
生物発光・蛍光プローブ用アンカーペプチド

【課題】従来の情報認識タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブ(蛍光・生物発光イメージングプローブ)の安定性を改善し、検出限界、検出範囲、選択性、感度などの性能を向上させる。
【解決手段】従来型の蛍光・生物発光プローブに、コリプレッサー活性化ペプチド、例えばコリプレッサー配列由来の「LXXIIXXXL」モチーフを含むペプチドをアンカーペプチドとして導入した。さらに、円順列置換(CP)を併用して各要素の結合順序を大幅に組み換える。
【解決手段】従来型の蛍光・生物発光プローブに、コリプレッサー活性化ペプチド、例えばコリプレッサー配列由来の「LXXIIXXXL」モチーフを含むペプチドをアンカーペプチドとして導入した。さらに、円順列置換(CP)を併用して各要素の結合順序を大幅に組み換える。
【発明の詳細な説明】
【技術分野】
【0001】
本願発明は、各種生物発光又は蛍光イメージングプローブを安定に保存するためのアンカーペプチドに関するものである。
【背景技術】
【0002】
生体試料を対象とした生物分析は、病気の早期診断などにおける欠かせない手段である。例えば、ストレスは生体内免疫系、内分泌系における様々な病気の原因として活発な研究が行われている。例えば、生体内ストレスホルモン濃度は、コルチゾールの過剰分泌に起因するクッシング症候群や、慢性副腎皮質機能不全(アジソン病)などを診断する上で指標となっている(非特許文献1)。したがって、多くの医学・薬学分野においての研究の中心は、生体試料を対象に病気の指標となる因子の同定、分析が重要な課題といえるが、実際に病院などの診断現場で用いられている診断法は、いまだに数十年前からの免疫染色法に基づいている。
まだ実用化には至っていないが、昨今、従来の免疫染色法に代わる可能性を秘めた診断試薬の研究開発が進んできている。生体試料を分析し、発色でイメージングする方法として、蛍光を利用したFRET(蛍光共鳴エネルギー転移)(特許文献1、非特許文献2,3)や発光を利用した2分子型タンパク質相補(protein complementation)法等が開発された(特許文献2)。また最近、本発明者によって一分子型生物発光プローブが開発され(非特許文献2〜6)、また発光酵素の円順列置換(circular permutation)による分子イメージングプローブも開発された(非特許文献6)。なお、これらについては本発明者らにより特許出願もされている(PCT/JP2008/050370、特願2007-202308、特願2008-116098)。
これらの発色プローブは、大きく分けて、蛍光タンパク質を利用する蛍光的な手法と、各種発光酵素を用いる発光的な手法の2種類に大別される。蛍光タンパク質を利用する場合には、自己蛍光によってバックグラウンドが非常に高くなることに加え、外部光源とフィルターシステムが必要となり、蛍光を測定するために比較的に巨大な蛍光顕微鏡が必要となるという問題がある。例えば、非特許文献7〜9に記載のYellow Camellonsを用いた場合、自己蛍光がバックグラウンドを必然的に高め、シグナルとノイズとのコントラスト(S/N比)を低下させる原因となっている。さらに、分析できる細胞数が制限されることにより、得られる結果は定量的というよりもむしろ、定性的となってしまう(非特許文献10)。
一方、発光的な手法の場合は、このような問題はほとんど生じないが(非特許文献10〜14)、蛍光に比べて光強度そのものが低い。光強度は、一分子内に全ての要素を集積した一分子型発光プローブの場合においても改善されない上、分子内非特異的な結合が起こりやすいため、プローブの計測性能(検出感度、選択性、S/N比)を損なう原因になっている。
このように、従来の融合蛋白質に基づく蛍光・生物発光可視化プローブの共通の問題点として、その潜在的な検出能が十分発揮できず、検出限界、検出範囲、選択性、感度が生体試料計測等には不十分であった。そしてさらに、容易に分解されたり、失活されてしまうという分子プローブそのものの安定性にも問題があった。
上記問題点を解決するために、従来の蛍光・発光プローブの安定性を増大させ、発光強度、検出限界を高め、検出能を向上させる技術開発が急務であった。
【先行技術文献】
【特許文献】
【0003】
【特許文献1】特開2007−49943号公報
【特許文献2】国際公開 WO2004/104222
【非特許文献】
【0004】
【非特許文献1】Burtis et al. Tietz, Tietz textbook of clinical chemistry, 3rd ed., W.B. Saunders, Philadelphia,1999.
【非特許文献2】Awais et al. Angew. Chem. Int. Ed. 2006, 45, 2707-2712.
【非特許文献3】Awais et al. Anal. Chem. 2004, 76, 2181-2186.
【非特許文献4】Kim et al. Anal. Chem. 2007, 79, 1874-1880.
【非特許文献5】Kim et al., ACS Chem. Biol. 2008, 3, 359-372.
【非特許文献6】Kim et al., Bioconju. Chem. 2008, 19(12), 2480.
【非特許文献7】Souslova et al. Biochemistry-Moscow 2007, 72, 683-697.
【非特許文献8】Truong et al. Nat. Struc. Biol. 2001, 8, 1069-1073.
【非特許文献9】Miyawaki et al. Nature 1997, 388,882-887.
【非特許文献10】Kim et al. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 11542-11547.
【非特許文献11】Paulmurugan et al. Anal. Chem. 2005, 77,1295-1302.
【非特許文献12】Paulmurugan et al. Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 15883-15888.
【非特許文献13】Remy et al. Nat. Meth. 2006, 3, 977-979.
【非特許文献14】Kim et al. ACS Chem. Biol. 2007, 2, 484-492.
【非特許文献15】Lee et al. Cell. Mol. Life Sci. 2003, 60, 1613-1622.
【非特許文献16】Maruvada et al. J. Biol. Chem. 2003, 278, 12425-12432.
【非特許文献17】Tamrazi et al. Mol. Endocrinol. 2002, 16, 2706-2719.
【非特許文献18】Perissi et al. Genes Dev. 1999, 13, 3198-3208.
【非特許文献19】Hu et al. Nature 1999, 402, 93-96.
【非特許文献20】Horlein et al. Nature 1995, 377, 397-404.
【非特許文献21】Wilson et al. Nat. Meth.,2004, 1(3), 255-262.
【非特許文献22】Ozawa et al. Anal. Chem., 2001, 73(24), 5866-5874.
【非特許文献23】Bracha-Drori et al. Plant J., 2004, 40(3), 419-427.
【非特許文献24】Fan et al. Biochem. Biophy. Res. Comm., 2008, 367(1), 47-53.
【非特許文献25】Galigniana et al. J. Biol. Chem., 1999, 274, 16222-16227.
【非特許文献26】Awais et al. Steroids 2007, 72, 949-954.
【非特許文献27】Kim et al. Anal. Biochem. , 2007, 362, 148-150.
【非特許文献28】Kim et al. Anal. Chem. 2005, 77, 6928-6934.
【発明の概要】
【発明が解決しようとする課題】
【0005】
従来の融合蛋白質に基づく蛍光・生物発光イメージングプローブの安定性を改善し、検出限界、検出範囲、選択性、感度などの検出能を向上させることを目的とする。
【課題を解決するための手段】
【0006】
本発明者らは、従来の蛍光・発光プローブの上述した問題点を鋭意検討した結果、従来型の蛍光・発光プローブは、分子認識タンパク質のみに注目し、生体内の分子認識メカニズムとしては、当該タンパク質以外に多くの共役因子(コアクチベーター、コリプレッサーなど)が関わっていることの配慮がなされなかったために、自然界での本来の分子認識をそのまま再現できていないことに思い至った。
そこで、本発明者らは、生体内の分子認識タンパク質が通常コリプレッサーとの結合によって安定化されている点に着目し、従来型の一分子発光プローブに、コリプレッサーのモチーフペプチドを導入することで、分子の安定化を飛躍的に増大させることができ、検出限界を格段に下げることに成功した。
【0007】
生体細胞内においては、核内受容体(NRs)は、多数のヒートショック蛋白質群及びコリプレッサー(転写共役制御因子)群と共にクラスターを形成する形で安定化されている。しかしながら、これら分子は分子量が数百キロダルトンに及ぶものもあるため、分子プローブに取り入れることが困難であり、生体内のメカニズムを正確に模すことはできない。本発明者らは、上記安定化メカニズムの中核部分としてコリプレッサー配列中の「LXXIIXXXL」モチーフを利用することを発想した。まず、典型的な核内受容体であるグルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)を、「LXXLL」モチーフに繋ぎ、その両端を発光酵素のガルシアルシフェラーゼ(GLuc)断片で挟み込んだものを、本分子組み込み型(IMF)プローブのプロトタイプとし、末端にコリプレッサー由来のモチーフを繋ぐ。なお、上記プロトタイプは、円順列置換法(CP)(非特許文献6)に基づき、各要素の結合順序を大幅に組み換えることができ、コリプレッサーのモチーフを置き換えることも可能である。このように作製された本発明のプローブは、ストレスホルモンの指示薬として用いたときに2相性の濃度―応答曲線を描くことを最大の特徴とし、円順列置換を併用することで(i)より低い検出限界とより長い検出範囲,(ii)蛍光の絶対量とリガンド選択性の拡大が可能となった。
具体的には、多種類のコリプレッサーペプチドを用いてそれぞれの分子安定化能を調べた結果、あるペプチドはプローブの検出限界を1000倍までさげる(図3、4、5)ことを確認し、さらに円順列置換法を併用することで、選択性が格段にあがり(図6)、直線性検出範囲も広がる(図5)という知見も得た。このように、各種のコリプレッサーペプチドを導入すれば、様々な従来の蛍光や生物発光融合蛋白質プローブにおける安定化効果と検出能を期待できることから、船の錨に因んで「アンカーペプチド」と名づけた。以下、本発明において「アンカーペプチド」というとき、従来型の生物発光又は蛍光により検知、測定する可視化プローブの末端に、安定化のためにさらに繋いだコリプレッサー活性ペプチドを指す。
【0008】
すなわち、本発明は、以上の知見を得て完成することができたものであり、具体的には以下の通りのものである。
〔1〕 情報認識タンパク質が当該タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブにおいて、当該プローブ分子末端に、前記情報認識タンパク質に対するコリプレッサー活性ペプチドがアンカーペプチドとして結合されており、当該ペプチドが、リガンドの非共存時に前記情報認識タンパク質と会合していることを特徴とする、可視化プローブ。
〔2〕 核内受容体のリガンド結合ドメインを含み、当該リガンド結合ドメインがリガンドの結合により活性化されて生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブにおいて、当該プローブ分子末端に、核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドがアンカーペプチドとして結合されており、当該ペプチドが、リガンドの非共存時に前記リガンド結合ドメインと会合していることを特徴とする、可視化プローブ。
〔3〕 前記核内受容体がグルココルチコイドレセプターであって、前記立体構造変化が、当該グルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)がリガンドとの結合により活性化され、プローブ分子内のコアクチベーター由来の「LXXLL」モチーフを含むペプチドと結合することにより生じる立体構造変化であり、前記生物発光又は蛍光が、プローブ分子中のそれぞれ別の位置に存在する、分割された生物発光酵素又は蛍光タンパク質のフラグメントがタンパク質相補して発する生物発光又は蛍光である、前記〔2〕に記載の可視化プローブ。
〔4〕 前記グルココルチコイドレセプター(GR LBD)のリガンド結合ドメインを含むポリペプチド部位が、リンカーペプチドを介して又は介さずに、コアクチベーター由来の「LXXLL」モチーフを含むペプチドに結合されており、同一のプローブ分子の他の位置に、前記分割された生物発光酵素又は蛍光タンパク質のフラグメントと共に、前記核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドが結合されている、前記〔3〕に記載の可視化プローブ。
〔5〕 前記分割された生物発光酵素又は蛍光タンパク質のN末側及びC末側フラグメントが、リンカーペプチドを介して又は介さずに逆向きに結合されており、前記グルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)を含むポリペプチド部位と、コアクチベーター由来の「LXXLL」モチーフを含むペプチド部位と共に、同一のプローブ分子の他の位置に、前記核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドが結合されている、前記〔3〕に記載の可視化プローブ。
〔6〕 前記グルココルチコイドレセプターのリガンド結合ドメインを含むポリペプチド部位が、コアクチベーター由来の「LXXLL」モチーフを含むペプチド部位と別のプローブ分子上に存在し、かつそれぞれの部位が分割された生物発光酵素又は蛍光タンパク質のN末側もしくはC末側フラグメントのいずれかと、リンカーペプチドを介して又は介さずに結合されており、前記グルココルチコイドレセプターのリガンド結合ドメインを含むプローブ分子末端には、さらに前記核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドが結合されている、前記〔3〕に記載のプローブ。
〔7〕 情報認識タンパク質が当該タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブに対して、当該プローブ分子の末端に、前記情報認識タンパク質に対するコリプレッサー活性ペプチドをアンカーペプチドとして結合させることにより、当該ペプチドが、リガンド非共存下においては前記情報認識タンパク質と会合していることを特徴とする、可視化プローブの安定化方法。
〔8〕 核内受容体のリガンド結合ドメインを含み、当該リガンド結合ドメインがリガンドの結合により活性化されて生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブに対して、当該プローブ分子の末端に、核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドをアンカーペプチドとして結合させることにより、当該ペプチドが、リガンド非共存下においては前記リガンド結合ドメインと会合していることを特徴とする、可視化プローブの安定化方法。
〔9〕 情報認識タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブ分子の末端に存在するアンカーペプチドであって、情報認識タンパク質に対するコリプレッサー活性ペプチドからなり、リガンドによる活性化前の可視化プローブを前記情報認識タンパク質と会合することで安定化させることを特徴とする、アンカーペプチド。
〔10〕 前記コリプレッサー活性ペプチドが、核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドからなる、前記〔9〕に記載のアンカーペプチド。
〔11〕 前記「LXXIIXXXL」モチーフ含有ペプチドが、配列番号5〜12又は配列番号23〜30のいずれかで表されるアミノ酸配列を含むペプチドである、前記〔10〕に記載のアンカーペプチド。
〔12〕 前記〔1〕〜〔6〕のいずれかに記載の可視化プローブをコードする核酸。
〔13〕 前記〔12〕に記載の核酸を含む、可視化プローブ発現用ベクター。
〔14〕 前記〔13〕に記載の発現ベクターを用いて形質転換され、可視化プローブを発現する形質転換細胞。
〔15〕 被検試料中のリガンドを検出又は定量するためのキットであって、前記〔1〕〜〔6〕のいずれかに記載の可視化プローブ又は前記〔14〕に記載の形質転換細胞を含むキット。
〔16〕 前記キットが、さらに発光酵素の基質を含み、紙ストリップ上に固定、乾燥されている前記〔15〕に記載のキット。
〔17〕 被検試料中のリガンドの検出方法又は定量方法であって、前記〔1〕〜〔6〕のいずれかに記載の可視化プローブ又は前記〔14〕に記載の形質転換細胞を用いることを特徴とする方法。
【発明の効果】
【0009】
本発明では、従来型の生物発光・蛍光プローブにアンカーペプチドを導入することで、生細胞内安定性を担保しながら、プローブの検出限界を顕著に下げることができ、特に円順列置換法と併用することで、選択性が格段にあがり、検出範囲も広げることができた。すなわち、本発明においては、生体や生細胞を対象とした計測能(選択性、信号強度、S/N比)が大きく改善された、安定な生物発光可視化プローブが提供できた。
また、本発明のプローブは、プローブ安定性が格段に高く、診断用紙片ストリップの製作にも適しているため、簡便な分析手段を提供できる。
【図面の簡単な説明】
【0010】
【図1】(A)本発明で使われた生物発光可視化プローブをコードする遺伝子構成物の概略図。省略名:GLuc−N,ガウシアルシフェラーゼのN−末側断片;GLuc−C,ガウシアルシフェラーゼのC−末側断片;GR LBD,グルココルチコイド受容体のリガンド結合ドメイン。(B)本発明で合成された生物発光プローブのおよその分子構造。(C)cPRESSO−C1のリガンド依存的な分子内構造変化の可視化戦略の模式図。アゴニスト無しの状態では,コリプレッサーモチーフと結合し安定化している。ところが,アゴニストによって刺激されれば,分子内発光酵素断片間相補が起こり,結果として発光酵素活性が回復する。
【図2】従来型プローブ(「SIMGR」系列)の分子認識能。(A)10-6 Mのコルチゾール添加に対する「SIMGR」系列のプローブの相対的な発光強度。挿入図は、蛋白質μgあたりの強度の絶対値を示している。(B)全タンパク量とプローブの発現レベルを示すウエスタンブロット分析。
【図3】PRESSOシリーズプローブ内のGLuc断片間相補によって回復した生物発光強度に基づいたコルチゾール濃度−応答曲線。(A)コルチゾール濃度依存的なPRESSO-C3、-C4の2相曲線。(B)コルチゾール濃度依存的なPRESSO-N2、-C2の2相曲線。
【図4】(A)本プローブの計測性における円順列変異とコリプレッサーモチーフの相乗効果。黒棒は10−5Mコルチゾール刺激による絶対発光値を示す。(B)コルチゾールとDHT濃度変化に対する,cPRESSO-C1とSIMGR3の濃度―応答曲線。
【図5】cPRESSO-C1,cPRESSO-C2,cPRESSO及びSIMGR3を発現するCOS-7細胞の絶対発光値を示す濃度−応答曲線。cPRESSO-C1による生物発光値が絶対発光強度と直線性領域等からもっとも優れている。
【図6】(A)cPRESSO-C1、PRESSO-C1、SIMGR3のリガンド選択性の比較。挿入図は10-6 M コルチゾール刺激前後の絶対発光値を示す。省略名:vehicle, 0.1% dimethyl sulfoxide; DHT,5α-dihydrotestosterone; T, testosterone; E2, 17β-estradiol; OHT, 4-hydroxytamoxifen; genis,genistein; ICI, ICI-182780。(B)cPRESSO-C1の反復したリガンド感受性測定。挿入図Aで示したようにcPRESSO-C1を持つCOS-7細胞に10-7Mコルチゾールの刺激と除去を繰り返した。
【図7】(A)リガンド刺激によるcPRESSO-C1からの発光値の経時変化。(B)非細胞系である生物発光紙片上でのリガンド活性の測定。ガラス基板上にニトロセルロース紙片を付着し,各紙片末端に細胞溶解液をスポットした。後にストレスホルモン有り,無しの状態で基質溶液を滴下して,発光値をスキャンした。
【発明を実施するための形態】
【0011】
1.本発明の生物発光又は蛍光プローブの構造
本発明は、従来の生物発光又は蛍光可視化プローブの安定化を図ることで、安定化と共に当初予期していなかった計測性の向上も可能にした改良発明に係るものであるといえるが、これらの可視化プローブにおいては、核内受容体などの情報認識タンパク質は、当該情報伝達物質(リガンド)との結合によって立体構造が変化したことを、生物発光又は蛍光により検知して測定するものである。当該可視化プローブで従来から用いられていた情報認識タンパク質は、いずれも生体内では1又は複数のコリプレッサータンパク質が結合した状態で安定して存在している。
つまり、生体内では、情報認識タンパク質は通常、コリプレッサータンパク質と結合して安定化されており、外部からの信号(リガンド)によって刺激されたときに初めてコリプレッサータンパク質と離れ、活性型に変貌する。
例えば、核内受容体は、生体内でコリプレッサータンパク質群と結合した形で安定化されているが、核内受容体と同様にその2量体形成メカニズムが可視化プローブに利用されているNuclear Factor-κB (NF-κB)(非特許文献27)の場合も、生体内では通常Inhibitor-κB(IκB)と結合した形で安定化されている。また、細胞内コレステロール濃度を司るsterol regulatory element-binding protein-2 (SREBP-2)は、サイト1分解酵素(site 1 protease; S1P)によって2分割されると2量体になることによって活性型に変わる。このSREBP-2の活性化メカニズムが、蛋白質スプライシングを介する生物発光可視化プローブ等に利用されているが(非特許文献28)、生体内では通常SREBP cleavage-activating protein(SCAP)と結合した形態で安定化されている。さらに、G-タンパク質は通常細胞内ではそのαとβユニットが結合した形態で安定化されており、外部信号が到来したときに、αユニットはβユニットから離れて細胞膜上受容体と結合して立体構造変化を起こす。このαユニットの立体構造変化は可視化プローブに利用でき、βサブユニット(システイン-リッチ配列)については、その安定化現象に対してアンカーペプチドとしての効果が期待できる。
本発明では、このように多くの情報認識タンパク質と生体内で通常結合しているコリプレッサー作用のあるペプチド配列を、可視化プローブ用アンカーペプチドとして用いようとする発明であるから、従来から可視化プローブでその立体構造変化が利用されていた情報認識タンパク質に対する公知コリプレッサーペプチドは、すべて利用可能である。本発明の実施態様では、これら情報認識タンパク質として典型的な核内受容体の場合を例として取り上げる。以下、核内受容体に対してコリプレッサーを連結して安定化させた生物発光プローブによって本発明を説明するが、生体内の情報認識タンパク質としての挙動はいずれもほぼ共通しているから、核内受容体に対するコリプレッサーの組み合わせと同様に、G-タンパク質αサブユニットに対するβサブユニットのシステイン-リッチ配列、NF-κBに対するIκBのreal homology domain (RHD)、及びSREBP-2に対するSCAPのTrp (W)-Asp (D)反復モチーフとの組み合わせも同様のアンカーペプチドとしての使用が可能であることは当然である。
なお、従来型の蛍光プローブ(FRET)において核内受容体を含むプローブ分子内にコリプレッサー活性ペプチドを有する例が報告されている(非特許文献26)が、当該ペプチドは、単にリガンドの結合による立体構造変化により蛍光が発せられるのを定性的に観察するための要素であり、アンカーペプチドとして用いるものではない。
【0012】
本発明の核内受容体のリガンド結合性を利用して、そのリガンド物質を検出する生物発光又は蛍光プローブにおいては、核内受容体のリガンド結合ドメインを含む従来型の生物発光プローブ又は蛍光プローブ(非特許文献2〜6、14、26)に対して、その分子中に、好ましくはN末側もしくはC末側に、コリプレッサー配列中の「LXXIIXXXL」モチーフをアンカーペプチドとして結合させることを特徴とするものである。
例えば、典型的な核内受容体であるグルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)を用いる場合についてみると、GR LBDのリガンド結合ドメインを含むポリペプチドを、そのコアクチベーターである「LXXLL」モチーフを含むペプチドに繋ぎ、その両端をガウシアルシフェラーゼ(GLuc)断片など、各種ルシフェラーゼ又はGFPで挟み込んだかたちで一分子に集積し、その末端にコリプレッサー由来モチーフを繋ぐ。ここで、上記プロトタイプは、(ア)円順列置換法(CP)に基づき、各要素の結合順序を大幅に組み換えることができ、(イ)コリプレッサーのモチーフを置き換えること、(ウ)各構成要素の間に挿入すること、(エ)またはさらに2種以上のコリプレッサーモチーフを繋げることも可能である。CPを適用した場合の典型的な各構成要素の順序は、図1に記載されるように、発光酵素、GFPの分割断片同士が隣り合って逆向きに繋がることとなる。
従来型の生物発光プローブ又は蛍光プローブが二分子型の場合、即ち、核内受容体のリガンド結合ドメインと、「LXXLL」モチーフとが別の分子中に存在する場合においても前者にアンカーペプチドを繋ぎ、各分子末端に発光酵素又はGFPの分割断片の片方ずつを繋げばよい。
それぞれの構成要素ペプチドは、適宜長さを調節したリンカーペプチドを介して繋がれる。リンカーペプチドとしては、Gly-Gly-Ser(配列番号17)を基本単位とする「GSリンカー」などを用いることができる。他に、柔軟なアミノ酸をいくつかが繋いだペプチドも適宜用いることができる。リンカーペプチドのアミノ酸配列長としては0〜15アミノ酸残基、好ましくは3〜6アミノ酸残基が一般に用いられ、GSリンカーの場合では1〜5単位、好ましくは1〜3単位で用いられる。
【0013】
2.核内受容体とコアクチベーター由来「LXXLL」モチーフ
核内受容体(nuclear receptor;NRs)は、細胞内にあり、リガンドが結合すると核内に移行し、DNAに直接結合してDNAの転写を制御する受容体であって、生体中の転写制御因子(transcriptional regulators)のうちでも最も多彩なメンバーを持つものの1つである(非特許文献15)。これら核内受容体には、恒常性維持、生殖、発生、及び代謝といった多様な機能が知られており、それぞれが持つリガンド認識能やリガンド刺激によって引き起こされる分子メカニズムは,新たな融合蛋白質プローブの設計に応用されてきた.例えば、α−ヘリカルペプチドと核内受容体のリガンド結合ドメイン(NR LBD)との相互作用を指標として、蛍光共鳴エネルギー移動(FRET)法(非特許文献2,3)と一分子型生物発光プローブ(IMF)が開発された(非特許文献4,5)。ここで、α−ヘリカルペプチドとは、核内受容体のリガンド結合ドメインを認識するコアクチベーター由来のペプチドであって、「LXXLL」モチーフを含むという配列上の特徴を有している(非特許文献2,3)。ただし、Lは他にもIのような疎水性アミノ酸に入れ替えしても同様の性質を保つが、ここでは通常の言い方に倣って「LXXLL」モチーフと称する。また、核内受容体の核内移行はGFP-修飾法(非特許文献16)と蛋白質スプライシング法(非特許文献10)によって観察できる。また、リガンドによる核内受容体間の2量化はFRET法を用いて以前観察できた(非特許文献17)。これらの研究例からも、生体内のような複雑系におけるリガンド活性を計測する上で、核内受容体の重要さを物語っている。
本発明のプローブには、これら核内受容体のうち少なくともリガンド結合ドメイン(NR LBD)を含む核内受容体タンパク質及び活性化された核内受容体タンパク質を認識するペプチドが用いられる。本発明のNR LBDとしては、コルチゾールなどのストレスホルモンをリガンドとするグルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)以外、どのような核受容体も置き換えて使用できるが、例えば、同じストレスホルモン受容体の一種であるミネラルコルチコイドレセプター(mineralocorticoid receptor; MR)、又は女性ホルモンレセプター(estrogen receptor; ER)、男性ホルモンレセプター(androgen receptor; AR)、黄体ホルモンレセプター(progesterone receptor; PR)、 ビタミンDレセプター(vitamine D receptor; VDR)のLBDなどを用いることができる。それぞれの活性化されたNR LBDを認識するペプチドは公知であり、GR LBDの場合においてもNR LBD共通のLXXLLモチーフを含むペプチド、また各種疎水性アミノ酸を含むペプチドが用いられる。
【0014】
3.コリプレッサー(転写抑制因子)
コリプレッサー(corepressor)は、コアクチベーター(coactivator)と同様に、核内受容体のリガンド結合ドメイン(NR LBD)と結合して転写活性を調節することによって、NR LBDの転写活性における中心的な役割を果たしている(非特許文献17,18)。最初、核内受容体のコリプレッサーは酵母の2−ハイブリッドスクリーニング法により単離されたが、その中には、核内受容体コリプレッサー(NcoR)(非特許文献19)及びレチノイン酸と甲状腺ホルモン受容体のサイレンシング・メディエーター(SMRT)(非特許文献20)等が含まれる。上記コリプレッサーに関する研究によれば、一般的なコリプレッサーの役割は、アゴニスト刺激がない時、ホスト分子を安定化し、その転写活性を抑制する作用をすることにある、といわれる(非特許文献19〜20)。これらのコリプレッサーの配列上の特徴としては、リガンド非存在条件下で核内受容体と結合能を持つ共通配列である「LXXIIXXXL」モチーフ(配列番号4)を共有している。
ここで、「LXXIIXXXL」モチーフとしては、NcoR由来の「LEDIIRKALMG:配列番号5」などの他、SMRT由来の「GMLAKRIIAELGMNTSA:配列番号6」など下記表2に示された配列が典型的であるが、他にSMRT ID2(RVVTLAQHISEVITQDYTR:配列番号7)やNCoR ID2(RLITLADHICQIITQDFAR:配列番号8)などがある(非特許文献19、20)。また、表1で示したように、最初のLは、Iであってもよく、4番目のアミノ酸はVでもよい。L, I, Vは、共に代表的な疎水性アミノ酸である特徴がある。また、Xは通常どのアミノ酸でもいいという意味ではあるが、本発明におけるコリプレッサーモチーフにおいては、E、R、K、D、S等のアミノ酸が好ましい。これらのアミノ酸は比較的に親水性アミノ酸である特徴がある(非特許文献19、20)。したがって、本発明において「LXXIIXXXL」モチーフというとき、「I、Lのような疎水性アミノ酸が一定間隔で規則的に反復される中で、その間にはアスパラギン酸(D)、グルタミン酸(E)のような親水性アミノ酸が入っているペプチド」の場合を好ましい条件とする。なお、モチーフを構成する一部のアミノ酸が欠損されても、そのレプレッサー性を完全には失わない。
核内受容体のコリプレッサーモチーフ以外にも、G-タンパク質αサブユニットに対するβサブユニットをアンカーペプチドとして用いる場合は、そのシステイン(C)−リッチモチーフ、NF-κBに対してIκBを用いる場合は、IκB のRel homology domain (RHD)、そしてSREBP-2に対してSCAPを用いる場合は、SCAPのTrp(W)-Asp(D)反復モチーフを含むペプチドが有効であり、本発明と同様に実施できる。
【0015】

【0016】
4.発光酵素(ルシフェラーゼ)又は蛍光色素蛋白質(FP)
本発明で2分割して用いるルシフェラーゼは、通常の発光プローブにおいて用いられるホタルルシフェラーゼ(FLuc)、ウミシイタケルシフェラーゼ(RLuc)、コメツキムシルシフェラーゼ(CBLuc)(非特許文献4,5,10)などいずれのルシフェラーゼであってもよいが、発光量の大きなガルシアルシフェラーゼ(Gluc)が特に好ましい(非特許文献6)。
また、蛍光色素としては、GFPが従来から用いられており(非特許文献21、22)、他にもYFP断片(非特許文献23)やVenus断片(非特許文献24)などが使われているが、本発明においても同様に用いることができる。
【0017】
5.本発明プローブの構築方法
本発明プローブは、その構成要素となるペプチドを直接もしくはリンカーペプチドを介して、化学的にペプチド結合で繋ぐこともできるが、それぞれのペプチドをコードするDNAを繋いで発現ベクター中に組み込み、その発現ベクターを導入した形質転換細胞により製造することができる。
既存の生物発光プローブ又は蛍光プローブ(非特許文献2〜6、9〜12,14)に対して、その末端に又は内部にアンカーペプチドを、直接又はリンカーペプチドを介して結合させてもよい。発現ベクターに組み込む場合も、既存の生物発光プローブ又は蛍光プローブをコードするDNAの5’末端又は3’末端に繋いで発現ベクターに挿入される。
また、当該発現ベクターのまま生体内の細胞中に組み込み、生細胞内でのイメージングが可能である。さらに当該発現ベクターが導入された形質転換細胞で産生させたプローブを用いて非細胞系分析用キットを作製することもできる。
【0018】
本発明プローブをコードする発現ベクターを導入する細胞としては、培養細胞(原核細胞、酵母細胞、昆虫細胞、哺乳類動物細胞など)のみならず、哺乳動物、昆虫など生体内の細胞も含み、典型的にはマウスなど実験動物もしくはその細胞である。
本発明で用いる「発現ベクター」としては、公知の真核または原核細胞発現ベクター(プラスミド、ウイルスベクターなど)を特段の制限なく使用することができ、公知の組織特異的、または特定刺激特異的プロモーター配列を組み込んでもよい。
本発明の発現ベクターの導入は、例えばマイクロインジェクション法やエレクトロポーレーション法、脂質試薬によるトランスフェクション法(TransIT, Mirus)により細胞内に導入することができる。また、これらの手法で導入される発現ベクターに単純にプローブの発現だけを託すのではなく、プローブの上流に発現制御をし得る制御配列を繋げることによって、プローブそのものの発現有無を指標に更なる生物分析系を組むことができる。
さらに、本発明プローブをコードするDNAを含む組換えDNAを、pMXのようなレトロウイルスベクターにつなげ、高力価のウイルス産生能を有するパッケイジング細胞PLATEに導入することによって、動物に感染するレトロウイルスの作成が可能である。このウイルスを実験動物の各臓器に感染させることによって、各臓器内におけるストレスホルモンなどのリアルタイムイメージングができる。
【0019】
6.本発明プローブの診断キットとしての使用方法
本発明のベクターを導入した形質転換細胞のまま、又は当該細胞からプローブを取り出して精製し、もしくは未精製のまま紙片上で用いることで、生体試料内の各種生理活性物質濃度、例えばストレスホルモン濃度を簡便に測定することができ、迅速にストレス量の診断ができる。
本発明のプローブは、細菌のような原核細胞で大量発現させることもできるが、哺乳類細胞の場合でも、適切な分泌シグナル配列(MGVKVLFALICIAVAEA:配列番号18など)を繋いだプローブを導入することによって、培地中に大量に分泌させることによって、精製工程なしでも分析に用いることのできる、大量なプローブ含有培養上清が得られる。また、さらにカラム精製用のタグ(例えば、His Tag; HHHHHH:配列番号19)をつけることで、大量に精製された本発明のプローブを得ることができる。
例えば、ストレスホルモンの定性定量分析用キットとする場合に、精製もしくは未精製発光プローブを持つ紙ストリップと発光酵素の基質を組み合わせたキット、又は本発明の発光プローブを発現している生細胞と発光酵素の基質を含有した培地とを組み合わせたキットを構成することができる。蛍光プローブの場合は、主に蛍光共鳴エネルギー移動(FRET)現象に基づいたプローブが作製されてきたが、バックグラウンドが高く信号蛍光は弱いのでフィルターを構える必要があったため、従来紙片上での蛍光断片を利用した例は報告されていないが、本発明により可能性となる。
【0020】
なお、診断用キットに用いる紙片や断片プレートの材質としては、ニトロセルロース(nitrocellulose)紙等の紙片、ナイロン66、polyvinylidene fluoride, hydrophobic
polymerなどが用いられ、特に、ニトロセルロース紙片を用いることで、リトマス試験紙のような取り扱いやすい発光プローブ付着型試験紙が提供できる。また、探針素子などの末端に本発光プローブを固定・乾燥した、発光型リガンドセンシング手段を提供できる。
【0021】
7.本発明の具体的な態様
本発明を具体的な態様で説明すると、以下の通りであるが、この方法には限定されない。
(1)本発明のプローブの構築方法
まず、グルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)を、GSリンカーを介して、α−ヘリカルコアクチベーターモチーフ(LXXLL:配列番号13)と繋ぐ。次いで、その融合蛋白質をQ105位置で分割したガウシアルシフェラーゼ(GLuc)のN-端側断片と、C-端側断片の間に挟み込み、さらに、このN-端及びC-端断片を両端に有する生物発光プローブをコリプレッサーモチーフで連結する。このようなコリプレッサーモチーフで修飾された生物発光プローブをコリプレッサーの英語スペルからとって「PRESSO」と命名した。但し、レプレッサーモチーフをプローブ構成要素の間に挟んでも構わない。
当該プローブでは、アゴニストに対して2相性の応答を示し、検出限界が最高で10-11 Mまで改善するという画期的な効果を示す。さらに、上記生物発光プローブプロトタイプにおいて円順列(circular permutation:CP)の手法で再構成することにより、さらに検出能を上げることができる。生物発光プローブ作製の際に、円順列手法は有効な再構成法であり、それが顕著に元のプローブの検出限界を改善する。上記実施態様においても、CP手法のみでコルチゾールで10-8 Mにまで検出できる。コリプレッサーモチーフの導入は、主に検量線における直線性範囲をのばすことに寄与している。
特にコリプレッサーモチーフはCP手法と組み合わせることで、相乗的に以下の効果が高まる。
(ア)発光強度の絶対値を高める。
(イ)蛍光強度のバックグラウンドを低く抑える。
(ウ)シグナル−ノイズ比(S/N比)を改善する。
(エ)リガンド選択性を改善する。
したがって、コリプレッサーモチーフを搭載した融合蛋白質プローブは、細胞内分子の事象を追跡するためにきわめて有用である。
特に、発光酵素(ルシフェラーゼ)としてGLucを使う場合は、他のルシフェラーゼと比較して100倍以上も明るいので、GFPを用いる場合と同様、生きたままの生細胞条件下でも明瞭で正確な分析ができる。なお、通常の発光酵素を用いる場合は、測定発光値が小さいため、最適緩衝溶液条件を作る必要があり、細胞を壊してLysateを作るのが一般的である。
【0022】
(2)本発明のプローブを用いた分析、スクリーニング方法
本発明の発光プローブを用いて生きた細胞内分子現象を分析、又は分子現象に影響を与えるリガンドのスクリーニングのためには、例えば以下の方法が適用できる。
(i)発光プローブをコードするcDNAを持つプラスミドを24穴プレート上の生細胞に導入し、さらに16時間培養する。
(ii)上記細胞の培養液を除去し、その変わりに基質(coelenterazine)を含んだ200μL緩衝溶液(例えば、HBSSバーファー)に細胞を浸す。
(iii)上記細胞を特定刺激物質で刺激し、その刺激前後の発光変化値を発光プレートリーダー(luminescence plate reader; 例えば、LB 941 Multimode Reader(Berthold))を用いて観測する。
また、発光プローブを用いた非細胞系実験(in vitro)としては、以下の分析、スクリーニング方法が適用できる。
(i)精製された発光プローブを直径1.2センチの十字架型紙ストリップの末端に垂らして乾燥させる。
(ii)上記十字架型紙ストリップの中央に刺激物質を含む基質溶液15μLを点滴し、即時に発光スキャナー(例えば、RAS-3000;FujiFilm)で発光値を観測する。
【0023】
(3)ストレスホルモンとしてのコルチゾールの測定
ストレスホルモンと呼ばれるコルチゾールの生体サンプル中の生理濃度はクッシング症候群とアジソン病のような病気の直接の指標となる(非特許論文1)。生体サンプル中の総コルチゾール濃度は、それぞれ0.8−6.4 x 10-7 M(血清)、0.6-7.5 x 10-7 M(小便)、0.2−2.8 x 10-8 M(唾液)となる。このような生理濃度範囲は、生体サンプル中の内分泌ストレスホルモンを検出するためには、感受性の高いセンサーが要求されることを意味する。従って、従来の測定法は主に放射性同位元素や非放射性免疫センシング法に依存してきた。このような従来法の代わりに、私たちはストレスホルモンを高感度で検出できるような一分子型生物発光プローブを合成した。このプローブの特色は、コリプレッサーモチーフを導入し、更に円順列変異と合わせることによって相乗効果を高めて、結果として、非放射性、非免疫学的、発色型の内分泌ステロイド計測手段といえる。
図3で示したような2相の濃度―反応曲線は、プローブ母体に与えるコリプレッサーペプチドの特質といえる。
【0024】
(4)本発明プローブを用いた測定のメカニズムに関する考察
本発明のプローブが図3などにおいて示すような2相濃度―反応曲線は一般的に、この系の中に2つの平行が存在することを意味する。本発明のプローブの構成成分を考慮すれば、第1平行はGR LBDとコリプレッサーモチーフ間の相互作用であり、第2平行はGR LBDとコアアクチベーターモチーフとの相互作用であると考えられるから、本発明のプローブのリガンドセンシングメカニズムは、以下のようになるといえる。
(ア)本発明のプローブ内のGR LBDは最初にはコリプレッサーモチーフによって安定化されている。この状態はHuらの文献(非特許論文19)の結果と一致する。
(イ)本発明のプローブは、低濃度のストレスホルモンに応答して、コリプレッサーモチーフを放し、この分離により、GR LBDとコアアクチベーターモチーフとが会合する確率を高める。
(ウ)本発明のプローブが、高濃度のストレスホルモンに応答して、GR LBDとコアアクチベーターモチーフとの完全結合によって最大発光値を出す。
【0025】
また、円順列法の適用により作製された、pcPressoの分子構造では、隣り合った2分割発光酵素の活性部位が反対方向に背向くことになる(図1(A))が、この分子構造では、分割された発光酵素の活性部位断片同士がたまたま衝突する確立は非常に低く抑えられる。この構造的利点により、プローブのバックグラウンド発光が劇的に減少し、信号―バックグラウンド比(S/N比)の改善に繋がったと考えられる。
また、本プローブにおけるコリプレッサーモチーフによる効果と円順列変異との相乗効果については、図4(B),図5におけるSIMGR3とcPRESSOの比較からみて、円順列変異によりバックグラウンド発光が抑えられることで検出限界が向上し、さらにNcoR由来のコリプレッサーモチーフを導入することによって、濃度―応答曲線での直線性応答領域が広くなり(cPRESSO-C1の場合)、絶対発光強度の増加も観測される(図4(A)と図5)、という相乗効果が観測される。すなわち、本プローブにおけるコリプレッサーモチーフの役割は、プローブ分子そのものを荒い海で船の錨のように硬くして細胞内などの測定環境の系において安定化させ、発光酵素の反応効率を上げることにあると考えられる。
【0026】
8.その他
本発明におけるその他の用語や概念は、発明の実施形態の説明や実施例において詳しく規定する。なお、用語は基本的にはIUPAC-IUB Commission on Biochemical Nomenclatureによるものであり、あるいは当該分野において慣用的に使用される用語の意味に基づくものである。また発明を実施するために使用する様々な技術は、特にその出典を明示した技術を除いては、公知の文献等に基づいて当業者であれば容易かつ確実に実施可能である。例えば、遺伝子工学および分子生物学的技術はJ. Sambrook, E. F. Fritsch & T. Maniatis, "Molecular Cloning: A Laboratory Manual (2nd edition)", Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York (1989); D. M. Glover et al. ed., "DNA Cloning", 2nd ed., Vol. 1 to 4, (The Practical Approach Series), IRL Press, Oxford University Press (1995); Ausubel, F. M. et al., Current Protocols in Molecular Biology, John Wiley & Sons, New York, N.Y, 1995;日本生化学会編、「続生化学実験講座1、遺伝子研究法II」、東京化学同人 (1986);日本生化学会編、「新生化学実験講座2、核酸 III(組換えDNA技術)」、東京化学同人 (1992); R. Wu ed., "Methods in Enzymology", Vol. 68 (Recombinant DNA), Academic Press, New York (1980); R. Wu et al. ed., "Methods in Enzymology", Vol. 100 (Recombinant DNA, Part B) & 101 (Recombinant DNA, Part C), Academic Press, New York (1983); R. Wu et al. ed., "Methods in Enzymology", Vol. 153 (Recombinant DNA, Part D), 154 (Recombinant DNA, Part E) & 155 (Recombinant DNA, Part F), Academic Press, New York (1987)などに記載の方法あるいはそこで引用された文献記載の方法またはそれらと実質的に同様な方法や改変法により行うことができる。また、本発明で使用する各種蛋白質やペプチド、あるいはそれらをコードするDNAについては、既存のデータベース(URL:http://www.ncbi.nlm.nih.gov/等)から入手することができる。
なお、本発明で引用した先行文献又は特許出願明細書の記載内容は、本明細書の記載として組み入れるものとする。
【実施例】
【0027】
以下、実施例を示して本発明をさらに詳細かつ具体的に説明するが、本発明は以下の例に限定されるものではない。
【0028】
(実施例1):プラスミドの構築
ガウシアルシフェラーゼ(GLuc)の全長をコードするcDNAのテンプレートは、Nanolight社から購入した。cDNAのN-端及びC-端側断片(GLuc-N (18-105 AA、配列番号1); GLuc-C (106-185 AA、配列番号2))はPCRにより合成した。この際、両断片の末端には特異的な制限酵素サイトHindIII/ KpnI (GLuc-N)及びBamHI/ XhoI (GLuc-C)を導入するために、好適なプライマーとその鋳型を用いてPCR反応を起こした。
グルココルチコイドレセプターのリガンド結合ドメイン(GR LBD; 527-777 AA、配列番号3)のcDNA末端に、PCR反応を用いて制限酵素サイトKpnI/ NotIを導入した。なお、両端のNotI/ BamHI制限酵素サイトを有する下記のペプチドのcDNAオリゴマーはExigen (Tokyo, Japan)社から発注・購入した。
(i)「Glucocorticoid Receptor Interacting Protein 1 Interaction domain 3」由来のα−ヘリカルLXXLLモチーフ(GRIP1 ID3;NALLRYLLDKD:配列番号14);
(ii)GRIP1 ID2 由来のLXXLLモチーフ(GRIP1 ID2: HKILHRLLQDS:配列番号15)
(iii) GRIP1 ID3のアラニン変異型LXXLLモチーフ(GRIP1 ID3m: NALARYALDKD:配列番号16).
上記cDNA断片は、図1に示されるとおりに繋がれ、pcDNA 3.1(+)ベクターにサブクローニングされている。
GRIP1 ID3、変異タイプのGRIP1 ID3及びGRIP1 ID2を含むプラスミドは、それぞれpSimgr3, pSimgr3m,及びpSimgr2と名付けた。この意味は、単一分子型のグルココルチコイドレセプター(GR LBD)搭載生物蛍光プローブ(SIngle-Molecule-format bioluminescent probe with Glucocorticoid Receptor)を意味しており、それぞれは、発現後のSIMGR3, SIMGR3M, 及び SIMGR2と呼ばれる融合蛋白質プローブに対応する。
さらに、pSimgr3のN-又はC-末端にコリプレッサー蛋白質のモチーフを付加する修飾を施した。つまり、そのN-末端にsilencing mediator for Retinoic acid and Thyroid hormone receptors Interaction Domain 1由来のLXXIIXXXLモチーフ(SMRT ID1; GMLAKRIIAELGMNTSA:配列番号11)を連結したもの、又はNuclear receptor CORepressor Interaction Domain 1由来のLXXIIXXXLモチーフ (NcoR ID1; ASNLGLEDIIRKALMG:配列番号9)を連結した。さらに、SMRT ID1の逆向きのLXXIIXXXL モチーフ(ASTNMGLEAIIRKALMG:配列番号12)やNcoR ID1の逆向きのLXXIIXXXL モチーフ(GMLAKRIIDELGLNSA:配列番号10)も上記プローブのN-末端に繋ぎ、GR LBDとの結合性を検討する。プローブのC-末端も、同様にSMRT ID1又はNcoR ID1由来のコリプレッサーペプチドを連結して結合特性を検討した。その構造は、模式的に図1に示した。その特異的なペプチド配列を〔表2〕に示す。
これらのプローブは、コリプレッサーモチーフ(corePRESSOr motif)を有する一分子集積型生物発光プローブ(integrated-molecule-format bioluminescent probe;IMF)、すなわちPRESSOと名付ける。そして、N-及びC-末端にコリプレッサーモチーフを有するプローブを、それぞれPRESSO-N 及びPRESSO-C系列指示薬とよぶ。
なお、これら各要素を連結する際には、リンカーペプチドとしてGly-Gly-Ser(配列番号17)を基本単位とするGSリンカーを適宜の長さに調節して用いた。
【0029】
さらに、PRESSOプローブの円順列バージョンを合成した。その合成理由としては、(i)リガンド感受性の増大、(ii)シグナルとバックグラウンドの比率(S/N比)の改善を見込んだものである。この具体的な手順は以下の通りである。最初に、GLucをQ105位置で2分割し、その5’末端と3’末端側断片を作成した。この新しい5’及び3’末端にそれぞれLXXLLモチーフ及びGR LBD (527-777 AA)を繋ぐ。一方、GLucの本来の5’及び3’末端は、3つのGSリンカーを繋いだ。このプラスミドはpcPresso系列と呼ぶ。ここで、‘c’は、円順列(circular permutation)を表す。pcPresso の3’末端には、更にコリプレッサーペプチドを連結した。これらのタイプのプラスミドは、pcPresso-C系列と呼ぶ。発現した後のそれぞれのプローブは、cPRESSO及びcPRESSO-Cと呼べる。
これらプラスミドの信頼性を確認するために、「BigDye Terminator Cycle sequencing kit」及び遺伝子分析機のABI Prism310 (PE Biosystems)を利用して、配列確認をした。本発明において合成された各プローブの構造は図1に模式的に示し、またその構成要素のリストは表2に示した。
【0030】
【0031】
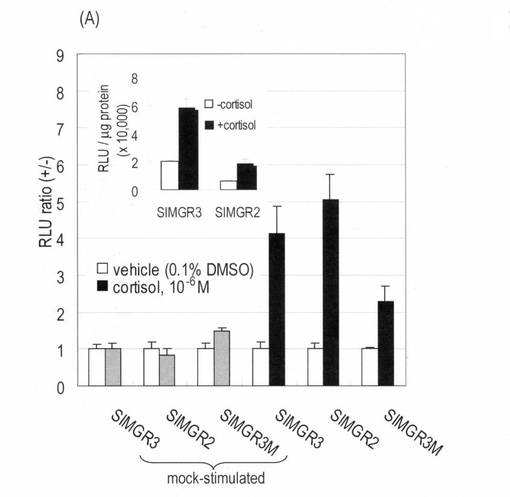
(実施例2)SIMGR系列プローブのストレスホルモン(コルチゾール)に対する感受性
COS-7細胞を12穴-プレート上で5%2酸化炭素培養機によって培養された。この時、10%ウシ胎児血漿(FBS)と1%ペニシリン-ストレプトマイシン(P/S)を添加したDulbeccoの改良Eagle培地(DMEM)を培地とした。プレート上のCOS-7細胞には、トランスフェクション試薬(TransIT-LT1;Mirus)を使ってpSimgr3,pSimgr2や pSimgr3m(1ウェルあたり0.2μg)をそれぞれ導入し、その後に16時間追加培養した。プレート上の細胞は、溶媒(0.1% DMSO)又はストレスホルモン(10-6 M cortisol)刺激を20分行い、その結果、回復されたルシフェラーゼ活性を特異的な基質キット(プロメガ)を使い、試薬マニュアル通りに測定した(図2(A)):簡単に説明すると、細胞をリン酸バーパーで一回洗浄し、キットから提供された細胞溶解バーパーで10分間溶解した。その溶解液を試験管に移し、40μLの基質溶液を加えて、ルミノメーター(Minilumat LB9506; Berthold)で発光値を最初15秒間測定した。
測定された発光強度は,以下の2つの何れかの方法で標準化された。(ア)細胞溶解液の単位蛋白質量で発光を換算する.この時の絶対発光値の単位は,RLU/μg of proteinになる.(イ)刺激有りの時の発光値(RLU(+))を、刺激無しの時の発光値(RLU(−))で割る。つまり、単位はRLU ratio (+/-)となる。ただし、この時のRLUは細胞溶解液の1μg蛋白質が出す発光値である。
【0032】
上述したように、一分子の中でストレスホルモン依存的な蛋白質間相互作用が起こるかどうかを検証するために、ストレスホルモン受容体(GR LBD)とコアクチベーター由来のLXXLLモチーフ間の結合を調べてみた(図2)。この結果、発光強度の回復がプローブ内に組み込まれたLXXLLモチーフの性質に依存していることが分かった。即ち、SIMGR3とSIMGR2共に10-6 Mストレスホルモン(cortisol)に応答したが、その絶対発光強度値はSIMGR3の方が強かった。LXXLLモチーフにアラニン(A)変異を導入したSIMGR3Mは、ストレスホルモンに弱い感度を示した。ウェスタンブロット結果によれば、上記プローブの何れも発現量の偏差は認められなかった(図2B)。
この結果は以下のように解析できる。
(ア)GR LBDは、以前X-線液晶構造解析結果(Protein Data Bank accession #: 1m2z)から予想されたように、LXXLLモチーフと相互作用する。
(イ)LXXLLモチーフの変異体による結果から予測されたように、LXXLLモチーフを構成する各アミノ酸が、GR LBDとLXXLLモチーフとの結合に決定的な役割をする。
【0033】
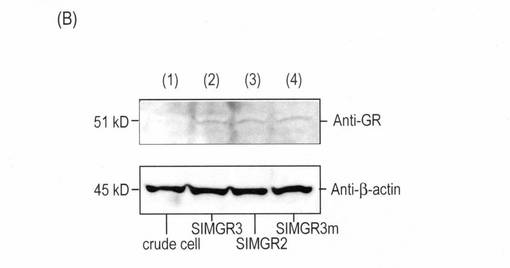
(実施例3)ウェスタンブロット分析
細胞溶解液中にある全蛋白質量と生物発光プローブの量をウェスタンブロットで検証した(図2(B))。まず、6ウェルプレート上で培養したCOS-7細胞にpSimgr3, pSimgr2, や pSimgr3mを導入し、更に16時間間培養した。その細胞をリン酸バーファー(PBS)で一回洗浄し、150μLの溶解バーファー(1% SDS/ 10% glycerol/ 10% 2-mercaptoethanol/ 0.001% bromophenol blue/ 50 mM Tris-HCl, pH 6.8)で溶解した。その後に細胞溶解液から5μLずつ取り、10%アクリルアミドゲル(TEFCO)上で電気泳動した。その後にニトロセルロース膜に移し、抗GR抗体や抗β-アクチン抗体で一次抗体処理をした。各汚点をhorseradish peroxidase (HRP)につないだ2次抗体処理し、最後に化学発光基質液(GE healthcare)で現象した。図4(A)のウェスタンブロットも上記記述と同様に行われた。
【0034】
(実施例4)PRESSO系列プローブのコルチゾールの用量応答曲線
PRESSOシリーズプローブのコルチゾール濃度―応答曲線(dose-response curve)を測定してみた(図3)。PRESSO-C3, -C4, や SIMGR3 の何れかを導入したCOS-7細胞に、一連の濃度のストレスホルモン(コルチゾール)で20分間刺激した。ほかにも同様にPRESSO-N2, -C2やSIMGR3の何れかを導入したCOS-7細胞に、一連のストレスホルモン刺激を20分間加え、蘇えた発光酵素活性を図2と同様のやり方でモニターした。
その結果、一番特徴的に見られる現象としては、プローブが描くストレスホルモン濃度―反応曲線が2相曲線を示す点である。つまり、コントロール(白丸)に比べて、PRESSO-C1と-C2は10-9から10-7 Mまでの濃度範囲において、発光値が増加する特徴を示す(図3(A))。
同様に、PRESSO-N1は10-11 から10-7 Mまでの領域において発光強度の上昇を示した(図3(B))。このような見かけ上の2相曲線を示すにも関わらず、そのシグナルーノイズ比(S/N比)は概ねコリプレッサーの影響を受けなかった。
【0035】
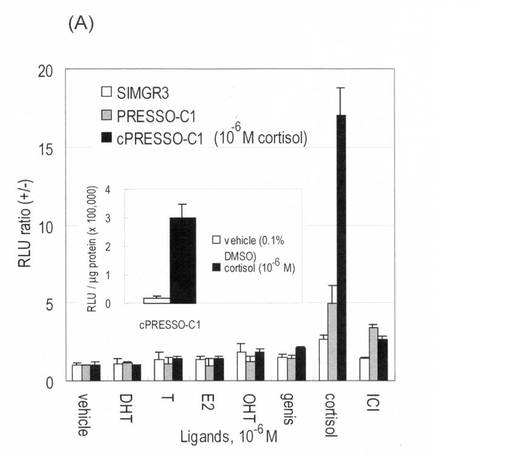
(実施例5)コリプレッサーモチーフと円順列変異(CP)との相乗効果
(5−1)COS-7細胞でのcPRESSO-C1, cPRESSO, cPRESSO-C2や SIMGR3のリガンド感受性
円順列変異とコリプレッサーモチーフを導入したプローブの相乗効果を調べた(図4,図5)。24ウェルプレートで培養したCOS-7細胞にpcPresso-c1, pcPresso, pcPresso-c2, や pSimgr3を導入した。その細胞を更に16時間間培養してから、溶媒か10-5 Mのコルチゾールで20分間刺激した。その発光強度を図2で記述したのと同様に評価した。図4(B)の濃度―応答曲線は図3で記述した方法で測定した。
その結果、cPRESSO-C1は、他のものに比べて、一番強い生物発光と直線性応答曲線を示した。一方、cPRESSOはテストしたプローブの中で一番高いS/N比を示した(例、10-6 M cortisolに対して約33倍)。
このようなコリプレッサーモチーフと円順列変異を導入することによって、およそ50-100倍程度の検出限界が下がる(改善する)ことを見出した。一番長い直線性応答範囲としては、cPRESSO-C1が10-8 から10-5 Mまでストレスホルモンに対して直線性応答を示した(図4(B),図5)。また、見かけ50%有効濃度(EC50)はおよそ2.5 x 10-7 M cortisolであった。その上、cPRESSO-C1のバックグラウンド発光値はコントロールに比べて、約1/3まで改善された。
【0036】
(5−2)コルチゾール濃度変化によるCOS-7細胞内PRESSO-C1のリガンド選択性
cPRESSO−C1のリガンド選択性をSIMGR3と比較してみた(図4(B))。24ウェルプレートで培養したCOS-7細胞にpcPresso-c1やpSimgr3を導入(transfection)した。
その細胞を10-6 M のステロイドや化学物質で20分間刺激した。その結果となる発光強度は図2で記述した方法と同様に測定した。
その結果、他のコントロールプローブに比べて、cPRESSO-C1は(ア)絶対発光強度の強さ、(イ)S/N比(約17倍)、(ウ)リガンド選択性において優れた結果を示した。
【0037】
(実施例6)コルチゾール検出におけるcPRESSO-C1の可逆性
ストレスホルモンセンシングにおけるcPRESSO-C1の可逆性をcortisolの刺激や除去を繰り返すことによって検証した(図6(B))。24ウェルプレート上に培養したCOS-7細胞にまずpcPresso-c1を導入し、更に16時間間培養した。次に細胞を溶媒(0.1% DMSO)又は10-7 M cortisolで20分間刺激した。その培養液をステロイドのない新鮮な培養液に置換することによってcortisolを一時除去した。図6(B)の挿入図で示したように、その細胞を更に10-7 M cortisolで刺激をする過程を繰り返した。毎段階における発光値の変化をその特異的な基質溶液キット(Promega)を使ってモニターした。
その結果は、10-7 Mのストレスホルモン刺激と洗浄の繰り返しにも関わらず、リガンド感度は衰えなかった。
この結果から、(ア)GR LBDとLXXLLモチーフとの結合は可逆的である。(イ)PRESSO-C1はストレスホルモンを繰り返して測定できるほど頑丈であり、この結果は以前のリガンド活性化されたGRの転写活性後リサイクルされる結論と一致する(非特許文献25)。
【0038】
(実施例7)PRESSO-C1からのリガンド応答に基づく蛍光強度の経時変化
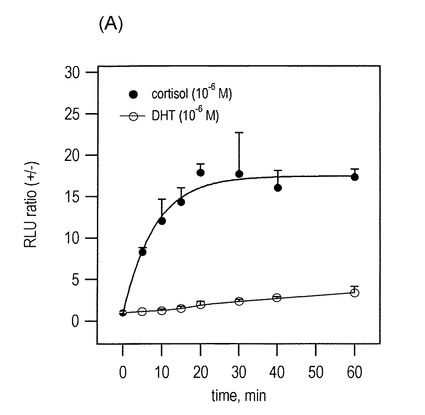
PRESSO-C1による発光強度の経時変化をリガンド刺激後1時間間観測した(図7(A))。24ウェルプレート上で培養したCOS-7細胞にトランスフェクション試薬(TransIT-LT, Mirus)を用いてpPresso-c1を導入した。その細胞を16時間間更に培養し、最後には10-6 Mストレスホルモンや男性ホルモン(5α-hydrotestosterone;DHT))で刺激した。刺激後5分、10分、15分、20分、30分、40分、60分にそれぞれ細胞を溶解・回収し、各々の発光強度をその特異的な基質溶液で現象した。この時のプロトコルは図2のものと同様である。
その結果、cPRESSO-C1による発光強度は、ストレスホルモンによる刺激後すぐ上がり始め、約20分ごろに最高値に達する。ところが、同一濃度の男性ホルモンに対しては、測定時間(1h)内に発光値の上昇は見られなかった。
この経時変化は以下のように結論できる。
(ア)cPRESSO-C1のストレスホルモンセンシングは約20分で完結される。
(イ)この20分の時間は、ホルモンの細胞膜透過、ホルモン-プローブ結合、分子内構造変化とタンパク質相補にかかる総時間を意味する。この反応時間は、以前GR LBD-LXXLLモチーフ間結合に基づくFRET研究結果と一致する(非特許文献26。)
【0039】
(実施例8)ガラス基板上に形成した生物発光紙stripを用いたストレスホルモン測定
非細胞系における本プローブの長所を披露するために、生物発光紙ストリップをガラス基板上(2 x 7 cm)に形成した(図7(B))。まず、Y字型紙片をニトロセルロース紙から切り出し、ガラススライド上に固定した。
一方、細胞溶解液そのもの、又はcPRESSO-C1を発現する細胞の溶解液をそれぞれ図2で記述した方法で準備した。それぞれの細胞溶解液12μLをY字型紙片の末端に塗布して、他に広がらないようにスポットを作った。後に溶媒のみ、又は10-6 Mコルチゾールを添加した基質溶液を紙片の真中に10μL滴下して、その発光強度を発光スキャナー(RAS-3000, FujiFilm)で10分間積算した。
その結果、図7(B)で示したように、紙片上ではストレスホルモン依存的に発光値が強化された。
この結果から以下の点がうかがえる。
(ア)ストレスホルモン活性度は紙片レベルでイメージングできる。
(イ)cPRESSO-C1は紙片上での乾燥にも耐えてホルモンをセンシングできるほど頑丈である。
(ウ)結果の発光強度がガラス基板上で従来の発光スキャナーで計測できるほど強い。
【図1A】
【図1BC】
【図3A】
【図3B】
【図4B】
【技術分野】
【0001】
本願発明は、各種生物発光又は蛍光イメージングプローブを安定に保存するためのアンカーペプチドに関するものである。
【背景技術】
【0002】
生体試料を対象とした生物分析は、病気の早期診断などにおける欠かせない手段である。例えば、ストレスは生体内免疫系、内分泌系における様々な病気の原因として活発な研究が行われている。例えば、生体内ストレスホルモン濃度は、コルチゾールの過剰分泌に起因するクッシング症候群や、慢性副腎皮質機能不全(アジソン病)などを診断する上で指標となっている(非特許文献1)。したがって、多くの医学・薬学分野においての研究の中心は、生体試料を対象に病気の指標となる因子の同定、分析が重要な課題といえるが、実際に病院などの診断現場で用いられている診断法は、いまだに数十年前からの免疫染色法に基づいている。
まだ実用化には至っていないが、昨今、従来の免疫染色法に代わる可能性を秘めた診断試薬の研究開発が進んできている。生体試料を分析し、発色でイメージングする方法として、蛍光を利用したFRET(蛍光共鳴エネルギー転移)(特許文献1、非特許文献2,3)や発光を利用した2分子型タンパク質相補(protein complementation)法等が開発された(特許文献2)。また最近、本発明者によって一分子型生物発光プローブが開発され(非特許文献2〜6)、また発光酵素の円順列置換(circular permutation)による分子イメージングプローブも開発された(非特許文献6)。なお、これらについては本発明者らにより特許出願もされている(PCT/JP2008/050370、特願2007-202308、特願2008-116098)。
これらの発色プローブは、大きく分けて、蛍光タンパク質を利用する蛍光的な手法と、各種発光酵素を用いる発光的な手法の2種類に大別される。蛍光タンパク質を利用する場合には、自己蛍光によってバックグラウンドが非常に高くなることに加え、外部光源とフィルターシステムが必要となり、蛍光を測定するために比較的に巨大な蛍光顕微鏡が必要となるという問題がある。例えば、非特許文献7〜9に記載のYellow Camellonsを用いた場合、自己蛍光がバックグラウンドを必然的に高め、シグナルとノイズとのコントラスト(S/N比)を低下させる原因となっている。さらに、分析できる細胞数が制限されることにより、得られる結果は定量的というよりもむしろ、定性的となってしまう(非特許文献10)。
一方、発光的な手法の場合は、このような問題はほとんど生じないが(非特許文献10〜14)、蛍光に比べて光強度そのものが低い。光強度は、一分子内に全ての要素を集積した一分子型発光プローブの場合においても改善されない上、分子内非特異的な結合が起こりやすいため、プローブの計測性能(検出感度、選択性、S/N比)を損なう原因になっている。
このように、従来の融合蛋白質に基づく蛍光・生物発光可視化プローブの共通の問題点として、その潜在的な検出能が十分発揮できず、検出限界、検出範囲、選択性、感度が生体試料計測等には不十分であった。そしてさらに、容易に分解されたり、失活されてしまうという分子プローブそのものの安定性にも問題があった。
上記問題点を解決するために、従来の蛍光・発光プローブの安定性を増大させ、発光強度、検出限界を高め、検出能を向上させる技術開発が急務であった。
【先行技術文献】
【特許文献】
【0003】
【特許文献1】特開2007−49943号公報
【特許文献2】国際公開 WO2004/104222
【非特許文献】
【0004】
【非特許文献1】Burtis et al. Tietz, Tietz textbook of clinical chemistry, 3rd ed., W.B. Saunders, Philadelphia,1999.
【非特許文献2】Awais et al. Angew. Chem. Int. Ed. 2006, 45, 2707-2712.
【非特許文献3】Awais et al. Anal. Chem. 2004, 76, 2181-2186.
【非特許文献4】Kim et al. Anal. Chem. 2007, 79, 1874-1880.
【非特許文献5】Kim et al., ACS Chem. Biol. 2008, 3, 359-372.
【非特許文献6】Kim et al., Bioconju. Chem. 2008, 19(12), 2480.
【非特許文献7】Souslova et al. Biochemistry-Moscow 2007, 72, 683-697.
【非特許文献8】Truong et al. Nat. Struc. Biol. 2001, 8, 1069-1073.
【非特許文献9】Miyawaki et al. Nature 1997, 388,882-887.
【非特許文献10】Kim et al. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 11542-11547.
【非特許文献11】Paulmurugan et al. Anal. Chem. 2005, 77,1295-1302.
【非特許文献12】Paulmurugan et al. Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 15883-15888.
【非特許文献13】Remy et al. Nat. Meth. 2006, 3, 977-979.
【非特許文献14】Kim et al. ACS Chem. Biol. 2007, 2, 484-492.
【非特許文献15】Lee et al. Cell. Mol. Life Sci. 2003, 60, 1613-1622.
【非特許文献16】Maruvada et al. J. Biol. Chem. 2003, 278, 12425-12432.
【非特許文献17】Tamrazi et al. Mol. Endocrinol. 2002, 16, 2706-2719.
【非特許文献18】Perissi et al. Genes Dev. 1999, 13, 3198-3208.
【非特許文献19】Hu et al. Nature 1999, 402, 93-96.
【非特許文献20】Horlein et al. Nature 1995, 377, 397-404.
【非特許文献21】Wilson et al. Nat. Meth.,2004, 1(3), 255-262.
【非特許文献22】Ozawa et al. Anal. Chem., 2001, 73(24), 5866-5874.
【非特許文献23】Bracha-Drori et al. Plant J., 2004, 40(3), 419-427.
【非特許文献24】Fan et al. Biochem. Biophy. Res. Comm., 2008, 367(1), 47-53.
【非特許文献25】Galigniana et al. J. Biol. Chem., 1999, 274, 16222-16227.
【非特許文献26】Awais et al. Steroids 2007, 72, 949-954.
【非特許文献27】Kim et al. Anal. Biochem. , 2007, 362, 148-150.
【非特許文献28】Kim et al. Anal. Chem. 2005, 77, 6928-6934.
【発明の概要】
【発明が解決しようとする課題】
【0005】
従来の融合蛋白質に基づく蛍光・生物発光イメージングプローブの安定性を改善し、検出限界、検出範囲、選択性、感度などの検出能を向上させることを目的とする。
【課題を解決するための手段】
【0006】
本発明者らは、従来の蛍光・発光プローブの上述した問題点を鋭意検討した結果、従来型の蛍光・発光プローブは、分子認識タンパク質のみに注目し、生体内の分子認識メカニズムとしては、当該タンパク質以外に多くの共役因子(コアクチベーター、コリプレッサーなど)が関わっていることの配慮がなされなかったために、自然界での本来の分子認識をそのまま再現できていないことに思い至った。
そこで、本発明者らは、生体内の分子認識タンパク質が通常コリプレッサーとの結合によって安定化されている点に着目し、従来型の一分子発光プローブに、コリプレッサーのモチーフペプチドを導入することで、分子の安定化を飛躍的に増大させることができ、検出限界を格段に下げることに成功した。
【0007】
生体細胞内においては、核内受容体(NRs)は、多数のヒートショック蛋白質群及びコリプレッサー(転写共役制御因子)群と共にクラスターを形成する形で安定化されている。しかしながら、これら分子は分子量が数百キロダルトンに及ぶものもあるため、分子プローブに取り入れることが困難であり、生体内のメカニズムを正確に模すことはできない。本発明者らは、上記安定化メカニズムの中核部分としてコリプレッサー配列中の「LXXIIXXXL」モチーフを利用することを発想した。まず、典型的な核内受容体であるグルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)を、「LXXLL」モチーフに繋ぎ、その両端を発光酵素のガルシアルシフェラーゼ(GLuc)断片で挟み込んだものを、本分子組み込み型(IMF)プローブのプロトタイプとし、末端にコリプレッサー由来のモチーフを繋ぐ。なお、上記プロトタイプは、円順列置換法(CP)(非特許文献6)に基づき、各要素の結合順序を大幅に組み換えることができ、コリプレッサーのモチーフを置き換えることも可能である。このように作製された本発明のプローブは、ストレスホルモンの指示薬として用いたときに2相性の濃度―応答曲線を描くことを最大の特徴とし、円順列置換を併用することで(i)より低い検出限界とより長い検出範囲,(ii)蛍光の絶対量とリガンド選択性の拡大が可能となった。
具体的には、多種類のコリプレッサーペプチドを用いてそれぞれの分子安定化能を調べた結果、あるペプチドはプローブの検出限界を1000倍までさげる(図3、4、5)ことを確認し、さらに円順列置換法を併用することで、選択性が格段にあがり(図6)、直線性検出範囲も広がる(図5)という知見も得た。このように、各種のコリプレッサーペプチドを導入すれば、様々な従来の蛍光や生物発光融合蛋白質プローブにおける安定化効果と検出能を期待できることから、船の錨に因んで「アンカーペプチド」と名づけた。以下、本発明において「アンカーペプチド」というとき、従来型の生物発光又は蛍光により検知、測定する可視化プローブの末端に、安定化のためにさらに繋いだコリプレッサー活性ペプチドを指す。
【0008】
すなわち、本発明は、以上の知見を得て完成することができたものであり、具体的には以下の通りのものである。
〔1〕 情報認識タンパク質が当該タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブにおいて、当該プローブ分子末端に、前記情報認識タンパク質に対するコリプレッサー活性ペプチドがアンカーペプチドとして結合されており、当該ペプチドが、リガンドの非共存時に前記情報認識タンパク質と会合していることを特徴とする、可視化プローブ。
〔2〕 核内受容体のリガンド結合ドメインを含み、当該リガンド結合ドメインがリガンドの結合により活性化されて生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブにおいて、当該プローブ分子末端に、核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドがアンカーペプチドとして結合されており、当該ペプチドが、リガンドの非共存時に前記リガンド結合ドメインと会合していることを特徴とする、可視化プローブ。
〔3〕 前記核内受容体がグルココルチコイドレセプターであって、前記立体構造変化が、当該グルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)がリガンドとの結合により活性化され、プローブ分子内のコアクチベーター由来の「LXXLL」モチーフを含むペプチドと結合することにより生じる立体構造変化であり、前記生物発光又は蛍光が、プローブ分子中のそれぞれ別の位置に存在する、分割された生物発光酵素又は蛍光タンパク質のフラグメントがタンパク質相補して発する生物発光又は蛍光である、前記〔2〕に記載の可視化プローブ。
〔4〕 前記グルココルチコイドレセプター(GR LBD)のリガンド結合ドメインを含むポリペプチド部位が、リンカーペプチドを介して又は介さずに、コアクチベーター由来の「LXXLL」モチーフを含むペプチドに結合されており、同一のプローブ分子の他の位置に、前記分割された生物発光酵素又は蛍光タンパク質のフラグメントと共に、前記核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドが結合されている、前記〔3〕に記載の可視化プローブ。
〔5〕 前記分割された生物発光酵素又は蛍光タンパク質のN末側及びC末側フラグメントが、リンカーペプチドを介して又は介さずに逆向きに結合されており、前記グルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)を含むポリペプチド部位と、コアクチベーター由来の「LXXLL」モチーフを含むペプチド部位と共に、同一のプローブ分子の他の位置に、前記核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドが結合されている、前記〔3〕に記載の可視化プローブ。
〔6〕 前記グルココルチコイドレセプターのリガンド結合ドメインを含むポリペプチド部位が、コアクチベーター由来の「LXXLL」モチーフを含むペプチド部位と別のプローブ分子上に存在し、かつそれぞれの部位が分割された生物発光酵素又は蛍光タンパク質のN末側もしくはC末側フラグメントのいずれかと、リンカーペプチドを介して又は介さずに結合されており、前記グルココルチコイドレセプターのリガンド結合ドメインを含むプローブ分子末端には、さらに前記核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドが結合されている、前記〔3〕に記載のプローブ。
〔7〕 情報認識タンパク質が当該タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブに対して、当該プローブ分子の末端に、前記情報認識タンパク質に対するコリプレッサー活性ペプチドをアンカーペプチドとして結合させることにより、当該ペプチドが、リガンド非共存下においては前記情報認識タンパク質と会合していることを特徴とする、可視化プローブの安定化方法。
〔8〕 核内受容体のリガンド結合ドメインを含み、当該リガンド結合ドメインがリガンドの結合により活性化されて生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブに対して、当該プローブ分子の末端に、核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドをアンカーペプチドとして結合させることにより、当該ペプチドが、リガンド非共存下においては前記リガンド結合ドメインと会合していることを特徴とする、可視化プローブの安定化方法。
〔9〕 情報認識タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブ分子の末端に存在するアンカーペプチドであって、情報認識タンパク質に対するコリプレッサー活性ペプチドからなり、リガンドによる活性化前の可視化プローブを前記情報認識タンパク質と会合することで安定化させることを特徴とする、アンカーペプチド。
〔10〕 前記コリプレッサー活性ペプチドが、核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドからなる、前記〔9〕に記載のアンカーペプチド。
〔11〕 前記「LXXIIXXXL」モチーフ含有ペプチドが、配列番号5〜12又は配列番号23〜30のいずれかで表されるアミノ酸配列を含むペプチドである、前記〔10〕に記載のアンカーペプチド。
〔12〕 前記〔1〕〜〔6〕のいずれかに記載の可視化プローブをコードする核酸。
〔13〕 前記〔12〕に記載の核酸を含む、可視化プローブ発現用ベクター。
〔14〕 前記〔13〕に記載の発現ベクターを用いて形質転換され、可視化プローブを発現する形質転換細胞。
〔15〕 被検試料中のリガンドを検出又は定量するためのキットであって、前記〔1〕〜〔6〕のいずれかに記載の可視化プローブ又は前記〔14〕に記載の形質転換細胞を含むキット。
〔16〕 前記キットが、さらに発光酵素の基質を含み、紙ストリップ上に固定、乾燥されている前記〔15〕に記載のキット。
〔17〕 被検試料中のリガンドの検出方法又は定量方法であって、前記〔1〕〜〔6〕のいずれかに記載の可視化プローブ又は前記〔14〕に記載の形質転換細胞を用いることを特徴とする方法。
【発明の効果】
【0009】
本発明では、従来型の生物発光・蛍光プローブにアンカーペプチドを導入することで、生細胞内安定性を担保しながら、プローブの検出限界を顕著に下げることができ、特に円順列置換法と併用することで、選択性が格段にあがり、検出範囲も広げることができた。すなわち、本発明においては、生体や生細胞を対象とした計測能(選択性、信号強度、S/N比)が大きく改善された、安定な生物発光可視化プローブが提供できた。
また、本発明のプローブは、プローブ安定性が格段に高く、診断用紙片ストリップの製作にも適しているため、簡便な分析手段を提供できる。
【図面の簡単な説明】
【0010】
【図1】(A)本発明で使われた生物発光可視化プローブをコードする遺伝子構成物の概略図。省略名:GLuc−N,ガウシアルシフェラーゼのN−末側断片;GLuc−C,ガウシアルシフェラーゼのC−末側断片;GR LBD,グルココルチコイド受容体のリガンド結合ドメイン。(B)本発明で合成された生物発光プローブのおよその分子構造。(C)cPRESSO−C1のリガンド依存的な分子内構造変化の可視化戦略の模式図。アゴニスト無しの状態では,コリプレッサーモチーフと結合し安定化している。ところが,アゴニストによって刺激されれば,分子内発光酵素断片間相補が起こり,結果として発光酵素活性が回復する。
【図2】従来型プローブ(「SIMGR」系列)の分子認識能。(A)10-6 Mのコルチゾール添加に対する「SIMGR」系列のプローブの相対的な発光強度。挿入図は、蛋白質μgあたりの強度の絶対値を示している。(B)全タンパク量とプローブの発現レベルを示すウエスタンブロット分析。
【図3】PRESSOシリーズプローブ内のGLuc断片間相補によって回復した生物発光強度に基づいたコルチゾール濃度−応答曲線。(A)コルチゾール濃度依存的なPRESSO-C3、-C4の2相曲線。(B)コルチゾール濃度依存的なPRESSO-N2、-C2の2相曲線。
【図4】(A)本プローブの計測性における円順列変異とコリプレッサーモチーフの相乗効果。黒棒は10−5Mコルチゾール刺激による絶対発光値を示す。(B)コルチゾールとDHT濃度変化に対する,cPRESSO-C1とSIMGR3の濃度―応答曲線。
【図5】cPRESSO-C1,cPRESSO-C2,cPRESSO及びSIMGR3を発現するCOS-7細胞の絶対発光値を示す濃度−応答曲線。cPRESSO-C1による生物発光値が絶対発光強度と直線性領域等からもっとも優れている。
【図6】(A)cPRESSO-C1、PRESSO-C1、SIMGR3のリガンド選択性の比較。挿入図は10-6 M コルチゾール刺激前後の絶対発光値を示す。省略名:vehicle, 0.1% dimethyl sulfoxide; DHT,5α-dihydrotestosterone; T, testosterone; E2, 17β-estradiol; OHT, 4-hydroxytamoxifen; genis,genistein; ICI, ICI-182780。(B)cPRESSO-C1の反復したリガンド感受性測定。挿入図Aで示したようにcPRESSO-C1を持つCOS-7細胞に10-7Mコルチゾールの刺激と除去を繰り返した。
【図7】(A)リガンド刺激によるcPRESSO-C1からの発光値の経時変化。(B)非細胞系である生物発光紙片上でのリガンド活性の測定。ガラス基板上にニトロセルロース紙片を付着し,各紙片末端に細胞溶解液をスポットした。後にストレスホルモン有り,無しの状態で基質溶液を滴下して,発光値をスキャンした。
【発明を実施するための形態】
【0011】
1.本発明の生物発光又は蛍光プローブの構造
本発明は、従来の生物発光又は蛍光可視化プローブの安定化を図ることで、安定化と共に当初予期していなかった計測性の向上も可能にした改良発明に係るものであるといえるが、これらの可視化プローブにおいては、核内受容体などの情報認識タンパク質は、当該情報伝達物質(リガンド)との結合によって立体構造が変化したことを、生物発光又は蛍光により検知して測定するものである。当該可視化プローブで従来から用いられていた情報認識タンパク質は、いずれも生体内では1又は複数のコリプレッサータンパク質が結合した状態で安定して存在している。
つまり、生体内では、情報認識タンパク質は通常、コリプレッサータンパク質と結合して安定化されており、外部からの信号(リガンド)によって刺激されたときに初めてコリプレッサータンパク質と離れ、活性型に変貌する。
例えば、核内受容体は、生体内でコリプレッサータンパク質群と結合した形で安定化されているが、核内受容体と同様にその2量体形成メカニズムが可視化プローブに利用されているNuclear Factor-κB (NF-κB)(非特許文献27)の場合も、生体内では通常Inhibitor-κB(IκB)と結合した形で安定化されている。また、細胞内コレステロール濃度を司るsterol regulatory element-binding protein-2 (SREBP-2)は、サイト1分解酵素(site 1 protease; S1P)によって2分割されると2量体になることによって活性型に変わる。このSREBP-2の活性化メカニズムが、蛋白質スプライシングを介する生物発光可視化プローブ等に利用されているが(非特許文献28)、生体内では通常SREBP cleavage-activating protein(SCAP)と結合した形態で安定化されている。さらに、G-タンパク質は通常細胞内ではそのαとβユニットが結合した形態で安定化されており、外部信号が到来したときに、αユニットはβユニットから離れて細胞膜上受容体と結合して立体構造変化を起こす。このαユニットの立体構造変化は可視化プローブに利用でき、βサブユニット(システイン-リッチ配列)については、その安定化現象に対してアンカーペプチドとしての効果が期待できる。
本発明では、このように多くの情報認識タンパク質と生体内で通常結合しているコリプレッサー作用のあるペプチド配列を、可視化プローブ用アンカーペプチドとして用いようとする発明であるから、従来から可視化プローブでその立体構造変化が利用されていた情報認識タンパク質に対する公知コリプレッサーペプチドは、すべて利用可能である。本発明の実施態様では、これら情報認識タンパク質として典型的な核内受容体の場合を例として取り上げる。以下、核内受容体に対してコリプレッサーを連結して安定化させた生物発光プローブによって本発明を説明するが、生体内の情報認識タンパク質としての挙動はいずれもほぼ共通しているから、核内受容体に対するコリプレッサーの組み合わせと同様に、G-タンパク質αサブユニットに対するβサブユニットのシステイン-リッチ配列、NF-κBに対するIκBのreal homology domain (RHD)、及びSREBP-2に対するSCAPのTrp (W)-Asp (D)反復モチーフとの組み合わせも同様のアンカーペプチドとしての使用が可能であることは当然である。
なお、従来型の蛍光プローブ(FRET)において核内受容体を含むプローブ分子内にコリプレッサー活性ペプチドを有する例が報告されている(非特許文献26)が、当該ペプチドは、単にリガンドの結合による立体構造変化により蛍光が発せられるのを定性的に観察するための要素であり、アンカーペプチドとして用いるものではない。
【0012】
本発明の核内受容体のリガンド結合性を利用して、そのリガンド物質を検出する生物発光又は蛍光プローブにおいては、核内受容体のリガンド結合ドメインを含む従来型の生物発光プローブ又は蛍光プローブ(非特許文献2〜6、14、26)に対して、その分子中に、好ましくはN末側もしくはC末側に、コリプレッサー配列中の「LXXIIXXXL」モチーフをアンカーペプチドとして結合させることを特徴とするものである。
例えば、典型的な核内受容体であるグルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)を用いる場合についてみると、GR LBDのリガンド結合ドメインを含むポリペプチドを、そのコアクチベーターである「LXXLL」モチーフを含むペプチドに繋ぎ、その両端をガウシアルシフェラーゼ(GLuc)断片など、各種ルシフェラーゼ又はGFPで挟み込んだかたちで一分子に集積し、その末端にコリプレッサー由来モチーフを繋ぐ。ここで、上記プロトタイプは、(ア)円順列置換法(CP)に基づき、各要素の結合順序を大幅に組み換えることができ、(イ)コリプレッサーのモチーフを置き換えること、(ウ)各構成要素の間に挿入すること、(エ)またはさらに2種以上のコリプレッサーモチーフを繋げることも可能である。CPを適用した場合の典型的な各構成要素の順序は、図1に記載されるように、発光酵素、GFPの分割断片同士が隣り合って逆向きに繋がることとなる。
従来型の生物発光プローブ又は蛍光プローブが二分子型の場合、即ち、核内受容体のリガンド結合ドメインと、「LXXLL」モチーフとが別の分子中に存在する場合においても前者にアンカーペプチドを繋ぎ、各分子末端に発光酵素又はGFPの分割断片の片方ずつを繋げばよい。
それぞれの構成要素ペプチドは、適宜長さを調節したリンカーペプチドを介して繋がれる。リンカーペプチドとしては、Gly-Gly-Ser(配列番号17)を基本単位とする「GSリンカー」などを用いることができる。他に、柔軟なアミノ酸をいくつかが繋いだペプチドも適宜用いることができる。リンカーペプチドのアミノ酸配列長としては0〜15アミノ酸残基、好ましくは3〜6アミノ酸残基が一般に用いられ、GSリンカーの場合では1〜5単位、好ましくは1〜3単位で用いられる。
【0013】
2.核内受容体とコアクチベーター由来「LXXLL」モチーフ
核内受容体(nuclear receptor;NRs)は、細胞内にあり、リガンドが結合すると核内に移行し、DNAに直接結合してDNAの転写を制御する受容体であって、生体中の転写制御因子(transcriptional regulators)のうちでも最も多彩なメンバーを持つものの1つである(非特許文献15)。これら核内受容体には、恒常性維持、生殖、発生、及び代謝といった多様な機能が知られており、それぞれが持つリガンド認識能やリガンド刺激によって引き起こされる分子メカニズムは,新たな融合蛋白質プローブの設計に応用されてきた.例えば、α−ヘリカルペプチドと核内受容体のリガンド結合ドメイン(NR LBD)との相互作用を指標として、蛍光共鳴エネルギー移動(FRET)法(非特許文献2,3)と一分子型生物発光プローブ(IMF)が開発された(非特許文献4,5)。ここで、α−ヘリカルペプチドとは、核内受容体のリガンド結合ドメインを認識するコアクチベーター由来のペプチドであって、「LXXLL」モチーフを含むという配列上の特徴を有している(非特許文献2,3)。ただし、Lは他にもIのような疎水性アミノ酸に入れ替えしても同様の性質を保つが、ここでは通常の言い方に倣って「LXXLL」モチーフと称する。また、核内受容体の核内移行はGFP-修飾法(非特許文献16)と蛋白質スプライシング法(非特許文献10)によって観察できる。また、リガンドによる核内受容体間の2量化はFRET法を用いて以前観察できた(非特許文献17)。これらの研究例からも、生体内のような複雑系におけるリガンド活性を計測する上で、核内受容体の重要さを物語っている。
本発明のプローブには、これら核内受容体のうち少なくともリガンド結合ドメイン(NR LBD)を含む核内受容体タンパク質及び活性化された核内受容体タンパク質を認識するペプチドが用いられる。本発明のNR LBDとしては、コルチゾールなどのストレスホルモンをリガンドとするグルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)以外、どのような核受容体も置き換えて使用できるが、例えば、同じストレスホルモン受容体の一種であるミネラルコルチコイドレセプター(mineralocorticoid receptor; MR)、又は女性ホルモンレセプター(estrogen receptor; ER)、男性ホルモンレセプター(androgen receptor; AR)、黄体ホルモンレセプター(progesterone receptor; PR)、 ビタミンDレセプター(vitamine D receptor; VDR)のLBDなどを用いることができる。それぞれの活性化されたNR LBDを認識するペプチドは公知であり、GR LBDの場合においてもNR LBD共通のLXXLLモチーフを含むペプチド、また各種疎水性アミノ酸を含むペプチドが用いられる。
【0014】
3.コリプレッサー(転写抑制因子)
コリプレッサー(corepressor)は、コアクチベーター(coactivator)と同様に、核内受容体のリガンド結合ドメイン(NR LBD)と結合して転写活性を調節することによって、NR LBDの転写活性における中心的な役割を果たしている(非特許文献17,18)。最初、核内受容体のコリプレッサーは酵母の2−ハイブリッドスクリーニング法により単離されたが、その中には、核内受容体コリプレッサー(NcoR)(非特許文献19)及びレチノイン酸と甲状腺ホルモン受容体のサイレンシング・メディエーター(SMRT)(非特許文献20)等が含まれる。上記コリプレッサーに関する研究によれば、一般的なコリプレッサーの役割は、アゴニスト刺激がない時、ホスト分子を安定化し、その転写活性を抑制する作用をすることにある、といわれる(非特許文献19〜20)。これらのコリプレッサーの配列上の特徴としては、リガンド非存在条件下で核内受容体と結合能を持つ共通配列である「LXXIIXXXL」モチーフ(配列番号4)を共有している。
ここで、「LXXIIXXXL」モチーフとしては、NcoR由来の「LEDIIRKALMG:配列番号5」などの他、SMRT由来の「GMLAKRIIAELGMNTSA:配列番号6」など下記表2に示された配列が典型的であるが、他にSMRT ID2(RVVTLAQHISEVITQDYTR:配列番号7)やNCoR ID2(RLITLADHICQIITQDFAR:配列番号8)などがある(非特許文献19、20)。また、表1で示したように、最初のLは、Iであってもよく、4番目のアミノ酸はVでもよい。L, I, Vは、共に代表的な疎水性アミノ酸である特徴がある。また、Xは通常どのアミノ酸でもいいという意味ではあるが、本発明におけるコリプレッサーモチーフにおいては、E、R、K、D、S等のアミノ酸が好ましい。これらのアミノ酸は比較的に親水性アミノ酸である特徴がある(非特許文献19、20)。したがって、本発明において「LXXIIXXXL」モチーフというとき、「I、Lのような疎水性アミノ酸が一定間隔で規則的に反復される中で、その間にはアスパラギン酸(D)、グルタミン酸(E)のような親水性アミノ酸が入っているペプチド」の場合を好ましい条件とする。なお、モチーフを構成する一部のアミノ酸が欠損されても、そのレプレッサー性を完全には失わない。
核内受容体のコリプレッサーモチーフ以外にも、G-タンパク質αサブユニットに対するβサブユニットをアンカーペプチドとして用いる場合は、そのシステイン(C)−リッチモチーフ、NF-κBに対してIκBを用いる場合は、IκB のRel homology domain (RHD)、そしてSREBP-2に対してSCAPを用いる場合は、SCAPのTrp(W)-Asp(D)反復モチーフを含むペプチドが有効であり、本発明と同様に実施できる。
【0015】
【0016】
4.発光酵素(ルシフェラーゼ)又は蛍光色素蛋白質(FP)
本発明で2分割して用いるルシフェラーゼは、通常の発光プローブにおいて用いられるホタルルシフェラーゼ(FLuc)、ウミシイタケルシフェラーゼ(RLuc)、コメツキムシルシフェラーゼ(CBLuc)(非特許文献4,5,10)などいずれのルシフェラーゼであってもよいが、発光量の大きなガルシアルシフェラーゼ(Gluc)が特に好ましい(非特許文献6)。
また、蛍光色素としては、GFPが従来から用いられており(非特許文献21、22)、他にもYFP断片(非特許文献23)やVenus断片(非特許文献24)などが使われているが、本発明においても同様に用いることができる。
【0017】
5.本発明プローブの構築方法
本発明プローブは、その構成要素となるペプチドを直接もしくはリンカーペプチドを介して、化学的にペプチド結合で繋ぐこともできるが、それぞれのペプチドをコードするDNAを繋いで発現ベクター中に組み込み、その発現ベクターを導入した形質転換細胞により製造することができる。
既存の生物発光プローブ又は蛍光プローブ(非特許文献2〜6、9〜12,14)に対して、その末端に又は内部にアンカーペプチドを、直接又はリンカーペプチドを介して結合させてもよい。発現ベクターに組み込む場合も、既存の生物発光プローブ又は蛍光プローブをコードするDNAの5’末端又は3’末端に繋いで発現ベクターに挿入される。
また、当該発現ベクターのまま生体内の細胞中に組み込み、生細胞内でのイメージングが可能である。さらに当該発現ベクターが導入された形質転換細胞で産生させたプローブを用いて非細胞系分析用キットを作製することもできる。
【0018】
本発明プローブをコードする発現ベクターを導入する細胞としては、培養細胞(原核細胞、酵母細胞、昆虫細胞、哺乳類動物細胞など)のみならず、哺乳動物、昆虫など生体内の細胞も含み、典型的にはマウスなど実験動物もしくはその細胞である。
本発明で用いる「発現ベクター」としては、公知の真核または原核細胞発現ベクター(プラスミド、ウイルスベクターなど)を特段の制限なく使用することができ、公知の組織特異的、または特定刺激特異的プロモーター配列を組み込んでもよい。
本発明の発現ベクターの導入は、例えばマイクロインジェクション法やエレクトロポーレーション法、脂質試薬によるトランスフェクション法(TransIT, Mirus)により細胞内に導入することができる。また、これらの手法で導入される発現ベクターに単純にプローブの発現だけを託すのではなく、プローブの上流に発現制御をし得る制御配列を繋げることによって、プローブそのものの発現有無を指標に更なる生物分析系を組むことができる。
さらに、本発明プローブをコードするDNAを含む組換えDNAを、pMXのようなレトロウイルスベクターにつなげ、高力価のウイルス産生能を有するパッケイジング細胞PLATEに導入することによって、動物に感染するレトロウイルスの作成が可能である。このウイルスを実験動物の各臓器に感染させることによって、各臓器内におけるストレスホルモンなどのリアルタイムイメージングができる。
【0019】
6.本発明プローブの診断キットとしての使用方法
本発明のベクターを導入した形質転換細胞のまま、又は当該細胞からプローブを取り出して精製し、もしくは未精製のまま紙片上で用いることで、生体試料内の各種生理活性物質濃度、例えばストレスホルモン濃度を簡便に測定することができ、迅速にストレス量の診断ができる。
本発明のプローブは、細菌のような原核細胞で大量発現させることもできるが、哺乳類細胞の場合でも、適切な分泌シグナル配列(MGVKVLFALICIAVAEA:配列番号18など)を繋いだプローブを導入することによって、培地中に大量に分泌させることによって、精製工程なしでも分析に用いることのできる、大量なプローブ含有培養上清が得られる。また、さらにカラム精製用のタグ(例えば、His Tag; HHHHHH:配列番号19)をつけることで、大量に精製された本発明のプローブを得ることができる。
例えば、ストレスホルモンの定性定量分析用キットとする場合に、精製もしくは未精製発光プローブを持つ紙ストリップと発光酵素の基質を組み合わせたキット、又は本発明の発光プローブを発現している生細胞と発光酵素の基質を含有した培地とを組み合わせたキットを構成することができる。蛍光プローブの場合は、主に蛍光共鳴エネルギー移動(FRET)現象に基づいたプローブが作製されてきたが、バックグラウンドが高く信号蛍光は弱いのでフィルターを構える必要があったため、従来紙片上での蛍光断片を利用した例は報告されていないが、本発明により可能性となる。
【0020】
なお、診断用キットに用いる紙片や断片プレートの材質としては、ニトロセルロース(nitrocellulose)紙等の紙片、ナイロン66、polyvinylidene fluoride, hydrophobic
polymerなどが用いられ、特に、ニトロセルロース紙片を用いることで、リトマス試験紙のような取り扱いやすい発光プローブ付着型試験紙が提供できる。また、探針素子などの末端に本発光プローブを固定・乾燥した、発光型リガンドセンシング手段を提供できる。
【0021】
7.本発明の具体的な態様
本発明を具体的な態様で説明すると、以下の通りであるが、この方法には限定されない。
(1)本発明のプローブの構築方法
まず、グルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)を、GSリンカーを介して、α−ヘリカルコアクチベーターモチーフ(LXXLL:配列番号13)と繋ぐ。次いで、その融合蛋白質をQ105位置で分割したガウシアルシフェラーゼ(GLuc)のN-端側断片と、C-端側断片の間に挟み込み、さらに、このN-端及びC-端断片を両端に有する生物発光プローブをコリプレッサーモチーフで連結する。このようなコリプレッサーモチーフで修飾された生物発光プローブをコリプレッサーの英語スペルからとって「PRESSO」と命名した。但し、レプレッサーモチーフをプローブ構成要素の間に挟んでも構わない。
当該プローブでは、アゴニストに対して2相性の応答を示し、検出限界が最高で10-11 Mまで改善するという画期的な効果を示す。さらに、上記生物発光プローブプロトタイプにおいて円順列(circular permutation:CP)の手法で再構成することにより、さらに検出能を上げることができる。生物発光プローブ作製の際に、円順列手法は有効な再構成法であり、それが顕著に元のプローブの検出限界を改善する。上記実施態様においても、CP手法のみでコルチゾールで10-8 Mにまで検出できる。コリプレッサーモチーフの導入は、主に検量線における直線性範囲をのばすことに寄与している。
特にコリプレッサーモチーフはCP手法と組み合わせることで、相乗的に以下の効果が高まる。
(ア)発光強度の絶対値を高める。
(イ)蛍光強度のバックグラウンドを低く抑える。
(ウ)シグナル−ノイズ比(S/N比)を改善する。
(エ)リガンド選択性を改善する。
したがって、コリプレッサーモチーフを搭載した融合蛋白質プローブは、細胞内分子の事象を追跡するためにきわめて有用である。
特に、発光酵素(ルシフェラーゼ)としてGLucを使う場合は、他のルシフェラーゼと比較して100倍以上も明るいので、GFPを用いる場合と同様、生きたままの生細胞条件下でも明瞭で正確な分析ができる。なお、通常の発光酵素を用いる場合は、測定発光値が小さいため、最適緩衝溶液条件を作る必要があり、細胞を壊してLysateを作るのが一般的である。
【0022】
(2)本発明のプローブを用いた分析、スクリーニング方法
本発明の発光プローブを用いて生きた細胞内分子現象を分析、又は分子現象に影響を与えるリガンドのスクリーニングのためには、例えば以下の方法が適用できる。
(i)発光プローブをコードするcDNAを持つプラスミドを24穴プレート上の生細胞に導入し、さらに16時間培養する。
(ii)上記細胞の培養液を除去し、その変わりに基質(coelenterazine)を含んだ200μL緩衝溶液(例えば、HBSSバーファー)に細胞を浸す。
(iii)上記細胞を特定刺激物質で刺激し、その刺激前後の発光変化値を発光プレートリーダー(luminescence plate reader; 例えば、LB 941 Multimode Reader(Berthold))を用いて観測する。
また、発光プローブを用いた非細胞系実験(in vitro)としては、以下の分析、スクリーニング方法が適用できる。
(i)精製された発光プローブを直径1.2センチの十字架型紙ストリップの末端に垂らして乾燥させる。
(ii)上記十字架型紙ストリップの中央に刺激物質を含む基質溶液15μLを点滴し、即時に発光スキャナー(例えば、RAS-3000;FujiFilm)で発光値を観測する。
【0023】
(3)ストレスホルモンとしてのコルチゾールの測定
ストレスホルモンと呼ばれるコルチゾールの生体サンプル中の生理濃度はクッシング症候群とアジソン病のような病気の直接の指標となる(非特許論文1)。生体サンプル中の総コルチゾール濃度は、それぞれ0.8−6.4 x 10-7 M(血清)、0.6-7.5 x 10-7 M(小便)、0.2−2.8 x 10-8 M(唾液)となる。このような生理濃度範囲は、生体サンプル中の内分泌ストレスホルモンを検出するためには、感受性の高いセンサーが要求されることを意味する。従って、従来の測定法は主に放射性同位元素や非放射性免疫センシング法に依存してきた。このような従来法の代わりに、私たちはストレスホルモンを高感度で検出できるような一分子型生物発光プローブを合成した。このプローブの特色は、コリプレッサーモチーフを導入し、更に円順列変異と合わせることによって相乗効果を高めて、結果として、非放射性、非免疫学的、発色型の内分泌ステロイド計測手段といえる。
図3で示したような2相の濃度―反応曲線は、プローブ母体に与えるコリプレッサーペプチドの特質といえる。
【0024】
(4)本発明プローブを用いた測定のメカニズムに関する考察
本発明のプローブが図3などにおいて示すような2相濃度―反応曲線は一般的に、この系の中に2つの平行が存在することを意味する。本発明のプローブの構成成分を考慮すれば、第1平行はGR LBDとコリプレッサーモチーフ間の相互作用であり、第2平行はGR LBDとコアアクチベーターモチーフとの相互作用であると考えられるから、本発明のプローブのリガンドセンシングメカニズムは、以下のようになるといえる。
(ア)本発明のプローブ内のGR LBDは最初にはコリプレッサーモチーフによって安定化されている。この状態はHuらの文献(非特許論文19)の結果と一致する。
(イ)本発明のプローブは、低濃度のストレスホルモンに応答して、コリプレッサーモチーフを放し、この分離により、GR LBDとコアアクチベーターモチーフとが会合する確率を高める。
(ウ)本発明のプローブが、高濃度のストレスホルモンに応答して、GR LBDとコアアクチベーターモチーフとの完全結合によって最大発光値を出す。
【0025】
また、円順列法の適用により作製された、pcPressoの分子構造では、隣り合った2分割発光酵素の活性部位が反対方向に背向くことになる(図1(A))が、この分子構造では、分割された発光酵素の活性部位断片同士がたまたま衝突する確立は非常に低く抑えられる。この構造的利点により、プローブのバックグラウンド発光が劇的に減少し、信号―バックグラウンド比(S/N比)の改善に繋がったと考えられる。
また、本プローブにおけるコリプレッサーモチーフによる効果と円順列変異との相乗効果については、図4(B),図5におけるSIMGR3とcPRESSOの比較からみて、円順列変異によりバックグラウンド発光が抑えられることで検出限界が向上し、さらにNcoR由来のコリプレッサーモチーフを導入することによって、濃度―応答曲線での直線性応答領域が広くなり(cPRESSO-C1の場合)、絶対発光強度の増加も観測される(図4(A)と図5)、という相乗効果が観測される。すなわち、本プローブにおけるコリプレッサーモチーフの役割は、プローブ分子そのものを荒い海で船の錨のように硬くして細胞内などの測定環境の系において安定化させ、発光酵素の反応効率を上げることにあると考えられる。
【0026】
8.その他
本発明におけるその他の用語や概念は、発明の実施形態の説明や実施例において詳しく規定する。なお、用語は基本的にはIUPAC-IUB Commission on Biochemical Nomenclatureによるものであり、あるいは当該分野において慣用的に使用される用語の意味に基づくものである。また発明を実施するために使用する様々な技術は、特にその出典を明示した技術を除いては、公知の文献等に基づいて当業者であれば容易かつ確実に実施可能である。例えば、遺伝子工学および分子生物学的技術はJ. Sambrook, E. F. Fritsch & T. Maniatis, "Molecular Cloning: A Laboratory Manual (2nd edition)", Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York (1989); D. M. Glover et al. ed., "DNA Cloning", 2nd ed., Vol. 1 to 4, (The Practical Approach Series), IRL Press, Oxford University Press (1995); Ausubel, F. M. et al., Current Protocols in Molecular Biology, John Wiley & Sons, New York, N.Y, 1995;日本生化学会編、「続生化学実験講座1、遺伝子研究法II」、東京化学同人 (1986);日本生化学会編、「新生化学実験講座2、核酸 III(組換えDNA技術)」、東京化学同人 (1992); R. Wu ed., "Methods in Enzymology", Vol. 68 (Recombinant DNA), Academic Press, New York (1980); R. Wu et al. ed., "Methods in Enzymology", Vol. 100 (Recombinant DNA, Part B) & 101 (Recombinant DNA, Part C), Academic Press, New York (1983); R. Wu et al. ed., "Methods in Enzymology", Vol. 153 (Recombinant DNA, Part D), 154 (Recombinant DNA, Part E) & 155 (Recombinant DNA, Part F), Academic Press, New York (1987)などに記載の方法あるいはそこで引用された文献記載の方法またはそれらと実質的に同様な方法や改変法により行うことができる。また、本発明で使用する各種蛋白質やペプチド、あるいはそれらをコードするDNAについては、既存のデータベース(URL:http://www.ncbi.nlm.nih.gov/等)から入手することができる。
なお、本発明で引用した先行文献又は特許出願明細書の記載内容は、本明細書の記載として組み入れるものとする。
【実施例】
【0027】
以下、実施例を示して本発明をさらに詳細かつ具体的に説明するが、本発明は以下の例に限定されるものではない。
【0028】
(実施例1):プラスミドの構築
ガウシアルシフェラーゼ(GLuc)の全長をコードするcDNAのテンプレートは、Nanolight社から購入した。cDNAのN-端及びC-端側断片(GLuc-N (18-105 AA、配列番号1); GLuc-C (106-185 AA、配列番号2))はPCRにより合成した。この際、両断片の末端には特異的な制限酵素サイトHindIII/ KpnI (GLuc-N)及びBamHI/ XhoI (GLuc-C)を導入するために、好適なプライマーとその鋳型を用いてPCR反応を起こした。
グルココルチコイドレセプターのリガンド結合ドメイン(GR LBD; 527-777 AA、配列番号3)のcDNA末端に、PCR反応を用いて制限酵素サイトKpnI/ NotIを導入した。なお、両端のNotI/ BamHI制限酵素サイトを有する下記のペプチドのcDNAオリゴマーはExigen (Tokyo, Japan)社から発注・購入した。
(i)「Glucocorticoid Receptor Interacting Protein 1 Interaction domain 3」由来のα−ヘリカルLXXLLモチーフ(GRIP1 ID3;NALLRYLLDKD:配列番号14);
(ii)GRIP1 ID2 由来のLXXLLモチーフ(GRIP1 ID2: HKILHRLLQDS:配列番号15)
(iii) GRIP1 ID3のアラニン変異型LXXLLモチーフ(GRIP1 ID3m: NALARYALDKD:配列番号16).
上記cDNA断片は、図1に示されるとおりに繋がれ、pcDNA 3.1(+)ベクターにサブクローニングされている。
GRIP1 ID3、変異タイプのGRIP1 ID3及びGRIP1 ID2を含むプラスミドは、それぞれpSimgr3, pSimgr3m,及びpSimgr2と名付けた。この意味は、単一分子型のグルココルチコイドレセプター(GR LBD)搭載生物蛍光プローブ(SIngle-Molecule-format bioluminescent probe with Glucocorticoid Receptor)を意味しており、それぞれは、発現後のSIMGR3, SIMGR3M, 及び SIMGR2と呼ばれる融合蛋白質プローブに対応する。
さらに、pSimgr3のN-又はC-末端にコリプレッサー蛋白質のモチーフを付加する修飾を施した。つまり、そのN-末端にsilencing mediator for Retinoic acid and Thyroid hormone receptors Interaction Domain 1由来のLXXIIXXXLモチーフ(SMRT ID1; GMLAKRIIAELGMNTSA:配列番号11)を連結したもの、又はNuclear receptor CORepressor Interaction Domain 1由来のLXXIIXXXLモチーフ (NcoR ID1; ASNLGLEDIIRKALMG:配列番号9)を連結した。さらに、SMRT ID1の逆向きのLXXIIXXXL モチーフ(ASTNMGLEAIIRKALMG:配列番号12)やNcoR ID1の逆向きのLXXIIXXXL モチーフ(GMLAKRIIDELGLNSA:配列番号10)も上記プローブのN-末端に繋ぎ、GR LBDとの結合性を検討する。プローブのC-末端も、同様にSMRT ID1又はNcoR ID1由来のコリプレッサーペプチドを連結して結合特性を検討した。その構造は、模式的に図1に示した。その特異的なペプチド配列を〔表2〕に示す。
これらのプローブは、コリプレッサーモチーフ(corePRESSOr motif)を有する一分子集積型生物発光プローブ(integrated-molecule-format bioluminescent probe;IMF)、すなわちPRESSOと名付ける。そして、N-及びC-末端にコリプレッサーモチーフを有するプローブを、それぞれPRESSO-N 及びPRESSO-C系列指示薬とよぶ。
なお、これら各要素を連結する際には、リンカーペプチドとしてGly-Gly-Ser(配列番号17)を基本単位とするGSリンカーを適宜の長さに調節して用いた。
【0029】
さらに、PRESSOプローブの円順列バージョンを合成した。その合成理由としては、(i)リガンド感受性の増大、(ii)シグナルとバックグラウンドの比率(S/N比)の改善を見込んだものである。この具体的な手順は以下の通りである。最初に、GLucをQ105位置で2分割し、その5’末端と3’末端側断片を作成した。この新しい5’及び3’末端にそれぞれLXXLLモチーフ及びGR LBD (527-777 AA)を繋ぐ。一方、GLucの本来の5’及び3’末端は、3つのGSリンカーを繋いだ。このプラスミドはpcPresso系列と呼ぶ。ここで、‘c’は、円順列(circular permutation)を表す。pcPresso の3’末端には、更にコリプレッサーペプチドを連結した。これらのタイプのプラスミドは、pcPresso-C系列と呼ぶ。発現した後のそれぞれのプローブは、cPRESSO及びcPRESSO-Cと呼べる。
これらプラスミドの信頼性を確認するために、「BigDye Terminator Cycle sequencing kit」及び遺伝子分析機のABI Prism310 (PE Biosystems)を利用して、配列確認をした。本発明において合成された各プローブの構造は図1に模式的に示し、またその構成要素のリストは表2に示した。
【0030】
【0031】
(実施例2)SIMGR系列プローブのストレスホルモン(コルチゾール)に対する感受性
COS-7細胞を12穴-プレート上で5%2酸化炭素培養機によって培養された。この時、10%ウシ胎児血漿(FBS)と1%ペニシリン-ストレプトマイシン(P/S)を添加したDulbeccoの改良Eagle培地(DMEM)を培地とした。プレート上のCOS-7細胞には、トランスフェクション試薬(TransIT-LT1;Mirus)を使ってpSimgr3,pSimgr2や pSimgr3m(1ウェルあたり0.2μg)をそれぞれ導入し、その後に16時間追加培養した。プレート上の細胞は、溶媒(0.1% DMSO)又はストレスホルモン(10-6 M cortisol)刺激を20分行い、その結果、回復されたルシフェラーゼ活性を特異的な基質キット(プロメガ)を使い、試薬マニュアル通りに測定した(図2(A)):簡単に説明すると、細胞をリン酸バーパーで一回洗浄し、キットから提供された細胞溶解バーパーで10分間溶解した。その溶解液を試験管に移し、40μLの基質溶液を加えて、ルミノメーター(Minilumat LB9506; Berthold)で発光値を最初15秒間測定した。
測定された発光強度は,以下の2つの何れかの方法で標準化された。(ア)細胞溶解液の単位蛋白質量で発光を換算する.この時の絶対発光値の単位は,RLU/μg of proteinになる.(イ)刺激有りの時の発光値(RLU(+))を、刺激無しの時の発光値(RLU(−))で割る。つまり、単位はRLU ratio (+/-)となる。ただし、この時のRLUは細胞溶解液の1μg蛋白質が出す発光値である。
【0032】
上述したように、一分子の中でストレスホルモン依存的な蛋白質間相互作用が起こるかどうかを検証するために、ストレスホルモン受容体(GR LBD)とコアクチベーター由来のLXXLLモチーフ間の結合を調べてみた(図2)。この結果、発光強度の回復がプローブ内に組み込まれたLXXLLモチーフの性質に依存していることが分かった。即ち、SIMGR3とSIMGR2共に10-6 Mストレスホルモン(cortisol)に応答したが、その絶対発光強度値はSIMGR3の方が強かった。LXXLLモチーフにアラニン(A)変異を導入したSIMGR3Mは、ストレスホルモンに弱い感度を示した。ウェスタンブロット結果によれば、上記プローブの何れも発現量の偏差は認められなかった(図2B)。
この結果は以下のように解析できる。
(ア)GR LBDは、以前X-線液晶構造解析結果(Protein Data Bank accession #: 1m2z)から予想されたように、LXXLLモチーフと相互作用する。
(イ)LXXLLモチーフの変異体による結果から予測されたように、LXXLLモチーフを構成する各アミノ酸が、GR LBDとLXXLLモチーフとの結合に決定的な役割をする。
【0033】
(実施例3)ウェスタンブロット分析
細胞溶解液中にある全蛋白質量と生物発光プローブの量をウェスタンブロットで検証した(図2(B))。まず、6ウェルプレート上で培養したCOS-7細胞にpSimgr3, pSimgr2, や pSimgr3mを導入し、更に16時間間培養した。その細胞をリン酸バーファー(PBS)で一回洗浄し、150μLの溶解バーファー(1% SDS/ 10% glycerol/ 10% 2-mercaptoethanol/ 0.001% bromophenol blue/ 50 mM Tris-HCl, pH 6.8)で溶解した。その後に細胞溶解液から5μLずつ取り、10%アクリルアミドゲル(TEFCO)上で電気泳動した。その後にニトロセルロース膜に移し、抗GR抗体や抗β-アクチン抗体で一次抗体処理をした。各汚点をhorseradish peroxidase (HRP)につないだ2次抗体処理し、最後に化学発光基質液(GE healthcare)で現象した。図4(A)のウェスタンブロットも上記記述と同様に行われた。
【0034】
(実施例4)PRESSO系列プローブのコルチゾールの用量応答曲線
PRESSOシリーズプローブのコルチゾール濃度―応答曲線(dose-response curve)を測定してみた(図3)。PRESSO-C3, -C4, や SIMGR3 の何れかを導入したCOS-7細胞に、一連の濃度のストレスホルモン(コルチゾール)で20分間刺激した。ほかにも同様にPRESSO-N2, -C2やSIMGR3の何れかを導入したCOS-7細胞に、一連のストレスホルモン刺激を20分間加え、蘇えた発光酵素活性を図2と同様のやり方でモニターした。
その結果、一番特徴的に見られる現象としては、プローブが描くストレスホルモン濃度―反応曲線が2相曲線を示す点である。つまり、コントロール(白丸)に比べて、PRESSO-C1と-C2は10-9から10-7 Mまでの濃度範囲において、発光値が増加する特徴を示す(図3(A))。
同様に、PRESSO-N1は10-11 から10-7 Mまでの領域において発光強度の上昇を示した(図3(B))。このような見かけ上の2相曲線を示すにも関わらず、そのシグナルーノイズ比(S/N比)は概ねコリプレッサーの影響を受けなかった。
【0035】
(実施例5)コリプレッサーモチーフと円順列変異(CP)との相乗効果
(5−1)COS-7細胞でのcPRESSO-C1, cPRESSO, cPRESSO-C2や SIMGR3のリガンド感受性
円順列変異とコリプレッサーモチーフを導入したプローブの相乗効果を調べた(図4,図5)。24ウェルプレートで培養したCOS-7細胞にpcPresso-c1, pcPresso, pcPresso-c2, や pSimgr3を導入した。その細胞を更に16時間間培養してから、溶媒か10-5 Mのコルチゾールで20分間刺激した。その発光強度を図2で記述したのと同様に評価した。図4(B)の濃度―応答曲線は図3で記述した方法で測定した。
その結果、cPRESSO-C1は、他のものに比べて、一番強い生物発光と直線性応答曲線を示した。一方、cPRESSOはテストしたプローブの中で一番高いS/N比を示した(例、10-6 M cortisolに対して約33倍)。
このようなコリプレッサーモチーフと円順列変異を導入することによって、およそ50-100倍程度の検出限界が下がる(改善する)ことを見出した。一番長い直線性応答範囲としては、cPRESSO-C1が10-8 から10-5 Mまでストレスホルモンに対して直線性応答を示した(図4(B),図5)。また、見かけ50%有効濃度(EC50)はおよそ2.5 x 10-7 M cortisolであった。その上、cPRESSO-C1のバックグラウンド発光値はコントロールに比べて、約1/3まで改善された。
【0036】
(5−2)コルチゾール濃度変化によるCOS-7細胞内PRESSO-C1のリガンド選択性
cPRESSO−C1のリガンド選択性をSIMGR3と比較してみた(図4(B))。24ウェルプレートで培養したCOS-7細胞にpcPresso-c1やpSimgr3を導入(transfection)した。
その細胞を10-6 M のステロイドや化学物質で20分間刺激した。その結果となる発光強度は図2で記述した方法と同様に測定した。
その結果、他のコントロールプローブに比べて、cPRESSO-C1は(ア)絶対発光強度の強さ、(イ)S/N比(約17倍)、(ウ)リガンド選択性において優れた結果を示した。
【0037】
(実施例6)コルチゾール検出におけるcPRESSO-C1の可逆性
ストレスホルモンセンシングにおけるcPRESSO-C1の可逆性をcortisolの刺激や除去を繰り返すことによって検証した(図6(B))。24ウェルプレート上に培養したCOS-7細胞にまずpcPresso-c1を導入し、更に16時間間培養した。次に細胞を溶媒(0.1% DMSO)又は10-7 M cortisolで20分間刺激した。その培養液をステロイドのない新鮮な培養液に置換することによってcortisolを一時除去した。図6(B)の挿入図で示したように、その細胞を更に10-7 M cortisolで刺激をする過程を繰り返した。毎段階における発光値の変化をその特異的な基質溶液キット(Promega)を使ってモニターした。
その結果は、10-7 Mのストレスホルモン刺激と洗浄の繰り返しにも関わらず、リガンド感度は衰えなかった。
この結果から、(ア)GR LBDとLXXLLモチーフとの結合は可逆的である。(イ)PRESSO-C1はストレスホルモンを繰り返して測定できるほど頑丈であり、この結果は以前のリガンド活性化されたGRの転写活性後リサイクルされる結論と一致する(非特許文献25)。
【0038】
(実施例7)PRESSO-C1からのリガンド応答に基づく蛍光強度の経時変化
PRESSO-C1による発光強度の経時変化をリガンド刺激後1時間間観測した(図7(A))。24ウェルプレート上で培養したCOS-7細胞にトランスフェクション試薬(TransIT-LT, Mirus)を用いてpPresso-c1を導入した。その細胞を16時間間更に培養し、最後には10-6 Mストレスホルモンや男性ホルモン(5α-hydrotestosterone;DHT))で刺激した。刺激後5分、10分、15分、20分、30分、40分、60分にそれぞれ細胞を溶解・回収し、各々の発光強度をその特異的な基質溶液で現象した。この時のプロトコルは図2のものと同様である。
その結果、cPRESSO-C1による発光強度は、ストレスホルモンによる刺激後すぐ上がり始め、約20分ごろに最高値に達する。ところが、同一濃度の男性ホルモンに対しては、測定時間(1h)内に発光値の上昇は見られなかった。
この経時変化は以下のように結論できる。
(ア)cPRESSO-C1のストレスホルモンセンシングは約20分で完結される。
(イ)この20分の時間は、ホルモンの細胞膜透過、ホルモン-プローブ結合、分子内構造変化とタンパク質相補にかかる総時間を意味する。この反応時間は、以前GR LBD-LXXLLモチーフ間結合に基づくFRET研究結果と一致する(非特許文献26。)
【0039】
(実施例8)ガラス基板上に形成した生物発光紙stripを用いたストレスホルモン測定
非細胞系における本プローブの長所を披露するために、生物発光紙ストリップをガラス基板上(2 x 7 cm)に形成した(図7(B))。まず、Y字型紙片をニトロセルロース紙から切り出し、ガラススライド上に固定した。
一方、細胞溶解液そのもの、又はcPRESSO-C1を発現する細胞の溶解液をそれぞれ図2で記述した方法で準備した。それぞれの細胞溶解液12μLをY字型紙片の末端に塗布して、他に広がらないようにスポットを作った。後に溶媒のみ、又は10-6 Mコルチゾールを添加した基質溶液を紙片の真中に10μL滴下して、その発光強度を発光スキャナー(RAS-3000, FujiFilm)で10分間積算した。
その結果、図7(B)で示したように、紙片上ではストレスホルモン依存的に発光値が強化された。
この結果から以下の点がうかがえる。
(ア)ストレスホルモン活性度は紙片レベルでイメージングできる。
(イ)cPRESSO-C1は紙片上での乾燥にも耐えてホルモンをセンシングできるほど頑丈である。
(ウ)結果の発光強度がガラス基板上で従来の発光スキャナーで計測できるほど強い。
【図1A】
【図1BC】
【図3A】
【図3B】
【図4B】
【特許請求の範囲】
【請求項1】
情報認識タンパク質が当該タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブにおいて、当該プローブ分子末端に、前記情報認識タンパク質に対するコリプレッサー活性ペプチドがアンカーペプチドとして結合されており、当該ペプチドが、リガンドの非共存時に前記情報認識タンパク質と会合していることを特徴とする、可視化プローブ。
【請求項2】
核内受容体のリガンド結合ドメインを含み、当該リガンド結合ドメインがリガンドの結合により活性化されて生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブにおいて、当該プローブ分子末端に、核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドがアンカーペプチドとして結合されており、当該ペプチドが、リガンドの非共存時に前記リガンド結合ドメインと会合していることを特徴とする、可視化プローブ。
【請求項3】
前記核内受容体がグルココルチコイドレセプターであって、前記立体構造変化が、当該グルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)がリガンドとの結合により活性化され、プローブ分子内のコアクチベーター由来の「LXXLL」モチーフを含むペプチドと結合することにより生じる立体構造変化であり、前記生物発光又は蛍光が、プローブ分子中のそれぞれ別の位置に存在する、分割された生物発光酵素又は蛍光タンパク質のフラグメントがタンパク質相補して発する生物発光又は蛍光である、請求項2に記載の可視化プローブ。
【請求項4】
前記グルココルチコイドレセプター(GR LBD)のリガンド結合ドメインを含むポリペプチド部位が、リンカーペプチドを介して又は介さずに、コアクチベーター由来の「LXXLL」モチーフを含むペプチドに結合されており、同一のプローブ分子の他の位置に、前記分割された生物発光酵素又は蛍光タンパク質のフラグメントと共に、前記核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドが結合されている、請求項3に記載の可視化プローブ。
【請求項5】
前記分割された生物発光酵素又は蛍光タンパク質のN末側及びC末側フラグメントが、リンカーペプチドを介して又は介さずに逆向きに結合されており、前記グルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)を含むポリペプチド部位と、コアクチベーター由来の「LXXLL」モチーフを含むペプチド部位と共に、同一のプローブ分子の他の位置に、前記核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドが結合されている、請求項3に記載の可視化プローブ。
【請求項6】
前記グルココルチコイドレセプターのリガンド結合ドメインを含むポリペプチド部位が、コアクチベーター由来の「LXXLL」モチーフを含むペプチド部位と別のプローブ分子上に存在し、かつそれぞれの部位が分割された生物発光酵素又は蛍光タンパク質のN末側もしくはC末側フラグメントのいずれかと、リンカーペプチドを介して又は介さずに結合されており、前記グルココルチコイドレセプターのリガンド結合ドメインを含むプローブ分子末端には、さらに前記核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドが結合されている、請求項3に記載のプローブ。
【請求項7】
情報認識タンパク質が当該タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブに対して、当該プローブ分子の末端に、前記情報認識タンパク質に対するコリプレッサー活性ペプチドをアンカーペプチドとして結合させることにより、当該ペプチドが、リガンド非共存下においては前記情報認識タンパク質と会合していることを特徴とする、可視化プローブの安定化方法。
【請求項8】
核内受容体のリガンド結合ドメインを含み、当該リガンド結合ドメインがリガンドの結合により活性化されて生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブに対して、当該プローブ分子の末端に、核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドをアンカーペプチドとして結合させることにより、当該ペプチドが、リガンド非共存下においては前記リガンド結合ドメインと会合していることを特徴とする、可視化プローブの安定化方法。
【請求項9】
情報認識タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブ分子の末端に存在するアンカーペプチドであって、情報認識タンパク質に対するコリプレッサー活性ペプチドからなり、リガンドによる活性化前の可視化プローブを前記情報認識タンパク質と会合することで安定化させることを特徴とする、アンカーペプチド。
【請求項10】
前記コリプレッサー活性ペプチドが、核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドからなる、請求項9に記載のアンカーペプチド。
【請求項11】
前記「LXXIIXXXL」モチーフ含有ペプチドが、配列番号5〜12又は配列番号23〜30のいずれかで表されるアミノ酸配列を含むペプチドである、請求項10に記載のアンカーペプチド。
【請求項12】
請求項1〜6のいずれかに記載の可視化プローブをコードする核酸。
【請求項13】
請求項12に記載の核酸を含む、可視化プローブ発現用ベクター。
【請求項14】
請求項13に記載の発現ベクターを用いて形質転換され、可視化プローブを発現する形質転換細胞。
【請求項15】
被検試料中のリガンドを検出又は定量するためのキットであって、請求項1〜6のいずれかに記載の可視化プローブ又は請求項14に記載の形質転換細胞を含むキット。
【請求項16】
前記キットが、さらに発光酵素の基質を含み、紙ストリップ上に固定、乾燥されている請求項15に記載のキット。
【請求項17】
被検試料中のリガンドの検出方法又は定量方法であって、請求項1〜6のいずれかに記載の可視化プローブ又は請求項14に記載の形質転換細胞を用いることを特徴とする方法。
【請求項1】
情報認識タンパク質が当該タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブにおいて、当該プローブ分子末端に、前記情報認識タンパク質に対するコリプレッサー活性ペプチドがアンカーペプチドとして結合されており、当該ペプチドが、リガンドの非共存時に前記情報認識タンパク質と会合していることを特徴とする、可視化プローブ。
【請求項2】
核内受容体のリガンド結合ドメインを含み、当該リガンド結合ドメインがリガンドの結合により活性化されて生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブにおいて、当該プローブ分子末端に、核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドがアンカーペプチドとして結合されており、当該ペプチドが、リガンドの非共存時に前記リガンド結合ドメインと会合していることを特徴とする、可視化プローブ。
【請求項3】
前記核内受容体がグルココルチコイドレセプターであって、前記立体構造変化が、当該グルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)がリガンドとの結合により活性化され、プローブ分子内のコアクチベーター由来の「LXXLL」モチーフを含むペプチドと結合することにより生じる立体構造変化であり、前記生物発光又は蛍光が、プローブ分子中のそれぞれ別の位置に存在する、分割された生物発光酵素又は蛍光タンパク質のフラグメントがタンパク質相補して発する生物発光又は蛍光である、請求項2に記載の可視化プローブ。
【請求項4】
前記グルココルチコイドレセプター(GR LBD)のリガンド結合ドメインを含むポリペプチド部位が、リンカーペプチドを介して又は介さずに、コアクチベーター由来の「LXXLL」モチーフを含むペプチドに結合されており、同一のプローブ分子の他の位置に、前記分割された生物発光酵素又は蛍光タンパク質のフラグメントと共に、前記核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドが結合されている、請求項3に記載の可視化プローブ。
【請求項5】
前記分割された生物発光酵素又は蛍光タンパク質のN末側及びC末側フラグメントが、リンカーペプチドを介して又は介さずに逆向きに結合されており、前記グルココルチコイドレセプターのリガンド結合ドメイン(GR LBD)を含むポリペプチド部位と、コアクチベーター由来の「LXXLL」モチーフを含むペプチド部位と共に、同一のプローブ分子の他の位置に、前記核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドが結合されている、請求項3に記載の可視化プローブ。
【請求項6】
前記グルココルチコイドレセプターのリガンド結合ドメインを含むポリペプチド部位が、コアクチベーター由来の「LXXLL」モチーフを含むペプチド部位と別のプローブ分子上に存在し、かつそれぞれの部位が分割された生物発光酵素又は蛍光タンパク質のN末側もしくはC末側フラグメントのいずれかと、リンカーペプチドを介して又は介さずに結合されており、前記グルココルチコイドレセプターのリガンド結合ドメインを含むプローブ分子末端には、さらに前記核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドが結合されている、請求項3に記載のプローブ。
【請求項7】
情報認識タンパク質が当該タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブに対して、当該プローブ分子の末端に、前記情報認識タンパク質に対するコリプレッサー活性ペプチドをアンカーペプチドとして結合させることにより、当該ペプチドが、リガンド非共存下においては前記情報認識タンパク質と会合していることを特徴とする、可視化プローブの安定化方法。
【請求項8】
核内受容体のリガンド結合ドメインを含み、当該リガンド結合ドメインがリガンドの結合により活性化されて生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブに対して、当該プローブ分子の末端に、核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドをアンカーペプチドとして結合させることにより、当該ペプチドが、リガンド非共存下においては前記リガンド結合ドメインと会合していることを特徴とする、可視化プローブの安定化方法。
【請求項9】
情報認識タンパク質のリガンドの結合により生じる立体構造変化を生物発光又は蛍光により検知し測定する可視化プローブ分子の末端に存在するアンカーペプチドであって、情報認識タンパク質に対するコリプレッサー活性ペプチドからなり、リガンドによる活性化前の可視化プローブを前記情報認識タンパク質と会合することで安定化させることを特徴とする、アンカーペプチド。
【請求項10】
前記コリプレッサー活性ペプチドが、核内受容体のコリプレッサー由来の「LXXIIXXXL」モチーフ含有ペプチドからなる、請求項9に記載のアンカーペプチド。
【請求項11】
前記「LXXIIXXXL」モチーフ含有ペプチドが、配列番号5〜12又は配列番号23〜30のいずれかで表されるアミノ酸配列を含むペプチドである、請求項10に記載のアンカーペプチド。
【請求項12】
請求項1〜6のいずれかに記載の可視化プローブをコードする核酸。
【請求項13】
請求項12に記載の核酸を含む、可視化プローブ発現用ベクター。
【請求項14】
請求項13に記載の発現ベクターを用いて形質転換され、可視化プローブを発現する形質転換細胞。
【請求項15】
被検試料中のリガンドを検出又は定量するためのキットであって、請求項1〜6のいずれかに記載の可視化プローブ又は請求項14に記載の形質転換細胞を含むキット。
【請求項16】
前記キットが、さらに発光酵素の基質を含み、紙ストリップ上に固定、乾燥されている請求項15に記載のキット。
【請求項17】
被検試料中のリガンドの検出方法又は定量方法であって、請求項1〜6のいずれかに記載の可視化プローブ又は請求項14に記載の形質転換細胞を用いることを特徴とする方法。
【図5】


【図6B】

【図7A】
【図7B】

【図2A】
【図2B】
【図4A】

【図6A】
【図6B】
【図7A】
【図7B】
【図2A】
【図2B】
【図4A】
【図6A】
【公開番号】特開2010−178676(P2010−178676A)
【公開日】平成22年8月19日(2010.8.19)
【国際特許分類】
【出願番号】特願2009−25229(P2009−25229)
【出願日】平成21年2月5日(2009.2.5)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成22年8月19日(2010.8.19)
【国際特許分類】
【出願日】平成21年2月5日(2009.2.5)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]