
ジョイントレス形質を有するトマトの作出方法

【課題】ジョイントレス形質を有する新規なトマト及びその作出方法を提供する。
【解決手段】トマトのDNA(LeMADS-MC遺伝子)の発現を抑制する;トマト由来の特定のアミノ酸配列からなるタンパク質をコードするDNA、該アミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
【解決手段】トマトのDNA(LeMADS-MC遺伝子)の発現を抑制する;トマト由来の特定のアミノ酸配列からなるタンパク質をコードするDNA、該アミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ジョイントレス形質を有するトマト及びその作出方法に関する。
【背景技術】
【0002】
果菜類や果樹の中には、果梗部分に離層が形成されるものがある。このような果実を収穫すると、果実は果梗部分の離層で離脱し、いわゆる「ヘタ」部分が果実と結合した状態で収穫される。
例えばトマトの場合、果実の状態で消費者に提供される生鮮トマトでは、「ヘタ」部分が果実に結合している事が、消費者に対して新鮮さのアピールに繋がる。
ところが加工用トマトの場合、加工後の製品の品質に影響しないように、この「ヘタ」部分を外してから加工工程へ供する必要があるが、離層が形成されるトマトを用いた場合、収穫後に果実から「ヘタ」を取り除く作業が必要であり、非常に重労働を強いることになる。
【0003】
トマトには、果梗部分の離層がない「ジョイントレス形質」を示す変異体(jointless、jointless2)が存在することが知られている。この変異体トマトは、収穫すると「ヘタ」の下から離脱するため、「ヘタ」のない果実が収穫できる。
これらの変異体のうち、jointlessに関してはジョイントレス形質の発現に関わる遺伝子が単離され、MADSボックスタンパク質をコードしていることが明らかになっている(非特許文献1)。この遺伝子は、11番染色体に座乗することが知られている。jointlessは、花房の先端に生長点が生じ、葉が発生するleafy形質を示すこともあり、実用的な品種の育成には利用されていない。
一方、jointless2に関してはマップベースドクローニングが進んでおり、転写因子遺伝子が候補として見つかっている(非特許文献2)。この遺伝子は、12番染色体に座乗していることが明らかにされている(非特許文献3)。また、jointless2に関しては他の形質に影響がないことが知られている。
よって最近では、加工へ供する前に「ヘタ」を取り除く作業を無くし、作業効率を格段に向上させるため、交配育種によってjointless2変異体由来の「ジョイントレス形質」を導入したトマトが、加工用品種として広く実用化されている。
【0004】
ところで、トマト(Solanum lycopersicum)のripening inhibitor (rin)遺伝子座には、MADSボックス転写因子をコードする二つの遺伝子が並んで配置されていることが明らかになっており(非特許文献4)、この二つの遺伝子の上流の遺伝子は成熟制御に関わるLeMADS-RIN遺伝子として、下流の遺伝子はがく片の大きさに関わるLeMADS-MC遺伝子として同定されている(非特許文献4)。LeMADS-MC遺伝子は、5番染色体に座乗していることが知られている。
一方、rin変異体は、LeMADS-RIN遺伝子の最終エクソン全体と転写終結シグナル部分を含む約1.7kbを欠失しており、LeMADS-RIN遺伝子が発現する際、mRNAの転写は転写終結シグナルがないために隣接するLeMADS-MC遺伝子全体を含んで進行し、したがってできあがったmRNAは最終エクソンを欠いたLeMADS-RIN遺伝子と第一エクソンを欠いたLeMADS-MC遺伝子の融合mRNAとなることが知られている(非特許文献5)。
このrin変異体は、1960年代にアメリカで自然突然変異体の中から見出され(非特許文献6)、(1)果実の肥大生長までは全く正常と同じである、(2)リコピンが全く生産されない、(3)軟化しない、(4)エチレン生産の上昇が全く見られないなどの特徴を有する(非特許文献7)。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】L. Mao et al., Nature 406, 910-913 (2000)
【非特許文献2】T. J. Yang et al., Chromosoma 114, 103-117 (2005)
【非特許文献3】Budiman et al., Theor. Appl. Genet., 108, 910-913 (2004)
【非特許文献4】J. Vrebalov et al., Science 296, 343-346 (2002)
【非特許文献5】J. J. Giovannoni, Plant Cell 16 Suppl., S170 (2004)
【非特許文献6】R. W. Robinson & M. L. Tomes, Rep. Tomato Genet. Coop. 18, 36-37 (1968)
【非特許文献7】E. C. Tigchelaar et al., HortScience 13, 508-513 (1978)
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、ジョイントレス形質を有する新規なトマト及びその作出方法を提供することを課題とする。また、本発明は、ジョイントレス形質を有するトマトを簡便に選抜する方法を提供することを課題とする。
【課題を解決するための手段】
【0007】
本発明者らは、日持ち性を向上させたトマトの開発において、上記のrin変異体に関し研究していく中で、正常型遺伝子(LeMADS-RIN遺伝子)及び変異型遺伝子(変異型rin遺伝子)の両方が発現しているrin F1植物体に、変異型rin遺伝子配列のうちLeMADS-MC遺伝子に由来する部分の配列に対し相補的なアンチセンスDNAを導入することで、変異型rin遺伝子の発現を抑制させた組換えトマトを作出した。
そして、この組換えトマトを観察したところ、全く予想外なことに果梗に離層が形成されていないこと、すなわち、トマトにジョイントレス形質が付与されていることを発見した。
【0008】
本発明者らは、この予想外の発見から、LeMADS-MC遺伝子がジョイントレス形質の発現に関与しているのではないかと推測し、正常型トマト品種、すなわち正常型遺伝子(LeMADS-RIN遺伝子)とLeMADS-MC遺伝子が発現しているトマト品種に、上記と同じアンチセンスDNAを導入してみたところ、このトマト品種にジョイントレス形質が付与されていることを確認した。このような経緯で、本発明者らは、トマトにおいてLeMADS-MC遺伝子の発現を抑制することにより、トマトの果梗部分の離層形成が阻害されることを見出し、本発明に至った。
【0009】
すなわち、本発明は、以下のとおりである。
〔1〕以下の(a)〜(d)の何れかのDNAの発現を抑制する工程を含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の作出方法;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
〔2〕 前記(a)〜(d)の何れかのDNAの転写産物に相補的なRNAをコードするDNAを細胞内に導入することにより、該DNAの発現を抑制する工程を含む、〔1〕に記載のトマトの作出方法。
〔3〕前記RNAをコードするDNAが、配列番号1に記載の塩基配列の182〜748番で示される塩基配列のうち少なくとも30以上の連続する塩基からなる塩基配列に対し相補的な塩基配列を含む、〔2〕に記載のトマトの作出方法。
〔4〕前記RNAをコードするDNAが、配列番号1に記載の塩基配列の211〜680番、182〜630番、又は296〜748番で示される塩基配列に対して相補的な塩基配列を含む、〔3〕に記載のトマトの作出方法。
【0010】
〔5〕以下の(a)〜(d)の何れかのDNAの発現が抑制された、ジョイントレス形質を有するトマト(Solanum lycopersicum);
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
〔6〕前記(a)〜(d)の何れかのDNAの転写産物に相補的なRNAをコードするDNAを細胞内に保持する、〔5〕に記載のトマト。
〔7〕前記RNAをコードするDNAが、配列番号1に記載の塩基配列の182〜748番で示される塩基配列のうち少なくとも30の連続する塩基からなる塩基配列に対し相補的な塩基配列を含む、〔6〕に記載のトマト。
〔8〕前記RNAをコードするDNAが、配列番号1に記載の塩基配列の211〜680番、182〜630番、又は296〜748番で示される塩基配列に対して相補的な塩基配列を含む、〔7〕に記載のトマト。
【0011】
〔9〕〔5〕〜〔8〕の何れかに記載のトマトから得られる種子。
〔10〕〔5〕〜〔8〕の何れかに記載のトマトを原料とする加工品。
【0012】
〔11〕〔9〕に記載の種子を栽培することを含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の作出方法。
【0013】
〔12〕以下の(a)〜(d)の何れかのDNAの転写産物に相補的なRNAをコードするDNA;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
〔13〕〔12〕に記載のDNAを含むベクター。
【0014】
〔14〕被検トマトにおける以下の(a)〜(d)の何れかのDNAの発現量を検出し、該DNAの発現量が正常型トマトに比して小さい被検トマトを選抜することを含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の選抜方法;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
〔15〕配列番号1に記載の塩基配列の167〜633番で示される塩基配列の相補配列
のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の192〜748番で示される塩基配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるリバースプライマーとからなるプライマーセットを用いて、被検トマトから調製したcDNAを鋳型としてPCRを行うことにより、前記(a)〜(d)の何れかのDNAの発現量を検出することを含む、〔14〕に記載のジョイントレス形質を有するトマトの選抜方法。
〔16〕被検トマトにおける以下の(a)〜(d)の何れかのDNAの存在を検出し、該DNAが存在しない被検トマトを選抜することを含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の選抜方法;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
〔17〕配列番号1に記載の塩基配列の14〜64番で示される塩基配列の相補配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の698〜748番で示される塩基配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるリバースプライマーとからなるプライマーセットを用いて、被検トマトから調製したゲノムDNAを鋳型としてPCRを行うことにより、前記(a)〜(d)の何れかのDNAの存在を検出することを含む、〔16〕に記載のジョイントレス形質を有するトマトの選抜方法。
〔18〕前記フォワードプライマーが、配列番号3に示す塩基配列を含むプライマー、又は該塩基配列と少なくとも90%の同一性を有する塩基配列からなるプライマーであり、前記リバースプライマーが、配列番号4に示す塩基配列を含むプライマー、又は該塩基配列と少なくとも90%の同一性を有する塩基配列からなるプライマーである、〔17〕に記載のジョイントレス形質を有するトマトの選抜方法。
【発明の効果】
【0015】
本発明により、ジョイントレス形質を有する種々の品種のトマトを簡便に作出又は製造し、新規なジョイントレス形質を有するトマトを提供することができる。
また、本発明により、ジョイントレス形質を有するトマトを簡便に選抜することができる。
【図面の簡単な説明】
【0016】
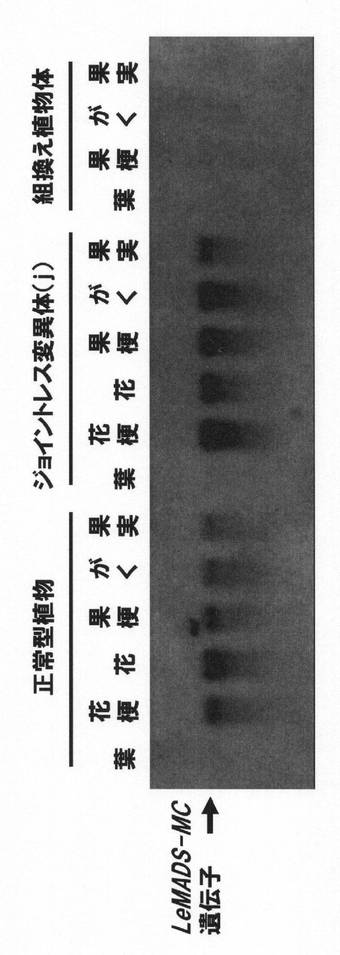
【図1】組換え植物におけるLeMADS-MC遺伝子の発現解析(電気泳動)の結果を示す写真である。左から、正常型トマトの葉・花梗・花・果梗・がく・果実、公知のジョイントレス変異体(jointless (j))の葉・花梗・花・果梗・がく・果実、本発明のジョイントレス形質が付与された組換えトマトの葉・果梗・がく・果実における発現解析の結果を示す。
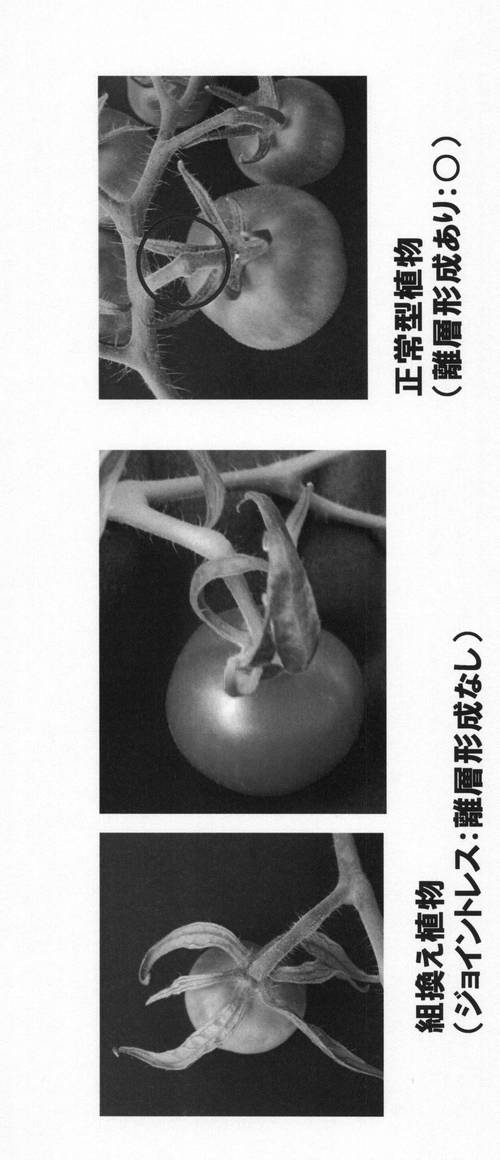
【図2】本発明のジョイントレス形質が付与された組換えトマト(左、中央)と正常型トマト(右)の果実の形態の写真である。
【図3】本発明のジョイントレス形質が付与された組換えトマトの後代における導入遺伝子の発現解析(電気泳動)の結果を示す写真である。左から、サイズマーカー(λHindIII)(M)、組換えトマト後代(4−ハ 1〜11)、正常型植物(W)。
【発明を実施するための形態】
【0017】
本発明のジョイントレス形質を有するトマトの作出方法は、果梗の離層形成を制御する活性を有するタンパク質をコードする遺伝子の発現を抑制する工程を含む。
本発明において、ジョイントレス形質とは、果梗に離層(ジョイント部分)が形成されない形質をいう。
【0018】
上記遺伝子のcDNAの塩基配列を配列番号1に示す。従来、上記遺伝子は、がく片の大きさの調節に関わるLeMADS-MC遺伝子として同定されていたが(『背景技術』の項を参照。)、LeMADS-MC遺伝子が果梗の離層形成に関わることは知られていなかった。なお、本明細書において、上記「果梗の離層形成を制御する活性を有するタンパク質をコードする遺伝子」を、便宜上「LeMADS-MC遺伝子」ということもある。
上記遺伝子のオープンリーディングフレームは、配列番号1に示す塩基配列における14〜748番の領域(735塩基)であり、配列番号2に示すアミノ酸配列(244残基)からなるタンパク質をコードする。
【0019】
本発明者らによって見出されたとおり、上記遺伝子(LeMADS-MC遺伝子)は果梗の離層形成を制御するため、その発現を抑制することによりトマトの果梗の離層形成を阻害し、ジョイントレス形質を有するトマトを作出することができる。
【0020】
具体的には、以下に示すDNAの発現を抑制する。
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA。
配列番号2のアミノ酸配列からなるタンパク質は、果梗の離層形成を制御する活性を有する。
【0021】
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
本発明において発現を抑制するターゲットDNAは、配列番号2のアミノ酸配列からなるタンパク質をコードするものに限定されない。すなわち、該DNAの変異体であっても、果梗の離層形成を制御する活性を有するタンパク質をコードするものである限り、ターゲットとなり得る。ここで、DNAの変異体が「果梗の離層形成を制御する活性を有するタンパク質をコードする」かどうかは、その変異体を発現しているトマトの果梗に離層が形成されているかどうか(ジョイントレス形質が付与されているかどうか)を観察することにより確認することができる。すなわち、あるDNAの変異体が発現するトマトの果梗に離層が形成されている場合には、その変異体は果梗の離層形成を制御する活性を有するタンパク質をコードするものと判断できる。
ここで、「数個」とは、通常2〜10個、好ましくは2〜7個、更に好ましくは2〜5個、より好ましくは2〜3個である。
【0022】
(c)配列番号1の塩基配列のコード領域を含むDNA。
前記配列番号2のアミノ酸配列からなるタンパク質をコードするDNAの一つとして、配列番号1の塩基配列のコード領域を含むDNAが挙げられる。
【0023】
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
本発明において発現を抑制するターゲットDNAは、配列番号1の塩基配列のコード領域を含むものに限定されない。すなわち、該DNAの変異体であっても、果梗の離層形成を制御する活性を有するタンパク質をコードするものである限り、ターゲットとなり得る。
ここで、「ストリンジェントな条件下」とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。例えば、0.1×SSC/0.1% SDS溶液/65℃や0.2×SSC/0.1% SDS溶液/68℃等の条件が挙げられる。
【0024】
また、(d)に示すDNAには、配列番号1の塩基配列に対して80%以上、より好ましくは90%以上、特に好ましくは95%以上の同一性を有する塩基配列からなり、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNAも含まれる。配列の同一性は、FASTA検索やBLAST検索により決定できる。
【0025】
本発明において、「DNAの発現を抑制する」とは、DNAの転写を抑制すること、及びタンパク質への翻訳を抑制することを含む。
DNAの発現を抑制する方法としては、該DNAの転写産物(mRNA)と相補的なRNAをコードするDNAを細胞に導入する方法(アンチセンス法、RNAi法)や、該転写産物を特異的に開裂するリボザイムをコードするDNAを細胞に導入する方法、遺伝子破壊法、共抑制法が挙げられる。
【0026】
前記転写産物と相補的なRNAは、該転写産物の全長に対して相補的である必要はなく、例えば、配列番号1の塩基配列のうち連続した少なくとも30、好ましくは少なくとも100、さらに好ましくは少なくとも400の塩基からなる塩基配列に対して相補的であればよい。
特に、配列番号1における182〜748番の領域は、果梗の離層形成を制御する活性を有するタンパク質をコードするDNAに特異的であるため、前記RNAは、この領域の連続した少なとも30、好ましくは少なくとも100、さらに好ましくは少なくとも400の塩基からなる塩基配列に相補的な塩基配列を含むことが好ましい。
このようなRNAとして、例えば、配列番号1の211〜680番、182〜630番、又は296〜748番に示される塩基からなる塩基配列に相補的な塩基配列を含むものが挙げられる。
ここで、「相補的」とは、DNAの発現を抑制することができる限り、上記転写産物に完全に相補的である場合のみならず、例えば、少なくとも90%程度、好ましくは少なくとも95%程度の相補性を有する場合も含む。
【0027】
前記転写産物と相補的なRNAには、アンチセンスRNA、dsRNAが含まれる。
【0028】
また、上記RNAをコードするDNAを含む組換えベクターは、トマトの形質転換に利用できる。
アンチセンスRNAをコードするDNAを導入するベクター(アンチセンスベクター)には、植物細胞において機能する適当なプロモーター及び好ましくはターミネーター等を含む通常トマトの形質転換に用いられるベクターを用いることができ、例えば、CaMV35Sプロモーター、NOSターミネーターを持つ「pBI121」などを用いることができる。
dsRNAコードするDNAをベクターに導入する場合には、一つのベクターにアンチセンスRNAをコードするDNA及びセンスRNAをコードするDNAを導入することもできるし、別のベクターにそれぞれアンチセンスRNAをコードするDNAとセンスRNAをコードするDNAを導入することもできる。
【0029】
アンチセンスベクターの作製は、例えば以下のようにして行うことができる。
配列番号1の塩基配列のコード領域の全長または一部の領域を、PCR法を用いて増幅しDNA断片を得る。アンチセンスRNAをコードするDNAの塩基配列の設計に好ましい領域は上述したとおりである。増幅したDNA断片を、制限酵素サイトを用いたリガーゼ反応により、ベクター内の適当なプロモーターの下流にアンチセンス方向に連結する。また、DNAの3’末端にはターミネーターを配置することが好ましい。
【0030】
組換えベクターを用いて、トマトの形質転換を行うには、アグロバクテリウム法、エレクトロポレーション法、ポリエチレングリコール法、リポソーム法、パーティクルガン法など、植物の形質転換の方法として公知の種々の方法を用いることができる。
形質転換を行う材料としては、トマトのプロトプラスト、培養細胞、カルス、根、茎、葉、種子等の組織を用いることができる。
【0031】
プロトプラストを用いる場合には、これを組換えベクターを保持するアグロバクテリウムと共存培養したり、スフェロプラスト化したアグロバクテリウムと融合したりすることによって、培養細胞を用いる場合は、組換えベクターを保持するアグロバクテリウムと共存培養することによって、組織を用いる場合には、葉の切片を組換えベクターを保持するアグロバクテリウム懸濁液に一定時間浸漬してアグロバクテリウムを組織に感染させ、共存培養することによって、形質転換することができる。
【0032】
このようにして形質転換した細胞、カルス、組織から公知の組織培養法により植物体を再生することができる。
【0033】
例えば、上記形質転換の植物材料に葉切片(リーフディスク)を用いる場合、葉切片にアグロバクテリウムを感染させた後、これらを無機塩類、ビタミン類、糖類、サイトカイニン等の植物ホルモンやカナマイシン等の選抜薬剤、殺菌剤等を加えた培地を用いて培養することで茎葉を形成させることができる。次に、上記培地よりサイトカイニン等の植物ホルモンを除いた培地を用いて茎葉を培養することにより発根を誘導することができる。
【0034】
本発明にいう「トマト」には、トマトの植物体全体、器官、組織、カルス、培養細胞の全てが含まれる。
【0035】
このようにして得られたトマトが形質転換されているか否かは、RT−PCR法、ノーザンハイブリダイゼーション法、ウェスタンブロッティング法により、導入遺伝子の発現量を検出したり、あるいは上記ターゲットDNAの発現の存在を検出したりすることで、確認することができる。
例えば、被検トマトのRNAを抽出し、配列番号1の塩基配列を基に設計したRNAプローブを用いてノーザンハイブリダイゼーションを行うことにより、ターゲットDNAの発現量を検出することもできる。
【0036】
また、本発明における「(a)〜(d)の何れかのDNAの発現が抑制された、ジョイントレス形質を有するトマト」には、該DNAの発現が抑制されている限り、形質転換操作を行った材料が再分化した当代トマトの後代トマトや、それらを片親にして交配したトマトやその後代トマト、さらに、当代トマトや後代トマトの細胞や組織を培養することにより得られたトマトも含まれる。
【0037】
また、本発明は、このようなジョイントレス形質を有するトマトから得られた種子、及びこの種子を栽培することを含むジョイントレス形質を有するトマトの製造方法を提供する。
また、本発明は、前記ジョイントレス形質を有するトマトを原料とする加工品、及び前記ジョイントレス形質を有するトマトを加工することを含むトマトを原料とする加工品の製造方法を提供する。このような加工品としては、例えばトマトの果実の加熱品、搾汁液及び抽出物、さらにはこれらを含む飲食品、食品添加物、飼料、医薬、化粧品等が挙げられる。
【0038】
本発明は、また被検トマトにおける前記(a)〜(d)の何れかのDNAの発現量を検出し、該DNAの発現量が正常型トマトに比して小さい被検トマトを選抜することを含む、ジョイントレス形質を有するトマトの選抜方法を提供する。
DNAの発現量の検出には、RT−PCR法、ノーザンハイブリダイゼーション法、ウェスタンブロッティング法を用いることができる。
【0039】
例えば、配列番号1の塩基配列に特異的なプライマーを設計し、被検トマトから調製したcDNAを鋳型としてPCRを行うことにより、上記DNAの発現量を検出することができる。
例えば、配列番号1における182〜748番の領域の100塩基以上、好ましくは少なくとも400の塩基からなる塩基配列を増幅することができるプライマーセットを設計し、これを用いてPCRを行うことができる。このようなプライマーセットとして、あくまでも一例ではあるが、配列番号1に記載の塩基配列の167〜633番で示される塩基配列の相補配列のうち連続した15以上、好ましくは20以上の塩基を含む塩基配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の192〜748番で示される塩基配列のうち連続した15以上、好ましくは20以上の塩基を含む塩基配列に相補的な塩基配列からなるリバースプライマーからなるプライマーセットが挙げられる。
【0040】
さらに具体例を挙げると、例えば以下のようなプライマーセットが挙げられる。
配列番号1に記載の塩基配列の211〜231番で示される塩基配列の相補配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の660〜680番で示される塩基配列に相補的な塩基配列からなるリバースプライマーからなるプライマーセット。
【0041】
配列番号1に記載の塩基配列の182〜202番で示される塩基配列の相補配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の610〜630番で示される塩基配列に相補的な塩基配列からなるリバースプライマーからなるプライマーセット。
【0042】
配列番号1に記載の塩基配列の296〜316番で示される塩基配列の相補配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の728〜748番で示される塩基配列に相補的な塩基配列からなるリバースプライマーからなるプライマーセット。
【0043】
ここで、「相補配列」とは、完全に相補的である配列をいう。
また、「相補的」とは、プライマーとして機能する限り、完全に相補的である場合のみならず、例えば、少なくとも90%程度、好ましくは少なくとも95%程度の相補性を有する場合も含む。
【0044】
本発明は、また被検トマトにおける前記(a)〜(d)の何れかのDNAの存在を検出し、該DNAが存在しない被検トマトを選抜することを含む、ジョイントレス形質を有するトマトの選抜方法を提供する。
DNAの存在の検出には、PCR法、サザンハイブリダイゼーション法を用いることができる。
【0045】
例えば、配列番号1の塩基配列に特異的なプライマーを設計し、被検トマトから調製したゲノムDNAを鋳型としてPCRを行うことにより、上記DNAの存在を検出することができる。
具体的には、配列番号1に記載の塩基配列の14〜64番で示される塩基配列の相補配列のうち連続した15以上、好ましくは20以上の塩基を含む塩基配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の698〜748番で示される塩基配列のうち連続した15以上、好ましくは20以上の塩基を含む塩基配列に相補的な塩基配列からなるリバースプライマーとからなるプライマーセットを用いて、PCRを行うことができる。
「相補配列」、「相補的」の定義は上述した通りである。
【0046】
フォワードプライマーとしては、例えば配列番号3に示す塩基配列を含むプライマー、又は該塩基配列と少なくとも90%程度、好ましくは少なくとも95%程度の同一性を有する塩基配列からなるプライマーが挙げられる。また、リバースプライマーとしては、例えば配列番号4に示す塩基配列を含むプライマー、又は該塩基配列と少なくとも90%程度、好ましくは少なくとも95%程度の同一性を有する塩基配列からなるプライマーが挙げられる。
また、このようにして選抜した被検トマトについて、上述した方法により該DNAの発現量を検出するなどして、ジョイントレス形質を有するトマトの選抜をさらに行うこともできる。
【実施例】
【0047】
以下、本発明を実施例を挙げてより詳細に説明する。
<1>バイナリーベクターの構築
DNA断片の加工やDNA断片相互の結合、DNA断片のクローニング、DNA配列の確認、大腸菌等の処理(形質転換、培養、プラスミドの抽出等)などの一般的操作の手順は、特に記載のない限り、Molecular Cloning 3rd editionに記載されている方法に従って行った。なお、以下の試験に用いるトマトは、何れも、Tomato Genetics Resource Center(カリフォルニア大学デービス校)(URL: http://tgrc.ucdavis.edu/)から分譲を受けた。
【0048】
1) LeMADS-MC遺伝子断片のクローニング
First-Strand cDNA Synthesis Kit (GE Healthcare Bio-Sciences社)を用いて正常型トマト(品種:LA2838A)の緑熟期果実の全RNA 2μgよりcDNAを合成した(配列番号1)。これを20倍希釈したものをテンプレートとしてGeneAmp(R)PCR System 9700 (登録商標、Applied Biosystems社)を用い、PCR反応を行った。PCR反応終了後は、5μLを1%アガロースゲルで電気泳動して増幅を確認した。尚、プライマーの配列、ならびにPCR反応条件は以下のとおりである。
増幅したPCR断片は、pBluescript SK(+)のEcoRV切断部位にクローニングし、DNAシークエンス解析により、PCRエラーがないことを確認した(このPCR断片をpBT-ASrin4と呼ぶ)。
【0049】
(i)LeMADS-MC(AF448521)プライマー配列
ベクター作製の便宜上、MC001プライマー5'末端にSacI認識部位を付加し、MC002プライマー5'末端にはBamHI認識部位を付加した。
MC001プライマー:5'-GAG CTC TTG AAC GAT ATG AAA G-3'(配列番号5)
MC002プライマー:5'-GG ATC CTC CTT GCT TCT GCT AC -3'(配列番号6)
【0050】
(ii)PCR反応条件
1サンプルあたり、表1に示した組成の反応液を使用し、表2に示したスケジュールで反応を行った。
【0051】
【表1】

【0052】
【表2】
【0053】
2) バイナリーベクターの構築
1)で得られたpBT-ASrin4 を、BamHIとSacIで制限酵素処理してLeMADS-MC cDNA断片を得た。これと、BamHIとSacIで制限酵素処理したpBI121ベクターとをライゲーション反応にて結合させて目的のバイナリーベクター(pBI-ASrin4)を作製した後、このバイナリーベクターで大腸菌を形質転換した。
【0054】
<2>アグロバクテリウムへのバイナリーベクターの導入
MicroPulserエレクトロポレーター(バイオ・ラッド ラボラトリーズ株式会社)を用いたエレクトロポレーション法でアグロバクテリウムEHA105株にpBI-ASrin4を導入した。アグロバクテリウムをYEBプレート(カナマイシン40 mg/mL入り)にまき、2、3日間暗黒下でインキュベートして、pBI-ASrin4が導入された株を選抜した。
【0055】
<3>トマトの形質転換
トマトの形質転換は、以下の論文に従って行った。
highly efficient transformation protocol for Micro-Tom, a model cultivar for tomato functional genomics.Sun HJ, Uchii S, Watanabe S, Ezura H. Plant Cell Physiol. 2006 Mar; 7(3):426-31.
【0056】
1) トマト種子の無菌播種
微粉ハイポネックス(登録商標、株式会社ハイポネックスジャパン)1.5g、シュクロース5.0gを水に溶かして1M NaOHでpH5.8に調整して1Lにした後、寒天8gを加えオートクレーブにて121℃、15分間滅菌し、300mL容プラントボックス(株式会社テックジャム)に30 m
Lずつ分注し、播種用培地を作製した。
一方で、トマト種子(品種:LA2838A)約100粒を5cm四方のガーゼに包んでホッチキスで留め、70%エタノール水溶液に2分間浸した。さらに10倍希釈したキッチンハイター(登録商標、花王株式会社)に45分間浸して消毒した。次に、ガーゼに包んだトマト種子を滅菌水で3回すすぎ、その後、滅菌水に浸して一晩吸水させた。
滅菌したキムタオル(登録商標、日本製紙クレシア株式会社)で、トマト種子に付着した余分な水分を取り、前記の播種用培地に16〜25粒ずつ播種し、25℃で16時間日長の部屋で発芽させた。
【0057】
2) 目的DNAを保持したアグロバクテリウムの培養
カナマイシン(100mg/L)を含むLBプレート培地にpBI-ASrin4を保持したアグロバクテリウムを植菌し、28℃でコロニーが十分大きくなるまで培養した。
LBプレート上に生育したコロニーのうち、1コロニーのアグロバクテリウムを、白金耳で別途調製したカナマイシン(100mg/L)を含む液体LB培地に接種し、28℃で24時間振とう培養した。
【0058】
3) アグロバクテリウム懸濁液の調製
24時間振とう培養した菌液(1mL)を遠心分離(5000rpm、5分間)して、アグロバクテリウムを集菌した。上清の液体LB培地を除去した後、同量のMS培地を添加して、アグロバクテリウムを懸濁した。それを、最終濃度100μMのアセトシリンゴンと10mMのメルカプトエタノールを添加したMS培地で40倍に希釈して、アグロバクテリウム懸濁液を調製した。
【0059】
4) アグロバクテリウムによる子葉切片の感染
発芽したトマトの子葉の真ん中を葉脈に対して垂直方向に切断し、一枚の子葉から2枚の子葉切片(リーフディスク)を作製した。子葉切片を前記のアグロバクテリウム懸濁液に10分間浸し、アグロバクテリウムを感染させた。
【0060】
5) 共存培養
感染処理後、滅菌したキムタオルに子葉切片をのせて懸濁液を吸い取り、共存培地に葉の表が下になるように、シャーレ1枚あたり15切片を並べ、アルミホイルで完全に包み、25℃の培養室で4日間共存培養を行った。
なお、共存培地は、MS培地1L、シュクロース30.0gを混合してpH5.8に調整し、ゲルライト(登録商標、和光純薬工業株式会社)3gを加えてオートクレーブにて121℃、15分間滅菌後、ゼアチン1.5mgを加え、シャーレに30 mLずつ分注して作製した。
【0061】
6) カルス誘導
子葉切片の周辺に菌が増殖している事が目視で確認できたら、カルス誘導培地に葉の表が上になるように移し替え、25℃、16時間日長で培養した。なお、子葉切片は2週間毎に新しいカルス誘導培地に移し替えた。
なお、カルス誘導培地は、MS培地1L、シュクロース30.0gを混合してpH5.8に調整し、ゲルライト(登録商標、和光純薬工業株式会社)3gを加えてオートクレーブにて121℃、15分間滅菌後、ゼアチン1.5mg、カナマイシン100mg、オーグメンチン(登録商標、グラクソ・スミスクライン株式会社)375mgを加え、シャーレに40 mLずつ分注して作製した。
【0062】
7) シュート誘導
子葉切片からカルスが形成され、そのカルスから茎及び葉(シュート)が見え始めたら茎及び葉(シュート)の生長を早めるため、茎及び葉(シュート)部分を子葉切片部分から切り落とし、シュート誘導培地に移した。
なお、シュート誘導培地は、MS培地1L、シュクロース30.0gを混合してpH5.8に調整し、ゲルライト(登録商標、和光純薬工業株式会社)3gを加えてオートクレーブにて121℃、1
5分間滅菌後、ゼアチン1.0mg、カナマイシン100mg、オーグメンチン(登録商標、グラクソ・スミスクライン株式会社)375mgを加え、シャーレに40 mLずつ分注して作製した。
【0063】
8) 発根誘導(発根選抜1)
茎及び葉の部分が1〜2cmの長さまで伸びてきたら、できるだけ根元で切り取り、発根培地に移した。
発根培地で2週間以内に発根した個体を形質転換体の候補として選抜した。一回目の発根培地で発根しなかったものは、切り口を薄く切り落とし、新しい発根培地へ移して発根誘導(発根選抜)を行った。
なお、発根培地は、1/2MS培地1L、シュクロース15gを混合してpH5.8に調整し、ゲルライト(登録商標、和光純薬工業株式会社)3gを加えてオートクレーブにて121℃、15分間滅菌後、カナマイシン50mg、オーグメンチン(登録商標、グラクソ・スミスクライン株式会社)375mgを加え、シャーレに40 mLずつ分注して作製した。
【0064】
9) 発根誘導(発根選抜2)
発根した個体は、発根培地を入れた300mL容プラントボックス(株式会社テックジャム)へ移した。その際、根を切り落とし、二度目の発根誘導(発根選抜)を行った。
【0065】
10) 順化
300mL容プラントボックス(株式会社テックジャム)で約1ヶ月間生長させた植物体は、土を入れた鉢に植え換え、ビニールで覆って湿度の急激な低下を避けながら、徐々に外気条件に慣らし、通常の鉢植え栽培へ移行した。
【0066】
<4>植物体における標的遺伝子(LeMADS-MC)の発現解析
1) 植物体からの全RNA抽出
上記で得られた組換え植物、正常型植物(品種:LA2838A)、及び既存のジョイントレス変異体(jointless (j))(品種:LA1806)の各植物体におけるLeMADS-MCの発現を解析した。
植物体からの全RNAの抽出には、SDS/フェノール法を用いた。植物体の凍結粉末サンプル1 gを乳鉢、乳棒を用いて液体窒素下でホモジナイズした後、10 mLの抽出バッファー(0.1 M Tris-HCl (pH 8.0)、0.1 M NaCl、1% SDS)、2 mLの2-メルカプトエタノール、10 mLのフェノールクロロホルム(フェノール:クロロホルム:イソアミルアルコール=25:24:1)を加え、5分間よく攪拌した。室温にて8,000 rpmで10分間遠心した後、上清を分取し、10 mLのフェノールクロロホルム(フェノール:クロロホルム:イソアミルアルコール=25:24:1)を加え、5分間よく攪拌した。再度、室温にて8,000 rpmで10分間遠心した後、上清を分取し、上清1 mL当り25μLの5 M NaClと0.8 mLの2-プロパノールを加え、-80℃で30分間静置した。その後、4℃にて8,000 rpmで30分間遠心した後、沈殿物を900μLの滅菌水に溶解し、300μLの10 M LiClを加え、-80℃で30分間静置した。さらに、4℃にて15,000 rpmで15分間遠心し、得られた沈殿物を70%エタノールでリンスした。リンス後の沈殿物を300μLの滅菌水に溶解し、30μLの3M酢酸ナトリウム、750μLの100%エタノールを加え、-80℃で30分間静置した後、4℃にて15,000 rpmで15分間遠心し、得られた沈殿物を70%エタノールで再びリンスした。沈殿物が完全に乾いたら、滅菌水を加えて溶解し、これを全RNAとし、-80℃で保存した。
【0067】
2)ノーザン解析
全RNAを1.2%アガロースゲル(1×MOPS、5%(w/v)フォルムアルデヒド)で、1レーン当り10μg泳動し、Hybond-N+メンブレン(GE Healthcare Bio-Sciences社)に転写した。Hybond-N+メンブレンをUVクロスリンクした後、ハイブリダイゼーションバッファー(5×SSC、50%フォルムアミド、0.02% SDS、0.1% N-ラウロイルサルコシン、2%ブロッキング試薬)で、68℃、30分間プレハイブリダイゼーションを行った。
LeMADS-MC遺伝子の翻訳開始点から数えて187番目のaから808番目のgまでを含むcDNA(配列番号1の塩基配列の200〜821番)をもとにDIG RNA Labeling Kit (Roche社)を用いてRNAプローブを合成し(配列番号7)、68℃で10分間熱変性させ、ハイブリダイゼーションバッファーに加え、68℃で一晩ハイブリダイゼーションを行った。その後、Hybond-N+メンブレンを2×SSC/0.1% SDS溶液で室温にて10分間、0.2×SSC/0.1% SDS溶液で室温にてさらに10分間、0.2×SSC/0.1% SDS溶液で68℃にて20分間×2回洗浄した。以降の操作はDIG検出マニュアル(Roche社)に従った。なお、シグナルの検出には化学発光基質としてCDP-Star(Roche社)を用いた。
【0068】
<5>形質転換体後代における導入DNAの解析
1) PCRによる導入DNAの確認
組換え遺伝子が、本当にジョイントレス形質を引き起こしているのかを明らかにするために、組換え植物から得た種子由来の複数の植物体について、ジョイントレス形質発現の有無と同時に組換え遺伝子が受け継がれているかどうかを確認した。
組換え遺伝子の導入によりLeMADS-MCの発現が抑制された組換え植物について、自殖後、着生した果実から種子を収集した。
収集した種子から、ランダムに十数個を選抜し、閉鎖系温室内において常法により栽培し、組換え植物から得た種子由来の植物体を得た。
組換え植物から得た種子由来の各植物体の葉から全DNAを抽出した。抽出にはGE Healthcare Nucleon Phytopure Genomic DNA Extraction Kits(GE Healthcare Bio-Sciences社)を使用した。次に得られたDNAに対してGeneAmp(R) PCR System 9700 (Applied Biosystems社)を用い、PCR反応を行った。PCR反応終了後、1%アガロースゲルで電気泳動を行い、導入DNAの増幅の有無を調べた。プライマー、PCR反応条件等は以下のとおりである。
【0069】
(i)導入遺伝子断片検出用プライマー配列
LeMADS-MC遺伝子断片上(MC001プライマー)と、pBI121由来のCaMV35Sプロモーター領域上(pBI121-Fプライマー)にプライマーを設計した。
MC001プライマー:5'-GAG CTC TTG AAC GAT ATG AAA G-3'(配列番号5)
PBI121-Fプライマー:5'-AGA CCC TTC CTC TAT ATA AG-3'(配列番号8)
【0070】
(ii)PCR反応条件
1サンプルあたり、表3に示した組成の反応液を使用し、表4に示したスケジュールで反応を行った。
【0071】
【表3】
【0072】
【表4】
【0073】
<6>結果
<4>のLeMADS-MC遺伝子の発現解析の結果を図1に示す。
正常型植物や既存のジョイントレス変異体(j)では、LeMADS-MC遺伝子の発現が認められるが、今回作出した組換え植物では、LeMADS-MC遺伝子の発現は認められない。このことから、正常型トマトが形質転換され、LeMADS-MC遺伝子の発現が抑制されたことが示された。また、j変異体ではLeMADS-MC遺伝子の発現が認められたことから、今回作出した組換え植物は、jと異なる遺伝子によってジョイントレス形質が付与されたものであることが示された。これは、jのジョイントレス形質の発現に関わる遺伝子が座乗している染色体(11番)とLeMADS-MC遺伝子が座乗している染色体(5番)が異なるという事実とも矛盾しないものである。
図2は、今回作出したジョイントレス形質が付与された組換え植物(左側)と正常型植物(右側)の果実の写真である。
右側の正常型植物は離層が形成されているのに対し、左側の組換え植物では離層が形成されていなかった。
なお、本試験では、既存のジョイントレス変異体(jointless2(j-2))のLeMADS-MC遺伝子の発現解析は行っていないが、j-2のジョイントレス形質の発現に関わる遺伝子は12番染色体に座乗しているという事実からみて、今回作出した組換え植物は、j-2とも異なる遺伝子によってジョイントレス形質が付与されたものであると考えられる。
【0074】
<5>で得られた組換え植物の後代における導入遺伝子の発現解析の結果を図3に示す。
表5は、今回作出したジョイントレス形質が付与された組換え植物の後代における、組換え遺伝子の有無と表現型(ジョイントレス形質)の相関性を示す。
組換え植物の後代において、組換え遺伝子が受け継がれた系統は離層が形成されていなかった(ジョイントレス形質を有していた)が、組換え遺伝子が受け継がれなかった系統では、離層が形成されていた。このことから、LeMADS-MC遺伝子がジョイントレス形質を付与していることが証明された。
【0075】
【表5】
【産業上の利用可能性】
【0076】
本発明によれば、さまざまな品種のトマトにジョイントレス形質を付与することができ、加工用トマトにバリエーションを与えることができる。
【技術分野】
【0001】
本発明は、ジョイントレス形質を有するトマト及びその作出方法に関する。
【背景技術】
【0002】
果菜類や果樹の中には、果梗部分に離層が形成されるものがある。このような果実を収穫すると、果実は果梗部分の離層で離脱し、いわゆる「ヘタ」部分が果実と結合した状態で収穫される。
例えばトマトの場合、果実の状態で消費者に提供される生鮮トマトでは、「ヘタ」部分が果実に結合している事が、消費者に対して新鮮さのアピールに繋がる。
ところが加工用トマトの場合、加工後の製品の品質に影響しないように、この「ヘタ」部分を外してから加工工程へ供する必要があるが、離層が形成されるトマトを用いた場合、収穫後に果実から「ヘタ」を取り除く作業が必要であり、非常に重労働を強いることになる。
【0003】
トマトには、果梗部分の離層がない「ジョイントレス形質」を示す変異体(jointless、jointless2)が存在することが知られている。この変異体トマトは、収穫すると「ヘタ」の下から離脱するため、「ヘタ」のない果実が収穫できる。
これらの変異体のうち、jointlessに関してはジョイントレス形質の発現に関わる遺伝子が単離され、MADSボックスタンパク質をコードしていることが明らかになっている(非特許文献1)。この遺伝子は、11番染色体に座乗することが知られている。jointlessは、花房の先端に生長点が生じ、葉が発生するleafy形質を示すこともあり、実用的な品種の育成には利用されていない。
一方、jointless2に関してはマップベースドクローニングが進んでおり、転写因子遺伝子が候補として見つかっている(非特許文献2)。この遺伝子は、12番染色体に座乗していることが明らかにされている(非特許文献3)。また、jointless2に関しては他の形質に影響がないことが知られている。
よって最近では、加工へ供する前に「ヘタ」を取り除く作業を無くし、作業効率を格段に向上させるため、交配育種によってjointless2変異体由来の「ジョイントレス形質」を導入したトマトが、加工用品種として広く実用化されている。
【0004】
ところで、トマト(Solanum lycopersicum)のripening inhibitor (rin)遺伝子座には、MADSボックス転写因子をコードする二つの遺伝子が並んで配置されていることが明らかになっており(非特許文献4)、この二つの遺伝子の上流の遺伝子は成熟制御に関わるLeMADS-RIN遺伝子として、下流の遺伝子はがく片の大きさに関わるLeMADS-MC遺伝子として同定されている(非特許文献4)。LeMADS-MC遺伝子は、5番染色体に座乗していることが知られている。
一方、rin変異体は、LeMADS-RIN遺伝子の最終エクソン全体と転写終結シグナル部分を含む約1.7kbを欠失しており、LeMADS-RIN遺伝子が発現する際、mRNAの転写は転写終結シグナルがないために隣接するLeMADS-MC遺伝子全体を含んで進行し、したがってできあがったmRNAは最終エクソンを欠いたLeMADS-RIN遺伝子と第一エクソンを欠いたLeMADS-MC遺伝子の融合mRNAとなることが知られている(非特許文献5)。
このrin変異体は、1960年代にアメリカで自然突然変異体の中から見出され(非特許文献6)、(1)果実の肥大生長までは全く正常と同じである、(2)リコピンが全く生産されない、(3)軟化しない、(4)エチレン生産の上昇が全く見られないなどの特徴を有する(非特許文献7)。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】L. Mao et al., Nature 406, 910-913 (2000)
【非特許文献2】T. J. Yang et al., Chromosoma 114, 103-117 (2005)
【非特許文献3】Budiman et al., Theor. Appl. Genet., 108, 910-913 (2004)
【非特許文献4】J. Vrebalov et al., Science 296, 343-346 (2002)
【非特許文献5】J. J. Giovannoni, Plant Cell 16 Suppl., S170 (2004)
【非特許文献6】R. W. Robinson & M. L. Tomes, Rep. Tomato Genet. Coop. 18, 36-37 (1968)
【非特許文献7】E. C. Tigchelaar et al., HortScience 13, 508-513 (1978)
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、ジョイントレス形質を有する新規なトマト及びその作出方法を提供することを課題とする。また、本発明は、ジョイントレス形質を有するトマトを簡便に選抜する方法を提供することを課題とする。
【課題を解決するための手段】
【0007】
本発明者らは、日持ち性を向上させたトマトの開発において、上記のrin変異体に関し研究していく中で、正常型遺伝子(LeMADS-RIN遺伝子)及び変異型遺伝子(変異型rin遺伝子)の両方が発現しているrin F1植物体に、変異型rin遺伝子配列のうちLeMADS-MC遺伝子に由来する部分の配列に対し相補的なアンチセンスDNAを導入することで、変異型rin遺伝子の発現を抑制させた組換えトマトを作出した。
そして、この組換えトマトを観察したところ、全く予想外なことに果梗に離層が形成されていないこと、すなわち、トマトにジョイントレス形質が付与されていることを発見した。
【0008】
本発明者らは、この予想外の発見から、LeMADS-MC遺伝子がジョイントレス形質の発現に関与しているのではないかと推測し、正常型トマト品種、すなわち正常型遺伝子(LeMADS-RIN遺伝子)とLeMADS-MC遺伝子が発現しているトマト品種に、上記と同じアンチセンスDNAを導入してみたところ、このトマト品種にジョイントレス形質が付与されていることを確認した。このような経緯で、本発明者らは、トマトにおいてLeMADS-MC遺伝子の発現を抑制することにより、トマトの果梗部分の離層形成が阻害されることを見出し、本発明に至った。
【0009】
すなわち、本発明は、以下のとおりである。
〔1〕以下の(a)〜(d)の何れかのDNAの発現を抑制する工程を含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の作出方法;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
〔2〕 前記(a)〜(d)の何れかのDNAの転写産物に相補的なRNAをコードするDNAを細胞内に導入することにより、該DNAの発現を抑制する工程を含む、〔1〕に記載のトマトの作出方法。
〔3〕前記RNAをコードするDNAが、配列番号1に記載の塩基配列の182〜748番で示される塩基配列のうち少なくとも30以上の連続する塩基からなる塩基配列に対し相補的な塩基配列を含む、〔2〕に記載のトマトの作出方法。
〔4〕前記RNAをコードするDNAが、配列番号1に記載の塩基配列の211〜680番、182〜630番、又は296〜748番で示される塩基配列に対して相補的な塩基配列を含む、〔3〕に記載のトマトの作出方法。
【0010】
〔5〕以下の(a)〜(d)の何れかのDNAの発現が抑制された、ジョイントレス形質を有するトマト(Solanum lycopersicum);
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
〔6〕前記(a)〜(d)の何れかのDNAの転写産物に相補的なRNAをコードするDNAを細胞内に保持する、〔5〕に記載のトマト。
〔7〕前記RNAをコードするDNAが、配列番号1に記載の塩基配列の182〜748番で示される塩基配列のうち少なくとも30の連続する塩基からなる塩基配列に対し相補的な塩基配列を含む、〔6〕に記載のトマト。
〔8〕前記RNAをコードするDNAが、配列番号1に記載の塩基配列の211〜680番、182〜630番、又は296〜748番で示される塩基配列に対して相補的な塩基配列を含む、〔7〕に記載のトマト。
【0011】
〔9〕〔5〕〜〔8〕の何れかに記載のトマトから得られる種子。
〔10〕〔5〕〜〔8〕の何れかに記載のトマトを原料とする加工品。
【0012】
〔11〕〔9〕に記載の種子を栽培することを含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の作出方法。
【0013】
〔12〕以下の(a)〜(d)の何れかのDNAの転写産物に相補的なRNAをコードするDNA;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
〔13〕〔12〕に記載のDNAを含むベクター。
【0014】
〔14〕被検トマトにおける以下の(a)〜(d)の何れかのDNAの発現量を検出し、該DNAの発現量が正常型トマトに比して小さい被検トマトを選抜することを含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の選抜方法;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
〔15〕配列番号1に記載の塩基配列の167〜633番で示される塩基配列の相補配列
のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の192〜748番で示される塩基配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるリバースプライマーとからなるプライマーセットを用いて、被検トマトから調製したcDNAを鋳型としてPCRを行うことにより、前記(a)〜(d)の何れかのDNAの発現量を検出することを含む、〔14〕に記載のジョイントレス形質を有するトマトの選抜方法。
〔16〕被検トマトにおける以下の(a)〜(d)の何れかのDNAの存在を検出し、該DNAが存在しない被検トマトを選抜することを含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の選抜方法;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
〔17〕配列番号1に記載の塩基配列の14〜64番で示される塩基配列の相補配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の698〜748番で示される塩基配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるリバースプライマーとからなるプライマーセットを用いて、被検トマトから調製したゲノムDNAを鋳型としてPCRを行うことにより、前記(a)〜(d)の何れかのDNAの存在を検出することを含む、〔16〕に記載のジョイントレス形質を有するトマトの選抜方法。
〔18〕前記フォワードプライマーが、配列番号3に示す塩基配列を含むプライマー、又は該塩基配列と少なくとも90%の同一性を有する塩基配列からなるプライマーであり、前記リバースプライマーが、配列番号4に示す塩基配列を含むプライマー、又は該塩基配列と少なくとも90%の同一性を有する塩基配列からなるプライマーである、〔17〕に記載のジョイントレス形質を有するトマトの選抜方法。
【発明の効果】
【0015】
本発明により、ジョイントレス形質を有する種々の品種のトマトを簡便に作出又は製造し、新規なジョイントレス形質を有するトマトを提供することができる。
また、本発明により、ジョイントレス形質を有するトマトを簡便に選抜することができる。
【図面の簡単な説明】
【0016】
【図1】組換え植物におけるLeMADS-MC遺伝子の発現解析(電気泳動)の結果を示す写真である。左から、正常型トマトの葉・花梗・花・果梗・がく・果実、公知のジョイントレス変異体(jointless (j))の葉・花梗・花・果梗・がく・果実、本発明のジョイントレス形質が付与された組換えトマトの葉・果梗・がく・果実における発現解析の結果を示す。
【図2】本発明のジョイントレス形質が付与された組換えトマト(左、中央)と正常型トマト(右)の果実の形態の写真である。
【図3】本発明のジョイントレス形質が付与された組換えトマトの後代における導入遺伝子の発現解析(電気泳動)の結果を示す写真である。左から、サイズマーカー(λHindIII)(M)、組換えトマト後代(4−ハ 1〜11)、正常型植物(W)。
【発明を実施するための形態】
【0017】
本発明のジョイントレス形質を有するトマトの作出方法は、果梗の離層形成を制御する活性を有するタンパク質をコードする遺伝子の発現を抑制する工程を含む。
本発明において、ジョイントレス形質とは、果梗に離層(ジョイント部分)が形成されない形質をいう。
【0018】
上記遺伝子のcDNAの塩基配列を配列番号1に示す。従来、上記遺伝子は、がく片の大きさの調節に関わるLeMADS-MC遺伝子として同定されていたが(『背景技術』の項を参照。)、LeMADS-MC遺伝子が果梗の離層形成に関わることは知られていなかった。なお、本明細書において、上記「果梗の離層形成を制御する活性を有するタンパク質をコードする遺伝子」を、便宜上「LeMADS-MC遺伝子」ということもある。
上記遺伝子のオープンリーディングフレームは、配列番号1に示す塩基配列における14〜748番の領域(735塩基)であり、配列番号2に示すアミノ酸配列(244残基)からなるタンパク質をコードする。
【0019】
本発明者らによって見出されたとおり、上記遺伝子(LeMADS-MC遺伝子)は果梗の離層形成を制御するため、その発現を抑制することによりトマトの果梗の離層形成を阻害し、ジョイントレス形質を有するトマトを作出することができる。
【0020】
具体的には、以下に示すDNAの発現を抑制する。
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA。
配列番号2のアミノ酸配列からなるタンパク質は、果梗の離層形成を制御する活性を有する。
【0021】
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
本発明において発現を抑制するターゲットDNAは、配列番号2のアミノ酸配列からなるタンパク質をコードするものに限定されない。すなわち、該DNAの変異体であっても、果梗の離層形成を制御する活性を有するタンパク質をコードするものである限り、ターゲットとなり得る。ここで、DNAの変異体が「果梗の離層形成を制御する活性を有するタンパク質をコードする」かどうかは、その変異体を発現しているトマトの果梗に離層が形成されているかどうか(ジョイントレス形質が付与されているかどうか)を観察することにより確認することができる。すなわち、あるDNAの変異体が発現するトマトの果梗に離層が形成されている場合には、その変異体は果梗の離層形成を制御する活性を有するタンパク質をコードするものと判断できる。
ここで、「数個」とは、通常2〜10個、好ましくは2〜7個、更に好ましくは2〜5個、より好ましくは2〜3個である。
【0022】
(c)配列番号1の塩基配列のコード領域を含むDNA。
前記配列番号2のアミノ酸配列からなるタンパク質をコードするDNAの一つとして、配列番号1の塩基配列のコード領域を含むDNAが挙げられる。
【0023】
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
本発明において発現を抑制するターゲットDNAは、配列番号1の塩基配列のコード領域を含むものに限定されない。すなわち、該DNAの変異体であっても、果梗の離層形成を制御する活性を有するタンパク質をコードするものである限り、ターゲットとなり得る。
ここで、「ストリンジェントな条件下」とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。例えば、0.1×SSC/0.1% SDS溶液/65℃や0.2×SSC/0.1% SDS溶液/68℃等の条件が挙げられる。
【0024】
また、(d)に示すDNAには、配列番号1の塩基配列に対して80%以上、より好ましくは90%以上、特に好ましくは95%以上の同一性を有する塩基配列からなり、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNAも含まれる。配列の同一性は、FASTA検索やBLAST検索により決定できる。
【0025】
本発明において、「DNAの発現を抑制する」とは、DNAの転写を抑制すること、及びタンパク質への翻訳を抑制することを含む。
DNAの発現を抑制する方法としては、該DNAの転写産物(mRNA)と相補的なRNAをコードするDNAを細胞に導入する方法(アンチセンス法、RNAi法)や、該転写産物を特異的に開裂するリボザイムをコードするDNAを細胞に導入する方法、遺伝子破壊法、共抑制法が挙げられる。
【0026】
前記転写産物と相補的なRNAは、該転写産物の全長に対して相補的である必要はなく、例えば、配列番号1の塩基配列のうち連続した少なくとも30、好ましくは少なくとも100、さらに好ましくは少なくとも400の塩基からなる塩基配列に対して相補的であればよい。
特に、配列番号1における182〜748番の領域は、果梗の離層形成を制御する活性を有するタンパク質をコードするDNAに特異的であるため、前記RNAは、この領域の連続した少なとも30、好ましくは少なくとも100、さらに好ましくは少なくとも400の塩基からなる塩基配列に相補的な塩基配列を含むことが好ましい。
このようなRNAとして、例えば、配列番号1の211〜680番、182〜630番、又は296〜748番に示される塩基からなる塩基配列に相補的な塩基配列を含むものが挙げられる。
ここで、「相補的」とは、DNAの発現を抑制することができる限り、上記転写産物に完全に相補的である場合のみならず、例えば、少なくとも90%程度、好ましくは少なくとも95%程度の相補性を有する場合も含む。
【0027】
前記転写産物と相補的なRNAには、アンチセンスRNA、dsRNAが含まれる。
【0028】
また、上記RNAをコードするDNAを含む組換えベクターは、トマトの形質転換に利用できる。
アンチセンスRNAをコードするDNAを導入するベクター(アンチセンスベクター)には、植物細胞において機能する適当なプロモーター及び好ましくはターミネーター等を含む通常トマトの形質転換に用いられるベクターを用いることができ、例えば、CaMV35Sプロモーター、NOSターミネーターを持つ「pBI121」などを用いることができる。
dsRNAコードするDNAをベクターに導入する場合には、一つのベクターにアンチセンスRNAをコードするDNA及びセンスRNAをコードするDNAを導入することもできるし、別のベクターにそれぞれアンチセンスRNAをコードするDNAとセンスRNAをコードするDNAを導入することもできる。
【0029】
アンチセンスベクターの作製は、例えば以下のようにして行うことができる。
配列番号1の塩基配列のコード領域の全長または一部の領域を、PCR法を用いて増幅しDNA断片を得る。アンチセンスRNAをコードするDNAの塩基配列の設計に好ましい領域は上述したとおりである。増幅したDNA断片を、制限酵素サイトを用いたリガーゼ反応により、ベクター内の適当なプロモーターの下流にアンチセンス方向に連結する。また、DNAの3’末端にはターミネーターを配置することが好ましい。
【0030】
組換えベクターを用いて、トマトの形質転換を行うには、アグロバクテリウム法、エレクトロポレーション法、ポリエチレングリコール法、リポソーム法、パーティクルガン法など、植物の形質転換の方法として公知の種々の方法を用いることができる。
形質転換を行う材料としては、トマトのプロトプラスト、培養細胞、カルス、根、茎、葉、種子等の組織を用いることができる。
【0031】
プロトプラストを用いる場合には、これを組換えベクターを保持するアグロバクテリウムと共存培養したり、スフェロプラスト化したアグロバクテリウムと融合したりすることによって、培養細胞を用いる場合は、組換えベクターを保持するアグロバクテリウムと共存培養することによって、組織を用いる場合には、葉の切片を組換えベクターを保持するアグロバクテリウム懸濁液に一定時間浸漬してアグロバクテリウムを組織に感染させ、共存培養することによって、形質転換することができる。
【0032】
このようにして形質転換した細胞、カルス、組織から公知の組織培養法により植物体を再生することができる。
【0033】
例えば、上記形質転換の植物材料に葉切片(リーフディスク)を用いる場合、葉切片にアグロバクテリウムを感染させた後、これらを無機塩類、ビタミン類、糖類、サイトカイニン等の植物ホルモンやカナマイシン等の選抜薬剤、殺菌剤等を加えた培地を用いて培養することで茎葉を形成させることができる。次に、上記培地よりサイトカイニン等の植物ホルモンを除いた培地を用いて茎葉を培養することにより発根を誘導することができる。
【0034】
本発明にいう「トマト」には、トマトの植物体全体、器官、組織、カルス、培養細胞の全てが含まれる。
【0035】
このようにして得られたトマトが形質転換されているか否かは、RT−PCR法、ノーザンハイブリダイゼーション法、ウェスタンブロッティング法により、導入遺伝子の発現量を検出したり、あるいは上記ターゲットDNAの発現の存在を検出したりすることで、確認することができる。
例えば、被検トマトのRNAを抽出し、配列番号1の塩基配列を基に設計したRNAプローブを用いてノーザンハイブリダイゼーションを行うことにより、ターゲットDNAの発現量を検出することもできる。
【0036】
また、本発明における「(a)〜(d)の何れかのDNAの発現が抑制された、ジョイントレス形質を有するトマト」には、該DNAの発現が抑制されている限り、形質転換操作を行った材料が再分化した当代トマトの後代トマトや、それらを片親にして交配したトマトやその後代トマト、さらに、当代トマトや後代トマトの細胞や組織を培養することにより得られたトマトも含まれる。
【0037】
また、本発明は、このようなジョイントレス形質を有するトマトから得られた種子、及びこの種子を栽培することを含むジョイントレス形質を有するトマトの製造方法を提供する。
また、本発明は、前記ジョイントレス形質を有するトマトを原料とする加工品、及び前記ジョイントレス形質を有するトマトを加工することを含むトマトを原料とする加工品の製造方法を提供する。このような加工品としては、例えばトマトの果実の加熱品、搾汁液及び抽出物、さらにはこれらを含む飲食品、食品添加物、飼料、医薬、化粧品等が挙げられる。
【0038】
本発明は、また被検トマトにおける前記(a)〜(d)の何れかのDNAの発現量を検出し、該DNAの発現量が正常型トマトに比して小さい被検トマトを選抜することを含む、ジョイントレス形質を有するトマトの選抜方法を提供する。
DNAの発現量の検出には、RT−PCR法、ノーザンハイブリダイゼーション法、ウェスタンブロッティング法を用いることができる。
【0039】
例えば、配列番号1の塩基配列に特異的なプライマーを設計し、被検トマトから調製したcDNAを鋳型としてPCRを行うことにより、上記DNAの発現量を検出することができる。
例えば、配列番号1における182〜748番の領域の100塩基以上、好ましくは少なくとも400の塩基からなる塩基配列を増幅することができるプライマーセットを設計し、これを用いてPCRを行うことができる。このようなプライマーセットとして、あくまでも一例ではあるが、配列番号1に記載の塩基配列の167〜633番で示される塩基配列の相補配列のうち連続した15以上、好ましくは20以上の塩基を含む塩基配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の192〜748番で示される塩基配列のうち連続した15以上、好ましくは20以上の塩基を含む塩基配列に相補的な塩基配列からなるリバースプライマーからなるプライマーセットが挙げられる。
【0040】
さらに具体例を挙げると、例えば以下のようなプライマーセットが挙げられる。
配列番号1に記載の塩基配列の211〜231番で示される塩基配列の相補配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の660〜680番で示される塩基配列に相補的な塩基配列からなるリバースプライマーからなるプライマーセット。
【0041】
配列番号1に記載の塩基配列の182〜202番で示される塩基配列の相補配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の610〜630番で示される塩基配列に相補的な塩基配列からなるリバースプライマーからなるプライマーセット。
【0042】
配列番号1に記載の塩基配列の296〜316番で示される塩基配列の相補配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の728〜748番で示される塩基配列に相補的な塩基配列からなるリバースプライマーからなるプライマーセット。
【0043】
ここで、「相補配列」とは、完全に相補的である配列をいう。
また、「相補的」とは、プライマーとして機能する限り、完全に相補的である場合のみならず、例えば、少なくとも90%程度、好ましくは少なくとも95%程度の相補性を有する場合も含む。
【0044】
本発明は、また被検トマトにおける前記(a)〜(d)の何れかのDNAの存在を検出し、該DNAが存在しない被検トマトを選抜することを含む、ジョイントレス形質を有するトマトの選抜方法を提供する。
DNAの存在の検出には、PCR法、サザンハイブリダイゼーション法を用いることができる。
【0045】
例えば、配列番号1の塩基配列に特異的なプライマーを設計し、被検トマトから調製したゲノムDNAを鋳型としてPCRを行うことにより、上記DNAの存在を検出することができる。
具体的には、配列番号1に記載の塩基配列の14〜64番で示される塩基配列の相補配列のうち連続した15以上、好ましくは20以上の塩基を含む塩基配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の698〜748番で示される塩基配列のうち連続した15以上、好ましくは20以上の塩基を含む塩基配列に相補的な塩基配列からなるリバースプライマーとからなるプライマーセットを用いて、PCRを行うことができる。
「相補配列」、「相補的」の定義は上述した通りである。
【0046】
フォワードプライマーとしては、例えば配列番号3に示す塩基配列を含むプライマー、又は該塩基配列と少なくとも90%程度、好ましくは少なくとも95%程度の同一性を有する塩基配列からなるプライマーが挙げられる。また、リバースプライマーとしては、例えば配列番号4に示す塩基配列を含むプライマー、又は該塩基配列と少なくとも90%程度、好ましくは少なくとも95%程度の同一性を有する塩基配列からなるプライマーが挙げられる。
また、このようにして選抜した被検トマトについて、上述した方法により該DNAの発現量を検出するなどして、ジョイントレス形質を有するトマトの選抜をさらに行うこともできる。
【実施例】
【0047】
以下、本発明を実施例を挙げてより詳細に説明する。
<1>バイナリーベクターの構築
DNA断片の加工やDNA断片相互の結合、DNA断片のクローニング、DNA配列の確認、大腸菌等の処理(形質転換、培養、プラスミドの抽出等)などの一般的操作の手順は、特に記載のない限り、Molecular Cloning 3rd editionに記載されている方法に従って行った。なお、以下の試験に用いるトマトは、何れも、Tomato Genetics Resource Center(カリフォルニア大学デービス校)(URL: http://tgrc.ucdavis.edu/)から分譲を受けた。
【0048】
1) LeMADS-MC遺伝子断片のクローニング
First-Strand cDNA Synthesis Kit (GE Healthcare Bio-Sciences社)を用いて正常型トマト(品種:LA2838A)の緑熟期果実の全RNA 2μgよりcDNAを合成した(配列番号1)。これを20倍希釈したものをテンプレートとしてGeneAmp(R)PCR System 9700 (登録商標、Applied Biosystems社)を用い、PCR反応を行った。PCR反応終了後は、5μLを1%アガロースゲルで電気泳動して増幅を確認した。尚、プライマーの配列、ならびにPCR反応条件は以下のとおりである。
増幅したPCR断片は、pBluescript SK(+)のEcoRV切断部位にクローニングし、DNAシークエンス解析により、PCRエラーがないことを確認した(このPCR断片をpBT-ASrin4と呼ぶ)。
【0049】
(i)LeMADS-MC(AF448521)プライマー配列
ベクター作製の便宜上、MC001プライマー5'末端にSacI認識部位を付加し、MC002プライマー5'末端にはBamHI認識部位を付加した。
MC001プライマー:5'-GAG CTC TTG AAC GAT ATG AAA G-3'(配列番号5)
MC002プライマー:5'-GG ATC CTC CTT GCT TCT GCT AC -3'(配列番号6)
【0050】
(ii)PCR反応条件
1サンプルあたり、表1に示した組成の反応液を使用し、表2に示したスケジュールで反応を行った。
【0051】
【表1】
【0052】
【表2】
【0053】
2) バイナリーベクターの構築
1)で得られたpBT-ASrin4 を、BamHIとSacIで制限酵素処理してLeMADS-MC cDNA断片を得た。これと、BamHIとSacIで制限酵素処理したpBI121ベクターとをライゲーション反応にて結合させて目的のバイナリーベクター(pBI-ASrin4)を作製した後、このバイナリーベクターで大腸菌を形質転換した。
【0054】
<2>アグロバクテリウムへのバイナリーベクターの導入
MicroPulserエレクトロポレーター(バイオ・ラッド ラボラトリーズ株式会社)を用いたエレクトロポレーション法でアグロバクテリウムEHA105株にpBI-ASrin4を導入した。アグロバクテリウムをYEBプレート(カナマイシン40 mg/mL入り)にまき、2、3日間暗黒下でインキュベートして、pBI-ASrin4が導入された株を選抜した。
【0055】
<3>トマトの形質転換
トマトの形質転換は、以下の論文に従って行った。
highly efficient transformation protocol for Micro-Tom, a model cultivar for tomato functional genomics.Sun HJ, Uchii S, Watanabe S, Ezura H. Plant Cell Physiol. 2006 Mar; 7(3):426-31.
【0056】
1) トマト種子の無菌播種
微粉ハイポネックス(登録商標、株式会社ハイポネックスジャパン)1.5g、シュクロース5.0gを水に溶かして1M NaOHでpH5.8に調整して1Lにした後、寒天8gを加えオートクレーブにて121℃、15分間滅菌し、300mL容プラントボックス(株式会社テックジャム)に30 m
Lずつ分注し、播種用培地を作製した。
一方で、トマト種子(品種:LA2838A)約100粒を5cm四方のガーゼに包んでホッチキスで留め、70%エタノール水溶液に2分間浸した。さらに10倍希釈したキッチンハイター(登録商標、花王株式会社)に45分間浸して消毒した。次に、ガーゼに包んだトマト種子を滅菌水で3回すすぎ、その後、滅菌水に浸して一晩吸水させた。
滅菌したキムタオル(登録商標、日本製紙クレシア株式会社)で、トマト種子に付着した余分な水分を取り、前記の播種用培地に16〜25粒ずつ播種し、25℃で16時間日長の部屋で発芽させた。
【0057】
2) 目的DNAを保持したアグロバクテリウムの培養
カナマイシン(100mg/L)を含むLBプレート培地にpBI-ASrin4を保持したアグロバクテリウムを植菌し、28℃でコロニーが十分大きくなるまで培養した。
LBプレート上に生育したコロニーのうち、1コロニーのアグロバクテリウムを、白金耳で別途調製したカナマイシン(100mg/L)を含む液体LB培地に接種し、28℃で24時間振とう培養した。
【0058】
3) アグロバクテリウム懸濁液の調製
24時間振とう培養した菌液(1mL)を遠心分離(5000rpm、5分間)して、アグロバクテリウムを集菌した。上清の液体LB培地を除去した後、同量のMS培地を添加して、アグロバクテリウムを懸濁した。それを、最終濃度100μMのアセトシリンゴンと10mMのメルカプトエタノールを添加したMS培地で40倍に希釈して、アグロバクテリウム懸濁液を調製した。
【0059】
4) アグロバクテリウムによる子葉切片の感染
発芽したトマトの子葉の真ん中を葉脈に対して垂直方向に切断し、一枚の子葉から2枚の子葉切片(リーフディスク)を作製した。子葉切片を前記のアグロバクテリウム懸濁液に10分間浸し、アグロバクテリウムを感染させた。
【0060】
5) 共存培養
感染処理後、滅菌したキムタオルに子葉切片をのせて懸濁液を吸い取り、共存培地に葉の表が下になるように、シャーレ1枚あたり15切片を並べ、アルミホイルで完全に包み、25℃の培養室で4日間共存培養を行った。
なお、共存培地は、MS培地1L、シュクロース30.0gを混合してpH5.8に調整し、ゲルライト(登録商標、和光純薬工業株式会社)3gを加えてオートクレーブにて121℃、15分間滅菌後、ゼアチン1.5mgを加え、シャーレに30 mLずつ分注して作製した。
【0061】
6) カルス誘導
子葉切片の周辺に菌が増殖している事が目視で確認できたら、カルス誘導培地に葉の表が上になるように移し替え、25℃、16時間日長で培養した。なお、子葉切片は2週間毎に新しいカルス誘導培地に移し替えた。
なお、カルス誘導培地は、MS培地1L、シュクロース30.0gを混合してpH5.8に調整し、ゲルライト(登録商標、和光純薬工業株式会社)3gを加えてオートクレーブにて121℃、15分間滅菌後、ゼアチン1.5mg、カナマイシン100mg、オーグメンチン(登録商標、グラクソ・スミスクライン株式会社)375mgを加え、シャーレに40 mLずつ分注して作製した。
【0062】
7) シュート誘導
子葉切片からカルスが形成され、そのカルスから茎及び葉(シュート)が見え始めたら茎及び葉(シュート)の生長を早めるため、茎及び葉(シュート)部分を子葉切片部分から切り落とし、シュート誘導培地に移した。
なお、シュート誘導培地は、MS培地1L、シュクロース30.0gを混合してpH5.8に調整し、ゲルライト(登録商標、和光純薬工業株式会社)3gを加えてオートクレーブにて121℃、1
5分間滅菌後、ゼアチン1.0mg、カナマイシン100mg、オーグメンチン(登録商標、グラクソ・スミスクライン株式会社)375mgを加え、シャーレに40 mLずつ分注して作製した。
【0063】
8) 発根誘導(発根選抜1)
茎及び葉の部分が1〜2cmの長さまで伸びてきたら、できるだけ根元で切り取り、発根培地に移した。
発根培地で2週間以内に発根した個体を形質転換体の候補として選抜した。一回目の発根培地で発根しなかったものは、切り口を薄く切り落とし、新しい発根培地へ移して発根誘導(発根選抜)を行った。
なお、発根培地は、1/2MS培地1L、シュクロース15gを混合してpH5.8に調整し、ゲルライト(登録商標、和光純薬工業株式会社)3gを加えてオートクレーブにて121℃、15分間滅菌後、カナマイシン50mg、オーグメンチン(登録商標、グラクソ・スミスクライン株式会社)375mgを加え、シャーレに40 mLずつ分注して作製した。
【0064】
9) 発根誘導(発根選抜2)
発根した個体は、発根培地を入れた300mL容プラントボックス(株式会社テックジャム)へ移した。その際、根を切り落とし、二度目の発根誘導(発根選抜)を行った。
【0065】
10) 順化
300mL容プラントボックス(株式会社テックジャム)で約1ヶ月間生長させた植物体は、土を入れた鉢に植え換え、ビニールで覆って湿度の急激な低下を避けながら、徐々に外気条件に慣らし、通常の鉢植え栽培へ移行した。
【0066】
<4>植物体における標的遺伝子(LeMADS-MC)の発現解析
1) 植物体からの全RNA抽出
上記で得られた組換え植物、正常型植物(品種:LA2838A)、及び既存のジョイントレス変異体(jointless (j))(品種:LA1806)の各植物体におけるLeMADS-MCの発現を解析した。
植物体からの全RNAの抽出には、SDS/フェノール法を用いた。植物体の凍結粉末サンプル1 gを乳鉢、乳棒を用いて液体窒素下でホモジナイズした後、10 mLの抽出バッファー(0.1 M Tris-HCl (pH 8.0)、0.1 M NaCl、1% SDS)、2 mLの2-メルカプトエタノール、10 mLのフェノールクロロホルム(フェノール:クロロホルム:イソアミルアルコール=25:24:1)を加え、5分間よく攪拌した。室温にて8,000 rpmで10分間遠心した後、上清を分取し、10 mLのフェノールクロロホルム(フェノール:クロロホルム:イソアミルアルコール=25:24:1)を加え、5分間よく攪拌した。再度、室温にて8,000 rpmで10分間遠心した後、上清を分取し、上清1 mL当り25μLの5 M NaClと0.8 mLの2-プロパノールを加え、-80℃で30分間静置した。その後、4℃にて8,000 rpmで30分間遠心した後、沈殿物を900μLの滅菌水に溶解し、300μLの10 M LiClを加え、-80℃で30分間静置した。さらに、4℃にて15,000 rpmで15分間遠心し、得られた沈殿物を70%エタノールでリンスした。リンス後の沈殿物を300μLの滅菌水に溶解し、30μLの3M酢酸ナトリウム、750μLの100%エタノールを加え、-80℃で30分間静置した後、4℃にて15,000 rpmで15分間遠心し、得られた沈殿物を70%エタノールで再びリンスした。沈殿物が完全に乾いたら、滅菌水を加えて溶解し、これを全RNAとし、-80℃で保存した。
【0067】
2)ノーザン解析
全RNAを1.2%アガロースゲル(1×MOPS、5%(w/v)フォルムアルデヒド)で、1レーン当り10μg泳動し、Hybond-N+メンブレン(GE Healthcare Bio-Sciences社)に転写した。Hybond-N+メンブレンをUVクロスリンクした後、ハイブリダイゼーションバッファー(5×SSC、50%フォルムアミド、0.02% SDS、0.1% N-ラウロイルサルコシン、2%ブロッキング試薬)で、68℃、30分間プレハイブリダイゼーションを行った。
LeMADS-MC遺伝子の翻訳開始点から数えて187番目のaから808番目のgまでを含むcDNA(配列番号1の塩基配列の200〜821番)をもとにDIG RNA Labeling Kit (Roche社)を用いてRNAプローブを合成し(配列番号7)、68℃で10分間熱変性させ、ハイブリダイゼーションバッファーに加え、68℃で一晩ハイブリダイゼーションを行った。その後、Hybond-N+メンブレンを2×SSC/0.1% SDS溶液で室温にて10分間、0.2×SSC/0.1% SDS溶液で室温にてさらに10分間、0.2×SSC/0.1% SDS溶液で68℃にて20分間×2回洗浄した。以降の操作はDIG検出マニュアル(Roche社)に従った。なお、シグナルの検出には化学発光基質としてCDP-Star(Roche社)を用いた。
【0068】
<5>形質転換体後代における導入DNAの解析
1) PCRによる導入DNAの確認
組換え遺伝子が、本当にジョイントレス形質を引き起こしているのかを明らかにするために、組換え植物から得た種子由来の複数の植物体について、ジョイントレス形質発現の有無と同時に組換え遺伝子が受け継がれているかどうかを確認した。
組換え遺伝子の導入によりLeMADS-MCの発現が抑制された組換え植物について、自殖後、着生した果実から種子を収集した。
収集した種子から、ランダムに十数個を選抜し、閉鎖系温室内において常法により栽培し、組換え植物から得た種子由来の植物体を得た。
組換え植物から得た種子由来の各植物体の葉から全DNAを抽出した。抽出にはGE Healthcare Nucleon Phytopure Genomic DNA Extraction Kits(GE Healthcare Bio-Sciences社)を使用した。次に得られたDNAに対してGeneAmp(R) PCR System 9700 (Applied Biosystems社)を用い、PCR反応を行った。PCR反応終了後、1%アガロースゲルで電気泳動を行い、導入DNAの増幅の有無を調べた。プライマー、PCR反応条件等は以下のとおりである。
【0069】
(i)導入遺伝子断片検出用プライマー配列
LeMADS-MC遺伝子断片上(MC001プライマー)と、pBI121由来のCaMV35Sプロモーター領域上(pBI121-Fプライマー)にプライマーを設計した。
MC001プライマー:5'-GAG CTC TTG AAC GAT ATG AAA G-3'(配列番号5)
PBI121-Fプライマー:5'-AGA CCC TTC CTC TAT ATA AG-3'(配列番号8)
【0070】
(ii)PCR反応条件
1サンプルあたり、表3に示した組成の反応液を使用し、表4に示したスケジュールで反応を行った。
【0071】
【表3】
【0072】
【表4】
【0073】
<6>結果
<4>のLeMADS-MC遺伝子の発現解析の結果を図1に示す。
正常型植物や既存のジョイントレス変異体(j)では、LeMADS-MC遺伝子の発現が認められるが、今回作出した組換え植物では、LeMADS-MC遺伝子の発現は認められない。このことから、正常型トマトが形質転換され、LeMADS-MC遺伝子の発現が抑制されたことが示された。また、j変異体ではLeMADS-MC遺伝子の発現が認められたことから、今回作出した組換え植物は、jと異なる遺伝子によってジョイントレス形質が付与されたものであることが示された。これは、jのジョイントレス形質の発現に関わる遺伝子が座乗している染色体(11番)とLeMADS-MC遺伝子が座乗している染色体(5番)が異なるという事実とも矛盾しないものである。
図2は、今回作出したジョイントレス形質が付与された組換え植物(左側)と正常型植物(右側)の果実の写真である。
右側の正常型植物は離層が形成されているのに対し、左側の組換え植物では離層が形成されていなかった。
なお、本試験では、既存のジョイントレス変異体(jointless2(j-2))のLeMADS-MC遺伝子の発現解析は行っていないが、j-2のジョイントレス形質の発現に関わる遺伝子は12番染色体に座乗しているという事実からみて、今回作出した組換え植物は、j-2とも異なる遺伝子によってジョイントレス形質が付与されたものであると考えられる。
【0074】
<5>で得られた組換え植物の後代における導入遺伝子の発現解析の結果を図3に示す。
表5は、今回作出したジョイントレス形質が付与された組換え植物の後代における、組換え遺伝子の有無と表現型(ジョイントレス形質)の相関性を示す。
組換え植物の後代において、組換え遺伝子が受け継がれた系統は離層が形成されていなかった(ジョイントレス形質を有していた)が、組換え遺伝子が受け継がれなかった系統では、離層が形成されていた。このことから、LeMADS-MC遺伝子がジョイントレス形質を付与していることが証明された。
【0075】
【表5】
【産業上の利用可能性】
【0076】
本発明によれば、さまざまな品種のトマトにジョイントレス形質を付与することができ、加工用トマトにバリエーションを与えることができる。
【特許請求の範囲】
【請求項1】
以下の(a)〜(d)の何れかのDNAの発現を抑制する工程を含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の作出方法;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
【請求項2】
前記(a)〜(d)の何れかのDNAの転写産物に相補的なRNAをコードするDNAを細胞内に導入することにより、該DNAの発現を抑制する工程を含む、請求項1に記載のトマトの作出方法。
【請求項3】
前記RNAをコードするDNAが、配列番号1に記載の塩基配列の182〜748番で示される塩基配列のうち少なくとも30以上の連続する塩基からなる塩基配列に対し相補的な塩基配列を含む、請求項2に記載のトマトの作出方法。
【請求項4】
前記RNAをコードするDNAが、配列番号1に記載の塩基配列の211〜680番、182〜630番、又は296〜748番で示される塩基配列に対して相補的な塩基配列を含む、請求項3に記載のトマトの作出方法。
【請求項5】
以下の(a)〜(d)の何れかのDNAの発現が抑制された、ジョイントレス形質を有するトマト(Solanum lycopersicum);
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
【請求項6】
前記(a)〜(d)の何れかのDNAの転写産物に相補的なRNAをコードするDNAを細胞内に保持する、請求項5に記載のトマト。
【請求項7】
前記RNAをコードするDNAが、配列番号1に記載の塩基配列の182〜748番で示される塩基配列のうち少なくとも30の連続する塩基からなる塩基配列に対し相補的な塩基配列を含む、請求項6に記載のトマト。
【請求項8】
前記RNAをコードするDNAが、配列番号1に記載の塩基配列の211〜680番、182〜630番、又は296〜748番で示される塩基配列に対して相補的な塩基配列を含む、請求項7に記載のトマト。
【請求項9】
請求項5〜8の何れか一項に記載のトマトから得られる種子。
【請求項10】
請求項5〜8の何れか一項に記載のトマトを原料とする加工品。
【請求項11】
請求項9に記載の種子を栽培することを含む、ジョイントレス形質を有するトマト(So
lanum lycopersicum)の作出方法。
【請求項12】
以下の(a)〜(d)の何れかのDNAの転写産物に相補的なRNAをコードするDNA;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
【請求項13】
請求項12に記載のDNAを含むベクター。
【請求項14】
被検トマトにおける以下の(a)〜(d)の何れかのDNAの発現量を検出し、該DNAの発現量が正常型トマトに比して小さい被検トマトを選抜することを含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の選抜方法;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
【請求項15】
配列番号1に記載の塩基配列の167〜633番で示される塩基配列の相補配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の192〜748番で示される塩基配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるリバースプライマーとからなるプライマーセットを用いて、被検トマトから調製したcDNAを鋳型としてPCRを行うことにより、前記(a)〜(d)の何れかのDNAの発現量を検出することを含む、請求項14に記載のジョイントレス形質を有するトマトの選抜方法。
【請求項16】
被検トマトにおける以下の(a)〜(d)の何れかのDNAの存在を検出し、該DNAが存在しない被検トマトを選抜することを含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の選抜方法;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
【請求項17】
配列番号1に記載の塩基配列の14〜64番で示される塩基配列の相補配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の698〜748番で示される塩基配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるリバースプライマーとからなるプライマーセットを用いて、被検トマトから調製したゲノムDNAを鋳型としてPCRを行うことにより、前記(a)〜(d)の何れかのDNAの存在を検出するこ
とを含む、請求項16に記載のジョイントレス形質を有するトマトの選抜方法。
【請求項18】
前記フォワードプライマーが、配列番号3に示す塩基配列を含むプライマー、又は該塩基配列と少なくとも90%の同一性を有する塩基配列からなるプライマーであり、前記リバースプライマーが、配列番号4に示す塩基配列を含むプライマー、又は該塩基配列と少なくとも90%の同一性を有する塩基配列からなるプライマーである、請求項17に記載のジョイントレス形質を有するトマトの選抜方法。
【請求項1】
以下の(a)〜(d)の何れかのDNAの発現を抑制する工程を含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の作出方法;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
【請求項2】
前記(a)〜(d)の何れかのDNAの転写産物に相補的なRNAをコードするDNAを細胞内に導入することにより、該DNAの発現を抑制する工程を含む、請求項1に記載のトマトの作出方法。
【請求項3】
前記RNAをコードするDNAが、配列番号1に記載の塩基配列の182〜748番で示される塩基配列のうち少なくとも30以上の連続する塩基からなる塩基配列に対し相補的な塩基配列を含む、請求項2に記載のトマトの作出方法。
【請求項4】
前記RNAをコードするDNAが、配列番号1に記載の塩基配列の211〜680番、182〜630番、又は296〜748番で示される塩基配列に対して相補的な塩基配列を含む、請求項3に記載のトマトの作出方法。
【請求項5】
以下の(a)〜(d)の何れかのDNAの発現が抑制された、ジョイントレス形質を有するトマト(Solanum lycopersicum);
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
【請求項6】
前記(a)〜(d)の何れかのDNAの転写産物に相補的なRNAをコードするDNAを細胞内に保持する、請求項5に記載のトマト。
【請求項7】
前記RNAをコードするDNAが、配列番号1に記載の塩基配列の182〜748番で示される塩基配列のうち少なくとも30の連続する塩基からなる塩基配列に対し相補的な塩基配列を含む、請求項6に記載のトマト。
【請求項8】
前記RNAをコードするDNAが、配列番号1に記載の塩基配列の211〜680番、182〜630番、又は296〜748番で示される塩基配列に対して相補的な塩基配列を含む、請求項7に記載のトマト。
【請求項9】
請求項5〜8の何れか一項に記載のトマトから得られる種子。
【請求項10】
請求項5〜8の何れか一項に記載のトマトを原料とする加工品。
【請求項11】
請求項9に記載の種子を栽培することを含む、ジョイントレス形質を有するトマト(So
lanum lycopersicum)の作出方法。
【請求項12】
以下の(a)〜(d)の何れかのDNAの転写産物に相補的なRNAをコードするDNA;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
【請求項13】
請求項12に記載のDNAを含むベクター。
【請求項14】
被検トマトにおける以下の(a)〜(d)の何れかのDNAの発現量を検出し、該DNAの発現量が正常型トマトに比して小さい被検トマトを選抜することを含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の選抜方法;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
【請求項15】
配列番号1に記載の塩基配列の167〜633番で示される塩基配列の相補配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の192〜748番で示される塩基配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるリバースプライマーとからなるプライマーセットを用いて、被検トマトから調製したcDNAを鋳型としてPCRを行うことにより、前記(a)〜(d)の何れかのDNAの発現量を検出することを含む、請求項14に記載のジョイントレス形質を有するトマトの選抜方法。
【請求項16】
被検トマトにおける以下の(a)〜(d)の何れかのDNAの存在を検出し、該DNAが存在しない被検トマトを選抜することを含む、ジョイントレス形質を有するトマト(Solanum lycopersicum)の選抜方法;
(a)配列番号2のアミノ酸配列からなるタンパク質をコードするDNA、
(b)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が置換、欠失、挿入、又は付加したアミノ酸配列を含み、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA、
(c)配列番号1の塩基配列のコード領域を含むDNA、
(d)配列番号1の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ果梗の離層形成を制御する活性を有するタンパク質をコードするDNA。
【請求項17】
配列番号1に記載の塩基配列の14〜64番で示される塩基配列の相補配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるフォワードプライマーと、配列番号1に記載の塩基配列の698〜748番で示される塩基配列のうち連続した少なくとも15の塩基を含む塩基配列に相補的な塩基配列からなるリバースプライマーとからなるプライマーセットを用いて、被検トマトから調製したゲノムDNAを鋳型としてPCRを行うことにより、前記(a)〜(d)の何れかのDNAの存在を検出するこ
とを含む、請求項16に記載のジョイントレス形質を有するトマトの選抜方法。
【請求項18】
前記フォワードプライマーが、配列番号3に示す塩基配列を含むプライマー、又は該塩基配列と少なくとも90%の同一性を有する塩基配列からなるプライマーであり、前記リバースプライマーが、配列番号4に示す塩基配列を含むプライマー、又は該塩基配列と少なくとも90%の同一性を有する塩基配列からなるプライマーである、請求項17に記載のジョイントレス形質を有するトマトの選抜方法。
【図1】

【図2】
【図3】

【図2】
【図3】
【公開番号】特開2011−130697(P2011−130697A)
【公開日】平成23年7月7日(2011.7.7)
【国際特許分類】
【出願番号】特願2009−292286(P2009−292286)
【出願日】平成21年12月24日(2009.12.24)
【出願人】(000104113)カゴメ株式会社 (50)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
【公開日】平成23年7月7日(2011.7.7)
【国際特許分類】
【出願日】平成21年12月24日(2009.12.24)
【出願人】(000104113)カゴメ株式会社 (50)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
[ Back to top ]