
インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子溶液を安定化させる方法
インフルエンザ抗原を含有するエンベロープウイルスベースのウイルス様粒子を伴う溶液を安定化させる方法、およびこのような安定化させた溶液について記載する。本開示の別の態様は、インフルエンザを治療または予防する方法であって、対象に、免疫原性量の、前出の態様およびその多様な実施形態のうちのいずれかによる、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物、または前出の方法の態様およびその多様な実施形態のうちのいずれかに従い安定化させた、免疫原性量の、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物を含有する溶液を投与するステップを含む方法を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、インフルエンザ抗原を包含する、エンベロープウイルスベースの安定化させたウイルス様粒子の分野に関する。本明細書では特に、このようなウイルス様粒子を安定化させる方法、およびこのようなウイルス様粒子を含む安定化させた組成物が開示される。
【背景技術】
【0002】
A型インフルエンザウイルスおよびB型インフルエンザウイルスは、流行性ヒト疾患を引き起こすインフルエンザウイルスの2つの型である(111)。A型インフルエンザウイルスは、2つの表面抗原:赤血球凝集素(HA)タンパク質およびノイラミニダーゼ(NA)タンパク質に基づく亜型にさらに類別される。B型インフルエンザウイルスは、亜型に類別されていないが、時間と共にウイルス株が分岐するドリフトを経る。1977年以来、A型(H1N1亜型)インフルエンザウイルス、A型(H3N2亜型)インフルエンザウイルス、およびB型インフルエンザウイルスが、世界中に蔓延している。おそらく、ヒトA型(H3N2亜型)ウイルスと、ヒトA型(H1N1亜型)ウイルスとが遺伝子再結合した後に出現したA型(H1N2亜型)インフルエンザウイルスは、近年多くの国々で検出されている。A型インフルエンザウイルスおよびB型インフルエンザウイルスのいずれもが、抗原特徴に基づき、さらに群分けされている。新型インフルエンザウイルスの変異型は、ウイルスの複製において生じる点突然変異の結果としてもたらされる、頻繁な抗原変化(すなわち、抗原ドリフト)から生じる。B型インフルエンザウイルスは、抗原ドリフトを経るのが、A型インフルエンザウイルスほど急速でない。抗原ドリフトによる抗原変異型の頻繁な発生が、例年の流行のウイルス学的基礎であり、毎年のインフルエンザワクチンに少なくとも1つの新たなウイルス株を組み込む理由でもある。
【0003】
表面抗原、とりわけ、赤血球凝集素タンパク質に対する対象者の免疫が、感染の可能性、および万一感染が生じた場合の疾患の重症度を低減する(112)。一般に、インフルエンザウイルスの1つの型または1つの亜型に対する抗体は、別の型もしくは別の亜型に対してもたらす防御が限定されたものであるか、または別の型もしくは別の亜型に対しては防御をもたらさないと考えられている。さらに、インフルエンザウイルスの1つの抗原変異型に対する抗体では、同じ型または同じ亜型の新たな抗原変異型に対する防御とならない可能性があることも、一般的に受け入れられている(113)。したがって、交差防御の実証は期待されない。
【0004】
1957年および1968年における、過去の二度にわたるインフルエンザ大流行の原因となったのは、ヒト−トリ間リアソータントのインフルエンザウイルスであった。1968年以後にH2亜型ウイルスがヒトにおいて蔓延しなくなって以来、H2亜型のリアソータントウイルスから抗原シフトが生じる可能性が、理論的にはいつでも存在する。しかし、高病原性トリインフルエンザ(HPAI)ウイルス(H5亜型およびH7亜型)が近年になって出現し、1997年以来、これらのウイルスが、散発的ながらも、トリからヒトに直接的に伝染するようになった(1〜5)ことから、H2亜型ウイルスに対する集団の感受性がかつてなく増大しているのに加えて、ヒトにおいて新たに大流行する脅威の可能性が高まっている。ヒトにおけるHPAI H5N1亜型の大発生が、異なる抗原によるものであったという事実は、大流行の脅威に対して、十分に適合させたワクチンの備蓄を前もって準備することをほとんど不可能にしている(5、6)。マウスにおけるH5亜型による免疫化および抗原曝露(challenge)のデータは、このモデルにおける各種のH5亜型単離物間に良好な交差反応性が見られることを示唆する(7)が、既存のワクチン法で、ヒトにおいても同様レベルの交差反応性が見られるかどうかは公知ではない。したがって、新たなウイルスの大発生に由来する抗原を包含するように迅速に適合させうるインフルエンザワクチンプラットフォームが必要とされている。
【0005】
現行の卵ベースの不活化ワクチン法では、卵内でHPAIウイルスを増殖させることが不可能であるために、新興の大流行による需要を満たし、バイオコンテインメントを増強する必要を満たすのには不十分である(6、8)。逆遺伝学法は、卵内で培養しうる所望のHA遺伝子組成およびNA遺伝子組成を伴う、低病原性のリアソータントを生成させる手段をもたらしている(7、9〜11)が、この手法により生成させるワクチンは、過去の知的財産および規制に関する問題のために、ようやく臨床段階に入りつつあるに過ぎない(8)。さらなる懸念は、ヒト臨床試験で評価されている、赤血球凝集素H5タンパク質と関連する免疫原性レベルが明らかに低いこと(12〜14)であり、これは、H5亜型に対して完全にナイーブな集団における防御を効率的に誘導するには、ワクチン、送達系、およびアジュバント使用の改善が必要とされることを明らかにしている。したがって、アジュバントと組み合わせたHPAI抗原の発現を可能とするインフルエンザワクチンプラットフォームが必要とされている。
【0006】
インフルエンザVLPは、インフルエンザワクチンを生成させるための代替的な技法を代表する。インフルエンザVLPとは、昆虫細胞内で発現させたインフルエンザのマトリックスタンパク質、HAタンパク質、およびNAタンパク質を用いて生成させたもので、鼻腔内送達後における免疫原性が顕著である(26:非特許文献1、27:非特許文献2)。実際、VLPは一般に、ロタウイルスVLP、ノロウイルスVLP、およびパピローマウイルスVLPについて示されている通り、鼻腔内送達後において、粘膜および全身における免疫を誘導するのに十分に適すると考えられる(28〜31:非特許文献3〜6)。インフルエンザVLPとは、真核細胞発現系において、インフルエンザのマトリックスタンパク質、HAタンパク質、およびNAタンパク質を発現させることにより生成させたものである。インフルエンザマトリックスタンパク質は、ウイルスの出芽を後押しする駆動力であり、インフルエンザNAタンパク質は、インフルエンザHAタンパク質も発現している場合、細胞表面のシアル酸とのHAタンパク質の会合により、出芽したVLPが生成細胞から放出されるのに必要とされる(51:非特許文献6)。また、マトリックスタンパク質と、HAタンパク質のC末端との相互作用が、出芽過程の一部として、マトリックスタンパク質を膜に方向づけるのに役割を果たすことを示唆するデータも存在する(51:非特許文献6)。昆虫細胞のバキュロウイルス発現系内で生成させたインフルエンザVLPは、動物試験において免疫原性であることが分かっており、将来の大流行に備えるための重要な戦略を代表する(26:非特許文献1、27:非特許文献2、47:非特許文献7)。加えて、インフルエンザVLPの鼻腔内送達は、非経口投与後に得られる抗体力価を超える抗体力価を結果としてもたらしうる。しかし、VLPは、膜内に包埋されるか、または膜と会合する、1または複数の異なる糖タンパク質と共に包埋される脂質膜の複合体構造である。実用性を有するために、VLPは、十分に長い保管寿命を有する必要がある。したがって、溶液中にインフルエンザ抗原を包含するウイルス様粒子を安定化させる方法が必要とされている。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】Latham, T., and J. M. Galarza. 2001. Formation of wild−type and chimeric influenza virus−like particles following simultaneous expression of only four structural proteins. J Virol 75:6154.
【非特許文献2】Galarza, J. M., T. Latham, and A. Cupo. 2005. Virus−like particle vaccine conferred complete protection against a lethal influenza virus challenge. Viral Immunol 18:365.
【非特許文献3】Fromantin, C., B. Jamot, J. Cohen, L. Piroth, P. Pothier, and E. Kohli. 2001. Rotavirus 2/6 virus−like particles administered intranasally in mice, with or without the mucosal adjuvants cholera toxin and Escherichia coli heat−labile toxin, induce a Th1/Th2−like immune response. J Virol 75:11010.
【非特許文献4】Harrington, P. R., B. Yount, R. E. Johnston, N. Davis, C. Moe, and R. S. Baric. 2002. Systemic, mucosal, and heterotypic immune induction in mice inoculated with Venezuelan equine encephalitis replicons expressing Norwalk virus−like particles. J Virol 76:730.
【非特許文献5】Shi, W., J. Liu, Y. Huang, and L. Qiao. 2001. Papillomavirus pseudovirus: a novel vaccine to induce mucosal and systemic cytotoxic T−lymphocyte responses. J Virol 75:10139.
【非特許文献6】Gomez−Puertas, P., C. Albo, E. Perez−Pastrana, A. Vivo, and A. Portela. 2000. Influenza virus matrix protein is the major driving force in virus budding. J Virol 74:11538.
【非特許文献7】Pushko, P., T. M. Tumpey, F. Bu, J. Knell, R. Robinson, and G. Smith. 2005. Influenza virus−like particles comprised of the HA, NA, and M1 proteins of H9N2 influenza virus induce protective immune responses in BALB/c mice. Vaccine 23:5751.
【発明の概要】
【課題を解決するための手段】
【0008】
本出願の開示は、インフルエンザ抗原を包含するウイルス様粒子を溶液中で安定化させる方法を、該安定化させた溶液、およびこのような安定化させた溶液を用いる方法と併せて提供することにより、この必要を満たす。
【0009】
開示の一態様は、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物を含有する溶液を安定化させる方法であって、(a)インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子を含有する溶液を用意するステップと、(b)(1)安定化量の、単糖、ソルビトール、二糖、トレハロース、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択される安定化剤を、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物に添加するステップ、(2)pHを、約pH6.5〜約pH8.0、約pH6.5〜約pH7.5、または約pH7とするように、該溶液を緩衝化するステップ、または(3)ステップ(1)およびステップ(2)の両方とを含み、ステップ(b)の後のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物が、以下の特徴:(i)光学濃度により測定した該ウイルス様粒子の凝集が、ステップ(b)の前のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していること;(ii)円偏光二色性またはANS結合により測定したインフルエンザ抗原が、ステップ(b)の前のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して安定化していること;および(iii)ラウルダン蛍光により測定した、該ウイルス様粒子の脂質二重層の、温度により誘導される水和が、ステップ(b)の前のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していることのうちの少なくとも1つを呈示する方法を提供する。ある実施形態では、緩衝化するステップを、リン酸バッファー、トリスバッファー、MESバッファー、クエン酸バッファー、および他のGRASバッファーからなる群から選択される緩衝剤を用いて実施する。前出の緩衝剤についての実施形態と組み合わせうるある実施形態では、安定化剤は、トレハロース、ソルビトール、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択され、該特徴は(i)である。前出の緩衝剤についての実施形態と組み合わせうるある実施形態では、安定化剤は、トレハロース、ソルビトール、および前出の安定化剤の組合せから選択され、該特徴は(ii)である。前出の緩衝剤についての実施形態と組み合わせうるある実施形態では、安定化剤は、トレハロースおよびグリシンから選択され、特徴は(iii)である。前出の緩衝剤についての実施形態と組み合わせうるある実施形態では、安定化剤はトレハロースであり、3つの特徴すべてが存在する。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチドを含む。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、gagポリペプチド、インフルエンザM1ポリペプチド、ニューカッスル病ウイルスマトリックスポリペプチド、エボラウイルスVP40ポリペプチド、およびマールブルグウイルスVP40ポリペプチドを含む群から選択される第2のポリペプチドを含む。gagポリペプチドを包含する前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該gagポリペプチドが、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス、ベータレトロウイルス、ガンマレトロウイルス、デルタレトロウイルス、およびレンチウイルスに由来しうる。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、ノイラミニダーゼポリペプチドをさらに含む。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化剤は、単糖、ソルビトール、二糖、およびトレハロースから選択され、安定化量は、10%(w/w)を超えるか、または少なくとも約20%(w/w)である。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化させるステップが、ガラス形成を必要としない。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化量は、凝固時のガラス形成に必要とされる量未満である。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化剤は、スクロースではない。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物は、該インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子と混合させたアジュバントをさらに含み、アジュバントは、前記ウイルス様粒子の内部に配置されてもよく、該ウイルス様粒子の外部に配置されてもよい。アジュバントおよび第2のポリペプチドを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、アジュバントを、該第2のポリペプチドに共有結合させて、共有結合を形成させる。アジュバントおよび赤血球凝集素ポリペプチドを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該アジュバントを、前記赤血球凝集素ポリペプチドに共有結合させて、共有結合を形成させる。アジュバントを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該アジュバントは、フラジェリンのアジュバント活性断片を含む。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該方法は、(c)溶液を、少なくとも2週間、少なくとも1カ月間、少なくとも2カ月間、少なくとも3カ月間、少なくとも4カ月間、少なくとも6カ月間、または少なくとも1年間にわたり液体形態で保存するステップをさらに含み、このような期間後のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物は、このような期間前のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物により誘導される免疫反応の少なくとも80パーセント、少なくとも90パーセント、または少なくとも95パーセントを誘導する。
【0010】
本開示の別の態様は、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子と、安定化量の、トレハロース、ソルビトール、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択される、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物に対する安定化剤とを含み、以下の特徴:(i)光学濃度により測定した該ウイルス様粒子の凝集が、該安定化剤を伴わない、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していること;(ii)円偏光二色性またはANS結合により測定したインフルエンザ抗原が、該安定化剤を伴わない、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して安定化していること;および(iii)ラウルダン蛍光により測定した、該ウイルス様粒子の脂質二重層の、温度により誘導される水和が、該安定化剤を伴わない、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していることのうちの少なくとも1つを呈示する、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物を提供する。ある実施形態では、緩衝化を、リン酸バッファー、トリスバッファー、MESバッファー、クエン酸バッファー、および他のGRASバッファーからなる群から選択される緩衝剤を用いて実施する。ある実施形態では、安定化剤は、トレハロース、ソルビトール、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択され、該特徴は(i)である。ある実施形態では、安定化剤は、トレハロース、ソルビトール、および前出の安定化剤の組合せから選択され、該特徴は(ii)である。ある実施形態では、安定化剤は、トレハロースおよびグリシンから選択され、該特徴は(iii)である。ある実施形態では、安定化剤はトレハロースであり、3つの特徴すべてが存在する。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子は、赤血球凝集素ポリペプチドを含む。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子は、gagポリペプチド、インフルエンザM1ポリペプチド、ニューカッスル病ウイルスマトリックスポリペプチド、エボラウイルスVP40ポリペプチド、およびマールブルグウイルスVP40ポリペプチドを含む群から選択される第2のポリペプチドを含む。gagポリペプチドを包含する前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該gagポリペプチドは、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス、ベータレトロウイルス、ガンマレトロウイルス、デルタレトロウイルス、およびレンチウイルスからなる群から選択されるレトロウイルスに由来する。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子は、ノイラミニダーゼポリペプチドをさらに含む。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化剤は、単糖、ソルビトール、二糖、およびトレハロースから選択され、安定化量は、10%(w/w)を超えるか、または少なくとも約20%(w/w)である。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化させるステップが、ガラス形成を必要としない。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化量は、凝固時のガラス形成に必要とされる量未満である。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化剤は、スクロースではない。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物は、該インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子と混合させたアジュバントをさらに含む。アジュバントを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該アジュバントは、前記ウイルス様粒子の内部に配置される。アジュバントを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該アジュバントは、前記ウイルス様粒子の外部に配置される。アジュバントおよび第2のポリペプチドを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該アジュバントを、前記第2のポリペプチドに共有結合させて、共有結合を形成させる。アジュバントを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、アジュバントを、前記赤血球凝集素ポリペプチドに共有結合させて、共有結合を形成させる。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、アジュバントは、フラジェリンのアジュバント活性断片を含む。
【0011】
本開示の別の態様は、インフルエンザを治療または予防する方法であって、対象に、免疫原性量の、前出の態様およびその多様な実施形態のうちのいずれかによる、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物、または前出の方法の態様およびその多様な実施形態のうちのいずれかに従い安定化させた、免疫原性量の、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物を含有する溶液を投与するステップを含む方法を提供する。ある実施形態では、投与するステップは、対象において防御的免疫反応を誘導する。前出の実施形態と組み合わせ得るある実施形態では、投与するステップは、皮下(subcutaneous)送達、経皮送達、皮内送達、皮下送達(subdermal)、筋肉内送達、経口(peroral)送達、経口(oral)送達、鼻腔内送達、頬(buccal)送達、舌下送達、腹腔内送達、膣内(intravaginal)送達、肛門(anal)送達、および頭蓋内送達からなる群から選択される。
【図面の簡単な説明】
【0012】
【図1】図1は、個別のGag遺伝子ベクター、HA遺伝子ベクター、または対照ベクターを感染させたSf9細胞、およびHA−gag−NA三重遺伝子ベクターを感染させたSf9細胞に由来する培地についてのウェスタンブロットを示す図である。(A)は、抗Gag抗体によりプローブしたものであり、(B)は、抗HA抗体によりプローブしたものである。
【図2】図2は、ペレット化させたHA−gag−NA VLPを、段階的なスクロース勾配上で再遠心分離することによる画分についてのウェスタンブロットを示す図である。(A)は、抗Gag抗体によりプローブしたものであり、(B)は、抗HA抗体によりプローブしたものである。
【図3】図3は、インフルエンザVLPによる動的光散乱を示す図である。有効直径(A)、静的光散乱の強度(B)、および試料の多分散性(C)を、温度の関数としてプロットする。各点は、3つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【図4】図4は、インフルエンザVLPの円偏光二色性スペクトルを示す図である。低温(10℃)では、4〜8の各単位pHにおけるスペクトル(A)を示し、pH7では、多様な温度におけるスペクトル(B)を示す。
【図5】図5は、熱ストレスに対する、インフルエンザVLPのタンパク質二次構造の反応を示す図である。227nmにおける標準化した(−1〜0の)CDを、温度の関数として示す。各点は、3つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【図6】図6は、インフルエンザVLPの内部蛍光のピーク位置を、温度の関数として示す図である。また、330nmにおける標準化した(0〜1の)蛍光強度も、温度の関数として示す(右下)。各点は、3つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【図7】図7は、インフルエンザVLPの物理的構造のプローブとしての、ANS蛍光を示す図である。ピーク発光の波長を、温度の関数として示す。また、485nmにおける標準化した(0〜1の)ANS蛍光強度も、温度の関数として示す(右下)。各点は、3つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【図8】図8は、インフルエンザVLPの存在下におけるラウルダン蛍光による一般化偏光(GP)を、温度の関数として示す図である。各点は、3つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【図9】図9は、インフルエンザVLPの生物物理的特徴づけに由来する実験相図(EPD)である。EPDは、温度に依存する有効直径、静的光散乱、多分散性、227nmにおけるCD、内部蛍光(ピーク位置、および330nmにおける相対強度)、ANS蛍光(ピーク位置、および485nmにおける相対強度)、および4〜8のpH範囲にわたり収集されたラウルダン蛍光データによるGPから作成した。
【図10】図10は、選択した安定化剤の存在下における、インフルエンザVLPによる内部蛍光を示す図である。ピーク発光の位置(波長)を、温度の関数として示す。各点は、2つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【図11】図11は、選択した安定化剤と共に調合されたインフルエンザVLPの存在下における、ラウルダン蛍光によるGPを示す図である。各点は、2つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【発明を実施するための形態】
【0013】
本発明は、インフルエンザ抗原のエンベロープウイルスベースのVLPを安定化させた製剤に基づく。例示的なエンベロープウイルスベースのVLPは、VLP製剤のベースとして、マウス白血病ウイルス(MLV)に由来するgagポリペプチドなどのgagポリペプチドを包含する。バキュロウイルス発現系において各種のレトロウイルスから得ることができるgag VLPの収量が大きいため、このようなgagタンパク質ベースのVLPを生成させる例示的な方法は、好ましくは、インフルエンザHAポリペプチド抗原およびインフルエンザNAポリペプチド抗原の共発現を含めた、昆虫細胞における発現を介する。
【0014】
安定化は、インフルエンザ抗原のウイルスベースのVLP調製物中に包含される安定化量の安定化剤を介する。例示的な安定化剤には、単糖(デキストロース、マンニトール、ソルビトールなど)、二糖(ラクトース、トレハロース、スクロースなど)、ジエタノールアミン、グリセロール、グリシン、またはこれらの組合せが含まれる。
【0015】
開示される方法およびプロトコールの実施は、別段に示唆されない限り、当業者の能力の範囲内にある、化学、分子生物学、微生物学、組換えDNA、および免疫学による従来の技法を使用する。このような技法は、文献において説明されている。例えば、J. Sambrook, E. F. Fritsch、およびT. Maniatis、1989年、Molecular Cloning: A Laboratory Manual、2版、1〜3巻、Cold Spring Harbor Laboratory Press;Ausubel, F. M.ら(1995年、および定期的な補遺、Current Protocols in Molecular Biology、9、13、および16章、John Wiley & Sons、New York、N.Y.);B. Roe、J. Crabtree、およびA. Kahn、1996年、DNA Isolation and Sequencing: Essential Techniques、John Wiley & Sons;J. M. PolakおよびJames O’D. McGee、1990年、In Situ Hybridization: Principles and Practice、Oxford University Press;M. J. Gait(編)、1984年、Oligonucleotide Synthesis: A Practical Approach、Irl Press;ならびにD. M. J. LilleyおよびJ. E. Dahlberg、1992年、Methods of Enzymology: DNA Structure、A部:Synthesis and Physical Analysis of DNA Methods in Enzymology、Academic Pressを参照されたい。これらの一般的な教科書の各々は、参照により本明細書に組み込まれる。
【0016】
定義
本明細書で用いられる「エンベロープウイルスベースのVLP」とは、エンベロープウイルスに由来する1または複数のコンポーネントを用いて形成されるウイルス様粒子を指す。限定なしに述べると、好ましい例には、インフルエンザ赤血球凝集素ポリペプチドと共にgagポリペプチドを用いて生成させたVLP、インフルエンザ赤血球凝集素ポリペプチドを用いて、または赤血球凝集素ポリペプチドと共にOrthomyxovirus属ウイルス(インフルエンザウイルスを含めた)M1ポリペプチドを用いて(場合によって、各場合にノイラミニダーゼポリペプチドを伴う)生成させたVLP、インフルエンザ赤血球凝集素ポリペプチドと共にParamyxovirus(Paromyxovirus)属ウイルス(ニューカッスル病ウイルスを含めた)マトリックスポリペプチドを用いて生成させたVLP、インフルエンザ赤血球凝集素ポリペプチドと共にFilovirus属ウイルス(エボラウイルスまたはマールブルグウイルスを含めた)VP40ポリペプチドを用いて生成させたVLPが含まれる。
【0017】
さらなる例には、エンベロープウイルスベースのVLPを形成するのに用いうる、エボラウイルスおよびマールブルグウイルスなどのフィロウイルス(例えば、細胞内でフィロウイルスに由来するウイルス性GPタンパク質およびウイルス性VP40タンパク質を共発現させ、脂質ラフト内でこれら2つのウイルス性タンパク質を会合させることにより、VLPを生成させる(米国特許公開第20060099225号を参照されたい));SARSウイルスなどのコロナウイルス(例えば、コロナウイルスのVLPを形成させるには、Eタンパク質およびMタンパク質で十分である(Fischerら、J. Virol.(1998年)、72巻:7885〜7894頁;およびVennemaら、EMBO J.(1996年)、15巻:2020〜2028頁を参照されたい));RS(respiratory syncytial)ウイルス(RSV)など、Paramyxoviridae(paramyxoviridae)科のウイルス(例えば、RSVウイルスのMタンパク質を発現させることにより、VLPを生成させる(例えば、米国特許公開第20080233150号を参照されたい));ならびに西ナイルウイルスなど、Flaviviridae(flaviviridae)科のウイルス(例えば、バキュロウイルス発現系内で西ナイルウイルスのprM遺伝子およびE遺伝子を含む構築物を発現させることにより、VLPを生成させる(例えば、米国特許公開第20080233150号を参照されたい))が含まれる。
【0018】
gagポリペプチドには、本明細書で説明するウイルス様粒子を形成する一因となる、レトロウイルス由来の構造ポリペプチドが含まれる。一部の実施形態では、RNAをパッケージングする傾向、または粒子形成および粒子出芽の効率など、一定の特徴に影響を及ぼす目的で、gagポリペプチドを意図的に突然変異させることができる。このような突然変異の一例をあげるならば、gag由来のVLPがRNAを組み込む能力に影響を及ぼすアミノ酸変化となろう。VLP出芽の効率を改善または改変する、他のこのようなアミノ酸変化ももたらしうるであろう。レトロウイルスのゲノムは、3つの主要な遺伝子産物:構造タンパク質をコードするgag遺伝子;逆転写酵素、ならびに関連するタンパク質分解ポリペプチド、ヌクレアーゼ、およびインテグラーゼに関連する機能をコードするpol遺伝子;それによりコードされる糖タンパク質である膜タンパク質が、感染細胞表面上で検出され、また、放出された成熟ウイルス粒子の表面上でも検出されるenv遺伝子による遺伝子産物をコードする。すべてのレトロウイルスのgag遺伝子は、全体的な構造類似性を有し、各レトロウイルス群内において、そのアミノ酸レベルが保存されている。gag遺伝子は、逆転写酵素を除いたコアタンパク質をもたらす。MLVでは、Gag前駆体であるポリタンパク質が、Pr65Gagであり、該前駆体におけるその順序がNH2−p15−pp12−p30−p10−COOHである4つのタンパク質に切断される。これらの切断は、ウイルス性プロテアーゼに媒介され、ウイルスに応じて、ウイルス放出の前に生じる場合もあり、ウイルス放出の後に生じる場合もある。MLV Gagタンパク質は、グリコシル化形態および非グリコシル化形態で存在する。グリコシル化形態は、非グリコシル化Pr65GagのAUGコドンより上流に位置する、異なるインフレームの開始コドンにより合成される、gPr80Gagから切断される。グリコシル化Gagを合成しないMLVの欠失突然変異体もなお感染性であり、該非グリコシル化Gagもなおウイルス様粒子を形成し得ることから、グリコシル化イベントの重要性について疑問が生じる。ウイルスによりコードされるプロテアーゼを介してHIV−1 Gag前駆体であるpr55Gagが翻訳後切断されると、N−ミリストイル化および内部リン酸化されたp17マトリックスタンパク質(p17MA)、リン酸化p24カプシドタンパク質(p24CA)、およびp15ヌクレオカプシドタンパク質(p15NC)(これはp9およびp6にさらに切断される)がもたらされる。
【0019】
構造的には、原型的なGagポリタンパク質が、レトロウイルスgag遺伝子では常に同じ順序で生じる3つの主要なタンパク質:マトリックスタンパク質(MA)(インフルエンザマトリックスタンパク質であるM1は、マトリックスという名称を共有しているが、MAとは異なるタンパク質であり、これとは区別する)、カプシドタンパク質(CA)、およびヌクレオカプシドタンパク質(NC)に分けられる。Gagポリタンパク質の成熟タンパク質へのプロセシングは、レトロウイルスによりコードされるプロテアーゼを介して触媒され、新たに出芽したウイルス粒子が成熟するときに生じる。機能的には、Gagポリタンパク質が、3つのドメイン:細胞膜をGagポリタンパク質の標的とする膜結合ドメイン;Gagの多量体化を促進する相互作用ドメイン;および宿主細胞から発生したビリオンの放出を促進する後期ドメインに分けられる。アセンブリーを媒介するGagタンパク質の形態は、ポリタンパク質である。したがって、アセンブリードメインは、後期に形成される切断産物のうちのいずれかの内部にもれなく収まる必要がない。こうして、本明細書に包含されるGagポリペプチドは、VLPの形成および放出に重要な機能的エレメントを包含する。当技術分野の現況は、これらの重要な機能エレメントに関して、極めて進展している。例えば、Hansenら、J. Virol.64巻、5306〜5316頁、1990年;Willら、AIDS 5巻、639〜654頁、1991年;Wangら、J. Virol.72巻、7950〜7959頁、1998年;McDonnellら、J. Mol. Biol.279巻、921〜928頁、1998年;SchultzおよびRein、J. Virol.63巻、2370〜2372頁、1989年;Accolaら、J. Virol.72巻、2072〜2078頁、1998年;Borsettiら、J. Virol.72巻、9313〜9317頁、1998年;Bowzardら、J. Virol.72巻、9034〜9044頁、1998年;Krishnaら、J. Virol.72巻、564〜577頁、1998年;Willsら、J. Virol.68巻、6605〜6618頁、1994年;Xiangら、J. Virol.70巻、5695〜5700頁、1996年;Garnierら、J. Virol.73巻、2309〜2320頁、1999年を参照されたい。
【0020】
例示的なレトロウイルスのGagポリペプチド供給源には、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス(ニワトリ白血病ウイルス、またはラウス肉腫ウイルスなど)、ベータレトロウイルス(マウス乳癌ウイルス、ヤーグジークテヒツジレトロウイルス、およびメーソン−ファイザーサルウイルスなど)、ガンマレトロウイルス(マウス白血病ウイルス、ネコ白血病ウイルス、細網内皮症ウイルス、およびテナガザル白血病ウイルスなど)、デルタレトロウイルス(ヒトTリンパ向性ウイルス、およびウシ白血病ウイルスなど)、イプシロンレトロウイルス(ウォールアイ皮膚肉腫ウイルスなど)、またはレンチウイルス(1型ヒト免疫不全ウイルス、HIV−2、サル免疫不全ウイルス、ネコ免疫不全ウイルス、ウマ感染性貧血ウイルス、およびヤギ関節炎脳炎ウイルスなど)が含まれる。
【0021】
本明細書で用いられる「赤血球凝集素ポリペプチド」は、インフルエンザウイルスが感染される細胞に結合することを媒介するインフルエンザウイルスタンパク質に由来する。赤血球凝集素タンパク質は、単一の膜貫通ドメインを介してインフルエンザウイルスの表面に係合されていることが見出される、抗原性の糖タンパク質である。インフルエンザウイルスの赤血球凝集素については、少なくとも16の亜型が同定され、H1〜H16と標識されている。ヒトインフルエンザウイルスにおいて見出されるのは、H1、H2、およびH3である。赤血球凝集素H5、赤血球凝集素H7または赤血球凝集素H9を伴う、高病原性トリインフルエンザウイルスがヒトに感染する割合は小さいことが判明している。赤血球凝集素H5がトリH5N1ウイルスの受容体特異性を著明に変化させることを可能とし、それらにヒト受容体に結合する能力をもたらすようにその受容体特異性を変化させる、トリインフルエンザウイルス株の赤血球凝集素H5型における単一のアミノ酸変化が、ヒト患者において見出されている(109および110)。この知見は、通常ではヒトに感染しないH5N1ウイルスが、どのようにして突然変異し、ヒト細胞に効率的に感染し得るようになるのかを説明する。
【0022】
赤血球凝集素は、ホモ三量体の内在性膜ポリペプチドである。その膜貫通ドメインは、天然でラフト脂質ドメインと会合し、それをgagポリペプチドと会合させて、VLPに組み込むことを可能とする。赤血球凝集素は、円筒様の形状であり、約135Åの長さである。HAを構成する3つの同一の単量体は、中央部のコイルドコイルおよびVLP表面上に曝露されているシアル酸結合部位を含有する球状ヘッドを形成する。HAの単量体は、グリコシル化されて2つの小型のポリペプチド:HA1サブユニットおよびHA2サブユニットに切断される、単一のポリペプチド前駆体として合成される。HA2サブユニットは、膜に係合する三量体のコイルドコイルを形成し、HA1サブユニットは、球状ヘッドを形成する。
【0023】
本発明のVLPにおいて用いられる赤血球凝集素ポリペプチドは、少なくとも膜アンカードメインおよび赤血球凝集素由来の少なくとも1つのエピトープを包含するものとする。赤血球凝集素ポリペプチドは、インフルエンザウイルスの任意の類型、亜型、株、または亜株、例えば、赤血球凝集素H1、赤血球凝集素H2、赤血球凝集素H3、赤血球凝集素H5、赤血球凝集素H7および赤血球凝集素H9に由来することが可能である。加えて、赤血球凝集素ポリペプチドは、異なるインフルエンザ赤血球凝集素のキメラ体でもあり得る。赤血球凝集素ポリペプチドは場合によって、1または複数のさらなるポリペプチドのコード配列を、そのコード配列にスプライシングすることにより生成させ得る、1または複数のさらなるポリペプチドを包含し得る。さらなるポリペプチドを赤血球凝集素ポリペプチドに挿入するための例示的な部位は、N末端である。
【0024】
本明細書で用いられる「ノイラミニダーゼポリペプチド」は、糖タンパク質から末端のシアル酸残基を切断することにより、細胞からのインフルエンザウイルスの放出を媒介する、インフルエンザウイルスタンパク質に由来する。ノイラミニダーゼの糖タンパク質は、ウイルス表面に発現する。ノイラミニダーゼタンパク質は、四量体であり、ベータ−ピンホイール構造を伴う球状ヘッド、細いストーク状領域、および単一の膜貫通ドメインを介してウイルス膜に該タンパク質を係合させる小型の疎水性領域からなる共通の構造を共有する。シアル酸残基切断の活性部位は、すべてのA型インフルエンザウイルスにおいて保存されている15の帯電アミノ酸により形成される各サブユニット表面においてポケット部分を包含する。インフルエンザノイラミニダーゼについては、少なくとも9つの亜型が同定されており、N1〜N9と標識されている。
【0025】
本明細書で開示されるVLPで用いられ得るノイラミニダーゼポリペプチドは、少なくとも膜アンカードメインおよび少なくともシアル酸残基切断活性を包含するものとする。機能領域に関する当技術分野の現況は、極めて高度である。例えば、Vargheseら、Nature 303巻、35〜40頁、1983年;Colmanら、Nature 303巻、41〜44頁、1983年;Lentzら、Biochem、26巻、5321〜5385頁、1987年;Websterら、Virol.135巻、30〜42頁、1984年を参照されたい。ノイラミニダーゼポリペプチドは、インフルエンザウイルスの任意の類型、亜型、株、または亜株、例えばノイラミニダーゼN1およびノイラミニダーゼN2に由来することが可能である。加えて、ノイラミニダーゼポリペプチドは、異なるインフルエンザノイラミニダーゼのキメラ体でもあり得る。ノイラミニダーゼポリペプチドは場合によって、1または複数のさらなるポリペプチドのコード配列を、赤血球凝集素ポリペプチドのコード配列にスプライシングすることにより生成させ得る、1または複数のさらなるポリペプチドを包含し得る。さらなるポリペプチドをノイラミニダーゼポリペプチドに挿入するための例示的な部位は、C末端である。
【0026】
本明細書で用いられる「GRASバッファー」とは、該当する政府の規制機関により公示されている通り、「一般に安全であると認識されている」バッファーを指す。GRASバッファーは、pH6〜pH8(例えば、pKa5〜9)の範囲内で緩衝作用をもたらすことが好ましい。化合物に有効な緩衝範囲は一般に、pKa±約1pH単位である、例えば、pKaが7.2であるH3PO4の緩衝能力は、約6.5〜8.0である。例示的なGRASバッファーには、トリス(ヒドロキシメチル)アミノメタン(トリス)、クエン酸水素またはクエン酸二水素、一塩基性リン酸カリウム、二塩基性リン酸カリウム、一塩基性リン酸ナトリウム、二塩基性リン酸ナトリウム、2−(N−モルホリノ)エタンスルホン酸(MES)、3−(N−モルホリノ)プロパンスルホン酸(MOPS)、N−(2−アセトアミド)イミノ二酢酸(ADA)、3−[N−トリス(ヒドロキシメチル)メチルアミノ]−2−ヒドロキシプロパンスルホン酸(TAPSO)、3−[N,N−ビス(2−ヒドロキシエチル)アミノ]−2−ヒドロキシプロパンスルホン酸(DIPSO)、ピペラジン−N,N’−ビス(2−ヒドロキシプロパンスルホン酸)(POPSO)、N−(2−ヒドロキシメチル)ピペラジン−N’−2−ヒドロキシプロパンスルホン酸(HEPPSO)、ピペラジン−N,N’−ビス(2−エタンスルホン酸)(PIPES)、N−(2−アセトアミド)−2−アミノエタンスルホン酸(ACES)、コラミンクロリド、N,N−ビス(2−ヒドロキシエチル)−2−アミノエタンスルホン酸(BES)、N−トリス(ヒドロキシメチル)メチル−2−アミノエタンスルホン酸(TES)、(4−(2−ヒドロキシエチル)−1−ピペラジンエタンスルホン酸)(HEPES)、アセトアミドグリシン、トリシン、グリシンアミド、ビシン、ヒスチジンなどのアミノ酸残基、酢酸、水酸化アンモニウム、イミダゾール、2−アミノ−2−メチル−1−プロパノール(AMP)、2−アミノ−2−メチル−1,3−プロパンジオール(AMPD)、2−[ビス(2−ヒドロキシエチル)アミノ]−2−(ヒドロキシメチル)−1,3−プロパンジオール(ビス−トリス)、1,3−ビス(トリス(ヒドロキシメチル)メチルアミノ)プロパン(ビス−トリスプロパン)、カルボン酸、クエン酸、エタノールアミン、グリシルグリシン、N−(2−ヒドロキシエチル)ピペラジン−N’(4−ブタンスルホン酸)(HEPBS)、およびマレイン酸が含まれる。適切な範囲で緩衝作用をもたらしうる例示的な官能基には、イミダゾール、カルボン酸、リン酸、ピペラジン、アミン、およびスルホン酸が含まれる。GRASは一般に、食品での使用について規定されているが、本明細書で用いられるGRASバッファーはまた、薬物製剤中に組み入れるのに薬学的に許容されるとみなされているバッファーも包含する。
【0027】
エンベロープウイルスベースのVLPを生成させる例示的な方法
エンベロープウイルスベースのVLPは、当業者に適用可能な任意の方法により生成させることができる。エンベロープウイルスベースのVLPは、インフルエンザ抗原ポリペプチドに加えて、VLPを形成する一因となる1または複数のポリペプチドを包含することが典型的であり、これらは、VLPを形成する一因となるポリペプチドが、それ自体、M1タンパク質などのインフルエンザ抗原であるか、または該インフルエンザ抗原ポリペプチドに連結される、キメラ形態を包含する。加えて、エンベロープウイルスベースのVLPは、第2のインフルエンザ抗原または別の抗原など、さらなる抗原をもたらすために、膜(脂質ラフトを含めた)会合ポリペプチドなど、1または複数のさらなるポリペプチドを包含することも可能である。ある実施形態では、ポリペプチドを、哺乳動物細胞による発現系および昆虫細胞による発現系など、細胞膜内に脂質ラフトドメインを包含する細胞ベースの系など、任意の適用可能なタンパク質発現系内で共発現させることができる。
【0028】
組換えによるVLP用ポリペプチドの発現は、該ポリペプチドのうちの1または複数をコードするポリヌクレオチドを含有する発現ベクターを伴う。ポリペプチドのうちの1または複数をコードするポリヌクレオチドが得られたら、該ポリペプチドを生成させるためのベクターを、当技術分野において周知の技法を用いる組換えDNA法により生成させることができる。このようにして、本明細書では、VLPポリペプチドをコードするヌクレオチド配列のうちのいずれかを含有するポリヌクレオチドを発現させることによりタンパク質を調製する方法が説明される。当業者に周知の方法を用いて、VLPポリペプチドのコード配列と、適切な転写制御シグナルおよび翻訳制御シグナルとを含有する発現ベクターを構築することができる。これらの方法には、例えば、in vitroにおける組換えDNA法、合成法、また、in vivoにおける遺伝子組換えが含まれる。したがって、本発明は、すべてを1または複数のプロモーターに作動可能に連結した、gagポリペプチドと、抗原に連結した脂質ラフト会合ポリペプチドとをコードするヌクレオチド配列を含む複製可能なベクターを提供する。
【0029】
発現ベクターを、従来の技法により宿主細胞に形質導入することができ、次いで、トランスフェクトされた細胞を、従来の技法により培養して、VLPポリペプチド(複数可)を生成させることができる。したがって、本発明は、異種プロモーターに作動可能に連結したVLPポリペプチドのうちの1または複数をコードするポリヌクレオチドを含有する宿主細胞を包含する。VLPを生成させるためのある実施形態では、以下で詳述する通り、gagポリペプチド、およびインフルエンザ抗原に連結した脂質ラフト会合ポリペプチド(またはインフルエンザ抗原自体が脂質ラフト会合ポリペプチドの場合もある)の両方をコードするベクターを、宿主細胞内で共発現させて、VLPを生成させる。
【0030】
各種の宿主発現ベクター系を使用して、VLPポリペプチドを発現させることができる。このような宿主発現系は、共発現などによりVLPを生成させる目的で、それを介してVLPポリペプチドを生成させうる媒体を表わす。広範囲にわたる宿主を、適切な発現ベクター構築物内において用いることができ、脂質ラフトベースのアセンブリーに依拠する場合、好ましい宿主発現系は、VLPのアセンブリーに適する脂質ラフトを有する宿主である。これらには、VLPポリペプチドコード配列を含有する組換えバクテリオファージDNA発現ベクター、組換えプラスミドDNA発現ベクター、もしくは組換えコスミドDNA発現ベクターにより形質転換した細菌(例えば、E. coli、B. subtilis);VLPポリペプチドコード配列を含有する組換え酵母発現ベクターにより形質転換した酵母(例えば、Saccharomyces属、Pichia属)などの微生物;VLPポリペプチドのコード配列を含有する組換えウイルス発現ベクター(例えば、バキュロウイルス)を感染させた昆虫細胞系;VLPポリペプチドのコード配列を含有する組換えウイルス発現ベクター(例えば、カリフラワーモザイクウイルス、CaMV;タバコモザイクウイルス、TMV)を感染させるか、もしくはVLPポリペプチドのコード配列を含有する組換えプラスミド発現ベクター(例えば、Tiプラスミド)により形質転換した植物細胞系;または、哺乳動物細胞のゲノムに由来するプロモーター(例えば、メタロチオネインプロモーター)、もしくは哺乳動物ウイルスに由来するプロモーター(例えば、アデノウイルス後期プロモーター;牛痘ウイルス7.5Kプロモーター)を含有する組換え発現構築物を保有する、哺乳動物細胞系(例えば、COS細胞、CHO細胞、BHK細胞、293細胞、3T3細胞)が含まれるがこれらに限定されない。VLPアセンブリーが、脂質ラフトの会合により駆動される場合は、哺乳動物細胞または昆虫細胞を用いて、VLPポリペプチドを発現させることができる。例えば、ヒトサイトメガロウイルスに由来する主要即初期(intermediate early)遺伝子プロモーターエレメントなどのベクターと併用される、MRC−5細胞、Vero細胞、PER.C6(TM)細胞、チャイニーズハムスター卵巣細胞(CHO)、およびHEK293細胞などの哺乳動物細胞は、VLPポリペプチドの有効な発現系である(Foeckingら、Gene、45巻:101頁(1986年); Cockettら、Bio/Technology、8巻:2頁(1990年))。
【0031】
昆虫系では、Autographa californica核多角体病ウイルス(AcNPV)を、外来遺伝子を発現するためのベクターとして用いることができる。該ウイルスは、Spodoptera frugiperda細胞内で増殖する。VLPポリペプチドのコード配列(複数可)は、ウイルスの非必須領域(例えば、ポリヘドリン遺伝子領域)内に個別にクローニングすることができ、また、AcNPVプロモーター(例えば、ポリヘドリンプロモーター)の制御下に置くことができる。
【0032】
哺乳動物の宿主細胞では、多くのウイルスベースの発現系を使用することができる。発現ベクターとしてアデノウイルスを用いる場合は、対象のVLPポリペプチド配列(複数可)を、アデノウイルス転写/翻訳制御複合体、例えば、後期プロモーターおよび三連リーダー配列にライゲーションすることができる。次いで、in vitroまたはin vivoにおける組換えにより、このキメラ遺伝子を、アデノウイルスゲノム内に挿入することができる。ウイルスゲノムの非必須領域(例えば、E1領域またはE3領域)内に挿入する結果、感染された宿主内において生存可能であり、また、VLPポリペプチド(複数可)を発現することが可能な組換えウイルスがもたらされる(例えば、LoganおよびShenk、Proc. Natl. Acad. Sci. USA、81巻:355〜359頁(1984年)を参照されたい)。挿入されたVLPポリペプチドのコード配列(複数可)を効率的に翻訳するには、特定の開始シグナルもまた必要とされうる。これらのシグナルには、ATG開始コドン、および隣接配列が含まれる。さらに、全挿入配列の翻訳を確保するには、該開始コドンが、所望されるコード配列のリーディングフレームと同フレームでなければならない。これらの外因性翻訳制御シグナルおよび開始コドンは、由来が多様であることが可能であり、天然および合成のいずれもが可能である。発現の効率は、適切な転写エンハンサーエレメント、転写ターミネーターなどを組み込むことにより、増強することができる(Bittnerら、Methods in Enzymol.、153巻:51〜544頁(1987年)を参照されたい)。一例をあげるならば、StratageneのAdEASY−XL(TM)システムなど、アデノウイルスベースのベクター系において用いられる、ヒトCMV即初期プロモーターであろう。
【0033】
加えて、挿入した配列の発現を調節するか、または該遺伝子産物を、所望される特定の形で修飾およびプロセシングする宿主細胞株も選択することができる。タンパク質産物に対するこのような修飾(例えば、グリコシル化)およびプロセシング(例えば、切断または膜への輸送)は、VLPを生成させるのに、またはVLPポリペプチド、もしくはアジュバントなどのさらなるポリペプチド、もしくはさらなる抗原が機能するのに重要でありうる。様々な宿主細胞が、タンパク質および遺伝子産物の翻訳後プロセシングおよび修飾の特徴的かつ特異的な機構を有する。発現させる外来タンパク質の適正な修飾およびプロセシングを確保するために、適切な細胞株または宿主系を選択することができる。この目的で、一次転写物を適正にプロセシングし、遺伝子産物を適正にグリコシル化およびリン酸化するための細胞機構を保有する真核宿主細胞を用いることができる。
【0034】
宿主細胞には、gagポリペプチドをコードする第1のベクターと、ウイルス性膜抗原、または抗原に連結した脂質ラフト関連ポリペプチドをコードする第2のベクターという、本発明の2つの発現ベクターを共トランスフェクトすることができる。2つのベクターは、各VLPポリペプチドの同等の発現を可能とする、同一の選択マーカーを含有しうる。代替的に、gagポリペプチドと、抗原に連結した脂質ラフト関連ポリペプチドとの両方をコードし、また、これらの両方を発現することが可能な単一のベクターを用いることもできる。
【0035】
宿主細胞によりVLPを生成させたら、ポリペプチドの精製について当技術分野で公知の任意の方法により、例えば、クロマトグラフィー(例えば、イオン交換クロマトグラフィー、特に、該ポリペプチドに付加した任意のアフィニティー精製タグによるアフィニティークロマトグラフィー、およびサイズ除外クロマトグラフィー)、遠心分離、吸収率較差溶解度法により、またはタンパク質もしくは他の高分子を精製するための他の任意の標準的な技法によりこれを精製することができる。加えて、VLPポリペプチドを、本明細書で説明されるか、またはこれ以外に当技術分野で公知の異種ポリペプチド配列に融合させ、VLPの精製を容易にすることもできる。精製後、VLPポリペプチドに対する共有結合を介して、または他の非共有結合機構により、さらなる抗原またはアジュバントなどのさらなるエレメントを、VLPに物理的に連結することができる。哺乳動物細胞および昆虫細胞など、脂質ラフトドメインを有する宿主細胞内においてVLPポリペプチドを共発現させるある実施形態では、VLPが自己アセンブリーして放出され、これにより、上記の方法のうちのいずれかによるVLPの精製が可能となる。VLPのある実施形態には、例えば、同種ウイルスのタンパク質から操作されたVLP、例えば、インフルエンザウイルスに由来するM1タンパク質、HAタンパク質、また、場合によってNAタンパク質から構築されたVLP、また、異種ウイルスのタンパク質から操作されたVLP、例えば、MLVもしくはHIVまたは他のレトロウイルスに由来するGagタンパク質を、異なるウイルスに由来する抗原、例えば、インフルエンザHAタンパク質およびインフルエンザNAタンパク質と共に操作することにより形成したVLPが含まれる。
【0036】
Gagタンパク質ベースのVLPを生成させる例示的な方法
当業者に適用可能な任意の方法により、VLPを容易にアセンブリーすることができ、この結果、gagポリペプチドと、インフルエンザ抗原ポリペプチドとを包含するVLPがアセンブリーされることが好ましい。ある実施形態では、哺乳動物細胞による発現系および昆虫細胞による発現系など、脂質内に脂質ラフトドメインを包含する細胞ベースの発現系など、任意の適用可能なタンパク質発現系において、ポリペプチドを共発現させることができる。
【0037】
gagポリペプチドを用いて形成されるVLP発現の例が数多く公表されており、これにより、VLPを生成させるのに適用可能な発現系の範囲が裏付けられている。複数のレトロウイルスによる研究は、他のウイルスコンポーネントの非存在下で発現させたGagポリペプチドが、細胞表面におけるVLPの形成および出芽に十分であることを裏付けている(WillsおよびCraven、AIDS、5巻、639〜654頁、1991年; Zhouら、J. Virol.、68巻、2556〜2569頁、1994年; Morikawaら、Virology、183巻、288〜297頁、1991年; Royerら、Virology、184巻、417〜422頁、1991年; Gheysenら、Cell、59巻、103〜112頁、1989年; Hughesら、Virology、193巻、242〜255頁、1993年; Yamshchikovら、Virology、214巻、50〜58頁、1995年)。バキュロウイルスベクターを用いる昆虫細胞において、Gag前駆体を発現させるとVLPが形成されることについては、複数の研究グループにより裏付けられている(Delchambreら、EMBO J.、8巻、2653〜2660頁、1989年; Luoら、Virology、179巻、874〜880頁、1990年; Royer ら、Virology、184巻、417〜422頁、1991年; Morikawa ら、Virology、183巻、288〜297頁、1991年; Zhouら、J. Virol.、68巻、2556〜2569頁、1994年; Gheysenら、Cell、59巻、103〜112頁、1989年; Hughesら、Virology、193巻、242〜255頁、1993年; Yamshchikovら、Virology、214巻、50〜58頁、1995年)。これらのVLPは、未成熟レンチウイルス粒子に類似しており、昆虫細胞の細胞膜から出芽することにより、効率的にアセンブリーされて放出される。
【0038】
Gag前駆体のアミノ末端領域は、ウイルスアセンブリーに必要とされる、細胞表面への輸送および膜結合を標的とするシグナル領域であることが報告されている(Yuら、J. Virol.、66巻、4966〜4971頁、1992年; Yuan, Xら、J. Virol.、67巻、6387〜6394頁、1993年; Zhouら、J. Virol.、68巻、2556〜2569頁、1994年; LeeおよびLinial、J. Virol.、68巻、6644〜6654頁、1994年; Dorfmanら、J. Virol.、68巻、1689〜1696頁、1994年; Fackeら、J. Virol.、67巻、4972〜4980頁、1993年)。牛痘ウイルスによる発現系を用いて、Gag構造タンパク質のほか、Env糖タンパク質であるgp120およびgp41も含有する、組換えHIVベースのVLPのアセンブリーが報告されている(Haffarら、J. Virol.、66巻、4279〜4287頁、1992年)。
【0039】
エンベロープウイルスベースのVLP調製物中における感染作用物質を不活化する例示的方法
電磁放射は、エンベロープウイルスベースのVLPの免疫原性を実質的に低下させることなく、感染作用物質を不活化することが可能なので、例示的な不活化の方法は、電磁放射を介する方法である。電磁放射の例示的な3つの方式のすべて(すなわち、光反応性化合物を伴うUV照射、UV照射単独、およびガンマ照射)には、血液、食品、ワクチンなど、多種多様な試料中における病原体を不活化するのに用いられてきた長い歴史があるため、不活化用電磁放射を適用するための多種多様な装置が市販されており、これらは、本明細書で開示される方法を実施するのにほとんど〜まったく改変せずに用いることができる。さらに、当技術分野では、波長および線量の最適化がルーチン化しており、したがって、当業者の能力範囲内で容易に実施される。
【0040】
光反応性化合物を伴うUV照射
電磁放射による例示的な不活化の方法は、感染作用物質中のポリヌクレオチドと反応する光反応性化合物などの、光反応性化合物の存在下における、UV−A照射などの紫外線照射の組合せである。
【0041】
例示的な光反応化合物には、アクチノマイシン、アントラサイクリノン、アントラマイシン、ベンゾジピロン、フルオレン、フルオレノン、フロクマリン、イソアロキサジン、マイトマイシン、Monostral Fast Blue、Norphillin A、フェナントリジン、フェナザチオニウム塩、フェナジン、フェノチアジン、フェニルアジド、キノリン、およびチオキサンテノン(thiaxanthenones)が含まれる。1つの分子種は、2つの主要なカテゴリーのうちの1つに属するフロクマリンである。第1のカテゴリーは、ソラーレン[7H−フロ(3,2−g)−(1)−ベンゾピラン−7−オン、または6−ヒドロキシ−5−ベンゾフランアクリル酸のデルタ−ラクトン]であり、これらは、直鎖状であり、中央の芳香環部分に付随する2つの酸素残基が1,3の配向性を有し、またさらに、フラン環部分が二環クマリン系の6位に結合している。第2のカテゴリーは、イソソラーレン[2H−フロ(2,3−h)−(1)−ベンゾピラン−2−オン、または4−ヒドロキシ−5−ベンゾフランアクリル酸のデルタ−ラクトン]であり、これらは、曲鎖状であり、中央の芳香環部分に付随する2つの酸素残基が1,3の配向性を有し、またさらに、フラン環部分が二環クマリン系の8位に結合している。3、4、5、8、4’、または5’位において直鎖状フロクマリンを置換することによりソラーレン誘導体を生成させ得る一方、3、4、5、6、4’、または5位において曲鎖状フロクマリンを置換することによりイソソラーレン誘導体を生成させ得る。ソラーレンは、二本鎖核酸の塩基対間に挿入することができ、長波長の紫外光(UVA)を吸収すると、ピリミジン塩基に対する共有結合付加体を形成する。例えば、G. D. Ciminoら、Ann. Rev. Biochem.54巻:1151頁(1985年);Hearstら、Quart. Rev. Biophys.17巻:1頁(1984頁)を参照されたい。
【0042】
例示的なUV(または場合によって、可視光)放射の波長は、適切な反応および/または光付加体が生成される波長に依存し、これは、光反応性化学物質の化学反応に依存する。例示を目的として述べると、多くのソラーレンには、320〜380nmの波長にあるUV放射が極めて有効であり、330〜360nmが最大の有効性を示す。また、同様のUV−A波長は、419nmなど、可視光とも併せて用いうる光反応性化合物であるリボフラビンと併用しても、病原体を不活化するのに極めて有効である。
【0043】
UV照射単独
光反応性化合物の存在下におけるUV照射に加え、感染作用物質は、UV照射単独でも不活化することができる。ある実施形態では、放射の波長が、約180〜320nm、または約225〜290nm、または約254nm(すなわち、ポリヌクレオチドの高吸光度ピークおよび低減したタンパク質吸収を有するスペクトル領域)であるUV−C放射である。UV−C放射は、エンベロープを形成する脂質二重層およびエンベロープ内のタンパク質など、本明細書で開示されるエンベロープウイルスベースのVLPのコンポーネントに対して、安定性および免疫原性の両面で有害性が低い一方で、感染作用物質を不活化するのに十分なエネルギーを保持するので、用いることができる。しかし、例えば、UV−AおよびUV−Bなど、他の種類のUV放射もまた用いることができる。
【0044】
ガンマ照射
本明細書で開示される方法を実施して組成物を生成させるには、ガンマ照射(すなわち、イオン化放射)もまた用いることができる。この実施形態では、10〜60kGyのガンマ照射線量が、病原体の不活化に有効である。ガンマ照射は、感染作用物質のゲノムをコードするポリヌクレオチド内において鎖の切断を導入することにより、感染作用物質を直接不活化することもでき、該ポリヌクレオチドを損傷させるフリーラジカルを生成することにより、感染作用物質を間接的に不活化することもできる。エンベロープVLPの脂質コンポーネントおよびタンパク質コンポーネントへの、ラジカルを介する損傷を阻止するには、ガンマ照射と共に、フリーラジカルスカベンジャーおよび低温を用いることができる。
【0045】
VLPを用いる例示的な方法
製剤
本明細書で説明されるエンベロープウイルスベースのVLPの例示的な使用は、ワクチン調製物としての使用である。典型的には、このようなワクチンは、溶液または懸濁液の注射剤として調製されるが、注射前に液体中に溶解させるか、または懸濁させるのに適する固体形態もまた調製することができる。このような調製物はまた、乳化させることもでき、乾燥粉末として生成させることもできる。免疫原性の有効成分は、薬学的に許容され、また、有効成分に適合する、賦形剤と混合することが多い。適切な賦形剤は、例えば、水、生理食塩液、デキストロース、スクロース、グリセロール、エタノールなどであり、また、これらの組合せである。加えて、所望の場合、ワクチンは、湿潤剤もしくは乳化剤、pH緩衝剤、または該ワクチンの有効性を増強するアジュバントを含有し得る。
【0046】
ワクチンは、注射、例えば、皮下(subcutaneously)注射、皮内注射、皮下(subdermally)注射、または筋肉内注射を介する、従来の非経口投与が可能である。他の投与方式に適する、さらなる製剤には坐剤が包含され、場合によっては、経口製剤、鼻腔内製剤、口腔内製剤、舌下製剤、腹腔内製剤、膣内製剤、経肛門製剤、および頭蓋内製剤も包含される。坐剤の場合、従来の結合剤および担体には、例えば、ポリアルキレングリコール(polyalkalene glycol)またはトリグリセリドが包含される場合があり、このような坐剤は、0.5%〜10%、または1〜2%の範囲でも、有効成分を含有する混合物から形成することができる。ある実施形態では、脂肪酸グリセリドまたはココアバターの混合物などの低温溶融ワックスをまず溶融させ、本明細書で説明されるエンベロープウイルスベースのVLPを、例えば、撹拌することにより、均一に分散させる。次いで、溶融した均一の混合物を、適当なサイズの鋳型に流し込み、冷却して固形化させる。
【0047】
鼻腔内送達に適する製剤には、液体(例えば、エアゾールまたは点鼻液として投与する水溶液)および乾燥粉末(例えば、鼻孔内への急速沈殿用)が包含される。製剤は、例えば、医薬品グレードのマンニトール、ラクトース、スクロース、トレハロース、キシリトール、およびキトサンなど、通常用いられる賦形剤を包含する。液体製剤または粉末製剤において、キトサンなどの粘膜付着剤を用いて、鼻腔内投与された製剤の粘液線毛クリアランスを遅延させることができる。マンニトールおよびスクロースなどの糖を、液体製剤では安定化剤として用いることができ、また、乾燥粉末製剤では安定化剤、充填剤、または粉末流動剤および粉末サイズ剤として用いることができる。加えて、モノホスホリル脂質A(MPL)またはCpGオリゴヌクレオチド(oligoneucleotides)などのアジュバントを、液体製剤および乾燥粉末製剤の両方において、免疫刺激性アジュバントとして用いることもできる。
【0048】
経口送達に適する製剤には、液体、固体、半固体、ゲル、錠剤、カプセル、トローチなどが包含される。経口送達に適する製剤には、錠剤、トローチ、カプセル、ゲル、液体、食品、飲料、栄養補助食品などが包含される。製剤は、例えば、医薬品グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなど、通常用いられる賦形剤を包含する。他のエンベロープウイルスベースのVLPワクチン組成物は、溶液、懸濁液、丸薬、持続放出製剤、または粉末の形態を取る場合があり、10〜95%の有効成分、または25〜70%の有効成分を含有する場合がある。経口製剤の場合、コレラ毒素が興味深い製剤パートナーである(また、可能なコンジュゲーションパートナーでもある)。
【0049】
膣内投与用に調合する場合、エンベロープウイルスベースのVLPワクチンは、ペッサリー、タンポン、クリーム、ゲル、ペースト、フォーム、またはスプレーの形態であり得る。前出の製剤のうちのいずれもが、エンベロープウイルスベースのVLPに加えて、担体など、当技術分野で適切であることが公知の薬剤を含有し得る。
【0050】
一部の実施形態では、エンベロープウイルスベースのVLPワクチンを、全身送達用に調合する場合もあり、局所送達用に調合する場合もある。このような製剤は、当技術分野において周知である。非経口媒体には、塩化ナトリウム溶液、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、乳酸加リンゲル液、または固定油が包含される。静脈内媒体には、流体および栄養物質補給剤、電解質補給剤(リンゲルデキストロースに基づく補給剤など)などが包含される。全身投与経路および局所投与経路には、例えば、皮内経路、局所適用経路、静脈内経路、筋肉内経路などが包含される。
【0051】
エンベロープウイルスベースのVLPは、中性製剤または塩ベースの製剤を包含するワクチンに調合することができる。薬学的に許容される塩には、酸添加塩(ペプチドの遊離アミノ基と共に形成され、また、例えば、塩酸もしくはリン酸などの無機酸、または酢酸、シュウ酸、酒石酸、マンデル酸などの有機酸と共に形成される)が包含される。遊離カルボキシル基と共に形成される塩はまた、例えば、水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、または水酸化鉄などの無機塩基、また、イソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジン、プロカインなどの有機塩基に由来し得る。
【0052】
ワクチンは、剤形に適合した形で、また、治療的に有効であり免疫原性となる量で投与することができる。投与量は、例えば、個体の免疫系が免疫反応を誘発する能力、および所望される防御の程度を包含する、治療される対象に依存する。適切な用量範囲は、1回のワクチン接種当たりの有効成分が数百マイクログラムのオーダーであり、例示的な範囲は、約0.5μg〜1000μgの範囲、1μg〜500μgの範囲、または、約10μg〜100μgの範囲など、約0.1μg〜2000μgの範囲である(1〜10mgの範囲にある高量も意図される)。初回投与および追加投与に適するレジメンもまた可変的であるが、初回投与にその後の接種または他の投与が続く形が典型的である。
【0053】
適用方式は多様に変化させることができる。従来のワクチン投与方法のうちのいずれもが適用可能である。これらには、固体の生理学的に許容されるベース上における経口適用、または注射など、生理学的に許容される分散剤による非経口適用が包含される。ワクチンの用量は、投与経路に依存し、ワクチン接種される人の年齢、および抗原製剤により変化する。
【0054】
ワクチン製剤の一部は、製剤自体でワクチンとして十分に免疫原性であるが、他の製剤では、該ワクチンがアジュバント物質をさらに含むと、免疫反応が増強される場合がある。
【0055】
とりわけ、鼻腔内ベース、経口ベース、または肺内ベースの送達製剤の場合、送達および免疫原性を改善するには、粘膜付着を改善する送達剤を用いることもできる。このような化合物である、キチンのN−脱アセチル化形態であるキトサンは、多くの医薬製剤で用いられる(32)。それが粘液線毛クリアランスを遅延させ、粘膜による抗原の取込みおよびプロセシングのためにより多くの時間を許容する能力のために、キトサンは、鼻腔内におけるワクチン送達にとって魅力的な粘膜付着剤である(33、34)。加えて、キトサンは、密着結合を一過性に開口させ、これにより、NALTへの抗原の経上皮輸送が増強され得る。近年のヒト試験では、キトサンを伴うが、さらなるアジュバントは伴わずに、三価の不活化インフルエンザワクチンを鼻腔内投与したところ、筋肉内接種後に得られたセロコンバージョンおよびHI力価をごくわずかに下回るセロコンバージョンおよびHI力価がもたらされた(33)。
【0056】
キトサンはまた、遺伝子的に解毒したE. coli易熱性エンテロトキシンの突然変異体であるLTK63など、鼻腔内において良好に機能するアジュバントと共に調合することもできる。これにより、キトサンにより付与される送達および付着の利益の上に、免疫刺激効果が付加され、その結果、粘膜反応および全身反応の増強をもたらす(35)。
【0057】
最後に、キトサン製剤はまた、ワクチンの安定性を改善し、さらに、粘液線毛クリアランスにおいて液体製剤を上回る遅延を結果としてもたらすことが示されている乾燥粉末フォーマットでも調製することができる(42)。これは、キトサンと共に調合された、鼻腔内用乾燥粉末によるジフテリア毒素ワクチンを伴う、近年のヒト臨床試験において見られ、ここで、鼻腔内経路は、従来の筋肉内経路と同程度に有効であり、分泌性IgA反応の利益が付加された(43)。該ワクチンはまた、忍容性も極めて良好であった。キトサンおよびMPLを含有する炭疽菌用の鼻腔内乾燥粉末ワクチンは、ウサギにおいて、筋肉内接種より強力な反応を誘導し、また、エアゾールによる胞子投与に対しても防御的であった(44)。
【0058】
鼻腔内ワクチンは、下気道に対してより良好に作用する非経口投与ワクチンに対して、上気道および下気道に作用し得るので、好ましい製剤を表わす。これは、アレルゲンベースのワクチンには忍容性を誘導し、また、病原体ベースのワクチンには免疫性を誘導する点で有益である。
【0059】
鼻腔内ワクチンは、上気道および下気道の両方において防御をもたらすのに加え、注射針による接種の煩雑さを回避し、また、粒子抗原および/または可溶性抗原が、鼻咽頭関連リンパ組織(NALT)と相互作用することを介して、粘膜および全身における体液性反応および細胞性反応の両方を誘導する手段をもたらす(16〜19)。歴史的に、鼻腔内経路は、非経口接種ほど有効ではなかったが、新規の送達製剤であるエンベロープウイルスベースのVLPおよびアジュバントを用いることにより、この枠組みが変わりつつある。実際、鼻腔粘膜内にはシアル酸を含有する受容体が豊富であるため、鼻腔内送達には、機能性の赤血球凝集素分子を含有するインフルエンザワクチンがとりわけよく適しており、その結果、HA抗原の結合が増強され、粘液線毛クリアランスが低減される可能性をもたらす。
【0060】
インフルエンザについては、並行する非経口投与の免疫原性が低く、ヘテロ亜型的防御が誘導されなかった実験の鼻腔内ワクチン送達後において、ヘテロ亜型的防御を含めた防御的免疫反応が報告されている(20〜22)。さらに、不活化インフルエンザは、鼻腔内投与されるサル免疫不全ウイルス(SIV)によるVLPワクチンと混合すると、全身および粘膜における体液性反応および細胞性反応に有効なアジュバントであることも示されている(23)。このアジュバント効果は、不活化インフルエンザビリオンが、VLPと共に凝集し、粘膜表面への結合を増強する能力に帰せられた。また、インフルエンザHAタンパク質をSIV VLPに直接に組み込んだところ、樹状細胞(DC)への結合およびDCの活性化が増強された場合にも、同様のアジュバント効果が見られた(24、25)。
【0061】
アジュバント
ワクチンに対するアジュバント効果を達成する各種の方法が公知であり、本明細書で開示されるエンベロープウイルスベースのVLPと共に用いることができる。一般的な原理および方法は、「The Theory and Practical Application of Adjuvants」、1995年、Duncan E. S. Stewart−Tull(編)、John Wiley & Sons Ltd.、ISBN 0−471−95170−6において詳述されており、また、「Vaccines: New Generation Immunological Adjuvants」、1995年、Gregoriadis Gら(編)、Plenum Press、New York、ISBN 0−306−45283−9においても詳述されており、どちらも参照により本明細書に組み込まれる。
【0062】
一部の実施形態では、VLPワクチンが、重量ベースの比率約10:1〜約1010:1のVLP:アジュバント、例えば、約10:1〜約100:1、約100:1〜約103:1、約103:1〜約104:1、約104:1〜約105:1、約105:1〜約106:1、約106:1〜約107:1、約107:1〜約108:1、約108:1〜約109:1、または約109:1〜約1010:1のVLP:アジュバントで、少なくとも1つのアジュバントと混合したエンベロープウイルスベースのVLPを含む。当業者は、アジュバントに関する情報、また、最適の比を決定する日常的実験に関する情報により、適切な比率を容易に決定することができる。当業者は、アジュバントに関する情報、および最適の比率を決定する日常的実験に関する情報により、適切な比率を容易に決定することができる。限定なしに述べると、本明細書で開示されるVLPとアジュバントとの混合物には、同じ溶液における個別のVLPとアジュバントとの混合物、共有結合させたVLPおよびアジュバント、イオン結合させたVLPおよびアジュバント、疎水性結合させたVLPおよびアジュバント(VLP膜内に部分的または完全に包埋させることを含めた)、親水性結合させたVLPおよびアジュバント、ならびに前出の任意の組合せを含め、当業者に利用可能な任意の形態の組合せが含まれうる。
【0063】
例示的なアジュバントには、toll様受容体(TLR)アゴニスト、モノホスホリル脂質A(MPL)、合成脂質A、脂質Aの模倣体または類似体、アルミニウム塩、サイトカイン、サポニン、ムラミルジペプチド(MDP)誘導体、CpGオリゴ、グラム陰性菌のリポ多糖(LPS)、ポリホスファゼン、エマルジョン、水中油エマルジョン、ビロソーム、コクリエート、ポリ(ラクチド−co−グリコリド)(PLG)マイクロ粒子、ポロキサマー粒子、マイクロ粒子、およびリポソームが包含され得るがこれらに限定されない。アジュバントは、細菌由来の外毒素ではないことが好ましい。好ましいアジュバントは、3DMPL、CpGオリゴヌクレオチド、またはQS21など、Th1型反応を刺激するアジュバントである。
【0064】
Salmonellaに由来する、脂質Aの非毒性誘導体であるモノホスホリル脂質A(MPL)は、ワクチンアジュバントとして開発された、強力なTLR−4アゴニストである(Evansら、2003年)。マウスによる前臨床研究では、鼻腔内MPLが、分泌性ならびに全身性の体液反応を増強することが示されている(Baldridgeら、2000年;Yangら、2002年)。MPLはまた、120,000例を超える患者による臨床研究でも、ワクチンアジュバントとして安全かつ有効であることが分かっている(Baldrickら、2002年;2004年)。MPLは、TLR−4受容体を介して自然免疫の誘導を刺激し、したがって、グラム陰性菌およびグラム陽性菌、ウイルス、および寄生虫を包含する、広範な範囲にわたる感染性病原体に対して、非特異的な免疫反応を誘発することが可能である(Baldrickら、2004年;Persingら、2002年)。鼻腔内製剤中にMPLを包含することにより、ウイルス感作に由来する非特異的な免疫反応を誘発する自然反応が迅速に誘導される一方で、該ワクチンの抗原成分により生成させる特異的反応が増強される。
【0065】
したがって、一実施形態では、本発明が、モノホスホリル脂質A(MPL(登録商標))または3−O−脱アシル化モノホスホリル脂質A(3 De−O−acylated monophosphoryl lipid A)(3D−MPL(登録商標))を、後天免疫および自然免疫の増強剤として含む組成物を提供する。化学的に述べると、3D−MPL(登録商標)とは、4位アシル化鎖、5位アシル化鎖、または6位アシル化鎖との3−O−脱アシル化モノホスホリル脂質Aの混合物である。3−O−脱アシル化モノホスホリル脂質Aの例示的な形態は、欧州特許第0689454B1号(SmithKline Beecham Biologicals SA)において開示されている。別の実施形態では、本発明が、合成脂質A、BioMiraによるPET Lipid Aなど、脂質Aの模倣体もしくは類似体、またはTLR−4アゴニストと同様に機能するようにデザインされている合成誘導体を含む組成物を提供する。
【0066】
例示的なアジュバントは、VLPポリペプチドと共発現させるか、またはVLPポリペプチドと融合させて、キメラポリペプチドを生成させることにより、本明細書で説明されるエンベロープウイルスベースのVLPに容易に添加し得るポリペプチドアジュバントである。鞭毛の主要なタンパク質成分である細菌性フラジェリンは、それが、toll様受容体であるTLR5を介する先天性免疫系により認識されるために、アジュバントタンパク質としてますます注目を集めているアジュバントである(65)。TLR5を介するフラジェリンシグナル伝達は、DCの成熟化および移動の他、マクロファージ、好中球、および腸上皮細胞の活性化も誘導することにより、先天性免疫機能および後天性免疫機能の両方に影響を及ぼし、その結果、炎症促進メディエーターを生成させる(66〜72)。
【0067】
TLR5は、免疫学的圧力に応答するその突然変異を除外し、このタンパク質に固有であり、また、鞭毛機能に必要とされる、フラジェリン単量体内における保存的構造を認識する(73)。該受容体は、100fMの濃度に対して感受性であるが、完全な線維は認識しない。鞭毛が単量体に解体されることが、結合および刺激に必要とされる。
【0068】
アジュバントとしてのフラジェリンは、非経口投与または鼻腔内投与された異種抗原に対する防御反応を誘導する強力な活性を有し(66、74〜77)、また、DNAワクチンに対するアジュバント効果もまた報告されている(78)。マウスまたはサルにおいてフラジェリンを用いると、Th2バイアス(これは、インフルエンザなどの呼吸器ウイルスには適切である)は観察されるが、IgE誘導の証拠は観察されていない。加えて、サルにおける鼻腔内投与または全身投与後における、局所炎症反応または全身炎症反応も報告されていない(74)。TLR5を介して、MyD88依存的な形でなされるフラジェリンシグナル、およびTLRを介する、他のすべてのMyD88依存的シグナルは、Th1バイアスを結果としてもたらすことが示されている(67、79)ため、フラジェリンを使用した後で誘発される反応におけるTh2の特徴は、ある意味で驚くべきことである。フラジェリンに対して予め存在する抗体が、アジュバントの有効性に対してさほどの影響を及ぼさない(74)ことから、フラジェリンは、複数回使用のためのアジュバントとして魅力的となっていることが重要である。
【0069】
近年における鼻腔内ワクチン試験における共通の主題は、ワクチンの有効性を改善するアジュバントおよび/または送達系の使用である。このような一研究では、遺伝子的に解毒したE. coli易熱性エンテロトキシンアジュバント(LT R192G)を含有するインフルエンザH3ワクチンが、H5による感作に対してヘテロ亜型的な防御を結果としてもたらしたが、これは、鼻腔内送達後に限られた。防御は、交差中和抗体の誘導に基づき、これにより、新たなインフルエンザワクチンの開発における鼻腔内経路についての重要な示唆が示された(22)。
【0070】
サイトカイン、コロニー刺激因子(例えば、GM−CSF、CSFなど);腫瘍壊死因子;インターロイキン−2、インターロイキン−7、インターロイキン−12、および他の同様の増殖因子は、VLPポリペプチドと混合または融合させることにより、エンベロープウイルスベースのVLPワクチン内に容易に包含され得るので、これらもまたアジュバントとして用いることができる。
【0071】
一部の実施形態では、本明細書で開示されるエンベロープウイルスベースのVLPワクチン組成物が、CpGオリゴヌクレオチドを含む、核酸のTLR9リガンド;イミダゾキノリンのTLR7リガンド;置換グアニンのTLR7/8リガンド;ロキソリビン、7−デアザデオキシグアノシン、7−チア−8−オキソデオキシグアノシン、二本鎖ポリ(I:C)、ポリイノシン酸、イミキモド(R−837)、およびレシキモド(R−848)などの他のTLR7リガンド;またはMPL(登録商標)もしくは合成誘導体などのTLR4アゴニストなど、Toll様受容体を介して作用する他のアジュバントを包含し得る。
【0072】
ある種のアジュバントは、樹状細胞などのAPCによるワクチン分子の取込みを促進し、これらを活性化する。非限定的な例は、免疫標的化アジュバント;毒素、サイトカイン、およびマイコバクテリア派生物などの免疫調節アジュバント;油製剤;ポリマー;ミセル形成アジュバント;サポニン;免疫刺激複合体マトリックス(ISCOMマトリックス);粒子;DDA;アルミニウムアジュバント;DNAアジュバント;MPL;および封入アジュバントからなる群から選択される。
【0073】
アジュバントのさらなる例には、緩衝生理食塩液中0.05〜0.1パーセントの溶液として一般的に用いられる水酸化物またはリン酸などのアルミニウム塩(ミョウバン)(例えば、Nicklas(1992年)Res. Immunol.143巻:489〜493頁を参照されたい)、0.25パーセントの溶液として用いられる糖の合成ポリマー(例えば、Carbopol(登録商標))との混合物、それぞれ、70℃〜101℃の範囲の温度で30秒間〜2分間にわたる熱処理によるワクチン中のタンパク質の凝集物などの薬剤が包含され、また、架橋形成剤による凝集物も可能である。ペプシン処理した、アルブミンに対する抗体(Fab断片)との再活性化による凝集物、C. parvumまたはグラム陰性菌の内毒素もしくはリポ多糖コンポーネントなどの細菌細胞との混合物、モノオレイン酸マンニド(Aracel A)などの生理学的に許容される油媒体中におけるエマルジョン、または保護置換基として用いられる、ペルフルオロカーボン(Fluosol−DA)の20パーセント溶液によるエマルジョンもまた用いることができる。スクアレンおよびIFAなどの油との混合物もまた用いることができる。
【0074】
DDA(ジメチルジオクタデシルアンモニウムブロミド)は、アジュバントの興味深い候補物質であるが、フロイントの完全アジュバントおよびフロイントの不完全アジュバントの他、QuilAおよびQS21などのキラヤサポニンもまた興味深い。さらなる可能性には、モノホスホリル脂質A(MPL(登録商標))、ムラミルジペプチド(MDP)、およびトレオニルムラミルジペプチド(tMDP)など、リポ多糖のポリ[ジ(カルボキシレートフェノキシ)ホスファゼン(poly[di(earboxylatophenoxy)phosphazene)(PCPP)誘導体が包含される。例えば、3−O−脱アシル化モノホスホリル脂質Aなどのモノホスホリル脂質Aをアルミニウム塩と併せた組合せを包含する、主にTh1型の反応をもたらすリポ多糖ベースのアジュバントを用いることができる。MPL(登録商標)アジュバントは、GlaxoSmithKlineから市販されている(例えば、それらの各々が、それらのリポ多糖に関する教示に対する具体的な言及を伴うそれらの全体において参照により組み込まれる、米国特許第4,436,727号;同第4,877,611号;同第4,866,034号;および同第4,912,094号を参照されたい)。
【0075】
リポソーム製剤もまた、アジュバント効果を付与することが知られており、したがって、リポソームアジュバントは、エンベロープウイルスベースのVLPと併用して用いることができる。
【0076】
免疫刺激複合体マトリックス型(ISCOM(登録商標)マトリックス)アジュバントは、とりわけ、この型のアジュバントが、APCによるMHCクラスII発現を上方制御することが可能であるため、エンベロープウイルスベースのVLPワクチンと共に用いることもできる。ISCOMマトリックスは、Quillaja saponariaに由来する(場合によって画分された)サポニン(トリテルペノイド)、コレステロール、およびリン脂質からなる。VLP中のものなどの免疫原性タンパク質と混合する場合、結果として得られる粒子状製剤は、サポニンが60〜70%w/wを占めることが可能であり、コレステロールおよびリン脂質が10〜15%w/w、およびタンパク質が10〜15%w/wを占めることが可能な、ISCOM粒子として公知の製剤である。免疫刺激複合体の組成および使用に関する詳細は、例えば、アジュバントを扱う上述の教科書中に見出し得るが、また、Morein Bら、1995年、Clin. Immunother.3巻:461〜475頁の他、Barr I GおよびMitchell G F、1996年、Immunol. and Cell Biol.74巻:8〜25頁(どちらも参照により本明細書に組み込まれる)も、完全な免疫刺激複合体を調製するのに有益な教示を提供している。
【0077】
本明細書で開示されるエンベロープウイルスベースのVLPワクチンとのアジュバントの組合せにおいて用い得るサポニンには、それらがISCOM形態にある場合であれ、そうでない場合であれ、米国特許第5,057,540号(Quil A画分、およびそれらを単離して用いる方法に対する具体的な言及を伴うその全体において参照により本明細書に組み込まれる)、および「Saponins as vaccine adjuvants」、Kensil, C. R.、Crit Rev Ther Drug Carrier Syst、1996年、12巻(1〜2号):1〜55頁;およびEP0362279B1で説明される、Quil Aと称する、Quillaja Saponaria Molinaの樹皮に由来するサポニン、ならびにそれらの画分が包含される。Quil Aの例示的な画分は、QS21、QS7、およびQS17である。
【0078】
β−エスシンは、本明細書で説明されるエンベロープウイルスベースのVLPワクチンのアジュバント組成物中で用いる別の溶血性サポニンである。エスシンは、メルクインデックス(第12版:登録番号3737)において、ラテン名:Aesculus hippocastanumであるトチノキの種子内で発生するサポニンの混合物として説明されている。クロマトグラフィーおよび精製(Fiedler、Arzneimittel−Forsch.4巻、213頁(1953年))による、およびイオン交換樹脂(Erbringら、米国特許第3,238,190号)によるその単離が、説明されている。エスシンの画分は精製されており、生物学的に活性であることが示されている(Yoshikawa Mら(Chem Pharm Bull(東京)、1996年8月;44巻(8号):1454〜1464頁))。β−エスシン(escin)はまた、エスシン(aescin)としても公知である。
【0079】
エンベロープウイルスベースのVLPワクチンと共に用いる別の溶血性サポニンは、ジギトニンである。ジギトニンは、メルクインデックス(第12版:登録番号3204)において、Digitalis purpureaの種子に由来し、また、Gisvoldら、J.Am.Pharm.Assoc.、1934年、23巻、664頁;およびRuhenstroth−Bauer、Physiol.Chem.、1955年、301巻、621頁において説明される手順により精製されるサポニンとして説明されている。その使用は、コレステロールを判定するための臨床試薬であると説明されている。
【0080】
アジュバント効果を達成する別の興味深い可能性は、Gosselinら、1992年(参照により本明細書に組み込まれる)において説明される技法を用いることである。略述すると、本明細書で説明されるVLPポリペプチドまたはさらなる抗原などの関与抗原の提示は、該抗原を、単球/マクロファージ上におけるFC受容体に対する抗体(または抗原に結合する抗体断片)にコンジュゲートすることにより増強することができる。とりわけ、抗原と抗FCRIとのコンジュゲートは、ワクチン接種のための免疫原性を増強することが示されている。該抗体は、VLPポリペプチドのうちのいずれか1つに対する融合体としての発現によることを包含する生成後において、または該生成の一部として、エンベロープウイルスベースのVLPにコンジュゲートすることができる。
【0081】
他の可能性は、標的化物質および免疫調節物質(すなわち、サイトカイン)の使用を伴う。加えて、ポリI:Cなど、合成のサイトカイン誘導物質もまた用いることができる。
【0082】
適切なマイコバクテリア派生物は、ムラミルジペプチド、フロイントの完全アジュバント、ならびに、TDMおよびTDEなど、トレハロースのジエステルからなる群から選択することができる。
【0083】
適切な免疫標的化アジュバントの例には、CD40リガンドおよびCD40抗体、またはこれらの特異的結合断片(上記の議論を参照されたい)、マンノース、Fab断片、ならびにCTLA−4が包含される。
【0084】
適切なポリマーアジュバントの例には、デキストランなどの炭水化物、PEG、デンプン、マンナン、およびマンノース;塑性ポリマー;ならびにラテックスビーズなどのラテックスが包含される。
【0085】
免疫反応を調節するさらに別の興味深い方法は、「仮想リンパ節」(VLN)(ImmunoTherapy、Inc.、360 Lexington Avenue, New York、N.Y. 10017−6501により開発された、特許品の医療デバイス)内に免疫原(場合によって、アジュバント、ならびに薬学的に許容される担体および媒体と併せて)を包含することである。VLN(細長型の管状デバイス)は、リンパ節の構造および機能を模倣する。VLNを皮下に挿入することにより、サイトカインおよびケモカインの急増による無菌的炎症部位が創出される。T細胞およびB細胞の他、APCも、危険シグナルに応答し、炎症部位に至り、VLNの多孔性マトリックスの内部に蓄積される。VLNを用いると、抗原に対する免疫反応を誘発するのに必要とされる抗原用量が低下し、VLNを用いるワクチン接種により付与される免疫的防御が、Ribiをアジュバントとして用いる従来の免疫化を凌駕することが示されている。この技法は、Gelber Cら、1998年、「Elicitation of Robust Cellular and Humoral Immune Responses to Small Amounts of Immunogens Using a Novel Medical Device Designated the Virtual Lymph Node」、「From the Laboratory to the Clinic、梗概、1998年10月12〜15日、Seascape Resort、Aptos Calif.」において略述されている。
【0086】
エンベロープウイルスベースのVLPと共に、オリゴヌクレオチドをアジュバントとして用いることができ、少なくとも3つ、または少なくとも6つ以上のヌクレオチドで隔てられた2つ以上のジヌクレオチドによるCpGモチーフを含有することができる。CpGを含有するオリゴヌクレオチド(ここで、CpGジヌクレオチドは、メチル化されていない)は、主にTh1反応を誘導する。このようなオリゴヌクレオチドは周知であり、例えば、それらの各々が、CpGオリゴヌクレオチドをアジュバントとして生成させて用いる方法に対する具体的な言及を伴うそれらの全体において参照により本明細書に組み込まれる、WO96/02555、WO99/33488、ならびに米国特許第6,008,200号および同第5,856,462号において説明されている。
【0087】
このようなオリゴヌクレオチドによるアジュバントは、デオキシヌクレオチドであり得る。ある実施形態では、該オリゴヌクレオチド内のヌクレオチド骨格は、ホスホロジチオエート結合であるか、またはホスホロチオエート結合であるが、骨格結合を混合したオリゴヌクレオチドを包含する、ホスホジエステル骨格、およびPNAなど他のヌクレオチド骨格をエンベロープウイルスベースのVLPワクチンと共に用いることができる。ホスホロチオエートオリゴヌクレオチドまたはホスホロジチオエートを生成させる方法は、それらの各々が、ホスホロチオエートおよびホスホロジチオエートについての教示に対する具体的な言及を伴うそれらの全体において参照により本明細書に組み込まれる、米国特許第5,666,153号、米国特許第5,278,302号、およびW095/26204において説明されている。
【0088】
例示的なオリゴヌクレオチドは、以下の配列を有する。配列は、ホスホロチオエート修飾されたヌクレオチド骨格を含有することができる。
【0089】
【化1】
代替的なCpGオリゴヌクレオチドには、これらに対する重要でない欠失または付加を伴う上記の配列が包含される。アジュバントとしてのCpGオリゴヌクレオチドは、当技術分野で公知である任意の方法(例えば、EP468520)により合成することができる。例えば、自動合成器を用いてこのようなオリゴヌクレオチドを合成し得る。このようなオリゴヌクレオチドによるアジュバントは、10〜50塩基の長さであり得る。別のアジュバント系は、CpGを含有するオリゴヌクレオチドと、サポニン誘導体との組合せを伴い、特に、CpGとQS21との組合せが、WO00/09159において開示されている。
【0090】
多くの単相または複相のエマルジョン系が説明されている。エマルジョンが、エンベロープウイルスベースのVLPの構造を破壊しないように、当業者は、エンベロープウイルスベースのVLPと共に用いられるこのようなエマルジョン系を容易に適合させることができる。水中油エマルジョンによるアジュバントは、それ自体、アジュバント組成物として有用であることが示唆されており(EPO399843B)、また、水中油エマルジョンと他の活性薬剤との組合せも、ワクチン用のアジュバントとして説明されている(WO95/17210;WO98/56414;WO99/12565;WO99/11241)。油中水エマルジョン(米国特許第5,422,109号;EP0480982B2)、および水中油中水エマルジョン(米国特許第5,424,067号;EP0480981B)など、他の油エマルジョンによるアジュバントが説明されている。
【0091】
本明細書で説明されるエンベロープウイルスベースのVLPと共に用いられる油エマルジョンによるアジュバントは、天然の場合もあり、合成の場合もあり、また、無機性の場合もあり、有機性の場合もある。鉱油および有機油の例は、当業者に容易に明らかであろう。
【0092】
任意の水中油組成物が、ヒトへの投与に適するためには、エマルジョン系の油相が、代謝性油を包含していてよい。代謝性油という用語の意味は、当技術分野において周知である。代謝性とは、「代謝により変換可能であること」と定義することができる(「Dorland’s Illustrated Medical Dictionary」、W.B. Sanders Company、第25版(1974年))。油は、受容者に毒性でなく、代謝により変換可能な、任意の植物油、魚油、動物油、または合成油であることが可能である。堅果(ラッカセイ油など)、種、穀類が、植物油の一般的な供給源である。合成油もエンベロープウイルスベースのVLPワクチンと共に用いることができ、これらには、NEOBEE(登録商標)および他の油など、市販の油が包含され得る。スクアレン(2,6,10,15,19,23−ヘキサメチル−2,6,10,14,18,22−テトラコサヘキサエン)は、サメの肝油中に高量で見出され、また、オリーブ油、コムギ胚種油、コメぬか油中、および酵母に低量で見出される不飽和油であり、本明細書で開示されるエンベロープウイルスベースのVLPワクチンと共に用いることができる。スクアレンは、コレステロールの生合成における中間体であるという事実により、代謝性油である(「メルクインデックス」、第10版、登録番号8619)。
【0093】
例示的な油エマルジョンは水中油エマルジョンであり、特に、水中スクアレンエマルジョンである。
【0094】
加えて、エンベロープウイルスベースのVLPワクチンにおいて用いる油エマルジョンによるアジュバントには、油であるα−トコフェロール(ビタミンE、EP0382271B1)などの、抗酸化剤が含まれ得る。
【0095】
WO95/17210およびWO99/11241は、場合によって、免疫刺激剤であるQS21および/または3D−MPLと共に調合される、スクアレン、α−トコフェロール、およびTWEEN 80(TM)に基づくエマルジョンによるアジュバントを開示している。WO99/12565は、油相へのステロールの添加による、これらのスクアレンエマルジョンに対する改善を開示している。加えて、エマルジョンを安定化させるために、トリカプリリン(C27H50O6)などのトリグリセリドを、油相に添加することができる(WO98/56414)。
【0096】
安定的な水中油エマルジョン中で見出される油滴のサイズは、光子相関分光分析により測定したときに、1ミクロン未満であってよく、実質的に30〜600nmの範囲内、実質的に直径約30〜500nm、実質的に直径150〜500nm、または直径約150nmであり得る。この点で、油滴数の80%がこれらの範囲内にあるものとし、油滴数の90%、または油滴数の95%超が規定のサイズ範囲内にある。油エマルジョン中に存在する成分の量は、従来、スクアレンなどの油が2〜10%の範囲内にあり;また、存在する場合、アルファトコフェロールが2〜10%であり、ポリオキシエチレンソルビタンモノオレエートなどの界面活性剤が0.3〜3%である。油:アルファトコフェロールの比は、より安定的なエマルジョンがもたらされるため、1以下であり得る。SPAN 85(TM)もまた、約1%のレベルで存在し得る。一部の場合には、本明細書で開示されるエンベロープウイルスベースのVLPワクチンが、安定化剤をさらに含有することが有利であり得る。
【0097】
水中油エマルジョンを生成させる方法は、当業者に周知である。一般に、該方法は、油相を、PBS/TWEEN80(登録商標)溶液などの界面活性剤と混合し、その後、ホモジナイザーを用いるホモジナイゼーションを包含するステップを含むが、該混合物に、シリンジ針内を2回流過させるステップを含む方法が、小容量の液体をホモジナイズするのには適切であることが、当業者には明らかであろう。同様に、当業者は、microfluidizer(M110S型マイクロ流体生成器、最大入力圧6バールで2分間にわたり、最大50回の流過(約850バールの出力圧))内で乳化過程を適合させれば、より低量または高量のエマルジョンを生成させることができるであろう。この適合は、必要とされる直径の油滴を有する調製物が達成されるまで、結果として得られるエマルジョンの測定を含む日常的な実験により達成し得るであろう。
【0098】
本明細書で開示されるエンベロープウイルスベースのVLPによるワクチン調製物は、前記ワクチンを、鼻腔内投与、筋肉内投与、腹腔内投与、皮内投与、経皮投与、静脈内投与、または皮下投与を介して投与することにより、ウイルス性インフルエンザを受けやすいか、またはこれを患う、哺乳動物またはトリを防御または治療するのに用いることができる。ワクチン調製物を全身投与する方法には、従来のシリンジおよび注射針、または固体ワクチンを弾道送達するためにデザインされたデバイス(WO99/27961)、または注射針なしの高圧液体ジェットデバイス(米国特許第4,596,556号;米国特許第5,993,412号)、または経皮パッチ(WO97/48440;WO98/28037)が包含され得る。エンベロープウイルスベースのVLPワクチンはまた、皮膚に貼付することもできる(経皮(transdermalまたはtranscutaneous)送達、WO98/20734;WO98/28037)。したがって、本明細書で開示されるエンベロープウイルスベースのVLPワクチンは、エンベロープウイルスベースのVLPワクチンまたはアジュバントによる組成物を予め充填した全身投与用の送達デバイスを包含し得る。したがって、哺乳動物またはトリなどの個体において免疫反応を誘導する方法であって、本明細書で説明されるエンベロープウイルスベースのVLP組成物のうちのいずれかを含み、また、場合によって、アジュバントおよび/または担体を包含するワクチンを該個体に投与するステップを含み、該ワクチンが、非経口経路または全身経路により投与される方法が提供される。
【0099】
本明細書で説明されるエンベロープウイルスベースのVLPワクチン調製物は、前記ワクチンを、経口/経消化管経路、または経鼻経路などの粘膜経路を介して投与することにより、ウイルス性インフルエンザを受けやすいか、またはこれを患う、哺乳動物またはトリを防御または治療するのに用い得る。代替的な粘膜経路は、膣内経路および直腸内経路である。例示的な粘膜投与経路は、経鼻経路を介し、鼻腔内ワクチン接種と称することができる。免疫化される個体の鼻咽頭内に、ワクチンの液滴形態、スプレー形態、または乾燥粉末形態を投与することを包含する、鼻腔内ワクチン接種の方法は、当技術分野において周知である。噴霧化またはエアゾール化されたワクチン製剤が、本明細書で開示されるエンベロープウイルスベースのVLPワクチンの例示的な形態である。経口投与用の胃液耐性のカプセルおよび顆粒などの腸溶製剤、直腸内投与または膣内投与用の坐剤もまた、本明細書で開示されるエンベロープウイルスベースのVLPワクチンの製剤である。
【0100】
本明細書で開示される、例示的なエンベロープウイルスベースのVLPによるワクチン組成物は、ヒトへの適用に適する経粘膜ワクチンのクラスを表わし、経粘膜ワクチン接種により全身ワクチン接種を代替する。
【0101】
エンベロープウイルスベースのVLPワクチンはまた、経口経路を介して投与することもできる。このような場合、薬学的に許容される賦形剤にはまた、アルカリ性バッファー、または腸溶性のカプセルもしくはマイクロ顆粒も包含され得る。エンベロープウイルスベースのVLPワクチンはまた、膣内経路によっても投与することができる。このような場合、薬学的に許容される賦形剤には、乳化剤、CARBOPOL(登録商標)などのポリマー、また、膣内クリームおよび膣内坐剤である他の公知の安定化剤も包含され得る。エンベロープウイルスベースのVLPワクチンはまた、直腸内経路によっても投与することができる。このような場合、賦形剤には、直腸内坐剤を形成するために当技術分野で公知のワックスおよびポリマーも包含され得る。
【0102】
代替的に、エンベロープウイルスベースのVLPによるワクチン製剤は、キトサン(上記で説明した)または他のポリカチオンポリマー、ポリラクチド粒子およびポリラクチド−コグリコリド粒子、ポリ−N−アセチルグルコサミンベースのポリマーマトリックス、多糖または化学修飾した多糖からなる粒子、リポソームおよび脂質ベースの粒子、グリセロールモノエステルからなる粒子などからなるワクチン媒体と組み合わせることができる。サポニンはまた、コレステロールの存在下において調合し、リポソームまたはISCOMなどの粒子構造を形成することもできる。さらに、サポニンは、非粒子状の溶液もしくは懸濁液中、または小ラメラリポソームもしくはISCOMなどの粒子状構造内において、ポリオキシエチレンエーテルまたはポリオキシエチレンエステルと併せて調合することができる。
【0103】
本明細書で説明されるエンベロープウイルスベースのVLPを用いる医薬組成物およびワクチン組成物中で用いられる、さらなる例示的なアジュバントには、SAF(Chiron、Calif.、United States)、MF−59(Chiron、例えば、Granoffら(1997年)Infect Immun.65巻(5号):1710〜1715頁を参照されたい)、SBASシリーズのアジュバント(例えば、SB−AS2(SmithKline Beechamのアジュバントシステム番号2;MPLおよびQS21を含有する水中油エマルジョン);SBAS−4(SmithKline Beechamのアジュバントシステム番号4;ミョウバンおよびMPLを含有する);SmithKline Beecham、Rixensart、Belgiumから市販されている)、Detox(Enhanzyn(登録商標))(GlaxoSmithKline)、RC−512、RC−522、RC−527、RC−529、RC−544、およびRC−560(GlaxoSmithKline)、ならびに、その開示全体が参照により本明細書に組み込まれる係属中の米国特許継続出願第08/853,826号、および同第09/074,720号において説明されるものなど、他のアミノアルキルグルコサミニド4−リン酸(AGP)が包含される。
【0104】
アジュバントの他の例には、HunterによるTiterMax(登録商標)アジュバント(CytRx Corp.、Norcross、Ga.);Gerbuアジュバント(Gerbu Biotechnik GmbH、Gaiberg、Germany);ニトロセルロース(NilssonおよびLarsson(1992年)Res. Immunol.143巻:553〜557頁);Seppic ISAシリーズのモンタニドアジュバント(例えば、ISA−51、ISA−57、ISA−720、ISA−151など;Seppic、Paris、France)など、鉱油エマルジョン、非鉱油エマルジョン、油中水エマルジョン、または水中油エマルジョンを包含する、ミョウバン(例えば、水酸化アルミニウム、リン酸アルミニウム)エマルジョンベースの製剤;およびPROVAX(登録商標)(IDEC Pharmaceuticals);OM−174(脂質Aと関連するグルコサミンジサッカリド);Leishmaniaの伸長因子;CRL 1005など、ミセルを形成する非イオン性のブロックコポリマー;ならびにSyntexアジュバント製剤が包含されるがこれらに限定されない。例えば、O’Haganら(2001年)Biomol Eng.18巻(3号):69〜85頁;および「Vaccine Adjuvants: Preparation Methods and Research Protocols」D. O’Hagan編(2000年)、Humana Pressを参照されたい。
【0105】
他のアジュバントには、一般式
HO(CH2CH2O)n−A−R (I)
[式中、nは1〜50であり、Aは結合または−C(O)−であり、RはC1〜50アルキル、またはフェニルC1〜50アルキルである]のアジュバント分子が包含される。
【0106】
本明細書で説明されるエンベロープウイルスベースのVLPワクチン製剤の一実施形態は、一般式(I)[式中、nは1〜50、4〜24、または9であり;R成分はC1〜50、C4〜C20アルキル、またはC12アルキルであり;また、Aは結合である]のポリオキシエチレンエーテルを包含する。ポリオキシエチレンエーテルの濃度は、0.1〜20%、0.1〜10%、または0.1〜1%の範囲であるものとする。例示的なポリオキシエチレンエーテルは、以下の群:ポリオキシエチレン−9−ラウリルエーテル、ポリオキシエチレン−9−ステアリルエーテル(polyoxyethylene−9−steoryl ether)、ポリオキシエチレン−8−ステアリルエーテル、ポリオキシエチレン−4−ラウリルエーテル、ポリオキシエチレン−35−ラウリルエーテル、およびポリオキシエチレン−23−ラウリルエーテルから選択される。ポリオキシエチレンラウリルエーテルなどのポリオキシエチレンエーテルは、メルクインデックス(第12版;登録番号7717)で説明されている。これらのアジュバント分子は、WO99/52549で説明されている。
【0107】
所望の場合、上記の一般式(I)に従うポリオキシエチレンエーテルを、別のアジュバントと組み合わせることができる。例えば、アジュバントの組合せは、上記で説明したCpGを包含し得る。
【0108】
本明細書で開示されるエンベロープウイルスベースのVLPワクチンと共に用いるのに適する、薬学的に許容される賦形剤のさらなる例には、水、リン酸緩衝生理食塩液、等張性緩衝液が包含される。
さらなるインフルエンザ抗原
本明細書で開示される安定化させたVLPは、インフルエンザウイルスの特定の株に対して、かつ/またはインフルエンザウイルスの複数の株にわたり免疫原性を増大させるために、インフルエンザウイルスに由来するさらなる抗原を包含しうる。
【0109】
例示的なさらなるインフルエンザ抗原は、M2ポリペプチド(インフルエンザBにおいてBM2とも呼ばれる)である。インフルエンザウイルスのM2ポリペプチドは、スプライシングイベント後におけるRNAのセグメント7(マトリックスセグメント)によりコードされる、97アミノ酸の低分子III型内在性膜タンパク質である(80、81)。ウイルス粒子上に存在するM2は極めてわずかであるが、感染細胞上においてより豊富に見出すことができる。M2は、ウイルスの侵入に必要な、プロトン選択性イオンチャネルとして用いられる(82、83)。感染時または従来のワクチン接種時における免疫原性は最小限であることから、その保存が説明されるが、代替的なフォーマットで存在する場合は、より免疫原性であり防御性である(84〜86)。これは、in vivoにおけるM2モノクローナル抗体の受動伝達により、ウイルスクリアランスが加速化され、結果として防御がもたらされるという観察(87)と符合する。M2の外部ドメインエピトープを、融合タンパク質として、HBVのコア粒子に連結すると、非経口接種および鼻腔内接種のいずれによっても、マウスにおいて防御性であり、3回のタンデムコピーを該コアタンパク質のN末端に融合させると、最も免疫原性となる(88〜90)。これは、エピトープ密度が増大すると免疫原性が増大することを示す他のキャリア−ハプテンデータ(91)と符合する。
【0110】
M2ワクチンを鼻腔内送達する場合、良好な防御を達成するにはアジュバントが必要とされる場合があり、LTR192G(88、90)およびCTA1−DD(89)により良好な結果が達成されている。ペプチドはまた、KLH、またはN. meningitidesの外膜タンパク質複合体、またはヒトパピローマウイルスのVLPなどのキャリアに化学的にコンジュゲートすることもでき、マウスおよび他の動物におけるワクチンとして防御性である(92、93)。
【0111】
M2タンパク質は、高度に保存性ではあるが、配列の相違をまったく示さないわけではない。一般的な株であるA/PR/8/34株(H1N1)およびA/Aichi/68株(H3N2)のM2外部ドメインエピトープは、A/Hong Kong/156/97株(H5N1)を除き、今日配列決定されている他のすべてのヒト株と、免疫学的に交差反応性であることが示された(92)。インフルエンザデータベースの配列の検討からも、A/Vietnam/1203/04株など、他のより近年の病原性H5N1ヒト単離物のM2配列内において、同様の相違が示されている。この知見は、M2エピトープを組み込む、有効なH5特異的な流行性ワクチンが、ヒトH1単離物およびH3単離物中において現在行きわたっているM2配列ではなく、病原性のトリ株に固有のM2配列を反映する必要があることを裏付ける。
【0112】
インフルエンザウイルスに由来するさらなるタンパク質(HA、NA、およびM2以外のタンパク質)も、共発現により、またはさらなる抗原の全部もしくは一部を、gagもしくはHAポリペプチドに連結することにより、VLPワクチン内に組み込むことができる。これらのさらなる抗原には、PB2、PB1、PA、核タンパク質、マトリックス(M1)、BM2、NS、NS1、およびNS2が包含される。A型インフルエンザウイルスの場合、例には、PB2タンパク質、PB1タンパク質、PAタンパク質、核タンパク質、マトリックス(M1)タンパク質、M2タンパク質、NS1タンパク質、およびNS2タンパク質が含まれる。B型インフルエンザウイルスの場合、例には、HAタンパク質、NAタンパク質、NPタンパク質、Mタンパク質、PB1タンパク質、PB2タンパク質、PAタンパク質、NSタンパク質、およびBM2タンパク質が含まれる。これら後者の抗原は、一般に、抗体反応を中和するための標的ではなく、T細胞により認識される重要なエピトープを含有し得る。このようなエピトープに対してVLPワクチンにより誘導されるT細胞反応が、防御的免疫性を促進するのに有益であることが判明し得る。
【実施例】
【0113】
(実施例1)
インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子の生成
MLV gagタンパク質のコード配列は、モロニーマウス白血病ウイルスの両種性プロウイルス配列全体を含有するプラスミドpAMS(ATCC)から、PCRにより得た。該gagタンパク質のコード配列を、pFastBac1(Invitrogen)のポリへドロンプロモーターの後に挿入し、結果として得られるプラスミドでDH10Bacコンピテント細胞を形質転換し、バキュロウイルスゲノムに組み換えた。次いで、高分子量のバクミドDNAを精製し、Sf9細胞にトランスフェクトし、gagタンパク質を発現する組換えバキュロウイルスを生成させた。ウイルスRNAに由来するHAタンパク質のコード配列およびNAタンパク質のコード配列をRT−PCRによりクローニングした後で、A/PR/8/34株(H1N1亜型)の赤血球凝集素タンパク質およびノイラミニダーゼタンパク質のそれぞれをコードする他の2つの組換えバキュロウイルスも、同様の形で生成させた。最後に、個別のpFastBac1プラスミドに由来する、HAタンパク質発現ユニット、gagタンパク質発現ユニット、およびNAタンパク質発現ユニット(ポリへドロンプロモーター−コード配列−ポリA部位)を、単一のpFastBac1ベクターに統合することにより、3つの産物すべて(HA−gag−NA)をコードする単一のバキュロウイルスベクターを生成させた。当初の解析のために、gagタンパク質またはHAタンパク質またはgag−HA−NAタンパク質をコードする組換えバキュロウイルスを、MOIを>1とする6ウェルプレート内のSf9細胞に感染させた。感染の3日後、培地上清から破砕物を除去し、次いで、20%のスクロースクッションを介する100,000×gでこれをペレット化させた。gagタンパク質特異的抗血清およびH1N1亜型特異的抗血清を用いるウェスタンブロット解析によりペレットを解析した(図1AおよびBを参照されたい)。
【0114】
図1AおよびBの各ブロットにおける左側の3つのレーンは、それぞれ、培地を収集する前の、Sf9細胞に、個別のgag遺伝子によるバキュロウイルスまたはHA遺伝子によるバキュロウイルスまたは対照(EV=空ベクター)のバキュロウイルスを感染させた結果を示す。予測される通り、gag遺伝子だけによるバキュロウイルスを感染させると、VLPの出芽により、高分子量の培地画分中に著明量のgag抗原が結果としてもたらされる(図1A;レーン「Gag」)。これに対し、HA遺伝子だけによるバキュロウイルスを感染させると、それ自体で培地中に放出される少量のHAタンパク質が結果としてもたらされる(図1B;レーン「HA」)。しかし、HA−gag−NA三重遺伝子ベクターをSf9細胞に感染させると、gagタンパク質およびHAタンパク質の両方が著明量で100,000×g画分中に結果として現れる(レーン1〜9;図1AおよびB)ことから、gagタンパク質の発現が、HAタンパク質を細胞から引き出しうることが示される。
【0115】
図2AおよびBは、ペレット化させたHA−gag−NA VLPを、20〜60%の段階的なスクロース勾配上で再遠心分離した後、個々の勾配画分に対してウェスタンブロット解析を行った結果を示す。gagタンパク質およびHAタンパク質の両方が、同じ画分中でピークに達することから、密度約1.16g/mlにおけるバンド形成の符合が裏付けられ、これにより、gagタンパク質およびHAタンパク質がVLP中に存在したことが示唆される。
【0116】
(実施例2)
VLPの安定性についての生物物理的特徴づけ:pHおよび温度
濃度は、モル濃度または容量当たりの重量百分率で報告する。6−ドデカノイル−2−ジメチルアミノナフタレン(ラウルダン)および8−アニリノ−1−ナフタレンスルホネート(ANS)は、Molecular Probes(Eugene、OR)から購入した。ジメチルスルホキシド(DMSO、Fisher Chemical)中に溶解させることにより、1.2mMのラウルダン原液および10mMのANS溶液を調製した。
【0117】
特徴づけされるVLPの調製
実施例1で示した通り、「三重遺伝子」による組換えバキュロウイルスを感染させた培養Sf9細胞内でVLPを生成させた。4〜8の各単位pHであるクエン酸/リン酸(CP)バッファーに透析することにより、特徴づけされるVLPを調製した。NaClを用いて、バッファーのイオン強度を0.1に維持した。透析カセット(10,000MWCO;Pierce、Rockford、IL)から回収した材料を、Amicon(登録商標)Ultra超遠心分離器(10,000MWCO;Millipore、Billerica、MA)により、4℃、3,150×gで濃縮した。残余物のタンパク質濃度は、BCA(ビシンコニン酸)比色法(Pierce、Rockford、IL)により推定した。別段に言及しない限り、残余物を適切なpHの20mM CPバッファーで希釈することにより、最終タンパク質濃度が90μg/mLの三連試料を調製した。
【0118】
表面赤血球凝集素タンパク質に対するトリプシン処理
トリプシン(Sigma;5μg/mLの最終濃度)をVLP原液(pH7.4の30%のスクロース/トリス緩衝生理食塩液中0.26mg/mLの全タンパク質)に添加し、2〜8℃の低温室で5分間にわたりインキュベートした。インキュベーション後、ダイズ(Fluka)に由来する、モル濃度を3倍としたトリプシン阻害剤を添加し、結果として得られる溶液に、0.45μmのシリンジフィルター(Millipore)を流過させた。次いで、上記で説明した通りに、試料を適切なCPバッファーに透析し、濃縮したが、100,000MWCOの透析チューブ(Spectrum Laboratories、Rancho Dominguez、CA)を用いた。HAタンパク質の切断、ならびにMLV gagタンパク質の非切断は、ウェスタンブロット解析により確認した。
【0119】
動的光散乱
動的光散乱(DLS)を用いて、温度上昇の関数としての、VLPの平均有効直径の変化を測定した。Brookhaven Instrument Corporation(Holtzville、NY)のシステムにより測定を行った。125mWのダイオード励起レーザーにより、532nmの入射光を生成させた。入射光に対して90°で散乱光を集光し、デジタル式自己相関計(BI−9000AT)を用いて、自己相関関数を創出した。10〜85℃の範囲にわたり、2.5℃おきに5回の測定を行った。キュムラント解析を用いて、相関関数から粒子の拡散係数を抽出し、ストークス−アインシュタインの式により、これらを粒子直径に変換した。この方法により計算される有効直径が正確なのは、直径が<1μmの粒子についてである(これより大きな粒子についての測定から得られる値は、定性的な比較だけに用いるべきである)ことに留意されたい。粒子サイズに加え、粒子拡散係数の分布についての第2キュムラントもまた、試料の多分散性の尺度としての相関関数から抽出した。
【0120】
実施例1のプロトコールを用いてなされた、VLP懸濁液についてのDLS測定値は、pHにより誘導される粒子サイズの変化および温度により誘導される粒子サイズの変化の両方についての証拠を示した(図3(A))。低温では、pH4または5における粒子サイズが、pH6〜8における粒子サイズの2〜3倍であったことから、酸性のpHにより、著明な凝集および/または膨張が誘導されたことが示唆される。pH4における試料は、約75℃まで、温度により誘導される粒子サイズの変化を示さないが、その後、有効直径の漸増が観察された。これに対し、pH5では、約50℃で粒子サイズが急激に増大したが、約75〜80℃でもさらなる増大が生じた可能性がある。6〜8の各pHの試料は、約58℃までの温度上昇に対して安定であったが、それより高温では、pH6および7の試料が、粒子サイズの顕著な増大を示した。また、pH8の試料も、約60℃でサイズ増大の証拠を示した。後者の場合におけるサイズの増大は、比較的小さく、VLPの凝集ではなく、VLPの膨張によった可能性がある。一般に、酸性度が増大するのに伴い、VLPの多分散性の増大が見られた(図3(C))。pH4および5の試料の多分散性が、温度範囲全体でほぼ一定を維持したのに対し、pH6以上の試料は、約60℃で多分散性の増大を示し、サイズデータで見られる変化と符合する。
【0121】
DLSの測定時にはまた、散乱光の強度も記録した(図3(B))。測定器の設定を各試料に応じて最適化させるために、標準化値を報告することから、試料間における有意義な直接的比較は不可能となる。一般に、粒子サイズまたは粒子の屈折率(溶媒と比べた)が増大すると、散乱光の強度が結果として増大する。しかし、粒子密度の減少(例えば、膨張による)の結果として屈折率が低下するならば、それは、散乱強度の低下として顕在化するであろう。これが、散乱光強度の滑らかで漸進的な低下が温度勾配の大半にわたって見られる、pH8の試料についてのデータに対する最良の説明である。60℃付近で曲線状の低下がわずかに乱れているが、これは、これらの試料について見られる有効直径の増大に対応する(図3(A))。また、pH5の試料による散乱光のプロットも、散乱光強度の急激な低下により顕在化する、60℃付近で生じる構造的変化の証拠を示す。pH6および7に対応するプロットは類似し、散乱光の低下が高温(約75℃)で生じる。これらの低下は、沈降する材料が入射光ビームの外で沈殿するために生じる可能性があり、上記で示したサイズデータについての解釈と符合する。pH4の加熱曲線は、強度の漸減だけを示し、粒子直径の測定された変化と相関する急激な変化は示さなかった(例外は、30〜35℃における散乱強度の一過性の低下であるが、以下のラウルダン蛍光についての結果を参照されたい)。
【0122】
円偏光二色性分光光度法
円偏光二色性分光光度法(CD)の測定は、100mdegの感度設定、2秒間の反応時間、および1nmのバンド幅を用いる、Jasco J−810分光光度計により行った。走査速度を20nm/秒、データピッチを0.5nm/秒として、VLPの複合スペクトル(3つ〜5つのスペクトルの集積)を得た。227nmにおけるCDシグナルをモニタリングする温度可変実験を実施して、全VLPタンパク質の二次構造の変化を、温度の関数として検出した。温度勾配を15℃/時間、遅延時間を2秒間として、10〜90℃の範囲にわたり、0.5℃おきに測定を行った。Origin(登録商標)データ解析ソフトウェアを用いて、温度に依存するデータをシグモイド関数に数学的に近似することにより、中点転移温度(Tm)値を決定した。スペクトルおよび加熱曲線では、3つの異なるタンパク質のうちで最も量の多いタンパク質からの寄与が優越すると推測されるが、スペクトルおよび加熱曲線のいずれもが、これらのタンパク質からの相加的寄与を反映する。これらのVLPでは、MLV gagタンパク質の量が、HAタンパク質の量より3〜4倍多いことが示されたが、HAタンパク質の量は、NAタンパク質の量をおそらく1桁上回る。
【0123】
pH4〜8では、インフルエンザVLPのCDスペクトルが、210nmおよび227nm付近で極小点を示したことは、対象のpH範囲にわたり著明な螺旋特徴が存在することを示唆する(図4(A))。高温ではシグナルが見られなくなる(図4(B))ことから、二次構造が温度に依存して見られなくなることが示唆された。この効果をさらに探索するため、227nmにおけるシグナルを、温度の関数としてモニタリングした(図5)。CDシグナルの急激な変化が観察されたことは、温度に依存するタンパク質構造の転移と符合した。pH6の試料は、55℃付近で最高のTmを示す。試料の酸性度を上昇させると、38℃(pH4)および47℃(pH5)という顕著に低いTm値がもたらされるのに対し、pH7または8の試料は、Tm値が、それぞれ、53℃および51℃で酷似している。溶融曲線の形状は、複数の成分を示唆し、系の異種性を反映すると推測される。
【0124】
蛍光分光光度法
別段に言及しない限り、Photon Technology Internationalによる蛍光測定器(Birmingham、NJ)を用いて、10〜85℃の範囲にわたり、2.5℃おきに蛍光発光スペクトルを収集した。15℃/時間の勾配で温度を上昇させ、すべての測定について、1nmのステップサイズおよび1秒間の積分時間を用いた。蛍光実験ではまた、第2の検出器(蛍光検出器から180°に配向させた)を用いることにより、静的光散乱もモニタリングした。Origin(登録商標)ソフトウェアパッケージを用いて、微分解析により発光ピークの位置を決定し、温度に依存するデータを、シグモイド関数に数学的に近似することにより、Tm値を決定した。
【0125】
芳香族アミノ酸であるトリプトファンおよびチロシンの内部蛍光を使用して、VLPタンパク質の三次構造の変化を、温度の関数として同定した。280nmで励起させた蛍光発光スペクトルを、300〜380nmで収集した。励起および発光のスリット幅は、それぞれ、3nmおよび4nmに設定した。
【0126】
VLP存在下における8−アニリノ−1−ナフタレンスルホネート(ANS)の蛍光発光を、VLPタンパク質の三次構造の安定性をモニタリングする代替的な方法として使用した。タンパク質の非極性領域に対してアフィニティーを有することが公知の低分子であるANSは、溶液中で示す蛍光は弱いが、結合すると、発光強度を増大させ、発光をシフト(通常は青色に)させる(114)。70μMのANSと共に調製したVLP試料を385nmで励起させた後で、その蛍光発光スペクトルを425〜550nmで収集した。ANS蛍光を測定するときの励起および発光のスリット幅はいずれも、4nmに設定した。
【0127】
熱により誘導されるVLP膜の流動性の変化を直接的にモニタリングするには、別の分子プローブである6−ドデカノイル−2−ジメチルアミノナフタレン(ラウルダン)を用いた。ラウルダンの化学構造は、誘導体化されたナフタレンに結合した長いアシル鎖を含有し、これにより、脂質二重層への容易な組込みが可能となる。膜における水和の増大は、二重層の流動性を、ゲル(流動性が低い)から液晶(流動性が高い)に転移させる可能性がある。340nmで励起させる場合、膜の水分含量が増大すると、ラウルダンの発光が約440nmから490nm付近にシフトする。有用なパラメータは、(115)において、GP=(I440−I480)/(I440+I480)[式中、Ix=波長xにおける強度]として定義されている、一般化偏光(GP)である。これにより、GP値が低下すると、膜における流動性の増大が示唆され、この逆も成り立つ。ラウルダンによる実験のスリット幅は、2nm(励起時)および5nm(発光時)に設定した。
【0128】
1.内部蛍光
すべての試料について、内部蛍光発光のピーク位置を、温度の関数として決定した(図6)。各場合において、約10〜40℃の温度範囲にわたり、ピーク位置がやや短波長となった後に、長波長への急激な遷移が生じた。タンパク質試料では、蛍光性アミノ酸側鎖を極性の大きな環境に曝露すると、ピーク極大点が熱に誘導されて赤方にシフトすることが観察された。これは、通常は少なくとも部分的に非極性のタンパク質コア内に包埋されているアミノ酸のフルオロフォアが水性溶媒に曝露される、アンフォールディングイベントと符合した。55〜65℃の温度では(pHに依存して)ピーク極大点が短波長に戻るが、これは、高温で観察されるVLPの凝集と符合する。330nmにおける(標準化された)発光強度も、温度の関数としてプロットした(図6)。構造転移の非存在下では、このようなプロットが、内部の温度効果に起因する温度の上昇と共に、発光強度の滑らかで曲線状の低下を呈示することが典型的である。45〜65℃の範囲にあるすべての試料について、曲線状のプロファイルからの逸脱が生じることから、加熱すると内部フルオロフォア環境が変化することが確認された。
【0129】
2.ANS蛍光
VLPの存在下におけるピーク位置(≧470nm)およびANS蛍光発光強度の増大から、低温では、これらの高分子複合体の非極性領域にANSが結合したことが示唆されるが、これは、脂質二重層の存在を踏まえれば予測されることであった。ANS発光のピーク位置を温度の関数としてプロットする(図7)と、すべての試料において、温度に依存する短波長へのシフトの後、場合によっては(すなわち、pH5、6、および7では)長波長へのシフトが生じることが示される。すべての試料において、ピーク位置の変化の度合い(2〜3nm)は、内部蛍光により観察される変化の度合いよりはるかに小さい。高温では、ピーク位置のばらつきおよびノイズが大きくなるために、これらのデータから実質的な結論を引き出すことは困難となる。485nmにおけるANSの相対強度(図7)は、温度により誘導されて非極性モチーフが曝露されることについてのより明確な証拠を顕示した。これは特に、予測される、蛍光に対する熱による非特異的な消光作用に対応する、発光の曲線状の低下の上に重ね合わせて、発光強度のわずかな増大(pH5では38℃付近で始まり、pH6〜8では43℃付近で始まる)が見られる、pH5〜8の試料についてのプロットに当てはまった。
【0130】
3.ラウルダン蛍光
一般化偏光(GP)を温度の関数としてプロットする(図8)ことにより、加熱すると、VLPの膜における水和(流動性)が漸増することが示唆される。VLP懸濁液のpHが、膜における水和の速度および程度に対して影響を及ぼしたことは、驚くべきことではない。低温では、すべての試料について、膜における水和の程度(すなわち、GP値)が酷似した。40℃を超えると、pH4の試料が示す膜における水和の変化が一貫してわずかであり、一般に、低いpHで調製された試料は、温度を上昇させても二重層内に水分子を取り込むのが遅かった(酸性度の高い試料では、GP=0となる温度が上昇した)。pH4、7、または8で調製した試料のGP値が、温度と共にシグモイド様に変化したのに対し、pH5および6の試料は、検討した温度範囲にわたり、GPの準直線的な低下を示した。pH5および6の試料では、膜における水和の程度が高温(すなわち、75℃を超える)で最大となった。また、ラウルダン蛍光による実験でも、340nmにおける静的光散乱をモニタリングした。これらのデータは、DLS実験で測定された静的光散乱と同様であったが、このさらなる実験では、pH4の試料について観察された散乱光強度の一過性の低下(図3(B)、30〜35℃)が観察されなかったことから、これはアーチファクトであった可能性が示唆される。
【0131】
実験相図
VLPの物理的な安定性についての包括的な視覚的表示を創出するために、上記で論じた各種の生物物理的方法に由来するデータを、多次元ベクトル空間の基底系に変換した。測定を行った温度およびpH(10〜85℃にわたる2.5℃おきの温度、4〜8の単位pH値)のすべての組合せについてn次元ベクトルを構築し、各ベクトルの成分を、適用したn種類の技法の各々による標準化された測定値とした。次いで、基底系におけるすべてのベクトルの射影演算子を足し合わせて、n個の固有ベクトルを伴うn×n密度行列を得た。次いで、データセットに対して最大の寄与(すなわち、最大の固有値)をもたらす3つの固有ベクトルを用いて、元のn次元ベクトル系を三次元に変換した。最後に、新たな三次元ベクトル各々の3つの成分に、3つの異なる色(赤色、緑色、青色)を割り当て、各個別のベクターに固有の色の組合せを得た。次いで、この方法により、n種類の測定を行った温度およびpHのすべての組合せに着色マーカーを割り当てることが可能となり、全データセットを温度およびpHの関数とする三色マップが得られた。このような相図の有用性は、各技法により検出される粒子構造の最も際立った変化を、VLP構造の見かけの「相」の差違と同時に視覚化しうることである。EPDの作成についてのより詳細な説明は、他所(116、117)において示されている。
【0132】
前出の節において示した、温度に依存するデータから、実験相図(図9)を作成した。実験空間にわたり、約10の異なる相を見ることができ、最大の相(pH6〜8、低温、青色)は、VLPの構造的破壊が最小となる状態に対応する。この相の上方に、pH6〜7では35〜55℃にわたり、かつ、pH8では35〜50℃にわたり転移領域が現れた(紫色)。pH6および7では60℃を超える色の変化する領域は、粒子の凝集に対応した。pH8では、著明な凝集がみられないことから、高温で、pH6または7で見られた相とは異なる相(暗赤色)がもたらされた。pH4および5では、低温で2つの異なる相が見られ、いずれも著明な構造の破壊を表わした(明青色)。pHが低い領域の35℃超では、温度に誘導されるさらなる立体構造変化により、複数の相がもたらされた(緑色/橙色)。pH5とpH6との間、および40℃超における見かけの相境界は、安定性が中程度である状態を表し、したがって、以下の実施例3の賦形剤スクリーニングアッセイを開発するための出発点を示す。
【0133】
結論
VLPの生物物理的特徴づけを組み合わせた結果に基づくと、凝集および溶解性に対する変化が、エンベロープウイルスベースのVLPの分解における重要な因子となる。VLPは、pH7〜pH8において最も安定的であった。安定性の理由および化学的な理由で、好ましい範囲は、約pH6.5〜約pH7.5である。この範囲で用いうる代表的なバッファーは、リン酸バッファー、トリスバッファー、MESバッファー、およびクエン酸バッファーである。
【0134】
(実施例3)
VLP賦形剤のスクリーニング
別段に言及しない限り、すべての潜在的な安定化剤は、Sigma−Aldrich(St.Louis、MO)から購入した。グアニジンHCl、塩化カルシウム二水和物、デキストロース、D−マンニトール、クエン酸、および二塩基性リン酸ナトリウムは、Fisher Chemical(Fair Lawn、NJ)から購入した。A型ブタゼラチンは、Dynagel(Calumet City、IL)から購入し、D−スクロースおよびD−トレハロースは、Ferro−Pfanstiehl Laboratories,Inc.(Waukegan、IL)から購入した。エクトイン(超純粋)は、Bitop AG(Witten、Germany)から購入し、NV10は、Expedeon(旧Novexin;Cambridge、UK)から購入した。適切なpHの20mMクエン酸/リン酸(CP)バッファーに溶解させることにより、濃縮賦形剤溶液を調製した。次いで、濃縮したNaOHまたはHClを用いて、pHを、標的pHに調整した(必要な場合)。0.22μmのDurapore(登録商標)(PVDF)膜によるシリンジフィルター(Millipore、Billerica、MA)により、最終原液を濾過した。
【0135】
賦形剤のスクリーニング
濁度(350nmにおける光学濃度;OD350)を時間の関数として測定することにより、pH6および60℃におけるVLPの凝集をモニタリングした。各種のGRAS(一般に安全と認識される)薬剤の存在下または非存在下において、濃縮した残余物(上記を参照されたい)を、20mMのCPバッファーおよび/または適切なpHの濃縮賦形剤溶液で希釈することにより、55μg/mLのタンパク質濃度で、二連のVLP試料を調製した。温度制御型Agilent 8453分光光度計(Palo Alto、CA)を用いて、2時間にわたり、30秒間ごとに測定を行った。
【0136】
溶液中のVLPに対する潜在的な安定化剤について、GRAS化合物のライブラリーをスクリーニングした。実施例2の特徴づけ研究から作成した実験相図(図9)を使用して、VLPの凝集(最も明らかな物理的分解過程)を防止する賦形剤を同定するためのスクリーニングアッセイを開発した。スクリーニングアッセイの温度およびpHについての初期条件の選択は、該相図(前出の実施例を参照されたい)を指針としたが、最終条件は、潜在的な安定化剤間のわずかな差違を増大させるように最適化した。対照試料の挙動に応じて、t=15分またはt=30分(凝集が最大となる時点を表わすいずれかの時点)において、凝集の阻止百分率(表1)を計算した。洗浄剤、ポリオール、アミノ酸、糖、および糖アルコールを含めた各種の分子クラスにおいて、最も有望な凝集阻止化合物が見出された。
【0137】
個々の安定化剤の効果
複数の分子クラス(すなわち、トレハロース、グリセロール、ソルビトール、リシン、およびジエタノールアミン(脂質二重層を破壊する洗浄剤の傾向を踏まえれば、凝集阻止剤としてのTween 20およびBrij 35の見かけの成功は、アーチファクトでありうる))に由来する、効能が最上位にある凝集阻止剤を用いて、潜在的な賦形剤の存在下におけるVLPのCDおよび蛍光の測定を実施した。実際のワクチン製剤をより緊密に近似するために、これらの実験では、溶液のpHを7に設定した。
【0138】
上記で説明した通り、温度可変CDの測定を使用して、選択された化合物のうちのいずれかが、VLPタンパク質(例示しない)の二次構造を安定化させるかどうかを決定した。調べたすべての化合物のうちで、ソルビトールだけが正の効果を示した;ソルビトールを含有する試料のTm(55℃)は、対照のTm(54℃)と比べてわずかに高かったが、他の賦形剤は、50℃(トレハロース)〜53℃(リシン)の範囲のTm値を示した。
【0139】
内部蛍光法を使用して、VLPタンパク質の三次構造に対する潜在的安定化剤の効果を測定した。発光のピーク位置を温度と対比させてプロットする(図10)と、調べた製剤の大半について、40℃またはその付近で始まる、約329nmから336nmへの遷移が示される。例外は、リシンを含有する製剤であり、これは、低温では344nm付近でその蛍光ピークを呈示し、43℃で始まる、わずかな長波長への可能な遷移(+1〜2nm)の証拠を示す(これらの測定値における誤差により、このシフトが統計学的に有意であるという結論は退けられる)。対照のTmは、51℃である。ジエタノールアミンは、49℃のTmを誘導するので有効な安定化剤ではないのに対し、グリセロール、トレハロース、およびソルビトールを含有する製剤はすべて、それぞれ、52、53、および54℃の、やや高温のTm値を示す。これらの実験において収集された静的光散乱(データは示さない)は、リシンを含有する製剤を除くすべての製剤間で同様の挙動を示唆した。リシンの存在下では、光散乱の強度が10分の1未満に低下したことから、VLPの構造破壊が示唆される。複数の化合物によるVLP膜の流動性に対する効果を、上昇させる温度の関数として測定するには、ラウルダン蛍光を用いた。これらの実験では、物理的安定性に対する正の効果が弱いかまたは正の効果が見られない賦形剤(すなわち、ジエタノールアミン、グリセロール、およびリシン)を、グリシン、エクトイン、およびNV10で置き換えた。グリシンは、インフルエンザHAタンパク質および/またはインフルエンザNAタンパク質を安定化させうるという私信により導入した。エクトイン(有機浸透圧調節物質)およびNV10(5kDaの直鎖状炭水化物ポリマー)は、それらが、高分子系を安定化させるのに一般に有効であるという報告(118)に基づき、潜在的な新規のVLP膜安定化剤として調べた。温度に依存するGPデータ(図11)を目視すると、調べた化合物のうちの1つまたは2つが、ゲルから液晶への転移を阻止すると考えられる。ここでもまた、データを近似するためにはシグモイド近似を用い、Tm値を抽出して、各安定化剤の効果を定量的に比較した。対照試料(Tm=52℃)と比較すると、ソルビトールまたはエクトインを含有する製剤は、54℃のやや高いTm値を示す。しかし、これらの測定に随伴する誤差の大きさを踏まえるなら、Tmの見かけの上昇は、いずれの場合にもおそらく有意ではない。他方、グリシンおよびトレハロースは、それぞれ、Tm値が59℃および60℃と高く、著明な安定化効果を及ぼしている。NV10は、20℃を超える温度でGP値の低下(膜における水和の増大)を誘導(対照と比べて)して、VLPエンベロープの安定性に対して負の効果を及ぼしている。NV10製剤について計算されたTmは、46℃である。
【0140】
結論
代表的な単糖(デキストロース、マンニトール、ソルビトール)および二糖(ラクトース、トレハロース、スクロース)を含めた、複数の炭水化物について調べた。デキストロースを除くすべての場合において、10%溶液では凝集が増大した。しかし、すべての場合において、20%溶液は、VLP溶液の凝集を阻止するのに有効であった(トレハロースが最も有効で、84%の阻止を示し、ソルビトールおよびラクトースがわずかの差でこれに続いた)。これに対し、オリゴ糖(シクロデキストランなど)は一般に、凝集を低減するのに有効ではなかった。グリセロール(ポリアルコールは、炭水化物と構造的な類似性を共有する)もまた、凝集を低減する(10%で82%の阻止)のに有効であった。
【0141】
4つの非イオン性界面活性剤を、0.01〜0.10パーセントの範囲の濃度で評価した。調べた4つの洗浄剤すべて(Brij 35、Tween 20、Tween 80、およびPluronic F−68)が、凝集を阻止するのに有効であった。調べたすべての洗浄剤が凝集の阻止を示したが、Tween 20が、他の洗浄剤に優ると考えられる。
【0142】
調べた一般に用いられる2つのタンパク質であるアルブミンおよびゼラチンは、凝集を阻止するのには有効でなかったが、この結果は、VLPではなく、アルブミンに関する問題に起因した可能性がある。
【0143】
また、代表的なアミノ酸についても調べた。300mMのジエタノールアミン、アルギニン、およびリシンは、凝集を70%阻止した。グアニジン(30%)、ヒスチジン(30%)、およびグリシン(12%)は、それほど有効でなく、アスパラギン酸は、凝集を加速化させた。しかし、凝集の阻止または加速化が、有機化合物により直接的に引き起こされたのか、またはpHの変化を介して間接的に引き起こされたのかは明確でない。リシンのさらなる特徴づけは、リシンを用いる結果として、VLP構造が破壊されることを明らかにし、したがって、濁度の低下はアーチファクトと考えられ、凝集を阻止する証拠ではないと考えられる。
【0144】
また、代表的な有機酸についても調べた。アスコルビン酸(150mM)が凝集を大幅に増大させたのに対し、同様の濃度の乳酸およびリンゴ酸は、わずかながら凝集を阻止した。ここでもまた、これらの観察が、pHの変化の結果であるのか、または有機酸がVLPと直接的に相互作用した結果であるのかは明確でない。
【0145】
調べた複数の炭水化物は、エンベロープウイルスベースのVLPを安定化させるのに有望である。炭水化物の濃度は重要であり、20%溶液では保護がもたらされるが、10%溶液または15%溶液では保護がもたらされない。しかし、これに対して、10%のグリセロールは、凝集を阻止するのに有効であった。以上に基づくなら、当業者は、安定化量のこれらの安定化剤を容易に選択することができる。
【0146】
最後に、トレハロースおよびソルビトールについての生物物理的研究により、炭水化物が、ウイルス性タンパク質の三次構造を安定化させることが示され、トレハロースはまた、温度により誘導される脂質二重層の水和を遅延させることも示された。
【0147】
したがって、この炭水化物は、高温で凝集を低減し、タンパク質の三次構造を安定化させ、膜における水和を最小化することが示されたので、VLPを安定化させるのに最も有望な候補物質は、20%のトレハロースであった。構造が類似し、凝集の阻止が濃度に依存するため、他の炭水化物も、同様の保護特性をもたらすと推測される。
【0148】
さらなる参考文献
【0149】
【数1】
【0150】
【数2】
【0151】
【数3】
【0152】
【数4】
【0153】
【数5】
【0154】
【数6】
【0155】
【数7】
【0156】
【数8】
【0157】
【数9】
【技術分野】
【0001】
本発明は、インフルエンザ抗原を包含する、エンベロープウイルスベースの安定化させたウイルス様粒子の分野に関する。本明細書では特に、このようなウイルス様粒子を安定化させる方法、およびこのようなウイルス様粒子を含む安定化させた組成物が開示される。
【背景技術】
【0002】
A型インフルエンザウイルスおよびB型インフルエンザウイルスは、流行性ヒト疾患を引き起こすインフルエンザウイルスの2つの型である(111)。A型インフルエンザウイルスは、2つの表面抗原:赤血球凝集素(HA)タンパク質およびノイラミニダーゼ(NA)タンパク質に基づく亜型にさらに類別される。B型インフルエンザウイルスは、亜型に類別されていないが、時間と共にウイルス株が分岐するドリフトを経る。1977年以来、A型(H1N1亜型)インフルエンザウイルス、A型(H3N2亜型)インフルエンザウイルス、およびB型インフルエンザウイルスが、世界中に蔓延している。おそらく、ヒトA型(H3N2亜型)ウイルスと、ヒトA型(H1N1亜型)ウイルスとが遺伝子再結合した後に出現したA型(H1N2亜型)インフルエンザウイルスは、近年多くの国々で検出されている。A型インフルエンザウイルスおよびB型インフルエンザウイルスのいずれもが、抗原特徴に基づき、さらに群分けされている。新型インフルエンザウイルスの変異型は、ウイルスの複製において生じる点突然変異の結果としてもたらされる、頻繁な抗原変化(すなわち、抗原ドリフト)から生じる。B型インフルエンザウイルスは、抗原ドリフトを経るのが、A型インフルエンザウイルスほど急速でない。抗原ドリフトによる抗原変異型の頻繁な発生が、例年の流行のウイルス学的基礎であり、毎年のインフルエンザワクチンに少なくとも1つの新たなウイルス株を組み込む理由でもある。
【0003】
表面抗原、とりわけ、赤血球凝集素タンパク質に対する対象者の免疫が、感染の可能性、および万一感染が生じた場合の疾患の重症度を低減する(112)。一般に、インフルエンザウイルスの1つの型または1つの亜型に対する抗体は、別の型もしくは別の亜型に対してもたらす防御が限定されたものであるか、または別の型もしくは別の亜型に対しては防御をもたらさないと考えられている。さらに、インフルエンザウイルスの1つの抗原変異型に対する抗体では、同じ型または同じ亜型の新たな抗原変異型に対する防御とならない可能性があることも、一般的に受け入れられている(113)。したがって、交差防御の実証は期待されない。
【0004】
1957年および1968年における、過去の二度にわたるインフルエンザ大流行の原因となったのは、ヒト−トリ間リアソータントのインフルエンザウイルスであった。1968年以後にH2亜型ウイルスがヒトにおいて蔓延しなくなって以来、H2亜型のリアソータントウイルスから抗原シフトが生じる可能性が、理論的にはいつでも存在する。しかし、高病原性トリインフルエンザ(HPAI)ウイルス(H5亜型およびH7亜型)が近年になって出現し、1997年以来、これらのウイルスが、散発的ながらも、トリからヒトに直接的に伝染するようになった(1〜5)ことから、H2亜型ウイルスに対する集団の感受性がかつてなく増大しているのに加えて、ヒトにおいて新たに大流行する脅威の可能性が高まっている。ヒトにおけるHPAI H5N1亜型の大発生が、異なる抗原によるものであったという事実は、大流行の脅威に対して、十分に適合させたワクチンの備蓄を前もって準備することをほとんど不可能にしている(5、6)。マウスにおけるH5亜型による免疫化および抗原曝露(challenge)のデータは、このモデルにおける各種のH5亜型単離物間に良好な交差反応性が見られることを示唆する(7)が、既存のワクチン法で、ヒトにおいても同様レベルの交差反応性が見られるかどうかは公知ではない。したがって、新たなウイルスの大発生に由来する抗原を包含するように迅速に適合させうるインフルエンザワクチンプラットフォームが必要とされている。
【0005】
現行の卵ベースの不活化ワクチン法では、卵内でHPAIウイルスを増殖させることが不可能であるために、新興の大流行による需要を満たし、バイオコンテインメントを増強する必要を満たすのには不十分である(6、8)。逆遺伝学法は、卵内で培養しうる所望のHA遺伝子組成およびNA遺伝子組成を伴う、低病原性のリアソータントを生成させる手段をもたらしている(7、9〜11)が、この手法により生成させるワクチンは、過去の知的財産および規制に関する問題のために、ようやく臨床段階に入りつつあるに過ぎない(8)。さらなる懸念は、ヒト臨床試験で評価されている、赤血球凝集素H5タンパク質と関連する免疫原性レベルが明らかに低いこと(12〜14)であり、これは、H5亜型に対して完全にナイーブな集団における防御を効率的に誘導するには、ワクチン、送達系、およびアジュバント使用の改善が必要とされることを明らかにしている。したがって、アジュバントと組み合わせたHPAI抗原の発現を可能とするインフルエンザワクチンプラットフォームが必要とされている。
【0006】
インフルエンザVLPは、インフルエンザワクチンを生成させるための代替的な技法を代表する。インフルエンザVLPとは、昆虫細胞内で発現させたインフルエンザのマトリックスタンパク質、HAタンパク質、およびNAタンパク質を用いて生成させたもので、鼻腔内送達後における免疫原性が顕著である(26:非特許文献1、27:非特許文献2)。実際、VLPは一般に、ロタウイルスVLP、ノロウイルスVLP、およびパピローマウイルスVLPについて示されている通り、鼻腔内送達後において、粘膜および全身における免疫を誘導するのに十分に適すると考えられる(28〜31:非特許文献3〜6)。インフルエンザVLPとは、真核細胞発現系において、インフルエンザのマトリックスタンパク質、HAタンパク質、およびNAタンパク質を発現させることにより生成させたものである。インフルエンザマトリックスタンパク質は、ウイルスの出芽を後押しする駆動力であり、インフルエンザNAタンパク質は、インフルエンザHAタンパク質も発現している場合、細胞表面のシアル酸とのHAタンパク質の会合により、出芽したVLPが生成細胞から放出されるのに必要とされる(51:非特許文献6)。また、マトリックスタンパク質と、HAタンパク質のC末端との相互作用が、出芽過程の一部として、マトリックスタンパク質を膜に方向づけるのに役割を果たすことを示唆するデータも存在する(51:非特許文献6)。昆虫細胞のバキュロウイルス発現系内で生成させたインフルエンザVLPは、動物試験において免疫原性であることが分かっており、将来の大流行に備えるための重要な戦略を代表する(26:非特許文献1、27:非特許文献2、47:非特許文献7)。加えて、インフルエンザVLPの鼻腔内送達は、非経口投与後に得られる抗体力価を超える抗体力価を結果としてもたらしうる。しかし、VLPは、膜内に包埋されるか、または膜と会合する、1または複数の異なる糖タンパク質と共に包埋される脂質膜の複合体構造である。実用性を有するために、VLPは、十分に長い保管寿命を有する必要がある。したがって、溶液中にインフルエンザ抗原を包含するウイルス様粒子を安定化させる方法が必要とされている。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】Latham, T., and J. M. Galarza. 2001. Formation of wild−type and chimeric influenza virus−like particles following simultaneous expression of only four structural proteins. J Virol 75:6154.
【非特許文献2】Galarza, J. M., T. Latham, and A. Cupo. 2005. Virus−like particle vaccine conferred complete protection against a lethal influenza virus challenge. Viral Immunol 18:365.
【非特許文献3】Fromantin, C., B. Jamot, J. Cohen, L. Piroth, P. Pothier, and E. Kohli. 2001. Rotavirus 2/6 virus−like particles administered intranasally in mice, with or without the mucosal adjuvants cholera toxin and Escherichia coli heat−labile toxin, induce a Th1/Th2−like immune response. J Virol 75:11010.
【非特許文献4】Harrington, P. R., B. Yount, R. E. Johnston, N. Davis, C. Moe, and R. S. Baric. 2002. Systemic, mucosal, and heterotypic immune induction in mice inoculated with Venezuelan equine encephalitis replicons expressing Norwalk virus−like particles. J Virol 76:730.
【非特許文献5】Shi, W., J. Liu, Y. Huang, and L. Qiao. 2001. Papillomavirus pseudovirus: a novel vaccine to induce mucosal and systemic cytotoxic T−lymphocyte responses. J Virol 75:10139.
【非特許文献6】Gomez−Puertas, P., C. Albo, E. Perez−Pastrana, A. Vivo, and A. Portela. 2000. Influenza virus matrix protein is the major driving force in virus budding. J Virol 74:11538.
【非特許文献7】Pushko, P., T. M. Tumpey, F. Bu, J. Knell, R. Robinson, and G. Smith. 2005. Influenza virus−like particles comprised of the HA, NA, and M1 proteins of H9N2 influenza virus induce protective immune responses in BALB/c mice. Vaccine 23:5751.
【発明の概要】
【課題を解決するための手段】
【0008】
本出願の開示は、インフルエンザ抗原を包含するウイルス様粒子を溶液中で安定化させる方法を、該安定化させた溶液、およびこのような安定化させた溶液を用いる方法と併せて提供することにより、この必要を満たす。
【0009】
開示の一態様は、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物を含有する溶液を安定化させる方法であって、(a)インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子を含有する溶液を用意するステップと、(b)(1)安定化量の、単糖、ソルビトール、二糖、トレハロース、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択される安定化剤を、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物に添加するステップ、(2)pHを、約pH6.5〜約pH8.0、約pH6.5〜約pH7.5、または約pH7とするように、該溶液を緩衝化するステップ、または(3)ステップ(1)およびステップ(2)の両方とを含み、ステップ(b)の後のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物が、以下の特徴:(i)光学濃度により測定した該ウイルス様粒子の凝集が、ステップ(b)の前のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していること;(ii)円偏光二色性またはANS結合により測定したインフルエンザ抗原が、ステップ(b)の前のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して安定化していること;および(iii)ラウルダン蛍光により測定した、該ウイルス様粒子の脂質二重層の、温度により誘導される水和が、ステップ(b)の前のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していることのうちの少なくとも1つを呈示する方法を提供する。ある実施形態では、緩衝化するステップを、リン酸バッファー、トリスバッファー、MESバッファー、クエン酸バッファー、および他のGRASバッファーからなる群から選択される緩衝剤を用いて実施する。前出の緩衝剤についての実施形態と組み合わせうるある実施形態では、安定化剤は、トレハロース、ソルビトール、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択され、該特徴は(i)である。前出の緩衝剤についての実施形態と組み合わせうるある実施形態では、安定化剤は、トレハロース、ソルビトール、および前出の安定化剤の組合せから選択され、該特徴は(ii)である。前出の緩衝剤についての実施形態と組み合わせうるある実施形態では、安定化剤は、トレハロースおよびグリシンから選択され、特徴は(iii)である。前出の緩衝剤についての実施形態と組み合わせうるある実施形態では、安定化剤はトレハロースであり、3つの特徴すべてが存在する。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチドを含む。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、gagポリペプチド、インフルエンザM1ポリペプチド、ニューカッスル病ウイルスマトリックスポリペプチド、エボラウイルスVP40ポリペプチド、およびマールブルグウイルスVP40ポリペプチドを含む群から選択される第2のポリペプチドを含む。gagポリペプチドを包含する前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該gagポリペプチドが、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス、ベータレトロウイルス、ガンマレトロウイルス、デルタレトロウイルス、およびレンチウイルスに由来しうる。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、ノイラミニダーゼポリペプチドをさらに含む。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化剤は、単糖、ソルビトール、二糖、およびトレハロースから選択され、安定化量は、10%(w/w)を超えるか、または少なくとも約20%(w/w)である。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化させるステップが、ガラス形成を必要としない。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化量は、凝固時のガラス形成に必要とされる量未満である。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化剤は、スクロースではない。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物は、該インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子と混合させたアジュバントをさらに含み、アジュバントは、前記ウイルス様粒子の内部に配置されてもよく、該ウイルス様粒子の外部に配置されてもよい。アジュバントおよび第2のポリペプチドを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、アジュバントを、該第2のポリペプチドに共有結合させて、共有結合を形成させる。アジュバントおよび赤血球凝集素ポリペプチドを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該アジュバントを、前記赤血球凝集素ポリペプチドに共有結合させて、共有結合を形成させる。アジュバントを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該アジュバントは、フラジェリンのアジュバント活性断片を含む。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該方法は、(c)溶液を、少なくとも2週間、少なくとも1カ月間、少なくとも2カ月間、少なくとも3カ月間、少なくとも4カ月間、少なくとも6カ月間、または少なくとも1年間にわたり液体形態で保存するステップをさらに含み、このような期間後のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物は、このような期間前のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物により誘導される免疫反応の少なくとも80パーセント、少なくとも90パーセント、または少なくとも95パーセントを誘導する。
【0010】
本開示の別の態様は、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子と、安定化量の、トレハロース、ソルビトール、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択される、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物に対する安定化剤とを含み、以下の特徴:(i)光学濃度により測定した該ウイルス様粒子の凝集が、該安定化剤を伴わない、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していること;(ii)円偏光二色性またはANS結合により測定したインフルエンザ抗原が、該安定化剤を伴わない、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して安定化していること;および(iii)ラウルダン蛍光により測定した、該ウイルス様粒子の脂質二重層の、温度により誘導される水和が、該安定化剤を伴わない、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していることのうちの少なくとも1つを呈示する、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物を提供する。ある実施形態では、緩衝化を、リン酸バッファー、トリスバッファー、MESバッファー、クエン酸バッファー、および他のGRASバッファーからなる群から選択される緩衝剤を用いて実施する。ある実施形態では、安定化剤は、トレハロース、ソルビトール、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択され、該特徴は(i)である。ある実施形態では、安定化剤は、トレハロース、ソルビトール、および前出の安定化剤の組合せから選択され、該特徴は(ii)である。ある実施形態では、安定化剤は、トレハロースおよびグリシンから選択され、該特徴は(iii)である。ある実施形態では、安定化剤はトレハロースであり、3つの特徴すべてが存在する。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子は、赤血球凝集素ポリペプチドを含む。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子は、gagポリペプチド、インフルエンザM1ポリペプチド、ニューカッスル病ウイルスマトリックスポリペプチド、エボラウイルスVP40ポリペプチド、およびマールブルグウイルスVP40ポリペプチドを含む群から選択される第2のポリペプチドを含む。gagポリペプチドを包含する前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該gagポリペプチドは、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス、ベータレトロウイルス、ガンマレトロウイルス、デルタレトロウイルス、およびレンチウイルスからなる群から選択されるレトロウイルスに由来する。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子は、ノイラミニダーゼポリペプチドをさらに含む。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化剤は、単糖、ソルビトール、二糖、およびトレハロースから選択され、安定化量は、10%(w/w)を超えるか、または少なくとも約20%(w/w)である。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化させるステップが、ガラス形成を必要としない。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化量は、凝固時のガラス形成に必要とされる量未満である。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、安定化剤は、スクロースではない。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物は、該インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子と混合させたアジュバントをさらに含む。アジュバントを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該アジュバントは、前記ウイルス様粒子の内部に配置される。アジュバントを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該アジュバントは、前記ウイルス様粒子の外部に配置される。アジュバントおよび第2のポリペプチドを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、該アジュバントを、前記第2のポリペプチドに共有結合させて、共有結合を形成させる。アジュバントを包含する、前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、アジュバントを、前記赤血球凝集素ポリペプチドに共有結合させて、共有結合を形成させる。前出の実施形態のうちのいずれかと組み合わせうるある実施形態では、アジュバントは、フラジェリンのアジュバント活性断片を含む。
【0011】
本開示の別の態様は、インフルエンザを治療または予防する方法であって、対象に、免疫原性量の、前出の態様およびその多様な実施形態のうちのいずれかによる、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物、または前出の方法の態様およびその多様な実施形態のうちのいずれかに従い安定化させた、免疫原性量の、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物を含有する溶液を投与するステップを含む方法を提供する。ある実施形態では、投与するステップは、対象において防御的免疫反応を誘導する。前出の実施形態と組み合わせ得るある実施形態では、投与するステップは、皮下(subcutaneous)送達、経皮送達、皮内送達、皮下送達(subdermal)、筋肉内送達、経口(peroral)送達、経口(oral)送達、鼻腔内送達、頬(buccal)送達、舌下送達、腹腔内送達、膣内(intravaginal)送達、肛門(anal)送達、および頭蓋内送達からなる群から選択される。
【図面の簡単な説明】
【0012】
【図1】図1は、個別のGag遺伝子ベクター、HA遺伝子ベクター、または対照ベクターを感染させたSf9細胞、およびHA−gag−NA三重遺伝子ベクターを感染させたSf9細胞に由来する培地についてのウェスタンブロットを示す図である。(A)は、抗Gag抗体によりプローブしたものであり、(B)は、抗HA抗体によりプローブしたものである。
【図2】図2は、ペレット化させたHA−gag−NA VLPを、段階的なスクロース勾配上で再遠心分離することによる画分についてのウェスタンブロットを示す図である。(A)は、抗Gag抗体によりプローブしたものであり、(B)は、抗HA抗体によりプローブしたものである。
【図3】図3は、インフルエンザVLPによる動的光散乱を示す図である。有効直径(A)、静的光散乱の強度(B)、および試料の多分散性(C)を、温度の関数としてプロットする。各点は、3つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【図4】図4は、インフルエンザVLPの円偏光二色性スペクトルを示す図である。低温(10℃)では、4〜8の各単位pHにおけるスペクトル(A)を示し、pH7では、多様な温度におけるスペクトル(B)を示す。
【図5】図5は、熱ストレスに対する、インフルエンザVLPのタンパク質二次構造の反応を示す図である。227nmにおける標準化した(−1〜0の)CDを、温度の関数として示す。各点は、3つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【図6】図6は、インフルエンザVLPの内部蛍光のピーク位置を、温度の関数として示す図である。また、330nmにおける標準化した(0〜1の)蛍光強度も、温度の関数として示す(右下)。各点は、3つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【図7】図7は、インフルエンザVLPの物理的構造のプローブとしての、ANS蛍光を示す図である。ピーク発光の波長を、温度の関数として示す。また、485nmにおける標準化した(0〜1の)ANS蛍光強度も、温度の関数として示す(右下)。各点は、3つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【図8】図8は、インフルエンザVLPの存在下におけるラウルダン蛍光による一般化偏光(GP)を、温度の関数として示す図である。各点は、3つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【図9】図9は、インフルエンザVLPの生物物理的特徴づけに由来する実験相図(EPD)である。EPDは、温度に依存する有効直径、静的光散乱、多分散性、227nmにおけるCD、内部蛍光(ピーク位置、および330nmにおける相対強度)、ANS蛍光(ピーク位置、および485nmにおける相対強度)、および4〜8のpH範囲にわたり収集されたラウルダン蛍光データによるGPから作成した。
【図10】図10は、選択した安定化剤の存在下における、インフルエンザVLPによる内部蛍光を示す図である。ピーク発光の位置(波長)を、温度の関数として示す。各点は、2つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【図11】図11は、選択した安定化剤と共に調合されたインフルエンザVLPの存在下における、ラウルダン蛍光によるGPを示す図である。各点は、2つの個別の試料の平均を表わし、誤差バーは、標準偏差を示す。
【発明を実施するための形態】
【0013】
本発明は、インフルエンザ抗原のエンベロープウイルスベースのVLPを安定化させた製剤に基づく。例示的なエンベロープウイルスベースのVLPは、VLP製剤のベースとして、マウス白血病ウイルス(MLV)に由来するgagポリペプチドなどのgagポリペプチドを包含する。バキュロウイルス発現系において各種のレトロウイルスから得ることができるgag VLPの収量が大きいため、このようなgagタンパク質ベースのVLPを生成させる例示的な方法は、好ましくは、インフルエンザHAポリペプチド抗原およびインフルエンザNAポリペプチド抗原の共発現を含めた、昆虫細胞における発現を介する。
【0014】
安定化は、インフルエンザ抗原のウイルスベースのVLP調製物中に包含される安定化量の安定化剤を介する。例示的な安定化剤には、単糖(デキストロース、マンニトール、ソルビトールなど)、二糖(ラクトース、トレハロース、スクロースなど)、ジエタノールアミン、グリセロール、グリシン、またはこれらの組合せが含まれる。
【0015】
開示される方法およびプロトコールの実施は、別段に示唆されない限り、当業者の能力の範囲内にある、化学、分子生物学、微生物学、組換えDNA、および免疫学による従来の技法を使用する。このような技法は、文献において説明されている。例えば、J. Sambrook, E. F. Fritsch、およびT. Maniatis、1989年、Molecular Cloning: A Laboratory Manual、2版、1〜3巻、Cold Spring Harbor Laboratory Press;Ausubel, F. M.ら(1995年、および定期的な補遺、Current Protocols in Molecular Biology、9、13、および16章、John Wiley & Sons、New York、N.Y.);B. Roe、J. Crabtree、およびA. Kahn、1996年、DNA Isolation and Sequencing: Essential Techniques、John Wiley & Sons;J. M. PolakおよびJames O’D. McGee、1990年、In Situ Hybridization: Principles and Practice、Oxford University Press;M. J. Gait(編)、1984年、Oligonucleotide Synthesis: A Practical Approach、Irl Press;ならびにD. M. J. LilleyおよびJ. E. Dahlberg、1992年、Methods of Enzymology: DNA Structure、A部:Synthesis and Physical Analysis of DNA Methods in Enzymology、Academic Pressを参照されたい。これらの一般的な教科書の各々は、参照により本明細書に組み込まれる。
【0016】
定義
本明細書で用いられる「エンベロープウイルスベースのVLP」とは、エンベロープウイルスに由来する1または複数のコンポーネントを用いて形成されるウイルス様粒子を指す。限定なしに述べると、好ましい例には、インフルエンザ赤血球凝集素ポリペプチドと共にgagポリペプチドを用いて生成させたVLP、インフルエンザ赤血球凝集素ポリペプチドを用いて、または赤血球凝集素ポリペプチドと共にOrthomyxovirus属ウイルス(インフルエンザウイルスを含めた)M1ポリペプチドを用いて(場合によって、各場合にノイラミニダーゼポリペプチドを伴う)生成させたVLP、インフルエンザ赤血球凝集素ポリペプチドと共にParamyxovirus(Paromyxovirus)属ウイルス(ニューカッスル病ウイルスを含めた)マトリックスポリペプチドを用いて生成させたVLP、インフルエンザ赤血球凝集素ポリペプチドと共にFilovirus属ウイルス(エボラウイルスまたはマールブルグウイルスを含めた)VP40ポリペプチドを用いて生成させたVLPが含まれる。
【0017】
さらなる例には、エンベロープウイルスベースのVLPを形成するのに用いうる、エボラウイルスおよびマールブルグウイルスなどのフィロウイルス(例えば、細胞内でフィロウイルスに由来するウイルス性GPタンパク質およびウイルス性VP40タンパク質を共発現させ、脂質ラフト内でこれら2つのウイルス性タンパク質を会合させることにより、VLPを生成させる(米国特許公開第20060099225号を参照されたい));SARSウイルスなどのコロナウイルス(例えば、コロナウイルスのVLPを形成させるには、Eタンパク質およびMタンパク質で十分である(Fischerら、J. Virol.(1998年)、72巻:7885〜7894頁;およびVennemaら、EMBO J.(1996年)、15巻:2020〜2028頁を参照されたい));RS(respiratory syncytial)ウイルス(RSV)など、Paramyxoviridae(paramyxoviridae)科のウイルス(例えば、RSVウイルスのMタンパク質を発現させることにより、VLPを生成させる(例えば、米国特許公開第20080233150号を参照されたい));ならびに西ナイルウイルスなど、Flaviviridae(flaviviridae)科のウイルス(例えば、バキュロウイルス発現系内で西ナイルウイルスのprM遺伝子およびE遺伝子を含む構築物を発現させることにより、VLPを生成させる(例えば、米国特許公開第20080233150号を参照されたい))が含まれる。
【0018】
gagポリペプチドには、本明細書で説明するウイルス様粒子を形成する一因となる、レトロウイルス由来の構造ポリペプチドが含まれる。一部の実施形態では、RNAをパッケージングする傾向、または粒子形成および粒子出芽の効率など、一定の特徴に影響を及ぼす目的で、gagポリペプチドを意図的に突然変異させることができる。このような突然変異の一例をあげるならば、gag由来のVLPがRNAを組み込む能力に影響を及ぼすアミノ酸変化となろう。VLP出芽の効率を改善または改変する、他のこのようなアミノ酸変化ももたらしうるであろう。レトロウイルスのゲノムは、3つの主要な遺伝子産物:構造タンパク質をコードするgag遺伝子;逆転写酵素、ならびに関連するタンパク質分解ポリペプチド、ヌクレアーゼ、およびインテグラーゼに関連する機能をコードするpol遺伝子;それによりコードされる糖タンパク質である膜タンパク質が、感染細胞表面上で検出され、また、放出された成熟ウイルス粒子の表面上でも検出されるenv遺伝子による遺伝子産物をコードする。すべてのレトロウイルスのgag遺伝子は、全体的な構造類似性を有し、各レトロウイルス群内において、そのアミノ酸レベルが保存されている。gag遺伝子は、逆転写酵素を除いたコアタンパク質をもたらす。MLVでは、Gag前駆体であるポリタンパク質が、Pr65Gagであり、該前駆体におけるその順序がNH2−p15−pp12−p30−p10−COOHである4つのタンパク質に切断される。これらの切断は、ウイルス性プロテアーゼに媒介され、ウイルスに応じて、ウイルス放出の前に生じる場合もあり、ウイルス放出の後に生じる場合もある。MLV Gagタンパク質は、グリコシル化形態および非グリコシル化形態で存在する。グリコシル化形態は、非グリコシル化Pr65GagのAUGコドンより上流に位置する、異なるインフレームの開始コドンにより合成される、gPr80Gagから切断される。グリコシル化Gagを合成しないMLVの欠失突然変異体もなお感染性であり、該非グリコシル化Gagもなおウイルス様粒子を形成し得ることから、グリコシル化イベントの重要性について疑問が生じる。ウイルスによりコードされるプロテアーゼを介してHIV−1 Gag前駆体であるpr55Gagが翻訳後切断されると、N−ミリストイル化および内部リン酸化されたp17マトリックスタンパク質(p17MA)、リン酸化p24カプシドタンパク質(p24CA)、およびp15ヌクレオカプシドタンパク質(p15NC)(これはp9およびp6にさらに切断される)がもたらされる。
【0019】
構造的には、原型的なGagポリタンパク質が、レトロウイルスgag遺伝子では常に同じ順序で生じる3つの主要なタンパク質:マトリックスタンパク質(MA)(インフルエンザマトリックスタンパク質であるM1は、マトリックスという名称を共有しているが、MAとは異なるタンパク質であり、これとは区別する)、カプシドタンパク質(CA)、およびヌクレオカプシドタンパク質(NC)に分けられる。Gagポリタンパク質の成熟タンパク質へのプロセシングは、レトロウイルスによりコードされるプロテアーゼを介して触媒され、新たに出芽したウイルス粒子が成熟するときに生じる。機能的には、Gagポリタンパク質が、3つのドメイン:細胞膜をGagポリタンパク質の標的とする膜結合ドメイン;Gagの多量体化を促進する相互作用ドメイン;および宿主細胞から発生したビリオンの放出を促進する後期ドメインに分けられる。アセンブリーを媒介するGagタンパク質の形態は、ポリタンパク質である。したがって、アセンブリードメインは、後期に形成される切断産物のうちのいずれかの内部にもれなく収まる必要がない。こうして、本明細書に包含されるGagポリペプチドは、VLPの形成および放出に重要な機能的エレメントを包含する。当技術分野の現況は、これらの重要な機能エレメントに関して、極めて進展している。例えば、Hansenら、J. Virol.64巻、5306〜5316頁、1990年;Willら、AIDS 5巻、639〜654頁、1991年;Wangら、J. Virol.72巻、7950〜7959頁、1998年;McDonnellら、J. Mol. Biol.279巻、921〜928頁、1998年;SchultzおよびRein、J. Virol.63巻、2370〜2372頁、1989年;Accolaら、J. Virol.72巻、2072〜2078頁、1998年;Borsettiら、J. Virol.72巻、9313〜9317頁、1998年;Bowzardら、J. Virol.72巻、9034〜9044頁、1998年;Krishnaら、J. Virol.72巻、564〜577頁、1998年;Willsら、J. Virol.68巻、6605〜6618頁、1994年;Xiangら、J. Virol.70巻、5695〜5700頁、1996年;Garnierら、J. Virol.73巻、2309〜2320頁、1999年を参照されたい。
【0020】
例示的なレトロウイルスのGagポリペプチド供給源には、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス(ニワトリ白血病ウイルス、またはラウス肉腫ウイルスなど)、ベータレトロウイルス(マウス乳癌ウイルス、ヤーグジークテヒツジレトロウイルス、およびメーソン−ファイザーサルウイルスなど)、ガンマレトロウイルス(マウス白血病ウイルス、ネコ白血病ウイルス、細網内皮症ウイルス、およびテナガザル白血病ウイルスなど)、デルタレトロウイルス(ヒトTリンパ向性ウイルス、およびウシ白血病ウイルスなど)、イプシロンレトロウイルス(ウォールアイ皮膚肉腫ウイルスなど)、またはレンチウイルス(1型ヒト免疫不全ウイルス、HIV−2、サル免疫不全ウイルス、ネコ免疫不全ウイルス、ウマ感染性貧血ウイルス、およびヤギ関節炎脳炎ウイルスなど)が含まれる。
【0021】
本明細書で用いられる「赤血球凝集素ポリペプチド」は、インフルエンザウイルスが感染される細胞に結合することを媒介するインフルエンザウイルスタンパク質に由来する。赤血球凝集素タンパク質は、単一の膜貫通ドメインを介してインフルエンザウイルスの表面に係合されていることが見出される、抗原性の糖タンパク質である。インフルエンザウイルスの赤血球凝集素については、少なくとも16の亜型が同定され、H1〜H16と標識されている。ヒトインフルエンザウイルスにおいて見出されるのは、H1、H2、およびH3である。赤血球凝集素H5、赤血球凝集素H7または赤血球凝集素H9を伴う、高病原性トリインフルエンザウイルスがヒトに感染する割合は小さいことが判明している。赤血球凝集素H5がトリH5N1ウイルスの受容体特異性を著明に変化させることを可能とし、それらにヒト受容体に結合する能力をもたらすようにその受容体特異性を変化させる、トリインフルエンザウイルス株の赤血球凝集素H5型における単一のアミノ酸変化が、ヒト患者において見出されている(109および110)。この知見は、通常ではヒトに感染しないH5N1ウイルスが、どのようにして突然変異し、ヒト細胞に効率的に感染し得るようになるのかを説明する。
【0022】
赤血球凝集素は、ホモ三量体の内在性膜ポリペプチドである。その膜貫通ドメインは、天然でラフト脂質ドメインと会合し、それをgagポリペプチドと会合させて、VLPに組み込むことを可能とする。赤血球凝集素は、円筒様の形状であり、約135Åの長さである。HAを構成する3つの同一の単量体は、中央部のコイルドコイルおよびVLP表面上に曝露されているシアル酸結合部位を含有する球状ヘッドを形成する。HAの単量体は、グリコシル化されて2つの小型のポリペプチド:HA1サブユニットおよびHA2サブユニットに切断される、単一のポリペプチド前駆体として合成される。HA2サブユニットは、膜に係合する三量体のコイルドコイルを形成し、HA1サブユニットは、球状ヘッドを形成する。
【0023】
本発明のVLPにおいて用いられる赤血球凝集素ポリペプチドは、少なくとも膜アンカードメインおよび赤血球凝集素由来の少なくとも1つのエピトープを包含するものとする。赤血球凝集素ポリペプチドは、インフルエンザウイルスの任意の類型、亜型、株、または亜株、例えば、赤血球凝集素H1、赤血球凝集素H2、赤血球凝集素H3、赤血球凝集素H5、赤血球凝集素H7および赤血球凝集素H9に由来することが可能である。加えて、赤血球凝集素ポリペプチドは、異なるインフルエンザ赤血球凝集素のキメラ体でもあり得る。赤血球凝集素ポリペプチドは場合によって、1または複数のさらなるポリペプチドのコード配列を、そのコード配列にスプライシングすることにより生成させ得る、1または複数のさらなるポリペプチドを包含し得る。さらなるポリペプチドを赤血球凝集素ポリペプチドに挿入するための例示的な部位は、N末端である。
【0024】
本明細書で用いられる「ノイラミニダーゼポリペプチド」は、糖タンパク質から末端のシアル酸残基を切断することにより、細胞からのインフルエンザウイルスの放出を媒介する、インフルエンザウイルスタンパク質に由来する。ノイラミニダーゼの糖タンパク質は、ウイルス表面に発現する。ノイラミニダーゼタンパク質は、四量体であり、ベータ−ピンホイール構造を伴う球状ヘッド、細いストーク状領域、および単一の膜貫通ドメインを介してウイルス膜に該タンパク質を係合させる小型の疎水性領域からなる共通の構造を共有する。シアル酸残基切断の活性部位は、すべてのA型インフルエンザウイルスにおいて保存されている15の帯電アミノ酸により形成される各サブユニット表面においてポケット部分を包含する。インフルエンザノイラミニダーゼについては、少なくとも9つの亜型が同定されており、N1〜N9と標識されている。
【0025】
本明細書で開示されるVLPで用いられ得るノイラミニダーゼポリペプチドは、少なくとも膜アンカードメインおよび少なくともシアル酸残基切断活性を包含するものとする。機能領域に関する当技術分野の現況は、極めて高度である。例えば、Vargheseら、Nature 303巻、35〜40頁、1983年;Colmanら、Nature 303巻、41〜44頁、1983年;Lentzら、Biochem、26巻、5321〜5385頁、1987年;Websterら、Virol.135巻、30〜42頁、1984年を参照されたい。ノイラミニダーゼポリペプチドは、インフルエンザウイルスの任意の類型、亜型、株、または亜株、例えばノイラミニダーゼN1およびノイラミニダーゼN2に由来することが可能である。加えて、ノイラミニダーゼポリペプチドは、異なるインフルエンザノイラミニダーゼのキメラ体でもあり得る。ノイラミニダーゼポリペプチドは場合によって、1または複数のさらなるポリペプチドのコード配列を、赤血球凝集素ポリペプチドのコード配列にスプライシングすることにより生成させ得る、1または複数のさらなるポリペプチドを包含し得る。さらなるポリペプチドをノイラミニダーゼポリペプチドに挿入するための例示的な部位は、C末端である。
【0026】
本明細書で用いられる「GRASバッファー」とは、該当する政府の規制機関により公示されている通り、「一般に安全であると認識されている」バッファーを指す。GRASバッファーは、pH6〜pH8(例えば、pKa5〜9)の範囲内で緩衝作用をもたらすことが好ましい。化合物に有効な緩衝範囲は一般に、pKa±約1pH単位である、例えば、pKaが7.2であるH3PO4の緩衝能力は、約6.5〜8.0である。例示的なGRASバッファーには、トリス(ヒドロキシメチル)アミノメタン(トリス)、クエン酸水素またはクエン酸二水素、一塩基性リン酸カリウム、二塩基性リン酸カリウム、一塩基性リン酸ナトリウム、二塩基性リン酸ナトリウム、2−(N−モルホリノ)エタンスルホン酸(MES)、3−(N−モルホリノ)プロパンスルホン酸(MOPS)、N−(2−アセトアミド)イミノ二酢酸(ADA)、3−[N−トリス(ヒドロキシメチル)メチルアミノ]−2−ヒドロキシプロパンスルホン酸(TAPSO)、3−[N,N−ビス(2−ヒドロキシエチル)アミノ]−2−ヒドロキシプロパンスルホン酸(DIPSO)、ピペラジン−N,N’−ビス(2−ヒドロキシプロパンスルホン酸)(POPSO)、N−(2−ヒドロキシメチル)ピペラジン−N’−2−ヒドロキシプロパンスルホン酸(HEPPSO)、ピペラジン−N,N’−ビス(2−エタンスルホン酸)(PIPES)、N−(2−アセトアミド)−2−アミノエタンスルホン酸(ACES)、コラミンクロリド、N,N−ビス(2−ヒドロキシエチル)−2−アミノエタンスルホン酸(BES)、N−トリス(ヒドロキシメチル)メチル−2−アミノエタンスルホン酸(TES)、(4−(2−ヒドロキシエチル)−1−ピペラジンエタンスルホン酸)(HEPES)、アセトアミドグリシン、トリシン、グリシンアミド、ビシン、ヒスチジンなどのアミノ酸残基、酢酸、水酸化アンモニウム、イミダゾール、2−アミノ−2−メチル−1−プロパノール(AMP)、2−アミノ−2−メチル−1,3−プロパンジオール(AMPD)、2−[ビス(2−ヒドロキシエチル)アミノ]−2−(ヒドロキシメチル)−1,3−プロパンジオール(ビス−トリス)、1,3−ビス(トリス(ヒドロキシメチル)メチルアミノ)プロパン(ビス−トリスプロパン)、カルボン酸、クエン酸、エタノールアミン、グリシルグリシン、N−(2−ヒドロキシエチル)ピペラジン−N’(4−ブタンスルホン酸)(HEPBS)、およびマレイン酸が含まれる。適切な範囲で緩衝作用をもたらしうる例示的な官能基には、イミダゾール、カルボン酸、リン酸、ピペラジン、アミン、およびスルホン酸が含まれる。GRASは一般に、食品での使用について規定されているが、本明細書で用いられるGRASバッファーはまた、薬物製剤中に組み入れるのに薬学的に許容されるとみなされているバッファーも包含する。
【0027】
エンベロープウイルスベースのVLPを生成させる例示的な方法
エンベロープウイルスベースのVLPは、当業者に適用可能な任意の方法により生成させることができる。エンベロープウイルスベースのVLPは、インフルエンザ抗原ポリペプチドに加えて、VLPを形成する一因となる1または複数のポリペプチドを包含することが典型的であり、これらは、VLPを形成する一因となるポリペプチドが、それ自体、M1タンパク質などのインフルエンザ抗原であるか、または該インフルエンザ抗原ポリペプチドに連結される、キメラ形態を包含する。加えて、エンベロープウイルスベースのVLPは、第2のインフルエンザ抗原または別の抗原など、さらなる抗原をもたらすために、膜(脂質ラフトを含めた)会合ポリペプチドなど、1または複数のさらなるポリペプチドを包含することも可能である。ある実施形態では、ポリペプチドを、哺乳動物細胞による発現系および昆虫細胞による発現系など、細胞膜内に脂質ラフトドメインを包含する細胞ベースの系など、任意の適用可能なタンパク質発現系内で共発現させることができる。
【0028】
組換えによるVLP用ポリペプチドの発現は、該ポリペプチドのうちの1または複数をコードするポリヌクレオチドを含有する発現ベクターを伴う。ポリペプチドのうちの1または複数をコードするポリヌクレオチドが得られたら、該ポリペプチドを生成させるためのベクターを、当技術分野において周知の技法を用いる組換えDNA法により生成させることができる。このようにして、本明細書では、VLPポリペプチドをコードするヌクレオチド配列のうちのいずれかを含有するポリヌクレオチドを発現させることによりタンパク質を調製する方法が説明される。当業者に周知の方法を用いて、VLPポリペプチドのコード配列と、適切な転写制御シグナルおよび翻訳制御シグナルとを含有する発現ベクターを構築することができる。これらの方法には、例えば、in vitroにおける組換えDNA法、合成法、また、in vivoにおける遺伝子組換えが含まれる。したがって、本発明は、すべてを1または複数のプロモーターに作動可能に連結した、gagポリペプチドと、抗原に連結した脂質ラフト会合ポリペプチドとをコードするヌクレオチド配列を含む複製可能なベクターを提供する。
【0029】
発現ベクターを、従来の技法により宿主細胞に形質導入することができ、次いで、トランスフェクトされた細胞を、従来の技法により培養して、VLPポリペプチド(複数可)を生成させることができる。したがって、本発明は、異種プロモーターに作動可能に連結したVLPポリペプチドのうちの1または複数をコードするポリヌクレオチドを含有する宿主細胞を包含する。VLPを生成させるためのある実施形態では、以下で詳述する通り、gagポリペプチド、およびインフルエンザ抗原に連結した脂質ラフト会合ポリペプチド(またはインフルエンザ抗原自体が脂質ラフト会合ポリペプチドの場合もある)の両方をコードするベクターを、宿主細胞内で共発現させて、VLPを生成させる。
【0030】
各種の宿主発現ベクター系を使用して、VLPポリペプチドを発現させることができる。このような宿主発現系は、共発現などによりVLPを生成させる目的で、それを介してVLPポリペプチドを生成させうる媒体を表わす。広範囲にわたる宿主を、適切な発現ベクター構築物内において用いることができ、脂質ラフトベースのアセンブリーに依拠する場合、好ましい宿主発現系は、VLPのアセンブリーに適する脂質ラフトを有する宿主である。これらには、VLPポリペプチドコード配列を含有する組換えバクテリオファージDNA発現ベクター、組換えプラスミドDNA発現ベクター、もしくは組換えコスミドDNA発現ベクターにより形質転換した細菌(例えば、E. coli、B. subtilis);VLPポリペプチドコード配列を含有する組換え酵母発現ベクターにより形質転換した酵母(例えば、Saccharomyces属、Pichia属)などの微生物;VLPポリペプチドのコード配列を含有する組換えウイルス発現ベクター(例えば、バキュロウイルス)を感染させた昆虫細胞系;VLPポリペプチドのコード配列を含有する組換えウイルス発現ベクター(例えば、カリフラワーモザイクウイルス、CaMV;タバコモザイクウイルス、TMV)を感染させるか、もしくはVLPポリペプチドのコード配列を含有する組換えプラスミド発現ベクター(例えば、Tiプラスミド)により形質転換した植物細胞系;または、哺乳動物細胞のゲノムに由来するプロモーター(例えば、メタロチオネインプロモーター)、もしくは哺乳動物ウイルスに由来するプロモーター(例えば、アデノウイルス後期プロモーター;牛痘ウイルス7.5Kプロモーター)を含有する組換え発現構築物を保有する、哺乳動物細胞系(例えば、COS細胞、CHO細胞、BHK細胞、293細胞、3T3細胞)が含まれるがこれらに限定されない。VLPアセンブリーが、脂質ラフトの会合により駆動される場合は、哺乳動物細胞または昆虫細胞を用いて、VLPポリペプチドを発現させることができる。例えば、ヒトサイトメガロウイルスに由来する主要即初期(intermediate early)遺伝子プロモーターエレメントなどのベクターと併用される、MRC−5細胞、Vero細胞、PER.C6(TM)細胞、チャイニーズハムスター卵巣細胞(CHO)、およびHEK293細胞などの哺乳動物細胞は、VLPポリペプチドの有効な発現系である(Foeckingら、Gene、45巻:101頁(1986年); Cockettら、Bio/Technology、8巻:2頁(1990年))。
【0031】
昆虫系では、Autographa californica核多角体病ウイルス(AcNPV)を、外来遺伝子を発現するためのベクターとして用いることができる。該ウイルスは、Spodoptera frugiperda細胞内で増殖する。VLPポリペプチドのコード配列(複数可)は、ウイルスの非必須領域(例えば、ポリヘドリン遺伝子領域)内に個別にクローニングすることができ、また、AcNPVプロモーター(例えば、ポリヘドリンプロモーター)の制御下に置くことができる。
【0032】
哺乳動物の宿主細胞では、多くのウイルスベースの発現系を使用することができる。発現ベクターとしてアデノウイルスを用いる場合は、対象のVLPポリペプチド配列(複数可)を、アデノウイルス転写/翻訳制御複合体、例えば、後期プロモーターおよび三連リーダー配列にライゲーションすることができる。次いで、in vitroまたはin vivoにおける組換えにより、このキメラ遺伝子を、アデノウイルスゲノム内に挿入することができる。ウイルスゲノムの非必須領域(例えば、E1領域またはE3領域)内に挿入する結果、感染された宿主内において生存可能であり、また、VLPポリペプチド(複数可)を発現することが可能な組換えウイルスがもたらされる(例えば、LoganおよびShenk、Proc. Natl. Acad. Sci. USA、81巻:355〜359頁(1984年)を参照されたい)。挿入されたVLPポリペプチドのコード配列(複数可)を効率的に翻訳するには、特定の開始シグナルもまた必要とされうる。これらのシグナルには、ATG開始コドン、および隣接配列が含まれる。さらに、全挿入配列の翻訳を確保するには、該開始コドンが、所望されるコード配列のリーディングフレームと同フレームでなければならない。これらの外因性翻訳制御シグナルおよび開始コドンは、由来が多様であることが可能であり、天然および合成のいずれもが可能である。発現の効率は、適切な転写エンハンサーエレメント、転写ターミネーターなどを組み込むことにより、増強することができる(Bittnerら、Methods in Enzymol.、153巻:51〜544頁(1987年)を参照されたい)。一例をあげるならば、StratageneのAdEASY−XL(TM)システムなど、アデノウイルスベースのベクター系において用いられる、ヒトCMV即初期プロモーターであろう。
【0033】
加えて、挿入した配列の発現を調節するか、または該遺伝子産物を、所望される特定の形で修飾およびプロセシングする宿主細胞株も選択することができる。タンパク質産物に対するこのような修飾(例えば、グリコシル化)およびプロセシング(例えば、切断または膜への輸送)は、VLPを生成させるのに、またはVLPポリペプチド、もしくはアジュバントなどのさらなるポリペプチド、もしくはさらなる抗原が機能するのに重要でありうる。様々な宿主細胞が、タンパク質および遺伝子産物の翻訳後プロセシングおよび修飾の特徴的かつ特異的な機構を有する。発現させる外来タンパク質の適正な修飾およびプロセシングを確保するために、適切な細胞株または宿主系を選択することができる。この目的で、一次転写物を適正にプロセシングし、遺伝子産物を適正にグリコシル化およびリン酸化するための細胞機構を保有する真核宿主細胞を用いることができる。
【0034】
宿主細胞には、gagポリペプチドをコードする第1のベクターと、ウイルス性膜抗原、または抗原に連結した脂質ラフト関連ポリペプチドをコードする第2のベクターという、本発明の2つの発現ベクターを共トランスフェクトすることができる。2つのベクターは、各VLPポリペプチドの同等の発現を可能とする、同一の選択マーカーを含有しうる。代替的に、gagポリペプチドと、抗原に連結した脂質ラフト関連ポリペプチドとの両方をコードし、また、これらの両方を発現することが可能な単一のベクターを用いることもできる。
【0035】
宿主細胞によりVLPを生成させたら、ポリペプチドの精製について当技術分野で公知の任意の方法により、例えば、クロマトグラフィー(例えば、イオン交換クロマトグラフィー、特に、該ポリペプチドに付加した任意のアフィニティー精製タグによるアフィニティークロマトグラフィー、およびサイズ除外クロマトグラフィー)、遠心分離、吸収率較差溶解度法により、またはタンパク質もしくは他の高分子を精製するための他の任意の標準的な技法によりこれを精製することができる。加えて、VLPポリペプチドを、本明細書で説明されるか、またはこれ以外に当技術分野で公知の異種ポリペプチド配列に融合させ、VLPの精製を容易にすることもできる。精製後、VLPポリペプチドに対する共有結合を介して、または他の非共有結合機構により、さらなる抗原またはアジュバントなどのさらなるエレメントを、VLPに物理的に連結することができる。哺乳動物細胞および昆虫細胞など、脂質ラフトドメインを有する宿主細胞内においてVLPポリペプチドを共発現させるある実施形態では、VLPが自己アセンブリーして放出され、これにより、上記の方法のうちのいずれかによるVLPの精製が可能となる。VLPのある実施形態には、例えば、同種ウイルスのタンパク質から操作されたVLP、例えば、インフルエンザウイルスに由来するM1タンパク質、HAタンパク質、また、場合によってNAタンパク質から構築されたVLP、また、異種ウイルスのタンパク質から操作されたVLP、例えば、MLVもしくはHIVまたは他のレトロウイルスに由来するGagタンパク質を、異なるウイルスに由来する抗原、例えば、インフルエンザHAタンパク質およびインフルエンザNAタンパク質と共に操作することにより形成したVLPが含まれる。
【0036】
Gagタンパク質ベースのVLPを生成させる例示的な方法
当業者に適用可能な任意の方法により、VLPを容易にアセンブリーすることができ、この結果、gagポリペプチドと、インフルエンザ抗原ポリペプチドとを包含するVLPがアセンブリーされることが好ましい。ある実施形態では、哺乳動物細胞による発現系および昆虫細胞による発現系など、脂質内に脂質ラフトドメインを包含する細胞ベースの発現系など、任意の適用可能なタンパク質発現系において、ポリペプチドを共発現させることができる。
【0037】
gagポリペプチドを用いて形成されるVLP発現の例が数多く公表されており、これにより、VLPを生成させるのに適用可能な発現系の範囲が裏付けられている。複数のレトロウイルスによる研究は、他のウイルスコンポーネントの非存在下で発現させたGagポリペプチドが、細胞表面におけるVLPの形成および出芽に十分であることを裏付けている(WillsおよびCraven、AIDS、5巻、639〜654頁、1991年; Zhouら、J. Virol.、68巻、2556〜2569頁、1994年; Morikawaら、Virology、183巻、288〜297頁、1991年; Royerら、Virology、184巻、417〜422頁、1991年; Gheysenら、Cell、59巻、103〜112頁、1989年; Hughesら、Virology、193巻、242〜255頁、1993年; Yamshchikovら、Virology、214巻、50〜58頁、1995年)。バキュロウイルスベクターを用いる昆虫細胞において、Gag前駆体を発現させるとVLPが形成されることについては、複数の研究グループにより裏付けられている(Delchambreら、EMBO J.、8巻、2653〜2660頁、1989年; Luoら、Virology、179巻、874〜880頁、1990年; Royer ら、Virology、184巻、417〜422頁、1991年; Morikawa ら、Virology、183巻、288〜297頁、1991年; Zhouら、J. Virol.、68巻、2556〜2569頁、1994年; Gheysenら、Cell、59巻、103〜112頁、1989年; Hughesら、Virology、193巻、242〜255頁、1993年; Yamshchikovら、Virology、214巻、50〜58頁、1995年)。これらのVLPは、未成熟レンチウイルス粒子に類似しており、昆虫細胞の細胞膜から出芽することにより、効率的にアセンブリーされて放出される。
【0038】
Gag前駆体のアミノ末端領域は、ウイルスアセンブリーに必要とされる、細胞表面への輸送および膜結合を標的とするシグナル領域であることが報告されている(Yuら、J. Virol.、66巻、4966〜4971頁、1992年; Yuan, Xら、J. Virol.、67巻、6387〜6394頁、1993年; Zhouら、J. Virol.、68巻、2556〜2569頁、1994年; LeeおよびLinial、J. Virol.、68巻、6644〜6654頁、1994年; Dorfmanら、J. Virol.、68巻、1689〜1696頁、1994年; Fackeら、J. Virol.、67巻、4972〜4980頁、1993年)。牛痘ウイルスによる発現系を用いて、Gag構造タンパク質のほか、Env糖タンパク質であるgp120およびgp41も含有する、組換えHIVベースのVLPのアセンブリーが報告されている(Haffarら、J. Virol.、66巻、4279〜4287頁、1992年)。
【0039】
エンベロープウイルスベースのVLP調製物中における感染作用物質を不活化する例示的方法
電磁放射は、エンベロープウイルスベースのVLPの免疫原性を実質的に低下させることなく、感染作用物質を不活化することが可能なので、例示的な不活化の方法は、電磁放射を介する方法である。電磁放射の例示的な3つの方式のすべて(すなわち、光反応性化合物を伴うUV照射、UV照射単独、およびガンマ照射)には、血液、食品、ワクチンなど、多種多様な試料中における病原体を不活化するのに用いられてきた長い歴史があるため、不活化用電磁放射を適用するための多種多様な装置が市販されており、これらは、本明細書で開示される方法を実施するのにほとんど〜まったく改変せずに用いることができる。さらに、当技術分野では、波長および線量の最適化がルーチン化しており、したがって、当業者の能力範囲内で容易に実施される。
【0040】
光反応性化合物を伴うUV照射
電磁放射による例示的な不活化の方法は、感染作用物質中のポリヌクレオチドと反応する光反応性化合物などの、光反応性化合物の存在下における、UV−A照射などの紫外線照射の組合せである。
【0041】
例示的な光反応化合物には、アクチノマイシン、アントラサイクリノン、アントラマイシン、ベンゾジピロン、フルオレン、フルオレノン、フロクマリン、イソアロキサジン、マイトマイシン、Monostral Fast Blue、Norphillin A、フェナントリジン、フェナザチオニウム塩、フェナジン、フェノチアジン、フェニルアジド、キノリン、およびチオキサンテノン(thiaxanthenones)が含まれる。1つの分子種は、2つの主要なカテゴリーのうちの1つに属するフロクマリンである。第1のカテゴリーは、ソラーレン[7H−フロ(3,2−g)−(1)−ベンゾピラン−7−オン、または6−ヒドロキシ−5−ベンゾフランアクリル酸のデルタ−ラクトン]であり、これらは、直鎖状であり、中央の芳香環部分に付随する2つの酸素残基が1,3の配向性を有し、またさらに、フラン環部分が二環クマリン系の6位に結合している。第2のカテゴリーは、イソソラーレン[2H−フロ(2,3−h)−(1)−ベンゾピラン−2−オン、または4−ヒドロキシ−5−ベンゾフランアクリル酸のデルタ−ラクトン]であり、これらは、曲鎖状であり、中央の芳香環部分に付随する2つの酸素残基が1,3の配向性を有し、またさらに、フラン環部分が二環クマリン系の8位に結合している。3、4、5、8、4’、または5’位において直鎖状フロクマリンを置換することによりソラーレン誘導体を生成させ得る一方、3、4、5、6、4’、または5位において曲鎖状フロクマリンを置換することによりイソソラーレン誘導体を生成させ得る。ソラーレンは、二本鎖核酸の塩基対間に挿入することができ、長波長の紫外光(UVA)を吸収すると、ピリミジン塩基に対する共有結合付加体を形成する。例えば、G. D. Ciminoら、Ann. Rev. Biochem.54巻:1151頁(1985年);Hearstら、Quart. Rev. Biophys.17巻:1頁(1984頁)を参照されたい。
【0042】
例示的なUV(または場合によって、可視光)放射の波長は、適切な反応および/または光付加体が生成される波長に依存し、これは、光反応性化学物質の化学反応に依存する。例示を目的として述べると、多くのソラーレンには、320〜380nmの波長にあるUV放射が極めて有効であり、330〜360nmが最大の有効性を示す。また、同様のUV−A波長は、419nmなど、可視光とも併せて用いうる光反応性化合物であるリボフラビンと併用しても、病原体を不活化するのに極めて有効である。
【0043】
UV照射単独
光反応性化合物の存在下におけるUV照射に加え、感染作用物質は、UV照射単独でも不活化することができる。ある実施形態では、放射の波長が、約180〜320nm、または約225〜290nm、または約254nm(すなわち、ポリヌクレオチドの高吸光度ピークおよび低減したタンパク質吸収を有するスペクトル領域)であるUV−C放射である。UV−C放射は、エンベロープを形成する脂質二重層およびエンベロープ内のタンパク質など、本明細書で開示されるエンベロープウイルスベースのVLPのコンポーネントに対して、安定性および免疫原性の両面で有害性が低い一方で、感染作用物質を不活化するのに十分なエネルギーを保持するので、用いることができる。しかし、例えば、UV−AおよびUV−Bなど、他の種類のUV放射もまた用いることができる。
【0044】
ガンマ照射
本明細書で開示される方法を実施して組成物を生成させるには、ガンマ照射(すなわち、イオン化放射)もまた用いることができる。この実施形態では、10〜60kGyのガンマ照射線量が、病原体の不活化に有効である。ガンマ照射は、感染作用物質のゲノムをコードするポリヌクレオチド内において鎖の切断を導入することにより、感染作用物質を直接不活化することもでき、該ポリヌクレオチドを損傷させるフリーラジカルを生成することにより、感染作用物質を間接的に不活化することもできる。エンベロープVLPの脂質コンポーネントおよびタンパク質コンポーネントへの、ラジカルを介する損傷を阻止するには、ガンマ照射と共に、フリーラジカルスカベンジャーおよび低温を用いることができる。
【0045】
VLPを用いる例示的な方法
製剤
本明細書で説明されるエンベロープウイルスベースのVLPの例示的な使用は、ワクチン調製物としての使用である。典型的には、このようなワクチンは、溶液または懸濁液の注射剤として調製されるが、注射前に液体中に溶解させるか、または懸濁させるのに適する固体形態もまた調製することができる。このような調製物はまた、乳化させることもでき、乾燥粉末として生成させることもできる。免疫原性の有効成分は、薬学的に許容され、また、有効成分に適合する、賦形剤と混合することが多い。適切な賦形剤は、例えば、水、生理食塩液、デキストロース、スクロース、グリセロール、エタノールなどであり、また、これらの組合せである。加えて、所望の場合、ワクチンは、湿潤剤もしくは乳化剤、pH緩衝剤、または該ワクチンの有効性を増強するアジュバントを含有し得る。
【0046】
ワクチンは、注射、例えば、皮下(subcutaneously)注射、皮内注射、皮下(subdermally)注射、または筋肉内注射を介する、従来の非経口投与が可能である。他の投与方式に適する、さらなる製剤には坐剤が包含され、場合によっては、経口製剤、鼻腔内製剤、口腔内製剤、舌下製剤、腹腔内製剤、膣内製剤、経肛門製剤、および頭蓋内製剤も包含される。坐剤の場合、従来の結合剤および担体には、例えば、ポリアルキレングリコール(polyalkalene glycol)またはトリグリセリドが包含される場合があり、このような坐剤は、0.5%〜10%、または1〜2%の範囲でも、有効成分を含有する混合物から形成することができる。ある実施形態では、脂肪酸グリセリドまたはココアバターの混合物などの低温溶融ワックスをまず溶融させ、本明細書で説明されるエンベロープウイルスベースのVLPを、例えば、撹拌することにより、均一に分散させる。次いで、溶融した均一の混合物を、適当なサイズの鋳型に流し込み、冷却して固形化させる。
【0047】
鼻腔内送達に適する製剤には、液体(例えば、エアゾールまたは点鼻液として投与する水溶液)および乾燥粉末(例えば、鼻孔内への急速沈殿用)が包含される。製剤は、例えば、医薬品グレードのマンニトール、ラクトース、スクロース、トレハロース、キシリトール、およびキトサンなど、通常用いられる賦形剤を包含する。液体製剤または粉末製剤において、キトサンなどの粘膜付着剤を用いて、鼻腔内投与された製剤の粘液線毛クリアランスを遅延させることができる。マンニトールおよびスクロースなどの糖を、液体製剤では安定化剤として用いることができ、また、乾燥粉末製剤では安定化剤、充填剤、または粉末流動剤および粉末サイズ剤として用いることができる。加えて、モノホスホリル脂質A(MPL)またはCpGオリゴヌクレオチド(oligoneucleotides)などのアジュバントを、液体製剤および乾燥粉末製剤の両方において、免疫刺激性アジュバントとして用いることもできる。
【0048】
経口送達に適する製剤には、液体、固体、半固体、ゲル、錠剤、カプセル、トローチなどが包含される。経口送達に適する製剤には、錠剤、トローチ、カプセル、ゲル、液体、食品、飲料、栄養補助食品などが包含される。製剤は、例えば、医薬品グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなど、通常用いられる賦形剤を包含する。他のエンベロープウイルスベースのVLPワクチン組成物は、溶液、懸濁液、丸薬、持続放出製剤、または粉末の形態を取る場合があり、10〜95%の有効成分、または25〜70%の有効成分を含有する場合がある。経口製剤の場合、コレラ毒素が興味深い製剤パートナーである(また、可能なコンジュゲーションパートナーでもある)。
【0049】
膣内投与用に調合する場合、エンベロープウイルスベースのVLPワクチンは、ペッサリー、タンポン、クリーム、ゲル、ペースト、フォーム、またはスプレーの形態であり得る。前出の製剤のうちのいずれもが、エンベロープウイルスベースのVLPに加えて、担体など、当技術分野で適切であることが公知の薬剤を含有し得る。
【0050】
一部の実施形態では、エンベロープウイルスベースのVLPワクチンを、全身送達用に調合する場合もあり、局所送達用に調合する場合もある。このような製剤は、当技術分野において周知である。非経口媒体には、塩化ナトリウム溶液、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、乳酸加リンゲル液、または固定油が包含される。静脈内媒体には、流体および栄養物質補給剤、電解質補給剤(リンゲルデキストロースに基づく補給剤など)などが包含される。全身投与経路および局所投与経路には、例えば、皮内経路、局所適用経路、静脈内経路、筋肉内経路などが包含される。
【0051】
エンベロープウイルスベースのVLPは、中性製剤または塩ベースの製剤を包含するワクチンに調合することができる。薬学的に許容される塩には、酸添加塩(ペプチドの遊離アミノ基と共に形成され、また、例えば、塩酸もしくはリン酸などの無機酸、または酢酸、シュウ酸、酒石酸、マンデル酸などの有機酸と共に形成される)が包含される。遊離カルボキシル基と共に形成される塩はまた、例えば、水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、または水酸化鉄などの無機塩基、また、イソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジン、プロカインなどの有機塩基に由来し得る。
【0052】
ワクチンは、剤形に適合した形で、また、治療的に有効であり免疫原性となる量で投与することができる。投与量は、例えば、個体の免疫系が免疫反応を誘発する能力、および所望される防御の程度を包含する、治療される対象に依存する。適切な用量範囲は、1回のワクチン接種当たりの有効成分が数百マイクログラムのオーダーであり、例示的な範囲は、約0.5μg〜1000μgの範囲、1μg〜500μgの範囲、または、約10μg〜100μgの範囲など、約0.1μg〜2000μgの範囲である(1〜10mgの範囲にある高量も意図される)。初回投与および追加投与に適するレジメンもまた可変的であるが、初回投与にその後の接種または他の投与が続く形が典型的である。
【0053】
適用方式は多様に変化させることができる。従来のワクチン投与方法のうちのいずれもが適用可能である。これらには、固体の生理学的に許容されるベース上における経口適用、または注射など、生理学的に許容される分散剤による非経口適用が包含される。ワクチンの用量は、投与経路に依存し、ワクチン接種される人の年齢、および抗原製剤により変化する。
【0054】
ワクチン製剤の一部は、製剤自体でワクチンとして十分に免疫原性であるが、他の製剤では、該ワクチンがアジュバント物質をさらに含むと、免疫反応が増強される場合がある。
【0055】
とりわけ、鼻腔内ベース、経口ベース、または肺内ベースの送達製剤の場合、送達および免疫原性を改善するには、粘膜付着を改善する送達剤を用いることもできる。このような化合物である、キチンのN−脱アセチル化形態であるキトサンは、多くの医薬製剤で用いられる(32)。それが粘液線毛クリアランスを遅延させ、粘膜による抗原の取込みおよびプロセシングのためにより多くの時間を許容する能力のために、キトサンは、鼻腔内におけるワクチン送達にとって魅力的な粘膜付着剤である(33、34)。加えて、キトサンは、密着結合を一過性に開口させ、これにより、NALTへの抗原の経上皮輸送が増強され得る。近年のヒト試験では、キトサンを伴うが、さらなるアジュバントは伴わずに、三価の不活化インフルエンザワクチンを鼻腔内投与したところ、筋肉内接種後に得られたセロコンバージョンおよびHI力価をごくわずかに下回るセロコンバージョンおよびHI力価がもたらされた(33)。
【0056】
キトサンはまた、遺伝子的に解毒したE. coli易熱性エンテロトキシンの突然変異体であるLTK63など、鼻腔内において良好に機能するアジュバントと共に調合することもできる。これにより、キトサンにより付与される送達および付着の利益の上に、免疫刺激効果が付加され、その結果、粘膜反応および全身反応の増強をもたらす(35)。
【0057】
最後に、キトサン製剤はまた、ワクチンの安定性を改善し、さらに、粘液線毛クリアランスにおいて液体製剤を上回る遅延を結果としてもたらすことが示されている乾燥粉末フォーマットでも調製することができる(42)。これは、キトサンと共に調合された、鼻腔内用乾燥粉末によるジフテリア毒素ワクチンを伴う、近年のヒト臨床試験において見られ、ここで、鼻腔内経路は、従来の筋肉内経路と同程度に有効であり、分泌性IgA反応の利益が付加された(43)。該ワクチンはまた、忍容性も極めて良好であった。キトサンおよびMPLを含有する炭疽菌用の鼻腔内乾燥粉末ワクチンは、ウサギにおいて、筋肉内接種より強力な反応を誘導し、また、エアゾールによる胞子投与に対しても防御的であった(44)。
【0058】
鼻腔内ワクチンは、下気道に対してより良好に作用する非経口投与ワクチンに対して、上気道および下気道に作用し得るので、好ましい製剤を表わす。これは、アレルゲンベースのワクチンには忍容性を誘導し、また、病原体ベースのワクチンには免疫性を誘導する点で有益である。
【0059】
鼻腔内ワクチンは、上気道および下気道の両方において防御をもたらすのに加え、注射針による接種の煩雑さを回避し、また、粒子抗原および/または可溶性抗原が、鼻咽頭関連リンパ組織(NALT)と相互作用することを介して、粘膜および全身における体液性反応および細胞性反応の両方を誘導する手段をもたらす(16〜19)。歴史的に、鼻腔内経路は、非経口接種ほど有効ではなかったが、新規の送達製剤であるエンベロープウイルスベースのVLPおよびアジュバントを用いることにより、この枠組みが変わりつつある。実際、鼻腔粘膜内にはシアル酸を含有する受容体が豊富であるため、鼻腔内送達には、機能性の赤血球凝集素分子を含有するインフルエンザワクチンがとりわけよく適しており、その結果、HA抗原の結合が増強され、粘液線毛クリアランスが低減される可能性をもたらす。
【0060】
インフルエンザについては、並行する非経口投与の免疫原性が低く、ヘテロ亜型的防御が誘導されなかった実験の鼻腔内ワクチン送達後において、ヘテロ亜型的防御を含めた防御的免疫反応が報告されている(20〜22)。さらに、不活化インフルエンザは、鼻腔内投与されるサル免疫不全ウイルス(SIV)によるVLPワクチンと混合すると、全身および粘膜における体液性反応および細胞性反応に有効なアジュバントであることも示されている(23)。このアジュバント効果は、不活化インフルエンザビリオンが、VLPと共に凝集し、粘膜表面への結合を増強する能力に帰せられた。また、インフルエンザHAタンパク質をSIV VLPに直接に組み込んだところ、樹状細胞(DC)への結合およびDCの活性化が増強された場合にも、同様のアジュバント効果が見られた(24、25)。
【0061】
アジュバント
ワクチンに対するアジュバント効果を達成する各種の方法が公知であり、本明細書で開示されるエンベロープウイルスベースのVLPと共に用いることができる。一般的な原理および方法は、「The Theory and Practical Application of Adjuvants」、1995年、Duncan E. S. Stewart−Tull(編)、John Wiley & Sons Ltd.、ISBN 0−471−95170−6において詳述されており、また、「Vaccines: New Generation Immunological Adjuvants」、1995年、Gregoriadis Gら(編)、Plenum Press、New York、ISBN 0−306−45283−9においても詳述されており、どちらも参照により本明細書に組み込まれる。
【0062】
一部の実施形態では、VLPワクチンが、重量ベースの比率約10:1〜約1010:1のVLP:アジュバント、例えば、約10:1〜約100:1、約100:1〜約103:1、約103:1〜約104:1、約104:1〜約105:1、約105:1〜約106:1、約106:1〜約107:1、約107:1〜約108:1、約108:1〜約109:1、または約109:1〜約1010:1のVLP:アジュバントで、少なくとも1つのアジュバントと混合したエンベロープウイルスベースのVLPを含む。当業者は、アジュバントに関する情報、また、最適の比を決定する日常的実験に関する情報により、適切な比率を容易に決定することができる。当業者は、アジュバントに関する情報、および最適の比率を決定する日常的実験に関する情報により、適切な比率を容易に決定することができる。限定なしに述べると、本明細書で開示されるVLPとアジュバントとの混合物には、同じ溶液における個別のVLPとアジュバントとの混合物、共有結合させたVLPおよびアジュバント、イオン結合させたVLPおよびアジュバント、疎水性結合させたVLPおよびアジュバント(VLP膜内に部分的または完全に包埋させることを含めた)、親水性結合させたVLPおよびアジュバント、ならびに前出の任意の組合せを含め、当業者に利用可能な任意の形態の組合せが含まれうる。
【0063】
例示的なアジュバントには、toll様受容体(TLR)アゴニスト、モノホスホリル脂質A(MPL)、合成脂質A、脂質Aの模倣体または類似体、アルミニウム塩、サイトカイン、サポニン、ムラミルジペプチド(MDP)誘導体、CpGオリゴ、グラム陰性菌のリポ多糖(LPS)、ポリホスファゼン、エマルジョン、水中油エマルジョン、ビロソーム、コクリエート、ポリ(ラクチド−co−グリコリド)(PLG)マイクロ粒子、ポロキサマー粒子、マイクロ粒子、およびリポソームが包含され得るがこれらに限定されない。アジュバントは、細菌由来の外毒素ではないことが好ましい。好ましいアジュバントは、3DMPL、CpGオリゴヌクレオチド、またはQS21など、Th1型反応を刺激するアジュバントである。
【0064】
Salmonellaに由来する、脂質Aの非毒性誘導体であるモノホスホリル脂質A(MPL)は、ワクチンアジュバントとして開発された、強力なTLR−4アゴニストである(Evansら、2003年)。マウスによる前臨床研究では、鼻腔内MPLが、分泌性ならびに全身性の体液反応を増強することが示されている(Baldridgeら、2000年;Yangら、2002年)。MPLはまた、120,000例を超える患者による臨床研究でも、ワクチンアジュバントとして安全かつ有効であることが分かっている(Baldrickら、2002年;2004年)。MPLは、TLR−4受容体を介して自然免疫の誘導を刺激し、したがって、グラム陰性菌およびグラム陽性菌、ウイルス、および寄生虫を包含する、広範な範囲にわたる感染性病原体に対して、非特異的な免疫反応を誘発することが可能である(Baldrickら、2004年;Persingら、2002年)。鼻腔内製剤中にMPLを包含することにより、ウイルス感作に由来する非特異的な免疫反応を誘発する自然反応が迅速に誘導される一方で、該ワクチンの抗原成分により生成させる特異的反応が増強される。
【0065】
したがって、一実施形態では、本発明が、モノホスホリル脂質A(MPL(登録商標))または3−O−脱アシル化モノホスホリル脂質A(3 De−O−acylated monophosphoryl lipid A)(3D−MPL(登録商標))を、後天免疫および自然免疫の増強剤として含む組成物を提供する。化学的に述べると、3D−MPL(登録商標)とは、4位アシル化鎖、5位アシル化鎖、または6位アシル化鎖との3−O−脱アシル化モノホスホリル脂質Aの混合物である。3−O−脱アシル化モノホスホリル脂質Aの例示的な形態は、欧州特許第0689454B1号(SmithKline Beecham Biologicals SA)において開示されている。別の実施形態では、本発明が、合成脂質A、BioMiraによるPET Lipid Aなど、脂質Aの模倣体もしくは類似体、またはTLR−4アゴニストと同様に機能するようにデザインされている合成誘導体を含む組成物を提供する。
【0066】
例示的なアジュバントは、VLPポリペプチドと共発現させるか、またはVLPポリペプチドと融合させて、キメラポリペプチドを生成させることにより、本明細書で説明されるエンベロープウイルスベースのVLPに容易に添加し得るポリペプチドアジュバントである。鞭毛の主要なタンパク質成分である細菌性フラジェリンは、それが、toll様受容体であるTLR5を介する先天性免疫系により認識されるために、アジュバントタンパク質としてますます注目を集めているアジュバントである(65)。TLR5を介するフラジェリンシグナル伝達は、DCの成熟化および移動の他、マクロファージ、好中球、および腸上皮細胞の活性化も誘導することにより、先天性免疫機能および後天性免疫機能の両方に影響を及ぼし、その結果、炎症促進メディエーターを生成させる(66〜72)。
【0067】
TLR5は、免疫学的圧力に応答するその突然変異を除外し、このタンパク質に固有であり、また、鞭毛機能に必要とされる、フラジェリン単量体内における保存的構造を認識する(73)。該受容体は、100fMの濃度に対して感受性であるが、完全な線維は認識しない。鞭毛が単量体に解体されることが、結合および刺激に必要とされる。
【0068】
アジュバントとしてのフラジェリンは、非経口投与または鼻腔内投与された異種抗原に対する防御反応を誘導する強力な活性を有し(66、74〜77)、また、DNAワクチンに対するアジュバント効果もまた報告されている(78)。マウスまたはサルにおいてフラジェリンを用いると、Th2バイアス(これは、インフルエンザなどの呼吸器ウイルスには適切である)は観察されるが、IgE誘導の証拠は観察されていない。加えて、サルにおける鼻腔内投与または全身投与後における、局所炎症反応または全身炎症反応も報告されていない(74)。TLR5を介して、MyD88依存的な形でなされるフラジェリンシグナル、およびTLRを介する、他のすべてのMyD88依存的シグナルは、Th1バイアスを結果としてもたらすことが示されている(67、79)ため、フラジェリンを使用した後で誘発される反応におけるTh2の特徴は、ある意味で驚くべきことである。フラジェリンに対して予め存在する抗体が、アジュバントの有効性に対してさほどの影響を及ぼさない(74)ことから、フラジェリンは、複数回使用のためのアジュバントとして魅力的となっていることが重要である。
【0069】
近年における鼻腔内ワクチン試験における共通の主題は、ワクチンの有効性を改善するアジュバントおよび/または送達系の使用である。このような一研究では、遺伝子的に解毒したE. coli易熱性エンテロトキシンアジュバント(LT R192G)を含有するインフルエンザH3ワクチンが、H5による感作に対してヘテロ亜型的な防御を結果としてもたらしたが、これは、鼻腔内送達後に限られた。防御は、交差中和抗体の誘導に基づき、これにより、新たなインフルエンザワクチンの開発における鼻腔内経路についての重要な示唆が示された(22)。
【0070】
サイトカイン、コロニー刺激因子(例えば、GM−CSF、CSFなど);腫瘍壊死因子;インターロイキン−2、インターロイキン−7、インターロイキン−12、および他の同様の増殖因子は、VLPポリペプチドと混合または融合させることにより、エンベロープウイルスベースのVLPワクチン内に容易に包含され得るので、これらもまたアジュバントとして用いることができる。
【0071】
一部の実施形態では、本明細書で開示されるエンベロープウイルスベースのVLPワクチン組成物が、CpGオリゴヌクレオチドを含む、核酸のTLR9リガンド;イミダゾキノリンのTLR7リガンド;置換グアニンのTLR7/8リガンド;ロキソリビン、7−デアザデオキシグアノシン、7−チア−8−オキソデオキシグアノシン、二本鎖ポリ(I:C)、ポリイノシン酸、イミキモド(R−837)、およびレシキモド(R−848)などの他のTLR7リガンド;またはMPL(登録商標)もしくは合成誘導体などのTLR4アゴニストなど、Toll様受容体を介して作用する他のアジュバントを包含し得る。
【0072】
ある種のアジュバントは、樹状細胞などのAPCによるワクチン分子の取込みを促進し、これらを活性化する。非限定的な例は、免疫標的化アジュバント;毒素、サイトカイン、およびマイコバクテリア派生物などの免疫調節アジュバント;油製剤;ポリマー;ミセル形成アジュバント;サポニン;免疫刺激複合体マトリックス(ISCOMマトリックス);粒子;DDA;アルミニウムアジュバント;DNAアジュバント;MPL;および封入アジュバントからなる群から選択される。
【0073】
アジュバントのさらなる例には、緩衝生理食塩液中0.05〜0.1パーセントの溶液として一般的に用いられる水酸化物またはリン酸などのアルミニウム塩(ミョウバン)(例えば、Nicklas(1992年)Res. Immunol.143巻:489〜493頁を参照されたい)、0.25パーセントの溶液として用いられる糖の合成ポリマー(例えば、Carbopol(登録商標))との混合物、それぞれ、70℃〜101℃の範囲の温度で30秒間〜2分間にわたる熱処理によるワクチン中のタンパク質の凝集物などの薬剤が包含され、また、架橋形成剤による凝集物も可能である。ペプシン処理した、アルブミンに対する抗体(Fab断片)との再活性化による凝集物、C. parvumまたはグラム陰性菌の内毒素もしくはリポ多糖コンポーネントなどの細菌細胞との混合物、モノオレイン酸マンニド(Aracel A)などの生理学的に許容される油媒体中におけるエマルジョン、または保護置換基として用いられる、ペルフルオロカーボン(Fluosol−DA)の20パーセント溶液によるエマルジョンもまた用いることができる。スクアレンおよびIFAなどの油との混合物もまた用いることができる。
【0074】
DDA(ジメチルジオクタデシルアンモニウムブロミド)は、アジュバントの興味深い候補物質であるが、フロイントの完全アジュバントおよびフロイントの不完全アジュバントの他、QuilAおよびQS21などのキラヤサポニンもまた興味深い。さらなる可能性には、モノホスホリル脂質A(MPL(登録商標))、ムラミルジペプチド(MDP)、およびトレオニルムラミルジペプチド(tMDP)など、リポ多糖のポリ[ジ(カルボキシレートフェノキシ)ホスファゼン(poly[di(earboxylatophenoxy)phosphazene)(PCPP)誘導体が包含される。例えば、3−O−脱アシル化モノホスホリル脂質Aなどのモノホスホリル脂質Aをアルミニウム塩と併せた組合せを包含する、主にTh1型の反応をもたらすリポ多糖ベースのアジュバントを用いることができる。MPL(登録商標)アジュバントは、GlaxoSmithKlineから市販されている(例えば、それらの各々が、それらのリポ多糖に関する教示に対する具体的な言及を伴うそれらの全体において参照により組み込まれる、米国特許第4,436,727号;同第4,877,611号;同第4,866,034号;および同第4,912,094号を参照されたい)。
【0075】
リポソーム製剤もまた、アジュバント効果を付与することが知られており、したがって、リポソームアジュバントは、エンベロープウイルスベースのVLPと併用して用いることができる。
【0076】
免疫刺激複合体マトリックス型(ISCOM(登録商標)マトリックス)アジュバントは、とりわけ、この型のアジュバントが、APCによるMHCクラスII発現を上方制御することが可能であるため、エンベロープウイルスベースのVLPワクチンと共に用いることもできる。ISCOMマトリックスは、Quillaja saponariaに由来する(場合によって画分された)サポニン(トリテルペノイド)、コレステロール、およびリン脂質からなる。VLP中のものなどの免疫原性タンパク質と混合する場合、結果として得られる粒子状製剤は、サポニンが60〜70%w/wを占めることが可能であり、コレステロールおよびリン脂質が10〜15%w/w、およびタンパク質が10〜15%w/wを占めることが可能な、ISCOM粒子として公知の製剤である。免疫刺激複合体の組成および使用に関する詳細は、例えば、アジュバントを扱う上述の教科書中に見出し得るが、また、Morein Bら、1995年、Clin. Immunother.3巻:461〜475頁の他、Barr I GおよびMitchell G F、1996年、Immunol. and Cell Biol.74巻:8〜25頁(どちらも参照により本明細書に組み込まれる)も、完全な免疫刺激複合体を調製するのに有益な教示を提供している。
【0077】
本明細書で開示されるエンベロープウイルスベースのVLPワクチンとのアジュバントの組合せにおいて用い得るサポニンには、それらがISCOM形態にある場合であれ、そうでない場合であれ、米国特許第5,057,540号(Quil A画分、およびそれらを単離して用いる方法に対する具体的な言及を伴うその全体において参照により本明細書に組み込まれる)、および「Saponins as vaccine adjuvants」、Kensil, C. R.、Crit Rev Ther Drug Carrier Syst、1996年、12巻(1〜2号):1〜55頁;およびEP0362279B1で説明される、Quil Aと称する、Quillaja Saponaria Molinaの樹皮に由来するサポニン、ならびにそれらの画分が包含される。Quil Aの例示的な画分は、QS21、QS7、およびQS17である。
【0078】
β−エスシンは、本明細書で説明されるエンベロープウイルスベースのVLPワクチンのアジュバント組成物中で用いる別の溶血性サポニンである。エスシンは、メルクインデックス(第12版:登録番号3737)において、ラテン名:Aesculus hippocastanumであるトチノキの種子内で発生するサポニンの混合物として説明されている。クロマトグラフィーおよび精製(Fiedler、Arzneimittel−Forsch.4巻、213頁(1953年))による、およびイオン交換樹脂(Erbringら、米国特許第3,238,190号)によるその単離が、説明されている。エスシンの画分は精製されており、生物学的に活性であることが示されている(Yoshikawa Mら(Chem Pharm Bull(東京)、1996年8月;44巻(8号):1454〜1464頁))。β−エスシン(escin)はまた、エスシン(aescin)としても公知である。
【0079】
エンベロープウイルスベースのVLPワクチンと共に用いる別の溶血性サポニンは、ジギトニンである。ジギトニンは、メルクインデックス(第12版:登録番号3204)において、Digitalis purpureaの種子に由来し、また、Gisvoldら、J.Am.Pharm.Assoc.、1934年、23巻、664頁;およびRuhenstroth−Bauer、Physiol.Chem.、1955年、301巻、621頁において説明される手順により精製されるサポニンとして説明されている。その使用は、コレステロールを判定するための臨床試薬であると説明されている。
【0080】
アジュバント効果を達成する別の興味深い可能性は、Gosselinら、1992年(参照により本明細書に組み込まれる)において説明される技法を用いることである。略述すると、本明細書で説明されるVLPポリペプチドまたはさらなる抗原などの関与抗原の提示は、該抗原を、単球/マクロファージ上におけるFC受容体に対する抗体(または抗原に結合する抗体断片)にコンジュゲートすることにより増強することができる。とりわけ、抗原と抗FCRIとのコンジュゲートは、ワクチン接種のための免疫原性を増強することが示されている。該抗体は、VLPポリペプチドのうちのいずれか1つに対する融合体としての発現によることを包含する生成後において、または該生成の一部として、エンベロープウイルスベースのVLPにコンジュゲートすることができる。
【0081】
他の可能性は、標的化物質および免疫調節物質(すなわち、サイトカイン)の使用を伴う。加えて、ポリI:Cなど、合成のサイトカイン誘導物質もまた用いることができる。
【0082】
適切なマイコバクテリア派生物は、ムラミルジペプチド、フロイントの完全アジュバント、ならびに、TDMおよびTDEなど、トレハロースのジエステルからなる群から選択することができる。
【0083】
適切な免疫標的化アジュバントの例には、CD40リガンドおよびCD40抗体、またはこれらの特異的結合断片(上記の議論を参照されたい)、マンノース、Fab断片、ならびにCTLA−4が包含される。
【0084】
適切なポリマーアジュバントの例には、デキストランなどの炭水化物、PEG、デンプン、マンナン、およびマンノース;塑性ポリマー;ならびにラテックスビーズなどのラテックスが包含される。
【0085】
免疫反応を調節するさらに別の興味深い方法は、「仮想リンパ節」(VLN)(ImmunoTherapy、Inc.、360 Lexington Avenue, New York、N.Y. 10017−6501により開発された、特許品の医療デバイス)内に免疫原(場合によって、アジュバント、ならびに薬学的に許容される担体および媒体と併せて)を包含することである。VLN(細長型の管状デバイス)は、リンパ節の構造および機能を模倣する。VLNを皮下に挿入することにより、サイトカインおよびケモカインの急増による無菌的炎症部位が創出される。T細胞およびB細胞の他、APCも、危険シグナルに応答し、炎症部位に至り、VLNの多孔性マトリックスの内部に蓄積される。VLNを用いると、抗原に対する免疫反応を誘発するのに必要とされる抗原用量が低下し、VLNを用いるワクチン接種により付与される免疫的防御が、Ribiをアジュバントとして用いる従来の免疫化を凌駕することが示されている。この技法は、Gelber Cら、1998年、「Elicitation of Robust Cellular and Humoral Immune Responses to Small Amounts of Immunogens Using a Novel Medical Device Designated the Virtual Lymph Node」、「From the Laboratory to the Clinic、梗概、1998年10月12〜15日、Seascape Resort、Aptos Calif.」において略述されている。
【0086】
エンベロープウイルスベースのVLPと共に、オリゴヌクレオチドをアジュバントとして用いることができ、少なくとも3つ、または少なくとも6つ以上のヌクレオチドで隔てられた2つ以上のジヌクレオチドによるCpGモチーフを含有することができる。CpGを含有するオリゴヌクレオチド(ここで、CpGジヌクレオチドは、メチル化されていない)は、主にTh1反応を誘導する。このようなオリゴヌクレオチドは周知であり、例えば、それらの各々が、CpGオリゴヌクレオチドをアジュバントとして生成させて用いる方法に対する具体的な言及を伴うそれらの全体において参照により本明細書に組み込まれる、WO96/02555、WO99/33488、ならびに米国特許第6,008,200号および同第5,856,462号において説明されている。
【0087】
このようなオリゴヌクレオチドによるアジュバントは、デオキシヌクレオチドであり得る。ある実施形態では、該オリゴヌクレオチド内のヌクレオチド骨格は、ホスホロジチオエート結合であるか、またはホスホロチオエート結合であるが、骨格結合を混合したオリゴヌクレオチドを包含する、ホスホジエステル骨格、およびPNAなど他のヌクレオチド骨格をエンベロープウイルスベースのVLPワクチンと共に用いることができる。ホスホロチオエートオリゴヌクレオチドまたはホスホロジチオエートを生成させる方法は、それらの各々が、ホスホロチオエートおよびホスホロジチオエートについての教示に対する具体的な言及を伴うそれらの全体において参照により本明細書に組み込まれる、米国特許第5,666,153号、米国特許第5,278,302号、およびW095/26204において説明されている。
【0088】
例示的なオリゴヌクレオチドは、以下の配列を有する。配列は、ホスホロチオエート修飾されたヌクレオチド骨格を含有することができる。
【0089】
【化1】
代替的なCpGオリゴヌクレオチドには、これらに対する重要でない欠失または付加を伴う上記の配列が包含される。アジュバントとしてのCpGオリゴヌクレオチドは、当技術分野で公知である任意の方法(例えば、EP468520)により合成することができる。例えば、自動合成器を用いてこのようなオリゴヌクレオチドを合成し得る。このようなオリゴヌクレオチドによるアジュバントは、10〜50塩基の長さであり得る。別のアジュバント系は、CpGを含有するオリゴヌクレオチドと、サポニン誘導体との組合せを伴い、特に、CpGとQS21との組合せが、WO00/09159において開示されている。
【0090】
多くの単相または複相のエマルジョン系が説明されている。エマルジョンが、エンベロープウイルスベースのVLPの構造を破壊しないように、当業者は、エンベロープウイルスベースのVLPと共に用いられるこのようなエマルジョン系を容易に適合させることができる。水中油エマルジョンによるアジュバントは、それ自体、アジュバント組成物として有用であることが示唆されており(EPO399843B)、また、水中油エマルジョンと他の活性薬剤との組合せも、ワクチン用のアジュバントとして説明されている(WO95/17210;WO98/56414;WO99/12565;WO99/11241)。油中水エマルジョン(米国特許第5,422,109号;EP0480982B2)、および水中油中水エマルジョン(米国特許第5,424,067号;EP0480981B)など、他の油エマルジョンによるアジュバントが説明されている。
【0091】
本明細書で説明されるエンベロープウイルスベースのVLPと共に用いられる油エマルジョンによるアジュバントは、天然の場合もあり、合成の場合もあり、また、無機性の場合もあり、有機性の場合もある。鉱油および有機油の例は、当業者に容易に明らかであろう。
【0092】
任意の水中油組成物が、ヒトへの投与に適するためには、エマルジョン系の油相が、代謝性油を包含していてよい。代謝性油という用語の意味は、当技術分野において周知である。代謝性とは、「代謝により変換可能であること」と定義することができる(「Dorland’s Illustrated Medical Dictionary」、W.B. Sanders Company、第25版(1974年))。油は、受容者に毒性でなく、代謝により変換可能な、任意の植物油、魚油、動物油、または合成油であることが可能である。堅果(ラッカセイ油など)、種、穀類が、植物油の一般的な供給源である。合成油もエンベロープウイルスベースのVLPワクチンと共に用いることができ、これらには、NEOBEE(登録商標)および他の油など、市販の油が包含され得る。スクアレン(2,6,10,15,19,23−ヘキサメチル−2,6,10,14,18,22−テトラコサヘキサエン)は、サメの肝油中に高量で見出され、また、オリーブ油、コムギ胚種油、コメぬか油中、および酵母に低量で見出される不飽和油であり、本明細書で開示されるエンベロープウイルスベースのVLPワクチンと共に用いることができる。スクアレンは、コレステロールの生合成における中間体であるという事実により、代謝性油である(「メルクインデックス」、第10版、登録番号8619)。
【0093】
例示的な油エマルジョンは水中油エマルジョンであり、特に、水中スクアレンエマルジョンである。
【0094】
加えて、エンベロープウイルスベースのVLPワクチンにおいて用いる油エマルジョンによるアジュバントには、油であるα−トコフェロール(ビタミンE、EP0382271B1)などの、抗酸化剤が含まれ得る。
【0095】
WO95/17210およびWO99/11241は、場合によって、免疫刺激剤であるQS21および/または3D−MPLと共に調合される、スクアレン、α−トコフェロール、およびTWEEN 80(TM)に基づくエマルジョンによるアジュバントを開示している。WO99/12565は、油相へのステロールの添加による、これらのスクアレンエマルジョンに対する改善を開示している。加えて、エマルジョンを安定化させるために、トリカプリリン(C27H50O6)などのトリグリセリドを、油相に添加することができる(WO98/56414)。
【0096】
安定的な水中油エマルジョン中で見出される油滴のサイズは、光子相関分光分析により測定したときに、1ミクロン未満であってよく、実質的に30〜600nmの範囲内、実質的に直径約30〜500nm、実質的に直径150〜500nm、または直径約150nmであり得る。この点で、油滴数の80%がこれらの範囲内にあるものとし、油滴数の90%、または油滴数の95%超が規定のサイズ範囲内にある。油エマルジョン中に存在する成分の量は、従来、スクアレンなどの油が2〜10%の範囲内にあり;また、存在する場合、アルファトコフェロールが2〜10%であり、ポリオキシエチレンソルビタンモノオレエートなどの界面活性剤が0.3〜3%である。油:アルファトコフェロールの比は、より安定的なエマルジョンがもたらされるため、1以下であり得る。SPAN 85(TM)もまた、約1%のレベルで存在し得る。一部の場合には、本明細書で開示されるエンベロープウイルスベースのVLPワクチンが、安定化剤をさらに含有することが有利であり得る。
【0097】
水中油エマルジョンを生成させる方法は、当業者に周知である。一般に、該方法は、油相を、PBS/TWEEN80(登録商標)溶液などの界面活性剤と混合し、その後、ホモジナイザーを用いるホモジナイゼーションを包含するステップを含むが、該混合物に、シリンジ針内を2回流過させるステップを含む方法が、小容量の液体をホモジナイズするのには適切であることが、当業者には明らかであろう。同様に、当業者は、microfluidizer(M110S型マイクロ流体生成器、最大入力圧6バールで2分間にわたり、最大50回の流過(約850バールの出力圧))内で乳化過程を適合させれば、より低量または高量のエマルジョンを生成させることができるであろう。この適合は、必要とされる直径の油滴を有する調製物が達成されるまで、結果として得られるエマルジョンの測定を含む日常的な実験により達成し得るであろう。
【0098】
本明細書で開示されるエンベロープウイルスベースのVLPによるワクチン調製物は、前記ワクチンを、鼻腔内投与、筋肉内投与、腹腔内投与、皮内投与、経皮投与、静脈内投与、または皮下投与を介して投与することにより、ウイルス性インフルエンザを受けやすいか、またはこれを患う、哺乳動物またはトリを防御または治療するのに用いることができる。ワクチン調製物を全身投与する方法には、従来のシリンジおよび注射針、または固体ワクチンを弾道送達するためにデザインされたデバイス(WO99/27961)、または注射針なしの高圧液体ジェットデバイス(米国特許第4,596,556号;米国特許第5,993,412号)、または経皮パッチ(WO97/48440;WO98/28037)が包含され得る。エンベロープウイルスベースのVLPワクチンはまた、皮膚に貼付することもできる(経皮(transdermalまたはtranscutaneous)送達、WO98/20734;WO98/28037)。したがって、本明細書で開示されるエンベロープウイルスベースのVLPワクチンは、エンベロープウイルスベースのVLPワクチンまたはアジュバントによる組成物を予め充填した全身投与用の送達デバイスを包含し得る。したがって、哺乳動物またはトリなどの個体において免疫反応を誘導する方法であって、本明細書で説明されるエンベロープウイルスベースのVLP組成物のうちのいずれかを含み、また、場合によって、アジュバントおよび/または担体を包含するワクチンを該個体に投与するステップを含み、該ワクチンが、非経口経路または全身経路により投与される方法が提供される。
【0099】
本明細書で説明されるエンベロープウイルスベースのVLPワクチン調製物は、前記ワクチンを、経口/経消化管経路、または経鼻経路などの粘膜経路を介して投与することにより、ウイルス性インフルエンザを受けやすいか、またはこれを患う、哺乳動物またはトリを防御または治療するのに用い得る。代替的な粘膜経路は、膣内経路および直腸内経路である。例示的な粘膜投与経路は、経鼻経路を介し、鼻腔内ワクチン接種と称することができる。免疫化される個体の鼻咽頭内に、ワクチンの液滴形態、スプレー形態、または乾燥粉末形態を投与することを包含する、鼻腔内ワクチン接種の方法は、当技術分野において周知である。噴霧化またはエアゾール化されたワクチン製剤が、本明細書で開示されるエンベロープウイルスベースのVLPワクチンの例示的な形態である。経口投与用の胃液耐性のカプセルおよび顆粒などの腸溶製剤、直腸内投与または膣内投与用の坐剤もまた、本明細書で開示されるエンベロープウイルスベースのVLPワクチンの製剤である。
【0100】
本明細書で開示される、例示的なエンベロープウイルスベースのVLPによるワクチン組成物は、ヒトへの適用に適する経粘膜ワクチンのクラスを表わし、経粘膜ワクチン接種により全身ワクチン接種を代替する。
【0101】
エンベロープウイルスベースのVLPワクチンはまた、経口経路を介して投与することもできる。このような場合、薬学的に許容される賦形剤にはまた、アルカリ性バッファー、または腸溶性のカプセルもしくはマイクロ顆粒も包含され得る。エンベロープウイルスベースのVLPワクチンはまた、膣内経路によっても投与することができる。このような場合、薬学的に許容される賦形剤には、乳化剤、CARBOPOL(登録商標)などのポリマー、また、膣内クリームおよび膣内坐剤である他の公知の安定化剤も包含され得る。エンベロープウイルスベースのVLPワクチンはまた、直腸内経路によっても投与することができる。このような場合、賦形剤には、直腸内坐剤を形成するために当技術分野で公知のワックスおよびポリマーも包含され得る。
【0102】
代替的に、エンベロープウイルスベースのVLPによるワクチン製剤は、キトサン(上記で説明した)または他のポリカチオンポリマー、ポリラクチド粒子およびポリラクチド−コグリコリド粒子、ポリ−N−アセチルグルコサミンベースのポリマーマトリックス、多糖または化学修飾した多糖からなる粒子、リポソームおよび脂質ベースの粒子、グリセロールモノエステルからなる粒子などからなるワクチン媒体と組み合わせることができる。サポニンはまた、コレステロールの存在下において調合し、リポソームまたはISCOMなどの粒子構造を形成することもできる。さらに、サポニンは、非粒子状の溶液もしくは懸濁液中、または小ラメラリポソームもしくはISCOMなどの粒子状構造内において、ポリオキシエチレンエーテルまたはポリオキシエチレンエステルと併せて調合することができる。
【0103】
本明細書で説明されるエンベロープウイルスベースのVLPを用いる医薬組成物およびワクチン組成物中で用いられる、さらなる例示的なアジュバントには、SAF(Chiron、Calif.、United States)、MF−59(Chiron、例えば、Granoffら(1997年)Infect Immun.65巻(5号):1710〜1715頁を参照されたい)、SBASシリーズのアジュバント(例えば、SB−AS2(SmithKline Beechamのアジュバントシステム番号2;MPLおよびQS21を含有する水中油エマルジョン);SBAS−4(SmithKline Beechamのアジュバントシステム番号4;ミョウバンおよびMPLを含有する);SmithKline Beecham、Rixensart、Belgiumから市販されている)、Detox(Enhanzyn(登録商標))(GlaxoSmithKline)、RC−512、RC−522、RC−527、RC−529、RC−544、およびRC−560(GlaxoSmithKline)、ならびに、その開示全体が参照により本明細書に組み込まれる係属中の米国特許継続出願第08/853,826号、および同第09/074,720号において説明されるものなど、他のアミノアルキルグルコサミニド4−リン酸(AGP)が包含される。
【0104】
アジュバントの他の例には、HunterによるTiterMax(登録商標)アジュバント(CytRx Corp.、Norcross、Ga.);Gerbuアジュバント(Gerbu Biotechnik GmbH、Gaiberg、Germany);ニトロセルロース(NilssonおよびLarsson(1992年)Res. Immunol.143巻:553〜557頁);Seppic ISAシリーズのモンタニドアジュバント(例えば、ISA−51、ISA−57、ISA−720、ISA−151など;Seppic、Paris、France)など、鉱油エマルジョン、非鉱油エマルジョン、油中水エマルジョン、または水中油エマルジョンを包含する、ミョウバン(例えば、水酸化アルミニウム、リン酸アルミニウム)エマルジョンベースの製剤;およびPROVAX(登録商標)(IDEC Pharmaceuticals);OM−174(脂質Aと関連するグルコサミンジサッカリド);Leishmaniaの伸長因子;CRL 1005など、ミセルを形成する非イオン性のブロックコポリマー;ならびにSyntexアジュバント製剤が包含されるがこれらに限定されない。例えば、O’Haganら(2001年)Biomol Eng.18巻(3号):69〜85頁;および「Vaccine Adjuvants: Preparation Methods and Research Protocols」D. O’Hagan編(2000年)、Humana Pressを参照されたい。
【0105】
他のアジュバントには、一般式
HO(CH2CH2O)n−A−R (I)
[式中、nは1〜50であり、Aは結合または−C(O)−であり、RはC1〜50アルキル、またはフェニルC1〜50アルキルである]のアジュバント分子が包含される。
【0106】
本明細書で説明されるエンベロープウイルスベースのVLPワクチン製剤の一実施形態は、一般式(I)[式中、nは1〜50、4〜24、または9であり;R成分はC1〜50、C4〜C20アルキル、またはC12アルキルであり;また、Aは結合である]のポリオキシエチレンエーテルを包含する。ポリオキシエチレンエーテルの濃度は、0.1〜20%、0.1〜10%、または0.1〜1%の範囲であるものとする。例示的なポリオキシエチレンエーテルは、以下の群:ポリオキシエチレン−9−ラウリルエーテル、ポリオキシエチレン−9−ステアリルエーテル(polyoxyethylene−9−steoryl ether)、ポリオキシエチレン−8−ステアリルエーテル、ポリオキシエチレン−4−ラウリルエーテル、ポリオキシエチレン−35−ラウリルエーテル、およびポリオキシエチレン−23−ラウリルエーテルから選択される。ポリオキシエチレンラウリルエーテルなどのポリオキシエチレンエーテルは、メルクインデックス(第12版;登録番号7717)で説明されている。これらのアジュバント分子は、WO99/52549で説明されている。
【0107】
所望の場合、上記の一般式(I)に従うポリオキシエチレンエーテルを、別のアジュバントと組み合わせることができる。例えば、アジュバントの組合せは、上記で説明したCpGを包含し得る。
【0108】
本明細書で開示されるエンベロープウイルスベースのVLPワクチンと共に用いるのに適する、薬学的に許容される賦形剤のさらなる例には、水、リン酸緩衝生理食塩液、等張性緩衝液が包含される。
さらなるインフルエンザ抗原
本明細書で開示される安定化させたVLPは、インフルエンザウイルスの特定の株に対して、かつ/またはインフルエンザウイルスの複数の株にわたり免疫原性を増大させるために、インフルエンザウイルスに由来するさらなる抗原を包含しうる。
【0109】
例示的なさらなるインフルエンザ抗原は、M2ポリペプチド(インフルエンザBにおいてBM2とも呼ばれる)である。インフルエンザウイルスのM2ポリペプチドは、スプライシングイベント後におけるRNAのセグメント7(マトリックスセグメント)によりコードされる、97アミノ酸の低分子III型内在性膜タンパク質である(80、81)。ウイルス粒子上に存在するM2は極めてわずかであるが、感染細胞上においてより豊富に見出すことができる。M2は、ウイルスの侵入に必要な、プロトン選択性イオンチャネルとして用いられる(82、83)。感染時または従来のワクチン接種時における免疫原性は最小限であることから、その保存が説明されるが、代替的なフォーマットで存在する場合は、より免疫原性であり防御性である(84〜86)。これは、in vivoにおけるM2モノクローナル抗体の受動伝達により、ウイルスクリアランスが加速化され、結果として防御がもたらされるという観察(87)と符合する。M2の外部ドメインエピトープを、融合タンパク質として、HBVのコア粒子に連結すると、非経口接種および鼻腔内接種のいずれによっても、マウスにおいて防御性であり、3回のタンデムコピーを該コアタンパク質のN末端に融合させると、最も免疫原性となる(88〜90)。これは、エピトープ密度が増大すると免疫原性が増大することを示す他のキャリア−ハプテンデータ(91)と符合する。
【0110】
M2ワクチンを鼻腔内送達する場合、良好な防御を達成するにはアジュバントが必要とされる場合があり、LTR192G(88、90)およびCTA1−DD(89)により良好な結果が達成されている。ペプチドはまた、KLH、またはN. meningitidesの外膜タンパク質複合体、またはヒトパピローマウイルスのVLPなどのキャリアに化学的にコンジュゲートすることもでき、マウスおよび他の動物におけるワクチンとして防御性である(92、93)。
【0111】
M2タンパク質は、高度に保存性ではあるが、配列の相違をまったく示さないわけではない。一般的な株であるA/PR/8/34株(H1N1)およびA/Aichi/68株(H3N2)のM2外部ドメインエピトープは、A/Hong Kong/156/97株(H5N1)を除き、今日配列決定されている他のすべてのヒト株と、免疫学的に交差反応性であることが示された(92)。インフルエンザデータベースの配列の検討からも、A/Vietnam/1203/04株など、他のより近年の病原性H5N1ヒト単離物のM2配列内において、同様の相違が示されている。この知見は、M2エピトープを組み込む、有効なH5特異的な流行性ワクチンが、ヒトH1単離物およびH3単離物中において現在行きわたっているM2配列ではなく、病原性のトリ株に固有のM2配列を反映する必要があることを裏付ける。
【0112】
インフルエンザウイルスに由来するさらなるタンパク質(HA、NA、およびM2以外のタンパク質)も、共発現により、またはさらなる抗原の全部もしくは一部を、gagもしくはHAポリペプチドに連結することにより、VLPワクチン内に組み込むことができる。これらのさらなる抗原には、PB2、PB1、PA、核タンパク質、マトリックス(M1)、BM2、NS、NS1、およびNS2が包含される。A型インフルエンザウイルスの場合、例には、PB2タンパク質、PB1タンパク質、PAタンパク質、核タンパク質、マトリックス(M1)タンパク質、M2タンパク質、NS1タンパク質、およびNS2タンパク質が含まれる。B型インフルエンザウイルスの場合、例には、HAタンパク質、NAタンパク質、NPタンパク質、Mタンパク質、PB1タンパク質、PB2タンパク質、PAタンパク質、NSタンパク質、およびBM2タンパク質が含まれる。これら後者の抗原は、一般に、抗体反応を中和するための標的ではなく、T細胞により認識される重要なエピトープを含有し得る。このようなエピトープに対してVLPワクチンにより誘導されるT細胞反応が、防御的免疫性を促進するのに有益であることが判明し得る。
【実施例】
【0113】
(実施例1)
インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子の生成
MLV gagタンパク質のコード配列は、モロニーマウス白血病ウイルスの両種性プロウイルス配列全体を含有するプラスミドpAMS(ATCC)から、PCRにより得た。該gagタンパク質のコード配列を、pFastBac1(Invitrogen)のポリへドロンプロモーターの後に挿入し、結果として得られるプラスミドでDH10Bacコンピテント細胞を形質転換し、バキュロウイルスゲノムに組み換えた。次いで、高分子量のバクミドDNAを精製し、Sf9細胞にトランスフェクトし、gagタンパク質を発現する組換えバキュロウイルスを生成させた。ウイルスRNAに由来するHAタンパク質のコード配列およびNAタンパク質のコード配列をRT−PCRによりクローニングした後で、A/PR/8/34株(H1N1亜型)の赤血球凝集素タンパク質およびノイラミニダーゼタンパク質のそれぞれをコードする他の2つの組換えバキュロウイルスも、同様の形で生成させた。最後に、個別のpFastBac1プラスミドに由来する、HAタンパク質発現ユニット、gagタンパク質発現ユニット、およびNAタンパク質発現ユニット(ポリへドロンプロモーター−コード配列−ポリA部位)を、単一のpFastBac1ベクターに統合することにより、3つの産物すべて(HA−gag−NA)をコードする単一のバキュロウイルスベクターを生成させた。当初の解析のために、gagタンパク質またはHAタンパク質またはgag−HA−NAタンパク質をコードする組換えバキュロウイルスを、MOIを>1とする6ウェルプレート内のSf9細胞に感染させた。感染の3日後、培地上清から破砕物を除去し、次いで、20%のスクロースクッションを介する100,000×gでこれをペレット化させた。gagタンパク質特異的抗血清およびH1N1亜型特異的抗血清を用いるウェスタンブロット解析によりペレットを解析した(図1AおよびBを参照されたい)。
【0114】
図1AおよびBの各ブロットにおける左側の3つのレーンは、それぞれ、培地を収集する前の、Sf9細胞に、個別のgag遺伝子によるバキュロウイルスまたはHA遺伝子によるバキュロウイルスまたは対照(EV=空ベクター)のバキュロウイルスを感染させた結果を示す。予測される通り、gag遺伝子だけによるバキュロウイルスを感染させると、VLPの出芽により、高分子量の培地画分中に著明量のgag抗原が結果としてもたらされる(図1A;レーン「Gag」)。これに対し、HA遺伝子だけによるバキュロウイルスを感染させると、それ自体で培地中に放出される少量のHAタンパク質が結果としてもたらされる(図1B;レーン「HA」)。しかし、HA−gag−NA三重遺伝子ベクターをSf9細胞に感染させると、gagタンパク質およびHAタンパク質の両方が著明量で100,000×g画分中に結果として現れる(レーン1〜9;図1AおよびB)ことから、gagタンパク質の発現が、HAタンパク質を細胞から引き出しうることが示される。
【0115】
図2AおよびBは、ペレット化させたHA−gag−NA VLPを、20〜60%の段階的なスクロース勾配上で再遠心分離した後、個々の勾配画分に対してウェスタンブロット解析を行った結果を示す。gagタンパク質およびHAタンパク質の両方が、同じ画分中でピークに達することから、密度約1.16g/mlにおけるバンド形成の符合が裏付けられ、これにより、gagタンパク質およびHAタンパク質がVLP中に存在したことが示唆される。
【0116】
(実施例2)
VLPの安定性についての生物物理的特徴づけ:pHおよび温度
濃度は、モル濃度または容量当たりの重量百分率で報告する。6−ドデカノイル−2−ジメチルアミノナフタレン(ラウルダン)および8−アニリノ−1−ナフタレンスルホネート(ANS)は、Molecular Probes(Eugene、OR)から購入した。ジメチルスルホキシド(DMSO、Fisher Chemical)中に溶解させることにより、1.2mMのラウルダン原液および10mMのANS溶液を調製した。
【0117】
特徴づけされるVLPの調製
実施例1で示した通り、「三重遺伝子」による組換えバキュロウイルスを感染させた培養Sf9細胞内でVLPを生成させた。4〜8の各単位pHであるクエン酸/リン酸(CP)バッファーに透析することにより、特徴づけされるVLPを調製した。NaClを用いて、バッファーのイオン強度を0.1に維持した。透析カセット(10,000MWCO;Pierce、Rockford、IL)から回収した材料を、Amicon(登録商標)Ultra超遠心分離器(10,000MWCO;Millipore、Billerica、MA)により、4℃、3,150×gで濃縮した。残余物のタンパク質濃度は、BCA(ビシンコニン酸)比色法(Pierce、Rockford、IL)により推定した。別段に言及しない限り、残余物を適切なpHの20mM CPバッファーで希釈することにより、最終タンパク質濃度が90μg/mLの三連試料を調製した。
【0118】
表面赤血球凝集素タンパク質に対するトリプシン処理
トリプシン(Sigma;5μg/mLの最終濃度)をVLP原液(pH7.4の30%のスクロース/トリス緩衝生理食塩液中0.26mg/mLの全タンパク質)に添加し、2〜8℃の低温室で5分間にわたりインキュベートした。インキュベーション後、ダイズ(Fluka)に由来する、モル濃度を3倍としたトリプシン阻害剤を添加し、結果として得られる溶液に、0.45μmのシリンジフィルター(Millipore)を流過させた。次いで、上記で説明した通りに、試料を適切なCPバッファーに透析し、濃縮したが、100,000MWCOの透析チューブ(Spectrum Laboratories、Rancho Dominguez、CA)を用いた。HAタンパク質の切断、ならびにMLV gagタンパク質の非切断は、ウェスタンブロット解析により確認した。
【0119】
動的光散乱
動的光散乱(DLS)を用いて、温度上昇の関数としての、VLPの平均有効直径の変化を測定した。Brookhaven Instrument Corporation(Holtzville、NY)のシステムにより測定を行った。125mWのダイオード励起レーザーにより、532nmの入射光を生成させた。入射光に対して90°で散乱光を集光し、デジタル式自己相関計(BI−9000AT)を用いて、自己相関関数を創出した。10〜85℃の範囲にわたり、2.5℃おきに5回の測定を行った。キュムラント解析を用いて、相関関数から粒子の拡散係数を抽出し、ストークス−アインシュタインの式により、これらを粒子直径に変換した。この方法により計算される有効直径が正確なのは、直径が<1μmの粒子についてである(これより大きな粒子についての測定から得られる値は、定性的な比較だけに用いるべきである)ことに留意されたい。粒子サイズに加え、粒子拡散係数の分布についての第2キュムラントもまた、試料の多分散性の尺度としての相関関数から抽出した。
【0120】
実施例1のプロトコールを用いてなされた、VLP懸濁液についてのDLS測定値は、pHにより誘導される粒子サイズの変化および温度により誘導される粒子サイズの変化の両方についての証拠を示した(図3(A))。低温では、pH4または5における粒子サイズが、pH6〜8における粒子サイズの2〜3倍であったことから、酸性のpHにより、著明な凝集および/または膨張が誘導されたことが示唆される。pH4における試料は、約75℃まで、温度により誘導される粒子サイズの変化を示さないが、その後、有効直径の漸増が観察された。これに対し、pH5では、約50℃で粒子サイズが急激に増大したが、約75〜80℃でもさらなる増大が生じた可能性がある。6〜8の各pHの試料は、約58℃までの温度上昇に対して安定であったが、それより高温では、pH6および7の試料が、粒子サイズの顕著な増大を示した。また、pH8の試料も、約60℃でサイズ増大の証拠を示した。後者の場合におけるサイズの増大は、比較的小さく、VLPの凝集ではなく、VLPの膨張によった可能性がある。一般に、酸性度が増大するのに伴い、VLPの多分散性の増大が見られた(図3(C))。pH4および5の試料の多分散性が、温度範囲全体でほぼ一定を維持したのに対し、pH6以上の試料は、約60℃で多分散性の増大を示し、サイズデータで見られる変化と符合する。
【0121】
DLSの測定時にはまた、散乱光の強度も記録した(図3(B))。測定器の設定を各試料に応じて最適化させるために、標準化値を報告することから、試料間における有意義な直接的比較は不可能となる。一般に、粒子サイズまたは粒子の屈折率(溶媒と比べた)が増大すると、散乱光の強度が結果として増大する。しかし、粒子密度の減少(例えば、膨張による)の結果として屈折率が低下するならば、それは、散乱強度の低下として顕在化するであろう。これが、散乱光強度の滑らかで漸進的な低下が温度勾配の大半にわたって見られる、pH8の試料についてのデータに対する最良の説明である。60℃付近で曲線状の低下がわずかに乱れているが、これは、これらの試料について見られる有効直径の増大に対応する(図3(A))。また、pH5の試料による散乱光のプロットも、散乱光強度の急激な低下により顕在化する、60℃付近で生じる構造的変化の証拠を示す。pH6および7に対応するプロットは類似し、散乱光の低下が高温(約75℃)で生じる。これらの低下は、沈降する材料が入射光ビームの外で沈殿するために生じる可能性があり、上記で示したサイズデータについての解釈と符合する。pH4の加熱曲線は、強度の漸減だけを示し、粒子直径の測定された変化と相関する急激な変化は示さなかった(例外は、30〜35℃における散乱強度の一過性の低下であるが、以下のラウルダン蛍光についての結果を参照されたい)。
【0122】
円偏光二色性分光光度法
円偏光二色性分光光度法(CD)の測定は、100mdegの感度設定、2秒間の反応時間、および1nmのバンド幅を用いる、Jasco J−810分光光度計により行った。走査速度を20nm/秒、データピッチを0.5nm/秒として、VLPの複合スペクトル(3つ〜5つのスペクトルの集積)を得た。227nmにおけるCDシグナルをモニタリングする温度可変実験を実施して、全VLPタンパク質の二次構造の変化を、温度の関数として検出した。温度勾配を15℃/時間、遅延時間を2秒間として、10〜90℃の範囲にわたり、0.5℃おきに測定を行った。Origin(登録商標)データ解析ソフトウェアを用いて、温度に依存するデータをシグモイド関数に数学的に近似することにより、中点転移温度(Tm)値を決定した。スペクトルおよび加熱曲線では、3つの異なるタンパク質のうちで最も量の多いタンパク質からの寄与が優越すると推測されるが、スペクトルおよび加熱曲線のいずれもが、これらのタンパク質からの相加的寄与を反映する。これらのVLPでは、MLV gagタンパク質の量が、HAタンパク質の量より3〜4倍多いことが示されたが、HAタンパク質の量は、NAタンパク質の量をおそらく1桁上回る。
【0123】
pH4〜8では、インフルエンザVLPのCDスペクトルが、210nmおよび227nm付近で極小点を示したことは、対象のpH範囲にわたり著明な螺旋特徴が存在することを示唆する(図4(A))。高温ではシグナルが見られなくなる(図4(B))ことから、二次構造が温度に依存して見られなくなることが示唆された。この効果をさらに探索するため、227nmにおけるシグナルを、温度の関数としてモニタリングした(図5)。CDシグナルの急激な変化が観察されたことは、温度に依存するタンパク質構造の転移と符合した。pH6の試料は、55℃付近で最高のTmを示す。試料の酸性度を上昇させると、38℃(pH4)および47℃(pH5)という顕著に低いTm値がもたらされるのに対し、pH7または8の試料は、Tm値が、それぞれ、53℃および51℃で酷似している。溶融曲線の形状は、複数の成分を示唆し、系の異種性を反映すると推測される。
【0124】
蛍光分光光度法
別段に言及しない限り、Photon Technology Internationalによる蛍光測定器(Birmingham、NJ)を用いて、10〜85℃の範囲にわたり、2.5℃おきに蛍光発光スペクトルを収集した。15℃/時間の勾配で温度を上昇させ、すべての測定について、1nmのステップサイズおよび1秒間の積分時間を用いた。蛍光実験ではまた、第2の検出器(蛍光検出器から180°に配向させた)を用いることにより、静的光散乱もモニタリングした。Origin(登録商標)ソフトウェアパッケージを用いて、微分解析により発光ピークの位置を決定し、温度に依存するデータを、シグモイド関数に数学的に近似することにより、Tm値を決定した。
【0125】
芳香族アミノ酸であるトリプトファンおよびチロシンの内部蛍光を使用して、VLPタンパク質の三次構造の変化を、温度の関数として同定した。280nmで励起させた蛍光発光スペクトルを、300〜380nmで収集した。励起および発光のスリット幅は、それぞれ、3nmおよび4nmに設定した。
【0126】
VLP存在下における8−アニリノ−1−ナフタレンスルホネート(ANS)の蛍光発光を、VLPタンパク質の三次構造の安定性をモニタリングする代替的な方法として使用した。タンパク質の非極性領域に対してアフィニティーを有することが公知の低分子であるANSは、溶液中で示す蛍光は弱いが、結合すると、発光強度を増大させ、発光をシフト(通常は青色に)させる(114)。70μMのANSと共に調製したVLP試料を385nmで励起させた後で、その蛍光発光スペクトルを425〜550nmで収集した。ANS蛍光を測定するときの励起および発光のスリット幅はいずれも、4nmに設定した。
【0127】
熱により誘導されるVLP膜の流動性の変化を直接的にモニタリングするには、別の分子プローブである6−ドデカノイル−2−ジメチルアミノナフタレン(ラウルダン)を用いた。ラウルダンの化学構造は、誘導体化されたナフタレンに結合した長いアシル鎖を含有し、これにより、脂質二重層への容易な組込みが可能となる。膜における水和の増大は、二重層の流動性を、ゲル(流動性が低い)から液晶(流動性が高い)に転移させる可能性がある。340nmで励起させる場合、膜の水分含量が増大すると、ラウルダンの発光が約440nmから490nm付近にシフトする。有用なパラメータは、(115)において、GP=(I440−I480)/(I440+I480)[式中、Ix=波長xにおける強度]として定義されている、一般化偏光(GP)である。これにより、GP値が低下すると、膜における流動性の増大が示唆され、この逆も成り立つ。ラウルダンによる実験のスリット幅は、2nm(励起時)および5nm(発光時)に設定した。
【0128】
1.内部蛍光
すべての試料について、内部蛍光発光のピーク位置を、温度の関数として決定した(図6)。各場合において、約10〜40℃の温度範囲にわたり、ピーク位置がやや短波長となった後に、長波長への急激な遷移が生じた。タンパク質試料では、蛍光性アミノ酸側鎖を極性の大きな環境に曝露すると、ピーク極大点が熱に誘導されて赤方にシフトすることが観察された。これは、通常は少なくとも部分的に非極性のタンパク質コア内に包埋されているアミノ酸のフルオロフォアが水性溶媒に曝露される、アンフォールディングイベントと符合した。55〜65℃の温度では(pHに依存して)ピーク極大点が短波長に戻るが、これは、高温で観察されるVLPの凝集と符合する。330nmにおける(標準化された)発光強度も、温度の関数としてプロットした(図6)。構造転移の非存在下では、このようなプロットが、内部の温度効果に起因する温度の上昇と共に、発光強度の滑らかで曲線状の低下を呈示することが典型的である。45〜65℃の範囲にあるすべての試料について、曲線状のプロファイルからの逸脱が生じることから、加熱すると内部フルオロフォア環境が変化することが確認された。
【0129】
2.ANS蛍光
VLPの存在下におけるピーク位置(≧470nm)およびANS蛍光発光強度の増大から、低温では、これらの高分子複合体の非極性領域にANSが結合したことが示唆されるが、これは、脂質二重層の存在を踏まえれば予測されることであった。ANS発光のピーク位置を温度の関数としてプロットする(図7)と、すべての試料において、温度に依存する短波長へのシフトの後、場合によっては(すなわち、pH5、6、および7では)長波長へのシフトが生じることが示される。すべての試料において、ピーク位置の変化の度合い(2〜3nm)は、内部蛍光により観察される変化の度合いよりはるかに小さい。高温では、ピーク位置のばらつきおよびノイズが大きくなるために、これらのデータから実質的な結論を引き出すことは困難となる。485nmにおけるANSの相対強度(図7)は、温度により誘導されて非極性モチーフが曝露されることについてのより明確な証拠を顕示した。これは特に、予測される、蛍光に対する熱による非特異的な消光作用に対応する、発光の曲線状の低下の上に重ね合わせて、発光強度のわずかな増大(pH5では38℃付近で始まり、pH6〜8では43℃付近で始まる)が見られる、pH5〜8の試料についてのプロットに当てはまった。
【0130】
3.ラウルダン蛍光
一般化偏光(GP)を温度の関数としてプロットする(図8)ことにより、加熱すると、VLPの膜における水和(流動性)が漸増することが示唆される。VLP懸濁液のpHが、膜における水和の速度および程度に対して影響を及ぼしたことは、驚くべきことではない。低温では、すべての試料について、膜における水和の程度(すなわち、GP値)が酷似した。40℃を超えると、pH4の試料が示す膜における水和の変化が一貫してわずかであり、一般に、低いpHで調製された試料は、温度を上昇させても二重層内に水分子を取り込むのが遅かった(酸性度の高い試料では、GP=0となる温度が上昇した)。pH4、7、または8で調製した試料のGP値が、温度と共にシグモイド様に変化したのに対し、pH5および6の試料は、検討した温度範囲にわたり、GPの準直線的な低下を示した。pH5および6の試料では、膜における水和の程度が高温(すなわち、75℃を超える)で最大となった。また、ラウルダン蛍光による実験でも、340nmにおける静的光散乱をモニタリングした。これらのデータは、DLS実験で測定された静的光散乱と同様であったが、このさらなる実験では、pH4の試料について観察された散乱光強度の一過性の低下(図3(B)、30〜35℃)が観察されなかったことから、これはアーチファクトであった可能性が示唆される。
【0131】
実験相図
VLPの物理的な安定性についての包括的な視覚的表示を創出するために、上記で論じた各種の生物物理的方法に由来するデータを、多次元ベクトル空間の基底系に変換した。測定を行った温度およびpH(10〜85℃にわたる2.5℃おきの温度、4〜8の単位pH値)のすべての組合せについてn次元ベクトルを構築し、各ベクトルの成分を、適用したn種類の技法の各々による標準化された測定値とした。次いで、基底系におけるすべてのベクトルの射影演算子を足し合わせて、n個の固有ベクトルを伴うn×n密度行列を得た。次いで、データセットに対して最大の寄与(すなわち、最大の固有値)をもたらす3つの固有ベクトルを用いて、元のn次元ベクトル系を三次元に変換した。最後に、新たな三次元ベクトル各々の3つの成分に、3つの異なる色(赤色、緑色、青色)を割り当て、各個別のベクターに固有の色の組合せを得た。次いで、この方法により、n種類の測定を行った温度およびpHのすべての組合せに着色マーカーを割り当てることが可能となり、全データセットを温度およびpHの関数とする三色マップが得られた。このような相図の有用性は、各技法により検出される粒子構造の最も際立った変化を、VLP構造の見かけの「相」の差違と同時に視覚化しうることである。EPDの作成についてのより詳細な説明は、他所(116、117)において示されている。
【0132】
前出の節において示した、温度に依存するデータから、実験相図(図9)を作成した。実験空間にわたり、約10の異なる相を見ることができ、最大の相(pH6〜8、低温、青色)は、VLPの構造的破壊が最小となる状態に対応する。この相の上方に、pH6〜7では35〜55℃にわたり、かつ、pH8では35〜50℃にわたり転移領域が現れた(紫色)。pH6および7では60℃を超える色の変化する領域は、粒子の凝集に対応した。pH8では、著明な凝集がみられないことから、高温で、pH6または7で見られた相とは異なる相(暗赤色)がもたらされた。pH4および5では、低温で2つの異なる相が見られ、いずれも著明な構造の破壊を表わした(明青色)。pHが低い領域の35℃超では、温度に誘導されるさらなる立体構造変化により、複数の相がもたらされた(緑色/橙色)。pH5とpH6との間、および40℃超における見かけの相境界は、安定性が中程度である状態を表し、したがって、以下の実施例3の賦形剤スクリーニングアッセイを開発するための出発点を示す。
【0133】
結論
VLPの生物物理的特徴づけを組み合わせた結果に基づくと、凝集および溶解性に対する変化が、エンベロープウイルスベースのVLPの分解における重要な因子となる。VLPは、pH7〜pH8において最も安定的であった。安定性の理由および化学的な理由で、好ましい範囲は、約pH6.5〜約pH7.5である。この範囲で用いうる代表的なバッファーは、リン酸バッファー、トリスバッファー、MESバッファー、およびクエン酸バッファーである。
【0134】
(実施例3)
VLP賦形剤のスクリーニング
別段に言及しない限り、すべての潜在的な安定化剤は、Sigma−Aldrich(St.Louis、MO)から購入した。グアニジンHCl、塩化カルシウム二水和物、デキストロース、D−マンニトール、クエン酸、および二塩基性リン酸ナトリウムは、Fisher Chemical(Fair Lawn、NJ)から購入した。A型ブタゼラチンは、Dynagel(Calumet City、IL)から購入し、D−スクロースおよびD−トレハロースは、Ferro−Pfanstiehl Laboratories,Inc.(Waukegan、IL)から購入した。エクトイン(超純粋)は、Bitop AG(Witten、Germany)から購入し、NV10は、Expedeon(旧Novexin;Cambridge、UK)から購入した。適切なpHの20mMクエン酸/リン酸(CP)バッファーに溶解させることにより、濃縮賦形剤溶液を調製した。次いで、濃縮したNaOHまたはHClを用いて、pHを、標的pHに調整した(必要な場合)。0.22μmのDurapore(登録商標)(PVDF)膜によるシリンジフィルター(Millipore、Billerica、MA)により、最終原液を濾過した。
【0135】
賦形剤のスクリーニング
濁度(350nmにおける光学濃度;OD350)を時間の関数として測定することにより、pH6および60℃におけるVLPの凝集をモニタリングした。各種のGRAS(一般に安全と認識される)薬剤の存在下または非存在下において、濃縮した残余物(上記を参照されたい)を、20mMのCPバッファーおよび/または適切なpHの濃縮賦形剤溶液で希釈することにより、55μg/mLのタンパク質濃度で、二連のVLP試料を調製した。温度制御型Agilent 8453分光光度計(Palo Alto、CA)を用いて、2時間にわたり、30秒間ごとに測定を行った。
【0136】
溶液中のVLPに対する潜在的な安定化剤について、GRAS化合物のライブラリーをスクリーニングした。実施例2の特徴づけ研究から作成した実験相図(図9)を使用して、VLPの凝集(最も明らかな物理的分解過程)を防止する賦形剤を同定するためのスクリーニングアッセイを開発した。スクリーニングアッセイの温度およびpHについての初期条件の選択は、該相図(前出の実施例を参照されたい)を指針としたが、最終条件は、潜在的な安定化剤間のわずかな差違を増大させるように最適化した。対照試料の挙動に応じて、t=15分またはt=30分(凝集が最大となる時点を表わすいずれかの時点)において、凝集の阻止百分率(表1)を計算した。洗浄剤、ポリオール、アミノ酸、糖、および糖アルコールを含めた各種の分子クラスにおいて、最も有望な凝集阻止化合物が見出された。
【0137】
個々の安定化剤の効果
複数の分子クラス(すなわち、トレハロース、グリセロール、ソルビトール、リシン、およびジエタノールアミン(脂質二重層を破壊する洗浄剤の傾向を踏まえれば、凝集阻止剤としてのTween 20およびBrij 35の見かけの成功は、アーチファクトでありうる))に由来する、効能が最上位にある凝集阻止剤を用いて、潜在的な賦形剤の存在下におけるVLPのCDおよび蛍光の測定を実施した。実際のワクチン製剤をより緊密に近似するために、これらの実験では、溶液のpHを7に設定した。
【0138】
上記で説明した通り、温度可変CDの測定を使用して、選択された化合物のうちのいずれかが、VLPタンパク質(例示しない)の二次構造を安定化させるかどうかを決定した。調べたすべての化合物のうちで、ソルビトールだけが正の効果を示した;ソルビトールを含有する試料のTm(55℃)は、対照のTm(54℃)と比べてわずかに高かったが、他の賦形剤は、50℃(トレハロース)〜53℃(リシン)の範囲のTm値を示した。
【0139】
内部蛍光法を使用して、VLPタンパク質の三次構造に対する潜在的安定化剤の効果を測定した。発光のピーク位置を温度と対比させてプロットする(図10)と、調べた製剤の大半について、40℃またはその付近で始まる、約329nmから336nmへの遷移が示される。例外は、リシンを含有する製剤であり、これは、低温では344nm付近でその蛍光ピークを呈示し、43℃で始まる、わずかな長波長への可能な遷移(+1〜2nm)の証拠を示す(これらの測定値における誤差により、このシフトが統計学的に有意であるという結論は退けられる)。対照のTmは、51℃である。ジエタノールアミンは、49℃のTmを誘導するので有効な安定化剤ではないのに対し、グリセロール、トレハロース、およびソルビトールを含有する製剤はすべて、それぞれ、52、53、および54℃の、やや高温のTm値を示す。これらの実験において収集された静的光散乱(データは示さない)は、リシンを含有する製剤を除くすべての製剤間で同様の挙動を示唆した。リシンの存在下では、光散乱の強度が10分の1未満に低下したことから、VLPの構造破壊が示唆される。複数の化合物によるVLP膜の流動性に対する効果を、上昇させる温度の関数として測定するには、ラウルダン蛍光を用いた。これらの実験では、物理的安定性に対する正の効果が弱いかまたは正の効果が見られない賦形剤(すなわち、ジエタノールアミン、グリセロール、およびリシン)を、グリシン、エクトイン、およびNV10で置き換えた。グリシンは、インフルエンザHAタンパク質および/またはインフルエンザNAタンパク質を安定化させうるという私信により導入した。エクトイン(有機浸透圧調節物質)およびNV10(5kDaの直鎖状炭水化物ポリマー)は、それらが、高分子系を安定化させるのに一般に有効であるという報告(118)に基づき、潜在的な新規のVLP膜安定化剤として調べた。温度に依存するGPデータ(図11)を目視すると、調べた化合物のうちの1つまたは2つが、ゲルから液晶への転移を阻止すると考えられる。ここでもまた、データを近似するためにはシグモイド近似を用い、Tm値を抽出して、各安定化剤の効果を定量的に比較した。対照試料(Tm=52℃)と比較すると、ソルビトールまたはエクトインを含有する製剤は、54℃のやや高いTm値を示す。しかし、これらの測定に随伴する誤差の大きさを踏まえるなら、Tmの見かけの上昇は、いずれの場合にもおそらく有意ではない。他方、グリシンおよびトレハロースは、それぞれ、Tm値が59℃および60℃と高く、著明な安定化効果を及ぼしている。NV10は、20℃を超える温度でGP値の低下(膜における水和の増大)を誘導(対照と比べて)して、VLPエンベロープの安定性に対して負の効果を及ぼしている。NV10製剤について計算されたTmは、46℃である。
【0140】
結論
代表的な単糖(デキストロース、マンニトール、ソルビトール)および二糖(ラクトース、トレハロース、スクロース)を含めた、複数の炭水化物について調べた。デキストロースを除くすべての場合において、10%溶液では凝集が増大した。しかし、すべての場合において、20%溶液は、VLP溶液の凝集を阻止するのに有効であった(トレハロースが最も有効で、84%の阻止を示し、ソルビトールおよびラクトースがわずかの差でこれに続いた)。これに対し、オリゴ糖(シクロデキストランなど)は一般に、凝集を低減するのに有効ではなかった。グリセロール(ポリアルコールは、炭水化物と構造的な類似性を共有する)もまた、凝集を低減する(10%で82%の阻止)のに有効であった。
【0141】
4つの非イオン性界面活性剤を、0.01〜0.10パーセントの範囲の濃度で評価した。調べた4つの洗浄剤すべて(Brij 35、Tween 20、Tween 80、およびPluronic F−68)が、凝集を阻止するのに有効であった。調べたすべての洗浄剤が凝集の阻止を示したが、Tween 20が、他の洗浄剤に優ると考えられる。
【0142】
調べた一般に用いられる2つのタンパク質であるアルブミンおよびゼラチンは、凝集を阻止するのには有効でなかったが、この結果は、VLPではなく、アルブミンに関する問題に起因した可能性がある。
【0143】
また、代表的なアミノ酸についても調べた。300mMのジエタノールアミン、アルギニン、およびリシンは、凝集を70%阻止した。グアニジン(30%)、ヒスチジン(30%)、およびグリシン(12%)は、それほど有効でなく、アスパラギン酸は、凝集を加速化させた。しかし、凝集の阻止または加速化が、有機化合物により直接的に引き起こされたのか、またはpHの変化を介して間接的に引き起こされたのかは明確でない。リシンのさらなる特徴づけは、リシンを用いる結果として、VLP構造が破壊されることを明らかにし、したがって、濁度の低下はアーチファクトと考えられ、凝集を阻止する証拠ではないと考えられる。
【0144】
また、代表的な有機酸についても調べた。アスコルビン酸(150mM)が凝集を大幅に増大させたのに対し、同様の濃度の乳酸およびリンゴ酸は、わずかながら凝集を阻止した。ここでもまた、これらの観察が、pHの変化の結果であるのか、または有機酸がVLPと直接的に相互作用した結果であるのかは明確でない。
【0145】
調べた複数の炭水化物は、エンベロープウイルスベースのVLPを安定化させるのに有望である。炭水化物の濃度は重要であり、20%溶液では保護がもたらされるが、10%溶液または15%溶液では保護がもたらされない。しかし、これに対して、10%のグリセロールは、凝集を阻止するのに有効であった。以上に基づくなら、当業者は、安定化量のこれらの安定化剤を容易に選択することができる。
【0146】
最後に、トレハロースおよびソルビトールについての生物物理的研究により、炭水化物が、ウイルス性タンパク質の三次構造を安定化させることが示され、トレハロースはまた、温度により誘導される脂質二重層の水和を遅延させることも示された。
【0147】
したがって、この炭水化物は、高温で凝集を低減し、タンパク質の三次構造を安定化させ、膜における水和を最小化することが示されたので、VLPを安定化させるのに最も有望な候補物質は、20%のトレハロースであった。構造が類似し、凝集の阻止が濃度に依存するため、他の炭水化物も、同様の保護特性をもたらすと推測される。
【0148】
さらなる参考文献
【0149】
【数1】
【0150】
【数2】
【0151】
【数3】
【0152】
【数4】
【0153】
【数5】
【0154】
【数6】
【0155】
【数7】
【0156】
【数8】
【0157】
【数9】
【特許請求の範囲】
【請求項1】
インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物を含有する溶液を安定化させる方法であって、
(a)前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子を含有する溶液を用意するステップと、
(b)(1)安定化量の、単糖、ソルビトール、二糖、トレハロース、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択される安定化剤を、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物に添加するステップ、(2)pHを、約pH6.5〜約pH8.0、約pH6.5〜約pH7.5、または約pH7とするように、前記溶液を緩衝化するステップ、または(3)ステップ(1)およびステップ(2)の両方と
を含み、
ステップ(b)の後の前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物が、以下の特徴:(i)光学濃度により測定した前記ウイルス様粒子の凝集が、ステップ(b)の前の前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していること;(ii)円偏光二色性またはANS結合により測定したインフルエンザ抗原が、ステップ(b)の前の前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して安定化していること;および(iii)ラウルダン蛍光により測定した、前記ウイルス様粒子の脂質二重層の、温度により誘導される水和が、ステップ(b)の前の前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していること
のうちの少なくとも1つを呈示する、方法。
【請求項2】
前記緩衝化するステップを、リン酸バッファー、トリスバッファー、MESバッファー、クエン酸バッファー、および他のGRASバッファーからなる群から選択される緩衝剤を用いて実施する、請求項1に記載の方法。
【請求項3】
前記安定化剤が、トレハロース、ソルビトール、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択され、そして前記特徴が(i)である、請求項1に記載の方法。
【請求項4】
前記安定化剤が、トレハロース、ソルビトール、および前出の安定化剤の組合せから選択され、そして前記特徴が(ii)である、請求項1に記載の方法。
【請求項5】
前記安定化剤が、トレハロースおよびグリシンから選択され、そして前記特徴が(iii)である、請求項1に記載の方法。
【請求項6】
前記安定化剤がトレハロースであり、そして3つの特徴すべてが存在する、請求項1に記載の方法。
【請求項7】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチドを含む、請求項1から6のうちのいずれか一項に記載の方法。
【請求項8】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、gagポリペプチド、インフルエンザM1ポリペプチド、ニューカッスル病ウイルスマトリックスポリペプチド、エボラウイルスVP40ポリペプチド、およびマールブルグウイルスVP40ポリペプチドを含む群から選択される第2のポリペプチドを含む、請求項1から7のうちのいずれか一項に記載の方法。
【請求項9】
前記gagポリペプチドが、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス、ベータレトロウイルス、ガンマレトロウイルス、デルタレトロウイルス、およびレンチウイルスからなる群から選択されるレトロウイルスに由来する、請求項8に記載の方法。
【請求項10】
前記gagポリペプチドが、マウス白血病ウイルスに由来する、請求項8に記載の方法。
【請求項11】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、ノイラミニダーゼポリペプチドをさらに含む、請求項1から10のうちのいずれか一項に記載の方法。
【請求項12】
前記安定化剤が、単糖、ソルビトール、二糖、およびトレハロースから選択され、そして前記安定化量が、10%(w/w)を超えるか、または少なくとも約20%(w/w)である、請求項1から11のうちのいずれか一項に記載の方法。
【請求項13】
前記安定化させるステップが、ガラス形成を必要としない、請求項1から12のうちのいずれか一項に記載の方法。
【請求項14】
前記安定化量が、凝固時のガラス形成に必要とされる量未満である、請求項1から13のうちのいずれか一項に記載の方法。
【請求項15】
前記安定化剤が、スクロースではない、請求項1から14のうちのいずれか一項に記載の方法。
【請求項16】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物が、前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子と混合させたアジュバントをさらに含む、請求項1から15のうちのいずれか一項に記載の方法。
【請求項17】
前記アジュバントが、前記ウイルス様粒子の内部に配置される、請求項16に記載の方法。
【請求項18】
前記アジュバントが、前記ウイルス様粒子の外部に配置される、請求項16に記載の方法。
【請求項19】
前記アジュバントを、前記第2のポリペプチドに共有結合させて、共有結合を形成させる、請求項16から18のうちのいずれか一項に記載の方法。
【請求項20】
前記アジュバントを、前記赤血球凝集素ポリペプチドに共有結合させて、共有結合を形成させる、請求項16から18のうちのいずれか一項に記載の方法。
【請求項21】
前記アジュバントが、フラジェリンのアジュバント活性断片を含む、請求項16から20のうちのいずれか一項に記載の方法。
【請求項22】
(c)前記溶液を、少なくとも2週間、少なくとも1カ月間、少なくとも2カ月間、少なくとも3カ月間、少なくとも4カ月間、少なくとも6カ月間、または少なくとも1年間にわたり液体形態で保存するステップをさらに含み、このような期間後の前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物が、このような期間前の前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物により誘導される免疫反応の少なくとも80パーセント、少なくとも90パーセント、または少なくとも95パーセントを誘導する、請求項1から21のうちのいずれか一項に記載の方法。
【請求項23】
インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子と、安定化量の、トレハロース、ソルビトール、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択される、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物に対する安定化剤とを含み、以下の特徴:(i)光学濃度により測定した前記ウイルス様粒子の凝集が、前記安定化剤を伴わない、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していること;(ii)円偏光二色性またはANS結合により測定したインフルエンザ抗原が、前記安定化剤を伴わない、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して安定化していること;および(iii)ラウルダン蛍光により測定した、前記ウイルス様粒子の脂質二重層の、温度により誘導される水和が、前記安定化剤を伴わない、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していること
のうちの少なくとも1つを呈示する、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項24】
前記緩衝化を、リン酸バッファー、トリスバッファー、MESバッファー、クエン酸バッファー、および他のGRASバッファーからなる群から選択される緩衝剤を用いて実施する、請求項23に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項25】
前記安定化剤が、トレハロース、ソルビトール、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択され、そして前記特徴が(i)である、請求項23に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項26】
前記安定化剤が、トレハロース、ソルビトール、および前出の安定化剤の組合せから選択され、そして前記特徴が(ii)である、請求項23に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項27】
前記安定化剤が、トレハロースおよびグリシンから選択され、そして前記特徴が(iii)である、請求項23に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項28】
前記安定化剤がトレハロースであり、そして3つの特徴すべてが存在する、請求項23に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項29】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチドを含む、請求項23から28のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項30】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、gagポリペプチド、インフルエンザM1ポリペプチド、ニューカッスル病ウイルスマトリックスポリペプチド、エボラウイルスVP40ポリペプチド、およびマールブルグウイルスVP40ポリペプチドを含む群から選択される第2のポリペプチドを含む、請求項23から29のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項31】
前記gagポリペプチドが、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス、ベータレトロウイルス、ガンマレトロウイルス、デルタレトロウイルス、およびレンチウイルスからなる群から選択されるレトロウイルスに由来する、請求項30に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項32】
前記gagポリペプチドが、マウス白血病ウイルスに由来する、請求項30に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項33】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、ノイラミニダーゼポリペプチドをさらに含む、請求項23から32のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項34】
前記安定化剤が、単糖、ソルビトール、二糖、およびトレハロースから選択され、そして前記安定化量が、10%(w/w)を超えるか、または少なくとも約20%(w/w)である、請求項23から33のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項35】
前記安定化させるステップが、ガラス形成を必要としない、請求項23から34のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項36】
前記安定化量が、凝固時のガラス形成に必要とされる量未満である、請求項23から35のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項37】
前記安定化剤が、スクロースではない、請求項23から36のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項38】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子と混合させたアジュバントをさらに含む、請求項23から37のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項39】
前記アジュバントが、前記ウイルス様粒子の内部に配置される、請求項38に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項40】
前記アジュバントが、前記ウイルス様粒子の外部に配置される、請求項38に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項41】
前記アジュバントを、前記第2のポリペプチドに共有結合させて、共有結合を形成させる、請求項38から40のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項42】
前記アジュバントを、前記赤血球凝集素ポリペプチドに共有結合させて、共有結合を形成させる、請求項38から40のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項43】
前記アジュバントが、フラジェリンのアジュバント活性断片を含む、請求項38から42のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項44】
インフルエンザを治療または予防する方法であって、対象に、免疫原性量の、請求項23から43のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物、または請求項1から21のいずれか一項に記載の方法に従い安定化させた、免疫原性量の、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物を含有する溶液を投与するステップを含む方法。
【請求項45】
前記投与するステップが、前記対象において防御的免疫反応を誘導する、請求項44に記載の方法。
【請求項46】
前記投与するステップが、皮下(subcutaneous)送達、経皮送達、皮内送達、皮下(subdermal)送達、筋肉内送達、経口(peroral)送達、経口(oral)送達、鼻腔内送達、頬送達、舌下送達、腹腔内送達、膣内送達、肛門送達、および頭蓋内送達からなる群から選択される、請求項44に記載の方法。
【請求項1】
インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物を含有する溶液を安定化させる方法であって、
(a)前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子を含有する溶液を用意するステップと、
(b)(1)安定化量の、単糖、ソルビトール、二糖、トレハロース、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択される安定化剤を、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物に添加するステップ、(2)pHを、約pH6.5〜約pH8.0、約pH6.5〜約pH7.5、または約pH7とするように、前記溶液を緩衝化するステップ、または(3)ステップ(1)およびステップ(2)の両方と
を含み、
ステップ(b)の後の前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物が、以下の特徴:(i)光学濃度により測定した前記ウイルス様粒子の凝集が、ステップ(b)の前の前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していること;(ii)円偏光二色性またはANS結合により測定したインフルエンザ抗原が、ステップ(b)の前の前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して安定化していること;および(iii)ラウルダン蛍光により測定した、前記ウイルス様粒子の脂質二重層の、温度により誘導される水和が、ステップ(b)の前の前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していること
のうちの少なくとも1つを呈示する、方法。
【請求項2】
前記緩衝化するステップを、リン酸バッファー、トリスバッファー、MESバッファー、クエン酸バッファー、および他のGRASバッファーからなる群から選択される緩衝剤を用いて実施する、請求項1に記載の方法。
【請求項3】
前記安定化剤が、トレハロース、ソルビトール、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択され、そして前記特徴が(i)である、請求項1に記載の方法。
【請求項4】
前記安定化剤が、トレハロース、ソルビトール、および前出の安定化剤の組合せから選択され、そして前記特徴が(ii)である、請求項1に記載の方法。
【請求項5】
前記安定化剤が、トレハロースおよびグリシンから選択され、そして前記特徴が(iii)である、請求項1に記載の方法。
【請求項6】
前記安定化剤がトレハロースであり、そして3つの特徴すべてが存在する、請求項1に記載の方法。
【請求項7】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチドを含む、請求項1から6のうちのいずれか一項に記載の方法。
【請求項8】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、gagポリペプチド、インフルエンザM1ポリペプチド、ニューカッスル病ウイルスマトリックスポリペプチド、エボラウイルスVP40ポリペプチド、およびマールブルグウイルスVP40ポリペプチドを含む群から選択される第2のポリペプチドを含む、請求項1から7のうちのいずれか一項に記載の方法。
【請求項9】
前記gagポリペプチドが、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス、ベータレトロウイルス、ガンマレトロウイルス、デルタレトロウイルス、およびレンチウイルスからなる群から選択されるレトロウイルスに由来する、請求項8に記載の方法。
【請求項10】
前記gagポリペプチドが、マウス白血病ウイルスに由来する、請求項8に記載の方法。
【請求項11】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、ノイラミニダーゼポリペプチドをさらに含む、請求項1から10のうちのいずれか一項に記載の方法。
【請求項12】
前記安定化剤が、単糖、ソルビトール、二糖、およびトレハロースから選択され、そして前記安定化量が、10%(w/w)を超えるか、または少なくとも約20%(w/w)である、請求項1から11のうちのいずれか一項に記載の方法。
【請求項13】
前記安定化させるステップが、ガラス形成を必要としない、請求項1から12のうちのいずれか一項に記載の方法。
【請求項14】
前記安定化量が、凝固時のガラス形成に必要とされる量未満である、請求項1から13のうちのいずれか一項に記載の方法。
【請求項15】
前記安定化剤が、スクロースではない、請求項1から14のうちのいずれか一項に記載の方法。
【請求項16】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物が、前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子と混合させたアジュバントをさらに含む、請求項1から15のうちのいずれか一項に記載の方法。
【請求項17】
前記アジュバントが、前記ウイルス様粒子の内部に配置される、請求項16に記載の方法。
【請求項18】
前記アジュバントが、前記ウイルス様粒子の外部に配置される、請求項16に記載の方法。
【請求項19】
前記アジュバントを、前記第2のポリペプチドに共有結合させて、共有結合を形成させる、請求項16から18のうちのいずれか一項に記載の方法。
【請求項20】
前記アジュバントを、前記赤血球凝集素ポリペプチドに共有結合させて、共有結合を形成させる、請求項16から18のうちのいずれか一項に記載の方法。
【請求項21】
前記アジュバントが、フラジェリンのアジュバント活性断片を含む、請求項16から20のうちのいずれか一項に記載の方法。
【請求項22】
(c)前記溶液を、少なくとも2週間、少なくとも1カ月間、少なくとも2カ月間、少なくとも3カ月間、少なくとも4カ月間、少なくとも6カ月間、または少なくとも1年間にわたり液体形態で保存するステップをさらに含み、このような期間後の前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物が、このような期間前の前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物により誘導される免疫反応の少なくとも80パーセント、少なくとも90パーセント、または少なくとも95パーセントを誘導する、請求項1から21のうちのいずれか一項に記載の方法。
【請求項23】
インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子と、安定化量の、トレハロース、ソルビトール、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択される、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物に対する安定化剤とを含み、以下の特徴:(i)光学濃度により測定した前記ウイルス様粒子の凝集が、前記安定化剤を伴わない、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していること;(ii)円偏光二色性またはANS結合により測定したインフルエンザ抗原が、前記安定化剤を伴わない、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して安定化していること;および(iii)ラウルダン蛍光により測定した、前記ウイルス様粒子の脂質二重層の、温度により誘導される水和が、前記安定化剤を伴わない、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物と比較して低減していること
のうちの少なくとも1つを呈示する、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項24】
前記緩衝化を、リン酸バッファー、トリスバッファー、MESバッファー、クエン酸バッファー、および他のGRASバッファーからなる群から選択される緩衝剤を用いて実施する、請求項23に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項25】
前記安定化剤が、トレハロース、ソルビトール、ジエタノールアミン、グリセロール、グリシン、および前出の安定化剤の組合せから選択され、そして前記特徴が(i)である、請求項23に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項26】
前記安定化剤が、トレハロース、ソルビトール、および前出の安定化剤の組合せから選択され、そして前記特徴が(ii)である、請求項23に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項27】
前記安定化剤が、トレハロースおよびグリシンから選択され、そして前記特徴が(iii)である、請求項23に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項28】
前記安定化剤がトレハロースであり、そして3つの特徴すべてが存在する、請求項23に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項29】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチドを含む、請求項23から28のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項30】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、gagポリペプチド、インフルエンザM1ポリペプチド、ニューカッスル病ウイルスマトリックスポリペプチド、エボラウイルスVP40ポリペプチド、およびマールブルグウイルスVP40ポリペプチドを含む群から選択される第2のポリペプチドを含む、請求項23から29のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項31】
前記gagポリペプチドが、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス、ベータレトロウイルス、ガンマレトロウイルス、デルタレトロウイルス、およびレンチウイルスからなる群から選択されるレトロウイルスに由来する、請求項30に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項32】
前記gagポリペプチドが、マウス白血病ウイルスに由来する、請求項30に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項33】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子が、ノイラミニダーゼポリペプチドをさらに含む、請求項23から32のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項34】
前記安定化剤が、単糖、ソルビトール、二糖、およびトレハロースから選択され、そして前記安定化量が、10%(w/w)を超えるか、または少なくとも約20%(w/w)である、請求項23から33のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項35】
前記安定化させるステップが、ガラス形成を必要としない、請求項23から34のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項36】
前記安定化量が、凝固時のガラス形成に必要とされる量未満である、請求項23から35のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項37】
前記安定化剤が、スクロースではない、請求項23から36のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項38】
前記インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子と混合させたアジュバントをさらに含む、請求項23から37のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項39】
前記アジュバントが、前記ウイルス様粒子の内部に配置される、請求項38に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項40】
前記アジュバントが、前記ウイルス様粒子の外部に配置される、請求項38に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項41】
前記アジュバントを、前記第2のポリペプチドに共有結合させて、共有結合を形成させる、請求項38から40のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項42】
前記アジュバントを、前記赤血球凝集素ポリペプチドに共有結合させて、共有結合を形成させる、請求項38から40のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項43】
前記アジュバントが、フラジェリンのアジュバント活性断片を含む、請求項38から42のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項44】
インフルエンザを治療または予防する方法であって、対象に、免疫原性量の、請求項23から43のいずれか一項に記載のインフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物、または請求項1から21のいずれか一項に記載の方法に従い安定化させた、免疫原性量の、インフルエンザ抗原のエンベロープウイルスベースのウイルス様粒子調製物を含有する溶液を投与するステップを含む方法。
【請求項45】
前記投与するステップが、前記対象において防御的免疫反応を誘導する、請求項44に記載の方法。
【請求項46】
前記投与するステップが、皮下(subcutaneous)送達、経皮送達、皮内送達、皮下(subdermal)送達、筋肉内送達、経口(peroral)送達、経口(oral)送達、鼻腔内送達、頬送達、舌下送達、腹腔内送達、膣内送達、肛門送達、および頭蓋内送達からなる群から選択される、請求項44に記載の方法。
【図1】

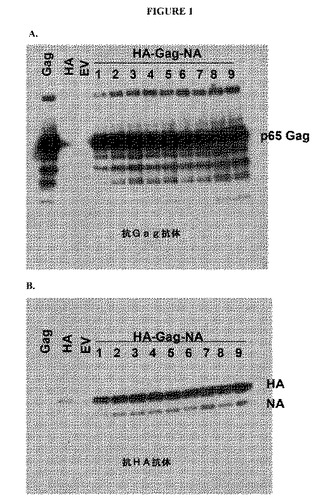
【図2】
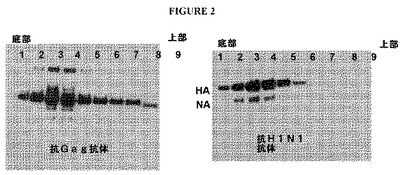
【図3】

【図4】
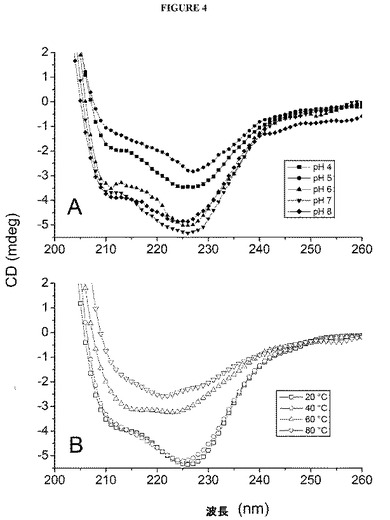
【図5】
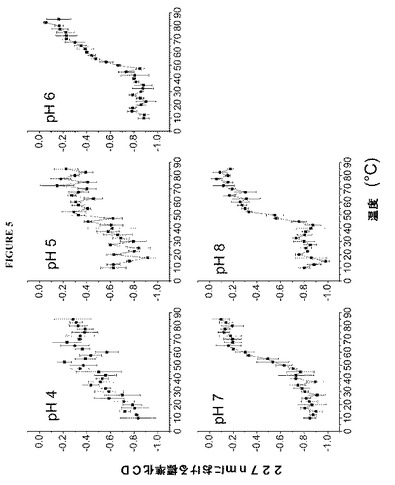
【図6】
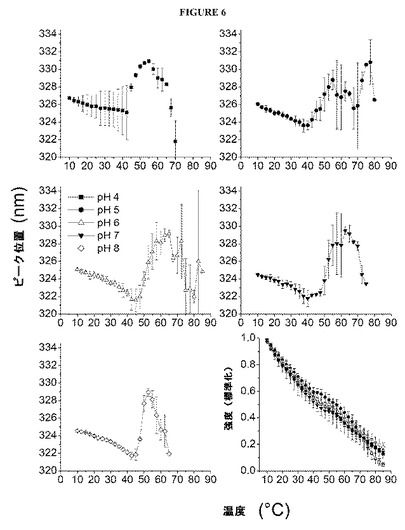
【図7】
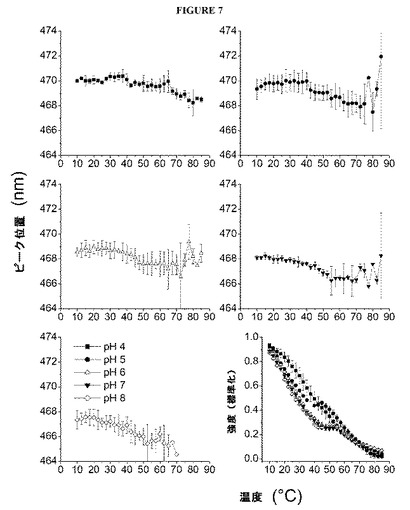
【図8】
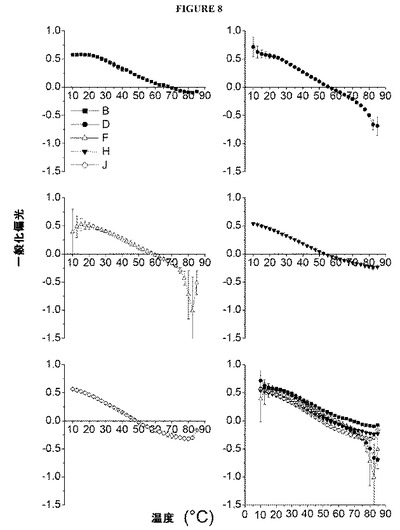
【図10】
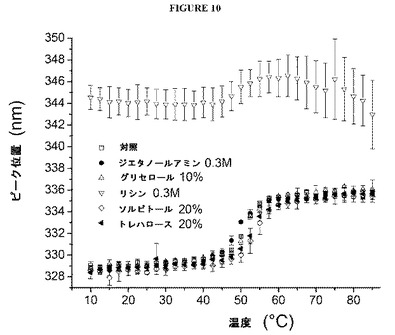
【図11】
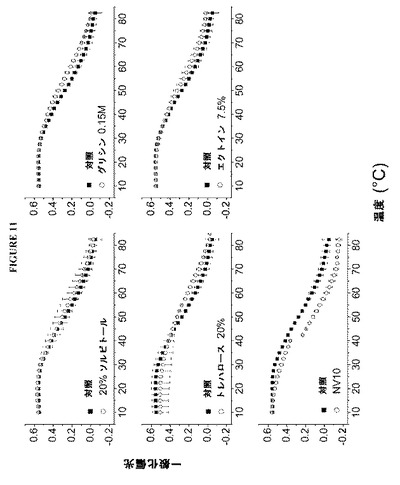
【図9】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図10】
【図11】
【図9】
【公表番号】特表2013−515748(P2013−515748A)
【公表日】平成25年5月9日(2013.5.9)
【国際特許分類】
【出願番号】特願2012−546257(P2012−546257)
【出願日】平成22年12月28日(2010.12.28)
【国際出願番号】PCT/US2010/062217
【国際公開番号】WO2011/090712
【国際公開日】平成23年7月28日(2011.7.28)
【出願人】(509024190)リゴサイト ファーマシューティカルズ インコーポレイテッド (6)
【氏名又は名称原語表記】LIGOCYTE PHARMACEUTICALS,INC.
【住所又は居所原語表記】2155 Analysis Drive,Bozeman,MT 59718−6831,United States of America
【Fターム(参考)】
【公表日】平成25年5月9日(2013.5.9)
【国際特許分類】
【出願日】平成22年12月28日(2010.12.28)
【国際出願番号】PCT/US2010/062217
【国際公開番号】WO2011/090712
【国際公開日】平成23年7月28日(2011.7.28)
【出願人】(509024190)リゴサイト ファーマシューティカルズ インコーポレイテッド (6)
【氏名又は名称原語表記】LIGOCYTE PHARMACEUTICALS,INC.
【住所又は居所原語表記】2155 Analysis Drive,Bozeman,MT 59718−6831,United States of America
【Fターム(参考)】
[ Back to top ]