
RNAi法を用いた有用アルカロイド生合成中間体の生産法
アルカロイドを産生する植物細胞、植物組織または植物体においてRNAi法を用いて、目的とするアルカロイド生合成中間体を基質とする酵素の発現を阻害することを含む、アルカロイド生合成中間体の生産方法および該方法に用いるRNAi遺伝子を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は有用アルカロイド生合成中間体の生産方法に関する。
【背景技術】
【0002】
アルカロイドは、植物に含まれる塩基性含窒素化合物の総称である。アルカロイドはその主要骨格に応じて、キノリン系、イソキノリン系、インドール系、トロパン系、キサンチン系アルカロイドなどに分類される。アルカロイドのなかには薬学的に有用なものが多く、たとえばコデインおよびモルヒネの鎮痛作用や商業的価値はよく知られている。ノスカピンは鎮咳作用化合物として有用である。パパベリンは平滑筋弛緩剤および脳血管拡張薬である。ベルベリンは、抗菌性、抗マラリア性および解熱性化合物として用いられてきた。
【0003】
イソキノリンアルカロイドは、イソキノリンを基本骨格とするアルカロイドの総称であり、分布も広く、構造も多様である。イソキノリンアルカロイドには例えば、モルヒネ(morphine)型(モルヒネなど)、プロトベルベリン(protoberberine)型(ベルベリンなど)、ベンゾフェナンスリジン(benzophenanthridine)型(サングイナリンなど)のアルカロイドが含まれる。イソキノリンアルカロイドを産生する植物としては、ケシ科(パパベラセアエ(Papaveraceae))、メギ科(ベルベリダセアエ(Berberidaceae))、キンポウゲ科(ラナンキュラセアエ(Ranunculaceae))、ツヅラフジ科(メニスペルマセアエ(Menispermaceae))、ミカン科(ルタセアエ(Rutaceae))などが挙げられる。
【0004】
イソキノリンアルカロイド生合成経路の中間体としては、レチクリンが多数の薬学的生成物への重要な前駆体として注目されてきた。例えば、特許文献1には、ケシにランダムに突然変異を導入し、レチクリン含量の高い植物体を選抜することを含む、レチクリンを製造する方法が開示されている。しかしこの方法は無作為に作成した突然変異体の中からレチクリン含量の高いものを選抜し、その植物体からレチクリンを抽出するという工程を含むため、煩雑なものであった(特許文献1を参照)。
【0005】
ベルベリン架橋酵素は、イソキノリンアルカロイド生合成経路の酵素の1つであり、レチクリンを基質とする。ベルベリン架橋酵素の発現を遺伝子操作を用いて低下させることにより、その基質であるレチクリン含量を増加させようとする試みがなされてきた。これらの試みは、アンチセンス法により、ベルベリン架橋酵素の発現を特異的に阻害しようとするものであった。しかし、その結果としてベルベリン架橋酵素の発現は阻害され、アルカロイド含量が全般的に著しく低下したものの、レチクリンのような中間代謝産物の特異的な蓄積は観察されなかった(例えば、非特許文献1および2を参照)。
【0006】
近年、アンチセンス法のほかに、遺伝子発現の抑制法としてRNAi法が利用されている。RNAi(RNA interference)とは、dsRNA(2本鎖RNA)の細胞内への導入によって、dsRNAと相同な配列を持つ遺伝子の発現が抑制される現象である。RNAiは様々な生物種における遺伝子機能解析に用いられている。例えば、非特許文献3では、ステロイド合成系に関与する酵素を特異的にRNAiによって阻害することにより、該酵素の基質となる中間体が蓄積することが報告された。しかし、この文献によると、目的の中間体から別の物質への変換がかなり起こっており、中間体を特異的に蓄積させるまでには至っていない。これはステロイドが細胞膜の必須成分であり細胞成長と関係しているため、完全には代謝経路の遮断が起こらないためと考えられる。また、この文献では植物細胞の成長が阻害されたことが報告されている(例えば、非特許文献3を参照)。
【0007】
RNAi法に用いられるRNAi用ベクターは二本鎖RNA(dsRNA)を植物体内で発現させることを目的として作製されるものである。RNAiベクターは、その構造に基づき、おおまかに2種類に分類される。
【0008】
一つは、別々に作製したセンスRNAを発現するプラスミドと、アンチセンスRNAを発現するプラスミドの組み合わせである。これら2つのプラスミドを、混合して細胞に形質転換し、細胞内で、dsRNAを形成させる。もう一つは、ヘアピン構造をとるRNAを発現するプラスミドである。後者については、線虫、ショウジョウバエ、植物、トリパノソーマなどで報告がある。植物については、導入遺伝子のヘアピン構造の中間にイントロンを導入したものとしないものの遺伝子抑制効力を比較した研究がある。この研究においては、イントロンを持った導入遺伝子のほうがより強力な発現抑制を引き起こすという結果が報告されている。
【0009】
これまでに、本発明者らは、内在遺伝子と80%程度の相同性をもつ100塩基対程度のdsRNAを発現させることにより、遺伝子抑制が起こることを観察している。この際dsRNAは、細胞内でRNaseによって分解を受け、約20塩基対のsiRNAとなり、siRNAがRISCと呼ばれる複合体に組み込まれ、この複合体が標的mRNAを分解すると考えられている。動物細胞においては、約20塩基対のsiRNAを導入することにより遺伝子発現の抑制がおこることが報告されている。
【特許文献1】特表2002−508947号公報
【非特許文献1】サング−ウン・パーク(Sang−Un Park)他著、「プラント・フィジオロジー(Plant Physiology)」第128巻、2002年2月、p.696−706
【非特許文献2】サング−ウン・パーク(Sang−Un Park)他著、「プラント・モレキュラー・バイオロジー(Plant Molecular Biology)」第51巻、2003年、p.153−164
【非特許文献3】セリン・バーガー(Celine Burger)他著、「ジャーナル・オブ・エクペリメンタル・ボタニー(Journal of Experimental Botany)」第54巻、第388号、2003年7月、p.1675−1683
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明は、特定のアルカロイド生合成中間体の生産方法を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明は、アルカロイドを産生する植物細胞、植物組織または植物体においてRNAi法を用いて、目的とするアルカロイド生合成中間体を基質とする酵素の発現を阻害することを含む、アルカロイド生合成中間体の生産方法を提供する。
【0012】
具体的には、後述のRNAi遺伝子を用いて目的とするアルカロイド生合成中間体を基質とする酵素の発現を阻害して、代謝の流れを遮断することにより、目的のアルカロイド生合成中間体が植物細胞内に蓄積する。
【0013】
本発明はまた、上記方法によって生産されたアルカロイド生合成中間体を提供する。
【0014】
さらに本発明は、上記方法に用いられる、
i)プロモーター、および、
ii)プロモーターの下流に、
a)目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列と相同的な正方向配列、および、
b)該正方向配列に相補的な逆方向配列
を含む遺伝子を提供する。本明細書においてこの遺伝子を「RNAi遺伝子」と称する。
【0015】
また、本発明は上記方法に用いられる、以下のAおよびBの遺伝子を含む遺伝子の組合せを提供する:
A.i)プロモーター、および、
ii)プロモーターの下流に、目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列と相同的な正方向配列を含む遺伝子、および、
B.i)プロモーター、および、
ii)プロモーターの下流に、該正方向配列に相補的な逆方向配列を含む遺伝子。
本明細書においてこの遺伝子の組合せを「RNAi遺伝子の組合せ」と称する。
【0016】
「目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列と相同的な正方向配列」とは、生産しようとするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列に対して相同性を有する配列であって、その長さが約100bp以上であり、ベクターなどのコンストラクトにおいて転写と同じ方向に挿入された配列のことを指す。
【0017】
ここで、「酵素の全体または一部をコードする配列」には、該酵素をコードする遺伝子の翻訳領域のみならず、非翻訳領域の配列も含まれる。
【0018】
「該正方向配列に相補的な逆方向配列」とは、上記の「目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列」に対して相補性を有する配列のことをいう。即ち、ベクターなどのコンストラクトにおいて転写と逆方向に挿入された正方向配列と相同性を有する配列を指す。
【0019】
RNAi遺伝子においては該正方向配列と逆方向配列はともにプロモーターの下流に位置する必要があるが、正方向配列と逆方向配列のどちらを上流に位置させてもよい。
【0020】
好ましくは、RNAi遺伝子においては、該正方向配列と逆方向配列の間にスペーサー配列を介在させる。スペーサーを介在させることによって、正方向配列と逆方向配列(以下両配列の繰り返しをインバーテッドリピートと称する)の対合を容易にする空間が生じる。スペーサー配列は特に限定されないが、通常数百bp〜1kbの長さの配列、例えばイントロン配列が好適に用いられる。
【0021】
正方向配列、スペーサー配列、および逆方向配列を含むRNAi遺伝子を植物細胞内で発現させることにより、植物細胞においてアルカロイド生合成酵素の発現が抑制される。
【0022】
該正方向配列、スペーサー配列および、該正方向配列に相補的な逆方向配列を含むDNAは、植物細胞においてプロモーターにより誘導されてmRNAへと転写される。正方向配列から転写された一本鎖RNA部分と逆方向配列から転写された一本鎖RNA部分は、相補的であるため互いに水素結合により結合する。好ましくはかかるRNAはスペーサー配列を介してヘアピン構造の二本鎖RNA(dsRNA)を形成する。このdsRNAがRNAi、即ち、標的であるアルカロイド生合成遺伝子の発現の抑制を引き起こすと考えられる。
【0023】
RNAi遺伝子の組合せを用いる場合は、プロモーターの下流に正方向配列を含むベクター(センスベクターとも呼ばれる)と、プロモーターの下流に逆方向配列を含むベクター(アンチセンスベクター)の両方を植物に導入する。該正方向配列および逆方向配列は、植物細胞においてプロモーターにより誘導されてmRNAへと転写され、正方向配列から転写された一本鎖RNAと逆方向配列から転写された一本鎖RNAが、互いに水素結合により結合して二本鎖RNA(dsRNA)を形成する。このdsRNAがRNAi、即ち、標的であるアルカロイド生合成遺伝子の発現の抑制を引き起こすと考えられる。
【0024】
このようにして、RNAi遺伝子あるいはRNAi遺伝子の組合せを用いることにより、二本鎖RNAが生じる結果、標的アルカロイド生合成遺伝子の発現が阻害され、該酵素の基質であるアルカロイド生合成中間体が細胞内に特異的に蓄積する。
【0025】
「目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列」は先に定義したように、必ずしも遺伝子のコード領域に含まれる必要はなく、5’UTR領域や3’UTR領域に位置する配列あるいはプロモーター領域でもよい。RNAiはこのような非コード領域の配列を用いることによっても起こる現象である。
【0026】
さらに本発明は上述したような遺伝子を含むベクターおよび該ベクターにより形質転換された植物細胞または植物組織あるいは植物体を提供する。
【発明の効果】
【0027】
これまでの研究では、生合成経路の最終産物の蓄積の抑制は報告されている。しかし、本発明のように中間代謝産物の特異的な蓄積を可能とし、中間体を大量に生産する、あるいは、さらに、新規な反応経路を追加することにより、新たな化合物を生産させようとする観点は無かった。また、かかる観点があったとしても、現実的には、そのようなことを可能とする技術はなかった。今回RNAi法によって、生合成経路の中間代謝産物が蓄積することが明らかになった。本発明は、様々な代謝系において、重要な代謝産物を蓄積させる可能性を示唆しており、その応用範囲は極めて広い。
【図面の簡単な説明】
【0028】
【図1】図1は、イソキノリンアルカロイド生合成経路と生合成酵素を示す図である。
【図2】図2は、インドールアルカロイド生合成経路と生合成酵素を示す図である。
【図3】図3は、中間体レチクリンから派生する化合物を示す図である。
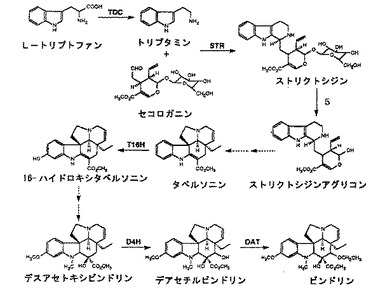
【図4】図4は、中間体ストリクトシジンから派生する化合物を示す図である。
【図5】図5は、ベルベリン架橋酵素(BBE)遺伝子に対するdsRNA発現ベクター、pART27−BBEir構築の手順を示す図である。
【図6】図6は、ハナビシソウBBEdsRNA形質転換体におけるレチクリン(m/z 330)の蓄積のLC/MS解析を示す図である。
【図7】図7は、コントロールおよびBBEdsRNA形質転換体における、BBE酵素活性を示す図である。
【図8】図8は、コントロールおよびBBEdsRNA形質転換体における、BBE酵素反応をLC/MS解析した結果を示す図である。
【図9】図9は、コントロールおよびBBEdsRNA形質転換体における、レチクリンと、サンギナリンの含量を示す図である。
【図10】図10は、BBEの阻害により、レチクリンからスコウレリンの反応が遮断されることを示す模式図である。
【符号の説明】
【0029】
1 ノルコクラウリン−6−O−メチルトランスフェラーゼ
2 コクラウリン−N−メチルトランスフェラーゼ
3 N−メチル−コクラウリン−3’−ヒドロキシラーゼ
4 ベルベリン架橋酵素(BBE)
5 グルコシダーゼI/II
【発明を実施するための最良の形態】
【0030】
本発明において標的とされる植物、即ちdsRNAを導入される植物は、アルカロイド生合成経路を有する植物であれば特に限定されない。アルカロイド生合成経路としてはイソキノリンアルカロイド生合成経路が好適であるが、その他にもインドールアルカロイド生合成経路などが好適な標的となりうる。
【0031】
具体的には、アルカロイド生合成経路を有する植物には以下のものが挙げられるがこれらに限定されるものではない。ベルベリンを産生する植物、例えば、オウレン(Coptis japonica,Coptis chinensis Franch,Coptis deltoides C.Y.Cheng et Hsiao等)、キハダ(Phellodendron amurense)、メギ(Berberis属)、ナンテン(Nandina domestica)、ヒイラギナンテン(Mahonia japonica)およびアキカラマツ(Thalictrum minus)、モルヒネやコデイン、パパベリンを産生する植物、例えば、ケシ(Papaver somniferum Linn)、アツミゲシ(Papaver setigerum DC)およびハカマオニゲシ(Papaver bracteatum)、モルヒネを生産しないが、類縁のアルカロイドを産生する植物、例えば、オニゲシ(Papaver orientale Linn)およびヒナゲシ(Papaver rhoeas)、サングイナリンを産生する植物、例えば、ハナビシソウ(Eschscholtzia californica)、サングイナリア(Sanguinarine canadensis L.)、コリダリンを産生する植物、例えば、エンゴサク(Corydalis属植物、例えば、Corydalis bulbosa DC.,Corydalis ternata Nakai,Corydalis nakaii Ishidoya,Corydalis decumbens Person)、コロンバミンを産生する植物、例えば、コロンボ(Jateorhiza columbaMiers)、セファランチン(cepharanthine)を産生する植物、例えば、タマサキツヅラフジ(Stephania capharantha)、シノメニン(sinomenine)を産生する植物、例えば、オオツヅラフジ(Sinomenium acutum Rehder et Wilson)、エメチン(emetine)を産生する植物、例えば、トコン(Cephaelis ipecacuanha)など。
【0032】
上記の植物の中ではイソキノリンアルカロイドを合成する植物が好適に用いられ、なかでもサングイナリンやベルベリンを産生する植物、例えば、ハナビシソウ、オウレン、キハダ、メギ、ナンテン、ヒイラギナンテンおよびアキカラマツが好ましく、特にハナビシソウが好ましい。
【0033】
RNAi遺伝子を導入される標的植物の起源と、導入するRNAi遺伝子の起源は、同じであっても異なっていてもよい。標的遺伝子と導入遺伝子の相同性を考慮すると、同じ植物種由来のものが特に好ましい。
【0034】
本発明の方法においては、目的とする生合成中間体を基質とする酵素をRNAiによる阻害の標的となる酵素とする。かかる酵素としては、例えば、イソキノリンアルカロイド合成系経路(図1を参照)のベルベリン架橋酵素(BBE)、ノルコクラウリン−6−O−メチルトランスフェラーゼ、コクラウリン−N−メチルトランスフェラーゼおよびN−メチルコクラウリン−3’−ヒドロキシラーゼが挙げられる。
【0035】
これらの酵素をRNAiにより阻害するとその基質であるアルカロイド生合成中間体が蓄積する。具体的には、ベルベリン架橋酵素を阻害するとレチクリン、ノルコクラウリン−6−O−メチルトランスフェラーゼを阻害するとノルコクラウリン、コクラウリン−N−メチルトランスフェラーゼを阻害するとコクラウリン、そしてN−メチルコクラウリン−3’−ヒドロキシラーゼを阻害するとN−メチルコクラウリンがそれぞれ蓄積する。
【0036】
イソキノリンアルカロイド生合成経路の酵素以外では例えば、インドールアルカロイド生合成経路(図2を参照)のグリコシダーゼI/IIを標的とすると、その基質であるストリクトシジンが蓄積する。
【0037】
好ましくは本発明の方法により生産されるアルカロイド生合成中間体は、レチクリン、ノルコクラウリン、コクラウリンおよびN−メチルコクラウリンからなる群から選択され、特に好ましくはレチクリンである。
【0038】
レチクリンを製造するには、標的酵素はベルベリン架橋酵素であり、その結果レチクリンが蓄積する。レチクリンの前駆体であるノルコクラウリン、コクラウリンおよびN−メチルコクラウリン(図1を参照)、ならびにレチクリンは、図3に示す様々なイソキノリンアルカロイドの前駆体として有用である。また、ストリクトシジンも図4に示す様々なインドールアルカロイドの前駆体として有用である。
【0039】
本発明のRNAiを引き起こす遺伝子は好ましくは、ベルベリン架橋酵素、ノルコクラウリン−6−O−メチルトランスフェラーゼ、コクラウリン−N−メチルトランスフェラーゼおよびN−メチルコクラウリン−3’−ヒドロキシラーゼからなる群から選択される酵素をコードする配列またはその一部である。RNAiを引き起こすためのdsRNAの長さは特に限定されないが、少なくとも23塩基以上、好ましくは100塩基対から2キロベースであり、最も好ましくは100塩基対〜800塩基対である。
【0040】
本発明においてRNAiを引き起こす遺伝子の発現を誘導するプロモーターとしては、植物へ導入した際に目的とする遺伝子を発現させることができるものであれば特に限定されない。かかるプロモーターは当業者によく知られており、例えば、カリフラワーモザイクウイルス35Sプロモーター、および、誘導性プロモーター、例えば、アルコールデヒドロゲナーゼプロモーターやテトラサイクリンリプレッサー/オペレーター制御系などが挙げられる。
【0041】
本発明において、正方向配列および逆方向配列と、標的の生合成酵素をコードする遺伝子の配列との相同性は、100%である必要はない。遺伝子の突然変異、多型、又は進化上の分岐による配列の変動は容認される。標的遺伝子に対する挿入、欠失、及び一塩基点突然変異を有するdsRNA鎖もまた、RNAiに有効である。具体的には、RNAiに用いる遺伝子は、標的遺伝子と完全に同一である必要はないが、少なくとも70%以上、好ましくは80%以上、さらに好ましくは90%以上、最も好ましくは95%以上の配列の同一性を有するものが好適である。
【0042】
また、正方向配列と逆方向配列は、転写された後に二本鎖RNAを形成し得るものであればよい。効率的にdsRNAを形成するためには、両者の相補性は、少なくとも70%、好ましくは少なくとも80%、さらに好ましくは少なくとも90%、最も好ましくは少なくとも95%である。
【0043】
本発明において標的とする植物へのベクターの導入には、ポリエチレングリコール法、電気穿孔法(エレクトロポレーション法)、アグロバクテリウムを介する方法、パーティクルガン法など、当業者に公知の種々の方法を用いることができる。それぞれの方法に適したベクターの調製、形質転換植物細胞からの植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である(Toki S,et al:Plant Physiol 100:1503,1995)。
【0044】
例えば、形質転換植物体を作出する手法については、ポリエチレングリコールを用いてプロトプラストへ遺伝子導入し、植物体を再生させる方法(Datta SK:In Gene Transfer To Plants(Potrykus I and Spangenberg,Eds)pp.66−74,1995)、電気パルスによりプロトプラストへ遺伝子導入し、植物体を再生させる方法(Toki S,et al:Plant Physiol 100:1503,1992)、パーティクルガン法により細胞へ遺伝子を直接導入し、植物体を再生させる方法(Christou P,et al:Biotechnology 9:957,1991)、およびアグロバクテリウムを介して遺伝子を導入し、植物体を再生させる方法(Hiei Y,et al:Plant J 6:271,1994)など、いくつかの技術が既に確立し、当該技術分野において広く用いられている。本発明においては、これらの方法を好適に用いることができる。
【0045】
本発明において、植物にRNAi遺伝子を導入する際に用いるベクターは、遺伝子導入手段に応じて適宜選択すればよく、特に限定されない。例えば、アグロバクテリウムによる遺伝子導入法を用いる場合は、バイナリーベクター、例えば、pART、pBI101、pBI121、pIG121Hm等が好適に用いられる。
【0046】
本発明のベクターの作成方法としては、特に限定されず、当業者に周知のいかなる方法を用いてもよい。本発明のベクターにおいては、3’末端側へターミネーターが挿入されるが、ターミネーターとしては公知のものがいずれも好適に用いられ、例えば、OCSターミネーター、nosターミネーター、35Sターミネーターなどが挙げられる。
【0047】
本明細書中、塩基配列の同一性は、カーリンおよびアルチュールによるアルゴリズムBLAST(Proc.Natl.Acad.Sei.USA 87:2264−2268,1990、Karlin S &Altschul SF:Proc Natl Acad Sci USA 90:5873,1993)を用いて決定できる。BLASTのアルゴリズムに基づいたBLASTNやBLASTXと呼ばれるプログラムが開発されている(Altschul SF,et al:J Mol Biol 215:403,1990)。これらの解析方法の具体的な手法は公知である(http://www.ncbi.nlm.nih.gov/)。
【実施例】
【0048】
以下に本発明を特定の実施例に言及して記載するがこれは本発明を例示する目的のものであり、限定する意図のものではない。
【0049】
(実施例の概要)
アルカロイド生合成遺伝子に対して特異的な二本鎖RNA(dsRNA)を合成する発現ベクターを構築し、その高い抑制効果により、標的遺伝子の発現および酵素活性を効果的に抑制し、中間体が蓄積生産できることを実証した。
【0050】
具体的には、重要な医薬品であるイソキノリンアルカロイドの生合成中間体を、同アルカロイドを生産する植物細胞の代謝経路を二本鎖RNA(dsRNA)を用いたRNA干渉(RNAi)法により遮断することにより、生産する方法を開示する。本実施例では、ベンゾフェナンスリジンアルカロイドを生産するハナビシソウ細胞に、その生合成経路の酵素の1つであるベルベリン架橋酵素(BBE)の配列の一部に対応するdsRNAを発現するベクターを導入した。その結果、かかるアルカロイド生合成系が遮断され、植物細胞における生合成中間体レチクリンの生産が可能となった。
【0051】
(実施例についての具体的説明)
形質転換が可能なイソキノリンアルカロイド生産性植物ハナビシソウから単離されている生合成酵素ベルベリン架橋酵素(BBE)遺伝子(配列番号1)に対して既知の配列情報によりプライマーを設計し、BBEのcDNAを単離した。このcDNAをもとにdsRNAを形成するようなBBEdsRNA発現ベクターを構築した。ついで、同ベクターをハナビシソウ細胞に導入し、BBEdsRNAを発現する形質転換体を得、ベルベリン架橋酵素の基質である生合成中間体であるレチクリンを蓄積する形質転換ハナビシソウ細胞を確立した。
【0052】
(実験材料および方法)
コンストラクトの作製には、インバーテッドリピート作成用ベクターとして広く用いられているpKANNIBALおよびpART27を用いた。pKANNIBALは、CaMV35Sプロモーター、その下流に両端に複数の制限酵素サイトをもつイントロン領域、さらにその下流にOCSターミネーターを有する。イントロンの両端に、正方向および逆方向配列を挿入する。かかる配列は、植物体内でmRNAへと転写され、スプライシングされる。該RNAは、インバーテッドリピート、すなわち、二本鎖RNA(dsRNA)を形成する。なお、本発明は、必ずしも、このベクターの使用に限定されるものではない。
【0053】
ハナビシソウBBE遺伝子の単離
データベースにすでに登録されている配列をもとに、3’arm(本明細書において逆方向配列と称される部分)にBamH I、Hind III、5’arm(本明細書において正方向配列と称される部分)にEcoR I、Xho Iの制限酵素サイトを持つハナビシソウBBE遺伝子のほぼ全長約1kb断片をPCRによりハナビシソウ培養細胞cDNAから単離した。
【0054】
用いたプライマーを以下に示す。
BBE−3’arm−フォワード(FM):ATG GAT CCG ATT CGG ACT CGG ATT TCA ACC
(配列番号2)
リバース(RV):ATT AAG CTT CCA CTT CGA TGA GGA AAC GG
(配列番号3)
5’arm−フォワード(FM):AAT CTC GAG ATT CGG ACT CGG ATT TCA ACC
(配列番号4)
リバース(RV):CGA ATT CCA CTT CGA TGA GGA AAC GG
(配列番号5)
【0055】
こうして単離した遺伝子をプラスミドpT7 Blue(Novagen)にサブクローニングしたのち、SIMADZU DSQ−2000Lにより塩基配列を確認した。
【0056】
dsRNA発現ベクターの作製(図5)
3’armの挿入
BBE−3’arm−FWとRVのプラーマーペアにより得られたPCR産物をpT7−Blueにサブクローニングし、配列確認したのち、BamH I、Hind IIIで切断した。ベクターpKANNIBALも同様に、BamH IとHind IIIで制限酵素で処理した。これらDNAを電気泳動後、フェノール処理、クロロフォルム抽出、エタノール沈殿し、10μlのTEに溶解させたのち、ライゲーション反応を行ない、XL1−Blueにトランスフォーメーションした。挿入の確認は制限酵素処理、およびOCSターミネーター配列に対するAS1を用いたシーケンシングによりおこなった。
【0057】
5’armの挿入
BBE5’arm FWとRVのプライマーペアにより得られたPCR産物をpT7−Blueにサブクローニングし、配列確認したのち、EcoRI、XhoIで切断した。タンデムな挿入を防ぐため断片にアルカリフォスファターゼ(Calf Intestine Alkaline Phosphatase:CIAP)を加えて脱リン酸処理した。CIAPを失活させるため、65℃で30分インキュベートした。その後エタノール沈殿により制限酵素を失活させ、20μlのTEに溶解した。先に、pKANNIBALに3’armを導入したベクターを同様にEcoRI、XhoIで切断したのち、ライゲーション反応をおこない、XL1−Blueにトランスフォーメーションした。挿入の確認は制限酵素処理およびOCSターミネーターに対するAS1プライマー、35Sプロモーターに対する−S1プライマー(35Spro−S1,GAG CTA CAC ATG CTC AGG TT(配列番号6)を用いたシーケンシングによりおこなった。得られたBBEの3’および5’armが挿入されたプラスミドをpKANNIBAL−BBEirと称する。
【0058】
バイナリ−ベクターpART27への導入
pART27ベクターを制限酵素NotIおよびアルカリフォスファターゼ(Calf Intestine Alkaline Phosphatase:CIAP)で処理した。また、先に作製したpKANNIBAL−BBEirをNotI処理することにより、インサートを調製した。こうして得られたベクターおよびインサートのそれぞれの反応液をフェノール抽出、クロロフォルム抽出、エタノール沈殿したのち、20μlTEバッファーに溶解させ、ライゲーションをおこなった。得られたコロニーからプラスミドを抽出し、制限酵素処理し、目的とするインサートが挿入されていることを確認した。さらに、プライマーとして35Spro−S1を用いた塩基配列決定をおこない、目的とするdsRNAを発現するベクターpART−27−BBEirが作製されていることを確認した。
【0059】
アグロバクテリウムへの導入
こうして作製したpART27−BBEirをエレクトロポーレーション法によりアグロバクテリウムLBA4404株へ導入した。形質転換は、出現してきたコロニーよりPromega SV Miniprepsを用いてプラスミド抽出し、制限酵素処理により確認した。
【0060】
ハナビシソウ細胞の形質転換
構築した発現ベクターの導入はProc.Nat.Acad.Sci.98:367−372(2001)7)に記載の方法に準じて行なった。ハナビシソウ(Eschscholtzia californica)の種子(カネコ種苗株式会社)をミラクロスに包み、1%オスバンで1分、70%エタノールで1分、1%アンチホルミンで14分除菌し、滅菌水で5分ずつ3回洗浄した。こうして除菌された種子を植物体用培地に播種し、25℃で培養した。発芽後2〜3週間後の植物体の、葉および葉柄をメスで切断し、5mm〜1cmの切片を得た。次に2昼夜25℃で振とう培養したAgrobacterium tumefaciens(pART27導入用(コントロール)およびpART27−BBEir導入用)を共存培養用培地で5倍希釈してシャーレに入れ、そこに植物体切片を10分浸した。その後植物体切片をキムタオルにのせ、培養液を除き、ろ紙をひいた共存用寒天培地に移植した。2日後植物体切片を選抜用寒天培地(アセトシリンゴン100μM、ナフタレン酢酸10μM、ベンジルアデニン1μM、ショ糖3%を含むLinsmaier−Skoog培地)で共存培養した。その後セフォタキシム200μg/ml、ハイグロマイシン20μg/ml、ナフタレン酢酸10μM、ベンジルアデニン1μM、ショ糖3%を含むLinsmaier−Skoog培地で選抜培養した。3週間ごとに培地を交換し、生育してくる細胞を選抜した。
【0061】
形質転換体の確認
形質転換体の確認は、導入した遺伝子の存在をゲノムDNAを用いてPCR解析した。
【0062】
アルカロイドの分析
細胞からのアルカロイドの抽出は、Proc.Nat.Acad.Sci.98:367−372(2001)7に記載のように行なった。具体的には細胞1gに対し、0.01N HClで酸性化したメタノール4mlを用いて一晩抽出し、その上清を遠心により分離した。その抽出物を島津製作所社HPLC SCL−10システム(移動相:50mM酒石酸/10mM SDS:アセトニトリル:メタノール=4:4:1、流速1.2ml/分,カラムTSK−GEL ODS−80)を用いて解析した。また、それぞれのアルカロイドの同定は、島津製作所社製液体クロマトグラフ質量分析計LC/MS−2010(移動相:水:アセトニトリル:メタノール:酢酸=391:400:100:9、流速0.5ml/分,カラムTSK−GEL ODS−80)を用い行なった(図6)。
【0063】
BBE酵素活性の測定
コントロールおよびBBE−dsRNAを導入したハナビシソウ形質転換体におけるBBEの活性測定を以下のように行った。まず、新鮮重1gの培養細胞に、2mlのグリシン緩衝液(50mMグリシン−NaOH、pH8.9)を加え、氷上にて磨砕した。次いで、細胞抽出液をPD−10カラムにて脱塩した後、500μlの粗酵素液あたり、1mM相当となるように基質であるレチクリンを加え、30℃にて反応させた。一定時間後、10μlの1N NaOHを加えて反応を停止させ、LC−MSにてBBEの代謝産物であるスコウレリンの生成を定量した(図7)。
【0064】
結果
BBE−dsRNAを導入したハナビシソウ形質転換体は、選抜後2ヶ月ほどでカルス化が観察され、その後、液体培地にて培養を行なった。コントロール(pART27導入株)で19ライン、BBE−dsRNAを導入したものでは20ラインの形質転換体を得た。これらコントロールとBBE−dsRNA導入株では表現型の違いが観察された。コントロールの細胞では細胞が赤色を帯びているのに対し、BBE−dsRNA導入株では白い細胞が多く観察された。実際にHPLCによりアルカロイド組成の変化を解析した結果、図6に示すように、BBEdsRNAを導入した形質転換体において、顕著なレチクリンの蓄積(新鮮物1g当たり約1.5mg)が認められた。
【0065】
BBE酵素活性については、コントロールと比較して、BBE−dsRNA導入株(図中、BBEirと示す)において顕著なBBE活性の低下が認められた。即ち、コントロールでは基質であるレチクリンが完全にスコウレリンに変換されているのに対し、BBE−dsRNA導入株ではスコウレリンへの転換はほとんど認められなかった(図7)。BBE活性のタイムコースを測定し、その活性の定量を行った結果、コントロール株C23の活性は、1.85±0.33pkat/mgタンパク質であるのに対して、BBE dsRNA導入株B14では、0.056±0.051pkat/mgタンパク質であった。即ち、BBE dsRNA導入株ではコントロール株の約3%程度にまでBBE活性が低下していることが認められた(図8)。
【0066】
なお、コントロール株とBBE dsRNA導入株のレチクリンおよびサンギナリン含量を調べたところ、図9に示すように、コントロールと比較して、BBE dsRNA導入株では総じてレチクリン含量が高かった。
【0067】
一方、ハナビシソウの根培養物にアンチセンスBBE RNA発現ベクターを導入した場合には、赤色色素が薄くなり、アルカロイド含量が全般的に著しく低下したという報告がなされているものの、本発明のような中間代謝産物の蓄積は観察されなかった(Plant Physiology,128,696−706(2002)およびPlant Molecular Biology 51:153−164(2003))。Plant Molecular Biology 51:153−164(2003)の結果によると、アンチセンス法における抑制によっても、0.6pkat/mgタンパク質程度の活性が残存しており、そのために代謝経路の遮断が不十分になったものと考えられる。
【0068】
これらの知見により、RNAi法が代謝系の遮断に極めて効果的であり、RNAi法を用いた効果的な遺伝子発現抑制によって、はじめて中間代謝産物の蓄積を可能とする代謝反応の抑制が可能になったと考えられる。
【0069】
このように,BBEに対するdsRNAの発現によって、その反応の基質となるレチクリンの蓄積が誘導されたことは、代謝系の遮断によって、様々な中間代謝産物の蓄積が可能となることを示唆している。
【0070】
アルカロイドの生合成経路の解明は進んでおり、生合成経路に関する酵素のいくつかは単離され、またその配列も知られている。
アルカロイド生合成経路についての情報は、例えば(P.J.Facchini,Alkaloid biosynthesis in plants:Annu.Rev.Plant Physiol.Plant Mol.Biol.2001,52:29−66を参照されたい。また、KEGG:http://www.genome.ad.jp/kegg/metabolism/htmlも参照されたい)から、また各酵素の配列についての情報は(例えば、DDBJ:http://www.ddbj.nig.ac.jp/welcome−j.html,およびGenbankTM:http://www/genome.ad.jp/dbget/debget.links.htmlから得ることができる。
したがって、本実施例と同様にして他のアルカロイド生合成中間体を植物体中に蓄積させることが可能である。
【0071】
例えば、図1に示すイソキノリンアルカロイド生合成系においては、ノルコクラウリン−6−O−メチルトランスフェラーゼ(1)(gb:029811)を抑制することにより、ノルコクラウリンが、コクラウリン−N−メチルトランスフェラーゼ(2)(gb:AB061863、gbu:AY217334)を抑制することにより、コクラウリンが、また、コクラウリン3’−ヒドロキシラーゼ(3)(gb:AF014801,gb:AB025030)を抑制することによりN−メチルコクラウリンが蓄積される。さらに、図2に示すインドールアルカロイド生合成系においては、グルコシダーゼI/II(5)(gb:AF112888)を抑制することによって、ストリクトシジンが蓄積される。これらの酵素の配列については、いずれも上記のデータバンクから得られる。
【産業上の利用可能性】
【0072】
本発明によりdsRNAを用いたRNAi法が、イソキノリンアルカロイドを含む有用物質生産経路の遮断に有効であることが明らかになり、有用代謝産物中間体の生産が初めて可能となった。本発明により確立した細胞株は、さらに、新しい生合成系の導入による新規な関連化合物生産の材料として、また、化学的変換の材料として、重要な医薬用アルカロイドを含む多様な関連化合物の生産開発に利用できる。
【技術分野】
【0001】
本発明は有用アルカロイド生合成中間体の生産方法に関する。
【背景技術】
【0002】
アルカロイドは、植物に含まれる塩基性含窒素化合物の総称である。アルカロイドはその主要骨格に応じて、キノリン系、イソキノリン系、インドール系、トロパン系、キサンチン系アルカロイドなどに分類される。アルカロイドのなかには薬学的に有用なものが多く、たとえばコデインおよびモルヒネの鎮痛作用や商業的価値はよく知られている。ノスカピンは鎮咳作用化合物として有用である。パパベリンは平滑筋弛緩剤および脳血管拡張薬である。ベルベリンは、抗菌性、抗マラリア性および解熱性化合物として用いられてきた。
【0003】
イソキノリンアルカロイドは、イソキノリンを基本骨格とするアルカロイドの総称であり、分布も広く、構造も多様である。イソキノリンアルカロイドには例えば、モルヒネ(morphine)型(モルヒネなど)、プロトベルベリン(protoberberine)型(ベルベリンなど)、ベンゾフェナンスリジン(benzophenanthridine)型(サングイナリンなど)のアルカロイドが含まれる。イソキノリンアルカロイドを産生する植物としては、ケシ科(パパベラセアエ(Papaveraceae))、メギ科(ベルベリダセアエ(Berberidaceae))、キンポウゲ科(ラナンキュラセアエ(Ranunculaceae))、ツヅラフジ科(メニスペルマセアエ(Menispermaceae))、ミカン科(ルタセアエ(Rutaceae))などが挙げられる。
【0004】
イソキノリンアルカロイド生合成経路の中間体としては、レチクリンが多数の薬学的生成物への重要な前駆体として注目されてきた。例えば、特許文献1には、ケシにランダムに突然変異を導入し、レチクリン含量の高い植物体を選抜することを含む、レチクリンを製造する方法が開示されている。しかしこの方法は無作為に作成した突然変異体の中からレチクリン含量の高いものを選抜し、その植物体からレチクリンを抽出するという工程を含むため、煩雑なものであった(特許文献1を参照)。
【0005】
ベルベリン架橋酵素は、イソキノリンアルカロイド生合成経路の酵素の1つであり、レチクリンを基質とする。ベルベリン架橋酵素の発現を遺伝子操作を用いて低下させることにより、その基質であるレチクリン含量を増加させようとする試みがなされてきた。これらの試みは、アンチセンス法により、ベルベリン架橋酵素の発現を特異的に阻害しようとするものであった。しかし、その結果としてベルベリン架橋酵素の発現は阻害され、アルカロイド含量が全般的に著しく低下したものの、レチクリンのような中間代謝産物の特異的な蓄積は観察されなかった(例えば、非特許文献1および2を参照)。
【0006】
近年、アンチセンス法のほかに、遺伝子発現の抑制法としてRNAi法が利用されている。RNAi(RNA interference)とは、dsRNA(2本鎖RNA)の細胞内への導入によって、dsRNAと相同な配列を持つ遺伝子の発現が抑制される現象である。RNAiは様々な生物種における遺伝子機能解析に用いられている。例えば、非特許文献3では、ステロイド合成系に関与する酵素を特異的にRNAiによって阻害することにより、該酵素の基質となる中間体が蓄積することが報告された。しかし、この文献によると、目的の中間体から別の物質への変換がかなり起こっており、中間体を特異的に蓄積させるまでには至っていない。これはステロイドが細胞膜の必須成分であり細胞成長と関係しているため、完全には代謝経路の遮断が起こらないためと考えられる。また、この文献では植物細胞の成長が阻害されたことが報告されている(例えば、非特許文献3を参照)。
【0007】
RNAi法に用いられるRNAi用ベクターは二本鎖RNA(dsRNA)を植物体内で発現させることを目的として作製されるものである。RNAiベクターは、その構造に基づき、おおまかに2種類に分類される。
【0008】
一つは、別々に作製したセンスRNAを発現するプラスミドと、アンチセンスRNAを発現するプラスミドの組み合わせである。これら2つのプラスミドを、混合して細胞に形質転換し、細胞内で、dsRNAを形成させる。もう一つは、ヘアピン構造をとるRNAを発現するプラスミドである。後者については、線虫、ショウジョウバエ、植物、トリパノソーマなどで報告がある。植物については、導入遺伝子のヘアピン構造の中間にイントロンを導入したものとしないものの遺伝子抑制効力を比較した研究がある。この研究においては、イントロンを持った導入遺伝子のほうがより強力な発現抑制を引き起こすという結果が報告されている。
【0009】
これまでに、本発明者らは、内在遺伝子と80%程度の相同性をもつ100塩基対程度のdsRNAを発現させることにより、遺伝子抑制が起こることを観察している。この際dsRNAは、細胞内でRNaseによって分解を受け、約20塩基対のsiRNAとなり、siRNAがRISCと呼ばれる複合体に組み込まれ、この複合体が標的mRNAを分解すると考えられている。動物細胞においては、約20塩基対のsiRNAを導入することにより遺伝子発現の抑制がおこることが報告されている。
【特許文献1】特表2002−508947号公報
【非特許文献1】サング−ウン・パーク(Sang−Un Park)他著、「プラント・フィジオロジー(Plant Physiology)」第128巻、2002年2月、p.696−706
【非特許文献2】サング−ウン・パーク(Sang−Un Park)他著、「プラント・モレキュラー・バイオロジー(Plant Molecular Biology)」第51巻、2003年、p.153−164
【非特許文献3】セリン・バーガー(Celine Burger)他著、「ジャーナル・オブ・エクペリメンタル・ボタニー(Journal of Experimental Botany)」第54巻、第388号、2003年7月、p.1675−1683
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明は、特定のアルカロイド生合成中間体の生産方法を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明は、アルカロイドを産生する植物細胞、植物組織または植物体においてRNAi法を用いて、目的とするアルカロイド生合成中間体を基質とする酵素の発現を阻害することを含む、アルカロイド生合成中間体の生産方法を提供する。
【0012】
具体的には、後述のRNAi遺伝子を用いて目的とするアルカロイド生合成中間体を基質とする酵素の発現を阻害して、代謝の流れを遮断することにより、目的のアルカロイド生合成中間体が植物細胞内に蓄積する。
【0013】
本発明はまた、上記方法によって生産されたアルカロイド生合成中間体を提供する。
【0014】
さらに本発明は、上記方法に用いられる、
i)プロモーター、および、
ii)プロモーターの下流に、
a)目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列と相同的な正方向配列、および、
b)該正方向配列に相補的な逆方向配列
を含む遺伝子を提供する。本明細書においてこの遺伝子を「RNAi遺伝子」と称する。
【0015】
また、本発明は上記方法に用いられる、以下のAおよびBの遺伝子を含む遺伝子の組合せを提供する:
A.i)プロモーター、および、
ii)プロモーターの下流に、目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列と相同的な正方向配列を含む遺伝子、および、
B.i)プロモーター、および、
ii)プロモーターの下流に、該正方向配列に相補的な逆方向配列を含む遺伝子。
本明細書においてこの遺伝子の組合せを「RNAi遺伝子の組合せ」と称する。
【0016】
「目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列と相同的な正方向配列」とは、生産しようとするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列に対して相同性を有する配列であって、その長さが約100bp以上であり、ベクターなどのコンストラクトにおいて転写と同じ方向に挿入された配列のことを指す。
【0017】
ここで、「酵素の全体または一部をコードする配列」には、該酵素をコードする遺伝子の翻訳領域のみならず、非翻訳領域の配列も含まれる。
【0018】
「該正方向配列に相補的な逆方向配列」とは、上記の「目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列」に対して相補性を有する配列のことをいう。即ち、ベクターなどのコンストラクトにおいて転写と逆方向に挿入された正方向配列と相同性を有する配列を指す。
【0019】
RNAi遺伝子においては該正方向配列と逆方向配列はともにプロモーターの下流に位置する必要があるが、正方向配列と逆方向配列のどちらを上流に位置させてもよい。
【0020】
好ましくは、RNAi遺伝子においては、該正方向配列と逆方向配列の間にスペーサー配列を介在させる。スペーサーを介在させることによって、正方向配列と逆方向配列(以下両配列の繰り返しをインバーテッドリピートと称する)の対合を容易にする空間が生じる。スペーサー配列は特に限定されないが、通常数百bp〜1kbの長さの配列、例えばイントロン配列が好適に用いられる。
【0021】
正方向配列、スペーサー配列、および逆方向配列を含むRNAi遺伝子を植物細胞内で発現させることにより、植物細胞においてアルカロイド生合成酵素の発現が抑制される。
【0022】
該正方向配列、スペーサー配列および、該正方向配列に相補的な逆方向配列を含むDNAは、植物細胞においてプロモーターにより誘導されてmRNAへと転写される。正方向配列から転写された一本鎖RNA部分と逆方向配列から転写された一本鎖RNA部分は、相補的であるため互いに水素結合により結合する。好ましくはかかるRNAはスペーサー配列を介してヘアピン構造の二本鎖RNA(dsRNA)を形成する。このdsRNAがRNAi、即ち、標的であるアルカロイド生合成遺伝子の発現の抑制を引き起こすと考えられる。
【0023】
RNAi遺伝子の組合せを用いる場合は、プロモーターの下流に正方向配列を含むベクター(センスベクターとも呼ばれる)と、プロモーターの下流に逆方向配列を含むベクター(アンチセンスベクター)の両方を植物に導入する。該正方向配列および逆方向配列は、植物細胞においてプロモーターにより誘導されてmRNAへと転写され、正方向配列から転写された一本鎖RNAと逆方向配列から転写された一本鎖RNAが、互いに水素結合により結合して二本鎖RNA(dsRNA)を形成する。このdsRNAがRNAi、即ち、標的であるアルカロイド生合成遺伝子の発現の抑制を引き起こすと考えられる。
【0024】
このようにして、RNAi遺伝子あるいはRNAi遺伝子の組合せを用いることにより、二本鎖RNAが生じる結果、標的アルカロイド生合成遺伝子の発現が阻害され、該酵素の基質であるアルカロイド生合成中間体が細胞内に特異的に蓄積する。
【0025】
「目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列」は先に定義したように、必ずしも遺伝子のコード領域に含まれる必要はなく、5’UTR領域や3’UTR領域に位置する配列あるいはプロモーター領域でもよい。RNAiはこのような非コード領域の配列を用いることによっても起こる現象である。
【0026】
さらに本発明は上述したような遺伝子を含むベクターおよび該ベクターにより形質転換された植物細胞または植物組織あるいは植物体を提供する。
【発明の効果】
【0027】
これまでの研究では、生合成経路の最終産物の蓄積の抑制は報告されている。しかし、本発明のように中間代謝産物の特異的な蓄積を可能とし、中間体を大量に生産する、あるいは、さらに、新規な反応経路を追加することにより、新たな化合物を生産させようとする観点は無かった。また、かかる観点があったとしても、現実的には、そのようなことを可能とする技術はなかった。今回RNAi法によって、生合成経路の中間代謝産物が蓄積することが明らかになった。本発明は、様々な代謝系において、重要な代謝産物を蓄積させる可能性を示唆しており、その応用範囲は極めて広い。
【図面の簡単な説明】
【0028】
【図1】図1は、イソキノリンアルカロイド生合成経路と生合成酵素を示す図である。
【図2】図2は、インドールアルカロイド生合成経路と生合成酵素を示す図である。
【図3】図3は、中間体レチクリンから派生する化合物を示す図である。
【図4】図4は、中間体ストリクトシジンから派生する化合物を示す図である。
【図5】図5は、ベルベリン架橋酵素(BBE)遺伝子に対するdsRNA発現ベクター、pART27−BBEir構築の手順を示す図である。
【図6】図6は、ハナビシソウBBEdsRNA形質転換体におけるレチクリン(m/z 330)の蓄積のLC/MS解析を示す図である。
【図7】図7は、コントロールおよびBBEdsRNA形質転換体における、BBE酵素活性を示す図である。
【図8】図8は、コントロールおよびBBEdsRNA形質転換体における、BBE酵素反応をLC/MS解析した結果を示す図である。
【図9】図9は、コントロールおよびBBEdsRNA形質転換体における、レチクリンと、サンギナリンの含量を示す図である。
【図10】図10は、BBEの阻害により、レチクリンからスコウレリンの反応が遮断されることを示す模式図である。
【符号の説明】
【0029】
1 ノルコクラウリン−6−O−メチルトランスフェラーゼ
2 コクラウリン−N−メチルトランスフェラーゼ
3 N−メチル−コクラウリン−3’−ヒドロキシラーゼ
4 ベルベリン架橋酵素(BBE)
5 グルコシダーゼI/II
【発明を実施するための最良の形態】
【0030】
本発明において標的とされる植物、即ちdsRNAを導入される植物は、アルカロイド生合成経路を有する植物であれば特に限定されない。アルカロイド生合成経路としてはイソキノリンアルカロイド生合成経路が好適であるが、その他にもインドールアルカロイド生合成経路などが好適な標的となりうる。
【0031】
具体的には、アルカロイド生合成経路を有する植物には以下のものが挙げられるがこれらに限定されるものではない。ベルベリンを産生する植物、例えば、オウレン(Coptis japonica,Coptis chinensis Franch,Coptis deltoides C.Y.Cheng et Hsiao等)、キハダ(Phellodendron amurense)、メギ(Berberis属)、ナンテン(Nandina domestica)、ヒイラギナンテン(Mahonia japonica)およびアキカラマツ(Thalictrum minus)、モルヒネやコデイン、パパベリンを産生する植物、例えば、ケシ(Papaver somniferum Linn)、アツミゲシ(Papaver setigerum DC)およびハカマオニゲシ(Papaver bracteatum)、モルヒネを生産しないが、類縁のアルカロイドを産生する植物、例えば、オニゲシ(Papaver orientale Linn)およびヒナゲシ(Papaver rhoeas)、サングイナリンを産生する植物、例えば、ハナビシソウ(Eschscholtzia californica)、サングイナリア(Sanguinarine canadensis L.)、コリダリンを産生する植物、例えば、エンゴサク(Corydalis属植物、例えば、Corydalis bulbosa DC.,Corydalis ternata Nakai,Corydalis nakaii Ishidoya,Corydalis decumbens Person)、コロンバミンを産生する植物、例えば、コロンボ(Jateorhiza columbaMiers)、セファランチン(cepharanthine)を産生する植物、例えば、タマサキツヅラフジ(Stephania capharantha)、シノメニン(sinomenine)を産生する植物、例えば、オオツヅラフジ(Sinomenium acutum Rehder et Wilson)、エメチン(emetine)を産生する植物、例えば、トコン(Cephaelis ipecacuanha)など。
【0032】
上記の植物の中ではイソキノリンアルカロイドを合成する植物が好適に用いられ、なかでもサングイナリンやベルベリンを産生する植物、例えば、ハナビシソウ、オウレン、キハダ、メギ、ナンテン、ヒイラギナンテンおよびアキカラマツが好ましく、特にハナビシソウが好ましい。
【0033】
RNAi遺伝子を導入される標的植物の起源と、導入するRNAi遺伝子の起源は、同じであっても異なっていてもよい。標的遺伝子と導入遺伝子の相同性を考慮すると、同じ植物種由来のものが特に好ましい。
【0034】
本発明の方法においては、目的とする生合成中間体を基質とする酵素をRNAiによる阻害の標的となる酵素とする。かかる酵素としては、例えば、イソキノリンアルカロイド合成系経路(図1を参照)のベルベリン架橋酵素(BBE)、ノルコクラウリン−6−O−メチルトランスフェラーゼ、コクラウリン−N−メチルトランスフェラーゼおよびN−メチルコクラウリン−3’−ヒドロキシラーゼが挙げられる。
【0035】
これらの酵素をRNAiにより阻害するとその基質であるアルカロイド生合成中間体が蓄積する。具体的には、ベルベリン架橋酵素を阻害するとレチクリン、ノルコクラウリン−6−O−メチルトランスフェラーゼを阻害するとノルコクラウリン、コクラウリン−N−メチルトランスフェラーゼを阻害するとコクラウリン、そしてN−メチルコクラウリン−3’−ヒドロキシラーゼを阻害するとN−メチルコクラウリンがそれぞれ蓄積する。
【0036】
イソキノリンアルカロイド生合成経路の酵素以外では例えば、インドールアルカロイド生合成経路(図2を参照)のグリコシダーゼI/IIを標的とすると、その基質であるストリクトシジンが蓄積する。
【0037】
好ましくは本発明の方法により生産されるアルカロイド生合成中間体は、レチクリン、ノルコクラウリン、コクラウリンおよびN−メチルコクラウリンからなる群から選択され、特に好ましくはレチクリンである。
【0038】
レチクリンを製造するには、標的酵素はベルベリン架橋酵素であり、その結果レチクリンが蓄積する。レチクリンの前駆体であるノルコクラウリン、コクラウリンおよびN−メチルコクラウリン(図1を参照)、ならびにレチクリンは、図3に示す様々なイソキノリンアルカロイドの前駆体として有用である。また、ストリクトシジンも図4に示す様々なインドールアルカロイドの前駆体として有用である。
【0039】
本発明のRNAiを引き起こす遺伝子は好ましくは、ベルベリン架橋酵素、ノルコクラウリン−6−O−メチルトランスフェラーゼ、コクラウリン−N−メチルトランスフェラーゼおよびN−メチルコクラウリン−3’−ヒドロキシラーゼからなる群から選択される酵素をコードする配列またはその一部である。RNAiを引き起こすためのdsRNAの長さは特に限定されないが、少なくとも23塩基以上、好ましくは100塩基対から2キロベースであり、最も好ましくは100塩基対〜800塩基対である。
【0040】
本発明においてRNAiを引き起こす遺伝子の発現を誘導するプロモーターとしては、植物へ導入した際に目的とする遺伝子を発現させることができるものであれば特に限定されない。かかるプロモーターは当業者によく知られており、例えば、カリフラワーモザイクウイルス35Sプロモーター、および、誘導性プロモーター、例えば、アルコールデヒドロゲナーゼプロモーターやテトラサイクリンリプレッサー/オペレーター制御系などが挙げられる。
【0041】
本発明において、正方向配列および逆方向配列と、標的の生合成酵素をコードする遺伝子の配列との相同性は、100%である必要はない。遺伝子の突然変異、多型、又は進化上の分岐による配列の変動は容認される。標的遺伝子に対する挿入、欠失、及び一塩基点突然変異を有するdsRNA鎖もまた、RNAiに有効である。具体的には、RNAiに用いる遺伝子は、標的遺伝子と完全に同一である必要はないが、少なくとも70%以上、好ましくは80%以上、さらに好ましくは90%以上、最も好ましくは95%以上の配列の同一性を有するものが好適である。
【0042】
また、正方向配列と逆方向配列は、転写された後に二本鎖RNAを形成し得るものであればよい。効率的にdsRNAを形成するためには、両者の相補性は、少なくとも70%、好ましくは少なくとも80%、さらに好ましくは少なくとも90%、最も好ましくは少なくとも95%である。
【0043】
本発明において標的とする植物へのベクターの導入には、ポリエチレングリコール法、電気穿孔法(エレクトロポレーション法)、アグロバクテリウムを介する方法、パーティクルガン法など、当業者に公知の種々の方法を用いることができる。それぞれの方法に適したベクターの調製、形質転換植物細胞からの植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である(Toki S,et al:Plant Physiol 100:1503,1995)。
【0044】
例えば、形質転換植物体を作出する手法については、ポリエチレングリコールを用いてプロトプラストへ遺伝子導入し、植物体を再生させる方法(Datta SK:In Gene Transfer To Plants(Potrykus I and Spangenberg,Eds)pp.66−74,1995)、電気パルスによりプロトプラストへ遺伝子導入し、植物体を再生させる方法(Toki S,et al:Plant Physiol 100:1503,1992)、パーティクルガン法により細胞へ遺伝子を直接導入し、植物体を再生させる方法(Christou P,et al:Biotechnology 9:957,1991)、およびアグロバクテリウムを介して遺伝子を導入し、植物体を再生させる方法(Hiei Y,et al:Plant J 6:271,1994)など、いくつかの技術が既に確立し、当該技術分野において広く用いられている。本発明においては、これらの方法を好適に用いることができる。
【0045】
本発明において、植物にRNAi遺伝子を導入する際に用いるベクターは、遺伝子導入手段に応じて適宜選択すればよく、特に限定されない。例えば、アグロバクテリウムによる遺伝子導入法を用いる場合は、バイナリーベクター、例えば、pART、pBI101、pBI121、pIG121Hm等が好適に用いられる。
【0046】
本発明のベクターの作成方法としては、特に限定されず、当業者に周知のいかなる方法を用いてもよい。本発明のベクターにおいては、3’末端側へターミネーターが挿入されるが、ターミネーターとしては公知のものがいずれも好適に用いられ、例えば、OCSターミネーター、nosターミネーター、35Sターミネーターなどが挙げられる。
【0047】
本明細書中、塩基配列の同一性は、カーリンおよびアルチュールによるアルゴリズムBLAST(Proc.Natl.Acad.Sei.USA 87:2264−2268,1990、Karlin S &Altschul SF:Proc Natl Acad Sci USA 90:5873,1993)を用いて決定できる。BLASTのアルゴリズムに基づいたBLASTNやBLASTXと呼ばれるプログラムが開発されている(Altschul SF,et al:J Mol Biol 215:403,1990)。これらの解析方法の具体的な手法は公知である(http://www.ncbi.nlm.nih.gov/)。
【実施例】
【0048】
以下に本発明を特定の実施例に言及して記載するがこれは本発明を例示する目的のものであり、限定する意図のものではない。
【0049】
(実施例の概要)
アルカロイド生合成遺伝子に対して特異的な二本鎖RNA(dsRNA)を合成する発現ベクターを構築し、その高い抑制効果により、標的遺伝子の発現および酵素活性を効果的に抑制し、中間体が蓄積生産できることを実証した。
【0050】
具体的には、重要な医薬品であるイソキノリンアルカロイドの生合成中間体を、同アルカロイドを生産する植物細胞の代謝経路を二本鎖RNA(dsRNA)を用いたRNA干渉(RNAi)法により遮断することにより、生産する方法を開示する。本実施例では、ベンゾフェナンスリジンアルカロイドを生産するハナビシソウ細胞に、その生合成経路の酵素の1つであるベルベリン架橋酵素(BBE)の配列の一部に対応するdsRNAを発現するベクターを導入した。その結果、かかるアルカロイド生合成系が遮断され、植物細胞における生合成中間体レチクリンの生産が可能となった。
【0051】
(実施例についての具体的説明)
形質転換が可能なイソキノリンアルカロイド生産性植物ハナビシソウから単離されている生合成酵素ベルベリン架橋酵素(BBE)遺伝子(配列番号1)に対して既知の配列情報によりプライマーを設計し、BBEのcDNAを単離した。このcDNAをもとにdsRNAを形成するようなBBEdsRNA発現ベクターを構築した。ついで、同ベクターをハナビシソウ細胞に導入し、BBEdsRNAを発現する形質転換体を得、ベルベリン架橋酵素の基質である生合成中間体であるレチクリンを蓄積する形質転換ハナビシソウ細胞を確立した。
【0052】
(実験材料および方法)
コンストラクトの作製には、インバーテッドリピート作成用ベクターとして広く用いられているpKANNIBALおよびpART27を用いた。pKANNIBALは、CaMV35Sプロモーター、その下流に両端に複数の制限酵素サイトをもつイントロン領域、さらにその下流にOCSターミネーターを有する。イントロンの両端に、正方向および逆方向配列を挿入する。かかる配列は、植物体内でmRNAへと転写され、スプライシングされる。該RNAは、インバーテッドリピート、すなわち、二本鎖RNA(dsRNA)を形成する。なお、本発明は、必ずしも、このベクターの使用に限定されるものではない。
【0053】
ハナビシソウBBE遺伝子の単離
データベースにすでに登録されている配列をもとに、3’arm(本明細書において逆方向配列と称される部分)にBamH I、Hind III、5’arm(本明細書において正方向配列と称される部分)にEcoR I、Xho Iの制限酵素サイトを持つハナビシソウBBE遺伝子のほぼ全長約1kb断片をPCRによりハナビシソウ培養細胞cDNAから単離した。
【0054】
用いたプライマーを以下に示す。
BBE−3’arm−フォワード(FM):ATG GAT CCG ATT CGG ACT CGG ATT TCA ACC
(配列番号2)
リバース(RV):ATT AAG CTT CCA CTT CGA TGA GGA AAC GG
(配列番号3)
5’arm−フォワード(FM):AAT CTC GAG ATT CGG ACT CGG ATT TCA ACC
(配列番号4)
リバース(RV):CGA ATT CCA CTT CGA TGA GGA AAC GG
(配列番号5)
【0055】
こうして単離した遺伝子をプラスミドpT7 Blue(Novagen)にサブクローニングしたのち、SIMADZU DSQ−2000Lにより塩基配列を確認した。
【0056】
dsRNA発現ベクターの作製(図5)
3’armの挿入
BBE−3’arm−FWとRVのプラーマーペアにより得られたPCR産物をpT7−Blueにサブクローニングし、配列確認したのち、BamH I、Hind IIIで切断した。ベクターpKANNIBALも同様に、BamH IとHind IIIで制限酵素で処理した。これらDNAを電気泳動後、フェノール処理、クロロフォルム抽出、エタノール沈殿し、10μlのTEに溶解させたのち、ライゲーション反応を行ない、XL1−Blueにトランスフォーメーションした。挿入の確認は制限酵素処理、およびOCSターミネーター配列に対するAS1を用いたシーケンシングによりおこなった。
【0057】
5’armの挿入
BBE5’arm FWとRVのプライマーペアにより得られたPCR産物をpT7−Blueにサブクローニングし、配列確認したのち、EcoRI、XhoIで切断した。タンデムな挿入を防ぐため断片にアルカリフォスファターゼ(Calf Intestine Alkaline Phosphatase:CIAP)を加えて脱リン酸処理した。CIAPを失活させるため、65℃で30分インキュベートした。その後エタノール沈殿により制限酵素を失活させ、20μlのTEに溶解した。先に、pKANNIBALに3’armを導入したベクターを同様にEcoRI、XhoIで切断したのち、ライゲーション反応をおこない、XL1−Blueにトランスフォーメーションした。挿入の確認は制限酵素処理およびOCSターミネーターに対するAS1プライマー、35Sプロモーターに対する−S1プライマー(35Spro−S1,GAG CTA CAC ATG CTC AGG TT(配列番号6)を用いたシーケンシングによりおこなった。得られたBBEの3’および5’armが挿入されたプラスミドをpKANNIBAL−BBEirと称する。
【0058】
バイナリ−ベクターpART27への導入
pART27ベクターを制限酵素NotIおよびアルカリフォスファターゼ(Calf Intestine Alkaline Phosphatase:CIAP)で処理した。また、先に作製したpKANNIBAL−BBEirをNotI処理することにより、インサートを調製した。こうして得られたベクターおよびインサートのそれぞれの反応液をフェノール抽出、クロロフォルム抽出、エタノール沈殿したのち、20μlTEバッファーに溶解させ、ライゲーションをおこなった。得られたコロニーからプラスミドを抽出し、制限酵素処理し、目的とするインサートが挿入されていることを確認した。さらに、プライマーとして35Spro−S1を用いた塩基配列決定をおこない、目的とするdsRNAを発現するベクターpART−27−BBEirが作製されていることを確認した。
【0059】
アグロバクテリウムへの導入
こうして作製したpART27−BBEirをエレクトロポーレーション法によりアグロバクテリウムLBA4404株へ導入した。形質転換は、出現してきたコロニーよりPromega SV Miniprepsを用いてプラスミド抽出し、制限酵素処理により確認した。
【0060】
ハナビシソウ細胞の形質転換
構築した発現ベクターの導入はProc.Nat.Acad.Sci.98:367−372(2001)7)に記載の方法に準じて行なった。ハナビシソウ(Eschscholtzia californica)の種子(カネコ種苗株式会社)をミラクロスに包み、1%オスバンで1分、70%エタノールで1分、1%アンチホルミンで14分除菌し、滅菌水で5分ずつ3回洗浄した。こうして除菌された種子を植物体用培地に播種し、25℃で培養した。発芽後2〜3週間後の植物体の、葉および葉柄をメスで切断し、5mm〜1cmの切片を得た。次に2昼夜25℃で振とう培養したAgrobacterium tumefaciens(pART27導入用(コントロール)およびpART27−BBEir導入用)を共存培養用培地で5倍希釈してシャーレに入れ、そこに植物体切片を10分浸した。その後植物体切片をキムタオルにのせ、培養液を除き、ろ紙をひいた共存用寒天培地に移植した。2日後植物体切片を選抜用寒天培地(アセトシリンゴン100μM、ナフタレン酢酸10μM、ベンジルアデニン1μM、ショ糖3%を含むLinsmaier−Skoog培地)で共存培養した。その後セフォタキシム200μg/ml、ハイグロマイシン20μg/ml、ナフタレン酢酸10μM、ベンジルアデニン1μM、ショ糖3%を含むLinsmaier−Skoog培地で選抜培養した。3週間ごとに培地を交換し、生育してくる細胞を選抜した。
【0061】
形質転換体の確認
形質転換体の確認は、導入した遺伝子の存在をゲノムDNAを用いてPCR解析した。
【0062】
アルカロイドの分析
細胞からのアルカロイドの抽出は、Proc.Nat.Acad.Sci.98:367−372(2001)7に記載のように行なった。具体的には細胞1gに対し、0.01N HClで酸性化したメタノール4mlを用いて一晩抽出し、その上清を遠心により分離した。その抽出物を島津製作所社HPLC SCL−10システム(移動相:50mM酒石酸/10mM SDS:アセトニトリル:メタノール=4:4:1、流速1.2ml/分,カラムTSK−GEL ODS−80)を用いて解析した。また、それぞれのアルカロイドの同定は、島津製作所社製液体クロマトグラフ質量分析計LC/MS−2010(移動相:水:アセトニトリル:メタノール:酢酸=391:400:100:9、流速0.5ml/分,カラムTSK−GEL ODS−80)を用い行なった(図6)。
【0063】
BBE酵素活性の測定
コントロールおよびBBE−dsRNAを導入したハナビシソウ形質転換体におけるBBEの活性測定を以下のように行った。まず、新鮮重1gの培養細胞に、2mlのグリシン緩衝液(50mMグリシン−NaOH、pH8.9)を加え、氷上にて磨砕した。次いで、細胞抽出液をPD−10カラムにて脱塩した後、500μlの粗酵素液あたり、1mM相当となるように基質であるレチクリンを加え、30℃にて反応させた。一定時間後、10μlの1N NaOHを加えて反応を停止させ、LC−MSにてBBEの代謝産物であるスコウレリンの生成を定量した(図7)。
【0064】
結果
BBE−dsRNAを導入したハナビシソウ形質転換体は、選抜後2ヶ月ほどでカルス化が観察され、その後、液体培地にて培養を行なった。コントロール(pART27導入株)で19ライン、BBE−dsRNAを導入したものでは20ラインの形質転換体を得た。これらコントロールとBBE−dsRNA導入株では表現型の違いが観察された。コントロールの細胞では細胞が赤色を帯びているのに対し、BBE−dsRNA導入株では白い細胞が多く観察された。実際にHPLCによりアルカロイド組成の変化を解析した結果、図6に示すように、BBEdsRNAを導入した形質転換体において、顕著なレチクリンの蓄積(新鮮物1g当たり約1.5mg)が認められた。
【0065】
BBE酵素活性については、コントロールと比較して、BBE−dsRNA導入株(図中、BBEirと示す)において顕著なBBE活性の低下が認められた。即ち、コントロールでは基質であるレチクリンが完全にスコウレリンに変換されているのに対し、BBE−dsRNA導入株ではスコウレリンへの転換はほとんど認められなかった(図7)。BBE活性のタイムコースを測定し、その活性の定量を行った結果、コントロール株C23の活性は、1.85±0.33pkat/mgタンパク質であるのに対して、BBE dsRNA導入株B14では、0.056±0.051pkat/mgタンパク質であった。即ち、BBE dsRNA導入株ではコントロール株の約3%程度にまでBBE活性が低下していることが認められた(図8)。
【0066】
なお、コントロール株とBBE dsRNA導入株のレチクリンおよびサンギナリン含量を調べたところ、図9に示すように、コントロールと比較して、BBE dsRNA導入株では総じてレチクリン含量が高かった。
【0067】
一方、ハナビシソウの根培養物にアンチセンスBBE RNA発現ベクターを導入した場合には、赤色色素が薄くなり、アルカロイド含量が全般的に著しく低下したという報告がなされているものの、本発明のような中間代謝産物の蓄積は観察されなかった(Plant Physiology,128,696−706(2002)およびPlant Molecular Biology 51:153−164(2003))。Plant Molecular Biology 51:153−164(2003)の結果によると、アンチセンス法における抑制によっても、0.6pkat/mgタンパク質程度の活性が残存しており、そのために代謝経路の遮断が不十分になったものと考えられる。
【0068】
これらの知見により、RNAi法が代謝系の遮断に極めて効果的であり、RNAi法を用いた効果的な遺伝子発現抑制によって、はじめて中間代謝産物の蓄積を可能とする代謝反応の抑制が可能になったと考えられる。
【0069】
このように,BBEに対するdsRNAの発現によって、その反応の基質となるレチクリンの蓄積が誘導されたことは、代謝系の遮断によって、様々な中間代謝産物の蓄積が可能となることを示唆している。
【0070】
アルカロイドの生合成経路の解明は進んでおり、生合成経路に関する酵素のいくつかは単離され、またその配列も知られている。
アルカロイド生合成経路についての情報は、例えば(P.J.Facchini,Alkaloid biosynthesis in plants:Annu.Rev.Plant Physiol.Plant Mol.Biol.2001,52:29−66を参照されたい。また、KEGG:http://www.genome.ad.jp/kegg/metabolism/htmlも参照されたい)から、また各酵素の配列についての情報は(例えば、DDBJ:http://www.ddbj.nig.ac.jp/welcome−j.html,およびGenbankTM:http://www/genome.ad.jp/dbget/debget.links.htmlから得ることができる。
したがって、本実施例と同様にして他のアルカロイド生合成中間体を植物体中に蓄積させることが可能である。
【0071】
例えば、図1に示すイソキノリンアルカロイド生合成系においては、ノルコクラウリン−6−O−メチルトランスフェラーゼ(1)(gb:029811)を抑制することにより、ノルコクラウリンが、コクラウリン−N−メチルトランスフェラーゼ(2)(gb:AB061863、gbu:AY217334)を抑制することにより、コクラウリンが、また、コクラウリン3’−ヒドロキシラーゼ(3)(gb:AF014801,gb:AB025030)を抑制することによりN−メチルコクラウリンが蓄積される。さらに、図2に示すインドールアルカロイド生合成系においては、グルコシダーゼI/II(5)(gb:AF112888)を抑制することによって、ストリクトシジンが蓄積される。これらの酵素の配列については、いずれも上記のデータバンクから得られる。
【産業上の利用可能性】
【0072】
本発明によりdsRNAを用いたRNAi法が、イソキノリンアルカロイドを含む有用物質生産経路の遮断に有効であることが明らかになり、有用代謝産物中間体の生産が初めて可能となった。本発明により確立した細胞株は、さらに、新しい生合成系の導入による新規な関連化合物生産の材料として、また、化学的変換の材料として、重要な医薬用アルカロイドを含む多様な関連化合物の生産開発に利用できる。
【特許請求の範囲】
【請求項1】
アルカロイドを産生する植物細胞、植物組織または植物体においてRNAi法を用いて、目的とするアルカロイド生合成中間体を基質とする酵素の発現を阻害することを含む、アルカロイド生合成中間体の生産方法。
【請求項2】
該アルカロイドがイソキノリンアルカロイドである請求項1に記載の方法。
【請求項3】
該酵素が、ベルベリン架橋酵素、ノルコクラウリン−6−O−メチルトランスフェラーゼ、コクラウリン−N−メチルトランスフェラーゼおよびN−メチルコクラウリン−3’−ヒドロキシラーゼからなる群から選択される請求項1に記載の方法。
【請求項4】
該酵素が、ベルベリン架橋酵素である請求項3に記載の方法。
【請求項5】
該アルカロイド生合成中間体がレチクリン、ノルコクラウリン、コクラウリンおよびN−メチルコクラウリンからなる群から選択される請求項1に記載の方法。
【請求項6】
該アルカロイド生合成中間体がレチクリンである請求項5に記載の方法。
【請求項7】
請求項1から6のいずれかに記載の方法によって生産されたアルカロイド生合成中間体。
【請求項8】
i)プロモーター、および、
ii)プロモーターの下流に、
a)目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列と相同的な正方向配列、および、
b)該正方向配列に相補的な逆方向配列
を含む、請求項1から7のいずれかに記載の方法に用いられる遺伝子。
【請求項9】
以下のAおよびBの遺伝子を含む、請求項1から7のいずれかに記載の方法に用いられる遺伝子の組合せ:
A.i)プロモーター、および、
ii)プロモーターの下流に、目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列と相同的な正方向配列を含む遺伝子、および、
B.i)プロモーター、および、
ii)プロモーターの下流に、該正方向配列に相補的な逆方向配列を含む遺伝子。
【請求項10】
該酵素が、ベルベリン架橋酵素、ノルコクラウリン−6−O−メチルトランスフェラーゼ、コクラウリン−N−メチルトランスフェラーゼおよびN−メチルコクラウリン−3’−ヒドロキシラーゼからなる群から選択される請求項8に記載の遺伝子。
【請求項11】
該酵素がベルベリン架橋酵素、ノルコクラウリン−6−O−メチルトランスフェラーゼ、コクラウリン−N−メチルトランスフェラーゼおよびN−メチルコクラウリン−3’−ヒドロキシラーゼからなる群から選択される請求項9に記載の遺伝子の組合せ。
【請求項12】
請求項8または10に記載の遺伝子を含むベクター。
【請求項13】
請求項9または11に記載の正方向配列を含む遺伝子を含むベクターおよび該正方向配列に相補的な逆方向配列を含む遺伝子を含むベクターの組合せ。
【請求項14】
請求項12に記載のベクターまたは請求項13に記載のベクターの組合せにより形質転換された植物細胞または植物組織あるいは植物体。
【請求項15】
植物がイソキノリンアルカロイド生産性植物である請求項14に記載の植物細胞または植物組織あるいは植物体。
【請求項16】
植物がハナビシソウである請求項15に記載の植物細胞または植物組織あるいは植物体。
【請求項1】
アルカロイドを産生する植物細胞、植物組織または植物体においてRNAi法を用いて、目的とするアルカロイド生合成中間体を基質とする酵素の発現を阻害することを含む、アルカロイド生合成中間体の生産方法。
【請求項2】
該アルカロイドがイソキノリンアルカロイドである請求項1に記載の方法。
【請求項3】
該酵素が、ベルベリン架橋酵素、ノルコクラウリン−6−O−メチルトランスフェラーゼ、コクラウリン−N−メチルトランスフェラーゼおよびN−メチルコクラウリン−3’−ヒドロキシラーゼからなる群から選択される請求項1に記載の方法。
【請求項4】
該酵素が、ベルベリン架橋酵素である請求項3に記載の方法。
【請求項5】
該アルカロイド生合成中間体がレチクリン、ノルコクラウリン、コクラウリンおよびN−メチルコクラウリンからなる群から選択される請求項1に記載の方法。
【請求項6】
該アルカロイド生合成中間体がレチクリンである請求項5に記載の方法。
【請求項7】
請求項1から6のいずれかに記載の方法によって生産されたアルカロイド生合成中間体。
【請求項8】
i)プロモーター、および、
ii)プロモーターの下流に、
a)目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列と相同的な正方向配列、および、
b)該正方向配列に相補的な逆方向配列
を含む、請求項1から7のいずれかに記載の方法に用いられる遺伝子。
【請求項9】
以下のAおよびBの遺伝子を含む、請求項1から7のいずれかに記載の方法に用いられる遺伝子の組合せ:
A.i)プロモーター、および、
ii)プロモーターの下流に、目的とするアルカロイド生合成中間体を基質とする酵素の全体または一部をコードする配列と相同的な正方向配列を含む遺伝子、および、
B.i)プロモーター、および、
ii)プロモーターの下流に、該正方向配列に相補的な逆方向配列を含む遺伝子。
【請求項10】
該酵素が、ベルベリン架橋酵素、ノルコクラウリン−6−O−メチルトランスフェラーゼ、コクラウリン−N−メチルトランスフェラーゼおよびN−メチルコクラウリン−3’−ヒドロキシラーゼからなる群から選択される請求項8に記載の遺伝子。
【請求項11】
該酵素がベルベリン架橋酵素、ノルコクラウリン−6−O−メチルトランスフェラーゼ、コクラウリン−N−メチルトランスフェラーゼおよびN−メチルコクラウリン−3’−ヒドロキシラーゼからなる群から選択される請求項9に記載の遺伝子の組合せ。
【請求項12】
請求項8または10に記載の遺伝子を含むベクター。
【請求項13】
請求項9または11に記載の正方向配列を含む遺伝子を含むベクターおよび該正方向配列に相補的な逆方向配列を含む遺伝子を含むベクターの組合せ。
【請求項14】
請求項12に記載のベクターまたは請求項13に記載のベクターの組合せにより形質転換された植物細胞または植物組織あるいは植物体。
【請求項15】
植物がイソキノリンアルカロイド生産性植物である請求項14に記載の植物細胞または植物組織あるいは植物体。
【請求項16】
植物がハナビシソウである請求項15に記載の植物細胞または植物組織あるいは植物体。
【図1】



【図2】
【図3】

【図4】

【図5】

【図6】

【図7】

【図8】

【図9】

【図10】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【国際公開番号】WO2005/033305
【国際公開日】平成17年4月14日(2005.4.14)
【発行日】平成19年11月29日(2007.11.29)
【国際特許分類】
【出願番号】特願2005−514384(P2005−514384)
【国際出願番号】PCT/JP2004/013449
【国際出願日】平成16年9月15日(2004.9.15)
【国等の委託研究の成果に係る記載事項】(出願人による申告)国等の委託研究の成果に係る特許出願(平成14年度独立行政法人新エネルギー・産業技術総合開発機構業務委託研究、産業活力再生特別措置法第30条の適用を受けるもの)
【出願人】(504132272)国立大学法人京都大学 (1,269)
【Fターム(参考)】
【国際公開日】平成17年4月14日(2005.4.14)
【発行日】平成19年11月29日(2007.11.29)
【国際特許分類】
【国際出願番号】PCT/JP2004/013449
【国際出願日】平成16年9月15日(2004.9.15)
【国等の委託研究の成果に係る記載事項】(出願人による申告)国等の委託研究の成果に係る特許出願(平成14年度独立行政法人新エネルギー・産業技術総合開発機構業務委託研究、産業活力再生特別措置法第30条の適用を受けるもの)
【出願人】(504132272)国立大学法人京都大学 (1,269)
【Fターム(参考)】
[ Back to top ]