
Fターム[4B024BA79]の内容
突然変異又は遺伝子工学 (218,933) | 目的とする生産物質 (23,930) | 食用又は観賞用植物 (394)
Fターム[4B024BA79]に分類される特許
381 - 394 / 394
SNOW1はICE1と結合し、CBF発現およびシロイヌナズナにおける凍結耐性を制御する

本発明は、タンパク質(Snow1)、その突然変異体、および前記タンパク質をコード化している核酸に関し、これはIce1と相互作用し、これはCBF3プロモーター活性を活性化し、このようにして植物における凍結耐性を制御する。 (もっと読む)
トマトhighpigment−1突然変異表現型(hp−1及びhp−1W)に関与する単離ヌクレオチド配列及びその使用
本発明は、トマト high pigment 1(hp−1)及びhigh pigment 1w(hp−1w)表現型に関与する単離ヌクレオチド配列(該配列は、改変トマトDDB1遺伝子配列又はその断片を含有し、該改変配列又は断片における該改変は、hp−1の場合は該DDB1遺伝子配列のヌクレオチド931でのAからTへの塩基転換を、hp−1wの場合は該DDB1遺伝子配列のヌクレオチド2392でのGからAへの転位を含有する。)を提供する。 (もっと読む)
植物において表皮特異的なトランスジェニック発現を行うためのプロモーター
本発明は、その制御下で、植物において表皮特異的に導入遺伝子を発現させることができるプロモーター領域に関する。本発明はまた、前記プロモーターを含む組換え核酸分子、これらの核酸分子で形質転換されたトランスジェニックな植物および植物細胞、ならびにそれらの製造方法に関する。本発明はさらに、本発明に係るプロモーターを含む核酸分子、病原体抵抗性を媒介する核酸配列または導入遺伝子、これらの核酸分子で形質転換された植物および植物細胞、ならびにそれらの製造方法に関する。 (もっと読む)
リグナンの生合成を触媒する酵素をコードする遺伝子、およびその利用
本発明は、ピノレジノールからピペリトールへの反応、およびピペリトールからセサミンへの反応を触媒する酵素、ならびに当該酵素をコードする遺伝子を提供する。本発明はさらに、上記酵素をコードする遺伝子を含むベクターおよび形質転換体、ならびに当該形質転換体を用いるタンパク質生産方法を提供する。 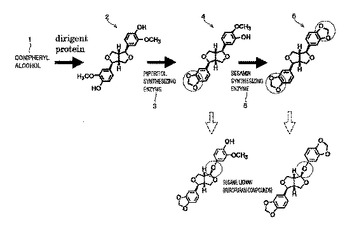 (もっと読む)
(もっと読む)
トランスジェニック構築体を用いる1を超える遺伝子の遺伝子発現の協調した低下および増加
本発明は、RNAの発現の同時の上方および下方調節と関連した核酸分子および核酸構築体ならびに他の剤に指向される。特に、それは、第2のRNAの発現を抑制すると同時に、第1のRNAの発現を同時に増強する方法を含む。また、本発明は、同時に第2のRNAの発現を抑制しつつ、第1のRNAの発現を同時に増強できる構築体、かかる剤を利用する方法、ならびにかかる剤を含む植物を特に提供する。また、本発明は、多シストロン性構築体を含む他の構築体を提供する。 (もっと読む)
バナナの抵抗性遺伝子およびそれらの使用法
本発明は、病原体抵抗性に関するものである。さらに詳細には本発明は、菌類病原体、詳細には土壌フザリウム菌類に対するバナナ植物の抵抗性メカニズムに関与するポリヌクレオチドおよびポリペプチド配列に関するものである。バナナの二つの抵抗性遺伝子RGA5およびRGA2が単離され、特徴づけされる。本発明は、植物の抵抗性を調整するため、および改変された病原体抵抗性の特性を有する遺伝子改変植物を産生させるためのこれらの配列の使用にも関するものでもある。 (もっと読む)
トランスジェニック生物における多不飽和脂肪酸の製造方法
本発明は、Δ5-エロンガーゼ活性を有するポリペプチドをコードする核酸を導入した生物において多不飽和脂肪酸を製造する方法に関する。該核酸配列は、場合によっては脂肪酸の生合成または脂質代謝のためのポリペプチドをコードする他の核酸配列と共に、該生物において有利に発現されうる。Δ6-デサチュラーゼ、Δ5-デサチュラーゼ、Δ4-デサチュラーゼおよび/またはΔ6-エロンガーゼ活性をコードする核酸配列が特に有利であり、有利には該デサチュラーゼおよびエロンガーゼはThalassiosira、EuglenaまたはOstreococcusに由来する。本発明は更に、長鎖多不飽和脂肪酸の含量が上昇した油および/またはトリアシルグリセリドの製造方法に関する。本発明の特定の実施形態は、不飽和ω3-脂肪酸の製造方法、および不飽和脂肪酸(特に、3個を超える二重結合を有するω3-脂肪酸)の含量が上昇したトリグリセリドの製造方法である。また、好ましくはω3-デサチュラーゼ、例えばPythiaceae科の真菌、例えばPhytophthora属、例えばPhytophthora infestans属・種からのω3-デサチュラーゼ、または藻類、例えばPrasinophyceae科、例えばOstreococcus属、特にOstreococcus tauri属・種、もしくはケイ藻類、例えばThalassiosira属、特にThalassiosira pseudonana属・種からのω3-デサチュラーゼと組み合わされた、前記方法において使用するエロンガーゼおよびデサチュラーゼの発現の結果として、ω3-脂肪酸、油または脂質(ω3-二重結合を有するもの)の含量が上昇したトランスジェニック生物、好ましくは、トランスジェニック植物またはトランスジェニック微生物の製造も開示する。本発明はまた、該核酸配列、核酸構築物、該核酸配列および/または核酸構築物を含有するベクターおよび生物、ならびに該核酸配列、核酸構築物および/またはベクターを含むトランスジェニック生物に関する。本発明のもう1つの態様は、前記方法により製造された油、脂質および/または脂肪酸、ならびにそれらの使用、さらには、不飽和脂肪酸、および不飽和脂肪酸の含量が上昇したトリグリセリド、ならびにそれらの使用に関する。 (もっと読む)
植物中の油レベルの上昇
本発明は、HOI001 GBSS対立遺伝子の発現によって、トウモロコシの穀穀粒組織中の油レベルを増加させるための方法を提供する。また、本発明は、HOI001 GBSSポリペプチドをコードする単離された核酸分子を提供する。 (もっと読む)
経口または経皮送達されたニコチンの有害な影響を低下させる方法
本発明は概して、従来のタバコ使用中断プログラムにおいて、経口または経皮送達されるニコチンの有害な影響を低下させることに関する。より詳細には、幾つかの実施形態は、低量のニコチンおよびタバコ特異的ニトロソアミン(TSNA)を含むタバコ製品を提供することによって、従来のタバコ使用中断プログラムと関係があるニコチン摂取の有害な影響を低下させる方法に関する。 (もっと読む)
eIF−5Aアイソフォーム:老化誘導eIF−5A;損傷誘導eIF−5A;成長eIF−5A;及びDHS
本発明は、真核生物開始因子5A(「eIF-5A」)の独自のアイソフォーム:老化-誘導eIF-5A;損傷-誘導eIF-5A;及び成長eIF-5A、並びにこれらの3種の因子をコードするポリヌクレオチドに関する。本発明はまた、これらの因子の発現を調節することを含む方法に関する。本発明はまた、デオキシハイプシン シンターゼ(「DHS」)、DHSをコードするポリヌクレオチド、及びDHSの発現を調節することを含む方法に関する。 (もっと読む)
1以上のアミノ酸レベルが増加した植物
本発明は、トレオニンデアミナーゼおよび/またはAHASをコードする外因性ポリヌクレオチドを含む構築体を提供する。また、該構築体で形質転換されたトランスジェニック植物ならびにこれらの植物から由来する種子および子孫を提供する。該トランスジェニック植物は、その同一種の非トランスジェニック植物に比較して増加したレベルの1以上のアミノ酸を有する。
 (もっと読む)
(もっと読む)
植物細胞における色素体標的タンパク質の生成方法
予め決定した植物の細胞で、対象のタンパク質を生成する方法であって、(i)融合タンパク質を色素体内にターゲティングする輸送ペプチド、及びそれに隣接して(ii)前記対象のタンパク質をN末端からC末端に含む前記融合タンパク質をコードするベクターを前記細胞内に導入することを含み、前記輸送ペプチドC末端の3個のアミノ酸X−3X−2X−1及び前記対象のタンパク質N末端のアミノ酸Zが、色素体で前記対象のタンパク質を放出するためにX−1とZとの間で前記融合タンパク質を開裂するための開裂部位X−3X−2X−1−Zを形成し、前記対象のタンパク質の予め決定したアミノ酸Zに対して、アミノ酸配列X−3X−2X−1が、植物の色素体を標的とする融合タンパク質で天然に存在するZに隣接するアミノ酸配列X−3X−2X−1の組から選択され、それにより前記開裂部位を形成する方法。 (もっと読む)
PUFAポリケチドシンターゼ系およびこれらの使用
本発明は概して、高度不飽和脂肪酸(PUFA)ポリケチドシンターゼ(PKS)系に関し、これらの相同体に関し、このようなPUFA PKS系の生物学的に活性なドメインをコードする単離された核酸分子および組換え核酸分子に関し、PUFA PKS系を含む遺伝子改変された生物に関し、関心対象の生理活性分子の産生のためにこのような系を作製および使用する方法に関し、かつこのようなPUFA PKS系を有する新たな非細菌性および細菌性の微生物を同定するための新規の方法に関する。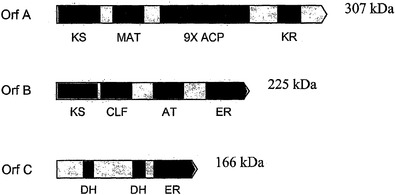
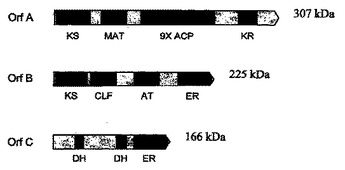 (もっと読む)
(もっと読む)
ワタアブラムシ耐性遺伝子
本発明は、ワタアブラムシおよび/または前記アブラムシによるウイルス伝搬に対する耐性に関与するVat遺伝子のクローニング、並びにその遺伝子の新しいマーカーの識別に関するものである。 (もっと読む)
381 - 394 / 394
[ Back to top ]