
新規なβ−フルクトフラノシダーゼ、その製造方法及びその利用
【課題】50〜65℃といった高温下において安定性の高い新規なβ−フルクトフラノシダーゼを提供する。
【解決手段】下記(A)〜(C)に記載の改変アミノ酸配列を含むβ−フルクトフラノシダーゼ。(A)特定のアミノ酸配列において、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンの少なくとも1種のアミノ酸が置換された改変アミノ酸配列。(B)前記改変アミノ酸配列Aから選ばれる少なくとも1種のアミノ酸の置換の他さらに1から数個のアミノ酸の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有しβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列B。(C)前記改変アミノ酸配列Aと70%以上の相同性を有し、β−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列C。
【解決手段】下記(A)〜(C)に記載の改変アミノ酸配列を含むβ−フルクトフラノシダーゼ。(A)特定のアミノ酸配列において、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンの少なくとも1種のアミノ酸が置換された改変アミノ酸配列。(B)前記改変アミノ酸配列Aから選ばれる少なくとも1種のアミノ酸の置換の他さらに1から数個のアミノ酸の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有しβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列B。(C)前記改変アミノ酸配列Aと70%以上の相同性を有し、β−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列C。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規なβ−フルクトフラノシダーゼ、該β−フルクトフラノシダーゼを製造するための核酸、組換えベクター及び形質転換体、該β−フルクトフラノシダーゼの製造方法、並びに該β−フルクトフラノシダーゼを利用したβ−フルクトフラノシル転移糖の製造方法に関する。
【背景技術】
【0002】
ラクトスクロースやフルクトオリゴ糖などのβ−フルクトフラノシル転移糖は、抗う蝕性及びミネラル吸収促進性などの性質を有し、さらに腸内細菌の栄養源として機能し得る。β−フルクトフラノシル転移糖を含む飲食料品は、虫歯になり難く、かつ整腸作用のある特定保健用食品として、全国で流通されている。さらに上記性質を有することから、β−フルクトフラノシル転移糖は、嗜好物、飼料、餌料、化粧品、医薬品、成形物、生活用品、農林水産用品、試薬、化学工業用品などの多岐用途への利用が期待されている。
【0003】
β−フルクトフラノシル転移糖を製造する方法には、酵素法がある。酵素法は、ショ糖などのβ−フルクトフラノシル基を有する供与体(以下、「β−フルクトフラノシル供与体」と呼ぶ場合もある)及び受容体(以下、「β−フルクトフラノシル受容体」と呼ぶ場合もある)を含有する溶液に、β−フルクトフラノシル供与体のβ−フルクトフラノシル基をβ−フルクトフラノシル受容体に転移させるβ−フルクトフラノシダーゼ活性を有する酵素を作用させることにより、β−フルクトフラノシル転移糖を製造する。β−フルクトフラノシル基を転移させる酵素としては、アスペルギルス・ニガー(Aspergillus niger FERM-P5886, ATCC20611;非特許文献1を参照)、ペニシリウム・ロッケフォルチ(Penicillium roqueforti IAM7254;特許文献1を参照)、スコプラリオプシス・ブレヴィカウリス(Scopulariopsis brevicaulis IFO4843;特許文献1を参照)などの真菌に由来するβ−フルクトフラノシダーゼ、並びにアルスロバクター・スピーシーズ(Arthrobacter sp. K-1 FERM P-10736/BP-3192;特許文献2を参照)、バチルス・メガテリウム(Bacillus megaterium IFO13498;特許文献3を参照)、バチルス・スピーシーズ(Bacillus sp. V230 FERM BP-5054;特許文献4を参照)などの細菌に由来するβ−フルクトフラノシダーゼなどが知られている。特に、細菌由来のβ−フルクトフラノシダーゼは、真菌由来の酵素と比較して、受容体糖質の選択の幅が広いことから、β−フルクトフラノシル転移糖であるラクトスクロースやキシロシルフラクトシドの製造に有用である。
【特許文献1】特許第3628336号公報
【特許文献2】特許第2781412号公報
【特許文献3】特開平4−200386号公報
【特許文献4】特開平9−224665号公報
【非特許文献1】H.Hidaka, M.Hirayama and N.Sumi:Agric. Biol. Chem.,52,1181 (1998)
【発明の開示】
【発明が解決しようとする課題】
【0004】
一般的に、酵素を利用した物の製造方法は、雑菌汚染を回避しつつ、反応速度を高めるために、高温条件下で行われることが望ましい。ところが、これまでに知られている細菌由来のβ−フルクトフラノシダーゼは、温度安定性が45℃程度までであり、50℃を超えると失活するという性質を有していた。その結果、従前の細菌由来のβ−フルクトフラノシダーゼを利用したβ−フルクトフラノシル転移糖の製造は、45℃以下の低温条件下で実施されており、雑菌汚染が発生し易く、かつ酵素活性が低いために酵素の添加量が多くなり経済性に欠けるという問題があった。
【0005】
そこで、本発明者らは、雑菌汚染の回避に有効な、例えば、50〜65℃といった高温下において安定性の高い新規なβ−フルクトフラノシダーゼ及び該β−フルクトフラノシダーゼの製造方法を提供することを、本発明の解決すべき課題とした。さらに、本発明者らは、該β−フルクトフラノシダーゼを利用することにより、雑菌汚染を回避しつつ、添加酵素量を低減させてβ−フルクトフラノシル転移糖を製造することができる方法を提供することを本発明の解決すべき課題とした。
【課題を解決するための手段】
【0006】
本発明者らは、上記課題を解決するために鋭意研究した結果、特許文献2に記載のアルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192に由来するβ−フルクトフラノシダーゼのアミノ酸配列において、酵素の耐熱性に寄与するアミノ酸残基を見出すことに成功した。さらに、本発明者らは、該アミノ酸残基を所定のアミノ酸に置換することにより、高温下で安定性のあるβ−フルクトフラノシダーゼを得ることに成功した。本発明は、かかる知見に基づいて完成されたものである。
【0007】
したがって、本発明によれば、下記(A)〜(C)に記載の改変アミノ酸配列A、改変アミノ酸配列B又は改変アミノ酸配列Cを含むβ−フルクトフラノシダーゼが提供される。
(A)配列表の配列番号1に記載のアミノ酸配列において、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸が下記[1]〜[5]に示す如く置換された改変アミノ酸配列A
[1]47位のスレオニンについて、セリン、グルタミン酸又はグリシンへの置換
[2]200位のセリンについて、スレオニン又はロイシンへの置換
[3]447位のフェニルアラニンについて、プロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインへの置換
[4]470位のフェニルアラニンについて、チロシン、アラニン、グルタミン酸又はバリンへの置換
[5]500位のプロリンについて、バリン、システイン、トリプトファン又はセリンへの置換
(B)前記改変アミノ酸配列Aにおいて、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸の置換の他さらに1から数個のアミノ酸の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列B
(C)前記改変アミノ酸配列Aと70%以上の相同性を有し、かつ前記改変アミノ酸配列Aの47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸の置換を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列C
【0008】
本発明のβ−フルクトフラノシダーゼの好ましい態様は、前記改変アミノ酸配列Aが、
200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び470位のフェニルアラニンがチロシンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;又は
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列である。
【0009】
本発明の別の側面によれば、本発明のβ−フルクトフラノシダーゼをコードする核酸が提供される。
【0010】
本発明の別の側面によれば、下記(a)〜(c)に記載の改変塩基配列a、改変塩基配列b又は改変塩基配列cを含むβ−フルクトフラノシダーゼをコードする核酸が提供される。
(a)配列表の配列番号2に記載の塩基配列において、139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の三塩基連鎖が下記[6]〜[10]に示す如く置換された改変塩基配列a
[6]139〜141位の三塩基連鎖について、セリン、グルタミン酸又はグリシンを規定するコドンに相補的な三塩基連鎖への置換
[7]598〜600位の三塩基連鎖について、スレオニン又はロイシンを規定するコドンに相補的な三塩基連鎖への置換
[8]1339〜1341位の三塩基連鎖について、プロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインを規定するコドンに相補的な三塩基連鎖への置換
[9]1408〜1410位の三塩基連鎖について、チロシン、アラニン、グルタミン酸又はバリンを規定するコドンに相補的な三塩基連鎖への置換
[10]1498〜1500位の三塩基連鎖について、バリン、システイン、トリプトファン又はセリンを規定するコドンに相補的な三塩基連鎖への置換
(b)前記改変塩基配列aにおいて、前記139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の三塩基連鎖の置換の他さらに1から数個の塩基の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードする改変塩基配列b
(c)前記改変塩基配列aとストリンジェントな条件下でハイブリダイズし、前記改変塩基配列aの139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位からなる群から選ばれる少なくとも1種の置換された三塩基連鎖と相補的な三塩基連鎖を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードする改変塩基配列c
【0011】
本発明の別の側面によれば、本発明の核酸を含む組換えベクターが提供される。
【0012】
本発明の別の側面によれば、本発明の核酸を導入してなる、又は本発明の組換えベクターを含む形質転換体が提供される。
【0013】
本発明の別の側面によれば、(a)本発明の形質転換体を培養して、β−フルクトフラノシダーゼを含む形質転換体培養物を得る工程、及び
(b)形質転換体培養物からβ−フルクトフラノシダーゼを得る工程
を含むβ−フルクトフラノシダーゼの製造方法が提供される。
【0014】
本発明の別の側面によれば、β−フルクトフラノシル供与体及びβ−フルクトフラノシル受容体を含有する溶液に、本発明のβ−フルクトフラノシダーゼ、又は本発明の形質転換体を作用させて、β−フルクトフラノシル転移糖を得ることを特徴とするβ−フルクトフラノシル転移糖の製造方法が提供される。
【0015】
本発明のβ−フルクトフラノシル転移糖の製造方法の好ましい態様は、β−フルクトフラノシル供与体が、ショ糖、ラフィノース及びエルロースからなる群から選ばれる少なくとも1種である。
【0016】
本発明のβ−フルクトフラノシル転移糖の製造方法の好ましい態様は、β−フルクトフラノシル受容体が、糖質、糖アルコール及びアルコールからなる群から選ばれる少なくとも1種である。
【0017】
本発明のβ−フルクトフラノシル転移糖の製造方法の好ましい態様は、β−フルクトフラノシル転移糖が、キシロシルフラクトシド、ガラクトシルフラクトシド、フコシルフラクトシド、ソルボシルフラクトシド、マルトオリゴシルフラクトシド、イソマルトシルフラクトシド、及びラクトシルフラクトシドからなる群から選ばれる少なくとも1種である。
【発明の効果】
【0018】
本発明によれば、工業的実施に有利な50〜65℃の高温下におけるβ−フルクトフラノシル転移糖の製造に利用可能なβ−フルクトフラノシダーゼを効率よく得ることができる。さらに、本発明によれば、雑菌汚染を回避しつつ、添加酵素量を低減させても、工業的規模で効率よくβ−フルクトフラノシル転移糖を製造することができる。したがって、本発明によれば、β−フルクトフラノシル転移糖を含む飲食物、化粧品、医薬品、成形物などの開発を促進することが期待できる。
【発明を実施するための最良の形態】
【0019】
以下、本発明の実施の形態について詳細に説明する。
(1)本発明の酵素
本発明者らは、特許文献2に記載されている、寄託番号FERM P-10736/BP-3192のアルスロバクター・スピーシーズ K-1(Arthrobacter sp. K-1 FERM P-10736/BP-3192)由来のβ−フルクトフラノシダーゼ遺伝子にランダム変異を導入し、得られた変異β−フルクトフラノシダーゼ遺伝子によって形質転換した約3000クローンから成る形質転換体ライブラリーを構築した。該形質転換体ライブラリーから、55℃の加熱処理後においても酵素活性が残存する変異β−フルクトフラノシダーゼの取得を目的として、該変異β−フルクトフラノシダーゼを発現する形質転換体をスクリーニングした。また、本発明者らは、スクリーニングにより得た変異β−フルクトフラノシダーゼのアミノ酸配列から、耐熱性に寄与するアミノ酸残基の部位と種類を特定し、本発明の酵素を構築することに成功した。
【0020】
本発明の酵素は、下記(A)〜(C)に記載の改変アミノ酸配列A、改変アミノ酸配列B又は改変アミノ酸配列Cを含むβ−フルクトフラノシダーゼである。
(A)配列表の配列番号1に記載のアミノ酸配列において、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸が下記[1]〜[5]に示す如く置換された改変アミノ酸配列A
[1]47位のスレオニンについて、セリン、グルタミン酸又はグリシンへの置換
[2]200位のセリンについて、スレオニン又はロイシンへの置換
[3]447位のフェニルアラニンについて、プロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインへの置換
[4]470位のフェニルアラニンについて、チロシン、アラニン、グルタミン酸又はバリンへの置換
[5]500位のプロリンについて、バリン、システイン、トリプトファン又はセリンへの置換
(B)前記改変アミノ酸配列Aにおいて、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸の置換の他さらに1から数個のアミノ酸の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列B
(C)前記改変アミノ酸配列Aと70%以上の相同性を有し、かつ前記改変アミノ酸配列Aの47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸の置換を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列C
【0021】
本発明の酵素は、pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有する。本発明の酵素のβ−フルクトフラノシダーゼ活性は、pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して、好ましくは70%以上、より好ましくは75%以上、さらに好ましくは80%以上、なおさらに好ましくは90%以上である。本発明の酵素のより好ましい態様として、pH6.5、60℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有する酵素;pH6.5、60℃、180分間の加熱処理後において、未処理の酵素に対して70%以上のβ−フルクトフラノシダーゼ活性を有する酵素;及び、pH6.5、60℃、180分間の加熱処理後において、未処理の酵素に対して75%以上のβ−フルクトフラノシダーゼ活性を有する酵素を挙げることができる。ただし、本発明の酵素は、上記好ましい態様に挙げた酵素に限定されるものではない。
【0022】
β−フルクトフラノシダーゼは、一般的に、ショ糖を加水分解してフルクトースを遊離させる活性(加水分解活性)、並びにショ糖のβ−フルクトフラノシル基を各種単糖、糖アルコール、アルキルアルコール、配糖体、オリゴ糖等の受容体に転移させる活性(β−フルクトフラノシル転移活性)を有する酵素である。
【0023】
本発明の酵素は、上記β−フルクトフラノシダーゼの触媒作用と同様の触媒作用を有することから、β−フルクトフラノシダーゼ(β-fructofuranosidase;EC 3.2.1.26)に属する酵素であると推測される。
【0024】
本発明の酵素を用いたβ−フルクトフラノシル転移反応に用いられるβ−フルクトフラノシル受容体は、特に限定されるものではなく、例えば、D−キシロース、D−ガラクトース、L−アラビノース、L−ソルボース、L−フコース、マルトース、スクロース、セロビオース、キシロビオース、イソマルトース、ラクトース、マルトトリオース、イソマルトトリオース、パノース、イソパノースなどを挙げることができる。本発明の酵素を用いたβ−フルクトフラノシル転移反応の結果として生成するβ−フルクトフラノシル転移糖としては、例えば、キシロシルフラクトシド、ガラクトシルフラクトシド、ラクトスクロース(別名、ラクトシルフラクトシド)、エルロース(別名、マルトシルフラクトシド)、イソマルトシルフラクトシドなどを挙げることができる。さらに、トレハロース、ネオトレハロースなどのグルコースからなる非還元性二糖類をβ−フルクトフラノシル受容体として、同様に本発明の酵素を作用させた場合、フラクトシルトレハロース、フラクトシルネオトレハロースなどのβ−フルクトフラノシル転移糖を生成し得る。単糖類、二糖類、三糖類のみならず、それ以上の重合度のキシロオリゴ糖、ガラクトオリゴ糖、マルトオリゴ糖、イソマルトオリゴ糖などのオリゴ糖類、ソルビトール、マルチトールなどの糖アルコール類もβ−フルクトフラノシル受容体となり得る。β−フルクトフラノシル受容体は、1種に限らず複数であってもよく、これらの混合物でもよく、澱粉、アラビノガラクタン、キシログルカンなどの多糖類の部分加水分解物でもよい。β−フルクトフラノシル供与体としては、ショ糖以外に、例えば、ラフィノース、エルロースなどのβ−フルクトフラノシド結合を有するオリゴ糖を用いることもできる。
【0025】
β−フルクトフラノシダーゼの加熱処理及びβ−フルクトフラノシダーゼ活性の測定は、以下の方法で実施される。
加熱処理は、β−フルクトフラノシダーゼを含む50mMのリン酸緩衝液(pH6.5)(以下、「酵素液」と呼ぶ場合がある)を45〜65℃で30〜180分間加熱することにより行う。β−フルクトフラノシダーゼ活性の測定は、50mMのリン酸緩衝液(pH6.5)に溶解させた10%(w/v)のショ糖30μlに加熱処理後の酵素液30μlを添加し、40℃で10分間反応させた後、100℃で15分間加熱して酵素反応を止める。続いて4℃に冷却した後、生成したグルコースをグルコースオキシダーゼ法(和光純薬工業(株)製、グルコースC−IIテスト・ワコー)で測定する。対照として、加熱処理に付していない未処理の酵素(β−フルクトフラノシダーゼ)のβ−フルクトフラノシダーゼ活性を上記の操作により測定する。ここで1単位の酵素活性は同条件下で1分間に1μmolのグルコースを生成する酵素量とする。
【0026】
β−フルクトフラノシダーゼの基質特異性は、以下の方法で測定される。
前記のβ−フルクトフラノシダーゼ活性の測定法において、使用する基質をショ糖の代わりに、エルロース、キシロシルフラクトシド、ラフィノース、ネオケストース、スタキオース、1−ケストース、ニストース、イヌロビオース、又はレバンビオースを基質としてこれらの基質の分解活性を調べることで測定する。
【0027】
β−フルクトフラノシダーゼの至適pHとpH安定性は、以下の方法で測定される。
至適pHは、前記のβ−フルクトフラノシダーゼ活性の測定法において、50mMリン酸緩衝液(pH6.5)の代わりに50mM Mcllvaine 緩衝液(pH3.0〜8.0)、50mM Kolthoff(pH8.0〜11.0)を用いて測定する。上記条件で最も高い酵素活性(以下、「最大活性」と略す)を有するpHを至適pHとする。
一方、pH安定性は、50mM Mcllvaine 緩衝液(pH3.0〜8.0)、50mM Kolthoff(pH8.0〜11.0)を用い、40℃で2時間処理した後、残存する酵素活性を求めて測定する。上記条件下における最大活性の値を基準(100%)として約80%以上の活性を示すpHを、pH安定性を示すpHまたは単に安定なpHという。
【0028】
β−フルクトフラノシダーゼの温度安定性は、以下の方法で測定される。
酵素液を各温度に設定した恒温槽に一定時間静置した後、4℃に冷却し、β−フルクトフラノシダーゼ活性を測定する。未処理の酵素活性の値を基準(100%)として残存活性(%)を求める。同条件下において60%以上の残存活性の値を示す温度を、温度安定性を示す温度、又は単に安定な温度という。
【0029】
β−フルクトフラノシダーゼの阻害剤は、各種化合物の存在下におけるβ−フルクトフラノシダーゼ活性を求めることで測定される。
【0030】
本発明の酵素のアミノ酸配列である改変アミノ酸配列は、配列表の配列番号1に記載のアミノ酸配列において、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる1種、好ましくは2種、より好ましくは3種、さらに好ましくは4種、なおさらに好ましくは5種のアミノ酸がそれぞれ別のアミノ酸に置換されたアミノ酸配列である。改変アミノ酸配列は、アミノ酸残基の置換の態様によって改変アミノ酸配列A〜Cに分類される。改変アミノ酸配列Aは、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる1種又は2種以上のアミノ酸がそれぞれ別のアミノ酸に置換されたアミノ酸配列である。具体的には、改変アミノ酸配列Aは、配列表の配列番号1に記載のアミノ酸配列において、47位のスレオニンについてセリン、グルタミン酸又はグリシンへの置換、200位のセリンについてスレオニン又はロイシンへの置換、447位のフェニルアラニンについてプロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインへの置換、470位のフェニルアラニンについてチロシン、アラニン、グルタミン酸又はバリンへの置換、及び500位のプロリンについてバリン、システイン、トリプトファン又はセリンへの置換からなる群から選ばれる1種又は2種以上の置換を有するものである。改変アミノ酸配列Aの好ましい態様としては、配列表の配列番号1に記載のアミノ酸配列において、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び470位のフェニルアラニンがチロシンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列などを挙げることができるが、これらに限定されるものではない。
【0031】
改変アミノ酸配列Bにおける「1から数個のアミノ酸の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異」における「1から数個」の範囲は、改変アミノ酸配列Bを有するタンパク質がpH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を維持できる程度の範囲であれば特に限定されないが、たとえば、1から20個、好ましくは1から10個、より好ましくは1から7個、さらに好ましくは1から5個、特に好ましくは1から3個程度を意味する。また、「アミノ酸の欠失」とは配列中のアミノ酸残基の欠落もしくは消失を意味し、「アミノ酸の置換」は配列中のアミノ酸残基が別のアミノ酸残基に置き換えられていること、「アミノ酸の逆位」とは隣り合う2以上のアミノ酸残基の位置が逆になっていること、「アミノ酸の付加」とはアミノ酸残基が付け加えられていること、「アミノ酸の挿入」とは配列中のアミノ酸残基の間に別のアミノ酸残基が挿し入れられていることをそれぞれ意味する。改変アミノ酸配列Bは、配列表の配列番号1に記載のアミノ酸配列おける47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種類のアミノ酸が別のアミノ酸に置換されており、さらにこれら以外に1から数個のアミノ酸の欠失等を有するアミノ酸配列を意味する。
【0032】
改変アミノ酸配列Bにおける「1から数個のアミノ酸の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異」の具体的な態様としては、1から数個のアミノ酸が別の化学的に類似したアミノ酸で置き換えらた態様がある。例えば、ある疎水性アミノ酸を別の疎水性アミノ酸に置換する場合、ある極性アミノ酸を同じ電荷を有する別の極性アミノ酸に置換する場合などを挙げることができる。斯く化学的に類似したアミノ酸は、アミノ酸毎に当該技術分野において知られている。具体例を挙げると、非極性(疎水性)アミノ酸としては、アラニン、バリン、イソロイシン、ロイシン、プロリン、トリプトファン、フェニルアラニン、メチオニンなどが挙げられる。極性(中性)アミノ酸としては、グリシン、セリン、スレオニン、チロシン、グルタミン、アスパラギン、システインなどが挙げられる。陽電荷をもつ(塩基性)アミノ酸としては、アルギニン、ヒスチジン、リジンなどが挙げられる。また、負電荷をもつ(酸性)アミノ酸としては、アスパラギン酸、グルタミン酸などが挙げられる。
【0033】
改変アミノ酸配列Cにおける「相同性」は、改変アミノ酸配列Cを有するタンパク質がpH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を維持できる程度の相同性であれば特に限定されないが、好ましくは70%以上が好ましく、より好ましくは80%以上、さらに好ましくは85%以上、なおさらに好ましくは90%以上、特に好ましくは95%以上である。ただし、改変アミノ酸配列Cは、配列表の配列番号1に記載のアミノ酸配列おける47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸が別のアミノ酸に置換されている。
【0034】
本発明の酵素を取得する方法は制限されるものではなく、本明細書に記載されている方法以外に、例えば、配列表の配列番号1の記載を参照して、物理化学的に合成してもよいし、本発明の酵素を発現する生物体から得てもよいし、遺伝子工学的に改変アミノ酸配列をコードする核酸から作成してもよいし、その他の通常知られる手段を用いた各種スクリーニングにより取得してもよい。
【0035】
(B)本発明の核酸
本発明の核酸は、本発明のβ−フルクトフラノシダーゼをコードする核酸である。本発明の核酸の好ましい態様は、例えば、下記(a)〜(c)に記載の改変塩基配列a、改変塩基配列b又は改変塩基配列cを含むβ−フルクトフラノシダーゼをコードする核酸である。
(a)配列表の配列番号2に記載の塩基配列において、139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の三塩基連鎖が下記[6]〜[10]に示す如く置換された改変塩基配列a
[6]139〜141位の三塩基連鎖について、セリン、グルタミン酸又はグリシンを規定するコドンに相補的な三塩基連鎖への置換
[7]598〜600位の三塩基連鎖について、スレオニン又はロイシンを規定するコドンに相補的な三塩基連鎖への置換
[8]1339〜1341位の三塩基連鎖について、プロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインを規定するコドンに相補的な三塩基連鎖への置換
[9]1408〜1410位の三塩基連鎖について、チロシン、アラニン、グルタミン酸又はバリンを規定するコドンに相補的な三塩基連鎖への置換
[10]1498〜1500位の三塩基連鎖について、バリン、システイン、トリプトファン又はセリンを規定するコドンに相補的な三塩基連鎖への置換
(b)前記改変塩基配列aにおいて、前記139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の三塩基連鎖の置換の他さらに1から数個の塩基の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードする改変塩基配列b
(c)前記改変塩基配列aとストリンジェントな条件下でハイブリダイズし、前記改変塩基配列aの139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の置換された三塩基連鎖と相補的な三塩基連鎖を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードする改変塩基配列c
【0036】
本発明の核酸の好ましい具体例としては、以下の改変アミノ酸配列をコードする塩基配列を含む核酸を挙げることができるが、これらに限定されるものではない。
配列表の配列番号1に記載のアミノ酸配列において、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び470位のフェニルアラニンがチロシンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列。
【0037】
改変塩基配列bの「1から数個の塩基の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異」における「1から数個」の範囲は、該改変塩基配列bがpH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードし得るものであれば特には限定されないが、例えば、1から40個、好ましくは1から30個、より好ましくは1から20個、より好ましくは1から10個、さらに好ましくは1から5個、特に好ましくは1から3個程度を意味する。また、「塩基の欠失」とは配列中の塩基の欠落もしくは消失を意味し、「塩基の置換」は配列中の塩基が別の塩基に置き換えられていること、「塩基の逆位」とは隣り合う2以上の塩基の位置が逆になっていること、「塩基の付加」とは塩基が付け加えられていること、「塩基の挿入」とは配列中の塩基の間に別の塩基が挿し入れられていることをそれぞれ意味する。改変塩基配列bは、配列表の配列番号2に記載の塩基配列おける139〜141位、598〜600位、1339〜1341位、1408〜1410位、及び1498〜1500位の塩基以外の塩基について欠失、置換、逆位、付加および/または挿入を有する。
【0038】
改変塩基配列cの「ストリンジェントな条件下でハイブリダイズする」とは、DNAをプローブとして使用し、コロニーハイブリダイゼーション法、プラークハイブリダイゼーション法、あるいはサザンブロットハイブリダイゼーション法等を用いることにより得られるDNAの塩基配列を意味し、例えば、コロニーあるいはプラーク由来のDNA又は該DNAの断片を固定化したフィルターを用いて、0.7〜1.0MのNaCl存在下、65℃でハイブリダイゼーションを行った後、0.1〜2×SSC溶液(1×SSC溶液は、150mM塩化ナトリウム、15mMクエン酸ナトリウム)を用い、65℃条件下でフィルターを洗浄することにより同定できるDNA等を挙げることができる。ハイブリダイゼーションは、Molecular Cloning: A laboratory Mannual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.,1989(以下、モレキュラークローニング第2版と略す)、Current Protocols in Molecular Biology, Supplement 1〜38, John Wiley & Sons (1987-1997)(以下、カレント・プロトコールズ・イン・モレキュラー・バイオロジーと略す)などに記載されている方法に準じて行うことができる。
【0039】
ストリンジェントな条件下でハイブリダイズするDNAとしては、プローブとして使用するDNAの塩基配列と一定以上の相同性を有するDNAが挙げられ、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは93%以上、特に好ましくは95%以上、最も好ましくは98%以上の相同性を有するDNAが挙げられる。ただし、ハイブリダイズする塩基配列は、配列表の配列番号2に記載の塩基配列おける139〜141位、598〜600位、1339〜1341位、1408〜1410位、及び1498〜1500位からなる群から選ばれる位置の1種又は2種以上の三塩基連鎖の置換を含む。
【0040】
本発明の核酸の取得方法は特に限定されない。たとえば、本明細書中の配列表の配列番号1に記載したアミノ酸配列および配列番号2に記載の塩基配列の情報に基づいて適当なプローブやプライマーを調製し、それらを用いて染色体DNAライブラリーから本発明の核酸を得ることができる。染色体DNAライブラリーは、例えば、本発明のβ−フルクトフラノシダーゼの野生型を発現するアルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192、本発明のβ−フルクトフラノシダーゼを発現するアルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192変異体などから得ることができる。アルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192変異体からβ−フルクトフラノシダーゼ遺伝子を含む核酸を取得する方法として、例えば、以下の方法を挙げることができる。
【0041】
アルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192変異体を栄養培地に植菌し、好気的条件下で約1〜3日間程度で培養した後、培養物から菌体を採取し、リゾチームやβ−グルカナーゼなどの細胞壁溶解酵素や超音波で処理することによりβ−フルクトフラノシダーゼ遺伝子を含む核酸を菌体外に溶出させる。このとき、細胞壁溶解酵素にプロテアーゼなどの蛋白質加水分解酵素を併用したり、菌体を超音波処理する際、SDSなどの界面活性剤を共存させたり、凍結融解してもよい。斯くして得られる処理物に、例えば、フェノール抽出、アルコール沈澱、遠心分離、プロテアーゼ処理、リボヌクレアーゼ処理などの当該技術分野において通常知られる方法を適用すればβ−フルクトフラノシダーゼ遺伝子を含む核酸が得られる。
【0042】
上記した染色体DNAライブラリーを得ることができれば、PCR法により本発明の核酸を取得することもできる。該染色体DNAライブラリーを鋳型として使用し、改変塩基配列a〜cを増幅できるように設計した1組又は2組以上のプライマーを用いてPCRを行う。PCRの反応条件は適宜設定することができ、例えば、94℃で15秒〜30秒(変性)、63℃で30秒〜1分間(アニーリング)、68℃で2〜3分間(伸長)からなる反応工程を1サイクルとして、例えば30〜50サイクル行った後、72℃で10分間反応させる条件などを挙げることができる。次いで、増幅されたDNA断片を、大腸菌等の宿主で増幅可能な適切なベクター中にクローニングすることができる。
【0043】
上記した各種操作、例えば、プローブ若しくはプライマーの調製、染色体DNAライブラリーの構築、DNAの抽出、cDNAライブラリーのスクリーニング、並びにβ−フルクトフラノシダーゼ遺伝子のクローニングなどの操作は当業者に既知であり、例えば、モレキュラークローニング第2版、またはカレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載の方法に準じて行うことができる。
【0044】
また、本発明のβ−フルクトフラノシダーゼや核酸は、本発明のβ−フルクトフラノシダーゼの野生型を発現しているアルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192由来のβ−フルクトフラノシダーゼ遺伝子から、改変アミノ酸配列Aおよび改変塩基配列aの情報に基づいて、化学合成、遺伝子工学的手法または突然変異誘発などの当業者に通常知られる任意の方法で作製することができる。具体的には、β−フルクトフラノシダーゼの野生型を発現しているアルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192由来のβ−フルクトフラノシダーゼ遺伝子に対し、変異原となる薬剤を接触作用させる方法、紫外線を照射する方法、遺伝子工学的手法を挙げることができる。遺伝子工学的手法の一つである部位特異的変異誘発法は対象となる位置に変異を導入できる手法であることから有用であり、サチュレーション変異法、ならびにモレキュラークローニング第2版およびカレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載の方法に準じて行うことができる。
【0045】
本発明の核酸は、通常、組換えDNAの形態で宿主に導入される。組換えDNAは、通常、DNAと自律増殖可能なベクターを含んでなり、本発明の核酸を入手できれば、通常知られる組換えDNA技術により比較的容易に調製することができる。例えば、本発明の核酸を自律増殖可能なベクターに挿入して組換えDNAとし、これを適宜宿主に導入して得られる形質転換体を培養し、培養物から菌体を採取し、この菌体から当該DNAを含むプラスミドを採取する。
【0046】
(C)本発明の組換えベクター
本発明の核酸は適当なベクター中に挿入して使用することができる。本発明で用いるベクターの種類は特に限定されず、例えば、自立的に複製することが可能なベクター(例えばプラスミド等)でもよいし、あるいは、宿主細胞に導入された際に宿主細胞の染色体に組み込まれ、染色体と共に複製されるものであってもよい。好ましくは、本発明で用いるベクターは発現ベクターである。発現ベクターにおいて本発明の遺伝子は、転写に必要な要素(例えば、プロモーター等)が機能的に連結されている。プロモーターは宿主細胞において転写活性を示すDNA配列であり、宿主の種類に応じて適宜選択することができる。
【0047】
また、自立的に複製することが可能なベクターの具体例としては、pET21a、pRSETA、pCR8、pBR322、pBluescript II SK(+)、pUC18、pCR2.1、pLEX、pJL3、pSW1、pSE280、pSE420、pHY300PLK、pTZ4、pC194、pUB110、pHV14、TRp7、YEp7、pBS7等のプラスミドベクターやλgt11、λZAP、λgt・λB、ρ11、φ1、φ105等のファージベクターが挙げられるが、大腸菌で発現させるには、pET21a、pRSETA、pCR8、pBR322、pBluescript II SK(+)、pUC18、pKK223−3、およびpCR2.1が好適であり、枯草菌で発現させるには、pUB110、pTZ4、pC194、ρ11、φ1及びφ105が好適である。pHY300PLK、pHV14、TRp7、YEp7及びpBS7は、組換えDNAを2種以上の宿主内で増殖させる場合に有用である。
【0048】
作動可能なプロモーターとしては、挿入断片に含まれる宿主中において機能することができるプロモーターであれば特に制限されないが、例えば、大腸菌においてはラクトースオペロン(lac)、トリプトファンオペロン(trp)などのプロモーターやtacプロモーターを挙げることができ;枯草菌においてはバチルス・ステアロテルモフィルス(Bacillus stearothermophilus)のマルトジェニックアミラーゼ遺伝子、バチルス・リケニホルミス(Bacillus licheniformis)のαアミラーゼ遺伝子、バチルス・アミロリケファチエンス(Bacillus amyloliquefaciens)のBANアミラーゼ遺伝子、バチルス・サブチリス(Bacillus subtilis)のアルカリプロテアーゼ遺伝子もしくはバチルス・プミルス(Bacillus pumilus)のキシロシダーゼ遺伝子などのプロモーターを挙げることができ;酵母ではアルコールデヒドロゲナーゼ遺伝子(ADH)、酸性フォスファターゼ遺伝子(PHO)、ガラクトシダーゼ遺伝子(GAL)、グリセロアルデヒド3リン酸デヒドロゲナーゼ遺伝子(GPD)などのプロモーター、酵母解糖系遺伝子由来のプロモーター、TPI1プロモーター、ADH2−4cプロモーターなどを挙げることができ;カビではα−アミラーゼ遺伝子(amy)、セロビオハイドロラーゼI遺伝子(CBHI)などのプロモーター、ADH3プロモーター、tpiAプロモーターなどを挙げることができ;哺乳動物細胞では、SV40プロモーター、MT−1(メタロチオネイン遺伝子)プロモーター、アデノウイルス2主後期プロモーターなどを挙げることができ;昆虫細胞では、ポリヘドリンプロモーター、P10プロモーター、オートグラファ・カリホルニカ・ポリヘドロシス塩基性タンパクプロモーター、バキュウロウイルス即時型初期遺伝子1プロモーター、バキュウロウイルス39K遅延型初期遺伝子プロモーターなどを挙げることでき、その他としてはファージ・ラムダのPRプロモーター、PLプロモーターなどを挙げることができる。
【0049】
本発明の組換えベクターはさらに選択マーカーを含有してもよい。選択マーカーとしては、例えば、薬剤耐性マーカー、栄養要求マーカーを使用することができる。選択マーカーの具体例としては、使用する宿主細胞が細菌の場合はアンピシリン耐性遺伝子、カナマイシン耐性遺伝子、テトラサイクリン耐性遺伝子、クロラムフェニコール耐性遺伝子、ネオマイシン耐性遺伝子、ヒグロマイシ耐性遺伝子などであり、酵母の場合はジヒドロ葉酸レダクターゼ(DHFR)遺伝子、シゾサッカロマイセス・ポンベTPI遺伝子、トリプトファン合成遺伝子(TRP1)、ウラシル合成遺伝子(URA3)、ロイシン合成遺伝子(LEU2)などがあり、カビの場合はハイグロマイシン耐性遺伝子(Hyg)、ビアラホス耐性遺伝子(Bar)、硝酸還元酵素遺伝子(niaD)などが挙げられる。
【0050】
本発明の組換えベクターは、本発明の核酸によってコードされるタンパク質の発現に必要なDNA配列、例えばプロモーター、エンハンサー、リボソーム結合部位、分泌シグナル配列、ターミネーターなどを適当な順で連結していることことが好ましい。本発明の組換えベクターを構築する方法や本発明の組換えベクターを宿主に挿入する方法は当業者に通常知られる。
【0051】
(D)本発明の形質転換体
本発明の核酸または組換えベクターを適当な宿主に導入することによって形質転換体を作製することができる。すなわち、本発明の形質転換体は、本発明の核酸を導入してなる、又は本発明の組換えベクターを含有する形質転換体である。
【0052】
本明細書でいう「核酸を導入」とは、バクテリオファージなどの通常知られる外来遺伝子を宿主細胞に導入する手段によって、本発明の核酸を宿主細胞へ導入すること、好ましくは、本発明の核酸が宿主細胞の染色体に組み込まれることを意味する。
【0053】
本発明の核酸または組換えベクターが導入される宿主細胞は、宿主−ベクター系が確立されているものであればいずれも利用可能であり、本発明のβ−フルクトフラノシダーゼ生産における形質転換体の培養条件において、基質であるβ−フルクトフラノシル供与体を分解する酵素を産生しないものが好ましく、細菌、酵母、真菌および高等真核細胞等が挙げられる。β−フルクトフラノシル供与体を分解する酵素としては、例えば、β−フルクトフラノシル供与体がショ糖である場合はスクラーゼを挙げることができる。
【0054】
細菌細胞の例としては、枯草菌または放線菌等のグラム陽性菌または大腸菌等のグラム陰性菌が挙げられる。これら細菌の形質転換は、プロトプラスト法、エレクトロポレーション法、カルシウムイオンを用いる方法等により行えばよい。哺乳類細胞の例としては、HEK293細胞、HeLa細胞、COS細胞、BHK細胞、CHL細胞またはCHO細胞等が挙げられる。哺乳類細胞を形質転換し、該細胞に導入されたDNA配列を発現させる方法も知られており、例えば、エレクトロポレーション法、リン酸カルシウム法、リポフェクション法等を用いることができる。
【0055】
酵母細胞の例としては、サッカロマイセスまたはシゾサッカロマイセスに属する細胞が挙げられ、例えば、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)またはサッカロマイセス・クルイベリ(Saccharomyces kluyveri)等が挙げられる。酵母宿主への組換えベクターの導入方法としては、例えば、エレクトロポレーション法、スフェロブラスト法、酢酸リチウム法等を挙げることができる。
【0056】
他の真菌細胞の例は、糸状菌、例えばアスペルギルス、ニューロスポラ、フザリウム、またはトリコデルマに属する細胞である。宿主細胞として糸状菌を用いる場合、DNA構築物を宿主染色体に組み込んで組換え宿主細胞を得ることにより形質転換を行うことができる。DNA構築物の宿主染色体への組み込みは、通常知られる方法に従い、例えば相同組換えまたは異種組換えにより行うことができる。
【0057】
昆虫細胞を宿主として用いる場合には、組換え遺伝子導入ベクターおよびバキュロウイルスを昆虫細胞に共導入して昆虫細胞培養上清中に組換えウイルスを得た後、さらに組換えウイルスを昆虫細胞に感染させ、タンパク質を発現させることができる。
【0058】
宿主細胞の形質転換により得られた形質転換体は、例えば、適当な条件で培養し、得られた培養液から一般的な方法によって酵素の分取や精製を行うことにより本発明のβ−フルクトフラノシダーゼを得ることができる。また、宿主が枯草菌、酵母、カビの場合には、分泌型ベクターを使用して、菌体外に本発明のβ−フルクトフラノシダーゼを分泌させることも有利である。
【0059】
(E)本発明の酵素の製造方法
本発明によれば、本発明の形質転換体を常法にしたがって適当な培地に接種して適切な条件下で培養し、得られた培養物から本発明の酵素を採取してなる、本発明の酵素の製造方法を包含する。すなわち、本発明の酵素の製造方法は、(a)本発明の形質転換体を培養して、β−フルクトフラノシダーゼを含む形質転換体培養物を得る工程、及び(b)形質転換体培養物からβ−フルクトフラノシダーゼを得る工程を含む。
【0060】
本発明の形質転換体の培養に用いる栄養培地としては、炭素源、窒素源、無機物、および必要に応じ使用菌株の必要とする微量栄養素を程よく含有するものであれば、天然培地、合成培地のいずれでもよい。
【0061】
本発明の形質転換体の培養に用いる栄養培地の炭素源としては、該形質転換体が資化しうる物であればよく、例えば、グルコース、マルトース、フルクトース、マンノース、トレハロース、スクロース、マンニトール、ソルビトール、デンプン、デキストリン、糖蜜などの糖質、またはクエン酸、コハク酸などの有機酸、またはグリセリンなどの多価アルコールや脂肪酸も使用することができる。
【0062】
本発明の形質転換体の培養に用いる栄養培地の窒素源としては、各種有機および無機の窒素化合物、さらに培地は各種の無機塩を含むことができる。たとえば、コーンスティープリカー、大豆粕、あるいは各種ペプトン類等の有機窒素源、そして塩化アンモニウム、硫酸アンモニウム、尿素、硝酸アンモニウム、硝酸ナトリウム、リン酸アンモニウム等の無機窒素源などの化合物が使用可能である。また、グルタミン酸などのアミノ酸および尿素などの有機窒素源が炭素源にもなることはいうまでもない。さらに、ペプトン、ポリペプトン、バクトペプトン、肉エキス、酵母エキス、コーンスティープリカー、大豆粉、大豆粕、乾燥酵母、カザミノ酸、ソリュブルベジタブルプロテイン等の窒素含有天然物も窒素源として使用できる。
【0063】
本発明の形質転換体の培養に用いる栄養培地の無機物としては、例えば、カルシウム塩、マグネシウム塩、カリウム塩、ナトリウム塩、リン酸塩、マンガン塩、亜鉛塩、鉄塩、銅塩、モリブデン塩、コバルト塩などが適宜用いられる。具体的には、リン酸二水素カリウム、リン酸水素二カリウム、硫酸マグネシウム、硫酸第一鉄、硫酸マンガン、硫酸亜鉛、塩化ナトリウム、塩化カリウム、塩化カルシウム等が用いられる。さらに、必要に応じて、アミノ酸ならびにビオチンおよびチアミンなどの微量栄養素ビタミンなども適宜用いられる。
【0064】
培養法としては液体培養法(振とう培養法もしくは通気攪拌培養法)がよく、工業的には通気攪拌培養法が好ましい。培養温度とpHは、使用する形質転換体の増殖に最も適した条件を選べばよい。たとえば、形質転換体が微生物の場合の培養は、通常、温度20〜45℃、好ましくは25〜42℃、pH5〜9、好ましくは6〜8から選ばれる条件で好気的に行われる。培養時間は微生物が増殖し始める時間以上の時間であればよく、好ましくは8〜120時間であり、さらに好ましくは本発明の酵素が最大に生成する時間までである。微生物の増殖を確認する方法は特に制限はないが、たとえば、培養物を採取して顕微鏡で観察してもよいし、吸光度で観察してもよい。また、培養液の溶存酸素濃度には特に制限はないが、通常は、0.5〜20ppmが好ましい。そのために、通気量を調節したり、撹拌したり、通気に酸素を追加したりすればよい。培養方式は、回分培養、流加培養、連続培養または灌流培養のいずれでもよい。
【0065】
本発明の形質転換体の培養において、組換えベクターに選択マーカーを含有させている場合などでは、選択マーカーに対応した抗生物質などを栄養培地とともに加える。たとえば、選択マーカーとしてアンピシリン耐性遺伝子およびクロラムフェニコール耐性遺伝子を含有する場合は、適当な濃度に調製したアンピシリン溶液およびクロラムフェニコール溶液をそれぞれ加える。また、必要であれば、本発明の核酸を有する遺伝子の発現を誘導する発現誘導剤を培養開始時または培養開始から形質転換体の増殖を確認した後に加える。たとえば、発現誘導剤としては、イソプロピル−β−D−チオガラクトピラノシド(IPTG)を用いる。
【0066】
このようにして得られた培養物から、本発明の酵素を採取する。本発明の核酸、組換えベクター、及び形質転換体の態様によって、本発明の酵素は細胞内外に蓄積される。そこで、細胞内若しくは細胞外に生成蓄積された本発明の酵素を採取する。
【0067】
本発明の酵素の採取法は、一般の酵素の採取の手段に準じて行うことができる。本発明の酵素が細胞外に生成蓄積された場合は、通常知られる手段によって細胞を除いた後に培養上清を粗酵素として用いることができる。細胞内に生成蓄積される場合は、例えば、培養して得られた培養物を固液分離操作によって細胞と培養上清に分離し、得られた細胞を破砕したものを、粗酵素として用いることができる。固液分離には、通常知られる方法を制限なく利用することができ、例えば、培養物そのものをそのまま遠心分離する方法、培養物に濾過助剤を加えることや濾過助剤をプレコートしたプレコートフィルターなどにより濾過分離する方法、平膜、中空糸膜などを用いる膜濾過分離する方法などが採用される。細胞の破砕は、通常知られる方法を制限なく利用することができ、例えば、有機溶剤やリゾチームなどの酵素によって細胞を溶解する方法、超音波破砕法、凍結融解法、フレンチプレス法、ガラスビーズ破砕法、ダイノミル破砕法などを採用することができる。細胞破砕物を固液分離して得た分離液を、粗酵素として用いることもできる。さらに、固液分離した細胞は、破砕することなく、そのまま粗酵素として用いられ得る。
【0068】
粗酵素は、そのままで使用することもできるが、精製して使用することもできる。例えば、ここに挙げるものに限定されるものではないが、得られた粗酵素を、熱処理の如く耐熱性の差を利用する方法、透析、限外ろ過、レジンカラム、ゲルろ過、ゲルろ過クロマトグラフィー及びSDS−ポリアクリルアミドゲル電気泳動の如く分子量の差を利用する方法、塩沈澱、硫安沈殿、アルコール沈殿及びその他の溶媒沈澱の如く溶解性の差を利用する方法、DEAE−トヨパール樹脂などを用いるイオン交換クロマトグラフィーの如く電荷の差を利用する方法、アフィニティークロマトグラフィーの如く特異的親和性を利用する方法、ブチルトヨパール樹脂などを用いる疎水クロマトグラフィー及び逆相クロマトグラフィーの如く疎水性の差を利用する方法、吸着クロマトグラフィーの如く物理化学的な吸着性の差を利用する方法、等電点電気泳動及び等電点クロマトグラフィーなどの如く等電点の差を利用する方法などの通常知られる方法を単独または組み合わせて供することにより、工業用途の精製酵素を調製できる。
【0069】
粗酵素及び精製酵素は、固定化することもできる。例えば、イオン交換体への結合法、樹脂及び膜などとの共有結合・吸着法、高分子物質を用いた包括法などを採用することができる。
【0070】
(F)本発明のβ−フルクトフラノシル転移糖の製造方法
β−フルクトフラノシル供与体とβ−フルクトフラノシル受容体とを含有する溶液に、本発明の酵素若しくは本発明の形質転換体(例えば、細菌)又はその培養物を作用させることにより、β−フルクトフラノシル転移糖を生成させることができる。
【0071】
本発明の酵素は、固体状または液体状の粗酵素および/または精製酵素として利用することができる。ただし、該粗酵素はスクラーゼ等のβ−フルクトフラノシル転移糖の製造に悪影響を及ぼす酵素を含まない粗酵素であることを必要とする。さらに、該β−フルクトフラノシダーゼの活性を有する菌体および該菌体を適当な担体に包括、吸着あるいは化学的に結合させた固定化菌体などをβ−フルクトフラノシル転移糖の製造に使用することができる。さらには、本発明の酵素は、通常知られる方法で固定化させた固定化酵素として使用することもできる。
【0072】
β−フルクトフラノシル供与体とβ−フルクトフラノシル受容体とを含有する溶液における水性媒体としては、β−フルクトフラノシル転移反応を妨げるものでなければ特に制限されないが、例えば、水、緩衝液などが挙げられる。緩衝液としては、例えば、酢酸緩衝液、リン酸緩衝液、クエン酸緩衝液、コハク酸緩衝液、トリス塩酸緩衝液などが採用される。
【0073】
本発明の酵素の使用量は特に制限はないが、β−フルクトフラノシル転移糖の収率及び経済性の観点から、β−フルクトフラノシド供与体1gに対して、好ましくは0.1から50単位であり、より好ましくは0.5から20単位であり、さらに好ましくは0.8から5単位が好適である。β−フルクトフラノシル受容体の濃度は、溶液に溶解し得る範囲であれば特に限定されないが、工業的には10w/w%以上が好ましく、20〜60w/w%がより好ましい。反応時のβ−フルクトフラノシル受容体分子とβ−フルクトフラノシル供与体分子のモル比は、特に限定されるものではないが、β−フルクトフラノシル転移糖の収率及び経済性の観点から、1:5から5:1が望ましい。
【0074】
前記反応は雑菌汚染を避けるとともに収率を挙げるために、好ましくは50〜70℃、より好ましくは60〜65℃で行う。pHは本発明の酵素の最適条件下で行うことが好ましく、例えば、6.0〜8.0で行うのが適当である。上記条件で十分なβ−フルクトフラノシル転移糖生成が見られた時点で反応を終了するが、反応は通常1〜100時間で終了する。β−フルクトフラノシル転移糖の生成率は、酵素反応液の基質濃度、用いる受容体の種類、反応条件などによって異なる。例えば、22w/w%ショ糖と18w/w%乳糖を基質として用いた場合、ラクトスクロースの生成率は、固形分当り最大約30%である。
【0075】
反応終了後、通常は、反応液を60〜135℃、好ましくは65〜100℃に加熱して酵素を失活させるか、又はpHの低下(塩酸等の酸の添加)などの適当な手段によって酵素を失活させて反応を停止する。次いで、常法により、濾過、遠心分離などして不溶物を除去した後、活性炭による脱色、H型、OH型イオン交換樹脂による脱塩などの単離・精製手段を適宜組み合わせてβ−フルクトフラノシル転移糖を得ることができる。さらに、必要であれば、活性炭カラムクロマトグラフィーなどのクロマト操作、公知の固定床方式、移動床方式、擬似移動床方式などを単独又は組み合わせることにより高純度に濃縮及び精製したβ−フルクトフラノシル転移糖精製品を得ることができる。β−フルクトフラノシル転移糖精製品はシラップ状であるが、必要に応じて、通常知られる乾燥工程を加えて粉末状製品とすることもできる。
【0076】
本発明の製造方法によって得られるβ−フルクトフラノシル転移糖は、味質良好な甘味を有し、また、浸透圧調節性、保湿性、照付与性、結晶防止性、澱粉老化防止性などの性質、また、抗う蝕性、ビフィズス菌増殖促進性、ミネラル吸収促進性などの機能をも有し得ることから、広く飲食物、嗜好物、飼料、餌料、化粧品、医薬品、成形物など、更には、生活用品、農林水産用品、試薬、化学工業用品などの各種組成物に有利に利用され得る。
【0077】
以下の実施例により本発明をより詳細に説明するが、本発明は以下の実施例に限定されるものではない。
【実施例1】
【0078】
耐熱性β−フルクトフラノシダーゼの取得
(1)変異遺伝子ライブラリーの構築
寄託番号FERM P−10736/BP−3192のアルスロバクター・スピーシーズ K−1(Arthrobacter sp. K−1)株由来のβ−フルクトフラノシダーゼ遺伝子を含む組換え体プラスミドpBFF1(特開2000−342273公報記載)を大腸菌HB101よりHigh Pure Plasmid Isolation Kit(ロシュ・ダイアグノスティックス株式会社)を用いることにより抽出、精製した。得られた組換え体プラスミドを鋳型として、配列表のプライマーAとプライマーBを用い、Diversify PCR Random Mutagenesis Kit(クロンテック社)を用い、キットのプロトコールにしたがって、β−フルクトフラノシダーゼ遺伝子にランダム変異を導入した。変異導入されたβ−フルクトフラノシダーゼ遺伝子を含む1.8k塩基対のDNA断片を精製した後、制限酵素NdeIおよびBamHIで切断した。これとプラスミドベクターpET21a(インビトロジェン社)を制限酵素NdeIおよびBamHIで切断したものを Ligation Kit ver 2.1(タカラバイオ社)を用いて、16℃で3時間ライゲーションをした後に、大腸菌BL21StarTM(DE3)pLysS[F- ompT hsdSB(rB-mB-)gal dcm rne131 (DE3)pLysS(CamR)]株へ形質転換を行うことによって、ランダム変異の導入された組換えプラスミドを有する組換え大腸菌ライブラリーを構築した。
【0079】
(2)耐熱性β−フルクトフラノシダーゼのスクリーニング
前記(1)の操作で得られた上記形質転換体をLB−Amp−Cm寒天培地(バクトペプトン1%、酵母エキス0.5%、塩化ナトリウム1%、バクトアガー1.5%、アンピシリン50μg/ml、クロラムフェニコール35μg/ml)上で37℃、一晩培養してコロニーを形成させた。各コロニーのうち、3000個を別々にLB−Amp−Cm培地(バクトペプトン1%、酵母エキス0.5%、塩化ナトリウム1%、アンピシリン50μg/ml、クロラムフェニコール35μg/ml)を1mlずつ入れた96穴ディープウェルプレートに接種し、20℃、8時間振とう培養を行った。その後、各ウェルに100mM イソプロピル−β−D−チオガラクトピラノシド(IPTG)10μlを添加し、さらに20℃で16時間振とう培養を行った。培養終了後、培養液を3000rpm、10分間遠心分離を行い、菌体を得た。得られた菌体に、BugBuster(ノバジェン社)を各ウェルに100μlずつ添加し、室温で5分間撹拌後、50mMのリン酸緩衝液(pH6.5)を900μlずつ添加し、さらに室温で5分間撹拌後、3000rpm、10分間遠心分離を行い、粗酵素抽出液を得た。得られた粗酵素抽出液に対して、55℃で30分間の熱処理を行った。対照として、野生型酵素遺伝子が挿入されたプラスミド(pBFF1)を有し、野生型酵素(BFF1)を生産する形質転換体についても同様の操作を行った。得られた酵素液について、熱処理前後のβ−フルクトフラノシダーゼの活性測定を行った。活性測定は50mMのリン酸緩衝液(pH6.5)に溶解させた10%(w/v)のショ糖30μlに酵素液30μlを添加し、40℃で10分間反応させた後、100℃で15分間加熱して酵素反応を止めた。続いて4℃に冷却した後、生成したグルコースをグルコースオキシダーゼ法(和光純薬工業(株)製、グルコースC−IIテスト・ワコー)で測定した。ここで1単位の酵素活性は同条件下で1分間に1μmolのグルコースを生成する酵素量とした。対照となる野生型酵素よりも高い残存活性を示す変異体酵素を選別した。その結果、酵素遺伝子に変異が導入されたプラスミドを含有する形質転換体のうち、4株が生産する変異体酵素(P494、P500、F285、およびF2139)が高い残存活性を有していた。
【0080】
(3)変異箇所の解析
上記の如く得られた優良変異株に含有されるβ−フルクトフラノシダーゼ遺伝子に対して、遺伝子のどの部位が変異操作により置換されたのか確認をする為に以下のような解析を行った。上記(2)で得た上記組換え体に含まれるプラスミド(pP494、pP500、pF285、およびpF2139)をHigh Pure Plasmid Isolation Kitを用いて精製した。このプラスミドの塩基配列はMegaBACE 1000(GEヘルスケアバイオサイエンス株式会社)を用いて決定した。結果は表1に示した。
【表1】

【0081】
(4)55℃における残存活性の測定
上記(2)で得られた4株の生産する変異体酵素(P494、P500、F285、およびF2139)および上記(2)記載の野生型酵素BFF1の粗酵素溶液を55℃の恒温槽に30分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。測定の結果は、熱処理前の酵素活性を100%とした場合の残存活性(%)として表2に示した。
【表2】
【実施例2】
【0082】
BFF1、P494、P500、F285、およびF2139の組み合わせによる変異体の作製および評価
(1)BFF1のN末端側のアミノ酸とP494、P500、F285、F2139の有するC末端側のアミノ酸の組み合わせによる変異体の作製および評価
pBFF1を鋳型とし、BFF1のN末端側のアミノ酸(37−407番目)をコードする遺伝子断片nbffNを、プライマーAおよびCを用いたPCRにより増幅した。また、P494、P500、F285、およびF2139のC末端側(395−579番目)のアミノ酸をコードする遺伝子断片、np494C、np500C、nf285C、nf2139CをpP494、pP500、pF285、pF2139を各々鋳型とし、プライマーBおよびDを用いたPCRにより増幅した。得られた遺伝子断片nbffNにnp494C、np500C、nf285Cまたはnf2139CをプライマーAおよびBを用いたリコンビナントPCR法により連結し、さらにこれを発現用プラスミドpET21aに連結した後に、大腸菌BL21StarTM(DE3)pLysSを形質転換した。得られたコロニーを培養し、実施例1(2)に記載の方法に従い粗酵素液を調製した。β−フルクトフラノシダーゼ活性を示す株から得られた粗酵素液を56℃の恒温槽に30分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。耐熱性の比較対照として、野生型酵素であるBFF1を同様の試験に供した。測定の結果は、熱処理前の酵素活性を100%とした場合の相対活性(%)として表3に示した。表3に示すように、P494、P500およびF2139の各変異体が有する395番アミノ酸よりC末端側の変異によって56℃、30分間の熱処理時における耐熱性が向上することが分かった。
【表3】
【0083】
(2)P494のN末端側のアミノ酸と、P494、P500、F285、F2139のC末端側のアミノ酸の組み合わせによる変異体の作製および評価
次にpP494を鋳型とし、P494のN末端側のアミノ酸(37−407番目)をコードする遺伝子断片np494Nを、プライマーAおよびCを用いたPCRにより増幅した。一方、P494、P500、F285およびF2139のC末端側(395−579番目)のアミノ酸をコードする遺伝子断片、np494C、np500C、nf285C、およびnf2139Cを、pP494、pP500、pF285およびpF2139を各々鋳型とし、プライマーBおよびDを用いたPCRにより増幅した。得られた遺伝子断片np494Nにnp494C、np500C、nf285C、またはnf2139CをプライマーAおよびBを用いたリコンビナントPCR法により連結し、さらにこれを発現用プラスミドpET21aに連結した後に、大腸菌BL21StarTM(DE3)pLysSを形質転換した。得られたコロニーを培養し、実施例1(2)に記載の方法に従い粗酵素液を調製した。β−フルクトフラノシダーゼ活性を示す株から得られた粗酵素液を55℃の恒温槽に30分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。耐熱性の比較対照として、野生型酵素であるBFF1を同様の試験に供した。測定の結果は、熱処理前の酵素活性を100%とした場合の残存活性(%)として表4に示した。表4に示すように、P494、P500およびF2139の各変異体が有する395番アミノ酸よりC末端側の変異によって55℃、30分間の熱処理時における耐熱性が向上した。
【表4】
【0084】
(3)F285のN末端側のアミノ酸と、P494、F285、F2139のC末端側のアミノ酸の組み合わせによる変異体の作製および評価
pF285を鋳型とし、F285のN末端側のアミノ酸(37−407番目)をコードする遺伝子断片nf285Nを、プライマーAおよびCを用いたPCRにより増幅した。一方、P494、F285およびF2139のC末端側(395−579番目)のアミノ酸をコードする遺伝子断片、np494C、nf285C、またはnf2139Cを、pP494、pF285およびpF2139を各鋳型とし、プライマーBおよびDを用いたPCRにより増幅した。得られた遺伝子断片nf285Nにnp494C、nf285C、またはnf2139CをプライマーAおよびBを用いたリコンビナントPCR法により連結し、さらにこれを発現用プラスミドpET21aに連結した後に、大腸菌BL21StarTM(DE3)pLysSを形質転換した。得られたコロニーを培養し、実施例1(2)に記載の方法に従い粗酵素液を調製した。β−フルクトフラノシダーゼ活性を示す株から得られた粗酵素液を56℃の恒温槽に30分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。測定の結果は、熱処理前の酵素活性を100%とした場合の残存活性(%)として表5に示した。表3と5を比較して、F285の有する395番のアミノ酸よりN末端側の変異が本酵素の耐熱性に寄与していることが分かった。表5に記載する77%の残存活性を示した変異体を24D、87%の残存活性を示した変異体を26F、84%の残存活性を示した変異体を26Iと命名した
【表5】
【実施例3】
【0085】
24D、26Fおよび26Iアミノ酸変異の組み合わせによる変異体24Yの作製および評価
24Dをコードする遺伝子配列を解析したところ、24Dにおけるアミノ酸変異箇所はT47S、S200T、F447V、およびP500Sの4カ所であった。また、26Fをコードする遺伝子配列を解析したところ、D42G、T47S、Q98R、S200T、N208D、R222C、F470Y、およびG530Dの8カ所であった。また、26Iをコードする遺伝子配列を解析したところ、T47S、S200T、F470Y、G530DおよびF552Lの5カ所であった。次に、24Dにおいては野生型配列と同じアミノ酸残基であり(F470)、26Fと26Iにおいて共通するアミノ酸変異であるF470Yを24Dに導入することとした。より具体的には24Dをコードするプラスミドp24Dを鋳型とし、24DのN末端アミノ酸から変位導入箇所近傍を含む476番目のアミノ酸をコードする遺伝子断片n24yNを、プライマーAおよび470番アミノ酸をチロシンに変異させる配列を含むアンチセンスプライマー(プライマーE)を用いたPCRにより増幅した。また、p24Dを鋳型とし、466番目のアミノ酸からC末端アミノ酸をコードする遺伝子断片n24yCを、プライマーBおよび470番アミノ酸をチロシンに変異させる配列を含むセンスプライマー(プライマーF)を用いたPCRにより増幅した。得られた遺伝子断片n24yNとn24yCをプライマーAおよびBを用いたリコンビナントPCR法により連結し、さらにこれを発現用プラスミドpET21aに連結した後に、大腸菌BL21StarTM(DE3)pLysSを形質転換した。得られたコロニーからプラスミドを抽出、精製した後、塩基配列の決定を行い、目的の変異が導入されていることを確認した。このようにして、5重変異体24Yをコードする遺伝子を含むプラスミドp24Y、および24Yを生産する組換え大腸菌を得た。得られたコロニーを培養し、実施例1(2)に記載の方法に従い粗酵素液を調製した。β−フルクトフラノシダーゼ活性を示す株から得られた粗酵素液を60℃の恒温槽に30分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。測定の結果は、熱処理前の酵素活性を100%とした場合の相対活性(%)として表6に示した。表6に示すように、24Yは24D、26Fよりも優れた耐熱性を示すことが分かった。
【表6】
【実施例4】
【0086】
47位、200位、447位、470位、および500位、アミノ酸のサチュレーション変異
47位、200位、447位、470位、および500位のアミノ酸について、サチュレーション変異を行った。所定の変異を起こさせるよう設計した5種類のセンスプライマー:Gfw、Hfw、Ifw、Jfw、Kfwと5種類のアンチセンスプライマー;Grv、Hrv、Irv、Jrv、Krvを用い、GfwとGrv、HfwとHrv、IfwとIrv、JfwとJrv、KfwとKrvをそれぞれ一組とし、p24Yを鋳型として、実施例3と同様な手順により、形質転換を行なった。得られた形質転換体のプラスミドが有する変異型β−フルクトフラノシダーゼ遺伝子をプライマーAおよびBを用いてPCRにより増幅した。増幅断片の遺伝子配列の解析はMegaBACE 1000を用いて行なった。各々、200クローンについて、実施例1(2)に記載の方法に従い粗酵素液を調製した。β−フルクトフラノシダーゼ活性を示す株から得られた粗酵素液を60℃の恒温槽に30分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。アミノ酸のサチュレーション変異により得られた変異酵素のうち、サチュレーション変異を施した各箇所が野生型アミノ酸に戻った4重変異酵素の示す残存活性を比較対照とした。対照よりも高い残存活性を示した変異体のアミノ酸配列を確認したところ、その内訳は、変異箇所47番アミノ酸では、S、E、Gであり、200番アミノ酸では、T、Lであり、447番アミノ酸では、P、I、V、K、Q、R、A、Y、H、T、S、W、Cであり、470番アミノ酸では、Y、A、E、Vであり、500番アミノ酸では、V、C、W、S、であった。これらの結果より、47位、200位、447位、470位、または500位の各アミノ酸が野生型から前述の各アミノ酸に置換されることにより、本酵素の耐熱性が向上することが明らかとなった。
【実施例5】
【0087】
24Y、24Y447P、24Y447I、24Y447FおよびBFF1の酵素溶液の作製
実施例3で得られたβ−フルクトフラノシダーゼ変異体である24Y、実施例4で得られた変異体酵素であり、24Yの447番目のアミノ酸がプロリン、イソロイシン、または野生型と同じフェニルアラニンに置換された変異体酵素(24Y447P、24Y447I、24Y447F)を生産する組換え大腸菌を各々培養し、大腸菌による組換え酵素の調製を行なった。より具体的な手順を以下に示す。培養は容量2Lの三角フラスコにLB培地(バクトペプトン1%、酵母エキス0.5%、塩化ナトリウム1%)0.5Lを入れて滅菌した後、アンピシリン溶液を終濃度で50μg/ml、クロラムフェニコール溶液を終濃度で35μg/mlになるように添加し、種培養液を1%になるように接種して、温度20℃、130rpmでOD600が0.5になるまで培養した。その後、IPTGを終濃度で0.5mMになるように添加し、さらに16時間培養を行った。培養終了後、培養液を5000g、10分間遠心分離を行い、菌体を得た。得られた大腸菌に50mM リン酸緩衝液(pH6.5) 20mlを添加し、細胞を超音波破砕した後に12000g、15分間遠心分離を行った。得られた粗抽出液をハイドロキシアパタイトカラム(生化学工業社製)によるクロマトグラフィー[(洗浄、50mM リン酸緩衝液(pH6.5);溶出、200mM リン酸緩衝液(pH6.5)]に供した。β−フルクトフラノシダーゼ活性を示す画分を集め、50mM リン酸緩衝液(pH6.5)で平衡化した陰イオン交換カラム(SuperQ 650Mカラム、東ソー社製)に供した。続いて、0−500mMのNaCl濃度勾配法により、酵素タンパクの溶出を行なった。β−フルクトフラノシダーゼ活性を示す画分を集め、限外ろ過膜(PM−10、Amicon社製)による濃縮を行なった後、50mM リン酸緩衝液(pH6.5)に緩衝液を置換し、酵素液を得た。比較対照として、野生型酵素であるBFF1についても、大腸菌による組換え酵素の調製を同様の手順で行なった。
【実施例6】
【0088】
酵素の半減期測定
実施例5の方法で得た本発明の酵素液を55℃または60℃の恒温槽に一定時間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。酵素の半減期を表7として示した。表7に示すように、447番目のアミノ酸は耐熱性向上に寄与していることが明らかとなった。さらに447番目のアミノ酸がプロリンの場合が最も高い耐熱性を示した。
【表7】
【実施例7】
【0089】
(1)24Y47T、24Y200S、24Y470F、24Y500P、24Y500C、24Y500Vの酵素溶液の作製
実施例4で得られた変異体酵素のうちから、24Yの47番目のアミノ酸が野生型と同じであるスレオニンに置換された変異体酵素(24Y47T)、200番目のアミノ酸が野生型と同じであるセリンに置換された変異体酵素(24Y200S)、470番目のアミノ酸が野生型と同じであるフェニルアラニンに置換された変異体酵素(24Y470F;実施例2(3)記載の24Dと同じアミノ酸配列を有する変異酵素)、500番目のアミノ酸が野生型と同じであるプロリンに置換された変異体酵素(24Y500P)、500番目のアミノ酸が野生型と同じであるシステインに置換された変異体酵素(24Y500C)、500番目のアミノ酸が野生型と同じであるバリンに置換された変異体酵素(24Y500V)を選択し、実施例5と同様の手順で大腸菌による組換え酵素の調製を行なった。
【0090】
(2)4重変異体、24Y200S447P500C、24Y200S447P500V、24Y200S447P500Wをコードするプラスミドと各酵素溶液の作製
実施例7(1)記載の4重変異体24Y200Sの447番目のアミノ酸であるバリン(野生型ではフェニルアラニン)がプロリンに置換し、且つ24Y200Sの500番目のアミノ酸であるセリン(野生型ではプロリン)がシステイン、バリンまたはトリプトファン置換した4重変異体を作製した。より具体的には24Y200Sをコードするプラスミドp24Y200Sを鋳型とし、24Y200SのN末端アミノ酸から変位導入箇所近傍を含む453番目のアミノ酸をコードする遺伝子断片n24y200SNを、プライマーAおよび447番アミノ酸をプロリンに変異させる配列を含むアンチセンスプライマー(プライマーL)を用いたPCRにより増幅した。また、p24Y500C、p24Y500Vまたはp24Y500Wを鋳型とし、441番目のアミノ酸からC末端アミノ酸をコードする遺伝子断片n24y500C、n24y500V、n24y500Wを、プライマーBおよび447番アミノ酸をプロリンに変異させる配列を含むセンスプライマー(プライマーM)を用いたPCRにより増幅した。得られた遺伝子断片n24y200SNに対して、n24y500C、n24y500Vまたはn24y500WをプライマーAおよびBを用いたリコンビナントPCR法により各々連結し、さらにこれを発現用プラスミドpET21aに連結した後に、大腸菌BL21StarTM(DE3)pLysSを形質転換した。得られたコロニーからプラスミドを抽出、精製した後、塩基配列の決定を行い、目的の変異が導入されていることを確認した。このようにして、4重変異体24Y200S447P500C、24Y200S447P500V、p24Y200S447P500Wをコードする遺伝子を含むプラスミドp24Y200S447P500C、p24Y200S447P500V、およびp24Y200S447P500Wを生産する組換え大腸菌を得た。得られた各組換え大腸菌より、実施例5と同様の手順で大腸菌による各組換え酵素の調製を行なった。
【0091】
(3)5重変異体の作製24Y447P500C、24Y447P500V、24Y447P500Wをコードするプラスミドと各酵素溶液の作製
実施例4で取得した5重変異体である24Y447Pの500番目のアミノ酸であるセリン(野生型ではプロリン)がシステイン、バリンまたはトリプトファンに置換した5重変異体3種を作製した。より具体的には24Yをコードするプラスミドp24Yを鋳型とし、24YのN末端アミノ酸から変位導入箇所近傍を含む453番目のアミノ酸をコードする遺伝子断片n24yN2を、プライマーAおよび447番アミノ酸をプロリンに変異させる配列を含むアンチセンスプライマー(プライマーL)を用いたPCRにより増幅した。また、p24Y500C、p24Y500Vまたはp24Y500Wを鋳型とし、441番目のアミノ酸からC末端アミノ酸をコードする遺伝子断片n24y500C、n24y500V、n24y500Wを、プライマーBおよび447番アミノ酸をプロリンに変異させる配列を含むセンスプライマー(プライマーM)を用いたPCRにより増幅した。得られた遺伝子断片n24yN2に対して、n24y500C、n24y500Vまたはn24y500WをプライマーAおよびBを用いたリコンビナントPCR法により各々連結し、さらにこれを発現用プラスミドpET21aに連結した後に、大腸菌BL21StarTM(DE3)pLysSを形質転換した。得られたコロニーからプラスミドを抽出、精製した後、塩基配列の決定を行い、目的の変異が導入されていることを確認した。このようにして、5重変異体24Y447P500C、24Y447P500V、p24Y447P500Wをコードする遺伝子を含むプラスミドp24Y447P500C、p24Y447P500V、およびp24Y447P500Wを生産する組換え大腸菌を得た。得られた各組換え大腸菌より、実施例5と同様の手順で大腸菌による各組換え酵素の調製を行なった。
【実施例8】
【0092】
酵素の耐熱性試験
実施例5および7(1)〜(3)の方法で得た本発明の酵素液(精製酵素)を45〜65度の各温度の恒温槽に180分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。測定の結果は、熱処理前の酵素活性を100%とした場合の残存活性(%)として図1に示した。図1に示す通り、本発明の変異体酵素24Y47T、24Y200S、24Y447F、24Y470F、24Y500P、24Y200S447P500C、24Y200S447P500V、24Y200S447P500W、24Y、24Y500C、24Y500V、24Y500W、24Y447P500C、24Y447P500V、および24Y447P500Wは野生型酵素BFF1と比較して耐熱性が著しく向上していることが明らかとなった。
【実施例9】
【0093】
糖転移試験
22%(W/W)ショ糖と18%(W/W)乳糖を含む水溶液(pH6.0、50mMリン酸緩衝液)に、ショ糖1gあたり1.0Uの実施例5および7(1)〜(3)の方法で得た本発明の酵素液(精製酵素)を添加し、50から75℃まで温度を変化させ、10時間転移反応を行った。反応液は100℃で10分間熱処理した後、HPLCに供し、生成したラクトスクロース(LS)を測定した(カラム:TSK−GEL Amide−80、移動相:70%アセトニトリル、流速:1.0mL/分、カラム温度:35℃、検出器:示差屈折計)。その結果、図2に示したように野生型酵素(BFF1)ではLS生成が急激に低下する60℃以上の高温域において耐熱性が向上した変異型酵素は高いLS生成量を維持することが明らかとなった。
【図面の簡単な説明】
【0094】
【図1】図1は、本発明の酵素及び野生型の酵素の耐熱性試験結果を示す。
【図2】図2は、本発明の酵素及び野生型の酵素の糖転移試験結果を示す。
【技術分野】
【0001】
本発明は、新規なβ−フルクトフラノシダーゼ、該β−フルクトフラノシダーゼを製造するための核酸、組換えベクター及び形質転換体、該β−フルクトフラノシダーゼの製造方法、並びに該β−フルクトフラノシダーゼを利用したβ−フルクトフラノシル転移糖の製造方法に関する。
【背景技術】
【0002】
ラクトスクロースやフルクトオリゴ糖などのβ−フルクトフラノシル転移糖は、抗う蝕性及びミネラル吸収促進性などの性質を有し、さらに腸内細菌の栄養源として機能し得る。β−フルクトフラノシル転移糖を含む飲食料品は、虫歯になり難く、かつ整腸作用のある特定保健用食品として、全国で流通されている。さらに上記性質を有することから、β−フルクトフラノシル転移糖は、嗜好物、飼料、餌料、化粧品、医薬品、成形物、生活用品、農林水産用品、試薬、化学工業用品などの多岐用途への利用が期待されている。
【0003】
β−フルクトフラノシル転移糖を製造する方法には、酵素法がある。酵素法は、ショ糖などのβ−フルクトフラノシル基を有する供与体(以下、「β−フルクトフラノシル供与体」と呼ぶ場合もある)及び受容体(以下、「β−フルクトフラノシル受容体」と呼ぶ場合もある)を含有する溶液に、β−フルクトフラノシル供与体のβ−フルクトフラノシル基をβ−フルクトフラノシル受容体に転移させるβ−フルクトフラノシダーゼ活性を有する酵素を作用させることにより、β−フルクトフラノシル転移糖を製造する。β−フルクトフラノシル基を転移させる酵素としては、アスペルギルス・ニガー(Aspergillus niger FERM-P5886, ATCC20611;非特許文献1を参照)、ペニシリウム・ロッケフォルチ(Penicillium roqueforti IAM7254;特許文献1を参照)、スコプラリオプシス・ブレヴィカウリス(Scopulariopsis brevicaulis IFO4843;特許文献1を参照)などの真菌に由来するβ−フルクトフラノシダーゼ、並びにアルスロバクター・スピーシーズ(Arthrobacter sp. K-1 FERM P-10736/BP-3192;特許文献2を参照)、バチルス・メガテリウム(Bacillus megaterium IFO13498;特許文献3を参照)、バチルス・スピーシーズ(Bacillus sp. V230 FERM BP-5054;特許文献4を参照)などの細菌に由来するβ−フルクトフラノシダーゼなどが知られている。特に、細菌由来のβ−フルクトフラノシダーゼは、真菌由来の酵素と比較して、受容体糖質の選択の幅が広いことから、β−フルクトフラノシル転移糖であるラクトスクロースやキシロシルフラクトシドの製造に有用である。
【特許文献1】特許第3628336号公報
【特許文献2】特許第2781412号公報
【特許文献3】特開平4−200386号公報
【特許文献4】特開平9−224665号公報
【非特許文献1】H.Hidaka, M.Hirayama and N.Sumi:Agric. Biol. Chem.,52,1181 (1998)
【発明の開示】
【発明が解決しようとする課題】
【0004】
一般的に、酵素を利用した物の製造方法は、雑菌汚染を回避しつつ、反応速度を高めるために、高温条件下で行われることが望ましい。ところが、これまでに知られている細菌由来のβ−フルクトフラノシダーゼは、温度安定性が45℃程度までであり、50℃を超えると失活するという性質を有していた。その結果、従前の細菌由来のβ−フルクトフラノシダーゼを利用したβ−フルクトフラノシル転移糖の製造は、45℃以下の低温条件下で実施されており、雑菌汚染が発生し易く、かつ酵素活性が低いために酵素の添加量が多くなり経済性に欠けるという問題があった。
【0005】
そこで、本発明者らは、雑菌汚染の回避に有効な、例えば、50〜65℃といった高温下において安定性の高い新規なβ−フルクトフラノシダーゼ及び該β−フルクトフラノシダーゼの製造方法を提供することを、本発明の解決すべき課題とした。さらに、本発明者らは、該β−フルクトフラノシダーゼを利用することにより、雑菌汚染を回避しつつ、添加酵素量を低減させてβ−フルクトフラノシル転移糖を製造することができる方法を提供することを本発明の解決すべき課題とした。
【課題を解決するための手段】
【0006】
本発明者らは、上記課題を解決するために鋭意研究した結果、特許文献2に記載のアルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192に由来するβ−フルクトフラノシダーゼのアミノ酸配列において、酵素の耐熱性に寄与するアミノ酸残基を見出すことに成功した。さらに、本発明者らは、該アミノ酸残基を所定のアミノ酸に置換することにより、高温下で安定性のあるβ−フルクトフラノシダーゼを得ることに成功した。本発明は、かかる知見に基づいて完成されたものである。
【0007】
したがって、本発明によれば、下記(A)〜(C)に記載の改変アミノ酸配列A、改変アミノ酸配列B又は改変アミノ酸配列Cを含むβ−フルクトフラノシダーゼが提供される。
(A)配列表の配列番号1に記載のアミノ酸配列において、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸が下記[1]〜[5]に示す如く置換された改変アミノ酸配列A
[1]47位のスレオニンについて、セリン、グルタミン酸又はグリシンへの置換
[2]200位のセリンについて、スレオニン又はロイシンへの置換
[3]447位のフェニルアラニンについて、プロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインへの置換
[4]470位のフェニルアラニンについて、チロシン、アラニン、グルタミン酸又はバリンへの置換
[5]500位のプロリンについて、バリン、システイン、トリプトファン又はセリンへの置換
(B)前記改変アミノ酸配列Aにおいて、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸の置換の他さらに1から数個のアミノ酸の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列B
(C)前記改変アミノ酸配列Aと70%以上の相同性を有し、かつ前記改変アミノ酸配列Aの47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸の置換を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列C
【0008】
本発明のβ−フルクトフラノシダーゼの好ましい態様は、前記改変アミノ酸配列Aが、
200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び470位のフェニルアラニンがチロシンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;又は
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列である。
【0009】
本発明の別の側面によれば、本発明のβ−フルクトフラノシダーゼをコードする核酸が提供される。
【0010】
本発明の別の側面によれば、下記(a)〜(c)に記載の改変塩基配列a、改変塩基配列b又は改変塩基配列cを含むβ−フルクトフラノシダーゼをコードする核酸が提供される。
(a)配列表の配列番号2に記載の塩基配列において、139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の三塩基連鎖が下記[6]〜[10]に示す如く置換された改変塩基配列a
[6]139〜141位の三塩基連鎖について、セリン、グルタミン酸又はグリシンを規定するコドンに相補的な三塩基連鎖への置換
[7]598〜600位の三塩基連鎖について、スレオニン又はロイシンを規定するコドンに相補的な三塩基連鎖への置換
[8]1339〜1341位の三塩基連鎖について、プロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインを規定するコドンに相補的な三塩基連鎖への置換
[9]1408〜1410位の三塩基連鎖について、チロシン、アラニン、グルタミン酸又はバリンを規定するコドンに相補的な三塩基連鎖への置換
[10]1498〜1500位の三塩基連鎖について、バリン、システイン、トリプトファン又はセリンを規定するコドンに相補的な三塩基連鎖への置換
(b)前記改変塩基配列aにおいて、前記139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の三塩基連鎖の置換の他さらに1から数個の塩基の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードする改変塩基配列b
(c)前記改変塩基配列aとストリンジェントな条件下でハイブリダイズし、前記改変塩基配列aの139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位からなる群から選ばれる少なくとも1種の置換された三塩基連鎖と相補的な三塩基連鎖を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードする改変塩基配列c
【0011】
本発明の別の側面によれば、本発明の核酸を含む組換えベクターが提供される。
【0012】
本発明の別の側面によれば、本発明の核酸を導入してなる、又は本発明の組換えベクターを含む形質転換体が提供される。
【0013】
本発明の別の側面によれば、(a)本発明の形質転換体を培養して、β−フルクトフラノシダーゼを含む形質転換体培養物を得る工程、及び
(b)形質転換体培養物からβ−フルクトフラノシダーゼを得る工程
を含むβ−フルクトフラノシダーゼの製造方法が提供される。
【0014】
本発明の別の側面によれば、β−フルクトフラノシル供与体及びβ−フルクトフラノシル受容体を含有する溶液に、本発明のβ−フルクトフラノシダーゼ、又は本発明の形質転換体を作用させて、β−フルクトフラノシル転移糖を得ることを特徴とするβ−フルクトフラノシル転移糖の製造方法が提供される。
【0015】
本発明のβ−フルクトフラノシル転移糖の製造方法の好ましい態様は、β−フルクトフラノシル供与体が、ショ糖、ラフィノース及びエルロースからなる群から選ばれる少なくとも1種である。
【0016】
本発明のβ−フルクトフラノシル転移糖の製造方法の好ましい態様は、β−フルクトフラノシル受容体が、糖質、糖アルコール及びアルコールからなる群から選ばれる少なくとも1種である。
【0017】
本発明のβ−フルクトフラノシル転移糖の製造方法の好ましい態様は、β−フルクトフラノシル転移糖が、キシロシルフラクトシド、ガラクトシルフラクトシド、フコシルフラクトシド、ソルボシルフラクトシド、マルトオリゴシルフラクトシド、イソマルトシルフラクトシド、及びラクトシルフラクトシドからなる群から選ばれる少なくとも1種である。
【発明の効果】
【0018】
本発明によれば、工業的実施に有利な50〜65℃の高温下におけるβ−フルクトフラノシル転移糖の製造に利用可能なβ−フルクトフラノシダーゼを効率よく得ることができる。さらに、本発明によれば、雑菌汚染を回避しつつ、添加酵素量を低減させても、工業的規模で効率よくβ−フルクトフラノシル転移糖を製造することができる。したがって、本発明によれば、β−フルクトフラノシル転移糖を含む飲食物、化粧品、医薬品、成形物などの開発を促進することが期待できる。
【発明を実施するための最良の形態】
【0019】
以下、本発明の実施の形態について詳細に説明する。
(1)本発明の酵素
本発明者らは、特許文献2に記載されている、寄託番号FERM P-10736/BP-3192のアルスロバクター・スピーシーズ K-1(Arthrobacter sp. K-1 FERM P-10736/BP-3192)由来のβ−フルクトフラノシダーゼ遺伝子にランダム変異を導入し、得られた変異β−フルクトフラノシダーゼ遺伝子によって形質転換した約3000クローンから成る形質転換体ライブラリーを構築した。該形質転換体ライブラリーから、55℃の加熱処理後においても酵素活性が残存する変異β−フルクトフラノシダーゼの取得を目的として、該変異β−フルクトフラノシダーゼを発現する形質転換体をスクリーニングした。また、本発明者らは、スクリーニングにより得た変異β−フルクトフラノシダーゼのアミノ酸配列から、耐熱性に寄与するアミノ酸残基の部位と種類を特定し、本発明の酵素を構築することに成功した。
【0020】
本発明の酵素は、下記(A)〜(C)に記載の改変アミノ酸配列A、改変アミノ酸配列B又は改変アミノ酸配列Cを含むβ−フルクトフラノシダーゼである。
(A)配列表の配列番号1に記載のアミノ酸配列において、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸が下記[1]〜[5]に示す如く置換された改変アミノ酸配列A
[1]47位のスレオニンについて、セリン、グルタミン酸又はグリシンへの置換
[2]200位のセリンについて、スレオニン又はロイシンへの置換
[3]447位のフェニルアラニンについて、プロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインへの置換
[4]470位のフェニルアラニンについて、チロシン、アラニン、グルタミン酸又はバリンへの置換
[5]500位のプロリンについて、バリン、システイン、トリプトファン又はセリンへの置換
(B)前記改変アミノ酸配列Aにおいて、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸の置換の他さらに1から数個のアミノ酸の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列B
(C)前記改変アミノ酸配列Aと70%以上の相同性を有し、かつ前記改変アミノ酸配列Aの47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸の置換を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列C
【0021】
本発明の酵素は、pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有する。本発明の酵素のβ−フルクトフラノシダーゼ活性は、pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して、好ましくは70%以上、より好ましくは75%以上、さらに好ましくは80%以上、なおさらに好ましくは90%以上である。本発明の酵素のより好ましい態様として、pH6.5、60℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有する酵素;pH6.5、60℃、180分間の加熱処理後において、未処理の酵素に対して70%以上のβ−フルクトフラノシダーゼ活性を有する酵素;及び、pH6.5、60℃、180分間の加熱処理後において、未処理の酵素に対して75%以上のβ−フルクトフラノシダーゼ活性を有する酵素を挙げることができる。ただし、本発明の酵素は、上記好ましい態様に挙げた酵素に限定されるものではない。
【0022】
β−フルクトフラノシダーゼは、一般的に、ショ糖を加水分解してフルクトースを遊離させる活性(加水分解活性)、並びにショ糖のβ−フルクトフラノシル基を各種単糖、糖アルコール、アルキルアルコール、配糖体、オリゴ糖等の受容体に転移させる活性(β−フルクトフラノシル転移活性)を有する酵素である。
【0023】
本発明の酵素は、上記β−フルクトフラノシダーゼの触媒作用と同様の触媒作用を有することから、β−フルクトフラノシダーゼ(β-fructofuranosidase;EC 3.2.1.26)に属する酵素であると推測される。
【0024】
本発明の酵素を用いたβ−フルクトフラノシル転移反応に用いられるβ−フルクトフラノシル受容体は、特に限定されるものではなく、例えば、D−キシロース、D−ガラクトース、L−アラビノース、L−ソルボース、L−フコース、マルトース、スクロース、セロビオース、キシロビオース、イソマルトース、ラクトース、マルトトリオース、イソマルトトリオース、パノース、イソパノースなどを挙げることができる。本発明の酵素を用いたβ−フルクトフラノシル転移反応の結果として生成するβ−フルクトフラノシル転移糖としては、例えば、キシロシルフラクトシド、ガラクトシルフラクトシド、ラクトスクロース(別名、ラクトシルフラクトシド)、エルロース(別名、マルトシルフラクトシド)、イソマルトシルフラクトシドなどを挙げることができる。さらに、トレハロース、ネオトレハロースなどのグルコースからなる非還元性二糖類をβ−フルクトフラノシル受容体として、同様に本発明の酵素を作用させた場合、フラクトシルトレハロース、フラクトシルネオトレハロースなどのβ−フルクトフラノシル転移糖を生成し得る。単糖類、二糖類、三糖類のみならず、それ以上の重合度のキシロオリゴ糖、ガラクトオリゴ糖、マルトオリゴ糖、イソマルトオリゴ糖などのオリゴ糖類、ソルビトール、マルチトールなどの糖アルコール類もβ−フルクトフラノシル受容体となり得る。β−フルクトフラノシル受容体は、1種に限らず複数であってもよく、これらの混合物でもよく、澱粉、アラビノガラクタン、キシログルカンなどの多糖類の部分加水分解物でもよい。β−フルクトフラノシル供与体としては、ショ糖以外に、例えば、ラフィノース、エルロースなどのβ−フルクトフラノシド結合を有するオリゴ糖を用いることもできる。
【0025】
β−フルクトフラノシダーゼの加熱処理及びβ−フルクトフラノシダーゼ活性の測定は、以下の方法で実施される。
加熱処理は、β−フルクトフラノシダーゼを含む50mMのリン酸緩衝液(pH6.5)(以下、「酵素液」と呼ぶ場合がある)を45〜65℃で30〜180分間加熱することにより行う。β−フルクトフラノシダーゼ活性の測定は、50mMのリン酸緩衝液(pH6.5)に溶解させた10%(w/v)のショ糖30μlに加熱処理後の酵素液30μlを添加し、40℃で10分間反応させた後、100℃で15分間加熱して酵素反応を止める。続いて4℃に冷却した後、生成したグルコースをグルコースオキシダーゼ法(和光純薬工業(株)製、グルコースC−IIテスト・ワコー)で測定する。対照として、加熱処理に付していない未処理の酵素(β−フルクトフラノシダーゼ)のβ−フルクトフラノシダーゼ活性を上記の操作により測定する。ここで1単位の酵素活性は同条件下で1分間に1μmolのグルコースを生成する酵素量とする。
【0026】
β−フルクトフラノシダーゼの基質特異性は、以下の方法で測定される。
前記のβ−フルクトフラノシダーゼ活性の測定法において、使用する基質をショ糖の代わりに、エルロース、キシロシルフラクトシド、ラフィノース、ネオケストース、スタキオース、1−ケストース、ニストース、イヌロビオース、又はレバンビオースを基質としてこれらの基質の分解活性を調べることで測定する。
【0027】
β−フルクトフラノシダーゼの至適pHとpH安定性は、以下の方法で測定される。
至適pHは、前記のβ−フルクトフラノシダーゼ活性の測定法において、50mMリン酸緩衝液(pH6.5)の代わりに50mM Mcllvaine 緩衝液(pH3.0〜8.0)、50mM Kolthoff(pH8.0〜11.0)を用いて測定する。上記条件で最も高い酵素活性(以下、「最大活性」と略す)を有するpHを至適pHとする。
一方、pH安定性は、50mM Mcllvaine 緩衝液(pH3.0〜8.0)、50mM Kolthoff(pH8.0〜11.0)を用い、40℃で2時間処理した後、残存する酵素活性を求めて測定する。上記条件下における最大活性の値を基準(100%)として約80%以上の活性を示すpHを、pH安定性を示すpHまたは単に安定なpHという。
【0028】
β−フルクトフラノシダーゼの温度安定性は、以下の方法で測定される。
酵素液を各温度に設定した恒温槽に一定時間静置した後、4℃に冷却し、β−フルクトフラノシダーゼ活性を測定する。未処理の酵素活性の値を基準(100%)として残存活性(%)を求める。同条件下において60%以上の残存活性の値を示す温度を、温度安定性を示す温度、又は単に安定な温度という。
【0029】
β−フルクトフラノシダーゼの阻害剤は、各種化合物の存在下におけるβ−フルクトフラノシダーゼ活性を求めることで測定される。
【0030】
本発明の酵素のアミノ酸配列である改変アミノ酸配列は、配列表の配列番号1に記載のアミノ酸配列において、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる1種、好ましくは2種、より好ましくは3種、さらに好ましくは4種、なおさらに好ましくは5種のアミノ酸がそれぞれ別のアミノ酸に置換されたアミノ酸配列である。改変アミノ酸配列は、アミノ酸残基の置換の態様によって改変アミノ酸配列A〜Cに分類される。改変アミノ酸配列Aは、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる1種又は2種以上のアミノ酸がそれぞれ別のアミノ酸に置換されたアミノ酸配列である。具体的には、改変アミノ酸配列Aは、配列表の配列番号1に記載のアミノ酸配列において、47位のスレオニンについてセリン、グルタミン酸又はグリシンへの置換、200位のセリンについてスレオニン又はロイシンへの置換、447位のフェニルアラニンについてプロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインへの置換、470位のフェニルアラニンについてチロシン、アラニン、グルタミン酸又はバリンへの置換、及び500位のプロリンについてバリン、システイン、トリプトファン又はセリンへの置換からなる群から選ばれる1種又は2種以上の置換を有するものである。改変アミノ酸配列Aの好ましい態様としては、配列表の配列番号1に記載のアミノ酸配列において、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び470位のフェニルアラニンがチロシンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列などを挙げることができるが、これらに限定されるものではない。
【0031】
改変アミノ酸配列Bにおける「1から数個のアミノ酸の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異」における「1から数個」の範囲は、改変アミノ酸配列Bを有するタンパク質がpH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を維持できる程度の範囲であれば特に限定されないが、たとえば、1から20個、好ましくは1から10個、より好ましくは1から7個、さらに好ましくは1から5個、特に好ましくは1から3個程度を意味する。また、「アミノ酸の欠失」とは配列中のアミノ酸残基の欠落もしくは消失を意味し、「アミノ酸の置換」は配列中のアミノ酸残基が別のアミノ酸残基に置き換えられていること、「アミノ酸の逆位」とは隣り合う2以上のアミノ酸残基の位置が逆になっていること、「アミノ酸の付加」とはアミノ酸残基が付け加えられていること、「アミノ酸の挿入」とは配列中のアミノ酸残基の間に別のアミノ酸残基が挿し入れられていることをそれぞれ意味する。改変アミノ酸配列Bは、配列表の配列番号1に記載のアミノ酸配列おける47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種類のアミノ酸が別のアミノ酸に置換されており、さらにこれら以外に1から数個のアミノ酸の欠失等を有するアミノ酸配列を意味する。
【0032】
改変アミノ酸配列Bにおける「1から数個のアミノ酸の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異」の具体的な態様としては、1から数個のアミノ酸が別の化学的に類似したアミノ酸で置き換えらた態様がある。例えば、ある疎水性アミノ酸を別の疎水性アミノ酸に置換する場合、ある極性アミノ酸を同じ電荷を有する別の極性アミノ酸に置換する場合などを挙げることができる。斯く化学的に類似したアミノ酸は、アミノ酸毎に当該技術分野において知られている。具体例を挙げると、非極性(疎水性)アミノ酸としては、アラニン、バリン、イソロイシン、ロイシン、プロリン、トリプトファン、フェニルアラニン、メチオニンなどが挙げられる。極性(中性)アミノ酸としては、グリシン、セリン、スレオニン、チロシン、グルタミン、アスパラギン、システインなどが挙げられる。陽電荷をもつ(塩基性)アミノ酸としては、アルギニン、ヒスチジン、リジンなどが挙げられる。また、負電荷をもつ(酸性)アミノ酸としては、アスパラギン酸、グルタミン酸などが挙げられる。
【0033】
改変アミノ酸配列Cにおける「相同性」は、改変アミノ酸配列Cを有するタンパク質がpH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を維持できる程度の相同性であれば特に限定されないが、好ましくは70%以上が好ましく、より好ましくは80%以上、さらに好ましくは85%以上、なおさらに好ましくは90%以上、特に好ましくは95%以上である。ただし、改変アミノ酸配列Cは、配列表の配列番号1に記載のアミノ酸配列おける47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸が別のアミノ酸に置換されている。
【0034】
本発明の酵素を取得する方法は制限されるものではなく、本明細書に記載されている方法以外に、例えば、配列表の配列番号1の記載を参照して、物理化学的に合成してもよいし、本発明の酵素を発現する生物体から得てもよいし、遺伝子工学的に改変アミノ酸配列をコードする核酸から作成してもよいし、その他の通常知られる手段を用いた各種スクリーニングにより取得してもよい。
【0035】
(B)本発明の核酸
本発明の核酸は、本発明のβ−フルクトフラノシダーゼをコードする核酸である。本発明の核酸の好ましい態様は、例えば、下記(a)〜(c)に記載の改変塩基配列a、改変塩基配列b又は改変塩基配列cを含むβ−フルクトフラノシダーゼをコードする核酸である。
(a)配列表の配列番号2に記載の塩基配列において、139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の三塩基連鎖が下記[6]〜[10]に示す如く置換された改変塩基配列a
[6]139〜141位の三塩基連鎖について、セリン、グルタミン酸又はグリシンを規定するコドンに相補的な三塩基連鎖への置換
[7]598〜600位の三塩基連鎖について、スレオニン又はロイシンを規定するコドンに相補的な三塩基連鎖への置換
[8]1339〜1341位の三塩基連鎖について、プロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインを規定するコドンに相補的な三塩基連鎖への置換
[9]1408〜1410位の三塩基連鎖について、チロシン、アラニン、グルタミン酸又はバリンを規定するコドンに相補的な三塩基連鎖への置換
[10]1498〜1500位の三塩基連鎖について、バリン、システイン、トリプトファン又はセリンを規定するコドンに相補的な三塩基連鎖への置換
(b)前記改変塩基配列aにおいて、前記139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の三塩基連鎖の置換の他さらに1から数個の塩基の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードする改変塩基配列b
(c)前記改変塩基配列aとストリンジェントな条件下でハイブリダイズし、前記改変塩基配列aの139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の置換された三塩基連鎖と相補的な三塩基連鎖を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードする改変塩基配列c
【0036】
本発明の核酸の好ましい具体例としては、以下の改変アミノ酸配列をコードする塩基配列を含む核酸を挙げることができるが、これらに限定されるものではない。
配列表の配列番号1に記載のアミノ酸配列において、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び470位のフェニルアラニンがチロシンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列。
【0037】
改変塩基配列bの「1から数個の塩基の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異」における「1から数個」の範囲は、該改変塩基配列bがpH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードし得るものであれば特には限定されないが、例えば、1から40個、好ましくは1から30個、より好ましくは1から20個、より好ましくは1から10個、さらに好ましくは1から5個、特に好ましくは1から3個程度を意味する。また、「塩基の欠失」とは配列中の塩基の欠落もしくは消失を意味し、「塩基の置換」は配列中の塩基が別の塩基に置き換えられていること、「塩基の逆位」とは隣り合う2以上の塩基の位置が逆になっていること、「塩基の付加」とは塩基が付け加えられていること、「塩基の挿入」とは配列中の塩基の間に別の塩基が挿し入れられていることをそれぞれ意味する。改変塩基配列bは、配列表の配列番号2に記載の塩基配列おける139〜141位、598〜600位、1339〜1341位、1408〜1410位、及び1498〜1500位の塩基以外の塩基について欠失、置換、逆位、付加および/または挿入を有する。
【0038】
改変塩基配列cの「ストリンジェントな条件下でハイブリダイズする」とは、DNAをプローブとして使用し、コロニーハイブリダイゼーション法、プラークハイブリダイゼーション法、あるいはサザンブロットハイブリダイゼーション法等を用いることにより得られるDNAの塩基配列を意味し、例えば、コロニーあるいはプラーク由来のDNA又は該DNAの断片を固定化したフィルターを用いて、0.7〜1.0MのNaCl存在下、65℃でハイブリダイゼーションを行った後、0.1〜2×SSC溶液(1×SSC溶液は、150mM塩化ナトリウム、15mMクエン酸ナトリウム)を用い、65℃条件下でフィルターを洗浄することにより同定できるDNA等を挙げることができる。ハイブリダイゼーションは、Molecular Cloning: A laboratory Mannual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.,1989(以下、モレキュラークローニング第2版と略す)、Current Protocols in Molecular Biology, Supplement 1〜38, John Wiley & Sons (1987-1997)(以下、カレント・プロトコールズ・イン・モレキュラー・バイオロジーと略す)などに記載されている方法に準じて行うことができる。
【0039】
ストリンジェントな条件下でハイブリダイズするDNAとしては、プローブとして使用するDNAの塩基配列と一定以上の相同性を有するDNAが挙げられ、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは93%以上、特に好ましくは95%以上、最も好ましくは98%以上の相同性を有するDNAが挙げられる。ただし、ハイブリダイズする塩基配列は、配列表の配列番号2に記載の塩基配列おける139〜141位、598〜600位、1339〜1341位、1408〜1410位、及び1498〜1500位からなる群から選ばれる位置の1種又は2種以上の三塩基連鎖の置換を含む。
【0040】
本発明の核酸の取得方法は特に限定されない。たとえば、本明細書中の配列表の配列番号1に記載したアミノ酸配列および配列番号2に記載の塩基配列の情報に基づいて適当なプローブやプライマーを調製し、それらを用いて染色体DNAライブラリーから本発明の核酸を得ることができる。染色体DNAライブラリーは、例えば、本発明のβ−フルクトフラノシダーゼの野生型を発現するアルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192、本発明のβ−フルクトフラノシダーゼを発現するアルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192変異体などから得ることができる。アルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192変異体からβ−フルクトフラノシダーゼ遺伝子を含む核酸を取得する方法として、例えば、以下の方法を挙げることができる。
【0041】
アルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192変異体を栄養培地に植菌し、好気的条件下で約1〜3日間程度で培養した後、培養物から菌体を採取し、リゾチームやβ−グルカナーゼなどの細胞壁溶解酵素や超音波で処理することによりβ−フルクトフラノシダーゼ遺伝子を含む核酸を菌体外に溶出させる。このとき、細胞壁溶解酵素にプロテアーゼなどの蛋白質加水分解酵素を併用したり、菌体を超音波処理する際、SDSなどの界面活性剤を共存させたり、凍結融解してもよい。斯くして得られる処理物に、例えば、フェノール抽出、アルコール沈澱、遠心分離、プロテアーゼ処理、リボヌクレアーゼ処理などの当該技術分野において通常知られる方法を適用すればβ−フルクトフラノシダーゼ遺伝子を含む核酸が得られる。
【0042】
上記した染色体DNAライブラリーを得ることができれば、PCR法により本発明の核酸を取得することもできる。該染色体DNAライブラリーを鋳型として使用し、改変塩基配列a〜cを増幅できるように設計した1組又は2組以上のプライマーを用いてPCRを行う。PCRの反応条件は適宜設定することができ、例えば、94℃で15秒〜30秒(変性)、63℃で30秒〜1分間(アニーリング)、68℃で2〜3分間(伸長)からなる反応工程を1サイクルとして、例えば30〜50サイクル行った後、72℃で10分間反応させる条件などを挙げることができる。次いで、増幅されたDNA断片を、大腸菌等の宿主で増幅可能な適切なベクター中にクローニングすることができる。
【0043】
上記した各種操作、例えば、プローブ若しくはプライマーの調製、染色体DNAライブラリーの構築、DNAの抽出、cDNAライブラリーのスクリーニング、並びにβ−フルクトフラノシダーゼ遺伝子のクローニングなどの操作は当業者に既知であり、例えば、モレキュラークローニング第2版、またはカレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載の方法に準じて行うことができる。
【0044】
また、本発明のβ−フルクトフラノシダーゼや核酸は、本発明のβ−フルクトフラノシダーゼの野生型を発現しているアルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192由来のβ−フルクトフラノシダーゼ遺伝子から、改変アミノ酸配列Aおよび改変塩基配列aの情報に基づいて、化学合成、遺伝子工学的手法または突然変異誘発などの当業者に通常知られる任意の方法で作製することができる。具体的には、β−フルクトフラノシダーゼの野生型を発現しているアルスロバクター・スピーシーズ K-1 FERM P-10736/BP-3192由来のβ−フルクトフラノシダーゼ遺伝子に対し、変異原となる薬剤を接触作用させる方法、紫外線を照射する方法、遺伝子工学的手法を挙げることができる。遺伝子工学的手法の一つである部位特異的変異誘発法は対象となる位置に変異を導入できる手法であることから有用であり、サチュレーション変異法、ならびにモレキュラークローニング第2版およびカレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載の方法に準じて行うことができる。
【0045】
本発明の核酸は、通常、組換えDNAの形態で宿主に導入される。組換えDNAは、通常、DNAと自律増殖可能なベクターを含んでなり、本発明の核酸を入手できれば、通常知られる組換えDNA技術により比較的容易に調製することができる。例えば、本発明の核酸を自律増殖可能なベクターに挿入して組換えDNAとし、これを適宜宿主に導入して得られる形質転換体を培養し、培養物から菌体を採取し、この菌体から当該DNAを含むプラスミドを採取する。
【0046】
(C)本発明の組換えベクター
本発明の核酸は適当なベクター中に挿入して使用することができる。本発明で用いるベクターの種類は特に限定されず、例えば、自立的に複製することが可能なベクター(例えばプラスミド等)でもよいし、あるいは、宿主細胞に導入された際に宿主細胞の染色体に組み込まれ、染色体と共に複製されるものであってもよい。好ましくは、本発明で用いるベクターは発現ベクターである。発現ベクターにおいて本発明の遺伝子は、転写に必要な要素(例えば、プロモーター等)が機能的に連結されている。プロモーターは宿主細胞において転写活性を示すDNA配列であり、宿主の種類に応じて適宜選択することができる。
【0047】
また、自立的に複製することが可能なベクターの具体例としては、pET21a、pRSETA、pCR8、pBR322、pBluescript II SK(+)、pUC18、pCR2.1、pLEX、pJL3、pSW1、pSE280、pSE420、pHY300PLK、pTZ4、pC194、pUB110、pHV14、TRp7、YEp7、pBS7等のプラスミドベクターやλgt11、λZAP、λgt・λB、ρ11、φ1、φ105等のファージベクターが挙げられるが、大腸菌で発現させるには、pET21a、pRSETA、pCR8、pBR322、pBluescript II SK(+)、pUC18、pKK223−3、およびpCR2.1が好適であり、枯草菌で発現させるには、pUB110、pTZ4、pC194、ρ11、φ1及びφ105が好適である。pHY300PLK、pHV14、TRp7、YEp7及びpBS7は、組換えDNAを2種以上の宿主内で増殖させる場合に有用である。
【0048】
作動可能なプロモーターとしては、挿入断片に含まれる宿主中において機能することができるプロモーターであれば特に制限されないが、例えば、大腸菌においてはラクトースオペロン(lac)、トリプトファンオペロン(trp)などのプロモーターやtacプロモーターを挙げることができ;枯草菌においてはバチルス・ステアロテルモフィルス(Bacillus stearothermophilus)のマルトジェニックアミラーゼ遺伝子、バチルス・リケニホルミス(Bacillus licheniformis)のαアミラーゼ遺伝子、バチルス・アミロリケファチエンス(Bacillus amyloliquefaciens)のBANアミラーゼ遺伝子、バチルス・サブチリス(Bacillus subtilis)のアルカリプロテアーゼ遺伝子もしくはバチルス・プミルス(Bacillus pumilus)のキシロシダーゼ遺伝子などのプロモーターを挙げることができ;酵母ではアルコールデヒドロゲナーゼ遺伝子(ADH)、酸性フォスファターゼ遺伝子(PHO)、ガラクトシダーゼ遺伝子(GAL)、グリセロアルデヒド3リン酸デヒドロゲナーゼ遺伝子(GPD)などのプロモーター、酵母解糖系遺伝子由来のプロモーター、TPI1プロモーター、ADH2−4cプロモーターなどを挙げることができ;カビではα−アミラーゼ遺伝子(amy)、セロビオハイドロラーゼI遺伝子(CBHI)などのプロモーター、ADH3プロモーター、tpiAプロモーターなどを挙げることができ;哺乳動物細胞では、SV40プロモーター、MT−1(メタロチオネイン遺伝子)プロモーター、アデノウイルス2主後期プロモーターなどを挙げることができ;昆虫細胞では、ポリヘドリンプロモーター、P10プロモーター、オートグラファ・カリホルニカ・ポリヘドロシス塩基性タンパクプロモーター、バキュウロウイルス即時型初期遺伝子1プロモーター、バキュウロウイルス39K遅延型初期遺伝子プロモーターなどを挙げることでき、その他としてはファージ・ラムダのPRプロモーター、PLプロモーターなどを挙げることができる。
【0049】
本発明の組換えベクターはさらに選択マーカーを含有してもよい。選択マーカーとしては、例えば、薬剤耐性マーカー、栄養要求マーカーを使用することができる。選択マーカーの具体例としては、使用する宿主細胞が細菌の場合はアンピシリン耐性遺伝子、カナマイシン耐性遺伝子、テトラサイクリン耐性遺伝子、クロラムフェニコール耐性遺伝子、ネオマイシン耐性遺伝子、ヒグロマイシ耐性遺伝子などであり、酵母の場合はジヒドロ葉酸レダクターゼ(DHFR)遺伝子、シゾサッカロマイセス・ポンベTPI遺伝子、トリプトファン合成遺伝子(TRP1)、ウラシル合成遺伝子(URA3)、ロイシン合成遺伝子(LEU2)などがあり、カビの場合はハイグロマイシン耐性遺伝子(Hyg)、ビアラホス耐性遺伝子(Bar)、硝酸還元酵素遺伝子(niaD)などが挙げられる。
【0050】
本発明の組換えベクターは、本発明の核酸によってコードされるタンパク質の発現に必要なDNA配列、例えばプロモーター、エンハンサー、リボソーム結合部位、分泌シグナル配列、ターミネーターなどを適当な順で連結していることことが好ましい。本発明の組換えベクターを構築する方法や本発明の組換えベクターを宿主に挿入する方法は当業者に通常知られる。
【0051】
(D)本発明の形質転換体
本発明の核酸または組換えベクターを適当な宿主に導入することによって形質転換体を作製することができる。すなわち、本発明の形質転換体は、本発明の核酸を導入してなる、又は本発明の組換えベクターを含有する形質転換体である。
【0052】
本明細書でいう「核酸を導入」とは、バクテリオファージなどの通常知られる外来遺伝子を宿主細胞に導入する手段によって、本発明の核酸を宿主細胞へ導入すること、好ましくは、本発明の核酸が宿主細胞の染色体に組み込まれることを意味する。
【0053】
本発明の核酸または組換えベクターが導入される宿主細胞は、宿主−ベクター系が確立されているものであればいずれも利用可能であり、本発明のβ−フルクトフラノシダーゼ生産における形質転換体の培養条件において、基質であるβ−フルクトフラノシル供与体を分解する酵素を産生しないものが好ましく、細菌、酵母、真菌および高等真核細胞等が挙げられる。β−フルクトフラノシル供与体を分解する酵素としては、例えば、β−フルクトフラノシル供与体がショ糖である場合はスクラーゼを挙げることができる。
【0054】
細菌細胞の例としては、枯草菌または放線菌等のグラム陽性菌または大腸菌等のグラム陰性菌が挙げられる。これら細菌の形質転換は、プロトプラスト法、エレクトロポレーション法、カルシウムイオンを用いる方法等により行えばよい。哺乳類細胞の例としては、HEK293細胞、HeLa細胞、COS細胞、BHK細胞、CHL細胞またはCHO細胞等が挙げられる。哺乳類細胞を形質転換し、該細胞に導入されたDNA配列を発現させる方法も知られており、例えば、エレクトロポレーション法、リン酸カルシウム法、リポフェクション法等を用いることができる。
【0055】
酵母細胞の例としては、サッカロマイセスまたはシゾサッカロマイセスに属する細胞が挙げられ、例えば、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)またはサッカロマイセス・クルイベリ(Saccharomyces kluyveri)等が挙げられる。酵母宿主への組換えベクターの導入方法としては、例えば、エレクトロポレーション法、スフェロブラスト法、酢酸リチウム法等を挙げることができる。
【0056】
他の真菌細胞の例は、糸状菌、例えばアスペルギルス、ニューロスポラ、フザリウム、またはトリコデルマに属する細胞である。宿主細胞として糸状菌を用いる場合、DNA構築物を宿主染色体に組み込んで組換え宿主細胞を得ることにより形質転換を行うことができる。DNA構築物の宿主染色体への組み込みは、通常知られる方法に従い、例えば相同組換えまたは異種組換えにより行うことができる。
【0057】
昆虫細胞を宿主として用いる場合には、組換え遺伝子導入ベクターおよびバキュロウイルスを昆虫細胞に共導入して昆虫細胞培養上清中に組換えウイルスを得た後、さらに組換えウイルスを昆虫細胞に感染させ、タンパク質を発現させることができる。
【0058】
宿主細胞の形質転換により得られた形質転換体は、例えば、適当な条件で培養し、得られた培養液から一般的な方法によって酵素の分取や精製を行うことにより本発明のβ−フルクトフラノシダーゼを得ることができる。また、宿主が枯草菌、酵母、カビの場合には、分泌型ベクターを使用して、菌体外に本発明のβ−フルクトフラノシダーゼを分泌させることも有利である。
【0059】
(E)本発明の酵素の製造方法
本発明によれば、本発明の形質転換体を常法にしたがって適当な培地に接種して適切な条件下で培養し、得られた培養物から本発明の酵素を採取してなる、本発明の酵素の製造方法を包含する。すなわち、本発明の酵素の製造方法は、(a)本発明の形質転換体を培養して、β−フルクトフラノシダーゼを含む形質転換体培養物を得る工程、及び(b)形質転換体培養物からβ−フルクトフラノシダーゼを得る工程を含む。
【0060】
本発明の形質転換体の培養に用いる栄養培地としては、炭素源、窒素源、無機物、および必要に応じ使用菌株の必要とする微量栄養素を程よく含有するものであれば、天然培地、合成培地のいずれでもよい。
【0061】
本発明の形質転換体の培養に用いる栄養培地の炭素源としては、該形質転換体が資化しうる物であればよく、例えば、グルコース、マルトース、フルクトース、マンノース、トレハロース、スクロース、マンニトール、ソルビトール、デンプン、デキストリン、糖蜜などの糖質、またはクエン酸、コハク酸などの有機酸、またはグリセリンなどの多価アルコールや脂肪酸も使用することができる。
【0062】
本発明の形質転換体の培養に用いる栄養培地の窒素源としては、各種有機および無機の窒素化合物、さらに培地は各種の無機塩を含むことができる。たとえば、コーンスティープリカー、大豆粕、あるいは各種ペプトン類等の有機窒素源、そして塩化アンモニウム、硫酸アンモニウム、尿素、硝酸アンモニウム、硝酸ナトリウム、リン酸アンモニウム等の無機窒素源などの化合物が使用可能である。また、グルタミン酸などのアミノ酸および尿素などの有機窒素源が炭素源にもなることはいうまでもない。さらに、ペプトン、ポリペプトン、バクトペプトン、肉エキス、酵母エキス、コーンスティープリカー、大豆粉、大豆粕、乾燥酵母、カザミノ酸、ソリュブルベジタブルプロテイン等の窒素含有天然物も窒素源として使用できる。
【0063】
本発明の形質転換体の培養に用いる栄養培地の無機物としては、例えば、カルシウム塩、マグネシウム塩、カリウム塩、ナトリウム塩、リン酸塩、マンガン塩、亜鉛塩、鉄塩、銅塩、モリブデン塩、コバルト塩などが適宜用いられる。具体的には、リン酸二水素カリウム、リン酸水素二カリウム、硫酸マグネシウム、硫酸第一鉄、硫酸マンガン、硫酸亜鉛、塩化ナトリウム、塩化カリウム、塩化カルシウム等が用いられる。さらに、必要に応じて、アミノ酸ならびにビオチンおよびチアミンなどの微量栄養素ビタミンなども適宜用いられる。
【0064】
培養法としては液体培養法(振とう培養法もしくは通気攪拌培養法)がよく、工業的には通気攪拌培養法が好ましい。培養温度とpHは、使用する形質転換体の増殖に最も適した条件を選べばよい。たとえば、形質転換体が微生物の場合の培養は、通常、温度20〜45℃、好ましくは25〜42℃、pH5〜9、好ましくは6〜8から選ばれる条件で好気的に行われる。培養時間は微生物が増殖し始める時間以上の時間であればよく、好ましくは8〜120時間であり、さらに好ましくは本発明の酵素が最大に生成する時間までである。微生物の増殖を確認する方法は特に制限はないが、たとえば、培養物を採取して顕微鏡で観察してもよいし、吸光度で観察してもよい。また、培養液の溶存酸素濃度には特に制限はないが、通常は、0.5〜20ppmが好ましい。そのために、通気量を調節したり、撹拌したり、通気に酸素を追加したりすればよい。培養方式は、回分培養、流加培養、連続培養または灌流培養のいずれでもよい。
【0065】
本発明の形質転換体の培養において、組換えベクターに選択マーカーを含有させている場合などでは、選択マーカーに対応した抗生物質などを栄養培地とともに加える。たとえば、選択マーカーとしてアンピシリン耐性遺伝子およびクロラムフェニコール耐性遺伝子を含有する場合は、適当な濃度に調製したアンピシリン溶液およびクロラムフェニコール溶液をそれぞれ加える。また、必要であれば、本発明の核酸を有する遺伝子の発現を誘導する発現誘導剤を培養開始時または培養開始から形質転換体の増殖を確認した後に加える。たとえば、発現誘導剤としては、イソプロピル−β−D−チオガラクトピラノシド(IPTG)を用いる。
【0066】
このようにして得られた培養物から、本発明の酵素を採取する。本発明の核酸、組換えベクター、及び形質転換体の態様によって、本発明の酵素は細胞内外に蓄積される。そこで、細胞内若しくは細胞外に生成蓄積された本発明の酵素を採取する。
【0067】
本発明の酵素の採取法は、一般の酵素の採取の手段に準じて行うことができる。本発明の酵素が細胞外に生成蓄積された場合は、通常知られる手段によって細胞を除いた後に培養上清を粗酵素として用いることができる。細胞内に生成蓄積される場合は、例えば、培養して得られた培養物を固液分離操作によって細胞と培養上清に分離し、得られた細胞を破砕したものを、粗酵素として用いることができる。固液分離には、通常知られる方法を制限なく利用することができ、例えば、培養物そのものをそのまま遠心分離する方法、培養物に濾過助剤を加えることや濾過助剤をプレコートしたプレコートフィルターなどにより濾過分離する方法、平膜、中空糸膜などを用いる膜濾過分離する方法などが採用される。細胞の破砕は、通常知られる方法を制限なく利用することができ、例えば、有機溶剤やリゾチームなどの酵素によって細胞を溶解する方法、超音波破砕法、凍結融解法、フレンチプレス法、ガラスビーズ破砕法、ダイノミル破砕法などを採用することができる。細胞破砕物を固液分離して得た分離液を、粗酵素として用いることもできる。さらに、固液分離した細胞は、破砕することなく、そのまま粗酵素として用いられ得る。
【0068】
粗酵素は、そのままで使用することもできるが、精製して使用することもできる。例えば、ここに挙げるものに限定されるものではないが、得られた粗酵素を、熱処理の如く耐熱性の差を利用する方法、透析、限外ろ過、レジンカラム、ゲルろ過、ゲルろ過クロマトグラフィー及びSDS−ポリアクリルアミドゲル電気泳動の如く分子量の差を利用する方法、塩沈澱、硫安沈殿、アルコール沈殿及びその他の溶媒沈澱の如く溶解性の差を利用する方法、DEAE−トヨパール樹脂などを用いるイオン交換クロマトグラフィーの如く電荷の差を利用する方法、アフィニティークロマトグラフィーの如く特異的親和性を利用する方法、ブチルトヨパール樹脂などを用いる疎水クロマトグラフィー及び逆相クロマトグラフィーの如く疎水性の差を利用する方法、吸着クロマトグラフィーの如く物理化学的な吸着性の差を利用する方法、等電点電気泳動及び等電点クロマトグラフィーなどの如く等電点の差を利用する方法などの通常知られる方法を単独または組み合わせて供することにより、工業用途の精製酵素を調製できる。
【0069】
粗酵素及び精製酵素は、固定化することもできる。例えば、イオン交換体への結合法、樹脂及び膜などとの共有結合・吸着法、高分子物質を用いた包括法などを採用することができる。
【0070】
(F)本発明のβ−フルクトフラノシル転移糖の製造方法
β−フルクトフラノシル供与体とβ−フルクトフラノシル受容体とを含有する溶液に、本発明の酵素若しくは本発明の形質転換体(例えば、細菌)又はその培養物を作用させることにより、β−フルクトフラノシル転移糖を生成させることができる。
【0071】
本発明の酵素は、固体状または液体状の粗酵素および/または精製酵素として利用することができる。ただし、該粗酵素はスクラーゼ等のβ−フルクトフラノシル転移糖の製造に悪影響を及ぼす酵素を含まない粗酵素であることを必要とする。さらに、該β−フルクトフラノシダーゼの活性を有する菌体および該菌体を適当な担体に包括、吸着あるいは化学的に結合させた固定化菌体などをβ−フルクトフラノシル転移糖の製造に使用することができる。さらには、本発明の酵素は、通常知られる方法で固定化させた固定化酵素として使用することもできる。
【0072】
β−フルクトフラノシル供与体とβ−フルクトフラノシル受容体とを含有する溶液における水性媒体としては、β−フルクトフラノシル転移反応を妨げるものでなければ特に制限されないが、例えば、水、緩衝液などが挙げられる。緩衝液としては、例えば、酢酸緩衝液、リン酸緩衝液、クエン酸緩衝液、コハク酸緩衝液、トリス塩酸緩衝液などが採用される。
【0073】
本発明の酵素の使用量は特に制限はないが、β−フルクトフラノシル転移糖の収率及び経済性の観点から、β−フルクトフラノシド供与体1gに対して、好ましくは0.1から50単位であり、より好ましくは0.5から20単位であり、さらに好ましくは0.8から5単位が好適である。β−フルクトフラノシル受容体の濃度は、溶液に溶解し得る範囲であれば特に限定されないが、工業的には10w/w%以上が好ましく、20〜60w/w%がより好ましい。反応時のβ−フルクトフラノシル受容体分子とβ−フルクトフラノシル供与体分子のモル比は、特に限定されるものではないが、β−フルクトフラノシル転移糖の収率及び経済性の観点から、1:5から5:1が望ましい。
【0074】
前記反応は雑菌汚染を避けるとともに収率を挙げるために、好ましくは50〜70℃、より好ましくは60〜65℃で行う。pHは本発明の酵素の最適条件下で行うことが好ましく、例えば、6.0〜8.0で行うのが適当である。上記条件で十分なβ−フルクトフラノシル転移糖生成が見られた時点で反応を終了するが、反応は通常1〜100時間で終了する。β−フルクトフラノシル転移糖の生成率は、酵素反応液の基質濃度、用いる受容体の種類、反応条件などによって異なる。例えば、22w/w%ショ糖と18w/w%乳糖を基質として用いた場合、ラクトスクロースの生成率は、固形分当り最大約30%である。
【0075】
反応終了後、通常は、反応液を60〜135℃、好ましくは65〜100℃に加熱して酵素を失活させるか、又はpHの低下(塩酸等の酸の添加)などの適当な手段によって酵素を失活させて反応を停止する。次いで、常法により、濾過、遠心分離などして不溶物を除去した後、活性炭による脱色、H型、OH型イオン交換樹脂による脱塩などの単離・精製手段を適宜組み合わせてβ−フルクトフラノシル転移糖を得ることができる。さらに、必要であれば、活性炭カラムクロマトグラフィーなどのクロマト操作、公知の固定床方式、移動床方式、擬似移動床方式などを単独又は組み合わせることにより高純度に濃縮及び精製したβ−フルクトフラノシル転移糖精製品を得ることができる。β−フルクトフラノシル転移糖精製品はシラップ状であるが、必要に応じて、通常知られる乾燥工程を加えて粉末状製品とすることもできる。
【0076】
本発明の製造方法によって得られるβ−フルクトフラノシル転移糖は、味質良好な甘味を有し、また、浸透圧調節性、保湿性、照付与性、結晶防止性、澱粉老化防止性などの性質、また、抗う蝕性、ビフィズス菌増殖促進性、ミネラル吸収促進性などの機能をも有し得ることから、広く飲食物、嗜好物、飼料、餌料、化粧品、医薬品、成形物など、更には、生活用品、農林水産用品、試薬、化学工業用品などの各種組成物に有利に利用され得る。
【0077】
以下の実施例により本発明をより詳細に説明するが、本発明は以下の実施例に限定されるものではない。
【実施例1】
【0078】
耐熱性β−フルクトフラノシダーゼの取得
(1)変異遺伝子ライブラリーの構築
寄託番号FERM P−10736/BP−3192のアルスロバクター・スピーシーズ K−1(Arthrobacter sp. K−1)株由来のβ−フルクトフラノシダーゼ遺伝子を含む組換え体プラスミドpBFF1(特開2000−342273公報記載)を大腸菌HB101よりHigh Pure Plasmid Isolation Kit(ロシュ・ダイアグノスティックス株式会社)を用いることにより抽出、精製した。得られた組換え体プラスミドを鋳型として、配列表のプライマーAとプライマーBを用い、Diversify PCR Random Mutagenesis Kit(クロンテック社)を用い、キットのプロトコールにしたがって、β−フルクトフラノシダーゼ遺伝子にランダム変異を導入した。変異導入されたβ−フルクトフラノシダーゼ遺伝子を含む1.8k塩基対のDNA断片を精製した後、制限酵素NdeIおよびBamHIで切断した。これとプラスミドベクターpET21a(インビトロジェン社)を制限酵素NdeIおよびBamHIで切断したものを Ligation Kit ver 2.1(タカラバイオ社)を用いて、16℃で3時間ライゲーションをした後に、大腸菌BL21StarTM(DE3)pLysS[F- ompT hsdSB(rB-mB-)gal dcm rne131 (DE3)pLysS(CamR)]株へ形質転換を行うことによって、ランダム変異の導入された組換えプラスミドを有する組換え大腸菌ライブラリーを構築した。
【0079】
(2)耐熱性β−フルクトフラノシダーゼのスクリーニング
前記(1)の操作で得られた上記形質転換体をLB−Amp−Cm寒天培地(バクトペプトン1%、酵母エキス0.5%、塩化ナトリウム1%、バクトアガー1.5%、アンピシリン50μg/ml、クロラムフェニコール35μg/ml)上で37℃、一晩培養してコロニーを形成させた。各コロニーのうち、3000個を別々にLB−Amp−Cm培地(バクトペプトン1%、酵母エキス0.5%、塩化ナトリウム1%、アンピシリン50μg/ml、クロラムフェニコール35μg/ml)を1mlずつ入れた96穴ディープウェルプレートに接種し、20℃、8時間振とう培養を行った。その後、各ウェルに100mM イソプロピル−β−D−チオガラクトピラノシド(IPTG)10μlを添加し、さらに20℃で16時間振とう培養を行った。培養終了後、培養液を3000rpm、10分間遠心分離を行い、菌体を得た。得られた菌体に、BugBuster(ノバジェン社)を各ウェルに100μlずつ添加し、室温で5分間撹拌後、50mMのリン酸緩衝液(pH6.5)を900μlずつ添加し、さらに室温で5分間撹拌後、3000rpm、10分間遠心分離を行い、粗酵素抽出液を得た。得られた粗酵素抽出液に対して、55℃で30分間の熱処理を行った。対照として、野生型酵素遺伝子が挿入されたプラスミド(pBFF1)を有し、野生型酵素(BFF1)を生産する形質転換体についても同様の操作を行った。得られた酵素液について、熱処理前後のβ−フルクトフラノシダーゼの活性測定を行った。活性測定は50mMのリン酸緩衝液(pH6.5)に溶解させた10%(w/v)のショ糖30μlに酵素液30μlを添加し、40℃で10分間反応させた後、100℃で15分間加熱して酵素反応を止めた。続いて4℃に冷却した後、生成したグルコースをグルコースオキシダーゼ法(和光純薬工業(株)製、グルコースC−IIテスト・ワコー)で測定した。ここで1単位の酵素活性は同条件下で1分間に1μmolのグルコースを生成する酵素量とした。対照となる野生型酵素よりも高い残存活性を示す変異体酵素を選別した。その結果、酵素遺伝子に変異が導入されたプラスミドを含有する形質転換体のうち、4株が生産する変異体酵素(P494、P500、F285、およびF2139)が高い残存活性を有していた。
【0080】
(3)変異箇所の解析
上記の如く得られた優良変異株に含有されるβ−フルクトフラノシダーゼ遺伝子に対して、遺伝子のどの部位が変異操作により置換されたのか確認をする為に以下のような解析を行った。上記(2)で得た上記組換え体に含まれるプラスミド(pP494、pP500、pF285、およびpF2139)をHigh Pure Plasmid Isolation Kitを用いて精製した。このプラスミドの塩基配列はMegaBACE 1000(GEヘルスケアバイオサイエンス株式会社)を用いて決定した。結果は表1に示した。
【表1】
【0081】
(4)55℃における残存活性の測定
上記(2)で得られた4株の生産する変異体酵素(P494、P500、F285、およびF2139)および上記(2)記載の野生型酵素BFF1の粗酵素溶液を55℃の恒温槽に30分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。測定の結果は、熱処理前の酵素活性を100%とした場合の残存活性(%)として表2に示した。
【表2】
【実施例2】
【0082】
BFF1、P494、P500、F285、およびF2139の組み合わせによる変異体の作製および評価
(1)BFF1のN末端側のアミノ酸とP494、P500、F285、F2139の有するC末端側のアミノ酸の組み合わせによる変異体の作製および評価
pBFF1を鋳型とし、BFF1のN末端側のアミノ酸(37−407番目)をコードする遺伝子断片nbffNを、プライマーAおよびCを用いたPCRにより増幅した。また、P494、P500、F285、およびF2139のC末端側(395−579番目)のアミノ酸をコードする遺伝子断片、np494C、np500C、nf285C、nf2139CをpP494、pP500、pF285、pF2139を各々鋳型とし、プライマーBおよびDを用いたPCRにより増幅した。得られた遺伝子断片nbffNにnp494C、np500C、nf285Cまたはnf2139CをプライマーAおよびBを用いたリコンビナントPCR法により連結し、さらにこれを発現用プラスミドpET21aに連結した後に、大腸菌BL21StarTM(DE3)pLysSを形質転換した。得られたコロニーを培養し、実施例1(2)に記載の方法に従い粗酵素液を調製した。β−フルクトフラノシダーゼ活性を示す株から得られた粗酵素液を56℃の恒温槽に30分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。耐熱性の比較対照として、野生型酵素であるBFF1を同様の試験に供した。測定の結果は、熱処理前の酵素活性を100%とした場合の相対活性(%)として表3に示した。表3に示すように、P494、P500およびF2139の各変異体が有する395番アミノ酸よりC末端側の変異によって56℃、30分間の熱処理時における耐熱性が向上することが分かった。
【表3】
【0083】
(2)P494のN末端側のアミノ酸と、P494、P500、F285、F2139のC末端側のアミノ酸の組み合わせによる変異体の作製および評価
次にpP494を鋳型とし、P494のN末端側のアミノ酸(37−407番目)をコードする遺伝子断片np494Nを、プライマーAおよびCを用いたPCRにより増幅した。一方、P494、P500、F285およびF2139のC末端側(395−579番目)のアミノ酸をコードする遺伝子断片、np494C、np500C、nf285C、およびnf2139Cを、pP494、pP500、pF285およびpF2139を各々鋳型とし、プライマーBおよびDを用いたPCRにより増幅した。得られた遺伝子断片np494Nにnp494C、np500C、nf285C、またはnf2139CをプライマーAおよびBを用いたリコンビナントPCR法により連結し、さらにこれを発現用プラスミドpET21aに連結した後に、大腸菌BL21StarTM(DE3)pLysSを形質転換した。得られたコロニーを培養し、実施例1(2)に記載の方法に従い粗酵素液を調製した。β−フルクトフラノシダーゼ活性を示す株から得られた粗酵素液を55℃の恒温槽に30分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。耐熱性の比較対照として、野生型酵素であるBFF1を同様の試験に供した。測定の結果は、熱処理前の酵素活性を100%とした場合の残存活性(%)として表4に示した。表4に示すように、P494、P500およびF2139の各変異体が有する395番アミノ酸よりC末端側の変異によって55℃、30分間の熱処理時における耐熱性が向上した。
【表4】
【0084】
(3)F285のN末端側のアミノ酸と、P494、F285、F2139のC末端側のアミノ酸の組み合わせによる変異体の作製および評価
pF285を鋳型とし、F285のN末端側のアミノ酸(37−407番目)をコードする遺伝子断片nf285Nを、プライマーAおよびCを用いたPCRにより増幅した。一方、P494、F285およびF2139のC末端側(395−579番目)のアミノ酸をコードする遺伝子断片、np494C、nf285C、またはnf2139Cを、pP494、pF285およびpF2139を各鋳型とし、プライマーBおよびDを用いたPCRにより増幅した。得られた遺伝子断片nf285Nにnp494C、nf285C、またはnf2139CをプライマーAおよびBを用いたリコンビナントPCR法により連結し、さらにこれを発現用プラスミドpET21aに連結した後に、大腸菌BL21StarTM(DE3)pLysSを形質転換した。得られたコロニーを培養し、実施例1(2)に記載の方法に従い粗酵素液を調製した。β−フルクトフラノシダーゼ活性を示す株から得られた粗酵素液を56℃の恒温槽に30分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。測定の結果は、熱処理前の酵素活性を100%とした場合の残存活性(%)として表5に示した。表3と5を比較して、F285の有する395番のアミノ酸よりN末端側の変異が本酵素の耐熱性に寄与していることが分かった。表5に記載する77%の残存活性を示した変異体を24D、87%の残存活性を示した変異体を26F、84%の残存活性を示した変異体を26Iと命名した
【表5】
【実施例3】
【0085】
24D、26Fおよび26Iアミノ酸変異の組み合わせによる変異体24Yの作製および評価
24Dをコードする遺伝子配列を解析したところ、24Dにおけるアミノ酸変異箇所はT47S、S200T、F447V、およびP500Sの4カ所であった。また、26Fをコードする遺伝子配列を解析したところ、D42G、T47S、Q98R、S200T、N208D、R222C、F470Y、およびG530Dの8カ所であった。また、26Iをコードする遺伝子配列を解析したところ、T47S、S200T、F470Y、G530DおよびF552Lの5カ所であった。次に、24Dにおいては野生型配列と同じアミノ酸残基であり(F470)、26Fと26Iにおいて共通するアミノ酸変異であるF470Yを24Dに導入することとした。より具体的には24Dをコードするプラスミドp24Dを鋳型とし、24DのN末端アミノ酸から変位導入箇所近傍を含む476番目のアミノ酸をコードする遺伝子断片n24yNを、プライマーAおよび470番アミノ酸をチロシンに変異させる配列を含むアンチセンスプライマー(プライマーE)を用いたPCRにより増幅した。また、p24Dを鋳型とし、466番目のアミノ酸からC末端アミノ酸をコードする遺伝子断片n24yCを、プライマーBおよび470番アミノ酸をチロシンに変異させる配列を含むセンスプライマー(プライマーF)を用いたPCRにより増幅した。得られた遺伝子断片n24yNとn24yCをプライマーAおよびBを用いたリコンビナントPCR法により連結し、さらにこれを発現用プラスミドpET21aに連結した後に、大腸菌BL21StarTM(DE3)pLysSを形質転換した。得られたコロニーからプラスミドを抽出、精製した後、塩基配列の決定を行い、目的の変異が導入されていることを確認した。このようにして、5重変異体24Yをコードする遺伝子を含むプラスミドp24Y、および24Yを生産する組換え大腸菌を得た。得られたコロニーを培養し、実施例1(2)に記載の方法に従い粗酵素液を調製した。β−フルクトフラノシダーゼ活性を示す株から得られた粗酵素液を60℃の恒温槽に30分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。測定の結果は、熱処理前の酵素活性を100%とした場合の相対活性(%)として表6に示した。表6に示すように、24Yは24D、26Fよりも優れた耐熱性を示すことが分かった。
【表6】
【実施例4】
【0086】
47位、200位、447位、470位、および500位、アミノ酸のサチュレーション変異
47位、200位、447位、470位、および500位のアミノ酸について、サチュレーション変異を行った。所定の変異を起こさせるよう設計した5種類のセンスプライマー:Gfw、Hfw、Ifw、Jfw、Kfwと5種類のアンチセンスプライマー;Grv、Hrv、Irv、Jrv、Krvを用い、GfwとGrv、HfwとHrv、IfwとIrv、JfwとJrv、KfwとKrvをそれぞれ一組とし、p24Yを鋳型として、実施例3と同様な手順により、形質転換を行なった。得られた形質転換体のプラスミドが有する変異型β−フルクトフラノシダーゼ遺伝子をプライマーAおよびBを用いてPCRにより増幅した。増幅断片の遺伝子配列の解析はMegaBACE 1000を用いて行なった。各々、200クローンについて、実施例1(2)に記載の方法に従い粗酵素液を調製した。β−フルクトフラノシダーゼ活性を示す株から得られた粗酵素液を60℃の恒温槽に30分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。アミノ酸のサチュレーション変異により得られた変異酵素のうち、サチュレーション変異を施した各箇所が野生型アミノ酸に戻った4重変異酵素の示す残存活性を比較対照とした。対照よりも高い残存活性を示した変異体のアミノ酸配列を確認したところ、その内訳は、変異箇所47番アミノ酸では、S、E、Gであり、200番アミノ酸では、T、Lであり、447番アミノ酸では、P、I、V、K、Q、R、A、Y、H、T、S、W、Cであり、470番アミノ酸では、Y、A、E、Vであり、500番アミノ酸では、V、C、W、S、であった。これらの結果より、47位、200位、447位、470位、または500位の各アミノ酸が野生型から前述の各アミノ酸に置換されることにより、本酵素の耐熱性が向上することが明らかとなった。
【実施例5】
【0087】
24Y、24Y447P、24Y447I、24Y447FおよびBFF1の酵素溶液の作製
実施例3で得られたβ−フルクトフラノシダーゼ変異体である24Y、実施例4で得られた変異体酵素であり、24Yの447番目のアミノ酸がプロリン、イソロイシン、または野生型と同じフェニルアラニンに置換された変異体酵素(24Y447P、24Y447I、24Y447F)を生産する組換え大腸菌を各々培養し、大腸菌による組換え酵素の調製を行なった。より具体的な手順を以下に示す。培養は容量2Lの三角フラスコにLB培地(バクトペプトン1%、酵母エキス0.5%、塩化ナトリウム1%)0.5Lを入れて滅菌した後、アンピシリン溶液を終濃度で50μg/ml、クロラムフェニコール溶液を終濃度で35μg/mlになるように添加し、種培養液を1%になるように接種して、温度20℃、130rpmでOD600が0.5になるまで培養した。その後、IPTGを終濃度で0.5mMになるように添加し、さらに16時間培養を行った。培養終了後、培養液を5000g、10分間遠心分離を行い、菌体を得た。得られた大腸菌に50mM リン酸緩衝液(pH6.5) 20mlを添加し、細胞を超音波破砕した後に12000g、15分間遠心分離を行った。得られた粗抽出液をハイドロキシアパタイトカラム(生化学工業社製)によるクロマトグラフィー[(洗浄、50mM リン酸緩衝液(pH6.5);溶出、200mM リン酸緩衝液(pH6.5)]に供した。β−フルクトフラノシダーゼ活性を示す画分を集め、50mM リン酸緩衝液(pH6.5)で平衡化した陰イオン交換カラム(SuperQ 650Mカラム、東ソー社製)に供した。続いて、0−500mMのNaCl濃度勾配法により、酵素タンパクの溶出を行なった。β−フルクトフラノシダーゼ活性を示す画分を集め、限外ろ過膜(PM−10、Amicon社製)による濃縮を行なった後、50mM リン酸緩衝液(pH6.5)に緩衝液を置換し、酵素液を得た。比較対照として、野生型酵素であるBFF1についても、大腸菌による組換え酵素の調製を同様の手順で行なった。
【実施例6】
【0088】
酵素の半減期測定
実施例5の方法で得た本発明の酵素液を55℃または60℃の恒温槽に一定時間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。酵素の半減期を表7として示した。表7に示すように、447番目のアミノ酸は耐熱性向上に寄与していることが明らかとなった。さらに447番目のアミノ酸がプロリンの場合が最も高い耐熱性を示した。
【表7】
【実施例7】
【0089】
(1)24Y47T、24Y200S、24Y470F、24Y500P、24Y500C、24Y500Vの酵素溶液の作製
実施例4で得られた変異体酵素のうちから、24Yの47番目のアミノ酸が野生型と同じであるスレオニンに置換された変異体酵素(24Y47T)、200番目のアミノ酸が野生型と同じであるセリンに置換された変異体酵素(24Y200S)、470番目のアミノ酸が野生型と同じであるフェニルアラニンに置換された変異体酵素(24Y470F;実施例2(3)記載の24Dと同じアミノ酸配列を有する変異酵素)、500番目のアミノ酸が野生型と同じであるプロリンに置換された変異体酵素(24Y500P)、500番目のアミノ酸が野生型と同じであるシステインに置換された変異体酵素(24Y500C)、500番目のアミノ酸が野生型と同じであるバリンに置換された変異体酵素(24Y500V)を選択し、実施例5と同様の手順で大腸菌による組換え酵素の調製を行なった。
【0090】
(2)4重変異体、24Y200S447P500C、24Y200S447P500V、24Y200S447P500Wをコードするプラスミドと各酵素溶液の作製
実施例7(1)記載の4重変異体24Y200Sの447番目のアミノ酸であるバリン(野生型ではフェニルアラニン)がプロリンに置換し、且つ24Y200Sの500番目のアミノ酸であるセリン(野生型ではプロリン)がシステイン、バリンまたはトリプトファン置換した4重変異体を作製した。より具体的には24Y200Sをコードするプラスミドp24Y200Sを鋳型とし、24Y200SのN末端アミノ酸から変位導入箇所近傍を含む453番目のアミノ酸をコードする遺伝子断片n24y200SNを、プライマーAおよび447番アミノ酸をプロリンに変異させる配列を含むアンチセンスプライマー(プライマーL)を用いたPCRにより増幅した。また、p24Y500C、p24Y500Vまたはp24Y500Wを鋳型とし、441番目のアミノ酸からC末端アミノ酸をコードする遺伝子断片n24y500C、n24y500V、n24y500Wを、プライマーBおよび447番アミノ酸をプロリンに変異させる配列を含むセンスプライマー(プライマーM)を用いたPCRにより増幅した。得られた遺伝子断片n24y200SNに対して、n24y500C、n24y500Vまたはn24y500WをプライマーAおよびBを用いたリコンビナントPCR法により各々連結し、さらにこれを発現用プラスミドpET21aに連結した後に、大腸菌BL21StarTM(DE3)pLysSを形質転換した。得られたコロニーからプラスミドを抽出、精製した後、塩基配列の決定を行い、目的の変異が導入されていることを確認した。このようにして、4重変異体24Y200S447P500C、24Y200S447P500V、p24Y200S447P500Wをコードする遺伝子を含むプラスミドp24Y200S447P500C、p24Y200S447P500V、およびp24Y200S447P500Wを生産する組換え大腸菌を得た。得られた各組換え大腸菌より、実施例5と同様の手順で大腸菌による各組換え酵素の調製を行なった。
【0091】
(3)5重変異体の作製24Y447P500C、24Y447P500V、24Y447P500Wをコードするプラスミドと各酵素溶液の作製
実施例4で取得した5重変異体である24Y447Pの500番目のアミノ酸であるセリン(野生型ではプロリン)がシステイン、バリンまたはトリプトファンに置換した5重変異体3種を作製した。より具体的には24Yをコードするプラスミドp24Yを鋳型とし、24YのN末端アミノ酸から変位導入箇所近傍を含む453番目のアミノ酸をコードする遺伝子断片n24yN2を、プライマーAおよび447番アミノ酸をプロリンに変異させる配列を含むアンチセンスプライマー(プライマーL)を用いたPCRにより増幅した。また、p24Y500C、p24Y500Vまたはp24Y500Wを鋳型とし、441番目のアミノ酸からC末端アミノ酸をコードする遺伝子断片n24y500C、n24y500V、n24y500Wを、プライマーBおよび447番アミノ酸をプロリンに変異させる配列を含むセンスプライマー(プライマーM)を用いたPCRにより増幅した。得られた遺伝子断片n24yN2に対して、n24y500C、n24y500Vまたはn24y500WをプライマーAおよびBを用いたリコンビナントPCR法により各々連結し、さらにこれを発現用プラスミドpET21aに連結した後に、大腸菌BL21StarTM(DE3)pLysSを形質転換した。得られたコロニーからプラスミドを抽出、精製した後、塩基配列の決定を行い、目的の変異が導入されていることを確認した。このようにして、5重変異体24Y447P500C、24Y447P500V、p24Y447P500Wをコードする遺伝子を含むプラスミドp24Y447P500C、p24Y447P500V、およびp24Y447P500Wを生産する組換え大腸菌を得た。得られた各組換え大腸菌より、実施例5と同様の手順で大腸菌による各組換え酵素の調製を行なった。
【実施例8】
【0092】
酵素の耐熱性試験
実施例5および7(1)〜(3)の方法で得た本発明の酵素液(精製酵素)を45〜65度の各温度の恒温槽に180分間静置した後、4℃に冷却し、酵素活性(残存活性)を測定した。測定の結果は、熱処理前の酵素活性を100%とした場合の残存活性(%)として図1に示した。図1に示す通り、本発明の変異体酵素24Y47T、24Y200S、24Y447F、24Y470F、24Y500P、24Y200S447P500C、24Y200S447P500V、24Y200S447P500W、24Y、24Y500C、24Y500V、24Y500W、24Y447P500C、24Y447P500V、および24Y447P500Wは野生型酵素BFF1と比較して耐熱性が著しく向上していることが明らかとなった。
【実施例9】
【0093】
糖転移試験
22%(W/W)ショ糖と18%(W/W)乳糖を含む水溶液(pH6.0、50mMリン酸緩衝液)に、ショ糖1gあたり1.0Uの実施例5および7(1)〜(3)の方法で得た本発明の酵素液(精製酵素)を添加し、50から75℃まで温度を変化させ、10時間転移反応を行った。反応液は100℃で10分間熱処理した後、HPLCに供し、生成したラクトスクロース(LS)を測定した(カラム:TSK−GEL Amide−80、移動相:70%アセトニトリル、流速:1.0mL/分、カラム温度:35℃、検出器:示差屈折計)。その結果、図2に示したように野生型酵素(BFF1)ではLS生成が急激に低下する60℃以上の高温域において耐熱性が向上した変異型酵素は高いLS生成量を維持することが明らかとなった。
【図面の簡単な説明】
【0094】
【図1】図1は、本発明の酵素及び野生型の酵素の耐熱性試験結果を示す。
【図2】図2は、本発明の酵素及び野生型の酵素の糖転移試験結果を示す。
【特許請求の範囲】
【請求項1】
下記(A)〜(C)に記載の改変アミノ酸配列A、改変アミノ酸配列B又は改変アミノ酸配列Cを含むβ−フルクトフラノシダーゼ。
(A)配列表の配列番号1に記載のアミノ酸配列において、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸が下記[1]〜[5]に示す如く置換された改変アミノ酸配列A
[1]47位のスレオニンについて、セリン、グルタミン酸又はグリシンへの置換
[2]200位のセリンについて、スレオニン又はロイシンへの置換
[3]447位のフェニルアラニンについて、プロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインへの置換
[4]470位のフェニルアラニンについて、チロシン、アラニン、グルタミン酸又はバリンへの置換
[5]500位のプロリンについて、バリン、システイン、トリプトファン又はセリンへの置換
(B)前記改変アミノ酸配列Aにおいて、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸の置換の他さらに1から数個のアミノ酸の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列B
(C)前記改変アミノ酸配列Aと70%以上の相同性を有し、かつ前記改変アミノ酸配列Aの47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸の置換を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列C
【請求項2】
前記改変アミノ酸配列Aが、
200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び470位のフェニルアラニンがチロシンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;又は
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列である、請求項1に記載のβ−フルクトフラノシダーゼ。
【請求項3】
請求項1又は2に記載のβ−フルクトフラノシダーゼをコードする核酸。
【請求項4】
下記(a)〜(c)に記載の改変塩基配列a、改変塩基配列b又は改変塩基配列cを含むβ−フルクトフラノシダーゼをコードする核酸。
(a)配列表の配列番号2に記載の塩基配列において、139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の三塩基連鎖が下記[6]〜[10]に示す如く置換された改変塩基配列a
[6]139〜141位の三塩基連鎖について、セリン、グルタミン酸又はグリシンを規定するコドンに相補的な三塩基連鎖への置換
[7]598〜600位の三塩基連鎖について、スレオニン又はロイシンを規定するコドンに相補的な三塩基連鎖への置換
[8]1339〜1341位の三塩基連鎖について、プロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインを規定するコドンに相補的な三塩基連鎖への置換
[9]1408〜1410位の三塩基連鎖について、チロシン、アラニン、グルタミン酸又はバリンを規定するコドンに相補的な三塩基連鎖への置換
[10]1498〜1500位の三塩基連鎖について、バリン、システイン、トリプトファン又はセリンを規定するコドンに相補的な三塩基連鎖への置換
(b)前記改変塩基配列aにおいて、前記139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の三塩基連鎖の置換の他さらに1から数個の塩基の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードする改変塩基配列b
(c)前記改変塩基配列aとストリンジェントな条件下でハイブリダイズし、前記改変塩基配列aの139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の置換された三塩基連鎖と相補的な三塩基連鎖を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードする改変塩基配列c
【請求項5】
請求項3又は4に記載の核酸を含む組換えベクター。
【請求項6】
請求項3若しくは4に記載の核酸を導入してなる、又は請求項5に記載の組換えベクターを含む形質転換体。
【請求項7】
(a)請求項6に記載の形質転換体を培養して、β−フルクトフラノシダーゼを含む形質転換体培養物を得る工程、及び
(b)形質転換体培養物からβ−フルクトフラノシダーゼを得る工程
を含むβ−フルクトフラノシダーゼの製造方法。
【請求項8】
β−フルクトフラノシル供与体及びβ−フルクトフラノシル受容体を含有する溶液に、請求項1若しくは2に記載のβ−フルクトフラノシダーゼ、又は請求項6に記載の形質転換体を作用させて、β−フルクトフラノシル転移糖を得ることを特徴とするβ−フルクトフラノシル転移糖の製造方法。
【請求項9】
β−フルクトフラノシル供与体が、ショ糖、ラフィノース及びエルロースからなる群から選ばれる少なくとも1種である、請求項8に記載の製造方法。
【請求項10】
β−フルクトフラノシル受容体が、糖質、糖アルコール及びアルコールからなる群から選ばれる少なくとも1種である、請求項8又は9に記載の製造方法。
【請求項11】
β−フルクトフラノシル転移糖が、キシロシルフラクトシド、ガラクトシルフラクトシド、フコシルフラクトシド、ソルボシルフラクトシド、マルトオリゴシルフラクトシド、イソマルトシルフラクトシド、及びラクトシルフラクトシドからなる群から選ばれる少なくとも1種である、請求項8〜10のいずれか1項に記載の製造方法。
【請求項1】
下記(A)〜(C)に記載の改変アミノ酸配列A、改変アミノ酸配列B又は改変アミノ酸配列Cを含むβ−フルクトフラノシダーゼ。
(A)配列表の配列番号1に記載のアミノ酸配列において、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸が下記[1]〜[5]に示す如く置換された改変アミノ酸配列A
[1]47位のスレオニンについて、セリン、グルタミン酸又はグリシンへの置換
[2]200位のセリンについて、スレオニン又はロイシンへの置換
[3]447位のフェニルアラニンについて、プロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインへの置換
[4]470位のフェニルアラニンについて、チロシン、アラニン、グルタミン酸又はバリンへの置換
[5]500位のプロリンについて、バリン、システイン、トリプトファン又はセリンへの置換
(B)前記改変アミノ酸配列Aにおいて、47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸の置換の他さらに1から数個のアミノ酸の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列B
(C)前記改変アミノ酸配列Aと70%以上の相同性を有し、かつ前記改変アミノ酸配列Aの47位のスレオニン、200位のセリン、447位のフェニルアラニン、470位のフェニルアラニン、及び500位のプロリンからなる群から選ばれる少なくとも1種のアミノ酸の置換を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質を構成する改変アミノ酸配列C
【請求項2】
前記改変アミノ酸配列Aが、
200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、及び470位のフェニルアラニンがチロシンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがバリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがセリンに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがシステインに置換された改変アミノ酸配列;
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがバリンに置換された改変アミノ酸配列;又は
47位のスレオニンがセリンに、200位のセリンがスレオニンに、447位のフェニルアラニンがプロリンに、470位のフェニルアラニンがチロシンに、及び500位のプロリンがトリプトファンに置換された改変アミノ酸配列である、請求項1に記載のβ−フルクトフラノシダーゼ。
【請求項3】
請求項1又は2に記載のβ−フルクトフラノシダーゼをコードする核酸。
【請求項4】
下記(a)〜(c)に記載の改変塩基配列a、改変塩基配列b又は改変塩基配列cを含むβ−フルクトフラノシダーゼをコードする核酸。
(a)配列表の配列番号2に記載の塩基配列において、139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の三塩基連鎖が下記[6]〜[10]に示す如く置換された改変塩基配列a
[6]139〜141位の三塩基連鎖について、セリン、グルタミン酸又はグリシンを規定するコドンに相補的な三塩基連鎖への置換
[7]598〜600位の三塩基連鎖について、スレオニン又はロイシンを規定するコドンに相補的な三塩基連鎖への置換
[8]1339〜1341位の三塩基連鎖について、プロリン、イソロイシン、バリン、リジン、グルタミン、アルギニン、アラニン、チロシン、ヒスチジン、スレオニン、セリン、トリプトファン又はシステインを規定するコドンに相補的な三塩基連鎖への置換
[9]1408〜1410位の三塩基連鎖について、チロシン、アラニン、グルタミン酸又はバリンを規定するコドンに相補的な三塩基連鎖への置換
[10]1498〜1500位の三塩基連鎖について、バリン、システイン、トリプトファン又はセリンを規定するコドンに相補的な三塩基連鎖への置換
(b)前記改変塩基配列aにおいて、前記139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の三塩基連鎖の置換の他さらに1から数個の塩基の欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも1種の変異を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードする改変塩基配列b
(c)前記改変塩基配列aとストリンジェントな条件下でハイブリダイズし、前記改変塩基配列aの139〜141位の三塩基連鎖、598〜600位の三塩基連鎖、1339〜1341位の三塩基連鎖、1408〜1410位の三塩基連鎖、及び1498〜1500位の三塩基連鎖からなる群から選ばれる少なくとも1種の置換された三塩基連鎖と相補的な三塩基連鎖を有し、
pH6.5、55℃、180分間の加熱処理後において、未処理の酵素に対して60%以上のβ−フルクトフラノシダーゼ活性を有するタンパク質をコードする改変塩基配列c
【請求項5】
請求項3又は4に記載の核酸を含む組換えベクター。
【請求項6】
請求項3若しくは4に記載の核酸を導入してなる、又は請求項5に記載の組換えベクターを含む形質転換体。
【請求項7】
(a)請求項6に記載の形質転換体を培養して、β−フルクトフラノシダーゼを含む形質転換体培養物を得る工程、及び
(b)形質転換体培養物からβ−フルクトフラノシダーゼを得る工程
を含むβ−フルクトフラノシダーゼの製造方法。
【請求項8】
β−フルクトフラノシル供与体及びβ−フルクトフラノシル受容体を含有する溶液に、請求項1若しくは2に記載のβ−フルクトフラノシダーゼ、又は請求項6に記載の形質転換体を作用させて、β−フルクトフラノシル転移糖を得ることを特徴とするβ−フルクトフラノシル転移糖の製造方法。
【請求項9】
β−フルクトフラノシル供与体が、ショ糖、ラフィノース及びエルロースからなる群から選ばれる少なくとも1種である、請求項8に記載の製造方法。
【請求項10】
β−フルクトフラノシル受容体が、糖質、糖アルコール及びアルコールからなる群から選ばれる少なくとも1種である、請求項8又は9に記載の製造方法。
【請求項11】
β−フルクトフラノシル転移糖が、キシロシルフラクトシド、ガラクトシルフラクトシド、フコシルフラクトシド、ソルボシルフラクトシド、マルトオリゴシルフラクトシド、イソマルトシルフラクトシド、及びラクトシルフラクトシドからなる群から選ばれる少なくとも1種である、請求項8〜10のいずれか1項に記載の製造方法。
【図1】

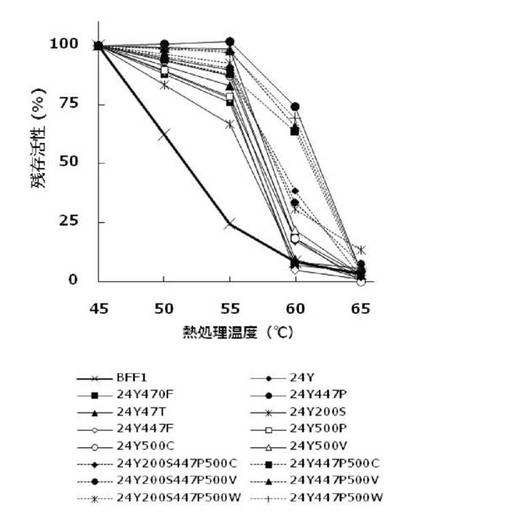
【図2】

【図2】
【公開番号】特開2010−148419(P2010−148419A)
【公開日】平成22年7月8日(2010.7.8)
【国際特許分類】
【出願番号】特願2008−329555(P2008−329555)
【出願日】平成20年12月25日(2008.12.25)
【出願人】(504194878)独立行政法人海洋研究開発機構 (110)
【出願人】(390021636)塩水港精糖株式会社 (11)
【Fターム(参考)】
【公開日】平成22年7月8日(2010.7.8)
【国際特許分類】
【出願日】平成20年12月25日(2008.12.25)
【出願人】(504194878)独立行政法人海洋研究開発機構 (110)
【出願人】(390021636)塩水港精糖株式会社 (11)
【Fターム(参考)】
[ Back to top ]