
哺乳動物中の発光複合体の非侵襲的局在化
【課題】哺乳動物などの生体内の病原体とその他の物体を検出し、位置決めし、追跡するための非侵襲的方法を提供すること。
【解決手段】生きている非ヒト哺乳類対象中の異種遺伝子の発現を検出するための非侵襲的方法であって、該方法が、以下:導入遺伝子を含む細胞を有する非ヒト哺乳類対象を提供する工程であって、ここで、(i)該導入遺伝子が、生物発光タンパク質または蛍光発生タンパク質をコードする異種遺伝子を含み、そして(ii)該対象は、不透明な組織を含む、工程;および不透明な組織を貫通する光子放射を光検出装置を用いて測定することによって、該異種遺伝子の発現を検出する工程であって、ここで、該細胞による該異種遺伝子の発現が許容される状況下に該対象が維持される、工程、を含む、方法。
【解決手段】生きている非ヒト哺乳類対象中の異種遺伝子の発現を検出するための非侵襲的方法であって、該方法が、以下:導入遺伝子を含む細胞を有する非ヒト哺乳類対象を提供する工程であって、ここで、(i)該導入遺伝子が、生物発光タンパク質または蛍光発生タンパク質をコードする異種遺伝子を含み、そして(ii)該対象は、不透明な組織を含む、工程;および不透明な組織を貫通する光子放射を光検出装置を用いて測定することによって、該異種遺伝子の発現を検出する工程であって、ここで、該細胞による該異種遺伝子の発現が許容される状況下に該対象が維持される、工程、を含む、方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、哺乳動物対象中の発光体および発光性生物事象を検出し、位置決めし、追跡するための非侵襲的な方法と組成物に関する。
【背景技術】
【0002】
組識内の感染性因子を検出し定量するための現行の生体外方法では、感染性疾患の進行をモニターする能力に限界がある。宿主における感染性因子の複製には、しばしば一次、二次および三次複製部位が関与する。複製部位と、感染性因子がそれらの部位を通過する過程は、接種経路、宿主がコードする因子、およびその感染性因子の決定基に基づいて観測される。
【0003】
考えられる複製部位と感染の進行は、経験から推定できる場合もある。しかし感染部位と疾患の速度は、知られないか、あるいは大雑把にしか推定できないことの方が多いだろう。また感染性疾患の進行は、マウスの同系交配系でさえ、個別的である場合が多く、実験的に感染させた宿主で疾患が平均してどのような経過をたどるのかを決定するために、数多くの感染動物の連続的な生体外分析を行なう必要がある。
【発明の概要】
【発明が解決しようとする課題】
【0004】
動物モデルにおける感染の進行を追跡する手段を持つことが望ましい。その追跡を非侵襲的に行なうことができ、一匹の動物を必要な頻度で有害な影響を与えずに評価できれば、理想的である。本発明は、哺乳動物などの生体内の病原体とその他の物体を検出し、位置決めし、追跡するための非侵襲的方法を提供することを目的とする。
【課題を解決するための手段】
【0005】
1つの局面において、本発明は、次の工程からなる哺乳類対象中の生物適合体の局在を検出する非侵襲的方法を提供する:(a)その対象に、その生物適合体と発光成分の複合体を投与し、(b)その複合体がその対象内で局在を達成しうる期間の後、その対象を光検出装置の検出フィールド内に固定し、(c)その対象を固定状態に維持し、(d)維持中に、その対象中に局在した発光成分からの光子放射を、光子放射の画像を構築することができるまで、その光検出装置で測定し、(e)その画像を構築する。
【0006】
1つの実施形態において、上記の方法は、工程(b)から(e)までを選択した間隔で反復することをさらに含み得、その反復が対象中の生物適合体の局在を経時的に追跡するのに有効であり得る。
【0007】
別の実施形態において、上記の方法は、その測定が増倍化電荷結合光検出素子を用いて行われ得る。
【0008】
別の実施形態において、上記の方法は、その投与が腫瘍細胞ターゲッティング成分を含有する複合体の投与を含み得る、哺乳類対象中の腫瘍細胞の局在を検出するための方法であり得る。
【0009】
別の実施形態において、上記の方法は、その投与が炎症ターゲッティング成分を含有する複合体の投与を含み得る、哺乳類対象中の炎症の局在を検出するための方法であり得る。
【0010】
別の実施形態において、上記の方法は、その投与が感染ターゲッティング成分を含有する複合体の投与を含み得る、哺乳類対象中の病原体による感染の局在を検出するための方法であり得る。
【0011】
別の実施形態において、上記の方法は、その投与が血栓性プラークターゲッティング成分を含有する複合体の投与を含み得る、哺乳類対象内の血栓性プラークの局在を検出するための方法であり得る。
【0012】
別の実施形態において、上記の方法は、その投与が発光成分を含有する粒子である複合体の投与を含み得る。
【0013】
別の実施形態において、上記の方法は、動物モデル中の病原体による感染症の局在を検出するための方法であり得、生物適合体がその病原体であり得る。好ましい実施形態において、その病原体がサルモネラであり得る。
【0014】
別の実施形態において、上記の発光成分が発光性タンパク質であり得る。好ましい実施形態において、上記タンパク質がルシフェラーゼ、黄色蛍光タンパク質およびフェレドキシンIVからなる群より選択され得る。別の好ましい実施形態において、その投与が抗体断片と発光性タンパク質の融合タンパク質である複合体の投与を含み得る。別の好ましい実施形態において、上記生物適合体が形質転換細胞であり得、上記発光成分がその細胞から発現する異種遺伝子の産物であり得る。さらに好ましい実施形態において、上記異種遺伝子の発現が活性化可能なプロモーターの制御下にあり得る。
【0015】
別の局面において、本発明は、哺乳類対象中の生物適合体のレベルを経時的に検出する非侵襲的方法であって、(a)その対象に、その生物適合体と発光成分の複合体を投与し、(b)その対象を光検出装置の検出フィールド内に置き、(c)その対象をその装置の検出フィールド中に維持し、(d)維持中に、その対象中の発光成分からの光子放射を、その光検出装置で測定し、(e)(b)から(d)までの工程を、選択した間隔で反復することからなり、その反復が、その対象中の生物適合体のレベルの経時変化を検出するのに有効である方法を提供する。
【0016】
さらなる局面において、本発明は、次の工程からなる哺乳類対象における導入遺伝子の組み込みを検出する非侵襲的方法を提供する:(a)その対象に、哺乳類細胞中に有効に組み込まれ、かつ、導入遺伝子、発光性タンパク質をコードする遺伝子および活性化可能なプロモーターを含有するベクター構築物(ただし、発光性タンパク質をコードする遺伝子はプロモーターの制御下にある)を投与し、(b)その構築物が組み込みを達成しうる期間の後、プロモーターを活性化し、(c)その対象を光検出装置の検出フィールド内に置き、(d)その対象をその光検出装置の検出フィールド内に維持し、(e)維持中に、その対象中の、発現した発光性タンパク質からの光子放射のレベルを、その光検出装置で測定し、(f)その光子放射のレベルがバックグラウンドより有意に高ければ、その導入遺伝子の組み込みを確認する。
【0017】
なおさらなる局面において、本発明は、次の工程からなる動物中のプロモーター誘導事象の局在を検出する非侵襲的方法を提供する:(a)その事象に反応する誘導性プロモーターとその制御下にある発光性タンパク質をコードする異種遺伝子とを持つトランスジェニック動物中で、その事象を誘発し、(b)その動物を光検出装置の検出フィールド内に置き、(c)その動物を固定状態に維持し、(d)維持中に、その動物内に局在する発現した発光性タンパク質からの光子放射を、光子放射の画像を構築することができるまで、その光検出装置で測定し、(e)その画像を構築する。
【0018】
なおさらなる別の局面において、本発明は、発光性タンパク質を発現させる遺伝子で形質転換されたサルモネラを提供する。
【0019】
一態様として、本発明は、哺乳類対象内の生物適合体の局在を検出する非侵襲的方法を包含する。その生物適合体は分子であってもよいし、巨大分子、細胞、微生物(病原体を含む)、あるいは粒子などであってもよい。
【0020】
この方法では、対象に当該物体と発光成分の複合体を投与する。発光成分は通例、光を放射する分子または高分子(巨大分子)である。この成分は、放射吸収の結果として発光したり(例えば蛍光性または燐光性分子)、あるいは化学反応の結果として発光することができる(例えば生物発光タンパク質)。発光成分の典型例は、ルシフェラーゼやエクオリンのような生物発光タンパク質および、黄色蛍光タンパク質やフェレドキシンIVのような有色または蛍光タンパク質である。
【0021】
この成分は、物体の合成中に組み込んだり(例えば化学的または遺伝子的組み込み、抗体断片と発光性タンパク質の融合タンパク質など)、合成後の化学的カップリング、非共有結合的会合(例えばリポソームによる封入)、物体内でのin situ合成(例えば形質転換細胞における異種生物発光タンパク質の発現)、またはプロモーター誘導物質で剌激されたトランスジェニック動物の細胞における生物発光タンパク質の活性化可能なプロモーター制御性のin situ発現(例えばウイルスの感染によって刺激されたインターフェロン活性化プロモーター)などといった様々な技術により、物体と複合させることができる。
【0022】
対象中で複合体が局在できた期間の後、対象を、十分な量の光子放射を(その光検出装置で)測定して画像を構築するのに有効な期間、光検出装置の検出フィールド内に固定する。典型的な光検出装置は、画像処理装置に接続した増倍化電荷結合素子(ICCD)カメラである。”固定されていない”対象が動く時間スケールに比べて短い時間内に画像を構築できるのであれば、対象は撮像中本質的に”固定されている”ことになり、特別な固定措置は必要ない。次に、光子放射データから画像を構築する。
【0023】
撮像工程を選択した間隔で繰り返し、各間隔に対応する画像を構築することにより、上述の方法で、対象内の物体の局在を経時的に追跡することができる。物体上にターゲッティング成分を結合したり、複合させたり、あるいは組み込むことにより、数多くの特定の応用に、上述の方法を使用することができる。ターゲッティング成分はその物体(例えば抗体または抗体断片)固有の特性であってもよいし、あるいはターゲッティング成分を物体に複合させたり、結合したりまたは組み込んでもよい(例えば抗体を含有するリポソーム)。ターゲッティング成分の例には、抗体、抗体断片、酵素阻害因子、レセプター結合分子、種々の毒素などがある。ターゲッティング成分の標的としては、炎症部位、感染部位、血栓性プラーク、腫瘍細胞などを挙げることができる。これらの標的を特徴づける、ターゲッティング成分による認識に適したマーカーは、よく知られている。
【0024】
さらに、発光成分と複合した病原体(例えばサルモネラ)を物体として使用すれば、この方法で、動物モデル内の病原体による感染部位を検出し、局在化(定位)することができる。
【0025】
関連する態様として、本発明は、哺乳類対象内の生体適合体のレベルを経時的に検出する非侵襲的方法を包含する。この方法は上述の方法に似ているが、必ずしも物体を画像の形で局在化するわけではなく、対象内での物体のレベルの変化を経時的に検出するよう設計される。この方法は、物体(例えば発光性細菌)のレベルに対する治療用物質(例えば抗生物質)の効果を経時的にモニターするのに、とりわけ有用である。
【0026】
もう1つの態様として、本発明は、哺乳類対象における導入遺伝子の組込みを検出する非侵襲的方法を包含する。この方法では、哺乳類細胞に導入遺伝子を組み込むのに有効なべクター構築物を、対象に投与する。そのような構築物は当技術分野ではよく知られている。この構築物は、効果的な組込みに必要な要素に加えて、導入遺伝子(例えば治療用遺伝子)と、発光性タンパク質をコードする遺伝子とを、選択した活性化可能なプロモーターの制御下に含有する。その構築物が組み込みを達成しうる期間の後、そのプロモーターを活性化する。例えば、インターフェロンプロモーターを使用した場合は、ポリイノシン−ポリシトシン二重鎖(ポリIC)を局所投与(例えば足蹠注射)することにより、インターフェロン産生を刺激することができる。次に、対象を光検出装置(例えば光増倍性”暗視”ゴーグルをつけた人)の検出フィールド内に置き、光子放射のレベルを測定または評価する。そのレベルがバックグラウンドを超える場合(すなわち光が”活性化した”領域に優先的に検出されうる場合)、その対象は当該導入遺伝子を組込んでいると記録される。
【0027】
関連する態様として、本発明は、発光性タンパク質をコードする遺伝子を誘導性プロモーターの制御下に含む構築物でトランスジェニックまたはキメラにした動物におけるプロモーター誘導事象の局在を検出する非侵襲的方法を包含する。プロモーター誘導事象には、そのプロモーターを直接活性化する物質の投与、内因性プロモーター活性化因子の産生を刺激する物質の投与(例えばRNAウイルス感染によるインターフェロン産生の剌激)、内因性プロモーター活性化因子の産生をもたらす状態に置くこと(例えば熱ショックまたはストレス)などがある。この事象を誘発し、上述のように動物を撮像する。
【0028】
さらなる態様として、本発明は、発光性タンパク質(例えばルシフェラーゼ)を発現させる遺伝子で形質転換された病原体(例えばサルモネラ)を包含する。
【0029】
またもう1つの側面として、本発明は、病原体による感染の蔓延を抑制するのに有効な治療用化合物を同定する方法をも包含する。この方法では、病原体と発光成分の複合体を対照動物と実験動物に投与し、その実験動物を治療用化合物候補で処置し、上述の方法によって対照動物内と実験動物内の発光性病原体を局在化し、その化合物が対照動物と比較して実験動物における病原体の蔓延または複製を有意に抑制する効力をもつのであれば、その化合物を治療上有効であると認める。複合体としては、蛍光標識された抗体、蛍光標識された粒子、蛍光標識された小分子などが挙げられる。
【0030】
さらなる側面として、本発明は、発光成分と共役した物体を、様々な不透明度を持つ媒質を通して局在化する方法を包含する。この方法では、光検出装置を用いて、その媒質を透過した光子を検出し、その光子を経時的に積分し、積分したその信号に基づいて画像を作成する。
【0031】
さらなる態様として、本発明は、ある生物内の特定の部位で、選択した物質(例えば溶存酸素またはカルシウム)の濃度を測定する方法を包含する。この方法には、濃度センサー(すなわち発光する能力が選択した物質の濃度に依存する発光性分子)を含有する物体(例えば細胞)が含まれる。その発光性分子を含有する物体を、動物内もしくは特定の組識または器官系(例えば脾臓)内に実質上均一に分布するように投与する。その生物を撮像する。光放射の強度と局在は、選択した物質の濃度と位置に相関する。別法として、当該物体は、第2の標識、例えば濃度センサーとは異なる波長で発光することができる分子を含有する。この第2標識を用いて宿主における物体の分布の非均一性を規格化すれば、選択した物質の濃度をより正確に決定することができる。
【0032】
もう1つの側面として、本発明は、腫瘍の増殖および/または転移性伝播を抑制するのに有効な治療用化合物を同定する方法を包含する。この方法では、(i)発光成分で標識した腫瘍細胞または発光成分を含有する腫瘍細胞を実験動物群と対照動物群に投与し、(ii)その実験群を選択した化合物で処置し、(iii)腫瘍細胞に付随する発光性分子からの光子放射を光検出装置で撮像することによって、両群の動物中の腫瘍細胞を局在化し、(iv)その化合物が対照群と比較して実験群における腫瘍の増殖および/または転移性伝播を有意に抑制できるならば、その化合物を治療上有効であると認める。
【0033】
本発明の上記その他の目的と特徴は、下記の発明の詳細な説明を添付の図面と合わせて読むことにより、より完全に理解されるだろう。
【0034】
すなわち、本発明により、哺乳動物などの生体内の病原体とその他の物体を検出し、位置決めし、追跡するための非侵襲的方法が提供される。
【図面の簡単な説明】
【0035】
【図1】図1は、サルモネラ(Salmonella)SL1344株、BJ66株およびLB5000株を形質転換してSL1344lux株、BJ66lux株およびLB5000lux株を作出するために使用したlux pCGLS1プラスミドのマップである。
【図2A】図2Aは、サルモネラSL1344lux株とBJ66lux株によるマクロファージとHEp−2細胞の接着および侵入を測定したアッセイの結果を示す。図2Aは、アッセイ皿のウェル内で局在化した発光性細菌細胞を示す。光子を1分間にわたって積分することによって得た擬似色像を、アッセイ皿のグレースケール像に重ね合せることにより、表記の”合成画像”を作成した。
【図2B】図2Bは、サルモネラSL1344lux株とBJ66lux株によるマクロファージとHEp−2細胞の接着および侵入を測定したアッセイの結果を示す。図2Bは、ゲンタマイシンで処理しなかったウェルの相対光強度を示す。
【図2C】図2Cは、サルモネラSL1344lux株とBJ66lux株によるマクロファージとHEp−2細胞の接着および侵入を測定したアッセイの結果を示す。図2Cは、図2Bで撮像したウェルと同じウェルから単離された1mlあたりのコロニー形成単位(CFU)の数を示す。
【図2D】図2Dは、サルモネラSL1344lux株とBJ66lux株によるマクロファージとHEp−2細胞の接着および侵入を測定したアッセイの結果を示す。図2Dは、ゲンタマイシンで処理したウェルの相対光強度を示す。
【図2E】図2Eは、サルモネラSL1344lux株とBJ66lux株によるマクロファージとHEp−2細胞の接着および侵入を測定したアッセイの結果を示す。図2Eは、図2Dで撮像したウェルと同じウェルから単離された1mlあたりのコロニー形成単位(CFU)の数を示す。
【図3A】図3Aは、LB5000lux細菌懸濁液の希釈液を含有する4つのガラス製毛細管の合成画像である。発光は30秒間の積分によって測定した。各毛細管内の懸濁液の両側には空気ポケットが存在する。
【図3B】図3Bは、マウスの腹腔内に野生型SL1344luxを接種した後の生物発光の分布を表わす。
【図4】図4は、LB5000luxが生成した光の透過を、動物組識越しに調べるために使用したバイアルの概略図である。
【図5A】図5Aは、低病原性LB5000luxサルモネラを経口接種し、図に表示した時間に撮像したBalb/cマウスの合成画像である。発光シグナルを5分間積分した。
【図5B】図5Bは、低病原性LB5000luxサルモネラを経口接種し、図に表示した時間に撮像したBalb/cマウスの合成画像である。発光シグナルを5分間積分した。
【図5C】図5Cは、非侵入性BJ66luxサルモネラを経口接種し、図に表示した時間に撮像したBalb/cマウスの合成画像である。発光シグナルを5分間積分した。
【図5D】図5Dは、非侵入性BJ66luxサルモネラを経口接種し、図に表示した時間に撮像したBalb/cマウスの合成画像である。発光シグナルを5分間積分した。
【図5E】図5Eは、病原性SL1344luxサルモネラを経口接種し、図に表示した時間に撮像したBalb/cマウスの合成画像である。発光シグナルを5分間積分した。
【図5F】図5Fは、病原性SL1344luxサルモネラを経口接種し、図に表示した時間に撮像したBalb/cマウスの合成画像である。発光シグナルを5分間積分した。
【図6】図6は、病原性SL1344lux細菌株(左の2匹)または低病原性SL5000lux細菌株(右の2匹)を腹腔内(i.p.)注射した32時間後のマウスにおけるサルモネラの分布を示す合成画像である。
【図7】図7Aと7Bは、全身性サルモネラ感染症に対して耐性なマウス(129XBalb/c,Ityr/s)における病原性サルモネラの分布を示す。図7Aは第1日、図7Bは第8日である。
【図8A】図8Aは、病原性が減少したサルモネラ突然変異体(BJ66lux)の経口接種7日後の分布を示す。図8Aは発光の外部非侵襲的撮像を示す。記号を付した器官は、C−盲腸、L−肝臓、I−小腸、Sp−脾臓である。
【図8B】図8Bは、病原性が減少したサルモネラ突然変異体(BJ66lux)の経口接種7日後の分布を示す。図8Bは図8Aと同じ動物を腹壁切開後に撮像したものである。記号を付した器官は、C−盲腸、L−肝臓、I−小腸、Sp−脾臓である。
【図8C】図8Cは、病原性が減少したサルモネラ突然変異体(BJ66lux)の経口接種7日後の分布を示す。記号を付した器官は、C−盲腸、L−肝臓、I−小腸、Sp−脾臓である。図8Cは、盲腸の前方と後方の腸管腔内に空気を注入した後に作成した腹壁切開後の画像を示す。
【図9A】図9Aは、SL1344luxの腹腔内接種後の感受性Balb/cマウスにおけるサルモネラSL1344luxの分布を示す。図9Aは腹腔を開く前に撮像した。
【図9B】図9Bは、SL1344luxの腹腔内接種後の感受性Balb/cマウスにおけるサルモネラSL1344luxの分布を示す。図9Bは腹腔を開いた後に撮像した。
【図9C】図9Cは、SL1344luxの腹腔内接種後の感受性Balb/cマウスにおけるサルモネラSL1344luxの分布を示す。図9Cは盲腸を左側に引き寄せた後に撮像した。
【図10A】図10Aは、経口接種マウス中のSL1344luxサルモネラからの生物発光に対するシプロフロキサシン処置の影響を表わす。図10Aは、処置動物と無処置動物について、腹部領域から測定された相対生物発光を、処置開始後の時間の関数として表わすグラフである。
【図10B】図10Bは、経口接種マウス中のSL1344luxサルモネラからの生物発光に対するシプロフロキサシン処置の影響を表わす。図10Bは、SL1344luxサルモネラを経口接種した8日後の、シプロフロキサシンで処置する前のマウスの合成画像である。
【図10C】図10Cは、経口接種マウス中のSL1344luxサルモネラからの生物発光に対するシプロフロキサシン処置の影響を表わす。図10Cは、同じマウスの対照の5.5時間後(処置なし)の合成画像である。
【図10D】図10Dは、経口接種マウス中のSL1344luxサルモネラからの生物発光に対するシプロフロキサシン処置の影響を表わす。図10Dは、SL1344luxサルモネラを経口接種した8日後の、シプロフロキサシンで処置する前のマウスの合成画像である。
【図10E】図10Eは、経口接種マウス中のSL1344luxサルモネラからの生物発光に対するシプロフロキサシン処置の影響を表わす。図10Eは、同じマウスの処置後5.5時間(図l0E)の合成画像である。
【発明を実施するための形態】
【0036】
(I.定義)
特に明示しない限り、本明細書で使用する用語はすべて、本発明の技術分野で使用される通常の意味を持つ。本明細書における不透明媒質とは「従来通りの」不透明な媒質を指し、必ずしも完全に不透明であるとは限らない。したがって不透明媒質とは、一般に透明でなく半透明でもないとみなされる媒質をいい、これには木片や哺乳動物の肉および皮膚などが含まれる。
【0037】
特に断わらない限り、ルシフェラーゼには原核生物ルシフェラーゼと真核生物ルシフェラーゼが含まれ、また、赤色領域の波長で発光するルシフェラーゼのような光学特性が変異または変化した変種も含まれる。
【0038】
生物適合体とは、哺乳動物に投与できる物体をいう。これには哺乳動物にとって有害でありうる病原体も含まれる。発光性タンパク質を発現させる導入遺伝子を含有する細胞を持つ動物の場合、生物適合体とは、その哺乳動物を構成する導入遺伝子含有細胞を指す。
【0039】
発光性とは、化学反応または放射線の吸収により光を発生しうることをいう。
【0040】
本明細書において光とは、特に断わらない限り、約300nmと約1100nmの間の波長を持つ電磁放射線をいう。
【0041】
感染の蔓延とは、通常、最初の感染部位以外の宿主部位の、病原体による伝播とコロニー形成を指す。ただしこの用語には、最初の感染部位における病原体の大きさおよび/または数の増大も含まれうる。
【0042】
lux−ルシフェラーゼおよび光子放射に関係する原核生物遺伝子。
【0043】
luc−ルシフェラーゼおよび光子放射に関係する真核生物遺伝子。
【0044】
プロモーター誘導事象とは、選択した誘導性プロモーターの直接的または間接的誘導をもたらす事象を指す。
【0045】
異種遺伝子とは、宿主生物中に形質移入された遺伝子を指す。通例、異種遺伝子とは、形質移入または形質転換された細胞のゲノムDNAに元来由来しない遺伝子を指す。
【0046】
(II.発明の一般的概観)
本発明は、哺乳類対象中の発光性複合体の非侵襲的撮像および/または検出に関する方法と組成物を包含する。この複合体は、生物適合体と発光成分とを含有する。生物適合体には、環状有機分子などの小分子;タンパク質などの高分子;ウイルス、細菌、酵母、カビなどの微生物;あらゆるタイプの病原体および病原性物質;ビーズやリポソームなどの粒子が含まれるが、これらに限らない。また、生物適合体は、撮像される哺乳類対象を構成する細胞の全部または一部であってもよい。
【0047】
発光能は、発光成分の複合体化によって物体に付与される。そのような成分には、蛍光分子、蛍光タンパク質、光子を放出する酵素反応、生物発光タンパク質などの発光物質がある。複合体化(共役)には、化学的カップリング、融合タンパク質の遺伝子操作、あるいは生物発光タンパク質を発現させる細胞、微生物または動物の形質転換が含まれうる。例えば、その物体が撮像する哺乳類対象を構成する細胞である場合、その発光成分はその細胞に「共役(複合)」させた生物発光タンパク質または蛍光タンパク質であってよく、それはトランスジェニック動物またはキメラ動物の作出によりその細胞内に導入されたべクター構築物からのプロモーター制御的発現の局在性による。
【0048】
通例、発光性複合体を様々な方法で対象に投与し、その対象内で局在させ、それを撮像する。撮像または対象からの光子放射の測定は数十分間続くことがあるので、常にそうであるとは限らないが、通常は撮像工程中、対象を固定しておく。
【0049】
発光体の撮像は、極めて低レベルの光(通常は単一光子事象)を検出し、画像の構築が可能になるまで光子放射を積分できる光検出器の使用を必要とする。そのような高感度光検出器の例には、単一光子事象を増幅した後、検出系に固有の背景ノイズに対して単一光子を検出できる(例えば液体窒素で冷却された)カメラまたはカメラ群で、その事象を検出する装置がある。
【0050】
光子放射画像を作成したら、通例、放射された光子の発生源に関して準拠座標系を与えるため(すなわち対象に関して発光性複合体を局在化するため)に、それをその対象の”通常の”反射光画像に重ね合わせる。次に、そのような”合成”画像を分析することにより、対象内の標的の位置および/または量を決定する。
【0051】
以下に、上述の段階と態様をより詳細に記述する。
【0052】
(III.発光体)
(A.発光成分)
本発明の実施に有用な発光成分(light−generating moiety;LGM)、発光分子または発光構築物は、その応用に応じて様々な形態のいずれをとってもよい。これらに共通する特徴は、それらが発光性であるということ、すなわちそれらが電子的励起状態から、より低いエネルギー状態(通常、基底状態)への遷移の結果として、原子または分子から紫外(UV)、可視および/または赤外(IR)領域の電磁放射線を放射するということである。
【0053】
発光成分の例には、蛍光分子、化学発光化合物、燐光化合物、生物発光化合物などの光冷光(フォトルミネセンス)分子がある。
【0054】
本発明との関連性が強いLGMの2つの特徴は、そのサイズとスペクトル特性である。これらについては、スペクトル特性に関する一般的議論の後、後述する発光成分の特定のタイプに関連して議論する。
【0055】
(1.スペクトル特性)
本発明の重要な側面は、非侵襲的に外部から検出できるように動物組識を貫通しうる光を生成する発光成分の選択である。光が動物組識(ほとんど水からなる)のような媒質を通過する能力は、主として、その光の強度と波長によって決まる。
【0056】
単位体積中の発光強度が強いほど、その光は検出しやすくなる。単位体積中に生成する光の強度は、後述する個々のLGMのスペクトル特性と、単位体積中の当該成分の濃度に依存する。したがって、通例、物体内または物体上に高濃度のLGMを配置する共役法(例えばリポソームの高効率負荷や、細胞内での生物発光タンパク質の高レベル発現など)により、例えば各物体にLGMを一つだけ共役させる方法よりも深い組識層を通して検出することが容易な、より明るい発光性複合体(light−emitting conjugate; LEC)を作成する。
【0057】
組識層越しのLGMの検出性を支配する第2の因子は、放射光の波長である。ほとんどの組織は主として水からなっているので、動物組識の吸収特性を近似するには水を使用することができる。水が長波長光(赤色領域の光)を短波長光より容易に透過させることはよく知られている。
【0058】
したがって、通例、黄色から赤色(550〜1100nm)の範囲の光を放射するLGMの方が、より短い波長で放射するLGMより好ましい。後述するLGMのいくつかはこの範囲で放射する。しかし、本発明の裏付けとして行なった後述の実験によれば、486nm域で放射するLGMで本発明を実施すると、それが最適な放射波長でないという事実にもかかわらず、良好な結果を得ることができる。これらの結果は、これらの実験で使用したLEC(形質転換サルモネラ細胞)内に比較的高濃度のLGM(ルシフェラーゼ分子)が存在することと、高感度な検出器を使用したことが、その一因だと考えられる。より最適な放射波長を持つLGMを使用すれば、より低濃度のLGMを持つLGEで同様の検出結果が得られることは、理解されるだろう。
【0059】
(2.蛍光系成分)
蛍光は、放射線源の除去後極めて短い持続時間を持つ単一の電子励起状態から生じる物質の発光である。励起光の一部はその蛍光分子によって熱に変換されるので、放射される蛍光の波長は励起光の波長より長い(ストークスの法則)。
【0060】
蛍光分子がルミネセンスを示すには光の入力が必要であるから、本発明における蛍光分子の使用は、生物発光分子の使用よりも複雑になるだろう。通例、対象から検出される蛍光光子シグナルを汚染しないように、励起光を遮蔽する予防措置がとられる。自明の予防措置としては、蛍光顕微鏡で使用されるような励起フィルターを放射線源に設置することが挙げられる。適切に選択された励起フィルターは、蛍光成分によって放射される光子の波長と類似する波長を持つ光子の大半を遮断する。また、蛍光光子の波長以外の波長を持つ光子の大半をふるい落とすために、障壁フィルターを検出器に使用する。上述のようなフィルターは、Omega Optical社(バーモント州ブラトルバロ)を含む様々な商業的供給源から入手できる。
【0061】
また、適切な励起波長に近く、蛍光放射波長には近くない高強度光を発生させるレーザーを使用して、蛍光成分を励起させることもできる。例えば共焦点顕微鏡のようにレーザーが対象を走査できるように、x−y変換機構を使用してもよい。
【0062】
もう1つの予防措置として、検出器の部位に到達した放射光子が、対象を完全に通過したものだけとなるように、放射線源を対象の背後に設置し、遮蔽することもできる。さらに、蛍光成分を励起するために使用する光の波長に対して低い感度を持つ検出器を選択してもよい。
【0063】
上述の措置を賢明に適用すれば、本発明の方法による蛍光性LGMの検出が可能である。
【0064】
蛍光成分としては、フルオレセインのような小さい蛍光分子と、緑色蛍光タンパク質(Chalfieら,1994,Science 263:802−805;MorinおよびHastings,1971,J.Cell.Physiol.77:313)やルマジンおよび黄色蛍光タンパク質(O’Kaneら,1991,PNAS 88:1100−1104;Daubnerら,1987,PNAS 84:8912−8916)のような蛍光タンパク質が挙げられる。また、ある種の有色タンパク質、例えばフェレドキシンIV(Grabauら,1991,J.Biol.Chem.266:3294−3299;その蛍光性はまだ評価されていない)などにも蛍光性があって、本発明で使用できるかもしれない。フェレドキシンIVは赤みがかった色を持ち、それは比較的長波長で蛍光または反射して組識を貫通するのに有効な光を生じる可能性を示すので、特に有望な候補である。さらに、この分子はタンパク質としては小さいので(95アミノ酸)、物体に共役させた時に、それらの機能に与える影響を最小限に抑えることができる。
【0065】
小さい蛍光分子の利点は、それを結合させる物体の生物活性を妨害する可能性が、より大きな発光成分よりも少ないということである。また、本発明での使用に適した種々の励起および放射スペクトルを持つ市販の蛍光分子を入手することもできる。例えばMolecular Probes社(オレゴン州ユージーン)は、ルシファーイエロー(428nmで吸収し、535nmで放射)やナイルレッド(551nmで吸収し、636nmで放射)を含む多数の発蛍光団を販売している。さらに、種々の結合法で使用できる様々な基で誘導体化された分子も(例えばMolecular Probes社から)入手することができる。
【0066】
(3.生物発光系成分)
化学発光(化学反応の結果としての発光)と生物発光(生体からの可視発光)は、多くの側面から詳細に研究されている(例えばCampbell,1988,Chemiluminescence.Principles and Applications in Biology and Medicine(英国チチェスター;Ellis Horwood社およびVCH出版社))。以下に顕著な特徴を簡単に要約する。
【0067】
生物発光分子は、光を放射するために放射エネルギーの入力を必要としない点で、蛍光分子とは異なる。むしろ生物発光分子は、ATPなどの化学エネルギーを利用して光を発生させる。蛍光成分に対する生物発光成分の利点は、そのシグナルに事実上バックグラウンドがないということである。検出される光は、外来の生物発光成分によって生成した光だけである。これに対して、蛍光分子を励起するために使用される光は、しばしば意図する標的以外の物質の蛍光をもたらす。これは”バックグラウンド”が生きている動物の内部環境のように複雑な場合は、とくにそうである。
【0068】
生物発光分子にはいくつかの種類が知られている。それらには、ルシフェラーゼファミリー(例えばWoodら,1989,Science 244:700−702)や、エクオリンファミリー(例えばPrasherら,Biochem.26:1326−1332)がある。ルシフェラーゼファミリーの要素は、種々の原核生物と真核生物で同定されている。原核発光(lux)系に関与するルシフェラーゼその他の酵素と、それに対応するlux遺伝子は、ビブリオ(Vibrio)属とフォトバクテリウム(Photobacterium)属の海生細菌およびキセノラブダス(Xenorhabdus)属の陸生細菌から単離されている。
【0069】
ルシフェラーゼ系(luc)を含有する代表的真核生物は、北アメリカホタルPhotinus pyralisである。ホタルルシフェラーゼは詳細に研究されており、ATPアッセイに広く使用されている。コメツキムシの一種 Pyrophorus plagiophthalamus由来のルシフェラーゼをコードするcDNAが、クローン化され、発現されている(Woodら,1989,Science 244:700−702)。この甲虫は、種の異なる要素が異なる色の生物発光を放射する点で珍しい。互いに95〜99%の相同性を持つ4種類のクローンが単離された。それらは546nm(緑色)、560nm(黄緑色)、578nm(黄色)、および593nm(橙色)の光を放射する。最後の種類(593nm)は、その放射光がそれより短い波長の光よりも容易に組識を貫通する波長を持つので、本発明で発光成分として使用するには特に好都合である。
【0070】
ルシフェラーゼ類とエクオリン様分子は、ATPやNAD(P)Hなどのエネルギー源と、ルシフェリンまたはコエレントリジン(coelentrizine)と酸素などの基質を必要とする。
【0071】
ルシフェラーゼ酵素を発光させるには、基質ルシフェリンをルシフェラーゼ酵素に供給しなければならない。ルシフェラーゼ酵素がluxルシフェラーゼをコードするcDNAを含有するベクターの発現産物として導入される場合、ルシフェリンを供給する便利な方法は、ルシフェラーゼを発現させるだけでなく、ルシフェリンを合成するための生合成酵素をも発現させることである。そのような構築物で形質転換された細胞では、生物発光に必要な外因性要素が酸素だけになる。そのような方法(実施例1に詳述)を用いてlux形質転換サルモネラを作成し、それを使って本発明を裏付ける後述の実験を行なう。
【0072】
土壌菌Xenorhabdus luminescensから得たluxオペロンをコードするプラスミド構築物(Frackmanら,1990,J.Bact.172:5767−5773)は、ヘテロ二量体ルシフェラーゼの2つのサブユニットと3つの補助タンパク質を発現させることにより、形質転換大腸菌に光子放射能を付与する(Frackmanら,1990)。X.luminescensのlux遺伝子を発現させる大腸菌の最適な生物発光は37℃で観察され(SzittnerおよびMeighen,1990,J.Biol.Chem.265:16581−16587;Xiら,1991,J.Bact.173:1399−1405)、これは真核発光生物や他の原核発光生物由来のルシフェラーゼの低い最適温度と対照的である(Campbell,1988,Chemiluminescence.Principles and Applications in Biology and Medicine(英国チチェスター:Ellis Horwood社およびVCH出版社))。したがってX.luminescens由来のルシフェラーゼは、動物中での研究にマーカーとして使用するのに適している。
【0073】
上述または実施例1に記述するようなルシフェラーゼベクター構築物は、大半の細菌と多くの真核細胞(luc構築物)を含む様々な宿主細胞の形質転換に使用できる。また、ヘルペスウイルスやワクシニアウイルスのようなある種のウイルスは、ルシフェラーゼを発現させるように遺伝子操作することができる。例えば、Kovacs Sz.およびMettenlieter,1991,J.Gen.Virol.,72:2999−3008には、ヘルペスウイルス内のホタルルシフェラーゼをコードする遺伝子の安定な発現が記述されている。BrasierおよびRon,1992,Meth.in Enzymol.216:386−396には、哺乳類細胞内でのルシフェラーゼ遺伝子構築物の使用が記述されている。培養哺乳類細胞からのルシフェラーゼ発現は、CCD撮像法で巨視的(IsraelおよびHonigman,1991,Gene 104:139−145)にも微視的(Hooperら,1990,J.Biolum.and Chemilum.5:123−130)にも研究されている。
【0074】
(B.物体)
本発明は、上述のような発光成分、発光性構築物または発光分子を含むように修飾したまたは共役させた物体を包含する。そのような共役体または修飾体を、発光体、発光性複合体(LEC)または単に複合体という。これらの物体自体は、例えば分子、高分子、粒子、微生物または細胞などの形態をとることができる。発光成分を物体に共役させる方法は、その成分と物体の性質に依存する。代表的な共役法については、後述の物体に関連して議論する。
【0075】
(1.小分子)
本発明の実施に有用であろう小分子物体としては、病原体もしくは内因性リガンドまたはレセプターと特異的に相互作用する化合物が挙げられる。そのような分子の例には、薬物や治療用化合物;ある種のクモ、ヘビ、サソリ、渦鞭毛藻類、海性巻貝、細菌を含む有毒生物の毒液中に存在するような毒素;NGF、PDGF、TGF、TNFなどの増殖因子;サイトカイン;生理活性ペプチドなどがあるが、これらに限るわけではない。
【0076】
これらの小分子は、その小分子の生物活性が仮に妨害されるとしてもそれが最小限であるような発光成分(例えば小さい蛍光分子(上述))と共役(結合)させることが好ましい。結合の性質は通例化学的であり、当業者の知る様々な方法のいずれで行なってもよい。
【0077】
小分子物体は、形式的な結合操作が必要ないように、発光成分を含有するよう合成することができる。また、発光成分と反応できる反応性基を持つ小分子物体を合成したり、その逆を行なうこともできる。
【0078】
本発明の発光成分と結合した小分子は、ヒトの疾患または状態の動物モデルで使用してもよいし、処置すべきヒト対象で直接使用してもよい。例えば、腫瘍細胞上に発現するレセプターに高い親和力で結合する小分子を動物モデルで使用することにより、腫瘍を局在化し、その大きさの推定値を得たり、治療薬候補による処置後の腫瘍の増殖または転移の変化をモニターすることができる。また、そのような分子を用いて、癌患者中の上述のような腫瘍の特徴をモニターすることもできる。
【0079】
(2.高分子)
ポリマーや生体ポリマーのような高分子は、本発明の実施に有用な物体のもう1つの例である。代表的な高分子には、抗体、抗体断片、融合タンパク質、ある種のベクター構築物がある。
【0080】
商業的供給源から購入したり、当技術分野で知られる方法(Harlowら,1988,Antibodies:A Laboratory Manual,第10章,402頁,Cold Spring Harbor Press)によって作成した抗体または抗体断片を用いて、それら抗体を発光成分と結合し、その複合体を対象に例えば注射などによって投与し、その複合体を抗原の部位に局在させ、その複合体を撮像することにより、哺乳類対象内の抗原を局在化することができる。
【0081】
抗体と抗体断片は、本発明での物体としての使用に関していくつかの利点を持つ。これらはその性質上、それら自体がターゲッティング成分を構成する。さらにその大きさゆえに、小さい蛍光分子、蛍光タンパク質および生物発光タンパク質を含む数タイプの発光成分との結合に馴染みやすく、また、例えば細胞やリポソームなどと比較して、迅速に拡散できる。
【0082】
発光成分は抗体または断片に直接結合させることもできるし、例えば蛍光性の二次抗体などを用いて間接的に結合させることもできる。直接的結合は、抗体または抗体断片に対する例えば発蛍光団の一般的な化学的結合によって、あるいは遺伝子操作によって行なうことができる。蛍光タンパク質または生物発光タンパク質に結合した抗体または抗体断片を含有するキメラまたは融合タンパク質を構築することができる。例えばCasadeiら,1990,PANS 87:2047−2051には、エクオリンと抗体の融合タンパク質遺伝子を哺乳類細胞内で発現させることができるベクター構築物の作成法が記述されている。
【0083】
抗体を含有する複合体は、本発明の多くの応用に使用できる。例えば炎症部位に発現するE−セレクチンに対する標識抗体を使用して、その炎症を局在化し、抗炎症薬候補の効果をモニターすることができる。
【0084】
ベクター構築物そのものも、本発明に応用できる高分子体となりうる。例えば、発光性分子をコードする遺伝子と治療用遺伝子とを、選択したプロモーター(すなわち当該治療用遺伝子が標的とする細胞内で発現するプロモーター)の制御下に含有する真核発現ベクターを構築することができる。本発明の方法を用いてアッセイされるその発光性分子の発現は、その治療用遺伝子の発現位置と発現レベルを決定するために使用できる。その治療用遺伝子の発現が、処置される人または動物モデル内で直接的な表現型を持たない場合に、この方法はとりわけ有用だろう。
【0085】
(3.ウイルス)
本発明のある種の側面で有用なもう1つの物体はウイルスである。多くのウイルスは哺乳類宿主に感染する病原体であるから、それらのウイルスを発光成分と共役させ、初期感染部位と感染の蔓延を研究するために使用することができる。また、発光成分で標識したウイルスは、感染もしくは感染の蔓延を抑制する薬物のスクリーニングに使用することもできる。
【0086】
ウイルスは、発光成分と結合した抗体を用いて間接的に標識したり、あるいは例えばウイルス粒子を(Dhawanら,1991,J.Immunol.147(1):102の方法などで)ビオチニル化した後、それらを検出可能成分(例えば蛍光分子)と結合したストレプトアビジンにさらすことにより、間接的に標識することができる。
【0087】
別法として、ウイルス粒子をローダミンのような発蛍光団で、例えばFanら,1992,J.Clin.Micro.30(4):905の方法を用いて直接標識してもよい。また、発光性タンパク質を発現させるようにウイルスを遺伝子操作することもできる。ヘルペスやワクシニアのようなある種のウイルスのゲノムは、本発明の裏付けとして行なった実験で使用したlux遺伝子やluc遺伝子のような大きさの遺伝子を十分に順応させうるほどに大きい。
【0088】
標識したウイルスを動物モデルで使用することにより、感染を局在化し、感染の進行をモニターすることができ、また感染の蔓延を抑制するのに有効な薬物をスクリーニングすることもできる。例えば、ヘルペスウイルス感染症は皮膚病変として現われるが、このウイルスはヘルペス脳炎をも引き起こしうる。上述の方法のいずれかで標識したウイルスを用いることにより、このような感染症を局在化し、モニターすることができると共に、様々な抗ウイルス剤を中枢神経系(CNS)感染症におけるその効力について調べることができる。
【0089】
(4.粒子)
ビーズ、リポソームなどを含む粒子は、本発明の実施に有用なもう1つの物体を構成する。これらはサイズが大きいので、粒子には、例えば小分子などの場合よりも多数の発光成分を共役させることができる。これは、より短い露光時間で、またはより厚い組識層を通して検出することができる、より高濃度の光放射をもたらす。また、本質的に純粋なターゲッティング成分またはリガンド(例えば抗原や抗体)をその表面に含むように、リポソームを構築することもできる。さらに、例えば生物発光タンパク質分子などを比較的高濃度に、リポソームに負荷することもできる(Campbell,1988,Chemiluminescence.Principles and Applications in Biology and Medicine(英国チチェスター:Ellis Horwood社およびVCH出版社))。
【0090】
さらに、2種類のリポソームを、その両方が存在する場合にのみ光が生成するように、同じ細胞タイプに向かわせることもできる。例えば一方のリポソームにルシフェラーゼを担持させ、他方にルシフェリンを担持させることができる。これらのリポソームはターゲッティング成分を保持してもよく、2つのリポソーム上のターゲッティング成分は同じであってもよいし、異なってもよい。感染細胞上のウイルスタンパク質は、感染した組識または器官の同定に使用できる。免疫系の細胞は、単一のまたは複数の細胞表面マーカーを用いて局在化することができる。
【0091】
血中循環時間を延長し、血流によるターゲッティングを増進するには、リポソームの表面を、例えばリン脂質−ポリエチレングリコール複合体の組み込みなどによってコーティングすることが好ましい。このタイプのリポソームはよく知られている。
【0092】
(5.細胞)
細胞は、原核、真核共に、本発明の実施に有用なもう1つの物体を構成する。細胞にも粒子と同様に比較的高濃度の発光成分を負荷できるが、細胞には、例えば異種遺伝子構築物で細胞をトランスフェクションすることによって、発光成分を供給しうるという利点がある。また、”ターゲッティング成分”もしくは細胞を対象内の所望の位置に導くのに有効な分子を発現させる細胞を選択することもできる。別法として、適当なターゲッティング成分を発現させるベクター構築物で細胞をトランスフェクションすることもできる。
【0093】
使用する細胞タイプは、その応用に依存する。例えば後に詳述するように、サルモネラなどの細菌細胞を使用することにより、高レベルな時間的空間的解像度でその感染過程を研究し、その感染過程に対する薬物または治療剤の効果を評価することができる。
【0094】
細菌細胞は有効な物体を構成する。例えば、それらをトランスフェクションすることにより、高レベルの発光成分や高レベルのターゲッティングタンパク質を容易に発現させることができる。また、表面結合型抗体を発現させる細菌を含む大腸菌ライブラリーを得て、それをスクリーニングすることにより、選択した抗原に対する抗体を発現させるコロニーを同定することも可能である(Stratagene社,カリフォルニア州ラホーヤ)。次に、このコロニーから得た細菌を、発光性タンパク質の遺伝子を含有する第2のプラスミドで形質転換し、形質転換体を本発明の方法で上述のように使用することにより、哺乳類宿主内の当該抗原を局在化できる。
【0095】
病原性細菌を発光成分と共役させ、それを動物モデル内で使用することにより、その感染過程を生体内で追跡したり、新しい抗生物質などの潜在的抗感染薬を、その感染の抑制に関するそれらの効力について評価することができる。本発明の裏付けとして行なった後述の実験によって、この応用の一例を説明する。
【0096】
真核細胞も本発明のある側面で物体として有用である。所望の調節要素を含有する適当な発現ベクターが市販されている。それらのベクターを用いて、初代培養細胞、体細胞、リンパ細胞などを含む種々の真核細胞中で所望の発光性タンパク質を発現させうる構築物を作成することができる。これらの細胞は、一過性発現試験にも使用できるし、細胞系の場合は安定な形質転換体として選択することもできる。
【0097】
形質転換細胞における発光性タンパク質の発現は、選択された様々なプロモーターのいずれかを用いて調節することができる。例えばその細胞を、発現したリガンドまたはレセプターによって対象内のある部位に向かわせる発光体として使用したい場合は、CMVプロモーターやSV40プロモーターなどの恒常的に活性なプロモーターを使用すればよい。そのような構築物で形質転換した細胞は、例えばその細胞を殺すことなどによって光の発生を阻害する化合物のアッセイにも使用できる。
【0098】
また、形質転換細胞が対象内に均一に分布し、ウイルスによる感染やサイトカインによる剌激を受けたときなど、一定の条件下でのみ発光性タンパク質が発現するように、形質転換細胞を投与してもよい。これらの剌激やその他の剌激に関係する因子に反応するプロモーターは、当技術分野で知られている。関連する側面として、Tet系(GossenおよびBujard,1992,PANS 89:5547−5551)のような誘導性プロモーターを使用して、発光性タンパク質の発現を一時的に活性化することもできる。
【0099】
例えばtat反応性HIV LTR要素を含有する構築物でCD4+リンパ細胞を形質転換し、それをHIV感染のアッセイとして使用することができる(IsraelおよびHonigman,1991,Gene 104:139−145)。そのような構築物で形質転換した細胞をSCID−huマウス(McCuneら,1988,Science 241:1632−1639)に導入し、それをヒトHIV感染症とAIDSのモデルとして使用することができる。
【0100】
例えば恒常的に活性なプロモーターを用いて上述のように形質転換した腫瘍細胞系は、腫瘍の増殖と転移をモニターするために使用できる。形質転換した腫瘍細胞を動物モデルに注射し、腫瘍塊を形成させ、増殖阻害剤候補または転移阻害剤候補で処置しながら、腫瘍の大きさと転移をモニターすることができる。
【0101】
腫瘍細胞は、様々な感染性因子または治療用化合物に感応する活性を持つ調節可能なプロモーターを含有する構築物で形質転換された細胞から作成することもできる。
【0102】
(6.細胞形質転換)
原核細胞と真核細胞の形質転換法は共に当技術分野でよく知られている(Sambrookら,1989,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory Press,第2巻)。適当な調節因子と多重クローニング部位を持つベクターは広く市販されている(例えばカリフォルニア州ラホーヤのStratagene社、カリフォルニア州パロアルトのClontech社)
(IV.発光性タンパク質をコードする遺伝子を含有するトランスジェニック動物)
もう1つの側面として、本発明は、発光性タンパク質または発光性のタンパク質複合体をコードする異種遺伝子構築物を含有するトランスジェニック動物を包含する。この構築物は選択したプロモーターによって駆動され、例えば発光性タンパク質の機能的発現に必要な種々の補助タンパク質や、選択マーカー、エンハンサー要素などを含みうる。
【0103】
プロモーターの活性化は、発光性分子と補助タンパク質をコードする遺伝子の発現量を増大させる。プロモーターの活性化は、選択した生物適合体またはその一部とプロモーター要素の相互作用によって達成される。活性化がその動物の一部でのみ起こる場合は、その部分の細胞のみが発光性タンパク質を発現させるだろう。
【0104】
例えばインターフェロン誘導性プロモーター(3’−5’ポリA合成酵素のプロモーターなど)を用いることにより、多数の異なるRNAウイルスによるトランスジェニック細胞の感染を検出できる。
【0105】
関連する側面として、ある種の疾患状態で発現するプロモーターを用いて、トランスジェニック動物内の患部を標識し、発光成分の発現を用いて、その疾患状態に対する処置の効果をモニターすることができる。例えば、E−セレクチンは生体内の炎症部位に発現する(PoberおよびCotran,1991,Lab.Invest.64:301−305)。したがってE−セレクチンプロモーターを単離し、それを用いてルシフェラーゼ遺伝子の発現を推進することができる。
【0106】
組識特異性プロモーターと共に本発明の方法を使用することも可能である。これは、例えば体内の特定の器官または組識の変性を引き起こす発病プロセスを阻害するのに有効な化合物のスクリーニングを可能にし、また発育中の動物などにおける細胞(例えばニューロン)の追跡を可能にする。
【0107】
当技術分野では、本発明で使用できるプロモーターが数多く知られている。また、プロモーター含有ゲノムDNAを単離するために、その遺伝子のcDNAから得られる情報を用いて、クローン化された遺伝子のプロモーターを単離する方法も知られている。
【0108】
(V.発光性複合体の撮像)
対象内の意図する部位に局在した発光性複合体は、いくつかの方法で撮像できる。そのような撮像の指針と具体例を以下に記述する。
【0109】
(A.発光性複合体の局在)
”標的型(targeted)”物体、すなわちターゲッティング成分(物体を対象または動物内の特定の一部位または複数部位に局在させるように設計された分子または特徴)を含有する物体の場合、局在とは、対象内で束縛された”局在型”物体と束縛されていない”遊離型”物体の間の平衡が本質的に成立した状態を指す。そのような平衡に到達する速度は投与経路に依存する。例えば血栓を局在化するために静脈内注射によって投与された複合体は、注射後数分以内にその血栓で局在または蓄積を達成するだろう。一方、腸内の感染を局在化するために経口投与された複合体は、局在を達成するのに数時間を要するだろう。
【0110】
また、局在という用語は、単に、物体を投与した後選択した時間におけるその物体の対象または動物内での位置をさす場合もある。例えば、本明細書に詳述する実験では、サルモネラを(例えば経口的に)投与し、その蔓延を時間の関数として追跡する。この場合、その物体は、それが投与した細菌の最初の位置を標識し、その後の蔓延または後退を撮像によって追跡できる限りは、経口投与後直ちに”局在型”となりうる。
【0111】
関連する側面として、例えば発光成分を発現させる、注射された腫瘍細胞の局在は、その動物内のある部位にそれらの細胞が定着し、腫瘍塊を形成することからなるだろう。
【0112】
もう1つの例として、局在は物体が投与後に分布した状態になった時に達成される。例えば、対象または動物じゅうの様々な器官における酸素濃度を測定するために投与された複合体の場合、その複合体は、それがその対象または動物内で本質的に定常的分布状態に到達した時に、”局在型”あるいは有益となる。
【0113】
上述のいずれの場合にも、当業者は、局在を達成するための時間を適切に見積もることができる。また、時間の関数としての局在の状態は、発光性複合体を本発明の方法に従って撮像することにより、追跡することができる。
【0114】
(B.光検出装置)
本発明の重要な側面は、哺乳動物内からの微弱な光を妥当な時間で(好ましくは約30分以内に)撮像することができ、その装置からの信号を使って画像を構築できるほど充分に感度の高い光検出装置の選択である。
【0115】
極めて明るい発光成分を使用できる場合および/または撮像する対象または動物の表面近くに局在した発光性複合体を検出することが可能な場合は、”暗視”ゴーグルや、シリコン増倍管(SIT)カメラ(例えば浜松ホトニクスシステムズ(Hamamatsu Photonic Systems)社,ニュージャージー州ブリッジウォーター)などの一般的高感度ビデオカメラを使用できる。しかし、より一般的には、もっと高感度な光検出法が必要である。
【0116】
本発明を実施する際に直面するような極めて低い光レベルでは、単位面積あたりの光子束があまりに低くなるので、撮影対象がもはや連続的でないように見える。その代わりに、それは時間的にも空間的にも互いに独立した個々の光子によって表される。モニターで見ると、そのような画像は、それぞれが検出された単一光子を表わす瞬く光の点のように見える。
【0117】
検出されたこれらの光子をデジタル画像処理装置で経時的に累積することにより、画像を獲得し、構築することができる。各画像点での信号に強度値を割り当てる従来のカメラとは対照的に、光子カウント撮像法では、信号の強度は意味を持たない。その目的は、単に信号(光子)の存在を検出し、その位置に関して信号の発生数を経時的に計数することである。
【0118】
後述する少なくとも2種類の光検出装置により、個々の光子を検出し、画像処理装置で分析できる信号を作成することができる。
【0119】
(1.ノイズ低減型光検出装置)
第1の種類は、光子信号の増幅とは対照的に、光子検出器内のバックグラウンドノイズを減少させることによって感度を得る装置である。ノイズは主として検出器アレイを冷却することによって低減される。この装置には、”バック薄型”冷却CCDカメラと呼ばれる電荷結合素子(CCD)カメラが含まれる。より高感度な装置では、例えばCCDアレイの温度を約−120℃まで下げる液体窒素を用いて、冷却が行われる。”バック薄型”とは、光子が検出されるためにたどる光路長を短くすることによって、量子効率を増大させる超薄型バックプレートを指す。とりわけ高感度なバック薄型低温CCDカメラは、Photometrics社(アリゾナ州トゥーソン)から入手できるシリーズ200カメラ”TECH 512”である。
【0120】
(2.光子増幅装置)
第2の種類の高感度光検出器には、光子が検出スクリーンに命中する前に光子を増幅する装置が含まれる。この種類には、マイクロチャネル増倍装置のような増倍装置を持つCCDカメラが含まれる。マイクロチャネル増倍装置は通例、カメラの検出スクリーンに対して垂直かつ同延的なチャネルの金属アレイを含有する。マイクロチャネルアレイは撮像しようとする試料、対象または動物とカメラの間に設置される。このアレイのチャネルに侵入する光子の大半は、チャネルを抜け出す前にチャネルの側面と接触する。アレイを横切ってかけられた電圧は、各光子衝突から多くの電子を放出させる。このような衝突から生じた電子は、それらが発生したチャネルから”散弾銃”式に抜け出して、カメラによって検出される。
【0121】
第1段階で生成した電子が、今度は第2段階で増幅された電子の信号をもたらすように、増倍マイクロチャネルアレイを直列に設置すれば、さらに高感度を得ることができる。しかし感度を増大させると、空間的解像度が犠牲になり、その空間的解像度は増幅段階を追加するごとに低下する。
【0122】
マイクロチャネル増倍装置型の単一光子検出装置の典型例は、Hamamatsu社から入手できるC2400シリーズである。
【0123】
(3.画像処理装置)
光子を計数する光検出装置によって生成した信号は、例えばモニターに表示したり、ビデオプリンターでプリントできる画像を構築するために、画像処理装置で処理する必要がある。そのような画像処理装置は、通例、上述の高感度光子計数カメラを含むシステムの一部として販売されており、したがって同じ供給者(例えばPhotometrics社やHamamatsu社)から入手できる。他の業者から入手した画像処理装置も使用できるが、一般的には、機能的なシステムを構築するのに、より多くの努力が必要になる。
【0124】
画像処理装置は通常、IBM互換PCやアップルマッキントッシュ(Apple Computer社,カリフォルニア州キューパーティーノ)のようなパーソナルコンピューターに接続する。このパーソナルコンピューターは購入した撮像システムの一部として含まれている場合もあるし、含まれていない場合もある。画像がデジタルファイルの形になったら、それを様々な画像処理プログラム(例えばAdobe Systems社(カリフォルニア州マウンテンビュー)の”ADOBE PHOTOSHOP”など)で操作し、印刷することができる。
【0125】
(C.装置の検出フィールドにおける対象の固定)
(1.装置の検出フィールド)
装置の検出フィールドとは、光子放射の一貫した測定値が得られる領域をいう。光学レンズを使用するカメラの場合、検出フィールドとは、単に、そのレンズがそのカメラに与える視野である。また、光検出装置が”暗視”ゴーグルである場合、検出フィールドとはそのゴーグルの視野である。
【0126】
また、検出フィールドは、密に充填されたアレイ状に配置した光ファイバーケーブルの末端が規定する表面であってもよい。そのアレイは、ケーブルの末端がカバーする領域(ケーブル間の空隙と相反する)が最大になるように構築され、対象に極めて近接して設置される。例えば、プレクシグラスなどの透明素材を対象に隣接して設置し、その透明素材に隣接して、対象とは反対側から、アレイを固定することができる。
【0127】
アレイの反対側にある光ファイバーケーブルの末端は、マイクロチャネル増倍装置の入力末端などの検出装置または増倍装置に直接接続することができ、そうすればレンズの必要がなくなる。
【0128】
この方法の利点は、対照と検出器の間の空隙の大半を排除することにより、かつ/または、レンズを排除することにより、光子の散乱および/または損失が減少することにある。本発明の裏付けとして行なった実験で使用した60mm AFニッコールマクロレンズのような高透過性レンズでさえ、フロントレンズ要素に到達する光の一部しか透過させない。
【0129】
より強度の強いLGMの場合、光ダイオードアレイを光子放射の測定に使用できる。光ダイオードアレイを比較的柔軟なシートに組み込めば、実施者は、そのアレイを対象のまわりに部分的に”巻き付ける”ことができる。この方法も光子の損失を最小限にし、さらに生物発光の三次元画像を得る手段にもなる。
【0130】
複数の検出器を対象の周囲に設置する方法や、走査型の検出器または検出器群などを含む他の方法を使用して、三次元画像を作成してもよい。
【0131】
動物または対象の全体が光検出装置の検出フィールド内にある必要はないということは理解されるだろう。例えば対象の特定の領域に局在することがわかっている発光性複合体を測定する場合、所望の情報を得るには、その領域とその周囲の十分な”暗”帯域からの光だけを測定すれば足りる。
【0132】
(2.対象の固定)
対象の2次元または3次元画像を作成したい場合、光子放射を測定している間は、対象を光検出装置の検出フィールド内に固定しておく場合がある。約20ミリ秒未満で測定した光子放射から画像を構築できるほどに信号が明るく、対象が特に動揺していない場合は、測定期間の開始時に対象が検出装置のフィールド内にあることを保証する以外に、特別な固定措置は必要ないだろう。
【0133】
これに対して、光子放射測定に約20msec以上を要し、対象が動揺している場合は、構築された画像に空間情報が保存されるように、その対象の動揺の程度に合わせて、光子放射測定中、対象の不動を保証するための予防措置を考慮する必要がある。例えば対象が人で、光子放射測定時間が数秒程度の場合は、その対象に光子放射測定(撮像)の間できるだけじっとしておいてくれるように頼むだけでよいだろう。これに対し、対象がマウスのような動物である場合は、その対象を例えば麻酔や機械的拘束具を用いて固定できる。
【0134】
様々な拘束具を作成することができる。例えば、プレクシグラスシートを発泡材クッション上に固定して、マウスを数十秒ないし数分間固定するのに有効な拘束具を作ることができる。そのクッションは、一端に、動物の頭部用のくぼみを持つ。動物は、その頭部がくぼみの上になり、息は自由にできるが、その体の動きは発泡材クッションによって束縛されるように、プレクシグラスの下に置かれる。
【0135】
対象または動物から放射され光の総量のみを測定したい場合は、光子放射測定期間が長くても、その対象を固定する必要はない。必要なことは、撮像の間、対象を光検出器の検出フィールドに閉じ込めておくことだけである。しかし、そのような測定中、対象を固定しておけば、検出される光子が通過する組識の厚さが動物間でより均一になるから、得られる結果の整合性が向上するであろうことは理解されるだろう。
【0136】
(D.撮像中のその他の問題)
(1.蛍光発生成分)
蛍光発生成分の可視化には、光検出器と共に、励起光源が必要である。またその励起光源が、発光成分からの光子放射を測定している間、点灯されることも理解されるだろう。
【0137】
発蛍光団の適切な選択、光源の設置、およびフィルターの選択と設置は、すべて情報に富む画像の構築を容易にするが、これらについては蛍光発生成分に関する項で上述した通りである。
【0138】
(2.高解像度画像)
組識による光散乱は、総光子放射の測定によるLGMの撮像で得ることができる解像度を制限する。本発明が、対象内の選択した点に集中させることができ、しかも組織内で有意に散乱しない外部源に、LGMの発光を同期させることによって、解像度のより高い画像を構築できる態様をも包含することは、理解されるだろう。例えば、集束させた超音波信号を使って、撮像する対象を三次元的に走査することができる。超音波の焦点内にある領域からの発光は、その超音波によって光に与えられた特徴的な振動により、他の光子放射から分離することができる(例えばHoustonおよびMoerner,米国特許第4,614,116号,1986年9月30日発行)
(E.光子放射画像の構築)
発光成分が例外的に明るいため、かつ/または、発光性複合体が対象の表面近くに局在するために、”暗視”ゴーグルまたは高感度ビデオカメラを使って画像を得た場合は、単に画像を見るか、ビデオモニターに表示するだけである。所望であれば、分析または印刷用に個々のビデオフレームをメモリーに保存でき、かつ/または、コンピューターで分析と印刷用に画像をデジタル化できる画像処理装置に、ビデオカメラからの信号を迂回させることができる。
【0139】
また、光子計数法を使用する場合は、光子放射の測定により、各ピクセル位置で検出した光子の数を表わす数字の配列が、画像処理装置内に生成する。これらの数字を用いて、通常は光子数を(一定の予め選択した値、またはいずれかのピクセルで検出された最大数に対して)規格化し、その規格化した数値を明るさ(グレースケール)または色(擬似色)に変換して、それをモニターに表示することにより、画像を作成する。擬似色表示の場合、典型的な色の割り当ては次の通りである。ゼロ光子カウントのピクセルには黒色を、低いカウントには青色を、カウントが増加するほど波長の長い色を割り当て、最高光子カウントには赤色を割り当てる。モニター上の色の位置は光子放射の分布を表わし、したがって発光性複合体の位置を表わす。
【0140】
複合体に対して準拠座標系を与えるために、通常は、光子放射を測定した(まだ固定されている)対象のグレースケール画像を構築する。このような画像は、例えばかすかなルームライトのなかで撮像室または撮像箱に向かって扉をあけ、反射した光子を(通例、光子放射を測定するのに要する時間のごく一部に相当する時間)測定することによって構築できる。このグレースケール像は、光子放射を測定する前に構築してもよいし、光子放射を測定した後に構築してもよい。
【0141】
通例、光子放射の画像をグレースケール像に重ね合せることにより、対象に関する光子放射の合成画像を作成する。
【0142】
例えば選択した生物適合成分の分布および/または局在に対するある処置の効果を記録するために、発光性複合体の局在および/または発光性複合体からの信号を経時的に追跡したい場合は、光子放射の測定または撮像を選択した時間間隔で反復することにより、一連の画像を構築することができる。間隔は数分程度の短いものであってもよいし、数日または数週間程度に長いものであってもよい。
【0143】
(VI.光子放射画像の分析)
本発明の方法および/または本発明の組成物を使用して作成した画像は、様々な方法で分析できる。それは、単純な視覚的検査、頭脳評価および/またはハードコピーの印刷から、複雑なデジタル画像分析まで様々である。分析によって得られる情報の解釈は、観察する現象と使用した物体に依存する。
【0144】
下記の実験は本発明の応用例の一つ(生体ラットにおけるサルモネラ感染の追跡)であり、本発明の方法で得られる画像を、どのように分析できるかを例示するものである。
【0145】
(VII.生体マウス内の発光性サルモネラの撮像)
本発明の裏付けとして行なった実験では、マウスにおけるネズミチフス菌(Salmonella typhimurium)感染(ヒトチフスの動物モデル)の分布を特徴づける。マウス病原性ネズミチフス菌SL1344(HoisethおよびStocker,1981,Nature 291:238−239)、SL1344の非侵入性突然変異体BJ66、およびサルモネラの低病原性LT−2株LB5000を、それぞれluxオペロンを含有するプラスミドで標識し、マウスにおけるサルモネラ感染を局在化する実験に使用した。
【0146】
(A.発光性サルモネラの構築)
(1.サルモネラ株)
マウスに対する経口および腹腔内接種によって明らかになる病原性表現型が異なる3株のネズミチフス菌を、形質転換のために選択した。
【0147】
ここで使用した最も病原性の高い表現型はSL1344であり、これはもともと子牛の致死的感染症から得られたマウス株である(HoisethおよびStocker,1981,Nature 291:238−239)。この株をマウスに経口接種すると、細菌はリンパ系を通って全身に転移し、肝臓、脾臓および骨髄のコロニー形成をもたらす(CarterおよびCollins,1974,J.Exper.Med.139:1189−1203;FinlayおよびFalkow,1989,Mol.Microbiol.3:1833−1841とHsu,1989,Microbiol.Rev.53:390−409の総説をも参照のこと)。
【0148】
SL1344の非侵入性突然変異体BJ66も評価する。BJ66の経口接種は通常、マウスにおける全身性感染を引き起こさないが、この株を腹腔内接種すると、全身性感染が起こる。
【0149】
サルモネラの低病原性LT−2株LB5000も調べる。LT−2株は、マウスに対する病原性が減少しているまたは変動することが知られている研究室株である。LB5000は複数の栄養要求性突然変異を含有し、ストレプトマイシン耐性であり、経口または腹腔内接種後、マウスから浄化される。
【0150】
(2.luxオペロンによるサルモネラ株の形質転換)
上記3株のそれぞれを、luxオペロンをコードするプラスミドで、実施例1に詳述するように形質転換する。土壌菌Xenorhabdus luminescens(Frackmanら,1990)から得られたこのプラスミドは、ヘテロ二量体ルシフェラーゼの2つのサブユニットと、3つの補助タンパク質luxC、luxDおよびluxEの発現によって、大腸菌に光子を放射する能力を与える。
【0151】
luxC、luxDおよびluxEが含まれるので、そのルシフェラーゼ発現細胞には、脂肪族アルデヒド基質ルシフェリンを与える必要がない。本明細書に記述するような生体系内の真核ルシフェラーゼに基質を供給することは困難だろうから、X.luminescensの全luxオペロンを使用する。このオペロンは、当該脂肪族アルデヒド基質を生合成するための酵素をもコードする。
【0152】
α−βヘテロ二量体混合機能オキシダーゼであるX.luminescensルシフェラーゼは、還元型フラビンと長鎖アルデヒドの酸化型フラビンと対応する長鎖脂肪酸への酸化を触媒する。アルデヒドの脂肪酸からの生成と再生には、脂肪酸レダクターゼ複合体が必要であり、NAD(P)H: フラビンオキシドレダクターゼが還元型フラビンを供給する。
【0153】
X.luminescensのlux遺伝子を発現させる大腸菌にとって最適な生物発光条件は37℃である(SzittnerおよびMeighen,1990,J.Biol.Chem.265:16581−16587,Xiら,1991,J.Bact.173:1399−1405)。これに対し、真核発光生物と他の原核発光生物に由来するルシフェラーゼ類は通例、より低い最適温度条件を持つ(Campbell,1988,Chemiluminescence.Principles and Applications in Biology and Medicine(英国チチェスター:Ellis Horwood社およびVCH出版社))。したがってX.luminescens由来のルシフェラーゼは、動物内での研究にマーカーとして使用するのに適している。
【0154】
全X.luminescens luxオペロンを含有し、かつ、サルモネラにアンピシリン耐性とカルベニシリン耐性を付与するプラスミドpGSL1(Frackmanら,1990)を用いて、上記3株をエレクトロポレーションによって形質転換する。このX.luminescens luxオペロンは、luxA、luxB、luxC、luxDおよびluxE遺伝子を含有する(Frackmanら,1990)。luxAとluxBはヘテロ二量体ルシフェラーゼの2つのサブユニットをコードし、luxCとluxDはルシフェラーゼ基質の生合成酵素をコードし、luxEは調節遺伝子である。培養中の細胞に外からルシフェリンを供給したり、動物を基質で処置するのとは対照的に、基質生合成用遺伝子の包含は、ルシフェラーゼに基質を提供する便利な手段である。
【0155】
(B.形質転換したサルモネラの試験管内での特徴づけ)
(1.付着性と侵入性)
上記luxプラスミドを含有する3つのサルモネラ株の付着性と侵入性を、FinlayおよびFalkow,1989,Mol.Microbiol.3:1833−1841に記述されているような一般的な侵入アッセイ(実施例2に詳述)により、培養中で互いに比較、またそれらの非発光性親株と比較する。
【0156】
このアッセイでは、上皮細胞系および腹腔マクロファージと共に培養した後、付着細菌と細胞内細菌を定量する。付着細菌と細胞内細菌は、生きている細胞からの光子の放出と、溶解してその細胞溶解液をカルベニシリン含有プレートで培養した後のコロニー形成単位との両者によって、検出、定量する。
【0157】
このアッセイの一部の結果を図2A〜2Eに示し、実施例8で考察する。lux発現プラスミドで形質転換した上記3株の表現型は、親サルモネラ株と比較して有意に変化しない。また、HEp−2細胞とマクロファージからの生物発光の強度とCFUの間には良好な相関が認められる。これらの結果は、細胞内細菌の指標としての発光が、培養内の細菌の侵入性をアッセイする迅速な方法であることを示している。
【0158】
BJ66のHEp−2に対する付着性はSL1344と比較して低かったが、マウス腹腔マクロファージの初代培養におけるこれら2株の付着性は同等だった。
【0159】
(2.光放射)
系の酸素要求性を調べるため、実施例3に詳述するように、細菌の10倍連続希釈液をガラス製毛細管内に入れ、撮像する。
【0160】
図3に、そのような実験の一つで作成した画像を示す。発光は、空気飽和培地中に少数の細菌を含む管でさえ、空気−液体界面でのみ検出される(51中に0.1mlの空気飽和緩衝液で最終O2濃度は5nMになる)。
【0161】
これらの結果から、酸素がおそらくは発光にとって制限因子であることは明らかである。
【0162】
(3.動物組識を貫通する光透過)
光が動物組識を貫通する程度を決定するために、発光性サルモネラから放射され組識を透過した光を、単一光子を検出するために高速同時検出器を切ったシンチレーション計数器を用いて定量する。このタイプの光電子増倍管の暗電流によるバックグラウンドは有意であるので、このアッセイは、比較的強い光子放射を伴う試料に限定される。
【0163】
透明度の異なる4タイプの組識:ヒヨコ胸部の筋肉、ヒヨコ胸部の皮膚、子ヒツジの腎臓、および子ヒツジの腎臓から得た腎臓髄質、をこの方法で比較する。組識を貫通して検出できる光子の数は、組識を通さない対照より約10倍少ない。
【0164】
(4.luxサルモネラの生体内での特徴づけ)
(a.経口投与)
経口接種はマウスやヒトにとって自然なサルモネラ感染経路であり、より遅延性の疾患進行をもたらす。この接種経路によるサルモネラ感染の進行を研究するために、2系統のマウスを3株のサルモネラに感染させる。耐性動物を用いて得た結果については、下記”耐性マウスの感染”という見出しの項で議論する。
【0165】
実施例5に記述するように、Balb/cマウスを病原性SL1344lux、非侵入性BL66luxおよび低病原性LB5000luxサルモネラに経口的に感染させる。その感染の進行を外部撮像法(”材料と方法”の項)によって8日間にわたって追跡する。
【0166】
典型的な画像を図5A〜Fに示す。接種後(p.i.)24時間の時点で、生物発光シグナルはすべての感染動物内で単一の病巣に局在する(図5A、5Cおよび5E)。接種後7日までには、低病原性LB5000luxに感染したすべての動物で、生物発光が消える(図5B)。これに対し、病原性SL1344luxに感染した動物の場合、蔓延が始まる時点は動物ごとにかなり変動するものの、しばしば腹腔の大半に蔓延する激しい感染を示す(図5F)。BJ66luxによる感染は通例、持続し、単一の部位に局在したままとなる(図5D)。
【0167】
(b.腹腔内接種)
サルモネラの複製部位にルシフェリンの酸化とそれに続く発光(Campbell,1988,Chemiluminescence.Principles and Applications in Biology and Medicine(英国チチェスター:Ellis Horwood社およびVCH出版社))に足るO2が存在するかどうかを評価するために、呼吸している動物の組識からの光放射を測定する。発光性のSL1344luxとLB5000luxを2群のBalb/cマウスの腹腔内に接種する。接種後(p.i.)32時間の時点で、透過した光子を撮像する(図6)。
【0168】
SL1344luxに感染したマウス(図の左側)では、透過した光子が大きな表面上に確認され、様々な強度の焦点が見える。これらの画像は播種性感染を示し、内臓(おそらくは肝臓と腸間膜リンパ節を含む)の広範なコロニー形成と合致する。これに対し、LB5000luxに感染した動物からの透過光子の分布は極めて制限されており、限定的感染を示す。
【0169】
LB5000lux感染マウスは接種後数週間にわたって健康を維持したが、SL1344lux感染マウスは接種後4日でほぼ瀕死状態になり、安楽死させた。
【0170】
これらの実験は、血中または組識中のO2レベルが、サルモネラから発現したluxルシフェラーゼの生物発光にとって十分であることを示している。また、これらの実験は、病原性が減少した研究室株LB5000と比較して病原性株SL1344の侵入性が強いこととも合致している。
【0171】
(c.耐性マウスの感染)
Ity遺伝子座がヘテロ接合性を示すマウス(Ity r/s)は、ネズミチフス菌による全身性感染症に対して耐性である(PlantおよびGlynn,1976,J.Infect.Dis.133:72−78)。Bcg(Grosら,1981,J.Immunol.127:2417−2421)またはLsh(Bradley,1977,Clin.and Exper.Immunol.30:130−140)とも呼ばれるこの遺伝子座は、鼡らい菌(Mycobacterium lepraemurium;Forgetら,1981,Infect.Immunol.32:42−47)、ウシ結核菌(M.bovis;Skameneら,1984,Immunogenet.19:117−120,SkameneおよびPietrangeli,1991,Nature 297:506−509)およびバテー杆菌(M.intracelluare;Gotoら,1989,Immunogenetics 30:218−221)のようなある種の細胞内病原体の発病プロセスを調節する。細胞内病原体に対する耐性と感受性の同様な遺伝子制御はヒトにも存在するようである(ヒト結核菌(M.tuberuculosis;Stead,1992,Annals of Intern.Med.116:937−941,Steadら,1990,New Eng.J.Med.322:422−427)とらい菌(M.leprae))。
【0172】
Ity遺伝子座はマウスの第1染色体に2つの対立遺伝子型、Ityr(耐性、優性)とItys(感受性、劣性)で存在する。Ity遺伝子座にコードされている遺伝子は、マクロファージが内部移行した病原体を破壊する能力に影響を与え(Blackwellら,1991,Immunol.Lett.30:241−248(1991)による総説がある;Skameneら,1984,Immunogenet.19:117−120とSkameneおよびPietrangeli,1991,Nature 297:506−509をも参照のこと)、それがマクロファージ媒介性とされる感染宿主内の他の部位への病原体の輸送という下流の機能に影響を及ぼすようである。Balb/cマウスはItys/sであり、129マウスはItyr/rである。ここに詳述する実験では、ヘテロ接合性Balb/cX129マウス(Ityr/s)を使用する。
【0173】
実施例7に詳述するように、耐性129XBalb/c(Ityr/s)生存マウスを、1X107個のSL1344luxサルモネラの胃内接種によって感染させる。そのマウスを注射後8日間(8 d.p.i)にわたって毎日撮像する。
【0174】
図7A(第1日)と図7B(第8日)に結果を示す。外部撮像法によって検出される発光は接種後24時間の時点で明白であり、全ての動物で単一の部位に局在しているようであった。その発光シグナルは試験期間(接種後8日まで)中常に存在する。発光の強度と発光源の位置は、一匹のマウス内で経時的にいくらか変動し、またマウスごとにも変動しうる。いずれの感染動物でも発光組識は盲腸であり(下記参照)、所在の変動性は(おそらくは強度の変動性も)齧歯類の内部器官が正しい位置に強く固定されていないという事実によるのだろう。
【0175】
これらの動物で観察された明らかな限定的感染は、このIty制限がマクロファージ輸送を遮断するという解釈を裏付けている。しかし10日間というこの感染の持続性は、腸粘膜に対する付着があることと、発光性の糞粒によって立証されるように、これらの動物の糞内に細菌の脱離が長期間続くことを示唆している。これらの結果は、生体内におけるこのサルモネラの発光性表現型が、Ity制限動物中で8日間にわたって保たれることと、経口接種後の局在が可能であることを示している。
【0176】
(d.経口接種後の内部撮像)
腹腔内の発光シグナルをさらに局在化するため、感染マウスを腹壁切開後に撮像する(図8)。経口経路で感染させた動物のいずれにおいても、主な疾患症状は盲腸の拡張である(図8A〜C)。”外部”画像(図8A)は限局的な発光を示し、それは腹壁切開後の画像(図8B)で盲腸にあたることがわかる。
【0177】
腸内に空気を注入すると、消化路の他の領域における細菌の存在が確認される。結腸と直腸内の細菌はルシフェラーゼを発現させているらしいが、それらの部位からの発光は、低い酸素濃度によって制限されているらしい。
【0178】
経口接種試験で得た画像は、接種後2日および接種後7日の発光シグナルが、LB5000luxに感染したマウスを除いて、ほとんど完全に各動物の盲腸に局在することを示している(Popeskoら,1990,A Colour Altas of Anatomy of Small Laboratory Animals Vol.Two:Rat Mouse Hamster(英国ロンドン:Wolfe))。発光は数匹の結腸にも見える。接種後7日までに、LB5000lux感染動物内に発光は認められなくなる。これらのマウスの器官中に存在するCFUを接種後2日と5日に測定する。
【0179】
侵入性株SL1344luxを胃内に感染させた動物では、盲腸内の発光が初期に現われ、その後、全身性の感染が起こる。これに対し、非侵入性BJ66lux株による感染は盲腸からの持続的な発光をもたらし、それが全試験期間(8日間)にわたって維持される動物もある。接種後8日までに、腹部表面の大半に発光が検出され、それはSL1344lux感染マウスにおける腹腔内接種後の光子の分布に似ている。
【0180】
SL1344luxによる感染は、予想通り、光子を放射する表面積が増大し、そこから放射される光子を累進的に増やしながら、全身性となるようである。いずれの株による感染でも、発光は腹部上に局在し、この領域の外側からの発光はほとんど検出されないようである。大量の透過光子が腹部上に単一の焦点として局在し、このことは、たとえ感染が全身性であっても、複製の大半が腸を取り巻く領域にあることを示唆している。
【0181】
盲腸上に発光が局在することは、腸のこの領域に大量の生物が存在することを示すだけでなく、発光するに足る酸素が得られるように、サルモネラがその粘膜の細胞に結合することをも示唆している。ルシフェラーゼからの光子の放射は酸素依存性であり、盲腸または小腸の管腔内の予想酸素レベルは、発光に必要なレベルより低い。ルシフェラーゼ反応は、細菌が腸上皮の細胞から酸素を得られない限り、腸内では機能しないと予想される。
【0182】
したがって全身性感染は侵入性表現型に関係し、腸の上皮細胞への単なる付着とは関係しないと思われる。これらの実験は、盲腸がこの発病プロセスの保菌状態で、あるいは播種部位として、何らかの役割を持つことを含意している。
【0183】
異なる組識への感染の進行をモニターすれば、発病プロセスにおけるこれらの段階についての理解を著しく深めることができ、選択した段階でその病原体を抑制するのに有効な化合物のスクリーニングが可能になるだろう。
【0184】
(e.腹腔内接種後の内部撮像)
SL1344luxを腹腔内に感染させたマウスを腹壁切開の前後に撮像する(実施例9)。その結果を図9に示す。その画像は、腹部の大部分に、複数の透過光子焦点を伴う発光を示す。盲腸は発光性サルモネラを含有しないようである。これらの実験から得られた結果は、すべてのサルモネラ株が、感染の初期相で発光するに足るO2を持つことを示している。しかし、その後の時点での全身性感染はSL1344lux感染マウスにのみ認められるので、サルモネラの粘膜の細胞への侵入と、その後の全身性感染は、侵入性表現型を持つ株に限られるようである。
【0185】
(f.サルモネラ感染に対するシプロフロキサシンの影響)
非侵襲的撮像法が薬物に対する感染症の反応を追跡するのに有用であることを示すために、実施例10に詳述する実験を行なう。マウスにSL1344luxを経口接種し、サルモネラ感染症に有効な抗生物質シプロフロキサシン100mgで処置する。処置後の選択した時期に、それらのマウスを撮像し、光子放射を測定することによって感染の程度を定量する。処置マウス内の光子放射を、処置開始前の値および感染したが処置していない対照マウスからの値と比較する。このような実験の一つで得た結果を図10A〜Eに示し、実施例10で考察する。抗生物質で処置したマウスにおける感染症は、処置マウスにおける0時間での病原体レベルおよび対照マウス内の病原体レベルと比較して、処置期間中常に低い。
【0186】
(g.カルベニシリン選択の効果)
Ducluzeauら,1970,Zeut.Bakt.5313:533−548は、抗生物質による動物の処置がサルモネラによる盲腸のコロニー形成を促進することを示した。本実験におけるマウスは、ルシフェラーゼクローンを含有するAmprサルモネラを選択するために、カルベニシリンの筋肉内注射という抗生物質措置をして維持される。この処置は、胃腸感染症の経過を変化させるかもしれないが、サルモネラが盲腸を裏打ちする細胞と結合できるという観察結果は、酸素を発光に利用できるということを示している。盲腸の管腔は一般に嫌気環境であると考えられているから、この観察結果は注目に値する。
【0187】
(VIII.応用)
生物発光技術は様々な宿主−病原体系に広く適用できると共に、例えば生きている哺乳動物内での腫瘍の進行や遺伝子発現などに関する他の生物学的事象の時間的空間的評価をも可能にするだろう。またこれは、医薬の開発とスクリーニングにも応用できる。病原体の生体内撮像を広範に使用すれば、病原および/またはリアルタイム試験抗菌剤に関する実験に必要な動物数と時間を減らすことができるだろう。また、発光性細菌は環境分析に使用されているが、生物発光性生物は生きている動物内のバイオセンサーとして有用だろう。例えばKorpelaらは、胃腸路の管腔内では酸素供給が制限されているため、酸素が(おそらくは上皮細胞または他の細胞タイプから)直接的にサルモネラに接近できる部位に、生物発光が限定されたことを示している。Korpelaら,1989,J.Biolum.Chemilum.4:551−554。この酸素要求性は、親密な細胞−細胞相互作用の指標として、あるいは生きている動物中の様々な部位における酸素濃度を調べるためのバイオセンサーとして利用できるだろう。以下、この技術の典型的応用例を例示のためにいくつか記述するが、これらは決して本発明の限定を意図するものではない。
【0188】
(A.酸素レベルの測定)
上に要約した実験で明示されたようにルシフェラーゼの発光に酸素が必要であるということは、本発明を、対象内の酸素濃度の空間的勾配の測定法として利用できるということを示している。10〜1mMの範囲の酸素レベルを測定するために、発光性細菌が使用されている。これらの研究によれば、0.1nMが検出下限だと予想される(Campbell,1988,Chemiluminescence.Pringiples and Applications in Biology and Medicine(英国チチェスター:Ellis Horwood社およびVCH出版社))。本明細書に記述する撮像法は、生きている動物内の様々な部位における酸素レベルを調べるのに利用できる。例えばO2中で、またはCa2+依存的に光を放射するように操作された微生物を、ちょうど発光性細菌が環境分析に使用されるように(Guzzoら,1992,Tox.Lett.64/65:687−693;Korpelaら,1989,J.Biolum.Chemilum.4:551−554;Jassimら,1990,J.Biolum.Chemilum.5:115−122)、対象内のバイオセンサーとして使用することができるだろう。O2濃度に関する発光のダイナミックレンジは、O2プローブよりはるかに広く、より低いO2濃度に達する(Campbell,1988,Chemiluminescence.Principles and Applications in Biology and Medicine(英国チチェスター:Ellis Horwood社およびVCH出版社))。さらに、O2濃度に比例する発光は、30nMから8mMまでの範囲にわたって直線的であり、1/2最大発光には9mM O2が必要である。
【0189】
(B.腫瘍細胞の局在)
対象内の腫瘍の増殖と転移的蔓延は、本発明の方法と組成物を用いてモニターできる。特にその個体が原発腫瘍を持つと診断される場合は、その腫瘍の細胞に対するLECを用いて、腫瘍の境界を明確にし、かつ、その原発腫瘍塊から細胞が離れた部位に移動し、定着したかどうかを決定することができる。
【0190】
例えば、腫瘍抗原に対する抗体を含有しかつLGMが添加されているリポソームなどのLECを、対象に投与し、対象内の腫瘍細胞に結合させ、撮像し、その光子放出の領域を腫瘍細胞の領域と相関させることができる。
【0191】
関連する側面として、上述のような腫瘍局在性LECを使用した画像を、選択した時間間隔で作成することにより、ある対象内で腫瘍の増殖、進行および転移を経時的にモニターすることができる。このようなモニタリングは、抗腫瘍療法の結果の記録に、あるいは腫瘍の増殖または転移を抑制するのに有用だと推定される治療用化合物のスクリーニングの一部として有用だろう。
【0192】
また、腫瘍細胞を恒常的に活性なプロモーターの制御下にあるルシフェラーゼ構築物で形質転換し、それを使って、上述のように、動物モデル中に発光性腫瘍を誘導することもできる。そのような動物モデルは、抗腫瘍化合物候補の効果を評価するのに使用できる。
【0193】
(C.炎症の局在)
上述と同様の方法で、本発明の組成物と方法を使用することにより、炎症部位を特定し、炎症を経時的にモニターし、かつ/または、抗炎症性化合物候補のスクリーニングを行なうことができる。炎症部位に向かわせるのに有用な分子には、セレクチンに結合するELANタンパク質ファミリーがある。ELAN分子を本発明の物体上にターゲッティング成分として組み込み、それを使って、炎症部位を標的にすることができる。
【0194】
また、E−セレクチンプロモーターの制御下にルシフェラーゼを持つよう遺伝子組換えした動物を作出することにより、抗炎症性物質候補を研究するための動物モデルを作出することができる。E−セレクチンは炎症部位に発現するので、炎症部位にあるトランスジェニック細胞はルシフェラーゼを発現させるだろう。
【0195】
この系は、抗炎症性物質のスクリーニングに使用できる。炎症性剌激を対照動物と実験動物に投与し、処置動物内で誘導される発光に対する抗炎症性化合物候補の影響を対照動物と比較することにより、それら抗炎症性化合物候補の効果を評価することができる。
【0196】
(D.感染の局在)
本発明を裏付けるために行なった上述の実験で例証されるように、LGCは、病原体による対象の感染の経過を追跡するのに有効に利用できる。本明細書に詳述する実験では、LGCは、ルシフェラーゼを発現させるように形質転換された病原性細胞(サルモネラ)である。このような系は、感染とその後の感染の蔓延をヒト疾患の動物モデルで研究するのに理想的である。このような系によれば、病因の研究に熱や腫脹などの伝統的な全身症状ではなく感染部位と疾患の進行部位を用いて、感染性疾患の進行をモニターすることができる。
【0197】
抗感染薬の効力をモニターするために外部撮像法を使用すれば、生きている個々の動物中で時間的空間的評価を行なうことができ、それによって病原および/または試験抗感染薬に関する実験に必要な動物の数を減らすことができる。
【0198】
以下、実施例により本発明を説明するが、これらは決して本発明の限定を意図するものではない。
【0199】
(材料と方法)
(A.細胞)
サルモネラ株SL1344とLB5000は B.A.D.Stocker(スタンフォード大学;HoisethおよびStocker,1981,Nature 291:238−239)から入手した。サルモネラ株BJ66はB.D.Jones(スタンフォード大学)から入手した。
【0200】
HEp−2細胞はAmerican Type Culture Collection(ATCC;メリーランド州ロックビル・パークローンドライブ12301;受託番号CCL−23)から入手した。
【0201】
マウス腹腔マクロファージは、安楽死させたBalb/cマウスの腹腔を7mlの生育培地で洗浄することによって得た(MaximowおよびBloom,1931,Textbook of Histology,Saunders,フィラデルフィア)。
【0202】
(B.静置培養)
100mg/mlのカルベニシリンを含むLBブロス3mlに、定常期培養から得た細菌懸濁液6μlを接種し、その細菌を7mlの静置培養管中37℃で終夜生育することにより、低酸素(静置)培養を調製した。
【0203】
(C.マウス)
Balb/c(Itys/s)マウスは、スタンフォード大学の腫瘍学科(the Department of Oncology)から入手した。129XBalb/c(Ityr/s)マウスはスタンフォードトランスジェニック動物施設(the Stanford Transgenic Animal Facility;カリフォルニア州スタンフォード)から入手した。動物はすべて、光周期、給餌法および温度条件を同一にして、スタンフォード大学研究動物施設(the Stanford University Research Animal Facility;カリフォルニア州スタンフォード)で飼育した。
【0204】
麻酔は動物の腹腔内に33μg/kg−体重のネンブタールを注射することによって行った。
【0205】
安楽死は、スタンフォード大学研究動物施設が推奨するプロトコールに従って、CO2による窒息または頚椎脱臼によって行った。頚椎脱臼は、窒息による生理学的変化が実験結果に影響を与えるかもしれない場合に使用した。
【0206】
lux形質転換サルモネラに感染したマウスには、luxオペロンを含有するAmprプラスミドが保持されるような選択圧を発光性サルモネラに与え続けるために、カルベニシリンの筋肉内(i.m.)注射(体重1kgあたり125mg)を毎日施した。
【0207】
(D.撮像)
撮像すべき動物または対象を、2段階マイクロチャネル増感ヘッドを持つ電荷結合素子(CCD)カメラ(モデルC2400−40,Hamamatsu社)と扉とを持つ光を通さない箱の中に固定した。そのカメラを、箱の外へ通じるケーブルを介して”ARGUS50”画像処理装置(Hamamatsu社)に接続した。
【0208】
上述のICCDシステムは、いったん10〜30光子の域値に到達すれば、単一光子を検出できる。このシステムの信号対雑音比は、信号強度に依存して2:1から1X104:1までに及んだ。
【0209】
グレースケール画像は、かすかなルームライトの中でライトボックスの扉をあけ、8〜64フレーム間積分することによって得た。グレースケール画像のゲインは、その画像が最適となるように設定した(通例、0〜10,000ボルトのスケールで3000ボルト)。
【0210】
生物発光データは、外部照射の不在下に得た。露出設定は次の通りとした:黒レベルはカメラ/画像処理装置によって自動的に設定、ゲインは増倍装置コントローラーで自動的に設定、Fストップは2.8に設定した。60mm”AFニッコール”マクロレンズを使用した(ニコン(Nikon)社,ニューヨーク州メルヴィル)。
【0211】
生物発光画像は、選択した時間(通例5分間)光子を積分することによって作成した。データは、いずれの動物についても、1ピクセルあたり0〜3ビットの最低ビットレンジ設定で表わす。生物発光シグナルの分解能を0〜3のビットレンジにできなかった他の物体(すなわち24ウェルプレート)の画像については、レンジを生物発光シグナルの局在化が可能な設定(通例1〜7)に増やした。5分間の撮像で付加的な情報が得られなかった場合は、物体をより短い時間で撮像した。
【0212】
外部撮像とは、動物の非侵襲的撮像を指す。内部撮像とは、動物の部分的切開(通例、腹壁切開)後の撮像を指す。内部撮像は、外部撮像で局在化した光子放射の光源を確認するために、選択した動物で行なう。
【0213】
生物発光画像データは、検出された光子の強度を表わす擬似色発光画像として表わす。通例、青色(低強度)から赤色(高強度)までの6つの強度レベルを使用する。
【0214】
ここに掲載する図を作成するために、グレースケール画像と生物発光画像を画像処理装置を使って重ね合せることにより、空間的準拠座標系を与える合成画像を作成した。
【0215】
合成画像はRGB CRT(赤、緑、青;ブラウン管)モニターに表示し、そのモニターを撮影してハードコピーを作った。また画像処理装置画像をデジタルファイルとして保存し、そのファイルをコンピューターに移し、それをそのコンピューターに接続したカラープリンターで印刷することによっても、ハードコピーを作成した。別法として、ビデオプリンターを使用してビデオ信号を直接印刷することにより、ハードコピーを作成してもよい。
【実施例】
【0216】
(実施例1)
(pCGLS1luxプラスミドによるサルモネラの形質転換)
サルモネラ株SL1344、BJ66およびLB5000を、Xenorhabdus luminescens由来のluxオペロン(Frackmanら,1990)をコードするpUC18系ベクターpCGLS1で形質転換した。
【0217】
(A.pCGLS1プラスミド)
pCGLS1プラスミドの概略を図1A、1Bおよび1Cに示す。このプラスミドは、土壌菌Xenorhabdus luminescens由来のlux遺伝子をコードする〜11kb領域(図1A;Frackmanら,1990)を、pUC18(図1C;Clontech社,カリフォルニア州パロアルト)のBam HI部位(図1B)にクローニングすることによって構築した。このべクターの構築はFrackmanらの報文(1990)に記述されている。
【0218】
図1A中の制限酵素部位は、次のように記載されている:Bs,Bst EII;C,Cla I;E,Eco RI;H,Hind III;M,Mlu I;S,Sca I;X,Xba I;B/Sa,Bam HIおよびSau 3A接合点。多重クローニング部位(MCS)を含む配列を図1Bに記載し、そのBam HI部位を太字で示す。
【0219】
挿入物を含まないpUC18ベクターの図解を図1Cに示す。印を付けた要素には、アンピシリン耐性遺伝子(Ap)、lac Z遺伝子(lac Z)および大腸菌の複製起点(Ori)が含まれる。非修飾pUC18ベクターのサイズは約2.7kbである。
【0220】
(B.サルモネラの形質転換)
サルモネラ株Sl1344、BJ66およびLB5000のエレクトロコンピテント細胞を一般的な方法(Sambrookら,1989,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory Press,第2巻)で作成し、使用直前まで−80℃で保存した。エレクトロポレーションは次のように行なった。1μlのプラスミド(0.2μg/ml)を、10%グリセロールに懸濁した40μlの氷冷エレクトロコンピテント細胞に加えた。その懸濁液を1分間穏やかに混合し、間隙1mmのエレクトロポレーションキュベットに入れ、Bio−Rad Gene−Pluser(Bio−Rad Laboratories社,カリフォルニア州ハーキュリーズ)を使って電気穿孔した。設定は、2.5キロボルト、400オームおよび25マイクロファラドとした。
【0221】
ルリア−べルチーニ(LB)ブロス中37℃で1時間攪拌培養した後、100μg/mlカルベニシリンを含む(LB)寒天上に細胞をまき、終夜生育した。
【0222】
標識されたサルモネラの生物発光を最大にするため、luxオペロンを高コピー数プラスミド上に維持し、単一コピー遺伝子として組み込まれないようにした。しかしプラスミドは、この研究で使用したSL1344やBJ66のようなrecA株では特に、細菌細胞による修飾を受ける。recA遺伝子座は、luxオペロンとβ−ラクタマーゼを含有するプラスミドの領域を欠失させうるリコンビナーゼをコードする。したがって培養細胞から回収したサルモネラをカルベニシリンの存在下と不在下の両方で平板培養し、生物発光が失われる頻度を決定するために撮像した。ゲンタマイシン処理し溶解したHEp−2細胞とマクロファージから回収されるコロニーはすべて、アンピシリン耐性(Ampr)かつ生物発光性だった。したがって、lux遺伝子は哺乳類細胞との同時培養中に失われないようだった。
【0223】
コロニーを暗室での視認によって発光について検査した。5つの形質転換体が高レベルの発光を持つと同定された。これらのうち3つ(SL1344、BJ66およびLB5000株からそれぞれ1つ)を、以降の実験のために選択した。それらをそれぞれSL1344lux、BJ66luxおよびLB5000luxと名づけた。
【0224】
(実施例2)
(通常のサルモネラおよび形質転換したサルモネラの侵入能力)
6株のサルモネラ(SL1344lux、LB5000lux、BJ66lux、SL1344、LB5000およびBJ66)の侵入能力を、2種類の細菌付着および侵入アッセイで測定した。コロニー形成単位(CFU)アッセイは、基本的に既に記述されている方法(FinlayおよびFalkow,1989,Mol.Microbiol.3:1833−1841)に変更(Leeら,1990,PNAS 87:4304−4308)を加えて行なった。生物発光アッセイは、細胞の数をCFUではなく生物発光を用いて定量する点以外は、基本的にCFUアッセイと同様に行なった。
【0225】
簡単に述べると、HE−p細胞と初代マウス腹腔マクロファージを24ウェル組識培養皿に、20mMグルタミン(Gibco/BRL社)および5%ウシ胎児血清(Hyclone社,ユタ州ローガン)を添加したRPMI(Gibco/BRL社,ニューヨーク州グランドアイランド)中、1X105細胞/ウェルの濃度で接種した。細胞を接種した24時間後(HEp−2)または数日間後(マクロファージ)に、静置培養からの細菌(上記”材料と方法”の項参照)を、1ウェルあたり1X106個(感染多重度(m.o.i)10)または1X107個(m.o.i=100,図2B〜Eの右側の柱)の密度で接種し、Beckman臨床用遠心器(Beckman Instruments社,メリーランド州コロンビア)を用いて、1000rpm(185Xg)で5分間、細胞単層上に遠心した。培地を、ゲンタマイシン(100mg/ml)を含む(侵入アッセイ)または含まない(付着アッセイ)RPMI培地(Gibco/BRL社)で置換した。その共存培養を5%CO2中35℃で合計3.5時間培養した。
【0226】
培養培地中のゲンタマイシンはHep−2細胞によって内部移行されなかった細菌を、HEp−2細胞の表面に付着しているものを含めて殺す。したがって、付着アッセイ(ゲンタマイシンを含まない)でのシグナルが、付着細菌と内部移行細菌の両方を表わすのに対し、侵入アッセイ(ゲンタマイシンを含む)でのシグナルは内部移行された細菌のみを表わす。
【0227】
付着と侵入を、3時点(接種の1.5時間後、3.0時間後および3.5時間後)で発光性細菌細胞を撮像することによりアッセイした。第1時点での撮像に先立って、付着していない細菌を除去するために細胞単層をリン酸緩衝食塩水(PBS)で3回洗浄し、新しいRPMIを加えた。発光は30秒間の露光時間で記録した。第2および第3時点での画像も同じ露光時間を用いて得たが、最初に細胞を洗うことはしなかった。
【0228】
図2Aは、最後の時点で記録したデータを、その培養皿ウェルのグレースケール画像に重ね合せた擬似色発光画像として表示したものである。細胞タイプ、サルモネラ株およびゲンタマイシンの用法を図中に示す。このデータを光子カウントの相対強度として図2Bと2Dのグラフにも要約する。
【0229】
3.5時間時点での撮像後、組識培養細胞をPBSで3回洗浄し、PBS中の0.2%”トリトン X−100”で溶解した。溶解によって放出された付着細菌および/または細胞内細菌をLB寒天プレートまたはLB−カルベニシリン寒天プレートにまき、35℃で18時間培養した。各ウェルから放出された細菌の数を、コロニー形成単位(CFU,FinlayおよびFalkow,1989,Mol.Microbiol.3:1833−1841;Leeら,1990,PNAS 87:4304−4309)の数を数えることによって決定した。これらのデータを、ゲンタマイシンと共にまたはゲンタマイシンなしで3.5時間培養した後の共存培養から回収された1mlあたりの総細菌コロニーとして表わし、図2Cおよび2Eのグラフに要約する。
【0230】
生物発光アッセイとCFUアッセイの両方から得られたデータは、(i)lux遺伝子で形質転換されたサルモネラが親系と類似する感染力を持つことと、(ii)発光検出とCFU決定がHEp−2細胞とマクロファージにおける2つのサルモネラ株の侵入力について同等な推定値を与えることを示している。CFUに対する生物発光の比率はマクロファージ培養の方が低く、これはおそらくサルモネラがマクロファージの細胞下区画に侵入するためだろう。
【0231】
(実施例3)
(形質転換したサルモネラの試験管内発光)
LB5000luxサルモネラの4つの10倍連続希釈液10μl(1mlあたり106細胞から103細胞まで)を4本の100μlガラス製毛細管(Becton Dickinson社のCaly−Adams事業部,ニュージャージー州パーシッパニー)に入れた。その細菌懸濁液は、毛細管中で両端に空気のポケットを持つ液体の柱を形成した。各毛細管の一端をクリトシール(critoseal;Clay−Adams社)で密封した。希釈液の作成に使用した培地は、空気に曝すことによりO2で飽和させた。
【0232】
その毛細管を透明なプラスチック製のラップで包み、上述のように30秒間撮像することにより、発光を測定した。典型的な画像を図3Aに示す。4本の毛細管を示す。これらは(上から下へ)106、105、104および103サルモネラ細胞/ml(104、103、102および10細胞/管)を含む。発光は、104細胞/ml(100細胞)しか含まない懸濁液でも検出できた。しかしその発光は空気/液体界面に限定され、この発光反応が比較的高レベルの酸素を必要とすることが示唆される。細胞の多くはおそらく液体柱の中にあり、空気/液体界面にはないだろうから、このデータは、図3Aに示す毛細管中の発光が、各管中の総細胞数よりかなり少ない細胞から生じることを示唆している。
【0233】
(実施例4)
(動物組識を貫通する発光の試験管内検出)
LB5000luxサルモネラの連続希釈液を含む内径3.5mmのガラス製毛細管で構成された微量試験管を、基本的に上記実施例3に記述したように調製した。ただし、本実施例では細菌懸濁液を毛細管の密封した末端に接触させ、上端のみを空気にさらした。その管を半透明のプラスチック製シンチレーションバイアルに入れ、次に挙げる動物組識の一つでそのまわりを囲んだ:ヒヨコ胸筋、ヒヨコ皮膚、子ヒツジ腎臓または子ヒツジ腎臓髄質。いずれの組識も地元のスーパーマーケット(Safeway社,カリフォルニア州マウンテンヴュー)の食肉部門から入手した。
【0234】
組識で囲まれた毛細管を含むバイアルの図を図4に示す。バイアル1は直径約1.4cmで、蓋2を含む。そのバイアルをその上部分3に沿って、不透明な素材(すなわち黒テープ)で覆う。動物組識4を、バイアルの底から不透明被覆3の下端のすぐ上までになるよう、バイアルに入れる。微量試験管5の底を栓7(すなわちクリトシール(crytoseal)栓)で密封し、それをバイアルの中心に、栓をした末端がバイアルの底に接触するか、きわめて接近するように置く。細菌懸濁液6は毛細管の底から上方約1cmに達する。
【0235】
組識を含むバイアルと組識を含まないバイアルおよび細菌を含むバイアルと細菌を含まないバイアルから放射された光子を、高速同時弁別器を無効にした液体シンチレーションカウンター(モデル1219 Rackbeta,LKB/Wallac社,メリーランド州ゲーサーズバーグ)でカウントした。
【0236】
組識を含まない対照は、細菌懸濁液をシンチレーションバイアルに直接入れてアッセイした。いずれの実験も3回ずつ行なった。
【0237】
各実験では、各カウントごとにバイアルを90℃回転させて、バイアルを2、3回カウントすることにより、考えうる組識の厚さの不一致の影響について検証した。有意差は検出されなかった。
【0238】
その結果を下記表1に要約する。
【0239】
【表1】

【0240】
腎臓組識内の1X103個のLB5000luxについてのシグナルは、シンチレーションカウンター中の光電子増倍管(PMT)でバックグランドレベルか、それに近かった。このタイプの検出におけるバックグラウンドはPMTの暗電流によるものであり、この試験はかなり強いシグナルの分析に限定される。
【0241】
約1X107個のLB5000luxからの生物発光は、0.5cmの鳥類筋肉、皮膚、ヒツジ腎臓髄質およびヒツジ腎臓を通して検出できた。これらの結果は、標識したサルモネラからの生物発光が様々な不透明度の動物組識を通して検出できることを示している。(図3Aで示したように)毛細管内では酸素が制限されていたと思われるので、このアッセイで表示している数よりも少ない数の生物発光性サルモネラが、組識を通して検出できると考えられる。
【0242】
(実施例5)
(生物発光性サルモネラの生体内検出)
感染中のサルモネラに対する酸素の利用性を評価するために、野生型SL1344 luxをBALB/cマウスの腹腔内(i.p.)に接種した。接種の24時間後、麻酔したラットで、細菌によって内部に放出され腹壁を透過した光子を、増倍化CCDカメラで外部から検出し、局在化した(図3B)。全身性サルモネラ感染には、リンパ節、脾臓、肝臓のコロニー形成が関与すると思われる。野生型SL1344 luxの腹腔内接種によって感染させたマウスの腹部画像は、腹部表面のほとんどに透過光子が様々な強度の焦点を作っていることを示した(図3B)。これらの結果は、おそらくは肝臓と腸間膜リンパ節を含む内臓の広範なコロニー形成と合致し、利用可能な酸素のレベルはいくつかの組識で標識病原体からの発光を外部から検出するのに十分でありうることを示している。
【0243】
(実施例6)
(Balb/cマウスに経口投与したluxサルモネラの検出)
病原性SL1344lux、非侵入性BJ66luxおよび低病原性LB5000luxサルモネラ1X107個の懸濁液50μlを経口的に与えることによって、Balb/cマウスを感染させた。感染時に4〜6週齢だったそれらのマウスを、5分間の積分時間で毎日撮像した(光子放射を5分間測定した)。撮像に先立って、マウスを33μg/kg−体重のネンブタールで麻酔した。
【0244】
典型的な画像を図5A〜Fに示す。接種後(p.i.)24時間の時点で、生物発光シグナルはいずれの感染動物でも単一の焦点に局在した(図5A、5Cおよび5E)。生物発光は、低病原性LB5000luxに感染したすべての動物で、接種後7日までに消失した(図5B)。野生型SL1344 luxに感染したBALB/cマウスでは、生物発光が試験期間中常に検出され、第8日では透過光子の焦点が複数認められた。これらの動物では、感染がしばしば腹腔のほとんどに広がった(図5F)。これらの動物の3分の1では、第8日に透過光子が腹部領域の大半で認められ、それは腹腔内接種後の光子の分布と似ていた(図3Bと5Fを参照のこと)。BJ66 luxによる感染の蔓延はかなり変動的だったが、通例、その感染は持続し、最初の部位に局在したままだった(図5D)。
【0245】
耐性BALB/cX129マウスを野生型SL1344 luxに感染させたところ、10匹のマウス群で、その生物発光シグナルはその試験期間中常に局在し、持続性を保っていた。この結果は、SL1344 luxに感染した感受性マウス(lytr/s)に観察された播種性生物発光とは対照的であった(実施例8と図7Aおよび7Bを参照のこと)が、侵入性の低いBJ66 luxによる感受性BALB/cマウスの持続性感染とは似ていた。対照として、持続的に感染した耐性BALB/cX129マウスから得たサルモネラを培養したところ、8日後に回収したコロニーの80〜90%はAmprだった。これらのうち90%以上は生物発光性であり、観察された相違がluxプラスミドの有意な減少によるのではなく、むしろ細菌株の病原性が真に相違するためであることが示唆された。
【0246】
(実施例7)
(サルモネラの病原性株および低病原性株の腹腔内接種後の感染の検出)
100μl懸濁液中の1X107細菌細胞を同時に空気を注入することなく腹腔内(i.p.)接種することによって、Balb/cマウスを病原性(SL1344lux)または低病原性(LB5000lux)サルモネラに感染させた。
【0247】
接種後(p.i.)32時間の時点で、上述のようにマウスを麻酔し、撮像した。その結果を図6に示す。この図の左側の2匹のマウス(病原性SL1344luxに感染)では広範な感染が明らかである。これに対して、低病原性LB5000lux株を注射した右側のマウスでは、発光はあったとしてもほとんど検出されない。
【0248】
(実施例8)
(サルモネラ経口接種後の耐性マウスにおける全身性感染の検出)
耐性129XBalb/c(Ityr/s)生存マウスを、1X107個のSL1344luxサルモネラの胃内接種によって感染させた。細菌は麻酔下に胃内給餌チューブを通して与えた。注入後8日間(8 d.p.i)、動物を毎日撮像した。
【0249】
結果を図7Aと7Bに示す。一組3匹のマウスを感染させ、8日間毎日撮像した。第1日の典型的画像(図7A)と第8日の典型的画像(図7B)を示す。これらのデータは、全身性サルモネラ感染症に対して耐性なマウスは盲腸に局在した慢性感染症を持つが、その感染は腹腔内に蔓延しないことを示している。
【0250】
(実施例9)
(サルモネラ経口接種後の腹壁切開後撮像)
腹腔内の発光シグナルの位置を正確に特定し、その位置を非侵襲的撮像法で得たものと比較するために、サルモネラの経口接種後に腹壁切開を行なった。実施例8に記述したように、動物に接種した。選択した期間(通例7日)の後、上述のようにマウスを麻酔し、外部から撮像した。典型的画像を図8Aに示す。外部撮像の後、腹腔を開き、図8Bに例示するように動物を再び撮像した。場合により、盲腸(C)の前後の腸管腔に空気を注入してから、3回目の撮像を行なった(図8C)。最後の撮像後、直ちにマウスを安楽死させた。
【0251】
経口接種後の感染初期に感受性マウス中で生物発光が限局的に観察された場合はいずれも、光子はほとんどもっぱら盲腸から生じており、限局的生物発光の正確な局在と強度の変動は、盲腸の位置が変動しやすいためであった。感染耐性BALB/cX129マウスに観察された限局的な生物発光も、同様に盲腸に局在した。これに対して、SL1344 luxに腹腔内感染させた動物では、そのような局在が観察されなかった(図3B)。野生型SL1344 luxを経口接種した感染感受性マウスの後期では、生物発光が多極性であったが、追加された発光の焦点は腹壁切開後に明白にならなかった。病原性の低いLB5000 luxに感染したマウスでは、第7日の時点で、生物発光が、いずれの組識または器官にも、皮膚および腹壁の除去後、限局的にさえ、検出できなかった。
【0252】
どの感染動物の脾臓または血流にも、生物発光は光学的に検出されなかった。肝臓からの生物発光は疾患後期にのみ認められた。また、胃腸路からの生物発光は疾患の初期には盲腸に限られた。このパターンは、異なる組識におけるサルモネラの数の相違もしくは利用できる酸素の欠如によるのだろう。経口感染したマウスの破砕した器官中に存在するAmpr cfuを定量することにより、標識サルモネラSL1344 luxの分布を調べた。第7日にSL1344 lux感染BALB/cマウスの肝臓、脾臓および肺から得たampr細菌コロニーは、いずれも、その90%以上が1.9X103〜>1.0X105の範囲の総cfuを示し、生体内では検出可能な光子放射を伴わなかった(表2)。これに対して、盲腸からの生物発光は検出可能であり、この組識は>1.0X108の総cfuを含有した。LB5000 lux感染マウスでは、どの組識にもcfuが検出されなかった。これらの結果は、現在の実験系を使用した場合、組識中1X106生物がほぼこの放射波長での検出限界であることを示唆している。
【0253】
酸素はルシフェラーゼ反応に必須の基質であるから、酸素化された微小環境中に存在するサルモネラだけが生物発光するはずである。したがって、胃腸路の管腔の嫌気環境にサルモネラからの生物発光がないことは予想できることであり、腸管腔を空気にさらせば、それまでは酸素がないために検出できなかった細菌の存在が明らかになるはずである。この見解の裏付けとして、盲腸だけに検出可能な生物発光を持つ一匹の動物が、空気にさらされたときに迅速に生物発光性になる糞粒を排泄した。腸管腔内に非発光性のルシフェラーゼ発現細菌が存在することを示すこの事実と、この組織内の好気地帯と嫌気地帯の明確な描写から、腸管腔に空気を注入すれば、さらなる細菌の存在が明らかになるであろうことが示唆された。同様の生物発光パターンを示す別の動物の回腸と結腸の管腔に空気を注入したところ、注射部位近くで光子が検出可能になった(図8)。最後に、盲腸生物発光をA示す第3のマウスを殺すと、生物発光は迅速に消滅した。明確な好気環境地帯と嫌気環境地帯を欠くため、他の組識部位に空気を注射した。
【0254】
【表2】
【0255】
(実施例10)
(サルモネラ腹腔内接種後の腹壁切開後撮像)
実施例7に記述したように、1X107個のサルモネラ(SL1344lux)を腹腔内接種することにより、Balb/cマウスを感染させた。そのような一匹の典型的画像を図9A、9Bおよび9Cに示す。
【0256】
注射後(p.i)24時間の時点で、動物を麻酔し、5分間撮像した(図9A)。腹腔を開き、そのマウスを再び5分間撮像した(図9B)。盲腸を左側に引き寄せ、その動物を再び5分間撮像した(図9A)。
【0257】
その結果から、非侵襲的撮像法で得られた感染部位の局在は、腹腔を開いて明らかになった部位とよく相関することがわかる。
【0258】
(実施例11)
(SL1344luxサルモネラからの生物発光に対するシプロフロキサシンの影響)
生体内撮像法の有用性を立証するために、感染した動物を、全身性サルモネラ感染症に有効であることが知られている抗生物質シプロフロキサシンで処置した。Magalianesら,1993,Antimicrobial Agents Chemo.37:2293。
【0259】
Balbcマウスの実験群と対照群にSL1344luxを経口接種した。接種の8日後に、実験群のマウスに100mgのシプロフロキサシン塩酸塩(3mg/kg−体重;Sigma Chemical社,ミズーリ州セントルイス)を腹腔内注射した。実験群の処置後、両群の動物を、処置後5.5時間にわたって数回(上述のように)撮像した。
【0260】
典型的画像を図10B〜Eに示す。図10Bと10Dは、それぞれ実験群の処置を開始する直前の対照群と処置群を代表する動物の合成画像である。図10Cと10Eは、処置の開始後5.5時間の同じ動物の合成画像である。対照の生物発光が7.5倍まで増大したのに対して、シプロフロキサシン処置動物の腹部上の生物発光はこの期間中に検出不能なレベルにまで減少した。腹部領域上に検出される光子の総数を決定し、t=0での値に対して規格化し、処置後の時間に関して図10Aにプロットした。
【0261】
このデータは、本発明の方法と組成物を、生体内感染の蔓延に対する薬物の効果の評価に使用できることを示している。
【0262】
特定の方法と態様に関して本発明を説明したが、本発明から逸脱することなく様々な変更や改変を施しうることは理解されるだろう。
【技術分野】
【0001】
本発明は、哺乳動物対象中の発光体および発光性生物事象を検出し、位置決めし、追跡するための非侵襲的な方法と組成物に関する。
【背景技術】
【0002】
組識内の感染性因子を検出し定量するための現行の生体外方法では、感染性疾患の進行をモニターする能力に限界がある。宿主における感染性因子の複製には、しばしば一次、二次および三次複製部位が関与する。複製部位と、感染性因子がそれらの部位を通過する過程は、接種経路、宿主がコードする因子、およびその感染性因子の決定基に基づいて観測される。
【0003】
考えられる複製部位と感染の進行は、経験から推定できる場合もある。しかし感染部位と疾患の速度は、知られないか、あるいは大雑把にしか推定できないことの方が多いだろう。また感染性疾患の進行は、マウスの同系交配系でさえ、個別的である場合が多く、実験的に感染させた宿主で疾患が平均してどのような経過をたどるのかを決定するために、数多くの感染動物の連続的な生体外分析を行なう必要がある。
【発明の概要】
【発明が解決しようとする課題】
【0004】
動物モデルにおける感染の進行を追跡する手段を持つことが望ましい。その追跡を非侵襲的に行なうことができ、一匹の動物を必要な頻度で有害な影響を与えずに評価できれば、理想的である。本発明は、哺乳動物などの生体内の病原体とその他の物体を検出し、位置決めし、追跡するための非侵襲的方法を提供することを目的とする。
【課題を解決するための手段】
【0005】
1つの局面において、本発明は、次の工程からなる哺乳類対象中の生物適合体の局在を検出する非侵襲的方法を提供する:(a)その対象に、その生物適合体と発光成分の複合体を投与し、(b)その複合体がその対象内で局在を達成しうる期間の後、その対象を光検出装置の検出フィールド内に固定し、(c)その対象を固定状態に維持し、(d)維持中に、その対象中に局在した発光成分からの光子放射を、光子放射の画像を構築することができるまで、その光検出装置で測定し、(e)その画像を構築する。
【0006】
1つの実施形態において、上記の方法は、工程(b)から(e)までを選択した間隔で反復することをさらに含み得、その反復が対象中の生物適合体の局在を経時的に追跡するのに有効であり得る。
【0007】
別の実施形態において、上記の方法は、その測定が増倍化電荷結合光検出素子を用いて行われ得る。
【0008】
別の実施形態において、上記の方法は、その投与が腫瘍細胞ターゲッティング成分を含有する複合体の投与を含み得る、哺乳類対象中の腫瘍細胞の局在を検出するための方法であり得る。
【0009】
別の実施形態において、上記の方法は、その投与が炎症ターゲッティング成分を含有する複合体の投与を含み得る、哺乳類対象中の炎症の局在を検出するための方法であり得る。
【0010】
別の実施形態において、上記の方法は、その投与が感染ターゲッティング成分を含有する複合体の投与を含み得る、哺乳類対象中の病原体による感染の局在を検出するための方法であり得る。
【0011】
別の実施形態において、上記の方法は、その投与が血栓性プラークターゲッティング成分を含有する複合体の投与を含み得る、哺乳類対象内の血栓性プラークの局在を検出するための方法であり得る。
【0012】
別の実施形態において、上記の方法は、その投与が発光成分を含有する粒子である複合体の投与を含み得る。
【0013】
別の実施形態において、上記の方法は、動物モデル中の病原体による感染症の局在を検出するための方法であり得、生物適合体がその病原体であり得る。好ましい実施形態において、その病原体がサルモネラであり得る。
【0014】
別の実施形態において、上記の発光成分が発光性タンパク質であり得る。好ましい実施形態において、上記タンパク質がルシフェラーゼ、黄色蛍光タンパク質およびフェレドキシンIVからなる群より選択され得る。別の好ましい実施形態において、その投与が抗体断片と発光性タンパク質の融合タンパク質である複合体の投与を含み得る。別の好ましい実施形態において、上記生物適合体が形質転換細胞であり得、上記発光成分がその細胞から発現する異種遺伝子の産物であり得る。さらに好ましい実施形態において、上記異種遺伝子の発現が活性化可能なプロモーターの制御下にあり得る。
【0015】
別の局面において、本発明は、哺乳類対象中の生物適合体のレベルを経時的に検出する非侵襲的方法であって、(a)その対象に、その生物適合体と発光成分の複合体を投与し、(b)その対象を光検出装置の検出フィールド内に置き、(c)その対象をその装置の検出フィールド中に維持し、(d)維持中に、その対象中の発光成分からの光子放射を、その光検出装置で測定し、(e)(b)から(d)までの工程を、選択した間隔で反復することからなり、その反復が、その対象中の生物適合体のレベルの経時変化を検出するのに有効である方法を提供する。
【0016】
さらなる局面において、本発明は、次の工程からなる哺乳類対象における導入遺伝子の組み込みを検出する非侵襲的方法を提供する:(a)その対象に、哺乳類細胞中に有効に組み込まれ、かつ、導入遺伝子、発光性タンパク質をコードする遺伝子および活性化可能なプロモーターを含有するベクター構築物(ただし、発光性タンパク質をコードする遺伝子はプロモーターの制御下にある)を投与し、(b)その構築物が組み込みを達成しうる期間の後、プロモーターを活性化し、(c)その対象を光検出装置の検出フィールド内に置き、(d)その対象をその光検出装置の検出フィールド内に維持し、(e)維持中に、その対象中の、発現した発光性タンパク質からの光子放射のレベルを、その光検出装置で測定し、(f)その光子放射のレベルがバックグラウンドより有意に高ければ、その導入遺伝子の組み込みを確認する。
【0017】
なおさらなる局面において、本発明は、次の工程からなる動物中のプロモーター誘導事象の局在を検出する非侵襲的方法を提供する:(a)その事象に反応する誘導性プロモーターとその制御下にある発光性タンパク質をコードする異種遺伝子とを持つトランスジェニック動物中で、その事象を誘発し、(b)その動物を光検出装置の検出フィールド内に置き、(c)その動物を固定状態に維持し、(d)維持中に、その動物内に局在する発現した発光性タンパク質からの光子放射を、光子放射の画像を構築することができるまで、その光検出装置で測定し、(e)その画像を構築する。
【0018】
なおさらなる別の局面において、本発明は、発光性タンパク質を発現させる遺伝子で形質転換されたサルモネラを提供する。
【0019】
一態様として、本発明は、哺乳類対象内の生物適合体の局在を検出する非侵襲的方法を包含する。その生物適合体は分子であってもよいし、巨大分子、細胞、微生物(病原体を含む)、あるいは粒子などであってもよい。
【0020】
この方法では、対象に当該物体と発光成分の複合体を投与する。発光成分は通例、光を放射する分子または高分子(巨大分子)である。この成分は、放射吸収の結果として発光したり(例えば蛍光性または燐光性分子)、あるいは化学反応の結果として発光することができる(例えば生物発光タンパク質)。発光成分の典型例は、ルシフェラーゼやエクオリンのような生物発光タンパク質および、黄色蛍光タンパク質やフェレドキシンIVのような有色または蛍光タンパク質である。
【0021】
この成分は、物体の合成中に組み込んだり(例えば化学的または遺伝子的組み込み、抗体断片と発光性タンパク質の融合タンパク質など)、合成後の化学的カップリング、非共有結合的会合(例えばリポソームによる封入)、物体内でのin situ合成(例えば形質転換細胞における異種生物発光タンパク質の発現)、またはプロモーター誘導物質で剌激されたトランスジェニック動物の細胞における生物発光タンパク質の活性化可能なプロモーター制御性のin situ発現(例えばウイルスの感染によって刺激されたインターフェロン活性化プロモーター)などといった様々な技術により、物体と複合させることができる。
【0022】
対象中で複合体が局在できた期間の後、対象を、十分な量の光子放射を(その光検出装置で)測定して画像を構築するのに有効な期間、光検出装置の検出フィールド内に固定する。典型的な光検出装置は、画像処理装置に接続した増倍化電荷結合素子(ICCD)カメラである。”固定されていない”対象が動く時間スケールに比べて短い時間内に画像を構築できるのであれば、対象は撮像中本質的に”固定されている”ことになり、特別な固定措置は必要ない。次に、光子放射データから画像を構築する。
【0023】
撮像工程を選択した間隔で繰り返し、各間隔に対応する画像を構築することにより、上述の方法で、対象内の物体の局在を経時的に追跡することができる。物体上にターゲッティング成分を結合したり、複合させたり、あるいは組み込むことにより、数多くの特定の応用に、上述の方法を使用することができる。ターゲッティング成分はその物体(例えば抗体または抗体断片)固有の特性であってもよいし、あるいはターゲッティング成分を物体に複合させたり、結合したりまたは組み込んでもよい(例えば抗体を含有するリポソーム)。ターゲッティング成分の例には、抗体、抗体断片、酵素阻害因子、レセプター結合分子、種々の毒素などがある。ターゲッティング成分の標的としては、炎症部位、感染部位、血栓性プラーク、腫瘍細胞などを挙げることができる。これらの標的を特徴づける、ターゲッティング成分による認識に適したマーカーは、よく知られている。
【0024】
さらに、発光成分と複合した病原体(例えばサルモネラ)を物体として使用すれば、この方法で、動物モデル内の病原体による感染部位を検出し、局在化(定位)することができる。
【0025】
関連する態様として、本発明は、哺乳類対象内の生体適合体のレベルを経時的に検出する非侵襲的方法を包含する。この方法は上述の方法に似ているが、必ずしも物体を画像の形で局在化するわけではなく、対象内での物体のレベルの変化を経時的に検出するよう設計される。この方法は、物体(例えば発光性細菌)のレベルに対する治療用物質(例えば抗生物質)の効果を経時的にモニターするのに、とりわけ有用である。
【0026】
もう1つの態様として、本発明は、哺乳類対象における導入遺伝子の組込みを検出する非侵襲的方法を包含する。この方法では、哺乳類細胞に導入遺伝子を組み込むのに有効なべクター構築物を、対象に投与する。そのような構築物は当技術分野ではよく知られている。この構築物は、効果的な組込みに必要な要素に加えて、導入遺伝子(例えば治療用遺伝子)と、発光性タンパク質をコードする遺伝子とを、選択した活性化可能なプロモーターの制御下に含有する。その構築物が組み込みを達成しうる期間の後、そのプロモーターを活性化する。例えば、インターフェロンプロモーターを使用した場合は、ポリイノシン−ポリシトシン二重鎖(ポリIC)を局所投与(例えば足蹠注射)することにより、インターフェロン産生を刺激することができる。次に、対象を光検出装置(例えば光増倍性”暗視”ゴーグルをつけた人)の検出フィールド内に置き、光子放射のレベルを測定または評価する。そのレベルがバックグラウンドを超える場合(すなわち光が”活性化した”領域に優先的に検出されうる場合)、その対象は当該導入遺伝子を組込んでいると記録される。
【0027】
関連する態様として、本発明は、発光性タンパク質をコードする遺伝子を誘導性プロモーターの制御下に含む構築物でトランスジェニックまたはキメラにした動物におけるプロモーター誘導事象の局在を検出する非侵襲的方法を包含する。プロモーター誘導事象には、そのプロモーターを直接活性化する物質の投与、内因性プロモーター活性化因子の産生を刺激する物質の投与(例えばRNAウイルス感染によるインターフェロン産生の剌激)、内因性プロモーター活性化因子の産生をもたらす状態に置くこと(例えば熱ショックまたはストレス)などがある。この事象を誘発し、上述のように動物を撮像する。
【0028】
さらなる態様として、本発明は、発光性タンパク質(例えばルシフェラーゼ)を発現させる遺伝子で形質転換された病原体(例えばサルモネラ)を包含する。
【0029】
またもう1つの側面として、本発明は、病原体による感染の蔓延を抑制するのに有効な治療用化合物を同定する方法をも包含する。この方法では、病原体と発光成分の複合体を対照動物と実験動物に投与し、その実験動物を治療用化合物候補で処置し、上述の方法によって対照動物内と実験動物内の発光性病原体を局在化し、その化合物が対照動物と比較して実験動物における病原体の蔓延または複製を有意に抑制する効力をもつのであれば、その化合物を治療上有効であると認める。複合体としては、蛍光標識された抗体、蛍光標識された粒子、蛍光標識された小分子などが挙げられる。
【0030】
さらなる側面として、本発明は、発光成分と共役した物体を、様々な不透明度を持つ媒質を通して局在化する方法を包含する。この方法では、光検出装置を用いて、その媒質を透過した光子を検出し、その光子を経時的に積分し、積分したその信号に基づいて画像を作成する。
【0031】
さらなる態様として、本発明は、ある生物内の特定の部位で、選択した物質(例えば溶存酸素またはカルシウム)の濃度を測定する方法を包含する。この方法には、濃度センサー(すなわち発光する能力が選択した物質の濃度に依存する発光性分子)を含有する物体(例えば細胞)が含まれる。その発光性分子を含有する物体を、動物内もしくは特定の組識または器官系(例えば脾臓)内に実質上均一に分布するように投与する。その生物を撮像する。光放射の強度と局在は、選択した物質の濃度と位置に相関する。別法として、当該物体は、第2の標識、例えば濃度センサーとは異なる波長で発光することができる分子を含有する。この第2標識を用いて宿主における物体の分布の非均一性を規格化すれば、選択した物質の濃度をより正確に決定することができる。
【0032】
もう1つの側面として、本発明は、腫瘍の増殖および/または転移性伝播を抑制するのに有効な治療用化合物を同定する方法を包含する。この方法では、(i)発光成分で標識した腫瘍細胞または発光成分を含有する腫瘍細胞を実験動物群と対照動物群に投与し、(ii)その実験群を選択した化合物で処置し、(iii)腫瘍細胞に付随する発光性分子からの光子放射を光検出装置で撮像することによって、両群の動物中の腫瘍細胞を局在化し、(iv)その化合物が対照群と比較して実験群における腫瘍の増殖および/または転移性伝播を有意に抑制できるならば、その化合物を治療上有効であると認める。
【0033】
本発明の上記その他の目的と特徴は、下記の発明の詳細な説明を添付の図面と合わせて読むことにより、より完全に理解されるだろう。
【0034】
すなわち、本発明により、哺乳動物などの生体内の病原体とその他の物体を検出し、位置決めし、追跡するための非侵襲的方法が提供される。
【図面の簡単な説明】
【0035】
【図1】図1は、サルモネラ(Salmonella)SL1344株、BJ66株およびLB5000株を形質転換してSL1344lux株、BJ66lux株およびLB5000lux株を作出するために使用したlux pCGLS1プラスミドのマップである。
【図2A】図2Aは、サルモネラSL1344lux株とBJ66lux株によるマクロファージとHEp−2細胞の接着および侵入を測定したアッセイの結果を示す。図2Aは、アッセイ皿のウェル内で局在化した発光性細菌細胞を示す。光子を1分間にわたって積分することによって得た擬似色像を、アッセイ皿のグレースケール像に重ね合せることにより、表記の”合成画像”を作成した。
【図2B】図2Bは、サルモネラSL1344lux株とBJ66lux株によるマクロファージとHEp−2細胞の接着および侵入を測定したアッセイの結果を示す。図2Bは、ゲンタマイシンで処理しなかったウェルの相対光強度を示す。
【図2C】図2Cは、サルモネラSL1344lux株とBJ66lux株によるマクロファージとHEp−2細胞の接着および侵入を測定したアッセイの結果を示す。図2Cは、図2Bで撮像したウェルと同じウェルから単離された1mlあたりのコロニー形成単位(CFU)の数を示す。
【図2D】図2Dは、サルモネラSL1344lux株とBJ66lux株によるマクロファージとHEp−2細胞の接着および侵入を測定したアッセイの結果を示す。図2Dは、ゲンタマイシンで処理したウェルの相対光強度を示す。
【図2E】図2Eは、サルモネラSL1344lux株とBJ66lux株によるマクロファージとHEp−2細胞の接着および侵入を測定したアッセイの結果を示す。図2Eは、図2Dで撮像したウェルと同じウェルから単離された1mlあたりのコロニー形成単位(CFU)の数を示す。
【図3A】図3Aは、LB5000lux細菌懸濁液の希釈液を含有する4つのガラス製毛細管の合成画像である。発光は30秒間の積分によって測定した。各毛細管内の懸濁液の両側には空気ポケットが存在する。
【図3B】図3Bは、マウスの腹腔内に野生型SL1344luxを接種した後の生物発光の分布を表わす。
【図4】図4は、LB5000luxが生成した光の透過を、動物組識越しに調べるために使用したバイアルの概略図である。
【図5A】図5Aは、低病原性LB5000luxサルモネラを経口接種し、図に表示した時間に撮像したBalb/cマウスの合成画像である。発光シグナルを5分間積分した。
【図5B】図5Bは、低病原性LB5000luxサルモネラを経口接種し、図に表示した時間に撮像したBalb/cマウスの合成画像である。発光シグナルを5分間積分した。
【図5C】図5Cは、非侵入性BJ66luxサルモネラを経口接種し、図に表示した時間に撮像したBalb/cマウスの合成画像である。発光シグナルを5分間積分した。
【図5D】図5Dは、非侵入性BJ66luxサルモネラを経口接種し、図に表示した時間に撮像したBalb/cマウスの合成画像である。発光シグナルを5分間積分した。
【図5E】図5Eは、病原性SL1344luxサルモネラを経口接種し、図に表示した時間に撮像したBalb/cマウスの合成画像である。発光シグナルを5分間積分した。
【図5F】図5Fは、病原性SL1344luxサルモネラを経口接種し、図に表示した時間に撮像したBalb/cマウスの合成画像である。発光シグナルを5分間積分した。
【図6】図6は、病原性SL1344lux細菌株(左の2匹)または低病原性SL5000lux細菌株(右の2匹)を腹腔内(i.p.)注射した32時間後のマウスにおけるサルモネラの分布を示す合成画像である。
【図7】図7Aと7Bは、全身性サルモネラ感染症に対して耐性なマウス(129XBalb/c,Ityr/s)における病原性サルモネラの分布を示す。図7Aは第1日、図7Bは第8日である。
【図8A】図8Aは、病原性が減少したサルモネラ突然変異体(BJ66lux)の経口接種7日後の分布を示す。図8Aは発光の外部非侵襲的撮像を示す。記号を付した器官は、C−盲腸、L−肝臓、I−小腸、Sp−脾臓である。
【図8B】図8Bは、病原性が減少したサルモネラ突然変異体(BJ66lux)の経口接種7日後の分布を示す。図8Bは図8Aと同じ動物を腹壁切開後に撮像したものである。記号を付した器官は、C−盲腸、L−肝臓、I−小腸、Sp−脾臓である。
【図8C】図8Cは、病原性が減少したサルモネラ突然変異体(BJ66lux)の経口接種7日後の分布を示す。記号を付した器官は、C−盲腸、L−肝臓、I−小腸、Sp−脾臓である。図8Cは、盲腸の前方と後方の腸管腔内に空気を注入した後に作成した腹壁切開後の画像を示す。
【図9A】図9Aは、SL1344luxの腹腔内接種後の感受性Balb/cマウスにおけるサルモネラSL1344luxの分布を示す。図9Aは腹腔を開く前に撮像した。
【図9B】図9Bは、SL1344luxの腹腔内接種後の感受性Balb/cマウスにおけるサルモネラSL1344luxの分布を示す。図9Bは腹腔を開いた後に撮像した。
【図9C】図9Cは、SL1344luxの腹腔内接種後の感受性Balb/cマウスにおけるサルモネラSL1344luxの分布を示す。図9Cは盲腸を左側に引き寄せた後に撮像した。
【図10A】図10Aは、経口接種マウス中のSL1344luxサルモネラからの生物発光に対するシプロフロキサシン処置の影響を表わす。図10Aは、処置動物と無処置動物について、腹部領域から測定された相対生物発光を、処置開始後の時間の関数として表わすグラフである。
【図10B】図10Bは、経口接種マウス中のSL1344luxサルモネラからの生物発光に対するシプロフロキサシン処置の影響を表わす。図10Bは、SL1344luxサルモネラを経口接種した8日後の、シプロフロキサシンで処置する前のマウスの合成画像である。
【図10C】図10Cは、経口接種マウス中のSL1344luxサルモネラからの生物発光に対するシプロフロキサシン処置の影響を表わす。図10Cは、同じマウスの対照の5.5時間後(処置なし)の合成画像である。
【図10D】図10Dは、経口接種マウス中のSL1344luxサルモネラからの生物発光に対するシプロフロキサシン処置の影響を表わす。図10Dは、SL1344luxサルモネラを経口接種した8日後の、シプロフロキサシンで処置する前のマウスの合成画像である。
【図10E】図10Eは、経口接種マウス中のSL1344luxサルモネラからの生物発光に対するシプロフロキサシン処置の影響を表わす。図10Eは、同じマウスの処置後5.5時間(図l0E)の合成画像である。
【発明を実施するための形態】
【0036】
(I.定義)
特に明示しない限り、本明細書で使用する用語はすべて、本発明の技術分野で使用される通常の意味を持つ。本明細書における不透明媒質とは「従来通りの」不透明な媒質を指し、必ずしも完全に不透明であるとは限らない。したがって不透明媒質とは、一般に透明でなく半透明でもないとみなされる媒質をいい、これには木片や哺乳動物の肉および皮膚などが含まれる。
【0037】
特に断わらない限り、ルシフェラーゼには原核生物ルシフェラーゼと真核生物ルシフェラーゼが含まれ、また、赤色領域の波長で発光するルシフェラーゼのような光学特性が変異または変化した変種も含まれる。
【0038】
生物適合体とは、哺乳動物に投与できる物体をいう。これには哺乳動物にとって有害でありうる病原体も含まれる。発光性タンパク質を発現させる導入遺伝子を含有する細胞を持つ動物の場合、生物適合体とは、その哺乳動物を構成する導入遺伝子含有細胞を指す。
【0039】
発光性とは、化学反応または放射線の吸収により光を発生しうることをいう。
【0040】
本明細書において光とは、特に断わらない限り、約300nmと約1100nmの間の波長を持つ電磁放射線をいう。
【0041】
感染の蔓延とは、通常、最初の感染部位以外の宿主部位の、病原体による伝播とコロニー形成を指す。ただしこの用語には、最初の感染部位における病原体の大きさおよび/または数の増大も含まれうる。
【0042】
lux−ルシフェラーゼおよび光子放射に関係する原核生物遺伝子。
【0043】
luc−ルシフェラーゼおよび光子放射に関係する真核生物遺伝子。
【0044】
プロモーター誘導事象とは、選択した誘導性プロモーターの直接的または間接的誘導をもたらす事象を指す。
【0045】
異種遺伝子とは、宿主生物中に形質移入された遺伝子を指す。通例、異種遺伝子とは、形質移入または形質転換された細胞のゲノムDNAに元来由来しない遺伝子を指す。
【0046】
(II.発明の一般的概観)
本発明は、哺乳類対象中の発光性複合体の非侵襲的撮像および/または検出に関する方法と組成物を包含する。この複合体は、生物適合体と発光成分とを含有する。生物適合体には、環状有機分子などの小分子;タンパク質などの高分子;ウイルス、細菌、酵母、カビなどの微生物;あらゆるタイプの病原体および病原性物質;ビーズやリポソームなどの粒子が含まれるが、これらに限らない。また、生物適合体は、撮像される哺乳類対象を構成する細胞の全部または一部であってもよい。
【0047】
発光能は、発光成分の複合体化によって物体に付与される。そのような成分には、蛍光分子、蛍光タンパク質、光子を放出する酵素反応、生物発光タンパク質などの発光物質がある。複合体化(共役)には、化学的カップリング、融合タンパク質の遺伝子操作、あるいは生物発光タンパク質を発現させる細胞、微生物または動物の形質転換が含まれうる。例えば、その物体が撮像する哺乳類対象を構成する細胞である場合、その発光成分はその細胞に「共役(複合)」させた生物発光タンパク質または蛍光タンパク質であってよく、それはトランスジェニック動物またはキメラ動物の作出によりその細胞内に導入されたべクター構築物からのプロモーター制御的発現の局在性による。
【0048】
通例、発光性複合体を様々な方法で対象に投与し、その対象内で局在させ、それを撮像する。撮像または対象からの光子放射の測定は数十分間続くことがあるので、常にそうであるとは限らないが、通常は撮像工程中、対象を固定しておく。
【0049】
発光体の撮像は、極めて低レベルの光(通常は単一光子事象)を検出し、画像の構築が可能になるまで光子放射を積分できる光検出器の使用を必要とする。そのような高感度光検出器の例には、単一光子事象を増幅した後、検出系に固有の背景ノイズに対して単一光子を検出できる(例えば液体窒素で冷却された)カメラまたはカメラ群で、その事象を検出する装置がある。
【0050】
光子放射画像を作成したら、通例、放射された光子の発生源に関して準拠座標系を与えるため(すなわち対象に関して発光性複合体を局在化するため)に、それをその対象の”通常の”反射光画像に重ね合わせる。次に、そのような”合成”画像を分析することにより、対象内の標的の位置および/または量を決定する。
【0051】
以下に、上述の段階と態様をより詳細に記述する。
【0052】
(III.発光体)
(A.発光成分)
本発明の実施に有用な発光成分(light−generating moiety;LGM)、発光分子または発光構築物は、その応用に応じて様々な形態のいずれをとってもよい。これらに共通する特徴は、それらが発光性であるということ、すなわちそれらが電子的励起状態から、より低いエネルギー状態(通常、基底状態)への遷移の結果として、原子または分子から紫外(UV)、可視および/または赤外(IR)領域の電磁放射線を放射するということである。
【0053】
発光成分の例には、蛍光分子、化学発光化合物、燐光化合物、生物発光化合物などの光冷光(フォトルミネセンス)分子がある。
【0054】
本発明との関連性が強いLGMの2つの特徴は、そのサイズとスペクトル特性である。これらについては、スペクトル特性に関する一般的議論の後、後述する発光成分の特定のタイプに関連して議論する。
【0055】
(1.スペクトル特性)
本発明の重要な側面は、非侵襲的に外部から検出できるように動物組識を貫通しうる光を生成する発光成分の選択である。光が動物組識(ほとんど水からなる)のような媒質を通過する能力は、主として、その光の強度と波長によって決まる。
【0056】
単位体積中の発光強度が強いほど、その光は検出しやすくなる。単位体積中に生成する光の強度は、後述する個々のLGMのスペクトル特性と、単位体積中の当該成分の濃度に依存する。したがって、通例、物体内または物体上に高濃度のLGMを配置する共役法(例えばリポソームの高効率負荷や、細胞内での生物発光タンパク質の高レベル発現など)により、例えば各物体にLGMを一つだけ共役させる方法よりも深い組識層を通して検出することが容易な、より明るい発光性複合体(light−emitting conjugate; LEC)を作成する。
【0057】
組識層越しのLGMの検出性を支配する第2の因子は、放射光の波長である。ほとんどの組織は主として水からなっているので、動物組識の吸収特性を近似するには水を使用することができる。水が長波長光(赤色領域の光)を短波長光より容易に透過させることはよく知られている。
【0058】
したがって、通例、黄色から赤色(550〜1100nm)の範囲の光を放射するLGMの方が、より短い波長で放射するLGMより好ましい。後述するLGMのいくつかはこの範囲で放射する。しかし、本発明の裏付けとして行なった後述の実験によれば、486nm域で放射するLGMで本発明を実施すると、それが最適な放射波長でないという事実にもかかわらず、良好な結果を得ることができる。これらの結果は、これらの実験で使用したLEC(形質転換サルモネラ細胞)内に比較的高濃度のLGM(ルシフェラーゼ分子)が存在することと、高感度な検出器を使用したことが、その一因だと考えられる。より最適な放射波長を持つLGMを使用すれば、より低濃度のLGMを持つLGEで同様の検出結果が得られることは、理解されるだろう。
【0059】
(2.蛍光系成分)
蛍光は、放射線源の除去後極めて短い持続時間を持つ単一の電子励起状態から生じる物質の発光である。励起光の一部はその蛍光分子によって熱に変換されるので、放射される蛍光の波長は励起光の波長より長い(ストークスの法則)。
【0060】
蛍光分子がルミネセンスを示すには光の入力が必要であるから、本発明における蛍光分子の使用は、生物発光分子の使用よりも複雑になるだろう。通例、対象から検出される蛍光光子シグナルを汚染しないように、励起光を遮蔽する予防措置がとられる。自明の予防措置としては、蛍光顕微鏡で使用されるような励起フィルターを放射線源に設置することが挙げられる。適切に選択された励起フィルターは、蛍光成分によって放射される光子の波長と類似する波長を持つ光子の大半を遮断する。また、蛍光光子の波長以外の波長を持つ光子の大半をふるい落とすために、障壁フィルターを検出器に使用する。上述のようなフィルターは、Omega Optical社(バーモント州ブラトルバロ)を含む様々な商業的供給源から入手できる。
【0061】
また、適切な励起波長に近く、蛍光放射波長には近くない高強度光を発生させるレーザーを使用して、蛍光成分を励起させることもできる。例えば共焦点顕微鏡のようにレーザーが対象を走査できるように、x−y変換機構を使用してもよい。
【0062】
もう1つの予防措置として、検出器の部位に到達した放射光子が、対象を完全に通過したものだけとなるように、放射線源を対象の背後に設置し、遮蔽することもできる。さらに、蛍光成分を励起するために使用する光の波長に対して低い感度を持つ検出器を選択してもよい。
【0063】
上述の措置を賢明に適用すれば、本発明の方法による蛍光性LGMの検出が可能である。
【0064】
蛍光成分としては、フルオレセインのような小さい蛍光分子と、緑色蛍光タンパク質(Chalfieら,1994,Science 263:802−805;MorinおよびHastings,1971,J.Cell.Physiol.77:313)やルマジンおよび黄色蛍光タンパク質(O’Kaneら,1991,PNAS 88:1100−1104;Daubnerら,1987,PNAS 84:8912−8916)のような蛍光タンパク質が挙げられる。また、ある種の有色タンパク質、例えばフェレドキシンIV(Grabauら,1991,J.Biol.Chem.266:3294−3299;その蛍光性はまだ評価されていない)などにも蛍光性があって、本発明で使用できるかもしれない。フェレドキシンIVは赤みがかった色を持ち、それは比較的長波長で蛍光または反射して組識を貫通するのに有効な光を生じる可能性を示すので、特に有望な候補である。さらに、この分子はタンパク質としては小さいので(95アミノ酸)、物体に共役させた時に、それらの機能に与える影響を最小限に抑えることができる。
【0065】
小さい蛍光分子の利点は、それを結合させる物体の生物活性を妨害する可能性が、より大きな発光成分よりも少ないということである。また、本発明での使用に適した種々の励起および放射スペクトルを持つ市販の蛍光分子を入手することもできる。例えばMolecular Probes社(オレゴン州ユージーン)は、ルシファーイエロー(428nmで吸収し、535nmで放射)やナイルレッド(551nmで吸収し、636nmで放射)を含む多数の発蛍光団を販売している。さらに、種々の結合法で使用できる様々な基で誘導体化された分子も(例えばMolecular Probes社から)入手することができる。
【0066】
(3.生物発光系成分)
化学発光(化学反応の結果としての発光)と生物発光(生体からの可視発光)は、多くの側面から詳細に研究されている(例えばCampbell,1988,Chemiluminescence.Principles and Applications in Biology and Medicine(英国チチェスター;Ellis Horwood社およびVCH出版社))。以下に顕著な特徴を簡単に要約する。
【0067】
生物発光分子は、光を放射するために放射エネルギーの入力を必要としない点で、蛍光分子とは異なる。むしろ生物発光分子は、ATPなどの化学エネルギーを利用して光を発生させる。蛍光成分に対する生物発光成分の利点は、そのシグナルに事実上バックグラウンドがないということである。検出される光は、外来の生物発光成分によって生成した光だけである。これに対して、蛍光分子を励起するために使用される光は、しばしば意図する標的以外の物質の蛍光をもたらす。これは”バックグラウンド”が生きている動物の内部環境のように複雑な場合は、とくにそうである。
【0068】
生物発光分子にはいくつかの種類が知られている。それらには、ルシフェラーゼファミリー(例えばWoodら,1989,Science 244:700−702)や、エクオリンファミリー(例えばPrasherら,Biochem.26:1326−1332)がある。ルシフェラーゼファミリーの要素は、種々の原核生物と真核生物で同定されている。原核発光(lux)系に関与するルシフェラーゼその他の酵素と、それに対応するlux遺伝子は、ビブリオ(Vibrio)属とフォトバクテリウム(Photobacterium)属の海生細菌およびキセノラブダス(Xenorhabdus)属の陸生細菌から単離されている。
【0069】
ルシフェラーゼ系(luc)を含有する代表的真核生物は、北アメリカホタルPhotinus pyralisである。ホタルルシフェラーゼは詳細に研究されており、ATPアッセイに広く使用されている。コメツキムシの一種 Pyrophorus plagiophthalamus由来のルシフェラーゼをコードするcDNAが、クローン化され、発現されている(Woodら,1989,Science 244:700−702)。この甲虫は、種の異なる要素が異なる色の生物発光を放射する点で珍しい。互いに95〜99%の相同性を持つ4種類のクローンが単離された。それらは546nm(緑色)、560nm(黄緑色)、578nm(黄色)、および593nm(橙色)の光を放射する。最後の種類(593nm)は、その放射光がそれより短い波長の光よりも容易に組識を貫通する波長を持つので、本発明で発光成分として使用するには特に好都合である。
【0070】
ルシフェラーゼ類とエクオリン様分子は、ATPやNAD(P)Hなどのエネルギー源と、ルシフェリンまたはコエレントリジン(coelentrizine)と酸素などの基質を必要とする。
【0071】
ルシフェラーゼ酵素を発光させるには、基質ルシフェリンをルシフェラーゼ酵素に供給しなければならない。ルシフェラーゼ酵素がluxルシフェラーゼをコードするcDNAを含有するベクターの発現産物として導入される場合、ルシフェリンを供給する便利な方法は、ルシフェラーゼを発現させるだけでなく、ルシフェリンを合成するための生合成酵素をも発現させることである。そのような構築物で形質転換された細胞では、生物発光に必要な外因性要素が酸素だけになる。そのような方法(実施例1に詳述)を用いてlux形質転換サルモネラを作成し、それを使って本発明を裏付ける後述の実験を行なう。
【0072】
土壌菌Xenorhabdus luminescensから得たluxオペロンをコードするプラスミド構築物(Frackmanら,1990,J.Bact.172:5767−5773)は、ヘテロ二量体ルシフェラーゼの2つのサブユニットと3つの補助タンパク質を発現させることにより、形質転換大腸菌に光子放射能を付与する(Frackmanら,1990)。X.luminescensのlux遺伝子を発現させる大腸菌の最適な生物発光は37℃で観察され(SzittnerおよびMeighen,1990,J.Biol.Chem.265:16581−16587;Xiら,1991,J.Bact.173:1399−1405)、これは真核発光生物や他の原核発光生物由来のルシフェラーゼの低い最適温度と対照的である(Campbell,1988,Chemiluminescence.Principles and Applications in Biology and Medicine(英国チチェスター:Ellis Horwood社およびVCH出版社))。したがってX.luminescens由来のルシフェラーゼは、動物中での研究にマーカーとして使用するのに適している。
【0073】
上述または実施例1に記述するようなルシフェラーゼベクター構築物は、大半の細菌と多くの真核細胞(luc構築物)を含む様々な宿主細胞の形質転換に使用できる。また、ヘルペスウイルスやワクシニアウイルスのようなある種のウイルスは、ルシフェラーゼを発現させるように遺伝子操作することができる。例えば、Kovacs Sz.およびMettenlieter,1991,J.Gen.Virol.,72:2999−3008には、ヘルペスウイルス内のホタルルシフェラーゼをコードする遺伝子の安定な発現が記述されている。BrasierおよびRon,1992,Meth.in Enzymol.216:386−396には、哺乳類細胞内でのルシフェラーゼ遺伝子構築物の使用が記述されている。培養哺乳類細胞からのルシフェラーゼ発現は、CCD撮像法で巨視的(IsraelおよびHonigman,1991,Gene 104:139−145)にも微視的(Hooperら,1990,J.Biolum.and Chemilum.5:123−130)にも研究されている。
【0074】
(B.物体)
本発明は、上述のような発光成分、発光性構築物または発光分子を含むように修飾したまたは共役させた物体を包含する。そのような共役体または修飾体を、発光体、発光性複合体(LEC)または単に複合体という。これらの物体自体は、例えば分子、高分子、粒子、微生物または細胞などの形態をとることができる。発光成分を物体に共役させる方法は、その成分と物体の性質に依存する。代表的な共役法については、後述の物体に関連して議論する。
【0075】
(1.小分子)
本発明の実施に有用であろう小分子物体としては、病原体もしくは内因性リガンドまたはレセプターと特異的に相互作用する化合物が挙げられる。そのような分子の例には、薬物や治療用化合物;ある種のクモ、ヘビ、サソリ、渦鞭毛藻類、海性巻貝、細菌を含む有毒生物の毒液中に存在するような毒素;NGF、PDGF、TGF、TNFなどの増殖因子;サイトカイン;生理活性ペプチドなどがあるが、これらに限るわけではない。
【0076】
これらの小分子は、その小分子の生物活性が仮に妨害されるとしてもそれが最小限であるような発光成分(例えば小さい蛍光分子(上述))と共役(結合)させることが好ましい。結合の性質は通例化学的であり、当業者の知る様々な方法のいずれで行なってもよい。
【0077】
小分子物体は、形式的な結合操作が必要ないように、発光成分を含有するよう合成することができる。また、発光成分と反応できる反応性基を持つ小分子物体を合成したり、その逆を行なうこともできる。
【0078】
本発明の発光成分と結合した小分子は、ヒトの疾患または状態の動物モデルで使用してもよいし、処置すべきヒト対象で直接使用してもよい。例えば、腫瘍細胞上に発現するレセプターに高い親和力で結合する小分子を動物モデルで使用することにより、腫瘍を局在化し、その大きさの推定値を得たり、治療薬候補による処置後の腫瘍の増殖または転移の変化をモニターすることができる。また、そのような分子を用いて、癌患者中の上述のような腫瘍の特徴をモニターすることもできる。
【0079】
(2.高分子)
ポリマーや生体ポリマーのような高分子は、本発明の実施に有用な物体のもう1つの例である。代表的な高分子には、抗体、抗体断片、融合タンパク質、ある種のベクター構築物がある。
【0080】
商業的供給源から購入したり、当技術分野で知られる方法(Harlowら,1988,Antibodies:A Laboratory Manual,第10章,402頁,Cold Spring Harbor Press)によって作成した抗体または抗体断片を用いて、それら抗体を発光成分と結合し、その複合体を対象に例えば注射などによって投与し、その複合体を抗原の部位に局在させ、その複合体を撮像することにより、哺乳類対象内の抗原を局在化することができる。
【0081】
抗体と抗体断片は、本発明での物体としての使用に関していくつかの利点を持つ。これらはその性質上、それら自体がターゲッティング成分を構成する。さらにその大きさゆえに、小さい蛍光分子、蛍光タンパク質および生物発光タンパク質を含む数タイプの発光成分との結合に馴染みやすく、また、例えば細胞やリポソームなどと比較して、迅速に拡散できる。
【0082】
発光成分は抗体または断片に直接結合させることもできるし、例えば蛍光性の二次抗体などを用いて間接的に結合させることもできる。直接的結合は、抗体または抗体断片に対する例えば発蛍光団の一般的な化学的結合によって、あるいは遺伝子操作によって行なうことができる。蛍光タンパク質または生物発光タンパク質に結合した抗体または抗体断片を含有するキメラまたは融合タンパク質を構築することができる。例えばCasadeiら,1990,PANS 87:2047−2051には、エクオリンと抗体の融合タンパク質遺伝子を哺乳類細胞内で発現させることができるベクター構築物の作成法が記述されている。
【0083】
抗体を含有する複合体は、本発明の多くの応用に使用できる。例えば炎症部位に発現するE−セレクチンに対する標識抗体を使用して、その炎症を局在化し、抗炎症薬候補の効果をモニターすることができる。
【0084】
ベクター構築物そのものも、本発明に応用できる高分子体となりうる。例えば、発光性分子をコードする遺伝子と治療用遺伝子とを、選択したプロモーター(すなわち当該治療用遺伝子が標的とする細胞内で発現するプロモーター)の制御下に含有する真核発現ベクターを構築することができる。本発明の方法を用いてアッセイされるその発光性分子の発現は、その治療用遺伝子の発現位置と発現レベルを決定するために使用できる。その治療用遺伝子の発現が、処置される人または動物モデル内で直接的な表現型を持たない場合に、この方法はとりわけ有用だろう。
【0085】
(3.ウイルス)
本発明のある種の側面で有用なもう1つの物体はウイルスである。多くのウイルスは哺乳類宿主に感染する病原体であるから、それらのウイルスを発光成分と共役させ、初期感染部位と感染の蔓延を研究するために使用することができる。また、発光成分で標識したウイルスは、感染もしくは感染の蔓延を抑制する薬物のスクリーニングに使用することもできる。
【0086】
ウイルスは、発光成分と結合した抗体を用いて間接的に標識したり、あるいは例えばウイルス粒子を(Dhawanら,1991,J.Immunol.147(1):102の方法などで)ビオチニル化した後、それらを検出可能成分(例えば蛍光分子)と結合したストレプトアビジンにさらすことにより、間接的に標識することができる。
【0087】
別法として、ウイルス粒子をローダミンのような発蛍光団で、例えばFanら,1992,J.Clin.Micro.30(4):905の方法を用いて直接標識してもよい。また、発光性タンパク質を発現させるようにウイルスを遺伝子操作することもできる。ヘルペスやワクシニアのようなある種のウイルスのゲノムは、本発明の裏付けとして行なった実験で使用したlux遺伝子やluc遺伝子のような大きさの遺伝子を十分に順応させうるほどに大きい。
【0088】
標識したウイルスを動物モデルで使用することにより、感染を局在化し、感染の進行をモニターすることができ、また感染の蔓延を抑制するのに有効な薬物をスクリーニングすることもできる。例えば、ヘルペスウイルス感染症は皮膚病変として現われるが、このウイルスはヘルペス脳炎をも引き起こしうる。上述の方法のいずれかで標識したウイルスを用いることにより、このような感染症を局在化し、モニターすることができると共に、様々な抗ウイルス剤を中枢神経系(CNS)感染症におけるその効力について調べることができる。
【0089】
(4.粒子)
ビーズ、リポソームなどを含む粒子は、本発明の実施に有用なもう1つの物体を構成する。これらはサイズが大きいので、粒子には、例えば小分子などの場合よりも多数の発光成分を共役させることができる。これは、より短い露光時間で、またはより厚い組識層を通して検出することができる、より高濃度の光放射をもたらす。また、本質的に純粋なターゲッティング成分またはリガンド(例えば抗原や抗体)をその表面に含むように、リポソームを構築することもできる。さらに、例えば生物発光タンパク質分子などを比較的高濃度に、リポソームに負荷することもできる(Campbell,1988,Chemiluminescence.Principles and Applications in Biology and Medicine(英国チチェスター:Ellis Horwood社およびVCH出版社))。
【0090】
さらに、2種類のリポソームを、その両方が存在する場合にのみ光が生成するように、同じ細胞タイプに向かわせることもできる。例えば一方のリポソームにルシフェラーゼを担持させ、他方にルシフェリンを担持させることができる。これらのリポソームはターゲッティング成分を保持してもよく、2つのリポソーム上のターゲッティング成分は同じであってもよいし、異なってもよい。感染細胞上のウイルスタンパク質は、感染した組識または器官の同定に使用できる。免疫系の細胞は、単一のまたは複数の細胞表面マーカーを用いて局在化することができる。
【0091】
血中循環時間を延長し、血流によるターゲッティングを増進するには、リポソームの表面を、例えばリン脂質−ポリエチレングリコール複合体の組み込みなどによってコーティングすることが好ましい。このタイプのリポソームはよく知られている。
【0092】
(5.細胞)
細胞は、原核、真核共に、本発明の実施に有用なもう1つの物体を構成する。細胞にも粒子と同様に比較的高濃度の発光成分を負荷できるが、細胞には、例えば異種遺伝子構築物で細胞をトランスフェクションすることによって、発光成分を供給しうるという利点がある。また、”ターゲッティング成分”もしくは細胞を対象内の所望の位置に導くのに有効な分子を発現させる細胞を選択することもできる。別法として、適当なターゲッティング成分を発現させるベクター構築物で細胞をトランスフェクションすることもできる。
【0093】
使用する細胞タイプは、その応用に依存する。例えば後に詳述するように、サルモネラなどの細菌細胞を使用することにより、高レベルな時間的空間的解像度でその感染過程を研究し、その感染過程に対する薬物または治療剤の効果を評価することができる。
【0094】
細菌細胞は有効な物体を構成する。例えば、それらをトランスフェクションすることにより、高レベルの発光成分や高レベルのターゲッティングタンパク質を容易に発現させることができる。また、表面結合型抗体を発現させる細菌を含む大腸菌ライブラリーを得て、それをスクリーニングすることにより、選択した抗原に対する抗体を発現させるコロニーを同定することも可能である(Stratagene社,カリフォルニア州ラホーヤ)。次に、このコロニーから得た細菌を、発光性タンパク質の遺伝子を含有する第2のプラスミドで形質転換し、形質転換体を本発明の方法で上述のように使用することにより、哺乳類宿主内の当該抗原を局在化できる。
【0095】
病原性細菌を発光成分と共役させ、それを動物モデル内で使用することにより、その感染過程を生体内で追跡したり、新しい抗生物質などの潜在的抗感染薬を、その感染の抑制に関するそれらの効力について評価することができる。本発明の裏付けとして行なった後述の実験によって、この応用の一例を説明する。
【0096】
真核細胞も本発明のある側面で物体として有用である。所望の調節要素を含有する適当な発現ベクターが市販されている。それらのベクターを用いて、初代培養細胞、体細胞、リンパ細胞などを含む種々の真核細胞中で所望の発光性タンパク質を発現させうる構築物を作成することができる。これらの細胞は、一過性発現試験にも使用できるし、細胞系の場合は安定な形質転換体として選択することもできる。
【0097】
形質転換細胞における発光性タンパク質の発現は、選択された様々なプロモーターのいずれかを用いて調節することができる。例えばその細胞を、発現したリガンドまたはレセプターによって対象内のある部位に向かわせる発光体として使用したい場合は、CMVプロモーターやSV40プロモーターなどの恒常的に活性なプロモーターを使用すればよい。そのような構築物で形質転換した細胞は、例えばその細胞を殺すことなどによって光の発生を阻害する化合物のアッセイにも使用できる。
【0098】
また、形質転換細胞が対象内に均一に分布し、ウイルスによる感染やサイトカインによる剌激を受けたときなど、一定の条件下でのみ発光性タンパク質が発現するように、形質転換細胞を投与してもよい。これらの剌激やその他の剌激に関係する因子に反応するプロモーターは、当技術分野で知られている。関連する側面として、Tet系(GossenおよびBujard,1992,PANS 89:5547−5551)のような誘導性プロモーターを使用して、発光性タンパク質の発現を一時的に活性化することもできる。
【0099】
例えばtat反応性HIV LTR要素を含有する構築物でCD4+リンパ細胞を形質転換し、それをHIV感染のアッセイとして使用することができる(IsraelおよびHonigman,1991,Gene 104:139−145)。そのような構築物で形質転換した細胞をSCID−huマウス(McCuneら,1988,Science 241:1632−1639)に導入し、それをヒトHIV感染症とAIDSのモデルとして使用することができる。
【0100】
例えば恒常的に活性なプロモーターを用いて上述のように形質転換した腫瘍細胞系は、腫瘍の増殖と転移をモニターするために使用できる。形質転換した腫瘍細胞を動物モデルに注射し、腫瘍塊を形成させ、増殖阻害剤候補または転移阻害剤候補で処置しながら、腫瘍の大きさと転移をモニターすることができる。
【0101】
腫瘍細胞は、様々な感染性因子または治療用化合物に感応する活性を持つ調節可能なプロモーターを含有する構築物で形質転換された細胞から作成することもできる。
【0102】
(6.細胞形質転換)
原核細胞と真核細胞の形質転換法は共に当技術分野でよく知られている(Sambrookら,1989,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory Press,第2巻)。適当な調節因子と多重クローニング部位を持つベクターは広く市販されている(例えばカリフォルニア州ラホーヤのStratagene社、カリフォルニア州パロアルトのClontech社)
(IV.発光性タンパク質をコードする遺伝子を含有するトランスジェニック動物)
もう1つの側面として、本発明は、発光性タンパク質または発光性のタンパク質複合体をコードする異種遺伝子構築物を含有するトランスジェニック動物を包含する。この構築物は選択したプロモーターによって駆動され、例えば発光性タンパク質の機能的発現に必要な種々の補助タンパク質や、選択マーカー、エンハンサー要素などを含みうる。
【0103】
プロモーターの活性化は、発光性分子と補助タンパク質をコードする遺伝子の発現量を増大させる。プロモーターの活性化は、選択した生物適合体またはその一部とプロモーター要素の相互作用によって達成される。活性化がその動物の一部でのみ起こる場合は、その部分の細胞のみが発光性タンパク質を発現させるだろう。
【0104】
例えばインターフェロン誘導性プロモーター(3’−5’ポリA合成酵素のプロモーターなど)を用いることにより、多数の異なるRNAウイルスによるトランスジェニック細胞の感染を検出できる。
【0105】
関連する側面として、ある種の疾患状態で発現するプロモーターを用いて、トランスジェニック動物内の患部を標識し、発光成分の発現を用いて、その疾患状態に対する処置の効果をモニターすることができる。例えば、E−セレクチンは生体内の炎症部位に発現する(PoberおよびCotran,1991,Lab.Invest.64:301−305)。したがってE−セレクチンプロモーターを単離し、それを用いてルシフェラーゼ遺伝子の発現を推進することができる。
【0106】
組識特異性プロモーターと共に本発明の方法を使用することも可能である。これは、例えば体内の特定の器官または組識の変性を引き起こす発病プロセスを阻害するのに有効な化合物のスクリーニングを可能にし、また発育中の動物などにおける細胞(例えばニューロン)の追跡を可能にする。
【0107】
当技術分野では、本発明で使用できるプロモーターが数多く知られている。また、プロモーター含有ゲノムDNAを単離するために、その遺伝子のcDNAから得られる情報を用いて、クローン化された遺伝子のプロモーターを単離する方法も知られている。
【0108】
(V.発光性複合体の撮像)
対象内の意図する部位に局在した発光性複合体は、いくつかの方法で撮像できる。そのような撮像の指針と具体例を以下に記述する。
【0109】
(A.発光性複合体の局在)
”標的型(targeted)”物体、すなわちターゲッティング成分(物体を対象または動物内の特定の一部位または複数部位に局在させるように設計された分子または特徴)を含有する物体の場合、局在とは、対象内で束縛された”局在型”物体と束縛されていない”遊離型”物体の間の平衡が本質的に成立した状態を指す。そのような平衡に到達する速度は投与経路に依存する。例えば血栓を局在化するために静脈内注射によって投与された複合体は、注射後数分以内にその血栓で局在または蓄積を達成するだろう。一方、腸内の感染を局在化するために経口投与された複合体は、局在を達成するのに数時間を要するだろう。
【0110】
また、局在という用語は、単に、物体を投与した後選択した時間におけるその物体の対象または動物内での位置をさす場合もある。例えば、本明細書に詳述する実験では、サルモネラを(例えば経口的に)投与し、その蔓延を時間の関数として追跡する。この場合、その物体は、それが投与した細菌の最初の位置を標識し、その後の蔓延または後退を撮像によって追跡できる限りは、経口投与後直ちに”局在型”となりうる。
【0111】
関連する側面として、例えば発光成分を発現させる、注射された腫瘍細胞の局在は、その動物内のある部位にそれらの細胞が定着し、腫瘍塊を形成することからなるだろう。
【0112】
もう1つの例として、局在は物体が投与後に分布した状態になった時に達成される。例えば、対象または動物じゅうの様々な器官における酸素濃度を測定するために投与された複合体の場合、その複合体は、それがその対象または動物内で本質的に定常的分布状態に到達した時に、”局在型”あるいは有益となる。
【0113】
上述のいずれの場合にも、当業者は、局在を達成するための時間を適切に見積もることができる。また、時間の関数としての局在の状態は、発光性複合体を本発明の方法に従って撮像することにより、追跡することができる。
【0114】
(B.光検出装置)
本発明の重要な側面は、哺乳動物内からの微弱な光を妥当な時間で(好ましくは約30分以内に)撮像することができ、その装置からの信号を使って画像を構築できるほど充分に感度の高い光検出装置の選択である。
【0115】
極めて明るい発光成分を使用できる場合および/または撮像する対象または動物の表面近くに局在した発光性複合体を検出することが可能な場合は、”暗視”ゴーグルや、シリコン増倍管(SIT)カメラ(例えば浜松ホトニクスシステムズ(Hamamatsu Photonic Systems)社,ニュージャージー州ブリッジウォーター)などの一般的高感度ビデオカメラを使用できる。しかし、より一般的には、もっと高感度な光検出法が必要である。
【0116】
本発明を実施する際に直面するような極めて低い光レベルでは、単位面積あたりの光子束があまりに低くなるので、撮影対象がもはや連続的でないように見える。その代わりに、それは時間的にも空間的にも互いに独立した個々の光子によって表される。モニターで見ると、そのような画像は、それぞれが検出された単一光子を表わす瞬く光の点のように見える。
【0117】
検出されたこれらの光子をデジタル画像処理装置で経時的に累積することにより、画像を獲得し、構築することができる。各画像点での信号に強度値を割り当てる従来のカメラとは対照的に、光子カウント撮像法では、信号の強度は意味を持たない。その目的は、単に信号(光子)の存在を検出し、その位置に関して信号の発生数を経時的に計数することである。
【0118】
後述する少なくとも2種類の光検出装置により、個々の光子を検出し、画像処理装置で分析できる信号を作成することができる。
【0119】
(1.ノイズ低減型光検出装置)
第1の種類は、光子信号の増幅とは対照的に、光子検出器内のバックグラウンドノイズを減少させることによって感度を得る装置である。ノイズは主として検出器アレイを冷却することによって低減される。この装置には、”バック薄型”冷却CCDカメラと呼ばれる電荷結合素子(CCD)カメラが含まれる。より高感度な装置では、例えばCCDアレイの温度を約−120℃まで下げる液体窒素を用いて、冷却が行われる。”バック薄型”とは、光子が検出されるためにたどる光路長を短くすることによって、量子効率を増大させる超薄型バックプレートを指す。とりわけ高感度なバック薄型低温CCDカメラは、Photometrics社(アリゾナ州トゥーソン)から入手できるシリーズ200カメラ”TECH 512”である。
【0120】
(2.光子増幅装置)
第2の種類の高感度光検出器には、光子が検出スクリーンに命中する前に光子を増幅する装置が含まれる。この種類には、マイクロチャネル増倍装置のような増倍装置を持つCCDカメラが含まれる。マイクロチャネル増倍装置は通例、カメラの検出スクリーンに対して垂直かつ同延的なチャネルの金属アレイを含有する。マイクロチャネルアレイは撮像しようとする試料、対象または動物とカメラの間に設置される。このアレイのチャネルに侵入する光子の大半は、チャネルを抜け出す前にチャネルの側面と接触する。アレイを横切ってかけられた電圧は、各光子衝突から多くの電子を放出させる。このような衝突から生じた電子は、それらが発生したチャネルから”散弾銃”式に抜け出して、カメラによって検出される。
【0121】
第1段階で生成した電子が、今度は第2段階で増幅された電子の信号をもたらすように、増倍マイクロチャネルアレイを直列に設置すれば、さらに高感度を得ることができる。しかし感度を増大させると、空間的解像度が犠牲になり、その空間的解像度は増幅段階を追加するごとに低下する。
【0122】
マイクロチャネル増倍装置型の単一光子検出装置の典型例は、Hamamatsu社から入手できるC2400シリーズである。
【0123】
(3.画像処理装置)
光子を計数する光検出装置によって生成した信号は、例えばモニターに表示したり、ビデオプリンターでプリントできる画像を構築するために、画像処理装置で処理する必要がある。そのような画像処理装置は、通例、上述の高感度光子計数カメラを含むシステムの一部として販売されており、したがって同じ供給者(例えばPhotometrics社やHamamatsu社)から入手できる。他の業者から入手した画像処理装置も使用できるが、一般的には、機能的なシステムを構築するのに、より多くの努力が必要になる。
【0124】
画像処理装置は通常、IBM互換PCやアップルマッキントッシュ(Apple Computer社,カリフォルニア州キューパーティーノ)のようなパーソナルコンピューターに接続する。このパーソナルコンピューターは購入した撮像システムの一部として含まれている場合もあるし、含まれていない場合もある。画像がデジタルファイルの形になったら、それを様々な画像処理プログラム(例えばAdobe Systems社(カリフォルニア州マウンテンビュー)の”ADOBE PHOTOSHOP”など)で操作し、印刷することができる。
【0125】
(C.装置の検出フィールドにおける対象の固定)
(1.装置の検出フィールド)
装置の検出フィールドとは、光子放射の一貫した測定値が得られる領域をいう。光学レンズを使用するカメラの場合、検出フィールドとは、単に、そのレンズがそのカメラに与える視野である。また、光検出装置が”暗視”ゴーグルである場合、検出フィールドとはそのゴーグルの視野である。
【0126】
また、検出フィールドは、密に充填されたアレイ状に配置した光ファイバーケーブルの末端が規定する表面であってもよい。そのアレイは、ケーブルの末端がカバーする領域(ケーブル間の空隙と相反する)が最大になるように構築され、対象に極めて近接して設置される。例えば、プレクシグラスなどの透明素材を対象に隣接して設置し、その透明素材に隣接して、対象とは反対側から、アレイを固定することができる。
【0127】
アレイの反対側にある光ファイバーケーブルの末端は、マイクロチャネル増倍装置の入力末端などの検出装置または増倍装置に直接接続することができ、そうすればレンズの必要がなくなる。
【0128】
この方法の利点は、対照と検出器の間の空隙の大半を排除することにより、かつ/または、レンズを排除することにより、光子の散乱および/または損失が減少することにある。本発明の裏付けとして行なった実験で使用した60mm AFニッコールマクロレンズのような高透過性レンズでさえ、フロントレンズ要素に到達する光の一部しか透過させない。
【0129】
より強度の強いLGMの場合、光ダイオードアレイを光子放射の測定に使用できる。光ダイオードアレイを比較的柔軟なシートに組み込めば、実施者は、そのアレイを対象のまわりに部分的に”巻き付ける”ことができる。この方法も光子の損失を最小限にし、さらに生物発光の三次元画像を得る手段にもなる。
【0130】
複数の検出器を対象の周囲に設置する方法や、走査型の検出器または検出器群などを含む他の方法を使用して、三次元画像を作成してもよい。
【0131】
動物または対象の全体が光検出装置の検出フィールド内にある必要はないということは理解されるだろう。例えば対象の特定の領域に局在することがわかっている発光性複合体を測定する場合、所望の情報を得るには、その領域とその周囲の十分な”暗”帯域からの光だけを測定すれば足りる。
【0132】
(2.対象の固定)
対象の2次元または3次元画像を作成したい場合、光子放射を測定している間は、対象を光検出装置の検出フィールド内に固定しておく場合がある。約20ミリ秒未満で測定した光子放射から画像を構築できるほどに信号が明るく、対象が特に動揺していない場合は、測定期間の開始時に対象が検出装置のフィールド内にあることを保証する以外に、特別な固定措置は必要ないだろう。
【0133】
これに対して、光子放射測定に約20msec以上を要し、対象が動揺している場合は、構築された画像に空間情報が保存されるように、その対象の動揺の程度に合わせて、光子放射測定中、対象の不動を保証するための予防措置を考慮する必要がある。例えば対象が人で、光子放射測定時間が数秒程度の場合は、その対象に光子放射測定(撮像)の間できるだけじっとしておいてくれるように頼むだけでよいだろう。これに対し、対象がマウスのような動物である場合は、その対象を例えば麻酔や機械的拘束具を用いて固定できる。
【0134】
様々な拘束具を作成することができる。例えば、プレクシグラスシートを発泡材クッション上に固定して、マウスを数十秒ないし数分間固定するのに有効な拘束具を作ることができる。そのクッションは、一端に、動物の頭部用のくぼみを持つ。動物は、その頭部がくぼみの上になり、息は自由にできるが、その体の動きは発泡材クッションによって束縛されるように、プレクシグラスの下に置かれる。
【0135】
対象または動物から放射され光の総量のみを測定したい場合は、光子放射測定期間が長くても、その対象を固定する必要はない。必要なことは、撮像の間、対象を光検出器の検出フィールドに閉じ込めておくことだけである。しかし、そのような測定中、対象を固定しておけば、検出される光子が通過する組識の厚さが動物間でより均一になるから、得られる結果の整合性が向上するであろうことは理解されるだろう。
【0136】
(D.撮像中のその他の問題)
(1.蛍光発生成分)
蛍光発生成分の可視化には、光検出器と共に、励起光源が必要である。またその励起光源が、発光成分からの光子放射を測定している間、点灯されることも理解されるだろう。
【0137】
発蛍光団の適切な選択、光源の設置、およびフィルターの選択と設置は、すべて情報に富む画像の構築を容易にするが、これらについては蛍光発生成分に関する項で上述した通りである。
【0138】
(2.高解像度画像)
組識による光散乱は、総光子放射の測定によるLGMの撮像で得ることができる解像度を制限する。本発明が、対象内の選択した点に集中させることができ、しかも組織内で有意に散乱しない外部源に、LGMの発光を同期させることによって、解像度のより高い画像を構築できる態様をも包含することは、理解されるだろう。例えば、集束させた超音波信号を使って、撮像する対象を三次元的に走査することができる。超音波の焦点内にある領域からの発光は、その超音波によって光に与えられた特徴的な振動により、他の光子放射から分離することができる(例えばHoustonおよびMoerner,米国特許第4,614,116号,1986年9月30日発行)
(E.光子放射画像の構築)
発光成分が例外的に明るいため、かつ/または、発光性複合体が対象の表面近くに局在するために、”暗視”ゴーグルまたは高感度ビデオカメラを使って画像を得た場合は、単に画像を見るか、ビデオモニターに表示するだけである。所望であれば、分析または印刷用に個々のビデオフレームをメモリーに保存でき、かつ/または、コンピューターで分析と印刷用に画像をデジタル化できる画像処理装置に、ビデオカメラからの信号を迂回させることができる。
【0139】
また、光子計数法を使用する場合は、光子放射の測定により、各ピクセル位置で検出した光子の数を表わす数字の配列が、画像処理装置内に生成する。これらの数字を用いて、通常は光子数を(一定の予め選択した値、またはいずれかのピクセルで検出された最大数に対して)規格化し、その規格化した数値を明るさ(グレースケール)または色(擬似色)に変換して、それをモニターに表示することにより、画像を作成する。擬似色表示の場合、典型的な色の割り当ては次の通りである。ゼロ光子カウントのピクセルには黒色を、低いカウントには青色を、カウントが増加するほど波長の長い色を割り当て、最高光子カウントには赤色を割り当てる。モニター上の色の位置は光子放射の分布を表わし、したがって発光性複合体の位置を表わす。
【0140】
複合体に対して準拠座標系を与えるために、通常は、光子放射を測定した(まだ固定されている)対象のグレースケール画像を構築する。このような画像は、例えばかすかなルームライトのなかで撮像室または撮像箱に向かって扉をあけ、反射した光子を(通例、光子放射を測定するのに要する時間のごく一部に相当する時間)測定することによって構築できる。このグレースケール像は、光子放射を測定する前に構築してもよいし、光子放射を測定した後に構築してもよい。
【0141】
通例、光子放射の画像をグレースケール像に重ね合せることにより、対象に関する光子放射の合成画像を作成する。
【0142】
例えば選択した生物適合成分の分布および/または局在に対するある処置の効果を記録するために、発光性複合体の局在および/または発光性複合体からの信号を経時的に追跡したい場合は、光子放射の測定または撮像を選択した時間間隔で反復することにより、一連の画像を構築することができる。間隔は数分程度の短いものであってもよいし、数日または数週間程度に長いものであってもよい。
【0143】
(VI.光子放射画像の分析)
本発明の方法および/または本発明の組成物を使用して作成した画像は、様々な方法で分析できる。それは、単純な視覚的検査、頭脳評価および/またはハードコピーの印刷から、複雑なデジタル画像分析まで様々である。分析によって得られる情報の解釈は、観察する現象と使用した物体に依存する。
【0144】
下記の実験は本発明の応用例の一つ(生体ラットにおけるサルモネラ感染の追跡)であり、本発明の方法で得られる画像を、どのように分析できるかを例示するものである。
【0145】
(VII.生体マウス内の発光性サルモネラの撮像)
本発明の裏付けとして行なった実験では、マウスにおけるネズミチフス菌(Salmonella typhimurium)感染(ヒトチフスの動物モデル)の分布を特徴づける。マウス病原性ネズミチフス菌SL1344(HoisethおよびStocker,1981,Nature 291:238−239)、SL1344の非侵入性突然変異体BJ66、およびサルモネラの低病原性LT−2株LB5000を、それぞれluxオペロンを含有するプラスミドで標識し、マウスにおけるサルモネラ感染を局在化する実験に使用した。
【0146】
(A.発光性サルモネラの構築)
(1.サルモネラ株)
マウスに対する経口および腹腔内接種によって明らかになる病原性表現型が異なる3株のネズミチフス菌を、形質転換のために選択した。
【0147】
ここで使用した最も病原性の高い表現型はSL1344であり、これはもともと子牛の致死的感染症から得られたマウス株である(HoisethおよびStocker,1981,Nature 291:238−239)。この株をマウスに経口接種すると、細菌はリンパ系を通って全身に転移し、肝臓、脾臓および骨髄のコロニー形成をもたらす(CarterおよびCollins,1974,J.Exper.Med.139:1189−1203;FinlayおよびFalkow,1989,Mol.Microbiol.3:1833−1841とHsu,1989,Microbiol.Rev.53:390−409の総説をも参照のこと)。
【0148】
SL1344の非侵入性突然変異体BJ66も評価する。BJ66の経口接種は通常、マウスにおける全身性感染を引き起こさないが、この株を腹腔内接種すると、全身性感染が起こる。
【0149】
サルモネラの低病原性LT−2株LB5000も調べる。LT−2株は、マウスに対する病原性が減少しているまたは変動することが知られている研究室株である。LB5000は複数の栄養要求性突然変異を含有し、ストレプトマイシン耐性であり、経口または腹腔内接種後、マウスから浄化される。
【0150】
(2.luxオペロンによるサルモネラ株の形質転換)
上記3株のそれぞれを、luxオペロンをコードするプラスミドで、実施例1に詳述するように形質転換する。土壌菌Xenorhabdus luminescens(Frackmanら,1990)から得られたこのプラスミドは、ヘテロ二量体ルシフェラーゼの2つのサブユニットと、3つの補助タンパク質luxC、luxDおよびluxEの発現によって、大腸菌に光子を放射する能力を与える。
【0151】
luxC、luxDおよびluxEが含まれるので、そのルシフェラーゼ発現細胞には、脂肪族アルデヒド基質ルシフェリンを与える必要がない。本明細書に記述するような生体系内の真核ルシフェラーゼに基質を供給することは困難だろうから、X.luminescensの全luxオペロンを使用する。このオペロンは、当該脂肪族アルデヒド基質を生合成するための酵素をもコードする。
【0152】
α−βヘテロ二量体混合機能オキシダーゼであるX.luminescensルシフェラーゼは、還元型フラビンと長鎖アルデヒドの酸化型フラビンと対応する長鎖脂肪酸への酸化を触媒する。アルデヒドの脂肪酸からの生成と再生には、脂肪酸レダクターゼ複合体が必要であり、NAD(P)H: フラビンオキシドレダクターゼが還元型フラビンを供給する。
【0153】
X.luminescensのlux遺伝子を発現させる大腸菌にとって最適な生物発光条件は37℃である(SzittnerおよびMeighen,1990,J.Biol.Chem.265:16581−16587,Xiら,1991,J.Bact.173:1399−1405)。これに対し、真核発光生物と他の原核発光生物に由来するルシフェラーゼ類は通例、より低い最適温度条件を持つ(Campbell,1988,Chemiluminescence.Principles and Applications in Biology and Medicine(英国チチェスター:Ellis Horwood社およびVCH出版社))。したがってX.luminescens由来のルシフェラーゼは、動物内での研究にマーカーとして使用するのに適している。
【0154】
全X.luminescens luxオペロンを含有し、かつ、サルモネラにアンピシリン耐性とカルベニシリン耐性を付与するプラスミドpGSL1(Frackmanら,1990)を用いて、上記3株をエレクトロポレーションによって形質転換する。このX.luminescens luxオペロンは、luxA、luxB、luxC、luxDおよびluxE遺伝子を含有する(Frackmanら,1990)。luxAとluxBはヘテロ二量体ルシフェラーゼの2つのサブユニットをコードし、luxCとluxDはルシフェラーゼ基質の生合成酵素をコードし、luxEは調節遺伝子である。培養中の細胞に外からルシフェリンを供給したり、動物を基質で処置するのとは対照的に、基質生合成用遺伝子の包含は、ルシフェラーゼに基質を提供する便利な手段である。
【0155】
(B.形質転換したサルモネラの試験管内での特徴づけ)
(1.付着性と侵入性)
上記luxプラスミドを含有する3つのサルモネラ株の付着性と侵入性を、FinlayおよびFalkow,1989,Mol.Microbiol.3:1833−1841に記述されているような一般的な侵入アッセイ(実施例2に詳述)により、培養中で互いに比較、またそれらの非発光性親株と比較する。
【0156】
このアッセイでは、上皮細胞系および腹腔マクロファージと共に培養した後、付着細菌と細胞内細菌を定量する。付着細菌と細胞内細菌は、生きている細胞からの光子の放出と、溶解してその細胞溶解液をカルベニシリン含有プレートで培養した後のコロニー形成単位との両者によって、検出、定量する。
【0157】
このアッセイの一部の結果を図2A〜2Eに示し、実施例8で考察する。lux発現プラスミドで形質転換した上記3株の表現型は、親サルモネラ株と比較して有意に変化しない。また、HEp−2細胞とマクロファージからの生物発光の強度とCFUの間には良好な相関が認められる。これらの結果は、細胞内細菌の指標としての発光が、培養内の細菌の侵入性をアッセイする迅速な方法であることを示している。
【0158】
BJ66のHEp−2に対する付着性はSL1344と比較して低かったが、マウス腹腔マクロファージの初代培養におけるこれら2株の付着性は同等だった。
【0159】
(2.光放射)
系の酸素要求性を調べるため、実施例3に詳述するように、細菌の10倍連続希釈液をガラス製毛細管内に入れ、撮像する。
【0160】
図3に、そのような実験の一つで作成した画像を示す。発光は、空気飽和培地中に少数の細菌を含む管でさえ、空気−液体界面でのみ検出される(51中に0.1mlの空気飽和緩衝液で最終O2濃度は5nMになる)。
【0161】
これらの結果から、酸素がおそらくは発光にとって制限因子であることは明らかである。
【0162】
(3.動物組識を貫通する光透過)
光が動物組識を貫通する程度を決定するために、発光性サルモネラから放射され組識を透過した光を、単一光子を検出するために高速同時検出器を切ったシンチレーション計数器を用いて定量する。このタイプの光電子増倍管の暗電流によるバックグラウンドは有意であるので、このアッセイは、比較的強い光子放射を伴う試料に限定される。
【0163】
透明度の異なる4タイプの組識:ヒヨコ胸部の筋肉、ヒヨコ胸部の皮膚、子ヒツジの腎臓、および子ヒツジの腎臓から得た腎臓髄質、をこの方法で比較する。組識を貫通して検出できる光子の数は、組識を通さない対照より約10倍少ない。
【0164】
(4.luxサルモネラの生体内での特徴づけ)
(a.経口投与)
経口接種はマウスやヒトにとって自然なサルモネラ感染経路であり、より遅延性の疾患進行をもたらす。この接種経路によるサルモネラ感染の進行を研究するために、2系統のマウスを3株のサルモネラに感染させる。耐性動物を用いて得た結果については、下記”耐性マウスの感染”という見出しの項で議論する。
【0165】
実施例5に記述するように、Balb/cマウスを病原性SL1344lux、非侵入性BL66luxおよび低病原性LB5000luxサルモネラに経口的に感染させる。その感染の進行を外部撮像法(”材料と方法”の項)によって8日間にわたって追跡する。
【0166】
典型的な画像を図5A〜Fに示す。接種後(p.i.)24時間の時点で、生物発光シグナルはすべての感染動物内で単一の病巣に局在する(図5A、5Cおよび5E)。接種後7日までには、低病原性LB5000luxに感染したすべての動物で、生物発光が消える(図5B)。これに対し、病原性SL1344luxに感染した動物の場合、蔓延が始まる時点は動物ごとにかなり変動するものの、しばしば腹腔の大半に蔓延する激しい感染を示す(図5F)。BJ66luxによる感染は通例、持続し、単一の部位に局在したままとなる(図5D)。
【0167】
(b.腹腔内接種)
サルモネラの複製部位にルシフェリンの酸化とそれに続く発光(Campbell,1988,Chemiluminescence.Principles and Applications in Biology and Medicine(英国チチェスター:Ellis Horwood社およびVCH出版社))に足るO2が存在するかどうかを評価するために、呼吸している動物の組識からの光放射を測定する。発光性のSL1344luxとLB5000luxを2群のBalb/cマウスの腹腔内に接種する。接種後(p.i.)32時間の時点で、透過した光子を撮像する(図6)。
【0168】
SL1344luxに感染したマウス(図の左側)では、透過した光子が大きな表面上に確認され、様々な強度の焦点が見える。これらの画像は播種性感染を示し、内臓(おそらくは肝臓と腸間膜リンパ節を含む)の広範なコロニー形成と合致する。これに対し、LB5000luxに感染した動物からの透過光子の分布は極めて制限されており、限定的感染を示す。
【0169】
LB5000lux感染マウスは接種後数週間にわたって健康を維持したが、SL1344lux感染マウスは接種後4日でほぼ瀕死状態になり、安楽死させた。
【0170】
これらの実験は、血中または組識中のO2レベルが、サルモネラから発現したluxルシフェラーゼの生物発光にとって十分であることを示している。また、これらの実験は、病原性が減少した研究室株LB5000と比較して病原性株SL1344の侵入性が強いこととも合致している。
【0171】
(c.耐性マウスの感染)
Ity遺伝子座がヘテロ接合性を示すマウス(Ity r/s)は、ネズミチフス菌による全身性感染症に対して耐性である(PlantおよびGlynn,1976,J.Infect.Dis.133:72−78)。Bcg(Grosら,1981,J.Immunol.127:2417−2421)またはLsh(Bradley,1977,Clin.and Exper.Immunol.30:130−140)とも呼ばれるこの遺伝子座は、鼡らい菌(Mycobacterium lepraemurium;Forgetら,1981,Infect.Immunol.32:42−47)、ウシ結核菌(M.bovis;Skameneら,1984,Immunogenet.19:117−120,SkameneおよびPietrangeli,1991,Nature 297:506−509)およびバテー杆菌(M.intracelluare;Gotoら,1989,Immunogenetics 30:218−221)のようなある種の細胞内病原体の発病プロセスを調節する。細胞内病原体に対する耐性と感受性の同様な遺伝子制御はヒトにも存在するようである(ヒト結核菌(M.tuberuculosis;Stead,1992,Annals of Intern.Med.116:937−941,Steadら,1990,New Eng.J.Med.322:422−427)とらい菌(M.leprae))。
【0172】
Ity遺伝子座はマウスの第1染色体に2つの対立遺伝子型、Ityr(耐性、優性)とItys(感受性、劣性)で存在する。Ity遺伝子座にコードされている遺伝子は、マクロファージが内部移行した病原体を破壊する能力に影響を与え(Blackwellら,1991,Immunol.Lett.30:241−248(1991)による総説がある;Skameneら,1984,Immunogenet.19:117−120とSkameneおよびPietrangeli,1991,Nature 297:506−509をも参照のこと)、それがマクロファージ媒介性とされる感染宿主内の他の部位への病原体の輸送という下流の機能に影響を及ぼすようである。Balb/cマウスはItys/sであり、129マウスはItyr/rである。ここに詳述する実験では、ヘテロ接合性Balb/cX129マウス(Ityr/s)を使用する。
【0173】
実施例7に詳述するように、耐性129XBalb/c(Ityr/s)生存マウスを、1X107個のSL1344luxサルモネラの胃内接種によって感染させる。そのマウスを注射後8日間(8 d.p.i)にわたって毎日撮像する。
【0174】
図7A(第1日)と図7B(第8日)に結果を示す。外部撮像法によって検出される発光は接種後24時間の時点で明白であり、全ての動物で単一の部位に局在しているようであった。その発光シグナルは試験期間(接種後8日まで)中常に存在する。発光の強度と発光源の位置は、一匹のマウス内で経時的にいくらか変動し、またマウスごとにも変動しうる。いずれの感染動物でも発光組識は盲腸であり(下記参照)、所在の変動性は(おそらくは強度の変動性も)齧歯類の内部器官が正しい位置に強く固定されていないという事実によるのだろう。
【0175】
これらの動物で観察された明らかな限定的感染は、このIty制限がマクロファージ輸送を遮断するという解釈を裏付けている。しかし10日間というこの感染の持続性は、腸粘膜に対する付着があることと、発光性の糞粒によって立証されるように、これらの動物の糞内に細菌の脱離が長期間続くことを示唆している。これらの結果は、生体内におけるこのサルモネラの発光性表現型が、Ity制限動物中で8日間にわたって保たれることと、経口接種後の局在が可能であることを示している。
【0176】
(d.経口接種後の内部撮像)
腹腔内の発光シグナルをさらに局在化するため、感染マウスを腹壁切開後に撮像する(図8)。経口経路で感染させた動物のいずれにおいても、主な疾患症状は盲腸の拡張である(図8A〜C)。”外部”画像(図8A)は限局的な発光を示し、それは腹壁切開後の画像(図8B)で盲腸にあたることがわかる。
【0177】
腸内に空気を注入すると、消化路の他の領域における細菌の存在が確認される。結腸と直腸内の細菌はルシフェラーゼを発現させているらしいが、それらの部位からの発光は、低い酸素濃度によって制限されているらしい。
【0178】
経口接種試験で得た画像は、接種後2日および接種後7日の発光シグナルが、LB5000luxに感染したマウスを除いて、ほとんど完全に各動物の盲腸に局在することを示している(Popeskoら,1990,A Colour Altas of Anatomy of Small Laboratory Animals Vol.Two:Rat Mouse Hamster(英国ロンドン:Wolfe))。発光は数匹の結腸にも見える。接種後7日までに、LB5000lux感染動物内に発光は認められなくなる。これらのマウスの器官中に存在するCFUを接種後2日と5日に測定する。
【0179】
侵入性株SL1344luxを胃内に感染させた動物では、盲腸内の発光が初期に現われ、その後、全身性の感染が起こる。これに対し、非侵入性BJ66lux株による感染は盲腸からの持続的な発光をもたらし、それが全試験期間(8日間)にわたって維持される動物もある。接種後8日までに、腹部表面の大半に発光が検出され、それはSL1344lux感染マウスにおける腹腔内接種後の光子の分布に似ている。
【0180】
SL1344luxによる感染は、予想通り、光子を放射する表面積が増大し、そこから放射される光子を累進的に増やしながら、全身性となるようである。いずれの株による感染でも、発光は腹部上に局在し、この領域の外側からの発光はほとんど検出されないようである。大量の透過光子が腹部上に単一の焦点として局在し、このことは、たとえ感染が全身性であっても、複製の大半が腸を取り巻く領域にあることを示唆している。
【0181】
盲腸上に発光が局在することは、腸のこの領域に大量の生物が存在することを示すだけでなく、発光するに足る酸素が得られるように、サルモネラがその粘膜の細胞に結合することをも示唆している。ルシフェラーゼからの光子の放射は酸素依存性であり、盲腸または小腸の管腔内の予想酸素レベルは、発光に必要なレベルより低い。ルシフェラーゼ反応は、細菌が腸上皮の細胞から酸素を得られない限り、腸内では機能しないと予想される。
【0182】
したがって全身性感染は侵入性表現型に関係し、腸の上皮細胞への単なる付着とは関係しないと思われる。これらの実験は、盲腸がこの発病プロセスの保菌状態で、あるいは播種部位として、何らかの役割を持つことを含意している。
【0183】
異なる組識への感染の進行をモニターすれば、発病プロセスにおけるこれらの段階についての理解を著しく深めることができ、選択した段階でその病原体を抑制するのに有効な化合物のスクリーニングが可能になるだろう。
【0184】
(e.腹腔内接種後の内部撮像)
SL1344luxを腹腔内に感染させたマウスを腹壁切開の前後に撮像する(実施例9)。その結果を図9に示す。その画像は、腹部の大部分に、複数の透過光子焦点を伴う発光を示す。盲腸は発光性サルモネラを含有しないようである。これらの実験から得られた結果は、すべてのサルモネラ株が、感染の初期相で発光するに足るO2を持つことを示している。しかし、その後の時点での全身性感染はSL1344lux感染マウスにのみ認められるので、サルモネラの粘膜の細胞への侵入と、その後の全身性感染は、侵入性表現型を持つ株に限られるようである。
【0185】
(f.サルモネラ感染に対するシプロフロキサシンの影響)
非侵襲的撮像法が薬物に対する感染症の反応を追跡するのに有用であることを示すために、実施例10に詳述する実験を行なう。マウスにSL1344luxを経口接種し、サルモネラ感染症に有効な抗生物質シプロフロキサシン100mgで処置する。処置後の選択した時期に、それらのマウスを撮像し、光子放射を測定することによって感染の程度を定量する。処置マウス内の光子放射を、処置開始前の値および感染したが処置していない対照マウスからの値と比較する。このような実験の一つで得た結果を図10A〜Eに示し、実施例10で考察する。抗生物質で処置したマウスにおける感染症は、処置マウスにおける0時間での病原体レベルおよび対照マウス内の病原体レベルと比較して、処置期間中常に低い。
【0186】
(g.カルベニシリン選択の効果)
Ducluzeauら,1970,Zeut.Bakt.5313:533−548は、抗生物質による動物の処置がサルモネラによる盲腸のコロニー形成を促進することを示した。本実験におけるマウスは、ルシフェラーゼクローンを含有するAmprサルモネラを選択するために、カルベニシリンの筋肉内注射という抗生物質措置をして維持される。この処置は、胃腸感染症の経過を変化させるかもしれないが、サルモネラが盲腸を裏打ちする細胞と結合できるという観察結果は、酸素を発光に利用できるということを示している。盲腸の管腔は一般に嫌気環境であると考えられているから、この観察結果は注目に値する。
【0187】
(VIII.応用)
生物発光技術は様々な宿主−病原体系に広く適用できると共に、例えば生きている哺乳動物内での腫瘍の進行や遺伝子発現などに関する他の生物学的事象の時間的空間的評価をも可能にするだろう。またこれは、医薬の開発とスクリーニングにも応用できる。病原体の生体内撮像を広範に使用すれば、病原および/またはリアルタイム試験抗菌剤に関する実験に必要な動物数と時間を減らすことができるだろう。また、発光性細菌は環境分析に使用されているが、生物発光性生物は生きている動物内のバイオセンサーとして有用だろう。例えばKorpelaらは、胃腸路の管腔内では酸素供給が制限されているため、酸素が(おそらくは上皮細胞または他の細胞タイプから)直接的にサルモネラに接近できる部位に、生物発光が限定されたことを示している。Korpelaら,1989,J.Biolum.Chemilum.4:551−554。この酸素要求性は、親密な細胞−細胞相互作用の指標として、あるいは生きている動物中の様々な部位における酸素濃度を調べるためのバイオセンサーとして利用できるだろう。以下、この技術の典型的応用例を例示のためにいくつか記述するが、これらは決して本発明の限定を意図するものではない。
【0188】
(A.酸素レベルの測定)
上に要約した実験で明示されたようにルシフェラーゼの発光に酸素が必要であるということは、本発明を、対象内の酸素濃度の空間的勾配の測定法として利用できるということを示している。10〜1mMの範囲の酸素レベルを測定するために、発光性細菌が使用されている。これらの研究によれば、0.1nMが検出下限だと予想される(Campbell,1988,Chemiluminescence.Pringiples and Applications in Biology and Medicine(英国チチェスター:Ellis Horwood社およびVCH出版社))。本明細書に記述する撮像法は、生きている動物内の様々な部位における酸素レベルを調べるのに利用できる。例えばO2中で、またはCa2+依存的に光を放射するように操作された微生物を、ちょうど発光性細菌が環境分析に使用されるように(Guzzoら,1992,Tox.Lett.64/65:687−693;Korpelaら,1989,J.Biolum.Chemilum.4:551−554;Jassimら,1990,J.Biolum.Chemilum.5:115−122)、対象内のバイオセンサーとして使用することができるだろう。O2濃度に関する発光のダイナミックレンジは、O2プローブよりはるかに広く、より低いO2濃度に達する(Campbell,1988,Chemiluminescence.Principles and Applications in Biology and Medicine(英国チチェスター:Ellis Horwood社およびVCH出版社))。さらに、O2濃度に比例する発光は、30nMから8mMまでの範囲にわたって直線的であり、1/2最大発光には9mM O2が必要である。
【0189】
(B.腫瘍細胞の局在)
対象内の腫瘍の増殖と転移的蔓延は、本発明の方法と組成物を用いてモニターできる。特にその個体が原発腫瘍を持つと診断される場合は、その腫瘍の細胞に対するLECを用いて、腫瘍の境界を明確にし、かつ、その原発腫瘍塊から細胞が離れた部位に移動し、定着したかどうかを決定することができる。
【0190】
例えば、腫瘍抗原に対する抗体を含有しかつLGMが添加されているリポソームなどのLECを、対象に投与し、対象内の腫瘍細胞に結合させ、撮像し、その光子放出の領域を腫瘍細胞の領域と相関させることができる。
【0191】
関連する側面として、上述のような腫瘍局在性LECを使用した画像を、選択した時間間隔で作成することにより、ある対象内で腫瘍の増殖、進行および転移を経時的にモニターすることができる。このようなモニタリングは、抗腫瘍療法の結果の記録に、あるいは腫瘍の増殖または転移を抑制するのに有用だと推定される治療用化合物のスクリーニングの一部として有用だろう。
【0192】
また、腫瘍細胞を恒常的に活性なプロモーターの制御下にあるルシフェラーゼ構築物で形質転換し、それを使って、上述のように、動物モデル中に発光性腫瘍を誘導することもできる。そのような動物モデルは、抗腫瘍化合物候補の効果を評価するのに使用できる。
【0193】
(C.炎症の局在)
上述と同様の方法で、本発明の組成物と方法を使用することにより、炎症部位を特定し、炎症を経時的にモニターし、かつ/または、抗炎症性化合物候補のスクリーニングを行なうことができる。炎症部位に向かわせるのに有用な分子には、セレクチンに結合するELANタンパク質ファミリーがある。ELAN分子を本発明の物体上にターゲッティング成分として組み込み、それを使って、炎症部位を標的にすることができる。
【0194】
また、E−セレクチンプロモーターの制御下にルシフェラーゼを持つよう遺伝子組換えした動物を作出することにより、抗炎症性物質候補を研究するための動物モデルを作出することができる。E−セレクチンは炎症部位に発現するので、炎症部位にあるトランスジェニック細胞はルシフェラーゼを発現させるだろう。
【0195】
この系は、抗炎症性物質のスクリーニングに使用できる。炎症性剌激を対照動物と実験動物に投与し、処置動物内で誘導される発光に対する抗炎症性化合物候補の影響を対照動物と比較することにより、それら抗炎症性化合物候補の効果を評価することができる。
【0196】
(D.感染の局在)
本発明を裏付けるために行なった上述の実験で例証されるように、LGCは、病原体による対象の感染の経過を追跡するのに有効に利用できる。本明細書に詳述する実験では、LGCは、ルシフェラーゼを発現させるように形質転換された病原性細胞(サルモネラ)である。このような系は、感染とその後の感染の蔓延をヒト疾患の動物モデルで研究するのに理想的である。このような系によれば、病因の研究に熱や腫脹などの伝統的な全身症状ではなく感染部位と疾患の進行部位を用いて、感染性疾患の進行をモニターすることができる。
【0197】
抗感染薬の効力をモニターするために外部撮像法を使用すれば、生きている個々の動物中で時間的空間的評価を行なうことができ、それによって病原および/または試験抗感染薬に関する実験に必要な動物の数を減らすことができる。
【0198】
以下、実施例により本発明を説明するが、これらは決して本発明の限定を意図するものではない。
【0199】
(材料と方法)
(A.細胞)
サルモネラ株SL1344とLB5000は B.A.D.Stocker(スタンフォード大学;HoisethおよびStocker,1981,Nature 291:238−239)から入手した。サルモネラ株BJ66はB.D.Jones(スタンフォード大学)から入手した。
【0200】
HEp−2細胞はAmerican Type Culture Collection(ATCC;メリーランド州ロックビル・パークローンドライブ12301;受託番号CCL−23)から入手した。
【0201】
マウス腹腔マクロファージは、安楽死させたBalb/cマウスの腹腔を7mlの生育培地で洗浄することによって得た(MaximowおよびBloom,1931,Textbook of Histology,Saunders,フィラデルフィア)。
【0202】
(B.静置培養)
100mg/mlのカルベニシリンを含むLBブロス3mlに、定常期培養から得た細菌懸濁液6μlを接種し、その細菌を7mlの静置培養管中37℃で終夜生育することにより、低酸素(静置)培養を調製した。
【0203】
(C.マウス)
Balb/c(Itys/s)マウスは、スタンフォード大学の腫瘍学科(the Department of Oncology)から入手した。129XBalb/c(Ityr/s)マウスはスタンフォードトランスジェニック動物施設(the Stanford Transgenic Animal Facility;カリフォルニア州スタンフォード)から入手した。動物はすべて、光周期、給餌法および温度条件を同一にして、スタンフォード大学研究動物施設(the Stanford University Research Animal Facility;カリフォルニア州スタンフォード)で飼育した。
【0204】
麻酔は動物の腹腔内に33μg/kg−体重のネンブタールを注射することによって行った。
【0205】
安楽死は、スタンフォード大学研究動物施設が推奨するプロトコールに従って、CO2による窒息または頚椎脱臼によって行った。頚椎脱臼は、窒息による生理学的変化が実験結果に影響を与えるかもしれない場合に使用した。
【0206】
lux形質転換サルモネラに感染したマウスには、luxオペロンを含有するAmprプラスミドが保持されるような選択圧を発光性サルモネラに与え続けるために、カルベニシリンの筋肉内(i.m.)注射(体重1kgあたり125mg)を毎日施した。
【0207】
(D.撮像)
撮像すべき動物または対象を、2段階マイクロチャネル増感ヘッドを持つ電荷結合素子(CCD)カメラ(モデルC2400−40,Hamamatsu社)と扉とを持つ光を通さない箱の中に固定した。そのカメラを、箱の外へ通じるケーブルを介して”ARGUS50”画像処理装置(Hamamatsu社)に接続した。
【0208】
上述のICCDシステムは、いったん10〜30光子の域値に到達すれば、単一光子を検出できる。このシステムの信号対雑音比は、信号強度に依存して2:1から1X104:1までに及んだ。
【0209】
グレースケール画像は、かすかなルームライトの中でライトボックスの扉をあけ、8〜64フレーム間積分することによって得た。グレースケール画像のゲインは、その画像が最適となるように設定した(通例、0〜10,000ボルトのスケールで3000ボルト)。
【0210】
生物発光データは、外部照射の不在下に得た。露出設定は次の通りとした:黒レベルはカメラ/画像処理装置によって自動的に設定、ゲインは増倍装置コントローラーで自動的に設定、Fストップは2.8に設定した。60mm”AFニッコール”マクロレンズを使用した(ニコン(Nikon)社,ニューヨーク州メルヴィル)。
【0211】
生物発光画像は、選択した時間(通例5分間)光子を積分することによって作成した。データは、いずれの動物についても、1ピクセルあたり0〜3ビットの最低ビットレンジ設定で表わす。生物発光シグナルの分解能を0〜3のビットレンジにできなかった他の物体(すなわち24ウェルプレート)の画像については、レンジを生物発光シグナルの局在化が可能な設定(通例1〜7)に増やした。5分間の撮像で付加的な情報が得られなかった場合は、物体をより短い時間で撮像した。
【0212】
外部撮像とは、動物の非侵襲的撮像を指す。内部撮像とは、動物の部分的切開(通例、腹壁切開)後の撮像を指す。内部撮像は、外部撮像で局在化した光子放射の光源を確認するために、選択した動物で行なう。
【0213】
生物発光画像データは、検出された光子の強度を表わす擬似色発光画像として表わす。通例、青色(低強度)から赤色(高強度)までの6つの強度レベルを使用する。
【0214】
ここに掲載する図を作成するために、グレースケール画像と生物発光画像を画像処理装置を使って重ね合せることにより、空間的準拠座標系を与える合成画像を作成した。
【0215】
合成画像はRGB CRT(赤、緑、青;ブラウン管)モニターに表示し、そのモニターを撮影してハードコピーを作った。また画像処理装置画像をデジタルファイルとして保存し、そのファイルをコンピューターに移し、それをそのコンピューターに接続したカラープリンターで印刷することによっても、ハードコピーを作成した。別法として、ビデオプリンターを使用してビデオ信号を直接印刷することにより、ハードコピーを作成してもよい。
【実施例】
【0216】
(実施例1)
(pCGLS1luxプラスミドによるサルモネラの形質転換)
サルモネラ株SL1344、BJ66およびLB5000を、Xenorhabdus luminescens由来のluxオペロン(Frackmanら,1990)をコードするpUC18系ベクターpCGLS1で形質転換した。
【0217】
(A.pCGLS1プラスミド)
pCGLS1プラスミドの概略を図1A、1Bおよび1Cに示す。このプラスミドは、土壌菌Xenorhabdus luminescens由来のlux遺伝子をコードする〜11kb領域(図1A;Frackmanら,1990)を、pUC18(図1C;Clontech社,カリフォルニア州パロアルト)のBam HI部位(図1B)にクローニングすることによって構築した。このべクターの構築はFrackmanらの報文(1990)に記述されている。
【0218】
図1A中の制限酵素部位は、次のように記載されている:Bs,Bst EII;C,Cla I;E,Eco RI;H,Hind III;M,Mlu I;S,Sca I;X,Xba I;B/Sa,Bam HIおよびSau 3A接合点。多重クローニング部位(MCS)を含む配列を図1Bに記載し、そのBam HI部位を太字で示す。
【0219】
挿入物を含まないpUC18ベクターの図解を図1Cに示す。印を付けた要素には、アンピシリン耐性遺伝子(Ap)、lac Z遺伝子(lac Z)および大腸菌の複製起点(Ori)が含まれる。非修飾pUC18ベクターのサイズは約2.7kbである。
【0220】
(B.サルモネラの形質転換)
サルモネラ株Sl1344、BJ66およびLB5000のエレクトロコンピテント細胞を一般的な方法(Sambrookら,1989,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory Press,第2巻)で作成し、使用直前まで−80℃で保存した。エレクトロポレーションは次のように行なった。1μlのプラスミド(0.2μg/ml)を、10%グリセロールに懸濁した40μlの氷冷エレクトロコンピテント細胞に加えた。その懸濁液を1分間穏やかに混合し、間隙1mmのエレクトロポレーションキュベットに入れ、Bio−Rad Gene−Pluser(Bio−Rad Laboratories社,カリフォルニア州ハーキュリーズ)を使って電気穿孔した。設定は、2.5キロボルト、400オームおよび25マイクロファラドとした。
【0221】
ルリア−べルチーニ(LB)ブロス中37℃で1時間攪拌培養した後、100μg/mlカルベニシリンを含む(LB)寒天上に細胞をまき、終夜生育した。
【0222】
標識されたサルモネラの生物発光を最大にするため、luxオペロンを高コピー数プラスミド上に維持し、単一コピー遺伝子として組み込まれないようにした。しかしプラスミドは、この研究で使用したSL1344やBJ66のようなrecA株では特に、細菌細胞による修飾を受ける。recA遺伝子座は、luxオペロンとβ−ラクタマーゼを含有するプラスミドの領域を欠失させうるリコンビナーゼをコードする。したがって培養細胞から回収したサルモネラをカルベニシリンの存在下と不在下の両方で平板培養し、生物発光が失われる頻度を決定するために撮像した。ゲンタマイシン処理し溶解したHEp−2細胞とマクロファージから回収されるコロニーはすべて、アンピシリン耐性(Ampr)かつ生物発光性だった。したがって、lux遺伝子は哺乳類細胞との同時培養中に失われないようだった。
【0223】
コロニーを暗室での視認によって発光について検査した。5つの形質転換体が高レベルの発光を持つと同定された。これらのうち3つ(SL1344、BJ66およびLB5000株からそれぞれ1つ)を、以降の実験のために選択した。それらをそれぞれSL1344lux、BJ66luxおよびLB5000luxと名づけた。
【0224】
(実施例2)
(通常のサルモネラおよび形質転換したサルモネラの侵入能力)
6株のサルモネラ(SL1344lux、LB5000lux、BJ66lux、SL1344、LB5000およびBJ66)の侵入能力を、2種類の細菌付着および侵入アッセイで測定した。コロニー形成単位(CFU)アッセイは、基本的に既に記述されている方法(FinlayおよびFalkow,1989,Mol.Microbiol.3:1833−1841)に変更(Leeら,1990,PNAS 87:4304−4308)を加えて行なった。生物発光アッセイは、細胞の数をCFUではなく生物発光を用いて定量する点以外は、基本的にCFUアッセイと同様に行なった。
【0225】
簡単に述べると、HE−p細胞と初代マウス腹腔マクロファージを24ウェル組識培養皿に、20mMグルタミン(Gibco/BRL社)および5%ウシ胎児血清(Hyclone社,ユタ州ローガン)を添加したRPMI(Gibco/BRL社,ニューヨーク州グランドアイランド)中、1X105細胞/ウェルの濃度で接種した。細胞を接種した24時間後(HEp−2)または数日間後(マクロファージ)に、静置培養からの細菌(上記”材料と方法”の項参照)を、1ウェルあたり1X106個(感染多重度(m.o.i)10)または1X107個(m.o.i=100,図2B〜Eの右側の柱)の密度で接種し、Beckman臨床用遠心器(Beckman Instruments社,メリーランド州コロンビア)を用いて、1000rpm(185Xg)で5分間、細胞単層上に遠心した。培地を、ゲンタマイシン(100mg/ml)を含む(侵入アッセイ)または含まない(付着アッセイ)RPMI培地(Gibco/BRL社)で置換した。その共存培養を5%CO2中35℃で合計3.5時間培養した。
【0226】
培養培地中のゲンタマイシンはHep−2細胞によって内部移行されなかった細菌を、HEp−2細胞の表面に付着しているものを含めて殺す。したがって、付着アッセイ(ゲンタマイシンを含まない)でのシグナルが、付着細菌と内部移行細菌の両方を表わすのに対し、侵入アッセイ(ゲンタマイシンを含む)でのシグナルは内部移行された細菌のみを表わす。
【0227】
付着と侵入を、3時点(接種の1.5時間後、3.0時間後および3.5時間後)で発光性細菌細胞を撮像することによりアッセイした。第1時点での撮像に先立って、付着していない細菌を除去するために細胞単層をリン酸緩衝食塩水(PBS)で3回洗浄し、新しいRPMIを加えた。発光は30秒間の露光時間で記録した。第2および第3時点での画像も同じ露光時間を用いて得たが、最初に細胞を洗うことはしなかった。
【0228】
図2Aは、最後の時点で記録したデータを、その培養皿ウェルのグレースケール画像に重ね合せた擬似色発光画像として表示したものである。細胞タイプ、サルモネラ株およびゲンタマイシンの用法を図中に示す。このデータを光子カウントの相対強度として図2Bと2Dのグラフにも要約する。
【0229】
3.5時間時点での撮像後、組識培養細胞をPBSで3回洗浄し、PBS中の0.2%”トリトン X−100”で溶解した。溶解によって放出された付着細菌および/または細胞内細菌をLB寒天プレートまたはLB−カルベニシリン寒天プレートにまき、35℃で18時間培養した。各ウェルから放出された細菌の数を、コロニー形成単位(CFU,FinlayおよびFalkow,1989,Mol.Microbiol.3:1833−1841;Leeら,1990,PNAS 87:4304−4309)の数を数えることによって決定した。これらのデータを、ゲンタマイシンと共にまたはゲンタマイシンなしで3.5時間培養した後の共存培養から回収された1mlあたりの総細菌コロニーとして表わし、図2Cおよび2Eのグラフに要約する。
【0230】
生物発光アッセイとCFUアッセイの両方から得られたデータは、(i)lux遺伝子で形質転換されたサルモネラが親系と類似する感染力を持つことと、(ii)発光検出とCFU決定がHEp−2細胞とマクロファージにおける2つのサルモネラ株の侵入力について同等な推定値を与えることを示している。CFUに対する生物発光の比率はマクロファージ培養の方が低く、これはおそらくサルモネラがマクロファージの細胞下区画に侵入するためだろう。
【0231】
(実施例3)
(形質転換したサルモネラの試験管内発光)
LB5000luxサルモネラの4つの10倍連続希釈液10μl(1mlあたり106細胞から103細胞まで)を4本の100μlガラス製毛細管(Becton Dickinson社のCaly−Adams事業部,ニュージャージー州パーシッパニー)に入れた。その細菌懸濁液は、毛細管中で両端に空気のポケットを持つ液体の柱を形成した。各毛細管の一端をクリトシール(critoseal;Clay−Adams社)で密封した。希釈液の作成に使用した培地は、空気に曝すことによりO2で飽和させた。
【0232】
その毛細管を透明なプラスチック製のラップで包み、上述のように30秒間撮像することにより、発光を測定した。典型的な画像を図3Aに示す。4本の毛細管を示す。これらは(上から下へ)106、105、104および103サルモネラ細胞/ml(104、103、102および10細胞/管)を含む。発光は、104細胞/ml(100細胞)しか含まない懸濁液でも検出できた。しかしその発光は空気/液体界面に限定され、この発光反応が比較的高レベルの酸素を必要とすることが示唆される。細胞の多くはおそらく液体柱の中にあり、空気/液体界面にはないだろうから、このデータは、図3Aに示す毛細管中の発光が、各管中の総細胞数よりかなり少ない細胞から生じることを示唆している。
【0233】
(実施例4)
(動物組識を貫通する発光の試験管内検出)
LB5000luxサルモネラの連続希釈液を含む内径3.5mmのガラス製毛細管で構成された微量試験管を、基本的に上記実施例3に記述したように調製した。ただし、本実施例では細菌懸濁液を毛細管の密封した末端に接触させ、上端のみを空気にさらした。その管を半透明のプラスチック製シンチレーションバイアルに入れ、次に挙げる動物組識の一つでそのまわりを囲んだ:ヒヨコ胸筋、ヒヨコ皮膚、子ヒツジ腎臓または子ヒツジ腎臓髄質。いずれの組識も地元のスーパーマーケット(Safeway社,カリフォルニア州マウンテンヴュー)の食肉部門から入手した。
【0234】
組識で囲まれた毛細管を含むバイアルの図を図4に示す。バイアル1は直径約1.4cmで、蓋2を含む。そのバイアルをその上部分3に沿って、不透明な素材(すなわち黒テープ)で覆う。動物組識4を、バイアルの底から不透明被覆3の下端のすぐ上までになるよう、バイアルに入れる。微量試験管5の底を栓7(すなわちクリトシール(crytoseal)栓)で密封し、それをバイアルの中心に、栓をした末端がバイアルの底に接触するか、きわめて接近するように置く。細菌懸濁液6は毛細管の底から上方約1cmに達する。
【0235】
組識を含むバイアルと組識を含まないバイアルおよび細菌を含むバイアルと細菌を含まないバイアルから放射された光子を、高速同時弁別器を無効にした液体シンチレーションカウンター(モデル1219 Rackbeta,LKB/Wallac社,メリーランド州ゲーサーズバーグ)でカウントした。
【0236】
組識を含まない対照は、細菌懸濁液をシンチレーションバイアルに直接入れてアッセイした。いずれの実験も3回ずつ行なった。
【0237】
各実験では、各カウントごとにバイアルを90℃回転させて、バイアルを2、3回カウントすることにより、考えうる組識の厚さの不一致の影響について検証した。有意差は検出されなかった。
【0238】
その結果を下記表1に要約する。
【0239】
【表1】
【0240】
腎臓組識内の1X103個のLB5000luxについてのシグナルは、シンチレーションカウンター中の光電子増倍管(PMT)でバックグランドレベルか、それに近かった。このタイプの検出におけるバックグラウンドはPMTの暗電流によるものであり、この試験はかなり強いシグナルの分析に限定される。
【0241】
約1X107個のLB5000luxからの生物発光は、0.5cmの鳥類筋肉、皮膚、ヒツジ腎臓髄質およびヒツジ腎臓を通して検出できた。これらの結果は、標識したサルモネラからの生物発光が様々な不透明度の動物組識を通して検出できることを示している。(図3Aで示したように)毛細管内では酸素が制限されていたと思われるので、このアッセイで表示している数よりも少ない数の生物発光性サルモネラが、組識を通して検出できると考えられる。
【0242】
(実施例5)
(生物発光性サルモネラの生体内検出)
感染中のサルモネラに対する酸素の利用性を評価するために、野生型SL1344 luxをBALB/cマウスの腹腔内(i.p.)に接種した。接種の24時間後、麻酔したラットで、細菌によって内部に放出され腹壁を透過した光子を、増倍化CCDカメラで外部から検出し、局在化した(図3B)。全身性サルモネラ感染には、リンパ節、脾臓、肝臓のコロニー形成が関与すると思われる。野生型SL1344 luxの腹腔内接種によって感染させたマウスの腹部画像は、腹部表面のほとんどに透過光子が様々な強度の焦点を作っていることを示した(図3B)。これらの結果は、おそらくは肝臓と腸間膜リンパ節を含む内臓の広範なコロニー形成と合致し、利用可能な酸素のレベルはいくつかの組識で標識病原体からの発光を外部から検出するのに十分でありうることを示している。
【0243】
(実施例6)
(Balb/cマウスに経口投与したluxサルモネラの検出)
病原性SL1344lux、非侵入性BJ66luxおよび低病原性LB5000luxサルモネラ1X107個の懸濁液50μlを経口的に与えることによって、Balb/cマウスを感染させた。感染時に4〜6週齢だったそれらのマウスを、5分間の積分時間で毎日撮像した(光子放射を5分間測定した)。撮像に先立って、マウスを33μg/kg−体重のネンブタールで麻酔した。
【0244】
典型的な画像を図5A〜Fに示す。接種後(p.i.)24時間の時点で、生物発光シグナルはいずれの感染動物でも単一の焦点に局在した(図5A、5Cおよび5E)。生物発光は、低病原性LB5000luxに感染したすべての動物で、接種後7日までに消失した(図5B)。野生型SL1344 luxに感染したBALB/cマウスでは、生物発光が試験期間中常に検出され、第8日では透過光子の焦点が複数認められた。これらの動物では、感染がしばしば腹腔のほとんどに広がった(図5F)。これらの動物の3分の1では、第8日に透過光子が腹部領域の大半で認められ、それは腹腔内接種後の光子の分布と似ていた(図3Bと5Fを参照のこと)。BJ66 luxによる感染の蔓延はかなり変動的だったが、通例、その感染は持続し、最初の部位に局在したままだった(図5D)。
【0245】
耐性BALB/cX129マウスを野生型SL1344 luxに感染させたところ、10匹のマウス群で、その生物発光シグナルはその試験期間中常に局在し、持続性を保っていた。この結果は、SL1344 luxに感染した感受性マウス(lytr/s)に観察された播種性生物発光とは対照的であった(実施例8と図7Aおよび7Bを参照のこと)が、侵入性の低いBJ66 luxによる感受性BALB/cマウスの持続性感染とは似ていた。対照として、持続的に感染した耐性BALB/cX129マウスから得たサルモネラを培養したところ、8日後に回収したコロニーの80〜90%はAmprだった。これらのうち90%以上は生物発光性であり、観察された相違がluxプラスミドの有意な減少によるのではなく、むしろ細菌株の病原性が真に相違するためであることが示唆された。
【0246】
(実施例7)
(サルモネラの病原性株および低病原性株の腹腔内接種後の感染の検出)
100μl懸濁液中の1X107細菌細胞を同時に空気を注入することなく腹腔内(i.p.)接種することによって、Balb/cマウスを病原性(SL1344lux)または低病原性(LB5000lux)サルモネラに感染させた。
【0247】
接種後(p.i.)32時間の時点で、上述のようにマウスを麻酔し、撮像した。その結果を図6に示す。この図の左側の2匹のマウス(病原性SL1344luxに感染)では広範な感染が明らかである。これに対して、低病原性LB5000lux株を注射した右側のマウスでは、発光はあったとしてもほとんど検出されない。
【0248】
(実施例8)
(サルモネラ経口接種後の耐性マウスにおける全身性感染の検出)
耐性129XBalb/c(Ityr/s)生存マウスを、1X107個のSL1344luxサルモネラの胃内接種によって感染させた。細菌は麻酔下に胃内給餌チューブを通して与えた。注入後8日間(8 d.p.i)、動物を毎日撮像した。
【0249】
結果を図7Aと7Bに示す。一組3匹のマウスを感染させ、8日間毎日撮像した。第1日の典型的画像(図7A)と第8日の典型的画像(図7B)を示す。これらのデータは、全身性サルモネラ感染症に対して耐性なマウスは盲腸に局在した慢性感染症を持つが、その感染は腹腔内に蔓延しないことを示している。
【0250】
(実施例9)
(サルモネラ経口接種後の腹壁切開後撮像)
腹腔内の発光シグナルの位置を正確に特定し、その位置を非侵襲的撮像法で得たものと比較するために、サルモネラの経口接種後に腹壁切開を行なった。実施例8に記述したように、動物に接種した。選択した期間(通例7日)の後、上述のようにマウスを麻酔し、外部から撮像した。典型的画像を図8Aに示す。外部撮像の後、腹腔を開き、図8Bに例示するように動物を再び撮像した。場合により、盲腸(C)の前後の腸管腔に空気を注入してから、3回目の撮像を行なった(図8C)。最後の撮像後、直ちにマウスを安楽死させた。
【0251】
経口接種後の感染初期に感受性マウス中で生物発光が限局的に観察された場合はいずれも、光子はほとんどもっぱら盲腸から生じており、限局的生物発光の正確な局在と強度の変動は、盲腸の位置が変動しやすいためであった。感染耐性BALB/cX129マウスに観察された限局的な生物発光も、同様に盲腸に局在した。これに対して、SL1344 luxに腹腔内感染させた動物では、そのような局在が観察されなかった(図3B)。野生型SL1344 luxを経口接種した感染感受性マウスの後期では、生物発光が多極性であったが、追加された発光の焦点は腹壁切開後に明白にならなかった。病原性の低いLB5000 luxに感染したマウスでは、第7日の時点で、生物発光が、いずれの組識または器官にも、皮膚および腹壁の除去後、限局的にさえ、検出できなかった。
【0252】
どの感染動物の脾臓または血流にも、生物発光は光学的に検出されなかった。肝臓からの生物発光は疾患後期にのみ認められた。また、胃腸路からの生物発光は疾患の初期には盲腸に限られた。このパターンは、異なる組識におけるサルモネラの数の相違もしくは利用できる酸素の欠如によるのだろう。経口感染したマウスの破砕した器官中に存在するAmpr cfuを定量することにより、標識サルモネラSL1344 luxの分布を調べた。第7日にSL1344 lux感染BALB/cマウスの肝臓、脾臓および肺から得たampr細菌コロニーは、いずれも、その90%以上が1.9X103〜>1.0X105の範囲の総cfuを示し、生体内では検出可能な光子放射を伴わなかった(表2)。これに対して、盲腸からの生物発光は検出可能であり、この組識は>1.0X108の総cfuを含有した。LB5000 lux感染マウスでは、どの組識にもcfuが検出されなかった。これらの結果は、現在の実験系を使用した場合、組識中1X106生物がほぼこの放射波長での検出限界であることを示唆している。
【0253】
酸素はルシフェラーゼ反応に必須の基質であるから、酸素化された微小環境中に存在するサルモネラだけが生物発光するはずである。したがって、胃腸路の管腔の嫌気環境にサルモネラからの生物発光がないことは予想できることであり、腸管腔を空気にさらせば、それまでは酸素がないために検出できなかった細菌の存在が明らかになるはずである。この見解の裏付けとして、盲腸だけに検出可能な生物発光を持つ一匹の動物が、空気にさらされたときに迅速に生物発光性になる糞粒を排泄した。腸管腔内に非発光性のルシフェラーゼ発現細菌が存在することを示すこの事実と、この組織内の好気地帯と嫌気地帯の明確な描写から、腸管腔に空気を注入すれば、さらなる細菌の存在が明らかになるであろうことが示唆された。同様の生物発光パターンを示す別の動物の回腸と結腸の管腔に空気を注入したところ、注射部位近くで光子が検出可能になった(図8)。最後に、盲腸生物発光をA示す第3のマウスを殺すと、生物発光は迅速に消滅した。明確な好気環境地帯と嫌気環境地帯を欠くため、他の組識部位に空気を注射した。
【0254】
【表2】
【0255】
(実施例10)
(サルモネラ腹腔内接種後の腹壁切開後撮像)
実施例7に記述したように、1X107個のサルモネラ(SL1344lux)を腹腔内接種することにより、Balb/cマウスを感染させた。そのような一匹の典型的画像を図9A、9Bおよび9Cに示す。
【0256】
注射後(p.i)24時間の時点で、動物を麻酔し、5分間撮像した(図9A)。腹腔を開き、そのマウスを再び5分間撮像した(図9B)。盲腸を左側に引き寄せ、その動物を再び5分間撮像した(図9A)。
【0257】
その結果から、非侵襲的撮像法で得られた感染部位の局在は、腹腔を開いて明らかになった部位とよく相関することがわかる。
【0258】
(実施例11)
(SL1344luxサルモネラからの生物発光に対するシプロフロキサシンの影響)
生体内撮像法の有用性を立証するために、感染した動物を、全身性サルモネラ感染症に有効であることが知られている抗生物質シプロフロキサシンで処置した。Magalianesら,1993,Antimicrobial Agents Chemo.37:2293。
【0259】
Balbcマウスの実験群と対照群にSL1344luxを経口接種した。接種の8日後に、実験群のマウスに100mgのシプロフロキサシン塩酸塩(3mg/kg−体重;Sigma Chemical社,ミズーリ州セントルイス)を腹腔内注射した。実験群の処置後、両群の動物を、処置後5.5時間にわたって数回(上述のように)撮像した。
【0260】
典型的画像を図10B〜Eに示す。図10Bと10Dは、それぞれ実験群の処置を開始する直前の対照群と処置群を代表する動物の合成画像である。図10Cと10Eは、処置の開始後5.5時間の同じ動物の合成画像である。対照の生物発光が7.5倍まで増大したのに対して、シプロフロキサシン処置動物の腹部上の生物発光はこの期間中に検出不能なレベルにまで減少した。腹部領域上に検出される光子の総数を決定し、t=0での値に対して規格化し、処置後の時間に関して図10Aにプロットした。
【0261】
このデータは、本発明の方法と組成物を、生体内感染の蔓延に対する薬物の効果の評価に使用できることを示している。
【0262】
特定の方法と態様に関して本発明を説明したが、本発明から逸脱することなく様々な変更や改変を施しうることは理解されるだろう。
【特許請求の範囲】
【請求項1】
本明細書に記載される発明。
【請求項1】
本明細書に記載される発明。
【図1】


【図2A】
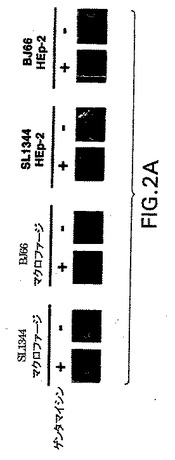
【図2B】

【図2C】
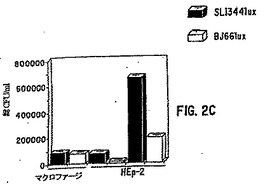
【図2D】

【図2E】

【図3A】

【図3B】

【図4】
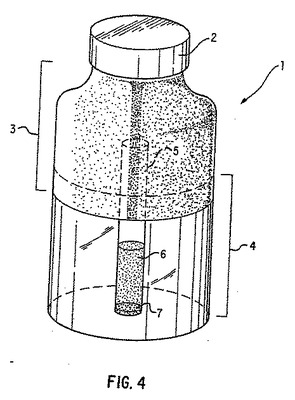
【図5A】

【図5B】

【図5C】

【図5D】

【図5E】

【図5F】
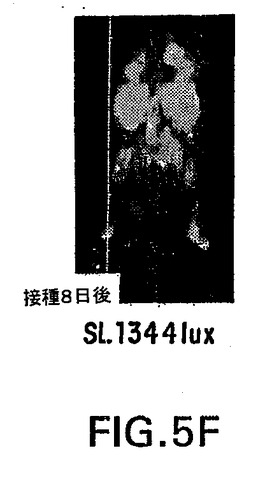
【図6】
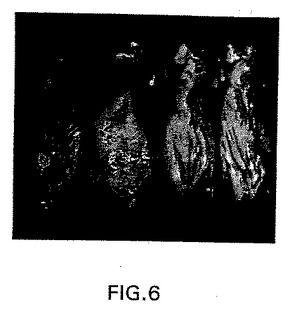
【図7】

【図8A】

【図8B】

【図8C】

【図9A】

【図9B】

【図9C】

【図10A】

【図10B】

【図10C】

【図10D】

【図10E】

【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図3A】
【図3B】
【図4】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図5F】
【図6】
【図7】
【図8A】
【図8B】
【図8C】
【図9A】
【図9B】
【図9C】
【図10A】
【図10B】
【図10C】
【図10D】
【図10E】
【公開番号】特開2011−160810(P2011−160810A)
【公開日】平成23年8月25日(2011.8.25)
【国際特許分類】
【出願番号】特願2011−111848(P2011−111848)
【出願日】平成23年5月18日(2011.5.18)
【分割の表示】特願2008−161037(P2008−161037)の分割
【原出願日】平成7年11月17日(1995.11.17)
【出願人】(503115205)ボード オブ トラスティーズ オブ ザ レランド スタンフォード ジュニア ユニバーシティ (69)
【Fターム(参考)】
【公開日】平成23年8月25日(2011.8.25)
【国際特許分類】
【出願日】平成23年5月18日(2011.5.18)
【分割の表示】特願2008−161037(P2008−161037)の分割
【原出願日】平成7年11月17日(1995.11.17)
【出願人】(503115205)ボード オブ トラスティーズ オブ ザ レランド スタンフォード ジュニア ユニバーシティ (69)
【Fターム(参考)】
[ Back to top ]