
新規なポリペプチドESDN、その製造方法、ESDNをコードするcDNA、そのcDNAからなるベクター、そのベクターで形質転換された宿主細胞、ESDNの抗体、およびESDNまたは抗体を含有する薬学的組成物
【構成】新規なポリペプチドESDN、その製造方法、ESDNをコードするcDNA、そのcDNAからなるベクター、そのベクターで形質転換された宿主細胞、ESDNの抗体、およびESDNまたは抗体を含有する薬学的組成物。
【効果】本発明のESDN蛋白は、冠動脈細胞ならびに平滑筋細胞で発現され、またバルーン傷害した動脈の血管平滑筋や総頸動脈の中膜で発現されることから、循環器領域におけるPTCA後の再狭窄や動脈硬化の治療に有用である。
【効果】本発明のESDN蛋白は、冠動脈細胞ならびに平滑筋細胞で発現され、またバルーン傷害した動脈の血管平滑筋や総頸動脈の中膜で発現されることから、循環器領域におけるPTCA後の再狭窄や動脈硬化の治療に有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規なポリペプチドである血管内皮細胞・平滑筋細胞由来ニューロピリン様分子(Endothelial and Smooth muscle cell-Derived Neuropilin-like molecule 、以下、ESDNと省略する。)、その製造方法、ESDNをコードするcDNA、そのcDNAからなるベクター、そのベクターで形質転換された宿主細胞、ESDNの抗体、ESDNまたは抗体を含有する薬学的組成物、ESDNを測定する方法、ESDNを測定する試薬およびESDNを用いたスクリーニング方法に関する。
【背景技術】
【0002】
血管内皮増殖因子(VEGF)のクローニング以来、血管形成を支配する複雑な細胞外シグナル伝達ネットワークの解明が画期的になされてきた。このリガンドやFlk−1/VEGFR2やFlt−1/VEGFR1といったチロシンキナーゼのノックアウトマウスの研究が、血管形成、発生の第一ステップに関する最初の重要点であった。
VEGF受容体−2(VEGFR2)は、内皮細胞形成において必要であり、またこの遺伝子の欠損は、血液細胞群あるいは組織化された血管の不足を招き発生9.5日目に胎生死につながる。しかしながら、RT−PCRを用いた初期マーカーの解析により初期の造血や内皮の前駆細胞が実際にVEGFR2非存在下において造られるという報告から、VEGFR2は血球芽細胞の形成においては必要不可欠ではないが、系統のその後の増加には必要であることを示唆している。
VEGF受容体−1(VEGFR1)ノックアウトマウスもまた発生9.5日目に死ぬが、それらの内皮細胞の分化は直接影響を受けていない。その代わりに、主に間充識の血球芽細胞の移行が不足していると考えられ、過密になった内皮前駆細胞が、脈管系の重篤な組織破壊を導く。
VEGFノックアウトマウスもまた発生9.5日目に著しい減少を伴って死ぬが、内皮細胞の分化を欠いていない。つまり、本質的にはVEGFR1ノックアウトマウスと同様であるが、より軽度なフェノタイプを示す。この遺伝子における目立った特長は、ヘテロ接合体の遺伝子欠失でも発生11.5日目に胎生致死になることである。このことは胎生期においてVEGFに対する著しく厳格な容量依存性が存在することを示している。
その他のレセプターチロシンキナーゼもまた、Tie−2やそのリガンドangiopoietin−1といったように血管内皮細胞に特異的に発現をしていて、血管形成における後期、血管新生のステップに関与している。これら遺伝子の破壊は、脈管形成には影響を及ぼさないが、血管のリモデリングに障害を与え、発生10.5日目に胎生死につながる。それらは、内皮細胞とそれを取り巻く血管平滑筋細胞(VSMC)あるいは間充識細胞間の相互作用において役割を担っている。これらの脈管特有のシステムに付け加えて、PDGF−BBあるいはTGF−βそしてそれらのレセプターもまた内皮細胞とそれを取り巻く細胞間で同様な役割を担っている。脈管システムに対するニューロピリン−1(NP−1;neuropilin-1)の関係の同定は、この分野の新局面を紹介した。NP−1は初め発達した神経系を特異的に認識するモノクローナル抗体によって認識される抗原としてクローニングされた。
そして、セマフォリン3A(semaphorin 3A;sema 3A)のレセプターとしての同定は、軸索形成において一躍注目を浴びるようになった。またその他のVEGFレセプターの探索がVEGFR2のレセプターでもあるNP−1の発見につながった。そしてそれは、一つのVEGFアイソフォーム(isoform)への結合の増加だけでなく、内皮細胞の遊走やおそらくマイトシス(mitosis)も同様に促進する。このことが同定される前は、NP−1を強制発現させたキメラマウスは、肥大した脈管を形成することが示された。NP−1ノックアウトマウスは、中枢神経系においては低い血管質を呈し、また大きな脈管の異形をも示した。一方sema 3Aノックアウトマウスでは血管の異常性の存在や非存在は報告されていないが、そのフェノタイプがNP−1強制発現マウスにおいても観察された薄い心筋層を呈することは興味深い。
Eph/ephrinシステムは、別の例であり、初めに神経科学において有名になった。しかし、後に現在までには唯一の動脈と静脈を識別する表面マーカーとして同定されている。それらもまた内皮細胞との接点である周囲の間充識細胞に発現を示している。これらの役割を持った分子以外にも、我々は更に多くの細胞外シグナル伝達分子が血管の分野においては存在すると想定した。そしてこの事の解明が、この複雑なシステムのさらなる理解に貢献できるであろうと考えた。
【0003】
従来、ある特定のポリペプチドまたはそれをコードするcDNAを得ようとする場合、組織や細胞培養液中に目的とする生物活性を確認し、次いでポリペプチドの単離精製を経て、遺伝子をクローニングするという方法、あるいはその生物活性を指標として遺伝子を発現クローニングする方法が一般的に用いられてきた。しかし、生体内生理活性ポリペプチドは、多様な生物活性を有している場合が多いので、あるひとつの活性を指標にして遺伝子をクローニングした結果、それが既知のポリペプチドと同一であることが後になって判明するという事例が増えている。また、微量しか産生されなかったり、特別な生理条件でのみ発現する因子も多く、そのことが単離、精製および生物活性の確認を困難なものとしている。
【0004】
本発明者らは、これまで造血系や免疫系で働く増殖分化因子の遺伝子のクローニングを研究してきた。そして、増殖分化因子(例えば、各種サイトカイン等)のような分泌蛋白質やそのレセプターのような膜蛋白質(以下、これらをまとめて分泌蛋白質等と呼ぶ。)の大部分がそのN末端にシグナルペプチドと呼ばれる配列を有していることに着目して、シグナルペプチドをコードする遺伝子を効率的かつ選択的にクローニングする方法を鋭意検討した。その結果、動物細胞を用いて、シグナルペプチドの有無を簡単に検索できる方法(シグナルシークエンストラップ(SST)法)を見出した(特許第3,229,590号参照)。さらに同じ概念のもとに、酵母を用いてさらに大量かつ簡便にシグナルペプチドをコードする遺伝子を単離する方法(酵母SST法)も開発された(米国特許No.5,536,637)。
【発明の開示】
【0005】
本発明者らは,循環器領域におけるPTCA後の再狭窄や動脈硬化での病的血管平滑筋で重要な役割を果たしている有益な新規因子(ポリペプチド)、特に分泌シグナルを有する分泌蛋白質および膜蛋白質に着目してそれを見出すべく、鋭意検討を行なった。
本発明者らは、血管細胞に対して新たに修正したSSTスクリーニングを行ない、新規トランスメンブラン(transmembrane)タンパクのクローニングに成功した。本発明の蛋白ESDNは、新規のI型transmembraneタンパク質であり、ニューロピリン(neuropilin)様の特徴的ドメインを持つものである。
【0006】
本発明のESDN蛋白は、後に詳述されるように、冠動脈細胞ならびに平滑筋細胞で発現され、またバルーン傷害した動脈の血管平滑筋や総頸動脈の中膜で発現されることから、循環器領域におけるPTCA後の再狭窄や動脈硬化の治療に有用であると考えられる。
【0007】
本発明が提供するcDNA配列は、配列番号2で示されるヒトESDNクローンとして同定され、冠動脈初代培養内皮細胞および平滑筋細胞から作製したcDNAライブラリーより、酵母SST法を使用して得た情報をもとに単離された。配列番号3で示されるヒトESDNクローンは分泌蛋白質(ここではヒトESDN蛋白として表される。)をコードする完全なcDNA配列を含む全長鎖cDNAである。
【0008】
本発明が提供するcDNA配列は、配列番号6で示されるマウスESDNクローンとして同定され、酵母SST法を使用して得た情報をもとに、マウスcDNAライブラリーより単離された。配列番号5で示されるマウスESDNクローンは分泌蛋白質(ここではマウスESDN蛋白として表される)をコードする完全なcDNA配列を含む全長鎖cDNAである。
【0009】
本発明が提供する配列番号9で示されるcDNA配列は、ラットESDNクローンとして同定され、酵母SST法を使用して得た情報をもとに、ラットcDNAライブラリーより単離された。配列番号8で示されるマウスESDNクローンは分泌蛋白質(ここではマウスESDN蛋白として表される)をコードする完全なcDNA配列を含む全長鎖cDNAである。
【0010】
核酸配列データベースに登録されている既知の核酸配列に対してジンバンク(GenBank)およびNCBIを用いたBLASTN、FASTAおよびUNIGENEなどにより、またアミノ酸配列データベースに登録されている既知のポリペプチドのアミノ酸配列に対してBLASTP、Fly Database、SwissProt などにより検索をした結果、本発明ポリペプチドであるラットESDNおよびそれらをコードする核酸配列と一致する配列はなかった。このことから、本発明のポリペプチドは新規な分泌蛋白質であることが判明した。
【0011】
本発明は、
1.実質的に純粋な形である配列番号2、5または8で示されるアミノ酸配列からなるポリペプチド、またはそのホモローグ、そのフラグメントまたはそのフラグメントのホモローグからなるポリペプチド。
2.配列番号2、5または8で示されるアミノ酸配列からなる前項1記載のポリペプチド、
3.前項1または2に記載されたポリペプチドをコードするcDNA、
4.配列番号3、6または9で示される塩基配列を有する前項3記載のcDNA、またはその配列に選択的にハイブリダイズするフラグメントからなるcDNA。
5.前項3または4に記載のcDNAからなる複製または発現ベクター、
6.前項5記載の複製または発現ベクターで形質転換された宿主細胞、
7.前項1または2に記載されたポリペプチドを発現させるための条件下、前項6記載の宿主細胞を培養することを特徴とするからなる前項1または2に記載のポリペプチドの製造方法、
8.前項1または2に記載されたポリペプチドのポリクローナルまたはモノクローナル抗体、
9.前項1または2に記載されたポリペプチドまたは前項8記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とする薬学的組成物、
10.前項1または2に記載されたポリペプチドまたは前項8記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とするPTCA後の再狭窄の治療に有効な薬学的組成物、
11.前項1または2に記載されたポリペプチドまたは前項8記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とする前項10記載の動脈硬化の治療に有効な薬学的組成物、
12.前項1または2に記載されたポリペプチドの測定方法、
13.前項8記載の抗体を用いることを特徴とする前項1または2に記載されたポリペプチドの免疫化学的測定方法、
14.前項12または13記載の方法を用いることを特徴とする前項1または2に記載されたポリペプチドを検出する試薬、
15.前項12または13記載の方法を用いることを特徴とするPTCA後の再狭窄を検査する試薬、
16.前項12または13記載の方法を用いることを特徴とする動脈硬化を検査する試薬、および
17.前項1または2に記載されたポリペプチドを用いることを特徴とする、該ポリペプチドに対するアンタゴニストまたはアゴニストとしての活性を有する化合物をスクリーニングする方法に関する。
【0012】
本発明者らは、該ポリペプチドが血管平滑筋の収縮形質よりも合成形質において発現が亢進することを確認した。そのため、該ポリペプチドを用いて循環器領域におけるPTCA後の再狭窄や動脈硬化の治療に有用であると考えられる。
また、本発明のポリペプチドは、実験に於いて、細胞増殖抑制作用を示したこと、VEGFと構造が類似であることから、以下の作用を有することが考えられる。
「サイトカイン活性および細胞増殖/分化活性」
本発明の蛋白は、強制発現系において細胞増殖を抑制したことから、サイトカイン活性および細胞増殖(誘導あるいは阻害)/分化活性(誘導あるいは阻害)を示す可能性、あるいはある細胞集団に他のサイトカインの産生を誘導あるいは抑制すると考えられる。
「免疫刺激/抑制活性」
本発明の蛋白は、免疫刺激活性および免疫抑制活性も示すと考えられる。例えば、Tリンパ球およびBリンパ球あるいはどちらか一方の成長および増殖を制御(刺激あるいは抑制)することや、同様にNK細胞や他の集団の細胞傷害性活性に影響を与えることによって、様々な免疫不全および疾患の治療に効果を示すと考えられる。特に腫瘍血管新生によって引き起こされるリンパ行性癌転移の抑制において効果を示すと考えられる。
「虚血性血管新生抑制活性」
本発明の蛋白は、糖尿病の合併症である網膜症を抑制すると考えられる。VEGFは虚血誘導によって強力な血管新生作用を持つことが知られている。糖尿病網膜症の血管新生も網膜無血管領域が形成された後に起こる虚血性血管新生である。本発明の蛋白は、ニューロピリン(neuropilin)との構造の類似性から新規のVEGF受容体として血管増殖を抑制するように働いている可能性が考えられる。このことから糖尿病網膜症において効果を示すと考えられる。
【発明を実施するための最良の形態】
【0013】
ハイブリダイズするcDNAには、上記配列の相補配列も含まれる。ハイブリダイズは、ストリンジェントな条件で行なうことが好ましい。
【0014】
実質的に純粋な形である配列番号2、5または8で示されるアミノ酸配列からなるポリペプチドとは、一般に、生産時のポリペプチドの90%以上、例えば、95、98または99%が配列番号2、5または8で示されるアミノ酸配列からなるポリペプチドであることを意味する。
【0015】
配列番号2、5または8で示されるアミノ酸配列からなるポリペプチドのホモローグとは、一般に少なくとも20個、好ましくは少なくとも30個、例えば40、60、80または100個の連続したアミノ酸領域で、少なくとも70%、好ましくは少なくとも80または90%、より好ましくは95%以上相同性であるものであり、そのようなホモローグは、以後本発明のポリペプチドとして記載される。
【0016】
さらに、配列番号2、5または8で示されるアミノ酸配列からなるポリペプチドのフラグメント、またはそれらのホモローグのフラグメントとは、少なくとも10アミノ酸、好ましくは少なくとも15アミノ酸、例えば20、25、30、40、50または60アミノ酸部分を意味する。
【0017】
配列番号3、6または9で示される塩基配列からなるcDNAに選択的にハイブリダイズするcDNAとは、一般に、少なくとも20個、好ましくは少なくとも30個、例えば40、60、80または100個の連続した塩基配列領域で、少なくとも70%、好ましくは少なくとも80または90%、より好ましくは95%以上相同性であるものであり、そのようなcDNAは、以後本発明のcDNAとして記載される。
【0018】
配列番号3、6または9で示される塩基配列からなるcDNAのフラグメントとは、少なくとも10塩基、好ましくは少なくとも15塩基、例えば20、25、30または40塩基部分を意味し、そのようなフラグメントも本発明のcDNAに含まれる。
【0019】
さらに、本発明には、本発明のcDNAからなる複製または発現ベクターが含まれる。ベクターとしては、例えば、ori領域と、必要により上記cDNAの発現のためのプロモーター、プロモーターの制御因子などからなるプラスミド、ウィルスまたはファージベクターが挙げられる。ベクターはひとつまたはそれ以上の選択的マーカー遺伝子、例えばアンピシリン耐性遺伝子を含んでいてもよい。ベクターは、イン・ビトロ(in vitro)において、例えばcDNAに対応するRNAの製造、宿主細胞の形質転換に用いることができる。
【0020】
さらに、本発明には、配列番号3、6また9で示される塩基配列、またはそれらのオープンリーディングフレームからなるcDNAを含む本発明のcDNAを複製または発現させるためのベクターで形質転換された宿主細胞も含まれる。細胞としては、例えば細菌、酵母、昆虫細胞または哺乳動物細胞が挙げられる。
【0021】
さらに、本発明には、本発明のポリペプチドを発現させるための条件下で、本発明の宿主細胞を培養することからなる本発明のポリペプチドの製造方法も含まれる。培養は、本発明のポリペプチドが発現し、宿主細胞より製造される条件下で行なわれることが好ましい。
【0022】
本発明のcDNAは、上記のようなベクターのアンチセンス領域に挿入することでアンチセンスRNAを製造することもできる。このようなアンチセンスRNAは、細胞中の本発明のポリペプチドのレベルを制御することに用いることもできる。
【0023】
本発明は、本発明におけるポリペプチドのモノクローナルまたはポリクローナル抗体も含む。さらに本発明におけるポリペプチドのモノクローナルまたはポリクローナル抗体の製造方法も含む。モノクローナル抗体は、本発明のペプチドまたは、その断片を抗原として用い、通常のハイブリドーマの技術により製造することができる。ポリクローナル抗体は、宿主動物(例えば、ラットやウサギ等)に本発明のポリペプチドを接種し、免疫血清を回収する、通常の方法により製造することができる。
【0024】
本発明には、本発明のポリペプチド、その抗体と薬学的に許容される賦形剤および/または担体を含有する薬学的組成物も含まれる。
【0025】
本発明のポリペプチドとしては、配列番号2、5または8で示されたアミノ酸配列からなるもの以外に、その一部が欠損したもの(例えば、配列番号2中、生物活性の発現に必須な部分だけからなる成熟ポリペプチド等)、その一部が他のアミノ酸と置換したもの(例えば、物性の類似したアミノ酸に置換したもの)、および上記本発明のポリペプチドに他のアミノ酸が付加または挿入されたものも含まれる。
【0026】
よく知られているように、ひとつのアミノ酸をコードするコドンは1〜6種類(例えば、Metは1種類、Leuは6種類)知られている。従って、ポリペプチドのアミノ酸配列を変えることなくcDNAの塩基配列を変えることができる。
【0027】
本発明のcDNAには、配列番号2、5または8で示されるポリペプチドをコードするすべての塩基配列群が含まれる。塩基配列を変えることによって、ポリペプチドの生産性が向上することがある。
【0028】
配列番号3、6または9で特定されるcDNAは、配列番号2、5または8で示されるポリペプチドをコードする塩基配列群の一態様であり、天然型配列を表わす。
【0029】
配列番号3、6または9で示される塩基配列からなるcDNAの作製は、以下の方法に従って行なわれる。
【0030】
はじめに酵母SST法(米国特許No.5,536,637に記載)の概要について説明する。
サッカロマイセス・セロヴィシエ(Saccharomyces cerevisiae)などの酵母がショ糖またはラフィノースをエネルギー源や炭素源として利用するためにはインベルターゼを培地中に分泌しなければならない(インベルターゼはラフィノースをショ糖とメリビオースに、ショ糖をフルクトースとグルコースに分解する酵素である。)。また数多くの既知の哺乳類のシグナルペプチドは酵母のインベルターゼを分泌させうることが知られている。
【0031】
これらの知見から、酵母のインベルターゼの分泌を可能にする新規のシグナルペプチドを哺乳類のcDNAライブラリーからラフィノース培地上での酵母の生育を指標にスクリーニングする方法として本方法は開発された。
【0032】
翻訳開始点ATGを削除した非分泌型のインベルターゼ遺伝子SUC2(GENBANK accession No.V01311) を酵母の発現ベクターに組み込んで酵母SST用ベクターpSUC2を作製した。発現ベクターには、AAH5プラスミド(Gammerer, Methods in Enzymol. 101,192-201,1983)由来の発現用プロモーター(ADHプロモーター)およびターミネーター(ADHターミネーター)が組み込まれ、酵母複製起点としては2μori、酵母選択マーカーにはTRP1、大腸菌複製起点としてはColE1 ori、大腸菌薬剤耐性マーカーにはアンピシリン耐性遺伝子がそれぞれ組み込まれている。そのSUC2遺伝子の上流に哺乳類のcDNAを組み込んで、酵母SSTcDNAライブラリーを調製した。このライブラリーを分泌型インベルターゼを欠損している酵母に形質転換した。
【0033】
組み込まれた哺乳類cDNAがシグナルペプチドをコードしている場合、酵母で発現されたインベルターゼに対しても分泌作用を持つと考えられ、その結果ラフィノース培地上での生育が可能となる。そこで出現したコロニーから酵母を培養してプラスミドを調製し、インサートcDNAの塩基配列を決定することによって、新規シグナルペプチドの検索を迅速かつ容易にした。
【0034】
酵母SSTcDNAライブラリーの作製は、
(1)対象となる細胞よりmRNAを単離し、特定の制限酵素(酵素I)サイトを連結したランダムプライマーを用いて二本鎖cDNAを合成し、
(2)酵素Iとは異なる特定の制限酵素(酵素II)サイトを含むアダプターを連結して、酵素Iで消化した後、適当なサイズで分画し、
(3)酵母発現ベクター内のシグナルペプチドを削除したインベルターゼ遺伝子の上流に得られたcDNA断片を連結し、形質転換する工程よりなる。
【0035】
各工程を詳しく説明すると、工程(1)では、対象となる哺乳類の臓器や細胞株などより、必要により適当な刺激剤で刺激した後、公知の方法(以下、公知の方法は特に記載がなければMolecular Cloning(Sambrook, J., Fritsch, E. F.および Maniatis, T. 著、Cold Spring Harbor Laboratory Press より1989年に発刊)またはCurrent Protocol in Molecular Biology (F.M.Ausubelら編、John Wiley & Sons, Inc. より発刊) に記載の方法に従って行なわれる。)に従ってmRNAの単離が行なわれる。
【0036】
対象となる組織としては、マウス胎児心臓が挙げられる。ランダムプライマーを用いる二本鎖cDNAの合成は公知の方法により行なわれる。
【0037】
アダプターに連結される制限酵素(酵素I)サイトと次の工程(2)で用いられる制限酵素(酵素II)サイトは、互いに異なるものであれば何を用いてもよい。好ましくは、酵素IとしてXhoI、酵素IIとしてはEcoRIが用いられる。
【0038】
工程(2)ではT4DNAポリメラーゼで末端を平滑化し、酵素IIアダプターを連結した後、酵素Iで消化し、アガロース電気泳動(AGE)により300〜800bpのcDNAを分画する。酵素IIは、前記したように酵素Iと異なるものなら何でもよい。
【0039】
工程(3)は、酵母発現用プラスミドベクターに連結されたシグナルペプチドを削除したインベルターゼの遺伝子の上流に(2)で得られたcDNA断片を組み込んで大腸菌に形質転換する工程である。ここで酵母発現用プラスミドベクターとしては、種々のものが知られているが、例えば、大腸菌内でも機能するYEp24などが用いられるが、好適には前述したプラスミドpSUC2が用いられる。
【0040】
形質転換のための宿主大腸菌株はすでに多くのものが知られており、好ましくはDH10Bのコンピテントセルである。また形質転換方法は公知の方法のいずれを用いてもよいが、好ましくはエレクトロポレーション法により行なわれる。形質転換体は公知の方法により培養され、酵母SST用のcDNAライブラリーが得られる。
【0041】
このcDNAライブラリーでは、すべてのクローンcDNA断片が導入されているわけではないし、またすべてが未知の(新規の)シグナルペプチドをコードする遺伝子断片とは限らない。そこで、次に該ライブラリーから未知のシグナルペプチドをコードする遺伝子断片をスクリーニングする必要がある。
【0042】
遺伝子をもたない酵母、サッカロマイセス・セロヴィシエ(Saccharomycs cerevisiae)(例えば、YT455株など)またはインベルターゼ遺伝子を人為的に欠損させた株(公知の方法に従い作製可能)に、該cDNAライブラリーを導入し、シグナルペプチドをコードする配列を有する断片のスクリーニングを行なう。酵母の形質転換は公知の方法、例えば酢酸リチウム法によって行なわれる。形質転換体を選択培地で生育後、ラフィノースを炭素源とする培地に移し、生育可能なコロニーを選択し、プラスミドを回収する。ラフィノースを炭素源として酵母が生育したということは、ライブラリー中に何らかの分泌蛋白質のシグナルペプチドが組み込まれていたことを示している。
【0043】
次に、単離した陽性クローンについて、塩基配列を決定し、未知の蛋白質をコードすることが明らかになったcDNAについては、それをプローブとして全長クローンを単離し、全長の塩基配列を決定することができる。これらの操作は、当業者にとってすべて公知の方法で行なわれる。
【0044】
配列番号3、6または9で示される塩基配列が、一部、好ましくは全てが確定されると哺乳類に存在する本発明の蛋白質をコードするcDNAもしくは本発明蛋白質のホモローグおよびサブセットをコードするcDNAを得ることができる。適当な塩基配列を有するオリゴヌクレオチドを合成し、それを用いて哺乳類由来のcDNAライブラリーあるいはmRNAからPCR法により、あるいは適当な塩基配列の断片をプローブとしてハイブリダイズさせることにより、他の哺乳類cDNAライブラリーあるいは該ゲノムライブラリーから、他の哺乳類型の当該蛋白質をコードするcDNAを得ることができる。
【0045】
このようにして得られたcDNAが、SSTで得られたcDNA断片の塩基配列(またはその相同配列)を含んでいるならばシグナルペプチドをコードしていることになるので、該cDNAが全長、またはほぼ全長であることは明らかである(シグナルペプチドは例外なく蛋白質のN末端に存在することから、cDNAのオープンリーディングフレームの5'末端にコードされている。)。
さらに公知の方法に従い、該cDNAをプローブとしてノザン(Northern)解析によって全長の確認をしてもよい。ハイブリダイズしたバンドから得られるmRNAのサイズと該cDNAのサイズを比較し、ほぼ同じであれば該cDNAはほぼ全長であると考えられる。
【0046】
本発明は、開示されたタンパクの全長型並びに成熟型の両方を提供する。それらのタンパクの全長型は、配列番号2、5または8で示される塩基配列の翻訳されるアミノ酸配列で同定される。それらの成熟タンパクは、適当な哺乳類の細胞あるいはその他の宿主細胞で開示された配列番号3、6または9で示される全長DNAを適当な哺乳類の細胞あるいはその他の宿主細胞で発現させることによって得られる。成熟型のタンパクの配列は、全長型のアミノ酸配列より予測可能である(図1参照)。
【0047】
配列番号3、6または9で示される塩基配列が一旦確定されると、その後は、化学合成によって、あるいは該塩基配列の断片を化学合成し、これをプローブとしてハイブリダイズさせることにより、本発明のcDNAを得ることができる。さらに、本cDNAを含有するベクターcDNAを適当な宿主に導入し、これを増殖させることによって、目的とするcDNAを必要量得ることができる。
【0048】
本発明のポリペプチドを取得する方法としては、
(1)生体または培養細胞から精製単離する方法、
(2)ペプチド合成する方法、または
(3)遺伝子組み換え技術を用いて生産する方法、
などが挙げられるが、工業的には(3)に記載した方法が好ましい。
【0049】
遺伝子組み換え技術を用いてポリペプチドを生産するための発現系(宿主−ベクター系)としては、例えば、細菌、酵母、昆虫細胞および哺乳動物細胞の発現系が挙げられる。
【0050】
例えば、大腸菌で発現させる場合には、成熟蛋白部分をコードするcDNAの5´末端に開始コドン(ATG)を付加し、得られたcDNAを、適当なプロモーター(例えば、trpプロモーター、lacプロモーター、λPLプロモーター、T7プロモーター等)の下流に接続し、大腸菌内で機能するベクター(例えば、pBR322、pUC18、pUC19等)に挿入して発現ベクターを作製する。
【0051】
次に、この発現ベクターで形質転換した大腸菌(例えば、E. Coli DH1、E. Coli JM109、E. Coli HB101株等)を適当な培地で培養して、その菌体より目的とするポリペプチドを得ることができる。また、バクテリアのシグナルペプチド(例えば、pelBのシグナルペプチド)を利用すれば、ペリプラズム中に目的とするポリペプチドを分泌することもできる。さらに、他のポリペプチドとのフュージョン・プロテイン(fusion protein)を生産することもできる。
【0052】
また、哺乳動物細胞で発現させる場合には、例えば、配列番号3、6または9で示される塩基配列を適当なベクター(例えば、レトロウイルスベクター、パピローマウイルスベクター、ワクシニアウイルスベクター、SV40系ベクター等)中の適当なプロモーター(例えば、SV40プロモーター、LTRプロモーター、メタロチオネインプロモーター等)の下流に挿入して発現ベクターを作製する。次に、得られた発現ベクターで適当な哺乳動物細胞(例えば,サルCOS−1細胞、COS−7細胞、チャイニーズハムスターCHO細胞、マウスL細胞等)を形質転換し、形質転換体を適当な培地で培養することによって、分泌蛋白である本発明の蛋白は、その細胞上清中に目的とするポリペプチドとして発現される。さらに、その他のポリペプチド、例えば抗体の定常領域(Fc portion)をコードするcDNA断片と連結することによって、フュージョン・プロテイン(fusion protein)を生産することもできる。以上のようにして得られたポリペプチドは、一般的な生化学的方法によって単離精製することができる。
【0053】
本発明の蛋白に関して記述される効果あるいは生物活性は、その蛋白の投与あるいは使用により、あるいは、その蛋白をコードするcDNAの投与あるいは使用(例えば、遺伝子療法(再生医療を含む。)やcDNA導入に適したベクター)により、提供される。
【0054】
また、該ポリペプチドのポリクローナル抗体またはモノクローナル抗体を用いて、生体における該ポリペプチドの定量が行なえ、これによって該ポリペプチドと疾患との関係の研究あるいは疾患の診断等に利用することができる。ポリクローナル抗体およびモノクローナル抗体は該ポリペプチドあるいはその断片を抗原として用いて公知の方法により作製することができる。
【0055】
また該ポリペプチドを用いることにより、例えばアフィニティーカラムを作製して、本ポリペプチドと結合する既知または未知の蛋白(リガンド)の同定、精製あるいはその遺伝子クローニングを行なうことができる。
【0056】
また該ポリペプチドを用いて、例えばウエスト−ウエスタン法により、または該cDNA(好ましくは該ポリペプチドをコードするcDNA)を用いて、例えば酵母2−ハイブリッド法により該ポリペプチドと相互作用する分子の同定、遺伝子クローニングを行なうこともできる。
【0057】
さらに本ポリペプチドを用いることによって、本ポリペプチドレセプターアゴニスト、アンタゴニストおよび受容体−シグナル伝達分子間の阻害剤等のスクリーニングを行なうこともできる。
【0058】
スクリーニングは、例えば、以下の方法により行なうことが出来る。すなわち、a)本発明のポリペプチド、スクリーニングすべき化合物、及び細胞を含む反応混合物を、細胞が該ペプチドにより正常に刺激される条件下に一緒にし(該反応混合物は細胞が増殖するに従い細胞中に導入される標識および該ペプチドの機能を効果的に観察させるための該ペプチド以外のペプチドを含む);ついで
b)細胞の増殖の程度を測定して、該化合物が有効なアンタゴニストまたはアゴニストであるかどうかを決定する。
【0059】
本発明のcDNAは、多大な有用性が期待される本発明のポリペプチドを生産する際の重要かつ必須の鋳型となるだけでなく、遺伝病の診断や治療(遺伝子欠損症の治療またはアンチセンスDNA(RNA)によって、ポリペプチドの発現を停止させることによる治療等)に利用できる。
また、本発明のcDNAをプローブとしてゲノミック(genomic)DNAを分離できる。
【0060】
[医薬品への適用]
前記の疾患に適応するために、本発明のポリペプチド、あるいは本発明のポリペプチドに対する抗体は通常、全身的又は局所的に、一般的には経口または非経口の形で投与される。好ましくは、経口投与、静脈内投与および脳室内投与である。
【0061】
投与量は、年齢、体重、症状、治療効果、投与方法、処理時間等により異なるが、通常、成人一人あたり、一回につき、100μgから100mgの範囲で、一日一回から数回経口投与されるかまたは、成人一人あたり、一回につき、10μgから100mgの範囲で、一日一回から数回非経口投与される。
【0062】
もちろん前記したように、投与量は、種々の条件により変動するので、上記投与量より少ない量で十分な場合もあるし、また範囲を越えて必要な場合もある。
【0063】
本発明化合物を投与する際には、経口投与のための固体組成物、液体組成物およびその他の組成物、非経口投与のための注射剤、外用剤、坐剤等として用いられる。
【0064】
経口投与のための固体組成物には、錠剤、丸剤、カプセル剤、散剤、顆粒剤等が含まれる。カプセルには、ソフトカプセルおよびハードカプセルが含まれる。
【0065】
このような固体組成物においては、一つまたはそれ以上の活性物質が、少なくとも一つの不活性な希釈剤(例えば、ラクトース、マンニトール、グルコース、ヒドロキシプロピルセルロース、微結晶セルロース、デンプン、ポリビニルピロリドン、メタケイ酸アルミン酸マグネシウム等)と混合される。組成物は、常法に従って、不活性な希釈剤以外の添加物、例えば、潤滑剤(ステアリン酸マグネシウム等)、崩壊剤(繊維素グリコール酸カルシウム等)、安定化剤(ヒト血清アルブミン、ラクトース等)、溶解補助剤(アルギニン、アスパラギン酸等)を含有していてもよい。
【0066】
錠剤または丸剤は、必要により白糖、ゼラチン、ヒドロキシプロピルセルロース、ヒドロキシプロピルメチルセルロースフタレート等の胃溶性あるいは腸溶性のフィルムで被膜してもよいし、また2以上の層で被膜してもよい。さらにゼラチンのような吸収されうる物質のカプセルも包含される。
【0067】
経口投与のための液体組成物は、薬学的に許容される乳濁剤、溶液剤、懸濁剤、シロップ剤、エリキシル剤等を含み、一般に用いられる不活性な希釈剤(例えば、精製水、エタノール等)を含んでいてもよい。この様な組成物は、不活性な希釈剤以外に湿潤剤、懸濁剤のような補助剤、甘味剤、風味剤、芳香剤、防腐剤を含有していてもよい。
【0068】
経口投与のためのその他の組成物としては、ひとつまたはそれ以上の活性物質を含み、それ自体公知の方法により処方されるスプレー剤が含まれる。この組成物は不活性な希釈剤以外に亜硫酸水素ナトリウムのような安定剤と等張性を与えるような安定化剤、塩化ナトリウム、クエン酸ナトリウムあるいはクエン酸のような等張剤を含有していてもよい。スプレー剤の製造方法は、例えば米国特許第2,868,691号および同第3,095,355号明細書に詳しく記載されている。
【0069】
本発明による非経口投与のための注射剤としては、無菌の水性または非水性の溶液剤、懸濁剤、乳濁剤を包含する。水性または非水性の溶液剤、懸濁剤としては、一つまたはそれ以上の活性物質が、少なくとも一つの不活性な希釈剤と混合される。水性の希釈剤としては、例えば注射用蒸留水および生理食塩水が挙げられる。非水性の希釈剤としては、例えばプロピレングリコール、ポリエチレングリコール、オリーブ油のような植物油、エタノールのようなアルコール類、ポリソルベート80(登録商標)等が挙げられる。
【0070】
このような組成物は、さらに防腐剤、湿潤剤、乳化剤、分散剤、安定化剤(例えば、ヒト血清アルブミン、ラクトース等)、溶解補助剤(例えば、アルギニン、アスパラギン酸等)のような補助剤を含んでいてもよい。
【実施例】
【0071】
以下に本発明のクローンESDNに関する実施例を挙げて本発明をより具体的に説明するが、これらは本発明の範囲を制限するものではない。
【0072】
実施例1:SSTによるスクリーニング
ヒト冠動脈初代培養内皮細胞(hCAEC)および平滑筋細胞(hCASMC)(Clontech)、そしてそれらの共培養(同じ細胞数のhCAECとhCASMCを混ぜ、EGM−2−MV(商品名、BioWhittakerより購入)を用いて2日間培養)をソースとしてcDNAライブラリーを構築した(特願平10-119731号参照)。次に酵母SST法(米国特許No.5,536,637を参照)によりスクリーニングを行なった結果、共培養をソースとしたcDNAライブラリーからCUBドメインを有するESDNを得た。
【0073】
実施例2:ヒト、マウス、ラットの全長cDNAクローニング
全長cDNAを得るためにマラソン・cDNA・アンプリフィケーション・キット(Marathon cDNA amplification kit)(商品名、CLONTECH社より購入)による5’−、3’−RACE(Rapid Amplification of cDNA End)法を用いて5’−、3’−末端cDNAのクローニングを行なった。同時にヒトの塩基配列情報に基づきプライマーを設計し、RT−PCRによりラット、マウスのESDNの単離も行なった。
マウスプライマー
5’−CTGCTCCAACTCCTCCTCCTTC−3’(配列番号10)
5’−CTGCTTCATTCCTTTCCACCAACCTG−3’(配列番号11)
ラットプライマー
5’−TGTGCTGGTCATGGTCCTCACTACTCTC−3’(配列番号12)
5’−TGTGCTTTAAAACGATGCTTTG−3’(配列番号13)
その結果、ESDNは、ヒト、マウス、ラット間において高い相同性を有していることが明らかとなったが、何れの種においても翻訳開始点ATGを含む5’末端配列を見出すことができなかった。この原因としてESDNの5’末端領域の高いGC含量が考えられる。そこでマウスphage genomic ライブラリー(Lambda FIXII library( 商品名、Strategene社より購入) )を用いたスクリーニングを行なった。その結果、翻訳開始点(Met)を含む2個の陽性クローンを得た。そこでこのMetを含む5’−側のプライマーを作成し、下流のエクソンを含む(genomic cloneには含まれていない)アンチセンスプライマー
Sense primer :5’−GCACTATGCGGGCGGATTGC−3’(配列番号14)
antisense primer :5’−GGATGTAAGGGTTCCACTCTCAGG−3’(配列番号15)
と共にRT−PCRを行なったところ、ヒト、ラットのカウンターパートを得た。得られたアミノ酸シークエンスよりヒトとげっ歯類は84〜5%の相同性、マウスとラットでは92%の相同性を表した。ヒト、ラットおよびマウスのESDNのアライメントを図1に示す。
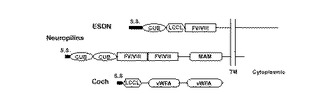
モチーフ検索の結果から、ESDNは、細胞外に1個のCUBドメインと1個のファクター(factor)V/VIII相同ドメインを有するタイプ(type)I膜貫通型タンパク質であることが判明した。また、この構造上の特徴からneuropilin(2個のCUB、2個のFV/VIIIドメイン、1個のMAMドメインを有する)との構造の類似性を指摘することができる。またその他の領域では、カブトガニのファクター(Factor)C、及びヒトにおける難聴の原因遺伝子の一つであるCochと4個のシステインが保存された相同性の高い領域がある。この領域はCochにおいては、DFNA9という遺伝性難聴においてこれまで発見されている4つの変異全てが局在している部分で、カブトガニのような進化系統樹で離れた生物にも見出されることから新しいドメイン構造と考えられる。図2および図3に示す。
【0074】
実施例3:哺乳動物細胞を用いたヒトESDN蛋白の発現と抗ペプチド抗体による同定
ヒト全長cDNAを哺乳動物細胞用発現ベクターのオリジナルV5エピトープをファッグ・タグ(FLAG tag)に交換し、pEF6V5−His(商品名、Invitrogen社より購入)にサブクローニングした。これらの発現ベクターは、293T細胞やCOS7細胞にセルフェクト(Cellphect)(商品名、Amersham Life Science社より購入)やリポフェクトアミン(Lipofectamine)(商品名、Life Technologies、社より購入)により細胞に導入した。なお目的の発現蛋白質の確認は、細胞のライセイト(lysate)を作成した後、各種抗体によるウエスタンブロッティングにより検出した。
anti-FLAG M2 monoclonal antibody( 商品名、Sigma社より購入)
GlyGluArgIleArgIleLysPheGlyAspGlyAspIleGluAspSerAsp(配列番号16)を用いて作成したウサギ抗CUBポリクローナル抗体
GlnAspLysIlePheGlnGlyAsnLysAspTyrHisLysAspValArgAsnAsn(配列番号17)を用いて作成した抗FV/VIII ポリクローナル抗体
は、それぞれKLHに連結させたポリペプチドをウサギに免疫することによって得た(Sawady Technology) 。
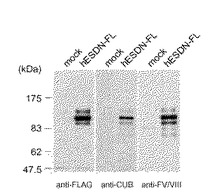
ウエスタンブロッティングは、ECL(商品名、Amersham Life Science社より購入)ルネッサンス(Renaissance)(NEN Life Science)のプロトコールに従った。その結果、発現ベクターのみを導入した細胞ライセイト(lysate)には認められないバンドが、127、106、93kDa付近に検出された(図6に示す。)。
【0075】
実施例4:シグナル配列の解析
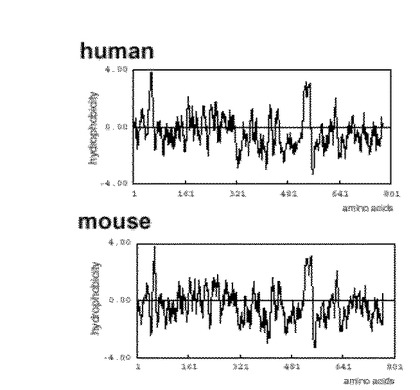
得られたアミノ酸配列の疎水性プロット(図4に示す。)から、ESDNのシグナル配列は、通常の配列よりも長く異型性であることが判明した。そこでヒト全長cDNAを導入したCOS7細胞を4%パラホルムアルデヒドにより固定した後、一次抗体により30分間室温で反応させた。次に30分間二次抗体、テキサス・レッド・アンチマウスIgG(Texas red anti-mouse IgG)(商品名、Vector Laboratories社より購入)やFITCアンチラビットIgG(FITC-anti-rabbit IgG)(商品名、Jackson Laboratories社より購入)で反応させた後、バイオ・ラッド・コンフォーカル・レーザー・スキャニング・マイクロスコープ(Bio-Rad confocal laser scanning microscope)(商品名、model MRC-1024)により解析した。その結果、同タンパク質は、細胞表面上に発現していることが判った(図7に示す。)。
次にシグナル配列の切断部位の確認を試みた。
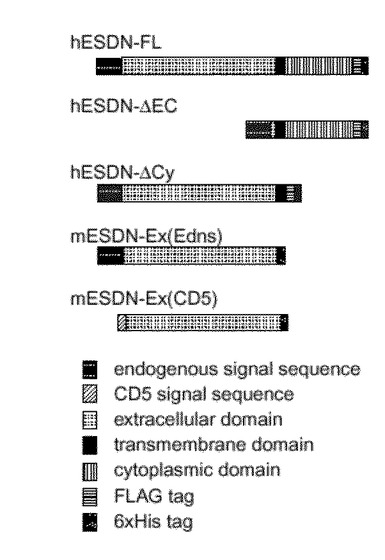
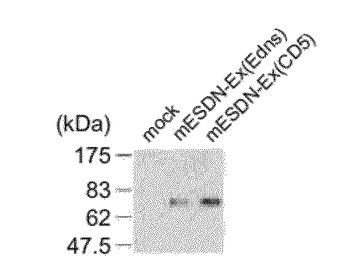
マウスESDNの細胞外領域のC端にHis−tagを付けたmESDN−Ex(Edns)やヒトCD5のシグナル配列:MetProMetGlySerLeuGlnProLeuAlaThrLeuTyrLeuLeuGlyMetLeuValAlaSerValLeuAla(配列番号18)に置換したmESDN−Ex(CD5)をpCAGGS(Science 261, 600-603 (1993))にサブクローニングした。293T細胞に導入後、培養上清に分泌された目的タンパク質を「His-probe H-15 polyclonal antibody」(商品名、Santa Cruz社より購入)によるウエスタンブロッティングにより検出した。その結果、mESDN−Ex(Edns)とmESDN−Ex(CD5)の両コンストラクト(図5に示す。)より同じサイズのタンパク質が確認された。ヒトCD5とマウスESDNのシグナル配列は、39アミノ酸(約4kDa)ヒトCD5の方が小さいが、得られたタンパク質のサイズが同じことから、同じ切断部位であることが予想された(図8に示す。)。
【0076】
実施例5:ESDN蛋白のサザーン・ズーブロット(southern zooblot)解析
[32P]dCTP−ラベルしたヒトESDNをプローブとして、哺乳類(マウス、ラット、ウサギ、ウシ、ヒト)、アフリカツメガエル(Xenopus)、ハエ(fly)、酵母(yeast)の種におけるサザーン・ズーブロット(southern zooblot)解析を行なった(37℃、1×SSCにより洗浄)。その結果、哺乳類においては強いバンドが確認され、ESDNが、哺乳類において高度に保存されていることが明らかとなった。一方、アフリカツメガエル(Xenopus)では弱いバンドが確認された。なお、ショウジョウバエ(fly)、酵母(yeast)においては、バンドは確認されなかった(図9Aに示す。)。
【0077】
実施例6:ヒトESDNの遺伝子座
ヒトにおいては、2個(stSG29921、sts-D29024) の独立したSTS(sequence-tagged site)の情報からラジエーション・ハイブリッド・マップ(radiation hybrid map)のD3S1603-D3S1271、D3S1552-D3S1603近傍(cytogenetic mapではchromosome 3q11.2に相当すると思われる部位)に遺伝子座があることが明らかとなった。このデータベースにより得られた結果を確認するために、マウスライン(Cell line)であるA9(Neo3)、A9(Neo12)(JCRB Cell Bank)を用いてゲノミック・サザン(genomic southern)ハイブリダイゼーションを行なった。その結果、ヒトESDNプローブは、マウスESDNも認識できるためA9(Neo3)、A9(Neo12)の両方においてマウスESDNが確認された。一方、ヒトESDNは、A9(Neo3)のみにおいて検出された。このことからヒトESDNは、クロモソーム(chromosome)3q11.2 に相当すると思われる部位に遺伝子座があることが明らかとなった(図9Bに示す。)。
【0078】
実施例7:ESDN mRNAの発現部位の解析
培養ヒト細胞のノザン(Northern)解析は、hCAEC、hCASMCそしてそれらの共培養からトータル(total)RNAをTRIzol(商品名、Life Technologies社より購入)により抽出した。ラット組織・細胞のノザン(Northern)解析は、トータル(total)RNAをTRIzol LS(全血に対して)、TRIzol(その他の組織や培養細胞に対して)により抽出した。次にpoly(A)RNAは、「OligotexTM-dT30 Super」(商品名、Roche Molecular Biochemicals社より購入)により精製した。次に[32P]dCTP−ラベルしたヒト、ラットESDNやGAPDH cDNAをプローブとして用い、ノザン・ブロット(Northern blot)解析(65℃、0.2 ×SSCにより洗浄)を行なった。その結果、hCASMC細胞において強い発現が認められた(6.4 、3kb)。一方hCAEC細胞においては、hCASMCに比べて弱い発現が認められた。なお、共培養することによる、ESDN mRNAの発現の変化は認められなかった(図10Aに示す。)。
ラット組織・細胞のノザン(Northern)解析の結果、全血細胞からESDN mRNAは確認されず、肝臓においても発現がかなり低いことが判った(図10Bに示す。)。
【0079】
実施例8:ヒト冠動脈平滑筋細胞を用いたESDN蛋白の発現
hCASMCを血清(serum)飢餓の状態のDMEM/2mMグルタミン(glutamine)で48時間培養した。次に指定濃度のPDGF−BB、AT−II、FCS(商品名、Sigma 、Life Technologies社より購入)を含む培地に置換し、刺激を行なった。トータル(total)RNAはTrizol(商品名、Life Technologies社より購入)により調製し、cDNA合成はスーパースクリプト・プレアンプリケーション・システム(SuperScript Preamplification System)(商品名、First Strand cDNA Synthesis :Life Technologies 社より購入)を用いて行なった。なおmRNAは、リアルタイム定量RT−PCR(real-time quantitative RT-PCR;PE Applied Biosystems Prism Model 7700 Sequence Detection System)により測定した。フォワード(Forward)リバース(Reverse)プライマーの配列は以下の通りである。
ESDN forward: 5’−CCCAGCAAGGTGATGGATG−3’(配列番号19)
ESDN reverse: 5’−CAAGAATCAGAATCTTCAATGTCAAAG−3’(配列番号20)
ESDN probe: 5’−(6−FAM)−CCTGAGAGTGGAACCCTTACATCCATAAAC−(TAMRA)−3’(配列番号21)
これらの配列はヒトを基準にしているが、げっ歯動物にも適応できることを確認した。
ヒトGAPDHのシークエンスは以下の通りである。
human GAPDH forward: 5’−GAAGGTGAAGGTCGGAGTC−3’(配列番号22)
human GAPDH reverse: 5’−GAAGATGGTGATGGGATTTC−3’(配列番号23)
human GAPDH probe: 5’−(VIC)−CAAGCTTCCCGTTCTCAGCC−(TAMRA)−3’(配列番号24)
「TaqMan Rodent GAPDH Control Reagents」(PE biosystems)をラット(rat)GAPDHの定量に用いた。ESDNとGAPDHのmRNAレベルはコピー数を意味している。そこで先方の発現レベルをGAPDHにより標準化した。このことは、スタンダードの連続希釈から作成した標準曲線によって成し遂げられた。なおスタンダードにはESDNあるいはGAPDHをpBlueScript SK(−)(商品名、Stratagene社より購入)にサブクローニングした既知量のプラスミドを準備した。
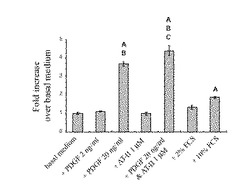
結果、ESDNはPDGF−BB刺激において濃度依存的に発現が増加するが、AT−II刺激においては発現の増加が認められなかった。またFCS刺激においてもESDNの発現増加が濃度依存的に認められたが、PDGF−BB刺激に比べてより少ない量であった(図11に示す。)。
【0080】
実施例9:ラットの総頸動脈におけるESDN蛋白の発現
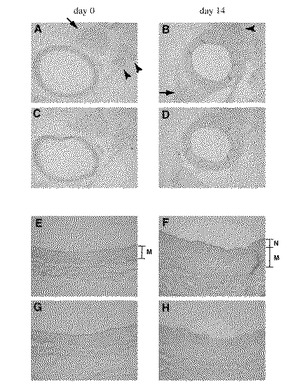
バルーン障害後(n=5,day 0,5,n=4,day 14)、0、5、14日後に頸動脈を採取した。トータル(total)RNAはTrizol(商品名、Life Technologies 社より購入)により調製し、cDNA合成は「SuperScript Preamplification System 」(商品名、First Strand cDNA Synthesis :Life Technologies 社より購入)を用いて行なった。ラットESDNやGAPDHのmRNAは「real-time quantitative RT-PCR 」(PE Applied Biosystems Prism Model 7700 Sequence Detection System)により測定した。その結果、5日目にESDNの増加の傾向が認められ、14日目には30%もの有意な増加が認められた。次に、このESDNの発現を免疫組織学的に解析した(図12に示す。)
処置あるいは未処置ラットを麻酔下にして、予め4℃に冷やした生理食塩水で組織を灌流した。そして局所灌流部分を4%パラホルムアルデヒドにより固定した。頸動脈を丁寧に採取し、ドライアイス−エタノール・バス上に置いた「Tissue-Tek O.C.T. Compound」(商品名、Sakura Finetechnical社より購入)により包埋した。組織を4μmの厚さにスライスし、「avidin-biotin-alkaline phosphatase complex」(商品名、Vector Laboratories 社より購入) 法により免疫組織解析を行なった。アルカリンホスファターゼ(Alkaline phosphatase)は「Vector Red」(商品名、Vector Laboratories 社より購入)により発色させた。切片はメチルグリーンにより対比染色を行なった。ウサギ抗ペプチドポリクローナル抗体(一次抗体)は、5〜10μg/mlの濃度で用いた。ネガティブコントロールには、正常ウサギIgG(DAKO)を同濃度で用いた。その結果、血管平滑筋の存在する大動脈や総頸動脈の中膜が染まるものの、むしろ脳・脊髄といった中枢神経系の一部(矢印)や迷走神経(アローヘッド)などの末梢神経が強く染色された(図13に示す。)。A、B、EおよびFは抗CUB抗体で染色した。C、D、GおよびHは、同じ濃度のウサギの抗IgG抗体を一次抗体として染色した(コントロール)。EおよびF中のMおよびNは、それぞれ血管中膜および血管内膜を示す。
【0081】
実施例10:293T細胞を用いたBrdUの取りこみ測定
全長ESDN、およびそのディレーション・ミュータント(deletion mutants)(hESDN−ΔEC、hESDN−ΔCy)を発現ベクター「QIAfilter Plasmid Midi Kit」(商品名、QIAGEN社より購入)により調整した。そしてさらに精製するためフェノール(phenol)/CIAAにより2回抽出し、CIAA抽出を1回行なった。セルフェクト(CellPhect)(商品名、Amersham Life Science 社より購入)を用いて293T細胞にトランスフェクトした。12時間後、メディウムを新鮮なDMEM/10%FCSに交換し、2時間インキュベーションした。次にトリプシンにより細胞を一旦回収した後、96穴プレート2枚に播き直した。一方のプレートは、2時間のBrdUの取り込みを24時間の培養後に行なった。BrdUの取り込みは、「Cell Proliferation ELISA、BrdU」(colorimetric)(商品名、Roche Diagnostics社より購入)により測定した。もう一方のプレートは、播きこみ後2 時間細胞数を「Premix WST-1 assay kit」(商品名、TaKaRa Biomedicals社より購入)により測定した。その結果、全長ESDNを発現させた細胞において、BrdUの取りこみが有意に抑制されることが明らかとなった。一方細胞外ドメインを欠いたものでは、この効果が弱められた。また細胞内ドメインを欠いたものでは全くこの効果が無くなることが確認された(図14に示す。)。
【図面の簡単な説明】
【0082】
【図1】ヒト、マウスおよびラットESDNのアライメントを表わす。
【図2】ESDN、ニューロピリンおよびCochにおけるドメイン構造を表わす。
【図3】ヒト、マウス、ラットESDN、リムラスファクターC、ラットLgl−1、ヒトCoch、マウスCochおよびラットCochのLCCL部分のアライメントを表わす。
【図4】ヒトおよびマウスESDNのハイドロフォビシティーを表わす。
【図5】発現ベクターの構成を表わす。
【図6】ヒト全長ESDNのウエスタンブロッティング解析を表わす。
【図7】細胞膜表面でESDNが発現されている様子を表わす。
【図8】開裂したシグナル配列のウエスタンブロッティング解析を表わす。
【図9】AはヒトESDNのcDNAを用いたサザンズーブロッティングを表わす。BはヒトESDNがクロモソーム3にあることを示す。
【図10】AおよびBは、ヒト頸動脈細胞のRNAおよびラット各臓器のRNAを用いたESDNのcDNAとのノザンブロッティング解析を示す。
【図11】ヒト平滑筋細胞において、PDGFまたはFCS刺激によりESDNのmRNAが誘導される様子を示す。
【図12】ラット頸動脈をバルーンで傷害することにより、ESDNのmRNAの発現が増加することを示す。
【図13】ラット頸動脈をバルーンで傷害することにより、ESDNの発現が増加することを示す。
【図14】ESDN過発現細胞においてBrdU取り込みが抑制されていることを示す。
【技術分野】
【0001】
本発明は、新規なポリペプチドである血管内皮細胞・平滑筋細胞由来ニューロピリン様分子(Endothelial and Smooth muscle cell-Derived Neuropilin-like molecule 、以下、ESDNと省略する。)、その製造方法、ESDNをコードするcDNA、そのcDNAからなるベクター、そのベクターで形質転換された宿主細胞、ESDNの抗体、ESDNまたは抗体を含有する薬学的組成物、ESDNを測定する方法、ESDNを測定する試薬およびESDNを用いたスクリーニング方法に関する。
【背景技術】
【0002】
血管内皮増殖因子(VEGF)のクローニング以来、血管形成を支配する複雑な細胞外シグナル伝達ネットワークの解明が画期的になされてきた。このリガンドやFlk−1/VEGFR2やFlt−1/VEGFR1といったチロシンキナーゼのノックアウトマウスの研究が、血管形成、発生の第一ステップに関する最初の重要点であった。
VEGF受容体−2(VEGFR2)は、内皮細胞形成において必要であり、またこの遺伝子の欠損は、血液細胞群あるいは組織化された血管の不足を招き発生9.5日目に胎生死につながる。しかしながら、RT−PCRを用いた初期マーカーの解析により初期の造血や内皮の前駆細胞が実際にVEGFR2非存在下において造られるという報告から、VEGFR2は血球芽細胞の形成においては必要不可欠ではないが、系統のその後の増加には必要であることを示唆している。
VEGF受容体−1(VEGFR1)ノックアウトマウスもまた発生9.5日目に死ぬが、それらの内皮細胞の分化は直接影響を受けていない。その代わりに、主に間充識の血球芽細胞の移行が不足していると考えられ、過密になった内皮前駆細胞が、脈管系の重篤な組織破壊を導く。
VEGFノックアウトマウスもまた発生9.5日目に著しい減少を伴って死ぬが、内皮細胞の分化を欠いていない。つまり、本質的にはVEGFR1ノックアウトマウスと同様であるが、より軽度なフェノタイプを示す。この遺伝子における目立った特長は、ヘテロ接合体の遺伝子欠失でも発生11.5日目に胎生致死になることである。このことは胎生期においてVEGFに対する著しく厳格な容量依存性が存在することを示している。
その他のレセプターチロシンキナーゼもまた、Tie−2やそのリガンドangiopoietin−1といったように血管内皮細胞に特異的に発現をしていて、血管形成における後期、血管新生のステップに関与している。これら遺伝子の破壊は、脈管形成には影響を及ぼさないが、血管のリモデリングに障害を与え、発生10.5日目に胎生死につながる。それらは、内皮細胞とそれを取り巻く血管平滑筋細胞(VSMC)あるいは間充識細胞間の相互作用において役割を担っている。これらの脈管特有のシステムに付け加えて、PDGF−BBあるいはTGF−βそしてそれらのレセプターもまた内皮細胞とそれを取り巻く細胞間で同様な役割を担っている。脈管システムに対するニューロピリン−1(NP−1;neuropilin-1)の関係の同定は、この分野の新局面を紹介した。NP−1は初め発達した神経系を特異的に認識するモノクローナル抗体によって認識される抗原としてクローニングされた。
そして、セマフォリン3A(semaphorin 3A;sema 3A)のレセプターとしての同定は、軸索形成において一躍注目を浴びるようになった。またその他のVEGFレセプターの探索がVEGFR2のレセプターでもあるNP−1の発見につながった。そしてそれは、一つのVEGFアイソフォーム(isoform)への結合の増加だけでなく、内皮細胞の遊走やおそらくマイトシス(mitosis)も同様に促進する。このことが同定される前は、NP−1を強制発現させたキメラマウスは、肥大した脈管を形成することが示された。NP−1ノックアウトマウスは、中枢神経系においては低い血管質を呈し、また大きな脈管の異形をも示した。一方sema 3Aノックアウトマウスでは血管の異常性の存在や非存在は報告されていないが、そのフェノタイプがNP−1強制発現マウスにおいても観察された薄い心筋層を呈することは興味深い。
Eph/ephrinシステムは、別の例であり、初めに神経科学において有名になった。しかし、後に現在までには唯一の動脈と静脈を識別する表面マーカーとして同定されている。それらもまた内皮細胞との接点である周囲の間充識細胞に発現を示している。これらの役割を持った分子以外にも、我々は更に多くの細胞外シグナル伝達分子が血管の分野においては存在すると想定した。そしてこの事の解明が、この複雑なシステムのさらなる理解に貢献できるであろうと考えた。
【0003】
従来、ある特定のポリペプチドまたはそれをコードするcDNAを得ようとする場合、組織や細胞培養液中に目的とする生物活性を確認し、次いでポリペプチドの単離精製を経て、遺伝子をクローニングするという方法、あるいはその生物活性を指標として遺伝子を発現クローニングする方法が一般的に用いられてきた。しかし、生体内生理活性ポリペプチドは、多様な生物活性を有している場合が多いので、あるひとつの活性を指標にして遺伝子をクローニングした結果、それが既知のポリペプチドと同一であることが後になって判明するという事例が増えている。また、微量しか産生されなかったり、特別な生理条件でのみ発現する因子も多く、そのことが単離、精製および生物活性の確認を困難なものとしている。
【0004】
本発明者らは、これまで造血系や免疫系で働く増殖分化因子の遺伝子のクローニングを研究してきた。そして、増殖分化因子(例えば、各種サイトカイン等)のような分泌蛋白質やそのレセプターのような膜蛋白質(以下、これらをまとめて分泌蛋白質等と呼ぶ。)の大部分がそのN末端にシグナルペプチドと呼ばれる配列を有していることに着目して、シグナルペプチドをコードする遺伝子を効率的かつ選択的にクローニングする方法を鋭意検討した。その結果、動物細胞を用いて、シグナルペプチドの有無を簡単に検索できる方法(シグナルシークエンストラップ(SST)法)を見出した(特許第3,229,590号参照)。さらに同じ概念のもとに、酵母を用いてさらに大量かつ簡便にシグナルペプチドをコードする遺伝子を単離する方法(酵母SST法)も開発された(米国特許No.5,536,637)。
【発明の開示】
【0005】
本発明者らは,循環器領域におけるPTCA後の再狭窄や動脈硬化での病的血管平滑筋で重要な役割を果たしている有益な新規因子(ポリペプチド)、特に分泌シグナルを有する分泌蛋白質および膜蛋白質に着目してそれを見出すべく、鋭意検討を行なった。
本発明者らは、血管細胞に対して新たに修正したSSTスクリーニングを行ない、新規トランスメンブラン(transmembrane)タンパクのクローニングに成功した。本発明の蛋白ESDNは、新規のI型transmembraneタンパク質であり、ニューロピリン(neuropilin)様の特徴的ドメインを持つものである。
【0006】
本発明のESDN蛋白は、後に詳述されるように、冠動脈細胞ならびに平滑筋細胞で発現され、またバルーン傷害した動脈の血管平滑筋や総頸動脈の中膜で発現されることから、循環器領域におけるPTCA後の再狭窄や動脈硬化の治療に有用であると考えられる。
【0007】
本発明が提供するcDNA配列は、配列番号2で示されるヒトESDNクローンとして同定され、冠動脈初代培養内皮細胞および平滑筋細胞から作製したcDNAライブラリーより、酵母SST法を使用して得た情報をもとに単離された。配列番号3で示されるヒトESDNクローンは分泌蛋白質(ここではヒトESDN蛋白として表される。)をコードする完全なcDNA配列を含む全長鎖cDNAである。
【0008】
本発明が提供するcDNA配列は、配列番号6で示されるマウスESDNクローンとして同定され、酵母SST法を使用して得た情報をもとに、マウスcDNAライブラリーより単離された。配列番号5で示されるマウスESDNクローンは分泌蛋白質(ここではマウスESDN蛋白として表される)をコードする完全なcDNA配列を含む全長鎖cDNAである。
【0009】
本発明が提供する配列番号9で示されるcDNA配列は、ラットESDNクローンとして同定され、酵母SST法を使用して得た情報をもとに、ラットcDNAライブラリーより単離された。配列番号8で示されるマウスESDNクローンは分泌蛋白質(ここではマウスESDN蛋白として表される)をコードする完全なcDNA配列を含む全長鎖cDNAである。
【0010】
核酸配列データベースに登録されている既知の核酸配列に対してジンバンク(GenBank)およびNCBIを用いたBLASTN、FASTAおよびUNIGENEなどにより、またアミノ酸配列データベースに登録されている既知のポリペプチドのアミノ酸配列に対してBLASTP、Fly Database、SwissProt などにより検索をした結果、本発明ポリペプチドであるラットESDNおよびそれらをコードする核酸配列と一致する配列はなかった。このことから、本発明のポリペプチドは新規な分泌蛋白質であることが判明した。
【0011】
本発明は、
1.実質的に純粋な形である配列番号2、5または8で示されるアミノ酸配列からなるポリペプチド、またはそのホモローグ、そのフラグメントまたはそのフラグメントのホモローグからなるポリペプチド。
2.配列番号2、5または8で示されるアミノ酸配列からなる前項1記載のポリペプチド、
3.前項1または2に記載されたポリペプチドをコードするcDNA、
4.配列番号3、6または9で示される塩基配列を有する前項3記載のcDNA、またはその配列に選択的にハイブリダイズするフラグメントからなるcDNA。
5.前項3または4に記載のcDNAからなる複製または発現ベクター、
6.前項5記載の複製または発現ベクターで形質転換された宿主細胞、
7.前項1または2に記載されたポリペプチドを発現させるための条件下、前項6記載の宿主細胞を培養することを特徴とするからなる前項1または2に記載のポリペプチドの製造方法、
8.前項1または2に記載されたポリペプチドのポリクローナルまたはモノクローナル抗体、
9.前項1または2に記載されたポリペプチドまたは前項8記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とする薬学的組成物、
10.前項1または2に記載されたポリペプチドまたは前項8記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とするPTCA後の再狭窄の治療に有効な薬学的組成物、
11.前項1または2に記載されたポリペプチドまたは前項8記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とする前項10記載の動脈硬化の治療に有効な薬学的組成物、
12.前項1または2に記載されたポリペプチドの測定方法、
13.前項8記載の抗体を用いることを特徴とする前項1または2に記載されたポリペプチドの免疫化学的測定方法、
14.前項12または13記載の方法を用いることを特徴とする前項1または2に記載されたポリペプチドを検出する試薬、
15.前項12または13記載の方法を用いることを特徴とするPTCA後の再狭窄を検査する試薬、
16.前項12または13記載の方法を用いることを特徴とする動脈硬化を検査する試薬、および
17.前項1または2に記載されたポリペプチドを用いることを特徴とする、該ポリペプチドに対するアンタゴニストまたはアゴニストとしての活性を有する化合物をスクリーニングする方法に関する。
【0012】
本発明者らは、該ポリペプチドが血管平滑筋の収縮形質よりも合成形質において発現が亢進することを確認した。そのため、該ポリペプチドを用いて循環器領域におけるPTCA後の再狭窄や動脈硬化の治療に有用であると考えられる。
また、本発明のポリペプチドは、実験に於いて、細胞増殖抑制作用を示したこと、VEGFと構造が類似であることから、以下の作用を有することが考えられる。
「サイトカイン活性および細胞増殖/分化活性」
本発明の蛋白は、強制発現系において細胞増殖を抑制したことから、サイトカイン活性および細胞増殖(誘導あるいは阻害)/分化活性(誘導あるいは阻害)を示す可能性、あるいはある細胞集団に他のサイトカインの産生を誘導あるいは抑制すると考えられる。
「免疫刺激/抑制活性」
本発明の蛋白は、免疫刺激活性および免疫抑制活性も示すと考えられる。例えば、Tリンパ球およびBリンパ球あるいはどちらか一方の成長および増殖を制御(刺激あるいは抑制)することや、同様にNK細胞や他の集団の細胞傷害性活性に影響を与えることによって、様々な免疫不全および疾患の治療に効果を示すと考えられる。特に腫瘍血管新生によって引き起こされるリンパ行性癌転移の抑制において効果を示すと考えられる。
「虚血性血管新生抑制活性」
本発明の蛋白は、糖尿病の合併症である網膜症を抑制すると考えられる。VEGFは虚血誘導によって強力な血管新生作用を持つことが知られている。糖尿病網膜症の血管新生も網膜無血管領域が形成された後に起こる虚血性血管新生である。本発明の蛋白は、ニューロピリン(neuropilin)との構造の類似性から新規のVEGF受容体として血管増殖を抑制するように働いている可能性が考えられる。このことから糖尿病網膜症において効果を示すと考えられる。
【発明を実施するための最良の形態】
【0013】
ハイブリダイズするcDNAには、上記配列の相補配列も含まれる。ハイブリダイズは、ストリンジェントな条件で行なうことが好ましい。
【0014】
実質的に純粋な形である配列番号2、5または8で示されるアミノ酸配列からなるポリペプチドとは、一般に、生産時のポリペプチドの90%以上、例えば、95、98または99%が配列番号2、5または8で示されるアミノ酸配列からなるポリペプチドであることを意味する。
【0015】
配列番号2、5または8で示されるアミノ酸配列からなるポリペプチドのホモローグとは、一般に少なくとも20個、好ましくは少なくとも30個、例えば40、60、80または100個の連続したアミノ酸領域で、少なくとも70%、好ましくは少なくとも80または90%、より好ましくは95%以上相同性であるものであり、そのようなホモローグは、以後本発明のポリペプチドとして記載される。
【0016】
さらに、配列番号2、5または8で示されるアミノ酸配列からなるポリペプチドのフラグメント、またはそれらのホモローグのフラグメントとは、少なくとも10アミノ酸、好ましくは少なくとも15アミノ酸、例えば20、25、30、40、50または60アミノ酸部分を意味する。
【0017】
配列番号3、6または9で示される塩基配列からなるcDNAに選択的にハイブリダイズするcDNAとは、一般に、少なくとも20個、好ましくは少なくとも30個、例えば40、60、80または100個の連続した塩基配列領域で、少なくとも70%、好ましくは少なくとも80または90%、より好ましくは95%以上相同性であるものであり、そのようなcDNAは、以後本発明のcDNAとして記載される。
【0018】
配列番号3、6または9で示される塩基配列からなるcDNAのフラグメントとは、少なくとも10塩基、好ましくは少なくとも15塩基、例えば20、25、30または40塩基部分を意味し、そのようなフラグメントも本発明のcDNAに含まれる。
【0019】
さらに、本発明には、本発明のcDNAからなる複製または発現ベクターが含まれる。ベクターとしては、例えば、ori領域と、必要により上記cDNAの発現のためのプロモーター、プロモーターの制御因子などからなるプラスミド、ウィルスまたはファージベクターが挙げられる。ベクターはひとつまたはそれ以上の選択的マーカー遺伝子、例えばアンピシリン耐性遺伝子を含んでいてもよい。ベクターは、イン・ビトロ(in vitro)において、例えばcDNAに対応するRNAの製造、宿主細胞の形質転換に用いることができる。
【0020】
さらに、本発明には、配列番号3、6また9で示される塩基配列、またはそれらのオープンリーディングフレームからなるcDNAを含む本発明のcDNAを複製または発現させるためのベクターで形質転換された宿主細胞も含まれる。細胞としては、例えば細菌、酵母、昆虫細胞または哺乳動物細胞が挙げられる。
【0021】
さらに、本発明には、本発明のポリペプチドを発現させるための条件下で、本発明の宿主細胞を培養することからなる本発明のポリペプチドの製造方法も含まれる。培養は、本発明のポリペプチドが発現し、宿主細胞より製造される条件下で行なわれることが好ましい。
【0022】
本発明のcDNAは、上記のようなベクターのアンチセンス領域に挿入することでアンチセンスRNAを製造することもできる。このようなアンチセンスRNAは、細胞中の本発明のポリペプチドのレベルを制御することに用いることもできる。
【0023】
本発明は、本発明におけるポリペプチドのモノクローナルまたはポリクローナル抗体も含む。さらに本発明におけるポリペプチドのモノクローナルまたはポリクローナル抗体の製造方法も含む。モノクローナル抗体は、本発明のペプチドまたは、その断片を抗原として用い、通常のハイブリドーマの技術により製造することができる。ポリクローナル抗体は、宿主動物(例えば、ラットやウサギ等)に本発明のポリペプチドを接種し、免疫血清を回収する、通常の方法により製造することができる。
【0024】
本発明には、本発明のポリペプチド、その抗体と薬学的に許容される賦形剤および/または担体を含有する薬学的組成物も含まれる。
【0025】
本発明のポリペプチドとしては、配列番号2、5または8で示されたアミノ酸配列からなるもの以外に、その一部が欠損したもの(例えば、配列番号2中、生物活性の発現に必須な部分だけからなる成熟ポリペプチド等)、その一部が他のアミノ酸と置換したもの(例えば、物性の類似したアミノ酸に置換したもの)、および上記本発明のポリペプチドに他のアミノ酸が付加または挿入されたものも含まれる。
【0026】
よく知られているように、ひとつのアミノ酸をコードするコドンは1〜6種類(例えば、Metは1種類、Leuは6種類)知られている。従って、ポリペプチドのアミノ酸配列を変えることなくcDNAの塩基配列を変えることができる。
【0027】
本発明のcDNAには、配列番号2、5または8で示されるポリペプチドをコードするすべての塩基配列群が含まれる。塩基配列を変えることによって、ポリペプチドの生産性が向上することがある。
【0028】
配列番号3、6または9で特定されるcDNAは、配列番号2、5または8で示されるポリペプチドをコードする塩基配列群の一態様であり、天然型配列を表わす。
【0029】
配列番号3、6または9で示される塩基配列からなるcDNAの作製は、以下の方法に従って行なわれる。
【0030】
はじめに酵母SST法(米国特許No.5,536,637に記載)の概要について説明する。
サッカロマイセス・セロヴィシエ(Saccharomyces cerevisiae)などの酵母がショ糖またはラフィノースをエネルギー源や炭素源として利用するためにはインベルターゼを培地中に分泌しなければならない(インベルターゼはラフィノースをショ糖とメリビオースに、ショ糖をフルクトースとグルコースに分解する酵素である。)。また数多くの既知の哺乳類のシグナルペプチドは酵母のインベルターゼを分泌させうることが知られている。
【0031】
これらの知見から、酵母のインベルターゼの分泌を可能にする新規のシグナルペプチドを哺乳類のcDNAライブラリーからラフィノース培地上での酵母の生育を指標にスクリーニングする方法として本方法は開発された。
【0032】
翻訳開始点ATGを削除した非分泌型のインベルターゼ遺伝子SUC2(GENBANK accession No.V01311) を酵母の発現ベクターに組み込んで酵母SST用ベクターpSUC2を作製した。発現ベクターには、AAH5プラスミド(Gammerer, Methods in Enzymol. 101,192-201,1983)由来の発現用プロモーター(ADHプロモーター)およびターミネーター(ADHターミネーター)が組み込まれ、酵母複製起点としては2μori、酵母選択マーカーにはTRP1、大腸菌複製起点としてはColE1 ori、大腸菌薬剤耐性マーカーにはアンピシリン耐性遺伝子がそれぞれ組み込まれている。そのSUC2遺伝子の上流に哺乳類のcDNAを組み込んで、酵母SSTcDNAライブラリーを調製した。このライブラリーを分泌型インベルターゼを欠損している酵母に形質転換した。
【0033】
組み込まれた哺乳類cDNAがシグナルペプチドをコードしている場合、酵母で発現されたインベルターゼに対しても分泌作用を持つと考えられ、その結果ラフィノース培地上での生育が可能となる。そこで出現したコロニーから酵母を培養してプラスミドを調製し、インサートcDNAの塩基配列を決定することによって、新規シグナルペプチドの検索を迅速かつ容易にした。
【0034】
酵母SSTcDNAライブラリーの作製は、
(1)対象となる細胞よりmRNAを単離し、特定の制限酵素(酵素I)サイトを連結したランダムプライマーを用いて二本鎖cDNAを合成し、
(2)酵素Iとは異なる特定の制限酵素(酵素II)サイトを含むアダプターを連結して、酵素Iで消化した後、適当なサイズで分画し、
(3)酵母発現ベクター内のシグナルペプチドを削除したインベルターゼ遺伝子の上流に得られたcDNA断片を連結し、形質転換する工程よりなる。
【0035】
各工程を詳しく説明すると、工程(1)では、対象となる哺乳類の臓器や細胞株などより、必要により適当な刺激剤で刺激した後、公知の方法(以下、公知の方法は特に記載がなければMolecular Cloning(Sambrook, J., Fritsch, E. F.および Maniatis, T. 著、Cold Spring Harbor Laboratory Press より1989年に発刊)またはCurrent Protocol in Molecular Biology (F.M.Ausubelら編、John Wiley & Sons, Inc. より発刊) に記載の方法に従って行なわれる。)に従ってmRNAの単離が行なわれる。
【0036】
対象となる組織としては、マウス胎児心臓が挙げられる。ランダムプライマーを用いる二本鎖cDNAの合成は公知の方法により行なわれる。
【0037】
アダプターに連結される制限酵素(酵素I)サイトと次の工程(2)で用いられる制限酵素(酵素II)サイトは、互いに異なるものであれば何を用いてもよい。好ましくは、酵素IとしてXhoI、酵素IIとしてはEcoRIが用いられる。
【0038】
工程(2)ではT4DNAポリメラーゼで末端を平滑化し、酵素IIアダプターを連結した後、酵素Iで消化し、アガロース電気泳動(AGE)により300〜800bpのcDNAを分画する。酵素IIは、前記したように酵素Iと異なるものなら何でもよい。
【0039】
工程(3)は、酵母発現用プラスミドベクターに連結されたシグナルペプチドを削除したインベルターゼの遺伝子の上流に(2)で得られたcDNA断片を組み込んで大腸菌に形質転換する工程である。ここで酵母発現用プラスミドベクターとしては、種々のものが知られているが、例えば、大腸菌内でも機能するYEp24などが用いられるが、好適には前述したプラスミドpSUC2が用いられる。
【0040】
形質転換のための宿主大腸菌株はすでに多くのものが知られており、好ましくはDH10Bのコンピテントセルである。また形質転換方法は公知の方法のいずれを用いてもよいが、好ましくはエレクトロポレーション法により行なわれる。形質転換体は公知の方法により培養され、酵母SST用のcDNAライブラリーが得られる。
【0041】
このcDNAライブラリーでは、すべてのクローンcDNA断片が導入されているわけではないし、またすべてが未知の(新規の)シグナルペプチドをコードする遺伝子断片とは限らない。そこで、次に該ライブラリーから未知のシグナルペプチドをコードする遺伝子断片をスクリーニングする必要がある。
【0042】
遺伝子をもたない酵母、サッカロマイセス・セロヴィシエ(Saccharomycs cerevisiae)(例えば、YT455株など)またはインベルターゼ遺伝子を人為的に欠損させた株(公知の方法に従い作製可能)に、該cDNAライブラリーを導入し、シグナルペプチドをコードする配列を有する断片のスクリーニングを行なう。酵母の形質転換は公知の方法、例えば酢酸リチウム法によって行なわれる。形質転換体を選択培地で生育後、ラフィノースを炭素源とする培地に移し、生育可能なコロニーを選択し、プラスミドを回収する。ラフィノースを炭素源として酵母が生育したということは、ライブラリー中に何らかの分泌蛋白質のシグナルペプチドが組み込まれていたことを示している。
【0043】
次に、単離した陽性クローンについて、塩基配列を決定し、未知の蛋白質をコードすることが明らかになったcDNAについては、それをプローブとして全長クローンを単離し、全長の塩基配列を決定することができる。これらの操作は、当業者にとってすべて公知の方法で行なわれる。
【0044】
配列番号3、6または9で示される塩基配列が、一部、好ましくは全てが確定されると哺乳類に存在する本発明の蛋白質をコードするcDNAもしくは本発明蛋白質のホモローグおよびサブセットをコードするcDNAを得ることができる。適当な塩基配列を有するオリゴヌクレオチドを合成し、それを用いて哺乳類由来のcDNAライブラリーあるいはmRNAからPCR法により、あるいは適当な塩基配列の断片をプローブとしてハイブリダイズさせることにより、他の哺乳類cDNAライブラリーあるいは該ゲノムライブラリーから、他の哺乳類型の当該蛋白質をコードするcDNAを得ることができる。
【0045】
このようにして得られたcDNAが、SSTで得られたcDNA断片の塩基配列(またはその相同配列)を含んでいるならばシグナルペプチドをコードしていることになるので、該cDNAが全長、またはほぼ全長であることは明らかである(シグナルペプチドは例外なく蛋白質のN末端に存在することから、cDNAのオープンリーディングフレームの5'末端にコードされている。)。
さらに公知の方法に従い、該cDNAをプローブとしてノザン(Northern)解析によって全長の確認をしてもよい。ハイブリダイズしたバンドから得られるmRNAのサイズと該cDNAのサイズを比較し、ほぼ同じであれば該cDNAはほぼ全長であると考えられる。
【0046】
本発明は、開示されたタンパクの全長型並びに成熟型の両方を提供する。それらのタンパクの全長型は、配列番号2、5または8で示される塩基配列の翻訳されるアミノ酸配列で同定される。それらの成熟タンパクは、適当な哺乳類の細胞あるいはその他の宿主細胞で開示された配列番号3、6または9で示される全長DNAを適当な哺乳類の細胞あるいはその他の宿主細胞で発現させることによって得られる。成熟型のタンパクの配列は、全長型のアミノ酸配列より予測可能である(図1参照)。
【0047】
配列番号3、6または9で示される塩基配列が一旦確定されると、その後は、化学合成によって、あるいは該塩基配列の断片を化学合成し、これをプローブとしてハイブリダイズさせることにより、本発明のcDNAを得ることができる。さらに、本cDNAを含有するベクターcDNAを適当な宿主に導入し、これを増殖させることによって、目的とするcDNAを必要量得ることができる。
【0048】
本発明のポリペプチドを取得する方法としては、
(1)生体または培養細胞から精製単離する方法、
(2)ペプチド合成する方法、または
(3)遺伝子組み換え技術を用いて生産する方法、
などが挙げられるが、工業的には(3)に記載した方法が好ましい。
【0049】
遺伝子組み換え技術を用いてポリペプチドを生産するための発現系(宿主−ベクター系)としては、例えば、細菌、酵母、昆虫細胞および哺乳動物細胞の発現系が挙げられる。
【0050】
例えば、大腸菌で発現させる場合には、成熟蛋白部分をコードするcDNAの5´末端に開始コドン(ATG)を付加し、得られたcDNAを、適当なプロモーター(例えば、trpプロモーター、lacプロモーター、λPLプロモーター、T7プロモーター等)の下流に接続し、大腸菌内で機能するベクター(例えば、pBR322、pUC18、pUC19等)に挿入して発現ベクターを作製する。
【0051】
次に、この発現ベクターで形質転換した大腸菌(例えば、E. Coli DH1、E. Coli JM109、E. Coli HB101株等)を適当な培地で培養して、その菌体より目的とするポリペプチドを得ることができる。また、バクテリアのシグナルペプチド(例えば、pelBのシグナルペプチド)を利用すれば、ペリプラズム中に目的とするポリペプチドを分泌することもできる。さらに、他のポリペプチドとのフュージョン・プロテイン(fusion protein)を生産することもできる。
【0052】
また、哺乳動物細胞で発現させる場合には、例えば、配列番号3、6または9で示される塩基配列を適当なベクター(例えば、レトロウイルスベクター、パピローマウイルスベクター、ワクシニアウイルスベクター、SV40系ベクター等)中の適当なプロモーター(例えば、SV40プロモーター、LTRプロモーター、メタロチオネインプロモーター等)の下流に挿入して発現ベクターを作製する。次に、得られた発現ベクターで適当な哺乳動物細胞(例えば,サルCOS−1細胞、COS−7細胞、チャイニーズハムスターCHO細胞、マウスL細胞等)を形質転換し、形質転換体を適当な培地で培養することによって、分泌蛋白である本発明の蛋白は、その細胞上清中に目的とするポリペプチドとして発現される。さらに、その他のポリペプチド、例えば抗体の定常領域(Fc portion)をコードするcDNA断片と連結することによって、フュージョン・プロテイン(fusion protein)を生産することもできる。以上のようにして得られたポリペプチドは、一般的な生化学的方法によって単離精製することができる。
【0053】
本発明の蛋白に関して記述される効果あるいは生物活性は、その蛋白の投与あるいは使用により、あるいは、その蛋白をコードするcDNAの投与あるいは使用(例えば、遺伝子療法(再生医療を含む。)やcDNA導入に適したベクター)により、提供される。
【0054】
また、該ポリペプチドのポリクローナル抗体またはモノクローナル抗体を用いて、生体における該ポリペプチドの定量が行なえ、これによって該ポリペプチドと疾患との関係の研究あるいは疾患の診断等に利用することができる。ポリクローナル抗体およびモノクローナル抗体は該ポリペプチドあるいはその断片を抗原として用いて公知の方法により作製することができる。
【0055】
また該ポリペプチドを用いることにより、例えばアフィニティーカラムを作製して、本ポリペプチドと結合する既知または未知の蛋白(リガンド)の同定、精製あるいはその遺伝子クローニングを行なうことができる。
【0056】
また該ポリペプチドを用いて、例えばウエスト−ウエスタン法により、または該cDNA(好ましくは該ポリペプチドをコードするcDNA)を用いて、例えば酵母2−ハイブリッド法により該ポリペプチドと相互作用する分子の同定、遺伝子クローニングを行なうこともできる。
【0057】
さらに本ポリペプチドを用いることによって、本ポリペプチドレセプターアゴニスト、アンタゴニストおよび受容体−シグナル伝達分子間の阻害剤等のスクリーニングを行なうこともできる。
【0058】
スクリーニングは、例えば、以下の方法により行なうことが出来る。すなわち、a)本発明のポリペプチド、スクリーニングすべき化合物、及び細胞を含む反応混合物を、細胞が該ペプチドにより正常に刺激される条件下に一緒にし(該反応混合物は細胞が増殖するに従い細胞中に導入される標識および該ペプチドの機能を効果的に観察させるための該ペプチド以外のペプチドを含む);ついで
b)細胞の増殖の程度を測定して、該化合物が有効なアンタゴニストまたはアゴニストであるかどうかを決定する。
【0059】
本発明のcDNAは、多大な有用性が期待される本発明のポリペプチドを生産する際の重要かつ必須の鋳型となるだけでなく、遺伝病の診断や治療(遺伝子欠損症の治療またはアンチセンスDNA(RNA)によって、ポリペプチドの発現を停止させることによる治療等)に利用できる。
また、本発明のcDNAをプローブとしてゲノミック(genomic)DNAを分離できる。
【0060】
[医薬品への適用]
前記の疾患に適応するために、本発明のポリペプチド、あるいは本発明のポリペプチドに対する抗体は通常、全身的又は局所的に、一般的には経口または非経口の形で投与される。好ましくは、経口投与、静脈内投与および脳室内投与である。
【0061】
投与量は、年齢、体重、症状、治療効果、投与方法、処理時間等により異なるが、通常、成人一人あたり、一回につき、100μgから100mgの範囲で、一日一回から数回経口投与されるかまたは、成人一人あたり、一回につき、10μgから100mgの範囲で、一日一回から数回非経口投与される。
【0062】
もちろん前記したように、投与量は、種々の条件により変動するので、上記投与量より少ない量で十分な場合もあるし、また範囲を越えて必要な場合もある。
【0063】
本発明化合物を投与する際には、経口投与のための固体組成物、液体組成物およびその他の組成物、非経口投与のための注射剤、外用剤、坐剤等として用いられる。
【0064】
経口投与のための固体組成物には、錠剤、丸剤、カプセル剤、散剤、顆粒剤等が含まれる。カプセルには、ソフトカプセルおよびハードカプセルが含まれる。
【0065】
このような固体組成物においては、一つまたはそれ以上の活性物質が、少なくとも一つの不活性な希釈剤(例えば、ラクトース、マンニトール、グルコース、ヒドロキシプロピルセルロース、微結晶セルロース、デンプン、ポリビニルピロリドン、メタケイ酸アルミン酸マグネシウム等)と混合される。組成物は、常法に従って、不活性な希釈剤以外の添加物、例えば、潤滑剤(ステアリン酸マグネシウム等)、崩壊剤(繊維素グリコール酸カルシウム等)、安定化剤(ヒト血清アルブミン、ラクトース等)、溶解補助剤(アルギニン、アスパラギン酸等)を含有していてもよい。
【0066】
錠剤または丸剤は、必要により白糖、ゼラチン、ヒドロキシプロピルセルロース、ヒドロキシプロピルメチルセルロースフタレート等の胃溶性あるいは腸溶性のフィルムで被膜してもよいし、また2以上の層で被膜してもよい。さらにゼラチンのような吸収されうる物質のカプセルも包含される。
【0067】
経口投与のための液体組成物は、薬学的に許容される乳濁剤、溶液剤、懸濁剤、シロップ剤、エリキシル剤等を含み、一般に用いられる不活性な希釈剤(例えば、精製水、エタノール等)を含んでいてもよい。この様な組成物は、不活性な希釈剤以外に湿潤剤、懸濁剤のような補助剤、甘味剤、風味剤、芳香剤、防腐剤を含有していてもよい。
【0068】
経口投与のためのその他の組成物としては、ひとつまたはそれ以上の活性物質を含み、それ自体公知の方法により処方されるスプレー剤が含まれる。この組成物は不活性な希釈剤以外に亜硫酸水素ナトリウムのような安定剤と等張性を与えるような安定化剤、塩化ナトリウム、クエン酸ナトリウムあるいはクエン酸のような等張剤を含有していてもよい。スプレー剤の製造方法は、例えば米国特許第2,868,691号および同第3,095,355号明細書に詳しく記載されている。
【0069】
本発明による非経口投与のための注射剤としては、無菌の水性または非水性の溶液剤、懸濁剤、乳濁剤を包含する。水性または非水性の溶液剤、懸濁剤としては、一つまたはそれ以上の活性物質が、少なくとも一つの不活性な希釈剤と混合される。水性の希釈剤としては、例えば注射用蒸留水および生理食塩水が挙げられる。非水性の希釈剤としては、例えばプロピレングリコール、ポリエチレングリコール、オリーブ油のような植物油、エタノールのようなアルコール類、ポリソルベート80(登録商標)等が挙げられる。
【0070】
このような組成物は、さらに防腐剤、湿潤剤、乳化剤、分散剤、安定化剤(例えば、ヒト血清アルブミン、ラクトース等)、溶解補助剤(例えば、アルギニン、アスパラギン酸等)のような補助剤を含んでいてもよい。
【実施例】
【0071】
以下に本発明のクローンESDNに関する実施例を挙げて本発明をより具体的に説明するが、これらは本発明の範囲を制限するものではない。
【0072】
実施例1:SSTによるスクリーニング
ヒト冠動脈初代培養内皮細胞(hCAEC)および平滑筋細胞(hCASMC)(Clontech)、そしてそれらの共培養(同じ細胞数のhCAECとhCASMCを混ぜ、EGM−2−MV(商品名、BioWhittakerより購入)を用いて2日間培養)をソースとしてcDNAライブラリーを構築した(特願平10-119731号参照)。次に酵母SST法(米国特許No.5,536,637を参照)によりスクリーニングを行なった結果、共培養をソースとしたcDNAライブラリーからCUBドメインを有するESDNを得た。
【0073】
実施例2:ヒト、マウス、ラットの全長cDNAクローニング
全長cDNAを得るためにマラソン・cDNA・アンプリフィケーション・キット(Marathon cDNA amplification kit)(商品名、CLONTECH社より購入)による5’−、3’−RACE(Rapid Amplification of cDNA End)法を用いて5’−、3’−末端cDNAのクローニングを行なった。同時にヒトの塩基配列情報に基づきプライマーを設計し、RT−PCRによりラット、マウスのESDNの単離も行なった。
マウスプライマー
5’−CTGCTCCAACTCCTCCTCCTTC−3’(配列番号10)
5’−CTGCTTCATTCCTTTCCACCAACCTG−3’(配列番号11)
ラットプライマー
5’−TGTGCTGGTCATGGTCCTCACTACTCTC−3’(配列番号12)
5’−TGTGCTTTAAAACGATGCTTTG−3’(配列番号13)
その結果、ESDNは、ヒト、マウス、ラット間において高い相同性を有していることが明らかとなったが、何れの種においても翻訳開始点ATGを含む5’末端配列を見出すことができなかった。この原因としてESDNの5’末端領域の高いGC含量が考えられる。そこでマウスphage genomic ライブラリー(Lambda FIXII library( 商品名、Strategene社より購入) )を用いたスクリーニングを行なった。その結果、翻訳開始点(Met)を含む2個の陽性クローンを得た。そこでこのMetを含む5’−側のプライマーを作成し、下流のエクソンを含む(genomic cloneには含まれていない)アンチセンスプライマー
Sense primer :5’−GCACTATGCGGGCGGATTGC−3’(配列番号14)
antisense primer :5’−GGATGTAAGGGTTCCACTCTCAGG−3’(配列番号15)
と共にRT−PCRを行なったところ、ヒト、ラットのカウンターパートを得た。得られたアミノ酸シークエンスよりヒトとげっ歯類は84〜5%の相同性、マウスとラットでは92%の相同性を表した。ヒト、ラットおよびマウスのESDNのアライメントを図1に示す。
モチーフ検索の結果から、ESDNは、細胞外に1個のCUBドメインと1個のファクター(factor)V/VIII相同ドメインを有するタイプ(type)I膜貫通型タンパク質であることが判明した。また、この構造上の特徴からneuropilin(2個のCUB、2個のFV/VIIIドメイン、1個のMAMドメインを有する)との構造の類似性を指摘することができる。またその他の領域では、カブトガニのファクター(Factor)C、及びヒトにおける難聴の原因遺伝子の一つであるCochと4個のシステインが保存された相同性の高い領域がある。この領域はCochにおいては、DFNA9という遺伝性難聴においてこれまで発見されている4つの変異全てが局在している部分で、カブトガニのような進化系統樹で離れた生物にも見出されることから新しいドメイン構造と考えられる。図2および図3に示す。
【0074】
実施例3:哺乳動物細胞を用いたヒトESDN蛋白の発現と抗ペプチド抗体による同定
ヒト全長cDNAを哺乳動物細胞用発現ベクターのオリジナルV5エピトープをファッグ・タグ(FLAG tag)に交換し、pEF6V5−His(商品名、Invitrogen社より購入)にサブクローニングした。これらの発現ベクターは、293T細胞やCOS7細胞にセルフェクト(Cellphect)(商品名、Amersham Life Science社より購入)やリポフェクトアミン(Lipofectamine)(商品名、Life Technologies、社より購入)により細胞に導入した。なお目的の発現蛋白質の確認は、細胞のライセイト(lysate)を作成した後、各種抗体によるウエスタンブロッティングにより検出した。
anti-FLAG M2 monoclonal antibody( 商品名、Sigma社より購入)
GlyGluArgIleArgIleLysPheGlyAspGlyAspIleGluAspSerAsp(配列番号16)を用いて作成したウサギ抗CUBポリクローナル抗体
GlnAspLysIlePheGlnGlyAsnLysAspTyrHisLysAspValArgAsnAsn(配列番号17)を用いて作成した抗FV/VIII ポリクローナル抗体
は、それぞれKLHに連結させたポリペプチドをウサギに免疫することによって得た(Sawady Technology) 。
ウエスタンブロッティングは、ECL(商品名、Amersham Life Science社より購入)ルネッサンス(Renaissance)(NEN Life Science)のプロトコールに従った。その結果、発現ベクターのみを導入した細胞ライセイト(lysate)には認められないバンドが、127、106、93kDa付近に検出された(図6に示す。)。
【0075】
実施例4:シグナル配列の解析
得られたアミノ酸配列の疎水性プロット(図4に示す。)から、ESDNのシグナル配列は、通常の配列よりも長く異型性であることが判明した。そこでヒト全長cDNAを導入したCOS7細胞を4%パラホルムアルデヒドにより固定した後、一次抗体により30分間室温で反応させた。次に30分間二次抗体、テキサス・レッド・アンチマウスIgG(Texas red anti-mouse IgG)(商品名、Vector Laboratories社より購入)やFITCアンチラビットIgG(FITC-anti-rabbit IgG)(商品名、Jackson Laboratories社より購入)で反応させた後、バイオ・ラッド・コンフォーカル・レーザー・スキャニング・マイクロスコープ(Bio-Rad confocal laser scanning microscope)(商品名、model MRC-1024)により解析した。その結果、同タンパク質は、細胞表面上に発現していることが判った(図7に示す。)。
次にシグナル配列の切断部位の確認を試みた。
マウスESDNの細胞外領域のC端にHis−tagを付けたmESDN−Ex(Edns)やヒトCD5のシグナル配列:MetProMetGlySerLeuGlnProLeuAlaThrLeuTyrLeuLeuGlyMetLeuValAlaSerValLeuAla(配列番号18)に置換したmESDN−Ex(CD5)をpCAGGS(Science 261, 600-603 (1993))にサブクローニングした。293T細胞に導入後、培養上清に分泌された目的タンパク質を「His-probe H-15 polyclonal antibody」(商品名、Santa Cruz社より購入)によるウエスタンブロッティングにより検出した。その結果、mESDN−Ex(Edns)とmESDN−Ex(CD5)の両コンストラクト(図5に示す。)より同じサイズのタンパク質が確認された。ヒトCD5とマウスESDNのシグナル配列は、39アミノ酸(約4kDa)ヒトCD5の方が小さいが、得られたタンパク質のサイズが同じことから、同じ切断部位であることが予想された(図8に示す。)。
【0076】
実施例5:ESDN蛋白のサザーン・ズーブロット(southern zooblot)解析
[32P]dCTP−ラベルしたヒトESDNをプローブとして、哺乳類(マウス、ラット、ウサギ、ウシ、ヒト)、アフリカツメガエル(Xenopus)、ハエ(fly)、酵母(yeast)の種におけるサザーン・ズーブロット(southern zooblot)解析を行なった(37℃、1×SSCにより洗浄)。その結果、哺乳類においては強いバンドが確認され、ESDNが、哺乳類において高度に保存されていることが明らかとなった。一方、アフリカツメガエル(Xenopus)では弱いバンドが確認された。なお、ショウジョウバエ(fly)、酵母(yeast)においては、バンドは確認されなかった(図9Aに示す。)。
【0077】
実施例6:ヒトESDNの遺伝子座
ヒトにおいては、2個(stSG29921、sts-D29024) の独立したSTS(sequence-tagged site)の情報からラジエーション・ハイブリッド・マップ(radiation hybrid map)のD3S1603-D3S1271、D3S1552-D3S1603近傍(cytogenetic mapではchromosome 3q11.2に相当すると思われる部位)に遺伝子座があることが明らかとなった。このデータベースにより得られた結果を確認するために、マウスライン(Cell line)であるA9(Neo3)、A9(Neo12)(JCRB Cell Bank)を用いてゲノミック・サザン(genomic southern)ハイブリダイゼーションを行なった。その結果、ヒトESDNプローブは、マウスESDNも認識できるためA9(Neo3)、A9(Neo12)の両方においてマウスESDNが確認された。一方、ヒトESDNは、A9(Neo3)のみにおいて検出された。このことからヒトESDNは、クロモソーム(chromosome)3q11.2 に相当すると思われる部位に遺伝子座があることが明らかとなった(図9Bに示す。)。
【0078】
実施例7:ESDN mRNAの発現部位の解析
培養ヒト細胞のノザン(Northern)解析は、hCAEC、hCASMCそしてそれらの共培養からトータル(total)RNAをTRIzol(商品名、Life Technologies社より購入)により抽出した。ラット組織・細胞のノザン(Northern)解析は、トータル(total)RNAをTRIzol LS(全血に対して)、TRIzol(その他の組織や培養細胞に対して)により抽出した。次にpoly(A)RNAは、「OligotexTM-dT30 Super」(商品名、Roche Molecular Biochemicals社より購入)により精製した。次に[32P]dCTP−ラベルしたヒト、ラットESDNやGAPDH cDNAをプローブとして用い、ノザン・ブロット(Northern blot)解析(65℃、0.2 ×SSCにより洗浄)を行なった。その結果、hCASMC細胞において強い発現が認められた(6.4 、3kb)。一方hCAEC細胞においては、hCASMCに比べて弱い発現が認められた。なお、共培養することによる、ESDN mRNAの発現の変化は認められなかった(図10Aに示す。)。
ラット組織・細胞のノザン(Northern)解析の結果、全血細胞からESDN mRNAは確認されず、肝臓においても発現がかなり低いことが判った(図10Bに示す。)。
【0079】
実施例8:ヒト冠動脈平滑筋細胞を用いたESDN蛋白の発現
hCASMCを血清(serum)飢餓の状態のDMEM/2mMグルタミン(glutamine)で48時間培養した。次に指定濃度のPDGF−BB、AT−II、FCS(商品名、Sigma 、Life Technologies社より購入)を含む培地に置換し、刺激を行なった。トータル(total)RNAはTrizol(商品名、Life Technologies社より購入)により調製し、cDNA合成はスーパースクリプト・プレアンプリケーション・システム(SuperScript Preamplification System)(商品名、First Strand cDNA Synthesis :Life Technologies 社より購入)を用いて行なった。なおmRNAは、リアルタイム定量RT−PCR(real-time quantitative RT-PCR;PE Applied Biosystems Prism Model 7700 Sequence Detection System)により測定した。フォワード(Forward)リバース(Reverse)プライマーの配列は以下の通りである。
ESDN forward: 5’−CCCAGCAAGGTGATGGATG−3’(配列番号19)
ESDN reverse: 5’−CAAGAATCAGAATCTTCAATGTCAAAG−3’(配列番号20)
ESDN probe: 5’−(6−FAM)−CCTGAGAGTGGAACCCTTACATCCATAAAC−(TAMRA)−3’(配列番号21)
これらの配列はヒトを基準にしているが、げっ歯動物にも適応できることを確認した。
ヒトGAPDHのシークエンスは以下の通りである。
human GAPDH forward: 5’−GAAGGTGAAGGTCGGAGTC−3’(配列番号22)
human GAPDH reverse: 5’−GAAGATGGTGATGGGATTTC−3’(配列番号23)
human GAPDH probe: 5’−(VIC)−CAAGCTTCCCGTTCTCAGCC−(TAMRA)−3’(配列番号24)
「TaqMan Rodent GAPDH Control Reagents」(PE biosystems)をラット(rat)GAPDHの定量に用いた。ESDNとGAPDHのmRNAレベルはコピー数を意味している。そこで先方の発現レベルをGAPDHにより標準化した。このことは、スタンダードの連続希釈から作成した標準曲線によって成し遂げられた。なおスタンダードにはESDNあるいはGAPDHをpBlueScript SK(−)(商品名、Stratagene社より購入)にサブクローニングした既知量のプラスミドを準備した。
結果、ESDNはPDGF−BB刺激において濃度依存的に発現が増加するが、AT−II刺激においては発現の増加が認められなかった。またFCS刺激においてもESDNの発現増加が濃度依存的に認められたが、PDGF−BB刺激に比べてより少ない量であった(図11に示す。)。
【0080】
実施例9:ラットの総頸動脈におけるESDN蛋白の発現
バルーン障害後(n=5,day 0,5,n=4,day 14)、0、5、14日後に頸動脈を採取した。トータル(total)RNAはTrizol(商品名、Life Technologies 社より購入)により調製し、cDNA合成は「SuperScript Preamplification System 」(商品名、First Strand cDNA Synthesis :Life Technologies 社より購入)を用いて行なった。ラットESDNやGAPDHのmRNAは「real-time quantitative RT-PCR 」(PE Applied Biosystems Prism Model 7700 Sequence Detection System)により測定した。その結果、5日目にESDNの増加の傾向が認められ、14日目には30%もの有意な増加が認められた。次に、このESDNの発現を免疫組織学的に解析した(図12に示す。)
処置あるいは未処置ラットを麻酔下にして、予め4℃に冷やした生理食塩水で組織を灌流した。そして局所灌流部分を4%パラホルムアルデヒドにより固定した。頸動脈を丁寧に採取し、ドライアイス−エタノール・バス上に置いた「Tissue-Tek O.C.T. Compound」(商品名、Sakura Finetechnical社より購入)により包埋した。組織を4μmの厚さにスライスし、「avidin-biotin-alkaline phosphatase complex」(商品名、Vector Laboratories 社より購入) 法により免疫組織解析を行なった。アルカリンホスファターゼ(Alkaline phosphatase)は「Vector Red」(商品名、Vector Laboratories 社より購入)により発色させた。切片はメチルグリーンにより対比染色を行なった。ウサギ抗ペプチドポリクローナル抗体(一次抗体)は、5〜10μg/mlの濃度で用いた。ネガティブコントロールには、正常ウサギIgG(DAKO)を同濃度で用いた。その結果、血管平滑筋の存在する大動脈や総頸動脈の中膜が染まるものの、むしろ脳・脊髄といった中枢神経系の一部(矢印)や迷走神経(アローヘッド)などの末梢神経が強く染色された(図13に示す。)。A、B、EおよびFは抗CUB抗体で染色した。C、D、GおよびHは、同じ濃度のウサギの抗IgG抗体を一次抗体として染色した(コントロール)。EおよびF中のMおよびNは、それぞれ血管中膜および血管内膜を示す。
【0081】
実施例10:293T細胞を用いたBrdUの取りこみ測定
全長ESDN、およびそのディレーション・ミュータント(deletion mutants)(hESDN−ΔEC、hESDN−ΔCy)を発現ベクター「QIAfilter Plasmid Midi Kit」(商品名、QIAGEN社より購入)により調整した。そしてさらに精製するためフェノール(phenol)/CIAAにより2回抽出し、CIAA抽出を1回行なった。セルフェクト(CellPhect)(商品名、Amersham Life Science 社より購入)を用いて293T細胞にトランスフェクトした。12時間後、メディウムを新鮮なDMEM/10%FCSに交換し、2時間インキュベーションした。次にトリプシンにより細胞を一旦回収した後、96穴プレート2枚に播き直した。一方のプレートは、2時間のBrdUの取り込みを24時間の培養後に行なった。BrdUの取り込みは、「Cell Proliferation ELISA、BrdU」(colorimetric)(商品名、Roche Diagnostics社より購入)により測定した。もう一方のプレートは、播きこみ後2 時間細胞数を「Premix WST-1 assay kit」(商品名、TaKaRa Biomedicals社より購入)により測定した。その結果、全長ESDNを発現させた細胞において、BrdUの取りこみが有意に抑制されることが明らかとなった。一方細胞外ドメインを欠いたものでは、この効果が弱められた。また細胞内ドメインを欠いたものでは全くこの効果が無くなることが確認された(図14に示す。)。
【図面の簡単な説明】
【0082】
【図1】ヒト、マウスおよびラットESDNのアライメントを表わす。
【図2】ESDN、ニューロピリンおよびCochにおけるドメイン構造を表わす。
【図3】ヒト、マウス、ラットESDN、リムラスファクターC、ラットLgl−1、ヒトCoch、マウスCochおよびラットCochのLCCL部分のアライメントを表わす。
【図4】ヒトおよびマウスESDNのハイドロフォビシティーを表わす。
【図5】発現ベクターの構成を表わす。
【図6】ヒト全長ESDNのウエスタンブロッティング解析を表わす。
【図7】細胞膜表面でESDNが発現されている様子を表わす。
【図8】開裂したシグナル配列のウエスタンブロッティング解析を表わす。
【図9】AはヒトESDNのcDNAを用いたサザンズーブロッティングを表わす。BはヒトESDNがクロモソーム3にあることを示す。
【図10】AおよびBは、ヒト頸動脈細胞のRNAおよびラット各臓器のRNAを用いたESDNのcDNAとのノザンブロッティング解析を示す。
【図11】ヒト平滑筋細胞において、PDGFまたはFCS刺激によりESDNのmRNAが誘導される様子を示す。
【図12】ラット頸動脈をバルーンで傷害することにより、ESDNのmRNAの発現が増加することを示す。
【図13】ラット頸動脈をバルーンで傷害することにより、ESDNの発現が増加することを示す。
【図14】ESDN過発現細胞においてBrdU取り込みが抑制されていることを示す。
【特許請求の範囲】
【請求項1】
実質的に純粋な形である配列番号2、5または8で示されるアミノ酸配列からなるポリペプチド。
【請求項2】
請求項1に記載されたポリペプチドをコードするcDNA。
【請求項3】
配列番号3、6または9で示される塩基配列を有する請求項2記載のcDNA。
【請求項4】
請求項3に記載のcDNAからなる複製または発現ベクター。
【請求項5】
請求項4記載の複製または発現ベクターで形質転換された宿主細胞。
【請求項6】
請求項1に記載されたポリペプチドを発現させるための条件下で請求項5記載の宿主細胞を培養することを特徴とする請求項1に記載のポリペプチドの製造方法。
【請求項7】
請求項1に記載されたポリペプチドのポリクローナルまたはモノクローナル抗体。
【請求項8】
請求項1に記載されたポリペプチドまたは請求項7記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とするPTCA後の再狭窄の治療に有効な薬学的組成物。
【請求項9】
請求項1に記載されたポリペプチドまたは請求項7記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とする動脈硬化の治療に有効な薬学的組成物。
【請求項1】
実質的に純粋な形である配列番号2、5または8で示されるアミノ酸配列からなるポリペプチド。
【請求項2】
請求項1に記載されたポリペプチドをコードするcDNA。
【請求項3】
配列番号3、6または9で示される塩基配列を有する請求項2記載のcDNA。
【請求項4】
請求項3に記載のcDNAからなる複製または発現ベクター。
【請求項5】
請求項4記載の複製または発現ベクターで形質転換された宿主細胞。
【請求項6】
請求項1に記載されたポリペプチドを発現させるための条件下で請求項5記載の宿主細胞を培養することを特徴とする請求項1に記載のポリペプチドの製造方法。
【請求項7】
請求項1に記載されたポリペプチドのポリクローナルまたはモノクローナル抗体。
【請求項8】
請求項1に記載されたポリペプチドまたは請求項7記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とするPTCA後の再狭窄の治療に有効な薬学的組成物。
【請求項9】
請求項1に記載されたポリペプチドまたは請求項7記載の抗体および薬学的に許容される賦形剤および/または担体を含有することを特徴とする動脈硬化の治療に有効な薬学的組成物。
【図1】



【図2】
【図3】

【図4】
【図5】
【図6】
【図7】

【図8】
【図9】

【図10】

【図11】
【図12】

【図13】
【図14】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公開番号】特開2007−130024(P2007−130024A)
【公開日】平成19年5月31日(2007.5.31)
【国際特許分類】
【出願番号】特願2006−344358(P2006−344358)
【出願日】平成18年12月21日(2006.12.21)
【分割の表示】特願2001−397725(P2001−397725)の分割
【原出願日】平成13年12月27日(2001.12.27)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2001年7月7日 発行の「The Journal of Biological Chemistry Vol.276,No.36,」に発表
【出願人】(000185983)小野薬品工業株式会社 (180)
【出願人】(396023812)
【Fターム(参考)】
【公開日】平成19年5月31日(2007.5.31)
【国際特許分類】
【出願日】平成18年12月21日(2006.12.21)
【分割の表示】特願2001−397725(P2001−397725)の分割
【原出願日】平成13年12月27日(2001.12.27)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2001年7月7日 発行の「The Journal of Biological Chemistry Vol.276,No.36,」に発表
【出願人】(000185983)小野薬品工業株式会社 (180)
【出願人】(396023812)
【Fターム(参考)】
[ Back to top ]