
ヒト4−1BB受容体タンパク質
【課題】新規ヒト4−1BB受容体タンパク質を提供すること。
【解決手段】ヒト4−1BB受容体タンパク質をコードする、特定のヌクレオチド配列を有するcDNAおよび特定のアミノ酸配列を有するヒト4−1BB受容体タンパク質を提供する。また、ヒト4−1BB受容体タンパク質を有する細胞外部分とアルカリホスファターゼからなる融合タンパク質を提供する。さらに、T細胞活性化を促進するために、4−1BB受容体タンパク質を有するT細胞をヒト4−1BB受容体タンパク質を認識するモノクローナル抗体で処理する方法を提供する。
【解決手段】ヒト4−1BB受容体タンパク質をコードする、特定のヌクレオチド配列を有するcDNAおよび特定のアミノ酸配列を有するヒト4−1BB受容体タンパク質を提供する。また、ヒト4−1BB受容体タンパク質を有する細胞外部分とアルカリホスファターゼからなる融合タンパク質を提供する。さらに、T細胞活性化を促進するために、4−1BB受容体タンパク質を有するT細胞をヒト4−1BB受容体タンパク質を認識するモノクローナル抗体で処理する方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、本発明者によるT細胞遺伝子の特異的発現によって分離および同定された類似のネズミ(マウス)受容体(レセプター)蛋白質4−1BBを用いた研究に基づいて分離および同定された以前に知られていないヒト受容体蛋白質H4−1BBに関する。
【背景技術】
【0002】
ヒトおよび他の種の免疫系は骨髄で作られる白血球を必要とし、その白血球には食細胞、リンパ球およびB細胞が含まれる。現在知られているように、食細胞にはこの系からウイルス蛋白質のような望ましくない物質を取り除くマクロファージ細胞が含まれる。リンパ球にはヘルパーT細胞、キラーT細胞およびB細胞ならびにサプレッサーT細胞に分類されるリンパ球を含む他の細胞が含まれる。キラーT細胞は細胞に物理的に突入し、またヘルパーT細胞は全過程を促進する。どのような事象においても、免疫過程はリンパ球によって促進される。
【0003】
リンホカインは免疫細胞が互いに連絡を取り合うための蛋白質である。科学者は免疫学的疾患に対して治療的に使用するためにそれらの蛋白質の充分量を製造する。多くのリンホカイン蛋白質が知られており、それらにはインターフェロン、インターロイキン1、2、3、4、5、6および7、コロニー刺激因子、リンホトキシン、腫瘍壊死因子、エリスロポイエチン、その他が含まれる。
【0004】
マクロファージから分泌されるインターロイキン1はヘルパーT細胞を活性化し、体温を上昇させて熱を生じさせ、これにより免疫細胞の活性を増強する。活性化ヘルパーT細胞はインターロイキン2を産生し、インターロイキン2はヘルパーおよびキラーT細胞を刺激して成長および分化させる。ヘルパーT細胞は、もうひとつのリンホカインである、B細胞の増殖をもたらすB細胞成長因子(BCGF)をも産生する。B細胞数の増加にしたがって、ヘルパーT細胞は、いくつかのB細胞に複製を停止し抗体産生を開始する指示を与える、B細胞分化因子(BCDF)として知られるもうひとつのリンホカインを産生する。T細胞はインターロイキン2様の増殖効果を有するリンホカインであるガンマインターフェロン(IF)も産生する。インターフェロンはキラーT細胞の活性化を助け、それらが侵入した微生物を攻撃することを可能にする。BCGF同様に、インターフェロンはB細胞の抗体産生能力を増大させる。インターフェロンはマクロファージにも影響を及ぼし、感染部位にマクロファージを保持し、マクロファージが包み込んだ細胞を消化するのを助ける。マクロファージとT細胞間の各種のリンホカイン信号によるはずみが次第に増加することによって、リンホカインは免疫系反応を増幅し、感染細胞のウイルス蛋白質や他の外来物質の上に覆いかぶさる。おそらく数100もしくはそれ以上の他の多くのリンホカインが存在し、免疫過程に関与している。多くのリンホカインが知られており、また知られていないものが多く存在する。
【0005】
リンホカインは時として細胞間ペプチドシグナルと呼ばれる。科学者の間で、リンホカイン産生者としてクローン化細胞株が広く使用されており、リンホカインmRNAの分離は一般的な技術になってきている。マウス受容体蛋白質4ー1BBは、刊行物(Proc.Natl.Acad.Sci.USA.84,2896-2900, 1987年5月, Immunology)において本発明者が示した技術を用いるT細胞遺伝子の特異的発現に基づいて分離および同定された(非特許文献1)。この刊行物に報告されたプロトコールを用いて、科学者は実質的にすべてのリンホカインを検出する事ができる。この方法は特異に発現した実質的にすべてのmRNAを検出するために計画され、また免疫細胞のmRNA配列は、発現レベルが低く、分泌されたリンホカイン蛋白質の量が少ない場合でも(それらがT細胞およびキラーT細胞に関連しているため)特異に発現する。生物学的に重要であるかまたは活性な分子は最も少ないメッセージによってコードされていることを多くの指摘があることから、本発明者は上記刊行物に記載の分析法によってリンホカインのような生物学的に重要な分子を明らかにすることができるものと考える。その一例は形質転換成長因子(TGF)であり、これは100万クローンにわずか1個しか発現しない。
【0006】
ほとんどのT細胞因子は、従来、アッセイにおいて生物活性を認識し、蛋白質情報を精製することによって同定されて来た。別のアプローチは、特異的発現に基づいて推定されたT細胞遺伝子を分離した後、その未知分子の機能を証明する事である。前記の改良分別スクリーニング法を用いて、本発明者はクローン化ヘルパーT細胞(HTL)L2およびクローン化細胞溶解性T細胞リンパ球(CTL)L3から一連のT細胞サブセット特異的cDNAをクローンした。
【0007】
一連のT細胞サブセット特異的cDNAは、改良分別スクリーニング法を用いてクローン化ネズミT細胞から分離した。cDNA群のいくつかのヌクレオチド配列および発現特性は報告されている。以前に特徴付けがなされていない、マウス受容体蛋白質4−1BBをコードしている遺伝子のひとつがさらに研究された。これらの研究はヒトにおける4−1BB類似体であるH4−1BBの分離をもたらした。
【非特許文献1】Proc. Natl. Acad. Sci. USA. 84, 2896-2900, 1987年5月, Immunology
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明はヒトの受容体蛋白質であるH4−1BBおよび該ヒト受容体蛋白質H4−1BBをコードするcDNAを提供する。
【0009】
本発明の主な目的は、本明細書中にその配列が示された、新規のヒト受容体であるH4−1BBを同定することである。
【0010】
別の本発明の目的は、H4−1BBの細胞外部分と検出蛋白質からなる融合蛋白質を提供することである。
【0011】
さらなる本発明の目的は、H4−1BBをコードするcDNA、受容体蛋白質H4−1BB、H4−1BBに対するモノクローナル抗体およびリガンドを用いる方法を提供することである。
【課題を解決するための手段】
【0012】
分離したcDNAのヌクレオチド配列は論理的に推理したアミノ酸配列と共に本明細書中に開示されている。pH4−1BBとして示されたcDNA遺伝子はAgricultural Research Service Culture Collectionに寄託された(受託番号:NRRL B21131)。
【0013】
cDNAおよびそのフラグメントならびに誘導体は、そのcDNAによってコードされた受容体蛋白質と同様な蛋白質をコードするDNA配列を分離するためのプローブとして用いることができる。ヒト受容体H4−1BBのcDNAはマウス4−1BBのcDNAと約65%相同であり、cDNA4−1BB由来のプローブを用いて分離された。p4−1BBで示されたcDNA遺伝子は、American Type Culture Collection(12301 Parlawn Drive,Rockville,Maryland 20852)に寄託された(ACTT No.67825)。
【0014】
ヒト受容体蛋白質H4−1BBは、
(1)H4−1BBのcDNAを適切な発現ベクターに挿入し、
(2)この発現ベクターを適切なトランスフェクション宿主中にトランスフェクトし、
(c)トランスフェクトさせた宿主を適切な培地中で増殖させ、
(d)培地から受容体蛋白質を精製する
ことによって製造することができる。
【0015】
本発明の蛋白質およびフラグメントならびに誘導体は、
1)ヒト受容体蛋白質H4−1BBに対するリガンドを分離するためのプローブとしてか、
2)H4−1BBリガンドを発現するB細胞の増殖を刺激するために、または
3)H4−1BBの結合をブロックするために用いることができる。
【0016】
B細胞の増殖は、受容体蛋白質H4−1BBに対するリガンドを発現しているB細胞を受容体蛋白質H4−1BBを発現している細胞で処理することによって誘導することができる。この種の出願のために、同様の共刺激免疫系経路が解析されつつある。「Mounting a Targeted Strike on Unwanted Immune Responses」(Jon Cohen,Science,257巻, 8-7-92);「Long Term Survival of Xenogeneic Pancreatic Islet Grafts Induced by CTLA4Ig」(Lenschowら, science,257巻, 7-8-92);および「Immunosuppresion in Vivo by a Soluble Form of the CTLA-4 T Cell Activation Molecule」(Linsleyら, Scienc, 257巻, 7-8-92)を参照。
【0017】
H4−1BBに対するモノクローナル抗体は、受容体蛋白質H4−1BBを発現しているT細胞を抗H4−1BBモノクローナル抗体で処理することによってT細胞の増殖を増強するために使用することができる。いくつかの腫瘍は、潜在的に免疫原性であるが、in vivoにおける有効な抗免疫反応を刺激しない。腫瘍は抗原特異的なシグナルをT細胞に供給できるかも知れないが、T細胞を完全に活性化するのに必要な共刺激シグナルを供給しないかも知れない。メラノーマ細胞における共刺激リガンドB7の発現は、in vivoにおけるネズミのメラノーマの拒絶を誘導しないことが明らかになった(「Tumor Rejection After Direact Co-Stimulation of CD8+T cells by B7-Transfected Melanoma Cells」(Sarah, E. TownsendおよびJames P. Allison, Science, 259巻, 1-5-93)。H4−1BBに対するモノクローナル抗体は、T細胞の増殖と活性化を誘導することが現在知られているのと同様の効果をもたらすことができるかも知れない。
【0018】
ヒト受容体蛋白質H4−1BBに対する細胞膜リガンドを検出するための融合蛋白質が開発された。この蛋白質は受容体蛋白質H4−1BBの細胞外部分と受容体蛋白質H4−1BBの一部分に結合した検出蛋白質(アルカリホスファターゼ)からなる。受容体蛋白質H4−1BBの一部分が細胞膜リガンドに結合し、結合は検出蛋白質に対する比活性アッセイによって検出することができる。融合蛋白質は受容体蛋白質B4−1BBを発現すると思われる細胞の存在下に置かれる。次に、この細胞を、いかなる融合蛋白質もその細胞膜リガンドに結合していないように洗浄する。一旦、洗浄した細胞を検出蛋白質の基質の存在下に置き、検出蛋白質の比活性を測定することができる。
【0019】
以下の詳細な説明では出願人の公開された研究と同様、既知の方法及び研究を引用する。これらの出版物は明確にするために引用して本願に組み込まれており、詳細な説明の最後に列挙されている。
【発明を実施するための最良の形態】
【0020】
マウスレセプター4−1BBの単離及び特性化
マウスレセプター4−1BBのヌクレオチド配列及び推定のアミノ酸配列は図1に示されている。メッセージ鎖のヌクレオチドは5’から3’側方向に番号付されており、番号は配列の両側に記載されている。ヌクレオチド残基1はATG開始コドンのAであり、残基1の5’側にあるヌクレオチドには負の番号が付されている。推定のアミノ酸配列はヌクレオチド配列の下方に記載されている。推定のシグナルペプチドには下線が引かれている。停止コドンは(−−−)で示されている。システイン残基は、点々で明示されている。4−1BB配列の通常でない特徴は、ヌクレオチド1158−1163にAATAAAで示される潜在的なポリアデニル化シグナルがあることである(図1、四角で囲んだ箇所)。遺伝子産物が少なくとも2つの異なる大きさのmRNAを産生することから、このシグナルは機能的であると信じられた。
【0021】
マウス脾細胞、T細胞クローン、及びハイブリドーマ内での4−1BBの転写はコンカナバリンAによって誘導可能であった。4−1BB転写物の発現はシクロスポリンAによって阻害された。4−1BBmRNAは抗原受容体刺激によって誘導可能であったが、クローン化T−細胞内でのIl−2刺激では誘導されなかった(1)。4−1BBcDNAは推定のリーダー配列、潜在的な膜アンカーセグメント、及び既知の受容体タンパク質の他の特徴を含む、256アミノ酸ペプチドをコードしている。従って、4−1BBの配列は受容体タンパク質のそれに一致しているように見えるが、発現パターンはリンホカインのそれに類似している。
【0022】
細胞表面における4−1BBの主な種は55ーkDaの2量体のようである。4−1BBはまた、30−kDaの単量体、そして恐らく110−kDaの4量体としても存在するようである。これらの4−1BB種は均質な細胞集団(T−細胞クローンF1)から免疫沈降されたので、いずれも各細胞上に共存している可能性がある。4−1BBが細胞表面でホモダイマーとして存在するか否かを決定するには、4−1BB単量体と2量体からのペプチド消化物を比較する必要があるだろう。インシュリン受容体(2)、B−細胞表面免疫グロブリン受容体(3)、T−細胞Ag受容体(4)、CD28共刺激(costimulatory)受容体(5)、及びCD27T細胞抗原(6)等の様々な細胞表面受容体はジスルフィド結合したサブユニットからなる。リガンドの結合及びその後の生化学的シグナル伝達(signaling)には、受容体の2量体化が必要かもしれない。
【0023】
4−1BBは休止期のT細胞には発現しないが、T細胞に対して完全な増殖刺激を与える活性化剤によって誘導可能である。PMAとイオノマイシンとの組み合わせは、T細胞の増殖に必要なこれらのシグナルをまねることができる。PMA又はイオノマイシン単独で4−1BBmRNAを誘導することができるが、PMAとイオノマイシンの併用により4−1BBの至適発現が得られた。しかも、4−1BBの発現は一過性でなかった。精製脾T細胞を固定化抗−CD3で刺激すると、4−1BBmRNAが発現され、この発現は刺激後96時間まで維持された。4−1BBが細胞周期の進行期間を通して発現されることを確認するためには、細胞周期の解析が必要であろう。
【0024】
4−1BBは構造的に神経成長因子受容体スーパーファミリーのメンバーに関連している。これらの受容体は構造上類似のリガンド−結合特性(システイン−豊富領域)を有するか、これらタンパク質の細胞質ドメインは非保存的であり、それがトランスメンブランシグナル伝達に多様性を与えているかもしれない。このファミリーには、T又はB細胞の活性化工程に関与しているメンバーもある。OX−40、CD40及びCD27抗原に関するインビトロ機能性データがある。OX−40に対する抗体は混合リンパ球反応におけるT細胞の応答性を増大し(7)、CD40に対する抗体はPMA又はCD20抗体のような共活性化剤(coactivator)の存在下でB−細胞の増殖を促進し、インビトロでIL−4と相乗的に作用してB−細胞の分化を誘導し長期正常B−細胞株を生成させる(8)。CD27分子上のエピトープを認識する1つのイノノクローナル(inonoclonal)抗体(抗−1A4)は、カルシウム動態化、IL−2分泌、ヘルパーT細胞機能、及びT細胞増殖を阻害した。一方、もう1つの抗CD27mAbであるCLB−CD27/1はPHA又は抗CD3mAbによって刺激されたヒトT細胞の増殖を促進した(6)。これらの結果はCD27分子がT細胞の活性化に重要な役割を果たしていることを示している。TNFR類、NCFR、及びCD40を除いて、スーパーファミリーのメンバーが結合するリガンド又は細胞表面分子はまだ同定されていない。受容体が結合するリガンドの同定及び特性化は4−1BBの生理学的な役割のより良い定義のために役立つであろう。
【0025】
細胞表面4−1BBがT細胞活性化に貢献しうるか否かを確かめるために、抗4−1BB53A2を4−1BBのアンタゴニストとして用いた。これらのデータは実際、4−1BBが、T細胞の活性化及び増殖の間、副シグナル分子として機能する能力を有することを示唆していた。固定化抗CD3で刺激した、精製脾由来T細胞に可溶性53A2を加えると、抗CD3単独で刺激した場合に比較して、3Hチミジンの取り込みが増大した。3回の独立実験における、この増大パターンの範囲は2〜10倍であった。
【0026】
Bretcher及びCohnの元々の2つのシグナルモデルで、彼らはシグナル1、T細胞抗原受容体(TCR)の占有、では補助細胞から供給されるシグナル2の不在下でT細胞が不活化されると提案した。これは以後様々な研究で確認された(9)。T細胞上の潜在的な共刺激受容体としての補助細胞CD28の同定は、T細胞の最適な増殖に必要な補助シグナル(群)の特性化を開始するのに重大な寄与であった(10)。他の細胞表面分子がこれらの共刺激活性化要求に寄与している可能性がある(11)。
【0027】
4−1BBを介して伝達される生化学的シグナルは、完全には分かっていない。考慮されている1つの可能性は、4−1BBが推定のp56kkチロシンキナーゼ結合ドメインをその細胞質尾部に含有しているとの観察であった。後に、p56kkチロシナーゼキナーゼは4−1BBに結合することが決定された。4−1BBを介するシグナル伝達が、その発現がT細胞の活性化及びその後の増殖に必要である、IL−2及びIL−2受容体のような遺伝子を調節できるかどうかを決定することも価値ある。
【0028】
神経成長因子受容体(NGFR)ファミリーのメンバーの正確な機能は多様であるように思われるが、これらの分子が様々な方法で、それらが発現される特定の細胞型の応答性又は生存可能性の維持に貢献しているかもしれない、というテーマが出現する。例えば、NGFはインビトロ及びインビボでのニューロンの生存に絶対的に必要である(12)。可溶性抗CD40モノクローナル抗体によるCD40のクロスリンキングは、胚中心中心細胞がインビトロでアポプトーシスを受けることを妨げる(13)。CD40を介して伝達されるシグナルもまた、分化因子に対する応答性の維持を補助しうる。CD40と抗−CD40F(ab’)2フラグメントのIL−4の存在下での結合(ライゲーション)はIgE合成の非常な増大を誘導した(14)。また、IL−10及びトランスフォーミング成長因子ーβで処理した、抗CD40活性化天然(固有)B細胞はIgA分泌を行うようになった(15)。
【0029】
4−1BBは、NGFRスーパーファミリーと共通する分子の特徴を有することに加えて、酵母e1F−2βタンパクの推定の亜鉛フィンガー構造を有することが分かった(16)。4−1BBはまたショウジョウバエのアプサンス(absentia)のsina sevenと、保存領域を共有しており、それは適正な光受容体細胞の発展に必要である(17)。その特定の領域はまた、その発現がcAMPによる凝集の期間に特異的に誘導される、DictyosteliumのDG17遺伝子のタンパク質生成物と類似している(18)。
【0030】
この領域はC−X2−C−X9−C−X3−H−X3−C−X−Cパターンを形成しており;システイン及びヒスチジンは4−1BB、sina及びDG17タンパク質内の、同様のスペースに保存されている。4−1BBタンパク質とsinaタンパク質との間で24アミノ酸の内、10アミノ酸が同一であり、24個の内3個は保存的置換である。保存のパターンはこれらのアミノ酸が機能的に重要であることを示している。sinaタンパク質は核に局在しており、これは、細胞内で調節機能を果たしていることを示唆している。4−1BBのアミノ酸配列が、亜鉛フィンガーモチーフ、核タンパク質、及び受容体ドメインの様な特徴を含有しているという事実は、4−1BBが細胞増殖及び分化の間に多様な役割を果たしていることを示している。
【0031】
4−1BBはT−細胞−APC相互作用に関連する他の細胞表面分子を表しているかもしれない。4−1BB−AP融合タンパク質は、成熟B−細胞株、抗μ−活性化初代B細胞、及び成熟マクロファージ細胞株と特異的に結合する。4−1BB−APは、未成熟な、B細胞株及びマクロファージ細胞株、T細胞クローン、T細胞株、T細胞の初代培養、及び種々の非リンパ様細胞株とは低レベル又は僅かなレベルで結合する。4−1BB−APは成熟B細胞及びマクロファージと結合するので、4−1BBの結合に際して伝達されるシグナルは何等かの方法でAPC機能を調節する可能性がある。この可能性は探索の余地がある。
【0032】
Chalupnyら(19)は4−1BBRg、4−1BBの細胞外領域とヒトIgGのFc領域からなる融合タンパク質、が細胞外マトリックス(ECM)に結合すると提案した。4−1BBRg結合はヒトビトロネクチンとの間で最高レベルであった。データは示されていないが、4−1BB−AP及びマイクロタイター上、1ウエル当たり0.007μg−10μgで固定化されたヒトビトロネクチンを用いてELISAが行われた[Yelios Pharmaceuticals/GIBCO-BRL, Grand Island, NY.)。AP活性に基づいて、4−1BB−APの結合は観察されなかった。細胞表面に外因的に結合しているタンパク質(細胞外マトリックス構成成分の可能性あり)に4−1BB−APが結合する可能性を排除するために、結合アッセイの前にB細胞リンパ腫を酸性条件で洗浄した。4−1BB−APは依然として成熟B−細胞に特異的に結合した。B細胞及びマクロファージ上で特異的に発現される4−1BB−リガンドが存在するか否か、及び4−1BB−APが特定の結合条件下でECMと結合するか否かはなお決定されるべきことである。ECMは4−1BBの特定の細胞表面リガンドへの結合を容易にしている可能性がある。
【0033】
B細胞とヘルパーT細胞とは、T細胞上の特異的な対応受容体に結合するB細胞上の受容体によって互いに相互作用する。この相互作用の結果、これら2つの細胞種間での生化学的シグナル伝達リレーのカスケードが起こると考えられている(20)。この相互作用が進行するにつれ、これら細胞は細胞周期のS期に入るようになる。TCRとT細胞上のCD4、およびB細胞上のプロセスされた抗原−MHCIIとの間の初期相互作用は、B細胞が細胞周期に入るという結果にはならない(21)。しかしながら、インビトロ系での研究は、T細胞が一旦刺激されると、B細胞を細胞周期に入らせうる新たに合成されたまたは修飾された細胞表面分子を発現することを示唆している(22、23)。このT細胞機能は抗原特異的でもMHCに限定されたものでもない(24)。加えて、B細胞活性化の活性化Th誘導には可溶性因子を必要としない(25)。B細胞が細胞周期に入ったら、IL−4がB細胞をG1期からS期への進行を誘導する。活性化T細胞またはT細胞膜がB細胞を細胞周期に入るのを促進する能力は、シクロヘキシミドかまたはシクロスポリンA処理により阻止することができる(26、27)。これらの新たに発現される膜タンパク質は、その誘導特性において「リンホカイン様」であると思われる。
【0034】
4−1BBは、B細胞のコスティミュレーターとしての要求を満たす発現特性を有する。4−1BBは抗CD3またはTCR媒体T細胞刺激によって誘導することができ、その発現はシクロスポリンA並びにシクロヘキシミド処理に感受性である(28)。興味深いことに、パラホルムアルデヒド固定したSF21−4−1BB細胞は、B細胞増殖の誘導において抗μと共同作用する(synergized)。SF21−4−1BBによる脾臓B細胞のコスティミュレーションは、抗μの最適用量(10μg/ml)および最適下限用量(1.0〜0.1μg/ml)で起こる。休止B細胞にSF21−4−1BB細胞を加えても有意のB細胞増殖とはならなかった。SF21−4−1BBは、B細胞増殖の誘導においてTPAまたはイオノマイシン、または最適下限濃度のLPSとは共同作用しなかった。
【0035】
大量の組換え可溶性タンパク質を発現させるためにバクロウイルス系が用いられてきているが、この系は組換え細胞表面タンパク質の発現のために用いることができる。バクロウイルス感染は、細胞当たりベースで均一な高レベルの組換えタンパク質を発現させるための都合の良い手段をもたらす。SF21細胞を単独で加えても有意レベルのコスティミュレーションとならなかったことは注目に値する。このことは、それ自身で強力なコスティミュレーター活性を示しうるcos−またはL−細胞株を用いるに際して潜在的な問題となりうる。
【0036】
NGFRスーパーファミリーの他の成員であるCD40はB細胞表面で発現され、活性化T細胞上で発現される分子であるgp39と相互作用する。マウスgp39タンパク質(29)およびヒトgp39タンパク質(30)をコードするcDNAはクローニングされている;この細胞表面分子は、腫瘍壊死因子と相同性を有するタイプIIの膜タンパク質である。ノエル(Noelle)ら(31)は、CD40−免疫グロブリン融合タンパク質がT細胞により誘導されたB細胞増殖および分化を用量に依存した仕方で阻止しうることを見いだした。アーミテージ(Armitage)らは、マウスgp39のcDNAを単離し、gp39がコスティミュレーター(co-stimuli)の不在下でB細胞増殖を誘導することができ、IL−4−の存在下でIgE産生という結果になることを示した。ホレンバウ(Hollenbaugh)ら(32)は、ヒトgp39でトランスフェクトしたCOS細胞がヒトB細胞増殖を誘導するうえでTPAかまたは抗CO20と共同作用しうること、およびコスティミュレーターなしでは低レベルでしかB細胞を刺激できないことを示している。これらデータは、CD40がT細胞との物理的接触の間にシグナルを伝達するB細胞表面分子の一つであることを示している。
【0037】
細胞表面受容体は、可溶性因子かまたは隣接細胞上に発現された他の細胞表面分子のいずれかと相互作用することによって外部環境と連絡している。細胞表面受容体と相互作用する可溶性因子によって送達される生化学的シグナルに対して細胞−細胞接触によって送達される生化学的シグナルの役割は明らかでない。NGFRが2以上のリガンドと結合するのと同様に、NGFRスーパーファミリーはTNFRIおよびIIに対して普通でない。TNFRIおよびIIはともにTNF−αおよびTNF−Rの両者に結合する(33)。NGFRは、NGF、脳由来神経栄養因子、およびニューロトロフィン−3に結合する(34)。
【0038】
加えて、一つのリガンドが細胞表面リガンドとしても可溶性リガンドとしても機能する。CD4−0リガンドであるgp39に関する最近の証拠は、このリガンドが膜結合したリガンドとしても可溶性リガンドとしても存在しうることを示唆している(35)。4−1BBが分泌され、B細胞と可溶性形態並びに膜結合形態で相互作用する可能性がある。NGFR受容体ファミリーの成員であるCD27は、T細胞上で発現され、分泌されるとともに細胞表面上に発現される(36)。1を越えるリガンド(可溶性および細胞表面)が4−1BBに結合する可能性もある。
【実施例】
【0039】
ヒト同族体、H4−1BBの単離
マウス4−1BBのヒト同族体(H4−1BB)を単離するため、2セットの複製連鎖反応(PCR)プライマーを設計した。PCRプライマーを設計するため、神経増殖因子受容体(NGFR)スーパーファミリーの成員間でアミノ酸配列を比較した。なぜなら4−1BBは該スーパーファミリーの成員であるからである(37)。使用したアミノ酸配列は、マウス4−1BB(38)、ヒトNGFR(39)、ヒト腫瘍壊死因子受容体(33)、ヒトCD40(40)、およびヒトCD27(6)であった。NGFRスーパーファミリー間での保存配列の領域を選択した。
【0040】
前(forward)プライマーI(H4−1BBFI)は、マウス4−1BBのアミノ酸36〜41の範囲、前プライマーII(HR−1BBFII)はアミノ酸52〜58の範囲にある。逆(reverse)プライマーI(H4−1BBRI)はマウス4−1BBのアミノ酸116〜121の範囲、逆プライマーII(H4−1BBRII)はアミノ酸122〜128の範囲にある。マウス4−1BBにおいてPCRプライマーとして用いた領域は図1に示してある。
【0041】
各プライマーのオリゴヌクレオチド配列は以下の通りである。
【表1】

【0042】
正常な健常者から末梢血リンパ球を単離し、PMA(10ng/ml)およびイオノマイシン(1μM)で活性化した。これらリンパ球からmRNAを単離した。逆転写酵素を用い、ヒトリンパ球mRNAを一本鎖cDNAに変換させた。ついで、一本鎖cDNAをTaqポリメラーゼと上記プライマーとの組わせにより増幅させた。プライマーの組わせは以下の通りであった:H4−1BBFIとH4−1BBRI;H4−1BBFIとH4−1BBRII;H4−1BBFIIとH4−1BBRI;およびH4−1BBFIIとH4−1BBRII。
【0043】
H4−1BBFIIとH4−1BBRIIとのプライマーセットは〜240bpの特異的バンドを生成した。この240bpは、ヒト同族体タンパク質がマウス4−1BBとサイズが同様であるとした場合にヒト4−1BBの予期されるサイズである。このPCR生成物(240bp)をPGEM3ベクター中にクローニングし、配列決定した。一つのオープンリーディングフレームはマウスの4−1BBと〜65%同一であった。それゆえ、この240bp PCR生成物はマウス4−1BBのヒト同族体であると結論した。この240bp PCR生成物を用いて活性化ヒトTリンパ球のλgt11 cDNAライブラリーをスクリーニングした。〜0.85kb cDNAが単離させた。このcDNAの配列を図2に示してあり、予測されるアミノ酸配列を図2bに示してある。同情報は本明細書中の配列表の配列1に示してある。
【0044】
H4−1BB−AP融合タンパク質を発現する発現プラスミドを構築した。シグナル配列および全細胞外ドメインをコードする配列を含むH4−1BB cDNAの5'部分をPCRにより増幅させた。正しい方向にクローニングするため、前プライマーの5'末端上にHindIII部位を、および逆プライマーの5'末端上にBglII部位をそれぞれ生成させた。
【0045】
HindIII−BglII H4−1BBフラグメントを哺乳動物発現ベクターAPtaq−1中、ヒト胎盤アルカリホスファターゼ(AP)をコードする配列の上流に挿入した。H4−1BBの5'部分の増幅に用いたオリゴヌクレオチドPCRプライマーは以下の通りである。
【表2】
【0046】
H4−1BB−APは、ヒト4−1BBのリガンド(すなわち、H4−1BBL)を発現する細胞および組織を同定するのに用いるであろう。マウス4−1BBを用いた研究は、4−1BBのリガンドが細胞表面上に存在することを示した。B細胞およびマクロファージが4−1BBを発現する主要な細胞である。H4−1BBLもまたヒトのB細胞およびマクロファージ上で発現されることが予想される。
【0047】
哺乳動物発現cDNAライブラリーは、H4−1BBLを発現するヒト細胞株から生成されるであろう。このライブラリーは[125]I−標識したH4−1BB−APによりスクリーニングされるであろう。ついで、H4−1BBLのcDNAが単離され、特徴付けられるであろう。ついで、可溶性組換えH4−1BBLが生成されるであろう。H4−1BB−APおよびH4−1BBLの両者とも、下記のように免疫応答を抑制または促進するために用いられるであろう。H4−1BBおよびH4−1BBLに対するモノクローナル抗体が産生されるであろう。 マウス4−1BBでの研究によれば、4−1BBはコスティミュラトリーシグナルとして働く。H4−1BBはT細胞活性化のコスティミュラトリーシグナルとして働くことが予想される。マウス4−1BBは、B細胞の増殖および分化を助ける。H4−1BBも同様であろうことが予想される。H4−1BB−AP、H4−1BBLおよびモノクローナル抗体は、ヒトの免疫応答を抑制または促進するために用いることができる。
【0048】
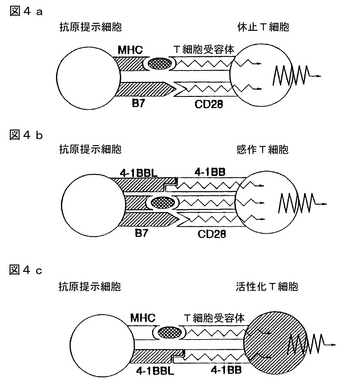
図3aおよび3bはT細胞活性化に関与する分子を示す。初期のT細胞活性化の間(認識(cognitive)期)、休止T細胞はTCR/CD3複合体および他の「アクセサリー(accessory)」分子を発現する。これら構成的に発現される分子のうち、CD4(またはCD3)、LFA−1およびCD28がおそらくコスティミュラトリーシグナルを受け取る分子である。これら「アクセサリー」コスティミュラトリーシグナルとともにTCR/CD3複合体と相互作用することにより、CD28、CTLA4、および4−1BBなどの他の受容体が引き続き発現される。これら新たに発現された分子は、おそらく、T細胞活性化の後期の段階で別の重要なコスティミュラトリーシグナルを受け取るであろう(クローン増殖)。
【0049】
免疫応答の抑制
図4a−cは正常なT−細胞活性経路を示す。図5a−cは4−1BBの可溶性キメラによる免疫応答阻害を示す。もし4−1BBがT細胞活性化になんらかの役割を果たすなら、抗原−提示細胞上のそのリガンドに対する相互作用の阻害がT−細胞依存免疫応答の抑制をもたらす筈である。カウンター−受容体B7に対するCD28の相互作用の阻害が、種々の程度でインビボの抗体産生および細胞−媒介免疫応答の両者を抑制することは十分に立証されている。両者の相互作用の阻害はより効果的な免疫抑制をもたらす。そのリガンドに対する4−1BBの相互作用の阻害はCD28/B7相互作用がもはや関連性がなくなった場合の活性化プロセスの後期の段階で重要になり得る。
【0050】
マウス受容体4−1BBおよびマウスリガンド4−1BBLについて、上記に詳述したように、H4−1BB−APの添加はH4−1BBL発現細胞を覆い、H4−1BBとH4−1BBL間の正常な相互作用を阻害する。これは免疫応答を抑制することになる。この種の免疫応答は抗原−特異的である。従って、antiCD3またはシクロスポリンA処置によって生じる一般化された免疫応答は回避される。H4−1BB−AP処置はある種の自己免疫疾患の処置に用いることができ、臓器移植を容易にする。
【0051】
免疫増強
H4−1BBはT−細胞活性化の後期の段階で機能し、T−細胞活性化の完成のための重要な細胞になり得る。多くの腫瘍は腫瘍−特異抗原を示す。しかしながら、なぜ免疫原性腫瘍が宿主免疫を免れ得るかの理由の1つは腫瘍−反応性T−細胞が不十分な共刺激(costimulation)を受けているからである。従って、H4−1BBなどの共刺激分子の腫瘍への導入は、細胞傷害性T細胞(CTL)の抗腫瘍免疫を増強し得た。H4−1BBLは細胞−特異様式で発現され得る。例えば、H4−1BBLは、例えば、チロシナーゼプロモーターなどのメラノサイト−特異プロモーターを用いる黒色腫中に発現され得る。H4−1BBL−発現黒色腫はH4−1BBを介して細胞傷害性T細胞を刺激し、黒色腫−特異CTLを活性化する。活性化された黒色腫−特異CTLは黒色腫を撲滅させ得る。
【0052】
引用により明細書の記載とされる文献の付録
1. Smith, C.A., Davis, T., Anderson, D., Solam, L., Beckmann, M.P., Jerzy, R., Dower, S.K., Cosman, D., and Goodwin, R.G. 1990. A receptor for tumor necrosis factor defines an unusual family of cellular and viral proteins.
Science 248:1019-1023.
2. Ebina, Y., L. Ellis, K. Jaruagin, M. Edery, L. Graf, E. Clauser, J. On, F. Marizrz, Y.W. Kan, J.D. Goldfine, R.A. Roth and W.J. Rutter, 1985, The human insulin receptor cDNA: the structural basis for hormone-activated transmembrane signalling, Cell 40:747.
3. Vassali, R., R. Tedghi, B. Listowska-Bernstein, A. Tartakoff and J.C. Jaton, 1979, Evidence for hydrophobic region within heavy chains of mouse B lymphocyte membranebound IgM, Proc. Natl. Acad. Sci. USA76:5515.
4. Haskins, K., R. Kubo, J. White, M. Pigeon, J. Kappler and P. Marrack, 1983, The major histocompatability complexrestricted antigen receptor on T cells I Isolation with monoclonal antibody, J. Exp. Med. 157:1149.
5. Lesslaver, W. and H.Gmunder, 1986, Biochemical characterization of the 9.3 antigens of human T-cells: stimultaneous expression of disulfide-bonded 90-Kiladalton dimers and free subunits at the cell surface, Mol. Immunol. 23:271.
6. Van Lier, R., J. Borst, T. Vroom, H. Klein, P. Mourik, W. Zeijlemaker and C. Melife, 1987, Tissue distribution and biochemical and functional properties of Tp55 (CD27) a novel T cell differentiation antigen, J. Immunol. 139:1589.
7. Mallett, S., S. Fossum and A. Barclay, 1990, Characterization of the MRC OX40 antigen of activated CD4 positive T lymphocytes-a molecule related to nerve growth factor receptor, EMBO J. 9:1603.
8. Banchereau, J., P. Paoli, A., Valle, E. Garcia and F. Roussel, 1991, Long-term human B cell lines dependent on interleukin-4 and antibody to CD40, Science 251:70.
9. Moeller, D.L., M.K. Jenkins and R.H. Schwartz, 1989, Clonal expansion versus functional colonal inactivation: a co-stimulatory signalling pathway determines the outcome of T cell antigen receptor occupancy, Ann. Rev. Immunol. 7:445.
10. June, D.H., J.A. Ledbetter, P.S. Linsley and C.B. Thompson, 1989, Role of CD28 receptor in T cell activation, Immunol. Today 11:211.
11. Yang, L., B. Jones, A. Aruffo, K.M. Sullivan, P.S. Linsley and C.A. Janeeway, Jr., 1992, Heat stable antigen is a co-stimulatory molecule for CD4 T cell growth, J. Exp. Med. 175:437.
12. Yamori, T., 1992, Molecular mechanisms for generation of neural diversity and specificity: foles of polypeptide factors in development of post-mitotic neurons, Neurosic. Res. 12:545.
13. Liu, Y.J., D.E. Joshua, G.T. Williams, C.A. Smith, J. Gordon and I.C. M. MacLennan, 1989, Mechanism of antigendriven selection in germinal centres, Nature, 342:929.
14. Jabara, H. H., s. M. Fu, R.S. Geha and D. Vercelli, 1990, CD40 and IfE: synergism between anti-CD40 momoclonal antibody and interleukin 4 in the induction of IgE synthesis by highly purified human B cells, J. Exp. Med. 172:1861.
15. Defrance, R., B. Vanbervliet, F. Briere, I. Durnad, F. Roussle and J. Banchereau, 1992, Interleukin 10 and transforming growth factor beta cooperate to induce anti-CD40 activated naive human B cells to secrete immunoglobulin A, J. Exp. Med. 175:671.
16. Donahue, T., Cigan, A., Pahich, E. and Valavicius, B., Mutations at a Zn(II) finger motif in the yeast elF-2beta gene alter ribosomal start-site selection during the scanning process, Cell 54 (1988) 621-632).
17. Carthew, R.W and Rubin, G.M., seven in absentia, a gene required for specification of R7 cell rate in the Drosophila eye, Cell, 63 (1990) 561-577.
18. Driscoll, D.M. and Williams, J.G., Two divergently transcribed genes of Dictyostelium discoideum are cyclic AMP-inducible and coregulated during development, Mol. and Cell. Biol. 7 (1987) 4482-4489.
19. Chalupny, N.J., Peach, R., Hollenbaugh, D., Ledbetter, J.A., Farr, A.G. and Aruffo, A., 1992, Proc. Natl. Acad. Sci USA 89:10360-10364.
20. Noelle, R.J., and Snow, E.C., 1991, The FASEB J. 5:2770-2776.
21. Noelle, R. and Snow, E., 1990, Immunol. Today 11:361-368.
22. Zurawski, G., Benedik, M., Kamb, B.J., Abrams, J.S., Zurawaki, S.M. and Lee, F.D. (1986) Science 232.772-775.
23. Kinachi, T. (1986) Nature 325,70-73.
24. Gershenfeld, H.K. and Weissman, I.L. Science (1986) 232.854-858.
25. Biggin, M., Gison, T. and Hung, G. (1983 Proc. Natl. Acad. Sci. USA 80,3963-3965.
26. Hodgkin, P.D., Yamashita, L.C., Coffman, R.L. and Kehry, M.R., 1990, J. Immunol. 145:2025-2034.
27. Barlett, W.C., McCann, J., Shephaer, D.M., Roy, M. and Noelle, R.J., 1990, J. Immunol. 145:3956-3962.
28. Kwon, B.S., Kestler, D.P., Eshhar, Z., Oh, K., and Wakulchik, M. 1989. Expression characteristics of two potential T cell mediator genes. Cell. Immunol. 121:414-422.
29. Armitage, R., Fanslow, W., Strockbine, L., Sato, T., Clifford, K., MacDuff, B., Anderson, D., Gimpel, S., Davis Smith, T., Maliszewski, C., Clark, E., Smith, C., Grabstein, K., Cosman, D. and Spriggs, M., 191, Nature 357:80-82.
30. Kwon, B., Kestler, D., Lee, E., Wakulchik, M. and Young J. (1988) J. Exp. Med) (1988) (In press).
31. Noelle, R.J., Roy, M., Shepherd, D.M., Stamenkovic, I., Ledbetter,J.A. and Aruffo, A., 1992, Proc. Natl. Acad. Sci. USA 89:6550-6554.
32. Hollenbaugh, D., Grosmaier, L.S., Kullas, C.D., Chalupny, N.J., Braesch-Andersen, S., Noelle, R.J., Stamenkovic, I., Ledbetter,J.A. and Aruffo, A., 1992, EMBO11:4314-4321.
33. Schall, T.J., M. Lewis, K.J. Koller, A. Lee, G.C. Rice, G.H. W. Wong, T. Gatanaga, G.A. Granger, R. Lentz, H. Raab, W.J. Kohr and D.V. Goeddel, 1990, Molecular cloning and expression of a receptor for human tumor necrosis factor, Cell 61:361.
34. Klein, R., Nanduri, V., Jing, S., Lamballe, F., Tapley, P., Bryant, S., Cordon-Cardo, C., Jones, K.R., Reichardt, L.F. and Barbacid, M., 1991, Cell 66:395-403.
35. Armitage, R.J., Sato, T.A., Macduff, B.M., Clifford, K.N., Alpert, A.R., Smith, C.A. and Fanslow, W.C., 1992, Eu8r. J. Immunol. 22:2071-2076.
36. Hintzen, R.Q., deJong, R., Hack, E.E., Chamuleau, M., de Vries, E.F.R., ten Berge, I.J.M., Borst, J. and van Lier, R.A.W., 1991, J. Immunol. 147:29-35.
37. Mallett, S., and Barclay, A.N. 1991. A new superfamily of cell surface proteins related to the nerve growth factor receptor. Immunol. Today. 12:220-223.
38. Kwon, B.S., and Weissman, S.M. 1989. cDNA sequences of two inducible T-cell genes. Proc. Natl. Acad. Sci. USA. 86:1963-1967.
39. Johnson, D., Lanahan, A., Buck C.R., Sehgal, A., Morgan, C., Mercer, E., Bothwell, M., and Chao, M. 1986. Expression and structure of the human NGF receptor. Cell 47:545-554.
40. Stamenkovic, I., Clark, E., and Seed, B. 1989. A Blymphocyte activation molecule related to the nerve growth factor receptor and induced by cytokines in carcinomas. EMBO. J. 8:1403-1408.
【0053】
上記の記載は、詳細な説明および解説の目的のために特許法の要件に従って、本発明の具体的な実施態様を述べたものである。しかしながら、本発明の範囲および要旨をはずれることなく、多くの修正および変更が可能であることは当業者にとって自明である。下記の特許請求の範囲はそのようなすべての修正を包含するものと理解されなければならない。
【図面の簡単な説明】
【0054】
【図1a】マウスレセプター(受容体)タンパク質4−1BBのcDNA配列及びヒト同族体H4−1BBを得るためのPCRプライマーとして用いた領域を示す。
【図1b】図1aの続き。マウスレセプター(受容体)タンパク質4−1BBのcDNA配列及びヒト同族体H4−1BBを得るためのPCRプライマーとして用いた領域を示す。
【図2a】ヒトレセプター(受容体)H4−1BBのヌクレオチド配列を示す。
【図2b】ヒトレセプター(受容体)H4−1BBの推定アミノ酸配列を示す。
【図3】T−細胞の活性化に関与する分子を示す。
【図4】正常なT−細胞活性化経路を示す。
【図5】T−細胞活性化経路における遮断工程における、(a)CTLA4−1g単独使用、(b)4−1BB/AP及びCTLA4−1g併用、および(c)4−1BB/AP単独使用を示す。
【技術分野】
【0001】
本発明は、本発明者によるT細胞遺伝子の特異的発現によって分離および同定された類似のネズミ(マウス)受容体(レセプター)蛋白質4−1BBを用いた研究に基づいて分離および同定された以前に知られていないヒト受容体蛋白質H4−1BBに関する。
【背景技術】
【0002】
ヒトおよび他の種の免疫系は骨髄で作られる白血球を必要とし、その白血球には食細胞、リンパ球およびB細胞が含まれる。現在知られているように、食細胞にはこの系からウイルス蛋白質のような望ましくない物質を取り除くマクロファージ細胞が含まれる。リンパ球にはヘルパーT細胞、キラーT細胞およびB細胞ならびにサプレッサーT細胞に分類されるリンパ球を含む他の細胞が含まれる。キラーT細胞は細胞に物理的に突入し、またヘルパーT細胞は全過程を促進する。どのような事象においても、免疫過程はリンパ球によって促進される。
【0003】
リンホカインは免疫細胞が互いに連絡を取り合うための蛋白質である。科学者は免疫学的疾患に対して治療的に使用するためにそれらの蛋白質の充分量を製造する。多くのリンホカイン蛋白質が知られており、それらにはインターフェロン、インターロイキン1、2、3、4、5、6および7、コロニー刺激因子、リンホトキシン、腫瘍壊死因子、エリスロポイエチン、その他が含まれる。
【0004】
マクロファージから分泌されるインターロイキン1はヘルパーT細胞を活性化し、体温を上昇させて熱を生じさせ、これにより免疫細胞の活性を増強する。活性化ヘルパーT細胞はインターロイキン2を産生し、インターロイキン2はヘルパーおよびキラーT細胞を刺激して成長および分化させる。ヘルパーT細胞は、もうひとつのリンホカインである、B細胞の増殖をもたらすB細胞成長因子(BCGF)をも産生する。B細胞数の増加にしたがって、ヘルパーT細胞は、いくつかのB細胞に複製を停止し抗体産生を開始する指示を与える、B細胞分化因子(BCDF)として知られるもうひとつのリンホカインを産生する。T細胞はインターロイキン2様の増殖効果を有するリンホカインであるガンマインターフェロン(IF)も産生する。インターフェロンはキラーT細胞の活性化を助け、それらが侵入した微生物を攻撃することを可能にする。BCGF同様に、インターフェロンはB細胞の抗体産生能力を増大させる。インターフェロンはマクロファージにも影響を及ぼし、感染部位にマクロファージを保持し、マクロファージが包み込んだ細胞を消化するのを助ける。マクロファージとT細胞間の各種のリンホカイン信号によるはずみが次第に増加することによって、リンホカインは免疫系反応を増幅し、感染細胞のウイルス蛋白質や他の外来物質の上に覆いかぶさる。おそらく数100もしくはそれ以上の他の多くのリンホカインが存在し、免疫過程に関与している。多くのリンホカインが知られており、また知られていないものが多く存在する。
【0005】
リンホカインは時として細胞間ペプチドシグナルと呼ばれる。科学者の間で、リンホカイン産生者としてクローン化細胞株が広く使用されており、リンホカインmRNAの分離は一般的な技術になってきている。マウス受容体蛋白質4ー1BBは、刊行物(Proc.Natl.Acad.Sci.USA.84,2896-2900, 1987年5月, Immunology)において本発明者が示した技術を用いるT細胞遺伝子の特異的発現に基づいて分離および同定された(非特許文献1)。この刊行物に報告されたプロトコールを用いて、科学者は実質的にすべてのリンホカインを検出する事ができる。この方法は特異に発現した実質的にすべてのmRNAを検出するために計画され、また免疫細胞のmRNA配列は、発現レベルが低く、分泌されたリンホカイン蛋白質の量が少ない場合でも(それらがT細胞およびキラーT細胞に関連しているため)特異に発現する。生物学的に重要であるかまたは活性な分子は最も少ないメッセージによってコードされていることを多くの指摘があることから、本発明者は上記刊行物に記載の分析法によってリンホカインのような生物学的に重要な分子を明らかにすることができるものと考える。その一例は形質転換成長因子(TGF)であり、これは100万クローンにわずか1個しか発現しない。
【0006】
ほとんどのT細胞因子は、従来、アッセイにおいて生物活性を認識し、蛋白質情報を精製することによって同定されて来た。別のアプローチは、特異的発現に基づいて推定されたT細胞遺伝子を分離した後、その未知分子の機能を証明する事である。前記の改良分別スクリーニング法を用いて、本発明者はクローン化ヘルパーT細胞(HTL)L2およびクローン化細胞溶解性T細胞リンパ球(CTL)L3から一連のT細胞サブセット特異的cDNAをクローンした。
【0007】
一連のT細胞サブセット特異的cDNAは、改良分別スクリーニング法を用いてクローン化ネズミT細胞から分離した。cDNA群のいくつかのヌクレオチド配列および発現特性は報告されている。以前に特徴付けがなされていない、マウス受容体蛋白質4−1BBをコードしている遺伝子のひとつがさらに研究された。これらの研究はヒトにおける4−1BB類似体であるH4−1BBの分離をもたらした。
【非特許文献1】Proc. Natl. Acad. Sci. USA. 84, 2896-2900, 1987年5月, Immunology
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明はヒトの受容体蛋白質であるH4−1BBおよび該ヒト受容体蛋白質H4−1BBをコードするcDNAを提供する。
【0009】
本発明の主な目的は、本明細書中にその配列が示された、新規のヒト受容体であるH4−1BBを同定することである。
【0010】
別の本発明の目的は、H4−1BBの細胞外部分と検出蛋白質からなる融合蛋白質を提供することである。
【0011】
さらなる本発明の目的は、H4−1BBをコードするcDNA、受容体蛋白質H4−1BB、H4−1BBに対するモノクローナル抗体およびリガンドを用いる方法を提供することである。
【課題を解決するための手段】
【0012】
分離したcDNAのヌクレオチド配列は論理的に推理したアミノ酸配列と共に本明細書中に開示されている。pH4−1BBとして示されたcDNA遺伝子はAgricultural Research Service Culture Collectionに寄託された(受託番号:NRRL B21131)。
【0013】
cDNAおよびそのフラグメントならびに誘導体は、そのcDNAによってコードされた受容体蛋白質と同様な蛋白質をコードするDNA配列を分離するためのプローブとして用いることができる。ヒト受容体H4−1BBのcDNAはマウス4−1BBのcDNAと約65%相同であり、cDNA4−1BB由来のプローブを用いて分離された。p4−1BBで示されたcDNA遺伝子は、American Type Culture Collection(12301 Parlawn Drive,Rockville,Maryland 20852)に寄託された(ACTT No.67825)。
【0014】
ヒト受容体蛋白質H4−1BBは、
(1)H4−1BBのcDNAを適切な発現ベクターに挿入し、
(2)この発現ベクターを適切なトランスフェクション宿主中にトランスフェクトし、
(c)トランスフェクトさせた宿主を適切な培地中で増殖させ、
(d)培地から受容体蛋白質を精製する
ことによって製造することができる。
【0015】
本発明の蛋白質およびフラグメントならびに誘導体は、
1)ヒト受容体蛋白質H4−1BBに対するリガンドを分離するためのプローブとしてか、
2)H4−1BBリガンドを発現するB細胞の増殖を刺激するために、または
3)H4−1BBの結合をブロックするために用いることができる。
【0016】
B細胞の増殖は、受容体蛋白質H4−1BBに対するリガンドを発現しているB細胞を受容体蛋白質H4−1BBを発現している細胞で処理することによって誘導することができる。この種の出願のために、同様の共刺激免疫系経路が解析されつつある。「Mounting a Targeted Strike on Unwanted Immune Responses」(Jon Cohen,Science,257巻, 8-7-92);「Long Term Survival of Xenogeneic Pancreatic Islet Grafts Induced by CTLA4Ig」(Lenschowら, science,257巻, 7-8-92);および「Immunosuppresion in Vivo by a Soluble Form of the CTLA-4 T Cell Activation Molecule」(Linsleyら, Scienc, 257巻, 7-8-92)を参照。
【0017】
H4−1BBに対するモノクローナル抗体は、受容体蛋白質H4−1BBを発現しているT細胞を抗H4−1BBモノクローナル抗体で処理することによってT細胞の増殖を増強するために使用することができる。いくつかの腫瘍は、潜在的に免疫原性であるが、in vivoにおける有効な抗免疫反応を刺激しない。腫瘍は抗原特異的なシグナルをT細胞に供給できるかも知れないが、T細胞を完全に活性化するのに必要な共刺激シグナルを供給しないかも知れない。メラノーマ細胞における共刺激リガンドB7の発現は、in vivoにおけるネズミのメラノーマの拒絶を誘導しないことが明らかになった(「Tumor Rejection After Direact Co-Stimulation of CD8+T cells by B7-Transfected Melanoma Cells」(Sarah, E. TownsendおよびJames P. Allison, Science, 259巻, 1-5-93)。H4−1BBに対するモノクローナル抗体は、T細胞の増殖と活性化を誘導することが現在知られているのと同様の効果をもたらすことができるかも知れない。
【0018】
ヒト受容体蛋白質H4−1BBに対する細胞膜リガンドを検出するための融合蛋白質が開発された。この蛋白質は受容体蛋白質H4−1BBの細胞外部分と受容体蛋白質H4−1BBの一部分に結合した検出蛋白質(アルカリホスファターゼ)からなる。受容体蛋白質H4−1BBの一部分が細胞膜リガンドに結合し、結合は検出蛋白質に対する比活性アッセイによって検出することができる。融合蛋白質は受容体蛋白質B4−1BBを発現すると思われる細胞の存在下に置かれる。次に、この細胞を、いかなる融合蛋白質もその細胞膜リガンドに結合していないように洗浄する。一旦、洗浄した細胞を検出蛋白質の基質の存在下に置き、検出蛋白質の比活性を測定することができる。
【0019】
以下の詳細な説明では出願人の公開された研究と同様、既知の方法及び研究を引用する。これらの出版物は明確にするために引用して本願に組み込まれており、詳細な説明の最後に列挙されている。
【発明を実施するための最良の形態】
【0020】
マウスレセプター4−1BBの単離及び特性化
マウスレセプター4−1BBのヌクレオチド配列及び推定のアミノ酸配列は図1に示されている。メッセージ鎖のヌクレオチドは5’から3’側方向に番号付されており、番号は配列の両側に記載されている。ヌクレオチド残基1はATG開始コドンのAであり、残基1の5’側にあるヌクレオチドには負の番号が付されている。推定のアミノ酸配列はヌクレオチド配列の下方に記載されている。推定のシグナルペプチドには下線が引かれている。停止コドンは(−−−)で示されている。システイン残基は、点々で明示されている。4−1BB配列の通常でない特徴は、ヌクレオチド1158−1163にAATAAAで示される潜在的なポリアデニル化シグナルがあることである(図1、四角で囲んだ箇所)。遺伝子産物が少なくとも2つの異なる大きさのmRNAを産生することから、このシグナルは機能的であると信じられた。
【0021】
マウス脾細胞、T細胞クローン、及びハイブリドーマ内での4−1BBの転写はコンカナバリンAによって誘導可能であった。4−1BB転写物の発現はシクロスポリンAによって阻害された。4−1BBmRNAは抗原受容体刺激によって誘導可能であったが、クローン化T−細胞内でのIl−2刺激では誘導されなかった(1)。4−1BBcDNAは推定のリーダー配列、潜在的な膜アンカーセグメント、及び既知の受容体タンパク質の他の特徴を含む、256アミノ酸ペプチドをコードしている。従って、4−1BBの配列は受容体タンパク質のそれに一致しているように見えるが、発現パターンはリンホカインのそれに類似している。
【0022】
細胞表面における4−1BBの主な種は55ーkDaの2量体のようである。4−1BBはまた、30−kDaの単量体、そして恐らく110−kDaの4量体としても存在するようである。これらの4−1BB種は均質な細胞集団(T−細胞クローンF1)から免疫沈降されたので、いずれも各細胞上に共存している可能性がある。4−1BBが細胞表面でホモダイマーとして存在するか否かを決定するには、4−1BB単量体と2量体からのペプチド消化物を比較する必要があるだろう。インシュリン受容体(2)、B−細胞表面免疫グロブリン受容体(3)、T−細胞Ag受容体(4)、CD28共刺激(costimulatory)受容体(5)、及びCD27T細胞抗原(6)等の様々な細胞表面受容体はジスルフィド結合したサブユニットからなる。リガンドの結合及びその後の生化学的シグナル伝達(signaling)には、受容体の2量体化が必要かもしれない。
【0023】
4−1BBは休止期のT細胞には発現しないが、T細胞に対して完全な増殖刺激を与える活性化剤によって誘導可能である。PMAとイオノマイシンとの組み合わせは、T細胞の増殖に必要なこれらのシグナルをまねることができる。PMA又はイオノマイシン単独で4−1BBmRNAを誘導することができるが、PMAとイオノマイシンの併用により4−1BBの至適発現が得られた。しかも、4−1BBの発現は一過性でなかった。精製脾T細胞を固定化抗−CD3で刺激すると、4−1BBmRNAが発現され、この発現は刺激後96時間まで維持された。4−1BBが細胞周期の進行期間を通して発現されることを確認するためには、細胞周期の解析が必要であろう。
【0024】
4−1BBは構造的に神経成長因子受容体スーパーファミリーのメンバーに関連している。これらの受容体は構造上類似のリガンド−結合特性(システイン−豊富領域)を有するか、これらタンパク質の細胞質ドメインは非保存的であり、それがトランスメンブランシグナル伝達に多様性を与えているかもしれない。このファミリーには、T又はB細胞の活性化工程に関与しているメンバーもある。OX−40、CD40及びCD27抗原に関するインビトロ機能性データがある。OX−40に対する抗体は混合リンパ球反応におけるT細胞の応答性を増大し(7)、CD40に対する抗体はPMA又はCD20抗体のような共活性化剤(coactivator)の存在下でB−細胞の増殖を促進し、インビトロでIL−4と相乗的に作用してB−細胞の分化を誘導し長期正常B−細胞株を生成させる(8)。CD27分子上のエピトープを認識する1つのイノノクローナル(inonoclonal)抗体(抗−1A4)は、カルシウム動態化、IL−2分泌、ヘルパーT細胞機能、及びT細胞増殖を阻害した。一方、もう1つの抗CD27mAbであるCLB−CD27/1はPHA又は抗CD3mAbによって刺激されたヒトT細胞の増殖を促進した(6)。これらの結果はCD27分子がT細胞の活性化に重要な役割を果たしていることを示している。TNFR類、NCFR、及びCD40を除いて、スーパーファミリーのメンバーが結合するリガンド又は細胞表面分子はまだ同定されていない。受容体が結合するリガンドの同定及び特性化は4−1BBの生理学的な役割のより良い定義のために役立つであろう。
【0025】
細胞表面4−1BBがT細胞活性化に貢献しうるか否かを確かめるために、抗4−1BB53A2を4−1BBのアンタゴニストとして用いた。これらのデータは実際、4−1BBが、T細胞の活性化及び増殖の間、副シグナル分子として機能する能力を有することを示唆していた。固定化抗CD3で刺激した、精製脾由来T細胞に可溶性53A2を加えると、抗CD3単独で刺激した場合に比較して、3Hチミジンの取り込みが増大した。3回の独立実験における、この増大パターンの範囲は2〜10倍であった。
【0026】
Bretcher及びCohnの元々の2つのシグナルモデルで、彼らはシグナル1、T細胞抗原受容体(TCR)の占有、では補助細胞から供給されるシグナル2の不在下でT細胞が不活化されると提案した。これは以後様々な研究で確認された(9)。T細胞上の潜在的な共刺激受容体としての補助細胞CD28の同定は、T細胞の最適な増殖に必要な補助シグナル(群)の特性化を開始するのに重大な寄与であった(10)。他の細胞表面分子がこれらの共刺激活性化要求に寄与している可能性がある(11)。
【0027】
4−1BBを介して伝達される生化学的シグナルは、完全には分かっていない。考慮されている1つの可能性は、4−1BBが推定のp56kkチロシンキナーゼ結合ドメインをその細胞質尾部に含有しているとの観察であった。後に、p56kkチロシナーゼキナーゼは4−1BBに結合することが決定された。4−1BBを介するシグナル伝達が、その発現がT細胞の活性化及びその後の増殖に必要である、IL−2及びIL−2受容体のような遺伝子を調節できるかどうかを決定することも価値ある。
【0028】
神経成長因子受容体(NGFR)ファミリーのメンバーの正確な機能は多様であるように思われるが、これらの分子が様々な方法で、それらが発現される特定の細胞型の応答性又は生存可能性の維持に貢献しているかもしれない、というテーマが出現する。例えば、NGFはインビトロ及びインビボでのニューロンの生存に絶対的に必要である(12)。可溶性抗CD40モノクローナル抗体によるCD40のクロスリンキングは、胚中心中心細胞がインビトロでアポプトーシスを受けることを妨げる(13)。CD40を介して伝達されるシグナルもまた、分化因子に対する応答性の維持を補助しうる。CD40と抗−CD40F(ab’)2フラグメントのIL−4の存在下での結合(ライゲーション)はIgE合成の非常な増大を誘導した(14)。また、IL−10及びトランスフォーミング成長因子ーβで処理した、抗CD40活性化天然(固有)B細胞はIgA分泌を行うようになった(15)。
【0029】
4−1BBは、NGFRスーパーファミリーと共通する分子の特徴を有することに加えて、酵母e1F−2βタンパクの推定の亜鉛フィンガー構造を有することが分かった(16)。4−1BBはまたショウジョウバエのアプサンス(absentia)のsina sevenと、保存領域を共有しており、それは適正な光受容体細胞の発展に必要である(17)。その特定の領域はまた、その発現がcAMPによる凝集の期間に特異的に誘導される、DictyosteliumのDG17遺伝子のタンパク質生成物と類似している(18)。
【0030】
この領域はC−X2−C−X9−C−X3−H−X3−C−X−Cパターンを形成しており;システイン及びヒスチジンは4−1BB、sina及びDG17タンパク質内の、同様のスペースに保存されている。4−1BBタンパク質とsinaタンパク質との間で24アミノ酸の内、10アミノ酸が同一であり、24個の内3個は保存的置換である。保存のパターンはこれらのアミノ酸が機能的に重要であることを示している。sinaタンパク質は核に局在しており、これは、細胞内で調節機能を果たしていることを示唆している。4−1BBのアミノ酸配列が、亜鉛フィンガーモチーフ、核タンパク質、及び受容体ドメインの様な特徴を含有しているという事実は、4−1BBが細胞増殖及び分化の間に多様な役割を果たしていることを示している。
【0031】
4−1BBはT−細胞−APC相互作用に関連する他の細胞表面分子を表しているかもしれない。4−1BB−AP融合タンパク質は、成熟B−細胞株、抗μ−活性化初代B細胞、及び成熟マクロファージ細胞株と特異的に結合する。4−1BB−APは、未成熟な、B細胞株及びマクロファージ細胞株、T細胞クローン、T細胞株、T細胞の初代培養、及び種々の非リンパ様細胞株とは低レベル又は僅かなレベルで結合する。4−1BB−APは成熟B細胞及びマクロファージと結合するので、4−1BBの結合に際して伝達されるシグナルは何等かの方法でAPC機能を調節する可能性がある。この可能性は探索の余地がある。
【0032】
Chalupnyら(19)は4−1BBRg、4−1BBの細胞外領域とヒトIgGのFc領域からなる融合タンパク質、が細胞外マトリックス(ECM)に結合すると提案した。4−1BBRg結合はヒトビトロネクチンとの間で最高レベルであった。データは示されていないが、4−1BB−AP及びマイクロタイター上、1ウエル当たり0.007μg−10μgで固定化されたヒトビトロネクチンを用いてELISAが行われた[Yelios Pharmaceuticals/GIBCO-BRL, Grand Island, NY.)。AP活性に基づいて、4−1BB−APの結合は観察されなかった。細胞表面に外因的に結合しているタンパク質(細胞外マトリックス構成成分の可能性あり)に4−1BB−APが結合する可能性を排除するために、結合アッセイの前にB細胞リンパ腫を酸性条件で洗浄した。4−1BB−APは依然として成熟B−細胞に特異的に結合した。B細胞及びマクロファージ上で特異的に発現される4−1BB−リガンドが存在するか否か、及び4−1BB−APが特定の結合条件下でECMと結合するか否かはなお決定されるべきことである。ECMは4−1BBの特定の細胞表面リガンドへの結合を容易にしている可能性がある。
【0033】
B細胞とヘルパーT細胞とは、T細胞上の特異的な対応受容体に結合するB細胞上の受容体によって互いに相互作用する。この相互作用の結果、これら2つの細胞種間での生化学的シグナル伝達リレーのカスケードが起こると考えられている(20)。この相互作用が進行するにつれ、これら細胞は細胞周期のS期に入るようになる。TCRとT細胞上のCD4、およびB細胞上のプロセスされた抗原−MHCIIとの間の初期相互作用は、B細胞が細胞周期に入るという結果にはならない(21)。しかしながら、インビトロ系での研究は、T細胞が一旦刺激されると、B細胞を細胞周期に入らせうる新たに合成されたまたは修飾された細胞表面分子を発現することを示唆している(22、23)。このT細胞機能は抗原特異的でもMHCに限定されたものでもない(24)。加えて、B細胞活性化の活性化Th誘導には可溶性因子を必要としない(25)。B細胞が細胞周期に入ったら、IL−4がB細胞をG1期からS期への進行を誘導する。活性化T細胞またはT細胞膜がB細胞を細胞周期に入るのを促進する能力は、シクロヘキシミドかまたはシクロスポリンA処理により阻止することができる(26、27)。これらの新たに発現される膜タンパク質は、その誘導特性において「リンホカイン様」であると思われる。
【0034】
4−1BBは、B細胞のコスティミュレーターとしての要求を満たす発現特性を有する。4−1BBは抗CD3またはTCR媒体T細胞刺激によって誘導することができ、その発現はシクロスポリンA並びにシクロヘキシミド処理に感受性である(28)。興味深いことに、パラホルムアルデヒド固定したSF21−4−1BB細胞は、B細胞増殖の誘導において抗μと共同作用する(synergized)。SF21−4−1BBによる脾臓B細胞のコスティミュレーションは、抗μの最適用量(10μg/ml)および最適下限用量(1.0〜0.1μg/ml)で起こる。休止B細胞にSF21−4−1BB細胞を加えても有意のB細胞増殖とはならなかった。SF21−4−1BBは、B細胞増殖の誘導においてTPAまたはイオノマイシン、または最適下限濃度のLPSとは共同作用しなかった。
【0035】
大量の組換え可溶性タンパク質を発現させるためにバクロウイルス系が用いられてきているが、この系は組換え細胞表面タンパク質の発現のために用いることができる。バクロウイルス感染は、細胞当たりベースで均一な高レベルの組換えタンパク質を発現させるための都合の良い手段をもたらす。SF21細胞を単独で加えても有意レベルのコスティミュレーションとならなかったことは注目に値する。このことは、それ自身で強力なコスティミュレーター活性を示しうるcos−またはL−細胞株を用いるに際して潜在的な問題となりうる。
【0036】
NGFRスーパーファミリーの他の成員であるCD40はB細胞表面で発現され、活性化T細胞上で発現される分子であるgp39と相互作用する。マウスgp39タンパク質(29)およびヒトgp39タンパク質(30)をコードするcDNAはクローニングされている;この細胞表面分子は、腫瘍壊死因子と相同性を有するタイプIIの膜タンパク質である。ノエル(Noelle)ら(31)は、CD40−免疫グロブリン融合タンパク質がT細胞により誘導されたB細胞増殖および分化を用量に依存した仕方で阻止しうることを見いだした。アーミテージ(Armitage)らは、マウスgp39のcDNAを単離し、gp39がコスティミュレーター(co-stimuli)の不在下でB細胞増殖を誘導することができ、IL−4−の存在下でIgE産生という結果になることを示した。ホレンバウ(Hollenbaugh)ら(32)は、ヒトgp39でトランスフェクトしたCOS細胞がヒトB細胞増殖を誘導するうえでTPAかまたは抗CO20と共同作用しうること、およびコスティミュレーターなしでは低レベルでしかB細胞を刺激できないことを示している。これらデータは、CD40がT細胞との物理的接触の間にシグナルを伝達するB細胞表面分子の一つであることを示している。
【0037】
細胞表面受容体は、可溶性因子かまたは隣接細胞上に発現された他の細胞表面分子のいずれかと相互作用することによって外部環境と連絡している。細胞表面受容体と相互作用する可溶性因子によって送達される生化学的シグナルに対して細胞−細胞接触によって送達される生化学的シグナルの役割は明らかでない。NGFRが2以上のリガンドと結合するのと同様に、NGFRスーパーファミリーはTNFRIおよびIIに対して普通でない。TNFRIおよびIIはともにTNF−αおよびTNF−Rの両者に結合する(33)。NGFRは、NGF、脳由来神経栄養因子、およびニューロトロフィン−3に結合する(34)。
【0038】
加えて、一つのリガンドが細胞表面リガンドとしても可溶性リガンドとしても機能する。CD4−0リガンドであるgp39に関する最近の証拠は、このリガンドが膜結合したリガンドとしても可溶性リガンドとしても存在しうることを示唆している(35)。4−1BBが分泌され、B細胞と可溶性形態並びに膜結合形態で相互作用する可能性がある。NGFR受容体ファミリーの成員であるCD27は、T細胞上で発現され、分泌されるとともに細胞表面上に発現される(36)。1を越えるリガンド(可溶性および細胞表面)が4−1BBに結合する可能性もある。
【実施例】
【0039】
ヒト同族体、H4−1BBの単離
マウス4−1BBのヒト同族体(H4−1BB)を単離するため、2セットの複製連鎖反応(PCR)プライマーを設計した。PCRプライマーを設計するため、神経増殖因子受容体(NGFR)スーパーファミリーの成員間でアミノ酸配列を比較した。なぜなら4−1BBは該スーパーファミリーの成員であるからである(37)。使用したアミノ酸配列は、マウス4−1BB(38)、ヒトNGFR(39)、ヒト腫瘍壊死因子受容体(33)、ヒトCD40(40)、およびヒトCD27(6)であった。NGFRスーパーファミリー間での保存配列の領域を選択した。
【0040】
前(forward)プライマーI(H4−1BBFI)は、マウス4−1BBのアミノ酸36〜41の範囲、前プライマーII(HR−1BBFII)はアミノ酸52〜58の範囲にある。逆(reverse)プライマーI(H4−1BBRI)はマウス4−1BBのアミノ酸116〜121の範囲、逆プライマーII(H4−1BBRII)はアミノ酸122〜128の範囲にある。マウス4−1BBにおいてPCRプライマーとして用いた領域は図1に示してある。
【0041】
各プライマーのオリゴヌクレオチド配列は以下の通りである。
【表1】
【0042】
正常な健常者から末梢血リンパ球を単離し、PMA(10ng/ml)およびイオノマイシン(1μM)で活性化した。これらリンパ球からmRNAを単離した。逆転写酵素を用い、ヒトリンパ球mRNAを一本鎖cDNAに変換させた。ついで、一本鎖cDNAをTaqポリメラーゼと上記プライマーとの組わせにより増幅させた。プライマーの組わせは以下の通りであった:H4−1BBFIとH4−1BBRI;H4−1BBFIとH4−1BBRII;H4−1BBFIIとH4−1BBRI;およびH4−1BBFIIとH4−1BBRII。
【0043】
H4−1BBFIIとH4−1BBRIIとのプライマーセットは〜240bpの特異的バンドを生成した。この240bpは、ヒト同族体タンパク質がマウス4−1BBとサイズが同様であるとした場合にヒト4−1BBの予期されるサイズである。このPCR生成物(240bp)をPGEM3ベクター中にクローニングし、配列決定した。一つのオープンリーディングフレームはマウスの4−1BBと〜65%同一であった。それゆえ、この240bp PCR生成物はマウス4−1BBのヒト同族体であると結論した。この240bp PCR生成物を用いて活性化ヒトTリンパ球のλgt11 cDNAライブラリーをスクリーニングした。〜0.85kb cDNAが単離させた。このcDNAの配列を図2に示してあり、予測されるアミノ酸配列を図2bに示してある。同情報は本明細書中の配列表の配列1に示してある。
【0044】
H4−1BB−AP融合タンパク質を発現する発現プラスミドを構築した。シグナル配列および全細胞外ドメインをコードする配列を含むH4−1BB cDNAの5'部分をPCRにより増幅させた。正しい方向にクローニングするため、前プライマーの5'末端上にHindIII部位を、および逆プライマーの5'末端上にBglII部位をそれぞれ生成させた。
【0045】
HindIII−BglII H4−1BBフラグメントを哺乳動物発現ベクターAPtaq−1中、ヒト胎盤アルカリホスファターゼ(AP)をコードする配列の上流に挿入した。H4−1BBの5'部分の増幅に用いたオリゴヌクレオチドPCRプライマーは以下の通りである。
【表2】
【0046】
H4−1BB−APは、ヒト4−1BBのリガンド(すなわち、H4−1BBL)を発現する細胞および組織を同定するのに用いるであろう。マウス4−1BBを用いた研究は、4−1BBのリガンドが細胞表面上に存在することを示した。B細胞およびマクロファージが4−1BBを発現する主要な細胞である。H4−1BBLもまたヒトのB細胞およびマクロファージ上で発現されることが予想される。
【0047】
哺乳動物発現cDNAライブラリーは、H4−1BBLを発現するヒト細胞株から生成されるであろう。このライブラリーは[125]I−標識したH4−1BB−APによりスクリーニングされるであろう。ついで、H4−1BBLのcDNAが単離され、特徴付けられるであろう。ついで、可溶性組換えH4−1BBLが生成されるであろう。H4−1BB−APおよびH4−1BBLの両者とも、下記のように免疫応答を抑制または促進するために用いられるであろう。H4−1BBおよびH4−1BBLに対するモノクローナル抗体が産生されるであろう。 マウス4−1BBでの研究によれば、4−1BBはコスティミュラトリーシグナルとして働く。H4−1BBはT細胞活性化のコスティミュラトリーシグナルとして働くことが予想される。マウス4−1BBは、B細胞の増殖および分化を助ける。H4−1BBも同様であろうことが予想される。H4−1BB−AP、H4−1BBLおよびモノクローナル抗体は、ヒトの免疫応答を抑制または促進するために用いることができる。
【0048】
図3aおよび3bはT細胞活性化に関与する分子を示す。初期のT細胞活性化の間(認識(cognitive)期)、休止T細胞はTCR/CD3複合体および他の「アクセサリー(accessory)」分子を発現する。これら構成的に発現される分子のうち、CD4(またはCD3)、LFA−1およびCD28がおそらくコスティミュラトリーシグナルを受け取る分子である。これら「アクセサリー」コスティミュラトリーシグナルとともにTCR/CD3複合体と相互作用することにより、CD28、CTLA4、および4−1BBなどの他の受容体が引き続き発現される。これら新たに発現された分子は、おそらく、T細胞活性化の後期の段階で別の重要なコスティミュラトリーシグナルを受け取るであろう(クローン増殖)。
【0049】
免疫応答の抑制
図4a−cは正常なT−細胞活性経路を示す。図5a−cは4−1BBの可溶性キメラによる免疫応答阻害を示す。もし4−1BBがT細胞活性化になんらかの役割を果たすなら、抗原−提示細胞上のそのリガンドに対する相互作用の阻害がT−細胞依存免疫応答の抑制をもたらす筈である。カウンター−受容体B7に対するCD28の相互作用の阻害が、種々の程度でインビボの抗体産生および細胞−媒介免疫応答の両者を抑制することは十分に立証されている。両者の相互作用の阻害はより効果的な免疫抑制をもたらす。そのリガンドに対する4−1BBの相互作用の阻害はCD28/B7相互作用がもはや関連性がなくなった場合の活性化プロセスの後期の段階で重要になり得る。
【0050】
マウス受容体4−1BBおよびマウスリガンド4−1BBLについて、上記に詳述したように、H4−1BB−APの添加はH4−1BBL発現細胞を覆い、H4−1BBとH4−1BBL間の正常な相互作用を阻害する。これは免疫応答を抑制することになる。この種の免疫応答は抗原−特異的である。従って、antiCD3またはシクロスポリンA処置によって生じる一般化された免疫応答は回避される。H4−1BB−AP処置はある種の自己免疫疾患の処置に用いることができ、臓器移植を容易にする。
【0051】
免疫増強
H4−1BBはT−細胞活性化の後期の段階で機能し、T−細胞活性化の完成のための重要な細胞になり得る。多くの腫瘍は腫瘍−特異抗原を示す。しかしながら、なぜ免疫原性腫瘍が宿主免疫を免れ得るかの理由の1つは腫瘍−反応性T−細胞が不十分な共刺激(costimulation)を受けているからである。従って、H4−1BBなどの共刺激分子の腫瘍への導入は、細胞傷害性T細胞(CTL)の抗腫瘍免疫を増強し得た。H4−1BBLは細胞−特異様式で発現され得る。例えば、H4−1BBLは、例えば、チロシナーゼプロモーターなどのメラノサイト−特異プロモーターを用いる黒色腫中に発現され得る。H4−1BBL−発現黒色腫はH4−1BBを介して細胞傷害性T細胞を刺激し、黒色腫−特異CTLを活性化する。活性化された黒色腫−特異CTLは黒色腫を撲滅させ得る。
【0052】
引用により明細書の記載とされる文献の付録
1. Smith, C.A., Davis, T., Anderson, D., Solam, L., Beckmann, M.P., Jerzy, R., Dower, S.K., Cosman, D., and Goodwin, R.G. 1990. A receptor for tumor necrosis factor defines an unusual family of cellular and viral proteins.
Science 248:1019-1023.
2. Ebina, Y., L. Ellis, K. Jaruagin, M. Edery, L. Graf, E. Clauser, J. On, F. Marizrz, Y.W. Kan, J.D. Goldfine, R.A. Roth and W.J. Rutter, 1985, The human insulin receptor cDNA: the structural basis for hormone-activated transmembrane signalling, Cell 40:747.
3. Vassali, R., R. Tedghi, B. Listowska-Bernstein, A. Tartakoff and J.C. Jaton, 1979, Evidence for hydrophobic region within heavy chains of mouse B lymphocyte membranebound IgM, Proc. Natl. Acad. Sci. USA76:5515.
4. Haskins, K., R. Kubo, J. White, M. Pigeon, J. Kappler and P. Marrack, 1983, The major histocompatability complexrestricted antigen receptor on T cells I Isolation with monoclonal antibody, J. Exp. Med. 157:1149.
5. Lesslaver, W. and H.Gmunder, 1986, Biochemical characterization of the 9.3 antigens of human T-cells: stimultaneous expression of disulfide-bonded 90-Kiladalton dimers and free subunits at the cell surface, Mol. Immunol. 23:271.
6. Van Lier, R., J. Borst, T. Vroom, H. Klein, P. Mourik, W. Zeijlemaker and C. Melife, 1987, Tissue distribution and biochemical and functional properties of Tp55 (CD27) a novel T cell differentiation antigen, J. Immunol. 139:1589.
7. Mallett, S., S. Fossum and A. Barclay, 1990, Characterization of the MRC OX40 antigen of activated CD4 positive T lymphocytes-a molecule related to nerve growth factor receptor, EMBO J. 9:1603.
8. Banchereau, J., P. Paoli, A., Valle, E. Garcia and F. Roussel, 1991, Long-term human B cell lines dependent on interleukin-4 and antibody to CD40, Science 251:70.
9. Moeller, D.L., M.K. Jenkins and R.H. Schwartz, 1989, Clonal expansion versus functional colonal inactivation: a co-stimulatory signalling pathway determines the outcome of T cell antigen receptor occupancy, Ann. Rev. Immunol. 7:445.
10. June, D.H., J.A. Ledbetter, P.S. Linsley and C.B. Thompson, 1989, Role of CD28 receptor in T cell activation, Immunol. Today 11:211.
11. Yang, L., B. Jones, A. Aruffo, K.M. Sullivan, P.S. Linsley and C.A. Janeeway, Jr., 1992, Heat stable antigen is a co-stimulatory molecule for CD4 T cell growth, J. Exp. Med. 175:437.
12. Yamori, T., 1992, Molecular mechanisms for generation of neural diversity and specificity: foles of polypeptide factors in development of post-mitotic neurons, Neurosic. Res. 12:545.
13. Liu, Y.J., D.E. Joshua, G.T. Williams, C.A. Smith, J. Gordon and I.C. M. MacLennan, 1989, Mechanism of antigendriven selection in germinal centres, Nature, 342:929.
14. Jabara, H. H., s. M. Fu, R.S. Geha and D. Vercelli, 1990, CD40 and IfE: synergism between anti-CD40 momoclonal antibody and interleukin 4 in the induction of IgE synthesis by highly purified human B cells, J. Exp. Med. 172:1861.
15. Defrance, R., B. Vanbervliet, F. Briere, I. Durnad, F. Roussle and J. Banchereau, 1992, Interleukin 10 and transforming growth factor beta cooperate to induce anti-CD40 activated naive human B cells to secrete immunoglobulin A, J. Exp. Med. 175:671.
16. Donahue, T., Cigan, A., Pahich, E. and Valavicius, B., Mutations at a Zn(II) finger motif in the yeast elF-2beta gene alter ribosomal start-site selection during the scanning process, Cell 54 (1988) 621-632).
17. Carthew, R.W and Rubin, G.M., seven in absentia, a gene required for specification of R7 cell rate in the Drosophila eye, Cell, 63 (1990) 561-577.
18. Driscoll, D.M. and Williams, J.G., Two divergently transcribed genes of Dictyostelium discoideum are cyclic AMP-inducible and coregulated during development, Mol. and Cell. Biol. 7 (1987) 4482-4489.
19. Chalupny, N.J., Peach, R., Hollenbaugh, D., Ledbetter, J.A., Farr, A.G. and Aruffo, A., 1992, Proc. Natl. Acad. Sci USA 89:10360-10364.
20. Noelle, R.J., and Snow, E.C., 1991, The FASEB J. 5:2770-2776.
21. Noelle, R. and Snow, E., 1990, Immunol. Today 11:361-368.
22. Zurawski, G., Benedik, M., Kamb, B.J., Abrams, J.S., Zurawaki, S.M. and Lee, F.D. (1986) Science 232.772-775.
23. Kinachi, T. (1986) Nature 325,70-73.
24. Gershenfeld, H.K. and Weissman, I.L. Science (1986) 232.854-858.
25. Biggin, M., Gison, T. and Hung, G. (1983 Proc. Natl. Acad. Sci. USA 80,3963-3965.
26. Hodgkin, P.D., Yamashita, L.C., Coffman, R.L. and Kehry, M.R., 1990, J. Immunol. 145:2025-2034.
27. Barlett, W.C., McCann, J., Shephaer, D.M., Roy, M. and Noelle, R.J., 1990, J. Immunol. 145:3956-3962.
28. Kwon, B.S., Kestler, D.P., Eshhar, Z., Oh, K., and Wakulchik, M. 1989. Expression characteristics of two potential T cell mediator genes. Cell. Immunol. 121:414-422.
29. Armitage, R., Fanslow, W., Strockbine, L., Sato, T., Clifford, K., MacDuff, B., Anderson, D., Gimpel, S., Davis Smith, T., Maliszewski, C., Clark, E., Smith, C., Grabstein, K., Cosman, D. and Spriggs, M., 191, Nature 357:80-82.
30. Kwon, B., Kestler, D., Lee, E., Wakulchik, M. and Young J. (1988) J. Exp. Med) (1988) (In press).
31. Noelle, R.J., Roy, M., Shepherd, D.M., Stamenkovic, I., Ledbetter,J.A. and Aruffo, A., 1992, Proc. Natl. Acad. Sci. USA 89:6550-6554.
32. Hollenbaugh, D., Grosmaier, L.S., Kullas, C.D., Chalupny, N.J., Braesch-Andersen, S., Noelle, R.J., Stamenkovic, I., Ledbetter,J.A. and Aruffo, A., 1992, EMBO11:4314-4321.
33. Schall, T.J., M. Lewis, K.J. Koller, A. Lee, G.C. Rice, G.H. W. Wong, T. Gatanaga, G.A. Granger, R. Lentz, H. Raab, W.J. Kohr and D.V. Goeddel, 1990, Molecular cloning and expression of a receptor for human tumor necrosis factor, Cell 61:361.
34. Klein, R., Nanduri, V., Jing, S., Lamballe, F., Tapley, P., Bryant, S., Cordon-Cardo, C., Jones, K.R., Reichardt, L.F. and Barbacid, M., 1991, Cell 66:395-403.
35. Armitage, R.J., Sato, T.A., Macduff, B.M., Clifford, K.N., Alpert, A.R., Smith, C.A. and Fanslow, W.C., 1992, Eu8r. J. Immunol. 22:2071-2076.
36. Hintzen, R.Q., deJong, R., Hack, E.E., Chamuleau, M., de Vries, E.F.R., ten Berge, I.J.M., Borst, J. and van Lier, R.A.W., 1991, J. Immunol. 147:29-35.
37. Mallett, S., and Barclay, A.N. 1991. A new superfamily of cell surface proteins related to the nerve growth factor receptor. Immunol. Today. 12:220-223.
38. Kwon, B.S., and Weissman, S.M. 1989. cDNA sequences of two inducible T-cell genes. Proc. Natl. Acad. Sci. USA. 86:1963-1967.
39. Johnson, D., Lanahan, A., Buck C.R., Sehgal, A., Morgan, C., Mercer, E., Bothwell, M., and Chao, M. 1986. Expression and structure of the human NGF receptor. Cell 47:545-554.
40. Stamenkovic, I., Clark, E., and Seed, B. 1989. A Blymphocyte activation molecule related to the nerve growth factor receptor and induced by cytokines in carcinomas. EMBO. J. 8:1403-1408.
【0053】
上記の記載は、詳細な説明および解説の目的のために特許法の要件に従って、本発明の具体的な実施態様を述べたものである。しかしながら、本発明の範囲および要旨をはずれることなく、多くの修正および変更が可能であることは当業者にとって自明である。下記の特許請求の範囲はそのようなすべての修正を包含するものと理解されなければならない。
【図面の簡単な説明】
【0054】
【図1a】マウスレセプター(受容体)タンパク質4−1BBのcDNA配列及びヒト同族体H4−1BBを得るためのPCRプライマーとして用いた領域を示す。
【図1b】図1aの続き。マウスレセプター(受容体)タンパク質4−1BBのcDNA配列及びヒト同族体H4−1BBを得るためのPCRプライマーとして用いた領域を示す。
【図2a】ヒトレセプター(受容体)H4−1BBのヌクレオチド配列を示す。
【図2b】ヒトレセプター(受容体)H4−1BBの推定アミノ酸配列を示す。
【図3】T−細胞の活性化に関与する分子を示す。
【図4】正常なT−細胞活性化経路を示す。
【図5】T−細胞活性化経路における遮断工程における、(a)CTLA4−1g単独使用、(b)4−1BB/AP及びCTLA4−1g併用、および(c)4−1BB/AP単独使用を示す。
【特許請求の範囲】
【請求項1】
ヒト4−1BBポリペプチドの少なくとも一部をコードする単離されたDNAであって、以下からなる群から選択されるヌクレオチド配列を有するDNA:
a)配列番号1のヌクレオチド41〜805;
b)配列番号1のヌクレオチド110〜805;
c) 配列番号2のアミノ酸1〜186を有する可溶性ヒト4−1BBポリペプチドまたはヒト4−1BBに対する細胞膜リガンドに結合可能なそのフラグメントをコードするヌクレオチド配列;
d)配列番号2のアミノ酸配列を有するヒト4−1BBをコードするヌクレオチド配列;および
e)前記可溶性ヒト4−1BBポリペプチドのC−末端に融合するポリペプチドをさらにコードする、c)記載のヌクレオチド配列。
【請求項2】
米国アグリカルチュラル・リサーチ・サービス・カルチャー・コレクションに受託番号NRRL B21131として寄託された、pH4−1BBとして特定される、請求項1に記載のDNA。
【請求項3】
a)適当なトランスフェクション宿主と、配列番号2を有するヒト4−1BBまたはヒト4−1BBに対する細胞膜リガンドに結合可能なそのフラグメントをコードするDNA配列を含んでなる発現ベクターとを接触させて、そのDNA配列によってコードされるタンパク質を発現するトラスフェクトされた宿主細胞を得;
b)ヒト4−1BBタンパク質を精製する
ことによって製造されるタンパク質。
【請求項4】
前記DNA配列が配列番号2のアミノ酸残基1〜186をコードする、請求項3に記載のタンパク質。
【請求項5】
前記DNA配列がヒト4−1BBと第2のポリペプチドからなる融合タンパク質をコードする請求項4に記載のタンパク質。
【請求項6】
前記第2のポリペプチドが検出タンパク質である、請求項5に記載のタンパク質。
【請求項7】
検出タンパク質がアルカリホスファターゼである、請求項6に記載のタンパク質。
【請求項8】
前記DNA配列が配列番号2をコードする、請求項3に記載のタンパク質。
【請求項9】
配列番号2のアミノ酸配列を有するヒト4−1BBを特異的に認識する モノクローナル抗体。
【請求項10】
配列番号2のアミノ酸配列を有するヒト4−1BBを特異的に認識する モノクローナル抗体を産生することが可能なハイブリドーマ。
【請求項11】
T細胞増殖を促進するために請求項9に記載のモノクローナル抗体を使用する方法であって、発現したヒト4−1BBを有するT細胞を該モノクローナル抗体によりインビトロで処理する工程を含む方法。
【請求項12】
前記処理をタンパク質チロシンキナーゼの存在下で行う工程をさらに含む、請求項11に記載の方法。
【請求項13】
T細胞活性化を促進するために請求項9に記載のモノクローナル抗体を使用する方法であって、発現したヒト4−1BBを有するT細胞を該モノクローナル抗体によりインビトロで処理する工程を含んでなる方法。
【請求項14】
前記処理をタンパク質チロシンキナーゼの存在下で行う工程をさらに含む、請求項13に記載の方法。
【請求項15】
a)ヒト4−1BBに対する細胞膜リガンドに結合する、配列番号2のアミノ酸配列を有するヒト4−1BBの細胞外ドメインに対応する、ヒト4−1BBの少なくとも一部;および
b)前記ヒト4−1BBの該一部に結合した検出タンパク質であって、リガンド結合が、該検出タンパク質の比活性アッセイによって検出できるような検出タンパク質
を含む融合タンパク質。
【請求項16】
前記検出タンパク質がアルカリホスファターゼである、請求項15に記載の融合タンパク質。
【請求項17】
ヒト4−1BBに対する細胞膜リガンドをインビトロで検出するための方法であって、
a)ヒト4−1BBに対する細胞膜リガンドを発現していると思われる細胞と、以下のものを含む融合タンパク質:
1)ヒト4−1BBの、前記細胞膜リガンドに結合する細胞外ドメインに対応する、配列番号2のアミノ酸配列を有する前記ヒト4−1BBの少なくとも一部分;および
2)ヒト4−1BBの該一部分に結合した検出タンパク質であって、該検出タンパク質の比活性アッセイによってリガンド結合が検出できるような検出タンパク質
とを接触させ;
b)前記細胞膜リガンドに結合しなかった融合タンパク質を前記細胞から洗い流し;そして
c)前記洗浄した細胞と、前記検出タンパク質に対する基質とを接触させて、前記検出タンパク質の比活性を測定する
ことを含む方法。
【請求項18】
前記検出タンパク質がアルカリホスファターゼである、請求項17に記載の方法。
【請求項19】
配列番号2のアミノ酸配列を有するヒト4−1BBまたはそのフラグメントをコードするDNA配列を含む組換え発現ベクターであって、宿主細胞中での該DNA配列の発現に適当な制御配列に作動可能なように結合し、該DNA配列が、
a)配列番号1のヌクレオチド41〜805;
b)配列番号1のヌクレオチド110〜805;
c)配列番号2のアミノ酸1〜186を有する可溶性ヒト4−1BBポリペプチドまたはヒト4−1BBに対する細胞膜リガンドに結合可能なそのフラグメントをコードするヌクレオチド配列;
d)配列番号2のアミノ酸配列を有するヒト4−1BBをコードするヌクレオチド配列;および
e)前記可溶性ヒト4−1BBポリペプチドのC−末端に融合するポリペプチドをさらにコードする、c)記載のヌクレオチド配列
からなる群から選択される組換え発現ベクター。
【請求項20】
e)におけるC−末端に融合するポリペプチドが検出タンパク質である、請求項1記載のDNA。
【請求項21】
検出タンパク質がアルカリホスファターゼである、請求項20に記載のDNA。
【請求項22】
前記e)におけるC−末端に融合するポリペプチドが検出タンパク質である、請求項19記載のベクター。
【請求項23】
検出タンパク質がアルカリホスファターゼである、請求項22に記載のベクター。
【請求項24】
a)配列番号2のアミノ酸配列またはヒト4−1BBに対する細胞膜リガンドを結合することが可能なそのフラグメント;
b)配列番号2のアミノ酸残基24〜255またはヒト4−1BBに対する細胞膜リガンドを結合することが可能なそのフラグメント;
c)配列番号2のアミノ酸残基24〜186またはヒト4−1BBに対する細胞膜リガンドを結合することが可能なそのフラグメント;および
d)N−末端アミノ酸配列が、Leu−Gln−Asp−Pro−Cys−Ser−Asn−Cys−Pro−Ala−Gly−Thrである配列番号2のアミノ酸配列
からなる群から選択されるアミノ酸配列を有するヒト4−1BBポリペプチド。
【請求項25】
ヒト4−1BBポリペプチドのC末端に融合した第2のポリペプチドをさらに含む、請求項24に記載のヒト4−1BBポリペプチド。
【請求項1】
ヒト4−1BBポリペプチドの少なくとも一部をコードする単離されたDNAであって、以下からなる群から選択されるヌクレオチド配列を有するDNA:
a)配列番号1のヌクレオチド41〜805;
b)配列番号1のヌクレオチド110〜805;
c) 配列番号2のアミノ酸1〜186を有する可溶性ヒト4−1BBポリペプチドまたはヒト4−1BBに対する細胞膜リガンドに結合可能なそのフラグメントをコードするヌクレオチド配列;
d)配列番号2のアミノ酸配列を有するヒト4−1BBをコードするヌクレオチド配列;および
e)前記可溶性ヒト4−1BBポリペプチドのC−末端に融合するポリペプチドをさらにコードする、c)記載のヌクレオチド配列。
【請求項2】
米国アグリカルチュラル・リサーチ・サービス・カルチャー・コレクションに受託番号NRRL B21131として寄託された、pH4−1BBとして特定される、請求項1に記載のDNA。
【請求項3】
a)適当なトランスフェクション宿主と、配列番号2を有するヒト4−1BBまたはヒト4−1BBに対する細胞膜リガンドに結合可能なそのフラグメントをコードするDNA配列を含んでなる発現ベクターとを接触させて、そのDNA配列によってコードされるタンパク質を発現するトラスフェクトされた宿主細胞を得;
b)ヒト4−1BBタンパク質を精製する
ことによって製造されるタンパク質。
【請求項4】
前記DNA配列が配列番号2のアミノ酸残基1〜186をコードする、請求項3に記載のタンパク質。
【請求項5】
前記DNA配列がヒト4−1BBと第2のポリペプチドからなる融合タンパク質をコードする請求項4に記載のタンパク質。
【請求項6】
前記第2のポリペプチドが検出タンパク質である、請求項5に記載のタンパク質。
【請求項7】
検出タンパク質がアルカリホスファターゼである、請求項6に記載のタンパク質。
【請求項8】
前記DNA配列が配列番号2をコードする、請求項3に記載のタンパク質。
【請求項9】
配列番号2のアミノ酸配列を有するヒト4−1BBを特異的に認識する モノクローナル抗体。
【請求項10】
配列番号2のアミノ酸配列を有するヒト4−1BBを特異的に認識する モノクローナル抗体を産生することが可能なハイブリドーマ。
【請求項11】
T細胞増殖を促進するために請求項9に記載のモノクローナル抗体を使用する方法であって、発現したヒト4−1BBを有するT細胞を該モノクローナル抗体によりインビトロで処理する工程を含む方法。
【請求項12】
前記処理をタンパク質チロシンキナーゼの存在下で行う工程をさらに含む、請求項11に記載の方法。
【請求項13】
T細胞活性化を促進するために請求項9に記載のモノクローナル抗体を使用する方法であって、発現したヒト4−1BBを有するT細胞を該モノクローナル抗体によりインビトロで処理する工程を含んでなる方法。
【請求項14】
前記処理をタンパク質チロシンキナーゼの存在下で行う工程をさらに含む、請求項13に記載の方法。
【請求項15】
a)ヒト4−1BBに対する細胞膜リガンドに結合する、配列番号2のアミノ酸配列を有するヒト4−1BBの細胞外ドメインに対応する、ヒト4−1BBの少なくとも一部;および
b)前記ヒト4−1BBの該一部に結合した検出タンパク質であって、リガンド結合が、該検出タンパク質の比活性アッセイによって検出できるような検出タンパク質
を含む融合タンパク質。
【請求項16】
前記検出タンパク質がアルカリホスファターゼである、請求項15に記載の融合タンパク質。
【請求項17】
ヒト4−1BBに対する細胞膜リガンドをインビトロで検出するための方法であって、
a)ヒト4−1BBに対する細胞膜リガンドを発現していると思われる細胞と、以下のものを含む融合タンパク質:
1)ヒト4−1BBの、前記細胞膜リガンドに結合する細胞外ドメインに対応する、配列番号2のアミノ酸配列を有する前記ヒト4−1BBの少なくとも一部分;および
2)ヒト4−1BBの該一部分に結合した検出タンパク質であって、該検出タンパク質の比活性アッセイによってリガンド結合が検出できるような検出タンパク質
とを接触させ;
b)前記細胞膜リガンドに結合しなかった融合タンパク質を前記細胞から洗い流し;そして
c)前記洗浄した細胞と、前記検出タンパク質に対する基質とを接触させて、前記検出タンパク質の比活性を測定する
ことを含む方法。
【請求項18】
前記検出タンパク質がアルカリホスファターゼである、請求項17に記載の方法。
【請求項19】
配列番号2のアミノ酸配列を有するヒト4−1BBまたはそのフラグメントをコードするDNA配列を含む組換え発現ベクターであって、宿主細胞中での該DNA配列の発現に適当な制御配列に作動可能なように結合し、該DNA配列が、
a)配列番号1のヌクレオチド41〜805;
b)配列番号1のヌクレオチド110〜805;
c)配列番号2のアミノ酸1〜186を有する可溶性ヒト4−1BBポリペプチドまたはヒト4−1BBに対する細胞膜リガンドに結合可能なそのフラグメントをコードするヌクレオチド配列;
d)配列番号2のアミノ酸配列を有するヒト4−1BBをコードするヌクレオチド配列;および
e)前記可溶性ヒト4−1BBポリペプチドのC−末端に融合するポリペプチドをさらにコードする、c)記載のヌクレオチド配列
からなる群から選択される組換え発現ベクター。
【請求項20】
e)におけるC−末端に融合するポリペプチドが検出タンパク質である、請求項1記載のDNA。
【請求項21】
検出タンパク質がアルカリホスファターゼである、請求項20に記載のDNA。
【請求項22】
前記e)におけるC−末端に融合するポリペプチドが検出タンパク質である、請求項19記載のベクター。
【請求項23】
検出タンパク質がアルカリホスファターゼである、請求項22に記載のベクター。
【請求項24】
a)配列番号2のアミノ酸配列またはヒト4−1BBに対する細胞膜リガンドを結合することが可能なそのフラグメント;
b)配列番号2のアミノ酸残基24〜255またはヒト4−1BBに対する細胞膜リガンドを結合することが可能なそのフラグメント;
c)配列番号2のアミノ酸残基24〜186またはヒト4−1BBに対する細胞膜リガンドを結合することが可能なそのフラグメント;および
d)N−末端アミノ酸配列が、Leu−Gln−Asp−Pro−Cys−Ser−Asn−Cys−Pro−Ala−Gly−Thrである配列番号2のアミノ酸配列
からなる群から選択されるアミノ酸配列を有するヒト4−1BBポリペプチド。
【請求項25】
ヒト4−1BBポリペプチドのC末端に融合した第2のポリペプチドをさらに含む、請求項24に記載のヒト4−1BBポリペプチド。
【図1a】


【図1b】

【図2a】

【図2b】

【図3】

【図4】
【図5】

【図1b】
【図2a】
【図2b】
【図3】
【図4】
【図5】
【公開番号】特開2006−149389(P2006−149389A)
【公開日】平成18年6月15日(2006.6.15)
【国際特許分類】
【出願番号】特願2005−368486(P2005−368486)
【出願日】平成17年12月21日(2005.12.21)
【分割の表示】特願平7−509364の分割
【原出願日】平成6年9月15日(1994.9.15)
【出願人】(500334014)インディアナ・ユニバーシティ・リサーチ・アンド・テクノロジー・コーポレイション (4)
【氏名又は名称原語表記】Indiana University Research and Technology Corporation
【Fターム(参考)】
【公開日】平成18年6月15日(2006.6.15)
【国際特許分類】
【出願日】平成17年12月21日(2005.12.21)
【分割の表示】特願平7−509364の分割
【原出願日】平成6年9月15日(1994.9.15)
【出願人】(500334014)インディアナ・ユニバーシティ・リサーチ・アンド・テクノロジー・コーポレイション (4)
【氏名又は名称原語表記】Indiana University Research and Technology Corporation
【Fターム(参考)】
[ Back to top ]