
基転移反応のアッセイ方法
本発明は、基転移反応における供与体生成物および該供与体生成物を産生させる触媒活性の検出、定量および高処理量スクリーニング方法に関する。本発明は、さらに、本発明の方法を実施するのに使用することができるイムノアッセイ法、抗体およびキットも提供する。

【発明の詳細な説明】
【技術分野】
【0001】
(関連出願との相互参照)
本出願は、2003年1月30日に出願された米国仮出願第60/443,746号に基づく利益を主張し、該米国出願は、その全体が引用により本明細書に取り込まれるものとする。
(技術分野)
本発明は、基転移反応方法論に関する。本発明は、基転移反応における供与体-生成物および該供与体-生成物を生成し得る触媒活性の検出および定量方法を提供する。また、本発明は、基転移反応を触媒する酵素の受容体基質、阻害因子または活性化因子の高処理量スクリーニング方法も提供する。さらに、本発明は、本発明を実施するためのイムノアッセイ法、抗体および関連キットも提供する。
【背景技術】
【0002】
基質が活性化供与体分子として知られる他の物質によって供与される化学基によって修飾される、多くの重要な生物学的反応が存在する。これらの生物学的反応は、“基転移反応”として広く認識されており、下記の一般反応式を有する:
供与体-X + 受容体 → 供与体生成物 + 受容体-X
典型的には、供与体-X、即ち、活性化供与体分子は、共有結合付加物に結合したヌクレオチドである。供与体-Xは、前記ヌクレオチド供与体中でホスホエステル結合の形成によって活性化される。また、受容体基質は、タンパク質または核酸のような小分子または巨大分子を含み得る。この反応の生成物は、修飾受容体、受容体-Xおよび供与体-生成物分子である。
基転移反応を触媒する、例えば、ATPを使用してリン酸基を供与するキナーゼ;ホスホアデノシン-ホスホスルフェート(PAPS)を使用してスルホネート基を供与するスルホトランスフェラーゼ(SULT);UDP-グルクロン酸を使用してグルクロン酸基を転移させるUDP-グルクロノシルトランスフェラーゼ(UGT);s-アデノシルトランスフェラーゼを使用してメチル基を供与するメチルトランスフェラーゼ;アセチル補酵素Aを使用してアセチル基を供与するアセチルトランスフェラーゼ;およびニコチンアミドアデニンジヌクレオチド(NAD)を使用してADP-リボース基を供与するADP-リボシルトランスフェラーゼのような多くの酵素が存在する。従って、基転移反応を触媒する多くの酵素は、製薬企業にとって興味あるものである。
【0003】
自動化高処理量スクリーニング(HTS)アッセイ法は、薬物発見環境における潜在的薬物分子とタンパク質との相互作用を同定するためのパラダイムであり、この方式は、好ましくは蛍光または化学発光読出しによる単純で強力な分子アッセイを必要とする。HTSに最も適する方式は、反応を開始させた後に何らの操作も必要とせず且つアッセイシグナルを連続的にモニターし得る均質アッセイ(例えば、“単回添加”または“混合および読取り”アッセイ)である。薬物発見の観点におけるこれらアッセイ法の重要性にもかかわらず、基転移酵素の製薬HTSプログラムへの取入れは、以下のようないくつかの理由により緩慢であり妨げられている:a)現在のところ、均質アッセイ法は利用可能ではない、b) 使用する検出方法が前記アッセイの使用に重大な制約をもたらしている、c) 前記アッセイ法の汎用的でない性質は、多様な受容体物質を試験するために多くの特異的な検出試薬の開発を必要とする。
スルホトランスフェラーゼ基質または阻害因子を同定するのに現在使用されている方法は、放射能の使用を必要とし、沈降またはクロマトグラフィーのような面倒な反応後分離工程を含む。例えば、35S-PAPSがスルホトランスフェラーゼ反応において使用され、標識化された生成物が、未反応35S-PAPSの選択的沈降後にシンチレーション計数によって定量される(Foldes, A. and Meek, J. L., Biochim Biophys Acta, 1973, 327:365-74)。この方法は、高処理量スクリーニング(HTS)法においては、高放射性廃棄物コスト故に、また、分離工程の取入れは自動化工程を複雑にすることから望ましくない。他のSULTアッセイ法が比色分析および蛍光手段を使用して開発されているが、これらのアッセイ法は、検出のために特定の受容体基質の使用に依存しており、従って、その使用は単一のSULTイソ型に制約され、これらのアッセイは多様な基質についてのスクリーニングに使用することはできない(Burkart, M. D. and Wong, C. H., Anal Biochem, 1999, 274:131-7;Frame, L. T. et al., Drug Metab Dispos, 2000, 28:1063-8)。結果として、SULT相互作用研究は、薬物の前臨床開発において、現在のところ含まれていない。また、UGTは、現在、放射性標識供与体を使用してアッセイされており、前臨床HTSプログラムを著しく阻害する薄層クロマトグラフィー(TLC)または高圧液体クロマトグラフィー(HPLC)のような反応後分離工程を必要とする(Ethell, B. T., et al., Anal Biochem, 1998, 255:142-7)。同様に、伝統的に、キナーゼは、32P-ATPまたは33P-ATPを供与体として使用して生成させた放射性標識ポリペプチド基質のフィルター捕捉または沈降によってアッセイされている。しかしながら、この方法は、濾過または遠心分離のような分離工程を必要とするので、自動化HTS方式に容易には適応させ得ない。表面近接アッセイ法(surface proximity assay) (SPA)は、マルチウェル方式での放射能アッセイを分離なしで可能にするが(Mallari, R., et al., J Biomol Screen, 2003, 8:198-204)、製薬企業によるその使用は、取扱い放射性同位元素の廃棄および法的コストのために減少してきている。
【0004】
キナーゼ阻害因子薬物開発における高レベルの興味故にこの酵素群の改良されたアッセイ方法を開発する多くの努力が科学者達によりなされている。高度に特異的な試薬を使用して、反応の他の成分の存在下での反応生成物を蛍光または化学発光のような光系読出しを使用して検出する均質アッセイ法が開発されている。キナーゼアッセイにおいて使用する最も一般的な均質方法は、種々の蛍光特性を示すホスホペプチド生成物の免疫検出である(Zaman, G. J., Comb Chem High Throughput Screen, 2003, 6:313-20)。この方法においては、基質ペプチドのリン酸化が抗-ホスホペプチドから蛍光ホスホペプチドトレーサーを外してこれを置換し、その蛍光特性の変化を生じさせる。この基本的方法は、蛍光偏光(FP)法 (Parker, G. J., et al., J Biomol Screen, 2000, 5:77-88);時間分解蛍光法(Xu, K., et al., J Biochem Mol Biol, 2003, 36:421-5);蛍光寿命識別法(Fowler, A., et al., Anal Biochem, 2002, 308:223-31);および化学発光法(Eglen, R. M. and Singh, R., Comb Chem High Throughput Screen, 2003, 6:381-7)のような拮抗イムノアッセイ法において使用する数種の読出し方式に適合されている。
この方法による欠点は、ホスホペプチド特異的抗体の必要性である。汎用的なホスホチロシン抗体が一般的であるが、ホスホセリン抗体およびホスホスレオニン抗体は、周知のとお作製するのが難しく、ホスホ-セリンまたは-スレオニンを特異的隣接アミノ酸との前後関係の中でしか認識しない(EglenおよびSingh、2003年)。ヒトにおいては400種を越えるキナーゼが存在し、リン酸化部位に対するそれらの特異性は広く変化する。即ち、多様な抗体を多様なキナーゼのアッセイまたは受容体基質のプロフィーリングにおいて必要とする。このことは、新たなキナーゼのHTSへの組入れを、とりわけそれらの基質特異性が良く明確にされていない場合に極めて複雑にする。また、このことは、アッセイの出力が使用する特定の抗体(Ab)-ホスホペプチド対に依存するので、多様な基質選択性を有するキナーゼ間でのデータ比較における解析問題も生ずる。汎用的なホスホセリン抗体を開発し且つより汎用的なキナーゼ基質を同定する努力は続いているものの(Sills, M. A., et al., J Biomol Screen, 2002, 7:191-214)、この方向性における研究は、今日まであまり成功していない。
【0005】
前後関係特異的Ab-ホスホペプチド相互作用の問題を回避するために多くの別の方法が開発されており、それらにはホスホペプチドを結合させるための金属複合体の使用(Scott, J. E. and Carpenter, J. W., Anal Biochem, 2003, 316:82-91)およびホスホペプチド生成物の共有結合タグ付けを可能にする修飾ATPアナログの使用(Allison Miller-Wing, E. G., Barbara Armstrong, Lindsey Yeats, Ram Bhatt, Frank Gonzales, and Steven Gessert., SBS 9th Annual Conference and Exhibition, Portland, OR., 2003)が含まれる。化学的リン酸結合試薬は、ヌクレオチドホスフェートへのバックグラウンド結合という問題があり、極めて低い非生理学的レベルのATPの使用を必要とし、これがアッセイの柔軟性を制約する。修飾ヌクレオチドは、該ATPアナログを供与体として使用する能力がキナーゼ間で変動するので汎用的な方式を提供せず、数多くの種々のアナログの開発を必要とする。また、キナーゼATP結合部位(キナーゼ阻害因子結合の最も頻繁な部位)での阻害因子と前記修飾ヌクレオチドとの拮抗も、生理学的状況を反映してない。ペプチドリン酸化によって生ずるプロテアーゼ感受性の差異が蛍光系キナーゼアッセイの開発において活用されているが(Kupcho, K., et al., Anal Biochem, 2003, 317:210-7)、これらのアッセイは、真に均質性ではない;即ち、これらのアッセイは、現像用プロテアーゼ試薬の反応後添加を必要とする。さらに、この方法の応用性は、キナーゼおよびプロテアーゼ特異性が重なり合うペプチドに限られる。
また、反応生成物と特異的マルチウェルプレート化学作用物との相互作用に依存する少数の方法も存在するが、これらの方法は、反応後の試薬添加および/または処理を必要とする点で真に均質性ではない。また、処理および/または検出のための特別仕様の機器を必要とすることも、殆どの製薬HTS基本構造に望まれるオープンアーキテクチャーに適合しない。
さらにまた、反応生成物の電気泳動分離に依存する微小流体工学系キナーゼアッセイも開発されてきている(Xue, Q., et al., Electrophoresis, 2001, 22:4000-7)。これらのアッセイ法においては、リン酸化ペプチド生成物をリン酸化されていない受容体基質から電気泳動により分離し、それによって特化された検出試薬の必要性を排除している。しかしながら、実際には、キナーゼアッセイは、多くの場合、マルチウェルプレートにおいて操作され、その後、生成物が分離用の微小流体装置に移される(HTS方式においては面倒な工程)。
【0006】
要するに、現在の基転移アッセイの汎用的でない性質は、個々の酵素またはそのファミリー内の下位群についてのアッセイを開発する必要がある故に、薬物発見におけるかなりの費用と遅れをもたらしている。また、現在のアッセイ法の多くは特異的なタグ付き受容体の修飾と検出に基づいていることから、多様な受容体基質を試験するには限られた能力しかない。多くの場合、使用するタグ付き化受容体基質は、生体内でリン酸化される基質とは異なり、従って、前記アッセイの生理学的適合性は疑わしい。さらに、製薬業界における大きな懸念事は、現在のアッセイ法の汎用的でない性質故に、場合によっては、研究者は異なるキナーゼに対しては異なる方法を使用することを余儀なくされることである。しかしながら、幾つかの研究において、異なるアッセイ方法を使用して同定した医薬ターゲットにおいて有意差があることが証明されており(Sills, M. A., et al., J Biomol Screen, 2002, 7:191-214)、これは、数種のキナーゼに亘る阻害因子選択性のプロファイリングにおいて大きな問題である。基転移反応についての現存するHTSアッセイ法のこれらの欠点は、製薬上の薬物発見プログラムにおける重要な酵素群の迅速な分析を妨げている。
基転移反応をアッセイする他の現存する方法は、供与体分子生成物を検出することによって、種々の化学物を基転移反応における基質としてスクリーニングすることを可能にしている;何故ならば、供与体分子生成物は、受容体が修飾されているかどうかにかかわらず、同じであるからである。供与体生成物の検出は汎用的アッセイ法の基本を提供するものと考えられており、ADPは常にキナーゼ反応の生成物であり、ホスホアデノシン-ホスフェート(PAP)は常にスルホトランスフェラーゼ反応の生成物である。しかしながら、例えば、ADPはATPと同じ蛍光および吸光特性を有する故に、開裂モノ-およびジ-ヌクレオチドを活性化供与体分子から吸光または蛍光特性に基づき識別することができず、これらの生成物の検出は複雑である。供与体生成物の供与体からの分離は、薄層クロマトグラフィーまたは高圧液体クロマトグラフィーのようなクロマトグラフィー法を使用して実施し得るが、これらの方法のHTS方式への取入れは面倒である。
【0007】
この困難性を回避するために、供与体生成物の検出を、さらなる酵素を使用して一次反応生成物(開裂モノ-およびジ-ヌクレオチド)から検出可能な生成物を生成させることによって達成している;これは、酵素共役反応として知られている。例えば、酵素および他の小分子をADP依存性のNADPH産生において使用でき、NADPHを340nmでの吸光または蛍光によって検出する(Walters, W. P. and Namchuk, M., Nat Rev Drug Discov, 2003, 2:259-66)。酵素共役反応は、もう1つのタイプの基転移酵素であるUGTにおいても開発されている(Mulder, G. J. and van Doorn, A. B., Biochem J, 1975, 151:131-40)。しかしながら、薬物化合物の吸光アッセイによる光干渉は、とりわけ紫外線において、この方法で広く認識された問題である。この方法のもう1つの欠点は、検出物と共役させるのに使用する酵素の全てがスクリーニングする化学物からの潜在的な阻害に供されることである。
もう1つの一般的な方法は、ルシフェラーゼをレポーターとして使用してATP消費量をモニターすることによってプロテインキナーゼ活性を検出することである。この方法の例は、Crouch等により、米国特許第6,599,711号において開示されている。該方法は、プロテインキナーゼによってリン酸化され得る基質をATPと試験すべきプロテインキナーゼを含む溶液、および試験すべきプロテインキナーゼの存在しないATPを含むもう1つの溶液に添加することによって、試験すべきプロテインキナーゼの活性を測定することを必要としていた。その後、ATPおよび/またはADPの濃度または時間変化速度を、生物発光を使用して測定していた。しかしながら、このアッセイ法は、高い初期シグナルの小さい低下に依存しているので最適ではない。バックグラウンドを最小にするためにATP濃度を低く保つ必要性があるので、アッセイ条件を注意深く制御しない限り非直線的な反応キネティクスが生じる。関連する方法においては、蛍光ATPアナログを使用する拮抗結合アッセイも開発されているが、酵素触媒活性の尺度を提供せず、従って、限られた利用性しかない。
【0008】
(発明の簡単な要約)
基転移反応を触媒する酵素のHTSアッセイへの使用は、製薬用の薬物ライブラリー中の分子からの干渉を受けない普遍的で均質性のアッセイ方法が無いために妨げられている。従って、HTS薬物発見に良好に適応する基転移反応に関連する酵素の普遍的なアッセイ方法を提供することが一般に望ましいであろう。
【発明の開示】
【0009】
重要な基転移酵素のHTSアッセイへの容易な組入れを可能にするために、出願人等は、前記供与体生成物の均質免疫検出に基づくアッセイ法を開発した。基転移反応の一般的方程式は、供与体-X + 受容体 → 供与体-生成物 + 受容体-Xであり、供与体-生成物を一般的検出反応:第1複合体 + 供与体-生成物 → 第2複合体 + 置換された検出可能タグによって検出する。前記供与体生成物は与えられた型の基転移反応を触媒する全ての酵素において同じであるので、同じ検出試薬を基転移酵素群内の全てのメンバーにおいてまた任意の受容体基質と一緒に使用し得る。アッセイ生成物は、製薬用の薬物ライブラリー中の分子からの有意の干渉またはバックグラウンドシグナルを受けない均質蛍光または化学発光法を使用して検出し得る。
【0010】
即ち、本発明は、基転移反応を触媒する酵素の活性を検出すること、および、これら酵素の受容体基質、阻害因子または活性化因子をスクリーニングしてより選択性で治療用の薬物の開発を容易にする方法およびその構成要素として要約される。このことは、基転移反応の供与体生成物と結合させるのに使用する高度に選択的な抗体によって達成される。抗体-抗原相互作用は、すでに他の人々によって記載されている多くの方法で検出し得る。出願人等が前記方法を実施するのに使用した検出方式は、拮抗蛍光偏光イムノアッセイ法(FPIA)(competitive fluorescence polarization immunoassay)であるが、この方法が製薬HTSアッセイに良好に適応するためである。この検出方式によれば、酵素により生成した供与体生成物は、トレーサーと呼ばれる該供与体生成物の蛍光誘導体を抗体から外して置き換わり(置換し)、トレーサー蛍光偏光の減少を生じさせる。このアッセイにおける主要試薬は、供与体生成物に高選択性でもって結合するが未開裂供与体分子には結合しない抗体、およびトレーサー(前記抗体に結合するのに十分にその構造を保持する前記供与体生成物の蛍光誘導体)である。本発明は、基転移反応を触媒する酵素の活性を種々の基質を使用して検出し定量する新規なアッセイ法を提供する。また、本発明は、基転移反応の基質、阻害因子または活性化因子のスクリーニング方法も提供する。
【0011】
1つの局面においては、本発明は、基転移反応の供与体-生成物の検出方法を提供し、該方法は、活性化型の供与体を触媒活性酵素の存在下に受容体と反応させること;供与体-生成物および受容体-Xを形成させること;前記供与体-生成物を、観測可能物理量を発生させ得る検出可能タグを含む第1複合体と接触させること;前記第1複合体の前記検出可能タグを前記供与体生成物によって拮抗的に置換して(外して)第2複合体および置換された(外された)検出可能タグを生成させること;および、前記第1複合体中の前記検出可能タグおよび前記置換された(外された)検出可能タグによって発生した観測可能物理量の変化を検出すること、を含む。
もう1つの局面においては、本発明は、基転移反応の供与体生成物に対して作製された抗体を提供し、該抗体は、供与体-生成物と供与体とを高供与体濃度の存在下に選択的に識別する能力を有する。
さらにもう1つの局面においては、本発明は、基転移反応の供与体-生成物の均質拮抗結合アッセイ法を提供し、該アッセイは、以下を含む:供与体-生成物をトレーサーおよび巨大分子と混合して混合物を調製することであって、前記巨大分子は前記供与体-生成物に対し特異的であり、前記トレーサーはフルオロフォアに結合させた前記供与体-生成物を含み、前記トレーサーは前記巨大分子と結合して蛍光偏光において検出可能な変化を生じ得る;前記混合物の蛍光偏光を測定して蛍光偏光測定値を得ること;および、前記蛍光偏光測定値を特性決定した蛍光偏光値と比較することであって、該特性決定した蛍光偏光値は既知の供与体生成物濃度に対応する。
さらにもう1つの局面においては、本発明は、基転移反応からの供与体生成物の特性決定用アッセイキットを提供し、該アッセイキットは、巨大分子とトレーサーの各々を、供与体生成物の少なくとも1回の蛍光偏光アッセイに適する量で含み、前記巨大分子は抗体または不活化酵素である。
他の利点、並びに特定の変形、組成的変更および全体的構成のより完全な認識は、特許請求の範囲と関連して行う以下の各種実施態様についての詳細な説明を検証するときに得られるであろう。
【0012】
本発明の実施態様を詳細に説明する前に、本発明は、その応用において、以下の説明において説明する詳細に限定されるものではないことを理解すべきである。本発明は、他の実施態様もあり得、種々の方法で実行または実施可能である。また、本明細書において使用する表現または用語は、説明目的のためであり、如何なる形でも限定するものとみなしてはならないことを理解すべきである。
【0013】
(発明の詳細な説明)
本発明は、広義には、供与体-生成物または基転移酵素に由来する前記供与体-生成物を生成させる触媒活性を、種々の基質を使用して検出し定量する新規なアッセイ方法に関する。また、本発明は、前記供与体生成物に対して特異的な抗体並びに本発明を高処理量スクリーニング方式において実施するためのアッセイキットも提供する。基転移反応の一般的等式は、供与体-X + 受容体 → 供与体-生成物 + 受容体-Xを含み、供与体生成物を一般的検出反応:第1複合体 + 供与体-生成物 → 第2複合体 + 置換された検出可能タグによって検出する。即ち、本発明によれば、高度に選択性の抗体を使用して前記基転移反応の供与体生成物に結合させ、この結合事象を、例えば、製薬HTSアッセイに良好に適応する拮抗結合FPIAのようなイムノアッセイ法を使用して検出する。この検出方式を使用することにより、酵素により生成した供与体生成物は抗体からトレーサーを外してこれに置き換わり、これによりトレーサー蛍光偏光の減少をもたらす。このアッセイにおける主要試薬は、前記供与体生成物に高選択性でもって結合するが未開裂供与体分子には結合しない抗体、およびトレーサー(前記抗体に結合するのに十分にその構造を保持する前記供与体生成物の蛍光誘導体)である。
【0014】
とりわけ、本発明は、単一系列の検出試薬を基転移酵素の所定群内の全ての酵素およびこの群に対する全ての受容体基質において使用し得る点で、普遍的なアッセイ方法を提供する。その普遍的性質故に、本発明は、SULT、プロテインキナーゼおよび他の基転移酵素のHTSスクリーニングへの組入れを促進するであろう。例えば、11種の既知のSULTイソ型が存在し、本発明の方法を使用すれば、11種のSULTイソ型の全てを、それらの種々の化合物を硫酸化能について、同じ試験において同じ検出試薬、プロトコールおよび器具を使用してスクリーニングすることができる。このことは、生体異物結合を触媒する酵素(例えば、SULTおよびUGT)が極めて広い受容体基質特異性を有することから、重要な能力である。キナーゼに関しては、酵素群内にさらにより多くの多様性が存在する。ヒトにおいて400種よりも多いプロテインキナーゼが存在し、それらの受容体基質特異性においては、生理学的タンパク質受容体基質並びに短ペプチドのいずれも使用し得るような大きな多様性が存在する。そのように、多数のプロテインキナーゼを、それらの多様な受容体基質を使用して、本発明のアッセイにおいて使用でき、同じ検出試薬、プロトコールおよび器具を使用して、阻害因子についてスクリーニングできる。これらの理由および以下で述べる他の理由により、SULT、UGTおよびキナーゼのような基転移反応における本発明のFPIA系供与体生成物検出アッセイ法は、なかんずく、自動化HTS用途に良好に適応する。
従って、本発明をそのような試みに関連して詳細に説明する;しかしながら、当業者であれば、本発明のそのような説明は、例示のみを意味し、本発明の範囲全体を限定するものとみなすべきでないことを理解されたい。
【0015】
定義
本明細書において使用するある種の用語は、以下の一般的定義を有するものとする:
本明細書において使用するときの用語“基転移反応”は、下記の一般的反応を称する:
供与体-X + 受容体 → 供与体-生成物 + 受容体-X
代表的な基転移反応は、以下のように示される:
キナーゼ反応:ATP + 受容体 → ADP + 受容体-PO4;
UGT反応:UDP-グルクロン酸 + 受容体 → UDP + 受容体-グルクロン酸;
SULT反応:受容体-XH + 3'-ホスホアデノシン5'-ホスホスルフェート →受容体-SO4 + 3'-ホスホアデノシン5'-スルフェート + H+;
メチルトランスフェラーゼ反応:s-アデノシルメチオニン + 受容体 → 受容体-CH3 + s-アデノシルホモシステイン;および、
アセチルトランスフェラーゼ反応:アセチル補酵素A + 受容体 → 受容体-COCH3 + 補酵素A。
SULTによる硫酸化、キナーゼによるリン酸化およびUGTによるUDP-グルクロン酸化のような部分的基転移反応は、それぞれ、PAP、ADPおよびUDPである供与体生成物に対する抗体/検出可能タグ対を単離し得るので、本発明の方法に適切に応用可能である。また、基転移反応は、ホルモン生体合成および機能;酵素調節および機能;および生体異物代謝のような多くの生物学的プロセスに一般的に関与する。
用語“普遍的アッセイ”および“汎用的アッセイ”は、互換的に使用され、基転移反応酵素群の全メンバーおよびそれらの受容体基質の全てを同じアッセイ試薬により検出し得る方法をいう。
用語“共有結合付加物”とは、基転移反応において供与体分子から受容体に転移される部分をいう;SULT、キナーゼおよびUGTにおける、それぞれ、スルホネート、ホスフェートおよびグルクロン酸。
【0016】
本明細書において用語“供与体-生成物”は、前記共有結合付加物が受容体に転移したときに生成する供与体分子のフラグメントである基転移反応の生成物をいう。多くの場合、供与体-生成物は、PAP、UDPまたはADPのようなヌクレオチド(天然または合成の);或いはs-アデノシルホモシステイン、ニコチンアミドまたは補酵素Aのような非ヌクレオチドである。供与体-生成物は、第1複合体 + 供与体-生成物 → 第2複合体 + 置換された(外された)検出可能タグのような一般的反応により検出する。
本明細書において用語“トレーサー”とは、特異的抗体と結合するのに十分にその構造を保持し、置換される、供与体生成物の検出可能な誘導体またはタグをいう。
本明細書において用語“供与体”とは、前記活性化共有結合付加物を担持する、基転移反応を触媒する酵素に対する基質を称する。適切な供与体の例としては、なかんずく、ヌクレオチドのみならずs-アデノシルメチオニンおよびアセチル-CoAがある。
用語“供与体-X”は(Xは前記共有結合付加物を表す)供与体分子のもう1つの用語である。
本明細書において用語“受容体”とは、前記共有結合付加物が共有結合することになる、基転移反応を触媒する酵素に対する基質を称する;該基質は、ポリペプチド、タンパク質、核酸、炭水化物、脂質、またはステロイドもしくはアミノ酸のような小分子基質である。
本明細書において使用するときの用語“受容体-X”は、Xが共有結合した共有結合付加物である反応生成物を称する;該共有結合付加物としては、ホスフェート、スルフェート、炭水化物、天然産アミノ酸、合成誘導アミノ酸、メチル、アセチルまたはグルタチオン成分の少なくとも1つ、およびこれらの組合せがある。前記共有結合付加物は、受容体基質の機能、安定性、または機能および安定性の双方を改変し得る。
【0017】
本明細書において用語“触媒活性酵素”とは、スルホトランスフェラーゼ、キナーゼ、UDP-グルクロノシルトランスフェラーゼ、メチルトランスフェラーゼ、アセチルトランスフェラーゼ、グルタチオントランスフェラーゼまたはADP-リボシルトランスフェラーゼの少なくとも1つをいう。
本明細書において用語“触媒活性”とは、化学触媒活性、酵素活性またはこれらの組合せをいう。
本明細書において用語“第1複合体”とは、巨大分子(即ち、抗体または不活化酵素)および検出可能タグを有する複合体をいう。
本明細書において用語“第2複合体”とは、巨大分子、および前記検出可能タグが前記供与体生成物によって拮抗的に置換された供与体生成物をいう。
本明細書において使用するときの用語“観測可能物理量”とは、前記第2複合体または前記転移検出可能タグの検出可能な蛍光の変化、蛍光強度、蛍光寿命、蛍光偏光、蛍光共鳴エネルギー移動(FRET)または化学発光、および測定観測可能物理量を得るためのこれらの組合せを称する。測定した観測可能物理量は特性決定されている観測可能物理量と比較されるが、この特性決定されている観測可能物理量は前記第1複合体に対応する。
本明細書において用語“検出可能タグ”とは、供与体生成物に結合させた蛍光または化学発光トレーサーをいう。蛍光が本発明における検出の好ましい形である。適切な検出可能タグは、例えば、化学発光タグまたはフルオロフォアタグをこれらのタグが抗体結合を有意に干渉しない方法で(即ち、もっともあり得るのは、ヌクレオチドのアデシン部分に結合するようにして)前記供与体生成物分子に結合させることによって調製し得る。本発明の方法に応用し得るフルオロフォアとしては、限定するものではないが、フルオレセイン、ローダミン、BODIPY、テキサスレッド、およびこれらの当該技術において公知の誘導体がある。ローダミン結合体および他の赤色結合体は、これらの高めの波長の発光がフルオレセインの緑色よりも自己蛍光からの干渉を少なく受けるので、検出可能タグとして合成して最適化し得る。
【0018】
本発明に応用し得る化学発光タグとしては、Lumigen TMA-6およびLumigen PS-3 (Lumigen社、ミシガン州サウスフィールド)があり、これらは適切な化学発光量子収率を有する。これらの試薬は、予測可能な発光時間経過を有する効率的な化学発光反応により、測定容易なシグナルを有する。さらにまた、化学発光タグは、生来のバックグラウンド化学発光に殆んどまたは全く寄与しない。また、光強度の測定も比較的簡単で、光電子倍増管またはフォトダイオードとシグナルを転換し記録するための関連電子機器とを必要とするだけである。さらに、化学発光シグナルは、イムノアッセイにおいて、酵素フラグメント相補法を使用して発生させることができる。この場合、検出可能タグはレポーター酵素のフラグメントAに結合させた供与体生成物であろうし、基転移反応において生成させた供与体生成物による抗体からのその転移はレポーター酵素のフラグメントBとの結合を可能にして、化学発光基質上で作用し得る触媒活性レポーター酵素の形成をもたらすであろう。この方法は、米国特許第4,708,929号に、レポーター酵素としてβ-ガラクトシダーゼを使用して記載されている。
本明細書において使用する用語“イムノアッセイ法”とは、例えば、均質アッセイ法、均質蛍光免疫測定法、均質蛍光強度免疫測定法、均質蛍光寿命免疫測定法、均質蛍光偏光免疫測定法(FPIA)、均質蛍光共鳴エネルギー移動(FRET)免疫測定法または均質化学発光免疫測定法或いは酵素結合免疫測定法(ELISA)のような非均質アッセイ法、並びにこれらの組合せのような、生成物を抗体によって検出する多くのアッセイ方法をいう。
【0019】
本明細書において使用する用語“蛍光偏光免疫測定法”または“FPIA”とは、基転移反応の生成物を検出するためのイムノアッセイ法をいう。蛍光偏光を使用し、分子相互作用を、蛍光標識化したまたは生来の蛍光分子の見掛けの大きさの変化をモニターすることによって調べる(Checovich, W. J., et al., Nature, 1995, 375:254-6;Owicki, J. C., J Biomol Screen, 2000, 5:297-306)。小蛍光分子(トレーサー)を平面偏光により励起させたとき、放出光は、該分子が蛍光事象中(励起と発光の間の時間)に溶液中で急速に回転するので、大きく偏光が解消される。しかしながら、前記トレーサーをはるかに大きいレセプターに結合させ、それによってその有効分子容積を増大させた場合、その回転は十分に低下して、トレーサーを励起させたのと同じ平面内で光を発出する。結合状態および遊離状態の蛍光分子は、各々、固有の偏光値を有し、結合状態で高い値を、遊離状態で低い値を有する。分子集団においては、測定偏光値は前記2つの値の加重平均であり、かくして、結合しているトレーサー分子の割合の直接の尺度を提供する。偏光値は、ミリ偏光単位(mP)として表し、500mPが最高理論値である。
実際には、フルオレセインのような小分子はおよそ20mPの偏光値を有し、抗体に結合させたとき、その偏光は100〜400mPまで増大する。拮抗FPIAにおいては、蛍光トレーサーは、図1に示すように、供与体生成物によって抗体への結合から外され得る。シグナルは結合対遊離のトレーサー画分の差に比例し、従って、該アッセイ法のダイナミックレンジおよび感度は、前記トレーサーおよび供与体生成物に対する抗体の親和性に依存する。FPIAにおける適切なダイナミックレンジを確立するには、約70〜80%のトレーサーを、拮抗因子の不存在下に、抗体に結合させなければならない。
例えば、図1は、SULT反応生成物PAPについての拮抗FPIAの図式を示しており;SULT反応から生成したPAPが、抗-PAP抗体への結合においてトレーサー(蛍光標識化PAP)と拮抗している。この方式においては、トレーサーの出発偏光は、トレーサーが殆んど全て抗体に結合しているので高く、反応が進行するにつれて低下し、トレーサーは抗体から外される。SULT反応において生成したPAPの量は、下記の等式を使用して定量し得る:
【0020】
log[PAP産物]=log[(最高偏光値-mP観測値)/(mP観測値-最低偏光値)]+ log IC50
【0021】
本発明に包含される酵素アッセイにおいては、SULTにおけるPAP、またはキナーゼにおけるADPは、それぞれ、硫酸化生成物またはリン酸化ペプチドと化学量論的量で生成する。即ち、PAPまたはADP、とりわけ供与体生成物における標準曲線の使用は、酵素代謝回転(enzyme turnover)の直接の尺度を与えるであろう。
本明細書において用語“高処理量スクリーニング”または“HTS”とは、何千もの分子(または試験化合物)をそのタンパク質の機能に対する作用について試験することを称する。基転移反応酵素の場合には、多くの分子をその触媒活性に対する作用について試験し得る。HTS法は当該技術において既知であり、該方法は、マルチウェルプレートにおいて、自動化液体操作および検出装置により一般に実施する;しかしながら、本発明の方法は、マイクロアレー上または微小流体装置内において実施し得ることも考慮される。
本明細書において用語“ライブラリー”または“薬物ライブラリー”は、複数の化学分子(試験化合物)、複数の核酸、複数のペプチドまたは複数のタンパク質、およびこれらの組合せをいう。そのスクリーニングを高処理量スクリーニング法で実施する場合、該方法は、マルチウェルプレートまたは微小流体装置を使用する。
本明細書において用語“結合性分子”または“巨大分子”とは、抗体または不活化酵素を称する。
本明細書において用語“抗体”とは、モノクローナル、ポリクローナルまたは組換え抗体をいう。抗体は、基転移反応の供与体生成物に対して生成させ、供与体生成物と供与体を高供与体濃度の存在において選択的に識別し得る。また、抗体は、ヌクレオチドの少なくとも1つのリン酸部分に対する特異性(即ち、5'-ホスフェート、5'-ホスホスルフェート、5'-ジホスフェートおよび5'-トリホスフェートを識別する能力)を示す。例えば、SULT反応においては、本発明の抗体は、PAPとPAPSを高厳格性でもって識別する。SULT反応における供与体-生成物分子のPAPSは、5'末端リン酸に結合した硫酸基の付加によってのみ異なる。これは、比較的小さい構造上の差異のように見え得るが、1個のリン酸基(スルフェート基に大きさおよび構造において極めて類似している)により異なる分子間を識別する抗体の実証された能力は、本発明の重要な特徴を提供している。例えば、分子のリボシルホスフェート部分に対して抗体を作製することが出来る。さらにまた、本発明のFPIA系アッセイ方法は、過剰のPAPSの存在下において反応生成物PAPに特異的に結合する抗体を適切に必要とする。
【0022】
キナーゼに関しての本発明の実施においては、担体タンパク質に結合させたADPを使用して、ADP分子のホスホ-リボシル部分は免疫反応性に暴露されるがアデニン部分は免疫反応性から大部分が隠蔽されるような方法で、動物内でまたはin vitro組換え法によりADPを特異的に認識するがATPは認識しない抗体を作製することが出来る。抗体の生成においては、アルファおよびベータホスフェート間を架橋するメチレンまたはイオウ基を含有するもののようなADPの非加水分解性アナログを使用して免疫動物中のヌクレオチダーゼまたはホスホターゼによる固定化ハプテンの加水分解を阻止するのが有益であり得る。また、メチルトランスフェラーゼおよびアセチルトランスフェラーゼに関しては、ヌクレオチド以外の小分子、それぞれ、s-アデノシルホモシステインおよび補酵素Aを特異的に認識する抗体を使用する。
本明細書において用語“不活化酵素”とは、供与体生成物に結合し得る結合性分子をいう。キナーゼの場合、ADPが供与体生成物であり、不活化ヌクレオシド二リン酸(NDP)キナーゼが結合性分子である。多くの酵素において、分離されたドメインが結合および触媒作用に関与することが知られている。遺伝子操作、アロステリック阻害、または共同因子、ヘミ基の除去のような他の化学的手段等によって結合特性を保持させながら触媒活性を選択的に破壊することは、酵素、とりわけ多サブユニット酵素がそれら基質に対する特異的な担体(もはやそれらの基質を化学的に改変することができない)として機能することを可能にする。
【0023】
本明細書において用語“抗体-検出可能タグ対”とは、基転移反応において生成した供与体生成物の検出を可能にする抗-供与体生成物抗体および検出可能タグ(蛍光供与体生成物)分子をいう。抗体/検出可能タグ対の結合における適切な解離定数は1x10-7Mまたはそれ以下であり、これにより未結合検出可能タグに対比して最少100mPの蛍光偏光変化をもたらし、且つ反応濃度で供与体分子に対し最少の交叉反応性を有する。最適の抗体-検出可能タグ対は、供与体分子への種々の結合部位、担体タンパク質へ種々のリンカーを使用して生成させ、且つフルオロフォアへの供与体分子結合の化学性を変化させることによって生成させた1群の検出可能タグとの相互作用のための数種の非加水分解性アナログを包含する数多くの種々の抗体を試験することによって同定し得る。例えば結合させたフルオロフォアを有し得る抗体との相互作用時の前記検出可能タグの蛍光または化学発光の変化をそれらの相互作用の尺度として使用し得る。
本明細書において使用するときの用語“リンカー”とは、スペーサーアーム構造を称する。アデニン環の接近性を立体的に最小にする短リンカー(即ち、カルバモイルおよびアミノエチル基)、および抗原のより柔軟性のある提示を可能にするより長めの炭素数6個のリンカーが存在する。リンカー分子は、検出可能タグ特性に、その抗原および蛍光特性の双方に影響を与える多くの重要な形で影響を及ぼす。一般に、抗原をフルオロフォアから十分に引き離して抗体との相互作用が妨害されないようにすることと、フルオロフォアにあまり自由過ぎる動きを生じさせないようにすることの間に注意しなければならない均衡が存在する。前者は親和性抗体結合性の低下およびフルオロフォアの消光をもたらし、一方、後者は抗体結合時の偏光変化を低下させ、それによってアッセイのダイナミックレンジを減じる。種々のリンカーは比較的単純な化学法を使用して取り替えることができる。即ち、リンカーは、検出可能タグの抗原および蛍光特性を最適にするために、多くの方法で変化され得る。さらにまた、ヘテロ二官能性リンカーを含む抗体のPAP結合について説明した方法と同様な方法を使用して、先ずスペーサーアームおよび/または芳香族置換基をPAP上に導入し、次いで反応性フルオレセイン誘導体と反応させることもできる。この方法は、可能性あるリンカー構造の範囲を大いに拡大する。
【0024】
方法および材料
本発明の以下の実験プロトコールは、特定の方法論、プロトコール、抗体、酵素、検出可能タグ、なかんずく、説明する試薬に限定されない;これらは、基転移反応によって変動し得るからである。また、本明細書において使用する用語は特定の実施態様を説明する目的のためだけであり、本発明の範囲を限定するものではないことを理解すべきである。
アッセイ試薬の開発
本発明においてHTSアッセイ法を成功裏に開発するには、アッセイ試薬相互作用の親和性および特異性、並びに得られる検出可能タグ蛍光特性の変化が、感度、ダイナミックレンジおよびシグナル対ノイズ比のようなアッセイの全体的性能を究極的に特徴付ける。それで、この相互作用を最適化するために、出願人等は、図4に示すような反復同時開発方法を提供して基転移反応生成物の検出、定量およびスクリーニングを最も効率的に達成している。
特に、図4は、SULT供与体生成物FPIA用の試薬、即ち、抗-PAP抗体および蛍光標識化PAP検出可能タグの反復同時開発戦略を説明する。図4は、ウサギの免疫用の種々の抗原を作製するためには、PAPを、以下で説明するような種々の長さのリンカー分子を使用し、アデニンのC2、C6およびN6アミノ基を介しての結合により、担体タンパク質(BSAまたはKLH)に結合させ得ることを示している。抗体結合において使用した同じPAP-リンカー-NH2中間体を使用して、検出可能タグとして試験するためのフルオレセイン-PAP結合体を合成することもできる。得られる抗体および検出可能タグのマトリックスは、FPアッセイ法を使用して、結合性について試験することができ、また、高親和性結合および最高FP変化を示す対は、トレーサー特性のさらなる最適化のための基礎として使用することができる。最適化した抗体-検出可能タグ対は、SULT1E1または他のSULTイソ型による硫酸化反応におけるPAP生成の検出について試験することができる。
また、数種のPAP抗原および検出可能タグを、PAP分子のリボシル-ホスフェート部分の抗体認識を最高にし、且つPAPSとの交叉反応性を最小にする全体的構造バイアスを保持させながら、ヌクレオチドを担体タンパク質およびフルオレセインに結合させることによって合成し得ることも考慮される。PAP抗原から生成した抗体は、検出可能タグとの相互作用について試験して、本発明に包含される新規なアッセイ法の最適結合性および蛍光特性を示す組合せを同定し得る。
【0025】
高親和性抗体の生成
本発明の方法に従い、当業者であれば、PAPのリボシル-ホスフェート部分のような抗原性ヌクレオチドまたは補酵素Aのような他の化学分子に対する種々の特異的抗体(即ち、ポリクローナル、モノクローナルまたは組換え)を容易に調製し得る。
一般に、ある範囲のPAP抗原に対するポリクローナル抗体は、ウサギにおいて適切に作製される。動物の注射および血清採集は、以下のプロトコールによって実施し得る。3羽のウサギを、開発する各抗原によって免疫し得る。力価、親和性に依存するが、1羽のウサギからの抗血清収量(約100ml)が数千回〜100万回のFPIAアッセイのために適当であり、ポリクローナル-抗原相互作用の多価性が極めて高い親和結合性を生じさせる。従って、これらの理由により、ポリクローナル抗体が多くの場合FPIAにおいて使用される(Nasir, M. S. and Jolley, M. E., Comb Chem High Throughput Screen, 1999, 2:177-90)。
また、以前の公表プロトコールに提示されている方法(Harlow, E. L., D., 1999)に従って作製し得るモノクローナル抗体も、本発明の範囲に属する。また、組換え単鎖抗体は、以前に公表されたin vitroコンビナトリアルエヴォリューションおよびディスプレイ法(Schaffitzel, C., et al., J Immunol Methods, 1999, 231:119-35;Breitling, F., Dubel, S., 1999)を使用して作製することが出来る。組換え抗体の調製例は、米国特許第5,693,780号、第5,658,570号、第5,876,961号(全て参考として本明細書に合体させる)に記載されている。
免疫応答の顕在化を助けるには、ウシ血清アルブミン(BSA)およびキーホールリンペットヘモシアニン(KLH)をPAP抗原への結合用担体タンパク質として使用し得る。KLH抗原は、一般に、哺乳類においてより強い免疫応答を引き出すが、BSA結合体よりも可溶性が低い傾向がある(Harlow, E. L., D., 1999)。PAPは、図7〜8に示すように、双方の担体タンパク質にアデニンのC2、C8およびN6アミノ基を介して結合させ得る。本発明の範囲において、FPIA系SULTアッセイ法の開発には、反応生成物PAPに過剰のPAPSの存在下において特異的に結合する抗体を必要とする。さらに適切には、前記抗体は、PAP分子のリボシルホスフェート部分を認識し得るものであり、PAPの5'-ホスフェートとPAPSの5'-ホスホスルフェートを識別し得る。
【0026】
結合方法
アデニンへの結合が、アデニン部分への結合により生じ得るPAPSとの抗体交叉反応を最小にすること(Goujon, L., et al., J Immunol Methods, 1998, 218:19-30;Horton, J. K., et al., 1992, J Immunol Methods, 155:31-40;Bredehorst, R., et al., Biochim Biophys Acta, 1981, 652:16-28)、さらにまた、得られる抗体の親和性にハプテン結合部位が影響を与え得ること(Crabbe, P., et al., J Agric Food Chem, 2000, 48:3633-8)が、当該技術において知られている。使用するハプテン結合化学法は、アデニンヌクレオチドの結合において広く使用されている詳しく記述された1または2工程合成方法である(Brodelius, P., et al., Eur J Biochem, 1974, 47:81-9;Lindberg, M. and Mosback, K., Eur J Biochem, 1975, 53:481-6;Camaioni, E., et al., J Med Chem, 1998, 41:183-90)。出発物質は全て商業的に入手可能であり、反応は良好な収率で進行し得ることが理解される。これらの結合方法を以下でさらに詳細に説明する。本発明の方法に従い、本発明は、以下のハプテン結合化学を包含するがそれに限定され得ないものとする
【0027】
N6アミノ基への結合
カルバモイル結合:N6-カルボキシメチル-PAPは、PAPをヨード酢酸によってアルキル化して1-カルボキシメチルアナログを形成させ、これを塩基中で転位させてN6-カルボキシメチルアナログを形成させることによって調製し得る(Lindberg, M. and Mosback, K., Eur J Biochem, 1975, 53:481-6)。N6-カルボニルメチル-PAPは、N-ヒドロキシスルホスクシンイミドにより促進させたカルボジイミドカップリングを使用して、担体タンパク質に結合させ得る。同様な方法を使用し、以前に公表されたプロトコールに記載されているようにして高特異的5'-AMP抗体を作製することが出来る(Bredehorst, R., et al., Biochim Biophys Acta, 1981, 652:16-28)。
C6結合:N6-アミノヘキシル-PAP (Sigma/Aldrich社、ミズーリ州セントルイス)は、a) N-スクシンイミジルS‐アセチルチオアセテート(Pierce社、イリノイ州ロックフォード)を使用して、タンパク質アミノ基(ほぼリジン)を反応性チオールに転換し、b) ヘテロ二官能性アミン-チオール反応性リンカー(Pierce社、イリノイ州ロックフォード)を使用して、PAPリンカー上の末端アミノ基を前記タンパク質チオールに結合させることによって、担体タンパク質に直接結合させ得る。特筆すべきは、アデノシンのN6アミノ基は実用目的上は反応性ではないことである;例えば、該アミノ基は、スクシンイミジルエステルとは極めて貧弱にしか反応しない。
【0028】
アデノシンC2への結合
2-クロロアデノシンをオキシ塩化リンと、次いで三塩化リンと反応させることによってリン酸化して、3',5'-ビスホスホ-クロロアデノシンを調製し得る。その後のエチレンジアミンおよびジアミノヘキサンとの反応並びにアニオン交換クロマトグラフィーによる精製を使用してアデニンの2位置におけるPAPの2-アミノエチル-および6-アミノヘキシル-アナログを調製し得る(Brodelius, P., et al., Eur J Biochem, 1974, 47:81-9)。これらの分子は、上述したようなヘテロ二官能性リンカーを使用して担体タンパク質と反応し得ることも意図する。
アデノシンC8への結合
PAPの8位置へのC2およびC6結合は、C2位置において説明したのと同じ方法を使用して生成することが出来るが、出発物質は8-ブロモアデノシン(Sigma/Aldrich社)であり得る。また、8-アジド-PAP (ICN社、カリフォルニア州コスタメサ)も、UV照射により、担体タンパク質に直接結合させ得る。
薄層クロマトグラフィー(TLC)は、アデニン環吸収スペクトル変化をモニターするのに使用し得る。アニオン交換(Dowex樹脂)および相分離は、抗原精製に使用し得る。高い抗原純度を確保するために、担体タンパク質に結合させるのに使用する最終PAP-リンカー分子をHPLCによって精製し、その存在性を質量スペクトルおよびNMR分析によって検証することができる。メチル化担体タンパク質の使用がリボース環とのカルボジイミド反応を阻止することが報告されているので(Bredehorst、 R., et al., Biochim Biophys Acta, 1981, 652:16-28)、N6-カルボキシメチル-PAPをメチル化BSAに結合させてもよい。
他のPAPアナログは、全てリンカー末端に反応性アミンを有する傾向にあり、これらは、より特異的なアミンおよびチオール化学物を使用して、BSAおよびKLHへの結合に使用することができ、それによってリボースとの反応を回避することができる。タンパク質アミンのチオールへの転化およびヘテロ二官能性リンカーを介しての結合は、担体タンパク質中にシステインよりも多くの反応性リジンが存在するので、システインへの直接結合よりも高密度の抗原結合をもたらし得る。担体タンパク質上のPAP密度はアデニン環についての質量分光分析によって測定でき、結合化学性を必要に応じて調整して高密度(タンパク質分子当り> 10のPAP)を得ることができ、それによって強い免疫応答が担保される。全部で約8〜10種の異なる抗原を合成することができ、幾つかはアデニン環の接近性を立体的に最小にするカルバモイルおよびアミノエチル基のような極めて短いリンカーを有し、また、抗原のより柔軟な提示を可能にする、より長めの炭素数6個のリンカーを有することが想定される。
【0029】
蛍光PAP分子の合成および精製
一般に、FPIA検出可能タグ分子は、3つの異なる構成部分、即ち、抗原、フルオロフォアおよびこれらを結合させるのに使用するリンカーに分けることができる;さらなる主要な構造的変数は、抗原へのリンカーの結合部位である。当業者であれば、適切な抗原、フルオロフォアおよびリンカーが本明細書において詳細に説明しているものに限定されないことを理解されたい。しかしながら、本発明の適切な局面においては、フルオロフォア、フルオレセインは、それを、担体タンパク質への抗原結合に使用したのと同じアデノシン上の部位に結合させるのに使用し得る。担体タンパク質への結合に使用するのと同じPAP-リンカー分子を、数種のアミン反応性フルオレセイン誘導体との結合に使用し得る。ある範囲の反応性フルオレセインの使用は、リンカー領域にさらなる構造的多様性を導入し得る。例えば、フルオレセインスクシンイミジルエステルは、種々の長さのスペーサーアームでもって利用でき、DTAF (4,6-ジクロロトリアジニルアミノフルオレセイン)は結合部位に隣接した嵩高で平面的な置換基を含有する。同様に、担体タンパク質結合部位近くでのAMPへの芳香環の導入(N6-ベンジルAMP)は、その抗体に対する親和性を未修飾AMPに対比して40倍増強する(Bredehorst, R., et al., Biochim Biophys Acta, 1981, 652:16-28)。簡単に言えば、アミノ末端を有する種々のPAP-リンカー分子はアミン反応性ローダミン、テキサスレッドまたは他の赤色フルオロフォアと反応させることができ、得られるPAP-フルオロフォア結合体は、抗体結合親和性および蛍光偏光の変化について試験することができる。
PAP結合体の精製は、薄層クロマトグラフィー(TLC)によって実施し得る。複数の分離を並行して操作することが可能であり、蛍光生成物の同定が目視により可能であり、さらに1回のクロマトグラフィー操作で数千もの結合性アッセイのための十分な量の蛍光化結合体が提供されるので、反応生成物の分離にはTLCが使用される。即ち、蛍光検出可能タグ分子のパイロット試験においては、TLCは、HPLCのような別の分離方法よりも優れている。PAP-フルオロフォア結合体のクロマトグラフィーおよび溶出が終了したならば、後述するようにして、強度、基底偏光(抗体なしでの)、安定性、抗-PAP抗体に対する結合性、並びに未標識PAPおよびPAPSによる拮抗性について結合体を分析することができる。
【0030】
抗体-検出可能タグ相互作用に影響するパラメーター
基転移方法の生成物を検出し、定量し、スクリーニングするための最も感度のあるアッセイ方法を創生するためには、抗体-検出可能タグ相互作用の親和性に影響するパラメーターを注意深くモニターする。このことは、例えば、ある種の受容体基質が示す強力な基質阻害によって、生成させ得る生成物の量が抑制されるSULT1E1の場合に重要である。本発明に属する新規なSULTアッセイにおいて必要な親和性は、高親和性で阻害性基質のエストラジオールの阻害例を考慮することによって推定する。一般に、エストラジオールのKiは80nMであり、Kmは約20nMであり(Zhang, H., et al., J Biol Chem, 1998, 273:10888-92)、従って、アッセイにおいては50nMのエストラジオールを使用し、酵素および反応時間を10%の基質が消費される(5nMのPAPの生成をもたらす)ように調整することが、有意の阻害を起すことなく初期速度の測定を可能にし得る良好な妥協点である。
5nMのPAPの検出を可能にするためには、蛍光-PAP(および未結合PAP)に対する抗体結合に必要な親和性は、およそ25nMであろう。殆どの抗体-抗原相互作用は、ポリクローナル抗体またはモノクローナル抗体のいずれであれ、1nM〜1μMのKdを有する(Harlow, E. L., D., 1999)。例えば、FPIA系キナーゼアッセイにおいて使用する抗-ホスホチロシン抗体は、リン酸化ペプチドおよびそれらの蛍光結合体に対しナノモル以下の親和性を示し(Parker, G. J., et al., J Biomol Screen, 2000, 5:77-88);上述した抗-AMPポリクローナル抗体は、100nMの親和性を有するヌクレオチドに結合した(Bredehorst, R., et al., Biochim Biophys Acta, 1981, 652:16-28)。抗原の構造が得られる抗体の親和性に影響を与えるかどうかを予測することは困難であるが、担体タンパク質および結合方式の双方は、大きな効果を有し得る(Crabbe, P., et al., J Agric Food Chem, 2000, 48:3633-8;Signorini, N., et al., Chem Res Toxcol, 1998, 11:1169-75;Oda, M. and Azuma, T., Mol Immunol, 2000, 37:1111-22)。従って、PAP上の異なる部位を介して2つの異なるタンパク質に結合させたPAPを使用して、さらに、異なる長さのリンカー領域を使用して、抗体を作製することが出来る。
【0031】
PAP検出の感度に影響を与えるもう1つの要因は、高濃度においてさえも極めて緩慢にしか硫酸化されない貧弱な基質である。これらの場合、必要とするSULT1E1酵素の量は、コスト性故に限界要因となる。エストラジオールによりVmaxの1%の速度で回転させる基質を使用すれば、5nMのPAP(約30分内での適切な検出限界)を生成させるのに30ngのSULT1E1を必要とするであろう。この量は、100mgの酵素バッチが300万回よりも多い反応に対処し得ることを意味し得るので、良好に許容範囲内にある。
さらにまた、遊離または結合状態のプローブの偏光の差異は、アッセイの全体的“広がり”即ちダイナミックレンジを特徴付ける。50mP未満の変化でSULT1E1活性の半定量検出に十分であり得るが、100mPまたはそれ以上の変化は、アッセイ方式の設計におけるはるかに大きい柔軟性およびより定量的な動的情報の双方を提供し得る。従って、偏光は分子容積に比例し、小分子量フルオロフォアへの抗体(150 kDa)結合時の有効容積の変化は、少なくとも300 mPの増加を生じさせることが予期し得る。しかしながら、フルオロフォアをPAPに可撓性スペーサーアームにより結合させることのような、遊離および結合検出可能タグの双方の観測偏光に影響を与え得る他の要因も存在する。即ち、それらの最高偏光は、抗体結合がスペーサーおよびフルオロフォアの移動性を低下させる度合の関数である;検出可能タグ“プロペラ効果”は、タンパク質結合フルオロフォアについてさえも比較的低い偏光値をもたらし得る。
さらなる考察点は、重複スペクトルを有するアミノ酸官能基からの消光を含む、レセプターへの結合時のフルオロフォアの強度の変化が存在し得るかどうかである。偏光は、蛍光強度に依存しない(Owicki, J. C., J Biomol Screen, 2000, 5:297-306);しかしながら、偏光は2つの平面で取った強度測定値の比であるので、有意の消光は、抗体検出可能タグ相互作用に影響を与え、アッセイ感度の低下をもたらし得る。抗体と結合したときの検出可能タグの蛍光特性の変化は、各相互作用分子双方の関数であり、予測するのは困難である。即ち、種々の抗体および検出可能タグ分子はマトリックスの形で調べられるであろうことが考慮される。さらに、検出可能タグの構造の最適化、或いは、結合させた検出可能タグの可動性を制限するためまたは消光を減じるためのアッセイ緩衝液の最適化も考慮される。
【0032】
最適抗体/検出可能タグ対(1種以上)の同定
最適抗体/検出可能タグ対を同定するためには、多様な組合せのフルオレセイン誘導体と結合化学物を一般に評価する。上述した合成および精製方法は、反応毎に数種の蛍光生成物の単離をもたらし、その結果、少なくとも100種の蛍光結合体を評価することができるが、数千の個々のFPアッセイを必要とする。FPアッセイの均質特性およびマルチウェル器具を利用できることは、スクリーニング操作を極めて迅速なものにしている;律速段階は、通常、合成と精製であり、抗体-検出可能タグ結合特性についての試験ではない。抗体/検出可能タグ試験は、Tecan UltraマルチウェルFPリーダーにおいて、Trisまたはリン酸緩衝液中で非特異相互作用を阻止するための低濃度の担体タンパク質(0.01%ウシガンマグロブリン)の存在下に実施し得る。初期スクリーニングは、以下の望ましい特性を見出すこと含む:抗体不存在での低偏光(<100mP);抗-PAP抗体への高親和結合性(Kd < 100nM);結合および遊離状態における偏光の最高差異(ΔmP >100mP);抗体に結合した場合の最少蛍光消失;PAPに対する選択性(即ち、PAPによる、PAPSまたは他のアデニンヌクレオチドによらない拮抗的置換);迅速な結合/解離速度;および、SULT1E1または他のアッセイ成分との相互作用の無いこと。
初期抗体結合性アッセイにおいては、精製した検出可能タグの各々を、結合フルオレセインの強度が消光されないことを想定して、蛍光強度を測定することによって定量し得る。常に該当するものではないが、有意の消光を有するものは、それらが低結合親和性を有するようなので、結合性分析によりそれらを排除することができる。見込みのある親和性(Kd < 100nM)および蛍光特性を有する抗体/検出可能タグ対は、未標識PAPおよびPAPSおよび他のヌクレオチドとの拮抗により、結合特異性について試験し得る。最も望ましいFPIA特性を有する抗体/検出可能タグ対は、さらに詳細に試験して、塩または変性剤の添加のような外因性条件における何らかの変化がその感度およびダイナミックレンジを改善し得るかどうかを評価し得る。さらなる検出可能タグを必要に応じて合成し、観測した構造-活性関係およびコンピュータ作成構造モデル予測に基づき試験し得る。
【0033】
本発明の応用性
本発明の方法は、後の実施例において説明するように、基転移反応における酵素活性の尺度として使用し得る。本発明の方法は、例えば、酵素モジュレーター、とりわけ阻害因子として作用し得る薬物の同定のため、また、潜在的薬物分子がどのように代謝されるかを決定するために、高処理量スクリーニング試験所における基転移酵素の分析を可能とするので、製薬工業においてとりわけの重要性を有する。
新たな治療薬の開発において目標とする基転移酵素の数が増大している。キナーゼは現在最高に興味あるものであるが、SULT、UGT、メチルトランスフェラーゼ、アセチルトランスフェラーゼおよびADP-リボシルトランスフェラーゼも対象とされている。このリストが増大するにつれ、各酵素についての或いは同じ受容体基質を改変する小下位群の酵素についての特異性試験を使用することは、現実性の無い極めて費用高となるであろう。本発明のアッセイ法とキットは、与えられた基転移酵素群の全ての酵素の検出を可能にし、これら酵素の全てについての一般的な評価項目検出系を提供する。このことは、とりわけ高処理量スクリーニング検査室においての使用の容易性を大きくし、ロボットに全ての検出試薬を備え付けることができ、各種基転移酵素および興味ある潜在的基質または阻害因子をマトリックス形式で試験し得る。
さらに、薬物および他の生体異物の共役的代謝における基転移反応の関与は、それらの薬力学、薬物動態学および副作用レベルの重要な決定因子である。この点、本発明のアッセイ法およびキットは、潜在的な薬物分子のSULT、UGT、メチルトランスフェラーゼおよびアセチルトランスフェラーゼによるスクリーニングをHTS方式で可能にする。多くの種々の生体異物共役イソ型が存在し、本発明方法の汎用的な性質故に、本発明のキットは、マトリックス形式のそれらの試験を大いに簡素化するであろう。
即ち、本発明の汎用的方法は、後述する統合化(integrated)薬物発見方法に貢献するために、ステロイドホルモン生体合成および機能、生体異物代謝、酵素レセプター調節およびシグナル伝達のような各種の異なる基転移関連酵素過程に応用し得る。
【0034】
薬物発見における硫酸化
硫酸化は、内在性ホルモンおよび生体異物のレベルおよび活性を調節する普遍的な共有結合性修飾である。ヒトにおいては、硫酸化は、各々異なるが重複した基質特異性と組織分布を有する1群の11種のスルホトランスフェラーゼ酵素(SULT)によって触媒されている(Strott, C. A., Endocr Rev, 2002, 23:703-32)。硫酸化における内在性基質としては、ステロイドホルモン、カテコールアミン、および甲状腺ホルモンのような多くの重要な情報伝達分子があり;硫酸化される生体異物としては、薬物、前駆型変異原(promutagen)、環境性内分泌撹乱物質がある(Strott, C. A., Endocr Rev, 2002, 23:703-32;Glatt, H., et al., Mutat Res, 2001, 482:27-40;Coughtrie, M. W. and Johnston, L. E., Drug Metab Dispos, 2001, 29:522-8)。硫酸化は、可逆性であり、情報伝達分子の活性を、多くの場合それら分子のレセプタータンパク質に対する親和性を改変することによって変化させ得る、即ち、硫酸化は、レセプターリガンドに対するオン-オフスイッチとして、リン酸化がその役割をタンパク質に対して果たすのと同程度に作動する。さらにまた、生体異物代謝における硫酸化の役割は、ホルモン情報伝達および細胞ホメオスタシスの調節におけるその関与と密接に絡み合っている。例えば、硫酸化は、うつ病、乳がんおよび心臓血管疾患用の薬物ターゲットとしての特異的神経伝達物質レセプター、核レセプターおよびプロテインキナーゼに対する内在性リガンドの活性を調節する(Plassart-Schiess, E. and Baulieu, E. E., Brain Res Rev, 2001, 37:133-40;Strott, C. A., Endocr Rev, 1996, 17:670-97;Kuroki, T., et al., Mutat Res, 2000, 462:189-95)。製薬企業によるこれらの障害に対するより選択性の治療薬の開発は、潜在的SLUT基質および阻害因子の高処理量スクリーニング(HTS)のための分子アッセイ法および精製SULTイソ型が無いために、現在のところ妨げられている。
【0035】
溶解性、大きさ、小細胞分布において異なり、20%未満の配列相同性を共有する2つの主要なクラスのヒトのSULTが存在する。膜結合酵素は、ゴルジ体内に存在し、ヘパリン、グリコサミノグリカンおよびタンパク質チロシンのような大内在性分子をスルホン化する(Strott, C. A., Endocr Rev, 2002, 23:703-32)。細胞質内スルホトランスフェラーゼ即ちSULTは、小分子量生体異物およびホルモンをスルホン化する。
11種の既知の細胞質内SULTイソ型が存在し、これらはその組織分布および特異性において異なる。使用されている命名法は、アミノ酸レベルでの相同性に基づいている:数字で表わされる同じ群内のSULTは少なくとも45%のアミノ酸相同性を共有し、文字で表される同じ下位群内のSULTは少なくとも60%の相同性を共有する。SULTは、ある程度は、基質特異性に基づいて分類することができるが、それらの基質プロフィールは、異なる群内の酵素間においてさえも、広範囲に重なり合う。
構造研究により、基質結合部位に柔軟性があること並びにリガンド結合性がより秩序立った構造への転換を引き出すことが明らかにされており、これらがSULTの広い基質特異性に寄与し得る(Bidwell, L.M., et al., J Mol Bil, 1999, 293:521-30)。こういう理由で、分子結合性のようなコンピュータ手法をSULT基質および阻害因子の同定に応用できるということはなさそうである;このことは、本発明に属するHTSアッセイ法の価値を増大させる。
【0036】
本発明のアッセイ法は、HTS方式において使用することができ、例えば、注目する化合物が1種以上のSULTイソ型と相互作用するかどうか、該化合物が基質または阻害因子であるかどうか、および相互作用における動的パラメーター(IC50、Km、Vmax)のようなSULT代謝情報を提供し得るということが考慮されている。本発明のHTSアッセイは、以下の2つの異なる方式でスクリーニングすることによって、前記情報の全てを提供することが想定されている:a)試験化合物の代謝回転の直接測定;およびb)プローブ基質硫酸化の化合物阻害の測定。直接代謝回転方式においては、化合物をSULTイソ型の集団に対してスクリーニングし、個々のSULTイソ酵素による化合物硫酸化をプロフィール化することができるであろう。イソ酵素同定後、適切なSULTイソ酵素によるより詳細なキネティクス試験を使用して生体内クリアランス速度を予測することができ(Obach, R. S., et al., J Pharmacol Exp Ther, 1997, 283:46-58)、それにより臨床試験おける化合物の不合格率を減じることができるであろう(Greco, G. N., E; Martin, YC, 1998, 219-245)。既知の基質(例えば、SULT1E1に対するエストラジオール)を使用し、試験化合物を阻害についてスクリーニングすることによって、SULT阻害因子を同定し得るであろう。イソ酵素特異的なキネティクスおよび阻害データは、特異的SULTによる代謝の知識を提供することにより、可能性のある薬物-薬物相互作用および薬理遺伝学問題を薬物発見チームに対して警告し、薬物発見をさらに改善するであろう。治療薬に対する種々の応答へのSULTにおける遺伝子的差異の寄与は、研究の活動的な領域である(Thomae, B. A., et al., Pharmacogenomics J, 2002, 2:48-56)。
薬物代謝に関与するSULTの同定は、潜在的な薬物-薬物相互作用の前臨床評価および毒物試験において使用する生体外アッセイ法または動物モデルの賢明な選択を促し、それによって不適切な或いは不必要な実験動物の使用を減じるであろう。代謝データは、合理的な薬物設計および先導的最適化の要素として使用し得る。種々のSULTイソ酵素に対する基質特異性を特徴付ける構造-活性関係のより良好な理解は、その代謝プロフィールを変化させる一次化合物の構造的修飾のための根拠を提供し得る(Greco, G. N., E; Martin, YC, 1998, 219-245)。従って、本発明の方法は、種々のSULTイソ酵素に対する基質特異性を規定する構造-活性関係のより良好な理解を可能にするであろう。
【0037】
薬物発見への統合化取り組み
本発明のアッセイ法がHTSアッセイとして使用できるということは、例えば、硫酸化が関連シグナル伝達生物学の要素である治療領域における薬物開発への合理的な統合化手段が可能になる。本発明のHTS方法を使用し得る幾つかの特定の領域としては、例えば、ステロイドホルモン系療法がある。硫酸化は、本発明によれば、乳腺腫瘍におけるエストロゲンレベル調節並びに前立腺がんにおけるアンドロゲンレベルへの関与にまで及ぶ。さらにまた、コルチゾール硫酸化は、よくは理解されてないものの、前記ホルモンをグルココルチコイドレセプターへの結合に対して不活化する。ステロイドリガンド-レセプター結合性についての高処理生化学アッセイおよび得られる転写調節補因子タンパク質結合性は、すでに商業的に利用可能であり、製薬HSTグループによって使用されてより選択性のステロイドホルモンモジュレーターが見出されている(Parker, G. J., et al., J Biomol Screen, 2000, 5:77-88;Spencer, T. A., et al., J Med Chem, 2001, 44:886-97)。従って、ステロイド硫酸化についての確固たるHTSアッセイ法を利用し得ることは、ステロイドシグナル伝達に絞った製薬グループにとって利用可能な分子手段備蓄への重要な追加物を提供し得る。例えば、ERをターゲットする化合物によってSULT1E1を阻害することは、腫瘍細胞中のエストロゲンレベルを上昇させることによる有害な効果を生じることがあった。SULT1E1阻害特性を有する分子は、本発明のアッセイ方法を使用した研究に基づいて選択或いは修飾することができるであろう。
神経ステロイドの調節は、神経興奮バランスを制御するための新規な薬理学的方法として研究されている(Malayev、 A., et al.,Br J Pharmacol, 2002, 135:901-9;Maurice, T., et al., Brain Res Res Brain Rev, 2001, 37:116-32;Park-Chung, M., et al., Brain Res, 1999,830:72-87)。本発明に包含される方法は、スルホ結合についての内在性および合成神経ステロイドの容易なスクリーニングを可能にし、基本的生物学に洞察力をもたらし、さらに、リード分子の同定および最適化のための手段を提供することによって、これらの努力を適切に促進させ得る。より良好な分子手段の必要性は、年齢関連の老化および記憶喪失を軽減することを目的とする“老化防止”栄養補助食品としてのDHEAのかなり大きい対抗市場がすでに存在するという事実によって強調される(Salek, F. S., et al., J Clin Pharmacol, 2002, 42)。
さらにまた、本発明の方法は、コレステロール流出、血小板凝集および心臓血管疾患の治療における皮膚発症、おそらくは、ある形状の皮膚がんの調節における硫酸コレステロールに関連する薬物ターゲットを適切に同定し得ることを意図する。この場合、スルホトランスフェラーゼ(おそらく、SULT2B1b)が薬物ターゲットとなり得、このイソ型を選択的に阻害する分子を同定する必要があり得る。ヒトSLUTの完全パネルおよび本発明の堅牢なHTSアッセイ法を利用し得ることは、そのような取組みにとって有益であり得る。
【0038】
薬物発見におけるグルクロン酸化
毒性代謝物の生成および好ましくない薬物動態のような薬物代謝問題は、臨床試験中に全ての薬物候補のほぼ半分の不合格をもたらしている(Obach, R.S., et al., J Pharmacol Exp Ther, 1997, 283:46-58)。大きな製薬企業は、全て、薬物動態学および薬理ゲノム学結果を薬物発見プロセスの早期に考察する必要性を認識しており、薬物代謝を評価するための高処理量in vitro方法の当面の必要性を生じている。P450依存性酸化に加えて、グルクロン酸化は、おそらく、肝臓薬物代謝の最も重要な経路である。非ステロイド系抗炎症剤、オピオイド、抗ヒスタミン剤、抗神経病薬および抗うつ薬のような広範囲の薬物は、グルクロン酸化によって排除または活性化される(Meech, R. and Mackenzie, P. I., Clin Exp Pharmacol Physiol, 1997, 24:907-15;Radominska-Pandya, A., et al., Drug Metab Rev, 1999, 31:817-99)。その重要性にもかかわらず、グルクロン酸化を触媒する肝臓UDP-グルクロノシルトランスフェラーゼ(UGT)の広く重なり合う基質特異性は、柔軟性のあるin vitroアッセイ方法が無いために、理解に乏しいままである。その主たる理由は、使用する触媒アッセイ法が基質特異性クロマトグラフィー工程を含む生成物からの反応物の分離を必要とし、従って、HTS方式においては現実的でないことである。
2種のUGTファミリー(例えば、UGT1およびUGT2)がヒトにおいて同定されている;これらのファミリーのメンバーは一次アミノ酸配列において50%未満の同一性しかないものの、これらは、基質特異性において有意の重なり合いを示す。生体異物代謝の大部分が起こるヒト肝臓内で発現するUGT1群のメンバーとしては、UGT 1A1、1A3、1A4、1A6および1A9がある。UGT2群は広くは研究されていないものの、UGT 2B4、2B7、2B10、2B11および2B15が肝臓内で発現することが知られている。UGTの変異は、5〜12%の頻度で発症し(Weber, W.、1997年)、神経毒症状、重篤な場合には、死に至る高ビリルビン血症のような有害作用を有することが知られている。P450のような他の薬物代謝性酵素における場合と同様に、UGT発現レベルの個人間の差が観察されており、薬物応答の差異に関連している(Iyer, L., et al., J Clin Invest, 1998, 101:874-54)。例えば、ギルバート症候群患者におけるようなUGT1A1の低発現は、有望な抗がん剤であるイリノテカン(Irinotecan)の毒性に関連している(Wasserman, E., et al., Ann Oncol, 1997, 8:1049-51)。さらに、腫瘍組織におけるUGTアップ調節は、抗がん薬抵抗性の可能性ある原因として同定されている(Franklin, T. J., et al., Cancer Res, 1996, 56:984-7;Takahashi, T., et al., Jpn J Cancer Res, 1997, 88:1211-7)。
【0039】
既知のUGTの全てが広い基質特異性を示しており、単一のイソ酵素が広範囲の構造的に関連の無い化合物のグルクロン酸化を触媒する;驚くに当たらないが、UGTイソ酵素の特異性においても大きな重なり合いが存在する(Radominska-Pandya, A., et al., Drug Metab Rev, 1999, 31:817-99)。内在性分子の生体転換に関しては、UGT1A1は、明らかに、テトラピロール、ビリルビンのグルクロン酸化に関与し、その排出をもたらす主要イソ型である。これ以上に特異性に関しての総括を行うことは、最近同定されたイソ型の殆どにおいて系統的な試験が無いために困難である。数多くの内在性ステロイドは、1A1、1A3、1A4、2B4、2B7および2B15のような肝臓イソ型の殆どにおいてアグリコンとして同定されている。脂質および胆汁酸は、2B4および2B7に対する基質として作用し、最近、レチノイドが双方の群に由来する幾つかのイソ型に対する基質として同定されている。既知の生体異物アグリコンの構造的多様性は、極めて広い;それには、イミプラミンのような第3級アミン、アセトミノフェンおよびナプロキセンのような非ステロイド系抗炎症剤(NSAID)、モルフィンおよびコデインのようなオピオイド、およびクロフィビリン酸のようなカルボン酸含有薬物を含む薬物および薬物様分子がある。
短期的には、製薬企業は、潜在的薬物候補が生体内でグルクロン酸化されるのかどうか、そうであれば、UGTイソ型によってグルクロン酸化されるかどうかを判定するためのより良好な方法の当面の必要性を有する。そして、長期的には、グルクロン酸化による薬物代謝を予測することができることの開発は、主要肝臓UGTの各々が認識する“化学空間”を十分に規定する系統的取組みを必要とするであろう。提案された第II段階研究は、大量の多様な化学物を分離したUGTイソ型による結合性と代謝について迅速にスクリーニングするのに使用し得るHTSアッセイ方法のようなこの取組みに必要な分子手段を産み出すであろう。
【0040】
医薬の研究と開発は、時間を消費するもので、費用が高く、非効率であり、高いコストまたは米国医療消費者における新治療薬の低利用性のいずれかをもたらす。現在、米国における新薬の開発には、10〜15年間と4億〜7.5億ドルの総研究開経費を要する。臨床試験は、典型的には総研究開発コストの30〜50%を占める最も費用の掛かる開発期間であるが、臨床試験において試験された全薬物候補の僅か10%しか、最終的には商業化されない(Obach, R.S., et al., J Pharmacol Exp Ther, 1997, 283:46-58)。さらにまた、臨床試験中の薬物候補脱落理由の分析によれば、化合物の成功または失敗の主要決定因子の幾つかは、クリアランス速度(薬物動態)を含むその代謝機能、他の薬物代謝による干渉の可能性および毒性代謝物生成の可能性であることが確認されている。貧弱な薬物動態(通常、過剰に速いクリアランス)と毒性との合計が全臨床失敗例の50%を上回る原因となっている。潜在的薬物候補が代謝されるかどうかについてのより良好な理解が発見段階の早期で得られることは、前臨床および臨床試験の成功率を改善し、より効率的な薬物開発並びに治療薬の経済性と利用性の増大をもたらすであろう。
UGTにおけるイムノアッセイ(即ち、FPIAベースの供与体生成物アッセイ)は、潜在的薬物候補が既知のUGTイソ型のいずれと相互作用するかどうかを判定するのに、SLUTについて説明した方法と極めて類似する方法で使用されることが意図されている。本発明の方法を使用することにより、注目する化合物が1種以上のUGTイソ型と相互作用するのかどうか、そうであれば、それは基質または阻害因子のいずれであるのかを判定することが可能であろう。また、注目する化合物と酵素イソ型間の相互作用における動的パラメーター(IC50、Km、Vmax)も同定し得る。組換え型の多くのUGTイソ型がすでに入手可能であることに留意すべきである(Invitrogen、Becton-Dickinson社)。これらのHTSアッセイにおいて得られた情報は、次のような方法で使用し得る、例えば、イソ酵素同定後、適切なUGTイソ酵素によるさらに詳細な動的試験を使用して生体内クリアランス速度を予測し、貧弱な薬物動態特性のために臨床試験で不合格となる化合物の数を減らし得る。また、UGTレベルにおける遺伝的相違が治療薬に対する種々の応答を変動させる重要な要因として認識されているので、特異的UGTによる代謝についての知識は、可能性のある薬物動態問題を薬物発見チームに警告する。さらにまた、薬物代謝に関与するUGTの同定は、薬物-薬物相互作用の前臨床評価および毒物試験において使用する生体外アッセイ法または動物モデルの賢明な選択を促し、それによって不適切な或いは不必要な実験動物の使用を減じるであろう。また、代謝データは、合理的な薬物設計の要素としても使用し得る。種々のUGTイソ酵素に対する基質特異性を規定する構造-活性関係のより良好な理解は、一次化合物の代謝プロフィールを変化させるその一次化合物構造的修飾のための根拠を提供するであろう。また、グルクロン酸化化合物の試験は、体内で活性型として代謝されるまでは不活性である価値あるプロドラッグの発見ももたらし得る。
【0041】
薬物発見におけるプロテインキナーゼ
ヒトゲノム中にコードされた400種を超える種々のキナーゼが存在する;疾病におけるそれらの役割を解明し、選択性の阻害因子を同定することは、大きな製薬上の主導事項である。キナーゼ機能不全は、がん、心臓血管疾病、炎症、神経変性疾患、および代謝障害のような最も重要な治療領域の全てに関連している。さらにまた、薬物ターゲットとしてのキナーゼの臨床上の検証は、それぞれ、乳がんおよび白血病に寄与する異所性チロシンキナーゼを阻害するHerceptinおよびGleevecの事例において最近示されている。高処理量スクリーニング(HTS) (薬物ターゲットによる相互作用についての何千もの化合物の並行試験)は、薬物発見の優勢な方式となってきている。2002年のHTSアッセイ試薬の総市場は4.74億ドルであり、およそ20%のスクリーニングがプロテインキナーゼにおいて実施された。高レベルの興味にもかかわらず、キナーゼをHTSプログラムに組み入れる製薬上の取組みは、前記アッセイ法による欠点により妨げられている。最も一般的に使用されるHTSキナーゼアッセイ法は、個々のキナーゼの基質特異性によって異なるリン酸化ペプチド反応生成物の蛍光系免疫検出に依存している。即ち、時間のかかる試薬開発が各キナーゼまたはキナーゼ群毎に必要となり、アッセイ間の結果の比較にも問題がある。この技術的障害を克服するために、本発明は、全てのキナーゼ反応の生成物であるアデノシンジホスフェート(ADP)の検出のためのFPIAを提供する。このアッセイ法は、如何なるキナーゼおよび如何なる受容体基質とも一緒に使用し得る普遍的な触媒アッセイを提供することによって、キナーゼ基質特異性を明らかにし新規な阻害因子を同定する取組みを促進させるであろう。
【0042】
プロテインキナーゼは、シグナル伝達において主要な役割を有する広くて多様なファミリーである。プロテインキナーゼは、ATPまたはGTPからの末端リン酸基を受容体タンパク質のセリン、スレオニンまたはチロシン残基への転移を触媒し、ヒトゲノムにおける最大のタンパク質ファミリーの1つである。最も広い意味においては、プロテインキナーゼは、セリン/スレオニンまたはチロシンキナーゼおよび可溶性酵素または膜貫通性レセプターとして分類し得る。最も最近の網羅的なゲノム分析においては、428種のヒトキナーゼが同定されており、これらは8種の異なる相同性群を含み、これらの相同性群は基質特異性、構造/局在化および/または調節の方式も反映したものである(Hanks, S. K., Genome Biol, 2003, 4:111)。例えば、EGFRおよびPDGRRのような膜貫通性成長因子レセプター並びにsrcキナーゼのような可溶性酵素の双方を含む84メンバーからなるチロシンキナーゼ群;61メンバーの環状ヌクレオチド依存性群、脂質依存性キナーゼ(PKCイソ型)を含むser/thrキナーゼ;および、45メンバーの有糸分裂促進性MAPキナーゼ情報伝達経路の成分を含む45メンバーの“STE”群が存在する。
キナーゼは、細胞内シグナル伝達経路の普遍的な調節因子であり、従って、広範囲の疾患および障害のより選択性の治療薬について研究中の製薬企業による強烈な関心の的となっている;キナーゼは、製薬優先順位付けの点でGプロテイン結合レセプターに次いで第2位である(Cohen, P., Nat Rev Drug Discov, 2002, 1:309-15)。リン酸化における細胞内ターゲットとしては、他のキナーゼ、転写因子;アクチンおよびチューブリンのような構造タンパク質;DNA複製および転写、およびタンパク質翻訳に関与する酵素;並びに代謝酵素がある(Cohen, P., Trends Biochem Sci, 2000, 25:596-601)。リン酸化は、タンパク質触媒活性、特異性、安定性、局在化および他の生体分子との結合に変化を生じさせることがある。タンパク質上の複数の部位での同時リン酸化は、異なる機能的結果を伴い、情報伝達経路の統合にとって一般的で中枢的である。
【0043】
リン酸化部位の多様性
各キナーゼは、1種以上の標的タンパク質を時には複数の部位でリン酸化することがあり、触媒活性または他の生体分子との相互作用を制御する1つ以上の調節ドメイン内で自己リン酸化することがある。健常および疾病状態からの細胞リン酸化プロフィールの機能的結果を明らかにすることはプロテオミクスの主たる第1歩である。しかしながら、この知識をどのキナーゼを薬物発見の標的とすべきかを決定するのに使用するには、その受容体基質に対する特異性も描き出さなければならない。キナーゼは、多くの場合ベータ屈曲部で生じているそれらの標的タンパク質の特異的直線配列を認識する。一般に、リン酸化残基の両側に隣接する3〜5個の残基のアミノ酸がリン酸化部位を決定している。既知のキナーゼリン酸化部位を蓄積しているPhophoBaseデータベースは、133種のヒトキナーゼについてのエントリーを収容しているが、これは全キナーゼの1/3に満たない。さらにまた、全部でないにしても殆どのこれらの特異性プロフィールは、各キナーゼについて、基質として同定されている1種または2種のペプチドしか示していないので不完全である。関連キナーゼ間で基質特異性の有意の重複は存在するものの、多数のキナーゼによってリン酸化されるコンセンサス配列は存在しない。この状況は、特異的リン酸化生成物の検出に依存しているHTSアッセイへの多様なまたは新規なキナーゼの組み入れを複雑にしている。
【0044】
表1
表1.抗がん剤としてのキナーゼ阻害因子の開発において選出した臨床試験(Dancey, J. and Sausville, E. A., Nat Rev Drug Discov, 2003, 2:296-313)。太字の薬物は、承認されている。
【0045】
がんにおけるプロテインキナーゼ
癌に干渉させるべくキナーゼを標的化する生物学的な合理的根拠は、ここで概説するにはあまりにも広過ぎる。しかしながら、1つの支配的なテーマは、細胞分裂速度(細胞サイクル進行)、細胞増殖(塊状)、および全てのがんにおいて撹乱されるプログラミングされた細胞死(アポトーシス)間の微妙な均衡の制御における数多くのキナーゼの関与である。増殖因子レセプターチロシンキナーゼ(RTK)は、ペプチド増殖因子シグナルを細胞外から細胞内経路に情報伝達する膜貫通タンパク質であり、これにより増殖促進(progrowth)および抗-アポトーシス遺伝子の活性化が生じる。ヒトにおける58種のRTKの大多数は優性腫瘍遺伝子であり、これは異常活性化または過発現が悪性細胞フェノタイプを生じさせることを意味する。驚くに当らないが、チロシンキナーゼは抗がん薬標的として積極的に追跡されており、そして、小分子阻害因子およびモノクローナル抗体阻害因子(それぞれ、GleevecおよびHerceptin)の双方が臨床的に承認されている。増殖因子レセプターからの下流シグナル伝達は、ser/thrおよびチロシンキナーゼが関与する複数の経路を介して生じる。支配的キナーゼの1つは、RafおよびMEKキナーゼを含む有糸分裂促進活性化プロテインキナーゼ(MAPK)経路である;これらキナーゼ全ての阻害因子は、臨床試験において現在試験中である(表1) (Dancey, J. and Sausville, E. A., Nat Rev Drug Discov, 2003, 2:296-313)。可溶性チロシンキナーゼ、とりわけ、Src群を含む11種の腫瘍遺伝子もRTKによって開始された有糸分裂促進シグナルを伝達し、これれは製薬者によりターゲットとされている(Warmuth, M., et al., Curr Pharm Des, 2003, 9:2043-59)。G1相へのエントリーを開始させる、RTKを介した有糸分裂促進シグナルに続いて、サイクリンタンパク質と関連する相特異的キナーゼの順次の活性化により細胞サイクルの進行が制御される。サイクリン依存性キナーゼは、製薬業者が悪性細胞増殖を抑制するという期待の下に追跡中であるさらにもう1つの重要なキナーゼ群を代表する(表1) (Elsayed, Y. A. and Sausville, E. A., Oncologist, 2001, 6:517-37)。
【0046】
他の疾病における標的としてのキナーゼ
キナーゼにおける製薬上の興味は、がんに最も強く的を絞っているが、主要治療領域の全てに及ぶ。表2は、広範囲の障害に対してのキナーゼ標的についての生物学的理論を説明する最近の論評を示している;それらの標的は、がんについて追跡中の標的と重複していることに留意されたい。全ての企業は、薬物発見におけるキナーゼ標的化に基づいて形成されてきた。これらには、Sugen (http://www.sugen.com、現在Pfizer社所有)、Signase (http://www.signase.com/index.htm)、およびProQinase (http://www.proqinase.com/index.html)がある。
表2
表2.プロテインキナーゼと各種疾病間の関連のリスト
【0047】
本FPIAベースの供与体生成物アッセイ法は、薬物ライブラリーをプロテインキナーゼの阻害因子または活性化因子をスクリーニングするために使用することを考慮する。また、本アッセイ法は、ペプチドまたはタンパク質をキナーゼに対する受容体基質としてスクリーニングするのにも有用である。これらの用途においては、本アッセイ法は、アッセイの普遍的性質、簡素化した均質アッセイ、放射活性の無いこと、および酵素代謝回転の定量能力のような、他の方法を上回る有意の利点を有する。
普遍的アッセイ方法
この方法は、真に汎用的であるので、プロテインキナーゼのHTSスクリーニングプログラムへの統合を促進させるであろう:検出試薬の単一の組を全てのキナーゼおよび全ての受容体基質に対して使用することができる。現行のアッセイ方法を上回る重要な利点は、生理学的タンパク質受容体基質および短ペプチドを使用することができる点である。
均質性
このアッセイ法は、単回添加、混合および読取り方式である。このことは、自動化高処理量環境におけるアッセイ法選択の決定を推進させる重要な要因である(High Tech Business Decisions, M., CA, Commisioned Market Analysis, 2002)。さらに、適切な結合速度を有する抗体を分離する場合は、このアッセイ法は、時間停止アッセイ法よりもより詳細な動的情報を提供する連続アッセイを可能にする。
蛍光検出
蛍光プローブを使用することにより、このFPIA方式は、放射線の取扱い、廃棄およびコストを排除する。最近の数年において、FPは、製薬者よって使用される主要HTSアッセイ基本骨格の1つになっていることに留意すべきである(Owicki, J. C., J Biomol Screen, 2000, 5:297-306)。2003年には、FPは一次スクリーニングアッセイ全体の約12%において製薬者に使用されることが予想される;これは、2001年のFP使用レベルの2倍である(High Tech Business Decisions, M., CA, Commisioned Market Analysis, 2002)。FPは、幾つかの市販HTSプレートリーダーにおける標準方式である。
定量性
提案した酵素アッセイにおいて、ADPは、リン酸化ペプチドまたはタンパク質と化学量論的量で生成され、従って、ADPについての標準曲線の使用は、酵素代謝回転の直接の測定を可能とするであろう。HTS用途におけるFPの使用は比較的最近の展開であるけれども、診断設定におけるホルモンおよび代謝物の定量検出のためのFPIAの使用は、極めて良く確立されている(Nasir, M. S. and Jolley, M. E., Comb Chem High Throughput Screen, 1999, 2:177-90)。
【0048】
(実施例)
実施例1
ウリジングルクロニドトランスフェラーゼアッセイ
本発明の1つの実施態様は、拮抗蛍光偏光イムノアッセイを使用するUGT反応の“供与体生成物”を検出することを含み、このイムノアッセイにおいては、抗体結合トレーサーが高偏光値を有し、この偏光値は、前記トレーサーがUDPのような分析物に置き換わったときに低下する(図2に示すように)。このアッセイにおいて必要とする主たる試薬は、UDPと高選択的に結合するが供与体UDP-グルクロン酸(UDPGA)に対しては無視し得る結合性を有する抗体の作製である。この高選択性抗体は、商業的に入手し得る蛍光UTP化合物と一緒に使用して前記アッセイ法を確立する。
UDPに対して作製したポリクローナル抗体は、担体タンパク質への共有結合を必要とした。トリホスフェートの反応性誘導体は容易に入手することができたが、ジホスフェートは入手できず、トリホスフェートの反応性誘導体が結合に使用することができたという理由のみにより、UTPをハプテンとして使用した。トリホスフェートの大多数が動物中でジ-およびモノ-ホスフェートに加水分解され得ると考えられた。UTPを担体タンパク質に結合するための数種の化学物を調査した。得られる抗体の特異性および抗原に対する親和性に結合の性質が深刻な影響を与え得るからである。注意を払って、リンカー分子がリボースまたはホスフェートよりはむしろウリジン環に結合するようにし、それによってUDP分子を供与体、UDPGAから識別し得るUDP分子の部分との免疫反応性を最高にした。
BSAに結合させたUTPおよびUDPの混合物、および商業的に入手し得るトレーサー分子、蛍光標識化UTP化合物(Alexa-UTP、分子プローブ)に対して生じさせたウサギ抗血清を、指定量のウリジンヌクレオチドを含有するブラックマルチウェルプレート(Thermo Labsystems Pt#7605)のウェルに添加した。Alexa-UTPをFPIA試験におけるトレーサーとして使用した。Tecan Ultraプレートリーダー内で、数時間の平衡後に蛍光偏光を読み取った。反応条件は、以下のとおりとした:50 mMのKPO4、pH 7.4、150 mMのNaCl、0.1 mg/mlのBGG、1 nMのChromaTide Alexa Fluor 488-5-UTP、1.25μlのウサギ血清、100μlの総体積。
【0049】
UGT反応からの実験結果は、図8A〜Bに示してある。図8は、UDP/UTPに対して生じさせた第1ポリクローナル抗体と商業的に入手し得るトレーサー分子(Alexa-UTP)を使用した、抗体-トレーサー複合体の各種ウリジンヌクレオチドによる滴定を示している。2つのグラフはX軸の尺度において異なること、および、UDP,即ち、供与体生成物による拮抗はおよそ10μMにおいて半最高であり、一方、UDPGA、即ち、供与体においては、半最高置換は1 mMよりも高く、これは、少なくとも100倍の選択性の差であることに注目されたい。
より関連性のあることは、UDPがトレーサーを低マイクロモル濃度で置換しているのに対し、100μMを越える濃度においてUDPGAとの検出し得る交叉反応性は存在しないことである。UTPとの交叉反応は、UTPがUGT酵素反応においては存在しないか或いは生成されないので、提案したアッセイにおいては問題ない。UDPがトレーサーを低マイクロモル濃度で置換できるということは、この抗体がUGT酵素反応において生成されたUDPの検出に適していることを意味している。さらにまた、予備結合速度試験により、UDPによるトレーサーの置換が極めて速い兆候が見えていたので、UGT代謝回転をリアルタイムでモニターするための抗体の使用、即ち、連続アッセイは、本発明に包含される。
【0050】
その重要性にもかかわらず、グルクロン酸化を触媒する肝臓UDP UGTの広くて重なり合う基質特異性は、柔軟性のある生体外アッセイ方法が無いために理解に乏しいままである。主たる問題は、使用する触媒アッセイが基質特異性クロマトグラフィー工程を含む生成物からの反応物の分離を必要とし、従って、HTS方式においては現実的でないということである。UGT反応生成物であるUDPを蛍光均質方式で測定する概念を確立することにより、本出願人等は、この問題を解決するための技術的基盤を提供する。本アッセイ法は、全てのUGT反応に共通する生成物を測定するので、あらゆるUGTイソ型およびあらゆる基質による単一方式の検出を可能にする。しかも、本方法は、生成物からの反応物の分離を必要とせず、本方法が比色分析ではなく蛍光検出を使用し、そのことが本方法を、製薬HTSの場(蛍光ベースの検出に大きく依存するようになってきている)においてより感度があってより望ましいものにしているので、他の関連アッセイ法を凌ぐ有意の改良法である。また、本方法は連続アッセイ方法であるので、本方法は、UGT酵素代謝回転についてのリアルタイムの動的データを提供し得る。さらに、抗体-抗原結合反応は、供与体生成物検出に従来使用されていた通常の結合酵素反応(Mulder, G.J. and A.B.D. Van Doorn, Biochem J., 1975, 151:p. 131-40)よりも試験サンプルからの干渉に対して影響を受けにくい。即ち、この新規なアッセイ方法は、単離したUGTイソ酵素の集団による多様な化合物の代謝についてのスクリーニングを可能にし、前臨床代謝試験を大いに増強し、臨床脱落率を強力に低下させる。
さらにまた、本出願人等が作製した抗体の性質は、他の重要なクラスの酵素についてのHTSアッセイ法の開発においてもより広い密接な関係を有する。図8に示すように、UDPがトレーサーを置換しているがUMPはそうではなく、このことはこの抗体が1個のリン酸基に基づき各ヌクレオチドを識別し得ることを例証している。即ち、反応生成物が供与体と1個のリン酸によって異なる、プロテインキナーゼ反応におけるADPのような他の“供与体生成物”に対する抗体を作製し得ることも包含される。
【0051】
実施例2
キナーゼアッセイ
本発明のアッセイ方法は、リン酸化ポリペプチドと化学量論的量で生成するキナーゼ反応生成物ADPのFPAI検出に依存する。上述したUGTアッセイと同様に、FPIAベースの供与体生成物キナーゼアッセイの構成要素は、供与体生成物に対する抗体、および検出可能タグと結合させた供与体生成物を含むトレーサーである。前記抗体は、ADPに対し高度に特異的である(即ち、前記抗体は、過剰のATPおよび蛍光トレーサーの存在下においてADPを認識し得る)。キナーゼアッセイにおける抗体とトレーサーを、アッセイの開始に最適の濃度でブラックマルチウェルプレートのウェルに加える。トレーサーの適切な濃度は1〜2nMであり、十分な抗体を使用してトレーサーに最高偏光変化のおよそ75%を生じさせる。受容体基質を、所望の、一般にKm値よりも2〜5倍高い濃度で、前記ウェルに加える。受容体基質は、ペプチドまたはインタクトタンパク質でよい。本発明の1つの利点は、他のキナーゼアッセイ法は、検出のためには特異的受容体を必要とするのに対し、任意の受容体基質を使用し得ることである。アッセイすべきキナーゼおよび抗体-トレーサー相互作用と適合する緩衝液、一般的には、およそ中性pHのTris-Clまたはリン酸緩衝液を使用する。この緩衝液と一緒に、100μM〜5mM濃度のATP、MgCl2およびキナーゼの活性化または安定化に必要な他の薬剤のような他の必要なアッセイ成分を添加する。その後、キナーゼ酵素を添加して反応を開始させ、偏光値をTecan Ultraのようなマルチウェルリーダーでモニターする。反応が進行するにつれて、リン酸化ペプチドまたはタンパク質と化学量論的量で生成したADPは、トレーサーを抗体から外して、偏光値の低下をもたらす。
阻害因子のスクリーニングにおいては、一般的には試験すべき化合物を何ら他のアッセイ成分を添加する前のウェル中に分配し、阻害因子を添加しない参照ウェルを比較のために含ませる。
各キナーゼは、1種以上のターゲットタンパク質を、時には複数部位でリン酸化させることがあり、触媒活性または他の生体分子との相互作用を制御する1個以上の調節ドメイン内で自己リン酸化することがある(Cohen, P., Trends Biochem Sci, 2000, 25:596-601)。健常および疾病状態における細胞リン酸化プロフィールの機能的結果を明らかにすることは、プロテオミクスの大きな第1歩であり、この知識は、薬物発見においてどのキナーゼを標的にするか、およびその受容体基質に対する特異性を決定するために使用し得る。関連キナーゼ間で基質特異性に有意の重なり合いは存在するものの、多数のキナーゼによってリン酸化されるコンセンサス配列は存在しない。この状況は、特異的リン酸化生成物の検出に依存しているHTSアッセイへの多様なまたは新規なキナーゼの組み入れを複雑にしている。
【0052】
実施例3
スルホトランスフェラーゼアッセイ
SULT1E1の発現および精製
スルホトランスフェラーゼHTSアッセイ法を確立するために、SULTイソ型であるSULT1E1を先ずC-末端6xヒスチジンタグを有する大腸菌発現ベクター中にサブクローニングし、発現タンパク質をアフィニティークロマトグラフィーにより精製し、その物理特性および酵素特性について特性決定した。
SDS-PAGE上でその算出分子量近くに移行した精製タンパク質は、予測される分子量と極めて緊密に一致しており、さらに重要には、質量分光分析において予測される分子量と極めて緊密に一致していた(それぞれ、36,161および36,374ダルトン)。このタンパク質は天然のN-末端を保持するように設計していたので、精製C-末端6xHis構築物をN-末端シークエンシングにも供した;15個のアミノ酸をシークエンシングしたところ、これらのアミノ酸は、GenBank中のヒトSULT1E1配列(NP005411)と同一であった。さらなる配列検証は、質量分光分析およびタンパク質シークエンシングにより得られた。
比較として使用するために、精製SULT1E1の酵素特性を試験した。エストラジオールおよびエストロンを生理学的基質として使用し、さらなる陽性(α-ナフトール)および陰性(ドーパミン)対照化合物も試験して特異性を評価した。2つのタイプの放射線アッセイ法をこれらの試験においては使用し、1つは35S-PAPS (Foldes, A. and Meek, J. L., Biochim Biophys Acta, 1973, 327:365-74)を使用し、他方は3H-エストラジオール(Zhang, H., et al., J Bio Chem, 1998, 273:10888-92)を使用した;両者は、SULT特性決定において広く使用されているものであり、以下に説明する。
【0053】
表4
【0054】
表4は、SULT1E1融合タンパク質の酵素特性を示している。KmおよびVmax測定を3H-エストラジオール放射線アッセイを使用して実施した。この測定では、放射活性硫酸化生成物を未反応3H-エストラジオールから有機/水性相抽出により分離し、液体シンチレーションカウンターで計数した。各キネティックパラメーターは、GraphPad/Prizmソフトウェアを使用して、非直線回帰によるV対S曲線から算出した。各種受容体による相対的反応速度は、35S-PAPS放射線アッセイにおいて、400nMの受容体基質を使用して測定した。この測定では、未反応35S-PAPSをバリウム金属錯体として沈降させ、35S-標識化スルホ結合体を含有する上清を液体シンチレーションカウンターで計数した。基本的反応条件は、双方の場合において、10 mMのKPO4、pH 6.5、10 mMのDTT、1.5 mMのMgCl2、10 mMのPAPS、0.5 ngのSULT1E1、0.0025〜15 mMの受容体であった。
精製SULT1E1において測定したVmaxおよびエストラジオールKm値は、精製組換えSULT1E1における公表された値(それぞれ、30〜40nモル/分/mgおよび5〜15nMである)に良好に匹敵していた。受容体基質特異性に関しての公表データはより変動しているが、その結果は、エストラジオールとエストロンが極めて良好な基質であり、α-ナフトールが中間であり、DHEAとドーパミンが極めて貧弱な基質であるという一般的傾向を反映している(表4参照)。即ち、出願人等が発現させたSULT1E1構築物の全てが、最高の公表値と同様な天然の基質特異性と触媒作用速度を示していた。
【0055】
抗原の合成および抗体作製
スルホトランスフェラーゼにおける蛍光系HTSアッセイ法の実際の開発に関しては、SULT触媒作用を検出するPAP FPIAを示す4つの要素が存在する:抗原の合成および抗体の生成;トレーサー分子の合成;相互作用および特異性についての抗体およびトレーサーの試験;および最適抗体-トレーサー対を使用したSULT1E1活性検出の実証。トレーサー合成の努力は、同じ反応性PAP誘導体を担体タンパク質およびフルオロフォアへの結合において使用するので、抗原合成の努力と有意に重なり合う。
提案したFPIA系SULTアッセイ法の開発は、SULT反応生成物のPAPと過剰のPAPSの存在下に特異的に結合する抗体を必要とする;即ち、単一の5'-リン酸基に基づき識別する抗体。リン酸基の数のみにおいて異なり、上述のような硫酸基と構造および大きさにおいて同様な各種ヌクレオチドを識別する抗体においては、十分な先例が存在する。しかしながら、PAPを特異的に認識する抗体の作製についての先例は存在しない。
PAPのような小分子は、免疫原として使用するためには担体タンパク質に結合させなければならない。適切には、担体タンパク質当り10〜20個の抗原密度が最適である。上述したように、本発明者らの抗原合成戦略の2つの要素は、a)数種の抗原の合成と試験(ヌクレオチドへの結合部位およびリンカー構造は得られる抗体の特性に有意に影響を与える得るから) (Crabbe, P., et al., J Agric Food Chem, 2000, 48:3633-8;Signorini, N., et al., Chem Res Toxicol, 1998, 11:1169-75;Oda, M. and Azuma, T., Mol Immunol, 2000, 37:1111-22)、および、b)アデニン環を介した結合(5-リン酸基の自由露出を可能にし、分子のアデニン部分との免疫反応性を阻害して、所望の特異性を有する抗原を生じるから)。アデニン環中にはタンパク質に直接または架橋性試薬を介して結合するのに十分に反応性である窒素はなく、従って、反応性PAP誘導体を開始点として使用した;唯一の商業的に入手可能な試薬は、N6-アミノヘキシルPAPであった。当初最も直接的な試みと思われた方法(N6-アミノヘキシルPAPを担体タンパク質とカルボジイミドカップリングまたはアミン反応性架橋剤により反応させる)は、かなりの実験努力にもかかわらず、許容し得ない低い抗原密度を生じた。
【0056】
別の化学物質を追求するために、出願人等は、光反応性分子である2-および8-アジド-PAP、およびC8-ヘキシルアミノ-PAP (図6)の合成を外部委託した。図6は、PAP抗原の合成を示す。ALT社(ケンタッキー州レキシントン)により特注合成された2-および8-アジド-PAPをBSAの存在下に照射した(254nm);未反応ヌクレオチドは、濾過および透析により除去した。7〜12PAP/BSAの最終抗原密度が、アデシン環の吸光度により測定したときに得られていた(270〜280nmに変化)。ウサギ(抗原毎に3羽)は、Lampire Biologicals社(ペンシルベニア州オッツビレ)により免疫化された。N6-アミノヘキシルPAP(Sigma社)は、グルタルアルデヒドを使用してKLHに結合させ、Biosynthesis社(テキサス州ルイスビラ)により3羽のウサギに注入された。免疫スケジュールは、全ての抗原において同様であり、4〜6週間に亘って3〜4回の注射を含んでいた。
前記2つの光反応性PAP誘導体からの抗原を作製し、抗体生成のために送った;KLHへのグルタルアルデヒド架橋を使用してのN6-アミノヘキシルPAPからの抗原調製および抗体産生は、別個の企業に外注した(図6)。3種の抗体すべてがELISAで測定したときにPAPに結合したことを外注製造者から報告された(データは示していない)。
さらに、出願人等は、反応性誘導体 N6-カルボキシメチルPAPおよび2'-O-スクシニル-PAPも合成した(図7);これらの誘導体からの抗体は、まだ産生させていない。拮抗FPIAを使用してのAb結合特性の詳細な説明は、トレーサー合成の説明に続いて後で説明する。
【0057】
PAP-蛍光体トレーサーの合成
FPIAトレーサー分子は、3種の異なる構成部分、即ち、抗原、蛍光体(fluor)およびこれらを結合させるのに使用するリンカーに分けることができる;さらなる主要な構造的変動要因は、抗原へのリンカーの結合部位である。トレーサーの同定は大いに経験的であるので、出願人等は、種々のリンカーを使用してPAPとフルオレセインを各分子上の種々の部位を介して結合させた;殆どの場合、最終リンカー領域は、使用した反応性フルオレセインとPAP分子の複合体であった。
抗体戦略は、PAPのリボシル-ホスフェート基に特異的に結合する抗体を作製するために、アデニン部分を介して結合させることであった。この部位は、所望のトレーサー分子の免疫反応性部分をAbを結合させるために遊離のまま残存させるための蛍光体結合用の自明な部位でもある。また、フルオレセインは、結合に使用した好ましい蛍光体であった。ローダミンのような赤色シフト蛍光体がHTS用途においてより望ましいが、フルオレセインによるFPトレーサーの開発は、フルオレセインが非特異結合を起こしにくく、多くの活性化誘導体を入手し得るので、通常はより簡単である。リンカー分子に関しては、該分子は、抗原性および蛍光特性の双方に影響を与える多くの重要な形でトレーサー特性に影響を及ぼす。一般に、抗原を蛍光体から十分に引き離して抗体との相互作用が妨害されないようにすることと、蛍光体にあまり自由過ぎる動きを生じさせないようにすることの間に注意しなければならない均衡が存在する。前者は親和性Ab結合性の低下および蛍光体の消光をもたらし、一方、後者はAb結合時の偏光変化を低下させ、それによってアッセイのダイナミックレンジを減じる。
N6-アミノヘキシルPAPは、トレーサー合成において有用である免疫原合成において使用した唯一の活性化PAP分子であった;前記2つのアジド-PAP誘導体を結合させるのに必要とした光活性化反応は、2つの小分子を結合するには不十分かもしれない。種々の位置で活性化させ且つ種々のリンカー領域を有するPAP分子を調製するために、出願人等は、C8-アミノヘキシル-PAPの調製を外部委託し、カルボキシ末端リンカーを含む2つのPAP誘導体、即ち、N6-カルボキシメチル-PAPと2'-O-スクシニル-PAPを自家合成した。後者の化合物はアデニンよりはむしろリボースヒドロキシルを介して結合させているが、この方法を使用して、cAMPに対して高度に特異的な抗体およびトレーサーが産生されている(Horton, J. K., et al., J Immunol Methods, 1992, 155:31-40)。
【0058】
以下は、活性化PAP分子の合成と精製並びにその各種活性化蛍光体への結合についての簡単な説明である;トレーサー合成成分は、図7および8に要約している。一般に、アミノ末端リンカーを含むPAP分子はスクシンイミジルエステル(または、1つの場合はイソシアネート)により活性化させた蛍光体に結合させ、カルボキシ末端リンカーを含むPAP分子は遊離アミノ基を含有するフルオレセイン誘導体とカルボジイミドカップリングを使用して反応させた。
図7は、トレーサー合成の各構成要素の構造を示す。図7は、左から右に、C8およびN6位置で結合させたアミノ末端リンカーを含むPAP分子;アミン-およびカルボキシ-反応性フルオレセイン誘導体;およびN6および2'-OHにおいてカルボキシ末端リンカーを含むPAP分子を示している。各PAP結合体をTLCにより分離し、抗-PAP抗体に対する結合性について試験した。本発明によれば、16の反応がPAPとフルオレセイン誘導体の種々の組合せを含有して進行し、各反応で、TLCで解像し得た1〜4種の蛍光生成物が得られた。全部で、40種以上のトレーサーを精製し、Abに対する結合性について試験した。N6-アミノヘキシルPAP(Sigma社)および反応性フルオレセイン誘導体の全て(Molecular Probes社)が商業的に入手可能である。2'-O-スクシニル-PAPおよびN6-カルボキシメチル-PAPは、以下で説明するようにして合成した;C8-アミノヘキシルPAPは、Jena Biosciences社(ドイツ国ジェナ)が合成した。図8は、代表的な最終トレーサー構造を示す。
【0059】
N6-カルボキシメチルPAPの調製
N6-カルボキシメチルPAPを調製するために、100mgのPAPを、0.3gのヨード酢酸と一緒に、LiOHでpH 6.5に調整した1.2mLの水中でインキュベートした。反応は、30℃で5〜7日間続き、定期的にpHを6.5に調整した。得られた1-カルボキシメチル-PAP生成物をエタノールで沈降させ、蒸留水中に再構成させ、pHをLiOHで8.5に調整した。この反応を90℃で1.5時間加熱してN6-カルボキシメチルPAPを得た。この生成物を、0.3 M LiCl、pH 2.75中で平衡化させたDowex 1-X2 (200〜400メッシュ)カラムで精製した。勾配を、0.5 M LiCl、pH 2.0を使用して10カラム体積に亘って適用した。N6-カルボキシメチルPAPはカラムから純生成物として溶出し、質量分光分析により確認した、約20%の収率。
2'-O-スクシニル-PAPの調製
2'-O-スクシニル-PAPを調製するために、10mg (0.024ミリモル)のPAPと無水コハク酸(43mg、0.426ミリモル)を、10%のトリエチルアミン含有する蒸留H2O (体積/体積、1mL)中に溶解させ、1.5時間振盪させ、反応の進行を、逆相薄層クロマトグラフィー(RP-TLC)を使用してモニターした。終了時に、反応を2回凍結乾燥させて残留トリエチルアミンが確実に残らないようにした。期待されたスクシネートを、50mLの250 mM NH4OAcで平衡化させたFast Flow Sepharose-Q樹脂(Pharmacia社)を使用してカラム精製し、500mMから750mMのNH4OAc(各々50mL)の直線勾配により溶出させた。生成物を含有する画分をロトバップ(回転エバポレーター)(rotovap)上で濃縮し、次いで凍結乾燥させて、吸光度測定によれば、8.4mg即ち収率67%を得た。ストック溶液を後の使用のために−20℃で保存した。
【0060】
アミノ活性化PAPを含む蛍光結合体の調製
アミノ活性化PAPを含む蛍光結合体を調製するために、N6-またはC8-アミノヘキシル-PAPの蒸留H2O中100mM溶液の10μLを、スクリューキャップ付きバイアル内で、モル当量のフルオレセインスクシンイミジルエステルと混合し、無水ジメチルスルホキシド(DMSO)を使用して反応物を200μLの最終体積とし、撹拌機(ボルテックス)(vortexer)上で24時間振盪させた。反応の進行を、展開用溶媒としてのH2Oを使用してRP-TLCにより追跡した。反応生成物を、分離用TLCを使用して、1:1のEtOH/0.5M NH4OAcを使用する正常相シリカゲル上で精製した。フルオレセイン標識化化合物(一般に、各反応において3〜5回生成させた)をUV光を使用して可視化し、TLCプレートからそぎ落とし、1:1のMeOH/0.5M NH4OAcを使用してシリカゲルから抽出した。個々の画分をボルテックス撹拌機上で1時間振盪させ、アルミニウムホイル内にラッピングし、4000RPMで8分間遠心分離した。上清を別のラベル付びんに移し、抽出工程を繰返した。集めた上清を後の使用のために50nM溶液に標準化し、褐色微量遠心用チューブ内で−20℃で凍結させた。
カルボキシ活性化PAPを含む蛍光結合体の調製
カルボキシ活性化PAPを含む蛍光結合体を調製するために、N6-カルボキシメチル-PAPまたは2'-O-スクシニル-PAPの蒸留水中100mM溶液の10mlを50当量の1-エチル-3-(ジメチルアミノプロピル)カルボジイミド(EDC) (無水DMSO中1M溶液の10μL)、次いで75当量のN-ヒドロキシスクシンイミド(無水DMSO中1000mM溶液の15μL)と混合し、反応物を無水DMSOにより180μLの中間体積とし、48時間振盪させた。その後、1モル当量(20ml)の遊離1級アミンを含むフルオレセイン誘導体を添加し、反応を攪拌させながら24時間インキュベートした。反応の進行を、展開用溶媒としてのH2Oを使用してRP-TLCにより追跡した。反応終了時に、蛍光生成物を、9:1 アセトニトリル/2mM NH4OAc、pH 5.5をTLC溶媒として使用し、8:2 アセトニトリル/2mM NH4OAc、pH 5.5を溶出用に使用した以外は、上述のようにして、正常相シリカTLCにより精製し、溶出させた。
全部で16組の活性化PAPとフルオレセイン分子の固有の組合せを反応させ、これらから、40種よりも多い蛍光生成物を単離し、抗体に対する結合性について試験した。この方法は“貧乏人の組合せ化学”の類とみることができるが、出願人等は、他の例において、FPトレーサーの開発において極めて成功裏であることを見出している。FPアッセイの均質性およびマルチウェル器具の利用可能性は、スクリーニング取組みを比較的迅速にすることに注目されたい。
【0061】
抗-PAP抗体およびトレーサーの特性決定
抗体/トレーサー相互作用の分析を蛍光偏光により実施した。蛍光偏光(上で簡単に説明した)を測定するには、垂直偏光励起光に対して平行および垂直である垂直面および水平面の双方において発光強度を測定する。放出光が偏光を規定する:P = (水平強度−垂直強度)/(水平強度−垂直強度)。偏光値は、mPとして報告され、これは、偏光値の1000倍である(30mP = 0.03P)である。フルオレセインは、pH 7.0のリン酸緩衝液中にて37℃でおよそ20mPの偏光を有する。数学的には、フルオレセインの限界偏光は、500mPである;実際には、フルオレセインに結合させた抗-フルオレセイン抗体は、約450mPの偏光を与える。Tecan Ultraのような現在入手可能なマルチウェルFPリーダーは、偏光値を高い精度で読取ることができる;即ち、<3mP未満の標準偏差での遊離および結合状態の1nMフルオレセインの検出。
拮抗FPIAにおいては、蛍光標識化抗原、即ち、トレーサーは、分析物により抗体との結合から外される。そのシグナルは、結合トレーサー画分対遊離トレーサー画分の差異に比例し、従って、アッセイのダイナミックレンジおよび感度の双方は、トレーサーおよび拮抗性サンプル分子に対する抗体親和性に依存している。FPIAの適切なダイナミックレンジを確立するためには、少なくとも70〜80%のトレーサーが拮抗因子の不存在において抗体に結合しなければならない。これは、Kdよりも2〜10倍低い濃度のトレーサーとKdよりも2〜3倍高い濃度の抗体を使用することによって達成される。その場合、アッセイの有用な濃度範囲は、Kd濃度の数倍の低さからKd濃度の約20倍の高さまでである。例えば、トレーサー/抗体複合体のKdが10nMである場合、サンプルの検出範囲は、約2nM〜約200nMであり得る。Kdが10倍低い場合(1nM)、アッセイ感度も約10倍改善する。
影響を受けるアッセイパラメーターの点で許容され得る抗-PAP抗体およびトレーサーの特性は、1) ダイナミックレンジ:Abの不存在下での低トレーサー偏光(<100mP)、および結合および遊離状態間での最大偏光差(ΔmP >100mP);2) 感度:トレーサーの抗-PAP抗体に対する高親和結合性(Kd <100nM)、同様なIC50を有する遊離PAPによる置換、および最小の結合によって生じる蛍光消失;3) シグナル/ノイズ:高Ab選択性;即ち、PAPによるが、PAPSまたは他のアデニンヌクレオチドにはよらないトレーサーの拮抗置換;また、SULT1E1または他のアッセイ成分とトレーサーとの相互作用の無いこと;および4) 連続アッセイ:迅速な結合/解離速度。
【0062】
Ab-トレーサー相互作用試験の結果
数種の濃度の各抗体をマルチウェルプレート中の1nmのトレーサーに添加し、トレーサー偏光の増加についてモニターすることによって、Ab-トレーサー結合性についての初期スクリーニングを実施した。抗体全てにおいて、固定化プロテインAまたはプロテインGを使用したIgG画分の精製が、非特異結合成分を除去するために必要であった。示したデータは、全て、プロテインG (Pierce NabTM Protein G Spin Cromatography Kit)で精製した抗体を使用している。初期スクリーニングにおいて相互作用を示したトレーサー/Ab組合せを、PAPによる拮抗置換およびPAPSに対するPAPの選択性についてさらに詳細に試験した。全部で9種の精製抗体(3種の抗原×3羽のウサギ)を44種のフルオレセイン標識化PAPトレーサーとの結合性について分析した。アッセイは、全て、ブラック96ウェルプレートまたはブラック低容積384ウェルプレート内で実施した。両者は本質的に同一の結果を与え、偏光値は蛍光フィルターセットを備えたTecan Ultraで読取った(485nmでの励起から535nmでの発光) (EX485/Em535)。
図9に示す結合等温線は、下記の表5に示す3種の抗体と2種のトレーサーを使用して作製しており、抗体およびトレーサーを結合性についてスクリーニングするのに使用した試験の代表的なものである。
【0063】
表5:代表的な抗体およびトレーサー
【0064】
また、図9は、抗-PAP抗体およびPAP-フルオレセイントレーサーにおける結合等温線を示している。Ab 1781○、1810◇および3642□に対するN6-PAP-F8トレーサー結合性、並びに同じ抗体1781●、1810◆、3642■によるC8-PAP-F14トレーサーを示している。各抗体は、96ウェルプレートにおいては100μLまたは384ウェルプレートにおいては12μlの総体積で、150mMのNaCl、0.1mg/mLのBGGおよび1nMのトレーサーを含有する50mMリン酸緩衝液(pH 7.4)中で連続2倍希釈し(2つのプレートにおける結果は同一である)、偏光値を、室温での1時間インキュベーション後に、Tecan Ultraで読取った。
異なる動物において同じ免疫原から生成させた抗体の結合特性は類似していたが同一ではなかった。この試験においては、3642抗体が1781よりも有意に強固に両トレーサーに結合し、1810は、試験した最高濃度においてさえもいずれのトレーサーとも結合しなかった。N6-PAP-F8およびC8-PAP-F14に結合する3642の最高偏光値は、それぞれ、140および300mPであった;未結合トレーサー偏光はおよそ20mPであった、従って、観測されたシフトはFPIAに使用するために充分以上である。結合が特異的であるかを試験するために、PAPによる拮抗置換を評価した。
図10は、Ab 3642および前記2つのトレーサー、PAPおよびPAPSに対するPAPおよびPAPSの拮抗曲線を示す。N6-PAP-F8トレーサーは、白抜き記号で示している:○(PAP)、□(PAPS)。C8-PAP-F14トレーサーは、黒塗り記号で示している:●(PAP)、■(PAPS)。PAPまたはPAPSを、12μlの総体積で、50mMリン酸緩衝液(pH 7.4)、150mMのNaCl、0.1mg/mLのBGG、1nMのC8-PAP-F14および1.5μLの精製Ab 3642または20μL体積中に1nMのN6-PAP-F8および0.5μLの3642 Abを含有する黒色マルチウェルプレート内で連続2倍希釈した。偏光値は、室温で1時間後に読取った。
C8-PAP-F14トレーサーによるPAPにおけるIC50は、SULT1E1活性をモニターするためのこれらの試薬の使用を可能にするのに十分に低い300nMである。より強固な結合性のN6-PAP-F8トレーサーを拮抗除去するにはかなり高濃度のPAP(およびPAPS)を必要とするに注目されたい。これは、この場合、トレーサーが抗体を生成させるのに使用した免疫原と同じリンカー基を有しており、抗体の集団がこのリンカーを認識してトレーサーが遊離PAPによって置換されるのをより困難にしていることにより得る。また、図10は、PAPSがPAPよりもトレーサーをAb 3642から外すのに有効性が低いことも示している。
【0065】
同様な結果は、図11に示すように、1781 Abによっても観察された。図11は、C8-PAP-F14トレーサーによる拮抗試験におけるPAP、PAPSおよび数種の同様な分子の完全曲線を含む。図11は、2つの抗-PAP抗体/トレーサー組合せによる拮抗曲線を示す。各拮抗因子を、12μlの総体積で、50mMリン酸緩衝液(pH 7.4)、150mMのNaCl、0.1mg/mLのBGG、1nMのトレーサーC8-PAP-F14および1.5μLのAb 3642 (A)または3μLのAb 1781 (B)のいずれかを含有する黒色384ウェルマイクロタイタープレート内で連続2倍希釈した。室温で1時間のインキュベーション後に、偏光値をTecan Ultraで読取った(EX485/Em535)。全てのデータ設定において複製の平均および標準偏差を示している。3642/トレーサーおよび1781/トレーサーによるPAPおよびPAPSのIC50は、それぞれ、0.3μM PAP、3.8μM PAPS;および0.3μM PAP、1.5μM PAPSであった。
1781および3642抗体は、それぞれ、PAPSに対してよりもPAPに対して5倍および13倍高い選択性を示し、試験した他のヌクレオチド全てに対してよりもはるかに高いPAPに対する選択性を示していた。PAPSとの交叉反応は、他のアデニンヌクレオチドとの交叉反応が無いことを考慮すれば、予期したよりも高い。この点、殆どの市販PAPS調製物は有意のPAP画分を含有しているが、出願人等は、95%よりも高い純度と分析されたHPLC精製調製物を購入し、その保存において予防策を講じて加水分解を阻止するのに使用したことに留意すべきである。いずれにしろ、これらの結果は、出願人等がPAPSの存在下でPAPに選択的に結合する抗体を作製し得ることを明らかに示しており、このことは、提案したSULTアッセイにおいて十分なシグナル:ノイズおよびダイナミックレンジを可能にする主要な実施可能性要素である。PAPSに対してよりもPAPに対するAb選択性がさらに10倍増加することは十分であり得、ポリクローナル抗体ではなくモノクローナル抗体を使用した場合の極めて合理的な期待事項である。
試験した他の抗体およびトレーサーにより観察した結果は類似していた。N6-アミノヘキシル-PAP-KLHおよび8-アジド-PAP-BSA免疫原から作製したさらなる抗体はトレーサーの多くと結合し、Ab/トレーサー複合体は全ての場合未標識PAPによって置換することができた。このことは、この相互作用が特異的であることを示唆している。1810抗体は、トレーサー結合性を極めて貧弱にしか或いは全く示さず、さらに試験しなかった。遊離トレーサーの偏光値は15〜40mPの範囲であり、Ab結合時に観測された増大は100mP未満からほぼ300mPまでの範囲であった。幾つかのトレーサーとの相互作用はAbとの飽和を可能にするには弱過ぎて、最高偏光変化は常には観測されなかった。試験したトレーサーの2/3よりも多くが両タイプのAbに結合したが、N6-カルボキシメチル-PAPから合成したトレーサーのうちで、いずれの免疫原から生成させた抗体とも結合したものはなかった。
【0066】
FPIAベースのSULTアッセイ
PAPとPAPSをより有効に識別する抗体を作製して高品質アッセイを開発する必要があるだろうが、出願人等は、3642 AbおよびC8-PAP-F14トレーサーを使用して精製SULT1E1-cHisを含有する反応において生成したPAPの検出をモニターすることができた。初期の試験において、出願人等は、最高シグナル:ノイズのための最適PAPS濃度を同定することを模索した。SULT1E1-cHisを、予め形成させたAb-トレーサー複合体の存在下に、エストラジオールおよび変動濃度のPAPSと一緒にインキュベートした;SULT1E1は、反応を完了まで迅速に推進させるに十分なレベルで存在していた(図12)。
図12は、酵素により産生させたPAPの検出におけるPAPS濃度の効果を示す。アッセイ混合物は200ngのSULT1E1-6xHisを含んでおり(□)、或いはアッセイ緩衝液(■)を、100μLの総アッセイ体積で、30mMのホスフェート(pH 7.4)、7mMのDTT、8mMのMgCl2、75mMのNaCl、0.5mg/mLのBGG、150nMのエストラジオール、1nMのC8-PAP-F14トレーサー、12.5μLのAb 3642および変動濃度のPAPSを含有するウェルに添加した。プレートを37℃で24時間インキュベートし、Tecan Ultraで読取った(EX485/Em535)。
典型的なFPIAにおけるように、トレーサーはKdよりもかなり低い濃度で使用し、Abは拮抗因子の不存在下で最高トレーサー偏光のおよそ85%を生じる濃度に調整した。図12は、酵素により産生させたPAPが約40mPのトレーサー偏光の減少(図12における白抜き四角と塗潰し四角との差)を生じさせるPAPS濃度範囲(約1〜5mM)が存在することを示している。
即ち、この抗体がPAPSと有意に交叉反応するとしても、この抗体を拮抗FPIA方式において使用して、飽和中のPAPS (SULT1E1によるPAPSのKmはおよそ50nMである)によるSULT反応において産生したPAPを40mPのダイナミックレンジでもって検出し得る。さらにまた、これらは、均質反応、即ち、反応および検出成分の全てを反応開始時に添加した単回添加反応であった;これは、HTSアッセイにおける好ましい方法である。同様な結果は、検出試薬を酵素反応が終了した後に添加する場合も達成される、従って、偏光シグナルは、酵素反応によって影響されない(データは示していない)。
【0067】
出願人等が作製したAb/トレーサー対の一つを、読取り前に完了まで進行させたSULT反応におけるPAPの検出においてどのように使用するかの実施態様を上述した。しかしながら、連続方法は、正確な酵素速度測定を可能にし且つクエンチング工程の必要性を排除するので、HTSにおける最も好ましい方式である。図13は、3642 AbおよびC8-PAP-F14トレーサーを使用してSULT1E1酵素活性を経時的に連続モニターして、種々の基質による酵素速度の測定を可能にし得ることを示す。
図13は、種々の基質によるSULT活性の連続FPIAに基づく検出を示すグラフである。受容体基質(200nM)を、100μLの総体積で、30mMのホスフェート(pH 7.4)、7mMのDTT、0.8mMのMgCl2、75mMのNaCl、0.5mg/mLのBGG、2μMのPAPS、200ngのSULT1E1-cHis、1nMのC8-PAP-F14トレーサーおよび12.5μLのAb 3642を含有するウェルに添加した。対照反応(各グラフの上の方のトレース)は、C-His-SULT1E1を含まず、全ての他の反応成分を含有していた。プレートを室温でインキュベートし、偏光を1分間隔で読取った。
各グラフの上の方のトレースはSULT1E1を欠く対照反応であるので偏光は有意には変化しないが、酵素を含有する各反応においては、偏光は時間につれて減少する。試験設定は、HTS設定においてなし得た設定と極めて類似のものである:抗体とトレーサーをSULT反応混合物に添加し、異なる受容体基質を含有する384ウェルプレート中に分配し、反応をSULT1E1-cHisの添加により開始させた。その後、各プレートを一定間隔で読取り、偏光値を時間の関数としてプロットした。既知のSULT基質による反応においては、偏光は、酵素を欠く対照反応と対比して経時的に減少していた;酵素により産生させたPAPがトレーサーをAbから外して置き換わるときに期待し得るように。受容体基質が存在しないとき(グラフf)或いはSULT1E1が存在しない場合(各グラフの上の方のトレース)、偏光の有意の変化はなく、PAPS分子が非生産性化学物または酵素加水分解に十分に抵抗性であることを示唆していることに注目されたい。
【0068】
これらの結果は、スルホトランスフェラーゼについて汎用的な蛍光系活性アッセイ法が技術的に実施可能であることを示している。これらの試験においては、3642 AbのPAPSとの交叉反応は有意なバックグラウンド(即ち、偏光の減少)に寄与して、アッセイのダイナミックレンジを制約しているが、酵素速度は、各速度曲線の直線部分から依然として得ることができている。図14の棒グラフは、図13の速度曲線の直線部分から算出した各基質の割合を示している。この時点では正確なアッセイでないものの、この基質の順位付けは、これら基質の公表されたKm値並びに出願人等が精製SULT1E1および35S-PAPS放射線アッセイを使用して測定した基質プロフィール(図14)と逆相関している。従って、図14は、FPIA系アッセイと35S-PAPS放射アッセイを使用して測定したSULT1E1受容体基質プロフィールの比較である。FPIA系反応の速度は、図13に示す曲線の直線部分から算出した。35S-放射線アッセイデータは表4に提示していることに留意されたい。受容体基質は、200nM (FPIA)および400nM (35S-放射線アッセイ)で使用した。
最後に、出願人等は、FPIA系アッセイを使用して阻害曲線を作成し得ることを示すために、既知のSULT阻害因子であるDCNPを使用した(図15)。詳細には、図15は、FPIA系アッセイにより測定した2,6ジクロロ-4-ニトロフェノール(DCNP)によるSULT1E1の阻害を示すグラフである。DCNPを、ウェル中で、46μLのリン酸アッセイ緩衝液(30mMのKPO4(pH 6.5)、0.5mg/mLのBGG、15mMのDTT、1.6mMのMgCl2、4μMのPAPS)中で連続2倍希釈し、次いで、50μlの2×抗体/トレーサー混合物(5μL 3642 Ab/2 nM トレーサーC8-PAP-F14)および200ngのSULT1Eを100μlの総体積中に添加した。プレートを室温で30分間インキュベートし、Tecan Ultraで読取った。ΔmP値を、SULT1E反応を無SULT1E対照から差し引くことによって算出した。全ての値は繰り返しの平均を表す。
DCNPに対する前記FPIAアッセイの応答を、35S-放射アッセイ(データは示していない)との比較により検証した;2つのアッセイ法で測定したKi値は、それぞれ、7.8mMおよび11mMであった。即ち、出願人等は、前記アッセイ法が基質および阻害因子の検出(共に意図する主要HTS用途)に使用し得たことを実証した。
要約するに、これらの結果は、提案したFPIA系SULTアッセイ法の構成要素、即ち、精製組換えSULT;PAPSの存在下においてPAPを選択的に認識する抗体;および、その結合およびAbからの拮抗的置換を偏光の有意の変化によって検出し得る蛍光PAPトレーサー、の全ての作製の実施可能性を明らかに立証するものである。さらに、出願人等は、前記アッセイ試薬を、アッセイをHTS設定において使用し得る方法に極めて類似した連続方式でSULT活性を検出するのに使用し得たこと;種々の基質に対する応答が標準の放射アッセイに類似していること;および前記アッセイ法は阻害因子の検出にも使用し得ることも実証することができた。
従って、商業的HTS用途において適切なダイナミックレンジおよびシグナル:ノイズを有するアッセイ法の開発を可能にする抗体、適切には、PAPに対しておよそ10倍高い親和性と選択性を有するモノクローナル抗体を産生させることが考慮されている。
【0069】
実施例4
アッセイ系
上述の方法は種々の異なるアッセイ系において使用し得るけれども、その最も簡単な形においては、本発明のアッセイ系は、アッセイする反応を実施するアッセイ容器、並びに反応の結果を検出するための検出器を含む。好ましい局面においては、前記アッセイ容器は、試験管、マルチウェルプレート内のウェルまたは他の同様な反応容器から選択する。そのような場合、各種試薬を容器中に導入し、蛍光偏光検出器のような上述した適切な検出装置を使用して典型的にアッセイする。容器以外に、ガラスまたはプラスチックのような平坦表面も使用することができ、反応成分はその表面にアレー状にスポットすることができる(マイクロアレーのように)。
別の等しく好ましいのは、反応容器が流体用チャネル、好ましくは微小流体用チャネルを含むことである。本明細書において、微小流体用なる用語は、約1ミクロン〜約500ミクロン範囲の少なくとも1つの断面寸法を有するチャネルまたは他の導管をいう。本明細書において説明する方法を実施するのに有用な微小流体装置の例としては、例えば、米国特許第5,942,443号、第5,779,868号、および国際特許出願 WO 98/46438号に記載されている装置があり、これら特許の開示は、引用により本明細書に取り込まれる。
上述の方法に従い、第1および第2反応物間の酵素介在カップリング反応を微小流体装置のチャネル内で実施することが考慮されている。そのように、微小流体工学基本骨格を使用することにより、真核細胞の区分化を模倣することが可能になり得る。この方法は、その場合、酵素活性に影響し得る他のタンパク質および細胞成分との関連において、より天然の環境において酵素により触媒作用される基転移反応の活性をモニターし得るであろう。従って、基転移反応を触媒する酵素の活性およびその活性の阻害結果に関するデータは、生体内環境をもっと正確に反映する条件設定において得ることができる。
【0070】
これらのアッセイ系は、興味ある酵素反応に影響を与える試験化合物をスクリーニングし得ることが考慮されている。必要に応じて、本発明に従って使用する装置は、例えば、米国特許第5,942,443号に記載されているような高処理量スクリーニング方式において操作するように形成される。とりわけ、潜在的試験化合物を装置本体と一体化させた貯蔵容器から反応領域に搬送する代りに、そのような試験化合物を、装置本体に装着し反応領域と流体的に連結させた外部サンプリングピペッターまたはキャピラリーにより、反応領域に導入する。そのようなピペッター装置は、例えば、米国特許第5,779,868号に記載されている(その全体を参考として本明細書に合体させる)。サンプリングピペッターは種々の試験化合物の源中に連続して浸漬され、それらの化合物は個別にまたは連続して反応領域に持ち込まれ、注目する反応に対してそれらの化合物の影響(あり得る場合の)が確認される。
これらの微小流体チャネルネットワークのチャネルを通る物質の移動は、電気動力学物質移動(例えば、米国特許第5,858,195号に記載されているような(該米国特許は引用により全てが本明細書に取り込まれる))、圧力系流動、軸流動、重力流動、またはこれらの任意のハイブリッド形のような任意の各種公知の方法によって実施する。
【0071】
実施例5
アッセイキット
本発明のもう1つの実施態様は、基転移反応の供与体生成物または基転移反応の供与体生成物を産生させる触媒活性を検出し定量するためのキットである。基転移反応の一般的等式は、供与体-X + 受容体 → 供与体-生成物 + 受容体-Xを含み、供与体-生成物を一般的検出反応:第1複合体 + 供与体-生成物 → 第2複合体 + 置換された検出可能タグによって検出する。最も単純な形において、FPIAイムノアッセイ用のキットは、巨大分子(即ち、抗体または不活化酵素)およびトレーサー(転移した)および、場合により注目する特異的基転移酵素を含み得る。前記巨大分子およびトレーサーは、別々または1つの溶液容器に混入させているかのいずれでもよいことに留意されたい。
また、前記キットは、不活化供与体、検出可能タグ、受容体基質、阻害因子、緩衝液、補因子、安定化剤、当該キットを使用するための使用説明書セットまたは包装材料、およびこれらの任意の組合せのような成分も含む。さらに、前記キットは、検出可能タグがその発生させる観測可能物理量に基づき識別され得る2以上の抗体/検出可能タグ対を使用することにより、多重検出用に定式化し得る。このイムノアッセイ法は、前記供与体生成物または前記供与体生成物を生成させる触媒活性を検出するのに使用し得る。
本発明の実施に当っては、前記キットを、酵素(該酵素は基転移反応において前記供与体生成物を産生させる)と接触し得る分子のライブラリーまたは分子群をスクリーニングするのに使用し得ることも考慮される。前記ライブラリーは、複数の化学分子、複数の核酸、複数のペプチド、およびこれらの組合せの少なくとも1つを含み得る;スクリーニングは、マルチウェルプレートまたは微小流体装置を使用する高処理量スクリーニング法により実施する。
さらに、前記キット内の巨大分子が、記供与体生成物と高親和性で接触し得る、抗体、ポリペプチド、タンパク質、核酸分子、不活化酵素、およびこれらの組合せの少なくとも1つを含むことが考慮される。さらに、前記キットが、スルホトランスフェラーゼ、キナーゼまたはUDP-グルクロノシルトランスフェラーゼ、メチルトランスフェラーゼ、アセチルトランスフェラーゼ、グルタチオントランスフェラーゼまたはADP-リボシルトランスフェラーゼ、およびこれらの組合せの少なくとも1つを含むことも考慮される。
【0072】
FPIAは適切な検出方式であるけれども、本発明の範囲には、均質アッセイ法、均質蛍光強度免疫測定法、均質蛍光寿命免疫測定法、均質蛍光共鳴エネルギー移動(FRET)免疫測定法または均質化学発光免疫測定法、或いは酵素結合免疫測定法(ELISA)のような非均質アッセイ法のような、供与体生成物または該供与体生成物を生成させる触媒活性を他の手段によって検出するのに使用するように設計されたキットも属する。均質蛍光強度免疫測定法または均質蛍光寿命免疫測定法の場合は、前記キットは、抗体と蛍光検出可能タグとから構成することができ、前記検出可能タグの強度および/または寿命は抗体と結合しているときには溶液中で遊離しているときと異なる。蛍光の差異、即ち、アッセイシグナルは、抗体の検出可能タグとの相互作用がその蛍光特性のさらなる変化、即ち、消光、増強または寿命の変化をもたらすような抗体の修飾によって増強させ得るであろう。均質FRET免疫測定法の場合は、検出可能タグと結合させた第1蛍光体と抗体に結合させた第2蛍光体(直接またはビオチンおよびストレプタビジンのような会合型結合性分子を介して)との相互作用が第2蛍光体の励起をもたらし(またはその逆)、それによって検出可能タグの波長とは異なる波長で蛍光放出を生じさせる。第2蛍光体は、小有機分子または発光ランタニドプローブであり得る。(ランタニド発光は、蛍光ではなく、発光系共鳴エネルギー移動、即ち、LRETと称することに留意されたい)。化学発光検出の場合は、検出可能タグは、β-ガラクトシダーゼのような化学発光検出において使用する酵素の1つのフラグメントに結合させた供与体生成物であり得るであろう。前記供与体生成物-フラグメント-1複合体は、供与体生成物により抗体から外されると、前記酵素のフラグメント2と結合して、適切な基質により化学発光を発生させ得るインタクト活性酵素を生成させる。ELISAの場合は、アッセイは均質ではなく、供与体生成物または抗体をマルチウェルプレートの表面に固定化させる必要があろう。検出酵素に結合させた二次抗体も、この方式に含ませ得るであろう。
【0073】
本明細書において引用した刊行物は、全て、その全体を本明細書に合体させる。本発明の開示と合体させた刊行物間で論争がある場合、本発明の開示が支配する。
今回、本発明を幾つかの具体例で説明し例示してきたが、当業者であれば、説明した範囲内でなし得る変更、付加および削除のような種々の修正を認識するであろう。従って、これらの修正も本発明に属し、そして、本発明の範囲は、特許請求の範囲に合法的に従う最も広い解釈によってのみ限定されるものとする。
【図面の簡単な説明】
【0074】
【図1】SULT反応生成物、即ち、PAPについての蛍光偏光免疫測定(FPIA)反応を例示する。
【図2】FPIAを使用してのUDPの生成、即ち、UGT反応の供与体生成物の検出と定量を例示する。
【図3】FPIAを使用してのADPの生成、即ち、キナーゼ反応の供与体生成物の検出と定量を例示する。
【図4】SULT1E1 FPIA用の試薬、即ち、抗-PAP抗体および蛍光標識化PAP検出可能タグの反復同時開発戦略を説明する。
【図5】図AおよびBは、UDP/UTPに対して産生させたポリクローナル抗体および商業的に入手し得るトレーサー分子(Alexa-UTP)を使用してのウリジンヌクレオチドの滴定または拮抗置換曲線を示す。
【図6】PAP抗原の合成を示す。
【図7】図A〜Cは、PAPトレーサー合成の諸ヨウ素を示す。
【図8】図A〜Cは、代表的な最終トレーサー構造を示す。
【図9】抗-PAP抗体およびPAP-フルオレセイントレーサーの結合等温線を示す。
【図10】Ab 3642に由来する2つの異なるトレーサーのPAPおよびPAPSによる拮抗置換を示す。
【図11】図AおよびBは、2つの抗-PAP抗体/トレーサー組合せによる拮抗曲線を示す。
【図12】酵素により産生させたPAPの検出におけるPAPS濃度の効果を示す。
【図13】図A〜Fは、種々の基質によるSULT活性の連続FPIA系検出を示す。
【図14】FPIA系アッセイおよび35S-PAPS放射アッセイを使用して測定したSULT1E1受容体基質プロフィールの比較を示す。
【図15】FPIA系アッセイにより測定した2,6ジクロロ-4-ニトロフェノール(DCNP)によるSULT1E1の阻害を示す。
【技術分野】
【0001】
(関連出願との相互参照)
本出願は、2003年1月30日に出願された米国仮出願第60/443,746号に基づく利益を主張し、該米国出願は、その全体が引用により本明細書に取り込まれるものとする。
(技術分野)
本発明は、基転移反応方法論に関する。本発明は、基転移反応における供与体-生成物および該供与体-生成物を生成し得る触媒活性の検出および定量方法を提供する。また、本発明は、基転移反応を触媒する酵素の受容体基質、阻害因子または活性化因子の高処理量スクリーニング方法も提供する。さらに、本発明は、本発明を実施するためのイムノアッセイ法、抗体および関連キットも提供する。
【背景技術】
【0002】
基質が活性化供与体分子として知られる他の物質によって供与される化学基によって修飾される、多くの重要な生物学的反応が存在する。これらの生物学的反応は、“基転移反応”として広く認識されており、下記の一般反応式を有する:
供与体-X + 受容体 → 供与体生成物 + 受容体-X
典型的には、供与体-X、即ち、活性化供与体分子は、共有結合付加物に結合したヌクレオチドである。供与体-Xは、前記ヌクレオチド供与体中でホスホエステル結合の形成によって活性化される。また、受容体基質は、タンパク質または核酸のような小分子または巨大分子を含み得る。この反応の生成物は、修飾受容体、受容体-Xおよび供与体-生成物分子である。
基転移反応を触媒する、例えば、ATPを使用してリン酸基を供与するキナーゼ;ホスホアデノシン-ホスホスルフェート(PAPS)を使用してスルホネート基を供与するスルホトランスフェラーゼ(SULT);UDP-グルクロン酸を使用してグルクロン酸基を転移させるUDP-グルクロノシルトランスフェラーゼ(UGT);s-アデノシルトランスフェラーゼを使用してメチル基を供与するメチルトランスフェラーゼ;アセチル補酵素Aを使用してアセチル基を供与するアセチルトランスフェラーゼ;およびニコチンアミドアデニンジヌクレオチド(NAD)を使用してADP-リボース基を供与するADP-リボシルトランスフェラーゼのような多くの酵素が存在する。従って、基転移反応を触媒する多くの酵素は、製薬企業にとって興味あるものである。
【0003】
自動化高処理量スクリーニング(HTS)アッセイ法は、薬物発見環境における潜在的薬物分子とタンパク質との相互作用を同定するためのパラダイムであり、この方式は、好ましくは蛍光または化学発光読出しによる単純で強力な分子アッセイを必要とする。HTSに最も適する方式は、反応を開始させた後に何らの操作も必要とせず且つアッセイシグナルを連続的にモニターし得る均質アッセイ(例えば、“単回添加”または“混合および読取り”アッセイ)である。薬物発見の観点におけるこれらアッセイ法の重要性にもかかわらず、基転移酵素の製薬HTSプログラムへの取入れは、以下のようないくつかの理由により緩慢であり妨げられている:a)現在のところ、均質アッセイ法は利用可能ではない、b) 使用する検出方法が前記アッセイの使用に重大な制約をもたらしている、c) 前記アッセイ法の汎用的でない性質は、多様な受容体物質を試験するために多くの特異的な検出試薬の開発を必要とする。
スルホトランスフェラーゼ基質または阻害因子を同定するのに現在使用されている方法は、放射能の使用を必要とし、沈降またはクロマトグラフィーのような面倒な反応後分離工程を含む。例えば、35S-PAPSがスルホトランスフェラーゼ反応において使用され、標識化された生成物が、未反応35S-PAPSの選択的沈降後にシンチレーション計数によって定量される(Foldes, A. and Meek, J. L., Biochim Biophys Acta, 1973, 327:365-74)。この方法は、高処理量スクリーニング(HTS)法においては、高放射性廃棄物コスト故に、また、分離工程の取入れは自動化工程を複雑にすることから望ましくない。他のSULTアッセイ法が比色分析および蛍光手段を使用して開発されているが、これらのアッセイ法は、検出のために特定の受容体基質の使用に依存しており、従って、その使用は単一のSULTイソ型に制約され、これらのアッセイは多様な基質についてのスクリーニングに使用することはできない(Burkart, M. D. and Wong, C. H., Anal Biochem, 1999, 274:131-7;Frame, L. T. et al., Drug Metab Dispos, 2000, 28:1063-8)。結果として、SULT相互作用研究は、薬物の前臨床開発において、現在のところ含まれていない。また、UGTは、現在、放射性標識供与体を使用してアッセイされており、前臨床HTSプログラムを著しく阻害する薄層クロマトグラフィー(TLC)または高圧液体クロマトグラフィー(HPLC)のような反応後分離工程を必要とする(Ethell, B. T., et al., Anal Biochem, 1998, 255:142-7)。同様に、伝統的に、キナーゼは、32P-ATPまたは33P-ATPを供与体として使用して生成させた放射性標識ポリペプチド基質のフィルター捕捉または沈降によってアッセイされている。しかしながら、この方法は、濾過または遠心分離のような分離工程を必要とするので、自動化HTS方式に容易には適応させ得ない。表面近接アッセイ法(surface proximity assay) (SPA)は、マルチウェル方式での放射能アッセイを分離なしで可能にするが(Mallari, R., et al., J Biomol Screen, 2003, 8:198-204)、製薬企業によるその使用は、取扱い放射性同位元素の廃棄および法的コストのために減少してきている。
【0004】
キナーゼ阻害因子薬物開発における高レベルの興味故にこの酵素群の改良されたアッセイ方法を開発する多くの努力が科学者達によりなされている。高度に特異的な試薬を使用して、反応の他の成分の存在下での反応生成物を蛍光または化学発光のような光系読出しを使用して検出する均質アッセイ法が開発されている。キナーゼアッセイにおいて使用する最も一般的な均質方法は、種々の蛍光特性を示すホスホペプチド生成物の免疫検出である(Zaman, G. J., Comb Chem High Throughput Screen, 2003, 6:313-20)。この方法においては、基質ペプチドのリン酸化が抗-ホスホペプチドから蛍光ホスホペプチドトレーサーを外してこれを置換し、その蛍光特性の変化を生じさせる。この基本的方法は、蛍光偏光(FP)法 (Parker, G. J., et al., J Biomol Screen, 2000, 5:77-88);時間分解蛍光法(Xu, K., et al., J Biochem Mol Biol, 2003, 36:421-5);蛍光寿命識別法(Fowler, A., et al., Anal Biochem, 2002, 308:223-31);および化学発光法(Eglen, R. M. and Singh, R., Comb Chem High Throughput Screen, 2003, 6:381-7)のような拮抗イムノアッセイ法において使用する数種の読出し方式に適合されている。
この方法による欠点は、ホスホペプチド特異的抗体の必要性である。汎用的なホスホチロシン抗体が一般的であるが、ホスホセリン抗体およびホスホスレオニン抗体は、周知のとお作製するのが難しく、ホスホ-セリンまたは-スレオニンを特異的隣接アミノ酸との前後関係の中でしか認識しない(EglenおよびSingh、2003年)。ヒトにおいては400種を越えるキナーゼが存在し、リン酸化部位に対するそれらの特異性は広く変化する。即ち、多様な抗体を多様なキナーゼのアッセイまたは受容体基質のプロフィーリングにおいて必要とする。このことは、新たなキナーゼのHTSへの組入れを、とりわけそれらの基質特異性が良く明確にされていない場合に極めて複雑にする。また、このことは、アッセイの出力が使用する特定の抗体(Ab)-ホスホペプチド対に依存するので、多様な基質選択性を有するキナーゼ間でのデータ比較における解析問題も生ずる。汎用的なホスホセリン抗体を開発し且つより汎用的なキナーゼ基質を同定する努力は続いているものの(Sills, M. A., et al., J Biomol Screen, 2002, 7:191-214)、この方向性における研究は、今日まであまり成功していない。
【0005】
前後関係特異的Ab-ホスホペプチド相互作用の問題を回避するために多くの別の方法が開発されており、それらにはホスホペプチドを結合させるための金属複合体の使用(Scott, J. E. and Carpenter, J. W., Anal Biochem, 2003, 316:82-91)およびホスホペプチド生成物の共有結合タグ付けを可能にする修飾ATPアナログの使用(Allison Miller-Wing, E. G., Barbara Armstrong, Lindsey Yeats, Ram Bhatt, Frank Gonzales, and Steven Gessert., SBS 9th Annual Conference and Exhibition, Portland, OR., 2003)が含まれる。化学的リン酸結合試薬は、ヌクレオチドホスフェートへのバックグラウンド結合という問題があり、極めて低い非生理学的レベルのATPの使用を必要とし、これがアッセイの柔軟性を制約する。修飾ヌクレオチドは、該ATPアナログを供与体として使用する能力がキナーゼ間で変動するので汎用的な方式を提供せず、数多くの種々のアナログの開発を必要とする。また、キナーゼATP結合部位(キナーゼ阻害因子結合の最も頻繁な部位)での阻害因子と前記修飾ヌクレオチドとの拮抗も、生理学的状況を反映してない。ペプチドリン酸化によって生ずるプロテアーゼ感受性の差異が蛍光系キナーゼアッセイの開発において活用されているが(Kupcho, K., et al., Anal Biochem, 2003, 317:210-7)、これらのアッセイは、真に均質性ではない;即ち、これらのアッセイは、現像用プロテアーゼ試薬の反応後添加を必要とする。さらに、この方法の応用性は、キナーゼおよびプロテアーゼ特異性が重なり合うペプチドに限られる。
また、反応生成物と特異的マルチウェルプレート化学作用物との相互作用に依存する少数の方法も存在するが、これらの方法は、反応後の試薬添加および/または処理を必要とする点で真に均質性ではない。また、処理および/または検出のための特別仕様の機器を必要とすることも、殆どの製薬HTS基本構造に望まれるオープンアーキテクチャーに適合しない。
さらにまた、反応生成物の電気泳動分離に依存する微小流体工学系キナーゼアッセイも開発されてきている(Xue, Q., et al., Electrophoresis, 2001, 22:4000-7)。これらのアッセイ法においては、リン酸化ペプチド生成物をリン酸化されていない受容体基質から電気泳動により分離し、それによって特化された検出試薬の必要性を排除している。しかしながら、実際には、キナーゼアッセイは、多くの場合、マルチウェルプレートにおいて操作され、その後、生成物が分離用の微小流体装置に移される(HTS方式においては面倒な工程)。
【0006】
要するに、現在の基転移アッセイの汎用的でない性質は、個々の酵素またはそのファミリー内の下位群についてのアッセイを開発する必要がある故に、薬物発見におけるかなりの費用と遅れをもたらしている。また、現在のアッセイ法の多くは特異的なタグ付き受容体の修飾と検出に基づいていることから、多様な受容体基質を試験するには限られた能力しかない。多くの場合、使用するタグ付き化受容体基質は、生体内でリン酸化される基質とは異なり、従って、前記アッセイの生理学的適合性は疑わしい。さらに、製薬業界における大きな懸念事は、現在のアッセイ法の汎用的でない性質故に、場合によっては、研究者は異なるキナーゼに対しては異なる方法を使用することを余儀なくされることである。しかしながら、幾つかの研究において、異なるアッセイ方法を使用して同定した医薬ターゲットにおいて有意差があることが証明されており(Sills, M. A., et al., J Biomol Screen, 2002, 7:191-214)、これは、数種のキナーゼに亘る阻害因子選択性のプロファイリングにおいて大きな問題である。基転移反応についての現存するHTSアッセイ法のこれらの欠点は、製薬上の薬物発見プログラムにおける重要な酵素群の迅速な分析を妨げている。
基転移反応をアッセイする他の現存する方法は、供与体分子生成物を検出することによって、種々の化学物を基転移反応における基質としてスクリーニングすることを可能にしている;何故ならば、供与体分子生成物は、受容体が修飾されているかどうかにかかわらず、同じであるからである。供与体生成物の検出は汎用的アッセイ法の基本を提供するものと考えられており、ADPは常にキナーゼ反応の生成物であり、ホスホアデノシン-ホスフェート(PAP)は常にスルホトランスフェラーゼ反応の生成物である。しかしながら、例えば、ADPはATPと同じ蛍光および吸光特性を有する故に、開裂モノ-およびジ-ヌクレオチドを活性化供与体分子から吸光または蛍光特性に基づき識別することができず、これらの生成物の検出は複雑である。供与体生成物の供与体からの分離は、薄層クロマトグラフィーまたは高圧液体クロマトグラフィーのようなクロマトグラフィー法を使用して実施し得るが、これらの方法のHTS方式への取入れは面倒である。
【0007】
この困難性を回避するために、供与体生成物の検出を、さらなる酵素を使用して一次反応生成物(開裂モノ-およびジ-ヌクレオチド)から検出可能な生成物を生成させることによって達成している;これは、酵素共役反応として知られている。例えば、酵素および他の小分子をADP依存性のNADPH産生において使用でき、NADPHを340nmでの吸光または蛍光によって検出する(Walters, W. P. and Namchuk, M., Nat Rev Drug Discov, 2003, 2:259-66)。酵素共役反応は、もう1つのタイプの基転移酵素であるUGTにおいても開発されている(Mulder, G. J. and van Doorn, A. B., Biochem J, 1975, 151:131-40)。しかしながら、薬物化合物の吸光アッセイによる光干渉は、とりわけ紫外線において、この方法で広く認識された問題である。この方法のもう1つの欠点は、検出物と共役させるのに使用する酵素の全てがスクリーニングする化学物からの潜在的な阻害に供されることである。
もう1つの一般的な方法は、ルシフェラーゼをレポーターとして使用してATP消費量をモニターすることによってプロテインキナーゼ活性を検出することである。この方法の例は、Crouch等により、米国特許第6,599,711号において開示されている。該方法は、プロテインキナーゼによってリン酸化され得る基質をATPと試験すべきプロテインキナーゼを含む溶液、および試験すべきプロテインキナーゼの存在しないATPを含むもう1つの溶液に添加することによって、試験すべきプロテインキナーゼの活性を測定することを必要としていた。その後、ATPおよび/またはADPの濃度または時間変化速度を、生物発光を使用して測定していた。しかしながら、このアッセイ法は、高い初期シグナルの小さい低下に依存しているので最適ではない。バックグラウンドを最小にするためにATP濃度を低く保つ必要性があるので、アッセイ条件を注意深く制御しない限り非直線的な反応キネティクスが生じる。関連する方法においては、蛍光ATPアナログを使用する拮抗結合アッセイも開発されているが、酵素触媒活性の尺度を提供せず、従って、限られた利用性しかない。
【0008】
(発明の簡単な要約)
基転移反応を触媒する酵素のHTSアッセイへの使用は、製薬用の薬物ライブラリー中の分子からの干渉を受けない普遍的で均質性のアッセイ方法が無いために妨げられている。従って、HTS薬物発見に良好に適応する基転移反応に関連する酵素の普遍的なアッセイ方法を提供することが一般に望ましいであろう。
【発明の開示】
【0009】
重要な基転移酵素のHTSアッセイへの容易な組入れを可能にするために、出願人等は、前記供与体生成物の均質免疫検出に基づくアッセイ法を開発した。基転移反応の一般的方程式は、供与体-X + 受容体 → 供与体-生成物 + 受容体-Xであり、供与体-生成物を一般的検出反応:第1複合体 + 供与体-生成物 → 第2複合体 + 置換された検出可能タグによって検出する。前記供与体生成物は与えられた型の基転移反応を触媒する全ての酵素において同じであるので、同じ検出試薬を基転移酵素群内の全てのメンバーにおいてまた任意の受容体基質と一緒に使用し得る。アッセイ生成物は、製薬用の薬物ライブラリー中の分子からの有意の干渉またはバックグラウンドシグナルを受けない均質蛍光または化学発光法を使用して検出し得る。
【0010】
即ち、本発明は、基転移反応を触媒する酵素の活性を検出すること、および、これら酵素の受容体基質、阻害因子または活性化因子をスクリーニングしてより選択性で治療用の薬物の開発を容易にする方法およびその構成要素として要約される。このことは、基転移反応の供与体生成物と結合させるのに使用する高度に選択的な抗体によって達成される。抗体-抗原相互作用は、すでに他の人々によって記載されている多くの方法で検出し得る。出願人等が前記方法を実施するのに使用した検出方式は、拮抗蛍光偏光イムノアッセイ法(FPIA)(competitive fluorescence polarization immunoassay)であるが、この方法が製薬HTSアッセイに良好に適応するためである。この検出方式によれば、酵素により生成した供与体生成物は、トレーサーと呼ばれる該供与体生成物の蛍光誘導体を抗体から外して置き換わり(置換し)、トレーサー蛍光偏光の減少を生じさせる。このアッセイにおける主要試薬は、供与体生成物に高選択性でもって結合するが未開裂供与体分子には結合しない抗体、およびトレーサー(前記抗体に結合するのに十分にその構造を保持する前記供与体生成物の蛍光誘導体)である。本発明は、基転移反応を触媒する酵素の活性を種々の基質を使用して検出し定量する新規なアッセイ法を提供する。また、本発明は、基転移反応の基質、阻害因子または活性化因子のスクリーニング方法も提供する。
【0011】
1つの局面においては、本発明は、基転移反応の供与体-生成物の検出方法を提供し、該方法は、活性化型の供与体を触媒活性酵素の存在下に受容体と反応させること;供与体-生成物および受容体-Xを形成させること;前記供与体-生成物を、観測可能物理量を発生させ得る検出可能タグを含む第1複合体と接触させること;前記第1複合体の前記検出可能タグを前記供与体生成物によって拮抗的に置換して(外して)第2複合体および置換された(外された)検出可能タグを生成させること;および、前記第1複合体中の前記検出可能タグおよび前記置換された(外された)検出可能タグによって発生した観測可能物理量の変化を検出すること、を含む。
もう1つの局面においては、本発明は、基転移反応の供与体生成物に対して作製された抗体を提供し、該抗体は、供与体-生成物と供与体とを高供与体濃度の存在下に選択的に識別する能力を有する。
さらにもう1つの局面においては、本発明は、基転移反応の供与体-生成物の均質拮抗結合アッセイ法を提供し、該アッセイは、以下を含む:供与体-生成物をトレーサーおよび巨大分子と混合して混合物を調製することであって、前記巨大分子は前記供与体-生成物に対し特異的であり、前記トレーサーはフルオロフォアに結合させた前記供与体-生成物を含み、前記トレーサーは前記巨大分子と結合して蛍光偏光において検出可能な変化を生じ得る;前記混合物の蛍光偏光を測定して蛍光偏光測定値を得ること;および、前記蛍光偏光測定値を特性決定した蛍光偏光値と比較することであって、該特性決定した蛍光偏光値は既知の供与体生成物濃度に対応する。
さらにもう1つの局面においては、本発明は、基転移反応からの供与体生成物の特性決定用アッセイキットを提供し、該アッセイキットは、巨大分子とトレーサーの各々を、供与体生成物の少なくとも1回の蛍光偏光アッセイに適する量で含み、前記巨大分子は抗体または不活化酵素である。
他の利点、並びに特定の変形、組成的変更および全体的構成のより完全な認識は、特許請求の範囲と関連して行う以下の各種実施態様についての詳細な説明を検証するときに得られるであろう。
【0012】
本発明の実施態様を詳細に説明する前に、本発明は、その応用において、以下の説明において説明する詳細に限定されるものではないことを理解すべきである。本発明は、他の実施態様もあり得、種々の方法で実行または実施可能である。また、本明細書において使用する表現または用語は、説明目的のためであり、如何なる形でも限定するものとみなしてはならないことを理解すべきである。
【0013】
(発明の詳細な説明)
本発明は、広義には、供与体-生成物または基転移酵素に由来する前記供与体-生成物を生成させる触媒活性を、種々の基質を使用して検出し定量する新規なアッセイ方法に関する。また、本発明は、前記供与体生成物に対して特異的な抗体並びに本発明を高処理量スクリーニング方式において実施するためのアッセイキットも提供する。基転移反応の一般的等式は、供与体-X + 受容体 → 供与体-生成物 + 受容体-Xを含み、供与体生成物を一般的検出反応:第1複合体 + 供与体-生成物 → 第2複合体 + 置換された検出可能タグによって検出する。即ち、本発明によれば、高度に選択性の抗体を使用して前記基転移反応の供与体生成物に結合させ、この結合事象を、例えば、製薬HTSアッセイに良好に適応する拮抗結合FPIAのようなイムノアッセイ法を使用して検出する。この検出方式を使用することにより、酵素により生成した供与体生成物は抗体からトレーサーを外してこれに置き換わり、これによりトレーサー蛍光偏光の減少をもたらす。このアッセイにおける主要試薬は、前記供与体生成物に高選択性でもって結合するが未開裂供与体分子には結合しない抗体、およびトレーサー(前記抗体に結合するのに十分にその構造を保持する前記供与体生成物の蛍光誘導体)である。
【0014】
とりわけ、本発明は、単一系列の検出試薬を基転移酵素の所定群内の全ての酵素およびこの群に対する全ての受容体基質において使用し得る点で、普遍的なアッセイ方法を提供する。その普遍的性質故に、本発明は、SULT、プロテインキナーゼおよび他の基転移酵素のHTSスクリーニングへの組入れを促進するであろう。例えば、11種の既知のSULTイソ型が存在し、本発明の方法を使用すれば、11種のSULTイソ型の全てを、それらの種々の化合物を硫酸化能について、同じ試験において同じ検出試薬、プロトコールおよび器具を使用してスクリーニングすることができる。このことは、生体異物結合を触媒する酵素(例えば、SULTおよびUGT)が極めて広い受容体基質特異性を有することから、重要な能力である。キナーゼに関しては、酵素群内にさらにより多くの多様性が存在する。ヒトにおいて400種よりも多いプロテインキナーゼが存在し、それらの受容体基質特異性においては、生理学的タンパク質受容体基質並びに短ペプチドのいずれも使用し得るような大きな多様性が存在する。そのように、多数のプロテインキナーゼを、それらの多様な受容体基質を使用して、本発明のアッセイにおいて使用でき、同じ検出試薬、プロトコールおよび器具を使用して、阻害因子についてスクリーニングできる。これらの理由および以下で述べる他の理由により、SULT、UGTおよびキナーゼのような基転移反応における本発明のFPIA系供与体生成物検出アッセイ法は、なかんずく、自動化HTS用途に良好に適応する。
従って、本発明をそのような試みに関連して詳細に説明する;しかしながら、当業者であれば、本発明のそのような説明は、例示のみを意味し、本発明の範囲全体を限定するものとみなすべきでないことを理解されたい。
【0015】
定義
本明細書において使用するある種の用語は、以下の一般的定義を有するものとする:
本明細書において使用するときの用語“基転移反応”は、下記の一般的反応を称する:
供与体-X + 受容体 → 供与体-生成物 + 受容体-X
代表的な基転移反応は、以下のように示される:
キナーゼ反応:ATP + 受容体 → ADP + 受容体-PO4;
UGT反応:UDP-グルクロン酸 + 受容体 → UDP + 受容体-グルクロン酸;
SULT反応:受容体-XH + 3'-ホスホアデノシン5'-ホスホスルフェート →受容体-SO4 + 3'-ホスホアデノシン5'-スルフェート + H+;
メチルトランスフェラーゼ反応:s-アデノシルメチオニン + 受容体 → 受容体-CH3 + s-アデノシルホモシステイン;および、
アセチルトランスフェラーゼ反応:アセチル補酵素A + 受容体 → 受容体-COCH3 + 補酵素A。
SULTによる硫酸化、キナーゼによるリン酸化およびUGTによるUDP-グルクロン酸化のような部分的基転移反応は、それぞれ、PAP、ADPおよびUDPである供与体生成物に対する抗体/検出可能タグ対を単離し得るので、本発明の方法に適切に応用可能である。また、基転移反応は、ホルモン生体合成および機能;酵素調節および機能;および生体異物代謝のような多くの生物学的プロセスに一般的に関与する。
用語“普遍的アッセイ”および“汎用的アッセイ”は、互換的に使用され、基転移反応酵素群の全メンバーおよびそれらの受容体基質の全てを同じアッセイ試薬により検出し得る方法をいう。
用語“共有結合付加物”とは、基転移反応において供与体分子から受容体に転移される部分をいう;SULT、キナーゼおよびUGTにおける、それぞれ、スルホネート、ホスフェートおよびグルクロン酸。
【0016】
本明細書において用語“供与体-生成物”は、前記共有結合付加物が受容体に転移したときに生成する供与体分子のフラグメントである基転移反応の生成物をいう。多くの場合、供与体-生成物は、PAP、UDPまたはADPのようなヌクレオチド(天然または合成の);或いはs-アデノシルホモシステイン、ニコチンアミドまたは補酵素Aのような非ヌクレオチドである。供与体-生成物は、第1複合体 + 供与体-生成物 → 第2複合体 + 置換された(外された)検出可能タグのような一般的反応により検出する。
本明細書において用語“トレーサー”とは、特異的抗体と結合するのに十分にその構造を保持し、置換される、供与体生成物の検出可能な誘導体またはタグをいう。
本明細書において用語“供与体”とは、前記活性化共有結合付加物を担持する、基転移反応を触媒する酵素に対する基質を称する。適切な供与体の例としては、なかんずく、ヌクレオチドのみならずs-アデノシルメチオニンおよびアセチル-CoAがある。
用語“供与体-X”は(Xは前記共有結合付加物を表す)供与体分子のもう1つの用語である。
本明細書において用語“受容体”とは、前記共有結合付加物が共有結合することになる、基転移反応を触媒する酵素に対する基質を称する;該基質は、ポリペプチド、タンパク質、核酸、炭水化物、脂質、またはステロイドもしくはアミノ酸のような小分子基質である。
本明細書において使用するときの用語“受容体-X”は、Xが共有結合した共有結合付加物である反応生成物を称する;該共有結合付加物としては、ホスフェート、スルフェート、炭水化物、天然産アミノ酸、合成誘導アミノ酸、メチル、アセチルまたはグルタチオン成分の少なくとも1つ、およびこれらの組合せがある。前記共有結合付加物は、受容体基質の機能、安定性、または機能および安定性の双方を改変し得る。
【0017】
本明細書において用語“触媒活性酵素”とは、スルホトランスフェラーゼ、キナーゼ、UDP-グルクロノシルトランスフェラーゼ、メチルトランスフェラーゼ、アセチルトランスフェラーゼ、グルタチオントランスフェラーゼまたはADP-リボシルトランスフェラーゼの少なくとも1つをいう。
本明細書において用語“触媒活性”とは、化学触媒活性、酵素活性またはこれらの組合せをいう。
本明細書において用語“第1複合体”とは、巨大分子(即ち、抗体または不活化酵素)および検出可能タグを有する複合体をいう。
本明細書において用語“第2複合体”とは、巨大分子、および前記検出可能タグが前記供与体生成物によって拮抗的に置換された供与体生成物をいう。
本明細書において使用するときの用語“観測可能物理量”とは、前記第2複合体または前記転移検出可能タグの検出可能な蛍光の変化、蛍光強度、蛍光寿命、蛍光偏光、蛍光共鳴エネルギー移動(FRET)または化学発光、および測定観測可能物理量を得るためのこれらの組合せを称する。測定した観測可能物理量は特性決定されている観測可能物理量と比較されるが、この特性決定されている観測可能物理量は前記第1複合体に対応する。
本明細書において用語“検出可能タグ”とは、供与体生成物に結合させた蛍光または化学発光トレーサーをいう。蛍光が本発明における検出の好ましい形である。適切な検出可能タグは、例えば、化学発光タグまたはフルオロフォアタグをこれらのタグが抗体結合を有意に干渉しない方法で(即ち、もっともあり得るのは、ヌクレオチドのアデシン部分に結合するようにして)前記供与体生成物分子に結合させることによって調製し得る。本発明の方法に応用し得るフルオロフォアとしては、限定するものではないが、フルオレセイン、ローダミン、BODIPY、テキサスレッド、およびこれらの当該技術において公知の誘導体がある。ローダミン結合体および他の赤色結合体は、これらの高めの波長の発光がフルオレセインの緑色よりも自己蛍光からの干渉を少なく受けるので、検出可能タグとして合成して最適化し得る。
【0018】
本発明に応用し得る化学発光タグとしては、Lumigen TMA-6およびLumigen PS-3 (Lumigen社、ミシガン州サウスフィールド)があり、これらは適切な化学発光量子収率を有する。これらの試薬は、予測可能な発光時間経過を有する効率的な化学発光反応により、測定容易なシグナルを有する。さらにまた、化学発光タグは、生来のバックグラウンド化学発光に殆んどまたは全く寄与しない。また、光強度の測定も比較的簡単で、光電子倍増管またはフォトダイオードとシグナルを転換し記録するための関連電子機器とを必要とするだけである。さらに、化学発光シグナルは、イムノアッセイにおいて、酵素フラグメント相補法を使用して発生させることができる。この場合、検出可能タグはレポーター酵素のフラグメントAに結合させた供与体生成物であろうし、基転移反応において生成させた供与体生成物による抗体からのその転移はレポーター酵素のフラグメントBとの結合を可能にして、化学発光基質上で作用し得る触媒活性レポーター酵素の形成をもたらすであろう。この方法は、米国特許第4,708,929号に、レポーター酵素としてβ-ガラクトシダーゼを使用して記載されている。
本明細書において使用する用語“イムノアッセイ法”とは、例えば、均質アッセイ法、均質蛍光免疫測定法、均質蛍光強度免疫測定法、均質蛍光寿命免疫測定法、均質蛍光偏光免疫測定法(FPIA)、均質蛍光共鳴エネルギー移動(FRET)免疫測定法または均質化学発光免疫測定法或いは酵素結合免疫測定法(ELISA)のような非均質アッセイ法、並びにこれらの組合せのような、生成物を抗体によって検出する多くのアッセイ方法をいう。
【0019】
本明細書において使用する用語“蛍光偏光免疫測定法”または“FPIA”とは、基転移反応の生成物を検出するためのイムノアッセイ法をいう。蛍光偏光を使用し、分子相互作用を、蛍光標識化したまたは生来の蛍光分子の見掛けの大きさの変化をモニターすることによって調べる(Checovich, W. J., et al., Nature, 1995, 375:254-6;Owicki, J. C., J Biomol Screen, 2000, 5:297-306)。小蛍光分子(トレーサー)を平面偏光により励起させたとき、放出光は、該分子が蛍光事象中(励起と発光の間の時間)に溶液中で急速に回転するので、大きく偏光が解消される。しかしながら、前記トレーサーをはるかに大きいレセプターに結合させ、それによってその有効分子容積を増大させた場合、その回転は十分に低下して、トレーサーを励起させたのと同じ平面内で光を発出する。結合状態および遊離状態の蛍光分子は、各々、固有の偏光値を有し、結合状態で高い値を、遊離状態で低い値を有する。分子集団においては、測定偏光値は前記2つの値の加重平均であり、かくして、結合しているトレーサー分子の割合の直接の尺度を提供する。偏光値は、ミリ偏光単位(mP)として表し、500mPが最高理論値である。
実際には、フルオレセインのような小分子はおよそ20mPの偏光値を有し、抗体に結合させたとき、その偏光は100〜400mPまで増大する。拮抗FPIAにおいては、蛍光トレーサーは、図1に示すように、供与体生成物によって抗体への結合から外され得る。シグナルは結合対遊離のトレーサー画分の差に比例し、従って、該アッセイ法のダイナミックレンジおよび感度は、前記トレーサーおよび供与体生成物に対する抗体の親和性に依存する。FPIAにおける適切なダイナミックレンジを確立するには、約70〜80%のトレーサーを、拮抗因子の不存在下に、抗体に結合させなければならない。
例えば、図1は、SULT反応生成物PAPについての拮抗FPIAの図式を示しており;SULT反応から生成したPAPが、抗-PAP抗体への結合においてトレーサー(蛍光標識化PAP)と拮抗している。この方式においては、トレーサーの出発偏光は、トレーサーが殆んど全て抗体に結合しているので高く、反応が進行するにつれて低下し、トレーサーは抗体から外される。SULT反応において生成したPAPの量は、下記の等式を使用して定量し得る:
【0020】
log[PAP産物]=log[(最高偏光値-mP観測値)/(mP観測値-最低偏光値)]+ log IC50
【0021】
本発明に包含される酵素アッセイにおいては、SULTにおけるPAP、またはキナーゼにおけるADPは、それぞれ、硫酸化生成物またはリン酸化ペプチドと化学量論的量で生成する。即ち、PAPまたはADP、とりわけ供与体生成物における標準曲線の使用は、酵素代謝回転(enzyme turnover)の直接の尺度を与えるであろう。
本明細書において用語“高処理量スクリーニング”または“HTS”とは、何千もの分子(または試験化合物)をそのタンパク質の機能に対する作用について試験することを称する。基転移反応酵素の場合には、多くの分子をその触媒活性に対する作用について試験し得る。HTS法は当該技術において既知であり、該方法は、マルチウェルプレートにおいて、自動化液体操作および検出装置により一般に実施する;しかしながら、本発明の方法は、マイクロアレー上または微小流体装置内において実施し得ることも考慮される。
本明細書において用語“ライブラリー”または“薬物ライブラリー”は、複数の化学分子(試験化合物)、複数の核酸、複数のペプチドまたは複数のタンパク質、およびこれらの組合せをいう。そのスクリーニングを高処理量スクリーニング法で実施する場合、該方法は、マルチウェルプレートまたは微小流体装置を使用する。
本明細書において用語“結合性分子”または“巨大分子”とは、抗体または不活化酵素を称する。
本明細書において用語“抗体”とは、モノクローナル、ポリクローナルまたは組換え抗体をいう。抗体は、基転移反応の供与体生成物に対して生成させ、供与体生成物と供与体を高供与体濃度の存在において選択的に識別し得る。また、抗体は、ヌクレオチドの少なくとも1つのリン酸部分に対する特異性(即ち、5'-ホスフェート、5'-ホスホスルフェート、5'-ジホスフェートおよび5'-トリホスフェートを識別する能力)を示す。例えば、SULT反応においては、本発明の抗体は、PAPとPAPSを高厳格性でもって識別する。SULT反応における供与体-生成物分子のPAPSは、5'末端リン酸に結合した硫酸基の付加によってのみ異なる。これは、比較的小さい構造上の差異のように見え得るが、1個のリン酸基(スルフェート基に大きさおよび構造において極めて類似している)により異なる分子間を識別する抗体の実証された能力は、本発明の重要な特徴を提供している。例えば、分子のリボシルホスフェート部分に対して抗体を作製することが出来る。さらにまた、本発明のFPIA系アッセイ方法は、過剰のPAPSの存在下において反応生成物PAPに特異的に結合する抗体を適切に必要とする。
【0022】
キナーゼに関しての本発明の実施においては、担体タンパク質に結合させたADPを使用して、ADP分子のホスホ-リボシル部分は免疫反応性に暴露されるがアデニン部分は免疫反応性から大部分が隠蔽されるような方法で、動物内でまたはin vitro組換え法によりADPを特異的に認識するがATPは認識しない抗体を作製することが出来る。抗体の生成においては、アルファおよびベータホスフェート間を架橋するメチレンまたはイオウ基を含有するもののようなADPの非加水分解性アナログを使用して免疫動物中のヌクレオチダーゼまたはホスホターゼによる固定化ハプテンの加水分解を阻止するのが有益であり得る。また、メチルトランスフェラーゼおよびアセチルトランスフェラーゼに関しては、ヌクレオチド以外の小分子、それぞれ、s-アデノシルホモシステインおよび補酵素Aを特異的に認識する抗体を使用する。
本明細書において用語“不活化酵素”とは、供与体生成物に結合し得る結合性分子をいう。キナーゼの場合、ADPが供与体生成物であり、不活化ヌクレオシド二リン酸(NDP)キナーゼが結合性分子である。多くの酵素において、分離されたドメインが結合および触媒作用に関与することが知られている。遺伝子操作、アロステリック阻害、または共同因子、ヘミ基の除去のような他の化学的手段等によって結合特性を保持させながら触媒活性を選択的に破壊することは、酵素、とりわけ多サブユニット酵素がそれら基質に対する特異的な担体(もはやそれらの基質を化学的に改変することができない)として機能することを可能にする。
【0023】
本明細書において用語“抗体-検出可能タグ対”とは、基転移反応において生成した供与体生成物の検出を可能にする抗-供与体生成物抗体および検出可能タグ(蛍光供与体生成物)分子をいう。抗体/検出可能タグ対の結合における適切な解離定数は1x10-7Mまたはそれ以下であり、これにより未結合検出可能タグに対比して最少100mPの蛍光偏光変化をもたらし、且つ反応濃度で供与体分子に対し最少の交叉反応性を有する。最適の抗体-検出可能タグ対は、供与体分子への種々の結合部位、担体タンパク質へ種々のリンカーを使用して生成させ、且つフルオロフォアへの供与体分子結合の化学性を変化させることによって生成させた1群の検出可能タグとの相互作用のための数種の非加水分解性アナログを包含する数多くの種々の抗体を試験することによって同定し得る。例えば結合させたフルオロフォアを有し得る抗体との相互作用時の前記検出可能タグの蛍光または化学発光の変化をそれらの相互作用の尺度として使用し得る。
本明細書において使用するときの用語“リンカー”とは、スペーサーアーム構造を称する。アデニン環の接近性を立体的に最小にする短リンカー(即ち、カルバモイルおよびアミノエチル基)、および抗原のより柔軟性のある提示を可能にするより長めの炭素数6個のリンカーが存在する。リンカー分子は、検出可能タグ特性に、その抗原および蛍光特性の双方に影響を与える多くの重要な形で影響を及ぼす。一般に、抗原をフルオロフォアから十分に引き離して抗体との相互作用が妨害されないようにすることと、フルオロフォアにあまり自由過ぎる動きを生じさせないようにすることの間に注意しなければならない均衡が存在する。前者は親和性抗体結合性の低下およびフルオロフォアの消光をもたらし、一方、後者は抗体結合時の偏光変化を低下させ、それによってアッセイのダイナミックレンジを減じる。種々のリンカーは比較的単純な化学法を使用して取り替えることができる。即ち、リンカーは、検出可能タグの抗原および蛍光特性を最適にするために、多くの方法で変化され得る。さらにまた、ヘテロ二官能性リンカーを含む抗体のPAP結合について説明した方法と同様な方法を使用して、先ずスペーサーアームおよび/または芳香族置換基をPAP上に導入し、次いで反応性フルオレセイン誘導体と反応させることもできる。この方法は、可能性あるリンカー構造の範囲を大いに拡大する。
【0024】
方法および材料
本発明の以下の実験プロトコールは、特定の方法論、プロトコール、抗体、酵素、検出可能タグ、なかんずく、説明する試薬に限定されない;これらは、基転移反応によって変動し得るからである。また、本明細書において使用する用語は特定の実施態様を説明する目的のためだけであり、本発明の範囲を限定するものではないことを理解すべきである。
アッセイ試薬の開発
本発明においてHTSアッセイ法を成功裏に開発するには、アッセイ試薬相互作用の親和性および特異性、並びに得られる検出可能タグ蛍光特性の変化が、感度、ダイナミックレンジおよびシグナル対ノイズ比のようなアッセイの全体的性能を究極的に特徴付ける。それで、この相互作用を最適化するために、出願人等は、図4に示すような反復同時開発方法を提供して基転移反応生成物の検出、定量およびスクリーニングを最も効率的に達成している。
特に、図4は、SULT供与体生成物FPIA用の試薬、即ち、抗-PAP抗体および蛍光標識化PAP検出可能タグの反復同時開発戦略を説明する。図4は、ウサギの免疫用の種々の抗原を作製するためには、PAPを、以下で説明するような種々の長さのリンカー分子を使用し、アデニンのC2、C6およびN6アミノ基を介しての結合により、担体タンパク質(BSAまたはKLH)に結合させ得ることを示している。抗体結合において使用した同じPAP-リンカー-NH2中間体を使用して、検出可能タグとして試験するためのフルオレセイン-PAP結合体を合成することもできる。得られる抗体および検出可能タグのマトリックスは、FPアッセイ法を使用して、結合性について試験することができ、また、高親和性結合および最高FP変化を示す対は、トレーサー特性のさらなる最適化のための基礎として使用することができる。最適化した抗体-検出可能タグ対は、SULT1E1または他のSULTイソ型による硫酸化反応におけるPAP生成の検出について試験することができる。
また、数種のPAP抗原および検出可能タグを、PAP分子のリボシル-ホスフェート部分の抗体認識を最高にし、且つPAPSとの交叉反応性を最小にする全体的構造バイアスを保持させながら、ヌクレオチドを担体タンパク質およびフルオレセインに結合させることによって合成し得ることも考慮される。PAP抗原から生成した抗体は、検出可能タグとの相互作用について試験して、本発明に包含される新規なアッセイ法の最適結合性および蛍光特性を示す組合せを同定し得る。
【0025】
高親和性抗体の生成
本発明の方法に従い、当業者であれば、PAPのリボシル-ホスフェート部分のような抗原性ヌクレオチドまたは補酵素Aのような他の化学分子に対する種々の特異的抗体(即ち、ポリクローナル、モノクローナルまたは組換え)を容易に調製し得る。
一般に、ある範囲のPAP抗原に対するポリクローナル抗体は、ウサギにおいて適切に作製される。動物の注射および血清採集は、以下のプロトコールによって実施し得る。3羽のウサギを、開発する各抗原によって免疫し得る。力価、親和性に依存するが、1羽のウサギからの抗血清収量(約100ml)が数千回〜100万回のFPIAアッセイのために適当であり、ポリクローナル-抗原相互作用の多価性が極めて高い親和結合性を生じさせる。従って、これらの理由により、ポリクローナル抗体が多くの場合FPIAにおいて使用される(Nasir, M. S. and Jolley, M. E., Comb Chem High Throughput Screen, 1999, 2:177-90)。
また、以前の公表プロトコールに提示されている方法(Harlow, E. L., D., 1999)に従って作製し得るモノクローナル抗体も、本発明の範囲に属する。また、組換え単鎖抗体は、以前に公表されたin vitroコンビナトリアルエヴォリューションおよびディスプレイ法(Schaffitzel, C., et al., J Immunol Methods, 1999, 231:119-35;Breitling, F., Dubel, S., 1999)を使用して作製することが出来る。組換え抗体の調製例は、米国特許第5,693,780号、第5,658,570号、第5,876,961号(全て参考として本明細書に合体させる)に記載されている。
免疫応答の顕在化を助けるには、ウシ血清アルブミン(BSA)およびキーホールリンペットヘモシアニン(KLH)をPAP抗原への結合用担体タンパク質として使用し得る。KLH抗原は、一般に、哺乳類においてより強い免疫応答を引き出すが、BSA結合体よりも可溶性が低い傾向がある(Harlow, E. L., D., 1999)。PAPは、図7〜8に示すように、双方の担体タンパク質にアデニンのC2、C8およびN6アミノ基を介して結合させ得る。本発明の範囲において、FPIA系SULTアッセイ法の開発には、反応生成物PAPに過剰のPAPSの存在下において特異的に結合する抗体を必要とする。さらに適切には、前記抗体は、PAP分子のリボシルホスフェート部分を認識し得るものであり、PAPの5'-ホスフェートとPAPSの5'-ホスホスルフェートを識別し得る。
【0026】
結合方法
アデニンへの結合が、アデニン部分への結合により生じ得るPAPSとの抗体交叉反応を最小にすること(Goujon, L., et al., J Immunol Methods, 1998, 218:19-30;Horton, J. K., et al., 1992, J Immunol Methods, 155:31-40;Bredehorst, R., et al., Biochim Biophys Acta, 1981, 652:16-28)、さらにまた、得られる抗体の親和性にハプテン結合部位が影響を与え得ること(Crabbe, P., et al., J Agric Food Chem, 2000, 48:3633-8)が、当該技術において知られている。使用するハプテン結合化学法は、アデニンヌクレオチドの結合において広く使用されている詳しく記述された1または2工程合成方法である(Brodelius, P., et al., Eur J Biochem, 1974, 47:81-9;Lindberg, M. and Mosback, K., Eur J Biochem, 1975, 53:481-6;Camaioni, E., et al., J Med Chem, 1998, 41:183-90)。出発物質は全て商業的に入手可能であり、反応は良好な収率で進行し得ることが理解される。これらの結合方法を以下でさらに詳細に説明する。本発明の方法に従い、本発明は、以下のハプテン結合化学を包含するがそれに限定され得ないものとする
【0027】
N6アミノ基への結合
カルバモイル結合:N6-カルボキシメチル-PAPは、PAPをヨード酢酸によってアルキル化して1-カルボキシメチルアナログを形成させ、これを塩基中で転位させてN6-カルボキシメチルアナログを形成させることによって調製し得る(Lindberg, M. and Mosback, K., Eur J Biochem, 1975, 53:481-6)。N6-カルボニルメチル-PAPは、N-ヒドロキシスルホスクシンイミドにより促進させたカルボジイミドカップリングを使用して、担体タンパク質に結合させ得る。同様な方法を使用し、以前に公表されたプロトコールに記載されているようにして高特異的5'-AMP抗体を作製することが出来る(Bredehorst, R., et al., Biochim Biophys Acta, 1981, 652:16-28)。
C6結合:N6-アミノヘキシル-PAP (Sigma/Aldrich社、ミズーリ州セントルイス)は、a) N-スクシンイミジルS‐アセチルチオアセテート(Pierce社、イリノイ州ロックフォード)を使用して、タンパク質アミノ基(ほぼリジン)を反応性チオールに転換し、b) ヘテロ二官能性アミン-チオール反応性リンカー(Pierce社、イリノイ州ロックフォード)を使用して、PAPリンカー上の末端アミノ基を前記タンパク質チオールに結合させることによって、担体タンパク質に直接結合させ得る。特筆すべきは、アデノシンのN6アミノ基は実用目的上は反応性ではないことである;例えば、該アミノ基は、スクシンイミジルエステルとは極めて貧弱にしか反応しない。
【0028】
アデノシンC2への結合
2-クロロアデノシンをオキシ塩化リンと、次いで三塩化リンと反応させることによってリン酸化して、3',5'-ビスホスホ-クロロアデノシンを調製し得る。その後のエチレンジアミンおよびジアミノヘキサンとの反応並びにアニオン交換クロマトグラフィーによる精製を使用してアデニンの2位置におけるPAPの2-アミノエチル-および6-アミノヘキシル-アナログを調製し得る(Brodelius, P., et al., Eur J Biochem, 1974, 47:81-9)。これらの分子は、上述したようなヘテロ二官能性リンカーを使用して担体タンパク質と反応し得ることも意図する。
アデノシンC8への結合
PAPの8位置へのC2およびC6結合は、C2位置において説明したのと同じ方法を使用して生成することが出来るが、出発物質は8-ブロモアデノシン(Sigma/Aldrich社)であり得る。また、8-アジド-PAP (ICN社、カリフォルニア州コスタメサ)も、UV照射により、担体タンパク質に直接結合させ得る。
薄層クロマトグラフィー(TLC)は、アデニン環吸収スペクトル変化をモニターするのに使用し得る。アニオン交換(Dowex樹脂)および相分離は、抗原精製に使用し得る。高い抗原純度を確保するために、担体タンパク質に結合させるのに使用する最終PAP-リンカー分子をHPLCによって精製し、その存在性を質量スペクトルおよびNMR分析によって検証することができる。メチル化担体タンパク質の使用がリボース環とのカルボジイミド反応を阻止することが報告されているので(Bredehorst、 R., et al., Biochim Biophys Acta, 1981, 652:16-28)、N6-カルボキシメチル-PAPをメチル化BSAに結合させてもよい。
他のPAPアナログは、全てリンカー末端に反応性アミンを有する傾向にあり、これらは、より特異的なアミンおよびチオール化学物を使用して、BSAおよびKLHへの結合に使用することができ、それによってリボースとの反応を回避することができる。タンパク質アミンのチオールへの転化およびヘテロ二官能性リンカーを介しての結合は、担体タンパク質中にシステインよりも多くの反応性リジンが存在するので、システインへの直接結合よりも高密度の抗原結合をもたらし得る。担体タンパク質上のPAP密度はアデニン環についての質量分光分析によって測定でき、結合化学性を必要に応じて調整して高密度(タンパク質分子当り> 10のPAP)を得ることができ、それによって強い免疫応答が担保される。全部で約8〜10種の異なる抗原を合成することができ、幾つかはアデニン環の接近性を立体的に最小にするカルバモイルおよびアミノエチル基のような極めて短いリンカーを有し、また、抗原のより柔軟な提示を可能にする、より長めの炭素数6個のリンカーを有することが想定される。
【0029】
蛍光PAP分子の合成および精製
一般に、FPIA検出可能タグ分子は、3つの異なる構成部分、即ち、抗原、フルオロフォアおよびこれらを結合させるのに使用するリンカーに分けることができる;さらなる主要な構造的変数は、抗原へのリンカーの結合部位である。当業者であれば、適切な抗原、フルオロフォアおよびリンカーが本明細書において詳細に説明しているものに限定されないことを理解されたい。しかしながら、本発明の適切な局面においては、フルオロフォア、フルオレセインは、それを、担体タンパク質への抗原結合に使用したのと同じアデノシン上の部位に結合させるのに使用し得る。担体タンパク質への結合に使用するのと同じPAP-リンカー分子を、数種のアミン反応性フルオレセイン誘導体との結合に使用し得る。ある範囲の反応性フルオレセインの使用は、リンカー領域にさらなる構造的多様性を導入し得る。例えば、フルオレセインスクシンイミジルエステルは、種々の長さのスペーサーアームでもって利用でき、DTAF (4,6-ジクロロトリアジニルアミノフルオレセイン)は結合部位に隣接した嵩高で平面的な置換基を含有する。同様に、担体タンパク質結合部位近くでのAMPへの芳香環の導入(N6-ベンジルAMP)は、その抗体に対する親和性を未修飾AMPに対比して40倍増強する(Bredehorst, R., et al., Biochim Biophys Acta, 1981, 652:16-28)。簡単に言えば、アミノ末端を有する種々のPAP-リンカー分子はアミン反応性ローダミン、テキサスレッドまたは他の赤色フルオロフォアと反応させることができ、得られるPAP-フルオロフォア結合体は、抗体結合親和性および蛍光偏光の変化について試験することができる。
PAP結合体の精製は、薄層クロマトグラフィー(TLC)によって実施し得る。複数の分離を並行して操作することが可能であり、蛍光生成物の同定が目視により可能であり、さらに1回のクロマトグラフィー操作で数千もの結合性アッセイのための十分な量の蛍光化結合体が提供されるので、反応生成物の分離にはTLCが使用される。即ち、蛍光検出可能タグ分子のパイロット試験においては、TLCは、HPLCのような別の分離方法よりも優れている。PAP-フルオロフォア結合体のクロマトグラフィーおよび溶出が終了したならば、後述するようにして、強度、基底偏光(抗体なしでの)、安定性、抗-PAP抗体に対する結合性、並びに未標識PAPおよびPAPSによる拮抗性について結合体を分析することができる。
【0030】
抗体-検出可能タグ相互作用に影響するパラメーター
基転移方法の生成物を検出し、定量し、スクリーニングするための最も感度のあるアッセイ方法を創生するためには、抗体-検出可能タグ相互作用の親和性に影響するパラメーターを注意深くモニターする。このことは、例えば、ある種の受容体基質が示す強力な基質阻害によって、生成させ得る生成物の量が抑制されるSULT1E1の場合に重要である。本発明に属する新規なSULTアッセイにおいて必要な親和性は、高親和性で阻害性基質のエストラジオールの阻害例を考慮することによって推定する。一般に、エストラジオールのKiは80nMであり、Kmは約20nMであり(Zhang, H., et al., J Biol Chem, 1998, 273:10888-92)、従って、アッセイにおいては50nMのエストラジオールを使用し、酵素および反応時間を10%の基質が消費される(5nMのPAPの生成をもたらす)ように調整することが、有意の阻害を起すことなく初期速度の測定を可能にし得る良好な妥協点である。
5nMのPAPの検出を可能にするためには、蛍光-PAP(および未結合PAP)に対する抗体結合に必要な親和性は、およそ25nMであろう。殆どの抗体-抗原相互作用は、ポリクローナル抗体またはモノクローナル抗体のいずれであれ、1nM〜1μMのKdを有する(Harlow, E. L., D., 1999)。例えば、FPIA系キナーゼアッセイにおいて使用する抗-ホスホチロシン抗体は、リン酸化ペプチドおよびそれらの蛍光結合体に対しナノモル以下の親和性を示し(Parker, G. J., et al., J Biomol Screen, 2000, 5:77-88);上述した抗-AMPポリクローナル抗体は、100nMの親和性を有するヌクレオチドに結合した(Bredehorst, R., et al., Biochim Biophys Acta, 1981, 652:16-28)。抗原の構造が得られる抗体の親和性に影響を与えるかどうかを予測することは困難であるが、担体タンパク質および結合方式の双方は、大きな効果を有し得る(Crabbe, P., et al., J Agric Food Chem, 2000, 48:3633-8;Signorini, N., et al., Chem Res Toxcol, 1998, 11:1169-75;Oda, M. and Azuma, T., Mol Immunol, 2000, 37:1111-22)。従って、PAP上の異なる部位を介して2つの異なるタンパク質に結合させたPAPを使用して、さらに、異なる長さのリンカー領域を使用して、抗体を作製することが出来る。
【0031】
PAP検出の感度に影響を与えるもう1つの要因は、高濃度においてさえも極めて緩慢にしか硫酸化されない貧弱な基質である。これらの場合、必要とするSULT1E1酵素の量は、コスト性故に限界要因となる。エストラジオールによりVmaxの1%の速度で回転させる基質を使用すれば、5nMのPAP(約30分内での適切な検出限界)を生成させるのに30ngのSULT1E1を必要とするであろう。この量は、100mgの酵素バッチが300万回よりも多い反応に対処し得ることを意味し得るので、良好に許容範囲内にある。
さらにまた、遊離または結合状態のプローブの偏光の差異は、アッセイの全体的“広がり”即ちダイナミックレンジを特徴付ける。50mP未満の変化でSULT1E1活性の半定量検出に十分であり得るが、100mPまたはそれ以上の変化は、アッセイ方式の設計におけるはるかに大きい柔軟性およびより定量的な動的情報の双方を提供し得る。従って、偏光は分子容積に比例し、小分子量フルオロフォアへの抗体(150 kDa)結合時の有効容積の変化は、少なくとも300 mPの増加を生じさせることが予期し得る。しかしながら、フルオロフォアをPAPに可撓性スペーサーアームにより結合させることのような、遊離および結合検出可能タグの双方の観測偏光に影響を与え得る他の要因も存在する。即ち、それらの最高偏光は、抗体結合がスペーサーおよびフルオロフォアの移動性を低下させる度合の関数である;検出可能タグ“プロペラ効果”は、タンパク質結合フルオロフォアについてさえも比較的低い偏光値をもたらし得る。
さらなる考察点は、重複スペクトルを有するアミノ酸官能基からの消光を含む、レセプターへの結合時のフルオロフォアの強度の変化が存在し得るかどうかである。偏光は、蛍光強度に依存しない(Owicki, J. C., J Biomol Screen, 2000, 5:297-306);しかしながら、偏光は2つの平面で取った強度測定値の比であるので、有意の消光は、抗体検出可能タグ相互作用に影響を与え、アッセイ感度の低下をもたらし得る。抗体と結合したときの検出可能タグの蛍光特性の変化は、各相互作用分子双方の関数であり、予測するのは困難である。即ち、種々の抗体および検出可能タグ分子はマトリックスの形で調べられるであろうことが考慮される。さらに、検出可能タグの構造の最適化、或いは、結合させた検出可能タグの可動性を制限するためまたは消光を減じるためのアッセイ緩衝液の最適化も考慮される。
【0032】
最適抗体/検出可能タグ対(1種以上)の同定
最適抗体/検出可能タグ対を同定するためには、多様な組合せのフルオレセイン誘導体と結合化学物を一般に評価する。上述した合成および精製方法は、反応毎に数種の蛍光生成物の単離をもたらし、その結果、少なくとも100種の蛍光結合体を評価することができるが、数千の個々のFPアッセイを必要とする。FPアッセイの均質特性およびマルチウェル器具を利用できることは、スクリーニング操作を極めて迅速なものにしている;律速段階は、通常、合成と精製であり、抗体-検出可能タグ結合特性についての試験ではない。抗体/検出可能タグ試験は、Tecan UltraマルチウェルFPリーダーにおいて、Trisまたはリン酸緩衝液中で非特異相互作用を阻止するための低濃度の担体タンパク質(0.01%ウシガンマグロブリン)の存在下に実施し得る。初期スクリーニングは、以下の望ましい特性を見出すこと含む:抗体不存在での低偏光(<100mP);抗-PAP抗体への高親和結合性(Kd < 100nM);結合および遊離状態における偏光の最高差異(ΔmP >100mP);抗体に結合した場合の最少蛍光消失;PAPに対する選択性(即ち、PAPによる、PAPSまたは他のアデニンヌクレオチドによらない拮抗的置換);迅速な結合/解離速度;および、SULT1E1または他のアッセイ成分との相互作用の無いこと。
初期抗体結合性アッセイにおいては、精製した検出可能タグの各々を、結合フルオレセインの強度が消光されないことを想定して、蛍光強度を測定することによって定量し得る。常に該当するものではないが、有意の消光を有するものは、それらが低結合親和性を有するようなので、結合性分析によりそれらを排除することができる。見込みのある親和性(Kd < 100nM)および蛍光特性を有する抗体/検出可能タグ対は、未標識PAPおよびPAPSおよび他のヌクレオチドとの拮抗により、結合特異性について試験し得る。最も望ましいFPIA特性を有する抗体/検出可能タグ対は、さらに詳細に試験して、塩または変性剤の添加のような外因性条件における何らかの変化がその感度およびダイナミックレンジを改善し得るかどうかを評価し得る。さらなる検出可能タグを必要に応じて合成し、観測した構造-活性関係およびコンピュータ作成構造モデル予測に基づき試験し得る。
【0033】
本発明の応用性
本発明の方法は、後の実施例において説明するように、基転移反応における酵素活性の尺度として使用し得る。本発明の方法は、例えば、酵素モジュレーター、とりわけ阻害因子として作用し得る薬物の同定のため、また、潜在的薬物分子がどのように代謝されるかを決定するために、高処理量スクリーニング試験所における基転移酵素の分析を可能とするので、製薬工業においてとりわけの重要性を有する。
新たな治療薬の開発において目標とする基転移酵素の数が増大している。キナーゼは現在最高に興味あるものであるが、SULT、UGT、メチルトランスフェラーゼ、アセチルトランスフェラーゼおよびADP-リボシルトランスフェラーゼも対象とされている。このリストが増大するにつれ、各酵素についての或いは同じ受容体基質を改変する小下位群の酵素についての特異性試験を使用することは、現実性の無い極めて費用高となるであろう。本発明のアッセイ法とキットは、与えられた基転移酵素群の全ての酵素の検出を可能にし、これら酵素の全てについての一般的な評価項目検出系を提供する。このことは、とりわけ高処理量スクリーニング検査室においての使用の容易性を大きくし、ロボットに全ての検出試薬を備え付けることができ、各種基転移酵素および興味ある潜在的基質または阻害因子をマトリックス形式で試験し得る。
さらに、薬物および他の生体異物の共役的代謝における基転移反応の関与は、それらの薬力学、薬物動態学および副作用レベルの重要な決定因子である。この点、本発明のアッセイ法およびキットは、潜在的な薬物分子のSULT、UGT、メチルトランスフェラーゼおよびアセチルトランスフェラーゼによるスクリーニングをHTS方式で可能にする。多くの種々の生体異物共役イソ型が存在し、本発明方法の汎用的な性質故に、本発明のキットは、マトリックス形式のそれらの試験を大いに簡素化するであろう。
即ち、本発明の汎用的方法は、後述する統合化(integrated)薬物発見方法に貢献するために、ステロイドホルモン生体合成および機能、生体異物代謝、酵素レセプター調節およびシグナル伝達のような各種の異なる基転移関連酵素過程に応用し得る。
【0034】
薬物発見における硫酸化
硫酸化は、内在性ホルモンおよび生体異物のレベルおよび活性を調節する普遍的な共有結合性修飾である。ヒトにおいては、硫酸化は、各々異なるが重複した基質特異性と組織分布を有する1群の11種のスルホトランスフェラーゼ酵素(SULT)によって触媒されている(Strott, C. A., Endocr Rev, 2002, 23:703-32)。硫酸化における内在性基質としては、ステロイドホルモン、カテコールアミン、および甲状腺ホルモンのような多くの重要な情報伝達分子があり;硫酸化される生体異物としては、薬物、前駆型変異原(promutagen)、環境性内分泌撹乱物質がある(Strott, C. A., Endocr Rev, 2002, 23:703-32;Glatt, H., et al., Mutat Res, 2001, 482:27-40;Coughtrie, M. W. and Johnston, L. E., Drug Metab Dispos, 2001, 29:522-8)。硫酸化は、可逆性であり、情報伝達分子の活性を、多くの場合それら分子のレセプタータンパク質に対する親和性を改変することによって変化させ得る、即ち、硫酸化は、レセプターリガンドに対するオン-オフスイッチとして、リン酸化がその役割をタンパク質に対して果たすのと同程度に作動する。さらにまた、生体異物代謝における硫酸化の役割は、ホルモン情報伝達および細胞ホメオスタシスの調節におけるその関与と密接に絡み合っている。例えば、硫酸化は、うつ病、乳がんおよび心臓血管疾患用の薬物ターゲットとしての特異的神経伝達物質レセプター、核レセプターおよびプロテインキナーゼに対する内在性リガンドの活性を調節する(Plassart-Schiess, E. and Baulieu, E. E., Brain Res Rev, 2001, 37:133-40;Strott, C. A., Endocr Rev, 1996, 17:670-97;Kuroki, T., et al., Mutat Res, 2000, 462:189-95)。製薬企業によるこれらの障害に対するより選択性の治療薬の開発は、潜在的SLUT基質および阻害因子の高処理量スクリーニング(HTS)のための分子アッセイ法および精製SULTイソ型が無いために、現在のところ妨げられている。
【0035】
溶解性、大きさ、小細胞分布において異なり、20%未満の配列相同性を共有する2つの主要なクラスのヒトのSULTが存在する。膜結合酵素は、ゴルジ体内に存在し、ヘパリン、グリコサミノグリカンおよびタンパク質チロシンのような大内在性分子をスルホン化する(Strott, C. A., Endocr Rev, 2002, 23:703-32)。細胞質内スルホトランスフェラーゼ即ちSULTは、小分子量生体異物およびホルモンをスルホン化する。
11種の既知の細胞質内SULTイソ型が存在し、これらはその組織分布および特異性において異なる。使用されている命名法は、アミノ酸レベルでの相同性に基づいている:数字で表わされる同じ群内のSULTは少なくとも45%のアミノ酸相同性を共有し、文字で表される同じ下位群内のSULTは少なくとも60%の相同性を共有する。SULTは、ある程度は、基質特異性に基づいて分類することができるが、それらの基質プロフィールは、異なる群内の酵素間においてさえも、広範囲に重なり合う。
構造研究により、基質結合部位に柔軟性があること並びにリガンド結合性がより秩序立った構造への転換を引き出すことが明らかにされており、これらがSULTの広い基質特異性に寄与し得る(Bidwell, L.M., et al., J Mol Bil, 1999, 293:521-30)。こういう理由で、分子結合性のようなコンピュータ手法をSULT基質および阻害因子の同定に応用できるということはなさそうである;このことは、本発明に属するHTSアッセイ法の価値を増大させる。
【0036】
本発明のアッセイ法は、HTS方式において使用することができ、例えば、注目する化合物が1種以上のSULTイソ型と相互作用するかどうか、該化合物が基質または阻害因子であるかどうか、および相互作用における動的パラメーター(IC50、Km、Vmax)のようなSULT代謝情報を提供し得るということが考慮されている。本発明のHTSアッセイは、以下の2つの異なる方式でスクリーニングすることによって、前記情報の全てを提供することが想定されている:a)試験化合物の代謝回転の直接測定;およびb)プローブ基質硫酸化の化合物阻害の測定。直接代謝回転方式においては、化合物をSULTイソ型の集団に対してスクリーニングし、個々のSULTイソ酵素による化合物硫酸化をプロフィール化することができるであろう。イソ酵素同定後、適切なSULTイソ酵素によるより詳細なキネティクス試験を使用して生体内クリアランス速度を予測することができ(Obach, R. S., et al., J Pharmacol Exp Ther, 1997, 283:46-58)、それにより臨床試験おける化合物の不合格率を減じることができるであろう(Greco, G. N., E; Martin, YC, 1998, 219-245)。既知の基質(例えば、SULT1E1に対するエストラジオール)を使用し、試験化合物を阻害についてスクリーニングすることによって、SULT阻害因子を同定し得るであろう。イソ酵素特異的なキネティクスおよび阻害データは、特異的SULTによる代謝の知識を提供することにより、可能性のある薬物-薬物相互作用および薬理遺伝学問題を薬物発見チームに対して警告し、薬物発見をさらに改善するであろう。治療薬に対する種々の応答へのSULTにおける遺伝子的差異の寄与は、研究の活動的な領域である(Thomae, B. A., et al., Pharmacogenomics J, 2002, 2:48-56)。
薬物代謝に関与するSULTの同定は、潜在的な薬物-薬物相互作用の前臨床評価および毒物試験において使用する生体外アッセイ法または動物モデルの賢明な選択を促し、それによって不適切な或いは不必要な実験動物の使用を減じるであろう。代謝データは、合理的な薬物設計および先導的最適化の要素として使用し得る。種々のSULTイソ酵素に対する基質特異性を特徴付ける構造-活性関係のより良好な理解は、その代謝プロフィールを変化させる一次化合物の構造的修飾のための根拠を提供し得る(Greco, G. N., E; Martin, YC, 1998, 219-245)。従って、本発明の方法は、種々のSULTイソ酵素に対する基質特異性を規定する構造-活性関係のより良好な理解を可能にするであろう。
【0037】
薬物発見への統合化取り組み
本発明のアッセイ法がHTSアッセイとして使用できるということは、例えば、硫酸化が関連シグナル伝達生物学の要素である治療領域における薬物開発への合理的な統合化手段が可能になる。本発明のHTS方法を使用し得る幾つかの特定の領域としては、例えば、ステロイドホルモン系療法がある。硫酸化は、本発明によれば、乳腺腫瘍におけるエストロゲンレベル調節並びに前立腺がんにおけるアンドロゲンレベルへの関与にまで及ぶ。さらにまた、コルチゾール硫酸化は、よくは理解されてないものの、前記ホルモンをグルココルチコイドレセプターへの結合に対して不活化する。ステロイドリガンド-レセプター結合性についての高処理生化学アッセイおよび得られる転写調節補因子タンパク質結合性は、すでに商業的に利用可能であり、製薬HSTグループによって使用されてより選択性のステロイドホルモンモジュレーターが見出されている(Parker, G. J., et al., J Biomol Screen, 2000, 5:77-88;Spencer, T. A., et al., J Med Chem, 2001, 44:886-97)。従って、ステロイド硫酸化についての確固たるHTSアッセイ法を利用し得ることは、ステロイドシグナル伝達に絞った製薬グループにとって利用可能な分子手段備蓄への重要な追加物を提供し得る。例えば、ERをターゲットする化合物によってSULT1E1を阻害することは、腫瘍細胞中のエストロゲンレベルを上昇させることによる有害な効果を生じることがあった。SULT1E1阻害特性を有する分子は、本発明のアッセイ方法を使用した研究に基づいて選択或いは修飾することができるであろう。
神経ステロイドの調節は、神経興奮バランスを制御するための新規な薬理学的方法として研究されている(Malayev、 A., et al.,Br J Pharmacol, 2002, 135:901-9;Maurice, T., et al., Brain Res Res Brain Rev, 2001, 37:116-32;Park-Chung, M., et al., Brain Res, 1999,830:72-87)。本発明に包含される方法は、スルホ結合についての内在性および合成神経ステロイドの容易なスクリーニングを可能にし、基本的生物学に洞察力をもたらし、さらに、リード分子の同定および最適化のための手段を提供することによって、これらの努力を適切に促進させ得る。より良好な分子手段の必要性は、年齢関連の老化および記憶喪失を軽減することを目的とする“老化防止”栄養補助食品としてのDHEAのかなり大きい対抗市場がすでに存在するという事実によって強調される(Salek, F. S., et al., J Clin Pharmacol, 2002, 42)。
さらにまた、本発明の方法は、コレステロール流出、血小板凝集および心臓血管疾患の治療における皮膚発症、おそらくは、ある形状の皮膚がんの調節における硫酸コレステロールに関連する薬物ターゲットを適切に同定し得ることを意図する。この場合、スルホトランスフェラーゼ(おそらく、SULT2B1b)が薬物ターゲットとなり得、このイソ型を選択的に阻害する分子を同定する必要があり得る。ヒトSLUTの完全パネルおよび本発明の堅牢なHTSアッセイ法を利用し得ることは、そのような取組みにとって有益であり得る。
【0038】
薬物発見におけるグルクロン酸化
毒性代謝物の生成および好ましくない薬物動態のような薬物代謝問題は、臨床試験中に全ての薬物候補のほぼ半分の不合格をもたらしている(Obach, R.S., et al., J Pharmacol Exp Ther, 1997, 283:46-58)。大きな製薬企業は、全て、薬物動態学および薬理ゲノム学結果を薬物発見プロセスの早期に考察する必要性を認識しており、薬物代謝を評価するための高処理量in vitro方法の当面の必要性を生じている。P450依存性酸化に加えて、グルクロン酸化は、おそらく、肝臓薬物代謝の最も重要な経路である。非ステロイド系抗炎症剤、オピオイド、抗ヒスタミン剤、抗神経病薬および抗うつ薬のような広範囲の薬物は、グルクロン酸化によって排除または活性化される(Meech, R. and Mackenzie, P. I., Clin Exp Pharmacol Physiol, 1997, 24:907-15;Radominska-Pandya, A., et al., Drug Metab Rev, 1999, 31:817-99)。その重要性にもかかわらず、グルクロン酸化を触媒する肝臓UDP-グルクロノシルトランスフェラーゼ(UGT)の広く重なり合う基質特異性は、柔軟性のあるin vitroアッセイ方法が無いために、理解に乏しいままである。その主たる理由は、使用する触媒アッセイ法が基質特異性クロマトグラフィー工程を含む生成物からの反応物の分離を必要とし、従って、HTS方式においては現実的でないことである。
2種のUGTファミリー(例えば、UGT1およびUGT2)がヒトにおいて同定されている;これらのファミリーのメンバーは一次アミノ酸配列において50%未満の同一性しかないものの、これらは、基質特異性において有意の重なり合いを示す。生体異物代謝の大部分が起こるヒト肝臓内で発現するUGT1群のメンバーとしては、UGT 1A1、1A3、1A4、1A6および1A9がある。UGT2群は広くは研究されていないものの、UGT 2B4、2B7、2B10、2B11および2B15が肝臓内で発現することが知られている。UGTの変異は、5〜12%の頻度で発症し(Weber, W.、1997年)、神経毒症状、重篤な場合には、死に至る高ビリルビン血症のような有害作用を有することが知られている。P450のような他の薬物代謝性酵素における場合と同様に、UGT発現レベルの個人間の差が観察されており、薬物応答の差異に関連している(Iyer, L., et al., J Clin Invest, 1998, 101:874-54)。例えば、ギルバート症候群患者におけるようなUGT1A1の低発現は、有望な抗がん剤であるイリノテカン(Irinotecan)の毒性に関連している(Wasserman, E., et al., Ann Oncol, 1997, 8:1049-51)。さらに、腫瘍組織におけるUGTアップ調節は、抗がん薬抵抗性の可能性ある原因として同定されている(Franklin, T. J., et al., Cancer Res, 1996, 56:984-7;Takahashi, T., et al., Jpn J Cancer Res, 1997, 88:1211-7)。
【0039】
既知のUGTの全てが広い基質特異性を示しており、単一のイソ酵素が広範囲の構造的に関連の無い化合物のグルクロン酸化を触媒する;驚くに当たらないが、UGTイソ酵素の特異性においても大きな重なり合いが存在する(Radominska-Pandya, A., et al., Drug Metab Rev, 1999, 31:817-99)。内在性分子の生体転換に関しては、UGT1A1は、明らかに、テトラピロール、ビリルビンのグルクロン酸化に関与し、その排出をもたらす主要イソ型である。これ以上に特異性に関しての総括を行うことは、最近同定されたイソ型の殆どにおいて系統的な試験が無いために困難である。数多くの内在性ステロイドは、1A1、1A3、1A4、2B4、2B7および2B15のような肝臓イソ型の殆どにおいてアグリコンとして同定されている。脂質および胆汁酸は、2B4および2B7に対する基質として作用し、最近、レチノイドが双方の群に由来する幾つかのイソ型に対する基質として同定されている。既知の生体異物アグリコンの構造的多様性は、極めて広い;それには、イミプラミンのような第3級アミン、アセトミノフェンおよびナプロキセンのような非ステロイド系抗炎症剤(NSAID)、モルフィンおよびコデインのようなオピオイド、およびクロフィビリン酸のようなカルボン酸含有薬物を含む薬物および薬物様分子がある。
短期的には、製薬企業は、潜在的薬物候補が生体内でグルクロン酸化されるのかどうか、そうであれば、UGTイソ型によってグルクロン酸化されるかどうかを判定するためのより良好な方法の当面の必要性を有する。そして、長期的には、グルクロン酸化による薬物代謝を予測することができることの開発は、主要肝臓UGTの各々が認識する“化学空間”を十分に規定する系統的取組みを必要とするであろう。提案された第II段階研究は、大量の多様な化学物を分離したUGTイソ型による結合性と代謝について迅速にスクリーニングするのに使用し得るHTSアッセイ方法のようなこの取組みに必要な分子手段を産み出すであろう。
【0040】
医薬の研究と開発は、時間を消費するもので、費用が高く、非効率であり、高いコストまたは米国医療消費者における新治療薬の低利用性のいずれかをもたらす。現在、米国における新薬の開発には、10〜15年間と4億〜7.5億ドルの総研究開経費を要する。臨床試験は、典型的には総研究開発コストの30〜50%を占める最も費用の掛かる開発期間であるが、臨床試験において試験された全薬物候補の僅か10%しか、最終的には商業化されない(Obach, R.S., et al., J Pharmacol Exp Ther, 1997, 283:46-58)。さらにまた、臨床試験中の薬物候補脱落理由の分析によれば、化合物の成功または失敗の主要決定因子の幾つかは、クリアランス速度(薬物動態)を含むその代謝機能、他の薬物代謝による干渉の可能性および毒性代謝物生成の可能性であることが確認されている。貧弱な薬物動態(通常、過剰に速いクリアランス)と毒性との合計が全臨床失敗例の50%を上回る原因となっている。潜在的薬物候補が代謝されるかどうかについてのより良好な理解が発見段階の早期で得られることは、前臨床および臨床試験の成功率を改善し、より効率的な薬物開発並びに治療薬の経済性と利用性の増大をもたらすであろう。
UGTにおけるイムノアッセイ(即ち、FPIAベースの供与体生成物アッセイ)は、潜在的薬物候補が既知のUGTイソ型のいずれと相互作用するかどうかを判定するのに、SLUTについて説明した方法と極めて類似する方法で使用されることが意図されている。本発明の方法を使用することにより、注目する化合物が1種以上のUGTイソ型と相互作用するのかどうか、そうであれば、それは基質または阻害因子のいずれであるのかを判定することが可能であろう。また、注目する化合物と酵素イソ型間の相互作用における動的パラメーター(IC50、Km、Vmax)も同定し得る。組換え型の多くのUGTイソ型がすでに入手可能であることに留意すべきである(Invitrogen、Becton-Dickinson社)。これらのHTSアッセイにおいて得られた情報は、次のような方法で使用し得る、例えば、イソ酵素同定後、適切なUGTイソ酵素によるさらに詳細な動的試験を使用して生体内クリアランス速度を予測し、貧弱な薬物動態特性のために臨床試験で不合格となる化合物の数を減らし得る。また、UGTレベルにおける遺伝的相違が治療薬に対する種々の応答を変動させる重要な要因として認識されているので、特異的UGTによる代謝についての知識は、可能性のある薬物動態問題を薬物発見チームに警告する。さらにまた、薬物代謝に関与するUGTの同定は、薬物-薬物相互作用の前臨床評価および毒物試験において使用する生体外アッセイ法または動物モデルの賢明な選択を促し、それによって不適切な或いは不必要な実験動物の使用を減じるであろう。また、代謝データは、合理的な薬物設計の要素としても使用し得る。種々のUGTイソ酵素に対する基質特異性を規定する構造-活性関係のより良好な理解は、一次化合物の代謝プロフィールを変化させるその一次化合物構造的修飾のための根拠を提供するであろう。また、グルクロン酸化化合物の試験は、体内で活性型として代謝されるまでは不活性である価値あるプロドラッグの発見ももたらし得る。
【0041】
薬物発見におけるプロテインキナーゼ
ヒトゲノム中にコードされた400種を超える種々のキナーゼが存在する;疾病におけるそれらの役割を解明し、選択性の阻害因子を同定することは、大きな製薬上の主導事項である。キナーゼ機能不全は、がん、心臓血管疾病、炎症、神経変性疾患、および代謝障害のような最も重要な治療領域の全てに関連している。さらにまた、薬物ターゲットとしてのキナーゼの臨床上の検証は、それぞれ、乳がんおよび白血病に寄与する異所性チロシンキナーゼを阻害するHerceptinおよびGleevecの事例において最近示されている。高処理量スクリーニング(HTS) (薬物ターゲットによる相互作用についての何千もの化合物の並行試験)は、薬物発見の優勢な方式となってきている。2002年のHTSアッセイ試薬の総市場は4.74億ドルであり、およそ20%のスクリーニングがプロテインキナーゼにおいて実施された。高レベルの興味にもかかわらず、キナーゼをHTSプログラムに組み入れる製薬上の取組みは、前記アッセイ法による欠点により妨げられている。最も一般的に使用されるHTSキナーゼアッセイ法は、個々のキナーゼの基質特異性によって異なるリン酸化ペプチド反応生成物の蛍光系免疫検出に依存している。即ち、時間のかかる試薬開発が各キナーゼまたはキナーゼ群毎に必要となり、アッセイ間の結果の比較にも問題がある。この技術的障害を克服するために、本発明は、全てのキナーゼ反応の生成物であるアデノシンジホスフェート(ADP)の検出のためのFPIAを提供する。このアッセイ法は、如何なるキナーゼおよび如何なる受容体基質とも一緒に使用し得る普遍的な触媒アッセイを提供することによって、キナーゼ基質特異性を明らかにし新規な阻害因子を同定する取組みを促進させるであろう。
【0042】
プロテインキナーゼは、シグナル伝達において主要な役割を有する広くて多様なファミリーである。プロテインキナーゼは、ATPまたはGTPからの末端リン酸基を受容体タンパク質のセリン、スレオニンまたはチロシン残基への転移を触媒し、ヒトゲノムにおける最大のタンパク質ファミリーの1つである。最も広い意味においては、プロテインキナーゼは、セリン/スレオニンまたはチロシンキナーゼおよび可溶性酵素または膜貫通性レセプターとして分類し得る。最も最近の網羅的なゲノム分析においては、428種のヒトキナーゼが同定されており、これらは8種の異なる相同性群を含み、これらの相同性群は基質特異性、構造/局在化および/または調節の方式も反映したものである(Hanks, S. K., Genome Biol, 2003, 4:111)。例えば、EGFRおよびPDGRRのような膜貫通性成長因子レセプター並びにsrcキナーゼのような可溶性酵素の双方を含む84メンバーからなるチロシンキナーゼ群;61メンバーの環状ヌクレオチド依存性群、脂質依存性キナーゼ(PKCイソ型)を含むser/thrキナーゼ;および、45メンバーの有糸分裂促進性MAPキナーゼ情報伝達経路の成分を含む45メンバーの“STE”群が存在する。
キナーゼは、細胞内シグナル伝達経路の普遍的な調節因子であり、従って、広範囲の疾患および障害のより選択性の治療薬について研究中の製薬企業による強烈な関心の的となっている;キナーゼは、製薬優先順位付けの点でGプロテイン結合レセプターに次いで第2位である(Cohen, P., Nat Rev Drug Discov, 2002, 1:309-15)。リン酸化における細胞内ターゲットとしては、他のキナーゼ、転写因子;アクチンおよびチューブリンのような構造タンパク質;DNA複製および転写、およびタンパク質翻訳に関与する酵素;並びに代謝酵素がある(Cohen, P., Trends Biochem Sci, 2000, 25:596-601)。リン酸化は、タンパク質触媒活性、特異性、安定性、局在化および他の生体分子との結合に変化を生じさせることがある。タンパク質上の複数の部位での同時リン酸化は、異なる機能的結果を伴い、情報伝達経路の統合にとって一般的で中枢的である。
【0043】
リン酸化部位の多様性
各キナーゼは、1種以上の標的タンパク質を時には複数の部位でリン酸化することがあり、触媒活性または他の生体分子との相互作用を制御する1つ以上の調節ドメイン内で自己リン酸化することがある。健常および疾病状態からの細胞リン酸化プロフィールの機能的結果を明らかにすることはプロテオミクスの主たる第1歩である。しかしながら、この知識をどのキナーゼを薬物発見の標的とすべきかを決定するのに使用するには、その受容体基質に対する特異性も描き出さなければならない。キナーゼは、多くの場合ベータ屈曲部で生じているそれらの標的タンパク質の特異的直線配列を認識する。一般に、リン酸化残基の両側に隣接する3〜5個の残基のアミノ酸がリン酸化部位を決定している。既知のキナーゼリン酸化部位を蓄積しているPhophoBaseデータベースは、133種のヒトキナーゼについてのエントリーを収容しているが、これは全キナーゼの1/3に満たない。さらにまた、全部でないにしても殆どのこれらの特異性プロフィールは、各キナーゼについて、基質として同定されている1種または2種のペプチドしか示していないので不完全である。関連キナーゼ間で基質特異性の有意の重複は存在するものの、多数のキナーゼによってリン酸化されるコンセンサス配列は存在しない。この状況は、特異的リン酸化生成物の検出に依存しているHTSアッセイへの多様なまたは新規なキナーゼの組み入れを複雑にしている。
【0044】
表1
表1.抗がん剤としてのキナーゼ阻害因子の開発において選出した臨床試験(Dancey, J. and Sausville, E. A., Nat Rev Drug Discov, 2003, 2:296-313)。太字の薬物は、承認されている。
【0045】
がんにおけるプロテインキナーゼ
癌に干渉させるべくキナーゼを標的化する生物学的な合理的根拠は、ここで概説するにはあまりにも広過ぎる。しかしながら、1つの支配的なテーマは、細胞分裂速度(細胞サイクル進行)、細胞増殖(塊状)、および全てのがんにおいて撹乱されるプログラミングされた細胞死(アポトーシス)間の微妙な均衡の制御における数多くのキナーゼの関与である。増殖因子レセプターチロシンキナーゼ(RTK)は、ペプチド増殖因子シグナルを細胞外から細胞内経路に情報伝達する膜貫通タンパク質であり、これにより増殖促進(progrowth)および抗-アポトーシス遺伝子の活性化が生じる。ヒトにおける58種のRTKの大多数は優性腫瘍遺伝子であり、これは異常活性化または過発現が悪性細胞フェノタイプを生じさせることを意味する。驚くに当らないが、チロシンキナーゼは抗がん薬標的として積極的に追跡されており、そして、小分子阻害因子およびモノクローナル抗体阻害因子(それぞれ、GleevecおよびHerceptin)の双方が臨床的に承認されている。増殖因子レセプターからの下流シグナル伝達は、ser/thrおよびチロシンキナーゼが関与する複数の経路を介して生じる。支配的キナーゼの1つは、RafおよびMEKキナーゼを含む有糸分裂促進活性化プロテインキナーゼ(MAPK)経路である;これらキナーゼ全ての阻害因子は、臨床試験において現在試験中である(表1) (Dancey, J. and Sausville, E. A., Nat Rev Drug Discov, 2003, 2:296-313)。可溶性チロシンキナーゼ、とりわけ、Src群を含む11種の腫瘍遺伝子もRTKによって開始された有糸分裂促進シグナルを伝達し、これれは製薬者によりターゲットとされている(Warmuth, M., et al., Curr Pharm Des, 2003, 9:2043-59)。G1相へのエントリーを開始させる、RTKを介した有糸分裂促進シグナルに続いて、サイクリンタンパク質と関連する相特異的キナーゼの順次の活性化により細胞サイクルの進行が制御される。サイクリン依存性キナーゼは、製薬業者が悪性細胞増殖を抑制するという期待の下に追跡中であるさらにもう1つの重要なキナーゼ群を代表する(表1) (Elsayed, Y. A. and Sausville, E. A., Oncologist, 2001, 6:517-37)。
【0046】
他の疾病における標的としてのキナーゼ
キナーゼにおける製薬上の興味は、がんに最も強く的を絞っているが、主要治療領域の全てに及ぶ。表2は、広範囲の障害に対してのキナーゼ標的についての生物学的理論を説明する最近の論評を示している;それらの標的は、がんについて追跡中の標的と重複していることに留意されたい。全ての企業は、薬物発見におけるキナーゼ標的化に基づいて形成されてきた。これらには、Sugen (http://www.sugen.com、現在Pfizer社所有)、Signase (http://www.signase.com/index.htm)、およびProQinase (http://www.proqinase.com/index.html)がある。
表2
表2.プロテインキナーゼと各種疾病間の関連のリスト
【0047】
本FPIAベースの供与体生成物アッセイ法は、薬物ライブラリーをプロテインキナーゼの阻害因子または活性化因子をスクリーニングするために使用することを考慮する。また、本アッセイ法は、ペプチドまたはタンパク質をキナーゼに対する受容体基質としてスクリーニングするのにも有用である。これらの用途においては、本アッセイ法は、アッセイの普遍的性質、簡素化した均質アッセイ、放射活性の無いこと、および酵素代謝回転の定量能力のような、他の方法を上回る有意の利点を有する。
普遍的アッセイ方法
この方法は、真に汎用的であるので、プロテインキナーゼのHTSスクリーニングプログラムへの統合を促進させるであろう:検出試薬の単一の組を全てのキナーゼおよび全ての受容体基質に対して使用することができる。現行のアッセイ方法を上回る重要な利点は、生理学的タンパク質受容体基質および短ペプチドを使用することができる点である。
均質性
このアッセイ法は、単回添加、混合および読取り方式である。このことは、自動化高処理量環境におけるアッセイ法選択の決定を推進させる重要な要因である(High Tech Business Decisions, M., CA, Commisioned Market Analysis, 2002)。さらに、適切な結合速度を有する抗体を分離する場合は、このアッセイ法は、時間停止アッセイ法よりもより詳細な動的情報を提供する連続アッセイを可能にする。
蛍光検出
蛍光プローブを使用することにより、このFPIA方式は、放射線の取扱い、廃棄およびコストを排除する。最近の数年において、FPは、製薬者よって使用される主要HTSアッセイ基本骨格の1つになっていることに留意すべきである(Owicki, J. C., J Biomol Screen, 2000, 5:297-306)。2003年には、FPは一次スクリーニングアッセイ全体の約12%において製薬者に使用されることが予想される;これは、2001年のFP使用レベルの2倍である(High Tech Business Decisions, M., CA, Commisioned Market Analysis, 2002)。FPは、幾つかの市販HTSプレートリーダーにおける標準方式である。
定量性
提案した酵素アッセイにおいて、ADPは、リン酸化ペプチドまたはタンパク質と化学量論的量で生成され、従って、ADPについての標準曲線の使用は、酵素代謝回転の直接の測定を可能とするであろう。HTS用途におけるFPの使用は比較的最近の展開であるけれども、診断設定におけるホルモンおよび代謝物の定量検出のためのFPIAの使用は、極めて良く確立されている(Nasir, M. S. and Jolley, M. E., Comb Chem High Throughput Screen, 1999, 2:177-90)。
【0048】
(実施例)
実施例1
ウリジングルクロニドトランスフェラーゼアッセイ
本発明の1つの実施態様は、拮抗蛍光偏光イムノアッセイを使用するUGT反応の“供与体生成物”を検出することを含み、このイムノアッセイにおいては、抗体結合トレーサーが高偏光値を有し、この偏光値は、前記トレーサーがUDPのような分析物に置き換わったときに低下する(図2に示すように)。このアッセイにおいて必要とする主たる試薬は、UDPと高選択的に結合するが供与体UDP-グルクロン酸(UDPGA)に対しては無視し得る結合性を有する抗体の作製である。この高選択性抗体は、商業的に入手し得る蛍光UTP化合物と一緒に使用して前記アッセイ法を確立する。
UDPに対して作製したポリクローナル抗体は、担体タンパク質への共有結合を必要とした。トリホスフェートの反応性誘導体は容易に入手することができたが、ジホスフェートは入手できず、トリホスフェートの反応性誘導体が結合に使用することができたという理由のみにより、UTPをハプテンとして使用した。トリホスフェートの大多数が動物中でジ-およびモノ-ホスフェートに加水分解され得ると考えられた。UTPを担体タンパク質に結合するための数種の化学物を調査した。得られる抗体の特異性および抗原に対する親和性に結合の性質が深刻な影響を与え得るからである。注意を払って、リンカー分子がリボースまたはホスフェートよりはむしろウリジン環に結合するようにし、それによってUDP分子を供与体、UDPGAから識別し得るUDP分子の部分との免疫反応性を最高にした。
BSAに結合させたUTPおよびUDPの混合物、および商業的に入手し得るトレーサー分子、蛍光標識化UTP化合物(Alexa-UTP、分子プローブ)に対して生じさせたウサギ抗血清を、指定量のウリジンヌクレオチドを含有するブラックマルチウェルプレート(Thermo Labsystems Pt#7605)のウェルに添加した。Alexa-UTPをFPIA試験におけるトレーサーとして使用した。Tecan Ultraプレートリーダー内で、数時間の平衡後に蛍光偏光を読み取った。反応条件は、以下のとおりとした:50 mMのKPO4、pH 7.4、150 mMのNaCl、0.1 mg/mlのBGG、1 nMのChromaTide Alexa Fluor 488-5-UTP、1.25μlのウサギ血清、100μlの総体積。
【0049】
UGT反応からの実験結果は、図8A〜Bに示してある。図8は、UDP/UTPに対して生じさせた第1ポリクローナル抗体と商業的に入手し得るトレーサー分子(Alexa-UTP)を使用した、抗体-トレーサー複合体の各種ウリジンヌクレオチドによる滴定を示している。2つのグラフはX軸の尺度において異なること、および、UDP,即ち、供与体生成物による拮抗はおよそ10μMにおいて半最高であり、一方、UDPGA、即ち、供与体においては、半最高置換は1 mMよりも高く、これは、少なくとも100倍の選択性の差であることに注目されたい。
より関連性のあることは、UDPがトレーサーを低マイクロモル濃度で置換しているのに対し、100μMを越える濃度においてUDPGAとの検出し得る交叉反応性は存在しないことである。UTPとの交叉反応は、UTPがUGT酵素反応においては存在しないか或いは生成されないので、提案したアッセイにおいては問題ない。UDPがトレーサーを低マイクロモル濃度で置換できるということは、この抗体がUGT酵素反応において生成されたUDPの検出に適していることを意味している。さらにまた、予備結合速度試験により、UDPによるトレーサーの置換が極めて速い兆候が見えていたので、UGT代謝回転をリアルタイムでモニターするための抗体の使用、即ち、連続アッセイは、本発明に包含される。
【0050】
その重要性にもかかわらず、グルクロン酸化を触媒する肝臓UDP UGTの広くて重なり合う基質特異性は、柔軟性のある生体外アッセイ方法が無いために理解に乏しいままである。主たる問題は、使用する触媒アッセイが基質特異性クロマトグラフィー工程を含む生成物からの反応物の分離を必要とし、従って、HTS方式においては現実的でないということである。UGT反応生成物であるUDPを蛍光均質方式で測定する概念を確立することにより、本出願人等は、この問題を解決するための技術的基盤を提供する。本アッセイ法は、全てのUGT反応に共通する生成物を測定するので、あらゆるUGTイソ型およびあらゆる基質による単一方式の検出を可能にする。しかも、本方法は、生成物からの反応物の分離を必要とせず、本方法が比色分析ではなく蛍光検出を使用し、そのことが本方法を、製薬HTSの場(蛍光ベースの検出に大きく依存するようになってきている)においてより感度があってより望ましいものにしているので、他の関連アッセイ法を凌ぐ有意の改良法である。また、本方法は連続アッセイ方法であるので、本方法は、UGT酵素代謝回転についてのリアルタイムの動的データを提供し得る。さらに、抗体-抗原結合反応は、供与体生成物検出に従来使用されていた通常の結合酵素反応(Mulder, G.J. and A.B.D. Van Doorn, Biochem J., 1975, 151:p. 131-40)よりも試験サンプルからの干渉に対して影響を受けにくい。即ち、この新規なアッセイ方法は、単離したUGTイソ酵素の集団による多様な化合物の代謝についてのスクリーニングを可能にし、前臨床代謝試験を大いに増強し、臨床脱落率を強力に低下させる。
さらにまた、本出願人等が作製した抗体の性質は、他の重要なクラスの酵素についてのHTSアッセイ法の開発においてもより広い密接な関係を有する。図8に示すように、UDPがトレーサーを置換しているがUMPはそうではなく、このことはこの抗体が1個のリン酸基に基づき各ヌクレオチドを識別し得ることを例証している。即ち、反応生成物が供与体と1個のリン酸によって異なる、プロテインキナーゼ反応におけるADPのような他の“供与体生成物”に対する抗体を作製し得ることも包含される。
【0051】
実施例2
キナーゼアッセイ
本発明のアッセイ方法は、リン酸化ポリペプチドと化学量論的量で生成するキナーゼ反応生成物ADPのFPAI検出に依存する。上述したUGTアッセイと同様に、FPIAベースの供与体生成物キナーゼアッセイの構成要素は、供与体生成物に対する抗体、および検出可能タグと結合させた供与体生成物を含むトレーサーである。前記抗体は、ADPに対し高度に特異的である(即ち、前記抗体は、過剰のATPおよび蛍光トレーサーの存在下においてADPを認識し得る)。キナーゼアッセイにおける抗体とトレーサーを、アッセイの開始に最適の濃度でブラックマルチウェルプレートのウェルに加える。トレーサーの適切な濃度は1〜2nMであり、十分な抗体を使用してトレーサーに最高偏光変化のおよそ75%を生じさせる。受容体基質を、所望の、一般にKm値よりも2〜5倍高い濃度で、前記ウェルに加える。受容体基質は、ペプチドまたはインタクトタンパク質でよい。本発明の1つの利点は、他のキナーゼアッセイ法は、検出のためには特異的受容体を必要とするのに対し、任意の受容体基質を使用し得ることである。アッセイすべきキナーゼおよび抗体-トレーサー相互作用と適合する緩衝液、一般的には、およそ中性pHのTris-Clまたはリン酸緩衝液を使用する。この緩衝液と一緒に、100μM〜5mM濃度のATP、MgCl2およびキナーゼの活性化または安定化に必要な他の薬剤のような他の必要なアッセイ成分を添加する。その後、キナーゼ酵素を添加して反応を開始させ、偏光値をTecan Ultraのようなマルチウェルリーダーでモニターする。反応が進行するにつれて、リン酸化ペプチドまたはタンパク質と化学量論的量で生成したADPは、トレーサーを抗体から外して、偏光値の低下をもたらす。
阻害因子のスクリーニングにおいては、一般的には試験すべき化合物を何ら他のアッセイ成分を添加する前のウェル中に分配し、阻害因子を添加しない参照ウェルを比較のために含ませる。
各キナーゼは、1種以上のターゲットタンパク質を、時には複数部位でリン酸化させることがあり、触媒活性または他の生体分子との相互作用を制御する1個以上の調節ドメイン内で自己リン酸化することがある(Cohen, P., Trends Biochem Sci, 2000, 25:596-601)。健常および疾病状態における細胞リン酸化プロフィールの機能的結果を明らかにすることは、プロテオミクスの大きな第1歩であり、この知識は、薬物発見においてどのキナーゼを標的にするか、およびその受容体基質に対する特異性を決定するために使用し得る。関連キナーゼ間で基質特異性に有意の重なり合いは存在するものの、多数のキナーゼによってリン酸化されるコンセンサス配列は存在しない。この状況は、特異的リン酸化生成物の検出に依存しているHTSアッセイへの多様なまたは新規なキナーゼの組み入れを複雑にしている。
【0052】
実施例3
スルホトランスフェラーゼアッセイ
SULT1E1の発現および精製
スルホトランスフェラーゼHTSアッセイ法を確立するために、SULTイソ型であるSULT1E1を先ずC-末端6xヒスチジンタグを有する大腸菌発現ベクター中にサブクローニングし、発現タンパク質をアフィニティークロマトグラフィーにより精製し、その物理特性および酵素特性について特性決定した。
SDS-PAGE上でその算出分子量近くに移行した精製タンパク質は、予測される分子量と極めて緊密に一致しており、さらに重要には、質量分光分析において予測される分子量と極めて緊密に一致していた(それぞれ、36,161および36,374ダルトン)。このタンパク質は天然のN-末端を保持するように設計していたので、精製C-末端6xHis構築物をN-末端シークエンシングにも供した;15個のアミノ酸をシークエンシングしたところ、これらのアミノ酸は、GenBank中のヒトSULT1E1配列(NP005411)と同一であった。さらなる配列検証は、質量分光分析およびタンパク質シークエンシングにより得られた。
比較として使用するために、精製SULT1E1の酵素特性を試験した。エストラジオールおよびエストロンを生理学的基質として使用し、さらなる陽性(α-ナフトール)および陰性(ドーパミン)対照化合物も試験して特異性を評価した。2つのタイプの放射線アッセイ法をこれらの試験においては使用し、1つは35S-PAPS (Foldes, A. and Meek, J. L., Biochim Biophys Acta, 1973, 327:365-74)を使用し、他方は3H-エストラジオール(Zhang, H., et al., J Bio Chem, 1998, 273:10888-92)を使用した;両者は、SULT特性決定において広く使用されているものであり、以下に説明する。
【0053】
表4
【0054】
表4は、SULT1E1融合タンパク質の酵素特性を示している。KmおよびVmax測定を3H-エストラジオール放射線アッセイを使用して実施した。この測定では、放射活性硫酸化生成物を未反応3H-エストラジオールから有機/水性相抽出により分離し、液体シンチレーションカウンターで計数した。各キネティックパラメーターは、GraphPad/Prizmソフトウェアを使用して、非直線回帰によるV対S曲線から算出した。各種受容体による相対的反応速度は、35S-PAPS放射線アッセイにおいて、400nMの受容体基質を使用して測定した。この測定では、未反応35S-PAPSをバリウム金属錯体として沈降させ、35S-標識化スルホ結合体を含有する上清を液体シンチレーションカウンターで計数した。基本的反応条件は、双方の場合において、10 mMのKPO4、pH 6.5、10 mMのDTT、1.5 mMのMgCl2、10 mMのPAPS、0.5 ngのSULT1E1、0.0025〜15 mMの受容体であった。
精製SULT1E1において測定したVmaxおよびエストラジオールKm値は、精製組換えSULT1E1における公表された値(それぞれ、30〜40nモル/分/mgおよび5〜15nMである)に良好に匹敵していた。受容体基質特異性に関しての公表データはより変動しているが、その結果は、エストラジオールとエストロンが極めて良好な基質であり、α-ナフトールが中間であり、DHEAとドーパミンが極めて貧弱な基質であるという一般的傾向を反映している(表4参照)。即ち、出願人等が発現させたSULT1E1構築物の全てが、最高の公表値と同様な天然の基質特異性と触媒作用速度を示していた。
【0055】
抗原の合成および抗体作製
スルホトランスフェラーゼにおける蛍光系HTSアッセイ法の実際の開発に関しては、SULT触媒作用を検出するPAP FPIAを示す4つの要素が存在する:抗原の合成および抗体の生成;トレーサー分子の合成;相互作用および特異性についての抗体およびトレーサーの試験;および最適抗体-トレーサー対を使用したSULT1E1活性検出の実証。トレーサー合成の努力は、同じ反応性PAP誘導体を担体タンパク質およびフルオロフォアへの結合において使用するので、抗原合成の努力と有意に重なり合う。
提案したFPIA系SULTアッセイ法の開発は、SULT反応生成物のPAPと過剰のPAPSの存在下に特異的に結合する抗体を必要とする;即ち、単一の5'-リン酸基に基づき識別する抗体。リン酸基の数のみにおいて異なり、上述のような硫酸基と構造および大きさにおいて同様な各種ヌクレオチドを識別する抗体においては、十分な先例が存在する。しかしながら、PAPを特異的に認識する抗体の作製についての先例は存在しない。
PAPのような小分子は、免疫原として使用するためには担体タンパク質に結合させなければならない。適切には、担体タンパク質当り10〜20個の抗原密度が最適である。上述したように、本発明者らの抗原合成戦略の2つの要素は、a)数種の抗原の合成と試験(ヌクレオチドへの結合部位およびリンカー構造は得られる抗体の特性に有意に影響を与える得るから) (Crabbe, P., et al., J Agric Food Chem, 2000, 48:3633-8;Signorini, N., et al., Chem Res Toxicol, 1998, 11:1169-75;Oda, M. and Azuma, T., Mol Immunol, 2000, 37:1111-22)、および、b)アデニン環を介した結合(5-リン酸基の自由露出を可能にし、分子のアデニン部分との免疫反応性を阻害して、所望の特異性を有する抗原を生じるから)。アデニン環中にはタンパク質に直接または架橋性試薬を介して結合するのに十分に反応性である窒素はなく、従って、反応性PAP誘導体を開始点として使用した;唯一の商業的に入手可能な試薬は、N6-アミノヘキシルPAPであった。当初最も直接的な試みと思われた方法(N6-アミノヘキシルPAPを担体タンパク質とカルボジイミドカップリングまたはアミン反応性架橋剤により反応させる)は、かなりの実験努力にもかかわらず、許容し得ない低い抗原密度を生じた。
【0056】
別の化学物質を追求するために、出願人等は、光反応性分子である2-および8-アジド-PAP、およびC8-ヘキシルアミノ-PAP (図6)の合成を外部委託した。図6は、PAP抗原の合成を示す。ALT社(ケンタッキー州レキシントン)により特注合成された2-および8-アジド-PAPをBSAの存在下に照射した(254nm);未反応ヌクレオチドは、濾過および透析により除去した。7〜12PAP/BSAの最終抗原密度が、アデシン環の吸光度により測定したときに得られていた(270〜280nmに変化)。ウサギ(抗原毎に3羽)は、Lampire Biologicals社(ペンシルベニア州オッツビレ)により免疫化された。N6-アミノヘキシルPAP(Sigma社)は、グルタルアルデヒドを使用してKLHに結合させ、Biosynthesis社(テキサス州ルイスビラ)により3羽のウサギに注入された。免疫スケジュールは、全ての抗原において同様であり、4〜6週間に亘って3〜4回の注射を含んでいた。
前記2つの光反応性PAP誘導体からの抗原を作製し、抗体生成のために送った;KLHへのグルタルアルデヒド架橋を使用してのN6-アミノヘキシルPAPからの抗原調製および抗体産生は、別個の企業に外注した(図6)。3種の抗体すべてがELISAで測定したときにPAPに結合したことを外注製造者から報告された(データは示していない)。
さらに、出願人等は、反応性誘導体 N6-カルボキシメチルPAPおよび2'-O-スクシニル-PAPも合成した(図7);これらの誘導体からの抗体は、まだ産生させていない。拮抗FPIAを使用してのAb結合特性の詳細な説明は、トレーサー合成の説明に続いて後で説明する。
【0057】
PAP-蛍光体トレーサーの合成
FPIAトレーサー分子は、3種の異なる構成部分、即ち、抗原、蛍光体(fluor)およびこれらを結合させるのに使用するリンカーに分けることができる;さらなる主要な構造的変動要因は、抗原へのリンカーの結合部位である。トレーサーの同定は大いに経験的であるので、出願人等は、種々のリンカーを使用してPAPとフルオレセインを各分子上の種々の部位を介して結合させた;殆どの場合、最終リンカー領域は、使用した反応性フルオレセインとPAP分子の複合体であった。
抗体戦略は、PAPのリボシル-ホスフェート基に特異的に結合する抗体を作製するために、アデニン部分を介して結合させることであった。この部位は、所望のトレーサー分子の免疫反応性部分をAbを結合させるために遊離のまま残存させるための蛍光体結合用の自明な部位でもある。また、フルオレセインは、結合に使用した好ましい蛍光体であった。ローダミンのような赤色シフト蛍光体がHTS用途においてより望ましいが、フルオレセインによるFPトレーサーの開発は、フルオレセインが非特異結合を起こしにくく、多くの活性化誘導体を入手し得るので、通常はより簡単である。リンカー分子に関しては、該分子は、抗原性および蛍光特性の双方に影響を与える多くの重要な形でトレーサー特性に影響を及ぼす。一般に、抗原を蛍光体から十分に引き離して抗体との相互作用が妨害されないようにすることと、蛍光体にあまり自由過ぎる動きを生じさせないようにすることの間に注意しなければならない均衡が存在する。前者は親和性Ab結合性の低下および蛍光体の消光をもたらし、一方、後者はAb結合時の偏光変化を低下させ、それによってアッセイのダイナミックレンジを減じる。
N6-アミノヘキシルPAPは、トレーサー合成において有用である免疫原合成において使用した唯一の活性化PAP分子であった;前記2つのアジド-PAP誘導体を結合させるのに必要とした光活性化反応は、2つの小分子を結合するには不十分かもしれない。種々の位置で活性化させ且つ種々のリンカー領域を有するPAP分子を調製するために、出願人等は、C8-アミノヘキシル-PAPの調製を外部委託し、カルボキシ末端リンカーを含む2つのPAP誘導体、即ち、N6-カルボキシメチル-PAPと2'-O-スクシニル-PAPを自家合成した。後者の化合物はアデニンよりはむしろリボースヒドロキシルを介して結合させているが、この方法を使用して、cAMPに対して高度に特異的な抗体およびトレーサーが産生されている(Horton, J. K., et al., J Immunol Methods, 1992, 155:31-40)。
【0058】
以下は、活性化PAP分子の合成と精製並びにその各種活性化蛍光体への結合についての簡単な説明である;トレーサー合成成分は、図7および8に要約している。一般に、アミノ末端リンカーを含むPAP分子はスクシンイミジルエステル(または、1つの場合はイソシアネート)により活性化させた蛍光体に結合させ、カルボキシ末端リンカーを含むPAP分子は遊離アミノ基を含有するフルオレセイン誘導体とカルボジイミドカップリングを使用して反応させた。
図7は、トレーサー合成の各構成要素の構造を示す。図7は、左から右に、C8およびN6位置で結合させたアミノ末端リンカーを含むPAP分子;アミン-およびカルボキシ-反応性フルオレセイン誘導体;およびN6および2'-OHにおいてカルボキシ末端リンカーを含むPAP分子を示している。各PAP結合体をTLCにより分離し、抗-PAP抗体に対する結合性について試験した。本発明によれば、16の反応がPAPとフルオレセイン誘導体の種々の組合せを含有して進行し、各反応で、TLCで解像し得た1〜4種の蛍光生成物が得られた。全部で、40種以上のトレーサーを精製し、Abに対する結合性について試験した。N6-アミノヘキシルPAP(Sigma社)および反応性フルオレセイン誘導体の全て(Molecular Probes社)が商業的に入手可能である。2'-O-スクシニル-PAPおよびN6-カルボキシメチル-PAPは、以下で説明するようにして合成した;C8-アミノヘキシルPAPは、Jena Biosciences社(ドイツ国ジェナ)が合成した。図8は、代表的な最終トレーサー構造を示す。
【0059】
N6-カルボキシメチルPAPの調製
N6-カルボキシメチルPAPを調製するために、100mgのPAPを、0.3gのヨード酢酸と一緒に、LiOHでpH 6.5に調整した1.2mLの水中でインキュベートした。反応は、30℃で5〜7日間続き、定期的にpHを6.5に調整した。得られた1-カルボキシメチル-PAP生成物をエタノールで沈降させ、蒸留水中に再構成させ、pHをLiOHで8.5に調整した。この反応を90℃で1.5時間加熱してN6-カルボキシメチルPAPを得た。この生成物を、0.3 M LiCl、pH 2.75中で平衡化させたDowex 1-X2 (200〜400メッシュ)カラムで精製した。勾配を、0.5 M LiCl、pH 2.0を使用して10カラム体積に亘って適用した。N6-カルボキシメチルPAPはカラムから純生成物として溶出し、質量分光分析により確認した、約20%の収率。
2'-O-スクシニル-PAPの調製
2'-O-スクシニル-PAPを調製するために、10mg (0.024ミリモル)のPAPと無水コハク酸(43mg、0.426ミリモル)を、10%のトリエチルアミン含有する蒸留H2O (体積/体積、1mL)中に溶解させ、1.5時間振盪させ、反応の進行を、逆相薄層クロマトグラフィー(RP-TLC)を使用してモニターした。終了時に、反応を2回凍結乾燥させて残留トリエチルアミンが確実に残らないようにした。期待されたスクシネートを、50mLの250 mM NH4OAcで平衡化させたFast Flow Sepharose-Q樹脂(Pharmacia社)を使用してカラム精製し、500mMから750mMのNH4OAc(各々50mL)の直線勾配により溶出させた。生成物を含有する画分をロトバップ(回転エバポレーター)(rotovap)上で濃縮し、次いで凍結乾燥させて、吸光度測定によれば、8.4mg即ち収率67%を得た。ストック溶液を後の使用のために−20℃で保存した。
【0060】
アミノ活性化PAPを含む蛍光結合体の調製
アミノ活性化PAPを含む蛍光結合体を調製するために、N6-またはC8-アミノヘキシル-PAPの蒸留H2O中100mM溶液の10μLを、スクリューキャップ付きバイアル内で、モル当量のフルオレセインスクシンイミジルエステルと混合し、無水ジメチルスルホキシド(DMSO)を使用して反応物を200μLの最終体積とし、撹拌機(ボルテックス)(vortexer)上で24時間振盪させた。反応の進行を、展開用溶媒としてのH2Oを使用してRP-TLCにより追跡した。反応生成物を、分離用TLCを使用して、1:1のEtOH/0.5M NH4OAcを使用する正常相シリカゲル上で精製した。フルオレセイン標識化化合物(一般に、各反応において3〜5回生成させた)をUV光を使用して可視化し、TLCプレートからそぎ落とし、1:1のMeOH/0.5M NH4OAcを使用してシリカゲルから抽出した。個々の画分をボルテックス撹拌機上で1時間振盪させ、アルミニウムホイル内にラッピングし、4000RPMで8分間遠心分離した。上清を別のラベル付びんに移し、抽出工程を繰返した。集めた上清を後の使用のために50nM溶液に標準化し、褐色微量遠心用チューブ内で−20℃で凍結させた。
カルボキシ活性化PAPを含む蛍光結合体の調製
カルボキシ活性化PAPを含む蛍光結合体を調製するために、N6-カルボキシメチル-PAPまたは2'-O-スクシニル-PAPの蒸留水中100mM溶液の10mlを50当量の1-エチル-3-(ジメチルアミノプロピル)カルボジイミド(EDC) (無水DMSO中1M溶液の10μL)、次いで75当量のN-ヒドロキシスクシンイミド(無水DMSO中1000mM溶液の15μL)と混合し、反応物を無水DMSOにより180μLの中間体積とし、48時間振盪させた。その後、1モル当量(20ml)の遊離1級アミンを含むフルオレセイン誘導体を添加し、反応を攪拌させながら24時間インキュベートした。反応の進行を、展開用溶媒としてのH2Oを使用してRP-TLCにより追跡した。反応終了時に、蛍光生成物を、9:1 アセトニトリル/2mM NH4OAc、pH 5.5をTLC溶媒として使用し、8:2 アセトニトリル/2mM NH4OAc、pH 5.5を溶出用に使用した以外は、上述のようにして、正常相シリカTLCにより精製し、溶出させた。
全部で16組の活性化PAPとフルオレセイン分子の固有の組合せを反応させ、これらから、40種よりも多い蛍光生成物を単離し、抗体に対する結合性について試験した。この方法は“貧乏人の組合せ化学”の類とみることができるが、出願人等は、他の例において、FPトレーサーの開発において極めて成功裏であることを見出している。FPアッセイの均質性およびマルチウェル器具の利用可能性は、スクリーニング取組みを比較的迅速にすることに注目されたい。
【0061】
抗-PAP抗体およびトレーサーの特性決定
抗体/トレーサー相互作用の分析を蛍光偏光により実施した。蛍光偏光(上で簡単に説明した)を測定するには、垂直偏光励起光に対して平行および垂直である垂直面および水平面の双方において発光強度を測定する。放出光が偏光を規定する:P = (水平強度−垂直強度)/(水平強度−垂直強度)。偏光値は、mPとして報告され、これは、偏光値の1000倍である(30mP = 0.03P)である。フルオレセインは、pH 7.0のリン酸緩衝液中にて37℃でおよそ20mPの偏光を有する。数学的には、フルオレセインの限界偏光は、500mPである;実際には、フルオレセインに結合させた抗-フルオレセイン抗体は、約450mPの偏光を与える。Tecan Ultraのような現在入手可能なマルチウェルFPリーダーは、偏光値を高い精度で読取ることができる;即ち、<3mP未満の標準偏差での遊離および結合状態の1nMフルオレセインの検出。
拮抗FPIAにおいては、蛍光標識化抗原、即ち、トレーサーは、分析物により抗体との結合から外される。そのシグナルは、結合トレーサー画分対遊離トレーサー画分の差異に比例し、従って、アッセイのダイナミックレンジおよび感度の双方は、トレーサーおよび拮抗性サンプル分子に対する抗体親和性に依存している。FPIAの適切なダイナミックレンジを確立するためには、少なくとも70〜80%のトレーサーが拮抗因子の不存在において抗体に結合しなければならない。これは、Kdよりも2〜10倍低い濃度のトレーサーとKdよりも2〜3倍高い濃度の抗体を使用することによって達成される。その場合、アッセイの有用な濃度範囲は、Kd濃度の数倍の低さからKd濃度の約20倍の高さまでである。例えば、トレーサー/抗体複合体のKdが10nMである場合、サンプルの検出範囲は、約2nM〜約200nMであり得る。Kdが10倍低い場合(1nM)、アッセイ感度も約10倍改善する。
影響を受けるアッセイパラメーターの点で許容され得る抗-PAP抗体およびトレーサーの特性は、1) ダイナミックレンジ:Abの不存在下での低トレーサー偏光(<100mP)、および結合および遊離状態間での最大偏光差(ΔmP >100mP);2) 感度:トレーサーの抗-PAP抗体に対する高親和結合性(Kd <100nM)、同様なIC50を有する遊離PAPによる置換、および最小の結合によって生じる蛍光消失;3) シグナル/ノイズ:高Ab選択性;即ち、PAPによるが、PAPSまたは他のアデニンヌクレオチドにはよらないトレーサーの拮抗置換;また、SULT1E1または他のアッセイ成分とトレーサーとの相互作用の無いこと;および4) 連続アッセイ:迅速な結合/解離速度。
【0062】
Ab-トレーサー相互作用試験の結果
数種の濃度の各抗体をマルチウェルプレート中の1nmのトレーサーに添加し、トレーサー偏光の増加についてモニターすることによって、Ab-トレーサー結合性についての初期スクリーニングを実施した。抗体全てにおいて、固定化プロテインAまたはプロテインGを使用したIgG画分の精製が、非特異結合成分を除去するために必要であった。示したデータは、全て、プロテインG (Pierce NabTM Protein G Spin Cromatography Kit)で精製した抗体を使用している。初期スクリーニングにおいて相互作用を示したトレーサー/Ab組合せを、PAPによる拮抗置換およびPAPSに対するPAPの選択性についてさらに詳細に試験した。全部で9種の精製抗体(3種の抗原×3羽のウサギ)を44種のフルオレセイン標識化PAPトレーサーとの結合性について分析した。アッセイは、全て、ブラック96ウェルプレートまたはブラック低容積384ウェルプレート内で実施した。両者は本質的に同一の結果を与え、偏光値は蛍光フィルターセットを備えたTecan Ultraで読取った(485nmでの励起から535nmでの発光) (EX485/Em535)。
図9に示す結合等温線は、下記の表5に示す3種の抗体と2種のトレーサーを使用して作製しており、抗体およびトレーサーを結合性についてスクリーニングするのに使用した試験の代表的なものである。
【0063】
表5:代表的な抗体およびトレーサー
【0064】
また、図9は、抗-PAP抗体およびPAP-フルオレセイントレーサーにおける結合等温線を示している。Ab 1781○、1810◇および3642□に対するN6-PAP-F8トレーサー結合性、並びに同じ抗体1781●、1810◆、3642■によるC8-PAP-F14トレーサーを示している。各抗体は、96ウェルプレートにおいては100μLまたは384ウェルプレートにおいては12μlの総体積で、150mMのNaCl、0.1mg/mLのBGGおよび1nMのトレーサーを含有する50mMリン酸緩衝液(pH 7.4)中で連続2倍希釈し(2つのプレートにおける結果は同一である)、偏光値を、室温での1時間インキュベーション後に、Tecan Ultraで読取った。
異なる動物において同じ免疫原から生成させた抗体の結合特性は類似していたが同一ではなかった。この試験においては、3642抗体が1781よりも有意に強固に両トレーサーに結合し、1810は、試験した最高濃度においてさえもいずれのトレーサーとも結合しなかった。N6-PAP-F8およびC8-PAP-F14に結合する3642の最高偏光値は、それぞれ、140および300mPであった;未結合トレーサー偏光はおよそ20mPであった、従って、観測されたシフトはFPIAに使用するために充分以上である。結合が特異的であるかを試験するために、PAPによる拮抗置換を評価した。
図10は、Ab 3642および前記2つのトレーサー、PAPおよびPAPSに対するPAPおよびPAPSの拮抗曲線を示す。N6-PAP-F8トレーサーは、白抜き記号で示している:○(PAP)、□(PAPS)。C8-PAP-F14トレーサーは、黒塗り記号で示している:●(PAP)、■(PAPS)。PAPまたはPAPSを、12μlの総体積で、50mMリン酸緩衝液(pH 7.4)、150mMのNaCl、0.1mg/mLのBGG、1nMのC8-PAP-F14および1.5μLの精製Ab 3642または20μL体積中に1nMのN6-PAP-F8および0.5μLの3642 Abを含有する黒色マルチウェルプレート内で連続2倍希釈した。偏光値は、室温で1時間後に読取った。
C8-PAP-F14トレーサーによるPAPにおけるIC50は、SULT1E1活性をモニターするためのこれらの試薬の使用を可能にするのに十分に低い300nMである。より強固な結合性のN6-PAP-F8トレーサーを拮抗除去するにはかなり高濃度のPAP(およびPAPS)を必要とするに注目されたい。これは、この場合、トレーサーが抗体を生成させるのに使用した免疫原と同じリンカー基を有しており、抗体の集団がこのリンカーを認識してトレーサーが遊離PAPによって置換されるのをより困難にしていることにより得る。また、図10は、PAPSがPAPよりもトレーサーをAb 3642から外すのに有効性が低いことも示している。
【0065】
同様な結果は、図11に示すように、1781 Abによっても観察された。図11は、C8-PAP-F14トレーサーによる拮抗試験におけるPAP、PAPSおよび数種の同様な分子の完全曲線を含む。図11は、2つの抗-PAP抗体/トレーサー組合せによる拮抗曲線を示す。各拮抗因子を、12μlの総体積で、50mMリン酸緩衝液(pH 7.4)、150mMのNaCl、0.1mg/mLのBGG、1nMのトレーサーC8-PAP-F14および1.5μLのAb 3642 (A)または3μLのAb 1781 (B)のいずれかを含有する黒色384ウェルマイクロタイタープレート内で連続2倍希釈した。室温で1時間のインキュベーション後に、偏光値をTecan Ultraで読取った(EX485/Em535)。全てのデータ設定において複製の平均および標準偏差を示している。3642/トレーサーおよび1781/トレーサーによるPAPおよびPAPSのIC50は、それぞれ、0.3μM PAP、3.8μM PAPS;および0.3μM PAP、1.5μM PAPSであった。
1781および3642抗体は、それぞれ、PAPSに対してよりもPAPに対して5倍および13倍高い選択性を示し、試験した他のヌクレオチド全てに対してよりもはるかに高いPAPに対する選択性を示していた。PAPSとの交叉反応は、他のアデニンヌクレオチドとの交叉反応が無いことを考慮すれば、予期したよりも高い。この点、殆どの市販PAPS調製物は有意のPAP画分を含有しているが、出願人等は、95%よりも高い純度と分析されたHPLC精製調製物を購入し、その保存において予防策を講じて加水分解を阻止するのに使用したことに留意すべきである。いずれにしろ、これらの結果は、出願人等がPAPSの存在下でPAPに選択的に結合する抗体を作製し得ることを明らかに示しており、このことは、提案したSULTアッセイにおいて十分なシグナル:ノイズおよびダイナミックレンジを可能にする主要な実施可能性要素である。PAPSに対してよりもPAPに対するAb選択性がさらに10倍増加することは十分であり得、ポリクローナル抗体ではなくモノクローナル抗体を使用した場合の極めて合理的な期待事項である。
試験した他の抗体およびトレーサーにより観察した結果は類似していた。N6-アミノヘキシル-PAP-KLHおよび8-アジド-PAP-BSA免疫原から作製したさらなる抗体はトレーサーの多くと結合し、Ab/トレーサー複合体は全ての場合未標識PAPによって置換することができた。このことは、この相互作用が特異的であることを示唆している。1810抗体は、トレーサー結合性を極めて貧弱にしか或いは全く示さず、さらに試験しなかった。遊離トレーサーの偏光値は15〜40mPの範囲であり、Ab結合時に観測された増大は100mP未満からほぼ300mPまでの範囲であった。幾つかのトレーサーとの相互作用はAbとの飽和を可能にするには弱過ぎて、最高偏光変化は常には観測されなかった。試験したトレーサーの2/3よりも多くが両タイプのAbに結合したが、N6-カルボキシメチル-PAPから合成したトレーサーのうちで、いずれの免疫原から生成させた抗体とも結合したものはなかった。
【0066】
FPIAベースのSULTアッセイ
PAPとPAPSをより有効に識別する抗体を作製して高品質アッセイを開発する必要があるだろうが、出願人等は、3642 AbおよびC8-PAP-F14トレーサーを使用して精製SULT1E1-cHisを含有する反応において生成したPAPの検出をモニターすることができた。初期の試験において、出願人等は、最高シグナル:ノイズのための最適PAPS濃度を同定することを模索した。SULT1E1-cHisを、予め形成させたAb-トレーサー複合体の存在下に、エストラジオールおよび変動濃度のPAPSと一緒にインキュベートした;SULT1E1は、反応を完了まで迅速に推進させるに十分なレベルで存在していた(図12)。
図12は、酵素により産生させたPAPの検出におけるPAPS濃度の効果を示す。アッセイ混合物は200ngのSULT1E1-6xHisを含んでおり(□)、或いはアッセイ緩衝液(■)を、100μLの総アッセイ体積で、30mMのホスフェート(pH 7.4)、7mMのDTT、8mMのMgCl2、75mMのNaCl、0.5mg/mLのBGG、150nMのエストラジオール、1nMのC8-PAP-F14トレーサー、12.5μLのAb 3642および変動濃度のPAPSを含有するウェルに添加した。プレートを37℃で24時間インキュベートし、Tecan Ultraで読取った(EX485/Em535)。
典型的なFPIAにおけるように、トレーサーはKdよりもかなり低い濃度で使用し、Abは拮抗因子の不存在下で最高トレーサー偏光のおよそ85%を生じる濃度に調整した。図12は、酵素により産生させたPAPが約40mPのトレーサー偏光の減少(図12における白抜き四角と塗潰し四角との差)を生じさせるPAPS濃度範囲(約1〜5mM)が存在することを示している。
即ち、この抗体がPAPSと有意に交叉反応するとしても、この抗体を拮抗FPIA方式において使用して、飽和中のPAPS (SULT1E1によるPAPSのKmはおよそ50nMである)によるSULT反応において産生したPAPを40mPのダイナミックレンジでもって検出し得る。さらにまた、これらは、均質反応、即ち、反応および検出成分の全てを反応開始時に添加した単回添加反応であった;これは、HTSアッセイにおける好ましい方法である。同様な結果は、検出試薬を酵素反応が終了した後に添加する場合も達成される、従って、偏光シグナルは、酵素反応によって影響されない(データは示していない)。
【0067】
出願人等が作製したAb/トレーサー対の一つを、読取り前に完了まで進行させたSULT反応におけるPAPの検出においてどのように使用するかの実施態様を上述した。しかしながら、連続方法は、正確な酵素速度測定を可能にし且つクエンチング工程の必要性を排除するので、HTSにおける最も好ましい方式である。図13は、3642 AbおよびC8-PAP-F14トレーサーを使用してSULT1E1酵素活性を経時的に連続モニターして、種々の基質による酵素速度の測定を可能にし得ることを示す。
図13は、種々の基質によるSULT活性の連続FPIAに基づく検出を示すグラフである。受容体基質(200nM)を、100μLの総体積で、30mMのホスフェート(pH 7.4)、7mMのDTT、0.8mMのMgCl2、75mMのNaCl、0.5mg/mLのBGG、2μMのPAPS、200ngのSULT1E1-cHis、1nMのC8-PAP-F14トレーサーおよび12.5μLのAb 3642を含有するウェルに添加した。対照反応(各グラフの上の方のトレース)は、C-His-SULT1E1を含まず、全ての他の反応成分を含有していた。プレートを室温でインキュベートし、偏光を1分間隔で読取った。
各グラフの上の方のトレースはSULT1E1を欠く対照反応であるので偏光は有意には変化しないが、酵素を含有する各反応においては、偏光は時間につれて減少する。試験設定は、HTS設定においてなし得た設定と極めて類似のものである:抗体とトレーサーをSULT反応混合物に添加し、異なる受容体基質を含有する384ウェルプレート中に分配し、反応をSULT1E1-cHisの添加により開始させた。その後、各プレートを一定間隔で読取り、偏光値を時間の関数としてプロットした。既知のSULT基質による反応においては、偏光は、酵素を欠く対照反応と対比して経時的に減少していた;酵素により産生させたPAPがトレーサーをAbから外して置き換わるときに期待し得るように。受容体基質が存在しないとき(グラフf)或いはSULT1E1が存在しない場合(各グラフの上の方のトレース)、偏光の有意の変化はなく、PAPS分子が非生産性化学物または酵素加水分解に十分に抵抗性であることを示唆していることに注目されたい。
【0068】
これらの結果は、スルホトランスフェラーゼについて汎用的な蛍光系活性アッセイ法が技術的に実施可能であることを示している。これらの試験においては、3642 AbのPAPSとの交叉反応は有意なバックグラウンド(即ち、偏光の減少)に寄与して、アッセイのダイナミックレンジを制約しているが、酵素速度は、各速度曲線の直線部分から依然として得ることができている。図14の棒グラフは、図13の速度曲線の直線部分から算出した各基質の割合を示している。この時点では正確なアッセイでないものの、この基質の順位付けは、これら基質の公表されたKm値並びに出願人等が精製SULT1E1および35S-PAPS放射線アッセイを使用して測定した基質プロフィール(図14)と逆相関している。従って、図14は、FPIA系アッセイと35S-PAPS放射アッセイを使用して測定したSULT1E1受容体基質プロフィールの比較である。FPIA系反応の速度は、図13に示す曲線の直線部分から算出した。35S-放射線アッセイデータは表4に提示していることに留意されたい。受容体基質は、200nM (FPIA)および400nM (35S-放射線アッセイ)で使用した。
最後に、出願人等は、FPIA系アッセイを使用して阻害曲線を作成し得ることを示すために、既知のSULT阻害因子であるDCNPを使用した(図15)。詳細には、図15は、FPIA系アッセイにより測定した2,6ジクロロ-4-ニトロフェノール(DCNP)によるSULT1E1の阻害を示すグラフである。DCNPを、ウェル中で、46μLのリン酸アッセイ緩衝液(30mMのKPO4(pH 6.5)、0.5mg/mLのBGG、15mMのDTT、1.6mMのMgCl2、4μMのPAPS)中で連続2倍希釈し、次いで、50μlの2×抗体/トレーサー混合物(5μL 3642 Ab/2 nM トレーサーC8-PAP-F14)および200ngのSULT1Eを100μlの総体積中に添加した。プレートを室温で30分間インキュベートし、Tecan Ultraで読取った。ΔmP値を、SULT1E反応を無SULT1E対照から差し引くことによって算出した。全ての値は繰り返しの平均を表す。
DCNPに対する前記FPIAアッセイの応答を、35S-放射アッセイ(データは示していない)との比較により検証した;2つのアッセイ法で測定したKi値は、それぞれ、7.8mMおよび11mMであった。即ち、出願人等は、前記アッセイ法が基質および阻害因子の検出(共に意図する主要HTS用途)に使用し得たことを実証した。
要約するに、これらの結果は、提案したFPIA系SULTアッセイ法の構成要素、即ち、精製組換えSULT;PAPSの存在下においてPAPを選択的に認識する抗体;および、その結合およびAbからの拮抗的置換を偏光の有意の変化によって検出し得る蛍光PAPトレーサー、の全ての作製の実施可能性を明らかに立証するものである。さらに、出願人等は、前記アッセイ試薬を、アッセイをHTS設定において使用し得る方法に極めて類似した連続方式でSULT活性を検出するのに使用し得たこと;種々の基質に対する応答が標準の放射アッセイに類似していること;および前記アッセイ法は阻害因子の検出にも使用し得ることも実証することができた。
従って、商業的HTS用途において適切なダイナミックレンジおよびシグナル:ノイズを有するアッセイ法の開発を可能にする抗体、適切には、PAPに対しておよそ10倍高い親和性と選択性を有するモノクローナル抗体を産生させることが考慮されている。
【0069】
実施例4
アッセイ系
上述の方法は種々の異なるアッセイ系において使用し得るけれども、その最も簡単な形においては、本発明のアッセイ系は、アッセイする反応を実施するアッセイ容器、並びに反応の結果を検出するための検出器を含む。好ましい局面においては、前記アッセイ容器は、試験管、マルチウェルプレート内のウェルまたは他の同様な反応容器から選択する。そのような場合、各種試薬を容器中に導入し、蛍光偏光検出器のような上述した適切な検出装置を使用して典型的にアッセイする。容器以外に、ガラスまたはプラスチックのような平坦表面も使用することができ、反応成分はその表面にアレー状にスポットすることができる(マイクロアレーのように)。
別の等しく好ましいのは、反応容器が流体用チャネル、好ましくは微小流体用チャネルを含むことである。本明細書において、微小流体用なる用語は、約1ミクロン〜約500ミクロン範囲の少なくとも1つの断面寸法を有するチャネルまたは他の導管をいう。本明細書において説明する方法を実施するのに有用な微小流体装置の例としては、例えば、米国特許第5,942,443号、第5,779,868号、および国際特許出願 WO 98/46438号に記載されている装置があり、これら特許の開示は、引用により本明細書に取り込まれる。
上述の方法に従い、第1および第2反応物間の酵素介在カップリング反応を微小流体装置のチャネル内で実施することが考慮されている。そのように、微小流体工学基本骨格を使用することにより、真核細胞の区分化を模倣することが可能になり得る。この方法は、その場合、酵素活性に影響し得る他のタンパク質および細胞成分との関連において、より天然の環境において酵素により触媒作用される基転移反応の活性をモニターし得るであろう。従って、基転移反応を触媒する酵素の活性およびその活性の阻害結果に関するデータは、生体内環境をもっと正確に反映する条件設定において得ることができる。
【0070】
これらのアッセイ系は、興味ある酵素反応に影響を与える試験化合物をスクリーニングし得ることが考慮されている。必要に応じて、本発明に従って使用する装置は、例えば、米国特許第5,942,443号に記載されているような高処理量スクリーニング方式において操作するように形成される。とりわけ、潜在的試験化合物を装置本体と一体化させた貯蔵容器から反応領域に搬送する代りに、そのような試験化合物を、装置本体に装着し反応領域と流体的に連結させた外部サンプリングピペッターまたはキャピラリーにより、反応領域に導入する。そのようなピペッター装置は、例えば、米国特許第5,779,868号に記載されている(その全体を参考として本明細書に合体させる)。サンプリングピペッターは種々の試験化合物の源中に連続して浸漬され、それらの化合物は個別にまたは連続して反応領域に持ち込まれ、注目する反応に対してそれらの化合物の影響(あり得る場合の)が確認される。
これらの微小流体チャネルネットワークのチャネルを通る物質の移動は、電気動力学物質移動(例えば、米国特許第5,858,195号に記載されているような(該米国特許は引用により全てが本明細書に取り込まれる))、圧力系流動、軸流動、重力流動、またはこれらの任意のハイブリッド形のような任意の各種公知の方法によって実施する。
【0071】
実施例5
アッセイキット
本発明のもう1つの実施態様は、基転移反応の供与体生成物または基転移反応の供与体生成物を産生させる触媒活性を検出し定量するためのキットである。基転移反応の一般的等式は、供与体-X + 受容体 → 供与体-生成物 + 受容体-Xを含み、供与体-生成物を一般的検出反応:第1複合体 + 供与体-生成物 → 第2複合体 + 置換された検出可能タグによって検出する。最も単純な形において、FPIAイムノアッセイ用のキットは、巨大分子(即ち、抗体または不活化酵素)およびトレーサー(転移した)および、場合により注目する特異的基転移酵素を含み得る。前記巨大分子およびトレーサーは、別々または1つの溶液容器に混入させているかのいずれでもよいことに留意されたい。
また、前記キットは、不活化供与体、検出可能タグ、受容体基質、阻害因子、緩衝液、補因子、安定化剤、当該キットを使用するための使用説明書セットまたは包装材料、およびこれらの任意の組合せのような成分も含む。さらに、前記キットは、検出可能タグがその発生させる観測可能物理量に基づき識別され得る2以上の抗体/検出可能タグ対を使用することにより、多重検出用に定式化し得る。このイムノアッセイ法は、前記供与体生成物または前記供与体生成物を生成させる触媒活性を検出するのに使用し得る。
本発明の実施に当っては、前記キットを、酵素(該酵素は基転移反応において前記供与体生成物を産生させる)と接触し得る分子のライブラリーまたは分子群をスクリーニングするのに使用し得ることも考慮される。前記ライブラリーは、複数の化学分子、複数の核酸、複数のペプチド、およびこれらの組合せの少なくとも1つを含み得る;スクリーニングは、マルチウェルプレートまたは微小流体装置を使用する高処理量スクリーニング法により実施する。
さらに、前記キット内の巨大分子が、記供与体生成物と高親和性で接触し得る、抗体、ポリペプチド、タンパク質、核酸分子、不活化酵素、およびこれらの組合せの少なくとも1つを含むことが考慮される。さらに、前記キットが、スルホトランスフェラーゼ、キナーゼまたはUDP-グルクロノシルトランスフェラーゼ、メチルトランスフェラーゼ、アセチルトランスフェラーゼ、グルタチオントランスフェラーゼまたはADP-リボシルトランスフェラーゼ、およびこれらの組合せの少なくとも1つを含むことも考慮される。
【0072】
FPIAは適切な検出方式であるけれども、本発明の範囲には、均質アッセイ法、均質蛍光強度免疫測定法、均質蛍光寿命免疫測定法、均質蛍光共鳴エネルギー移動(FRET)免疫測定法または均質化学発光免疫測定法、或いは酵素結合免疫測定法(ELISA)のような非均質アッセイ法のような、供与体生成物または該供与体生成物を生成させる触媒活性を他の手段によって検出するのに使用するように設計されたキットも属する。均質蛍光強度免疫測定法または均質蛍光寿命免疫測定法の場合は、前記キットは、抗体と蛍光検出可能タグとから構成することができ、前記検出可能タグの強度および/または寿命は抗体と結合しているときには溶液中で遊離しているときと異なる。蛍光の差異、即ち、アッセイシグナルは、抗体の検出可能タグとの相互作用がその蛍光特性のさらなる変化、即ち、消光、増強または寿命の変化をもたらすような抗体の修飾によって増強させ得るであろう。均質FRET免疫測定法の場合は、検出可能タグと結合させた第1蛍光体と抗体に結合させた第2蛍光体(直接またはビオチンおよびストレプタビジンのような会合型結合性分子を介して)との相互作用が第2蛍光体の励起をもたらし(またはその逆)、それによって検出可能タグの波長とは異なる波長で蛍光放出を生じさせる。第2蛍光体は、小有機分子または発光ランタニドプローブであり得る。(ランタニド発光は、蛍光ではなく、発光系共鳴エネルギー移動、即ち、LRETと称することに留意されたい)。化学発光検出の場合は、検出可能タグは、β-ガラクトシダーゼのような化学発光検出において使用する酵素の1つのフラグメントに結合させた供与体生成物であり得るであろう。前記供与体生成物-フラグメント-1複合体は、供与体生成物により抗体から外されると、前記酵素のフラグメント2と結合して、適切な基質により化学発光を発生させ得るインタクト活性酵素を生成させる。ELISAの場合は、アッセイは均質ではなく、供与体生成物または抗体をマルチウェルプレートの表面に固定化させる必要があろう。検出酵素に結合させた二次抗体も、この方式に含ませ得るであろう。
【0073】
本明細書において引用した刊行物は、全て、その全体を本明細書に合体させる。本発明の開示と合体させた刊行物間で論争がある場合、本発明の開示が支配する。
今回、本発明を幾つかの具体例で説明し例示してきたが、当業者であれば、説明した範囲内でなし得る変更、付加および削除のような種々の修正を認識するであろう。従って、これらの修正も本発明に属し、そして、本発明の範囲は、特許請求の範囲に合法的に従う最も広い解釈によってのみ限定されるものとする。
【図面の簡単な説明】
【0074】
【図1】SULT反応生成物、即ち、PAPについての蛍光偏光免疫測定(FPIA)反応を例示する。
【図2】FPIAを使用してのUDPの生成、即ち、UGT反応の供与体生成物の検出と定量を例示する。
【図3】FPIAを使用してのADPの生成、即ち、キナーゼ反応の供与体生成物の検出と定量を例示する。
【図4】SULT1E1 FPIA用の試薬、即ち、抗-PAP抗体および蛍光標識化PAP検出可能タグの反復同時開発戦略を説明する。
【図5】図AおよびBは、UDP/UTPに対して産生させたポリクローナル抗体および商業的に入手し得るトレーサー分子(Alexa-UTP)を使用してのウリジンヌクレオチドの滴定または拮抗置換曲線を示す。
【図6】PAP抗原の合成を示す。
【図7】図A〜Cは、PAPトレーサー合成の諸ヨウ素を示す。
【図8】図A〜Cは、代表的な最終トレーサー構造を示す。
【図9】抗-PAP抗体およびPAP-フルオレセイントレーサーの結合等温線を示す。
【図10】Ab 3642に由来する2つの異なるトレーサーのPAPおよびPAPSによる拮抗置換を示す。
【図11】図AおよびBは、2つの抗-PAP抗体/トレーサー組合せによる拮抗曲線を示す。
【図12】酵素により産生させたPAPの検出におけるPAPS濃度の効果を示す。
【図13】図A〜Fは、種々の基質によるSULT活性の連続FPIA系検出を示す。
【図14】FPIA系アッセイおよび35S-PAPS放射アッセイを使用して測定したSULT1E1受容体基質プロフィールの比較を示す。
【図15】FPIA系アッセイにより測定した2,6ジクロロ-4-ニトロフェノール(DCNP)によるSULT1E1の阻害を示す。
【特許請求の範囲】
【請求項1】
a) 活性化型の供与体を触媒活性酵素の存在下に受容体と反応させること;
b) 供与体-生成物および受容体-Xを形成させること;
c) 前記供与体-生成物を、観測可能物理量を生じさせ得る検出可能タグを含む第1複合体と接触させること;
d) 前記第1複合体の検出可能タグを前記供与体-生成物によって拮抗的に置換して第2の複合体および外された検出可能タグを生成させること;および、
e) 前記第1複合体中の検出可能タグおよび前記外された検出可能タグによって発生した観測可能物理量の変化を検出すること;
を含む、基転移反応の供与体生成物の検出方法。
【請求項2】
(f) 工程(e)の観測可能物理量を定量することをさらに含む、請求項1記載の方法。
【請求項3】
a) 供与体の活性化型が供与体-Xを含み;
b) 受容体が触媒活性酵素に対する基質を含み、前記基質が、ポリペプチド、タンパク質、核酸、脂質、炭水化物および小分子基質からなる群から選ばれ;
c) 供与体-生成物がヌクレオチドまたは非ヌクレオチドを含み、前記非ヌクレオチドが、s-アデノシルホモシステイン、ニコチンアミドまたは補酵素Aからなる群から選ばれる代謝中間体であり;
d) 受容体-Xが、Xが共有結合付加物である反応生成物を含み;前記共有結合付加物が、ホスフェート、スルフェート、炭水化物、天然に存在するアミノ酸、合成誘導アミノ酸、ADP-リボース、ヌクレオチド、メチル、アセチルおよびグルタチオン部分からなる群から選ばれ;前記共有結合付加物は、必要に応じて、前記受容体の機能、安定性または機能と安定性の双方を改変することができ;
e) 第1複合体が巨大分子と検出可能タグを含み;かつ、
f) 第2複合体が前記巨大分子を含み、前記検出可能タグが前記供与体-生成物により拮抗的に置換されて観測可能物理量を生じさせる、請求項1記載の方法。
【請求項4】
巨大分子が、供与体-生成物と高親和性で接触可能である、抗体、ポリペプチド、タンパク質、核酸分子、および不活化酵素からなる群から選ばれる、請求項3記載の方法。
【請求項5】
抗体が、モノクローナル抗体、ポリクローナル抗体または組換え抗体である、請求項4記載の方法。
【請求項6】
抗体が供与体-生成物に対して特異的であり、供与体-Xと交叉反応する抗体のレベルが、前期抗体が前期供与体-生成物に対して示す特異性のレベルよりも低い、請求項4記載の方法。
【請求項7】
検出可能タグがトレーサーであり、前記トレーサーがヌクレオチドまたは非ヌクレオチドに結合させた蛍光または化学発光分子である、請求項1記載の方法。
【請求項8】
触媒活性を検出することをさらに含み、前記触媒活性が基転移反応において供与体-生成物を生成させる活性である、請求項1記載の方法。
【請求項9】
触媒活性が化学触媒活性、酵素活性またはこれらの組合せを含み、前記酵素活性が、スルホトランスフェラーゼ、キナーゼ、UDP-グルクロノシルトランスフェラーゼ、メチルトランスフェラーゼ、アセチルトランスフェラーゼ、グルタチオントランスフェラーゼおよびADP-リボシルトランスフェラーゼの活性を含む、請求項8記載の方法。
【請求項10】
イムノアッセイ法である、請求項1記載の方法。
【請求項11】
イムノアッセイ法が、蛍光偏光免疫測定法(FPIA)、蛍光共鳴エネルギー移動法(FRET)、酵素結合免疫吸着測定法(ELISA)、化学発光免疫測定法からなる群から選ばれる、請求項10記載の方法。
【請求項12】
基転移反応酵素を活性化または阻害し得る分子を同定するために、化学ライブラリーのスクリーニングにおいて使用される、請求項1記載の方法。
【請求項13】
分子が、受容体の機能、安定性または機能と安定性の双方を改変し得る、請求項12記載の方法。
【請求項14】
分子が治療作用を示し得る、請求項12記載の方法。
【請求項15】
ライブラリーを、マルチウェルプレート、マイクロアレーまたは微小流体装置を含む高処理量スクリーニング法を使用してスクリーニングする、請求項12記載の方法。
【請求項16】
供与体-生成物と供与体を高供与体濃度の存在下に選択的に識別する能力を有する、基転移反応の供与体-生成物に対して生成された抗体。
【請求項17】
供与体生成物が、ヌクレオチドまたは非ヌクレオチドからなる群から選ばれる、請求項16記載の抗体。
【請求項18】
ヌクレオチドのリン酸部分に対して特異的であり、5'-ホスフェート、5'-ホスホスルフェート、5'-ジホスフェートおよび5'-トリホスフェート間を識別する能力を有する、請求項16記載の抗体。
【請求項19】
下記の工程を含む、基転移反応の供与体-生成物の均質拮抗結合アッセイ法:
a) 供与体-生成物をトレーサーおよび巨大分子と混合して混合物を調製する工程であって、前記巨大分子は前記供与体生成物に対し特異的であり、前記トレーサーはフルオロフォアに結合させた前記供与体生成物を含み、前記トレーサーは前記巨大分子と結合して蛍光偏光において検出可能な変化を生じさせることのできる、前記工程;
b) 前記混合物の蛍光偏光を測定して蛍光偏光測定値を得る工程;および、
c) 前記蛍光偏光測定値を特性決定した蛍光偏光値と比較する工程であって、前記特性決定した蛍光偏光値が既知の供与体生成物濃度に対応する、前記工程。
【請求項20】
基転移反応が酵素によって触媒作用される、請求項19記載のアッセイ法。
【請求項21】
酵素が、キナーゼ、スルホトランスフェラーゼ、メチルトランスフェラーゼ、UDP-グルクロノシルトランスフェラーゼ、アセチルトランスフェラーゼ、グルタチオントランスフェラーゼおよびADP-リボシルトランスフェラーゼからなる群から選ばれる、請求項19記載のアッセイ法。
【請求項22】
供与体-生成物が、ホスホアデノシン-ホスホスルフェート(PAP)、アデノシンジホスフェート(ADP)、ウリジンジホスフェート(UDP)、s-アデノシルホモシステイン、ニコチンアミドおよび補酵素Aからなる群から選ばれる、請求項19記載のアッセイ法。
【請求項23】
フルオロフォアが、フルオレセイン、ローダミン、テキサスレッドおよびこれらの誘導体からなる群から選ばれる、請求項19記載のアッセイ法。
【請求項24】
化学ライブラリーをスクリーニングして基転移反応酵素を阻害または活性化し得る分子を同定するための、請求項19記載のアッセイ法の使用方法。
【請求項25】
巨大分子とトレーサーの各々を、供与体生成物の少なくとも1回の均質蛍光偏光アッセイ法に適する量で含み、前記巨大分子が抗体または不活化酵素であり、前記巨大分子と前記トレーサーが容器内に別々または一緒に存在し得る、基転移反応からの供与体-生成物の特性決定用アッセイキット。
【請求項26】
こん包材と、抗体およびトレーサーの均質蛍光偏光アッセイ法における使用に関する使用説明書とをさらに含み、前記抗体が供与体-生成物に対して特異的であり、前記トレーサーがフルオロフォアに結合させた供与体-生成物を含み、前記トレーサーが前記抗体に結合して蛍光偏光において検出可能な変化を発生させ得る、請求項25記載のアッセイキット。
【請求項27】
フルオロフォアが、フルオレセイン、ローダミン、テキサスレッドおよびこれらの誘導体からなる群から選ばれる、請求項26記載のアッセイキット。
【請求項1】
a) 活性化型の供与体を触媒活性酵素の存在下に受容体と反応させること;
b) 供与体-生成物および受容体-Xを形成させること;
c) 前記供与体-生成物を、観測可能物理量を生じさせ得る検出可能タグを含む第1複合体と接触させること;
d) 前記第1複合体の検出可能タグを前記供与体-生成物によって拮抗的に置換して第2の複合体および外された検出可能タグを生成させること;および、
e) 前記第1複合体中の検出可能タグおよび前記外された検出可能タグによって発生した観測可能物理量の変化を検出すること;
を含む、基転移反応の供与体生成物の検出方法。
【請求項2】
(f) 工程(e)の観測可能物理量を定量することをさらに含む、請求項1記載の方法。
【請求項3】
a) 供与体の活性化型が供与体-Xを含み;
b) 受容体が触媒活性酵素に対する基質を含み、前記基質が、ポリペプチド、タンパク質、核酸、脂質、炭水化物および小分子基質からなる群から選ばれ;
c) 供与体-生成物がヌクレオチドまたは非ヌクレオチドを含み、前記非ヌクレオチドが、s-アデノシルホモシステイン、ニコチンアミドまたは補酵素Aからなる群から選ばれる代謝中間体であり;
d) 受容体-Xが、Xが共有結合付加物である反応生成物を含み;前記共有結合付加物が、ホスフェート、スルフェート、炭水化物、天然に存在するアミノ酸、合成誘導アミノ酸、ADP-リボース、ヌクレオチド、メチル、アセチルおよびグルタチオン部分からなる群から選ばれ;前記共有結合付加物は、必要に応じて、前記受容体の機能、安定性または機能と安定性の双方を改変することができ;
e) 第1複合体が巨大分子と検出可能タグを含み;かつ、
f) 第2複合体が前記巨大分子を含み、前記検出可能タグが前記供与体-生成物により拮抗的に置換されて観測可能物理量を生じさせる、請求項1記載の方法。
【請求項4】
巨大分子が、供与体-生成物と高親和性で接触可能である、抗体、ポリペプチド、タンパク質、核酸分子、および不活化酵素からなる群から選ばれる、請求項3記載の方法。
【請求項5】
抗体が、モノクローナル抗体、ポリクローナル抗体または組換え抗体である、請求項4記載の方法。
【請求項6】
抗体が供与体-生成物に対して特異的であり、供与体-Xと交叉反応する抗体のレベルが、前期抗体が前期供与体-生成物に対して示す特異性のレベルよりも低い、請求項4記載の方法。
【請求項7】
検出可能タグがトレーサーであり、前記トレーサーがヌクレオチドまたは非ヌクレオチドに結合させた蛍光または化学発光分子である、請求項1記載の方法。
【請求項8】
触媒活性を検出することをさらに含み、前記触媒活性が基転移反応において供与体-生成物を生成させる活性である、請求項1記載の方法。
【請求項9】
触媒活性が化学触媒活性、酵素活性またはこれらの組合せを含み、前記酵素活性が、スルホトランスフェラーゼ、キナーゼ、UDP-グルクロノシルトランスフェラーゼ、メチルトランスフェラーゼ、アセチルトランスフェラーゼ、グルタチオントランスフェラーゼおよびADP-リボシルトランスフェラーゼの活性を含む、請求項8記載の方法。
【請求項10】
イムノアッセイ法である、請求項1記載の方法。
【請求項11】
イムノアッセイ法が、蛍光偏光免疫測定法(FPIA)、蛍光共鳴エネルギー移動法(FRET)、酵素結合免疫吸着測定法(ELISA)、化学発光免疫測定法からなる群から選ばれる、請求項10記載の方法。
【請求項12】
基転移反応酵素を活性化または阻害し得る分子を同定するために、化学ライブラリーのスクリーニングにおいて使用される、請求項1記載の方法。
【請求項13】
分子が、受容体の機能、安定性または機能と安定性の双方を改変し得る、請求項12記載の方法。
【請求項14】
分子が治療作用を示し得る、請求項12記載の方法。
【請求項15】
ライブラリーを、マルチウェルプレート、マイクロアレーまたは微小流体装置を含む高処理量スクリーニング法を使用してスクリーニングする、請求項12記載の方法。
【請求項16】
供与体-生成物と供与体を高供与体濃度の存在下に選択的に識別する能力を有する、基転移反応の供与体-生成物に対して生成された抗体。
【請求項17】
供与体生成物が、ヌクレオチドまたは非ヌクレオチドからなる群から選ばれる、請求項16記載の抗体。
【請求項18】
ヌクレオチドのリン酸部分に対して特異的であり、5'-ホスフェート、5'-ホスホスルフェート、5'-ジホスフェートおよび5'-トリホスフェート間を識別する能力を有する、請求項16記載の抗体。
【請求項19】
下記の工程を含む、基転移反応の供与体-生成物の均質拮抗結合アッセイ法:
a) 供与体-生成物をトレーサーおよび巨大分子と混合して混合物を調製する工程であって、前記巨大分子は前記供与体生成物に対し特異的であり、前記トレーサーはフルオロフォアに結合させた前記供与体生成物を含み、前記トレーサーは前記巨大分子と結合して蛍光偏光において検出可能な変化を生じさせることのできる、前記工程;
b) 前記混合物の蛍光偏光を測定して蛍光偏光測定値を得る工程;および、
c) 前記蛍光偏光測定値を特性決定した蛍光偏光値と比較する工程であって、前記特性決定した蛍光偏光値が既知の供与体生成物濃度に対応する、前記工程。
【請求項20】
基転移反応が酵素によって触媒作用される、請求項19記載のアッセイ法。
【請求項21】
酵素が、キナーゼ、スルホトランスフェラーゼ、メチルトランスフェラーゼ、UDP-グルクロノシルトランスフェラーゼ、アセチルトランスフェラーゼ、グルタチオントランスフェラーゼおよびADP-リボシルトランスフェラーゼからなる群から選ばれる、請求項19記載のアッセイ法。
【請求項22】
供与体-生成物が、ホスホアデノシン-ホスホスルフェート(PAP)、アデノシンジホスフェート(ADP)、ウリジンジホスフェート(UDP)、s-アデノシルホモシステイン、ニコチンアミドおよび補酵素Aからなる群から選ばれる、請求項19記載のアッセイ法。
【請求項23】
フルオロフォアが、フルオレセイン、ローダミン、テキサスレッドおよびこれらの誘導体からなる群から選ばれる、請求項19記載のアッセイ法。
【請求項24】
化学ライブラリーをスクリーニングして基転移反応酵素を阻害または活性化し得る分子を同定するための、請求項19記載のアッセイ法の使用方法。
【請求項25】
巨大分子とトレーサーの各々を、供与体生成物の少なくとも1回の均質蛍光偏光アッセイ法に適する量で含み、前記巨大分子が抗体または不活化酵素であり、前記巨大分子と前記トレーサーが容器内に別々または一緒に存在し得る、基転移反応からの供与体-生成物の特性決定用アッセイキット。
【請求項26】
こん包材と、抗体およびトレーサーの均質蛍光偏光アッセイ法における使用に関する使用説明書とをさらに含み、前記抗体が供与体-生成物に対して特異的であり、前記トレーサーがフルオロフォアに結合させた供与体-生成物を含み、前記トレーサーが前記抗体に結合して蛍光偏光において検出可能な変化を発生させ得る、請求項25記載のアッセイキット。
【請求項27】
フルオロフォアが、フルオレセイン、ローダミン、テキサスレッドおよびこれらの誘導体からなる群から選ばれる、請求項26記載のアッセイキット。
【図1】


【図2】

【図3】
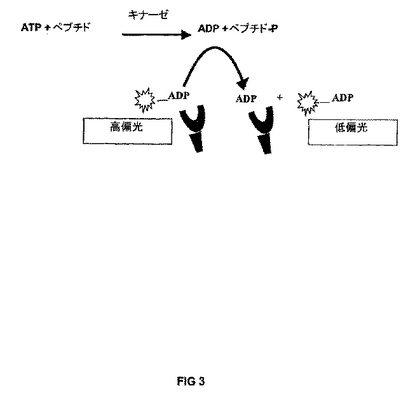
【図4】
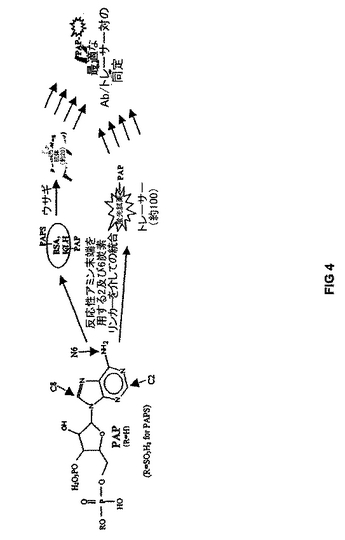
【図5】

【図6】

【図7】

【図8】

【図9】

【図10】

【図11】

【図12】

【図13】

【図14】

【図15】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公表番号】特表2006−519013(P2006−519013A)
【公表日】平成18年8月24日(2006.8.24)
【国際特許分類】
【出願番号】特願2006−503179(P2006−503179)
【出願日】平成16年1月30日(2004.1.30)
【国際出願番号】PCT/US2004/002618
【国際公開番号】WO2004/068115
【国際公開日】平成16年8月12日(2004.8.12)
【出願人】(505289443)ベル ブルック ラブズ リミテッド ライアビリティ カンパニー (2)
【Fターム(参考)】
【公表日】平成18年8月24日(2006.8.24)
【国際特許分類】
【出願日】平成16年1月30日(2004.1.30)
【国際出願番号】PCT/US2004/002618
【国際公開番号】WO2004/068115
【国際公開日】平成16年8月12日(2004.8.12)
【出願人】(505289443)ベル ブルック ラブズ リミテッド ライアビリティ カンパニー (2)
【Fターム(参考)】
[ Back to top ]