
微生物宿主を迅速にスクリーニングして、異種タンパク質発現の収率および/または質が改善されている特定の株を同定する方法

本発明は、改善された収率および/または質で異種タンパク質を産生できる宿主細胞集団を迅速に同定するためのアレイを提供する。このアレイは、タンパク質産生に関与する1つもしくは複数の標的遺伝子の発現が増大するように、タンパク質分解に関与する1つもしくは複数の標的遺伝子の発現が低下するように、またはそれらの両方となるように遺伝子改変されている1つまたは複数の宿主細胞集団を含む。このアレイ内の1つまたは複数の株は、対象とする異種タンパク質をペリプラズム区画に発現することもでき、または外側細胞壁を通して細胞外に異種タンパク質を分泌することもできる。株アレイは、治療用タンパク質、ホルモン、成長因子、細胞外受容体またはリガンド、プロテアーゼ、キナーゼ、血液タンパク質、ケモカイン、サイトカインおよび抗体などを含めた、対象とするいかなるタンパク質の発現改善のスクリーニングにも有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、タンパク質生産(production)の分野に属し、詳細には、適切にプロセシングされた異種タンパク質の大規模生産に最適な宿主細胞の同定に関する。
【背景技術】
【0002】
バイオテクノロジー薬物およびワクチンとしての使用について、150種を超える、組換えにより産生されたタンパク質およびポリペプチドが米国食品医薬品局(FDA)によって承認されており、さらに370種が臨床試験中である。化学合成を介して生産される小分子治療薬と異なり、タンパク質およびポリペプチドは、生細胞内で最も効率的に産生される。しかし、細菌内における組換えタンパク質産生の現在の方法はしばしば、不適切にフォールディングされたタンパク質、凝集したタンパク質または不活性なタンパク質を産生し、そして多くのタイプのタンパク質が二次修飾を必要とするが、それらの修飾は、既知の方法を用いて行うと非効率的である。
【0003】
組換え系における、適切にフォールディングされたタンパク質の産生を増大させるために、多数の試みが開発されている。例えば、研究者たちは、発酵条件を変えるか(Schein(1989)、Bio/Technology、7:1141〜1149)、プロモーター強度を変えるか、または過剰発現させたシャペロンタンパク質を使用しており(Hockney(1994)、Trends Biotechnol.12:456〜463)、これらは、封入体の形成を防止するのに役立ち得る。
【0004】
細胞から上清中にタンパク質を排出させるために、複数の方法(strategies)が開発されている。例えば、米国特許第5,348,867号、米国特許第6,329,172号、WO96/17943、WO02/40696および米国特許出願公開第2003/0013150号を参照。増大した発現を得るための他の方法は、タンパク質をペリプラズム(periplasm、周辺質)にターゲティングすることに向けられている。一部の研究は、非Sec型の分泌に焦点をおいている(例えば、WO03/079007、米国特許出願公開第2003/0180937号、米国特許出願公開第2003/0064435号およびWO00/59537を参照)。しかし、大部分の研究は、Sec型分泌系を用いた外因性タンパク質の分泌に焦点をおいてきた。
【0005】
組換えポリペプチドまたはタンパク質の発現に使用するために、多くの分泌シグナルが記載されている。例えば、米国特許第5,914,254号、米国特許第4,963,495号、欧州特許第0 177 343号、米国特許第5,082,783号、WO89/10971、米国特許第6,156,552号、米国特許第6,495,357号、第6,509,181号、第6,524,827号、第6,528,298号、第6,558,939号、第6,608,018号、第6,617,143号、米国特許第5,595,898号、第5,698,435号および第6,204,023号、米国特許第6,258,560号、WO01/21662、WO02/068660および米国特許出願公開第2003/0044906号、米国特許第5,641,671号ならびに欧州特許第0 121 352号を参照。
【発明の概要】
【発明が解決しようとする課題】
【0006】
異種タンパク質産生はしばしば、不溶性タンパク質または不適切にフォールディングされたタンパク質の形成をもたらし、それらは、回収が困難であり、かつ不活性であり得る。さらに、特定の宿主細胞プロテアーゼの存在によって、対象とするタンパク質(protein of interest)が分解され、それによって最終収率が低下し得る。全ての異種タンパク質の産生を改善する単一の因子は存在しない。結果として、当技術分野では、組換えポリペプチドを分泌し、その適切なプロセシングを行って、適切にプロセシングされた形態でトランスジェニックタンパク質を産生することができる、改善された大規模発現系を同定する必要性が存在する。
【課題を解決するための手段】
【0007】
本発明は、収率および/または質が改善された望ましい仕様(desired specification)に従って、少なくとも1種の異種ポリペプチドを産生できる宿主細胞集団を迅速に同定するための組成物および方法を提供する。組成物は、タンパク質産生(production)に関与する1つまたは複数の標的遺伝子の発現が増大するように、タンパク質分解(degradation)に関与する1つまたは複数の標的遺伝子の発現が低下するように、またはタンパク質産物に影響を与える異種遺伝子を発現するように、またはその組合せとなるように遺伝子改変されている2つ以上の宿主細胞集団を含む。対象とするポリペプチドを様々な改変宿主細胞で発現する能力は、対象とするポリペプチドに最適な宿主細胞を決定するための迅速かつ効率的な手段を提供する。望ましい仕様は、対象とするポリペプチドに応じて異なるが、それには、収率、質および活性などが含まれる。
【0008】
対象とするポリペプチドの発現を促進する内因性配列および/または外因性配列の発現レベルに影響を与える核酸配列の多数の組合せを発現するように宿主細胞集団を改変できることが認識されている。一実施形態では、2つ以上の宿主細胞集団が、対象とする異種タンパク質の適切な発現、プロセシングおよび/または移行のうち1つまたは複数に関与する1つまたは複数の標的遺伝子の発現が増大するように遺伝子改変されている。別の実施形態では、標的遺伝子はタンパク質フォールディング調節因子である。別の実施形態では、タンパク質フォールディング調節因子は表1のリストから選択される。
【0009】
別の実施形態では、1つまたは複数の宿主細胞集団が、タンパク質分解に関与する1つまたは複数の標的遺伝子の発現が低下するように遺伝子改変されている。別の実施形態では、標的遺伝子はプロテアーゼである。別の実施形態では、プロテアーゼは表2のリストから選択される。
【0010】
一実施形態では、対象とするタンパク質をコードするヌクレオチド配列が、本明細書に記載の、P.fluorescensのSec系分泌シグナルに作動可能に連結(operably linked)されている。アレイ内の株のうち1つまたは複数が、対象とする異種タンパク質をペリプラズム区画に発現し得る。特定の実施形態では、1つまたは複数の株が、外側細胞壁を通して細胞外に異種タンパク質を分泌し得る。
【0011】
宿主細胞には、酵母細胞、昆虫細胞、哺乳動物細胞および植物細胞などを含めた真核細胞、ならびにP.fluorescensおよびE.coliなどの細菌細胞を含めた原核細胞が含まれる。
【0012】
示されているように、異種発現されたタンパク質の、改善された収率および/または質を有する(1つまたは複数の)特定の株を同定するために、宿主細胞集団のライブラリーを迅速にスクリーニングできる。株アレイは、治療用タンパク質、ホルモン、成長因子、細胞外受容体またはリガンド、プロテアーゼ、キナーゼ、血液タンパク質、ケモカイン、サイトカインおよび抗体などを含めた、対象とするいかなるタンパク質の発現改善のスクリーニングにも有用である。
【図面の簡単な説明】
【0013】
【図1A】図1Aは、P.fluorescensにおけるゲノム欠失を遺伝子工学処理するために使用したプラスミドpDOW1261−2を示す図である。
【図1B】図1Bは、遺伝子X欠失の構築の概略図である。
【図2】図2は、Δprc1、ΔdegP2、ΔLa2およびgrpEdnaKJ同時発現菌株(実施例6)において誘導後0および24時間で調製した可溶性細胞分画(それぞれI0およびI24)のウエスタンブロット分析の図である。上部矢印は、対照(DC440)ではなく同時発現菌株において完全に構築されたモノクローナル抗体を指す。r=組換え体(recombinant);n−r=非組換え体(nonreconbinant)。
【発明を実施するための形態】
【0014】
これより、本発明の全ての実施形態ではなく、それらの一部が示されている添付図面を参照して、以下に本発明をより完全に説明する。実際、これらの発明は、多くの異なった形態で具体化でき、本明細書に示す実施形態に限定して解釈するべきではなく、これらの実施形態は、この開示が適用される(applicable)法的必要条件を満たすように提示されている。
【0015】
以上の説明と添付図面とに提示されている教示の恩恵により、これらの発明が属する分野の当業者ならば、本明細書に示す本発明の多くの変更形態および他の実施形態が想起されよう。
【0016】
したがって、本発明は、開示されている特定の実施形態に限定されるものではなく、変更形態および他の実施形態も本発明の範囲に含まれるものであることを理解されたい。本明細書では特定の用語が用いられているが、それらは包括的および説明的な意味でのみで用いられており、限定を目的としたものではない。
【0017】
概要(Overview)
宿主細胞内で適切にプロセシングされた高レベルの異種ポリペプチドを産生するのに最適な宿主株(optimal host strain)、例えば、Pseudomonas fluorescens宿主株を同定するための組成物および方法を提供する。詳細には、ライブラリー内の各株(または「宿主細胞の集団」)が、宿主細胞内の1つまたは複数の標的遺伝子の発現を調節するように遺伝子改変されている宿主株のライブラリー(または「アレイ」)を提供する。「最適な宿主株」は、アレイ内の、表現型が異なる宿主細胞の他の集団と比較した、対象とする発現タンパク質の量、質および/または位置に基づいて同定または選択できる。したがって、最適な宿主株は、対象とするポリペプチドを望ましい仕様に従って産生する株である。望ましい仕様は、産生されるポリペプチドに応じて異なるであろうが、仕様には、タンパク質の質および/または量、そのタンパク質が隔離(sequestered)されるか、または分泌されるか、ならびにタンパク質フォールディングが含まれる。
【0018】
「異種(heterologous)」、「異種発現された(heterologously expressed)」または「組換え(recombinant)」は通常、宿主細胞で内因的に生じたのではないか、または、それがその中に存在している天然ゲノムにおける位置で内因的に生じたのではなく、感染、形質移入、マイクロインジェクション、エレクトロポレーション、マイクロプロジェクション、または同様の方法によって細胞に添加された遺伝子またはタンパク質を指す。
【0019】
アレイ内の宿主細胞集団のうち1つまたは複数は、宿主細胞内の1つまたは複数の標的遺伝子の発現を調節するように改変されている。「標的遺伝子(target gene)」とは、宿主細胞内の異種タンパク質産生に影響を与える遺伝子を意味する。異種タンパク質産生に影響を与える標的遺伝子には、異種タンパク質の発現、活性、溶解性、移行、タンパク質分解および/または切断を調節するタンパク質をコードする遺伝子が含まれる。例えば、標的遺伝子は、宿主細胞プロテアーゼ、タンパク質フォールディング調節因子、転写因子、翻訳因子および分泌調節因子のうち少なくとも1つをコードするものでもよく、または対象とする異種タンパク質の適切な転写、翻訳、プロセシングおよび/または移行に関与する他の任意のタンパク質でもよい。「標的タンパク質(target protein)」は、標的遺伝子の発現の結果として生じるタンパク質またはポリペプチドを指す。1つまたは複数の標的遺伝子の発現および/または活性は、標的遺伝子またはタンパク質の機能に応じて、増大または低下する。例えば、1つまたは複数の宿主細胞プロテアーゼの発現が低下し、一方、1つまたは複数のタンパク質フォールディング調節因子の発現が増大してもよい。
【0020】
本明細書に記載のアレイは、異種タンパク質または対象とするペプチドの産生に最適な宿主細胞を迅速に同定するのに有用である。異種タンパク質産生はしばしば、不溶性タンパク質または不適切にフォールディングされたタンパク質の形成をもたらし、それらは、回収が困難であり、かつ不活性であり得る。さらに、特定の宿主細胞プロテアーゼの存在によって、対象とするタンパク質が分解され、それによって最終収率が低下し得る。対象とする全てのポリペプチドまたはタンパク質を最適に産生する単一の宿主細胞集団は存在しない。したがって、本発明の組成物および方法を用いて、改変細胞集団のライブラリーから、最適な宿主細胞を迅速かつ効率的に同定できる。その後、対象とするタンパク質を十分な量で産生するために、または商業的生産のために、その最適な宿主株を使用できる。同様に、最適な宿主株に基づいて、対象とするタンパク質の発現用に宿主株を改変できる。
【0021】
一実施形態では、この方法は、P.fluorescens細胞の少なくとも第1および第2の集団を含むアレイであって、各集団が、(i)タンパク質分解に関与する少なくとも標的遺伝子の発現が低減するように遺伝子改変されているP.fluorescens細胞集団、(ii)タンパク質産生に関与する少なくとも1つの標的遺伝子の発現が増大するように遺伝子改変されているP.fluorescens細胞集団、ならびに(iii)タンパク質分解に関与する少なくとも1つの標的遺伝子の発現が低減するように、かつタンパク質産生に関与する少なくとも標的遺伝子の発現が増大するように遺伝子改変されている少なくとも1つのP.fluorescens細胞集団からなる群から選択されるアレイを得るステップと、対象とする少なくとも1つの異種タンパク質をコードする少なくとも1つの遺伝子を含む発現コンストラクトを、各集団の少なくとも1つの細胞に導入するステップと;少なくとも1つの細胞集団における前記対象とするタンパク質の発現に十分な条件下で前記細胞を維持するステップと、対象とする異種タンパク質が産生される最適な細胞集団を選択するステップとを含み、アレイ内の各集団が同一でなく、かつ各集団が物理的に相互に離れており、最適な細胞集団では、アレイ内の他の集団と比較して、対象とする異種タンパク質が、改善された発現、改善された活性、改善された溶解性、改善された移行、または低減したタンパク質分解もしくは切断のうち1つまたは複数を示す。
【0022】
アレイは、宿主細胞プロテアーゼまたはタンパク質フォールディング調節因子の発現を改変するように遺伝子改変されていない宿主細胞(例えばP.fluorescens宿主細胞)の集団をさらに含んでもよい。この集団は、野生型株でもよく、または、タンパク質産生、プロセシングまたは移行に関与しない、またはもしくは複数の遺伝子(or or more genes)の発現を変えるように遺伝子改変されている株でもよい(例えば、選択マーカー遺伝子を発現するように遺伝子改変されたものでもよい)。
【0023】
一実施形態では、P.fluorescens宿主細胞の各集団は、相互に表現型が異なる(phenotypically distinct)(すなわち「同一でない(non-identical)」)。「表現型が異なる」とは、各集団が、1つまたは複数の標的遺伝子を測定可能に異なった量で産生することを意味する。この実施形態では、1つまたは複数の異なった標的遺伝子の発現を変えるように各株が遺伝子改変されている。宿主細胞の一集団内で複数の標的遺伝子の発現が調節される場合、標的遺伝子の組合せが、ライブラリー内の他の集団とは表現型上異なる。本発明による、表現型が異なる複数の宿主細胞集団を含むアレイは、異種タンパク質または対象とするペプチドを産生するのに有用な1つまたは複数の株をそれから選択する多様な集団を提供するものである。当業者ならば、そのようなアレイが、宿主細胞の任意の1つまたは複数の集団の複製(replicates)(例えば二つ組(duplicates)、三つ組(triplicates)など)も含むことを理解するであろう。
【0024】
アレイ(Arrays)
迅速にスクリーニングして、異種タンパク質の収率および/または質が改善されている(1つまたは複数の)特定の株を同定できる宿主細胞集団のアレイ(すなわち「株アレイ(strain array)」)をここに提供する。本明細書で使用される場合、「株アレイ」という用語は、アドレス特定されているか、アドレス特定可能な複数の位置を指す(例えば、ディープウェルまたはマイクロウェルなどのウェル)。通常、アレイ内のマイクロウェルまたはマイクロウェル群のそれぞれの位置は既知であり、それによって、対象とする異種タンパク質の発現に最適な宿主細胞の同定が可能となっている。
【0025】
株アレイは、表現型が異なる複数の宿主株を含む。アレイは、低密度アレイでも、高密度アレイでもよく、約2以上、約4以上、約8以上、約12以上、約16以上、約20以上、約24以上、約32以上、約40以上、約48以上、約64以上、約72以上、約80以上、約96以上、約192または約384以上の宿主細胞集団を含有し得る。
【0026】
本発明の宿主細胞集団は、マルチウェルまたはディープウェル容器内で維持および/またはスクリーニングできる。この容器は、任意の望ましい数のウェルを含有し得るが、最小限の試薬と比較的少ない数の細胞とを用いて、宿主細胞の各集団を個別かつ同時にスクリーニングするには、小型化された細胞培養マイクロアレイプラットフォームが有用である。このアッセイで有用な典型的なマルチウェルマイクロタイター容器は、限定されるものではないが、10ウェルプレート、28ウェルプレート、96ウェルプレート、384ウェルプレート、および384超のウェルを有するプレートを含めたマルチウェルプレートである。代替として、所望の容積に応じて、チューブ、ホルダー、カートリッジ、ミニチューブ、微量遠心チューブ、クライオバイアル、角型ウェルプレートチューブ、プレート、スラントまたは培養フラスコのアレイを用いてもよい。
【0027】
容器(vessel)は、対象とする宿主細胞、例えば、シュードモナスを培養および/またはスクリーニングするのに適したいかなる材質で作製されたものでもよい。例えば、容器は、その材質が生体適合性である限り、プラスチックまたは他の人工高分子材料など、容易に殺菌できる材質であり得る。限定されるものではないが、ポリスチレン、ポリプロピレン、ポリビニル化合物(例えばポリ塩化ビニル)、ポリカーボネート(ポリ塩化ビニル)、ポリテトラフルオロエチレン(PTFE)、ポリグリコール酸(PGA)、セルロース、ガラス、フッ素重合体、フッ素化エチレンプロピレン、ポリビニリデン、ポリジメチルシロキサンおよびシリコンなどを含めた多くの物質が使用できる。
【0028】
細胞の自動形質転換および自動コロニーピッカーは、所望の細胞の迅速なスクリーニングを容易にする。アレイは、当技術分野で知られているスポッター装置(例えば自動ロボット装置)を用いて作製および/またはスクリーニングできる。
【0029】
標的遺伝子(Target genes)
本発明の株アレイは、表現型および遺伝子型が異なる複数の宿主細胞集団を含み、アレイ内の各集団は、宿主細胞内の1つまたは複数の標的遺伝子の発現を調節するように遺伝子改変されている。「標的遺伝子」とは、宿主細胞内の異種タンパク質産生に影響を与える遺伝子を意味する。標的遺伝子は、宿主細胞プロテアーゼまたは内因性もしくは外因性タンパク質フォールディング調節因子、転写因子、翻訳因子、分泌調節因子、または対象とする異種タンパク質の適切な転写、翻訳、プロセシングおよび/または移行に関与する他の任意のタンパク質でもよい。「標的タンパク質」は、標的遺伝子の発現の結果として生じるタンパク質またはポリペプチドを指す。1つまたは複数の標的遺伝子の発現および/または活性は、標的遺伝子またはタンパク質の機能に応じて、増大または低下する。標的遺伝子は、宿主細胞で内因的に生じたものでもよく、またはアレイ内の宿主細胞集団のそれぞれで異種発現された遺伝子でもよい。
【0030】
一実施形態では、1つまたは複数の標的遺伝子が、タンパク質フォールディング調節因子、推定上のタンパク質フォールディング調節因子、またはフォールディング調節因子の補因子もしくはサブユニットのうち少なくとも1つである。一部の実施形態では、1つまたは複数の標的遺伝子が、シャペロンタンパク質、フォルダーゼ、ペプチジルプロリルイソメラーゼおよびジスルフィド結合イソメラーゼから選択できる。一部の実施形態では、1つまたは複数の標的遺伝子が、htpG、cbpA、dnaJ、dnaKおよびfkbPから選択できる。P.fluorescens由来の例示的タンパク質フォールディング調節因子を表1に列挙する。
【0031】
他の実施形態では、標的遺伝子が、推定上のプロテアーゼ、プロテアーゼ様タンパク質、プロテアーゼの補因子またはサブユニットのうち少なくとも1つを含む。例えば、上記1つまたは複数の標的遺伝子が、セリン、スレオニン、システイン、アスパラギン酸またはメタロペプチダーゼであり得る。一実施形態では、1つまたは複数の標的遺伝子が、hslV、hslU、clpA、clpBおよびclpXから選択できる。標的遺伝子はプロテアーゼの補因子でもあり得る。P.fluorescens由来の例示的プロテアーゼを表2に列挙する。様々な生物由来のプロテアーゼは、Wellcome Trust Sanger Institute、Cambridge、UKによって維持されているMEROPSペプチダーゼデータベースに見出すことができる(ウェブサイトアドレスmerops.sanger.ac.uk/を参照)。
【0032】
タンパク質フォールディング調節因子(Protein folding modulators)
宿主細胞での異種タンパク質産生における別の重大な障害は、しばしば細胞が、可溶性または活性のタンパク質を産生するように適切に装備されていないことである。タンパク質の一次構造はアミノ酸配列によって規定(defined)されるが、二次構造はαへリックスまたはβシートの存在によって規定され、三次構造はジスルフィド結合などの隣接するタンパク質ストレッチ間の共有結合によって規定される。異種タンパク質を発現する場合、とりわけ大規模産生では、そのタンパク質それ自体の二次構造および三次構造が極めて重要である。タンパク質構造におけるいかなる有意な(significant)変化も機能的に不活性な分子または生物活性が有意に低下したタンパク質を生じ得る。多くの場合、宿主細胞は、活性な異種タンパク質の適切な産生に必要なタンパク質フォールディング調節因子(protein folding modulators)(PFMs)を発現している。しかし、使用可能かつ経済的に満足できるバイオテクノロジー製品を生産するのに通常必要な高レベルでの発現では、細胞は、しばしば、異種発現されたタンパク質をプロセシングするのに十分な天然の1つまたは複数のタンパク質フォールディング調節因子を産生できない。
【0033】
特定の発現系では、異種タンパク質の過剰産生に、それらのミスフォールディングおよび不溶性凝集体への隔離(segregation)が伴い得る。細菌細胞では、これらの凝集体は封入体として知られている。E.coliでは、フォールディング調節因子/シャペロンのネットワークに、Hsp70ファミリーが含まれる。主要なHsp70シャペロンであるDnaKは、タンパク質凝集を効率的に防止し、損傷タンパク質のリフォールディングを補助する。タンパク質凝集体への熱ショックタンパク質の取込みによって脱凝集が促進され得る。しかし、特定の場合には、不溶性画分の追加プロセシングを介して、封入体内にプロセシングされたタンパク質が回収され得る。封入体内に見出されたタンパク質は通常、変性(denaturation)および復元(renaturation)を含めた複数のステップを経て精製されなければならない。封入体にターゲティングされたタンパク質の典型的な復元方法は、高濃度の変成剤中に凝集体を溶解する試みと、それに続く、希釈による変成剤の除去とを含む。しばしばこの段階で、凝集体が再形成される。追加の処理はコストを増大させ、in vitroのリフォールディングが生理的に活性のある生成物を産生するであろうという保証がなく、回収されるタンパク質は大量の断片不純物を含み得る。
【0034】
分子シャペロンおよびフォルダーゼによって、in vivoでのタンパク質フォールディングが補助されており、分子シャペロンはフォールディング中間体と一時的に相互作用することによって他のポリペプチドの適切な異性化および細胞ターゲッティングを促進し、フォルダーゼはフォールディング経路における律速段階を加速するという最近の認識によって、封入体形成の問題と戦うための追加アプローチが提示されている(例えば、Thomas J Gら(1997)、Appl Biochem Biotechnol 66:197〜238を参照)。
【0035】
特定の場合には、シャペロンの過剰発現が、凝集しやすいタンパク質(aggregation-prone proteins)の可溶性収率を増大させることが見出されている(Baneyx,F.(1999)、Curr.Opin.Biotech、10:411−421およびその中の参考文献を参照)。これらのシャペロンの細胞内濃度の増大に伴う有益効果は、過剰産生されるタンパク質の性質に強く依存しているようであり、同じ(1つまたは複数の)タンパク質フォールディング調節因子の過剰発現が全ての異種タンパク質に必要であるとは限らない。
【0036】
シャペロン、ジスルフィド結合イソメラーゼ、およびペプチジルプロリルシス−トランスイソメラーゼ(PPIase)を含めたタンパク質フォールディング調節因子は、新生ポリペプチドのフォールディング、アンフォールディングおよび分解を補助する、全ての細胞に存在する1クラスのタンパク質である。
【0037】
シャペロンは、新生ポリペプチドに結合し、それらを安定化し、それらが適切にフォールディングするのを可能にすることによって作用する。タンパク質は、疎水性残基および親水性残基の両方を有し、前者は通常表面に露出しており、一方、後者は構造内に埋められており、それらは構造内で、その分子を取り囲んでいる水ではなく、他の親水性残基と相互作用している。しかし、フォールディング中のポリペプチド鎖では、タンパク質が部分的にフォールディングされた状態またはミスフォールディングされた状態で存在しているので、しばしば、親水性残基がしばらくの間露出する。この間に、生成中のポリペプチドが恒久的にミスフォールディングされた状態になるか、または他のミスフォールディングされたタンパク質と相互作用して、細胞内に大きな凝集体または封入体を形成し得る。シャペロンは通常、部分的にフォールディングされた鎖の疎水性領域に結合し、それらが完全にミスフォールディングするか、他のタンパク質と凝集するのを防止することによって作用する。シャペロンは、封入体内のタンパク質にも結合し、それらが脱凝集するのを可能にできる(Ransonら、1998)。フォールディング調節因子のGroES/EL、DnaKJ、Clp、Hsp90およびSecBファミリーは全てシャペロン様活性を有するタンパク質の例である。
【0038】
フォールディング調節因子の別の重要なタイプはジスルフィド結合イソメラーゼである。これらのタンパク質は、フォールディングポリペプチドが適切なタンパク質内ジスルフィド結合を形成するのを助ける極めて特殊な1セットの反応を触媒する。2残基以上のシステインを有するいかなるタンパク質も、誤った残基間でジスルフィド結合を形成する危険性がある。ジスルフィド結合生成ファミリーは、Dsbタンパク質からなり、Dsbタンパク質は、ペリプラズムの非還元環境におけるジスルフィド結合の形成を触媒する。ペリプラズムポリペプチドがミスフォールディングした場合、ジスルフィド結合イソメラーゼであるDsbCは、ジスルフィド結合を再編成して、そのタンパク質が正しい結合を再形成するのを可能にできる。
【0039】
プロリン残基は、そのすぐ前のペプチジル結合がシス配置(conformation)またはトランス配置をとり得るという点で、アミノ酸のなかで独特である。他の全てのアミノ酸について、これは、立体障害のために好ましくない。ペプチジルプロリルシスートランスイソメラーゼ(PPIase)は、この結合の一方の形態からもう一方への変換を触媒する。この異性化は、細胞内におけるタンパク質のフォールディング、リフォールディング、サブユニット集合および輸送の補助となり得る(Dolinskiら、1997)。
【0040】
一般的なシャペロンは非特異的な様式でタンパク質と相互作用するように見えるが、それに加えて、特定の標的のフォールディングを補助するシャペロンも存在する。これらのタンパク質特異的なシャペロンは、それらの標的と複合体を形成し、凝集および分解を防止し、それらが集合して、多重サブユニット構造になるための時間を与える。PapDシャペロンは、このタイプのよく知られている一例である(Lombardoら、1997)。
【0041】
フォールディング調節因子には、例えばHSP70タンパク質、HSP110/SSEタンパク質、HSP40(DNAJ関連)のタンパク質、GRPE様タンパク質、HSP90タンパク質、CPN60およびCPN10タンパク質、サイトゾルシャペロン、HSP100タンパク質、低分子量HSP、カルネキシンおよびカルレティキュリン、PDIおよびチオレドキシン関連タンパク質、ペプチジルプロリルイソメラーゼ、シクロフィリンPPIase、FK−506結合タンパク質、パルブリンPPIase、個別のシャペロニング、タンパク質特異的シャペロンまたは分子内シャペロンも含まれる。フォールディング調節因子については、概ね、「Guidebook to Molecular Chaperones and Protein−Folding Catalysts」(1997)、M.Gething編集、Melbourne University,Australiaに記載されている。
【0042】
E.coli細胞質中で最も詳細に特徴付けられている分子シャペロンは、ATP依存性のDnaK−DnaJ−GrpE系およびGroEL−GroES系である。in vitro研究および相同性考察に基づいて、多くの追加の細胞質タンパク質が、E.coliで分子シャペロンとして機能していると提唱されている。これらには、ClpB、HtpGおよびIbpA/Bが含まれ、それらは、DnaK−DnaJ−GrpEおよびGroEL−GroESのように、ストレスレギュロンに属する熱ショックタンパク質(Hsp)である。新生タンパク質鎖では、X−Pro結合のトランス配座がエネルギー的に支持されるが、天然タンパク質では、全てのプロリルペプチド結合の約5%がシス配座で見出されている。X−Pro結合のトランスからシスへの異性化は、多くのポリペプチドのフォールディングで律速となっており、in vivoでは、ペプチジルプロリルシス/トランスイソメラーゼ(PPIase)によって触媒されている。SlyD、SlpAおよびトリガーファクター(TF)という3種の細胞質PPIaseがこれまでにE.coliで同定されている。TFは、50Sリボソームサブユニットに随伴する48kDaタンパク質であり、E.coliでシャペロンと協力して、新規合成タンパク質の適切なフォールディングを保証すると仮定(postulated)されている。少なくとも5種のタンパク質(それぞれtrxA、trxC、grxA、grxBおよびgrxC遺伝子の産物である、チオレドキシン1および2、ならびにグルタレドキシン1、2および3)が、細胞質酵素内に一時的に生じるジスルフィド架橋の還元(reduction)に関与している。したがって、標的遺伝子は、適切なジスルフィド結合形成を可能にするジスルフィド結合形成タンパク質またはシャペロンであり得る。
【0043】
【表1】

【0044】
プロテアーゼ(Protease)
異種発現されたタンパク質の好ましくない分解は、特定の発現系の効率的な使用に障害を与える。多量の標的タンパク質を産生するように細胞が改変されている場合、細胞はストレス下に置かれ、しばしば他のタンパク質を誘導または抑制することによって反応する。異種タンパク質産生中に宿主細胞が経験するストレスは、例えば、過剰発現した異種タンパク質の分解を引き起こす特定のタンパク質または補因子の発現を増大し得る。代償性の(compensatory)タンパク質の発現増大は、高レベルの活性かつ完全長の異種タンパク質を発現するという目標にとって反生産的であり得る。他のタンパク質の発現低下または適切な発現の不足は、異種発現されたタンパク質のミスフォールディングおよび凝集を引き起こし得る。ストレス下の細胞はタンパク質発現のプロファイルを変えることが知られているが、異種発現された全てのタンパク質が特定の宿主細胞内で同じタンパク質の発現を調節するわけではない。
【0045】
したがって、1つまたは複数のプロテアーゼ酵素の発現が低下するように遺伝子操作された複数の宿主細胞集団を含むアレイを用いて、最適な宿主株、例えばP.fluorescens宿主株を同定できる。一実施形態では、そのゲノムから、少なくとも1つのプロテアーゼの発現を低下させること、阻害すること、または除去することによって、1つまたは複数の宿主細胞集団が改変される。改変は、複数のプロテアーゼに施すこともできる。関連の実施形態では、プロテアーゼ補因子またはプロテアーゼタンパク質の発現を低下させることによって、細胞が改変されている。別の実施形態では、プロテアーゼまたは関連タンパク質のプロモーターの阻害によって、宿主細胞が改変されており、プロモーターは天然のプロモーターであり得る。代替として、遺伝子改変は、標的遺伝子に相同なタンパク質を調節するためのものでもあり得る。
【0046】
改変宿主株を含むアレイは、下記に論じる通り、対象とする(1つまたは複数の)異種タンパク質を発現させ、タンパク質産生の質および/または量を評価することによってスクリーニングできる。代替として、対象とする異種タンパク質の単離株は、プロテアーゼ欠失宿主細胞集団それぞれから収集された溶解物と共に、独立にインキュベートすることもでき、タンパク分解のレベルを、最適な宿主細胞を同定するのに用いることができる。この実施形態では、最適な宿主細胞集団は、最小量の異種タンパク質分解をもたらすものである。したがって、一実施形態では、最適な宿主細胞集団の溶解物が、約50%未満、約45%未満、約40%未満、約35%未満、約30%未満、約25%未満、約20%未満、約10%未満、約5%未満、約4%未満、約3%未満、約2%未満、約1%未満の異種タンパク質、またはさらに少ないタンパク質によって分解され得る。
【0047】
例示的な標的プロテアーゼ遺伝子には、アミノペプチダーゼ、ジペプチダーゼ、ジペプチジルペプチダーゼおよびトリペプチジルペプチダーゼ、ペプチジルジペプチダーゼ、セリン型カルボキシペプチダーゼ、メタロカルボキシペプチダーゼ、システイン型カルボキシペプチダーゼ、オメガペプチダーゼ、セリンプロテイナーゼ、システインプロテイナーゼ、アスパラギン酸プロテイナーゼ、メタロプロテイナーゼまたは作用機序未知のプロテイナーゼと分類されるプロテアーゼが含まれる。
【0048】
アミノペプチダーゼには、サイトゾルアミノペプチダーゼ(ロイシルアミノペプチダーゼ)、膜アラニルアミノペプチダーゼ、シスチニルアミノペプチダーゼ、トリペプチドアミノペプチダーゼ、プロリルアミノペプチダーゼ、アルギニルアミノペプチダーゼ、グルタミルアミノペプチダーゼ、x−proアミノペプチダーゼ、細菌性ロイシルアミノペプチダーゼ、好熱性アミノペプチダーゼ、クロストリジウムアミノペプチダーゼ、サイトゾルアラニルアミノペプチダーゼ、リジルアミノペプチダーゼ、x−trpアミノペプチダーゼ、トリプトファニルアミノペプチダーゼ、メチオニルアミノペプチダーゼ、d−立体特異的アミノペプチダーゼ、アミノペプチダーゼEyが含まれる。ジペプチダーゼには、x−his、x−argジペプチダーゼ、x−メチル−hisジペプチダーゼ、cys−glyジペプチダーゼ、glu−gluジペプチダーゼ、pro−xジペプチダーゼ、x−proジペプチダーゼ、met−xジペプチダーゼ、非立体特異的ジペプチダーゼ、サイトゾル非特異的ジペプチダーゼ、膜ジペプチダーゼ、β−ala−hisジペプチダーゼが含まれる。ジペプチジルペプチダーゼおよびトリペプチジルペプチダーゼには、ジペプチジルペプチダーゼi、ジペプチジルペプチダーゼii、ジペプチジルペプチダーゼiii、ジペプチジルペプチダーゼiv、ジペプチジル−ジペプチダーゼ、トリペプチジルペプチダーゼI、トリペプチジルペプチダーゼIIが含まれる。ペプチジルジペプチダーゼには、ペプチジルジペプチダーゼaおよびペプチジルジペプチダーゼbが含まれる。セリンカルボキシペプチダーゼには、リソソームpro−xカルボキシペプチダーゼ、セリン型D−ala−D−alaカルボキシペプチダーゼ、カルボキシペプチダーゼC、カルボキシペプチダーゼDが含まれる。メタロカルボキシペプチダーゼには、カルボキシペプチダーゼa、カルボキシペプチダーゼB、リジン(アルギニン)カルボキシペプチダーゼ、gly−Xカルボキシペプチダーゼ、アラニンカルボキシペプチダーゼ、ムラモイルペンタペプチドカルボキシペプチダーゼ、カルボキシペプチダーゼh、グルタミン酸カルボキシペプチダーゼ、カルボキシペプチダーゼM、ムラモイルテトラペプチドカルボキシペプチダーゼ、亜鉛d−ala−d−alaカルボキシペプチダーゼ、カルボキシペプチダーゼA2、膜pro−xカルボキシペプチダーゼ、チューブリン−tyrカルボキシペプチダーゼ、カルボキシペプチダーゼtが含まれる。オメガペプチダーゼには、アシルアミノアシルペプチダーゼ、ペプチジルグリシンアミダーゼ、ピログルタミルペプチダーゼI、βアスパラチルペプチダーゼ、ピログルタミルペプチダーゼII、nホルミルメチオニルペプチダーゼ、プテロイルポリ−γ−グルタミン酸カルボキシペプチダーゼ、γ−glu−Xカルボキシペプチダーゼ、アシルムラモイル−alaペプチダーゼが含まれる。セリンプロテイナーゼには、キモトリプシン、キモトリプシンc、メトリジン、トリプシン、トロンビン、凝固因子Xa、プラスミン、エンテロペプチダーゼ、アクロシン、α分解プロテアーゼ、グルタミル、エンドペプチダーゼ、カテプシンG、凝血因子viia、凝固因子ixa、ククミシ(cucumisi)、プロリルオリゴペプチダーゼ、凝固因子xia、ブラキウリン、血漿カリクレイン、組織カリクレイン、膵臓エラスターゼ、白血球エラスターゼ、凝固因子xiia、キマーゼ、補体成分c1r55、補体成分c1s55、古典的補体経路c3/c5コンバターゼ、補体因子I、補体因子D、第二補体経路c3/c5コンバターゼ、セレビシン、ヒポデルミンC、リジルエンドペプチダーゼ、エンドペプチダーゼ1a、γ−レニ(gamma-reni)、ベノムビンab、ロイシルエンドペプチダーゼ、トリプターゼ、スクテラリン、ケキシン、ズブチリシン、オリジン(oryzin)、エンドペプチダーゼk、サーモミコリン、テルミターゼ、エンドペプチダーゼSO、T−プラスミノーゲンアクチベータ、プロテインC、膵臓エンドペプチダーゼE、膵臓エラスターゼii、IGA特異的セリンエンドペプチダーゼ、U−プラスミノーゲン, 活性化因子(U-plasminogen, activator)、ベノムビンA、フリン、ミエロブラスチン、セメノゲラーゼ、グランザイムAすなわち傷害性Tリンパ細胞プロテイナーゼ1、グランザイムBすなわち傷害性Tリンパ細胞プロテイナーゼ2、ストレプトグリシンA、トレプトグリシンB(trestgrisin B)、グルタミルエンドペプチダーゼII、オリゴペプチダーゼB、リムルス凝固因子c、リムルス凝固因子、リムルス凝血酵素、オンプチン、レプレッサーlexa、細菌リーダーペプチダーゼI、トガビリン、フラビリンが含まれる。システインプロテイナーゼには、カテプシンB、パパイン、フィシン、キモパパイン、アスクレパイン、クロストリパイン、ストレプトパイン、アクチニド、カテプシン1、カテプシンH、カルパイン、カテプシンt、グリシル, エンドペプチダーゼ(glycyl, endopeptidase)、癌凝血原、カテプシンS、ピコルナイン3C、ピコルナイン2A、カリカイン、アナナイン、幹ブロメライン、果実ブロメライン、レグミン、ヒストリサイン、インターロイキン1−β変換酵素が含まれる。アスパラギン酸プロテイナーゼには、ペプシンA、ペプシンB、ガストリクシン、キモシン、カテプシンD、ネオペンテシン、レニン、レトロペプシン、プロオピオメラノコルチン変換酵素、アスペルギロペプシンI、アスペルギロペプシンII、ペニシロペプシン、リゾプスペプシン、エンドチアペプシン、ムコロペプシン、カンジダペプシン、サッカロペプシン、ロドトルラペプシン、フィサロペプシン、アクロシリンドロペプシン、ポリポロペプシン、ピクノポロペプシン、シタリドペプシンa、シタリドペプシンb、キサントモノナペプシン、カテプシンe、バリヤーペプシン、細菌性リーダーペプチダーゼI、シュードモナペプシン、プラスメプシンが含まれる。メタロプロテイナーゼには、アトロリシンa、微生物コラゲナーゼ、ロイコリシン、間質コラゲナーゼ、ネプリリシン、エンベリシン、iga特異的メタロエンドペプチダーゼ、プロコラーゲンN−エンドペプチダーゼ、チメットオリゴペプチダーゼ、神経溶解素、ストロメライシン1、メプリンA、プロコラーゲンC−エンドペプチダーゼ、ペプチジル−Lysメタロエンドペプチダーゼ、アスタシン、ストロメライシン, 2(stromelysin, 2)、マトリリジンゼラチナーゼ、エアロモノリシン、プソイドリシン、サーモリシン、バシロリシン、アウレオリシン、ココリシン、ミコリシン、β−分解メタロエンドペプチダーゼ、ペプチジル−Aspメタロエンドペプチダーゼ、好中球コラゲナーゼ、ゲラチナーゼB、リーシュマノリシン、サッカロリシン、自己溶菌酵素、デウテロリシン、セラリシン、アトロリシンB、アトロリシンC、アトロキサーゼ、アトロリシンE、アトロリシンF、アダマリシン、ホリリシン、ラバリシン、ボソロパシン、ボソロリシン、オフィオリシン、トリメレリシンI、トリメレリシンII、ムクロリシン、ピトリリシン、インスリシン、O−シアロ糖タンパク質エンドペプチダーゼ、ルスセリリシン、ミトコンドリア, 間質性, ペプチダーゼ(mitochondrial intermediate peptidase)、ダクチリシン、ナルジリシン、マグノリシン、メプリンB、ミトコンドリアプロセシングペプチダーゼ、マクロファージエラスターゼ、コリオリシン、トキシリシンが含まれる。作用機序未知のプロテイナーゼには、サーモプシンおよび多触媒エンドペプチダーゼ複合体が含まれる。
【0049】
【表2】
【0050】
その他のタンパク質修飾酵素(Additional protein modification enzymes)
別の実施形態では、標的遺伝子が、適切なタンパク質プロセシングおよび/または修飾に関与する遺伝子を含む。一般的な修飾には、ジスルフィド結合形成、グリコシル化、アセチル化、アシル化、リン酸化およびγ−カルボキシ化が含まれ、これらは全て、タンパク質フォールディングおよび生物活性を調節し得る。タンパク質プロセシングに関与する酵素のいくつかのクラスの網羅的ではないリストが表3に示されている。当業者ならば、アレイ用に選択された宿主細胞内で有用であるか、または対象とする異種タンパク質と併用して有用である標的遺伝子を、どのようにして表3に列挙したタンパク質修飾酵素のクラスの中から同定するか認識するであろう。標的遺伝子は、使用される宿主細胞に内因的なものでもよく、または対象とする異種タンパク質が由来する生物に内因的なものもよく、または対象とする異種発現タンパク質の適切なプロセシングを促進することが知られているものでもよい。タンパク質産生に関与するいかなる遺伝子も、対象とする異種タンパク質の望ましい仕様に従ってターゲッティングできることも認識されている。
【0051】
【表3】
【0052】
標的遺伝子の発現を調節する方法(Methods for modulating the expression of target genes)
当技術分野で知られている任意の技法によって、アレイ内の1つまたは複数の宿主細胞集団を改変でき、例えば、ゲノム内の少なくとも1つの標的遺伝子がノックアウトされる技法によって、または少なくとも1つの標的遺伝子の発現が低下するように、遺伝子に変異導入することによって、または少なくとも1つの標的遺伝子の発現が低下するように、標的遺伝子の少なくとも1つのプロモーターを改変することによって、または宿主ゲノム内の標的遺伝子または標的遺伝子の阻害因子を(対象とする異種タンパク質またはポリペプチドと共に)同時発現させることによって改変できる。上記に論じたように、標的遺伝子はアレイ内の宿主細胞集団に内因的なものでもよく、または宿主細胞集団それぞれで異種発現されるものでもよい。
【0053】
標的遺伝子の発現は、例えば、タンパク質産生に関与する1つまたは複数の標的遺伝子を含む発現ベクターを、宿主集団内の少なくとも1つの細胞に導入することによって増大できる。標的遺伝子発現は、例えば、標的遺伝子のプロモーターに変異導入することによっても増大できる。異種タンパク質を発現する宿主細胞または生物は、タンパク質産生に関与する少なくとも1つの標的遺伝子の発現が増大し、かつタンパク質分解に関与する少なくとも1つの標的遺伝子の発現が低下するように、遺伝子改変することもできる。
【0054】
ゲノムは、外因性遺伝子またはプロモーターエレメントをゲノム内に、もしくは発現ベクターで宿主内に含めることによって、または特定の標的遺伝子の、mRNAもしくはタンパク質産生能力を高めることによって、または標的遺伝子もしくはプロモーターエレメントを除去もしくは破壊することによって、または標的遺伝子の、mRNAもしくはタンパク質産生能力を低下させることによって、1つまたは複数の標的遺伝子の発現を調節するように改変できる。例えば、置換、欠失(「ノックアウト」)、同時発現または挿入(「ノックイン」)技法によって、遺伝暗号を改変し、それによって、標的遺伝子の転写および/または翻訳に影響を与えることができる。既存の標的配列の転写を調節する所望のタンパク質または調節配列の追加遺伝子を挿入することもできる。
【0055】
ゲノム改変(Genome modification)
宿主細胞のゲノムは、遺伝的ターゲッティング事象を介して改変することができ、それは、挿入または組換え、例えば相同組換えによるものであり得る。相同組換えは、配列相同性に基づいたDNA組換え過程を指す。相同組換えは、内因性遺伝子内における部位特異的な改変を可能にし、それゆえ、ゲノム内の新規な改変が構築できる(例えば、Radding(1982)、Ann.Rev.Genet.16:405;米国特許第4,888,274号参照)。
【0056】
標的遺伝子座の相同組換え用には、様々なコンストラクトが調製できる。通常、コンストラクトは、同定された遺伝子座と相同な少なくとも10bp、20bp、30bp、40bp、50bp、70bp、100bp、500bp、1kbp、2kbp、4kbp、5kbp、10kbp、15kbp、20kbpまたは50kbpの配列を含み得る。標的遺伝子配列の相同性の程度を決定するには、例えば、標的遺伝子座のサイズ、配列の利用可能性、標的遺伝子座における二重交差事象の相対効率および他の配列との標的配列の類似性など、様々な要件が関与し得る。
【0057】
改変遺伝子には、改変されるべきゲノム内の、対応する標的配列と実質的に同質遺伝子的なDNAによって、所望の配列改変が挟まれている配列が含まれ得る。「改変遺伝子(modified gene)」は、ゲノム内に導入されて、宿主細胞内のプロテアーゼまたはタンパク質フォールディング調節因子の発現を改変する配列である。「標的遺伝子(target gene)」は、改変遺伝子によって置換される配列である。実質的に同質遺伝子的な配列は、対応する標的配列に、(所望の配列改変を除いて)少なくとも約95%、97〜98%、99.0〜99.5%、99.6〜99.9%または100%同一であり得る。改変遺伝子および標的遺伝子は、100%同一である少なくとも約10、20、30、50、75、150または500塩基対のDNAストレッチを共有し得る。
【0058】
ヌクレオチドコンストラクトは、内因性の標的遺伝子産物を改変するように設計できる。改変遺伝子配列は、結果として生じる遺伝子産物の機能を破壊するように設計された1つまたは複数の欠失、挿入、置換またはこれらの組合せを有し得る。一実施形態では、上記改変が、標的遺伝子の上流配列と、リーディングフレーム内で融合している選択マーカー遺伝子の挿入であり得る。
【0059】
ゲノムは、挿入不活化を用いて改変することもできる。この実施形態では、遺伝子産物の生成を阻害する遺伝子内の配列の組換えによってゲノムが改変される。この挿入は、別個のエレメントを挿入することによって遺伝子を破壊するか、遺伝子の必須部分を除去することができる。一実施形態では、抗生物質などの特定のストレッサーに対する耐性、または特定の培地中での増殖、例えば必須アミノ酸の産生をコードする遺伝子の挿入も挿入欠失に含まれる。
【0060】
ゲノムは、トランスポゾンの使用によって改変することもできる。トランスポゾンは、相同組換えとは独立した作用機序によって、原核生物ゲノム内の複数の部位に挿入され得る遺伝的エレメントである。トランスポゾンには、例えば、E.coliにおけるTn7、Tn5またはTn10、S.aureusにおけるTn554、M.paratuberculosis由来のIS900、Pseudomonas atlantica由来のIS492、ストレプトミセス由来のIS116ならびにM.paratuberculosis由来のIS900が含まれ得る。遺伝子転位に関与すると考えられているステップには、3’OHを生じる、トランスポゾン末端の切断と、トランスポザーゼによってトランスポゾンの3’OH露出末端と同定された配列とが引き合わされる鎖転移と、同定されたDNAへのトランスポゾンの共有結合を生じる1ステップエステル交換反応とが含まれる。トランスポザーゼによって行われる、鍵となる反応は、ニック形成または鎖交換であり、残りの過程は宿主酵素によって行われると一般的に考えられている。
【0061】
一実施形態では、標的タンパク質またはその相同体をコードする遺伝子の配列を、組換えによってゲノム内に組込むことによって、標的遺伝子またはタンパク質の発現または活性を増大させる。別の実施形態では、ゲノム内にプロモーターを挿入して、標的遺伝子または相同体の発現を亢進する。別の実施形態では、不活性遺伝子を用いた組換えによって、標的遺伝子またはその相同体の発現または活性を低下させる。別の実施形態では、細胞内で別個の機能を有し得る、または耐性マーカーなどのレポーター遺伝子であるか、もしくは他の方法で検出可能なマーカー遺伝子であり得る異なった遺伝子をコードする配列を、組換えによってゲノム内に挿入できる。さらに別の実施形態では、1つまたは複数の位置で変異導入されている標的遺伝子の少なくとも一部の1コピーを、組換えによってゲノム内に挿入する。この変異バージョンの標的遺伝子は、タンパク質をコードしていなくてもよく、または変異遺伝子によってコードされているタンパク質が不活性でもよく、活性が調節(増大または低下)されていてもよく、または天然タンパク質と比較した際に、変異体タンパクが異なった活性を有することもできる。
【0062】
細菌で遺伝子をノックアウトするための戦略があり、それらは概ね、E.coliで例示されている。1つの経路は、抗生物質耐性の遺伝子(例えばアンピシリン)を含有するベクターに遺伝子内DNA断片をクローニングするものである。接合伝達、化学的形質転換またはエレクトロポレーション(Puehlerら(1984)、Advanced Molecular Genetics、New York、Heidelberg、Berlin、Tokyo、Springer Verlag)を介して細胞を形質転換する前に、栄養プラスミド複製(oriV遺伝子座)などの複製開始点を切除し、残りのDNA断片を再連結および精製する(Sambrookら(2000)、Molecular cloning:A laboratory manual、第3版、Cold Spring Harbor、N.Y.、Cold Spring Harbor Laboratory Press)。代替として、DNA複製開始点を有する抗生物質抵抗性のプラスミドを用いることもできる。形質転換後に、適切な抗生物質(例えば200μg/mLのアンピシリン)を含有する、例えばLBアガープレート上に細胞をプレーティングする。抗生物質を含有するプレート上で増殖するコロニーは、おそらく、相同遺伝子座における、ゲノム内への全DNA断片の組込みをもたらす単一の組換え事象を経たものである(Snyder,L.、W.Champnessら(1997)、Molecular Genetics of Bacteria、Washington DC、ASM Press)。所望の遺伝子ノックアウトが所望の遺伝子座で起こったことを確かめる抗生物質耐性細胞のさらなる分析は、例えばPCR診断による(McPherson,M.J.、P.Quirkeら(1991)、PCR:A Practical Approach、New York、Oxford University Press)。ここでは、少なくとも2通りのPCRプライマーが設計される。ひとつは、遺伝子ノックアウトの構築に使用されるDNA領域の外側でハイブリッド形成するものであり、ひとつは、残りのプラスミドバックボーン内でハイブリッド形成するものである。正しいサイズを有するDNA断片のPCR増幅に成功して、それに続いてDNA配列分析を行うことによって、細菌染色体内の正しい位置で遺伝子ノックアウトが起こったことが確認されるであろう。新規に構築された変異体株の表現型は、その後、例えばSDSポリアクリルアミドゲル電気泳動によって分析できる(Simpson,R.J.(2003)、Proteins and Proteomics−A Laboratory Manual、Cold Spring Harbor、N.Y.、Cold Spring Harbor Laboratory Press)。
【0063】
遺伝子ノックアウトを生成する代替の経路は、pSC101レプリコンなどの温度感受性レプリコンを使用して、遺伝子置換を促進するものである(Hamiltonら(1989)、Journal of Bacteriology 171(9):4617〜22)。この過程は、染色体上の遺伝子と、DNA複製が温度感受性であるプラスミド上に保持されている相同配列との間の相同組換えによって進行する。このプラスミドを適切な宿主内に導入して形質転換した後、プラスミドが染色体内に組込まれているものを44℃で選択することが可能である。それに続いてこれらの共組換え体を30℃で増殖させることによって、第2の組換え事象がもたらされ、その結果、それらの解離が起こる。第2の組換え事象がどこで起こるかに応じて、その染色体が、遺伝子置換を経たものになるか、またはその遺伝子の元のコピーを保持することになる。
【0064】
特定の遺伝子産物の発現を阻害する他の戦略も開発されている。例えば、特定遺伝子産物の発現の低下または消失さえもたらすために、RNA干渉(RNAi)、とりわけ低分子干渉RNA(siRNA)を用いたものが、広範に開発されている。siRNAは、相補的なmRNAを、分解するためにターゲッティングできる短い二本鎖RNA分子である。RNAiは、二本鎖RNAの導入によって相同遺伝子の発現が抑制される現象である。dsRNA分子は、in vivoで、RNAi効果の媒介因子である21〜23ntのsiRNAにされる。2本鎖RNAは、導入された際に、Dicerと呼ばれるRNaseIII様の酵素によって、20〜25ヌクレオチドのsiRNAにプロセシングされる(開始段階)。その後、siRNAは、RNA誘導サイレンシング複合体(RISC)として知られる、エンドリボヌクレアーゼを含有する複合体の中に組み込まれ、この過程で巻き戻される。続いて、siRNA鎖は、RISCを相補的なRNA分子へと導き、そこで、それらは同族のRNAを切断して、破壊する(エフェクター段階)。同族のRNAの切断は、siRNA鎖が結合した領域の中央近くで行われる。RNAiは、ゼブラフィッシュ、線虫(C.elegans)、昆虫(Drosophila melanogaster)、プラナリア、刺胞動物、トリパノソーマ、マウス、および哺乳動物細胞を含めた様々な生物で、遺伝子発現を低減するのに使用され、成功している。
【0065】
標的遺伝子をコードするオープンリーディングフレーム内の1または複数ヌクレオチドを変異させることによっても、ゲノムを改変できる。遺伝的変異のため技法、例えば部位特異的変異誘発は、当技術分野でよく知られている。一部のアプローチは、X線および化学物質によって誘導されるものなど、染色体DNAにおけるランダム変異の生成に焦点をおいている。
【0066】
同時発現(Coexpression)
一実施形態では、(1つまたは複数の)標的遺伝子をコードする1または複数種のベクターを含めて、異種タンパク質またはペプチドとの標的遺伝子の同時発現を促進することによって、宿主細胞内の1つまたは複数の標的遺伝子を改変できる。別の実施形態では、宿主細胞ゲノムへの外因性プロモーターの付加を含めた、標的遺伝子のプロモーターの亢進によって、宿主細胞を改変する。
【0067】
別の実施形態では、プロテアーゼ阻害物質など、標的遺伝子の阻害物質をコードする1または複数種のベクターを含めて、標的プロテアーゼの活性を阻害することによって、宿主細胞内の1つまたは複数の標的遺伝子を改変する。そのような阻害物質は、標的遺伝子、標的遺伝子の補因子または標的遺伝子の相同体の発現を制限するアンチセンス分子であり得る。通常、アンチセンスは、標的遺伝子の少なくとも一部に相補的な配列を有する核酸分子を指すのに使用される。加えて、阻害物質は、干渉(interfering)RNAまたは干渉RNAをコードする遺伝子であり得る。例えば、Fire,A.ら(1998)、Nature 391:806〜11、Elbashirら(2001)、Genes&Development 15(2):188〜200、Elbashirら(2001)、Nature 411(6836):494〜8、Carnegie Instituteの米国特許第6,506,559号、Benitecの第6,573,099号、the Whitehead Inst.の米国特許出願第2003/0108923号、ならびに米国特許出願第2003/0114409号、WO03/006477、WO03/012052、WO03/023015、WO03/056022、WO03/064621およびWO03/070966に記載されている通り、真核生物では、そのような干渉RNAが、低分子干渉(small interfering)RNAまたはリボザイムであり得る。
【0068】
阻害物質は、別のタンパク質またはペプチドであり得る。阻害物質は、例えば、標的タンパク質のコンセンサス配列を有するペプチドであり得る。阻害物質は、標的タンパク質の直接的または間接的阻害分子を宿主で産生できるタンパク質またはペプチドであり得る。例えば、プロテアーゼ阻害物質には、アマスタチン、E−64、アンチパイン、エラスタチナール、APMSF、ロイペプチン、ベスタチン、ペプスタチン、ベンズアミジン、1,10−フェナントロリン、キモスタチン、ホスホラミドン、3,4−ジクロロイソクマリン、TLCK、DFP、TPCKが含まれ得る。これまでに、100を超える天然存在タンパク質プロテアーゼ阻害物質が同定されている。それらは、細菌から動植物までの様々な生物で単離されている。それらは、立体障害を介して活性部位への基質接近を防止する、プロテアーゼの強結合可逆阻害物質または疑似不可逆阻害物質として働く。それらのサイズも、50残基(例えばBPTI:ウシ膵臓トリプシン阻害物質)から、最大400残基(例えばα−1PI:α−1プロテイナーゼ阻害物質)まで極めて可変的である。それらは、α−マクログロブリンファミリーのタンパク質(例えばα−2マクログロブリン)を除いて、厳密にクラス特異的である。α−マクログロブリンファミリーのタンパク質は、分子トラップ作用機序を介して、ほとんどのプロテアーゼに結合し、それらを阻害する。
【0069】
外因性のベクターまたはDNAコンストラクトを宿主細胞内に導入して、形質移入(transfected)または形質転換(transformed)することができる。外因性の核酸で真核細胞および原核細胞をそれぞれ形質移入および形質転換する技法は、当技術分野でよく知られている。これらには、脂質小胞媒介の取り込み、リン酸カルシウム媒介の形質移入(リン酸カルシウム/DNA共沈)、ウイルス感染、とりわけ、例えば改変アデノウイルスなどの改変ウイルスを用いたもの、微量注入およびエレクトロポレーションが含まれ得る。原核細胞の形質転換には、熱ショック媒介の取り込み、無傷細胞を用いた細菌プロトプラスト融合、微量注入およびエレクトロポレーションが技法に含まれ得る。植物を形質転換する技法には、A.tumefaciensなどのアグロバクテリウム媒介の導入、高速推進のタングステンまたは金マイクロプロジェクタイル、エレクトロポレーション、微量注入、およびポリエチレングリコール媒介の取り込みが含まれる。DNAは、1本鎖または2本鎖、直鎖状または環状、リラックス型またはスーパーコイルド型のDNAであり得る。哺乳動物細胞に形質移入する様々な技法に関しては、例えば、Keownら(1990)、Processes in Enzymology、第185巻、527〜537ページを参照のこと。
【0070】
下記の通り、対象とする異種タンパク質またはポリペプチドを含む発現コンストラクト用に、標的遺伝子またはそのエンハンサーもしくは阻害物質をコードする発現コンストラクトを構築できる。例えば、コンストラクトは、1箇所または複数箇所の内部リボソーム進入部位(IRES)を含有できる。コンストラクトは、標的遺伝子の少なくとも一部、または標的遺伝子の補因子、変異体バージョンの標的遺伝子の少なくとも一部、もしくは一部の実施形態では標的遺伝子の阻害物質をコードする核酸配列に作用可能に連結したプロモーターをも含有できる。代替として、コンストラクトは、プロモーターの無いものであり得る。コンストラクトが細胞DNA/ゲノムに組み込まれるように設計されていない場合には、ベクターは通常、少なくとも1つのプロモーターエレメントを含有する。上記核酸配列に加えて、発現ベクターは、選択マーカー配列を含有し得る。発現コンストラクトは、転写開始、終止および/または、リボソーム結合部位のための部位をさらに含有し得る。特定されたコンストラクトは、限定されるものではないが、P.fluorescensもしくはE.coliなどの細菌細胞、酵母細胞、CHO細胞などの哺乳動物細胞または植物細胞を含めた、いかなる原核細胞または真核細胞にも挿入でき、発現させることができる。
【0071】
コンストラクトは、当技術分野で知られている方法に従って調製できる。望ましいコンストラクトが得られるまで、様々な断片を集め、適切なベクター内に導入し、クローニングし、分析し、その後さらに操作することができる。制限分析、切断、プローブの同定などを可能にする様々な改変を配列に加えることができる。必要に応じて、サイレント突然変異を導入できる。様々な段階で、制限分析、配列決定、ポリメラーゼ連鎖反応を用いた増幅、プライマー修復、in vitro突然変異誘発などを利用できる。抗生物質耐性遺伝子およびネガティブ選択因子を組込む方法は、当業者ならばよく知っているであろう(例えば、国際公開第99/15650号、米国特許第6,080,576号、米国特許第6,136,566号、Niwaら、J.Biochem.113:343〜349(1993);およびYoshidaら、Transgenic Research,4:277〜287(1995)を参照)。
【0072】
原核細胞複製系、例えばP.fluorescensまたはE.coliなどの原核細胞によって認識可能なオリジンを含有する細菌ベクターを用いて、コンストラクトを調製できる。マーカーを利用することができ、それは挿入に使用されるべきマーカーと同じか、または異なっており、宿主細胞に導入する前に除去できる。ひとたびコンストラクトを含有するベクターが完成したならば、特定の配列を除去するか、直線化するか、相同配列中に変異、欠失または他の配列を導入することなどによって、それをさらに操作できる。一実施形態では、標的遺伝子コンストラクトおよび異種タンパク質コンストラクトが同じ発現ベクターの一部であり、かつ同じプロモーターエレメントの制御下でも、そうでなくてもよい。別の実施形態では、それは別々の発現ベクター上にある。最終操作の後、コンストラクトを細胞に導入することができる。
【0073】
細胞増殖条件(Cell growth conditions)
本明細書に記載の宿主細胞の細胞増殖条件には、対象とするタンパク質の発現をアレイ内の少なくとも1つの株(または少なくともそれらの細胞の一部)で促進する条件および/または発現された、対象とするタンパク質の発酵を促進する条件が含まれる。本明細書で使用される場合、「発酵」という用語には、文字通りの発酵が用いられる実施形態と、他の、非発酵性の培養モードが用いられる実施形態との両方が含まれる。アレイ内の宿主細胞集団の増殖、維持および/または発酵は、いかなるスケールで行ってもよい。しかし、宿主細胞の複数集団が同時にスクリーニングされる場合には、異なった集団の数ならびに宿主細胞の複数集団を増殖および試験する能力によってスケールが限定されるであろう。一実施形態では、発酵培地が、富栄養培地、最小培地および無機塩培地の中から選択され得る。別の実施形態では、最少培地または無機塩培地のいずれかが選択される。さらに別の実施形態では、最少培地が選択される。さらに別の実施形態では、無機塩培地が選択される。
【0074】
無機塩培地は、無機塩と、例えば、グルコース、ショ糖またはグリセロールなどの炭素源とからなる。無機塩培地の例には、例えば、M9培地、シュードモナス培地(ATCC179)、ならびにDavisおよびMingioli培地(BD DavisおよびES Mingioli(1950)、J.Bact.60:17〜28を参照)が含まれる。無機塩培地を作製するのに使用される無機塩には、例えば、リン酸カリウム、硫酸もしくは塩化アンモニウム、硫酸もしくは塩化マグネシウム、ならびに微量無機塩、すなわち鉄、銅、マンガンおよび亜鉛の塩化カルシウム塩、ホウ酸塩および硫酸塩などの中から選択されたものが含まれる。有機窒素源、すなわち、ペプトン、トリプトン、アミノ酸または酵母抽出物などは、無機塩培地に含まれていない。代わりに、無機窒素源が使用され、これは、例えば、アンモニウム塩、アンモニア水および気体アンモニアの中から選択され得る。好ましい無機塩培地は、炭素源としてグルコースを含有する。無機塩培地と比較すると、最小培地も、無機質塩および炭素源を含有し得るが、例えば、低レベルのアミノ酸、ビタミン、ペプトンまたは他の成分が捕捉されていてもよい。但し、これらは極めて最小レベルで添加される。
【0075】
一実施形態では、下記表4に示す成分を用いて、培地を調製できる。それらの成分は、以下の順序で添加できる。すなわち、最初に、約30リットルの蒸留水に(NH4)HPO4、KH2PO4およびクエン酸を溶解することができる。その後、微量元素の溶液を添加し、それに続いて、Ucolub N115などの消泡剤を添加することができる。次に、乾熱滅菌(約121℃など)の後、グルコース、MgSO4およびチアミン−HCLの無菌液を添加できる。約6.8のpH調節は、アンモニア水を用いて実現できる。その後、滅菌蒸留水を添加して、371からグリセロールストック(123mL)を引いた容積に、初期容積を調整することができる。これらの化学物質は、Merckなどの様々な供給業者から市販されている。
【0076】
【表4】
【0077】
本発明では、形質転換された宿主細胞の増殖、培養および/または発酵を、宿主細胞の生存を許容する温度範囲内、好ましくは、包括的に、約4℃から約55℃の範囲内の温度で行う。したがって、例えば、「増殖」(および「増殖する」、「増殖すること」)、「培養すること」(および「培養」)」ならびに「発酵」(および「発酵させる」、「発酵させること」)という用語は、本明細書で使用される場合、本発明の宿主細胞に関して、本質的に、包括的に約4℃から約55℃の温度範囲内での「増殖」、「培養すること」および「発酵」を意味する。加えて、「増殖」は、活動的な細胞分裂および/または拡大の生物学的状態、ならびに無分裂および/または無拡大の細胞が代謝によって維持されている生物学的状態の両方を示すのに使用され、「増殖」という用語の後者の使用は、「維持」という用語と同義である。
【0078】
アレイの宿主細胞は、その細胞型の正常な増殖に適した温度で培養および維持するべきである。そのような正常増殖温度は、選択された宿主細胞の既知の増殖必要条件に基づいて容易に選択できる。培養を確立している間、とりわけスクリーニングの過程の間は、対象とする異種タンパク質またはポリペプチドで形質転換させる前後に、選択された細胞の増殖に適したCO2/N2湿度に制御された中で、細胞培養物をインキュベートすることが好ましい。インキュベーションの湿度は、培養器からの蒸発を最小限にし、より小さい容積の使用が可能となるように制御する。湿度の制御の代替として、またはそれに加えて、蒸発を最小限にするために、容器をフタで覆ってもよい。培養温度の選択は、主として、使用される宿主細胞のアイデンティティによる。蒸発を制御する湿度(パーセント)の選択は、選択された容器容積、ならびに容器内の細胞培養物の濃度および容積、ならびに培養温度に基づく。したがって、湿度は、約10%から約80%まで変動し得る。適した状態の選択は、十分に当技術分野の技術範囲内であることを理解するべきである。
【0079】
スクリーニング(Screening)
対象とする異種タンパク質を発現するのに最適な宿主細胞集団を求めて、本明細書に記載の株アレイをスクリーニングすることができる。最適な宿主細胞集団は、発現された、対象とするタンパク質の量、質および/または局在に基づいて同定または選択できる。一実施形態では、最適な宿主細胞集団は、アレイ内の、表現型が異なる宿主細胞の他の集団と比較して、宿主細胞内における、対象とするタンパク質またはポリペプチドの収率増大をもたらすものである。
【0080】
産生の増大は、代替として、産生されたタンパク質1グラムあたり、または宿主タンパク質1グラムあたりの、適切にプロセシングされたタンパク質またはポリペプチドのレベルの増大であり得る。産生の増大は、異種タンパク質1グラムあたり、または宿主細胞タンパク質1グラムあたりに産生された、回収可能なタンパク質またはポリペプチドのレベルの増大でもあり得る。産生の増大は、総タンパク質レベルの増大、適切にプロセシングされたタンパク質もしくは適切にフォールディングされたタンパク質のレベルの増大、または、活性もしくは可溶性タンパク質のレベルの増大の任意な組合せであり得る。この実施形態では、「増大」または「改善」という用語は、対象とするタンパク質またはポリペプチドを、アレイ内の、宿主細胞の他の1つまたは複数の集団で発現させた場合に、産生される、適切にプロセシングされる、可溶性である、かつ/または回収可能であるタンパク質またはポリペプチドのレベルに対して相対的なものである。産生の増大は、例えば、エネルギー消費量を低下させることによって、利用可能資源の使用を増大させることによって、または増殖培地中の増殖補助物質の必要性を低下させることによって、細胞または生物の効率を最適化し得る。産生の増大は、発現されたタンパク質のタンパク質分解の低下の結果であってもよい。
【0081】
一実施形態では、アレイ内の少なくとも1つの株が、少なくとも0.1mg/mlの正しくプロセシングされたタンパク質を産生する。正しくプロセシングされたタンパク質は、天然タンパク質のアミノ末端を有する。別の実施形態では、少なくとも1つの株が、0.1〜10mg/mlの正しくプロセシングされたタンパク質を細胞内に産生し、これには、少なくとも約0.2、約0.3、約0.4、約0.5、約0.6、約0.7、約0.8、約0.9または少なくとも約1.0mg/mlの正しくプロセシングされたタンパク質が含まれる。別の実施形態では、アレイ内の少なくとも1つの株によって産生される、正しくプロセシングされた、対象とするタンパク質またはポリペプチドの総タンパク質量が、少なくとも1.0mg/ml、少なくとも約2mg/ml、少なくとも約3mg/ml、約4mg/ml、約5mg/ml、約6mg/ml、約7mg/ml、約8mg/ml、約10mg/ml、約15mg/ml、約20mg/ml、約25mg/ml、約30mg/ml、約35mg/ml、約40mg/ml、約45mg/mlもしくは少なくとも約50mg/mlであるか、またはそれより多い。一部の実施形態では、産生される、正しくプロセシングされたタンパク質の量が、正しくプロセシングされた形態の異種タンパク質の総タンパク質量の少なくとも約5%、約10%、約15%、約20%、約25%、約30%、約40%、約50%、約60%、約70%、約80%、約90%、約95%、約96%、約97%、約98%もしくは少なくとも約99%であるか、またはそれより多い。
【0082】
対象とするタンパク質またはポリペプチドの改善された発現は、タンパク質の溶解性の増大も指し得る。対象とするタンパク質またはポリペプチドを宿主細胞の細胞質、ペリプラズムまたは細胞外の培地中に産生させて、そこから回収することができる。タンパク質またはポリペプチドは、不溶性または可溶性であり得る。タンパク質またはポリペプチドは、上記に論じた通り、1つまたは複数の標的(例えばシグナルまたはリーダー)配列または精製に役立つ配列を含有し得る。
【0083】
「可溶性(soluble)」という用語は、本明細書で使用される場合、生理的条件下にある緩衝液中で10〜30分間遠心処理された場合に、約5000〜20000×重力の遠心処理によって、そのタンパク質が沈殿しないことを意味する。可溶性タンパク質は、封入体または他の沈殿物質塊の一部でない。同様に、「不溶性(insoluble)」は、生理的条件下にある緩衝液中で10〜30分間遠心処理された場合に、5000〜20000×重力の遠心処理によって、そのタンパク質またはポリペプチドが沈殿し得ることを意味する。不溶性タンパク質またはポリペプチドは、封入体または他の沈殿物質塊の一部であり得る。「封入体(inclusion body)」という用語には、細胞内に含有されている、タンパク質またはポリペプチドの凝集体が隔離されている任意の細胞内物体が含まれるものとする。
【0084】
別の実施形態では、最適な宿主細胞集団は、ペリプラズムに輸送されるか、または宿主細胞の細胞外空間に分泌される、対象とするタンパク質を、増大した量で産生する。一実施形態では、アレイ内の少なくとも1つの株が、少なくとも0.1mg/mlのタンパク質をペリプラズム区画に産生する。別の実施形態では、少なくとも1つの株が、0.1〜10mg/mlのペリプラズムタンパク質、または少なくとも0.2ml、約0.3ml、約0.4ml、約0.5ml、約0.6ml、約0.7ml、約0.8ml、約0.9mlもしくは少なくとも約1.0mg/mlのペリプラズムタンパク質を細胞内で産生する。一実施形態では、アレイ内の少なくとも1つの株によって産生される、対象とするタンパク質またはポリペプチドの総タンパク質量が、少なくとも1.0mg/ml、少なくとも約2mg/ml、少なくとも約3mg/ml、約4mg/ml、約5mg/ml、約6mg/ml、約7mg/ml、約8mg/ml、約10mg/ml、約15mg/ml、約20mg/mlもしくは少なくとも約25mg/mlであるか、またはそれより多い。一部の実施形態では、産生されるペリプラズムタンパク質の量が、産生される対象とするタンパク質またはポリペプチドの少なくとも約5%、約10%、約15%、約20%、約25%、約30%、約40%、約50%、約60%、約70%、約80%、約90%、約95%、約96%、約97%、約98%もしくは約99%であるか、またはそれより多い。
【0085】
本発明のアレイの少なくとも1つの株は、対象とするタンパク質またはポリペプチドの収率の増大ももたらし得る。一実施形態では、少なくと1つの株が、対象とするタンパク質またはポリペプチドを、全細胞タンパク質(total cell protein)(tcp)の少なくとも約5%、少なくとも約10%、約15%、約20%、約25%、約30%、約40%、約45%、約50%、約55%、約60%、約65%、約70%もしくは約75%、またはそれより多い量で産生する。「パーセント全細胞タンパク質(percent total cell protein)」は、総計細胞タンパク質のパーセントとしての、宿主細胞内のタンパク質またはポリペプチドの量である。パーセント全細胞タンパク質を測定する方法は、当技術分野でよく知られている。
【0086】
特定の実施形態では、無機塩培地中で増殖(すなわち、約10℃、約15℃、約20℃、約25℃、約30℃、約35℃、約40℃、約45℃および約50℃を含めた、約4℃〜約55℃の温度範囲内で増殖)させた場合、アレイ内の少なくとも1つの宿主細胞集団が、少なくとも1%tcpの異種タンパク質産生レベルおよび少なくとも40mg/mlの細胞密度を有し得る。特に好ましい実施形態では、無機塩培地中で増殖(すなわち、包括的に約4℃〜約55℃の温度範囲内で増殖)させた場合、発現系が、少なくとも5%tcpのタンパク質またはポリペプチド発現レベルおよび少なくとも40g/Lの細胞密度を有するであろう。
【0087】
実際、ペリプラズムにターゲッティングされた異種タンパク質は、おそらく外側細胞膜への損傷、またはその流動性の増大から、しばしば、ブロス(broth)中に見出される(欧州特許第EP0 288 451号を参照)。この「受動的な」分泌の速度は、コリシン(Mikschら(1997)、Arch.Microbiol.167:143〜150)、増殖速度(Shokriら(2002)、App Miocrobiol Biotechnol 58:386〜392)、TolIII過剰発現(WanおよびBaneyx(1998)、Protein Expression Purif.14:13〜22)、バクテリオシン放出タンパク質(Hsiungら(1989)、Bio/Technology7:267〜71)、コリシンA溶解タンパク質(Lloubesら(1993)、Biochimie 75:451〜8)、周辺質(periplasmic)タンパク質を漏出する変異体(FurlongおよびSundstrom(1989)、Developments in Indus.Microbio.30:141〜8)、融合パートナー(JeongおよびLee(2002)、Appl.Environ.Microbio.68:4979〜4985)または浸透圧ショックによる回収(Taguchiら(1990)、Biochimica Biophysica Acta 1049:278〜85)を含めた、外側細胞膜を透過処理する様々な作用機序を用いることによって増大し得る。E.coliで適切にフォールディングされた活性タンパク質を産生するのに、周辺腔(periplasmic space)への改変タンパク質の輸送、およびそれに続くブロス中への局在化が使用されている(WanおよびBaneyx(1998)、Protein Expression Purif.14:13〜22;Simmonsら(2002)、J.Immun.Meth.263:133〜147;Lundellら(1990)、J.Indust.Microbio.5:215〜27)。
【0088】
この方法は、対象とするタンパク質またはポリペプチドをペリプラズムから、または細胞外の培地から精製するステップも含み得る。異種タンパク質またはポリペプチドがタグタンパク質に連結されるような方法で、それを発現させることができ、「タグ付き」のタンパク質を細胞または細胞外の培地から精製することができる。
【0089】
一部の実施形態では、対象とするタンパク質またはポリペプチドを、活性形態でアレイの少なくとも1つの株によって産生することもできる。「活性」という用語は、生物活性の存在を意味し、生物活性は、対応する天然タンパク質またはポリペプチドの生物活性に匹敵するか、それに実質的に相当する。タンパク質に関する文脈では、通常これは、標準的なパラメータを用いて、対応する天然タンパク質またはポリペプチドと比較した場合、ポリヌクレオチドまたはポリペプチドが、少なくとも約20%、約50%、好ましくは少なくとも約60〜80%、そして最も好ましくは少なくとも約90〜95%の活性を有する生物学的機能または作用を含むことを意味する。しかし、一部の実施形態では、天然タンパク質と比較して、改変または改善された活性を有するポリペプチド(例えば免疫応答性および基質の特異性などを改変または改善したもの)を産生することが望ましい可能性がある。改変または改善されたポリペプチドは、アレイ内の1つまたは複数の宿主細胞集団によって生成された特定の立体配座の結果として生じたものであり得る。
【0090】
タンパク質またはポリペプチドの活性の測定は、特定のタンパク質またはポリペプチドを標的とした、対応する標準的な生物学的比較アッセイを用いて行うことができ、それらのアッセイを用いて、生物活性を評価することができる。
【0091】
対象とする活性なタンパク質またはポリペプチドの回収も、本発明のアレイ内の1つまたは複数の他の株と比較して、最適な宿主株で改善されている可能性がある。活性タンパク質は、その配列が由来する天然タンパク質またはポリペプチドの活性の少なくとも約20%、少なくとも約30%、少なくとも約40%、約50%、約60%、少なくとも約70%、約80%、約90%、または少なくとも95約%の比活性を有し得る。さらに、基質特異性(kcat/Km)が天然タンパク質またはポリペプチドと実質的に同様であってもよい。通常、kcat/Kmは、少なくとも約30%、約40%、約50%、約60%、約70%、約80%、少なくとも約90%もしくは少なくとも約95%、またはそれより大きいであろう。タンパク質およびポリペプチドの活性および基質特異性(kcat/Km)の尺度をアッセイおよび定量化する方法は、当業者によく知られている。
【0092】
タンパク質活性の測定(Measurement of protein activity)
対象とする異種発現タンパク質またはポリペプチドの活性は、事前に確立されている天然タンパク質またはポリペプチドの標準的な活性と比較できる。代替として、対象のタンパク質またはポリペプチドの活性は、同時または実質的に同時に行われる、天然タンパク質またはポリペプチドを用いた比較アッセイで測定できる。例えば、発現された酵素と基質との間、発現されたホルモンとホルモン受容体との間、および発現された抗体と抗原との間の相互作用など、例えば、対象とするタンパク質またはポリペプチドと標的との間の検出可能な相互作用を測定するのに、in vitroアッセイを用いることができる。そのような検出には、熱量変化の測定、増殖変化、細胞死、細胞忌避(cell repelling)、放射能の変化、溶解性の変化、ゲル電気泳動および/またはゲル濾過法で測定した分子量の変化、リン酸化能、ELISAアッセイなどの抗体特異性アッセイなどが含まれ得る。加えて、in vivoアッセイには、天然タンパク質またはポリペプチドの生理作用と比較して、異種発現されたタンパク質またはポリペプチドの生理作用を検出するアッセイ、例えば、質量増大、電解質バランスの変化、血液凝固時間の変化、血餅融解の変化、および抗原性反応の誘導を検出するアッセイが含まれるが、これらに限定されない。通常、対象とするタンパク質またはポリペプチドの活性特性を、そのような活性がアッセイ可能である限り、天然タンパク質またはポリペプチドとの比較分析を可能にする任意のin vitroまたはin vivoアッセイを用いて決定できる。代替として、本発明のアレイ内の少なくとも1つの株で産生されたタンパク質またはポリペプチドを、そのタンパク質またはポリペプチドと、通常、そのタンパク質またはポリペプチドと相互作用する分子、例えば天然タンパク質が通常相互作用するシグナル経路の基質または構成要素との間の相互作用を刺激または抑制する能力についてアッセイできる。そのようなアッセイは、通常、タンパク質またはポリペプチドが標的分子と相互作用するのを可能にする条件下で、タンパク質を基質分子と混合し、上記タンパク質と標的分子との相互作用の生化学的帰結を検出するステップを含み得る。
【0093】
タンパク質またはポリペプチドの活性を測定するのに利用できるアッセイは、例えば、Ralph,P.J.ら(1984)、J.Immunol.132:1858もしくはSaikiら(1981)、J.Immunol.127:1044、Steward,W.E.II(1980)、「The Interferon Systems」、Springer−Verlag,ViennaおよびNew York、Broxmeyer,H.E.ら(1982)、Blood 60:595、「Molecular Cloning:A Laboratory Manua」、第2版、Cold Spring Harbor Laboratory Press、Sambrook,J.、E.F.FritschおよびT.Maniatis編集、1989、および「Methods in Enzymology:Guide to Molecular Cloning Techniques」、Academic Press、Berger,S.L.およびA.R.Kimmel編集、1987、A K Patraら、Protein Expr Purif,18(2):182〜92ページ(2000)、Kodamaら、J.Biochem.99:1465〜1472(1986)、Stewartら、Proc.Nat’l Acad.Sci.USA 90:5209〜5213(1993)、(Lombilloら、J.Cell Biol.128:107〜115(1995);(Valeら、Cell 42:39〜50(1985)に記載されている。アレイ内の1つまたは複数の他の宿主細胞集団に由来する、異種発現されたタンパク質の試料間で活性を比較することも、天然タンパク質の活性と比較することも、またはそれらを両方行うこともできる。活性測定は、単離されたタンパク質で行うこともでき、または宿主細胞内でin vitroで行うこともできる。
【0094】
別の実施形態では、例えば、従来の顕微鏡、ルミノメーターまたはプレートリーダーにおける蛍光測定または分光測定によって、タンパク質産生および/または活性を培養物中で直接モニターすることができる。対象とするタンパク質が、その基質が既知の酵素である場合、基質を培地に添加することができ、基質が酵素によって産物に変換された際に蛍光シグナルが放出される。一実施形態では、対象とする異種タンパク質またはポリペプチドをコードする発現コンストラクトが、レポータータンパク質をさらにコードする。「レポータータンパク質」は、細胞内または細胞表面にそれが存在することによって、または培地中に分泌された場合に、レポータータンパク質を含有しない細胞からその細胞を識別するのを可能にするタンパク質を意味する。対象とする異種タンパク質の産生は、宿主細胞集団の検出可能な変化をもたらす。レポーター分子は、ホタルルシフェラーゼおよびGFP、または他の任意の蛍光分子でもよく、β−ガラクトシダーゼ遺伝子(beta.gal)、クロラムフェニコールおよびアセチルトランスフェラーゼ遺伝子(CAT)でもよい。これらのレポーター遺伝子エレメントそれぞれと連動して産生される発現のアッセイは、当業者によく知られている。
【0095】
レポーター遺伝子は、検出可能なタンパク質もしくは間接的に検出可能なタンパク質をコードするものであり得、またはレポーター遺伝子が生存遺伝子であり得る。好ましい実施形態では、レポータータンパク質が検出可能なタンパク質である。(検出可能な遺伝子または検出遺伝子によってコードされている)「検出可能なタンパク質」または「検出タンパク質」は、直接標識として使用できるタンパク質であり、すなわち、このタンパク質は、さらなる操作なしに検出可能(好ましくは、上記検出可能なタンパク質を含む細胞が検出可能)である。したがって、この実施形態では、レポーター遺伝子のタンパク質産物それ自体が、検出可能な遺伝子を発現している細胞を識別するのに役立ち得る。この実施形態では、適した検出可能遺伝子に、自己蛍光タンパク質をコードするものが含まれる。
【0096】
当技術分野で知られている通り、様々な既知の自己蛍光タンパク質が存在しており、これらは通常、Aequoreaの緑色蛍光タンパク質(GFP)およびその変種をベースにしており、これらには、GFP(Chalfieら(1994)、Science 263(5148):802〜805)、高感度GFP(EGFP;Clontech−Genbank受託番号U55762)、青色蛍光タンパク質(BFP;Quantum Biotechnologies,Inc.、Montreal,Canada);Stauber(1998)、Biotechniques 24(3):462〜471;HeimおよびTsien(1996)、Curr.Biol.6:178〜182)、高感度黄色蛍光タンパク質(EYFP;Clontech Laboratories,Inc.、Palo Alto,CA)、および赤色蛍光タンパク質が含まれるが、これらに限定されない。加えて、RenillaおよびPtilosarcus種由来の自己蛍光タンパク質が最近報告されている。全てが参照により明示的に本明細書に組み込まれている、WO92/15673、WO95/07463、WO98/14605、WO98/26277、WO99/49019、米国特許第5,292,658号、米国特許第5,418,155号、米国特許第5,683,888号、米国特許第5,741,668号、米国特許第5,777,079号、米国特許第5,804,387号、米国特許第5,874,304号、米国特許第5,876,995号および米国特許第5,925,558を参照のこと。
【0097】
対象とするタンパク質またはポリペプチドの単離(Isolation of Protein or Polypeptide of Interest)
対象とするタンパク質の収率、溶解性、立体配座および/または活性を測定するために、アレイ内の1つまたは複数の株からタンパク質を単離することが望ましい可能性がある。適切な測定を行うのに用いたアッセイの必要条件に応じて、単離は、粗単離でも、準粗単離でも、純粋単離でもよい。タンパク質は、細胞質で産生されるものでも、ペリプラズムにターゲッティングされるものでも、または培養もしくは発酵培地中に分泌されるものでもよい。ペリプラズムにターゲッティングされたタンパク質を遊離させるには、クロロホルム(Amesら(1984)、J.Bacteriol.、160:1181〜1183)、グアニジン−HClおよびトリトンX−100(NaglakおよびWang(1990)、Enzyme Microb.Technol.、12:603〜611)などの化学物質を用いた処置が使用されている。しかし、これらの化学物質は不活性ではなく、多くの異種タンパク質産物またはその後の精製方法に対する有害効果を有し得る。E.coliのグリシン処理は、外膜の透過化を引き起こし、ペリプラズム内容物を放出させることも報告されている(Arigaら(1989)、J.Ferm.Bioeng.68:243〜246)。異種タンパク質をペリプラズムから放出させる最も広範に使用されている方法は、浸透圧ショック(NosalおよびHeppel(1966)、J.Biol.Chem.241:3055〜3062;NeuおよびHeppel(1965)、J.Biol.Chem.240:3685〜3692)、雌鳥卵白(HEW)−リゾチーム/エチレンジアミン四酢酸(EDTA)処理(NeuおよびHeppel(1964)、J.Biol.Chem、239:3893〜3900;Witholtら(1976)、Biochim.Biophys.Acta,443:534〜544;Pierceら(1995)、ICheme Research.Event,2:995〜997)、およびHEW−リゾチーム/浸透圧ショック併用処理(Frenchら(1996)、Enzyme and Microb.Tech.19:332〜338)である。French法は、分画緩衝液中の細胞の再懸濁、およびそれに続くペリプラズム画分の回収を行ものであり、その際、浸透圧ショックはリゾチーム処理のすぐ後に行う。
【0098】
通常、これらの手順は、浸透圧的に安定化されている培地中での初期破壊と、それに続く、非安定化培地中における選択的放出とを含む。これらの培地の組成(pH、保護剤)および使用される破壊方法(クロロホルム、HEW−リゾチーム、EDTA、超音波処理)は、報告されている特定な手順相互で異なっている。EDTAの代わりに双極性イオン界面活性剤を用いるHEW−リゾチーム/EDTA処理の変法が、Stabelら(1994)、Veterinary Microbiol.38:307〜314に論じられている。E.coliを破壊するための細胞内溶菌酵素系の使用に関する一般的概説には、DaboraおよびCooney(1990)、Advances in Biochemical Engineering/Biotechnology、第43巻、A.Fiechter編集(Springer−Verlag:Berlin)、11〜30ページを参照のこと。
【0099】
対象とするタンパク質またはポリペプチドを、可溶性タンパク質または屈折性粒子として細胞質から回収する従来の方法は、機械的な切断による細菌細胞の崩壊を用いる。機械的破壊には、通常、液体懸濁液中に局所的なキャビテーションの産生、強固なビーズを用いた高速撹拌、超音波処理または細胞懸濁液のグラインディングを用いる(「Bacterial Cell Surface Techniques,Hancock and Poxton」(John Wiley&Sons Ltd,1988)、第3章、55ページ)。
【0100】
HEW−リゾチームは、細胞壁のペプチドグリカンバックボーンを加水分解するように生化学的に作用する。この方法は、最初、ZinderおよびArndt(1956)、Proc.Natl.Acad.Sci.USA、42:586〜590によって開発されたが、ZinderおよびArndtは、卵アルブミン(HEW−リゾチームを含有する)でE.coliを処理して、後にスフェロプラストとして知られる丸い細胞球を産生させた。これらの構造は、一部の細胞壁成分を保持していたが、細胞質膜が露出した大きな表面積を有していた。米国特許第5,169,772号は、浸透圧的に安定化された培地中、例えば20%ショ糖液中で、例えば、EDTA、リゾチーム、または有機化合物を用いて細菌のエンベロープを破壊するステップと、細菌を低イオン強度緩衝液に暴露することによって、破壊した細菌の周辺腔から非ヘパリナーゼ様タンパク質を遊離させるステップと、低イオン強度で洗浄された細菌を緩衝化塩溶液に暴露することによって、ヘパリナーゼ様タンパク質を遊離させるステップとを含む、細菌からヘパリナーゼを精製する方法を開示する。
【0101】
これらの方法の多くの異なった変法が、広範な発現系で使用されており、様々な程度に成功している(Joseph−Liazunら(1990)、Gene、86:291〜295;Carterら(1992)、Bio/Technology、10:163〜167)。組換え細胞培養物を誘導して、リゾチームを産生させる試みが報告されている。EP0 155 189は、組換え細胞培養物を誘導して、リゾチームを産生させる手段を開示し、この手段は、通常、細胞壁構造を破壊または溶解することによって、そのような宿主細胞を死滅させると予測される。
【0102】
米国特許第4,595,658号は、細菌の周辺腔に輸送されたタンパク質の外在化を促進する方法を開示する。この方法は、細胞のリゾチーム処理、機械的粉砕または浸透圧ショック処理を必要とせずに、ペリプラズム中に局在するタンパク質の選択的単離を可能にする。米国特許第4,637,980号は、産物を直接的または間接的にコードするDNA分子で温度感受性溶原菌を形質転換させ、形質転換体を許容条件下で培養して、細胞内で遺伝子産物を発現させ、温度を上げることによって産物を外在化させ、ファージにコードされている機能を誘導することによって、細菌産物を産生させる方法を開示する。Asamiら(1997)、J.Ferment and Bioeng、83:511〜516は、T4ファージ感染によるE.coli細胞の同調破壊を開示し、Tanjiら(1998)、J.Ferment.and Bioeng,85:74〜78は、E.coli細胞を穏やかに破壊するための、T4ファージコードされている溶菌遺伝子の調節発現を開示する。
【0103】
細胞溶解の際には、ゲノムDNAが細胞質から培地中に漏出し、その結果、流体粘度の著しい増大がもたらされる。これは、遠心力場内における固形物の沈降を妨害し得る。DNA重合体を破壊するための機械的な破壊の間に作用するものなどのせん断力の不在下では、粘性流体を通る固形物の沈降速度が遅延する結果、遠心処理中の固形物と液体との分離が悪くなる。機械的せん断力の他に、DNA重合体を分解する核酸分解酵素が存在する。E.coliでは、内因性遺伝子であるendAが、エンドヌクレアーゼ(成熟タンパク質の分子量は約24.5kD)をコードしている。エンドヌクレアーゼは、通常、ペリプラズムに分泌され、ヌクレオチド鎖切断の様式でDNAをオリゴデオキシリボヌクレオチドへと切断する。E.coliによって、endAが比較的弱く発現されると示唆されている(Wackemagelら(1995)、Gene 154:55〜59)。
【0104】
望ましい場合には、本発明のアレイ内の1つまたは複数の株を用いて産生されたタンパク質を、当技術分野でよく知られている標準的な技法によって、実質的な純度にまで単離および精製することができ、これらの技法には、硫酸アンモニウムもしくはエタノール沈殿、酸抽出、陰イオンもしくは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ニッケルクロマトグラフィー、ヒドロキシアパタイトクロマトグラフィー、逆相クロマトグラフィー、レクチンクロマトグラフィー、分取電気泳動、界面活性剤可溶化、カラムクロマトグラフィーのような物質を用いた選択的沈殿、および免疫精製法などが含まれるが、これらに限定されない。例えば、確立された分子付着特性を有するタンパク質をリガンドに可逆的に融合させることができる。適切なリガンドを用いて、タンパク質を精製カラムに選択的に吸着させ、その後、比較的純粋な形態でカラムから遊離させることができる。融合タンパク質は、その後、酵素活性によって除去する。加えて、イムノアフィニティーカラムまたはNi−NTAカラムを用いて、タンパク質を精製することができる。一般技法については、例えば、R.Scopes、「Protein Purification:Principles and Practice」、Springer−Verlag:N.Y.(1982);Deutscher、「Guide to Protein Purification」Academic Press(1990);米国特許第4,511,503号;S.Roe、「Protein Purification Techniques:A Practical Approach(Practical Approach Series)」、Oxford Press(2001);D.Bollagら、「Protein Methods」、Wiley−Lisa,Inc.(1996);AK Patraら、Protein Expr Purif;18(2)、182〜92ページ(2000);およびR.Mukhijaら、Gene 165(2):303〜6ページ(1995)にさらに記載されている。例えば、Ausubelら(1987年および定期増補);Deutscher(1990)、「Guide to Protein Purification」、Methods in Enzymology、第182巻およびこのシリーズの他の巻;Coliganら(1996年および定期増補)、「Current Protocols in Protein Science」、Wiley/Greene,NY;ならびにタンパク質精製製品の使用に関する製造会社、例えば、Pharmacia、Piscataway,N.J.またはBio−Rad、Richmond,Calif.の文献も参照のこと。組換え技法との併用によって、プロテアーゼで除去可能な配列を介して融合させることのできる、例えばFLAG配列またはその等価物に、適したセグメントを融合させることが可能となる。例えば、Hochuli(1989)、Chemische Industrie 12:69〜70;Hochuli(1990)、「Purification of Recombinant Proteins with Metal Chelate Absorbent」、Setlow(編集)、Genetic Engineering,Principle and Methods 12:87−98、Plenum Press,NY;およびCroweら(1992)、QIAexpress:The High Level Expression&Protein Purification System、QUIAGEN,Inc、Chatsworth,Calif.も参照のこと。
【0105】
発現されたタンパク質の検出は、当技術分野で知られている方法で行われ、それには、例えば、ラジオイムノアッセイ、ウエスタンブロット技法または免疫沈降反応が含まれる。
【0106】
本発明のアレイ内の株によって発現される特定のタンパク質は、不溶性の凝集体(「封入体」)を形成し得る。いくつかのプロトコールが、封入体からタンパク質を精製するのに適している。例えば、封入体の精製は通常、例えば、50mM TRIS/HCL、pH7.5、50mM NaCl、5mM MgCl2、1mM DTT、0.1mM ATPおよび1mM PMSFという緩衝液中でのインキュベーションによって、宿主細胞を破壊することによる、封入体の抽出、分離および/または精製を要する。細胞懸濁液は、通常、フレンチプレスに通した2〜3回の通過を用いて溶解する。細胞懸濁液は、Polytron(Brinkman Instruments)を用いてホモジナイズすることも、氷上で超音波処理することもできる。細菌を溶解する代替法は、当業者には明らかである(例えば、Sambrookら、同上;Ausubelら、同上を参照)。
【0107】
必要な場合には、封入体は可溶化させることができ、通常は、好ましくない不溶物を除去するために、溶解細胞懸濁液を遠心処理することができる。封入体を形成したタンパク質は、適合した緩衝液で希釈または透析することによって再生させることができる。適した溶剤には、尿素(約4M〜約8M)、ホルムアミド(少なくとも約80%、容積/容積ベース)および塩酸グアニジン(約4M〜約8M)が含まれるが、これらに限定されない。塩酸グアニジンおよび類似の薬剤は、変性剤であるが、この変性は不可逆的でなく、変成剤を除去(例えば透析によって)または希釈した際に、再生(renaturation)が起こり得る。これにより、免疫学的および/または生物学的に活性なタンパク質の再形成(re-formation)が可能となる。他の適した緩衝液も、当業者に知られている。
【0108】
当業者によく知られている標準的な分離技法によって、上清中に存在する異種発現タンパク質を宿主タンパク質から分離することができる。例えば、最初の塩分画によって、多くの好ましくない宿主細胞タンパク質(細胞培地由来のタンパク質)を、対象とするタンパク質またはポリペプチドから分離し得る。その一例が硫酸アンモニウムであり得る。硫酸アンモニウムは、タンパク質混合物中の水の量を効果的に低下させることによって、タンパク質を沈殿させる。その際、タンパク質は、それらの溶解性に基づいて沈殿する。タンパク質が疎水性であるほど、より低い濃度の硫酸アンモニウムで沈殿する可能性が大きい。典型的なプロトコールは、その結果の硫酸アンモニウム濃度が20〜30%となるように、飽和硫酸アンモニウムをタンパク質溶液に添加するステップを含む。この濃度は、ほとんどの疎水性タンパク質を沈殿させるであろう。その後、(対象とするタンパク質が疎水性でない限り)沈殿を廃棄し、対象とするタンパク質を沈殿させることが既知の濃度まで上清に硫酸アンモニウムを添加する。その後、緩衝液中で沈殿を可溶化し、必要に応じて、透析(dialysis)またはダイアフィルトレーション(diafiltration)を介して、過剰な塩を除去する。冷エタノール沈殿など、タンパク質の溶解性に依存した他の方法も、当業者にはよく知られており、複雑なタンパク質混合物を分画するのに使用できる。
【0109】
対象とするタンパク質またはポリペプチドの分子量を用いて、それを、より大きなサイズのタンパク質およびより小さなサイズのタンパク質から、異なった孔径の膜(例えばAmiconまたはMillipore膜)を通す限外濾過を用いて単離することができる。最初のステップとして、対象とするタンパク質の分子量より小さい分子量のカットオフを有する孔径を備えた膜を通して、タンパク質混合物を限外濾過にかけることができる。その後、対象とするタンパク質の分子量より大きな分子カットオフを有する膜に対して、保持液の限外濾過を行うことができる。対象とするタンパク質またはポリペプチドは、膜を通り抜けて、濾液中に入る。その後、以下に記載の通り、濾液をクロマトグラフィーにかけることができる。
【0110】
発現された、対象とするタンパク質またはポリペプチドを、そのサイズ、正味の表面電荷、疎水性度、およびリガンドへのアフィニティーに基づいて、他のタンパク質から分離することができる。加えて、タンパク質に対して産生された抗体をカラムマトリクスに結合させ、そのタンパク質を免疫精製することができる。これらの方法は全て、当技術分野でよく知られている。任意のスケールで、かつ多くの異なった製造会社(例えばPharmacia Biotech)の機器を用いて、クロマトグラフィー技法を実施できることは、当業者には明らかであろう。
【0111】
再生およびリフォールディング(Renaturation and Refolding)
異種発現されたタンパク質が変性形態で産生される場合、二次および三次タンパク質構造を生成するように、不溶性タンパク質を再生またはリフォールディングさせることができる。異種産物の立体配座を完成させるのに、タンパク質リフォールディングステップを、必要に応じて用いることができる。当技術分野で知られている、タンパク質の解離/会合を促進する薬剤を用いて、リフォールディングおよび再生を実現できる。例えば、タンパク質をジチオスレイトールと共にインキュベートし、それに続いて、酸化グルタチオン二ナトリウム塩と共にインキュベートし、それに続いて、尿素などのリフォールディング剤を含有する緩衝液と共にインキュベートすることができる。
【0112】
例えば、リン酸緩衝食塩水(PBS)または50mM Na−酢酸、pH6緩衝液に200mM NaClを加えたものに対して、対象とするタンパク質またはポリペプチドを透析することによって、それを再生させることもできる。代替として、プロテアーゼ阻害物質を含有する、500mM NaCl、20%グリセロール、20mM Tris/HCl pH7.4中における直線的な6Mから1Mの尿素勾配を用いることによって、Ni NTAカラムなどのカラムに固定化しながら、タンパク質をリフォールディングさせることができる。1.5時間を超える時間にわたって、再生を行うことができる。再生の後、250mMイミダゾールを添加することによって、タンパク質を溶出できる。イミダゾールは、PBSまたは50mM酢酸ナトリウムpH6緩衝液に200mM NaClを加えたものに対する最終の透析ステップによって除去できる。精製されたタンパク質は、4℃で保存するか、または−80℃で冷凍できる。
【0113】
他の方法には、例えば、M H Leeら、Protein Expr.Purif、25(1):166〜73ページ(2002)、W.K.Choら、J.Biotechnology,77(2〜3):169〜78ページ(2000)、Ausubelら(1987年および定期的な増補)、Deutscher(1990)「Guide to Protein Purification」、Methods in Enzymology、第182巻、およびこのシリーズの他の巻、Coliganら(1996年および定期的な増補)、「Current Protocols in Protein Science」、Wiley/Greene,NY、S.Roe、「Protein Purification Techniques:A Practical Approach(Practical Approach Series)」、Oxford Press(2001年);D.Bollagら、「Protein Methods」、Wiley−Lisa,Inc.(1996)に記載されている可能性のあるものが含まれる。
【0114】
発現ベクター(Expression vectors)
対象とする異種タンパク質は、それぞれの菌株に対象とする異種タンパク質をコードする発現ベクターを導入することによって、本明細書に開示する1つまたは複数の宿主細胞において産生することができる。一実施形態では、ベクターは、選択した宿主細胞において働くことができるプロモーター、ならびに全ての他の必要とされる転写および翻訳調節エレメントと作用可能に連結した対象とするタンパク質をコードするポリヌクレオチド配列を含む。
【0115】
用語「作用可能に連結した(Operably linked)」は、転写および任意の翻訳調節エレメントが、コード配列に対して、宿主細胞中またはその作用によって、調節エレメントがコード配列の発現を誘導することができるような配置で、コード配列と共有結合した任意の形態を指す。
【0116】
対象とする異種タンパク質は、異種ポリペプチドコード配列が転写および翻訳調節エレメントと作用可能に連結して、宿主細胞がそこからタンパク質またはポリペプチドを発現することができる機能遺伝子を形成する、ポリヌクレオチドから発現させることが可能である。コード配列は異種ポリペプチドの天然コード配列であってよく、あるいは、例えば、宿主種のコドン使用の偏向を反映するために遺伝子を合成することによって、選択した発現宿主細胞中で使用するために選択、改良、または最適化されたコード配列であってよい。本発明の一実施形態では、宿主種はP.fluorescensであり、ポリペプチドコード配列の設計時には、P.fluorescensのコドンの偏向を考慮に入れる。(1つまたは複数の)遺伝子を1つまたは複数のベクター内に構築し、または1つまたは複数のベクターに挿入し、次いで発現宿主細胞に形質転換することができる。
【0117】
他の調節エレメントを、(「発現コンストラクト(expression construct)」とも呼ばれる)ベクター中に含めることができる。ベクターは典型的には、1つまたは複数の表現型選択可能マーカーおよび1つの複製起点を含んで、ベクターの維持を確実にし、および望ましい場合、宿主内の増幅をもたらす。追加のエレメントには、発現したポリペプチドの同定、分離、精製、および/または単離を容易にする、例えば転写エンハンサー配列、翻訳エンハンサー配列、他のプロモーター、アクチベーター、翻訳開始および停止シグナル、転写ターミネーター、シストロン性レギュレーター、ポリシストロン性レギュレーター、またはヌクレオチド配列「タグ」および「タグ」ポリペプチドコード配列などのタグ配列があるが、これらだけには限られない。
【0118】
別の実施形態では、発現ベクターは、対象とするタンパク質またはポリペプチドのコード配列と隣接したタグ配列をさらに含む。一実施形態では、このタグ配列はタンパク質の精製を可能にする。タグ配列は、ヘキサ−ヒスチジン親和性タグなどの親和性タグであってよい。別の実施形態では、親和性タグはグルタチオン−S−トランスフェラーゼ分子であってよい。タグは、YFPまたはGFPなどの蛍光分子、またはこのような蛍光タンパク質の類似体であってもよい。タグは、抗体分子、または既知の抗原もしくは精製に有用な既知の結合パートナーのリガンドの一部分であってもよい。
【0119】
本発明によるタンパク質コード遺伝子は、タンパク質コード配列に加えて、そこに作用可能に連結した以下の調節エレメント:プロモーター、リボソーム結合部位(ribosome binding site)(RBS)、転写ターミネーター、翻訳開始および停止シグナルを含むことができる。有用なRBSは、本発明による発現系中で宿主細胞として有用な任意の種から、好ましくは選択した宿主細胞から得ることができる。多くの特異的および様々なコンセンサスRBS、例えばD.Frishmanら、Gene234(2):257〜65ページ(1999年7月8日);およびB.E.Suzekら、Bioinformatics17(12):1123〜30(2001年12月)中に記載されそれらによって言及されたRBSが知られている。さらに、天然または合成RBSのいずれか、例えばEP0207459(合成RBS);O.Ikehataら、Eur.J.Biochem.181(3):563〜70ページ(1989)(AAGGAAGの天然RBS配列)中に記載されたRBSを使用することができる。本発明中で有用な方法、ベクター、および翻訳および転写エレメント、および他のエレメントのさらなる例は、例えばGilroyへの米国特許第5,055,294号、およびGilroyらへの米国特許第5,128,130号、Rammlerらへの米国特許第5,281,532号、Barnesらへの米国特許第4,695,455号および米国特許第4,861,595号、Grayらへの米国特許第4,755,465号、およびWilcoxへの米国特許第5,169,760号中に記載されている。
【0120】
対象とする異種タンパク質をコードするDNAの転写は、エンハンサー配列をベクターまたはプラスミドに挿入することによって増大する。典型的なエンハンサーは、プロモーターに作用してその転写を増大させる、通常約10〜300bpの大きさのDNAのシス作用性エレメントである。例には様々なシュードモナスのエンハンサーがある。
【0121】
一般に、異種発現ベクターは、宿主細胞の形質転換を可能にする複製起点および選択可能マーカー、および下流構造配列の転写を誘導するための高発現遺伝子由来のプロモーターを含む。このようなプロモーターは、特に3−ホスホグリセレートキナーゼ(PGK)、酸性ホスファターゼ、または熱ショックタンパク質などの酵素をコードするオペロンから誘導することができる。シグナル配列を使用する場合、異種コード配列は翻訳開始および停止配列と適切に一致して構築され、かつシグナル配列は翻訳されるタンパク質の部分的蓄積または分泌を誘導することができる。場合によっては異種配列は、望ましい性質、例えば発現される異種産物の安定性または精製の単純化をもたらすN末端同定ポリペプチドを含む融合酵素をコードすることができる。前に論じたように、融合ポリペプチドは、1つまたは複数の標的タンパク質またはその阻害剤もしくはエンハンサーも含むことができる。
【0122】
宿主細胞中で異種タンパク質を発現させるためのベクターが当技術分野で知られており、任意のこれらのベクターは、本発明による遺伝子を発現させるために使用することができる。このようなベクターには、例えばプラスミド、コスミド、およびファージ発現ベクターがある。有用なプラスミドベクターの例には、発現プラスミドpBBR1MCS、pDSK519、pKT240、pML122、pPS10、RK2、RK6、pRO1600、およびRSF1010があるが、これらだけには限られない。このような有用なベクターの他の例には、例えば:N.Hayase、Appl.Envir.Microbiol.60(9):3336〜42ページ(1994年9月)中、A.A.Lushnikovら、Basic Life Sci.30:657〜62ページ(1985)中、S.Graupner & W.Wackemagel、Biomolec.Eng.17(1):11〜16ページ(2000年10月)中、H.P.Schweizer、Curr.Opin.Biotech.12(5):439〜45ページ(2001年10月)中、M.Bagdasarian & K.N.Timmis、Curr.Topics Microbiol.Immunol.96:47〜67ページ(1982)中、T.Ishiiら、FEMS Microbiol.Lett.116(3):307〜13ページ(1994年3月1日)中、I.N.Olekhnovich & Y.K.Fomichev、Gene140(1):63〜65ページ(1994年3月11日)中、M.Tsuda & T.Nakazawa、Gene136(1〜2):257〜62ページ(1993年12月22日)中、C.Nietoら、Gene87(1):145〜49ページ(1990年3月1日)中、J.D.Jones & N.Gutterson、Gene61(3):299〜306ページ(1987)中、M.Bagdasarianら、Gene16(1〜3):237〜47ページ(1981年12月)中、H.P.Schweizerら、Genet.Eng.(NY)23:69〜81ページ(2001年)中、P.Mukhopadhyayら、J.Bact.172(1):477〜80ページ(1990年1月)中、D.O.Woodら、J.Bact.145(3):1448〜51ページ(1981年3月)中、およびR.Holtwickら、Microbiology147(Pt2):337〜44ページ(2001年2月)中によって記載されたベクターがある。
【0123】
本発明の宿主細胞において有用であり得る発現ベクターのさらなる例には、示したレプリコン由来の表5中に挙げたベクターがある。
【0124】
【表5】
【0125】
発現プラスミド、RSF1010は、例えばF.Heffronらによって、Proc.Nat’l Acad.Sci.USA72(9):3623〜27ページ(1975年9月)中、およびK.Nagahari & K.Sakaguchiによって、J.Bact.133(3):1527〜29ページ(1978年3月)中に記載されている。プラスミドRSF1010およびその誘導体は、本発明中で特に有用なベクターである。当技術分野で知られているRSF1010の例示的で有用な誘導体には、例えばpKT212、pKT214、pKT231および関連プラスミド、およびpMYC1050および関連プラスミド(例えば、Thompsonらへの米国特許第5,527,883号および米国特許第5,840,554号を参照)、例えばpMYC1803などがある。プラスミドpMYC1803は、調節型テトラサイクリン耐性マーカーならびにRSF1010プラスミド由来の複製および可動性遺伝子座を有する、RSF1010系プラスミドpTJS260に由来する(Wilcoxへの米国特許第5,169,760号を参照)。他の例示的で有用なベクターには、Puhlerらへ米国特許第4,680,264号の中に記載されたベクターがある。
【0126】
一実施形態では、発現プラスミドは発現ベクターとして使用する。別の実施形態では、RSF1010またはその誘導体を発現ベクターとして使用する。さらに別の実施形態では、pMYC1050またはその誘導体、またはpMYC4803またはその誘導体を発現ベクターとして使用する。
【0127】
プラスミド中に選択マーカー遺伝子を封入することによって、プラスミドを宿主細胞中に維持することができる。これは(1つまたは複数の)抗生物質耐性遺伝子であってよく、この場合対応する(1つまたは複数の)抗生物質、または当技術分野で知られている任意の他型の選択マーカー遺伝子、例えば原栄養体復元遺伝子を発酵培地に加え、この場合プラスミドは、対応する形質、例えばアミノ酸生合成またはヌクレオチド生合成形質、または炭素源利用形質などの生体触媒形質の栄養要求性である宿主細胞において使用する。
【0128】
本発明に従って使用するプロモーターは、構成的プロモーター(constitutive promoters)または調節型プロモーター(regulated promoters)であってよい。有用な調節型プロモーターの一般例には、lacプロモーター由来のファミリーのプロモーター(すなわち、lacZプロモーター)、特にDeBoerへの米国特許第4,551,433号中に記載されたtacおよびtrcプロモーター、ならびにPtac16、Ptac17、PtacII、PlacUV5、およびT71acプロモーターがある。一実施形態では、プロモーターは宿主細胞生物に由来しない。特定の実施形態では、プロモーターはE.coli生物に由来する。
【0129】
本発明による発現系中で有用な非lac型プロモーターの一般例には、例えば表6中に挙げたプロモーターがある。
【0130】
【表6】
【0131】
例えば、J.Sanchez−Romero & V.De Lorenzo(1999)Manual of Industrial Microbiology and Biotechnology(A.Demain & J.Davies、eds.)460〜74ページ(ASM Press、Washington、D.C.);H.Schweizer(2001)Current Opinion in Biotechnology、12:439〜445ページ;およびR.Slater & R.Williams(2000 Molecular Biology and Biotechnology(J.Walker & R.Rapley、eds.)125〜54ページ(The Royal Society of Chemistry、Cambridge、UK))を参照。選択した細菌宿主細胞に由来するプロモーターのヌクレオチド配列を有するプロモーター、例えばPseudomonasアントラニル酸塩または安息香酸オペロンプロモーター(Pant、Pben)を使用して、標的ポリペプチドをコードするトランス遺伝子の発現を制御することもできる。配列が同じであろうと異なろうと複数のプロモーターが互いに共有結合しているタンデムプロモーター、または同じ生物または異なる生物のいずれに由来しようと、例えばPant−Pbenタンデムプロモーター(プロモーター間ハイブリッド)またはPlac−Placタンデムプロモーターを使用することもできる。
【0132】
調節型プロモーターはプロモーター調節タンパク質を利用して、プロモーターがその一部分である遺伝子の転写を制御する。調節型プロモーターを本明細書において使用する場合、対応するプロモーター調節タンパク質も本発明による発現系の一部分であり得る。プロモーター調節タンパク質の例には、活性化タンパク質、例えばE.coliカタボライト活性化タンパク質、MalTタンパク質、AraCファミリー転写活性化因子、リプレッサータンパク質、例えば、E.coliLacIタンパク質、および二重機能調節タンパク質、例えばE.coliNagCタンパク質がある。多くの調節型プロモーター/プロモーター調節タンパク質のペアが当技術分野で知られている。一実施形態では、(1つまたは複数の)標的タンパク質の発現コンストラクトと対象とする異種タンパク質は、同じ調節エレメントの制御下にある。
【0133】
プロモーター調節タンパク質は、エフェクター化合物、すなわち可逆的または不可逆的に調節タンパク質と会合して、タンパク質の放出、またはプロモーターの制御下にある遺伝子の少なくとも1つのDNA転写調節領域との結合のいずれかを可能にし、それによって遺伝子の転写を開始する際のトランスクリプターゼ酵素の作用を許容または阻害する化合物と相互作用する。エフェクター化合物はインデューサーまたはコリプレッサーのいずれかとして分類され、かつこれらの化合物は、天然エフェクター化合物および余分なインデューサー化合物を含む。多くの調節型プロモーター/プロモーター調節タンパク質/エフェクター化合物のトリオが当技術分野で知られている。エフェクター化合物は細胞培養または発酵中を通して使用することができるが、調節型プロモーターを使用する好ましい実施形態では、望ましい量または密度の宿主細胞バイオマスの増殖後、適切なエフェクター化合物を培養物に直接的または間接的に加え、対象とするタンパク質またはポリペプチドをコードする(1つまたは複数の)望ましい遺伝子の発現をもたらす。
【0134】
例えば、lacファミリーのプロモーターを利用する場合、lacI遺伝子が系中に存在してもよい。(通常)構成的に発現される遺伝子であるlacI遺伝子は、これらのプロモーターのlacオペレーターと結合するLacリプレッサータンパク質(LacDタンパク質)をコードする。したがって、lacファミリーのプロモーターを利用する場合、lacI遺伝子を発現系中に含め発現させることが可能である。lacプロモーターファミリーのメンバー、例えばtacプロモーターの場合、エフェクター化合物はインデューサー、好ましくはIPTG(「イソプロピルチオガラクトシド」とも呼ばれるイソプロピル−D−1−チオガラクトピラノシドなどの余分なインデューサーである。
【0135】
対象とするタンパク質またはポリペプチドの発現用に、任意の植物プロモーターを使用することもできる。プロモーターは植物RNAポリメラーゼIIプロモーターであってよい。植物プロモーター中に含まれるエレメントは、転写開始部位の約25〜35塩基対上流(5’)に典型的に位置するTATAボックスまたはGoldberg−Hognessボックス、および70と100塩基対上流の間に位置するCCAATボックスであってよい。植物において、CCAATボックスは、哺乳動物プロモーターの機能的に類似した配列と異なるコンセンサス配列を有し得る(Messingら、(1983)In:Genetic Engineering of Plants、Kosugeら、eds.、211〜227ページ)。さらに、ほぼ全てのプロモーターは、転写開始部位の約−100bp〜−1,000bp以上上流に広がる追加の上流活性化配列またはエンハンサーを含む(Benoist and Chambon(1981)Nature290:304〜310ページ;Grussら(1981)Proc.Nat.Acad.Sci.78:943〜947ページ;およびKhoury and Gruss(1983)Cell27:313〜314ページ)。
【0136】
発現系(Expression Systems)
対象とするタンパク質またはポリペプチドを、アレイ内の1つまたは複数の宿主細胞集団のペリプラズム、または細胞外空間に標的化することは望ましい可能性がある。一実施形態では、発現ベクターは、対象とするタンパク質またはポリペプチドをコードするヌクレオチド配列と作用可能に連結した、分泌シグナル配列ポリペプチドをコードするヌクレオチド配列をさらに含む。いくつかの実施形態では、シグナル配列と対象とするタンパク質またはポリペプチドの間に修飾を施さない。しかしながら、特定の実施形態では、追加の切断シグナルを取り込んでポリペプチドのアミノ末端の正確なプロセシングを促進する。
【0137】
ベクターは、前に記載した特性のいずれかを有し得る。一実施形態では、対象とするタンパク質またはポリペプチドのコード配列を含むベクターは、シグナル配列、例えば分泌シグナル配列をさらに含む。
【0138】
したがって一実施形態では、この単離したポリペプチドは、分泌シグナルと対象とするタンパク質またはポリペプチドの融合タンパク質である。しかしながら、タンパク質がペリプラズムを標的化するとき、分泌シグナルをタンパク質から切断することもできる。一実施形態では、Sec系分泌シグナルとタンパク質またはポリペプチドの間の結合を修飾して分泌シグナルの切断を増大する。
【0139】
CHAMPION(商標)pET発現系は高レベルのタンパク質産生をもたらす。発現は強力なT7lacプロモーターから誘導される。この系は、対象とする遺伝子の高レベルの転写に関するバクテリオファージT7RNAポリメラーゼの高い活性および特異性を利用する。プロモーター領域に位置するlacオペレーターは伝統的なT7系ベクターより強い調節をもたらし、プラスミドの安定性および細胞の生存を改善する(Studier and Moffatt(1986)J Molecular Biology189(1):113〜30ページ;Rosenbergら(1987)Gene56(1):125〜35ページ)。T7発現系は、対象とする遺伝子の高レベルの転写にT7プロモーターおよびT7RNAポリメラーゼ(T7RNAP)を使用する。T7RNAPは天然E.coliRNAPよりプロセシングされやすく、対象とする遺伝子の転写に限られるので、高レベルの発現がT7発現系において得られる。同定した遺伝子の発現は、宿主細胞中にT7RNAPの供給源を提供することによって誘導する。これはT7RNAP遺伝子の染色体コピーを含有するBL21E.coli宿主を使用することによって実施する。T7RNAP遺伝子は、IPTGによって誘導することができるlacUV5プロモーターの制御下にある。T7RNAPは誘導によって発現され、対象とする遺伝子を転写する。
【0140】
pBAD発現系は、グルコース、グリセロールおよびアラビノースなどの特異的炭素源の存在によって、対象とするタンパク質またはポリペプチドの厳重に制御された、滴定可能な発現を可能にする(Guzmanら、(1995)J Bacteriology 177(14):4121〜30ページ)。pBADベクターは独自に設計して発現レベルに対する正確な制御をもたらす。pBADベクターからの異種遺伝子の発現はaraBADプロモーターで始まる。このプロモーターは、araC遺伝子の産物によって正と負の両方に制御される。AraCは、L−アラビノースと複合体を形成する転写レギュレーターである。L−アラビノースの不在下では、AraC二量体は転写を阻害する。最大の転写活性化のためには、2つの事象、(i)L−アラビノースがAraCと結合して転写の開始を可能にすること、および(ii)cAMP活性化タンパク質(CAP)−cAMP複合体がDNAと結合してプロモーター領域の正確な位置へのAraCの結合を刺激することが必要とされる。
【0141】
trc発現系は、E.coli中でのtrcプロモーターからの、高レベルの制御された発現を可能にする。trc発現ベクターは、E.coli中での真核生物遺伝子の発現用に最適化されている。trcプロモーターは、トリプトファン(trp)およびラクトース(lac)プロモーター由来の強力なハイブリッドプロモーターである。それはlacOオペレーターおよびlacIQ遺伝子の産物によって調節される(Brosius、J.(1984)Gene27(2):161〜72ページ)。
【0142】
本明細書に開示する(1つまたは複数の)ベクターを用いた宿主細胞の形質転換は、当技術分野で知られている任意の形質転換法を使用して実施することができ、かつ細菌宿主細胞は無傷細胞(intact cells)として、またはプロトプラスト(すなわち、サイトプラスト含む)として形質転換することができる。例示的な形質転換法にはポレーション法、例えばエレクトロポレーション、プロトプラスト融合、細菌接合、および二価カチオン処理、例えば塩化カルシウム処理またはCaCl/Mg2+処理、または当技術分野でよく知られている他の方法がある。例えば、Morrison、J Bact.、132:349〜351ページ(1977年);Clark−Curtiss & Curtiss、Methods in Enzymology、101:347〜362ページ(Wuら、eds、1983年)、Sambrookら、Molecular Cloning、A Laboratory Manual(1989年第2版);Kriegler、Gene Transfer and Expression:A Laboratory Manual(1990年);およびCurrent Protocols in Molecular Biology(Ausubelら、eds.、1994年))を参照。
【0143】
対象とするタンパク質(Proteins of interst)
本発明の方法および組成物は、高レベルの適切にプロセシングされた対象とするタンパク質またはポリペプチドを産生するのに最適であるP.fluorescens菌株を同定するのに有用である。任意の種および任意の大きさの対象とするタンパク質またはポリペプチドの産生をスクリーニングするのに、アレイは有用である。しかしながら、特定の実施形態では、対象とするタンパク質またはポリペプチドは、治療上有用なタンパク質またはポリペプチドである。いくつかの実施形態では、タンパク質は哺乳動物タンパク質、例えばヒトタンパク質であってよく、かつ例えば成長因子、サイトイン、ケモカインまたは血中タンパク質であってよい。対象とするタンパク質またはポリペプチドは、天然タンパク質またはポリペプチドと同様の方法でプロセシングすることができる。特定の実施形態では、対象とするタンパク質またはポリペプチドは、100kD未満、50kD未満、または30kD未満の大きさである。特定の実施形態では、対象とするタンパク質またはポリペプチドは、少なくとも約5、10、15、20、30、40、50または100またはそれより多くのアミノ酸のポリペプチドである。
【0144】
対象とするタンパク質またはポリペプチドのコード配列は、ポリペプチドの天然コード配列であってよく、利用可能である場合、但し、例えば、P.fluorescensなどのシュードモナス種または他の適切な生物のコドン使用の偏向を反映するために遺伝子を最適化することによって、アレイの菌株において発現可能な形で使用するために選択、改良、または最適化されたコード配列であることがより好ましい。遺伝子最適化のために、1つまたは複数の稀なコドンを除去してリボゾーム消失を避け、アミノ酸の取り込み違いを最小にすることができる。1つまたは複数の遺伝子内リボソーム結合部位を除去して、切断型タンパク質産物を避けることもできる。CおよびGヌクレオチドの長い延長部分を除去して、フレームシフトをもたらす可能性があるRNAポリメラーゼのスリップを避けることができる。強力な遺伝子内ステムループ構造、特にリボソーム結合部位を覆う構造を除去することもできる。
【0145】
他の実施形態では、タンパク質は産生時に、追加の標的配列、例えばタンパク質をペリプラズムまたは細胞外培地に標的化する配列も含む。一実施形態では、その追加の標的配列は、タンパク質のカルボキシ末端と作用可能に連結する。別の実施形態では、タンパク質は、オートトランスポーターの分泌シグナル、2つのパートナー分泌系、主要末端分岐系または線毛外膜ポーリンを含む。
【0146】
生成する(1つまたは複数の)遺伝子は1つまたは複数のベクター内で構築されるか、またはその中に挿入され、次いでアレイ内の宿主細胞集団のそれぞれに形質転換される。「発現可能な形」で提供されると言われる核酸またはポリヌクレオチドは、本発明の1つまたは複数の宿主細胞集団によって発現され得る少なくとも1つの遺伝子を含有する、核酸またはポリヌクレオチドを意味する。
【0147】
分子遺伝学および遺伝子工学技法に必要とされる広範囲の配列情報が、広く公に利用されている。哺乳動物の完全なヌクレオチド配列、ならびにヒト、遺伝子、cDNA配列、アミノ酸配列およびゲノムへのアクセスは、ウェブサイトwww.ncbi.nlm.nih.gov/EntrezでGenBankから得ることができる。追加の情報はGeneCards、遺伝子およびそれらの産物に関する情報を統合した電子百科事典、およびWeizmann Institute of Science Genome and Bioinformatics(bioinformatics.weizmann.ac.il/cards)からの生物医学的応用から得ることもでき、ヌクレオチド配列情報は、EMBLヌクレオチド配列データベース(www.ebi.ac.uk/embl/)またはDNAデータバンクまたは日本(the DNA Databank or Japan)(DDBJ、www.ddbi.nig.ac.ii/;アミノ酸配列に関する情報の追加のサイトには、Georgetownのタンパク質情報リソースウェブサイト(www−nbrf.Reorgetown.edu/pirl)およびSwiss−Prot(au.expasy.org/sprot/sprot−top.html)がある)から得ることもできる。
【0148】
本発明において発現させることが可能であるタンパク質の例には、例えばレニン、ヒト成長ホルモン、ウシ成長ホルモン、成長ホルモン放出因子を含めた成長ホルモン、副甲状腺ホルモン、甲状腺刺激ホルモン、リポタンパク質、α−1−アンチトリプシン、インシュリンA鎖、インシュリンB鎖、プロインシュリン、トロンボポエチン、卵胞刺激ホルモン、カルシトニン、黄体形成ホルモン、グルカゴン、因子VIIIC、因子IX、組織因子、およびフォンウィルブランド因子などの凝固因子、プロテインCなどの抗凝固因子、心房性ナトリウム利尿因子、肺表面活性剤、ウロキナーゼまたはヒト尿または組織型プラスミノーゲン活性化因子(t−PA)などのプラスミノーゲン活性化因子、ボムベシン、トロンビン、造血成長因子、腫瘍壊死因子−αおよび−β、エンケファリナーゼ、ヒト血清アルブミンなどの血清アルブミン、ミュラー管抑制物質、レラキシンA鎖、レラキシンB鎖、プロレラキシン、マウスゴナドトロピン関連ポリペプチド、β−ラクタマーゼなどの微生物タンパク質、デオキシリボヌクレアーゼ、インヒビン、アクチビン、血管内皮増殖因子(VEGF)、ホルモンまたは増殖因子に対する受容体、インテグリン、プロテインAまたはD、リウマチ因子、脳由来神経栄養因子(BDNF)、ニューロトロフィン−3、−4、−5、または−6(NT−3、NT−4、NT−5、またはNT−6)、またはNGF−βなどの神経成長因子などの神経栄養因子、カルジオトロフィン−1(CT−1)などのカルジオトロフィン(心肥大因子)、血小板由来成長因子(PDGF)、aFGFおよびbFGFなどの線維芽細胞成長因子、表皮成長因子(EGF)、TGF−αおよびTGF−β1、TGF−β2、TGF−β3、TGF−β4、またはTGF−β5を含めたTGF−βなどの形質転換成長因子(TGF)、インシュリン様成長因子−Iおよび−II(IGF−IおよびIGF−II);des(1〜3)−IGF−I(脳IGF−I)、インシュリン様成長因子結合タンパク質、CD−3、CD−4、CD−8、およびCD−19などのCDタンパク質、エリスロポエチン、骨誘導因子、免疫毒素、骨形成タンパク質(BMP)、インターフェロン−α、−β、および−γなどのインターフェロン、コロニー刺激因子(CSF)、例えばM−CSF、GM−CSF、およびG−CSF、インターロイキン(IL)、例えばIL−1〜IL−10、抗HER−2抗体、スーパーオキシドジスムターゼ、T細胞受容体、表面膜タンパク質、分解促進因子、例えばAIDSエンベロープの一部分などのウイルス抗原、輸送タンパク質、ホーミング受容体、アドレシン、調節タンパク質、抗体、および前に挙げたポリペプチドのいずれかの断片などの分子がある。
【0149】
特定の実施形態では、タンパク質またはポリペプチドは、IL−1、IL−1a、IL−1b、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12、IL−12elasti、IL−13、IL−15、IL−16、IL−18、IL−18BPa、IL−23、IL−24、VIP、エリスロポエチン、GM−CSF、G−CSF、M−CSF、血小板由来成長因子(PDGF)、MSF、FLT−3リガンド、EGF、線維芽細胞成長因子(FGF;例えば、α−FGF(FGF−1)、β−FGF(FGF−2)、FGF−3、FGF−4、FGF−5、FGF−6、またはFGF−7)、インシュリン様成長因子(例えば、IGF−1、IGF−2)、腫瘍壊死因子(例えば、TNF、リンホトキシン)、神経成長因子(例えば、NGF)、血管内皮増殖因子(VEGF)、インターフェロン(例えば、IFN−α、IFN−β、IFN−γ)、白血病抑制因子(LIF)、絨毛神経栄養因子(CNTF)、オンコスタチンM、幹細胞因子(SCF)、形質転換成長因子(例えば、TGF−α、TGF−β1、TGF−β2、TGF−β3)、TNFスーパーファミリー(例えば、LIGHT/TNFSF14、STALL−l/TNFSF13B(BLy5、BAFF、THANK)、TNFα/TNFSF2およびTWEAK/TNFSF12)、またはケモカイン(BCA−1/BLC−1、BRAK/Kec、CXCL16、CXCR3、ENA−78/LIX、エオタキシン−1、エオタキシン−2/MPIF−2、エクソダス(Exodus)−2/SLC、フラクタルカイン/ニューロタクチン、GROα/MGSA、HCC−1、I−TAC、リンホタクチン/ATAC/SCM、MCP−1/MCAF、MCP−3、MCP−4、MDC/STCP−1/ABCD−1、MIP−1.クアドラチュア(quadrature)、MIP−1.クアドラチュア、MIP−2.クアドラチュア/GRO.クアドラチュア、MIP−3.クアドラチュア./エクソダス/LARC、MIP−3/エクソダス−3/ELC、MIP−4/PARC/DC−CK1、PF−4、RANTES、SDF1、TARC、TECK、微生物毒素、ADPリボシル化毒素、微生物またはウイルス抗原)から選択することができる。
【0150】
本発明の一実施形態では、対象とするタンパク質はマルチサブユニットタンパク質またはポリペプチドであってよい。発現させることが可能であるマルチサブユニットタンパク質には、同種および異種タンパク質がある。マルチサブユニットタンパク質は、同じであるかまたは異なってよい2個以上のサブユニットを含むことができる。例えばタンパク質は、2、3、4、5、6、7、8、9、10、11、12個またはそれより多くのサブユニットを含む同種タンパク質であってよい。タンパク質は、2、3、4、5、6、7、8、9、10、11、12個、またはそれより多くのサブユニットを含む異種タンパク質であってもよい。例示的なマルチサブユニットタンパク質には、イオンチャネル受容体を含めた受容体、コンドロイチン、コラーゲンを含めた細胞外マトリクスタンパク質、MHCタンパク質、完全鎖抗体、および抗体断片を含めた免疫修飾物質、RNAポリメラーゼ、およびDNAポリメラーゼを含めた酵素、および膜タンパク質がある。
【0151】
別の実施形態では、対象とするタンパク質は血中タンパク質であってよい。この実施形態において発現される血中タンパク質には、担体タンパク質、ヒトおよびウシアルブミンを含めたアルブミンなど、トランスフェリン、組換え体トランスフェリン半分子、ハプトグロビン、フィブリノゲンおよび他の凝固因子、補体成分、免疫グロブリン、酵素阻害剤、アンギオテンシンおよびブラジキニン、インシュリン、エンドセリン、およびα、β、およびγ−グロブリンを含めたグロブリンなどの物質の前駆体、および哺乳動物の血液中で主に見られる他の型のタンパク質、ポリペプチド、およびそれらの断片があるが、これらだけには限られない。ヒト血清アルブミンに関するアミノ酸配列(Lawn、L.Mら、(1981年)Nucleic Acids Research、9:6103〜6114ページ)およびヒト血清トランスフェリンに関するアミノ酸配列(Yang、Fら、(1984年)Proc.Natl.Acad.Sci.USA81:2752〜2756ページ)を含めた、多数の血中タンパク質に関するアミノ酸配列が報告されてきている(S.S.Baldwin(1993年)Comp.Biochem Physiol.106b:203〜218ページ参照)。
【0152】
別の実施形態では、対象とするタンパク質は酵素または補因子であってよい。この実施形態において発現される酵素および補因子には、アルドラーゼ、アミンオキシダーゼ、アミノ酸オキシダーゼ、アスパルターゼ、B12依存性酵素、カルボキシペプチダーゼ、カルボキシエステラーゼ、カルボキシリアーゼ、ケモトリプシン、CoA要求酵素、シアノヒドリンシンテターゼ、シスタチオンシンターゼ、デカルボキシラーゼ、デヒドロゲナーゼ、アルコールデヒドロゲナーゼ、デヒドラターゼ、ジアホラーゼ、ジオキシゲナーゼ、エノアートリダクターゼ、エポキシドヒドラーゼ、フメラーゼ、ガラクトースオキシダ−ゼ、グルコースイソメラーゼ、グルコースオキシダーゼ、グリコシルトランスフェラーゼ、メチルトランスフェラーゼ、ニトリルヒドラーゼ、ヌクレオシドホスホリラーゼ、オキシドリダクターゼ、オキシニチラーゼ、ペプチダーゼ、グリコシルトランスフェラーゼ、ペロキシダーゼ、治療活性ポリペプチドと融合した酵素、組織型プラスミノーゲン活性化因子、ウロキナーゼ、レプチラーゼ、ストレプトキナーゼ、カタラーゼ、スーパーオキシドジスムターゼ、デオキシリボヌクレアーゼ、アミノ酸加水分解酵素(例えば、アスパラギナーゼ、アミド加水分解酵素)、カルボキシペプチダーゼ、プロテアーゼ、トリプシン、ペプシン、キモトリプシン、パパイン、ブロメライン、コラゲナーゼ、ノイラミニダーゼ、ラクターゼ、マルターゼ、スクラーゼ、およびアラビノフラノシダーゼがあるが、これらだけには限られない。
【0153】
別の実施形態では、対象とするタンパク質は単鎖、Fab断片および/または完全鎖抗体、またはその断片もしくは一部分であってよい。単鎖抗体は、1つの安定的にフォールディングしたポリペプチド鎖上の抗体の抗原結合領域を含むことができる。Fab断片は特定の抗体の一片であってよい。Fab断片は抗原結合部位を含有することができる。Fab断片は2本の鎖、軽鎖および重鎖断片を含有することができる。これらの断片は、リンカーまたはジスルフィド結合によって連結させることが可能である。
【0154】
他の実施形態では、対象とするタンパク質は、約20〜約42℃の温度で活性があるタンパク質である。一実施形態では、タンパク質は生理的温度で活性があり、高温または極温(high or extreme temperatures)、65℃を超える温度に加熱すると不活性である。
【0155】
一実施形態では、対象とするタンパク質は約20〜約42℃の温度で活性があるタンパク質であり、および/または高温または極温、65℃を超える温度に加熱すると不活性であり、あるいは天然哺乳動物またはヒトタンパク質などの天然タンパク質と実質的に相同的であり、コンカタマー型の核酸から発現されず、この場合プロモーターはアレイ内で使用する宿主細胞における天然プロモーターではなく、E.coliなどの他の生物に由来する。
【0156】
宿主細胞(Host cell)
一実施形態では本発明は、そこから対象とする異種タンパク質またはペプチドを場合によっては産生するための、P.fluorescens宿主細胞のアレイを提供する。P.fluorescensは様々なタンパク質を産生するための改良された基盤であることが実証されており、いくつかの有効な分泌シグナルがこの生物から同定されている(その全容が参照によって本明細書に組み込まれる、米国出願公開第2006/0008877号を参照)。
【0157】
シュードモナス系は、他の細菌発現系と比較して、ポリペプチドおよび酵素の工業的発現に関して利点をもたらす。特に、P.fluorescensは有利な発現系として同定されている。P.fluorescensは、土壌、水および植物表面環境にコロニー形成する、一群の一般的な、非病原性の腐生菌を含む。洗浄添加剤として、および立体選択的加水分解用に、P.fluorescens由来の市販の酵素が使用されて、環境汚染物質が減らされている。P.fluorescensは病原体を制御するために農業用にも使用されている。米国特許第4,695,462号は、P.fluorescensにおける組換え体細菌タンパク質の発現を記載している。
【0158】
しかしながら、代替宿主細胞、またはさらに多様な異なる宿主細胞を使用して、前に論じたように1つまたは複数の標的遺伝子の発現を調節するために遺伝子改変された複数の表現型が異なる宿主細胞を含む、アレイを作製することができることは企図(comtemplated)される。宿主細胞は、その中で標的遺伝子を改変することができる任意の生物であってよい。表1および2中に挙げた標的遺伝子と相同的な標的遺伝子を同定する方法は、当技術分野で知られている。さらに、当業者は、対象とする宿主細胞に由来するかまたはその中で有用である標的遺伝子の同定の仕方を理解しているはずである。これらのタンパク質の多くは当技術分野でよく知られている。例えば、米国特許出願公開第2006/0110747号)を参照。
【0159】
宿主細胞は「グラム陰性プロテオバクテリア亜群18」から選択することができる。「グラム陰性プロテオバクテリア亜群18」は、例えば以下の(丸括弧中に示すATCCまたは他の寄託番号の(1つまたは複数の)例示的菌株):次亜種1または次亜種Iとも呼ばれる、Pseudomonas fluorescensバイオタイプA(ATCC13525);次亜種2または次亜種IIとも呼ばれる、Pseudomonas fluorescensバイオタイプB(ATCC17816);次亜種3または次亜種IIIとも呼ばれる、Pseudomonas fluorescensバイオタイプC(ATCC17400);次亜種4または次亜種IVとも呼ばれる、Pseudomonas fluorescensバイオタイプF(ATCC12983);次亜種5または次亜種Vとも呼ばれる、Pseudomonas fluorescensバイオタイプG(ATCC17518);Pseudomonas fluorescensバイオタイプVI;Pseudomonas fluorescensPf0−1;Pseudomonas fluorescensPf−5(ATCC BAA−477);Pseudomonas fluorescensSBW25;およびPseudomonas fluorescens亜種cellulosa(NCIMB10462)に属するものを含めた、Pseudomonas fluorescens種の全ての亜種、変種、菌株、および他の下位専門単位の群として定義する。
【0160】
宿主細胞は「グラム陰性プロテオバクテリア亜群19」から選択することができる。「グラム陰性プロテオバクテリア亜群19」は、Pseudomonas fluorescensバイオタイプAの全ての菌株の群として定義する。このバイオタイプの特に好ましい菌株は、P.fluorescens菌株MB101(Wilcoxへの米国特許第5,169,760号を参照)、およびその誘導体である。その好ましい誘導体の一例は、MB101染色体asd(アスパラギン酸デヒドロゲナーゼ遺伝子)遺伝子座に挿入することによって構築されたP.fluorescens菌株MB214、天然E.coliPlacI−lacI−lacZYAコンストラクト(すなわち、その中でPlacZが欠失した)である。
【0161】
本発明において使用することができる追加のP.fluorescens菌株には、以下のATCC指定を有する、Pseudomonas fluorescens MigulaおよびPseudomonas fluorescens Loitokitokがある:[NCIB 8286];NRRL B−1244;NCIB 8865株CO1;NCIB 8866株CO2;1291[ATCC 17458;IFO 15837;NCIB 8917;LA;NRRL B−1864;ピロリジン;PW2[ICMP 3966;NCPPB 967;NRRL B−899];13475;NCTC 10038;NRRL B−1603[6;IFO 15840];52−1C;CCEB 488−A[BU 140];CCEB 553[EM 15/47];IAM 1008[AHH−27];IAM 1055[AHH−23];I[IFO 15842];12[ATCC 25323;NIH 11;den Dooren de Jong 216];18[IFO 15833;WRRL P−7];93[TR−10];108[52−22;IFO 15832];143[IFO 15836;PL];149[2−40−40;IFO 15838];182[IFO 3081;PJ 73];184[IFO 15830];185[W2 L−1];186[IFO 15829;PJ 79];187[NCPPB 263];188[NCPPB 316];189[PJ227;1208];191[IFO 15834;PJ 236;22/1];194[Klinge R−60;PJ 253];196[PJ 288];197[PJ 290];198[PJ 302];201[PJ 368];202[PJ 372];203[PJ 376];204[IFO 15835;PJ 682];205[PJ 686];206[PJ 692];207[PJ 693];208[PJ 722];212.[PJ 832];215[PJ 849];216[PJ 885];267[B−9];271[B−1612];401[C71A;IFO 15831;PJ 187];NRRL B−3178[4;IFO.15841];KY 8521;3081;30−21;[IFO 3081];N;PYR;PW;D946−B83[BU 2183;FERM−P 3328];P−2563[FERM−P 2894;IFO 13658];IAM−1126[43F];M−1;A506[A5−06];A505[A5−05−1];A526[A5−26];B69;72;NRRL B−4290;PMW6[NCIB 11615];SC 12936;Al[IFO 15839];F 1847[CDC−EB];F 1848[CDC 93];NCIB 10586;P17;F−12;AmMS 257;PRA25;6133D02;6519E01;Ni;SC15208;BNL−WVC;NCTC 2583[NCIB 8194];H13;1013[ATCC 11251;CCEB 295];IFO 3903;1062;またはPf−5。
【0162】
一実施形態では、宿主細胞は、前に記載したP.fluorescens細胞を含めた、対象とするタンパク質またはポリペプチドを産生することができる任意の細胞であってよい。対象とするタンパク質またはポリペプチドを産生するために最も一般的に使用される系は、大規模なバッチ式培養においてタンパク質を産生するための、それらの比較的安価な増殖要件および潜在能力のために、特定の細菌細胞、特にE.coliを含む。酵母菌をさらに使用して、特に研究目的で、生物学的関連があるタンパク質およびポリペプチドを発現させる。系にはSaccharomyces cerevisiaeまたはPichia pastorisがある。これらの系は十分特徴付けられており、一般に認められるレベルの全タンパク質産生をもたらし、比較的迅速および安価である。昆虫細胞発現系も、生物活性型の組換えタンパク質を発現させるための代替として浮上している。いくつかの場合、翻訳後修飾された、正確にフォールディングされたタンパク質を産生することができる。チャイニーズハムスター卵巣細胞などの哺乳動物細胞発現系も、対象とするタンパク質またはポリペプチドの発現用に使用されている。小規模では、これらの発現系は有効であることが多い。特定の生物製剤は、特に動物またはヒトの健康用途で、タンパク質から誘導することができる。別の実施形態では、宿主細胞は、これらだけには限られないが、タバコ細胞、コーン細胞、シロイヌナズナ種由来の細胞、ジャガイモまたはコメ細胞を含めた植物細胞である。
【0163】
別の実施形態では、宿主細胞は、これらだけには限られないが、エシェリキアまたはシュードモナス種を含めた細菌細胞などの原核細胞であってよい。典型的な細菌細胞は、ウェブサイトwww.emc.maricotpa.edu/faculty/farabee/BIOBK/BioBookDiversityで、Estrella Mountain Community College、Arizona、USAのDr M J Farabeeによって提供された、例えば「Biological Diversity:Bacteria and Archaeans」、the On−Line Biology Bookの一章中に記載されている。特定の実施形態では、宿主細胞はシュードモナス細胞であってよく、かつ典型的にはP.fluorescens細胞であってよい。他の実施形態では、宿主細胞はE.coli細胞であってもよい。別の実施形態では、宿主細胞は真核細胞、例えばこれらだけには限られないがスポドプテラ、トリコプルシア、ショウジョウバエまたはエスティグメン種由来の細胞を含めた昆虫細胞、またはこれらだけには限られないがネズミ細胞、ハムスター細胞、サル、霊長類またはヒト細胞を含めた哺乳動物細胞であってよい。
【0164】
一実施形態では、宿主細胞は細菌分類群のいずれかのメンバーであってよい。細胞は例えば、真正細菌の任意の種のメンバーであってよい。宿主は、分類群:アシドバクテリア、アクチノバクテリア、アクイフェックス、バクテロイデス、クロロビウム、クラミジア、コロフレキシ、クリシオジェネス、シアノバクテリア、デフェリバクター、デイノコッカス、ディクチオグロムス、フィブロバクター、ファーミキューテス、フソバクテリア、ゲマティモナス、レンチスファエラ、ニトロスピラ、プランクトミセス、プロテオバクテリア、スピロヘータ、サーモデスルフォバクテリア、サーモミクロビア、サーモトガ、サーマス(サーマレス)、またはベルコミクロビアのいずれか1つのメンバーであってよい。真正細菌宿主細胞の一実施形態では、細胞は、シアノバクテリアを除く真正細菌の任意の種のメンバーであってよい。
【0165】
細菌宿主は、プロテオバクテリアの任意の種のメンバーであってもよい。プロテオバクテリア宿主細胞は、分類群アルファプロテオバクテリア、ベータプロテオバクテリア、ガンマプロテオバクテリア、デルタプロテオバクテリア、またはイプシロンプロテオバクテリアのいずれか1つのメンバーであってよい。さらに宿主は、分類群アルファプロテオバクテリア、ベータプロテオバクテリア、またはガンマプロテオバクテリアのいずれか1つのメンバー、およびガンマプロテオバクテリアの任意の種のメンバーであってよい。
【0166】
ガンマプロテオバクテリア宿主の一実施形態では、宿主は、分類群エロモナス目、アルテロモナス目、腸内細菌目、シュードモナス目、またはキサントモナス目のいずれか1つのメンバー、または腸内細菌目もしくはシュードモナスの目の任意の種のメンバーであり得る。一実施形態では、宿主細胞は腸内細菌の目であってよく、宿主細胞は腸内細菌科のメンバーであり得る、またはエルビニア、エシェリキア、またはセラチア属のいずれか1つのメンバー、またはエシェリキア属のメンバーであってよい。宿主細胞がシュードモナスの目である場合、宿主細胞は、シュードモナス属を含めたシュードモナス科のメンバーであってよい。ガンマプロテオバクテリア宿主には、Escherichaia.coli種のメンバーおよびPseudomonas fluorescens種のメンバーがある。
【0167】
他のシュードモナス生物も有用である可能性がある。シュードモナスおよび近縁の種には、R.E.Buchanan and N.E.Gibbons(eds.)、Bergey’s Manual of Determinative Bacteriology、217〜289ページ(第8版、1974年)(The Williams & Wilkins Co.、Baltimore、Md.、USA)(本明細書では以後「Bergey(1974)」)によって「Gram−Negative Aerobic Rods and Cocci」として記載された、科および/または属に属するプロテオバクテリアの群を含む、グラム陰性プロテオバクテリア亜群1がある。表7は、これらの生物の科および属を表す。
【0168】
【表7】
【0169】
「グラム陰性プロテオバクテリア亜群1」は、分類において使用する基準に従ってこの表題に分類され得るプロテオバクテリアも含む。この表題は、この項で前に分類したがそれ以上は分類されない群、アシドボラックス、ブレブンジモナス、バークホルデアリア、好気性水素酸化細菌、オーシャンイモナス、ラルストニア、およびステノトロフォモナス属、(以前に命名された種の)キサントモナス属に属する生物を再グループ化することによって生成したスフィンゴモナス属(およびそれに由来するブラストモナス属)、Bergey(1974)において定義されたアセトバクター属に属する生物を再グループ化することによって生成したアシドモナス属なども含む。さらに宿主は、それぞれAlteromonas haloplanktis、Alteromonas nigrifaciens、およびAlteromonas putrefaciensとして再分類された、シュードモナス属、Pseudomonas enalia(ATCC14393)、Pseudomonas nigrifaciensi(ATCC19375)、およびPseudomonas putrefaciens(ATCC8071)由来の細胞を含むことができる。同様に、例えば、Pseudomonas acidovorans(ATCC15668)およびPseudomonas testosteroni(ATCC11996)が、それぞれComamonas acidovoransおよびComamonas testosteroniとして以後再分類されており、かつPseudomonas nigrifaciens(ATCC19375)およびPseudomonas piscicida(ATCC15057)が、それぞれPseudoalteromonas nigrifaciensおよびPseudoalteromonas piscicidaとして再分類されている。「グラム陰性プロテオバクテリア亜群1」は、科:シュードモナス科、アゾトバクター科(現在、同義語、「シュードモナス科のアゾトバクター族」によって呼ばれることが多い)、リゾビウム科、およびメチロモナス科(現在、同義語、「メチロコッカス科」によって呼ばれることが多い)のいずれかに属するとして分類されたプロテオバクテリアも含む。したがって、本明細書で他に記載する属に加えて、「グラム陰性プロテオバクテリア亜群1」の範疇のさらなるプロテオバクテリア属には、1)アゾリゾフィルス属のアゾトバクター群の細菌、2)セルビブリオ、オリゲラ、およびテレジニバクター属のシュードモナス科の細菌、3)ケラトバクター、エンシファー、リベリバクター(「カンジダタスリベリバクター」とも呼ばれる)、およびシノリゾビウム属のリゾビウム科の細菌、および4)メチロバクター、メチロカルダム、メチロミクロビウム、メチロサルシナ、およびメチロスファエラ属のメチロコッカス科の細菌がある。
【0170】
別の実施形態では、宿主細胞は「グラム陰性プロテオバクテリア亜群2」から選択される。「グラム陰性プロテオバクテリア亜群2」は、以下の属(および全数のカタログに挙げられ、公に入手可能な、他に示した場合を除きいずれもATCCで寄託された、丸括弧中に示すその寄託菌株):アシドモナス(2);アセトバクター(93);グルコノバクター(37);ブレブンジモナス(23);ベイエリンキア(13);デルキシア(2);ブルセラ(4);アグロバクテリウム(79);ケラトバクター(2);エンシファー(3);リゾビウム(144);シノリゾビウム(24);ブラストモナス(1);スフィンゴモナス(27);アルカリゲネス(88);ボルデテラ(43);バークホルデアリア(73);ラルストニア(33);アシドボラックス(20);好気性水素酸化細菌(9);ズーグレア(9);メチロバクター(2);メチロカルダム(NCIMBで1);メチロコッカス(2);メチロミクロビウム(2);メチロモナス(9);メチロサルシナ(1);メチロスファエラ、アゾモナス(9);アゾリゾフィルス(5);アゾトバクター(64);セルビブリオ(3);オリゲラ(5);シュードモナス(1139);フランシセラ(4);キサントモナス(229);ステノトロフォモナス(50);およびオーシャンイモナス(4)のプロテオバクテリアの群として定義する。
【0171】
「グラム陰性プロテオバクテリア亜群2」の例示的な宿主細胞種には、以下の細菌(および丸括弧中に示すATCCまたは他の寄託番号の(1つまたは複数の)例示的菌株)があるが、これらだけには限られない:Acidomonas methanolica(ATCC 43581);Acetobacter aceti(ATCC 15973);Gluconobacter oxydans(ATCC 19357);Brevundimonas diminuta(ATCC 11568);Beijerinckia indica(ATCC 9039 およびATCC 19361);Derxia gummosa(ATCC 15994);Brucella melitensis(ATCC 23456),Brucella abortus(ATCC 23448);Agrobacterium tumefaciens(ATCC 23308),Agrobacterium radiobacter(ATCC 19358),Agrobacterium rhizogenes(ATCC 11325);Chelatobacter heintzii(ATCC 29600);Ensifer adhaerens(ATCC 33212);Rhizobium leguminosarum(ATCC 10004);Sinorhizobium frediii(ATCC 35423);Blastomonas natatoria(ATCC 35951);Sphingomonas paucimobilis(ATCC 29837);Alcaligenes faecalis(ATCC 8750);Bordetella pertussis(ATCC 9797);Burkholderia cepacia(ATCC 25416);Ralstonia pickettii(ATCC 27511);Acidovorax facilis(ATCC 11228);Hydrogenophagaflava(ATCC 33667);Zoogloea ramigera(ATCC 19544);Methylobacter luteus(ATCC 49878);Methylocaldum gracile(NCIMB 11912);Methylococcus capsulatus(ATCC 19069);Methylomicrobium agile(ATCC 35068);Methylomonas methanica(ATCC 35067);Methylosarcina fibrata(ATCC 700909);Methylosphaera hansonii(ACAM 549);Azomonas agilis(ATCC 7494);Azorhizophilus paspali(ATCC 23833);Azotobacter chroococcum(ATCC 9043);Cellvibrio mixtus(UQM 2601);Oligella urethralis(ATCC 17960);Pseudomonas aeruginosa(ATCC 10145),Pseudomonas fluorescens(ATCC 35858);Francisella tularensis(ATCC 6223);Stenotrophomonas maltophilia(ATCC 13637);Xanthomonas campestris(ATCC 33913);およびOceanimonas doudoroffii(ATCC 27123)。
【0172】
別の実施形態では、宿主細胞は「グラム陰性プロテオバクテリア亜群3」から選択される。「グラム陰性プロテオバクテリア亜群3」は、以下の属:ブレブンジモナス、アグロバクテリウム、リゾビウム、シノリゾビウム、ブラストモナス、スフィンゴモナス、アルカリゲネス、バークホルデアリア、ラルストニア、アシドボラックス、好気性水素酸化細菌、メチロバクター、メチロカルダム、メチロコッカス、メチロミクロビウム、メチロモナス、メチロサルシナ、メチロスファエラ、アゾモナス、アゾリゾフィルス、アゾトバクター、セルビブリオ、オリゲラ、シュードモナス、テレジニバクター、フランシセラ、ステノトロフォモナス、キサントモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0173】
別の実施形態では、宿主細胞は「グラム陰性プロテオバクテリア亜群4」から選択される。「グラム陰性プロテオバクテリア亜群4」は、以下の属:ブレブンジモナス、ブラストモナス、スフィンゴモナス、バークホルデアリア、ラルストニア、アシドボラックス、好気性水素酸化細菌、メチロバクター、メチロカルダム、メチロコッカス、メチロミクロビウム、メチロモナス、メチロサルシナ、メチロスファエラ、アゾモナス、アゾリゾフィルス、アゾトバクター、セルビブリオ、オリゲラ、シュードモナス、テレジニバクター、フランシセラ、ステノトロフォモナス、キサントモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0174】
別の実施形態では、宿主細胞は「グラム陰性プロテオバクテリア亜群5」から選択される。「グラム陰性プロテオバクテリア亜群5」は、以下の属:メチロバクター、メチロカルダム、メチロコッカス、メチロミクロビウム、メチロモナス、メチロサルシナ、メチロスファエラ、アゾモナス、アゾリゾフィルス、アゾトバクター、セルビブリオ、オリゲラ、シュードモナス、テレジニバクター、フランシセラ、ステノトロフォモナス、キサントモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0175】
宿主細胞は、「グラム陰性プロテオバクテリア亜群6」から選択することができる。「グラム陰性プロテオバクテリア亜群6」は、以下の属:ブレブンジモナス、ブラストモナス、スフィンゴモナス、バークホルデアリア、ラルストニア、アシドボラックス、好気性水素酸化細菌、アゾモナス、アゾリゾフィルス、アゾトバクター、セルビブリオ、オリゲラ、シュードモナス、テレジニバクター、ステノトロフォモナス、キサントモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0176】
宿主細胞は、「グラム陰性プロテオバクテリア亜群7」から選択することができる。「グラム陰性プロテオバクテリア亜群7」は、以下の属:アゾモナス、アゾリゾフィルス、アゾトバクター、セルビブリオ、オリゲラ、シュードモナス、テレジニバクター、ステノトロフォモナス、キサントモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0177】
宿主細胞は、「グラム陰性プロテオバクテリア亜群8」から選択することができる。「グラム陰性プロテオバクテリア亜群8」は、以下の属:ブレブンジモナス、ブラストモナス、スフィンゴモナス、バークホルデアリア、ラルストニア、アシドボラックス、好気性水素酸化細菌、シュードモナス、ステノトロフォモナス、キサントモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0178】
宿主細胞は、「グラム陰性プロテオバクテリア亜群9」から選択することができる。「グラム陰性プロテオバクテリア亜群9」は、以下の属:ブレブンジモナス、バークホルデアリア、ラルストニア、アシドボラックス、好気性水素酸化細菌、シュードモナス、ステノトロフォモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0179】
宿主細胞は、「グラム陰性プロテオバクテリア亜群10」から選択することができる。「グラム陰性プロテオバクテリア亜群10」は、以下の属:バークホルデアリア、ラルストニア、シュードモナス、ステノトロフォモナス、およびキサントモナスのプロテオバクテリアの群として定義する。
【0180】
宿主細胞は、「グラム陰性プロテオバクテリア亜群11」から選択することができる。「グラム陰性プロテオバクテリア亜群11」は、シュードモナス、ステノトロフォモナス、およびキサントモナス属のプロテオバクテリアの群として定義する。
【0181】
宿主細胞は、「グラム陰性プロテオバクテリア亜群12」から選択することができる。「グラム陰性プロテオバクテリア亜群12」は、以下の属:バークホルデアリア、ラルストニア、シュードモナスのプロテオバクテリアの群として定義する。宿主細胞は、「グラム陰性プロテオバクテリア亜群13」から選択することができる。「グラム陰性プロテオバクテリア亜群13」は、以下の属:バークホルデアリア、ラルストニア、シュードモナス、およびキサントモナスのプロテオバクテリアの群として定義する。宿主細胞は、「グラム陰性プロテオバクテリア亜群14」から選択することができる。「グラム陰性プロテオバクテリア亜群14」は、以下の属:シュードモナスおよびキサントモナスのプロテオバクテリアの群として定義する。宿主細胞は、「グラム陰性プロテオバクテリア亜群15」から選択することができる。「グラム陰性プロテオバクテリア亜群15」は、シュードモナス属のプロテオバクテリアの群として定義する。
【0182】
宿主細胞は、「グラム陰性プロテオバクテリア亜群16」から選択することができる。「グラム陰性プロテオバクテリア亜群16」は、以下のシュードモナス種(および丸括弧中に示すATCCまたは他の寄託番号の(1つまたは複数の)例示的菌株)のプロテオバクテリアの群として定義する:Pseudomonas abietaniphila(ATCC 700689);Pseudomonas aeruginosa(ATCC 10145);Pseudomonas alcaligenes(ATCC 14909);Pseudomonas anguilliseptica(ATCC 33660);Pseudomonas citronellolis(ATCC 13674);Pseudomonas flavescens(ATCC 51555);Pseudomonas mendocina(ATCC 25411);Pseudomonas nitroreducens(ATCC 33634);Pseudomonas oleovorans(ATCC 8062);Pseudomonas pseudoalcaligenes(ATCC 17440);Pseudomonas resinovorans(ATCC 14235);Pseudomonas straminea(ATCC 33636);Pseudomonas agarici(ATCC 25941);Pseudomonas alcaliphila;Pseudomonas alginovora;Pseudomonas andersonii;Pseudomonas asplenii(ATCC 23835);Pseudomonas azelaica(ATCC 27162);Pseudomonas beyerinckii(ATCC 19372);Pseudomonas borealis;Pseudomonas boreopolis(ATCC 33662);Pseudomonas brassicacearum;Pseudomonas butanovora(ATCC 43655);Pseudomonas cellulosa(ATCC 55703);Pseudomonas aurantiaca(ATCC 33663);Pseudomonas chlororaphis(ATCC 9446,ATCC 13985,ATCC 17418,ATCC 17461);Pseudomonas fragi(ATCC 4973);Pseudomonas lundensis(ATCC 49968);Pseudomonas taetrolens(ATCC 4683);Pseudomonas cissicola(ATCC 33616);Pseudomonas coronafaciens;Pseudomonas diterpeniphila;Pseudomonas elongata(ATCC 10144);Pseudomonasflectens(ATCC 12775);Pseudomonas azotoformans;Pseudomonas brenneri;Pseudomonas cedrella;Pseudomonas corrugata(ATCC 29736);Pseudomonas extremorientalis;Pseudomonas fluorescens(ATCC 35858);Pseudomonas gessardii;Pseudomonas libanensis;Pseudomonas mandelii(ATCC 700871);Pseudomonas marginalis(ATCC 10844);Pseudomonas migulae;Pseudomonas mucidolens(ATCC 4685);Pseudomonas orientalis;Pseudomonas rhodesiae;Pseudomonas synxantha(ATCC 9890);Pseudomonas tolaasii(ATCC 33618);Pseudomonas veronii(ATCC 700474);Pseudomonas frederiksbergensis;Pseudomonas geniculata(ATCC 19374);Pseudomonas gingeri;Pseudomonas graminis;Pseudomonas grimontii;Pseudomonas halodenitrificans;Pseudomonas halophila;Pseudomonas hibiscicola(ATCC 19867);Pseudomonas huttiensis(ATCC 14670);Pseudomonas hydrogenovora;Pseudomonas jessenii(ATCC 700870);Pseudomonas kilonensis;Pseudomonas lanceolata(ATCC 14669);Pseudomonas lini;Pseudomonas marginata(ATCC 25417);Pseudomonas mephitica(ATCC 33665);Pseudomonas denitrificans(ATCC 19244);Pseudomonas pertucinogena(ATCC 190);Pseudomonas pictorum(ATCC 23328);Pseudomonas psychrophila;Pseudomonas filva(ATCC 31418);Pseudomonas monteilii(ATCC 700476);Pseudomonas mosselii;Pseudomonas oryzihabitans(ATCC 43272);Pseudomonas plecoglossicida(ATCC 700383);Pseudomonas putida(ATCC 12633);Pseudomonas reactans;Pseudomonas spinosa(ATCC 14606);Pseudomonas balearica;Pseudomonas luteola(ATCC 43273);Pseudomonas stutzeri(ATCC 17588);Pseudomonas amygdali(ATCC 33614);Pseudomonas avellanae(ATCC 700331);Pseudomonas caricapapayae(ATCC 33615);Pseudomonas cichorii(ATCC 10857);Pseudomonas ficuserectae(ATCC 35104);Pseudomonas fuscovaginae;Pseudomonas meliae(ATCC 33050);Pseudomonas syringae(ATCC 19310);Pseudomonas viridiflava(ATCC 13223);Pseudomonas thermocarboxydovorans(ATCC 35961);Pseudomonas thermotolerans;Pseudomonas thivervalensis;Pseudomonas vancouverensis(ATCC 700688);Pseudomonas wisconsinensis;およびPseudomonas xiamenensis。
【0183】
宿主細胞は、「グラム陰性プロテオバクテリア亜群17」から選択することができる。「グラム陰性プロテオバクテリア亜群17」は、例えば以下のシュードモナス種に属する種を含めた「fluorescent Pseudomonads」として当技術分野で知られているプロテオバクテリアの群として定義する:Pseudomonas azotoformans;Pseudomonas brenneri;Pseudomonas cedrella;Pseudomonas corrugata;Pseudomonas extremorientalis;Pseudomonas fluorescens;Pseudomonas gessardii;Pseudomonas libanensis;Pseudomonas mandelii;Pseudomonas marginalis;Pseudomonas migulae;Pseudomonas mucidolens;Pseudomonas orientalis;Pseudomonas rhodesiae;Pseudomonas synxantha;Pseudomonas tolaasii;およびPseudomonas veronii。
【0184】
他の適切な宿主には、グラム(+)プロテオバクテリアなどの参照の他の部分で分類した宿主がある。一実施形態では、宿主細胞はE.coliである。E.coliに関するゲノム配列は、E.coliMG1655に関して確定している(Blattnerら、(1997年)。E.coliK−12の完全なゲノム配列、Science277(5331):1453〜74ページ)およびDNAマイクロアレイが、E.coliK12に関して市販されている(MWG Inc、High Point、N.C.)。E.coliは、1%グルコースなどの適切な炭素源と共に、Luria−Bertani(LB)(10g/Lのトリプトン、5g/LのNaCl、5g/Lの酵母エキス)などの富栄養培地中、またはM9(6g/LのNa2HPO4、3g/LのKH2PO4、1g/LのNH4Cl、0.5g/LのNaCl、pH7.4)などの定義した最小培地中のいずれかで培養することができる。通常、E.coli細胞の一晩培養物を希釈し、振とうフラスコまたは発酵槽中のいずれかの新たな富栄養または最小培地に接種し、37℃で増殖する。
【0185】
宿主細胞は、任意のヒトを含めた哺乳動物または非ヒト哺乳動物由来の細胞などの、哺乳動物起源の細胞であってもよい。哺乳動物は、霊長類、サル、ブタ、ヒツジ類、ウシ、げっ歯類、有蹄動物、家畜ブタ、ブタ類、ヒツジ、ラム、ヤギ、畜牛、シカ、ラバ、ウマ、サル、類人猿、イヌ、ネコ、ラット、およびマウスを含み得るが、これらだけには限られない。
【0186】
宿主細胞は、植物起源の細胞であってもよい。対象とする異種タンパク質の産生を求めてスクリーニングするために任意の植物由来の細胞を選択することができる。適切な植物の例には、アルファルファ、リンゴ、アプリコット、シロイヌナズナ、チョウセンアザミ、ルッコラ、アスパラガス、アボカド、バナナ、オオムギ、マメ、ビート、ブラックチェリー、ブルーベリー、ブロッコリ、芽キャベツ、キャベツ、キャノーラ、マスクメロン、ニンジン、キャッサバ、トウゴマの実、カリフラワー、セロリ、チェリー、チコリ、シラントロ、シトラス、クレメンタイン、クローバー、ココナッツ、コーヒー、コーン、コットン、クランベリー、キュウリ、ダグラスファー、ナス、エンダイブ、キクチシャ、ユーカリ、フェンネル、イチジク、ニンニク、ウリ類、グレープ、グレープフルーツ、甘露、クズイモ、キウィフルーツ、レタス、リーキ、レモン、ライム、テーダマツ、リンシード、マンゴー、メロン、マッシュルーム、ネクタリン、ナッツ、オートムギ、油ヤシ、ナタネ、オクラ、オリーブ、オニオン、オレンジ、観賞植物、ヤシ、パパイヤ、パセリ、パースニップ、エンドウマメ、モモ、ピーナッツ、ナシ、コショウ、カキ、パイン、パインアップル、プランテイン、プラム、ザクロ、ポプラ、ジャガイモ、パンプキン、マルメロ、ラジアタパイン、ラディッキオ、ラディッシュ、ナタネ、ラズベリー、コメ、ライムギ、ソルガム、サザンパイン、ダイズ、ホウレンソウ、カボチャ、イチゴ、テンサイ、サトウキビ、ヒマワリ、サツマイモ、モミジバフウ、タンジェリン、茶、タバコ、トマト、ライコムギ、シバ、カブ、ブドウの木、スイカ、コムギ、ヤムイモ、およびズッキーニがあるが、これらだけには限られない。いくつかの実施形態では、方法中で有用な植物は、シロイヌナズナ、コーン、コムギ、ダイズ、およびコットンである。
【0187】
キット(Kits)
本発明は、対象とする異種タンパク質またはポリペプチドの産生に最適な宿主菌株、例えばP.fluorescens宿主菌株を同定するのに有用なキットも提供する。キットは複数の表現型が異なる宿主細胞を含み、各集団は、タンパク質産生に関与する1つもしくは複数の標的遺伝子の発現が増大するように、タンパク質分解に関与する1つもしくは複数の標的遺伝子の発現が低下するように、またはそれらの両方となるように遺伝子改変されている。アレイは、タンパク質産生に関与する遺伝子またはタンパク質分解に関与する遺伝子のいずれかの発現を調節するように遺伝子改変されていない1つまたは複数の細胞集団をさらに含むことができる。これらのキットは、細胞集団の増殖および維持を容易にするのに十分な試薬、ならびに対象とする異種タンパク質またはポリペプチドの発現用の試薬および/またはコンストラクトも含むことができる。宿主細胞の集団は、細胞集団の保存、輸送、および再構築に適した任意の方法でキットにおいて提供することができる。細胞集団はチューブ中、プレート上、またはスラント上に生きた状態で提供することができ、あるいはチューブまたはバイアル中に凍結乾燥または乾燥のいずれかの状態で保存することができる。細胞集団は、グリセロール、スクロース、アルブミン、または他の適切な保護剤もしくは保存剤などの追加の構成要素を保存培地中に含有することができる。
【0188】
以下の実施例は、限定の目的ではなく例示の目的で提供される。
【0189】
実験例(EXPERIMENTAL EXAMPLES)
概説(Overview)
異種タンパク質産生はしばしば、不溶性タンパク質または不適切にフォールディングされたタンパク質の形成をもたらし、それらは、回収が困難であり、かつ不活性であり得る。さらに、特定の宿主細胞プロテアーゼの存在によって、対象とするタンパク質が分解され、それによって最終収率が低下し得る。全ての異種タンパク質の産生を改善する単一の因子は存在しない。したがって、考えられる候補の集合由来の特定の異種タンパク質に特異的な因子を同定するための方法を探究した。
【0190】
Systems Biologyのツールを使用して、P.fluorescensのゲノムを調べて、宿主細胞のタンパク質フォールディング調節因子およびプロテアーゼ遺伝子を同定した。次いで、全体的な遺伝子発現分析を実施して上方制御された標的を優先し、およびその後、新規なタンパク質産生菌株を構築した。結果として、宿主細胞プロテアーゼが欠損しているかまたはタンパク質フォールディング調節因子の同時過剰発現を可能にする、複数の表現型が異なるP.fluorescens宿主菌株からなる、「Pfenex菌株アレイ」を構築した。この菌株アレイを使用して、特定の異種タンパク質の収率または質を特異的に高める因子を求めてスクリーニングすることができる。複数の表現型が異なる宿主菌株を提供することによって、対象とする任意の個々の異種タンパク質の産生を増大させ得る宿主菌株を同定する成功の機会が増大する。
【0191】
本発明は、Pseudomonas fluorescensにおける異種タンパク質の産生の改善をもたらす。同じ遺伝的背景の宿主菌株の利用可能なライブラリーを有することによって、異種発現したタンパク質の収率および/または質を高める因子の、迅速なスクリーニングおよび同定が可能である。P.fluorescensのゲノム配列には注釈付けしており、標的宿主細胞のフォールディング調節因子およびプロテアーゼを同定している。フォールディング調節因子はタンパク質の適切なフォールディングを手助けし、シャペロン、シャペロニン、ペプチジル−プロリンイソメラーゼ(PPIase)、およびジスルフィド結合形成タンパク質を含む。プロテアーゼは対象とするタンパク質を分解し、したがって異種タンパク質の収率および質に影響を与える可能性がある。考えられる標的を同定するために文献からの背景知識およびDNAマイクロアレイ分析を使用して、約80の標的遺伝子の一覧を構築した。同じ遺伝的背景を有する宿主細胞において、これらの遺伝子をゲノムから除去、またはプラスミドにクローニングして、異種タンパク質との同時過剰発現を可能にした。生成した菌株は96ウェルフォーマットに並べ、および、対象とする異種タンパク質を発現するプラスミドの形質転換後、改善されたタンパク質の収率および/または質を求めてスクリーニングした。
【実施例1】
【0192】
P.fluorescens菌株MB214のゲノム中のフォールディング調節因子遺伝子の同定
フォールディング調節因子は、新生および異種ポリペプチドのフォールディング、アンフォールディングおよび分解を助長する、全ての細胞中に存在するクラスのタンパク質である。フォールディング調節因子には、シャペロン、シャペロニン、ペプチジル−プロリルシス−トランスイソメラーゼ、およびタンパク質ジスルフィド結合形成に関与するタンパク質がある。異種タンパク質のフォールディングに助力する能力を有する新規な産生菌株を構築するための第一ステップとして、P.fluorescensのゲノムを調べて、宿主細胞のフォールディング調節因子遺伝子を同定した。
【0193】
P.fluorescens MB214のゲノムの6,433の予想ORFのそれぞれを、以下の方法を使用して、それらがフォールディング調節因子をコードしていた確率に関して分析した。対象とするいくつかのフォールディング調節因子は、ゲノム注釈の分析によってDowの研究者により既に同定されている(Ramseierら、2001)。これらの出発タンパク質の相同体は、クエリーとして出発タンパク質およびサブジェクトとして全てのMB214翻訳ORFのデータベースを用いて、タンパク質/タンパク質BLASTを使用して同定した。有意な相同性でクエリータンパク質と適合した翻訳ORFを、さらなる分析用のリストに加えた。有意な相同性は、アラインメントの長さおよび質に基づく人間による判断を考慮に入れてle−30以下のe−値を有するものとしてここでは定義する。この試験の目的は、全ての考えられるフォールディング調節因子を同定し得る機会を最大にすることを全体に含めることであった。
【0194】
より多くのORFを、キーワード「シャペロン」を含有する前の注釈からの、それらの管理された機能に基づいてリストに加えた。最終的に、InterProデータベースバージョン7.0(Mulderら、2005年)に対するタンパク質シグネチャーファミリー検索プログラムInterProScan(Quevillonら、2005年)によってORFを分析した。これらのORFは、InterProScanソフトウェアおよびこれらのファミリーと関係があるGene Ontology(GO)カテゴリー(Gene Ontology Consortium.2004年)によって割り当てられたタンパク質ファミリーであった。これらの自動GO割り当てを使用して、GO用語「GO:0006457生物学的過程:タンパク質フォールディング」または「GO:0003754分子機能:シャペロン活性」を割り当てた全てのORFを、さらなる分析用のリストに加えた。
【0195】
次いでリストを分析して、フォールディング調節因子をコードする低い確率を有していたORFを除去した。再度、この試験の目的は非常に包括的であることであったが、これらの半自動的方法によってリストに割り当てたORFの多くは、限られた基準および人間による判断に基づいて、フォールディング調節因子をコードしていないとして容易に同定することができた。
【0196】
特定のORFを除外した最も一般的な理由は、このORFが実際にフォールディング調節因子である根拠が乏しいため、すなわち、フォールディング調節因子としてのORFの注釈付けの理由が不明確または矛盾のいずれかであった、前の注釈に基づいてリストに割り当てたORFであるためであった。InterProScanは実際に異なるプログラムの集合体であり、これらのプログラムのいくつかは他のプログラムより信頼できると考えられる。ScanRegExpまたはProfileScanコンポーネントのアウトプットのみに基づいてORFをリストに割り当てた場合、したがってそれは除去した。P.fluorescensのフォールディング調節因子の最終的なリストは43のメンバーを有し、それは表1中に示す。
【実施例2】
【0197】
P.fluorescens菌株MB214のゲノム中のプロテアーゼ遺伝子の同定
プロテアーゼは、ペプチド結合を加水分解し全ての生物の生存に必要な酵素である。しかしながら、細胞中でのそれらの役割は、Pfenex Expression Technology(商標)も含めた任意の異種タンパク質発現系中で、プロテアーゼは組換えタンパク質の収率および/または質に弊害をもたらす可能性があることを意味する。ゲノムから除去されたプロテアーゼ遺伝子を有する新規な産生菌株を構築するための第一ステップとして、P.fluorescensのゲノムを調べて、宿主細胞のプロテアーゼ遺伝子を同定した。
【0198】
P.fluorescens MB214のゲノムの6,433の予想ORFのそれぞれを、以下の方法を使用して、それらがプロテアーゼをコードしていた確率に関して分析した。MEROPSデータベースは、Wellcome Trust Sanger Institute、Cambridge、UK(Rawlingsら、2006年、http://merops.sanger.ac.uk)で研究者によって手作業で管理されている。それは、研究室での実験と既知のプロテアーゼファミリーとの相同性の両方によって発見された、プロテアーゼの総合目録である。データベースの長所の1つは、MEROPS階層的分類スキームである。このシステムでは、同じ機能を共有する相同体を複数のファミリーに一緒にグループ分けする。複数のファミリーは、ここでも類似の構造特性に基づく進化的関係に基づいて複数の仲間にグループ分けする。この方法はこのデータベースを大いに利用して、P.fluorescensゲノム内のプロテアーゼ相同体を同定する。
【0199】
MEROPSデータベースとの相同体は、クエリーとしてそれぞれのMB214翻訳ORFおよびサブジェクトとして全てのMEROPSタンパク質のデータベースを用いて、タンパク質/タンパク質BLASTを使用して同定した。有意な相同性でクエリータンパク質と適合した翻訳ORFを、さらなる分析用のリストに加えた。この場合の有意な相同性は、アラインメントの長さおよび質に基づく人間による判断を考慮に入れてle−−60以下のe−値を有するものとしてここでは定義する。このステップによって、リスト用の109の考えられるプロテアーゼを得た。
【0200】
InterProデータベースバージョン7.0(Mulderら、2005年)に対するタンパク質シグネチャーファミリー検索プログラムInterProScan(Quevillonら、2005年)によってもORFを分析した。これらのORFは、InterProScanソフトウェアおよびこれらのファミリーと関係があるGene Ontology(GO)カテゴリー(Gene Ontology Consortium.2004年)によって割り当てられたタンパク質ファミリーであった。これらの自動GO割り当てを使用して、文字列「ペプチダーゼ」、「プロテアーゼ」または「タンパク質分解」を含有したGO名を割り当てた全てのORFを、さらなる分析用のリストに加えた。このステップによって、前のステップで同定されなかった追加の70の考えられるプロテアーゼを得た。
【0201】
より多くのORFを、キーワード「ペプチダーゼ」または「プロテアーゼ」を含有する前の注釈(annotation)(Ramseierら、2001年)からの、それらの管理された機能に基づいてリストに加えた。このステップによって、前のステップで再度同定しなかった32の考えられるプロテアーゼを得た。
【0202】
次いでリストを分析して、プロテアーゼをコードする低い確率を有していたORFを除去した。再度、この試験の目的は非常に包括的であることであったが、これらの半自動的方法によってリストに割り当てたORFの多くは、限られた基準および人間による判断に基づいて、プロテアーゼをコードしていないとして容易に同定することができた。複数の遺伝子を除外した2つの一般的な理由は、特定のORFが実際にプロテアーゼであることの根拠が乏しいため、または特定の遺伝子が、プロテアーゼ自体ではなくプロテアーゼ相同体であることが知られている他のタンパク質と最大の相同性を示したことであった。P.fluorescensのプロテアーゼの最終的なリストは90のメンバーを有し、それは表2中に示す。
【実施例3】
【0203】
in silicoでのフォールディング調節因子およびプロテアーゼタンパク質の細胞内位置の予想
Pfenex Expression Technology(商標)の長所の1つは、特定の異種タンパク質を分離することができる細胞内区画(cellular compartment)を制御するその能力である。したがって、同定した宿主細胞のフォールディング調節因子およびプロテアーゼタンパク質が位置する細胞内区画を予想した。これらの予想を行うために、2つのプログラムを選択した。PsortB2.0は、所与のペプチドの細胞内位置を予想する12の別個のアルゴリズムの結果を組み合わせる。大部分のこれらのアルゴリズムは、クエリータンパク質と既知の細胞内位置のタンパク質の間の相同性の検出に依存する。PsortBは、Hidden Markov Models(HMM)を使用して膜貫通フォールディングドメインまたはI型分泌シグナル配列の存在をそれぞれ検出する、HMMTOPおよびSignalPなどのアルゴリズムも含む。PsortBの結果に加えて、SignalP HMMを使用して、I型分泌シグナル配列の存在を予想した。シグナル配列を検出するが細胞内位置を示す他の具体的情報が得られないとき、PsortBのアウトプットは曖昧である可能性があるので、これは必要であった。これらの場合、PsortBはタンパク質の細胞内局在が知られていないことを示す、何故ならそれは実際、細胞質膜、ペリプラズム、外膜または細胞外区画のいずれか1つに分離する可能性があったからである。しかしながら、タンパク質はおそらく細胞質中に位置しないことを知り、表中においてそれに注目することは十分有益である。したがって表2は、その結果が知られていない場合以外のPsortBアルゴリズムの結果を挙げる。これらの場合、SignalP HMM単独の結果は、シグナルペプチドが検出されたことを示す「シグナルペプチド(Signal Peptide)」、およびシグナルペプチドが検出されなかったことを示す「非分泌性(Non Secretory)」を用いて示す。
【実施例4】
【0204】
フォールディング調節因子の同時過剰発現を可能にするプラスミドの構築
フォールディング調節因子遺伝子を、対象とする異種タンパク質を発現させるために通常使用される別のプラスミドと適合性がある、pCNのプラスミド誘導体(Nietoら、1990年)にクローニングした(Squiresら、2004年;Chewら、2005年)。マンニトール誘導性grpE−dnaKJ含有プラスミドの構築を例示する。他のフォールディング調節因子を、1つの遺伝子または多数の遺伝子としてオペロンで編成されるとき、以下に概説したのと同様にクローニングした。
【0205】
鋳型およびプライマーRC199(5−ATATACTAGTAGGAGGTAACTTATGGCTGACGAACAGACGCA−3’)(配列番号:1)およびRC200(5’ATATTCTAGATTACAGGTCGCCGAAGAAGC−3’)(配列番号:2)として、P.fluorescens MB214(DNeasy;Qiagen、Valencia、CA)から単離したゲノムDNAを利用して、grpE−dnaKJ遺伝子を、製造者の推奨に従いPfuTurbo(Stratagene、La Jolla、CA)を使用して増幅した。生成した4kbのPCR産物はSpeIおよびXbaI(前述のプライマー中で下線を引いた制限部位)で消化し、pDOW1306−6(Schneiderら、2005b)の誘導体であるpDOW2236に連結させて、tacプロモーターの制御下でgrpE−dnaKJオペロンを含有するpDOW2240を作製した。次いでプラスミドpDOW2240をSpeIおよびHindIIIで消化し、生成したgrpE−dnaKJ含有4.0kbのDNA断片はQiaquick(Qiagen、Valencia、CA)を使用してゲル精製し、P.fluorescensマンニトール調節型プロモーター(Schneiderら、2005a)を有するpCNの誘導体であるpDOW2247に連結させ、それをさらにSpeIおよびHindIIIで消化した。生成したプラスミド、pDOW3501は、マンニトールプロモーターの制御下でgrpE−dnaKJオペロンを含有していた。次いでプラスミドpDOW3501を、250ug/mlのウラシルを補ったM9グルコースプレート上での選択によって、DC388および他のウラシル要求性菌株に形質転換した。
【実施例5】
【0206】
プロテアーゼ遺伝子のゲノムが欠失したP.fluorescens菌株の構築
ゲノム欠失の生成を可能にしたプラスミドは、欠失する遺伝子の5’と3’の両方の500〜1000bpのDNA断片の増幅によって構築した。生成する5’PCR産物は典型的には欠失する遺伝子の翻訳開始コドン(ATGまたはGTGまたはTGT)で終わり、一方3’PCR産物は典型的には欠失する遺伝子の停止コドン(TAAまたはTGAまたはTAG)で始まる。これら2つのPCR産物は追加の増幅ステップによって1つに融合し、次いでSOE PCR(Hortonら、1990年)を使用してpDOW1261(図1)(Chewら、2005年)にクローニングした。
【実施例6】
【0207】
P.fluorescens菌株におけるハイスループット増殖および異種タンパク質発現の分析
プラスミドpDOW2787はモノクローナル抗体(mAb)gal2をコードし、その重鎖はPbp分泌リーダーおよびtacプロモーターの制御下で発現する。その軽鎖はOprF分泌リーダーおよびマンニトールプロモーターの制御下で発現する。プラスミドは、対象遺伝子の欠失または同時発現用のフォールディング調節因子を有するpDOW2247のいずれかを有する63菌株、および野生型菌株を含有する5つの対照菌株のコンピテント細胞にエレクトロポレーションした。300rpmで振とうすることによって、グリセロールを含む増殖培地を含有する同一の深さのウェル単位で、細胞を培養した。0.1mMのイソプロピルβ−D−チオガラクトピラノシド(IPTG)および1%マンニトールを用いて、24時間でタンパク質発現を誘導した。誘導後24時間で、アリコートを溶かし、抗原の抗原結合を測定して活性抗体の量を定量化した。その値をOD600で割って細胞特異的活性を測定した。菌株Δprc1、ΔdegP2、ΔLa2、ΔclpP、およびΔprc2、Δprc2、grpEdnaKJ同時発現菌株、Δtig、ΔclpX、およびΔlonはいずれも対照菌株より2.4倍以上高く、これは統計上有意であった(p<0.5)。可溶性細胞分画をΔprc1、ΔdegP2、ΔLa2およびgrpEdnaKJ同時発現菌株から調製し、ウエスタン分析にかけた(図2)。完全構築抗体と一致した大きさを有するバンドを4つの試験菌株において検出したが、対照においては検出しなかった。
(参考文献)
【表8】
【0208】
【表9】
【0209】
本明細書中に述べる全ての刊行物および特許出願は、本発明が関係する技術分野の当業者のレベルを示す。全ての刊行物および特許出願は、それぞれ個々の刊行物または特許出願が参照によって組み込まれることが具体的および個々に示されるのと同じ程度で、参照によって本明細書に組み込まれる。
【0210】
前述の本発明を、理解を明確にする目的で例示および実施例によってある程度詳細に説明してきたが、特定の変化および変更形態を添付の特許請求の範囲内で実施することができることは明らかであろう。
【技術分野】
【0001】
本発明は、タンパク質生産(production)の分野に属し、詳細には、適切にプロセシングされた異種タンパク質の大規模生産に最適な宿主細胞の同定に関する。
【背景技術】
【0002】
バイオテクノロジー薬物およびワクチンとしての使用について、150種を超える、組換えにより産生されたタンパク質およびポリペプチドが米国食品医薬品局(FDA)によって承認されており、さらに370種が臨床試験中である。化学合成を介して生産される小分子治療薬と異なり、タンパク質およびポリペプチドは、生細胞内で最も効率的に産生される。しかし、細菌内における組換えタンパク質産生の現在の方法はしばしば、不適切にフォールディングされたタンパク質、凝集したタンパク質または不活性なタンパク質を産生し、そして多くのタイプのタンパク質が二次修飾を必要とするが、それらの修飾は、既知の方法を用いて行うと非効率的である。
【0003】
組換え系における、適切にフォールディングされたタンパク質の産生を増大させるために、多数の試みが開発されている。例えば、研究者たちは、発酵条件を変えるか(Schein(1989)、Bio/Technology、7:1141〜1149)、プロモーター強度を変えるか、または過剰発現させたシャペロンタンパク質を使用しており(Hockney(1994)、Trends Biotechnol.12:456〜463)、これらは、封入体の形成を防止するのに役立ち得る。
【0004】
細胞から上清中にタンパク質を排出させるために、複数の方法(strategies)が開発されている。例えば、米国特許第5,348,867号、米国特許第6,329,172号、WO96/17943、WO02/40696および米国特許出願公開第2003/0013150号を参照。増大した発現を得るための他の方法は、タンパク質をペリプラズム(periplasm、周辺質)にターゲティングすることに向けられている。一部の研究は、非Sec型の分泌に焦点をおいている(例えば、WO03/079007、米国特許出願公開第2003/0180937号、米国特許出願公開第2003/0064435号およびWO00/59537を参照)。しかし、大部分の研究は、Sec型分泌系を用いた外因性タンパク質の分泌に焦点をおいてきた。
【0005】
組換えポリペプチドまたはタンパク質の発現に使用するために、多くの分泌シグナルが記載されている。例えば、米国特許第5,914,254号、米国特許第4,963,495号、欧州特許第0 177 343号、米国特許第5,082,783号、WO89/10971、米国特許第6,156,552号、米国特許第6,495,357号、第6,509,181号、第6,524,827号、第6,528,298号、第6,558,939号、第6,608,018号、第6,617,143号、米国特許第5,595,898号、第5,698,435号および第6,204,023号、米国特許第6,258,560号、WO01/21662、WO02/068660および米国特許出願公開第2003/0044906号、米国特許第5,641,671号ならびに欧州特許第0 121 352号を参照。
【発明の概要】
【発明が解決しようとする課題】
【0006】
異種タンパク質産生はしばしば、不溶性タンパク質または不適切にフォールディングされたタンパク質の形成をもたらし、それらは、回収が困難であり、かつ不活性であり得る。さらに、特定の宿主細胞プロテアーゼの存在によって、対象とするタンパク質(protein of interest)が分解され、それによって最終収率が低下し得る。全ての異種タンパク質の産生を改善する単一の因子は存在しない。結果として、当技術分野では、組換えポリペプチドを分泌し、その適切なプロセシングを行って、適切にプロセシングされた形態でトランスジェニックタンパク質を産生することができる、改善された大規模発現系を同定する必要性が存在する。
【課題を解決するための手段】
【0007】
本発明は、収率および/または質が改善された望ましい仕様(desired specification)に従って、少なくとも1種の異種ポリペプチドを産生できる宿主細胞集団を迅速に同定するための組成物および方法を提供する。組成物は、タンパク質産生(production)に関与する1つまたは複数の標的遺伝子の発現が増大するように、タンパク質分解(degradation)に関与する1つまたは複数の標的遺伝子の発現が低下するように、またはタンパク質産物に影響を与える異種遺伝子を発現するように、またはその組合せとなるように遺伝子改変されている2つ以上の宿主細胞集団を含む。対象とするポリペプチドを様々な改変宿主細胞で発現する能力は、対象とするポリペプチドに最適な宿主細胞を決定するための迅速かつ効率的な手段を提供する。望ましい仕様は、対象とするポリペプチドに応じて異なるが、それには、収率、質および活性などが含まれる。
【0008】
対象とするポリペプチドの発現を促進する内因性配列および/または外因性配列の発現レベルに影響を与える核酸配列の多数の組合せを発現するように宿主細胞集団を改変できることが認識されている。一実施形態では、2つ以上の宿主細胞集団が、対象とする異種タンパク質の適切な発現、プロセシングおよび/または移行のうち1つまたは複数に関与する1つまたは複数の標的遺伝子の発現が増大するように遺伝子改変されている。別の実施形態では、標的遺伝子はタンパク質フォールディング調節因子である。別の実施形態では、タンパク質フォールディング調節因子は表1のリストから選択される。
【0009】
別の実施形態では、1つまたは複数の宿主細胞集団が、タンパク質分解に関与する1つまたは複数の標的遺伝子の発現が低下するように遺伝子改変されている。別の実施形態では、標的遺伝子はプロテアーゼである。別の実施形態では、プロテアーゼは表2のリストから選択される。
【0010】
一実施形態では、対象とするタンパク質をコードするヌクレオチド配列が、本明細書に記載の、P.fluorescensのSec系分泌シグナルに作動可能に連結(operably linked)されている。アレイ内の株のうち1つまたは複数が、対象とする異種タンパク質をペリプラズム区画に発現し得る。特定の実施形態では、1つまたは複数の株が、外側細胞壁を通して細胞外に異種タンパク質を分泌し得る。
【0011】
宿主細胞には、酵母細胞、昆虫細胞、哺乳動物細胞および植物細胞などを含めた真核細胞、ならびにP.fluorescensおよびE.coliなどの細菌細胞を含めた原核細胞が含まれる。
【0012】
示されているように、異種発現されたタンパク質の、改善された収率および/または質を有する(1つまたは複数の)特定の株を同定するために、宿主細胞集団のライブラリーを迅速にスクリーニングできる。株アレイは、治療用タンパク質、ホルモン、成長因子、細胞外受容体またはリガンド、プロテアーゼ、キナーゼ、血液タンパク質、ケモカイン、サイトカインおよび抗体などを含めた、対象とするいかなるタンパク質の発現改善のスクリーニングにも有用である。
【図面の簡単な説明】
【0013】
【図1A】図1Aは、P.fluorescensにおけるゲノム欠失を遺伝子工学処理するために使用したプラスミドpDOW1261−2を示す図である。
【図1B】図1Bは、遺伝子X欠失の構築の概略図である。
【図2】図2は、Δprc1、ΔdegP2、ΔLa2およびgrpEdnaKJ同時発現菌株(実施例6)において誘導後0および24時間で調製した可溶性細胞分画(それぞれI0およびI24)のウエスタンブロット分析の図である。上部矢印は、対照(DC440)ではなく同時発現菌株において完全に構築されたモノクローナル抗体を指す。r=組換え体(recombinant);n−r=非組換え体(nonreconbinant)。
【発明を実施するための形態】
【0014】
これより、本発明の全ての実施形態ではなく、それらの一部が示されている添付図面を参照して、以下に本発明をより完全に説明する。実際、これらの発明は、多くの異なった形態で具体化でき、本明細書に示す実施形態に限定して解釈するべきではなく、これらの実施形態は、この開示が適用される(applicable)法的必要条件を満たすように提示されている。
【0015】
以上の説明と添付図面とに提示されている教示の恩恵により、これらの発明が属する分野の当業者ならば、本明細書に示す本発明の多くの変更形態および他の実施形態が想起されよう。
【0016】
したがって、本発明は、開示されている特定の実施形態に限定されるものではなく、変更形態および他の実施形態も本発明の範囲に含まれるものであることを理解されたい。本明細書では特定の用語が用いられているが、それらは包括的および説明的な意味でのみで用いられており、限定を目的としたものではない。
【0017】
概要(Overview)
宿主細胞内で適切にプロセシングされた高レベルの異種ポリペプチドを産生するのに最適な宿主株(optimal host strain)、例えば、Pseudomonas fluorescens宿主株を同定するための組成物および方法を提供する。詳細には、ライブラリー内の各株(または「宿主細胞の集団」)が、宿主細胞内の1つまたは複数の標的遺伝子の発現を調節するように遺伝子改変されている宿主株のライブラリー(または「アレイ」)を提供する。「最適な宿主株」は、アレイ内の、表現型が異なる宿主細胞の他の集団と比較した、対象とする発現タンパク質の量、質および/または位置に基づいて同定または選択できる。したがって、最適な宿主株は、対象とするポリペプチドを望ましい仕様に従って産生する株である。望ましい仕様は、産生されるポリペプチドに応じて異なるであろうが、仕様には、タンパク質の質および/または量、そのタンパク質が隔離(sequestered)されるか、または分泌されるか、ならびにタンパク質フォールディングが含まれる。
【0018】
「異種(heterologous)」、「異種発現された(heterologously expressed)」または「組換え(recombinant)」は通常、宿主細胞で内因的に生じたのではないか、または、それがその中に存在している天然ゲノムにおける位置で内因的に生じたのではなく、感染、形質移入、マイクロインジェクション、エレクトロポレーション、マイクロプロジェクション、または同様の方法によって細胞に添加された遺伝子またはタンパク質を指す。
【0019】
アレイ内の宿主細胞集団のうち1つまたは複数は、宿主細胞内の1つまたは複数の標的遺伝子の発現を調節するように改変されている。「標的遺伝子(target gene)」とは、宿主細胞内の異種タンパク質産生に影響を与える遺伝子を意味する。異種タンパク質産生に影響を与える標的遺伝子には、異種タンパク質の発現、活性、溶解性、移行、タンパク質分解および/または切断を調節するタンパク質をコードする遺伝子が含まれる。例えば、標的遺伝子は、宿主細胞プロテアーゼ、タンパク質フォールディング調節因子、転写因子、翻訳因子および分泌調節因子のうち少なくとも1つをコードするものでもよく、または対象とする異種タンパク質の適切な転写、翻訳、プロセシングおよび/または移行に関与する他の任意のタンパク質でもよい。「標的タンパク質(target protein)」は、標的遺伝子の発現の結果として生じるタンパク質またはポリペプチドを指す。1つまたは複数の標的遺伝子の発現および/または活性は、標的遺伝子またはタンパク質の機能に応じて、増大または低下する。例えば、1つまたは複数の宿主細胞プロテアーゼの発現が低下し、一方、1つまたは複数のタンパク質フォールディング調節因子の発現が増大してもよい。
【0020】
本明細書に記載のアレイは、異種タンパク質または対象とするペプチドの産生に最適な宿主細胞を迅速に同定するのに有用である。異種タンパク質産生はしばしば、不溶性タンパク質または不適切にフォールディングされたタンパク質の形成をもたらし、それらは、回収が困難であり、かつ不活性であり得る。さらに、特定の宿主細胞プロテアーゼの存在によって、対象とするタンパク質が分解され、それによって最終収率が低下し得る。対象とする全てのポリペプチドまたはタンパク質を最適に産生する単一の宿主細胞集団は存在しない。したがって、本発明の組成物および方法を用いて、改変細胞集団のライブラリーから、最適な宿主細胞を迅速かつ効率的に同定できる。その後、対象とするタンパク質を十分な量で産生するために、または商業的生産のために、その最適な宿主株を使用できる。同様に、最適な宿主株に基づいて、対象とするタンパク質の発現用に宿主株を改変できる。
【0021】
一実施形態では、この方法は、P.fluorescens細胞の少なくとも第1および第2の集団を含むアレイであって、各集団が、(i)タンパク質分解に関与する少なくとも標的遺伝子の発現が低減するように遺伝子改変されているP.fluorescens細胞集団、(ii)タンパク質産生に関与する少なくとも1つの標的遺伝子の発現が増大するように遺伝子改変されているP.fluorescens細胞集団、ならびに(iii)タンパク質分解に関与する少なくとも1つの標的遺伝子の発現が低減するように、かつタンパク質産生に関与する少なくとも標的遺伝子の発現が増大するように遺伝子改変されている少なくとも1つのP.fluorescens細胞集団からなる群から選択されるアレイを得るステップと、対象とする少なくとも1つの異種タンパク質をコードする少なくとも1つの遺伝子を含む発現コンストラクトを、各集団の少なくとも1つの細胞に導入するステップと;少なくとも1つの細胞集団における前記対象とするタンパク質の発現に十分な条件下で前記細胞を維持するステップと、対象とする異種タンパク質が産生される最適な細胞集団を選択するステップとを含み、アレイ内の各集団が同一でなく、かつ各集団が物理的に相互に離れており、最適な細胞集団では、アレイ内の他の集団と比較して、対象とする異種タンパク質が、改善された発現、改善された活性、改善された溶解性、改善された移行、または低減したタンパク質分解もしくは切断のうち1つまたは複数を示す。
【0022】
アレイは、宿主細胞プロテアーゼまたはタンパク質フォールディング調節因子の発現を改変するように遺伝子改変されていない宿主細胞(例えばP.fluorescens宿主細胞)の集団をさらに含んでもよい。この集団は、野生型株でもよく、または、タンパク質産生、プロセシングまたは移行に関与しない、またはもしくは複数の遺伝子(or or more genes)の発現を変えるように遺伝子改変されている株でもよい(例えば、選択マーカー遺伝子を発現するように遺伝子改変されたものでもよい)。
【0023】
一実施形態では、P.fluorescens宿主細胞の各集団は、相互に表現型が異なる(phenotypically distinct)(すなわち「同一でない(non-identical)」)。「表現型が異なる」とは、各集団が、1つまたは複数の標的遺伝子を測定可能に異なった量で産生することを意味する。この実施形態では、1つまたは複数の異なった標的遺伝子の発現を変えるように各株が遺伝子改変されている。宿主細胞の一集団内で複数の標的遺伝子の発現が調節される場合、標的遺伝子の組合せが、ライブラリー内の他の集団とは表現型上異なる。本発明による、表現型が異なる複数の宿主細胞集団を含むアレイは、異種タンパク質または対象とするペプチドを産生するのに有用な1つまたは複数の株をそれから選択する多様な集団を提供するものである。当業者ならば、そのようなアレイが、宿主細胞の任意の1つまたは複数の集団の複製(replicates)(例えば二つ組(duplicates)、三つ組(triplicates)など)も含むことを理解するであろう。
【0024】
アレイ(Arrays)
迅速にスクリーニングして、異種タンパク質の収率および/または質が改善されている(1つまたは複数の)特定の株を同定できる宿主細胞集団のアレイ(すなわち「株アレイ(strain array)」)をここに提供する。本明細書で使用される場合、「株アレイ」という用語は、アドレス特定されているか、アドレス特定可能な複数の位置を指す(例えば、ディープウェルまたはマイクロウェルなどのウェル)。通常、アレイ内のマイクロウェルまたはマイクロウェル群のそれぞれの位置は既知であり、それによって、対象とする異種タンパク質の発現に最適な宿主細胞の同定が可能となっている。
【0025】
株アレイは、表現型が異なる複数の宿主株を含む。アレイは、低密度アレイでも、高密度アレイでもよく、約2以上、約4以上、約8以上、約12以上、約16以上、約20以上、約24以上、約32以上、約40以上、約48以上、約64以上、約72以上、約80以上、約96以上、約192または約384以上の宿主細胞集団を含有し得る。
【0026】
本発明の宿主細胞集団は、マルチウェルまたはディープウェル容器内で維持および/またはスクリーニングできる。この容器は、任意の望ましい数のウェルを含有し得るが、最小限の試薬と比較的少ない数の細胞とを用いて、宿主細胞の各集団を個別かつ同時にスクリーニングするには、小型化された細胞培養マイクロアレイプラットフォームが有用である。このアッセイで有用な典型的なマルチウェルマイクロタイター容器は、限定されるものではないが、10ウェルプレート、28ウェルプレート、96ウェルプレート、384ウェルプレート、および384超のウェルを有するプレートを含めたマルチウェルプレートである。代替として、所望の容積に応じて、チューブ、ホルダー、カートリッジ、ミニチューブ、微量遠心チューブ、クライオバイアル、角型ウェルプレートチューブ、プレート、スラントまたは培養フラスコのアレイを用いてもよい。
【0027】
容器(vessel)は、対象とする宿主細胞、例えば、シュードモナスを培養および/またはスクリーニングするのに適したいかなる材質で作製されたものでもよい。例えば、容器は、その材質が生体適合性である限り、プラスチックまたは他の人工高分子材料など、容易に殺菌できる材質であり得る。限定されるものではないが、ポリスチレン、ポリプロピレン、ポリビニル化合物(例えばポリ塩化ビニル)、ポリカーボネート(ポリ塩化ビニル)、ポリテトラフルオロエチレン(PTFE)、ポリグリコール酸(PGA)、セルロース、ガラス、フッ素重合体、フッ素化エチレンプロピレン、ポリビニリデン、ポリジメチルシロキサンおよびシリコンなどを含めた多くの物質が使用できる。
【0028】
細胞の自動形質転換および自動コロニーピッカーは、所望の細胞の迅速なスクリーニングを容易にする。アレイは、当技術分野で知られているスポッター装置(例えば自動ロボット装置)を用いて作製および/またはスクリーニングできる。
【0029】
標的遺伝子(Target genes)
本発明の株アレイは、表現型および遺伝子型が異なる複数の宿主細胞集団を含み、アレイ内の各集団は、宿主細胞内の1つまたは複数の標的遺伝子の発現を調節するように遺伝子改変されている。「標的遺伝子」とは、宿主細胞内の異種タンパク質産生に影響を与える遺伝子を意味する。標的遺伝子は、宿主細胞プロテアーゼまたは内因性もしくは外因性タンパク質フォールディング調節因子、転写因子、翻訳因子、分泌調節因子、または対象とする異種タンパク質の適切な転写、翻訳、プロセシングおよび/または移行に関与する他の任意のタンパク質でもよい。「標的タンパク質」は、標的遺伝子の発現の結果として生じるタンパク質またはポリペプチドを指す。1つまたは複数の標的遺伝子の発現および/または活性は、標的遺伝子またはタンパク質の機能に応じて、増大または低下する。標的遺伝子は、宿主細胞で内因的に生じたものでもよく、またはアレイ内の宿主細胞集団のそれぞれで異種発現された遺伝子でもよい。
【0030】
一実施形態では、1つまたは複数の標的遺伝子が、タンパク質フォールディング調節因子、推定上のタンパク質フォールディング調節因子、またはフォールディング調節因子の補因子もしくはサブユニットのうち少なくとも1つである。一部の実施形態では、1つまたは複数の標的遺伝子が、シャペロンタンパク質、フォルダーゼ、ペプチジルプロリルイソメラーゼおよびジスルフィド結合イソメラーゼから選択できる。一部の実施形態では、1つまたは複数の標的遺伝子が、htpG、cbpA、dnaJ、dnaKおよびfkbPから選択できる。P.fluorescens由来の例示的タンパク質フォールディング調節因子を表1に列挙する。
【0031】
他の実施形態では、標的遺伝子が、推定上のプロテアーゼ、プロテアーゼ様タンパク質、プロテアーゼの補因子またはサブユニットのうち少なくとも1つを含む。例えば、上記1つまたは複数の標的遺伝子が、セリン、スレオニン、システイン、アスパラギン酸またはメタロペプチダーゼであり得る。一実施形態では、1つまたは複数の標的遺伝子が、hslV、hslU、clpA、clpBおよびclpXから選択できる。標的遺伝子はプロテアーゼの補因子でもあり得る。P.fluorescens由来の例示的プロテアーゼを表2に列挙する。様々な生物由来のプロテアーゼは、Wellcome Trust Sanger Institute、Cambridge、UKによって維持されているMEROPSペプチダーゼデータベースに見出すことができる(ウェブサイトアドレスmerops.sanger.ac.uk/を参照)。
【0032】
タンパク質フォールディング調節因子(Protein folding modulators)
宿主細胞での異種タンパク質産生における別の重大な障害は、しばしば細胞が、可溶性または活性のタンパク質を産生するように適切に装備されていないことである。タンパク質の一次構造はアミノ酸配列によって規定(defined)されるが、二次構造はαへリックスまたはβシートの存在によって規定され、三次構造はジスルフィド結合などの隣接するタンパク質ストレッチ間の共有結合によって規定される。異種タンパク質を発現する場合、とりわけ大規模産生では、そのタンパク質それ自体の二次構造および三次構造が極めて重要である。タンパク質構造におけるいかなる有意な(significant)変化も機能的に不活性な分子または生物活性が有意に低下したタンパク質を生じ得る。多くの場合、宿主細胞は、活性な異種タンパク質の適切な産生に必要なタンパク質フォールディング調節因子(protein folding modulators)(PFMs)を発現している。しかし、使用可能かつ経済的に満足できるバイオテクノロジー製品を生産するのに通常必要な高レベルでの発現では、細胞は、しばしば、異種発現されたタンパク質をプロセシングするのに十分な天然の1つまたは複数のタンパク質フォールディング調節因子を産生できない。
【0033】
特定の発現系では、異種タンパク質の過剰産生に、それらのミスフォールディングおよび不溶性凝集体への隔離(segregation)が伴い得る。細菌細胞では、これらの凝集体は封入体として知られている。E.coliでは、フォールディング調節因子/シャペロンのネットワークに、Hsp70ファミリーが含まれる。主要なHsp70シャペロンであるDnaKは、タンパク質凝集を効率的に防止し、損傷タンパク質のリフォールディングを補助する。タンパク質凝集体への熱ショックタンパク質の取込みによって脱凝集が促進され得る。しかし、特定の場合には、不溶性画分の追加プロセシングを介して、封入体内にプロセシングされたタンパク質が回収され得る。封入体内に見出されたタンパク質は通常、変性(denaturation)および復元(renaturation)を含めた複数のステップを経て精製されなければならない。封入体にターゲティングされたタンパク質の典型的な復元方法は、高濃度の変成剤中に凝集体を溶解する試みと、それに続く、希釈による変成剤の除去とを含む。しばしばこの段階で、凝集体が再形成される。追加の処理はコストを増大させ、in vitroのリフォールディングが生理的に活性のある生成物を産生するであろうという保証がなく、回収されるタンパク質は大量の断片不純物を含み得る。
【0034】
分子シャペロンおよびフォルダーゼによって、in vivoでのタンパク質フォールディングが補助されており、分子シャペロンはフォールディング中間体と一時的に相互作用することによって他のポリペプチドの適切な異性化および細胞ターゲッティングを促進し、フォルダーゼはフォールディング経路における律速段階を加速するという最近の認識によって、封入体形成の問題と戦うための追加アプローチが提示されている(例えば、Thomas J Gら(1997)、Appl Biochem Biotechnol 66:197〜238を参照)。
【0035】
特定の場合には、シャペロンの過剰発現が、凝集しやすいタンパク質(aggregation-prone proteins)の可溶性収率を増大させることが見出されている(Baneyx,F.(1999)、Curr.Opin.Biotech、10:411−421およびその中の参考文献を参照)。これらのシャペロンの細胞内濃度の増大に伴う有益効果は、過剰産生されるタンパク質の性質に強く依存しているようであり、同じ(1つまたは複数の)タンパク質フォールディング調節因子の過剰発現が全ての異種タンパク質に必要であるとは限らない。
【0036】
シャペロン、ジスルフィド結合イソメラーゼ、およびペプチジルプロリルシス−トランスイソメラーゼ(PPIase)を含めたタンパク質フォールディング調節因子は、新生ポリペプチドのフォールディング、アンフォールディングおよび分解を補助する、全ての細胞に存在する1クラスのタンパク質である。
【0037】
シャペロンは、新生ポリペプチドに結合し、それらを安定化し、それらが適切にフォールディングするのを可能にすることによって作用する。タンパク質は、疎水性残基および親水性残基の両方を有し、前者は通常表面に露出しており、一方、後者は構造内に埋められており、それらは構造内で、その分子を取り囲んでいる水ではなく、他の親水性残基と相互作用している。しかし、フォールディング中のポリペプチド鎖では、タンパク質が部分的にフォールディングされた状態またはミスフォールディングされた状態で存在しているので、しばしば、親水性残基がしばらくの間露出する。この間に、生成中のポリペプチドが恒久的にミスフォールディングされた状態になるか、または他のミスフォールディングされたタンパク質と相互作用して、細胞内に大きな凝集体または封入体を形成し得る。シャペロンは通常、部分的にフォールディングされた鎖の疎水性領域に結合し、それらが完全にミスフォールディングするか、他のタンパク質と凝集するのを防止することによって作用する。シャペロンは、封入体内のタンパク質にも結合し、それらが脱凝集するのを可能にできる(Ransonら、1998)。フォールディング調節因子のGroES/EL、DnaKJ、Clp、Hsp90およびSecBファミリーは全てシャペロン様活性を有するタンパク質の例である。
【0038】
フォールディング調節因子の別の重要なタイプはジスルフィド結合イソメラーゼである。これらのタンパク質は、フォールディングポリペプチドが適切なタンパク質内ジスルフィド結合を形成するのを助ける極めて特殊な1セットの反応を触媒する。2残基以上のシステインを有するいかなるタンパク質も、誤った残基間でジスルフィド結合を形成する危険性がある。ジスルフィド結合生成ファミリーは、Dsbタンパク質からなり、Dsbタンパク質は、ペリプラズムの非還元環境におけるジスルフィド結合の形成を触媒する。ペリプラズムポリペプチドがミスフォールディングした場合、ジスルフィド結合イソメラーゼであるDsbCは、ジスルフィド結合を再編成して、そのタンパク質が正しい結合を再形成するのを可能にできる。
【0039】
プロリン残基は、そのすぐ前のペプチジル結合がシス配置(conformation)またはトランス配置をとり得るという点で、アミノ酸のなかで独特である。他の全てのアミノ酸について、これは、立体障害のために好ましくない。ペプチジルプロリルシスートランスイソメラーゼ(PPIase)は、この結合の一方の形態からもう一方への変換を触媒する。この異性化は、細胞内におけるタンパク質のフォールディング、リフォールディング、サブユニット集合および輸送の補助となり得る(Dolinskiら、1997)。
【0040】
一般的なシャペロンは非特異的な様式でタンパク質と相互作用するように見えるが、それに加えて、特定の標的のフォールディングを補助するシャペロンも存在する。これらのタンパク質特異的なシャペロンは、それらの標的と複合体を形成し、凝集および分解を防止し、それらが集合して、多重サブユニット構造になるための時間を与える。PapDシャペロンは、このタイプのよく知られている一例である(Lombardoら、1997)。
【0041】
フォールディング調節因子には、例えばHSP70タンパク質、HSP110/SSEタンパク質、HSP40(DNAJ関連)のタンパク質、GRPE様タンパク質、HSP90タンパク質、CPN60およびCPN10タンパク質、サイトゾルシャペロン、HSP100タンパク質、低分子量HSP、カルネキシンおよびカルレティキュリン、PDIおよびチオレドキシン関連タンパク質、ペプチジルプロリルイソメラーゼ、シクロフィリンPPIase、FK−506結合タンパク質、パルブリンPPIase、個別のシャペロニング、タンパク質特異的シャペロンまたは分子内シャペロンも含まれる。フォールディング調節因子については、概ね、「Guidebook to Molecular Chaperones and Protein−Folding Catalysts」(1997)、M.Gething編集、Melbourne University,Australiaに記載されている。
【0042】
E.coli細胞質中で最も詳細に特徴付けられている分子シャペロンは、ATP依存性のDnaK−DnaJ−GrpE系およびGroEL−GroES系である。in vitro研究および相同性考察に基づいて、多くの追加の細胞質タンパク質が、E.coliで分子シャペロンとして機能していると提唱されている。これらには、ClpB、HtpGおよびIbpA/Bが含まれ、それらは、DnaK−DnaJ−GrpEおよびGroEL−GroESのように、ストレスレギュロンに属する熱ショックタンパク質(Hsp)である。新生タンパク質鎖では、X−Pro結合のトランス配座がエネルギー的に支持されるが、天然タンパク質では、全てのプロリルペプチド結合の約5%がシス配座で見出されている。X−Pro結合のトランスからシスへの異性化は、多くのポリペプチドのフォールディングで律速となっており、in vivoでは、ペプチジルプロリルシス/トランスイソメラーゼ(PPIase)によって触媒されている。SlyD、SlpAおよびトリガーファクター(TF)という3種の細胞質PPIaseがこれまでにE.coliで同定されている。TFは、50Sリボソームサブユニットに随伴する48kDaタンパク質であり、E.coliでシャペロンと協力して、新規合成タンパク質の適切なフォールディングを保証すると仮定(postulated)されている。少なくとも5種のタンパク質(それぞれtrxA、trxC、grxA、grxBおよびgrxC遺伝子の産物である、チオレドキシン1および2、ならびにグルタレドキシン1、2および3)が、細胞質酵素内に一時的に生じるジスルフィド架橋の還元(reduction)に関与している。したがって、標的遺伝子は、適切なジスルフィド結合形成を可能にするジスルフィド結合形成タンパク質またはシャペロンであり得る。
【0043】
【表1】
【0044】
プロテアーゼ(Protease)
異種発現されたタンパク質の好ましくない分解は、特定の発現系の効率的な使用に障害を与える。多量の標的タンパク質を産生するように細胞が改変されている場合、細胞はストレス下に置かれ、しばしば他のタンパク質を誘導または抑制することによって反応する。異種タンパク質産生中に宿主細胞が経験するストレスは、例えば、過剰発現した異種タンパク質の分解を引き起こす特定のタンパク質または補因子の発現を増大し得る。代償性の(compensatory)タンパク質の発現増大は、高レベルの活性かつ完全長の異種タンパク質を発現するという目標にとって反生産的であり得る。他のタンパク質の発現低下または適切な発現の不足は、異種発現されたタンパク質のミスフォールディングおよび凝集を引き起こし得る。ストレス下の細胞はタンパク質発現のプロファイルを変えることが知られているが、異種発現された全てのタンパク質が特定の宿主細胞内で同じタンパク質の発現を調節するわけではない。
【0045】
したがって、1つまたは複数のプロテアーゼ酵素の発現が低下するように遺伝子操作された複数の宿主細胞集団を含むアレイを用いて、最適な宿主株、例えばP.fluorescens宿主株を同定できる。一実施形態では、そのゲノムから、少なくとも1つのプロテアーゼの発現を低下させること、阻害すること、または除去することによって、1つまたは複数の宿主細胞集団が改変される。改変は、複数のプロテアーゼに施すこともできる。関連の実施形態では、プロテアーゼ補因子またはプロテアーゼタンパク質の発現を低下させることによって、細胞が改変されている。別の実施形態では、プロテアーゼまたは関連タンパク質のプロモーターの阻害によって、宿主細胞が改変されており、プロモーターは天然のプロモーターであり得る。代替として、遺伝子改変は、標的遺伝子に相同なタンパク質を調節するためのものでもあり得る。
【0046】
改変宿主株を含むアレイは、下記に論じる通り、対象とする(1つまたは複数の)異種タンパク質を発現させ、タンパク質産生の質および/または量を評価することによってスクリーニングできる。代替として、対象とする異種タンパク質の単離株は、プロテアーゼ欠失宿主細胞集団それぞれから収集された溶解物と共に、独立にインキュベートすることもでき、タンパク分解のレベルを、最適な宿主細胞を同定するのに用いることができる。この実施形態では、最適な宿主細胞集団は、最小量の異種タンパク質分解をもたらすものである。したがって、一実施形態では、最適な宿主細胞集団の溶解物が、約50%未満、約45%未満、約40%未満、約35%未満、約30%未満、約25%未満、約20%未満、約10%未満、約5%未満、約4%未満、約3%未満、約2%未満、約1%未満の異種タンパク質、またはさらに少ないタンパク質によって分解され得る。
【0047】
例示的な標的プロテアーゼ遺伝子には、アミノペプチダーゼ、ジペプチダーゼ、ジペプチジルペプチダーゼおよびトリペプチジルペプチダーゼ、ペプチジルジペプチダーゼ、セリン型カルボキシペプチダーゼ、メタロカルボキシペプチダーゼ、システイン型カルボキシペプチダーゼ、オメガペプチダーゼ、セリンプロテイナーゼ、システインプロテイナーゼ、アスパラギン酸プロテイナーゼ、メタロプロテイナーゼまたは作用機序未知のプロテイナーゼと分類されるプロテアーゼが含まれる。
【0048】
アミノペプチダーゼには、サイトゾルアミノペプチダーゼ(ロイシルアミノペプチダーゼ)、膜アラニルアミノペプチダーゼ、シスチニルアミノペプチダーゼ、トリペプチドアミノペプチダーゼ、プロリルアミノペプチダーゼ、アルギニルアミノペプチダーゼ、グルタミルアミノペプチダーゼ、x−proアミノペプチダーゼ、細菌性ロイシルアミノペプチダーゼ、好熱性アミノペプチダーゼ、クロストリジウムアミノペプチダーゼ、サイトゾルアラニルアミノペプチダーゼ、リジルアミノペプチダーゼ、x−trpアミノペプチダーゼ、トリプトファニルアミノペプチダーゼ、メチオニルアミノペプチダーゼ、d−立体特異的アミノペプチダーゼ、アミノペプチダーゼEyが含まれる。ジペプチダーゼには、x−his、x−argジペプチダーゼ、x−メチル−hisジペプチダーゼ、cys−glyジペプチダーゼ、glu−gluジペプチダーゼ、pro−xジペプチダーゼ、x−proジペプチダーゼ、met−xジペプチダーゼ、非立体特異的ジペプチダーゼ、サイトゾル非特異的ジペプチダーゼ、膜ジペプチダーゼ、β−ala−hisジペプチダーゼが含まれる。ジペプチジルペプチダーゼおよびトリペプチジルペプチダーゼには、ジペプチジルペプチダーゼi、ジペプチジルペプチダーゼii、ジペプチジルペプチダーゼiii、ジペプチジルペプチダーゼiv、ジペプチジル−ジペプチダーゼ、トリペプチジルペプチダーゼI、トリペプチジルペプチダーゼIIが含まれる。ペプチジルジペプチダーゼには、ペプチジルジペプチダーゼaおよびペプチジルジペプチダーゼbが含まれる。セリンカルボキシペプチダーゼには、リソソームpro−xカルボキシペプチダーゼ、セリン型D−ala−D−alaカルボキシペプチダーゼ、カルボキシペプチダーゼC、カルボキシペプチダーゼDが含まれる。メタロカルボキシペプチダーゼには、カルボキシペプチダーゼa、カルボキシペプチダーゼB、リジン(アルギニン)カルボキシペプチダーゼ、gly−Xカルボキシペプチダーゼ、アラニンカルボキシペプチダーゼ、ムラモイルペンタペプチドカルボキシペプチダーゼ、カルボキシペプチダーゼh、グルタミン酸カルボキシペプチダーゼ、カルボキシペプチダーゼM、ムラモイルテトラペプチドカルボキシペプチダーゼ、亜鉛d−ala−d−alaカルボキシペプチダーゼ、カルボキシペプチダーゼA2、膜pro−xカルボキシペプチダーゼ、チューブリン−tyrカルボキシペプチダーゼ、カルボキシペプチダーゼtが含まれる。オメガペプチダーゼには、アシルアミノアシルペプチダーゼ、ペプチジルグリシンアミダーゼ、ピログルタミルペプチダーゼI、βアスパラチルペプチダーゼ、ピログルタミルペプチダーゼII、nホルミルメチオニルペプチダーゼ、プテロイルポリ−γ−グルタミン酸カルボキシペプチダーゼ、γ−glu−Xカルボキシペプチダーゼ、アシルムラモイル−alaペプチダーゼが含まれる。セリンプロテイナーゼには、キモトリプシン、キモトリプシンc、メトリジン、トリプシン、トロンビン、凝固因子Xa、プラスミン、エンテロペプチダーゼ、アクロシン、α分解プロテアーゼ、グルタミル、エンドペプチダーゼ、カテプシンG、凝血因子viia、凝固因子ixa、ククミシ(cucumisi)、プロリルオリゴペプチダーゼ、凝固因子xia、ブラキウリン、血漿カリクレイン、組織カリクレイン、膵臓エラスターゼ、白血球エラスターゼ、凝固因子xiia、キマーゼ、補体成分c1r55、補体成分c1s55、古典的補体経路c3/c5コンバターゼ、補体因子I、補体因子D、第二補体経路c3/c5コンバターゼ、セレビシン、ヒポデルミンC、リジルエンドペプチダーゼ、エンドペプチダーゼ1a、γ−レニ(gamma-reni)、ベノムビンab、ロイシルエンドペプチダーゼ、トリプターゼ、スクテラリン、ケキシン、ズブチリシン、オリジン(oryzin)、エンドペプチダーゼk、サーモミコリン、テルミターゼ、エンドペプチダーゼSO、T−プラスミノーゲンアクチベータ、プロテインC、膵臓エンドペプチダーゼE、膵臓エラスターゼii、IGA特異的セリンエンドペプチダーゼ、U−プラスミノーゲン, 活性化因子(U-plasminogen, activator)、ベノムビンA、フリン、ミエロブラスチン、セメノゲラーゼ、グランザイムAすなわち傷害性Tリンパ細胞プロテイナーゼ1、グランザイムBすなわち傷害性Tリンパ細胞プロテイナーゼ2、ストレプトグリシンA、トレプトグリシンB(trestgrisin B)、グルタミルエンドペプチダーゼII、オリゴペプチダーゼB、リムルス凝固因子c、リムルス凝固因子、リムルス凝血酵素、オンプチン、レプレッサーlexa、細菌リーダーペプチダーゼI、トガビリン、フラビリンが含まれる。システインプロテイナーゼには、カテプシンB、パパイン、フィシン、キモパパイン、アスクレパイン、クロストリパイン、ストレプトパイン、アクチニド、カテプシン1、カテプシンH、カルパイン、カテプシンt、グリシル, エンドペプチダーゼ(glycyl, endopeptidase)、癌凝血原、カテプシンS、ピコルナイン3C、ピコルナイン2A、カリカイン、アナナイン、幹ブロメライン、果実ブロメライン、レグミン、ヒストリサイン、インターロイキン1−β変換酵素が含まれる。アスパラギン酸プロテイナーゼには、ペプシンA、ペプシンB、ガストリクシン、キモシン、カテプシンD、ネオペンテシン、レニン、レトロペプシン、プロオピオメラノコルチン変換酵素、アスペルギロペプシンI、アスペルギロペプシンII、ペニシロペプシン、リゾプスペプシン、エンドチアペプシン、ムコロペプシン、カンジダペプシン、サッカロペプシン、ロドトルラペプシン、フィサロペプシン、アクロシリンドロペプシン、ポリポロペプシン、ピクノポロペプシン、シタリドペプシンa、シタリドペプシンb、キサントモノナペプシン、カテプシンe、バリヤーペプシン、細菌性リーダーペプチダーゼI、シュードモナペプシン、プラスメプシンが含まれる。メタロプロテイナーゼには、アトロリシンa、微生物コラゲナーゼ、ロイコリシン、間質コラゲナーゼ、ネプリリシン、エンベリシン、iga特異的メタロエンドペプチダーゼ、プロコラーゲンN−エンドペプチダーゼ、チメットオリゴペプチダーゼ、神経溶解素、ストロメライシン1、メプリンA、プロコラーゲンC−エンドペプチダーゼ、ペプチジル−Lysメタロエンドペプチダーゼ、アスタシン、ストロメライシン, 2(stromelysin, 2)、マトリリジンゼラチナーゼ、エアロモノリシン、プソイドリシン、サーモリシン、バシロリシン、アウレオリシン、ココリシン、ミコリシン、β−分解メタロエンドペプチダーゼ、ペプチジル−Aspメタロエンドペプチダーゼ、好中球コラゲナーゼ、ゲラチナーゼB、リーシュマノリシン、サッカロリシン、自己溶菌酵素、デウテロリシン、セラリシン、アトロリシンB、アトロリシンC、アトロキサーゼ、アトロリシンE、アトロリシンF、アダマリシン、ホリリシン、ラバリシン、ボソロパシン、ボソロリシン、オフィオリシン、トリメレリシンI、トリメレリシンII、ムクロリシン、ピトリリシン、インスリシン、O−シアロ糖タンパク質エンドペプチダーゼ、ルスセリリシン、ミトコンドリア, 間質性, ペプチダーゼ(mitochondrial intermediate peptidase)、ダクチリシン、ナルジリシン、マグノリシン、メプリンB、ミトコンドリアプロセシングペプチダーゼ、マクロファージエラスターゼ、コリオリシン、トキシリシンが含まれる。作用機序未知のプロテイナーゼには、サーモプシンおよび多触媒エンドペプチダーゼ複合体が含まれる。
【0049】
【表2】
【0050】
その他のタンパク質修飾酵素(Additional protein modification enzymes)
別の実施形態では、標的遺伝子が、適切なタンパク質プロセシングおよび/または修飾に関与する遺伝子を含む。一般的な修飾には、ジスルフィド結合形成、グリコシル化、アセチル化、アシル化、リン酸化およびγ−カルボキシ化が含まれ、これらは全て、タンパク質フォールディングおよび生物活性を調節し得る。タンパク質プロセシングに関与する酵素のいくつかのクラスの網羅的ではないリストが表3に示されている。当業者ならば、アレイ用に選択された宿主細胞内で有用であるか、または対象とする異種タンパク質と併用して有用である標的遺伝子を、どのようにして表3に列挙したタンパク質修飾酵素のクラスの中から同定するか認識するであろう。標的遺伝子は、使用される宿主細胞に内因的なものでもよく、または対象とする異種タンパク質が由来する生物に内因的なものもよく、または対象とする異種発現タンパク質の適切なプロセシングを促進することが知られているものでもよい。タンパク質産生に関与するいかなる遺伝子も、対象とする異種タンパク質の望ましい仕様に従ってターゲッティングできることも認識されている。
【0051】
【表3】
【0052】
標的遺伝子の発現を調節する方法(Methods for modulating the expression of target genes)
当技術分野で知られている任意の技法によって、アレイ内の1つまたは複数の宿主細胞集団を改変でき、例えば、ゲノム内の少なくとも1つの標的遺伝子がノックアウトされる技法によって、または少なくとも1つの標的遺伝子の発現が低下するように、遺伝子に変異導入することによって、または少なくとも1つの標的遺伝子の発現が低下するように、標的遺伝子の少なくとも1つのプロモーターを改変することによって、または宿主ゲノム内の標的遺伝子または標的遺伝子の阻害因子を(対象とする異種タンパク質またはポリペプチドと共に)同時発現させることによって改変できる。上記に論じたように、標的遺伝子はアレイ内の宿主細胞集団に内因的なものでもよく、または宿主細胞集団それぞれで異種発現されるものでもよい。
【0053】
標的遺伝子の発現は、例えば、タンパク質産生に関与する1つまたは複数の標的遺伝子を含む発現ベクターを、宿主集団内の少なくとも1つの細胞に導入することによって増大できる。標的遺伝子発現は、例えば、標的遺伝子のプロモーターに変異導入することによっても増大できる。異種タンパク質を発現する宿主細胞または生物は、タンパク質産生に関与する少なくとも1つの標的遺伝子の発現が増大し、かつタンパク質分解に関与する少なくとも1つの標的遺伝子の発現が低下するように、遺伝子改変することもできる。
【0054】
ゲノムは、外因性遺伝子またはプロモーターエレメントをゲノム内に、もしくは発現ベクターで宿主内に含めることによって、または特定の標的遺伝子の、mRNAもしくはタンパク質産生能力を高めることによって、または標的遺伝子もしくはプロモーターエレメントを除去もしくは破壊することによって、または標的遺伝子の、mRNAもしくはタンパク質産生能力を低下させることによって、1つまたは複数の標的遺伝子の発現を調節するように改変できる。例えば、置換、欠失(「ノックアウト」)、同時発現または挿入(「ノックイン」)技法によって、遺伝暗号を改変し、それによって、標的遺伝子の転写および/または翻訳に影響を与えることができる。既存の標的配列の転写を調節する所望のタンパク質または調節配列の追加遺伝子を挿入することもできる。
【0055】
ゲノム改変(Genome modification)
宿主細胞のゲノムは、遺伝的ターゲッティング事象を介して改変することができ、それは、挿入または組換え、例えば相同組換えによるものであり得る。相同組換えは、配列相同性に基づいたDNA組換え過程を指す。相同組換えは、内因性遺伝子内における部位特異的な改変を可能にし、それゆえ、ゲノム内の新規な改変が構築できる(例えば、Radding(1982)、Ann.Rev.Genet.16:405;米国特許第4,888,274号参照)。
【0056】
標的遺伝子座の相同組換え用には、様々なコンストラクトが調製できる。通常、コンストラクトは、同定された遺伝子座と相同な少なくとも10bp、20bp、30bp、40bp、50bp、70bp、100bp、500bp、1kbp、2kbp、4kbp、5kbp、10kbp、15kbp、20kbpまたは50kbpの配列を含み得る。標的遺伝子配列の相同性の程度を決定するには、例えば、標的遺伝子座のサイズ、配列の利用可能性、標的遺伝子座における二重交差事象の相対効率および他の配列との標的配列の類似性など、様々な要件が関与し得る。
【0057】
改変遺伝子には、改変されるべきゲノム内の、対応する標的配列と実質的に同質遺伝子的なDNAによって、所望の配列改変が挟まれている配列が含まれ得る。「改変遺伝子(modified gene)」は、ゲノム内に導入されて、宿主細胞内のプロテアーゼまたはタンパク質フォールディング調節因子の発現を改変する配列である。「標的遺伝子(target gene)」は、改変遺伝子によって置換される配列である。実質的に同質遺伝子的な配列は、対応する標的配列に、(所望の配列改変を除いて)少なくとも約95%、97〜98%、99.0〜99.5%、99.6〜99.9%または100%同一であり得る。改変遺伝子および標的遺伝子は、100%同一である少なくとも約10、20、30、50、75、150または500塩基対のDNAストレッチを共有し得る。
【0058】
ヌクレオチドコンストラクトは、内因性の標的遺伝子産物を改変するように設計できる。改変遺伝子配列は、結果として生じる遺伝子産物の機能を破壊するように設計された1つまたは複数の欠失、挿入、置換またはこれらの組合せを有し得る。一実施形態では、上記改変が、標的遺伝子の上流配列と、リーディングフレーム内で融合している選択マーカー遺伝子の挿入であり得る。
【0059】
ゲノムは、挿入不活化を用いて改変することもできる。この実施形態では、遺伝子産物の生成を阻害する遺伝子内の配列の組換えによってゲノムが改変される。この挿入は、別個のエレメントを挿入することによって遺伝子を破壊するか、遺伝子の必須部分を除去することができる。一実施形態では、抗生物質などの特定のストレッサーに対する耐性、または特定の培地中での増殖、例えば必須アミノ酸の産生をコードする遺伝子の挿入も挿入欠失に含まれる。
【0060】
ゲノムは、トランスポゾンの使用によって改変することもできる。トランスポゾンは、相同組換えとは独立した作用機序によって、原核生物ゲノム内の複数の部位に挿入され得る遺伝的エレメントである。トランスポゾンには、例えば、E.coliにおけるTn7、Tn5またはTn10、S.aureusにおけるTn554、M.paratuberculosis由来のIS900、Pseudomonas atlantica由来のIS492、ストレプトミセス由来のIS116ならびにM.paratuberculosis由来のIS900が含まれ得る。遺伝子転位に関与すると考えられているステップには、3’OHを生じる、トランスポゾン末端の切断と、トランスポザーゼによってトランスポゾンの3’OH露出末端と同定された配列とが引き合わされる鎖転移と、同定されたDNAへのトランスポゾンの共有結合を生じる1ステップエステル交換反応とが含まれる。トランスポザーゼによって行われる、鍵となる反応は、ニック形成または鎖交換であり、残りの過程は宿主酵素によって行われると一般的に考えられている。
【0061】
一実施形態では、標的タンパク質またはその相同体をコードする遺伝子の配列を、組換えによってゲノム内に組込むことによって、標的遺伝子またはタンパク質の発現または活性を増大させる。別の実施形態では、ゲノム内にプロモーターを挿入して、標的遺伝子または相同体の発現を亢進する。別の実施形態では、不活性遺伝子を用いた組換えによって、標的遺伝子またはその相同体の発現または活性を低下させる。別の実施形態では、細胞内で別個の機能を有し得る、または耐性マーカーなどのレポーター遺伝子であるか、もしくは他の方法で検出可能なマーカー遺伝子であり得る異なった遺伝子をコードする配列を、組換えによってゲノム内に挿入できる。さらに別の実施形態では、1つまたは複数の位置で変異導入されている標的遺伝子の少なくとも一部の1コピーを、組換えによってゲノム内に挿入する。この変異バージョンの標的遺伝子は、タンパク質をコードしていなくてもよく、または変異遺伝子によってコードされているタンパク質が不活性でもよく、活性が調節(増大または低下)されていてもよく、または天然タンパク質と比較した際に、変異体タンパクが異なった活性を有することもできる。
【0062】
細菌で遺伝子をノックアウトするための戦略があり、それらは概ね、E.coliで例示されている。1つの経路は、抗生物質耐性の遺伝子(例えばアンピシリン)を含有するベクターに遺伝子内DNA断片をクローニングするものである。接合伝達、化学的形質転換またはエレクトロポレーション(Puehlerら(1984)、Advanced Molecular Genetics、New York、Heidelberg、Berlin、Tokyo、Springer Verlag)を介して細胞を形質転換する前に、栄養プラスミド複製(oriV遺伝子座)などの複製開始点を切除し、残りのDNA断片を再連結および精製する(Sambrookら(2000)、Molecular cloning:A laboratory manual、第3版、Cold Spring Harbor、N.Y.、Cold Spring Harbor Laboratory Press)。代替として、DNA複製開始点を有する抗生物質抵抗性のプラスミドを用いることもできる。形質転換後に、適切な抗生物質(例えば200μg/mLのアンピシリン)を含有する、例えばLBアガープレート上に細胞をプレーティングする。抗生物質を含有するプレート上で増殖するコロニーは、おそらく、相同遺伝子座における、ゲノム内への全DNA断片の組込みをもたらす単一の組換え事象を経たものである(Snyder,L.、W.Champnessら(1997)、Molecular Genetics of Bacteria、Washington DC、ASM Press)。所望の遺伝子ノックアウトが所望の遺伝子座で起こったことを確かめる抗生物質耐性細胞のさらなる分析は、例えばPCR診断による(McPherson,M.J.、P.Quirkeら(1991)、PCR:A Practical Approach、New York、Oxford University Press)。ここでは、少なくとも2通りのPCRプライマーが設計される。ひとつは、遺伝子ノックアウトの構築に使用されるDNA領域の外側でハイブリッド形成するものであり、ひとつは、残りのプラスミドバックボーン内でハイブリッド形成するものである。正しいサイズを有するDNA断片のPCR増幅に成功して、それに続いてDNA配列分析を行うことによって、細菌染色体内の正しい位置で遺伝子ノックアウトが起こったことが確認されるであろう。新規に構築された変異体株の表現型は、その後、例えばSDSポリアクリルアミドゲル電気泳動によって分析できる(Simpson,R.J.(2003)、Proteins and Proteomics−A Laboratory Manual、Cold Spring Harbor、N.Y.、Cold Spring Harbor Laboratory Press)。
【0063】
遺伝子ノックアウトを生成する代替の経路は、pSC101レプリコンなどの温度感受性レプリコンを使用して、遺伝子置換を促進するものである(Hamiltonら(1989)、Journal of Bacteriology 171(9):4617〜22)。この過程は、染色体上の遺伝子と、DNA複製が温度感受性であるプラスミド上に保持されている相同配列との間の相同組換えによって進行する。このプラスミドを適切な宿主内に導入して形質転換した後、プラスミドが染色体内に組込まれているものを44℃で選択することが可能である。それに続いてこれらの共組換え体を30℃で増殖させることによって、第2の組換え事象がもたらされ、その結果、それらの解離が起こる。第2の組換え事象がどこで起こるかに応じて、その染色体が、遺伝子置換を経たものになるか、またはその遺伝子の元のコピーを保持することになる。
【0064】
特定の遺伝子産物の発現を阻害する他の戦略も開発されている。例えば、特定遺伝子産物の発現の低下または消失さえもたらすために、RNA干渉(RNAi)、とりわけ低分子干渉RNA(siRNA)を用いたものが、広範に開発されている。siRNAは、相補的なmRNAを、分解するためにターゲッティングできる短い二本鎖RNA分子である。RNAiは、二本鎖RNAの導入によって相同遺伝子の発現が抑制される現象である。dsRNA分子は、in vivoで、RNAi効果の媒介因子である21〜23ntのsiRNAにされる。2本鎖RNAは、導入された際に、Dicerと呼ばれるRNaseIII様の酵素によって、20〜25ヌクレオチドのsiRNAにプロセシングされる(開始段階)。その後、siRNAは、RNA誘導サイレンシング複合体(RISC)として知られる、エンドリボヌクレアーゼを含有する複合体の中に組み込まれ、この過程で巻き戻される。続いて、siRNA鎖は、RISCを相補的なRNA分子へと導き、そこで、それらは同族のRNAを切断して、破壊する(エフェクター段階)。同族のRNAの切断は、siRNA鎖が結合した領域の中央近くで行われる。RNAiは、ゼブラフィッシュ、線虫(C.elegans)、昆虫(Drosophila melanogaster)、プラナリア、刺胞動物、トリパノソーマ、マウス、および哺乳動物細胞を含めた様々な生物で、遺伝子発現を低減するのに使用され、成功している。
【0065】
標的遺伝子をコードするオープンリーディングフレーム内の1または複数ヌクレオチドを変異させることによっても、ゲノムを改変できる。遺伝的変異のため技法、例えば部位特異的変異誘発は、当技術分野でよく知られている。一部のアプローチは、X線および化学物質によって誘導されるものなど、染色体DNAにおけるランダム変異の生成に焦点をおいている。
【0066】
同時発現(Coexpression)
一実施形態では、(1つまたは複数の)標的遺伝子をコードする1または複数種のベクターを含めて、異種タンパク質またはペプチドとの標的遺伝子の同時発現を促進することによって、宿主細胞内の1つまたは複数の標的遺伝子を改変できる。別の実施形態では、宿主細胞ゲノムへの外因性プロモーターの付加を含めた、標的遺伝子のプロモーターの亢進によって、宿主細胞を改変する。
【0067】
別の実施形態では、プロテアーゼ阻害物質など、標的遺伝子の阻害物質をコードする1または複数種のベクターを含めて、標的プロテアーゼの活性を阻害することによって、宿主細胞内の1つまたは複数の標的遺伝子を改変する。そのような阻害物質は、標的遺伝子、標的遺伝子の補因子または標的遺伝子の相同体の発現を制限するアンチセンス分子であり得る。通常、アンチセンスは、標的遺伝子の少なくとも一部に相補的な配列を有する核酸分子を指すのに使用される。加えて、阻害物質は、干渉(interfering)RNAまたは干渉RNAをコードする遺伝子であり得る。例えば、Fire,A.ら(1998)、Nature 391:806〜11、Elbashirら(2001)、Genes&Development 15(2):188〜200、Elbashirら(2001)、Nature 411(6836):494〜8、Carnegie Instituteの米国特許第6,506,559号、Benitecの第6,573,099号、the Whitehead Inst.の米国特許出願第2003/0108923号、ならびに米国特許出願第2003/0114409号、WO03/006477、WO03/012052、WO03/023015、WO03/056022、WO03/064621およびWO03/070966に記載されている通り、真核生物では、そのような干渉RNAが、低分子干渉(small interfering)RNAまたはリボザイムであり得る。
【0068】
阻害物質は、別のタンパク質またはペプチドであり得る。阻害物質は、例えば、標的タンパク質のコンセンサス配列を有するペプチドであり得る。阻害物質は、標的タンパク質の直接的または間接的阻害分子を宿主で産生できるタンパク質またはペプチドであり得る。例えば、プロテアーゼ阻害物質には、アマスタチン、E−64、アンチパイン、エラスタチナール、APMSF、ロイペプチン、ベスタチン、ペプスタチン、ベンズアミジン、1,10−フェナントロリン、キモスタチン、ホスホラミドン、3,4−ジクロロイソクマリン、TLCK、DFP、TPCKが含まれ得る。これまでに、100を超える天然存在タンパク質プロテアーゼ阻害物質が同定されている。それらは、細菌から動植物までの様々な生物で単離されている。それらは、立体障害を介して活性部位への基質接近を防止する、プロテアーゼの強結合可逆阻害物質または疑似不可逆阻害物質として働く。それらのサイズも、50残基(例えばBPTI:ウシ膵臓トリプシン阻害物質)から、最大400残基(例えばα−1PI:α−1プロテイナーゼ阻害物質)まで極めて可変的である。それらは、α−マクログロブリンファミリーのタンパク質(例えばα−2マクログロブリン)を除いて、厳密にクラス特異的である。α−マクログロブリンファミリーのタンパク質は、分子トラップ作用機序を介して、ほとんどのプロテアーゼに結合し、それらを阻害する。
【0069】
外因性のベクターまたはDNAコンストラクトを宿主細胞内に導入して、形質移入(transfected)または形質転換(transformed)することができる。外因性の核酸で真核細胞および原核細胞をそれぞれ形質移入および形質転換する技法は、当技術分野でよく知られている。これらには、脂質小胞媒介の取り込み、リン酸カルシウム媒介の形質移入(リン酸カルシウム/DNA共沈)、ウイルス感染、とりわけ、例えば改変アデノウイルスなどの改変ウイルスを用いたもの、微量注入およびエレクトロポレーションが含まれ得る。原核細胞の形質転換には、熱ショック媒介の取り込み、無傷細胞を用いた細菌プロトプラスト融合、微量注入およびエレクトロポレーションが技法に含まれ得る。植物を形質転換する技法には、A.tumefaciensなどのアグロバクテリウム媒介の導入、高速推進のタングステンまたは金マイクロプロジェクタイル、エレクトロポレーション、微量注入、およびポリエチレングリコール媒介の取り込みが含まれる。DNAは、1本鎖または2本鎖、直鎖状または環状、リラックス型またはスーパーコイルド型のDNAであり得る。哺乳動物細胞に形質移入する様々な技法に関しては、例えば、Keownら(1990)、Processes in Enzymology、第185巻、527〜537ページを参照のこと。
【0070】
下記の通り、対象とする異種タンパク質またはポリペプチドを含む発現コンストラクト用に、標的遺伝子またはそのエンハンサーもしくは阻害物質をコードする発現コンストラクトを構築できる。例えば、コンストラクトは、1箇所または複数箇所の内部リボソーム進入部位(IRES)を含有できる。コンストラクトは、標的遺伝子の少なくとも一部、または標的遺伝子の補因子、変異体バージョンの標的遺伝子の少なくとも一部、もしくは一部の実施形態では標的遺伝子の阻害物質をコードする核酸配列に作用可能に連結したプロモーターをも含有できる。代替として、コンストラクトは、プロモーターの無いものであり得る。コンストラクトが細胞DNA/ゲノムに組み込まれるように設計されていない場合には、ベクターは通常、少なくとも1つのプロモーターエレメントを含有する。上記核酸配列に加えて、発現ベクターは、選択マーカー配列を含有し得る。発現コンストラクトは、転写開始、終止および/または、リボソーム結合部位のための部位をさらに含有し得る。特定されたコンストラクトは、限定されるものではないが、P.fluorescensもしくはE.coliなどの細菌細胞、酵母細胞、CHO細胞などの哺乳動物細胞または植物細胞を含めた、いかなる原核細胞または真核細胞にも挿入でき、発現させることができる。
【0071】
コンストラクトは、当技術分野で知られている方法に従って調製できる。望ましいコンストラクトが得られるまで、様々な断片を集め、適切なベクター内に導入し、クローニングし、分析し、その後さらに操作することができる。制限分析、切断、プローブの同定などを可能にする様々な改変を配列に加えることができる。必要に応じて、サイレント突然変異を導入できる。様々な段階で、制限分析、配列決定、ポリメラーゼ連鎖反応を用いた増幅、プライマー修復、in vitro突然変異誘発などを利用できる。抗生物質耐性遺伝子およびネガティブ選択因子を組込む方法は、当業者ならばよく知っているであろう(例えば、国際公開第99/15650号、米国特許第6,080,576号、米国特許第6,136,566号、Niwaら、J.Biochem.113:343〜349(1993);およびYoshidaら、Transgenic Research,4:277〜287(1995)を参照)。
【0072】
原核細胞複製系、例えばP.fluorescensまたはE.coliなどの原核細胞によって認識可能なオリジンを含有する細菌ベクターを用いて、コンストラクトを調製できる。マーカーを利用することができ、それは挿入に使用されるべきマーカーと同じか、または異なっており、宿主細胞に導入する前に除去できる。ひとたびコンストラクトを含有するベクターが完成したならば、特定の配列を除去するか、直線化するか、相同配列中に変異、欠失または他の配列を導入することなどによって、それをさらに操作できる。一実施形態では、標的遺伝子コンストラクトおよび異種タンパク質コンストラクトが同じ発現ベクターの一部であり、かつ同じプロモーターエレメントの制御下でも、そうでなくてもよい。別の実施形態では、それは別々の発現ベクター上にある。最終操作の後、コンストラクトを細胞に導入することができる。
【0073】
細胞増殖条件(Cell growth conditions)
本明細書に記載の宿主細胞の細胞増殖条件には、対象とするタンパク質の発現をアレイ内の少なくとも1つの株(または少なくともそれらの細胞の一部)で促進する条件および/または発現された、対象とするタンパク質の発酵を促進する条件が含まれる。本明細書で使用される場合、「発酵」という用語には、文字通りの発酵が用いられる実施形態と、他の、非発酵性の培養モードが用いられる実施形態との両方が含まれる。アレイ内の宿主細胞集団の増殖、維持および/または発酵は、いかなるスケールで行ってもよい。しかし、宿主細胞の複数集団が同時にスクリーニングされる場合には、異なった集団の数ならびに宿主細胞の複数集団を増殖および試験する能力によってスケールが限定されるであろう。一実施形態では、発酵培地が、富栄養培地、最小培地および無機塩培地の中から選択され得る。別の実施形態では、最少培地または無機塩培地のいずれかが選択される。さらに別の実施形態では、最少培地が選択される。さらに別の実施形態では、無機塩培地が選択される。
【0074】
無機塩培地は、無機塩と、例えば、グルコース、ショ糖またはグリセロールなどの炭素源とからなる。無機塩培地の例には、例えば、M9培地、シュードモナス培地(ATCC179)、ならびにDavisおよびMingioli培地(BD DavisおよびES Mingioli(1950)、J.Bact.60:17〜28を参照)が含まれる。無機塩培地を作製するのに使用される無機塩には、例えば、リン酸カリウム、硫酸もしくは塩化アンモニウム、硫酸もしくは塩化マグネシウム、ならびに微量無機塩、すなわち鉄、銅、マンガンおよび亜鉛の塩化カルシウム塩、ホウ酸塩および硫酸塩などの中から選択されたものが含まれる。有機窒素源、すなわち、ペプトン、トリプトン、アミノ酸または酵母抽出物などは、無機塩培地に含まれていない。代わりに、無機窒素源が使用され、これは、例えば、アンモニウム塩、アンモニア水および気体アンモニアの中から選択され得る。好ましい無機塩培地は、炭素源としてグルコースを含有する。無機塩培地と比較すると、最小培地も、無機質塩および炭素源を含有し得るが、例えば、低レベルのアミノ酸、ビタミン、ペプトンまたは他の成分が捕捉されていてもよい。但し、これらは極めて最小レベルで添加される。
【0075】
一実施形態では、下記表4に示す成分を用いて、培地を調製できる。それらの成分は、以下の順序で添加できる。すなわち、最初に、約30リットルの蒸留水に(NH4)HPO4、KH2PO4およびクエン酸を溶解することができる。その後、微量元素の溶液を添加し、それに続いて、Ucolub N115などの消泡剤を添加することができる。次に、乾熱滅菌(約121℃など)の後、グルコース、MgSO4およびチアミン−HCLの無菌液を添加できる。約6.8のpH調節は、アンモニア水を用いて実現できる。その後、滅菌蒸留水を添加して、371からグリセロールストック(123mL)を引いた容積に、初期容積を調整することができる。これらの化学物質は、Merckなどの様々な供給業者から市販されている。
【0076】
【表4】
【0077】
本発明では、形質転換された宿主細胞の増殖、培養および/または発酵を、宿主細胞の生存を許容する温度範囲内、好ましくは、包括的に、約4℃から約55℃の範囲内の温度で行う。したがって、例えば、「増殖」(および「増殖する」、「増殖すること」)、「培養すること」(および「培養」)」ならびに「発酵」(および「発酵させる」、「発酵させること」)という用語は、本明細書で使用される場合、本発明の宿主細胞に関して、本質的に、包括的に約4℃から約55℃の温度範囲内での「増殖」、「培養すること」および「発酵」を意味する。加えて、「増殖」は、活動的な細胞分裂および/または拡大の生物学的状態、ならびに無分裂および/または無拡大の細胞が代謝によって維持されている生物学的状態の両方を示すのに使用され、「増殖」という用語の後者の使用は、「維持」という用語と同義である。
【0078】
アレイの宿主細胞は、その細胞型の正常な増殖に適した温度で培養および維持するべきである。そのような正常増殖温度は、選択された宿主細胞の既知の増殖必要条件に基づいて容易に選択できる。培養を確立している間、とりわけスクリーニングの過程の間は、対象とする異種タンパク質またはポリペプチドで形質転換させる前後に、選択された細胞の増殖に適したCO2/N2湿度に制御された中で、細胞培養物をインキュベートすることが好ましい。インキュベーションの湿度は、培養器からの蒸発を最小限にし、より小さい容積の使用が可能となるように制御する。湿度の制御の代替として、またはそれに加えて、蒸発を最小限にするために、容器をフタで覆ってもよい。培養温度の選択は、主として、使用される宿主細胞のアイデンティティによる。蒸発を制御する湿度(パーセント)の選択は、選択された容器容積、ならびに容器内の細胞培養物の濃度および容積、ならびに培養温度に基づく。したがって、湿度は、約10%から約80%まで変動し得る。適した状態の選択は、十分に当技術分野の技術範囲内であることを理解するべきである。
【0079】
スクリーニング(Screening)
対象とする異種タンパク質を発現するのに最適な宿主細胞集団を求めて、本明細書に記載の株アレイをスクリーニングすることができる。最適な宿主細胞集団は、発現された、対象とするタンパク質の量、質および/または局在に基づいて同定または選択できる。一実施形態では、最適な宿主細胞集団は、アレイ内の、表現型が異なる宿主細胞の他の集団と比較して、宿主細胞内における、対象とするタンパク質またはポリペプチドの収率増大をもたらすものである。
【0080】
産生の増大は、代替として、産生されたタンパク質1グラムあたり、または宿主タンパク質1グラムあたりの、適切にプロセシングされたタンパク質またはポリペプチドのレベルの増大であり得る。産生の増大は、異種タンパク質1グラムあたり、または宿主細胞タンパク質1グラムあたりに産生された、回収可能なタンパク質またはポリペプチドのレベルの増大でもあり得る。産生の増大は、総タンパク質レベルの増大、適切にプロセシングされたタンパク質もしくは適切にフォールディングされたタンパク質のレベルの増大、または、活性もしくは可溶性タンパク質のレベルの増大の任意な組合せであり得る。この実施形態では、「増大」または「改善」という用語は、対象とするタンパク質またはポリペプチドを、アレイ内の、宿主細胞の他の1つまたは複数の集団で発現させた場合に、産生される、適切にプロセシングされる、可溶性である、かつ/または回収可能であるタンパク質またはポリペプチドのレベルに対して相対的なものである。産生の増大は、例えば、エネルギー消費量を低下させることによって、利用可能資源の使用を増大させることによって、または増殖培地中の増殖補助物質の必要性を低下させることによって、細胞または生物の効率を最適化し得る。産生の増大は、発現されたタンパク質のタンパク質分解の低下の結果であってもよい。
【0081】
一実施形態では、アレイ内の少なくとも1つの株が、少なくとも0.1mg/mlの正しくプロセシングされたタンパク質を産生する。正しくプロセシングされたタンパク質は、天然タンパク質のアミノ末端を有する。別の実施形態では、少なくとも1つの株が、0.1〜10mg/mlの正しくプロセシングされたタンパク質を細胞内に産生し、これには、少なくとも約0.2、約0.3、約0.4、約0.5、約0.6、約0.7、約0.8、約0.9または少なくとも約1.0mg/mlの正しくプロセシングされたタンパク質が含まれる。別の実施形態では、アレイ内の少なくとも1つの株によって産生される、正しくプロセシングされた、対象とするタンパク質またはポリペプチドの総タンパク質量が、少なくとも1.0mg/ml、少なくとも約2mg/ml、少なくとも約3mg/ml、約4mg/ml、約5mg/ml、約6mg/ml、約7mg/ml、約8mg/ml、約10mg/ml、約15mg/ml、約20mg/ml、約25mg/ml、約30mg/ml、約35mg/ml、約40mg/ml、約45mg/mlもしくは少なくとも約50mg/mlであるか、またはそれより多い。一部の実施形態では、産生される、正しくプロセシングされたタンパク質の量が、正しくプロセシングされた形態の異種タンパク質の総タンパク質量の少なくとも約5%、約10%、約15%、約20%、約25%、約30%、約40%、約50%、約60%、約70%、約80%、約90%、約95%、約96%、約97%、約98%もしくは少なくとも約99%であるか、またはそれより多い。
【0082】
対象とするタンパク質またはポリペプチドの改善された発現は、タンパク質の溶解性の増大も指し得る。対象とするタンパク質またはポリペプチドを宿主細胞の細胞質、ペリプラズムまたは細胞外の培地中に産生させて、そこから回収することができる。タンパク質またはポリペプチドは、不溶性または可溶性であり得る。タンパク質またはポリペプチドは、上記に論じた通り、1つまたは複数の標的(例えばシグナルまたはリーダー)配列または精製に役立つ配列を含有し得る。
【0083】
「可溶性(soluble)」という用語は、本明細書で使用される場合、生理的条件下にある緩衝液中で10〜30分間遠心処理された場合に、約5000〜20000×重力の遠心処理によって、そのタンパク質が沈殿しないことを意味する。可溶性タンパク質は、封入体または他の沈殿物質塊の一部でない。同様に、「不溶性(insoluble)」は、生理的条件下にある緩衝液中で10〜30分間遠心処理された場合に、5000〜20000×重力の遠心処理によって、そのタンパク質またはポリペプチドが沈殿し得ることを意味する。不溶性タンパク質またはポリペプチドは、封入体または他の沈殿物質塊の一部であり得る。「封入体(inclusion body)」という用語には、細胞内に含有されている、タンパク質またはポリペプチドの凝集体が隔離されている任意の細胞内物体が含まれるものとする。
【0084】
別の実施形態では、最適な宿主細胞集団は、ペリプラズムに輸送されるか、または宿主細胞の細胞外空間に分泌される、対象とするタンパク質を、増大した量で産生する。一実施形態では、アレイ内の少なくとも1つの株が、少なくとも0.1mg/mlのタンパク質をペリプラズム区画に産生する。別の実施形態では、少なくとも1つの株が、0.1〜10mg/mlのペリプラズムタンパク質、または少なくとも0.2ml、約0.3ml、約0.4ml、約0.5ml、約0.6ml、約0.7ml、約0.8ml、約0.9mlもしくは少なくとも約1.0mg/mlのペリプラズムタンパク質を細胞内で産生する。一実施形態では、アレイ内の少なくとも1つの株によって産生される、対象とするタンパク質またはポリペプチドの総タンパク質量が、少なくとも1.0mg/ml、少なくとも約2mg/ml、少なくとも約3mg/ml、約4mg/ml、約5mg/ml、約6mg/ml、約7mg/ml、約8mg/ml、約10mg/ml、約15mg/ml、約20mg/mlもしくは少なくとも約25mg/mlであるか、またはそれより多い。一部の実施形態では、産生されるペリプラズムタンパク質の量が、産生される対象とするタンパク質またはポリペプチドの少なくとも約5%、約10%、約15%、約20%、約25%、約30%、約40%、約50%、約60%、約70%、約80%、約90%、約95%、約96%、約97%、約98%もしくは約99%であるか、またはそれより多い。
【0085】
本発明のアレイの少なくとも1つの株は、対象とするタンパク質またはポリペプチドの収率の増大ももたらし得る。一実施形態では、少なくと1つの株が、対象とするタンパク質またはポリペプチドを、全細胞タンパク質(total cell protein)(tcp)の少なくとも約5%、少なくとも約10%、約15%、約20%、約25%、約30%、約40%、約45%、約50%、約55%、約60%、約65%、約70%もしくは約75%、またはそれより多い量で産生する。「パーセント全細胞タンパク質(percent total cell protein)」は、総計細胞タンパク質のパーセントとしての、宿主細胞内のタンパク質またはポリペプチドの量である。パーセント全細胞タンパク質を測定する方法は、当技術分野でよく知られている。
【0086】
特定の実施形態では、無機塩培地中で増殖(すなわち、約10℃、約15℃、約20℃、約25℃、約30℃、約35℃、約40℃、約45℃および約50℃を含めた、約4℃〜約55℃の温度範囲内で増殖)させた場合、アレイ内の少なくとも1つの宿主細胞集団が、少なくとも1%tcpの異種タンパク質産生レベルおよび少なくとも40mg/mlの細胞密度を有し得る。特に好ましい実施形態では、無機塩培地中で増殖(すなわち、包括的に約4℃〜約55℃の温度範囲内で増殖)させた場合、発現系が、少なくとも5%tcpのタンパク質またはポリペプチド発現レベルおよび少なくとも40g/Lの細胞密度を有するであろう。
【0087】
実際、ペリプラズムにターゲッティングされた異種タンパク質は、おそらく外側細胞膜への損傷、またはその流動性の増大から、しばしば、ブロス(broth)中に見出される(欧州特許第EP0 288 451号を参照)。この「受動的な」分泌の速度は、コリシン(Mikschら(1997)、Arch.Microbiol.167:143〜150)、増殖速度(Shokriら(2002)、App Miocrobiol Biotechnol 58:386〜392)、TolIII過剰発現(WanおよびBaneyx(1998)、Protein Expression Purif.14:13〜22)、バクテリオシン放出タンパク質(Hsiungら(1989)、Bio/Technology7:267〜71)、コリシンA溶解タンパク質(Lloubesら(1993)、Biochimie 75:451〜8)、周辺質(periplasmic)タンパク質を漏出する変異体(FurlongおよびSundstrom(1989)、Developments in Indus.Microbio.30:141〜8)、融合パートナー(JeongおよびLee(2002)、Appl.Environ.Microbio.68:4979〜4985)または浸透圧ショックによる回収(Taguchiら(1990)、Biochimica Biophysica Acta 1049:278〜85)を含めた、外側細胞膜を透過処理する様々な作用機序を用いることによって増大し得る。E.coliで適切にフォールディングされた活性タンパク質を産生するのに、周辺腔(periplasmic space)への改変タンパク質の輸送、およびそれに続くブロス中への局在化が使用されている(WanおよびBaneyx(1998)、Protein Expression Purif.14:13〜22;Simmonsら(2002)、J.Immun.Meth.263:133〜147;Lundellら(1990)、J.Indust.Microbio.5:215〜27)。
【0088】
この方法は、対象とするタンパク質またはポリペプチドをペリプラズムから、または細胞外の培地から精製するステップも含み得る。異種タンパク質またはポリペプチドがタグタンパク質に連結されるような方法で、それを発現させることができ、「タグ付き」のタンパク質を細胞または細胞外の培地から精製することができる。
【0089】
一部の実施形態では、対象とするタンパク質またはポリペプチドを、活性形態でアレイの少なくとも1つの株によって産生することもできる。「活性」という用語は、生物活性の存在を意味し、生物活性は、対応する天然タンパク質またはポリペプチドの生物活性に匹敵するか、それに実質的に相当する。タンパク質に関する文脈では、通常これは、標準的なパラメータを用いて、対応する天然タンパク質またはポリペプチドと比較した場合、ポリヌクレオチドまたはポリペプチドが、少なくとも約20%、約50%、好ましくは少なくとも約60〜80%、そして最も好ましくは少なくとも約90〜95%の活性を有する生物学的機能または作用を含むことを意味する。しかし、一部の実施形態では、天然タンパク質と比較して、改変または改善された活性を有するポリペプチド(例えば免疫応答性および基質の特異性などを改変または改善したもの)を産生することが望ましい可能性がある。改変または改善されたポリペプチドは、アレイ内の1つまたは複数の宿主細胞集団によって生成された特定の立体配座の結果として生じたものであり得る。
【0090】
タンパク質またはポリペプチドの活性の測定は、特定のタンパク質またはポリペプチドを標的とした、対応する標準的な生物学的比較アッセイを用いて行うことができ、それらのアッセイを用いて、生物活性を評価することができる。
【0091】
対象とする活性なタンパク質またはポリペプチドの回収も、本発明のアレイ内の1つまたは複数の他の株と比較して、最適な宿主株で改善されている可能性がある。活性タンパク質は、その配列が由来する天然タンパク質またはポリペプチドの活性の少なくとも約20%、少なくとも約30%、少なくとも約40%、約50%、約60%、少なくとも約70%、約80%、約90%、または少なくとも95約%の比活性を有し得る。さらに、基質特異性(kcat/Km)が天然タンパク質またはポリペプチドと実質的に同様であってもよい。通常、kcat/Kmは、少なくとも約30%、約40%、約50%、約60%、約70%、約80%、少なくとも約90%もしくは少なくとも約95%、またはそれより大きいであろう。タンパク質およびポリペプチドの活性および基質特異性(kcat/Km)の尺度をアッセイおよび定量化する方法は、当業者によく知られている。
【0092】
タンパク質活性の測定(Measurement of protein activity)
対象とする異種発現タンパク質またはポリペプチドの活性は、事前に確立されている天然タンパク質またはポリペプチドの標準的な活性と比較できる。代替として、対象のタンパク質またはポリペプチドの活性は、同時または実質的に同時に行われる、天然タンパク質またはポリペプチドを用いた比較アッセイで測定できる。例えば、発現された酵素と基質との間、発現されたホルモンとホルモン受容体との間、および発現された抗体と抗原との間の相互作用など、例えば、対象とするタンパク質またはポリペプチドと標的との間の検出可能な相互作用を測定するのに、in vitroアッセイを用いることができる。そのような検出には、熱量変化の測定、増殖変化、細胞死、細胞忌避(cell repelling)、放射能の変化、溶解性の変化、ゲル電気泳動および/またはゲル濾過法で測定した分子量の変化、リン酸化能、ELISAアッセイなどの抗体特異性アッセイなどが含まれ得る。加えて、in vivoアッセイには、天然タンパク質またはポリペプチドの生理作用と比較して、異種発現されたタンパク質またはポリペプチドの生理作用を検出するアッセイ、例えば、質量増大、電解質バランスの変化、血液凝固時間の変化、血餅融解の変化、および抗原性反応の誘導を検出するアッセイが含まれるが、これらに限定されない。通常、対象とするタンパク質またはポリペプチドの活性特性を、そのような活性がアッセイ可能である限り、天然タンパク質またはポリペプチドとの比較分析を可能にする任意のin vitroまたはin vivoアッセイを用いて決定できる。代替として、本発明のアレイ内の少なくとも1つの株で産生されたタンパク質またはポリペプチドを、そのタンパク質またはポリペプチドと、通常、そのタンパク質またはポリペプチドと相互作用する分子、例えば天然タンパク質が通常相互作用するシグナル経路の基質または構成要素との間の相互作用を刺激または抑制する能力についてアッセイできる。そのようなアッセイは、通常、タンパク質またはポリペプチドが標的分子と相互作用するのを可能にする条件下で、タンパク質を基質分子と混合し、上記タンパク質と標的分子との相互作用の生化学的帰結を検出するステップを含み得る。
【0093】
タンパク質またはポリペプチドの活性を測定するのに利用できるアッセイは、例えば、Ralph,P.J.ら(1984)、J.Immunol.132:1858もしくはSaikiら(1981)、J.Immunol.127:1044、Steward,W.E.II(1980)、「The Interferon Systems」、Springer−Verlag,ViennaおよびNew York、Broxmeyer,H.E.ら(1982)、Blood 60:595、「Molecular Cloning:A Laboratory Manua」、第2版、Cold Spring Harbor Laboratory Press、Sambrook,J.、E.F.FritschおよびT.Maniatis編集、1989、および「Methods in Enzymology:Guide to Molecular Cloning Techniques」、Academic Press、Berger,S.L.およびA.R.Kimmel編集、1987、A K Patraら、Protein Expr Purif,18(2):182〜92ページ(2000)、Kodamaら、J.Biochem.99:1465〜1472(1986)、Stewartら、Proc.Nat’l Acad.Sci.USA 90:5209〜5213(1993)、(Lombilloら、J.Cell Biol.128:107〜115(1995);(Valeら、Cell 42:39〜50(1985)に記載されている。アレイ内の1つまたは複数の他の宿主細胞集団に由来する、異種発現されたタンパク質の試料間で活性を比較することも、天然タンパク質の活性と比較することも、またはそれらを両方行うこともできる。活性測定は、単離されたタンパク質で行うこともでき、または宿主細胞内でin vitroで行うこともできる。
【0094】
別の実施形態では、例えば、従来の顕微鏡、ルミノメーターまたはプレートリーダーにおける蛍光測定または分光測定によって、タンパク質産生および/または活性を培養物中で直接モニターすることができる。対象とするタンパク質が、その基質が既知の酵素である場合、基質を培地に添加することができ、基質が酵素によって産物に変換された際に蛍光シグナルが放出される。一実施形態では、対象とする異種タンパク質またはポリペプチドをコードする発現コンストラクトが、レポータータンパク質をさらにコードする。「レポータータンパク質」は、細胞内または細胞表面にそれが存在することによって、または培地中に分泌された場合に、レポータータンパク質を含有しない細胞からその細胞を識別するのを可能にするタンパク質を意味する。対象とする異種タンパク質の産生は、宿主細胞集団の検出可能な変化をもたらす。レポーター分子は、ホタルルシフェラーゼおよびGFP、または他の任意の蛍光分子でもよく、β−ガラクトシダーゼ遺伝子(beta.gal)、クロラムフェニコールおよびアセチルトランスフェラーゼ遺伝子(CAT)でもよい。これらのレポーター遺伝子エレメントそれぞれと連動して産生される発現のアッセイは、当業者によく知られている。
【0095】
レポーター遺伝子は、検出可能なタンパク質もしくは間接的に検出可能なタンパク質をコードするものであり得、またはレポーター遺伝子が生存遺伝子であり得る。好ましい実施形態では、レポータータンパク質が検出可能なタンパク質である。(検出可能な遺伝子または検出遺伝子によってコードされている)「検出可能なタンパク質」または「検出タンパク質」は、直接標識として使用できるタンパク質であり、すなわち、このタンパク質は、さらなる操作なしに検出可能(好ましくは、上記検出可能なタンパク質を含む細胞が検出可能)である。したがって、この実施形態では、レポーター遺伝子のタンパク質産物それ自体が、検出可能な遺伝子を発現している細胞を識別するのに役立ち得る。この実施形態では、適した検出可能遺伝子に、自己蛍光タンパク質をコードするものが含まれる。
【0096】
当技術分野で知られている通り、様々な既知の自己蛍光タンパク質が存在しており、これらは通常、Aequoreaの緑色蛍光タンパク質(GFP)およびその変種をベースにしており、これらには、GFP(Chalfieら(1994)、Science 263(5148):802〜805)、高感度GFP(EGFP;Clontech−Genbank受託番号U55762)、青色蛍光タンパク質(BFP;Quantum Biotechnologies,Inc.、Montreal,Canada);Stauber(1998)、Biotechniques 24(3):462〜471;HeimおよびTsien(1996)、Curr.Biol.6:178〜182)、高感度黄色蛍光タンパク質(EYFP;Clontech Laboratories,Inc.、Palo Alto,CA)、および赤色蛍光タンパク質が含まれるが、これらに限定されない。加えて、RenillaおよびPtilosarcus種由来の自己蛍光タンパク質が最近報告されている。全てが参照により明示的に本明細書に組み込まれている、WO92/15673、WO95/07463、WO98/14605、WO98/26277、WO99/49019、米国特許第5,292,658号、米国特許第5,418,155号、米国特許第5,683,888号、米国特許第5,741,668号、米国特許第5,777,079号、米国特許第5,804,387号、米国特許第5,874,304号、米国特許第5,876,995号および米国特許第5,925,558を参照のこと。
【0097】
対象とするタンパク質またはポリペプチドの単離(Isolation of Protein or Polypeptide of Interest)
対象とするタンパク質の収率、溶解性、立体配座および/または活性を測定するために、アレイ内の1つまたは複数の株からタンパク質を単離することが望ましい可能性がある。適切な測定を行うのに用いたアッセイの必要条件に応じて、単離は、粗単離でも、準粗単離でも、純粋単離でもよい。タンパク質は、細胞質で産生されるものでも、ペリプラズムにターゲッティングされるものでも、または培養もしくは発酵培地中に分泌されるものでもよい。ペリプラズムにターゲッティングされたタンパク質を遊離させるには、クロロホルム(Amesら(1984)、J.Bacteriol.、160:1181〜1183)、グアニジン−HClおよびトリトンX−100(NaglakおよびWang(1990)、Enzyme Microb.Technol.、12:603〜611)などの化学物質を用いた処置が使用されている。しかし、これらの化学物質は不活性ではなく、多くの異種タンパク質産物またはその後の精製方法に対する有害効果を有し得る。E.coliのグリシン処理は、外膜の透過化を引き起こし、ペリプラズム内容物を放出させることも報告されている(Arigaら(1989)、J.Ferm.Bioeng.68:243〜246)。異種タンパク質をペリプラズムから放出させる最も広範に使用されている方法は、浸透圧ショック(NosalおよびHeppel(1966)、J.Biol.Chem.241:3055〜3062;NeuおよびHeppel(1965)、J.Biol.Chem.240:3685〜3692)、雌鳥卵白(HEW)−リゾチーム/エチレンジアミン四酢酸(EDTA)処理(NeuおよびHeppel(1964)、J.Biol.Chem、239:3893〜3900;Witholtら(1976)、Biochim.Biophys.Acta,443:534〜544;Pierceら(1995)、ICheme Research.Event,2:995〜997)、およびHEW−リゾチーム/浸透圧ショック併用処理(Frenchら(1996)、Enzyme and Microb.Tech.19:332〜338)である。French法は、分画緩衝液中の細胞の再懸濁、およびそれに続くペリプラズム画分の回収を行ものであり、その際、浸透圧ショックはリゾチーム処理のすぐ後に行う。
【0098】
通常、これらの手順は、浸透圧的に安定化されている培地中での初期破壊と、それに続く、非安定化培地中における選択的放出とを含む。これらの培地の組成(pH、保護剤)および使用される破壊方法(クロロホルム、HEW−リゾチーム、EDTA、超音波処理)は、報告されている特定な手順相互で異なっている。EDTAの代わりに双極性イオン界面活性剤を用いるHEW−リゾチーム/EDTA処理の変法が、Stabelら(1994)、Veterinary Microbiol.38:307〜314に論じられている。E.coliを破壊するための細胞内溶菌酵素系の使用に関する一般的概説には、DaboraおよびCooney(1990)、Advances in Biochemical Engineering/Biotechnology、第43巻、A.Fiechter編集(Springer−Verlag:Berlin)、11〜30ページを参照のこと。
【0099】
対象とするタンパク質またはポリペプチドを、可溶性タンパク質または屈折性粒子として細胞質から回収する従来の方法は、機械的な切断による細菌細胞の崩壊を用いる。機械的破壊には、通常、液体懸濁液中に局所的なキャビテーションの産生、強固なビーズを用いた高速撹拌、超音波処理または細胞懸濁液のグラインディングを用いる(「Bacterial Cell Surface Techniques,Hancock and Poxton」(John Wiley&Sons Ltd,1988)、第3章、55ページ)。
【0100】
HEW−リゾチームは、細胞壁のペプチドグリカンバックボーンを加水分解するように生化学的に作用する。この方法は、最初、ZinderおよびArndt(1956)、Proc.Natl.Acad.Sci.USA、42:586〜590によって開発されたが、ZinderおよびArndtは、卵アルブミン(HEW−リゾチームを含有する)でE.coliを処理して、後にスフェロプラストとして知られる丸い細胞球を産生させた。これらの構造は、一部の細胞壁成分を保持していたが、細胞質膜が露出した大きな表面積を有していた。米国特許第5,169,772号は、浸透圧的に安定化された培地中、例えば20%ショ糖液中で、例えば、EDTA、リゾチーム、または有機化合物を用いて細菌のエンベロープを破壊するステップと、細菌を低イオン強度緩衝液に暴露することによって、破壊した細菌の周辺腔から非ヘパリナーゼ様タンパク質を遊離させるステップと、低イオン強度で洗浄された細菌を緩衝化塩溶液に暴露することによって、ヘパリナーゼ様タンパク質を遊離させるステップとを含む、細菌からヘパリナーゼを精製する方法を開示する。
【0101】
これらの方法の多くの異なった変法が、広範な発現系で使用されており、様々な程度に成功している(Joseph−Liazunら(1990)、Gene、86:291〜295;Carterら(1992)、Bio/Technology、10:163〜167)。組換え細胞培養物を誘導して、リゾチームを産生させる試みが報告されている。EP0 155 189は、組換え細胞培養物を誘導して、リゾチームを産生させる手段を開示し、この手段は、通常、細胞壁構造を破壊または溶解することによって、そのような宿主細胞を死滅させると予測される。
【0102】
米国特許第4,595,658号は、細菌の周辺腔に輸送されたタンパク質の外在化を促進する方法を開示する。この方法は、細胞のリゾチーム処理、機械的粉砕または浸透圧ショック処理を必要とせずに、ペリプラズム中に局在するタンパク質の選択的単離を可能にする。米国特許第4,637,980号は、産物を直接的または間接的にコードするDNA分子で温度感受性溶原菌を形質転換させ、形質転換体を許容条件下で培養して、細胞内で遺伝子産物を発現させ、温度を上げることによって産物を外在化させ、ファージにコードされている機能を誘導することによって、細菌産物を産生させる方法を開示する。Asamiら(1997)、J.Ferment and Bioeng、83:511〜516は、T4ファージ感染によるE.coli細胞の同調破壊を開示し、Tanjiら(1998)、J.Ferment.and Bioeng,85:74〜78は、E.coli細胞を穏やかに破壊するための、T4ファージコードされている溶菌遺伝子の調節発現を開示する。
【0103】
細胞溶解の際には、ゲノムDNAが細胞質から培地中に漏出し、その結果、流体粘度の著しい増大がもたらされる。これは、遠心力場内における固形物の沈降を妨害し得る。DNA重合体を破壊するための機械的な破壊の間に作用するものなどのせん断力の不在下では、粘性流体を通る固形物の沈降速度が遅延する結果、遠心処理中の固形物と液体との分離が悪くなる。機械的せん断力の他に、DNA重合体を分解する核酸分解酵素が存在する。E.coliでは、内因性遺伝子であるendAが、エンドヌクレアーゼ(成熟タンパク質の分子量は約24.5kD)をコードしている。エンドヌクレアーゼは、通常、ペリプラズムに分泌され、ヌクレオチド鎖切断の様式でDNAをオリゴデオキシリボヌクレオチドへと切断する。E.coliによって、endAが比較的弱く発現されると示唆されている(Wackemagelら(1995)、Gene 154:55〜59)。
【0104】
望ましい場合には、本発明のアレイ内の1つまたは複数の株を用いて産生されたタンパク質を、当技術分野でよく知られている標準的な技法によって、実質的な純度にまで単離および精製することができ、これらの技法には、硫酸アンモニウムもしくはエタノール沈殿、酸抽出、陰イオンもしくは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ニッケルクロマトグラフィー、ヒドロキシアパタイトクロマトグラフィー、逆相クロマトグラフィー、レクチンクロマトグラフィー、分取電気泳動、界面活性剤可溶化、カラムクロマトグラフィーのような物質を用いた選択的沈殿、および免疫精製法などが含まれるが、これらに限定されない。例えば、確立された分子付着特性を有するタンパク質をリガンドに可逆的に融合させることができる。適切なリガンドを用いて、タンパク質を精製カラムに選択的に吸着させ、その後、比較的純粋な形態でカラムから遊離させることができる。融合タンパク質は、その後、酵素活性によって除去する。加えて、イムノアフィニティーカラムまたはNi−NTAカラムを用いて、タンパク質を精製することができる。一般技法については、例えば、R.Scopes、「Protein Purification:Principles and Practice」、Springer−Verlag:N.Y.(1982);Deutscher、「Guide to Protein Purification」Academic Press(1990);米国特許第4,511,503号;S.Roe、「Protein Purification Techniques:A Practical Approach(Practical Approach Series)」、Oxford Press(2001);D.Bollagら、「Protein Methods」、Wiley−Lisa,Inc.(1996);AK Patraら、Protein Expr Purif;18(2)、182〜92ページ(2000);およびR.Mukhijaら、Gene 165(2):303〜6ページ(1995)にさらに記載されている。例えば、Ausubelら(1987年および定期増補);Deutscher(1990)、「Guide to Protein Purification」、Methods in Enzymology、第182巻およびこのシリーズの他の巻;Coliganら(1996年および定期増補)、「Current Protocols in Protein Science」、Wiley/Greene,NY;ならびにタンパク質精製製品の使用に関する製造会社、例えば、Pharmacia、Piscataway,N.J.またはBio−Rad、Richmond,Calif.の文献も参照のこと。組換え技法との併用によって、プロテアーゼで除去可能な配列を介して融合させることのできる、例えばFLAG配列またはその等価物に、適したセグメントを融合させることが可能となる。例えば、Hochuli(1989)、Chemische Industrie 12:69〜70;Hochuli(1990)、「Purification of Recombinant Proteins with Metal Chelate Absorbent」、Setlow(編集)、Genetic Engineering,Principle and Methods 12:87−98、Plenum Press,NY;およびCroweら(1992)、QIAexpress:The High Level Expression&Protein Purification System、QUIAGEN,Inc、Chatsworth,Calif.も参照のこと。
【0105】
発現されたタンパク質の検出は、当技術分野で知られている方法で行われ、それには、例えば、ラジオイムノアッセイ、ウエスタンブロット技法または免疫沈降反応が含まれる。
【0106】
本発明のアレイ内の株によって発現される特定のタンパク質は、不溶性の凝集体(「封入体」)を形成し得る。いくつかのプロトコールが、封入体からタンパク質を精製するのに適している。例えば、封入体の精製は通常、例えば、50mM TRIS/HCL、pH7.5、50mM NaCl、5mM MgCl2、1mM DTT、0.1mM ATPおよび1mM PMSFという緩衝液中でのインキュベーションによって、宿主細胞を破壊することによる、封入体の抽出、分離および/または精製を要する。細胞懸濁液は、通常、フレンチプレスに通した2〜3回の通過を用いて溶解する。細胞懸濁液は、Polytron(Brinkman Instruments)を用いてホモジナイズすることも、氷上で超音波処理することもできる。細菌を溶解する代替法は、当業者には明らかである(例えば、Sambrookら、同上;Ausubelら、同上を参照)。
【0107】
必要な場合には、封入体は可溶化させることができ、通常は、好ましくない不溶物を除去するために、溶解細胞懸濁液を遠心処理することができる。封入体を形成したタンパク質は、適合した緩衝液で希釈または透析することによって再生させることができる。適した溶剤には、尿素(約4M〜約8M)、ホルムアミド(少なくとも約80%、容積/容積ベース)および塩酸グアニジン(約4M〜約8M)が含まれるが、これらに限定されない。塩酸グアニジンおよび類似の薬剤は、変性剤であるが、この変性は不可逆的でなく、変成剤を除去(例えば透析によって)または希釈した際に、再生(renaturation)が起こり得る。これにより、免疫学的および/または生物学的に活性なタンパク質の再形成(re-formation)が可能となる。他の適した緩衝液も、当業者に知られている。
【0108】
当業者によく知られている標準的な分離技法によって、上清中に存在する異種発現タンパク質を宿主タンパク質から分離することができる。例えば、最初の塩分画によって、多くの好ましくない宿主細胞タンパク質(細胞培地由来のタンパク質)を、対象とするタンパク質またはポリペプチドから分離し得る。その一例が硫酸アンモニウムであり得る。硫酸アンモニウムは、タンパク質混合物中の水の量を効果的に低下させることによって、タンパク質を沈殿させる。その際、タンパク質は、それらの溶解性に基づいて沈殿する。タンパク質が疎水性であるほど、より低い濃度の硫酸アンモニウムで沈殿する可能性が大きい。典型的なプロトコールは、その結果の硫酸アンモニウム濃度が20〜30%となるように、飽和硫酸アンモニウムをタンパク質溶液に添加するステップを含む。この濃度は、ほとんどの疎水性タンパク質を沈殿させるであろう。その後、(対象とするタンパク質が疎水性でない限り)沈殿を廃棄し、対象とするタンパク質を沈殿させることが既知の濃度まで上清に硫酸アンモニウムを添加する。その後、緩衝液中で沈殿を可溶化し、必要に応じて、透析(dialysis)またはダイアフィルトレーション(diafiltration)を介して、過剰な塩を除去する。冷エタノール沈殿など、タンパク質の溶解性に依存した他の方法も、当業者にはよく知られており、複雑なタンパク質混合物を分画するのに使用できる。
【0109】
対象とするタンパク質またはポリペプチドの分子量を用いて、それを、より大きなサイズのタンパク質およびより小さなサイズのタンパク質から、異なった孔径の膜(例えばAmiconまたはMillipore膜)を通す限外濾過を用いて単離することができる。最初のステップとして、対象とするタンパク質の分子量より小さい分子量のカットオフを有する孔径を備えた膜を通して、タンパク質混合物を限外濾過にかけることができる。その後、対象とするタンパク質の分子量より大きな分子カットオフを有する膜に対して、保持液の限外濾過を行うことができる。対象とするタンパク質またはポリペプチドは、膜を通り抜けて、濾液中に入る。その後、以下に記載の通り、濾液をクロマトグラフィーにかけることができる。
【0110】
発現された、対象とするタンパク質またはポリペプチドを、そのサイズ、正味の表面電荷、疎水性度、およびリガンドへのアフィニティーに基づいて、他のタンパク質から分離することができる。加えて、タンパク質に対して産生された抗体をカラムマトリクスに結合させ、そのタンパク質を免疫精製することができる。これらの方法は全て、当技術分野でよく知られている。任意のスケールで、かつ多くの異なった製造会社(例えばPharmacia Biotech)の機器を用いて、クロマトグラフィー技法を実施できることは、当業者には明らかであろう。
【0111】
再生およびリフォールディング(Renaturation and Refolding)
異種発現されたタンパク質が変性形態で産生される場合、二次および三次タンパク質構造を生成するように、不溶性タンパク質を再生またはリフォールディングさせることができる。異種産物の立体配座を完成させるのに、タンパク質リフォールディングステップを、必要に応じて用いることができる。当技術分野で知られている、タンパク質の解離/会合を促進する薬剤を用いて、リフォールディングおよび再生を実現できる。例えば、タンパク質をジチオスレイトールと共にインキュベートし、それに続いて、酸化グルタチオン二ナトリウム塩と共にインキュベートし、それに続いて、尿素などのリフォールディング剤を含有する緩衝液と共にインキュベートすることができる。
【0112】
例えば、リン酸緩衝食塩水(PBS)または50mM Na−酢酸、pH6緩衝液に200mM NaClを加えたものに対して、対象とするタンパク質またはポリペプチドを透析することによって、それを再生させることもできる。代替として、プロテアーゼ阻害物質を含有する、500mM NaCl、20%グリセロール、20mM Tris/HCl pH7.4中における直線的な6Mから1Mの尿素勾配を用いることによって、Ni NTAカラムなどのカラムに固定化しながら、タンパク質をリフォールディングさせることができる。1.5時間を超える時間にわたって、再生を行うことができる。再生の後、250mMイミダゾールを添加することによって、タンパク質を溶出できる。イミダゾールは、PBSまたは50mM酢酸ナトリウムpH6緩衝液に200mM NaClを加えたものに対する最終の透析ステップによって除去できる。精製されたタンパク質は、4℃で保存するか、または−80℃で冷凍できる。
【0113】
他の方法には、例えば、M H Leeら、Protein Expr.Purif、25(1):166〜73ページ(2002)、W.K.Choら、J.Biotechnology,77(2〜3):169〜78ページ(2000)、Ausubelら(1987年および定期的な増補)、Deutscher(1990)「Guide to Protein Purification」、Methods in Enzymology、第182巻、およびこのシリーズの他の巻、Coliganら(1996年および定期的な増補)、「Current Protocols in Protein Science」、Wiley/Greene,NY、S.Roe、「Protein Purification Techniques:A Practical Approach(Practical Approach Series)」、Oxford Press(2001年);D.Bollagら、「Protein Methods」、Wiley−Lisa,Inc.(1996)に記載されている可能性のあるものが含まれる。
【0114】
発現ベクター(Expression vectors)
対象とする異種タンパク質は、それぞれの菌株に対象とする異種タンパク質をコードする発現ベクターを導入することによって、本明細書に開示する1つまたは複数の宿主細胞において産生することができる。一実施形態では、ベクターは、選択した宿主細胞において働くことができるプロモーター、ならびに全ての他の必要とされる転写および翻訳調節エレメントと作用可能に連結した対象とするタンパク質をコードするポリヌクレオチド配列を含む。
【0115】
用語「作用可能に連結した(Operably linked)」は、転写および任意の翻訳調節エレメントが、コード配列に対して、宿主細胞中またはその作用によって、調節エレメントがコード配列の発現を誘導することができるような配置で、コード配列と共有結合した任意の形態を指す。
【0116】
対象とする異種タンパク質は、異種ポリペプチドコード配列が転写および翻訳調節エレメントと作用可能に連結して、宿主細胞がそこからタンパク質またはポリペプチドを発現することができる機能遺伝子を形成する、ポリヌクレオチドから発現させることが可能である。コード配列は異種ポリペプチドの天然コード配列であってよく、あるいは、例えば、宿主種のコドン使用の偏向を反映するために遺伝子を合成することによって、選択した発現宿主細胞中で使用するために選択、改良、または最適化されたコード配列であってよい。本発明の一実施形態では、宿主種はP.fluorescensであり、ポリペプチドコード配列の設計時には、P.fluorescensのコドンの偏向を考慮に入れる。(1つまたは複数の)遺伝子を1つまたは複数のベクター内に構築し、または1つまたは複数のベクターに挿入し、次いで発現宿主細胞に形質転換することができる。
【0117】
他の調節エレメントを、(「発現コンストラクト(expression construct)」とも呼ばれる)ベクター中に含めることができる。ベクターは典型的には、1つまたは複数の表現型選択可能マーカーおよび1つの複製起点を含んで、ベクターの維持を確実にし、および望ましい場合、宿主内の増幅をもたらす。追加のエレメントには、発現したポリペプチドの同定、分離、精製、および/または単離を容易にする、例えば転写エンハンサー配列、翻訳エンハンサー配列、他のプロモーター、アクチベーター、翻訳開始および停止シグナル、転写ターミネーター、シストロン性レギュレーター、ポリシストロン性レギュレーター、またはヌクレオチド配列「タグ」および「タグ」ポリペプチドコード配列などのタグ配列があるが、これらだけには限られない。
【0118】
別の実施形態では、発現ベクターは、対象とするタンパク質またはポリペプチドのコード配列と隣接したタグ配列をさらに含む。一実施形態では、このタグ配列はタンパク質の精製を可能にする。タグ配列は、ヘキサ−ヒスチジン親和性タグなどの親和性タグであってよい。別の実施形態では、親和性タグはグルタチオン−S−トランスフェラーゼ分子であってよい。タグは、YFPまたはGFPなどの蛍光分子、またはこのような蛍光タンパク質の類似体であってもよい。タグは、抗体分子、または既知の抗原もしくは精製に有用な既知の結合パートナーのリガンドの一部分であってもよい。
【0119】
本発明によるタンパク質コード遺伝子は、タンパク質コード配列に加えて、そこに作用可能に連結した以下の調節エレメント:プロモーター、リボソーム結合部位(ribosome binding site)(RBS)、転写ターミネーター、翻訳開始および停止シグナルを含むことができる。有用なRBSは、本発明による発現系中で宿主細胞として有用な任意の種から、好ましくは選択した宿主細胞から得ることができる。多くの特異的および様々なコンセンサスRBS、例えばD.Frishmanら、Gene234(2):257〜65ページ(1999年7月8日);およびB.E.Suzekら、Bioinformatics17(12):1123〜30(2001年12月)中に記載されそれらによって言及されたRBSが知られている。さらに、天然または合成RBSのいずれか、例えばEP0207459(合成RBS);O.Ikehataら、Eur.J.Biochem.181(3):563〜70ページ(1989)(AAGGAAGの天然RBS配列)中に記載されたRBSを使用することができる。本発明中で有用な方法、ベクター、および翻訳および転写エレメント、および他のエレメントのさらなる例は、例えばGilroyへの米国特許第5,055,294号、およびGilroyらへの米国特許第5,128,130号、Rammlerらへの米国特許第5,281,532号、Barnesらへの米国特許第4,695,455号および米国特許第4,861,595号、Grayらへの米国特許第4,755,465号、およびWilcoxへの米国特許第5,169,760号中に記載されている。
【0120】
対象とする異種タンパク質をコードするDNAの転写は、エンハンサー配列をベクターまたはプラスミドに挿入することによって増大する。典型的なエンハンサーは、プロモーターに作用してその転写を増大させる、通常約10〜300bpの大きさのDNAのシス作用性エレメントである。例には様々なシュードモナスのエンハンサーがある。
【0121】
一般に、異種発現ベクターは、宿主細胞の形質転換を可能にする複製起点および選択可能マーカー、および下流構造配列の転写を誘導するための高発現遺伝子由来のプロモーターを含む。このようなプロモーターは、特に3−ホスホグリセレートキナーゼ(PGK)、酸性ホスファターゼ、または熱ショックタンパク質などの酵素をコードするオペロンから誘導することができる。シグナル配列を使用する場合、異種コード配列は翻訳開始および停止配列と適切に一致して構築され、かつシグナル配列は翻訳されるタンパク質の部分的蓄積または分泌を誘導することができる。場合によっては異種配列は、望ましい性質、例えば発現される異種産物の安定性または精製の単純化をもたらすN末端同定ポリペプチドを含む融合酵素をコードすることができる。前に論じたように、融合ポリペプチドは、1つまたは複数の標的タンパク質またはその阻害剤もしくはエンハンサーも含むことができる。
【0122】
宿主細胞中で異種タンパク質を発現させるためのベクターが当技術分野で知られており、任意のこれらのベクターは、本発明による遺伝子を発現させるために使用することができる。このようなベクターには、例えばプラスミド、コスミド、およびファージ発現ベクターがある。有用なプラスミドベクターの例には、発現プラスミドpBBR1MCS、pDSK519、pKT240、pML122、pPS10、RK2、RK6、pRO1600、およびRSF1010があるが、これらだけには限られない。このような有用なベクターの他の例には、例えば:N.Hayase、Appl.Envir.Microbiol.60(9):3336〜42ページ(1994年9月)中、A.A.Lushnikovら、Basic Life Sci.30:657〜62ページ(1985)中、S.Graupner & W.Wackemagel、Biomolec.Eng.17(1):11〜16ページ(2000年10月)中、H.P.Schweizer、Curr.Opin.Biotech.12(5):439〜45ページ(2001年10月)中、M.Bagdasarian & K.N.Timmis、Curr.Topics Microbiol.Immunol.96:47〜67ページ(1982)中、T.Ishiiら、FEMS Microbiol.Lett.116(3):307〜13ページ(1994年3月1日)中、I.N.Olekhnovich & Y.K.Fomichev、Gene140(1):63〜65ページ(1994年3月11日)中、M.Tsuda & T.Nakazawa、Gene136(1〜2):257〜62ページ(1993年12月22日)中、C.Nietoら、Gene87(1):145〜49ページ(1990年3月1日)中、J.D.Jones & N.Gutterson、Gene61(3):299〜306ページ(1987)中、M.Bagdasarianら、Gene16(1〜3):237〜47ページ(1981年12月)中、H.P.Schweizerら、Genet.Eng.(NY)23:69〜81ページ(2001年)中、P.Mukhopadhyayら、J.Bact.172(1):477〜80ページ(1990年1月)中、D.O.Woodら、J.Bact.145(3):1448〜51ページ(1981年3月)中、およびR.Holtwickら、Microbiology147(Pt2):337〜44ページ(2001年2月)中によって記載されたベクターがある。
【0123】
本発明の宿主細胞において有用であり得る発現ベクターのさらなる例には、示したレプリコン由来の表5中に挙げたベクターがある。
【0124】
【表5】
【0125】
発現プラスミド、RSF1010は、例えばF.Heffronらによって、Proc.Nat’l Acad.Sci.USA72(9):3623〜27ページ(1975年9月)中、およびK.Nagahari & K.Sakaguchiによって、J.Bact.133(3):1527〜29ページ(1978年3月)中に記載されている。プラスミドRSF1010およびその誘導体は、本発明中で特に有用なベクターである。当技術分野で知られているRSF1010の例示的で有用な誘導体には、例えばpKT212、pKT214、pKT231および関連プラスミド、およびpMYC1050および関連プラスミド(例えば、Thompsonらへの米国特許第5,527,883号および米国特許第5,840,554号を参照)、例えばpMYC1803などがある。プラスミドpMYC1803は、調節型テトラサイクリン耐性マーカーならびにRSF1010プラスミド由来の複製および可動性遺伝子座を有する、RSF1010系プラスミドpTJS260に由来する(Wilcoxへの米国特許第5,169,760号を参照)。他の例示的で有用なベクターには、Puhlerらへ米国特許第4,680,264号の中に記載されたベクターがある。
【0126】
一実施形態では、発現プラスミドは発現ベクターとして使用する。別の実施形態では、RSF1010またはその誘導体を発現ベクターとして使用する。さらに別の実施形態では、pMYC1050またはその誘導体、またはpMYC4803またはその誘導体を発現ベクターとして使用する。
【0127】
プラスミド中に選択マーカー遺伝子を封入することによって、プラスミドを宿主細胞中に維持することができる。これは(1つまたは複数の)抗生物質耐性遺伝子であってよく、この場合対応する(1つまたは複数の)抗生物質、または当技術分野で知られている任意の他型の選択マーカー遺伝子、例えば原栄養体復元遺伝子を発酵培地に加え、この場合プラスミドは、対応する形質、例えばアミノ酸生合成またはヌクレオチド生合成形質、または炭素源利用形質などの生体触媒形質の栄養要求性である宿主細胞において使用する。
【0128】
本発明に従って使用するプロモーターは、構成的プロモーター(constitutive promoters)または調節型プロモーター(regulated promoters)であってよい。有用な調節型プロモーターの一般例には、lacプロモーター由来のファミリーのプロモーター(すなわち、lacZプロモーター)、特にDeBoerへの米国特許第4,551,433号中に記載されたtacおよびtrcプロモーター、ならびにPtac16、Ptac17、PtacII、PlacUV5、およびT71acプロモーターがある。一実施形態では、プロモーターは宿主細胞生物に由来しない。特定の実施形態では、プロモーターはE.coli生物に由来する。
【0129】
本発明による発現系中で有用な非lac型プロモーターの一般例には、例えば表6中に挙げたプロモーターがある。
【0130】
【表6】
【0131】
例えば、J.Sanchez−Romero & V.De Lorenzo(1999)Manual of Industrial Microbiology and Biotechnology(A.Demain & J.Davies、eds.)460〜74ページ(ASM Press、Washington、D.C.);H.Schweizer(2001)Current Opinion in Biotechnology、12:439〜445ページ;およびR.Slater & R.Williams(2000 Molecular Biology and Biotechnology(J.Walker & R.Rapley、eds.)125〜54ページ(The Royal Society of Chemistry、Cambridge、UK))を参照。選択した細菌宿主細胞に由来するプロモーターのヌクレオチド配列を有するプロモーター、例えばPseudomonasアントラニル酸塩または安息香酸オペロンプロモーター(Pant、Pben)を使用して、標的ポリペプチドをコードするトランス遺伝子の発現を制御することもできる。配列が同じであろうと異なろうと複数のプロモーターが互いに共有結合しているタンデムプロモーター、または同じ生物または異なる生物のいずれに由来しようと、例えばPant−Pbenタンデムプロモーター(プロモーター間ハイブリッド)またはPlac−Placタンデムプロモーターを使用することもできる。
【0132】
調節型プロモーターはプロモーター調節タンパク質を利用して、プロモーターがその一部分である遺伝子の転写を制御する。調節型プロモーターを本明細書において使用する場合、対応するプロモーター調節タンパク質も本発明による発現系の一部分であり得る。プロモーター調節タンパク質の例には、活性化タンパク質、例えばE.coliカタボライト活性化タンパク質、MalTタンパク質、AraCファミリー転写活性化因子、リプレッサータンパク質、例えば、E.coliLacIタンパク質、および二重機能調節タンパク質、例えばE.coliNagCタンパク質がある。多くの調節型プロモーター/プロモーター調節タンパク質のペアが当技術分野で知られている。一実施形態では、(1つまたは複数の)標的タンパク質の発現コンストラクトと対象とする異種タンパク質は、同じ調節エレメントの制御下にある。
【0133】
プロモーター調節タンパク質は、エフェクター化合物、すなわち可逆的または不可逆的に調節タンパク質と会合して、タンパク質の放出、またはプロモーターの制御下にある遺伝子の少なくとも1つのDNA転写調節領域との結合のいずれかを可能にし、それによって遺伝子の転写を開始する際のトランスクリプターゼ酵素の作用を許容または阻害する化合物と相互作用する。エフェクター化合物はインデューサーまたはコリプレッサーのいずれかとして分類され、かつこれらの化合物は、天然エフェクター化合物および余分なインデューサー化合物を含む。多くの調節型プロモーター/プロモーター調節タンパク質/エフェクター化合物のトリオが当技術分野で知られている。エフェクター化合物は細胞培養または発酵中を通して使用することができるが、調節型プロモーターを使用する好ましい実施形態では、望ましい量または密度の宿主細胞バイオマスの増殖後、適切なエフェクター化合物を培養物に直接的または間接的に加え、対象とするタンパク質またはポリペプチドをコードする(1つまたは複数の)望ましい遺伝子の発現をもたらす。
【0134】
例えば、lacファミリーのプロモーターを利用する場合、lacI遺伝子が系中に存在してもよい。(通常)構成的に発現される遺伝子であるlacI遺伝子は、これらのプロモーターのlacオペレーターと結合するLacリプレッサータンパク質(LacDタンパク質)をコードする。したがって、lacファミリーのプロモーターを利用する場合、lacI遺伝子を発現系中に含め発現させることが可能である。lacプロモーターファミリーのメンバー、例えばtacプロモーターの場合、エフェクター化合物はインデューサー、好ましくはIPTG(「イソプロピルチオガラクトシド」とも呼ばれるイソプロピル−D−1−チオガラクトピラノシドなどの余分なインデューサーである。
【0135】
対象とするタンパク質またはポリペプチドの発現用に、任意の植物プロモーターを使用することもできる。プロモーターは植物RNAポリメラーゼIIプロモーターであってよい。植物プロモーター中に含まれるエレメントは、転写開始部位の約25〜35塩基対上流(5’)に典型的に位置するTATAボックスまたはGoldberg−Hognessボックス、および70と100塩基対上流の間に位置するCCAATボックスであってよい。植物において、CCAATボックスは、哺乳動物プロモーターの機能的に類似した配列と異なるコンセンサス配列を有し得る(Messingら、(1983)In:Genetic Engineering of Plants、Kosugeら、eds.、211〜227ページ)。さらに、ほぼ全てのプロモーターは、転写開始部位の約−100bp〜−1,000bp以上上流に広がる追加の上流活性化配列またはエンハンサーを含む(Benoist and Chambon(1981)Nature290:304〜310ページ;Grussら(1981)Proc.Nat.Acad.Sci.78:943〜947ページ;およびKhoury and Gruss(1983)Cell27:313〜314ページ)。
【0136】
発現系(Expression Systems)
対象とするタンパク質またはポリペプチドを、アレイ内の1つまたは複数の宿主細胞集団のペリプラズム、または細胞外空間に標的化することは望ましい可能性がある。一実施形態では、発現ベクターは、対象とするタンパク質またはポリペプチドをコードするヌクレオチド配列と作用可能に連結した、分泌シグナル配列ポリペプチドをコードするヌクレオチド配列をさらに含む。いくつかの実施形態では、シグナル配列と対象とするタンパク質またはポリペプチドの間に修飾を施さない。しかしながら、特定の実施形態では、追加の切断シグナルを取り込んでポリペプチドのアミノ末端の正確なプロセシングを促進する。
【0137】
ベクターは、前に記載した特性のいずれかを有し得る。一実施形態では、対象とするタンパク質またはポリペプチドのコード配列を含むベクターは、シグナル配列、例えば分泌シグナル配列をさらに含む。
【0138】
したがって一実施形態では、この単離したポリペプチドは、分泌シグナルと対象とするタンパク質またはポリペプチドの融合タンパク質である。しかしながら、タンパク質がペリプラズムを標的化するとき、分泌シグナルをタンパク質から切断することもできる。一実施形態では、Sec系分泌シグナルとタンパク質またはポリペプチドの間の結合を修飾して分泌シグナルの切断を増大する。
【0139】
CHAMPION(商標)pET発現系は高レベルのタンパク質産生をもたらす。発現は強力なT7lacプロモーターから誘導される。この系は、対象とする遺伝子の高レベルの転写に関するバクテリオファージT7RNAポリメラーゼの高い活性および特異性を利用する。プロモーター領域に位置するlacオペレーターは伝統的なT7系ベクターより強い調節をもたらし、プラスミドの安定性および細胞の生存を改善する(Studier and Moffatt(1986)J Molecular Biology189(1):113〜30ページ;Rosenbergら(1987)Gene56(1):125〜35ページ)。T7発現系は、対象とする遺伝子の高レベルの転写にT7プロモーターおよびT7RNAポリメラーゼ(T7RNAP)を使用する。T7RNAPは天然E.coliRNAPよりプロセシングされやすく、対象とする遺伝子の転写に限られるので、高レベルの発現がT7発現系において得られる。同定した遺伝子の発現は、宿主細胞中にT7RNAPの供給源を提供することによって誘導する。これはT7RNAP遺伝子の染色体コピーを含有するBL21E.coli宿主を使用することによって実施する。T7RNAP遺伝子は、IPTGによって誘導することができるlacUV5プロモーターの制御下にある。T7RNAPは誘導によって発現され、対象とする遺伝子を転写する。
【0140】
pBAD発現系は、グルコース、グリセロールおよびアラビノースなどの特異的炭素源の存在によって、対象とするタンパク質またはポリペプチドの厳重に制御された、滴定可能な発現を可能にする(Guzmanら、(1995)J Bacteriology 177(14):4121〜30ページ)。pBADベクターは独自に設計して発現レベルに対する正確な制御をもたらす。pBADベクターからの異種遺伝子の発現はaraBADプロモーターで始まる。このプロモーターは、araC遺伝子の産物によって正と負の両方に制御される。AraCは、L−アラビノースと複合体を形成する転写レギュレーターである。L−アラビノースの不在下では、AraC二量体は転写を阻害する。最大の転写活性化のためには、2つの事象、(i)L−アラビノースがAraCと結合して転写の開始を可能にすること、および(ii)cAMP活性化タンパク質(CAP)−cAMP複合体がDNAと結合してプロモーター領域の正確な位置へのAraCの結合を刺激することが必要とされる。
【0141】
trc発現系は、E.coli中でのtrcプロモーターからの、高レベルの制御された発現を可能にする。trc発現ベクターは、E.coli中での真核生物遺伝子の発現用に最適化されている。trcプロモーターは、トリプトファン(trp)およびラクトース(lac)プロモーター由来の強力なハイブリッドプロモーターである。それはlacOオペレーターおよびlacIQ遺伝子の産物によって調節される(Brosius、J.(1984)Gene27(2):161〜72ページ)。
【0142】
本明細書に開示する(1つまたは複数の)ベクターを用いた宿主細胞の形質転換は、当技術分野で知られている任意の形質転換法を使用して実施することができ、かつ細菌宿主細胞は無傷細胞(intact cells)として、またはプロトプラスト(すなわち、サイトプラスト含む)として形質転換することができる。例示的な形質転換法にはポレーション法、例えばエレクトロポレーション、プロトプラスト融合、細菌接合、および二価カチオン処理、例えば塩化カルシウム処理またはCaCl/Mg2+処理、または当技術分野でよく知られている他の方法がある。例えば、Morrison、J Bact.、132:349〜351ページ(1977年);Clark−Curtiss & Curtiss、Methods in Enzymology、101:347〜362ページ(Wuら、eds、1983年)、Sambrookら、Molecular Cloning、A Laboratory Manual(1989年第2版);Kriegler、Gene Transfer and Expression:A Laboratory Manual(1990年);およびCurrent Protocols in Molecular Biology(Ausubelら、eds.、1994年))を参照。
【0143】
対象とするタンパク質(Proteins of interst)
本発明の方法および組成物は、高レベルの適切にプロセシングされた対象とするタンパク質またはポリペプチドを産生するのに最適であるP.fluorescens菌株を同定するのに有用である。任意の種および任意の大きさの対象とするタンパク質またはポリペプチドの産生をスクリーニングするのに、アレイは有用である。しかしながら、特定の実施形態では、対象とするタンパク質またはポリペプチドは、治療上有用なタンパク質またはポリペプチドである。いくつかの実施形態では、タンパク質は哺乳動物タンパク質、例えばヒトタンパク質であってよく、かつ例えば成長因子、サイトイン、ケモカインまたは血中タンパク質であってよい。対象とするタンパク質またはポリペプチドは、天然タンパク質またはポリペプチドと同様の方法でプロセシングすることができる。特定の実施形態では、対象とするタンパク質またはポリペプチドは、100kD未満、50kD未満、または30kD未満の大きさである。特定の実施形態では、対象とするタンパク質またはポリペプチドは、少なくとも約5、10、15、20、30、40、50または100またはそれより多くのアミノ酸のポリペプチドである。
【0144】
対象とするタンパク質またはポリペプチドのコード配列は、ポリペプチドの天然コード配列であってよく、利用可能である場合、但し、例えば、P.fluorescensなどのシュードモナス種または他の適切な生物のコドン使用の偏向を反映するために遺伝子を最適化することによって、アレイの菌株において発現可能な形で使用するために選択、改良、または最適化されたコード配列であることがより好ましい。遺伝子最適化のために、1つまたは複数の稀なコドンを除去してリボゾーム消失を避け、アミノ酸の取り込み違いを最小にすることができる。1つまたは複数の遺伝子内リボソーム結合部位を除去して、切断型タンパク質産物を避けることもできる。CおよびGヌクレオチドの長い延長部分を除去して、フレームシフトをもたらす可能性があるRNAポリメラーゼのスリップを避けることができる。強力な遺伝子内ステムループ構造、特にリボソーム結合部位を覆う構造を除去することもできる。
【0145】
他の実施形態では、タンパク質は産生時に、追加の標的配列、例えばタンパク質をペリプラズムまたは細胞外培地に標的化する配列も含む。一実施形態では、その追加の標的配列は、タンパク質のカルボキシ末端と作用可能に連結する。別の実施形態では、タンパク質は、オートトランスポーターの分泌シグナル、2つのパートナー分泌系、主要末端分岐系または線毛外膜ポーリンを含む。
【0146】
生成する(1つまたは複数の)遺伝子は1つまたは複数のベクター内で構築されるか、またはその中に挿入され、次いでアレイ内の宿主細胞集団のそれぞれに形質転換される。「発現可能な形」で提供されると言われる核酸またはポリヌクレオチドは、本発明の1つまたは複数の宿主細胞集団によって発現され得る少なくとも1つの遺伝子を含有する、核酸またはポリヌクレオチドを意味する。
【0147】
分子遺伝学および遺伝子工学技法に必要とされる広範囲の配列情報が、広く公に利用されている。哺乳動物の完全なヌクレオチド配列、ならびにヒト、遺伝子、cDNA配列、アミノ酸配列およびゲノムへのアクセスは、ウェブサイトwww.ncbi.nlm.nih.gov/EntrezでGenBankから得ることができる。追加の情報はGeneCards、遺伝子およびそれらの産物に関する情報を統合した電子百科事典、およびWeizmann Institute of Science Genome and Bioinformatics(bioinformatics.weizmann.ac.il/cards)からの生物医学的応用から得ることもでき、ヌクレオチド配列情報は、EMBLヌクレオチド配列データベース(www.ebi.ac.uk/embl/)またはDNAデータバンクまたは日本(the DNA Databank or Japan)(DDBJ、www.ddbi.nig.ac.ii/;アミノ酸配列に関する情報の追加のサイトには、Georgetownのタンパク質情報リソースウェブサイト(www−nbrf.Reorgetown.edu/pirl)およびSwiss−Prot(au.expasy.org/sprot/sprot−top.html)がある)から得ることもできる。
【0148】
本発明において発現させることが可能であるタンパク質の例には、例えばレニン、ヒト成長ホルモン、ウシ成長ホルモン、成長ホルモン放出因子を含めた成長ホルモン、副甲状腺ホルモン、甲状腺刺激ホルモン、リポタンパク質、α−1−アンチトリプシン、インシュリンA鎖、インシュリンB鎖、プロインシュリン、トロンボポエチン、卵胞刺激ホルモン、カルシトニン、黄体形成ホルモン、グルカゴン、因子VIIIC、因子IX、組織因子、およびフォンウィルブランド因子などの凝固因子、プロテインCなどの抗凝固因子、心房性ナトリウム利尿因子、肺表面活性剤、ウロキナーゼまたはヒト尿または組織型プラスミノーゲン活性化因子(t−PA)などのプラスミノーゲン活性化因子、ボムベシン、トロンビン、造血成長因子、腫瘍壊死因子−αおよび−β、エンケファリナーゼ、ヒト血清アルブミンなどの血清アルブミン、ミュラー管抑制物質、レラキシンA鎖、レラキシンB鎖、プロレラキシン、マウスゴナドトロピン関連ポリペプチド、β−ラクタマーゼなどの微生物タンパク質、デオキシリボヌクレアーゼ、インヒビン、アクチビン、血管内皮増殖因子(VEGF)、ホルモンまたは増殖因子に対する受容体、インテグリン、プロテインAまたはD、リウマチ因子、脳由来神経栄養因子(BDNF)、ニューロトロフィン−3、−4、−5、または−6(NT−3、NT−4、NT−5、またはNT−6)、またはNGF−βなどの神経成長因子などの神経栄養因子、カルジオトロフィン−1(CT−1)などのカルジオトロフィン(心肥大因子)、血小板由来成長因子(PDGF)、aFGFおよびbFGFなどの線維芽細胞成長因子、表皮成長因子(EGF)、TGF−αおよびTGF−β1、TGF−β2、TGF−β3、TGF−β4、またはTGF−β5を含めたTGF−βなどの形質転換成長因子(TGF)、インシュリン様成長因子−Iおよび−II(IGF−IおよびIGF−II);des(1〜3)−IGF−I(脳IGF−I)、インシュリン様成長因子結合タンパク質、CD−3、CD−4、CD−8、およびCD−19などのCDタンパク質、エリスロポエチン、骨誘導因子、免疫毒素、骨形成タンパク質(BMP)、インターフェロン−α、−β、および−γなどのインターフェロン、コロニー刺激因子(CSF)、例えばM−CSF、GM−CSF、およびG−CSF、インターロイキン(IL)、例えばIL−1〜IL−10、抗HER−2抗体、スーパーオキシドジスムターゼ、T細胞受容体、表面膜タンパク質、分解促進因子、例えばAIDSエンベロープの一部分などのウイルス抗原、輸送タンパク質、ホーミング受容体、アドレシン、調節タンパク質、抗体、および前に挙げたポリペプチドのいずれかの断片などの分子がある。
【0149】
特定の実施形態では、タンパク質またはポリペプチドは、IL−1、IL−1a、IL−1b、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12、IL−12elasti、IL−13、IL−15、IL−16、IL−18、IL−18BPa、IL−23、IL−24、VIP、エリスロポエチン、GM−CSF、G−CSF、M−CSF、血小板由来成長因子(PDGF)、MSF、FLT−3リガンド、EGF、線維芽細胞成長因子(FGF;例えば、α−FGF(FGF−1)、β−FGF(FGF−2)、FGF−3、FGF−4、FGF−5、FGF−6、またはFGF−7)、インシュリン様成長因子(例えば、IGF−1、IGF−2)、腫瘍壊死因子(例えば、TNF、リンホトキシン)、神経成長因子(例えば、NGF)、血管内皮増殖因子(VEGF)、インターフェロン(例えば、IFN−α、IFN−β、IFN−γ)、白血病抑制因子(LIF)、絨毛神経栄養因子(CNTF)、オンコスタチンM、幹細胞因子(SCF)、形質転換成長因子(例えば、TGF−α、TGF−β1、TGF−β2、TGF−β3)、TNFスーパーファミリー(例えば、LIGHT/TNFSF14、STALL−l/TNFSF13B(BLy5、BAFF、THANK)、TNFα/TNFSF2およびTWEAK/TNFSF12)、またはケモカイン(BCA−1/BLC−1、BRAK/Kec、CXCL16、CXCR3、ENA−78/LIX、エオタキシン−1、エオタキシン−2/MPIF−2、エクソダス(Exodus)−2/SLC、フラクタルカイン/ニューロタクチン、GROα/MGSA、HCC−1、I−TAC、リンホタクチン/ATAC/SCM、MCP−1/MCAF、MCP−3、MCP−4、MDC/STCP−1/ABCD−1、MIP−1.クアドラチュア(quadrature)、MIP−1.クアドラチュア、MIP−2.クアドラチュア/GRO.クアドラチュア、MIP−3.クアドラチュア./エクソダス/LARC、MIP−3/エクソダス−3/ELC、MIP−4/PARC/DC−CK1、PF−4、RANTES、SDF1、TARC、TECK、微生物毒素、ADPリボシル化毒素、微生物またはウイルス抗原)から選択することができる。
【0150】
本発明の一実施形態では、対象とするタンパク質はマルチサブユニットタンパク質またはポリペプチドであってよい。発現させることが可能であるマルチサブユニットタンパク質には、同種および異種タンパク質がある。マルチサブユニットタンパク質は、同じであるかまたは異なってよい2個以上のサブユニットを含むことができる。例えばタンパク質は、2、3、4、5、6、7、8、9、10、11、12個またはそれより多くのサブユニットを含む同種タンパク質であってよい。タンパク質は、2、3、4、5、6、7、8、9、10、11、12個、またはそれより多くのサブユニットを含む異種タンパク質であってもよい。例示的なマルチサブユニットタンパク質には、イオンチャネル受容体を含めた受容体、コンドロイチン、コラーゲンを含めた細胞外マトリクスタンパク質、MHCタンパク質、完全鎖抗体、および抗体断片を含めた免疫修飾物質、RNAポリメラーゼ、およびDNAポリメラーゼを含めた酵素、および膜タンパク質がある。
【0151】
別の実施形態では、対象とするタンパク質は血中タンパク質であってよい。この実施形態において発現される血中タンパク質には、担体タンパク質、ヒトおよびウシアルブミンを含めたアルブミンなど、トランスフェリン、組換え体トランスフェリン半分子、ハプトグロビン、フィブリノゲンおよび他の凝固因子、補体成分、免疫グロブリン、酵素阻害剤、アンギオテンシンおよびブラジキニン、インシュリン、エンドセリン、およびα、β、およびγ−グロブリンを含めたグロブリンなどの物質の前駆体、および哺乳動物の血液中で主に見られる他の型のタンパク質、ポリペプチド、およびそれらの断片があるが、これらだけには限られない。ヒト血清アルブミンに関するアミノ酸配列(Lawn、L.Mら、(1981年)Nucleic Acids Research、9:6103〜6114ページ)およびヒト血清トランスフェリンに関するアミノ酸配列(Yang、Fら、(1984年)Proc.Natl.Acad.Sci.USA81:2752〜2756ページ)を含めた、多数の血中タンパク質に関するアミノ酸配列が報告されてきている(S.S.Baldwin(1993年)Comp.Biochem Physiol.106b:203〜218ページ参照)。
【0152】
別の実施形態では、対象とするタンパク質は酵素または補因子であってよい。この実施形態において発現される酵素および補因子には、アルドラーゼ、アミンオキシダーゼ、アミノ酸オキシダーゼ、アスパルターゼ、B12依存性酵素、カルボキシペプチダーゼ、カルボキシエステラーゼ、カルボキシリアーゼ、ケモトリプシン、CoA要求酵素、シアノヒドリンシンテターゼ、シスタチオンシンターゼ、デカルボキシラーゼ、デヒドロゲナーゼ、アルコールデヒドロゲナーゼ、デヒドラターゼ、ジアホラーゼ、ジオキシゲナーゼ、エノアートリダクターゼ、エポキシドヒドラーゼ、フメラーゼ、ガラクトースオキシダ−ゼ、グルコースイソメラーゼ、グルコースオキシダーゼ、グリコシルトランスフェラーゼ、メチルトランスフェラーゼ、ニトリルヒドラーゼ、ヌクレオシドホスホリラーゼ、オキシドリダクターゼ、オキシニチラーゼ、ペプチダーゼ、グリコシルトランスフェラーゼ、ペロキシダーゼ、治療活性ポリペプチドと融合した酵素、組織型プラスミノーゲン活性化因子、ウロキナーゼ、レプチラーゼ、ストレプトキナーゼ、カタラーゼ、スーパーオキシドジスムターゼ、デオキシリボヌクレアーゼ、アミノ酸加水分解酵素(例えば、アスパラギナーゼ、アミド加水分解酵素)、カルボキシペプチダーゼ、プロテアーゼ、トリプシン、ペプシン、キモトリプシン、パパイン、ブロメライン、コラゲナーゼ、ノイラミニダーゼ、ラクターゼ、マルターゼ、スクラーゼ、およびアラビノフラノシダーゼがあるが、これらだけには限られない。
【0153】
別の実施形態では、対象とするタンパク質は単鎖、Fab断片および/または完全鎖抗体、またはその断片もしくは一部分であってよい。単鎖抗体は、1つの安定的にフォールディングしたポリペプチド鎖上の抗体の抗原結合領域を含むことができる。Fab断片は特定の抗体の一片であってよい。Fab断片は抗原結合部位を含有することができる。Fab断片は2本の鎖、軽鎖および重鎖断片を含有することができる。これらの断片は、リンカーまたはジスルフィド結合によって連結させることが可能である。
【0154】
他の実施形態では、対象とするタンパク質は、約20〜約42℃の温度で活性があるタンパク質である。一実施形態では、タンパク質は生理的温度で活性があり、高温または極温(high or extreme temperatures)、65℃を超える温度に加熱すると不活性である。
【0155】
一実施形態では、対象とするタンパク質は約20〜約42℃の温度で活性があるタンパク質であり、および/または高温または極温、65℃を超える温度に加熱すると不活性であり、あるいは天然哺乳動物またはヒトタンパク質などの天然タンパク質と実質的に相同的であり、コンカタマー型の核酸から発現されず、この場合プロモーターはアレイ内で使用する宿主細胞における天然プロモーターではなく、E.coliなどの他の生物に由来する。
【0156】
宿主細胞(Host cell)
一実施形態では本発明は、そこから対象とする異種タンパク質またはペプチドを場合によっては産生するための、P.fluorescens宿主細胞のアレイを提供する。P.fluorescensは様々なタンパク質を産生するための改良された基盤であることが実証されており、いくつかの有効な分泌シグナルがこの生物から同定されている(その全容が参照によって本明細書に組み込まれる、米国出願公開第2006/0008877号を参照)。
【0157】
シュードモナス系は、他の細菌発現系と比較して、ポリペプチドおよび酵素の工業的発現に関して利点をもたらす。特に、P.fluorescensは有利な発現系として同定されている。P.fluorescensは、土壌、水および植物表面環境にコロニー形成する、一群の一般的な、非病原性の腐生菌を含む。洗浄添加剤として、および立体選択的加水分解用に、P.fluorescens由来の市販の酵素が使用されて、環境汚染物質が減らされている。P.fluorescensは病原体を制御するために農業用にも使用されている。米国特許第4,695,462号は、P.fluorescensにおける組換え体細菌タンパク質の発現を記載している。
【0158】
しかしながら、代替宿主細胞、またはさらに多様な異なる宿主細胞を使用して、前に論じたように1つまたは複数の標的遺伝子の発現を調節するために遺伝子改変された複数の表現型が異なる宿主細胞を含む、アレイを作製することができることは企図(comtemplated)される。宿主細胞は、その中で標的遺伝子を改変することができる任意の生物であってよい。表1および2中に挙げた標的遺伝子と相同的な標的遺伝子を同定する方法は、当技術分野で知られている。さらに、当業者は、対象とする宿主細胞に由来するかまたはその中で有用である標的遺伝子の同定の仕方を理解しているはずである。これらのタンパク質の多くは当技術分野でよく知られている。例えば、米国特許出願公開第2006/0110747号)を参照。
【0159】
宿主細胞は「グラム陰性プロテオバクテリア亜群18」から選択することができる。「グラム陰性プロテオバクテリア亜群18」は、例えば以下の(丸括弧中に示すATCCまたは他の寄託番号の(1つまたは複数の)例示的菌株):次亜種1または次亜種Iとも呼ばれる、Pseudomonas fluorescensバイオタイプA(ATCC13525);次亜種2または次亜種IIとも呼ばれる、Pseudomonas fluorescensバイオタイプB(ATCC17816);次亜種3または次亜種IIIとも呼ばれる、Pseudomonas fluorescensバイオタイプC(ATCC17400);次亜種4または次亜種IVとも呼ばれる、Pseudomonas fluorescensバイオタイプF(ATCC12983);次亜種5または次亜種Vとも呼ばれる、Pseudomonas fluorescensバイオタイプG(ATCC17518);Pseudomonas fluorescensバイオタイプVI;Pseudomonas fluorescensPf0−1;Pseudomonas fluorescensPf−5(ATCC BAA−477);Pseudomonas fluorescensSBW25;およびPseudomonas fluorescens亜種cellulosa(NCIMB10462)に属するものを含めた、Pseudomonas fluorescens種の全ての亜種、変種、菌株、および他の下位専門単位の群として定義する。
【0160】
宿主細胞は「グラム陰性プロテオバクテリア亜群19」から選択することができる。「グラム陰性プロテオバクテリア亜群19」は、Pseudomonas fluorescensバイオタイプAの全ての菌株の群として定義する。このバイオタイプの特に好ましい菌株は、P.fluorescens菌株MB101(Wilcoxへの米国特許第5,169,760号を参照)、およびその誘導体である。その好ましい誘導体の一例は、MB101染色体asd(アスパラギン酸デヒドロゲナーゼ遺伝子)遺伝子座に挿入することによって構築されたP.fluorescens菌株MB214、天然E.coliPlacI−lacI−lacZYAコンストラクト(すなわち、その中でPlacZが欠失した)である。
【0161】
本発明において使用することができる追加のP.fluorescens菌株には、以下のATCC指定を有する、Pseudomonas fluorescens MigulaおよびPseudomonas fluorescens Loitokitokがある:[NCIB 8286];NRRL B−1244;NCIB 8865株CO1;NCIB 8866株CO2;1291[ATCC 17458;IFO 15837;NCIB 8917;LA;NRRL B−1864;ピロリジン;PW2[ICMP 3966;NCPPB 967;NRRL B−899];13475;NCTC 10038;NRRL B−1603[6;IFO 15840];52−1C;CCEB 488−A[BU 140];CCEB 553[EM 15/47];IAM 1008[AHH−27];IAM 1055[AHH−23];I[IFO 15842];12[ATCC 25323;NIH 11;den Dooren de Jong 216];18[IFO 15833;WRRL P−7];93[TR−10];108[52−22;IFO 15832];143[IFO 15836;PL];149[2−40−40;IFO 15838];182[IFO 3081;PJ 73];184[IFO 15830];185[W2 L−1];186[IFO 15829;PJ 79];187[NCPPB 263];188[NCPPB 316];189[PJ227;1208];191[IFO 15834;PJ 236;22/1];194[Klinge R−60;PJ 253];196[PJ 288];197[PJ 290];198[PJ 302];201[PJ 368];202[PJ 372];203[PJ 376];204[IFO 15835;PJ 682];205[PJ 686];206[PJ 692];207[PJ 693];208[PJ 722];212.[PJ 832];215[PJ 849];216[PJ 885];267[B−9];271[B−1612];401[C71A;IFO 15831;PJ 187];NRRL B−3178[4;IFO.15841];KY 8521;3081;30−21;[IFO 3081];N;PYR;PW;D946−B83[BU 2183;FERM−P 3328];P−2563[FERM−P 2894;IFO 13658];IAM−1126[43F];M−1;A506[A5−06];A505[A5−05−1];A526[A5−26];B69;72;NRRL B−4290;PMW6[NCIB 11615];SC 12936;Al[IFO 15839];F 1847[CDC−EB];F 1848[CDC 93];NCIB 10586;P17;F−12;AmMS 257;PRA25;6133D02;6519E01;Ni;SC15208;BNL−WVC;NCTC 2583[NCIB 8194];H13;1013[ATCC 11251;CCEB 295];IFO 3903;1062;またはPf−5。
【0162】
一実施形態では、宿主細胞は、前に記載したP.fluorescens細胞を含めた、対象とするタンパク質またはポリペプチドを産生することができる任意の細胞であってよい。対象とするタンパク質またはポリペプチドを産生するために最も一般的に使用される系は、大規模なバッチ式培養においてタンパク質を産生するための、それらの比較的安価な増殖要件および潜在能力のために、特定の細菌細胞、特にE.coliを含む。酵母菌をさらに使用して、特に研究目的で、生物学的関連があるタンパク質およびポリペプチドを発現させる。系にはSaccharomyces cerevisiaeまたはPichia pastorisがある。これらの系は十分特徴付けられており、一般に認められるレベルの全タンパク質産生をもたらし、比較的迅速および安価である。昆虫細胞発現系も、生物活性型の組換えタンパク質を発現させるための代替として浮上している。いくつかの場合、翻訳後修飾された、正確にフォールディングされたタンパク質を産生することができる。チャイニーズハムスター卵巣細胞などの哺乳動物細胞発現系も、対象とするタンパク質またはポリペプチドの発現用に使用されている。小規模では、これらの発現系は有効であることが多い。特定の生物製剤は、特に動物またはヒトの健康用途で、タンパク質から誘導することができる。別の実施形態では、宿主細胞は、これらだけには限られないが、タバコ細胞、コーン細胞、シロイヌナズナ種由来の細胞、ジャガイモまたはコメ細胞を含めた植物細胞である。
【0163】
別の実施形態では、宿主細胞は、これらだけには限られないが、エシェリキアまたはシュードモナス種を含めた細菌細胞などの原核細胞であってよい。典型的な細菌細胞は、ウェブサイトwww.emc.maricotpa.edu/faculty/farabee/BIOBK/BioBookDiversityで、Estrella Mountain Community College、Arizona、USAのDr M J Farabeeによって提供された、例えば「Biological Diversity:Bacteria and Archaeans」、the On−Line Biology Bookの一章中に記載されている。特定の実施形態では、宿主細胞はシュードモナス細胞であってよく、かつ典型的にはP.fluorescens細胞であってよい。他の実施形態では、宿主細胞はE.coli細胞であってもよい。別の実施形態では、宿主細胞は真核細胞、例えばこれらだけには限られないがスポドプテラ、トリコプルシア、ショウジョウバエまたはエスティグメン種由来の細胞を含めた昆虫細胞、またはこれらだけには限られないがネズミ細胞、ハムスター細胞、サル、霊長類またはヒト細胞を含めた哺乳動物細胞であってよい。
【0164】
一実施形態では、宿主細胞は細菌分類群のいずれかのメンバーであってよい。細胞は例えば、真正細菌の任意の種のメンバーであってよい。宿主は、分類群:アシドバクテリア、アクチノバクテリア、アクイフェックス、バクテロイデス、クロロビウム、クラミジア、コロフレキシ、クリシオジェネス、シアノバクテリア、デフェリバクター、デイノコッカス、ディクチオグロムス、フィブロバクター、ファーミキューテス、フソバクテリア、ゲマティモナス、レンチスファエラ、ニトロスピラ、プランクトミセス、プロテオバクテリア、スピロヘータ、サーモデスルフォバクテリア、サーモミクロビア、サーモトガ、サーマス(サーマレス)、またはベルコミクロビアのいずれか1つのメンバーであってよい。真正細菌宿主細胞の一実施形態では、細胞は、シアノバクテリアを除く真正細菌の任意の種のメンバーであってよい。
【0165】
細菌宿主は、プロテオバクテリアの任意の種のメンバーであってもよい。プロテオバクテリア宿主細胞は、分類群アルファプロテオバクテリア、ベータプロテオバクテリア、ガンマプロテオバクテリア、デルタプロテオバクテリア、またはイプシロンプロテオバクテリアのいずれか1つのメンバーであってよい。さらに宿主は、分類群アルファプロテオバクテリア、ベータプロテオバクテリア、またはガンマプロテオバクテリアのいずれか1つのメンバー、およびガンマプロテオバクテリアの任意の種のメンバーであってよい。
【0166】
ガンマプロテオバクテリア宿主の一実施形態では、宿主は、分類群エロモナス目、アルテロモナス目、腸内細菌目、シュードモナス目、またはキサントモナス目のいずれか1つのメンバー、または腸内細菌目もしくはシュードモナスの目の任意の種のメンバーであり得る。一実施形態では、宿主細胞は腸内細菌の目であってよく、宿主細胞は腸内細菌科のメンバーであり得る、またはエルビニア、エシェリキア、またはセラチア属のいずれか1つのメンバー、またはエシェリキア属のメンバーであってよい。宿主細胞がシュードモナスの目である場合、宿主細胞は、シュードモナス属を含めたシュードモナス科のメンバーであってよい。ガンマプロテオバクテリア宿主には、Escherichaia.coli種のメンバーおよびPseudomonas fluorescens種のメンバーがある。
【0167】
他のシュードモナス生物も有用である可能性がある。シュードモナスおよび近縁の種には、R.E.Buchanan and N.E.Gibbons(eds.)、Bergey’s Manual of Determinative Bacteriology、217〜289ページ(第8版、1974年)(The Williams & Wilkins Co.、Baltimore、Md.、USA)(本明細書では以後「Bergey(1974)」)によって「Gram−Negative Aerobic Rods and Cocci」として記載された、科および/または属に属するプロテオバクテリアの群を含む、グラム陰性プロテオバクテリア亜群1がある。表7は、これらの生物の科および属を表す。
【0168】
【表7】
【0169】
「グラム陰性プロテオバクテリア亜群1」は、分類において使用する基準に従ってこの表題に分類され得るプロテオバクテリアも含む。この表題は、この項で前に分類したがそれ以上は分類されない群、アシドボラックス、ブレブンジモナス、バークホルデアリア、好気性水素酸化細菌、オーシャンイモナス、ラルストニア、およびステノトロフォモナス属、(以前に命名された種の)キサントモナス属に属する生物を再グループ化することによって生成したスフィンゴモナス属(およびそれに由来するブラストモナス属)、Bergey(1974)において定義されたアセトバクター属に属する生物を再グループ化することによって生成したアシドモナス属なども含む。さらに宿主は、それぞれAlteromonas haloplanktis、Alteromonas nigrifaciens、およびAlteromonas putrefaciensとして再分類された、シュードモナス属、Pseudomonas enalia(ATCC14393)、Pseudomonas nigrifaciensi(ATCC19375)、およびPseudomonas putrefaciens(ATCC8071)由来の細胞を含むことができる。同様に、例えば、Pseudomonas acidovorans(ATCC15668)およびPseudomonas testosteroni(ATCC11996)が、それぞれComamonas acidovoransおよびComamonas testosteroniとして以後再分類されており、かつPseudomonas nigrifaciens(ATCC19375)およびPseudomonas piscicida(ATCC15057)が、それぞれPseudoalteromonas nigrifaciensおよびPseudoalteromonas piscicidaとして再分類されている。「グラム陰性プロテオバクテリア亜群1」は、科:シュードモナス科、アゾトバクター科(現在、同義語、「シュードモナス科のアゾトバクター族」によって呼ばれることが多い)、リゾビウム科、およびメチロモナス科(現在、同義語、「メチロコッカス科」によって呼ばれることが多い)のいずれかに属するとして分類されたプロテオバクテリアも含む。したがって、本明細書で他に記載する属に加えて、「グラム陰性プロテオバクテリア亜群1」の範疇のさらなるプロテオバクテリア属には、1)アゾリゾフィルス属のアゾトバクター群の細菌、2)セルビブリオ、オリゲラ、およびテレジニバクター属のシュードモナス科の細菌、3)ケラトバクター、エンシファー、リベリバクター(「カンジダタスリベリバクター」とも呼ばれる)、およびシノリゾビウム属のリゾビウム科の細菌、および4)メチロバクター、メチロカルダム、メチロミクロビウム、メチロサルシナ、およびメチロスファエラ属のメチロコッカス科の細菌がある。
【0170】
別の実施形態では、宿主細胞は「グラム陰性プロテオバクテリア亜群2」から選択される。「グラム陰性プロテオバクテリア亜群2」は、以下の属(および全数のカタログに挙げられ、公に入手可能な、他に示した場合を除きいずれもATCCで寄託された、丸括弧中に示すその寄託菌株):アシドモナス(2);アセトバクター(93);グルコノバクター(37);ブレブンジモナス(23);ベイエリンキア(13);デルキシア(2);ブルセラ(4);アグロバクテリウム(79);ケラトバクター(2);エンシファー(3);リゾビウム(144);シノリゾビウム(24);ブラストモナス(1);スフィンゴモナス(27);アルカリゲネス(88);ボルデテラ(43);バークホルデアリア(73);ラルストニア(33);アシドボラックス(20);好気性水素酸化細菌(9);ズーグレア(9);メチロバクター(2);メチロカルダム(NCIMBで1);メチロコッカス(2);メチロミクロビウム(2);メチロモナス(9);メチロサルシナ(1);メチロスファエラ、アゾモナス(9);アゾリゾフィルス(5);アゾトバクター(64);セルビブリオ(3);オリゲラ(5);シュードモナス(1139);フランシセラ(4);キサントモナス(229);ステノトロフォモナス(50);およびオーシャンイモナス(4)のプロテオバクテリアの群として定義する。
【0171】
「グラム陰性プロテオバクテリア亜群2」の例示的な宿主細胞種には、以下の細菌(および丸括弧中に示すATCCまたは他の寄託番号の(1つまたは複数の)例示的菌株)があるが、これらだけには限られない:Acidomonas methanolica(ATCC 43581);Acetobacter aceti(ATCC 15973);Gluconobacter oxydans(ATCC 19357);Brevundimonas diminuta(ATCC 11568);Beijerinckia indica(ATCC 9039 およびATCC 19361);Derxia gummosa(ATCC 15994);Brucella melitensis(ATCC 23456),Brucella abortus(ATCC 23448);Agrobacterium tumefaciens(ATCC 23308),Agrobacterium radiobacter(ATCC 19358),Agrobacterium rhizogenes(ATCC 11325);Chelatobacter heintzii(ATCC 29600);Ensifer adhaerens(ATCC 33212);Rhizobium leguminosarum(ATCC 10004);Sinorhizobium frediii(ATCC 35423);Blastomonas natatoria(ATCC 35951);Sphingomonas paucimobilis(ATCC 29837);Alcaligenes faecalis(ATCC 8750);Bordetella pertussis(ATCC 9797);Burkholderia cepacia(ATCC 25416);Ralstonia pickettii(ATCC 27511);Acidovorax facilis(ATCC 11228);Hydrogenophagaflava(ATCC 33667);Zoogloea ramigera(ATCC 19544);Methylobacter luteus(ATCC 49878);Methylocaldum gracile(NCIMB 11912);Methylococcus capsulatus(ATCC 19069);Methylomicrobium agile(ATCC 35068);Methylomonas methanica(ATCC 35067);Methylosarcina fibrata(ATCC 700909);Methylosphaera hansonii(ACAM 549);Azomonas agilis(ATCC 7494);Azorhizophilus paspali(ATCC 23833);Azotobacter chroococcum(ATCC 9043);Cellvibrio mixtus(UQM 2601);Oligella urethralis(ATCC 17960);Pseudomonas aeruginosa(ATCC 10145),Pseudomonas fluorescens(ATCC 35858);Francisella tularensis(ATCC 6223);Stenotrophomonas maltophilia(ATCC 13637);Xanthomonas campestris(ATCC 33913);およびOceanimonas doudoroffii(ATCC 27123)。
【0172】
別の実施形態では、宿主細胞は「グラム陰性プロテオバクテリア亜群3」から選択される。「グラム陰性プロテオバクテリア亜群3」は、以下の属:ブレブンジモナス、アグロバクテリウム、リゾビウム、シノリゾビウム、ブラストモナス、スフィンゴモナス、アルカリゲネス、バークホルデアリア、ラルストニア、アシドボラックス、好気性水素酸化細菌、メチロバクター、メチロカルダム、メチロコッカス、メチロミクロビウム、メチロモナス、メチロサルシナ、メチロスファエラ、アゾモナス、アゾリゾフィルス、アゾトバクター、セルビブリオ、オリゲラ、シュードモナス、テレジニバクター、フランシセラ、ステノトロフォモナス、キサントモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0173】
別の実施形態では、宿主細胞は「グラム陰性プロテオバクテリア亜群4」から選択される。「グラム陰性プロテオバクテリア亜群4」は、以下の属:ブレブンジモナス、ブラストモナス、スフィンゴモナス、バークホルデアリア、ラルストニア、アシドボラックス、好気性水素酸化細菌、メチロバクター、メチロカルダム、メチロコッカス、メチロミクロビウム、メチロモナス、メチロサルシナ、メチロスファエラ、アゾモナス、アゾリゾフィルス、アゾトバクター、セルビブリオ、オリゲラ、シュードモナス、テレジニバクター、フランシセラ、ステノトロフォモナス、キサントモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0174】
別の実施形態では、宿主細胞は「グラム陰性プロテオバクテリア亜群5」から選択される。「グラム陰性プロテオバクテリア亜群5」は、以下の属:メチロバクター、メチロカルダム、メチロコッカス、メチロミクロビウム、メチロモナス、メチロサルシナ、メチロスファエラ、アゾモナス、アゾリゾフィルス、アゾトバクター、セルビブリオ、オリゲラ、シュードモナス、テレジニバクター、フランシセラ、ステノトロフォモナス、キサントモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0175】
宿主細胞は、「グラム陰性プロテオバクテリア亜群6」から選択することができる。「グラム陰性プロテオバクテリア亜群6」は、以下の属:ブレブンジモナス、ブラストモナス、スフィンゴモナス、バークホルデアリア、ラルストニア、アシドボラックス、好気性水素酸化細菌、アゾモナス、アゾリゾフィルス、アゾトバクター、セルビブリオ、オリゲラ、シュードモナス、テレジニバクター、ステノトロフォモナス、キサントモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0176】
宿主細胞は、「グラム陰性プロテオバクテリア亜群7」から選択することができる。「グラム陰性プロテオバクテリア亜群7」は、以下の属:アゾモナス、アゾリゾフィルス、アゾトバクター、セルビブリオ、オリゲラ、シュードモナス、テレジニバクター、ステノトロフォモナス、キサントモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0177】
宿主細胞は、「グラム陰性プロテオバクテリア亜群8」から選択することができる。「グラム陰性プロテオバクテリア亜群8」は、以下の属:ブレブンジモナス、ブラストモナス、スフィンゴモナス、バークホルデアリア、ラルストニア、アシドボラックス、好気性水素酸化細菌、シュードモナス、ステノトロフォモナス、キサントモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0178】
宿主細胞は、「グラム陰性プロテオバクテリア亜群9」から選択することができる。「グラム陰性プロテオバクテリア亜群9」は、以下の属:ブレブンジモナス、バークホルデアリア、ラルストニア、アシドボラックス、好気性水素酸化細菌、シュードモナス、ステノトロフォモナス、およびオーシャンイモナスのプロテオバクテリアの群として定義する。
【0179】
宿主細胞は、「グラム陰性プロテオバクテリア亜群10」から選択することができる。「グラム陰性プロテオバクテリア亜群10」は、以下の属:バークホルデアリア、ラルストニア、シュードモナス、ステノトロフォモナス、およびキサントモナスのプロテオバクテリアの群として定義する。
【0180】
宿主細胞は、「グラム陰性プロテオバクテリア亜群11」から選択することができる。「グラム陰性プロテオバクテリア亜群11」は、シュードモナス、ステノトロフォモナス、およびキサントモナス属のプロテオバクテリアの群として定義する。
【0181】
宿主細胞は、「グラム陰性プロテオバクテリア亜群12」から選択することができる。「グラム陰性プロテオバクテリア亜群12」は、以下の属:バークホルデアリア、ラルストニア、シュードモナスのプロテオバクテリアの群として定義する。宿主細胞は、「グラム陰性プロテオバクテリア亜群13」から選択することができる。「グラム陰性プロテオバクテリア亜群13」は、以下の属:バークホルデアリア、ラルストニア、シュードモナス、およびキサントモナスのプロテオバクテリアの群として定義する。宿主細胞は、「グラム陰性プロテオバクテリア亜群14」から選択することができる。「グラム陰性プロテオバクテリア亜群14」は、以下の属:シュードモナスおよびキサントモナスのプロテオバクテリアの群として定義する。宿主細胞は、「グラム陰性プロテオバクテリア亜群15」から選択することができる。「グラム陰性プロテオバクテリア亜群15」は、シュードモナス属のプロテオバクテリアの群として定義する。
【0182】
宿主細胞は、「グラム陰性プロテオバクテリア亜群16」から選択することができる。「グラム陰性プロテオバクテリア亜群16」は、以下のシュードモナス種(および丸括弧中に示すATCCまたは他の寄託番号の(1つまたは複数の)例示的菌株)のプロテオバクテリアの群として定義する:Pseudomonas abietaniphila(ATCC 700689);Pseudomonas aeruginosa(ATCC 10145);Pseudomonas alcaligenes(ATCC 14909);Pseudomonas anguilliseptica(ATCC 33660);Pseudomonas citronellolis(ATCC 13674);Pseudomonas flavescens(ATCC 51555);Pseudomonas mendocina(ATCC 25411);Pseudomonas nitroreducens(ATCC 33634);Pseudomonas oleovorans(ATCC 8062);Pseudomonas pseudoalcaligenes(ATCC 17440);Pseudomonas resinovorans(ATCC 14235);Pseudomonas straminea(ATCC 33636);Pseudomonas agarici(ATCC 25941);Pseudomonas alcaliphila;Pseudomonas alginovora;Pseudomonas andersonii;Pseudomonas asplenii(ATCC 23835);Pseudomonas azelaica(ATCC 27162);Pseudomonas beyerinckii(ATCC 19372);Pseudomonas borealis;Pseudomonas boreopolis(ATCC 33662);Pseudomonas brassicacearum;Pseudomonas butanovora(ATCC 43655);Pseudomonas cellulosa(ATCC 55703);Pseudomonas aurantiaca(ATCC 33663);Pseudomonas chlororaphis(ATCC 9446,ATCC 13985,ATCC 17418,ATCC 17461);Pseudomonas fragi(ATCC 4973);Pseudomonas lundensis(ATCC 49968);Pseudomonas taetrolens(ATCC 4683);Pseudomonas cissicola(ATCC 33616);Pseudomonas coronafaciens;Pseudomonas diterpeniphila;Pseudomonas elongata(ATCC 10144);Pseudomonasflectens(ATCC 12775);Pseudomonas azotoformans;Pseudomonas brenneri;Pseudomonas cedrella;Pseudomonas corrugata(ATCC 29736);Pseudomonas extremorientalis;Pseudomonas fluorescens(ATCC 35858);Pseudomonas gessardii;Pseudomonas libanensis;Pseudomonas mandelii(ATCC 700871);Pseudomonas marginalis(ATCC 10844);Pseudomonas migulae;Pseudomonas mucidolens(ATCC 4685);Pseudomonas orientalis;Pseudomonas rhodesiae;Pseudomonas synxantha(ATCC 9890);Pseudomonas tolaasii(ATCC 33618);Pseudomonas veronii(ATCC 700474);Pseudomonas frederiksbergensis;Pseudomonas geniculata(ATCC 19374);Pseudomonas gingeri;Pseudomonas graminis;Pseudomonas grimontii;Pseudomonas halodenitrificans;Pseudomonas halophila;Pseudomonas hibiscicola(ATCC 19867);Pseudomonas huttiensis(ATCC 14670);Pseudomonas hydrogenovora;Pseudomonas jessenii(ATCC 700870);Pseudomonas kilonensis;Pseudomonas lanceolata(ATCC 14669);Pseudomonas lini;Pseudomonas marginata(ATCC 25417);Pseudomonas mephitica(ATCC 33665);Pseudomonas denitrificans(ATCC 19244);Pseudomonas pertucinogena(ATCC 190);Pseudomonas pictorum(ATCC 23328);Pseudomonas psychrophila;Pseudomonas filva(ATCC 31418);Pseudomonas monteilii(ATCC 700476);Pseudomonas mosselii;Pseudomonas oryzihabitans(ATCC 43272);Pseudomonas plecoglossicida(ATCC 700383);Pseudomonas putida(ATCC 12633);Pseudomonas reactans;Pseudomonas spinosa(ATCC 14606);Pseudomonas balearica;Pseudomonas luteola(ATCC 43273);Pseudomonas stutzeri(ATCC 17588);Pseudomonas amygdali(ATCC 33614);Pseudomonas avellanae(ATCC 700331);Pseudomonas caricapapayae(ATCC 33615);Pseudomonas cichorii(ATCC 10857);Pseudomonas ficuserectae(ATCC 35104);Pseudomonas fuscovaginae;Pseudomonas meliae(ATCC 33050);Pseudomonas syringae(ATCC 19310);Pseudomonas viridiflava(ATCC 13223);Pseudomonas thermocarboxydovorans(ATCC 35961);Pseudomonas thermotolerans;Pseudomonas thivervalensis;Pseudomonas vancouverensis(ATCC 700688);Pseudomonas wisconsinensis;およびPseudomonas xiamenensis。
【0183】
宿主細胞は、「グラム陰性プロテオバクテリア亜群17」から選択することができる。「グラム陰性プロテオバクテリア亜群17」は、例えば以下のシュードモナス種に属する種を含めた「fluorescent Pseudomonads」として当技術分野で知られているプロテオバクテリアの群として定義する:Pseudomonas azotoformans;Pseudomonas brenneri;Pseudomonas cedrella;Pseudomonas corrugata;Pseudomonas extremorientalis;Pseudomonas fluorescens;Pseudomonas gessardii;Pseudomonas libanensis;Pseudomonas mandelii;Pseudomonas marginalis;Pseudomonas migulae;Pseudomonas mucidolens;Pseudomonas orientalis;Pseudomonas rhodesiae;Pseudomonas synxantha;Pseudomonas tolaasii;およびPseudomonas veronii。
【0184】
他の適切な宿主には、グラム(+)プロテオバクテリアなどの参照の他の部分で分類した宿主がある。一実施形態では、宿主細胞はE.coliである。E.coliに関するゲノム配列は、E.coliMG1655に関して確定している(Blattnerら、(1997年)。E.coliK−12の完全なゲノム配列、Science277(5331):1453〜74ページ)およびDNAマイクロアレイが、E.coliK12に関して市販されている(MWG Inc、High Point、N.C.)。E.coliは、1%グルコースなどの適切な炭素源と共に、Luria−Bertani(LB)(10g/Lのトリプトン、5g/LのNaCl、5g/Lの酵母エキス)などの富栄養培地中、またはM9(6g/LのNa2HPO4、3g/LのKH2PO4、1g/LのNH4Cl、0.5g/LのNaCl、pH7.4)などの定義した最小培地中のいずれかで培養することができる。通常、E.coli細胞の一晩培養物を希釈し、振とうフラスコまたは発酵槽中のいずれかの新たな富栄養または最小培地に接種し、37℃で増殖する。
【0185】
宿主細胞は、任意のヒトを含めた哺乳動物または非ヒト哺乳動物由来の細胞などの、哺乳動物起源の細胞であってもよい。哺乳動物は、霊長類、サル、ブタ、ヒツジ類、ウシ、げっ歯類、有蹄動物、家畜ブタ、ブタ類、ヒツジ、ラム、ヤギ、畜牛、シカ、ラバ、ウマ、サル、類人猿、イヌ、ネコ、ラット、およびマウスを含み得るが、これらだけには限られない。
【0186】
宿主細胞は、植物起源の細胞であってもよい。対象とする異種タンパク質の産生を求めてスクリーニングするために任意の植物由来の細胞を選択することができる。適切な植物の例には、アルファルファ、リンゴ、アプリコット、シロイヌナズナ、チョウセンアザミ、ルッコラ、アスパラガス、アボカド、バナナ、オオムギ、マメ、ビート、ブラックチェリー、ブルーベリー、ブロッコリ、芽キャベツ、キャベツ、キャノーラ、マスクメロン、ニンジン、キャッサバ、トウゴマの実、カリフラワー、セロリ、チェリー、チコリ、シラントロ、シトラス、クレメンタイン、クローバー、ココナッツ、コーヒー、コーン、コットン、クランベリー、キュウリ、ダグラスファー、ナス、エンダイブ、キクチシャ、ユーカリ、フェンネル、イチジク、ニンニク、ウリ類、グレープ、グレープフルーツ、甘露、クズイモ、キウィフルーツ、レタス、リーキ、レモン、ライム、テーダマツ、リンシード、マンゴー、メロン、マッシュルーム、ネクタリン、ナッツ、オートムギ、油ヤシ、ナタネ、オクラ、オリーブ、オニオン、オレンジ、観賞植物、ヤシ、パパイヤ、パセリ、パースニップ、エンドウマメ、モモ、ピーナッツ、ナシ、コショウ、カキ、パイン、パインアップル、プランテイン、プラム、ザクロ、ポプラ、ジャガイモ、パンプキン、マルメロ、ラジアタパイン、ラディッキオ、ラディッシュ、ナタネ、ラズベリー、コメ、ライムギ、ソルガム、サザンパイン、ダイズ、ホウレンソウ、カボチャ、イチゴ、テンサイ、サトウキビ、ヒマワリ、サツマイモ、モミジバフウ、タンジェリン、茶、タバコ、トマト、ライコムギ、シバ、カブ、ブドウの木、スイカ、コムギ、ヤムイモ、およびズッキーニがあるが、これらだけには限られない。いくつかの実施形態では、方法中で有用な植物は、シロイヌナズナ、コーン、コムギ、ダイズ、およびコットンである。
【0187】
キット(Kits)
本発明は、対象とする異種タンパク質またはポリペプチドの産生に最適な宿主菌株、例えばP.fluorescens宿主菌株を同定するのに有用なキットも提供する。キットは複数の表現型が異なる宿主細胞を含み、各集団は、タンパク質産生に関与する1つもしくは複数の標的遺伝子の発現が増大するように、タンパク質分解に関与する1つもしくは複数の標的遺伝子の発現が低下するように、またはそれらの両方となるように遺伝子改変されている。アレイは、タンパク質産生に関与する遺伝子またはタンパク質分解に関与する遺伝子のいずれかの発現を調節するように遺伝子改変されていない1つまたは複数の細胞集団をさらに含むことができる。これらのキットは、細胞集団の増殖および維持を容易にするのに十分な試薬、ならびに対象とする異種タンパク質またはポリペプチドの発現用の試薬および/またはコンストラクトも含むことができる。宿主細胞の集団は、細胞集団の保存、輸送、および再構築に適した任意の方法でキットにおいて提供することができる。細胞集団はチューブ中、プレート上、またはスラント上に生きた状態で提供することができ、あるいはチューブまたはバイアル中に凍結乾燥または乾燥のいずれかの状態で保存することができる。細胞集団は、グリセロール、スクロース、アルブミン、または他の適切な保護剤もしくは保存剤などの追加の構成要素を保存培地中に含有することができる。
【0188】
以下の実施例は、限定の目的ではなく例示の目的で提供される。
【0189】
実験例(EXPERIMENTAL EXAMPLES)
概説(Overview)
異種タンパク質産生はしばしば、不溶性タンパク質または不適切にフォールディングされたタンパク質の形成をもたらし、それらは、回収が困難であり、かつ不活性であり得る。さらに、特定の宿主細胞プロテアーゼの存在によって、対象とするタンパク質が分解され、それによって最終収率が低下し得る。全ての異種タンパク質の産生を改善する単一の因子は存在しない。したがって、考えられる候補の集合由来の特定の異種タンパク質に特異的な因子を同定するための方法を探究した。
【0190】
Systems Biologyのツールを使用して、P.fluorescensのゲノムを調べて、宿主細胞のタンパク質フォールディング調節因子およびプロテアーゼ遺伝子を同定した。次いで、全体的な遺伝子発現分析を実施して上方制御された標的を優先し、およびその後、新規なタンパク質産生菌株を構築した。結果として、宿主細胞プロテアーゼが欠損しているかまたはタンパク質フォールディング調節因子の同時過剰発現を可能にする、複数の表現型が異なるP.fluorescens宿主菌株からなる、「Pfenex菌株アレイ」を構築した。この菌株アレイを使用して、特定の異種タンパク質の収率または質を特異的に高める因子を求めてスクリーニングすることができる。複数の表現型が異なる宿主菌株を提供することによって、対象とする任意の個々の異種タンパク質の産生を増大させ得る宿主菌株を同定する成功の機会が増大する。
【0191】
本発明は、Pseudomonas fluorescensにおける異種タンパク質の産生の改善をもたらす。同じ遺伝的背景の宿主菌株の利用可能なライブラリーを有することによって、異種発現したタンパク質の収率および/または質を高める因子の、迅速なスクリーニングおよび同定が可能である。P.fluorescensのゲノム配列には注釈付けしており、標的宿主細胞のフォールディング調節因子およびプロテアーゼを同定している。フォールディング調節因子はタンパク質の適切なフォールディングを手助けし、シャペロン、シャペロニン、ペプチジル−プロリンイソメラーゼ(PPIase)、およびジスルフィド結合形成タンパク質を含む。プロテアーゼは対象とするタンパク質を分解し、したがって異種タンパク質の収率および質に影響を与える可能性がある。考えられる標的を同定するために文献からの背景知識およびDNAマイクロアレイ分析を使用して、約80の標的遺伝子の一覧を構築した。同じ遺伝的背景を有する宿主細胞において、これらの遺伝子をゲノムから除去、またはプラスミドにクローニングして、異種タンパク質との同時過剰発現を可能にした。生成した菌株は96ウェルフォーマットに並べ、および、対象とする異種タンパク質を発現するプラスミドの形質転換後、改善されたタンパク質の収率および/または質を求めてスクリーニングした。
【実施例1】
【0192】
P.fluorescens菌株MB214のゲノム中のフォールディング調節因子遺伝子の同定
フォールディング調節因子は、新生および異種ポリペプチドのフォールディング、アンフォールディングおよび分解を助長する、全ての細胞中に存在するクラスのタンパク質である。フォールディング調節因子には、シャペロン、シャペロニン、ペプチジル−プロリルシス−トランスイソメラーゼ、およびタンパク質ジスルフィド結合形成に関与するタンパク質がある。異種タンパク質のフォールディングに助力する能力を有する新規な産生菌株を構築するための第一ステップとして、P.fluorescensのゲノムを調べて、宿主細胞のフォールディング調節因子遺伝子を同定した。
【0193】
P.fluorescens MB214のゲノムの6,433の予想ORFのそれぞれを、以下の方法を使用して、それらがフォールディング調節因子をコードしていた確率に関して分析した。対象とするいくつかのフォールディング調節因子は、ゲノム注釈の分析によってDowの研究者により既に同定されている(Ramseierら、2001)。これらの出発タンパク質の相同体は、クエリーとして出発タンパク質およびサブジェクトとして全てのMB214翻訳ORFのデータベースを用いて、タンパク質/タンパク質BLASTを使用して同定した。有意な相同性でクエリータンパク質と適合した翻訳ORFを、さらなる分析用のリストに加えた。有意な相同性は、アラインメントの長さおよび質に基づく人間による判断を考慮に入れてle−30以下のe−値を有するものとしてここでは定義する。この試験の目的は、全ての考えられるフォールディング調節因子を同定し得る機会を最大にすることを全体に含めることであった。
【0194】
より多くのORFを、キーワード「シャペロン」を含有する前の注釈からの、それらの管理された機能に基づいてリストに加えた。最終的に、InterProデータベースバージョン7.0(Mulderら、2005年)に対するタンパク質シグネチャーファミリー検索プログラムInterProScan(Quevillonら、2005年)によってORFを分析した。これらのORFは、InterProScanソフトウェアおよびこれらのファミリーと関係があるGene Ontology(GO)カテゴリー(Gene Ontology Consortium.2004年)によって割り当てられたタンパク質ファミリーであった。これらの自動GO割り当てを使用して、GO用語「GO:0006457生物学的過程:タンパク質フォールディング」または「GO:0003754分子機能:シャペロン活性」を割り当てた全てのORFを、さらなる分析用のリストに加えた。
【0195】
次いでリストを分析して、フォールディング調節因子をコードする低い確率を有していたORFを除去した。再度、この試験の目的は非常に包括的であることであったが、これらの半自動的方法によってリストに割り当てたORFの多くは、限られた基準および人間による判断に基づいて、フォールディング調節因子をコードしていないとして容易に同定することができた。
【0196】
特定のORFを除外した最も一般的な理由は、このORFが実際にフォールディング調節因子である根拠が乏しいため、すなわち、フォールディング調節因子としてのORFの注釈付けの理由が不明確または矛盾のいずれかであった、前の注釈に基づいてリストに割り当てたORFであるためであった。InterProScanは実際に異なるプログラムの集合体であり、これらのプログラムのいくつかは他のプログラムより信頼できると考えられる。ScanRegExpまたはProfileScanコンポーネントのアウトプットのみに基づいてORFをリストに割り当てた場合、したがってそれは除去した。P.fluorescensのフォールディング調節因子の最終的なリストは43のメンバーを有し、それは表1中に示す。
【実施例2】
【0197】
P.fluorescens菌株MB214のゲノム中のプロテアーゼ遺伝子の同定
プロテアーゼは、ペプチド結合を加水分解し全ての生物の生存に必要な酵素である。しかしながら、細胞中でのそれらの役割は、Pfenex Expression Technology(商標)も含めた任意の異種タンパク質発現系中で、プロテアーゼは組換えタンパク質の収率および/または質に弊害をもたらす可能性があることを意味する。ゲノムから除去されたプロテアーゼ遺伝子を有する新規な産生菌株を構築するための第一ステップとして、P.fluorescensのゲノムを調べて、宿主細胞のプロテアーゼ遺伝子を同定した。
【0198】
P.fluorescens MB214のゲノムの6,433の予想ORFのそれぞれを、以下の方法を使用して、それらがプロテアーゼをコードしていた確率に関して分析した。MEROPSデータベースは、Wellcome Trust Sanger Institute、Cambridge、UK(Rawlingsら、2006年、http://merops.sanger.ac.uk)で研究者によって手作業で管理されている。それは、研究室での実験と既知のプロテアーゼファミリーとの相同性の両方によって発見された、プロテアーゼの総合目録である。データベースの長所の1つは、MEROPS階層的分類スキームである。このシステムでは、同じ機能を共有する相同体を複数のファミリーに一緒にグループ分けする。複数のファミリーは、ここでも類似の構造特性に基づく進化的関係に基づいて複数の仲間にグループ分けする。この方法はこのデータベースを大いに利用して、P.fluorescensゲノム内のプロテアーゼ相同体を同定する。
【0199】
MEROPSデータベースとの相同体は、クエリーとしてそれぞれのMB214翻訳ORFおよびサブジェクトとして全てのMEROPSタンパク質のデータベースを用いて、タンパク質/タンパク質BLASTを使用して同定した。有意な相同性でクエリータンパク質と適合した翻訳ORFを、さらなる分析用のリストに加えた。この場合の有意な相同性は、アラインメントの長さおよび質に基づく人間による判断を考慮に入れてle−−60以下のe−値を有するものとしてここでは定義する。このステップによって、リスト用の109の考えられるプロテアーゼを得た。
【0200】
InterProデータベースバージョン7.0(Mulderら、2005年)に対するタンパク質シグネチャーファミリー検索プログラムInterProScan(Quevillonら、2005年)によってもORFを分析した。これらのORFは、InterProScanソフトウェアおよびこれらのファミリーと関係があるGene Ontology(GO)カテゴリー(Gene Ontology Consortium.2004年)によって割り当てられたタンパク質ファミリーであった。これらの自動GO割り当てを使用して、文字列「ペプチダーゼ」、「プロテアーゼ」または「タンパク質分解」を含有したGO名を割り当てた全てのORFを、さらなる分析用のリストに加えた。このステップによって、前のステップで同定されなかった追加の70の考えられるプロテアーゼを得た。
【0201】
より多くのORFを、キーワード「ペプチダーゼ」または「プロテアーゼ」を含有する前の注釈(annotation)(Ramseierら、2001年)からの、それらの管理された機能に基づいてリストに加えた。このステップによって、前のステップで再度同定しなかった32の考えられるプロテアーゼを得た。
【0202】
次いでリストを分析して、プロテアーゼをコードする低い確率を有していたORFを除去した。再度、この試験の目的は非常に包括的であることであったが、これらの半自動的方法によってリストに割り当てたORFの多くは、限られた基準および人間による判断に基づいて、プロテアーゼをコードしていないとして容易に同定することができた。複数の遺伝子を除外した2つの一般的な理由は、特定のORFが実際にプロテアーゼであることの根拠が乏しいため、または特定の遺伝子が、プロテアーゼ自体ではなくプロテアーゼ相同体であることが知られている他のタンパク質と最大の相同性を示したことであった。P.fluorescensのプロテアーゼの最終的なリストは90のメンバーを有し、それは表2中に示す。
【実施例3】
【0203】
in silicoでのフォールディング調節因子およびプロテアーゼタンパク質の細胞内位置の予想
Pfenex Expression Technology(商標)の長所の1つは、特定の異種タンパク質を分離することができる細胞内区画(cellular compartment)を制御するその能力である。したがって、同定した宿主細胞のフォールディング調節因子およびプロテアーゼタンパク質が位置する細胞内区画を予想した。これらの予想を行うために、2つのプログラムを選択した。PsortB2.0は、所与のペプチドの細胞内位置を予想する12の別個のアルゴリズムの結果を組み合わせる。大部分のこれらのアルゴリズムは、クエリータンパク質と既知の細胞内位置のタンパク質の間の相同性の検出に依存する。PsortBは、Hidden Markov Models(HMM)を使用して膜貫通フォールディングドメインまたはI型分泌シグナル配列の存在をそれぞれ検出する、HMMTOPおよびSignalPなどのアルゴリズムも含む。PsortBの結果に加えて、SignalP HMMを使用して、I型分泌シグナル配列の存在を予想した。シグナル配列を検出するが細胞内位置を示す他の具体的情報が得られないとき、PsortBのアウトプットは曖昧である可能性があるので、これは必要であった。これらの場合、PsortBはタンパク質の細胞内局在が知られていないことを示す、何故ならそれは実際、細胞質膜、ペリプラズム、外膜または細胞外区画のいずれか1つに分離する可能性があったからである。しかしながら、タンパク質はおそらく細胞質中に位置しないことを知り、表中においてそれに注目することは十分有益である。したがって表2は、その結果が知られていない場合以外のPsortBアルゴリズムの結果を挙げる。これらの場合、SignalP HMM単独の結果は、シグナルペプチドが検出されたことを示す「シグナルペプチド(Signal Peptide)」、およびシグナルペプチドが検出されなかったことを示す「非分泌性(Non Secretory)」を用いて示す。
【実施例4】
【0204】
フォールディング調節因子の同時過剰発現を可能にするプラスミドの構築
フォールディング調節因子遺伝子を、対象とする異種タンパク質を発現させるために通常使用される別のプラスミドと適合性がある、pCNのプラスミド誘導体(Nietoら、1990年)にクローニングした(Squiresら、2004年;Chewら、2005年)。マンニトール誘導性grpE−dnaKJ含有プラスミドの構築を例示する。他のフォールディング調節因子を、1つの遺伝子または多数の遺伝子としてオペロンで編成されるとき、以下に概説したのと同様にクローニングした。
【0205】
鋳型およびプライマーRC199(5−ATATACTAGTAGGAGGTAACTTATGGCTGACGAACAGACGCA−3’)(配列番号:1)およびRC200(5’ATATTCTAGATTACAGGTCGCCGAAGAAGC−3’)(配列番号:2)として、P.fluorescens MB214(DNeasy;Qiagen、Valencia、CA)から単離したゲノムDNAを利用して、grpE−dnaKJ遺伝子を、製造者の推奨に従いPfuTurbo(Stratagene、La Jolla、CA)を使用して増幅した。生成した4kbのPCR産物はSpeIおよびXbaI(前述のプライマー中で下線を引いた制限部位)で消化し、pDOW1306−6(Schneiderら、2005b)の誘導体であるpDOW2236に連結させて、tacプロモーターの制御下でgrpE−dnaKJオペロンを含有するpDOW2240を作製した。次いでプラスミドpDOW2240をSpeIおよびHindIIIで消化し、生成したgrpE−dnaKJ含有4.0kbのDNA断片はQiaquick(Qiagen、Valencia、CA)を使用してゲル精製し、P.fluorescensマンニトール調節型プロモーター(Schneiderら、2005a)を有するpCNの誘導体であるpDOW2247に連結させ、それをさらにSpeIおよびHindIIIで消化した。生成したプラスミド、pDOW3501は、マンニトールプロモーターの制御下でgrpE−dnaKJオペロンを含有していた。次いでプラスミドpDOW3501を、250ug/mlのウラシルを補ったM9グルコースプレート上での選択によって、DC388および他のウラシル要求性菌株に形質転換した。
【実施例5】
【0206】
プロテアーゼ遺伝子のゲノムが欠失したP.fluorescens菌株の構築
ゲノム欠失の生成を可能にしたプラスミドは、欠失する遺伝子の5’と3’の両方の500〜1000bpのDNA断片の増幅によって構築した。生成する5’PCR産物は典型的には欠失する遺伝子の翻訳開始コドン(ATGまたはGTGまたはTGT)で終わり、一方3’PCR産物は典型的には欠失する遺伝子の停止コドン(TAAまたはTGAまたはTAG)で始まる。これら2つのPCR産物は追加の増幅ステップによって1つに融合し、次いでSOE PCR(Hortonら、1990年)を使用してpDOW1261(図1)(Chewら、2005年)にクローニングした。
【実施例6】
【0207】
P.fluorescens菌株におけるハイスループット増殖および異種タンパク質発現の分析
プラスミドpDOW2787はモノクローナル抗体(mAb)gal2をコードし、その重鎖はPbp分泌リーダーおよびtacプロモーターの制御下で発現する。その軽鎖はOprF分泌リーダーおよびマンニトールプロモーターの制御下で発現する。プラスミドは、対象遺伝子の欠失または同時発現用のフォールディング調節因子を有するpDOW2247のいずれかを有する63菌株、および野生型菌株を含有する5つの対照菌株のコンピテント細胞にエレクトロポレーションした。300rpmで振とうすることによって、グリセロールを含む増殖培地を含有する同一の深さのウェル単位で、細胞を培養した。0.1mMのイソプロピルβ−D−チオガラクトピラノシド(IPTG)および1%マンニトールを用いて、24時間でタンパク質発現を誘導した。誘導後24時間で、アリコートを溶かし、抗原の抗原結合を測定して活性抗体の量を定量化した。その値をOD600で割って細胞特異的活性を測定した。菌株Δprc1、ΔdegP2、ΔLa2、ΔclpP、およびΔprc2、Δprc2、grpEdnaKJ同時発現菌株、Δtig、ΔclpX、およびΔlonはいずれも対照菌株より2.4倍以上高く、これは統計上有意であった(p<0.5)。可溶性細胞分画をΔprc1、ΔdegP2、ΔLa2およびgrpEdnaKJ同時発現菌株から調製し、ウエスタン分析にかけた(図2)。完全構築抗体と一致した大きさを有するバンドを4つの試験菌株において検出したが、対照においては検出しなかった。
(参考文献)
【表8】
【0208】
【表9】
【0209】
本明細書中に述べる全ての刊行物および特許出願は、本発明が関係する技術分野の当業者のレベルを示す。全ての刊行物および特許出願は、それぞれ個々の刊行物または特許出願が参照によって組み込まれることが具体的および個々に示されるのと同じ程度で、参照によって本明細書に組み込まれる。
【0210】
前述の本発明を、理解を明確にする目的で例示および実施例によってある程度詳細に説明してきたが、特定の変化および変更形態を添付の特許請求の範囲内で実施することができることは明らかであろう。
【特許請求の範囲】
【請求項1】
Pseudomonas fluorescens(P.fluorescens)細胞の少なくとも第1および第2の集団を含むアレイであって、各集団が、
a)タンパク質分解に関与する少なくとも1つの標的遺伝子の発現が低減するように遺伝子改変されている少なくとも1つのP.fluorescens細胞集団、
b)タンパク質産生に関与する少なくとも1つの標的遺伝子の発現が増大するように遺伝子改変されている少なくとも1つのP.fluorescens細胞集団、ならびに
c)(a)および(b)に従って遺伝子改変されている少なくとも1つのP.fluorescens細胞集団
からなる群から選択され、アレイ内の各集団が同一でなく、各集団が物理的に相互に離れているアレイ。
【請求項2】
前記タンパク質分解に関与する少なくとも1つの標的遺伝子がプロテアーゼである、請求項1に記載のアレイ。
【請求項3】
前記少なくとも1つのプロテアーゼが表2に記載のプロテアーゼから選択される、請求項2に記載のアレイ。
【請求項4】
前記タンパク質産生に関与する少なくとも1つの標的遺伝子が異種タンパク質の適切なタンパク質転写、翻訳、プロセシングまたは移行に関与する遺伝子である、請求項1に記載のアレイ。
【請求項5】
前記少なくとも1つの標的遺伝子がタンパク質フォールディング調節因子である、請求項4に記載のアレイ。
【請求項6】
前記タンパク質フォールディング調節因子が表1に記載のタンパク質フォールディング調節因子から選択される、請求項5に記載のアレイ。
【請求項7】
前記タンパク質産生に関与する標的遺伝子または前記タンパク質分解に関与する標的遺伝子の発現が増大するように遺伝子改変されていない少なくとも1つのP.fluorescens細胞集団をさらに含む、請求項1から6のいずれか一項に記載のアレイ。
【請求項8】
少なくとも1つの集団が、タンパク質産生に関与する2つ以上の標的遺伝子の発現が増大するように遺伝子改変されている、請求項1から7のいずれか一項に記載のアレイ。
【請求項9】
少なくとも1つの集団が、タンパク質分解に関与する2つ以上の標的遺伝子の発現が低減するように遺伝子改変されている、請求項1から7のいずれか一項に記載のアレイ。
【請求項10】
前記アレイが、前記アレイのハイスループットスクリーニングに役立つフォーマットである、請求項1から9のいずれか一項に記載のアレイ。
【請求項11】
前記フォーマットが96ウェルフォーマットである、請求項10に記載のアレイ。
【請求項12】
対象とする少なくとも1つの異種タンパク質をコードする少なくとも1つの遺伝子を含む発現コンストラクトで形質転換された場合、少なくとも1つの集団が、前記対象とする少なくとも1つの異種タンパク質を発現できる、請求項1から11のいずれか一項に記載のアレイ。
【請求項13】
請求項1から12のいずれか一項に記載のアレイを含むキット。
【請求項14】
対象とする少なくとも1つの異種タンパク質を発現するのに最適なPseudomonas fluorescens(P.fluorescens)宿主細胞を同定する方法であって、
a)P.fluorescens細胞の少なくとも第1および第2の集団を含むアレイであって、各集団が、
i)タンパク質分解に関与する少なくとも1つの標的遺伝子の発現が低減するように遺伝子改変されている少なくとも1つのP.fluorescens細胞集団、
ii)タンパク質産生に関与する少なくとも1つの標的遺伝子の発現が増大するように遺伝子改変されている少なくとも1つのP.fluorescens細胞集団、ならびに
iii)(i)および(ii)に従って遺伝子改変されている少なくとも1つのP.fluorescens細胞集団
からなる群から選択され、アレイ内の各集団が同一でなく、各集団が物理的に相互に離れているアレイを得るステップと、
b)対象とする少なくとも1つの異種タンパク質をコードする少なくとも1つの遺伝子を含む発現カセットを、各集団の少なくとも1つの細胞に導入するステップと、
c)少なくとも1つの細胞集団における前記対象とするタンパク質の発現に十分な条件下で前記細胞を維持するステップと、
d)前記対象とする異種タンパク質がその中で産生される最適な細胞集団を選択するステップと
を含み、前記対象とする異種タンパク質が、前記最適な細胞集団で、アレイにおける他の集団と比較して改善された発現、改善された活性、改善された溶解性、改善された移行、または低減したタンパク分解のうち1つまたは複数を示す方法。
【請求項15】
前記タンパク質分解に関与する少なくとも1つの標的遺伝子がプロテアーゼである、請求項14に記載の方法。
【請求項16】
前記プロテアーゼが表2に記載のプロテアーゼから選択される、請求項15に記載の方法。
【請求項17】
前記タンパク質産生に関与する少なくとも1つの標的遺伝子が、異種タンパク質の適切なタンパク質転写、翻訳、プロセシングまたは移行に関与する遺伝子である、請求項14に記載の方法。
【請求項18】
前記少なくとも1つの標的遺伝子がタンパク質フォールディング調節因子である、請求項17に記載の方法。
【請求項19】
前記タンパク質フォールディング調節因子が表1に記載のタンパク質フォールディング調節因子から選択される、請求項18に記載の方法。
【請求項20】
アレイが、前記タンパク質産生に関与する標的遺伝子または前記タンパク質分解に関与する標的遺伝子の発現が増大するように遺伝子改変されていない少なくとも1つのP.fluorescens細胞集団をさらに含む、請求項14から19のいずれか一項に記載の方法。
【請求項21】
少なくとも1つの集団が、タンパク質産生に関与する2つ以上の標的遺伝子の発現が増大するように遺伝子改変されている、請求項14から20のいずれか一項に記載の方法。
【請求項22】
少なくとも1つの集団が、タンパク質分解に関与する2つ以上の標的遺伝子の発現が低下するように遺伝子改変されている、請求項14から20のいずれか一項に記載の方法。
【請求項23】
前記アレイが、前記アレイのハイスループットスクリーニングに役立つフォーマットである、請求項14から22のいずれか一項に記載の方法。
【請求項24】
前記フォーマットが96ウェルフォーマットである、請求項23に記載の方法。
【請求項25】
前記発現カセットが、前記対象とする異種タンパク質に作動可能に連結したシグナリングペプチドをさらに含む、請求項14に記載の方法。
【請求項26】
前記シグナリングペプチドが分泌シグナルペプチドである、請求項19に記載の方法。
【請求項27】
前記シグナリングペプチドがPseudomonas fluorescens宿主細胞に由来する、請求項19に記載の方法。
【請求項28】
対象とする異種タンパク質の発現に最適なPseudomonas fluorescens(P.fluorescens)宿主細胞を同定する方法であって、
a)P.fluorescens細胞の少なくとも第1および第2の集団を含むアレイであって、各集団が、タンパク質分解に関与する少なくとも1つのタンパク質の発現が低減するように遺伝子改変されており、アレイ内の各集団が同一でなく、各集団が物理的に相互に離れているアレイを得るステップと、
b)細胞を溶解するステップと、
c)各集団の細胞の溶解物を、対象とする異種タンパク質と接触させるステップと、
d)最適な細胞集団を選択するステップと
を含み、前記最適な細胞集団が、アレイにおける他の集団と比較して、ステップ(c)において対象とする異種タンパク質の分解レベルが低下した集団である方法。
【請求項1】
Pseudomonas fluorescens(P.fluorescens)細胞の少なくとも第1および第2の集団を含むアレイであって、各集団が、
a)タンパク質分解に関与する少なくとも1つの標的遺伝子の発現が低減するように遺伝子改変されている少なくとも1つのP.fluorescens細胞集団、
b)タンパク質産生に関与する少なくとも1つの標的遺伝子の発現が増大するように遺伝子改変されている少なくとも1つのP.fluorescens細胞集団、ならびに
c)(a)および(b)に従って遺伝子改変されている少なくとも1つのP.fluorescens細胞集団
からなる群から選択され、アレイ内の各集団が同一でなく、各集団が物理的に相互に離れているアレイ。
【請求項2】
前記タンパク質分解に関与する少なくとも1つの標的遺伝子がプロテアーゼである、請求項1に記載のアレイ。
【請求項3】
前記少なくとも1つのプロテアーゼが表2に記載のプロテアーゼから選択される、請求項2に記載のアレイ。
【請求項4】
前記タンパク質産生に関与する少なくとも1つの標的遺伝子が異種タンパク質の適切なタンパク質転写、翻訳、プロセシングまたは移行に関与する遺伝子である、請求項1に記載のアレイ。
【請求項5】
前記少なくとも1つの標的遺伝子がタンパク質フォールディング調節因子である、請求項4に記載のアレイ。
【請求項6】
前記タンパク質フォールディング調節因子が表1に記載のタンパク質フォールディング調節因子から選択される、請求項5に記載のアレイ。
【請求項7】
前記タンパク質産生に関与する標的遺伝子または前記タンパク質分解に関与する標的遺伝子の発現が増大するように遺伝子改変されていない少なくとも1つのP.fluorescens細胞集団をさらに含む、請求項1から6のいずれか一項に記載のアレイ。
【請求項8】
少なくとも1つの集団が、タンパク質産生に関与する2つ以上の標的遺伝子の発現が増大するように遺伝子改変されている、請求項1から7のいずれか一項に記載のアレイ。
【請求項9】
少なくとも1つの集団が、タンパク質分解に関与する2つ以上の標的遺伝子の発現が低減するように遺伝子改変されている、請求項1から7のいずれか一項に記載のアレイ。
【請求項10】
前記アレイが、前記アレイのハイスループットスクリーニングに役立つフォーマットである、請求項1から9のいずれか一項に記載のアレイ。
【請求項11】
前記フォーマットが96ウェルフォーマットである、請求項10に記載のアレイ。
【請求項12】
対象とする少なくとも1つの異種タンパク質をコードする少なくとも1つの遺伝子を含む発現コンストラクトで形質転換された場合、少なくとも1つの集団が、前記対象とする少なくとも1つの異種タンパク質を発現できる、請求項1から11のいずれか一項に記載のアレイ。
【請求項13】
請求項1から12のいずれか一項に記載のアレイを含むキット。
【請求項14】
対象とする少なくとも1つの異種タンパク質を発現するのに最適なPseudomonas fluorescens(P.fluorescens)宿主細胞を同定する方法であって、
a)P.fluorescens細胞の少なくとも第1および第2の集団を含むアレイであって、各集団が、
i)タンパク質分解に関与する少なくとも1つの標的遺伝子の発現が低減するように遺伝子改変されている少なくとも1つのP.fluorescens細胞集団、
ii)タンパク質産生に関与する少なくとも1つの標的遺伝子の発現が増大するように遺伝子改変されている少なくとも1つのP.fluorescens細胞集団、ならびに
iii)(i)および(ii)に従って遺伝子改変されている少なくとも1つのP.fluorescens細胞集団
からなる群から選択され、アレイ内の各集団が同一でなく、各集団が物理的に相互に離れているアレイを得るステップと、
b)対象とする少なくとも1つの異種タンパク質をコードする少なくとも1つの遺伝子を含む発現カセットを、各集団の少なくとも1つの細胞に導入するステップと、
c)少なくとも1つの細胞集団における前記対象とするタンパク質の発現に十分な条件下で前記細胞を維持するステップと、
d)前記対象とする異種タンパク質がその中で産生される最適な細胞集団を選択するステップと
を含み、前記対象とする異種タンパク質が、前記最適な細胞集団で、アレイにおける他の集団と比較して改善された発現、改善された活性、改善された溶解性、改善された移行、または低減したタンパク分解のうち1つまたは複数を示す方法。
【請求項15】
前記タンパク質分解に関与する少なくとも1つの標的遺伝子がプロテアーゼである、請求項14に記載の方法。
【請求項16】
前記プロテアーゼが表2に記載のプロテアーゼから選択される、請求項15に記載の方法。
【請求項17】
前記タンパク質産生に関与する少なくとも1つの標的遺伝子が、異種タンパク質の適切なタンパク質転写、翻訳、プロセシングまたは移行に関与する遺伝子である、請求項14に記載の方法。
【請求項18】
前記少なくとも1つの標的遺伝子がタンパク質フォールディング調節因子である、請求項17に記載の方法。
【請求項19】
前記タンパク質フォールディング調節因子が表1に記載のタンパク質フォールディング調節因子から選択される、請求項18に記載の方法。
【請求項20】
アレイが、前記タンパク質産生に関与する標的遺伝子または前記タンパク質分解に関与する標的遺伝子の発現が増大するように遺伝子改変されていない少なくとも1つのP.fluorescens細胞集団をさらに含む、請求項14から19のいずれか一項に記載の方法。
【請求項21】
少なくとも1つの集団が、タンパク質産生に関与する2つ以上の標的遺伝子の発現が増大するように遺伝子改変されている、請求項14から20のいずれか一項に記載の方法。
【請求項22】
少なくとも1つの集団が、タンパク質分解に関与する2つ以上の標的遺伝子の発現が低下するように遺伝子改変されている、請求項14から20のいずれか一項に記載の方法。
【請求項23】
前記アレイが、前記アレイのハイスループットスクリーニングに役立つフォーマットである、請求項14から22のいずれか一項に記載の方法。
【請求項24】
前記フォーマットが96ウェルフォーマットである、請求項23に記載の方法。
【請求項25】
前記発現カセットが、前記対象とする異種タンパク質に作動可能に連結したシグナリングペプチドをさらに含む、請求項14に記載の方法。
【請求項26】
前記シグナリングペプチドが分泌シグナルペプチドである、請求項19に記載の方法。
【請求項27】
前記シグナリングペプチドがPseudomonas fluorescens宿主細胞に由来する、請求項19に記載の方法。
【請求項28】
対象とする異種タンパク質の発現に最適なPseudomonas fluorescens(P.fluorescens)宿主細胞を同定する方法であって、
a)P.fluorescens細胞の少なくとも第1および第2の集団を含むアレイであって、各集団が、タンパク質分解に関与する少なくとも1つのタンパク質の発現が低減するように遺伝子改変されており、アレイ内の各集団が同一でなく、各集団が物理的に相互に離れているアレイを得るステップと、
b)細胞を溶解するステップと、
c)各集団の細胞の溶解物を、対象とする異種タンパク質と接触させるステップと、
d)最適な細胞集団を選択するステップと
を含み、前記最適な細胞集団が、アレイにおける他の集団と比較して、ステップ(c)において対象とする異種タンパク質の分解レベルが低下した集団である方法。
【図1A】


【図1B】

【図2】
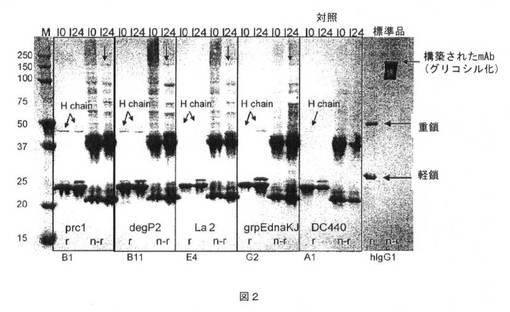
【図1B】
【図2】
【公表番号】特表2010−524506(P2010−524506A)
【公表日】平成22年7月22日(2010.7.22)
【国際特許分類】
【出願番号】特願2010−506503(P2010−506503)
【出願日】平成20年4月25日(2008.4.25)
【国際出願番号】PCT/US2008/061483
【国際公開番号】WO2008/134461
【国際公開日】平成20年11月6日(2008.11.6)
【出願人】(502141050)ダウ グローバル テクノロジーズ インコーポレイティド (1,383)
【Fターム(参考)】
【公表日】平成22年7月22日(2010.7.22)
【国際特許分類】
【出願日】平成20年4月25日(2008.4.25)
【国際出願番号】PCT/US2008/061483
【国際公開番号】WO2008/134461
【国際公開日】平成20年11月6日(2008.11.6)
【出願人】(502141050)ダウ グローバル テクノロジーズ インコーポレイティド (1,383)
【Fターム(参考)】
[ Back to top ]