
雑種ラクツカ・サティバ(Lactucasativa)種子の作製
本発明は、雑種ラクツカ・サティバ(Lactuca sativa)種子であって、雄性不稔遺伝子型であるとともに、特徴的な表現型である当該種子に由来する植物を提供する、少なくとも1つの特徴的遺伝子の異種接合体である種子に関する。本発明はまた、雑種ラクツカ・サティバ種子の作製のための方法であって、一定環境中で表現型的に雄性不稔植物及び表現型的に雄性不稔性植物の栽培のステップ、植物の開花の際に一定環境中に導入される双翅目による授粉のステップ、及び雄性不稔植物により作製される種子の収穫のステップ、を含んでなる方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、雑種ラクツカ・サティバ(Lactuca sativa)種子及び当該種子の作製のための方法に関する。本発明はまた、雑種ラクツカ・サティバ植物、及び雑種ラクツカ・サティバ植物由来の細胞に関する。
【背景技術】
【0002】
雑種強勢(Heterosis又はhybrid vigour)、恒常性(異なる環境における植物の安定性)、虫、真菌、細菌又はウイルスへの耐性、又は非生物ストレスへの適応、例えば極限温度、すなわち5℃未満又は30℃超の温度への耐性、又は微光レベルへの耐性を有する遺伝子蓄積の可能性は、雑種植物由来の種子を大スケールで作製するための、いくつかの理由を構成する。
【0003】
特に、ラクツカ・サティバ(栽培品種レタス)において、問題の遺伝子が、同じ遺伝子座(各対立遺伝子の一方)又は非常に近接する遺伝子座に位置するため、同型接合系統における特定の耐性遺伝子を集積することが、不可能又は非常に困難であることが示されている。
【0004】
雑種F1栽培レタスを得ること、特に同じ遺伝子座の様々な対立遺伝子、又は非常に近接する遺伝子座によりもたらされる、農業で注目の複数の相互優性(co-dominant)又は優性遺伝子を蓄積する、大スケールの雑種作製をさせることについては、大きな関心を集めるであろう。
【0005】
ラクツカ(Lactuca)属は100種以上が含まれており、例えばラクツカ・サティバ(レタス、Lactuca sativa)(栽培品種)、ラクツカ・サリグナ(Lactuca saligna)(野生型種)、ラクツカ・セリオラ(Lactuca serriola)(野生型種)及びラクツカ・ビロサ(Lactuca virosa)(野生型種)がある。
【0006】
ラクツカ・サティバ(レタス)は、種子作製は、98%が自己授粉による、ほとんどが自家生殖の二倍性種(2n=18)である。雄蕊(雄器官)は集団を形成し、葯の花粉嚢を開く雄蕊管を形成する。雌蕊(雌器官)は子房、花柱、及び二分された柱頭から構成される。開花時は、雄蕊管内部に花柱が伸びて、柱頭が花粉で覆われ、植物が自己受精する。一般的に1つのラクツカ・サティバ植物は、栽培条件次第で、0.5〜6グラム又は最大で10グラムもの種子を作製し、その1グラムには一般的に600〜1000個の種子が含まれる。
【0007】
雑種植物の大スケールでの作製のための従来技術は、互いに近接する「親」として使用される2つの変種を栽培し、授粉のための虫を使用することからなり、雄性不稔性をもたらす一方の親が、自己授粉による汚染(contamination)を防止する。その後、雄性不稔性親から種子を収穫する。
【0008】
この技術は、その組(parcel)の均一性を維持しながら、各親において異種接合状態で存在する注目の遺伝子を蓄積でき、そのF1世代は収穫段階で(開花前に)100%異種接合体の表現型である。
【0009】
前記技術を用いて大スケールで栽培雑種レタス(ラクツカ・サティバ)を作製することは非常に困難であることが観察されている。これはラクツカ・サティバの花が、通常の授粉する虫、例えば特定種のハチ(ミツバチ)又はバンブルビー(bumble bee)(Bombus種)がまだ活動しない日中(早朝)の時間帯に、一度だけ、たった数時間、例えば1〜4時間しか開花しないことが原因である。
ラクツカ・サティバはまた、風による授粉はしない。
【0010】
すなわち、雑種レタスの作製のために使用できる授粉する虫を確認することは、多数の問題に直面する。具体的には:
開花の期間が短く且つ早朝であるために、虫が花を訪れることの確認を困難にする。
開花が1度しかないために、最大数の雄性不稔花の受粉を確実にするためには、選択された授粉する虫が花を毎日訪れる虫でなければならない。
【0011】
さらに、F1雑種の作製は、「雄」系及び「雌」系の供給が必要である。「雌」系は、当該植物の手作業で去葯することにより得ることができる。ただし、レタスの場合は、開花が1回で且つ非常に短期間(数時間)なので、この方法はかなりの労力を要するため、流通スケールでの実施は困難である。
【0012】
「雌」系はまた、葯の不存在、空の葯又は生育不能花粉により、それ自身が発現する不稔性の遺伝子型(又は核型)又は細胞質型を誘導することにより得ることができる。
【0013】
その作業において、Goubara及びTakasakiは、雑種ラクツカ・サティバレタスの作製のために使用できる、可能性ある授粉する虫を確認する目的で、野外及び閉鎖条件で実験を行った(Appl Entomo Zool 38(4), 571-581, 2003 [1])。22種の虫が観察され(21種のハチ及び1種の吸蜜(nectar-feeding)ハエ種、シルフィダエ・エリスタリス・テナクス(syrphidae eristalis tenax))、そのうちハチのラジオグロッサム・ビロスラム・トリコプセ(lagioglossum villosulum trichopse)は最も可能性ある授粉する虫であることがわかった。
【0014】
Goubara及びTakasakiはまた、閉鎖環境中にハチのラジオグロッサム・ビロスラム・トリコプセを存在させて、雄性不稔性遺伝子を有するレタスと雄性稔性レタスとの間で小スケールのハイブリダイゼーション試験を行った。その著者等は、雑種F1レタスは得られたが収率が非常に低かったことを報告している。
【0015】
現時点では、授粉する虫を使用する雑種ラクツカ・サティバの作製実験で他に知られるものはない。
今日、F1雑種レタス変種で、流通スケールで作製されているものはない。
【0016】
本発明との関連において、以下の用語は以下の意味を有する。
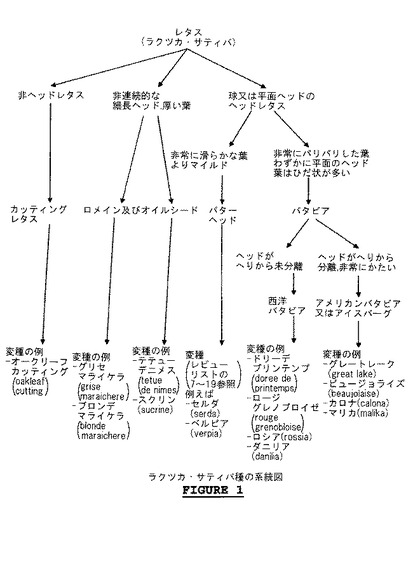
「栽培レタス」は、ラクツカ・サティバ種を意味する。基本的な栽培レタスの5つの栽培品種群(cultigroups)が存在する(図1を参照):ラクツカ・サティバ変種 アングスタナ(angustana)(アスパラガス(asparagus)レタス); ラクツカ・サティバ変種 カピタタ(capitata)(バターヘッド(butterhead)レタス); ラクツカ・サティバ変種 クリスパ(crispa)(バタビア(batavia)又はアイスバーグ(Iceberg)レタス); ラクツカ・サティバ変種 ロンギフォリア(longifolia)(ロマイネ(Romaine)レタス)及びラクツカ・サティバ変種 アセファラ(acephala)(フリシー(frisee)レタス、カッティング(cutting)レタス)。本発明は、これらの様々な型のレタスの各々の使用を包含する。
【0017】
「授粉(受粉)(pollinization)」は、葯からの花粉を、同じ花又は別の花の柱頭に移転することを意味する。この生殖系は花を有する植物の繁殖の好ましい態様である(被子植物及び裸子植物)。花粉の粒を柱頭に到達させ、その後、それを受精させるために、花柱を通って、胚珠に到達する花粉管を形成させる。
【0018】
「自己授粉」は、自身の持つ花粉を用いる個体又は生物型の授粉を意味し、その結果当該個体は自己授粉したと言われる。
「自家生殖の」なる用語は、2つの配偶子が同じ個体に由来する、植物の自己授粉する能力を意味する。
「他殖性の」は、ある個体の花が他の1又は複数の個体からの花粉で受精する表現型を意味する。
「授粉する虫」は、花の研究において(例えば蜜を研究する場合)、雄蕊を摺ることにより花粉の数粒を収穫し、その後これを別の花に置いてくる虫を意味する。
【0019】
「遺伝子座」は、染色体上での遺伝子又は対立遺伝子により占められる場所を意味する。
「対立遺伝子」は、所与の染色体位置(遺伝子座)に位置する特定の遺伝子内での変異である。遺伝子の異なる対立遺伝子は、異なる形質の発現をもたらす。
【0020】
「クラスター」は、同じ染色体上に互いに近接して位置する2以上の遺伝子を意味する。レタスにおいて確認された耐性遺伝子のクラスターは、特にKesseli等により報告されている[12]。
「優性遺伝子」は、1対の2つの染色体に存在するか1つだけに存在するかにかかわらず、表現型を提供する遺伝子を意味する。
「劣性遺伝子」は、それが2つの相同染色体のうちの各々に存在する場合、1の表現型のみを提供する遺伝子を意味する。
「相互優性(co-dominanace)」は、2つの対立遺伝子が決定する表現型的特徴の、一方及び他方を発現する遺伝子特性を意味する。異種接合体において、物質が2つの相互優性の対立遺伝子を有すると、遺伝子型はこれらの遺伝子によりもたらされた情報に従ってその表現型で完全に発現する。
「異種接合体」は、染色体の同じ対の既定の遺伝子座上の2つの異なる対立遺伝子を有する細胞又は個体を意味する。
「同型接合体」は、染色体の同じ対の規定の遺伝子座上の2つの同一の対立遺伝子を有する細胞又は個体を意味する。
「雑種」なる用語は、異なる遺伝子構成の、好ましくは同一種由来の個体間の交配産物を意味する。
【0021】
「F1雑種」なる用語は、異なる遺伝子構成の個体間の交配から得られる第一世代を意味する。結果としてF1雑種は、少なくとも1の遺伝子の異種接合体である。
「戻し交配」は、雑種とその親の一方との間の交配を意味する。
「栽培品種」は、ある変種を意味する。
「遺伝子型」は、個体からもたらされた遺伝物質であって、その遺伝形質を構成するものの全てを意味する。
「表現型」は、遺伝子型の発現比率と外部環境により決定される現象の両方に対応する、個体の、明らかな形態的又は機能的特徴の組を意味する。
「農業で注目の表現型」なる用語は、例えば2つの同型接合遺伝子型間での交配から得られた表現型であって、農業的観点から注目の特性、例えば様々な病原菌又は虫への耐性の蓄積、雑種強勢(すなわち、雑種形質の平均は、2つの親のものの平均より高い)、恒常性、非生物ストレスに適応する能力、形態学的特徴、例えば色、形、葉の柔軟性又は剛性の性質、栄養組成物又は植物の味質等を伴う表現型を意味する。
「雑種強勢(heterosis又はhybrid vigour)」とは、F1雑種が、一又は複数の特徴の点で、特に強勢(vigour)に関して、その最良の親よりも顕著に優勢である表現型を意味する。
「恒常性」とは、環境又は複数の環境的特徴まで適応する、植物の能力を意味する。
「不稔性雄」とは、花の雄的要素の不稔性が原因で、自己授粉により繁殖できない植物を意味する。例としては、花粉が非機能的である、又は雄性繁殖器官、例えばタペータム構造的異常がある可能性がある。
「細胞質雄性不稔性」とは、細胞質型の遺伝により、その母から均一に遺伝する不稔性を意味する。
「核(又は遺伝子的)雄性不稔性」とは、劣性遺伝又は優性遺伝のいずれに依存してもよいが、核のDNAによりもたらされる、メンデル遺伝による不稔性を意味する。
「一遺伝子的雄性不稔性」とは、単一の遺伝子によりもたらされる雄性不稔性を意味する。
「多遺伝子的雄性不稔性」とは、複数の遺伝子によりもたらされる雄性不稔性を意味する。
「耐性」とは、病原又は有害生物にとって、類似する環境且つ圧力条件下で、感受性の高い変種と比較して、既定の病原又は有害生物及び/又はそれが引き起こす障害の増殖及び発展を制限する様々な能力を意味する。ただしこれらの変種は、病原又は有害生物の圧力が高い場合、病気のいくつかの症状又はいくつかの障害を発現する可能性がある。
「標準的又は高い耐性」とは、感受性の高い変種と比較して、通常の圧力条件下で既定の病原又は有害生物の増殖又は発展を強力に制限する様々な能力を意味する。ただし、これらの変種は、病原又は有害生物の圧力が高い場合、症状又は障害を発現することもある。
「中程度又は調整的な耐性」とは、既定の病原又は有害生物の増殖及び発展を制限するが、高い/標準的な耐性変種と比較すると、症状又は障害をより発現し得る能力を意味する。中程度の耐性変種は、類似する環境及び/又は病原又は有害生物にとっての圧力条件下で、感受性の高い変種で観察されたものより、重篤性の低い症状又は障害を示す。
「分子マーカー」とは、個体の全ゲノムにおいて確認され得る、及び注目の遺伝子を局在化させるために、又は親生物からのある特定の特徴が遺伝した個体であるかを検証するために使用できる、DNAの特定の断片を意味する。それはコード配列であってもそうでなくてもよい。遺伝的交配において、注目の遺伝子は、一般的に分子マーカーと連結した状態である。つまり、分子マーカーの検出とは、注目の遺伝子が存在する個体が、その遺伝子の配列を知る必要なく、セレクションできることを意味する。
【0022】
農学分野において、分子マーカーの使用は、植物が所望の特徴を保持することを、セレクションの際、迅速に試験できることを意味する。特定の場合において、形質に関連するマーカーの存在により、特定の試験又は表現型の観察が不必要になる。具体的には、雄性不稔性遺伝子のための、好ましくは雄性不稔性の優性遺伝子のための、特定の分子マーカーを使用することで、開花前の雄性不稔性植物を早期にセレクションできる。このセレクションは、例えば雄性不稔性の優性遺伝子を使用する場合、雑種種子の作製のための、「雌」(雄性不稔性)と推測される親として使用される植物の集団の一部を形成し得る、雄性稔性植物を除去することができ、その結果、これらの雄性稔性植物の自己受精に結びつく汚染のリスクを低減できる。
【発明の概要】
【課題を解決するための手段】
【0023】
本発明は、雄性不稔性遺伝子型を有するとともに、雄性不稔性に関与せず、且つその植物に検出可能な表現型を提供する少なくとも1つの遺伝子との異種接合体であることを特徴とする、雑種ラクツカ・サティバ植物由来の種子、植物及び細胞に関する。
【図面の簡単な説明】
【0024】
【図1】ラクツカ・サティバの系統図。
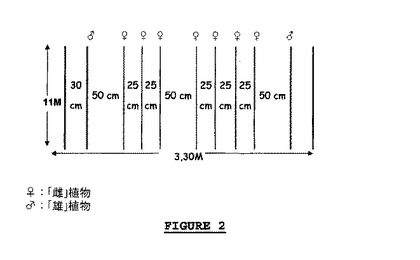
【図2】試行1で「雄」及び「雌」親として使用されたレタスの性質。
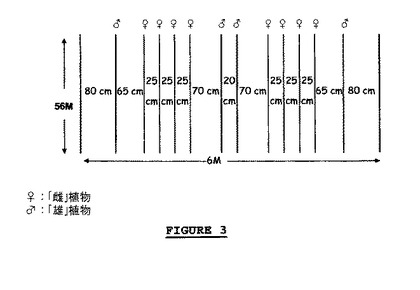
【図3】試行2で「雄」及び「雌」親として使用されたレタスの性質。
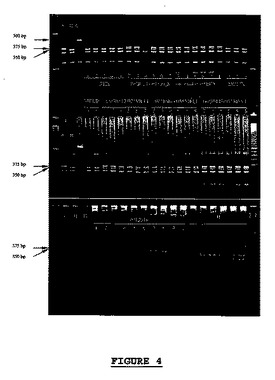
【図4】試行10から得られた50個の雑種植物の遺伝子型決定。TagIとの消化処理後の、分子マーカーSCW09の増幅プロファイル。SCW09マーカーは、ブレミア・ラクツカエに耐性のあるDm6及びDm8を標識する。A=Dm18+/Dm18+コントロール、B=Dm18−/Dm18−コントロール、C=Dm6+コントロール、D=TaqIでの消化処理なし。
【図5】試行10から得られた50個の雑種植物の遺伝子型決定。ブレミア・ラクツカエに耐性のあるDm3遺伝子を標識する分子マーカーB1の増幅プロファイル。A=Dm3+コントロール、B=Dm3−コントロール。
【図6】マーカーBA05−675(矢印)を伴うOPBA05プライマーの電気泳動プロファイル。1.0%アガロースゲル電気泳動、190Vで1時間45分。M:100bpラダー分子量マーカー(Pharmasica Biotech、27.4001.01参照)。
【図7】レタス、ラクツカ・サティバにおけるMs7遺伝子座に関連するBA05−675分子マーカーのマッピング(Kosambi関数を用いて計算した距離)。
【図8】レタス、ラクツカ・サティバにおけるMs7遺伝子座に関連するBA05−675分子マーカーの5'〜3'ヌクレオチド配列(配列番号2)。
【発明を実施するための形態】
【0025】
ラクツカ・サティバ植物由来の、種子、植物又は細胞によりもたらされた雄性不稔性は、核又は細胞質起源であってよい。それが細胞質性である場合、その不稔性は雌親から遺伝する。これは細胞質に存在するミトコンドリアと核遺伝子との間の相互作用に起因することが多い。細胞質雄性不稔性は、雄親のゲノムで稔性を回復する遺伝子の不存在下で、100%雄性不稔性の表現型を有する子孫の出現により特徴付けられる。雄性不稔性は、「雄」親のゲノムにおける稔性回復遺伝子 (restoring gene (Rf))の存在によって、その子孫で回復してもよい。
【0026】
より具体的には、本発明は雄性不稔性遺伝子型が、少なくとも1の核遺伝子を含み、且つその種子、細胞又は植物が、雄性不稔性の原因であるその1又は複数の遺伝子の異種接合体である場合に関する。
核雄性不稔性は、その核のDNAから遺伝した雄性不稔性の、1又は複数の遺伝子により引き起こされ、この1又は複数の遺伝子は、優性でも劣性でもあり得る。
【0027】
これら雄性不稔性の遺伝子には、Ryderによる(J Am Soc Hort Sci 96(6), 826-828, 1971 [8])に記載の、優性核遺伝子Ms7がある。当該雄性不稔性植物は、異種接合体Ms7ms7である。稔性植物は、同種接合体ms7ms7である。優性のMs7Ms7同種接合体の獲得は、不可能とは言わないまでもかなり困難である。さらに、Ryderによればこの雄性不稔性の優性形質は、雑種レタスの作製のためにこの遺伝子を使用することについて問題がある。本発明の方法を使用して獲得されるラクツカ・サティバ種子のサンプルで、NCIMB(NCIMB Ltd., Ferguson Building, Craibstone Estate, Bucksburn, Aberdeen, A21 9YA, スコットランド、英国)に2007年2月13日に、受託番号NCIMB 41470で寄託されたサンプルを、Ms7遺伝子の源とした。さらに、本発明者等により開発された分子マーカーRAPD BA05-675 (配列番号2)は、次の交配のために使用されるはずの雄性不稔性植物を確認するための、開花前の初期段階で実施されるべき分子試験を可能にするので、雄性不稔性レタス植物Ms7のセレクションを補助の点で特に注目される。この図8に示される配列のマーカーの検出は、全てのレタス類型を共に考慮すると、平均で96%の事例において雄性不稔性植物の確認ができる(表18)。
【0028】
雄性不稔性のための3つの劣性遺伝子、ms1、ms2及びms3はまた、Lindqvist の(Heriditas 46 :387- 470, 1960 [3])により、他の3つ、ms4、ms5及びms6は、Ryder (Proc. Am. Soc. Hort . Sci 83 : 585-595, 1963 [6], Proc . Am. Soc. Hort. Sci 91 : 366-368, 1967 [7])により確認されている。ただし、1979年に公表された彼の研究(Leafy Salad Vegetables, p 30 [9])で、Ryderは、不稔性遺伝子ms1、ms2、ms3、ms4、ms5、ms6及びMs7は全て核性であるため、F1雑種を作製するために使用できることが全く見込めない。これにもかかわらず、本発明者等は、核起源の雄性不稔体を使用する、雑種の種子、植物及び細胞を獲得することに成功した。
【0029】
核雄性不稔性は、多遺伝子的であっても一遺伝子的であってもよい。多遺伝子的である場合、例えば上記記載のms1、ms2及びms3遺伝子の集積による、劣性遺伝子の複合体を使用して獲得してもよい。好ましくは、本発明は、雄性不稔性が一遺伝子的である場合に関する。
核雄性不稔性は、優性であっても劣性であってもよい。具体的には、本発明は雄性不稔性が優性の一遺伝子的である場合に関する。より具体的には、本発明は雄性不稔性が、上記記載の優性のMs7遺伝子により提供される場合に関する。
【0030】
好ましくは、本発明の種子、植物又は細胞のゲノムは、650〜700ヌクレオチドの、例えば655〜695、660〜690、665〜685、670〜680、673〜677ヌクレオチドの、又は674、675又は676ヌクレオチドの二本鎖DNA配列を含んでなり、ここで、各2つの5'末端は、配列「5' TGCGTTCCAC 3'」(配列番号1)から開始する。好ましい実施態様によれば、本発明の種子、植物又は細胞のゲノムは、図8(配列番号2)に示したヌクレオチド配列を含んでなるか、又は1〜10個の、好ましくは1〜5個の又はより好ましくは1〜3個のヌクレオチドが他のものにより置換、削除又は付加される、この配列に由来する配列を含んでなる。
【0031】
好ましくは、本発明は、1又は複数の異種接合遺伝子により提供された検出可能な表現型が、農業で注目の表現型、例えば様々な病原又は虫に対する耐性、雑種強勢、恒常性(異なる環境での植物の安定性)、非生物ストレス、例えば極限温度又は微光レベルへの適応、雑種由来の変種のものより高い収率、形態的特長、例えば色、形、大きさ、葉の柔軟性もしくは剛性の性質、栄養組成物又は植物の味質等である場合に関する。
【0032】
さらに、驚くべきことに本発明者等は、異なるレタス間での様々な交配から得られる本発明の種子由来の植物は、冬期に且つ温暖な地域での栽培で、同じ条件下で栽培した親植物より平均で7〜10日早く成熟に到達し、より早く生育することを観察した。F1雑種の組の中で(intraparcel)発育の均一性(1組当たりおよそ30植物の株でのもの)は、組系列(親植物)のものより高いことも観察された。言い換えると、同じ組由来の全ての植物と比較して遅延又は向上した発育を示す植物の数は、変種系列由来の組よりも、F1雑種の組の方がかなり少なかった。結果として、これらのF1雑種の特徴は、レタス作製周期を短縮できるだけでなく、収穫を短期間に集中させることができる。
【0033】
農業で注目の表現型は、ウイルス、細菌、虫又は真菌による感染に対して、より具体的には以下の真菌:ブレミア・ラクツカエ(Bremia lactucae)、フサリウム・オキシスポラム(Fusarium oxysporum)、スクレロチニア・マイナー(Sclerotinia minor)もしくはスクレロトラム(sclerotorum)、又はボトリチス・シネレア(Botrytis cinerea)、リジクトニア・ソラニ(Rhizictonia solani)、ミクロドシウム・パナットニアナム(Microdochium panattonianum)、ベルチシウリウム・ダリアエ(Verticiulium dahliae)又はエリシフェ・チコセアラム(Erysiphe chicocearum)又はピチウム・トラケイフィラム(Pithium tracheiphilum)の1種に対して、以下の虫:ナソノビア・リビスニグリ(Nasonovia ribisnigri)、マイザス・ペルシカエ(Myzus persicae)、マクロシファム・ユーフォルビア(Macrosiphum euphorbia)、ネマトドス・プラチレンカス(Nematodes pratylenchus)、又はネコブ線虫(meloidogyne)、ハモグリバエ(leafminers):リリオミザ・フリドブレンシス(Liriomyza huidobrensis)又はペンフィガス・ブサリウス(Pemphigus busarius)の1種に対して、以下の細菌:シュードモナス(pseudomonas)、キサントモナス(xanthomonas)又はリゾモナス(rhizomonas)の1種に対して、又は以下のウイルス:LMW(レタスモザイクウイルス)、TSWV(鉢植えトマト立ち枯れ病ウイルス)、「ビッグ・ベイン」(LBVV(レタスビッグベインウイルス)及びMILV(ミラフィオリ(mirafiori)レタスウイルス)からなる)、TBSV(トマト茂み(bushy)発育阻害ウイルス)、LNSV(レタス壊死発育阻害ウイルス)、TuMV(カブモザイクウイルス)、CMV(カボチャモザイクウイルス)又はBMYV(ビート西洋黄色(beet western yellows)ウイルス)の1種に対する、標準的又は中程度の耐性である1又は複数の遺伝子から提供されてもよい。
【0034】
農業で注目の表現型は、ウイルス、細菌、虫又は真菌による感染に対して、標準的又は中程度の耐性のある1又は複数の遺伝子から提供され、この表現型を提供する当該又は前記の標準的又は中程度の耐性のある遺伝子は、具体的に、ブレミア Dm10、R17m、Dm5、Dm8、R36、R37(ラクツカ・サティバのクラスター1上に位置する遺伝子)、Dm1、Dm2、Dm3、Dm6、Dm14、Dm15、Dm16、Dm18、(ラクツカ・サティバのクラスター2上に位置する遺伝子)、Dm4、Dm7、Dm11、R38(ラクツカ・サティバのクラスター4上に位置する遺伝子)への耐性遺伝子;又はクラスター1上に位置するTuMVに耐性のあるTu遺伝子;クラスター2上に位置するナソノビアに耐性のあるNr遺伝子;又はクラスター4上に位置するLMVに耐性のあるmol.1及びmol.2遺伝子、から選択してもよい。上記のクラスター1、2及び4は、Michelmore R.W.による(Plant Pathol, 1987, vol. 36, no4 : 499-514 [4], Theor . Appl. Genet., 1993, vol. 85, No 8 : 985-993 [5])で定義されている。
【0035】
より具体的には、農業で注目の表現型は、レタスを侵す病気、ブレミア・ラクツカエ、白色を引き起こす真菌、枝内面に落下粉末(powdery down on the inner face of the limb)の1つに対し、標準的又は中程度の耐性のある、1又は複数の遺伝子により提供されてもよい。さらに、ブレミアは、高度に適応でき、植物繁殖者によってその変種に既に導入された耐性を征服できる、新規な品種の出現をもたらす。
【0036】
より好ましくは、本発明は、ラクツカ・サティバ植物の種子、植物又は細胞が、雄性不稔性に関与せず、且つ検出可能な表現型を提供する、少なくとも2つの遺伝子の異種接合体である。さらにより好ましくは、検出可能な表現型を植物に提供する2以上の遺伝子は、同じクラスターに、例えばラクツカ・サティバのクラスター1、2又は4の耐性遺伝子のクラスター上に位置する。
【0037】
本発明はまた、雄性不稔性遺伝子型であるとともに、雄性不稔性に関与せず、且つ当該種子由来の植物を検出可能な、少なくとも1つの遺伝子の異種接合体であり、任意に上記の種子の追加の特徴を有する、雑種ラクツカ・サティバ種子集団であって、少なくとも105個の種子、好ましくは少なくとも106、及びより好ましくは少なくとも107個の種子を含んでなるような、種子集団に関する。
【0038】
種子を2つのラクツカ・サティバ植物の交配により得る場合、このうちの1つ(「雌」親)は1又は複数の優性遺伝子により提供される核雄性不稔性をもたらし、「雌」親として使用される植物は、不稔性遺伝子の異種接合体であることが必要である。実際は、1又は複数の優性の雄性不稔性遺伝子の同種接合体となるものとして、交配により得られる「雌」親として使用される植物由来の当該2つの植物は、自身が雄性不稔性の遺伝子のキャリアとなるはずである。ただし、その場合はこれらの2つの植物間の交配は不可能である。
【0039】
結果として、一方(その「雌」親は)は1又は複数の優性遺伝子から提供される核雄性不稔性のキャリアである、2つの型のラクツカ・サティバ植物の間の交配から得られる本発明の種子集団は、減数分裂中の染色体の分離現象が原因で、1又は複数の雄性不稔性の優性の遺伝子を保有するラクツカ・サティバ種子、及び当該1又は複数の遺伝子を保有しない種子からなる。一般的に「雌」親によりもたらされる雄性不稔性が一遺伝子的且つ優性である場合、不稔性雄性種子の割合は少なくとも40%である。
【0040】
本発明は、発明者等による以下の観察により生まれた。すなわち双翅目はラクツカ・サティバの花に定常的に授粉する虫ではないが、さらにその蜜を餌とすることは知られておらず、これらの虫、具体的な種としては、カリホラ・ボミタリア(Calliphora vomitaria)、カリホラ・エリトロセファラ(Calliphora erythrocephala)、及びルシリア・カエサル(Lucilia Caesar)は、閉鎖環境に過剰数を導入すると、ラクツカ・サティバの花粉授粉者としてふるまうという観察である。
【0041】
さらに本発明は、具体的には雑種ラクツカ・サティバ植物を得る観点で、閉鎖環境で雄性稔性植物により雄性不稔性ラクツカ・サティバ植物の授粉を行うための、双翅目の使用に関する。好ましくは、使用される雄性不稔性ラクツカ・サティバ植物は、単一の優性遺伝子により、好ましくは上記の雄性不稔性Ms7によりもたらされる雄性不稔性を有する。使用される稔性雄性植物は、好ましくはラクツカ・サティバ植物であり、より好ましくは栽培品種である。
【0042】
本発明の閉鎖環境は、具体的には温室、ケージ又はトンネルであってもよく、好ましくは30m2以上の、より好ましくは300m2以上の、例えば30〜1500m2又は50〜1000m2の表面積である。閉鎖環境の高さは、通常、2m〜4mの範囲であり、好ましくは2.5m〜3.5m、例えば3mである。具体的には、換気、散水、温度制御及び光レベル制御のための手段があってもよい。好ましくは、当該閉鎖環境は、虫に対して密閉するように封じられる囲いである。
【0043】
使用される双翅目は、好ましくは1m2当たり少なくとも100匹、好ましくは1m2当たり少なくとも250匹、例えば1m2当たり100〜1000匹の濃度で存在させるべきである。この濃度はまた、1m3当たりの双翅目の数として決定してもよく、好ましくは、1m3当たり少なくとも25匹、より好ましくは1m3当たり少なくとも50匹、さらに好ましくは1m3当たり少なくとも75匹、例えば1m3当たり25〜500匹、1m3当たり75〜250匹でもよい。
【0044】
本発明はまた、雑種ラクツカ・サティバ種子を獲得するための方法であって、
表現型的に「雌」親として使用される雄性不稔性ラクツカ・サティバ植物、及び表現型的に「雄」親として使用される雄性稔性ラクツカ・サティバ植物を、互いに近接した状態で、閉鎖環境中で栽培するステップであって、2つの親の一方が、雄性不稔性以外の検出可能な表現型をそれに提供する遺伝子の同種接合体であるという追加的な特徴を有し、他方の親はこの遺伝子を保有しないものであるステップ、
植物の開花の際に、1m2当たり100匹以上の濃度で、好ましくは1m2当たり少なくとも250匹の濃度で、閉鎖環境へ導入する双翅目を用いて授粉するステップ、及び
雄性不稔性植物により作られる種子を収穫するステップ、
を含んでなる方法に関する。
【0045】
好ましくは、第二の親はまた、他方の親からはもたらされない、雄性不稔性以外の検出可能な表現型をそれに提供する少なくとも1つの遺伝子の異種接合体である。好ましくは、当該閉鎖環境は、虫に対して密閉するように封じられる囲いである。
【0046】
最良の結果は双翅目が過剰の場合に観察され、さらにその濃度は、好ましくは1m2当たり400匹以上、より好ましくは1m2当たり500匹以上である。
【0047】
双翅目は卵期、幼虫期、蛹期又は成虫期に導入してよい。好ましくは、これらを蛹の状態で導入する。
使用する双翅目は好ましくは短角類(brachycera)であり、例えばシクロラファ・ブラキセラ(cyclorhapha brachycera)、より好ましくはイエバエ科(Muscidae)又はクロバエ科(Calliphoride)由来の短角類、例えばカリホラ・ボミタリア(Calliphora vomitaria)、カリホラ・エリトロセファラ(Calliphora erythrocephala)又はルシリア・カエサル(Lucilia Caesar)である。
【0048】
双翅目の導入が、少なくとも1週間に1回、好ましくは1週間に2回、少なくとも3〜4週にわたって繰り返されると、通常より良好な収率が得られる。
同様に、閉鎖環境中での雄性不稔性植物の数が、雄性稔性植物の数より大きくなると、通常より良好な収率が得られる。閉鎖環境中の雄性不稔性植物の数は、例えば少なくとも200、及び雄性稔性植物の数は、例えば少なくとも1000である。
【0049】
本発明の方法の実施において、2つの親の開花が同時に起こることが必要である。これはこの形質のためのセレクションにより、又は適切な栽培技術を使用することにより達成できる。
【0050】
好ましくは、雌親として使用する植物の雄性不稔性は、一遺伝子的、優性及び核性であり、より好ましくは遺伝子Ms7により提供される。
好ましくは、雌親として使用する植物のゲノムは、650〜700ヌクレオチドの、例えば655〜695、660〜690、665〜685、670〜680、673〜677ヌクレオチドの、又は674、675又は676ヌクレオチドの二本鎖DNA配列を含んでなり、ここで、各2つの5'末端は、配列「5' TGCGTTCCAC 3'」(配列番号1)から開始する。好ましい実施態様に従うと、「雌」親として使用する植物のゲノムは、図8に示すヌクレオチド配列(配列番号2)を含んでなるか、又は1〜10個の、好ましくは1〜5個又はより好ましくは1〜3個のヌクレオチドが他のものにより置換、削除又は付加される前記配列に由来する配列を含んでなる。
【0051】
上記の説明の通り、減数分裂中の染色体の分離現象が原因で、従来の交配技術では、雄性不稔性の優性遺伝子を保有し、「雌」植物として使用される植物の同種接合体集団を得ることはできない。結果として、雄性不稔性の優性遺伝子を使用する場合、雌親として使用される植物は、
核雄性不稔性の優性遺伝子の異種接合体であるラクツカ・サティバ植物と、不稔性遺伝子を保有しない雄性稔性のラクツカ・サティバ植物との間で交配するステップ、
当該交配から得られる種子を栽培するステップ、及び
雄性稔性表現型を有する植物を除去するステップ、
を含んでなる方法により得ることができる。
【0052】
この雄性稔性表現型を有する植物を除去するステップは、例えば、雄性不稔性ラクツカ・サティバ植物を、雄性稔性ラクツカ・サティバ植物から識別可能とする可視的な特徴に基づき、手作業で行ってもよい。例としては、Ms7遺伝子に関連する不稔性の場合、雄性不稔性植物の花頭は、雄性稔性植物のそれより長時間開いており、その雄性不稔性植物は花粉を持たないという事実を有効に用いることにより、分類を行ってもよい。
【0053】
代替の実施態様に従えば、雄性稔性表現型の植物を除去するステップは、各植物のサンプルにおける雄性不稔性の優性遺伝子に特異的な分子マーカーの不存在を検出することにより行う。例えばそれは、約675塩基対(例えば650〜700、655〜695、660〜690、665〜685、670〜680、673〜677、674、675又は676塩基対)の長さの、2本鎖の各々の5'末端は、配列「5' TGCGTTCCAC 3'」(配列番号1)で開始する、RAPD(多形DNAのランダム増幅(random amplification of polymorphic))マーカーであって、例えば発明者等により開発されたマーカーBA05−675(配列番号2)又はSCAR(配列特徴付け増幅領域(sequence characterized amplified region))マーカー、CAPS(開裂増幅多形配列(cleaved amplified polymorphic sequence))マーカー、又はこのRAPDマーカーを用いて開発したSTS(配列タグ化部位(sequence tagged site))として一般的に設計された任意の他のマーカーがある。
【0054】
好ましい実施態様によれば、雄性不稔性の優性遺伝子の特異的分子マーカーの存在又は不存在の検出は、雌親として使用される植物の開花前に、好ましくは植物の生育の初期段階に、例えば1〜5葉期に、又はより好ましくは1〜2葉期に行う。
【0055】
さらに好ましくは、使用される分子マーカーは、少なくとも70%、75%、80%、85%、95%、98%又は99%、又は100%もの感受性のある雄性不稔性植物を検出でき、その感受性は、分子マーカーを有する雄性不稔性植物(真陽性)の数と、真陽性の数と分子マーカーを有さない雄性不稔性植物(偽陰性)の数の総和との間の比率として定義される(表17を参照)。
【0056】
一般的に、本発明の方法を用いて得られた種子のTSW(1000種子質量(thousand seed weight))は、「雄」親として使用される雄性稔性ラクツカ・サティバ植物の自己受精により得られた種子のTSWより、少なくとも10%、又は少なくとも20%又は少なくとも30%大きい。
【0057】
本発明はまた、上記記載の方法を用いて得ることができる種子集団で、少なくとも105個の種子、好ましくは106個の種子、及びより好ましくは107個の種子含んでなる集団に関する。
【実施例】
【0058】
実施例1:雑種レタス種子の獲得
「雌」親として以下に記載の種々の試行で使用される雄性不稔性ラクツカ・サティバ植物は、Ms7不稔性遺伝子を保有した。当該植物は、INRA、USDA等の種々の研究組織から入手できるMs7遺伝子(X Girelle 94-9538-1)を保有する温室バターヘッドレタスの第5代戻し交配(BC5)由来であった。
【0059】
Ms7遺伝子は優性であるため、Ms7遺伝子を保有する2つの植物間の交配は不可能である。結果として、雄性不稔性表現型を有する植物は、Ms7遺伝子の異種接合体である必要があり、Ms7/ms7(不稔性)とms7/ms7(稔性)植物との間で交配することによってのみ、得ることができる。
【0060】
結果として、雄性稔性植物を除去するための精製ステップが必要となる。この後者のステップは、Ms7/ms7植物が花粉を持たず、且つMs7/ms7植物の花頭はms7/ms7植物のそれよりも長期間開いたままであるため、可能となった。
【0061】
試行1
第一の試行の環境としては、
Nacre/Cambria Dm18/R38栽培品種(クラスター2からのDm18耐性遺伝子及びクラスター4からのR38耐性遺伝子)由来のバターヘッドレタスとの、Ms7バターヘッドレタスの第3代戻し交配(BC3)由来の450個の温室バターヘッドレタスラクツカ・サティバ植物は、ブレミア・ラクツカエ品種24(Bl24)に感受性の高い組み合わせであり、これが「雌」親として使用され(その約半分は不稔性雄);及び
BRA Dm18/R37温室バターヘッド栽培品種(クラスター2からのDm18耐性遺伝子及びクラスター1からのR37)由来の100個のラクツカ・サティバ植物は、B124に対して耐性のある組み合わせであり、「雄」親として使用された。
この2つの遺伝的父はまさに同時に開花し、これは受精を可能な限り均一にできることを意味する。
【0062】
植物を36m2の虫を密閉して封じる囲いの中に(湿度及び温度制御を備えるダッチ型温室で、11m×3.30m)、以下の通りに(図2を参照)配置した。
「雄」親として使用される植物を、密封囲いの端に沿って2列に配置し、植物は20cmずつ間隔をあけ、及び
「雌」親として使用される植物を、「雄」植物の2列の間であって、そこから50cmの距離の点に、7列で配列し、植物は15cmずつ、且つ列は25cmずつ間隔をあけた。
【0063】
播種は13週目に、植え付けは16週目に行った。
Ms7遺伝子の優性性質のための、雄性稔性表現型を持つ雌親として使用される植物の精製(除去)のためのステップが必要であった。本ステップは、Ms7/ms7植物が花粉を持たず、Ms7/ms7(雄性不稔性)植物の花頭が、ms7/ms7植物(雄性稔性)のものより長期間開いた状態であるという事実を利用して、主に26週と27週の間で行った。
雌親として使用される450植物のうち、217は雄性不稔性であり、効率的に使用された。
【0064】
2種類の双翅目クロバエ科の虫を授粉する虫として使用した。
「ウジ虫(maggots)」として知られる(キンバエ(bluebottle)又はニクバエ(blowflie)としても知られる)、カリホラ・ボミタリア及びカリホラ・エリトロセファラハエは、15〜20℃で取り扱い、及び
「ピンキーズ(pinkies)」として知られるラクツカ・カエサーハエ(キンバエ(greenbottle)としても知られる)は、20〜25℃で取り扱った。
授粉する虫は、28週〜31週の間に隔週で、ハエが1m2当たり300匹となる濃度で添加した。
自己授粉制御として使用される、雄性不稔性植物(「雌」親)及び雄性稔性植物(「雄」親)も、33週で収穫した。植物を乾燥させ、脱穀し、その後種子の洗浄を完了させるとともに残存した軽量ごみを除去するため、パルスエアのカラムを通し、重さと幅を調整した。
【0065】
試行1で得られた2500種子のサンプルを、2007年2月13日にNCIMB(NCIMB Ltd., Ferguson Building, Craibstone Estate, Bucksburn, Aberdeen, A21 9YA,スコットランド、英国)に、受託番号NCIMB 41470で寄託した。
【0066】
試行2
同様に第二の試行を、336m2の虫を密閉して封じる囲いの中で(長さ:56m、幅:6m、高さ:3m)、より大きなスケールで実施した。
【0067】
第二の試行の環境としては、
Nacre/Cambria Dm18/R38栽培品種との、Ms7バターヘッドレタスの第4代戻し交配(BC4)由来の30000個の温室バターヘッドレタスラクツカ・サティバ植物、ブレミア(Bl24に感受性の高い組み合わせ)を、「雌」親として使用し(その約半分は雄性不稔性である);及び
B124に対して耐性のある、BRA Dm18/R37温室バターヘッド栽培品種由来の1500個の対応する(parallel)植物を、「雄」親として使用した。
【0068】
植物を以下の通りに密封囲い中に配列した(図3を参照)。
「雄」親として使用される植物は4列に配置し、密封囲いの各端に沿って(80cmのところに)1列、中央に2列を(20cmの間隔をあけて)配置し、配置において植物はねじれており、15cmの間隔をあけ;及び
「雌」親として使用される植物を、稔性雄親の列の間にそこから65と70cmの距離の点に8列で配置し、植物は15cmずつ、列は25cmずつ間隔をあけた。
播種は12週目に、植え付けは16週目に行った。
【0069】
試行1と同様の方法で、雄性稔性表現型を持つ雌親として使用される植物の精製(除去)のためのステップが必要であった。本ステップは、Ms7/ms7植物が花粉を持たず、Ms7/ms7(雄性不稔性)植物の花頭が、ms7/ms7植物(雄性稔性)のものより長期間開いた状態であるという事実を利用して、主に26週と27週の間で行った。
雌親として使用される3000植物のうち、1250は雄性不稔性であり、効率的に使用された。
16のms7/ms7植物(雄性稔性)を保存したが、精製の間に、自己受精コントロールとして使用されるための、虫を通過させないようなネットで構成された、密封して封じられた囲いの中で自己受精した。
【0070】
使用した授粉する虫は試行1で使用したものと同じである。
授粉する虫は、25週〜29週の間に毎週、ハエが1m2当たり1100匹となる濃度で添加した。
【0071】
同時に、精製のステップで保存した、10の稔性植物(雄親)及び16の稔性ms7/ms7植物を、自己受精コントロールとして作用するための、虫を通過させないようなネットで構成された、密封して封じられた囲いの中で自己受精した。
【0072】
雄性不稔性植物(「雌」親)、並びに自己授粉制御として使用した精製ステップで保存した10の雄性稔性植物(雄親)及び16の稔性ms7/ms7植物を、33週で収穫した。
植物を乾燥させ、脱穀し、その後種子の洗浄を完了させるとともに残存した軽量ごみを除去するため、パルスエアのカラムを通し、重さと幅を調整した。
【0073】
得られた種子のサンプルを播種し、得られた植物のB124に対する耐性を、公式のGEVESプロトコル(Groupe d’Etude et de controle des Varietes et des Semences [Varieties and Seeds Study and Monitoring Group])に従って試験した。当該植物がB124に耐性であることが明らかになった。
【0074】
試行1及び2の結果
試行1及び2の結果を以下の表に示す。
表1:試行1及び2で収穫された種子の質量の比較
【表1】

【0075】
表2:試行1及び2で作成した、雑種種子及び自己受精コントロールに由来する種子の計測の比較
【表2】
【0076】
表3:試行1及び2で作成した、雑種種子及び自己受精コントロールに由来する種子のTSW(1000種子質量)の比較
【表3】
【0077】
考察
第一の試行では、217の雄性不稔性ラクツカ・サティバ植物(雌親)から730グラムの種子、すなわち1植物当たり3.36グラムの収率で得られ、自己受精コントロールの15.30%の比率に相当した(雄親)。
第二の試行では、1170の雄性不稔性ラクツカ・サティバ植物(雌親)から7.730kgの種子、すなわち植物あたり6.60グラムの収率で得られ、自己受精コントロールの2つ(「雄」親とms/ms雄性稔性「雌」親、精製の間保存され、虫密閉の下で受精した)の平均の50%の比率に相当した。
【0078】
第二の試行では、F1雑種を計測する有用種子の量が、自己受精したコントロールの10%以上であった。
2つの試行では、F1雑種のTSW(1000種子質量)が、自己受精コントロールの30%以上であった。
第二の試行で得られた種子由来の植物は、B124に耐性であった。
【0079】
試行3〜8
異なる種類のバターヘッド及びバタビアのレタスを用い、(Ms7遺伝子を保有するラクツカ・サティバ植物と稔性ラクツカ・サティバ植物との間の戻し交配に由来する50%の雄性不稔性である)「雌」親として使用される8つのラクツカ・サティバ植物と、「雄」親として使用される4つの雄性稔性ラクツカ・サティバ植物を各々含有する、換気された温室に置いた、虫試験用の3.3m×1.1mケージ中で、6つの同様の試行を同時に開始した。
播種を22週目に、植え付けを27週目に行い、31週目の開花の兆候で雄性稔性「雌」植物から雄性不稔性を分類し、ハエを32、33及び34週の3週間にわたり1週間当たりのある量、すなわちハエ約100匹/m2を添加し、37週目に収穫を行った。
【0080】
試行3〜8の結果
ケージ1: CHARLIN(68/12624)* CHARLIN (バターヘッド)
1種の雌性植物:9.20 g
1種の雄性植物:28.30 g
ケージ2:CHARLIN(BC/77)* CHARLIN (バターヘッド)
1種の雌性植物:5.45 g
1種の雄性植物:19.40 g
ケージ3:BRA 68/12588 (温室リンゴレタス)* BRA 68/12588
6種の雌性植物:
植物1:7.00 g
植物2:2.00 g
植物3:5.20 g
植物4:3.40 g
植物5:9.60 g
植物6:8.40 g
平均:5.9 g/植物
1種の雄性植物:34.40 g
ケージ4: BRA 68/12588 * BRA 68/12588
4種の雌性植物:
植物1:11.95 g
植物2:7.15 g
植物3:15.90 g
植物4:11.90 g
平均:11.25 g/植物
1種の雄性植物:29.00 g
ケージ5:BVA 68/12553 (温室バタビアレタス)* BVA 68/12553
5種の雌性植物:
植物1:.50 g
植物2:1.30 g
植物3:2.20 g
植物4:1.70 g
植物5:4.20 g
平均:3.30 g/植物
1種の雄性植物:3.60 g
ケージ6:BVA 68/12553 * BVA 68/12553
3種の雌性植物:
植物1:4.60 g
植物2:3.20 g
植物3:3.70 g
平均:3.80 g/植物
1種の雄性植物:23.40 g
【0081】
考察
「雌」親の産物の平均は、3.3g〜11.2g/植物であり、バタビア類型物(BVA 68/12553)は、約8.5g/植物のバターヘッド類型物(Charlin, BRA 68/12588)と比較して、かなり低い収率(約3.5g/植物)であり、当該類型物はその最終収率に寄与すると考えられる。
【0082】
試行9
新規な作製試行を、開放地面上の換気された温室内に置いた、直径0.40mの測定用の虫小さな試験用ケージ中で行った。
播種を22週目で、再植え付けを25週目で行い、30週目の開花の兆候で雄性稔性植物から「雌」植物を分類し、31週目で植え付けし、ハエを32、33及び34週の3週間にわたり1週間当たりのある量、すなわちハエ約100匹/m2を添加し、35週目に収穫を行った。
【0083】
結果
得られた結果を以下の表4及び5にまとめた。
表4:「雄」及び「雌」親、Charlin (温室バターヘッド)による種子の作製
【表4】
【0084】
表5:「雄」及び「雌」親、BVA 68/12553 (バタビア)による種子の作製
【表5】
【0085】
考察
バタビアレタス(BVA 68/12553)で得られた産物の平均は、自己受精コントロールの1.7g/植物に対して約1 g/植物であり、且つ温室バターヘッドレタス(Charlin)で得られた産物の平均は、約1 g/植物であり自己受精コントロールとほぼ同じであった点で、結果は満足のいくものであった。
【0086】
試行10:雑種の組み合わせによるDm3/Dm18蓄積
2つの優性遺伝子、Dm3及びDm18であって、クラスター2に属し同じ遺伝子座に位置する品種24(B124)及び品種23(B123)の、ブレミア・ラクツカエ耐性用遺伝子を蓄積するF1雑種レタスは、上記記載の試行で記載された方法を用いて得た。
【0087】
「雌」親として使用される植物(06/30443: MS7/21NACRExDEVONIAxCAMBRIA)は、Dm18及びR38耐性遺伝子のキャリアである。「雄」親として使用される植物は、Dm3耐性遺伝子を保有する温室バターヘッドレタスである(BC: 06/30443 * Rex 又は BC:06/30443 * Melina)。
以下の5つの交配を行った。
06/30443/01 (MS7/21xNACRExDEVONIAxCAMBRIA-01) Dml8/R38 * Rex Dm3、
06/30443/10 (MS7/21xNACRExDEVONIAxCAMBRIA-10) Dml8/R38 * Rex Dm3、
06/30443/02 (MS7/21xNACRExDEVONIAxCAMBRIA-02) Dml8/R38 * Melina Dm3、
06/30443/03 (MS7/21xNACRExDEVONIAxCAMBRIA-03) Dml8/R38 * Melina Dm3、
06/30443/04 (MS7/21xNACRExDEVONIAxCAMBRIA-04) Dml8/R38 * Melina Dm3。
【0088】
当該5つの交配由来の種子は播種され、葉ディスク上でのインキュベーション試験及び分子標識をF1植物で行うとともに、「雄」(Rex/Melina)及び「雌」親で行った。
【0089】
I 葉ディスク上でのインキュベーション試験
装置:
植物原料は、1植物当たり及び1品種当たり5ディスクの数で、1起源当たり10植物及び2つのブレミア・ラクツカエ品種で(Dm18/R38蓄積を攻撃し、且つDm3がそれに対する耐性を与えるB124、及びDm3/R38蓄積を攻撃し、且つDm18がそれに対する耐性を与えるB123)、1.5cm直径の葉ディスクからなる。すなわち、総数で80の植物を2つの品種で試験した。
【0090】
技術:
葉ディスクに、ブレミアB123及びB124を独立に伴ったスポロシストを噴霧することにより植菌を行った。
【0091】
1/葉ディスクの除去:
1植物当たり5つの葉ディスクを除去し、2重の厚みの湿ったブロット紙の上に配列させた。各皿には、6列の試験するディスク及び1列の感受性の高いコントロールを入れた。
2/植菌:
6皿の葉ディスクを植菌するために必要な植菌材料の量は11mlであった。100 mlの脱塩水に4滴のTween 80を使用することにより、Tween 80溶液を調製した。植菌材料の容積のための抽出溶液の量は、100mlの植菌材料当たり5 mlのTweenストック溶液であった。植菌材料は、5%のTween 80ストック溶液を補充した水道水中で、スポロシストを分離することにより作製した。スポロシストを、108胞子/ml近傍のスポロシスト濃度を得るため、1植物/mlの植菌材料量で除去した。250mlビーカーに、ガーゼを通して溶液を通過させて濾過を行った。
3/噴霧:
植菌は、葉ディスクを採取した日に行った。植菌材料溶液の容積を測定し、その後配置した皿に植菌した。植菌材料を、各ディスクの表面全体に、微細な液滴で均一に噴霧した。この皿を密封し、15℃で、日光14時間、夜間10時間の単位で7日間インキュベートした。
【0092】
結果
植菌の結果を、以下の表6〜13に示す(S=感受性、R=耐性)。種々の交配から得られる雑種は2つの品種B123及びB124に耐性であり、クラスター2由来の2つの遺伝子Dm18及びDm3の存在を示すものである。
【0093】
表6:「雌」30443 (Dm18/R38)植物: MS7/21xNACRExDEVONIAxCAMBRIA:B123耐性及びB124感受性。
【表6】
表7:「雄」Melina (Dm3) 植物:B123感受性及びB124耐性(10植物での試験)
【表7】
表8: 06/30443/02 * Melina (Dml8/R38*Dm3) 交配=MS7/2IxNACRExDEVONIAxCAMBRIA-02*MELINA
【表8】
表9: 06/30443/03 * Melina (Dml8/R38*Dm3) 交配=MS7/2IxNACRExDEVONIAxCAMBRIA-03*MELINA
【表9】
表10: 06/30443/04 * Melina (Dml8/R38*Dm3) 交配=MS7/2IxNACRExDEVONIAxCAMBRIA-03*MELINA
【表10】
表11:「雄」Rex (Dm3) 植物:B123感受性及びB124耐性(10植物での試験)
【表11】
表12:06/30443/01 * Rex (Dml8/R38*Dm3) 交配=MS7/21xNACRExDEVONIAxCAMBRIA-01*REX
【表12】
表13: 06/30443/10 * Rex (Dml8/R38*Dm3) 交配=MS7/21xNACRExDEVONIAxCAMBRIA-10*REX
【表13】
【0094】
II 分子標識
2つの分子マーカーを用いることで、ラクツカ・サティバにおいて、優性であり且つ2つのブレミア・ラクツカエ種に耐性である、2つの密接に関連した遺伝子(同じ遺伝子座に位置する)の蓄積を証明することができる。
【0095】
方法:
1/植物原料:
植物原料は、1植物当たり1ディスクで、1.5cm直径の葉ディスクからなる。遺伝子型当たり5〜10植物(すなわち総数で50植物)由来のDNAを、CTAB/クロロホルム抽出により得て、その後5ng/μlの濃度で、TE 0.1×溶液中で懸濁として溶解させた。
【0096】
2/遺伝子型決定:
Maisonneuve等による[10]及びKuang等による[11]で報告された、SCW09及びB1マーカーを前記50植物の遺伝子型決定のために使用した(表14)。PCR条件は表15に記載する。
SCW09及びB1のPCRサイクルはそれぞれ以下の通りである:
94℃/30秒−94℃/1分、60℃/1分、72℃/2分;40サイクル−72℃/5分−4℃;
94℃/30秒−94℃/1分、63℃/1分、72℃/2分;40サイクル−72℃/5分−8℃。
SCW09マーカーは、移行ステップの前にTaqI酵素での酵素的消化を受けた(バッファ1×、BSA1×及びTaqI 1U)。電気泳動を、2%(SCW09)又は1.5%(B1)のアガロースゲルを用い、TBE1×バッファ中で、1時間(B1)又は1時間30分(SCW09)、定電圧220Vで泳動した。電気泳動プロファイルは、UVランプ下、エチジウムブロミド(ETB)での染色により明らかになった。
【0097】
表14:SCW09及びB1マーカーの特徴
【表14】
表15:SCW09及びB1マーカーのPCR条件
【表15】
【0098】
結果
結果を表16並びに図4及び5に示す。REX及びMELINAの遺伝子型は、Dm18−/Dm18−且つDm3+プロファイルであった。06/30443遺伝子型はDm18+/Dm18+且つDm3−プロファイルであった。親(REX又はMELINA)の一方又は他方との交配の組は、Dm18+/Dm18−及びDm3+プロファイル、すなわちDm18及びDm3に耐性であった。
【0099】
表16:SCW09及びB1マーカーでの遺伝子型決定の結果(S=感受性、R=耐性)。
【表16】
【表17】
【0100】
実施例2:Ms7遺伝子用分子マーカーの同定
以下に記載のラクツカ・サティバ(レタス)中の雄性不稔性Ms7に関連する分子マーカーを得るための方法を、指示する目的で提供する。
1−レタス(ラクツカ・サティバ)中で核雄性不稔性Ms7と関連するマーカーの開発用の群の構成
Ms7雄性不稔性であるレタス用の異なる類型の2つの群を、戻し交配で作製した。結果として得られた、200植物の反復第4代群(BC4)の2つを雄性不稔性表現型に対応する「生花粉の不存在」形質で表現型検査を行った。2つの表現型グループを、雄性稔性(F)又は雄性不稔性(S)として1:1の比率で同定した。
この比率は、Ms7遺伝子座で異種接合体の親により提供される、雄性不稔性の優性一遺伝子的性質を裏付けるものであった。
【0101】
2−レタス植物由来のDNAの抽出
DNAは、修正CTABプロトコル(Tomas等, 1989 [20]; Doyle 等, 1990 [15]; Edwards 等, 1991 [16])を用い、新鮮な幼葉を使用して抽出した。
1.5ml管を用いて得た新鮮な葉ディスクを、500μlの抽出バッファ(Tris−HCl 0.1M、NaCl 0.7 M、EDTA 10 mM、CTAB 1%、βメルカプトエタノール 1%)中に置いた。この置いた原料を65℃で1時間インキュベートし、インキュベーション中に2〜3回反転させて混合した。
その後、200μlの24:1のクロロホルム:イソアミル溶液を添加し、反転させて混合した。6000rpmで10分間、20℃で遠心分離後に、400μlの上澄みを回収し、400μlのイソプロパノールと混合した。
−20℃で1時間後、混合物を6000rpmで10分間、4℃で遠心分離した。管の中身を出した。管底に付着したDNA残基は12時間空気中で乾燥させた。
このDNAを、200μlのTE 0.1×溶液(tris−HCl 1 mM、EDTA 0.1 mM)中で懸濁として溶解させた。終濃度を、1%アガロースゲル上で、Lambda DNA-HindIII消化アッセイにより測定した。
【0102】
3−BSA(bulk segregant analysis)法を用いる、ラクツカ・サティバにおけるMs7雄性不稔性に関連するRAPDマーカーの研究
雄性不稔性に関連する分子マーカーMs7を同定する目的で、Operon Technologies Inc.(Huntsville, AL 35805、米国)製のRAPD(登録商標)10merキット及びBuIk Segregant Analysis法(Michelmore 等, 1991 [17] 及び Paran 等, 1991 [18])を使用した。
上記記載の2つのBC4群の表現型検査から開始するため、10植物の混合物の2つのサンプルを、2つのグループ「雄性不稔性」及び「雄性稔性」用として作製した。OPA-01〜OPBH-20の1200個のプライマー(Operon Biotechnologies Inc. (Huntsville, AL 35805、米国))を、William 等, 1990 [22] 及び Welsh 等, 1990 [21]により報告されたRAPD技術を用いて、1群当たり4サンプルを試験した。「雄性不稔性」サンプルに特異的なバンドを表し且つ読みやすい電気泳動プロファイルを有するプライマーを選択した。
【0103】
PCR反応は、PCR1×バッファ、3 mM MgCl2、200μMのdNTP、400nMのプライマー、1単位のAmpliTaq (Perkin-Elmer cetus)DNAポリメラーゼで組成した、総反応量25μlで実施した。複数サイクルからなるPCR反応を以下に記載した。94℃30秒間で1ステップ、94℃1分、その後35℃1分で最終的に72℃にして2分、及び最後の伸長ステップを72℃5分間を45サイクルで実施し、その後反応物を4℃で保存した。
増幅産物を、2%アガロースゲルで、以下の電気泳動条件下で分離した:移動バッファTBE 1×(tris−ホウ酸塩−EDTA)、190Vで2時間15分。
【0104】
残存するRAPDプライマーを確認する(validate)ために、2つのBC4群由来の200植物を、以下の配列:5' TGCGTTCCAC 3'(配列番号1)(これは675bpのマーカー、BA05-675(配列番号2)と対応する)を有し、Ms7を同時分離する、OPBA05プライマーで個別に試験した。このマーカーから得られた電気泳動プロファイルを図6に示す。
【0105】
このマーカーを12の群からの約25植物で、種々の類型のレタス(バタビア、バターヘッド、アイスバーグ(iceberg)、ロメイン(romaine)、オークリーフ(oakleaf)及びロロロッサ(lollo rossa))で確認した。試験条件は、抽出、増幅及び電気泳動のステップで既に記載したものと同じであった。
RAPD BA05-675マーカーの感受性及び特異性の計算からは、類型によって異なる値が得られ、それぞれ平均値は96%及び94%であった(表17及び18を参照)。
【0106】
表17:保有する分子マーカーの感受性及び特異性の予測される値の、計算の詳細。
【表18】
表18:レタスの類型と、Ms7と関連する分子マーカーBA05-675への感受性(S*)及び特異性(F**)の平均値の結果。
【表19】
【0107】
4−ラクツカ・サティバ種におけるMs7雄性不稔性に関連するBA05-675の遺伝子マッピング
上記のms7のために解体されたBC4の群由来の200植物を基に、Ms7に関連するBA05-675マーカーの遺伝子マッピング(William 等, 1993 [23])を、JoinMap(登録商標)4 (Stam 等, 1996 [19]) 及び CarteBlanche (Keygene N P, P O Box 216, 6700 A E Wageningen, The Netherlands)を用いて実施した。優性のBA05-675マーカーについて、メンデル型1:1比率のゼロ仮定を、X2統計的検定で確認した(検定はp > 0.05で有意性がない)(表19を参照)。3.0以上の可能性ある比率(又はLODスコア)での結合試験により、Ms7の1.8 cMマーカーをマップできた(図6)。
【0108】
表19:MS7に関連する分子分離(segregation)マーカーBA05-675の比率及び比率1:1でのメンデル型分離のゼロ仮説を用いる1ddlでのX2検定の結果(F:稔性優性植物の数、S:不稔性雄性植物の数)。
【表20】
【0109】
5−ラクツカ・サティバレタスにおける雄性不稔性に関連するRAPD BA05-675分子マーカーのクローニング及び配列決定
アガロースゲル電気泳動プロファイルから、675bpのDNA断片を単離し、TEバッファ(10 mM tris-HCl、1 mM EDTA)中で懸濁物として溶解させ、その後上記のPCR条件下で再増幅させた。この再増幅した単離断片を、市販のPCR(登録商標)4-TOPO(登録商標)キット(Invitrogen, Carlsbad, California 92008, 米国)を用いてクローン化した。クローニングの確認後、DNA抽出(Midiprep system)は、Promegaカラム精製キット((Madison, WI, 米国)を用いて行った。
75 ng/μlに濃縮した精製クローンを、Cogenics (38944 Meylan, フランス)により配列決定した。すなわち、図8に記載のBA05-675マーカーの配列が得られた(配列番号2)。
【0110】
6−結果
本発明者等が実施した研究により、RAPDマーカー、BA05-675(配列番号2)の特徴づけが可能となった。その配列は、図8に示され、Ms7から1.8cMの距離で(図7)、且つレタスの類型について個別にそれぞれ平均96%及び94%の感受性及び特異性を有する(表18)。2つのBC4群由来の個別の植物でのこのマーカーの評価から、不稔性の雄性植物を同定するための平均予測値(Altman 1994a [13])が97%近傍であるという推定ができる。
【0111】
参考文献
1. Goubara & Takasaki (2003) Flower visitors of lettuce under field and enclosure conditions, Appl . Entomo. Zool . 38(4) : 571-581.
2. Goubara & Takasaki (2004) Pollination effects of the sweat bee Lasioglossum vilosulum trichopse on genic-male lettuce, Appl. Entomo. Zool. 39(1) : 163-169, 2004.
3. Lindqvist, K. (1960) Inheritance studies in lettuce, Heriditas 46 : 387-470.
4. Michelmore, R.W. et al. (1987) Genetic analysis of factors for resistance to downy mildew in lettuce, Plant Pathol., vol. 36, no 4 : 499-514.
5. Michelmore, R.W. et al. (1993) Development of reliable PCR-based markers linked to downy mildew resistance genes in lettuce, Theor. Appl . Genet., vol. 85, No 8 : 985-993.
6. Ryder, E. J. (1963) An epistatically controlled pollen sterile in lettuce (Lactuca sativa L.), Proc. Am. Soc. Hort. Sci. 96 : 826-828.
7. Ryder, E.J. (1967) A recessive male sterile gene in lettuce, Proc. Am. Soc. Hort. Sci 91 : 366-368.
8. Ryder, E.J. (1971) Genetic Studies in Lettuce {Lactuca sativa L.), J.Amer.Soc .Hort Sci 96(6) 826-828.
9. Ryder, E.J. (1979) Leafy Salad Vegetables, Avi . Pub. Co. , page 30.
10. Maisonneuve, B. et al. (1994) Rapid mapping of two genes for resistance to downy mildew from Lactuca serriola to existing clusters of resistance genes, Theor. Appl. Genet. 89 : 96-104.
11. Kuang H., et al. (2004) Multiple genetic processes result in heterogeneous rates of evolution within the major cluster disease resistance genes in lettuce, The plant cell, Vol. 16, 2870-2894.
12. Kesseli R.V., et al. (1994) Analysis of a detailed linkage map of Lactuca sativa (lettuce) constructed from RFLP and RAPD markers, Genetics 136: 1435-1446.
13. Altman D. G., Bland J.M., 1994a. Statistics Notes: Diagnostic tests 1: sensitivity and specificity. British Medical Journal, 308:1552.
14. Altman D. G., Bland J.M., 1994b. Statistics Notes: Diagnostic tests 2: predictive values. British Medical Journal, 30:102.
15. Doyle J. J., and Doyle J. L., 1990. Isolation of plant DNA from fresh tissue. Focus 12, 13-15.
16. Edwards K., Johnstone C. and Thompson C, 1991. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acid Res 19:1349.
17. Michelmore R.W., Paran I., Kesseli R.V., 1991. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA 88:9828- 9832.
18. Paran I., Kesseli R., Michelmore R.W., 1991. Identification of restriction fragment length polymorphism and random amplified polymorphic DNA markers linked to downy mildew genes in lettuce, using near-isogenic lines Genome 34:1021-1027.
19. Stam P., & J.W. van Ooijen, 1996. JoinMap, version 2.0. Software for the calculation of genetic linkage maps . Ed. CPRO-DLO, 60 pp.
20. Tomas H.T., and Tanksley S.D., 1989. A rapid and inexpensive method for isolation of total DNA from dehydrated plant tissue. Plant Mol Biol Rep 12:106-109.
21. Welsh J, McClelland M., 1990. Fingerprinting genomes using PCR with arbitrary primers . Nucleic Acids Res, 18:7213-7218.
22. Williams J.K.G., Kubelik A.R., Livak K. J., Rafalsky J.A., Tynger S.V., 1990. DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res, 18:6531-6535.
23. Williams J. K. G., Reiter R. S., Young R.M., Scolnik P.A., 1993. Genetic mapping of mutations using phenotypic pools and mapped RAPD markers. Nucleic Acids Res., 21(11): 2697- 2702.
【技術分野】
【0001】
本発明は、雑種ラクツカ・サティバ(Lactuca sativa)種子及び当該種子の作製のための方法に関する。本発明はまた、雑種ラクツカ・サティバ植物、及び雑種ラクツカ・サティバ植物由来の細胞に関する。
【背景技術】
【0002】
雑種強勢(Heterosis又はhybrid vigour)、恒常性(異なる環境における植物の安定性)、虫、真菌、細菌又はウイルスへの耐性、又は非生物ストレスへの適応、例えば極限温度、すなわち5℃未満又は30℃超の温度への耐性、又は微光レベルへの耐性を有する遺伝子蓄積の可能性は、雑種植物由来の種子を大スケールで作製するための、いくつかの理由を構成する。
【0003】
特に、ラクツカ・サティバ(栽培品種レタス)において、問題の遺伝子が、同じ遺伝子座(各対立遺伝子の一方)又は非常に近接する遺伝子座に位置するため、同型接合系統における特定の耐性遺伝子を集積することが、不可能又は非常に困難であることが示されている。
【0004】
雑種F1栽培レタスを得ること、特に同じ遺伝子座の様々な対立遺伝子、又は非常に近接する遺伝子座によりもたらされる、農業で注目の複数の相互優性(co-dominant)又は優性遺伝子を蓄積する、大スケールの雑種作製をさせることについては、大きな関心を集めるであろう。
【0005】
ラクツカ(Lactuca)属は100種以上が含まれており、例えばラクツカ・サティバ(レタス、Lactuca sativa)(栽培品種)、ラクツカ・サリグナ(Lactuca saligna)(野生型種)、ラクツカ・セリオラ(Lactuca serriola)(野生型種)及びラクツカ・ビロサ(Lactuca virosa)(野生型種)がある。
【0006】
ラクツカ・サティバ(レタス)は、種子作製は、98%が自己授粉による、ほとんどが自家生殖の二倍性種(2n=18)である。雄蕊(雄器官)は集団を形成し、葯の花粉嚢を開く雄蕊管を形成する。雌蕊(雌器官)は子房、花柱、及び二分された柱頭から構成される。開花時は、雄蕊管内部に花柱が伸びて、柱頭が花粉で覆われ、植物が自己受精する。一般的に1つのラクツカ・サティバ植物は、栽培条件次第で、0.5〜6グラム又は最大で10グラムもの種子を作製し、その1グラムには一般的に600〜1000個の種子が含まれる。
【0007】
雑種植物の大スケールでの作製のための従来技術は、互いに近接する「親」として使用される2つの変種を栽培し、授粉のための虫を使用することからなり、雄性不稔性をもたらす一方の親が、自己授粉による汚染(contamination)を防止する。その後、雄性不稔性親から種子を収穫する。
【0008】
この技術は、その組(parcel)の均一性を維持しながら、各親において異種接合状態で存在する注目の遺伝子を蓄積でき、そのF1世代は収穫段階で(開花前に)100%異種接合体の表現型である。
【0009】
前記技術を用いて大スケールで栽培雑種レタス(ラクツカ・サティバ)を作製することは非常に困難であることが観察されている。これはラクツカ・サティバの花が、通常の授粉する虫、例えば特定種のハチ(ミツバチ)又はバンブルビー(bumble bee)(Bombus種)がまだ活動しない日中(早朝)の時間帯に、一度だけ、たった数時間、例えば1〜4時間しか開花しないことが原因である。
ラクツカ・サティバはまた、風による授粉はしない。
【0010】
すなわち、雑種レタスの作製のために使用できる授粉する虫を確認することは、多数の問題に直面する。具体的には:
開花の期間が短く且つ早朝であるために、虫が花を訪れることの確認を困難にする。
開花が1度しかないために、最大数の雄性不稔花の受粉を確実にするためには、選択された授粉する虫が花を毎日訪れる虫でなければならない。
【0011】
さらに、F1雑種の作製は、「雄」系及び「雌」系の供給が必要である。「雌」系は、当該植物の手作業で去葯することにより得ることができる。ただし、レタスの場合は、開花が1回で且つ非常に短期間(数時間)なので、この方法はかなりの労力を要するため、流通スケールでの実施は困難である。
【0012】
「雌」系はまた、葯の不存在、空の葯又は生育不能花粉により、それ自身が発現する不稔性の遺伝子型(又は核型)又は細胞質型を誘導することにより得ることができる。
【0013】
その作業において、Goubara及びTakasakiは、雑種ラクツカ・サティバレタスの作製のために使用できる、可能性ある授粉する虫を確認する目的で、野外及び閉鎖条件で実験を行った(Appl Entomo Zool 38(4), 571-581, 2003 [1])。22種の虫が観察され(21種のハチ及び1種の吸蜜(nectar-feeding)ハエ種、シルフィダエ・エリスタリス・テナクス(syrphidae eristalis tenax))、そのうちハチのラジオグロッサム・ビロスラム・トリコプセ(lagioglossum villosulum trichopse)は最も可能性ある授粉する虫であることがわかった。
【0014】
Goubara及びTakasakiはまた、閉鎖環境中にハチのラジオグロッサム・ビロスラム・トリコプセを存在させて、雄性不稔性遺伝子を有するレタスと雄性稔性レタスとの間で小スケールのハイブリダイゼーション試験を行った。その著者等は、雑種F1レタスは得られたが収率が非常に低かったことを報告している。
【0015】
現時点では、授粉する虫を使用する雑種ラクツカ・サティバの作製実験で他に知られるものはない。
今日、F1雑種レタス変種で、流通スケールで作製されているものはない。
【0016】
本発明との関連において、以下の用語は以下の意味を有する。
「栽培レタス」は、ラクツカ・サティバ種を意味する。基本的な栽培レタスの5つの栽培品種群(cultigroups)が存在する(図1を参照):ラクツカ・サティバ変種 アングスタナ(angustana)(アスパラガス(asparagus)レタス); ラクツカ・サティバ変種 カピタタ(capitata)(バターヘッド(butterhead)レタス); ラクツカ・サティバ変種 クリスパ(crispa)(バタビア(batavia)又はアイスバーグ(Iceberg)レタス); ラクツカ・サティバ変種 ロンギフォリア(longifolia)(ロマイネ(Romaine)レタス)及びラクツカ・サティバ変種 アセファラ(acephala)(フリシー(frisee)レタス、カッティング(cutting)レタス)。本発明は、これらの様々な型のレタスの各々の使用を包含する。
【0017】
「授粉(受粉)(pollinization)」は、葯からの花粉を、同じ花又は別の花の柱頭に移転することを意味する。この生殖系は花を有する植物の繁殖の好ましい態様である(被子植物及び裸子植物)。花粉の粒を柱頭に到達させ、その後、それを受精させるために、花柱を通って、胚珠に到達する花粉管を形成させる。
【0018】
「自己授粉」は、自身の持つ花粉を用いる個体又は生物型の授粉を意味し、その結果当該個体は自己授粉したと言われる。
「自家生殖の」なる用語は、2つの配偶子が同じ個体に由来する、植物の自己授粉する能力を意味する。
「他殖性の」は、ある個体の花が他の1又は複数の個体からの花粉で受精する表現型を意味する。
「授粉する虫」は、花の研究において(例えば蜜を研究する場合)、雄蕊を摺ることにより花粉の数粒を収穫し、その後これを別の花に置いてくる虫を意味する。
【0019】
「遺伝子座」は、染色体上での遺伝子又は対立遺伝子により占められる場所を意味する。
「対立遺伝子」は、所与の染色体位置(遺伝子座)に位置する特定の遺伝子内での変異である。遺伝子の異なる対立遺伝子は、異なる形質の発現をもたらす。
【0020】
「クラスター」は、同じ染色体上に互いに近接して位置する2以上の遺伝子を意味する。レタスにおいて確認された耐性遺伝子のクラスターは、特にKesseli等により報告されている[12]。
「優性遺伝子」は、1対の2つの染色体に存在するか1つだけに存在するかにかかわらず、表現型を提供する遺伝子を意味する。
「劣性遺伝子」は、それが2つの相同染色体のうちの各々に存在する場合、1の表現型のみを提供する遺伝子を意味する。
「相互優性(co-dominanace)」は、2つの対立遺伝子が決定する表現型的特徴の、一方及び他方を発現する遺伝子特性を意味する。異種接合体において、物質が2つの相互優性の対立遺伝子を有すると、遺伝子型はこれらの遺伝子によりもたらされた情報に従ってその表現型で完全に発現する。
「異種接合体」は、染色体の同じ対の既定の遺伝子座上の2つの異なる対立遺伝子を有する細胞又は個体を意味する。
「同型接合体」は、染色体の同じ対の規定の遺伝子座上の2つの同一の対立遺伝子を有する細胞又は個体を意味する。
「雑種」なる用語は、異なる遺伝子構成の、好ましくは同一種由来の個体間の交配産物を意味する。
【0021】
「F1雑種」なる用語は、異なる遺伝子構成の個体間の交配から得られる第一世代を意味する。結果としてF1雑種は、少なくとも1の遺伝子の異種接合体である。
「戻し交配」は、雑種とその親の一方との間の交配を意味する。
「栽培品種」は、ある変種を意味する。
「遺伝子型」は、個体からもたらされた遺伝物質であって、その遺伝形質を構成するものの全てを意味する。
「表現型」は、遺伝子型の発現比率と外部環境により決定される現象の両方に対応する、個体の、明らかな形態的又は機能的特徴の組を意味する。
「農業で注目の表現型」なる用語は、例えば2つの同型接合遺伝子型間での交配から得られた表現型であって、農業的観点から注目の特性、例えば様々な病原菌又は虫への耐性の蓄積、雑種強勢(すなわち、雑種形質の平均は、2つの親のものの平均より高い)、恒常性、非生物ストレスに適応する能力、形態学的特徴、例えば色、形、葉の柔軟性又は剛性の性質、栄養組成物又は植物の味質等を伴う表現型を意味する。
「雑種強勢(heterosis又はhybrid vigour)」とは、F1雑種が、一又は複数の特徴の点で、特に強勢(vigour)に関して、その最良の親よりも顕著に優勢である表現型を意味する。
「恒常性」とは、環境又は複数の環境的特徴まで適応する、植物の能力を意味する。
「不稔性雄」とは、花の雄的要素の不稔性が原因で、自己授粉により繁殖できない植物を意味する。例としては、花粉が非機能的である、又は雄性繁殖器官、例えばタペータム構造的異常がある可能性がある。
「細胞質雄性不稔性」とは、細胞質型の遺伝により、その母から均一に遺伝する不稔性を意味する。
「核(又は遺伝子的)雄性不稔性」とは、劣性遺伝又は優性遺伝のいずれに依存してもよいが、核のDNAによりもたらされる、メンデル遺伝による不稔性を意味する。
「一遺伝子的雄性不稔性」とは、単一の遺伝子によりもたらされる雄性不稔性を意味する。
「多遺伝子的雄性不稔性」とは、複数の遺伝子によりもたらされる雄性不稔性を意味する。
「耐性」とは、病原又は有害生物にとって、類似する環境且つ圧力条件下で、感受性の高い変種と比較して、既定の病原又は有害生物及び/又はそれが引き起こす障害の増殖及び発展を制限する様々な能力を意味する。ただしこれらの変種は、病原又は有害生物の圧力が高い場合、病気のいくつかの症状又はいくつかの障害を発現する可能性がある。
「標準的又は高い耐性」とは、感受性の高い変種と比較して、通常の圧力条件下で既定の病原又は有害生物の増殖又は発展を強力に制限する様々な能力を意味する。ただし、これらの変種は、病原又は有害生物の圧力が高い場合、症状又は障害を発現することもある。
「中程度又は調整的な耐性」とは、既定の病原又は有害生物の増殖及び発展を制限するが、高い/標準的な耐性変種と比較すると、症状又は障害をより発現し得る能力を意味する。中程度の耐性変種は、類似する環境及び/又は病原又は有害生物にとっての圧力条件下で、感受性の高い変種で観察されたものより、重篤性の低い症状又は障害を示す。
「分子マーカー」とは、個体の全ゲノムにおいて確認され得る、及び注目の遺伝子を局在化させるために、又は親生物からのある特定の特徴が遺伝した個体であるかを検証するために使用できる、DNAの特定の断片を意味する。それはコード配列であってもそうでなくてもよい。遺伝的交配において、注目の遺伝子は、一般的に分子マーカーと連結した状態である。つまり、分子マーカーの検出とは、注目の遺伝子が存在する個体が、その遺伝子の配列を知る必要なく、セレクションできることを意味する。
【0022】
農学分野において、分子マーカーの使用は、植物が所望の特徴を保持することを、セレクションの際、迅速に試験できることを意味する。特定の場合において、形質に関連するマーカーの存在により、特定の試験又は表現型の観察が不必要になる。具体的には、雄性不稔性遺伝子のための、好ましくは雄性不稔性の優性遺伝子のための、特定の分子マーカーを使用することで、開花前の雄性不稔性植物を早期にセレクションできる。このセレクションは、例えば雄性不稔性の優性遺伝子を使用する場合、雑種種子の作製のための、「雌」(雄性不稔性)と推測される親として使用される植物の集団の一部を形成し得る、雄性稔性植物を除去することができ、その結果、これらの雄性稔性植物の自己受精に結びつく汚染のリスクを低減できる。
【発明の概要】
【課題を解決するための手段】
【0023】
本発明は、雄性不稔性遺伝子型を有するとともに、雄性不稔性に関与せず、且つその植物に検出可能な表現型を提供する少なくとも1つの遺伝子との異種接合体であることを特徴とする、雑種ラクツカ・サティバ植物由来の種子、植物及び細胞に関する。
【図面の簡単な説明】
【0024】
【図1】ラクツカ・サティバの系統図。
【図2】試行1で「雄」及び「雌」親として使用されたレタスの性質。
【図3】試行2で「雄」及び「雌」親として使用されたレタスの性質。
【図4】試行10から得られた50個の雑種植物の遺伝子型決定。TagIとの消化処理後の、分子マーカーSCW09の増幅プロファイル。SCW09マーカーは、ブレミア・ラクツカエに耐性のあるDm6及びDm8を標識する。A=Dm18+/Dm18+コントロール、B=Dm18−/Dm18−コントロール、C=Dm6+コントロール、D=TaqIでの消化処理なし。
【図5】試行10から得られた50個の雑種植物の遺伝子型決定。ブレミア・ラクツカエに耐性のあるDm3遺伝子を標識する分子マーカーB1の増幅プロファイル。A=Dm3+コントロール、B=Dm3−コントロール。
【図6】マーカーBA05−675(矢印)を伴うOPBA05プライマーの電気泳動プロファイル。1.0%アガロースゲル電気泳動、190Vで1時間45分。M:100bpラダー分子量マーカー(Pharmasica Biotech、27.4001.01参照)。
【図7】レタス、ラクツカ・サティバにおけるMs7遺伝子座に関連するBA05−675分子マーカーのマッピング(Kosambi関数を用いて計算した距離)。
【図8】レタス、ラクツカ・サティバにおけるMs7遺伝子座に関連するBA05−675分子マーカーの5'〜3'ヌクレオチド配列(配列番号2)。
【発明を実施するための形態】
【0025】
ラクツカ・サティバ植物由来の、種子、植物又は細胞によりもたらされた雄性不稔性は、核又は細胞質起源であってよい。それが細胞質性である場合、その不稔性は雌親から遺伝する。これは細胞質に存在するミトコンドリアと核遺伝子との間の相互作用に起因することが多い。細胞質雄性不稔性は、雄親のゲノムで稔性を回復する遺伝子の不存在下で、100%雄性不稔性の表現型を有する子孫の出現により特徴付けられる。雄性不稔性は、「雄」親のゲノムにおける稔性回復遺伝子 (restoring gene (Rf))の存在によって、その子孫で回復してもよい。
【0026】
より具体的には、本発明は雄性不稔性遺伝子型が、少なくとも1の核遺伝子を含み、且つその種子、細胞又は植物が、雄性不稔性の原因であるその1又は複数の遺伝子の異種接合体である場合に関する。
核雄性不稔性は、その核のDNAから遺伝した雄性不稔性の、1又は複数の遺伝子により引き起こされ、この1又は複数の遺伝子は、優性でも劣性でもあり得る。
【0027】
これら雄性不稔性の遺伝子には、Ryderによる(J Am Soc Hort Sci 96(6), 826-828, 1971 [8])に記載の、優性核遺伝子Ms7がある。当該雄性不稔性植物は、異種接合体Ms7ms7である。稔性植物は、同種接合体ms7ms7である。優性のMs7Ms7同種接合体の獲得は、不可能とは言わないまでもかなり困難である。さらに、Ryderによればこの雄性不稔性の優性形質は、雑種レタスの作製のためにこの遺伝子を使用することについて問題がある。本発明の方法を使用して獲得されるラクツカ・サティバ種子のサンプルで、NCIMB(NCIMB Ltd., Ferguson Building, Craibstone Estate, Bucksburn, Aberdeen, A21 9YA, スコットランド、英国)に2007年2月13日に、受託番号NCIMB 41470で寄託されたサンプルを、Ms7遺伝子の源とした。さらに、本発明者等により開発された分子マーカーRAPD BA05-675 (配列番号2)は、次の交配のために使用されるはずの雄性不稔性植物を確認するための、開花前の初期段階で実施されるべき分子試験を可能にするので、雄性不稔性レタス植物Ms7のセレクションを補助の点で特に注目される。この図8に示される配列のマーカーの検出は、全てのレタス類型を共に考慮すると、平均で96%の事例において雄性不稔性植物の確認ができる(表18)。
【0028】
雄性不稔性のための3つの劣性遺伝子、ms1、ms2及びms3はまた、Lindqvist の(Heriditas 46 :387- 470, 1960 [3])により、他の3つ、ms4、ms5及びms6は、Ryder (Proc. Am. Soc. Hort . Sci 83 : 585-595, 1963 [6], Proc . Am. Soc. Hort. Sci 91 : 366-368, 1967 [7])により確認されている。ただし、1979年に公表された彼の研究(Leafy Salad Vegetables, p 30 [9])で、Ryderは、不稔性遺伝子ms1、ms2、ms3、ms4、ms5、ms6及びMs7は全て核性であるため、F1雑種を作製するために使用できることが全く見込めない。これにもかかわらず、本発明者等は、核起源の雄性不稔体を使用する、雑種の種子、植物及び細胞を獲得することに成功した。
【0029】
核雄性不稔性は、多遺伝子的であっても一遺伝子的であってもよい。多遺伝子的である場合、例えば上記記載のms1、ms2及びms3遺伝子の集積による、劣性遺伝子の複合体を使用して獲得してもよい。好ましくは、本発明は、雄性不稔性が一遺伝子的である場合に関する。
核雄性不稔性は、優性であっても劣性であってもよい。具体的には、本発明は雄性不稔性が優性の一遺伝子的である場合に関する。より具体的には、本発明は雄性不稔性が、上記記載の優性のMs7遺伝子により提供される場合に関する。
【0030】
好ましくは、本発明の種子、植物又は細胞のゲノムは、650〜700ヌクレオチドの、例えば655〜695、660〜690、665〜685、670〜680、673〜677ヌクレオチドの、又は674、675又は676ヌクレオチドの二本鎖DNA配列を含んでなり、ここで、各2つの5'末端は、配列「5' TGCGTTCCAC 3'」(配列番号1)から開始する。好ましい実施態様によれば、本発明の種子、植物又は細胞のゲノムは、図8(配列番号2)に示したヌクレオチド配列を含んでなるか、又は1〜10個の、好ましくは1〜5個の又はより好ましくは1〜3個のヌクレオチドが他のものにより置換、削除又は付加される、この配列に由来する配列を含んでなる。
【0031】
好ましくは、本発明は、1又は複数の異種接合遺伝子により提供された検出可能な表現型が、農業で注目の表現型、例えば様々な病原又は虫に対する耐性、雑種強勢、恒常性(異なる環境での植物の安定性)、非生物ストレス、例えば極限温度又は微光レベルへの適応、雑種由来の変種のものより高い収率、形態的特長、例えば色、形、大きさ、葉の柔軟性もしくは剛性の性質、栄養組成物又は植物の味質等である場合に関する。
【0032】
さらに、驚くべきことに本発明者等は、異なるレタス間での様々な交配から得られる本発明の種子由来の植物は、冬期に且つ温暖な地域での栽培で、同じ条件下で栽培した親植物より平均で7〜10日早く成熟に到達し、より早く生育することを観察した。F1雑種の組の中で(intraparcel)発育の均一性(1組当たりおよそ30植物の株でのもの)は、組系列(親植物)のものより高いことも観察された。言い換えると、同じ組由来の全ての植物と比較して遅延又は向上した発育を示す植物の数は、変種系列由来の組よりも、F1雑種の組の方がかなり少なかった。結果として、これらのF1雑種の特徴は、レタス作製周期を短縮できるだけでなく、収穫を短期間に集中させることができる。
【0033】
農業で注目の表現型は、ウイルス、細菌、虫又は真菌による感染に対して、より具体的には以下の真菌:ブレミア・ラクツカエ(Bremia lactucae)、フサリウム・オキシスポラム(Fusarium oxysporum)、スクレロチニア・マイナー(Sclerotinia minor)もしくはスクレロトラム(sclerotorum)、又はボトリチス・シネレア(Botrytis cinerea)、リジクトニア・ソラニ(Rhizictonia solani)、ミクロドシウム・パナットニアナム(Microdochium panattonianum)、ベルチシウリウム・ダリアエ(Verticiulium dahliae)又はエリシフェ・チコセアラム(Erysiphe chicocearum)又はピチウム・トラケイフィラム(Pithium tracheiphilum)の1種に対して、以下の虫:ナソノビア・リビスニグリ(Nasonovia ribisnigri)、マイザス・ペルシカエ(Myzus persicae)、マクロシファム・ユーフォルビア(Macrosiphum euphorbia)、ネマトドス・プラチレンカス(Nematodes pratylenchus)、又はネコブ線虫(meloidogyne)、ハモグリバエ(leafminers):リリオミザ・フリドブレンシス(Liriomyza huidobrensis)又はペンフィガス・ブサリウス(Pemphigus busarius)の1種に対して、以下の細菌:シュードモナス(pseudomonas)、キサントモナス(xanthomonas)又はリゾモナス(rhizomonas)の1種に対して、又は以下のウイルス:LMW(レタスモザイクウイルス)、TSWV(鉢植えトマト立ち枯れ病ウイルス)、「ビッグ・ベイン」(LBVV(レタスビッグベインウイルス)及びMILV(ミラフィオリ(mirafiori)レタスウイルス)からなる)、TBSV(トマト茂み(bushy)発育阻害ウイルス)、LNSV(レタス壊死発育阻害ウイルス)、TuMV(カブモザイクウイルス)、CMV(カボチャモザイクウイルス)又はBMYV(ビート西洋黄色(beet western yellows)ウイルス)の1種に対する、標準的又は中程度の耐性である1又は複数の遺伝子から提供されてもよい。
【0034】
農業で注目の表現型は、ウイルス、細菌、虫又は真菌による感染に対して、標準的又は中程度の耐性のある1又は複数の遺伝子から提供され、この表現型を提供する当該又は前記の標準的又は中程度の耐性のある遺伝子は、具体的に、ブレミア Dm10、R17m、Dm5、Dm8、R36、R37(ラクツカ・サティバのクラスター1上に位置する遺伝子)、Dm1、Dm2、Dm3、Dm6、Dm14、Dm15、Dm16、Dm18、(ラクツカ・サティバのクラスター2上に位置する遺伝子)、Dm4、Dm7、Dm11、R38(ラクツカ・サティバのクラスター4上に位置する遺伝子)への耐性遺伝子;又はクラスター1上に位置するTuMVに耐性のあるTu遺伝子;クラスター2上に位置するナソノビアに耐性のあるNr遺伝子;又はクラスター4上に位置するLMVに耐性のあるmol.1及びmol.2遺伝子、から選択してもよい。上記のクラスター1、2及び4は、Michelmore R.W.による(Plant Pathol, 1987, vol. 36, no4 : 499-514 [4], Theor . Appl. Genet., 1993, vol. 85, No 8 : 985-993 [5])で定義されている。
【0035】
より具体的には、農業で注目の表現型は、レタスを侵す病気、ブレミア・ラクツカエ、白色を引き起こす真菌、枝内面に落下粉末(powdery down on the inner face of the limb)の1つに対し、標準的又は中程度の耐性のある、1又は複数の遺伝子により提供されてもよい。さらに、ブレミアは、高度に適応でき、植物繁殖者によってその変種に既に導入された耐性を征服できる、新規な品種の出現をもたらす。
【0036】
より好ましくは、本発明は、ラクツカ・サティバ植物の種子、植物又は細胞が、雄性不稔性に関与せず、且つ検出可能な表現型を提供する、少なくとも2つの遺伝子の異種接合体である。さらにより好ましくは、検出可能な表現型を植物に提供する2以上の遺伝子は、同じクラスターに、例えばラクツカ・サティバのクラスター1、2又は4の耐性遺伝子のクラスター上に位置する。
【0037】
本発明はまた、雄性不稔性遺伝子型であるとともに、雄性不稔性に関与せず、且つ当該種子由来の植物を検出可能な、少なくとも1つの遺伝子の異種接合体であり、任意に上記の種子の追加の特徴を有する、雑種ラクツカ・サティバ種子集団であって、少なくとも105個の種子、好ましくは少なくとも106、及びより好ましくは少なくとも107個の種子を含んでなるような、種子集団に関する。
【0038】
種子を2つのラクツカ・サティバ植物の交配により得る場合、このうちの1つ(「雌」親)は1又は複数の優性遺伝子により提供される核雄性不稔性をもたらし、「雌」親として使用される植物は、不稔性遺伝子の異種接合体であることが必要である。実際は、1又は複数の優性の雄性不稔性遺伝子の同種接合体となるものとして、交配により得られる「雌」親として使用される植物由来の当該2つの植物は、自身が雄性不稔性の遺伝子のキャリアとなるはずである。ただし、その場合はこれらの2つの植物間の交配は不可能である。
【0039】
結果として、一方(その「雌」親は)は1又は複数の優性遺伝子から提供される核雄性不稔性のキャリアである、2つの型のラクツカ・サティバ植物の間の交配から得られる本発明の種子集団は、減数分裂中の染色体の分離現象が原因で、1又は複数の雄性不稔性の優性の遺伝子を保有するラクツカ・サティバ種子、及び当該1又は複数の遺伝子を保有しない種子からなる。一般的に「雌」親によりもたらされる雄性不稔性が一遺伝子的且つ優性である場合、不稔性雄性種子の割合は少なくとも40%である。
【0040】
本発明は、発明者等による以下の観察により生まれた。すなわち双翅目はラクツカ・サティバの花に定常的に授粉する虫ではないが、さらにその蜜を餌とすることは知られておらず、これらの虫、具体的な種としては、カリホラ・ボミタリア(Calliphora vomitaria)、カリホラ・エリトロセファラ(Calliphora erythrocephala)、及びルシリア・カエサル(Lucilia Caesar)は、閉鎖環境に過剰数を導入すると、ラクツカ・サティバの花粉授粉者としてふるまうという観察である。
【0041】
さらに本発明は、具体的には雑種ラクツカ・サティバ植物を得る観点で、閉鎖環境で雄性稔性植物により雄性不稔性ラクツカ・サティバ植物の授粉を行うための、双翅目の使用に関する。好ましくは、使用される雄性不稔性ラクツカ・サティバ植物は、単一の優性遺伝子により、好ましくは上記の雄性不稔性Ms7によりもたらされる雄性不稔性を有する。使用される稔性雄性植物は、好ましくはラクツカ・サティバ植物であり、より好ましくは栽培品種である。
【0042】
本発明の閉鎖環境は、具体的には温室、ケージ又はトンネルであってもよく、好ましくは30m2以上の、より好ましくは300m2以上の、例えば30〜1500m2又は50〜1000m2の表面積である。閉鎖環境の高さは、通常、2m〜4mの範囲であり、好ましくは2.5m〜3.5m、例えば3mである。具体的には、換気、散水、温度制御及び光レベル制御のための手段があってもよい。好ましくは、当該閉鎖環境は、虫に対して密閉するように封じられる囲いである。
【0043】
使用される双翅目は、好ましくは1m2当たり少なくとも100匹、好ましくは1m2当たり少なくとも250匹、例えば1m2当たり100〜1000匹の濃度で存在させるべきである。この濃度はまた、1m3当たりの双翅目の数として決定してもよく、好ましくは、1m3当たり少なくとも25匹、より好ましくは1m3当たり少なくとも50匹、さらに好ましくは1m3当たり少なくとも75匹、例えば1m3当たり25〜500匹、1m3当たり75〜250匹でもよい。
【0044】
本発明はまた、雑種ラクツカ・サティバ種子を獲得するための方法であって、
表現型的に「雌」親として使用される雄性不稔性ラクツカ・サティバ植物、及び表現型的に「雄」親として使用される雄性稔性ラクツカ・サティバ植物を、互いに近接した状態で、閉鎖環境中で栽培するステップであって、2つの親の一方が、雄性不稔性以外の検出可能な表現型をそれに提供する遺伝子の同種接合体であるという追加的な特徴を有し、他方の親はこの遺伝子を保有しないものであるステップ、
植物の開花の際に、1m2当たり100匹以上の濃度で、好ましくは1m2当たり少なくとも250匹の濃度で、閉鎖環境へ導入する双翅目を用いて授粉するステップ、及び
雄性不稔性植物により作られる種子を収穫するステップ、
を含んでなる方法に関する。
【0045】
好ましくは、第二の親はまた、他方の親からはもたらされない、雄性不稔性以外の検出可能な表現型をそれに提供する少なくとも1つの遺伝子の異種接合体である。好ましくは、当該閉鎖環境は、虫に対して密閉するように封じられる囲いである。
【0046】
最良の結果は双翅目が過剰の場合に観察され、さらにその濃度は、好ましくは1m2当たり400匹以上、より好ましくは1m2当たり500匹以上である。
【0047】
双翅目は卵期、幼虫期、蛹期又は成虫期に導入してよい。好ましくは、これらを蛹の状態で導入する。
使用する双翅目は好ましくは短角類(brachycera)であり、例えばシクロラファ・ブラキセラ(cyclorhapha brachycera)、より好ましくはイエバエ科(Muscidae)又はクロバエ科(Calliphoride)由来の短角類、例えばカリホラ・ボミタリア(Calliphora vomitaria)、カリホラ・エリトロセファラ(Calliphora erythrocephala)又はルシリア・カエサル(Lucilia Caesar)である。
【0048】
双翅目の導入が、少なくとも1週間に1回、好ましくは1週間に2回、少なくとも3〜4週にわたって繰り返されると、通常より良好な収率が得られる。
同様に、閉鎖環境中での雄性不稔性植物の数が、雄性稔性植物の数より大きくなると、通常より良好な収率が得られる。閉鎖環境中の雄性不稔性植物の数は、例えば少なくとも200、及び雄性稔性植物の数は、例えば少なくとも1000である。
【0049】
本発明の方法の実施において、2つの親の開花が同時に起こることが必要である。これはこの形質のためのセレクションにより、又は適切な栽培技術を使用することにより達成できる。
【0050】
好ましくは、雌親として使用する植物の雄性不稔性は、一遺伝子的、優性及び核性であり、より好ましくは遺伝子Ms7により提供される。
好ましくは、雌親として使用する植物のゲノムは、650〜700ヌクレオチドの、例えば655〜695、660〜690、665〜685、670〜680、673〜677ヌクレオチドの、又は674、675又は676ヌクレオチドの二本鎖DNA配列を含んでなり、ここで、各2つの5'末端は、配列「5' TGCGTTCCAC 3'」(配列番号1)から開始する。好ましい実施態様に従うと、「雌」親として使用する植物のゲノムは、図8に示すヌクレオチド配列(配列番号2)を含んでなるか、又は1〜10個の、好ましくは1〜5個又はより好ましくは1〜3個のヌクレオチドが他のものにより置換、削除又は付加される前記配列に由来する配列を含んでなる。
【0051】
上記の説明の通り、減数分裂中の染色体の分離現象が原因で、従来の交配技術では、雄性不稔性の優性遺伝子を保有し、「雌」植物として使用される植物の同種接合体集団を得ることはできない。結果として、雄性不稔性の優性遺伝子を使用する場合、雌親として使用される植物は、
核雄性不稔性の優性遺伝子の異種接合体であるラクツカ・サティバ植物と、不稔性遺伝子を保有しない雄性稔性のラクツカ・サティバ植物との間で交配するステップ、
当該交配から得られる種子を栽培するステップ、及び
雄性稔性表現型を有する植物を除去するステップ、
を含んでなる方法により得ることができる。
【0052】
この雄性稔性表現型を有する植物を除去するステップは、例えば、雄性不稔性ラクツカ・サティバ植物を、雄性稔性ラクツカ・サティバ植物から識別可能とする可視的な特徴に基づき、手作業で行ってもよい。例としては、Ms7遺伝子に関連する不稔性の場合、雄性不稔性植物の花頭は、雄性稔性植物のそれより長時間開いており、その雄性不稔性植物は花粉を持たないという事実を有効に用いることにより、分類を行ってもよい。
【0053】
代替の実施態様に従えば、雄性稔性表現型の植物を除去するステップは、各植物のサンプルにおける雄性不稔性の優性遺伝子に特異的な分子マーカーの不存在を検出することにより行う。例えばそれは、約675塩基対(例えば650〜700、655〜695、660〜690、665〜685、670〜680、673〜677、674、675又は676塩基対)の長さの、2本鎖の各々の5'末端は、配列「5' TGCGTTCCAC 3'」(配列番号1)で開始する、RAPD(多形DNAのランダム増幅(random amplification of polymorphic))マーカーであって、例えば発明者等により開発されたマーカーBA05−675(配列番号2)又はSCAR(配列特徴付け増幅領域(sequence characterized amplified region))マーカー、CAPS(開裂増幅多形配列(cleaved amplified polymorphic sequence))マーカー、又はこのRAPDマーカーを用いて開発したSTS(配列タグ化部位(sequence tagged site))として一般的に設計された任意の他のマーカーがある。
【0054】
好ましい実施態様によれば、雄性不稔性の優性遺伝子の特異的分子マーカーの存在又は不存在の検出は、雌親として使用される植物の開花前に、好ましくは植物の生育の初期段階に、例えば1〜5葉期に、又はより好ましくは1〜2葉期に行う。
【0055】
さらに好ましくは、使用される分子マーカーは、少なくとも70%、75%、80%、85%、95%、98%又は99%、又は100%もの感受性のある雄性不稔性植物を検出でき、その感受性は、分子マーカーを有する雄性不稔性植物(真陽性)の数と、真陽性の数と分子マーカーを有さない雄性不稔性植物(偽陰性)の数の総和との間の比率として定義される(表17を参照)。
【0056】
一般的に、本発明の方法を用いて得られた種子のTSW(1000種子質量(thousand seed weight))は、「雄」親として使用される雄性稔性ラクツカ・サティバ植物の自己受精により得られた種子のTSWより、少なくとも10%、又は少なくとも20%又は少なくとも30%大きい。
【0057】
本発明はまた、上記記載の方法を用いて得ることができる種子集団で、少なくとも105個の種子、好ましくは106個の種子、及びより好ましくは107個の種子含んでなる集団に関する。
【実施例】
【0058】
実施例1:雑種レタス種子の獲得
「雌」親として以下に記載の種々の試行で使用される雄性不稔性ラクツカ・サティバ植物は、Ms7不稔性遺伝子を保有した。当該植物は、INRA、USDA等の種々の研究組織から入手できるMs7遺伝子(X Girelle 94-9538-1)を保有する温室バターヘッドレタスの第5代戻し交配(BC5)由来であった。
【0059】
Ms7遺伝子は優性であるため、Ms7遺伝子を保有する2つの植物間の交配は不可能である。結果として、雄性不稔性表現型を有する植物は、Ms7遺伝子の異種接合体である必要があり、Ms7/ms7(不稔性)とms7/ms7(稔性)植物との間で交配することによってのみ、得ることができる。
【0060】
結果として、雄性稔性植物を除去するための精製ステップが必要となる。この後者のステップは、Ms7/ms7植物が花粉を持たず、且つMs7/ms7植物の花頭はms7/ms7植物のそれよりも長期間開いたままであるため、可能となった。
【0061】
試行1
第一の試行の環境としては、
Nacre/Cambria Dm18/R38栽培品種(クラスター2からのDm18耐性遺伝子及びクラスター4からのR38耐性遺伝子)由来のバターヘッドレタスとの、Ms7バターヘッドレタスの第3代戻し交配(BC3)由来の450個の温室バターヘッドレタスラクツカ・サティバ植物は、ブレミア・ラクツカエ品種24(Bl24)に感受性の高い組み合わせであり、これが「雌」親として使用され(その約半分は不稔性雄);及び
BRA Dm18/R37温室バターヘッド栽培品種(クラスター2からのDm18耐性遺伝子及びクラスター1からのR37)由来の100個のラクツカ・サティバ植物は、B124に対して耐性のある組み合わせであり、「雄」親として使用された。
この2つの遺伝的父はまさに同時に開花し、これは受精を可能な限り均一にできることを意味する。
【0062】
植物を36m2の虫を密閉して封じる囲いの中に(湿度及び温度制御を備えるダッチ型温室で、11m×3.30m)、以下の通りに(図2を参照)配置した。
「雄」親として使用される植物を、密封囲いの端に沿って2列に配置し、植物は20cmずつ間隔をあけ、及び
「雌」親として使用される植物を、「雄」植物の2列の間であって、そこから50cmの距離の点に、7列で配列し、植物は15cmずつ、且つ列は25cmずつ間隔をあけた。
【0063】
播種は13週目に、植え付けは16週目に行った。
Ms7遺伝子の優性性質のための、雄性稔性表現型を持つ雌親として使用される植物の精製(除去)のためのステップが必要であった。本ステップは、Ms7/ms7植物が花粉を持たず、Ms7/ms7(雄性不稔性)植物の花頭が、ms7/ms7植物(雄性稔性)のものより長期間開いた状態であるという事実を利用して、主に26週と27週の間で行った。
雌親として使用される450植物のうち、217は雄性不稔性であり、効率的に使用された。
【0064】
2種類の双翅目クロバエ科の虫を授粉する虫として使用した。
「ウジ虫(maggots)」として知られる(キンバエ(bluebottle)又はニクバエ(blowflie)としても知られる)、カリホラ・ボミタリア及びカリホラ・エリトロセファラハエは、15〜20℃で取り扱い、及び
「ピンキーズ(pinkies)」として知られるラクツカ・カエサーハエ(キンバエ(greenbottle)としても知られる)は、20〜25℃で取り扱った。
授粉する虫は、28週〜31週の間に隔週で、ハエが1m2当たり300匹となる濃度で添加した。
自己授粉制御として使用される、雄性不稔性植物(「雌」親)及び雄性稔性植物(「雄」親)も、33週で収穫した。植物を乾燥させ、脱穀し、その後種子の洗浄を完了させるとともに残存した軽量ごみを除去するため、パルスエアのカラムを通し、重さと幅を調整した。
【0065】
試行1で得られた2500種子のサンプルを、2007年2月13日にNCIMB(NCIMB Ltd., Ferguson Building, Craibstone Estate, Bucksburn, Aberdeen, A21 9YA,スコットランド、英国)に、受託番号NCIMB 41470で寄託した。
【0066】
試行2
同様に第二の試行を、336m2の虫を密閉して封じる囲いの中で(長さ:56m、幅:6m、高さ:3m)、より大きなスケールで実施した。
【0067】
第二の試行の環境としては、
Nacre/Cambria Dm18/R38栽培品種との、Ms7バターヘッドレタスの第4代戻し交配(BC4)由来の30000個の温室バターヘッドレタスラクツカ・サティバ植物、ブレミア(Bl24に感受性の高い組み合わせ)を、「雌」親として使用し(その約半分は雄性不稔性である);及び
B124に対して耐性のある、BRA Dm18/R37温室バターヘッド栽培品種由来の1500個の対応する(parallel)植物を、「雄」親として使用した。
【0068】
植物を以下の通りに密封囲い中に配列した(図3を参照)。
「雄」親として使用される植物は4列に配置し、密封囲いの各端に沿って(80cmのところに)1列、中央に2列を(20cmの間隔をあけて)配置し、配置において植物はねじれており、15cmの間隔をあけ;及び
「雌」親として使用される植物を、稔性雄親の列の間にそこから65と70cmの距離の点に8列で配置し、植物は15cmずつ、列は25cmずつ間隔をあけた。
播種は12週目に、植え付けは16週目に行った。
【0069】
試行1と同様の方法で、雄性稔性表現型を持つ雌親として使用される植物の精製(除去)のためのステップが必要であった。本ステップは、Ms7/ms7植物が花粉を持たず、Ms7/ms7(雄性不稔性)植物の花頭が、ms7/ms7植物(雄性稔性)のものより長期間開いた状態であるという事実を利用して、主に26週と27週の間で行った。
雌親として使用される3000植物のうち、1250は雄性不稔性であり、効率的に使用された。
16のms7/ms7植物(雄性稔性)を保存したが、精製の間に、自己受精コントロールとして使用されるための、虫を通過させないようなネットで構成された、密封して封じられた囲いの中で自己受精した。
【0070】
使用した授粉する虫は試行1で使用したものと同じである。
授粉する虫は、25週〜29週の間に毎週、ハエが1m2当たり1100匹となる濃度で添加した。
【0071】
同時に、精製のステップで保存した、10の稔性植物(雄親)及び16の稔性ms7/ms7植物を、自己受精コントロールとして作用するための、虫を通過させないようなネットで構成された、密封して封じられた囲いの中で自己受精した。
【0072】
雄性不稔性植物(「雌」親)、並びに自己授粉制御として使用した精製ステップで保存した10の雄性稔性植物(雄親)及び16の稔性ms7/ms7植物を、33週で収穫した。
植物を乾燥させ、脱穀し、その後種子の洗浄を完了させるとともに残存した軽量ごみを除去するため、パルスエアのカラムを通し、重さと幅を調整した。
【0073】
得られた種子のサンプルを播種し、得られた植物のB124に対する耐性を、公式のGEVESプロトコル(Groupe d’Etude et de controle des Varietes et des Semences [Varieties and Seeds Study and Monitoring Group])に従って試験した。当該植物がB124に耐性であることが明らかになった。
【0074】
試行1及び2の結果
試行1及び2の結果を以下の表に示す。
表1:試行1及び2で収穫された種子の質量の比較
【表1】
【0075】
表2:試行1及び2で作成した、雑種種子及び自己受精コントロールに由来する種子の計測の比較
【表2】
【0076】
表3:試行1及び2で作成した、雑種種子及び自己受精コントロールに由来する種子のTSW(1000種子質量)の比較
【表3】
【0077】
考察
第一の試行では、217の雄性不稔性ラクツカ・サティバ植物(雌親)から730グラムの種子、すなわち1植物当たり3.36グラムの収率で得られ、自己受精コントロールの15.30%の比率に相当した(雄親)。
第二の試行では、1170の雄性不稔性ラクツカ・サティバ植物(雌親)から7.730kgの種子、すなわち植物あたり6.60グラムの収率で得られ、自己受精コントロールの2つ(「雄」親とms/ms雄性稔性「雌」親、精製の間保存され、虫密閉の下で受精した)の平均の50%の比率に相当した。
【0078】
第二の試行では、F1雑種を計測する有用種子の量が、自己受精したコントロールの10%以上であった。
2つの試行では、F1雑種のTSW(1000種子質量)が、自己受精コントロールの30%以上であった。
第二の試行で得られた種子由来の植物は、B124に耐性であった。
【0079】
試行3〜8
異なる種類のバターヘッド及びバタビアのレタスを用い、(Ms7遺伝子を保有するラクツカ・サティバ植物と稔性ラクツカ・サティバ植物との間の戻し交配に由来する50%の雄性不稔性である)「雌」親として使用される8つのラクツカ・サティバ植物と、「雄」親として使用される4つの雄性稔性ラクツカ・サティバ植物を各々含有する、換気された温室に置いた、虫試験用の3.3m×1.1mケージ中で、6つの同様の試行を同時に開始した。
播種を22週目に、植え付けを27週目に行い、31週目の開花の兆候で雄性稔性「雌」植物から雄性不稔性を分類し、ハエを32、33及び34週の3週間にわたり1週間当たりのある量、すなわちハエ約100匹/m2を添加し、37週目に収穫を行った。
【0080】
試行3〜8の結果
ケージ1: CHARLIN(68/12624)* CHARLIN (バターヘッド)
1種の雌性植物:9.20 g
1種の雄性植物:28.30 g
ケージ2:CHARLIN(BC/77)* CHARLIN (バターヘッド)
1種の雌性植物:5.45 g
1種の雄性植物:19.40 g
ケージ3:BRA 68/12588 (温室リンゴレタス)* BRA 68/12588
6種の雌性植物:
植物1:7.00 g
植物2:2.00 g
植物3:5.20 g
植物4:3.40 g
植物5:9.60 g
植物6:8.40 g
平均:5.9 g/植物
1種の雄性植物:34.40 g
ケージ4: BRA 68/12588 * BRA 68/12588
4種の雌性植物:
植物1:11.95 g
植物2:7.15 g
植物3:15.90 g
植物4:11.90 g
平均:11.25 g/植物
1種の雄性植物:29.00 g
ケージ5:BVA 68/12553 (温室バタビアレタス)* BVA 68/12553
5種の雌性植物:
植物1:.50 g
植物2:1.30 g
植物3:2.20 g
植物4:1.70 g
植物5:4.20 g
平均:3.30 g/植物
1種の雄性植物:3.60 g
ケージ6:BVA 68/12553 * BVA 68/12553
3種の雌性植物:
植物1:4.60 g
植物2:3.20 g
植物3:3.70 g
平均:3.80 g/植物
1種の雄性植物:23.40 g
【0081】
考察
「雌」親の産物の平均は、3.3g〜11.2g/植物であり、バタビア類型物(BVA 68/12553)は、約8.5g/植物のバターヘッド類型物(Charlin, BRA 68/12588)と比較して、かなり低い収率(約3.5g/植物)であり、当該類型物はその最終収率に寄与すると考えられる。
【0082】
試行9
新規な作製試行を、開放地面上の換気された温室内に置いた、直径0.40mの測定用の虫小さな試験用ケージ中で行った。
播種を22週目で、再植え付けを25週目で行い、30週目の開花の兆候で雄性稔性植物から「雌」植物を分類し、31週目で植え付けし、ハエを32、33及び34週の3週間にわたり1週間当たりのある量、すなわちハエ約100匹/m2を添加し、35週目に収穫を行った。
【0083】
結果
得られた結果を以下の表4及び5にまとめた。
表4:「雄」及び「雌」親、Charlin (温室バターヘッド)による種子の作製
【表4】
【0084】
表5:「雄」及び「雌」親、BVA 68/12553 (バタビア)による種子の作製
【表5】
【0085】
考察
バタビアレタス(BVA 68/12553)で得られた産物の平均は、自己受精コントロールの1.7g/植物に対して約1 g/植物であり、且つ温室バターヘッドレタス(Charlin)で得られた産物の平均は、約1 g/植物であり自己受精コントロールとほぼ同じであった点で、結果は満足のいくものであった。
【0086】
試行10:雑種の組み合わせによるDm3/Dm18蓄積
2つの優性遺伝子、Dm3及びDm18であって、クラスター2に属し同じ遺伝子座に位置する品種24(B124)及び品種23(B123)の、ブレミア・ラクツカエ耐性用遺伝子を蓄積するF1雑種レタスは、上記記載の試行で記載された方法を用いて得た。
【0087】
「雌」親として使用される植物(06/30443: MS7/21NACRExDEVONIAxCAMBRIA)は、Dm18及びR38耐性遺伝子のキャリアである。「雄」親として使用される植物は、Dm3耐性遺伝子を保有する温室バターヘッドレタスである(BC: 06/30443 * Rex 又は BC:06/30443 * Melina)。
以下の5つの交配を行った。
06/30443/01 (MS7/21xNACRExDEVONIAxCAMBRIA-01) Dml8/R38 * Rex Dm3、
06/30443/10 (MS7/21xNACRExDEVONIAxCAMBRIA-10) Dml8/R38 * Rex Dm3、
06/30443/02 (MS7/21xNACRExDEVONIAxCAMBRIA-02) Dml8/R38 * Melina Dm3、
06/30443/03 (MS7/21xNACRExDEVONIAxCAMBRIA-03) Dml8/R38 * Melina Dm3、
06/30443/04 (MS7/21xNACRExDEVONIAxCAMBRIA-04) Dml8/R38 * Melina Dm3。
【0088】
当該5つの交配由来の種子は播種され、葉ディスク上でのインキュベーション試験及び分子標識をF1植物で行うとともに、「雄」(Rex/Melina)及び「雌」親で行った。
【0089】
I 葉ディスク上でのインキュベーション試験
装置:
植物原料は、1植物当たり及び1品種当たり5ディスクの数で、1起源当たり10植物及び2つのブレミア・ラクツカエ品種で(Dm18/R38蓄積を攻撃し、且つDm3がそれに対する耐性を与えるB124、及びDm3/R38蓄積を攻撃し、且つDm18がそれに対する耐性を与えるB123)、1.5cm直径の葉ディスクからなる。すなわち、総数で80の植物を2つの品種で試験した。
【0090】
技術:
葉ディスクに、ブレミアB123及びB124を独立に伴ったスポロシストを噴霧することにより植菌を行った。
【0091】
1/葉ディスクの除去:
1植物当たり5つの葉ディスクを除去し、2重の厚みの湿ったブロット紙の上に配列させた。各皿には、6列の試験するディスク及び1列の感受性の高いコントロールを入れた。
2/植菌:
6皿の葉ディスクを植菌するために必要な植菌材料の量は11mlであった。100 mlの脱塩水に4滴のTween 80を使用することにより、Tween 80溶液を調製した。植菌材料の容積のための抽出溶液の量は、100mlの植菌材料当たり5 mlのTweenストック溶液であった。植菌材料は、5%のTween 80ストック溶液を補充した水道水中で、スポロシストを分離することにより作製した。スポロシストを、108胞子/ml近傍のスポロシスト濃度を得るため、1植物/mlの植菌材料量で除去した。250mlビーカーに、ガーゼを通して溶液を通過させて濾過を行った。
3/噴霧:
植菌は、葉ディスクを採取した日に行った。植菌材料溶液の容積を測定し、その後配置した皿に植菌した。植菌材料を、各ディスクの表面全体に、微細な液滴で均一に噴霧した。この皿を密封し、15℃で、日光14時間、夜間10時間の単位で7日間インキュベートした。
【0092】
結果
植菌の結果を、以下の表6〜13に示す(S=感受性、R=耐性)。種々の交配から得られる雑種は2つの品種B123及びB124に耐性であり、クラスター2由来の2つの遺伝子Dm18及びDm3の存在を示すものである。
【0093】
表6:「雌」30443 (Dm18/R38)植物: MS7/21xNACRExDEVONIAxCAMBRIA:B123耐性及びB124感受性。
【表6】
表7:「雄」Melina (Dm3) 植物:B123感受性及びB124耐性(10植物での試験)
【表7】
表8: 06/30443/02 * Melina (Dml8/R38*Dm3) 交配=MS7/2IxNACRExDEVONIAxCAMBRIA-02*MELINA
【表8】
表9: 06/30443/03 * Melina (Dml8/R38*Dm3) 交配=MS7/2IxNACRExDEVONIAxCAMBRIA-03*MELINA
【表9】
表10: 06/30443/04 * Melina (Dml8/R38*Dm3) 交配=MS7/2IxNACRExDEVONIAxCAMBRIA-03*MELINA
【表10】
表11:「雄」Rex (Dm3) 植物:B123感受性及びB124耐性(10植物での試験)
【表11】
表12:06/30443/01 * Rex (Dml8/R38*Dm3) 交配=MS7/21xNACRExDEVONIAxCAMBRIA-01*REX
【表12】
表13: 06/30443/10 * Rex (Dml8/R38*Dm3) 交配=MS7/21xNACRExDEVONIAxCAMBRIA-10*REX
【表13】
【0094】
II 分子標識
2つの分子マーカーを用いることで、ラクツカ・サティバにおいて、優性であり且つ2つのブレミア・ラクツカエ種に耐性である、2つの密接に関連した遺伝子(同じ遺伝子座に位置する)の蓄積を証明することができる。
【0095】
方法:
1/植物原料:
植物原料は、1植物当たり1ディスクで、1.5cm直径の葉ディスクからなる。遺伝子型当たり5〜10植物(すなわち総数で50植物)由来のDNAを、CTAB/クロロホルム抽出により得て、その後5ng/μlの濃度で、TE 0.1×溶液中で懸濁として溶解させた。
【0096】
2/遺伝子型決定:
Maisonneuve等による[10]及びKuang等による[11]で報告された、SCW09及びB1マーカーを前記50植物の遺伝子型決定のために使用した(表14)。PCR条件は表15に記載する。
SCW09及びB1のPCRサイクルはそれぞれ以下の通りである:
94℃/30秒−94℃/1分、60℃/1分、72℃/2分;40サイクル−72℃/5分−4℃;
94℃/30秒−94℃/1分、63℃/1分、72℃/2分;40サイクル−72℃/5分−8℃。
SCW09マーカーは、移行ステップの前にTaqI酵素での酵素的消化を受けた(バッファ1×、BSA1×及びTaqI 1U)。電気泳動を、2%(SCW09)又は1.5%(B1)のアガロースゲルを用い、TBE1×バッファ中で、1時間(B1)又は1時間30分(SCW09)、定電圧220Vで泳動した。電気泳動プロファイルは、UVランプ下、エチジウムブロミド(ETB)での染色により明らかになった。
【0097】
表14:SCW09及びB1マーカーの特徴
【表14】
表15:SCW09及びB1マーカーのPCR条件
【表15】
【0098】
結果
結果を表16並びに図4及び5に示す。REX及びMELINAの遺伝子型は、Dm18−/Dm18−且つDm3+プロファイルであった。06/30443遺伝子型はDm18+/Dm18+且つDm3−プロファイルであった。親(REX又はMELINA)の一方又は他方との交配の組は、Dm18+/Dm18−及びDm3+プロファイル、すなわちDm18及びDm3に耐性であった。
【0099】
表16:SCW09及びB1マーカーでの遺伝子型決定の結果(S=感受性、R=耐性)。
【表16】
【表17】
【0100】
実施例2:Ms7遺伝子用分子マーカーの同定
以下に記載のラクツカ・サティバ(レタス)中の雄性不稔性Ms7に関連する分子マーカーを得るための方法を、指示する目的で提供する。
1−レタス(ラクツカ・サティバ)中で核雄性不稔性Ms7と関連するマーカーの開発用の群の構成
Ms7雄性不稔性であるレタス用の異なる類型の2つの群を、戻し交配で作製した。結果として得られた、200植物の反復第4代群(BC4)の2つを雄性不稔性表現型に対応する「生花粉の不存在」形質で表現型検査を行った。2つの表現型グループを、雄性稔性(F)又は雄性不稔性(S)として1:1の比率で同定した。
この比率は、Ms7遺伝子座で異種接合体の親により提供される、雄性不稔性の優性一遺伝子的性質を裏付けるものであった。
【0101】
2−レタス植物由来のDNAの抽出
DNAは、修正CTABプロトコル(Tomas等, 1989 [20]; Doyle 等, 1990 [15]; Edwards 等, 1991 [16])を用い、新鮮な幼葉を使用して抽出した。
1.5ml管を用いて得た新鮮な葉ディスクを、500μlの抽出バッファ(Tris−HCl 0.1M、NaCl 0.7 M、EDTA 10 mM、CTAB 1%、βメルカプトエタノール 1%)中に置いた。この置いた原料を65℃で1時間インキュベートし、インキュベーション中に2〜3回反転させて混合した。
その後、200μlの24:1のクロロホルム:イソアミル溶液を添加し、反転させて混合した。6000rpmで10分間、20℃で遠心分離後に、400μlの上澄みを回収し、400μlのイソプロパノールと混合した。
−20℃で1時間後、混合物を6000rpmで10分間、4℃で遠心分離した。管の中身を出した。管底に付着したDNA残基は12時間空気中で乾燥させた。
このDNAを、200μlのTE 0.1×溶液(tris−HCl 1 mM、EDTA 0.1 mM)中で懸濁として溶解させた。終濃度を、1%アガロースゲル上で、Lambda DNA-HindIII消化アッセイにより測定した。
【0102】
3−BSA(bulk segregant analysis)法を用いる、ラクツカ・サティバにおけるMs7雄性不稔性に関連するRAPDマーカーの研究
雄性不稔性に関連する分子マーカーMs7を同定する目的で、Operon Technologies Inc.(Huntsville, AL 35805、米国)製のRAPD(登録商標)10merキット及びBuIk Segregant Analysis法(Michelmore 等, 1991 [17] 及び Paran 等, 1991 [18])を使用した。
上記記載の2つのBC4群の表現型検査から開始するため、10植物の混合物の2つのサンプルを、2つのグループ「雄性不稔性」及び「雄性稔性」用として作製した。OPA-01〜OPBH-20の1200個のプライマー(Operon Biotechnologies Inc. (Huntsville, AL 35805、米国))を、William 等, 1990 [22] 及び Welsh 等, 1990 [21]により報告されたRAPD技術を用いて、1群当たり4サンプルを試験した。「雄性不稔性」サンプルに特異的なバンドを表し且つ読みやすい電気泳動プロファイルを有するプライマーを選択した。
【0103】
PCR反応は、PCR1×バッファ、3 mM MgCl2、200μMのdNTP、400nMのプライマー、1単位のAmpliTaq (Perkin-Elmer cetus)DNAポリメラーゼで組成した、総反応量25μlで実施した。複数サイクルからなるPCR反応を以下に記載した。94℃30秒間で1ステップ、94℃1分、その後35℃1分で最終的に72℃にして2分、及び最後の伸長ステップを72℃5分間を45サイクルで実施し、その後反応物を4℃で保存した。
増幅産物を、2%アガロースゲルで、以下の電気泳動条件下で分離した:移動バッファTBE 1×(tris−ホウ酸塩−EDTA)、190Vで2時間15分。
【0104】
残存するRAPDプライマーを確認する(validate)ために、2つのBC4群由来の200植物を、以下の配列:5' TGCGTTCCAC 3'(配列番号1)(これは675bpのマーカー、BA05-675(配列番号2)と対応する)を有し、Ms7を同時分離する、OPBA05プライマーで個別に試験した。このマーカーから得られた電気泳動プロファイルを図6に示す。
【0105】
このマーカーを12の群からの約25植物で、種々の類型のレタス(バタビア、バターヘッド、アイスバーグ(iceberg)、ロメイン(romaine)、オークリーフ(oakleaf)及びロロロッサ(lollo rossa))で確認した。試験条件は、抽出、増幅及び電気泳動のステップで既に記載したものと同じであった。
RAPD BA05-675マーカーの感受性及び特異性の計算からは、類型によって異なる値が得られ、それぞれ平均値は96%及び94%であった(表17及び18を参照)。
【0106】
表17:保有する分子マーカーの感受性及び特異性の予測される値の、計算の詳細。
【表18】
表18:レタスの類型と、Ms7と関連する分子マーカーBA05-675への感受性(S*)及び特異性(F**)の平均値の結果。
【表19】
【0107】
4−ラクツカ・サティバ種におけるMs7雄性不稔性に関連するBA05-675の遺伝子マッピング
上記のms7のために解体されたBC4の群由来の200植物を基に、Ms7に関連するBA05-675マーカーの遺伝子マッピング(William 等, 1993 [23])を、JoinMap(登録商標)4 (Stam 等, 1996 [19]) 及び CarteBlanche (Keygene N P, P O Box 216, 6700 A E Wageningen, The Netherlands)を用いて実施した。優性のBA05-675マーカーについて、メンデル型1:1比率のゼロ仮定を、X2統計的検定で確認した(検定はp > 0.05で有意性がない)(表19を参照)。3.0以上の可能性ある比率(又はLODスコア)での結合試験により、Ms7の1.8 cMマーカーをマップできた(図6)。
【0108】
表19:MS7に関連する分子分離(segregation)マーカーBA05-675の比率及び比率1:1でのメンデル型分離のゼロ仮説を用いる1ddlでのX2検定の結果(F:稔性優性植物の数、S:不稔性雄性植物の数)。
【表20】
【0109】
5−ラクツカ・サティバレタスにおける雄性不稔性に関連するRAPD BA05-675分子マーカーのクローニング及び配列決定
アガロースゲル電気泳動プロファイルから、675bpのDNA断片を単離し、TEバッファ(10 mM tris-HCl、1 mM EDTA)中で懸濁物として溶解させ、その後上記のPCR条件下で再増幅させた。この再増幅した単離断片を、市販のPCR(登録商標)4-TOPO(登録商標)キット(Invitrogen, Carlsbad, California 92008, 米国)を用いてクローン化した。クローニングの確認後、DNA抽出(Midiprep system)は、Promegaカラム精製キット((Madison, WI, 米国)を用いて行った。
75 ng/μlに濃縮した精製クローンを、Cogenics (38944 Meylan, フランス)により配列決定した。すなわち、図8に記載のBA05-675マーカーの配列が得られた(配列番号2)。
【0110】
6−結果
本発明者等が実施した研究により、RAPDマーカー、BA05-675(配列番号2)の特徴づけが可能となった。その配列は、図8に示され、Ms7から1.8cMの距離で(図7)、且つレタスの類型について個別にそれぞれ平均96%及び94%の感受性及び特異性を有する(表18)。2つのBC4群由来の個別の植物でのこのマーカーの評価から、不稔性の雄性植物を同定するための平均予測値(Altman 1994a [13])が97%近傍であるという推定ができる。
【0111】
参考文献
1. Goubara & Takasaki (2003) Flower visitors of lettuce under field and enclosure conditions, Appl . Entomo. Zool . 38(4) : 571-581.
2. Goubara & Takasaki (2004) Pollination effects of the sweat bee Lasioglossum vilosulum trichopse on genic-male lettuce, Appl. Entomo. Zool. 39(1) : 163-169, 2004.
3. Lindqvist, K. (1960) Inheritance studies in lettuce, Heriditas 46 : 387-470.
4. Michelmore, R.W. et al. (1987) Genetic analysis of factors for resistance to downy mildew in lettuce, Plant Pathol., vol. 36, no 4 : 499-514.
5. Michelmore, R.W. et al. (1993) Development of reliable PCR-based markers linked to downy mildew resistance genes in lettuce, Theor. Appl . Genet., vol. 85, No 8 : 985-993.
6. Ryder, E. J. (1963) An epistatically controlled pollen sterile in lettuce (Lactuca sativa L.), Proc. Am. Soc. Hort. Sci. 96 : 826-828.
7. Ryder, E.J. (1967) A recessive male sterile gene in lettuce, Proc. Am. Soc. Hort. Sci 91 : 366-368.
8. Ryder, E.J. (1971) Genetic Studies in Lettuce {Lactuca sativa L.), J.Amer.Soc .Hort Sci 96(6) 826-828.
9. Ryder, E.J. (1979) Leafy Salad Vegetables, Avi . Pub. Co. , page 30.
10. Maisonneuve, B. et al. (1994) Rapid mapping of two genes for resistance to downy mildew from Lactuca serriola to existing clusters of resistance genes, Theor. Appl. Genet. 89 : 96-104.
11. Kuang H., et al. (2004) Multiple genetic processes result in heterogeneous rates of evolution within the major cluster disease resistance genes in lettuce, The plant cell, Vol. 16, 2870-2894.
12. Kesseli R.V., et al. (1994) Analysis of a detailed linkage map of Lactuca sativa (lettuce) constructed from RFLP and RAPD markers, Genetics 136: 1435-1446.
13. Altman D. G., Bland J.M., 1994a. Statistics Notes: Diagnostic tests 1: sensitivity and specificity. British Medical Journal, 308:1552.
14. Altman D. G., Bland J.M., 1994b. Statistics Notes: Diagnostic tests 2: predictive values. British Medical Journal, 30:102.
15. Doyle J. J., and Doyle J. L., 1990. Isolation of plant DNA from fresh tissue. Focus 12, 13-15.
16. Edwards K., Johnstone C. and Thompson C, 1991. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acid Res 19:1349.
17. Michelmore R.W., Paran I., Kesseli R.V., 1991. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA 88:9828- 9832.
18. Paran I., Kesseli R., Michelmore R.W., 1991. Identification of restriction fragment length polymorphism and random amplified polymorphic DNA markers linked to downy mildew genes in lettuce, using near-isogenic lines Genome 34:1021-1027.
19. Stam P., & J.W. van Ooijen, 1996. JoinMap, version 2.0. Software for the calculation of genetic linkage maps . Ed. CPRO-DLO, 60 pp.
20. Tomas H.T., and Tanksley S.D., 1989. A rapid and inexpensive method for isolation of total DNA from dehydrated plant tissue. Plant Mol Biol Rep 12:106-109.
21. Welsh J, McClelland M., 1990. Fingerprinting genomes using PCR with arbitrary primers . Nucleic Acids Res, 18:7213-7218.
22. Williams J.K.G., Kubelik A.R., Livak K. J., Rafalsky J.A., Tynger S.V., 1990. DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res, 18:6531-6535.
23. Williams J. K. G., Reiter R. S., Young R.M., Scolnik P.A., 1993. Genetic mapping of mutations using phenotypic pools and mapped RAPD markers. Nucleic Acids Res., 21(11): 2697- 2702.
【特許請求の範囲】
【請求項1】
雑種ラクツカ・サティバ(Lactuca sativa)種子であって、それが
雄性不稔性遺伝子型を有するとともに、
検出可能な表現型を当該種子に由来する植物に提供する、少なくとも1つの特徴的遺伝子の異種接合体であること、
を特徴とする種子。
【請求項2】
前記雄性不稔性遺伝子型が、少なくとも1の核遺伝子を含有し、且つ前記種子が雄性不稔性の原因である1又は複数の遺伝子の異種接合体である、請求項1に記載の種子。
【請求項3】
前記雄性不稔性が一遺伝子的である、請求項2に記載の種子。
【請求項4】
前記雄性不稔性の遺伝子が優性である、請求項3に記載の種子。
【請求項5】
前記雄性不稔性遺伝子型がMs7不稔性遺伝子により提供される、請求項1に記載の種子。
【請求項6】
ゲノムが、650〜700ヌクレオチドの二本鎖DNA配列を含んでなり、ここでその2本の鎖の各々の5'末端が配列 5' TGCGTTCCAC 3'(配列番号1)で開始することを特徴とする、請求項1〜5のいずれか1項に記載の種子。
【請求項7】
前記表現型が農業で注目の表現型である、請求項1〜6のいずれか1項に記載の種子。
【請求項8】
前記表現型が、ウイルス、細菌、虫又は真菌による感染に対して、標準的又は中程度に耐性である遺伝子により提供される、請求項1〜7のいずれか1項に記載の種子。
【請求項9】
前記表現型が、
以下の真菌:ブレミア・ラクツカエ(Bremia lactucae)、フサリウム・オキシスポラム(Fusarium oxysporum)、スクレロティニア・マイナー(Sclerotinia minor)もしくはスクレロトラム(sclerotorum)、ボトリチス・シネレア(Botrytis cinerea)、リゾクトニア・ソラニ(Rhizictonia solani)、ミクロドチウム・パナットニアナム(Microdochium panattonianum)、ベルチシリウム・ダーリエ(Verticiulium dahliae)、エリシフェ・チコセアラム(Erysiphe chicocearum)又はピチウム・トラケイフィルム(Pithium tracheiphilum)の1種に対して、以下の虫:ナソノビア・リビスニグリ(Nasonovia ribisnigri)、マイザス・ペルシカエ(Myzus persicae)、マクロシファム・エウフォルビア(Macrosiphum euphorbia)、ネマトデス・プラチレクス(Nematodes pratylenchus)又はネコブ線虫(meloidogyne)、ハモグリバエ(leafminers)リリオマイザ・フイドブレンシス(liriomyza huidobrensis)又はペンフィガス・ブサリウス(Pemphigus busarius)の1種に対して、以下の細菌:シュードモナス(pseudomonas)、キサントモナス(xanthomonas)又はリゾモナス(rhizomonas)の1種に対して、又は以下のウイルス:LMV、TSWV、「ビッグベイン(Big vein)」、TBSV、LNSV、TuMV、CMV又はBWYVの1種に対する、標準的又は中程度の耐性である、請求項1〜8のいずれか1項に記載の種子。
【請求項10】
前記種子が、異種接合状態で、標準的又は中程度の耐性である、第二の特徴的遺伝子を提示する、請求項8又は9に記載の種子。
【請求項11】
前記標準的又は中程度の耐性遺伝子が、以下の、ブレミア(bremia)耐性遺伝子:Dm10、R17、Dm5/Dm8、R36/R37(クラスター1)、Dm1、Dm2、Dm3、Dm6、Dm14、Dml5、Dml6、Dml8 (クラスター2)、又はDm4、Dm7、Dm11、R38 (クラスター4);又はTuMV耐性のTu遺伝子(クラスター1);ナソノビア(Nasonovia)耐性のNr遺伝子(クラスター2);又はLMV耐性の遺伝子mol1、mol2(クラスター4)から選択される、請求項8又は10に記載の種子。
【請求項12】
雑種ラクツカ・サティバ植物であって、それが
雄性不稔性遺伝子型の異種接合体であるとともに、
検出可能な表現型を提供する、少なくとも1つの特徴的遺伝子の異種接合体であること、
を特徴とする植物。
【請求項13】
前記雄性不稔性遺伝子型が核由来であり、且つ前記植物が雄性不稔性の原因である1又は複数の遺伝子の異種接合体である、請求項12に記載の植物。
【請求項14】
前記雄性不稔性が一遺伝子的である、請求項13に記載の植物。
【請求項15】
前記雄性不稔性の遺伝子が優性である、請求項14に記載の植物。
【請求項16】
前記雄性不稔性遺伝子型が不稔性遺伝子Ms7により提供される、請求項12に記載の遺伝子。
【請求項17】
雑種ラクツカ・サティバ植物由来の細胞であって、それが
雄性不稔性遺伝子型の異種接合体であるとともに、
植物中で検出可能な表現型を提供する、少なくとも1つの特徴的遺伝子の異種接合体であること、
を特徴とする細胞。
【請求項18】
前記雄性不稔遺伝型が核由来であり、且つ前記植物が雄性不稔性の原因である1又は複数の遺伝子の異種接合体である、請求項17に記載の細胞。
【請求項19】
前記雄性不稔性が一遺伝子的である、請求項18に記載の細胞。
【請求項20】
前記雄性不稔性遺伝子が優性である、請求項19に記載の細胞。
【請求項21】
前記雄性不稔性遺伝子型が不稔性遺伝子Ms7により提供される、請求項17に記載の細胞。
【請求項22】
少なくとも105個の種子、好ましくは少なくとも106個の種子を含んでなる、請求項1〜11のいずれか1項に記載の種子の集団。
【請求項23】
少なくとも107種子を含んでなる、請求項22に記載の集団。
【請求項24】
請求項1〜11のいずれか1項に記載の種子を少なくとも40%含んでなる、ラクツカ・サティバ種子の集団。
【請求項25】
その他の種子が雄性不稔性遺伝子を含まない、請求項24に記載の集団。
【請求項26】
雄性稔性植物による雄性不稔性ラクツカ・サティバ植物の閉鎖環境中での授粉を実施するための、1m2当たり100匹以上、又は1m3当たり25匹以上の濃度での双翅目の虫の使用。
【請求項27】
前記双翅目が、1m2当たり少なくとも250匹、又は1m3当たり少なくとも75匹の濃度で存在する、請求項26に記載の使用。
【請求項28】
雑種ラクツカ・サティバ植物を得るための、請求項26に記載の使用。
【請求項29】
前記雄性不稔性ラクツカ・サティバ植物が、単一の優性遺伝子により、好ましくはMs7雄性不稔性遺伝子によりもたらされる雄性不稔性を有する、請求項26〜28のいずれか1項に記載の使用。
【請求項30】
前記雄性稔性植物がまた、ラクツカ・サティバ植物、好ましくは栽培品種である、請求項26〜29のいずれか1項に記載の使用。
【請求項31】
雑種ラクツカ・サティバ種子を得るための方法であって、
表現型的に「雌」親として使用される雄性不稔性ラクツカ・サティバ植物、及び表現型的に「雄」親として使用される雄性稔性ラクツカ・サティバ植物を、互いに近接した状態で、閉鎖環境中で栽培するステップであって、2つの親の一方が、雄性不稔性以外の検出可能な表現型をそれに提供する遺伝子の同種接合体であるという追加的な特徴を有し、他方の親はこの遺伝子を保有しないものであるステップ、
植物の開花の際に、1m2当たり100匹以上又は1m3当たり25匹以上の濃度で、好ましくは1m2当たり少なくとも250匹の又は1m3当たり少なくとも75匹の濃度で、閉鎖環境へ導入する双翅目を用いて授粉するステップ、及び
雄性不稔性植物により作られる種子を収穫するステップ
を含んでなる方法。
【請求項32】
第二の親も、他方の親が有しない、雄性不稔性以外の検出可能な表現型をそれに提供する少なくとも1の遺伝子の異種接合体である、請求項31に記載の方法。
【請求項33】
前記導入する双翅目の濃度が、1m2当たり400匹以上、好ましくは1m2当たり50匹以上である、請求項31又は32に記載の方法。
【請求項34】
前記双翅目を蛹の状態で導入する、請求項31〜33のいずれか1項に記載の方法。
【請求項35】
前記閉鎖環境が虫に対して密閉されている、請求項31〜34のいずれか1項に記載の方法。
【請求項36】
前記双翅目が、短角類(ブラキセラ、brachycera)、好ましくはシクロラファ・ブラキセラ(cyclorhapha brachycera)である、請求項31〜35のいずれか1項に記載の方法。
【請求項37】
前記双翅目がクロバエ(Calliphorides)又はイヌバエ(Muscidae)ファミリーに属する、請求項36に記載の方法。
【請求項38】
前記双翅目が、カリホラ・ボミタリア(Calliphora vomitaria)、カリホラ・エリトロセファラ(Calliphora erythrocephala)又はルシリア・カエサル(Lucilia Caesar)である、請求項37に記載の方法。
【請求項39】
閉鎖環境中で、雄性不稔性植物の数が雄性稔性植物の数より多い、請求項31〜38のいずれか1項に記載の方法。
【請求項40】
閉鎖環境中で、雄性不稔性植物の数が少なくとも2000であり、雄性稔性植物の数が少なくとも1000である、請求項31〜39のいずれか1項に記載の方法。
【請求項41】
双翅目の導入を、少なくとも1週間に1度、3〜4週間にわたって繰り返す、請求項31〜40のいずれか1項に記載の方法。
【請求項42】
雌親として使用される植物の雄性不稔性が一遺伝子起源、優性且つ核性であるとともに、雌親として使用される植物が以下の、
優性の核雄性不稔性遺伝子の異種接合体であるラクツカ・サティバ植物と、不稔性遺伝子を保有しない雄性稔性のラクツカ・サティバ植物との間で交配するステップ、
前記交配から得られる種子を栽培するステップ、及び
雄性稔性表現型を有する植物を除去するステップ、
を含んでなる方法により得られる、
請求項31〜41のいずれか1項に記載の方法。
【請求項43】
前記雄性稔性表現型を有する植物を除去するステップが、植物の各サンプルにおいて、雄性不稔優性遺伝子に特異的な分子マーカーの不存在を検出することにより実施される、請求項42に記載の方法。
【請求項44】
雌親として使用される植物の雄性不稔性がMs7遺伝子により提供される、請求項42又は43に記載の方法。
【請求項45】
雌親として使用される植物のゲノムが、650〜700ヌクレオチドの二本鎖DNA配列を含んでなり、ここでその2本の鎖の各々の5'末端が配列 5' TGCGTTCCAC 3'(配列番号1)で開始する、請求項31〜44のいずれか1項に記載の方法。
【請求項46】
少なくとも105個の種子、好ましくは106個の種子及びより好ましくは107個の種子を含んでなる、請求項31〜45のいずれか1項に記載の方法を使用して得られる種子の集団。
【請求項47】
TSW(1000種子質量、thousand seed weight)が、「雄」親として使用される雄性稔性ラクツカ・サティバ植物の自己受精により得られる植物のTSWより、少なくとも10%、好ましくは少なくとも20%及びより好ましくは少なくとも30%大きい、請求項46に記載の集団。
【請求項1】
雑種ラクツカ・サティバ(Lactuca sativa)種子であって、それが
雄性不稔性遺伝子型を有するとともに、
検出可能な表現型を当該種子に由来する植物に提供する、少なくとも1つの特徴的遺伝子の異種接合体であること、
を特徴とする種子。
【請求項2】
前記雄性不稔性遺伝子型が、少なくとも1の核遺伝子を含有し、且つ前記種子が雄性不稔性の原因である1又は複数の遺伝子の異種接合体である、請求項1に記載の種子。
【請求項3】
前記雄性不稔性が一遺伝子的である、請求項2に記載の種子。
【請求項4】
前記雄性不稔性の遺伝子が優性である、請求項3に記載の種子。
【請求項5】
前記雄性不稔性遺伝子型がMs7不稔性遺伝子により提供される、請求項1に記載の種子。
【請求項6】
ゲノムが、650〜700ヌクレオチドの二本鎖DNA配列を含んでなり、ここでその2本の鎖の各々の5'末端が配列 5' TGCGTTCCAC 3'(配列番号1)で開始することを特徴とする、請求項1〜5のいずれか1項に記載の種子。
【請求項7】
前記表現型が農業で注目の表現型である、請求項1〜6のいずれか1項に記載の種子。
【請求項8】
前記表現型が、ウイルス、細菌、虫又は真菌による感染に対して、標準的又は中程度に耐性である遺伝子により提供される、請求項1〜7のいずれか1項に記載の種子。
【請求項9】
前記表現型が、
以下の真菌:ブレミア・ラクツカエ(Bremia lactucae)、フサリウム・オキシスポラム(Fusarium oxysporum)、スクレロティニア・マイナー(Sclerotinia minor)もしくはスクレロトラム(sclerotorum)、ボトリチス・シネレア(Botrytis cinerea)、リゾクトニア・ソラニ(Rhizictonia solani)、ミクロドチウム・パナットニアナム(Microdochium panattonianum)、ベルチシリウム・ダーリエ(Verticiulium dahliae)、エリシフェ・チコセアラム(Erysiphe chicocearum)又はピチウム・トラケイフィルム(Pithium tracheiphilum)の1種に対して、以下の虫:ナソノビア・リビスニグリ(Nasonovia ribisnigri)、マイザス・ペルシカエ(Myzus persicae)、マクロシファム・エウフォルビア(Macrosiphum euphorbia)、ネマトデス・プラチレクス(Nematodes pratylenchus)又はネコブ線虫(meloidogyne)、ハモグリバエ(leafminers)リリオマイザ・フイドブレンシス(liriomyza huidobrensis)又はペンフィガス・ブサリウス(Pemphigus busarius)の1種に対して、以下の細菌:シュードモナス(pseudomonas)、キサントモナス(xanthomonas)又はリゾモナス(rhizomonas)の1種に対して、又は以下のウイルス:LMV、TSWV、「ビッグベイン(Big vein)」、TBSV、LNSV、TuMV、CMV又はBWYVの1種に対する、標準的又は中程度の耐性である、請求項1〜8のいずれか1項に記載の種子。
【請求項10】
前記種子が、異種接合状態で、標準的又は中程度の耐性である、第二の特徴的遺伝子を提示する、請求項8又は9に記載の種子。
【請求項11】
前記標準的又は中程度の耐性遺伝子が、以下の、ブレミア(bremia)耐性遺伝子:Dm10、R17、Dm5/Dm8、R36/R37(クラスター1)、Dm1、Dm2、Dm3、Dm6、Dm14、Dml5、Dml6、Dml8 (クラスター2)、又はDm4、Dm7、Dm11、R38 (クラスター4);又はTuMV耐性のTu遺伝子(クラスター1);ナソノビア(Nasonovia)耐性のNr遺伝子(クラスター2);又はLMV耐性の遺伝子mol1、mol2(クラスター4)から選択される、請求項8又は10に記載の種子。
【請求項12】
雑種ラクツカ・サティバ植物であって、それが
雄性不稔性遺伝子型の異種接合体であるとともに、
検出可能な表現型を提供する、少なくとも1つの特徴的遺伝子の異種接合体であること、
を特徴とする植物。
【請求項13】
前記雄性不稔性遺伝子型が核由来であり、且つ前記植物が雄性不稔性の原因である1又は複数の遺伝子の異種接合体である、請求項12に記載の植物。
【請求項14】
前記雄性不稔性が一遺伝子的である、請求項13に記載の植物。
【請求項15】
前記雄性不稔性の遺伝子が優性である、請求項14に記載の植物。
【請求項16】
前記雄性不稔性遺伝子型が不稔性遺伝子Ms7により提供される、請求項12に記載の遺伝子。
【請求項17】
雑種ラクツカ・サティバ植物由来の細胞であって、それが
雄性不稔性遺伝子型の異種接合体であるとともに、
植物中で検出可能な表現型を提供する、少なくとも1つの特徴的遺伝子の異種接合体であること、
を特徴とする細胞。
【請求項18】
前記雄性不稔遺伝型が核由来であり、且つ前記植物が雄性不稔性の原因である1又は複数の遺伝子の異種接合体である、請求項17に記載の細胞。
【請求項19】
前記雄性不稔性が一遺伝子的である、請求項18に記載の細胞。
【請求項20】
前記雄性不稔性遺伝子が優性である、請求項19に記載の細胞。
【請求項21】
前記雄性不稔性遺伝子型が不稔性遺伝子Ms7により提供される、請求項17に記載の細胞。
【請求項22】
少なくとも105個の種子、好ましくは少なくとも106個の種子を含んでなる、請求項1〜11のいずれか1項に記載の種子の集団。
【請求項23】
少なくとも107種子を含んでなる、請求項22に記載の集団。
【請求項24】
請求項1〜11のいずれか1項に記載の種子を少なくとも40%含んでなる、ラクツカ・サティバ種子の集団。
【請求項25】
その他の種子が雄性不稔性遺伝子を含まない、請求項24に記載の集団。
【請求項26】
雄性稔性植物による雄性不稔性ラクツカ・サティバ植物の閉鎖環境中での授粉を実施するための、1m2当たり100匹以上、又は1m3当たり25匹以上の濃度での双翅目の虫の使用。
【請求項27】
前記双翅目が、1m2当たり少なくとも250匹、又は1m3当たり少なくとも75匹の濃度で存在する、請求項26に記載の使用。
【請求項28】
雑種ラクツカ・サティバ植物を得るための、請求項26に記載の使用。
【請求項29】
前記雄性不稔性ラクツカ・サティバ植物が、単一の優性遺伝子により、好ましくはMs7雄性不稔性遺伝子によりもたらされる雄性不稔性を有する、請求項26〜28のいずれか1項に記載の使用。
【請求項30】
前記雄性稔性植物がまた、ラクツカ・サティバ植物、好ましくは栽培品種である、請求項26〜29のいずれか1項に記載の使用。
【請求項31】
雑種ラクツカ・サティバ種子を得るための方法であって、
表現型的に「雌」親として使用される雄性不稔性ラクツカ・サティバ植物、及び表現型的に「雄」親として使用される雄性稔性ラクツカ・サティバ植物を、互いに近接した状態で、閉鎖環境中で栽培するステップであって、2つの親の一方が、雄性不稔性以外の検出可能な表現型をそれに提供する遺伝子の同種接合体であるという追加的な特徴を有し、他方の親はこの遺伝子を保有しないものであるステップ、
植物の開花の際に、1m2当たり100匹以上又は1m3当たり25匹以上の濃度で、好ましくは1m2当たり少なくとも250匹の又は1m3当たり少なくとも75匹の濃度で、閉鎖環境へ導入する双翅目を用いて授粉するステップ、及び
雄性不稔性植物により作られる種子を収穫するステップ
を含んでなる方法。
【請求項32】
第二の親も、他方の親が有しない、雄性不稔性以外の検出可能な表現型をそれに提供する少なくとも1の遺伝子の異種接合体である、請求項31に記載の方法。
【請求項33】
前記導入する双翅目の濃度が、1m2当たり400匹以上、好ましくは1m2当たり50匹以上である、請求項31又は32に記載の方法。
【請求項34】
前記双翅目を蛹の状態で導入する、請求項31〜33のいずれか1項に記載の方法。
【請求項35】
前記閉鎖環境が虫に対して密閉されている、請求項31〜34のいずれか1項に記載の方法。
【請求項36】
前記双翅目が、短角類(ブラキセラ、brachycera)、好ましくはシクロラファ・ブラキセラ(cyclorhapha brachycera)である、請求項31〜35のいずれか1項に記載の方法。
【請求項37】
前記双翅目がクロバエ(Calliphorides)又はイヌバエ(Muscidae)ファミリーに属する、請求項36に記載の方法。
【請求項38】
前記双翅目が、カリホラ・ボミタリア(Calliphora vomitaria)、カリホラ・エリトロセファラ(Calliphora erythrocephala)又はルシリア・カエサル(Lucilia Caesar)である、請求項37に記載の方法。
【請求項39】
閉鎖環境中で、雄性不稔性植物の数が雄性稔性植物の数より多い、請求項31〜38のいずれか1項に記載の方法。
【請求項40】
閉鎖環境中で、雄性不稔性植物の数が少なくとも2000であり、雄性稔性植物の数が少なくとも1000である、請求項31〜39のいずれか1項に記載の方法。
【請求項41】
双翅目の導入を、少なくとも1週間に1度、3〜4週間にわたって繰り返す、請求項31〜40のいずれか1項に記載の方法。
【請求項42】
雌親として使用される植物の雄性不稔性が一遺伝子起源、優性且つ核性であるとともに、雌親として使用される植物が以下の、
優性の核雄性不稔性遺伝子の異種接合体であるラクツカ・サティバ植物と、不稔性遺伝子を保有しない雄性稔性のラクツカ・サティバ植物との間で交配するステップ、
前記交配から得られる種子を栽培するステップ、及び
雄性稔性表現型を有する植物を除去するステップ、
を含んでなる方法により得られる、
請求項31〜41のいずれか1項に記載の方法。
【請求項43】
前記雄性稔性表現型を有する植物を除去するステップが、植物の各サンプルにおいて、雄性不稔優性遺伝子に特異的な分子マーカーの不存在を検出することにより実施される、請求項42に記載の方法。
【請求項44】
雌親として使用される植物の雄性不稔性がMs7遺伝子により提供される、請求項42又は43に記載の方法。
【請求項45】
雌親として使用される植物のゲノムが、650〜700ヌクレオチドの二本鎖DNA配列を含んでなり、ここでその2本の鎖の各々の5'末端が配列 5' TGCGTTCCAC 3'(配列番号1)で開始する、請求項31〜44のいずれか1項に記載の方法。
【請求項46】
少なくとも105個の種子、好ましくは106個の種子及びより好ましくは107個の種子を含んでなる、請求項31〜45のいずれか1項に記載の方法を使用して得られる種子の集団。
【請求項47】
TSW(1000種子質量、thousand seed weight)が、「雄」親として使用される雄性稔性ラクツカ・サティバ植物の自己受精により得られる植物のTSWより、少なくとも10%、好ましくは少なくとも20%及びより好ましくは少なくとも30%大きい、請求項46に記載の集団。
【図1】

【図2】
【図3】
【図4】
【図5】

【図6】

【図7】

【図8】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公表番号】特表2010−519923(P2010−519923A)
【公表日】平成22年6月10日(2010.6.10)
【国際特許分類】
【出願番号】特願2009−552247(P2009−552247)
【出願日】平成20年3月5日(2008.3.5)
【国際出願番号】PCT/FR2008/000289
【国際公開番号】WO2008/132310
【国際公開日】平成20年11月6日(2008.11.6)
【出願人】(509248198)
【Fターム(参考)】
【公表日】平成22年6月10日(2010.6.10)
【国際特許分類】
【出願日】平成20年3月5日(2008.3.5)
【国際出願番号】PCT/FR2008/000289
【国際公開番号】WO2008/132310
【国際公開日】平成20年11月6日(2008.11.6)
【出願人】(509248198)
【Fターム(参考)】
[ Back to top ]