 抗CD3, 抗Gr1, 抗MAC1, 抗Sca1抗体、Phamingen社)を用い、FACS Vantage(BectonDickinson社)にてソーティングしたTrizol試薬(Invitrogen社)にてRNAを抽出し、ランダムヘキサマーを用いてRT反応を行った上で、同様にPCR反応を行った。
【0044】
その結果、第14図に示される様に全てのSDHF遺伝子はOP9細胞での発現が確認された。特にSDHF−4は発現が脳と骨髄に限局して発現しており、また骨髄内においてはストローマ細胞に限局して発現しているなど興味深い。
【0045】
Long Term Culture-Initiating Cells Assay(LTC−IC法)による幹細胞支持機能の検定
上記のCAFC法では比較的短期間の造血支持機能を反映しているにすぎないため、より長期の幹細胞支持機能評価法としてLTC−IC法を行った。上記と同様に単離した遺伝子をレトロウィルス発現ベクターpMX−puroを用いてOP9細胞に高発現させLTC−IC法を行った。
【0046】
まず造血幹細胞を純化する。6週齢から8週齢のC57BL/6マウスの大腿骨より骨髄細胞を採取し、Ficoll法(比重1.100) にて単核細胞を遠心分離した。PBSにて洗浄後、ビオチン標識した1次抗体(抗Lineage抗体カクテル:抗CD3, 抗CD4, 抗CD8, 抗B220, 抗Ter119, 抗Gr1,抗 MAC1抗体、以上Phamingen社)およびストレプトアビジン標識磁気ビーズにそれぞれ4℃15分間反応させた。MACS法にてカラムを通過する分画(LIN(-)細胞)を回収し、遠心洗浄後、さらにPE標識抗Sca1抗体、FITC標識抗c−kit抗体、Per−CP−Cy5.5標識ストレプトアビジン(以上Phamingen社)にて4℃15分間反応させ、FACS Vantage(Becton Dickinson社)にてLIN(−),Sca1(+),c−kit(+)分画(以上にて得られた細胞をKSL細胞と定義する。)SDHF−1−6のカルボキシル末端にFLAGペプチドを付加したcDNAをレトロウィルスベクター(pMX-puro) にてOP9細胞に遺伝子導入して樹立した安定発現株を6穴ディッシュに2.5x105ずつ蒔き、一晩培養した後、KSL細胞を100個ずつまき、3週間培養した。その後細胞を回収し、γ線を20Gy照射したOP9細胞を1穴に3000個ずつまいておいた96穴ディッシュに、8穴ずつペアで2分の1ずつ薄めて蒔いた。5週間後、各穴から細胞を回収しそれぞれメチルセルロース培地(IMDM, メチルセルロース1%、牛胎児血清15%、牛血清アルブミン 1%、ヒトエリスロポイエチン3U/ml、ヒトIL-6 10ng/ml、マウスIL-3 10ng/ml、マウスSCF 100ng/ml牛インスリン10μg/ml)に蒔いて12日後に血球コロニー(CFU-C)の有無で判定した。幹細胞の頻度はL−Calcソフトウェア(Stemcell社)にて行った。
【0047】
その結果、第15図に示されるようにSDHF−4遺伝子に著明な造血支持能が観察された。この遺伝子の造血支持機能が示された。
【0048】
SDHF−4細胞外領域・リコンビナントタンパク質によるマウス造血幹細胞の増幅効果
SDHF−4の細胞膜貫通領域をPSORT IIプログラム(http://psort.ims.u-tokyo.ac.jp/form2.html)で予測し(アミノ酸524-540)、細胞外領域(アミノ酸1-523)にヒト免疫グロブリンFc領域と融合タンパク質(SDHF-4-ΔTM-Fc)を作製、発現ベクターpSSRaに組み込み、CHO−k1細胞に安定的に遺伝子導入した。無血清培地CD−CHO(Invitrogen社)にて培養することで、上清から融合リコンビナントタンパク質をProteinAカラムにて精製した(第16図)。マウス幹細胞はLTC−IC法の項に記載した様にマウス骨髄LIN(−),Sca1(+),c−kit(+)分画(KSL細胞)100個ずつ、SCF(50ng/ml),IL−3(10ng/ml),IL−6(10ng/ml)及びSDHF−4−ΔTM−Fc(100ng/ml)を添加したStem Span培地(StemCell社)にて培養した。7日後、細胞を回収し、FACS CaliburにてLIN,Sca1,c−kitの発現を解析した。
【0049】
その結果、第17図に示されるように、SCF,IL−3,IL−6を添加したStemSpan培地にて培養したLSK細胞はSDHF−4−ΔTM−Fc100ng/mlの存在下において非存在下と比較してLIN(−)細胞の割合が増加するだけでなく、LIN(−),Sca1(+),c−kit(+)分画の割合も増加することが判明した。この結果は、SDHF−4細胞外領域・リコンビナントタンパク質にマウス造血幹細胞の未分化状態を維持する作用があることを示している。
【0050】
脂肪細胞分化の検定とオイルレッドO染色法
SDHF−6遺伝子をレトロウィルスベクター(pMX-puro)導入した細胞とベクターのみを導入した細胞をそれぞれAlpha−MEM+20%牛胎児血清の条件にて4週間以上継代せずにConfluentの状態で培養し続けた。細胞は10%ホルマリンにて固定後オイルレッドOにて染色した。
その結果、第18図に示されるように、SDHF−6をレトロウィルス発現ベクターpMX−puroを用いてOP9細胞に高発現させると約4週間の培養の後、効率に脂肪細胞に分化させる効果があることが判明した。この遺伝子が、造血幹細胞よりもむしろ間葉系の細胞に作用して脂肪細胞への分化に関与していると考えられた。
【0051】
SDHF−4に対するポリクローナル抗体の作製及びそれによる骨髄内造血支持細胞の単離
SDHF−4のアミノ酸配列をもとに、合成ペプチド(GYMAKDKFRRMNEGQVY(第32-48番目のアミノ酸に相当する))を設計しラビットに免疫してポリクローナル抗体を作製した。
このペプチドを Epoxy-activated Sepharose 6B(ファルマシア社)に結合させ、抗原カラムを作製、これにより、得られたポリクローナル抗体をアフィニティー精製した。この抗体を用い、CHO−k1細胞にpSSRα−bsrベクターを用いてSDHF−4を発現させた細胞(SDHF4-cl12)とベクターだけ導入した細胞(mock)を比較してウェスタン解析を施行したところ、非常に特異的にSDHF−4遺伝子産物を認識できることがわかった(第19図)。また、得られた精製抗体1mgを LinKit Fluoro-Link (ISL社) によりFITC標識し、同じ細胞をFACS calibur(Becton Dickinson社)にて解析したところ、この抗体はFlowcytemetryにも使用可能であることがわかった(第20図; ○線がmock、■線がSDHF4-cl12)。この抗体を用いてC57BL/6Jマウスの骨髄細胞を染色し、FACS Vantage(Becton Dickinson社)によりSDHF4を発現している細胞をソーティングして回収したところ(第21図)、これらの細胞は培養ディッシュに付着し、形態学的にも骨髄造血支持細胞に相当する間葉系の細胞であった(第22図)。
【産業上の利用可能性】
【0052】
本発明によって、ストローマ細胞に由来する、新規な造血関連遺伝子の単離を行うことに成功し、それらが造血幹細胞増殖調節活性を有することが実証された。これによって、造血幹細胞の生体外増幅、造血器悪性腫瘍の移植治療、幹細胞による再生医学、遺伝子治療、免疫療法、細胞移植、遺伝子治療、神経疾患(アルツハイマー病、脳梗塞、変性神経疾患等)、肝疾患(肝硬変等)、肺疾患、心筋疾患、糖尿病、骨疾患、及び慢性腎疾患等の広範囲な応用を可能としたものである。
【図面の簡単な説明】
【0053】
【図1】第1図は、SDHF−1のアミノ酸配列を示す。
【図2】第2図は、SDHF−2のアミノ酸配列を示す。
【図3】第3図は、SDHF−3のアミノ酸配列を示す。
【図4】第4図は、SDHF−4のアミノ酸配列を示す。
【図5】第5図は、SDHF−5のアミノ酸配列を示す。
【図6】第6図は、SDHF−6のアミノ酸配列を示す。
【図7】第7図は、マイクロアレイ法により得られた結果を示す。
【図8−1】第8図は、ノーザンブロッティングにより得られた結果を示す。
【図8−2】第8図は、ノーザンブロッティングにより得られた結果を示す。
【図9】第9図は、ウェスタン解析により得られた結果を示す。
【図10】第10図は、免疫染色法により得られた結果を示す。
【図11】第11図は、CAFC解析の結果を示す。
【図12】第12図は、ウェスタン解析により検出された本発明遺伝子SDHF−1、SDHF−5の蛋白質翻訳後修飾の結果を示す。★は糖鎖が付加されて分子量が大きくなった成熟型の蛋白質を示し、▲は糖鎖の付加される以前の生成過程の蛋白質を示し、▼は切断後の残存蛋白質を示し、●は分泌型蛋白質を示す。
【図13】第13図は、細胞培養上清の分泌蛋白質のメチオニン標識による検出の結果を示す。●は分泌型蛋白質を示し、→は切断後培養上清に分泌された蛋白質を示す。
【図14−1】第14図は、RT−PCR法による本発明遺伝子の発現部位の検定の結果を示す、電気泳動の写真である。
【図14−2】第14図は、RT−PCR法による本発明遺伝子の発現部位の検定の結果を示す、電気泳動の写真である。
【図15】第15図は、LTC−IC法による本発明遺伝子の幹細胞支持機能の検定の結果を示す。
【図16】第16図は、SDHF−4細胞外領域・リコンビナントタンパク質をProteinAカラムにて精製した様子を示す、電気泳動の写真である。
【図17】第17図は、SDHF−4細胞外領域・リコンビナントタンパク質がマウス造血幹細胞の未分化状態を維持する作用を有することを示すグラフである。
【図18−1】第18図は、SDHF−6をレトロウィルス発現ベクターpMX−puroを用いてOP9細胞に高発現させると約4週間の培養の後、効率に脂肪細胞に分化させる効果があることを示す、顕微鏡写真(倍率:×40)である。
【図18−2】第18図は、SDHF−6をレトロウィルス発現ベクターpMX−puroを用いてOP9細胞に高発現させると約4週間の培養の後、効率に脂肪細胞に分化させる効果があることを示す、顕微鏡写真(倍率:×40)である。
【図19】第19図は、SDHF−4の部分ペプチドに対する抗体を用い、CHO−k1細胞にpSSRα−bsrベクターを用いてSDHF−4を発現させた結果を示す、ウェスタン解析による写真である。
【図20】第20図は、上記の精製抗体をFITC標識し、SDHF−4を発現させたCHO−k1細胞をFACS calibur (BectonDickinson社)にて解析した結果を示す。
【図21】第21図は、同抗体を用いてC57BL/6Jマウスの骨髄細胞を染色し、FACS Vantage (BectonDickinson社) によりSDHF4を発現している細胞をソーティングした結果を示す。
【図22】第22図は、ソーティングにより選択された上記細胞の顕微鏡写真(倍率:×200倍)である。
【図23】第23図は、SDHF−1からSDHF−6の分子構造を模式的に表したものである。
【特許請求の範囲】
【請求項1】
以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号10に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【請求項2】
以下の(a)又は(b)のDNAを含む遺伝子:
(a)配列番号9に示される塩基配列からなるDNA
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNA。
【請求項3】
以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号12に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【請求項4】
以下の(a)又は(b)のDNAを含む遺伝子:
(a)配列番号11に示される塩基配列からなるDNA
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNA。
【請求項5】
以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号2に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【請求項6】
以下の(a)又は(b)のDNAを含む遺伝子:
(a)配列番号1に示される塩基配列からなるDNA
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNA。
【請求項7】
以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号4に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【請求項8】
以下の(a)又は(b)のDNAを含む遺伝子:
(a)配列番号3に示される塩基配列からなるDNA
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNA。
【請求項9】
以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号6に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【請求項10】
以下の(a)又は(b)のDNAを含む遺伝子:
(a)配列番号5に示される塩基配列からなるDNA
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNA。
【請求項11】
請求項1ないし10のいずれか一項に記載の遺伝子にコードされる蛋白質。
【請求項12】
請求項1ないし10のいずれか一項に記載の少なくとも一種の遺伝子を含有する組換え発現ビークル。
【請求項13】
組換えプラスミドベクターである請求項12に記載の組換え発現ビークル。
【請求項14】
組換えレトロウィルスベクターである請求項12に記載の組換え発現ビークル。
【請求項15】
請求項12、13又は14に記載の発現ビークルによって形質転換された形質転換体。
【請求項16】
形質転換体がCOS−7細胞である請求項15に記載の形質転換体。
【請求項17】
形質転換体がストローマ細胞である請求項15に記載の形質転換体。
【請求項18】
請求項15又は16に記載の形質転換体を培養することを含む、請求項11に記載の蛋白質の製造方法。
【請求項19】
請求項1ないし10のいずれか一項に記載の少なくとも一種の遺伝子によりストローマ細胞を形質転換することから成る、該ストローマ細胞の造血幹細胞増殖活性の調節方法。
【請求項20】
造血幹細胞増殖活性を増幅することを特徴とする、請求項19に記載の調節方法。
【請求項21】
該ストローマ細胞がマウス・ストローマ細胞である、請求項19又は20に記載の造血幹細胞増殖活性の調節方法。
【請求項22】
以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号13に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質、
でストローマ細胞を形質転換することから成る、該ストローマ細胞の造血幹細胞増殖調節活性の修飾方法。
【請求項23】
造血幹細胞増殖活性を増幅することを特徴とする、請求項22に記載の調節方法。
【請求項24】
該ストローマ細胞がマウス・ストローマ細胞である、請求項21、22又は23に記載の造血幹細胞増殖活性の調節方法。
【請求項25】
請求項13に記載の少なくとも一種の蛋白質、及び/又は、
(a)配列番号13に示されるアミノ酸配列、若しくは
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質、
を有効成分として含有する、造血幹細胞増殖調節用組成物。
【請求項26】
請求項1ないし10のいずれか一項に記載の遺伝子にコードされる蛋白質又はその部分ペプチドに対する抗体。
【請求項27】
ポリクローナル抗体であることを特徴とする、請求項26に記載の抗体。
【請求項28】
請求項26ないし27のいずれ一項に記載の抗体を用いて、ストローマ細胞を単離する方法。
【請求項29】
ストローマ細胞が骨髄造血支持細胞である、請求項28に記載の方法。
【請求項1】
以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号10に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【請求項2】
以下の(a)又は(b)のDNAを含む遺伝子:
(a)配列番号9に示される塩基配列からなるDNA
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNA。
【請求項3】
以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号12に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【請求項4】
以下の(a)又は(b)のDNAを含む遺伝子:
(a)配列番号11に示される塩基配列からなるDNA
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNA。
【請求項5】
以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号2に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【請求項6】
以下の(a)又は(b)のDNAを含む遺伝子:
(a)配列番号1に示される塩基配列からなるDNA
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNA。
【請求項7】
以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号4に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【請求項8】
以下の(a)又は(b)のDNAを含む遺伝子:
(a)配列番号3に示される塩基配列からなるDNA
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNA。
【請求項9】
以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号6に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質。
【請求項10】
以下の(a)又は(b)のDNAを含む遺伝子:
(a)配列番号5に示される塩基配列からなるDNA
(b)塩基配列(a)からなるDNAとストリンジェントな条件下でハイブリダイズし、且つ、造血幹細胞増殖調節活性を有する蛋白質をコードするDNA。
【請求項11】
請求項1ないし10のいずれか一項に記載の遺伝子にコードされる蛋白質。
【請求項12】
請求項1ないし10のいずれか一項に記載の少なくとも一種の遺伝子を含有する組換え発現ビークル。
【請求項13】
組換えプラスミドベクターである請求項12に記載の組換え発現ビークル。
【請求項14】
組換えレトロウィルスベクターである請求項12に記載の組換え発現ビークル。
【請求項15】
請求項12、13又は14に記載の発現ビークルによって形質転換された形質転換体。
【請求項16】
形質転換体がCOS−7細胞である請求項15に記載の形質転換体。
【請求項17】
形質転換体がストローマ細胞である請求項15に記載の形質転換体。
【請求項18】
請求項15又は16に記載の形質転換体を培養することを含む、請求項11に記載の蛋白質の製造方法。
【請求項19】
請求項1ないし10のいずれか一項に記載の少なくとも一種の遺伝子によりストローマ細胞を形質転換することから成る、該ストローマ細胞の造血幹細胞増殖活性の調節方法。
【請求項20】
造血幹細胞増殖活性を増幅することを特徴とする、請求項19に記載の調節方法。
【請求項21】
該ストローマ細胞がマウス・ストローマ細胞である、請求項19又は20に記載の造血幹細胞増殖活性の調節方法。
【請求項22】
以下の(a)又は(b)の蛋白質をコードする遺伝子:
(a)配列番号13に示されるアミノ酸配列、
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質、
でストローマ細胞を形質転換することから成る、該ストローマ細胞の造血幹細胞増殖調節活性の修飾方法。
【請求項23】
造血幹細胞増殖活性を増幅することを特徴とする、請求項22に記載の調節方法。
【請求項24】
該ストローマ細胞がマウス・ストローマ細胞である、請求項21、22又は23に記載の造血幹細胞増殖活性の調節方法。
【請求項25】
請求項13に記載の少なくとも一種の蛋白質、及び/又は、
(a)配列番号13に示されるアミノ酸配列、若しくは
(b)アミノ酸配列(a)において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、造血幹細胞増殖調節活性を有する蛋白質、
を有効成分として含有する、造血幹細胞増殖調節用組成物。
【請求項26】
請求項1ないし10のいずれか一項に記載の遺伝子にコードされる蛋白質又はその部分ペプチドに対する抗体。
【請求項27】
ポリクローナル抗体であることを特徴とする、請求項26に記載の抗体。
【請求項28】
請求項26ないし27のいずれ一項に記載の抗体を用いて、ストローマ細胞を単離する方法。
【請求項29】
ストローマ細胞が骨髄造血支持細胞である、請求項28に記載の方法。
【図1】


【図2】

【図3】

【図4】

【図5】

【図6】

【図7】

【図11】

【図15】

【図16】

【図17】

【図19】

【図20】

【図21】

【図8−1】

【図8−2】

【図9】

【図10】

【図12】

【図13】

【図14−1】

【図14−2】

【図18−1】

【図18−2】

【図22】

【図23】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図11】
【図15】
【図16】
【図17】
【図19】
【図20】
【図21】
【図8−1】
【図8−2】
【図9】
【図10】
【図12】
【図13】
【図14−1】
【図14−2】
【図18−1】
【図18−2】
【図22】
【図23】
【公開番号】特開2007−143556(P2007−143556A)
【公開日】平成19年6月14日(2007.6.14)
【国際特許分類】
【出願番号】特願2007−25808(P2007−25808)
【出願日】平成19年2月5日(2007.2.5)
【分割の表示】特願2003−523654(P2003−523654)の分割
【原出願日】平成14年8月22日(2002.8.22)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【出願人】(590001452)国立がんセンター総長 (80)
【Fターム(参考)】
【公開日】平成19年6月14日(2007.6.14)
【国際特許分類】
【出願日】平成19年2月5日(2007.2.5)
【分割の表示】特願2003−523654(P2003−523654)の分割
【原出願日】平成14年8月22日(2002.8.22)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【出願人】(590001452)国立がんセンター総長 (80)
【Fターム(参考)】
[ Back to top ]