
Aequoreacoerulescens由来の新規な蛍光タンパク質およびその使用方法

【課題】標識化は、関心のあるタンパク質、細胞または生物に印をつけるためのツールであり、生化学的な応用、分子生物学的な応用および医学的診断への応用の多くで突出した役割を果たしている。しかしながら、新しい標識の開発が引き続き注目されている。特に注目されるものは、蛍光タンパク質標識を含む新しいタンパク質標識の開発である。
【解決手段】Aequorea coerulescensから得られる新規な無色のGFP様タンパク質(acGFP)、ならびにその蛍光性および非蛍光性の変異体および誘導体をコードする核酸組成物、そしてまた、これらの核酸組成物によってコードされるペプチドおよびタンパク質を提供する。該タンパク質および核酸組成物は有色であり、かつ/または蛍光性であり、かつ/または光活性化が可能であり、従って、様々な異なる生物学的適用において、特に標識化のために使用することができる。
【解決手段】Aequorea coerulescensから得られる新規な無色のGFP様タンパク質(acGFP)、ならびにその蛍光性および非蛍光性の変異体および誘導体をコードする核酸組成物、そしてまた、これらの核酸組成物によってコードされるペプチドおよびタンパク質を提供する。該タンパク質および核酸組成物は有色であり、かつ/または蛍光性であり、かつ/または光活性化が可能であり、従って、様々な異なる生物学的適用において、特に標識化のために使用することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、蛍光タンパク質に関する。
【背景技術】
【0002】
標識化は、関心のあるタンパク質、細胞または生物に印をつけるためのツールであり、生化学的な応用、分子生物学的な応用および医学的診断への応用の多くで突出した役割を果たしている。様々な異なる標識がこの分野で開発および使用されており、これらには、様々な性質および最適な用途を有する放射性標識、発色性標識、蛍光性標識、化学発光性標識などが含まれる。しかしながら、新しい標識の開発が引き続き注目されている。特に注目されるものは、蛍光タンパク質標識を含む新しいタンパク質標識の開発である。
【0003】
注目される米国特許には、米国特許第6,066,476号、同第6,020,192号、同第5,985,577号、同第5,976,796号、同第5,968,750号、同第5,968,738号、同第5,958,713号、同第5,919,445号、同第5,874,304号および同第5,491,084号が含まれる。注目される国際特許出願公開には、WO00/46233、WO99/49019、およびドイツ国特許出願公開DE19718640Aが含まれる。また、Anderluh等、Biochemical and Biophysical Research Communications(1996)、220:437〜442;Dove等、Biological Bulletin(1995)、189:288〜297;Fradkov等、FEBS Lett.(2000)、479(3):127〜30;Gurskaya等、FEBS Lett.(2001)、507(1):16〜20;Gurskaya等、BMC Biochem.(2001)、2:6;Lukyanov,K.等(2000)、J.Biol.Chem.、275(34):25879〜25882;Macek等、Eur.J.Biochem.(1995)、234:329〜335;Martynov等、J.Biol.Chem.(2001)、276:21012〜6;Matz,M.V.等(1999)、Nature Biotechnol.、17:969〜973;Tershikh等、Science(2000)、290:1585〜8;Tsien、Annual Rev.of Biochemistry(1998)、67:509〜544;Tsien、Nat.Biotech.(1999)、17:956〜957;Ward等、J.Biol.Chem.(1979)、254:781〜788;Wiedermann等、Jarhrestagung der Deutschen Gesellschact fur Tropenokologie−gto.Ulm.、1999年2月17日〜19日、ポスターP−4.20;Yarbrough等、Proceedings of the National Academy of Sciences(2001)、98:462〜7も注目される。
【発明の概要】
【発明が解決しようとする課題】
【0004】
本発明は、Aequorea coerulescens由来の他に類のない無色のタンパク質ならびにその蛍光性変異体および非蛍光性変異体をコードする核酸組成物を提供し、また、そのような核酸によってコードされるタンパク質およびペプチドも提供する。本発明のタンパク質は有色であり、かつ/または蛍光性であり、かつ/または光活性化が可能であり、この場合、この光学的特徴はタンパク質の2つ以上のアミノ酸残基の相互作用から生じている。また、本発明のペプチドが組み込まれた融合タンパク質を含む上記の特定のタンパク質に実質的に類似するタンパク質やその誘導体または変異体に注目され、また、これらのタンパク質に対する抗体も注目される。本発明のタンパク質および核酸組成物は様々な異なる応用への用途がある。最後に、本発明は、標識化適用において使用されるキットを提供する。
【課題を解決するための手段】
【0005】
本発明は、蛍光タンパク質acGFP、その変異体またはその誘導体をコードする核酸を提供する。ある実施形態においては、核酸は単離されているか、操作されているか、またはその天然の環境とは異なる環境に存在する。ある実施形態においては、核酸は、配列番号1、3、5、7、9、11、13、15、17、19、21または23に由来する少なくとも10残基の長さのヌクレオチド配列と実質的に同じであるか、または同一である残基の配列を有する。
【0006】
ある実施形態においては、本発明の核酸は、配列番号1、3、5、7、9、11、13、15、17、19、21または23の配列と少なくとも約60%の配列類似性を有し、長さが少なくとも10残基である。ある実施形態においては、本発明の核酸は、Aequorea coerulescensの蛍光タンパク質の変異体タンパク質または誘導体タンパク質をコードする、配列番号2、4、6、8、10、12、14、16、18、20、22または24からなる群から選択されるアミノ酸配列を有するタンパク質をコードする。
【0007】
本発明の核酸の断片もまた提供される。更に、ストリンジェントな条件のもとで本発明の核酸にハイブリダイゼーションする核酸またはそのミメティック(mimetic)が提供される。ベクターおよび本発明の核酸を含む構築物もまた提供される。更に、本発明は発現カセットを提供する。ここで、発現カセットは、発現宿主において機能し得る転写開始領域と、本発明の核酸と、細胞内への発現カセットの導入の結果、染色体外エレメントの一部としての発現カセットにおいて、または細胞のゲノムに組み込まれる発現カセットにおいて機能し得る転写終結領域とを含む。
【0008】
色素タンパク質および/または蛍光タンパク質を製造する方法もまた提供され、この方法は、本発明の細胞を増殖させること、上記タンパク質を細胞において発現させること、および他のタンパク質を実質的に含まない上記タンパク質を単離することを含む。
【0009】
更に、本発明の核酸によってコードされるタンパク質、断片またはペプチドが提供され、また、本発明のタンパク質またはペプチドに対して特異的に結合する抗体も提供される。
【0010】
更に、本発明の核酸を含む遺伝子組換え細胞(またはその子孫)が提供され、また、本発明の核酸を含む遺伝子組換え生物も提供される。
【0011】
また、本発明の色素タンパク質または蛍光タンパク質を用いる方法、あるいは、本発明の色素タンパク質または蛍光タンパク質をコードする核酸を用いる方法が提供される。
【0012】
更に、本発明の核酸またはタンパク質と、その使用説明書とを含むキットが提供される。
【0013】
本発明に従って、この分野の技術に含まれる、従来の、分子生物学、微生物学および組換えDNA技術を用いることができる。そのような技術は文献に詳しく説明されている。例えば、Maniatis、Fritsch&Sambrook、Molecular Cloning:A Laboratory Manual(1982年);DNA Cloning:A Practical Approach、第1巻および第2巻(D.N.Glover編、1985年);Oligonucleotide Synthesis(M.J.Gait編、1984年);Nucleic Acid Hybridization(B.D.Hames&S.J.Higgins編(1984年));Animal Cell Culture(R.I.Freshney編(1986年));Immobilized Cells and Enzymes(IRL Press(1986年));およびB.Perbal、A Practical Guide to Molecular Cloning(1984年)を参照のこと。
【0014】
「ベクター」は、別のDNAセグメントを結合させることができる、プラスミド、ファージまたはコスミドなどのレプリコンである。
【0015】
「DNA分子」は、デオキシリボヌクレオチド(アデニン、グアニン、チミンまたはシトシン)のポリマー形態を示し、一本鎖形態または二本鎖らせんのどちらでもよい。この用語は、分子の一次構造および二次構造を示すだけであり、分子を何らかの特定の三次構造に限定しない。従って、「DNA分子」には、とりわけ、線状DNA分子(例えば、制限酵素断片)、ウイルス、プラスミドおよび染色体に見出される二本鎖DNAが含まれる。
【0016】
DNAの「コード配列」は、適切な制御配列の制御下に置かれたとき、インビボで転写され、ポリペプチドに翻訳されるDNA配列である。コード配列の境界は、5’(アミノ)末端における開始コドンと、3’(カルボキシル)末端における翻訳終止コドンとによって決定される。コード配列には、原核生物配列、真核生物mRNA由来のcDNA、真核生物(例えば、哺乳動物)DNA由来のゲノムDNA配列、および合成DNA配列が含まれ得るが、これらに限定されない。ポリアデニル化シグナルおよび転写終結配列がコード配列の3’に存在し得る。
【0017】
本明細書中で使用される用語「ハイブリダイゼーション」は、2つの核酸鎖が会合して、対になる核酸鎖の残基間の水素結合によって安定化される反平行の二重鎖を形成するプロセスを示す。
【0018】
用語「オリゴヌクレオチド」は、短い(100塩基未満の長さの)核酸分子を示す。
【0019】
本明細書中で使用される「DNA制御配列」は、宿主細胞におけるコード配列の発現を規定および/または調節する転写制御配列および翻訳制御配列(例えば、プロモーター、エンハンサー、ポリアデニル化シグナルおよびターミネーターなど)である。
【0020】
「プロモーター配列」は、細胞内のRNAポリメラーゼと結合し、コード配列の転写を開始させることができるDNA制御領域である。例えば、プロモーター配列は、転写開始部位をその3’末端での境界とすることができ、そして、上流(5’方向)に広がり、バックグラウンドを上回る検出可能なレベルで転写を開始させるために必要な最小数の塩基またはエレメントを含むことができる。プロモーター配列内には、転写開始部位、ならびに、RNAポリメラーゼの結合に関わるタンパク質結合ドメインが見出され得る。真核生物のプロモーターは、常にというわけではないが、多くの場合、「TATA」ボックスおよび「CAT」ボックスを含有する。誘導性プロモーターを含む様々なプロモーターを、本発明の様々なベクターを駆動させるために使用することができる。
【0021】
本明細書中で使用される用語「制限エンドヌクレアーゼ」および用語「制限酵素」は、特定のヌクレオチド配列でまたはその近くで二本鎖DNAを切断する細菌酵素を示す。
【0022】
細胞は、外因性または異種のDNAが細胞の内部に導入されているとき、外因性または異種のDNAによって「形質転換」または「トランスフェクション(形質移入)」されている。形質転換DNAは細胞のゲノムに組み込まれ(共有結合的に連結され)てもよく、または組み込まれなくてもよい。例えば、原核生物、酵母細胞および哺乳動物細胞では、形質転換DNAは、プラスミドなどのエピソームエレメントに維持され得る。真核細胞に関して、安定に形質転換された細胞は、形質転換DNAが宿主細胞の染色体に組み込まれているか、または染色体外に維持され、その結果、形質転換DNAが細胞複製のときに娘細胞によって受け継がれる細胞である。そのような安定に形質転換された真核生物細胞は、形質転換DNAを含有する娘細胞の集団から構成される細胞株またはクローンを樹立することができる。「クローン」は、単一細胞または共通の祖先から有糸分裂によって得られる細胞の集団である。「細胞株」は、多世代にわたってインビトロで安定に成長可能なクローンまたは細胞である。
【0023】
DNA構築物の「異種」領域は、より大きなDNA分子における同定可能なDNAセグメントであって、そのより大きな分子に関連して天然に見出されないDNAセグメントであり、例えば、非哺乳類生物のゲノムにおいて哺乳類のゲノムDNAを異種領域がコードするときのDNAセグメントである。別の例では、異種DNAには、2つの異なる供給源に由来する遺伝子の一部が一緒になり、その結果、融合タンパク質産物が産生されるようになっている構築物におけるコード配列が含まれる。
【0024】
本明細書中で使用される用語「レポーター遺伝子」は、異種のプロモーターエレメントまたはエンハンサーエレメントに結合されたコード配列で、構築物が組織または細胞に導入されたとき、その産物を容易かつ定量的にアッセイすることができるコード配列を示す。
【0025】
本明細書中に記載されるアミノ酸は「L」型であることが好ましい。アミノ酸配列は一文字記号で示される(A:アラニン、C:システイン、D:アスパラギン酸、E:グルタミン酸、F:フェニルアラニン、G:グリシン、H:ヒスチジン、I:イソロイシン、K:リシン、L:ロイシン、M:メチオニン、N:アスパラギン、P:プロリン、Q:グルタミン、R:アルギニン、S:セリン、T:トレオニン、V:バリン、W:トリプトファン、Y:チロシン、X:任意の残基)。NH2は、ポリペプチドのアミノ末端に存在する遊離型アミノ基を示す。COOHは、ポリペプチドのカルボキシ末端に存在する遊離型カルボキシ基を示す。標準的なポリペプチド命名法(J.Biol.Chem.、243、3552〜59(1969)を参照のこと)が使用されている。
【0026】
用語「免疫学的に活性な」によって、天然型、組換え型または合成型の色素タンパク質もしくは蛍光タンパク質またはそれらの任意のオリゴペプチドが特異的な免疫応答を適切な動物または細胞において誘導し、特異的な抗体と結合する能力が規定される。本明細書中で使用される「抗原性アミノ酸配列」は、単独でまたはキャリア分子と一緒になって、哺乳動物において抗体応答を誘発することができるアミノ鎖配列を意味する。抗体が抗原に結合するという状況における、用語「特異的に結合する」は、この分野で広く理解されている用語であり、ある抗体がその抗体を産生させた抗原には結合するが、関連性のない他の抗原には結合しないことを示す。
【0027】
本明細書中で使用される用語「単離された」は、ポリヌクレオチド、ポリペプチド、抗体、または宿主細胞が天然に存在する環境とは異なる環境にあるポリヌクレオチド、ポリペプチド、抗体または宿主細胞を記載することが意味される。
【0028】
生物発光は、暗所で見ることができる、生物による光の放射である(例えば、Harvey、Bioluminescence、New York:Academic Press(1952);Cell Physiology(Speralakis編)のHastings、「生物発光」の項、New York、Academic Press、651頁〜81頁(1995);WilsonおよびHastings、「生物発光」、Annu.Rev.Cell.Dev.Biol.、14、197頁〜230頁(1998)を参照のこと)。生物発光には、高感度な光度測定装置を使用して事実上すべての生体構造体において検出され得るいわゆる超微弱な光放射は含まれず(MurphyおよびSies、「生物学システムにおける可視範囲の低レベル化学発光」、Meth.Enzymol.、186、595頁〜610頁(1990);Radotic等、「植物における自発的な超微弱な生物発光:起源、機構および性質」、Gen.Physiol.Biophys.、17、289頁〜308頁(1998))、また、生物発光は、例えば、タケの成長円錐の発光など、生態学的役割をおそらくは果たしていない弱い光放射から生じるものではなく(Totsune等、「筍切断体からの化学発光」、Biochem.Biophys.Res.Comm.、194、1025頁〜1029頁(1993))、動物の卵の受精時における光放射から生じるものでもない(Klebanoff等、「受精と食作用との間における代謝的類似性」、J.Exp.Med.、149、938頁〜53頁(1979);SchomerおよびEpel、「海洋無脊椎動物の卵の受精および成熟化の際の酸化還元変化」、Dev.Biol.、2003、1頁〜11頁(1998))。
【0029】
本明細書中で使用される用語「GFP様タンパク質」は、Aequorea victoria由来の緑色蛍光タンパク質(GFP)に類似するタンパク質を記載することが意味される。
【発明の効果】
【0030】
Aequorea coerulescens由来の無色のGFP様タンパク質であるacGFP、その蛍光性誘導体および非蛍光性誘導体、またはそれらの変異体をコードする核酸組成物、そしてまた、これらの核酸組成物によってコードされるタンパク質およびペプチドが提供される。注目されるタンパク質は、有色であり、かつ/または蛍光性であり、かつ/または光活性化され得るタンパク質であり、この場合、色、蛍光または光活性化の特徴は、タンパク質の2つ以上のアミノ酸残基の相互作用から生じている。また、上記の特定のタンパク質に実質的に類似するタンパク質、その誘導体タンパク質またはその変異体タンパク質も注目される。また、核酸断片、およびそれによってコードされるペプチドが提供され、また、本発明のタンパク質およびペプチドに対する抗体が提供される。更に、遺伝子組換え細胞および遺伝子組換え生物が提供される。本発明のタンパク質および核酸組成物は、様々な異なる応用および方法における用途があり、特に、タンパク質標識への応用がある。最後に、そのような方法および適用において使用されるキットが提供される。
【図面の簡単な説明】
【0031】
【図1】本明細書中でacGFPと表す、Aequorea coerulescens由来の野生型GFP様タンパク質をコードするアミノ酸配列および核酸配列である。
【図2】Aequorea victoria GFPおよびAequorea coerulescenssc GFPのアミノ酸配列の比較である。
【図3】acGFP変異体Z1をコードするアミノ酸配列および核酸配列である。
【図4】変異体Z1の励起−発光スペクトルである。
【図5】acGFP変異体Z2をコードするアミノ酸配列および核酸配列である。
【図6】acGFP変異体G1をコードするアミノ酸配列および核酸配列である。
【図7】acGFP変異体G2をコードするアミノ酸配列および核酸配列である。
【図8】変異体G2の励起−発光スペクトルである。
【図9】acGFP変異体G22をコードするアミノ酸配列および核酸配列である。
【図10】変異体G22の励起−発光スペクトルである。
【図11a】A.victoriaのGFPに対する抗体を使用した、A.coerulescensから得られた可溶性タンパク質抽出物のウエスタンブロット分析である。レーン1は、精製された組換えA.victoria GFPであり、レーン2はA.coerulescens抽出物である。
【図11b】加熱処理(レーン1〜3)対非加熱(レーン4〜6)のタンパク質サンプルの移動度が比較される。レーン1およびレーン4はA.victoriaのGFPであり、レーン2およびレーン5はG22変異体であり、レーン3およびレーン6はG22−G222E変異体である。クーマシーブルー染色が示される。
【図11c】UV光のもとでの非加熱タンパク質の蛍光が示される。
【図12】acGFP変異体G22−G222Eをコードするアミノ酸配列および核酸配列である。
【図13a】変異体G22−G222Eの吸収スペクトルである。
【図13b】変異体G22−G222Eの励起−発光スペクトルである。
【図14】G22−G222EのUV誘導の光変換に対するスペクトルを示す。
【図15】acGFP変異体G22−G222E/Y220Lをコードするアミノ酸配列および核酸配列である。
【図16】変異体G22−G222E/Y220Lの励起−発光スペクトルである。
【図17】acGFP変異体220−II−5をコードするアミノ酸配列および核酸配列である。
【図18a】変異体220−II−5のスペクトル特性を示す。
【図18b】変異体220−II−5のスペクトル特性を示す。
【図19】acGFP変異体CFP−rand3をコードするアミノ酸配列および核酸配列である。
【図20】変異体CFP−rand3に対する励起−発光スペクトルである。
【図21】acGFP変異体CFP−3をコードするアミノ酸配列および核酸配列である。
【図22a】変異体CFP−3のスペクトル特性を示す。
【図22b】変異体CFP−3のスペクトル特性を示す。
【図23】変異体G22のヒト化体のアミノ酸配列および核酸配列である。
【図24a】G22−h変異体(293T)を発現する哺乳動物細胞の顕微鏡写真を示す。
【図24b】G22−h変異体(vero)を発現する哺乳動物細胞の顕微鏡写真を示す。
【図24c】G22−h変異体(3T3)を発現する哺乳動物細胞の顕微鏡写真を示す。
【図24d】G22−h変異体(L929)を発現する哺乳動物細胞の顕微鏡写真を示す。
【図24e】G22−h変異体(COS1)を発現する哺乳動物細胞の顕微鏡写真を示す。
【図24f】G22−β−アクチン融合タンパク質(3T3)を発現する哺乳動物細胞の顕微鏡写真を示す。
【図25】大腸菌コロニーにおける変異体220−11−5の光活性化を示す。
【発明を実施するための形態】
【0032】
本発明を更に記載する前に、本発明は、下記に記載される本発明の特定の実施形態に限定されないことを理解しなければならない。これは、そのような特定の実施形態の様々な変化を行うことができ、そして、そのような変化は依然として本発明の範囲に含まれるからである。用いられている用語は、特定の実施形態を記載するためであり、限定的であることを意図しないこともまた理解しなければならない。
【0033】
本明細書において、単数形態(“a”、“an”および“the”)は、文脈により、別のことが明らかに示されない限り、複数である参照体を含む。別途定義されない限り、本明細書中で使用されている技術的用語および科学的用語はすべて、本発明が属する分野の当業者に一般に理解されているのと同じ意味を有する。本明細書中に記載されているものと類似している、または同等である、方法、デバイスおよび材料はどれも、本発明の実施または試験において使用することができるが、好ましい方法、デバイスおよび材料が下記に記載される。本明細書中で言及される刊行物はすべて、本発明を記載および開示する目的のために参考として本明細書中に組み入れる。
【0034】
本発明を記載するにあたり、核酸組成物が最初に記載し、その後、タンパク質組成物、抗体組成物、そして、遺伝子組換え細胞および遺伝子組換え生物を議論する。次に、本発明のタンパク質が使用される例示的な方法を概説する。
【0035】
核酸組成物
【0036】
上で要約したように、本発明は、Aequorea coerulescens由来の無色のタンパク質であるacGFPをコードする核酸組成物、または、acGFPの蛍光性および非蛍光性の変異体または誘導体をコードする核酸組成物を提供し、それらの核酸組成物の断片およびホモログも提供する。「蛍光タンパク質」という用語は、蛍光性であるタンパク質を意味する。例えば、蛍光タンパク質は、適切な励起波長の光で照射されたとき、弱い、中程度の、または強い蛍光を示すことができる。本発明のタンパク質は、蛍光特性が、1つだけのアミノ酸残基からではなく、タンパク質の2つ以上のアミノ酸残基の相互作用から生じる特性であるタンパク質である。そのため、本発明の蛍光タンパク質には、固有的な蛍光体として単独で作用する残基(すなわち、トリプトファン、チロシンおよびフェニルアラニン)のみに由来する蛍光を示すタンパク質は含まれない。従って、本発明の蛍光タンパク質は、上記に示された単一アミノ酸残基とは異なるタンパク質内の何らかの構造からその蛍光が生じる蛍光タンパク質である。例えば、蛍光は2つ以上のアミノ酸残基の相互作用から生じている。
【0037】
本発明の核酸組成物の1つは、本発明のポリペプチドをコードするオープンリーディングフレーム(翻訳領域)を有するDNA配列を含む組成物(すなわち、蛍光タンパク質遺伝子)である。そのような核酸組成物は、適切な条件のもとで蛍光タンパク質として発現させることができる。核酸組成物の用語にはまた、本発明の核酸に対して相同的な核酸、実質的に類似する核酸、もしくは同一である核酸、または本発明の核酸のミメティックが包含される。本発明の核酸は、その天然の環境とは異なる環境に存在する。例えば、本発明の核酸は単離されているか、濃縮された量で存在するか、あるいは、インビトロにおいてまたは天然に存在する環境とは異なる細胞もしくは生物において存在するか、または発現させられる。
【0038】
本発明の別の実施形態において、核酸は、配列番号1、3、5、7、9、11、13、15、17、19、21または23によりコードされ得るか、そのような核酸に由来する核酸であるか、またはそのような核酸のホモログである。
【0039】
上記の特定の核酸組成物に加えて、上記配列のホモログもまた注目される。本発明の核酸のホモログに関して、相同的な遺伝子の供給源は植物または動物の任意の種であり得るか、あるいは、配列は核酸ミメティックを取り込む配列を含む、全体または一部が合成されたものであり得る。ある実施形態において、ホモログ間の配列類似性は少なくとも約40%であり、50%、60%、70%またはそれ以上であってもよく、これには、75%、80%、85%、90%および95%またはそれ以上が含まれる。配列類似性は、より大きな配列のサブセット(例えば、保存モチーフ、翻訳領域、隣接領域など)であり得る基準配列に基づいて計算される。基準配列は、通常、長さが少なくとも約18ヌクレオチドであり、より一般的には、長さが少なくとも約30ヌクレオチドであり、また、比較されている完全な配列にまで及び得る。配列解析のための様々なアルゴリズムがこの分野では知られており、例えば、Altschul等、J.Mol.Biol.、215、403頁〜10頁(1990)に記載されるBLAST(例えば、デフォルト条件、すなわち、w=4およびT=17のパラメーターを使用する)などがある。
【0040】
ホモログが多数の方法のいずれかによって同定される。本発明のcDNAのフラグメントは、目的とする標的生物から得られたcDNAライブラリーに対するハイブリダイゼーションプローブとして使用することができ、この場合には、低いストリンジェンシーの条件が使用される。プローブは、大きいフラグメントであり得るか、または1つ以上の短い縮重プライマーであり得る。配列類似性を有する核酸が、低いストリンジェンシーの条件のもとでのハイブリダイゼーションによって、例えば、50℃および6×SSC(0.9M塩化ナトリウム/0.09Mクエン酸ナトリウム)におけるハイブリダイゼーションによって検出され、そして、1×SSC(0.15M塩化ナトリウム/0.015Mクエン酸ナトリウム)における55℃での洗浄に供されたとき、結合したままである。配列同一性を、ストリンジェントな条件のもとでのハイブリダイゼーションによって、例えば、50℃以上および0.1×SSC(15mM塩化ナトリウム/1.5mMクエン酸ナトリウム)におけるハイブリダイゼーションによって決定することができる。示された配列に対して実質的な同一性を有する領域を有する核酸、例えば、対立遺伝子変異体、遺伝子の遺伝的に変化した形態などは、ストリンジェントなハイブリダイゼーション条件のもとで、示された配列に結合する。プローブ(特に、DNA配列の標識されたプローブ)を使用することによって、ホモログまたは関連遺伝子を単離することができる。
【0041】
本発明の特定の実施形態において特に注目されるものは、配列番号1、3、5、7、9、11、13、15、17、19、21または23として同定された核酸と実質的に同じ長さの核酸である。この場合、「実質的に同じ長さ」とは、長さの差が約20%を超えず、一般的には約10%を超えず、より一般的には約5%を超えないことを意味する。好ましい実施形態において、実質的に同じ長さのヌクレオチドは、配列番号1、3、5、7、9、11、13、15、17、19、21または23に対する配列同一性が、核酸の全長について、少なくとも約90%(例えば、少なくとも約92%、93%、94%)であり、通常は少なくとも約95%、96%、97%、または98%、または約99%でさえある。「実質的に類似する」とは、配列同一性が、一般には少なくとも約60%であり、通常は少なくとも約75%であり、多くの場合には、少なくとも約80%、85%、90%(例えば、92%、93%、94%)、または95%でさえあり、例えば、96%、98%、98%、99%または99.5%以上であるという意味である。
【0042】
更に、本発明には、先に記載された核酸によってコードされるタンパク質をコードするが、遺伝暗号の縮重性のために、先に記載された核酸とは配列が異なる核酸が含まれる。
【0043】
また、先に記載された核酸にストリンジェントな条件のもとでハイブリダイゼーションする核酸(すなわち、先に記載された核酸の相補体)も提供される。ストリンジェントなハイブリダイゼーション条件の例には、50℃以上および0.1×SSC(15mM塩化ナトリウム/1.5mMクエン酸ナトリウム)におけるハイブリダイゼーションがある。ストリンジェントなハイブリダイゼーション条件の別の例には、50%ホルムアミド、5×SSC(150mM塩化ナトリウム、15mMクエン酸ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハルト溶液、10%デキストラン硫酸および20μg/mlの変性剪断サケ精子DNAからなる溶液における42℃での一晩のインキュベーション、その後、0.1×SSCにおいて約65℃での洗浄がある。ストリンジェントなハイブリダイゼーション条件は、上記の代表的な条件と少なくとも80%のストリンジェンシーであるハイブリダイゼーション条件である。他の様々なストリンジェントなハイブリダイゼーション条件がこの分野では知られており、これらもまた、本発明のこの特定の実施形態の核酸を同定するために用いることができる。
【0044】
本発明のタンパク質の変異体または誘導体をコードする核酸もまた提供される。変異型核酸は、この分野で広く知られている技術を使用して、ランダム変異誘発または標的化変異誘発によって作製することができる。目的とする変異には、欠失、付加および置換が含まれる。ある実施形態においては、ホモログまたは変異体をコードする核酸によってコードされる蛍光タンパク質は、野生型の蛍光タンパク質と同じ蛍光特性を有する。別の実施形態においては、変異体のacGFPタンパク質について本明細書中でより詳しく記載されるように、ホモログまたは変異体の核酸は、変化したスペクトル特性を有する蛍光タンパク質をコードする。
【0045】
本発明の核酸はcDNA、ゲノムDNAまたはそれらの断片であり得る。特定の実施形態において、本発明の核酸は、特定の蛍光タンパク質およびポリペプチドをコードするオープンリーディングフレーム、そしてイントロンを含む、また、翻訳領域を超えて約20kbまでの配列(しかし、おそらくは更なる配列)をいずれかの方向で含む、発現の制御に関与する隣接した5’及び3’の非翻訳ヌクレオチド配列の1つ以上を含む。本発明の核酸は、染色体外での維持のために、または宿主ゲノム内への組み込みのために、適切なベクターに導入することができ、より詳しくは以下に記載される。
【0046】
本明細書中で使用される用語「cDNA」は、エキソンならびに5’及び3’非翻訳領域である、天然の成熟mRNA種に見出される配列エレメントの配置を有する核酸を包含することが意図される。通常、mRNA種は、タンパク質をコードする連続したオープンリーディングフレームを生じさせるために、切れ目なく連続したエキソンを有しており、介在するイントロンは、存在するときには、核でのRNAスプライシングによって除かれている。
【0047】
注目されるゲノム配列は、天然の染色体に通常存在するイントロンのすべてを含む、示された配列において定義される開始コドンと終止コドンとの間に存在する核酸を含むことができる。注目されるゲノム配列は更に、成熟mRNAに見出される5’非翻訳領域および3’非翻訳領域を含んでいてもよく、また、転写領域の5’末端または3’末端のいずれかでの隣接ゲノムDNAの約1kb(しかし、おそらくは更に多く)を含む特異的な転写制御配列および翻訳制御配列(例えば、プロモーター、エンハンサーなど)を含むことができる。ゲノムDNAは、100kb以下のフラグメントとして単離することができ、従って、隣接する染色体配列を実質的に含まない。翻訳領域と3’または5’のいずれかで隣接するゲノムDNA、あるいはイントロン内で時々見出されることがある内部の制御配列は、適切な組織特異的な発現および段階特異的な発現のために要求される配列を含有することがある。
【0048】
本発明の核酸組成物は本発明のタンパク質の全体または一部をコードすることができる。二本鎖フラグメントまたは一本鎖フラグメントは、従来の方法に従ってオリゴヌクレオチドを化学合成することによって、制限酵素消化によって、PCR増幅によって、そして他の方法によって、DNA配列から得られる。たいていの場合、DNA断片は長さが少なくとも約15ヌクレオチドであり、一般的には長さが少なくとも約18ヌクレオチドであるか、または長さが約25ヌクレオチドであり、そして長さが少なくとも約50ヌクレオチドであり得る。ある実施形態においては、本発明の核酸分子は、長さが、約100ヌクレオチド、約200ヌクレオチド、約300ヌクレオチド、約400ヌクレオチド、約500ヌクレオチド、約600ヌクレオチド、約700ヌクレオチドまたはそれ以上であり得る。本発明の核酸は、本発明のタンパク質の断片または全長のタンパク質をコードし得る。例えば、本発明の核酸は、約25アミノ酸、約50アミノ酸、約75アミノ酸、約100アミノ酸、約125アミノ酸、約150アミノ酸、約200アミノ酸のポリペプチドから、全長のタンパク質までをコードし得る。
【0049】
本発明の核酸は、一般には、無傷の(インタクト)染色体としてではなく、他の核酸として、実質的な純度で単離され、得ることができる。通常、本発明の核酸またはそのフラグメントを含まない核酸配列が実質的に存在しないDNAが得られる。実質的な純度とは、核酸が少なくとも約50%の純度であり、一般的には少なくとも約90%の純度であり、典型的には「組換え」であること、すなわち、核酸には、その天然の宿主生物での天然に存在する染色体において通常の場合には関連しないヌクレオチドが1つ以上隣接することを意味する。
【0050】
本発明のポリヌクレオチド、例えば、配列番号1、3、5、7、9、11、13、15、17、19、21または23の配列を有するポリヌクレオチド、対応するcDNA、全長遺伝子および構築物を、当業者に知られている多数の異なるプロトコルによって合成的に作製することができる。適切なポリヌクレオチド構築物は、例えば、Sambrook等、Molecular Cloning:A Laboratory Manual(第2版)、Cold Spring Harbor Press、Cold Spring Harbor、NY(1989)に記載されるような標準的な組換えDNA技術を、例えば、米国保健社会福祉省、国立衛生研究所(NIH)組換えDNA研究指針に記載される規制のもとで使用して精製される。
【0051】
また、第2のペプチドまたはタンパク質に融合された本発明の対象タンパク質またはペプチドの融合タンパク質またはそのフラグメントをコードする核酸が提供される。第2のタンパク質は、例えば、分解配列、シグナルペプチド、または注目される任意のタンパク質であり得る。acGFPポリペプチドまたは変異体acGFPポリペプチドと、acGFPポリペプチドのN末端および/またはC末端においてインフレームで(翻訳領域内部に)融合された第2のポリペプチド(「融合パートナー」)とを、例えば、融合タンパク質は含むことができる。融合パートナーには、融合パートナーに対して特異的な抗体と結合することができるポリペプチド(例えば、エピトープタグ)、抗体またはその結合性フラグメント、触媒機能を提供するポリペプチドまたは細胞応答を誘導するポリペプチド、そしてリガンドまたは受容体またはそのミメティックなどが含まれるが、これらに限定されない。そのような融合タンパク質において、融合パートナーは、一般に天然では融合タンパク質のacGFP部分と会合せず、典型的にはAequorea coerulescensのタンパク質またはその誘導体/断片ではない。すなわち、融合パートナーはAequorea属の種において見出されない。
【0052】
本発明の核酸を含むベクターおよび他の核酸構築物もまた提供され、この場合、そのような構築物は、増殖、タンパク質製造などを含む数多くの応用のために使用することができる。ウイルスベクターおよび非ウイルスベクターを調製し、使用することができ、これには、プラスミドが含まれる。ベクターの選択は、増殖が所望される細胞のタイプ、および増殖の目的に依存する。ある種のベクターが、多量の所望するDNA配列を増幅し、作製するために有用である。他の様々なベクターが、培養での細胞における発現のために好適である。更に他の様々なベクターが、動物での細胞における移入および発現のために好適である。適切なベクターの選択は十分にこの分野の技術の範囲内であり、多くのそのようなベクターが市販されている。構築物を調製するために、部分ポリヌクレオチドまたは全長ポリヌクレオチドが、典型的にはベクター内の切断された制限酵素部位へのDNAリガーゼ結合によってベクター内に挿入される。あるいは、所望されるヌクレオチド配列を、インビボでの相同的組換えによって、典型的には、相同性の領域を所望するヌクレオチド配列の両側においてベクターに結合することによって挿入することができる。相同性の領域は、例えば、オリゴヌクレオチドの連結によって、または、相同性の領域と、所望するヌクレオチド配列の一部との両方を含むプライマーを使用するポリメラーゼ連鎖反応によって付加される。
【0053】
また、他の適用の中でも、本発明の色素タンパク質または蛍光タンパク質またはその融合タンパク質の合成において使用される発現カセットまたは発現系も提供される。発現のために、本発明のポリヌクレオチドによってコードされる遺伝子産物が、例えば、細菌、酵母、昆虫、両生類および哺乳類の系を含む任意の好都合な発現系において発現させられる。そのようなベクターおよび宿主細胞が米国特許第5,654,173号に記載される。発現ベクターにおいて、本発明のポリヌクレオチド、例えば、配列番号1、3、5、7、9、11、13、15、17、19、21または23に示されるようなポリヌクレオチドが、所望する発現特性を得るために適するように制御配列に連結される。これらの制御配列には、プロモーター(センス鎖の5’端またはアンチセンス鎖の3’端のいずれかに結合している)、エンハンサー、ターミネーター、オペレーター、リプレッサーおよびインデューサーが含まれ得る。プロモーターは制御型または構成型であり得る。状況により、例えば、組織特異的プロモーターまたは発達段階特異的プロモーターなどの条件的に活性なプロモーターを使用することが望ましい場合がある。これらは、ベクターへの連結について上記に記載される技術を使用して、所望するヌクレオチド配列に連結される。この分野で知られている技術はどれも使用することができる。すなわち、発現ベクターは、転写開始領域および翻訳開始領域(これらは誘導型でも構成型でもよい)を提供するが、この場合、転写開始領域、転写終結領域および翻訳終結領域の転写制御下に機能的に連結される。これらの制御領域は、問題とする核酸が得られる問題とする種に対して生来的であり得るか、または外因性の供給源に由来し得る。
【0054】
発現ベクターは、一般には、異種タンパク質をコードする核酸配列の挿入をもたらすために、好都合な制限部位がプロモーター配列の近くに存在する。発現宿主において機能的な選択マーカーを存在させることができる。発現ベクターは、とりわけ、上述のように、融合タンパク質を産生させるために使用することができる。
【0055】
転写開始領域、遺伝子またはその断片、および転写終結領域を含む発現カセットを調製することができる。特に注目されるものは、一般的には長さが少なくとも約8アミノ酸であり、より一般的には長さが少なくとも約15アミノ酸であり、また、約25アミノ酸までであり、そして遺伝子の完全なオープンリーディングフレームまでである、機能的なエピトープまたはドメインの発現を可能にする、配列の使用である。DNAの導入後、構築物を含有する細胞を選択マーカーによって選択することができ、その後、拡大培養された細胞を発現のために使用することができる。
【0056】
先に記載された発現系は、発現させる目的に依存して、従来の方法に従って原核生物または真核生物と一緒に用いることができる。タンパク質の大規模な生産の場合、単細胞生物(E.Coli(大腸菌)、B.subtilis(枯草菌)、S.cerevisiaeなど)、バキュロウイルスベクターと組合せた昆虫細胞、または脊椎動物などの高等生物の細胞(例えば、COS7細胞、HEK293、CHO、アフリカツメガエル卵母細胞など)を発現宿主細胞として使用することができる。状況により、発現タンパク質が本来の折畳みおよび翻訳後修飾から利益を受けられる真核生物細胞において、遺伝子を発現させることが望ましい。小ペプチドは実験室で合成することもできる。完全なタンパク質配列のサブセットであるポリペプチドは、機能について重要なタンパク質の一部分を同定し、調べるために使用することができる。
【0057】
注目される特異的な発現系には、細菌、酵母、昆虫細胞および哺乳動物細胞に由来する発現系が含まれる。これらのカテゴリーのそれぞれから代表的なシステムに対して引かれる参考文献が下記に示される。
【0058】
細菌における発現系には、Chang等、Nature、275:615(1978);Goeddel等、Nature、281:544(1979);Goeddel等、Nucleic Acids Res.、8:4057(1980);欧州特許第0036,776号、米国特許第4,551,433号;DeBoer等、Proc.Natl.Acad.Sci.(USA)、80:21〜25(1983);Siebenlist等、Cell、20:269(1980)に記載される発現系が含まれる。
【0059】
酵母における発現系には、Hinnen等、Proc.Natl.Acad.Sci.(USA)、75:1929(1978);Ito等、J.Bacteriol.、153:163(1983);Kurtz等、Mol.Cell Biol.、6:142(1986);Kunze等、J.Basic Microbiol.、25:141(1985);Gleeson等、J.Gen.Microbiol.、132:3459(1986);Roggenkamp等、Mol.Gen.Genet.、202:302(1986);Das等、J.Bacteriol.、158:1165(1984);De Louvencount等、J.Bacteriol.、154:737(1983);Van den Berg等、Bio/Technology、8:135(1990);Kunze等、J.Basic Microbiol.、25:141(1985);Cregg等、Mol.Cell.Biol.、5:3376(1985);米国特許第4,837,148号および同第4,929,555号;BeachおよびNurse、Nature、300:706(1981);Davidow等、Curr.Genet.、10:380(1985);Gaillardin等、Curr.Genet.、10:49(1985);Ballance等、Biochem.Biophys.Res.Commun.、112:284〜289(1983);Tilburn等、Gene、26:205〜221(1983);Yelton等、Proc.Natl.Acad.Sci.(81:1470〜1474(1984);KellyおよびHynes、EMBO J.、4:475479(1985);欧州特許第0244,234号;国際特許出願公開WO91/00357に記載される発現系が含まれる。
【0060】
昆虫における異種遺伝子の発現は、米国特許第4,745,051号;Friesen等、「バキュロウイルス遺伝子発現における制御」、The Molecular Biology of Baculoviruses(W.Doerfler編)(1986);欧州特許第0127,839号;欧州特許第0155,476号;Vlak等、J.Gen.Virol.、69:765〜776(1988);Miller等、Ann.Rev.Microbiol.、42:177(1988);Carbonell等、Gene、73:409(1988);Maeda等、Nature、315:592〜594(1985);Labacq−Verheyden等、Mol.Cell.Biol.、8:3129(1988);Smith等、Proc.Natl.Acad.Sci.、82:8844(1985);Miyajima等、Gene、58:273(1987);Martin等、DNA、7:99(1988)に記載されるように達成される。宿主に由来する数多くのバキュロウイルス株および変異株ならびに対応する許容性の昆虫宿主細胞が、Luckow等、Bio/Technology、6:47〜55(1988);Miller等、Genetic Engineering、8:277〜279(1986);Maeda等、Nature、315:592〜594(1985)に記載される。
【0061】
哺乳動物発現は、Dijkema等、EMBO J.、4:761(1985);Gorman等、Proc.Natl.Acad.Sci.(USA)、79:6777(1982);Boshart等、Cell、41:521(1985);米国特許第4,399,216号に記載されるように達成される。哺乳動物の発現の他の特徴が、HammおよびWallace、Meth.Enz.、58:44(1979);BarnesおよびSata、Anal.Biochem.、102:255(1980);米国特許第4,767,704号、同第4,657,866号、同第4,927,762号、同第4,560,655号;国際特許出願公開WO91/103430、同WO87/00195;米国再発行特許第30,985号に記載されるように容易に行われる。
【0062】
上記の宿主細胞のいずれか、あるいは他の適切な宿主細胞または宿主生物を使用して、本発明のポリヌクレオチドまたは核酸を複製し、かつ/または発現させるとき、得られる複製された核酸、RNA、発現タンパク質またはポリペプチドは、宿主細胞または宿主生物の産物として本発明の範囲内である。産物は、この分野で知られている適切な手段によって回収することができる。
【0063】
選択されたポリヌクレオチドに対応する遺伝子が同定されると、遺伝子が生来的である細胞において、その発現を調節することができる。例えば、細胞の内因性遺伝子を、細胞における遺伝子の発現を少なくとも増強するために十分な位置において細胞のゲノム内に挿入された外因性の制御配列によって制御することができる。制御配列は、米国特許第5,641,670号および同第5,733,761号(これらの開示は参考として本明細書中に組み入れる)に開示されるような相同的な組換えによってゲノム内に組み込まれるように設計することができ、または、国際特許出願公開WO99/15650(この開示もまた参考として本明細書中に組み入れる)に開示されるような非相同的な組換えによってゲノム内に組み込まれるように設計することができる。そのため、コード核酸自体の操作を伴わず、しかし、その代わり、所望するタンパク質をコードする遺伝子を既に含む細胞のゲノムに調節配列を組み込むことによるタンパク質の産生もまた本発明に包含される。
【0064】
本発明のゲノム配列のプロモーター配列もまた注目される。この場合、5’隣接領域の配列を、例えば、本発明のタンパク質遺伝子が発現する細胞/組織における発現の制御をもたらす、エンハンサー結合部位を含むプロモーターエレメントのために利用することができる。
【0065】
本発明の核酸の小さいDNA断片もまた注目され、これらは、PCR用プライマー、ハイブリダイゼーションスクリーニングプローブなどとして有用である。より大きいDNA断片は、先に記載されたように、コードされるポリペプチドを産生させるために有用である。しかしながら、幾何級数的PCRなどの幾何級数的増幅反応において使用される場合、一対の小さいDNAフラグメント(すなわち、プライマー)が使用される。プライマー配列の正確な組成は本発明にとって重要ではなく、しかし、ほとんどの適用の場合、プライマーは、この分野で知られているように、ストリンジェントな条件のもとで、対象とする配列にハイブリダイゼーションする。少なくとも約50ヌクレオチド(好ましくは少なくとも約100ヌクレオチド)の増幅産物を生じさせる一対のプライマーを選ぶことが好ましい。プライマー配列を選択するための様々なアルゴリズムが一般に知られており、市販のソフトウエアパッケージで入手可能である。増幅プライマーはDNAの相補鎖にハイブリダイゼーションし、互いに向かって伸張を開始する。
【0066】
本発明の核酸組成物はまた、生物学的試料における遺伝子の発現を確認するために使用することができる。特定のヌクレオチド配列(例えば、ゲノムDNAまたはRNAなど)の存在について細胞をプローブする様式はこの分野では十分に確立されている。簡単に記載すると、DNAまたはmRNAが細胞サンプルから単離される。mRNAは、相補的DNA鎖を形成させるために逆転写酵素を使用し、その後、対象DNA配列に対して特異的なプライマーを使用するポリメラーゼ連鎖反応増幅を行うRT−PCRによって増幅することができる。あるいは、mRNAサンプルはゲル電気泳動によって分離され、好適な担体(例えば、ニトロセルロース、ナイロンなど)に移され、その後、対象DNAの断片をプローブとして用いてプローブされる。他の技術、例えば、オリゴヌクレオチドライゲーションアッセイ、インサイチュハイブリダイゼーション、および固体チップ上に配列されたDNAプローブに対するハイブリダイゼーションなどもまた使用することができる。対象配列にハイブリダイゼーションしているmRNAの検出により、サンプルにおける遺伝子発現が示される。
【0067】
本発明の核酸(これは隣接プロモーター領域および翻訳領域を含む)は、プロモーター強度の目的とする変化を生じさせるために、あるいはコードされるタンパク質の配列またはコードされるタンパク質の性質(これには、コードされるタンパク質の蛍光特性が含まれる)を変化させるために、この分野で知られている様々な方法で変異させることができる。そのような変異を有するDNA配列またはタンパク質産物は、本明細書中に示される配列番号1〜24に対して実質的に類似している。これらの配列の配列変化は、置換、挿入、欠失またはそれらの組合せであり得る。欠失は更に、ドメインまたはエキソンの欠失などの大きな変化、例えば、10、20、50、75、100、150アミノ酸残基またはそれ以上のアミノ酸残基の領域の欠失を含むことがある。インビトロ変異誘発のための様々な技術を、Gustin等、Biotechniques、14:22(1993);Barany、Gene、37:111〜23(1985);Colicelli等、Mol.Gen.Genet.199:537〜9(1985)に見出すことができる。部位特異的変異誘発のための様々な方法を、Sambrook等、Molecular Cloning:A Laboratory Manual、CHS Press、15.3頁〜15.108頁(1986)に見出すことができる。そのような変異させられた核酸誘導体は、特定の色素性蛍光タンパク質の構造−機能の関係を研究するために、あるいはその機能または制御に影響を及ぼすタンパク質の性質を変化させるために使用することができる。
【0068】
また、本明細書中に記載されるacGFPのhG22変異体などの、本発明の核酸のヒト化体も注目される。本明細書中で使用される用語「ヒト化(された)」は、ヒト細胞におけるタンパク質の発現のためにコドンを最適化するために核酸配列になされた変化を示す(Yang等、Nucleic Acids Research、24:4592〜93(1996))。タンパク質のヒト化を記載する米国特許第5,795,737号もまた参照のこと(この開示は参考として本明細書中に組み入れる)。
【0069】
ペプチド組成物
【0070】
本発明は、蛍光タンパク質acGFPおよびその誘導体、ならびに関連するポリペプチド断片を提供する。本明細書中で使用される用語の蛍光タンパク質は、光を照射されたとき、例えば、白色光または特定の波長の光(または励起波長などの狭い波長バンド)で照射されたとき、蛍光を発するタンパク質を示す。本明細書中で使用される用語のポリペプチドは、全長タンパク質、ならびにタンパク質の一部分または断片の両方を示す。また、この用語には、天然に存在するタンパク質の変異体が含まれ、この場合、そのような変異体は、より詳細には下に記載されるように、天然に存在するタンパク質に対して相同的または実質的に類似し、そして天然に存在するタンパク質の変異体である。本発明のポリペプチドは、その天然の環境とは異なる環境に存在する。
【0071】
多くの実施形態において、本発明のタンパク質は、約300nm〜700nm、一般的には約350nm〜550nm、より一般的には約400nm〜500nm、多くの場合には約450nm〜490nm、例えば、470nm〜490nm、の範囲に及ぶ吸収極大を有し、一方で、本発明のタンパク質の発光スペクトルは、典型的には、約400nm〜700nm、一般的には約450nm〜650nm、より一般的には約500nm〜600nm、の範囲にあり、その一方で、多くの実施形態において、発光スペクトルは約500nm〜550nm、例えば、500nm〜525nm、または500nm〜510nm、の範囲にある。本発明のタンパク質は、一般に、約25,000〜150,000、一般的には約45,000〜120,000、例えば、50,000〜100,000、の範囲に及ぶ最大吸光係数を有する。本発明のタンパク質は、典型的には、長さが約150アミノ酸〜300アミノ酸、一般的には約200アミノ酸残基〜300アミノ酸残基、の範囲にあり、そして一般に、分子量が約15kDa〜35kDa、一般的には約17.5kDa〜32.5kDa、の範囲にある。
【0072】
特定の実施形態において、本発明のタンパク質は鮮明(bright)である。この場合、「鮮明」とは、色素タンパク質およびその蛍光性誘導体が一般的な方法(例えば、目視スクリーニング、分光測定法、分光蛍光法、蛍光顕微鏡観察、FACS計測など)によって検出され得ることを意味する。特定の蛍光タンパク質の蛍光輝度は、最大吸光係数を乗じたその量子収量によって決定される。色素タンパク質の輝度はその最大吸光係数によって表されることがある。
【0073】
特定の実施形態において、本発明のタンパク質は、宿主細胞で発現した後、迅速に折りたたまれる。「迅速に折りたたまれる」とは、タンパク質が、短い時間で、その発色性または蛍光性を生じさせる三次構造を達成することを意味する。これらの実施形態において、タンパク質は、一般には約3日を超えない期間で、一般的には約2日を超えない期間で、より一般的には約1日を超えない期間で折りたたまれる。
【0074】
注目される具体的なタンパク質には、野生型acGFP蛍光タンパク質およびその変異体が含まれるが、それらは、例えば、acGFP、Z1、Z2、G1、G2、G22、G22−G22E、G22−G22E/Y220L、220−11−5、CFP−rand3およびCFP−3ならびにヒト化G22をコードする、配列番号2、4、6、8、10、12、14、16、18、20、22または24で提供される。
【0075】
上記に提供された具体的なアミノ酸配列、すなわち、配列番号2、4、6、8、10、12、14、16、18、20、22または24、とは配列が異なるタンパク質のホモログ(またはその断片)もまた提供される。「ホモログ」は、D.G.HigginsおよびP.M.Sharp、「マイクロコンピューターによる迅速かつ高感度な多重配列アラインメント」、CABIOS、5、151頁〜3頁(1989)に記載されるようなMegAlignのDNAstarクラスターアルゴリズム(ktuple=1、gap penalty=3、window=5およびdiagonals saved=5のパラメーターを使用する)を使用して決定されたとき、配列番号2、4、6、8、10、12、14、16、18、20、22または24のアミノ酸配列に対する少なくとも約35%、一般的には少なくとも約40%、より一般的には少なくとも約60%、のアミノ酸配列同一性を有するタンパク質を意味する。多くの実施形態において、注目されるホモログは、特にタンパク質の機能的な領域を提供するアミノ酸配列について、はるかにより大きい配列同一性を有し、例えば、65%、70%、75%、80%、85%、90%(例えば、92%、93%、94%)またはそれ以上、例えば、95%、96%、97%、98%、99%、99.5%の配列同一性を有する。
【0076】
野生型タンパク質に対して実質的に同一であるタンパク質もまた提供される。この場合、「実質的に同一」は、タンパク質が、野生型タンパク質の配列に対して少なくとも約60%、一般的には少なくとも約65%、より一般的には少なくとも約70%、のアミノ酸配列同一性を有すること、そして、場合により、同一性がはるかにより大きく、例えば、75%、80%、85%、90%(例えば、92%、93%、94%)、95%またはそれ以上、例えば、95%、96%、97%、98%、99%、99.5%であり得ることを意味する。
【0077】
先に記載された天然に存在するタンパク質の誘導体または変異体であるタンパク質もまた提供される。変異体は、野生型(例えば、天然に存在する)タンパク質の生物学的性質を保持し得るか、または野生型タンパク質とは異なる生物学的性質を有し得る。本発明のタンパク質の用語「生物学的性質」は、(例えば、野生型タンパク質、またはA.victoria由来の緑色蛍光タンパク質(GFP)などの別の基準タンパク質と比較されるような)スペクトル特性、例えば、吸収極大、発光極大、最大吸光係数、輝度など;インビボおよび/またはインビトロでの安定性(例えば、半減期);ならびに他のそのような性質を示すが、これらに限定されない。変異には、単一アミノ酸変化、1つ以上のアミノ酸の欠失、N末端切断(トランケーション)、C末端切断、挿入などが含まれる。
【0078】
変異体タンパク質は、分子生物学の標準的な技術を使用して、例えば、先に記載されたようなランダム変異誘発および標的化変異誘発を使用して作製することができる。いくつかの変異体が本明細書中に記載される。実施例に示される指針に従い、そして標準的な技術を使用して、当業者は、広範囲の様々な更なる変異体を容易に作製し、そして、生物学的性質が変化しているかどうかを調べることができる。例えば、蛍光強度を、分光光度計を様々な励起波長で使用して測定することができる。
【0079】
天然に存在するタンパク質である本発明のそのようなタンパク質は、天然に存在しない環境に存在し、例えば、その天然に存在する環境から分離される。特定の実施形態において、本発明のタンパク質は、天然に存在する環境と比較して、本発明のタンパク質について濃縮された組成物に存在する。例えば、精製されたタンパク質が提供され、この場合、「精製された」は、タンパク質が、目的とする色素タンパク質または蛍光タンパク質でないタンパク質を実質的に含まない組成物に存在することを意味し、この場合、「実質的に含まない」は、組成物の90%未満、一般的に60%未満、より一般的には50%未満、が非色素タンパク質または非蛍光タンパク質またはその変異体から構成されることを意味する。本発明のタンパク質はまた、単離体として存在し得る。この場合、単離体により、タンパク質が、他のタンパク質および他の天然に存在する生物学的分子(例えば、オリゴ糖、ポリヌクレオチドおよびその断片など)を実質的に含まないことが意味され、ただし、この場合の用語「実質的に含まない」は、単離されたタンパク質を含有する組成物の70%未満、一般的には60%未満、より一般的には50%未満、が何らかの他の天然に存在する生物学的分子であることを意味する。特定の実施形態において、タンパク質は実質的に純粋な形態で存在する。この場合、「実質的に純粋な形態」により、純度が少なくとも95%、一般的には少なくとも97%、より一般的には少なくとも99%、であることが意味される。
【0080】
天然に存在するタンパク質に加えて、天然に存在するタンパク質とは異なるポリペプチド、例えば、先に記載された変異体タンパク質もまた提供される、一般に、そのようなポリペプチドは、本発明の野生型タンパク質をコードする遺伝子のオープンリーディングフレーム(ORF)によってコードされるアミノ酸配列を含み、これらには、全長タンパク質およびその断片、特に、生物学的に活性な断片、および/または機能的ドメインに対応する断片などが含まれ、また、他のタンパク質またはペプチドに対する本発明のポリペプチドの融合体が含まれる。注目される断片は、典型的には、長さが少なくとも約10アミノ酸であり、一般的には長さが少なくとも約50アミノ酸であり、そして、長さが300アミノ酸またはそれ以上もの長さである場合があるが、一般的には約250アミノ酸の長さを超えない。この場合、断片は、少なくとも約10アミノ酸、一般的には少なくとも約15アミノ酸、多くの実施形態では少なくとも約50アミノ酸、の長さである本発明のタンパク質に対して同一であるアミノ酸領域を有する。いくつかの実施形態では、本発明のポリペプチドは、長さが、約25アミノ酸、約50アミノ酸、約75アミノ酸、約100アミノ酸、約125アミノ酸、約150アミノ酸、約200アミノ酸または約250アミノ酸から、タンパク質の全長までである。いくつかの実施形態において、タンパク質断片は、野生型タンパク質の特異的な性質のすべてまたは実質的にすべてを保持する。
【0081】
本発明のタンパク質およびポリペプチドは、天然に存在する供給源から得ることができ、または合成的に作製することができる。例えば、野生型タンパク質を、そのタンパク質を発現する生物学的供給源から、例えば、Aequorea coerulescensから得ることができる。本発明のタンパク質はまた、合成的手段から、例えば、先に記載されたように、目的とするタンパク質をコードする組換え遺伝子または核酸コード配列を好適な宿主において発現させることによって得ることができる。任意の好都合なタンパク質精製手法を用いることができ、様々な好適なタンパク質精製方法論が、Guide to Protein Purification(Deuthser編)(Academic Press、1990)に記載される。例えば、可溶化液(ライセート)を元の供給源から調製し、そして、HPLC、排除クロマトグラフィー、ゲル電気泳動、アフィニティークロマトグラフィーなどを使用して精製することができる。
【0082】
抗体組成物
【0083】
本発明の蛍光タンパク質に対して特異的に結合する抗体もまた提供される。好適な抗体が、タンパク質の全体または一部を含むペプチドで宿主動物を免疫化することによって得られる。好適な宿主動物は、マウス、ラット、ヒツジ、ヤギ、ハムスター、ウサギなどが含まれる。免疫源は、完全なタンパク質、またはその断片および誘導体を含み得る。好ましい免疫源はタンパク質の全体または一部を含み、この場合、タンパク質は、天然の標的タンパク質において見出される翻訳後修飾を含む。免疫源は、この分野で知られている様々な方法で作製され、そのような方法には、例えば、従来の組換え方法を使用するクローン化遺伝子の発現、または、直接的にAequorea coerulescensからの単離がある。
【0084】
ポリクローナル抗体を調製する場合、最初の工程には、ペプチド免疫源による宿主動物の免疫化が伴い、この場合、ペプチドタンパク質免疫源は、好ましくは、約1%未満の混入物を含む実質的に純粋な形態である。免疫源は、完全なタンパク質あるいはその断片または誘導体を含み得る。宿主動物の免疫応答を増大させるために、標的タンパク質はアジュバントと組み合わせることができ、この場合、好適なアジュバントには、ミョウバン、デキストラン硫酸、大きなポリマーアニオン、油および水のエマルション、フロイントアジュバント、フロイント完全アジュバントなどが含まれる。ペプチド免疫源はまた、合成キャリアタンパク質または合成抗原にコンジュゲート化することができる。
【0085】
ペプチド免疫源は、一般的には皮内投与で宿主に投与されるが、初期投与の
後、1回以上、一般的には少なくとも2回、の更なる追加免疫投与が行われる。免疫後、宿主からの血液が採取され、この場合、血清が血液細胞から分離される。得られた抗血清に存在する免疫グロブリンを、知られている方法を使用して、例えば、アンモニウム塩分画化、DEAEクロマトグラフィーなどを使用して精製することができる。
【0086】
あるいは、モノクローナル抗体を従来の技術によって作製することができる。一般には、免疫化された宿主動物の脾臓および/またはリンパ節が形質細胞の供給源となる。形質細胞はミエローマ細胞との融合によって不死化され、ハイブリドーマ細胞が作製される。個々のハイブリドーマから得られる培養上清が、所望する特異性を有する抗体を産生するハイブリドーマを同定するために、標準的な技術を使用してスクリーニングされる。ヒトタンパク質に対するモノクローナル抗体を作製するための好適な動物には、マウス、ラット、ハムスターなどが含まれる。マウスタンパク質に対する抗体を産生するためには、動物は、一般に、ハムスター、モルモットまたはウサギである。抗体は、従来の技術によって、例えば、プロテインAセファロースのような不溶性担体に結合させたタンパク質を使用するアフィニティークロマトグラフィーなどによってハイブリドーマ細胞上清または腹水から精製することができる。
【0087】
抗体は、通常の多量体構造の代わりに、単鎖体(single chain)として作製することができる。単鎖抗体が、Jost等、J.B.C.、269:26267〜73(1994)などに記載される。重鎖の可変領域および軽鎖の可変領域をコードするDNA配列を、グリシンおよび/またはセリンを含む小さい中性アミノ酸の少なくとも4個のアミノ酸をコードするスペーサーに連結することができる。この融合体によってコードされるタンパク質は、元の抗体の特異性および親和性を保持する機能的な可変領域の組み立てを可能にする。
【0088】
特定の実施形態では、ヒト化抗体もまた注目される。抗体をヒト化する様々な方法がこの分野では知られている。ヒト化抗体は、遺伝子組換えされたヒト免疫グロブリン定常領域遺伝子を有する動物の産物であり得る(例えば、国際特許出願公開WO90/10077および同第WO90/04036を参照のこと)。あるいは、注目される抗体は、CH1、CH2、CH3、ヒンジドメインおよび/またはフレームワークドメインを対応するヒト配列で置換するために組換えDNA技術によって操作することができる(国際特許出願公開WO92/02190を参照のこと)。
【0089】
キメラな免疫グロブリン遺伝子を構築するために免疫グロブリンcDNAを使用することがこの分野では知られている(Liu等、Proceedings of the National Academy of Sciences、84:3439(1987)、およびJ.Immunol.、139:3521(1987))。本質的には、mRNAが、抗体を産生するハイブリドーマまたは他の細胞から単離され、cDNAを作製するために使用される。目的とするcDNAを、特異的なプライマーを使用するポリメラーゼ連鎖反応によって増幅することができる(米国特許第4,683,195号および同第4,683,202号)。あるいは、目的とする配列を単離するために、ライブラリーが作製され、スクリーニングされる。その後、抗体の可変領域をコードするDNA配列がヒト定常領域配列に融合される。ヒト定常領域遺伝子の配列は、Kabat等、「免疫学的に注目されるタンパク質の配列」、N.I.H.刊行物番号91−3242(1991)に見出され得る。ヒトC領域遺伝子を知られているクローンから容易に入手することができる。アイソタイプの選択は、補体固定、または抗体依存的な細胞の細胞毒性における活性などの所望するエフェクター機能によって導かれる。好ましいアイソタイプは、IgG1、IgG3およびIgG4である。ヒト軽鎖定常領域のカッパまたはラムダのいずれかを使用することができる。キメラなヒト化抗体は、その後、従来の方法によって発現される。
【0090】
抗体断片、例えば、Fv、F(ab’)2およびF(ab)などを、インタクトなタンパク質を、例えば、プロテアーゼまたは化学的切断により切断することによって調製することができる。あるいは、切断された(truncated)遺伝子を設計することができる。例えば、F(ab’)2断片の一部をコードするキメラな遺伝子により、H鎖のCH1ドメインおよびヒンジ領域、その後に翻訳終止コドンをコードして、切断型分子を生じさせるDNA配列が示される。
【0091】
H鎖およびL鎖のJ領域のコンセンサス配列を使用して、V領域セグメントをヒトC領域セグメントにその後で連結するために有用な制限部位をJ領域に導入するためのプライマーとして使用されるオリゴヌクレオチドを設計することができる。C領域のcDNAを、制限部位をヒト配列において類似する位置に設置するために部位特異的変異誘発によって改変することができる。
【0092】
発現ベクターには、プラスミド、レトロウイルス、YAC、EBV由来エピソームなどが含まれる。好都合なベクターは、機能的に完全なヒトCH免疫グロブリン配列またはCL免疫グロブリン配列を、任意のVH配列またはVL配列が容易に挿入および発現され得るように設計された適切な制限部位とともにコードするベクターである。そのようなベクターでは、スプライシングが、通常、挿入されたJ領域内のスプライスドナー部位と、ヒトC領域に先行するスプライスアクセプター部位との間において、そしてまた、ヒトCHエキソンの中に存在するスプライス領域において生じる。ポリアデニル化および転写終結が、翻訳領域の下流にある生来的な染色体部位において生じる。得られるキメラな抗体は、レトロウイルスLTRを含む任意の強いプロモーターに、例えば、SV−40初期プロモーター(Okayama等、Mol.Cell.Bio.、3:280(1983))、ラウス肉腫ウイルスLTR(Gorman等、Proceedings of the National Academy of Sciences、79:6777(1982))、またはモロニーマウス白血病ウイルスLTR(Grosschedl等、Cell、41:885(1985))、あるいは生来的なIgプロモーターなどに結合することができる。
【0093】
遺伝子組換え体
【0094】
本発明の核酸は、遺伝子組み換えされた非ヒトの植物または動物または部位特異的遺伝子改変体を細胞株において作製するために使用することができる。本発明の遺伝子組換え細胞は、導入遺伝子として存在する本発明による1つ以上の核酸を含む。この場合、この定義には、導入遺伝子を含むように形質転換された親細胞およびその子孫が含まれる。多くの実施形態において、遺伝子組換え細胞は、本発明による核酸を通常の場合には保有または含有しない細胞である。遺伝子組換え細胞が本発明の核酸を通常の場合には含有しないそのような実施形態において、核酸は、細胞内では、非天然の位置において細胞のゲノム物質に組み込まれるなどの、その天然の位置とは異なる位置に存在する。遺伝子組換え動物は、内因性遺伝子座が変化する相同的組換えによって作製することができる。あるいは、核酸構築物をゲノム内にランダムに組み込むことができる。安定な組み込みのためのベクターには、プラスミド、レトロウイルスおよび他の動物ウイルス、YACなどが含まれる。
【0095】
注目される遺伝子組換え生物には、タンパク質またはその変異体が、通常の場合には発現しない細胞または組織で発現し、かつ/あるいはそのような細胞または組織に通常の場合には存在しないレベルで発現する細胞および多細胞生物(植物および動物の両方)が含まれる。
【0096】
相同的組換えのためのDNA構築物は本発明の核酸の一部を少なくとも含む。この場合、遺伝子は、所望する遺伝子修飾を有し、そして標的遺伝子座に対する相同性の領域を含む。ランダム組み込みのためのDNA構築物は、組換えを媒介するための相同性の領域を含む必要はない。好都合なことに、正の選択および負の選択を行うためのマーカーを含むことができる。標的化された遺伝子修飾を相同的組換えにより有する細胞を作製するための様々な方法がこの分野では知られている。哺乳動物細胞をトランスフェクションするための技術については、Keown等、Meth.Enzymol.、185:527〜37(1990)を参照のこと。
【0097】
胚性幹(ES)細胞の場合、ES細胞株を用いることができ、または胚細胞を、マウス、ラット、モルモットなどの宿主から新たに得ることができる。そのような細胞は、適切な繊維芽細胞フェーダー層上で成長し、または白血病阻害因子(LIF)の存在下で成長する。ES細胞または胚細胞が形質転換されているとき、それらを使用して、遺伝子組換え動物を作製することができる。形質転換後、細胞は適切な培地においてフィーダー層に置床される。構築物を含有する細胞を、選択培地を用いることによって検出することができる。十分な時間が、コロニーが成長するために与えられた後、コロニーは選ばれ、構築物の相同的組換えまたは組み込みが生じているかどうか解析される。陽性であるそのようなコロニーは、その後、胚操作および胚盤胞への注入のために使用することができる。胚盤胞は、4週齢〜6週齢の過排卵処理されたメスから得られる。ES細胞はトリプシン処理され、改変された細胞が胚盤胞の胞胚腔に注入される。注入後、胚盤胞は、偽妊娠メスのそれぞれの子宮角に戻される。その後、メスは出産させられ、得られた子が、構築物についてスクリーニングされる。キメラな子孫は、形質転換された細胞の表現型が、天然に存在する細胞と何らかの点で異なる場合(例えば、蛍光を示すことなど)、容易に検出することができる。
【0098】
キメラな動物は、改変された遺伝子の存在についてスクリーニングされ、その改変を有するオスおよびメスが、ホモ接合の子孫を作製するためにかけ合わせられる。特に本発明の融合タンパク質に関しては可能性があるが、遺伝子の変化が発達における何らかのところで致死性を生じさせる場合、組織または器官を、同種(アロジェニック)または類似遺伝子(コンジェニック)の移植片または移植体として維持することができ、あるいはインビボ培養で維持することができる。遺伝子組換え動物は、任意の非ヒト哺乳動物であり、例えば、実験室動物、家畜などであり、機能的研究、薬物スクリーニングなどにおいて使用することができる。遺伝子組換え動物の使用の代表的な例には、下に記載される例が含まれる。
【0099】
遺伝子組換え植物もまた作製することができる。遺伝子組換えされた植物細胞および植物を調製する様々な方法が、米国特許第5,767,367号、同第5,570,870号、同第5,739,409号、同第5,689,049号、同第5,689,045号、同第5,674,731号、同第5,656,466号、同第5,633,155号、同第5,629,470号、同第5,595,896号、同第5,576,198号、同第5,538,879号、同第5,484,956号(これらの開示は参考として本明細書中に組み入れる)に記載される。遺伝子組換え植物を作製する様々な方法がまた、Plant Biochemistry and Molecular Biology(LeaおよびLeegood編、John Wiley&Sons)、275頁〜295頁(1993)に総説される。簡単に記載すると、好適な植物細胞または植物組織が、植物種の特性に依存して集められる。そのため、特定の場合には、プロトプラストが単離され、この場合、そのようなプロトプラストを様々な異なる植物組織(例えば、葉、胚軸、根など)から単離することができる。プロトプラストを単離する場合、集められた細胞は、細胞壁を除くためにセルラーゼの存在下でインキュベーションされる。この場合、正確なインキュベーション条件は、細胞が得られる植物および/または組織のタイプに依存して変化する。得られたプロトプラストは、その後、生じた細胞破片からふるい分けおよび遠心分離によって分離される。
【0100】
あるいは、体細胞を含む胚性外植片を、遺伝子組換え宿主を調製するために使用することができる。細胞または組織を集めた後、目的とする外因性DNAが植物細胞に導入される。この場合、様々な異なる技術をそのような導入のために利用することができる。単離されたプロトプラストの場合、プロトプラストを裸のDNA(例えば、目的とする外因性のコード配列を含むプラスミドなど)を多価カチオン(例えば、PEGまたはPLO)の存在下でインキュベーションすること、または、目的とする外因性のコード配列を含む裸のDNAの存在下でプロトプラストをエレクトポレーションすることを含むDNA媒介遺伝子移入プロトコルによる導入を行うことができる。外因性DNAを取り込むことに成功したプロトプラストは、その後、選択され、カルスに成長させられ、そして最終的には、適切な量および比率の刺激因子(例えば、オーキシンおよびサイトカイニンなど)との接触によって遺伝子組換え植物に成長させられる。
【0101】
胚性外植片の場合、外因性DNAを標的の体細胞に導入する好都合な方法は、粒子加速プロトコル、すなわち、「遺伝子銃」プロトコルの使用による。得られる外植片は、その後、キメラな植物に成長させられ、交配され、その後、遺伝子組換え子孫が得られる。
【0102】
先に記載された裸のDNA法の代わりに、遺伝子組換え植物を作製する別の方法には、アグロバクテリウム媒介形質転換による方法がある。アグロバクテリウム媒介形質転換の場合、外因性DNAを含む同時組み込みベクターまたはバイナリーベクターが調製され、その後、適切なアグロバクテリウム株(例えば、A.tumefaciens)に導入される。得られた細菌が、その後、調製されたプロトプラストまたは組織外植片(例えば、リーフディスクなど)とインキュベーションされ、カルスが作製される。その後、カルスは、選択条件のもとで成長させられ、選択され、そして根および苗条(シュート)の成長を誘導し、最終的には遺伝子組換え植物を得るために成長培地に供される。
【0103】
使用方法
【0104】
本発明の蛍光タンパク質およびペプチドは様々な異なる応用において使用される。これらのタイプのタンパク質のそれぞれについて代表的な使用が以下に記載されるが、この場合、本明細書中に記載される使用は、単なる例示にすぎず、本発明のタンパク質の使用を記載される方法に限定することを決して意図していない。
【0105】
注目される第1の応用は、本発明のタンパク質を蛍光共鳴エネルギー転移(FRET)法において使用することである。これらの方法では、本発明のタンパク質は、下記のような第2の蛍光タンパク質または蛍光色素と組合せてドナーおよび/またはアクセプターとして役立つ:例えば、Matz等、Nature Biotechnology、17:969〜973(1999)に記載されるような蛍光タンパク質;例えば、米国特許第6,066,476号、同第6,020,192号、同第5,985,577号、同第5,976,796号、同第5,968,750号、同第5,968,738号、同第5,958,713号、同第5,919,445号、同第5,874,304号(これらの開示は参考して本明細書中に組み入れる)に記載される、Aequorea victoria由来の緑色蛍光タンパク質またはその蛍光変異体、;他の蛍光性色素、例えば、クマリンおよびその誘導体(7−アミノ−4−メチルクマリンおよびアミノクマリン)など;bodipy色素;カスケードブルー;またはフルオレセインおよびその誘導体、例えば、フルオレセインイソチオシアナートおよびオレゴングリーンなど;ローダミン色素、例えば、テキサスレッド、テトラメチルローダミン、エオシン類およびエリトロシン類など;Cy3およびCy5などのシアニン色素;ランタニドイオンの大環状キレート、例えば、量子色素など;そして、ルシフェラーゼなどの化学発光色素、これには、米国特許第5,843,746号、同第5,700,673号、同第5,674,713号、同第5,618,722号、同第5,418,155号、同第5,330,906号、同第5,229,285号、同第5,221,623号、同第5,182,202号(これらの開示は参考して本明細書中に組み入れる)に記載される色素が含まれる。
【0106】
本発明の蛍光タンパク質を用いるFRETアッセイが使用され得る具体的な例には、下記の例が含まれるが、これらに限定されない。例えば、哺乳動物ツーハイブリッドシステム、転写因子の二量体化、膜タンパク質の多量体化、多タンパク質複合体形成などにおけるタンパク質−タンパク質相互作用の検出;多数の異なる事象に対するバイオセンサーとして、この場合、ペプチドまたはタンパク質により、本発明の蛍光タンパク質を含むFRET蛍光性組合せが共有結合的に連結され、連結用のペプチドまたはタンパク質には、例えば、カスパーゼが媒介する切断のためのプロテアーゼ特異的基質、FRETを増大または減少させるシグナルを受け取ったときに立体配座的変化を受けるペプチド、例えば、PKA調節ドメイン(cAMPセンサー)、リン酸化部位(例えば、リン酸化部位がペプチドに存在する場合、または、ペプチドが別のタンパク質のリン酸化/脱リン酸化ドメインに対する結合特異性を有する場合、あるいは、ペプチドがCa2+結合ドメインを有する場合)がある。更に、本発明のタンパク質が使用される蛍光共鳴エネルギー転移適用、すなわち、FRETの応用には、米国特許第6,008,373号、同第5,998,146号、同第5,981,200号、同第5,945,526号、同第5,945,283号、同第5,911,952号、同第5,869,255号、同第5,866,336号、同第5,863,727号、同第5,728,528号、同第5,707,804号、同第5,688,648号、同第5,439,797号(これらの開示は参考して本明細書中に組み入れる)に記載される適用が含まれるが、それらに限定されない。
【0107】
本発明の蛍光タンパク質はまた、原核生物細胞および真核生物細胞におけるバイオセンサーとして、例えば、Ca2+イオン指示薬、pH指示薬、リン酸化指示薬、または他のイオン(例えば、マグネシウム、ナトリウム、カリウム、塩化物およびハロゲン化物など)の指示薬などとして使用される。例えば、Ca2+イオンを検出する場合、EFハンドモチーフを含有するタンパク質は、Ca2+が結合したとき、細胞質ゾルから膜に移動することが知られている。これらのタンパク質は、タンパク質の他の領域との疎水性相互作用により分子内に埋もれているミリストイル基を含有する。Ca2+の結合は、ミリストイル基を露出させる立体配座の変化を誘導し、その後、ミリストイル基は、脂質二重層内への挿入のために使用され得る(「Ca2+−ミリストイルスイッチ」と呼ばれる)。そのようなEFハンド含有タンパク質の蛍光タンパク質への融合は、細胞質ゾルから形質膜への移動を共焦点顕微鏡観察によりモニターすることによって、EFハンド含有タンパク質を細胞内Ca2+の指示薬にする。このシステムにおいて使用される好適なEFハンドタンパク質には、リカバリン、カルシニューリンB、トロポニンC、ビシニン、ニューロカルシン、カルモジュリン、パルブアルブミンなどが含まれるが、これらに限定されない。
【0108】
pH指示の場合、ヒサクトフィリン類(hisactophilins)に基づくシステムを用いることができる。ヒサクトフィリン類は、ジクチオステリウム(Dyctyostelium)属に存在することが知られているミリストイル化されたヒスチジンリッチなタンパク質である。アクチンおよび酸性脂質に対するそれらの結合は、細胞質のpH変化の範囲内において鋭いpH依存性を有する。生細胞では、膜に結合することにより、ヒサクトフィリン類はアクチンフィラメントと相互作用しなくなるようである。6.5以下のpHにおいて、ヒサクトフィリン類は形質膜および核に移動する。対照的に、7.5のpHでは、ヒサクトフィリン類は細胞質空間全体に一様に分布する。分布のこの変化は可逆的であり、分子表面にループ状に露出したヒスチジンクラスターに起因する。細胞質のpH変化の範囲における細胞内分布の反転は、ヒスチジン残基の6.5のpKと一致している。細胞分布はタンパク質のミリストイル化とは無関係である。蛍光タンパク質をヒサクトフィリンに融合することによって、融合タンパク質の細胞内分布をレーザー走査共焦点顕微鏡観察または標準的な蛍光顕微鏡観察によって追跡することができる。
【0109】
そのような研究の場合、定量的な蛍光分析を、細胞を通るライン操作(レーザー走査共焦点顕微鏡観察)または他の電子的データ分析(例えば、メタモルフソフトウエア(Universal Imaging Corp)を使用する)を行い、細胞集団において集められたデータを平均化することによって行うことができる。細胞質ゾルから形質膜へのヒサクトフィリン/蛍光タンパク質の実質的なpH依存的再分布は1分〜2分のうちに生じ、5分〜10分の後に定常状態レベルに達する。逆反応が、類似する時間スケールで生じる。そのため、類似した様式で作用するヒサクトフィリン−蛍光タンパク質の融合タンパク質は、生きた哺乳動物細胞においてリアルタイムで細胞質ゾルのpH変化をモニターするために使用することができる。そのような方法は、ハイスループットへの応用において、例えば、増殖因子受容体活性化(例えば、上皮増殖因子または血小板由来増殖因子)、走化性刺激/細胞移動の結果としてのpH変化の測定において、二次メッセンジャーとしての細胞内pH変化の検出において、pH操作実験における細胞内pHのモニターリングなどにおいて使用される。
【0110】
PKC活性を検出する場合、レポーターシステムでは、MARCKS(ミリストイル化アラニンリッチCキナーゼ基質)と呼ばれる分子がPKCの基質であるという事実が利用される。MARCKSは、ミリストイル化と、静電的相互作用により負にチャージした形質膜に結合する正にチャージしたアミノ酸の領域(EDドメイン)と、を介して形質膜に固定される。PKCが活性化されると、EDドメインがPKCによってリン酸化され、それにより、負荷電となり、そして、静電的反発の結果として、MARCKSが形質膜から細胞質に移動する(これは「ミリストイル−静電的スイッチ」と呼ばれる)。MARCKSのミリストイル化モチーフからEDドメインまでのMARCKSのN末端を本発明の蛍光タンパク質に融合することにより、PKC活性に対する検出器システムが提供される。PKCによってリン酸化されたとき、融合タンパク質は形質膜から細胞質ゾルに移動する。この移動は、例えば、Cellomics Inc.の技術、または他のハイコンテントスクリーニングシステム(例えば、Universal Imaging Corp.またはBecton Dickinsonから得られるシステムなど)を使用することにより、標準的な蛍光顕微鏡観察または共焦点顕微鏡観察によって追跡することができる。上記のレポーターシステムは、PKC阻害剤に対するハイコンテントスクリーニングに応用され、また、このシグナル伝達経路を妨害する潜在的な試薬についてのスクリーニングアッセイにおけるPKC活性に対する指示薬として適用される。蛍光タンパク質をバイオセンサーとして使用する方法にはまた、米国特許第5,972,638号、同第5,824,485号および同第5,650,135号(ならびにそれらにおいて引用される参考文献)(これらの開示は参考して本明細書中に組み入れる)に記載される方法が含まれる。
【0111】
本発明の蛍光タンパク質はまた、顕微鏡画像化および電子的解析を使用することによる、蛍光性のレポーター基を発現する細胞のアレイの自動化されたスクリーニングを伴う適用において使用される。スクリーニングは、薬物探索のために、また、機能的ゲノミクスの分野で使用することができ、この場合、本発明のタンパク質は、多細胞性の再編成および遊走における変化、例えば、内皮細胞による多細胞細管の形成(血管形成)、Fluoroblok Insertシステム(Becton Dickinson Co.)を介する細胞の遊走、創傷治癒、または神経突起の伸長における変化を検出するために細胞全体のマーカーとして使用される。本発明のタンパク質が、例えば、シグナル伝達における細胞活性についての指示薬として細胞内位置の変化(刺激に対するキナーゼおよび転写因子の移動など)を検出するペプチド(例えば、標的配列など)またはタンパク質に融合されたマーカーとして使用されるスクリーニングもまた用いることができる。例には、プロテインキナーゼC、プロテインキナーゼA、転写因子NFκBおよびNFAT;細胞周期タンパク質、例えば、サイクリンA、サイクリンB1およびサイクリンEなど;引き続いて切断基質が移動するプロテアーゼ切断;リン脂質が、細胞内構造体(例えば、小胞体、ゴルジ装置、ミトコンドリア、ペルオキシソーム、核、核小体、形質膜、ヒストン、エンドソーム、リソソームまたは微小管など)に対するマーカーとともに含まれる。
【0112】
本発明のタンパク質は、局在化マーカーを細胞内蛍光タンパク質/ペプチドの移動の指示薬として、またはマーカー単独として他の蛍光性融合タンパク質の同時局在化を検出するためのハイコンテントスクリーニングにおいて使用することもできる。本発明の蛍光タンパク質が使用される細胞アレイの自動化されたスクリーニングを伴う適用の例には、米国特許第5,989,835号、ならびに、国際特許出願公開WO0017624、同WO00/26408、同WO00/17643および同00/03246(これらの開示は参考して本明細書中に組み入れる)が含まれる。
【0113】
本発明の蛍光タンパク質は、ハイスループットスクリーニングアッセイにおいて使用される。本発明の蛍光タンパク質は、24時間を超える半減期を有する安定なタンパク質である。また、薬物探索のための転写リポーターとして使用することができる、低下した半減期を有する本発明の蛍光タンパク質の脱安定化体が提供される。例えば、本発明によるタンパク質は、より短い半減期を有するタンパク質に由来する推定的なタンパク質分解シグナル配列(例えば、マウスオルニチンデカルボキシラーゼ遺伝子に由来するPEST配列など)、マウスサイクリンB1破壊ボックスまたはユビキチンなどと融合させることができる。脱安定化されたタンパク質、およびそのようなタンパク質を作製するために用いられ得るベクターの記載については、例えば、米国特許第6,130,313号(この開示は参考して本明細書中に組み入れる)を参照のこと。本発明の蛍光タンパク質の脱安定化体を薬物スクリーニングのために使用して、シグナル伝達経路におけるプロモーターを検出することができ、例えば、AP1、NFAT、NFκB、Smad、STAT、p53、E2F、Rb、myc、CRE、ER、GRおよびTREなどである。
【0114】
本発明のタンパク質は、例えば、PattersonおよびLippincott−Scott、Science、13:1873〜77(2002);Ando等、Proc.Natl.Acad.Sci.USA、99:12651〜56(2002)に記載されるように、タンパク質、オルガネラまたは細胞の正確なインビボ光標識、そして、その輸送を追跡するための光活性化標識として使用することができる。
【0115】
更に、本発明のタンパク質は、本発明のタンパク質を特異的なドメイン(例えば、PKCγのCa結合ドメイン、PKCγのDAG結合ドメイン、SH2ドメインまたはSH3ドメインなど)に融合することによって二次メッセンジャー検出器として使用することができる。
【0116】
分泌リーディング配列(secreted leading sequence)を本発明のタンパク質に融合して、本発明のタンパク質の分泌型形態を構築することによって、本発明のタンパク質の分泌型形態を調製することができ、そのような分泌型形態は次いで様々な異なる適用において使用することができる。
【0117】
本発明のタンパク質は、蛍光活性化細胞分取(FACS)適用において使用される。そのような適用において、本発明の蛍光タンパク質は、細胞集団に標識をつけるための標識として使用され、そして、得られる標識された細胞集団が、その後、この分野で知られているように、蛍光活性化細胞分取装置で分取される。様々なFACS法が米国特許第5,968,738号および同第5,804,387号(これらの開示は参考して本明細書中に組み入れる)に記載される。
【0118】
本発明のタンパク質はまた、遺伝子組換え動物におけるインビボマーカーとしても使用される。例えば、本発明のタンパク質の発現を組織特異的なプロモーターによって行わせることができる。この場合、そのような方法は、他の適用の中でも、導入遺伝子の発現効率を調べることなどの、遺伝子治療のための研究において使用される。そのような適用を例示する、遺伝子組換え動物における蛍光タンパク質の代表的な適用が、国際特許出願公開WO00/02997(この開示は参考して本明細書中に組み入れる)に見出される。
【0119】
本発明のタンパク質の更なる適用には、細胞または動物に注入された後、定量的測定のための校正におけるマーカーとしての使用;細胞生存性をモニターするための酸素バイオセンサーデバイスにおけるマーカーまたはレポーターとしての使用;動物、ペット、玩具、食物などに対するマーカーまたは標識としての使用が含まれる。
【0120】
本発明のタンパク質はまた、プロテアーゼ切断アッセイにおいて使用される。例えば、切断不活性型蛍光アッセイを、本発明のタンパク質を使用して開発することができ、この場合、本発明のタンパク質は、タンパク質の蛍光性を破壊することなく、プロテアーゼ特異的切断配列を含むために設計される。活性化されたプロテアーゼによって蛍光タンパク質が切断されたとき、機能的な発色団が破壊されることにより、蛍光が急激に減少する。あるいは、切断により活性化される蛍光を、本発明のタンパク質を使用して生じさせることができ、この場合、タンパク質は、発色団のすぐ近くおよび/または内部に更なるスペーサー配列を含有するために設計される。この変異体は、機能的な発色団の一部がスペーサーによって分割されているので、その蛍光活性が著しく減少する。スペーサーは2つの同一のプロテアーゼ特異的切断部位によって組み立てられる。活性化されたプロテアーゼによって切断されると、スペーサーは切り離され、蛍光タンパク質の2つの残った「サブユニット」が再び会合して、機能的な蛍光タンパク質を生じさせることができる。上記適用はともに、様々な異なるタイプのプロテアーゼ(例えば、カスパーゼなど)に対するアッセイにおいて開発することができる。
【0121】
本発明のタンパク質は、生物学的膜におけるリン脂質組成を明らかにするためのアッセイにおいて使用することもできる。例えば、特異的なリン脂質に結合して、生物学的膜におけるリン脂質分布のパターンを局在化/可視化することを可能し、その一方で、特定のリン脂質領域における膜タンパク質の同時局在化を可能にする、本発明のタンパク質の融合タンパク質(または本発明のタンパク質の任意の他の種類の共有結合的修飾もしくは非共有結合的修飾)を、本発明のタンパク質を用いて達成することができる。例えば、GRP1のPHドメインは、大きい親和性をホスファチジルイノシトール三リン酸(PIP3)に対して有するが、PIP2に対しては有していない。そのため、GRP1のPHドメインと本発明のタンパク質との融合タンパク質を、生物学的膜におけるPIP3リッチ領域を特異的に標識するために構築することができる。
【0122】
本発明のタンパク質の更に別の適用は、蛍光性タイマーとしてであり、この場合、蛍光タンパク質の経時変化に付随するある蛍光色から別の蛍光色への変化(例えば、緑色から赤色への変化)が、発生上の遺伝子発現、細胞周期に依存する遺伝子発現、および概日リズム特異的な遺伝子発現などの遺伝子発現の活性化/不活化を明らかにするために使用される。
【0123】
上術の本発明の抗体はまた、本発明のタンパク質を他の蛍光タンパク質から区別することを含む多数の適用において使用される。
【0124】
キット
【0125】
先に記載された適用の1つ以上を実施する際に使用されるキットもまた本発明によって提供される。この場合、キットは、典型的には、本発明のタンパク質を発現させるための要素、例えば、本発明のタンパク質に対する翻訳領域を含むベクターを構成する構築物を含む。キットの構成要素は、典型的には好適な容器において、典型的には、好適な貯蔵媒体(例えば、緩衝化溶液など)に存在する。また、キットには、提供されたタンパク質に対する抗体を存在させることができる。特定の実施形態において、キットは、本発明のタンパク質をそれぞれがコードする多数の異なるベクターを含む。この場合、ベクターは、異なる環境および/または異なる条件下での発現のために、例えば、ベクターが哺乳動物細胞における発現のために強力なプロモーターを含む構成的発現のために設計されているか、あるいは、プロモーターを独自に挿入するための多数のクローニング部位を有し、発現が独自に調節される無プロモーターベクターなどである。
【0126】
上記の構成要素に加えて、本発明のキットは更に、本発明の方法を実施するための説明書を含む。このような説明書は、キットにはそのうちの1つ以上が存在し得る様々な形態で本発明のキットに存在させることができる。これらの説明書が存在し得る1つの形態は、キットを包装する際に含められる印刷情報(例えば、包装添付文書など)としてである。更に別の手段には、情報が記録されているコンピューター読み取り可能な媒体(例えば、ディスケット、CDなど)がある。存在させることができる更に別の手段には、離れた場所でインターネットを介して情報を入手するために使用され得るウェブサイトアドレスがある。好都合な手段はどれも、キットに存在させることができる。
【実施例】
【0127】
下記の実施例は、限定としてではなく、例示として提供される。
大きいhydromedusaの数個の試料が、2001年8月に、ウラジオストック近くの日本海のロシア沿岸で集められた。1組の特徴により、これらのクラゲをAequorea coerulescensとして同定することが可能である(Kramp、Dana.Rept.、72:201〜202(1968);PogodinおよびYakovlev、Rus.J.Mar.Biol.、25:417〜419(1999))。A.coerulescensおよびA.victoria(A.forscalea、A.aequoreaの異名)は類似しているが、それらの特徴のいくつかは非常に異なる。最も明白な差は触手の数である。すなわち、A.victoriaは放射状導管1つあたり1つの触手を有するだけであり、一方、A.coerulescensは、隣接する放射状導管の各対の間に4つ〜6つの触手を有する。
【0128】
捕まえられたA.coerulescens試料は生物発光性であった。A.victoriaとは対照的に、それらは、緑色ではなく、青色の発光を示した。検出可能な蛍光は、UV光において、または蛍光顕微鏡を使用したとき、A.coerulescensクラゲでは観測されなかった。それにもかかわらず、A.victoriaのGFPに対するモノクローナル抗体により、A.coerulescensから得られたタンパク質抽出物においてGFP様タンパク質が検出された。図11は、acGFPのタンパク質ゲル電気泳動分析を示す。図11aは、A.victoriaのGFPに対する抗体を使用した、A.coerulescensから得られた可溶性タンパク質抽出物のウエスタンブロット分析である。レーン1は、精製された組換えA.victoria GFPであり、レーン2はA.coerulescens抽出物である。
【0129】
GFP様タンパク質をクローン化するために、保存されたアミノ酸配列に対応する縮重プライマーを用いてPCRを行った。GFP様タンパク質をコードするcDNAがクローン化された。野生型acGFPタンパク質のヌクレオチド配列およびアミノ酸配列が図1に示される。このacGFPタンパク質は、GFPに対して非常に大きい類似性を示し、92%のアミノ酸配列類似性を有した(図2参照)。すべての知られている重要な残基(これらには、発色団を形成するSer65、Tyr66およびGly67、進化的に変化していないArg96およびGlu222、そして発色団に対して空間的に近い残基のHis148、Phe165、Ile167およびThr203が含まれる)が、acGFPでは変化していないことが見出された。3個の内部アミノ酸のみが、これらのタンパク質の間では異なっていた。
【0130】
非常に大きい配列類似性を考慮に入れると、acGFPのスペクトル特性はGFPのスペクトル特性と非常に類似していることが予想された。それにもかかわらず、acGFPを発現する大腸菌コロニーは蛍光または着色のいずれも示さなかった。この事実の最も単純な説明、すなわち、大腸菌ではacGFPの折りたたみが完全ではないということは、ほんの部分的には正しく、それは更なる実験で明らかにされた。
【0131】
クラゲの様々な部分におけるacGFP cDNAの存在を、特異的なプライマーを使用するPCRによって調べた。クラゲの傘縁、放射状導管および口盤に対応する3つのcDNAサンプルが調べられた。acGFP cDNAが傘縁において明瞭に検出されたが、2つの他のサンプルには存在しなかった。従って、A.coerulescensにおけるacGFPの分布は、A.victoriaの傘縁において蛍光性の環を形成するA.victoriaにおけるGFPの分布と類似している。
【0132】
acGFPのランダム変異誘発により、多数の緑色蛍光クローンが得られ、そのうちの一部が特徴づけられた。それらの性質および考えられる適用は、A.victoriaの増強型GFP(EGFP)についての性質および適用と類似する。
【0133】
変異体Z1は、図3に示されるように、1つのアミノ酸置換E222G(222位のグルタミン酸の代わりにグリシン)を含有した。変異体Z1タンパク質は、低い輝度、非常に遅い折りたたみを有し、20℃未満の温度が成熟化のために必要であった。30℃で成長させた後、Z1を発現する大腸菌コロニーは、蛍光が見えるようになるためには、室温または4℃で3日間〜5日間貯蔵しなければならない。Z1変異体に対する励起スペクトルおよび発光スペクトルは、それぞれ、480nmおよび504nmに極大を有する(図4参照)。
【0134】
変異体Z2は、図5に示されるように、2つのアミノ酸置換、具体的には、N19D(19位のアスパラギンの代わりにアスパラギン酸)およびE222G(222位のグルタミン酸の代わりにグリシン)を含有した。変異体Z2は、低い輝度、非常に遅い折りたたみ効率を有し、20℃未満の温度が成熟化のために必要であった。37℃で一晩成長させた後、Z2を発現する大腸菌コロニーは、蛍光が見えるようになるためには、室温または4℃で3日間〜5日間貯蔵しなければならない。Z2変異体に対する励起スペクトルおよび発光スペクトルは変異体Z1の励起スペクトルおよび発光スペクトルと非常に類似している。
【0135】
変異体G1は、図6に示されるように、V11I(11位のバリンの代わりにイソロイシン)、K101E(101位のリシンの代わりにグルタミン酸)およびE222G(222位のグルタミン酸の代わりにグリシン)の置換を有する。変異体G1は別のランダム変異誘発処理で作製された。置換E222Gがこれらの変異体の3つで見出されたので、この変異はこれらの変異体の緑色蛍光のために重要であり得ることが考えられた。変異体G1はかなり低い輝度を有する。この変異体の蛍光は、大腸菌を一晩成長させた後の第1日目に見えるようになる。変異体G1の励起スペクトルおよび発光スペクトルは変異体Z1の励起スペクトルおよび発光スペクトルと非常に類似している。
【0136】
変異体G2は、図7に示されるように、V11I(11位のバリンの代わりにイソロイシン)、F64L(64位のフェニルアラニンの代わりにロイシン)、K101E(101位のリシンの代わりにグルタミン酸)およびE222G(222位のグルタミン酸の代わりにグリシン)の置換を有する。変異体G2は、2回目のランダム変異誘発を使用して変異体G1に基づいて作製された。G1と比較して、G2変異体タンパク質は、輝度およびタンパク質折りたたみ速度の改善された特徴を有する。興味深いことに、G2は、増強型GFP(EGFP)についてもまた特徴づけられている置換F64Lを含有する(Cormack等、Gene、173:33〜38(1996);Yang等、Nuc.Acids Res.、24:4592〜4593(1996))。G2のスペクトル特性が下記の表1に示される。G2に対する励起−発光スペクトルが図8に示される。
【0137】
変異体G22は、図9に示されるように、V11I(11位のバリンの代わりにイソロイシン)、F64L(64位のフェニルアラニンの代わりにロイシン)、K101E(101位のリシンの代わりにグルタミン酸)、T206A(206位のトレオニンの代わりにアラニン)およびE222G(222位のグルタミン酸の代わりにグリシン)の置換を有する。変異体G22は、3回目のランダム変異誘発を使用して変異体G2から作製された。G2と比較して、G22は、更に大きく改善された輝度を有する。G22のスペクトル特性が表1に報告される。G22に対する励起−発光スペクトルが図10に示される。50,000M−1cm−1の吸光係数および0.55の量子収量により、このタンパク質は、広く使用されている増強型GFPとほぼ同じくらい明るくなっている。非加熱の変異体G22タンパク質のゲルろ過試験ならびにSDS−PAGEは、G22が単量体であることを明らかにした。図11Bでは、加熱処理(レーン1〜3)対非加熱(レーン4〜6)のタンパク質サンプルの移動度が比較される。レーン1およびレーン4はA.victoriaのGFPであり、レーン2およびレーン5はG22変異体であり、レーン3およびレーン6はG22−G222E変異体である。クーマシーブルー染色が図11bに示され、UV光のもとでの非加熱タンパク質の蛍光が図11cに示される。
【0138】
上述のacGFPの蛍光性変異体のすべてが、470nm〜480nmおよび500nm〜510nmをそれぞれピークとした、類似した励起スペクトルおよび発光スペクトルを有する。それらの励起スペクトルの形状は、野生型A.victoria GFPではなく、増強型GFPの励起スペクトル形状と類似している。acGFP変異体の蛍光団は、増強型GFPについて示されているように、常に脱プロトン化形態であると考えられる。考えられる説明は、Glu222の非存在であり、このことはプロトン移動のために重要であると考えられ得る(Ehrig等、FEBS Lett.、367:163〜167(1995))。
【0139】
蛍光に対するE222G置換の重要性を明らかにするために、逆のG222E置換を変異体G22(V11I、F64L、K101E、T206Aの置換)に対して行った。この逆変異体G22−E222Gの核酸配列およびアミノ酸配列が図12に示される。この逆変異はG22−E222G変異体タンパク質を無色の状態に容易に変換した。G22−G222Eを発現する大腸菌コロニーは着色または検出可能な蛍光のいずれも示さなかった。
【0140】
G22−G222Eは、問題なく、37℃で大腸菌において発現し、折りたたまれるが、これは可溶性組換えタンパク質の収量が大きいこと(総タンパク質の約20%)によって明白に示される。精製されたG22−G222Eは、280nmでの主ピークおよび390nmでの副ピークを有する吸収スペクトルを示した(図13a参照)。アルカリ変性処理されたG22−G222Eタンパク質は、GFP発色団のアニオン型に明らかに対応する吸収ピークを446nmに示した。発色団について44,000M−1cm−1の吸光係数を仮定すると、390nmにおける未変性G22−G222Eの吸光は33,000M−1cm−1であると推定された。280nmピークと390nmピークとの測定された比(280nmにおけるモル吸光係数は23,500M−1cm−1であると計算された)は、可溶性G22−G222Eの約3%のみが成熟形態で存在することを示した。390nmでの励起は、0.07の量子収量で460nmおよび505nmにピークを有する弱い二色蛍光をもたらした(図13b参照)。
【0141】
GFP様タンパク質は、タンパク質サンプルがゲルへの負荷前に加熱されない限り、そのスペクトル特性およびオリゴマー化状態を一般的なSDS−PAGEの条件のもとで保持することが広く知られている。この試験が、G22−G222Eの折りたたみ状態を調べるために使用された。ゲル電気泳動により、明瞭な違いが非変性タンパク質および変性タンパク質の移動度において明らかにされた(図11参照)。更に、非加熱G22−G222Eタンパク質のバンドはUV光のもとで非常に弱い蛍光をもたらした(再度、図11参照)。このタンパク質の約97%が非吸収性形態で存在するので、これらの結果は、この非吸収性形態の立体配座が、変性状態にではなく、未変性状態に近いことを示している。
【0142】
これらの結果により勢いづけられ、組換え野生型acGFPを得て、特徴づける試みを繰り返した。acGFPを発現する大腸菌を、誘導することなく、室温で成長させ、その後、4℃で数日間インキュベーションすることにより、可溶性acGFPの小さい画分の出現がもたらされた(総acGFPの約5%)。可溶性の野生型acGFPに対する吸収スペクトルおよび蛍光スペクトルの形状は、G22−G222Eのスペクトル形状と非常に類似していた。G22−G222E変異体は、大腸菌で発現させられたとき、天然型acGFPの性質を反映し、しかし、改善されたタンパク質折りたたみおよび温度安定性を有すると結論された。
【0143】
これらのデータは、可溶性G22−G222Eが、野生型acGFPと同様に、2つの形態で存在することを示していた。これらのタンパク質の大部分は、スペクトル的に検出可能な発色団を有しない、折りたたまれたが、未熟な形態で存在する。少ない330nm吸収性形態は、中性状態ではGFP様発色団を含有し、弱い二色蛍光を有する。
【0144】
新規なタイプの光変換が変異体G22−G222Eで観測された。G22−G222Eタンパク質サンプルに250nm〜300nmのUV光を照射すると、480nmのピークが吸収/励起スペクトルに出現した。図14において、照射前のG22−G222Eの励起スペクトル(線1)、および250nm〜300nmの光をタンパク質サンプルに照射したことによる曲線のゆっくりした変化に留意すること。番号のない線は光変換後の発光スペクトルを表す(480nmでの励起)。これは、390nmの吸収ピークが、この光変換の間、低下しなかったので、タンパク質のスペクトル的に検出できない未成熟な形態から生じていると考えられる。480nmのピークにおける励起は、大きい量子収量(0.45)で、505nmでの緑色蛍光をもたらした。1000倍を超える、緑色蛍光強度のUV誘導増強が達成された(480nmでの励起)。
【0145】
変異体G22−G222E/Y220Lは、図15に示されるように、野生型scGFPと比較して、V11I(11位のバリンの代わりにイソロイシン)、F64L(64位のフェニルアラニンの代わりにロイシン)、V68A(68位のバリンの代わりにアラニン)、K101E(101位のリシンの代わりにグルタミン酸)、T206A(206位のトレオニンの代わりにアラニン)およびY220L(220位のチロシンの代わりにロイシン)の置換を有する。この変異体は37℃でタンパク質の折りたたみを示し、508nmでの明瞭な緑色蛍光を有した。G22−G222E/Y220Lに対する励起スペクトルは396nmでの主ピークおよび493nmでの副ピークを有した(10:1の比率)(励起スペクトルが点線であり、発光スペクトルが実線である図16を参照のこと)。この変異体のスペクトル特性は、Aequorea victoria由来の野生型GFPのスペクトル特性と非常に類似していた。
【0146】
変異体G22−G222E/Y220Lを基礎として使用したとき、V11I(11位のバリンの代わりにイソロイシン)、F64L(64位のフェニルアラニンの代わりにロイシン)、K101E(101位のリシンの代わりにグルタミン酸)、E115K(115位のグルタミン酸の代わりにリシン)、H148Q(148位のヒスチジンの代わりにグルタミン)、T206A(206位のトレオニンの代わりにアラニン)、Y220L(220位のチロシンの代わりにロイシン)、F221L(221位のフェニルアラニンの代わりにロイシン)およびK238Q(238位のリシンの代わりにグルタミン)の置換を有する変異体220−II−5が得られた。この変異体に対する核酸配列およびアミノ酸配列が図17に示される。この変異体は、主励起ピークを395nmに有するが、約480nmには励起ピークを有さず、512nmに発光ピークを有する(図18a参照)。長波長側の励起ピークの抑制は、発色団の荷電部分の消失を生じさせる置換H148Qによって説明できると考えられる。蛍光顕微鏡下で約400nmの光による比較的強い照射を数分間行った後、変異体G22−G222E/Y220Lの励起スペクトルが変化した。同時に、395nmのピークが低下し、480nmの励起ピークが出現した(図18b参照)。結果として、510nmでの発光の蛍光輝度における100倍を超えるコントラストを、400nmの波長の強い光を照射する前と、光を照射した後とを比較したとき、480nmの励起光で得ることができる。図25には、変異体220−11−5を発現する2つの大腸菌コロニーが蛍光顕微鏡下で示される。上部コロニーにおける2つの領域が、強い400nmの光によって予備的に光活性化された。従って、G22−G222E/Y220L変異体は、PattersonおよびLippincott−Schwartz、Science、13:1873〜1877(2002)におけるPA−GFP変異体の使用に関する最近発表された方法と類似する、生物を光標識するための光活性化蛍光マーカーとして使用することができる。
【0147】
シアン蛍光タンパク質変異体CFP−rand3は、V11I(11位のバリンの代わりにイソロイシン)、T62A(62位のトレオニンの代わりにアラニン)、F64L(64位のフェニルアラニンの代わりにロイシン)、K101E(101位のリシンの代わりにグルタミン酸)、N121S(121位のアスパラギンの代わりにセリン)、H148T(148位のヒスチジンの代わりにトレオニン)、E172K(172位のグルタミン酸の代わりにリシン)およびT206A(206位のトレオニンの代わりにアラニン)における置換を有する。CFP−rand3変異体のアミノ酸配列および核酸配列が図19に示される。この変異体は励起ピークを402nmに有し、1つだけの発光ピークを467nmに有する(図20参照)。
【0148】
別の変異体シアン蛍光タンパク質CFP−3が作製されたが、これは、V11I(11位のバリンの代わりにイソロイシン)、F64L(64位のフェニルアラニンの代わりにロイシン)、K101E(101位のリシンの代わりにグルタミン酸)、H148S(148位のヒスチジンの代わりにセリン)、F165L(165位のフェニルアラニンの代わりにロイシン)、E172A(172位のグルタミン酸の代わりにアラニン)およびT206A(206位のトレオニンの代わりにアラニン)の置換を有する。この変異体に対するアミノ酸配列および核酸配列が図21に示される。この変異体は励起ピークを390nmに有し、1つだけの発光ピークを470nmに有する(図22a)。400nmでの光による比較的強い照射を数分間行った後、励起スペクトルおよび発光スペクトルの下記の変化が観測された:(i)390nmの励起ピークのかなり大きい低下、および(ii)505nmでの発光を伴う480nmの励起ピークの出現(図22b参照)。
【0149】
結果として、505nmでの発光の蛍光輝度における30倍を超えるコントラストが、405nmでの強い照射の前後でスペクトルを比較したとき、480nmでの励起光を使用して得ることができる。励起パラメーターおよび発光パラメーターの同時変化により、シアン変異体CFP−3は、強い400nmの照射に応答して緑色蛍光タンパク質に変換させられる。従って、CFP−3変異体は、生物を光標識するための、光活性化または「光スイッチ」の蛍光性マーカーとして使用することができる。
【0150】
1つの例示的な実験において、G22変異体が、哺乳動物細胞におけるタンパク質発現を調べるための蛍光性タグとして使用された。しかしながら、予想外ではあったが、変異体G22はヒト細胞株において非常に弱い蛍光シグナルをもたらした。これは、最適でないコドン使用頻度またはG22変異体遺伝子における潜在性イントロンの存在のいずれかによって説明され得ると考えられる。これらの問題の両方を克服するために、哺乳動物に最適化されたコドン使用頻度を取り込むG22−h変異体遺伝子を合成した。このヒト化変異体に対するアミノ酸配列および核酸配列が図23に示される。種々の細胞株におけるG22−hの一過性発現は、凝集を伴わない明るい緑色シグナルを示した。
【0151】
図24には、種々の哺乳動物細胞株におけるG22−h変異体(パネルA〜E)およびG22−β−アクチン融合タンパク質(パネルF)の一過性発現が示される。パネルA−293T;パネルB−vero;パネルC−3T3;パネルD−L929;パネルE−COS1;パネルF−3T3。蛍光がトランスフェクションの24時間後に明瞭に検出された。毒性は観測されなかった。タンパク質を標識するG22−hの能力が、細胞質β−アクチンとの融合タンパク質を構築することによって明らかにされた。3T3細胞におけるこの融合体の一過性発現は、明るい正確なアクチン標識化を示した(図24f参照)。ストレスファイバー、限局的接着域および細胞集団を明瞭に認めることができた。細胞の接着性または生存生に対して有害であることは観測されず、また、何らかの非特異的なタンパク質凝集は観測されなかった。
【0152】
方法
【0153】
トータルRNAを、傘縁および放射状導管を含むAequorea coerulescens生物の小さい生体解剖物から、NucleoSpin RNA IIキット(Clontech)を使用して単離した。cDNAが、SMART PCR cDNA合成キット(Clontech)を用いて合成され、また増幅された。新規な蛍光タンパク質遺伝子の断片が、縮重プライマーを用いたPCRによって得られた。ステップアウトPCR RACE法を使用して、標的cDNAの5’端断片をクローン化した。新規な蛍光タンパク質acGFPをコードするcDNAのヌクレオチド配列はアクセション番号AY151052でGenBankに提出されている。acGFPの細菌発現の場合、全長の翻訳領域が、特異的なプライマーを使用して増幅され、pQE30ベクター(Qiagen)にクローン化された。
【0154】
ダイバーシティPCRランダム変異誘発キット(CLONTECH)が、1000塩基対あたり5つ〜6つの変異のために最適な条件で、acGFPのランダム変異誘発のために使用された。変異体タンパク質を発現する大腸菌コロニーは、蛍光実体顕微鏡SZX−12(Olympus)を用いて目視によりスクリーニングされた。最も明るい変異体が選択され、更なるランダム変異誘発処理に供された。部位特異的変異誘発が、適切な標的置換を含有するプライマーを用いて重複伸張法を使用するPCRによって行われた(例えば、Ho等、Gene、77:51〜59(1989)を参照のこと)。
【0155】
N末端の6ヒスチジンタグに融合されたタンパク質を大腸菌XL1 blue株(Invitrogen)において発現させ、TALON金属アフィニティー樹脂(Clontech)を使用して精製した。吸収スペクトルはBeckman DU520UV/VIS分光光度計により記録された。Varian Cary Eclipse蛍光分光光度計が、励起−発光スペクトルを測定するために使用された。
【0156】
モル吸光係数を測定する場合、成熟発色団の濃度の推定が使用された。タンパク質を等量の2M NaOHでアルカリ変性した。これらの条件のもとで、A.victoriaのGFP発色団は446nmで吸収し、そのモル吸光係数は44,000M−1cm−1に等しい(Ward等、Photochem.Photobiol.、31:611〜615(1980))。未変性タンパク質およびアルカリ変性タンパク質に対する吸収スペクトルを測定した。未変性状態に対するモル吸光係数を、基礎としての変性タンパク質の吸収を使用して推定した。量子収量を測定する場合、変異体の蛍光が増強型GFPの量子収量(0.60の蛍光収量)と比較された。
【0157】
acGFP−G222EのUV誘導による光変換を、Cary Eclipse蛍光分光光度計を使用して行った。タンパク質サンプルには、走査モードで250nm〜300nmの波長の光が数時間照射された(励起スリット:20nm、走査速度:30nm/分、平均化時間:1秒、サイクルモード)。
【0158】
精製されたタンパク質のサンプル(〜1mg/ml)をSephadex−100カラム(0.7×60cm)に負荷し、100mMのNaClを含む50mMリン酸塩緩衝液(pH7.0)で溶出した。EGFP、HcRed1およびDsRed2(Clontech)が、それぞれ、単量体、二量体および四量体の標準物として使用された。
【0159】
タンパク質のゲル分析の場合、加熱サンプルおよび非加熱サンプルを一般的な12%SDS−PAGEに負荷し、電気泳動を15mA/ゲルで行った。ウエスタンブロッティングの場合、タンパク質を、標準的な手法を使用してHydond Cメンブラン(Amersham)に転写した。メンブランを、GFPに対するマウス抗体(Clontech)(1:2500)でプローブし、その後、HRPコンジュゲート化抗マウス抗体(Amersham)で1:2500にてプローブした。染色パターンを発色させるために、ECLウエスタンブロティング分析システム(Amersham Pharmacia Biotech)が使用された。これには、検出試薬1および2、そしてHyperfilm ECLが含まれる。
【0160】
真核生物における発現の場合、acGFPが、AgeIおよびBglIIの制限部位の間で(EGFP翻訳領域の代わりに)、pEGFP−C1およびpEGFP−Actinの両ベクター(CLONTECH)にクローン化された。下記の細胞株が使用された:ヒト腎臓上皮細胞293T、マウス胚繊維芽細胞3T3、マウス皮下繊維芽細胞L929、アフリカミドリサル腎臓上皮細胞Vero、およびアフリカミドリサル腎臓繊維芽細胞COS1。細胞は、LipofectAMINE試薬(Invitrogen)を用いてトランスフェクションされ、トランスフェクションの20時間後に試験された。CCDカメラDP−50(Olympus)を備えたOlympus CK40蛍光顕微鏡が細胞画像化のために使用された。
【0161】
【表1】

【0162】
本明細書に引用されたすべての刊行物および特許明細書は、それぞれの個々の刊行物または特許明細書が、参考として組み入れられるために具体的かつ個々に示されていたかのように、参考として本明細書中に組み入れる。何らかの刊行物の引用は、出願日前におけるこの開示のためであり、本発明は、先行発明によるそのような公開がより以前であったとする権利を有しないという承認として解釈してはならない。
【0163】
前述の発明は理解の明確化のための例示および例として幾分詳しく記載されているが、ある種の様々な変化および改変が、添付された請求項の精神または範囲から逸脱することなく本発明に対してなされ得ることが、本発明の教示に照らして当業者には容易に明らかである。
【産業上の利用可能性】
【0164】
Aequorea coerulescens由来の無色のGFP様タンパク質であるacGFP、その蛍光性誘導体および非蛍光性誘導体、またはそれらの変異体をコードする核酸組成物、そしてまた、これらの核酸組成物によってコードされるタンパク質およびペプチドが提供される。注目されるタンパク質は、有色であり、かつ/または蛍光性であり、かつ/または光活性化され得るタンパク質であり、この場合、色、蛍光または光活性化の特徴は、タンパク質の2つ以上のアミノ酸残基の相互作用から生じている。また、上記の特定のタンパク質に実質的に類似するタンパク質、その誘導体タンパク質またはその変異体タンパク質も注目される。また、核酸断片、およびそれによってコードされるペプチドが提供され、また、本発明のタンパク質およびペプチドに対する抗体が提供される。更に、遺伝子組換え細胞および遺伝子組換え生物が提供される。本発明のタンパク質および核酸組成物は、様々な異なる応用および方法における用途があり、特に、タンパク質標識への応用がある。最後に、そのような方法および適用において使用されるキットが提供される。
【技術分野】
【0001】
本発明は、蛍光タンパク質に関する。
【背景技術】
【0002】
標識化は、関心のあるタンパク質、細胞または生物に印をつけるためのツールであり、生化学的な応用、分子生物学的な応用および医学的診断への応用の多くで突出した役割を果たしている。様々な異なる標識がこの分野で開発および使用されており、これらには、様々な性質および最適な用途を有する放射性標識、発色性標識、蛍光性標識、化学発光性標識などが含まれる。しかしながら、新しい標識の開発が引き続き注目されている。特に注目されるものは、蛍光タンパク質標識を含む新しいタンパク質標識の開発である。
【0003】
注目される米国特許には、米国特許第6,066,476号、同第6,020,192号、同第5,985,577号、同第5,976,796号、同第5,968,750号、同第5,968,738号、同第5,958,713号、同第5,919,445号、同第5,874,304号および同第5,491,084号が含まれる。注目される国際特許出願公開には、WO00/46233、WO99/49019、およびドイツ国特許出願公開DE19718640Aが含まれる。また、Anderluh等、Biochemical and Biophysical Research Communications(1996)、220:437〜442;Dove等、Biological Bulletin(1995)、189:288〜297;Fradkov等、FEBS Lett.(2000)、479(3):127〜30;Gurskaya等、FEBS Lett.(2001)、507(1):16〜20;Gurskaya等、BMC Biochem.(2001)、2:6;Lukyanov,K.等(2000)、J.Biol.Chem.、275(34):25879〜25882;Macek等、Eur.J.Biochem.(1995)、234:329〜335;Martynov等、J.Biol.Chem.(2001)、276:21012〜6;Matz,M.V.等(1999)、Nature Biotechnol.、17:969〜973;Tershikh等、Science(2000)、290:1585〜8;Tsien、Annual Rev.of Biochemistry(1998)、67:509〜544;Tsien、Nat.Biotech.(1999)、17:956〜957;Ward等、J.Biol.Chem.(1979)、254:781〜788;Wiedermann等、Jarhrestagung der Deutschen Gesellschact fur Tropenokologie−gto.Ulm.、1999年2月17日〜19日、ポスターP−4.20;Yarbrough等、Proceedings of the National Academy of Sciences(2001)、98:462〜7も注目される。
【発明の概要】
【発明が解決しようとする課題】
【0004】
本発明は、Aequorea coerulescens由来の他に類のない無色のタンパク質ならびにその蛍光性変異体および非蛍光性変異体をコードする核酸組成物を提供し、また、そのような核酸によってコードされるタンパク質およびペプチドも提供する。本発明のタンパク質は有色であり、かつ/または蛍光性であり、かつ/または光活性化が可能であり、この場合、この光学的特徴はタンパク質の2つ以上のアミノ酸残基の相互作用から生じている。また、本発明のペプチドが組み込まれた融合タンパク質を含む上記の特定のタンパク質に実質的に類似するタンパク質やその誘導体または変異体に注目され、また、これらのタンパク質に対する抗体も注目される。本発明のタンパク質および核酸組成物は様々な異なる応用への用途がある。最後に、本発明は、標識化適用において使用されるキットを提供する。
【課題を解決するための手段】
【0005】
本発明は、蛍光タンパク質acGFP、その変異体またはその誘導体をコードする核酸を提供する。ある実施形態においては、核酸は単離されているか、操作されているか、またはその天然の環境とは異なる環境に存在する。ある実施形態においては、核酸は、配列番号1、3、5、7、9、11、13、15、17、19、21または23に由来する少なくとも10残基の長さのヌクレオチド配列と実質的に同じであるか、または同一である残基の配列を有する。
【0006】
ある実施形態においては、本発明の核酸は、配列番号1、3、5、7、9、11、13、15、17、19、21または23の配列と少なくとも約60%の配列類似性を有し、長さが少なくとも10残基である。ある実施形態においては、本発明の核酸は、Aequorea coerulescensの蛍光タンパク質の変異体タンパク質または誘導体タンパク質をコードする、配列番号2、4、6、8、10、12、14、16、18、20、22または24からなる群から選択されるアミノ酸配列を有するタンパク質をコードする。
【0007】
本発明の核酸の断片もまた提供される。更に、ストリンジェントな条件のもとで本発明の核酸にハイブリダイゼーションする核酸またはそのミメティック(mimetic)が提供される。ベクターおよび本発明の核酸を含む構築物もまた提供される。更に、本発明は発現カセットを提供する。ここで、発現カセットは、発現宿主において機能し得る転写開始領域と、本発明の核酸と、細胞内への発現カセットの導入の結果、染色体外エレメントの一部としての発現カセットにおいて、または細胞のゲノムに組み込まれる発現カセットにおいて機能し得る転写終結領域とを含む。
【0008】
色素タンパク質および/または蛍光タンパク質を製造する方法もまた提供され、この方法は、本発明の細胞を増殖させること、上記タンパク質を細胞において発現させること、および他のタンパク質を実質的に含まない上記タンパク質を単離することを含む。
【0009】
更に、本発明の核酸によってコードされるタンパク質、断片またはペプチドが提供され、また、本発明のタンパク質またはペプチドに対して特異的に結合する抗体も提供される。
【0010】
更に、本発明の核酸を含む遺伝子組換え細胞(またはその子孫)が提供され、また、本発明の核酸を含む遺伝子組換え生物も提供される。
【0011】
また、本発明の色素タンパク質または蛍光タンパク質を用いる方法、あるいは、本発明の色素タンパク質または蛍光タンパク質をコードする核酸を用いる方法が提供される。
【0012】
更に、本発明の核酸またはタンパク質と、その使用説明書とを含むキットが提供される。
【0013】
本発明に従って、この分野の技術に含まれる、従来の、分子生物学、微生物学および組換えDNA技術を用いることができる。そのような技術は文献に詳しく説明されている。例えば、Maniatis、Fritsch&Sambrook、Molecular Cloning:A Laboratory Manual(1982年);DNA Cloning:A Practical Approach、第1巻および第2巻(D.N.Glover編、1985年);Oligonucleotide Synthesis(M.J.Gait編、1984年);Nucleic Acid Hybridization(B.D.Hames&S.J.Higgins編(1984年));Animal Cell Culture(R.I.Freshney編(1986年));Immobilized Cells and Enzymes(IRL Press(1986年));およびB.Perbal、A Practical Guide to Molecular Cloning(1984年)を参照のこと。
【0014】
「ベクター」は、別のDNAセグメントを結合させることができる、プラスミド、ファージまたはコスミドなどのレプリコンである。
【0015】
「DNA分子」は、デオキシリボヌクレオチド(アデニン、グアニン、チミンまたはシトシン)のポリマー形態を示し、一本鎖形態または二本鎖らせんのどちらでもよい。この用語は、分子の一次構造および二次構造を示すだけであり、分子を何らかの特定の三次構造に限定しない。従って、「DNA分子」には、とりわけ、線状DNA分子(例えば、制限酵素断片)、ウイルス、プラスミドおよび染色体に見出される二本鎖DNAが含まれる。
【0016】
DNAの「コード配列」は、適切な制御配列の制御下に置かれたとき、インビボで転写され、ポリペプチドに翻訳されるDNA配列である。コード配列の境界は、5’(アミノ)末端における開始コドンと、3’(カルボキシル)末端における翻訳終止コドンとによって決定される。コード配列には、原核生物配列、真核生物mRNA由来のcDNA、真核生物(例えば、哺乳動物)DNA由来のゲノムDNA配列、および合成DNA配列が含まれ得るが、これらに限定されない。ポリアデニル化シグナルおよび転写終結配列がコード配列の3’に存在し得る。
【0017】
本明細書中で使用される用語「ハイブリダイゼーション」は、2つの核酸鎖が会合して、対になる核酸鎖の残基間の水素結合によって安定化される反平行の二重鎖を形成するプロセスを示す。
【0018】
用語「オリゴヌクレオチド」は、短い(100塩基未満の長さの)核酸分子を示す。
【0019】
本明細書中で使用される「DNA制御配列」は、宿主細胞におけるコード配列の発現を規定および/または調節する転写制御配列および翻訳制御配列(例えば、プロモーター、エンハンサー、ポリアデニル化シグナルおよびターミネーターなど)である。
【0020】
「プロモーター配列」は、細胞内のRNAポリメラーゼと結合し、コード配列の転写を開始させることができるDNA制御領域である。例えば、プロモーター配列は、転写開始部位をその3’末端での境界とすることができ、そして、上流(5’方向)に広がり、バックグラウンドを上回る検出可能なレベルで転写を開始させるために必要な最小数の塩基またはエレメントを含むことができる。プロモーター配列内には、転写開始部位、ならびに、RNAポリメラーゼの結合に関わるタンパク質結合ドメインが見出され得る。真核生物のプロモーターは、常にというわけではないが、多くの場合、「TATA」ボックスおよび「CAT」ボックスを含有する。誘導性プロモーターを含む様々なプロモーターを、本発明の様々なベクターを駆動させるために使用することができる。
【0021】
本明細書中で使用される用語「制限エンドヌクレアーゼ」および用語「制限酵素」は、特定のヌクレオチド配列でまたはその近くで二本鎖DNAを切断する細菌酵素を示す。
【0022】
細胞は、外因性または異種のDNAが細胞の内部に導入されているとき、外因性または異種のDNAによって「形質転換」または「トランスフェクション(形質移入)」されている。形質転換DNAは細胞のゲノムに組み込まれ(共有結合的に連結され)てもよく、または組み込まれなくてもよい。例えば、原核生物、酵母細胞および哺乳動物細胞では、形質転換DNAは、プラスミドなどのエピソームエレメントに維持され得る。真核細胞に関して、安定に形質転換された細胞は、形質転換DNAが宿主細胞の染色体に組み込まれているか、または染色体外に維持され、その結果、形質転換DNAが細胞複製のときに娘細胞によって受け継がれる細胞である。そのような安定に形質転換された真核生物細胞は、形質転換DNAを含有する娘細胞の集団から構成される細胞株またはクローンを樹立することができる。「クローン」は、単一細胞または共通の祖先から有糸分裂によって得られる細胞の集団である。「細胞株」は、多世代にわたってインビトロで安定に成長可能なクローンまたは細胞である。
【0023】
DNA構築物の「異種」領域は、より大きなDNA分子における同定可能なDNAセグメントであって、そのより大きな分子に関連して天然に見出されないDNAセグメントであり、例えば、非哺乳類生物のゲノムにおいて哺乳類のゲノムDNAを異種領域がコードするときのDNAセグメントである。別の例では、異種DNAには、2つの異なる供給源に由来する遺伝子の一部が一緒になり、その結果、融合タンパク質産物が産生されるようになっている構築物におけるコード配列が含まれる。
【0024】
本明細書中で使用される用語「レポーター遺伝子」は、異種のプロモーターエレメントまたはエンハンサーエレメントに結合されたコード配列で、構築物が組織または細胞に導入されたとき、その産物を容易かつ定量的にアッセイすることができるコード配列を示す。
【0025】
本明細書中に記載されるアミノ酸は「L」型であることが好ましい。アミノ酸配列は一文字記号で示される(A:アラニン、C:システイン、D:アスパラギン酸、E:グルタミン酸、F:フェニルアラニン、G:グリシン、H:ヒスチジン、I:イソロイシン、K:リシン、L:ロイシン、M:メチオニン、N:アスパラギン、P:プロリン、Q:グルタミン、R:アルギニン、S:セリン、T:トレオニン、V:バリン、W:トリプトファン、Y:チロシン、X:任意の残基)。NH2は、ポリペプチドのアミノ末端に存在する遊離型アミノ基を示す。COOHは、ポリペプチドのカルボキシ末端に存在する遊離型カルボキシ基を示す。標準的なポリペプチド命名法(J.Biol.Chem.、243、3552〜59(1969)を参照のこと)が使用されている。
【0026】
用語「免疫学的に活性な」によって、天然型、組換え型または合成型の色素タンパク質もしくは蛍光タンパク質またはそれらの任意のオリゴペプチドが特異的な免疫応答を適切な動物または細胞において誘導し、特異的な抗体と結合する能力が規定される。本明細書中で使用される「抗原性アミノ酸配列」は、単独でまたはキャリア分子と一緒になって、哺乳動物において抗体応答を誘発することができるアミノ鎖配列を意味する。抗体が抗原に結合するという状況における、用語「特異的に結合する」は、この分野で広く理解されている用語であり、ある抗体がその抗体を産生させた抗原には結合するが、関連性のない他の抗原には結合しないことを示す。
【0027】
本明細書中で使用される用語「単離された」は、ポリヌクレオチド、ポリペプチド、抗体、または宿主細胞が天然に存在する環境とは異なる環境にあるポリヌクレオチド、ポリペプチド、抗体または宿主細胞を記載することが意味される。
【0028】
生物発光は、暗所で見ることができる、生物による光の放射である(例えば、Harvey、Bioluminescence、New York:Academic Press(1952);Cell Physiology(Speralakis編)のHastings、「生物発光」の項、New York、Academic Press、651頁〜81頁(1995);WilsonおよびHastings、「生物発光」、Annu.Rev.Cell.Dev.Biol.、14、197頁〜230頁(1998)を参照のこと)。生物発光には、高感度な光度測定装置を使用して事実上すべての生体構造体において検出され得るいわゆる超微弱な光放射は含まれず(MurphyおよびSies、「生物学システムにおける可視範囲の低レベル化学発光」、Meth.Enzymol.、186、595頁〜610頁(1990);Radotic等、「植物における自発的な超微弱な生物発光:起源、機構および性質」、Gen.Physiol.Biophys.、17、289頁〜308頁(1998))、また、生物発光は、例えば、タケの成長円錐の発光など、生態学的役割をおそらくは果たしていない弱い光放射から生じるものではなく(Totsune等、「筍切断体からの化学発光」、Biochem.Biophys.Res.Comm.、194、1025頁〜1029頁(1993))、動物の卵の受精時における光放射から生じるものでもない(Klebanoff等、「受精と食作用との間における代謝的類似性」、J.Exp.Med.、149、938頁〜53頁(1979);SchomerおよびEpel、「海洋無脊椎動物の卵の受精および成熟化の際の酸化還元変化」、Dev.Biol.、2003、1頁〜11頁(1998))。
【0029】
本明細書中で使用される用語「GFP様タンパク質」は、Aequorea victoria由来の緑色蛍光タンパク質(GFP)に類似するタンパク質を記載することが意味される。
【発明の効果】
【0030】
Aequorea coerulescens由来の無色のGFP様タンパク質であるacGFP、その蛍光性誘導体および非蛍光性誘導体、またはそれらの変異体をコードする核酸組成物、そしてまた、これらの核酸組成物によってコードされるタンパク質およびペプチドが提供される。注目されるタンパク質は、有色であり、かつ/または蛍光性であり、かつ/または光活性化され得るタンパク質であり、この場合、色、蛍光または光活性化の特徴は、タンパク質の2つ以上のアミノ酸残基の相互作用から生じている。また、上記の特定のタンパク質に実質的に類似するタンパク質、その誘導体タンパク質またはその変異体タンパク質も注目される。また、核酸断片、およびそれによってコードされるペプチドが提供され、また、本発明のタンパク質およびペプチドに対する抗体が提供される。更に、遺伝子組換え細胞および遺伝子組換え生物が提供される。本発明のタンパク質および核酸組成物は、様々な異なる応用および方法における用途があり、特に、タンパク質標識への応用がある。最後に、そのような方法および適用において使用されるキットが提供される。
【図面の簡単な説明】
【0031】
【図1】本明細書中でacGFPと表す、Aequorea coerulescens由来の野生型GFP様タンパク質をコードするアミノ酸配列および核酸配列である。
【図2】Aequorea victoria GFPおよびAequorea coerulescenssc GFPのアミノ酸配列の比較である。
【図3】acGFP変異体Z1をコードするアミノ酸配列および核酸配列である。
【図4】変異体Z1の励起−発光スペクトルである。
【図5】acGFP変異体Z2をコードするアミノ酸配列および核酸配列である。
【図6】acGFP変異体G1をコードするアミノ酸配列および核酸配列である。
【図7】acGFP変異体G2をコードするアミノ酸配列および核酸配列である。
【図8】変異体G2の励起−発光スペクトルである。
【図9】acGFP変異体G22をコードするアミノ酸配列および核酸配列である。
【図10】変異体G22の励起−発光スペクトルである。
【図11a】A.victoriaのGFPに対する抗体を使用した、A.coerulescensから得られた可溶性タンパク質抽出物のウエスタンブロット分析である。レーン1は、精製された組換えA.victoria GFPであり、レーン2はA.coerulescens抽出物である。
【図11b】加熱処理(レーン1〜3)対非加熱(レーン4〜6)のタンパク質サンプルの移動度が比較される。レーン1およびレーン4はA.victoriaのGFPであり、レーン2およびレーン5はG22変異体であり、レーン3およびレーン6はG22−G222E変異体である。クーマシーブルー染色が示される。
【図11c】UV光のもとでの非加熱タンパク質の蛍光が示される。
【図12】acGFP変異体G22−G222Eをコードするアミノ酸配列および核酸配列である。
【図13a】変異体G22−G222Eの吸収スペクトルである。
【図13b】変異体G22−G222Eの励起−発光スペクトルである。
【図14】G22−G222EのUV誘導の光変換に対するスペクトルを示す。
【図15】acGFP変異体G22−G222E/Y220Lをコードするアミノ酸配列および核酸配列である。
【図16】変異体G22−G222E/Y220Lの励起−発光スペクトルである。
【図17】acGFP変異体220−II−5をコードするアミノ酸配列および核酸配列である。
【図18a】変異体220−II−5のスペクトル特性を示す。
【図18b】変異体220−II−5のスペクトル特性を示す。
【図19】acGFP変異体CFP−rand3をコードするアミノ酸配列および核酸配列である。
【図20】変異体CFP−rand3に対する励起−発光スペクトルである。
【図21】acGFP変異体CFP−3をコードするアミノ酸配列および核酸配列である。
【図22a】変異体CFP−3のスペクトル特性を示す。
【図22b】変異体CFP−3のスペクトル特性を示す。
【図23】変異体G22のヒト化体のアミノ酸配列および核酸配列である。
【図24a】G22−h変異体(293T)を発現する哺乳動物細胞の顕微鏡写真を示す。
【図24b】G22−h変異体(vero)を発現する哺乳動物細胞の顕微鏡写真を示す。
【図24c】G22−h変異体(3T3)を発現する哺乳動物細胞の顕微鏡写真を示す。
【図24d】G22−h変異体(L929)を発現する哺乳動物細胞の顕微鏡写真を示す。
【図24e】G22−h変異体(COS1)を発現する哺乳動物細胞の顕微鏡写真を示す。
【図24f】G22−β−アクチン融合タンパク質(3T3)を発現する哺乳動物細胞の顕微鏡写真を示す。
【図25】大腸菌コロニーにおける変異体220−11−5の光活性化を示す。
【発明を実施するための形態】
【0032】
本発明を更に記載する前に、本発明は、下記に記載される本発明の特定の実施形態に限定されないことを理解しなければならない。これは、そのような特定の実施形態の様々な変化を行うことができ、そして、そのような変化は依然として本発明の範囲に含まれるからである。用いられている用語は、特定の実施形態を記載するためであり、限定的であることを意図しないこともまた理解しなければならない。
【0033】
本明細書において、単数形態(“a”、“an”および“the”)は、文脈により、別のことが明らかに示されない限り、複数である参照体を含む。別途定義されない限り、本明細書中で使用されている技術的用語および科学的用語はすべて、本発明が属する分野の当業者に一般に理解されているのと同じ意味を有する。本明細書中に記載されているものと類似している、または同等である、方法、デバイスおよび材料はどれも、本発明の実施または試験において使用することができるが、好ましい方法、デバイスおよび材料が下記に記載される。本明細書中で言及される刊行物はすべて、本発明を記載および開示する目的のために参考として本明細書中に組み入れる。
【0034】
本発明を記載するにあたり、核酸組成物が最初に記載し、その後、タンパク質組成物、抗体組成物、そして、遺伝子組換え細胞および遺伝子組換え生物を議論する。次に、本発明のタンパク質が使用される例示的な方法を概説する。
【0035】
核酸組成物
【0036】
上で要約したように、本発明は、Aequorea coerulescens由来の無色のタンパク質であるacGFPをコードする核酸組成物、または、acGFPの蛍光性および非蛍光性の変異体または誘導体をコードする核酸組成物を提供し、それらの核酸組成物の断片およびホモログも提供する。「蛍光タンパク質」という用語は、蛍光性であるタンパク質を意味する。例えば、蛍光タンパク質は、適切な励起波長の光で照射されたとき、弱い、中程度の、または強い蛍光を示すことができる。本発明のタンパク質は、蛍光特性が、1つだけのアミノ酸残基からではなく、タンパク質の2つ以上のアミノ酸残基の相互作用から生じる特性であるタンパク質である。そのため、本発明の蛍光タンパク質には、固有的な蛍光体として単独で作用する残基(すなわち、トリプトファン、チロシンおよびフェニルアラニン)のみに由来する蛍光を示すタンパク質は含まれない。従って、本発明の蛍光タンパク質は、上記に示された単一アミノ酸残基とは異なるタンパク質内の何らかの構造からその蛍光が生じる蛍光タンパク質である。例えば、蛍光は2つ以上のアミノ酸残基の相互作用から生じている。
【0037】
本発明の核酸組成物の1つは、本発明のポリペプチドをコードするオープンリーディングフレーム(翻訳領域)を有するDNA配列を含む組成物(すなわち、蛍光タンパク質遺伝子)である。そのような核酸組成物は、適切な条件のもとで蛍光タンパク質として発現させることができる。核酸組成物の用語にはまた、本発明の核酸に対して相同的な核酸、実質的に類似する核酸、もしくは同一である核酸、または本発明の核酸のミメティックが包含される。本発明の核酸は、その天然の環境とは異なる環境に存在する。例えば、本発明の核酸は単離されているか、濃縮された量で存在するか、あるいは、インビトロにおいてまたは天然に存在する環境とは異なる細胞もしくは生物において存在するか、または発現させられる。
【0038】
本発明の別の実施形態において、核酸は、配列番号1、3、5、7、9、11、13、15、17、19、21または23によりコードされ得るか、そのような核酸に由来する核酸であるか、またはそのような核酸のホモログである。
【0039】
上記の特定の核酸組成物に加えて、上記配列のホモログもまた注目される。本発明の核酸のホモログに関して、相同的な遺伝子の供給源は植物または動物の任意の種であり得るか、あるいは、配列は核酸ミメティックを取り込む配列を含む、全体または一部が合成されたものであり得る。ある実施形態において、ホモログ間の配列類似性は少なくとも約40%であり、50%、60%、70%またはそれ以上であってもよく、これには、75%、80%、85%、90%および95%またはそれ以上が含まれる。配列類似性は、より大きな配列のサブセット(例えば、保存モチーフ、翻訳領域、隣接領域など)であり得る基準配列に基づいて計算される。基準配列は、通常、長さが少なくとも約18ヌクレオチドであり、より一般的には、長さが少なくとも約30ヌクレオチドであり、また、比較されている完全な配列にまで及び得る。配列解析のための様々なアルゴリズムがこの分野では知られており、例えば、Altschul等、J.Mol.Biol.、215、403頁〜10頁(1990)に記載されるBLAST(例えば、デフォルト条件、すなわち、w=4およびT=17のパラメーターを使用する)などがある。
【0040】
ホモログが多数の方法のいずれかによって同定される。本発明のcDNAのフラグメントは、目的とする標的生物から得られたcDNAライブラリーに対するハイブリダイゼーションプローブとして使用することができ、この場合には、低いストリンジェンシーの条件が使用される。プローブは、大きいフラグメントであり得るか、または1つ以上の短い縮重プライマーであり得る。配列類似性を有する核酸が、低いストリンジェンシーの条件のもとでのハイブリダイゼーションによって、例えば、50℃および6×SSC(0.9M塩化ナトリウム/0.09Mクエン酸ナトリウム)におけるハイブリダイゼーションによって検出され、そして、1×SSC(0.15M塩化ナトリウム/0.015Mクエン酸ナトリウム)における55℃での洗浄に供されたとき、結合したままである。配列同一性を、ストリンジェントな条件のもとでのハイブリダイゼーションによって、例えば、50℃以上および0.1×SSC(15mM塩化ナトリウム/1.5mMクエン酸ナトリウム)におけるハイブリダイゼーションによって決定することができる。示された配列に対して実質的な同一性を有する領域を有する核酸、例えば、対立遺伝子変異体、遺伝子の遺伝的に変化した形態などは、ストリンジェントなハイブリダイゼーション条件のもとで、示された配列に結合する。プローブ(特に、DNA配列の標識されたプローブ)を使用することによって、ホモログまたは関連遺伝子を単離することができる。
【0041】
本発明の特定の実施形態において特に注目されるものは、配列番号1、3、5、7、9、11、13、15、17、19、21または23として同定された核酸と実質的に同じ長さの核酸である。この場合、「実質的に同じ長さ」とは、長さの差が約20%を超えず、一般的には約10%を超えず、より一般的には約5%を超えないことを意味する。好ましい実施形態において、実質的に同じ長さのヌクレオチドは、配列番号1、3、5、7、9、11、13、15、17、19、21または23に対する配列同一性が、核酸の全長について、少なくとも約90%(例えば、少なくとも約92%、93%、94%)であり、通常は少なくとも約95%、96%、97%、または98%、または約99%でさえある。「実質的に類似する」とは、配列同一性が、一般には少なくとも約60%であり、通常は少なくとも約75%であり、多くの場合には、少なくとも約80%、85%、90%(例えば、92%、93%、94%)、または95%でさえあり、例えば、96%、98%、98%、99%または99.5%以上であるという意味である。
【0042】
更に、本発明には、先に記載された核酸によってコードされるタンパク質をコードするが、遺伝暗号の縮重性のために、先に記載された核酸とは配列が異なる核酸が含まれる。
【0043】
また、先に記載された核酸にストリンジェントな条件のもとでハイブリダイゼーションする核酸(すなわち、先に記載された核酸の相補体)も提供される。ストリンジェントなハイブリダイゼーション条件の例には、50℃以上および0.1×SSC(15mM塩化ナトリウム/1.5mMクエン酸ナトリウム)におけるハイブリダイゼーションがある。ストリンジェントなハイブリダイゼーション条件の別の例には、50%ホルムアミド、5×SSC(150mM塩化ナトリウム、15mMクエン酸ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハルト溶液、10%デキストラン硫酸および20μg/mlの変性剪断サケ精子DNAからなる溶液における42℃での一晩のインキュベーション、その後、0.1×SSCにおいて約65℃での洗浄がある。ストリンジェントなハイブリダイゼーション条件は、上記の代表的な条件と少なくとも80%のストリンジェンシーであるハイブリダイゼーション条件である。他の様々なストリンジェントなハイブリダイゼーション条件がこの分野では知られており、これらもまた、本発明のこの特定の実施形態の核酸を同定するために用いることができる。
【0044】
本発明のタンパク質の変異体または誘導体をコードする核酸もまた提供される。変異型核酸は、この分野で広く知られている技術を使用して、ランダム変異誘発または標的化変異誘発によって作製することができる。目的とする変異には、欠失、付加および置換が含まれる。ある実施形態においては、ホモログまたは変異体をコードする核酸によってコードされる蛍光タンパク質は、野生型の蛍光タンパク質と同じ蛍光特性を有する。別の実施形態においては、変異体のacGFPタンパク質について本明細書中でより詳しく記載されるように、ホモログまたは変異体の核酸は、変化したスペクトル特性を有する蛍光タンパク質をコードする。
【0045】
本発明の核酸はcDNA、ゲノムDNAまたはそれらの断片であり得る。特定の実施形態において、本発明の核酸は、特定の蛍光タンパク質およびポリペプチドをコードするオープンリーディングフレーム、そしてイントロンを含む、また、翻訳領域を超えて約20kbまでの配列(しかし、おそらくは更なる配列)をいずれかの方向で含む、発現の制御に関与する隣接した5’及び3’の非翻訳ヌクレオチド配列の1つ以上を含む。本発明の核酸は、染色体外での維持のために、または宿主ゲノム内への組み込みのために、適切なベクターに導入することができ、より詳しくは以下に記載される。
【0046】
本明細書中で使用される用語「cDNA」は、エキソンならびに5’及び3’非翻訳領域である、天然の成熟mRNA種に見出される配列エレメントの配置を有する核酸を包含することが意図される。通常、mRNA種は、タンパク質をコードする連続したオープンリーディングフレームを生じさせるために、切れ目なく連続したエキソンを有しており、介在するイントロンは、存在するときには、核でのRNAスプライシングによって除かれている。
【0047】
注目されるゲノム配列は、天然の染色体に通常存在するイントロンのすべてを含む、示された配列において定義される開始コドンと終止コドンとの間に存在する核酸を含むことができる。注目されるゲノム配列は更に、成熟mRNAに見出される5’非翻訳領域および3’非翻訳領域を含んでいてもよく、また、転写領域の5’末端または3’末端のいずれかでの隣接ゲノムDNAの約1kb(しかし、おそらくは更に多く)を含む特異的な転写制御配列および翻訳制御配列(例えば、プロモーター、エンハンサーなど)を含むことができる。ゲノムDNAは、100kb以下のフラグメントとして単離することができ、従って、隣接する染色体配列を実質的に含まない。翻訳領域と3’または5’のいずれかで隣接するゲノムDNA、あるいはイントロン内で時々見出されることがある内部の制御配列は、適切な組織特異的な発現および段階特異的な発現のために要求される配列を含有することがある。
【0048】
本発明の核酸組成物は本発明のタンパク質の全体または一部をコードすることができる。二本鎖フラグメントまたは一本鎖フラグメントは、従来の方法に従ってオリゴヌクレオチドを化学合成することによって、制限酵素消化によって、PCR増幅によって、そして他の方法によって、DNA配列から得られる。たいていの場合、DNA断片は長さが少なくとも約15ヌクレオチドであり、一般的には長さが少なくとも約18ヌクレオチドであるか、または長さが約25ヌクレオチドであり、そして長さが少なくとも約50ヌクレオチドであり得る。ある実施形態においては、本発明の核酸分子は、長さが、約100ヌクレオチド、約200ヌクレオチド、約300ヌクレオチド、約400ヌクレオチド、約500ヌクレオチド、約600ヌクレオチド、約700ヌクレオチドまたはそれ以上であり得る。本発明の核酸は、本発明のタンパク質の断片または全長のタンパク質をコードし得る。例えば、本発明の核酸は、約25アミノ酸、約50アミノ酸、約75アミノ酸、約100アミノ酸、約125アミノ酸、約150アミノ酸、約200アミノ酸のポリペプチドから、全長のタンパク質までをコードし得る。
【0049】
本発明の核酸は、一般には、無傷の(インタクト)染色体としてではなく、他の核酸として、実質的な純度で単離され、得ることができる。通常、本発明の核酸またはそのフラグメントを含まない核酸配列が実質的に存在しないDNAが得られる。実質的な純度とは、核酸が少なくとも約50%の純度であり、一般的には少なくとも約90%の純度であり、典型的には「組換え」であること、すなわち、核酸には、その天然の宿主生物での天然に存在する染色体において通常の場合には関連しないヌクレオチドが1つ以上隣接することを意味する。
【0050】
本発明のポリヌクレオチド、例えば、配列番号1、3、5、7、9、11、13、15、17、19、21または23の配列を有するポリヌクレオチド、対応するcDNA、全長遺伝子および構築物を、当業者に知られている多数の異なるプロトコルによって合成的に作製することができる。適切なポリヌクレオチド構築物は、例えば、Sambrook等、Molecular Cloning:A Laboratory Manual(第2版)、Cold Spring Harbor Press、Cold Spring Harbor、NY(1989)に記載されるような標準的な組換えDNA技術を、例えば、米国保健社会福祉省、国立衛生研究所(NIH)組換えDNA研究指針に記載される規制のもとで使用して精製される。
【0051】
また、第2のペプチドまたはタンパク質に融合された本発明の対象タンパク質またはペプチドの融合タンパク質またはそのフラグメントをコードする核酸が提供される。第2のタンパク質は、例えば、分解配列、シグナルペプチド、または注目される任意のタンパク質であり得る。acGFPポリペプチドまたは変異体acGFPポリペプチドと、acGFPポリペプチドのN末端および/またはC末端においてインフレームで(翻訳領域内部に)融合された第2のポリペプチド(「融合パートナー」)とを、例えば、融合タンパク質は含むことができる。融合パートナーには、融合パートナーに対して特異的な抗体と結合することができるポリペプチド(例えば、エピトープタグ)、抗体またはその結合性フラグメント、触媒機能を提供するポリペプチドまたは細胞応答を誘導するポリペプチド、そしてリガンドまたは受容体またはそのミメティックなどが含まれるが、これらに限定されない。そのような融合タンパク質において、融合パートナーは、一般に天然では融合タンパク質のacGFP部分と会合せず、典型的にはAequorea coerulescensのタンパク質またはその誘導体/断片ではない。すなわち、融合パートナーはAequorea属の種において見出されない。
【0052】
本発明の核酸を含むベクターおよび他の核酸構築物もまた提供され、この場合、そのような構築物は、増殖、タンパク質製造などを含む数多くの応用のために使用することができる。ウイルスベクターおよび非ウイルスベクターを調製し、使用することができ、これには、プラスミドが含まれる。ベクターの選択は、増殖が所望される細胞のタイプ、および増殖の目的に依存する。ある種のベクターが、多量の所望するDNA配列を増幅し、作製するために有用である。他の様々なベクターが、培養での細胞における発現のために好適である。更に他の様々なベクターが、動物での細胞における移入および発現のために好適である。適切なベクターの選択は十分にこの分野の技術の範囲内であり、多くのそのようなベクターが市販されている。構築物を調製するために、部分ポリヌクレオチドまたは全長ポリヌクレオチドが、典型的にはベクター内の切断された制限酵素部位へのDNAリガーゼ結合によってベクター内に挿入される。あるいは、所望されるヌクレオチド配列を、インビボでの相同的組換えによって、典型的には、相同性の領域を所望するヌクレオチド配列の両側においてベクターに結合することによって挿入することができる。相同性の領域は、例えば、オリゴヌクレオチドの連結によって、または、相同性の領域と、所望するヌクレオチド配列の一部との両方を含むプライマーを使用するポリメラーゼ連鎖反応によって付加される。
【0053】
また、他の適用の中でも、本発明の色素タンパク質または蛍光タンパク質またはその融合タンパク質の合成において使用される発現カセットまたは発現系も提供される。発現のために、本発明のポリヌクレオチドによってコードされる遺伝子産物が、例えば、細菌、酵母、昆虫、両生類および哺乳類の系を含む任意の好都合な発現系において発現させられる。そのようなベクターおよび宿主細胞が米国特許第5,654,173号に記載される。発現ベクターにおいて、本発明のポリヌクレオチド、例えば、配列番号1、3、5、7、9、11、13、15、17、19、21または23に示されるようなポリヌクレオチドが、所望する発現特性を得るために適するように制御配列に連結される。これらの制御配列には、プロモーター(センス鎖の5’端またはアンチセンス鎖の3’端のいずれかに結合している)、エンハンサー、ターミネーター、オペレーター、リプレッサーおよびインデューサーが含まれ得る。プロモーターは制御型または構成型であり得る。状況により、例えば、組織特異的プロモーターまたは発達段階特異的プロモーターなどの条件的に活性なプロモーターを使用することが望ましい場合がある。これらは、ベクターへの連結について上記に記載される技術を使用して、所望するヌクレオチド配列に連結される。この分野で知られている技術はどれも使用することができる。すなわち、発現ベクターは、転写開始領域および翻訳開始領域(これらは誘導型でも構成型でもよい)を提供するが、この場合、転写開始領域、転写終結領域および翻訳終結領域の転写制御下に機能的に連結される。これらの制御領域は、問題とする核酸が得られる問題とする種に対して生来的であり得るか、または外因性の供給源に由来し得る。
【0054】
発現ベクターは、一般には、異種タンパク質をコードする核酸配列の挿入をもたらすために、好都合な制限部位がプロモーター配列の近くに存在する。発現宿主において機能的な選択マーカーを存在させることができる。発現ベクターは、とりわけ、上述のように、融合タンパク質を産生させるために使用することができる。
【0055】
転写開始領域、遺伝子またはその断片、および転写終結領域を含む発現カセットを調製することができる。特に注目されるものは、一般的には長さが少なくとも約8アミノ酸であり、より一般的には長さが少なくとも約15アミノ酸であり、また、約25アミノ酸までであり、そして遺伝子の完全なオープンリーディングフレームまでである、機能的なエピトープまたはドメインの発現を可能にする、配列の使用である。DNAの導入後、構築物を含有する細胞を選択マーカーによって選択することができ、その後、拡大培養された細胞を発現のために使用することができる。
【0056】
先に記載された発現系は、発現させる目的に依存して、従来の方法に従って原核生物または真核生物と一緒に用いることができる。タンパク質の大規模な生産の場合、単細胞生物(E.Coli(大腸菌)、B.subtilis(枯草菌)、S.cerevisiaeなど)、バキュロウイルスベクターと組合せた昆虫細胞、または脊椎動物などの高等生物の細胞(例えば、COS7細胞、HEK293、CHO、アフリカツメガエル卵母細胞など)を発現宿主細胞として使用することができる。状況により、発現タンパク質が本来の折畳みおよび翻訳後修飾から利益を受けられる真核生物細胞において、遺伝子を発現させることが望ましい。小ペプチドは実験室で合成することもできる。完全なタンパク質配列のサブセットであるポリペプチドは、機能について重要なタンパク質の一部分を同定し、調べるために使用することができる。
【0057】
注目される特異的な発現系には、細菌、酵母、昆虫細胞および哺乳動物細胞に由来する発現系が含まれる。これらのカテゴリーのそれぞれから代表的なシステムに対して引かれる参考文献が下記に示される。
【0058】
細菌における発現系には、Chang等、Nature、275:615(1978);Goeddel等、Nature、281:544(1979);Goeddel等、Nucleic Acids Res.、8:4057(1980);欧州特許第0036,776号、米国特許第4,551,433号;DeBoer等、Proc.Natl.Acad.Sci.(USA)、80:21〜25(1983);Siebenlist等、Cell、20:269(1980)に記載される発現系が含まれる。
【0059】
酵母における発現系には、Hinnen等、Proc.Natl.Acad.Sci.(USA)、75:1929(1978);Ito等、J.Bacteriol.、153:163(1983);Kurtz等、Mol.Cell Biol.、6:142(1986);Kunze等、J.Basic Microbiol.、25:141(1985);Gleeson等、J.Gen.Microbiol.、132:3459(1986);Roggenkamp等、Mol.Gen.Genet.、202:302(1986);Das等、J.Bacteriol.、158:1165(1984);De Louvencount等、J.Bacteriol.、154:737(1983);Van den Berg等、Bio/Technology、8:135(1990);Kunze等、J.Basic Microbiol.、25:141(1985);Cregg等、Mol.Cell.Biol.、5:3376(1985);米国特許第4,837,148号および同第4,929,555号;BeachおよびNurse、Nature、300:706(1981);Davidow等、Curr.Genet.、10:380(1985);Gaillardin等、Curr.Genet.、10:49(1985);Ballance等、Biochem.Biophys.Res.Commun.、112:284〜289(1983);Tilburn等、Gene、26:205〜221(1983);Yelton等、Proc.Natl.Acad.Sci.(81:1470〜1474(1984);KellyおよびHynes、EMBO J.、4:475479(1985);欧州特許第0244,234号;国際特許出願公開WO91/00357に記載される発現系が含まれる。
【0060】
昆虫における異種遺伝子の発現は、米国特許第4,745,051号;Friesen等、「バキュロウイルス遺伝子発現における制御」、The Molecular Biology of Baculoviruses(W.Doerfler編)(1986);欧州特許第0127,839号;欧州特許第0155,476号;Vlak等、J.Gen.Virol.、69:765〜776(1988);Miller等、Ann.Rev.Microbiol.、42:177(1988);Carbonell等、Gene、73:409(1988);Maeda等、Nature、315:592〜594(1985);Labacq−Verheyden等、Mol.Cell.Biol.、8:3129(1988);Smith等、Proc.Natl.Acad.Sci.、82:8844(1985);Miyajima等、Gene、58:273(1987);Martin等、DNA、7:99(1988)に記載されるように達成される。宿主に由来する数多くのバキュロウイルス株および変異株ならびに対応する許容性の昆虫宿主細胞が、Luckow等、Bio/Technology、6:47〜55(1988);Miller等、Genetic Engineering、8:277〜279(1986);Maeda等、Nature、315:592〜594(1985)に記載される。
【0061】
哺乳動物発現は、Dijkema等、EMBO J.、4:761(1985);Gorman等、Proc.Natl.Acad.Sci.(USA)、79:6777(1982);Boshart等、Cell、41:521(1985);米国特許第4,399,216号に記載されるように達成される。哺乳動物の発現の他の特徴が、HammおよびWallace、Meth.Enz.、58:44(1979);BarnesおよびSata、Anal.Biochem.、102:255(1980);米国特許第4,767,704号、同第4,657,866号、同第4,927,762号、同第4,560,655号;国際特許出願公開WO91/103430、同WO87/00195;米国再発行特許第30,985号に記載されるように容易に行われる。
【0062】
上記の宿主細胞のいずれか、あるいは他の適切な宿主細胞または宿主生物を使用して、本発明のポリヌクレオチドまたは核酸を複製し、かつ/または発現させるとき、得られる複製された核酸、RNA、発現タンパク質またはポリペプチドは、宿主細胞または宿主生物の産物として本発明の範囲内である。産物は、この分野で知られている適切な手段によって回収することができる。
【0063】
選択されたポリヌクレオチドに対応する遺伝子が同定されると、遺伝子が生来的である細胞において、その発現を調節することができる。例えば、細胞の内因性遺伝子を、細胞における遺伝子の発現を少なくとも増強するために十分な位置において細胞のゲノム内に挿入された外因性の制御配列によって制御することができる。制御配列は、米国特許第5,641,670号および同第5,733,761号(これらの開示は参考として本明細書中に組み入れる)に開示されるような相同的な組換えによってゲノム内に組み込まれるように設計することができ、または、国際特許出願公開WO99/15650(この開示もまた参考として本明細書中に組み入れる)に開示されるような非相同的な組換えによってゲノム内に組み込まれるように設計することができる。そのため、コード核酸自体の操作を伴わず、しかし、その代わり、所望するタンパク質をコードする遺伝子を既に含む細胞のゲノムに調節配列を組み込むことによるタンパク質の産生もまた本発明に包含される。
【0064】
本発明のゲノム配列のプロモーター配列もまた注目される。この場合、5’隣接領域の配列を、例えば、本発明のタンパク質遺伝子が発現する細胞/組織における発現の制御をもたらす、エンハンサー結合部位を含むプロモーターエレメントのために利用することができる。
【0065】
本発明の核酸の小さいDNA断片もまた注目され、これらは、PCR用プライマー、ハイブリダイゼーションスクリーニングプローブなどとして有用である。より大きいDNA断片は、先に記載されたように、コードされるポリペプチドを産生させるために有用である。しかしながら、幾何級数的PCRなどの幾何級数的増幅反応において使用される場合、一対の小さいDNAフラグメント(すなわち、プライマー)が使用される。プライマー配列の正確な組成は本発明にとって重要ではなく、しかし、ほとんどの適用の場合、プライマーは、この分野で知られているように、ストリンジェントな条件のもとで、対象とする配列にハイブリダイゼーションする。少なくとも約50ヌクレオチド(好ましくは少なくとも約100ヌクレオチド)の増幅産物を生じさせる一対のプライマーを選ぶことが好ましい。プライマー配列を選択するための様々なアルゴリズムが一般に知られており、市販のソフトウエアパッケージで入手可能である。増幅プライマーはDNAの相補鎖にハイブリダイゼーションし、互いに向かって伸張を開始する。
【0066】
本発明の核酸組成物はまた、生物学的試料における遺伝子の発現を確認するために使用することができる。特定のヌクレオチド配列(例えば、ゲノムDNAまたはRNAなど)の存在について細胞をプローブする様式はこの分野では十分に確立されている。簡単に記載すると、DNAまたはmRNAが細胞サンプルから単離される。mRNAは、相補的DNA鎖を形成させるために逆転写酵素を使用し、その後、対象DNA配列に対して特異的なプライマーを使用するポリメラーゼ連鎖反応増幅を行うRT−PCRによって増幅することができる。あるいは、mRNAサンプルはゲル電気泳動によって分離され、好適な担体(例えば、ニトロセルロース、ナイロンなど)に移され、その後、対象DNAの断片をプローブとして用いてプローブされる。他の技術、例えば、オリゴヌクレオチドライゲーションアッセイ、インサイチュハイブリダイゼーション、および固体チップ上に配列されたDNAプローブに対するハイブリダイゼーションなどもまた使用することができる。対象配列にハイブリダイゼーションしているmRNAの検出により、サンプルにおける遺伝子発現が示される。
【0067】
本発明の核酸(これは隣接プロモーター領域および翻訳領域を含む)は、プロモーター強度の目的とする変化を生じさせるために、あるいはコードされるタンパク質の配列またはコードされるタンパク質の性質(これには、コードされるタンパク質の蛍光特性が含まれる)を変化させるために、この分野で知られている様々な方法で変異させることができる。そのような変異を有するDNA配列またはタンパク質産物は、本明細書中に示される配列番号1〜24に対して実質的に類似している。これらの配列の配列変化は、置換、挿入、欠失またはそれらの組合せであり得る。欠失は更に、ドメインまたはエキソンの欠失などの大きな変化、例えば、10、20、50、75、100、150アミノ酸残基またはそれ以上のアミノ酸残基の領域の欠失を含むことがある。インビトロ変異誘発のための様々な技術を、Gustin等、Biotechniques、14:22(1993);Barany、Gene、37:111〜23(1985);Colicelli等、Mol.Gen.Genet.199:537〜9(1985)に見出すことができる。部位特異的変異誘発のための様々な方法を、Sambrook等、Molecular Cloning:A Laboratory Manual、CHS Press、15.3頁〜15.108頁(1986)に見出すことができる。そのような変異させられた核酸誘導体は、特定の色素性蛍光タンパク質の構造−機能の関係を研究するために、あるいはその機能または制御に影響を及ぼすタンパク質の性質を変化させるために使用することができる。
【0068】
また、本明細書中に記載されるacGFPのhG22変異体などの、本発明の核酸のヒト化体も注目される。本明細書中で使用される用語「ヒト化(された)」は、ヒト細胞におけるタンパク質の発現のためにコドンを最適化するために核酸配列になされた変化を示す(Yang等、Nucleic Acids Research、24:4592〜93(1996))。タンパク質のヒト化を記載する米国特許第5,795,737号もまた参照のこと(この開示は参考として本明細書中に組み入れる)。
【0069】
ペプチド組成物
【0070】
本発明は、蛍光タンパク質acGFPおよびその誘導体、ならびに関連するポリペプチド断片を提供する。本明細書中で使用される用語の蛍光タンパク質は、光を照射されたとき、例えば、白色光または特定の波長の光(または励起波長などの狭い波長バンド)で照射されたとき、蛍光を発するタンパク質を示す。本明細書中で使用される用語のポリペプチドは、全長タンパク質、ならびにタンパク質の一部分または断片の両方を示す。また、この用語には、天然に存在するタンパク質の変異体が含まれ、この場合、そのような変異体は、より詳細には下に記載されるように、天然に存在するタンパク質に対して相同的または実質的に類似し、そして天然に存在するタンパク質の変異体である。本発明のポリペプチドは、その天然の環境とは異なる環境に存在する。
【0071】
多くの実施形態において、本発明のタンパク質は、約300nm〜700nm、一般的には約350nm〜550nm、より一般的には約400nm〜500nm、多くの場合には約450nm〜490nm、例えば、470nm〜490nm、の範囲に及ぶ吸収極大を有し、一方で、本発明のタンパク質の発光スペクトルは、典型的には、約400nm〜700nm、一般的には約450nm〜650nm、より一般的には約500nm〜600nm、の範囲にあり、その一方で、多くの実施形態において、発光スペクトルは約500nm〜550nm、例えば、500nm〜525nm、または500nm〜510nm、の範囲にある。本発明のタンパク質は、一般に、約25,000〜150,000、一般的には約45,000〜120,000、例えば、50,000〜100,000、の範囲に及ぶ最大吸光係数を有する。本発明のタンパク質は、典型的には、長さが約150アミノ酸〜300アミノ酸、一般的には約200アミノ酸残基〜300アミノ酸残基、の範囲にあり、そして一般に、分子量が約15kDa〜35kDa、一般的には約17.5kDa〜32.5kDa、の範囲にある。
【0072】
特定の実施形態において、本発明のタンパク質は鮮明(bright)である。この場合、「鮮明」とは、色素タンパク質およびその蛍光性誘導体が一般的な方法(例えば、目視スクリーニング、分光測定法、分光蛍光法、蛍光顕微鏡観察、FACS計測など)によって検出され得ることを意味する。特定の蛍光タンパク質の蛍光輝度は、最大吸光係数を乗じたその量子収量によって決定される。色素タンパク質の輝度はその最大吸光係数によって表されることがある。
【0073】
特定の実施形態において、本発明のタンパク質は、宿主細胞で発現した後、迅速に折りたたまれる。「迅速に折りたたまれる」とは、タンパク質が、短い時間で、その発色性または蛍光性を生じさせる三次構造を達成することを意味する。これらの実施形態において、タンパク質は、一般には約3日を超えない期間で、一般的には約2日を超えない期間で、より一般的には約1日を超えない期間で折りたたまれる。
【0074】
注目される具体的なタンパク質には、野生型acGFP蛍光タンパク質およびその変異体が含まれるが、それらは、例えば、acGFP、Z1、Z2、G1、G2、G22、G22−G22E、G22−G22E/Y220L、220−11−5、CFP−rand3およびCFP−3ならびにヒト化G22をコードする、配列番号2、4、6、8、10、12、14、16、18、20、22または24で提供される。
【0075】
上記に提供された具体的なアミノ酸配列、すなわち、配列番号2、4、6、8、10、12、14、16、18、20、22または24、とは配列が異なるタンパク質のホモログ(またはその断片)もまた提供される。「ホモログ」は、D.G.HigginsおよびP.M.Sharp、「マイクロコンピューターによる迅速かつ高感度な多重配列アラインメント」、CABIOS、5、151頁〜3頁(1989)に記載されるようなMegAlignのDNAstarクラスターアルゴリズム(ktuple=1、gap penalty=3、window=5およびdiagonals saved=5のパラメーターを使用する)を使用して決定されたとき、配列番号2、4、6、8、10、12、14、16、18、20、22または24のアミノ酸配列に対する少なくとも約35%、一般的には少なくとも約40%、より一般的には少なくとも約60%、のアミノ酸配列同一性を有するタンパク質を意味する。多くの実施形態において、注目されるホモログは、特にタンパク質の機能的な領域を提供するアミノ酸配列について、はるかにより大きい配列同一性を有し、例えば、65%、70%、75%、80%、85%、90%(例えば、92%、93%、94%)またはそれ以上、例えば、95%、96%、97%、98%、99%、99.5%の配列同一性を有する。
【0076】
野生型タンパク質に対して実質的に同一であるタンパク質もまた提供される。この場合、「実質的に同一」は、タンパク質が、野生型タンパク質の配列に対して少なくとも約60%、一般的には少なくとも約65%、より一般的には少なくとも約70%、のアミノ酸配列同一性を有すること、そして、場合により、同一性がはるかにより大きく、例えば、75%、80%、85%、90%(例えば、92%、93%、94%)、95%またはそれ以上、例えば、95%、96%、97%、98%、99%、99.5%であり得ることを意味する。
【0077】
先に記載された天然に存在するタンパク質の誘導体または変異体であるタンパク質もまた提供される。変異体は、野生型(例えば、天然に存在する)タンパク質の生物学的性質を保持し得るか、または野生型タンパク質とは異なる生物学的性質を有し得る。本発明のタンパク質の用語「生物学的性質」は、(例えば、野生型タンパク質、またはA.victoria由来の緑色蛍光タンパク質(GFP)などの別の基準タンパク質と比較されるような)スペクトル特性、例えば、吸収極大、発光極大、最大吸光係数、輝度など;インビボおよび/またはインビトロでの安定性(例えば、半減期);ならびに他のそのような性質を示すが、これらに限定されない。変異には、単一アミノ酸変化、1つ以上のアミノ酸の欠失、N末端切断(トランケーション)、C末端切断、挿入などが含まれる。
【0078】
変異体タンパク質は、分子生物学の標準的な技術を使用して、例えば、先に記載されたようなランダム変異誘発および標的化変異誘発を使用して作製することができる。いくつかの変異体が本明細書中に記載される。実施例に示される指針に従い、そして標準的な技術を使用して、当業者は、広範囲の様々な更なる変異体を容易に作製し、そして、生物学的性質が変化しているかどうかを調べることができる。例えば、蛍光強度を、分光光度計を様々な励起波長で使用して測定することができる。
【0079】
天然に存在するタンパク質である本発明のそのようなタンパク質は、天然に存在しない環境に存在し、例えば、その天然に存在する環境から分離される。特定の実施形態において、本発明のタンパク質は、天然に存在する環境と比較して、本発明のタンパク質について濃縮された組成物に存在する。例えば、精製されたタンパク質が提供され、この場合、「精製された」は、タンパク質が、目的とする色素タンパク質または蛍光タンパク質でないタンパク質を実質的に含まない組成物に存在することを意味し、この場合、「実質的に含まない」は、組成物の90%未満、一般的に60%未満、より一般的には50%未満、が非色素タンパク質または非蛍光タンパク質またはその変異体から構成されることを意味する。本発明のタンパク質はまた、単離体として存在し得る。この場合、単離体により、タンパク質が、他のタンパク質および他の天然に存在する生物学的分子(例えば、オリゴ糖、ポリヌクレオチドおよびその断片など)を実質的に含まないことが意味され、ただし、この場合の用語「実質的に含まない」は、単離されたタンパク質を含有する組成物の70%未満、一般的には60%未満、より一般的には50%未満、が何らかの他の天然に存在する生物学的分子であることを意味する。特定の実施形態において、タンパク質は実質的に純粋な形態で存在する。この場合、「実質的に純粋な形態」により、純度が少なくとも95%、一般的には少なくとも97%、より一般的には少なくとも99%、であることが意味される。
【0080】
天然に存在するタンパク質に加えて、天然に存在するタンパク質とは異なるポリペプチド、例えば、先に記載された変異体タンパク質もまた提供される、一般に、そのようなポリペプチドは、本発明の野生型タンパク質をコードする遺伝子のオープンリーディングフレーム(ORF)によってコードされるアミノ酸配列を含み、これらには、全長タンパク質およびその断片、特に、生物学的に活性な断片、および/または機能的ドメインに対応する断片などが含まれ、また、他のタンパク質またはペプチドに対する本発明のポリペプチドの融合体が含まれる。注目される断片は、典型的には、長さが少なくとも約10アミノ酸であり、一般的には長さが少なくとも約50アミノ酸であり、そして、長さが300アミノ酸またはそれ以上もの長さである場合があるが、一般的には約250アミノ酸の長さを超えない。この場合、断片は、少なくとも約10アミノ酸、一般的には少なくとも約15アミノ酸、多くの実施形態では少なくとも約50アミノ酸、の長さである本発明のタンパク質に対して同一であるアミノ酸領域を有する。いくつかの実施形態では、本発明のポリペプチドは、長さが、約25アミノ酸、約50アミノ酸、約75アミノ酸、約100アミノ酸、約125アミノ酸、約150アミノ酸、約200アミノ酸または約250アミノ酸から、タンパク質の全長までである。いくつかの実施形態において、タンパク質断片は、野生型タンパク質の特異的な性質のすべてまたは実質的にすべてを保持する。
【0081】
本発明のタンパク質およびポリペプチドは、天然に存在する供給源から得ることができ、または合成的に作製することができる。例えば、野生型タンパク質を、そのタンパク質を発現する生物学的供給源から、例えば、Aequorea coerulescensから得ることができる。本発明のタンパク質はまた、合成的手段から、例えば、先に記載されたように、目的とするタンパク質をコードする組換え遺伝子または核酸コード配列を好適な宿主において発現させることによって得ることができる。任意の好都合なタンパク質精製手法を用いることができ、様々な好適なタンパク質精製方法論が、Guide to Protein Purification(Deuthser編)(Academic Press、1990)に記載される。例えば、可溶化液(ライセート)を元の供給源から調製し、そして、HPLC、排除クロマトグラフィー、ゲル電気泳動、アフィニティークロマトグラフィーなどを使用して精製することができる。
【0082】
抗体組成物
【0083】
本発明の蛍光タンパク質に対して特異的に結合する抗体もまた提供される。好適な抗体が、タンパク質の全体または一部を含むペプチドで宿主動物を免疫化することによって得られる。好適な宿主動物は、マウス、ラット、ヒツジ、ヤギ、ハムスター、ウサギなどが含まれる。免疫源は、完全なタンパク質、またはその断片および誘導体を含み得る。好ましい免疫源はタンパク質の全体または一部を含み、この場合、タンパク質は、天然の標的タンパク質において見出される翻訳後修飾を含む。免疫源は、この分野で知られている様々な方法で作製され、そのような方法には、例えば、従来の組換え方法を使用するクローン化遺伝子の発現、または、直接的にAequorea coerulescensからの単離がある。
【0084】
ポリクローナル抗体を調製する場合、最初の工程には、ペプチド免疫源による宿主動物の免疫化が伴い、この場合、ペプチドタンパク質免疫源は、好ましくは、約1%未満の混入物を含む実質的に純粋な形態である。免疫源は、完全なタンパク質あるいはその断片または誘導体を含み得る。宿主動物の免疫応答を増大させるために、標的タンパク質はアジュバントと組み合わせることができ、この場合、好適なアジュバントには、ミョウバン、デキストラン硫酸、大きなポリマーアニオン、油および水のエマルション、フロイントアジュバント、フロイント完全アジュバントなどが含まれる。ペプチド免疫源はまた、合成キャリアタンパク質または合成抗原にコンジュゲート化することができる。
【0085】
ペプチド免疫源は、一般的には皮内投与で宿主に投与されるが、初期投与の
後、1回以上、一般的には少なくとも2回、の更なる追加免疫投与が行われる。免疫後、宿主からの血液が採取され、この場合、血清が血液細胞から分離される。得られた抗血清に存在する免疫グロブリンを、知られている方法を使用して、例えば、アンモニウム塩分画化、DEAEクロマトグラフィーなどを使用して精製することができる。
【0086】
あるいは、モノクローナル抗体を従来の技術によって作製することができる。一般には、免疫化された宿主動物の脾臓および/またはリンパ節が形質細胞の供給源となる。形質細胞はミエローマ細胞との融合によって不死化され、ハイブリドーマ細胞が作製される。個々のハイブリドーマから得られる培養上清が、所望する特異性を有する抗体を産生するハイブリドーマを同定するために、標準的な技術を使用してスクリーニングされる。ヒトタンパク質に対するモノクローナル抗体を作製するための好適な動物には、マウス、ラット、ハムスターなどが含まれる。マウスタンパク質に対する抗体を産生するためには、動物は、一般に、ハムスター、モルモットまたはウサギである。抗体は、従来の技術によって、例えば、プロテインAセファロースのような不溶性担体に結合させたタンパク質を使用するアフィニティークロマトグラフィーなどによってハイブリドーマ細胞上清または腹水から精製することができる。
【0087】
抗体は、通常の多量体構造の代わりに、単鎖体(single chain)として作製することができる。単鎖抗体が、Jost等、J.B.C.、269:26267〜73(1994)などに記載される。重鎖の可変領域および軽鎖の可変領域をコードするDNA配列を、グリシンおよび/またはセリンを含む小さい中性アミノ酸の少なくとも4個のアミノ酸をコードするスペーサーに連結することができる。この融合体によってコードされるタンパク質は、元の抗体の特異性および親和性を保持する機能的な可変領域の組み立てを可能にする。
【0088】
特定の実施形態では、ヒト化抗体もまた注目される。抗体をヒト化する様々な方法がこの分野では知られている。ヒト化抗体は、遺伝子組換えされたヒト免疫グロブリン定常領域遺伝子を有する動物の産物であり得る(例えば、国際特許出願公開WO90/10077および同第WO90/04036を参照のこと)。あるいは、注目される抗体は、CH1、CH2、CH3、ヒンジドメインおよび/またはフレームワークドメインを対応するヒト配列で置換するために組換えDNA技術によって操作することができる(国際特許出願公開WO92/02190を参照のこと)。
【0089】
キメラな免疫グロブリン遺伝子を構築するために免疫グロブリンcDNAを使用することがこの分野では知られている(Liu等、Proceedings of the National Academy of Sciences、84:3439(1987)、およびJ.Immunol.、139:3521(1987))。本質的には、mRNAが、抗体を産生するハイブリドーマまたは他の細胞から単離され、cDNAを作製するために使用される。目的とするcDNAを、特異的なプライマーを使用するポリメラーゼ連鎖反応によって増幅することができる(米国特許第4,683,195号および同第4,683,202号)。あるいは、目的とする配列を単離するために、ライブラリーが作製され、スクリーニングされる。その後、抗体の可変領域をコードするDNA配列がヒト定常領域配列に融合される。ヒト定常領域遺伝子の配列は、Kabat等、「免疫学的に注目されるタンパク質の配列」、N.I.H.刊行物番号91−3242(1991)に見出され得る。ヒトC領域遺伝子を知られているクローンから容易に入手することができる。アイソタイプの選択は、補体固定、または抗体依存的な細胞の細胞毒性における活性などの所望するエフェクター機能によって導かれる。好ましいアイソタイプは、IgG1、IgG3およびIgG4である。ヒト軽鎖定常領域のカッパまたはラムダのいずれかを使用することができる。キメラなヒト化抗体は、その後、従来の方法によって発現される。
【0090】
抗体断片、例えば、Fv、F(ab’)2およびF(ab)などを、インタクトなタンパク質を、例えば、プロテアーゼまたは化学的切断により切断することによって調製することができる。あるいは、切断された(truncated)遺伝子を設計することができる。例えば、F(ab’)2断片の一部をコードするキメラな遺伝子により、H鎖のCH1ドメインおよびヒンジ領域、その後に翻訳終止コドンをコードして、切断型分子を生じさせるDNA配列が示される。
【0091】
H鎖およびL鎖のJ領域のコンセンサス配列を使用して、V領域セグメントをヒトC領域セグメントにその後で連結するために有用な制限部位をJ領域に導入するためのプライマーとして使用されるオリゴヌクレオチドを設計することができる。C領域のcDNAを、制限部位をヒト配列において類似する位置に設置するために部位特異的変異誘発によって改変することができる。
【0092】
発現ベクターには、プラスミド、レトロウイルス、YAC、EBV由来エピソームなどが含まれる。好都合なベクターは、機能的に完全なヒトCH免疫グロブリン配列またはCL免疫グロブリン配列を、任意のVH配列またはVL配列が容易に挿入および発現され得るように設計された適切な制限部位とともにコードするベクターである。そのようなベクターでは、スプライシングが、通常、挿入されたJ領域内のスプライスドナー部位と、ヒトC領域に先行するスプライスアクセプター部位との間において、そしてまた、ヒトCHエキソンの中に存在するスプライス領域において生じる。ポリアデニル化および転写終結が、翻訳領域の下流にある生来的な染色体部位において生じる。得られるキメラな抗体は、レトロウイルスLTRを含む任意の強いプロモーターに、例えば、SV−40初期プロモーター(Okayama等、Mol.Cell.Bio.、3:280(1983))、ラウス肉腫ウイルスLTR(Gorman等、Proceedings of the National Academy of Sciences、79:6777(1982))、またはモロニーマウス白血病ウイルスLTR(Grosschedl等、Cell、41:885(1985))、あるいは生来的なIgプロモーターなどに結合することができる。
【0093】
遺伝子組換え体
【0094】
本発明の核酸は、遺伝子組み換えされた非ヒトの植物または動物または部位特異的遺伝子改変体を細胞株において作製するために使用することができる。本発明の遺伝子組換え細胞は、導入遺伝子として存在する本発明による1つ以上の核酸を含む。この場合、この定義には、導入遺伝子を含むように形質転換された親細胞およびその子孫が含まれる。多くの実施形態において、遺伝子組換え細胞は、本発明による核酸を通常の場合には保有または含有しない細胞である。遺伝子組換え細胞が本発明の核酸を通常の場合には含有しないそのような実施形態において、核酸は、細胞内では、非天然の位置において細胞のゲノム物質に組み込まれるなどの、その天然の位置とは異なる位置に存在する。遺伝子組換え動物は、内因性遺伝子座が変化する相同的組換えによって作製することができる。あるいは、核酸構築物をゲノム内にランダムに組み込むことができる。安定な組み込みのためのベクターには、プラスミド、レトロウイルスおよび他の動物ウイルス、YACなどが含まれる。
【0095】
注目される遺伝子組換え生物には、タンパク質またはその変異体が、通常の場合には発現しない細胞または組織で発現し、かつ/あるいはそのような細胞または組織に通常の場合には存在しないレベルで発現する細胞および多細胞生物(植物および動物の両方)が含まれる。
【0096】
相同的組換えのためのDNA構築物は本発明の核酸の一部を少なくとも含む。この場合、遺伝子は、所望する遺伝子修飾を有し、そして標的遺伝子座に対する相同性の領域を含む。ランダム組み込みのためのDNA構築物は、組換えを媒介するための相同性の領域を含む必要はない。好都合なことに、正の選択および負の選択を行うためのマーカーを含むことができる。標的化された遺伝子修飾を相同的組換えにより有する細胞を作製するための様々な方法がこの分野では知られている。哺乳動物細胞をトランスフェクションするための技術については、Keown等、Meth.Enzymol.、185:527〜37(1990)を参照のこと。
【0097】
胚性幹(ES)細胞の場合、ES細胞株を用いることができ、または胚細胞を、マウス、ラット、モルモットなどの宿主から新たに得ることができる。そのような細胞は、適切な繊維芽細胞フェーダー層上で成長し、または白血病阻害因子(LIF)の存在下で成長する。ES細胞または胚細胞が形質転換されているとき、それらを使用して、遺伝子組換え動物を作製することができる。形質転換後、細胞は適切な培地においてフィーダー層に置床される。構築物を含有する細胞を、選択培地を用いることによって検出することができる。十分な時間が、コロニーが成長するために与えられた後、コロニーは選ばれ、構築物の相同的組換えまたは組み込みが生じているかどうか解析される。陽性であるそのようなコロニーは、その後、胚操作および胚盤胞への注入のために使用することができる。胚盤胞は、4週齢〜6週齢の過排卵処理されたメスから得られる。ES細胞はトリプシン処理され、改変された細胞が胚盤胞の胞胚腔に注入される。注入後、胚盤胞は、偽妊娠メスのそれぞれの子宮角に戻される。その後、メスは出産させられ、得られた子が、構築物についてスクリーニングされる。キメラな子孫は、形質転換された細胞の表現型が、天然に存在する細胞と何らかの点で異なる場合(例えば、蛍光を示すことなど)、容易に検出することができる。
【0098】
キメラな動物は、改変された遺伝子の存在についてスクリーニングされ、その改変を有するオスおよびメスが、ホモ接合の子孫を作製するためにかけ合わせられる。特に本発明の融合タンパク質に関しては可能性があるが、遺伝子の変化が発達における何らかのところで致死性を生じさせる場合、組織または器官を、同種(アロジェニック)または類似遺伝子(コンジェニック)の移植片または移植体として維持することができ、あるいはインビボ培養で維持することができる。遺伝子組換え動物は、任意の非ヒト哺乳動物であり、例えば、実験室動物、家畜などであり、機能的研究、薬物スクリーニングなどにおいて使用することができる。遺伝子組換え動物の使用の代表的な例には、下に記載される例が含まれる。
【0099】
遺伝子組換え植物もまた作製することができる。遺伝子組換えされた植物細胞および植物を調製する様々な方法が、米国特許第5,767,367号、同第5,570,870号、同第5,739,409号、同第5,689,049号、同第5,689,045号、同第5,674,731号、同第5,656,466号、同第5,633,155号、同第5,629,470号、同第5,595,896号、同第5,576,198号、同第5,538,879号、同第5,484,956号(これらの開示は参考として本明細書中に組み入れる)に記載される。遺伝子組換え植物を作製する様々な方法がまた、Plant Biochemistry and Molecular Biology(LeaおよびLeegood編、John Wiley&Sons)、275頁〜295頁(1993)に総説される。簡単に記載すると、好適な植物細胞または植物組織が、植物種の特性に依存して集められる。そのため、特定の場合には、プロトプラストが単離され、この場合、そのようなプロトプラストを様々な異なる植物組織(例えば、葉、胚軸、根など)から単離することができる。プロトプラストを単離する場合、集められた細胞は、細胞壁を除くためにセルラーゼの存在下でインキュベーションされる。この場合、正確なインキュベーション条件は、細胞が得られる植物および/または組織のタイプに依存して変化する。得られたプロトプラストは、その後、生じた細胞破片からふるい分けおよび遠心分離によって分離される。
【0100】
あるいは、体細胞を含む胚性外植片を、遺伝子組換え宿主を調製するために使用することができる。細胞または組織を集めた後、目的とする外因性DNAが植物細胞に導入される。この場合、様々な異なる技術をそのような導入のために利用することができる。単離されたプロトプラストの場合、プロトプラストを裸のDNA(例えば、目的とする外因性のコード配列を含むプラスミドなど)を多価カチオン(例えば、PEGまたはPLO)の存在下でインキュベーションすること、または、目的とする外因性のコード配列を含む裸のDNAの存在下でプロトプラストをエレクトポレーションすることを含むDNA媒介遺伝子移入プロトコルによる導入を行うことができる。外因性DNAを取り込むことに成功したプロトプラストは、その後、選択され、カルスに成長させられ、そして最終的には、適切な量および比率の刺激因子(例えば、オーキシンおよびサイトカイニンなど)との接触によって遺伝子組換え植物に成長させられる。
【0101】
胚性外植片の場合、外因性DNAを標的の体細胞に導入する好都合な方法は、粒子加速プロトコル、すなわち、「遺伝子銃」プロトコルの使用による。得られる外植片は、その後、キメラな植物に成長させられ、交配され、その後、遺伝子組換え子孫が得られる。
【0102】
先に記載された裸のDNA法の代わりに、遺伝子組換え植物を作製する別の方法には、アグロバクテリウム媒介形質転換による方法がある。アグロバクテリウム媒介形質転換の場合、外因性DNAを含む同時組み込みベクターまたはバイナリーベクターが調製され、その後、適切なアグロバクテリウム株(例えば、A.tumefaciens)に導入される。得られた細菌が、その後、調製されたプロトプラストまたは組織外植片(例えば、リーフディスクなど)とインキュベーションされ、カルスが作製される。その後、カルスは、選択条件のもとで成長させられ、選択され、そして根および苗条(シュート)の成長を誘導し、最終的には遺伝子組換え植物を得るために成長培地に供される。
【0103】
使用方法
【0104】
本発明の蛍光タンパク質およびペプチドは様々な異なる応用において使用される。これらのタイプのタンパク質のそれぞれについて代表的な使用が以下に記載されるが、この場合、本明細書中に記載される使用は、単なる例示にすぎず、本発明のタンパク質の使用を記載される方法に限定することを決して意図していない。
【0105】
注目される第1の応用は、本発明のタンパク質を蛍光共鳴エネルギー転移(FRET)法において使用することである。これらの方法では、本発明のタンパク質は、下記のような第2の蛍光タンパク質または蛍光色素と組合せてドナーおよび/またはアクセプターとして役立つ:例えば、Matz等、Nature Biotechnology、17:969〜973(1999)に記載されるような蛍光タンパク質;例えば、米国特許第6,066,476号、同第6,020,192号、同第5,985,577号、同第5,976,796号、同第5,968,750号、同第5,968,738号、同第5,958,713号、同第5,919,445号、同第5,874,304号(これらの開示は参考して本明細書中に組み入れる)に記載される、Aequorea victoria由来の緑色蛍光タンパク質またはその蛍光変異体、;他の蛍光性色素、例えば、クマリンおよびその誘導体(7−アミノ−4−メチルクマリンおよびアミノクマリン)など;bodipy色素;カスケードブルー;またはフルオレセインおよびその誘導体、例えば、フルオレセインイソチオシアナートおよびオレゴングリーンなど;ローダミン色素、例えば、テキサスレッド、テトラメチルローダミン、エオシン類およびエリトロシン類など;Cy3およびCy5などのシアニン色素;ランタニドイオンの大環状キレート、例えば、量子色素など;そして、ルシフェラーゼなどの化学発光色素、これには、米国特許第5,843,746号、同第5,700,673号、同第5,674,713号、同第5,618,722号、同第5,418,155号、同第5,330,906号、同第5,229,285号、同第5,221,623号、同第5,182,202号(これらの開示は参考して本明細書中に組み入れる)に記載される色素が含まれる。
【0106】
本発明の蛍光タンパク質を用いるFRETアッセイが使用され得る具体的な例には、下記の例が含まれるが、これらに限定されない。例えば、哺乳動物ツーハイブリッドシステム、転写因子の二量体化、膜タンパク質の多量体化、多タンパク質複合体形成などにおけるタンパク質−タンパク質相互作用の検出;多数の異なる事象に対するバイオセンサーとして、この場合、ペプチドまたはタンパク質により、本発明の蛍光タンパク質を含むFRET蛍光性組合せが共有結合的に連結され、連結用のペプチドまたはタンパク質には、例えば、カスパーゼが媒介する切断のためのプロテアーゼ特異的基質、FRETを増大または減少させるシグナルを受け取ったときに立体配座的変化を受けるペプチド、例えば、PKA調節ドメイン(cAMPセンサー)、リン酸化部位(例えば、リン酸化部位がペプチドに存在する場合、または、ペプチドが別のタンパク質のリン酸化/脱リン酸化ドメインに対する結合特異性を有する場合、あるいは、ペプチドがCa2+結合ドメインを有する場合)がある。更に、本発明のタンパク質が使用される蛍光共鳴エネルギー転移適用、すなわち、FRETの応用には、米国特許第6,008,373号、同第5,998,146号、同第5,981,200号、同第5,945,526号、同第5,945,283号、同第5,911,952号、同第5,869,255号、同第5,866,336号、同第5,863,727号、同第5,728,528号、同第5,707,804号、同第5,688,648号、同第5,439,797号(これらの開示は参考して本明細書中に組み入れる)に記載される適用が含まれるが、それらに限定されない。
【0107】
本発明の蛍光タンパク質はまた、原核生物細胞および真核生物細胞におけるバイオセンサーとして、例えば、Ca2+イオン指示薬、pH指示薬、リン酸化指示薬、または他のイオン(例えば、マグネシウム、ナトリウム、カリウム、塩化物およびハロゲン化物など)の指示薬などとして使用される。例えば、Ca2+イオンを検出する場合、EFハンドモチーフを含有するタンパク質は、Ca2+が結合したとき、細胞質ゾルから膜に移動することが知られている。これらのタンパク質は、タンパク質の他の領域との疎水性相互作用により分子内に埋もれているミリストイル基を含有する。Ca2+の結合は、ミリストイル基を露出させる立体配座の変化を誘導し、その後、ミリストイル基は、脂質二重層内への挿入のために使用され得る(「Ca2+−ミリストイルスイッチ」と呼ばれる)。そのようなEFハンド含有タンパク質の蛍光タンパク質への融合は、細胞質ゾルから形質膜への移動を共焦点顕微鏡観察によりモニターすることによって、EFハンド含有タンパク質を細胞内Ca2+の指示薬にする。このシステムにおいて使用される好適なEFハンドタンパク質には、リカバリン、カルシニューリンB、トロポニンC、ビシニン、ニューロカルシン、カルモジュリン、パルブアルブミンなどが含まれるが、これらに限定されない。
【0108】
pH指示の場合、ヒサクトフィリン類(hisactophilins)に基づくシステムを用いることができる。ヒサクトフィリン類は、ジクチオステリウム(Dyctyostelium)属に存在することが知られているミリストイル化されたヒスチジンリッチなタンパク質である。アクチンおよび酸性脂質に対するそれらの結合は、細胞質のpH変化の範囲内において鋭いpH依存性を有する。生細胞では、膜に結合することにより、ヒサクトフィリン類はアクチンフィラメントと相互作用しなくなるようである。6.5以下のpHにおいて、ヒサクトフィリン類は形質膜および核に移動する。対照的に、7.5のpHでは、ヒサクトフィリン類は細胞質空間全体に一様に分布する。分布のこの変化は可逆的であり、分子表面にループ状に露出したヒスチジンクラスターに起因する。細胞質のpH変化の範囲における細胞内分布の反転は、ヒスチジン残基の6.5のpKと一致している。細胞分布はタンパク質のミリストイル化とは無関係である。蛍光タンパク質をヒサクトフィリンに融合することによって、融合タンパク質の細胞内分布をレーザー走査共焦点顕微鏡観察または標準的な蛍光顕微鏡観察によって追跡することができる。
【0109】
そのような研究の場合、定量的な蛍光分析を、細胞を通るライン操作(レーザー走査共焦点顕微鏡観察)または他の電子的データ分析(例えば、メタモルフソフトウエア(Universal Imaging Corp)を使用する)を行い、細胞集団において集められたデータを平均化することによって行うことができる。細胞質ゾルから形質膜へのヒサクトフィリン/蛍光タンパク質の実質的なpH依存的再分布は1分〜2分のうちに生じ、5分〜10分の後に定常状態レベルに達する。逆反応が、類似する時間スケールで生じる。そのため、類似した様式で作用するヒサクトフィリン−蛍光タンパク質の融合タンパク質は、生きた哺乳動物細胞においてリアルタイムで細胞質ゾルのpH変化をモニターするために使用することができる。そのような方法は、ハイスループットへの応用において、例えば、増殖因子受容体活性化(例えば、上皮増殖因子または血小板由来増殖因子)、走化性刺激/細胞移動の結果としてのpH変化の測定において、二次メッセンジャーとしての細胞内pH変化の検出において、pH操作実験における細胞内pHのモニターリングなどにおいて使用される。
【0110】
PKC活性を検出する場合、レポーターシステムでは、MARCKS(ミリストイル化アラニンリッチCキナーゼ基質)と呼ばれる分子がPKCの基質であるという事実が利用される。MARCKSは、ミリストイル化と、静電的相互作用により負にチャージした形質膜に結合する正にチャージしたアミノ酸の領域(EDドメイン)と、を介して形質膜に固定される。PKCが活性化されると、EDドメインがPKCによってリン酸化され、それにより、負荷電となり、そして、静電的反発の結果として、MARCKSが形質膜から細胞質に移動する(これは「ミリストイル−静電的スイッチ」と呼ばれる)。MARCKSのミリストイル化モチーフからEDドメインまでのMARCKSのN末端を本発明の蛍光タンパク質に融合することにより、PKC活性に対する検出器システムが提供される。PKCによってリン酸化されたとき、融合タンパク質は形質膜から細胞質ゾルに移動する。この移動は、例えば、Cellomics Inc.の技術、または他のハイコンテントスクリーニングシステム(例えば、Universal Imaging Corp.またはBecton Dickinsonから得られるシステムなど)を使用することにより、標準的な蛍光顕微鏡観察または共焦点顕微鏡観察によって追跡することができる。上記のレポーターシステムは、PKC阻害剤に対するハイコンテントスクリーニングに応用され、また、このシグナル伝達経路を妨害する潜在的な試薬についてのスクリーニングアッセイにおけるPKC活性に対する指示薬として適用される。蛍光タンパク質をバイオセンサーとして使用する方法にはまた、米国特許第5,972,638号、同第5,824,485号および同第5,650,135号(ならびにそれらにおいて引用される参考文献)(これらの開示は参考して本明細書中に組み入れる)に記載される方法が含まれる。
【0111】
本発明の蛍光タンパク質はまた、顕微鏡画像化および電子的解析を使用することによる、蛍光性のレポーター基を発現する細胞のアレイの自動化されたスクリーニングを伴う適用において使用される。スクリーニングは、薬物探索のために、また、機能的ゲノミクスの分野で使用することができ、この場合、本発明のタンパク質は、多細胞性の再編成および遊走における変化、例えば、内皮細胞による多細胞細管の形成(血管形成)、Fluoroblok Insertシステム(Becton Dickinson Co.)を介する細胞の遊走、創傷治癒、または神経突起の伸長における変化を検出するために細胞全体のマーカーとして使用される。本発明のタンパク質が、例えば、シグナル伝達における細胞活性についての指示薬として細胞内位置の変化(刺激に対するキナーゼおよび転写因子の移動など)を検出するペプチド(例えば、標的配列など)またはタンパク質に融合されたマーカーとして使用されるスクリーニングもまた用いることができる。例には、プロテインキナーゼC、プロテインキナーゼA、転写因子NFκBおよびNFAT;細胞周期タンパク質、例えば、サイクリンA、サイクリンB1およびサイクリンEなど;引き続いて切断基質が移動するプロテアーゼ切断;リン脂質が、細胞内構造体(例えば、小胞体、ゴルジ装置、ミトコンドリア、ペルオキシソーム、核、核小体、形質膜、ヒストン、エンドソーム、リソソームまたは微小管など)に対するマーカーとともに含まれる。
【0112】
本発明のタンパク質は、局在化マーカーを細胞内蛍光タンパク質/ペプチドの移動の指示薬として、またはマーカー単独として他の蛍光性融合タンパク質の同時局在化を検出するためのハイコンテントスクリーニングにおいて使用することもできる。本発明の蛍光タンパク質が使用される細胞アレイの自動化されたスクリーニングを伴う適用の例には、米国特許第5,989,835号、ならびに、国際特許出願公開WO0017624、同WO00/26408、同WO00/17643および同00/03246(これらの開示は参考して本明細書中に組み入れる)が含まれる。
【0113】
本発明の蛍光タンパク質は、ハイスループットスクリーニングアッセイにおいて使用される。本発明の蛍光タンパク質は、24時間を超える半減期を有する安定なタンパク質である。また、薬物探索のための転写リポーターとして使用することができる、低下した半減期を有する本発明の蛍光タンパク質の脱安定化体が提供される。例えば、本発明によるタンパク質は、より短い半減期を有するタンパク質に由来する推定的なタンパク質分解シグナル配列(例えば、マウスオルニチンデカルボキシラーゼ遺伝子に由来するPEST配列など)、マウスサイクリンB1破壊ボックスまたはユビキチンなどと融合させることができる。脱安定化されたタンパク質、およびそのようなタンパク質を作製するために用いられ得るベクターの記載については、例えば、米国特許第6,130,313号(この開示は参考して本明細書中に組み入れる)を参照のこと。本発明の蛍光タンパク質の脱安定化体を薬物スクリーニングのために使用して、シグナル伝達経路におけるプロモーターを検出することができ、例えば、AP1、NFAT、NFκB、Smad、STAT、p53、E2F、Rb、myc、CRE、ER、GRおよびTREなどである。
【0114】
本発明のタンパク質は、例えば、PattersonおよびLippincott−Scott、Science、13:1873〜77(2002);Ando等、Proc.Natl.Acad.Sci.USA、99:12651〜56(2002)に記載されるように、タンパク質、オルガネラまたは細胞の正確なインビボ光標識、そして、その輸送を追跡するための光活性化標識として使用することができる。
【0115】
更に、本発明のタンパク質は、本発明のタンパク質を特異的なドメイン(例えば、PKCγのCa結合ドメイン、PKCγのDAG結合ドメイン、SH2ドメインまたはSH3ドメインなど)に融合することによって二次メッセンジャー検出器として使用することができる。
【0116】
分泌リーディング配列(secreted leading sequence)を本発明のタンパク質に融合して、本発明のタンパク質の分泌型形態を構築することによって、本発明のタンパク質の分泌型形態を調製することができ、そのような分泌型形態は次いで様々な異なる適用において使用することができる。
【0117】
本発明のタンパク質は、蛍光活性化細胞分取(FACS)適用において使用される。そのような適用において、本発明の蛍光タンパク質は、細胞集団に標識をつけるための標識として使用され、そして、得られる標識された細胞集団が、その後、この分野で知られているように、蛍光活性化細胞分取装置で分取される。様々なFACS法が米国特許第5,968,738号および同第5,804,387号(これらの開示は参考して本明細書中に組み入れる)に記載される。
【0118】
本発明のタンパク質はまた、遺伝子組換え動物におけるインビボマーカーとしても使用される。例えば、本発明のタンパク質の発現を組織特異的なプロモーターによって行わせることができる。この場合、そのような方法は、他の適用の中でも、導入遺伝子の発現効率を調べることなどの、遺伝子治療のための研究において使用される。そのような適用を例示する、遺伝子組換え動物における蛍光タンパク質の代表的な適用が、国際特許出願公開WO00/02997(この開示は参考して本明細書中に組み入れる)に見出される。
【0119】
本発明のタンパク質の更なる適用には、細胞または動物に注入された後、定量的測定のための校正におけるマーカーとしての使用;細胞生存性をモニターするための酸素バイオセンサーデバイスにおけるマーカーまたはレポーターとしての使用;動物、ペット、玩具、食物などに対するマーカーまたは標識としての使用が含まれる。
【0120】
本発明のタンパク質はまた、プロテアーゼ切断アッセイにおいて使用される。例えば、切断不活性型蛍光アッセイを、本発明のタンパク質を使用して開発することができ、この場合、本発明のタンパク質は、タンパク質の蛍光性を破壊することなく、プロテアーゼ特異的切断配列を含むために設計される。活性化されたプロテアーゼによって蛍光タンパク質が切断されたとき、機能的な発色団が破壊されることにより、蛍光が急激に減少する。あるいは、切断により活性化される蛍光を、本発明のタンパク質を使用して生じさせることができ、この場合、タンパク質は、発色団のすぐ近くおよび/または内部に更なるスペーサー配列を含有するために設計される。この変異体は、機能的な発色団の一部がスペーサーによって分割されているので、その蛍光活性が著しく減少する。スペーサーは2つの同一のプロテアーゼ特異的切断部位によって組み立てられる。活性化されたプロテアーゼによって切断されると、スペーサーは切り離され、蛍光タンパク質の2つの残った「サブユニット」が再び会合して、機能的な蛍光タンパク質を生じさせることができる。上記適用はともに、様々な異なるタイプのプロテアーゼ(例えば、カスパーゼなど)に対するアッセイにおいて開発することができる。
【0121】
本発明のタンパク質は、生物学的膜におけるリン脂質組成を明らかにするためのアッセイにおいて使用することもできる。例えば、特異的なリン脂質に結合して、生物学的膜におけるリン脂質分布のパターンを局在化/可視化することを可能し、その一方で、特定のリン脂質領域における膜タンパク質の同時局在化を可能にする、本発明のタンパク質の融合タンパク質(または本発明のタンパク質の任意の他の種類の共有結合的修飾もしくは非共有結合的修飾)を、本発明のタンパク質を用いて達成することができる。例えば、GRP1のPHドメインは、大きい親和性をホスファチジルイノシトール三リン酸(PIP3)に対して有するが、PIP2に対しては有していない。そのため、GRP1のPHドメインと本発明のタンパク質との融合タンパク質を、生物学的膜におけるPIP3リッチ領域を特異的に標識するために構築することができる。
【0122】
本発明のタンパク質の更に別の適用は、蛍光性タイマーとしてであり、この場合、蛍光タンパク質の経時変化に付随するある蛍光色から別の蛍光色への変化(例えば、緑色から赤色への変化)が、発生上の遺伝子発現、細胞周期に依存する遺伝子発現、および概日リズム特異的な遺伝子発現などの遺伝子発現の活性化/不活化を明らかにするために使用される。
【0123】
上術の本発明の抗体はまた、本発明のタンパク質を他の蛍光タンパク質から区別することを含む多数の適用において使用される。
【0124】
キット
【0125】
先に記載された適用の1つ以上を実施する際に使用されるキットもまた本発明によって提供される。この場合、キットは、典型的には、本発明のタンパク質を発現させるための要素、例えば、本発明のタンパク質に対する翻訳領域を含むベクターを構成する構築物を含む。キットの構成要素は、典型的には好適な容器において、典型的には、好適な貯蔵媒体(例えば、緩衝化溶液など)に存在する。また、キットには、提供されたタンパク質に対する抗体を存在させることができる。特定の実施形態において、キットは、本発明のタンパク質をそれぞれがコードする多数の異なるベクターを含む。この場合、ベクターは、異なる環境および/または異なる条件下での発現のために、例えば、ベクターが哺乳動物細胞における発現のために強力なプロモーターを含む構成的発現のために設計されているか、あるいは、プロモーターを独自に挿入するための多数のクローニング部位を有し、発現が独自に調節される無プロモーターベクターなどである。
【0126】
上記の構成要素に加えて、本発明のキットは更に、本発明の方法を実施するための説明書を含む。このような説明書は、キットにはそのうちの1つ以上が存在し得る様々な形態で本発明のキットに存在させることができる。これらの説明書が存在し得る1つの形態は、キットを包装する際に含められる印刷情報(例えば、包装添付文書など)としてである。更に別の手段には、情報が記録されているコンピューター読み取り可能な媒体(例えば、ディスケット、CDなど)がある。存在させることができる更に別の手段には、離れた場所でインターネットを介して情報を入手するために使用され得るウェブサイトアドレスがある。好都合な手段はどれも、キットに存在させることができる。
【実施例】
【0127】
下記の実施例は、限定としてではなく、例示として提供される。
大きいhydromedusaの数個の試料が、2001年8月に、ウラジオストック近くの日本海のロシア沿岸で集められた。1組の特徴により、これらのクラゲをAequorea coerulescensとして同定することが可能である(Kramp、Dana.Rept.、72:201〜202(1968);PogodinおよびYakovlev、Rus.J.Mar.Biol.、25:417〜419(1999))。A.coerulescensおよびA.victoria(A.forscalea、A.aequoreaの異名)は類似しているが、それらの特徴のいくつかは非常に異なる。最も明白な差は触手の数である。すなわち、A.victoriaは放射状導管1つあたり1つの触手を有するだけであり、一方、A.coerulescensは、隣接する放射状導管の各対の間に4つ〜6つの触手を有する。
【0128】
捕まえられたA.coerulescens試料は生物発光性であった。A.victoriaとは対照的に、それらは、緑色ではなく、青色の発光を示した。検出可能な蛍光は、UV光において、または蛍光顕微鏡を使用したとき、A.coerulescensクラゲでは観測されなかった。それにもかかわらず、A.victoriaのGFPに対するモノクローナル抗体により、A.coerulescensから得られたタンパク質抽出物においてGFP様タンパク質が検出された。図11は、acGFPのタンパク質ゲル電気泳動分析を示す。図11aは、A.victoriaのGFPに対する抗体を使用した、A.coerulescensから得られた可溶性タンパク質抽出物のウエスタンブロット分析である。レーン1は、精製された組換えA.victoria GFPであり、レーン2はA.coerulescens抽出物である。
【0129】
GFP様タンパク質をクローン化するために、保存されたアミノ酸配列に対応する縮重プライマーを用いてPCRを行った。GFP様タンパク質をコードするcDNAがクローン化された。野生型acGFPタンパク質のヌクレオチド配列およびアミノ酸配列が図1に示される。このacGFPタンパク質は、GFPに対して非常に大きい類似性を示し、92%のアミノ酸配列類似性を有した(図2参照)。すべての知られている重要な残基(これらには、発色団を形成するSer65、Tyr66およびGly67、進化的に変化していないArg96およびGlu222、そして発色団に対して空間的に近い残基のHis148、Phe165、Ile167およびThr203が含まれる)が、acGFPでは変化していないことが見出された。3個の内部アミノ酸のみが、これらのタンパク質の間では異なっていた。
【0130】
非常に大きい配列類似性を考慮に入れると、acGFPのスペクトル特性はGFPのスペクトル特性と非常に類似していることが予想された。それにもかかわらず、acGFPを発現する大腸菌コロニーは蛍光または着色のいずれも示さなかった。この事実の最も単純な説明、すなわち、大腸菌ではacGFPの折りたたみが完全ではないということは、ほんの部分的には正しく、それは更なる実験で明らかにされた。
【0131】
クラゲの様々な部分におけるacGFP cDNAの存在を、特異的なプライマーを使用するPCRによって調べた。クラゲの傘縁、放射状導管および口盤に対応する3つのcDNAサンプルが調べられた。acGFP cDNAが傘縁において明瞭に検出されたが、2つの他のサンプルには存在しなかった。従って、A.coerulescensにおけるacGFPの分布は、A.victoriaの傘縁において蛍光性の環を形成するA.victoriaにおけるGFPの分布と類似している。
【0132】
acGFPのランダム変異誘発により、多数の緑色蛍光クローンが得られ、そのうちの一部が特徴づけられた。それらの性質および考えられる適用は、A.victoriaの増強型GFP(EGFP)についての性質および適用と類似する。
【0133】
変異体Z1は、図3に示されるように、1つのアミノ酸置換E222G(222位のグルタミン酸の代わりにグリシン)を含有した。変異体Z1タンパク質は、低い輝度、非常に遅い折りたたみを有し、20℃未満の温度が成熟化のために必要であった。30℃で成長させた後、Z1を発現する大腸菌コロニーは、蛍光が見えるようになるためには、室温または4℃で3日間〜5日間貯蔵しなければならない。Z1変異体に対する励起スペクトルおよび発光スペクトルは、それぞれ、480nmおよび504nmに極大を有する(図4参照)。
【0134】
変異体Z2は、図5に示されるように、2つのアミノ酸置換、具体的には、N19D(19位のアスパラギンの代わりにアスパラギン酸)およびE222G(222位のグルタミン酸の代わりにグリシン)を含有した。変異体Z2は、低い輝度、非常に遅い折りたたみ効率を有し、20℃未満の温度が成熟化のために必要であった。37℃で一晩成長させた後、Z2を発現する大腸菌コロニーは、蛍光が見えるようになるためには、室温または4℃で3日間〜5日間貯蔵しなければならない。Z2変異体に対する励起スペクトルおよび発光スペクトルは変異体Z1の励起スペクトルおよび発光スペクトルと非常に類似している。
【0135】
変異体G1は、図6に示されるように、V11I(11位のバリンの代わりにイソロイシン)、K101E(101位のリシンの代わりにグルタミン酸)およびE222G(222位のグルタミン酸の代わりにグリシン)の置換を有する。変異体G1は別のランダム変異誘発処理で作製された。置換E222Gがこれらの変異体の3つで見出されたので、この変異はこれらの変異体の緑色蛍光のために重要であり得ることが考えられた。変異体G1はかなり低い輝度を有する。この変異体の蛍光は、大腸菌を一晩成長させた後の第1日目に見えるようになる。変異体G1の励起スペクトルおよび発光スペクトルは変異体Z1の励起スペクトルおよび発光スペクトルと非常に類似している。
【0136】
変異体G2は、図7に示されるように、V11I(11位のバリンの代わりにイソロイシン)、F64L(64位のフェニルアラニンの代わりにロイシン)、K101E(101位のリシンの代わりにグルタミン酸)およびE222G(222位のグルタミン酸の代わりにグリシン)の置換を有する。変異体G2は、2回目のランダム変異誘発を使用して変異体G1に基づいて作製された。G1と比較して、G2変異体タンパク質は、輝度およびタンパク質折りたたみ速度の改善された特徴を有する。興味深いことに、G2は、増強型GFP(EGFP)についてもまた特徴づけられている置換F64Lを含有する(Cormack等、Gene、173:33〜38(1996);Yang等、Nuc.Acids Res.、24:4592〜4593(1996))。G2のスペクトル特性が下記の表1に示される。G2に対する励起−発光スペクトルが図8に示される。
【0137】
変異体G22は、図9に示されるように、V11I(11位のバリンの代わりにイソロイシン)、F64L(64位のフェニルアラニンの代わりにロイシン)、K101E(101位のリシンの代わりにグルタミン酸)、T206A(206位のトレオニンの代わりにアラニン)およびE222G(222位のグルタミン酸の代わりにグリシン)の置換を有する。変異体G22は、3回目のランダム変異誘発を使用して変異体G2から作製された。G2と比較して、G22は、更に大きく改善された輝度を有する。G22のスペクトル特性が表1に報告される。G22に対する励起−発光スペクトルが図10に示される。50,000M−1cm−1の吸光係数および0.55の量子収量により、このタンパク質は、広く使用されている増強型GFPとほぼ同じくらい明るくなっている。非加熱の変異体G22タンパク質のゲルろ過試験ならびにSDS−PAGEは、G22が単量体であることを明らかにした。図11Bでは、加熱処理(レーン1〜3)対非加熱(レーン4〜6)のタンパク質サンプルの移動度が比較される。レーン1およびレーン4はA.victoriaのGFPであり、レーン2およびレーン5はG22変異体であり、レーン3およびレーン6はG22−G222E変異体である。クーマシーブルー染色が図11bに示され、UV光のもとでの非加熱タンパク質の蛍光が図11cに示される。
【0138】
上述のacGFPの蛍光性変異体のすべてが、470nm〜480nmおよび500nm〜510nmをそれぞれピークとした、類似した励起スペクトルおよび発光スペクトルを有する。それらの励起スペクトルの形状は、野生型A.victoria GFPではなく、増強型GFPの励起スペクトル形状と類似している。acGFP変異体の蛍光団は、増強型GFPについて示されているように、常に脱プロトン化形態であると考えられる。考えられる説明は、Glu222の非存在であり、このことはプロトン移動のために重要であると考えられ得る(Ehrig等、FEBS Lett.、367:163〜167(1995))。
【0139】
蛍光に対するE222G置換の重要性を明らかにするために、逆のG222E置換を変異体G22(V11I、F64L、K101E、T206Aの置換)に対して行った。この逆変異体G22−E222Gの核酸配列およびアミノ酸配列が図12に示される。この逆変異はG22−E222G変異体タンパク質を無色の状態に容易に変換した。G22−G222Eを発現する大腸菌コロニーは着色または検出可能な蛍光のいずれも示さなかった。
【0140】
G22−G222Eは、問題なく、37℃で大腸菌において発現し、折りたたまれるが、これは可溶性組換えタンパク質の収量が大きいこと(総タンパク質の約20%)によって明白に示される。精製されたG22−G222Eは、280nmでの主ピークおよび390nmでの副ピークを有する吸収スペクトルを示した(図13a参照)。アルカリ変性処理されたG22−G222Eタンパク質は、GFP発色団のアニオン型に明らかに対応する吸収ピークを446nmに示した。発色団について44,000M−1cm−1の吸光係数を仮定すると、390nmにおける未変性G22−G222Eの吸光は33,000M−1cm−1であると推定された。280nmピークと390nmピークとの測定された比(280nmにおけるモル吸光係数は23,500M−1cm−1であると計算された)は、可溶性G22−G222Eの約3%のみが成熟形態で存在することを示した。390nmでの励起は、0.07の量子収量で460nmおよび505nmにピークを有する弱い二色蛍光をもたらした(図13b参照)。
【0141】
GFP様タンパク質は、タンパク質サンプルがゲルへの負荷前に加熱されない限り、そのスペクトル特性およびオリゴマー化状態を一般的なSDS−PAGEの条件のもとで保持することが広く知られている。この試験が、G22−G222Eの折りたたみ状態を調べるために使用された。ゲル電気泳動により、明瞭な違いが非変性タンパク質および変性タンパク質の移動度において明らかにされた(図11参照)。更に、非加熱G22−G222Eタンパク質のバンドはUV光のもとで非常に弱い蛍光をもたらした(再度、図11参照)。このタンパク質の約97%が非吸収性形態で存在するので、これらの結果は、この非吸収性形態の立体配座が、変性状態にではなく、未変性状態に近いことを示している。
【0142】
これらの結果により勢いづけられ、組換え野生型acGFPを得て、特徴づける試みを繰り返した。acGFPを発現する大腸菌を、誘導することなく、室温で成長させ、その後、4℃で数日間インキュベーションすることにより、可溶性acGFPの小さい画分の出現がもたらされた(総acGFPの約5%)。可溶性の野生型acGFPに対する吸収スペクトルおよび蛍光スペクトルの形状は、G22−G222Eのスペクトル形状と非常に類似していた。G22−G222E変異体は、大腸菌で発現させられたとき、天然型acGFPの性質を反映し、しかし、改善されたタンパク質折りたたみおよび温度安定性を有すると結論された。
【0143】
これらのデータは、可溶性G22−G222Eが、野生型acGFPと同様に、2つの形態で存在することを示していた。これらのタンパク質の大部分は、スペクトル的に検出可能な発色団を有しない、折りたたまれたが、未熟な形態で存在する。少ない330nm吸収性形態は、中性状態ではGFP様発色団を含有し、弱い二色蛍光を有する。
【0144】
新規なタイプの光変換が変異体G22−G222Eで観測された。G22−G222Eタンパク質サンプルに250nm〜300nmのUV光を照射すると、480nmのピークが吸収/励起スペクトルに出現した。図14において、照射前のG22−G222Eの励起スペクトル(線1)、および250nm〜300nmの光をタンパク質サンプルに照射したことによる曲線のゆっくりした変化に留意すること。番号のない線は光変換後の発光スペクトルを表す(480nmでの励起)。これは、390nmの吸収ピークが、この光変換の間、低下しなかったので、タンパク質のスペクトル的に検出できない未成熟な形態から生じていると考えられる。480nmのピークにおける励起は、大きい量子収量(0.45)で、505nmでの緑色蛍光をもたらした。1000倍を超える、緑色蛍光強度のUV誘導増強が達成された(480nmでの励起)。
【0145】
変異体G22−G222E/Y220Lは、図15に示されるように、野生型scGFPと比較して、V11I(11位のバリンの代わりにイソロイシン)、F64L(64位のフェニルアラニンの代わりにロイシン)、V68A(68位のバリンの代わりにアラニン)、K101E(101位のリシンの代わりにグルタミン酸)、T206A(206位のトレオニンの代わりにアラニン)およびY220L(220位のチロシンの代わりにロイシン)の置換を有する。この変異体は37℃でタンパク質の折りたたみを示し、508nmでの明瞭な緑色蛍光を有した。G22−G222E/Y220Lに対する励起スペクトルは396nmでの主ピークおよび493nmでの副ピークを有した(10:1の比率)(励起スペクトルが点線であり、発光スペクトルが実線である図16を参照のこと)。この変異体のスペクトル特性は、Aequorea victoria由来の野生型GFPのスペクトル特性と非常に類似していた。
【0146】
変異体G22−G222E/Y220Lを基礎として使用したとき、V11I(11位のバリンの代わりにイソロイシン)、F64L(64位のフェニルアラニンの代わりにロイシン)、K101E(101位のリシンの代わりにグルタミン酸)、E115K(115位のグルタミン酸の代わりにリシン)、H148Q(148位のヒスチジンの代わりにグルタミン)、T206A(206位のトレオニンの代わりにアラニン)、Y220L(220位のチロシンの代わりにロイシン)、F221L(221位のフェニルアラニンの代わりにロイシン)およびK238Q(238位のリシンの代わりにグルタミン)の置換を有する変異体220−II−5が得られた。この変異体に対する核酸配列およびアミノ酸配列が図17に示される。この変異体は、主励起ピークを395nmに有するが、約480nmには励起ピークを有さず、512nmに発光ピークを有する(図18a参照)。長波長側の励起ピークの抑制は、発色団の荷電部分の消失を生じさせる置換H148Qによって説明できると考えられる。蛍光顕微鏡下で約400nmの光による比較的強い照射を数分間行った後、変異体G22−G222E/Y220Lの励起スペクトルが変化した。同時に、395nmのピークが低下し、480nmの励起ピークが出現した(図18b参照)。結果として、510nmでの発光の蛍光輝度における100倍を超えるコントラストを、400nmの波長の強い光を照射する前と、光を照射した後とを比較したとき、480nmの励起光で得ることができる。図25には、変異体220−11−5を発現する2つの大腸菌コロニーが蛍光顕微鏡下で示される。上部コロニーにおける2つの領域が、強い400nmの光によって予備的に光活性化された。従って、G22−G222E/Y220L変異体は、PattersonおよびLippincott−Schwartz、Science、13:1873〜1877(2002)におけるPA−GFP変異体の使用に関する最近発表された方法と類似する、生物を光標識するための光活性化蛍光マーカーとして使用することができる。
【0147】
シアン蛍光タンパク質変異体CFP−rand3は、V11I(11位のバリンの代わりにイソロイシン)、T62A(62位のトレオニンの代わりにアラニン)、F64L(64位のフェニルアラニンの代わりにロイシン)、K101E(101位のリシンの代わりにグルタミン酸)、N121S(121位のアスパラギンの代わりにセリン)、H148T(148位のヒスチジンの代わりにトレオニン)、E172K(172位のグルタミン酸の代わりにリシン)およびT206A(206位のトレオニンの代わりにアラニン)における置換を有する。CFP−rand3変異体のアミノ酸配列および核酸配列が図19に示される。この変異体は励起ピークを402nmに有し、1つだけの発光ピークを467nmに有する(図20参照)。
【0148】
別の変異体シアン蛍光タンパク質CFP−3が作製されたが、これは、V11I(11位のバリンの代わりにイソロイシン)、F64L(64位のフェニルアラニンの代わりにロイシン)、K101E(101位のリシンの代わりにグルタミン酸)、H148S(148位のヒスチジンの代わりにセリン)、F165L(165位のフェニルアラニンの代わりにロイシン)、E172A(172位のグルタミン酸の代わりにアラニン)およびT206A(206位のトレオニンの代わりにアラニン)の置換を有する。この変異体に対するアミノ酸配列および核酸配列が図21に示される。この変異体は励起ピークを390nmに有し、1つだけの発光ピークを470nmに有する(図22a)。400nmでの光による比較的強い照射を数分間行った後、励起スペクトルおよび発光スペクトルの下記の変化が観測された:(i)390nmの励起ピークのかなり大きい低下、および(ii)505nmでの発光を伴う480nmの励起ピークの出現(図22b参照)。
【0149】
結果として、505nmでの発光の蛍光輝度における30倍を超えるコントラストが、405nmでの強い照射の前後でスペクトルを比較したとき、480nmでの励起光を使用して得ることができる。励起パラメーターおよび発光パラメーターの同時変化により、シアン変異体CFP−3は、強い400nmの照射に応答して緑色蛍光タンパク質に変換させられる。従って、CFP−3変異体は、生物を光標識するための、光活性化または「光スイッチ」の蛍光性マーカーとして使用することができる。
【0150】
1つの例示的な実験において、G22変異体が、哺乳動物細胞におけるタンパク質発現を調べるための蛍光性タグとして使用された。しかしながら、予想外ではあったが、変異体G22はヒト細胞株において非常に弱い蛍光シグナルをもたらした。これは、最適でないコドン使用頻度またはG22変異体遺伝子における潜在性イントロンの存在のいずれかによって説明され得ると考えられる。これらの問題の両方を克服するために、哺乳動物に最適化されたコドン使用頻度を取り込むG22−h変異体遺伝子を合成した。このヒト化変異体に対するアミノ酸配列および核酸配列が図23に示される。種々の細胞株におけるG22−hの一過性発現は、凝集を伴わない明るい緑色シグナルを示した。
【0151】
図24には、種々の哺乳動物細胞株におけるG22−h変異体(パネルA〜E)およびG22−β−アクチン融合タンパク質(パネルF)の一過性発現が示される。パネルA−293T;パネルB−vero;パネルC−3T3;パネルD−L929;パネルE−COS1;パネルF−3T3。蛍光がトランスフェクションの24時間後に明瞭に検出された。毒性は観測されなかった。タンパク質を標識するG22−hの能力が、細胞質β−アクチンとの融合タンパク質を構築することによって明らかにされた。3T3細胞におけるこの融合体の一過性発現は、明るい正確なアクチン標識化を示した(図24f参照)。ストレスファイバー、限局的接着域および細胞集団を明瞭に認めることができた。細胞の接着性または生存生に対して有害であることは観測されず、また、何らかの非特異的なタンパク質凝集は観測されなかった。
【0152】
方法
【0153】
トータルRNAを、傘縁および放射状導管を含むAequorea coerulescens生物の小さい生体解剖物から、NucleoSpin RNA IIキット(Clontech)を使用して単離した。cDNAが、SMART PCR cDNA合成キット(Clontech)を用いて合成され、また増幅された。新規な蛍光タンパク質遺伝子の断片が、縮重プライマーを用いたPCRによって得られた。ステップアウトPCR RACE法を使用して、標的cDNAの5’端断片をクローン化した。新規な蛍光タンパク質acGFPをコードするcDNAのヌクレオチド配列はアクセション番号AY151052でGenBankに提出されている。acGFPの細菌発現の場合、全長の翻訳領域が、特異的なプライマーを使用して増幅され、pQE30ベクター(Qiagen)にクローン化された。
【0154】
ダイバーシティPCRランダム変異誘発キット(CLONTECH)が、1000塩基対あたり5つ〜6つの変異のために最適な条件で、acGFPのランダム変異誘発のために使用された。変異体タンパク質を発現する大腸菌コロニーは、蛍光実体顕微鏡SZX−12(Olympus)を用いて目視によりスクリーニングされた。最も明るい変異体が選択され、更なるランダム変異誘発処理に供された。部位特異的変異誘発が、適切な標的置換を含有するプライマーを用いて重複伸張法を使用するPCRによって行われた(例えば、Ho等、Gene、77:51〜59(1989)を参照のこと)。
【0155】
N末端の6ヒスチジンタグに融合されたタンパク質を大腸菌XL1 blue株(Invitrogen)において発現させ、TALON金属アフィニティー樹脂(Clontech)を使用して精製した。吸収スペクトルはBeckman DU520UV/VIS分光光度計により記録された。Varian Cary Eclipse蛍光分光光度計が、励起−発光スペクトルを測定するために使用された。
【0156】
モル吸光係数を測定する場合、成熟発色団の濃度の推定が使用された。タンパク質を等量の2M NaOHでアルカリ変性した。これらの条件のもとで、A.victoriaのGFP発色団は446nmで吸収し、そのモル吸光係数は44,000M−1cm−1に等しい(Ward等、Photochem.Photobiol.、31:611〜615(1980))。未変性タンパク質およびアルカリ変性タンパク質に対する吸収スペクトルを測定した。未変性状態に対するモル吸光係数を、基礎としての変性タンパク質の吸収を使用して推定した。量子収量を測定する場合、変異体の蛍光が増強型GFPの量子収量(0.60の蛍光収量)と比較された。
【0157】
acGFP−G222EのUV誘導による光変換を、Cary Eclipse蛍光分光光度計を使用して行った。タンパク質サンプルには、走査モードで250nm〜300nmの波長の光が数時間照射された(励起スリット:20nm、走査速度:30nm/分、平均化時間:1秒、サイクルモード)。
【0158】
精製されたタンパク質のサンプル(〜1mg/ml)をSephadex−100カラム(0.7×60cm)に負荷し、100mMのNaClを含む50mMリン酸塩緩衝液(pH7.0)で溶出した。EGFP、HcRed1およびDsRed2(Clontech)が、それぞれ、単量体、二量体および四量体の標準物として使用された。
【0159】
タンパク質のゲル分析の場合、加熱サンプルおよび非加熱サンプルを一般的な12%SDS−PAGEに負荷し、電気泳動を15mA/ゲルで行った。ウエスタンブロッティングの場合、タンパク質を、標準的な手法を使用してHydond Cメンブラン(Amersham)に転写した。メンブランを、GFPに対するマウス抗体(Clontech)(1:2500)でプローブし、その後、HRPコンジュゲート化抗マウス抗体(Amersham)で1:2500にてプローブした。染色パターンを発色させるために、ECLウエスタンブロティング分析システム(Amersham Pharmacia Biotech)が使用された。これには、検出試薬1および2、そしてHyperfilm ECLが含まれる。
【0160】
真核生物における発現の場合、acGFPが、AgeIおよびBglIIの制限部位の間で(EGFP翻訳領域の代わりに)、pEGFP−C1およびpEGFP−Actinの両ベクター(CLONTECH)にクローン化された。下記の細胞株が使用された:ヒト腎臓上皮細胞293T、マウス胚繊維芽細胞3T3、マウス皮下繊維芽細胞L929、アフリカミドリサル腎臓上皮細胞Vero、およびアフリカミドリサル腎臓繊維芽細胞COS1。細胞は、LipofectAMINE試薬(Invitrogen)を用いてトランスフェクションされ、トランスフェクションの20時間後に試験された。CCDカメラDP−50(Olympus)を備えたOlympus CK40蛍光顕微鏡が細胞画像化のために使用された。
【0161】
【表1】
【0162】
本明細書に引用されたすべての刊行物および特許明細書は、それぞれの個々の刊行物または特許明細書が、参考として組み入れられるために具体的かつ個々に示されていたかのように、参考として本明細書中に組み入れる。何らかの刊行物の引用は、出願日前におけるこの開示のためであり、本発明は、先行発明によるそのような公開がより以前であったとする権利を有しないという承認として解釈してはならない。
【0163】
前述の発明は理解の明確化のための例示および例として幾分詳しく記載されているが、ある種の様々な変化および改変が、添付された請求項の精神または範囲から逸脱することなく本発明に対してなされ得ることが、本発明の教示に照らして当業者には容易に明らかである。
【産業上の利用可能性】
【0164】
Aequorea coerulescens由来の無色のGFP様タンパク質であるacGFP、その蛍光性誘導体および非蛍光性誘導体、またはそれらの変異体をコードする核酸組成物、そしてまた、これらの核酸組成物によってコードされるタンパク質およびペプチドが提供される。注目されるタンパク質は、有色であり、かつ/または蛍光性であり、かつ/または光活性化され得るタンパク質であり、この場合、色、蛍光または光活性化の特徴は、タンパク質の2つ以上のアミノ酸残基の相互作用から生じている。また、上記の特定のタンパク質に実質的に類似するタンパク質、その誘導体タンパク質またはその変異体タンパク質も注目される。また、核酸断片、およびそれによってコードされるペプチドが提供され、また、本発明のタンパク質およびペプチドに対する抗体が提供される。更に、遺伝子組換え細胞および遺伝子組換え生物が提供される。本発明のタンパク質および核酸組成物は、様々な異なる応用および方法における用途があり、特に、タンパク質標識への応用がある。最後に、そのような方法および適用において使用されるキットが提供される。
【特許請求の範囲】
【請求項1】
Aequorea coerulescens由来の蛍光タンパク質をコードする、天然の環境と異なる環境に存在する、核酸分子。
【請求項2】
単離されている、請求項1に記載の核酸。
【請求項3】
前記蛍光タンパク質が、配列番号2、4、6、8、10、12、14、16、18、20、22または24からなる群から選択されるアミノ酸配列を有する、請求項1に記載の核酸。
【請求項4】
配列番号1、3、5、7、9、11、13、15、17、19、21または23からなる配列群から選ばれる少なくとも10残基の長さの配列に対して少なくとも約70%の配列類似性を有する、請求項3に記載の核酸。
【請求項5】
変異体蛍光タンパク質をコードする、請求項1に記載の核酸。
【請求項6】
前記変異体タンパク質が、野生型タンパク質と比較して少なくとも1つの点変異を含む、請求項5に記載の核酸。
【請求項7】
前記変異体タンパク質が、野生型タンパク質と比較して少なくとも1つの欠失変異を含む、請求項5に記載の核酸。
【請求項8】
配列番号1、3、5、7、9、11、13、15、17、19、21または23から選ばれる少なくとも10残基の長さのヌクレオチド配列に対して実質的に類似または同一である配列を有する、核酸分子。
【請求項9】
ストリンジェントな条件のもとで、
(a)Aequorea coerulescens由来の蛍光タンパク質をコードする単離された核酸;
(b)配列番号1、3、5、7、9、11、13、15、17、19、21または23に由来する少なくとも10残基の長さのヌクレオチド配列に対して実質的に類似または同一である配列を有する核酸;
(c)Aequorea coerulescens由来の変異体蛍光タンパク質をコードする単離された核酸;
(d)核酸(a)〜(c)の相補体;または
(e)核酸(a)〜(c)の断片
からなる群から選択される核酸にハイブリダイゼーションする、単離された核酸またはそのミメティック。
【請求項10】
ベクターと、請求項9に記載の核酸とを含む、構築物。
【請求項11】
(a)発現宿主において機能し得る転写開始領域;
(b)請求項9に記載の核酸;および
(c)発現宿主において機能し得る転写終結領域
を含む、発現カセット。
【請求項12】
請求項11に記載の発現カセットを含む、細胞またはその子孫。
【請求項13】
色素タンパク質または蛍光タンパク質を製造する方法であって、色素タンパク質または蛍光タンパク質が発現する条件のもとで請求項12に記載の細胞を増殖させることを含む、方法。
【請求項14】
他のタンパク質を実質的に含まない色素タンパク質または蛍光タンパク質を単離する工程を更に含む、請求項13に記載の方法。
【請求項15】
請求項9に記載の核酸によってコードされる、タンパク質またはその断片。
【請求項16】
請求項15に記載のタンパク質または断片に対して少なくとも約95%の配列類似性を有する、タンパク質またはその断片。
【請求項17】
請求項15に記載のタンパク質または断片を組み込んだ、融合タンパク質。
【請求項18】
請求項15に記載のタンパク質に対して特異的に結合する抗体。
【請求項19】
請求項9に記載の核酸を含む、遺伝子組換え生物。
【請求項20】
請求項9に記載の核酸と、前記核酸を使用するための説明書とを含む、キット。
【請求項1】
Aequorea coerulescens由来の蛍光タンパク質をコードする、天然の環境と異なる環境に存在する、核酸分子。
【請求項2】
単離されている、請求項1に記載の核酸。
【請求項3】
前記蛍光タンパク質が、配列番号2、4、6、8、10、12、14、16、18、20、22または24からなる群から選択されるアミノ酸配列を有する、請求項1に記載の核酸。
【請求項4】
配列番号1、3、5、7、9、11、13、15、17、19、21または23からなる配列群から選ばれる少なくとも10残基の長さの配列に対して少なくとも約70%の配列類似性を有する、請求項3に記載の核酸。
【請求項5】
変異体蛍光タンパク質をコードする、請求項1に記載の核酸。
【請求項6】
前記変異体タンパク質が、野生型タンパク質と比較して少なくとも1つの点変異を含む、請求項5に記載の核酸。
【請求項7】
前記変異体タンパク質が、野生型タンパク質と比較して少なくとも1つの欠失変異を含む、請求項5に記載の核酸。
【請求項8】
配列番号1、3、5、7、9、11、13、15、17、19、21または23から選ばれる少なくとも10残基の長さのヌクレオチド配列に対して実質的に類似または同一である配列を有する、核酸分子。
【請求項9】
ストリンジェントな条件のもとで、
(a)Aequorea coerulescens由来の蛍光タンパク質をコードする単離された核酸;
(b)配列番号1、3、5、7、9、11、13、15、17、19、21または23に由来する少なくとも10残基の長さのヌクレオチド配列に対して実質的に類似または同一である配列を有する核酸;
(c)Aequorea coerulescens由来の変異体蛍光タンパク質をコードする単離された核酸;
(d)核酸(a)〜(c)の相補体;または
(e)核酸(a)〜(c)の断片
からなる群から選択される核酸にハイブリダイゼーションする、単離された核酸またはそのミメティック。
【請求項10】
ベクターと、請求項9に記載の核酸とを含む、構築物。
【請求項11】
(a)発現宿主において機能し得る転写開始領域;
(b)請求項9に記載の核酸;および
(c)発現宿主において機能し得る転写終結領域
を含む、発現カセット。
【請求項12】
請求項11に記載の発現カセットを含む、細胞またはその子孫。
【請求項13】
色素タンパク質または蛍光タンパク質を製造する方法であって、色素タンパク質または蛍光タンパク質が発現する条件のもとで請求項12に記載の細胞を増殖させることを含む、方法。
【請求項14】
他のタンパク質を実質的に含まない色素タンパク質または蛍光タンパク質を単離する工程を更に含む、請求項13に記載の方法。
【請求項15】
請求項9に記載の核酸によってコードされる、タンパク質またはその断片。
【請求項16】
請求項15に記載のタンパク質または断片に対して少なくとも約95%の配列類似性を有する、タンパク質またはその断片。
【請求項17】
請求項15に記載のタンパク質または断片を組み込んだ、融合タンパク質。
【請求項18】
請求項15に記載のタンパク質に対して特異的に結合する抗体。
【請求項19】
請求項9に記載の核酸を含む、遺伝子組換え生物。
【請求項20】
請求項9に記載の核酸と、前記核酸を使用するための説明書とを含む、キット。
【図1】


【図2】

【図3】

【図4】

【図5】
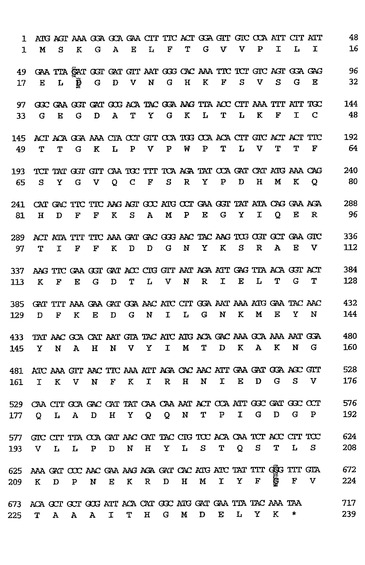
【図6】

【図7】

【図8】
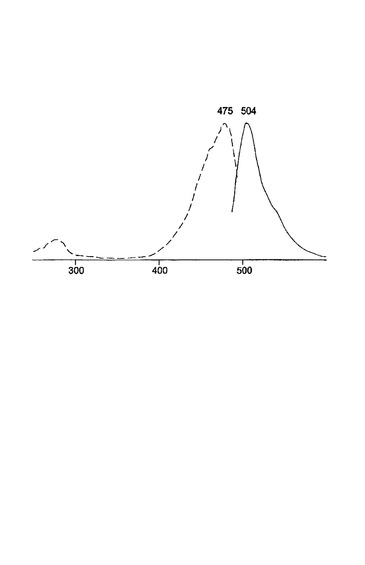
【図9】

【図10】
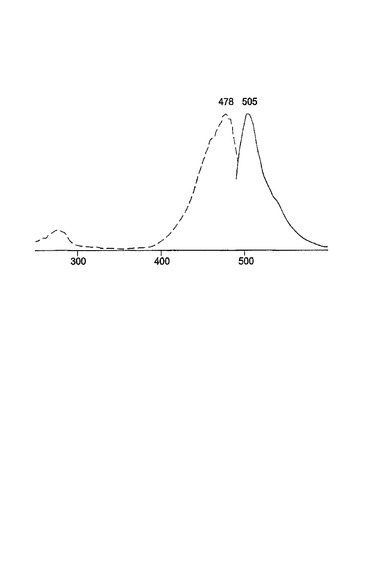
【図11a】
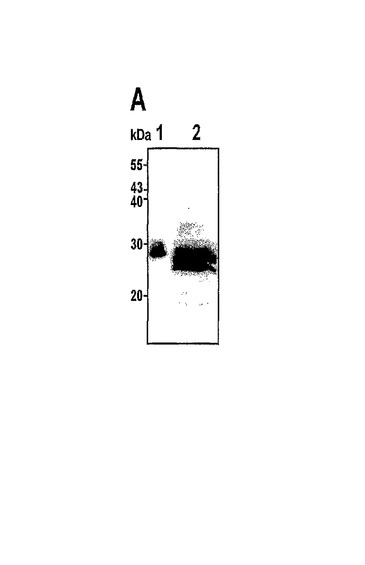
【図11b】
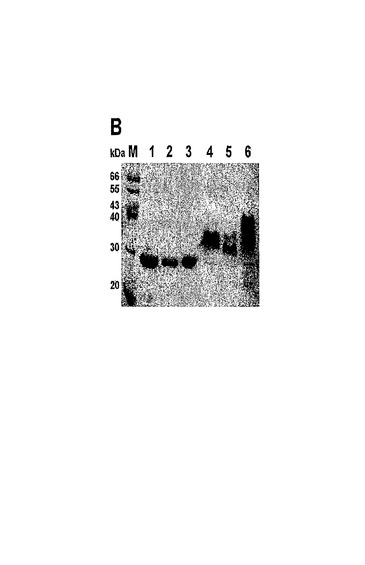
【図11c】

【図12】

【図13a】

【図13b】
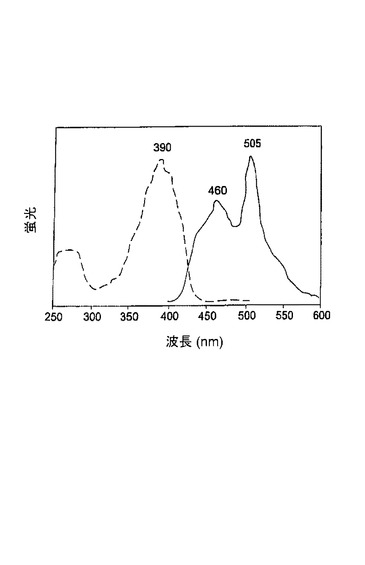
【図14】

【図15】

【図16】
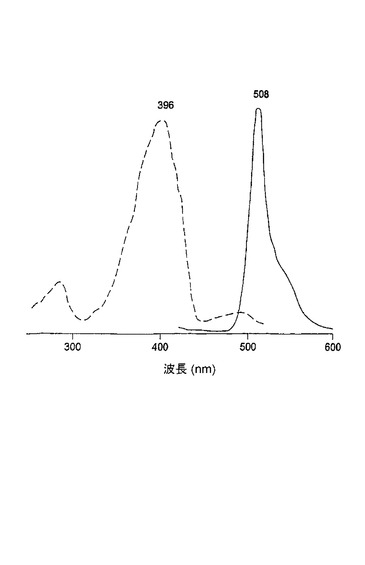
【図17】

【図18a】
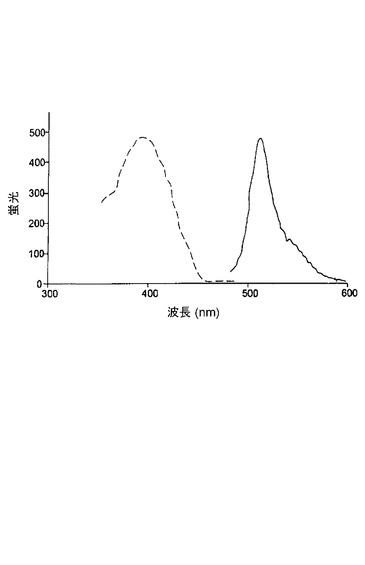
【図18b】
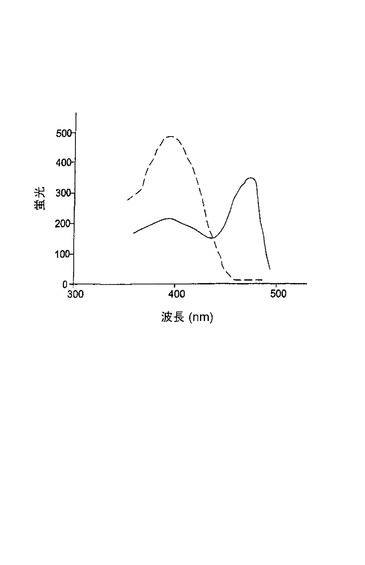
【図19】

【図20】

【図21】

【図22a】

【図22b】

【図23】

【図24a】

【図24b】

【図24c】
【図24d】

【図24e】

【図24f】

【図25】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11a】
【図11b】
【図11c】
【図12】
【図13a】
【図13b】
【図14】
【図15】
【図16】
【図17】
【図18a】
【図18b】
【図19】
【図20】
【図21】
【図22a】
【図22b】
【図23】
【図24a】
【図24b】
【図24c】
【図24d】
【図24e】
【図24f】
【図25】
【公開番号】特開2010−46084(P2010−46084A)
【公開日】平成22年3月4日(2010.3.4)
【国際特許分類】
【出願番号】特願2009−243614(P2009−243614)
【出願日】平成21年10月22日(2009.10.22)
【分割の表示】特願2003−562147(P2003−562147)の分割
【原出願日】平成15年1月17日(2003.1.17)
【出願人】(505009818)
【Fターム(参考)】
【公開日】平成22年3月4日(2010.3.4)
【国際特許分類】
【出願日】平成21年10月22日(2009.10.22)
【分割の表示】特願2003−562147(P2003−562147)の分割
【原出願日】平成15年1月17日(2003.1.17)
【出願人】(505009818)
【Fターム(参考)】
[ Back to top ]