
感染症の処置のためのIL−23アンタゴニストの使用
【課題】慢性の細菌感染症、ウイルス感染症、および真菌感染症などの感染症を処置するための、IL−23アンタゴニストを含む方法および組成物を提供すること。
【解決手段】一実施形態では、アンタゴニストは、IL−23またはそのp19サブユニットに結合する抗体またはその結合性断片などの結合性化合物である。いくつかの実施形態では、抗体の結合は、IL−23またはそのp19サブユニットの、IL−23受容体またはそのIL−23Rサブユニットに対する結合を阻止する。別の実施形態では、IL−23アンタゴニストは、IL−23受容体またはそのIL−23Rサブユニットに結合する。
【解決手段】一実施形態では、アンタゴニストは、IL−23またはそのp19サブユニットに結合する抗体またはその結合性断片などの結合性化合物である。いくつかの実施形態では、抗体の結合は、IL−23またはそのp19サブユニットの、IL−23受容体またはそのIL−23Rサブユニットに対する結合を阻止する。別の実施形態では、IL−23アンタゴニストは、IL−23受容体またはそのIL−23Rサブユニットに結合する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一般的に感染症の処置に関する。具体的には、本発明は、慢性の細菌感染症、真菌感染症、またはウイルス感染症などの感染症を示す被験体に、IL−23アンタゴニスト、たとえば抗体を投与することに関する。
【背景技術】
【0002】
慢性感染症は、多くの病原体により引き起こされる。種々のウイルス、真菌、および細菌は、たとえば、消散させることができない持続的感染症を引き起こす場合がある。
【0003】
重篤な真菌感染症数が増加し続けているため、それらを処置するための方法および組成物の必要性がより差し迫っている。主要な真菌病原体には、Candida albicans、Aspergillus fumigatus、およびCryptococcus neoformansが含まれ、これらの病原体に関連する侵襲性真菌症の推定年間発生率は、米国の人口100万人当たり72〜228件(Candida種属の場合)、12〜34件(Aspergillus種属の場合)、および30〜66件(C.neoformansの場合)の感染である。非特許文献1。真菌感染症の増加は、医学の進歩(移植および化学療法)の結果として、およびAIDS患者人口の増加の結果として、主として免疫無防備状態の患者数の増加に起因する。免疫無防備状態の患者における真菌感染症の80%超は、Candida種属により引き起こされる。クリプトコックス症は、カンジダ症に次いで、AIDS患者で2番目に蔓延している真菌感染症である。アスペルギルス症は、癌患者および臓器移植患者における感染症の少なくとも30%に関与しており、高い死亡率を示す。
【0004】
フルコナゾールは、長年の間、真菌病原体に対して有効な薬物であるが、耐性は増加している。アムホテリシンBなどの代替薬は、発熱、腎臓障害、貧血、低血圧、頭痛、悪心、嘔吐、および静脈炎などの副作用を含む深刻な欠陥を有している。
【0005】
細菌感染症は、免疫無防備状態の個体人口の増加および抗生物質耐性細菌株の広範な発生に照らすと、抗生物質の普及にもかかわらず重要な課題のままである。免疫無防備状態の個体には、高齢者、移植レシピエント、化学療法患者、および後天性免疫不全症候群(AIDS)の個体が含まれる。米国では、毎年ほぼ200万人の患者が病院で感染症に罹り、それらの感染症の原因である細菌の70%は、少なくとも1種の抗生物質に耐性である。NIAID概況報告書(fact sheet)、「The Problem of Antimicrobial Resistance」、2006年4月。近年米国では、約90,000人が感染症で死亡し、1992年の13,300人から増加している。ほとんどの細菌感染症は、少なくとも1種の抗生物質の長期的治療過程(たとえば、バンコマイシンの持続的静脈内投与)に感受性のままであるが、これが今後の病原性細菌について当てはまるという保証はない。メチシリン耐性Stapholococcus aureus(MRSA)は、重要な公衆衛生上の課題を代表する多抗生物質耐性微生物の主要な例である。2002年には、疾病予防管理センター(CDC)は、ミシガン州の患者でバンコマイシンに対して完全に耐性であるS.aureus(VRSAと呼ばれる)感染症の最初の症例を報告した。持続感染性の病原細菌には、Salmonella種属、Brucella種属、およびChlamydia種属も含まれる。
【0006】
マイコバクテリアとは、高い細胞壁脂質含有量および低い増殖率を示す、好気性で非胞子形成性の非運動性桿菌の広く分布する多様な集団である。Mycobacterium属のメンバーは、たとえば、無害から重大な病原性を有する種、たとえば結核(TB)の原因因子であるM.tuberculosisまでと、毒力が様々である。TBは、世界で2番目に多い感染性の死因である。約20億人、すなわち世界人口の3分の1が、M.tuberculosisに感染していると推定される。800万件の新しい症例およびほぼ300万人の死亡が、毎年発生している。TBは世界中の全死亡の7%の直接的原因であり、この世界的な流行は、薬剤耐性生物の拡散および進行中のHIV流行の結果、悪化する可能性が高い。たとえば、Dale and Federman(編)(2002年)WebMD Scientific American Medicine、WebMD Professional Publishing、ニューヨーク、米国ニューヨーク州を参照されたい。
【0007】
TBを処置する最新の方法は、イソニアジド、リファンピン、ピラジナミド、エタンブトール、ストレプトマイシン、シプロフロキサシン、およびオフロキサシンなどの広域スペクトルの抗感染症薬の使用を含む。しかしながら、そのような作用剤は種々の器官で毒性を引き起こす場合があり、いくつかの抗生物質耐性TB株の増大と共に、有効性を失いつつある。多様な非抗生物質薬を使用して、結核患者の肺におけるマイコバクテリア負荷を低減すると、疾患形成、伝播、および死亡が防止できる。
【0008】
慢性のウイルス感染症も、公衆衛生に対する大きな脅威を代表する。C型肝炎ウイルス(HCV)またはヒト免疫不全ウイルス(HIV)などのウイルス感染症を完全に根絶できないと、それぞれ、その後の再活性化および肝臓癌または後天性免疫不全症候群(AIDS)などの合併症に結び付く場合がある。非特許文献2。さらに、ヒトパピローマウイルス(HPV)遺伝子型16、18、31、33、45、および56は、子宮頚癌の症例の95%超を占める。非特許文献3。事実上100%のHIV感染症例、55〜85%のHCV感染症例、および30%を超えるHPV症例で、慢性感染症が発生すると推定されている。Berzofskyら(2004年)。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】Pfallerら(2006年)Clin. Infect. Dis. 第43巻:S3〜14頁
【非特許文献2】Robertson & Hasenkrug(2006年)Springer Semin. Immun. 第28巻:51頁
【非特許文献3】Berzofskyら(2004年)J. Clin. Invest. 第114巻:450頁
【発明の概要】
【発明が解決しようとする課題】
【0010】
細菌感染症、ウイルス感染症、および真菌感染症の処置および/または予防のための方法および組成物の改良に対する必要性が存在する。そのような方法および組成物は、好ましくは、既存の処置方法および組成物より毒性が低くおよび/またはより効果的である。
【課題を解決するための手段】
【0011】
本発明は、細菌感染症、ウイルス感染症、および真菌感染症と戦うために、IL−23アンタゴニストを使用する組成物、薬剤、および方法を提供することにより、これらの必要性およびその他を満たす。
【0012】
一局面では、本発明は、IL−23アンタゴニストの投与を伴う、感染症を有する被験体、感染症を有する疑いのある被験体、または感染症に罹るリスクのある被験体を処置するための方法に関する。一実施形態では、アンタゴニストは、IL−23またはそのp19サブユニットに結合する抗体またはその結合性断片などの結合性化合物である。いくつかの実施形態では、抗体の結合は、IL−23またはそのp19サブユニットの、IL−23受容体またはそのIL−23Rサブユニットに対する結合を阻止する。別の実施形態では、IL−23アンタゴニストは、IL−23受容体またはそのIL−23Rサブユニットに結合する。いくつかの実施形態では、アンタゴニストは、IL−23受容体またはそのIL−23Rサブユニットに結合し、IL−23またはそのp19サブユニットの結合を阻止する。別の局面では、本発明は、前記処置方法で使用するための組成物に関する。
【0013】
いくつかの実施形態では、感染性障害には、細菌感染症、マイコバクテリア感染症、ウイルス感染症、または真菌感染症などの感染性疾患が含まれる。一実施形態では、感染性障害は、M.bovis、M.leprae、またはM.tuberculosisにより引き起こされるマイコバクテリア感染症である。一実施形態では、感染性障害はTBである。別の実施形態では、感染性障害は、爪甲真菌症、カンジダ症、アスペルギルス症、クリプトコックス症からなる群から選択される真菌感染症である。さらに別の実施形態では、感染性障害は、C.albicans(たとえば、慢性皮膚粘膜カンジダ症、鵞口瘡)、C.neoformans、またはA.fumigatusにより引き起こされる真菌感染症である。さらなる実施形態では、感染性障害は、ウイルス感染症、たとえばヒト免疫不全ウイルス(HIV)、B型肝炎ウイルス(HBV)、C型肝炎ウイルス(HCV)、またはヒトパピローマウイルス(HPV)により引き起こされるウイルス感染症である。
【0014】
他の実施形態では、感染性障害は慢性感染症である。種々の実施形態では、慢性感染症は、感染を消散するための少なくとも1つの過去の試みにもかかわらず、1、2、4、6、9、12、18、24、36または48カ月以上の間持続する。種々の実施形態では、慢性感染症を消散しようとする過去の試みは、抗菌物質、抗生物質、抗ウイルス物質、または抗真菌物質を用いた処置を伴う。
【0015】
一実施形態では、本発明は、IL−23アンタゴニストの投与を、別の治療薬などの少なくとも1つの他の治療様式と組み合わせた併用治療に関する。種々の実施形態では、他の治療薬は、IL−17Aアンタゴニスト、IL−17Fアンタゴニスト、IL−12アゴニスト(IL−12を含む)、TGF−βアンタゴニスト、またはIL−6アンタゴニストである。別の実施形態では、他の治療薬は、ポサコナゾール、フルコナゾール、ボリコナゾール、イトラコナゾール、ケトコナゾール、リアロゾール(liarozole)、イルテマゾール(irtemazol)、クロトリマゾール、ミコナゾール、エコナゾール、ブトコナゾール、オキシコナゾール、スルコナゾール、チオコナゾール、およびテルコナゾール、置換チアゾール、チアジアゾール、オキサジアゾール、カスポファンギン、アムホテリシンB、ナイスタチン、ピマリシン、フルシトシン(5−フルオロシトシン)、ナフチフィン、テルビナフィン、ブテナフィン、チオカーボネートトルナフテート(thiocarbonate tolnaftate)、グリセオフルビン、アミオダロン、シクロピロックス、スルベンチン、アモロルフィン、クリオキノール、ゲンチアナバイオレット、ヨウ化カリウム、チオ硫酸ナトリウム、石炭酸フクシン溶液、およびエキノキャンディン(たとえば、カスポファンギンアセテート(caspofungin acetate)、ミカファンギン、およびアニデュラファンギン)からなる群から選択される1つまたは複数の抗真菌物質である。
【0016】
別の実施形態では、他の治療薬は、イソニアジド、リファンピン、ピラジナミド、エタンブトール、ストレプトマイシン、シプロフロキサシン、バンコマイシン、またはオフロキサシンからなる群から選択される1つまたは複数の抗菌物質である。
【0017】
別の実施形態では、他の治療薬は、アバカビル、アシクロビル、アマンタジン、アンプレナビル、デラビルジン、ジダノシン、エファビレンツ、ファムシクロビル、インジナビル、インターフェロンアルファ(IFN−α)、リバビリン、ラミブジン、ネルフィナビル、ネビラピン、オセルタミビル、ペンシクロビル、リバビリン、リトナビル、サキナビル、スタブジン、バラシクロビル、ザルシタビン、ザナミビル、ジドブジン(アジドデオキシチミジン、AZT)からなる群から選択される1つまたは複数の抗ウイルス物質である。
【0018】
一局面では、本発明は、感染症を有する被験体、または感染症を有する疑いのある被験体の1型(Th1)免疫応答を増強するための方法に関する。種々の実施形態では、Th1応答の増強は、IL−23アンタゴニストで処置する前のT細胞の割合と比較した場合の、IFN−γ発現CD4+T細胞の割合の増加、IL−17A発現CD4+T細胞の割合の減少、またはその両方により反映される。種々の実施形態では、増加は、1.5倍、2倍、3倍、5倍、10倍、20倍、もしくは50倍以上であり、または減少は、3分の2、2分の1、3分の1、5分の1、10分の1、20分の1、もしくは50分の1以下である。別の局面では、本発明は、前記Th1応答を増強する方法で使用するための組成物に関する。
【0019】
種々の実施形態では、他の治療薬は、IL−23アンタゴニストを投与する前、および/または投与と同時に、および/または投与した後で投与される。一実施形態では、IL−17Aアンタゴニストは、IL−23アンタゴニストの前および/または同時に投与される。別の実施形態では、抗菌、抗真菌、または抗ウイルス物質は、IL−23アンタゴニストと同時に投与される。
【0020】
別の局面では、本発明は、サイトカイン自体に対する、またはそれらのそれぞれの受容体もしくは受容体サブユニットに対するアンタゴニスト抗体などの、IL−17Aおよび/またはIL−17Fのアンタゴニストの投与を伴う、感染症を有する被験体、感染症を有する疑いのある被験体、または感染症に罹るリスクのある被験体の処置方法に関する。
【0021】
他の実施形態では、IL−23アンタゴニストはポリヌクレオチドを含む。種々の実施形態では、ポリヌクレオチドは、アンチセンスポリヌクレオチド(たとえば、アンチセンスRNA)または低分子干渉RNA(siRNA)である。一実施形態では、IL−23のポリヌクレオチドアンタゴニストは、アデノウイルス、レンチウイルス、レトロウイルス、またはアデノ随伴ウイルスベクターなどの遺伝子治療用ベクター中で送達される。別の実施形態では、IL−23のポリヌクレオチドアンタゴニストは、治療薬として送達される。
【0022】
さらに別の実施形態では、IL−23アンタゴニストは、可溶性受容体ポリペプチドを含む。一実施形態では、IL−23アンタゴニストは、IL−23Rの細胞外ドメインに由来する可溶性断片である。
【0023】
種々の実施形態では、IL−23アンタゴニストは、抗体またはその抗原結合性断片である。種々の実施形態では、抗体またはその抗原結合性断片は、ポリクローナル抗体、モノクローナル抗体、ヒト化抗体、完全ヒト抗体;抗体断片(たとえば、Fab、Fab’、Fab’−SH、Fv、scFv、F(ab’)2、およびダイアボディ(diabody))を含む。他の実施形態では、アンタゴニストは、抗体のペプチド模倣物質を含む。いっそうさらなる実施形態では、抗体またはその抗原結合性断片は、検出可能なように標識される。一実施形態では、IL−23アンタゴニストは、補体活性化の低減、抗体依存性細胞傷害(ADCC)の低減、またはその両方を示す抗体またはその抗原結合性断片である。一実施形態では、エフェクター機能が低減されたIL−23アンタゴニスト抗体またはその断片は、抗IL−23受容体(たとえば、抗IL−23R)抗体または断片である。種々の実施形態では、エフェクター機能が低減された抗体は、抗体断片(たとえば、Fab、Fab’、Fab’−SH、Fv、scFv、F(ab’)2)、IgG4であるか、または改変グリコシル化を有する。
【0024】
一実施形態では、本発明は、IL−23p19、IL−23R、IL−17A、IL−17F、IL−17RA、IL−17RC、IL−6、およびTGF−βからなる群から選択される任意の2つのタンパク質に特異的に結合する有効量の二重特異性抗体の投与により、感染症、たとえば慢性の真菌感染症、細菌感染症、またはウイルス感染症を処置することに関する。一実施形態では、タンパク質はヒトタンパク質である。
【0025】
一実施形態では、IL−23アンタゴニストは、IL−23(またはその受容体)に特異的であり、IL−12(またはその受容体)に拮抗しない。種々の実施形態では、拮抗作用は、in vitro結合アッセイ(たとえば、ELISA)、またはバイオアッセイ(たとえば、BaF3細胞増殖またはTh17細胞産生の促進)により測定される。種々の実施形態では、IL−23とその受容体の結合の阻害に関するIC50に対する、IL−12とその受容体の結合の阻害に関するIC50の比率(IC50IL−12/IC50IL−23)は、1.5、2、3、4、5、7、10、15、20、50、または100以上である。
【0026】
一実施形態では、本発明の方法および組成物は、TBを処置するために使用され、処置の成功は、細菌負荷の低減により測定される。種々の実施形態では、マイコバクテリア負荷は、ツベルクリン検査、マントー試験(Mantoux test)、または臨床試料中のマイコバクテリアDNAまたはRNAの存在により測定される。
【0027】
本発明のいくつかの実施形態では、感染症を有する被験体は、他の方法または組成物を使用して以前に感染症の処置を受けたことがある被験体である。一実施形態では、以前の処置は、感染の除去に有効ではなかった。別の実施形態では、感染症を有する被験体、感染症を有する疑いのある被験体、または感染症に罹るリスクのある被験体は、たとえばAIDS、移植、または化学療法の結果として免疫無防備状態である。
【0028】
本発明は、真菌感染症、持続性真菌感染症、カンジダ症、慢性皮膚粘膜カンジダ症(CMC)、アスペルギルス症、クリプトコックス症、ウイルス感染症、持続性ウイルス感染症、HIV感染、HBV感染、HCV感染、持続性細菌感染症、マイコバクテリア感染症、M.tuberculosis感染、M.bovis感染、およびM.leprae感染からなる群から選択される1つまたは複数の感染症を処置するための薬剤の製造におけるIL−23アンタゴニストの使用をさらに包含する。いくつかの実施形態では、薬剤は、1つまたは複数の追加的な治療薬を含み得る。他の実施形態では、本発明の薬剤は、1つまたは複数の他の治療薬と併せて使用できる。
例えば、本発明は以下の項目を提供する。
(項目1)
感染症を有する被験体を処置するための方法であって、有効量のIL−23アンタゴニストを投与する工程を含む方法。
(項目2)
前記感染症は、ウイルス感染症、真菌感染症、および細菌感染症からなる群から選択される慢性感染症である、項目1に記載の方法。
(項目3)
前記慢性感染症は真菌感染症である、項目2に記載の方法。
(項目4)
前記真菌感染症は、カンジダ症、アスペルギルス症、クリプトコックス症、および爪甲真菌症からなる群から選択される、項目3に記載の方法。
(項目5)
前記真菌感染症はカンジダ症である、項目4に記載の方法。
(項目6)
前記真菌感染症は慢性皮膚粘膜カンジダ症(CMC)である、項目3に記載の方法。
(項目7)
前記被験体に、ポサコナゾール、フルコナゾール、ボリコナゾール、イトラコナゾール、ケトコナゾール、リアロゾール、イルテマゾール、クロトリマゾール、ミコナゾール、エコナゾール、ブトコナゾール、オキシコナゾール、スルコナゾール、チオコナゾール、およびテルコナゾール、チアジアゾール、オキサジアゾール、カスポファンギン、アムホテリシンB、ナイスタチン、ピマリシン、フルシトシン(5−フルオロシトシン)、ナフチフィン、テルビナフィン、ブテナフィン、チオカーボネートトルナフテート、グリセオフルビン、アミオダロン、シクロピロックス、スルベンチン、アモロルフィン、クリオキノール、ゲンチアナバイオレット、ヨウ化カリウム、チオ硫酸ナトリウム、石炭酸フクシン溶液、カスポファンギンアセテート、ミカファンギン、およびアニデュラファンギンからなる群から選択される1つまたは複数の抗真菌物質を投与する工程をさらに含む、項目3に記載の方法。
(項目8)
前記1つまたは複数の抗真菌物質は、アムホテリシンB、フルシトシン、イトラコナゾール、ボリコナゾール、フルコナゾール、またはポサコナゾールである、項目7に記載の方法。
(項目9)
前記慢性感染症は細菌感染症である、項目2に記載の方法。
(項目10)
前記細菌感染症はマイコバクテリア感染症である、項目9に記載の方法。
(項目11)
前記マイコバクテリア感染症は、M.bovis、M.leprae、およびM.tuberculosisからなる群から選択されるマイコバクテリアにより引き起こされる、項目10に記載の方法。
(項目12)
前記マイコバクテリア感染症はTBである、項目10に記載の方法。
(項目13)
前記被験体に、イソニアジド、リファンピン、ピラジナミド、エタンブトール、ストレプトマイシン、シプロフロキサシン、バンコマイシン、およびオフロキサシンからなる群から選択される1つまたは複数の抗菌物質を投与する工程をさらに含む、項目9に記載の方法。
(項目14)
前記抗菌物質はバンコマイシンである、項目13に記載の方法。
(項目15)
前記慢性感染症はウイルス感染症である、項目2に記載の方法。
(項目16)
前記ウイルス感染症は、HIV、HPV、HBV、およびHCVからなる群から選択されるウイルスにより引き起こされる、項目15に記載の方法。
(項目17)
前記ウイルス感染症はHCVにより引き起こされる、項目16に記載の方法。
(項目18)
前記ウイルス感染症はHIVにより引き起こされる、項目16に記載の方法。
(項目19)
前記被験体に、アバカビル、アシクロビル、アマンタジン、アンプレナビル、デラビルジン、ジダノシン、エファビレンツ、ファムシクロビル、インジナビル、インターフェロンアルファ、リバビリン、ラミブジン、ネルフィナビル、ネビラピン、オセルタミビル、ペンシクロビル、リバビリン、リトナビル、サキナビル、スタブジン、バラシクロビル、ザルシタビン、ザナミビル、およびジドブジンからなる群から選択される1つまたは複数の抗ウイルス物質を投与する工程をさらに含む、項目15に記載の方法。
(項目20)
前記抗ウイルス物質はジドブジンである、項目19に記載の方法。
(項目21)
感染症を有する被験体においてTh1免疫応答を増強するための方法であって、IL−23アンタゴニストを投与する工程を含む方法。
(項目22)
前記増強されたTh1免疫応答は、前記IL−23アンタゴニストを投与する前のIFN−γ発現CD4+T細胞の割合と比較して、IFN−γ発現CD4+T細胞の割合が2倍以上に増加する工程を含む、項目21に記載の方法。
(項目23)
前記増強されたTh1免疫応答は、前記IL−23アンタゴニストを投与する前のIL−17発現CD4+T細胞の割合と比較して、IL−17発現CD4+T細胞の割合が2分の1以下に減少する工程を含む、項目21に記載の方法。
(項目24)
前記感染症は慢性真菌感染症である、項目21に記載の方法。
(項目25)
IL−17Aアンタゴニストを投与する工程をさらに含む、前記項目のいずれかに記載の方法。
(項目26)
IL−6アンタゴニストを投与する工程をさらに含む、前記項目のいずれかに記載の方法。
(項目27)
TGF−βアンタゴニストを投与する工程をさらに含む、前記項目のいずれかに記載の方法。
(項目28)
IL−12またはIL−12アゴニストを投与する工程をさらに含む、前記項目のいずれかに記載の方法。
(項目29)
前記IL−23アンタゴニストは、IL−23p19に結合する結合性化合物である、前記項目のいずれかに記載の方法。
(項目30)
前記IL−23アンタゴニストは、IL−23Rに結合する結合性化合物である、前記項目のいずれかに記載の方法。
(項目31)
前記結合性化合物は抗体またはその抗原結合性断片である、項目29または項目30に記載の方法。
(項目32)
IL−23とその受容体との結合の阻害に関するIC50に対する、IL−12とその受容体との結合の阻害に関するIC50の比率(IC50IL−12/IC50IL−23)は、ELISAにより測定された場合に5以上である、項目29または項目30に記載の方法。
(項目33)
前記結合性化合物は、Fab、Fab’、Fab’−SH、Fv、scFv、F(ab’)2、およびダイアボディからなる群から選択される抗体断片である、項目31に記載の方法。
(項目34)
前記抗体はヒト化抗体である、項目31に記載の方法。
(項目35)
前記抗体は完全ヒト抗体である、項目31に記載の方法。
(項目36)
前記結合性化合物は単鎖抗体である、項目29または項目30に記載の方法。
(項目37)
前記抗体は、補体活性化、ADCC、またはその両方を低減するエフェクター機能を変更した抗体である、項目31に記載の方法。
(項目38)
前記結合性化合物は、IL−23Rに由来する可溶性受容体断片である、項目29に記載の方法。
(項目39)
前記IL−23アンタゴニストはsiRNAである、項目1から28のいずれかに記載の方法。
(項目40)
前記IL−23アンタゴニストはアンチセンス核酸である、項目1から28のいずれかに記載の方法。
(項目41)
前記感染症を有する前記被験体は、他の方法または組成物を使用して以前に該感染症の処置を受けたことがある被験体である、前記項目のいずれかに記載の方法。
(項目42)
前記感染症を有する前記被験体は免疫無防備状態である、前記項目のいずれかに記載の方法。
(項目43)
感染症を処置するための医薬の製造におけるIL−23アンタゴニストの使用。
(項目44)
前記感染症は慢性真菌感染症である、項目43に記載の使用。
(項目45)
感染症を有する疑いのある被験体を処置するための方法であって、有効量のIL−23アンタゴニストを投与する工程を含む方法。
(項目46)
感染症に罹るリスクのある被験体を処置するための方法であって、有効量のIL−23アンタゴニストを投与する工程を含む方法。
(項目47)
感染症を有する被験体を処置するための方法であって、有効量のIL−17AもしくはIL−17Fのアンタゴニスト、またはその両方を投与することを含む方法。
(項目48)
感染症を処置するための方法であって、IL−23p19、IL−23R、IL−17A、IL−17F、IL−17RA、IL−17RC、IL−6、およびTGF−βからなる群から選択される任意の2つのタンパク質に特異的に結合する有効量の二重特異性抗体を投与する工程を含む方法。
(項目49)
前記タンパク質はヒトタンパク質である、項目48に記載の方法。
(項目50)
本明細書に記載の任意の発明。
【図面の簡単な説明】
【0029】
【図1−1】カンジダ症に対する感受性におけるIL−23/IL−17依存性経路の役割に関する実験の結果を示す図である。マウスに108個の毒性Candidaを胃内注入した。結果は3回の実験から集める(1回の実験につき1群当たり6匹のマウス)。図1Aは、p19−/−、p35−/−、p40−/−およびC57BL/6(野生型)マウスの経時的な生存率(%)を示す図である。図1Bは、感染3日後および感染10日後の胃中の真菌増殖(CFU)を示す図である。3日後と10日後の両時点でC57BL/6マウスと比較したとき、p19−/−、p35−/−またはp40−/−マウスに関して結果は統計的差異があった(p<0.05、*によって示す)。図1Cは、MLNにおけるp35およびp19のmRNA発現(感染翌日)ならびにIL−12β2RおよびIL−23RのmRNA発現(感染3日後)を示す図である。mRNA発現はリアルタイムRT−PCRによって測定した。図1Dは、感染1週間後のIFN−γ、IL−17またはIL−4産生MLN CD4+細胞の頻度を示す図である。サイトカイン産生細胞の頻度はELISPOTアッセイにより測定し、値は細胞105個当たりのサイトカイン産生細胞の平均数(±SE)である。図1Eは、感染3日後の胃ホモジネート中の炎症性サイトカイン(IL−17、IL−23、IFN−γ、IL−12)のレベルを示す図である。サイトカインはELISAによって測定した(pg/ml)。図1C〜1Eにおいて、図に示されるように、感染マウス(+)と非感染マウス(−)を比較したとき(*)、およびp19−/−またはp35−/−マウスとC57BL/6マウスを比較したとき(**)、統計的有意差があった(p<0.05)。
【図1−2】カンジダ症に対する感受性におけるIL−23/IL−17依存性経路の役割に関する実験の結果を示す図である。マウスに108個の毒性Candidaを胃内注入した。結果は3回の実験から集める(1回の実験につき1群当たり6匹のマウス)。図1Aは、p19−/−、p35−/−、p40−/−およびC57BL/6(野生型)マウスの経時的な生存率(%)を示す図である。図1Bは、感染3日後および感染10日後の胃中の真菌増殖(CFU)を示す図である。3日後と10日後の両時点でC57BL/6マウスと比較したとき、p19−/−、p35−/−またはp40−/−マウスに関して結果は統計的差異があった(p<0.05、*によって示す)。図1Cは、MLNにおけるp35およびp19のmRNA発現(感染翌日)ならびにIL−12β2RおよびIL−23RのmRNA発現(感染3日後)を示す図である。mRNA発現はリアルタイムRT−PCRによって測定した。図1Dは、感染1週間後のIFN−γ、IL−17またはIL−4産生MLN CD4+細胞の頻度を示す図である。サイトカイン産生細胞の頻度はELISPOTアッセイにより測定し、値は細胞105個当たりのサイトカイン産生細胞の平均数(±SE)である。図1Eは、感染3日後の胃ホモジネート中の炎症性サイトカイン(IL−17、IL−23、IFN−γ、IL−12)のレベルを示す図である。サイトカインはELISAによって測定した(pg/ml)。図1C〜1Eにおいて、図に示されるように、感染マウス(+)と非感染マウス(−)を比較したとき(*)、およびp19−/−またはp35−/−マウスとC57BL/6マウスを比較したとき(**)、統計的有意差があった(p<0.05)。
【図2】アスペルギルス症に対する感受性におけるIL−23/IL−17依存性経路の役割に関する実験の結果を示す図である。マウスに2×107個のAspergillus静止期分生子を鼻腔内感染させた。図2Aおよび2B中に示す結果は4回の実験から集める(動物6匹/群)。図2Aは、感染3日後の肺中の真菌増殖(キチン含有量は、器官当たりのグルコサミンのμg数として表す)を示す図である。p19−/−、p35−/−またはp40−/−マウスとC57BL/6マウスを比較したとき(*)、統計的有意差があった(p<0.05)。図2Bは、TLNにおけるp35/p19のmRNA発現(感染翌日)ならびにIL−12β2R/IL−23RのmRNA発現(感染3日後)を示す図である。メッセンジャーRNAの発現はRT−PCRによって測定した。図に示されるように、感染マウス(+)と非感染マウス(−)を比較したとき(*)、およびp19−/−またはp35−/−マウスとC57BL/6マウスを比較したとき(**)、統計的有意差があった(p<0.05)。
【図3】真菌感染症に対する感受性におけるIL−23/IL−17依存性経路の重要性に関する実験の結果を示す図である。マウスは図1および2と同様に感染させ、感染5時間後に200μgのp19またはIL−17中和抗体、または感染5時間後と24時間後に1mgのTGF−β中和抗体で処置した。図3Aは、感染3日後のカンジダ症(C.albicans)またはアスペルギルス症(A.fumigatus)を有するマウスの胃または肺中での真菌増殖を示す図である。図に示されるように、処置マウス(+)と未処置マウス(−)を比較したとき(*)、統計的有意差があった(p<0.05)。図3Bは、ELISPOTアッセイにより測定した、それぞれカンジダ症またはアスペルギルス症を有するマウス由来のMLNまたはTLN由来のIFN−γまたはIL−17産生CD4+細胞の頻度を示す図である。値は細胞105個当たりのサイトカイン産生細胞の平均数(±SE)である。図3Bは、抗原刺激した未分画MLNまたはTLNの培養物上清における実際のIL−17産生(感染1週間後)をさらに示す。図に示されるように、感染マウスと非感染(Ct)マウスを比較したとき(*)、および処置マウス(+)と未処置マウス(−)を比較したとき(**)、統計的有意差があった(p<0.05)。図3Cは、感染3日後の、前述のp19中和抗体で処置した、カンジダ症を有するマウスの胃中での真菌増殖を示す図である。図に示されるように、処置(+)マウスと未処置(−)マウスを比較したとき(*)、およびIL−4−/−、IFN−γ−/−、p35−/−またはIFN−γ−/−/p35−/−マウスとBALB/cマウスを比較したとき(**)、統計的有意差があった(p<0.05)。
【図4】真菌に応答したDCサブセットにおけるIL−23およびIL−12産生に関する実験の結果を示す図である。GM−CSF+IL−4(GM−DC)またはFLT3−L(FL−DC)の存在下で得た骨髄DCは真菌で刺激し、サイトカインの発現を評価した。図4Aは、サイトカインmRNA発現のリアルタイムRT−PCR分析を示す図である。図4Bは、ELISAによって測定したサイトカインの発現(pg/ml)を示す図である。ザイモザン、LPS(10μg/ml)またはCpG−ODN2006(0.06μM)を陽性対照として使用した。DCは10:1の比で酵母に曝露した。図に示されるように、曝露DCと非曝露(「曝露なし」)DCを比較したとき(*)、統計的有意差があった(p<0.05)。図4Cは、p19−/−またはp35−/−マウス由来の脾性CD11c+DCにおけるIL−12およびIL−23産生を示す図である。培養物上清中のサイトカインの測定前に真菌でマウスを刺激した。図4Dは、図に示されるように、IL−12またはIL−23(10ng/ml)の存在(+)または不在(−)、または中和抗IL−12または抗IL−23抗体(10μg/ml)の存在下で、12時間真菌に曝露したC57BL/6マウス由来の脾性CD11c+DCにおけるIL−12およびIL−23産生を示す図である。
【図5】真菌に応答した炎症性DCによるIL−23産生に関する実験の結果、および特にこのような産生がTLRおよびT細胞依存性であるかどうかを示す図である。図5Aは、12時間前に真菌に曝露した異なる型のマウス由来の脾性CD11c+DCにおけるIL−23産生(pg/ml)を示す図である。4回の実験から集めた結果を示す。図に示されるように、曝露DCと非曝露(「曝露なし」)DCを比較したとき(*)、統計的有意差があった(p<0.05)。図5Bは、様々な細胞培養物および同時培養物におけるサイトカインの発現を示す図である。C57BL/6(野生型)またはp35−/−マウス由来の脾性CD4+T細胞を、パルスしていない(群2および5)またはカンジダ酵母(Ag)でパルスした(群3および6)対応する脾性DCの存在下で培養した。サイトカイン(IL−12、IL−23、IFN−γ、IL−17)を、パルス5日後にELISAによって測定した。群1および4は、真菌で刺激しT細胞では刺激しなかったC57BL/6またはp35−/−DCである。群7および8は、真菌の存在下でそれぞれC57BL/6またはp35−/−DCと共に培養した、p35−/−またはC57BL/6CD4+T細胞である。図に示されるように、群3および7をIFN−γ産生に関して群1と比較したとき、および群6および8をIL−23およびIL−17産生に関して群4と比較したとき、統計的有意差がある(p<0.05、*によって示す)。図5Cは、サンプルのいくつかが抗IL−23または抗TGF−β抗体を含むこと以外、図5B中に示したデータと同様のデータを示す図である。C57BL/6(野生型)(群1〜3)またはp35−/−(群4〜6)マウス由来の脾性CD4+T細胞を、対応する脾性CDの存在下で培養した。培養物は10μg/mlのIL−23またはTGF−β中和抗体の存在下で5日間カンジダ酵母(Ag)でパルスし、かつサイトカイン(IFN−γ、IL−17)は、ELISAによって培養物上清において定量化した。図に示されるように、群2および3をIFN−γおよびIL−17産生に関して群1と比較したとき、および群5をIL−17産生に関して群4と比較したとき、統計的有意差がある(p<0.05、*によって示す)。
【図6】PMNの、抗真菌エフェクター機能を障害し抗炎症性プログラムを破壊するIL−23およびIL−17の能力に関する実験の結果を示す図である。図6Aは、37℃における5:1のエフェクターと真菌細胞の比での非オプソニン化酵母(30分間)または分生子(60分間)とのインキュベーション後の、C57BL/6(野生型)、p19−/−またはp35−/−マウス由来のPMNにおける抗真菌活性を示す図である。結果はコロニー形成単位阻害の割合(平均±SE)としてプロットする。結果は3回の実験から集めたデータを反映する。図に示されるように、p19−/−またはp35−/−PMNとC57BL/6(野生型)PMNを比較したとき(*)、統計的有意差があった(p<0.05)。図6Bは、示した濃度でIL−23またはIL−17に曝露したC57BL/6(野生型)マウス由来のPMNの抗真菌活性を示す図である。サイトカイン曝露PMNと非曝露PMNを比較したとき(*)、統計的有意差があった(p<0.05)。図6Cおよび6Dは、60分間IFN−γ(50ng/ml)、IL−23(100ng/ml)およびIL−17(100ng/ml)の様々な組合せに曝露したC57BL/6(野生型)マウス由来のPMNの抗真菌活性を示す図である。カンジダ酵母またはAspergillus分生子に対する抗真菌活性を測定した(図6C)。MMP9/MPO産生も測定した(図6D)。ゼラチナーゼおよびミエロペロキシダーゼの産生はゼラチンザイモグラフィーによって評価し、かつウエスタンブロット分析を培養物上清に実施した。ゲルは、活性のある92kDaのMMP9および60kDaのMPOに対応するバンドを示す。図に示されるように、サイトカイン曝露PMNと非曝露PMNを比較したとき(*)、および(IFN−γ+IL−23)または(IFN−γ+IL−17)曝露PMNとIFN−γ曝露PMNを比較したとき(**)、統計的有意差があった(p<0.05)。図6Eは、ウエスタンブロット上のバンドを示す図である。PMNは12時間インビトロでIFN−γ、IL−23およびIL−17の様々な組合せに曝露した。次いでIDOタンパク質の発現をウエスタンブロッティングによって決定した。IDO発現MC24トランスフェクタントおよびモックトランスフェクトMC22細胞は、それぞれ陽性対照および陰性対照として働いた。β−チューブリンはローディング対照として働く。
【発明を実施するための形態】
【0030】
添付の特許請求の範囲を含む本明細書で使用される場合、「a」、「an」、および「the」などの単数形の単語は、文脈上明確に別段の記載がない限り、それらの対応する複数の参照を含む。特に明記のない限り、本明細書で提供された典型的な実施形態は、本発明の範囲を制限するとは見なされない。「たとえば(e.g.)」、「たとえば(for example)」、「一実施形態では」などの語句または他のそのような非限定的な語が、そのような典型的な実施形態に先行してもよく、またはそれらの典型的な性質が、文脈(たとえば「実施例」)から明白であってもよい。特に明記のない限り、「阻害しない」などの用語は、絶対的というよりむしろ相対的であると意図される。たとえば、IL−23を阻害するがIL−12は阻害「しない」作用剤は、2つのサイトカインの比較可能なアッセイにおいて作用剤が所定の濃度で存在する場合に、IL−23よりIL−12の阻害に効果が低い作用剤を指す。
【0031】
本明細書で引用されるすべての参考文献は、あたかも各個々の刊行物、データベースエントリー、特許出願、または特許が、参照によって具体的におよび個別に組み込まれるのと同程度に、参照によって全体が組み込まれる。
【0032】
I.定義。
【0033】
「活性化」、「刺激」、および「処置」は、細胞または受容体に適用される場合、文脈によりまたは明示的に特に明記のない限り、同じ意味を有していてもよく、たとえば細胞または受容体をリガンドで活性化、刺激、または処置することである。「リガンド」は、天然および合成リガンド、たとえばサイトカイン、サイトカイン改変体、類似体、ムテイン(mutein)、および抗体に由来する結合性組成物を包含する。「リガンド」は、小分子(small molecule)、たとえばサイトカインのペプチド模倣物質および抗体のペプチド模倣物質も包含する。「活性化」は、内部機構によりならびに外部的または環境的要因により制御されるような細胞活性化を指すことができる。たとえば、細胞、組織、器官、または生物の「応答」は、生化学的なもしくは生理学的な挙動、たとえば濃度、密度、接着、もしくは生物学的区画内での遊走、遺伝子発現の速さ、または分化の状態における変化を包含し、この変化は、活性化、刺激、もしくは処置と関連があるか、または遺伝プログラミングなどの内部機構と関連がある。
【0034】
分子の「活性」は、リガンドまたは受容体への分子の結合、触媒活性;遺伝子発現もしくは細胞シグナル伝達、分化、または成熟化を刺激する能力;抗原活性、他の分子の活性の調節などを記述するかまたはそれらを指すことができる。分子の「活性」は、細胞間相互作用、たとえば接着を調節または維持する活性、または細胞の構造、たとえば細胞膜または細胞骨格を維持する活性を指すこともできる。「活性」は、比活性、たとえば、[触媒活性]/[mgタンパク質]、もしくは[免疫学的活性]/[mgタンパク質]、または生物学的区画における濃度などを意味することもできる。「増殖活性」は、たとえば、正常な細胞分裂ならびに癌、腫瘍、異形成、細胞の形質転換、転移、および新脈管形成を促進するまたはそれに必要であるまたはそれに特異的に関連する活性を包含する。
【0035】
「投与」および「処置」は、動物、ヒト、実験被験体、細胞、組織、器官、または生物学的液体に適用される場合、外因性の医薬品、治療剤、診断用薬、または組成物の、動物、ヒト、被験体、細胞、組織、器官、または生物学的液体に対する接触を指す。「投与」および「処置」は、たとえば、治療方法、薬物動態学的方法、診断方法、研究方法、および実験方法を指すことができる。細胞の処置は、細胞に対する試薬の接触および細胞と接触している体液に対する試薬の接触を包含する。「投与」および「処置」はまた、たとえば、試薬、診断薬、結合性組成物による、または他の細胞による細胞のインビトロおよび生体外(ex vivo)での処置をも意味する。「処置」は、ヒト被験体、獣医学的被験体、または研究被験体に適用される場合、治療処置、予防的または防止的手段、研究への応用、および診断への応用を指す。「処置」は、ヒト被験体、獣医学的被験体、もしくは研究被験体または細胞、組織、もしくは器官に適用される場合、IL−23またはIL−23Rアンタゴニストの、ヒト被験体または動物被験体、細胞、組織、生理的コンパートメント、または生理的液体に対する接触を包含する。「細胞の処置」はまた、IL−23またはIL−23Rアンタゴニストが、たとえば体液相またはコロイド相中でIL−23R複合体(IL−23R/IL−12Rベータ1ヘテロ二量体)と接触する状況だけではなく、アンタゴニストが細胞または受容体と接触しない状況をも包含する。
【0036】
「結合性組成物」は、標的に結合することができる分子、小分子、高分子、抗体、その断片もしくは類似体、または可溶性受容体を指す。「結合性組成物」はまた、標的に結合することができる、分子の複合体、たとえば非共有結合複合体、イオン化分子、および共有結合でまたは非共有結合で改変された分子、たとえばリン酸化、アシル化、架橋、環化、または限定切断で改変された分子を指すものであってもよい。「結合性組成物」はまた、安定剤、賦形剤、塩、緩衝剤、溶媒、または添加剤と組み合わせた、標的に結合することができる分子を指すものであってもよい。「結合」は、結合性組成物の標的との関連性として定義されてもよく、その関連性は、結合性組成物が溶液中に溶解するまたは懸濁することができる場合、結合性組成物の通常のブラウン運動における低下をもたらす。
【0037】
本発明の結合性化合物は、二重特異性抗体を含むことができる。本明細書に使用される場合、用語「二重特異性抗体」は、少なくとも2つの異なる抗原エピトープに対する結合特異性を有する抗体、典型的にはモノクローナル抗体を指す。一実施形態では、エピトープは、同じ抗原に由来する。他の実施形態では、エピトープは、2つの異なる抗原に由来する。二重特異性抗体を作製するための方法は、当技術分野で知られている。たとえば、二重特異性抗体は、2つの免疫グロブリン重鎖/軽鎖対の同時発現を使用して組換えで産生することができる。たとえばMilsteinら(1983年)Nature 305:537〜39頁を参照されたい。あるいは、二重特異性抗体は、化学的連結を用いて調製することができる。たとえばBrennanら(1985年)Science 229:81頁を参照されたい。二重特異性抗体は、二重特異性抗体の断片を含む。たとえばHolligerら(1993年)Proc. Natl. Acad. Sci. U.S.A. 90:6444〜48頁、Gruberら、J. Immunol. 152:5368頁(1994年)を参照されたい。
【0038】
「古典的TH1型T細胞」とは、IL−4、IL−5、またはIL−13の各々の発現を超える程度まで、インターフェロン−ガンマ(IFNγ)を発現するT細胞であり、一方で「古典的TH2型T細胞」とは、各々がIFNγの発現を超える程度まで、IL−4、IL−5、またはIL−13を発現するT細胞である。「程度」とは、古典的TH2型細胞の、典型的には4倍以上、より典型的には8倍以上、および最も典型的には16倍以上である。
【0039】
本明細書で定義される「記憶T細胞」とは、所定の抗原に以前曝露されている長寿命のT細胞のサブセットである。記憶T細胞は、生物中に何年間も存在することができ、同じ抗原によるその後の曝露に対する迅速な応答を可能にする。マウス記憶T細胞の表現型は、CD4+highCD45RBlowと定義される。ヒト記憶T細胞の表現型は、CD45RAneg/lowCD45R0highと定義される。これらの記憶T細胞のIL−23処置は、増殖およびIL−17の発現に帰着する。特に明記のない限り、「IL−17」とは、本明細書で使用される場合、IL−17Aを指す。たとえば、Moseleyら(2003年)Cytokine & Growth Factor Rev. 第14巻:155頁を参照されたい。
【0040】
「保存的修飾改変体」とは、アミノ酸配列および核酸配列の両方に適用される。特定の核酸配列に関して、保存的修飾改変体とは、同一または本質的に同一なアミノ酸配列をコードするこれらの核酸を指すか、または核酸がアミノ酸配列をコードしない場合は、本質的に同一の核酸配列を指す。遺伝コードの縮重のため、多数の機能的に同一な核酸が、任意の所定のタンパク質をコードできる。
【0041】
アミノ酸配列に関しては、当業者であれば、コードされた配列において1つのアミノ酸またはわずかな割合のアミノ酸を、保存されたアミノ酸に置換する、核酸、ペプチド、ポリペプチド、またはタンパク質配列における変化が、「保存的修飾改変体」であることを認識する。機能的に類似のアミノ酸を提供する保存的置換表は、当技術分野で周知である。保存的置換の一例は、以下の群のもののうち1つのアミノ酸を同じ群の別のアミノ酸に交換することである:(Leeらに交付された米国特許第5,767,063号の明細書;Kyte and Doolittle(1982年)J. Mol. Biol. 第157巻:105〜132頁):
(1)疎水性:ノルロイシン、Ile、Val、Leu、Phe、Cys、またはMet;
(2)中性親水性:Cys、Ser、Thr;
(3)酸性:Asp、Glu;
(4)塩基性:Asn、Gln、His、Lys、Arg;
(5)鎖の配向に影響を及ぼす残基:Gly、Pro;
(6)芳香族:Trp、Tyr、Phe;
(7)小型アミノ酸:Gly、Ala、Ser。
【0042】
「有効量」は、病状の症候または徴候を改善または予防するのに十分な量を包含する。有効量は、診断を可能または容易にするのに十分な量も意味する。特定の患者または獣医学的被験体のための有効量は、処置されている状態、患者の健康全般、投与の方法、経路、および用量、ならびに副作用の重症度などの要因に依存して変動し得る。たとえば米国特許第5,888,530号明細書を参照されたい。有効量は、著しい副作用または毒性作用を回避する最大の用量または投薬プロトコルであり得る。効果は、診断の尺度またはパラメーターが、正常被験体により示される診断パラメーターを100%と定義した場合、少なくとも5%、通常は少なくとも10%、より通常は少なくとも20%、最も通常は少なくとも30%、好ましくは少なくとも40%、より好ましくは少なくとも50%、最も好ましくは少なくとも60%、理想的には少なくとも70%、より理想的には少なくとも80%、および最も理想的には少なくとも90%改善することに帰着する。たとえば、Maynardら(1996年)A Handbook of SOPs for Good Clinical Practice、Interpharm Press、ボーカラトーン、米国フロリダ州;Dent(2001年)Good Laboratory
and Good Clinical Practice、Urch Publ.、ロンドン、英国を参照されたい。
【0043】
「外因性」とは、文脈に依存して、生物、細胞、または人体の外部で産生される物質を指す。「内因性」とは、文脈に依存して、細胞、生物、または人体の内部で産生される物質を指す。
【0044】
「感染性疾患」とは、生物、器官、組織、または細胞の微生物感染症、たとえば、細菌感染症、ウイルス感染症、および/または真菌感染症を指す。
【0045】
「IL−17産生細胞」とは、古典的TH1型T細胞または古典的TH2型T細胞ではないT細胞を意味する。「IL−17産生細胞」は、遺伝子またはポリペプチド(たとえば、分裂促進因子応答性P−タンパク質;ケモカインリガンド2;インターロイキン−17(IL−17);転写因子ROR−γT関連;サイトカインシグナル伝達抑制因子3など)を発現するT細胞も意味し、IL−23アゴニストで処置した発現は、IL−12アゴニストでの処置より高く、「より高い」とは以下の通り定義される。IL−23アゴニストでの発現は、IL−12処置よりも、普通は少なくとも5倍高く、典型的には少なくとも10倍高く、より典型的には少なくとも15倍高く、最も典型的には少なくとも20倍高く、好ましくは少なくとも25倍高く、および最も好ましくは少なくとも30倍高い。発現は、たとえば、実質的に純粋なIL−17産生細胞の集団を処置して測定できる。
【0046】
さらに、「IL−17産生細胞」は、細胞発生または細胞分化の経路において、上記で定義されたIL−17産生細胞への分化に決定された前駆体または前駆細胞を含む。IL−17産生細胞への前駆体または前駆細胞は、流入領域リンパ節(DLN)に見出すことができる。さらに、「IL−17産生細胞」は、たとえばホルボールエステル、イオノフォア、および/または発癌物質により、たとえば活性化され、さらには分化され、保管され、冷凍され、乾燥され、不活性化され、たとえばアポトーシス、タンパク質分解、または脂質酸化により部分的に分解され、またはたとえば組換え技術により改変された、上記で定義されたIL−17産生細胞を包含する。
【0047】
「阻害剤」および「アンタゴニスト」とは、たとえばリガンド、受容体、補助因子、遺伝子、細胞、組織、または器官を阻害する阻害分子を指す。遺伝子、受容体、リガンド、または細胞の調節因子とは、遺伝子、受容体、リガンド、または細胞の活性を変化させる分子であり、活性は、その制御特性において活性化、阻害、または変化させることができる。調節因子は、単独で機能してもよく、または補助因子、たとえばタンパク質、金属イオン、もしくは小分子を使用してもよい。阻害剤とは、たとえば遺伝子、タンパク質、リガンド、受容体、または細胞を減少、阻止、防止、活性化遅延、不活性化、除感作、または下方制御する化合物である。活性化剤とは、たとえば遺伝子、タンパク質、リガンド、受容体、または細胞を増加、活性化、促進、活性化増強、感作、または上方制御する化合物である。阻害剤は、恒常的活性を低減、阻止、または不活性化する組成物とも定義できる。「アンタゴニスト」とは、アゴニストの作用に対抗する化合物である。アンタゴニストは、アゴニストの活性を防止、低減、阻害、または中和する。アンタゴニストは、同定されたアゴニストが存在しない場合でさえ、標的、たとえば標的受容体の恒常的活性を防止、阻害、または低減することもできる。
【0048】
IL−23アンタゴニストは、たとえば、以下でより詳細に記述するTh17細胞の増幅および生存などの、IL−23の生物活性を混乱させるあらゆる作用剤を含む。IL−23受容体およびIL−23Rのアンタゴニストは、IL−23シグナル伝達を阻止することによりIL−23の活性を阻止する役目を果たすため、IL−23アンタゴニストのサブセットである。
【0049】
阻害の程度を検討するためには、たとえば、所定のタンパク質、遺伝子、細胞、または生物を含む試料またはアッセイを、有望な活性化剤または阻害剤で処理し、阻害剤無しの対照試料と比較する。対照試料、つまりアンタゴニストで処理されなかった試料には、100%の相対的活性値が割り当てられる。対照と比較した活性値が、約90%以下、典型的には85%以下、より典型的には80%以下、最も典型的には75%以下、一般的には70%以下、より一般的には65%以下、最も一般的には60%以下、典型的には55%以下、通常は50%以下、より通常は45%以下、最も通常は40%以下、好ましくは35%以下、より好ましくは30%以下、さらにより好ましくは25%以下、および最も好ましくは25%未満である場合、阻害は達成される。対照と比較した活性値が、約110%、一般的には少なくとも120%、より一般的には少なくとも140%、より一般的には少なくとも160%、多くの場合少なくとも180%、より多くの場合少なくとも2倍、最も多くの場合少なくとも2.5倍、通常は少なくとも5倍、より通常は少なくとも10倍、好ましくは少なくとも20倍、より好ましくは少なくとも40倍、および最も好ましくは40倍を超えてより高い場合、活性化は達成される。
【0050】
活性化または阻害の終点(endpoint)は、以下のようにモニターできる。たとえば細胞、生理的液体、組織、器官、および動物被験体またはヒト被験体の活性化、阻害、および処置に対する応答は、終点によりモニターできる。終点は、あらかじめ定められた量または割合の、たとえば細菌負荷低減、腫瘍原性低減、もしくは細胞脱顆粒低減、またはサイトカイン、毒性酸素、もしくはプロテアーゼの放出などの分泌低減の徴候を含み得る。終点は、たとえば、あらかじめ定められた量のイオンフラックス(flux)または輸送;細胞遊走;細胞接着;細胞増殖;転移能;細胞分化;および表現型の変化、たとえば炎症、アポトーシス、形質転換、細胞周期、または転移に関連する遺伝子発現の変化を含み得る。たとえば、Knight(2000年)Ann. Clin. Lab. Sci. 第30巻:145〜158頁;Hood and Cheresh(2002年)Nature Rev. Cancer 第2巻:91〜100頁;Timmeら(2003年)Curr. Drug Targets 第4巻:251〜261頁; Robbins and Itzkowitz(2002年)Med. Clin. North Am. 第86巻:1467〜1495頁;Grady and Markowitz(2002年)Annu. Rev. Genomics Hum. Genet. 第3巻:101〜128頁;Bauerら(2001年)Glia 第36巻:235〜243頁;Stanimirovic and Satoh(2000年)Brain Pathol. 第10巻:113〜126頁を参照されたい。
【0051】
阻害の終点は、一般的には対照の75%以下、好ましくは対照の50%以下、より好ましくは対照の25%以下、最も好ましくは対照の10%以下である。一般的には、活性化の終点は、少なくとも150%対照、好ましくは少なくとも2倍対照、より好ましくは少なくとも4倍対照、および最も好ましくは少なくとも10倍対照である。
【0052】
「ノックアウト」(KO)とは、遺伝子によりコードされたポリペプチド、たとえばIL−23またはIL−23受容体のサブユニットをコードするポリペプチドの少なくとも一部の発現の部分的または完全な低減を指し、その遺伝子は、哺乳類の単一の細胞、選択された細胞、またはすべての細胞に内因性である。KOは、生物学的な機能は低減されるが発現は必ずしも低減されない実施形態、たとえば挿入された不活性化ペプチドを含有するポリペプチドも包含する。コード配列または制御配列の破壊は、ノックアウト技術により包含される。細胞または哺乳動物は、内因性遺伝子の1つの対立遺伝子が破壊された「ヘテロ接合性ノックアウト」であり得る。あるいは、細胞または哺乳動物は、内因性遺伝子の両対立遺伝子が破壊された「ホモ接合性ノックアウト」であり得る。「ホモ接合性ノックアウト」は、両対立遺伝子の破壊を同一技術またはゲノムでの同一結果に限定するとは意図されていない。
【0053】
「標識された」組成物は、分光法、光化学法、生化学法、免疫化学法、同位体法、または化学的方法により、直接的または間接的のいずれかで検出可能である。たとえば、有用な標識には、32P、33P、35S、14C、3H、125I、安定同位体、蛍光色素、高電子密度の試薬、基質、エピトープタグ、またはたとえば酵素結合免疫測定(enzyme−linked immunoassay)もしくはフルオレッテ(fluorette)で使用されるような酵素が含まれる。たとえば、Rozinov and Nolan(1998年)Chem. Biol. 第5巻:713〜728頁を参照されたい。
【0054】
「リガンド」とは、受容体のアゴニストまたはアンタゴニストとして作用し得る、たとえば、小分子、ペプチド、ポリペプチド、および膜関連もしくは膜結合分子、またはそれらの複合体を指す。「リガンド」は、アゴニストまたはアンタゴニストではないが受容体に結合できる作用剤も包含する。さらに、「リガンド」は、たとえば化学的方法または組換え法により、可溶型の膜結合リガンドに変更された膜結合リガンドを含む。通常は、リガンドが第1の細胞に膜結合している場合、通常第2の細胞に受容体が生じる。第2の細胞は、第1の細胞と同じまたは異なる同一性を有していてよい。リガンドまたは受容体は、完全に細胞内性であってもよく、すなわち、細胞質ゾル、核、またはいくつかの他の細胞内区画に存在していてもよい。リガンドまたは受容体は、たとえば、細胞内区画から原形質膜の外部面へとその位置を変化させることができる。リガンドおよび受容体の複合体は、「リガンド受容体複合体」と名付けられる。リガンドおよび受容体がシグナル伝達経路に関与する場合、リガンドはシグナル伝達経路の上流位置で生じ、受容体は下流位置で生じる。
【0055】
「マーカー」は、細胞、組織、器官、動物の表現型、たとえばIL−17産生細胞の表現型に関する。マーカーは、たとえば、細胞の精製中、定量化中、遊走中、活性化中、成熟中、または発生中に細胞を検出するために使用され、in vitroおよびin vivo研究の両方で使用できる。活性化マーカーとは、細胞活性化に関連するマーカーである。
【0056】
「精製された細胞」は、たとえば、他のタイプの細胞が実質的に存在しない、たとえば他のタイプのT細胞による汚染が実質的に存在しない1つまたは複数の「IL−17産生細胞」を包含する。純度は、幾何学的な座標により、またはたとえばフラスコ、チューブ、またはバイアルを含む区画により定義される容積を使用することにより算定できる。「精製されたIL−17産生細胞」は、たとえば、「IL−17産生細胞」が、通常は全細胞の少なくとも20%、より通常には全細胞の少なくとも30%、最も通常には全細胞の少なくとも40%、一般的には全細胞の少なくとも50%、より一般的には全細胞の少なくとも60%、最も一般的には全細胞の少なくとも70%、好ましくは全細胞の少なくとも80%、より好ましくは全細胞の少なくとも90%;最も好ましくは全細胞の少なくとも95%を占める区画により定義できる。
【0057】
「小分子」は、10kD未満、典型的には2kD未満、および好ましくは1kD未満の分子量を有する分子として定義される。小分子には、それらに限定されないが、無機分子、有機分子、無機成分を含有する有機分子、放射性原子を含む分子、合成分子、ペプチド模倣物質、および抗体模倣物質が含まれる。治療薬として、小分子は、大分子より細胞に対してより透過性があり、分解を受けにくく、免疫応答を誘発する傾向が少ない場合がある。抗体およびサイトカインのペプチド模倣物質などの小分子ならびに小分子トキシンが、当技術分野で知られている。たとえば、Cassetら(2003年)Biochem. Biophys. Res. Commun. 第307巻:198〜205頁;Muyldermans(2001年)J. Biotechnol. 第74巻:277〜302頁;Li(2000年)Nat. Biotechnol. 第18巻:1251〜1256頁;Apostolopoulosら(2002年)Curr. Med.
Chem. 第9巻:411〜420頁;Monfardiniら(2002年)Curr. Pharm. Des. 第8巻:2185〜2199頁;Dominguesら(1999年)Nat. Struct. Biol. 第6巻:652〜656頁;Sato and Sone(2003年)Biochem. J. 第371巻:603〜608頁;米国特許第6,326,482号明細書を参照されたい。
【0058】
リガンド/受容体、抗体/抗原、または他の結合対に関する場合、「特異的に」または「選択的に」結合するとは、タンパク質および他の生物製剤の不均一集団におけるタンパク質の存在の決定要因である結合反応を示す。したがって、指定された条件下で、特定のリガンドは特定の受容体に結合し、試料中に存在する他のタンパク質に対しては相当量は結合しない。意図された方法の抗体、または抗体の抗原結合部位に由来する結合性組成物は、任意の他の抗体またはそれに由来する結合性組成物の親和性より、少なくとも2倍高い、好ましくは少なくとも10倍高い、より好ましくは少なくとも20倍高い、および最も好ましくは少なくとも100倍高い親和性で、その抗原またはその改変体もしくはムテインに結合する。好ましい実施形態では、抗体は、たとえばスキャチャード解析により決定されるように、約109リットル/molより高い、所望の標的に対する親和性を有す。Munsenら(1980年)Analyt. Biochem. 第107巻:220〜239頁。
【0059】
IL−23またはIL−23受容体に「特異的に結合する」抗体は、IL−23由来配列を含まないタンパク質には結合せず、つまり本明細書で使用される場合、「特異性」とはIL−23特異性に関し、問題のタンパク質に存在し得る任意の他の配列には関しない。たとえば、本明細書で使用される場合、IL−23に「特異的に結合する」抗体は、典型的には、IL−23およびFLAG(登録商標)ペプチドタグを含む融合タンパク質であるFLAG−hIL−23に結合するが、FLAG(登録商標)ペプチドタグ単独には、またはそれがIL−23以外のタンパク質に融合されている場合には、結合しない。文脈に依存して、IL−23に対する特異性は、IL−23(またはその受容体)には結合するが、IL−12(またはその受容体)などの他のタンパク質には結合しないという能力を指すこともできる。
【0060】
II.一般。
【0061】
IL−23およびIL−12は両方とも、共通のサブユニットおよび共通の受容体サブユニットを共有するヘテロ二量体サイトカインであるが、最近の結果によると、炎症および宿主防御におけるそれらの役割は、重複的というより拮抗的であることが示唆されている。インターロイキン23(IL−23)は、2つのサブユニット、つまりp19およびp40で構成されたヘテロ二量体サイトカインである。p19サブユニットは、IL−6、顆粒球コロニー刺激因子(G−CSF)、およびIL−12のp35サブユニットと構造的に関連している。p40サブユニットは、p35およびp40で構成されるサイトカインIL−12の一部である。ヘテロ二量体IL−12は、多くの場合IL−12p70と呼ばれる。IL−23は、IL−23RおよびIL−12Rβ1で構成されるヘテロ二量体受容体に結合することによりシグナル伝達を媒介する。IL−12Rβ1サブユニットは、IL−12Rβ1およびIL−12Rβ2で構成されるIL−12受容体により共有されている。IL−23およびIL−23受容体、ならびにそれぞれのIL−23特異的サブユニットは、国際公開第99/05280号パンフレット、国際公開第01/18051号パンフレット、国際公開第00/73451号パンフレット、および国際公開第01/85790号パンフレットに開示されている。
【0062】
IL−I2に関する多くの初期研究では、p40の遺伝的欠損(p40ノックアウトマウス;p40KOマウス)が必要とされたが、その後IL−23の発見と共に、そのようなマウスはIL−12およびIL−23の両方を欠損していたことが判明した。Oppmannら(2000年)Immunity 第13巻:715〜725頁;Wiekowskiら(2001年)J. Immunol. 第166巻:7563〜7570頁;Parhamら(2002年)J. Immunol. 第168巻:5699〜708頁;Frucht(2002年)Sci STKE 2002、E1〜E3頁;Elkinsら(2002年)Infection Immunity 第70巻:1936〜1948頁。これらの結果により、IL−12およびTh1応答に関係すると当初は考えられていた初期観察の多数の解釈が変更された。
【0063】
最近の研究によると、CD4+T細胞エフェクターレパートリーの多様性は、Th1/Th2パラダイムにより包含される多様性より大きいことが示唆されている。Th17細胞は、以前はTh1系列に帰されていた免疫病原性に寄与するエフェクターTh細胞の異なる系列であると今では考えられている。Th17分化に結び付く経路はまだ不明確であるが(Dong(2006年)Nat Rev Immunol 第6巻:329頁)、IL−23は、この系列の産生および維持のための重要なサイトカインである(Trinchieriら(2003年)Immunity 第19巻:641頁)。IL−12およびIL−23の両方がCD4+T細胞でIFN−γの発現を誘導できる一方で、IL−23は単独で、Th細胞による炎症促進性サイトカインIL−17の産生を促進する。
【0064】
多数の類似性にもかかわらず、IL−12およびIL−23が、多岐にわたる免疫経路を駆動するという証拠が増加している。IL−17で開始された(primed)Th細胞(Th17細胞)は、実験的自己免疫性脳脊髄炎(EAE)(Langrishら(2005年)J. Exp. Med. 第201巻:233頁)、関節炎(Murphyら(2003年)J. Exp. Med. 第198巻:1951頁)、大腸炎(Yenら(2006年)J. Clin. Invest. 第116巻:1310頁)、および自己免疫性心筋炎(Sondereggerら(2006年)Eur. J. Immunol. 第36巻:2844頁)を含む、種々の臓器関連自己免疫性疾患(Harringtonら(2006年)Curr. Opin. Immunol. 第18巻:349頁)に関与している。さらに、それほど明確ではないが、高レベルIL−23/IL−17の産生は、IL−12/IFN−γより、多様な感染症における疾患重症度および免疫病理とより良好に相関する。Hunter(2005年)Nat. Rev. Immunol. 第5巻:521頁;Rutitzky(2005年)J. Immunol. 第175巻:3920頁。他の研究によると、IL−12およびIL−23は、Klebsiella pneumoniae(Happelら(2005年)J. Exp. Med. 第202巻:761頁)およびCitrobacter rodentium(Manganら(2006年)Nature 第441巻:231頁)に対する宿主防御において別個の役割を有することが示唆されている。これらの研究によると、IL−12およびIL−23は、in vivoで、抗菌性の免疫応答および疾患の促進に別個の役割を有することが示唆されている。
【0065】
炎症および宿主防御におけるIL−12およびIL−23の役割が異なることは、慢性真菌感染症などの慢性感染症にとって重要な意味合いを有する。炎症は、真菌感染症を迅速に制御するために必要であるが、炎症の消散は、感染症および関連疾患における防御と免疫病理との間のバランスを維持するために不可欠である。Han and Ulevitch(2005年)Nat. Immunol. 第6巻:1198頁。持続性炎症は、広範な慢性疾患および自己免疫の特徴である。Han & Ulevitch(2005年)。Candidaの場合、炎症消散の失敗は、不完全な真菌除去と関係する。この未消散のCandida感染症は、慢性皮膚粘膜カンジダ症(CMC)に帰着する。CMCは、自己免疫性多発性内分泌腺症−カンジダ症−外胚葉ジストロフィー、機能不全性T細胞活性の状態と関係する。Ryanら(2005年)J. Allergy Clin. Immunol. 第116巻:1158頁。CMCは、未知の免疫病原性の多様な臨床的障害も包含する。Lilic(2002年)Curr. Opin. Infect. Dis. 第15巻:143頁。Aspergillusの場合、感染が難治性である持続性炎症は、同種造血幹細胞移植後の非好中球減少患者(Ortegaら(2006年)Bone Marrow Transplant 第37巻:499頁)、ならびにアレルギー性真菌疾患(Schubert(2006年)Clin. Rev. Allergy Immunol. 第30巻:205頁)において一般的である。この20年間、真菌感染症および関連炎症性疾患の免疫病原性は、様々なタイプの制御性T細胞(Treg)の組合せにより影響されるようなTh1/Th2バランスの点から、主として説明されている。Romani(2004年)Nat. Rev. Immunol. 第4巻:1頁;Romani and Puccetti(2006年)Trends Microbiol. 第14巻:183頁。
【0066】
炎症は、真菌に対する防御応答の不可欠な構成要素であるが、その調節障害は、真菌疾患を著しく悪化させる場合がある。本明細書で開示されたように、IL−23およびIL−17は、真菌に対するIL−12/Th1媒介性免疫耐性を負に制御し、以前は抑制不良のTh1細胞応答に帰されていた起炎性の役割を果たす。IL−23は、Th17細胞(IL−17を産生し、炎症を引き起こす)の生存を促進し、IL−12媒介性Th1応答(インターフェロン−γ(IFN−γ)の産生を伴う)に拮抗することが知られている。Langrishら(2004年)Immunol. Rev. 第202巻:96頁。本明細書で実証されたように、IL−12産生およびTh1応答のIL−23対抗制御は、2つの主要なヒト真菌病原体であるCandida albicansおよびAspergillus fumigatusの制御不良の炎症および増殖に帰着する。IL−23およびIL−17の両方は、好中球の炎症性プログラムおよび抗真菌活性を破壊し、感染に伴う重篤な組織炎症性病理に帰着する。要約すると、IL−23が駆動する炎症は、感染を促進し、抗真菌免疫耐性を損なう。Zelanteら(2007年)Eur. J. Immunol. 第37巻:2695頁、およびCooper(2007年)Eur. J. Immunol. 第37巻:2680頁の関連解説も参照されたい。IL−23に対する拮抗作用による炎症性応答の調節は、真菌に対する防御免疫応答を刺激するための有望な戦略を代表する。
【0067】
本発明は、IL−23および/またはIL−17の活性を阻止して、Th17細胞の効果を低減し、頑健なTh1応答を出現させ、感染した細胞または生物を除去することにより、慢性感染症などの感染症を処置するための組成物および方法を提供する。最適な場合、結果は、感染が完全に消散される無菌治癒である(つまり、処置を中止できるが感染は再発しない)。
【0068】
望ましくない結果を生む炎症状態の維持におけるTh17細胞の役割に関する同じ論拠が、結核(TB)などの慢性のウイルスおよび細菌感染症の場合に当てはまる。サイトカインは、免疫細胞の可溶性媒介物質である。以下のサイトカインが、TBに感染した患者の胸腔洗浄液または気管支肺胞洗浄(BAL)液で検出されている:IL−1β、TNFα、IFNγ、TGFβ、およびIL−12。たとえば、Crystalら(編)(1997年)The Lung; Scientific Foundations、Lippincott−Raven、ニューヨーク、米国ニューヨーク州、2381〜2394頁を参照されたい。IFN−γおよびTNFαは、マイコバクテリア感染症の制御に重要な役割を果たすことが示されている。たとえば、Cooperら(1993年)J. Exp. Med. 第178巻:2243頁;Flynnら(1993年)J. Exp. Med. 第178巻:2249頁;Kindlerら(1989年)Cell 第56巻:731頁;Cheuseら(1994年)Am. J. Pathol. 第145巻:1105頁を参照されたい。IL−12駆動のIFN−γ産生を低減するTh17細胞の生存をIL−23が促進する範囲内において、IL−23活性に対する拮抗作用が細菌感染症の消散を増強すると期待できる。
【0069】
文献によると、IL−23アンタゴニストでの処置は、たとえば自己免疫障害または慢性感染症の処置において、IL−12アンタゴニストでの処置より安全であり得ることが示唆される。Chackerianらは、p19−/−ノックアウト(KO)マウスにおける抗体中和または遺伝子除去のいずれかによるIL−23活性の除去が、マイコバクテリア(BCG)感染に対する免疫を損なわなかったという実験を記述している。Chackerianら(2006年)J. Exp. Med. 第74巻:6092頁。IL−23p19KOマウスにおける感染の経過は、野生型マウスにおける経過と区別がつかず、抗IL−23p19処置マウスにおける細菌コロニー形成単位数は、イソタイプ対照処置マウスにおける単位数と変わらなかった。対照的に、IL−12欠損KOマウスはBCGの増殖を制御できず、IL−12の抗体阻止は、イソタイプ対照処置マウスと比較して、脾臓、肝臓、および肺において著しくより高いCFU数と相関した。これらの結果により、IL−23は、IL−12の存在下ではマイコバクテリアに対する宿主防御に重要な役割を果たさず、したがってIL−23の選択的阻害は、IL−12の中和を伴う処置(IL−23を同時中和して、またはしないでのいずれかで)より安全であり得ることが示唆される。本明細書で提示された結果は、これらの結果を進展させ、IL−23アンタゴニストは、宿主防御を損なわないという点でより安全であるだけでなく、IL−23/IL−17炎症の調節障害により引き起こされる慢性感染症の消散を支援する点で実際に有益であり得ることを示唆する。
【0070】
Chackerianらの実験(2006年)は、IL−23の阻止が、既存の慢性マイコバクテリア感染の除去を増強するか否かの問題に取り組むようには設計されていなかった。対照マウス(WTマウス、未処置であるかまたはイソタイプ対照抗体のみで処置された)は、慢性感染症を発症せず、感染を効果的に除去できた。Chackerianら(2006年)のKOマウスおよび抗体阻止実験の両方で、IL−23活性は、感染後ではなく、静脈内BCGによる感染の前に除去された。本明細書に記載の実験は、マイコバクテリア感染症ではなく、真菌感染症に関していた。さらに、本明細書の実施例4および5に記載の実験は、静脈内投与ではなく、真菌病原体の胃内および鼻腔内投与を使用した実験を含む。これらの真菌病原体を肺および胃に直接送達すると、静脈内送達と比べてより生理学的に関係性のある疾患モデルがもたらされる。本明細書に開示の実験において真菌病原体に感染した組織は、Th17応答がその最も重要な生理学的役割を有し得る組織、つまり肺および腸の粘膜関門であると示唆されている。Cua and Kastelein(2006年)Nature Immunol. 第7巻:557頁。さらに、本明細書に記載の実験においては、抗IL−23p19および抗IL−17抗体は、感染前ではなく、感染の5時間後に投与された。
【0071】
III.真菌感染症の実験結果
持続性炎症は、広範な慢性疾患および自己免疫の特徴である。Han & Ulevitch(2005年)。IL−23が発見され、自己免疫炎症におけるその役割が近年になって文献化される前(Cuaら(2003年)Nature 第421巻:744頁;Langrishら(2005年)J. Exp. Med. 第201巻:233頁)は、IL−12が、Th1応答を開始および維持することにより、過剰反応免疫および自己免疫障害に関与すると考えられていた。これは、真菌感染症、ならびに炎症および制御されないTh1/Th2抗真菌反応性の微調整に免疫制御が不可欠であると証明された疾患にも当てはまった。Ryanら(2005年);Romani(2004年);Romani & Puccetti(2006年)。
【0072】
本研究の結果により、制御されないTh1応答ではなくIL−23/IL−17軸が、不完全な病原体除去、炎症消散の失敗、ならびにCandidaおよびAspergillusに対する防御免疫応答の開始失敗と関係していることが示される。したがって、この新しい知見は、進行中の炎症に直面して真菌の残留が生じる難治形態の真菌感染症と慢性炎症反応との矛盾した関連性を調整する役割を果たすことができる。
【0073】
IL−23およびIL−17の両方は、IFN−γの存在下でさえPMNの抗真菌エフェクター活性を損ない、この知見はTh17エフェクター経路がTh1経路より優勢であることを示唆する。さらに、両サイトカインは、真菌に対するPMNの炎症性状態を限定することが知られている、インドールアミン2,3−ジオキシゲナーゼ(IDO)のIFN−γ依存性活性化に対抗することにより(Bozzaら(2005年)J. Immunol. 第174巻:2910頁)、ならびにTh17細胞活性化に伴う高い炎症性病理および組織破壊の原因である可能性が高いMMP9およびMPOの放出を誘導することにより、PMNの炎症性プログラムを活性化した。
【0074】
IDOに対する作用は興味深い。IDOは、C.albicansで発現され、真菌発芽のトリプトファン栄養要求依存性阻害に関与する。Bozzaら(2005年)。IDO遮断と同様におよびIFN−γと対向して(Kalo−Kleinら(1990年)Infect. Immun. 第58巻:260頁)、IL−17は真菌発芽を促進したが(データ非表示)、この知見は、哺乳類宿主免疫系からのシグナルに高度に反応する酵素である真菌IDOに対する作用を示唆する。Mellor and Munn(2004年)Nat. Rev. Immunol. 第4巻:762頁。したがって、他の感染症について既に記述されているように(McKenzieら(2006年)Trends Immunol. 第27巻:17頁)、Th17経路の機能は、炎症を促進し抗菌免疫を破壊するその能力の範囲を超えて、真菌の形態および毒性に対する作用を含み得る。これは、高レベルの真菌増殖に厳密に依存することが知られている共動性IL−4+Th2細胞活性化に変形する場合があり(Mencacciら(1996年)Infect. Immun. 第64巻:4907頁)、Th1が機能することをさらに防止する。
【0075】
他の感染症について既に記述されているように(Cruzら(2006年)J. Immunol. 第177巻:1416頁;Parkら(2005年)Nat. Immunol. 第6巻:1133頁)、Th1またはTh17経路は、両真菌感染症において相互に制御された。この知見は、真菌に応答するどちらか一方の経路の出現が、厳密な環境制御下にあることを示唆する。制御は、様々な段階で起こり得る。1つの明白なレベルの制御は、Th17細胞の誘導を制御することが知られているIFN−γにより代表される。Cruzら(2006年);Parkら(2005年)。IL−23/IL−17軸は、IFN−γ欠損の場合、両感染症において実際に亢進され、IFN−γ産生細胞数はIL−17中和に際して増加した。これらのデータは、IFN−γが、カンジダ症を有するマウスのIL−12応答に必要であるという概念と一致している。Cenciら(1998年)J. Immunol. 第161巻:3543頁。
【0076】
より重要なことだが、IL−12の産生はp19−/−DCにおいてより高く、IL−23の産生はp35−/−DCにおいてより高く、両サイトカインはWTDCにおいて交差制御された。これらの知見は、これらのサイトカインが、DC産生のレベルで相互に制御されることを示唆する。Beckerら(2006年)J. Immunol. 第177巻:2760頁。しかしながら、炎症性DCは、真菌に応答して免疫寛容原誘発性DCより多くのIL−23を産生すると考えられるため、これは、Th1/Th17のバランスが、様々な体内部位のDCサブセット間の相互制御にも依存するように現れる。
【0077】
危険性の高い炎症の状態において、真菌に応答して、すなわちTLR−/MyD88経路を介する高酵母数に応答した炎症性DCにより、IL−23が産生されるという知見は、重要な意味合いを有する。それは、炎症性の過程および真菌毒性間の重要な分子的結び付きとしてIL−23を指摘するだけでなく、悪循環がそれにより作用し得るというシナリオも確立する。p19−/−マウスがより少ないIL−17を産生し、TGF−βが真菌に対するTh17活性化および/または維持に非不可欠な役割を示したため、IL−23は、IL−17の近位媒介物質として作用することが考えられる。このシナリオでは、制御されない真菌増殖は、非防御的Th2細胞の同時活性化と関係する病原性Th17細胞の活性化を永続化する場合がある。
【0078】
この研究における1つの興味深い所見は、微生物刺激がIL−23分泌の主要な誘発因子であり得るが、適応免疫過程はその産生を調節することもできることであった。これを支持するものであるが、本発明者らは、DCによるIL−23分泌がT細胞の存在下で劇的に増加したという証拠を提供したが、この知見は、活性化T細胞がIL−23のさらなる誘導に正のフィードバックループを提供できることを示唆した。
【0079】
上記の考察は、共生生物または遍在性のいずれの真菌であっても、免疫恒常性およびその調節障害内に適応させるのに役立ち得る。IL−23/IL−17軸の活性化を介した炎症性プログラムを破壊する能力が、最終的に免疫調節障害に結び付き得る場合、CandidaまたはAspergillusのいずれかに対する防御免疫の不可欠で本質的な構成要素であるTreg細胞を活性化するそれらの能力(Romani & Puccetti(2006年))は、それにより制御されない免疫が防止される機序を代表し得る。この点に関しては、in vivoでのIL−17産生細胞の発生におけるIL−10の阻害的役割を含む、Th17細胞およびTreg細胞間の機能的拮抗作用が記述されている(Bettelli & Kuchroo(2005年)J. Exp. Med. 第201巻:169頁)。Kullbergら(2006年)J. Exp. Med. 第203巻:2485頁。したがって、Th17細胞およびTreg細胞の産生のための相互的経路は、真菌感染症でも起こり得る可能性がある。本発明者らは、感染後のp35−/−マウスにおけるCD4+CD25+Treg細胞活性化の証拠を見出しておらず、この知見は、Th17細胞およびTreg細胞が相互に排他的であることを示唆する。その代わりに、IL−10産生の著しい減少にかかわらず、CD4+CD25+Treg細胞がp19−/−マウスで観察され、それはT細胞によるIL−10産生を上方制御するIL−23の能力と一致する。Vanden Eijndenら(2005年)Eur. J. Immunol. 第35巻:469〜475頁。
【0080】
本研究の別の重要な所見は、IL−23/IL−17依存性経路が、IFN−γ欠損の場合、p35依存性経路を介していくつかの抗真菌耐性を提供できるということである。IL−12欠損の場合IL−23が防御的役割を果たすことができることは、慢性クリプトコックス症(Kleinschekら(2006年)J. Immunol. 第176巻:1098頁)で、マイコバクテリア感染症で(Khaderら(2005年)J. Immunol. 第175巻:788頁)で、および急性肺性Klebsiella pneumoniae感染症(Happelら(2005年)Infect. Immun. 第73巻:5782頁)で既に報告されており、防御は、IL−12p70と独立して抗原特異的IFN−γ産生CD4+T細胞を活性化するIL−23の能力、および病原体除去を媒介するPMNを動員するIL−23の能力と相関していた。Happelら(2005年)J. Exp. Med. 第202巻:761頁。実際のところ、実験的Helicobacter hepaticus誘導性大腸炎では、IL−23が、IFN−γ産生細胞およびIL−17産生細胞の両方を駆動することが明らかに示された。Kullbergら(2006年)。本発明者らの結果は、IL−23の作用におけるp35依存性経路を包含する、感染に際したTh1経路およびTh17経路間のさらなるレベルの交差制御を示唆すると考えられる。最終的には、IL−12依存性軸により支配された適切な免疫エフェクター機能の開始前に、初期炎症の危険シグナルを処理するIL−23の能力(McKenzieら(2006年))は、アンタゴニストだけでなくこのサイトカイン対間の協同的活性とも一致する。
【0081】
まとめると、この研究で提示されたデータは、真菌に対する宿主防御、免疫恒常性、および免疫の機序に重要な意味合いを有する、以前は未決定だったIL−23依存性Th17系列の真菌感染症における役割を実証する。さらに、それらは、炎症消散の失敗と抗真菌免疫耐性の欠如との間の分子的関連性を示す。この最新の結果は、炎症を限定して効果的な免疫応答を刺激することを試みる、真菌感染症の免疫治療の戦略を示唆する。
【0082】
IV.IL−23アンタゴニスト。
【0083】
IL−23アンタゴニストは、IL−23の1つまたは複数の生物活性を阻害可能な任意の物質または方法を含む。そのような活性には、IL−23(p19およびp40サブユニットを含む)、IL−23受容体(IL−23RおよびIL−12Rβ1サブユニットを含む)への結合、ならびにTh17細胞の促進および維持が含まれる。アンタゴニストには、たとえば小分子、抗体または抗体断片、ペプチド模倣物質、アプタマー(たとえば米国特許出願公開第2006−0193821号明細書に開示されているような)、IL−23受容体サブユニットの細胞外領域上に由来する可溶性受容体、および核酸に基づくアンタゴニストが含まれ得る。
【0084】
IL−23の核酸に基づくアンタゴニストには、アンチセンス核酸、およびIL−23p19遺伝子またはIL−23R遺伝子に対するsiRNAが含まれる。一般的なsiRNA方法論については、国際公開第2006/06060598号パンフレットを参照されたい。Arenz and Schepers(2003年)Naturwissenschaften第90巻:345頁;Sazani and Kole(2003年)J. Clin. Invest. 第112巻:481頁;Pirolloら(2003年)Pharmacol. Therapeutics第99巻:55頁;Wangら(2003年)Antisense Nucl. Acid Drug Devel. 第13巻:169頁も参照されたい。アンチセンスおよびsiRNA分子は、ヒトIL−23p19およびIL−23R mRNAの既知の配列に基づいて設計できる。ヒトIL−23p19のmRNAおよびアミノ酸配列は、それぞれ、GenBankアクセッション番号NM_016584およびNP_057668に見出される。ヒトIL−23RのcDNAおよびアミノ酸配列は、それぞれ、GenBankアクセッション番号AF461422およびAAM44229に見出される。本発明は、RNA干渉用の組成物も提供する。
【0085】
siRNAを生成および使用する方法は、たとえば米国特許第6,506,559号明細書(国際公開第99/32619号パンフレット);第6,673,611号明細書(国際公開第99/054459号パンフレット);第7,078,196号明細書(国際公開第01/75164号パンフレット);第7,071,311号明細書、およびPCT公報、国際公開第03/70914号パンフレット;国際公開第03/70918号パンフレット;国際公開第03/70966号パンフレット;国際公開第03/74654号パンフレット;国際公開第04/14312号パンフレット;国際公開第04/13280号パンフレット;国際公開第04/13355号パンフレット;国際公開第04/58940号パンフレット;国際公開第04/93788号パンフレット;国際公開第05/19453号パンフレット;国際公開第05/44981号パンフレット;国際公開第03/78097号パンフレットに開示されている。米国特許は、関連PCT公報と共に列挙されている。遺伝子抑制(gene silencing)および治療的処置にsiRNAを使用する典型的な方法は、PCT公報、国際公開第02/096927号パンフレット(VEGFおよびVEGF受容体);国際公開第03/70742号パンフレット(テロメラーゼ);国際公開第03/70886号パンフレット(タンパク質チロシンホスファターゼIVA型(Prl3));国際公開第03/70888号パンフレット(Chk1);国際公開第03/70895号パンフレット、および国際公開第05/03350号パンフレット(アルツハイマー病);国際公開第03/70983号パンフレット(タンパク質キナーゼCアルファ);国際公開第03/72590号パンフレット(MAPキナーゼ);国際公開第03/72705号パンフレット(サイクリンD);国際公開第05/45034号パンフレット(パーキンソン病)に開示されている。siRNAの治療的使用に関する典型的な実験は、Zenderら(2003年)Proc. Nat’l. Acad. Sci. (USA) 第100巻:7797頁;Paddisonら(2002年)Proc. Nat’l. Acad. Sci. (USA) 第99巻:1443頁;およびSah(2006年)Life Sci. 第79巻:1773頁にも開示されている。siRNA分子は、たとえば慢性骨髄性白血病(CML)(ClinicalTrials.gov識別番号:NCT00257647)および加齢黄斑変性(AMD)(ClinicalTrials.gov識別番号:NCT00363714)の臨床試験でも使用されている。
【0086】
「siRNA」という用語は、RNA干渉経路を介して遺伝子抑制を誘導するために使用される分子(Fireら(1998年)Nature 第391巻:806頁)を参照するために本明細書で使用されるが、そのようなsiRNA分子は、厳密にポリリボヌクレオチドである必要はなく、その代わり、治療薬としてのその特性を改良するために、核酸に対する1つまたは複数の改変を含有できる。そのような作用剤は、短い干渉核酸(short interfering nucleic acid)の代わりに「siNA」と呼ばれることがある。そのような変更により、この分子は、「リボ」ヌクレオチドの定義から正式には外れる場合があるが、それにもかかわらず本明細書では、そのような分子を「siRNA」分子と呼ぶ。RNA干渉経路を介して遺伝子抑制を誘導するために使用される他の核酸改変体には、たとえば、米国特許出願公開第20060115453号明細書で開示されているような短いヘアピン型RNA(short hairpin RNA)(「shRNA」)が含まれる。核酸に基づく阻害剤は、たとえば、プラスミドまたはウイルス(たとえば裸の(naked)DNAとして)などの組換えベクターを用いた形質転換により、または知られている遺伝子治療用ベクター(たとえばアデノ随伴ウイルス(AAV)、アデノウイルス、レトロウイルス、またはレンチウイルス)のいずれかを用いた遺伝子治療により送達できる。核酸は、形質転換、エレクトロポレーション、遺伝子銃、または当技術分野で知られている他の方法により送達できる。
【0087】
本発明の組成物および方法で使用するためのIL−23の抗体アンタゴニストには、IL−23に対する抗体およびIL−23受容体に対する抗体が含まれる。IL−23に対する典型的なアンタゴニスト抗体には、同一人に譲渡された米国特許仮出願第60/891,409号明細書および第60/891,413号明細書(両方とも2007年2月23日に出願された)、米国特許出願公開第2007−0009526号明細書および第2007−0048315号明細書、ならびに国際公開第2007/076524号パンフレット、国際公開第2007/024846号パンフレット、および国際公開第2007/147019号パンフレットに開示されているような、抗ヒトIL−23p19抗体およびその断片が含まれる。IL−23に対する抗体アンタゴニストには、そのサブユニットがIL−23p19に結合する場合にはIL−12p40サブユニットに結合する抗体も含まれるが、IL−12p35に結合する場合には含まれない。たとえば、米国特許出願公開第2005−0137385号明細書、米国特許第7,252,971号明細書を参照されたい。IL−23に対する典型的なアンタゴニスト抗体には、抗ヒトIL−23受容体抗体、たとえば抗IL−23R抗体およびその断片が含まれる。IL−23Rに対する典型的なアンタゴニスト抗体は、同一人に譲渡された米国特許仮出願第60/892,104号明細書(2007年2月28日に出願された)および第60/945,183号明細書(2007年6月20日に出願された)に開示されている。IL−23アンタゴニストには、二重特異性抗体も含まれる。
【0088】
抗原性の増加した領域は、抗体産生に使用できる。ヒトp19の抗原性が増加した領域は、たとえば、GenBank AAQ89442(gi:37183284)のアミノ酸16〜28位;57〜87位;110〜114位;136〜154位;および182〜186位に生じている。ヒトIL−23Rの抗原性が増加した領域は、たとえば、GenBank AAM44229(gi:21239252)のアミノ酸22〜33位;57〜63位;68〜74位;101〜112位;117〜133位;164〜177位;244〜264位;294〜302位;315〜326位;347〜354位;444〜473位;510〜530位;および554〜558位に生じている。解析は、Vector NTI(登録商標)Suite(Informax社、ベテスダ、米国メリーランド州)を使用したパーカープロット(Parker plot)により行った。本発明は、つまりIL−23Rの細胞外領域、たとえばGenBank AAM44229のアミノ酸1〜353位またはその断片を含む可溶性受容体であり、この細胞外領域またはその断片がIL−23に特異的に結合するIL−23アンタゴニストも提供する。マウスIL−23Rは、GenBank NP_653131(gi:21362353)である。ムテインおよび改変体、たとえばペグ化、または脱アミドするアスパラギン残基を欠失もしくは置換する変異誘発が企図される。
【0089】
本発明の方法および組成物で使用するための、IL−23活性に拮抗する追加的な考え得る方法には、線維状赤血球凝集素(FHA)に投与する工程(国際公開第2006/109195号パンフレット)、およびIL−23に対する免疫応答を生じさせるために予防接種する工程(国際公開第2005/058349号パンフレット)が含まれる。
【0090】
一実施形態では、IL−17産生(Th17)細胞のアンタゴニストは、たとえばナイーブT細胞、Th1型T細胞、TH2型T細胞、上皮細胞、および/または内皮細胞の活性に、たとえば実質的に影響を与えずに、Th17細胞の活性を特異的に調節する試薬を包含する。この試薬は、たとえば、IL−17産生細胞の転写因子(たとえばRORγt)または接着タンパク質の発現または活性を調節できる。さらに、IL−23、TGF−β、またはIL−6のアンタゴニストは、Th17細胞の生成および生存を減少でき、IL−17のアンタゴニストは、そのような細胞の炎症性効果(たとえば、好中球動員)を減少できる。
【0091】
モノクローナル抗体、ポリクローナル抗体、およびヒト化抗体は、調製可能である(たとえば、Sheperd and Dean(編)(2000年)Monoclonal Antibodies、Oxford Univ. Press、ニューヨーク、米国ニューヨーク州);Kontermann and Dubel(編)(2001年)Antibody Engineering、Springer−Verlag、ニューヨーク;Harlow and Lane(1988年)Antibodies A Laboratory Manual、Cold Spring Harbor Laboratory Press、コールドスプリングハーバー、米国ニューヨーク州、139〜243頁;Carpenterら(2000年)J. Immunol. 第165巻:6205頁;Heら(1998年)J. Immunol. 第160巻:1029頁;Tangら(1999年)J. Biol. Chem. 第274巻:27371頁;Bacaら(1997年)J. Biol. Chem. 第272巻:10678頁;Chothiaら(1989年)Nature 第342巻:877頁;Foote and Winter(1992年)J. Mol. Biol. 第224巻:487頁;米国特許第6,329,511号明細書を参照)。抗体配列の全体がヒト生殖細胞系列(germline)配列に由来する完全ヒト抗体も調製できる。そのような完全ヒト抗体は、ヒト免疫グロブリン遺伝子を発現するように遺伝子操作されたトランスジェニック動物から、またはファージディスプレイなどの方法により、調製できる。たとえば、Lonberg(2005年)Nature Biotechnol. 第23巻:1117頁;Vaughanら(1998年)Nature Biotechnol. 第16巻:535頁を参照されたい。抗体断片には、Fab、Fab’、Fab’−SH、Fv、単鎖Fv(scFv)、F(ab’)2、およびダイアボディが含まれる。Pluckthun(1994年)THE PHARMACOLOGY OF MONOCLONAL ANTIBODIES、第113巻、Rosenburg and Moore編、Springer−Verlag、ニューヨーク、269〜315頁;Holliger and Hudson(2005年)Nature Biotechnol. 第23巻:1126〜1136頁。
【0092】
本発明の抗体はまた、変化したエフェクター機能を提供するために改変(または遮断)Fc領域を有する抗体を含む。たとえば米国特許第5,624,821号;国際公開第2003/086310号;国際公開第2005/120571号;国際公開第2006/0057702号;Presta(2006年)Adv. Drug Delivery
Rev. 58:640〜656頁を参照されたい。そのような改変は、診断および治療での可能性のある有益な効果を伴って、免疫系の多様な反応を増強するまたは抑制するために使用することができる。Fc領域の改変は、アミノ酸変化(置換、欠失、および挿入)、糖化または糖鎖除去、ならびに複数のFcの付加を含む。Fcに対する変更はまた、治療用抗体中の抗体の半減期を変化させることもでき、より長い半減期により、投薬がそれほど頻繁ではなくなり、同時に利便性が増加し、材料の使用が減少する。エフェクター機能の変更は、IgG1のFc部分に特定の変異を導入することにより、たとえば、Asn297をたとえばAlaまたはGlnに変更すること(N297AまたはN297Q)により達成できる。Presta(2005年)J. Allergy Clin. Immunol. 第116巻:731頁の734〜35頁を参照されたい。エフェクター機能は、様々な定常ドメインの選択により変更することもできる。たとえば、本発明の抗体(または断片)の特定の使用目的が、エフェクター機能の変更を要求した場合、IgG1以外の重鎖定常ドメインが使用できる。IgG1抗体は、長期の半減期ならびに補体活性化および抗体依存性細胞傷害活性(ADCC)などのエフェクター機能を備えるが、そのような活性は、抗体のすべての用途にとって望ましいとは限らない場合がある。そのような場合には、たとえばIgG4定常ドメインが使用できる。一実施形態(唯一の実施形態ではない)では、目的がIL−23受容体を発現する細胞の殺滅を誘導することではなく、その代わり単にそのような細胞におけるIL−23シグナル伝達を阻止することであるため、エフェクター機能の変更は、IL−23受容体に対する(たとえばIL−23Rに対する)抗体の場合、特に関連性がある。この実施形態では、Th17系列からTh1系列へとTh細胞を転換することが目的であり、その場合には、細胞殺滅は非産生的である。
【0093】
抗原の精製は、抗体の産生に必要ではない。免疫化は、DNAベクター免疫化により実施でき、たとえば、Wangら(1997年)Virology 第228巻:278頁を参照されたい。あるいは、目的の抗原を保持する細胞で動物を免疫できる。その後、免疫された動物から脾細胞を単離することができ、脾細胞を骨髄腫細胞株と融合させてハイブリドーマを産生することができる(Meyaardら(1997年)Immunity 第7巻:283頁;Wrightら(2000年)Immunity 第13巻:233頁;Prestonら(1997年)Eur. J. Immunol. 第27巻:1911頁)。その結果生じるハイブリドーマは、機能アッセイまたはバイオアッセイ、すなわち精製された抗原の所有に依存しないアッセイにより、所望の抗体の産生のためにスクリーニングできる。細胞による免疫化は、精製された抗原を用いた免疫化より抗体産生に優れていることが証明できる(Kaithamanaら(1999年)J. Immunol. 第163巻:5157頁)。
【0094】
抗体−抗原結合特性およびリガンド−受容体結合特性は、たとえば、表面プラズモン共鳴(Karlssonら(1991年)J. Immunol. Methods 第145巻:229頁;Neriら(1997年)Nat. Biotechnol. 第15巻:1271頁;Jonssonら(1991年)Biotechniques 第11巻:620頁)、または競合ELISA(Friguetら(1985年)J. Immunol. Methods 第77巻:305頁;Hubble(1997年)Immunol. Today 第18巻:305頁)により、測定できる。抗体は、抗体の標的抗原および関連結合タンパク質を単離するために、親和性精製(affinity purification)用に使用できる。たとえば、Wilchekら(1984年)Meth. Enzymol. 第104巻:3頁を参照されたい。
【0095】
抗体は、通常は少なくとも約10−6M、典型的には少なくとも10−7M、より典型的には少なくとも10−8M、好ましくは少なくとも約10−9M、およびより好ましくは少なくとも10−10M、および最も好ましくは少なくとも10−11MのKDで結合する。たとえば、Prestaら(2001年)Thromb. Haemost. 第85巻:379頁;Yangら(2001年)Crit. Rev. Oncol. Hematol. 第38巻:17頁;Carnahanら(2003年)Clin. Cancer Res. (Suppl.) 第9巻:3982頁を参照されたい。
【0096】
IL−23Rの細胞外ドメインを含む可溶性受容体は、本発明の組成物および方法に有用である。可溶性受容体は、標準的方法により調製および使用できる。たとえば、Jonesら(2002年)Biochim. Biophys. Acta 第1592巻:251頁;Prudhommeら(2001年)Expert Opinion Biol. Ther. 第1巻:359頁;Fernandez−Botran(1999年)Crit. Rev. Clin. Lab Sci. 第36巻:165〜224頁を参照されたい。
【0097】
一実施形態では、本発明の組成物および方法は、IL−23に対する拮抗作用を必要とするが、IL−12に対する拮抗作用は必要としない。IL−23およびIL−12の活性を両方とも阻止するIL−12およびIL−23両方のIL−12p40サブユニットを標的とするいくつかの有望な治療薬が現在開発中である。そのような作用剤は、本発明が促進することを目的とする頑健なIL−12媒介性Th1応答を阻害するため、本発明の組成物および方法のこの実施形態における使用に好適ではない。IL−12との関連ではなくIL−23との関連においてのみIL−12p40に結合する作用剤を開発することは、原理的には可能であるが(米国特許出願公開第2005−0137385号明細書および米国特許第7,252,971号明細書を参照)、IL−12p40を標的とする大多数の作用剤はIL−12を阻害する可能性が高く、したがって本発明には好適ではない。同じ議論が、IL−23およびIL−12の共通受容体サブユニット、IL−12Rβ1に当てはまる。IL−12受容体との関連ではなくIL−23受容体との関連においてのみIL−12Rβ1に結合する作用剤を開発することは、原理的には可能であるが、IL−12Rβ1を標的とする大多数の作用剤はIL−12受容体を阻害する可能性が高く、したがって本発明には好適ではない。対照的に、IL−23またはその受容体に特異的なサブユニット、つまりそれぞれp19およびIL−23Rに結合し拮抗する作用剤は、IL−12ではなくIL−23の特異的阻害剤である可能性が高く、したがって本発明の組成物および方法における使用に、より好適である。
【0098】
有望な治療薬がIL−12ではなくIL−23を特異的に阻害するかどうかは、当技術分野で知られている任意の方法により決定できる。たとえば、有望なIL−23特異的アンタゴニストについては、その受容体に対するIL−23の結合またはその受容体に対するIL−12の結合を阻止するその能力を試験できる。そのような阻止アッセイは、溶液中で(たとえば、蛍光活性化セルソーティング(cell sorting)により)、または固体支持体上で(たとえば、酵素結合免疫吸着測定法−ELISAにより)実施できる。IL−23およびIL−12受容体阻止は、Ba/F3細胞増殖アッセイなどのバイオアッセイで測定することもできる。たとえば、Hoら(1995年)Mol. Cell. Biol.(1995年)第15巻:5043頁を参照されたい。そのような結合アッセイでは、有望なIL−23アンタゴニストの効力および特異性は、IC50として、すなわち所定の一組のアッセイ条件下で、IL−23結合(または結合に依存する生物活性)の50%低減を達成するのに必要なこの有望なアンタゴニストの濃度として表すことができる。より低いIC50は、より効果的なアンタゴニストを示す。有望なアンタゴニストのIL−23特異性は、IL−23とその受容体の結合の阻害に関するIC50に対する、IL−12とその受容体の結合の阻害に関するIC50の比率(IC50IL−12/IC50IL−23)として表すことができる。種々の実施形態では、有望なIL−23特異的アンタゴニストは、この比率(IC50IL−12/IC50IL−23)が、1.5、2、3、4、5、7、10、15、20、50、または100以上である場合、IL−23特異的と見なされる。好ましい実施形態では、阻害アッセイで使用されるIL−23およびIL−12のレベルは、IL−23およびIL−12アッセイの少なくとも1つ、好ましくはその両方が、直線的用量応答濃度範囲中で実施されることを保証するように調整される。
【0099】
IL−23およびIL−12は、拮抗作用の特異性を決定するために使用できる様々な生物学的機能も有する。IL−12とは対照的に、IL−23は、ヒトおよびマウスの両方のナイーブT細胞集団に対して優先的に記憶を刺激する。IL−23は、多数の細胞内細胞シグナル伝達分子、たとえばJak2、Tyk2、Stat1、Stat2、Stat3、およびStat4を活性化する。IL−12は、この同じ群の分子を活性化するが、IL−23に対するStat4応答は比較的弱く、その一方でIL−12に対するStat4応答は強い。Oppmannら(2000年);Parhamら(2002年)J. Immunol. 第168巻:5699頁。
【0100】
有望なIL−23特異的アンタゴニストについては、IL−12およびIL−23によりTh1およびTh17細胞の増幅および生存を阻害するその能力を試験することもできる。IL−23特異的アンタゴニストは、Th1細胞のIL−12媒介性増幅および生存ではなく、Th17細胞のIL−23媒介性増幅および生存を優先的に阻害する。Th17細胞はIL−17を特徴的に分泌する一方で、Th1細胞はIFN−γを特徴的に分泌する。典型的なTh1/Th17アッセイのデータは、Langrishら(2005年)J. Exp. Med. 第201巻:233頁の図2に見出され、IL−23がIL−17産生CD4+T細胞の増幅および生存を促進する一方で、IL−12はIFN−γ産生CD4+T細胞の増幅および生存を促進することを実証する。本発明の一実施形態では、作用剤は、Th17細胞のIL−23媒介性増幅および生存を阻害できるが、Th1細胞のIL−12媒介性増幅および生存を阻害しない場合に、「IL−23特異的」アンタゴニスト(IL−12に対して)であると見なされる。Th17/Th1細胞増殖の阻害は、IC50として、すなわち所定の一組のアッセイ条件下で、IL−17またはIFN−γを産生する特定のT細胞サブセットの増幅および生存の促進におけるIL−23活性の50%低減を達成するのに必要な作用剤の濃度として表すことができる。典型的なアッセイは、実施例13で提供される。実施例13に記載したアンタゴニストのように、バイオアッセイにおけるIL−23アンタゴニストの効力は、IC50IL−23として、つまりIL−23活性をその非阻害値の50%に低減するのに必要なアンタゴニストの濃度として表すことができる。類似したIC50IL−12は、IFN−γ産生細胞の産生促進におけるIL−12およびその活性で決定できる。その後、アンタゴニストのIL−23特異性は、比率IC50IL−12/IC50IL−23として表すことができる。種々の実施形態では、確認した(validated)IL−23特異的アンタゴニストのIC50IL−12/IC50IL−23比率は、1.5、2、3、4、5、7、10、15、20、50、または100以上である。
【0101】
Langrishら(2005年)に本質的に記載されているように、IL−17AおよびIFN−γの産生は、サイトカインに結合する蛍光性試薬を用いた蛍光活性化セルソーティング(FACS(登録商標)解析)による細胞内サイトカインの流動細胞計測法(flow cytometry)により測定できる。その細胞が「IL−17産生」または「IFN−γ産生」と見なされるための、生のCD4+T細胞中のIL−17AまたはIFN−γの閾値レベルを定義することが重要である。一実施形態では、閾値レベルは、生のCD4+T細胞の5%が、未処置細胞の対照試料中で「IL−17産生」または「IFN−γ産生」であるレベルと定義される。典型的な未処置細胞には、プロテオリピドタンパク質(PLP)で免疫されたSJLマウス(The Jackson Laboratories社、バーハーバー、メーン州、米国)から単離され、PLPの存在下で培養された流入領域リンパ節(DLN)細胞が含まれる。
【0102】
V.組成物および方法。
【0103】
IL−23のアンタゴニストを含む医薬組成物または無菌組成物を調製するために、アンタゴニストは、薬学的に許容可能な担体または賦形剤と混合される。たとえばRemington’s Pharmaceutical Sciences and U.S. Pharmacopeia: National Formulary、Mack Publishing Company、Easton、PA(1984年)を参照されたい。治療薬の処方は、生理学的に許容可能な担体、賦形剤、または安定剤と混ぜることによって、たとえば凍結乾燥粉末、スラリー、水性溶液、または懸濁液の形状で調製されてもよい(たとえばHardmanら(2001年)Goodman and Gilman’s The Pharmacological Basis of Therapeutics、McGraw−Hill、New York、NY;Gennaro(2000年)Remington: The Science and Practice of Pharmacy、Lippincott, Williams, and Wilkins、New York、NY;Avisら(編)(1993年)Pharmaceutical Dosage Forms: Parenteral Medications、Marcel Dekker、NY;Liebermanら(編)(1990年)Pharmaceutical Dosage Forms: Tablets、Marcel Dekker、NY;Liebermanら(編)(1990年)Pharmaceutical Dosage Forms: Disperse Systems、Marcel Dekker、NY;WeinerおよびKotkoskie(2000年)Excipient Toxicity and Safety、Marcel
Dekker, Inc.、New York、NYを参照されたい)。
【0104】
投与経路は、たとえば局所的もしくは皮膚塗布、静脈内注射または注入、腹腔内、脳内、筋肉内、眼内、動脈内、脳脊髄内、病巣内、もしくは肺性の経路によるか、または徐放性システムもしくは埋没物(implant)による。遺伝子移入ベクターの中枢神経系への注射は記述されている。たとえば、Cuaら(2001年)J. Immunol.
第166巻:602頁;Sidmanら(1983年)Biopolymers 第22巻:547頁;Langerら(1981年)J. Biomed. Mater. Res. 第15巻:167頁;Langer(1982年)Chem. Tech. 第12巻:98頁;Epsteinら(1985年)Proc. Natl. Acad. Sci. USA 第82巻:3688頁;Hwangら(1980年)Proc. Natl. Acad. Sci. USA 第77巻:4030頁;米国特許第6,350,466号明細書および第6,316,024号明細書を参照されたい。
【0105】
治療剤に対する投与計画の選択は、作用物質の血清代謝回転速度または組織代謝回転速度、症状のレベル、作用物質の免疫原性、および生物学的マトリックス中での標的細胞の到達性を含むいくつかの要因に依存する。好ましくは、投与計画は、許容可能なレベルの副作用と調和した、患者に送達される治療剤の量を最大限にする。したがって、送達される作用物質の量は、部分的に、特定の実体および処置されている状態の重症度に依存する。抗体、サイトカイン、および小分子の適正用量の選択の手引きが入手可能である。たとえばWawrzynczak(1996年)Antibody Therapy、Bios Scientific Pub. Ltd、Oxfordshire、UK;Kresina(編)(1991年)Monoclonal Antibodies, Cytokines and Arthritis、Marcel Dekker、New York、NY;Bach(編)(1993年)Monoclonal Antibodies and Peptide Therapy in Autoimmune Diseases、Marcel Dekker、New York、NY;Baertら(2003年)New Engl. J. Med. 348:601頁;Milgromら(1999年)New Engl. J. Med. 341:1966頁;Slamonら(2001年)New Engl. J. Med. 344:783頁;Beniaminovitzら(2000年)New Engl. J. Med. 342:613頁;Ghoshら(2003年)New Engl. J. Med. 348:24頁;Lipskyら(2000年)New Engl. J. Med. 343:1594頁を参照されたい。
【0106】
抗体、抗体断片、およびサイトカインは、持続注入により、またはたとえば1日、1週、または1週当たり1〜7回の間隔での投与により提供できる。投与は、静脈内に、皮下に、局所的に、経口的に、経鼻的に、直腸的に、筋肉内に、脳内に、脊髄内に、または吸入により提供できる。種々の実施形態では、投与方法は、たとえば肺または胃腸管(GI
tract)など、感染の原発部位に基づいて選択される。
【0107】
好ましい投与プロトコルは、著しい望ましくない副作用を回避する最大用量または投与頻度を含むプロトコルである。週用量の合計は、一般的に少なくとも約0.05μg/kg、0.2μg/kg、0.5μg/kg、1μg/kg、10μg/kg、100μg/kg、0.2mg/kg、1.0mg/kg、2.0mg/kg、10mg/kg、25mg/kg、または50mg/kgである。たとえば、Yangら(2003年)New Engl. J. Med. 第349巻:427頁;Heroldら(2002年)New Engl. J. Med. 第346巻:1692頁;Liuら(1999年)J. Neurol. Neurosurg. Psych. 第67巻:451頁;Portieljiら(2003年)Cancer Immunol. Immunother. 第52巻:133頁を参照されたい。小分子治療薬、たとえばペプチド模倣物質、天然産物、または有機化学物質の望ましい用量は、抗体またはポリペプチドとほぼ同じで、モル/kg(mole/kg)程度である。
【0108】
特定の患者についての有効量は、処置されている状態、患者の健康全般、投与の方法、経路、および用量、ならびに副作用の重症度などの要因に依存して変動し得る。たとえば、Maynardら(1996年)A Handbook of SOPs for Good Clinical Practice、Interpharm Press、ボーカラトーン、米国フロリダ州;Dent(2001年)Good Laboratory and Good Clinical Practice、Urch Publ.、ロンドン、英国を参照されたい。
【0109】
典型的な獣医学的被験体、実験被験体、または研究被験体は、サル、イヌ、ネコ、ラット、マウス、ウサギ、モルモット、ウマ、およびヒトを含む。
【0110】
適正用量の決定は、たとえば、処置に影響することが当技術分野で知られているもしくは疑われるまたは処置に影響することが予測されるパラメーターまたは要因を使用して、臨床医によってなされる。一般に、用量は、やや最適用量未満の量から始まり、あらゆる負の副作用と比較して、所望または最適の効果が達成されるまで、用量は、その後、少量ずつ増加させることによって増加する。重要な診断尺度には、たとえば感染または感染レベルの徴候の尺度が含まれる。好ましくは、使用される生物製剤は、処置目標の動物と同一種に由来するか、または同一種に由来するタンパク質を模倣するように改変されており(たとえば、ヒト化抗体)、それにより試薬に対する体液性応答を最小限に抑える。
【0111】
第2の治療薬、たとえばサイトカイン、ステロイド、化学療法剤、抗生物質、または放射線との同時投与または同時処置のための方法は、当技術分野でよく知られている。たとえばHardmanら(編)(2001年)Goodman and Gilman’s
The Pharmacological Basis of Therapeutics、第10版、McGraw−Hill、New York、NY;PooleおよびPeterson(編)(2001年)Pharmacotherapeutics for Advanced Practice: A Practical Approach、Lippincott, Williams & Wilkins、Phila.、PA;ChabnerおよびLongo(編)(2001年)Cancer Chemotherapy and Biotherapy、Lippincott, Williams & Wilkins、Phila.、PAを参照されたい。治療剤の有効量は、典型的には少なくとも10%、一般に少なくとも20%、好ましくは、少なくとも約30%、より好ましくは、少なくとも40%、および最も好ましくは、少なくとも50%、症状を減少させる。
【0112】
本発明は、それらに限定されないが、真菌感染症、持続性真菌感染症、カンジダ症、CMC、アスペルギルス症、クリプトコックス症、ウイルス感染症、持続性ウイルス感染症、HIV感染、HBV感染、HCV感染、細菌感染症、マイコバクテリア感染症、M.tuberculosis感染、M.bovis感染、およびM.leprae感染からなる群から選択される症状を含む感染症を処置するための薬剤の製造における、IL−23またはIL−23R、あるいはその両方のアンタゴニストの使用をさらに提供する。いくつかの実施形態では、薬剤は、1つまたは複数の追加的な治療薬を含むことができる。他の実施形態では、本発明の薬剤は、1つまたは複数の他の治療薬と併せて使用できる。
【0113】
VI.抗イディオタイプ抗体。
【0114】
本発明は、本発明の治療用抗IL−23または抗IL−23R抗体に対する抗イディオタイプ抗体をさらに提供する。抗イディオタイプ抗体とは、一般的には別の抗体の抗原結合領域に結合する、固有の決定基を認識する抗体である。抗イディオタイプ抗体は、抗IL−23抗体またはそのCDR含有領域を有する元の抗IL−23抗体の供給源と同一の種および遺伝子型(たとえば、マウス株)の動物を免疫することにより調製できる。その後、免疫された動物は、免疫する抗体のイディオタイプ決定基に対する抗体を生成して、抗イディオタイプ抗体を産生する。抗イディオタイプ抗体は、さらに別の動物に免疫応答を誘導する免疫原としても使用でき、いわゆる「抗抗Id抗体(anti−anti−Id antibody)」を産生する。
【0115】
たとえば、抗イディオタイプ抗体は、被験体における、たとえば抗IL−23治療を受けている被験体の体液(たとえば血液)における、治療用抗IL−23(または抗IL−23R)抗体のレベルを決定するために使用できる。被験体における抗IL−23(または抗IL−23R)抗体レベルの決定は、そのような決定に応じて用量が変更され得るため、被験体における抗IL−23抗体の望ましいレベルを維持するのに有用であり得る。望ましい範囲の値内の抗IL−23抗体の循環レベルを得るために、用量を増加または減少させることができる(頻度および/または1回の投与当たりの量において)。望ましい範囲は、当技術分野で典型的な方法により医師によって決定することができ、抗IL−23(または抗IL−23R)抗体またはその断片に関する治療指数に依存する場合がある。
【0116】
抗イディオタイプ抗体は、ポリペプチドタグ(たとえば、FLAG(登録商標)タグ)を有する抗体、または色素、同位体、酵素、および金属に結合された抗体を含む、簡易検出に好適な形状で供給できる。たとえば、Le Doussalら(1991年)New Engl. J. Med. 第146巻:169頁;Gibelliniら(1998年)J. Immunol. 第160巻:3891頁;Hsing and Bishop(1999年)New Engl. J. Med. 第162巻:2804頁;Evertsら(2002年)New Engl. J. Med. 第168巻:883頁を参照されたい。放射性免疫測定法(RIA)、ELISA、およびラボオンチップ(lab on a chip)などの種々のアッセイ形式が存在する。米国特許第6,176,962号明細書および第6,517,234号明細書。
【0117】
VII.キット。
【0118】
さらに、本発明は、慢性の細菌感染症、マイコバクテリア感染症、ウイルス感染症、および真菌感染症などの感染症を患っている被験体(ヒトまたは非ヒト)の処置に使用するためのキット中にIL−23アンタゴニストを提供する。一実施形態では、キットは、IL−23アンタゴニストを含有するための区画、IL−23アンタゴニスト自体(抗体など)、および自由選択的に使用説明書、1つまたは複数の追加的な治療薬(複数可)、および1つまたは複数の投与用医療機器(たとえば、RedipenTMインジェクターデバイスなどの注射器または使い捨てインジェクター)を含む。IL−23アンタゴニストは、本明細書に記載された作用剤のいずれかであってよく、それらに限定されないが、抗p19抗体またはそのp19結合性断片、抗IL−23R抗体またはそのIL−23R結合性断片、または可溶性IL−23R断片を含む。
【0119】
1つまたは複数の追加的な治療薬には、それらに限定されないが、非ステロイド系抗炎症剤(NSAID)、ステロイド、IL−12またはそのアゴニスト、およびIL−17A、IL−17F、TGF−β、IL−6などのサイトカインまたはそれらのそれぞれの受容体のアンタゴニストが含まれる。サイトカインのアンタゴニストには、サイトカイン、そのサブユニット、またはその受容体に結合する抗体が含まれる。サイトカインまたはそれらの受容体に結合する抗体は、すべてが必ずしもアンタゴニストとは限らないが、そのようなアンタゴニスト活性は、バイオアッセイまたは受容体結合アッセイなどの当技術分野で一般的に知られている技術により容易にアッセイできる。IL−17A(NM_002190、NP_002181)、IL−17F(NM_052872、NP_443104);IL−17RA(NM_014339、NP_055154);IL−17RC(転写改変体NM_153461、NM_153460、NM_032732、およびそれらのそれぞれの翻訳体)を含む、種々の(ヒト)サイトカインおよび受容体の核酸およびアミノ酸配列は知られている。
【0120】
本発明は、本発明の治療用抗IL−23(または抗IL−23R)抗体に対する抗イディオタイプ抗体を含むキットをさらに提供する。一実施形態では、キットは、抗イディオタイプ抗体を含有するための区画、抗イディオタイプ抗体自体、および自由選択的に使用説明書、1つまたは複数の検出用試薬、抗イディオタイプ抗体検出用の1つまたは複数の機器(マイクロタイタープレートなど)、および検出する抗IL−23抗体の1つまたは複数の試料(または他の陽性対照)を含む。
【0121】
VIII.使用
結核の発症前には、長期の無症候性の症状発症前の期間が存在することが多い。したがって、IL−23およびIL−23Rアンタゴニスト治療は、種々のTB診断マーカーの解析のときに開始できる。正常な非感染患者と比較して、陽性のツベルクリン検査またはマントー試験(たとえば、Dale and Federman(2002年)を参照)を示す患者には、IL−23またはIL−23Rアンタゴニスト治療を実施して、マイコバクテリアのさらなる増殖を防止または既存の非病理学的感染を除去することができる。生体試料、たとえばBALに高レベルのマイコバクテリアを有する患者も、肺中のマイコバクテリアのさらなる増殖を防止し細菌負荷を除去するためのIL−23およびIL−23Rアンタゴニスト治療から恩恵を受けることができる。同様の処置は、臨床試料中のマイコバクテリアDNAまたはRNAのレベルが高い患者または培養物中のナイアシン検査が陽性である患者に使用できる。細菌負荷を軽減または除去するために、病理学的に症候的なTB感染と併せたIL−23およびIL−23Rアンタゴニストの使用も企図される。
【0122】
本発明の方法および組成物を使用して処置できる細菌感染症には、それらに限定されないが、以下により引き起こされる感染症が含まれる:
【0123】
【化1】

【0124】
【化2】
本発明の方法および組成物を使用して処置できるマイコバクテリア感染症には、それらに限定されないが、以下により引き起こされる感染症が含まれる:
【0125】
【化3】
本発明の方法および組成物は、これらに限定されないが、ヒストプラスマ症、コクシジオイデス真菌症、ブラストミセス症、アスペルギルス症、ペニシリウム症、カンジダ症、およびクリプトコックス症を含む真菌症状を処置するためにも使用できる。真菌症のリスク因子には、血液および骨髄移植、実体のある臓器移植、大手術(特に胃腸管手術)、AIDS、新生物疾患、高齢、免疫抑制治療、ならびに乳幼児の未成熟が含まれる。
【0126】
本発明の方法および組成物を使用して処置できる感染症(および臨床症候群(clinical syndrome))を引き起こす真菌病原体には、これらに限定されないが、Candida albicans(鵞口瘡、膣カンジダ症、食道カンジダ症)、Cryptococcus neoformans(髄膜炎)、Histoplasma capsulatum(発熱および体重喪失を伴う播種性感染症)、Coccidioides immitis(散在性および局限性肺疾患)、Blastomyces dermatitidis(髄膜炎を含む、局所的肺疾患および播種性感染症)、Aspergillus fumigatus(発熱、咳、および喀血を伴う肺疾患)およびPenicillium marneffei(発熱のみ、または肺浸潤、リンパ節症、もしくは皮膚病変を伴う)が含まれる。本発明の方法および組成物は、Candida種属、C.glabrata、C.parapsilosis、C.tropicalis、C.krusei、C.lusitaniae、C.guilliermondii、およびC.rugosaによる感染症を処置するためにも使用できる。前述の真菌病原体(および臨床症候群)は、一般的にHIV感染に付随する。
【0127】
本発明の方法および組成物は、C.glabrata、C.parapsilosis、C.tropicalis、C.krusei、C.lusitaniae、C.guilliermondii、およびC.rugosaなどのCandida種属による感染症を処置するためにも使用できる。本発明の方法および組成物は、A.flavus、A.niger、A.ustus、およびA.terreusなどのAspergillus種属による感染症を処置するためにも使用できる。追加的な真菌病原体には、Fusarium種属(たとえば、F.moniliforme、F.solani、F.oxysporum)およびScedosporium種属(たとえば、S.apiosperum、S.prolificans)が含まれる。追加的な真菌疾患には、Rhizopus(たとえば、R.oryzae、R.arrhizus)、Rhizomucor、Absidia、Cunninghamellaの種属により引き起こされる接合真菌症が含まれる。
【0128】
IL−23およびIL−23Rのアンタゴニストは、単独で、あるいはTh1応答を増強する目的の作用剤(たとえば、IL−12またはそのアゴニスト)、もしくはTh17応答を阻害する目的の作用剤(たとえば、TGF−βアンタゴニスト;IL−6アンタゴニスト;IL−17Aおよび/またはIL−17Fアンタゴニスト)、またはその両方の作用剤と共に使用できる。これらのサイトカインの受容体のアゴニストおよびアンタゴニストも使用できる。そのような作用剤には、抗体およびその抗原結合性断片、小分子、siRNA、およびアンチセンス核酸が含まれ得る。IL−23およびIL−23Rのアンタゴニストは、コルチコステロイド、たとえばプレドニゾンなどの抗炎症薬と共に使用することもできる。
【0129】
IL−17アンタゴニストは、IL−17A、IL−17F、IL−17RA、またはIL−17RCの発現を阻害することができるか、または機能的なリガンド−受容体相互作用を防止するために、これらのポリペプチドの1つまたは複数と直接的にまたは間接的に相互作用することによりIL−17シグナル伝達を阻害できる。いくつかの好ましい実施形態では、IL−17アンタゴニストは、IL−17A、IL−17F、IL−17RA、またはIL−17RCのいずれかに結合してその活性を阻害する抗体または抗体断片である。1つの特に好ましい実施形態では、IL−17アンタゴニストは、IL−17Aに特異的に結合するモノクローナル抗体である。IL−17Aに対する典型的なアンタゴニスト抗体には、同一人に譲渡された米国特許出願第11/836,318号明細書(2007年8月9日に出願された)、および国際公開第2006/013107号パンフレットおよび国際公開第2006/054059号パンフレットに開示されている抗ヒトIL−17A抗体およびそれらの断片が含まれる。別の実施形態では、IL−17アンタゴニストには二重特異性抗体が含まれる。
【0130】
一実施形態では、IL−23アンタゴニストには、IL−23に結合しその活性を阻害する二重特異性抗体が含まれる。そのような二重特異性抗体は、IL−23p19またはIL−23Rに結合でき、IL−17A、IL−17F、IL−17RA、IL−17RCにも結合できる。他の実施形態では、IL−23アンタゴニストは、IL−23p19およびIL−17に結合しIL−23およびIL−17の活性を阻害する二重特異性抗体である。たとえば、国際公開第2007/147019号パンフレットを参照されたい。あるいは、IL−23およびIL−17アンタゴニスト二重特異性抗体は、アンタゴニスト抗体である限り、それぞれIL−23受容体(たとえばIL−23R)またはIL−17受容体(IL−17RAまたはIL−17RC)のいずれかに結合できる。IL−17およびIL−23活性の両方に拮抗する二重特異性抗体は、当技術分野で知られている任意の技術により産生することができる。たとえば、二重特異性抗体は、2つの免疫グロブリン重鎖/軽鎖対の共発現を使用して組換え的に産生することができる。たとえば、Milsteinら(1983年)Nature 第305巻:537〜39頁を参照されたい。あるいは、二重特異性抗体は、化学的結合を使用して調製することができる。たとえば、Brennanら(1985年)Science 第229巻:81頁を参照されたい。二機能性抗体(bifunctional antibody)は、ジスルフィド交換、ハイブリッドハイブリドーマ(クアドローマ(quadroma))の産生により、二重特異性抗体を具現化する単一のポリペプチド鎖を産生するための転写および翻訳により、または二重特異性抗体を産生するために共有結合で結合され得る複数のポリペプチド鎖を産生するための転写および翻訳によっても調製できる。企図された二重特異性抗体は、全化学合成により作製することもできる。二重特異性抗体は、2つの異なる可変領域、2つの異なる定常領域、可変領域および定常領域、または他の改変を含むことができる。
【0131】
IL−23およびIL−23Rのアンタゴニストは、単独で使用し、またはイソニアジド、リファンピン、ピラジナミド、エタンブトール、ストレプトマイシン、シプロフロキサシン、およびオフロキサシンなどの既知の抗菌物質と共投与することができる。追加的な抗菌物質には、これらに限定されないが、アラトロフロキサシン(alatrofloxacin)、アジスロマイシン、バクロフェン、ベンザチンペニシリン、シノキサシン、クラリスロマイシン、クロファジミン、クロキサシリン、デメクロサイクリン、ジリスロマイシン、ドキシサイクリン、エリスロマイシン、エチオナミド、フラゾリドン、グレパフロキサシン、イミペネム、レボフロキサシン、ロレフロキサシン(lorefloxacin)、モキシフロキサシンHCl、ナリジキシン酸、ニトロフラントイン、ノルフロキサシン、オフロキサシン、リファブチン、リファペンチン、スパルフロキサシン、スピラマイシン、スルファベンズアミド(sulphabenzamide)、スルファドキシン、スルファメラジン、ウルファセタミド(ulphacetamide)、スルファジアジン、スルファフラゾール(sulphafurazole)、スルファメトキサゾール、スルファピリジン、テトラサイクリン、トリメトプリム、トロバフロキサシン、およびバンコマイシンが含まれる。
【0132】
本発明の方法および組成物は、これらに限定されないが、HBV、HCV、HIV、ヒトパピローマウイルス(HPV)により引き起こされる感染症を含む、持続性のウイルス感染症を処置するために使用できる。そのような慢性感染症は、感染を根絶するための免疫応答の失敗を表す。IL−23およびIL−23Rのアンタゴニストは、単独で、またはこれらに限定されないが、アバカビル、アシクロビル、アマンタジン、アンプレナビル、デラビルジン、ジダノシン、エファビレンツ、ファムシクロビル、インジナビル、インターフェロンアルファ、リバビリン、ラミブジン、ネルフィナビル、ネビラピン、オセルタミビル、ペンシクロビル、リバビリン、リトナビル、サキナビル、スタブジン、バラシクロビル、ザルシタビン、ザナミビル、ジドブジン(アジドデオキシチミジン、AZT)を含む他の抗ウイルス物質と併せて使用できる。好ましいインターフェロンアルファ剤には、ペグ化インターフェロンアルファ2aおよびペグ化インターフェロン2bが含まれる。インターフェロンアルファの典型的な形状は、米国特許第6,923,966号明細書で考察されている。IL−23アンタゴニストは、慢性HCV感染用のHCVプロテアーゼまたはHCVポリメラーゼ阻害剤、および慢性HIV感染用のCCR5アンタゴニストなどのウイルス特異的作用剤と組み合わせて使用することもできる。
【0133】
IL−23およびIL−23Rのアンタゴニストは、治療用ワクチン、たとえばHIV感染用のgp120枯渇全殺滅ウイルス、HCV感染用の組換えE1タンパク質、ならびにHPV感染用のウイルスE6およびE7腫瘍性タンパク質と併せて使用することもできる。Berzofskyら(2004年)を参照されたい。そのような治療用ワクチンには、DNAワクチンまたはウイルスベクターが含まれ、自由選択的には、ウイルスベクターワクチンがDNAワクチン後になされる異種性の初回抗原刺激および追加免疫投与計画で投与される。Berzofskyら(2004年)。
【0134】
IL−23およびIL−23Rのアンタゴニストは、単独で、またはこれらに限定されないが、ポサコナゾール、フルコナゾール(米国特許第4,404,216号明細書)、ボリコナゾール、イトラコナゾール(米国特許第4,267,179号明細書)、ケトコナゾール(米国特許第4,144,346号明細書および第4,223,036号明細書)、リアロゾール、イルテマゾール、クロトリマゾール、ミコナゾール、エコナゾール、ブトコナゾール、オキシコナゾール、スルコナゾール、チオコナゾール、およびテルコナゾール、置換チアゾール、チアジアゾール、オキサジアゾール、カスポファンギン、アムホテリシンB、ナイスタチン、ピマリシン、フルシトシン(5−フルオロシトシン)、ナフチフィン、テルビナフィン、ブテナフィン、チオカーボネートトルナフテート(thiocarbonate tolnaftate)、グリセオフルビン、アミオダロン、シクロピロックス、スルベンチン、アモロルフィン、クリオキノール、ゲンチアナバイオレット、ヨウ化カリウム、チオ硫酸ナトリウム、石炭酸フクシン溶液、およびエキノキャンディン(たとえば、カスポファンギンアセテート、ミカファンギン、およびアニデュラファンギン)を含む他の抗真菌物質と併せて使用できる。
【0135】
本発明のIL−23およびIL−23Rアンタゴニストは、単一の作用剤として使用される場合はそれらの通常用量で、または薬物が併用される場合は効果に何らかの相乗的増強があればより低用量で、標準的抗真菌物質と組み合わせて使用できる。フルコナゾールは、たとえば、400〜800mg/日で投与できる。ボリコナゾールは、1日2回4mg/kgで投与できる。イトラコナゾールは、200〜600mg/日で投与できる。アムホテリシンBデゾキシコレート(D−AmB)は、0.5〜1mg/kg/日で投与できる。本発明の組成物および方法と組み合わせ得る作用剤および処置計画のタイプに関する一般的なガイダンスは、米国感染症学会(IDSA)により刊行された診療ガイドラインのPappasら(2004年)Clin. Infect. Dis. 第38巻:161頁(カンジダ症)、Stevensら(2000年)Clin. Infect. Dis. 第30巻:696頁(アスペルギルス症)に見出すことができる。結核処置の診療ガイドラインは、2006年3月22日に刊行され、IDSAにより承認された結核医療の国際基準(International Standards for Tuberculosis Care)に見出される。
【0136】
本発明のいくつかの実施形態では、感染症を有する被験体、または感染症を有する疑いのある被験体は、他の方法または組成物(つまり、本発明の方法または組成物ではない)を使用して以前に感染症の処置を受けたことがある被験体である。以前の処置には、本明細書に開示された抗菌物質、抗生物質、抗真菌物質、抗ウイルス物質、または任意の他の処置方法もしくは組成物のいずれかでの処置が含まれ得る。
【0137】
いくつかの実施形態では、被験体は感染症の正式な診断を受け、自由選択的には病因物質が同定されているが、他の実施形態では、被験体は、正式な診断を受けていないか、または病因物質が限定されたが完全には同定されていない部分診断を受けている場合がある。他の実施形態では、被験体は、感染症を有する疑いのあるだけである。他の実施形態では、被験体は、感染症を有するまたは感染症に罹るリスクがあり、たとえば被験体は、免疫抑制治療を受けており、AIDSなどが原因で真菌感染症に罹るリスクがある。いくつかの実施形態では、感染症を有する被験体、感染症を有する疑いのある被験体、または感染症を有するもしくは感染症に罹るリスクのある被験体は、たとえばAIDS、化学療法、移植、高齢のため免疫無防備状態のになっている。
【0138】
当業者であれば明白であるが、本発明の趣旨および範囲から逸脱することなく、本発明の改変および変更を多数行うことができる。本明細書に記載された特定の実施形態は例示目的でのみ提示されており、本発明は、添付の特許請求の範囲の用語により制限されると共に、そのような特許請求の範囲に与えられる等価物の包括的範囲により制限され、本発明は、例示目的で本明細書に提示された特定の実施形態により制限されるものではない。
【実施例】
【0139】
(実施例1)
一般的な方法
分子生物学における標準的な方法を記載する(Maniatisら、(1982年)Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、NY;Sambrook and Russell(2001年)Molecular Cloning、第3版、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、NY;Wu(1993年)Recombinant DNA、第217巻、Academic Press、San Diego、CA)。標準的な方法はAusbelら、(2001年)Current Protocols in Molecular Biology、第1〜4巻、John Wiley and Sons、Inc.New York、NY中にも載っており、これは細菌細胞におけるクローニングおよびDNA変異誘発(第1巻)、哺乳動物細胞および酵母におけるクローニング(第2巻)、複合糖質およびタンパク質発現(第3巻)、およびバイオインフォマティクス(第4巻)を記載している。
【0140】
免疫沈降、クロマトグラフィー、電気泳動、遠心分離、および結晶化を含めたタンパク質精製のための方法を記載する。Coliganら、(2000年)Current Protocols in Protein Science、第1巻、John Wiley and Sons、Inc.、New York。化学分析、化学改変、翻訳後改変、融合タンパク質の生成、タンパク質のグリコシル化を記載する。たとえば、Coliganら、(2000年)Current Protocols in Protein Science、第2巻、John Wiley and Sons、Inc.、New York;Ausubelら、(2001年)Current Protocols in Molecular Biology、第3巻、John Wiley and Sons、Inc.、NY、NY、16.0.5〜16.22.17頁;Sigma−Aldrich、Co.(2001年)Products for Life Science Research、St.Louis、MO;45〜89頁;Amersham Pharmacia Biotech(2001年)BioDirectory、Piscataway、N.J.384〜391頁を参照。ポリクローナルおよびモノクローナル抗体の産生、精製、および断片化を記載する。Coliganら、(2001年)Current Protocols in Immunology、第1巻、John Wiley and Sons、Inc.、New York;Harlow and Lane(1999年)Using Antibodies、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、NY; Harlow and Lane(1998年)。リガンド/受容体相互作用を特徴付けするための標準的な技法が利用可能である。たとえば、Coliganら、(2001年)Current Protocols in Immunology、第4巻、John Wiley Inc.、New Yorkを参照。
【0141】
蛍光活性化セルソーティング(FACS)を含めた、フローサイトメトリーのための方法が利用可能である(たとえば、Owensら、(1994年)Flow Cytometry Principles for Clinical Laboratory Practice、John Wiley and Sons、Hoboken、NJ;Givan(2001年)Flow Cytometry、第2版;Wiley−Liss、Hoboken、NJ;Shapiro(2003年)Practical Flow Cytometry、John Wiley and Sons、Hoboken、NJ中を参照)。たとえば診断用試薬として使用するための、核酸プライマーおよびプローブ、ポリペプチド、および抗体を含めた核酸を改変するのに適した蛍光試薬が利用可能である(Molecular Probes(2003年)Catalogue、Molecular Probes、Inc.、Eugene、OR;Sigma−Aldrich(2003年)Catalogue、St.Louis、MO)。
【0142】
免疫系の組織学の標準的な方法を記載する。たとえば、Muller−Harmelink(ed.)(1986年)Human Thymus:Histopathology and Pathology、Springer Verlag、New York、NY;Hiattら、(2000年)Color Atlas of Histology、Lippincott、Williams、and Wilkins、Phila、PA;Louisら、(2002年)Basic Histology:Text and Atlas、McGraw−Hill、New York、NYを参照。
【0143】
たとえば抗原断片、リーダー配列、タンパク質フォールディング、機能性ドメイン、グリコシル化部位、および配列アラインメントを決定するための、ソフトウェアパッケージおよびデータベースが利用可能である。たとえば、GenBank、Vector NTI(登録商標)Suite(Informax、Inc、Bethesda、MD);GCG Wisconsin Package(Accelrys、Inc.、San Diego、CA);DeCypher(登録商標)(TimeLogicCrop.、Crystal Bay、Nevada);Menneら、(2000年)Bioinformatics 16:741〜742頁;Menneら、(2000年)Bioinformatics Applications Note 16:741頁;Wrenら、(2002年)Comput.Methods Programs Biomed. 68:177頁;von Heijne(1983年)Eur.J.Biochem. 133:17頁;von Heijne(1986年)Nucleic Acids Res. 14:4683頁を参照。
【0144】
(実施例2)
真菌感染モデル
真菌感染の試験用のマウス系統は以下のように得た。雌性C57BL/6およびBALB/cマウス、8〜10週齢はCharles River(Calco、イタリア)から購入した。同型IL−12p35、IL−23pl9またはIL−12p40欠損マウス(ここでは以後、それぞれp35−/−、p19−/−およびp40−/−と呼ぶ)、TLR−2−、TLR−4、MyD88またはTRIF欠損マウス(ここでは以後、TLR−2−/−、TLR−4−/−、MyD88−/−またはTRIF−/−と呼ぶ)C57BL/6バックグラウンドのマウスを、ペルージャ大学、ペルージャ、イタリアの動物施設において、特定病原体未感染の条件下で飼育した。BALB/cバックグラウンドのIFN−γ−/−p35−/−マウスのつがいは、Dr.M.Colombo(Istituto Tumori、Milan、イタリア)によって提供された。BALB/cバックグラウンドのIFN−γ−/−およびIL−4−/−マウスも、ペルージャ大学の動物施設で飼育した。Italian Approved Animal Welfare Assurance A−3143−01に従い実験を実施した。
【0145】
真菌感染およびそれらの処置は以下のように試験した。この試験で使用したC.albicans系統の起源および特徴は既に記載されている。Bacciら、(2002年)J. Immunol. 168:2904頁。胃腸感染用に、108個のCandida細胞を胃内注射し、かつ真菌増殖の定量化は、記載したように器官当たりのCFU(平均±SE)として表した。Bacciら、(2002年)。静脈内感染用に、マウスには静脈内に0.5ml当たり異なる量の真菌を与えた。A.fumigatusの系統および培養条件は記載した通りであった。Montagnoliら、(2006年)J. Immunol. 176:1712頁。マウスには2回用量の2×107個のAspergillus静止期分生子を鼻腔内に与えた。Limulus amebocyte血球抽出成分(LAL)の方法により決定して、1.0EU/ml未満でエンドトキシンフリー溶液(Detoxi−gel、Pierce、Rockford、IL)中に真菌を懸濁させた。真菌増殖はキチンアッセイ(chitin assay)によって定量化し、結果は器官当たりのグルコサミンのマイクログラム数として表した。組織検査用に、組織を切除し、すぐにホルマリン中に固定し、かつパラフィン包埋組織の切片(3〜4μm)を過ヨウ素酸シッフ試薬で染色し調べた。Bacciら、(2002年);Montagnoliら、(2006年)。感染動物は200μgのp19中和抗体(Belladonnaら、(2006年)Cytokine 34:161頁)またはIL−17A中和モノクローナル抗体(TC11−18H10、PharMingen、San Diego、CA)で処置し、感染5時間後に腹腔内投与した。合計1mgの精製抗TGF−β1、β2、β3モノクローナル抗体(2G7)(Lucasら、(1990年)J. Immunol. 145:1415頁)を、感染5時間後および24時間後に腹腔内投与した。対照マウスにはPBSを注射した、何故なら、PBS処置動物とアイソタイプ対照処置(各処置)動物(各群に関してn>6)の間で差異を観察しなかったからである。
【0146】
細胞は以下のように精製した。Gr−1+CD11b+多形核好中球(PMN、FACS分析において98%を超えて純粋)を、Ly−6G MicroBeadsおよびMidiMacs(Miltenyi Biotech、Bergisch Gladbach、ドイツ)を使用した磁気活性化ソーティング(magnetic−activating sorting)によってマウスの腹膜腔から単離した。CD4+T細胞は、CD4MicroBeadsおよびMidiMacs(Miltenyi Biotech)を使用した磁気活性化ソーティングによって腸間膜リンパ節(MLN)、胸部リンパ節(TLN)および脾臓から精製した。DCは、CD11b+DCを得るために7日間150U/m1のマウスrGM−CSF(Sigma−Aldrich、St.Louis、MO)および75U/mlのrIL−4(R&D Systems、Minneapolis、MN)、またはFL−DCを得るために9日間200ng/mlのFLT3−L(R&D Systems)の存在下において、Iscoveの改変培地中で培養した骨髄細胞から得た。Romaniら、(2006年)Blood 108:2265頁。90〜95%のCD8−、5〜10%のCD8+、および1〜5%のB220+細胞)からなる脾性DC(>99%CD11c+および<0.1%CD3+)は、CD11c MicroBeadsおよびMidiMacs(Miltenyi Biotech)を使用した磁気活性化ソーティングによって精製した。Saccharomyces cerevisiae由来のザイモザン(10μg/ml、Sigma−Aldrich)、Salmonella minnesota Re595由来の超高純度(ultra−pure)LPS(10μg/ml、Labogen、Rho、Milan、イタリア)およびCpGオリゴヌクレオチド2006(CpGODN、0.06μM)を、記載したように使用した。Bellocchioら、(2004年)J. Immunol、173:7406頁。
【0147】
DC細胞を以下のようにパルスし培養した。DCは、記載したように1:1の細胞:真菌比で、生きた非オプソニン化真菌に、(R&D Systems;Space Import−Export srl、Milan、イタリア;およびBD Biosciences−PharMingen、San Diego、CAからの)10ng/mlのサイトカイン有りおよび無しで、または中和抗体(10μg/ml)有りおよび無しで曝露した。Bacciら、(2002年);Montagnoliら、(2006年)。培養12時間でRT−PCR用に細胞を採取し、かつ上清はELISAによってサイトカイン含有量に関して評価した。脾性CD4+T細胞(106/ml)は、培養物上清におけるサイトカイン定量化前に、中和抗体(10μg/ml)有りおよび無しで、5日間5×105個のCandidaでパルスした脾性DCの存在下において平底96ウェルプレート中で培養した。非分画MLNまたはTLN細胞は、5日後の培養物上清におけるサイトカイン測定前に、記載したように(Montagnoliら、(2006年);Montagnoliら、(2002年)J. Immunol. 169:6298頁)不活化真菌と共に培養した。
【0148】
(実施例3)
抗真菌活性アッセイ
非オプソニン化Candida酵母またはAspergillus分生子のPMN食作用、および抗真菌活性のアッセイを、記載したように実施した。Bellocchioら、(2004年)。結果はCFU阻害の割合(平均±SE)として表す。PMNは、IDOのウエスタンブロッティング前に12時間、または抗真菌活性およびMMP9/MPO(マウスミエロペロキシダーゼ)測定の試験用にさらに60分間真菌を加える前に60分間、様々な濃度のIL−17もしくはIL−23または50ng/mlのIFN−γ±IL−23/IL−17(100ng/ml)に曝露した。ゼラチンザイモグラフィーを記載したように実施した。Bellocchioら、(2004年)。マトリクスメタロプロテイナーゼ9(MMP9)のゼラチン分解活性は、72−kD領域中の溶解バンドをスキャンすることによって測定した。MPO測定用に、サンプルはウサギポリクローナル抗ヒトMPO抗体(Calbiochem、San Diego、CA)でプローブ処理し、電気化学発光(ECL)(Amersham Pharmacia Biotech、Piscataway、NJ)を使用して目に見える状態にした。
【0149】
インドールアミン2,3−ジオキシゲナーゼ(IDO)は、記載したようにウサギポリクローナルIDO特異的抗体を用いたイムノブロッティングによって検出した。Bozzaら、(2005年)。陽性対照はIDO発現MC24トランスフェクタントからなっており、かつ陰性対照はモックトランスフェクト(mock−transfectes)MC22細胞であった。
【0150】
サイトカインは以下のようにリアルタイムRT−PCR、ELISAおよびELISPOTアッセイによって定量化した。リアルタイムRT−PCRは、iCycler iQ(登録商標)検出システム(Bio−Rad、Hercules、CA)およびSYBR(登録商標)Green chemistry(Finnzymes Oy、Espoo、フィンランド)を使用して実施した。細胞を溶解し、すべてのRNAはRNeasy Miniキット(QIAGEN S.p.A.、Milan、イタリア)を使用して抽出し、製造者の指示に従いSensiscript逆転写酵素(QIAGEN)を用いて逆転写した。PCRプライマーはInvitrogen(Carlsbad、CA)から得た。使用したPCRプライマーは以下の通りであった:
マウスIL−12p35の場合、フォワードプライマー、
【0151】
【化4】
、およびリバースプライマー、
【0152】
【化5】
、;マウスIL−23p19の場合、フォワードプライマー、
【0153】
【化6】
、およびリバースプライマー
【0154】
【化7】
、;IL−12Rβ2の場合、フォワードプライマー、
【0155】
【化8】
、およびリバースプライマー
【0156】
【化9】
;IL−23Rの場合、フォワードプライマー、
【0157】
【化10】
、およびリバースプライマー
【0158】
【化11】
;マウスガンマ−アクチンの場合、フォワードプライマー、
【0159】
【化12】
、およびリバースプライマー、
【0160】
【化13】
。
【0161】
製造者の説明書(Applied Biosystems、Foster City、CA)に従い、ハウスキーピングγ−アクチン遺伝子のPCR増幅を、それぞれのサンプルに実施して(三連)、サンプルローディングを調節し、サンプル間の標準化を可能にした。水対照を含めて特異性を確実にした。SYBR(登録商標)GreenリアルタイムPCRに関する温度プロファイルは、95℃で3分間、次に95℃で15秒間の変性および60℃で1分間のアニーリング/延長ステップの40サイクルであった。増幅プロットの分析によって、完全性に関してそれぞれのデータ地点を調べた。mRNAの標準化データは、モック感染細胞のそれと比較した処置細胞中の相対的サイトカインmRNAとして表した。サイトカイン含有量は、培養細胞の組織ホモジネートまたは上清に関する酵素結合免疫吸着アッセイ(R&D SystemsおよびIL−23に関しては、eBioscience、Societa Italiana Chimici、Roma、イタリア)によって評価した。アッセイの検出限界(pg/ml)は、IL−12p70に関して<16、IL−23に関して<30、IFN−γに関して<10、IL−10に関して<3、IL−17に関して<10およびTGF−β1に関して<4,6であった。AID EliSpotアッセイキット(Amplimedical、Buttigliera Alta、Turin、イタリア)を、3日間CandidaでパルスしたDCと同時培養した精製MLN CD4+T細胞に使用して、サイトカイン産生細胞を計数した。
【0162】
データの統計分析は以下のように実施した。対数順位検定を、カプランマイヤー生存曲線の対データ分析に使用した。スチューデントのt検定または分散分析(ANOVA)およびボンフェロニの検定を使用して、臓器クリアランスおよびインビトロアッセイにおける統計的有意差を決定した。有意はp<0.05として定義した。記載されたデータは、3回の別個の実験の中の1回の代表的な実験からのデータ、または3回〜5回の実験から集めたデータのいずれかである。インビボ群は群当たり6〜8匹のマウスからなっていた。
【0163】
(実施例4)
カンジダ症に対する感受性におけるIL−23/IL−17の役割
C.albicans感染に対するIL−23/IL−17経路の貢献を評価するために、本発明者らは、生存、真菌増殖、および組織病理学の点で胃腸感染に対する感受性に関して、ならびに炎症および適応Th1/Th17免疫のパラメーターに関して、p19−/−、p35−/−、p40−/−およびC57BL/6マウスを比較した。結果(図1A〜E)は、カンジダ症に対する耐性はp35−/−マウス中では著しく低下し、それらは50%を超えて感染に屈し(図1A)、胃中では真菌増殖が高まった(図1B)ことを示した。対照的に、真菌増殖を制限する能力は、感染3日後および10日後にC57BL/6マウスと比較して、p19−/−マウス中で大幅に増大した。特に、IL−12とIL−23の両方が欠損したp40−/−マウスはp35−/−マウスよりカンジダ症に対する感受性が低く、かつp19−/−マウスより感受性が高く、Candidaの調節に関するIL−12の差次的役割が強調された。Mencacciら、(1998年)J. Immunol. 161:6228頁。同様の結果をp35−/−およびp19−/−マウスの静脈内感染後に観察し、それぞれ6±2対20±3日(5×105個の真菌細胞接種)、および4±2対15±3日(106個の真菌細胞接種)の平均生存時間(MST)であった。胃の組織病理学的検査によって、C57BL/6、p19−/−またはp40−/−マウス中での錯角化、表皮肥厚および限られた炎症反応の存在が明らかになったが、p40−/−、および特にp19−/−マウスでは単核細胞の浸潤を示した。対照的に、p35−/−マウスの胃中のPMNの多量浸潤、上皮壊死および顕著な表皮肥厚の徴候と併せて、多数の菌糸が角質化層中に存在した。これらの結果は、IL−23経路とIL−12経路はカンジダ症において相違する役割を有することを示唆する。
【0164】
これらの発見とIL−12/Th1およびIL−23/Th17免疫応答を関連付けるために、マウスは、感染翌日または3日後にMLNにおけるp35、p19、IL−12Rβ2およびIL−23RのmRNA発現(図1C)、および感染7日後のMLNにおけるIFN−γ、IL−4またはIL−17産生CD4+細胞の頻度に関して評価した(図1D)。本発明者らは、C57BL/6マウスと比較して、p19−/−マウス中での増大したレベルのp35およびIL−12Rβ2、およびIFN−γ+細胞の数を発見し、IL−23の不在下での増強したIL−I2/Th1応答を実証した。対照的に、p19およびIL−23RのレベルおよびIL−17産生細胞の数は、IL−12を欠くマウス(p35−/−)において増大した。予想通り、IL−4産生細胞の数もp35−/−マウスにおいて相当増大した。これらのデータは、野生型C57BL/6マウス中でIL−12によって促進されIL−23によって制限される主要なTh1応答を実証する。IL−12はIL−23およびIL−17産生を抑制し、および逆にIL−23はIL−12およびIFN−γ産生を阻害し、IL−23/Th17とIL−12/Th1経路の交差制御を示す。これらのデータは、高いIL−23/Th17応答はマウスをカンジダ症に対して非常に敏感にすることを示唆する。
【0165】
(実施例5)
アスペルギルス症に対する感受性におけるIL−23/IL−17の役割
カンジダ症と同様に、IL−23/IL−17経路の活性化がアスペルギルス症に対する感受性と関係があるかどうか決定するために、肺アスペルギルス症に対する感受性、ならびに炎症および適応Th1/Th17免疫のパラメーターに関してp19−/−、p35−/−、p40−/−またはC57BL/6マウスを評価した。結果(図2A)は、真菌負荷はp35−/−マウス中で、およびp19−/−マウス中ではさらに高い程度で低下したことを示し、IL−12、および特にIL−23はAspergillus感染の調節(すなわち促進)を阻害することを示唆する。肺の組織病理学的検査によって、他の完全な肺実質中に散乱した炎症性単核細胞のわずかな浸潤によって特徴付けられる、C57BL/6、p40−/−またはp19−/−マウスにおける軽度の炎症性病理の存在が明らかになった。浸潤単核細胞の数はp19−/−マウス中でより多かったが、実質破壊の徴候は観察しなかった。対照的に、PMNの多量浸潤(Gr1+CD11c−PMNの約8〜10倍の増大)が、広範囲の間質性肺炎の徴候を伴うp35−/−マウスの肺において存在した。Candidaによる感染と同様に、IL−12およびIL−23およびそれらの各々の受容体の発現は交差制御され、C57BL/6マウスと比較して、p19−/−マウスのTLNにおいてp35およびIL−12Rβ2は上方制御され、かつp35−/−マウスのTLNにおいてp19およびIL−23Rは上方制御された(図2B)。対照的に、p40−/−マウス中でのIL−12とIL−23の両方の不在が、p35およびp19またはそれらの受容体IL−12Rβ2およびIL−23Rの発現を有意に変えることはなかった。さらに、IFN−γ+およびIL−17+産生CD4+T細胞の数は、感染7日後にそれぞれp19−/−およびp35−/−マウスにおいて増大した(データ示さず)。肺中では、IL−12p70のレベルはC57BL/6マウス中(68±8pg/ml)よりp19−/−中ではるかに高かった(554±44pg/ml)、かつIL−23はp35−/−マウス中のみで検出することができた(79±11)。IL−17はC57BL/6マウス(37±7pg/ml)と比較してp35−/−マウス中で増大した(246±17pg/ml)。これらのデータは、増大したIL−23/IL−17依存性炎症性応答はアスペルギルス症に対する感受性とも関係があることを示唆する。
【0166】
(実施例6)
真菌感染に対するIL−23/IL−17仲介感受性におけるTGF−βの役割
真菌感染に対する感受性におけるIL−17の役割を試験するために、本発明者らは、真菌感染直後に抗IL−17中和抗体でマウスを処置した。IL−17の遮断は、低下した真菌増殖(図3A)、関連標的器官中の組織炎症およびPMN浸潤(データ示さず)により判断して、C.albicansとA.fumigatusの両方に対する耐性を大幅に増大した。耐性はIFN−γ+Th1細胞の頻度の増大およびTh17細胞の頻度の低下を伴い、MLN細胞によって分泌されるIL−17の量の低下をもたらした(図3B)。同様に、抗体によるIL−23の中和によって真菌感染およびTh1発生に対する耐性が増大し、p19−/−マウスにおいて得た本発明者らのデータを確認する(図3B)。これらの結果は、IL−23/IL−17経路は防御Th1免疫の阻害によって真菌感染に対する感受性をもたらすことを明らかに実証する。
【0167】
最新の結果は、TGF−βはIL−6と一緒にTh17発生を促進することを示唆する。Bettelli and Kuchroo(2005年)J. Exp. Med.
201:169頁;Manganら、(2006年)Nature 441:231頁;Veldhoenら、(2006年)Immunity 24:179頁。本発明者らは、TGF−β中和抗体を用いた処置によって、マウス中でのTh細胞の発生および真菌調節に対するTGF−βの影響を評価した。特に、TGF−β阻害はC.albicansとA.fumigatusの両方の感染中にIL−17産生細胞の発生に影響を与えず(図3B)、真菌負荷のわずかではあるが有意な低下はCandidaを有するマウスではなくAspergillusを有するマウスのみで観察したが(図3A)、いずれの感染においても処置によって影響を受けるCD4+Th17細胞の活性化はなかった。TGF−β中和は感染中のTGF−β産生を効果的に低下させたので(胃中では46pg/mlから24pg/ml、および肺中では36pg/mlから15pg/ml)、本発明者らは、TGF−βは真菌感染に対するTh17仲介の感受性においてわずかな役割を果たすと結論付ける。
【0168】
(実施例7)
IL−12の不在下での真菌感染におけるIL−23/IL−17の役割
上記のデータは、それによってIL−23/IL−17軸が真菌感染に対する感受性を決定する1つの可能性のある機構は、防御Th1応答を抑制する相対能力に頼ることを示唆する。それを正式に証明するために、IL−23の遮断を増大(IL−4−/−マウス)または欠損(IFN−γ−/−マウス)Th1反応性のいずれかの条件下で行った。マウスにはC.albicansを胃内感染させ、中和抗体によってIL−23遮断を施した。以前の刊行物(Romaniら、(1992年)J. Exp. Med. 176:19頁;Cenciら、(1998年)J. Immunol. 161:3543頁)と一致して、真菌負荷はBALB/cマウスと比較してIL−4−/−マウス中で低く、かつIFN−γ−/−マウス中で高く、感染の調節に関するIFN−γの重要性を実証した。野生型マウスと同様に、IL−23の遮断はIL−4−/−マウスの胃中の真菌負荷を大幅に低下させ(図3C)、および同時にMLNにおいてIL−12p70/IFN−γ産生を増大させ(データ示さず)、Th2とIL−23/Th17経路の両方が防御性抗真菌応答に相加的に拮抗することを示唆した。驚くことに、IFN−γ−/−マウスにおける高い真菌負荷はIL−23の中和によってさらに増大し(図3C)、これは抗IL−23処置マウスにおいて低いIL−23産生(229対21pg/ml)IL−17産生(279対95pg/ml)をもたらした。したがって、IL−23はIFN−γの不在下で真菌感染において防御的役割を有し得る。しかしながら、p35−/−または二重欠損IFN−γ−/−/p35−/−マウスにおけるIL−23の中和によって低下した真菌負荷により実証されたように、IL−23はIL−12p70の不在下、またはIL−12p70とIFN−γの両方の不在下では反対の影響を有する(図3C)。これらのデータは、IFN−γの不在下でのIL−23の防御的役割はIL−12p70によって仲介されることを示唆する。特に、IL−12p70の不在下でのIL−23の適度な防御的役割は結核においても観察されており、この場合IL−23は、防御的IFN−γ産生CD4+T細胞の誘導においてIL−12p70に部分的に置き換わった。Khaderら、(2005年)。
【0169】
(実施例8)
真菌感染に応答した樹状細胞中でのIL−23およびIL−12の産生
IL−23はin vitroではAspergillusに応答してヒトDCによって産生されることは、既に示されている。Gafaら、(2006年)Infect. Immun. 74:1480頁。本発明者らは、IL−23がC.albicansに応答してDCによって産生されるかどうか、およびそれがIL−12およびIL−10、真菌に対する防御耐性の誘導に本来必要とされる2つのサイトカインの産生とどのような関係があるかをここで評価した。Romani & Puccetti(2006年)。
【0170】
この目的のために、本発明者らは、それぞれ骨髄性DCおよび形質細胞様(plasmacytoid)DCの特徴を共有するGM−CSF(GM−DC)またはFlt3−L(FL−DC)のいずれかの存在下で、骨髄由来のDCを作製した。FL−DCは新たに採取した脾性CD8+、CD8−およびB220+LyC6+形質細胞様DCの混合物と同等の集団を包含するが(Naikら、(2005年)J. Immunol. 174:6592頁)、本発明者らは、FL−DCの機能活性は形質細胞様DC中またはCD8−とCD8+DCの組合せ中に存在することを近年実証している。Romaniら、(2006年)Blood 108:2265頁。DCはin vitroにおいて酵母または真菌の菌糸で刺激し、サイトカインmRNA発現および産生に関して評価した。ザイモザンおよびLPSはGM−DCの陽性対照として、およびCpG−ODNはFL−DCの陽性対照として使用した。
【0171】
これらの結果は、真菌に応答したDCサブセットの2つのサブセットによるサイトカイン発現および産生において二分(dichotomy)を示した。RT−PCR分析は、p19mRNA発現のみが菌糸より酵母に応答してGM−DCにおいて増大し、p35mRNA発現は酵母に応答してGM−DCにおいてわずかに増大したが、IL−10と同様に、菌糸に曝露したFL−DCにおいて大幅に増大したことを明らかにした(図4A)。培養物上清における実際のサイトカイン産生の測定によって、IL−23が酵母、特に高い真菌:DC比、およびザイモザンまたはLPSに応答してGM−DCによって産生されたことを確認した(図4B)。IL−23産生の最高レベルはインキュベーションの12時間で観察し(図4B)、その後低下した(データ示さず)。逆に、IL−12p70とIL−10の両方は、12時間Candida菌糸、LPSまたはCpG−ODNで刺激したFL−DCによって主に産生され(図4B)、その後上昇し続けた(データ示さず)。一緒にするとこれらのデータは、IL−23は真菌に応答して、特に高レベル真菌増殖の条件で、かつ他の誘導サイトカインより早く骨髄性DCによって産生されることを示唆する。Candidaに応答して誘導サイトカインを産生する異なるDCサブセットの能力は、したがってin vivoでそれらの抗真菌免疫を調節することができる。実際のところ、Aspergillusに関して既に示されているように(Romaniら、(2006年)Blood 108:2265頁)、CandidaでパルスしたFL−DCは防御をもたらし、かつCandidaでパルスしたGM−DCはカンジダ症を有するレシピエントマウスへの養子移入によって感染を悪化させた。
【0172】
(実施例9)
IL−23とIL−12の交差制御
IL−12p70およびIL−23産生が真菌に応答して交差制御されるかどうかを検証するために、本発明者らは、IL−12p70またはIL−23、または対応する中和抗体のいずれかへの曝露後にp19−/−、p35−/−、およびC57BL/6対照マウス由来の脾性DCによるIL−12p70およびIL−23の分泌を測定した。図4Cは、野生型DCと比較してIL−12p70の産生がp19−/−DC中で高く、かつIL−23の産生がp35−/−DC中で高かったので、IL−12p70とIL−23は実際交差制御されることを示す。さらに、IL−12p70またはIL−23のいずれかへの曝露は、野生型DCによるIL−23またはIL−12p70の分泌をそれぞれ有意に低下させ、かつIL−12またはIL−23中和の状態において逆も真であった(図4D)。RT−PCRによって無刺激DCは両方のサイトカイン受容体を発現することを明らかにしたので(データ示さず)、これらのデータは、それによってDCによるIL−12p70およびIL−23産生が相互に制御されるパラクリンループ(paracrine loop)の存在を示唆する。
【0173】
(実施例10)
樹状細胞によるIL−23産生におけるTLRの役割
真菌に応答したIL−23産生のTLR依存性の可能性を明確にするために、本発明者らは、TLR−2−/−またはTLR−4−/−マウスから、ならびにMyD88−/−およびTRIF−/−マウスから作製したGM−DCによる酵母または分生子に応答したIL−23産生を測定した。Akira and Takeda(2004年)Nat. Rev. Immunol. 4:499頁。図5Aは、TLR2とTLR4の両方が、TRIFではなくMyD88を介したシグナル伝達によるIL−23産生に必要不可欠であることを示す。特に、TRIFの不在下でさえIL−23は促進されたように現れた。したがってIL−23は、TLR/MyD88依存性炎症性経路を介して、真菌に応答して通常のDCによって産生される。
【0174】
T細胞もIL−23産生を制御することができるかどうかを明確にするために、本発明者らは、CD4+T細胞と共に培養したDCの上清に産生されたIL−23のレベルを評価した。これらの結果は、C57BL/6および特にp35−/−マウス(群3対群6、図5B)由来のCandidaでパルスしたDCを用いて刺激したT細胞の培養物においてIL−23産生が上方制御されたことを明らかに示し、この発見により、活性化T細胞がIL−23産生の増幅のための正のフィードバックループをもたらし得ることを示唆した。さらに、十字交差実験の結果によって、IL−23産生DCはIL−17産生細胞を活性化するのに必要かつ十分であったことを確認した(群7および8)。さらに、DCとT細胞の同時培養物に加えたモノクローナル抗体によるIL−23の中和はIL−l7産生を阻害し(群4対群5、図5C)、一方TGF−β中和はIL−l7(群4対群6)ではなくIFN−γ産生に影響を与えた(群1対群3)。
【0175】
(実施例11)
多形核好中球の抗真菌エフェクター機能に対するIL−23およびIL−17の影響
PMNは真菌に対する急性炎症性応答の開始および遂行において必要不可欠である。Romani(2004年)。p35−/−マウスにおける初期の真菌増殖とともに、PMNが感染部位に多量に動員されたという発見により、本発明者らは、IL−23/IL−17依存性経路はPMNの抗真菌エフェクター機能に悪影響を与え得ると仮定するに至った。したがって本発明者らは、p19−/−マウスまたはp35−/−マウスのいずれか由来、およびIFN−γの不在または存在下で組換えIL−23またはIL−17と共に培養した野生型マウス由来のPMNの抗真菌活性を評価した。殺滅活性は、C57BL/6 PMNと比較してp19−/−PMNにおいて有意に増大し、かつp35−/−マウスにおいて低下した(図6A)。PMNをこれらのサイトカインに曝露する前に、本発明者らは、IL−17Rと同様に(Yaoら、(1995年)Immunity 3:811〜821頁)、IL−23RはマウスPMNでも発現されたかどうかを検証した。定量RT−PCRは、無刺激PMNがIL−23Rを発現し、その発現はLPSで刺激した後さらに増大したこと(データ示さず)を明らかにし、この発見により、PMNはIL−23にも応答性があることを示唆した。
【0176】
いずれかのサイトカインへの曝露は、用量依存的様式で(図6B)IFN−γの不在および存在下で(図6C)野生型PMNの殺滅活性を弱めた。したがって、IL−23およびIL−17はPMNの抗真菌エフェクター機能を負に制御し、これはp35−/−マウスが真菌増殖を効率良く制限できない原因である可能性がある。したがって、感染器官への末梢PMNの流入低下がカンジダ症に対するIL−17AR欠損マウスの高い感受性の原因であったように(Huangら、(2004年)J. Infect. Dis. 190:624頁)、IL−17はPMNの強力な化学誘引物質であるが(Yeら、(2001年)J. Exp. Med. 194:519頁)、本発明者らの結果は、PMN機能に対するIL−17の有害な影響も指摘する。
【0177】
(実施例12)
多形核好中球のIDO依存性抗炎症性プログラムに対するIL−23およびIL−17の影響
本発明者らは、IDO遮断がPMNの炎症性状態の促進をもたらしたように、IFN−γ仲介IDO活性化がCandidaに対するPMNの炎症性プログラムを負に制御することを既に示している。Bozzaら、(2005年)。MMP−9およびMPOは、IL−17によって活性化されることが示されている典型的な炎症性マーカーである。Kolls and Linden(2004年)Immunity 21:467頁。したがって本発明者らは、C.albicansに対するIL−23とIL−17の両方の影響が、MMP−9、MPOおよびIFN−γ仲介IDO産生を誘導したと評価した。IL−23と、特にIL−17の両方が、MMP−9およびMPOを相当増大させた(図6D)。野生型PMNのIDO発現および炎症性応答。対照的に、両方のサイトカインがIFN−γによるIDOの誘導に完全に拮抗した(図6E)。興味深いことに、アポトーシスPMNの数はIL−23とIL−17の両方への曝露によって有意に減少し(データ示さず)、これらのサイトカインはPMNの生存能力も高めることが示唆された。これは炎症がTh17経路によって持続するさらなる機構である可能性がある。したがって、PMNの炎症性プログラムを妨害する能力、および炎症組織中の増大した実質的タンパク質分解負荷は、真菌感染中のTh17細胞活性化と関係がある炎症性病理の原因である可能性がある。
【0178】
(実施例13)
IL−17産生に基づくIL−23特異的アンタゴニストのアッセイ
マウス流入領域リンパ節(DLN)細胞を使用するin vitro試験は、蛍光活性化細胞ソーティング(FACS(登録商標))分析により測定したとき、IL−23の除去はIL−17産生細胞を阻害または除去し、一方IL−23の添加はIL−17分泌を発生または刺激することを実証した。WO2004/071517、Langrishら、(2005年)J. Exp. Med. 201:233頁を参照。Aggarwalら、(2003年)J. Biol. Chem. 278:1910頁も参照。これらの実験では、5日間サイトカインまたは抗体でDLN細胞を処置した。細胞は抗原刺激した正常野生型マウスから単離し、rIL−12またはrIL−23のいずれかの存在下で培養した。DLN培養物中のCD4+T細胞の分析は、IL−17産生集団の消失を伴いながら、IL−12がIFN−γ産生細胞の発生を促進したことを実証した。対照的に、IFN−γ産生集団の消失を伴いながら、IL−23はIL−17産生細胞の発生を促進した。抗p19抗体はIL−17産生を低下させたがIFN−γレベルには影響を与えず、一方抗p35抗体はIL−17産生を変えなかった。総合するとこれらの結果は、IL−23がIL−17産生CD4+T細胞の発生を選択的に促進することを示した。
【0179】
IL−23とIL−12の生物活性のこの差異を使用して、以下のようにIL−12と比較して、可能性のあるIL−23アンタゴニストの効力および特異性を評価する。
【0180】
可能性のあるIL−23特異的アンタゴニストの不在下でのIL−23およびIL−12活性に関する基線データ(baseline data)を、以下のようにして得る。正常野生型SJLマウスは、フロイント完全アジュバントで乳化したプロテオリピドペプチド(PLP)で(皮下)、および百日咳毒素で免疫処置する(静脈内)。流入領域リンパ節を免疫処置の9日後に除去し、単核細胞は細胞内IFN−γおよびIL−17産生に関してすぐに評価する(以下に記載するように)、または単離しおよび5日間PLPに加えrIL−12もしくはrIL−23のどちらかの存在下で単離および培養するかのいずれかである。細胞はPMA(50ng/ml)で3時間/ゴルジ−プラグ(Golgi−plug)の存在下において4時間イオノマイシン(500ng/ml)で刺激し、次いで表面をCD4に関して染色し、透過処理し、かつ細胞内をIFN−γおよびIL−17に関して染色する。フローサイトメトリーのプロットを生存CD4+T細胞において得る。
【0181】
IL−23およびIL−12の影響を、サイトカイン処置しなかった対照細胞と比較して評価する。典型的には、IL−23処置細胞は増大した割合のIL−17産生細胞を示し、IFN−γ産生細胞の増大は示さず、一方IL−12処置細胞は増大した割合のIFN−γ産生細胞を示し、IL−12産生細胞の増大(またはさらに減少)は示さない。
【0182】
可能性のあるIL−23特異的アンタゴニストの効力および特異性は、アンタゴニスト、または好ましくは一連の濃度のアンタゴニストの存在下で同じ実験を実施することによって決定する。IL−23特異的アンタゴニストはIL−23の活性を阻害するが(すなわち、このアンタゴニストは、他の場合IL−23によって誘導されるIL−17産生細胞の割合を減少させる)、IL−12の活性は実質的に低下させない。IL−12またはIL−12とIL−23の両方の活性を阻害する作用物質は、IL−23特異的アンタゴニストではない。
【0183】
場合によっては、陽性対照を含めることができ、その中で既知の抗p19アンタゴニスト抗体を使用して、IL−23の活性を特異的に阻害する。
【0184】
(実施例14)
マイコバクテリア感染症
マイコバクテリア感染症の処置における本発明の組成物および方法の有効性を実証する方法を提供する。C57BL/6マウスに以下のようにマイコバクテリアを感染させる。Theracys−BCG Live(Aventis Pasteur、Inc.、Swiftwater、PA)、Bacille CabmetteおよびGuerinのConnaught系統ならびにM.bovisの弱毒系統の凍結乾燥調製物を、製造者によって推奨されるように元の状態に戻す。元の状態に戻した(reconstituted)細菌は、10%グリセロール生理食塩液中に約6×107cfu/mLの濃度にする。マウスへの注射前に、0.02%Tween−80/0.9%生理食塩液中にアリコートを適切な濃度に希釈する。
【0185】
6〜8週齢の雌性C57BL/6マウスに、約3.5×105cfuのBCGを側方の尾静脈を介して静脈内感染させる。マウスには1mgの適切なモノクローナル抗体を(たとえば、アイソタイプ対照、抗IL−23p19、または抗IL−23R)0.9%生理食塩液中で与え、マイコバクテリア感染前第1日、およびマイコバクテリア感染1〜2週間後に再度皮下投与する。CO2ナルコーシスによって、感染後の適切な時間地点でマウスを屠殺する。
【0186】
屠殺したBCG感染マウスは以下のように分析する。下大静脈を切断した後に心臓の右心室を介してRPMI 1640を灌流させることによって、肺から血液を取り除く。左肺、肝臓右半部下葉(lower right liver lobe)、および脾臓の半分を無菌状態で除去する。これらの組織を、Mini−Bead Beater−8ホモジェナイザー(BioSpec Products、Bartlesville、OK)を用いて0.9%NaCl/0.02%Tween80中に均質化する。生存マイコバクテリアは、10倍連続希釈の臓器ホモジネートを7H10Middlebrook寒天プレート(Becton Dickinson、Sparks、MD)に平板培養することによって定量化する。コロニー形成単位(CFU)は、37℃で2週間のインキュベーションの後に手作業で計数する。対照マウス(たとえば、アイソタイプ対照)と比較した、抗IL−23抗体(たとえば、抗IL−23p19抗体または抗IL−23R抗体)で処置した動物における(CFUによって測定した)細菌負荷の統計的に有意な低下は、マイコバクテリア感染症の有効な処置の証拠である。
【技術分野】
【0001】
本発明は、一般的に感染症の処置に関する。具体的には、本発明は、慢性の細菌感染症、真菌感染症、またはウイルス感染症などの感染症を示す被験体に、IL−23アンタゴニスト、たとえば抗体を投与することに関する。
【背景技術】
【0002】
慢性感染症は、多くの病原体により引き起こされる。種々のウイルス、真菌、および細菌は、たとえば、消散させることができない持続的感染症を引き起こす場合がある。
【0003】
重篤な真菌感染症数が増加し続けているため、それらを処置するための方法および組成物の必要性がより差し迫っている。主要な真菌病原体には、Candida albicans、Aspergillus fumigatus、およびCryptococcus neoformansが含まれ、これらの病原体に関連する侵襲性真菌症の推定年間発生率は、米国の人口100万人当たり72〜228件(Candida種属の場合)、12〜34件(Aspergillus種属の場合)、および30〜66件(C.neoformansの場合)の感染である。非特許文献1。真菌感染症の増加は、医学の進歩(移植および化学療法)の結果として、およびAIDS患者人口の増加の結果として、主として免疫無防備状態の患者数の増加に起因する。免疫無防備状態の患者における真菌感染症の80%超は、Candida種属により引き起こされる。クリプトコックス症は、カンジダ症に次いで、AIDS患者で2番目に蔓延している真菌感染症である。アスペルギルス症は、癌患者および臓器移植患者における感染症の少なくとも30%に関与しており、高い死亡率を示す。
【0004】
フルコナゾールは、長年の間、真菌病原体に対して有効な薬物であるが、耐性は増加している。アムホテリシンBなどの代替薬は、発熱、腎臓障害、貧血、低血圧、頭痛、悪心、嘔吐、および静脈炎などの副作用を含む深刻な欠陥を有している。
【0005】
細菌感染症は、免疫無防備状態の個体人口の増加および抗生物質耐性細菌株の広範な発生に照らすと、抗生物質の普及にもかかわらず重要な課題のままである。免疫無防備状態の個体には、高齢者、移植レシピエント、化学療法患者、および後天性免疫不全症候群(AIDS)の個体が含まれる。米国では、毎年ほぼ200万人の患者が病院で感染症に罹り、それらの感染症の原因である細菌の70%は、少なくとも1種の抗生物質に耐性である。NIAID概況報告書(fact sheet)、「The Problem of Antimicrobial Resistance」、2006年4月。近年米国では、約90,000人が感染症で死亡し、1992年の13,300人から増加している。ほとんどの細菌感染症は、少なくとも1種の抗生物質の長期的治療過程(たとえば、バンコマイシンの持続的静脈内投与)に感受性のままであるが、これが今後の病原性細菌について当てはまるという保証はない。メチシリン耐性Stapholococcus aureus(MRSA)は、重要な公衆衛生上の課題を代表する多抗生物質耐性微生物の主要な例である。2002年には、疾病予防管理センター(CDC)は、ミシガン州の患者でバンコマイシンに対して完全に耐性であるS.aureus(VRSAと呼ばれる)感染症の最初の症例を報告した。持続感染性の病原細菌には、Salmonella種属、Brucella種属、およびChlamydia種属も含まれる。
【0006】
マイコバクテリアとは、高い細胞壁脂質含有量および低い増殖率を示す、好気性で非胞子形成性の非運動性桿菌の広く分布する多様な集団である。Mycobacterium属のメンバーは、たとえば、無害から重大な病原性を有する種、たとえば結核(TB)の原因因子であるM.tuberculosisまでと、毒力が様々である。TBは、世界で2番目に多い感染性の死因である。約20億人、すなわち世界人口の3分の1が、M.tuberculosisに感染していると推定される。800万件の新しい症例およびほぼ300万人の死亡が、毎年発生している。TBは世界中の全死亡の7%の直接的原因であり、この世界的な流行は、薬剤耐性生物の拡散および進行中のHIV流行の結果、悪化する可能性が高い。たとえば、Dale and Federman(編)(2002年)WebMD Scientific American Medicine、WebMD Professional Publishing、ニューヨーク、米国ニューヨーク州を参照されたい。
【0007】
TBを処置する最新の方法は、イソニアジド、リファンピン、ピラジナミド、エタンブトール、ストレプトマイシン、シプロフロキサシン、およびオフロキサシンなどの広域スペクトルの抗感染症薬の使用を含む。しかしながら、そのような作用剤は種々の器官で毒性を引き起こす場合があり、いくつかの抗生物質耐性TB株の増大と共に、有効性を失いつつある。多様な非抗生物質薬を使用して、結核患者の肺におけるマイコバクテリア負荷を低減すると、疾患形成、伝播、および死亡が防止できる。
【0008】
慢性のウイルス感染症も、公衆衛生に対する大きな脅威を代表する。C型肝炎ウイルス(HCV)またはヒト免疫不全ウイルス(HIV)などのウイルス感染症を完全に根絶できないと、それぞれ、その後の再活性化および肝臓癌または後天性免疫不全症候群(AIDS)などの合併症に結び付く場合がある。非特許文献2。さらに、ヒトパピローマウイルス(HPV)遺伝子型16、18、31、33、45、および56は、子宮頚癌の症例の95%超を占める。非特許文献3。事実上100%のHIV感染症例、55〜85%のHCV感染症例、および30%を超えるHPV症例で、慢性感染症が発生すると推定されている。Berzofskyら(2004年)。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】Pfallerら(2006年)Clin. Infect. Dis. 第43巻:S3〜14頁
【非特許文献2】Robertson & Hasenkrug(2006年)Springer Semin. Immun. 第28巻:51頁
【非特許文献3】Berzofskyら(2004年)J. Clin. Invest. 第114巻:450頁
【発明の概要】
【発明が解決しようとする課題】
【0010】
細菌感染症、ウイルス感染症、および真菌感染症の処置および/または予防のための方法および組成物の改良に対する必要性が存在する。そのような方法および組成物は、好ましくは、既存の処置方法および組成物より毒性が低くおよび/またはより効果的である。
【課題を解決するための手段】
【0011】
本発明は、細菌感染症、ウイルス感染症、および真菌感染症と戦うために、IL−23アンタゴニストを使用する組成物、薬剤、および方法を提供することにより、これらの必要性およびその他を満たす。
【0012】
一局面では、本発明は、IL−23アンタゴニストの投与を伴う、感染症を有する被験体、感染症を有する疑いのある被験体、または感染症に罹るリスクのある被験体を処置するための方法に関する。一実施形態では、アンタゴニストは、IL−23またはそのp19サブユニットに結合する抗体またはその結合性断片などの結合性化合物である。いくつかの実施形態では、抗体の結合は、IL−23またはそのp19サブユニットの、IL−23受容体またはそのIL−23Rサブユニットに対する結合を阻止する。別の実施形態では、IL−23アンタゴニストは、IL−23受容体またはそのIL−23Rサブユニットに結合する。いくつかの実施形態では、アンタゴニストは、IL−23受容体またはそのIL−23Rサブユニットに結合し、IL−23またはそのp19サブユニットの結合を阻止する。別の局面では、本発明は、前記処置方法で使用するための組成物に関する。
【0013】
いくつかの実施形態では、感染性障害には、細菌感染症、マイコバクテリア感染症、ウイルス感染症、または真菌感染症などの感染性疾患が含まれる。一実施形態では、感染性障害は、M.bovis、M.leprae、またはM.tuberculosisにより引き起こされるマイコバクテリア感染症である。一実施形態では、感染性障害はTBである。別の実施形態では、感染性障害は、爪甲真菌症、カンジダ症、アスペルギルス症、クリプトコックス症からなる群から選択される真菌感染症である。さらに別の実施形態では、感染性障害は、C.albicans(たとえば、慢性皮膚粘膜カンジダ症、鵞口瘡)、C.neoformans、またはA.fumigatusにより引き起こされる真菌感染症である。さらなる実施形態では、感染性障害は、ウイルス感染症、たとえばヒト免疫不全ウイルス(HIV)、B型肝炎ウイルス(HBV)、C型肝炎ウイルス(HCV)、またはヒトパピローマウイルス(HPV)により引き起こされるウイルス感染症である。
【0014】
他の実施形態では、感染性障害は慢性感染症である。種々の実施形態では、慢性感染症は、感染を消散するための少なくとも1つの過去の試みにもかかわらず、1、2、4、6、9、12、18、24、36または48カ月以上の間持続する。種々の実施形態では、慢性感染症を消散しようとする過去の試みは、抗菌物質、抗生物質、抗ウイルス物質、または抗真菌物質を用いた処置を伴う。
【0015】
一実施形態では、本発明は、IL−23アンタゴニストの投与を、別の治療薬などの少なくとも1つの他の治療様式と組み合わせた併用治療に関する。種々の実施形態では、他の治療薬は、IL−17Aアンタゴニスト、IL−17Fアンタゴニスト、IL−12アゴニスト(IL−12を含む)、TGF−βアンタゴニスト、またはIL−6アンタゴニストである。別の実施形態では、他の治療薬は、ポサコナゾール、フルコナゾール、ボリコナゾール、イトラコナゾール、ケトコナゾール、リアロゾール(liarozole)、イルテマゾール(irtemazol)、クロトリマゾール、ミコナゾール、エコナゾール、ブトコナゾール、オキシコナゾール、スルコナゾール、チオコナゾール、およびテルコナゾール、置換チアゾール、チアジアゾール、オキサジアゾール、カスポファンギン、アムホテリシンB、ナイスタチン、ピマリシン、フルシトシン(5−フルオロシトシン)、ナフチフィン、テルビナフィン、ブテナフィン、チオカーボネートトルナフテート(thiocarbonate tolnaftate)、グリセオフルビン、アミオダロン、シクロピロックス、スルベンチン、アモロルフィン、クリオキノール、ゲンチアナバイオレット、ヨウ化カリウム、チオ硫酸ナトリウム、石炭酸フクシン溶液、およびエキノキャンディン(たとえば、カスポファンギンアセテート(caspofungin acetate)、ミカファンギン、およびアニデュラファンギン)からなる群から選択される1つまたは複数の抗真菌物質である。
【0016】
別の実施形態では、他の治療薬は、イソニアジド、リファンピン、ピラジナミド、エタンブトール、ストレプトマイシン、シプロフロキサシン、バンコマイシン、またはオフロキサシンからなる群から選択される1つまたは複数の抗菌物質である。
【0017】
別の実施形態では、他の治療薬は、アバカビル、アシクロビル、アマンタジン、アンプレナビル、デラビルジン、ジダノシン、エファビレンツ、ファムシクロビル、インジナビル、インターフェロンアルファ(IFN−α)、リバビリン、ラミブジン、ネルフィナビル、ネビラピン、オセルタミビル、ペンシクロビル、リバビリン、リトナビル、サキナビル、スタブジン、バラシクロビル、ザルシタビン、ザナミビル、ジドブジン(アジドデオキシチミジン、AZT)からなる群から選択される1つまたは複数の抗ウイルス物質である。
【0018】
一局面では、本発明は、感染症を有する被験体、または感染症を有する疑いのある被験体の1型(Th1)免疫応答を増強するための方法に関する。種々の実施形態では、Th1応答の増強は、IL−23アンタゴニストで処置する前のT細胞の割合と比較した場合の、IFN−γ発現CD4+T細胞の割合の増加、IL−17A発現CD4+T細胞の割合の減少、またはその両方により反映される。種々の実施形態では、増加は、1.5倍、2倍、3倍、5倍、10倍、20倍、もしくは50倍以上であり、または減少は、3分の2、2分の1、3分の1、5分の1、10分の1、20分の1、もしくは50分の1以下である。別の局面では、本発明は、前記Th1応答を増強する方法で使用するための組成物に関する。
【0019】
種々の実施形態では、他の治療薬は、IL−23アンタゴニストを投与する前、および/または投与と同時に、および/または投与した後で投与される。一実施形態では、IL−17Aアンタゴニストは、IL−23アンタゴニストの前および/または同時に投与される。別の実施形態では、抗菌、抗真菌、または抗ウイルス物質は、IL−23アンタゴニストと同時に投与される。
【0020】
別の局面では、本発明は、サイトカイン自体に対する、またはそれらのそれぞれの受容体もしくは受容体サブユニットに対するアンタゴニスト抗体などの、IL−17Aおよび/またはIL−17Fのアンタゴニストの投与を伴う、感染症を有する被験体、感染症を有する疑いのある被験体、または感染症に罹るリスクのある被験体の処置方法に関する。
【0021】
他の実施形態では、IL−23アンタゴニストはポリヌクレオチドを含む。種々の実施形態では、ポリヌクレオチドは、アンチセンスポリヌクレオチド(たとえば、アンチセンスRNA)または低分子干渉RNA(siRNA)である。一実施形態では、IL−23のポリヌクレオチドアンタゴニストは、アデノウイルス、レンチウイルス、レトロウイルス、またはアデノ随伴ウイルスベクターなどの遺伝子治療用ベクター中で送達される。別の実施形態では、IL−23のポリヌクレオチドアンタゴニストは、治療薬として送達される。
【0022】
さらに別の実施形態では、IL−23アンタゴニストは、可溶性受容体ポリペプチドを含む。一実施形態では、IL−23アンタゴニストは、IL−23Rの細胞外ドメインに由来する可溶性断片である。
【0023】
種々の実施形態では、IL−23アンタゴニストは、抗体またはその抗原結合性断片である。種々の実施形態では、抗体またはその抗原結合性断片は、ポリクローナル抗体、モノクローナル抗体、ヒト化抗体、完全ヒト抗体;抗体断片(たとえば、Fab、Fab’、Fab’−SH、Fv、scFv、F(ab’)2、およびダイアボディ(diabody))を含む。他の実施形態では、アンタゴニストは、抗体のペプチド模倣物質を含む。いっそうさらなる実施形態では、抗体またはその抗原結合性断片は、検出可能なように標識される。一実施形態では、IL−23アンタゴニストは、補体活性化の低減、抗体依存性細胞傷害(ADCC)の低減、またはその両方を示す抗体またはその抗原結合性断片である。一実施形態では、エフェクター機能が低減されたIL−23アンタゴニスト抗体またはその断片は、抗IL−23受容体(たとえば、抗IL−23R)抗体または断片である。種々の実施形態では、エフェクター機能が低減された抗体は、抗体断片(たとえば、Fab、Fab’、Fab’−SH、Fv、scFv、F(ab’)2)、IgG4であるか、または改変グリコシル化を有する。
【0024】
一実施形態では、本発明は、IL−23p19、IL−23R、IL−17A、IL−17F、IL−17RA、IL−17RC、IL−6、およびTGF−βからなる群から選択される任意の2つのタンパク質に特異的に結合する有効量の二重特異性抗体の投与により、感染症、たとえば慢性の真菌感染症、細菌感染症、またはウイルス感染症を処置することに関する。一実施形態では、タンパク質はヒトタンパク質である。
【0025】
一実施形態では、IL−23アンタゴニストは、IL−23(またはその受容体)に特異的であり、IL−12(またはその受容体)に拮抗しない。種々の実施形態では、拮抗作用は、in vitro結合アッセイ(たとえば、ELISA)、またはバイオアッセイ(たとえば、BaF3細胞増殖またはTh17細胞産生の促進)により測定される。種々の実施形態では、IL−23とその受容体の結合の阻害に関するIC50に対する、IL−12とその受容体の結合の阻害に関するIC50の比率(IC50IL−12/IC50IL−23)は、1.5、2、3、4、5、7、10、15、20、50、または100以上である。
【0026】
一実施形態では、本発明の方法および組成物は、TBを処置するために使用され、処置の成功は、細菌負荷の低減により測定される。種々の実施形態では、マイコバクテリア負荷は、ツベルクリン検査、マントー試験(Mantoux test)、または臨床試料中のマイコバクテリアDNAまたはRNAの存在により測定される。
【0027】
本発明のいくつかの実施形態では、感染症を有する被験体は、他の方法または組成物を使用して以前に感染症の処置を受けたことがある被験体である。一実施形態では、以前の処置は、感染の除去に有効ではなかった。別の実施形態では、感染症を有する被験体、感染症を有する疑いのある被験体、または感染症に罹るリスクのある被験体は、たとえばAIDS、移植、または化学療法の結果として免疫無防備状態である。
【0028】
本発明は、真菌感染症、持続性真菌感染症、カンジダ症、慢性皮膚粘膜カンジダ症(CMC)、アスペルギルス症、クリプトコックス症、ウイルス感染症、持続性ウイルス感染症、HIV感染、HBV感染、HCV感染、持続性細菌感染症、マイコバクテリア感染症、M.tuberculosis感染、M.bovis感染、およびM.leprae感染からなる群から選択される1つまたは複数の感染症を処置するための薬剤の製造におけるIL−23アンタゴニストの使用をさらに包含する。いくつかの実施形態では、薬剤は、1つまたは複数の追加的な治療薬を含み得る。他の実施形態では、本発明の薬剤は、1つまたは複数の他の治療薬と併せて使用できる。
例えば、本発明は以下の項目を提供する。
(項目1)
感染症を有する被験体を処置するための方法であって、有効量のIL−23アンタゴニストを投与する工程を含む方法。
(項目2)
前記感染症は、ウイルス感染症、真菌感染症、および細菌感染症からなる群から選択される慢性感染症である、項目1に記載の方法。
(項目3)
前記慢性感染症は真菌感染症である、項目2に記載の方法。
(項目4)
前記真菌感染症は、カンジダ症、アスペルギルス症、クリプトコックス症、および爪甲真菌症からなる群から選択される、項目3に記載の方法。
(項目5)
前記真菌感染症はカンジダ症である、項目4に記載の方法。
(項目6)
前記真菌感染症は慢性皮膚粘膜カンジダ症(CMC)である、項目3に記載の方法。
(項目7)
前記被験体に、ポサコナゾール、フルコナゾール、ボリコナゾール、イトラコナゾール、ケトコナゾール、リアロゾール、イルテマゾール、クロトリマゾール、ミコナゾール、エコナゾール、ブトコナゾール、オキシコナゾール、スルコナゾール、チオコナゾール、およびテルコナゾール、チアジアゾール、オキサジアゾール、カスポファンギン、アムホテリシンB、ナイスタチン、ピマリシン、フルシトシン(5−フルオロシトシン)、ナフチフィン、テルビナフィン、ブテナフィン、チオカーボネートトルナフテート、グリセオフルビン、アミオダロン、シクロピロックス、スルベンチン、アモロルフィン、クリオキノール、ゲンチアナバイオレット、ヨウ化カリウム、チオ硫酸ナトリウム、石炭酸フクシン溶液、カスポファンギンアセテート、ミカファンギン、およびアニデュラファンギンからなる群から選択される1つまたは複数の抗真菌物質を投与する工程をさらに含む、項目3に記載の方法。
(項目8)
前記1つまたは複数の抗真菌物質は、アムホテリシンB、フルシトシン、イトラコナゾール、ボリコナゾール、フルコナゾール、またはポサコナゾールである、項目7に記載の方法。
(項目9)
前記慢性感染症は細菌感染症である、項目2に記載の方法。
(項目10)
前記細菌感染症はマイコバクテリア感染症である、項目9に記載の方法。
(項目11)
前記マイコバクテリア感染症は、M.bovis、M.leprae、およびM.tuberculosisからなる群から選択されるマイコバクテリアにより引き起こされる、項目10に記載の方法。
(項目12)
前記マイコバクテリア感染症はTBである、項目10に記載の方法。
(項目13)
前記被験体に、イソニアジド、リファンピン、ピラジナミド、エタンブトール、ストレプトマイシン、シプロフロキサシン、バンコマイシン、およびオフロキサシンからなる群から選択される1つまたは複数の抗菌物質を投与する工程をさらに含む、項目9に記載の方法。
(項目14)
前記抗菌物質はバンコマイシンである、項目13に記載の方法。
(項目15)
前記慢性感染症はウイルス感染症である、項目2に記載の方法。
(項目16)
前記ウイルス感染症は、HIV、HPV、HBV、およびHCVからなる群から選択されるウイルスにより引き起こされる、項目15に記載の方法。
(項目17)
前記ウイルス感染症はHCVにより引き起こされる、項目16に記載の方法。
(項目18)
前記ウイルス感染症はHIVにより引き起こされる、項目16に記載の方法。
(項目19)
前記被験体に、アバカビル、アシクロビル、アマンタジン、アンプレナビル、デラビルジン、ジダノシン、エファビレンツ、ファムシクロビル、インジナビル、インターフェロンアルファ、リバビリン、ラミブジン、ネルフィナビル、ネビラピン、オセルタミビル、ペンシクロビル、リバビリン、リトナビル、サキナビル、スタブジン、バラシクロビル、ザルシタビン、ザナミビル、およびジドブジンからなる群から選択される1つまたは複数の抗ウイルス物質を投与する工程をさらに含む、項目15に記載の方法。
(項目20)
前記抗ウイルス物質はジドブジンである、項目19に記載の方法。
(項目21)
感染症を有する被験体においてTh1免疫応答を増強するための方法であって、IL−23アンタゴニストを投与する工程を含む方法。
(項目22)
前記増強されたTh1免疫応答は、前記IL−23アンタゴニストを投与する前のIFN−γ発現CD4+T細胞の割合と比較して、IFN−γ発現CD4+T細胞の割合が2倍以上に増加する工程を含む、項目21に記載の方法。
(項目23)
前記増強されたTh1免疫応答は、前記IL−23アンタゴニストを投与する前のIL−17発現CD4+T細胞の割合と比較して、IL−17発現CD4+T細胞の割合が2分の1以下に減少する工程を含む、項目21に記載の方法。
(項目24)
前記感染症は慢性真菌感染症である、項目21に記載の方法。
(項目25)
IL−17Aアンタゴニストを投与する工程をさらに含む、前記項目のいずれかに記載の方法。
(項目26)
IL−6アンタゴニストを投与する工程をさらに含む、前記項目のいずれかに記載の方法。
(項目27)
TGF−βアンタゴニストを投与する工程をさらに含む、前記項目のいずれかに記載の方法。
(項目28)
IL−12またはIL−12アゴニストを投与する工程をさらに含む、前記項目のいずれかに記載の方法。
(項目29)
前記IL−23アンタゴニストは、IL−23p19に結合する結合性化合物である、前記項目のいずれかに記載の方法。
(項目30)
前記IL−23アンタゴニストは、IL−23Rに結合する結合性化合物である、前記項目のいずれかに記載の方法。
(項目31)
前記結合性化合物は抗体またはその抗原結合性断片である、項目29または項目30に記載の方法。
(項目32)
IL−23とその受容体との結合の阻害に関するIC50に対する、IL−12とその受容体との結合の阻害に関するIC50の比率(IC50IL−12/IC50IL−23)は、ELISAにより測定された場合に5以上である、項目29または項目30に記載の方法。
(項目33)
前記結合性化合物は、Fab、Fab’、Fab’−SH、Fv、scFv、F(ab’)2、およびダイアボディからなる群から選択される抗体断片である、項目31に記載の方法。
(項目34)
前記抗体はヒト化抗体である、項目31に記載の方法。
(項目35)
前記抗体は完全ヒト抗体である、項目31に記載の方法。
(項目36)
前記結合性化合物は単鎖抗体である、項目29または項目30に記載の方法。
(項目37)
前記抗体は、補体活性化、ADCC、またはその両方を低減するエフェクター機能を変更した抗体である、項目31に記載の方法。
(項目38)
前記結合性化合物は、IL−23Rに由来する可溶性受容体断片である、項目29に記載の方法。
(項目39)
前記IL−23アンタゴニストはsiRNAである、項目1から28のいずれかに記載の方法。
(項目40)
前記IL−23アンタゴニストはアンチセンス核酸である、項目1から28のいずれかに記載の方法。
(項目41)
前記感染症を有する前記被験体は、他の方法または組成物を使用して以前に該感染症の処置を受けたことがある被験体である、前記項目のいずれかに記載の方法。
(項目42)
前記感染症を有する前記被験体は免疫無防備状態である、前記項目のいずれかに記載の方法。
(項目43)
感染症を処置するための医薬の製造におけるIL−23アンタゴニストの使用。
(項目44)
前記感染症は慢性真菌感染症である、項目43に記載の使用。
(項目45)
感染症を有する疑いのある被験体を処置するための方法であって、有効量のIL−23アンタゴニストを投与する工程を含む方法。
(項目46)
感染症に罹るリスクのある被験体を処置するための方法であって、有効量のIL−23アンタゴニストを投与する工程を含む方法。
(項目47)
感染症を有する被験体を処置するための方法であって、有効量のIL−17AもしくはIL−17Fのアンタゴニスト、またはその両方を投与することを含む方法。
(項目48)
感染症を処置するための方法であって、IL−23p19、IL−23R、IL−17A、IL−17F、IL−17RA、IL−17RC、IL−6、およびTGF−βからなる群から選択される任意の2つのタンパク質に特異的に結合する有効量の二重特異性抗体を投与する工程を含む方法。
(項目49)
前記タンパク質はヒトタンパク質である、項目48に記載の方法。
(項目50)
本明細書に記載の任意の発明。
【図面の簡単な説明】
【0029】
【図1−1】カンジダ症に対する感受性におけるIL−23/IL−17依存性経路の役割に関する実験の結果を示す図である。マウスに108個の毒性Candidaを胃内注入した。結果は3回の実験から集める(1回の実験につき1群当たり6匹のマウス)。図1Aは、p19−/−、p35−/−、p40−/−およびC57BL/6(野生型)マウスの経時的な生存率(%)を示す図である。図1Bは、感染3日後および感染10日後の胃中の真菌増殖(CFU)を示す図である。3日後と10日後の両時点でC57BL/6マウスと比較したとき、p19−/−、p35−/−またはp40−/−マウスに関して結果は統計的差異があった(p<0.05、*によって示す)。図1Cは、MLNにおけるp35およびp19のmRNA発現(感染翌日)ならびにIL−12β2RおよびIL−23RのmRNA発現(感染3日後)を示す図である。mRNA発現はリアルタイムRT−PCRによって測定した。図1Dは、感染1週間後のIFN−γ、IL−17またはIL−4産生MLN CD4+細胞の頻度を示す図である。サイトカイン産生細胞の頻度はELISPOTアッセイにより測定し、値は細胞105個当たりのサイトカイン産生細胞の平均数(±SE)である。図1Eは、感染3日後の胃ホモジネート中の炎症性サイトカイン(IL−17、IL−23、IFN−γ、IL−12)のレベルを示す図である。サイトカインはELISAによって測定した(pg/ml)。図1C〜1Eにおいて、図に示されるように、感染マウス(+)と非感染マウス(−)を比較したとき(*)、およびp19−/−またはp35−/−マウスとC57BL/6マウスを比較したとき(**)、統計的有意差があった(p<0.05)。
【図1−2】カンジダ症に対する感受性におけるIL−23/IL−17依存性経路の役割に関する実験の結果を示す図である。マウスに108個の毒性Candidaを胃内注入した。結果は3回の実験から集める(1回の実験につき1群当たり6匹のマウス)。図1Aは、p19−/−、p35−/−、p40−/−およびC57BL/6(野生型)マウスの経時的な生存率(%)を示す図である。図1Bは、感染3日後および感染10日後の胃中の真菌増殖(CFU)を示す図である。3日後と10日後の両時点でC57BL/6マウスと比較したとき、p19−/−、p35−/−またはp40−/−マウスに関して結果は統計的差異があった(p<0.05、*によって示す)。図1Cは、MLNにおけるp35およびp19のmRNA発現(感染翌日)ならびにIL−12β2RおよびIL−23RのmRNA発現(感染3日後)を示す図である。mRNA発現はリアルタイムRT−PCRによって測定した。図1Dは、感染1週間後のIFN−γ、IL−17またはIL−4産生MLN CD4+細胞の頻度を示す図である。サイトカイン産生細胞の頻度はELISPOTアッセイにより測定し、値は細胞105個当たりのサイトカイン産生細胞の平均数(±SE)である。図1Eは、感染3日後の胃ホモジネート中の炎症性サイトカイン(IL−17、IL−23、IFN−γ、IL−12)のレベルを示す図である。サイトカインはELISAによって測定した(pg/ml)。図1C〜1Eにおいて、図に示されるように、感染マウス(+)と非感染マウス(−)を比較したとき(*)、およびp19−/−またはp35−/−マウスとC57BL/6マウスを比較したとき(**)、統計的有意差があった(p<0.05)。
【図2】アスペルギルス症に対する感受性におけるIL−23/IL−17依存性経路の役割に関する実験の結果を示す図である。マウスに2×107個のAspergillus静止期分生子を鼻腔内感染させた。図2Aおよび2B中に示す結果は4回の実験から集める(動物6匹/群)。図2Aは、感染3日後の肺中の真菌増殖(キチン含有量は、器官当たりのグルコサミンのμg数として表す)を示す図である。p19−/−、p35−/−またはp40−/−マウスとC57BL/6マウスを比較したとき(*)、統計的有意差があった(p<0.05)。図2Bは、TLNにおけるp35/p19のmRNA発現(感染翌日)ならびにIL−12β2R/IL−23RのmRNA発現(感染3日後)を示す図である。メッセンジャーRNAの発現はRT−PCRによって測定した。図に示されるように、感染マウス(+)と非感染マウス(−)を比較したとき(*)、およびp19−/−またはp35−/−マウスとC57BL/6マウスを比較したとき(**)、統計的有意差があった(p<0.05)。
【図3】真菌感染症に対する感受性におけるIL−23/IL−17依存性経路の重要性に関する実験の結果を示す図である。マウスは図1および2と同様に感染させ、感染5時間後に200μgのp19またはIL−17中和抗体、または感染5時間後と24時間後に1mgのTGF−β中和抗体で処置した。図3Aは、感染3日後のカンジダ症(C.albicans)またはアスペルギルス症(A.fumigatus)を有するマウスの胃または肺中での真菌増殖を示す図である。図に示されるように、処置マウス(+)と未処置マウス(−)を比較したとき(*)、統計的有意差があった(p<0.05)。図3Bは、ELISPOTアッセイにより測定した、それぞれカンジダ症またはアスペルギルス症を有するマウス由来のMLNまたはTLN由来のIFN−γまたはIL−17産生CD4+細胞の頻度を示す図である。値は細胞105個当たりのサイトカイン産生細胞の平均数(±SE)である。図3Bは、抗原刺激した未分画MLNまたはTLNの培養物上清における実際のIL−17産生(感染1週間後)をさらに示す。図に示されるように、感染マウスと非感染(Ct)マウスを比較したとき(*)、および処置マウス(+)と未処置マウス(−)を比較したとき(**)、統計的有意差があった(p<0.05)。図3Cは、感染3日後の、前述のp19中和抗体で処置した、カンジダ症を有するマウスの胃中での真菌増殖を示す図である。図に示されるように、処置(+)マウスと未処置(−)マウスを比較したとき(*)、およびIL−4−/−、IFN−γ−/−、p35−/−またはIFN−γ−/−/p35−/−マウスとBALB/cマウスを比較したとき(**)、統計的有意差があった(p<0.05)。
【図4】真菌に応答したDCサブセットにおけるIL−23およびIL−12産生に関する実験の結果を示す図である。GM−CSF+IL−4(GM−DC)またはFLT3−L(FL−DC)の存在下で得た骨髄DCは真菌で刺激し、サイトカインの発現を評価した。図4Aは、サイトカインmRNA発現のリアルタイムRT−PCR分析を示す図である。図4Bは、ELISAによって測定したサイトカインの発現(pg/ml)を示す図である。ザイモザン、LPS(10μg/ml)またはCpG−ODN2006(0.06μM)を陽性対照として使用した。DCは10:1の比で酵母に曝露した。図に示されるように、曝露DCと非曝露(「曝露なし」)DCを比較したとき(*)、統計的有意差があった(p<0.05)。図4Cは、p19−/−またはp35−/−マウス由来の脾性CD11c+DCにおけるIL−12およびIL−23産生を示す図である。培養物上清中のサイトカインの測定前に真菌でマウスを刺激した。図4Dは、図に示されるように、IL−12またはIL−23(10ng/ml)の存在(+)または不在(−)、または中和抗IL−12または抗IL−23抗体(10μg/ml)の存在下で、12時間真菌に曝露したC57BL/6マウス由来の脾性CD11c+DCにおけるIL−12およびIL−23産生を示す図である。
【図5】真菌に応答した炎症性DCによるIL−23産生に関する実験の結果、および特にこのような産生がTLRおよびT細胞依存性であるかどうかを示す図である。図5Aは、12時間前に真菌に曝露した異なる型のマウス由来の脾性CD11c+DCにおけるIL−23産生(pg/ml)を示す図である。4回の実験から集めた結果を示す。図に示されるように、曝露DCと非曝露(「曝露なし」)DCを比較したとき(*)、統計的有意差があった(p<0.05)。図5Bは、様々な細胞培養物および同時培養物におけるサイトカインの発現を示す図である。C57BL/6(野生型)またはp35−/−マウス由来の脾性CD4+T細胞を、パルスしていない(群2および5)またはカンジダ酵母(Ag)でパルスした(群3および6)対応する脾性DCの存在下で培養した。サイトカイン(IL−12、IL−23、IFN−γ、IL−17)を、パルス5日後にELISAによって測定した。群1および4は、真菌で刺激しT細胞では刺激しなかったC57BL/6またはp35−/−DCである。群7および8は、真菌の存在下でそれぞれC57BL/6またはp35−/−DCと共に培養した、p35−/−またはC57BL/6CD4+T細胞である。図に示されるように、群3および7をIFN−γ産生に関して群1と比較したとき、および群6および8をIL−23およびIL−17産生に関して群4と比較したとき、統計的有意差がある(p<0.05、*によって示す)。図5Cは、サンプルのいくつかが抗IL−23または抗TGF−β抗体を含むこと以外、図5B中に示したデータと同様のデータを示す図である。C57BL/6(野生型)(群1〜3)またはp35−/−(群4〜6)マウス由来の脾性CD4+T細胞を、対応する脾性CDの存在下で培養した。培養物は10μg/mlのIL−23またはTGF−β中和抗体の存在下で5日間カンジダ酵母(Ag)でパルスし、かつサイトカイン(IFN−γ、IL−17)は、ELISAによって培養物上清において定量化した。図に示されるように、群2および3をIFN−γおよびIL−17産生に関して群1と比較したとき、および群5をIL−17産生に関して群4と比較したとき、統計的有意差がある(p<0.05、*によって示す)。
【図6】PMNの、抗真菌エフェクター機能を障害し抗炎症性プログラムを破壊するIL−23およびIL−17の能力に関する実験の結果を示す図である。図6Aは、37℃における5:1のエフェクターと真菌細胞の比での非オプソニン化酵母(30分間)または分生子(60分間)とのインキュベーション後の、C57BL/6(野生型)、p19−/−またはp35−/−マウス由来のPMNにおける抗真菌活性を示す図である。結果はコロニー形成単位阻害の割合(平均±SE)としてプロットする。結果は3回の実験から集めたデータを反映する。図に示されるように、p19−/−またはp35−/−PMNとC57BL/6(野生型)PMNを比較したとき(*)、統計的有意差があった(p<0.05)。図6Bは、示した濃度でIL−23またはIL−17に曝露したC57BL/6(野生型)マウス由来のPMNの抗真菌活性を示す図である。サイトカイン曝露PMNと非曝露PMNを比較したとき(*)、統計的有意差があった(p<0.05)。図6Cおよび6Dは、60分間IFN−γ(50ng/ml)、IL−23(100ng/ml)およびIL−17(100ng/ml)の様々な組合せに曝露したC57BL/6(野生型)マウス由来のPMNの抗真菌活性を示す図である。カンジダ酵母またはAspergillus分生子に対する抗真菌活性を測定した(図6C)。MMP9/MPO産生も測定した(図6D)。ゼラチナーゼおよびミエロペロキシダーゼの産生はゼラチンザイモグラフィーによって評価し、かつウエスタンブロット分析を培養物上清に実施した。ゲルは、活性のある92kDaのMMP9および60kDaのMPOに対応するバンドを示す。図に示されるように、サイトカイン曝露PMNと非曝露PMNを比較したとき(*)、および(IFN−γ+IL−23)または(IFN−γ+IL−17)曝露PMNとIFN−γ曝露PMNを比較したとき(**)、統計的有意差があった(p<0.05)。図6Eは、ウエスタンブロット上のバンドを示す図である。PMNは12時間インビトロでIFN−γ、IL−23およびIL−17の様々な組合せに曝露した。次いでIDOタンパク質の発現をウエスタンブロッティングによって決定した。IDO発現MC24トランスフェクタントおよびモックトランスフェクトMC22細胞は、それぞれ陽性対照および陰性対照として働いた。β−チューブリンはローディング対照として働く。
【発明を実施するための形態】
【0030】
添付の特許請求の範囲を含む本明細書で使用される場合、「a」、「an」、および「the」などの単数形の単語は、文脈上明確に別段の記載がない限り、それらの対応する複数の参照を含む。特に明記のない限り、本明細書で提供された典型的な実施形態は、本発明の範囲を制限するとは見なされない。「たとえば(e.g.)」、「たとえば(for example)」、「一実施形態では」などの語句または他のそのような非限定的な語が、そのような典型的な実施形態に先行してもよく、またはそれらの典型的な性質が、文脈(たとえば「実施例」)から明白であってもよい。特に明記のない限り、「阻害しない」などの用語は、絶対的というよりむしろ相対的であると意図される。たとえば、IL−23を阻害するがIL−12は阻害「しない」作用剤は、2つのサイトカインの比較可能なアッセイにおいて作用剤が所定の濃度で存在する場合に、IL−23よりIL−12の阻害に効果が低い作用剤を指す。
【0031】
本明細書で引用されるすべての参考文献は、あたかも各個々の刊行物、データベースエントリー、特許出願、または特許が、参照によって具体的におよび個別に組み込まれるのと同程度に、参照によって全体が組み込まれる。
【0032】
I.定義。
【0033】
「活性化」、「刺激」、および「処置」は、細胞または受容体に適用される場合、文脈によりまたは明示的に特に明記のない限り、同じ意味を有していてもよく、たとえば細胞または受容体をリガンドで活性化、刺激、または処置することである。「リガンド」は、天然および合成リガンド、たとえばサイトカイン、サイトカイン改変体、類似体、ムテイン(mutein)、および抗体に由来する結合性組成物を包含する。「リガンド」は、小分子(small molecule)、たとえばサイトカインのペプチド模倣物質および抗体のペプチド模倣物質も包含する。「活性化」は、内部機構によりならびに外部的または環境的要因により制御されるような細胞活性化を指すことができる。たとえば、細胞、組織、器官、または生物の「応答」は、生化学的なもしくは生理学的な挙動、たとえば濃度、密度、接着、もしくは生物学的区画内での遊走、遺伝子発現の速さ、または分化の状態における変化を包含し、この変化は、活性化、刺激、もしくは処置と関連があるか、または遺伝プログラミングなどの内部機構と関連がある。
【0034】
分子の「活性」は、リガンドまたは受容体への分子の結合、触媒活性;遺伝子発現もしくは細胞シグナル伝達、分化、または成熟化を刺激する能力;抗原活性、他の分子の活性の調節などを記述するかまたはそれらを指すことができる。分子の「活性」は、細胞間相互作用、たとえば接着を調節または維持する活性、または細胞の構造、たとえば細胞膜または細胞骨格を維持する活性を指すこともできる。「活性」は、比活性、たとえば、[触媒活性]/[mgタンパク質]、もしくは[免疫学的活性]/[mgタンパク質]、または生物学的区画における濃度などを意味することもできる。「増殖活性」は、たとえば、正常な細胞分裂ならびに癌、腫瘍、異形成、細胞の形質転換、転移、および新脈管形成を促進するまたはそれに必要であるまたはそれに特異的に関連する活性を包含する。
【0035】
「投与」および「処置」は、動物、ヒト、実験被験体、細胞、組織、器官、または生物学的液体に適用される場合、外因性の医薬品、治療剤、診断用薬、または組成物の、動物、ヒト、被験体、細胞、組織、器官、または生物学的液体に対する接触を指す。「投与」および「処置」は、たとえば、治療方法、薬物動態学的方法、診断方法、研究方法、および実験方法を指すことができる。細胞の処置は、細胞に対する試薬の接触および細胞と接触している体液に対する試薬の接触を包含する。「投与」および「処置」はまた、たとえば、試薬、診断薬、結合性組成物による、または他の細胞による細胞のインビトロおよび生体外(ex vivo)での処置をも意味する。「処置」は、ヒト被験体、獣医学的被験体、または研究被験体に適用される場合、治療処置、予防的または防止的手段、研究への応用、および診断への応用を指す。「処置」は、ヒト被験体、獣医学的被験体、もしくは研究被験体または細胞、組織、もしくは器官に適用される場合、IL−23またはIL−23Rアンタゴニストの、ヒト被験体または動物被験体、細胞、組織、生理的コンパートメント、または生理的液体に対する接触を包含する。「細胞の処置」はまた、IL−23またはIL−23Rアンタゴニストが、たとえば体液相またはコロイド相中でIL−23R複合体(IL−23R/IL−12Rベータ1ヘテロ二量体)と接触する状況だけではなく、アンタゴニストが細胞または受容体と接触しない状況をも包含する。
【0036】
「結合性組成物」は、標的に結合することができる分子、小分子、高分子、抗体、その断片もしくは類似体、または可溶性受容体を指す。「結合性組成物」はまた、標的に結合することができる、分子の複合体、たとえば非共有結合複合体、イオン化分子、および共有結合でまたは非共有結合で改変された分子、たとえばリン酸化、アシル化、架橋、環化、または限定切断で改変された分子を指すものであってもよい。「結合性組成物」はまた、安定剤、賦形剤、塩、緩衝剤、溶媒、または添加剤と組み合わせた、標的に結合することができる分子を指すものであってもよい。「結合」は、結合性組成物の標的との関連性として定義されてもよく、その関連性は、結合性組成物が溶液中に溶解するまたは懸濁することができる場合、結合性組成物の通常のブラウン運動における低下をもたらす。
【0037】
本発明の結合性化合物は、二重特異性抗体を含むことができる。本明細書に使用される場合、用語「二重特異性抗体」は、少なくとも2つの異なる抗原エピトープに対する結合特異性を有する抗体、典型的にはモノクローナル抗体を指す。一実施形態では、エピトープは、同じ抗原に由来する。他の実施形態では、エピトープは、2つの異なる抗原に由来する。二重特異性抗体を作製するための方法は、当技術分野で知られている。たとえば、二重特異性抗体は、2つの免疫グロブリン重鎖/軽鎖対の同時発現を使用して組換えで産生することができる。たとえばMilsteinら(1983年)Nature 305:537〜39頁を参照されたい。あるいは、二重特異性抗体は、化学的連結を用いて調製することができる。たとえばBrennanら(1985年)Science 229:81頁を参照されたい。二重特異性抗体は、二重特異性抗体の断片を含む。たとえばHolligerら(1993年)Proc. Natl. Acad. Sci. U.S.A. 90:6444〜48頁、Gruberら、J. Immunol. 152:5368頁(1994年)を参照されたい。
【0038】
「古典的TH1型T細胞」とは、IL−4、IL−5、またはIL−13の各々の発現を超える程度まで、インターフェロン−ガンマ(IFNγ)を発現するT細胞であり、一方で「古典的TH2型T細胞」とは、各々がIFNγの発現を超える程度まで、IL−4、IL−5、またはIL−13を発現するT細胞である。「程度」とは、古典的TH2型細胞の、典型的には4倍以上、より典型的には8倍以上、および最も典型的には16倍以上である。
【0039】
本明細書で定義される「記憶T細胞」とは、所定の抗原に以前曝露されている長寿命のT細胞のサブセットである。記憶T細胞は、生物中に何年間も存在することができ、同じ抗原によるその後の曝露に対する迅速な応答を可能にする。マウス記憶T細胞の表現型は、CD4+highCD45RBlowと定義される。ヒト記憶T細胞の表現型は、CD45RAneg/lowCD45R0highと定義される。これらの記憶T細胞のIL−23処置は、増殖およびIL−17の発現に帰着する。特に明記のない限り、「IL−17」とは、本明細書で使用される場合、IL−17Aを指す。たとえば、Moseleyら(2003年)Cytokine & Growth Factor Rev. 第14巻:155頁を参照されたい。
【0040】
「保存的修飾改変体」とは、アミノ酸配列および核酸配列の両方に適用される。特定の核酸配列に関して、保存的修飾改変体とは、同一または本質的に同一なアミノ酸配列をコードするこれらの核酸を指すか、または核酸がアミノ酸配列をコードしない場合は、本質的に同一の核酸配列を指す。遺伝コードの縮重のため、多数の機能的に同一な核酸が、任意の所定のタンパク質をコードできる。
【0041】
アミノ酸配列に関しては、当業者であれば、コードされた配列において1つのアミノ酸またはわずかな割合のアミノ酸を、保存されたアミノ酸に置換する、核酸、ペプチド、ポリペプチド、またはタンパク質配列における変化が、「保存的修飾改変体」であることを認識する。機能的に類似のアミノ酸を提供する保存的置換表は、当技術分野で周知である。保存的置換の一例は、以下の群のもののうち1つのアミノ酸を同じ群の別のアミノ酸に交換することである:(Leeらに交付された米国特許第5,767,063号の明細書;Kyte and Doolittle(1982年)J. Mol. Biol. 第157巻:105〜132頁):
(1)疎水性:ノルロイシン、Ile、Val、Leu、Phe、Cys、またはMet;
(2)中性親水性:Cys、Ser、Thr;
(3)酸性:Asp、Glu;
(4)塩基性:Asn、Gln、His、Lys、Arg;
(5)鎖の配向に影響を及ぼす残基:Gly、Pro;
(6)芳香族:Trp、Tyr、Phe;
(7)小型アミノ酸:Gly、Ala、Ser。
【0042】
「有効量」は、病状の症候または徴候を改善または予防するのに十分な量を包含する。有効量は、診断を可能または容易にするのに十分な量も意味する。特定の患者または獣医学的被験体のための有効量は、処置されている状態、患者の健康全般、投与の方法、経路、および用量、ならびに副作用の重症度などの要因に依存して変動し得る。たとえば米国特許第5,888,530号明細書を参照されたい。有効量は、著しい副作用または毒性作用を回避する最大の用量または投薬プロトコルであり得る。効果は、診断の尺度またはパラメーターが、正常被験体により示される診断パラメーターを100%と定義した場合、少なくとも5%、通常は少なくとも10%、より通常は少なくとも20%、最も通常は少なくとも30%、好ましくは少なくとも40%、より好ましくは少なくとも50%、最も好ましくは少なくとも60%、理想的には少なくとも70%、より理想的には少なくとも80%、および最も理想的には少なくとも90%改善することに帰着する。たとえば、Maynardら(1996年)A Handbook of SOPs for Good Clinical Practice、Interpharm Press、ボーカラトーン、米国フロリダ州;Dent(2001年)Good Laboratory
and Good Clinical Practice、Urch Publ.、ロンドン、英国を参照されたい。
【0043】
「外因性」とは、文脈に依存して、生物、細胞、または人体の外部で産生される物質を指す。「内因性」とは、文脈に依存して、細胞、生物、または人体の内部で産生される物質を指す。
【0044】
「感染性疾患」とは、生物、器官、組織、または細胞の微生物感染症、たとえば、細菌感染症、ウイルス感染症、および/または真菌感染症を指す。
【0045】
「IL−17産生細胞」とは、古典的TH1型T細胞または古典的TH2型T細胞ではないT細胞を意味する。「IL−17産生細胞」は、遺伝子またはポリペプチド(たとえば、分裂促進因子応答性P−タンパク質;ケモカインリガンド2;インターロイキン−17(IL−17);転写因子ROR−γT関連;サイトカインシグナル伝達抑制因子3など)を発現するT細胞も意味し、IL−23アゴニストで処置した発現は、IL−12アゴニストでの処置より高く、「より高い」とは以下の通り定義される。IL−23アゴニストでの発現は、IL−12処置よりも、普通は少なくとも5倍高く、典型的には少なくとも10倍高く、より典型的には少なくとも15倍高く、最も典型的には少なくとも20倍高く、好ましくは少なくとも25倍高く、および最も好ましくは少なくとも30倍高い。発現は、たとえば、実質的に純粋なIL−17産生細胞の集団を処置して測定できる。
【0046】
さらに、「IL−17産生細胞」は、細胞発生または細胞分化の経路において、上記で定義されたIL−17産生細胞への分化に決定された前駆体または前駆細胞を含む。IL−17産生細胞への前駆体または前駆細胞は、流入領域リンパ節(DLN)に見出すことができる。さらに、「IL−17産生細胞」は、たとえばホルボールエステル、イオノフォア、および/または発癌物質により、たとえば活性化され、さらには分化され、保管され、冷凍され、乾燥され、不活性化され、たとえばアポトーシス、タンパク質分解、または脂質酸化により部分的に分解され、またはたとえば組換え技術により改変された、上記で定義されたIL−17産生細胞を包含する。
【0047】
「阻害剤」および「アンタゴニスト」とは、たとえばリガンド、受容体、補助因子、遺伝子、細胞、組織、または器官を阻害する阻害分子を指す。遺伝子、受容体、リガンド、または細胞の調節因子とは、遺伝子、受容体、リガンド、または細胞の活性を変化させる分子であり、活性は、その制御特性において活性化、阻害、または変化させることができる。調節因子は、単独で機能してもよく、または補助因子、たとえばタンパク質、金属イオン、もしくは小分子を使用してもよい。阻害剤とは、たとえば遺伝子、タンパク質、リガンド、受容体、または細胞を減少、阻止、防止、活性化遅延、不活性化、除感作、または下方制御する化合物である。活性化剤とは、たとえば遺伝子、タンパク質、リガンド、受容体、または細胞を増加、活性化、促進、活性化増強、感作、または上方制御する化合物である。阻害剤は、恒常的活性を低減、阻止、または不活性化する組成物とも定義できる。「アンタゴニスト」とは、アゴニストの作用に対抗する化合物である。アンタゴニストは、アゴニストの活性を防止、低減、阻害、または中和する。アンタゴニストは、同定されたアゴニストが存在しない場合でさえ、標的、たとえば標的受容体の恒常的活性を防止、阻害、または低減することもできる。
【0048】
IL−23アンタゴニストは、たとえば、以下でより詳細に記述するTh17細胞の増幅および生存などの、IL−23の生物活性を混乱させるあらゆる作用剤を含む。IL−23受容体およびIL−23Rのアンタゴニストは、IL−23シグナル伝達を阻止することによりIL−23の活性を阻止する役目を果たすため、IL−23アンタゴニストのサブセットである。
【0049】
阻害の程度を検討するためには、たとえば、所定のタンパク質、遺伝子、細胞、または生物を含む試料またはアッセイを、有望な活性化剤または阻害剤で処理し、阻害剤無しの対照試料と比較する。対照試料、つまりアンタゴニストで処理されなかった試料には、100%の相対的活性値が割り当てられる。対照と比較した活性値が、約90%以下、典型的には85%以下、より典型的には80%以下、最も典型的には75%以下、一般的には70%以下、より一般的には65%以下、最も一般的には60%以下、典型的には55%以下、通常は50%以下、より通常は45%以下、最も通常は40%以下、好ましくは35%以下、より好ましくは30%以下、さらにより好ましくは25%以下、および最も好ましくは25%未満である場合、阻害は達成される。対照と比較した活性値が、約110%、一般的には少なくとも120%、より一般的には少なくとも140%、より一般的には少なくとも160%、多くの場合少なくとも180%、より多くの場合少なくとも2倍、最も多くの場合少なくとも2.5倍、通常は少なくとも5倍、より通常は少なくとも10倍、好ましくは少なくとも20倍、より好ましくは少なくとも40倍、および最も好ましくは40倍を超えてより高い場合、活性化は達成される。
【0050】
活性化または阻害の終点(endpoint)は、以下のようにモニターできる。たとえば細胞、生理的液体、組織、器官、および動物被験体またはヒト被験体の活性化、阻害、および処置に対する応答は、終点によりモニターできる。終点は、あらかじめ定められた量または割合の、たとえば細菌負荷低減、腫瘍原性低減、もしくは細胞脱顆粒低減、またはサイトカイン、毒性酸素、もしくはプロテアーゼの放出などの分泌低減の徴候を含み得る。終点は、たとえば、あらかじめ定められた量のイオンフラックス(flux)または輸送;細胞遊走;細胞接着;細胞増殖;転移能;細胞分化;および表現型の変化、たとえば炎症、アポトーシス、形質転換、細胞周期、または転移に関連する遺伝子発現の変化を含み得る。たとえば、Knight(2000年)Ann. Clin. Lab. Sci. 第30巻:145〜158頁;Hood and Cheresh(2002年)Nature Rev. Cancer 第2巻:91〜100頁;Timmeら(2003年)Curr. Drug Targets 第4巻:251〜261頁; Robbins and Itzkowitz(2002年)Med. Clin. North Am. 第86巻:1467〜1495頁;Grady and Markowitz(2002年)Annu. Rev. Genomics Hum. Genet. 第3巻:101〜128頁;Bauerら(2001年)Glia 第36巻:235〜243頁;Stanimirovic and Satoh(2000年)Brain Pathol. 第10巻:113〜126頁を参照されたい。
【0051】
阻害の終点は、一般的には対照の75%以下、好ましくは対照の50%以下、より好ましくは対照の25%以下、最も好ましくは対照の10%以下である。一般的には、活性化の終点は、少なくとも150%対照、好ましくは少なくとも2倍対照、より好ましくは少なくとも4倍対照、および最も好ましくは少なくとも10倍対照である。
【0052】
「ノックアウト」(KO)とは、遺伝子によりコードされたポリペプチド、たとえばIL−23またはIL−23受容体のサブユニットをコードするポリペプチドの少なくとも一部の発現の部分的または完全な低減を指し、その遺伝子は、哺乳類の単一の細胞、選択された細胞、またはすべての細胞に内因性である。KOは、生物学的な機能は低減されるが発現は必ずしも低減されない実施形態、たとえば挿入された不活性化ペプチドを含有するポリペプチドも包含する。コード配列または制御配列の破壊は、ノックアウト技術により包含される。細胞または哺乳動物は、内因性遺伝子の1つの対立遺伝子が破壊された「ヘテロ接合性ノックアウト」であり得る。あるいは、細胞または哺乳動物は、内因性遺伝子の両対立遺伝子が破壊された「ホモ接合性ノックアウト」であり得る。「ホモ接合性ノックアウト」は、両対立遺伝子の破壊を同一技術またはゲノムでの同一結果に限定するとは意図されていない。
【0053】
「標識された」組成物は、分光法、光化学法、生化学法、免疫化学法、同位体法、または化学的方法により、直接的または間接的のいずれかで検出可能である。たとえば、有用な標識には、32P、33P、35S、14C、3H、125I、安定同位体、蛍光色素、高電子密度の試薬、基質、エピトープタグ、またはたとえば酵素結合免疫測定(enzyme−linked immunoassay)もしくはフルオレッテ(fluorette)で使用されるような酵素が含まれる。たとえば、Rozinov and Nolan(1998年)Chem. Biol. 第5巻:713〜728頁を参照されたい。
【0054】
「リガンド」とは、受容体のアゴニストまたはアンタゴニストとして作用し得る、たとえば、小分子、ペプチド、ポリペプチド、および膜関連もしくは膜結合分子、またはそれらの複合体を指す。「リガンド」は、アゴニストまたはアンタゴニストではないが受容体に結合できる作用剤も包含する。さらに、「リガンド」は、たとえば化学的方法または組換え法により、可溶型の膜結合リガンドに変更された膜結合リガンドを含む。通常は、リガンドが第1の細胞に膜結合している場合、通常第2の細胞に受容体が生じる。第2の細胞は、第1の細胞と同じまたは異なる同一性を有していてよい。リガンドまたは受容体は、完全に細胞内性であってもよく、すなわち、細胞質ゾル、核、またはいくつかの他の細胞内区画に存在していてもよい。リガンドまたは受容体は、たとえば、細胞内区画から原形質膜の外部面へとその位置を変化させることができる。リガンドおよび受容体の複合体は、「リガンド受容体複合体」と名付けられる。リガンドおよび受容体がシグナル伝達経路に関与する場合、リガンドはシグナル伝達経路の上流位置で生じ、受容体は下流位置で生じる。
【0055】
「マーカー」は、細胞、組織、器官、動物の表現型、たとえばIL−17産生細胞の表現型に関する。マーカーは、たとえば、細胞の精製中、定量化中、遊走中、活性化中、成熟中、または発生中に細胞を検出するために使用され、in vitroおよびin vivo研究の両方で使用できる。活性化マーカーとは、細胞活性化に関連するマーカーである。
【0056】
「精製された細胞」は、たとえば、他のタイプの細胞が実質的に存在しない、たとえば他のタイプのT細胞による汚染が実質的に存在しない1つまたは複数の「IL−17産生細胞」を包含する。純度は、幾何学的な座標により、またはたとえばフラスコ、チューブ、またはバイアルを含む区画により定義される容積を使用することにより算定できる。「精製されたIL−17産生細胞」は、たとえば、「IL−17産生細胞」が、通常は全細胞の少なくとも20%、より通常には全細胞の少なくとも30%、最も通常には全細胞の少なくとも40%、一般的には全細胞の少なくとも50%、より一般的には全細胞の少なくとも60%、最も一般的には全細胞の少なくとも70%、好ましくは全細胞の少なくとも80%、より好ましくは全細胞の少なくとも90%;最も好ましくは全細胞の少なくとも95%を占める区画により定義できる。
【0057】
「小分子」は、10kD未満、典型的には2kD未満、および好ましくは1kD未満の分子量を有する分子として定義される。小分子には、それらに限定されないが、無機分子、有機分子、無機成分を含有する有機分子、放射性原子を含む分子、合成分子、ペプチド模倣物質、および抗体模倣物質が含まれる。治療薬として、小分子は、大分子より細胞に対してより透過性があり、分解を受けにくく、免疫応答を誘発する傾向が少ない場合がある。抗体およびサイトカインのペプチド模倣物質などの小分子ならびに小分子トキシンが、当技術分野で知られている。たとえば、Cassetら(2003年)Biochem. Biophys. Res. Commun. 第307巻:198〜205頁;Muyldermans(2001年)J. Biotechnol. 第74巻:277〜302頁;Li(2000年)Nat. Biotechnol. 第18巻:1251〜1256頁;Apostolopoulosら(2002年)Curr. Med.
Chem. 第9巻:411〜420頁;Monfardiniら(2002年)Curr. Pharm. Des. 第8巻:2185〜2199頁;Dominguesら(1999年)Nat. Struct. Biol. 第6巻:652〜656頁;Sato and Sone(2003年)Biochem. J. 第371巻:603〜608頁;米国特許第6,326,482号明細書を参照されたい。
【0058】
リガンド/受容体、抗体/抗原、または他の結合対に関する場合、「特異的に」または「選択的に」結合するとは、タンパク質および他の生物製剤の不均一集団におけるタンパク質の存在の決定要因である結合反応を示す。したがって、指定された条件下で、特定のリガンドは特定の受容体に結合し、試料中に存在する他のタンパク質に対しては相当量は結合しない。意図された方法の抗体、または抗体の抗原結合部位に由来する結合性組成物は、任意の他の抗体またはそれに由来する結合性組成物の親和性より、少なくとも2倍高い、好ましくは少なくとも10倍高い、より好ましくは少なくとも20倍高い、および最も好ましくは少なくとも100倍高い親和性で、その抗原またはその改変体もしくはムテインに結合する。好ましい実施形態では、抗体は、たとえばスキャチャード解析により決定されるように、約109リットル/molより高い、所望の標的に対する親和性を有す。Munsenら(1980年)Analyt. Biochem. 第107巻:220〜239頁。
【0059】
IL−23またはIL−23受容体に「特異的に結合する」抗体は、IL−23由来配列を含まないタンパク質には結合せず、つまり本明細書で使用される場合、「特異性」とはIL−23特異性に関し、問題のタンパク質に存在し得る任意の他の配列には関しない。たとえば、本明細書で使用される場合、IL−23に「特異的に結合する」抗体は、典型的には、IL−23およびFLAG(登録商標)ペプチドタグを含む融合タンパク質であるFLAG−hIL−23に結合するが、FLAG(登録商標)ペプチドタグ単独には、またはそれがIL−23以外のタンパク質に融合されている場合には、結合しない。文脈に依存して、IL−23に対する特異性は、IL−23(またはその受容体)には結合するが、IL−12(またはその受容体)などの他のタンパク質には結合しないという能力を指すこともできる。
【0060】
II.一般。
【0061】
IL−23およびIL−12は両方とも、共通のサブユニットおよび共通の受容体サブユニットを共有するヘテロ二量体サイトカインであるが、最近の結果によると、炎症および宿主防御におけるそれらの役割は、重複的というより拮抗的であることが示唆されている。インターロイキン23(IL−23)は、2つのサブユニット、つまりp19およびp40で構成されたヘテロ二量体サイトカインである。p19サブユニットは、IL−6、顆粒球コロニー刺激因子(G−CSF)、およびIL−12のp35サブユニットと構造的に関連している。p40サブユニットは、p35およびp40で構成されるサイトカインIL−12の一部である。ヘテロ二量体IL−12は、多くの場合IL−12p70と呼ばれる。IL−23は、IL−23RおよびIL−12Rβ1で構成されるヘテロ二量体受容体に結合することによりシグナル伝達を媒介する。IL−12Rβ1サブユニットは、IL−12Rβ1およびIL−12Rβ2で構成されるIL−12受容体により共有されている。IL−23およびIL−23受容体、ならびにそれぞれのIL−23特異的サブユニットは、国際公開第99/05280号パンフレット、国際公開第01/18051号パンフレット、国際公開第00/73451号パンフレット、および国際公開第01/85790号パンフレットに開示されている。
【0062】
IL−I2に関する多くの初期研究では、p40の遺伝的欠損(p40ノックアウトマウス;p40KOマウス)が必要とされたが、その後IL−23の発見と共に、そのようなマウスはIL−12およびIL−23の両方を欠損していたことが判明した。Oppmannら(2000年)Immunity 第13巻:715〜725頁;Wiekowskiら(2001年)J. Immunol. 第166巻:7563〜7570頁;Parhamら(2002年)J. Immunol. 第168巻:5699〜708頁;Frucht(2002年)Sci STKE 2002、E1〜E3頁;Elkinsら(2002年)Infection Immunity 第70巻:1936〜1948頁。これらの結果により、IL−12およびTh1応答に関係すると当初は考えられていた初期観察の多数の解釈が変更された。
【0063】
最近の研究によると、CD4+T細胞エフェクターレパートリーの多様性は、Th1/Th2パラダイムにより包含される多様性より大きいことが示唆されている。Th17細胞は、以前はTh1系列に帰されていた免疫病原性に寄与するエフェクターTh細胞の異なる系列であると今では考えられている。Th17分化に結び付く経路はまだ不明確であるが(Dong(2006年)Nat Rev Immunol 第6巻:329頁)、IL−23は、この系列の産生および維持のための重要なサイトカインである(Trinchieriら(2003年)Immunity 第19巻:641頁)。IL−12およびIL−23の両方がCD4+T細胞でIFN−γの発現を誘導できる一方で、IL−23は単独で、Th細胞による炎症促進性サイトカインIL−17の産生を促進する。
【0064】
多数の類似性にもかかわらず、IL−12およびIL−23が、多岐にわたる免疫経路を駆動するという証拠が増加している。IL−17で開始された(primed)Th細胞(Th17細胞)は、実験的自己免疫性脳脊髄炎(EAE)(Langrishら(2005年)J. Exp. Med. 第201巻:233頁)、関節炎(Murphyら(2003年)J. Exp. Med. 第198巻:1951頁)、大腸炎(Yenら(2006年)J. Clin. Invest. 第116巻:1310頁)、および自己免疫性心筋炎(Sondereggerら(2006年)Eur. J. Immunol. 第36巻:2844頁)を含む、種々の臓器関連自己免疫性疾患(Harringtonら(2006年)Curr. Opin. Immunol. 第18巻:349頁)に関与している。さらに、それほど明確ではないが、高レベルIL−23/IL−17の産生は、IL−12/IFN−γより、多様な感染症における疾患重症度および免疫病理とより良好に相関する。Hunter(2005年)Nat. Rev. Immunol. 第5巻:521頁;Rutitzky(2005年)J. Immunol. 第175巻:3920頁。他の研究によると、IL−12およびIL−23は、Klebsiella pneumoniae(Happelら(2005年)J. Exp. Med. 第202巻:761頁)およびCitrobacter rodentium(Manganら(2006年)Nature 第441巻:231頁)に対する宿主防御において別個の役割を有することが示唆されている。これらの研究によると、IL−12およびIL−23は、in vivoで、抗菌性の免疫応答および疾患の促進に別個の役割を有することが示唆されている。
【0065】
炎症および宿主防御におけるIL−12およびIL−23の役割が異なることは、慢性真菌感染症などの慢性感染症にとって重要な意味合いを有する。炎症は、真菌感染症を迅速に制御するために必要であるが、炎症の消散は、感染症および関連疾患における防御と免疫病理との間のバランスを維持するために不可欠である。Han and Ulevitch(2005年)Nat. Immunol. 第6巻:1198頁。持続性炎症は、広範な慢性疾患および自己免疫の特徴である。Han & Ulevitch(2005年)。Candidaの場合、炎症消散の失敗は、不完全な真菌除去と関係する。この未消散のCandida感染症は、慢性皮膚粘膜カンジダ症(CMC)に帰着する。CMCは、自己免疫性多発性内分泌腺症−カンジダ症−外胚葉ジストロフィー、機能不全性T細胞活性の状態と関係する。Ryanら(2005年)J. Allergy Clin. Immunol. 第116巻:1158頁。CMCは、未知の免疫病原性の多様な臨床的障害も包含する。Lilic(2002年)Curr. Opin. Infect. Dis. 第15巻:143頁。Aspergillusの場合、感染が難治性である持続性炎症は、同種造血幹細胞移植後の非好中球減少患者(Ortegaら(2006年)Bone Marrow Transplant 第37巻:499頁)、ならびにアレルギー性真菌疾患(Schubert(2006年)Clin. Rev. Allergy Immunol. 第30巻:205頁)において一般的である。この20年間、真菌感染症および関連炎症性疾患の免疫病原性は、様々なタイプの制御性T細胞(Treg)の組合せにより影響されるようなTh1/Th2バランスの点から、主として説明されている。Romani(2004年)Nat. Rev. Immunol. 第4巻:1頁;Romani and Puccetti(2006年)Trends Microbiol. 第14巻:183頁。
【0066】
炎症は、真菌に対する防御応答の不可欠な構成要素であるが、その調節障害は、真菌疾患を著しく悪化させる場合がある。本明細書で開示されたように、IL−23およびIL−17は、真菌に対するIL−12/Th1媒介性免疫耐性を負に制御し、以前は抑制不良のTh1細胞応答に帰されていた起炎性の役割を果たす。IL−23は、Th17細胞(IL−17を産生し、炎症を引き起こす)の生存を促進し、IL−12媒介性Th1応答(インターフェロン−γ(IFN−γ)の産生を伴う)に拮抗することが知られている。Langrishら(2004年)Immunol. Rev. 第202巻:96頁。本明細書で実証されたように、IL−12産生およびTh1応答のIL−23対抗制御は、2つの主要なヒト真菌病原体であるCandida albicansおよびAspergillus fumigatusの制御不良の炎症および増殖に帰着する。IL−23およびIL−17の両方は、好中球の炎症性プログラムおよび抗真菌活性を破壊し、感染に伴う重篤な組織炎症性病理に帰着する。要約すると、IL−23が駆動する炎症は、感染を促進し、抗真菌免疫耐性を損なう。Zelanteら(2007年)Eur. J. Immunol. 第37巻:2695頁、およびCooper(2007年)Eur. J. Immunol. 第37巻:2680頁の関連解説も参照されたい。IL−23に対する拮抗作用による炎症性応答の調節は、真菌に対する防御免疫応答を刺激するための有望な戦略を代表する。
【0067】
本発明は、IL−23および/またはIL−17の活性を阻止して、Th17細胞の効果を低減し、頑健なTh1応答を出現させ、感染した細胞または生物を除去することにより、慢性感染症などの感染症を処置するための組成物および方法を提供する。最適な場合、結果は、感染が完全に消散される無菌治癒である(つまり、処置を中止できるが感染は再発しない)。
【0068】
望ましくない結果を生む炎症状態の維持におけるTh17細胞の役割に関する同じ論拠が、結核(TB)などの慢性のウイルスおよび細菌感染症の場合に当てはまる。サイトカインは、免疫細胞の可溶性媒介物質である。以下のサイトカインが、TBに感染した患者の胸腔洗浄液または気管支肺胞洗浄(BAL)液で検出されている:IL−1β、TNFα、IFNγ、TGFβ、およびIL−12。たとえば、Crystalら(編)(1997年)The Lung; Scientific Foundations、Lippincott−Raven、ニューヨーク、米国ニューヨーク州、2381〜2394頁を参照されたい。IFN−γおよびTNFαは、マイコバクテリア感染症の制御に重要な役割を果たすことが示されている。たとえば、Cooperら(1993年)J. Exp. Med. 第178巻:2243頁;Flynnら(1993年)J. Exp. Med. 第178巻:2249頁;Kindlerら(1989年)Cell 第56巻:731頁;Cheuseら(1994年)Am. J. Pathol. 第145巻:1105頁を参照されたい。IL−12駆動のIFN−γ産生を低減するTh17細胞の生存をIL−23が促進する範囲内において、IL−23活性に対する拮抗作用が細菌感染症の消散を増強すると期待できる。
【0069】
文献によると、IL−23アンタゴニストでの処置は、たとえば自己免疫障害または慢性感染症の処置において、IL−12アンタゴニストでの処置より安全であり得ることが示唆される。Chackerianらは、p19−/−ノックアウト(KO)マウスにおける抗体中和または遺伝子除去のいずれかによるIL−23活性の除去が、マイコバクテリア(BCG)感染に対する免疫を損なわなかったという実験を記述している。Chackerianら(2006年)J. Exp. Med. 第74巻:6092頁。IL−23p19KOマウスにおける感染の経過は、野生型マウスにおける経過と区別がつかず、抗IL−23p19処置マウスにおける細菌コロニー形成単位数は、イソタイプ対照処置マウスにおける単位数と変わらなかった。対照的に、IL−12欠損KOマウスはBCGの増殖を制御できず、IL−12の抗体阻止は、イソタイプ対照処置マウスと比較して、脾臓、肝臓、および肺において著しくより高いCFU数と相関した。これらの結果により、IL−23は、IL−12の存在下ではマイコバクテリアに対する宿主防御に重要な役割を果たさず、したがってIL−23の選択的阻害は、IL−12の中和を伴う処置(IL−23を同時中和して、またはしないでのいずれかで)より安全であり得ることが示唆される。本明細書で提示された結果は、これらの結果を進展させ、IL−23アンタゴニストは、宿主防御を損なわないという点でより安全であるだけでなく、IL−23/IL−17炎症の調節障害により引き起こされる慢性感染症の消散を支援する点で実際に有益であり得ることを示唆する。
【0070】
Chackerianらの実験(2006年)は、IL−23の阻止が、既存の慢性マイコバクテリア感染の除去を増強するか否かの問題に取り組むようには設計されていなかった。対照マウス(WTマウス、未処置であるかまたはイソタイプ対照抗体のみで処置された)は、慢性感染症を発症せず、感染を効果的に除去できた。Chackerianら(2006年)のKOマウスおよび抗体阻止実験の両方で、IL−23活性は、感染後ではなく、静脈内BCGによる感染の前に除去された。本明細書に記載の実験は、マイコバクテリア感染症ではなく、真菌感染症に関していた。さらに、本明細書の実施例4および5に記載の実験は、静脈内投与ではなく、真菌病原体の胃内および鼻腔内投与を使用した実験を含む。これらの真菌病原体を肺および胃に直接送達すると、静脈内送達と比べてより生理学的に関係性のある疾患モデルがもたらされる。本明細書に開示の実験において真菌病原体に感染した組織は、Th17応答がその最も重要な生理学的役割を有し得る組織、つまり肺および腸の粘膜関門であると示唆されている。Cua and Kastelein(2006年)Nature Immunol. 第7巻:557頁。さらに、本明細書に記載の実験においては、抗IL−23p19および抗IL−17抗体は、感染前ではなく、感染の5時間後に投与された。
【0071】
III.真菌感染症の実験結果
持続性炎症は、広範な慢性疾患および自己免疫の特徴である。Han & Ulevitch(2005年)。IL−23が発見され、自己免疫炎症におけるその役割が近年になって文献化される前(Cuaら(2003年)Nature 第421巻:744頁;Langrishら(2005年)J. Exp. Med. 第201巻:233頁)は、IL−12が、Th1応答を開始および維持することにより、過剰反応免疫および自己免疫障害に関与すると考えられていた。これは、真菌感染症、ならびに炎症および制御されないTh1/Th2抗真菌反応性の微調整に免疫制御が不可欠であると証明された疾患にも当てはまった。Ryanら(2005年);Romani(2004年);Romani & Puccetti(2006年)。
【0072】
本研究の結果により、制御されないTh1応答ではなくIL−23/IL−17軸が、不完全な病原体除去、炎症消散の失敗、ならびにCandidaおよびAspergillusに対する防御免疫応答の開始失敗と関係していることが示される。したがって、この新しい知見は、進行中の炎症に直面して真菌の残留が生じる難治形態の真菌感染症と慢性炎症反応との矛盾した関連性を調整する役割を果たすことができる。
【0073】
IL−23およびIL−17の両方は、IFN−γの存在下でさえPMNの抗真菌エフェクター活性を損ない、この知見はTh17エフェクター経路がTh1経路より優勢であることを示唆する。さらに、両サイトカインは、真菌に対するPMNの炎症性状態を限定することが知られている、インドールアミン2,3−ジオキシゲナーゼ(IDO)のIFN−γ依存性活性化に対抗することにより(Bozzaら(2005年)J. Immunol. 第174巻:2910頁)、ならびにTh17細胞活性化に伴う高い炎症性病理および組織破壊の原因である可能性が高いMMP9およびMPOの放出を誘導することにより、PMNの炎症性プログラムを活性化した。
【0074】
IDOに対する作用は興味深い。IDOは、C.albicansで発現され、真菌発芽のトリプトファン栄養要求依存性阻害に関与する。Bozzaら(2005年)。IDO遮断と同様におよびIFN−γと対向して(Kalo−Kleinら(1990年)Infect. Immun. 第58巻:260頁)、IL−17は真菌発芽を促進したが(データ非表示)、この知見は、哺乳類宿主免疫系からのシグナルに高度に反応する酵素である真菌IDOに対する作用を示唆する。Mellor and Munn(2004年)Nat. Rev. Immunol. 第4巻:762頁。したがって、他の感染症について既に記述されているように(McKenzieら(2006年)Trends Immunol. 第27巻:17頁)、Th17経路の機能は、炎症を促進し抗菌免疫を破壊するその能力の範囲を超えて、真菌の形態および毒性に対する作用を含み得る。これは、高レベルの真菌増殖に厳密に依存することが知られている共動性IL−4+Th2細胞活性化に変形する場合があり(Mencacciら(1996年)Infect. Immun. 第64巻:4907頁)、Th1が機能することをさらに防止する。
【0075】
他の感染症について既に記述されているように(Cruzら(2006年)J. Immunol. 第177巻:1416頁;Parkら(2005年)Nat. Immunol. 第6巻:1133頁)、Th1またはTh17経路は、両真菌感染症において相互に制御された。この知見は、真菌に応答するどちらか一方の経路の出現が、厳密な環境制御下にあることを示唆する。制御は、様々な段階で起こり得る。1つの明白なレベルの制御は、Th17細胞の誘導を制御することが知られているIFN−γにより代表される。Cruzら(2006年);Parkら(2005年)。IL−23/IL−17軸は、IFN−γ欠損の場合、両感染症において実際に亢進され、IFN−γ産生細胞数はIL−17中和に際して増加した。これらのデータは、IFN−γが、カンジダ症を有するマウスのIL−12応答に必要であるという概念と一致している。Cenciら(1998年)J. Immunol. 第161巻:3543頁。
【0076】
より重要なことだが、IL−12の産生はp19−/−DCにおいてより高く、IL−23の産生はp35−/−DCにおいてより高く、両サイトカインはWTDCにおいて交差制御された。これらの知見は、これらのサイトカインが、DC産生のレベルで相互に制御されることを示唆する。Beckerら(2006年)J. Immunol. 第177巻:2760頁。しかしながら、炎症性DCは、真菌に応答して免疫寛容原誘発性DCより多くのIL−23を産生すると考えられるため、これは、Th1/Th17のバランスが、様々な体内部位のDCサブセット間の相互制御にも依存するように現れる。
【0077】
危険性の高い炎症の状態において、真菌に応答して、すなわちTLR−/MyD88経路を介する高酵母数に応答した炎症性DCにより、IL−23が産生されるという知見は、重要な意味合いを有する。それは、炎症性の過程および真菌毒性間の重要な分子的結び付きとしてIL−23を指摘するだけでなく、悪循環がそれにより作用し得るというシナリオも確立する。p19−/−マウスがより少ないIL−17を産生し、TGF−βが真菌に対するTh17活性化および/または維持に非不可欠な役割を示したため、IL−23は、IL−17の近位媒介物質として作用することが考えられる。このシナリオでは、制御されない真菌増殖は、非防御的Th2細胞の同時活性化と関係する病原性Th17細胞の活性化を永続化する場合がある。
【0078】
この研究における1つの興味深い所見は、微生物刺激がIL−23分泌の主要な誘発因子であり得るが、適応免疫過程はその産生を調節することもできることであった。これを支持するものであるが、本発明者らは、DCによるIL−23分泌がT細胞の存在下で劇的に増加したという証拠を提供したが、この知見は、活性化T細胞がIL−23のさらなる誘導に正のフィードバックループを提供できることを示唆した。
【0079】
上記の考察は、共生生物または遍在性のいずれの真菌であっても、免疫恒常性およびその調節障害内に適応させるのに役立ち得る。IL−23/IL−17軸の活性化を介した炎症性プログラムを破壊する能力が、最終的に免疫調節障害に結び付き得る場合、CandidaまたはAspergillusのいずれかに対する防御免疫の不可欠で本質的な構成要素であるTreg細胞を活性化するそれらの能力(Romani & Puccetti(2006年))は、それにより制御されない免疫が防止される機序を代表し得る。この点に関しては、in vivoでのIL−17産生細胞の発生におけるIL−10の阻害的役割を含む、Th17細胞およびTreg細胞間の機能的拮抗作用が記述されている(Bettelli & Kuchroo(2005年)J. Exp. Med. 第201巻:169頁)。Kullbergら(2006年)J. Exp. Med. 第203巻:2485頁。したがって、Th17細胞およびTreg細胞の産生のための相互的経路は、真菌感染症でも起こり得る可能性がある。本発明者らは、感染後のp35−/−マウスにおけるCD4+CD25+Treg細胞活性化の証拠を見出しておらず、この知見は、Th17細胞およびTreg細胞が相互に排他的であることを示唆する。その代わりに、IL−10産生の著しい減少にかかわらず、CD4+CD25+Treg細胞がp19−/−マウスで観察され、それはT細胞によるIL−10産生を上方制御するIL−23の能力と一致する。Vanden Eijndenら(2005年)Eur. J. Immunol. 第35巻:469〜475頁。
【0080】
本研究の別の重要な所見は、IL−23/IL−17依存性経路が、IFN−γ欠損の場合、p35依存性経路を介していくつかの抗真菌耐性を提供できるということである。IL−12欠損の場合IL−23が防御的役割を果たすことができることは、慢性クリプトコックス症(Kleinschekら(2006年)J. Immunol. 第176巻:1098頁)で、マイコバクテリア感染症で(Khaderら(2005年)J. Immunol. 第175巻:788頁)で、および急性肺性Klebsiella pneumoniae感染症(Happelら(2005年)Infect. Immun. 第73巻:5782頁)で既に報告されており、防御は、IL−12p70と独立して抗原特異的IFN−γ産生CD4+T細胞を活性化するIL−23の能力、および病原体除去を媒介するPMNを動員するIL−23の能力と相関していた。Happelら(2005年)J. Exp. Med. 第202巻:761頁。実際のところ、実験的Helicobacter hepaticus誘導性大腸炎では、IL−23が、IFN−γ産生細胞およびIL−17産生細胞の両方を駆動することが明らかに示された。Kullbergら(2006年)。本発明者らの結果は、IL−23の作用におけるp35依存性経路を包含する、感染に際したTh1経路およびTh17経路間のさらなるレベルの交差制御を示唆すると考えられる。最終的には、IL−12依存性軸により支配された適切な免疫エフェクター機能の開始前に、初期炎症の危険シグナルを処理するIL−23の能力(McKenzieら(2006年))は、アンタゴニストだけでなくこのサイトカイン対間の協同的活性とも一致する。
【0081】
まとめると、この研究で提示されたデータは、真菌に対する宿主防御、免疫恒常性、および免疫の機序に重要な意味合いを有する、以前は未決定だったIL−23依存性Th17系列の真菌感染症における役割を実証する。さらに、それらは、炎症消散の失敗と抗真菌免疫耐性の欠如との間の分子的関連性を示す。この最新の結果は、炎症を限定して効果的な免疫応答を刺激することを試みる、真菌感染症の免疫治療の戦略を示唆する。
【0082】
IV.IL−23アンタゴニスト。
【0083】
IL−23アンタゴニストは、IL−23の1つまたは複数の生物活性を阻害可能な任意の物質または方法を含む。そのような活性には、IL−23(p19およびp40サブユニットを含む)、IL−23受容体(IL−23RおよびIL−12Rβ1サブユニットを含む)への結合、ならびにTh17細胞の促進および維持が含まれる。アンタゴニストには、たとえば小分子、抗体または抗体断片、ペプチド模倣物質、アプタマー(たとえば米国特許出願公開第2006−0193821号明細書に開示されているような)、IL−23受容体サブユニットの細胞外領域上に由来する可溶性受容体、および核酸に基づくアンタゴニストが含まれ得る。
【0084】
IL−23の核酸に基づくアンタゴニストには、アンチセンス核酸、およびIL−23p19遺伝子またはIL−23R遺伝子に対するsiRNAが含まれる。一般的なsiRNA方法論については、国際公開第2006/06060598号パンフレットを参照されたい。Arenz and Schepers(2003年)Naturwissenschaften第90巻:345頁;Sazani and Kole(2003年)J. Clin. Invest. 第112巻:481頁;Pirolloら(2003年)Pharmacol. Therapeutics第99巻:55頁;Wangら(2003年)Antisense Nucl. Acid Drug Devel. 第13巻:169頁も参照されたい。アンチセンスおよびsiRNA分子は、ヒトIL−23p19およびIL−23R mRNAの既知の配列に基づいて設計できる。ヒトIL−23p19のmRNAおよびアミノ酸配列は、それぞれ、GenBankアクセッション番号NM_016584およびNP_057668に見出される。ヒトIL−23RのcDNAおよびアミノ酸配列は、それぞれ、GenBankアクセッション番号AF461422およびAAM44229に見出される。本発明は、RNA干渉用の組成物も提供する。
【0085】
siRNAを生成および使用する方法は、たとえば米国特許第6,506,559号明細書(国際公開第99/32619号パンフレット);第6,673,611号明細書(国際公開第99/054459号パンフレット);第7,078,196号明細書(国際公開第01/75164号パンフレット);第7,071,311号明細書、およびPCT公報、国際公開第03/70914号パンフレット;国際公開第03/70918号パンフレット;国際公開第03/70966号パンフレット;国際公開第03/74654号パンフレット;国際公開第04/14312号パンフレット;国際公開第04/13280号パンフレット;国際公開第04/13355号パンフレット;国際公開第04/58940号パンフレット;国際公開第04/93788号パンフレット;国際公開第05/19453号パンフレット;国際公開第05/44981号パンフレット;国際公開第03/78097号パンフレットに開示されている。米国特許は、関連PCT公報と共に列挙されている。遺伝子抑制(gene silencing)および治療的処置にsiRNAを使用する典型的な方法は、PCT公報、国際公開第02/096927号パンフレット(VEGFおよびVEGF受容体);国際公開第03/70742号パンフレット(テロメラーゼ);国際公開第03/70886号パンフレット(タンパク質チロシンホスファターゼIVA型(Prl3));国際公開第03/70888号パンフレット(Chk1);国際公開第03/70895号パンフレット、および国際公開第05/03350号パンフレット(アルツハイマー病);国際公開第03/70983号パンフレット(タンパク質キナーゼCアルファ);国際公開第03/72590号パンフレット(MAPキナーゼ);国際公開第03/72705号パンフレット(サイクリンD);国際公開第05/45034号パンフレット(パーキンソン病)に開示されている。siRNAの治療的使用に関する典型的な実験は、Zenderら(2003年)Proc. Nat’l. Acad. Sci. (USA) 第100巻:7797頁;Paddisonら(2002年)Proc. Nat’l. Acad. Sci. (USA) 第99巻:1443頁;およびSah(2006年)Life Sci. 第79巻:1773頁にも開示されている。siRNA分子は、たとえば慢性骨髄性白血病(CML)(ClinicalTrials.gov識別番号:NCT00257647)および加齢黄斑変性(AMD)(ClinicalTrials.gov識別番号:NCT00363714)の臨床試験でも使用されている。
【0086】
「siRNA」という用語は、RNA干渉経路を介して遺伝子抑制を誘導するために使用される分子(Fireら(1998年)Nature 第391巻:806頁)を参照するために本明細書で使用されるが、そのようなsiRNA分子は、厳密にポリリボヌクレオチドである必要はなく、その代わり、治療薬としてのその特性を改良するために、核酸に対する1つまたは複数の改変を含有できる。そのような作用剤は、短い干渉核酸(short interfering nucleic acid)の代わりに「siNA」と呼ばれることがある。そのような変更により、この分子は、「リボ」ヌクレオチドの定義から正式には外れる場合があるが、それにもかかわらず本明細書では、そのような分子を「siRNA」分子と呼ぶ。RNA干渉経路を介して遺伝子抑制を誘導するために使用される他の核酸改変体には、たとえば、米国特許出願公開第20060115453号明細書で開示されているような短いヘアピン型RNA(short hairpin RNA)(「shRNA」)が含まれる。核酸に基づく阻害剤は、たとえば、プラスミドまたはウイルス(たとえば裸の(naked)DNAとして)などの組換えベクターを用いた形質転換により、または知られている遺伝子治療用ベクター(たとえばアデノ随伴ウイルス(AAV)、アデノウイルス、レトロウイルス、またはレンチウイルス)のいずれかを用いた遺伝子治療により送達できる。核酸は、形質転換、エレクトロポレーション、遺伝子銃、または当技術分野で知られている他の方法により送達できる。
【0087】
本発明の組成物および方法で使用するためのIL−23の抗体アンタゴニストには、IL−23に対する抗体およびIL−23受容体に対する抗体が含まれる。IL−23に対する典型的なアンタゴニスト抗体には、同一人に譲渡された米国特許仮出願第60/891,409号明細書および第60/891,413号明細書(両方とも2007年2月23日に出願された)、米国特許出願公開第2007−0009526号明細書および第2007−0048315号明細書、ならびに国際公開第2007/076524号パンフレット、国際公開第2007/024846号パンフレット、および国際公開第2007/147019号パンフレットに開示されているような、抗ヒトIL−23p19抗体およびその断片が含まれる。IL−23に対する抗体アンタゴニストには、そのサブユニットがIL−23p19に結合する場合にはIL−12p40サブユニットに結合する抗体も含まれるが、IL−12p35に結合する場合には含まれない。たとえば、米国特許出願公開第2005−0137385号明細書、米国特許第7,252,971号明細書を参照されたい。IL−23に対する典型的なアンタゴニスト抗体には、抗ヒトIL−23受容体抗体、たとえば抗IL−23R抗体およびその断片が含まれる。IL−23Rに対する典型的なアンタゴニスト抗体は、同一人に譲渡された米国特許仮出願第60/892,104号明細書(2007年2月28日に出願された)および第60/945,183号明細書(2007年6月20日に出願された)に開示されている。IL−23アンタゴニストには、二重特異性抗体も含まれる。
【0088】
抗原性の増加した領域は、抗体産生に使用できる。ヒトp19の抗原性が増加した領域は、たとえば、GenBank AAQ89442(gi:37183284)のアミノ酸16〜28位;57〜87位;110〜114位;136〜154位;および182〜186位に生じている。ヒトIL−23Rの抗原性が増加した領域は、たとえば、GenBank AAM44229(gi:21239252)のアミノ酸22〜33位;57〜63位;68〜74位;101〜112位;117〜133位;164〜177位;244〜264位;294〜302位;315〜326位;347〜354位;444〜473位;510〜530位;および554〜558位に生じている。解析は、Vector NTI(登録商標)Suite(Informax社、ベテスダ、米国メリーランド州)を使用したパーカープロット(Parker plot)により行った。本発明は、つまりIL−23Rの細胞外領域、たとえばGenBank AAM44229のアミノ酸1〜353位またはその断片を含む可溶性受容体であり、この細胞外領域またはその断片がIL−23に特異的に結合するIL−23アンタゴニストも提供する。マウスIL−23Rは、GenBank NP_653131(gi:21362353)である。ムテインおよび改変体、たとえばペグ化、または脱アミドするアスパラギン残基を欠失もしくは置換する変異誘発が企図される。
【0089】
本発明の方法および組成物で使用するための、IL−23活性に拮抗する追加的な考え得る方法には、線維状赤血球凝集素(FHA)に投与する工程(国際公開第2006/109195号パンフレット)、およびIL−23に対する免疫応答を生じさせるために予防接種する工程(国際公開第2005/058349号パンフレット)が含まれる。
【0090】
一実施形態では、IL−17産生(Th17)細胞のアンタゴニストは、たとえばナイーブT細胞、Th1型T細胞、TH2型T細胞、上皮細胞、および/または内皮細胞の活性に、たとえば実質的に影響を与えずに、Th17細胞の活性を特異的に調節する試薬を包含する。この試薬は、たとえば、IL−17産生細胞の転写因子(たとえばRORγt)または接着タンパク質の発現または活性を調節できる。さらに、IL−23、TGF−β、またはIL−6のアンタゴニストは、Th17細胞の生成および生存を減少でき、IL−17のアンタゴニストは、そのような細胞の炎症性効果(たとえば、好中球動員)を減少できる。
【0091】
モノクローナル抗体、ポリクローナル抗体、およびヒト化抗体は、調製可能である(たとえば、Sheperd and Dean(編)(2000年)Monoclonal Antibodies、Oxford Univ. Press、ニューヨーク、米国ニューヨーク州);Kontermann and Dubel(編)(2001年)Antibody Engineering、Springer−Verlag、ニューヨーク;Harlow and Lane(1988年)Antibodies A Laboratory Manual、Cold Spring Harbor Laboratory Press、コールドスプリングハーバー、米国ニューヨーク州、139〜243頁;Carpenterら(2000年)J. Immunol. 第165巻:6205頁;Heら(1998年)J. Immunol. 第160巻:1029頁;Tangら(1999年)J. Biol. Chem. 第274巻:27371頁;Bacaら(1997年)J. Biol. Chem. 第272巻:10678頁;Chothiaら(1989年)Nature 第342巻:877頁;Foote and Winter(1992年)J. Mol. Biol. 第224巻:487頁;米国特許第6,329,511号明細書を参照)。抗体配列の全体がヒト生殖細胞系列(germline)配列に由来する完全ヒト抗体も調製できる。そのような完全ヒト抗体は、ヒト免疫グロブリン遺伝子を発現するように遺伝子操作されたトランスジェニック動物から、またはファージディスプレイなどの方法により、調製できる。たとえば、Lonberg(2005年)Nature Biotechnol. 第23巻:1117頁;Vaughanら(1998年)Nature Biotechnol. 第16巻:535頁を参照されたい。抗体断片には、Fab、Fab’、Fab’−SH、Fv、単鎖Fv(scFv)、F(ab’)2、およびダイアボディが含まれる。Pluckthun(1994年)THE PHARMACOLOGY OF MONOCLONAL ANTIBODIES、第113巻、Rosenburg and Moore編、Springer−Verlag、ニューヨーク、269〜315頁;Holliger and Hudson(2005年)Nature Biotechnol. 第23巻:1126〜1136頁。
【0092】
本発明の抗体はまた、変化したエフェクター機能を提供するために改変(または遮断)Fc領域を有する抗体を含む。たとえば米国特許第5,624,821号;国際公開第2003/086310号;国際公開第2005/120571号;国際公開第2006/0057702号;Presta(2006年)Adv. Drug Delivery
Rev. 58:640〜656頁を参照されたい。そのような改変は、診断および治療での可能性のある有益な効果を伴って、免疫系の多様な反応を増強するまたは抑制するために使用することができる。Fc領域の改変は、アミノ酸変化(置換、欠失、および挿入)、糖化または糖鎖除去、ならびに複数のFcの付加を含む。Fcに対する変更はまた、治療用抗体中の抗体の半減期を変化させることもでき、より長い半減期により、投薬がそれほど頻繁ではなくなり、同時に利便性が増加し、材料の使用が減少する。エフェクター機能の変更は、IgG1のFc部分に特定の変異を導入することにより、たとえば、Asn297をたとえばAlaまたはGlnに変更すること(N297AまたはN297Q)により達成できる。Presta(2005年)J. Allergy Clin. Immunol. 第116巻:731頁の734〜35頁を参照されたい。エフェクター機能は、様々な定常ドメインの選択により変更することもできる。たとえば、本発明の抗体(または断片)の特定の使用目的が、エフェクター機能の変更を要求した場合、IgG1以外の重鎖定常ドメインが使用できる。IgG1抗体は、長期の半減期ならびに補体活性化および抗体依存性細胞傷害活性(ADCC)などのエフェクター機能を備えるが、そのような活性は、抗体のすべての用途にとって望ましいとは限らない場合がある。そのような場合には、たとえばIgG4定常ドメインが使用できる。一実施形態(唯一の実施形態ではない)では、目的がIL−23受容体を発現する細胞の殺滅を誘導することではなく、その代わり単にそのような細胞におけるIL−23シグナル伝達を阻止することであるため、エフェクター機能の変更は、IL−23受容体に対する(たとえばIL−23Rに対する)抗体の場合、特に関連性がある。この実施形態では、Th17系列からTh1系列へとTh細胞を転換することが目的であり、その場合には、細胞殺滅は非産生的である。
【0093】
抗原の精製は、抗体の産生に必要ではない。免疫化は、DNAベクター免疫化により実施でき、たとえば、Wangら(1997年)Virology 第228巻:278頁を参照されたい。あるいは、目的の抗原を保持する細胞で動物を免疫できる。その後、免疫された動物から脾細胞を単離することができ、脾細胞を骨髄腫細胞株と融合させてハイブリドーマを産生することができる(Meyaardら(1997年)Immunity 第7巻:283頁;Wrightら(2000年)Immunity 第13巻:233頁;Prestonら(1997年)Eur. J. Immunol. 第27巻:1911頁)。その結果生じるハイブリドーマは、機能アッセイまたはバイオアッセイ、すなわち精製された抗原の所有に依存しないアッセイにより、所望の抗体の産生のためにスクリーニングできる。細胞による免疫化は、精製された抗原を用いた免疫化より抗体産生に優れていることが証明できる(Kaithamanaら(1999年)J. Immunol. 第163巻:5157頁)。
【0094】
抗体−抗原結合特性およびリガンド−受容体結合特性は、たとえば、表面プラズモン共鳴(Karlssonら(1991年)J. Immunol. Methods 第145巻:229頁;Neriら(1997年)Nat. Biotechnol. 第15巻:1271頁;Jonssonら(1991年)Biotechniques 第11巻:620頁)、または競合ELISA(Friguetら(1985年)J. Immunol. Methods 第77巻:305頁;Hubble(1997年)Immunol. Today 第18巻:305頁)により、測定できる。抗体は、抗体の標的抗原および関連結合タンパク質を単離するために、親和性精製(affinity purification)用に使用できる。たとえば、Wilchekら(1984年)Meth. Enzymol. 第104巻:3頁を参照されたい。
【0095】
抗体は、通常は少なくとも約10−6M、典型的には少なくとも10−7M、より典型的には少なくとも10−8M、好ましくは少なくとも約10−9M、およびより好ましくは少なくとも10−10M、および最も好ましくは少なくとも10−11MのKDで結合する。たとえば、Prestaら(2001年)Thromb. Haemost. 第85巻:379頁;Yangら(2001年)Crit. Rev. Oncol. Hematol. 第38巻:17頁;Carnahanら(2003年)Clin. Cancer Res. (Suppl.) 第9巻:3982頁を参照されたい。
【0096】
IL−23Rの細胞外ドメインを含む可溶性受容体は、本発明の組成物および方法に有用である。可溶性受容体は、標準的方法により調製および使用できる。たとえば、Jonesら(2002年)Biochim. Biophys. Acta 第1592巻:251頁;Prudhommeら(2001年)Expert Opinion Biol. Ther. 第1巻:359頁;Fernandez−Botran(1999年)Crit. Rev. Clin. Lab Sci. 第36巻:165〜224頁を参照されたい。
【0097】
一実施形態では、本発明の組成物および方法は、IL−23に対する拮抗作用を必要とするが、IL−12に対する拮抗作用は必要としない。IL−23およびIL−12の活性を両方とも阻止するIL−12およびIL−23両方のIL−12p40サブユニットを標的とするいくつかの有望な治療薬が現在開発中である。そのような作用剤は、本発明が促進することを目的とする頑健なIL−12媒介性Th1応答を阻害するため、本発明の組成物および方法のこの実施形態における使用に好適ではない。IL−12との関連ではなくIL−23との関連においてのみIL−12p40に結合する作用剤を開発することは、原理的には可能であるが(米国特許出願公開第2005−0137385号明細書および米国特許第7,252,971号明細書を参照)、IL−12p40を標的とする大多数の作用剤はIL−12を阻害する可能性が高く、したがって本発明には好適ではない。同じ議論が、IL−23およびIL−12の共通受容体サブユニット、IL−12Rβ1に当てはまる。IL−12受容体との関連ではなくIL−23受容体との関連においてのみIL−12Rβ1に結合する作用剤を開発することは、原理的には可能であるが、IL−12Rβ1を標的とする大多数の作用剤はIL−12受容体を阻害する可能性が高く、したがって本発明には好適ではない。対照的に、IL−23またはその受容体に特異的なサブユニット、つまりそれぞれp19およびIL−23Rに結合し拮抗する作用剤は、IL−12ではなくIL−23の特異的阻害剤である可能性が高く、したがって本発明の組成物および方法における使用に、より好適である。
【0098】
有望な治療薬がIL−12ではなくIL−23を特異的に阻害するかどうかは、当技術分野で知られている任意の方法により決定できる。たとえば、有望なIL−23特異的アンタゴニストについては、その受容体に対するIL−23の結合またはその受容体に対するIL−12の結合を阻止するその能力を試験できる。そのような阻止アッセイは、溶液中で(たとえば、蛍光活性化セルソーティング(cell sorting)により)、または固体支持体上で(たとえば、酵素結合免疫吸着測定法−ELISAにより)実施できる。IL−23およびIL−12受容体阻止は、Ba/F3細胞増殖アッセイなどのバイオアッセイで測定することもできる。たとえば、Hoら(1995年)Mol. Cell. Biol.(1995年)第15巻:5043頁を参照されたい。そのような結合アッセイでは、有望なIL−23アンタゴニストの効力および特異性は、IC50として、すなわち所定の一組のアッセイ条件下で、IL−23結合(または結合に依存する生物活性)の50%低減を達成するのに必要なこの有望なアンタゴニストの濃度として表すことができる。より低いIC50は、より効果的なアンタゴニストを示す。有望なアンタゴニストのIL−23特異性は、IL−23とその受容体の結合の阻害に関するIC50に対する、IL−12とその受容体の結合の阻害に関するIC50の比率(IC50IL−12/IC50IL−23)として表すことができる。種々の実施形態では、有望なIL−23特異的アンタゴニストは、この比率(IC50IL−12/IC50IL−23)が、1.5、2、3、4、5、7、10、15、20、50、または100以上である場合、IL−23特異的と見なされる。好ましい実施形態では、阻害アッセイで使用されるIL−23およびIL−12のレベルは、IL−23およびIL−12アッセイの少なくとも1つ、好ましくはその両方が、直線的用量応答濃度範囲中で実施されることを保証するように調整される。
【0099】
IL−23およびIL−12は、拮抗作用の特異性を決定するために使用できる様々な生物学的機能も有する。IL−12とは対照的に、IL−23は、ヒトおよびマウスの両方のナイーブT細胞集団に対して優先的に記憶を刺激する。IL−23は、多数の細胞内細胞シグナル伝達分子、たとえばJak2、Tyk2、Stat1、Stat2、Stat3、およびStat4を活性化する。IL−12は、この同じ群の分子を活性化するが、IL−23に対するStat4応答は比較的弱く、その一方でIL−12に対するStat4応答は強い。Oppmannら(2000年);Parhamら(2002年)J. Immunol. 第168巻:5699頁。
【0100】
有望なIL−23特異的アンタゴニストについては、IL−12およびIL−23によりTh1およびTh17細胞の増幅および生存を阻害するその能力を試験することもできる。IL−23特異的アンタゴニストは、Th1細胞のIL−12媒介性増幅および生存ではなく、Th17細胞のIL−23媒介性増幅および生存を優先的に阻害する。Th17細胞はIL−17を特徴的に分泌する一方で、Th1細胞はIFN−γを特徴的に分泌する。典型的なTh1/Th17アッセイのデータは、Langrishら(2005年)J. Exp. Med. 第201巻:233頁の図2に見出され、IL−23がIL−17産生CD4+T細胞の増幅および生存を促進する一方で、IL−12はIFN−γ産生CD4+T細胞の増幅および生存を促進することを実証する。本発明の一実施形態では、作用剤は、Th17細胞のIL−23媒介性増幅および生存を阻害できるが、Th1細胞のIL−12媒介性増幅および生存を阻害しない場合に、「IL−23特異的」アンタゴニスト(IL−12に対して)であると見なされる。Th17/Th1細胞増殖の阻害は、IC50として、すなわち所定の一組のアッセイ条件下で、IL−17またはIFN−γを産生する特定のT細胞サブセットの増幅および生存の促進におけるIL−23活性の50%低減を達成するのに必要な作用剤の濃度として表すことができる。典型的なアッセイは、実施例13で提供される。実施例13に記載したアンタゴニストのように、バイオアッセイにおけるIL−23アンタゴニストの効力は、IC50IL−23として、つまりIL−23活性をその非阻害値の50%に低減するのに必要なアンタゴニストの濃度として表すことができる。類似したIC50IL−12は、IFN−γ産生細胞の産生促進におけるIL−12およびその活性で決定できる。その後、アンタゴニストのIL−23特異性は、比率IC50IL−12/IC50IL−23として表すことができる。種々の実施形態では、確認した(validated)IL−23特異的アンタゴニストのIC50IL−12/IC50IL−23比率は、1.5、2、3、4、5、7、10、15、20、50、または100以上である。
【0101】
Langrishら(2005年)に本質的に記載されているように、IL−17AおよびIFN−γの産生は、サイトカインに結合する蛍光性試薬を用いた蛍光活性化セルソーティング(FACS(登録商標)解析)による細胞内サイトカインの流動細胞計測法(flow cytometry)により測定できる。その細胞が「IL−17産生」または「IFN−γ産生」と見なされるための、生のCD4+T細胞中のIL−17AまたはIFN−γの閾値レベルを定義することが重要である。一実施形態では、閾値レベルは、生のCD4+T細胞の5%が、未処置細胞の対照試料中で「IL−17産生」または「IFN−γ産生」であるレベルと定義される。典型的な未処置細胞には、プロテオリピドタンパク質(PLP)で免疫されたSJLマウス(The Jackson Laboratories社、バーハーバー、メーン州、米国)から単離され、PLPの存在下で培養された流入領域リンパ節(DLN)細胞が含まれる。
【0102】
V.組成物および方法。
【0103】
IL−23のアンタゴニストを含む医薬組成物または無菌組成物を調製するために、アンタゴニストは、薬学的に許容可能な担体または賦形剤と混合される。たとえばRemington’s Pharmaceutical Sciences and U.S. Pharmacopeia: National Formulary、Mack Publishing Company、Easton、PA(1984年)を参照されたい。治療薬の処方は、生理学的に許容可能な担体、賦形剤、または安定剤と混ぜることによって、たとえば凍結乾燥粉末、スラリー、水性溶液、または懸濁液の形状で調製されてもよい(たとえばHardmanら(2001年)Goodman and Gilman’s The Pharmacological Basis of Therapeutics、McGraw−Hill、New York、NY;Gennaro(2000年)Remington: The Science and Practice of Pharmacy、Lippincott, Williams, and Wilkins、New York、NY;Avisら(編)(1993年)Pharmaceutical Dosage Forms: Parenteral Medications、Marcel Dekker、NY;Liebermanら(編)(1990年)Pharmaceutical Dosage Forms: Tablets、Marcel Dekker、NY;Liebermanら(編)(1990年)Pharmaceutical Dosage Forms: Disperse Systems、Marcel Dekker、NY;WeinerおよびKotkoskie(2000年)Excipient Toxicity and Safety、Marcel
Dekker, Inc.、New York、NYを参照されたい)。
【0104】
投与経路は、たとえば局所的もしくは皮膚塗布、静脈内注射または注入、腹腔内、脳内、筋肉内、眼内、動脈内、脳脊髄内、病巣内、もしくは肺性の経路によるか、または徐放性システムもしくは埋没物(implant)による。遺伝子移入ベクターの中枢神経系への注射は記述されている。たとえば、Cuaら(2001年)J. Immunol.
第166巻:602頁;Sidmanら(1983年)Biopolymers 第22巻:547頁;Langerら(1981年)J. Biomed. Mater. Res. 第15巻:167頁;Langer(1982年)Chem. Tech. 第12巻:98頁;Epsteinら(1985年)Proc. Natl. Acad. Sci. USA 第82巻:3688頁;Hwangら(1980年)Proc. Natl. Acad. Sci. USA 第77巻:4030頁;米国特許第6,350,466号明細書および第6,316,024号明細書を参照されたい。
【0105】
治療剤に対する投与計画の選択は、作用物質の血清代謝回転速度または組織代謝回転速度、症状のレベル、作用物質の免疫原性、および生物学的マトリックス中での標的細胞の到達性を含むいくつかの要因に依存する。好ましくは、投与計画は、許容可能なレベルの副作用と調和した、患者に送達される治療剤の量を最大限にする。したがって、送達される作用物質の量は、部分的に、特定の実体および処置されている状態の重症度に依存する。抗体、サイトカイン、および小分子の適正用量の選択の手引きが入手可能である。たとえばWawrzynczak(1996年)Antibody Therapy、Bios Scientific Pub. Ltd、Oxfordshire、UK;Kresina(編)(1991年)Monoclonal Antibodies, Cytokines and Arthritis、Marcel Dekker、New York、NY;Bach(編)(1993年)Monoclonal Antibodies and Peptide Therapy in Autoimmune Diseases、Marcel Dekker、New York、NY;Baertら(2003年)New Engl. J. Med. 348:601頁;Milgromら(1999年)New Engl. J. Med. 341:1966頁;Slamonら(2001年)New Engl. J. Med. 344:783頁;Beniaminovitzら(2000年)New Engl. J. Med. 342:613頁;Ghoshら(2003年)New Engl. J. Med. 348:24頁;Lipskyら(2000年)New Engl. J. Med. 343:1594頁を参照されたい。
【0106】
抗体、抗体断片、およびサイトカインは、持続注入により、またはたとえば1日、1週、または1週当たり1〜7回の間隔での投与により提供できる。投与は、静脈内に、皮下に、局所的に、経口的に、経鼻的に、直腸的に、筋肉内に、脳内に、脊髄内に、または吸入により提供できる。種々の実施形態では、投与方法は、たとえば肺または胃腸管(GI
tract)など、感染の原発部位に基づいて選択される。
【0107】
好ましい投与プロトコルは、著しい望ましくない副作用を回避する最大用量または投与頻度を含むプロトコルである。週用量の合計は、一般的に少なくとも約0.05μg/kg、0.2μg/kg、0.5μg/kg、1μg/kg、10μg/kg、100μg/kg、0.2mg/kg、1.0mg/kg、2.0mg/kg、10mg/kg、25mg/kg、または50mg/kgである。たとえば、Yangら(2003年)New Engl. J. Med. 第349巻:427頁;Heroldら(2002年)New Engl. J. Med. 第346巻:1692頁;Liuら(1999年)J. Neurol. Neurosurg. Psych. 第67巻:451頁;Portieljiら(2003年)Cancer Immunol. Immunother. 第52巻:133頁を参照されたい。小分子治療薬、たとえばペプチド模倣物質、天然産物、または有機化学物質の望ましい用量は、抗体またはポリペプチドとほぼ同じで、モル/kg(mole/kg)程度である。
【0108】
特定の患者についての有効量は、処置されている状態、患者の健康全般、投与の方法、経路、および用量、ならびに副作用の重症度などの要因に依存して変動し得る。たとえば、Maynardら(1996年)A Handbook of SOPs for Good Clinical Practice、Interpharm Press、ボーカラトーン、米国フロリダ州;Dent(2001年)Good Laboratory and Good Clinical Practice、Urch Publ.、ロンドン、英国を参照されたい。
【0109】
典型的な獣医学的被験体、実験被験体、または研究被験体は、サル、イヌ、ネコ、ラット、マウス、ウサギ、モルモット、ウマ、およびヒトを含む。
【0110】
適正用量の決定は、たとえば、処置に影響することが当技術分野で知られているもしくは疑われるまたは処置に影響することが予測されるパラメーターまたは要因を使用して、臨床医によってなされる。一般に、用量は、やや最適用量未満の量から始まり、あらゆる負の副作用と比較して、所望または最適の効果が達成されるまで、用量は、その後、少量ずつ増加させることによって増加する。重要な診断尺度には、たとえば感染または感染レベルの徴候の尺度が含まれる。好ましくは、使用される生物製剤は、処置目標の動物と同一種に由来するか、または同一種に由来するタンパク質を模倣するように改変されており(たとえば、ヒト化抗体)、それにより試薬に対する体液性応答を最小限に抑える。
【0111】
第2の治療薬、たとえばサイトカイン、ステロイド、化学療法剤、抗生物質、または放射線との同時投与または同時処置のための方法は、当技術分野でよく知られている。たとえばHardmanら(編)(2001年)Goodman and Gilman’s
The Pharmacological Basis of Therapeutics、第10版、McGraw−Hill、New York、NY;PooleおよびPeterson(編)(2001年)Pharmacotherapeutics for Advanced Practice: A Practical Approach、Lippincott, Williams & Wilkins、Phila.、PA;ChabnerおよびLongo(編)(2001年)Cancer Chemotherapy and Biotherapy、Lippincott, Williams & Wilkins、Phila.、PAを参照されたい。治療剤の有効量は、典型的には少なくとも10%、一般に少なくとも20%、好ましくは、少なくとも約30%、より好ましくは、少なくとも40%、および最も好ましくは、少なくとも50%、症状を減少させる。
【0112】
本発明は、それらに限定されないが、真菌感染症、持続性真菌感染症、カンジダ症、CMC、アスペルギルス症、クリプトコックス症、ウイルス感染症、持続性ウイルス感染症、HIV感染、HBV感染、HCV感染、細菌感染症、マイコバクテリア感染症、M.tuberculosis感染、M.bovis感染、およびM.leprae感染からなる群から選択される症状を含む感染症を処置するための薬剤の製造における、IL−23またはIL−23R、あるいはその両方のアンタゴニストの使用をさらに提供する。いくつかの実施形態では、薬剤は、1つまたは複数の追加的な治療薬を含むことができる。他の実施形態では、本発明の薬剤は、1つまたは複数の他の治療薬と併せて使用できる。
【0113】
VI.抗イディオタイプ抗体。
【0114】
本発明は、本発明の治療用抗IL−23または抗IL−23R抗体に対する抗イディオタイプ抗体をさらに提供する。抗イディオタイプ抗体とは、一般的には別の抗体の抗原結合領域に結合する、固有の決定基を認識する抗体である。抗イディオタイプ抗体は、抗IL−23抗体またはそのCDR含有領域を有する元の抗IL−23抗体の供給源と同一の種および遺伝子型(たとえば、マウス株)の動物を免疫することにより調製できる。その後、免疫された動物は、免疫する抗体のイディオタイプ決定基に対する抗体を生成して、抗イディオタイプ抗体を産生する。抗イディオタイプ抗体は、さらに別の動物に免疫応答を誘導する免疫原としても使用でき、いわゆる「抗抗Id抗体(anti−anti−Id antibody)」を産生する。
【0115】
たとえば、抗イディオタイプ抗体は、被験体における、たとえば抗IL−23治療を受けている被験体の体液(たとえば血液)における、治療用抗IL−23(または抗IL−23R)抗体のレベルを決定するために使用できる。被験体における抗IL−23(または抗IL−23R)抗体レベルの決定は、そのような決定に応じて用量が変更され得るため、被験体における抗IL−23抗体の望ましいレベルを維持するのに有用であり得る。望ましい範囲の値内の抗IL−23抗体の循環レベルを得るために、用量を増加または減少させることができる(頻度および/または1回の投与当たりの量において)。望ましい範囲は、当技術分野で典型的な方法により医師によって決定することができ、抗IL−23(または抗IL−23R)抗体またはその断片に関する治療指数に依存する場合がある。
【0116】
抗イディオタイプ抗体は、ポリペプチドタグ(たとえば、FLAG(登録商標)タグ)を有する抗体、または色素、同位体、酵素、および金属に結合された抗体を含む、簡易検出に好適な形状で供給できる。たとえば、Le Doussalら(1991年)New Engl. J. Med. 第146巻:169頁;Gibelliniら(1998年)J. Immunol. 第160巻:3891頁;Hsing and Bishop(1999年)New Engl. J. Med. 第162巻:2804頁;Evertsら(2002年)New Engl. J. Med. 第168巻:883頁を参照されたい。放射性免疫測定法(RIA)、ELISA、およびラボオンチップ(lab on a chip)などの種々のアッセイ形式が存在する。米国特許第6,176,962号明細書および第6,517,234号明細書。
【0117】
VII.キット。
【0118】
さらに、本発明は、慢性の細菌感染症、マイコバクテリア感染症、ウイルス感染症、および真菌感染症などの感染症を患っている被験体(ヒトまたは非ヒト)の処置に使用するためのキット中にIL−23アンタゴニストを提供する。一実施形態では、キットは、IL−23アンタゴニストを含有するための区画、IL−23アンタゴニスト自体(抗体など)、および自由選択的に使用説明書、1つまたは複数の追加的な治療薬(複数可)、および1つまたは複数の投与用医療機器(たとえば、RedipenTMインジェクターデバイスなどの注射器または使い捨てインジェクター)を含む。IL−23アンタゴニストは、本明細書に記載された作用剤のいずれかであってよく、それらに限定されないが、抗p19抗体またはそのp19結合性断片、抗IL−23R抗体またはそのIL−23R結合性断片、または可溶性IL−23R断片を含む。
【0119】
1つまたは複数の追加的な治療薬には、それらに限定されないが、非ステロイド系抗炎症剤(NSAID)、ステロイド、IL−12またはそのアゴニスト、およびIL−17A、IL−17F、TGF−β、IL−6などのサイトカインまたはそれらのそれぞれの受容体のアンタゴニストが含まれる。サイトカインのアンタゴニストには、サイトカイン、そのサブユニット、またはその受容体に結合する抗体が含まれる。サイトカインまたはそれらの受容体に結合する抗体は、すべてが必ずしもアンタゴニストとは限らないが、そのようなアンタゴニスト活性は、バイオアッセイまたは受容体結合アッセイなどの当技術分野で一般的に知られている技術により容易にアッセイできる。IL−17A(NM_002190、NP_002181)、IL−17F(NM_052872、NP_443104);IL−17RA(NM_014339、NP_055154);IL−17RC(転写改変体NM_153461、NM_153460、NM_032732、およびそれらのそれぞれの翻訳体)を含む、種々の(ヒト)サイトカインおよび受容体の核酸およびアミノ酸配列は知られている。
【0120】
本発明は、本発明の治療用抗IL−23(または抗IL−23R)抗体に対する抗イディオタイプ抗体を含むキットをさらに提供する。一実施形態では、キットは、抗イディオタイプ抗体を含有するための区画、抗イディオタイプ抗体自体、および自由選択的に使用説明書、1つまたは複数の検出用試薬、抗イディオタイプ抗体検出用の1つまたは複数の機器(マイクロタイタープレートなど)、および検出する抗IL−23抗体の1つまたは複数の試料(または他の陽性対照)を含む。
【0121】
VIII.使用
結核の発症前には、長期の無症候性の症状発症前の期間が存在することが多い。したがって、IL−23およびIL−23Rアンタゴニスト治療は、種々のTB診断マーカーの解析のときに開始できる。正常な非感染患者と比較して、陽性のツベルクリン検査またはマントー試験(たとえば、Dale and Federman(2002年)を参照)を示す患者には、IL−23またはIL−23Rアンタゴニスト治療を実施して、マイコバクテリアのさらなる増殖を防止または既存の非病理学的感染を除去することができる。生体試料、たとえばBALに高レベルのマイコバクテリアを有する患者も、肺中のマイコバクテリアのさらなる増殖を防止し細菌負荷を除去するためのIL−23およびIL−23Rアンタゴニスト治療から恩恵を受けることができる。同様の処置は、臨床試料中のマイコバクテリアDNAまたはRNAのレベルが高い患者または培養物中のナイアシン検査が陽性である患者に使用できる。細菌負荷を軽減または除去するために、病理学的に症候的なTB感染と併せたIL−23およびIL−23Rアンタゴニストの使用も企図される。
【0122】
本発明の方法および組成物を使用して処置できる細菌感染症には、それらに限定されないが、以下により引き起こされる感染症が含まれる:
【0123】
【化1】
【0124】
【化2】
本発明の方法および組成物を使用して処置できるマイコバクテリア感染症には、それらに限定されないが、以下により引き起こされる感染症が含まれる:
【0125】
【化3】
本発明の方法および組成物は、これらに限定されないが、ヒストプラスマ症、コクシジオイデス真菌症、ブラストミセス症、アスペルギルス症、ペニシリウム症、カンジダ症、およびクリプトコックス症を含む真菌症状を処置するためにも使用できる。真菌症のリスク因子には、血液および骨髄移植、実体のある臓器移植、大手術(特に胃腸管手術)、AIDS、新生物疾患、高齢、免疫抑制治療、ならびに乳幼児の未成熟が含まれる。
【0126】
本発明の方法および組成物を使用して処置できる感染症(および臨床症候群(clinical syndrome))を引き起こす真菌病原体には、これらに限定されないが、Candida albicans(鵞口瘡、膣カンジダ症、食道カンジダ症)、Cryptococcus neoformans(髄膜炎)、Histoplasma capsulatum(発熱および体重喪失を伴う播種性感染症)、Coccidioides immitis(散在性および局限性肺疾患)、Blastomyces dermatitidis(髄膜炎を含む、局所的肺疾患および播種性感染症)、Aspergillus fumigatus(発熱、咳、および喀血を伴う肺疾患)およびPenicillium marneffei(発熱のみ、または肺浸潤、リンパ節症、もしくは皮膚病変を伴う)が含まれる。本発明の方法および組成物は、Candida種属、C.glabrata、C.parapsilosis、C.tropicalis、C.krusei、C.lusitaniae、C.guilliermondii、およびC.rugosaによる感染症を処置するためにも使用できる。前述の真菌病原体(および臨床症候群)は、一般的にHIV感染に付随する。
【0127】
本発明の方法および組成物は、C.glabrata、C.parapsilosis、C.tropicalis、C.krusei、C.lusitaniae、C.guilliermondii、およびC.rugosaなどのCandida種属による感染症を処置するためにも使用できる。本発明の方法および組成物は、A.flavus、A.niger、A.ustus、およびA.terreusなどのAspergillus種属による感染症を処置するためにも使用できる。追加的な真菌病原体には、Fusarium種属(たとえば、F.moniliforme、F.solani、F.oxysporum)およびScedosporium種属(たとえば、S.apiosperum、S.prolificans)が含まれる。追加的な真菌疾患には、Rhizopus(たとえば、R.oryzae、R.arrhizus)、Rhizomucor、Absidia、Cunninghamellaの種属により引き起こされる接合真菌症が含まれる。
【0128】
IL−23およびIL−23Rのアンタゴニストは、単独で、あるいはTh1応答を増強する目的の作用剤(たとえば、IL−12またはそのアゴニスト)、もしくはTh17応答を阻害する目的の作用剤(たとえば、TGF−βアンタゴニスト;IL−6アンタゴニスト;IL−17Aおよび/またはIL−17Fアンタゴニスト)、またはその両方の作用剤と共に使用できる。これらのサイトカインの受容体のアゴニストおよびアンタゴニストも使用できる。そのような作用剤には、抗体およびその抗原結合性断片、小分子、siRNA、およびアンチセンス核酸が含まれ得る。IL−23およびIL−23Rのアンタゴニストは、コルチコステロイド、たとえばプレドニゾンなどの抗炎症薬と共に使用することもできる。
【0129】
IL−17アンタゴニストは、IL−17A、IL−17F、IL−17RA、またはIL−17RCの発現を阻害することができるか、または機能的なリガンド−受容体相互作用を防止するために、これらのポリペプチドの1つまたは複数と直接的にまたは間接的に相互作用することによりIL−17シグナル伝達を阻害できる。いくつかの好ましい実施形態では、IL−17アンタゴニストは、IL−17A、IL−17F、IL−17RA、またはIL−17RCのいずれかに結合してその活性を阻害する抗体または抗体断片である。1つの特に好ましい実施形態では、IL−17アンタゴニストは、IL−17Aに特異的に結合するモノクローナル抗体である。IL−17Aに対する典型的なアンタゴニスト抗体には、同一人に譲渡された米国特許出願第11/836,318号明細書(2007年8月9日に出願された)、および国際公開第2006/013107号パンフレットおよび国際公開第2006/054059号パンフレットに開示されている抗ヒトIL−17A抗体およびそれらの断片が含まれる。別の実施形態では、IL−17アンタゴニストには二重特異性抗体が含まれる。
【0130】
一実施形態では、IL−23アンタゴニストには、IL−23に結合しその活性を阻害する二重特異性抗体が含まれる。そのような二重特異性抗体は、IL−23p19またはIL−23Rに結合でき、IL−17A、IL−17F、IL−17RA、IL−17RCにも結合できる。他の実施形態では、IL−23アンタゴニストは、IL−23p19およびIL−17に結合しIL−23およびIL−17の活性を阻害する二重特異性抗体である。たとえば、国際公開第2007/147019号パンフレットを参照されたい。あるいは、IL−23およびIL−17アンタゴニスト二重特異性抗体は、アンタゴニスト抗体である限り、それぞれIL−23受容体(たとえばIL−23R)またはIL−17受容体(IL−17RAまたはIL−17RC)のいずれかに結合できる。IL−17およびIL−23活性の両方に拮抗する二重特異性抗体は、当技術分野で知られている任意の技術により産生することができる。たとえば、二重特異性抗体は、2つの免疫グロブリン重鎖/軽鎖対の共発現を使用して組換え的に産生することができる。たとえば、Milsteinら(1983年)Nature 第305巻:537〜39頁を参照されたい。あるいは、二重特異性抗体は、化学的結合を使用して調製することができる。たとえば、Brennanら(1985年)Science 第229巻:81頁を参照されたい。二機能性抗体(bifunctional antibody)は、ジスルフィド交換、ハイブリッドハイブリドーマ(クアドローマ(quadroma))の産生により、二重特異性抗体を具現化する単一のポリペプチド鎖を産生するための転写および翻訳により、または二重特異性抗体を産生するために共有結合で結合され得る複数のポリペプチド鎖を産生するための転写および翻訳によっても調製できる。企図された二重特異性抗体は、全化学合成により作製することもできる。二重特異性抗体は、2つの異なる可変領域、2つの異なる定常領域、可変領域および定常領域、または他の改変を含むことができる。
【0131】
IL−23およびIL−23Rのアンタゴニストは、単独で使用し、またはイソニアジド、リファンピン、ピラジナミド、エタンブトール、ストレプトマイシン、シプロフロキサシン、およびオフロキサシンなどの既知の抗菌物質と共投与することができる。追加的な抗菌物質には、これらに限定されないが、アラトロフロキサシン(alatrofloxacin)、アジスロマイシン、バクロフェン、ベンザチンペニシリン、シノキサシン、クラリスロマイシン、クロファジミン、クロキサシリン、デメクロサイクリン、ジリスロマイシン、ドキシサイクリン、エリスロマイシン、エチオナミド、フラゾリドン、グレパフロキサシン、イミペネム、レボフロキサシン、ロレフロキサシン(lorefloxacin)、モキシフロキサシンHCl、ナリジキシン酸、ニトロフラントイン、ノルフロキサシン、オフロキサシン、リファブチン、リファペンチン、スパルフロキサシン、スピラマイシン、スルファベンズアミド(sulphabenzamide)、スルファドキシン、スルファメラジン、ウルファセタミド(ulphacetamide)、スルファジアジン、スルファフラゾール(sulphafurazole)、スルファメトキサゾール、スルファピリジン、テトラサイクリン、トリメトプリム、トロバフロキサシン、およびバンコマイシンが含まれる。
【0132】
本発明の方法および組成物は、これらに限定されないが、HBV、HCV、HIV、ヒトパピローマウイルス(HPV)により引き起こされる感染症を含む、持続性のウイルス感染症を処置するために使用できる。そのような慢性感染症は、感染を根絶するための免疫応答の失敗を表す。IL−23およびIL−23Rのアンタゴニストは、単独で、またはこれらに限定されないが、アバカビル、アシクロビル、アマンタジン、アンプレナビル、デラビルジン、ジダノシン、エファビレンツ、ファムシクロビル、インジナビル、インターフェロンアルファ、リバビリン、ラミブジン、ネルフィナビル、ネビラピン、オセルタミビル、ペンシクロビル、リバビリン、リトナビル、サキナビル、スタブジン、バラシクロビル、ザルシタビン、ザナミビル、ジドブジン(アジドデオキシチミジン、AZT)を含む他の抗ウイルス物質と併せて使用できる。好ましいインターフェロンアルファ剤には、ペグ化インターフェロンアルファ2aおよびペグ化インターフェロン2bが含まれる。インターフェロンアルファの典型的な形状は、米国特許第6,923,966号明細書で考察されている。IL−23アンタゴニストは、慢性HCV感染用のHCVプロテアーゼまたはHCVポリメラーゼ阻害剤、および慢性HIV感染用のCCR5アンタゴニストなどのウイルス特異的作用剤と組み合わせて使用することもできる。
【0133】
IL−23およびIL−23Rのアンタゴニストは、治療用ワクチン、たとえばHIV感染用のgp120枯渇全殺滅ウイルス、HCV感染用の組換えE1タンパク質、ならびにHPV感染用のウイルスE6およびE7腫瘍性タンパク質と併せて使用することもできる。Berzofskyら(2004年)を参照されたい。そのような治療用ワクチンには、DNAワクチンまたはウイルスベクターが含まれ、自由選択的には、ウイルスベクターワクチンがDNAワクチン後になされる異種性の初回抗原刺激および追加免疫投与計画で投与される。Berzofskyら(2004年)。
【0134】
IL−23およびIL−23Rのアンタゴニストは、単独で、またはこれらに限定されないが、ポサコナゾール、フルコナゾール(米国特許第4,404,216号明細書)、ボリコナゾール、イトラコナゾール(米国特許第4,267,179号明細書)、ケトコナゾール(米国特許第4,144,346号明細書および第4,223,036号明細書)、リアロゾール、イルテマゾール、クロトリマゾール、ミコナゾール、エコナゾール、ブトコナゾール、オキシコナゾール、スルコナゾール、チオコナゾール、およびテルコナゾール、置換チアゾール、チアジアゾール、オキサジアゾール、カスポファンギン、アムホテリシンB、ナイスタチン、ピマリシン、フルシトシン(5−フルオロシトシン)、ナフチフィン、テルビナフィン、ブテナフィン、チオカーボネートトルナフテート(thiocarbonate tolnaftate)、グリセオフルビン、アミオダロン、シクロピロックス、スルベンチン、アモロルフィン、クリオキノール、ゲンチアナバイオレット、ヨウ化カリウム、チオ硫酸ナトリウム、石炭酸フクシン溶液、およびエキノキャンディン(たとえば、カスポファンギンアセテート、ミカファンギン、およびアニデュラファンギン)を含む他の抗真菌物質と併せて使用できる。
【0135】
本発明のIL−23およびIL−23Rアンタゴニストは、単一の作用剤として使用される場合はそれらの通常用量で、または薬物が併用される場合は効果に何らかの相乗的増強があればより低用量で、標準的抗真菌物質と組み合わせて使用できる。フルコナゾールは、たとえば、400〜800mg/日で投与できる。ボリコナゾールは、1日2回4mg/kgで投与できる。イトラコナゾールは、200〜600mg/日で投与できる。アムホテリシンBデゾキシコレート(D−AmB)は、0.5〜1mg/kg/日で投与できる。本発明の組成物および方法と組み合わせ得る作用剤および処置計画のタイプに関する一般的なガイダンスは、米国感染症学会(IDSA)により刊行された診療ガイドラインのPappasら(2004年)Clin. Infect. Dis. 第38巻:161頁(カンジダ症)、Stevensら(2000年)Clin. Infect. Dis. 第30巻:696頁(アスペルギルス症)に見出すことができる。結核処置の診療ガイドラインは、2006年3月22日に刊行され、IDSAにより承認された結核医療の国際基準(International Standards for Tuberculosis Care)に見出される。
【0136】
本発明のいくつかの実施形態では、感染症を有する被験体、または感染症を有する疑いのある被験体は、他の方法または組成物(つまり、本発明の方法または組成物ではない)を使用して以前に感染症の処置を受けたことがある被験体である。以前の処置には、本明細書に開示された抗菌物質、抗生物質、抗真菌物質、抗ウイルス物質、または任意の他の処置方法もしくは組成物のいずれかでの処置が含まれ得る。
【0137】
いくつかの実施形態では、被験体は感染症の正式な診断を受け、自由選択的には病因物質が同定されているが、他の実施形態では、被験体は、正式な診断を受けていないか、または病因物質が限定されたが完全には同定されていない部分診断を受けている場合がある。他の実施形態では、被験体は、感染症を有する疑いのあるだけである。他の実施形態では、被験体は、感染症を有するまたは感染症に罹るリスクがあり、たとえば被験体は、免疫抑制治療を受けており、AIDSなどが原因で真菌感染症に罹るリスクがある。いくつかの実施形態では、感染症を有する被験体、感染症を有する疑いのある被験体、または感染症を有するもしくは感染症に罹るリスクのある被験体は、たとえばAIDS、化学療法、移植、高齢のため免疫無防備状態のになっている。
【0138】
当業者であれば明白であるが、本発明の趣旨および範囲から逸脱することなく、本発明の改変および変更を多数行うことができる。本明細書に記載された特定の実施形態は例示目的でのみ提示されており、本発明は、添付の特許請求の範囲の用語により制限されると共に、そのような特許請求の範囲に与えられる等価物の包括的範囲により制限され、本発明は、例示目的で本明細書に提示された特定の実施形態により制限されるものではない。
【実施例】
【0139】
(実施例1)
一般的な方法
分子生物学における標準的な方法を記載する(Maniatisら、(1982年)Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、NY;Sambrook and Russell(2001年)Molecular Cloning、第3版、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、NY;Wu(1993年)Recombinant DNA、第217巻、Academic Press、San Diego、CA)。標準的な方法はAusbelら、(2001年)Current Protocols in Molecular Biology、第1〜4巻、John Wiley and Sons、Inc.New York、NY中にも載っており、これは細菌細胞におけるクローニングおよびDNA変異誘発(第1巻)、哺乳動物細胞および酵母におけるクローニング(第2巻)、複合糖質およびタンパク質発現(第3巻)、およびバイオインフォマティクス(第4巻)を記載している。
【0140】
免疫沈降、クロマトグラフィー、電気泳動、遠心分離、および結晶化を含めたタンパク質精製のための方法を記載する。Coliganら、(2000年)Current Protocols in Protein Science、第1巻、John Wiley and Sons、Inc.、New York。化学分析、化学改変、翻訳後改変、融合タンパク質の生成、タンパク質のグリコシル化を記載する。たとえば、Coliganら、(2000年)Current Protocols in Protein Science、第2巻、John Wiley and Sons、Inc.、New York;Ausubelら、(2001年)Current Protocols in Molecular Biology、第3巻、John Wiley and Sons、Inc.、NY、NY、16.0.5〜16.22.17頁;Sigma−Aldrich、Co.(2001年)Products for Life Science Research、St.Louis、MO;45〜89頁;Amersham Pharmacia Biotech(2001年)BioDirectory、Piscataway、N.J.384〜391頁を参照。ポリクローナルおよびモノクローナル抗体の産生、精製、および断片化を記載する。Coliganら、(2001年)Current Protocols in Immunology、第1巻、John Wiley and Sons、Inc.、New York;Harlow and Lane(1999年)Using Antibodies、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、NY; Harlow and Lane(1998年)。リガンド/受容体相互作用を特徴付けするための標準的な技法が利用可能である。たとえば、Coliganら、(2001年)Current Protocols in Immunology、第4巻、John Wiley Inc.、New Yorkを参照。
【0141】
蛍光活性化セルソーティング(FACS)を含めた、フローサイトメトリーのための方法が利用可能である(たとえば、Owensら、(1994年)Flow Cytometry Principles for Clinical Laboratory Practice、John Wiley and Sons、Hoboken、NJ;Givan(2001年)Flow Cytometry、第2版;Wiley−Liss、Hoboken、NJ;Shapiro(2003年)Practical Flow Cytometry、John Wiley and Sons、Hoboken、NJ中を参照)。たとえば診断用試薬として使用するための、核酸プライマーおよびプローブ、ポリペプチド、および抗体を含めた核酸を改変するのに適した蛍光試薬が利用可能である(Molecular Probes(2003年)Catalogue、Molecular Probes、Inc.、Eugene、OR;Sigma−Aldrich(2003年)Catalogue、St.Louis、MO)。
【0142】
免疫系の組織学の標準的な方法を記載する。たとえば、Muller−Harmelink(ed.)(1986年)Human Thymus:Histopathology and Pathology、Springer Verlag、New York、NY;Hiattら、(2000年)Color Atlas of Histology、Lippincott、Williams、and Wilkins、Phila、PA;Louisら、(2002年)Basic Histology:Text and Atlas、McGraw−Hill、New York、NYを参照。
【0143】
たとえば抗原断片、リーダー配列、タンパク質フォールディング、機能性ドメイン、グリコシル化部位、および配列アラインメントを決定するための、ソフトウェアパッケージおよびデータベースが利用可能である。たとえば、GenBank、Vector NTI(登録商標)Suite(Informax、Inc、Bethesda、MD);GCG Wisconsin Package(Accelrys、Inc.、San Diego、CA);DeCypher(登録商標)(TimeLogicCrop.、Crystal Bay、Nevada);Menneら、(2000年)Bioinformatics 16:741〜742頁;Menneら、(2000年)Bioinformatics Applications Note 16:741頁;Wrenら、(2002年)Comput.Methods Programs Biomed. 68:177頁;von Heijne(1983年)Eur.J.Biochem. 133:17頁;von Heijne(1986年)Nucleic Acids Res. 14:4683頁を参照。
【0144】
(実施例2)
真菌感染モデル
真菌感染の試験用のマウス系統は以下のように得た。雌性C57BL/6およびBALB/cマウス、8〜10週齢はCharles River(Calco、イタリア)から購入した。同型IL−12p35、IL−23pl9またはIL−12p40欠損マウス(ここでは以後、それぞれp35−/−、p19−/−およびp40−/−と呼ぶ)、TLR−2−、TLR−4、MyD88またはTRIF欠損マウス(ここでは以後、TLR−2−/−、TLR−4−/−、MyD88−/−またはTRIF−/−と呼ぶ)C57BL/6バックグラウンドのマウスを、ペルージャ大学、ペルージャ、イタリアの動物施設において、特定病原体未感染の条件下で飼育した。BALB/cバックグラウンドのIFN−γ−/−p35−/−マウスのつがいは、Dr.M.Colombo(Istituto Tumori、Milan、イタリア)によって提供された。BALB/cバックグラウンドのIFN−γ−/−およびIL−4−/−マウスも、ペルージャ大学の動物施設で飼育した。Italian Approved Animal Welfare Assurance A−3143−01に従い実験を実施した。
【0145】
真菌感染およびそれらの処置は以下のように試験した。この試験で使用したC.albicans系統の起源および特徴は既に記載されている。Bacciら、(2002年)J. Immunol. 168:2904頁。胃腸感染用に、108個のCandida細胞を胃内注射し、かつ真菌増殖の定量化は、記載したように器官当たりのCFU(平均±SE)として表した。Bacciら、(2002年)。静脈内感染用に、マウスには静脈内に0.5ml当たり異なる量の真菌を与えた。A.fumigatusの系統および培養条件は記載した通りであった。Montagnoliら、(2006年)J. Immunol. 176:1712頁。マウスには2回用量の2×107個のAspergillus静止期分生子を鼻腔内に与えた。Limulus amebocyte血球抽出成分(LAL)の方法により決定して、1.0EU/ml未満でエンドトキシンフリー溶液(Detoxi−gel、Pierce、Rockford、IL)中に真菌を懸濁させた。真菌増殖はキチンアッセイ(chitin assay)によって定量化し、結果は器官当たりのグルコサミンのマイクログラム数として表した。組織検査用に、組織を切除し、すぐにホルマリン中に固定し、かつパラフィン包埋組織の切片(3〜4μm)を過ヨウ素酸シッフ試薬で染色し調べた。Bacciら、(2002年);Montagnoliら、(2006年)。感染動物は200μgのp19中和抗体(Belladonnaら、(2006年)Cytokine 34:161頁)またはIL−17A中和モノクローナル抗体(TC11−18H10、PharMingen、San Diego、CA)で処置し、感染5時間後に腹腔内投与した。合計1mgの精製抗TGF−β1、β2、β3モノクローナル抗体(2G7)(Lucasら、(1990年)J. Immunol. 145:1415頁)を、感染5時間後および24時間後に腹腔内投与した。対照マウスにはPBSを注射した、何故なら、PBS処置動物とアイソタイプ対照処置(各処置)動物(各群に関してn>6)の間で差異を観察しなかったからである。
【0146】
細胞は以下のように精製した。Gr−1+CD11b+多形核好中球(PMN、FACS分析において98%を超えて純粋)を、Ly−6G MicroBeadsおよびMidiMacs(Miltenyi Biotech、Bergisch Gladbach、ドイツ)を使用した磁気活性化ソーティング(magnetic−activating sorting)によってマウスの腹膜腔から単離した。CD4+T細胞は、CD4MicroBeadsおよびMidiMacs(Miltenyi Biotech)を使用した磁気活性化ソーティングによって腸間膜リンパ節(MLN)、胸部リンパ節(TLN)および脾臓から精製した。DCは、CD11b+DCを得るために7日間150U/m1のマウスrGM−CSF(Sigma−Aldrich、St.Louis、MO)および75U/mlのrIL−4(R&D Systems、Minneapolis、MN)、またはFL−DCを得るために9日間200ng/mlのFLT3−L(R&D Systems)の存在下において、Iscoveの改変培地中で培養した骨髄細胞から得た。Romaniら、(2006年)Blood 108:2265頁。90〜95%のCD8−、5〜10%のCD8+、および1〜5%のB220+細胞)からなる脾性DC(>99%CD11c+および<0.1%CD3+)は、CD11c MicroBeadsおよびMidiMacs(Miltenyi Biotech)を使用した磁気活性化ソーティングによって精製した。Saccharomyces cerevisiae由来のザイモザン(10μg/ml、Sigma−Aldrich)、Salmonella minnesota Re595由来の超高純度(ultra−pure)LPS(10μg/ml、Labogen、Rho、Milan、イタリア)およびCpGオリゴヌクレオチド2006(CpGODN、0.06μM)を、記載したように使用した。Bellocchioら、(2004年)J. Immunol、173:7406頁。
【0147】
DC細胞を以下のようにパルスし培養した。DCは、記載したように1:1の細胞:真菌比で、生きた非オプソニン化真菌に、(R&D Systems;Space Import−Export srl、Milan、イタリア;およびBD Biosciences−PharMingen、San Diego、CAからの)10ng/mlのサイトカイン有りおよび無しで、または中和抗体(10μg/ml)有りおよび無しで曝露した。Bacciら、(2002年);Montagnoliら、(2006年)。培養12時間でRT−PCR用に細胞を採取し、かつ上清はELISAによってサイトカイン含有量に関して評価した。脾性CD4+T細胞(106/ml)は、培養物上清におけるサイトカイン定量化前に、中和抗体(10μg/ml)有りおよび無しで、5日間5×105個のCandidaでパルスした脾性DCの存在下において平底96ウェルプレート中で培養した。非分画MLNまたはTLN細胞は、5日後の培養物上清におけるサイトカイン測定前に、記載したように(Montagnoliら、(2006年);Montagnoliら、(2002年)J. Immunol. 169:6298頁)不活化真菌と共に培養した。
【0148】
(実施例3)
抗真菌活性アッセイ
非オプソニン化Candida酵母またはAspergillus分生子のPMN食作用、および抗真菌活性のアッセイを、記載したように実施した。Bellocchioら、(2004年)。結果はCFU阻害の割合(平均±SE)として表す。PMNは、IDOのウエスタンブロッティング前に12時間、または抗真菌活性およびMMP9/MPO(マウスミエロペロキシダーゼ)測定の試験用にさらに60分間真菌を加える前に60分間、様々な濃度のIL−17もしくはIL−23または50ng/mlのIFN−γ±IL−23/IL−17(100ng/ml)に曝露した。ゼラチンザイモグラフィーを記載したように実施した。Bellocchioら、(2004年)。マトリクスメタロプロテイナーゼ9(MMP9)のゼラチン分解活性は、72−kD領域中の溶解バンドをスキャンすることによって測定した。MPO測定用に、サンプルはウサギポリクローナル抗ヒトMPO抗体(Calbiochem、San Diego、CA)でプローブ処理し、電気化学発光(ECL)(Amersham Pharmacia Biotech、Piscataway、NJ)を使用して目に見える状態にした。
【0149】
インドールアミン2,3−ジオキシゲナーゼ(IDO)は、記載したようにウサギポリクローナルIDO特異的抗体を用いたイムノブロッティングによって検出した。Bozzaら、(2005年)。陽性対照はIDO発現MC24トランスフェクタントからなっており、かつ陰性対照はモックトランスフェクト(mock−transfectes)MC22細胞であった。
【0150】
サイトカインは以下のようにリアルタイムRT−PCR、ELISAおよびELISPOTアッセイによって定量化した。リアルタイムRT−PCRは、iCycler iQ(登録商標)検出システム(Bio−Rad、Hercules、CA)およびSYBR(登録商標)Green chemistry(Finnzymes Oy、Espoo、フィンランド)を使用して実施した。細胞を溶解し、すべてのRNAはRNeasy Miniキット(QIAGEN S.p.A.、Milan、イタリア)を使用して抽出し、製造者の指示に従いSensiscript逆転写酵素(QIAGEN)を用いて逆転写した。PCRプライマーはInvitrogen(Carlsbad、CA)から得た。使用したPCRプライマーは以下の通りであった:
マウスIL−12p35の場合、フォワードプライマー、
【0151】
【化4】
、およびリバースプライマー、
【0152】
【化5】
、;マウスIL−23p19の場合、フォワードプライマー、
【0153】
【化6】
、およびリバースプライマー
【0154】
【化7】
、;IL−12Rβ2の場合、フォワードプライマー、
【0155】
【化8】
、およびリバースプライマー
【0156】
【化9】
;IL−23Rの場合、フォワードプライマー、
【0157】
【化10】
、およびリバースプライマー
【0158】
【化11】
;マウスガンマ−アクチンの場合、フォワードプライマー、
【0159】
【化12】
、およびリバースプライマー、
【0160】
【化13】
。
【0161】
製造者の説明書(Applied Biosystems、Foster City、CA)に従い、ハウスキーピングγ−アクチン遺伝子のPCR増幅を、それぞれのサンプルに実施して(三連)、サンプルローディングを調節し、サンプル間の標準化を可能にした。水対照を含めて特異性を確実にした。SYBR(登録商標)GreenリアルタイムPCRに関する温度プロファイルは、95℃で3分間、次に95℃で15秒間の変性および60℃で1分間のアニーリング/延長ステップの40サイクルであった。増幅プロットの分析によって、完全性に関してそれぞれのデータ地点を調べた。mRNAの標準化データは、モック感染細胞のそれと比較した処置細胞中の相対的サイトカインmRNAとして表した。サイトカイン含有量は、培養細胞の組織ホモジネートまたは上清に関する酵素結合免疫吸着アッセイ(R&D SystemsおよびIL−23に関しては、eBioscience、Societa Italiana Chimici、Roma、イタリア)によって評価した。アッセイの検出限界(pg/ml)は、IL−12p70に関して<16、IL−23に関して<30、IFN−γに関して<10、IL−10に関して<3、IL−17に関して<10およびTGF−β1に関して<4,6であった。AID EliSpotアッセイキット(Amplimedical、Buttigliera Alta、Turin、イタリア)を、3日間CandidaでパルスしたDCと同時培養した精製MLN CD4+T細胞に使用して、サイトカイン産生細胞を計数した。
【0162】
データの統計分析は以下のように実施した。対数順位検定を、カプランマイヤー生存曲線の対データ分析に使用した。スチューデントのt検定または分散分析(ANOVA)およびボンフェロニの検定を使用して、臓器クリアランスおよびインビトロアッセイにおける統計的有意差を決定した。有意はp<0.05として定義した。記載されたデータは、3回の別個の実験の中の1回の代表的な実験からのデータ、または3回〜5回の実験から集めたデータのいずれかである。インビボ群は群当たり6〜8匹のマウスからなっていた。
【0163】
(実施例4)
カンジダ症に対する感受性におけるIL−23/IL−17の役割
C.albicans感染に対するIL−23/IL−17経路の貢献を評価するために、本発明者らは、生存、真菌増殖、および組織病理学の点で胃腸感染に対する感受性に関して、ならびに炎症および適応Th1/Th17免疫のパラメーターに関して、p19−/−、p35−/−、p40−/−およびC57BL/6マウスを比較した。結果(図1A〜E)は、カンジダ症に対する耐性はp35−/−マウス中では著しく低下し、それらは50%を超えて感染に屈し(図1A)、胃中では真菌増殖が高まった(図1B)ことを示した。対照的に、真菌増殖を制限する能力は、感染3日後および10日後にC57BL/6マウスと比較して、p19−/−マウス中で大幅に増大した。特に、IL−12とIL−23の両方が欠損したp40−/−マウスはp35−/−マウスよりカンジダ症に対する感受性が低く、かつp19−/−マウスより感受性が高く、Candidaの調節に関するIL−12の差次的役割が強調された。Mencacciら、(1998年)J. Immunol. 161:6228頁。同様の結果をp35−/−およびp19−/−マウスの静脈内感染後に観察し、それぞれ6±2対20±3日(5×105個の真菌細胞接種)、および4±2対15±3日(106個の真菌細胞接種)の平均生存時間(MST)であった。胃の組織病理学的検査によって、C57BL/6、p19−/−またはp40−/−マウス中での錯角化、表皮肥厚および限られた炎症反応の存在が明らかになったが、p40−/−、および特にp19−/−マウスでは単核細胞の浸潤を示した。対照的に、p35−/−マウスの胃中のPMNの多量浸潤、上皮壊死および顕著な表皮肥厚の徴候と併せて、多数の菌糸が角質化層中に存在した。これらの結果は、IL−23経路とIL−12経路はカンジダ症において相違する役割を有することを示唆する。
【0164】
これらの発見とIL−12/Th1およびIL−23/Th17免疫応答を関連付けるために、マウスは、感染翌日または3日後にMLNにおけるp35、p19、IL−12Rβ2およびIL−23RのmRNA発現(図1C)、および感染7日後のMLNにおけるIFN−γ、IL−4またはIL−17産生CD4+細胞の頻度に関して評価した(図1D)。本発明者らは、C57BL/6マウスと比較して、p19−/−マウス中での増大したレベルのp35およびIL−12Rβ2、およびIFN−γ+細胞の数を発見し、IL−23の不在下での増強したIL−I2/Th1応答を実証した。対照的に、p19およびIL−23RのレベルおよびIL−17産生細胞の数は、IL−12を欠くマウス(p35−/−)において増大した。予想通り、IL−4産生細胞の数もp35−/−マウスにおいて相当増大した。これらのデータは、野生型C57BL/6マウス中でIL−12によって促進されIL−23によって制限される主要なTh1応答を実証する。IL−12はIL−23およびIL−17産生を抑制し、および逆にIL−23はIL−12およびIFN−γ産生を阻害し、IL−23/Th17とIL−12/Th1経路の交差制御を示す。これらのデータは、高いIL−23/Th17応答はマウスをカンジダ症に対して非常に敏感にすることを示唆する。
【0165】
(実施例5)
アスペルギルス症に対する感受性におけるIL−23/IL−17の役割
カンジダ症と同様に、IL−23/IL−17経路の活性化がアスペルギルス症に対する感受性と関係があるかどうか決定するために、肺アスペルギルス症に対する感受性、ならびに炎症および適応Th1/Th17免疫のパラメーターに関してp19−/−、p35−/−、p40−/−またはC57BL/6マウスを評価した。結果(図2A)は、真菌負荷はp35−/−マウス中で、およびp19−/−マウス中ではさらに高い程度で低下したことを示し、IL−12、および特にIL−23はAspergillus感染の調節(すなわち促進)を阻害することを示唆する。肺の組織病理学的検査によって、他の完全な肺実質中に散乱した炎症性単核細胞のわずかな浸潤によって特徴付けられる、C57BL/6、p40−/−またはp19−/−マウスにおける軽度の炎症性病理の存在が明らかになった。浸潤単核細胞の数はp19−/−マウス中でより多かったが、実質破壊の徴候は観察しなかった。対照的に、PMNの多量浸潤(Gr1+CD11c−PMNの約8〜10倍の増大)が、広範囲の間質性肺炎の徴候を伴うp35−/−マウスの肺において存在した。Candidaによる感染と同様に、IL−12およびIL−23およびそれらの各々の受容体の発現は交差制御され、C57BL/6マウスと比較して、p19−/−マウスのTLNにおいてp35およびIL−12Rβ2は上方制御され、かつp35−/−マウスのTLNにおいてp19およびIL−23Rは上方制御された(図2B)。対照的に、p40−/−マウス中でのIL−12とIL−23の両方の不在が、p35およびp19またはそれらの受容体IL−12Rβ2およびIL−23Rの発現を有意に変えることはなかった。さらに、IFN−γ+およびIL−17+産生CD4+T細胞の数は、感染7日後にそれぞれp19−/−およびp35−/−マウスにおいて増大した(データ示さず)。肺中では、IL−12p70のレベルはC57BL/6マウス中(68±8pg/ml)よりp19−/−中ではるかに高かった(554±44pg/ml)、かつIL−23はp35−/−マウス中のみで検出することができた(79±11)。IL−17はC57BL/6マウス(37±7pg/ml)と比較してp35−/−マウス中で増大した(246±17pg/ml)。これらのデータは、増大したIL−23/IL−17依存性炎症性応答はアスペルギルス症に対する感受性とも関係があることを示唆する。
【0166】
(実施例6)
真菌感染に対するIL−23/IL−17仲介感受性におけるTGF−βの役割
真菌感染に対する感受性におけるIL−17の役割を試験するために、本発明者らは、真菌感染直後に抗IL−17中和抗体でマウスを処置した。IL−17の遮断は、低下した真菌増殖(図3A)、関連標的器官中の組織炎症およびPMN浸潤(データ示さず)により判断して、C.albicansとA.fumigatusの両方に対する耐性を大幅に増大した。耐性はIFN−γ+Th1細胞の頻度の増大およびTh17細胞の頻度の低下を伴い、MLN細胞によって分泌されるIL−17の量の低下をもたらした(図3B)。同様に、抗体によるIL−23の中和によって真菌感染およびTh1発生に対する耐性が増大し、p19−/−マウスにおいて得た本発明者らのデータを確認する(図3B)。これらの結果は、IL−23/IL−17経路は防御Th1免疫の阻害によって真菌感染に対する感受性をもたらすことを明らかに実証する。
【0167】
最新の結果は、TGF−βはIL−6と一緒にTh17発生を促進することを示唆する。Bettelli and Kuchroo(2005年)J. Exp. Med.
201:169頁;Manganら、(2006年)Nature 441:231頁;Veldhoenら、(2006年)Immunity 24:179頁。本発明者らは、TGF−β中和抗体を用いた処置によって、マウス中でのTh細胞の発生および真菌調節に対するTGF−βの影響を評価した。特に、TGF−β阻害はC.albicansとA.fumigatusの両方の感染中にIL−17産生細胞の発生に影響を与えず(図3B)、真菌負荷のわずかではあるが有意な低下はCandidaを有するマウスではなくAspergillusを有するマウスのみで観察したが(図3A)、いずれの感染においても処置によって影響を受けるCD4+Th17細胞の活性化はなかった。TGF−β中和は感染中のTGF−β産生を効果的に低下させたので(胃中では46pg/mlから24pg/ml、および肺中では36pg/mlから15pg/ml)、本発明者らは、TGF−βは真菌感染に対するTh17仲介の感受性においてわずかな役割を果たすと結論付ける。
【0168】
(実施例7)
IL−12の不在下での真菌感染におけるIL−23/IL−17の役割
上記のデータは、それによってIL−23/IL−17軸が真菌感染に対する感受性を決定する1つの可能性のある機構は、防御Th1応答を抑制する相対能力に頼ることを示唆する。それを正式に証明するために、IL−23の遮断を増大(IL−4−/−マウス)または欠損(IFN−γ−/−マウス)Th1反応性のいずれかの条件下で行った。マウスにはC.albicansを胃内感染させ、中和抗体によってIL−23遮断を施した。以前の刊行物(Romaniら、(1992年)J. Exp. Med. 176:19頁;Cenciら、(1998年)J. Immunol. 161:3543頁)と一致して、真菌負荷はBALB/cマウスと比較してIL−4−/−マウス中で低く、かつIFN−γ−/−マウス中で高く、感染の調節に関するIFN−γの重要性を実証した。野生型マウスと同様に、IL−23の遮断はIL−4−/−マウスの胃中の真菌負荷を大幅に低下させ(図3C)、および同時にMLNにおいてIL−12p70/IFN−γ産生を増大させ(データ示さず)、Th2とIL−23/Th17経路の両方が防御性抗真菌応答に相加的に拮抗することを示唆した。驚くことに、IFN−γ−/−マウスにおける高い真菌負荷はIL−23の中和によってさらに増大し(図3C)、これは抗IL−23処置マウスにおいて低いIL−23産生(229対21pg/ml)IL−17産生(279対95pg/ml)をもたらした。したがって、IL−23はIFN−γの不在下で真菌感染において防御的役割を有し得る。しかしながら、p35−/−または二重欠損IFN−γ−/−/p35−/−マウスにおけるIL−23の中和によって低下した真菌負荷により実証されたように、IL−23はIL−12p70の不在下、またはIL−12p70とIFN−γの両方の不在下では反対の影響を有する(図3C)。これらのデータは、IFN−γの不在下でのIL−23の防御的役割はIL−12p70によって仲介されることを示唆する。特に、IL−12p70の不在下でのIL−23の適度な防御的役割は結核においても観察されており、この場合IL−23は、防御的IFN−γ産生CD4+T細胞の誘導においてIL−12p70に部分的に置き換わった。Khaderら、(2005年)。
【0169】
(実施例8)
真菌感染に応答した樹状細胞中でのIL−23およびIL−12の産生
IL−23はin vitroではAspergillusに応答してヒトDCによって産生されることは、既に示されている。Gafaら、(2006年)Infect. Immun. 74:1480頁。本発明者らは、IL−23がC.albicansに応答してDCによって産生されるかどうか、およびそれがIL−12およびIL−10、真菌に対する防御耐性の誘導に本来必要とされる2つのサイトカインの産生とどのような関係があるかをここで評価した。Romani & Puccetti(2006年)。
【0170】
この目的のために、本発明者らは、それぞれ骨髄性DCおよび形質細胞様(plasmacytoid)DCの特徴を共有するGM−CSF(GM−DC)またはFlt3−L(FL−DC)のいずれかの存在下で、骨髄由来のDCを作製した。FL−DCは新たに採取した脾性CD8+、CD8−およびB220+LyC6+形質細胞様DCの混合物と同等の集団を包含するが(Naikら、(2005年)J. Immunol. 174:6592頁)、本発明者らは、FL−DCの機能活性は形質細胞様DC中またはCD8−とCD8+DCの組合せ中に存在することを近年実証している。Romaniら、(2006年)Blood 108:2265頁。DCはin vitroにおいて酵母または真菌の菌糸で刺激し、サイトカインmRNA発現および産生に関して評価した。ザイモザンおよびLPSはGM−DCの陽性対照として、およびCpG−ODNはFL−DCの陽性対照として使用した。
【0171】
これらの結果は、真菌に応答したDCサブセットの2つのサブセットによるサイトカイン発現および産生において二分(dichotomy)を示した。RT−PCR分析は、p19mRNA発現のみが菌糸より酵母に応答してGM−DCにおいて増大し、p35mRNA発現は酵母に応答してGM−DCにおいてわずかに増大したが、IL−10と同様に、菌糸に曝露したFL−DCにおいて大幅に増大したことを明らかにした(図4A)。培養物上清における実際のサイトカイン産生の測定によって、IL−23が酵母、特に高い真菌:DC比、およびザイモザンまたはLPSに応答してGM−DCによって産生されたことを確認した(図4B)。IL−23産生の最高レベルはインキュベーションの12時間で観察し(図4B)、その後低下した(データ示さず)。逆に、IL−12p70とIL−10の両方は、12時間Candida菌糸、LPSまたはCpG−ODNで刺激したFL−DCによって主に産生され(図4B)、その後上昇し続けた(データ示さず)。一緒にするとこれらのデータは、IL−23は真菌に応答して、特に高レベル真菌増殖の条件で、かつ他の誘導サイトカインより早く骨髄性DCによって産生されることを示唆する。Candidaに応答して誘導サイトカインを産生する異なるDCサブセットの能力は、したがってin vivoでそれらの抗真菌免疫を調節することができる。実際のところ、Aspergillusに関して既に示されているように(Romaniら、(2006年)Blood 108:2265頁)、CandidaでパルスしたFL−DCは防御をもたらし、かつCandidaでパルスしたGM−DCはカンジダ症を有するレシピエントマウスへの養子移入によって感染を悪化させた。
【0172】
(実施例9)
IL−23とIL−12の交差制御
IL−12p70およびIL−23産生が真菌に応答して交差制御されるかどうかを検証するために、本発明者らは、IL−12p70またはIL−23、または対応する中和抗体のいずれかへの曝露後にp19−/−、p35−/−、およびC57BL/6対照マウス由来の脾性DCによるIL−12p70およびIL−23の分泌を測定した。図4Cは、野生型DCと比較してIL−12p70の産生がp19−/−DC中で高く、かつIL−23の産生がp35−/−DC中で高かったので、IL−12p70とIL−23は実際交差制御されることを示す。さらに、IL−12p70またはIL−23のいずれかへの曝露は、野生型DCによるIL−23またはIL−12p70の分泌をそれぞれ有意に低下させ、かつIL−12またはIL−23中和の状態において逆も真であった(図4D)。RT−PCRによって無刺激DCは両方のサイトカイン受容体を発現することを明らかにしたので(データ示さず)、これらのデータは、それによってDCによるIL−12p70およびIL−23産生が相互に制御されるパラクリンループ(paracrine loop)の存在を示唆する。
【0173】
(実施例10)
樹状細胞によるIL−23産生におけるTLRの役割
真菌に応答したIL−23産生のTLR依存性の可能性を明確にするために、本発明者らは、TLR−2−/−またはTLR−4−/−マウスから、ならびにMyD88−/−およびTRIF−/−マウスから作製したGM−DCによる酵母または分生子に応答したIL−23産生を測定した。Akira and Takeda(2004年)Nat. Rev. Immunol. 4:499頁。図5Aは、TLR2とTLR4の両方が、TRIFではなくMyD88を介したシグナル伝達によるIL−23産生に必要不可欠であることを示す。特に、TRIFの不在下でさえIL−23は促進されたように現れた。したがってIL−23は、TLR/MyD88依存性炎症性経路を介して、真菌に応答して通常のDCによって産生される。
【0174】
T細胞もIL−23産生を制御することができるかどうかを明確にするために、本発明者らは、CD4+T細胞と共に培養したDCの上清に産生されたIL−23のレベルを評価した。これらの結果は、C57BL/6および特にp35−/−マウス(群3対群6、図5B)由来のCandidaでパルスしたDCを用いて刺激したT細胞の培養物においてIL−23産生が上方制御されたことを明らかに示し、この発見により、活性化T細胞がIL−23産生の増幅のための正のフィードバックループをもたらし得ることを示唆した。さらに、十字交差実験の結果によって、IL−23産生DCはIL−17産生細胞を活性化するのに必要かつ十分であったことを確認した(群7および8)。さらに、DCとT細胞の同時培養物に加えたモノクローナル抗体によるIL−23の中和はIL−l7産生を阻害し(群4対群5、図5C)、一方TGF−β中和はIL−l7(群4対群6)ではなくIFN−γ産生に影響を与えた(群1対群3)。
【0175】
(実施例11)
多形核好中球の抗真菌エフェクター機能に対するIL−23およびIL−17の影響
PMNは真菌に対する急性炎症性応答の開始および遂行において必要不可欠である。Romani(2004年)。p35−/−マウスにおける初期の真菌増殖とともに、PMNが感染部位に多量に動員されたという発見により、本発明者らは、IL−23/IL−17依存性経路はPMNの抗真菌エフェクター機能に悪影響を与え得ると仮定するに至った。したがって本発明者らは、p19−/−マウスまたはp35−/−マウスのいずれか由来、およびIFN−γの不在または存在下で組換えIL−23またはIL−17と共に培養した野生型マウス由来のPMNの抗真菌活性を評価した。殺滅活性は、C57BL/6 PMNと比較してp19−/−PMNにおいて有意に増大し、かつp35−/−マウスにおいて低下した(図6A)。PMNをこれらのサイトカインに曝露する前に、本発明者らは、IL−17Rと同様に(Yaoら、(1995年)Immunity 3:811〜821頁)、IL−23RはマウスPMNでも発現されたかどうかを検証した。定量RT−PCRは、無刺激PMNがIL−23Rを発現し、その発現はLPSで刺激した後さらに増大したこと(データ示さず)を明らかにし、この発見により、PMNはIL−23にも応答性があることを示唆した。
【0176】
いずれかのサイトカインへの曝露は、用量依存的様式で(図6B)IFN−γの不在および存在下で(図6C)野生型PMNの殺滅活性を弱めた。したがって、IL−23およびIL−17はPMNの抗真菌エフェクター機能を負に制御し、これはp35−/−マウスが真菌増殖を効率良く制限できない原因である可能性がある。したがって、感染器官への末梢PMNの流入低下がカンジダ症に対するIL−17AR欠損マウスの高い感受性の原因であったように(Huangら、(2004年)J. Infect. Dis. 190:624頁)、IL−17はPMNの強力な化学誘引物質であるが(Yeら、(2001年)J. Exp. Med. 194:519頁)、本発明者らの結果は、PMN機能に対するIL−17の有害な影響も指摘する。
【0177】
(実施例12)
多形核好中球のIDO依存性抗炎症性プログラムに対するIL−23およびIL−17の影響
本発明者らは、IDO遮断がPMNの炎症性状態の促進をもたらしたように、IFN−γ仲介IDO活性化がCandidaに対するPMNの炎症性プログラムを負に制御することを既に示している。Bozzaら、(2005年)。MMP−9およびMPOは、IL−17によって活性化されることが示されている典型的な炎症性マーカーである。Kolls and Linden(2004年)Immunity 21:467頁。したがって本発明者らは、C.albicansに対するIL−23とIL−17の両方の影響が、MMP−9、MPOおよびIFN−γ仲介IDO産生を誘導したと評価した。IL−23と、特にIL−17の両方が、MMP−9およびMPOを相当増大させた(図6D)。野生型PMNのIDO発現および炎症性応答。対照的に、両方のサイトカインがIFN−γによるIDOの誘導に完全に拮抗した(図6E)。興味深いことに、アポトーシスPMNの数はIL−23とIL−17の両方への曝露によって有意に減少し(データ示さず)、これらのサイトカインはPMNの生存能力も高めることが示唆された。これは炎症がTh17経路によって持続するさらなる機構である可能性がある。したがって、PMNの炎症性プログラムを妨害する能力、および炎症組織中の増大した実質的タンパク質分解負荷は、真菌感染中のTh17細胞活性化と関係がある炎症性病理の原因である可能性がある。
【0178】
(実施例13)
IL−17産生に基づくIL−23特異的アンタゴニストのアッセイ
マウス流入領域リンパ節(DLN)細胞を使用するin vitro試験は、蛍光活性化細胞ソーティング(FACS(登録商標))分析により測定したとき、IL−23の除去はIL−17産生細胞を阻害または除去し、一方IL−23の添加はIL−17分泌を発生または刺激することを実証した。WO2004/071517、Langrishら、(2005年)J. Exp. Med. 201:233頁を参照。Aggarwalら、(2003年)J. Biol. Chem. 278:1910頁も参照。これらの実験では、5日間サイトカインまたは抗体でDLN細胞を処置した。細胞は抗原刺激した正常野生型マウスから単離し、rIL−12またはrIL−23のいずれかの存在下で培養した。DLN培養物中のCD4+T細胞の分析は、IL−17産生集団の消失を伴いながら、IL−12がIFN−γ産生細胞の発生を促進したことを実証した。対照的に、IFN−γ産生集団の消失を伴いながら、IL−23はIL−17産生細胞の発生を促進した。抗p19抗体はIL−17産生を低下させたがIFN−γレベルには影響を与えず、一方抗p35抗体はIL−17産生を変えなかった。総合するとこれらの結果は、IL−23がIL−17産生CD4+T細胞の発生を選択的に促進することを示した。
【0179】
IL−23とIL−12の生物活性のこの差異を使用して、以下のようにIL−12と比較して、可能性のあるIL−23アンタゴニストの効力および特異性を評価する。
【0180】
可能性のあるIL−23特異的アンタゴニストの不在下でのIL−23およびIL−12活性に関する基線データ(baseline data)を、以下のようにして得る。正常野生型SJLマウスは、フロイント完全アジュバントで乳化したプロテオリピドペプチド(PLP)で(皮下)、および百日咳毒素で免疫処置する(静脈内)。流入領域リンパ節を免疫処置の9日後に除去し、単核細胞は細胞内IFN−γおよびIL−17産生に関してすぐに評価する(以下に記載するように)、または単離しおよび5日間PLPに加えrIL−12もしくはrIL−23のどちらかの存在下で単離および培養するかのいずれかである。細胞はPMA(50ng/ml)で3時間/ゴルジ−プラグ(Golgi−plug)の存在下において4時間イオノマイシン(500ng/ml)で刺激し、次いで表面をCD4に関して染色し、透過処理し、かつ細胞内をIFN−γおよびIL−17に関して染色する。フローサイトメトリーのプロットを生存CD4+T細胞において得る。
【0181】
IL−23およびIL−12の影響を、サイトカイン処置しなかった対照細胞と比較して評価する。典型的には、IL−23処置細胞は増大した割合のIL−17産生細胞を示し、IFN−γ産生細胞の増大は示さず、一方IL−12処置細胞は増大した割合のIFN−γ産生細胞を示し、IL−12産生細胞の増大(またはさらに減少)は示さない。
【0182】
可能性のあるIL−23特異的アンタゴニストの効力および特異性は、アンタゴニスト、または好ましくは一連の濃度のアンタゴニストの存在下で同じ実験を実施することによって決定する。IL−23特異的アンタゴニストはIL−23の活性を阻害するが(すなわち、このアンタゴニストは、他の場合IL−23によって誘導されるIL−17産生細胞の割合を減少させる)、IL−12の活性は実質的に低下させない。IL−12またはIL−12とIL−23の両方の活性を阻害する作用物質は、IL−23特異的アンタゴニストではない。
【0183】
場合によっては、陽性対照を含めることができ、その中で既知の抗p19アンタゴニスト抗体を使用して、IL−23の活性を特異的に阻害する。
【0184】
(実施例14)
マイコバクテリア感染症
マイコバクテリア感染症の処置における本発明の組成物および方法の有効性を実証する方法を提供する。C57BL/6マウスに以下のようにマイコバクテリアを感染させる。Theracys−BCG Live(Aventis Pasteur、Inc.、Swiftwater、PA)、Bacille CabmetteおよびGuerinのConnaught系統ならびにM.bovisの弱毒系統の凍結乾燥調製物を、製造者によって推奨されるように元の状態に戻す。元の状態に戻した(reconstituted)細菌は、10%グリセロール生理食塩液中に約6×107cfu/mLの濃度にする。マウスへの注射前に、0.02%Tween−80/0.9%生理食塩液中にアリコートを適切な濃度に希釈する。
【0185】
6〜8週齢の雌性C57BL/6マウスに、約3.5×105cfuのBCGを側方の尾静脈を介して静脈内感染させる。マウスには1mgの適切なモノクローナル抗体を(たとえば、アイソタイプ対照、抗IL−23p19、または抗IL−23R)0.9%生理食塩液中で与え、マイコバクテリア感染前第1日、およびマイコバクテリア感染1〜2週間後に再度皮下投与する。CO2ナルコーシスによって、感染後の適切な時間地点でマウスを屠殺する。
【0186】
屠殺したBCG感染マウスは以下のように分析する。下大静脈を切断した後に心臓の右心室を介してRPMI 1640を灌流させることによって、肺から血液を取り除く。左肺、肝臓右半部下葉(lower right liver lobe)、および脾臓の半分を無菌状態で除去する。これらの組織を、Mini−Bead Beater−8ホモジェナイザー(BioSpec Products、Bartlesville、OK)を用いて0.9%NaCl/0.02%Tween80中に均質化する。生存マイコバクテリアは、10倍連続希釈の臓器ホモジネートを7H10Middlebrook寒天プレート(Becton Dickinson、Sparks、MD)に平板培養することによって定量化する。コロニー形成単位(CFU)は、37℃で2週間のインキュベーションの後に手作業で計数する。対照マウス(たとえば、アイソタイプ対照)と比較した、抗IL−23抗体(たとえば、抗IL−23p19抗体または抗IL−23R抗体)で処置した動物における(CFUによって測定した)細菌負荷の統計的に有意な低下は、マイコバクテリア感染症の有効な処置の証拠である。
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1−1】

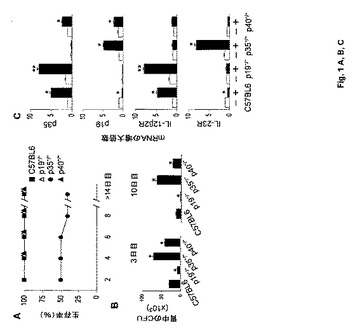
【図1−2】
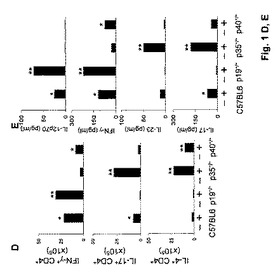
【図2】
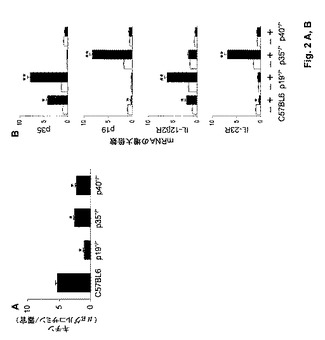
【図3】

【図4】

【図5】

【図6】
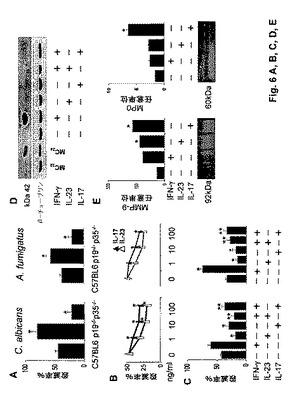
【図1−2】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2013−14624(P2013−14624A)
【公開日】平成25年1月24日(2013.1.24)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−230578(P2012−230578)
【出願日】平成24年10月18日(2012.10.18)
【分割の表示】特願2009−549119(P2009−549119)の分割
【原出願日】平成20年2月8日(2008.2.8)
【出願人】(596129215)メルク・シャープ・アンド・ドーム・コーポレーション (785)
【氏名又は名称原語表記】Merck Sharp & Dohme Corp.
【住所又は居所原語表記】One Merck Drive,Whitehouse Station,New Jersey 08889,U.S.A.
【Fターム(参考)】
【公開日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願番号】特願2012−230578(P2012−230578)
【出願日】平成24年10月18日(2012.10.18)
【分割の表示】特願2009−549119(P2009−549119)の分割
【原出願日】平成20年2月8日(2008.2.8)
【出願人】(596129215)メルク・シャープ・アンド・ドーム・コーポレーション (785)
【氏名又は名称原語表記】Merck Sharp & Dohme Corp.
【住所又は居所原語表記】One Merck Drive,Whitehouse Station,New Jersey 08889,U.S.A.
【Fターム(参考)】
[ Back to top ]