
新生児仮死を治療するための低体温療法におけるキセノンの使用
本発明は、対象である新生児において新生児仮死を治療する薬剤であって、低体温療法と組み合わせて使用する薬剤の調製におけるキセノンの使用に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新生児仮死の治療方法に関する。
【背景技術】
【0002】
低酸素性虚血症(HI)としても知られる新生児(又は周生期)仮死は、陣痛時、分娩時、又は出生直後の時期に、充分な酸素が産児の体内に取り込まれないことが原因で生じる状態である。新生児仮死は、依然として新生児における慢性神経疾患罹患率及び急性死亡率の主要な原因となっており(Balduini et al, 2000; Vannucci et al, 1997)、低酸素性虚血性脳症に至る場合が多い。
【0003】
6分間という短時間であっても、新生児仮死(低酸素症)によって永久的な神経損傷が生じる恐れがあることが、これまでの研究によって示されている。仮死状態の霊長類新生児では脳組織の喪失が認められ、脳組織の喪失は記憶障害及び痙攣性麻痺と相関がある(Windle, WF, 1969)。
【0004】
出生時における全死亡数の約14.6%は、新生児仮死によるものである。西洋においては、新生児の約0.9%(すなわち、100〜130,000)に、新生児仮死が発生している。約15〜20%は死亡し、命を取り留めた新生児の25%が、精神遅滞、脳性麻痺、痙攣、学習障害及び/又はてんかんなどの長期にわたる合併症による重度の障害を背負っている(Law et al, 1993; Perlman et al, 1999)。また、軽度の仮死状態で合併症も起こさずに回復したと当初は思われていた子どもが、この新生児期の障害に由来すると思われるような問題行動を小児期に示すという認識も増えてきている。新生児仮死は、罹患率が住人10000人当たり5人未満であるため希少疾病医薬品としての基準を満たしているとともに、生命を脅かす、確立した治療法のない重篤な消耗性疾患である。
【0005】
HIモデル動物の新生児におけるこの種の脳障害に関与する細胞死の機序には、グルタミン受容体、特に、シナプス形成期における神経毒性に対する感受性が最も高いことから、N−メチル−D−アスパラギン酸(NMDA)受容体の過剰活性化(Jevtovic-Todorovic and Olney, 2003)によって、及びアポトーシス性神経変性(Ikonomidou et al, 1989; Pohl et al, 1999)によって生じる興奮毒性損傷(又は壊死)の組み合わせが含まれることが、これまでに示されている。この種の損傷は、低酸素性障害の重篤度と関連しており(Jevtovic-Todorovic and Olney, 2003)、また、別の脳領域の脆弱性におけるばらつきにも関連している(Northington et al, 2001)。現時点では、HIによって生じた急性神経細胞死に対処する効果的な治療法は存在しないが、多種多様な薬理学的及び非薬理学的介入の実験的研究が進行中である(Vannucci and Perlman, 1997)。
【0006】
本発明は、新生児仮死の治療方法の提供を目指すものである。
【0007】
【非特許文献1】Balduini et al, 2000
【非特許文献2】Vannucci et al, 1997
【非特許文献3】Windle, WF, 1969
【非特許文献4】Law et al, 1993
【非特許文献5】Perlman et al, 1999
【非特許文献6】Jevtovic-Todorovic and Olney, 2003
【非特許文献7】Ikonomidou et al, 1989
【非特許文献8】Pohl et al, 1999
【非特許文献9】Northington et al, 2001
【非特許文献10】Vannucci and Perlman, 1997
【発明の開示】
【0008】
本発明の第1の態様は、対象である新生児において新生児仮死を治療する薬剤であって、低体温療法と組み合わせて使用する薬剤の調製におけるキセノンの使用に関する。
【0009】
本発明の第2の態様は、新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、
(a)治療上有効な量のキセノンを前記哺乳動物に投与するステップ、及び
(b)前記哺乳動物に低体温療法を施すステップ
を含む方法に関する。
【0010】
本発明の第3の態様は、新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、治療上有効な量のキセノンを、低体温療法と組み合わせて前記哺乳動物に投与するステップを含む方法に関する。
【0011】
本発明の第4の態様は、新生児仮死治療用薬剤の調製におけるキセノンの使用であって、前記治療が、低体温療法と組み合わせて、同時、順次、又は別個にキセノンを対象に投与するステップを含む使用に関する。
【0012】
本発明の第5の態様は、新生児仮死を治療するための、低体温療法との組み合わせによる、キセノンの使用に関する。
【発明を実施するための最良の形態】
【0013】
未成熟CNSの通常の生理機能
興奮性アミノ酸(EAA)のグルタミン酸及びアスパラギン酸は、成熟中枢神経系(CNS)における興奮性シナプス伝達の主要な仲介役となっている(Dingledine and McBain,1999)。それらはまた、未成熟CNSにおいて、シナプス形成、ニューロンの生存、シナプス可塑性、並びに樹状及び軸索構造など、多くの生理学的プロセスに関与しており、未成熟CNSの発達にきわめて重要な役割を演じている。しかし、発達中にこれらのアミノ酸受容体が過剰に活性化すると、ニューロンの損傷及び死につながる恐れがある。これを「興奮毒性」という。
【0014】
EAAのうち、最も豊富に存在するのがグルタミン酸である(Dingledine and McBain,1999)。グルタミン酸はシナプス小胞に保存され、シナプス前終末から放出されると、シナプス後膜のカルシウム依存性膜脱分極を引き起こす。グルタミン酸は、N−メチル−D−アスパラギン酸(NMDA)型と非NMDA型に分けることができる各種受容体サブタイプにてその興奮性作用を発揮するが、発達中のCNSにおいては、HIに付随する脳障害で主要な役割を果たすのはNMDA受容体サブタイプであることが判明している(Ikonomidou et al, 1989; Komuro, 1993; MacDonald et al, 1986)。
【0015】
NMDA受容体はグルタミン酸受容体の主要なサブクラスであり、グルタミン酸は、哺乳類の中枢神経系における最も重要な興奮性神経伝達物質だと考えられている。重要なことに、NMDA受容体の活性化は、頭部外傷、卒中及びその後の心停止に続いて起こる低酸素症及び虚血の結果として生じるものであることに加えて、数多くの病態における興奮毒性及びニューロン死につながる中心的な事象であることが判明している。
【0016】
NMDA受容体はCNS全体に偏在するイオンチャネル型受容体であり、シナプス後膜及びシナプス外膜両方の表面に位置している(Riccio and Ginty, 2002; Sattler et al, 2000)。NMDA受容体はNa+イオンとCa2+イオンの両方に透過性のあるカチオンチャネルと結合し、通常の生理学的状態では、負の静止膜電位でMg2+によってブロックされる。細胞膜の脱分極によってそのブロックは外され、チャネルを通じてCa2+の流入が可能になり、受容体がその細胞内作用を発揮できるようになる(Hardingham and Bading, 2003)。
【0017】
NMDA受容体は正常な脳機能に不可欠であり、正常な生理機能におけるその重要性は、記憶と学習におけるその中心的役割によって実証されている(Bliss and Collingridge, 1993)。これとは逆に、過剰なグルタミン酸によるNMDA受容体の病的活性化は、細胞内Ca2+の制御が乱されるため、脳に対する虚血性障害の後に起こるニューロン細胞死の主要な原因である。これは、HIにおいてNMDA受容体が演じる中心的役割を強調するものである。
【0018】
新生児における低酸素性虚血性障害
脳が機能するためには、酸素と糖の継続的な供給が必要であるため、脳の機能は血液の充分な供給に依存している(Choi and Rothman, 1990)。新生児仮死の場合のように、血液の供給が妨げられると、その後数分以内に下流部位に低酸素性虚血性損傷が発生する。こうした酸素欠乏条件下では、細胞代謝が、好気的代謝から、細胞のエネルギー必要量を満たす効果が低い嫌気的代謝へと変化する(Vannucci and Perlman, 1997)。これによりエネルギーの貯蔵が減少し、特に、ニューロン及びグリア細胞区画におけるATPなどの高エネルギーリン酸の備蓄に影響する(Dingledine and McBain, 1999)。アシドーシスにつながるH+イオンの蓄積と、さらなる細胞損傷の一因となる遊離基の放出とが同時に発生する。
【0019】
生理学的状態下では、ニューロン細胞に位置するグルタミン酸トランスポーターの作用により、グルタミン酸の細胞外濃度は低いレベルに維持されているが、グリア細胞では優先的に発現している(Dingledine and McBain, 1999)。グルタミン酸を取り込む担体は数種類存在するが、基本的に、同様の方式でその機能を発揮する。すなわち、Na+カチオン2個及びグルタミン酸アニオン1個を細胞内へ輸送し、K+カチオン1個及びOH−アニオン1個を細胞内から細胞外空間へ輸送する(Dingledine and McBain, 1999)。これらのイオンポンプは電気化学勾配に逆らって作用するため、正しく機能するには、ATPの形態のエネルギーに依存することになる。よって、HIの結果ATPの濃度が下がることにより、これらのポンプの静止膜電位維持能力が低下する。その結果として、ATP依存性ポンプの不全が、膜の脱分極及びポンピング方向の逆転につながっていく(Eilers and Bickler, 1996; Kauppinen et al, 1988)。このようにして、グルタミン酸は細胞から運び出され、過剰な濃度のグルタミン酸が細胞外空間に蓄積する。取り込みの減少によりグルタミン酸の濃度が上昇するだけではなく、膜脱分極により活動電位が生じるため、シナプス前終末からのグルタミン酸の放出も増加する(Dingledine and McBain, 1999)。過剰な細胞外グルタミン酸をもたらすこうしたプロセスの例は、インビトロ(Bosley et al, 1983; Hauptman et al, 1984; Pellegrino-Giampietro et al, 1990)及びインビボ(Erecinska et al, 1984; Graham et al,1990; Ikeda et al, 1989)の両方で見受けられる。
【0020】
興奮毒性は、過剰な細胞外グルタミン酸が継続的にシナプス後受容体(特にNMDA受容体)を活性化する場合に発生し、結果として生じたカルシウムの流入が浸透圧勾配を作り、それに基づいて水分が移動して細胞の膨張を引き起こす。カルシウム依存性酵素系も細胞内で活性化され、これら2つのプロセスの結果、急性ニューロン細胞死が起こる(Choi and Rothman, 1990)。
【0021】
細胞死の機序
Wyllie et al(Wyllie et al, 1980)が仮定したように、ニューロン細胞死は常に壊死とアポトーシスという2つの機序のいずれかによって起こると考えられてきた。しかし、最近はこれらのカテゴリーが疑問視されている。なぜなら、細胞死は興奮毒性細胞死とアポトーシスというカテゴリーに分けるべきだと示唆する多くの証拠が明らかになってきたためである(Olney, 2003)。興奮毒性細胞死は、壊死プロセス(Gwag et al, 1997; Katja and Green, 2001)、アポトーシスプロセス、及びそれら2つの連続体(Leist and Nicotera, 1998; Nakajima et al, 2000)として説明されてきた。通常、アポトーシスと壊死とは、そのはっきりと異なる形態学的外観によって区別されている。アポトーシスはATPと新規タンパク質合成を必要とし、細胞収縮、辺縁化を伴うクロマチン凝集、及び膜に囲まれたアポトーシス小体の形成によって確認される。一方、壊死は、核崩壊及び核濃縮という変化を伴う核収縮によって認識される(Hill et al, 1995)。
【0022】
HIによる細胞死では、壊死という初期段階の後に、アポトーシス細胞死という波が遅れて発生することがわかっている(Northington etal, 2001)。その後に生じる障害のタイプは時間依存的かつ位置依存的であると思われ、HIモデル新生児ラットの同側前脳に限局する初期の壊死障害、及び、視床に発生する遅延性アポトーシス障害を伴う(Northington etal, 2001)。このことは、異なる脳領域がHI後の異なる時期の各タイプの細胞死に対して差別的脆弱性を示すことを示唆している。
【0023】
正常な発達において、アポトーシスは、不要な又は損傷したニューロンがそれによって「自殺を図る」定期的な事象である(Ikonomidou et al,2001)。HIでは、初期の興奮毒性細胞死はNMDA受容体の過剰な活性化に仲介され、その結果、グルタミン酸が無制御状態で放出され、周辺のニューロンに損傷をもたらす。ニューロンがプログラムされた細胞死を開始することは、シナプス形成中の損傷に対する自然な反応であり(Olney, 2003)、これは周辺組織を保護するために活性化する機序であると考えられる(Leist and Nicotera, 1998)。
【0024】
神経保護剤としてのキセノン
NMDA受容体が、ある侵害受容経路及び痛みの知覚における場合と同様に、記憶や学習など多くの高度な認知機能の根底にあるシナプス可塑性においても主要な役割を果たしていることは、当該技術で周知である(Collingridge et al, The NMDA Receptor, Oxford University Press, 1994)。また、NMDA受容体のある特質は、意識それ自体の根底にある脳内情報処理にNMDA受容体が関与している可能性を示唆している。
【0025】
NMDA受容体アンタゴニストは、多くの理由により、治療上有用である。第一に、NMDA受容体アンタゴニストによって、全身麻酔及び鎮静の極めて望ましい構成要素である深い無痛覚がもたらされる。第二に、NMDA受容体アンタゴニストは、臨床的に関連する多くの状況(虚血症、脳外傷、神経因性疼痛、及びある種のけいれんを含む)下において神経保護剤となる。第三に、NMDA受容体アンタゴニストにより、有益な程度の健忘症がもたらされる。
【0026】
HIの発症機序におけるNMDA受容体の重要性を考えると、神経保護剤の候補としてNMDAアンタゴニストを研究してきたのは適切なことであった。MK−801やケタミンなどの多くのNMDAアンタゴニストは、インビボモデル及びインビトロモデルの両方で神経保護剤となることがわかっている(Albers et al, 1989; Arias et al, 1999; Choi et al, 1988; Kudo et al, 2001)。しかし、こうした心強い結果にもかかわらず、NMDA受容体アンタゴニストは、ヒトにおける精神異常発現という副作用も有すること(Krystal et al, 1994)、並びに後帯状皮質(PC)及び後板状皮質(RS)に損傷を引き起こすこと(Olney et al, 1991)がわかっている。さらに、従来の多くのNMDA受容体アンタゴニストは、不随意運動の生起、交感神経系の刺激、高用量時の神経毒性の誘発(NMDA受容体アンタゴニストは全身麻酔剤としては効力が弱いため、これはもっともなことである)、心筋の機能低下、及びいくつかのてんかん発生例におけるけいれん前症状(proconvulsions)、例えば、「燃え上がり現象(kindling)」をもたらす(Wlaz P et al, Eur. J. Neurosci. 1994; 6: 1710-1719)。また、血液脳関門を通過できる新規NMDA受容体アンタゴニストの開発には、これまでかなりの困難を伴っている。
【0027】
キセノンは、無極性の不活性ガスで、強力なNMDAアンタゴニストである(Franks et al, 1998)。他のNMDAアンタゴニストと同様に、インビトロ(Petzelt et al, 2003)及びインビボ(Homi et al, 2003; Wilhelm et al, 2002)の両方で多くの形態のニューロン損傷に対する神経保護剤となることが判明している。しかし、他の多くのNMDA受容体アンタゴニストとは異なり、キセノンには神経毒性がない(Ma et al, 2002)。キセノンをNMDAアンタゴニストとして使用することのさらなる利点は、その分子が不活性で、呼吸により速やかに排出可能な揮発性のガスである点にある。
【0028】
キセノンは他にも好ましい性質を数多く有している。外科で最初に用いられて以来(Cullen SC et al, Science 1951; 113: 580-582)、代謝副産物がないこと、深い無痛覚、薬効の発現及び薬効からの回復が速やかであること、心血管系への影響が最小であることなどの優れた薬理学的性質がキセノンにあることを、多数の研究グループが示してきた(Lachmann B et al, Lancet 1990; 335: 1413-1415; Kennedy RR et al, Anaesth. Intens. Care 1992; 20: 66-70; Luttropp HH et al, Acta Anaesthesiol. Scand. 1994; 38: 121-125; Goto T et al, Anesthesiology 1997; 86: 1273-1278; Marx T et al, Br. J. Anaesth. 1997; 78: 326-327)。また、キセノンは小型で電荷のない原子であるため、容易に血液脳関門を通過することが可能であり、したがって、作用が速やかに現れる(Nakata et al, 2001)。血液ガス分配計数も非常に低く、この計数の低さがキセノン麻酔からの迅速な覚醒に役立っている(Goto et al, 1997)。これらの利点に加え、キセノンは非爆発性、無毒及び非反応性である(Shichino et al, 2002)ため、キセノンは新生児の神経保護剤としての使用に理想的な候補物質となっている。
【0029】
ここで使用する「神経保護剤」との語は、神経保護作用をもたらすこと、すなわち、ニューロンなどの神経に関わる構成要素を、例えば虚血性障害又は外傷性障害に由来する、進行中の障害から保護することが可能な作因を意味する。
【0030】
神経保護剤としての低体温療法
1941年、手術で用いる低体温療法の神経保護性を、Talbotが初めて実証した(Talbot, 1941)。現在、低体温療法が決まって用いられるのは、心肺バイパス手術のみであり、脳の術中虚血を防止することを目的としている。しかし、他の脳障害モデルにおける低体温療法の治療効果を実証する発表がこれまでに何件かなされている。例えば、新生児仮死のインビトロモデル(Onitsuka et al, 1998)及びインビボモデルの両方について、低体温療法のよい効果を示す多くの発表がなされている(Debillon et al, 2003; Treschera et al, 1997)。組織の障害と脳の冷却程度との間には直接的な相関が存在することが示されており(Towfighi et al,1994)、正常酸素状態では、体温が1℃低下するごとに大脳の代謝速度が5%低下する(Yager and Asselin, 1996)。
【0031】
低体温療法がその神経保護的効果を発揮する機序はまだ解明されていないが、多くの理論が想定されている。低体温療法が保護作用を示す機序は温度及び時間依存的であり、HI障害につながる一連の事象に沿った複数のポイントで作用すると思われることが、これまでの研究で示唆されている(Yager and Asselin, 1996)。このことは、31℃という中程度の体温は大脳のエネルギー代謝を低下させることにより神経保護作用を示す一方で、34℃という軽度の低体温療法は、同じく神経保護作用を示すもののエネルギー代謝には影響を及ぼさないため、異なる機序を介して作用しているに違いない、という事実によって裏付けられている(Yager and Asselin, 1996)。Taylor et alによる他の研究(Taylor et al, 2002)では、HI障害後に開始した低体温療法は、虚血中の低体温療法よりも効果的であると示しており、これは回復期に発生する有害な作用を減少させることによるものであろうと示唆している。そうした機序の一例として、再灌流中に続いて起こる興奮毒性損傷が低体温療法によって減少することが挙げられるかもしれない(Taylor et al, 2002)。活性酸素種の減少(Taylor et al, 2002)、組織アシドーシスの減少(Chopp et al, 1989)、及びHI後におけるニューロンのアポトーシスの緩和(Xu et al, 2002)を含め、低体温療法による保護機序は他にも多く示唆されている。
【0032】
キセノンと低体温療法の組み合わせ
前述のとおり、本発明の第1の態様は、対象である新生児において新生児仮死を治療する薬剤であって、低体温療法と組み合わせて使用する薬剤の調製におけるキセノンの使用に関する。
【0033】
ここで使用されている「低体温療法」との語は、ある特定の対象(この場合は新生児の対象)を、例えば受動的又は能動的な技法によって体温を好ましくは3〜5℃下げることにより、低体温状態にすることに関するものである。一般的には、低体温状態にすると対象の体組織代謝が減少し、それにより、酸素要求量も減少する。
【0034】
前述のとおり、新生児仮死の治療における低体温療法の使用は、当該技術において文書で充分に裏付けられている(例えば、Volpe, 2001; Gunn et al, 2000を参照のこと)。しかし、低体温療法をキセノン投与と組み合わせて使用できるかもしれないということについては、今日まで当該技術において教示も示唆もされてこなかった。そのような組み合わせ療法によって、結果として得られる神経保護作用が驚異的かつ予想もしない程度に増強されるだろうとの示唆も皆無であった。
【0035】
本出願人による以前の研究により、キセノンに神経保護性があることが明らかになった。特にここに参照としてその内容を組み込んでいるWO01/08692は、神経保護剤として、及び/又はシナプス可塑性の阻害剤としてのキセノンの使用に関するものである。しかし、本出願で請求項に記載されている発明との関連においてキセノンが神経保護剤として効果的であることは、先行技術には教示も示唆もされていない。
【0036】
本発明のある好ましい実施形態において、キセノンは薬学的に許容可能な希釈剤、賦形剤又は担体と混合されている。
【0037】
本明細書に記載した種々様々な形態の医薬組成物に適切な賦形剤の例は、Handbook of Pharmaceutical Excipients, 2nd Edition, (1994), Edited by A Wade and PJ Wellerに掲載されている。
【0038】
治療用として許容可能な担体又は希釈剤は薬学分野において周知であり、例えば、Remington's Pharmaceutical Sciences, Mack Publishing Co. (A. R. Gennaro edit. 1985) に記載されている。適切な担体の例としては、ラクトース、澱粉、グルコース、メチルセルロース、ステアリン酸マグネシウム、マンニトール、ソルビトールなどが挙げられる。適切な希釈剤の例としては、エタノール、グリセロール及び水が挙げられる。
【0039】
薬剤用の担体、賦形剤、又は希釈剤の選択は、目的の投与経路及び薬学の標準的技法を参照しながら行うことができる。医薬組成物には、担体、賦形剤又は希釈剤が含まれていてもよく、あるいはそれらに加えて、適切な結合剤、滑沢剤、懸濁剤、コーティング剤、可溶化剤が含まれていてもよい。
【0040】
適切な結合剤の例としては、澱粉、ゼラチン、並びにグルコース、無水ラクトース、フリーフローラクトース、ベータラクトース、コーンシロップなどの天然糖、並びにアカシア、トラガカント又はアルギン酸ナトリウムなどの天然及び合成ゴム、カルボキシメチルセルロース及びポリエチレングリコールが挙げられる。
【0041】
適切な滑沢剤の例としては、オレイン酸ナトリウム、ステアリン酸ナトリウム、ステアリン酸マグネシウム、安息香酸ナトリウム、酢酸ナトリウム、塩化ナトリウムなどが挙げられる。
【0042】
防腐剤、安定剤及び着色剤を医薬組成物に添加してもよい。防腐剤の例として、安息香酸ナトリウム、ソルビン酸、及びp−ヒドロキシ安息香酸エステルが挙げられる。抗酸化剤及び懸濁剤も用いてもよい。
【0043】
本発明は、動物の治療にも適用することができる。この点において本発明はさらに、獣医学的に許容可能な希釈剤、賦形剤又は担体と組み合わせたキセノンの使用にも関するものである。
【0044】
獣医学用としては、獣医学における通常の技法にしたがってキセノンを投与するのが一般的であり、ある特定の動物に最も適切と思われる投与計画及び投与経路の決定は、獣医が行う。
【0045】
他の薬学的活性物質と組み合わせてキセノンを使用してもよい。そのような物質としては、GABA性活性を促進する麻酔剤又は鎮静剤を含め、どのような薬学的活性物質であってもよい。かかるGABA性物質の例としては、イソフルラン、プロポフォール及びベンゾジアゼピンが挙げられる。
【0046】
ある好ましい実施形態において、キセノンは、揮発性麻酔剤、好ましくはイソフルラン、セボフルラン又はデスフルランと組み合わせて投与される。
【0047】
また、キセノンを、その他の有効成分、例えばL型カルシウムチャネル遮断薬、N型カルシウムチャネル遮断薬、物質Pアンタゴニスト、ナトリウムチャネル遮断薬、プリン受容体遮断薬、又はそれらを組み合わせたものなどと組み合わせて投与してもよい。
【0048】
単数又は複数の適切な送達機構によって、キセノンを投与してもよい。
【0049】
ある特に好ましい実施形態においては、灌流によりキセノンを投与する。本発明との関連では、「灌流」との語は、特殊な心肺装置を用いて、患者に酸素/キセノン混合体を導入し、患者から二酸化炭素を除去することを示す。一般的な用語では、心肺装置は心臓と肺の機能を代替するものであり、無血で不動の手術野を外科医に提供する。灌流技師は、患者の血液に酸素を供給し、酸素と二酸化炭素の量を制御する。本発明との関連では、灌流技師はキセノンも患者の血液に導入する。その後灌流技師は血液を動脈系に戻し、栄養血流を手術中に患者の全重要臓器及び組織に供給する。
【0050】
ある特に好ましい実施形態では、薬剤はガス状である。
【0051】
別の非常に好ましい実施形態では、キセノンは吸入により投与される。より好ましくは、キセノンは70〜30%v/vのキセノン・酸素混合体の吸入により投与される。
【0052】
より好ましくは、キセノンは20〜70%v/vのキセノン・空気混合体の形態で投与される。
【0053】
本発明のさらに別の好ましい実施形態においては、薬剤は、液体又は溶液の形態である。
【0054】
好ましくは液体は、静脈内、動脈内、髄腔内、皮下、皮内、腹腔内又は筋肉内に注入可能な、溶液の形態、又は無菌若しくは無菌処理可能な溶液から調製された乳剤の形態で投与される。
【0055】
ある特に好ましい実施形態においては、キセノンは、脂肪乳剤の形態で投与される。静脈用製剤は、一般に、キセノンの溶解性を充分に高めて所望の臨床効果を達成する脂肪乳剤(市販されているイントラリピッド(登録商標)10、イントラリピッド(登録商標)20、イントラファット(登録商標)、リポファンジン(Lipofundin)(登録商標)S若しくはリポシン(Liposyn)(登録商標)の乳剤、又は溶解性を最大にするために特別に製剤されたもの)を含んでいる。この種の脂肪乳剤に関する詳しい情報は、G. Kleinberger and H. Pamper, Infusionstherapie, 108-117 (1983) 3に掲載されている。
【0056】
ガスを溶解又は分散させる本発明の脂質相は、一般的に、8〜30個の炭素原子を含む飽和及び不飽和長鎖及び中鎖脂肪酸エステルから形成される。これらの脂質は水性溶液中でリポソームを形成する。例としては、魚油、及び大豆油、アザミ油又は綿実油などの植物油を挙げることができる。一般的に、本発明の脂肪乳剤は、乳剤中の脂肪の割合が従来的に5〜30重量%、好ましくは10〜20重量%の水中油型乳剤である。この種の水中油型乳剤は、大豆リン脂質などの乳化剤の存在下で調製されることが多い。
【0057】
本発明のリポソームを形成する脂質は、天然物でも人工物でもよく、コレステロール、グリコリピッド、スフィンゴミエリン、グルコリピッド、グリコスフィンゴリピッド、ホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルセリン、ホスファチジルグリセロール、ホスファチジルイノシトールなどが例示できる。
【0058】
本発明の脂肪乳剤は、付加的な成分を含んでいてもよい。そうした成分としては、抗酸化剤、脂質相の周囲の水相の浸透圧を血液と等張にする添加剤、又はリポソーム表面を改変するポリマーが挙げられる。
【0059】
相当量のキセノンを脂肪乳剤に添加してもよいことは、すでに立証されている。20℃で通常圧力という最も単純な場合でさえ、キセノンは、乳剤1ml当たり0.2〜10ml以上の濃度で溶解又は分散できる。溶解したガスの濃度は、温度、圧力及び脂質濃度を含めた多くの要因によって左右される。
【0060】
本発明の脂肪乳剤に、ガス状のキセノンを入れておいてもよい。一般に、乳剤と、その乳剤に浸漬されている焼結ガラス気泡管を通過するガス状又は蒸気状の麻酔剤とを装置に充填する。選択した分圧で乳剤を麻酔ガス又は蒸気と平衡させる。気密性容器に保存する場合、これらの脂肪乳剤は、従来の貯蔵期間中はガスとして放出されないという、麻酔剤として十分な安定性を示す。
【0061】
本発明の脂肪乳剤に、飽和レベルになるようにキセノンを入れてもよい。あるいは、より低濃度のキセノンを存在させてもよいが、それは、例えば、その乳剤を投与することにより所望の薬学的活性が得られるというような場合に限られる。
【0062】
本発明で用いるキセノンの濃度は、所望の臨床効果を得るのに必要な最低濃度でよい。通常は、医者が患者個人に最適と思われる実際の投与量を判断し、この投与量はその特定の患者の年齢、体重、及び反応によって異なる。もちろん、個々の事例によっては、高め又は低めの用量域が適切な場合もありうる。それらもまた、本発明の範囲内である。
【0063】
好ましくは、薬剤は、静脈内、軸索内、又は経皮的デリバリーに適切な形態である。
【0064】
好ましくは、キセノンは、低体温療法と同時に、組み合わせて、順次に、又は別個に投与される。
【0065】
本明細書では、「同時」は、キセノンが低体温療法と時を同じくして投与されることを意味するのに用いており、一方「組み合わせて」は、同時ではないにせよ、キセノンが投与されることを意味するのに用いている。「順次」とは、キセノンと低体温療法が共に治療効果を示す一定の時間枠内のこと、すなわち、それらが共に同一の時間枠内で治療効果を示すことができるということである。したがって、キセノンの循環半減期が、対象の新生児が低体温状態に置かれている時点でキセノンが治療上有効な量で存在するようなものであれば、低体温療法の5分前、10分前、又は何時間も前にキセノンを投与しても、「順次」投与となりうる。
【0066】
本発明の別の好ましい実施形態においては、新生児はキセノンでの治療に先立って、低体温療法を施される。
【0067】
「組み合わせて」又は「順次」に対し、「別個」は、キセノンの投与と、新生児への低体温療法の実施との間の間隔がかなり広い、すなわち、対象の新生児が低体温状態に置かれている時点では、もはやキセノンは治療上有効な量で血流中に存在しないことを意味するために、本明細書で用いられている。
【0068】
ある好ましい実施形態においては、キセノンは低体温療法と順次に投与される。
【0069】
より好ましくは、キセノンは低体温療法に先立って、順次に投与される。
【0070】
別の好ましい実施形態においては、キセノンは低体温療法に先立って、別個に投与される。
【0071】
ある好ましい実施形態においては、キセノンは低体温療法の後に、順次に投与される。
【0072】
別の好ましい実施形態においては、キセノンは低体温療法の後に、別個に投与される。
【0073】
より好ましくは、キセノンは低体温療法と順次又は同時に投与され、さらに好ましくは、同時に投与される。
【0074】
本発明のある好ましい実施形態においては、キセノンは、治療上有効な量で投与される。
【0075】
別の好ましい実施形態においては、キセノンは治療上有効な量以下で投与される。すなわち、キセノンは、低体温ではない状態で投与された場合、所望の治療効果を得るには不十分な量で投与される。
【0076】
さらに好ましくは、キセノンと低体温療法の組み合わせにより相乗効果が得られる、すなわち、この組み合わせは相乗的である。
【0077】
ある特に好ましい実施形態においては、キセノンは低酸素性障害の前に投与される。したがって、ある好ましい実施形態においては、キセノンは、新生児が出生する前に、例えば、陣痛の前又は陣痛中に母親に投与することによって、母親を通じて新生児に投与される。好ましくは、出産前に約48〜24時間、より好ましくは約12時間、さらに好ましくは、約6時間又は3時間又は1時間、母親に投与される。出生後、新生児は低体温状態におかれる。
【0078】
本発明の別の態様は、新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、
(a)治療上有効な量のキセノンを前記哺乳動物の母親に、陣痛前及び/又は陣痛中に投与するステップ、及び
(b)前記哺乳動物に出生後に低体温療法を施すステップ
を含む方法に関する。
【0079】
好ましくは、低体温療法は、低酸素性虚血性(HI)障害後、少なくとも約6時間、より好ましくは約12時間続けられる。
【0080】
ある好ましい実施形態では、低体温療法が、低酸素性虚血性(HI)障害後、約6〜24時間続けられる。
【0081】
好ましくは、低体温療法が、出生後、少なくとも約6時間、より好ましくは少なくとも約12時間続けられる。
【0082】
ある好ましい実施形態においては、低体温療法が、出生後、約6〜約24時間続けられる。
【0083】
好ましくは、本発明の方法に準拠する治療は、低酸素性虚血性(HI)障害から6時間以内に開始され、より好ましくは、低酸素性虚血性障害から2時間以内に開始される。
【0084】
体温が低下するにまかせて、意図的に体温を維持しようとしないことで、受動的に低体温療法の状態を作り出してもよい。新生児は変温性であるため、体温が急速にその周囲の温度に近づいていく。あるいは、故意に周囲の温度を下げることにより、患者を能動的に低体温状態にしてもよい。
【0085】
本発明の第二の態様は、新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、
(a)治療上有効な量のキセノンを前記哺乳動物に投与するステップ、及び
(b)前記哺乳動物に低体温療法を施す又は低体温状態にするステップ
を含む方法に関する。
【0086】
好ましい実施形態においては、哺乳動物は新生児である対象で、生後4週間以内である。より好ましくは、哺乳動物は生後2週間以内、さらに好ましくは、生後1週間以内である。
【0087】
好ましくは、かかる哺乳動物はヒトである。
【0088】
好ましくは、哺乳動物は軽度の低体温状態に置かれる。本明細書においては、「軽度の低体温療法」とは、一般に、中核体温が37℃から約33℃へ低下することを示すものである。
【0089】
ある好ましい実施形態においては、哺乳動物の体温は、約31℃〜約36℃の温度で維持される。
【0090】
より好ましくは、哺乳動物の体温は、約32℃〜約36℃の温度で維持され、さらに好ましくは約32℃〜約35℃、さらにまた好ましくは約33℃〜約35℃に維持される。
【0091】
第二の態様の好ましい実施形態は、第一の態様に関して上述したものと同一である。
【0092】
本発明の別の態様は、新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、治療上有効な量のキセノンを、低体温療法と組み合わせて前記哺乳動物に投与するステップを含む方法に関する。
本発明のさらに別の態様は、新生児仮死治療用薬剤の調製におけるキセノンの使用であって、前記治療が、低体温療法と組み合わせて、同時、順次、又は別個にキセノンを対象に投与するステップを含む使用に関する。
【0093】
本発明のさらなる態様は、新生児仮死を治療するための、低体温療法との組み合わせによる、キセノンの使用に関する。
【0094】
インビトロ研究
HIのモデル動物を用いて、新生児ラットに対し、キセノンと低体温療法による処置を一匹ずつ個別に施した。アポトーシス細胞死の量を減少させることで、キセノンが新生児のHIに対して神経保護作用を示すことが示された。一方、低体温療法の効果はそれよりも低いようであった。キセノンと低体温療法は、組み合わせることにより、抗アポトーシス機構を介して神経保護作用を示すようになる(図17)。それらを組み合わせた場合の効果は、相乗的であることがわかった。
【0095】
新生児ラットHIモデルは確固として確立されており、これまでの多くの研究で使用が妥当であることが確認されている(Levine, 1960; Rice et al, 1981)。このモデルで使用されるラットの年齢は、ヒト新生児の脳成熟期間に相当することがわかっており(Clancy et al, 2001; Ikonimidou et al, 1989)、したがって、これらの二者について、かなり正確な比較を行うことができる。
【0096】
低体温療法実験の間、1匹のラットの皮質に挿入したプローブを用いてラット仔の体温をモニターした。プローブが平衡に達するまで約15分かかったが、この時間を考慮して、プローブが正確な体温を記録し始めるまで実験開始時刻を遅らせた。平均値前後で体温のばらつきがあったが、持続的モニタリングと必要に応じて手動で水槽を調節することにより、それらを制御した。ラットに生じる外傷及びプローブによって皮質に与えられる損傷を最小限にするために、1グループにつき1匹のラットのみ、体温をモニターした。プローブを挿入したラットを組織学的分析にかけることは不可能であった。
【0097】
麻酔ガスのキセノンは、数例の成体神経損傷モデルにおいて神経保護作用を示すことがわかっている。現時点では、キセノンが新生児において同様の神経保護的効果を有することを確認した公開済のデータはない。本研究の結果は、キセノンにはかなりの神経保護性があるというこれまでの知見を裏付けるものであり、また、この神経保護作用が低酸素性虚血により誘導される脳損傷新生児モデルにまで拡大することを示唆している。
【0098】
グルタミン受容体のNMDAサブタイプの活性化がHIにおける進行中のニューロンの損傷及び死を持続させるために必要であることは、かなり以前から周知であり、キセノンがこれらの受容体を遮断することによってその無痛覚及び麻酔効果を示すことは、文書による充分な裏付けがなされている。したがって、キセノンの神経保護性はこのアンタゴニズムの結果と考えられる。以前に、インビトロ研究で神経保護作用をはっきりと示した他のNMDAアンタゴニストがいくつか存在したが、その後、臨床設定において利用された際には不首尾に終わった(Muir and Lees, 1995)。臨床でのこうした不首尾の理由は不明だが、グルタミン酸受容体サブタイプの遮断が、障害を防止するには不十分なものであった可能性がある。これは、キセノンがその神経保護的効果を他の機構を通じて発揮していることを暗示している。
【0099】
本研究において、キセノンが、抗アポトーシス機構を介して、新生児HIを有意に防止していることが示された。アポトーシスと壊死は共にHI障害後のニューロン喪失の重要な要素だが、アポトーシスは、新生児の転帰を決定する点においてより重要なタイプの細胞死と思われる(Taylor et al, 1999)。アポトーシス死が起こる前には、アポトーシス促進性又は抗アポトーシス性のいずれかの、多くの遺伝子(転写因子も含む)が活性化する場合が多い。キセノンはアポトーシス細胞死に干渉すると思われるため、その作用をこれら遺伝子のうちの1つに発揮する可能性、又はアポトーシス経路のある点で発揮する可能性がある。現在、外因性経路及び内在性経路という異なる2つのアポトーシス経路について証拠がある。外因性経路(「死受容体経路」とも称される)は、カスパーゼ−8を活性化する死受容体へのサイトカインの結合を伴うものであり、続いてそのカスパーゼ−8が、アポトーシス細胞死の誘導を開始する「執行カスパーゼ」であるカスパーゼ−3を活性化する(Mehmet, 2000)。内在性経路はミトコンドリアに多大に依存しており、アポトーシス促進性タンパク質baxによって生じるミトコンドリア膜透過性の増大を伴う。これはサイトクロムcの放出、サイトクロムc、Apaf−1(アポトーシスプロテアーゼ活性化因子−1)及びカスパーゼ−9との複合体の形成、並びにその後のカスパーゼ−3の活性化につながる。これらの経路のいずれか1つに対してキセノンが作用するのは全く可能なことであるが、HIに誘導されたアポトーシス性神経変性はミトコンドリア経路及びbax依存性ミトコンドリア変化の開始に仲介されることを示唆する証拠がある(Taylor et al, 1999)。これに加え、NMDAアンタゴニストのケタミンは、アポトーシス促進性タンパク質と抗アポトーシス性タンパク質とのバランスを早期に調節することにより、すなわち、HI誘導性bax増大を阻害することにより、不完全な大脳虚血及び再灌流を防止することがわかっている(Engelhard et al, 2003)。このように、キセノンは、baxをダウンレギュレートすることによりアポトーシスを阻害できる、という可能性がある。Bcl−2は、ミトコンドリア膜の透過性を減少させる作用を有する抗アポトーシス性タンパク質であり、そのため、サイトクロムcの放出を阻害する。その過剰発現は、アレチネズミにおいて一過性全脳虚血により生じる神経損傷を減少させることがわかっている(Engelhard et al, 2003)。したがって、bcl−2のアップレギュレーションは、キセノンの別の潜在的な標的である。キセノンは無極性で脂溶性であることから、身体全体に広く分布させることができる。膜を通過することができ、その結果、遺伝子転写を変化させて生存経路をアップレギュレートすることにより、又はアポトーシス促進性分子のRNA及びタンパク質合成を阻害することにより、核内にも作用を及ぼす場合がある。
【0100】
キセノンによる抗壊死作用は、皮質の48時間群において、統計的に有意であることが判明したが、脳回ではそうではなかった(図16)。他の全ての時間群では、キセノンは抗壊死作用を示さなかった。これに関して考えられる解釈が1つ挙げられる。以前の研究によれば(Northington et al, 2001)、皮質だけに見られる壊死性細胞死の第二の波が48時間群で存在するとのことである。これは、48時間群のポジティブコントロールに存在する壊死細胞の比率が、16時間群及び24時間群と比較して増加していることを説明するものである。皮質の48時間群でキセノンがどのようにして抗壊死作用を発揮するのかは明らかではないが、キセノンは、(16時間群及び24時間群の場合と同様に)キセノン投与前に生じた壊死を防止することは不可能である一方で、キセノン投与後に生じた第二の壊死の波に何らかの方法で対抗することができる、ということだと思われる。最初の壊死は、HI障害の3時間後に早くも発生しており(Northington et al, 2001)、この時点でキセノンはまだ投与されていなかった。したがって、すでに発生したプロセスを停止又は逆行させることはできるとは考えにくい。しかし、第二の壊死の波はキセノンが脳内に48時間存在した時点で発生し、これは、壊死出現時にキセノンが存在することによって、このタイプの細胞死を減少させることが可能な場合があることを示唆している。この相互作用の正確な機序を確認するには、さらなる研究を完成させなくてはならない。
【0101】
33℃という軽度の低体温療法は、虚血性神経損傷に対する神経保護作用があることが、以前の研究で示されている(Busto et al, 1987)。他の研究では、この神経保護作用が抗アポトーシス機構を介して達成されていることが示唆されている(Xu et al, 2002)。16時間群又は24時間群(それぞれ図13及び15)では神経保護作用が皆無であることが、実験で示されている。
【0102】
しかし、48時間群では、異なる機構によってではあるが、皮質と脳回の両方で有意な神経保護作用が達成されていた。低体温療法は、皮質においては抗壊死性であり、脳回においては抗アポトーシス性である(図16)。本研究のデータはこの作用を説明するものではないが、考えられる理由を1つ挙げるとすれば、異なる脳領域が差別的な脆弱性を発現するということがあるのかもしれない(Northington et al, 2001)。皮質においては、低体温療法がすでに行われた時点で第二の壊死の波(上述)が発生するため、効果が増大すると思われる。しかし脳回では遅発性の壊死は存在しないため、抗壊死作用も観察されない。この領域では抗アポトーシスが神経保護的機構であると思われ、時間間隔が短い場合にははっきりしない低体温療法の予期された抗アポトーシス性神経保護的効果が、より長い時間を経て露になった可能性がある。
【0103】
こうした結果により、20%のキセノンと35℃の低体温療法を組み合わせて使用すると、驚異的なレベルの神経保護作用がもたらされることが示された。これらの数値は、それぞれの作因を単独で使用した場合には全く神経保護作用をもたらさなかったため、こうした結果は付加的な機構によって説明できるものではなかったが、これら2つの作因の相乗効果的な相互作用によるものにちがいなかった。
【0104】
すなわち、本研究は、キセノンが新生児において神経保護作用を示し、低酸素性虚血性障害に誘導されたアポトーシスを有意に防止することを示すため、インビボラットモデルを用いて示したものである。本研究のデータは、キセノンと低体温療法とを同一モデルで組み合わせて使用すると、相乗的な相互作用が発生して、アポトーシス性細胞死を劇的に減少させることを示唆している。したがって、この組み合わせは、新生児仮死の破壊的な神経学的結果を防止する効果的治療法の代表となるかもしれない。
【0105】
本発明を、実施例により、また、後述する図面により、詳細に説明する。
【実施例1】
【0106】
新生児仮死モデル
手術用麻酔(1〜1.5%のイソフルランを含む純粋酸素)下で、生後7日のSprague-Dawleyラットの右総頚動脈を結紮した。結紮後、それらの動物を母親のもとへ戻し、室温(23℃)及び湿度(48%)を一定にした特別設計の区画に収容した。手術の1時間後、8%の酸素と0、20、40、60、又は70%のキセノンとを組み合わせたもの(残りは窒素で補充した)を入れた特別設計のチャンバーに、新生児ラットを収用した。収容時間は90分間で、その際の温度は37℃であった(チャンバーの外側に設置した水槽で温度を維持した)。実験の7日後、ラット(日齢14)を殺処分し、脳を摘出した。左半球に対する右半球の重量比(R/L比)を計算した。数群のラット仔については、生後30日まで生存させ、その時点でそれらの神経運動機能及び協調を、確立されたプロトコルによって査定した(神経運動試験及びロータロッド試験)。
【0107】
低酸素状態の時間を延長したところ、損傷(脳重量の減少として測定)が明瞭に認められるのは、低酸素状態が90分を越えた場合のみであることが結果として示された(図1)。そのため、低酸素障害の標準時間を90分に設定した。
【0108】
90分間の低酸素性虚血性障害を受けた動物の脳切片を図2に示す。詳細には、図2(中央)は、巨視的な解剖学的損傷(障害を受けた側の脳――この図では左側)を、コントロール動物(左)と比較して示す。右側の脳スライスは、同じく低酸素性虚血を経験したものの、低酸素状態の間に70%のキセノンを呼吸した動物に由来するものである。これらの脳は正常のものとよく似ており、キセノンによってもたらされた驚異的神経保護作用を示している。
【0109】
90分間の低酸素性虚血の24時間後に明瞭に認められる多大な細胞損傷を図3に示す。
【0110】
キセノンの神経保護作用の濃度依存性(左半球に対する右半球の重量比と、キセノンの濃度)を図4に示す。詳細には、図4は、生後7日で行った、各種濃度のキセノンを組み合わせた又は組み合わせない低酸素性虚血の後の、日齢14のラット脳の同側/対側半球重量比を示す。麻酔域下濃度の場合でさえ、神経保護作用は明瞭に認められた。コントロール動物には頚動脈結紮を行ったが、低酸素状態にはしなかった。結果は、平均値±SEM(n=5−8)である。8%のO2に対して、*P<0.01。
【0111】
低酸素性虚血性(HI)障害後にかなりの時間をおいて査定した、70%のキセノンの神経機能に対する作用を図5に示す。生後7日で右頚動脈を結紮し、ラット仔を低酸素環境(8%の酸素+70%のキセノン、残りは窒素で補充した)に90分間暴露させた。障害から30日後、(A)把握牽引(prehensile traction)、強度、及び平均台パフォーマンス(0〜9のレベルに採点)の検定を含むパネルの使用により、及び(B)バランスと神経運動機能の標準的なテストであるロータロッドでのバランスにより、ラットの神経運動機能を評価した。ラット1個体からのデータポイントは、三回のテストの合計である。水平のバーは、各群の中央値を示す。
【0112】
HI障害の2時間後にキセノンが投与される場合の、N2とキセノンそれぞれについて観察された神経保護的効果(右半球/左の比)を図6に示す。より詳細には、低酸素状態の時間が終了してから2時間後にキセノンが投与された場合でさえ、キセノンが神経保護作用を効果的に発揮していることを、データは示している。生後7日で、90分間の低酸素性虚血性障害後に、70%のN2又は70%のキセノン+30%のO2に90分間暴露させてから2時間の回復期を与えた、日齢14のラット脳の同側/対側半球重量比。結果は、平均値±SEM(n=6)である。
【0113】
キセノンの神経保護的効果に対する軽度の低体温療法の作用(LDH放出と、キセノン濃度(%原子))を図7に示す。穏当な低体温療法は、キセノンの神経保護作用に、非常に大型で予想しない増強をもたらす。体温が4℃下がると、LDH放出のブロッキングにおけるキセノンの効能が大幅に増強する。より詳細には、この図は、酸素−グルコース欠乏(OGD)が誘導する乳酸脱水素酵素(LDH)放出に対し、キセノンと低体温療法との組み合わせが及ぼす作用を示す。図7は、濃度を増大させたキセノンの存在下、37℃(赤)又は33℃(青)のいずれかで、ニューロン培養物をOGDに75分間暴露させた結果を示す。37℃でのキセノン、33℃でのキセノンに対するED50値は、それぞれ35.9+/−2.15%、11.5+/−2.0%(平均値+/−SEM)であった。ニューロンの損傷は、キセノン又は低体温療法いずれかの不在下での、75分間のOGD及び6時間の回復期後におけるLDHの最大放出量のパーセンテージで表した。点は平均値を示す。エラーバーは、標準誤差を示す。
【0114】
かかるプロセスの温度依存性の程度を図8に示す。図8には、絶対温度の逆数に対してプロットされたLDH放出の自然対数のファント・ホッフプロットが示されている。そのようなプロットのスロープから、かかるプロセスのエンタルピーの変化が計算でき、その大きさは温度依存性の尺度となる。赤のデータは、キセノン不在下でのLDH放出に対する温度の作用を示す。温度の低下に伴う放出の減少が予想されるが、穏当なものである。12.5%のキセノンが存在する場合、温度依存性は非常に高く、予想不可能である。そのため、低体温療法は、キセノンの神経保護的効果を大幅に増強させると思われる。したがって、こうした結果は、低体温療法とキセノンが神経保護剤として相乗的に作用することを示唆している。
【0115】
より詳細な研究を下記の実施例2において概述する。
【実施例2】
【0116】
材料と方法
本研究は、1986年の英国動物(科学的処置)法に準拠するものであり、内務省(英国)により認可されている。
【0117】
低酸素性虚血症のモデル動物
本研究では、脳障害のパターンが満期産のヒト新生児における低酸素性虚血性障害のものと類似している日齢7(p7)の局所的HIモデルラットを使用する(Johnston, 1983)。
【0118】
英国Harlan社のp7のSprague-Dawleyラット仔(体重は10〜14g)に、以前記載されたモデルのHI障害を受けさせた(Levine, 1960; Rice et al,1981)。すなわち、2%のイソフルランでラット仔に麻酔をかけた後、頸部正中切開及び5.0絹縫合糸による右総頚動脈の永久的な片側結紮を行った。手術が完了すると、ラット仔は麻酔から覚醒し、その後実験時まで母親のもとへ戻された。
【0119】
手術の1時間後、一部が37℃の水槽に沈められている、専用の気密チャンバーにラット仔を収容することにより、ラット仔を低酸素状態に暴露させた(図9)。90分間という低酸素時間は、半球の重量として測定された低酸素性虚血性損傷が、この時間の後に最大となることが予備実験で示されたために、選択された。8%の加湿酸素に残りを窒素で補充して、それを継続的に流し入れることで、低酸素状態を誘導した。この混合気体を、15分ごとにDatex Ohmeda(英国、ブラッドフォード)でモニターした(気体はすべてBOC社より供給された)。
【0120】
実験的処置
HIの後、ラット仔を回復のために4時間母親のもとへ戻し、その後、下記の実験的介入の1つを90分間受けさせた。予備実験のデータにより、介入を適用する最適の時期は、HIと同時かその4時間後であることが示された。この2つの時期については有意な差異は存在しなかったため、臨床的に最適と思われたとの理由により、介入の適用時間として障害の4時間後を選択した。
【0121】
ラット仔は、処置の16、24、及び48時間後に殺処分されるまで、母親のもとに戻された(Northington et al, 2001)(図10)。
【0122】
コントロール
コントロール群は、(a)ネガティブコントロールとして使用される、切開は行ったが結紮は行っていない無処置の同腹子(すなわち、擬似手術動物)と、(b)ポジティブコントロールとしての役割を果たす、HIの措置は行ったが実験的介入の措置は行っていない同腹子とからなる。これらの動物は、25%の酸素に残りを窒素で補充した混合気体に90分間、37℃でさらされた。
【0123】
実験用ラット
HI及び回復期の後、実験用ラットを下記の5つの実験的介入の1つで90分間処置した。5つの処置のそれぞれは、別個の群のラットに対して行われた。
【0124】
低体温療法での処置
ラット仔に、軽度の低体温療法(33℃)での処置を90分間施した。無作為に1匹のラット仔を選別し、イソフルラン及び局所麻酔下で、温度プローブ(Mini-Mitter Co. Inc.社製、米国、オレゴン州、ベンド)を皮質に挿入し、強力接着剤でその場所に固定した。その後、すべてのラット仔を気密チャンバー(前述のとおり)に収容し、25%の酸素に残りを窒素で補充した混合気体をポンプで送った。温度プローブとVital Viewコンピュータプログラムによって測定されたラットの皮質の温度を正確に33℃に保つように維持された水槽に、チャンバーを部分的に水没させた。この温度を選択したのは、それが「軽度の(mild)」低体温療法を示すものであること、したがって、臨床的に適していると考えられ、副作用と効能との間のバランスがよいという理由によるものである。処置の90分後、ラット仔を母親のもとへ戻し、殺処分時まで回復させた。温度プローブを設置されたラット仔は、実験後直ちに殺処分され、その脳を分析に使うことはなかった。
【0125】
キセノンでの処置
キセノンでの処置にも同じ実験用パラダイムが遵守されたが、低体温療法の代わりに、水槽を37℃に維持し、混合気体を25%の酸素と75%のキセノンに変えて、90分間行った。キセノンの漏出を最小限にするため、専用の閉鎖システムにガスを送達した(図9)。ここでも、殺処分時までラット仔を母親のもとへ戻した。
【0126】
組み合わせプロトコル
組み合わせパラダイムにおいて、ラットに低体温療法とキセノンの両方を同時に90分間施した。再びラット仔を気密チャンバーに収容したが、この場合は、その温度を35℃に維持し、混合気体の構成は、25%の酸素にキセノンは20%のみで、残りを窒素で補充したものであった。この温度及びキセノン濃度は、別々に使用しても発達途中の脳に神経保護的な効能を与えないことが、予備実験で示されていたものである。したがって、これらの数値を用いることにより神経保護的効能が少しでも生じれば、それはこの2つの作因間の相乗作用を示すことになる。処置後、ラット仔を殺処分時まで母親のもとへ戻した。
【0127】
組み合わせ群で使用した数値が、別個に使用された場合にはまったく神経保護作用を及ぼさないことを示すため、実験ラット群をもう2群使用した。一方の群には35℃での低体温療法(前述のとおり)を施し、他方の群には濃度20%のキセノンへの暴露を行った。
【0128】
組織の調製
脳の採取
実験プロトコルの終了から16、24、及び48時間後に、脳を摘出した。
【0129】
腹腔内へペントバルビタールを過剰投与(100mg/kg)してラットを殺処分し、その後、2.5u/mlのヘパリンを含むPBSを用いて左心室からの経心臓的灌流を行って血液を抜いた。引き続き、4%のパラホルムアルデヒドを含むPBSを20ml使って灌流した後、脳を除去した。その後、同じ固定剤で脳を一晩後固定した。各時間群について、コントロールの数又は実験用脳を均等に分配し、免疫組織化学用の凍結切片としてスライスするか、組織学的分析用にパラフィン包埋するかのいずれかを行った。同側半球と対側半球とを区別するため、スライスする前に、対側の影響を受けていない(左)半球にペーパークリップで穴をあけた。
【0130】
パラフィン包埋
後固定後、組織学用の脳を、Histokinette 2000組織包埋プロセッサー(Leica U. K. Ltd.社製、英国、ミルトン・ケインズ)で乾燥させ、その後、パラフィンワックスブロックに包埋した。パラフィン包埋した脳を、ミクロトーム(Leica社製、ドイツ)を用いて、厚さ5μmの冠状切片にカットした。図10.3は、薄片が採取された脳部位を示す。各脳の、ブレグマから約−3.6mmにある海馬部位(この部位がHI障害に最も脆弱な部位であるため)から、約20の薄片を採取した。
【0131】
凍結切片
脳を一晩後固定した後、30%のスクロースを含むPBS(これには、2mg/mlのアジ化ナトリウムも含まれている)中で凍結防止処理を施し、冷蔵庫で48時間又は脳がチューブの底に沈むまで保管した。その後、脳をO.C.T.化合物(BDH社製、英国、プール)中で−22℃にて凍結させ、冠状切片をスライディング・クライオスタット(Bright Instrument Company Ltd社製、英国、ハンチントン)上で30μmにカットした。スライスした脳を、0.1MのPBS及び1mg/mlのアジ化ナトリウムを含むウェルに入れて、冷蔵庫にて保管した後、免疫組織化学用に染色した(下記参照)。
【0132】
染色の手順
組織学
パラフィン包埋した切片を、スライドに載せ、以前の記載のとおり、組織学用にクレシルバイオレットで染色した(Wilhelm et al, 2002)。
【0133】
壊死及びアポトーシスの神経病理学的分析
組織学的顕微鏡検査
クレシルバイオレットで染色したパラフィン脳薄片の組織学的分析により、ニューロン損傷を査定した。クレシルバイオレットは、RNAを多く含むリボソームのようなニューロン細胞質の酸性成分、並びに神経細胞の核及び核小体に結合する基本的な染色液である。この技法を用いて、細胞の生存、及び生育不能な細胞が示すものがアポトーシスであるか、又は壊死であるかを、有効な形態学的基準(Nakajima et al, 2000)に基づいて査定した。
【0134】
各実験群は、3つの時間群(16、24、48時間)からなり、各時間群は、3匹の動物を擁していた(したがって、各実験群には、合計9匹の動物が存在した)。ブレグマから−3.6mmの脳部位から、1匹あたり3枚のスライドが選ばれた(図11)。その後、スライドを時間群に分けて、試験者にはその介入がわからないようにした。
【0135】
BX60光学顕微鏡(Olympus社製、英国、サウスオール)を用いて各脳の同側の2つの部位を分析した――1つは皮質内の、もう1つは海馬の脳回内の部位である(図11)。グリッド付きの40倍の対物レンズを用いて、グリッド上に認められる細胞の総数を計数した。細胞に対し、その形態学的外観に基づき、生存、アポトーシス、又は壊死のいずれかとしてスコアをつけ、各細胞型のパーセンテージを、それぞれの脳部位について書き記した。Axiocamデジタルカメラ(Zeiss社製、ドイツ、ゲッチンゲン)を顕微鏡と併用して、脳薄片の顕微鏡写真を撮影した。
【0136】
細胞をそれぞれのカテゴリーに割り当てる際の基準は、以下のとおりである(図12):
1.いずれかのタイプの細胞死(アポトーシス又は壊死)を起こしている細胞は、通常の形態を有するとともに細胞質が淡色で核が明瞭に視認可能な濃色である生存細胞よりも、クレシルバイオレット染色液の取り込みが強かった。
2.アポトーシスであると分類された細胞は、非常に濃く染色され、縮退している球状の核及び完全な細胞膜を有しており、周囲に空胞化が見られる場合が多い。
3.一方、壊死細胞は、同じく強く染色されるが、不規則な形状の、拡大した核を有している。
【0137】
アポトーシス細胞、壊死細胞及び生存細胞の数を、総細胞数のパーセンテージとして示し、それぞれのスライドに書き記した。その後、これら3つのスライドから各動物について平均パーセンテージを計算して、1つの時間群における3匹の動物それぞれの数値を1つにまとめた。各時間群についての数値を1つにまとめるため、これら3匹の動物から再び平均値を取り、標準偏差を記録した。
【0138】
統計学的分析
データ分析は、一元配置分散分析を用いて行い、適切とされる場合には、その後に、Student-Newman-Keulsを用いた。p<0.05は、統計学的に有意であると考えられた。
【0139】
結果
別個の作因としてのキセノン及び低体温療法
キセノンは、新生児において抗アポトーシス機構による神経保護作用を有する
脳の皮質及び海馬領域の顕微鏡分析を行ったところ、擬似手術脳と比較してキセノン処置脳の形態学的外観が類似していること、及びキセノンで処置しなかったラットの脳と比較して外観が相違していることから、キセノンの神経保護性が実証された(図14)。新生児ラットにおける低酸素性虚血性障害に対する強い神経保護作用は、キセノンをその最大濃度(75%)で使用したことにより達成され、これは、クレシルバイオレットで染色した脳薄片の組織化学的分析により定量化されている。この濃度のキセノンを別個に使用すると、アポトーシス細胞死を有意に減少させ、生存細胞数を増大させる。16時間群では、皮質のアポトーシスは、ポジティブコントロールにおいて、36.5%±2.5%から8.5%±1.6%(p<0.001)に減少しており、生存細胞数は、ポジティブコントロールにおいて、52.9%±2.3%から80.6%±0.2%(p<0.001)に増大している(図13A)。脳回においては、アポトーシスは、ポジティブコントロールにおいて、33.6%±1.8%から13.9%±2.4%(p<0.01)に減少しており、生存細胞数は、ポジティブコントロールにおいて、56.5%±2.6%から77.1%±3.3%(p<0.01)に増大している(図13B)。24時間群及び48時間群(それぞれ、図15及び図16)では、16時間群と類似した結果が示されており、キセノンは、皮質と脳回の両方において、ポジティブコントロール動物と比較して、統計学的に有意な抗アポトーシス作用を示している。
【0140】
キセノンによる抗壊死作用は、ポジティブコントロールにおいて壊死が16.6%±0.2%から10.7%±0.4%(p<0.01)に減少していた皮質の48時間群で統計学的に有意であることが示された(図16A)。しかし、キセノンは、脳回の48時間群においては抗壊死作用を示さなかった(図16B)。他のすべての時間群(16及び24時間)では、キセノンは抗壊死作用を示さなかった。
【0141】
中程度のHIの後に、33℃での低体温療法を90分間行っても効果はない
16時間群又は24時間群(それぞれ、図13及び15)では、33℃での低体温療法については神経保護作用が観察されなかった。16時間群では、低体温療法は皮質において有意な抗アポトーシス作用を有するようであったが、生存細胞数についてはポジティブコントロール動物と統計学的な差異がなかったため、この介入により神経保護作用はもたらされなかったと結論づけることができる。しかし、48時間群では、低体温療法は皮質の抗壊死機構を介して神経保護作用を示し、壊死細胞数をポジティブコントロールにおいて、16.6%±0.2%から12%±2.3%に減少させ、生存細胞数を43%±3.4%から52.3%±3.1%に増大させた(図16A)。脳回の48時間群においては、低体温療法は、抗アポトーシス性の神経保護作用を示した(図16B)。
【0142】
キセノンと低体温療法との組み合わせ
20%のキセノンのみでの処置では、神経保護作用は示されない
75%のキセノンを用いて得られた結果とは異なり、20%のキセノンはまったく神経保護的作用を発揮しない。図17を見ると、皮質の16時間群で認められたアポトーシスのパーセンテージが、ポジティブコントロール動物(p>0.05)の37%±2.5%と比較して20%のキセノン群では36%±5.7%であること、生存細胞数のパーセンテージが、ポジティブコントロール動物(p>0.05)の53%±2.3%と比較して20%のキセノン群では51%±7.8%であることがわかる(すなわち、20%群とポジティブコントロール群との間に統計学的な差異はない)。脳回のデータからも、非常に良く似た結果が出た。
【0143】
35℃での低体温療法のみでの処置では、神経保護作用は示されない
35℃の低体温療法を単独で使用してもHIに対して効果がなく、ポジティブコントロールと比較して、どの脳部位においても統計学的な差異は示されない。アポトーシスのパーセンテージは、ポジティブコントロールの37%±2.5%に対して48%±10.1%であり、細胞生存のパーセンテージは、53%±2.3%に対して44%±10.3%である。
【0144】
20%のキセノンと35℃での低体温療法とを組み合わせた処置では、抗アポトーシス機構を介した相乗的神経保護作用が示される。キセノン(20%)又は低体温療法(35℃)のいずれかという、すでに効果がないことが証明された介入を組み合わせて使うことにより、抗アポトーシス機構を介して、3つすべての時間群(16、24、48時間)を通じて、脳の両方の部位で強い相乗的な神経保護作用が示された。かかる組み合わせ処置を虚血後に適用すると、アポトーシス細胞死の程度を有意に低下させ、生存細胞の割合を増加させた(図17参照)。皮質の16時間群では、組み合わせ療法によるアポトーシスの減少は、20%のキセノン群と35℃での低体温療法群ではそれぞれ35.8%±5.7%と47.6%±10.1%であったのが、組み合わせ群(それぞれ、p<0.01及びp<0.001)においてはわずか7.2%±2%に減少しているのがわかった。一方、生存細胞は、51%±7.8%と43.7%±10.3%であったのが、82.3%±4.9%(両方の群において、p<0.01)に増加していた。脳回のデータからも、抗アポトーシス性保護作用に加えて抗壊死性保護作用も示されると思われる24時間群以外は、良く似た結果が出た(図17B)。
【0145】
個々の作因からはまったく神経保護作用がもたらされないという事実に鑑みて、かかる組み合わせの結果は、驚くべきものであり、予想をはるかに上回るものであることは確かである。別々では効果のない2つの介入を組み合わせたことによりもたらされる神経保護作用の程度は、キセノンと低体温療法との間に、インビボでの相乗性が存在することを示している。
【0146】
本発明の範囲及び精神から逸脱することのない、ここに記載した本発明の方法の様々な修正及び変更は、当該分野の熟練者には自明なことであろう。本発明を特定の好ましい実施形態に関して記載したが、関連分野の熟練者にとっては自明なことである、ここに記載した本発明の実施形態の様々な修正は、添付した請求の範囲の範囲内であると解釈される。
【0147】
[参考文献]
Albers, G., Goldberg, M.P., Choi, D.W., 1989. N-methyl-D-aspartate antagonists: ready for clinical trial in brain ischemia? Ann Neurol 25:398-403.
Ankarcrona, M., 1995. Glutamate-induced neuronal death: A succession of necrosis or apoptosis depending on mitochondrial function. Neuron 15:961-973.
Arias, R.L., Tasse, J.R.P., Bowlby, M.R., 1999. Neuroprotective interaction effects of NMDA and AMPA receptor antagonists in an in vitro model of cerebral ischemia. Brain Research 816:299-308.
Balduini, W., De Angelis, V., Mazzoni, E., Cimino, M., 2000. Long-lasting behavioural alterations following a hypoxic/ischemic brain injury in neonatal rats. Brain Research 859:318-325.
Bliss, T.V.P., Collingridge, G.L., 1993. A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361:31-38.
Bosley, T.M., Woodhams, P.L., Gordon, R.D., Belazs, R., 1983. Effects of anoxia on the stimulated release of amino acid neurotransmitters in the cerebellum in vitro. J. Neurochem 40:189-201.
Busto, R., Dietrich, W.D., Globus, M.Y.T., 1987. Small differences in intraischemic brain temperature critically determine the extent of ischemic neuronal injury. Blood Flow Metab 7:729-738.
Choi, D.W., Koh, J-Y., Peters, S., 1988. Pharmacology of glutamate neurotoxicity in cortical cell culture: attenuation by NMDA antagonists. J. Neurosci 8:185-196.
Choi, D.W., Rothman, S.M., 1990. The role of glutamate neurotoxicity in hypoxic-ischemic neuronal death. Annu Rev Neurosci 13:171-182.
Chopp, M., Knight, R., Tidwell, C.D., Helpern, J.A., Brown, E., Welch, K.M., 1989. The metabolic effects of mild hypothermia on global cerebral ischemia and recirculation in the cat: comparison to normothermia and hypothermia. J Cereb Blood Flow Metab 9:141-148.
Chow, A., Ma, D., Hossain, M., Franks, N.P., Maze, M., 2003. Combined neuroprotection by xenon and hypothermia. Society Neuroscience Abstract 893(1).
Clancy, B., Darlington, R.B., Finlay, B.L., 2001. Translating developmental time across mammalian species. Neurosci 105(1):7-17.
Cullen, S.C., Gross, E.G., 1951. The anaesthetic properties of xenon in animals and human beings, with additional observations on krypton. Science 113:580-581.
Debillon, T., Daoud, P., Durand, P., Cantagrel, S., Jouvet, P., Saizou, C., Zupan, V., 2003. Whole-body cooling after perinatal asphyxia: a pilot study in term neonates. Developmental Medicine and Child Neurology 45:17-23.
Dingledine, R., McBain, C.J., 1999. Glutamate and aspartate in: Siegel, S.J., Agranoff, B.W., Albers, R.W., Fisher, S.K., Uhler, M.D., (Eds), Basic neurochemistry: Molecular, cellular and medical aspects, 6th edition, Lippincott-Raven, pp316-333.
Eilers, H., Bickler, P.E., 1996. Hypothermia and Isoflurane similarly inhibit glutamate release evoked by chemical anoxia in rat cortical brain slices. Anesthesiology 85:600-607.
Engelhard, K., Werner, C., Ebersp臘her, E., Bachl, M., Blobner, M., Eberhard, H., Hutzler, P., Eberhard, K., 2003. The effect of the 2-agonist dexmedetomidine and the N-methyl-D-aspartate antagonist S(+)-ketamine on the expression of apoptosis-regulating proteins after incomplete cerebral ischemia and reperfusion in rats. Anesth Analg 96:524-531.
Erecinska, M., Nelson, D., Wilson, F., Silver, I.A., 1984. Neurotransmitter amino acid levels in rat brain during ischemia and reperfusion. Brain Research 304:9-22.
Franks, N.P., Dickinson, R., de Sousa, S.L.M., Hall, A.C., Lieb, W.R., 1998. How does xenon produce anaesthesia? Nature 396(6709):324.
Goto, T., Nakata, Y., Morita, S., 2003. Will xenon be a stranger or a friend? Anesthesiology 98:1-2.
Goto, T., Saito, H., Nakata, Y., Uezono, S., Ichinose, F., Morita, S., 1997. Emergence times from xenon anaesthesia are independent of the duration of anaesthesia. British Journal of Anaesthesia 79:595-599.
Graham, S.H., Shiraishi, K., Panter, S.S., Simon, E.P., Faden, A.I., 1990. Changes in extracellular amino acid neurotransmitters produced by focal cerebral ischemia. Neurosci Lett 110:124-130.
Gunn et al, 2000. Curr Opin Pediatr. Apr;12(2):111-5.
Gwag, B.J., Koh, J.Y., Demaro, J.A., Ying, H.S., Jacquin, M., Choi, D.W., 1997. Slowly triggered excitotoxicity occurs by necrosis in cortical cultures. Neurosci 77:393-401.
Hardingham, G.E., Bading, H., 2003. The yin and yang of NMDA receptor signalling. Trends in Neuroscience 26(2):81-89.
Hauptman, M., Nelson, D., Wilson, D.F., Erecinska, M., 1984. Some changes in amino acid levels in rat brain synaptosomes during and after in vitro anoxia and simulated ischemia. Brain Research 304:23-35.
Hill, I.E., MacManus, J.P., Rasquinha, I., Tuor, U.I., 1995. DNA fragmentation indicative of apoptosis following unilateral cerebral hypoxia-ischemia in the neonatal rat. Brain Res 676:398-403.
Homi, H.M., Yokoo, N., Ma, D., Warner, D.S., Franks, N.P., Maze, M., Grocott, H.P., 2003. The neuroprotective effect of xenon administration during transient middle cerebral artery occlusion in mice. Anesthesiology 99:876-881.
Ikeda, M., Nakazawa, T., Abe, K., Kaneko, T., Yamatsu, K., 1989. Extracellular accumulation of glutamate in the hippocampus induced by ischemia is not calcium dependent in vitro and in vivo evidence. Neurosci Lett 96:202-206.
Ikonomidou, C., Bittigau, P., Koch, C., Genz, K., Hoerster, F., Felderhoff-Mueser, U., Tenkova, T., Dikranian, K., Olney, J.W., 2001. Neurotransmitters and apoptosis in the developing brain. Biochemical Pharmacology 62:401-405.
Ikonomidou, C., Mosinger, J.L., Olney, J.W., 1989. Hypothermia enhances protective effect of MK-801 against hypoxic/ischemic brain damage in infant rats. Brain Research 487:184-187.
Ikonomidou, C., Mosinger, J.L., Salles, K.S., 1989. Sensitivity of the developing rat brain to hypobaric/ischemic damage parallels sensitivity to N-methyl-D-aspartate neurotoxicity. J Neurosci 9:2809.
Ikonomidou, C., Price, M.T., Mosinger, J.L., 1989. Hypobaric-ischemic conditions produce glutamate-like cytopathology in infant rat brain. J Neurosci 9:1693.
Jevtovic-Todorovic, V., Olney, J.W., 2003. Neuroprotective agents in: Evers, A.S., Maze, M., (Eds), Anesthetic Pharmacology: Physiological principles and clinical practice, Churchill Livingstone, pp557-572.
Johnston, M.V., 1983. Neurotransmitter alterations in a model of perinatal hypoxic-ischemic brain injury. Ann Neurol 13:511-518.
Katja, C.Z., Green, D.R., 2001. How cells die: Apoptosis pathways. J Allergy Clin Immunol 108(4):S99-S103.
Kauppinen, R.A., McMahon, H., Nicholls, D.G., 1988. Ca2+-dependent and Ca2+-independent glutamate release, energy status and cytosolic free Ca2+-concentration in isolated nerve terminals following in vitro hypoglycaemia and anoxia. Neuroscience 27:175-182.
Komuro, R.P., 1993. Modulation of neuronal migration by NMDA receptors. Science 260:95.
Krystal, J.H., Karper, L.P., Seibyl, J.P., Freeman, R., Delaney, R., Bremner, J.D., Heninger, G.R., Bowers, M.B., Charney, D.S., 1994. Subanaesthetic effects of the non-competitive NMDA antagonist, ketamine, in humans: psychotomimetic, perceptual, cognitive and neuroendocrine responses. Arch Gen Psychiatry 51:199-214.
Kudo, M., Aono, M., Lee, Y., Massey, G., Pearlstein, R.D., Warner, D.S., 2001. Effects of volatile anesthetics on N-methyl-D-aspartate excitotoxicity in primary rat neuronal-glial cultures. Anesthesiology 95:756-765.
Leist, M., Nicotera, P., 1998. Apoptosis, excitotoxicity, and neuropathology. Experimental Cell Research 239:183-201.
Levine, S., 1960. Anoxic-ischemic encephalopathy in rats. Am J Pathol 36:1-17.
Low, J.A., 1993. The relationship of asphyxia in the mature fetus to long-term neurologic function. Clinical Obstetrics and Gynaecology 36(1):82-90.
Ma, D., Wilhelm, S., Maze, M., Franks, N.P., 2002. Neuroprotective and neurotoxic properties of the ‘inert’ gas, xenon. British Journal of Anaesthesia 89:739-746.
MacDonald, J.F., Schneiderman, J.H., Miljkovic, Z., 1986. Excitatory amino acids and regenerative activity in cultured neurons. Adv Exp Med Biol 203:425.
Mehmet, H., 2000. Caspases find a new place to hide. Nature 403:29-30.
Muir, K.W., Lees, K.R., 1995. Clinical experience with excitatory amino acid antagonist drugs. Stroke 26:503-513.
Nakajima, W., Ishida, A., Lange, M.S., Gabrielson, K.L., Wilson, M.A., Martin, L.J., Blue, M.E., Johnston, M.V., 2000. Apoptosis has a prolonged role in the neurodegeneration after hypoxic ischemia in the newborn rat. J Neurosci 20(2):7994-8004.
Nakata, Y., Goto, T., Morito, S., 2001. Clinical pharmacology of xenon. Int Anesthiol Clin 39:63-75.
Nicholls, D.G., Budd, S.L., 2000. Mithochondria and neuronal survival. Physiol Rev 80:315-360.
Northington, F.J., Ferriero, D.M., Graham, E.M., Traystman, R.J., Martin, L.J., 2001. Early neurodegeneration after hypoxia-ischemia in neonatal rat is necrosis while delayed neuronal death is apoptosis. Neurobiology of Disease 8:207-219.
Olney, J.W., 2003. Excitotoxicity, apoptosis and neuropsychiatric disorders. Current Opinion in Pharmacology 3:101-109.
Olney, J.W., Labruyere, J., Wang, G., Wozniak, D.F., Price, M.T., Sesma, M.A., 1991. NMDA antagonist neurotoxicity: mechanism and prevention. Science 254:1515-1518.
Onitsuka, M., Satoshi, M., Inokuchi, H., Shigemori, M., Higashi, H., 1998. Mild hypothermia protects rat hippocampal CA1 neurons from irreversible membrane dysfunction induced by experimental ischemia. Neuroscience Research 30:1-6.
Pellegrino-Giampietro, D.E., Cherici, G., Alesiana, M., Carla, V., Moroni, F., 1990. Excitatory amino acid release and free radical formation may cooperate in the genesis of ischemia-induced damage. J. Neurosci 10:1035-1041.
Perlman, J.M., 1999. Markers of asphyxia and neonatal brain injury. NEJM 341(5):364-365.
Petzelt, C., Blom, P., Schmehl, W., Muller, J., Kox, W.J., 2003. Prevention of neurotoxicity in hypoxic cortical neurons by the noble gas xenon. Life Sciences 72:1909-1918.
Pohl, D., Bittigau, P., Ishimaru, M.J., 1999. N-methyl-D-aspartate antagonists and apoptotic cell death triggered by head trauma in developing rat brain. Proc Natl Acad Sci 96:2508.
Reinelt, H., Marx, T., Schirmer, U., Schmidt, M., 2001. Xenon expenditure and nitrogen accumulation in closed-circuit anaesthesia. Anaesthesia 56(4):309-311.
Riccio, A., Ginty, D.D., 2002. Nature Neurosci 5(5):389-390.
Rice, J.E., Vannucci, R.C., Brierley, J.B., 1981. The influence of immaturity on hypoxic-ischemic brain damage in the rat. Ann Neurol 9:131-141.
Rothman, S.M., Olney, J.W., 1986. Glutamate and the pathophysiology of hypoxic-ischemic brain damage. Ann Neurol 19:105-111.
Sanders, R.D., Franks, N.P., Maze, M., 2003. Xenon: no stranger to anaesthesia. BJA 91(5):709-717.
Sattler, R., Xiong, Z., Lu, W., MacDonald, J.F., Tymianski, M., 2000. Distinct roles of synaptic and extrasynaptic NMDA receptors in excitotoxicity. J Neurosci 20(1):22-33.
Shichino, T., Murakawa, M., Adachi, T., Miyazaki, Y., Segawa, H., Fukuda, K., Mori, K., 2002. Effects of xenon on acetlycholine release in the rat cerebral cortex in vivo. British Journal of Anaesthesia 88(6):866-868.
Talbot, J.H., 1941. The physiologic and therapeutic effects of hypothermia. N Eng J Med 224:281.
Taylor, D.L., Mehmet, H., Cady, E.B., Edwards, A.D., 2002. Improved neuroprotection with hypothermia delayed by 6 hours following cerebral hypoxia-ischemia in the 14-day-old rat. Ped Res 51(1):13-19.
Towfighi, J., Housman, C., Heitjan, D.F., Vannucci, R.C., Yager, J.Y., 1994. The effect of focal cerebral cooling on perinatal hypoxic-ischemic brain damage. Acta Neuropathol (Berl.) 87:598-604.
Treschera, W.H., Ishiwac, S., Johnston, M.V., 1997. Brief post-hypoxic-ischemic hypothermia markedly delays neonatal brain injury. Brain and Development 19:326-338.
Vannucci, R.C., 1997. Hypoxia-ischemia: Clinical Aspects in: Fanaroff, A.A., Martin, R.J., (Eds), Neonatal-perinatal medicine IV, Mosby-Yearbook Inc, Philadelphia, pp877-891.
Vannucci, R.C., Connor, J.R., Mauger, D.T., Palmer, C., Smith, M.B., Towfighi, J., Vannucci, S.J., 1999. Mini-Review: Rat model of perinatal hypoxic-ischemic brain damage. J Neuro Res 55:158-163.
Vannucci, R.C., Perlman, J.M., 1997. Interventions for perinatal hypoxic-ischemic encephalopathy. Pediatrics 100(6):1004-1014.
Volpe, 2001. Mental Retardation and Developmental Disabilities Research Reviews 7: 56-64.
Wilhelm, S., Ma, D., Maze, M., Franks, N.P., 2002. Effects of xenon on in vitro and in vivo models of neuronal injury. Anesthesiology 96:1485-1491.
Wyllie, A.H., Kerr, J.F.R., Currie, A.R., 1980. Cell death: the significance of apoptosis. Int Rev Cytol 68:251-306.
Windle, WF, 1969. Brain Damage by Asphyxia at Birth. Scientific American Oct; 221(4):76-84.
Xu, L., Yenari, M.A., Steinberg, G.K., Giffard, R.G., 2002. Mild hypothermia reduces apoptosis of mouse neurons in vitro early in the cascade. J Cereb Blood Flow Metab 22:21-28.
Yager, J.Y., Asselin, J., 1996. Effect of mild hypothermia on cerebral energy metabolism during the evolution of hypoxic-ischemic brain damage in the immature rat. Stroke 27:919-926.
【図面の簡単な説明】
【0148】
【図1】図1は、Sprague-Dawleyラットにおける、脳重量(左半球に対する右半球の比)の減少により測定した損傷と、低酸素状態の持続時間(分)との間の関係を示す。
【図2】図2は、90分間の低酸素性虚血性障害を経験したSprague-Dawleyラットの脳切片を示す。
【図3】図3は、90分間の低酸素性虚血の24時間後に、Sprague-Dawleyラットにおいて明瞭に認められる多大な細胞損傷を示す。
【図4】図4は、キセノンによる神経保護作用の濃度依存性(左半球に対する右半球の重量比と、キセノンの濃度)を示す。
【図5】図5は、低酸素性虚血性(HI)障害後にかなりの時間をおいて査定した、神経学的機能に対する70%キセノンの作用を示す。
【図6】図6は、キセノンがHI障害の2時間後に投与された場合の、N2及びキセノンそれぞれに観察される神経保護的効果(左半球に対する右半球の比)を比較するものである。
【図7】図7は、キセノンの神経保護的効果に対する軽度の低体温療法の作用を示す(LDH放出と、キセノン濃度(%原子))。
【図8】図8は、絶対温度の逆数に対してプロットされたLDH放出の自然対数のファント・ホッフプロットを示す。
【図9】図9は、ガスの供給に用いる専用の気密チャンバーを示す。水槽及び閉回路キセノン供給システムも表されている。
【図10】図10は、使用した方法のスケジュールの概略である。60分とは、n=12のラット仔の手術に要した時間である。回復期は母親のもとで過ごさせた。使用した介入措置は、擬似手術動物、ポジティブコントロール、75%のキセノン(残りは酸素)、33℃での低体温療法、20%のキセノン(25%の酸素、55%の窒素)、35℃での低体温療法、及び35℃での低体温療法と20%のキセノンの組み合わせであった。特に記載のない限り、動物は37℃に保たれ、25%の酸素と残りは窒素からなる混合気体を呼吸した。擬似手術動物には切開を施したが結紮やHIは行わず、ポジティブコントロールには、手術とHIの両方を行った。各群の動物は、変更可能な回復期(16、24及び48時間)に均等に分けられ、その後処分された。略語:HI、低酸素性虚血。
【図11】図11は、(A)ウェブサイトfaculty.virginia.edu/…/RatBrainLabels.jpgから修正したラット脳の矢状断面を示す。破線は、ブレグマから約−3.6mmに位置する、冠状断面(B)を採取した脳部位を表す。(B)はクレシルバイオレットで染色した冠状断面を示す。ボックスは、計算盤を設置して細胞を分析した部位を示す。上のボックスは皮質を示し、下のボックスは脳回を示す。Xは、非結紮(対側)半球を示すために安全ピンで故意に作られた穴の上につけられている。
【図12】図12は、アポトーシス細胞、壊死細胞及び生存細胞間の形態学的外観の違いが明示されている、100x油浸レンズとAxiocamデジタルカメラで撮影された皮質(クレシルバイオレット染色)の顕微鏡写真を示す。生存細胞の染色程度は、いずれのタイプの細胞死よりも弱く、そのため、細胞質の色が薄い。その一方、死亡細胞はより濃く染色されている。壊死細胞及びアポトーシス細胞は、それらの核の外観における相違点に基づいて、区別される。壊死核は大きく不規則な形をしており、一方、アポトーシス核は小さく、縮退し、球形である。
【図13】図13は、キセノンが16時間群で抗アポトーシス機構を介した神経保護作用を有することを示す。より具体的には、図13は、16時間群での、(A)皮質、(B)脳回における、低酸素性虚血に誘導されたアポトーシス性及び壊死性ニューロン死のグラフ、及びそのような細胞死に及ぼす75%のキセノンと33℃での低体温療法の作用を示す。両方の脳部位において、キセノンは、ポジティブコントロール動物と比較して、アポトーシス細胞の比率を低下させただけではなく、生存細胞の比率を有意に上昇させた。皮質においては、低体温療法はアポトーシス細胞の比率を減少させたものの、生存細胞数を増加させることはなかったため、神経保護作用を有するとは考えられない。結果は平均値±SD(n=3)である。ポジティブコントロールに対して、*P<0.05、**P<0.01、***P<0.001。
【図14】図14は、16時間群の、擬似手術動物、ポジティブコントロール動物、及び75%のキセノンで処置した動物における皮質及び脳回を表す顕微鏡写真を示す。75%群は、外観に関しては、ポジティブコントロール群よりも擬似手術群に似ている。これは、16時間群でのキセノンの神経保護的効果を確認するものである。コントロール群の脳回は、細胞死及び空胞化の量が増大したことにより、形にゆがみが見られる。
【図15】図15は、キセノンが24時間群で抗アポトーシス機構を介した神経保護作用を有することを示す。より具体的には、図15は、24時間群での、(A)皮質、(B)脳回における、低酸素性虚血に誘導されたアポトーシス性及び壊死性ニューロン死のグラフ、及びそのような細胞死に及ぼす75%のキセノンと33℃での低体温療法の作用を示す。両方の脳部位において、キセノンは、壊死細胞数の減少によって生存細胞の比率を有意に上昇させた。結果は平均値±SD(n=3)である。ポジティブコントロールに対して、*P<0.05。
【図16】図16は、キセノンが48時間群で抗アポトーシス機構を介した神経保護作用を有することを示す。より具体的には、図16は、48時間群での、(A)皮質、(B)脳回における、低酸素性虚血に誘導されたアポトーシス性及び壊死性ニューロン死のグラフ、及びそのような細胞死に及ぼす75%のキセノンと33℃での低体温療法の作用を示す。皮質と脳回の両方において、キセノンは、抗アポトーシス機構を介して神経保護作用を示している。また、キセノンは、皮質において抗壊死作用を有している。33℃での低体温療法は両方の脳部位において神経保護作用を有しているようだが、その機序は異なる――皮質では抗壊死性であり、脳回では抗アポトーシス性である。結果は平均値±SD(n=3)である。ポジティブコントロールに対して、*P<0.05、**P<0.01、***P<0.001。
【図17】図17は、キセノンと低体温療法との組み合わせにより相乗的な相互作用が発生して抗アポトーシス性の神経保護作用が生じることを示す。より具体的には、図17は、16、24.及び48時間群での、(A)皮質、(B)脳回における、低酸素性虚血症に誘導されたアポトーシス性及び壊死性ニューロン死のグラフ、及びそのような細胞死に及ぼす20%のキセノンと35℃での低体温療法との組み合わせの作用を示す。20%のキセノン群及び35℃での低体温療法群をポジティブコントロール群と比較した際には何の相違点も見られなかった。したがって、これらの数値では、神経保護作用は存在しない。しかし、これらの数値が組み合わされて使用された場合には、ポジティブコントロール動物と比較してアポトーシス細胞数が有意に減少することにより、生存細胞の比率が劇的に上昇する。脳回においてその組み合わせは、さらに24時間群で抗壊死作用ももたらす。結果は平均値±SD(n=3)である。ポジティブコントロールに対して、*P<0.05、**P<0.01、***P<0.001。これらの図に関するより詳細な論述が、実施例の項目に記載されている。
【技術分野】
【0001】
本発明は、新生児仮死の治療方法に関する。
【背景技術】
【0002】
低酸素性虚血症(HI)としても知られる新生児(又は周生期)仮死は、陣痛時、分娩時、又は出生直後の時期に、充分な酸素が産児の体内に取り込まれないことが原因で生じる状態である。新生児仮死は、依然として新生児における慢性神経疾患罹患率及び急性死亡率の主要な原因となっており(Balduini et al, 2000; Vannucci et al, 1997)、低酸素性虚血性脳症に至る場合が多い。
【0003】
6分間という短時間であっても、新生児仮死(低酸素症)によって永久的な神経損傷が生じる恐れがあることが、これまでの研究によって示されている。仮死状態の霊長類新生児では脳組織の喪失が認められ、脳組織の喪失は記憶障害及び痙攣性麻痺と相関がある(Windle, WF, 1969)。
【0004】
出生時における全死亡数の約14.6%は、新生児仮死によるものである。西洋においては、新生児の約0.9%(すなわち、100〜130,000)に、新生児仮死が発生している。約15〜20%は死亡し、命を取り留めた新生児の25%が、精神遅滞、脳性麻痺、痙攣、学習障害及び/又はてんかんなどの長期にわたる合併症による重度の障害を背負っている(Law et al, 1993; Perlman et al, 1999)。また、軽度の仮死状態で合併症も起こさずに回復したと当初は思われていた子どもが、この新生児期の障害に由来すると思われるような問題行動を小児期に示すという認識も増えてきている。新生児仮死は、罹患率が住人10000人当たり5人未満であるため希少疾病医薬品としての基準を満たしているとともに、生命を脅かす、確立した治療法のない重篤な消耗性疾患である。
【0005】
HIモデル動物の新生児におけるこの種の脳障害に関与する細胞死の機序には、グルタミン受容体、特に、シナプス形成期における神経毒性に対する感受性が最も高いことから、N−メチル−D−アスパラギン酸(NMDA)受容体の過剰活性化(Jevtovic-Todorovic and Olney, 2003)によって、及びアポトーシス性神経変性(Ikonomidou et al, 1989; Pohl et al, 1999)によって生じる興奮毒性損傷(又は壊死)の組み合わせが含まれることが、これまでに示されている。この種の損傷は、低酸素性障害の重篤度と関連しており(Jevtovic-Todorovic and Olney, 2003)、また、別の脳領域の脆弱性におけるばらつきにも関連している(Northington et al, 2001)。現時点では、HIによって生じた急性神経細胞死に対処する効果的な治療法は存在しないが、多種多様な薬理学的及び非薬理学的介入の実験的研究が進行中である(Vannucci and Perlman, 1997)。
【0006】
本発明は、新生児仮死の治療方法の提供を目指すものである。
【0007】
【非特許文献1】Balduini et al, 2000
【非特許文献2】Vannucci et al, 1997
【非特許文献3】Windle, WF, 1969
【非特許文献4】Law et al, 1993
【非特許文献5】Perlman et al, 1999
【非特許文献6】Jevtovic-Todorovic and Olney, 2003
【非特許文献7】Ikonomidou et al, 1989
【非特許文献8】Pohl et al, 1999
【非特許文献9】Northington et al, 2001
【非特許文献10】Vannucci and Perlman, 1997
【発明の開示】
【0008】
本発明の第1の態様は、対象である新生児において新生児仮死を治療する薬剤であって、低体温療法と組み合わせて使用する薬剤の調製におけるキセノンの使用に関する。
【0009】
本発明の第2の態様は、新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、
(a)治療上有効な量のキセノンを前記哺乳動物に投与するステップ、及び
(b)前記哺乳動物に低体温療法を施すステップ
を含む方法に関する。
【0010】
本発明の第3の態様は、新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、治療上有効な量のキセノンを、低体温療法と組み合わせて前記哺乳動物に投与するステップを含む方法に関する。
【0011】
本発明の第4の態様は、新生児仮死治療用薬剤の調製におけるキセノンの使用であって、前記治療が、低体温療法と組み合わせて、同時、順次、又は別個にキセノンを対象に投与するステップを含む使用に関する。
【0012】
本発明の第5の態様は、新生児仮死を治療するための、低体温療法との組み合わせによる、キセノンの使用に関する。
【発明を実施するための最良の形態】
【0013】
未成熟CNSの通常の生理機能
興奮性アミノ酸(EAA)のグルタミン酸及びアスパラギン酸は、成熟中枢神経系(CNS)における興奮性シナプス伝達の主要な仲介役となっている(Dingledine and McBain,1999)。それらはまた、未成熟CNSにおいて、シナプス形成、ニューロンの生存、シナプス可塑性、並びに樹状及び軸索構造など、多くの生理学的プロセスに関与しており、未成熟CNSの発達にきわめて重要な役割を演じている。しかし、発達中にこれらのアミノ酸受容体が過剰に活性化すると、ニューロンの損傷及び死につながる恐れがある。これを「興奮毒性」という。
【0014】
EAAのうち、最も豊富に存在するのがグルタミン酸である(Dingledine and McBain,1999)。グルタミン酸はシナプス小胞に保存され、シナプス前終末から放出されると、シナプス後膜のカルシウム依存性膜脱分極を引き起こす。グルタミン酸は、N−メチル−D−アスパラギン酸(NMDA)型と非NMDA型に分けることができる各種受容体サブタイプにてその興奮性作用を発揮するが、発達中のCNSにおいては、HIに付随する脳障害で主要な役割を果たすのはNMDA受容体サブタイプであることが判明している(Ikonomidou et al, 1989; Komuro, 1993; MacDonald et al, 1986)。
【0015】
NMDA受容体はグルタミン酸受容体の主要なサブクラスであり、グルタミン酸は、哺乳類の中枢神経系における最も重要な興奮性神経伝達物質だと考えられている。重要なことに、NMDA受容体の活性化は、頭部外傷、卒中及びその後の心停止に続いて起こる低酸素症及び虚血の結果として生じるものであることに加えて、数多くの病態における興奮毒性及びニューロン死につながる中心的な事象であることが判明している。
【0016】
NMDA受容体はCNS全体に偏在するイオンチャネル型受容体であり、シナプス後膜及びシナプス外膜両方の表面に位置している(Riccio and Ginty, 2002; Sattler et al, 2000)。NMDA受容体はNa+イオンとCa2+イオンの両方に透過性のあるカチオンチャネルと結合し、通常の生理学的状態では、負の静止膜電位でMg2+によってブロックされる。細胞膜の脱分極によってそのブロックは外され、チャネルを通じてCa2+の流入が可能になり、受容体がその細胞内作用を発揮できるようになる(Hardingham and Bading, 2003)。
【0017】
NMDA受容体は正常な脳機能に不可欠であり、正常な生理機能におけるその重要性は、記憶と学習におけるその中心的役割によって実証されている(Bliss and Collingridge, 1993)。これとは逆に、過剰なグルタミン酸によるNMDA受容体の病的活性化は、細胞内Ca2+の制御が乱されるため、脳に対する虚血性障害の後に起こるニューロン細胞死の主要な原因である。これは、HIにおいてNMDA受容体が演じる中心的役割を強調するものである。
【0018】
新生児における低酸素性虚血性障害
脳が機能するためには、酸素と糖の継続的な供給が必要であるため、脳の機能は血液の充分な供給に依存している(Choi and Rothman, 1990)。新生児仮死の場合のように、血液の供給が妨げられると、その後数分以内に下流部位に低酸素性虚血性損傷が発生する。こうした酸素欠乏条件下では、細胞代謝が、好気的代謝から、細胞のエネルギー必要量を満たす効果が低い嫌気的代謝へと変化する(Vannucci and Perlman, 1997)。これによりエネルギーの貯蔵が減少し、特に、ニューロン及びグリア細胞区画におけるATPなどの高エネルギーリン酸の備蓄に影響する(Dingledine and McBain, 1999)。アシドーシスにつながるH+イオンの蓄積と、さらなる細胞損傷の一因となる遊離基の放出とが同時に発生する。
【0019】
生理学的状態下では、ニューロン細胞に位置するグルタミン酸トランスポーターの作用により、グルタミン酸の細胞外濃度は低いレベルに維持されているが、グリア細胞では優先的に発現している(Dingledine and McBain, 1999)。グルタミン酸を取り込む担体は数種類存在するが、基本的に、同様の方式でその機能を発揮する。すなわち、Na+カチオン2個及びグルタミン酸アニオン1個を細胞内へ輸送し、K+カチオン1個及びOH−アニオン1個を細胞内から細胞外空間へ輸送する(Dingledine and McBain, 1999)。これらのイオンポンプは電気化学勾配に逆らって作用するため、正しく機能するには、ATPの形態のエネルギーに依存することになる。よって、HIの結果ATPの濃度が下がることにより、これらのポンプの静止膜電位維持能力が低下する。その結果として、ATP依存性ポンプの不全が、膜の脱分極及びポンピング方向の逆転につながっていく(Eilers and Bickler, 1996; Kauppinen et al, 1988)。このようにして、グルタミン酸は細胞から運び出され、過剰な濃度のグルタミン酸が細胞外空間に蓄積する。取り込みの減少によりグルタミン酸の濃度が上昇するだけではなく、膜脱分極により活動電位が生じるため、シナプス前終末からのグルタミン酸の放出も増加する(Dingledine and McBain, 1999)。過剰な細胞外グルタミン酸をもたらすこうしたプロセスの例は、インビトロ(Bosley et al, 1983; Hauptman et al, 1984; Pellegrino-Giampietro et al, 1990)及びインビボ(Erecinska et al, 1984; Graham et al,1990; Ikeda et al, 1989)の両方で見受けられる。
【0020】
興奮毒性は、過剰な細胞外グルタミン酸が継続的にシナプス後受容体(特にNMDA受容体)を活性化する場合に発生し、結果として生じたカルシウムの流入が浸透圧勾配を作り、それに基づいて水分が移動して細胞の膨張を引き起こす。カルシウム依存性酵素系も細胞内で活性化され、これら2つのプロセスの結果、急性ニューロン細胞死が起こる(Choi and Rothman, 1990)。
【0021】
細胞死の機序
Wyllie et al(Wyllie et al, 1980)が仮定したように、ニューロン細胞死は常に壊死とアポトーシスという2つの機序のいずれかによって起こると考えられてきた。しかし、最近はこれらのカテゴリーが疑問視されている。なぜなら、細胞死は興奮毒性細胞死とアポトーシスというカテゴリーに分けるべきだと示唆する多くの証拠が明らかになってきたためである(Olney, 2003)。興奮毒性細胞死は、壊死プロセス(Gwag et al, 1997; Katja and Green, 2001)、アポトーシスプロセス、及びそれら2つの連続体(Leist and Nicotera, 1998; Nakajima et al, 2000)として説明されてきた。通常、アポトーシスと壊死とは、そのはっきりと異なる形態学的外観によって区別されている。アポトーシスはATPと新規タンパク質合成を必要とし、細胞収縮、辺縁化を伴うクロマチン凝集、及び膜に囲まれたアポトーシス小体の形成によって確認される。一方、壊死は、核崩壊及び核濃縮という変化を伴う核収縮によって認識される(Hill et al, 1995)。
【0022】
HIによる細胞死では、壊死という初期段階の後に、アポトーシス細胞死という波が遅れて発生することがわかっている(Northington etal, 2001)。その後に生じる障害のタイプは時間依存的かつ位置依存的であると思われ、HIモデル新生児ラットの同側前脳に限局する初期の壊死障害、及び、視床に発生する遅延性アポトーシス障害を伴う(Northington etal, 2001)。このことは、異なる脳領域がHI後の異なる時期の各タイプの細胞死に対して差別的脆弱性を示すことを示唆している。
【0023】
正常な発達において、アポトーシスは、不要な又は損傷したニューロンがそれによって「自殺を図る」定期的な事象である(Ikonomidou et al,2001)。HIでは、初期の興奮毒性細胞死はNMDA受容体の過剰な活性化に仲介され、その結果、グルタミン酸が無制御状態で放出され、周辺のニューロンに損傷をもたらす。ニューロンがプログラムされた細胞死を開始することは、シナプス形成中の損傷に対する自然な反応であり(Olney, 2003)、これは周辺組織を保護するために活性化する機序であると考えられる(Leist and Nicotera, 1998)。
【0024】
神経保護剤としてのキセノン
NMDA受容体が、ある侵害受容経路及び痛みの知覚における場合と同様に、記憶や学習など多くの高度な認知機能の根底にあるシナプス可塑性においても主要な役割を果たしていることは、当該技術で周知である(Collingridge et al, The NMDA Receptor, Oxford University Press, 1994)。また、NMDA受容体のある特質は、意識それ自体の根底にある脳内情報処理にNMDA受容体が関与している可能性を示唆している。
【0025】
NMDA受容体アンタゴニストは、多くの理由により、治療上有用である。第一に、NMDA受容体アンタゴニストによって、全身麻酔及び鎮静の極めて望ましい構成要素である深い無痛覚がもたらされる。第二に、NMDA受容体アンタゴニストは、臨床的に関連する多くの状況(虚血症、脳外傷、神経因性疼痛、及びある種のけいれんを含む)下において神経保護剤となる。第三に、NMDA受容体アンタゴニストにより、有益な程度の健忘症がもたらされる。
【0026】
HIの発症機序におけるNMDA受容体の重要性を考えると、神経保護剤の候補としてNMDAアンタゴニストを研究してきたのは適切なことであった。MK−801やケタミンなどの多くのNMDAアンタゴニストは、インビボモデル及びインビトロモデルの両方で神経保護剤となることがわかっている(Albers et al, 1989; Arias et al, 1999; Choi et al, 1988; Kudo et al, 2001)。しかし、こうした心強い結果にもかかわらず、NMDA受容体アンタゴニストは、ヒトにおける精神異常発現という副作用も有すること(Krystal et al, 1994)、並びに後帯状皮質(PC)及び後板状皮質(RS)に損傷を引き起こすこと(Olney et al, 1991)がわかっている。さらに、従来の多くのNMDA受容体アンタゴニストは、不随意運動の生起、交感神経系の刺激、高用量時の神経毒性の誘発(NMDA受容体アンタゴニストは全身麻酔剤としては効力が弱いため、これはもっともなことである)、心筋の機能低下、及びいくつかのてんかん発生例におけるけいれん前症状(proconvulsions)、例えば、「燃え上がり現象(kindling)」をもたらす(Wlaz P et al, Eur. J. Neurosci. 1994; 6: 1710-1719)。また、血液脳関門を通過できる新規NMDA受容体アンタゴニストの開発には、これまでかなりの困難を伴っている。
【0027】
キセノンは、無極性の不活性ガスで、強力なNMDAアンタゴニストである(Franks et al, 1998)。他のNMDAアンタゴニストと同様に、インビトロ(Petzelt et al, 2003)及びインビボ(Homi et al, 2003; Wilhelm et al, 2002)の両方で多くの形態のニューロン損傷に対する神経保護剤となることが判明している。しかし、他の多くのNMDA受容体アンタゴニストとは異なり、キセノンには神経毒性がない(Ma et al, 2002)。キセノンをNMDAアンタゴニストとして使用することのさらなる利点は、その分子が不活性で、呼吸により速やかに排出可能な揮発性のガスである点にある。
【0028】
キセノンは他にも好ましい性質を数多く有している。外科で最初に用いられて以来(Cullen SC et al, Science 1951; 113: 580-582)、代謝副産物がないこと、深い無痛覚、薬効の発現及び薬効からの回復が速やかであること、心血管系への影響が最小であることなどの優れた薬理学的性質がキセノンにあることを、多数の研究グループが示してきた(Lachmann B et al, Lancet 1990; 335: 1413-1415; Kennedy RR et al, Anaesth. Intens. Care 1992; 20: 66-70; Luttropp HH et al, Acta Anaesthesiol. Scand. 1994; 38: 121-125; Goto T et al, Anesthesiology 1997; 86: 1273-1278; Marx T et al, Br. J. Anaesth. 1997; 78: 326-327)。また、キセノンは小型で電荷のない原子であるため、容易に血液脳関門を通過することが可能であり、したがって、作用が速やかに現れる(Nakata et al, 2001)。血液ガス分配計数も非常に低く、この計数の低さがキセノン麻酔からの迅速な覚醒に役立っている(Goto et al, 1997)。これらの利点に加え、キセノンは非爆発性、無毒及び非反応性である(Shichino et al, 2002)ため、キセノンは新生児の神経保護剤としての使用に理想的な候補物質となっている。
【0029】
ここで使用する「神経保護剤」との語は、神経保護作用をもたらすこと、すなわち、ニューロンなどの神経に関わる構成要素を、例えば虚血性障害又は外傷性障害に由来する、進行中の障害から保護することが可能な作因を意味する。
【0030】
神経保護剤としての低体温療法
1941年、手術で用いる低体温療法の神経保護性を、Talbotが初めて実証した(Talbot, 1941)。現在、低体温療法が決まって用いられるのは、心肺バイパス手術のみであり、脳の術中虚血を防止することを目的としている。しかし、他の脳障害モデルにおける低体温療法の治療効果を実証する発表がこれまでに何件かなされている。例えば、新生児仮死のインビトロモデル(Onitsuka et al, 1998)及びインビボモデルの両方について、低体温療法のよい効果を示す多くの発表がなされている(Debillon et al, 2003; Treschera et al, 1997)。組織の障害と脳の冷却程度との間には直接的な相関が存在することが示されており(Towfighi et al,1994)、正常酸素状態では、体温が1℃低下するごとに大脳の代謝速度が5%低下する(Yager and Asselin, 1996)。
【0031】
低体温療法がその神経保護的効果を発揮する機序はまだ解明されていないが、多くの理論が想定されている。低体温療法が保護作用を示す機序は温度及び時間依存的であり、HI障害につながる一連の事象に沿った複数のポイントで作用すると思われることが、これまでの研究で示唆されている(Yager and Asselin, 1996)。このことは、31℃という中程度の体温は大脳のエネルギー代謝を低下させることにより神経保護作用を示す一方で、34℃という軽度の低体温療法は、同じく神経保護作用を示すもののエネルギー代謝には影響を及ぼさないため、異なる機序を介して作用しているに違いない、という事実によって裏付けられている(Yager and Asselin, 1996)。Taylor et alによる他の研究(Taylor et al, 2002)では、HI障害後に開始した低体温療法は、虚血中の低体温療法よりも効果的であると示しており、これは回復期に発生する有害な作用を減少させることによるものであろうと示唆している。そうした機序の一例として、再灌流中に続いて起こる興奮毒性損傷が低体温療法によって減少することが挙げられるかもしれない(Taylor et al, 2002)。活性酸素種の減少(Taylor et al, 2002)、組織アシドーシスの減少(Chopp et al, 1989)、及びHI後におけるニューロンのアポトーシスの緩和(Xu et al, 2002)を含め、低体温療法による保護機序は他にも多く示唆されている。
【0032】
キセノンと低体温療法の組み合わせ
前述のとおり、本発明の第1の態様は、対象である新生児において新生児仮死を治療する薬剤であって、低体温療法と組み合わせて使用する薬剤の調製におけるキセノンの使用に関する。
【0033】
ここで使用されている「低体温療法」との語は、ある特定の対象(この場合は新生児の対象)を、例えば受動的又は能動的な技法によって体温を好ましくは3〜5℃下げることにより、低体温状態にすることに関するものである。一般的には、低体温状態にすると対象の体組織代謝が減少し、それにより、酸素要求量も減少する。
【0034】
前述のとおり、新生児仮死の治療における低体温療法の使用は、当該技術において文書で充分に裏付けられている(例えば、Volpe, 2001; Gunn et al, 2000を参照のこと)。しかし、低体温療法をキセノン投与と組み合わせて使用できるかもしれないということについては、今日まで当該技術において教示も示唆もされてこなかった。そのような組み合わせ療法によって、結果として得られる神経保護作用が驚異的かつ予想もしない程度に増強されるだろうとの示唆も皆無であった。
【0035】
本出願人による以前の研究により、キセノンに神経保護性があることが明らかになった。特にここに参照としてその内容を組み込んでいるWO01/08692は、神経保護剤として、及び/又はシナプス可塑性の阻害剤としてのキセノンの使用に関するものである。しかし、本出願で請求項に記載されている発明との関連においてキセノンが神経保護剤として効果的であることは、先行技術には教示も示唆もされていない。
【0036】
本発明のある好ましい実施形態において、キセノンは薬学的に許容可能な希釈剤、賦形剤又は担体と混合されている。
【0037】
本明細書に記載した種々様々な形態の医薬組成物に適切な賦形剤の例は、Handbook of Pharmaceutical Excipients, 2nd Edition, (1994), Edited by A Wade and PJ Wellerに掲載されている。
【0038】
治療用として許容可能な担体又は希釈剤は薬学分野において周知であり、例えば、Remington's Pharmaceutical Sciences, Mack Publishing Co. (A. R. Gennaro edit. 1985) に記載されている。適切な担体の例としては、ラクトース、澱粉、グルコース、メチルセルロース、ステアリン酸マグネシウム、マンニトール、ソルビトールなどが挙げられる。適切な希釈剤の例としては、エタノール、グリセロール及び水が挙げられる。
【0039】
薬剤用の担体、賦形剤、又は希釈剤の選択は、目的の投与経路及び薬学の標準的技法を参照しながら行うことができる。医薬組成物には、担体、賦形剤又は希釈剤が含まれていてもよく、あるいはそれらに加えて、適切な結合剤、滑沢剤、懸濁剤、コーティング剤、可溶化剤が含まれていてもよい。
【0040】
適切な結合剤の例としては、澱粉、ゼラチン、並びにグルコース、無水ラクトース、フリーフローラクトース、ベータラクトース、コーンシロップなどの天然糖、並びにアカシア、トラガカント又はアルギン酸ナトリウムなどの天然及び合成ゴム、カルボキシメチルセルロース及びポリエチレングリコールが挙げられる。
【0041】
適切な滑沢剤の例としては、オレイン酸ナトリウム、ステアリン酸ナトリウム、ステアリン酸マグネシウム、安息香酸ナトリウム、酢酸ナトリウム、塩化ナトリウムなどが挙げられる。
【0042】
防腐剤、安定剤及び着色剤を医薬組成物に添加してもよい。防腐剤の例として、安息香酸ナトリウム、ソルビン酸、及びp−ヒドロキシ安息香酸エステルが挙げられる。抗酸化剤及び懸濁剤も用いてもよい。
【0043】
本発明は、動物の治療にも適用することができる。この点において本発明はさらに、獣医学的に許容可能な希釈剤、賦形剤又は担体と組み合わせたキセノンの使用にも関するものである。
【0044】
獣医学用としては、獣医学における通常の技法にしたがってキセノンを投与するのが一般的であり、ある特定の動物に最も適切と思われる投与計画及び投与経路の決定は、獣医が行う。
【0045】
他の薬学的活性物質と組み合わせてキセノンを使用してもよい。そのような物質としては、GABA性活性を促進する麻酔剤又は鎮静剤を含め、どのような薬学的活性物質であってもよい。かかるGABA性物質の例としては、イソフルラン、プロポフォール及びベンゾジアゼピンが挙げられる。
【0046】
ある好ましい実施形態において、キセノンは、揮発性麻酔剤、好ましくはイソフルラン、セボフルラン又はデスフルランと組み合わせて投与される。
【0047】
また、キセノンを、その他の有効成分、例えばL型カルシウムチャネル遮断薬、N型カルシウムチャネル遮断薬、物質Pアンタゴニスト、ナトリウムチャネル遮断薬、プリン受容体遮断薬、又はそれらを組み合わせたものなどと組み合わせて投与してもよい。
【0048】
単数又は複数の適切な送達機構によって、キセノンを投与してもよい。
【0049】
ある特に好ましい実施形態においては、灌流によりキセノンを投与する。本発明との関連では、「灌流」との語は、特殊な心肺装置を用いて、患者に酸素/キセノン混合体を導入し、患者から二酸化炭素を除去することを示す。一般的な用語では、心肺装置は心臓と肺の機能を代替するものであり、無血で不動の手術野を外科医に提供する。灌流技師は、患者の血液に酸素を供給し、酸素と二酸化炭素の量を制御する。本発明との関連では、灌流技師はキセノンも患者の血液に導入する。その後灌流技師は血液を動脈系に戻し、栄養血流を手術中に患者の全重要臓器及び組織に供給する。
【0050】
ある特に好ましい実施形態では、薬剤はガス状である。
【0051】
別の非常に好ましい実施形態では、キセノンは吸入により投与される。より好ましくは、キセノンは70〜30%v/vのキセノン・酸素混合体の吸入により投与される。
【0052】
より好ましくは、キセノンは20〜70%v/vのキセノン・空気混合体の形態で投与される。
【0053】
本発明のさらに別の好ましい実施形態においては、薬剤は、液体又は溶液の形態である。
【0054】
好ましくは液体は、静脈内、動脈内、髄腔内、皮下、皮内、腹腔内又は筋肉内に注入可能な、溶液の形態、又は無菌若しくは無菌処理可能な溶液から調製された乳剤の形態で投与される。
【0055】
ある特に好ましい実施形態においては、キセノンは、脂肪乳剤の形態で投与される。静脈用製剤は、一般に、キセノンの溶解性を充分に高めて所望の臨床効果を達成する脂肪乳剤(市販されているイントラリピッド(登録商標)10、イントラリピッド(登録商標)20、イントラファット(登録商標)、リポファンジン(Lipofundin)(登録商標)S若しくはリポシン(Liposyn)(登録商標)の乳剤、又は溶解性を最大にするために特別に製剤されたもの)を含んでいる。この種の脂肪乳剤に関する詳しい情報は、G. Kleinberger and H. Pamper, Infusionstherapie, 108-117 (1983) 3に掲載されている。
【0056】
ガスを溶解又は分散させる本発明の脂質相は、一般的に、8〜30個の炭素原子を含む飽和及び不飽和長鎖及び中鎖脂肪酸エステルから形成される。これらの脂質は水性溶液中でリポソームを形成する。例としては、魚油、及び大豆油、アザミ油又は綿実油などの植物油を挙げることができる。一般的に、本発明の脂肪乳剤は、乳剤中の脂肪の割合が従来的に5〜30重量%、好ましくは10〜20重量%の水中油型乳剤である。この種の水中油型乳剤は、大豆リン脂質などの乳化剤の存在下で調製されることが多い。
【0057】
本発明のリポソームを形成する脂質は、天然物でも人工物でもよく、コレステロール、グリコリピッド、スフィンゴミエリン、グルコリピッド、グリコスフィンゴリピッド、ホスファチジルコリン、ホスファチジルエタノールアミン、ホスファチジルセリン、ホスファチジルグリセロール、ホスファチジルイノシトールなどが例示できる。
【0058】
本発明の脂肪乳剤は、付加的な成分を含んでいてもよい。そうした成分としては、抗酸化剤、脂質相の周囲の水相の浸透圧を血液と等張にする添加剤、又はリポソーム表面を改変するポリマーが挙げられる。
【0059】
相当量のキセノンを脂肪乳剤に添加してもよいことは、すでに立証されている。20℃で通常圧力という最も単純な場合でさえ、キセノンは、乳剤1ml当たり0.2〜10ml以上の濃度で溶解又は分散できる。溶解したガスの濃度は、温度、圧力及び脂質濃度を含めた多くの要因によって左右される。
【0060】
本発明の脂肪乳剤に、ガス状のキセノンを入れておいてもよい。一般に、乳剤と、その乳剤に浸漬されている焼結ガラス気泡管を通過するガス状又は蒸気状の麻酔剤とを装置に充填する。選択した分圧で乳剤を麻酔ガス又は蒸気と平衡させる。気密性容器に保存する場合、これらの脂肪乳剤は、従来の貯蔵期間中はガスとして放出されないという、麻酔剤として十分な安定性を示す。
【0061】
本発明の脂肪乳剤に、飽和レベルになるようにキセノンを入れてもよい。あるいは、より低濃度のキセノンを存在させてもよいが、それは、例えば、その乳剤を投与することにより所望の薬学的活性が得られるというような場合に限られる。
【0062】
本発明で用いるキセノンの濃度は、所望の臨床効果を得るのに必要な最低濃度でよい。通常は、医者が患者個人に最適と思われる実際の投与量を判断し、この投与量はその特定の患者の年齢、体重、及び反応によって異なる。もちろん、個々の事例によっては、高め又は低めの用量域が適切な場合もありうる。それらもまた、本発明の範囲内である。
【0063】
好ましくは、薬剤は、静脈内、軸索内、又は経皮的デリバリーに適切な形態である。
【0064】
好ましくは、キセノンは、低体温療法と同時に、組み合わせて、順次に、又は別個に投与される。
【0065】
本明細書では、「同時」は、キセノンが低体温療法と時を同じくして投与されることを意味するのに用いており、一方「組み合わせて」は、同時ではないにせよ、キセノンが投与されることを意味するのに用いている。「順次」とは、キセノンと低体温療法が共に治療効果を示す一定の時間枠内のこと、すなわち、それらが共に同一の時間枠内で治療効果を示すことができるということである。したがって、キセノンの循環半減期が、対象の新生児が低体温状態に置かれている時点でキセノンが治療上有効な量で存在するようなものであれば、低体温療法の5分前、10分前、又は何時間も前にキセノンを投与しても、「順次」投与となりうる。
【0066】
本発明の別の好ましい実施形態においては、新生児はキセノンでの治療に先立って、低体温療法を施される。
【0067】
「組み合わせて」又は「順次」に対し、「別個」は、キセノンの投与と、新生児への低体温療法の実施との間の間隔がかなり広い、すなわち、対象の新生児が低体温状態に置かれている時点では、もはやキセノンは治療上有効な量で血流中に存在しないことを意味するために、本明細書で用いられている。
【0068】
ある好ましい実施形態においては、キセノンは低体温療法と順次に投与される。
【0069】
より好ましくは、キセノンは低体温療法に先立って、順次に投与される。
【0070】
別の好ましい実施形態においては、キセノンは低体温療法に先立って、別個に投与される。
【0071】
ある好ましい実施形態においては、キセノンは低体温療法の後に、順次に投与される。
【0072】
別の好ましい実施形態においては、キセノンは低体温療法の後に、別個に投与される。
【0073】
より好ましくは、キセノンは低体温療法と順次又は同時に投与され、さらに好ましくは、同時に投与される。
【0074】
本発明のある好ましい実施形態においては、キセノンは、治療上有効な量で投与される。
【0075】
別の好ましい実施形態においては、キセノンは治療上有効な量以下で投与される。すなわち、キセノンは、低体温ではない状態で投与された場合、所望の治療効果を得るには不十分な量で投与される。
【0076】
さらに好ましくは、キセノンと低体温療法の組み合わせにより相乗効果が得られる、すなわち、この組み合わせは相乗的である。
【0077】
ある特に好ましい実施形態においては、キセノンは低酸素性障害の前に投与される。したがって、ある好ましい実施形態においては、キセノンは、新生児が出生する前に、例えば、陣痛の前又は陣痛中に母親に投与することによって、母親を通じて新生児に投与される。好ましくは、出産前に約48〜24時間、より好ましくは約12時間、さらに好ましくは、約6時間又は3時間又は1時間、母親に投与される。出生後、新生児は低体温状態におかれる。
【0078】
本発明の別の態様は、新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、
(a)治療上有効な量のキセノンを前記哺乳動物の母親に、陣痛前及び/又は陣痛中に投与するステップ、及び
(b)前記哺乳動物に出生後に低体温療法を施すステップ
を含む方法に関する。
【0079】
好ましくは、低体温療法は、低酸素性虚血性(HI)障害後、少なくとも約6時間、より好ましくは約12時間続けられる。
【0080】
ある好ましい実施形態では、低体温療法が、低酸素性虚血性(HI)障害後、約6〜24時間続けられる。
【0081】
好ましくは、低体温療法が、出生後、少なくとも約6時間、より好ましくは少なくとも約12時間続けられる。
【0082】
ある好ましい実施形態においては、低体温療法が、出生後、約6〜約24時間続けられる。
【0083】
好ましくは、本発明の方法に準拠する治療は、低酸素性虚血性(HI)障害から6時間以内に開始され、より好ましくは、低酸素性虚血性障害から2時間以内に開始される。
【0084】
体温が低下するにまかせて、意図的に体温を維持しようとしないことで、受動的に低体温療法の状態を作り出してもよい。新生児は変温性であるため、体温が急速にその周囲の温度に近づいていく。あるいは、故意に周囲の温度を下げることにより、患者を能動的に低体温状態にしてもよい。
【0085】
本発明の第二の態様は、新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、
(a)治療上有効な量のキセノンを前記哺乳動物に投与するステップ、及び
(b)前記哺乳動物に低体温療法を施す又は低体温状態にするステップ
を含む方法に関する。
【0086】
好ましい実施形態においては、哺乳動物は新生児である対象で、生後4週間以内である。より好ましくは、哺乳動物は生後2週間以内、さらに好ましくは、生後1週間以内である。
【0087】
好ましくは、かかる哺乳動物はヒトである。
【0088】
好ましくは、哺乳動物は軽度の低体温状態に置かれる。本明細書においては、「軽度の低体温療法」とは、一般に、中核体温が37℃から約33℃へ低下することを示すものである。
【0089】
ある好ましい実施形態においては、哺乳動物の体温は、約31℃〜約36℃の温度で維持される。
【0090】
より好ましくは、哺乳動物の体温は、約32℃〜約36℃の温度で維持され、さらに好ましくは約32℃〜約35℃、さらにまた好ましくは約33℃〜約35℃に維持される。
【0091】
第二の態様の好ましい実施形態は、第一の態様に関して上述したものと同一である。
【0092】
本発明の別の態様は、新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、治療上有効な量のキセノンを、低体温療法と組み合わせて前記哺乳動物に投与するステップを含む方法に関する。
本発明のさらに別の態様は、新生児仮死治療用薬剤の調製におけるキセノンの使用であって、前記治療が、低体温療法と組み合わせて、同時、順次、又は別個にキセノンを対象に投与するステップを含む使用に関する。
【0093】
本発明のさらなる態様は、新生児仮死を治療するための、低体温療法との組み合わせによる、キセノンの使用に関する。
【0094】
インビトロ研究
HIのモデル動物を用いて、新生児ラットに対し、キセノンと低体温療法による処置を一匹ずつ個別に施した。アポトーシス細胞死の量を減少させることで、キセノンが新生児のHIに対して神経保護作用を示すことが示された。一方、低体温療法の効果はそれよりも低いようであった。キセノンと低体温療法は、組み合わせることにより、抗アポトーシス機構を介して神経保護作用を示すようになる(図17)。それらを組み合わせた場合の効果は、相乗的であることがわかった。
【0095】
新生児ラットHIモデルは確固として確立されており、これまでの多くの研究で使用が妥当であることが確認されている(Levine, 1960; Rice et al, 1981)。このモデルで使用されるラットの年齢は、ヒト新生児の脳成熟期間に相当することがわかっており(Clancy et al, 2001; Ikonimidou et al, 1989)、したがって、これらの二者について、かなり正確な比較を行うことができる。
【0096】
低体温療法実験の間、1匹のラットの皮質に挿入したプローブを用いてラット仔の体温をモニターした。プローブが平衡に達するまで約15分かかったが、この時間を考慮して、プローブが正確な体温を記録し始めるまで実験開始時刻を遅らせた。平均値前後で体温のばらつきがあったが、持続的モニタリングと必要に応じて手動で水槽を調節することにより、それらを制御した。ラットに生じる外傷及びプローブによって皮質に与えられる損傷を最小限にするために、1グループにつき1匹のラットのみ、体温をモニターした。プローブを挿入したラットを組織学的分析にかけることは不可能であった。
【0097】
麻酔ガスのキセノンは、数例の成体神経損傷モデルにおいて神経保護作用を示すことがわかっている。現時点では、キセノンが新生児において同様の神経保護的効果を有することを確認した公開済のデータはない。本研究の結果は、キセノンにはかなりの神経保護性があるというこれまでの知見を裏付けるものであり、また、この神経保護作用が低酸素性虚血により誘導される脳損傷新生児モデルにまで拡大することを示唆している。
【0098】
グルタミン受容体のNMDAサブタイプの活性化がHIにおける進行中のニューロンの損傷及び死を持続させるために必要であることは、かなり以前から周知であり、キセノンがこれらの受容体を遮断することによってその無痛覚及び麻酔効果を示すことは、文書による充分な裏付けがなされている。したがって、キセノンの神経保護性はこのアンタゴニズムの結果と考えられる。以前に、インビトロ研究で神経保護作用をはっきりと示した他のNMDAアンタゴニストがいくつか存在したが、その後、臨床設定において利用された際には不首尾に終わった(Muir and Lees, 1995)。臨床でのこうした不首尾の理由は不明だが、グルタミン酸受容体サブタイプの遮断が、障害を防止するには不十分なものであった可能性がある。これは、キセノンがその神経保護的効果を他の機構を通じて発揮していることを暗示している。
【0099】
本研究において、キセノンが、抗アポトーシス機構を介して、新生児HIを有意に防止していることが示された。アポトーシスと壊死は共にHI障害後のニューロン喪失の重要な要素だが、アポトーシスは、新生児の転帰を決定する点においてより重要なタイプの細胞死と思われる(Taylor et al, 1999)。アポトーシス死が起こる前には、アポトーシス促進性又は抗アポトーシス性のいずれかの、多くの遺伝子(転写因子も含む)が活性化する場合が多い。キセノンはアポトーシス細胞死に干渉すると思われるため、その作用をこれら遺伝子のうちの1つに発揮する可能性、又はアポトーシス経路のある点で発揮する可能性がある。現在、外因性経路及び内在性経路という異なる2つのアポトーシス経路について証拠がある。外因性経路(「死受容体経路」とも称される)は、カスパーゼ−8を活性化する死受容体へのサイトカインの結合を伴うものであり、続いてそのカスパーゼ−8が、アポトーシス細胞死の誘導を開始する「執行カスパーゼ」であるカスパーゼ−3を活性化する(Mehmet, 2000)。内在性経路はミトコンドリアに多大に依存しており、アポトーシス促進性タンパク質baxによって生じるミトコンドリア膜透過性の増大を伴う。これはサイトクロムcの放出、サイトクロムc、Apaf−1(アポトーシスプロテアーゼ活性化因子−1)及びカスパーゼ−9との複合体の形成、並びにその後のカスパーゼ−3の活性化につながる。これらの経路のいずれか1つに対してキセノンが作用するのは全く可能なことであるが、HIに誘導されたアポトーシス性神経変性はミトコンドリア経路及びbax依存性ミトコンドリア変化の開始に仲介されることを示唆する証拠がある(Taylor et al, 1999)。これに加え、NMDAアンタゴニストのケタミンは、アポトーシス促進性タンパク質と抗アポトーシス性タンパク質とのバランスを早期に調節することにより、すなわち、HI誘導性bax増大を阻害することにより、不完全な大脳虚血及び再灌流を防止することがわかっている(Engelhard et al, 2003)。このように、キセノンは、baxをダウンレギュレートすることによりアポトーシスを阻害できる、という可能性がある。Bcl−2は、ミトコンドリア膜の透過性を減少させる作用を有する抗アポトーシス性タンパク質であり、そのため、サイトクロムcの放出を阻害する。その過剰発現は、アレチネズミにおいて一過性全脳虚血により生じる神経損傷を減少させることがわかっている(Engelhard et al, 2003)。したがって、bcl−2のアップレギュレーションは、キセノンの別の潜在的な標的である。キセノンは無極性で脂溶性であることから、身体全体に広く分布させることができる。膜を通過することができ、その結果、遺伝子転写を変化させて生存経路をアップレギュレートすることにより、又はアポトーシス促進性分子のRNA及びタンパク質合成を阻害することにより、核内にも作用を及ぼす場合がある。
【0100】
キセノンによる抗壊死作用は、皮質の48時間群において、統計的に有意であることが判明したが、脳回ではそうではなかった(図16)。他の全ての時間群では、キセノンは抗壊死作用を示さなかった。これに関して考えられる解釈が1つ挙げられる。以前の研究によれば(Northington et al, 2001)、皮質だけに見られる壊死性細胞死の第二の波が48時間群で存在するとのことである。これは、48時間群のポジティブコントロールに存在する壊死細胞の比率が、16時間群及び24時間群と比較して増加していることを説明するものである。皮質の48時間群でキセノンがどのようにして抗壊死作用を発揮するのかは明らかではないが、キセノンは、(16時間群及び24時間群の場合と同様に)キセノン投与前に生じた壊死を防止することは不可能である一方で、キセノン投与後に生じた第二の壊死の波に何らかの方法で対抗することができる、ということだと思われる。最初の壊死は、HI障害の3時間後に早くも発生しており(Northington et al, 2001)、この時点でキセノンはまだ投与されていなかった。したがって、すでに発生したプロセスを停止又は逆行させることはできるとは考えにくい。しかし、第二の壊死の波はキセノンが脳内に48時間存在した時点で発生し、これは、壊死出現時にキセノンが存在することによって、このタイプの細胞死を減少させることが可能な場合があることを示唆している。この相互作用の正確な機序を確認するには、さらなる研究を完成させなくてはならない。
【0101】
33℃という軽度の低体温療法は、虚血性神経損傷に対する神経保護作用があることが、以前の研究で示されている(Busto et al, 1987)。他の研究では、この神経保護作用が抗アポトーシス機構を介して達成されていることが示唆されている(Xu et al, 2002)。16時間群又は24時間群(それぞれ図13及び15)では神経保護作用が皆無であることが、実験で示されている。
【0102】
しかし、48時間群では、異なる機構によってではあるが、皮質と脳回の両方で有意な神経保護作用が達成されていた。低体温療法は、皮質においては抗壊死性であり、脳回においては抗アポトーシス性である(図16)。本研究のデータはこの作用を説明するものではないが、考えられる理由を1つ挙げるとすれば、異なる脳領域が差別的な脆弱性を発現するということがあるのかもしれない(Northington et al, 2001)。皮質においては、低体温療法がすでに行われた時点で第二の壊死の波(上述)が発生するため、効果が増大すると思われる。しかし脳回では遅発性の壊死は存在しないため、抗壊死作用も観察されない。この領域では抗アポトーシスが神経保護的機構であると思われ、時間間隔が短い場合にははっきりしない低体温療法の予期された抗アポトーシス性神経保護的効果が、より長い時間を経て露になった可能性がある。
【0103】
こうした結果により、20%のキセノンと35℃の低体温療法を組み合わせて使用すると、驚異的なレベルの神経保護作用がもたらされることが示された。これらの数値は、それぞれの作因を単独で使用した場合には全く神経保護作用をもたらさなかったため、こうした結果は付加的な機構によって説明できるものではなかったが、これら2つの作因の相乗効果的な相互作用によるものにちがいなかった。
【0104】
すなわち、本研究は、キセノンが新生児において神経保護作用を示し、低酸素性虚血性障害に誘導されたアポトーシスを有意に防止することを示すため、インビボラットモデルを用いて示したものである。本研究のデータは、キセノンと低体温療法とを同一モデルで組み合わせて使用すると、相乗的な相互作用が発生して、アポトーシス性細胞死を劇的に減少させることを示唆している。したがって、この組み合わせは、新生児仮死の破壊的な神経学的結果を防止する効果的治療法の代表となるかもしれない。
【0105】
本発明を、実施例により、また、後述する図面により、詳細に説明する。
【実施例1】
【0106】
新生児仮死モデル
手術用麻酔(1〜1.5%のイソフルランを含む純粋酸素)下で、生後7日のSprague-Dawleyラットの右総頚動脈を結紮した。結紮後、それらの動物を母親のもとへ戻し、室温(23℃)及び湿度(48%)を一定にした特別設計の区画に収容した。手術の1時間後、8%の酸素と0、20、40、60、又は70%のキセノンとを組み合わせたもの(残りは窒素で補充した)を入れた特別設計のチャンバーに、新生児ラットを収用した。収容時間は90分間で、その際の温度は37℃であった(チャンバーの外側に設置した水槽で温度を維持した)。実験の7日後、ラット(日齢14)を殺処分し、脳を摘出した。左半球に対する右半球の重量比(R/L比)を計算した。数群のラット仔については、生後30日まで生存させ、その時点でそれらの神経運動機能及び協調を、確立されたプロトコルによって査定した(神経運動試験及びロータロッド試験)。
【0107】
低酸素状態の時間を延長したところ、損傷(脳重量の減少として測定)が明瞭に認められるのは、低酸素状態が90分を越えた場合のみであることが結果として示された(図1)。そのため、低酸素障害の標準時間を90分に設定した。
【0108】
90分間の低酸素性虚血性障害を受けた動物の脳切片を図2に示す。詳細には、図2(中央)は、巨視的な解剖学的損傷(障害を受けた側の脳――この図では左側)を、コントロール動物(左)と比較して示す。右側の脳スライスは、同じく低酸素性虚血を経験したものの、低酸素状態の間に70%のキセノンを呼吸した動物に由来するものである。これらの脳は正常のものとよく似ており、キセノンによってもたらされた驚異的神経保護作用を示している。
【0109】
90分間の低酸素性虚血の24時間後に明瞭に認められる多大な細胞損傷を図3に示す。
【0110】
キセノンの神経保護作用の濃度依存性(左半球に対する右半球の重量比と、キセノンの濃度)を図4に示す。詳細には、図4は、生後7日で行った、各種濃度のキセノンを組み合わせた又は組み合わせない低酸素性虚血の後の、日齢14のラット脳の同側/対側半球重量比を示す。麻酔域下濃度の場合でさえ、神経保護作用は明瞭に認められた。コントロール動物には頚動脈結紮を行ったが、低酸素状態にはしなかった。結果は、平均値±SEM(n=5−8)である。8%のO2に対して、*P<0.01。
【0111】
低酸素性虚血性(HI)障害後にかなりの時間をおいて査定した、70%のキセノンの神経機能に対する作用を図5に示す。生後7日で右頚動脈を結紮し、ラット仔を低酸素環境(8%の酸素+70%のキセノン、残りは窒素で補充した)に90分間暴露させた。障害から30日後、(A)把握牽引(prehensile traction)、強度、及び平均台パフォーマンス(0〜9のレベルに採点)の検定を含むパネルの使用により、及び(B)バランスと神経運動機能の標準的なテストであるロータロッドでのバランスにより、ラットの神経運動機能を評価した。ラット1個体からのデータポイントは、三回のテストの合計である。水平のバーは、各群の中央値を示す。
【0112】
HI障害の2時間後にキセノンが投与される場合の、N2とキセノンそれぞれについて観察された神経保護的効果(右半球/左の比)を図6に示す。より詳細には、低酸素状態の時間が終了してから2時間後にキセノンが投与された場合でさえ、キセノンが神経保護作用を効果的に発揮していることを、データは示している。生後7日で、90分間の低酸素性虚血性障害後に、70%のN2又は70%のキセノン+30%のO2に90分間暴露させてから2時間の回復期を与えた、日齢14のラット脳の同側/対側半球重量比。結果は、平均値±SEM(n=6)である。
【0113】
キセノンの神経保護的効果に対する軽度の低体温療法の作用(LDH放出と、キセノン濃度(%原子))を図7に示す。穏当な低体温療法は、キセノンの神経保護作用に、非常に大型で予想しない増強をもたらす。体温が4℃下がると、LDH放出のブロッキングにおけるキセノンの効能が大幅に増強する。より詳細には、この図は、酸素−グルコース欠乏(OGD)が誘導する乳酸脱水素酵素(LDH)放出に対し、キセノンと低体温療法との組み合わせが及ぼす作用を示す。図7は、濃度を増大させたキセノンの存在下、37℃(赤)又は33℃(青)のいずれかで、ニューロン培養物をOGDに75分間暴露させた結果を示す。37℃でのキセノン、33℃でのキセノンに対するED50値は、それぞれ35.9+/−2.15%、11.5+/−2.0%(平均値+/−SEM)であった。ニューロンの損傷は、キセノン又は低体温療法いずれかの不在下での、75分間のOGD及び6時間の回復期後におけるLDHの最大放出量のパーセンテージで表した。点は平均値を示す。エラーバーは、標準誤差を示す。
【0114】
かかるプロセスの温度依存性の程度を図8に示す。図8には、絶対温度の逆数に対してプロットされたLDH放出の自然対数のファント・ホッフプロットが示されている。そのようなプロットのスロープから、かかるプロセスのエンタルピーの変化が計算でき、その大きさは温度依存性の尺度となる。赤のデータは、キセノン不在下でのLDH放出に対する温度の作用を示す。温度の低下に伴う放出の減少が予想されるが、穏当なものである。12.5%のキセノンが存在する場合、温度依存性は非常に高く、予想不可能である。そのため、低体温療法は、キセノンの神経保護的効果を大幅に増強させると思われる。したがって、こうした結果は、低体温療法とキセノンが神経保護剤として相乗的に作用することを示唆している。
【0115】
より詳細な研究を下記の実施例2において概述する。
【実施例2】
【0116】
材料と方法
本研究は、1986年の英国動物(科学的処置)法に準拠するものであり、内務省(英国)により認可されている。
【0117】
低酸素性虚血症のモデル動物
本研究では、脳障害のパターンが満期産のヒト新生児における低酸素性虚血性障害のものと類似している日齢7(p7)の局所的HIモデルラットを使用する(Johnston, 1983)。
【0118】
英国Harlan社のp7のSprague-Dawleyラット仔(体重は10〜14g)に、以前記載されたモデルのHI障害を受けさせた(Levine, 1960; Rice et al,1981)。すなわち、2%のイソフルランでラット仔に麻酔をかけた後、頸部正中切開及び5.0絹縫合糸による右総頚動脈の永久的な片側結紮を行った。手術が完了すると、ラット仔は麻酔から覚醒し、その後実験時まで母親のもとへ戻された。
【0119】
手術の1時間後、一部が37℃の水槽に沈められている、専用の気密チャンバーにラット仔を収容することにより、ラット仔を低酸素状態に暴露させた(図9)。90分間という低酸素時間は、半球の重量として測定された低酸素性虚血性損傷が、この時間の後に最大となることが予備実験で示されたために、選択された。8%の加湿酸素に残りを窒素で補充して、それを継続的に流し入れることで、低酸素状態を誘導した。この混合気体を、15分ごとにDatex Ohmeda(英国、ブラッドフォード)でモニターした(気体はすべてBOC社より供給された)。
【0120】
実験的処置
HIの後、ラット仔を回復のために4時間母親のもとへ戻し、その後、下記の実験的介入の1つを90分間受けさせた。予備実験のデータにより、介入を適用する最適の時期は、HIと同時かその4時間後であることが示された。この2つの時期については有意な差異は存在しなかったため、臨床的に最適と思われたとの理由により、介入の適用時間として障害の4時間後を選択した。
【0121】
ラット仔は、処置の16、24、及び48時間後に殺処分されるまで、母親のもとに戻された(Northington et al, 2001)(図10)。
【0122】
コントロール
コントロール群は、(a)ネガティブコントロールとして使用される、切開は行ったが結紮は行っていない無処置の同腹子(すなわち、擬似手術動物)と、(b)ポジティブコントロールとしての役割を果たす、HIの措置は行ったが実験的介入の措置は行っていない同腹子とからなる。これらの動物は、25%の酸素に残りを窒素で補充した混合気体に90分間、37℃でさらされた。
【0123】
実験用ラット
HI及び回復期の後、実験用ラットを下記の5つの実験的介入の1つで90分間処置した。5つの処置のそれぞれは、別個の群のラットに対して行われた。
【0124】
低体温療法での処置
ラット仔に、軽度の低体温療法(33℃)での処置を90分間施した。無作為に1匹のラット仔を選別し、イソフルラン及び局所麻酔下で、温度プローブ(Mini-Mitter Co. Inc.社製、米国、オレゴン州、ベンド)を皮質に挿入し、強力接着剤でその場所に固定した。その後、すべてのラット仔を気密チャンバー(前述のとおり)に収容し、25%の酸素に残りを窒素で補充した混合気体をポンプで送った。温度プローブとVital Viewコンピュータプログラムによって測定されたラットの皮質の温度を正確に33℃に保つように維持された水槽に、チャンバーを部分的に水没させた。この温度を選択したのは、それが「軽度の(mild)」低体温療法を示すものであること、したがって、臨床的に適していると考えられ、副作用と効能との間のバランスがよいという理由によるものである。処置の90分後、ラット仔を母親のもとへ戻し、殺処分時まで回復させた。温度プローブを設置されたラット仔は、実験後直ちに殺処分され、その脳を分析に使うことはなかった。
【0125】
キセノンでの処置
キセノンでの処置にも同じ実験用パラダイムが遵守されたが、低体温療法の代わりに、水槽を37℃に維持し、混合気体を25%の酸素と75%のキセノンに変えて、90分間行った。キセノンの漏出を最小限にするため、専用の閉鎖システムにガスを送達した(図9)。ここでも、殺処分時までラット仔を母親のもとへ戻した。
【0126】
組み合わせプロトコル
組み合わせパラダイムにおいて、ラットに低体温療法とキセノンの両方を同時に90分間施した。再びラット仔を気密チャンバーに収容したが、この場合は、その温度を35℃に維持し、混合気体の構成は、25%の酸素にキセノンは20%のみで、残りを窒素で補充したものであった。この温度及びキセノン濃度は、別々に使用しても発達途中の脳に神経保護的な効能を与えないことが、予備実験で示されていたものである。したがって、これらの数値を用いることにより神経保護的効能が少しでも生じれば、それはこの2つの作因間の相乗作用を示すことになる。処置後、ラット仔を殺処分時まで母親のもとへ戻した。
【0127】
組み合わせ群で使用した数値が、別個に使用された場合にはまったく神経保護作用を及ぼさないことを示すため、実験ラット群をもう2群使用した。一方の群には35℃での低体温療法(前述のとおり)を施し、他方の群には濃度20%のキセノンへの暴露を行った。
【0128】
組織の調製
脳の採取
実験プロトコルの終了から16、24、及び48時間後に、脳を摘出した。
【0129】
腹腔内へペントバルビタールを過剰投与(100mg/kg)してラットを殺処分し、その後、2.5u/mlのヘパリンを含むPBSを用いて左心室からの経心臓的灌流を行って血液を抜いた。引き続き、4%のパラホルムアルデヒドを含むPBSを20ml使って灌流した後、脳を除去した。その後、同じ固定剤で脳を一晩後固定した。各時間群について、コントロールの数又は実験用脳を均等に分配し、免疫組織化学用の凍結切片としてスライスするか、組織学的分析用にパラフィン包埋するかのいずれかを行った。同側半球と対側半球とを区別するため、スライスする前に、対側の影響を受けていない(左)半球にペーパークリップで穴をあけた。
【0130】
パラフィン包埋
後固定後、組織学用の脳を、Histokinette 2000組織包埋プロセッサー(Leica U. K. Ltd.社製、英国、ミルトン・ケインズ)で乾燥させ、その後、パラフィンワックスブロックに包埋した。パラフィン包埋した脳を、ミクロトーム(Leica社製、ドイツ)を用いて、厚さ5μmの冠状切片にカットした。図10.3は、薄片が採取された脳部位を示す。各脳の、ブレグマから約−3.6mmにある海馬部位(この部位がHI障害に最も脆弱な部位であるため)から、約20の薄片を採取した。
【0131】
凍結切片
脳を一晩後固定した後、30%のスクロースを含むPBS(これには、2mg/mlのアジ化ナトリウムも含まれている)中で凍結防止処理を施し、冷蔵庫で48時間又は脳がチューブの底に沈むまで保管した。その後、脳をO.C.T.化合物(BDH社製、英国、プール)中で−22℃にて凍結させ、冠状切片をスライディング・クライオスタット(Bright Instrument Company Ltd社製、英国、ハンチントン)上で30μmにカットした。スライスした脳を、0.1MのPBS及び1mg/mlのアジ化ナトリウムを含むウェルに入れて、冷蔵庫にて保管した後、免疫組織化学用に染色した(下記参照)。
【0132】
染色の手順
組織学
パラフィン包埋した切片を、スライドに載せ、以前の記載のとおり、組織学用にクレシルバイオレットで染色した(Wilhelm et al, 2002)。
【0133】
壊死及びアポトーシスの神経病理学的分析
組織学的顕微鏡検査
クレシルバイオレットで染色したパラフィン脳薄片の組織学的分析により、ニューロン損傷を査定した。クレシルバイオレットは、RNAを多く含むリボソームのようなニューロン細胞質の酸性成分、並びに神経細胞の核及び核小体に結合する基本的な染色液である。この技法を用いて、細胞の生存、及び生育不能な細胞が示すものがアポトーシスであるか、又は壊死であるかを、有効な形態学的基準(Nakajima et al, 2000)に基づいて査定した。
【0134】
各実験群は、3つの時間群(16、24、48時間)からなり、各時間群は、3匹の動物を擁していた(したがって、各実験群には、合計9匹の動物が存在した)。ブレグマから−3.6mmの脳部位から、1匹あたり3枚のスライドが選ばれた(図11)。その後、スライドを時間群に分けて、試験者にはその介入がわからないようにした。
【0135】
BX60光学顕微鏡(Olympus社製、英国、サウスオール)を用いて各脳の同側の2つの部位を分析した――1つは皮質内の、もう1つは海馬の脳回内の部位である(図11)。グリッド付きの40倍の対物レンズを用いて、グリッド上に認められる細胞の総数を計数した。細胞に対し、その形態学的外観に基づき、生存、アポトーシス、又は壊死のいずれかとしてスコアをつけ、各細胞型のパーセンテージを、それぞれの脳部位について書き記した。Axiocamデジタルカメラ(Zeiss社製、ドイツ、ゲッチンゲン)を顕微鏡と併用して、脳薄片の顕微鏡写真を撮影した。
【0136】
細胞をそれぞれのカテゴリーに割り当てる際の基準は、以下のとおりである(図12):
1.いずれかのタイプの細胞死(アポトーシス又は壊死)を起こしている細胞は、通常の形態を有するとともに細胞質が淡色で核が明瞭に視認可能な濃色である生存細胞よりも、クレシルバイオレット染色液の取り込みが強かった。
2.アポトーシスであると分類された細胞は、非常に濃く染色され、縮退している球状の核及び完全な細胞膜を有しており、周囲に空胞化が見られる場合が多い。
3.一方、壊死細胞は、同じく強く染色されるが、不規則な形状の、拡大した核を有している。
【0137】
アポトーシス細胞、壊死細胞及び生存細胞の数を、総細胞数のパーセンテージとして示し、それぞれのスライドに書き記した。その後、これら3つのスライドから各動物について平均パーセンテージを計算して、1つの時間群における3匹の動物それぞれの数値を1つにまとめた。各時間群についての数値を1つにまとめるため、これら3匹の動物から再び平均値を取り、標準偏差を記録した。
【0138】
統計学的分析
データ分析は、一元配置分散分析を用いて行い、適切とされる場合には、その後に、Student-Newman-Keulsを用いた。p<0.05は、統計学的に有意であると考えられた。
【0139】
結果
別個の作因としてのキセノン及び低体温療法
キセノンは、新生児において抗アポトーシス機構による神経保護作用を有する
脳の皮質及び海馬領域の顕微鏡分析を行ったところ、擬似手術脳と比較してキセノン処置脳の形態学的外観が類似していること、及びキセノンで処置しなかったラットの脳と比較して外観が相違していることから、キセノンの神経保護性が実証された(図14)。新生児ラットにおける低酸素性虚血性障害に対する強い神経保護作用は、キセノンをその最大濃度(75%)で使用したことにより達成され、これは、クレシルバイオレットで染色した脳薄片の組織化学的分析により定量化されている。この濃度のキセノンを別個に使用すると、アポトーシス細胞死を有意に減少させ、生存細胞数を増大させる。16時間群では、皮質のアポトーシスは、ポジティブコントロールにおいて、36.5%±2.5%から8.5%±1.6%(p<0.001)に減少しており、生存細胞数は、ポジティブコントロールにおいて、52.9%±2.3%から80.6%±0.2%(p<0.001)に増大している(図13A)。脳回においては、アポトーシスは、ポジティブコントロールにおいて、33.6%±1.8%から13.9%±2.4%(p<0.01)に減少しており、生存細胞数は、ポジティブコントロールにおいて、56.5%±2.6%から77.1%±3.3%(p<0.01)に増大している(図13B)。24時間群及び48時間群(それぞれ、図15及び図16)では、16時間群と類似した結果が示されており、キセノンは、皮質と脳回の両方において、ポジティブコントロール動物と比較して、統計学的に有意な抗アポトーシス作用を示している。
【0140】
キセノンによる抗壊死作用は、ポジティブコントロールにおいて壊死が16.6%±0.2%から10.7%±0.4%(p<0.01)に減少していた皮質の48時間群で統計学的に有意であることが示された(図16A)。しかし、キセノンは、脳回の48時間群においては抗壊死作用を示さなかった(図16B)。他のすべての時間群(16及び24時間)では、キセノンは抗壊死作用を示さなかった。
【0141】
中程度のHIの後に、33℃での低体温療法を90分間行っても効果はない
16時間群又は24時間群(それぞれ、図13及び15)では、33℃での低体温療法については神経保護作用が観察されなかった。16時間群では、低体温療法は皮質において有意な抗アポトーシス作用を有するようであったが、生存細胞数についてはポジティブコントロール動物と統計学的な差異がなかったため、この介入により神経保護作用はもたらされなかったと結論づけることができる。しかし、48時間群では、低体温療法は皮質の抗壊死機構を介して神経保護作用を示し、壊死細胞数をポジティブコントロールにおいて、16.6%±0.2%から12%±2.3%に減少させ、生存細胞数を43%±3.4%から52.3%±3.1%に増大させた(図16A)。脳回の48時間群においては、低体温療法は、抗アポトーシス性の神経保護作用を示した(図16B)。
【0142】
キセノンと低体温療法との組み合わせ
20%のキセノンのみでの処置では、神経保護作用は示されない
75%のキセノンを用いて得られた結果とは異なり、20%のキセノンはまったく神経保護的作用を発揮しない。図17を見ると、皮質の16時間群で認められたアポトーシスのパーセンテージが、ポジティブコントロール動物(p>0.05)の37%±2.5%と比較して20%のキセノン群では36%±5.7%であること、生存細胞数のパーセンテージが、ポジティブコントロール動物(p>0.05)の53%±2.3%と比較して20%のキセノン群では51%±7.8%であることがわかる(すなわち、20%群とポジティブコントロール群との間に統計学的な差異はない)。脳回のデータからも、非常に良く似た結果が出た。
【0143】
35℃での低体温療法のみでの処置では、神経保護作用は示されない
35℃の低体温療法を単独で使用してもHIに対して効果がなく、ポジティブコントロールと比較して、どの脳部位においても統計学的な差異は示されない。アポトーシスのパーセンテージは、ポジティブコントロールの37%±2.5%に対して48%±10.1%であり、細胞生存のパーセンテージは、53%±2.3%に対して44%±10.3%である。
【0144】
20%のキセノンと35℃での低体温療法とを組み合わせた処置では、抗アポトーシス機構を介した相乗的神経保護作用が示される。キセノン(20%)又は低体温療法(35℃)のいずれかという、すでに効果がないことが証明された介入を組み合わせて使うことにより、抗アポトーシス機構を介して、3つすべての時間群(16、24、48時間)を通じて、脳の両方の部位で強い相乗的な神経保護作用が示された。かかる組み合わせ処置を虚血後に適用すると、アポトーシス細胞死の程度を有意に低下させ、生存細胞の割合を増加させた(図17参照)。皮質の16時間群では、組み合わせ療法によるアポトーシスの減少は、20%のキセノン群と35℃での低体温療法群ではそれぞれ35.8%±5.7%と47.6%±10.1%であったのが、組み合わせ群(それぞれ、p<0.01及びp<0.001)においてはわずか7.2%±2%に減少しているのがわかった。一方、生存細胞は、51%±7.8%と43.7%±10.3%であったのが、82.3%±4.9%(両方の群において、p<0.01)に増加していた。脳回のデータからも、抗アポトーシス性保護作用に加えて抗壊死性保護作用も示されると思われる24時間群以外は、良く似た結果が出た(図17B)。
【0145】
個々の作因からはまったく神経保護作用がもたらされないという事実に鑑みて、かかる組み合わせの結果は、驚くべきものであり、予想をはるかに上回るものであることは確かである。別々では効果のない2つの介入を組み合わせたことによりもたらされる神経保護作用の程度は、キセノンと低体温療法との間に、インビボでの相乗性が存在することを示している。
【0146】
本発明の範囲及び精神から逸脱することのない、ここに記載した本発明の方法の様々な修正及び変更は、当該分野の熟練者には自明なことであろう。本発明を特定の好ましい実施形態に関して記載したが、関連分野の熟練者にとっては自明なことである、ここに記載した本発明の実施形態の様々な修正は、添付した請求の範囲の範囲内であると解釈される。
【0147】
[参考文献]
Albers, G., Goldberg, M.P., Choi, D.W., 1989. N-methyl-D-aspartate antagonists: ready for clinical trial in brain ischemia? Ann Neurol 25:398-403.
Ankarcrona, M., 1995. Glutamate-induced neuronal death: A succession of necrosis or apoptosis depending on mitochondrial function. Neuron 15:961-973.
Arias, R.L., Tasse, J.R.P., Bowlby, M.R., 1999. Neuroprotective interaction effects of NMDA and AMPA receptor antagonists in an in vitro model of cerebral ischemia. Brain Research 816:299-308.
Balduini, W., De Angelis, V., Mazzoni, E., Cimino, M., 2000. Long-lasting behavioural alterations following a hypoxic/ischemic brain injury in neonatal rats. Brain Research 859:318-325.
Bliss, T.V.P., Collingridge, G.L., 1993. A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361:31-38.
Bosley, T.M., Woodhams, P.L., Gordon, R.D., Belazs, R., 1983. Effects of anoxia on the stimulated release of amino acid neurotransmitters in the cerebellum in vitro. J. Neurochem 40:189-201.
Busto, R., Dietrich, W.D., Globus, M.Y.T., 1987. Small differences in intraischemic brain temperature critically determine the extent of ischemic neuronal injury. Blood Flow Metab 7:729-738.
Choi, D.W., Koh, J-Y., Peters, S., 1988. Pharmacology of glutamate neurotoxicity in cortical cell culture: attenuation by NMDA antagonists. J. Neurosci 8:185-196.
Choi, D.W., Rothman, S.M., 1990. The role of glutamate neurotoxicity in hypoxic-ischemic neuronal death. Annu Rev Neurosci 13:171-182.
Chopp, M., Knight, R., Tidwell, C.D., Helpern, J.A., Brown, E., Welch, K.M., 1989. The metabolic effects of mild hypothermia on global cerebral ischemia and recirculation in the cat: comparison to normothermia and hypothermia. J Cereb Blood Flow Metab 9:141-148.
Chow, A., Ma, D., Hossain, M., Franks, N.P., Maze, M., 2003. Combined neuroprotection by xenon and hypothermia. Society Neuroscience Abstract 893(1).
Clancy, B., Darlington, R.B., Finlay, B.L., 2001. Translating developmental time across mammalian species. Neurosci 105(1):7-17.
Cullen, S.C., Gross, E.G., 1951. The anaesthetic properties of xenon in animals and human beings, with additional observations on krypton. Science 113:580-581.
Debillon, T., Daoud, P., Durand, P., Cantagrel, S., Jouvet, P., Saizou, C., Zupan, V., 2003. Whole-body cooling after perinatal asphyxia: a pilot study in term neonates. Developmental Medicine and Child Neurology 45:17-23.
Dingledine, R., McBain, C.J., 1999. Glutamate and aspartate in: Siegel, S.J., Agranoff, B.W., Albers, R.W., Fisher, S.K., Uhler, M.D., (Eds), Basic neurochemistry: Molecular, cellular and medical aspects, 6th edition, Lippincott-Raven, pp316-333.
Eilers, H., Bickler, P.E., 1996. Hypothermia and Isoflurane similarly inhibit glutamate release evoked by chemical anoxia in rat cortical brain slices. Anesthesiology 85:600-607.
Engelhard, K., Werner, C., Ebersp臘her, E., Bachl, M., Blobner, M., Eberhard, H., Hutzler, P., Eberhard, K., 2003. The effect of the 2-agonist dexmedetomidine and the N-methyl-D-aspartate antagonist S(+)-ketamine on the expression of apoptosis-regulating proteins after incomplete cerebral ischemia and reperfusion in rats. Anesth Analg 96:524-531.
Erecinska, M., Nelson, D., Wilson, F., Silver, I.A., 1984. Neurotransmitter amino acid levels in rat brain during ischemia and reperfusion. Brain Research 304:9-22.
Franks, N.P., Dickinson, R., de Sousa, S.L.M., Hall, A.C., Lieb, W.R., 1998. How does xenon produce anaesthesia? Nature 396(6709):324.
Goto, T., Nakata, Y., Morita, S., 2003. Will xenon be a stranger or a friend? Anesthesiology 98:1-2.
Goto, T., Saito, H., Nakata, Y., Uezono, S., Ichinose, F., Morita, S., 1997. Emergence times from xenon anaesthesia are independent of the duration of anaesthesia. British Journal of Anaesthesia 79:595-599.
Graham, S.H., Shiraishi, K., Panter, S.S., Simon, E.P., Faden, A.I., 1990. Changes in extracellular amino acid neurotransmitters produced by focal cerebral ischemia. Neurosci Lett 110:124-130.
Gunn et al, 2000. Curr Opin Pediatr. Apr;12(2):111-5.
Gwag, B.J., Koh, J.Y., Demaro, J.A., Ying, H.S., Jacquin, M., Choi, D.W., 1997. Slowly triggered excitotoxicity occurs by necrosis in cortical cultures. Neurosci 77:393-401.
Hardingham, G.E., Bading, H., 2003. The yin and yang of NMDA receptor signalling. Trends in Neuroscience 26(2):81-89.
Hauptman, M., Nelson, D., Wilson, D.F., Erecinska, M., 1984. Some changes in amino acid levels in rat brain synaptosomes during and after in vitro anoxia and simulated ischemia. Brain Research 304:23-35.
Hill, I.E., MacManus, J.P., Rasquinha, I., Tuor, U.I., 1995. DNA fragmentation indicative of apoptosis following unilateral cerebral hypoxia-ischemia in the neonatal rat. Brain Res 676:398-403.
Homi, H.M., Yokoo, N., Ma, D., Warner, D.S., Franks, N.P., Maze, M., Grocott, H.P., 2003. The neuroprotective effect of xenon administration during transient middle cerebral artery occlusion in mice. Anesthesiology 99:876-881.
Ikeda, M., Nakazawa, T., Abe, K., Kaneko, T., Yamatsu, K., 1989. Extracellular accumulation of glutamate in the hippocampus induced by ischemia is not calcium dependent in vitro and in vivo evidence. Neurosci Lett 96:202-206.
Ikonomidou, C., Bittigau, P., Koch, C., Genz, K., Hoerster, F., Felderhoff-Mueser, U., Tenkova, T., Dikranian, K., Olney, J.W., 2001. Neurotransmitters and apoptosis in the developing brain. Biochemical Pharmacology 62:401-405.
Ikonomidou, C., Mosinger, J.L., Olney, J.W., 1989. Hypothermia enhances protective effect of MK-801 against hypoxic/ischemic brain damage in infant rats. Brain Research 487:184-187.
Ikonomidou, C., Mosinger, J.L., Salles, K.S., 1989. Sensitivity of the developing rat brain to hypobaric/ischemic damage parallels sensitivity to N-methyl-D-aspartate neurotoxicity. J Neurosci 9:2809.
Ikonomidou, C., Price, M.T., Mosinger, J.L., 1989. Hypobaric-ischemic conditions produce glutamate-like cytopathology in infant rat brain. J Neurosci 9:1693.
Jevtovic-Todorovic, V., Olney, J.W., 2003. Neuroprotective agents in: Evers, A.S., Maze, M., (Eds), Anesthetic Pharmacology: Physiological principles and clinical practice, Churchill Livingstone, pp557-572.
Johnston, M.V., 1983. Neurotransmitter alterations in a model of perinatal hypoxic-ischemic brain injury. Ann Neurol 13:511-518.
Katja, C.Z., Green, D.R., 2001. How cells die: Apoptosis pathways. J Allergy Clin Immunol 108(4):S99-S103.
Kauppinen, R.A., McMahon, H., Nicholls, D.G., 1988. Ca2+-dependent and Ca2+-independent glutamate release, energy status and cytosolic free Ca2+-concentration in isolated nerve terminals following in vitro hypoglycaemia and anoxia. Neuroscience 27:175-182.
Komuro, R.P., 1993. Modulation of neuronal migration by NMDA receptors. Science 260:95.
Krystal, J.H., Karper, L.P., Seibyl, J.P., Freeman, R., Delaney, R., Bremner, J.D., Heninger, G.R., Bowers, M.B., Charney, D.S., 1994. Subanaesthetic effects of the non-competitive NMDA antagonist, ketamine, in humans: psychotomimetic, perceptual, cognitive and neuroendocrine responses. Arch Gen Psychiatry 51:199-214.
Kudo, M., Aono, M., Lee, Y., Massey, G., Pearlstein, R.D., Warner, D.S., 2001. Effects of volatile anesthetics on N-methyl-D-aspartate excitotoxicity in primary rat neuronal-glial cultures. Anesthesiology 95:756-765.
Leist, M., Nicotera, P., 1998. Apoptosis, excitotoxicity, and neuropathology. Experimental Cell Research 239:183-201.
Levine, S., 1960. Anoxic-ischemic encephalopathy in rats. Am J Pathol 36:1-17.
Low, J.A., 1993. The relationship of asphyxia in the mature fetus to long-term neurologic function. Clinical Obstetrics and Gynaecology 36(1):82-90.
Ma, D., Wilhelm, S., Maze, M., Franks, N.P., 2002. Neuroprotective and neurotoxic properties of the ‘inert’ gas, xenon. British Journal of Anaesthesia 89:739-746.
MacDonald, J.F., Schneiderman, J.H., Miljkovic, Z., 1986. Excitatory amino acids and regenerative activity in cultured neurons. Adv Exp Med Biol 203:425.
Mehmet, H., 2000. Caspases find a new place to hide. Nature 403:29-30.
Muir, K.W., Lees, K.R., 1995. Clinical experience with excitatory amino acid antagonist drugs. Stroke 26:503-513.
Nakajima, W., Ishida, A., Lange, M.S., Gabrielson, K.L., Wilson, M.A., Martin, L.J., Blue, M.E., Johnston, M.V., 2000. Apoptosis has a prolonged role in the neurodegeneration after hypoxic ischemia in the newborn rat. J Neurosci 20(2):7994-8004.
Nakata, Y., Goto, T., Morito, S., 2001. Clinical pharmacology of xenon. Int Anesthiol Clin 39:63-75.
Nicholls, D.G., Budd, S.L., 2000. Mithochondria and neuronal survival. Physiol Rev 80:315-360.
Northington, F.J., Ferriero, D.M., Graham, E.M., Traystman, R.J., Martin, L.J., 2001. Early neurodegeneration after hypoxia-ischemia in neonatal rat is necrosis while delayed neuronal death is apoptosis. Neurobiology of Disease 8:207-219.
Olney, J.W., 2003. Excitotoxicity, apoptosis and neuropsychiatric disorders. Current Opinion in Pharmacology 3:101-109.
Olney, J.W., Labruyere, J., Wang, G., Wozniak, D.F., Price, M.T., Sesma, M.A., 1991. NMDA antagonist neurotoxicity: mechanism and prevention. Science 254:1515-1518.
Onitsuka, M., Satoshi, M., Inokuchi, H., Shigemori, M., Higashi, H., 1998. Mild hypothermia protects rat hippocampal CA1 neurons from irreversible membrane dysfunction induced by experimental ischemia. Neuroscience Research 30:1-6.
Pellegrino-Giampietro, D.E., Cherici, G., Alesiana, M., Carla, V., Moroni, F., 1990. Excitatory amino acid release and free radical formation may cooperate in the genesis of ischemia-induced damage. J. Neurosci 10:1035-1041.
Perlman, J.M., 1999. Markers of asphyxia and neonatal brain injury. NEJM 341(5):364-365.
Petzelt, C., Blom, P., Schmehl, W., Muller, J., Kox, W.J., 2003. Prevention of neurotoxicity in hypoxic cortical neurons by the noble gas xenon. Life Sciences 72:1909-1918.
Pohl, D., Bittigau, P., Ishimaru, M.J., 1999. N-methyl-D-aspartate antagonists and apoptotic cell death triggered by head trauma in developing rat brain. Proc Natl Acad Sci 96:2508.
Reinelt, H., Marx, T., Schirmer, U., Schmidt, M., 2001. Xenon expenditure and nitrogen accumulation in closed-circuit anaesthesia. Anaesthesia 56(4):309-311.
Riccio, A., Ginty, D.D., 2002. Nature Neurosci 5(5):389-390.
Rice, J.E., Vannucci, R.C., Brierley, J.B., 1981. The influence of immaturity on hypoxic-ischemic brain damage in the rat. Ann Neurol 9:131-141.
Rothman, S.M., Olney, J.W., 1986. Glutamate and the pathophysiology of hypoxic-ischemic brain damage. Ann Neurol 19:105-111.
Sanders, R.D., Franks, N.P., Maze, M., 2003. Xenon: no stranger to anaesthesia. BJA 91(5):709-717.
Sattler, R., Xiong, Z., Lu, W., MacDonald, J.F., Tymianski, M., 2000. Distinct roles of synaptic and extrasynaptic NMDA receptors in excitotoxicity. J Neurosci 20(1):22-33.
Shichino, T., Murakawa, M., Adachi, T., Miyazaki, Y., Segawa, H., Fukuda, K., Mori, K., 2002. Effects of xenon on acetlycholine release in the rat cerebral cortex in vivo. British Journal of Anaesthesia 88(6):866-868.
Talbot, J.H., 1941. The physiologic and therapeutic effects of hypothermia. N Eng J Med 224:281.
Taylor, D.L., Mehmet, H., Cady, E.B., Edwards, A.D., 2002. Improved neuroprotection with hypothermia delayed by 6 hours following cerebral hypoxia-ischemia in the 14-day-old rat. Ped Res 51(1):13-19.
Towfighi, J., Housman, C., Heitjan, D.F., Vannucci, R.C., Yager, J.Y., 1994. The effect of focal cerebral cooling on perinatal hypoxic-ischemic brain damage. Acta Neuropathol (Berl.) 87:598-604.
Treschera, W.H., Ishiwac, S., Johnston, M.V., 1997. Brief post-hypoxic-ischemic hypothermia markedly delays neonatal brain injury. Brain and Development 19:326-338.
Vannucci, R.C., 1997. Hypoxia-ischemia: Clinical Aspects in: Fanaroff, A.A., Martin, R.J., (Eds), Neonatal-perinatal medicine IV, Mosby-Yearbook Inc, Philadelphia, pp877-891.
Vannucci, R.C., Connor, J.R., Mauger, D.T., Palmer, C., Smith, M.B., Towfighi, J., Vannucci, S.J., 1999. Mini-Review: Rat model of perinatal hypoxic-ischemic brain damage. J Neuro Res 55:158-163.
Vannucci, R.C., Perlman, J.M., 1997. Interventions for perinatal hypoxic-ischemic encephalopathy. Pediatrics 100(6):1004-1014.
Volpe, 2001. Mental Retardation and Developmental Disabilities Research Reviews 7: 56-64.
Wilhelm, S., Ma, D., Maze, M., Franks, N.P., 2002. Effects of xenon on in vitro and in vivo models of neuronal injury. Anesthesiology 96:1485-1491.
Wyllie, A.H., Kerr, J.F.R., Currie, A.R., 1980. Cell death: the significance of apoptosis. Int Rev Cytol 68:251-306.
Windle, WF, 1969. Brain Damage by Asphyxia at Birth. Scientific American Oct; 221(4):76-84.
Xu, L., Yenari, M.A., Steinberg, G.K., Giffard, R.G., 2002. Mild hypothermia reduces apoptosis of mouse neurons in vitro early in the cascade. J Cereb Blood Flow Metab 22:21-28.
Yager, J.Y., Asselin, J., 1996. Effect of mild hypothermia on cerebral energy metabolism during the evolution of hypoxic-ischemic brain damage in the immature rat. Stroke 27:919-926.
【図面の簡単な説明】
【0148】
【図1】図1は、Sprague-Dawleyラットにおける、脳重量(左半球に対する右半球の比)の減少により測定した損傷と、低酸素状態の持続時間(分)との間の関係を示す。
【図2】図2は、90分間の低酸素性虚血性障害を経験したSprague-Dawleyラットの脳切片を示す。
【図3】図3は、90分間の低酸素性虚血の24時間後に、Sprague-Dawleyラットにおいて明瞭に認められる多大な細胞損傷を示す。
【図4】図4は、キセノンによる神経保護作用の濃度依存性(左半球に対する右半球の重量比と、キセノンの濃度)を示す。
【図5】図5は、低酸素性虚血性(HI)障害後にかなりの時間をおいて査定した、神経学的機能に対する70%キセノンの作用を示す。
【図6】図6は、キセノンがHI障害の2時間後に投与された場合の、N2及びキセノンそれぞれに観察される神経保護的効果(左半球に対する右半球の比)を比較するものである。
【図7】図7は、キセノンの神経保護的効果に対する軽度の低体温療法の作用を示す(LDH放出と、キセノン濃度(%原子))。
【図8】図8は、絶対温度の逆数に対してプロットされたLDH放出の自然対数のファント・ホッフプロットを示す。
【図9】図9は、ガスの供給に用いる専用の気密チャンバーを示す。水槽及び閉回路キセノン供給システムも表されている。
【図10】図10は、使用した方法のスケジュールの概略である。60分とは、n=12のラット仔の手術に要した時間である。回復期は母親のもとで過ごさせた。使用した介入措置は、擬似手術動物、ポジティブコントロール、75%のキセノン(残りは酸素)、33℃での低体温療法、20%のキセノン(25%の酸素、55%の窒素)、35℃での低体温療法、及び35℃での低体温療法と20%のキセノンの組み合わせであった。特に記載のない限り、動物は37℃に保たれ、25%の酸素と残りは窒素からなる混合気体を呼吸した。擬似手術動物には切開を施したが結紮やHIは行わず、ポジティブコントロールには、手術とHIの両方を行った。各群の動物は、変更可能な回復期(16、24及び48時間)に均等に分けられ、その後処分された。略語:HI、低酸素性虚血。
【図11】図11は、(A)ウェブサイトfaculty.virginia.edu/…/RatBrainLabels.jpgから修正したラット脳の矢状断面を示す。破線は、ブレグマから約−3.6mmに位置する、冠状断面(B)を採取した脳部位を表す。(B)はクレシルバイオレットで染色した冠状断面を示す。ボックスは、計算盤を設置して細胞を分析した部位を示す。上のボックスは皮質を示し、下のボックスは脳回を示す。Xは、非結紮(対側)半球を示すために安全ピンで故意に作られた穴の上につけられている。
【図12】図12は、アポトーシス細胞、壊死細胞及び生存細胞間の形態学的外観の違いが明示されている、100x油浸レンズとAxiocamデジタルカメラで撮影された皮質(クレシルバイオレット染色)の顕微鏡写真を示す。生存細胞の染色程度は、いずれのタイプの細胞死よりも弱く、そのため、細胞質の色が薄い。その一方、死亡細胞はより濃く染色されている。壊死細胞及びアポトーシス細胞は、それらの核の外観における相違点に基づいて、区別される。壊死核は大きく不規則な形をしており、一方、アポトーシス核は小さく、縮退し、球形である。
【図13】図13は、キセノンが16時間群で抗アポトーシス機構を介した神経保護作用を有することを示す。より具体的には、図13は、16時間群での、(A)皮質、(B)脳回における、低酸素性虚血に誘導されたアポトーシス性及び壊死性ニューロン死のグラフ、及びそのような細胞死に及ぼす75%のキセノンと33℃での低体温療法の作用を示す。両方の脳部位において、キセノンは、ポジティブコントロール動物と比較して、アポトーシス細胞の比率を低下させただけではなく、生存細胞の比率を有意に上昇させた。皮質においては、低体温療法はアポトーシス細胞の比率を減少させたものの、生存細胞数を増加させることはなかったため、神経保護作用を有するとは考えられない。結果は平均値±SD(n=3)である。ポジティブコントロールに対して、*P<0.05、**P<0.01、***P<0.001。
【図14】図14は、16時間群の、擬似手術動物、ポジティブコントロール動物、及び75%のキセノンで処置した動物における皮質及び脳回を表す顕微鏡写真を示す。75%群は、外観に関しては、ポジティブコントロール群よりも擬似手術群に似ている。これは、16時間群でのキセノンの神経保護的効果を確認するものである。コントロール群の脳回は、細胞死及び空胞化の量が増大したことにより、形にゆがみが見られる。
【図15】図15は、キセノンが24時間群で抗アポトーシス機構を介した神経保護作用を有することを示す。より具体的には、図15は、24時間群での、(A)皮質、(B)脳回における、低酸素性虚血に誘導されたアポトーシス性及び壊死性ニューロン死のグラフ、及びそのような細胞死に及ぼす75%のキセノンと33℃での低体温療法の作用を示す。両方の脳部位において、キセノンは、壊死細胞数の減少によって生存細胞の比率を有意に上昇させた。結果は平均値±SD(n=3)である。ポジティブコントロールに対して、*P<0.05。
【図16】図16は、キセノンが48時間群で抗アポトーシス機構を介した神経保護作用を有することを示す。より具体的には、図16は、48時間群での、(A)皮質、(B)脳回における、低酸素性虚血に誘導されたアポトーシス性及び壊死性ニューロン死のグラフ、及びそのような細胞死に及ぼす75%のキセノンと33℃での低体温療法の作用を示す。皮質と脳回の両方において、キセノンは、抗アポトーシス機構を介して神経保護作用を示している。また、キセノンは、皮質において抗壊死作用を有している。33℃での低体温療法は両方の脳部位において神経保護作用を有しているようだが、その機序は異なる――皮質では抗壊死性であり、脳回では抗アポトーシス性である。結果は平均値±SD(n=3)である。ポジティブコントロールに対して、*P<0.05、**P<0.01、***P<0.001。
【図17】図17は、キセノンと低体温療法との組み合わせにより相乗的な相互作用が発生して抗アポトーシス性の神経保護作用が生じることを示す。より具体的には、図17は、16、24.及び48時間群での、(A)皮質、(B)脳回における、低酸素性虚血症に誘導されたアポトーシス性及び壊死性ニューロン死のグラフ、及びそのような細胞死に及ぼす20%のキセノンと35℃での低体温療法との組み合わせの作用を示す。20%のキセノン群及び35℃での低体温療法群をポジティブコントロール群と比較した際には何の相違点も見られなかった。したがって、これらの数値では、神経保護作用は存在しない。しかし、これらの数値が組み合わされて使用された場合には、ポジティブコントロール動物と比較してアポトーシス細胞数が有意に減少することにより、生存細胞の比率が劇的に上昇する。脳回においてその組み合わせは、さらに24時間群で抗壊死作用ももたらす。結果は平均値±SD(n=3)である。ポジティブコントロールに対して、*P<0.05、**P<0.01、***P<0.001。これらの図に関するより詳細な論述が、実施例の項目に記載されている。
【特許請求の範囲】
【請求項1】
対象である新生児において新生児仮死を治療する薬剤であって、低体温療法と組み合わせて使用する薬剤の調製におけるキセノンの使用。
【請求項2】
キセノンが、薬学上許容可能な希釈剤、賦形剤又は担体と混合されていることを特徴とする請求項1記載の使用。
【請求項3】
薬剤が、ガス状であることを特徴とする請求項1又は2記載の使用。
【請求項4】
薬剤が、吸入により投与されることを特徴とする請求項3記載の使用。
【請求項5】
キセノンが、20〜70%v/vのキセノン・空気混合体の形態で投与されることを特徴とする請求項1〜4のいずれか記載の使用。
【請求項6】
キセノンが、灌流により投与されることを特徴とする請求項1又は2記載の使用。
【請求項7】
薬剤が、液体又は溶液の形態であることを特徴とする請求項1又は2記載の使用。
【請求項8】
薬剤が、脂肪乳剤の形態であることを特徴とする請求項7記載の使用。
【請求項9】
薬剤が、静脈内、軸索内又は経皮的デリバリーに好適な形態であることを特徴とする請求項7又は8記載の使用。
【請求項10】
キセノンの投与が、低体温療法と同時、順次、又は別個に行われることを特徴とする請求項1〜9のいずれか記載の使用。
【請求項11】
キセノンの投与が、低体温療法と同時に行われることを特徴とする請求項10記載の使用。
【請求項12】
キセノンが、対象である新生児の母親に、出産前に投与されることを特徴とする請求項1〜11のいずれか記載の使用。
【請求項13】
キセノンが、対象である新生児の母親に、陣痛前又は陣痛中に投与されることを特徴とする請求項12記載の使用。
【請求項14】
キセノンが、対象である新生児の母親に、出産前に最長約24時間投与されることを特徴とする請求項13記載の使用。
【請求項15】
低体温療法が、低酸素性虚血性(HI)障害後、少なくとも約6時間続けられることを特徴とする請求項1〜14のいずれか記載の使用。
【請求項16】
低体温療法が、低酸素性虚血性(HI)障害後、約6〜約24時間続けられることを特徴とする請求項1〜15のいずれか記載の使用。
【請求項17】
キセノンが、治療上有効な量で投与されることを特徴とする請求項1〜16のいずれか記載の使用。
【請求項18】
キセノンが、治療上有効な量以下で投与されることを特徴とする請求項1〜16のいずれか記載の使用。
【請求項19】
キセノンが、イソフルラン、セボフルラン及びデスフルランから選択される麻酔剤と組み合わせて投与されることを特徴とする請求項1〜18のいずれか記載の使用。
【請求項20】
新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、
(a)治療上有効な量のキセノンを前記哺乳動物に投与するステップ、及び
(b)前記哺乳動物に低体温療法を施すステップ
を含む方法。
【請求項21】
哺乳動物がヒトであることを特徴とする請求項20記載の方法。
【請求項22】
キセノンが、薬学上許容可能な担体、希釈剤又は賦形剤と組み合わせて投与されることを特徴とする請求項20又は21記載の方法。
【請求項23】
キセノンが、吸入により投与されることを特徴とする請求項20〜22のいずれか記載の方法。
【請求項24】
キセノンが、20〜70%v/vのキセノン・空気混合体の形態で投与されることを特徴とする請求項23記載の方法。
【請求項25】
キセノンが、灌流により投与されることを特徴とする請求項20〜22のいずれか記載の方法。
【請求項26】
キセノンが、溶液又は乳剤の形態で投与されることを特徴とする請求項20〜22のいずれか記載の方法。
【請求項27】
キセノンが、脂肪乳剤の形態で投与されることを特徴とする請求項26記載の方法。
【請求項28】
キセノンが、静脈内、軸索内又は経皮的に投与されることを特徴とする請求項26又は27のいずれか記載の方法。
【請求項29】
キセノンの投与が、低体温療法と同時、順次、又は別個に行われることを特徴とする請求項20〜28のいずれか記載の方法。
【請求項30】
キセノンの投与が、低体温療法と同時に行われることを特徴とする請求項29記載の方法。
【請求項31】
哺乳動物の体温が、約32℃〜約36℃に維持されることを特徴とする請求項20〜30のいずれか記載の方法。
【請求項32】
哺乳動物の体温が、約33℃〜約35℃に維持されることを特徴とする請求項31記載の方法。
【請求項33】
低体温療法が、低酸素性虚血性(HI)障害後、少なくとも6時間続けられることを特徴とする請求項20〜32のいずれか記載の方法。
【請求項34】
低体温療法が、低酸素性虚血性(HI)障害後、約6〜約24時間続けられることを特徴とする請求項20〜33のいずれか記載の方法。
【請求項35】
キセノンが、哺乳動物の母親に、出産前に投与されることを特徴とする請求項20〜34のいずれか記載の方法。
【請求項36】
キセノンが、哺乳動物の母親に、陣痛前又は陣痛中に投与されることを特徴とする請求項35記載の方法。
【請求項37】
キセノンが、哺乳動物の母親に、出産前に最長約24時間投与されることを特徴とする請求項35又は36記載の方法。
【請求項38】
キセノンが、治療上有効な量で投与されることを特徴とする請求項20〜37のいずれか記載の方法。
【請求項39】
キセノンが、治療上有効な量以下で投与されることを特徴とする請求項20〜37のいずれか記載の方法。
【請求項40】
キセノンが、イソフルラン、セボフルラン及びデスフルランから選択される麻酔剤と組み合わせて投与されることを特徴とする請求項20〜38のいずれか記載の方法。
【請求項41】
新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、治療上有効な量のキセノンを、低体温療法と組み合わせて前記哺乳動物に投与するステップを含む方法。
【請求項42】
新生児仮死治療用薬剤の調製におけるキセノンの使用であって、前記治療が、低体温療法と組み合わせて、同時、順次、又は別個にキセノンを対象に投与するステップを含む使用。
【請求項43】
新生児仮死を治療するための、低体温療法との組み合わせによる、キセノンの使用。
【請求項44】
本発明の別の態様は、新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、
(a)治療上有効な量のキセノンを前記哺乳動物の母親に、陣痛前及び/又は陣痛中に投与するステップ、及び
(b)前記哺乳動物に出生後に低体温療法を施すステップ
を含む方法に関する。
【請求項45】
本明細書中において、及び添付の図面を参照して、実質的に記載された方法又は使用。
【請求項1】
対象である新生児において新生児仮死を治療する薬剤であって、低体温療法と組み合わせて使用する薬剤の調製におけるキセノンの使用。
【請求項2】
キセノンが、薬学上許容可能な希釈剤、賦形剤又は担体と混合されていることを特徴とする請求項1記載の使用。
【請求項3】
薬剤が、ガス状であることを特徴とする請求項1又は2記載の使用。
【請求項4】
薬剤が、吸入により投与されることを特徴とする請求項3記載の使用。
【請求項5】
キセノンが、20〜70%v/vのキセノン・空気混合体の形態で投与されることを特徴とする請求項1〜4のいずれか記載の使用。
【請求項6】
キセノンが、灌流により投与されることを特徴とする請求項1又は2記載の使用。
【請求項7】
薬剤が、液体又は溶液の形態であることを特徴とする請求項1又は2記載の使用。
【請求項8】
薬剤が、脂肪乳剤の形態であることを特徴とする請求項7記載の使用。
【請求項9】
薬剤が、静脈内、軸索内又は経皮的デリバリーに好適な形態であることを特徴とする請求項7又は8記載の使用。
【請求項10】
キセノンの投与が、低体温療法と同時、順次、又は別個に行われることを特徴とする請求項1〜9のいずれか記載の使用。
【請求項11】
キセノンの投与が、低体温療法と同時に行われることを特徴とする請求項10記載の使用。
【請求項12】
キセノンが、対象である新生児の母親に、出産前に投与されることを特徴とする請求項1〜11のいずれか記載の使用。
【請求項13】
キセノンが、対象である新生児の母親に、陣痛前又は陣痛中に投与されることを特徴とする請求項12記載の使用。
【請求項14】
キセノンが、対象である新生児の母親に、出産前に最長約24時間投与されることを特徴とする請求項13記載の使用。
【請求項15】
低体温療法が、低酸素性虚血性(HI)障害後、少なくとも約6時間続けられることを特徴とする請求項1〜14のいずれか記載の使用。
【請求項16】
低体温療法が、低酸素性虚血性(HI)障害後、約6〜約24時間続けられることを特徴とする請求項1〜15のいずれか記載の使用。
【請求項17】
キセノンが、治療上有効な量で投与されることを特徴とする請求項1〜16のいずれか記載の使用。
【請求項18】
キセノンが、治療上有効な量以下で投与されることを特徴とする請求項1〜16のいずれか記載の使用。
【請求項19】
キセノンが、イソフルラン、セボフルラン及びデスフルランから選択される麻酔剤と組み合わせて投与されることを特徴とする請求項1〜18のいずれか記載の使用。
【請求項20】
新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、
(a)治療上有効な量のキセノンを前記哺乳動物に投与するステップ、及び
(b)前記哺乳動物に低体温療法を施すステップ
を含む方法。
【請求項21】
哺乳動物がヒトであることを特徴とする請求項20記載の方法。
【請求項22】
キセノンが、薬学上許容可能な担体、希釈剤又は賦形剤と組み合わせて投与されることを特徴とする請求項20又は21記載の方法。
【請求項23】
キセノンが、吸入により投与されることを特徴とする請求項20〜22のいずれか記載の方法。
【請求項24】
キセノンが、20〜70%v/vのキセノン・空気混合体の形態で投与されることを特徴とする請求項23記載の方法。
【請求項25】
キセノンが、灌流により投与されることを特徴とする請求項20〜22のいずれか記載の方法。
【請求項26】
キセノンが、溶液又は乳剤の形態で投与されることを特徴とする請求項20〜22のいずれか記載の方法。
【請求項27】
キセノンが、脂肪乳剤の形態で投与されることを特徴とする請求項26記載の方法。
【請求項28】
キセノンが、静脈内、軸索内又は経皮的に投与されることを特徴とする請求項26又は27のいずれか記載の方法。
【請求項29】
キセノンの投与が、低体温療法と同時、順次、又は別個に行われることを特徴とする請求項20〜28のいずれか記載の方法。
【請求項30】
キセノンの投与が、低体温療法と同時に行われることを特徴とする請求項29記載の方法。
【請求項31】
哺乳動物の体温が、約32℃〜約36℃に維持されることを特徴とする請求項20〜30のいずれか記載の方法。
【請求項32】
哺乳動物の体温が、約33℃〜約35℃に維持されることを特徴とする請求項31記載の方法。
【請求項33】
低体温療法が、低酸素性虚血性(HI)障害後、少なくとも6時間続けられることを特徴とする請求項20〜32のいずれか記載の方法。
【請求項34】
低体温療法が、低酸素性虚血性(HI)障害後、約6〜約24時間続けられることを特徴とする請求項20〜33のいずれか記載の方法。
【請求項35】
キセノンが、哺乳動物の母親に、出産前に投与されることを特徴とする請求項20〜34のいずれか記載の方法。
【請求項36】
キセノンが、哺乳動物の母親に、陣痛前又は陣痛中に投与されることを特徴とする請求項35記載の方法。
【請求項37】
キセノンが、哺乳動物の母親に、出産前に最長約24時間投与されることを特徴とする請求項35又は36記載の方法。
【請求項38】
キセノンが、治療上有効な量で投与されることを特徴とする請求項20〜37のいずれか記載の方法。
【請求項39】
キセノンが、治療上有効な量以下で投与されることを特徴とする請求項20〜37のいずれか記載の方法。
【請求項40】
キセノンが、イソフルラン、セボフルラン及びデスフルランから選択される麻酔剤と組み合わせて投与されることを特徴とする請求項20〜38のいずれか記載の方法。
【請求項41】
新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、治療上有効な量のキセノンを、低体温療法と組み合わせて前記哺乳動物に投与するステップを含む方法。
【請求項42】
新生児仮死治療用薬剤の調製におけるキセノンの使用であって、前記治療が、低体温療法と組み合わせて、同時、順次、又は別個にキセノンを対象に投与するステップを含む使用。
【請求項43】
新生児仮死を治療するための、低体温療法との組み合わせによる、キセノンの使用。
【請求項44】
本発明の別の態様は、新生児仮死の治療を必要とする哺乳動物における新生児仮死の治療方法であって、
(a)治療上有効な量のキセノンを前記哺乳動物の母親に、陣痛前及び/又は陣痛中に投与するステップ、及び
(b)前記哺乳動物に出生後に低体温療法を施すステップ
を含む方法に関する。
【請求項45】
本明細書中において、及び添付の図面を参照して、実質的に記載された方法又は使用。
【図1】



【図4】

【図5】

【図6】

【図7】

【図8】

【図14】

【図15】

【図16】
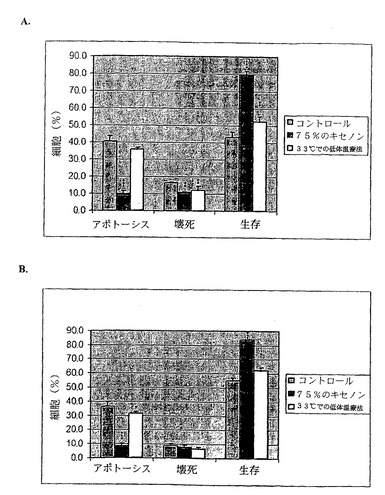
【図17】

【図2】

【図3】

【図9】

【図10】

【図11】

【図12】

【図13】

【図4】
【図5】
【図6】
【図7】
【図8】
【図14】
【図15】
【図16】
【図17】
【図2】
【図3】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2007−508284(P2007−508284A)
【公表日】平成19年4月5日(2007.4.5)
【国際特許分類】
【出願番号】特願2006−530602(P2006−530602)
【出願日】平成16年10月11日(2004.10.11)
【国際出願番号】PCT/GB2004/004298
【国際公開番号】WO2005/034966
【国際公開日】平成17年4月21日(2005.4.21)
【出願人】(504405707)プロテクソン リミテッド (3)
【Fターム(参考)】
【公表日】平成19年4月5日(2007.4.5)
【国際特許分類】
【出願日】平成16年10月11日(2004.10.11)
【国際出願番号】PCT/GB2004/004298
【国際公開番号】WO2005/034966
【国際公開日】平成17年4月21日(2005.4.21)
【出願人】(504405707)プロテクソン リミテッド (3)
【Fターム(参考)】
[ Back to top ]