
マウスのSCURFYの表現型を引き起こす遺伝子およびそのヒトのオルソログの同定
【課題】Fkhsfをコードする単離された核酸分子およびその変異形態を提供すること。
【解決手段】本発明によって、上記課題が解決された。本発明にしたがって、Fkhsfをコードする単離された核酸分子およびその変異形態が提供される。このような核酸分子の発現のために適切な発現ベクター、およびこのような発現ベクターを含む宿主細胞もまた、提供される。本明細書中で開示される核酸配列(およびその変異体形態)に基づくアッセイを利用して、免疫系を調節する多数の分子が同定され得る。
【解決手段】本発明によって、上記課題が解決された。本発明にしたがって、Fkhsfをコードする単離された核酸分子およびその変異形態が提供される。このような核酸分子の発現のために適切な発現ベクター、およびこのような発現ベクターを含む宿主細胞もまた、提供される。本明細書中で開示される核酸配列(およびその変異体形態)に基づくアッセイを利用して、免疫系を調節する多数の分子が同定され得る。
【発明の詳細な説明】
【技術分野】
【0001】
(技術分野)
本発明は、一般的には、薬学的な生成物および方法、ならびにより詳細には、scurfy関連疾患を診断するために有用な方法および組成物に関し、そして、免疫系を調節し得る化合物を同定するための方法に関する。
【背景技術】
【0002】
(発明の背景)
マウスの免疫系に影響を与える遺伝した変異は、免疫系の調節に対して重要な新規の遺伝子の豊富な供給源であることが証明されており、そしてヒトの免疫学的な障害の重要な動物モデルを提供した。これらとしては、xid(X連鎖無ガンマグロブリン血症のマウスの等価物)(非特許文献1および2)、beige(チェディアック‐東症候群の等価物)(Barbosaら、Nature、382:262、1996)、lprおよびgld(fasおよびfasリガンドの欠損)、X連鎖重症複合免疫不全(Sugamuraら、Annu.Rev.Immunol.14:179、1996)、および造血細胞ホスファターゼ変異体motheaten(SHP−1)(BignonおよびSiminovitch、Clin Immunol Immunopathol、73:168、1994)が挙げられる。
【0003】
特定の目的の1つのマウスの変異体は、いまだなおクローン化されていないX連鎖マウス変異体、scurfy(sf)である。簡潔には、scurfy変異について半接合性のマウスは、重篤なリンパ増殖障害を示す。詳細には、scurfy変異について半接合性の雌性(Xsf/Y)は、リンパ節、脾臓、肝臓、および皮膚の進行性のリンパ球浸潤を発症し、それによって巨脾腫、肝腫、大いに肥大したリンパ節、矮小、離脱性皮膚炎、および厚くなった奇形の耳を含む、肉眼的な形態学的な症状を生じる(Godfreyら、Amer.J.Pathol.138:1379、1991;Godfreyら、Proc.Natl.Acad.Sci.USA 88:5528、1991)。他の臨床的な症状としては、増大した白血球数、高ガンマグロブリン血症、および重篤な貧血が挙げられる(Lyonら、Proc.Natl.Acad.Sci.USA 87:2433、1990);罹患した雄性の死亡は、通常は3週齢までに生じる。sf遺伝子座は、X染色体の極めて近位の領域に、sparse−fur(spf)(Lyonら、Proc.Natl.Acad.Sci.USA 87:2433、1990;Blairら、Mamm.Genome 5:652、1994)(それ自体が、オルニチントランスカルバミラーゼ遺伝子(Otc)内の点変異(Veresら、Science 237:415、1987))についての遺伝子座からおよそ0.7センチモルガンに、マップされている。sf遺伝子座はまた、マウスのGata1、Tcfe3、およびWasp遺伝子座と密接に連鎖している(Blairら、Mamm.Genome 5:652、1994;Derryら、Genomics 29:471、1995)。scurfyおよびヒトのWiscott−Aldrich症候群(WAS)との間の類似性が報告されており(Lyonら、Proc.Natl.Acad.Sci.USA 87、2433、1990)、そしてマウスのWasp遺伝子が、scurfyについての候補として提案されている(Lyonら、Proc.Natl.Acad.Sci.USA 87:2433、1990;Derryら、Genomics 29:471、1995)。より最近の生物学的な実験は、WASとscurfyとの間の有意な差異を明らかにするが、しかし、この2つの遺伝子座は、対立遺伝子ではないことが実証されている(JefferyおよびBrunkow、公開されていないデータ)。従って、出願人らの発明の以前には、scurfy遺伝子の正体は、未決定のままであった。
本発明は、scurfyに関連する疾患の診断のために有用な方法および組成物、ならびに、免疫系を調節し得る化合物を同定するための方法を開示し、そしてさらに他の関連する利点を提供する。
この出願の発明に関連する先行技術文献情報としては、次のものがある。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Thomasら、Science、261:355、1993
【非特許文献2】Rawlingsら、Science、261:358、1993
【発明の概要】
【課題を解決するための手段】
【0005】
(発明の要旨)
本発明は、以下を提供する:
(項目1) Fkhsfをコードする、単離された核酸分子。
(項目2) 前記FkhsfがマウスのFkhsfである、項目1に記載の単離された核酸分子。
(項目3) 前記FkhsfがヒトのFKHsfである、項目1に記載の単離された核酸分子。
(項目4) 前記核酸分子が、(a)配列番号2または4を含むアミノ酸配列をコードする核酸分子、(b)配列番号1もしくは3のヌクレオチド配列を有する核酸分子、またはその相補物に対してストリンジェントな条件下でハイブリダイズする核酸分子、および(c)(a)または(b)のいずれかによってコードされるポリペプチドの機能的なフラグメントをコードする核酸分子、
からなる群より選択される、項目1に記載の単離された核酸分子。
(項目5) 前記核酸分子が配列番号2のアミノ酸配列をコードする、項目1に記載の単離された核酸分子。
(項目6) 前記核酸分子が配列番号1のヌクレオチド配列を含む、項目5に記載の単離された核酸分子。
(項目7) 項目1に記載の単離された核酸分子を含む、ベクター。
(項目8) 前記ベクターがウイルスベクターである、項目7に記載のベクター。
(項目9) 前記ウイルスベクターが、レトロウイルス、アデノウイルス、ヘルペスウイルス、アデノ随伴ウイルス、およびアルファウイルスからなる群より選択されるウイルスから生成される、項目8に記載のベクター。
(項目10) 項目1に記載の単離された核酸分子およびプロモーターを含む発現ベクターであって、ここで、該プロモーターが該核酸分子と作動可能に連結されている、発現ベクター。
(項目11) 項目10に記載の発現ベクターを含む、組換え宿主細胞。
(項目12) Fkhsfタンパク質を調製するために項目10に記載の発現ベクターを使用する方法であって、該方法が以下の工程:
(a)該発現ベクターを含み、そして該タンパク質を産生する、組換え宿主細胞を培養する工程、および
(b)該培養した組換え宿主細胞から該タンパク質を単離する工程、
を包含する、方法。
(項目13) 項目1から6のいずれか1項に記載の核酸分子によってコードされるアミノ酸配列を含む、単離されたポリペプチド。
(項目14) 項目1に記載の核酸分子によってコードされるポリペプチドと特異的に結合する、抗体または抗体フラグメント。
(項目15) 前記抗体が、以下:
(a)ポリクローナル抗体、
(b)マウスのモノクローナル抗体、
(c)(b)から誘導されるヒト化抗体、および
(d)ヒトのモノクローナル抗体、
からなる群より選択される、項目13に記載の抗体。
(項目16) 前記抗体フラグメントが、F(ab’)2、F(ab)2、Fab’、Fab、Fv、sFv、および最小認識ユニットからなる群より選択される、項目14に記載の抗体フラグメント。
(項目17) 項目13に記載のポリペプチドを含む、融合タンパク質。
(項目18) 被験体に由来する生物学的サンプル中のFkhsf核酸配列の存在を検出する方法であって、以下の工程:
(a)Fkhsf特異的核酸プローブと、(i)該生物学的サンプルから単離した試験核酸分子、または(ii)RNA分子から合成した核酸分子のいずれかとを、ハイブリダイゼーション条件下で接触させる工程であって、ここで、該プローブが、項目1に記載のヌクレオチド配列の少なくとも一部を認識する、工程、および
(b)該核酸プローブと(i)または(ii)とのハイブリッドの形成を検出する工程、
を包含する、方法。
(項目19) 前記試験核酸分子が、RT−PCRによって得られる、項目18に記載の方法。
(項目20) 生物学的サンプル中のFkhsfまたはその変異形態の存在を検出する方法であって、以下の工程:
(a)該生物学的サンプルを、抗Fkhsf抗体または抗体フラグメントと接触させる工程であって、ここで、該接触工程が、該生物学的サンプルに対する該抗体または抗体フラグメントの結合を可能にする条件下で行われる、工程、および
(b)任意の該結合した抗体または結合した抗体フラグメントを検出する工程、
を包含する、方法。
(項目21) 前記抗体または前記抗体フラグメントが、以下:
(a)ポリクローナル抗体、
(b)マウスのモノクローナル抗体、
(c)(b)から誘導されるヒト化抗体、
(d)ヒトのモノクローナル抗体、および
(e)(b)、(c)、または(d)から誘導される抗体フラグメント、
からなる群より選択される、項目20に記載の方法。
(項目22) 前記抗体フラグメントが、F(ab’)2、F(ab)2、Fab’、Fab、Fv、sFv、および最小認識ユニットからなる群より選択される、項目20に記載の方法。
(項目23) 前記抗体または前記抗体フラグメントが、放射性同位元素、蛍光標識、化学発光標識、酵素標識、生体発光標識、およびコロイド状の金からなる群より選択される検出可能な標識をさらに含む、項目20に記載の方法。
(項目24) 項目1に記載の核酸分子にハイブリダイズし得る、単離されたオリゴヌクレオチド。
(項目25) 検出可能な標識をさらに含む、項目24に記載のオリゴヌクレオチド。
(項目26) 動物に対して項目1に記載のFkhsf核酸分子を投与する工程を包含する、動物に対してFkhsf核酸分子を導入する方法。
(項目27) 前記核酸分子がウイルスベクターによって発現される、項目26に記載の方法。
(項目28) 前記核酸分子がプラスミドベクターによって発現される、項目26に記載の方法。
(項目29) 前記核酸分子がインビボで動物に投与される、項目26に記載の方法。
(項目30) 前記核酸分子がエキソビボで細胞に投与され、次いで該細胞が前記動物に投与される、項目26に記載の方法。
(項目31) 前記細胞が造血細胞である、項目26に記載の方法。
(項目32) 前記造血細胞がT細胞である、項目26に記載の方法。
(項目33) 前記動物が、ヒト、サル、イヌ、ネコ、ラット、およびマウスからなる群より選択される、項目26に記載の方法。
(項目34) その細胞がFkhsfタンパク質をコードする配列を含むトランスジーンを発現する、ヒト以外のトランスジェニック動物。
本発明は、一般的には、変異した場合に、重大なリンパ増殖障害を生じる新規の遺伝子の発見に関する。詳細には、変異マウス(「Scurfy」と命名された)が、戻し交配分析、物理的なマッピング、および大規模なDNAの配列決定を通じたこの障害の原因遺伝子の同定のために使用された。この遺伝子の配列の分析は、この遺伝子が、関連の遺伝子ファミリー(全てが、翼状(winged)へリックスDNA結合ドメインを含有する)に属することを、示した。
【0006】
従って、本発明の1つの局面においては、FKHsfまたはFkhsfをコードする単離された核酸分子(それらの変異形態を含む)が提供される。特定の実施態様においては、任意の型のFkhsfは、温血動物(例えば、マウスまたはヒト)に由来し得る。さらなる実施態様においては、単離された核酸分子が提供され、ここで、この核酸分子は、以下からなる群より選択される:(a)配列番号2または配列番号4を含むアミノ酸配列をコードする核酸分子、(b)配列番号1もしくは配列番号3のヌクレオチド配列、またはその相補物を有する核酸分子に対してストリンジェントな条件下でハイブリダイズする核酸分子、および(c)(a)または(b)のいずれかによってコードされるポリペプチドの機能的なフラグメントをコードする核酸分子。好ましくは、核酸分子は、JM2ではない。関連する局面においては、ベクター(発現ベクターを含む)および組換え宿主細胞、ならびに上記の核酸分子によってコードされるタンパク質もまた、提供される。さらに、別のタンパク質のコード領域と上記の核酸分子の少なくとも一部とを組み合わせた融合タンパク質もまた、提供される。上記の配列に基づくオリゴヌクレオチドフラグメント(プローブおよびプライマーを含む)もまた、提供される。このようなフラグメントは、少なくとも8、10、12、15、20、または25ヌクレオチド長であり、そして100、200、500、1000、1500、または2000ヌクレオチド長にまで及び得る。
【0007】
他の局面においては、Fkhsfタンパク質(任意の型)を産生するために上記の発現ベクターを使用する方法が提供される。この方法は、(a)この発現ベクターを含みそしてFkhsfタンパク質を産生する組換え宿主細胞を培養する工程、および(b)培養した組換え宿主細胞からタンパク質を単離する工程、という一般的な工程を包含する。
【0008】
Fkhsfタンパク質に特異的に結合する抗体および抗体フラグメントもまた、提供される。このような抗体の代表的な例としては、ポリクローナル抗体およびモノクローナル抗体(マウスのハイブリドーマから得られたかまたはヒトの形態に誘導されたかにはかかわらず)の両方が挙げられる。抗体フラグメントの代表的な例としては、F(ab’)2、F(ab)2、Fab’、Fab、Fv、sFv、および最小認識ユニットまたは相補性決定領域が挙げられる。
【0009】
なお他の局面においては、被験体に由来する生物学的サンプル中のFkhsf核酸配列の存在を検出するための方法が、提供される。この方法は、以下の工程を包含する:(a)Fkhsf特異的核酸プローブと、(i)上記の生物学的サンプルから単離された試験核酸分子、または(ii)RNA分子から合成された核酸分子のいずれかとを、ハイブリダイゼーション条件下で接触させる工程(ここで、上記のプローブは、請求項1に記載のヌクレオチド配列の少なくとも一部を認識する)、および(b)上記の核酸プローブと(i)または(ii)とのハイブリッドの形成を検出する工程。
【0010】
別の関連する実施態様においては、生物学的サンプル中の、Fkhsfまたはその変異形態の存在を検出するための方法が提供される。この方法は、以下の工程を包含する:(a)生物学的サンプルを、抗Fkhsf抗体または抗体フラグメントと接触させる工程(ここで、上記の接触工程は、上記の生物学的サンプルに対する上記の抗体または抗体フラグメントの結合を可能にする条件下で行われる)、および(b)任意の上記の結合した抗体または結合した抗体フラグメントを検出する工程。
【0011】
本発明の他の局面においては、動物に対してFkhsf核酸分子を導入する方法が提供される。この方法は、動物(例えば、ヒト、サル、イヌ、ネコ、ラット、またはマウス)に対して、本明細書中に記載されているようなFkhsf核酸分子を投与する工程を包含する。1つの実施態様においては、核酸分子は、ウイルスベクター(例えば、レトロウイルス、アデノウイルス、アデノ随伴ウイルス、ヘルペスウイルスまたはアルファウイルスから少なくとも一部生成されたベクター)中に含まれ、そしてそれによって発現される。別の実施態様においては、核酸分子は、プラスミドベクターによって発現されるか、またはその中に含まれる。このようなベクターは、インビボまたはエキソビボのいずれかで、(例えば、T細胞のような造血細胞に)投与され得る。
【0012】
他の実施態様においては、ヒト以外のトランスジェニック動物が提供される。ここで、この動物の細胞は、Fkhsfタンパク質をコードする配列を含むトランスジーンを発現する。
本発明のこれらおよび他の局面は、以下の詳細な説明および添付の図面を参照して明らかとなる。さらに、特定の手順または組成物(例えば、プラスミドなど)をより詳細に記載する種々の参考文献が、本明細書中に示されており、そしてそれゆえそれらの全体が、参考として援用されている。
【図面の簡単な説明】
【0013】
【図1A】図1は、マウスのFkhsf cDNAのヌクレオチド配列(配列番号1)を示す;翻訳は、259位で始まり、そして1546位で終わると推定される。
【図1B】図1は、マウスのFkhsf cDNAのヌクレオチド配列(配列番号1)を示す;翻訳は、259位で始まり、そして1546位で終わると推定される。
【図2】図2は、マウスのFkhsfのアミノ酸配列(配列番号2)を示す。
【図3A】図3は、ヒトのFKHsf cDNAに対応する1735bpのヌクレオチド配列(配列番号3、1293bpのコード領域を含む)を示す;翻訳は、55位で始まり、そして1348位で終わると推定される。
【図3B】図3は、ヒトのFKHsf cDNAに対応する1735bpのヌクレオチド配列(配列番号3、1293bpのコード領域を含む)を示す;翻訳は、55位で始まり、そして1348位で終わると推定される。
【図4】図4は、ヒトのFKHsfタンパク質の431アミノ酸の配列(配列番号4)を示す。
【図5】図5は、FKHsfトランスジェニックマウスの生成のためのベクターを模式的に示す。
【図6】図6は、FKHsfトランスジーンが、scurfy動物における欠損を修復することを示す写真である。
【図7】図7は、FKHsf tgマウスが、正常な細胞と比較して、減少したリンパ節細胞を有することを示すグラフである。
【図8】図8は、FKHsfトランスジェニックマウスがインビトロでの刺激にほとんど応答しないことを示すグラフである。
【図9】図9は、FKHsfとJM2とのcDNAの比較である。
【図10】図10は、ヒトのFKHsfとマウスのFkhsfとの種々の領域における相同性を比較する。
【発明を実施するための形態】
【0014】
(発明の詳細な説明)
(定義)
本発明を詳細に示す前に、特定の用語の定義を示すこと、および本明細書中で以降に使用される略号を列挙しそしてそれらを定義することは、本発明の理解に役立ち得る。
【0015】
「Scurfy」は、重篤なリンパ増殖障害を示すマウスにおける遺伝した疾患をいう(例えば、Lyonら、Proc.Natl.Acad.Sci.USA 87:2433、1990を参照のこと)。原因遺伝子(その変異形態が、この疾患の原因である)を、配列番号1および3に示す。
【0016】
「分子」は、タンパク質またはペプチド(例えば、抗体、組換え結合パートナー、所望される結合親和性を有するペプチド)、核酸(例えば、DNA、RNA、キメラ核酸分子、およびPNAのような核酸アナログ)、および有機化合物または無機化合物を含むと理解されるはずである。
【0017】
「核酸」または「核酸分子」は、任意のデオキシリボ核酸(DNA)、リボ核酸(RNA)、オリゴヌクレオチド、ポリメラーゼ連鎖反応(PCR)によって生成されたフラグメント、ならびに任意の連結、切断、エンドヌクレアーゼ作用、およびエキソヌクレアーゼ作用によって生成されたフラグメントをいう。核酸は、天然に存在するヌクレオチド(例えば、デオキシリボヌクレオチドおよびリボヌクレオチド)、または天然に存在するヌクレオチドのアナログ(例えば、天然に存在するヌクレオチドのα−エナンチオマー形態)であるモノマー、あるいは両方の組み合わせから構成され得る。改変されたヌクレオチドは、糖部分、および/またはピリミジン塩基部分もしくはプリン塩基部分において修飾を有し得る。糖の修飾としては、例えば、ハロゲン、アルキル基、アミン、およびアジド基での1つ以上のヒドロキシル基の置換が挙げられ、または糖はエーテルまたはエステルとして官能化され得る。さらに、全体的な糖部分は、立体構造的に、および電気的に類似の構造(例えば、アザ糖、および炭素環式糖アナログ)で置きかえられ得る。塩基部分における修飾の例としては、アルキル化されたプリンおよびピリミジン、アシル化されたプリンまたはピリミジン、あるいは他の周知の複素環式置換基が挙げられる。核酸のモノマーは、ホスホジエステル結合またはこのような連結のアナログによって連結され得る。ホスホジエステル結合のアナログとしては、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホルアニリデート、ホスホルアミデートなどが挙げられる。用語「核酸」はまた、いわゆる「ペプチド核酸」を含む。これは、ポリアミド骨格に対して結合した、天然に存在するかまたは修飾された核酸塩基を含む。核酸は、一本鎖または二本鎖のいずれかであり得る。
【0018】
「単離された核酸分子」は、生物体のゲノムDNA中には組み込まれていない核酸分子である。例えば、真核生物細胞のゲノムDNAから分離されている遺伝子をコードするDNA分子は、単離されたDNA分子である。単離された核酸分子の別の例は、生物体のゲノム中には組み込まれていない、化学的に合成された核酸分子である。
【0019】
「プロモーター」は、構造遺伝子の転写を指向するヌクレオチド配列である。代表的には、プロモーターは、構造遺伝子の転写開始部位の近位の、遺伝子の5’領域中に配置される。プロモーターが誘導性プロモーターである場合は、転写速度は、誘導剤に応答して増大する。対照的に、プロモーターが構成的プロモーターである場合は、転写の速度は、誘導剤によっては調節されない。
【0020】
「ベクター」は、所望されるタンパク質の発現を指向し得るアセンブリをいう。ベクターは、目的の遺伝子に対して作動可能に連結された転写プロモーターエレメントを含まなければならない。ベクターは、デオキシリボ核酸(「DNA」)、リボ核酸(「RNA」)、またはこの2つの組み合わせ(例えば、DNA−RNAキメラ)のいずれかから構成され得る。必要に応じて、ベクターは、ポリアデニル化配列、1つ以上の制限部位、ならびに1つ以上の選択マーカー(例えば、ネオマイシンホスホトランスフェラーゼまたはハイグロマイシンホスホトランスフェラーゼ)を含み得る。さらに、選択される宿主細胞および使用されるベクターに依存して、他の遺伝的なエレメント(例えば、複製起点、さらなる核酸制限部位、エンハンサー、転写誘導性を付与する配列、および選択マーカー)もまた、本明細書中に記載されているベクター中に取りこまれ得る。
【0021】
「単離された」は、タンパク質またはポリペプチドの場合においては、他の生物学的巨大分子の実質的非存在下に存在し、そしてクマシーブルー染色を用いてSDS−PAGEゲル上に名目上単一のバンドとして出現する分子をいう。「単離された」は、有機分子について言及する場合は、その化合物が、当該分野で周知の方法(例えば、NMR、融点)を利用して90%よりも純粋であることを意味する。
【0022】
「クローニングベクター」は、宿主細胞中で自律複製する能力を有する、プラスミド、コスミド、またはバクテリオファージのような核酸分子をいう。クローニングベクターは、代表的には、1または少数の制限エンドヌクレアーゼ認識部位(ここでは、外来のヌクレオチド配列が、ベクターの本質的な生物学的な機能の損失を伴わずに、決定可能な様式で挿入され得る)およびクローニングベクターで形質転換された細胞の同定および選択における使用に適切であるマーカー遺伝子をコードするヌクレオチド配列を含む。マーカー遺伝子としては、代表的には、テトラサイクリン耐性またはアンピシリン耐性を提供する遺伝子が挙げられる。
【0023】
「発現ベクター」は、宿主細胞中で発現される遺伝子をコードする核酸分子をいう。代表的には、遺伝子の発現は、プロモーターの制御下に配置され、そして必要に応じて、少なくとも1つの調節エレメントの制御下に配置される。このような遺伝子は、プロモーター「に対して作動可能に連結された」といわれる。同様に、調節エレメントおよびプロモーターは、調節エレメントがプロモーターの活性を調節する場合は、作動可能に連結される。
【0024】
「組換え宿主」は、クローニングベクターまたは発現ベクターのいずれかを含む、任意の原核生物細胞または真核生物細胞をいう。この用語はまた、宿主細胞の染色体またはゲノム中のクローン化された遺伝子を含むように遺伝子操作されている、原核生物細胞または真核生物細胞を含む。
【0025】
真核生物においては、RMAポリメラーゼIIは、mRNAを産生するように、構造遺伝子の転写を触媒する。核酸分子は、RNAポリメラーゼIIの鋳型を含むように設計され得、ここで、RNA転写物は、特異的なmRNAの配列に対して相補的である配列を有する。このRNA転写物は、「アンチセンスRNA」と呼ばれ、そしてアンチセンスRNAをコードする核酸分子は、「アンチセンス遺伝子」と呼ばれる。アンチセンスRNA分子は、mRNA分子に結合し得、それによってmRNAの翻訳の阻害を生じる。
【0026】
「Fkhsfに特異的なアンチセンスオリゴヌクレオチド」または「Fkhsfアンチセンスオリゴヌクレオチド」は、(a)遺伝子の一部と安定な三重鎖を形成し得るか、または(b)mRNA転写物の一部と安定な二重鎖を形成し得る配列を有するオリゴヌクレオチドである。同様に、「Fkhsfに特異的なアンチセンスオリゴヌクレオチド」または「Fkhsfアンチセンスオリゴヌクレオチド」は、(a)Fkhsf遺伝子の一部と安定な三重鎖を形成し得るか、または(b)Fkhsf遺伝子のmRNA転写物の一部と安定な二重鎖を形成し得る配列を有するオリゴヌクレオチドである。
【0027】
「リボザイム」は、触媒中心を含む核酸分子である。この用語は、RNA酵素、自己スプライシングRNA、自己切断RNA、およびこれらの触媒機能を行う核酸分子を含む。リボザイムをコードする核酸分子は、「リボザイム遺伝子」と呼ばれる。
【0028】
略号:YAC、酵母人工染色体;PCR、ポリメラーゼ連鎖反応;RT−PCR、逆転写酵素(RT)を使用して最初の工程でRNAが最初にDNAに転写されるPCRプロセス;cDNA、DNA形態にRNA配列をコピーすることによって作成される、任意のDNA。本明細書中で利用される場合は、「Fkhsf」は、(この遺伝子が、ヒト、哺乳動物、または任意の他の温血動物から得られたかどうかにはかかわらず)Fkhsf遺伝子の遺伝子産物をいう。「FKHsf」と大文字になっている場合は、この遺伝子産物(および遺伝子)は、ヒトに由来すると理解されるべきである。
【0029】
上記のように、本発明は、一般的には、薬学的な生成物および方法、そしてより詳細には、scurfyに関連する疾患を診断するために有用な方法および組成物、ならびに免疫系を調節し得る化合物を同定するための方法に関する。
【0030】
従って、以下により詳細に議論されるように、この発見は、免疫系のアゴニストまたは、あるいはアンタゴニストとして作用し得る分子を選択するために利用され得るアッセイの開発を導いた。さらに、このようなアッセイは、免疫系を調節することにおいて同様に活性である、他の遺伝子および遺伝子産物を同定するために利用され得る。
【0031】
(SCURFY) 簡潔には、本発明は、Fkhsfをコードする遺伝子における変異が稀な状態(scurfy)を生じるという予想外の発見に基づく。この状態は、リンパ節、脾臓、肝臓、および皮膚の進行性のリンパ球浸潤によって特徴付けられ、それによって巨脾腫、肝腫、大いに肥大したリンパ節、矮小、脱離性皮膚炎、および厚くなった奇形の耳を含む、肉眼的な形態学的な症状を生じる(Godfreyら、Amer.J.Pathol.138:1379、1991;Godfreyら、Proc.Natl.Acad.Sci.USA、88:5528、1991)。この翼状ヘリックスのファミリーの新規のメンバーは、免疫系の新規の成分を示す。
【0032】
scurfyの原因遺伝子を発見するために利用された方法は、以下の実施例1に提供される。マウスのscurfyの原因遺伝子、およびヒトのオルソログをクローニングするための方法は、実施例2および3において以下に提供される。トランスジェニックマウスを使用して決定されるような、遺伝子の正体および遺伝子機能との相関の確認のための方法もまた、実施例において提供される。
【0033】
Fkhsf遺伝子および遺伝子産物の存在を決定するための方法もまた、本発明によって提供される。1つの実施態様においては、このような方法は、以下の一般的な工程を包含する:(a)Fkhsf特異的核酸プローブと、(i)生物学的サンプルから単離された試験核酸分子、または(ii)RNA分子から合成された核酸分子のいずれかとを、ハイブリダイゼーション条件下で接触させる工程(ここで、プローブは、Fkhsfのヌクレオチド配列の少なくとも一部を認識する)、および(b)上記の核酸プローブと(i)または(ii)とのハイブリッドの形成を検出する工程。種々の方法(例えば、RNA増幅(Lizardiら、Bio/Technology 6:1197−1202、1988;Kramerら、Nature、339:401−402、1989;Lomeliら、Clinical Chem.35(9):1826−1831、1989;米国特許第4,786,600号を参照のこと)、およびポリメラーゼ連鎖反応(「PCR」)を利用する核酸の増幅(米国特許第4,683,195号、同第4,683,202号、および同第4,800,159号を参照のこと)、逆転写酵素PCRおよびCPT(米国特許第4,876,187号および同第5,011,769号を参照のこと)を含む)が、選択された配列を増幅するために利用され得る。
【0034】
あるいは、抗体は、Fkhsf遺伝子産物の存在を検出するために利用され得る。より詳細には、1つの実施態様においては、生物学的サンプル中のFkhsfペプチドまたはその変異体形態の存在を検出するための方法が提供される。この方法は、以下の工程を包含する:(a)抗Fkhsf抗体または抗体フラグメントと生物学的サンプルとを接触させる工程(ここで、上記の接触工程は、生物学的サンプルに対する上記の抗体または抗体フラグメントの結合を可能にする条件下で行われる)、および(b)任意の結合した抗体または結合した抗体フラグメントを検出する工程。
【0035】
このような方法は、例えば、向流免疫電気泳動(Countercurrent Immuno−Electrophoresis、CIEP)、放射免疫アッセイ、放射免疫沈降、酵素結合免疫吸着アッセイ(ELISA)、ドットブロットアッセイ、阻害または競合アッセイ、およびサンドイッチアッセイを含む、広範な種々のアッセイ形式において達成され得る(米国特許第4,376,110号および同第4,486,530号を参照のこと;Antibodies:A Laboratory Manual、前出もまた参照のこと)。
【0036】
(核酸分子、タンパク質、およびタンパク質の産生方法) 種々のFKHsfまたはFkhsfのタンパク質および核酸分子(またはその一部)が本明細書中で提供されているが、本発明の状況においては、1つ以上のこれらのタンパク質に関する言及は、実質的に類似の活性のタンパク質を含むと理解されるべきであると理解されるべきである。本明細書中で使用される場合は、タンパク質は、(a)これらがこのタンパク質をコードする遺伝子のコード領域に由来するヌクレオチド配列(例えば、この配列の一部、またはこの配列の対立遺伝子改変体を含む)によってコードされる;(b)このヌクレオチド配列が、中程度、高い、または非常に高いストリンジェンシー(Sambrookら、Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor Laboratory Press、NY、1989を参照のこと)の下で本発明のヌクレオチド配列に対してハイブリダイゼーションし得るか、または本明細書中で開示されている配列に対して、少なくとも50%、60%、70%、75%、80%、90%、95%、もしくはそれより高い相同性を有する;あるいは(c)このDNA配列が、(a)または(b)において定義されるDNA配列に対する遺伝コードの結果として縮重している場合に、「実質的に類似」であると考えられる。さらに、本明細書中で開示されている核酸分子は、配列が、他の点では本明細書中に示される基準を満たす限りは、相補配列および非相補配列の両方を含む。本発明の状況においては、高いストリンジェンシーは、標準的なハイブリダイゼーション条件(例えば、65℃にて、5×SSPE、0.5%のSDS、またはそれに等価なもの)を意味する。ハイブリダイゼーションの目的のためには、アミノ末端ドメイン、ジンクフィンガードメイン、中央ドメイン、またはフォークヘッドドメイン(実施例10を参照のこと)をコードする核酸分子が、利用され得る。
【0037】
本明細書中に記載されている核酸分子によってコードされるタンパク質の構造は、例えば、P/C遺伝子またはIntelligenetics Suite(Intelligenetics,Mountain View、California)の疎水性プロット関数を使用して、またはKyteおよびDoolittle(J.Mol.Biol.157:105−132、1982)によって記載されている方法に従って、一次翻訳産物から推定され得る。
【0038】
本発明のタンパク質は、酸の塩もしくは塩基の塩の形態で、または中性の形態で、調製され得る。さらに、個々のアミノ酸残基は、酸化または還元によって改変され得る。さらに、種々の置換、欠失、または付加が、アミノ酸または核酸の配列に対して行われ得、その正味の影響は、変異体または野生型のタンパク質の生物学的な活性を維持するか、またはさらに増強もしくは減少させる。さらに、遺伝コードの縮重に起因して、例えば、同じアミノ酸配列をコードするヌクレオチド配列におけるかなりのバリエーションが存在し得る。
【0039】
本明細書中に開示されているタンパク質の他の誘導体としては、他のタンパク質またはポリペプチドとのタンパク質の結合体が挙げられる。これは、例えば、タンパク質の精製または同定を容易にするために付加され得る、N末端融合タンパク質またはC末端融合タンパク質の合成によって達成され得る(米国特許第4,851,341号を参照のこと、また、Hoppら、Bio/Technology 6:1204、1988をもまた参照のこと)。あるいは、融合タンパク質(例えば、FKHもしくはFkh−ルシフェラーゼ、またはFKHもしくはFkh−GFP)が、タンパク質の同定、発現、および分析を補助するために構築され得る。
【0040】
本発明のタンパク質は、本明細書中に記載されている広範な種々の技術を使用して構築され得る。さらに、変異は、天然の配列のフラグメントに対する連結を可能にする制限部位に隣接した、変異体配列を含むオリゴヌクレオチドを合成することによって、特定の遺伝子座に導入され得る。連結後、得られた再構築された配列は、所望のアミノ酸の挿入、置換、または欠失を有する誘導体をコードする。
【0041】
あるいは、オリゴヌクレオチドによって指向される部位特異的(またはセグメント特異的)変異誘発手順が、要求される置換、欠失、または挿入に従って変更された特定のコドンを有する変更された遺伝子を提供するために使用され得る。上記に示される変更を作成する例示的な方法は、Walderら(Gene 42:133、1986);Bauerら(Gene 37:73、1985);Craik(BioTechniques、1985年1月、12−19);Smithら(Genetic Engineering:Principles and Methods、Plenum Press、1981):およびSambrookら(前出)によって開示されている。タンパク質の欠失誘導体または短縮誘導体(例えば、可溶性の細胞外部分)もまた、所望される欠失に対して隣接する便利な制限エンドヌクレアーゼ部位を利用することによって、構築され得る。制限消化に続いて、突出がフィルインされ得、そしてDNAが再連結され得る。上記に示される変更を作成する例示的な方法が、Sambrookら(Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor Laboratory Press、1989)によって開示されている。
【0042】
本発明の核酸分子中に作成される変異は、好ましくは、コード配列のリーディングフレームを保存する。さらに、変異は、好ましくは、mRNAの翻訳に有害な影響を与える二次的なmRNA構造(例えば、ループまたはヘアピン)を生じるようにハイブリダイズし得る相補性領域を作成しない。変異部位は予め決定され得るが、変異の性質自体は予め決定される必要はない。例えば、所定の部位で変異体の最適の特徴について選択するためには、ランダムな変異誘発が標的のコドンで行われ得、そして発現された変異体が表示生物学的活性についてスクリーニングされ得る。あるいは、変異は、天然の配列のフラグメントへの連結を可能にする制限部位に隣接した変異体配列を含むオリゴヌクレオチドを合成することによって、特定の遺伝子座に導入され得る。連結後、得られた再構築された配列は、所望のアミノ酸の挿入、置換、または欠失を有する誘導体をコードする。変異は、タンパク質の活性を保存するかもしくは増大させる、またはタンパク質(例えば、変異体Fkh)を減少させるかもしくは不能にする目的のために導入され得る。
【0043】
本発明のタンパク質をコードする核酸分子もまた、PCR変異誘発、化学的変異誘発の技術(DrinkwaterおよびKlinedinst、PNAS 83:3402−3406、1986)を利用して、ヌクレオチドの誤った取りこみを強制することによって(例えば、LiaoおよびWise、Gene 88:107−111、1990)、またはランダムに変異誘発したオリゴヌクレオチドの使用によって(Horwitzら、Genome 3:112−117、1989)、構築され得る。
【0044】
本発明はまた、上記に記載されている遺伝子を発現し得るベクターを含有する宿主細胞を培養することによって、上記に記載されている遺伝子の操作および発現を提供する。このようなベクターまたはベクター構築物は、適切な転写調節エレメントまたは翻訳調節エレメントに対して作動可能に連結された所望のタンパク質をコードする、合成核酸分子またはcDNAから誘導される核酸分子のいずれかを含む。適切な調節エレメントは、細菌、真菌、ウイルス、哺乳動物、昆虫、または植物の遺伝子を含む種々の供給源から誘導され得る。適切な調節エレメントの選択は、選択される宿主細胞に依存し、そして当業者によって容易に達成され得る。調節エレメントの例としては、以下が挙げられる:転写プロモーターおよびエンハンサー、またはRNAポリメラーゼ結合配列、転写ターミネーター、およびリボソーム結合配列(翻訳開始シグナルを含む)。
【0045】
上記に記載されている任意のタンパク質をコードする核酸分子は、細菌、哺乳動物、酵母もしくは他の真菌、ウイルス、または植物細胞を含む、広範な種々の原核生物宿主細胞および真核生物宿主細胞によって容易に発現され得る。このような細胞を外来DNAを発現するように形質転換またはトランスフェクトするための方法は、当該分野で周知である(例えば、Itakuraら、米国特許第4,704,362号;Hinnenら、Proc.Natl.Acad.Sci.USA 75:1929−1933、1978;Murrayら、米国特許第4,801,542号;Upshallら、米国特許第4,935,349号;Hagenら、米国特許第4,784,950号;Axelら、米国特許第4,399,216号;Goeddelら、米国特許第4,766,075号;およびSambrookら、Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor Laboratory Press、1989を参照のこと;植物細胞については、CzakoおよびMarton、Plant Physiol.、104:1067−1071、1994;およびPaszkowskiら、Biotech.24:387−392、1992を参照のこと)。
【0046】
本発明を実行するために適切な細菌宿主細胞としては、E.coli、B.subtilis、Salmonella typhimurium、ならびにPseudomonas属、Streptomyces属、およびStaphylococcus属内の種々の種、ならびに当業者に周知の多くの他の細菌種が挙げられる。細菌宿主細胞の代表的な例としては、DH5αが挙げられる(Stratagene、LaJolla、California)。
【0047】
細菌の発現ベクターは、好ましくは、宿主細胞中で機能するプロモーター、1つ以上の選択可能な表現型マーカー、および細菌の複製起点を含む。代表的なプロモーターとしては、β−ラクタマーゼ(ペニシリナーゼ)およびラクトースのプロモーターシステム(Changら、Nature 275:615、1978を参照のこと)、T7 RNAポリメラーゼプロモーター(Studierら、Meth.Enzymol.185:60−89、1990)、λプロモーター(Elvinら、Gene 87:123−126、1990)、trpプロモーター(NicholsおよびYanofsky、Meth.in Enzymology 101:155、1983)、およびtacプロモーター(Russellら、Gene 20:231、1982)が挙げられる。代表的な選択マーカーとしては、種々の抗生物質耐性マーカー(例えば、カナマイシン耐性遺伝子またはアンピシリン耐性遺伝子)が挙げられる。宿主細胞を形質転換するために適切な多くのプラスミドが、当該分野で周知であり、中でも、pBR322(Bolivarら、Gene 2:95、1977を参照のこと)、pUCプラスミドpUC18、pUC19、pUC118、pUC119(Messing、Meth.in Enzymology 101:20−77、1983、ならびにVieiraおよびMessing、Gene 19:259−268、1982を参照のこと)、ならびにpNH8A、pNH16a、pNH18a、ならびにBluescript M13(Stratagene、La Jolla、California)が挙げられる。
【0048】
本発明を実行するために適切な酵母宿主細胞および真菌宿主細胞としては、中でも、Saccharomyces pombe、Saccharomyces cerevisiae、Pichia属またはKluyveromyces属、およびAspergillus属の種々の種(McKnightら、米国特許第4,935,349号)が挙げられる。酵母および真菌についての適切な発現ベクターとしては、中でも、酵母についてはYCp50(ATCC第37419号)、およびamdSクローニングベクターpV3(Turnbull、Bio/Technology 7:169、1989)、YRp7(Struhlら、Proc.Natl.Acad.Sci.USA 76:1035−1039、1978)、YEp13(Broachら、Gene 8:121−133、1979)、pJDB249およびpJDB219(Beggs、Nature 275:104−108、1978)およびそれらの誘導体が挙げられる。
【0049】
酵母中での使用に好ましいプロモーターとしては、酵母の解糖遺伝子に由来するプロモーター(Hitzemanら、J.Biol.Chem.255:12073−12080、1980;AlberおよびKawasaki,J.Mol.Appl.Genet.1:419−434、1982)またはアルコールデヒドロゲナーゼ遺伝子に由来するプロモーター(Youngら、Genetic Engineering of Microorganisms for Chemicals、Hollaenderら(編)、355頁、Plenum,New York、1982;Ammerer、Meth Enzymol.101:192−201、1983)が挙げられる。真菌ベクターについての有用なプロモーターの例としては、Aspergillus nidulans解糖遺伝子に由来するプロモーター(例えば、adh3プロモーター(McKnightら、EMBO J.4:2093−2099、1985)が挙げられる。発現ユニットはまた、転写ターミネーターを含み得る。適切なターミネーターの例は、adh3ターミネーターである(McKnightら、同書、1985)。
【0050】
細菌のベクターを用いる場合は、酵母のベクターは、一般的には、表現型アッセイが存在して形質転換体が選択されることを可能にする優性表現型を示す任意の数の遺伝子の1つであり得る、選択マーカーを含む。好ましい選択マーカーは、宿主細胞の栄養要求性を相補するか、抗生物質耐性を提供するか、または細胞が特定の炭素源を利用することを可能にするものであり、そしてleu2(Broachら、同書)、ura3(Botsteinら、Gene 8:17、1979)、またはhis3(Struhlら、同書)を含む。別の適切な選択マーカーは、酵母細胞に対してクロラムフェニコール耐性を付与する、cat遺伝子である。
【0051】
真菌を形質転換するための技術は、文献において周知であり、そして例えば、Beggs(同書)、Hinnenら(Proc.Natl.Acad.Sci.USA 75:1929−1933、1978)、Yeltonら(Proc.Natl.Acad.Sci.USA 81:1740−1747、1984)、およびRussell(Nature 301:167−169、1983)によって記載されている。宿主細胞の遺伝子型は、発現ベクター上に存在する選択マーカーによって相補される遺伝的な欠損を含み得る。特定の宿主および選択マーカーの選択は、十分に当業者のレベルの範囲内である。
【0052】
酵母を形質転換するためのプロトコールもまた、当業者に周知である。例えば、形質転換は、DNAを用いた酵母スフェロプラストの調製によるか(Hinnenら、PNAS USA 75:1929、1978を参照のこと)、またはLiClのようなアルカリ塩での処理による(Itohら、J.Bacteriology 153:163、1983を参照のこと)かのいずれかによって、容易に達成され得る。真菌の形質転換はまた、Cullenら(Bio/Technology 5:369、1987)によって記載されているようにポリエチレングリコールを使用して行われ得る。
【0053】
ウイルスベクターとしては、上記に記載されているような所望のタンパク質をコードする単離された核酸分子の発現を指向するプロモーターを含むベクターが挙げられる。以下を含む広範な種々のプロモーターが、本発明の状況において利用され得る:例えば、MoMLV LTR、RSV LTR、Friend MuLV LTR、アデノウイルスプロモーター(Ohnoら、Science 265:781−784、1994)、ネオマイシンホスホトランスフェラーゼプロモーター/エンハンサー、後期パルボウイルスプロモーター(Koeringら、Hum.Gene Therap.5:457−463、1994)、Herpes TKプロモーター、SV40プロモーター、メタロチオネインIIa遺伝子エンハンサー/プロモーター、サイトメガロウイルス即時初期プロモーター、およびサイトメガロウイルス即時後期プロモーターのようなプロモーター。本発明の特定の好ましい実施態様においては、プロモーターは、組織特異的プロモーターである(例えば、第WO91/02805号;第EP 0,415,731号;および第WO90/07936号を参照のこと)。適切な組織特異的プロモーターの代表的な例としては、神経特異的エノラーゼプロモーター、血小板由来増殖因子βプロモーター、ヒトα1−キマエリン(chimaerin)プロモーター、シナプシンIプロモーター、およびシナプシンIIプロモーターが挙げられる。上記のプロモーターに加えて、他のウイルス特異的プロモーター(例えば、レトロウイルスプロモーター(上記のプロモーターおよびHIVプロモーターのような他のプロモーターを含む)、肝炎、ヘルペス(例えば、EBV)、および細菌、真菌、または寄生生物(例えば、マラリア)特異的プロモーター)が、ウイルス、細菌、真菌、または寄生生物に感染した特異的な細胞または組織を標的化するために利用され得る。
【0054】
本発明を実行するために適切な哺乳動物細胞としては、中でも以下が挙げられる:PC12(ATCC第CRL1721号)、N1E−115神経芽細胞腫、SK−N−BE(2)C神経芽細胞腫、SHSY5アドレナリン作動性神経芽細胞腫、NS20YおよびNG108−15マウスコリン作用性細胞株、またはラットのF2後根神経節株、COS(例えば、ATCC第CRL1650号または同第1651号)、BHK(例えば、ATCC第CRL 6281号;BHK570細胞株(登録番号第CRL10314号のもとで、アメリカンタイプカルチャーコレクションに寄託された))、CHO(ATCC第CCL61号)、HeLa(例えば、ATCC第CCL2号)、293(ATCC第1573号;Grahamら、J.Gen.Virol.36:59−72、1977)、およびNS−1細胞。以下を含む他の哺乳動物細胞株が、本発明において使用され得る:ラットのHep1(ATCC第CRL1600号)、ラットのHepII(ATCC第CRL1548号)、TCMK(ATCC第CCL139号)、ヒトの肺(ATCC第CCL75.1号)、ヒトの肝ガン(ATCC第HTB−52号)、Hep G2(ATCC第HB8065号)、マウスの肝臓(ATCC第CCL29.1号)、NCTC 1469(ATCC第CCL9.1号)、SP2/0−Ag14(ATCC第1581号)、HIT−T15(ATCC第CRL1777号)、Jurkat(ATCC第Tib 152号)、およびRINm 5AHT2B(OrskovおよびNielsen、FEBS 22
9(1):175−178、1988)。
【0055】
本発明を実行することにおける使用のための哺乳動物の発現ベクターとしては、クローン化された遺伝子またはcDNAの転写を指向し得るプロモーターが挙げられる。好ましいプロモーターとしては、ウイルスプロモーターおよび細胞性のプロモーターが挙げられる。ウイルスプロモーターとしては、サイトメガロウイルス即時初期プロモーター(Boshartら、Cell 41:521−530、1985)、サイトメガロウイルス即時後期プロモーター、SV40プロモーター(Subramaniら、Mol.Cell.Biol.1:854−864、1981)、MMTV LTR、RSV LTR、メタロチオネイン−1、アデノウイルスE1aが挙げられる。細胞性のプロモーターとしては、マウスのメタロチオネイン−1プロモーター(Palmiterら、米国特許第4,579,821号)、マウスVKプロモーター(Bergmanら、Proc.Natl
.Acad.Sci.USA、81:7041−7045、1983);Grantら、Nucl.Acids Res.15:5496、1987)、およびマウスVHプロモ
ーター(Lohら、Cell 33:85−93、1983)が挙げられる。プロモーターの選択は、少なくとも一部、所望される発現のレベル、またはトランスフェクトされるレシピエント細胞株に依存する。
【0056】
このような発現ベクターはまた、プロモーターの下流、および目的のペプチドまたはタンパク質をコードするDNA配列の上流に配置された、RNAスプライス部位のセットを含み得る。好ましいRNAスプライス部位は、アデノウイルスおよび/または免疫グロブリン遺伝子から得られ得る。目的のコード配列の下流に配置されたポリアデニル化シグナルもまた、発現ベクター中に含まれる。適切なポリアデニル化シグナルとしては、SV40に由来する初期または後期ポリアデニル化シグナル(KaufmanおよびSharp、同書)、アデノウイルス5 E1B領域に由来するポリアデニル化シグナルおよびヒト成長ホルモン遺伝子ターミネーター(DeNotoら、Nuc.Acids Res.9:3719−3730、1981)が挙げられる。発現ベクターは、プロモーターとRNAスプライス部位との間に配置された、アデノウイルス2 三分節(tripartile)リーダーのような、非コードウイルスリーダー配列を含み得る。好ましいベクターはまた、SV40エンハンサーのようなエンハンサー配列を含み得る。発現ベクターはまた、アデノウイルスのVA RNAをコードする配列を含み得る。適切な発現ベクターは、商業的供給源(例えば、Stratagene、La Jolla、California)から得ることができる。
【0057】
クローン化されたDNA配列を含むベクター構築物は、例えば、リン酸カルシウムによって媒介されるトランスフェクション(Wiglerら、Cell 14:725、1978;CorsaroおよびPearson、Somatic Cell Genetics 7:603、1981;GrahamおよびVan der Eb.Virology 52:456、1973)、エレクトロポレーション(Neumannら、EMBO J.1:841−845、1982)、またはDEAE−デキストラン媒介性トランスフェクション(Ausubelら(編)、Current Protocols in Molecular Biology、John Wiley and Sons,Inc.,NY,1987)によって、培養された哺乳動物細胞中に導入され得る。クローン化されたDNAを安定に組み込んだ細胞を同定するために、選択マーカーが一般的には、目的の遺伝子またはcDNAとともに細胞中に導入される。培養された哺乳動物細胞における使用に好ましい選択マーカーとしては、ネオマイシン、ハイグロマイシン、およびメトトレキセートのような薬物に対する耐性を付与する遺伝子が挙げられる。他の選択マーカーとしては、蛍光タンパク質(例えば、GFP(緑色蛍光タンパク質)またはBFP(青色蛍光タンパク質))が挙げられる。選択マーカーは、増幅可能な選択マーカーであり得る。好ましい増幅可能な選択マーカーは、DHFR遺伝子およびネオマイシン耐性遺伝子である。適切なマーカーは、Thilly(Mammalian Cell Technology、Butterworth Publishers、Stoneham、MA、これは、本明細書中で参考として援用されている)によって概説されている。
【0058】
適切なベクターを含有する哺乳動物細胞は、目的のDNA配列(単数または複数)の発現を開始するために、一定の期間(代表的には、1〜2日間)増殖させることが可能である。次いで、薬物選択が、安定な様式で選択マーカーを発現する細胞の増殖について選択するために適用される。増幅可能な選択マーカーでトランスフェクトされている細胞については、薬物濃度が、クローン化された配列の増大したコピー数について選択するために、段階的な様式で増大させられ得、それによって発現レベルを増大させる。導入された配列を発現する細胞が選択され、そして所望の形態または所望のレベルでの目的のタンパク質の産生についてスクリーニングされる。次いで、これらの基準を満たす細胞がクローン化され得、そして産生のためにスケールアップされ得る。細胞はまた、フローサイトメーターを使用してGFP陽性細胞について分類することによって、それらのGFP発現に基づいて、トランスフェクションについて選択され得る。
【0059】
哺乳動物細胞のトランスフェクションのためのプロトコールは、当業者に周知である。代表的な方法としては、リン酸カルシウムによって媒介されるトランスフェクション、エレクトロポレーション、リポフェクション、レトロウイルス、アデノウイルス、およびプロトプラスト融合によって媒介されるトランスフェクション(Sambrookら、前出を参照のこと)が挙げられる。裸のベクター構築物もまた、筋肉細胞または他の適切な細胞によって取りこまれ得、続いて、哺乳動物(または他の動物)の筋肉中に注射され得る。
【0060】
当該分野で公知の多数の昆虫宿主細胞もまた、本明細書を参照して本発明において有用であり得る。例えば、昆虫細胞中で異種DNA配列を発現するためのベクターとしてのバキュロウイルスの使用が、Atkinsonら(Pestic.Sci.28:215−224、1990)によって概説されている。
【0061】
当該分野で公知の多数の植物宿主細胞もまた、本明細書を参照して本発明において有用であり得る。例えば、植物細胞中での遺伝子の発現のためのベクターとしてのAgrobacterium rhizogenesの使用が、Sinkarら(J.Biosci.(Bangalore)11:47−58、1987)によって概説されている。
【0062】
本発明の関連する局面においては、所望のタンパク質をコードし、そして遺伝子の発現に有効なプロモーターに対して作動可能に連結されている遺伝子をその生殖細胞および体細胞が含むトランスジェニック動物中で本発明のタンパク質は発現され得る。あるいは、類似の様式においては、所望の遺伝子を欠失しているトランスジェニック動物が、調製され得る(例えば、「ノックアウト」マウス)。このようなトランスジェニックは、ヒト以外の種々の動物(マウス、ラット、ウサギ、ヒツジ、イヌ、ヤギ、およびブタを含む)において調製され得る(Hammerら、Nature 315:680−683、1985、Palmiterら、Science 222:809−814、1983、Brinsterら、Proc.Natl.Acad.Sci.USA 82:4438−4442、1985、PalmiterおよびBrinster、Cell 41:343−345、1985、ならびに米国特許第5,175,383号、同第5,087,571号、同第4,736,866号、同第5,387,742号、同第5,347,075号、同第5,221,778号、および同第5,175,384号を参照のこと)。簡潔には、発現される核酸分子を、適切に配置された発現制御配列とともに含む発現ベクターは、例えば、マイクロインジェクションによって、受精卵の前核中に導入される。注入されたDNAの組込みは、組織サンプルから、DNAのブロット分析によって検出される。導入されたDNAが、動物の生殖列系中に取りこまれ、その結果、その動物の子孫に伝達されることが、好ましい。組織特異的な発現は、組織特異的プロモーターの使用を通じて、またはトランスジーンの調節された発現を可能にする誘導性プロモーター(例えば、メタロチオネイン遺伝子のプロモーター(Palmiterら、1983、同書))の使用を通じて、達成され得る。
【0063】
天然に存在するscurfy変異体(「sf」)以外のFkhsfの変異体形態を産生するか、または天然に存在する変異体とは異なる遺伝的なバックグラウンドにある動物は、本明細書中に提供される開示を考慮して容易に産生され得る。
【0064】
タンパク質は、数ある方法の中でも、本発明の組換え翻訳産物を産生するために適切な宿主およびベクターシステムを培養することによって単離され得る。次いで、このような細胞株に由来する上清、またはタンパク質が上清中には分泌されない場合はタンパク質封入物もしくは細胞全体が、所望のタンパク質を単離するために種々の精製手順によって処理され得る。例えば、上清は、市販のタンパク質濃縮フィルター(例えば、AmiconまたはMillipore Pellicon限外濾過ユニット)を使用して最初に濃縮され得る。濃縮後、濃縮物は、適切な精製マトリックス(例えば、適切な支持体に結合させられた抗タンパク質抗体のような)に対して適用され得る。あるいは、陰イオン交換樹脂または陽イオン交換樹脂が、タンパク質を精製するために使用され得る。さらに別のものとしては、1つ以上の逆相高速液体クロマトグラフィー(RP−HPLC)工程が、タンパク質をさらに精製するために使用され得る。本発明のタンパク質を単離する他の方法は、当該分野で周知である。
【0065】
タンパク質は、他の(所望されない)タンパク質が、SDS−PAGE分析、続いてクマシーブルーでの染色によって検出されない場合は、本発明の状況において「単離された」と考えられる。他の実施態様においては、所望されるタンパク質は、従って、他の(所望されない)タンパク質が、SDS−AGE分析、続いて銀染色によって検出されないように、単離され得る。
【0066】
(免疫系を調節する分子を選択するためのアッセイ) 上記のように、本発明は、免疫系を調節し得る分子を選択および/または単離するための方法を提供する。適切なアッセイの代表的な例としては、酵母および哺乳動物の2−ハイブリッドシステム(例えば、Dangら、Mol.Cell.Biol.11:954、1991;Fearonら、Proc.Natl.Acad.Sci.USA、89:7958、1992)、DNA結合アッセイ、アンチセンスアッセイ、伝統的なタンパク質結合アッセイ(例えば、125Iまたは時間消散蛍光を利用する)、ゲル電気泳動と組み合わせた免疫沈降、および直接的なタンパク質配列決定、Fkhsfによって調節される遺伝子の転写分析、サイトカイン産生、ならびに増殖アッセイが挙げられる。
【0067】
例えば、1つの実施態様においては、Fkhsfと直接相互作用するタンパク質は、酵母の2−ハイブリッド結合システム(例えば、米国特許第5,283,173号、同第5,468,614号、同第5,610,015号、および同第5,667,973号を参照のこと)のようなアッセイによって検出され得る。簡潔には、2−ハイブリッドシステムにおいては、DNA−結合ドメイン−Fkhsfタンパク質の融合体(例えば、GAL4−Fkhsf融合体)が構築され、そして選択マーカー遺伝子に連結されたGAL4結合部位を含む細胞中にトランスフェクトされる。全Fkhsfタンパク質またはFkhsfのサブ領域が、使用され得る。GAL4活性化ドメインに対して融合されたcDNAのライブラリーもまた、構築され、そして同時トランスフェクトされる、cDNA−GAL4活性化ドメイン融合体中のcDNAがFkhsfと相互作用するタンパク質をコードする場合は、選択マーカーが発現される、次いで、cDNAを含有する細胞が増殖させられ、構築物が単離され、そして特徴付けられる。他のアッセイもまた、相互作用するタンパク質を同定するために使用され得る。このようなアッセイとしては、ELISA、ウェスタンブロッティング、同時免疫沈降、インビトロでの転写/翻訳分析などが挙げられる。
【0068】
本発明の別の局面においては、選択された分子が免疫系を調節し得るかどうかを決定するための方法が提供される。この方法は、以下の工程を包含する:(a)FkhsfまたはFkhsfの変異体を発現する細胞に対して選択された候補分子を曝露する工程、および(b)この分子がFkhsfの活性を調節するかどうかを決定する工程、およびそれによって上記の分子が免疫系を調節するかどうかを決定する工程。このような試験のための細胞は、(a)正常なリンパ球、(b)FKHsf(もしくはFkhsf)タンパク質(またはその変異形態)を過剰発現するように操作された細胞株、あるいは(c)上記のタンパク質を発現するように操作されたトランスジェニック動物に由来し得る。このようなトランスジェニックマウスに由来する細胞は、減少した細胞数および種々の刺激に対する減少した応答性を含む、低応答性(hyporesponsive)状態によって、一部特徴付けられる(例えば、実施例8)。
【0069】
本明細書中に記載されている方法は、個々の試験分子の分析を言及し得るが、本発明がそのようには限定されるべきでないことが、注意されるべきである。特に、選択された分子は、化合物の混合物中に含まれ得る。従って、記載されている方法は、さらに、所望される分子を単離する工程を包含し得る。さらに、候補分子が、多数のパラメーター(例えば、T細胞の増殖、サイトカインの産生などを含む)によって、免疫系を調節するそれらの能力について評価され得る。
【0070】
(候補分子)
広範な種々の分子が、免疫系を調節するそれらの能力についてアッセイされ得る。より詳細に以下で議論される代表的な例としては、有機分子、タンパク質またはペプチド、および核酸分子が挙げられる。
【0071】
(1.有機分子)
多数の有機分子が、免疫系を調節するそれらの能力についてアッセイされ得る。例えば、本発明の1つの実施態様においては、適切な有機分子が、化学的なライブラリー(ここでは、化合物が別々にアッセイされる)から、またはコンビナトリアル化学ライブラリー(ここでは、複数の化合物が一度にアッセイされ、次いで、最も活性な化合物を決定および単離するために取り組まれる)から、のいずれかから選択され得る。
【0072】
このようなコンビナトリアル化学ライブラリーの代表的な例としては、以下に記載されているものが挙げられる:Agrafiotisら「System and method of automatically generating chemical compounds with desired properties」、米国特許第5,463,564号;Armstrong,R.W.、「Synthesis of combinatorial arrays of organic compounds through the use of multiple component combinatorial array syntheses」、第WO95/02566号;Baldwin,J.J.ら、「Sulfonamide derivatives and their use」、第WO95/24186号;Baldwin,J.J.ら、「Combinatorial dehydrobenzopyran library」、第WO95/30642号;Brenner,S.、「New kit for preparing combinatorial libraries」、第WO95/16918号;Chenera,B.ら、「Preparation of library of resin−bound aromatic carbocyclic compounds」、第WO95/16712号;Ellman,J.A.、「Solid phase and combinatorial synthesis of benzodiazepine compounds on a solid support」、米国特許第5,288,514号;Felder,E.ら、「Novel combinatorial compound libraries」、第WO95/16209号:Lerner,R.ら、「Encoded combinatorial chemical libraries」、第WO93/20242号;Pavia,M.R.ら、「A method for preparing and selecting,pharmaceutically useful non−peptide compounds from a structurally diverse universal library」、第WO95/04277号;Summerton,J.E.およびD.D.Weller、「Morpholino−subunit combinatorial library and method」、米国特許第5,506,337号;Holmes,C.、「Methods for the Solid Phase Synthesis of Tiazolidinones、Metathiazanones,and Derivatives thereof」、第WO96/00148号;Phillips,G.B.およびG.P.Wei、「Solid−phase Synthesis of Benzimidazoles」、Tet.Letters 37:4887−90、1996;Ruhland,B.ら、「Solid−supported Combinatorial Synthesis of
Structurally Diverse β−Lactams」J.Amer.Chem.Soc.111:253−4、1996;Look,G.C.ら、「The Indentification of Cyclooxygenase−1 Inhibitors from 4−Tiazolidinone Combinatorial Libraries」、Bioorg and Med.Chem.Letters 6:707−12、1996。
【0073】
(2.タンパク質およびペプチド)
広範なタンパク質およびペプチドは、同様に、免疫系を調節するための候補分子として利用される。
【0074】
(a.コンビナトリアルペプチドライブラリー)
免疫系を調節するペプチド分子は、コンビナトリアルペプチドライブラリーのスクリーニングを通じて得られ得る。このようなライブラリーは、当業者によって調製され得る(例えば、米国特許第4,528,266号、および同第4,359,535号、ならびにPCT公開番号第WO92/15679号、同第WO92/15677号、同第WO90/07862号、同第WO90/02809号を参照のこと)か、または市販の供給源(例えば、New England Biolabs Ph.D.TM Pharge Display Peptide Library Kit)から購入され得るかのいずれかであり得る。
【0075】
(b.抗体)
免疫系を調節する抗体は、本明細書中に提供されている開示を考慮すれば、容易に調製され得る。本発明の状況においては、抗体は、モノクローナル抗体、ポリクローナル抗体、抗イディオタイプ抗体、抗体フラグメント(例えば、FabおよびF(ab’)2、Fv可変領域、または相補性決定領域)を含むことが理解される。上記で議論されているように、抗体は、それらが、107Mよりも大きいかまたはそれと同等である、好ましくは、108Mよりも大きいかまたはそれと同等であるKaを有して結合する場合は、Fkhsfに対して特異的であると理解される。モノクローナル抗体または結合パートナーの親和性、ならびに結合の阻害は、当業者によって容易に決定され得る(Scatchard,Ann.N.Y.Acad.Sci.51:660−672,1949を参照のこと)。
【0076】
簡潔には、ポリクローナル抗体は、ウマ、ウシ、種々の家禽、ウサギ、マウス、またはラットのような種々の温血動物から、当業者によって容易に生成され得る。代表的には、Fkhsfまたは13−20アミノ酸のその特有のペプチド(好ましくは、グルタルアルデヒドでの架橋によってキーホルリンペットヘモシアニンに対して結合体化される)が、フロイト完全アジュバントまたはフロイント不完全アジュバントのようなアジュバントと組み合わせて、腹腔内、筋肉内、眼内、または皮下注射を通じて動物を免疫するために利用される。数回の追加免疫後、血清のサンプルが回収され、そしてタンパク質またはペプチドに対する反応性について試験される。特に好ましいポリクローナル抗血清は、少なくともバックグラウンドよりも3倍大きい、これらのアッセイの1つに対するシグナルを生じる。一旦、タンパク質に対するその反応性に関して、動物の力価がプラトーに到達すると、より大量の抗血清が、毎週の採血によるか、または動物を瀉血するかのいずれかによって、容易に得られ得る。
【0077】
モノクローナル抗体はまた、従来技術を使用して容易に生成され得る(米国特許第RE32,011号、同第4,902,614号、同第4,543,439号、および同第4,411,993号(これらは、本明細書中で参考として援用されている)を参照のこと;Monoclonal Antibodies,Hybridomas:A New Dimension in Biological Analyses,Plenum Press,Kennett、McKearnおよびBechtol(編)、1980、およびAntibodies:A Laboratory Manual,HarlowおよびLane(編)、Cold Spring Harbor Laboratory Press,1988(これもまた、本明細書中で参考として援用されている)もまた参照のこと)。
【0078】
他の技術もまた、モノクローナル抗体を構築するために利用され得る(William D.Huseら、「Generation of a Large Combinatorial Library of the Immunoglobulin Repertoire in Phage Lambda」、Science、246:1275−1281、1989年12月を参照のこと;L.Sastryら、「Cloning of the Immunological Repertoire in Eschrichia coli for Generation of Monoclonal Catalytic Antibodies:Construction of a Heavy Chain Variable Region−Specific cDNA Library」、Proc.Natl.Acad.Sci.USA、86:5728−5732、1989年8月もまた参照のこと;Michelle Alting−Meesら、「Monoclonal Antibody Expression Libraries:A Rapid Alternative to Hybridomas」、Strategies in Molecular Biology 3:1−9、1990年1月もまた参照のこと)。
【0079】
以下を含む広範な種々のアッセイが、Fkhsf(または本明細書中に記載されているFkhsfの変異形態)に対して反応性である抗体の存在を決定するために利用され得る:例えば、向流免疫電気泳動、放射免疫アッセイ、放射免疫沈降、酵素結合免疫吸着アッセイ(ELISA)、ドットブロットアッセイ、ウェスタンブロット、免疫沈降、阻害または競合アッセイ、およびサンドイッチアッセイ(米国特許第4,376,110号および同第4,486,530号を参照のこと;Antibodies:A Laboratory Manual、HarlowおよびLane(編)、Cold Spring Harbor Laboratory Press、1988もまた参照のこと)。
【0080】
一旦、適切な抗体が得られると、それらは、当業者に周知の多くの技術によって単離または精製され得る(Antibodies:A Laboratory Manual、HarlowおよびLane(編)、Cold Spring Harbor Laboratory Press、1988を参照のこと)。適切な技術としては、ペプチドまたはタンパク質アフィニティーカラム、HPLCまたはRP−HPLC、プロテインAもしくはプロテインGカラム上での精製、または任意のこれらの技術の組み合わせが挙げられる。
【0081】
本発明の抗体は、免疫系を調節するためだけではなく、診断試験のため(例えば、FKHsfまたはFkhsfのタンパク質またはペプチドの存在を決定するため)、治療目的のため、またはタンパク質の精製のためにも利用され得る。
【0082】
(c.変異体Fkhsf)
本明細書中および以下の実施例において記載されているように、Fkhsfの改変されたバージョンが、Fkhsfの正常な活性を阻害し、それによって免疫系を調節するために利用され得る(一般的には、上記の核酸分子およびタンパク質を参照のこと)。
【0083】
FKHsfまたはFkhsfのさらなる変異体または改変された形態は、広範な種々のインビトロでのアッセイのため(例えば、種々のモデルにおいてこのようなタンパク質の影響を試験するため)、または抗体の開発のために、利用され得る。
【0084】
(3.核酸分子)
本発明の他の局面においては、免疫系を調節し得る核酸分子が提供される。例えば、1つの実施態様においては、FKHsfもしくはFkhsfの核酸配列、または変異体FKHsfもしくはFkhsfの発現を特異的に阻害するアンチセンスオリゴヌクレオチド分子が、提供される(一般的には、Hirashimaら、Molecular Biology of RNA:New Perspectives(M.InouyeおよびB.S.Dudock.編、1987 Academic Press,San Diego、401頁);Oligonucleotides:Antisense Inhibitors of Gene Expression(J.S.Cohen編、1989、MacMillan Press、London);SteinおよびCheng、Science、261:1004−1012、1993;第WO95/10607号;米国特許第5,359,051号;同第WO92/06693号;およびEP−A2−612844号を参照のこと)。簡潔には、このような分子は、それらが、転写されるFkhsfのmRNA配列の領域と相補的であり、そしてワトソン−クリック塩基対を形成し得るように、構築される。得られた二本鎖の核酸は、mRNAの続くプロセシングを妨害し、それによってタンパク質の合成を妨げる。
【0085】
本発明の他の局面においては、FKHsfもしくはFkhsf、またはFKHsfもしくはFkhsfの変異形態を阻害し得るリボザイムが提供される。本明細書中で使用される場合、「リボザイム」は、特異的認識のためのアンチセンス配列およびRNA−切断酵素活性を含むRNA分子を含むことが意図される。触媒的な鎖は、化学量論的な濃度を超える濃度で、標的RNAの特異的な部位を切断する。広範な種々のリボザイムが、本発明の状況において利用され得る。これは例えば、ハンマーヘッドリボザイム(例えば、ForsterおよびSymons、Cell 48:211−220、1987;HaseloffおよびGerlach、Nature、328:596−600、1988;WalbotおよびBruening、Nature、334:196、1988、HaseloffおよびGerlach、Nature、334:585、1988によって記載されている);ヘアピンリボザイム(例えば、Haseloffら、1993年10月19日に発行された米国特許第5,254,678号、Hempelら、1990年3月26日に公開された、欧州特許公開番号第0 360 257号によって記載されている);およびTetrahymenaのリボソームRNAに基づくリボザイム(Cechら、米国特許第4,987,071号を参照のこと)を含む。本発明のリボザイムは、代表的には、RNAから構成されるが、DNA、核酸アナログ(例えば、ホスホロチオエート)、またはそれらのキメラ(例えば、DNA/RNA/RNA)からも構成され得る。
【0086】
(4.標識)
FKHsfもしくはFkhsf(ならびにそれらの変異形態)、または上記または下記の任意の候補分子が、以下を含む種々の化合物で標識され得る:例えば、蛍光分子、毒素、および放射性核種。蛍光分子の代表的な例としては、フルオレセイン、Phycobiliタンパク質(例えば、フィコエリトリン)、ローダミン、テキサスレッド、およびルシフェラーゼが挙げられる。毒素の代表的な例としては、リシン、アブリン、ジフテリア毒素、コレラ毒素、ゲロニン、アメリカヤマゴボウ抗ウイルスタンパク質、トリチン、Shigella毒素、およびPseudomonas外毒素Aが挙げられる。放射性核種の代表的な例としては、Cu−64、Ga−67、Ga−68、Zr−89、Ru−97、Tc−99m、Rh−105、Pd−109、In−111、I−123、I−125、I−131、Re−186、Re−188,Au−198、Au−199,Pb−203、At−211、Pb−212、およびBi−212が挙げられる。さらに、上記の抗体もまた、標識され得るか、またはリガンド結合対の1つのパートナーに対して結合体化され得る。代表的な例としては、アビジン−ビオチン、およびリボフラビン−リボフラビン結合タンパク質が挙げられる。
【0087】
上記の代表的な標識と本明細書中に記載されている分子とを結合体化または標識するための方法は、当業者によって容易に達成され得る(Trichothecene Antibody Conjugate、米国特許第4,744,981号;Antibody conjugate、米国特許第5,106,951号;Florogenic Materials and Labeling Techniques、米国特許第4,018,884号;Metal Radionuclide Labeled Proteins for Diagnosis and Therapy、米国特許第4,897,255号;およびMetal Radionuclide Chelating Compounds for Improved Chelation Kinetics、米国特許第4,988,496号を参照のこと;Inman.Methods In Enzymology、第34巻、Affinity Techniques、Enzyme Purification:Part B、JakobyおよびWilchek(編)、Academic Press,New York、30頁、1974をもまた参照のこと;WilchekおよびBayer、「The Avidin−Biotin Complex in Bioanalytical Applications」Anal.Biochem.171:1−32、1988をもまた参照のこと)。
【0088】
(薬学的組成物)
上記のように、本発明はまた、薬学的または生理学的に受容可能なキャリア、賦形剤、または希釈剤とともに、免疫系を調節する上記の分子の1つを含む、種々の薬学的組成物を提供する。一般的には、このようなキャリアは、使用される投与量および濃度では、レシピエントに対しては非毒性である。通常は、このような組成物の調製は、緩衝液、抗酸化剤(例えば、アスコルビン酸、低分子量(約10残基未満)のポリペプチド、タンパク質、アミノ酸、炭水化物(グルコース、スクロース、またはデキストリンを含む))、キレート化剤(例えば、EDTA、グルタチオンなど)、安定剤、および賦形剤と、治療薬との混合を含む。中性の緩衝化生理食塩水または非特異的な血清アルブミンと混合した生理食塩水は、例示的な適切な希釈剤である。好ましくは、薬学的組成物(または「医薬品」)は、滅菌の、発熱物質を含まない形態で提供される。
【0089】
さらに、本発明の薬学的組成物は、種々の異なる経路による投与のために調製され得る。さらに、本発明の薬学的組成物は、このような薬学的組成物の使用に関する説明書を提供するパッケージング材料とともに、容器内に配置され得る。一般的には、このような説明者は、試薬の濃度、ならびに特定の実施態様においては、薬学的組成物の再構成のために必要とされ得る賦形剤成分または希釈剤(例えば、水、生理食塩水、もしくはPBS)の相対的な量を記載する具体的な表現を含む。
【0090】
(処置方法)
本発明はまた、免疫系を調節するための方法を提供する。免疫系を調節する、本明細書中に記載されている分子の使用を通じて、温血動物中の広範な種々の状態が、容易に処置され得るかまたは予防され得る。処置され得る温血動物の例としては、脊椎動物および哺乳動物(例えば、ヒト、ウマ、ウシ、ブタ、ヒツジ、イヌ、ネコ、ラット、およびマウスを含む)の両方が挙げられる。このような方法は、変更された免疫系を有する患者において治療的な価値があり得る。これは、種々の免疫不全症候群、ならびにT細胞媒介自己免疫疾患を有する患者の、化学療法を受けている患者を含む。治療的な価値はまた、ワクチンアジュバントとしての有用性によって認識され得る。
【0091】
治療用分子は、分子の型に依存して、種々の処方物において種々の経路を介して投与され得る。例えば、1つの実施態様においては、有機分子が、経口もしくは鼻内経路によって送達され得るか、または注射(例えば、筋肉内、静脈内など)によって送達され得る。
【0092】
1つの局面においては、免疫系を調節するための方法が提供される。この方法は、免疫系を調節する分子の発現を指向するベクターを、リンパ系細胞中に導入する工程、および温血動物にベクターを含有する細胞を投与する工程を包含する。他の関連する実施態様においては、このベクターは、所望される標的の位置(例えば、骨髄)に直接投与され得る。
【0093】
ウイルスベクターおよび非ウイルスベクターの両方を含む、広範な種々のベクターが、このような治療目的のために利用され得る。適切なウイルスベクターの代表的な例としては、ヘルペスウイルスベクター(例えば、米国特許第5,288,641号)、アデノウイルスベクター(例えば、第WO94/26914号、第WO93/9191号、第WO99/20778号、第WO99/20773号;第WO99/20779号;Kollsら、PNAS 91(1):215−219、1994;Kass−Eislerら、PNAS 90(24):11498−502、1993;Guzmanら、Circulation 88(6):2838−48、1993;Guzmanら、Cir.Res.73(6):1202−1207、1993;Zabnerら、Cell、75(2):207−216、1993;Liら、Hum.Gene Ther.4(4):403−409、1993:Caillaudら、Eur.J.Neurosci.5(10):1287−1291、1993;Vincentら、Nat.Genet.5(2):130−134、1993;Jaffeら、Nat.Genet.1(5):372−378、1992;およびLevreroら、Gene、101(2):195−202、1991)、アデノ随伴ウイルスベクター(第WO95/13365号;Flotteら、PNAS 90(22):10613−10617、1993)、バキュロウイルスベクター、パルボウイルスベクター(Koeringら、Hum.Gene Therap.5:457−463、1994)、ポックスウイルスベクター(PanicaliおよびPaoletti、PNAS、79:4927−4931,1982;およびOzakiら、Biochem.Biophys.Res.Comm.193(2):653−660、1993)、およびレトロウイルス(例えば、第EP0,415,731号;第WO90/07936号;第WO91/0285号、第WO94/03622号;第WO93/25698号;第WO93/25234号;米国特許第5,219,740号;第WO93/11230号;第WO93/10218号)が挙げられる。同様に、種々のウイルスまたは非ウイルス供給源に由来する種々のエレメント(例えば、プロモーター、エンベロープ配列など)の混合物を含むウイルスベクターが、構築され得る。種々の実施態様においては、ウイルスベクター自体、またはウイルスベクターを含むウイルス粒子のいずれかが、以下に記載されている方法および組成物において利用され得る。
【0094】
本発明の他の実施態様においては、免疫系を調節する分子(例えば、変異体Fkhsf、またはアンチセンスもしくはFkhsfを切断するリボザイム分子)をコードする核酸分子が、以下を含む種々の別の技術によって投与され得る:例えば、ポリ−L−リジンDNA複合体と結合体化したアシアロオソムコイド(ASOR)の投与(Cristanoら、PNAS 92122−92126,1993)、殺傷したアデノウイルスに連結されたDNA(Curielら、Hum.Gene Ther.3(2):147−154、1992)、サイトフェクチンによって媒介される導入(DMRIE−DOPE、Vical、California)、直接的なDNA注入(Acsadiら、Nature、352:815−818、1991);DNAリガンド(Wuら、J.of Biol.Chem.264;16985−16987、1989);リポフェクション(Felgnerら、Proc.Natl.Acad.Sci.USA 84:7413−7417、1989);リポソーム(Pickeringら、Circ、89(1):13−21、1994;およびWangら、PNAS、84:7851−7855、1987);マイクロプロジェクタイルボンバードメント(Williamsら、PNAS、88:2726−2730、1991);およびタンパク質自体をコードする核酸分子の、単独(VileおよびHart、Cancer Res.53:3860−3864、1993)またはPEG−核酸複合体を利用するかのいずれかである直接的な送達。
【0095】
本発明のベクターによって発現され得る分子の代表的な例としては、リボザイムおよびアンチセンス分子(これらのそれぞれは、以下により詳細に議論されている)が挙げられる。
【0096】
当業者に明らかであるように、投与の量および頻度は、もちろん、処置される徴候の性質および重篤度、所望される応答、患者の状態のような因子に依存する。代表的には、組成物は、上記のような種々の技術によって投与され得る。
【0097】
以下の実施例は、例示によって付与され,そして限定によっては与えられない。
【0098】
(実施例)
(実施例1)
(SCURFY変異体の原因である遺伝子の同定)
(A.SCURFY遺伝子のクローニング)
もともとのscurfy変異は、Oak Ridge National Laboratory(ORNL)に1949年にストックされた一部近交系のMRにおいて自発的に生じた。戻し交配分析を、マウスScurfy変異を含むX染色体の動原体周辺領域(peri−centromeric region)を詳細にマップするために使用した。同じ領域をカバーする物理的なマップを、重複している酵母および細菌の人工染色体(YACおよびBAC)の単離を通じて並行して生成した。一旦、候補領域が、約500キロ塩基対(kb)にまで狭められると、大規模なDNA配列決定を、4個の重複するBACクローンについて行った。この500kbの領域内の全ての転写ユニットを、配列データベースの検索とコンピューターによるエキソン予想プログラムの適用との組合せにより同定した。次いで、候補遺伝子を、正常なおよびScurfyに由来するRNAサンプルからの逆転写ポリメラーゼ連鎖反応(RT−PCR)手順によって得たcDNAの配列を比較することによって、Scurfy特異的変異についてスクリーニングした。Fkhsfと本明細書中で呼ばれる1つの遺伝子においては、2塩基対(bp)の挿入が、正常なcDNAと比較してScurfy cDNAのコード領域中に見出された。挿入を、いくつかのマウスの株(Scurfy変異体を含む)のゲノムDNAに由来するPCR産物のDNA配列を比較することによって確認した。再び、2bpの挿入を、Scurfyサンプル中でのみ見出し、これによって、これをScurfy欠失の原因である可能性があると確定した。
【0099】
マウスのFkhsf遺伝子は、BACクローン8C22中に含まれ、そして完全に配列決定されている。これは約14kbにまたがり、そして11個のコードエキソンを含む。エキソン中断の位置は、GenScanエキソン推定プログラムを使用して、ゲノムDNA配列のコンピューターによる分析によって最初に同定した;次いで、エキソンの位置を、ゲノム配列に対する正常なマウスの組織に由来するcDNA配列の直接的な比較によって確認した。
【0100】
得られたcDNAの長さは、2160bpである;コード領域は、その1287bpにまたがり、429アミノ酸のタンパク質をコードする。図1はマウスのFkhsf cDNAのヌクレオチド配列を示す;翻訳は、259位で開始し、そして1546位で終結すると推定される。図2は、マウスのFkhsfのアミノ酸配列を示す。
【0101】
(b.Fkhsfトランスジェニックマウスの生成)
Scurfy表現型の真の原因としてのFkhsf遺伝子の実態を、トランスジェニックマウスにおいて確認した。簡潔には、Fkhsf遺伝子の約7kbのコード領域、ならびに隣接する配列の上流の約20kb、および下流の配列の約4kb(図5)を含む正常なゲノムDNAの30kbのフラグメントを、正常なマウスの1細胞胚中にマイクロインジェクションした。それぞれが別々のインテグリンを有する5匹の個々の崩壊させた動物を生成し、そして各トランスジェニック株に由来する雄性の動物を、雌性のsfキャリアと交配した。トランスジーン(正常なFkhsf)およびsf変異(変異Fkhsf)の両方を有する雄性の子孫を、分析した。
【0102】
分析は、倭小(runting)、鱗状の皮膚、毛皮の異常、およびscurfy表現型の他の特徴についての動物の試験からなる。さらに、リンパ系組織(胸腺、脾臓、および結節)を回収し、そしてそれらの大きさおよび細胞数を試験し、そして正常な動物およびscurfyマウスの両方について比較した。全ての5個のトランスジェニック株について、トランスジーンを保有した雄性のsf子孫は、大きさおよび重量において正常であり、そして全ての点で健常であるように見えた。これらのトランスジェニックマウスにおけるリンパ節の大きさは、正常な動物にものと類似(またはそれよりも小さい)であり(図6)、そして活性化されたT細胞の徴候はなかった。これらのパラメーターは、sfマウスとは極端に異なり、そして正常なFkhsf遺伝子の添加が、scurfyマウス中に見出される欠損を克服し得ることを示す。従って、これによって、Fkhsf遺伝子中の変異がScurfy疾患の原因であることを確認する。
【0103】
(実施例2)
(FKHsf cDNAの生成)
完全なマウスのFkhsfタンパク質をコードする相補的DNA(cDNA)は、逆転写酵素ポリメラーゼ連鎖反応(RT−PCR)手順によって得られ得る。より詳細には、第1鎖のcDNAが、適切な供給源(例えば、マウスの脾臓)に由来する5μgの全RNAを、オリゴdTプライミングすること、および標準的な条件下で逆転写酵素を用いて伸張すること(例えば、Gibco/BRL SuperScriptキット)によって、生成する。次いで、第1鎖のcDNAのアリコートを、正方向および逆方向プライマー(正方向プライマー:GCAGATCTCC TGACTCTGCC TTC;逆方向プライマー:GCAGATCTGACAAGCTGTGT CTG)(0.2mMの最終濃度))、60mMのTris−HCl、15mMの硫酸アンモニウム、1.5mMの塩化マグネシウム、0.2mMの各dNTP、および1ユニットのTaqポリメラーゼの存在下での、35サイクルのPCR(94℃にて30秒間、60℃にて30秒間、72℃にて2分間)に供する。
【0104】
(実施例3)
(マウスのFkhsfに対するヒトのオルソログの生成)
完全なFKHsfタンパク質をコードするヒトFKHsf cDNAは、実施例2に記載されているものと本質的に同じ手順によって得ることができる。詳細には、全脾臓RNAを用いて開始し、そして以下のオリゴヌクレオチドプライマー(正方向プライマー:AGCCTGCCCT TGGACAAGGA C;逆方向プライマー:GCAAGACAGT GGAAACCTCA C)、および60℃のアニーリング温度を除いて上記と同じPCR条件を利用する。
【0105】
図4は、今日までに得られた1869bpのcDNAのヌクレオチド配列(1293bpのコード領域を含む)を示す;翻訳は、189位で開始し、そして1482位で終結すると推定される。図4は、431アミノ酸のヒトFKHsfタンパク質の配列を示す。マウスcDNA配列に対するヒト遺伝子の推定されるコード領域の比較は、ほぼ同一であるエキソン構造、および全タンパク質にわたる86.1%のアミノ酸配列同一性を明らかにする。
【0106】
(実施例4)
(SCURFY変異体を検出するための方法)
上記のように、Scurfy変異体を、最初に、sfのRT−PCRによって誘導されたcDNAおよび正常なマウスのRNAサンプルを直接配列決定することによって発見し、そしてゲノムDNAに由来する同じ領域を配列決定することによって確認した。変異の性質(すなわち、2bpの挿入)は、それ自体を、多数の種々の変異検出アッセイに導く。これは、最初に、オリゴヌクレオチドプローブのディファレンシャルハイブリダイゼーションに基づく。このような、ハイブリダイゼーションに基づくアッセイは、対立遺伝子特異的発現の定量的な分析を可能にし得る。
【0107】
一例としては、360bpのDNAフラグメントを、以下のオリゴを使用して、第1鎖のcDNAから増幅する:
DMO5985(正方向):CTACCCACTGCTGGCAAATG(図1のntd.825−844)
DMO6724(逆方向):GAAGGAACTATTGCCATGGCTTC(ntd 1221−1199)。
【0108】
PCR産物を、1.8%のアガロースゲル上で泳動し、ナイロン膜に移し、そしてScurfy特異的な2bpの挿入部位に対応する領域に相補的である末端標識オリゴヌクレオチドで標識する。2つの別々のハイブリダイゼーション反応を、正常なおよびScurfy PCR産物を検出するために、以下のオリゴヌクレオチド(2bpの挿入部位を太字で示す)を使用して行う:
正常:ATGCAGCAAGAGCTCTTGTCCATTGAGG
DMO7439
Scurfy:GCAGCAAGAGCTCTTTTGTCCATTGAGG
DMO6919。
【0109】
Scurfy変異はまた、冷却一本鎖立体構造多形性(cSSCP)アッセイによっても検出され得る。このアッセイにおいては、上記と同じPCR産物を、20%のアクリルアミド(TBE)ゲル上で、鎖の変性後に泳動する。Scurfyの挿入は、正常な配列と比較して鎖の移動においてシフトを生じ、そして別の鎖が、エチジウムブロマイドでの染色後に検出される。
【0110】
(実施例5)
(FKHSF遺伝子の発現)
半定量的なRT−PCRが、広範な種々の組織および細胞株中におけるマウスおよびヒトのFkhsf遺伝子の発現パターンを分析するために使用されている。発現のレベルは、偏在して発現されるDAD−1遺伝子について正規化される。簡潔には、Fkhsf遺伝子は、非常に低いレベルではあるが、これまでに試験されたほぼ全ての組織(胸腺、脾臓、分類されたCD4+およびCD4−CD8−、Tリンパ球、ならびに腎臓、脳、ならびに種々のマウスおよびヒトのT細胞株、ならびにヒト腫瘍を含む)中で発現される。しかし、発現がないことが、新たに分類したマウスのB細胞において注目された。
【0111】
予想されるように、RT−PCRアッセイにおける正常な組織対Scurfy組織において、発現レベルにおける差異は観察されなかった。
【0112】
(実施例6)
(インビトロでのFKHSFの発現)
全長のマウスおよびヒトのFkhsf cDNA、ならびに種々のcDNAのサブ領域を、哺乳動物細胞(たとえば、ヒトのJurkat T細胞株)、E.coli、または酵母中での発現が可能であるベクター中にクローン化した。E.coli系または酵母系を、Fkhsf特異的抗体を惹起する目的のためのタンパク質の産生に使用し得る(以下を参照のこと)。
【0113】
(実施例7)
(抗FKHSF抗体の生成)
実施例6に記載されているベクターから発現されるタンパク質を、FKHsf特異的抗体の産生のために適切な動物を免疫化するために使用する。全長または短縮されたタンパク質のいずれかを、この目的のために使用し得る。タンパク質を、例えば、E.coliのような細菌、昆虫細胞、または哺乳動物細胞から、入手し得る。動物種としては、マウス、ウサギ、モルモット、ニワトリ、または他のものが挙げられ得る。FKHsfに対して特異的なウサギ抗血清は、生化学的な特徴によって決定されるように、生成されている(免疫沈降およびウェスタンブロッティング)。
【0114】
(実施例8)
(FKHSF遺伝子の機能についてのアッセイ)
FKHsfタンパク質の機能の欠損が、scurfy動物において観察される表現型(るいそう、敏感な免疫応答、および死)を生じるので、FKHsfタンパク質の過剰な発現を評価するためのアッセイを記載する。トランスジェニック動物(実施例1に記載されている)を、それらの免疫応答能力の状態について、いくつかの異なるパラメーターを使用して試験する。動物を、リンパ節および胸腺中に存在するリンパ系細胞の数について(図7)、およびインビトロでの刺激に対するT細胞の応答性について(図8)試験する。
【0115】
Scurfy変異動物は、正常な動物のそれらのリンパ節中の細胞よりもほぼ2倍多い細胞を有する。しかし、正常なFKHsfタンパク質の過剰なレベルを発現するマウスは、ほぼ3倍多い細胞を含む(図7)。さらに、標準的な抗血清を使用するフローサイトメトリーによって評価される場合に、それらの細胞表面の表現型である場合(示さない)は、胸腺細胞の数は正常であり(図7)、このことは、過剰のFKHsfタンパク質に関連する発達的欠損が存在しないことを示す。
【0116】
正常なscurfy動物およびトランスジェニック動物はさらに、T細胞の刺激に対するそれらの増殖性応答について試験される。CD4+T細胞はCD3およびCD28に対する抗体と反応し、そしてそれらの増殖性応答が、放射活性チミジンの取りこみを使用して測定される。scurfy細胞は、刺激の非存在下でのみ分裂するが、正常な細胞は、刺激の後に十分応答する。FKHsfトランスジェニック細胞もまた刺激に対して応答するが、応答は、正常な細胞のものよりも有意に少ない(図8)。このことは、過剰のFKHsfを発現するCD4+T細胞が、刺激に対して応答する能力を低下させられたことを示す。
【0117】
(実施例9)
(JM2に関連するヒトのFKHSF cDNA配列)
ヒトFKHsf cDNA配列の改変されたバージョンが、GenBankによって公開されている配列データベース中に存在する。この配列は、JM2と呼ばれ(GenBank登録番号第AJ005891号)、そしてFKHsf遺伝子を含むゲノム配列に対するエキソン推定プログラムの適用の結果である(Strom,T.M.ら、未公開−GenBank登録番号第AJ005891を参照のこと)。対称的に、FKHsf cDNAの構造は、実験によって決定された。Genetics Computer Group(GCG;Madison,USA)、Wisconsin配列分析パッケージのGAPプログラムを、2つの配列を比較するために使用し、そして差異を図9に示す。2つの配列の5’末端は、ゲノムDNA配列の状況におけるそれらの位置において異なり、FKHsfの第2のコードエキソンがJM2から欠落しており、そしてFKHsf遺伝子の最後のイントロンが、JM2配列中ではスプライシングされない。これらの差異は、FKHsfと比較して、より短いアミノ末端ドメイン、カルボキシ末端でのフォークヘッドドメイン(以下を参照のこと)中への大きい挿入を有するJM2タンパク質を生じる。
【0118】
(実施例10)
(FKHsfタンパク質は種を超えて保存されている)
FKHsfタンパク質は、機能的ドメインを示し得る配列モチーフに基づいて、サブ領域に分けられ得る。FKHsf中の2つの重要なモチーフは、タンパク質の中央部分中のC2
II2クラスの単一のジンクフィンガー(ZNF)、およびタンパク質の最もカルボキシ
末端での二股の矢じりまたは翼状のへリックスドメインである。FKHsfと他のタンパク質との間での相同性の程度を特徴付ける目的のために、本発明者らは、タンパク質を以下の4つの領域に分けた:
アミノ末端ドメイン :図2の残基1−197
図4の残基1−198
ジンクフィンガードメイン :図2の残基198−221
図4の残基199−222
中央ドメイン :図2の残基222−336
図4の残基223−336
フォークヘッドドメイン :図2の残基337−429
図4の残基337−431。
【0119】
DNAStar配列分析パッケージによるMultiple Sequence Alignmentプログラムを使用して、Lipman−Pearsonアルゴリズムを、これらの4つのドメインにわたる、ヒトFKHsfとマウスFkhsfタンパク質との間での類似性の程度を決定するために使用した。結果を、図10に示す。類似性は、82.8%から96.4%までの範囲を示し、このことは、このタンパク質が非常に高度に種をこえて保存されていることを示す。
【0120】
(実施例11)
(新規のFKHSFに関連する遺伝子の同定)
FKHsf遺伝子配列の特有の特徴を利用して、二股の矢じりを含む分子の同じサブクラスに入る他の新規の遺伝子(およびタンパク質)を同定し得る。FKHsfタンパク質は、それが、フォークヘッドドメインに対して単一のジンクフィンガードメインのアミノ末端を、そしてフォークヘッドドメインの最もカルボキシ末端に有する点で、特有である。縮重PCRアプローチを、フォークヘッドドメインの上流にジンクフィンガー配列を含む新規の遺伝子を単離するために行い得る。例えば、以下の縮重プライマーを合成した(縮重の位置を括弧で示し、そして「I」はヌクレオシドイノシンを示す):
正方向プライマー:CA(TC)GGIGA(GA)TG(CT)AA(GA)TGG 逆方向プライマー:(GA)AACCA(GA)TT(AG)TA(AGT)AT(CT)TC(GA)TT。
【0121】
正方向プライマーは、ジンクフィンガー配列中の領域に対応し、そして逆方向プライマーは、フォークヘッドドメインの中央の領域に対応する。これらのプライマーを使用して、種々のヒト組織(肝臓、脾臓、脳、肺、腎臓などを含む)から実施例2のように産生した第1鎖のcDNAを増幅した。以下のPCR条件:0.2mMの最終濃度の正方向プライマーおよび逆方向プライマー、60mMのTris−HCl、15mMの硫酸アンモニウム、1.5mMの塩化マグネシウム、0.2mMの各dNTP、および1ユニットのTaqポリメラーゼ、を使用し35サイクル(94℃にて30秒間、50℃にて30秒間、72℃にて2分間)に供した。PCR産物を、1.8%のアガロースゲル(1×TAE中で泳動)上で可視化し、そしてTAクローニングベクター(Invitrogen、Carlsbad、CA)中にサブクローン化した;個々のクローンを配列決定し、そして全長cDNAのさらなる特徴付けのために使用した。
【0122】
あるいは、FKHsf遺伝子の特有の領域(すなわち、「アミノ末端」ドメインおよび「中央」ドメイン)を、ハイブリダイゼーションによるcDNAライブラリーのスクリーニングのために使用し得る。種々のヒトおよび/またはマウスの組織に由来し、そしてλファージベクター(例えば、λgt11)中で増殖させたcDNAライブラリーをアガロース上にプレートし、プラークをナイロン膜に移し、そしてFKHsf遺伝子の特有の領域に由来するフラグメントを用いてプローブした。高ストリンジェンシーの条件(例えば、65℃にて、5×SSPE、5×Denhardt溶液、0.5%のSDS中でのハイブリダイゼーション、65℃にて、0.1×SSPE、0.1%のSDS中での洗浄)下では、密接に関連するごくわずかな配列のみが、ハイブリダイズすると予想した(すなわち、90−100%の相同性)。上記と同じ緩衝液中での45−55℃でのこのようなハイブリダイゼーションおよび洗浄のような低いストリンジェンシーの下では、FKHsfに関連する遺伝子(65−90%の相同性)が同定され得る。FKHsfの特有の配列を用いて公開されているデータベースの検索によって得られる結果に基づくと、低いストリンジェンシーから中程度のストリンジェンシーまでのハイブダイゼーション実験を通じて同定される任意の遺伝子が、「FKHsfファミリー」の新規のメンバーを提示すると予想される。
【0123】
上記から、本発明の特定の実施態様が、例示の目的のために本明細書中で記載されているが、種々の改変が本発明の精神および範囲を逸脱することなく行われ得ることが、理解される。従って、本発明は、添付の特許請求の範囲による場合を除いて、限定されない。
【技術分野】
【0001】
(技術分野)
本発明は、一般的には、薬学的な生成物および方法、ならびにより詳細には、scurfy関連疾患を診断するために有用な方法および組成物に関し、そして、免疫系を調節し得る化合物を同定するための方法に関する。
【背景技術】
【0002】
(発明の背景)
マウスの免疫系に影響を与える遺伝した変異は、免疫系の調節に対して重要な新規の遺伝子の豊富な供給源であることが証明されており、そしてヒトの免疫学的な障害の重要な動物モデルを提供した。これらとしては、xid(X連鎖無ガンマグロブリン血症のマウスの等価物)(非特許文献1および2)、beige(チェディアック‐東症候群の等価物)(Barbosaら、Nature、382:262、1996)、lprおよびgld(fasおよびfasリガンドの欠損)、X連鎖重症複合免疫不全(Sugamuraら、Annu.Rev.Immunol.14:179、1996)、および造血細胞ホスファターゼ変異体motheaten(SHP−1)(BignonおよびSiminovitch、Clin Immunol Immunopathol、73:168、1994)が挙げられる。
【0003】
特定の目的の1つのマウスの変異体は、いまだなおクローン化されていないX連鎖マウス変異体、scurfy(sf)である。簡潔には、scurfy変異について半接合性のマウスは、重篤なリンパ増殖障害を示す。詳細には、scurfy変異について半接合性の雌性(Xsf/Y)は、リンパ節、脾臓、肝臓、および皮膚の進行性のリンパ球浸潤を発症し、それによって巨脾腫、肝腫、大いに肥大したリンパ節、矮小、離脱性皮膚炎、および厚くなった奇形の耳を含む、肉眼的な形態学的な症状を生じる(Godfreyら、Amer.J.Pathol.138:1379、1991;Godfreyら、Proc.Natl.Acad.Sci.USA 88:5528、1991)。他の臨床的な症状としては、増大した白血球数、高ガンマグロブリン血症、および重篤な貧血が挙げられる(Lyonら、Proc.Natl.Acad.Sci.USA 87:2433、1990);罹患した雄性の死亡は、通常は3週齢までに生じる。sf遺伝子座は、X染色体の極めて近位の領域に、sparse−fur(spf)(Lyonら、Proc.Natl.Acad.Sci.USA 87:2433、1990;Blairら、Mamm.Genome 5:652、1994)(それ自体が、オルニチントランスカルバミラーゼ遺伝子(Otc)内の点変異(Veresら、Science 237:415、1987))についての遺伝子座からおよそ0.7センチモルガンに、マップされている。sf遺伝子座はまた、マウスのGata1、Tcfe3、およびWasp遺伝子座と密接に連鎖している(Blairら、Mamm.Genome 5:652、1994;Derryら、Genomics 29:471、1995)。scurfyおよびヒトのWiscott−Aldrich症候群(WAS)との間の類似性が報告されており(Lyonら、Proc.Natl.Acad.Sci.USA 87、2433、1990)、そしてマウスのWasp遺伝子が、scurfyについての候補として提案されている(Lyonら、Proc.Natl.Acad.Sci.USA 87:2433、1990;Derryら、Genomics 29:471、1995)。より最近の生物学的な実験は、WASとscurfyとの間の有意な差異を明らかにするが、しかし、この2つの遺伝子座は、対立遺伝子ではないことが実証されている(JefferyおよびBrunkow、公開されていないデータ)。従って、出願人らの発明の以前には、scurfy遺伝子の正体は、未決定のままであった。
本発明は、scurfyに関連する疾患の診断のために有用な方法および組成物、ならびに、免疫系を調節し得る化合物を同定するための方法を開示し、そしてさらに他の関連する利点を提供する。
この出願の発明に関連する先行技術文献情報としては、次のものがある。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Thomasら、Science、261:355、1993
【非特許文献2】Rawlingsら、Science、261:358、1993
【発明の概要】
【課題を解決するための手段】
【0005】
(発明の要旨)
本発明は、以下を提供する:
(項目1) Fkhsfをコードする、単離された核酸分子。
(項目2) 前記FkhsfがマウスのFkhsfである、項目1に記載の単離された核酸分子。
(項目3) 前記FkhsfがヒトのFKHsfである、項目1に記載の単離された核酸分子。
(項目4) 前記核酸分子が、(a)配列番号2または4を含むアミノ酸配列をコードする核酸分子、(b)配列番号1もしくは3のヌクレオチド配列を有する核酸分子、またはその相補物に対してストリンジェントな条件下でハイブリダイズする核酸分子、および(c)(a)または(b)のいずれかによってコードされるポリペプチドの機能的なフラグメントをコードする核酸分子、
からなる群より選択される、項目1に記載の単離された核酸分子。
(項目5) 前記核酸分子が配列番号2のアミノ酸配列をコードする、項目1に記載の単離された核酸分子。
(項目6) 前記核酸分子が配列番号1のヌクレオチド配列を含む、項目5に記載の単離された核酸分子。
(項目7) 項目1に記載の単離された核酸分子を含む、ベクター。
(項目8) 前記ベクターがウイルスベクターである、項目7に記載のベクター。
(項目9) 前記ウイルスベクターが、レトロウイルス、アデノウイルス、ヘルペスウイルス、アデノ随伴ウイルス、およびアルファウイルスからなる群より選択されるウイルスから生成される、項目8に記載のベクター。
(項目10) 項目1に記載の単離された核酸分子およびプロモーターを含む発現ベクターであって、ここで、該プロモーターが該核酸分子と作動可能に連結されている、発現ベクター。
(項目11) 項目10に記載の発現ベクターを含む、組換え宿主細胞。
(項目12) Fkhsfタンパク質を調製するために項目10に記載の発現ベクターを使用する方法であって、該方法が以下の工程:
(a)該発現ベクターを含み、そして該タンパク質を産生する、組換え宿主細胞を培養する工程、および
(b)該培養した組換え宿主細胞から該タンパク質を単離する工程、
を包含する、方法。
(項目13) 項目1から6のいずれか1項に記載の核酸分子によってコードされるアミノ酸配列を含む、単離されたポリペプチド。
(項目14) 項目1に記載の核酸分子によってコードされるポリペプチドと特異的に結合する、抗体または抗体フラグメント。
(項目15) 前記抗体が、以下:
(a)ポリクローナル抗体、
(b)マウスのモノクローナル抗体、
(c)(b)から誘導されるヒト化抗体、および
(d)ヒトのモノクローナル抗体、
からなる群より選択される、項目13に記載の抗体。
(項目16) 前記抗体フラグメントが、F(ab’)2、F(ab)2、Fab’、Fab、Fv、sFv、および最小認識ユニットからなる群より選択される、項目14に記載の抗体フラグメント。
(項目17) 項目13に記載のポリペプチドを含む、融合タンパク質。
(項目18) 被験体に由来する生物学的サンプル中のFkhsf核酸配列の存在を検出する方法であって、以下の工程:
(a)Fkhsf特異的核酸プローブと、(i)該生物学的サンプルから単離した試験核酸分子、または(ii)RNA分子から合成した核酸分子のいずれかとを、ハイブリダイゼーション条件下で接触させる工程であって、ここで、該プローブが、項目1に記載のヌクレオチド配列の少なくとも一部を認識する、工程、および
(b)該核酸プローブと(i)または(ii)とのハイブリッドの形成を検出する工程、
を包含する、方法。
(項目19) 前記試験核酸分子が、RT−PCRによって得られる、項目18に記載の方法。
(項目20) 生物学的サンプル中のFkhsfまたはその変異形態の存在を検出する方法であって、以下の工程:
(a)該生物学的サンプルを、抗Fkhsf抗体または抗体フラグメントと接触させる工程であって、ここで、該接触工程が、該生物学的サンプルに対する該抗体または抗体フラグメントの結合を可能にする条件下で行われる、工程、および
(b)任意の該結合した抗体または結合した抗体フラグメントを検出する工程、
を包含する、方法。
(項目21) 前記抗体または前記抗体フラグメントが、以下:
(a)ポリクローナル抗体、
(b)マウスのモノクローナル抗体、
(c)(b)から誘導されるヒト化抗体、
(d)ヒトのモノクローナル抗体、および
(e)(b)、(c)、または(d)から誘導される抗体フラグメント、
からなる群より選択される、項目20に記載の方法。
(項目22) 前記抗体フラグメントが、F(ab’)2、F(ab)2、Fab’、Fab、Fv、sFv、および最小認識ユニットからなる群より選択される、項目20に記載の方法。
(項目23) 前記抗体または前記抗体フラグメントが、放射性同位元素、蛍光標識、化学発光標識、酵素標識、生体発光標識、およびコロイド状の金からなる群より選択される検出可能な標識をさらに含む、項目20に記載の方法。
(項目24) 項目1に記載の核酸分子にハイブリダイズし得る、単離されたオリゴヌクレオチド。
(項目25) 検出可能な標識をさらに含む、項目24に記載のオリゴヌクレオチド。
(項目26) 動物に対して項目1に記載のFkhsf核酸分子を投与する工程を包含する、動物に対してFkhsf核酸分子を導入する方法。
(項目27) 前記核酸分子がウイルスベクターによって発現される、項目26に記載の方法。
(項目28) 前記核酸分子がプラスミドベクターによって発現される、項目26に記載の方法。
(項目29) 前記核酸分子がインビボで動物に投与される、項目26に記載の方法。
(項目30) 前記核酸分子がエキソビボで細胞に投与され、次いで該細胞が前記動物に投与される、項目26に記載の方法。
(項目31) 前記細胞が造血細胞である、項目26に記載の方法。
(項目32) 前記造血細胞がT細胞である、項目26に記載の方法。
(項目33) 前記動物が、ヒト、サル、イヌ、ネコ、ラット、およびマウスからなる群より選択される、項目26に記載の方法。
(項目34) その細胞がFkhsfタンパク質をコードする配列を含むトランスジーンを発現する、ヒト以外のトランスジェニック動物。
本発明は、一般的には、変異した場合に、重大なリンパ増殖障害を生じる新規の遺伝子の発見に関する。詳細には、変異マウス(「Scurfy」と命名された)が、戻し交配分析、物理的なマッピング、および大規模なDNAの配列決定を通じたこの障害の原因遺伝子の同定のために使用された。この遺伝子の配列の分析は、この遺伝子が、関連の遺伝子ファミリー(全てが、翼状(winged)へリックスDNA結合ドメインを含有する)に属することを、示した。
【0006】
従って、本発明の1つの局面においては、FKHsfまたはFkhsfをコードする単離された核酸分子(それらの変異形態を含む)が提供される。特定の実施態様においては、任意の型のFkhsfは、温血動物(例えば、マウスまたはヒト)に由来し得る。さらなる実施態様においては、単離された核酸分子が提供され、ここで、この核酸分子は、以下からなる群より選択される:(a)配列番号2または配列番号4を含むアミノ酸配列をコードする核酸分子、(b)配列番号1もしくは配列番号3のヌクレオチド配列、またはその相補物を有する核酸分子に対してストリンジェントな条件下でハイブリダイズする核酸分子、および(c)(a)または(b)のいずれかによってコードされるポリペプチドの機能的なフラグメントをコードする核酸分子。好ましくは、核酸分子は、JM2ではない。関連する局面においては、ベクター(発現ベクターを含む)および組換え宿主細胞、ならびに上記の核酸分子によってコードされるタンパク質もまた、提供される。さらに、別のタンパク質のコード領域と上記の核酸分子の少なくとも一部とを組み合わせた融合タンパク質もまた、提供される。上記の配列に基づくオリゴヌクレオチドフラグメント(プローブおよびプライマーを含む)もまた、提供される。このようなフラグメントは、少なくとも8、10、12、15、20、または25ヌクレオチド長であり、そして100、200、500、1000、1500、または2000ヌクレオチド長にまで及び得る。
【0007】
他の局面においては、Fkhsfタンパク質(任意の型)を産生するために上記の発現ベクターを使用する方法が提供される。この方法は、(a)この発現ベクターを含みそしてFkhsfタンパク質を産生する組換え宿主細胞を培養する工程、および(b)培養した組換え宿主細胞からタンパク質を単離する工程、という一般的な工程を包含する。
【0008】
Fkhsfタンパク質に特異的に結合する抗体および抗体フラグメントもまた、提供される。このような抗体の代表的な例としては、ポリクローナル抗体およびモノクローナル抗体(マウスのハイブリドーマから得られたかまたはヒトの形態に誘導されたかにはかかわらず)の両方が挙げられる。抗体フラグメントの代表的な例としては、F(ab’)2、F(ab)2、Fab’、Fab、Fv、sFv、および最小認識ユニットまたは相補性決定領域が挙げられる。
【0009】
なお他の局面においては、被験体に由来する生物学的サンプル中のFkhsf核酸配列の存在を検出するための方法が、提供される。この方法は、以下の工程を包含する:(a)Fkhsf特異的核酸プローブと、(i)上記の生物学的サンプルから単離された試験核酸分子、または(ii)RNA分子から合成された核酸分子のいずれかとを、ハイブリダイゼーション条件下で接触させる工程(ここで、上記のプローブは、請求項1に記載のヌクレオチド配列の少なくとも一部を認識する)、および(b)上記の核酸プローブと(i)または(ii)とのハイブリッドの形成を検出する工程。
【0010】
別の関連する実施態様においては、生物学的サンプル中の、Fkhsfまたはその変異形態の存在を検出するための方法が提供される。この方法は、以下の工程を包含する:(a)生物学的サンプルを、抗Fkhsf抗体または抗体フラグメントと接触させる工程(ここで、上記の接触工程は、上記の生物学的サンプルに対する上記の抗体または抗体フラグメントの結合を可能にする条件下で行われる)、および(b)任意の上記の結合した抗体または結合した抗体フラグメントを検出する工程。
【0011】
本発明の他の局面においては、動物に対してFkhsf核酸分子を導入する方法が提供される。この方法は、動物(例えば、ヒト、サル、イヌ、ネコ、ラット、またはマウス)に対して、本明細書中に記載されているようなFkhsf核酸分子を投与する工程を包含する。1つの実施態様においては、核酸分子は、ウイルスベクター(例えば、レトロウイルス、アデノウイルス、アデノ随伴ウイルス、ヘルペスウイルスまたはアルファウイルスから少なくとも一部生成されたベクター)中に含まれ、そしてそれによって発現される。別の実施態様においては、核酸分子は、プラスミドベクターによって発現されるか、またはその中に含まれる。このようなベクターは、インビボまたはエキソビボのいずれかで、(例えば、T細胞のような造血細胞に)投与され得る。
【0012】
他の実施態様においては、ヒト以外のトランスジェニック動物が提供される。ここで、この動物の細胞は、Fkhsfタンパク質をコードする配列を含むトランスジーンを発現する。
本発明のこれらおよび他の局面は、以下の詳細な説明および添付の図面を参照して明らかとなる。さらに、特定の手順または組成物(例えば、プラスミドなど)をより詳細に記載する種々の参考文献が、本明細書中に示されており、そしてそれゆえそれらの全体が、参考として援用されている。
【図面の簡単な説明】
【0013】
【図1A】図1は、マウスのFkhsf cDNAのヌクレオチド配列(配列番号1)を示す;翻訳は、259位で始まり、そして1546位で終わると推定される。
【図1B】図1は、マウスのFkhsf cDNAのヌクレオチド配列(配列番号1)を示す;翻訳は、259位で始まり、そして1546位で終わると推定される。
【図2】図2は、マウスのFkhsfのアミノ酸配列(配列番号2)を示す。
【図3A】図3は、ヒトのFKHsf cDNAに対応する1735bpのヌクレオチド配列(配列番号3、1293bpのコード領域を含む)を示す;翻訳は、55位で始まり、そして1348位で終わると推定される。
【図3B】図3は、ヒトのFKHsf cDNAに対応する1735bpのヌクレオチド配列(配列番号3、1293bpのコード領域を含む)を示す;翻訳は、55位で始まり、そして1348位で終わると推定される。
【図4】図4は、ヒトのFKHsfタンパク質の431アミノ酸の配列(配列番号4)を示す。
【図5】図5は、FKHsfトランスジェニックマウスの生成のためのベクターを模式的に示す。
【図6】図6は、FKHsfトランスジーンが、scurfy動物における欠損を修復することを示す写真である。
【図7】図7は、FKHsf tgマウスが、正常な細胞と比較して、減少したリンパ節細胞を有することを示すグラフである。
【図8】図8は、FKHsfトランスジェニックマウスがインビトロでの刺激にほとんど応答しないことを示すグラフである。
【図9】図9は、FKHsfとJM2とのcDNAの比較である。
【図10】図10は、ヒトのFKHsfとマウスのFkhsfとの種々の領域における相同性を比較する。
【発明を実施するための形態】
【0014】
(発明の詳細な説明)
(定義)
本発明を詳細に示す前に、特定の用語の定義を示すこと、および本明細書中で以降に使用される略号を列挙しそしてそれらを定義することは、本発明の理解に役立ち得る。
【0015】
「Scurfy」は、重篤なリンパ増殖障害を示すマウスにおける遺伝した疾患をいう(例えば、Lyonら、Proc.Natl.Acad.Sci.USA 87:2433、1990を参照のこと)。原因遺伝子(その変異形態が、この疾患の原因である)を、配列番号1および3に示す。
【0016】
「分子」は、タンパク質またはペプチド(例えば、抗体、組換え結合パートナー、所望される結合親和性を有するペプチド)、核酸(例えば、DNA、RNA、キメラ核酸分子、およびPNAのような核酸アナログ)、および有機化合物または無機化合物を含むと理解されるはずである。
【0017】
「核酸」または「核酸分子」は、任意のデオキシリボ核酸(DNA)、リボ核酸(RNA)、オリゴヌクレオチド、ポリメラーゼ連鎖反応(PCR)によって生成されたフラグメント、ならびに任意の連結、切断、エンドヌクレアーゼ作用、およびエキソヌクレアーゼ作用によって生成されたフラグメントをいう。核酸は、天然に存在するヌクレオチド(例えば、デオキシリボヌクレオチドおよびリボヌクレオチド)、または天然に存在するヌクレオチドのアナログ(例えば、天然に存在するヌクレオチドのα−エナンチオマー形態)であるモノマー、あるいは両方の組み合わせから構成され得る。改変されたヌクレオチドは、糖部分、および/またはピリミジン塩基部分もしくはプリン塩基部分において修飾を有し得る。糖の修飾としては、例えば、ハロゲン、アルキル基、アミン、およびアジド基での1つ以上のヒドロキシル基の置換が挙げられ、または糖はエーテルまたはエステルとして官能化され得る。さらに、全体的な糖部分は、立体構造的に、および電気的に類似の構造(例えば、アザ糖、および炭素環式糖アナログ)で置きかえられ得る。塩基部分における修飾の例としては、アルキル化されたプリンおよびピリミジン、アシル化されたプリンまたはピリミジン、あるいは他の周知の複素環式置換基が挙げられる。核酸のモノマーは、ホスホジエステル結合またはこのような連結のアナログによって連結され得る。ホスホジエステル結合のアナログとしては、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホルアニリデート、ホスホルアミデートなどが挙げられる。用語「核酸」はまた、いわゆる「ペプチド核酸」を含む。これは、ポリアミド骨格に対して結合した、天然に存在するかまたは修飾された核酸塩基を含む。核酸は、一本鎖または二本鎖のいずれかであり得る。
【0018】
「単離された核酸分子」は、生物体のゲノムDNA中には組み込まれていない核酸分子である。例えば、真核生物細胞のゲノムDNAから分離されている遺伝子をコードするDNA分子は、単離されたDNA分子である。単離された核酸分子の別の例は、生物体のゲノム中には組み込まれていない、化学的に合成された核酸分子である。
【0019】
「プロモーター」は、構造遺伝子の転写を指向するヌクレオチド配列である。代表的には、プロモーターは、構造遺伝子の転写開始部位の近位の、遺伝子の5’領域中に配置される。プロモーターが誘導性プロモーターである場合は、転写速度は、誘導剤に応答して増大する。対照的に、プロモーターが構成的プロモーターである場合は、転写の速度は、誘導剤によっては調節されない。
【0020】
「ベクター」は、所望されるタンパク質の発現を指向し得るアセンブリをいう。ベクターは、目的の遺伝子に対して作動可能に連結された転写プロモーターエレメントを含まなければならない。ベクターは、デオキシリボ核酸(「DNA」)、リボ核酸(「RNA」)、またはこの2つの組み合わせ(例えば、DNA−RNAキメラ)のいずれかから構成され得る。必要に応じて、ベクターは、ポリアデニル化配列、1つ以上の制限部位、ならびに1つ以上の選択マーカー(例えば、ネオマイシンホスホトランスフェラーゼまたはハイグロマイシンホスホトランスフェラーゼ)を含み得る。さらに、選択される宿主細胞および使用されるベクターに依存して、他の遺伝的なエレメント(例えば、複製起点、さらなる核酸制限部位、エンハンサー、転写誘導性を付与する配列、および選択マーカー)もまた、本明細書中に記載されているベクター中に取りこまれ得る。
【0021】
「単離された」は、タンパク質またはポリペプチドの場合においては、他の生物学的巨大分子の実質的非存在下に存在し、そしてクマシーブルー染色を用いてSDS−PAGEゲル上に名目上単一のバンドとして出現する分子をいう。「単離された」は、有機分子について言及する場合は、その化合物が、当該分野で周知の方法(例えば、NMR、融点)を利用して90%よりも純粋であることを意味する。
【0022】
「クローニングベクター」は、宿主細胞中で自律複製する能力を有する、プラスミド、コスミド、またはバクテリオファージのような核酸分子をいう。クローニングベクターは、代表的には、1または少数の制限エンドヌクレアーゼ認識部位(ここでは、外来のヌクレオチド配列が、ベクターの本質的な生物学的な機能の損失を伴わずに、決定可能な様式で挿入され得る)およびクローニングベクターで形質転換された細胞の同定および選択における使用に適切であるマーカー遺伝子をコードするヌクレオチド配列を含む。マーカー遺伝子としては、代表的には、テトラサイクリン耐性またはアンピシリン耐性を提供する遺伝子が挙げられる。
【0023】
「発現ベクター」は、宿主細胞中で発現される遺伝子をコードする核酸分子をいう。代表的には、遺伝子の発現は、プロモーターの制御下に配置され、そして必要に応じて、少なくとも1つの調節エレメントの制御下に配置される。このような遺伝子は、プロモーター「に対して作動可能に連結された」といわれる。同様に、調節エレメントおよびプロモーターは、調節エレメントがプロモーターの活性を調節する場合は、作動可能に連結される。
【0024】
「組換え宿主」は、クローニングベクターまたは発現ベクターのいずれかを含む、任意の原核生物細胞または真核生物細胞をいう。この用語はまた、宿主細胞の染色体またはゲノム中のクローン化された遺伝子を含むように遺伝子操作されている、原核生物細胞または真核生物細胞を含む。
【0025】
真核生物においては、RMAポリメラーゼIIは、mRNAを産生するように、構造遺伝子の転写を触媒する。核酸分子は、RNAポリメラーゼIIの鋳型を含むように設計され得、ここで、RNA転写物は、特異的なmRNAの配列に対して相補的である配列を有する。このRNA転写物は、「アンチセンスRNA」と呼ばれ、そしてアンチセンスRNAをコードする核酸分子は、「アンチセンス遺伝子」と呼ばれる。アンチセンスRNA分子は、mRNA分子に結合し得、それによってmRNAの翻訳の阻害を生じる。
【0026】
「Fkhsfに特異的なアンチセンスオリゴヌクレオチド」または「Fkhsfアンチセンスオリゴヌクレオチド」は、(a)遺伝子の一部と安定な三重鎖を形成し得るか、または(b)mRNA転写物の一部と安定な二重鎖を形成し得る配列を有するオリゴヌクレオチドである。同様に、「Fkhsfに特異的なアンチセンスオリゴヌクレオチド」または「Fkhsfアンチセンスオリゴヌクレオチド」は、(a)Fkhsf遺伝子の一部と安定な三重鎖を形成し得るか、または(b)Fkhsf遺伝子のmRNA転写物の一部と安定な二重鎖を形成し得る配列を有するオリゴヌクレオチドである。
【0027】
「リボザイム」は、触媒中心を含む核酸分子である。この用語は、RNA酵素、自己スプライシングRNA、自己切断RNA、およびこれらの触媒機能を行う核酸分子を含む。リボザイムをコードする核酸分子は、「リボザイム遺伝子」と呼ばれる。
【0028】
略号:YAC、酵母人工染色体;PCR、ポリメラーゼ連鎖反応;RT−PCR、逆転写酵素(RT)を使用して最初の工程でRNAが最初にDNAに転写されるPCRプロセス;cDNA、DNA形態にRNA配列をコピーすることによって作成される、任意のDNA。本明細書中で利用される場合は、「Fkhsf」は、(この遺伝子が、ヒト、哺乳動物、または任意の他の温血動物から得られたかどうかにはかかわらず)Fkhsf遺伝子の遺伝子産物をいう。「FKHsf」と大文字になっている場合は、この遺伝子産物(および遺伝子)は、ヒトに由来すると理解されるべきである。
【0029】
上記のように、本発明は、一般的には、薬学的な生成物および方法、そしてより詳細には、scurfyに関連する疾患を診断するために有用な方法および組成物、ならびに免疫系を調節し得る化合物を同定するための方法に関する。
【0030】
従って、以下により詳細に議論されるように、この発見は、免疫系のアゴニストまたは、あるいはアンタゴニストとして作用し得る分子を選択するために利用され得るアッセイの開発を導いた。さらに、このようなアッセイは、免疫系を調節することにおいて同様に活性である、他の遺伝子および遺伝子産物を同定するために利用され得る。
【0031】
(SCURFY) 簡潔には、本発明は、Fkhsfをコードする遺伝子における変異が稀な状態(scurfy)を生じるという予想外の発見に基づく。この状態は、リンパ節、脾臓、肝臓、および皮膚の進行性のリンパ球浸潤によって特徴付けられ、それによって巨脾腫、肝腫、大いに肥大したリンパ節、矮小、脱離性皮膚炎、および厚くなった奇形の耳を含む、肉眼的な形態学的な症状を生じる(Godfreyら、Amer.J.Pathol.138:1379、1991;Godfreyら、Proc.Natl.Acad.Sci.USA、88:5528、1991)。この翼状ヘリックスのファミリーの新規のメンバーは、免疫系の新規の成分を示す。
【0032】
scurfyの原因遺伝子を発見するために利用された方法は、以下の実施例1に提供される。マウスのscurfyの原因遺伝子、およびヒトのオルソログをクローニングするための方法は、実施例2および3において以下に提供される。トランスジェニックマウスを使用して決定されるような、遺伝子の正体および遺伝子機能との相関の確認のための方法もまた、実施例において提供される。
【0033】
Fkhsf遺伝子および遺伝子産物の存在を決定するための方法もまた、本発明によって提供される。1つの実施態様においては、このような方法は、以下の一般的な工程を包含する:(a)Fkhsf特異的核酸プローブと、(i)生物学的サンプルから単離された試験核酸分子、または(ii)RNA分子から合成された核酸分子のいずれかとを、ハイブリダイゼーション条件下で接触させる工程(ここで、プローブは、Fkhsfのヌクレオチド配列の少なくとも一部を認識する)、および(b)上記の核酸プローブと(i)または(ii)とのハイブリッドの形成を検出する工程。種々の方法(例えば、RNA増幅(Lizardiら、Bio/Technology 6:1197−1202、1988;Kramerら、Nature、339:401−402、1989;Lomeliら、Clinical Chem.35(9):1826−1831、1989;米国特許第4,786,600号を参照のこと)、およびポリメラーゼ連鎖反応(「PCR」)を利用する核酸の増幅(米国特許第4,683,195号、同第4,683,202号、および同第4,800,159号を参照のこと)、逆転写酵素PCRおよびCPT(米国特許第4,876,187号および同第5,011,769号を参照のこと)を含む)が、選択された配列を増幅するために利用され得る。
【0034】
あるいは、抗体は、Fkhsf遺伝子産物の存在を検出するために利用され得る。より詳細には、1つの実施態様においては、生物学的サンプル中のFkhsfペプチドまたはその変異体形態の存在を検出するための方法が提供される。この方法は、以下の工程を包含する:(a)抗Fkhsf抗体または抗体フラグメントと生物学的サンプルとを接触させる工程(ここで、上記の接触工程は、生物学的サンプルに対する上記の抗体または抗体フラグメントの結合を可能にする条件下で行われる)、および(b)任意の結合した抗体または結合した抗体フラグメントを検出する工程。
【0035】
このような方法は、例えば、向流免疫電気泳動(Countercurrent Immuno−Electrophoresis、CIEP)、放射免疫アッセイ、放射免疫沈降、酵素結合免疫吸着アッセイ(ELISA)、ドットブロットアッセイ、阻害または競合アッセイ、およびサンドイッチアッセイを含む、広範な種々のアッセイ形式において達成され得る(米国特許第4,376,110号および同第4,486,530号を参照のこと;Antibodies:A Laboratory Manual、前出もまた参照のこと)。
【0036】
(核酸分子、タンパク質、およびタンパク質の産生方法) 種々のFKHsfまたはFkhsfのタンパク質および核酸分子(またはその一部)が本明細書中で提供されているが、本発明の状況においては、1つ以上のこれらのタンパク質に関する言及は、実質的に類似の活性のタンパク質を含むと理解されるべきであると理解されるべきである。本明細書中で使用される場合は、タンパク質は、(a)これらがこのタンパク質をコードする遺伝子のコード領域に由来するヌクレオチド配列(例えば、この配列の一部、またはこの配列の対立遺伝子改変体を含む)によってコードされる;(b)このヌクレオチド配列が、中程度、高い、または非常に高いストリンジェンシー(Sambrookら、Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor Laboratory Press、NY、1989を参照のこと)の下で本発明のヌクレオチド配列に対してハイブリダイゼーションし得るか、または本明細書中で開示されている配列に対して、少なくとも50%、60%、70%、75%、80%、90%、95%、もしくはそれより高い相同性を有する;あるいは(c)このDNA配列が、(a)または(b)において定義されるDNA配列に対する遺伝コードの結果として縮重している場合に、「実質的に類似」であると考えられる。さらに、本明細書中で開示されている核酸分子は、配列が、他の点では本明細書中に示される基準を満たす限りは、相補配列および非相補配列の両方を含む。本発明の状況においては、高いストリンジェンシーは、標準的なハイブリダイゼーション条件(例えば、65℃にて、5×SSPE、0.5%のSDS、またはそれに等価なもの)を意味する。ハイブリダイゼーションの目的のためには、アミノ末端ドメイン、ジンクフィンガードメイン、中央ドメイン、またはフォークヘッドドメイン(実施例10を参照のこと)をコードする核酸分子が、利用され得る。
【0037】
本明細書中に記載されている核酸分子によってコードされるタンパク質の構造は、例えば、P/C遺伝子またはIntelligenetics Suite(Intelligenetics,Mountain View、California)の疎水性プロット関数を使用して、またはKyteおよびDoolittle(J.Mol.Biol.157:105−132、1982)によって記載されている方法に従って、一次翻訳産物から推定され得る。
【0038】
本発明のタンパク質は、酸の塩もしくは塩基の塩の形態で、または中性の形態で、調製され得る。さらに、個々のアミノ酸残基は、酸化または還元によって改変され得る。さらに、種々の置換、欠失、または付加が、アミノ酸または核酸の配列に対して行われ得、その正味の影響は、変異体または野生型のタンパク質の生物学的な活性を維持するか、またはさらに増強もしくは減少させる。さらに、遺伝コードの縮重に起因して、例えば、同じアミノ酸配列をコードするヌクレオチド配列におけるかなりのバリエーションが存在し得る。
【0039】
本明細書中に開示されているタンパク質の他の誘導体としては、他のタンパク質またはポリペプチドとのタンパク質の結合体が挙げられる。これは、例えば、タンパク質の精製または同定を容易にするために付加され得る、N末端融合タンパク質またはC末端融合タンパク質の合成によって達成され得る(米国特許第4,851,341号を参照のこと、また、Hoppら、Bio/Technology 6:1204、1988をもまた参照のこと)。あるいは、融合タンパク質(例えば、FKHもしくはFkh−ルシフェラーゼ、またはFKHもしくはFkh−GFP)が、タンパク質の同定、発現、および分析を補助するために構築され得る。
【0040】
本発明のタンパク質は、本明細書中に記載されている広範な種々の技術を使用して構築され得る。さらに、変異は、天然の配列のフラグメントに対する連結を可能にする制限部位に隣接した、変異体配列を含むオリゴヌクレオチドを合成することによって、特定の遺伝子座に導入され得る。連結後、得られた再構築された配列は、所望のアミノ酸の挿入、置換、または欠失を有する誘導体をコードする。
【0041】
あるいは、オリゴヌクレオチドによって指向される部位特異的(またはセグメント特異的)変異誘発手順が、要求される置換、欠失、または挿入に従って変更された特定のコドンを有する変更された遺伝子を提供するために使用され得る。上記に示される変更を作成する例示的な方法は、Walderら(Gene 42:133、1986);Bauerら(Gene 37:73、1985);Craik(BioTechniques、1985年1月、12−19);Smithら(Genetic Engineering:Principles and Methods、Plenum Press、1981):およびSambrookら(前出)によって開示されている。タンパク質の欠失誘導体または短縮誘導体(例えば、可溶性の細胞外部分)もまた、所望される欠失に対して隣接する便利な制限エンドヌクレアーゼ部位を利用することによって、構築され得る。制限消化に続いて、突出がフィルインされ得、そしてDNAが再連結され得る。上記に示される変更を作成する例示的な方法が、Sambrookら(Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor Laboratory Press、1989)によって開示されている。
【0042】
本発明の核酸分子中に作成される変異は、好ましくは、コード配列のリーディングフレームを保存する。さらに、変異は、好ましくは、mRNAの翻訳に有害な影響を与える二次的なmRNA構造(例えば、ループまたはヘアピン)を生じるようにハイブリダイズし得る相補性領域を作成しない。変異部位は予め決定され得るが、変異の性質自体は予め決定される必要はない。例えば、所定の部位で変異体の最適の特徴について選択するためには、ランダムな変異誘発が標的のコドンで行われ得、そして発現された変異体が表示生物学的活性についてスクリーニングされ得る。あるいは、変異は、天然の配列のフラグメントへの連結を可能にする制限部位に隣接した変異体配列を含むオリゴヌクレオチドを合成することによって、特定の遺伝子座に導入され得る。連結後、得られた再構築された配列は、所望のアミノ酸の挿入、置換、または欠失を有する誘導体をコードする。変異は、タンパク質の活性を保存するかもしくは増大させる、またはタンパク質(例えば、変異体Fkh)を減少させるかもしくは不能にする目的のために導入され得る。
【0043】
本発明のタンパク質をコードする核酸分子もまた、PCR変異誘発、化学的変異誘発の技術(DrinkwaterおよびKlinedinst、PNAS 83:3402−3406、1986)を利用して、ヌクレオチドの誤った取りこみを強制することによって(例えば、LiaoおよびWise、Gene 88:107−111、1990)、またはランダムに変異誘発したオリゴヌクレオチドの使用によって(Horwitzら、Genome 3:112−117、1989)、構築され得る。
【0044】
本発明はまた、上記に記載されている遺伝子を発現し得るベクターを含有する宿主細胞を培養することによって、上記に記載されている遺伝子の操作および発現を提供する。このようなベクターまたはベクター構築物は、適切な転写調節エレメントまたは翻訳調節エレメントに対して作動可能に連結された所望のタンパク質をコードする、合成核酸分子またはcDNAから誘導される核酸分子のいずれかを含む。適切な調節エレメントは、細菌、真菌、ウイルス、哺乳動物、昆虫、または植物の遺伝子を含む種々の供給源から誘導され得る。適切な調節エレメントの選択は、選択される宿主細胞に依存し、そして当業者によって容易に達成され得る。調節エレメントの例としては、以下が挙げられる:転写プロモーターおよびエンハンサー、またはRNAポリメラーゼ結合配列、転写ターミネーター、およびリボソーム結合配列(翻訳開始シグナルを含む)。
【0045】
上記に記載されている任意のタンパク質をコードする核酸分子は、細菌、哺乳動物、酵母もしくは他の真菌、ウイルス、または植物細胞を含む、広範な種々の原核生物宿主細胞および真核生物宿主細胞によって容易に発現され得る。このような細胞を外来DNAを発現するように形質転換またはトランスフェクトするための方法は、当該分野で周知である(例えば、Itakuraら、米国特許第4,704,362号;Hinnenら、Proc.Natl.Acad.Sci.USA 75:1929−1933、1978;Murrayら、米国特許第4,801,542号;Upshallら、米国特許第4,935,349号;Hagenら、米国特許第4,784,950号;Axelら、米国特許第4,399,216号;Goeddelら、米国特許第4,766,075号;およびSambrookら、Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor Laboratory Press、1989を参照のこと;植物細胞については、CzakoおよびMarton、Plant Physiol.、104:1067−1071、1994;およびPaszkowskiら、Biotech.24:387−392、1992を参照のこと)。
【0046】
本発明を実行するために適切な細菌宿主細胞としては、E.coli、B.subtilis、Salmonella typhimurium、ならびにPseudomonas属、Streptomyces属、およびStaphylococcus属内の種々の種、ならびに当業者に周知の多くの他の細菌種が挙げられる。細菌宿主細胞の代表的な例としては、DH5αが挙げられる(Stratagene、LaJolla、California)。
【0047】
細菌の発現ベクターは、好ましくは、宿主細胞中で機能するプロモーター、1つ以上の選択可能な表現型マーカー、および細菌の複製起点を含む。代表的なプロモーターとしては、β−ラクタマーゼ(ペニシリナーゼ)およびラクトースのプロモーターシステム(Changら、Nature 275:615、1978を参照のこと)、T7 RNAポリメラーゼプロモーター(Studierら、Meth.Enzymol.185:60−89、1990)、λプロモーター(Elvinら、Gene 87:123−126、1990)、trpプロモーター(NicholsおよびYanofsky、Meth.in Enzymology 101:155、1983)、およびtacプロモーター(Russellら、Gene 20:231、1982)が挙げられる。代表的な選択マーカーとしては、種々の抗生物質耐性マーカー(例えば、カナマイシン耐性遺伝子またはアンピシリン耐性遺伝子)が挙げられる。宿主細胞を形質転換するために適切な多くのプラスミドが、当該分野で周知であり、中でも、pBR322(Bolivarら、Gene 2:95、1977を参照のこと)、pUCプラスミドpUC18、pUC19、pUC118、pUC119(Messing、Meth.in Enzymology 101:20−77、1983、ならびにVieiraおよびMessing、Gene 19:259−268、1982を参照のこと)、ならびにpNH8A、pNH16a、pNH18a、ならびにBluescript M13(Stratagene、La Jolla、California)が挙げられる。
【0048】
本発明を実行するために適切な酵母宿主細胞および真菌宿主細胞としては、中でも、Saccharomyces pombe、Saccharomyces cerevisiae、Pichia属またはKluyveromyces属、およびAspergillus属の種々の種(McKnightら、米国特許第4,935,349号)が挙げられる。酵母および真菌についての適切な発現ベクターとしては、中でも、酵母についてはYCp50(ATCC第37419号)、およびamdSクローニングベクターpV3(Turnbull、Bio/Technology 7:169、1989)、YRp7(Struhlら、Proc.Natl.Acad.Sci.USA 76:1035−1039、1978)、YEp13(Broachら、Gene 8:121−133、1979)、pJDB249およびpJDB219(Beggs、Nature 275:104−108、1978)およびそれらの誘導体が挙げられる。
【0049】
酵母中での使用に好ましいプロモーターとしては、酵母の解糖遺伝子に由来するプロモーター(Hitzemanら、J.Biol.Chem.255:12073−12080、1980;AlberおよびKawasaki,J.Mol.Appl.Genet.1:419−434、1982)またはアルコールデヒドロゲナーゼ遺伝子に由来するプロモーター(Youngら、Genetic Engineering of Microorganisms for Chemicals、Hollaenderら(編)、355頁、Plenum,New York、1982;Ammerer、Meth Enzymol.101:192−201、1983)が挙げられる。真菌ベクターについての有用なプロモーターの例としては、Aspergillus nidulans解糖遺伝子に由来するプロモーター(例えば、adh3プロモーター(McKnightら、EMBO J.4:2093−2099、1985)が挙げられる。発現ユニットはまた、転写ターミネーターを含み得る。適切なターミネーターの例は、adh3ターミネーターである(McKnightら、同書、1985)。
【0050】
細菌のベクターを用いる場合は、酵母のベクターは、一般的には、表現型アッセイが存在して形質転換体が選択されることを可能にする優性表現型を示す任意の数の遺伝子の1つであり得る、選択マーカーを含む。好ましい選択マーカーは、宿主細胞の栄養要求性を相補するか、抗生物質耐性を提供するか、または細胞が特定の炭素源を利用することを可能にするものであり、そしてleu2(Broachら、同書)、ura3(Botsteinら、Gene 8:17、1979)、またはhis3(Struhlら、同書)を含む。別の適切な選択マーカーは、酵母細胞に対してクロラムフェニコール耐性を付与する、cat遺伝子である。
【0051】
真菌を形質転換するための技術は、文献において周知であり、そして例えば、Beggs(同書)、Hinnenら(Proc.Natl.Acad.Sci.USA 75:1929−1933、1978)、Yeltonら(Proc.Natl.Acad.Sci.USA 81:1740−1747、1984)、およびRussell(Nature 301:167−169、1983)によって記載されている。宿主細胞の遺伝子型は、発現ベクター上に存在する選択マーカーによって相補される遺伝的な欠損を含み得る。特定の宿主および選択マーカーの選択は、十分に当業者のレベルの範囲内である。
【0052】
酵母を形質転換するためのプロトコールもまた、当業者に周知である。例えば、形質転換は、DNAを用いた酵母スフェロプラストの調製によるか(Hinnenら、PNAS USA 75:1929、1978を参照のこと)、またはLiClのようなアルカリ塩での処理による(Itohら、J.Bacteriology 153:163、1983を参照のこと)かのいずれかによって、容易に達成され得る。真菌の形質転換はまた、Cullenら(Bio/Technology 5:369、1987)によって記載されているようにポリエチレングリコールを使用して行われ得る。
【0053】
ウイルスベクターとしては、上記に記載されているような所望のタンパク質をコードする単離された核酸分子の発現を指向するプロモーターを含むベクターが挙げられる。以下を含む広範な種々のプロモーターが、本発明の状況において利用され得る:例えば、MoMLV LTR、RSV LTR、Friend MuLV LTR、アデノウイルスプロモーター(Ohnoら、Science 265:781−784、1994)、ネオマイシンホスホトランスフェラーゼプロモーター/エンハンサー、後期パルボウイルスプロモーター(Koeringら、Hum.Gene Therap.5:457−463、1994)、Herpes TKプロモーター、SV40プロモーター、メタロチオネインIIa遺伝子エンハンサー/プロモーター、サイトメガロウイルス即時初期プロモーター、およびサイトメガロウイルス即時後期プロモーターのようなプロモーター。本発明の特定の好ましい実施態様においては、プロモーターは、組織特異的プロモーターである(例えば、第WO91/02805号;第EP 0,415,731号;および第WO90/07936号を参照のこと)。適切な組織特異的プロモーターの代表的な例としては、神経特異的エノラーゼプロモーター、血小板由来増殖因子βプロモーター、ヒトα1−キマエリン(chimaerin)プロモーター、シナプシンIプロモーター、およびシナプシンIIプロモーターが挙げられる。上記のプロモーターに加えて、他のウイルス特異的プロモーター(例えば、レトロウイルスプロモーター(上記のプロモーターおよびHIVプロモーターのような他のプロモーターを含む)、肝炎、ヘルペス(例えば、EBV)、および細菌、真菌、または寄生生物(例えば、マラリア)特異的プロモーター)が、ウイルス、細菌、真菌、または寄生生物に感染した特異的な細胞または組織を標的化するために利用され得る。
【0054】
本発明を実行するために適切な哺乳動物細胞としては、中でも以下が挙げられる:PC12(ATCC第CRL1721号)、N1E−115神経芽細胞腫、SK−N−BE(2)C神経芽細胞腫、SHSY5アドレナリン作動性神経芽細胞腫、NS20YおよびNG108−15マウスコリン作用性細胞株、またはラットのF2後根神経節株、COS(例えば、ATCC第CRL1650号または同第1651号)、BHK(例えば、ATCC第CRL 6281号;BHK570細胞株(登録番号第CRL10314号のもとで、アメリカンタイプカルチャーコレクションに寄託された))、CHO(ATCC第CCL61号)、HeLa(例えば、ATCC第CCL2号)、293(ATCC第1573号;Grahamら、J.Gen.Virol.36:59−72、1977)、およびNS−1細胞。以下を含む他の哺乳動物細胞株が、本発明において使用され得る:ラットのHep1(ATCC第CRL1600号)、ラットのHepII(ATCC第CRL1548号)、TCMK(ATCC第CCL139号)、ヒトの肺(ATCC第CCL75.1号)、ヒトの肝ガン(ATCC第HTB−52号)、Hep G2(ATCC第HB8065号)、マウスの肝臓(ATCC第CCL29.1号)、NCTC 1469(ATCC第CCL9.1号)、SP2/0−Ag14(ATCC第1581号)、HIT−T15(ATCC第CRL1777号)、Jurkat(ATCC第Tib 152号)、およびRINm 5AHT2B(OrskovおよびNielsen、FEBS 22
9(1):175−178、1988)。
【0055】
本発明を実行することにおける使用のための哺乳動物の発現ベクターとしては、クローン化された遺伝子またはcDNAの転写を指向し得るプロモーターが挙げられる。好ましいプロモーターとしては、ウイルスプロモーターおよび細胞性のプロモーターが挙げられる。ウイルスプロモーターとしては、サイトメガロウイルス即時初期プロモーター(Boshartら、Cell 41:521−530、1985)、サイトメガロウイルス即時後期プロモーター、SV40プロモーター(Subramaniら、Mol.Cell.Biol.1:854−864、1981)、MMTV LTR、RSV LTR、メタロチオネイン−1、アデノウイルスE1aが挙げられる。細胞性のプロモーターとしては、マウスのメタロチオネイン−1プロモーター(Palmiterら、米国特許第4,579,821号)、マウスVKプロモーター(Bergmanら、Proc.Natl
.Acad.Sci.USA、81:7041−7045、1983);Grantら、Nucl.Acids Res.15:5496、1987)、およびマウスVHプロモ
ーター(Lohら、Cell 33:85−93、1983)が挙げられる。プロモーターの選択は、少なくとも一部、所望される発現のレベル、またはトランスフェクトされるレシピエント細胞株に依存する。
【0056】
このような発現ベクターはまた、プロモーターの下流、および目的のペプチドまたはタンパク質をコードするDNA配列の上流に配置された、RNAスプライス部位のセットを含み得る。好ましいRNAスプライス部位は、アデノウイルスおよび/または免疫グロブリン遺伝子から得られ得る。目的のコード配列の下流に配置されたポリアデニル化シグナルもまた、発現ベクター中に含まれる。適切なポリアデニル化シグナルとしては、SV40に由来する初期または後期ポリアデニル化シグナル(KaufmanおよびSharp、同書)、アデノウイルス5 E1B領域に由来するポリアデニル化シグナルおよびヒト成長ホルモン遺伝子ターミネーター(DeNotoら、Nuc.Acids Res.9:3719−3730、1981)が挙げられる。発現ベクターは、プロモーターとRNAスプライス部位との間に配置された、アデノウイルス2 三分節(tripartile)リーダーのような、非コードウイルスリーダー配列を含み得る。好ましいベクターはまた、SV40エンハンサーのようなエンハンサー配列を含み得る。発現ベクターはまた、アデノウイルスのVA RNAをコードする配列を含み得る。適切な発現ベクターは、商業的供給源(例えば、Stratagene、La Jolla、California)から得ることができる。
【0057】
クローン化されたDNA配列を含むベクター構築物は、例えば、リン酸カルシウムによって媒介されるトランスフェクション(Wiglerら、Cell 14:725、1978;CorsaroおよびPearson、Somatic Cell Genetics 7:603、1981;GrahamおよびVan der Eb.Virology 52:456、1973)、エレクトロポレーション(Neumannら、EMBO J.1:841−845、1982)、またはDEAE−デキストラン媒介性トランスフェクション(Ausubelら(編)、Current Protocols in Molecular Biology、John Wiley and Sons,Inc.,NY,1987)によって、培養された哺乳動物細胞中に導入され得る。クローン化されたDNAを安定に組み込んだ細胞を同定するために、選択マーカーが一般的には、目的の遺伝子またはcDNAとともに細胞中に導入される。培養された哺乳動物細胞における使用に好ましい選択マーカーとしては、ネオマイシン、ハイグロマイシン、およびメトトレキセートのような薬物に対する耐性を付与する遺伝子が挙げられる。他の選択マーカーとしては、蛍光タンパク質(例えば、GFP(緑色蛍光タンパク質)またはBFP(青色蛍光タンパク質))が挙げられる。選択マーカーは、増幅可能な選択マーカーであり得る。好ましい増幅可能な選択マーカーは、DHFR遺伝子およびネオマイシン耐性遺伝子である。適切なマーカーは、Thilly(Mammalian Cell Technology、Butterworth Publishers、Stoneham、MA、これは、本明細書中で参考として援用されている)によって概説されている。
【0058】
適切なベクターを含有する哺乳動物細胞は、目的のDNA配列(単数または複数)の発現を開始するために、一定の期間(代表的には、1〜2日間)増殖させることが可能である。次いで、薬物選択が、安定な様式で選択マーカーを発現する細胞の増殖について選択するために適用される。増幅可能な選択マーカーでトランスフェクトされている細胞については、薬物濃度が、クローン化された配列の増大したコピー数について選択するために、段階的な様式で増大させられ得、それによって発現レベルを増大させる。導入された配列を発現する細胞が選択され、そして所望の形態または所望のレベルでの目的のタンパク質の産生についてスクリーニングされる。次いで、これらの基準を満たす細胞がクローン化され得、そして産生のためにスケールアップされ得る。細胞はまた、フローサイトメーターを使用してGFP陽性細胞について分類することによって、それらのGFP発現に基づいて、トランスフェクションについて選択され得る。
【0059】
哺乳動物細胞のトランスフェクションのためのプロトコールは、当業者に周知である。代表的な方法としては、リン酸カルシウムによって媒介されるトランスフェクション、エレクトロポレーション、リポフェクション、レトロウイルス、アデノウイルス、およびプロトプラスト融合によって媒介されるトランスフェクション(Sambrookら、前出を参照のこと)が挙げられる。裸のベクター構築物もまた、筋肉細胞または他の適切な細胞によって取りこまれ得、続いて、哺乳動物(または他の動物)の筋肉中に注射され得る。
【0060】
当該分野で公知の多数の昆虫宿主細胞もまた、本明細書を参照して本発明において有用であり得る。例えば、昆虫細胞中で異種DNA配列を発現するためのベクターとしてのバキュロウイルスの使用が、Atkinsonら(Pestic.Sci.28:215−224、1990)によって概説されている。
【0061】
当該分野で公知の多数の植物宿主細胞もまた、本明細書を参照して本発明において有用であり得る。例えば、植物細胞中での遺伝子の発現のためのベクターとしてのAgrobacterium rhizogenesの使用が、Sinkarら(J.Biosci.(Bangalore)11:47−58、1987)によって概説されている。
【0062】
本発明の関連する局面においては、所望のタンパク質をコードし、そして遺伝子の発現に有効なプロモーターに対して作動可能に連結されている遺伝子をその生殖細胞および体細胞が含むトランスジェニック動物中で本発明のタンパク質は発現され得る。あるいは、類似の様式においては、所望の遺伝子を欠失しているトランスジェニック動物が、調製され得る(例えば、「ノックアウト」マウス)。このようなトランスジェニックは、ヒト以外の種々の動物(マウス、ラット、ウサギ、ヒツジ、イヌ、ヤギ、およびブタを含む)において調製され得る(Hammerら、Nature 315:680−683、1985、Palmiterら、Science 222:809−814、1983、Brinsterら、Proc.Natl.Acad.Sci.USA 82:4438−4442、1985、PalmiterおよびBrinster、Cell 41:343−345、1985、ならびに米国特許第5,175,383号、同第5,087,571号、同第4,736,866号、同第5,387,742号、同第5,347,075号、同第5,221,778号、および同第5,175,384号を参照のこと)。簡潔には、発現される核酸分子を、適切に配置された発現制御配列とともに含む発現ベクターは、例えば、マイクロインジェクションによって、受精卵の前核中に導入される。注入されたDNAの組込みは、組織サンプルから、DNAのブロット分析によって検出される。導入されたDNAが、動物の生殖列系中に取りこまれ、その結果、その動物の子孫に伝達されることが、好ましい。組織特異的な発現は、組織特異的プロモーターの使用を通じて、またはトランスジーンの調節された発現を可能にする誘導性プロモーター(例えば、メタロチオネイン遺伝子のプロモーター(Palmiterら、1983、同書))の使用を通じて、達成され得る。
【0063】
天然に存在するscurfy変異体(「sf」)以外のFkhsfの変異体形態を産生するか、または天然に存在する変異体とは異なる遺伝的なバックグラウンドにある動物は、本明細書中に提供される開示を考慮して容易に産生され得る。
【0064】
タンパク質は、数ある方法の中でも、本発明の組換え翻訳産物を産生するために適切な宿主およびベクターシステムを培養することによって単離され得る。次いで、このような細胞株に由来する上清、またはタンパク質が上清中には分泌されない場合はタンパク質封入物もしくは細胞全体が、所望のタンパク質を単離するために種々の精製手順によって処理され得る。例えば、上清は、市販のタンパク質濃縮フィルター(例えば、AmiconまたはMillipore Pellicon限外濾過ユニット)を使用して最初に濃縮され得る。濃縮後、濃縮物は、適切な精製マトリックス(例えば、適切な支持体に結合させられた抗タンパク質抗体のような)に対して適用され得る。あるいは、陰イオン交換樹脂または陽イオン交換樹脂が、タンパク質を精製するために使用され得る。さらに別のものとしては、1つ以上の逆相高速液体クロマトグラフィー(RP−HPLC)工程が、タンパク質をさらに精製するために使用され得る。本発明のタンパク質を単離する他の方法は、当該分野で周知である。
【0065】
タンパク質は、他の(所望されない)タンパク質が、SDS−PAGE分析、続いてクマシーブルーでの染色によって検出されない場合は、本発明の状況において「単離された」と考えられる。他の実施態様においては、所望されるタンパク質は、従って、他の(所望されない)タンパク質が、SDS−AGE分析、続いて銀染色によって検出されないように、単離され得る。
【0066】
(免疫系を調節する分子を選択するためのアッセイ) 上記のように、本発明は、免疫系を調節し得る分子を選択および/または単離するための方法を提供する。適切なアッセイの代表的な例としては、酵母および哺乳動物の2−ハイブリッドシステム(例えば、Dangら、Mol.Cell.Biol.11:954、1991;Fearonら、Proc.Natl.Acad.Sci.USA、89:7958、1992)、DNA結合アッセイ、アンチセンスアッセイ、伝統的なタンパク質結合アッセイ(例えば、125Iまたは時間消散蛍光を利用する)、ゲル電気泳動と組み合わせた免疫沈降、および直接的なタンパク質配列決定、Fkhsfによって調節される遺伝子の転写分析、サイトカイン産生、ならびに増殖アッセイが挙げられる。
【0067】
例えば、1つの実施態様においては、Fkhsfと直接相互作用するタンパク質は、酵母の2−ハイブリッド結合システム(例えば、米国特許第5,283,173号、同第5,468,614号、同第5,610,015号、および同第5,667,973号を参照のこと)のようなアッセイによって検出され得る。簡潔には、2−ハイブリッドシステムにおいては、DNA−結合ドメイン−Fkhsfタンパク質の融合体(例えば、GAL4−Fkhsf融合体)が構築され、そして選択マーカー遺伝子に連結されたGAL4結合部位を含む細胞中にトランスフェクトされる。全Fkhsfタンパク質またはFkhsfのサブ領域が、使用され得る。GAL4活性化ドメインに対して融合されたcDNAのライブラリーもまた、構築され、そして同時トランスフェクトされる、cDNA−GAL4活性化ドメイン融合体中のcDNAがFkhsfと相互作用するタンパク質をコードする場合は、選択マーカーが発現される、次いで、cDNAを含有する細胞が増殖させられ、構築物が単離され、そして特徴付けられる。他のアッセイもまた、相互作用するタンパク質を同定するために使用され得る。このようなアッセイとしては、ELISA、ウェスタンブロッティング、同時免疫沈降、インビトロでの転写/翻訳分析などが挙げられる。
【0068】
本発明の別の局面においては、選択された分子が免疫系を調節し得るかどうかを決定するための方法が提供される。この方法は、以下の工程を包含する:(a)FkhsfまたはFkhsfの変異体を発現する細胞に対して選択された候補分子を曝露する工程、および(b)この分子がFkhsfの活性を調節するかどうかを決定する工程、およびそれによって上記の分子が免疫系を調節するかどうかを決定する工程。このような試験のための細胞は、(a)正常なリンパ球、(b)FKHsf(もしくはFkhsf)タンパク質(またはその変異形態)を過剰発現するように操作された細胞株、あるいは(c)上記のタンパク質を発現するように操作されたトランスジェニック動物に由来し得る。このようなトランスジェニックマウスに由来する細胞は、減少した細胞数および種々の刺激に対する減少した応答性を含む、低応答性(hyporesponsive)状態によって、一部特徴付けられる(例えば、実施例8)。
【0069】
本明細書中に記載されている方法は、個々の試験分子の分析を言及し得るが、本発明がそのようには限定されるべきでないことが、注意されるべきである。特に、選択された分子は、化合物の混合物中に含まれ得る。従って、記載されている方法は、さらに、所望される分子を単離する工程を包含し得る。さらに、候補分子が、多数のパラメーター(例えば、T細胞の増殖、サイトカインの産生などを含む)によって、免疫系を調節するそれらの能力について評価され得る。
【0070】
(候補分子)
広範な種々の分子が、免疫系を調節するそれらの能力についてアッセイされ得る。より詳細に以下で議論される代表的な例としては、有機分子、タンパク質またはペプチド、および核酸分子が挙げられる。
【0071】
(1.有機分子)
多数の有機分子が、免疫系を調節するそれらの能力についてアッセイされ得る。例えば、本発明の1つの実施態様においては、適切な有機分子が、化学的なライブラリー(ここでは、化合物が別々にアッセイされる)から、またはコンビナトリアル化学ライブラリー(ここでは、複数の化合物が一度にアッセイされ、次いで、最も活性な化合物を決定および単離するために取り組まれる)から、のいずれかから選択され得る。
【0072】
このようなコンビナトリアル化学ライブラリーの代表的な例としては、以下に記載されているものが挙げられる:Agrafiotisら「System and method of automatically generating chemical compounds with desired properties」、米国特許第5,463,564号;Armstrong,R.W.、「Synthesis of combinatorial arrays of organic compounds through the use of multiple component combinatorial array syntheses」、第WO95/02566号;Baldwin,J.J.ら、「Sulfonamide derivatives and their use」、第WO95/24186号;Baldwin,J.J.ら、「Combinatorial dehydrobenzopyran library」、第WO95/30642号;Brenner,S.、「New kit for preparing combinatorial libraries」、第WO95/16918号;Chenera,B.ら、「Preparation of library of resin−bound aromatic carbocyclic compounds」、第WO95/16712号;Ellman,J.A.、「Solid phase and combinatorial synthesis of benzodiazepine compounds on a solid support」、米国特許第5,288,514号;Felder,E.ら、「Novel combinatorial compound libraries」、第WO95/16209号:Lerner,R.ら、「Encoded combinatorial chemical libraries」、第WO93/20242号;Pavia,M.R.ら、「A method for preparing and selecting,pharmaceutically useful non−peptide compounds from a structurally diverse universal library」、第WO95/04277号;Summerton,J.E.およびD.D.Weller、「Morpholino−subunit combinatorial library and method」、米国特許第5,506,337号;Holmes,C.、「Methods for the Solid Phase Synthesis of Tiazolidinones、Metathiazanones,and Derivatives thereof」、第WO96/00148号;Phillips,G.B.およびG.P.Wei、「Solid−phase Synthesis of Benzimidazoles」、Tet.Letters 37:4887−90、1996;Ruhland,B.ら、「Solid−supported Combinatorial Synthesis of
Structurally Diverse β−Lactams」J.Amer.Chem.Soc.111:253−4、1996;Look,G.C.ら、「The Indentification of Cyclooxygenase−1 Inhibitors from 4−Tiazolidinone Combinatorial Libraries」、Bioorg and Med.Chem.Letters 6:707−12、1996。
【0073】
(2.タンパク質およびペプチド)
広範なタンパク質およびペプチドは、同様に、免疫系を調節するための候補分子として利用される。
【0074】
(a.コンビナトリアルペプチドライブラリー)
免疫系を調節するペプチド分子は、コンビナトリアルペプチドライブラリーのスクリーニングを通じて得られ得る。このようなライブラリーは、当業者によって調製され得る(例えば、米国特許第4,528,266号、および同第4,359,535号、ならびにPCT公開番号第WO92/15679号、同第WO92/15677号、同第WO90/07862号、同第WO90/02809号を参照のこと)か、または市販の供給源(例えば、New England Biolabs Ph.D.TM Pharge Display Peptide Library Kit)から購入され得るかのいずれかであり得る。
【0075】
(b.抗体)
免疫系を調節する抗体は、本明細書中に提供されている開示を考慮すれば、容易に調製され得る。本発明の状況においては、抗体は、モノクローナル抗体、ポリクローナル抗体、抗イディオタイプ抗体、抗体フラグメント(例えば、FabおよびF(ab’)2、Fv可変領域、または相補性決定領域)を含むことが理解される。上記で議論されているように、抗体は、それらが、107Mよりも大きいかまたはそれと同等である、好ましくは、108Mよりも大きいかまたはそれと同等であるKaを有して結合する場合は、Fkhsfに対して特異的であると理解される。モノクローナル抗体または結合パートナーの親和性、ならびに結合の阻害は、当業者によって容易に決定され得る(Scatchard,Ann.N.Y.Acad.Sci.51:660−672,1949を参照のこと)。
【0076】
簡潔には、ポリクローナル抗体は、ウマ、ウシ、種々の家禽、ウサギ、マウス、またはラットのような種々の温血動物から、当業者によって容易に生成され得る。代表的には、Fkhsfまたは13−20アミノ酸のその特有のペプチド(好ましくは、グルタルアルデヒドでの架橋によってキーホルリンペットヘモシアニンに対して結合体化される)が、フロイト完全アジュバントまたはフロイント不完全アジュバントのようなアジュバントと組み合わせて、腹腔内、筋肉内、眼内、または皮下注射を通じて動物を免疫するために利用される。数回の追加免疫後、血清のサンプルが回収され、そしてタンパク質またはペプチドに対する反応性について試験される。特に好ましいポリクローナル抗血清は、少なくともバックグラウンドよりも3倍大きい、これらのアッセイの1つに対するシグナルを生じる。一旦、タンパク質に対するその反応性に関して、動物の力価がプラトーに到達すると、より大量の抗血清が、毎週の採血によるか、または動物を瀉血するかのいずれかによって、容易に得られ得る。
【0077】
モノクローナル抗体はまた、従来技術を使用して容易に生成され得る(米国特許第RE32,011号、同第4,902,614号、同第4,543,439号、および同第4,411,993号(これらは、本明細書中で参考として援用されている)を参照のこと;Monoclonal Antibodies,Hybridomas:A New Dimension in Biological Analyses,Plenum Press,Kennett、McKearnおよびBechtol(編)、1980、およびAntibodies:A Laboratory Manual,HarlowおよびLane(編)、Cold Spring Harbor Laboratory Press,1988(これもまた、本明細書中で参考として援用されている)もまた参照のこと)。
【0078】
他の技術もまた、モノクローナル抗体を構築するために利用され得る(William D.Huseら、「Generation of a Large Combinatorial Library of the Immunoglobulin Repertoire in Phage Lambda」、Science、246:1275−1281、1989年12月を参照のこと;L.Sastryら、「Cloning of the Immunological Repertoire in Eschrichia coli for Generation of Monoclonal Catalytic Antibodies:Construction of a Heavy Chain Variable Region−Specific cDNA Library」、Proc.Natl.Acad.Sci.USA、86:5728−5732、1989年8月もまた参照のこと;Michelle Alting−Meesら、「Monoclonal Antibody Expression Libraries:A Rapid Alternative to Hybridomas」、Strategies in Molecular Biology 3:1−9、1990年1月もまた参照のこと)。
【0079】
以下を含む広範な種々のアッセイが、Fkhsf(または本明細書中に記載されているFkhsfの変異形態)に対して反応性である抗体の存在を決定するために利用され得る:例えば、向流免疫電気泳動、放射免疫アッセイ、放射免疫沈降、酵素結合免疫吸着アッセイ(ELISA)、ドットブロットアッセイ、ウェスタンブロット、免疫沈降、阻害または競合アッセイ、およびサンドイッチアッセイ(米国特許第4,376,110号および同第4,486,530号を参照のこと;Antibodies:A Laboratory Manual、HarlowおよびLane(編)、Cold Spring Harbor Laboratory Press、1988もまた参照のこと)。
【0080】
一旦、適切な抗体が得られると、それらは、当業者に周知の多くの技術によって単離または精製され得る(Antibodies:A Laboratory Manual、HarlowおよびLane(編)、Cold Spring Harbor Laboratory Press、1988を参照のこと)。適切な技術としては、ペプチドまたはタンパク質アフィニティーカラム、HPLCまたはRP−HPLC、プロテインAもしくはプロテインGカラム上での精製、または任意のこれらの技術の組み合わせが挙げられる。
【0081】
本発明の抗体は、免疫系を調節するためだけではなく、診断試験のため(例えば、FKHsfまたはFkhsfのタンパク質またはペプチドの存在を決定するため)、治療目的のため、またはタンパク質の精製のためにも利用され得る。
【0082】
(c.変異体Fkhsf)
本明細書中および以下の実施例において記載されているように、Fkhsfの改変されたバージョンが、Fkhsfの正常な活性を阻害し、それによって免疫系を調節するために利用され得る(一般的には、上記の核酸分子およびタンパク質を参照のこと)。
【0083】
FKHsfまたはFkhsfのさらなる変異体または改変された形態は、広範な種々のインビトロでのアッセイのため(例えば、種々のモデルにおいてこのようなタンパク質の影響を試験するため)、または抗体の開発のために、利用され得る。
【0084】
(3.核酸分子)
本発明の他の局面においては、免疫系を調節し得る核酸分子が提供される。例えば、1つの実施態様においては、FKHsfもしくはFkhsfの核酸配列、または変異体FKHsfもしくはFkhsfの発現を特異的に阻害するアンチセンスオリゴヌクレオチド分子が、提供される(一般的には、Hirashimaら、Molecular Biology of RNA:New Perspectives(M.InouyeおよびB.S.Dudock.編、1987 Academic Press,San Diego、401頁);Oligonucleotides:Antisense Inhibitors of Gene Expression(J.S.Cohen編、1989、MacMillan Press、London);SteinおよびCheng、Science、261:1004−1012、1993;第WO95/10607号;米国特許第5,359,051号;同第WO92/06693号;およびEP−A2−612844号を参照のこと)。簡潔には、このような分子は、それらが、転写されるFkhsfのmRNA配列の領域と相補的であり、そしてワトソン−クリック塩基対を形成し得るように、構築される。得られた二本鎖の核酸は、mRNAの続くプロセシングを妨害し、それによってタンパク質の合成を妨げる。
【0085】
本発明の他の局面においては、FKHsfもしくはFkhsf、またはFKHsfもしくはFkhsfの変異形態を阻害し得るリボザイムが提供される。本明細書中で使用される場合、「リボザイム」は、特異的認識のためのアンチセンス配列およびRNA−切断酵素活性を含むRNA分子を含むことが意図される。触媒的な鎖は、化学量論的な濃度を超える濃度で、標的RNAの特異的な部位を切断する。広範な種々のリボザイムが、本発明の状況において利用され得る。これは例えば、ハンマーヘッドリボザイム(例えば、ForsterおよびSymons、Cell 48:211−220、1987;HaseloffおよびGerlach、Nature、328:596−600、1988;WalbotおよびBruening、Nature、334:196、1988、HaseloffおよびGerlach、Nature、334:585、1988によって記載されている);ヘアピンリボザイム(例えば、Haseloffら、1993年10月19日に発行された米国特許第5,254,678号、Hempelら、1990年3月26日に公開された、欧州特許公開番号第0 360 257号によって記載されている);およびTetrahymenaのリボソームRNAに基づくリボザイム(Cechら、米国特許第4,987,071号を参照のこと)を含む。本発明のリボザイムは、代表的には、RNAから構成されるが、DNA、核酸アナログ(例えば、ホスホロチオエート)、またはそれらのキメラ(例えば、DNA/RNA/RNA)からも構成され得る。
【0086】
(4.標識)
FKHsfもしくはFkhsf(ならびにそれらの変異形態)、または上記または下記の任意の候補分子が、以下を含む種々の化合物で標識され得る:例えば、蛍光分子、毒素、および放射性核種。蛍光分子の代表的な例としては、フルオレセイン、Phycobiliタンパク質(例えば、フィコエリトリン)、ローダミン、テキサスレッド、およびルシフェラーゼが挙げられる。毒素の代表的な例としては、リシン、アブリン、ジフテリア毒素、コレラ毒素、ゲロニン、アメリカヤマゴボウ抗ウイルスタンパク質、トリチン、Shigella毒素、およびPseudomonas外毒素Aが挙げられる。放射性核種の代表的な例としては、Cu−64、Ga−67、Ga−68、Zr−89、Ru−97、Tc−99m、Rh−105、Pd−109、In−111、I−123、I−125、I−131、Re−186、Re−188,Au−198、Au−199,Pb−203、At−211、Pb−212、およびBi−212が挙げられる。さらに、上記の抗体もまた、標識され得るか、またはリガンド結合対の1つのパートナーに対して結合体化され得る。代表的な例としては、アビジン−ビオチン、およびリボフラビン−リボフラビン結合タンパク質が挙げられる。
【0087】
上記の代表的な標識と本明細書中に記載されている分子とを結合体化または標識するための方法は、当業者によって容易に達成され得る(Trichothecene Antibody Conjugate、米国特許第4,744,981号;Antibody conjugate、米国特許第5,106,951号;Florogenic Materials and Labeling Techniques、米国特許第4,018,884号;Metal Radionuclide Labeled Proteins for Diagnosis and Therapy、米国特許第4,897,255号;およびMetal Radionuclide Chelating Compounds for Improved Chelation Kinetics、米国特許第4,988,496号を参照のこと;Inman.Methods In Enzymology、第34巻、Affinity Techniques、Enzyme Purification:Part B、JakobyおよびWilchek(編)、Academic Press,New York、30頁、1974をもまた参照のこと;WilchekおよびBayer、「The Avidin−Biotin Complex in Bioanalytical Applications」Anal.Biochem.171:1−32、1988をもまた参照のこと)。
【0088】
(薬学的組成物)
上記のように、本発明はまた、薬学的または生理学的に受容可能なキャリア、賦形剤、または希釈剤とともに、免疫系を調節する上記の分子の1つを含む、種々の薬学的組成物を提供する。一般的には、このようなキャリアは、使用される投与量および濃度では、レシピエントに対しては非毒性である。通常は、このような組成物の調製は、緩衝液、抗酸化剤(例えば、アスコルビン酸、低分子量(約10残基未満)のポリペプチド、タンパク質、アミノ酸、炭水化物(グルコース、スクロース、またはデキストリンを含む))、キレート化剤(例えば、EDTA、グルタチオンなど)、安定剤、および賦形剤と、治療薬との混合を含む。中性の緩衝化生理食塩水または非特異的な血清アルブミンと混合した生理食塩水は、例示的な適切な希釈剤である。好ましくは、薬学的組成物(または「医薬品」)は、滅菌の、発熱物質を含まない形態で提供される。
【0089】
さらに、本発明の薬学的組成物は、種々の異なる経路による投与のために調製され得る。さらに、本発明の薬学的組成物は、このような薬学的組成物の使用に関する説明書を提供するパッケージング材料とともに、容器内に配置され得る。一般的には、このような説明者は、試薬の濃度、ならびに特定の実施態様においては、薬学的組成物の再構成のために必要とされ得る賦形剤成分または希釈剤(例えば、水、生理食塩水、もしくはPBS)の相対的な量を記載する具体的な表現を含む。
【0090】
(処置方法)
本発明はまた、免疫系を調節するための方法を提供する。免疫系を調節する、本明細書中に記載されている分子の使用を通じて、温血動物中の広範な種々の状態が、容易に処置され得るかまたは予防され得る。処置され得る温血動物の例としては、脊椎動物および哺乳動物(例えば、ヒト、ウマ、ウシ、ブタ、ヒツジ、イヌ、ネコ、ラット、およびマウスを含む)の両方が挙げられる。このような方法は、変更された免疫系を有する患者において治療的な価値があり得る。これは、種々の免疫不全症候群、ならびにT細胞媒介自己免疫疾患を有する患者の、化学療法を受けている患者を含む。治療的な価値はまた、ワクチンアジュバントとしての有用性によって認識され得る。
【0091】
治療用分子は、分子の型に依存して、種々の処方物において種々の経路を介して投与され得る。例えば、1つの実施態様においては、有機分子が、経口もしくは鼻内経路によって送達され得るか、または注射(例えば、筋肉内、静脈内など)によって送達され得る。
【0092】
1つの局面においては、免疫系を調節するための方法が提供される。この方法は、免疫系を調節する分子の発現を指向するベクターを、リンパ系細胞中に導入する工程、および温血動物にベクターを含有する細胞を投与する工程を包含する。他の関連する実施態様においては、このベクターは、所望される標的の位置(例えば、骨髄)に直接投与され得る。
【0093】
ウイルスベクターおよび非ウイルスベクターの両方を含む、広範な種々のベクターが、このような治療目的のために利用され得る。適切なウイルスベクターの代表的な例としては、ヘルペスウイルスベクター(例えば、米国特許第5,288,641号)、アデノウイルスベクター(例えば、第WO94/26914号、第WO93/9191号、第WO99/20778号、第WO99/20773号;第WO99/20779号;Kollsら、PNAS 91(1):215−219、1994;Kass−Eislerら、PNAS 90(24):11498−502、1993;Guzmanら、Circulation 88(6):2838−48、1993;Guzmanら、Cir.Res.73(6):1202−1207、1993;Zabnerら、Cell、75(2):207−216、1993;Liら、Hum.Gene Ther.4(4):403−409、1993:Caillaudら、Eur.J.Neurosci.5(10):1287−1291、1993;Vincentら、Nat.Genet.5(2):130−134、1993;Jaffeら、Nat.Genet.1(5):372−378、1992;およびLevreroら、Gene、101(2):195−202、1991)、アデノ随伴ウイルスベクター(第WO95/13365号;Flotteら、PNAS 90(22):10613−10617、1993)、バキュロウイルスベクター、パルボウイルスベクター(Koeringら、Hum.Gene Therap.5:457−463、1994)、ポックスウイルスベクター(PanicaliおよびPaoletti、PNAS、79:4927−4931,1982;およびOzakiら、Biochem.Biophys.Res.Comm.193(2):653−660、1993)、およびレトロウイルス(例えば、第EP0,415,731号;第WO90/07936号;第WO91/0285号、第WO94/03622号;第WO93/25698号;第WO93/25234号;米国特許第5,219,740号;第WO93/11230号;第WO93/10218号)が挙げられる。同様に、種々のウイルスまたは非ウイルス供給源に由来する種々のエレメント(例えば、プロモーター、エンベロープ配列など)の混合物を含むウイルスベクターが、構築され得る。種々の実施態様においては、ウイルスベクター自体、またはウイルスベクターを含むウイルス粒子のいずれかが、以下に記載されている方法および組成物において利用され得る。
【0094】
本発明の他の実施態様においては、免疫系を調節する分子(例えば、変異体Fkhsf、またはアンチセンスもしくはFkhsfを切断するリボザイム分子)をコードする核酸分子が、以下を含む種々の別の技術によって投与され得る:例えば、ポリ−L−リジンDNA複合体と結合体化したアシアロオソムコイド(ASOR)の投与(Cristanoら、PNAS 92122−92126,1993)、殺傷したアデノウイルスに連結されたDNA(Curielら、Hum.Gene Ther.3(2):147−154、1992)、サイトフェクチンによって媒介される導入(DMRIE−DOPE、Vical、California)、直接的なDNA注入(Acsadiら、Nature、352:815−818、1991);DNAリガンド(Wuら、J.of Biol.Chem.264;16985−16987、1989);リポフェクション(Felgnerら、Proc.Natl.Acad.Sci.USA 84:7413−7417、1989);リポソーム(Pickeringら、Circ、89(1):13−21、1994;およびWangら、PNAS、84:7851−7855、1987);マイクロプロジェクタイルボンバードメント(Williamsら、PNAS、88:2726−2730、1991);およびタンパク質自体をコードする核酸分子の、単独(VileおよびHart、Cancer Res.53:3860−3864、1993)またはPEG−核酸複合体を利用するかのいずれかである直接的な送達。
【0095】
本発明のベクターによって発現され得る分子の代表的な例としては、リボザイムおよびアンチセンス分子(これらのそれぞれは、以下により詳細に議論されている)が挙げられる。
【0096】
当業者に明らかであるように、投与の量および頻度は、もちろん、処置される徴候の性質および重篤度、所望される応答、患者の状態のような因子に依存する。代表的には、組成物は、上記のような種々の技術によって投与され得る。
【0097】
以下の実施例は、例示によって付与され,そして限定によっては与えられない。
【0098】
(実施例)
(実施例1)
(SCURFY変異体の原因である遺伝子の同定)
(A.SCURFY遺伝子のクローニング)
もともとのscurfy変異は、Oak Ridge National Laboratory(ORNL)に1949年にストックされた一部近交系のMRにおいて自発的に生じた。戻し交配分析を、マウスScurfy変異を含むX染色体の動原体周辺領域(peri−centromeric region)を詳細にマップするために使用した。同じ領域をカバーする物理的なマップを、重複している酵母および細菌の人工染色体(YACおよびBAC)の単離を通じて並行して生成した。一旦、候補領域が、約500キロ塩基対(kb)にまで狭められると、大規模なDNA配列決定を、4個の重複するBACクローンについて行った。この500kbの領域内の全ての転写ユニットを、配列データベースの検索とコンピューターによるエキソン予想プログラムの適用との組合せにより同定した。次いで、候補遺伝子を、正常なおよびScurfyに由来するRNAサンプルからの逆転写ポリメラーゼ連鎖反応(RT−PCR)手順によって得たcDNAの配列を比較することによって、Scurfy特異的変異についてスクリーニングした。Fkhsfと本明細書中で呼ばれる1つの遺伝子においては、2塩基対(bp)の挿入が、正常なcDNAと比較してScurfy cDNAのコード領域中に見出された。挿入を、いくつかのマウスの株(Scurfy変異体を含む)のゲノムDNAに由来するPCR産物のDNA配列を比較することによって確認した。再び、2bpの挿入を、Scurfyサンプル中でのみ見出し、これによって、これをScurfy欠失の原因である可能性があると確定した。
【0099】
マウスのFkhsf遺伝子は、BACクローン8C22中に含まれ、そして完全に配列決定されている。これは約14kbにまたがり、そして11個のコードエキソンを含む。エキソン中断の位置は、GenScanエキソン推定プログラムを使用して、ゲノムDNA配列のコンピューターによる分析によって最初に同定した;次いで、エキソンの位置を、ゲノム配列に対する正常なマウスの組織に由来するcDNA配列の直接的な比較によって確認した。
【0100】
得られたcDNAの長さは、2160bpである;コード領域は、その1287bpにまたがり、429アミノ酸のタンパク質をコードする。図1はマウスのFkhsf cDNAのヌクレオチド配列を示す;翻訳は、259位で開始し、そして1546位で終結すると推定される。図2は、マウスのFkhsfのアミノ酸配列を示す。
【0101】
(b.Fkhsfトランスジェニックマウスの生成)
Scurfy表現型の真の原因としてのFkhsf遺伝子の実態を、トランスジェニックマウスにおいて確認した。簡潔には、Fkhsf遺伝子の約7kbのコード領域、ならびに隣接する配列の上流の約20kb、および下流の配列の約4kb(図5)を含む正常なゲノムDNAの30kbのフラグメントを、正常なマウスの1細胞胚中にマイクロインジェクションした。それぞれが別々のインテグリンを有する5匹の個々の崩壊させた動物を生成し、そして各トランスジェニック株に由来する雄性の動物を、雌性のsfキャリアと交配した。トランスジーン(正常なFkhsf)およびsf変異(変異Fkhsf)の両方を有する雄性の子孫を、分析した。
【0102】
分析は、倭小(runting)、鱗状の皮膚、毛皮の異常、およびscurfy表現型の他の特徴についての動物の試験からなる。さらに、リンパ系組織(胸腺、脾臓、および結節)を回収し、そしてそれらの大きさおよび細胞数を試験し、そして正常な動物およびscurfyマウスの両方について比較した。全ての5個のトランスジェニック株について、トランスジーンを保有した雄性のsf子孫は、大きさおよび重量において正常であり、そして全ての点で健常であるように見えた。これらのトランスジェニックマウスにおけるリンパ節の大きさは、正常な動物にものと類似(またはそれよりも小さい)であり(図6)、そして活性化されたT細胞の徴候はなかった。これらのパラメーターは、sfマウスとは極端に異なり、そして正常なFkhsf遺伝子の添加が、scurfyマウス中に見出される欠損を克服し得ることを示す。従って、これによって、Fkhsf遺伝子中の変異がScurfy疾患の原因であることを確認する。
【0103】
(実施例2)
(FKHsf cDNAの生成)
完全なマウスのFkhsfタンパク質をコードする相補的DNA(cDNA)は、逆転写酵素ポリメラーゼ連鎖反応(RT−PCR)手順によって得られ得る。より詳細には、第1鎖のcDNAが、適切な供給源(例えば、マウスの脾臓)に由来する5μgの全RNAを、オリゴdTプライミングすること、および標準的な条件下で逆転写酵素を用いて伸張すること(例えば、Gibco/BRL SuperScriptキット)によって、生成する。次いで、第1鎖のcDNAのアリコートを、正方向および逆方向プライマー(正方向プライマー:GCAGATCTCC TGACTCTGCC TTC;逆方向プライマー:GCAGATCTGACAAGCTGTGT CTG)(0.2mMの最終濃度))、60mMのTris−HCl、15mMの硫酸アンモニウム、1.5mMの塩化マグネシウム、0.2mMの各dNTP、および1ユニットのTaqポリメラーゼの存在下での、35サイクルのPCR(94℃にて30秒間、60℃にて30秒間、72℃にて2分間)に供する。
【0104】
(実施例3)
(マウスのFkhsfに対するヒトのオルソログの生成)
完全なFKHsfタンパク質をコードするヒトFKHsf cDNAは、実施例2に記載されているものと本質的に同じ手順によって得ることができる。詳細には、全脾臓RNAを用いて開始し、そして以下のオリゴヌクレオチドプライマー(正方向プライマー:AGCCTGCCCT TGGACAAGGA C;逆方向プライマー:GCAAGACAGT GGAAACCTCA C)、および60℃のアニーリング温度を除いて上記と同じPCR条件を利用する。
【0105】
図4は、今日までに得られた1869bpのcDNAのヌクレオチド配列(1293bpのコード領域を含む)を示す;翻訳は、189位で開始し、そして1482位で終結すると推定される。図4は、431アミノ酸のヒトFKHsfタンパク質の配列を示す。マウスcDNA配列に対するヒト遺伝子の推定されるコード領域の比較は、ほぼ同一であるエキソン構造、および全タンパク質にわたる86.1%のアミノ酸配列同一性を明らかにする。
【0106】
(実施例4)
(SCURFY変異体を検出するための方法)
上記のように、Scurfy変異体を、最初に、sfのRT−PCRによって誘導されたcDNAおよび正常なマウスのRNAサンプルを直接配列決定することによって発見し、そしてゲノムDNAに由来する同じ領域を配列決定することによって確認した。変異の性質(すなわち、2bpの挿入)は、それ自体を、多数の種々の変異検出アッセイに導く。これは、最初に、オリゴヌクレオチドプローブのディファレンシャルハイブリダイゼーションに基づく。このような、ハイブリダイゼーションに基づくアッセイは、対立遺伝子特異的発現の定量的な分析を可能にし得る。
【0107】
一例としては、360bpのDNAフラグメントを、以下のオリゴを使用して、第1鎖のcDNAから増幅する:
DMO5985(正方向):CTACCCACTGCTGGCAAATG(図1のntd.825−844)
DMO6724(逆方向):GAAGGAACTATTGCCATGGCTTC(ntd 1221−1199)。
【0108】
PCR産物を、1.8%のアガロースゲル上で泳動し、ナイロン膜に移し、そしてScurfy特異的な2bpの挿入部位に対応する領域に相補的である末端標識オリゴヌクレオチドで標識する。2つの別々のハイブリダイゼーション反応を、正常なおよびScurfy PCR産物を検出するために、以下のオリゴヌクレオチド(2bpの挿入部位を太字で示す)を使用して行う:
正常:ATGCAGCAAGAGCTCTTGTCCATTGAGG
DMO7439
Scurfy:GCAGCAAGAGCTCTTTTGTCCATTGAGG
DMO6919。
【0109】
Scurfy変異はまた、冷却一本鎖立体構造多形性(cSSCP)アッセイによっても検出され得る。このアッセイにおいては、上記と同じPCR産物を、20%のアクリルアミド(TBE)ゲル上で、鎖の変性後に泳動する。Scurfyの挿入は、正常な配列と比較して鎖の移動においてシフトを生じ、そして別の鎖が、エチジウムブロマイドでの染色後に検出される。
【0110】
(実施例5)
(FKHSF遺伝子の発現)
半定量的なRT−PCRが、広範な種々の組織および細胞株中におけるマウスおよびヒトのFkhsf遺伝子の発現パターンを分析するために使用されている。発現のレベルは、偏在して発現されるDAD−1遺伝子について正規化される。簡潔には、Fkhsf遺伝子は、非常に低いレベルではあるが、これまでに試験されたほぼ全ての組織(胸腺、脾臓、分類されたCD4+およびCD4−CD8−、Tリンパ球、ならびに腎臓、脳、ならびに種々のマウスおよびヒトのT細胞株、ならびにヒト腫瘍を含む)中で発現される。しかし、発現がないことが、新たに分類したマウスのB細胞において注目された。
【0111】
予想されるように、RT−PCRアッセイにおける正常な組織対Scurfy組織において、発現レベルにおける差異は観察されなかった。
【0112】
(実施例6)
(インビトロでのFKHSFの発現)
全長のマウスおよびヒトのFkhsf cDNA、ならびに種々のcDNAのサブ領域を、哺乳動物細胞(たとえば、ヒトのJurkat T細胞株)、E.coli、または酵母中での発現が可能であるベクター中にクローン化した。E.coli系または酵母系を、Fkhsf特異的抗体を惹起する目的のためのタンパク質の産生に使用し得る(以下を参照のこと)。
【0113】
(実施例7)
(抗FKHSF抗体の生成)
実施例6に記載されているベクターから発現されるタンパク質を、FKHsf特異的抗体の産生のために適切な動物を免疫化するために使用する。全長または短縮されたタンパク質のいずれかを、この目的のために使用し得る。タンパク質を、例えば、E.coliのような細菌、昆虫細胞、または哺乳動物細胞から、入手し得る。動物種としては、マウス、ウサギ、モルモット、ニワトリ、または他のものが挙げられ得る。FKHsfに対して特異的なウサギ抗血清は、生化学的な特徴によって決定されるように、生成されている(免疫沈降およびウェスタンブロッティング)。
【0114】
(実施例8)
(FKHSF遺伝子の機能についてのアッセイ)
FKHsfタンパク質の機能の欠損が、scurfy動物において観察される表現型(るいそう、敏感な免疫応答、および死)を生じるので、FKHsfタンパク質の過剰な発現を評価するためのアッセイを記載する。トランスジェニック動物(実施例1に記載されている)を、それらの免疫応答能力の状態について、いくつかの異なるパラメーターを使用して試験する。動物を、リンパ節および胸腺中に存在するリンパ系細胞の数について(図7)、およびインビトロでの刺激に対するT細胞の応答性について(図8)試験する。
【0115】
Scurfy変異動物は、正常な動物のそれらのリンパ節中の細胞よりもほぼ2倍多い細胞を有する。しかし、正常なFKHsfタンパク質の過剰なレベルを発現するマウスは、ほぼ3倍多い細胞を含む(図7)。さらに、標準的な抗血清を使用するフローサイトメトリーによって評価される場合に、それらの細胞表面の表現型である場合(示さない)は、胸腺細胞の数は正常であり(図7)、このことは、過剰のFKHsfタンパク質に関連する発達的欠損が存在しないことを示す。
【0116】
正常なscurfy動物およびトランスジェニック動物はさらに、T細胞の刺激に対するそれらの増殖性応答について試験される。CD4+T細胞はCD3およびCD28に対する抗体と反応し、そしてそれらの増殖性応答が、放射活性チミジンの取りこみを使用して測定される。scurfy細胞は、刺激の非存在下でのみ分裂するが、正常な細胞は、刺激の後に十分応答する。FKHsfトランスジェニック細胞もまた刺激に対して応答するが、応答は、正常な細胞のものよりも有意に少ない(図8)。このことは、過剰のFKHsfを発現するCD4+T細胞が、刺激に対して応答する能力を低下させられたことを示す。
【0117】
(実施例9)
(JM2に関連するヒトのFKHSF cDNA配列)
ヒトFKHsf cDNA配列の改変されたバージョンが、GenBankによって公開されている配列データベース中に存在する。この配列は、JM2と呼ばれ(GenBank登録番号第AJ005891号)、そしてFKHsf遺伝子を含むゲノム配列に対するエキソン推定プログラムの適用の結果である(Strom,T.M.ら、未公開−GenBank登録番号第AJ005891を参照のこと)。対称的に、FKHsf cDNAの構造は、実験によって決定された。Genetics Computer Group(GCG;Madison,USA)、Wisconsin配列分析パッケージのGAPプログラムを、2つの配列を比較するために使用し、そして差異を図9に示す。2つの配列の5’末端は、ゲノムDNA配列の状況におけるそれらの位置において異なり、FKHsfの第2のコードエキソンがJM2から欠落しており、そしてFKHsf遺伝子の最後のイントロンが、JM2配列中ではスプライシングされない。これらの差異は、FKHsfと比較して、より短いアミノ末端ドメイン、カルボキシ末端でのフォークヘッドドメイン(以下を参照のこと)中への大きい挿入を有するJM2タンパク質を生じる。
【0118】
(実施例10)
(FKHsfタンパク質は種を超えて保存されている)
FKHsfタンパク質は、機能的ドメインを示し得る配列モチーフに基づいて、サブ領域に分けられ得る。FKHsf中の2つの重要なモチーフは、タンパク質の中央部分中のC2
II2クラスの単一のジンクフィンガー(ZNF)、およびタンパク質の最もカルボキシ
末端での二股の矢じりまたは翼状のへリックスドメインである。FKHsfと他のタンパク質との間での相同性の程度を特徴付ける目的のために、本発明者らは、タンパク質を以下の4つの領域に分けた:
アミノ末端ドメイン :図2の残基1−197
図4の残基1−198
ジンクフィンガードメイン :図2の残基198−221
図4の残基199−222
中央ドメイン :図2の残基222−336
図4の残基223−336
フォークヘッドドメイン :図2の残基337−429
図4の残基337−431。
【0119】
DNAStar配列分析パッケージによるMultiple Sequence Alignmentプログラムを使用して、Lipman−Pearsonアルゴリズムを、これらの4つのドメインにわたる、ヒトFKHsfとマウスFkhsfタンパク質との間での類似性の程度を決定するために使用した。結果を、図10に示す。類似性は、82.8%から96.4%までの範囲を示し、このことは、このタンパク質が非常に高度に種をこえて保存されていることを示す。
【0120】
(実施例11)
(新規のFKHSFに関連する遺伝子の同定)
FKHsf遺伝子配列の特有の特徴を利用して、二股の矢じりを含む分子の同じサブクラスに入る他の新規の遺伝子(およびタンパク質)を同定し得る。FKHsfタンパク質は、それが、フォークヘッドドメインに対して単一のジンクフィンガードメインのアミノ末端を、そしてフォークヘッドドメインの最もカルボキシ末端に有する点で、特有である。縮重PCRアプローチを、フォークヘッドドメインの上流にジンクフィンガー配列を含む新規の遺伝子を単離するために行い得る。例えば、以下の縮重プライマーを合成した(縮重の位置を括弧で示し、そして「I」はヌクレオシドイノシンを示す):
正方向プライマー:CA(TC)GGIGA(GA)TG(CT)AA(GA)TGG 逆方向プライマー:(GA)AACCA(GA)TT(AG)TA(AGT)AT(CT)TC(GA)TT。
【0121】
正方向プライマーは、ジンクフィンガー配列中の領域に対応し、そして逆方向プライマーは、フォークヘッドドメインの中央の領域に対応する。これらのプライマーを使用して、種々のヒト組織(肝臓、脾臓、脳、肺、腎臓などを含む)から実施例2のように産生した第1鎖のcDNAを増幅した。以下のPCR条件:0.2mMの最終濃度の正方向プライマーおよび逆方向プライマー、60mMのTris−HCl、15mMの硫酸アンモニウム、1.5mMの塩化マグネシウム、0.2mMの各dNTP、および1ユニットのTaqポリメラーゼ、を使用し35サイクル(94℃にて30秒間、50℃にて30秒間、72℃にて2分間)に供した。PCR産物を、1.8%のアガロースゲル(1×TAE中で泳動)上で可視化し、そしてTAクローニングベクター(Invitrogen、Carlsbad、CA)中にサブクローン化した;個々のクローンを配列決定し、そして全長cDNAのさらなる特徴付けのために使用した。
【0122】
あるいは、FKHsf遺伝子の特有の領域(すなわち、「アミノ末端」ドメインおよび「中央」ドメイン)を、ハイブリダイゼーションによるcDNAライブラリーのスクリーニングのために使用し得る。種々のヒトおよび/またはマウスの組織に由来し、そしてλファージベクター(例えば、λgt11)中で増殖させたcDNAライブラリーをアガロース上にプレートし、プラークをナイロン膜に移し、そしてFKHsf遺伝子の特有の領域に由来するフラグメントを用いてプローブした。高ストリンジェンシーの条件(例えば、65℃にて、5×SSPE、5×Denhardt溶液、0.5%のSDS中でのハイブリダイゼーション、65℃にて、0.1×SSPE、0.1%のSDS中での洗浄)下では、密接に関連するごくわずかな配列のみが、ハイブリダイズすると予想した(すなわち、90−100%の相同性)。上記と同じ緩衝液中での45−55℃でのこのようなハイブリダイゼーションおよび洗浄のような低いストリンジェンシーの下では、FKHsfに関連する遺伝子(65−90%の相同性)が同定され得る。FKHsfの特有の配列を用いて公開されているデータベースの検索によって得られる結果に基づくと、低いストリンジェンシーから中程度のストリンジェンシーまでのハイブダイゼーション実験を通じて同定される任意の遺伝子が、「FKHsfファミリー」の新規のメンバーを提示すると予想される。
【0123】
上記から、本発明の特定の実施態様が、例示の目的のために本明細書中で記載されているが、種々の改変が本発明の精神および範囲を逸脱することなく行われ得ることが、理解される。従って、本発明は、添付の特許請求の範囲による場合を除いて、限定されない。
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1A】



【図1B】

【図2】

【図3A】

【図3B】

【図4】

【図5】
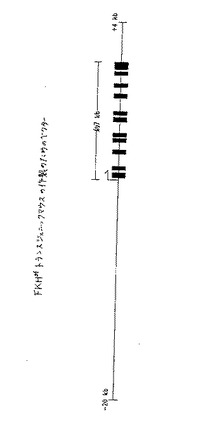
【図6】

【図7】

【図8】
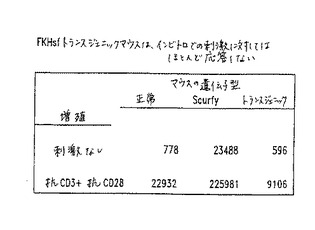
【図9】
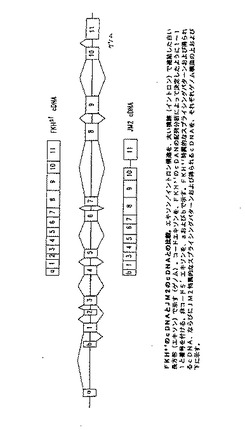
【図10】

【図1B】
【図2】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2012−191949(P2012−191949A)
【公開日】平成24年10月11日(2012.10.11)
【国際特許分類】
【出願番号】特願2012−150555(P2012−150555)
【出願日】平成24年7月4日(2012.7.4)
【分割の表示】特願2009−251526(P2009−251526)の分割
【原出願日】平成11年8月11日(1999.8.11)
【出願人】(398057477)ダーウィン・ディスカバリー・リミテッド (4)
【氏名又は名称原語表記】DARWIN DISCOVERY LIMITED
【Fターム(参考)】
【公開日】平成24年10月11日(2012.10.11)
【国際特許分類】
【出願日】平成24年7月4日(2012.7.4)
【分割の表示】特願2009−251526(P2009−251526)の分割
【原出願日】平成11年8月11日(1999.8.11)
【出願人】(398057477)ダーウィン・ディスカバリー・リミテッド (4)
【氏名又は名称原語表記】DARWIN DISCOVERY LIMITED
【Fターム(参考)】
[ Back to top ]