
Toll様レセプター強制発現細胞の利用
【課題】Toll様レセプター強制発現細胞の利用法を提供することを課題とする。
【解決手段】Toll様レセプター9(TLR9)遺伝子およびToll様レセプター2(TLR2)遺伝子をブタ腸管パイエル板よりクローニングし、ブタTLR9またはブタTLR2を強制発現させた細胞を作製した。ブタTLR9トランスフェクタントを用いたCpG DNAに対する機能性解析を行なった結果、ブタTLR9はマウス特異的CpG DNAモチーフ(CpG1826)よりもヒトのCpG DNAモチーフ(CpG2006)に対する認識性が高いことが判明した。さらにReal-time PCR法によりmRNAの発現量を各種組織において比較した結果、腸管免疫系で中心的な役割を果たすパイエル板および腸管膜リンパ節において、脾臓の3倍以上のmRNAが発現していることが判明した。また、ブタTLR2トランスフェクタントが酵母細胞壁成分からのザイモサンのみならず、intactな乳酸菌を認識することができ、それによって転写因子(NF-κB)活性化を誘導できることを示した。よって、TLR9やTLR2等の腸管組織において発現しているTLRを強制発現させた細胞は、腸管免疫系を活性化する試料の同定に利用できる。
【解決手段】Toll様レセプター9(TLR9)遺伝子およびToll様レセプター2(TLR2)遺伝子をブタ腸管パイエル板よりクローニングし、ブタTLR9またはブタTLR2を強制発現させた細胞を作製した。ブタTLR9トランスフェクタントを用いたCpG DNAに対する機能性解析を行なった結果、ブタTLR9はマウス特異的CpG DNAモチーフ(CpG1826)よりもヒトのCpG DNAモチーフ(CpG2006)に対する認識性が高いことが判明した。さらにReal-time PCR法によりmRNAの発現量を各種組織において比較した結果、腸管免疫系で中心的な役割を果たすパイエル板および腸管膜リンパ節において、脾臓の3倍以上のmRNAが発現していることが判明した。また、ブタTLR2トランスフェクタントが酵母細胞壁成分からのザイモサンのみならず、intactな乳酸菌を認識することができ、それによって転写因子(NF-κB)活性化を誘導できることを示した。よって、TLR9やTLR2等の腸管組織において発現しているTLRを強制発現させた細胞は、腸管免疫系を活性化する試料の同定に利用できる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、Toll様レセプター強制発現細胞の利用法に関する。
【背景技術】
【0002】
生体の免疫システムには先天的な自然免疫と後天的に生じる獲得免疫の2種類がある。ヒト等の高等生物にしか存在しない獲得免疫に対して、自然免疫は昆虫からヒトまで広く保存された免疫機構である。自然免疫を担当する細胞(マクロファージや樹状細胞等)は外界からの病原細菌等が侵入すると、すばやくこれを探知し、食作用等の直接的な攻撃に加え、サイトカイン等の警戒シグナルを発して獲得免疫系を活性化するといった感染防御の第一段階として機能している。この様な一連の免疫反応システムにおいて最初のきっかけとなる細菌を発見する役目を果たしている受容体タンパクがToll様レセプター(Toll-like receptor;TLR)である。TLRは現在までにヒトにおいて10種類が同定されており、それぞれのTLRは認識する分子構造が異なるものと考えられている(非特許文献1)。その中で2000年に審良らの研究グループによって報告されたTLR9は細菌のDNA、特にCpGモチーフを認識するレセプターとして同定された (非特許文献2、特許文献1)。また、TLR2は、ペプチドグリカン(PGN)、リポテイコ酸、微生物のリポタンパク質、およびザイモサンのような微生物の細胞壁成分に反応した細胞シグナル伝達を媒介することが認識されている(非特許文献3)。
【0003】
すでに、リポペプチドがヒトTLR2を強制発現させたCHO細胞におけるNF-κBの発現活性に与える影響について、また、病原性大腸菌由来CpG DNAの様々なモチーフがヒトTLR9を強制発現させたHEK293細胞におけるサイトカイン(IL-8)の産生量に与える影響について報告されている(非特許文献4〜6)。
【0004】
尚、本出願の発明に関連する先行技術文献情報を以下に示す。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2002-34565
【非特許文献】
【0006】
【非特許文献1】O. Takeuchi, S. Akira, International Immunopharmacology 1 (2001) 625-635
【非特許文献2】H. Hemmi et al, Nature 408 (2000) 740-745
【非特許文献3】Wetzler, L.M. (2003) The role of Toll-like receptor 2 in microbial disease and immunity. Vaccine 21, S55-S60.
【非特許文献4】Yoshimura A. Takada H. Kaneko T. Kato I. Golenbock D. Hara Y. Structural requirements of muramylpeptides for induction of Toll-like receptor 2-mediated NF-kappaB activation in CHO cells. Journal of Endotoxin Research. 6(5):407-10, 2000.
【非特許文献5】Klinman DM. Takeshita F. Gursel I. Leifer C. Ishii KJ. Verthelyi D. Gursel M. CpG DNA: recognition by and activation of monocytes. Microbes & Infection. 4(9):897-901, 2002 Jul.
【非特許文献6】Takeshita F. Leifer CA. Gursel I. Ishii KJ. Takeshita S. Gursel M. Klinman DM. Cutting edge: Role of Toll-like receptor 9 in CpG DNA-induced activation of human cells. Journal of Immunology. 167(7):3555-8, 2001 Oct 1.
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明者らはこれまでにプロバイオティック乳酸菌を含む乳業用乳酸菌由来のDNAモチーフが腸管免疫系において免疫賦活化機能を有することを明らかにしてきた(H. Kitazawa et al, Int. J. Food Microbiol. 65 (2001) 149-162、H. Kitazawa et al, Int. J. Food Microbiol, (2003) in press)。本発明者らは、この知見からTLR9が病原細菌由来DNAのみならず、食餌性乳酸菌由来のDNAに対しても認識性を持ち、免疫の活性化に寄与する可能性を考えた。多様なDNAモチーフに対する認識性および活性評価系の構築が可能となれば、今後、病原性細菌の負の評価のためのみならず、乳業用乳酸菌を用いた機能性食品開発の上で重要なツールとなる。
【0008】
また、最近、臨床および実験的試験において、乳酸菌は代表的なイムノバイオティクスとして用いられており、その細胞壁成分は、以下の手段によって宿主に対して有用な作用を付与することが報告されている:分裂促進活性(Takeda, K., et al. (1997) Milchwissenshaft 52, 21-25.; Kirjavainen, P.V., et al. (1999) FEMS Immunol. Med. Microbiol. 26, 131-135.)、抗癌活性(Bogdanov, I.G., et al. (1975) FEBS Lett. 57, 259-261.)、抗変異原活性(Bodana, A.R. and Rao, D.R. (1990) J. Dairy Sci. 73, 3379-3384.)、抗感染作用(Perdigon, G., et al. (1986) Infect. Immun. 53, 404-410.; Popova, P., et al. (1993) Int. J. Immunopharmacol. 15, 25-37.)、およびサイトカイン産生の誘導(Popova, P., et al. (1993) Int. J. Immunopharmacol. 15, 25-37.; Pereyra, B.S., et al. (1991) Eur. Cytokine Netw. 2, 299-303.; De Simone, C., et al. (1993) Int. J. Immunother. 9, 23-28.)。
【0009】
機能性食品の開発を考える場合、最終的にヒトに対する効果を評価する必要があるが、その基礎的知見を得るためには、動物細胞および実験動物を用いた検討が必要不可欠となる。そこで本発明では、実験対象動物として、臓器移植等の観点からヒトモデル系としてその利用性が期待され、また食品産業の面から大きな価値を持つブタに着目し、ブタのTLR遺伝子のクローニングと、その遺伝子を導入し強制発現させた動物細胞を作製し、機能性DNAや機能性微生物等の評価系のためのTLR強制発現細胞を構築することを考えた。
本発明は、このような状況に鑑みてなされたものであり、その目的は、TLR強制発現細胞の利用法を提供することを目的とする。
【課題を解決するための手段】
【0010】
本発明者らは、病原細菌由来のCpG DNAモチーフを認識する受容体タンパクToll様レセプター9遺伝子をブタ腸管パイエル板よりクローニングし、ブタTLR9(sTLR9)を強制発現させた動物細胞(トランスフェクタント)を作製した。動物細胞におけるsTLR9タンパクの発現の有無は、sTLR9に対するポリクローナル抗体を作成し、同抗体を用いて確認した。さらに、sTLR9トランスフェクタントのCpG DNAに対する機能性を解析し、乳酸菌DNAの活性評価系への応用を目指した。
【0011】
具体的には、下記(1)〜(5)を行った。
(1)ブタ腸管パイエル板よりTotal RNAを抽出し、ヒトおよびマウスTLR9遺伝子における保存性の高い領域より作製したプライマーを用いてRT-PCR法、RACE法によりブタTLR9遺伝子をクローニングし、その全遺伝子配列を決定した。
(2)遺伝子情報から得られたブタTLR9全アミノ酸配列から抗原決定基部位を探索し、選抜した領域をペプチド合成しブタTLR9ポリクローナル抗体作製用抗原とした。抗原は化学合成後、定法によりウサギに免疫し、ブタTLR9に対するポリクローナル抗体を作成した。
(3)ブタTLR9遺伝子をHEK293T細胞(ヒト胎児腎細胞)に導入し、ブタTLR9遺伝子導入細胞(トランスフェクタント)を作製した。
(4)ブタTLR9のHEK293Tにおける発現については、ブタTLR9 mRNAの発現をRT-PCR法により、また、ブタTLR9膜タンパクの発現をブタTLR9ポリクローナル抗体による免疫染色法によりレーザー顕微鏡およびフローサイトメーターを用いて確認した。
(5)ヒトおよびマウス細胞を強く刺激する特異的CpG DNAモチーフを含むオリゴDNA(それぞれCpG2006,CpG1826)に対する反応性を解析した。
【0012】
検討の結果、ブタTLR9遺伝子は1029残基(MW:115.8)のアミノ酸をコードする3090塩基からなり、その遺伝子を含め3145塩基のcDNA塩基配列を決定することができた。ブタTLR9のアミノ酸配列はヒトTLR9に対して82.9%と非常に高く、マウスTLR9には74.9%の相同性を示したことから、ヒトTLR9に対してマウスよりも比較的高い相同性を示すことが明らかとなった。ブタTLR9トランスフェクタントにおけるブタTLR9タンパク質はRT-PCR法およびブタTLR9ポリクローナル抗体の免疫染色から確実に発現し、膜タンパク質として発現したことを確認できた。このことは、ブタTLR9トランスフェクタントの作製に成功したことを意味する。本トランスフェクタントを用いたCpG DNAに対する機能性解析よりブタTLR9はCpG1826よりもCpG2006に対して強い反応性を示したことから、ブタTLR9はマウス特異的CpG DNAモチーフよりも比較的ヒトのモチーフに対する認識性が高いことが判明した。Real-time PCR法によりmRNAの発現量を各種組織において比較した結果、驚くべきことに、腸管免疫系で中心的な役割を果たすパイエル板および腸管膜リンパ節において、脾臓の3倍以上のmRNAが発現していることが判明した。
【0013】
また、本発明者らは回腸のパイエル板からブタTLR2(sTLR2)cDNAを単離し、その後sTLR2発現トランスフェクタントを構築することによって、イムノバイオティック微生物の免疫応答に関する評価法の開発を行った。
【0014】
即ち、本発明は、以下の〔1〕〜〔21〕を提供するものである。
〔1〕 被験試料が腸管免疫系を活性化するか否かを評価する方法であって、
(a)腸管組織において発現しているToll様レセプターを強制発現させた細胞に、被験試料を接触させる工程
(b)該細胞におけるシグナル伝達を指標に、該Toll様レセプターの活性を測定する工程
を含み、上記Toll様レセプターの活性が、上記被験試料を接触させないときに比べ上昇する場合に、被験試料が腸管免疫系を活性化すると判定される方法。
〔2〕 以下の(a)および(b)の工程を含む、腸管免疫系を活性化する試料のスクリーニング方法。
(a)〔1〕に記載の評価方法により、複数の被験試料について、腸管免疫系を活性化するか否かを評価する工程
(b)複数の被験試料から、腸管免疫系を活性化すると評価された試料を選択する工程
〔3〕 〔2〕に記載の工程に、さらに腸管免疫系を活性化すると評価された試料と医薬上許容される担体とを混合する工程を含む、腸管免疫系を活性化する医薬組成物の製造方法。
〔4〕 被験微生物が腸管免疫系を活性化する微生物であるか否かを評価する方法であって、
(a)被験微生物から抽出物を調製する工程
(b)腸管組織において発現しているToll様レセプターを強制発現させた細胞に、該抽出物を接触させる工程
(c)該細胞におけるシグナル伝達を指標に、該Toll様レセプターの活性を測定する工程
を含み、上記Toll様レセプターの活性が、上記抽出物を接触させないときに比べ上昇する場合に、被験微生物が腸管免疫系を活性化する微生物であると判定される方法。
〔5〕 以下の(a)および(b)の工程を含む、腸管免疫系を活性化する微生物のスクリーニング方法。
(a)〔4〕に記載の評価方法により、複数の被験微生物について、腸管免疫系を活性化する微生物であるか否かを評価する工程
(b)複数の被験微生物から、腸管免疫系を活性化する微生物であると評価された微生物を選択する工程
〔6〕 〔5〕に記載の工程に、さらに腸管免疫系を活性化する微生物であると評価された微生物と食品上許容される担体とを混合する工程を含む、腸管免疫系を活性化する食品組成物の製造方法。
〔7〕 微生物が乳酸菌であり、食品組成物が乳製品である、〔6〕に記載の方法。
〔8〕 微生物が細菌である、〔4〕〜〔6〕のいずれかに記載の方法。
〔9〕 細菌が乳酸菌である、〔8〕に記載の方法。
〔10〕 腸管組織において発現しているToll様レセプターをコードするDNAを有する発現ベクターを細胞に導入する工程を含む、腸管免疫担当細胞のモデル細胞の製造方法。
〔11〕 腸管組織において発現しているToll様レセプターを強制発現させた細胞を、腸管免疫担当細胞のモデル細胞として使用する方法。
〔12〕 腸管組織が腸管リンパ系組織である、〔1〕〜〔11〕のいずれかに記載の方法。
〔13〕 腸管リンパ系組織がパイエル板または腸管リンパ節である、〔12〕に記載の方法。
〔14〕 Toll様レセプターがブタ由来である、〔1〕〜〔13〕のいずれかに記載の方法。
〔15〕 Toll様レセプターがToll様レセプター9および/または2である、〔1〕〜〔13〕のいずれかに記載の方法。
〔16〕 〔1〕〜〔9〕のいずれかに記載の方法に用いるための、腸管組織において発現しているToll様レセプターを強制発現させた細胞。
〔17〕 腸管組織において発現しているToll様レセプターをコードするDNAを有する発現ベクターを細胞に導入することで製造される、腸管免疫担当細胞のモデル細胞。
〔18〕 腸管組織が腸管リンパ系組織である、〔16〕または〔17〕に記載の細胞。
〔19〕 腸管リンパ系組織がパイエル板または腸管リンパ節である、〔18〕に記載の細胞。
〔20〕 Toll様レセプターがブタ由来である、〔16〕〜〔19〕のいずれかに記載の細胞。
〔21〕 Toll様レセプターがToll様レセプター9および/または2である、〔16〕〜〔19〕のいずれかに記載の細胞。
【発明の効果】
【0015】
本発明においてTLRトランスフェクタントの利用法が提供された。TLRトランスフェクタントは、腸管免疫担当細胞のモデル細胞として利用できる。また、TLRトランスフェクタントを利用することで、腸管免疫系を活性化する試料および微生物を同定でき、腸管免疫系を活性化する医薬組成物および食品組成物を製造できる。該試料、微生物、医薬組成物、および食品組成物は、例えば、アレルギー、癌、感染症等の治療または予防に使用できる。
【0016】
また、本発明により、同じ菌種の異なる株の差をも区別できるTLRトランスフェクタントが提供された。TLRによって免疫調節作用を発揮するイムノバイオティック微生物を用いて機能的食品を開発する場合、TLR-発現トランスフェクタントは、イムノバイオティック微生物をも評価することができる免疫スクリーニング系の貴重なツールとして、TLR−イムノバイオティック微生物間の関係の評価に用いることができる。
【図面の簡単な説明】
【0017】
【図1】ブタTLR9 cDNA塩基配列を示す図である。下線はシグナルペプチド領域を示す。
【図2】ブタTLR9 cDNA塩基配列を示す図である。図1の続きを示す。下線は膜貫通領域を示す。
【図3】ブタTLR9のSMARTによるドメイン解析を示す図である。
【図4】TLR9アミノ酸配列のアライメントを示す図である。
【図5】TLR9アミノ酸配列のアライメントを示す図である。図4の続きを示す。
【図6】トランスフェクタントにおけるブタTLR9のRT-PCR法による発現解析を示す写真である。
【図7】トランスフェクタントにおけるブタTLR9の発現とCpG DNAの取り込み解析を示す図である。a;抗FLAG抗体による解析を示す。b;抗ブタTLR9抗体による解析を示す。c;CpG DNAの取り込み解析を示す。図中の矢印(1)はコントロール細胞のCpG1826および2006の取り込みを示し、矢印(2)はトランスフェクタントにおけるCpG1826を示し、矢印(3)はトランスフェクタントにおけるCpG2006を示す。
【図8】トランスフェクタントの共焦点レーザー顕微鏡による解析を示す写真である。a,b,c;対照細胞、d,e,f;ブタTLR9トランスフェクタントを示す。a,d;抗FLAG抗体による解析を示す。b,e;抗ブタTLR9抗体による解析を示す。c,f;CpG DNAの取り込み解析を示す。
【図9】real-time定量PCRによるブタTLR9の各種組織における発現解析を示す図である。
【図10】CHO K-1sTLR2trans細胞上でのsTLR2 mRNA発現をRT-PCRによって分析した結果を示す写真である。逆転写された総RNA 1 mgからのPCR(30サイクル)増幅産物をアガロースゲル上で検出した。
【図11】(a,b)CHO K-1sTLR2trans細胞上でのsTLR2タンパク質の発現をフローサイトメトリーにより検出した結果を示す図である。(a)抗FLAGモノクローナル抗体(PE)および(b)抗sTLR2ポリクローナル抗体(アレクサ488)によって検出した。線(2)および線(4)は、CHO K-1sTLR2trans細胞を表す。線(1),(3)は、ベクター対照細胞を表す。(c)CHO K-1sTLR2trans細胞によるFITC-標識ザイモサン(6)の取り込みを、フローサイトメトリーによって観察した結果を示す図である((5):ベクター対照細胞によるザイモサンの取り込み)。
【図12】sTLR2の細胞局在を共焦点レーザー顕微鏡によって観察した写真である。CHO K-1sTLR2trans細胞(d、e、f)またはCHO K-1Cont.細胞(a、b、c)を抗FLAG M5 モノクローナル抗体によって染色した後、アレクサ660結合抗マウスIgG(青色;d、f)または抗sTLR2ポリクローナルAbによって染色した後、アレクサ488結合抗ウサギIgG(緑色;e)によって染色した。細胞をFITC標識ザイモサンによって37℃で1時間処置した(c、f)。核はヨウ化プロピジウム(PI)(赤またはオレンジ色)によって染色した。矢印は、同時局在を示すより大きいザイモサン小胞を示す(f)。6回の独立した実験の一つからの代表的な切片を示す(尺度のバー;10 μm)。
【図13】プラスミドベクターのみをトランスフェクトしたCHO K-1細胞(b)およびCHO K-1sTLR2trans細胞(a,c)の表面を、EMによって観察した写真である。パネル(a)および(b,c)の倍率はそれぞれ、1,000倍および10,000倍である。
【図14】四つのLAB株におけるNF-κB依存的ルシフェラーゼ活性の比較を示す図である。1 μM D25、10 μg/mlザイモサン、10 μg/ml PGN、および100 μg/mlのintactなLABによる処置後24時間でのトランスフェクト細胞のルシフェラーゼ活性を示す。結果は、独立した実験3回〜4回のS.I.の平均値±SDを表す;培地のみにおいて培養した同一に処置した細胞と比較して*:P<0.05、**:P<0.01
【発明を実施するための形態】
【0018】
免疫活性化に関与するToll様レセプター9(Toll-like receptor 9;TLR9)は、脾臓において強く発現していることが知られていた(Zarember KA. and Godowski PJ. Journal of Immunology. 168 (2002) 554-561)。これに対し、本発明者らは、TLR9が腸管リンパ系組織、特にパイエル板や腸管膜リンパ節において強く発現していることを初めて見出した。
【0019】
腸管は体内と体外の境界組織として、経口摂取された栄養成分の吸収のみならず、外界からの刺激(細菌やウイルスなどの微生物、薬剤や食品添加物、食物に付着した農薬や環境汚染物質など)に常に接触しており、これら異物の受容、伝達、応答、および排除など種々の第一線生体防御機構(first defense line)が存在している(Mantis NJ. et al, J. Immunol. 169 (2002) 1844-1851)。さらに防御機構として特有のリンパ組織、腸管関連リンパ組織(GALT; gut-associated lymphoid tissue)が存在し、GALTは非集合性組成と集合性組成から構成され、非集合性組成には腸管上皮間リンパ球と粘膜固有層リンパ球が含まれ、集合性組成にはパイエル板、リンパ濾胞および腸管膜リンパ節が含まれる(Spahn TW. et al, Eur. J. Immunol. 32 (2002) :1109-1113)。パイエル板はリンパ組織関連上皮(Follicle-associated epithelium; FAE)に覆われ、腸管絨毛の存在しない部位でドーム形態をなしている。また、胚中心を持ちB細胞の存在する濾胞域(follicular area)と、ヘルパーT細胞を含む傍濾胞域(parafollicular area)を備えている(Owen RL. Sem. Immunol. 11 (1999) 157-163)。腸管の局所免疫機構の最初の役割を果たす細胞として、FAEには特殊な上皮細胞であるMembranous epithelial cells(M cells;M細胞)が散在し、細胞質を通って基底表層へとつながる抗原のトンネルとしてリンパ球や樹状細胞、マクロファージなど抗原提示細胞(antigen presenting cell)との接触のためM細胞膜は深く陥入している。気管上皮粘膜においてもM細胞の存在は確認されており、結核菌などの病原体の入り口として貢献していることが報告されている(Teitelbaum R. et al, Immunity. 10 (1999) 641-650)。また、食品中に含まれる機能性因子のみならず微生物や食物抗原の入り口としても存在することが認められている。腸管内の抗原(特に高分子)はM細胞に取り込まれた後、パイエル板の内部へと運ばれ、パイエル板内部の樹状細胞、マクロファージなどのMHC(major histocompatibility complex)クラスII陽性の抗原提示細胞に接触する(Kaneko K. et al, J. Veterinary. Med. Sci. 61 (1999) 1175-1177、Gebert A. et al, American J. Pathology. 154 (1999) 1573-1582、Jensen VB. et al, Infection & Immunity. 66 (1998) 3758-3766、Penheiter KL. et al, Mol. Microbiol. 24 (1997) 697-709、Debard N. et al, Gastroenterology. 120 (2001) 1173-1182、Gebert A. et al, Int. Rev. Cytology. 167 (1996) 91-159)。抗原刺激を受けたヘルパーT細胞はFcレセプターや抗体結合因子(IBF)、IL-2、IL-4、IL-5、IL-6を産生し、抗原提示を受け活性化したT細胞およびB細胞は循環帰巣(homing)という現象を開始し、腸管膜リンパ節を経て、胸腺に入り、体内循環して実行組織である腸管粘膜固有層、乳腺、涙腺、唾液腺、泌尿生殖器などに送られ、さらにB細胞は形質細胞となってIgAを産生する。分泌型IgAは、腸管およびその他の粘膜組織に侵入したウイルス、細菌、細菌毒素、アレルゲンなどを排除する役割を果たしている(Vaerman JP. et al, Immunology. 54 (1985) 601-603、Machtinger S. and Moss R. J. Allergy. Clinical. Immunol. 77 (1986) 341-347、Mathewson JJ. et al, J. Infectious Diseases. 169 (1994) 614-617)。さらにパイエル板の下には腸管膜リンパ節が発達し、パイエル板を越える多くのリンパ球、樹状細胞、マクロファージが待機している。このように、パイエル板や腸管膜リンパ節は、腸管免疫系(上記のような腸管に存在する免疫系)において中心的な役割を果たすことが知られている。
【0020】
一方、TLR4抗体、IRAK抗体を用いて、サルの腸管上皮からのFITC標識したTLR4のリガンドとして知られるLPS(リポポリサッカライド)の取り込みと腸管粘膜上皮細胞との関係について解析した結果、LPSが腸管上皮細胞中のTLR4とIRAKを発現する細胞により取り込まれ、粘膜固有層にまで運ばれる様子が観察された(Imaeda H. et al, Histochemical Cell Biology. 118 (2002) 381-388)。以上を考慮すると、TLR9やTLR2等の腸管組織において発現しているTLRは、腸管免疫系に関与していると考えられる。
【0021】
本発明は、以上の知見に基づき、被験試料が腸管免疫系を活性化するか否かを評価する方法を提供する。該評価方法としては、まず、腸管組織において発現しているTLRを強制発現させた細胞(TLRトランスフェクタント)に、被験試料を接触させる。この工程により、被験試料を、トランスフェクタント表面のTLRに接触させる。次いで、該TLRトランスフェクタントにおけるシグナル伝達を指標に、該TLRの活性を測定する。該評価方法においては、該TLRの活性が、被験試料を接触させないときに比べ上昇する場合に、被験試料が腸管免疫系を活性化すると判定される。
【0022】
本発明において、被験試料としては、例えば、DNA、DNA断片、天然化合物、有機化合物、無機化合物、タンパク質、ペプチド等の単一化合物、並びに、化合物ライブラリー、遺伝子ライブラリーの発現産物、非哺乳動物細胞培養上清、非哺乳動物細胞抽出物、発酵微生物産生物、微生物培養上清、微生物抽出物、海洋生物抽出物、植物抽出物、微生物等を挙げることができるが、これらに限定されるものではない。上記DNA断片としては、由来する生物に制限はないが、CpG モチーフ、ATモチーフまたはCpG 様モチーフを有するDNA断片が好ましい。また、上記微生物抽出物としては、例えば、細胞壁、細胞膜、DNA、RNA、鞭毛等が挙げられる。また、該微生物としては、細菌、酵母等を例示できる。また、該細菌としては、病原性細菌、乳酸菌等を挙げることができる。上記被験試料は必要に応じて適宜標識して用いることができる。標識としては、例えば、放射標識、蛍光標識等を挙げることができる。
【0023】
また、本発明における腸管としては、十二指腸、空腸、回腸等が挙げられるが、これらに限定されるものではない。また、本発明における腸管組織としては、特に制限はないが、好ましくは腸管リンパ系組織、より好ましくはパイエル板または腸管リンパ節、さらに好ましくは回腸由来パイエル板または回腸由来腸管リンパ節が挙げられる。
【0024】
また、腸管組織において発現しているTLRには、全てのタイプのTLRが包含され、例えばTLR1〜TLR10が挙げられる。現在までに10種類のTLRの存在が報告されており、TLRファミリーはそれぞれ認識分子すなわち細菌性モデュリンが異なると考えられている。細菌性モデュリンとは、細菌に特異的な分子パターン(pathogen associated molecular patterns; PAMPs)で、宿主に対しサイトカイン誘導能を示し、免疫応答を制御するものと定義されている。また、TLRは、細胞外にロイシンリッチリピート(LRRs)、細胞内にインターロイキン1受容体と相同性のある領域(TIRドメイン)を持つことが知られている。また、腸管組織において発現しているTLRが由来する生物としては、ブタ、ヒト、マウス、ネコ等、無脊椎、脊椎動物全般あるいは生物一般が挙げられる。
【0025】
本発明におけるTLR9としては、例えば、配列番号:2、4、6または8に記載のアミノ酸配列を含むタンパク質が挙げられる。また、本発明におけるTLR2としては、例えば、配列番号:36に記載のアミノ酸配列を含むタンパク質が挙げられる。配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質をコードするDNAは、例えば、胸腺、肺臓、脾臓、十二指腸、パイエル板および腸管膜リンパ節よりRNAを調製し、逆転写酵素によりcDNAを合成した後、配列番号:1、3、5、7または35に基づいてオリゴDNAを合成し、これをプライマーとして用いてPCR反応を行い、上記タンパク質をコードするcDNAを増幅させることにより調製できる。
【0026】
また、本発明におけるTLR9には、配列番号:2、4、6または8に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質、TLR2には、配列番号:36に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質も含まれる。このようなタンパク質には、例えば、配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質の変異体、アレル、バリアント、ホモログ等が含まれる。ここで「機能的に同等」とは、対象となるタンパク質が、配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質と同等の生物学的機能(生物学的役割)、生化学的機能(生化学的活性)を有することを指す。
【0027】
TLRは菌体成分を認識すると細胞内シグナル伝達系路を活性化し、共通のアダプター分子MyD88を介してIL-1 receptor associated kinase (IRAK)、TNF receptor associated factor 6 (TRAF6)、そして転写因子であるNF-κBの核移行を促し、最終的にtumor necrosis factor α(TNF-α)、interleukin(IL)-6、IL-12、IL-18、IFN-γなどといった様々な炎症性サイトカインの産生や、細胞表面共刺激因子(co-stimulatory molecule)の発現を誘導する(Kaisho T. and Akira S. Trends in Immunology. 22 (2001) 78-83)。TLRは主に病原体の糖質、脂質や核酸を認識することから、タンパクを認識する獲得免疫と相補的であるといえる。
【0028】
すなわち、配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質の生物学的機能、生化学的機能としては、微生物成分認識能、細胞内シグナル伝達系路の活性化能、炎症性サイトカイン発現誘導能、細胞表面共刺激因子発現誘導能などが挙げられる。
【0029】
あるタンパク質と機能的に同等なタンパク質をコードするDNAを調製する方法としては、ハイブリダイゼーション技術(Sambrook,J et al., Molecular Cloning 2nd ed., 9.47-9.58, Cold Spring Harbor Lab. press, 1989)を利用する方法が挙げられる。即ち、配列番号:1、3、5、7または35に記載の塩基配列もしくはその一部を利用して、配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質をコードするDNAを単離することができる。
【0030】
配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質をコードするDNAを単離するためのハイブリダイゼーションの条件は、当業者であれば適宜選択することができる。ハイブリダイゼーションの条件としては、例えば、低ストリンジェントな条件が挙げられる。低ストリンジェントな条件とは、ハイブリダイゼーション後の洗浄において、例えば42℃、5xSSC、0.1%SDSの条件であり、好ましくは50℃、5xSSC、0.1%SDSの条件である。より好ましいハイブリダイゼーションの条件としては、高ストリンジェントな条件が挙げられる。高ストリンジェントな条件とは、例えば65℃、0.1xSSC及び0.1%SDSの条件である。これらの条件において、温度を上げる程に高い相同性を有するDNAが効率的に得られると考えられる。但し、ハイブリダイゼーションのストリンジェンシーに影響する要素としては温度や塩濃度等複数の要素が考えられ、当業者であればこれら要素を適宜選択することで同様のストリンジェンシーを実現することが可能である。
【0031】
また、配列番号:1、3、5、7または35に記載の配列情報を基に合成したプライマーを用いる遺伝子増幅法、例えば、ポリメラーゼ連鎖反応(PCR)法を利用して、配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質をコードするDNAを単離することも可能である。
【0032】
これらハイブリダイゼーション技術や遺伝子増幅技術により単離されるDNAがコードする、配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質は、通常、配列番号:2、4、6、8または36のアミノ酸配列を含むタンパク質とアミノ酸配列レベルにおいて高い相同性を有する。高い相同性とは、アミノ酸レベルにおいて、少なくとも60%以上の同一性、好ましくは70%以上の同一性、より好ましくは80%以上の同一性、さらに好ましくは90%、よりさらに好ましくは95%以上、最も好ましくは98%以上の同一性を指す。
【0033】
アミノ酸配列や塩基配列の同一性は、Karlin and AltschulによるアルゴリズムBLAST(Proc. Natl. Acad. Sci. USA 90:5873-5877, 1993)によって決定することができる。このアルゴリズムに基づいて、BLASTNやBLASTXと呼ばれるプログラムが開発されている(Altschul et al. J. Mol. Biol.215:403-410, 1990)。BLASTに基づいてBLASTNによって塩基配列を解析する場合には、パラメーターはたとえばscore = 100、wordlength = 12とする。また、BLASTに基づいてBLASTXによってアミノ酸配列を解析する場合には、パラメーターはたとえばscore = 50、wordlength = 3とする。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である(http://www.ncbi.nlm.nih.gov.)。
【0034】
また、配列番号:2、4、6、8または36に記載のアミノ酸配列において1もしくは複数のアミノ酸が変異したアミノ酸配列からなるタンパク質も本発明に含まれる。このようなアミノ酸の変異は自然界においても生じうる。変異するアミノ酸数は、通常、30アミノ酸以内であり、好ましくは15アミノ酸以内であり、より好ましくは5アミノ酸以内であり、さらに好ましくは2アミノ酸以内である。
【0035】
本発明の方法においては、上記TLRトランスフェクタントが使用される。該TLRトランスフェクタントは、腸管組織において発現しているTLRをコードするDNAを含む発現ベクターを細胞に導入することで製造することができる。該発現ベクターとしては、例えば、哺乳動物由来の発現ベクター(例えば、pcDNA3 (インビトロゲン社製)や、pEGF-BOS (Nucleic Acids. Res.1990, 18(17),p5322)、pEF、pCDM8)、昆虫細胞由来の発現ベクター(例えば「Bac-to-BAC baculovairus expression system」(インビトロゲン社製)、pBacPAK8)、動物ウィルス由来の発現ベクター(例えば、pHSV、pMV、pAdexLcw)、レトロウィルス由来の発現ベクター(例えば、pZIPneo)が挙げられる。
【0036】
CHO細胞、COS細胞、NIH3T3細胞等の動物細胞での発現を目的とした場合には、細胞内で発現させるために必要なプロモーター、例えばSV40プロモーター(Mulligan et al., Nature (1979) 277, 108)、MMLV-LTRプロモーター、EF1αプロモーター(Mizushima et al., Nucleic Acids Res. (1990) 18, 5322)、CMVプロモーター等を持っていることが不可欠であり、細胞への形質転換を選抜するための遺伝子(例えば、薬剤(ネオマイシン、G418等)により判別できるような薬剤耐性遺伝子)を有すればさらに好ましい。
【0037】
さらに、遺伝子を安定的に発現させ、かつ、細胞内での遺伝子のコピー数の増幅を目的とする場合には、核酸合成経路が欠損したCHO細胞にそれを相補するジヒドロ葉酸還元酵素遺伝子(dhfr遺伝子)を有するベクター(例えば、pCHOI等)を導入し、メトトレキセート(MTX)により増幅させる方法が挙げられ、また、遺伝子の一過性の発現を目的とする場合には、SV40 T抗原を発現する遺伝子を染色体上に持つCOS細胞を用いてSV40の複製起点を持つベクター(pcD等)で形質転換する方法が挙げられる。複製開始点としては、また、ポリオーマウィルス、アデノウィルス、ウシパピローマウィルス(BPV)等の由来のものを用いることもできる。さらに、宿主細胞系で遺伝子コピー数増幅のため、発現ベクターは選択マーカーとして、アミノグリコシドトランスフェラーゼ(APH)遺伝子、チミジンキナーゼ(TK)遺伝子、dhfr遺伝子等を含むことができる。
【0038】
また、宿主細胞としては、例えば、哺乳類細胞、昆虫類細胞等が挙げられるが、これらに限定されるものではない。哺乳類細胞としては、例えば、HEK293T細胞、CHO(J. Exp. Med. (1995) 108, 945)、COS、NIH3T3、ミエローマ、BHK(baby hamster kidney)、HeLa、Vero、昆虫類細胞としては、例えば、Sf9、Sf21、Tn5が知られている。
【0039】
CHO細胞としては、特に、dhfr遺伝子を欠損したCHO細胞であるdhfr-CHO(Proc. Natl. Acad. Sci. USA (1980) 77, 4216-4220)やCHO K-1(Proc. Natl. Acad. Sci. USA (1968) 60, 1275)を好適に使用することができる。動物細胞において、大量発現を目的とする場合には特にCHO細胞が好ましい。また、宿主細胞としては、不死化した細胞株が好ましい。
【0040】
宿主細胞へのベクターの導入は、例えば、リン酸カルシウム法、DEAEデキストラン法、カチオニックリボソームDOTAP(ロッシュダイアグノスティックス社製)を用いた方法、エレクトロポーレーション法、リポフェクション等当業者に周知の方法で行うことが可能である。
【0041】
本発明においては、上記トランスフェクタントにおけるシグナル伝達を指標に、TLRの活性を測定する。TLRの活性は、例えば、サイトカイン(例えばIL-6、IL-12、IFN-γ、TNF-α等)の発現、シグナル伝達経路における分子(例えばNF-κB、JNK、IRAK等)の活性化等を指標に測定できる。サイトカインの発現は、mRNAレベルでもタンパク質レベルでも行うことが可能である。例えば、サイトカインのタンパク質レベルでの発現は、ヒトIL-6, IL-12, TNF-α, IFN-γELISA kit(株式会社テイエフビー(TFB,INC.))等の既存のヒト用キットを使用することで測定できる。また、シグナル伝達経路における分子の活性上昇は、ルシフェラーゼアッセイで行うことが可能である。例えば、ブタTLRトランスフェクタントにNF-κBおよびルシフェラーゼ遺伝子を含むプラスミドベクター(pGLM-ENH)をトランスフェクションし、18時間後にリガンド(DNA等)で刺激する。刺激24時間後に細胞を溶解後、細胞溶解液を回収し測定まで−80℃で保存する。ルシフェラーゼ活性は、細胞溶解液にルシフェリンを含む反応液を添加後、2秒後から8秒間にわたる発光の変化を測定する。各試料で3回同様の操作を行い、その平均をとる。以上の測定で発光が強い程、NF-κBの活性が強いことを示す。
【0042】
本発明はまた、腸管免疫系を活性化する試料のスクリーニング方法を提供する。本発明におけるスクリーニング方法としては、上記評価方法を利用して、複数の被験試料について、腸管免疫系を活性化するか否かを評価し、腸管免疫系を活性化すると評価された試料を選択する。
【0043】
また、腸管組織において発現しているTLRに結合する試料を予めスクリーニングし、これにより得られた試料を被験試料として用いてもよい。腸管組織において発現しているTLRに結合する試料のスクリーニングに用いられるTLRは、組換えタンパク質であっても、天然由来のタンパク質であってもよい。また、スクリーニングに用いられるTLRは部分ペプチドであってもよい。腸管組織において発現しているTLRに結合する試料のスクリーニング方法としては、まず、複数の被験試料を腸管組織において発現しているTLRに接触させる。次いで、該TLRと被験試料との結合を検出する。次いで、該TLRと結合する被験試料を選択する。TLRと被験試料との結合は、当業者に周知の方法で検出できる。
上記評価方法またはスクリーニング方法によって得られた試料は、免疫賦活化機能を有する試料として、例えば、アレルギー、癌、感染症等の治療または予防に使用できる。
【0044】
さらに、本発明においては、腸管免疫系を活性化する医薬組成物の製造方法を提供する。本発明の医薬組成物は、免疫賦活化機能を有する医薬組成物として、例えば、アレルギー、癌、感染症等の治療または予防に使用できる。また、本発明の医薬組成物は、好ましくはワクチンとして使用される。
【0045】
本発明の医薬組成物の製造方法においては、上記スクリーニング方法によって腸管免疫系を活性化すると評価された試料と医薬上許容される担体とを混合する。医薬上許容される担体としては、例えばアジュバント(抗体産生増強剤)が挙げられる。また、界面活性剤、賦形剤、着色料、着香料、保存料、安定剤、緩衝剤、懸濁剤、等張化剤、結合剤、崩壊剤、滑沢剤、流動性促進剤、矯味剤等も医薬上許容される担体として挙げられるが、その他常用の担体を適宜使用することができる。具体的には、軽質無水ケイ酸、乳糖、結晶セルロース、マンニトール、デンプン、カルメロースカルシウム、カルメロースナトリウム、ヒドロキシプロピルセルロース、ヒドロキシプロピルメチルセルロース、ポリビニルアセタールジエチルアミノアセテート、ポリビニルピロリドン、ゼラチン、中鎖脂肪酸トリグリセライド、ポリオキシエチレン硬化ヒマシ油60、白糖、カルボキシメチルセルロース、コーンスターチ、無機塩類等を挙げることができる。このようにして製造された腸管免疫系を活性化する医薬組成物は、経口剤または注射剤として使用できる。
【0046】
また、本発明は、被験微生物が腸管免疫系を活性化する微生物であるか否かを評価する方法も提供する。本発明における微生物としては、細菌、酵母等が例示できるが、これらに限定されるものではない。また、細菌としては、特に制限はないが、乳酸菌(例えば腸管常在乳酸菌や乳業用乳酸菌)が挙げられる。
【0047】
該評価方法においては、まず、被験微生物から抽出物を調製する。微生物抽出物としては、例えば、細胞壁、細胞膜、DNA、RNA、鞭毛等が挙げられる。また、該DNAとしては、CpG モチーフ、ATモチーフまたはCpG 様モチーフを有する断片が好適である。このような抽出物は、当業者に周知の方法によって微生物から調製できる。
以下に、乳業用乳酸菌からCpG モチーフ、ATモチーフまたはCpG 様モチーフを有する断片の調整方法を例示するが、本発明の方法はこれに限定されない。
【0048】
LactobacilliはLactobacilli MRS Broth(Difco laboratories, Detroit, MI, USA)で、StreptococciはElliker brothでそれぞれ3回継代培養(37℃, 24時間)後、50mlの培地に1%接種し、37℃で16時間培養する。菌体を遠心(3,000xg, 4℃, 20分間)集菌し、TE緩衝液(10mM Tris-Cl,1mM EDTA pH7.5)で2回遠心(4,000xg, 4℃, 20分間)洗浄後、5.0mlのTE緩衝液に再懸濁する。この菌液に、2.5mlのリゾチーム(生化学工業、東京、30mg/ml)、20μlのN-アセチルムラミダーゼSG(生化学工業、東京、250μg/ml)を加え、37℃で10〜30分間反応する。反応後、10mlの0.1M Tris-1% SDS溶液を添加し、穏やかに撹拌する。これにプロテイナーゼK(TaKaRa,京都20mg/ml,150μl)溶液を加え、37℃で一晩反応させる。反応後、5M NaCl溶液(2.5ml)を加え撹拌し、滅菌したビーカーに移す。100%エタノール(50ml)を加えることにより核酸を沈殿させ、生じた沈殿を滅菌したガラス棒を用いて巻き取り、70%エタノールで洗浄後、TE緩衝液(10ml)に溶解し、一晩4℃下で完全に溶解させる。RNaseA(SIGMA,10mg/ml,100μl)を加え、37℃で60分間インキュベートした後、1/10量の5M NaCl(1ml)及び等量の100%エタノールを加え、核酸を沈殿させる。生じた沈殿を滅菌したガラス棒を用いて巻き取り、70%エタノールで洗浄後、TE緩衝液(2ml)に溶解する。以上の操作でDNAが精製され、使用までの間4℃下で保存する。染色体DNAはSau 3A1 (制限酵素)により切断処理し、3%アガロースゲル電気泳動に供する。アガロースゲルよりDNAを回収し、プラスミドベクターにライゲーション後、クローニングしDNA配列をシークエンサーにて決定し、CpG モチーフ、ATモチーフまたはCpG 様モチーフを有する断片を調製する。
【0049】
該評価方法においては、次いで、該抽出物をTLRトランスフェクタントに接触させる。次いで、該形質転換細胞におけるシグナル伝達を指標に、腸管組織において発現しているTLRの活性を測定する。該評価方法においては、該TLRの活性が、該抽出物を接触させないときに比べ上昇する場合に、被験微生物が腸管免疫系を活性化する微生物であると判定される。
【0050】
また、本発明は、腸管免疫系を活性化する微生物のスクリーニング方法も提供する。該スクリーニング方法としては、上記評価方法を利用して、複数の被験微生物について、腸管免疫系を活性化するか否かを評価し、腸管免疫系を活性化すると評価された微生物を選択する。
上記評価方法またはスクリーニング方法によって得られた微生物は、免疫賦活化機能を有する微生物として、例えば、アレルギー、癌、感染症等の治療または予防に使用できる。
【0051】
腸管免疫系を活性化する微生物を使用することで、腸管免疫系を活性化する食品組成物を製造できる。本発明は、このような腸管免疫系を活性化する食品組成物の製造方法もまた提供する。本発明の食品組成物の製造方法においては、上記スクリーニング方法によって腸管免疫系を活性化する微生物であると評価された微生物と食品上許容される担体とを混合する。食品上許容される担体としては、例えば安定化剤、保存剤、着色料、香料等が挙げられる。
【0052】
本発明の食品組成物の好ましい態様としては、乳酸菌や酵母を含む食品、ドリンク等が挙げられる。乳酸菌や酵母を含む食品、ドリンクとしては、例えば乳製品が挙げられる。本発明の乳製品としては、発酵乳、チーズ、発酵食品(乳酸菌を含む食品、キムチ等)が挙げられる。これらは、当業者に周知の方法で製造できる。
このようにして製造された食品組成物は、免疫賦活化機能を有する食品組成物(例えば機能性食品、健康食品、特定保健用食品等)として、アレルギー、癌、感染症等の治療または予防に使用できる。
【0053】
本発明者らは、TLRと腸管免疫との関連を初めて見出した。また、TLRは、自然免疫を担当する細胞(例えばマクロファージや樹状細胞等)に発現していることが知られている(Gordon S. Cell. 111 (2002) 927-930、Akira S. et al, Nature Immunology. 2 (2001) 675-680)。よって、本発明のTLRトランスフェクタントは、腸管免疫担当細胞のモデル細胞として、例えば上記本発明の方法に使用できる。また、該TLRトランスフェクタントは、本発明の方法に用いる他の成分とともに、本発明の方法用のキットとして利用することもできる。
【0054】
本発明は、上記本発明の方法に利用できるTLRトランスフェクタント、該TLRトランスフェクタントを腸管免疫担当細胞のモデル細胞として使用する方法、該腸管免疫担当細胞のモデル細胞の製造方法、および該製造方法によって製造されるモデル細胞もまた提供するものである。
【実施例】
【0055】
以下、本発明を実施例により、さらに具体的に説明するが本発明はこれら実施例に制限されるものではない。
〔実施例1〕
【0056】
1.1 ブタ組織
フナコシ株式会社より購入した。
【0057】
1.2 ブタTLR9遺伝子のクローニングと塩基配列決定
DDBJ/EMBL/GenBankにおいて公開されているヒトおよびマウスTLR9(それぞれAccession Number AB045180およびAF348140)遺伝子配列をもとに保存性の高い領域を探索しプライマーを作製した。そのプライマーを用いてブタ腸管パイエル板由来total RNAからRT-PCR法によりブタTLR9遺伝子断片を得た。得られた遺伝子断片はpGEM-T-Easy vectorにライゲーションしE.coli JM109コンピテントセルにトランスフォーメーションしサブクローニングした。DNA配列はDNA sequencer Model 4000L(Li-Cor, Lincoln, NE, USA)により決定した。塩基配列、アミノ酸配列解析にはGENETYX-SV/RC Ver.11.0.3.1を用いた。残りのブタTLR9遺伝子配列はブタTLR9遺伝子断片から設計したプライマーを用いてRACE法により得た。全TLR9遺伝子をPCR法により増幅しクローニングした。
【0058】
1.3 ブタTLR9特異的ポリクローナル抗体の作製
GENETYX-SV/RC Ver.11.0.3.1を用いた抗原決定基(エピトープ)解析とタンパク2次構造解析よりブタTLR9アミノ酸配列中より268から284番目の領域が高い抗原性を持つことを見出した。その領域のペプチド合成および合成ペプチドを抗原としたポリクローナル抗体作製をサワディテクノロジー株式会社に依頼した。
【0059】
1.4 ブタTLR9遺伝子を導入したトランスフェクタントの構築
ブタTLR9遺伝子を導入する宿主細胞としては、ヒトの細胞系で遺伝子導入のホスト細胞として広く利用されているHEK293T細胞(ヒト胎児腎細胞)を選択した。ヒトTLR9特異的プライマーを用いてHEK293T細胞がTLR9遺伝子を発現していないことを解析し、シグナルペプチドを除くブタTLR9遺伝子をライゲーションしたpCXN2.1-FLAG 遺伝子発現ベクター (H. Niwa et al, Gene, 108 (1991) 193-199 )(大阪大学大学院医学研究科宮崎純一氏より分譲) をリポフェクション法によりHEK293T細胞にトランスフェクションした。抗生物質にはG418ネオマイシン(SIGMA)を用い、BECKMAN COULTER社のEPICSセルソーターシステムによりブタTLR9発現細胞をセレクションした。
【0060】
1.5 RT-PCR法によるトランスフェクタントにおけるブタTLR9発現解析
トランスフェクタントよりTRIzol(インビトロジェン)を用いてtotal RNAを抽出し、ブタTLR9特異的プライマー(本発明で設計)、ヒトTLR9特異的プライマーおよびポジティブコントロールとしてヒトGAPDHプライマー(K. A. Zarember, P. J. Godowski, J. Immunology, 168 (2002) 554-561) を用いてRT-PCR法によるTLR9 mRNA遺伝子発現解析を行った。
【0061】
1.6 フローサイトメトリー法および共焦点レーザー顕微鏡によるブタTLR9のトランスフェクタントにおける発現解析
細胞を1次抗体としてanti-FLAG mouse IgGモノクローナル抗体(SIGMA)で4℃、1時間処理後、2次抗体としてanti-mouse IgG-PerCP 標識抗体で4℃、30分間染色した。核染色はPropidium iodideを4℃、10分間処理で行った。解析にはFACSCaliburTM(日本BECTON DICKINSON株式会社)を用いた。ブタTLR9ポリクローナル抗体による免疫染色は、上記のように1次抗体染色後、2次抗体として、anti-rabbit IgG-Alexa 488標識抗体で4℃、30分間染色し、核染後解析した。また、細胞をI型コラーゲンコートDisk(IWAKI)に播き1次抗体としてanti-FLAG mouse IgGビオチン標識抗体(SIGMA)で4℃、1時間処理後、2次抗体としてストレプトアビジン-PE-Cy5抗体で4℃、30分間染色した。核染色にはPropidium iodide 4℃、10分間処理した。解析には共焦点レーザー顕微鏡(BIO-RAD社)を用いた。
【0062】
1.7 トランスフェクタントを用いたCpG DNA取り込みの解析
CpG DNAは、既に報告のあるヒト免疫細胞を強く刺激する大腸菌ゲノムDNA由来CpG2006型(5’-TCGTCGTTTTGTCGTTTTGTCGTT-3’(配列番号:9))とマウス免疫細胞を強く刺激するCpG1826型(5’-TCCATGACGTTCCTGACGTT-3’(配列番号:10)) (S. Pichyangkul et al, J. Immunological Methods, 247 (2001) 83-94) を用い、1Mの濃度のCpG DNAで、37℃にて1時間反応後、FACSCaliburTMおよび共焦点レーザー顕微鏡を用いて解析を行なった。
【0063】
1.8 ブタ各種組織におけるTLR9のreal-time PCR法による発現解析
ブタ各種組織(心臓、胸腺、肺臓、脾臓、肝臓、腎臓、骨格筋、十二指腸、空腸、回腸、回腸由来パイエル板、回腸由来腸管膜リンパ節)よりtotal RNAを抽出し、total RNA 1μgからoligo-d(T)18プライマーを用いてcDNAを合成後精製した。ブタTLR9特異的プライマーおよび精製cDNAからLightCycler (Roche)を用いて real-time quantitative PCRに供した。リアクションキットとしてLight Cycler-Fast Start DNA Master SYBR Green (Roche)を用いた。検量線から求めたブタTLR9遺伝子量とハウスキーピング遺伝子であるβアクチン遺伝子量との比から、ブタTLR9 mRNA量を算出した。結果は、脾臓中のTLR9発現量を1.000とした場合の各種組織に含まれるブタTLR9 mRNA量を数値化し比較した。
【0064】
本発明で用いたプライマーの塩基配列情報を表1にまとめた。
【表1】

【0065】
2. ブタTLR9遺伝子配列の決定と他種TLR9との相同性
本発明により決定したブタTLR9 cDNA塩基配列は3090塩基の構造遺伝子(ORF)を含む3145塩基(5’側に54塩基の非転写領域を含む)であった。ORFは1029残基のアミノ酸をコードしており、分子量は115.8kDaであった(図1および2)。多種TLR9に対するアライメントを図4および5に示した。ブタTLR9アミノ酸配列とヒト、マウス、ネコTLR9に対してそれぞれ82.0%、74.9%および86.6%の相同性が認められた(表2)。
【0066】
〔表2〕
−−−−−−−−−−−−−−−−−−−−−
TLR9 塩基配列a アミノ酸配列a
相同性(%) 相同性(%)
−−−−−−−−−−−−−−−−−−−−−
ヒト 84.9 82.0
マウス 78.2 74.9
ネコ 86.6 86.6
−−−−−−−−−−−−−−−−−−−−−
aヒト、マウスおよびネコTLR9の配列情報は、DDBJより以下のaccession No.(それぞれAB045180、AF348140およびAY137581)で入手した。
【0067】
3. ブタTLR9トランスフェクタントの発現解析
コントロール細胞とトランスフェクタント由来total RNAを鋳型としたブタTLR9 mRNAとヒトTLR9 mRNAのRT-PCR発現解析によりトランスフェクタントにおけるブタTLR9 mRNAの強い発現が認められた。また、両者ともにヒトTLR9 mRNAの発現は認められなかった(図6)。
フローサイトメトリーによる発現解析から、1次抗体として抗-FLAG抗体を用いた場合、コントロール細胞と比較してポジティブ側への大きなシフトがみられた(図7-a)。またブタTLR9抗体を用いた場合もポジティブ側へのシフトがみられた(図7-b)。レーザー顕微鏡による解析からも同様に発現が確認された(図8‐a、c、b、d)。
【0068】
4. CpG DNA取り込み解析
共焦点レーザー顕微鏡による解析ではCpG DNAの違いによる取り込みの顕著な差は判断できなかったが、フローサイトメトリーによるCpG DNAの取り込み解析によりブタTLR9はマウス型CpG 1826よりもヒト型CpG2006を比較的多く取り込んでいることが明らかとなった(図7‐c、8‐e、f)。
【0069】
5. 各種組織におけるブタTLR9 mRNAの発現解析
real-time PCR法による各種組織におけるブタTLR9 mRNAの発現解析の結果、ブタTLR9は、腸管リンパ系組織において、特にパイエル板と腸管膜リンパ節において強く発現していることが明らかとなった(図9)。
本実施例によりブタTLR9はマウスに比べヒトやネコTLR9と高い相同性を示すことが明らかとなった。また、real-time PCRによる発現解析から、これまでTLR9は脾臓において強い発現が認められるという報告がされていたが、経口的に病原細菌に曝される可能性が最も高い腸管粘膜系の内、腸管免疫の中心的な役割を果たすパイエル板や腸管膜リンパ節において、脾臓の約3倍以上のmRNA発現が認められた点は大変興味深い新知見である。また、本実施例はブタTLR9トランスフェクタントを構築することに成功し、本トランスフェクタントを用いたCpG DNAの取り込み解析から、ブタTLR9がマウス型に比べてヒト型CpG DNAに対する反応性が高かったことから、今後、このトランスフェクタントを用いた機能性乳酸菌DNAの認識性の解析が飛躍的に進み、ブタを実験動物としたヒトへのモデル系の計画的解析を通して、機能性食品開発の基礎研究が分子間の反応としてより詳細に検討できる。
【0070】
TLR9により認識される細菌DNAはマクロファージや樹状細胞等を刺激し、サイトカイン等のシグナル分子の産生を促すことが知られている(M. Bauer et al , J. Immunol. 166 (2001) 5000-5007)。本実施例の成果は細菌DNAによる刺激とTLR9による認識およびシグナル伝達システムを応用したワクチンの開発につながるものであり(R. L. Modlin, Nature 408 (2000) 659-660)、感染を抑制するだけの低い免疫応答から、細菌を殺す有効性の高い応答へと切り換えることができるため、結核等の感染症の治療、さらには癌、アレルギー等への幅広い応用が期待できる。最近、腸管まで届く生きた抗原運搬役として乳酸菌が注目を浴びているが、それに加えて乳酸菌由来DNAもまた免疫賦活化能があることが本発明者らの研究で明らかになったことから乳酸菌に対する期待が大いに高まっている。本発明でTLR9がパイエル板および腸管膜リンパ節で強い発現が認められたことから、TLR9が腸管免疫系において重要な役割を担っていることが強く示唆され、腸管に存在する乳酸菌や食餌性の乳業用乳酸菌のDNAがTLR9を刺激し、免疫を活性化する可能性が考えられる。それゆえ、乳酸菌DNAのTLR9を介した腸管免疫系を明らかにすることは、腸管常在乳酸菌や乳業用乳酸菌を用いたDNAワクチンの開発に向けた解決すべき課題である。
【0071】
また、パイエル板および腸管膜リンパ節におけるTLR9の強い発現は、これら腸管免疫における自然免疫系の顕著な発達を意味しているかもしれない。近年、医学、免疫学の分野において腸管免疫系が極めて重要な領域として注目を集めるようになってきた。比較的新しい研究分野であるため基本的な免疫機構についての知見が少なく、その機構解明が大いに期待されている。
本発明で得られた知見は、今後腸管におけるTLR9を介したシグナル伝達経路についてその認識メカニズムを分子レベルで明らかするための糸口となり、自然免疫という基本的な免疫システムがより発展的に解明されるための原動力ともなる。
【0072】
〔実施例2〕
1.1 細胞株
チャイニーズハムスター卵巣(CHO)K-1細胞は、TKG細胞バンク(東北大学、加齢医学研究所)から得て、10%仔ウシ胎児血清、50 mg/mlペニシリン/ストレプトマイシン、2 mM L-グルタミン、10 mM HEPES(2-[4-(2-ヒドロキシエチル)-1-ピペラジニル]エタンスルホン酸)緩衝液、0.11 mg/mlピルビン酸ナトリウム、および0.5 mM 2-メルカプトエタノール(シグマ(Sigma)、東京、日本)を添加した完全なダルベッコ改変イーグル培地において維持した。
【0073】
1.2 乳酸菌(LAB)調製物
2種の乳酸桿菌、デルブリュック菌亜種ブルガリクス(L. delbrueckii ssp.bulgaricus)とL.ガセリ(L. gasseri)を用いた。L.ブルガリクスNIAI B6は、国立動物産業研究所(つくば市、日本)から得た。L.ブルガリクス JCM 1002TおよびL.ガセリJCM 1131Tは、日本微生物コレクション(JCM、埼玉県、日本)から購入した。L. ガセリOLL2716は、明治乳業(小田原市、神奈川県、日本)の食品機能性研究所から供与された。株は全てMRSブロス(ディフコ(Difco)、デトロイト、アメリカ)において37℃で16時間培養して、凍結乾燥し、NF-κBレポーターアッセイのために用いた。
【0074】
1.3 プラスミドの構築
sTLR2のコード領域を、ブタパイエル板から単離した総RNAからRT-PCRによって増幅した。N-末端FLAGタグ-sTLR2発現ベクターは、ブタTLR2のコード領域を、単離されたsTLR2 cDNAからPCRによって増幅して、N-末端FLAGエピトープをコードするpCXN2.1-FLAG発現ベクターのNotI-EcoRI部位にクローニングした(Niwa, H., et al. (1991) Gene 108, 193-199.)。構築物の構造は、制限酵素マッピングおよびDNA配列分析によって確認した。
【0075】
1.4 安定なトランスフェクタントの単離
トランスフェクションの前日に、CHO K-1細胞4×105個/ウェルを6ウェルプレートに播種した。培養1日後、リポフェクタミンTM試薬(インビトロジェン(Invitrogen Corp.)、カールスバッド、カリフォルニア州、アメリカ)を用いて、sTLR2をコードするpCXN2.1-FLAG発現ベクター(1μg/ウェル)を細胞にトランスフェクトした。sTLR2発現トランスフェクタントをG418(インビトロジェン)(1.0 mg/ml)によって10日間陽性選択して、これをCHO K-1sTLR2 transと命名した。プラスミドベクターのみをトランスフェクトしたCHO K-1細胞は、CHO K-1Cont.と命名した。sTLR2の発現は、sTLR2に結合させたFLAGタンパク質の細胞外染色によって確認した(1.7 タンパク質発現分析参照)。
【0076】
1.5 RT-PCR分析
cDNAを総RNAから合成して、トランスフェクタントおよび非トランスフェクタントにおいてTRIZOL試薬(ライフテクノロジーズ(Life Technologies)、ロックビル、メリーランド州)によって調製して、rTaqポリメラーゼ(タカラバイオインク(TaKaRa BIO Inc)、大津、日本)を用いて増幅した。外因性のsTLR2を検出するために、以下のプライマーを用いてDNAを増幅した。
センス:5'-TTC AGG CCA AGG ATT TCC AG-3'(配列番号:37)
アンチセンス:5'-TCA CTG TGC TGG GTT CAT TG-3'(配列番号:38)
【0077】
βアクチンプライマーの実際の配列は、以下のようにチャイニーズハムスターのβ-アクチン(GenBank アクセッション番号U20114)のDNA配列に基づいて、以下のようにデザインして、ハウスキーピング遺伝子対照として用いた。
センス:5'-TCT ACA ACG SGC TGC GTG TG-3'(配列番号39)
アンチセンス:5'-GCT TCT CTT TGA TGT CAC GC-3'(配列番号:40)
PCRサイクル条件は、95℃で5分の後95℃で30秒、55℃で30秒、および72℃で30秒を30サイクルであった。PCR産物は、1.5%アガロースゲル上で、エチジウムブロマイドによって可視化して分析した。
【0078】
1.6 sTLR2に対するポリクローナル抗体(Ab)の作製
sTLR2タンパク質は、二次構造、疎水性、および抗原性を予測するためにジェネティクス-SV/RC(11.0.3.1)を用いて分析した後、細胞外ドメインにおけるアミノ酸76〜90個を含む領域(CVNLRALRLGANSIH)(配列番号:41)をペプチド合成のために選択した。免疫抗原としての合成ポリペプチドを、フロイントの完全アジュバントと共に1:1の割合で乳化した。抗sTLR2ポリクローナルAbは、0.3 mg/ウサギの用量で皮下注射した合成ポリペプチドによってウサギ(日本白色種)を免疫することによって産生した。1ヶ月間隔で追加免疫を2回行った後、ウサギから採血して抗血清を回収した後、エピトープのアフィニティクロマトグラフィーによる精製を行った。最終的に、直接的な酵素結合イムノソルベントアッセイ(データは示していない)によって、sTLR2の高い抗体価が得られた。
【0079】
1.7 タンパク質発現分析
細胞を洗浄して、固定し、透過性にして、N-末端FLAG-sTLR2タンパク質に関して抗FLAG M5マウスモノクローナルAb(シグマ)および第一抗体としてウサギ抗sTLR2ポリクローナルAbを用いて染色した。次に、細胞をそれぞれ、フィコエリスリン(PE)またはアレクサ660結合抗マウスIgGおよびアレクサ488結合抗ウサギIgGによって処置した。PEおよびアレクサ488シグナルは、フローサイトメトリー(FACSキャリバーTM、ベクトンディッキンソン(BECTON DICKINSON)、東京)によって決定した。アレクサ660およびアレクサ488シグナルは、共焦点レーザー顕微鏡(バイオラド・ラボラトリーズインク(Bio-Rad Laboratories Inc.)、ハーキュリーズ、カリフォルニア州)によって決定した。
【0080】
1.8 走査型電子顕微鏡分析
10 μg/mlザイモサン(モレキュラープローブス(Molecular Probes)、ユージーン、オレゴン州)によって刺激して一晩培養した細胞を、2%グルタルアルデヒドを含む0.2 M燐酸緩衝生理食塩液(pH 7.4)において30分間固定した。段階的な濃度のエタノール(50%、60%、70%、80%、90%および99%)を用いて細胞を脱水した後、無水エタノールによって2回洗浄した。試料を臨界点乾燥させて、金-パラジウムによってコーティングしてからS-4200走査型電子顕微鏡(ヒタチ(HITACHI)、東京、日本)によって観察した。
【0081】
1.9 ザイモサンの取り込み試験
細胞を、最終濃度20 μg/mlのFITC-結合ザイモサンによって37℃で1時間処置した。細胞を固定した後、ザイモサンの取り込みをフローサイトメトリーおよび共焦点レーザー顕微鏡によって検出した。
【0082】
1.10 NF-κBレポーターアッセイ
細胞(24ウェルプレートにおいて8×104個/ウェル)を、リポフェクタミンTM試薬と共にヒトNF-κB遺伝子A1およびA2部位を含むpGLM-ENH-luciベクター(Ueda, A., et al. (1997) J. Biol. Chem. 272, 31092-31099.; Inadera, H., et al. (2000) Endocrinology 141, 50-59.)0.5 μgを用いて18時間トランスフェクトさせた。様々なリガンドによる24時間の刺激の後、製造元(プロメガ(Promega)、東京、日本)が推奨する条件を用いてルシフェラーゼアッセイを行った。刺激指数(S.I.)は、以下の等式によって推定した:[(CHO K-1sTLR2trans.細胞における共鳴単位(RU)−(バックグラウンドのRU)]/[(CHO K-1Cont.細胞における共鳴単位(RU)−(バックグラウンドのRU)]。アッセイは全て、異なる3つの培養ウェルを用いて少なくとも3回の異なる実験によって行った。
【0083】
2. ブタTLR2遺伝子配列の決定
腸関連リンパ様組織に属する回腸パイエル板から単離したブタmRNAを用いて、Horton R. H.らにより報告されているオーバーラップPCR伸長法によってsTLR2の完全長のcDNAを作製した(Horton, R.H., et al. (1989) Gene 77, 61-68.)。sTLR2の完全長のcDNAは、アミノ酸785個をコードする2358 bpであり、計算分子量は89.6 kDaであった。得られたcDNAは、Munetaら(Muneta, Y., et al. (2003) J. Interferon Cytokine Res. 23, 583-589.)によって最近報告された、リポ多糖類によって刺激したブタ肺胞マクロファージから得られたcDNAと同一であった。ブタTLR2のヌクレオチド配列は、アクセッション番号AB072190としてDDBJ、EMBL、およびGenBankヌクレオチドデータベースに提出されている。
【0084】
3. ブタTLR2発現トランスフェクタントの発現および機能的分析
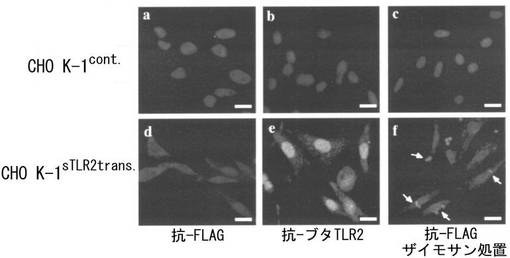
CHO K-1細胞は、チャイニーズハムスターの機能的TLR2を発現しないことから、sTLR2をコードするpCXN-FLAGベクターによるリポフェクションによってCHO K-1sTLR2trans細胞を確立した(Heine, H., et al. (1999) J. Immunol. 162, 6971-6975.)。RT-PCR分析で、sTLR2のmRNAがCHO K-1sTLR2trans細胞において発現されたことから、外因性のsTLR2 mRNAがCHO K-1sTLR2trans細胞において発現されたことが示された(図10)。プラスミドベクターのみをトランスフェクトしたCHO K-1Cont.細胞とCHO K-1sTLR2trans細胞とをマウス抗FLAG mAbおよび抗-sTLR2ポリクローナル抗体によって染色すると、FLAGおよびsTLR2陽性細胞は、フローサイトメトリー(図11-a、b)および共焦点レーザー顕微鏡(図12-a、b、d、e)によってCHO K-1sTLR2trans細胞でのみ検出された。さらに、走査型電子顕微鏡写真から、TLR2-特異的リガンドとして同定されているザイモサンによって細胞を処置した場合、CHO K-1sTLR2trans細胞の表面は劇的に粗くなったが、CHO K-1Cont.細胞では粗くならなかったことが示された(図13)。これらの結果により、sTLR2がCHO K-1sTLR2trans細胞において確実に発現していることが確認された。CHO K-1Cont.細胞およびCHO K-1sTLR2trans細胞をザイモサンによって1時間処置すると、ザイモサンを取り込んだ細胞は、CHO K-1sTLR2trans細胞では強く検出されたが、CHO K-1Cont.細胞では検出されなかった(図11-c)。さらに、FITC標識ザイモサン(緑色)およびFLAG-sTLR2(青色)が同時に局在することは、CHO K-1sTLR2trans細胞でのみ示された(図12-c、f)。これらは、微生物の細胞壁成分によって誘導されたシグナル伝達が、sTLR2を通して核に達し、トランスフェクタントの生物機能の活性化が起こった可能性を示している。
【0085】
4. ブタTLR2トランスフェクタントのLAB株におけるNF-κB依存的ルシフェラーゼ活性
Underhillらにより、TLR2が、5分以内にザイモサンを含むマクロファージファゴソームに特異的に補充されることが示されている(Underhill, D., et al. (1999) Nature 401, 811-815.)。粒子のインターナリゼーションの全段階において、TLR2は液胞の内容物を認識し、特定の生物に対する防御に対して適当な炎症反応を誘発した。したがって、CHO K-1sTLR2trans細胞におけるTLR2による活性化を評価するために、細胞にNF-κB依存的ルシフェラーゼレポーター(pGLM-ENH-luci)ベクターを一過性に同時トランスフェクトした。ザイモサンおよびPGNによって刺激したCHO K-1sTLR2trans細胞は、NF-κB依存的ルシフェラーゼ活性を有意に増加させたが、sTLR9のリガンドDNAであるD25には反応しなかった(図14)。これらの知見は、CHO K-1sTLR2trans細胞が微生物の細胞壁成分を選択的に認識して、それによってNF-κB活性化が誘導されうることを示唆する。
【0086】
次に、本発明者らは、CHO K-1sTLR2trans細胞がNF-κB依存的ルシフェラーゼ活性を発揮した後にintactなLABと共に細胞活性成分に反応するか否かを確認しようと試みた。CHO K-1sTLR2trans細胞をL.ブルガリクス(NIAI B6、LCM1002T)およびL.ガセリ(JCM1131T、OLL2716)の四つの株によって刺激すると、TLR2を通しての細胞活性化はCHO K-1sTLR2trans細胞において検出され、この細胞はPGNおよびリポテイコ酸のような細胞壁成分を含むLABの細胞表面を認識する可能性がある。さらに、intactなLABの刺激は、個々の細胞壁成分と比較してCHO K-1sTLR2trans細胞活性化を有意に増加させることから、CHO K-1sTLR2trans細胞の活性化が、intactなLABを構成する様々な細胞壁成分によって減弱したことを意味している。興味深いことに、ルシフェラーゼ活性の差は、類似の遺伝的背景にもかかわらず、それぞれのLABの二つの個々の株においても示された。本発明者らのこれまでの研究および他の報告から、細胞表面構成成分における構造的、化学的な違い、およびコンフォメーションの違いが、遺伝的に近縁のLAB株においても認められたことが示されている(Schar-Zammaretti, P. and Ubbink, J. (2003) Biophys. J. 85, 4076-4092.; Takeda, K., et al. (1997) Milchwissenshaft 52, 21-25.)。この証拠は、LABの細胞壁によって誘導される様々な免疫調節作用が、細胞壁特性の差に反映される可能性があるという考え方を支持する。このように、本発明者らは、同じ種の異なる株における活性のわずかな差も区別できるCHO K-1sTLR2trans細胞の構築に成功した。
【0087】
腸の菌叢は、免疫コンピテント細胞の産生およびTヘルパー1(TH1)とTヘルパー2(TH2)免疫のあいだのバランスの調節に密接に関与している(Neurath, M.F., et al. (2002) Nat. Med. 8, 567-573.)。ほとんどのアレルギー疾患は、アレルゲン性の分子に対する免疫応答がTH2型に向けて明らかに偏った、リンパ球が支配する免疫における不均衡によって反映される。in vitroおよびin vivoにおいて、細菌のリポ多糖類およびCpGオリゴヌクレオチドは、免疫学的環境において樹状細胞による炎症性サイトカインの産生を通してTH1型T細胞発達を増強する(Trinchieri, G. (2003) Nat. Rev. Immunol. 3, 133-46.)。これまで、本発明者らは、マウスB細胞における変異原性を誘導するLABからの免疫刺激オリゴヌクレオチドの発見に成功した(Kitazawa, H., et al. (2001) Int. J. Food Microbiol. 65, 149-162.; Kitazawa, H., et al. (2003) Int. J. Food Microbiol. 85, 11-21.)。本発明者らの最近の研究において、トランスフェクタント実験により、粘膜免疫において重要な役割を果たす可能性があるTLR9が、LABからのCpGのみならずATモチーフを認識することが示された(Shimosato, T., et al. (2003) Biochim. Biophys. Acta 1627, 56-61.; Shimosato, T., et al. (2004) Animal Science Journal 75, In press.)。LABからのオリゴヌクレオチドの他に、細胞壁は、LABによるインターフェロン誘導能を決定するための重要な要因である(Solis-Pereya, B., et al. (1997) Am. J. Clin. Nutr. 66, 521S-525S.)。本実施例において示した結果は、intactな共生LABと共に細胞壁成分がTLR2を通して免疫応答の誘導能を有することを示し、活性は、CHO K-1sTLR2trans細胞によって十分に検出可能となりうることを示している。本発明者らの研究において、ブタがヒトモデルとなると予想され、場合によってはin vivo実験をヒトの代わりにブタにおいて行うことができることから、sTLR2を用いたアッセイ系を開発した。このように、本研究により、sTLR2発現トランスフェクタントを利用することで、LABのような特定のイムノバイオティック微生物を評価するための主な免疫アッセイ系を提供できることが示された。これはTH1/TH2極性の維持のような免疫調節能を有する新規機能性食品の生産に貢献する可能性がある。
【技術分野】
【0001】
本発明は、Toll様レセプター強制発現細胞の利用法に関する。
【背景技術】
【0002】
生体の免疫システムには先天的な自然免疫と後天的に生じる獲得免疫の2種類がある。ヒト等の高等生物にしか存在しない獲得免疫に対して、自然免疫は昆虫からヒトまで広く保存された免疫機構である。自然免疫を担当する細胞(マクロファージや樹状細胞等)は外界からの病原細菌等が侵入すると、すばやくこれを探知し、食作用等の直接的な攻撃に加え、サイトカイン等の警戒シグナルを発して獲得免疫系を活性化するといった感染防御の第一段階として機能している。この様な一連の免疫反応システムにおいて最初のきっかけとなる細菌を発見する役目を果たしている受容体タンパクがToll様レセプター(Toll-like receptor;TLR)である。TLRは現在までにヒトにおいて10種類が同定されており、それぞれのTLRは認識する分子構造が異なるものと考えられている(非特許文献1)。その中で2000年に審良らの研究グループによって報告されたTLR9は細菌のDNA、特にCpGモチーフを認識するレセプターとして同定された (非特許文献2、特許文献1)。また、TLR2は、ペプチドグリカン(PGN)、リポテイコ酸、微生物のリポタンパク質、およびザイモサンのような微生物の細胞壁成分に反応した細胞シグナル伝達を媒介することが認識されている(非特許文献3)。
【0003】
すでに、リポペプチドがヒトTLR2を強制発現させたCHO細胞におけるNF-κBの発現活性に与える影響について、また、病原性大腸菌由来CpG DNAの様々なモチーフがヒトTLR9を強制発現させたHEK293細胞におけるサイトカイン(IL-8)の産生量に与える影響について報告されている(非特許文献4〜6)。
【0004】
尚、本出願の発明に関連する先行技術文献情報を以下に示す。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2002-34565
【非特許文献】
【0006】
【非特許文献1】O. Takeuchi, S. Akira, International Immunopharmacology 1 (2001) 625-635
【非特許文献2】H. Hemmi et al, Nature 408 (2000) 740-745
【非特許文献3】Wetzler, L.M. (2003) The role of Toll-like receptor 2 in microbial disease and immunity. Vaccine 21, S55-S60.
【非特許文献4】Yoshimura A. Takada H. Kaneko T. Kato I. Golenbock D. Hara Y. Structural requirements of muramylpeptides for induction of Toll-like receptor 2-mediated NF-kappaB activation in CHO cells. Journal of Endotoxin Research. 6(5):407-10, 2000.
【非特許文献5】Klinman DM. Takeshita F. Gursel I. Leifer C. Ishii KJ. Verthelyi D. Gursel M. CpG DNA: recognition by and activation of monocytes. Microbes & Infection. 4(9):897-901, 2002 Jul.
【非特許文献6】Takeshita F. Leifer CA. Gursel I. Ishii KJ. Takeshita S. Gursel M. Klinman DM. Cutting edge: Role of Toll-like receptor 9 in CpG DNA-induced activation of human cells. Journal of Immunology. 167(7):3555-8, 2001 Oct 1.
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明者らはこれまでにプロバイオティック乳酸菌を含む乳業用乳酸菌由来のDNAモチーフが腸管免疫系において免疫賦活化機能を有することを明らかにしてきた(H. Kitazawa et al, Int. J. Food Microbiol. 65 (2001) 149-162、H. Kitazawa et al, Int. J. Food Microbiol, (2003) in press)。本発明者らは、この知見からTLR9が病原細菌由来DNAのみならず、食餌性乳酸菌由来のDNAに対しても認識性を持ち、免疫の活性化に寄与する可能性を考えた。多様なDNAモチーフに対する認識性および活性評価系の構築が可能となれば、今後、病原性細菌の負の評価のためのみならず、乳業用乳酸菌を用いた機能性食品開発の上で重要なツールとなる。
【0008】
また、最近、臨床および実験的試験において、乳酸菌は代表的なイムノバイオティクスとして用いられており、その細胞壁成分は、以下の手段によって宿主に対して有用な作用を付与することが報告されている:分裂促進活性(Takeda, K., et al. (1997) Milchwissenshaft 52, 21-25.; Kirjavainen, P.V., et al. (1999) FEMS Immunol. Med. Microbiol. 26, 131-135.)、抗癌活性(Bogdanov, I.G., et al. (1975) FEBS Lett. 57, 259-261.)、抗変異原活性(Bodana, A.R. and Rao, D.R. (1990) J. Dairy Sci. 73, 3379-3384.)、抗感染作用(Perdigon, G., et al. (1986) Infect. Immun. 53, 404-410.; Popova, P., et al. (1993) Int. J. Immunopharmacol. 15, 25-37.)、およびサイトカイン産生の誘導(Popova, P., et al. (1993) Int. J. Immunopharmacol. 15, 25-37.; Pereyra, B.S., et al. (1991) Eur. Cytokine Netw. 2, 299-303.; De Simone, C., et al. (1993) Int. J. Immunother. 9, 23-28.)。
【0009】
機能性食品の開発を考える場合、最終的にヒトに対する効果を評価する必要があるが、その基礎的知見を得るためには、動物細胞および実験動物を用いた検討が必要不可欠となる。そこで本発明では、実験対象動物として、臓器移植等の観点からヒトモデル系としてその利用性が期待され、また食品産業の面から大きな価値を持つブタに着目し、ブタのTLR遺伝子のクローニングと、その遺伝子を導入し強制発現させた動物細胞を作製し、機能性DNAや機能性微生物等の評価系のためのTLR強制発現細胞を構築することを考えた。
本発明は、このような状況に鑑みてなされたものであり、その目的は、TLR強制発現細胞の利用法を提供することを目的とする。
【課題を解決するための手段】
【0010】
本発明者らは、病原細菌由来のCpG DNAモチーフを認識する受容体タンパクToll様レセプター9遺伝子をブタ腸管パイエル板よりクローニングし、ブタTLR9(sTLR9)を強制発現させた動物細胞(トランスフェクタント)を作製した。動物細胞におけるsTLR9タンパクの発現の有無は、sTLR9に対するポリクローナル抗体を作成し、同抗体を用いて確認した。さらに、sTLR9トランスフェクタントのCpG DNAに対する機能性を解析し、乳酸菌DNAの活性評価系への応用を目指した。
【0011】
具体的には、下記(1)〜(5)を行った。
(1)ブタ腸管パイエル板よりTotal RNAを抽出し、ヒトおよびマウスTLR9遺伝子における保存性の高い領域より作製したプライマーを用いてRT-PCR法、RACE法によりブタTLR9遺伝子をクローニングし、その全遺伝子配列を決定した。
(2)遺伝子情報から得られたブタTLR9全アミノ酸配列から抗原決定基部位を探索し、選抜した領域をペプチド合成しブタTLR9ポリクローナル抗体作製用抗原とした。抗原は化学合成後、定法によりウサギに免疫し、ブタTLR9に対するポリクローナル抗体を作成した。
(3)ブタTLR9遺伝子をHEK293T細胞(ヒト胎児腎細胞)に導入し、ブタTLR9遺伝子導入細胞(トランスフェクタント)を作製した。
(4)ブタTLR9のHEK293Tにおける発現については、ブタTLR9 mRNAの発現をRT-PCR法により、また、ブタTLR9膜タンパクの発現をブタTLR9ポリクローナル抗体による免疫染色法によりレーザー顕微鏡およびフローサイトメーターを用いて確認した。
(5)ヒトおよびマウス細胞を強く刺激する特異的CpG DNAモチーフを含むオリゴDNA(それぞれCpG2006,CpG1826)に対する反応性を解析した。
【0012】
検討の結果、ブタTLR9遺伝子は1029残基(MW:115.8)のアミノ酸をコードする3090塩基からなり、その遺伝子を含め3145塩基のcDNA塩基配列を決定することができた。ブタTLR9のアミノ酸配列はヒトTLR9に対して82.9%と非常に高く、マウスTLR9には74.9%の相同性を示したことから、ヒトTLR9に対してマウスよりも比較的高い相同性を示すことが明らかとなった。ブタTLR9トランスフェクタントにおけるブタTLR9タンパク質はRT-PCR法およびブタTLR9ポリクローナル抗体の免疫染色から確実に発現し、膜タンパク質として発現したことを確認できた。このことは、ブタTLR9トランスフェクタントの作製に成功したことを意味する。本トランスフェクタントを用いたCpG DNAに対する機能性解析よりブタTLR9はCpG1826よりもCpG2006に対して強い反応性を示したことから、ブタTLR9はマウス特異的CpG DNAモチーフよりも比較的ヒトのモチーフに対する認識性が高いことが判明した。Real-time PCR法によりmRNAの発現量を各種組織において比較した結果、驚くべきことに、腸管免疫系で中心的な役割を果たすパイエル板および腸管膜リンパ節において、脾臓の3倍以上のmRNAが発現していることが判明した。
【0013】
また、本発明者らは回腸のパイエル板からブタTLR2(sTLR2)cDNAを単離し、その後sTLR2発現トランスフェクタントを構築することによって、イムノバイオティック微生物の免疫応答に関する評価法の開発を行った。
【0014】
即ち、本発明は、以下の〔1〕〜〔21〕を提供するものである。
〔1〕 被験試料が腸管免疫系を活性化するか否かを評価する方法であって、
(a)腸管組織において発現しているToll様レセプターを強制発現させた細胞に、被験試料を接触させる工程
(b)該細胞におけるシグナル伝達を指標に、該Toll様レセプターの活性を測定する工程
を含み、上記Toll様レセプターの活性が、上記被験試料を接触させないときに比べ上昇する場合に、被験試料が腸管免疫系を活性化すると判定される方法。
〔2〕 以下の(a)および(b)の工程を含む、腸管免疫系を活性化する試料のスクリーニング方法。
(a)〔1〕に記載の評価方法により、複数の被験試料について、腸管免疫系を活性化するか否かを評価する工程
(b)複数の被験試料から、腸管免疫系を活性化すると評価された試料を選択する工程
〔3〕 〔2〕に記載の工程に、さらに腸管免疫系を活性化すると評価された試料と医薬上許容される担体とを混合する工程を含む、腸管免疫系を活性化する医薬組成物の製造方法。
〔4〕 被験微生物が腸管免疫系を活性化する微生物であるか否かを評価する方法であって、
(a)被験微生物から抽出物を調製する工程
(b)腸管組織において発現しているToll様レセプターを強制発現させた細胞に、該抽出物を接触させる工程
(c)該細胞におけるシグナル伝達を指標に、該Toll様レセプターの活性を測定する工程
を含み、上記Toll様レセプターの活性が、上記抽出物を接触させないときに比べ上昇する場合に、被験微生物が腸管免疫系を活性化する微生物であると判定される方法。
〔5〕 以下の(a)および(b)の工程を含む、腸管免疫系を活性化する微生物のスクリーニング方法。
(a)〔4〕に記載の評価方法により、複数の被験微生物について、腸管免疫系を活性化する微生物であるか否かを評価する工程
(b)複数の被験微生物から、腸管免疫系を活性化する微生物であると評価された微生物を選択する工程
〔6〕 〔5〕に記載の工程に、さらに腸管免疫系を活性化する微生物であると評価された微生物と食品上許容される担体とを混合する工程を含む、腸管免疫系を活性化する食品組成物の製造方法。
〔7〕 微生物が乳酸菌であり、食品組成物が乳製品である、〔6〕に記載の方法。
〔8〕 微生物が細菌である、〔4〕〜〔6〕のいずれかに記載の方法。
〔9〕 細菌が乳酸菌である、〔8〕に記載の方法。
〔10〕 腸管組織において発現しているToll様レセプターをコードするDNAを有する発現ベクターを細胞に導入する工程を含む、腸管免疫担当細胞のモデル細胞の製造方法。
〔11〕 腸管組織において発現しているToll様レセプターを強制発現させた細胞を、腸管免疫担当細胞のモデル細胞として使用する方法。
〔12〕 腸管組織が腸管リンパ系組織である、〔1〕〜〔11〕のいずれかに記載の方法。
〔13〕 腸管リンパ系組織がパイエル板または腸管リンパ節である、〔12〕に記載の方法。
〔14〕 Toll様レセプターがブタ由来である、〔1〕〜〔13〕のいずれかに記載の方法。
〔15〕 Toll様レセプターがToll様レセプター9および/または2である、〔1〕〜〔13〕のいずれかに記載の方法。
〔16〕 〔1〕〜〔9〕のいずれかに記載の方法に用いるための、腸管組織において発現しているToll様レセプターを強制発現させた細胞。
〔17〕 腸管組織において発現しているToll様レセプターをコードするDNAを有する発現ベクターを細胞に導入することで製造される、腸管免疫担当細胞のモデル細胞。
〔18〕 腸管組織が腸管リンパ系組織である、〔16〕または〔17〕に記載の細胞。
〔19〕 腸管リンパ系組織がパイエル板または腸管リンパ節である、〔18〕に記載の細胞。
〔20〕 Toll様レセプターがブタ由来である、〔16〕〜〔19〕のいずれかに記載の細胞。
〔21〕 Toll様レセプターがToll様レセプター9および/または2である、〔16〕〜〔19〕のいずれかに記載の細胞。
【発明の効果】
【0015】
本発明においてTLRトランスフェクタントの利用法が提供された。TLRトランスフェクタントは、腸管免疫担当細胞のモデル細胞として利用できる。また、TLRトランスフェクタントを利用することで、腸管免疫系を活性化する試料および微生物を同定でき、腸管免疫系を活性化する医薬組成物および食品組成物を製造できる。該試料、微生物、医薬組成物、および食品組成物は、例えば、アレルギー、癌、感染症等の治療または予防に使用できる。
【0016】
また、本発明により、同じ菌種の異なる株の差をも区別できるTLRトランスフェクタントが提供された。TLRによって免疫調節作用を発揮するイムノバイオティック微生物を用いて機能的食品を開発する場合、TLR-発現トランスフェクタントは、イムノバイオティック微生物をも評価することができる免疫スクリーニング系の貴重なツールとして、TLR−イムノバイオティック微生物間の関係の評価に用いることができる。
【図面の簡単な説明】
【0017】
【図1】ブタTLR9 cDNA塩基配列を示す図である。下線はシグナルペプチド領域を示す。
【図2】ブタTLR9 cDNA塩基配列を示す図である。図1の続きを示す。下線は膜貫通領域を示す。
【図3】ブタTLR9のSMARTによるドメイン解析を示す図である。
【図4】TLR9アミノ酸配列のアライメントを示す図である。
【図5】TLR9アミノ酸配列のアライメントを示す図である。図4の続きを示す。
【図6】トランスフェクタントにおけるブタTLR9のRT-PCR法による発現解析を示す写真である。
【図7】トランスフェクタントにおけるブタTLR9の発現とCpG DNAの取り込み解析を示す図である。a;抗FLAG抗体による解析を示す。b;抗ブタTLR9抗体による解析を示す。c;CpG DNAの取り込み解析を示す。図中の矢印(1)はコントロール細胞のCpG1826および2006の取り込みを示し、矢印(2)はトランスフェクタントにおけるCpG1826を示し、矢印(3)はトランスフェクタントにおけるCpG2006を示す。
【図8】トランスフェクタントの共焦点レーザー顕微鏡による解析を示す写真である。a,b,c;対照細胞、d,e,f;ブタTLR9トランスフェクタントを示す。a,d;抗FLAG抗体による解析を示す。b,e;抗ブタTLR9抗体による解析を示す。c,f;CpG DNAの取り込み解析を示す。
【図9】real-time定量PCRによるブタTLR9の各種組織における発現解析を示す図である。
【図10】CHO K-1sTLR2trans細胞上でのsTLR2 mRNA発現をRT-PCRによって分析した結果を示す写真である。逆転写された総RNA 1 mgからのPCR(30サイクル)増幅産物をアガロースゲル上で検出した。
【図11】(a,b)CHO K-1sTLR2trans細胞上でのsTLR2タンパク質の発現をフローサイトメトリーにより検出した結果を示す図である。(a)抗FLAGモノクローナル抗体(PE)および(b)抗sTLR2ポリクローナル抗体(アレクサ488)によって検出した。線(2)および線(4)は、CHO K-1sTLR2trans細胞を表す。線(1),(3)は、ベクター対照細胞を表す。(c)CHO K-1sTLR2trans細胞によるFITC-標識ザイモサン(6)の取り込みを、フローサイトメトリーによって観察した結果を示す図である((5):ベクター対照細胞によるザイモサンの取り込み)。
【図12】sTLR2の細胞局在を共焦点レーザー顕微鏡によって観察した写真である。CHO K-1sTLR2trans細胞(d、e、f)またはCHO K-1Cont.細胞(a、b、c)を抗FLAG M5 モノクローナル抗体によって染色した後、アレクサ660結合抗マウスIgG(青色;d、f)または抗sTLR2ポリクローナルAbによって染色した後、アレクサ488結合抗ウサギIgG(緑色;e)によって染色した。細胞をFITC標識ザイモサンによって37℃で1時間処置した(c、f)。核はヨウ化プロピジウム(PI)(赤またはオレンジ色)によって染色した。矢印は、同時局在を示すより大きいザイモサン小胞を示す(f)。6回の独立した実験の一つからの代表的な切片を示す(尺度のバー;10 μm)。
【図13】プラスミドベクターのみをトランスフェクトしたCHO K-1細胞(b)およびCHO K-1sTLR2trans細胞(a,c)の表面を、EMによって観察した写真である。パネル(a)および(b,c)の倍率はそれぞれ、1,000倍および10,000倍である。
【図14】四つのLAB株におけるNF-κB依存的ルシフェラーゼ活性の比較を示す図である。1 μM D25、10 μg/mlザイモサン、10 μg/ml PGN、および100 μg/mlのintactなLABによる処置後24時間でのトランスフェクト細胞のルシフェラーゼ活性を示す。結果は、独立した実験3回〜4回のS.I.の平均値±SDを表す;培地のみにおいて培養した同一に処置した細胞と比較して*:P<0.05、**:P<0.01
【発明を実施するための形態】
【0018】
免疫活性化に関与するToll様レセプター9(Toll-like receptor 9;TLR9)は、脾臓において強く発現していることが知られていた(Zarember KA. and Godowski PJ. Journal of Immunology. 168 (2002) 554-561)。これに対し、本発明者らは、TLR9が腸管リンパ系組織、特にパイエル板や腸管膜リンパ節において強く発現していることを初めて見出した。
【0019】
腸管は体内と体外の境界組織として、経口摂取された栄養成分の吸収のみならず、外界からの刺激(細菌やウイルスなどの微生物、薬剤や食品添加物、食物に付着した農薬や環境汚染物質など)に常に接触しており、これら異物の受容、伝達、応答、および排除など種々の第一線生体防御機構(first defense line)が存在している(Mantis NJ. et al, J. Immunol. 169 (2002) 1844-1851)。さらに防御機構として特有のリンパ組織、腸管関連リンパ組織(GALT; gut-associated lymphoid tissue)が存在し、GALTは非集合性組成と集合性組成から構成され、非集合性組成には腸管上皮間リンパ球と粘膜固有層リンパ球が含まれ、集合性組成にはパイエル板、リンパ濾胞および腸管膜リンパ節が含まれる(Spahn TW. et al, Eur. J. Immunol. 32 (2002) :1109-1113)。パイエル板はリンパ組織関連上皮(Follicle-associated epithelium; FAE)に覆われ、腸管絨毛の存在しない部位でドーム形態をなしている。また、胚中心を持ちB細胞の存在する濾胞域(follicular area)と、ヘルパーT細胞を含む傍濾胞域(parafollicular area)を備えている(Owen RL. Sem. Immunol. 11 (1999) 157-163)。腸管の局所免疫機構の最初の役割を果たす細胞として、FAEには特殊な上皮細胞であるMembranous epithelial cells(M cells;M細胞)が散在し、細胞質を通って基底表層へとつながる抗原のトンネルとしてリンパ球や樹状細胞、マクロファージなど抗原提示細胞(antigen presenting cell)との接触のためM細胞膜は深く陥入している。気管上皮粘膜においてもM細胞の存在は確認されており、結核菌などの病原体の入り口として貢献していることが報告されている(Teitelbaum R. et al, Immunity. 10 (1999) 641-650)。また、食品中に含まれる機能性因子のみならず微生物や食物抗原の入り口としても存在することが認められている。腸管内の抗原(特に高分子)はM細胞に取り込まれた後、パイエル板の内部へと運ばれ、パイエル板内部の樹状細胞、マクロファージなどのMHC(major histocompatibility complex)クラスII陽性の抗原提示細胞に接触する(Kaneko K. et al, J. Veterinary. Med. Sci. 61 (1999) 1175-1177、Gebert A. et al, American J. Pathology. 154 (1999) 1573-1582、Jensen VB. et al, Infection & Immunity. 66 (1998) 3758-3766、Penheiter KL. et al, Mol. Microbiol. 24 (1997) 697-709、Debard N. et al, Gastroenterology. 120 (2001) 1173-1182、Gebert A. et al, Int. Rev. Cytology. 167 (1996) 91-159)。抗原刺激を受けたヘルパーT細胞はFcレセプターや抗体結合因子(IBF)、IL-2、IL-4、IL-5、IL-6を産生し、抗原提示を受け活性化したT細胞およびB細胞は循環帰巣(homing)という現象を開始し、腸管膜リンパ節を経て、胸腺に入り、体内循環して実行組織である腸管粘膜固有層、乳腺、涙腺、唾液腺、泌尿生殖器などに送られ、さらにB細胞は形質細胞となってIgAを産生する。分泌型IgAは、腸管およびその他の粘膜組織に侵入したウイルス、細菌、細菌毒素、アレルゲンなどを排除する役割を果たしている(Vaerman JP. et al, Immunology. 54 (1985) 601-603、Machtinger S. and Moss R. J. Allergy. Clinical. Immunol. 77 (1986) 341-347、Mathewson JJ. et al, J. Infectious Diseases. 169 (1994) 614-617)。さらにパイエル板の下には腸管膜リンパ節が発達し、パイエル板を越える多くのリンパ球、樹状細胞、マクロファージが待機している。このように、パイエル板や腸管膜リンパ節は、腸管免疫系(上記のような腸管に存在する免疫系)において中心的な役割を果たすことが知られている。
【0020】
一方、TLR4抗体、IRAK抗体を用いて、サルの腸管上皮からのFITC標識したTLR4のリガンドとして知られるLPS(リポポリサッカライド)の取り込みと腸管粘膜上皮細胞との関係について解析した結果、LPSが腸管上皮細胞中のTLR4とIRAKを発現する細胞により取り込まれ、粘膜固有層にまで運ばれる様子が観察された(Imaeda H. et al, Histochemical Cell Biology. 118 (2002) 381-388)。以上を考慮すると、TLR9やTLR2等の腸管組織において発現しているTLRは、腸管免疫系に関与していると考えられる。
【0021】
本発明は、以上の知見に基づき、被験試料が腸管免疫系を活性化するか否かを評価する方法を提供する。該評価方法としては、まず、腸管組織において発現しているTLRを強制発現させた細胞(TLRトランスフェクタント)に、被験試料を接触させる。この工程により、被験試料を、トランスフェクタント表面のTLRに接触させる。次いで、該TLRトランスフェクタントにおけるシグナル伝達を指標に、該TLRの活性を測定する。該評価方法においては、該TLRの活性が、被験試料を接触させないときに比べ上昇する場合に、被験試料が腸管免疫系を活性化すると判定される。
【0022】
本発明において、被験試料としては、例えば、DNA、DNA断片、天然化合物、有機化合物、無機化合物、タンパク質、ペプチド等の単一化合物、並びに、化合物ライブラリー、遺伝子ライブラリーの発現産物、非哺乳動物細胞培養上清、非哺乳動物細胞抽出物、発酵微生物産生物、微生物培養上清、微生物抽出物、海洋生物抽出物、植物抽出物、微生物等を挙げることができるが、これらに限定されるものではない。上記DNA断片としては、由来する生物に制限はないが、CpG モチーフ、ATモチーフまたはCpG 様モチーフを有するDNA断片が好ましい。また、上記微生物抽出物としては、例えば、細胞壁、細胞膜、DNA、RNA、鞭毛等が挙げられる。また、該微生物としては、細菌、酵母等を例示できる。また、該細菌としては、病原性細菌、乳酸菌等を挙げることができる。上記被験試料は必要に応じて適宜標識して用いることができる。標識としては、例えば、放射標識、蛍光標識等を挙げることができる。
【0023】
また、本発明における腸管としては、十二指腸、空腸、回腸等が挙げられるが、これらに限定されるものではない。また、本発明における腸管組織としては、特に制限はないが、好ましくは腸管リンパ系組織、より好ましくはパイエル板または腸管リンパ節、さらに好ましくは回腸由来パイエル板または回腸由来腸管リンパ節が挙げられる。
【0024】
また、腸管組織において発現しているTLRには、全てのタイプのTLRが包含され、例えばTLR1〜TLR10が挙げられる。現在までに10種類のTLRの存在が報告されており、TLRファミリーはそれぞれ認識分子すなわち細菌性モデュリンが異なると考えられている。細菌性モデュリンとは、細菌に特異的な分子パターン(pathogen associated molecular patterns; PAMPs)で、宿主に対しサイトカイン誘導能を示し、免疫応答を制御するものと定義されている。また、TLRは、細胞外にロイシンリッチリピート(LRRs)、細胞内にインターロイキン1受容体と相同性のある領域(TIRドメイン)を持つことが知られている。また、腸管組織において発現しているTLRが由来する生物としては、ブタ、ヒト、マウス、ネコ等、無脊椎、脊椎動物全般あるいは生物一般が挙げられる。
【0025】
本発明におけるTLR9としては、例えば、配列番号:2、4、6または8に記載のアミノ酸配列を含むタンパク質が挙げられる。また、本発明におけるTLR2としては、例えば、配列番号:36に記載のアミノ酸配列を含むタンパク質が挙げられる。配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質をコードするDNAは、例えば、胸腺、肺臓、脾臓、十二指腸、パイエル板および腸管膜リンパ節よりRNAを調製し、逆転写酵素によりcDNAを合成した後、配列番号:1、3、5、7または35に基づいてオリゴDNAを合成し、これをプライマーとして用いてPCR反応を行い、上記タンパク質をコードするcDNAを増幅させることにより調製できる。
【0026】
また、本発明におけるTLR9には、配列番号:2、4、6または8に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質、TLR2には、配列番号:36に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質も含まれる。このようなタンパク質には、例えば、配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質の変異体、アレル、バリアント、ホモログ等が含まれる。ここで「機能的に同等」とは、対象となるタンパク質が、配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質と同等の生物学的機能(生物学的役割)、生化学的機能(生化学的活性)を有することを指す。
【0027】
TLRは菌体成分を認識すると細胞内シグナル伝達系路を活性化し、共通のアダプター分子MyD88を介してIL-1 receptor associated kinase (IRAK)、TNF receptor associated factor 6 (TRAF6)、そして転写因子であるNF-κBの核移行を促し、最終的にtumor necrosis factor α(TNF-α)、interleukin(IL)-6、IL-12、IL-18、IFN-γなどといった様々な炎症性サイトカインの産生や、細胞表面共刺激因子(co-stimulatory molecule)の発現を誘導する(Kaisho T. and Akira S. Trends in Immunology. 22 (2001) 78-83)。TLRは主に病原体の糖質、脂質や核酸を認識することから、タンパクを認識する獲得免疫と相補的であるといえる。
【0028】
すなわち、配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質の生物学的機能、生化学的機能としては、微生物成分認識能、細胞内シグナル伝達系路の活性化能、炎症性サイトカイン発現誘導能、細胞表面共刺激因子発現誘導能などが挙げられる。
【0029】
あるタンパク質と機能的に同等なタンパク質をコードするDNAを調製する方法としては、ハイブリダイゼーション技術(Sambrook,J et al., Molecular Cloning 2nd ed., 9.47-9.58, Cold Spring Harbor Lab. press, 1989)を利用する方法が挙げられる。即ち、配列番号:1、3、5、7または35に記載の塩基配列もしくはその一部を利用して、配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質をコードするDNAを単離することができる。
【0030】
配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質をコードするDNAを単離するためのハイブリダイゼーションの条件は、当業者であれば適宜選択することができる。ハイブリダイゼーションの条件としては、例えば、低ストリンジェントな条件が挙げられる。低ストリンジェントな条件とは、ハイブリダイゼーション後の洗浄において、例えば42℃、5xSSC、0.1%SDSの条件であり、好ましくは50℃、5xSSC、0.1%SDSの条件である。より好ましいハイブリダイゼーションの条件としては、高ストリンジェントな条件が挙げられる。高ストリンジェントな条件とは、例えば65℃、0.1xSSC及び0.1%SDSの条件である。これらの条件において、温度を上げる程に高い相同性を有するDNAが効率的に得られると考えられる。但し、ハイブリダイゼーションのストリンジェンシーに影響する要素としては温度や塩濃度等複数の要素が考えられ、当業者であればこれら要素を適宜選択することで同様のストリンジェンシーを実現することが可能である。
【0031】
また、配列番号:1、3、5、7または35に記載の配列情報を基に合成したプライマーを用いる遺伝子増幅法、例えば、ポリメラーゼ連鎖反応(PCR)法を利用して、配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質をコードするDNAを単離することも可能である。
【0032】
これらハイブリダイゼーション技術や遺伝子増幅技術により単離されるDNAがコードする、配列番号:2、4、6、8または36に記載のアミノ酸配列を含むタンパク質と機能的に同等なタンパク質は、通常、配列番号:2、4、6、8または36のアミノ酸配列を含むタンパク質とアミノ酸配列レベルにおいて高い相同性を有する。高い相同性とは、アミノ酸レベルにおいて、少なくとも60%以上の同一性、好ましくは70%以上の同一性、より好ましくは80%以上の同一性、さらに好ましくは90%、よりさらに好ましくは95%以上、最も好ましくは98%以上の同一性を指す。
【0033】
アミノ酸配列や塩基配列の同一性は、Karlin and AltschulによるアルゴリズムBLAST(Proc. Natl. Acad. Sci. USA 90:5873-5877, 1993)によって決定することができる。このアルゴリズムに基づいて、BLASTNやBLASTXと呼ばれるプログラムが開発されている(Altschul et al. J. Mol. Biol.215:403-410, 1990)。BLASTに基づいてBLASTNによって塩基配列を解析する場合には、パラメーターはたとえばscore = 100、wordlength = 12とする。また、BLASTに基づいてBLASTXによってアミノ酸配列を解析する場合には、パラメーターはたとえばscore = 50、wordlength = 3とする。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である(http://www.ncbi.nlm.nih.gov.)。
【0034】
また、配列番号:2、4、6、8または36に記載のアミノ酸配列において1もしくは複数のアミノ酸が変異したアミノ酸配列からなるタンパク質も本発明に含まれる。このようなアミノ酸の変異は自然界においても生じうる。変異するアミノ酸数は、通常、30アミノ酸以内であり、好ましくは15アミノ酸以内であり、より好ましくは5アミノ酸以内であり、さらに好ましくは2アミノ酸以内である。
【0035】
本発明の方法においては、上記TLRトランスフェクタントが使用される。該TLRトランスフェクタントは、腸管組織において発現しているTLRをコードするDNAを含む発現ベクターを細胞に導入することで製造することができる。該発現ベクターとしては、例えば、哺乳動物由来の発現ベクター(例えば、pcDNA3 (インビトロゲン社製)や、pEGF-BOS (Nucleic Acids. Res.1990, 18(17),p5322)、pEF、pCDM8)、昆虫細胞由来の発現ベクター(例えば「Bac-to-BAC baculovairus expression system」(インビトロゲン社製)、pBacPAK8)、動物ウィルス由来の発現ベクター(例えば、pHSV、pMV、pAdexLcw)、レトロウィルス由来の発現ベクター(例えば、pZIPneo)が挙げられる。
【0036】
CHO細胞、COS細胞、NIH3T3細胞等の動物細胞での発現を目的とした場合には、細胞内で発現させるために必要なプロモーター、例えばSV40プロモーター(Mulligan et al., Nature (1979) 277, 108)、MMLV-LTRプロモーター、EF1αプロモーター(Mizushima et al., Nucleic Acids Res. (1990) 18, 5322)、CMVプロモーター等を持っていることが不可欠であり、細胞への形質転換を選抜するための遺伝子(例えば、薬剤(ネオマイシン、G418等)により判別できるような薬剤耐性遺伝子)を有すればさらに好ましい。
【0037】
さらに、遺伝子を安定的に発現させ、かつ、細胞内での遺伝子のコピー数の増幅を目的とする場合には、核酸合成経路が欠損したCHO細胞にそれを相補するジヒドロ葉酸還元酵素遺伝子(dhfr遺伝子)を有するベクター(例えば、pCHOI等)を導入し、メトトレキセート(MTX)により増幅させる方法が挙げられ、また、遺伝子の一過性の発現を目的とする場合には、SV40 T抗原を発現する遺伝子を染色体上に持つCOS細胞を用いてSV40の複製起点を持つベクター(pcD等)で形質転換する方法が挙げられる。複製開始点としては、また、ポリオーマウィルス、アデノウィルス、ウシパピローマウィルス(BPV)等の由来のものを用いることもできる。さらに、宿主細胞系で遺伝子コピー数増幅のため、発現ベクターは選択マーカーとして、アミノグリコシドトランスフェラーゼ(APH)遺伝子、チミジンキナーゼ(TK)遺伝子、dhfr遺伝子等を含むことができる。
【0038】
また、宿主細胞としては、例えば、哺乳類細胞、昆虫類細胞等が挙げられるが、これらに限定されるものではない。哺乳類細胞としては、例えば、HEK293T細胞、CHO(J. Exp. Med. (1995) 108, 945)、COS、NIH3T3、ミエローマ、BHK(baby hamster kidney)、HeLa、Vero、昆虫類細胞としては、例えば、Sf9、Sf21、Tn5が知られている。
【0039】
CHO細胞としては、特に、dhfr遺伝子を欠損したCHO細胞であるdhfr-CHO(Proc. Natl. Acad. Sci. USA (1980) 77, 4216-4220)やCHO K-1(Proc. Natl. Acad. Sci. USA (1968) 60, 1275)を好適に使用することができる。動物細胞において、大量発現を目的とする場合には特にCHO細胞が好ましい。また、宿主細胞としては、不死化した細胞株が好ましい。
【0040】
宿主細胞へのベクターの導入は、例えば、リン酸カルシウム法、DEAEデキストラン法、カチオニックリボソームDOTAP(ロッシュダイアグノスティックス社製)を用いた方法、エレクトロポーレーション法、リポフェクション等当業者に周知の方法で行うことが可能である。
【0041】
本発明においては、上記トランスフェクタントにおけるシグナル伝達を指標に、TLRの活性を測定する。TLRの活性は、例えば、サイトカイン(例えばIL-6、IL-12、IFN-γ、TNF-α等)の発現、シグナル伝達経路における分子(例えばNF-κB、JNK、IRAK等)の活性化等を指標に測定できる。サイトカインの発現は、mRNAレベルでもタンパク質レベルでも行うことが可能である。例えば、サイトカインのタンパク質レベルでの発現は、ヒトIL-6, IL-12, TNF-α, IFN-γELISA kit(株式会社テイエフビー(TFB,INC.))等の既存のヒト用キットを使用することで測定できる。また、シグナル伝達経路における分子の活性上昇は、ルシフェラーゼアッセイで行うことが可能である。例えば、ブタTLRトランスフェクタントにNF-κBおよびルシフェラーゼ遺伝子を含むプラスミドベクター(pGLM-ENH)をトランスフェクションし、18時間後にリガンド(DNA等)で刺激する。刺激24時間後に細胞を溶解後、細胞溶解液を回収し測定まで−80℃で保存する。ルシフェラーゼ活性は、細胞溶解液にルシフェリンを含む反応液を添加後、2秒後から8秒間にわたる発光の変化を測定する。各試料で3回同様の操作を行い、その平均をとる。以上の測定で発光が強い程、NF-κBの活性が強いことを示す。
【0042】
本発明はまた、腸管免疫系を活性化する試料のスクリーニング方法を提供する。本発明におけるスクリーニング方法としては、上記評価方法を利用して、複数の被験試料について、腸管免疫系を活性化するか否かを評価し、腸管免疫系を活性化すると評価された試料を選択する。
【0043】
また、腸管組織において発現しているTLRに結合する試料を予めスクリーニングし、これにより得られた試料を被験試料として用いてもよい。腸管組織において発現しているTLRに結合する試料のスクリーニングに用いられるTLRは、組換えタンパク質であっても、天然由来のタンパク質であってもよい。また、スクリーニングに用いられるTLRは部分ペプチドであってもよい。腸管組織において発現しているTLRに結合する試料のスクリーニング方法としては、まず、複数の被験試料を腸管組織において発現しているTLRに接触させる。次いで、該TLRと被験試料との結合を検出する。次いで、該TLRと結合する被験試料を選択する。TLRと被験試料との結合は、当業者に周知の方法で検出できる。
上記評価方法またはスクリーニング方法によって得られた試料は、免疫賦活化機能を有する試料として、例えば、アレルギー、癌、感染症等の治療または予防に使用できる。
【0044】
さらに、本発明においては、腸管免疫系を活性化する医薬組成物の製造方法を提供する。本発明の医薬組成物は、免疫賦活化機能を有する医薬組成物として、例えば、アレルギー、癌、感染症等の治療または予防に使用できる。また、本発明の医薬組成物は、好ましくはワクチンとして使用される。
【0045】
本発明の医薬組成物の製造方法においては、上記スクリーニング方法によって腸管免疫系を活性化すると評価された試料と医薬上許容される担体とを混合する。医薬上許容される担体としては、例えばアジュバント(抗体産生増強剤)が挙げられる。また、界面活性剤、賦形剤、着色料、着香料、保存料、安定剤、緩衝剤、懸濁剤、等張化剤、結合剤、崩壊剤、滑沢剤、流動性促進剤、矯味剤等も医薬上許容される担体として挙げられるが、その他常用の担体を適宜使用することができる。具体的には、軽質無水ケイ酸、乳糖、結晶セルロース、マンニトール、デンプン、カルメロースカルシウム、カルメロースナトリウム、ヒドロキシプロピルセルロース、ヒドロキシプロピルメチルセルロース、ポリビニルアセタールジエチルアミノアセテート、ポリビニルピロリドン、ゼラチン、中鎖脂肪酸トリグリセライド、ポリオキシエチレン硬化ヒマシ油60、白糖、カルボキシメチルセルロース、コーンスターチ、無機塩類等を挙げることができる。このようにして製造された腸管免疫系を活性化する医薬組成物は、経口剤または注射剤として使用できる。
【0046】
また、本発明は、被験微生物が腸管免疫系を活性化する微生物であるか否かを評価する方法も提供する。本発明における微生物としては、細菌、酵母等が例示できるが、これらに限定されるものではない。また、細菌としては、特に制限はないが、乳酸菌(例えば腸管常在乳酸菌や乳業用乳酸菌)が挙げられる。
【0047】
該評価方法においては、まず、被験微生物から抽出物を調製する。微生物抽出物としては、例えば、細胞壁、細胞膜、DNA、RNA、鞭毛等が挙げられる。また、該DNAとしては、CpG モチーフ、ATモチーフまたはCpG 様モチーフを有する断片が好適である。このような抽出物は、当業者に周知の方法によって微生物から調製できる。
以下に、乳業用乳酸菌からCpG モチーフ、ATモチーフまたはCpG 様モチーフを有する断片の調整方法を例示するが、本発明の方法はこれに限定されない。
【0048】
LactobacilliはLactobacilli MRS Broth(Difco laboratories, Detroit, MI, USA)で、StreptococciはElliker brothでそれぞれ3回継代培養(37℃, 24時間)後、50mlの培地に1%接種し、37℃で16時間培養する。菌体を遠心(3,000xg, 4℃, 20分間)集菌し、TE緩衝液(10mM Tris-Cl,1mM EDTA pH7.5)で2回遠心(4,000xg, 4℃, 20分間)洗浄後、5.0mlのTE緩衝液に再懸濁する。この菌液に、2.5mlのリゾチーム(生化学工業、東京、30mg/ml)、20μlのN-アセチルムラミダーゼSG(生化学工業、東京、250μg/ml)を加え、37℃で10〜30分間反応する。反応後、10mlの0.1M Tris-1% SDS溶液を添加し、穏やかに撹拌する。これにプロテイナーゼK(TaKaRa,京都20mg/ml,150μl)溶液を加え、37℃で一晩反応させる。反応後、5M NaCl溶液(2.5ml)を加え撹拌し、滅菌したビーカーに移す。100%エタノール(50ml)を加えることにより核酸を沈殿させ、生じた沈殿を滅菌したガラス棒を用いて巻き取り、70%エタノールで洗浄後、TE緩衝液(10ml)に溶解し、一晩4℃下で完全に溶解させる。RNaseA(SIGMA,10mg/ml,100μl)を加え、37℃で60分間インキュベートした後、1/10量の5M NaCl(1ml)及び等量の100%エタノールを加え、核酸を沈殿させる。生じた沈殿を滅菌したガラス棒を用いて巻き取り、70%エタノールで洗浄後、TE緩衝液(2ml)に溶解する。以上の操作でDNAが精製され、使用までの間4℃下で保存する。染色体DNAはSau 3A1 (制限酵素)により切断処理し、3%アガロースゲル電気泳動に供する。アガロースゲルよりDNAを回収し、プラスミドベクターにライゲーション後、クローニングしDNA配列をシークエンサーにて決定し、CpG モチーフ、ATモチーフまたはCpG 様モチーフを有する断片を調製する。
【0049】
該評価方法においては、次いで、該抽出物をTLRトランスフェクタントに接触させる。次いで、該形質転換細胞におけるシグナル伝達を指標に、腸管組織において発現しているTLRの活性を測定する。該評価方法においては、該TLRの活性が、該抽出物を接触させないときに比べ上昇する場合に、被験微生物が腸管免疫系を活性化する微生物であると判定される。
【0050】
また、本発明は、腸管免疫系を活性化する微生物のスクリーニング方法も提供する。該スクリーニング方法としては、上記評価方法を利用して、複数の被験微生物について、腸管免疫系を活性化するか否かを評価し、腸管免疫系を活性化すると評価された微生物を選択する。
上記評価方法またはスクリーニング方法によって得られた微生物は、免疫賦活化機能を有する微生物として、例えば、アレルギー、癌、感染症等の治療または予防に使用できる。
【0051】
腸管免疫系を活性化する微生物を使用することで、腸管免疫系を活性化する食品組成物を製造できる。本発明は、このような腸管免疫系を活性化する食品組成物の製造方法もまた提供する。本発明の食品組成物の製造方法においては、上記スクリーニング方法によって腸管免疫系を活性化する微生物であると評価された微生物と食品上許容される担体とを混合する。食品上許容される担体としては、例えば安定化剤、保存剤、着色料、香料等が挙げられる。
【0052】
本発明の食品組成物の好ましい態様としては、乳酸菌や酵母を含む食品、ドリンク等が挙げられる。乳酸菌や酵母を含む食品、ドリンクとしては、例えば乳製品が挙げられる。本発明の乳製品としては、発酵乳、チーズ、発酵食品(乳酸菌を含む食品、キムチ等)が挙げられる。これらは、当業者に周知の方法で製造できる。
このようにして製造された食品組成物は、免疫賦活化機能を有する食品組成物(例えば機能性食品、健康食品、特定保健用食品等)として、アレルギー、癌、感染症等の治療または予防に使用できる。
【0053】
本発明者らは、TLRと腸管免疫との関連を初めて見出した。また、TLRは、自然免疫を担当する細胞(例えばマクロファージや樹状細胞等)に発現していることが知られている(Gordon S. Cell. 111 (2002) 927-930、Akira S. et al, Nature Immunology. 2 (2001) 675-680)。よって、本発明のTLRトランスフェクタントは、腸管免疫担当細胞のモデル細胞として、例えば上記本発明の方法に使用できる。また、該TLRトランスフェクタントは、本発明の方法に用いる他の成分とともに、本発明の方法用のキットとして利用することもできる。
【0054】
本発明は、上記本発明の方法に利用できるTLRトランスフェクタント、該TLRトランスフェクタントを腸管免疫担当細胞のモデル細胞として使用する方法、該腸管免疫担当細胞のモデル細胞の製造方法、および該製造方法によって製造されるモデル細胞もまた提供するものである。
【実施例】
【0055】
以下、本発明を実施例により、さらに具体的に説明するが本発明はこれら実施例に制限されるものではない。
〔実施例1〕
【0056】
1.1 ブタ組織
フナコシ株式会社より購入した。
【0057】
1.2 ブタTLR9遺伝子のクローニングと塩基配列決定
DDBJ/EMBL/GenBankにおいて公開されているヒトおよびマウスTLR9(それぞれAccession Number AB045180およびAF348140)遺伝子配列をもとに保存性の高い領域を探索しプライマーを作製した。そのプライマーを用いてブタ腸管パイエル板由来total RNAからRT-PCR法によりブタTLR9遺伝子断片を得た。得られた遺伝子断片はpGEM-T-Easy vectorにライゲーションしE.coli JM109コンピテントセルにトランスフォーメーションしサブクローニングした。DNA配列はDNA sequencer Model 4000L(Li-Cor, Lincoln, NE, USA)により決定した。塩基配列、アミノ酸配列解析にはGENETYX-SV/RC Ver.11.0.3.1を用いた。残りのブタTLR9遺伝子配列はブタTLR9遺伝子断片から設計したプライマーを用いてRACE法により得た。全TLR9遺伝子をPCR法により増幅しクローニングした。
【0058】
1.3 ブタTLR9特異的ポリクローナル抗体の作製
GENETYX-SV/RC Ver.11.0.3.1を用いた抗原決定基(エピトープ)解析とタンパク2次構造解析よりブタTLR9アミノ酸配列中より268から284番目の領域が高い抗原性を持つことを見出した。その領域のペプチド合成および合成ペプチドを抗原としたポリクローナル抗体作製をサワディテクノロジー株式会社に依頼した。
【0059】
1.4 ブタTLR9遺伝子を導入したトランスフェクタントの構築
ブタTLR9遺伝子を導入する宿主細胞としては、ヒトの細胞系で遺伝子導入のホスト細胞として広く利用されているHEK293T細胞(ヒト胎児腎細胞)を選択した。ヒトTLR9特異的プライマーを用いてHEK293T細胞がTLR9遺伝子を発現していないことを解析し、シグナルペプチドを除くブタTLR9遺伝子をライゲーションしたpCXN2.1-FLAG 遺伝子発現ベクター (H. Niwa et al, Gene, 108 (1991) 193-199 )(大阪大学大学院医学研究科宮崎純一氏より分譲) をリポフェクション法によりHEK293T細胞にトランスフェクションした。抗生物質にはG418ネオマイシン(SIGMA)を用い、BECKMAN COULTER社のEPICSセルソーターシステムによりブタTLR9発現細胞をセレクションした。
【0060】
1.5 RT-PCR法によるトランスフェクタントにおけるブタTLR9発現解析
トランスフェクタントよりTRIzol(インビトロジェン)を用いてtotal RNAを抽出し、ブタTLR9特異的プライマー(本発明で設計)、ヒトTLR9特異的プライマーおよびポジティブコントロールとしてヒトGAPDHプライマー(K. A. Zarember, P. J. Godowski, J. Immunology, 168 (2002) 554-561) を用いてRT-PCR法によるTLR9 mRNA遺伝子発現解析を行った。
【0061】
1.6 フローサイトメトリー法および共焦点レーザー顕微鏡によるブタTLR9のトランスフェクタントにおける発現解析
細胞を1次抗体としてanti-FLAG mouse IgGモノクローナル抗体(SIGMA)で4℃、1時間処理後、2次抗体としてanti-mouse IgG-PerCP 標識抗体で4℃、30分間染色した。核染色はPropidium iodideを4℃、10分間処理で行った。解析にはFACSCaliburTM(日本BECTON DICKINSON株式会社)を用いた。ブタTLR9ポリクローナル抗体による免疫染色は、上記のように1次抗体染色後、2次抗体として、anti-rabbit IgG-Alexa 488標識抗体で4℃、30分間染色し、核染後解析した。また、細胞をI型コラーゲンコートDisk(IWAKI)に播き1次抗体としてanti-FLAG mouse IgGビオチン標識抗体(SIGMA)で4℃、1時間処理後、2次抗体としてストレプトアビジン-PE-Cy5抗体で4℃、30分間染色した。核染色にはPropidium iodide 4℃、10分間処理した。解析には共焦点レーザー顕微鏡(BIO-RAD社)を用いた。
【0062】
1.7 トランスフェクタントを用いたCpG DNA取り込みの解析
CpG DNAは、既に報告のあるヒト免疫細胞を強く刺激する大腸菌ゲノムDNA由来CpG2006型(5’-TCGTCGTTTTGTCGTTTTGTCGTT-3’(配列番号:9))とマウス免疫細胞を強く刺激するCpG1826型(5’-TCCATGACGTTCCTGACGTT-3’(配列番号:10)) (S. Pichyangkul et al, J. Immunological Methods, 247 (2001) 83-94) を用い、1Mの濃度のCpG DNAで、37℃にて1時間反応後、FACSCaliburTMおよび共焦点レーザー顕微鏡を用いて解析を行なった。
【0063】
1.8 ブタ各種組織におけるTLR9のreal-time PCR法による発現解析
ブタ各種組織(心臓、胸腺、肺臓、脾臓、肝臓、腎臓、骨格筋、十二指腸、空腸、回腸、回腸由来パイエル板、回腸由来腸管膜リンパ節)よりtotal RNAを抽出し、total RNA 1μgからoligo-d(T)18プライマーを用いてcDNAを合成後精製した。ブタTLR9特異的プライマーおよび精製cDNAからLightCycler (Roche)を用いて real-time quantitative PCRに供した。リアクションキットとしてLight Cycler-Fast Start DNA Master SYBR Green (Roche)を用いた。検量線から求めたブタTLR9遺伝子量とハウスキーピング遺伝子であるβアクチン遺伝子量との比から、ブタTLR9 mRNA量を算出した。結果は、脾臓中のTLR9発現量を1.000とした場合の各種組織に含まれるブタTLR9 mRNA量を数値化し比較した。
【0064】
本発明で用いたプライマーの塩基配列情報を表1にまとめた。
【表1】
【0065】
2. ブタTLR9遺伝子配列の決定と他種TLR9との相同性
本発明により決定したブタTLR9 cDNA塩基配列は3090塩基の構造遺伝子(ORF)を含む3145塩基(5’側に54塩基の非転写領域を含む)であった。ORFは1029残基のアミノ酸をコードしており、分子量は115.8kDaであった(図1および2)。多種TLR9に対するアライメントを図4および5に示した。ブタTLR9アミノ酸配列とヒト、マウス、ネコTLR9に対してそれぞれ82.0%、74.9%および86.6%の相同性が認められた(表2)。
【0066】
〔表2〕
−−−−−−−−−−−−−−−−−−−−−
TLR9 塩基配列a アミノ酸配列a
相同性(%) 相同性(%)
−−−−−−−−−−−−−−−−−−−−−
ヒト 84.9 82.0
マウス 78.2 74.9
ネコ 86.6 86.6
−−−−−−−−−−−−−−−−−−−−−
aヒト、マウスおよびネコTLR9の配列情報は、DDBJより以下のaccession No.(それぞれAB045180、AF348140およびAY137581)で入手した。
【0067】
3. ブタTLR9トランスフェクタントの発現解析
コントロール細胞とトランスフェクタント由来total RNAを鋳型としたブタTLR9 mRNAとヒトTLR9 mRNAのRT-PCR発現解析によりトランスフェクタントにおけるブタTLR9 mRNAの強い発現が認められた。また、両者ともにヒトTLR9 mRNAの発現は認められなかった(図6)。
フローサイトメトリーによる発現解析から、1次抗体として抗-FLAG抗体を用いた場合、コントロール細胞と比較してポジティブ側への大きなシフトがみられた(図7-a)。またブタTLR9抗体を用いた場合もポジティブ側へのシフトがみられた(図7-b)。レーザー顕微鏡による解析からも同様に発現が確認された(図8‐a、c、b、d)。
【0068】
4. CpG DNA取り込み解析
共焦点レーザー顕微鏡による解析ではCpG DNAの違いによる取り込みの顕著な差は判断できなかったが、フローサイトメトリーによるCpG DNAの取り込み解析によりブタTLR9はマウス型CpG 1826よりもヒト型CpG2006を比較的多く取り込んでいることが明らかとなった(図7‐c、8‐e、f)。
【0069】
5. 各種組織におけるブタTLR9 mRNAの発現解析
real-time PCR法による各種組織におけるブタTLR9 mRNAの発現解析の結果、ブタTLR9は、腸管リンパ系組織において、特にパイエル板と腸管膜リンパ節において強く発現していることが明らかとなった(図9)。
本実施例によりブタTLR9はマウスに比べヒトやネコTLR9と高い相同性を示すことが明らかとなった。また、real-time PCRによる発現解析から、これまでTLR9は脾臓において強い発現が認められるという報告がされていたが、経口的に病原細菌に曝される可能性が最も高い腸管粘膜系の内、腸管免疫の中心的な役割を果たすパイエル板や腸管膜リンパ節において、脾臓の約3倍以上のmRNA発現が認められた点は大変興味深い新知見である。また、本実施例はブタTLR9トランスフェクタントを構築することに成功し、本トランスフェクタントを用いたCpG DNAの取り込み解析から、ブタTLR9がマウス型に比べてヒト型CpG DNAに対する反応性が高かったことから、今後、このトランスフェクタントを用いた機能性乳酸菌DNAの認識性の解析が飛躍的に進み、ブタを実験動物としたヒトへのモデル系の計画的解析を通して、機能性食品開発の基礎研究が分子間の反応としてより詳細に検討できる。
【0070】
TLR9により認識される細菌DNAはマクロファージや樹状細胞等を刺激し、サイトカイン等のシグナル分子の産生を促すことが知られている(M. Bauer et al , J. Immunol. 166 (2001) 5000-5007)。本実施例の成果は細菌DNAによる刺激とTLR9による認識およびシグナル伝達システムを応用したワクチンの開発につながるものであり(R. L. Modlin, Nature 408 (2000) 659-660)、感染を抑制するだけの低い免疫応答から、細菌を殺す有効性の高い応答へと切り換えることができるため、結核等の感染症の治療、さらには癌、アレルギー等への幅広い応用が期待できる。最近、腸管まで届く生きた抗原運搬役として乳酸菌が注目を浴びているが、それに加えて乳酸菌由来DNAもまた免疫賦活化能があることが本発明者らの研究で明らかになったことから乳酸菌に対する期待が大いに高まっている。本発明でTLR9がパイエル板および腸管膜リンパ節で強い発現が認められたことから、TLR9が腸管免疫系において重要な役割を担っていることが強く示唆され、腸管に存在する乳酸菌や食餌性の乳業用乳酸菌のDNAがTLR9を刺激し、免疫を活性化する可能性が考えられる。それゆえ、乳酸菌DNAのTLR9を介した腸管免疫系を明らかにすることは、腸管常在乳酸菌や乳業用乳酸菌を用いたDNAワクチンの開発に向けた解決すべき課題である。
【0071】
また、パイエル板および腸管膜リンパ節におけるTLR9の強い発現は、これら腸管免疫における自然免疫系の顕著な発達を意味しているかもしれない。近年、医学、免疫学の分野において腸管免疫系が極めて重要な領域として注目を集めるようになってきた。比較的新しい研究分野であるため基本的な免疫機構についての知見が少なく、その機構解明が大いに期待されている。
本発明で得られた知見は、今後腸管におけるTLR9を介したシグナル伝達経路についてその認識メカニズムを分子レベルで明らかするための糸口となり、自然免疫という基本的な免疫システムがより発展的に解明されるための原動力ともなる。
【0072】
〔実施例2〕
1.1 細胞株
チャイニーズハムスター卵巣(CHO)K-1細胞は、TKG細胞バンク(東北大学、加齢医学研究所)から得て、10%仔ウシ胎児血清、50 mg/mlペニシリン/ストレプトマイシン、2 mM L-グルタミン、10 mM HEPES(2-[4-(2-ヒドロキシエチル)-1-ピペラジニル]エタンスルホン酸)緩衝液、0.11 mg/mlピルビン酸ナトリウム、および0.5 mM 2-メルカプトエタノール(シグマ(Sigma)、東京、日本)を添加した完全なダルベッコ改変イーグル培地において維持した。
【0073】
1.2 乳酸菌(LAB)調製物
2種の乳酸桿菌、デルブリュック菌亜種ブルガリクス(L. delbrueckii ssp.bulgaricus)とL.ガセリ(L. gasseri)を用いた。L.ブルガリクスNIAI B6は、国立動物産業研究所(つくば市、日本)から得た。L.ブルガリクス JCM 1002TおよびL.ガセリJCM 1131Tは、日本微生物コレクション(JCM、埼玉県、日本)から購入した。L. ガセリOLL2716は、明治乳業(小田原市、神奈川県、日本)の食品機能性研究所から供与された。株は全てMRSブロス(ディフコ(Difco)、デトロイト、アメリカ)において37℃で16時間培養して、凍結乾燥し、NF-κBレポーターアッセイのために用いた。
【0074】
1.3 プラスミドの構築
sTLR2のコード領域を、ブタパイエル板から単離した総RNAからRT-PCRによって増幅した。N-末端FLAGタグ-sTLR2発現ベクターは、ブタTLR2のコード領域を、単離されたsTLR2 cDNAからPCRによって増幅して、N-末端FLAGエピトープをコードするpCXN2.1-FLAG発現ベクターのNotI-EcoRI部位にクローニングした(Niwa, H., et al. (1991) Gene 108, 193-199.)。構築物の構造は、制限酵素マッピングおよびDNA配列分析によって確認した。
【0075】
1.4 安定なトランスフェクタントの単離
トランスフェクションの前日に、CHO K-1細胞4×105個/ウェルを6ウェルプレートに播種した。培養1日後、リポフェクタミンTM試薬(インビトロジェン(Invitrogen Corp.)、カールスバッド、カリフォルニア州、アメリカ)を用いて、sTLR2をコードするpCXN2.1-FLAG発現ベクター(1μg/ウェル)を細胞にトランスフェクトした。sTLR2発現トランスフェクタントをG418(インビトロジェン)(1.0 mg/ml)によって10日間陽性選択して、これをCHO K-1sTLR2 transと命名した。プラスミドベクターのみをトランスフェクトしたCHO K-1細胞は、CHO K-1Cont.と命名した。sTLR2の発現は、sTLR2に結合させたFLAGタンパク質の細胞外染色によって確認した(1.7 タンパク質発現分析参照)。
【0076】
1.5 RT-PCR分析
cDNAを総RNAから合成して、トランスフェクタントおよび非トランスフェクタントにおいてTRIZOL試薬(ライフテクノロジーズ(Life Technologies)、ロックビル、メリーランド州)によって調製して、rTaqポリメラーゼ(タカラバイオインク(TaKaRa BIO Inc)、大津、日本)を用いて増幅した。外因性のsTLR2を検出するために、以下のプライマーを用いてDNAを増幅した。
センス:5'-TTC AGG CCA AGG ATT TCC AG-3'(配列番号:37)
アンチセンス:5'-TCA CTG TGC TGG GTT CAT TG-3'(配列番号:38)
【0077】
βアクチンプライマーの実際の配列は、以下のようにチャイニーズハムスターのβ-アクチン(GenBank アクセッション番号U20114)のDNA配列に基づいて、以下のようにデザインして、ハウスキーピング遺伝子対照として用いた。
センス:5'-TCT ACA ACG SGC TGC GTG TG-3'(配列番号39)
アンチセンス:5'-GCT TCT CTT TGA TGT CAC GC-3'(配列番号:40)
PCRサイクル条件は、95℃で5分の後95℃で30秒、55℃で30秒、および72℃で30秒を30サイクルであった。PCR産物は、1.5%アガロースゲル上で、エチジウムブロマイドによって可視化して分析した。
【0078】
1.6 sTLR2に対するポリクローナル抗体(Ab)の作製
sTLR2タンパク質は、二次構造、疎水性、および抗原性を予測するためにジェネティクス-SV/RC(11.0.3.1)を用いて分析した後、細胞外ドメインにおけるアミノ酸76〜90個を含む領域(CVNLRALRLGANSIH)(配列番号:41)をペプチド合成のために選択した。免疫抗原としての合成ポリペプチドを、フロイントの完全アジュバントと共に1:1の割合で乳化した。抗sTLR2ポリクローナルAbは、0.3 mg/ウサギの用量で皮下注射した合成ポリペプチドによってウサギ(日本白色種)を免疫することによって産生した。1ヶ月間隔で追加免疫を2回行った後、ウサギから採血して抗血清を回収した後、エピトープのアフィニティクロマトグラフィーによる精製を行った。最終的に、直接的な酵素結合イムノソルベントアッセイ(データは示していない)によって、sTLR2の高い抗体価が得られた。
【0079】
1.7 タンパク質発現分析
細胞を洗浄して、固定し、透過性にして、N-末端FLAG-sTLR2タンパク質に関して抗FLAG M5マウスモノクローナルAb(シグマ)および第一抗体としてウサギ抗sTLR2ポリクローナルAbを用いて染色した。次に、細胞をそれぞれ、フィコエリスリン(PE)またはアレクサ660結合抗マウスIgGおよびアレクサ488結合抗ウサギIgGによって処置した。PEおよびアレクサ488シグナルは、フローサイトメトリー(FACSキャリバーTM、ベクトンディッキンソン(BECTON DICKINSON)、東京)によって決定した。アレクサ660およびアレクサ488シグナルは、共焦点レーザー顕微鏡(バイオラド・ラボラトリーズインク(Bio-Rad Laboratories Inc.)、ハーキュリーズ、カリフォルニア州)によって決定した。
【0080】
1.8 走査型電子顕微鏡分析
10 μg/mlザイモサン(モレキュラープローブス(Molecular Probes)、ユージーン、オレゴン州)によって刺激して一晩培養した細胞を、2%グルタルアルデヒドを含む0.2 M燐酸緩衝生理食塩液(pH 7.4)において30分間固定した。段階的な濃度のエタノール(50%、60%、70%、80%、90%および99%)を用いて細胞を脱水した後、無水エタノールによって2回洗浄した。試料を臨界点乾燥させて、金-パラジウムによってコーティングしてからS-4200走査型電子顕微鏡(ヒタチ(HITACHI)、東京、日本)によって観察した。
【0081】
1.9 ザイモサンの取り込み試験
細胞を、最終濃度20 μg/mlのFITC-結合ザイモサンによって37℃で1時間処置した。細胞を固定した後、ザイモサンの取り込みをフローサイトメトリーおよび共焦点レーザー顕微鏡によって検出した。
【0082】
1.10 NF-κBレポーターアッセイ
細胞(24ウェルプレートにおいて8×104個/ウェル)を、リポフェクタミンTM試薬と共にヒトNF-κB遺伝子A1およびA2部位を含むpGLM-ENH-luciベクター(Ueda, A., et al. (1997) J. Biol. Chem. 272, 31092-31099.; Inadera, H., et al. (2000) Endocrinology 141, 50-59.)0.5 μgを用いて18時間トランスフェクトさせた。様々なリガンドによる24時間の刺激の後、製造元(プロメガ(Promega)、東京、日本)が推奨する条件を用いてルシフェラーゼアッセイを行った。刺激指数(S.I.)は、以下の等式によって推定した:[(CHO K-1sTLR2trans.細胞における共鳴単位(RU)−(バックグラウンドのRU)]/[(CHO K-1Cont.細胞における共鳴単位(RU)−(バックグラウンドのRU)]。アッセイは全て、異なる3つの培養ウェルを用いて少なくとも3回の異なる実験によって行った。
【0083】
2. ブタTLR2遺伝子配列の決定
腸関連リンパ様組織に属する回腸パイエル板から単離したブタmRNAを用いて、Horton R. H.らにより報告されているオーバーラップPCR伸長法によってsTLR2の完全長のcDNAを作製した(Horton, R.H., et al. (1989) Gene 77, 61-68.)。sTLR2の完全長のcDNAは、アミノ酸785個をコードする2358 bpであり、計算分子量は89.6 kDaであった。得られたcDNAは、Munetaら(Muneta, Y., et al. (2003) J. Interferon Cytokine Res. 23, 583-589.)によって最近報告された、リポ多糖類によって刺激したブタ肺胞マクロファージから得られたcDNAと同一であった。ブタTLR2のヌクレオチド配列は、アクセッション番号AB072190としてDDBJ、EMBL、およびGenBankヌクレオチドデータベースに提出されている。
【0084】
3. ブタTLR2発現トランスフェクタントの発現および機能的分析
CHO K-1細胞は、チャイニーズハムスターの機能的TLR2を発現しないことから、sTLR2をコードするpCXN-FLAGベクターによるリポフェクションによってCHO K-1sTLR2trans細胞を確立した(Heine, H., et al. (1999) J. Immunol. 162, 6971-6975.)。RT-PCR分析で、sTLR2のmRNAがCHO K-1sTLR2trans細胞において発現されたことから、外因性のsTLR2 mRNAがCHO K-1sTLR2trans細胞において発現されたことが示された(図10)。プラスミドベクターのみをトランスフェクトしたCHO K-1Cont.細胞とCHO K-1sTLR2trans細胞とをマウス抗FLAG mAbおよび抗-sTLR2ポリクローナル抗体によって染色すると、FLAGおよびsTLR2陽性細胞は、フローサイトメトリー(図11-a、b)および共焦点レーザー顕微鏡(図12-a、b、d、e)によってCHO K-1sTLR2trans細胞でのみ検出された。さらに、走査型電子顕微鏡写真から、TLR2-特異的リガンドとして同定されているザイモサンによって細胞を処置した場合、CHO K-1sTLR2trans細胞の表面は劇的に粗くなったが、CHO K-1Cont.細胞では粗くならなかったことが示された(図13)。これらの結果により、sTLR2がCHO K-1sTLR2trans細胞において確実に発現していることが確認された。CHO K-1Cont.細胞およびCHO K-1sTLR2trans細胞をザイモサンによって1時間処置すると、ザイモサンを取り込んだ細胞は、CHO K-1sTLR2trans細胞では強く検出されたが、CHO K-1Cont.細胞では検出されなかった(図11-c)。さらに、FITC標識ザイモサン(緑色)およびFLAG-sTLR2(青色)が同時に局在することは、CHO K-1sTLR2trans細胞でのみ示された(図12-c、f)。これらは、微生物の細胞壁成分によって誘導されたシグナル伝達が、sTLR2を通して核に達し、トランスフェクタントの生物機能の活性化が起こった可能性を示している。
【0085】
4. ブタTLR2トランスフェクタントのLAB株におけるNF-κB依存的ルシフェラーゼ活性
Underhillらにより、TLR2が、5分以内にザイモサンを含むマクロファージファゴソームに特異的に補充されることが示されている(Underhill, D., et al. (1999) Nature 401, 811-815.)。粒子のインターナリゼーションの全段階において、TLR2は液胞の内容物を認識し、特定の生物に対する防御に対して適当な炎症反応を誘発した。したがって、CHO K-1sTLR2trans細胞におけるTLR2による活性化を評価するために、細胞にNF-κB依存的ルシフェラーゼレポーター(pGLM-ENH-luci)ベクターを一過性に同時トランスフェクトした。ザイモサンおよびPGNによって刺激したCHO K-1sTLR2trans細胞は、NF-κB依存的ルシフェラーゼ活性を有意に増加させたが、sTLR9のリガンドDNAであるD25には反応しなかった(図14)。これらの知見は、CHO K-1sTLR2trans細胞が微生物の細胞壁成分を選択的に認識して、それによってNF-κB活性化が誘導されうることを示唆する。
【0086】
次に、本発明者らは、CHO K-1sTLR2trans細胞がNF-κB依存的ルシフェラーゼ活性を発揮した後にintactなLABと共に細胞活性成分に反応するか否かを確認しようと試みた。CHO K-1sTLR2trans細胞をL.ブルガリクス(NIAI B6、LCM1002T)およびL.ガセリ(JCM1131T、OLL2716)の四つの株によって刺激すると、TLR2を通しての細胞活性化はCHO K-1sTLR2trans細胞において検出され、この細胞はPGNおよびリポテイコ酸のような細胞壁成分を含むLABの細胞表面を認識する可能性がある。さらに、intactなLABの刺激は、個々の細胞壁成分と比較してCHO K-1sTLR2trans細胞活性化を有意に増加させることから、CHO K-1sTLR2trans細胞の活性化が、intactなLABを構成する様々な細胞壁成分によって減弱したことを意味している。興味深いことに、ルシフェラーゼ活性の差は、類似の遺伝的背景にもかかわらず、それぞれのLABの二つの個々の株においても示された。本発明者らのこれまでの研究および他の報告から、細胞表面構成成分における構造的、化学的な違い、およびコンフォメーションの違いが、遺伝的に近縁のLAB株においても認められたことが示されている(Schar-Zammaretti, P. and Ubbink, J. (2003) Biophys. J. 85, 4076-4092.; Takeda, K., et al. (1997) Milchwissenshaft 52, 21-25.)。この証拠は、LABの細胞壁によって誘導される様々な免疫調節作用が、細胞壁特性の差に反映される可能性があるという考え方を支持する。このように、本発明者らは、同じ種の異なる株における活性のわずかな差も区別できるCHO K-1sTLR2trans細胞の構築に成功した。
【0087】
腸の菌叢は、免疫コンピテント細胞の産生およびTヘルパー1(TH1)とTヘルパー2(TH2)免疫のあいだのバランスの調節に密接に関与している(Neurath, M.F., et al. (2002) Nat. Med. 8, 567-573.)。ほとんどのアレルギー疾患は、アレルゲン性の分子に対する免疫応答がTH2型に向けて明らかに偏った、リンパ球が支配する免疫における不均衡によって反映される。in vitroおよびin vivoにおいて、細菌のリポ多糖類およびCpGオリゴヌクレオチドは、免疫学的環境において樹状細胞による炎症性サイトカインの産生を通してTH1型T細胞発達を増強する(Trinchieri, G. (2003) Nat. Rev. Immunol. 3, 133-46.)。これまで、本発明者らは、マウスB細胞における変異原性を誘導するLABからの免疫刺激オリゴヌクレオチドの発見に成功した(Kitazawa, H., et al. (2001) Int. J. Food Microbiol. 65, 149-162.; Kitazawa, H., et al. (2003) Int. J. Food Microbiol. 85, 11-21.)。本発明者らの最近の研究において、トランスフェクタント実験により、粘膜免疫において重要な役割を果たす可能性があるTLR9が、LABからのCpGのみならずATモチーフを認識することが示された(Shimosato, T., et al. (2003) Biochim. Biophys. Acta 1627, 56-61.; Shimosato, T., et al. (2004) Animal Science Journal 75, In press.)。LABからのオリゴヌクレオチドの他に、細胞壁は、LABによるインターフェロン誘導能を決定するための重要な要因である(Solis-Pereya, B., et al. (1997) Am. J. Clin. Nutr. 66, 521S-525S.)。本実施例において示した結果は、intactな共生LABと共に細胞壁成分がTLR2を通して免疫応答の誘導能を有することを示し、活性は、CHO K-1sTLR2trans細胞によって十分に検出可能となりうることを示している。本発明者らの研究において、ブタがヒトモデルとなると予想され、場合によってはin vivo実験をヒトの代わりにブタにおいて行うことができることから、sTLR2を用いたアッセイ系を開発した。このように、本研究により、sTLR2発現トランスフェクタントを利用することで、LABのような特定のイムノバイオティック微生物を評価するための主な免疫アッセイ系を提供できることが示された。これはTH1/TH2極性の維持のような免疫調節能を有する新規機能性食品の生産に貢献する可能性がある。
【特許請求の範囲】
【請求項1】
被験試料が腸管免疫系を活性化するか否かを評価する方法であって、
(a)腸管組織において発現しているToll様レセプターを強制発現させた細胞に、被験試料を接触させる工程
(b)該細胞におけるシグナル伝達を指標に、該Toll様レセプターの活性を測定する工程
を含み、上記Toll様レセプターの活性が、上記被験試料を接触させないときに比べ上昇する場合に、被験試料が腸管免疫系を活性化すると判定される方法。
【請求項2】
以下の(a)および(b)の工程を含む、腸管免疫系を活性化する試料のスクリーニング方法。
(a)請求項1に記載の評価方法により、複数の被験試料について、腸管免疫系を活性化するか否かを評価する工程
(b)複数の被験試料から、腸管免疫系を活性化すると評価された試料を選択する工程
【請求項3】
請求項2に記載の工程に、さらに腸管免疫系を活性化すると評価された試料と医薬上許容される担体とを混合する工程を含む、腸管免疫系を活性化する医薬組成物の製造方法。
【請求項4】
被験微生物が腸管免疫系を活性化する微生物であるか否かを評価する方法であって、
(a)被験微生物から抽出物を調製する工程
(b)腸管組織において発現しているToll様レセプターを強制発現させた細胞に、該抽出物を接触させる工程
(c)該細胞におけるシグナル伝達を指標に、該Toll様レセプターの活性を測定する工程
を含み、上記Toll様レセプターの活性が、上記抽出物を接触させないときに比べ上昇する場合に、被験微生物が腸管免疫系を活性化する微生物であると判定される方法。
【請求項5】
以下の(a)および(b)の工程を含む、腸管免疫系を活性化する微生物のスクリーニング方法。
(a)請求項4に記載の評価方法により、複数の被験微生物について、腸管免疫系を活性化する微生物であるか否かを評価する工程
(b)複数の被験微生物から、腸管免疫系を活性化する微生物であると評価された微生物を選択する工程
【請求項6】
請求項5に記載の工程に、さらに腸管免疫系を活性化する微生物であると評価された微生物と食品上許容される担体とを混合する工程を含む、腸管免疫系を活性化する食品組成物の製造方法。
【請求項7】
微生物が乳酸菌であり、食品組成物が乳製品である、請求項6に記載の方法。
【請求項8】
微生物が細菌である、請求項4〜6のいずれかに記載の方法。
【請求項9】
細菌が乳酸菌である、請求項8に記載の方法。
【請求項10】
腸管組織において発現しているToll様レセプターをコードするDNAを有する発現ベクターを細胞に導入する工程を含む、腸管免疫担当細胞のモデル細胞の製造方法。
【請求項11】
腸管組織において発現しているToll様レセプターを強制発現させた細胞を、腸管免疫担当細胞のモデル細胞として使用する方法。
【請求項12】
腸管組織が腸管リンパ系組織である、請求項1〜11のいずれかに記載の方法。
【請求項13】
腸管リンパ系組織がパイエル板または腸管リンパ節である、請求項12に記載の方法。
【請求項14】
Toll様レセプターがブタ由来である、請求項1〜13のいずれかに記載の方法。
【請求項15】
Toll様レセプターがToll様レセプター9および/または2である、請求項1〜13のいずれかに記載の方法。
【請求項16】
請求項1〜9のいずれかに記載の方法に用いるための、腸管組織において発現しているToll様レセプターを強制発現させた細胞。
【請求項17】
腸管組織において発現しているToll様レセプターをコードするDNAを有する発現ベクターを細胞に導入することで製造される、腸管免疫担当細胞のモデル細胞。
【請求項18】
腸管組織が腸管リンパ系組織である、請求項16または17に記載の細胞。
【請求項19】
腸管リンパ系組織がパイエル板または腸管リンパ節である、請求項18に記載の細胞。
【請求項20】
Toll様レセプターがブタ由来である、請求項16〜19のいずれかに記載の細胞。
【請求項21】
Toll様レセプターがToll様レセプター9および/または2である、請求項16〜19のいずれかに記載の細胞。
【請求項1】
被験試料が腸管免疫系を活性化するか否かを評価する方法であって、
(a)腸管組織において発現しているToll様レセプターを強制発現させた細胞に、被験試料を接触させる工程
(b)該細胞におけるシグナル伝達を指標に、該Toll様レセプターの活性を測定する工程
を含み、上記Toll様レセプターの活性が、上記被験試料を接触させないときに比べ上昇する場合に、被験試料が腸管免疫系を活性化すると判定される方法。
【請求項2】
以下の(a)および(b)の工程を含む、腸管免疫系を活性化する試料のスクリーニング方法。
(a)請求項1に記載の評価方法により、複数の被験試料について、腸管免疫系を活性化するか否かを評価する工程
(b)複数の被験試料から、腸管免疫系を活性化すると評価された試料を選択する工程
【請求項3】
請求項2に記載の工程に、さらに腸管免疫系を活性化すると評価された試料と医薬上許容される担体とを混合する工程を含む、腸管免疫系を活性化する医薬組成物の製造方法。
【請求項4】
被験微生物が腸管免疫系を活性化する微生物であるか否かを評価する方法であって、
(a)被験微生物から抽出物を調製する工程
(b)腸管組織において発現しているToll様レセプターを強制発現させた細胞に、該抽出物を接触させる工程
(c)該細胞におけるシグナル伝達を指標に、該Toll様レセプターの活性を測定する工程
を含み、上記Toll様レセプターの活性が、上記抽出物を接触させないときに比べ上昇する場合に、被験微生物が腸管免疫系を活性化する微生物であると判定される方法。
【請求項5】
以下の(a)および(b)の工程を含む、腸管免疫系を活性化する微生物のスクリーニング方法。
(a)請求項4に記載の評価方法により、複数の被験微生物について、腸管免疫系を活性化する微生物であるか否かを評価する工程
(b)複数の被験微生物から、腸管免疫系を活性化する微生物であると評価された微生物を選択する工程
【請求項6】
請求項5に記載の工程に、さらに腸管免疫系を活性化する微生物であると評価された微生物と食品上許容される担体とを混合する工程を含む、腸管免疫系を活性化する食品組成物の製造方法。
【請求項7】
微生物が乳酸菌であり、食品組成物が乳製品である、請求項6に記載の方法。
【請求項8】
微生物が細菌である、請求項4〜6のいずれかに記載の方法。
【請求項9】
細菌が乳酸菌である、請求項8に記載の方法。
【請求項10】
腸管組織において発現しているToll様レセプターをコードするDNAを有する発現ベクターを細胞に導入する工程を含む、腸管免疫担当細胞のモデル細胞の製造方法。
【請求項11】
腸管組織において発現しているToll様レセプターを強制発現させた細胞を、腸管免疫担当細胞のモデル細胞として使用する方法。
【請求項12】
腸管組織が腸管リンパ系組織である、請求項1〜11のいずれかに記載の方法。
【請求項13】
腸管リンパ系組織がパイエル板または腸管リンパ節である、請求項12に記載の方法。
【請求項14】
Toll様レセプターがブタ由来である、請求項1〜13のいずれかに記載の方法。
【請求項15】
Toll様レセプターがToll様レセプター9および/または2である、請求項1〜13のいずれかに記載の方法。
【請求項16】
請求項1〜9のいずれかに記載の方法に用いるための、腸管組織において発現しているToll様レセプターを強制発現させた細胞。
【請求項17】
腸管組織において発現しているToll様レセプターをコードするDNAを有する発現ベクターを細胞に導入することで製造される、腸管免疫担当細胞のモデル細胞。
【請求項18】
腸管組織が腸管リンパ系組織である、請求項16または17に記載の細胞。
【請求項19】
腸管リンパ系組織がパイエル板または腸管リンパ節である、請求項18に記載の細胞。
【請求項20】
Toll様レセプターがブタ由来である、請求項16〜19のいずれかに記載の細胞。
【請求項21】
Toll様レセプターがToll様レセプター9および/または2である、請求項16〜19のいずれかに記載の細胞。
【図1】


【図2】

【図3】

【図4】

【図5】

【図7】

【図9】

【図11】

【図14】

【図6】

【図8】

【図10】

【図12】
【図13】

【図2】
【図3】
【図4】
【図5】
【図7】
【図9】
【図11】
【図14】
【図6】
【図8】
【図10】
【図12】
【図13】
【公開番号】特開2010−252800(P2010−252800A)
【公開日】平成22年11月11日(2010.11.11)
【国際特許分類】
【出願番号】特願2010−138149(P2010−138149)
【出願日】平成22年6月17日(2010.6.17)
【分割の表示】特願2004−178266(P2004−178266)の分割
【原出願日】平成16年6月16日(2004.6.16)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成15年3月5日 社団法人日本農芸化学会発行の「大会講演要旨集■2003年度(平成15年度)大会[東京]■」に発表
【出願人】(000006138)明治乳業株式会社 (265)
【Fターム(参考)】
【公開日】平成22年11月11日(2010.11.11)
【国際特許分類】
【出願日】平成22年6月17日(2010.6.17)
【分割の表示】特願2004−178266(P2004−178266)の分割
【原出願日】平成16年6月16日(2004.6.16)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成15年3月5日 社団法人日本農芸化学会発行の「大会講演要旨集■2003年度(平成15年度)大会[東京]■」に発表
【出願人】(000006138)明治乳業株式会社 (265)
【Fターム(参考)】
[ Back to top ]