
核酸を検出するためのプライマ伸長法
【課題】異質性生体試料の多数のゲノム突然変異に向けた、選択性および感受性のある核酸検出法ならびに信頼性のある大規模スクリーニング法を提供すること。
【解決手段】本発明は、高い感受性と高い選択性を有する突然変異検出法を提供する。一般的実施例においては、本発明は、少なくとも1回反復する単一塩基伸長反応を含む。本発明の方法は、異質性生体試料において、遺伝子突然変異または疾患を引き起こす微生物の存在を検出および同定する際に有用である。
【解決手段】本発明は、高い感受性と高い選択性を有する突然変異検出法を提供する。一般的実施例においては、本発明は、少なくとも1回反復する単一塩基伸長反応を含む。本発明の方法は、異質性生体試料において、遺伝子突然変異または疾患を引き起こす微生物の存在を検出および同定する際に有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本特許出願は、1997年6月16日に提出されたU.S.Serial Number 08/877,333の一部継続出願であり、その開示は本明細書に引例として包含する。
【0002】
[発明の属する技術分野]
本発明は、一般に、核酸試料中の単一ヌクレオチドを同定するためのオリゴヌクレオチドプライマ伸長法に関する。本発明の方法は、生体試料中の遺伝子突然変異の存在、または疾患を引き起こす微生物の存在を検出および同定することにより、疾患診断に有用となる。
【背景技術】
【0003】
[発明の背景]
遺伝性疾患等の疾患を引き起こす分子欠損の知見は、急速に増加している。遺伝子突然変異が引き起こすと考えられる遺伝性疾患には、鎌型赤血球貧血症、α−およびβ−地中海貧血、フェニルケトン尿症、血友病、α1−アンチトリプシン欠損症、ならびに膵嚢胞性繊維症が挙げられる。例えば、鎌型赤血球貧血症は、β−グロブリン遺伝子の6番目のコドンでの単一塩基対置換がもたらす同型接合性によるものと報告されている。Antonarakis,New England J.Med.320:153−163(1989)。インシュリン受容体遺伝子およびインシュリン応答性グルコース輸送体遺伝子の突然変異が、インシュリン抵抗性糖尿病で検出されている。非特許文献1。
【0004】
癌は、多数の癌遺伝子および腫瘍抑圧遺伝子の遺伝子突然変異に関連してきた。Duffy,Clin.Chem.,41:1410−1413(1993)。例えば、そのras遺伝子の点突然変異が、その遺伝子を形質転換癌遺伝子に転換させることが示されている。Bos et al.,Nature,315:725−730。p53腫瘍抑圧遺伝子座での突然変異および異型接合性の喪失は、種々の癌に関連してきた。Ridanpaa et al.,Path.Res.Pract,191:399−402(1995);Hollstein et al.,Science,253:49−53(1991)。加えて、apoおよびdoc腫瘍抑圧遺伝子の欠損またはその他の突然変異もまた、腫瘍発生に関連してきた。Blum,Europ.J.Cancer,31A:1369−1372(1995)。それらの突然変異は、疾患の初期ステップおよびそれの素因に対しマーカの役割を果たす可能性がある。初期診断は、治療の成功に重要なだけでなく、慢性的徴候が生じる前に予防または治療を導入することも可能にする。
【0005】
結腸直腸癌は、初期に検出されれば治癒される可能性が高い疾患の一例である。結腸癌は、初期の検出により、例えば癌にかかった組織の外科的除去で、効果的に治療し得る。結腸癌は、結腸上皮の細胞に始まり、初期ステップでは大循環(general circulation)から分離されるため、初期ステップの結腸癌の外科的除去は通常は成功する。こうして、結腸直腸細胞の初期突然変異の検出は、一般に、生存率を大幅に上昇させるはずである。結腸直腸癌の現行の検出法は、糞便潜血の存在または血清中を循環する癌胎児性抗原の存在等、癌の存在を示す細胞外証拠に集中している。そのような細胞外証拠は、典型的には、癌が浸潤性になった後にのみ生じる。この意味で、結腸直腸癌は治療が非常に困難である。
【0006】
疾患関連遺伝子内の突然変異の存在を検出するために、方法が改正されてきた。そのような方法の1つは、試料のゲノム領域の完全なヌクレオチド配列を、相当する野生型領域と比較することである。例えば、Engelke et al.,Proc.Acad.Sci,U.S.A.85:544−548(1988)、Wong et al.,Nature,330:384−386(1988)を参照されたい。しかし、そのような方法では費用と時間がかかり、低頻度突然変異を明白に検出するために、標的遺伝子の多重クローン(multiple clones)を分析する必要がある。そのため、遺伝子突然変異の大規模スクリーニングに、広範囲の塩基配列決定を利用するのは実用的ではない。
【0007】
酵素的および化学的切断技術を用いてDNAの配列変異を活用する様々な検出方法が開発されてきた。DNA多型のために一般的に用いられるスクリーニングは、Botstein et al.,Am.J.Hum.Genet.,32:314−331(1980)とWhite et al.,Sci.Am.,258:40−48(1988)により報告されるように、制限エンドヌクレアーゼでDNAを消化することと、生成した断片をサザンブロット法で分析することからなる。エンドヌクレアーゼの認識配列に影響を与える突然変異は、その部位での酵素的切断を妨害し、それによりDNAの切断様式を変化させる。制限断片長の差を捜索することにより、配列を比較する。この方法(制限断片長多型マッピングもしくはRFLPマッピングと呼ばれる)での問題は、制限エンドヌクレアーゼによる切断に影響を与えない突然変異の検出が不可能なことである。ある試験は、ヒトDNAの40,000塩基対領域での存在が推定される突然変異体で、RFLP分析により検出されるのがわすか0.7%であることを報告した。Jeffrey,Cell,18:1−18(1979)。
【0008】
単一塩基突然変異は、対立遺伝子特異性オリゴヌクレオチド(ASO)プローブを用いた差分ハイブリッド形成法により検出されてきた。Saiki et al.,Proc.Natl.Acad.Sci.USA.56:6230−6234(1989)。突然変異は、ミスマッチしたプローブと比較して、完全にマッチしたプローブのより高い熱安定性を基に同定する。突然変異分析のこの方法の欠点には、:(1)各プローブでハイブリッド形成を最適化する必要があることと、(2)プローブの判別度でのミスマッチおよび局所配列の適用制限(local sequence impose limitations)の性質とが挙げられる。実際に、検査試料の配列複雑性が高い場合(例えば異質性生体試料)には、核酸ハイブリッド形成のパラメータのみに基づく検査は機能性が乏しくなる。これは、幾分かは、単一ヌクレオチドの変化により生じるハイブリッド安定性の小さな熱力学的差異による。つまり、核酸ハイブリッド形成は、一般に、分析および診断目的で、その他幾つかの選択または強化手順を組み込んでいる。
【0009】
酵素を介したライゲーション法では、突然変異は、標的DNA分子またはRNA分子上で互いに直接隣接してアニーリングすることが可能な2つのオリゴヌクレオチドにより識別され、オリゴヌクレオチドの1つは、点突然変異に相補的な3’末端を有する。隣接するオリゴヌクレオチド配列は、両方のヌクレオチドが正しくの塩基対を形成する場合には、共有結合のみの結合である。こうして、点突然変異の存在は、2つの隣接するオリゴヌクレオチドのライゲーションにより示される。Grossman et al.,Nucleic Acid Research,22:4527−4534(1994)。しかし、この検出方法の有用性は、あるヌクレオチドミスマッチの許容性から、または非鋳型指向性ライゲーション反応(non−template directed ligation reactions)から生じる高いバックグランドにより損なわれる。Barringer et al.,Gene,89:117−122(1990)。
【0010】
鋳型依存性プライマ伸長反応に基づき、検出法が多数開発されてきた。これらの方法は、主に2つのカテゴリに分類される:(1)突然変異を識別する領域をつなぐプライマを用いた方法、および(2)突然変異を識別する領域の近傍と上流でハイブリッド形成するプライマを用いた方法。
【0011】
最初のカテゴリでは、ミスマッチのあるものに比べ完全にマッチしたプライマの結合に有利なハイブリッド形成条件下で、標的核酸の単一塩基突然変異が拮抗的オリゴヌクレオチドプライミングにより検出されるような方法を、CaskeyとGibbs(U.S.Patent No.5,578,458)が報告している。VaryとDiamond(U.S.Patent No.4,851,331)は、プライマの3’末端ヌクレオチドが、当該の変異体ヌクレオチドに相当する、同様の方法を記載している。プライマの3’末端におけるプライマと鋳型とのミスマッチが延長を阻害するために、通常のプライマ伸長条件下では、トレーサヌクレオチドの取り込み量に有意差が生じる。
【0012】
プライマ依存性DNAポリメラーゼが、一般に、低い複製エラー率を有するとは知られて久しい。この特性は、後代への障害作用を有する遺伝的誤りを防ぐ上で重要である。2番目のカテゴリの方法は、この酵素学的反応に特有の高い忠実度を活用している。突然変異の検出は、プライマ伸長と検出可能な鎖終結ヌクレオチド三リン酸の取り込みに基づいている。DNAポリメラーゼの高い忠実度により、レポータ分子を標識した正しい塩基の特異的取り込みが確実に行われる。そのような単一ヌクレオチドプライマ誘導性伸長測定は、アスパルチルグリコサミン尿症、血友病B、および膵嚢胞性繊維症を検出するために、さらにレーバー遺伝性視神経萎縮(LHON)に関連する点突然変異を定量するために用いられてきた。例えば、Kuppuswamy et al.,Proc.Natl.Acad.Sci.USA,88:1143−1147(1991)、Syvanen et al.,Genomics,8:684−692(1990)、Juvonen et al.,Human Genetics,93:16−20(1994)、Ikonen et al.,PCR Meth.Applications,1:234−240(1992)、Ikonen et al.,Proc.Natl.Acad.Sci.USA,88:11222−11226(1991)、Nikiforov et al.,Nucleic Acids Research,22:4167−4175(1994)を参照されたい。分子量に基づき伸長したプライマの分解能を高めるために、鎖終結ヌクレオチドの前に幾つかのヌクレオチドを添加することを含む別のプライマ伸長法も提案されている。例えば、Fahy et al.,WO/96/30545(1996)を参照されたい。
【0013】
プライマ伸長に基づく方策には、完全にアニーリングしたオリゴヌクレオチドのみが、伸長反応のプライマとして確実に機能するためにかなりの最適化が必要となる。ポリメラーゼの高い忠実度が与える利点は、プライマの鋳型へのハイブリッド形成におけるヌクレオチドミスマッチの許容性により損なわれる可能性がある。あらゆる「不正確な」プライミングを、真の陽性シグナルと区別することは困難である。
【0014】
オリゴヌクレオチドプライマ伸長測定の選択性および感受性は、オリゴヌクレオチドプライマの長さと反応条件とに関係する。一般に、高い選択性に有利なプライマ長と反応条件は、低い感受性をもたらす。これとは逆に、高い感受性に有利なプライマ長と反応条件は、低い選択性をもたらす。
【0015】
典型的な反応条件下では、短いプライマ(即ち約15マー未満)は一時的で不安定なハイブリッド形成を呈する。そのため、一時的で不安定なオリゴヌクレオチドハイブリッドは、伸長反応を容易に開始せず、伸長したオリゴヌクレオチドの低い収率をもたらすため、短いプライマを使用する場合には、プライマ伸長測定の感受性が低くなる。その上、複雑な異質性生体試料では、短いプライマは、様々な種類の完全にマッチした相補的配列への非特異的結合を呈する。こうして、それらの低い安定性と高い非特異的結合のために、周知の位置にある突然変異の信頼性のある同定において、短いプライマはさほど有用ではない。そのため、プライマ伸長測定に基づいた検出法は、15マー〜25マーの長さ範囲のオリゴヌクレオチドプライマを使用する。例えば、特許文献1;特許文献2および特許文献3を参照されたい。しかし、安定性を増大させるプローブの延長は、選択性を低減させる作用を有する。ミスマッチしたプライマと完全にマッチしたプライマとの熱力学的差異は比較的小さいため、単一塩基ミスマッチは通常、より長いオリゴヌクレオチドプライマの結合効率に、より短いプライマに対してよりも低い作用を有する。より長いプライマの鋳型へのハイブリッド形成で、ヌクレオチドのスマッチの許容性が高い程、複雑な異質性生体試料での非特異的で「不正確な」プライミングが高レベルでもたらされる可能性がある。
【0016】
プライマ伸長反応の反応条件は、ミスマッチしたオリゴヌクレオチドによる「不正確な」プライミングを減少させるよう最適化することが可能である。しかし、最適化は、過酷で費用のかかる労働であり、伸長したプライマの収率減少により低い感受性をもたらすことが多々ある。その上、完全にアニーリングされたオリゴヌクレオチドのみが伸長反応のプライマとして確実に機能するためには、かなりの最適化が必要とされるため、プライマ伸長測定の限定されたマルチプレックシングのみが可能となる。Krook et al.,supra(非特許文献1)は、2つの別個の単一ヌクレオチド取り込み反応において、異なる長さのプライマを用いることにより、および各突然変異部位の野生型ヌクレオチドと突然変異ヌクレオチドとをモニタリングすることにより、マルチプレックシングが達成し得ることを報告している。しかし、オリゴヌクレオチドプライマ伸長測定の選択性および安定性が、オリゴヌクレオチドプライマの長さと反応条件により決定されるとすれば、所与の反応混合物において同時に検査し得るプライマ数は、非常に限定されてしまう。
【0017】
当該技術分野の方法は、検査試料の配列複雑性を減少させることにより、不正確なプライミングの可能性を低下させる。こうして、ゲノムDNAは、生体試料から分離され、かつ/または識別される領域の側面に作用する(flank)プライマを用いたPCRにより増幅される。その後、プライマ伸長分析は、精製したPCR生成物上で実施される。特許文献1、特許文献2、および特許文献3を参照されたい。しかし、これらの方法は試料の処理という追加的ステップを含むため、時間と費用がかかる。その上、これらの方法は、単一試料における多重プライマ伸長反応(multiple primer extension reaction)には適応しない。
【先行技術文献】
【特許文献】
【0018】
【特許文献1】国際公開第91/13075号パンフレット
【特許文献2】国際公開第92/15712号パンフレット
【特許文献3】国際公開第96/30545号パンフレット
【非特許文献】
【0019】
【非特許文献1】Krool et al.,Human Molecular Genetics,1:391−396(1992)
【発明の概要】
【発明が解決しようとする課題】
【0020】
そのため、当該技術分野では、異質性生体試料の多数のゲノム突然変異に向けた、選択性および感受性のある核酸検出法ならびに信頼性のある大規模スクリーニング法が求められている。そのような方法を本明細書において提供する。
【課題を解決するための手段】
【0021】
[発明の概要]
本発明は、高い感受性と高い選択性を有する突然変異検出法を提供する。一般的実施例においては、本発明は、少なくとも1回反復する単一塩基伸長反応を含む。本発明の方法は、異質性生体試料において、遺伝子突然変異または疾患を引き起こす微生物の存在を検出および同定する際に有用である。
本発明の方法は、単一塩基伸長反応を複数のサイクルで実施し、それにより選択性を損なうことなくプライマ伸長測定の感受性を増大させることを含む。好ましい実施例においては、本発明の方法は、2〜100サイクルのプライマ伸長を含む。より好ましくは、10〜50サイクル実施する。最も好ましくは、約30サイクル実施する。
好ましい実施例においては、過剰のプライマを用いて、各伸長サイクルにおいて伸長したプライマを確実に生成する。オリゴヌクレオチドのプライマ長は、好ましくは約10〜約100ヌクレオチド、より好ましくは約15〜約35ヌクレオチド、最も好ましくは約25ヌクレオチドである。
【0022】
好ましい実施例においては、各伸長反応は、完全な相補的配列を備える核酸のみへのプライマのハイブリッド形成を促進させる条件を含む(即ち、ミスマッチ塩基対は許容されない)。1つの実施例においては、ハイブリッド形成は、測定でのプライマのほぼTmで実施される。より好ましい実施例においては、ハイブリッド形成は、プライマのTmより高温で実施される。
【0023】
1つの実施例においては、ハイブリッド形成されたオリゴヌクレオチドプライマは、標識された末端ヌクレオチドにより伸長される。標識ddNTPおよびdNTPは、好ましくは伸長プライマの検出を容易にする「検出部分」を含む。検出部分は、蛍光、発光または放射性標識、酵素、ハプテン、および、例えば分光光度法により、標識伸長生成物を容易に検出させる、ビオチン等のその他の化学的識別票からなる群から選択される。
【0024】
好ましい実施例においては、ハイブリッド形成して伸長したプライマを変性し、伸長していないプライマをアニーリングして、新たにハイブリッド形成したプライマを伸長することにより、プライマ伸長のさらなるサイクルを開始する。反応での過剰なプライマの存在は、反応の各サイクルの伸長していないプライマのアニーリングを促進させる。
【0025】
さらなる実施例においては、本発明の方法は、分断したプライマを用いて、少なくとも2サイクルの単一塩基伸長反応を実施することを含む。本発明の方法は、推測される突然変異の部位に隣接する2つのプローブをハイブリッド形成することを含み、ここでいずれのプローブも単独では鋳型依存性伸長のプライマにはなり得ないが、プローブが互いに隣接してハイブリッド形成する場合には、伸長を開始することが可能である。好ましい実施例においては、本発明の方法は、約5塩基〜約10塩基の長さを有するプローブを標的核酸へハイブリッド形成することを含み、ここでプローブは推測される突然変異の直ぐ上流でハイブリッド形成する。本発明の方法は、さらに第一プローブの上流にある第二プローブをハイブリッド形成することを含み、その第二プローブは約15〜約100ヌクレオチドの長さを有し、3’末端の伸長不能なヌクレオチドを有する。第二プローブは、第一プローブと実質的に近接している。好ましくは、実質的に近接するプローブは、0〜約1ヌクレオチド隔てている。リンカが核酸伸長反応を妨害しないとすれば、好ましくは第一および第二プローブが2以上のヌクレオチドにより分離されるようにリンカが使用される。そのようなリンカは、当該技術分野で周知であり、例えば、ペプチド核酸、DNA結合蛋白質、およびライゲーションが挙げられる。
【0026】
別の実施例においては、分断したプライマは、第一オリゴヌクレオチドプローブ系列(a series of first oligonucleotide probe)を含む。第一プローブ系列の各構成員が、標的核酸の実質的な近接部分に同時にハイブリッド形成し、それにより近接するプライマを形成する場合を除けば、前記系列の構成員はいずれも、核酸ポリメライゼーションのプライマにはなり得ない。1つの実施例においては、分断したプライマは、3つの8マー第一プローブを含む。別の実施例においては、分断したプライマは4つの6マー第一プローブを含む。
【0027】
伸長測定の各サイクルにおいて、伸長反応は、鋳型依存法による上記プローブの共同ハイブリッド形成(co−hybridization)からもたらされる分断したプライマにヌクレオチドを付加させる。好ましい実施例においては、標的核酸へハイブリッド形成された第一プローブは、標識した末端ヌクレオチドにより伸長するが、野生型核酸または非標的核酸へハイブリッド形成された第一プローブは、非標識の末端ヌクレオチドで伸長する。標識ddNTPまたはdNTPは、好ましくは標識した末端ヌクレオチドで伸長した短いプローブの検出を容易にする「検出部分」を含む。検出部分は、蛍光、発光または放射性標識、酵素、ハプテン、および、例えば分光光度法により、標識伸長生成物を容易に検出させる、ビオチン等のその他の化学的識別票からなる群から選択される。
【0028】
好ましい実施例においては、測定シグナルを増幅させるために、数サイクルの伸長反応が実施される。伸長反応は、過剰な第一及び第二プローブ、標識dNTPまたはdNTP、ならびに熱安定ポリメラーゼの存在下で実施される。一旦伸長反応が完了すると、標的核酸に結合した第一および第二プローブは、反応混合物をハイブリッドの融解温度を超える温度で加熱することにより分解される。その後、反応混合物は、ハイブリッドの融解温度未満に冷却され、第一および第二プローブはもう1つの伸長反応のために標的核酸と会合させる。好ましい実施例においては、10〜50サイクルの伸長反応が実施される。最も好ましい実施例においては、30サイクルの伸長反応が実施される。
【0029】
本明細書に開示される方法を用いて、単一ヌクレオチド多型(SNP)、つまり挿入、欠失、および置換などの突然変異を検出してもよい。本明細書の方法でスクリーニング可能な核酸試料は、ヒト核酸試料を含む。プライマ(もしくは分断したプライマ)は、ハイブリッド形成されたプライマの3’末端が、測定されるヌクレオチド位置の相補的位置の直ぐ上流にあるように設計される。測定されるヌクレオチド位置は、単一塩基プライマ伸長反応で取り込まれるヌクレオチドに相補的なヌクレオチドとして同定される。例えば、Gがその反応で取り込まれる場合には、Cは生体試料中の核酸の相補的位置に存在する。好ましい実施例においては、プライマ伸長反応は、4つのヌクレオチド、好ましくは鎖終結ヌクレオチド(例えばジデオキシヌクレオチドの、ddATP、ddCTP、ddGTP、およびddTTP)の存在下で実施される。より好ましい実施例においては、ヌクレオチドは検出可能に標識され、好ましくは判別的に標識される。別の実施例においては、伸長反応は、1、2、または3の異なるヌクレオチドの存在下で実施される。測定されるヌクレオチド位置で、生体試料が異質性であれば、プライマ伸長測定では、相補的ヌクレオチドが取り込まれる(それらがプライマ伸長反応に含まれる場合)。
【0030】
本明細書に開示される方法を用いて、癌等の疾患に関連する突然変異を検出してもよい。加えて、本発明の方法を用いて、酵素活性の完全喪失または部分的喪失など、代謝エラーを引き起こす欠失または塩基置換突然変異を検出してもよい。
【0031】
別の実施例においては、特異的核酸配列は、病的症状または症候に関与することが知られるヒトゲノム核酸中の特定遺伝子または遺伝子座の一部を含む。非限定的例には、膵嚢胞性繊維症、テイ−サックス病、鎌型赤血球貧血症、β−地中海貧血、およびゴーシェ病が挙げられる。
【0032】
さらに別の実施例においては、特異的核酸配列は、特定疾患に関連することが知られていないが、多型が知られるまたは推測される、特定遺伝子または遺伝子座の一部を含む。
【0033】
さらに別の実施例においては、特異的核酸配列は、例えば侵入した微生物のゲノム等、外来遺伝子配列の一部を含む。非限定的例には、細菌およびそのファージ、ウイルス、菌類、原生動物等が挙げられる。適切な治療的介入を選択するために、微生物の異なる変異種または菌株を区別することが望ましい場合に、本発明の方法は特に適用できる。
【0034】
ゲノム核酸試料は、生体試料から分離される。一旦分離された核酸は、さらに操作することなく本発明に使用してもよい。別法として、核酸に存在する1以上の特異的領域を、例えばPCRにより増幅してもよい。このステップでの増幅は、標的核酸配列集団内で特異的核酸配列の濃度を増加させるという利益をもたらす。別の実施例においては、ゲノム核酸は、さらに分析される前に切断される。
【0035】
別の実施例においては、核酸は固相支持体に結合させる。これにより、多数の試料の同時処理およびスクリーニングが可能となる。本発明での使用に適した支持体の非限定的例には、ニトロセルロースまたはナイロンのフィルタ、ガラスビーズ、親和性捕獲用の薬剤をコーティングした磁気ビーズ、処理したまたは未処理のミクロタイタープレート等が挙げられる。好ましい実施例においては、支持体は、多数のウェルを有するミクロタイターディッシュである。そのような支持体の使用により、多数の検体および対照の同時測定が可能となり、こうして分析が促進される。その上、自動化装置を使用して、そのようなミクロタイターディッシュへ試薬を提供することも可能である。別の実施例においては、本発明の方法は水相において実施される。
【0036】
本発明の1つの実施例においては、伸長プライマまたはプローブが算出される。プライマまたはプローブは、好ましくはインピーダンスビーズで標識したヌクレオチドで伸長し、インピーダンスビーズの数が計数される(例えばコールターカウンタを用いる)。その後、標識プライマの数は、インピーダンスビーズの数から決定される。標識は、より好ましくは放射性同位体であり、標識プライマまたはプローブに関連する放射性崩壊の量を測定する。標識プライマまたはプローブの数は、放射性崩壊の量から計算する。伸長したプライマまたはプローブの数は、循環した伸長反応の統計解析に有用である。
【0037】
最後に、本発明の方法はさらに、伸長したプライマまたは第一プローブを分離することおよび塩基配列決定することを含む。プライマまたは第一プローブは、好ましくはそれらの分離を容易にする「分離部分」を含む。分離部分の非限定的例には、ハプテン、ビオチン、およびジゴキシゲニンが挙げられる。好ましい実施例においては、分離部分を含むプライマまたは第一プローブは、分離部分に対し親和性を有する(例えば、抗ハプテン、アビジン、ストレプトアビジンまたは抗ジゴキシゲニンでコーティングされた)固形支持体に固定されている。固形支持体は、ガラス、プラスチック、および紙からなる群から選択される。支持体は、円柱、ビーズ、棒、または試験管として形づくられる。好ましい実施例においては、分離部分は、標識ddNTPまたはdNTPに取り込まれて、標識ddNTPまたはdNTPで伸長した第一プローブのみが支持体に固定される。そのことにより、標識したプライマまたは第一プローブが、伸長していないプライマまたは第一プローブ、および第二プローブから分離される。別の好ましい実施例においては、鋳型とハイブリッド形成して伸長する第一プローブの能力を分離部分が妨害しないならば、分離部分は第一プローブ全てに取り込まれる。分離部分を取り込むことにより、第一プローブの全てが固形支持体に固定される。第一プローブは、1回以上の洗浄ステップにより第2プローブから分離される。
【0038】
その後、標識したプライマまたは第一プローブは、突然変異または疾患を引き起こす微生物を同定するために塩基配列決定する。1つの実施例においては、固定したプライマまたはプローブを、例えば当該技術分野で標準的な化学的方法を用いて、塩基配列決定に直接供試される。その他の実施例においては、標識した第一プローブを固形支持体から除去して、水性溶液中で、標識した第一プローブの塩基配列決定を実施する。分離した第一プローブを複数の相補的オリゴヌクレオチドと接触させる。1つの実施例においては、分離した第一プローブをプライマとして、相補的オリゴヌクレオチドを鋳型として用いて、酵素的塩基配列決定を実施する。別の実施例においては、分離した第一プローブをプライマとして、相補的オリゴヌクレオチドを鋳型として用いて、単一塩基伸長反応を実施する。酵素的塩基配列決定により、伸長生成物の配列を決定する。伸長した標識第一プローブの配列が、遺伝突然変異または試料中に存在する、疾患を引き起こす微生物を同定する。
本発明は、さらに、例えば以下の項目を提供する。
(項目1)核酸試料中の単一ヌクレオチドを同定するための方法であって、
(a)オリゴヌクレオチドプライマと核酸試料中の核酸の一部の間の厳密な相補的ハイブリッド形成を促進する条件下で、前記プライマを前記試料へアニーリングするステップと、
(b)単一塩基により前記プライマを伸長するステップと、
(c)前記伸長したプライマを前記部分から分離するステップと、
(d)ステップ(a)〜ステップ(c)を反復するステップと、
(e)前記伸長したプライマに取り込まれた塩基を同定し、それにより前記単一ヌクレオチドを同定するステップと、を含む方法。
(項目2)ステップ(b)の前記塩基が、鎖終結ヌクレオチドである、項目1に記載の方法。
(項目3)ステップ(b)の前記伸長反応が、1以上の鎖終結ヌクレオチドの付加により実施される、項目1または2に記載の方法。
(項目4)前記単一塩基が、検出可能に標識される、項目1〜3のいずれか1項に記載の方法。
(項目5)前記単一ヌクレオチドが、多型変異体である、項目1〜4のいずれか1項に記載の方法。
(項目6)前記試料が、異なる個体からの複数の組織または体液検体から集められた核酸を含む、項目1〜5のいずれか1項に記載の方法。
(項目7)前記プライマが、分断したプライマである、項目1〜6のいずれか1項に記載の方法。
(項目8)前記分断したプライマが、15未満のヌクレオチドを有する第一プライマと、15を超えるヌクレオチドを有する第二プライマとを含む、項目1〜7のいずれか1項に記載の方法。
(項目9)周知の単一ヌクレオチド多型変異体の存在を検出するための方法であって、
(a)オリゴヌクレオチドプライマを、単一ヌクレオチド多型変異体の直ぐ上流にあると推測される核酸の一部へアニーリングするステップと、
(b)単一塩基により前記プライマを伸長するステップと、
(c)前記プライマを前記核酸から分離するステップと、
(d)ステップ(a)〜ステップ(c)を少なくとも1回反復するステップと、
(e)前記単一塩基を同定し、それにより前記多型変異体を同定するステップと、を含む方法。
(項目10)集団中の単一ヌクレオチド多型変異体を同定するための方法であって、
(a)集団の複数の個体の各構成員から核酸を含む試料を得るステップと、
(b)推測される単一ヌクレオチド多型変異体の直ぐ上流の前記核酸の一部に相補的なオリゴヌクレオチドプライマを、各試料中の前記核酸の一部へアニーリングするステップと、
(c)単一塩基により各試料中の前記プライマを伸長するステップと、
(d)前記プライマを各試料中の前記核酸から分離するステップと、
(e)ステップ(b)〜ステップ(d)を少なくとも1回反復するステップと、
(f)各試料中の前記単一塩基を同定するステップと、
(g)単一ヌクレオチド多型変異体を、ステップ(f)で同定された1以上の塩基の存在として同定するステップと、を含む方法。
(項目11)前記試料が混合されている、項目10に記載の方法。
(項目12)単一ヌクレオチド多型変異体が疾患の証拠である、項目9〜11のいずれか1項に記載の方法。
(項目13)前記疾患が、癌、膵嚢胞性繊維症、テイ−サックス病、鎌型赤血球貧血症、ゴーシェ病、β−地中海貧血、および寄生虫感染からなる群から選択される、項目9〜12のいずれか1項に記載の方法。
本発明の更なる様相および利益は、以下に詳述するそれの説明を検討することにより明白となる。
【図面の簡単な説明】
【0039】
【図1】単一塩基多型を検出するための単一塩基伸長反応において、分断したプライマの使用を記述する線図である。白線は鋳型を表し、濃灰色は第一プローブ(薄灰色)に実質的に近接する鋳型の領域へハイブリッド形成する第二プローブを表す。単一塩基突然変異が推測される部位はAと標識する。検出可能な標識をBと印字する。
【図2A】統計確率の低い領域を示すガウス分布モデルである。
【図2B】統計確率の低い領域を示すガウス分布モデルである。
【図3】細胞の異質性集団で細胞の1%が突然変異している確率値Nを示すグラフである。
【発明を実施するための形態】
【0040】
[発明の詳細な説明]
本発明は、第一に、高い選択性と高い感受性の両者を備える単一塩基伸長測定を提供することである。本発明は、高い感受性と高い選択性の両者を備える生体試料の特異的核酸を検出する方法を提供する。本発明は、伸長生成物の低い収率により感受性を喪失することなく、厳しいハイブリッド形成条件の高い選択性を提供する。一般に、本発明の方法は、生体試料中で複数サイクルの単一塩基伸長反応を実施することを含む。循環により、伸長生成物の収率は高くなり、プライマのハイブリッド形成条件が、単一塩基伸長反応中に典型的に適用されるものに比べ厳しく保たれるために、選択性の有意な喪失がない。本発明の方法は、癌等の疾患に関連する突然変異、酵素活性の完全喪失または部分喪失等、代謝エラーを引き起こす欠失または塩基置換突然変異、病的症状または症候に関与することが知られる患者のゲノム核酸中の特定遺伝子または遺伝子座の一部、単一ヌクレオチド多型(SNP)、あるいは、例えば侵入した疾患を引き起こす微生物のゲノムの、外来遺伝子配列の一部を検出および同定するのに有用である。
【0041】
単一塩基プライマ伸長反応は、オリゴヌクレオチドプライマを相補的核酸へアニーリングすることにより、およびアニーリングされたプライマの3’末端を、DNAポリメラーゼで触媒する鋳型指向性反応で付加された鎖終結ヌクレオチドで伸長することにより実施される。単一塩基プライマ伸長反応の選択性および感受性は、オリゴヌクレオチドプライマ長と反応条件(例えば、アニーリング温度、塩濃度)とに影響される。
【0042】
プライマ伸長反応の選択性は、試料中でのオリゴヌクレオチドプライマと核酸との厳密な相補的ハイブリッド形成量に反映される。選択性の高い反応は、厳密な相補的配列(即ち、ハイブリッド形成されたプライマと核酸との間に塩基ミスマッチがない)を備える核酸へのプライマハイブリッド形成のみを促進する。これに反して、非選択的反応では、プライマは、部分的な相補的配列(即ち、ハイブリッド形成されたプライマと核酸との間に塩基ミスマッチがある)を備える核酸にもハイブリッド形成する。一般に、選択的なプライマハイブリッド形成に有利なパラメータ(例えば、より短いプライマおよびより高いアニーリング温度)は、ハイブリッド形成されたプライマレベルの低下をもたらす。そのため、選択的な単一塩基プライマ伸長反応測定に有利なパラメータは、測定の感受性低下をもたらす。
【0043】
本発明の方法は、少なくとも2サイクルの単一塩基伸長反応を実施することを含む。本発明の方法は、単一塩基伸長反応を反復することにより、測定の選択性を低下させることなく、単一塩基プライマ伸長測定のシグナルを増強させる。循環によりシグナルが増強し、そのため伸長反応が選択性の高い条件下(例えば、プライマがほぼTmかそれを上回る温度でアニーリングされる)で実施されることが可能である。
【0044】
好ましい実施例においては、本発明の方法は、厳密なハイブリッド形成に有利な条件下で過剰のプライマをアニーリングすること、ハイブリッド形成したプライマを伸長すること、伸長したプライマを変性すること、およびアニーリングと伸長反応を少なくとも1回反復すること、により実施される。最も好ましい実施例においては、反応サイクルは熱変性のステップを含み、ポリメラーゼは温度安定性がある(例えば、TqポリメラーゼまたはVentポリメラーゼ)。
好ましいプライマ長は、10〜100ヌクレオチドであり、より好ましくは10〜50ヌクレオチドであり、最も好ましくは約30ヌクレオチドである。有用なプライマは、プライマの3’末端の単一塩基伸長が、鋳型に存在する突然変異ヌクレオチドに相補的なヌクレオチドを取り込むよう、推測される突然変異部位に隣接してハイブリッド形成するものである。
【0045】
好ましいハイブリッド形成条件は、反応でのオリゴヌクレオチドのほぼTmかそれを上回るアニーリング温度を含む。オリゴヌクレオチドプライマのTmは、その長さとGC含量により測定され、当該技術分野で周知の複数の式の1つを用いて計算される。標準的アニーリング条件では、約25ヌクレオチド長のプライマの好ましい式は、Tm(℃)=4x(G数+C数)+2x(A数+T数)。
好ましい反応では、アニーリングと変性のステップは、反応温度を変化させることにより実施される。本発明の1つの実施例においては、プライマは、そのプライマのほぼTmでアニーリングされ、その温度を伸長の最適温度まで昇温させ、その後その温度を変性温度にまで昇温させる。アニーリング、伸長、および変性の温度と時間との例を、例2に記載する。本発明のより好ましい実施例においては、反応は、アニーリング温度と変性温度の間で循環され、単一塩基伸長は、アニーリング条件から変性条件までの変移の間に生じる。
【0046】
本発明の好ましい実施例においては、2サイクル以上の伸長を実施する。より好ましい実施例においては、5〜100サイクル実施される。これ以外の実施例においては10〜50サイクル、最も好ましくは約30サイクル実施される。
【0047】
本発明の好ましい実施例においては、鋳型依存性反応のプライマの3’末端に付加されるヌクレオチドは、例えばジデオキシヌクレオチドの、鎖終結ヌクレオチドである。より好ましい実施例においては、ヌクレオチドは論じたinfraと同様に検出可能に標識される。
【0048】
I.分割プライマを用いる循環伸長反応
一実施例において、本発明の方法は、分割プライマを用い少なくとも二つの単一基伸長を行うステップを含む。好適実施例において、分割プライマは、ターゲット核酸の実質的連続部分にハイブリッド形成の能力がある短い第一プローブ及び長い第二プローブを含む。二つのプローブは、長い第二プローブがない状態では短い第一プローブのハイブリッド形成を利さないとの条件下で試料に露出される。ハイブリッド形成に影響する要因は当該技術で周知であり、温度、イオン濃度、pH、プローブ長さ、及びプローブGC含有量などが含まれる。第一プローブは、大きさが小さいので、平均的ゲノムの多くの場所でハイブリッド形成する。例えば、人ゲノムにおいて任意の所与の8−merは約65,000回起こる。しかし、一つの8−merは融解温度(Tm)が低く、単一基不整合がこれの不安定度を大きく誇張する。他方第二プローブは、第一プローブより大きいので融解温度Tmが高い。例えば、20−mer第二プローブは、一般的に8−merより良い安定度でハイブリッド形成する。しかし、単一ヌクレオチド変化によりハイブリッド安定度に生じる熱力学的相違が小さいため、長いプローブは安定なハイブリッドを形成するけれども、ヌクレオチド不整合を許容するので、選択性が低い。したがって、第一プローブに不利なハイブリッド形成条件の下では(例えば、第一プローブの上10−40度C)、第一プローブは高い選択性でハイブリッドを形成する(即ち、単一の不整合があるだけでも配列にほとんどハイブリッド形成しない)が、単独で(即ち、第二プローブが存在しなくて)ハイブリッド形成するときは、不安定なハイブリッドを形成する。第二プローブは、安定なハイブリッドを形成するが、不整合に対する許容性のため選択性が低い。
【0049】
本発明における伸長反応は、第一及び第二のプローブの連続ハイブリッド形成が無いと、起こらない。第一(近接)プローブ単独では、分析に用いられる反応条件の下で安定ハイブリッドを形成しないので、鋳型に基く核酸伸長に関しプライマにならない。第一プローブは約5個から10個の間のヌクレオチドを含むのが好適である。第一プローブは、突然変異が疑われる核酸に隣接してハイブリッド形成をする。本発明の突然変異判定方法における第二(遠隔)プローブは、第一プローブの上流側でターゲット(鋳型)のほぼ連続領域においてハイブリッド形成をする。第二プローブ単独では、3’伸長不能ヌクレオチドから構成されるので、鋳型に基く核酸伸長のプライマにはならない。第二プローブは、第一プローブより大きく、長さがヌクレオチド約15個と約100個の間であるのが好適である。
【0050】
本発明の方法にしたがうと、鋳型依存の伸長は、第一プローブが第二プローブの次ぎにハイブリッド形成するときにのみ起こる。これが起こるとき、短い第一プローブは、単一基突然変異の疑われるサイトの直ぐ隣りにハイブリッド形成する。第二プローブは第一プローブの5’終端近傍にハイブリッド形成する。二つのプローブの存在は、共同束縛効果により、一緒になって安定性を増す。二つのプローブは一緒に、ポリメラーゼによりプライマとして認識される。このシステムは、短いプローブの選択性と長いプローブの安定性とを用いてハイブリッド形成するプライマを生じるため、短いプローブの高い選択性と、長いプローブが与えるハイブリッド形成安定性と利用する。したがって、本質的に、分割プライマに伴う偽始動がない。長い第二プローブによる不整合許容範囲が偽信号を発生しないので、対応する長い第二プローブを欠いた短い第一プローブの伸長をハイブリッド形成条件が許さない限り、幾つかの分割プライマを同一反応において分析することが出来る。さらに、ターゲットに関する選択性が増加するので、本発明の方法は、検出のため試料中で少量しか利用出来ず、通常は例えばPCRなどを用いて拡大しなければならないターゲット核酸を、検出し同定するのに用いることができる。
【0051】
プローブ二つのハイブリッド形成を要することにより、偽陽性信号が減少又は消滅する。このように、分割オリゴヌクレオチドの使用により、現在必要とされる個々のプローブについてのハイブリッド形成条件の慎重な最適化が不要となり、広範な多重化が出来るようになる。対応する長い第二プローブを欠いた短い第一プローブの安定ハイブリッド形成をハイブリッド形成条件が許さない限り、幾つかのオリゴヌクレオチドを使用して、同一反応で分析される幾つかのターゲット配列をプローブすることが出来る。
第一及び第二プローブは、ターゲットの実質的連続部分にハイブリッド形成する。本発明の目的に関し、実質的連続部分とは、ハイブリッド形成された第一及び第二プローブが単一プローブとして(例えば核酸伸長のプライマとして)機能することが出来るのに十分近い部分である。実質的連続部分は、ヌクレオチド0個分(即ち、部分間に間隔がなく正確に連続)とヌクレオチド約1個分の間だけ離れているのが好適である。第一及び第二プローブがヌクレオチド2個分以上離れているときは、リンカーが分析(例えば、核酸伸長反応)を妨害しないとの条件で、リンカーを使用するのが好適である。このようなリンカーは当該技術では周知で、例えば、ペプチド核酸、DNA結合蛋白、及び結紮などが含まれる。隣接プローブは、長い第二プローブが短い第一プローブに安定性を与えるよう、共同的に結びつくことを明らかした。しかし、第二プローブが与える安定性は、第一プローブの選択性(即ち、不整合に対する非許容性)にうち勝たない。したがって、本発明の方法は、短い第一プローブの高い選択性と長い第二プローブが与える安定性との利点を利用する。
【0052】
こうして、好適実施例においては、第一及び第二プローブがターゲットの実質的連続部分にハイブリッド形成され、このとき第一プローブは突然変異、例えば単一基突然変異、の疑われるサイトの直ぐ隣で上流側にある。このとき試料は、疑わしいサイトで可能な突然変異の相補体であるジデオキシ核酸に露出される。例えば、野生型核酸が既知のサイトでアデニンであれば、ジデオキシ・アデニン、ジデオキシ・シトシン及びジデオキシ・グアニンが試料内に置かれる。ジデオキシ核酸にはラベルを付けるのが好適である。単一ヌクレオチド添加後、反応が停止するときは、代わりにデオキシ核酸を使ってもよい。ポリメラーゼを内生的又は外生的に供給すると、ジデオキシ基の第一プローブ上への組み込みが促進される。ラベルの検出は、非野生型(即ち突然変異体)基の組み込みを示すので、第一プローブに隣接するサイトに突然変異がある。代わりに、本発明の方法は、野生型配列が未知であるときに常用される。この場合、四つの共通ジデオキシ・ヌクレオチドに別々のラベルを付ける。上述の分析における一つ以上のラベルの出現は、突然変異の存在を示す。
代替好適実施例においては、分割オリゴヌクレオチドが一連の第一プローブを構成する。このとき、分割オリゴヌクレオチドの構成要員すべてが同時に核酸の実質的連続部分にハイブリッド形成するとき十分な安定性が得られる。ここで、短いプローブは過渡的で不安定なハイブリッド形成を示すけれども、隣接する短いプローブは共同的に結びついて個々のプローブそれぞれより大きい安定性を持つことが明らかになった。隣接してハイブリッド形成された一連の第一プローブは纏まって、個々のプローブ又は一連のプローブのサブセットより大きい安定性を持つ。例えば、三個の一連の第一プローブから構成される分割プライマ(即ち、ターゲット核酸上流核酸の実質的連続部分にハイブリッド形成能力のある終端ヌクレオチドのない三個の短いプローブ)を用いる伸長反応において、三個のプローブの同時ハイブリッド形成が、核酸伸長始動のための三個のプローブについて十分な共同安定性を生じ、疑わしい突然変異に直ちに隣接する短いプローブが伸長される。こうして一連の短い第一プローブから構成される分割プローブは、短いプローブの高い選択性(即ち不整合の非許容性)と長いプローブの安定性を提供する。
【0053】
好適実施例においては、分析信号を拡大するため数サイクルの伸長反応を実行する。伸長反応は、dNTP又はddNTPラベル付きの余分な第一及び第二プローブ、及び熱安定ポリメラーゼを存在させて行う。伸長反応が完了すると、ターゲット核酸に束縛された第一及び第二プローブは、反応混合物をハイブリッドの融解温度以上に加熱することにより解離する。次いで反応混合物をハイブリッドの融解温度以下に冷却すると、第一及び第二プローブは別の伸長反応のためターゲット核酸に会合出来るようになる。好適実施例においては、10乃至50サイクルの伸長反応を行った。最好適実施例においては、30サイクルの伸長反応を行った。
【0054】
II.伸長プライマの検出
ラベル付きddNTP又はdNTPは、分割プライマ反応において伸長したプライマ又は伸長した短い第一プローブの検出を容易にする「検出部分」を含むのが好適である。検出部分は、蛍光性、発光性又は放射性ラベル、酵素、ハプテン及びその他の化学的タグ、ラベル付き伸長生成物を容易に検出出来るビオチンなどを含むグループから選ぶ。例えば、ダンシル基のような蛍光ラベル、フルオレスセイン及び置換フルオレスセイン、アクリジン誘導体、カウマリン誘導体、フタロシアニン、テトラメチルローダミン、テキサスレッド、9−(カルボキシエチル)−3−ヒドロキシ−6−オキサ−6H−キサンテン、DABCYL、およびBODIPY(分子プローブ、オイゲン、OR)などは、ここに記述する方法に特に有利である。このようなラベルは、多数試料の同時高生産分析用自動装置に日常的に使用されている。
好適実施例においては、プライマ即ち第一プローブは「分離部分」を含む。このような分離部分は、例えば、ハプテン、ビオチン、又はジゴキシゲニンである。これら分離部分を含むプライマ即ち第一プローブは、(例えば、抗ハプテン、アビジン、ステプタビジン又は抗ジゴキシゲニンで被覆された)分離部分に関し親和性を有する固体相マトリクスに固定して、反応混合物から隔離される。本発明での利用に適するマトリクスの非限定例には、ニトロセルローズ又はナイロンフィルタ、親和性キャプチャ用薬剤で被覆されたガラスビーズ、磁性ビーズ、処理又は未処理ミクロタイタープレート、及び類似物が含まれる。
【0055】
好適実施例において、分離部分はラベル付きddNTP又はdNTPに組み込まれる。ハイブリッド形成されたプライマ又はプローブの変性、及びラベル付きddNTP又はdNTPと共に伸長したプライマ又は第一プローブの固定マトリクスへの固定化により、ラベル付きプライマ又はラベル付き第一プローブが伸長しないプライマ又は伸長しない第一プローブ及び第二プローブ、及びラベル無しddNTPと共に伸長したプライマ又は第一プローブから、1回以上の洗滌ステップにより分離される。
代替好適実施例において分離部分は、分離部分が第一プライマの又はプローブの鋳型にハイブリッド形成して伸長する能力を妨害しない限り、プライマ又は第一プローブに組み込まれる。溶離プライマ又は第一プローブは固体支持体に固定化されるので、溶離第二プローブから1回以上の洗滌ステップにより分離することが出来る。
【0056】
代案として、ラベル付き終端ヌクレオチドと共に伸長したプライマ又は第一プローブの存在は、ハイブリッド形成したプライマ又はプローブを溶離しないで判定できる。検出方法は、プライマ又は第一プローブに組み込まれたラベル又はタグに左右される。例えば、ターゲット核酸に結合された放射性ラベル付き又は化学発光第一プローブは、フィルタをX線フィルムに露出して検出することが出来る。代わりに、蛍光ラベルを含むプライマ又は第一プローブは、レーザー又はランプに基づくシステムを用い蛍光物質を固有吸収波長で励起して検出することが出来る。
代替実施例においては、束縛されたプライマ又は第一及び第二プローブが、マトリクス固定ターゲット核酸から溶離される(下記を参照)。溶離は、当該技術で周知の、核酸ハイブリッドを不安定化する任意の手段(即ち、塩の低下、温度上昇、フォルムアミド、アルキルへの露出など)を用いて行われる。好適実施例において、束縛されたオリゴヌクレオチド・プローブは、ターゲット核酸−分割プライマ複合体を水中培養し、反応物をハイブリッドの融解温度以上に加熱して、溶離した。
【0057】
上記反応のいずれにおいても検出可能単一伸長基として、デオキシヌクレオチドを使用することが出来る。しかし、このような方法では単一デオキシヌクレオチド付加後、伸長反応を停止しなければならない。このような方法は、固有突然変異が既知(即ち、C→G)であるか否かに関わりなく採用できる。さらに、単一基突然変異検出用試料内でデオキシヌクレオチドのラベル付き種を一つだけ利用出来るときは、デオキシヌクレオチド一つだけを付加した後、伸長反応を停止する必要はない。この方法は、疑問の単一基位置を含むヌクレオチド反復があるとき、実際に信号を強化する。
好適実施例においては、ターゲット核酸をプライマ又は分割プライマに露出して伸長反応を行う前に、ターゲット核酸を固定支持体に固定化する。核酸試料を一旦固定化したら、試料を洗滌して固定化されなかった物質を洗い流す。次いで核酸試料を、本発明にしたがってプライマ又は分割プライマの一つ以上の組に露出する。一旦単一基伸長反応が完了したら、ラベル付きddNTP又はdNTPと共に伸長したプライマ又は第一プローブを、伸長しなかったプローブ及びラベル無しddNTP又はdNTPと共に伸長したプローブから隔離するのが好適である。束縛されたプライマ又は第一及び第二プローブは支持体に束縛されたターゲット核酸から溶離される。溶離は、当該技術に周知の、核酸ハイブリッドを不安定化する任意の手段(即ち、塩の低下、温度上昇、フォルマミド、アルキルへの露出など)を用いて行われる。好適実施例において、第一及び第二プローブは、ターゲット核酸−分割プライマ複合体を水中培養して解離し、反応をハイブリッドの融解温度以上に加熱して伸長した第一プローブを隔離した。代替好適実施例において伸長反応は、水様溶液中で実行した。単一基伸長反応が一旦完了すると、オリゴヌクレオチド・プローブをターゲット核酸から解離して伸長した第一プローブを隔離する。代替実施例においては、核酸が水様相中に止まる。
【0058】
最後に、本発明の方法は、伸長した第一プローブを隔離するステップと配列するステップとを含む。伸長した第一プローブの隔離には、例えば、ハプテン、ビオチン又はジゴキシゲニンのような「分離部分」を用いる。好適実施例において、分離部分を含む第一プローブは、分離部分に関し親和性を有する(例えば、抗ハプテン、アビジン、ステプタビジン又は抗ジゴキシゲニンで被覆された)固体支持体に固定化される。本発明での利用に適する支持体の非限定例には、ニトロセルローズ又はナイロンフィルタ、親和性キャプチャ用薬剤で被覆されたガラスビーズ、磁性ビーズ、処理又は未処理ミクロタイタープレート、及び類似物が含まれる。
好適実施例において、分離部分はラベル付きddNTP又はdNTPに組み込まれる。ラベル付きddNTP又はdNTPと共に伸長した溶離プライマ又は第一プローブの固体支持体への固定化により、ラベル付きプライマ又は第一プローブが伸長しない第一プローブ及び第二プローブ、及びラベル無しddNTPと共に伸長したプライマ又は第一プローブから、1回以上の洗滌ステップにより分離される。
【0059】
代替好適実施例において分離部分は、分離部分が第一プライマの又はプローブの鋳型にハイブリッド形成して伸長する能力を妨害しない限り、プライマ又は第一プローブに組み込まれる。溶離プライマ又は第一プローブは固体支持体に固定化されるので、溶離第二プローブから1回以上の洗滌ステップにより隔離することが出来る。
次いでラベル付きプライマ又は第一プローブを配列して、検出された突然変異又は病原微生物を同定する。一実施例においては、標準技術の化学的方法を用いて、固定化プローブを直接配列する。別の実施例においては、ラベル付き固定化第一プローブを固体支持体から移動してラベル付き第一プローブの配列は水様溶液中で行う。
【0060】
III.列挙的検出方法
本発明の方法は、核酸列挙が必要又は好ましいいずれの場合にも、有用である。第一に、上述の検出方法は、生体試料におけるヌクレオチド突然変異検出のため有用である。したがって、本発明の方法は、疾病に関連又は関連疑惑の核酸列挙(例えば対立因子、単一ヌクレオチド多形性又は突然変異)のため有用である。患者試料中で一旦ターゲット核酸数が決定されると、その数を、試料が健康人から得られたのであれば存在すると期待される数と比較する。患者試料中の核酸数と健康患者に期待される数(これはプールされた健康人試料から決定される)との間に統計的有意の差が存在するとき、その患者は疾病又はその傾向を有すると診断される。本発明の方法はまた、生体試料中の核酸検出にも有用である。これらは不均質であることが多く、突然変異核酸が野生型核酸に対し少量で存在する。例えば、スツール試料において、スツール上に落ちた変形細胞からの突然変異体核酸は、スツール上に落ちた通常細胞からの野生型核酸に対し、特に結腸癌の早期ステップで、滅多にない。本発明の方法は、本発明の単一基伸長分析からの結果が、生体試料中の突然変異体核酸存在を示すか否かを判定する統計的解析を含む。好適実施例において、本発明の方法は、単一基伸長したプライマ又はプローブの列挙を含む。さらに好適な実施例において、伸長したプライマ又はプローブの数を解析して、統計的有意量の突然変異体核酸配列が生物学的試料中に存在するか否かを判定する。
【0061】
本発明の一実施例において、ここで論じたように、プライマ又はプローブはラベル付きヌクレオチド共に伸長させるのが好ましい。このときはラベル付きプライマの数を測定する。ラベルを放射性同位元素とし、ラベル付きプライマ又はプローブに関連する放射性崩壊の量を測定するのが好ましい。ラベル付きプライマ又はプローブの数は、放射性崩壊の量から計算する。分子数は(例えば、所定期間中の全計測数の測定又は所定計数を得るに要する時間の測定など)ラベル付きプライマ又はプローブに固有結合した放射性崩壊現象の数Xを測定して計数する。数Xを、ラベル付きプライマ又はプローブに固有結合した放射性核種の数X1の計算に用いる。数X1は、参照としてここに共有、共出願の特許出願 号(代理人名簿番号EXT−005)で開示したように、分析中の各ラベル付き分子に結合した放射性核種分子の数を知った上で、ラベル付きプライマ又はプローブ分子の数X2の計算に用いる。分析に存在する伸長したプライマ又はプローブの数は引き続く統計的解析に有用である。
本発明の方法は、不純細胞母集団中の小数細胞における異型接合性喪失検出のため有用である。このような方法は精密な欠失終末点を知ることに依存せず、このような方法は試料中の異質DNA存在により影響されないからである。例えば、異型接合性喪失に際しては、ゲノムの大部分にわたって欠失が起こり、全染色体アームが無くなる。本発明の方法は、欠失の疑われるターゲット核酸の分子数計数及びそれらの基準数との比較を含む。好適実施例において基準数は、同一試料内で欠失していないと思われる核酸の分子数である。知る必要のあるすべては、欠失したと思われるターゲット核酸配列の少なくとも一部及び、欠失していないと思われる基準核酸配列の少なくとも一部である。本発明の方法は、突然変異多重検出を受け入れられるけれども、異質試料内の癌徴候を検出するためには突然変異多重検出を必要としない。
【0062】
したがって、本発明の方法は試料中の細胞副母集団又はそれからの断片における異型接合性喪失の検出に有用である。異型接合性喪失は、一般的に細胞副母集団における少なくとも一つの野生型対立因子配列の欠失として生じる。腫瘍疑惑遺伝子の場合、欠失は一般的に、異型接合性喪失に特有の大量欠失の形を取る。一定の形の癌の場合のように、発病性欠失は最初単一細胞中で起こることが多く、これが次いで突然変異体細胞の小さい副母集団を作る。突然変異の臨床的明徴が検出された時には、疾病は治療不能のステップに進行している。本発明の方法は、欠失が、試料中に全細胞又は細胞断片の小さい百分比でのみ存在するとき、その検出を可能にする。
【0063】
本発明の方法は、試料中に正常(突然変異していない)細胞と同数存在すると期待される二つの核酸の分子数比較を含む。好適実施例において、比較は(1)試料細胞内で突然変異していないと判明又は推定される(「基準」)ゲノム的ポリヌクレオチド断片の量と(2)試料(「ターゲット」)内で突然変異している推定される野生型(突然変異体でない)ゲノム的ポリヌクレオチド断片の量との間でなされる。二つのゲノム的ポリヌクレオチド断片の間の統計的有意差は、突然変異が起こったことを示す。
好適実施例において、基準及びターゲットの核酸は、同一因子座の対立因子である。対立因子は、一つの対立因子を別の対立因子から区別する配列相違があるとき、本発明の方法において有用である。好適実施例において、因子座は腫瘍抑制因子の上又は近くにある。異型接合性喪失は対立因子いずれかの喪失を生じるので、対立因子のいずれかを基準対立因子として役立てることが出来る。重要情報は、試料中の各対立因子の分子数間に統計的有意差が有るか無いかである。好適実施例においてはまた、基準及びターゲットの核酸は異なる因子座、例えば、異なる遺伝子である。好適実施例において、基準核酸は基準因子座の両対立因子を含み、ターゲット核酸はターゲット因子座、例えば腫瘍抑制遺伝子、の両対立因子を含む。明細には、腫瘍抑制遺伝子における欠失の場合、基準因子の検出量はターゲット因子の検出量より著しく大きい。特定の腫瘍因子突然変異の場合のように、ターゲット配列が拡大されると、ターゲットの検出量が基準因子の検出量より十分有意な余裕を持って大きくなる。
【0064】
当該技術で知られた方法は、欠失又は点突然変異を検出するため、通常はPCRプローブ及び/又はハイブリッド形成プローブの形で数多のプローブの使用を必要とする。しかし、本発明の方法は、ヌクレオチド配列の列挙的検出及び安定と知られた配列と不安定と思われる配列との間の列挙的比較を含むので、癌危険性を正確に分析するため僅か数個のプローブを使わなければならないだけである。事実、単一大型欠失検出に必要なすべては、プライマ又はプローブのセットたった一つである。癌の危険性は、腫瘍発生に関わると知られた又は思われる因子領域における突然変異の存在により示される。本発明の方法にしたがって実施された試験に基づき危険性があると同定された患者は、疾病の確認及び/又は治療のため、他の一般的には侵襲的処理に回される。
【0065】
本発明の方法にしたがうと、ターゲット及び基準核酸は、循環単一基伸長反応を用いて別様にラベル付けされる。その循環単一基伸長反応は、別様にラベル付けされたヌクレオチドを、ターゲット及び基準核酸に選択的にハイブリッド形成されるプライマ又はプローブの3’端に組み込む。例えば、プライマ又はプローブは、ターゲット核酸にハイブリッド形成されたプライマ又はプローブの単一基伸長に向けられた鋳型がTの添加を生じ、基準核酸にハイブリッド形成されたプライマ又はプローブの単一基伸長に向けられた鋳型がGの添加を生じるように、設計される。伸長反応は、例えば、35Sでラベルされた終端T及び32Pでラベルされた終端Gの存在下で行われる。代わりに、二つの終端核酸をそれぞれ、大きいインピーダンスと小さいインピーダンスのビーズでラベル付けする。これらの終端核酸は、ここで論じたように、伸長したプライマ又はプローブの列挙が出来るようにする検出可能な任意の各種マーカーを用いてラベル付けすることが出来る。
生体試料中で一様に分布する核酸配列の列挙的試料採取は、一般的にポアソン分布にしたがう。生体試料中にあるゲノムのポリヌクレオチド断片の典型数のような、大型母集団について、ポアソン分布は、平均値N及び、Nの二乗平方根で近似される標準偏差を有する正規(ガウス)曲線と同様である。
生体試料から得られたターゲット及び基準の遺伝子の数の間の統計的有意性は、任意の適切な方法を用いて決定される。例えば、本書の開示を参照によりここに組み込んだ、スチール他「統計の原理と手順、生物統計学的方法」(マクグローヒル社、1980年)を参照。例示的方法は、特異性(偽陽性の許容範囲)と感度(偽陰性の許容範囲)の好ましい水準に基づき、選択した信頼性水準の範囲内で、選択した統計的有意水準に達するよう、得るべきターゲット及び基準の遺伝子の数の間の差を決定することである。このような決定におけるしきい値問題は、統計的に有意な判定が出来るため母集団中で利用出来なければならない(ターゲット及び基準についての)遺伝子の最小数Nである。数Nは、突然変異体対立因子を含む試料の中の突然変異体対立因子最小数推定値(ここでは最小1%と推定)及び正規試料は突然変異体対立因子を含まないとの追加推定に依存する。ターゲット及び基準の遺伝子数の間の差のしきい値は、試料中の細胞副母集団に突然変異体が存在するとの診断に関し、少なくとも0.5%でなければならないこともまた仮定する。前述の仮定に基づいて、突然変異体数と基準対立因子数との間の差0.5%以下が検出されたとき真に陰性(即ち、試料中に突然変異体副母集団がない)である結果が回数の99.9%で生じるよう、Nがどれだけ大きくなければならないかを決定することが可能である。
【0066】
特異性に関するNの計算は、このとき、一つの試料測定が母集団の最低部分3.16%を覆うガウス分布の一部(図2Aで「A」とマークした部分)であるとの確率及び、他の試料測定が母集団の最高部分3.16%を覆うガウス分布の一部(図2Bで「B」とマークした部分)であるとの確率に基づく。二つの試料測定は独立事象なので、両事象が単一試料中で同時に起こる確率は、約0.001即ち0.1%である。こうして、ガウス分布の93.68%(100%−2×3.16%)が図3でA及びBとマークした領域の間に入る。統計表は、当該領域が標準偏差3.72に等しいことを示す。したがって、0.5%Nは3.72シグマに等しいと設定される。シグマ(標準偏差)は√Nに等しいので、方程式はNについて553,536と解ける。これは、基準及びターゲットを示す二つの数字の低い方が553,536以上であって、患者が真に正常であるときは、数字の間の差は回数の約99.9%で0.5%以下であることを意味する。
99%感度に必要な最小Nを決定するため同様の解析を行う。今度は一端を切り取ったガウス分布表が、平均値からの1.28標準偏差(シグマ)がガウス分布の90%を覆うことを示す。さらに、数字の一つ(基準又はターゲット)が図3で「A」とマークした面積又は図3で「B」とマークした面積に入る10%(1%の二乗平方根)の確率がある。二つの母集団平均が全体で1%の相違であり、ターゲットと基準因子の数字の間に0.5%の差がある筈とすると、いずれかの平均値から統計的有意に関するしきい値までの差は、99%感度に関し0.25%Nに等しい(図3参照)。図3に示すように、0.25%Nはガウス分布片側の約40%に相当する。統計表は、ガウス分布の40%が平均からの標準偏差1.28に相当することを明らかにする。したがって、1.28シグマは0.0025Nに等しく、Nは262.144に等しい。こうして、異常試料については、二つの数字の低い方が262,144以上であるとき、回数の99%以上において、差が0.5%を超える。逆に陰性誤診断は、これら条件の下で回数の僅か1%になる。
【0067】
99.9%特異性(偽陽性の回避)と99%感度(偽陰性の回避)の両方を持つためには、DNAを有する553,536個以上(大凡550,000個以上)の細胞から誘導したDNAを持つ試料を計数しなければならない。得られた数の間の0.5%以上の差が感度に関し99.0%信頼度水準で有意であり、数の間の0.5%未満の差が特異性に関し99.9%信頼度水準で有意である。上に注記したように、統計的有意性判定のため他の標準統計的試験を使用してもよく、前記は当該試験の一つをあらわすに過ぎない。
前記説明に基づいて、本発明の方法が任意の生体試料内のポリヌクレオチド副母集団にある突然変異検出に有用であることを、当業者は理解するであろう。例えば、ここに開示した方法は、癌のような疾病に伴う対立因子喪失(異型接合性喪失)の検出に使用される。加えて、本発明の方法は、酵素活動の完全又は部分的喪失のような、物質交代エラーを引き起こす欠失又は基置換突然変異の検出に使用される。例示目的で、以下に結腸癌検出における本発明の方法の利用法詳細を述べる。発明の方法は腫瘍抑制因子における突然変異(及び特に異型接合性喪失に典型的な大型欠失)の早期発見に特に有用である。したがって、以下の方法で例示したが、本発明はそのように制限されるものではなく、それらの考察に当たって当業者はその広範な利用性を理解するであろう。
【0068】
本発明の方法は、突然変異したと既知又は推定されるターゲット・ポリヌクレオチドの数を突然変異していないと既知又は推定される基準ポリヌクレオチドの数と比較するステップを含むのが好適である。対立因子又は因子座のいずれかを基準及びターゲット核酸として利用する代替実施例に加えて、本発明は正常対立因子におけるミクロサテライト繰り返し領域を突然変異が既知又は推定される対立因子における対応ミクロサテライト繰り返し領域と比較するステップを含む。本発明の例示的検出方法は、測定される各核酸の計数の間に差が存在するか否かを決定するステップを含む。統計的有意差の存在は、測定された核酸の一つに突然変異が起こったことを示す。
【実施例】
【0069】
使用例
例示目的で、以下に結腸癌検出における本発明の方法の利用法詳細を述べる。発明の方法は突然変異の早期発見に特に有用である。したがって、以下の方法で例示したが、本発明はそのように制限されるものではなく、それらの考察に当たって当業者はその広範な利用性を理解するであろう。
【0070】
結腸の癌又は前癌状態の検出に関する例示的方法
使用例1.試料準備
本発明にしたがって、ターゲット核酸は患者から隔離した核酸試料をあらわす。この核酸は任意の細胞源又は体液から採取することができる。臨床行為で得られる細胞源の非限定例には、血液細胞、頬面細胞、膣頚部細胞、尿からの上皮細胞、胎児細胞、又は生検で得られた組織内に存在する任意の細胞が含まれる。体液には、血液、尿、脳脊髄液、及び感染又は炎症部位からの組織滲出液が含まれる。
好適実施例において、試料はスツールの横断面又は周辺部分である。スツールの横断面又は周辺部分作成に好適な方法は、参照によりここに組み込んだ共有米国特許5,741650号及び、共有共出願の特許 号(代理人名簿番号EXT−015)で提供されている。スツールが結腸を通過するとき、結腸上皮細胞から脱け落ちた細胞及び細胞断片を付着する。同様に、細胞及び細胞断片は結腸ポリープ(突然変異DNAを含む)が脱ぎ落とす。しかし、ポリープと接触するスツールの一部だけが脱落細胞を付着する。したがって、スツール試料が、推定癌細胞(例えばポリープ)の落としたものを含む脱落細胞すべての混合物を確実に含むためには、少なくともスツールの横断面又は周辺部分を得ることが必要である。
【0071】
試料作成の後、NaCl又はKCl200mMのような塩及び1−10%SDS又はトリトンTMのような洗浄剤、及び/又はプロティナーゼKのようなプロティナーゼを含むリン酸塩緩衝サリンのような、適切なバッファの中で試料を均質化する。特に好適なバッファは、参照によりここに組み込んだ共有共出願の米国特許出願 号(代理人名簿番号EXT−006)で開示したような、Tris−EDTA−NaClバッファである。このバッファはまた、DNA及びRNA劣化酵素の抑制剤をも含むことがある。試料中の二重撚りDNAは周知の方法で融解(変性して単一撚りDNAを形成)する。例えば、参照によりここに組み込んだギレンステン他、組み換えDNAにおける方法論II、565−578頁(Wu版、1995年)を参照。DNAは次いで、当該技術で標準の多数の方法のいずれかを用いて、細胞源又は体液から分離される。参照によりここに組み込んだスミスーレビン他、Gut、36,81−86頁(1995年)を参照。DNA抽出に用いる特定の方法が供給源の性質に左右されることは、理解されるであろう。
一旦抽出したら、ターゲット核酸は追加処理することなく本発明に採用してよい。代わりに、ターゲット核酸中に存在する一つ以上の特定領域をPCRにより拡大してもよい。この場合、拡大される領域は、プライマとして使用のため特定の側面配列の選択により、規定される。このステップでの拡大は、ターゲット核酸配列母集団の中で特定核酸配列の濃度を増加するとの利点を備える。
一実施例において、特定配列先行拡大を伴う又は伴わないターゲット核酸が、固定相支持物に束縛される。これにより多数の患者試料の同時処理及び篩い分けが出来る。本発明における使用に適切な支持物の非限定的例示には、ニトロセルローズ又はナイロンフィルタ、親和性キャプチャ用薬剤で被覆されたガラスビーズ、磁性ビーズ、処理又は未処理ミクロタイタープレート、及び類似物が含まれる。診断試験室及び組織培養に用いられる在来の96穴ミクロタイターディッシュは好適支持物である。好適実施例において、支持物は多数の穴を有するミクロタイターディッシュである。このような支持物の使用により、多数の試料及び対照の同時判定が出来、その結果、解析を促進する。さらに、自動システムを使用してこのようなミクロタイターディッシュに試薬を提供することが出来る。ターゲット核酸を支持物に束縛する方法が、使用する特定マトリクスに左右されることは、当業者に理解されるであろう。例えば、DNAのニトロセルローズへの束縛は、フィルタにDNAを単に吸収させ、次いでフィルタを75−80度C真空下で15分ー2時間焼くことにより、達成される。代わりに、帯電ナイロン皮膜を使用することが出来、これは束縛核酸の追加処理を何も必要としない。アビジン又はストレプタビジンで被覆したビーズ及びミクロタイターを、ビオチンが(例えば、ビオチン抱合PCRプライマを使って)付着されたターゲット核酸の束縛に使うことが出来る。加えて、ターゲット核酸表面を抗体で覆い抗体特有付着素をターゲット核酸に組み込むことにより、抗体を用いてターゲット核酸を任意の上記固体支持体に付着することが出来る。またエポキシド/アミン結合化学作用を用いて、ターゲット核酸を任意の上記固体支持体に直接付着することが出来る。エガース他、DNA配列技術の進歩、SPIE会議議事録(1993)を参照。一旦核酸試料が固定されたら、試料を洗滌して非固定物質を除去する。次いで核酸試料を本発明にしたがって一つ以上の分割プライマに露出する。代替実施例において、核酸は水様相に止まる。
【0072】
使用例2.単一基伸長反応の多重循環
a)プライマ選択
一つ以上の突然変異を含むと思われる遺伝子領域は、GenBank、EMBLのようなヌクレオチド・テータベース若しくは適切なデータベース又は刊行物を参照して、又は配列により同定する。癌検出については、数多の腫瘍発生遺伝因子及び腫瘍抑制因子における遺伝子突然変異が知られている。デュフィ、臨床化学、41巻、1410−1413頁(1933年)。本発明の突然変異検出法での使用に好適な遺伝子には、一つ以上の腫瘍発生遺伝因子及び/又は一つ以上の腫瘍抑制因子が含まれる。特に好適な遺伝子には、ras腫瘍発生遺伝因子、p53、dcc、apc、mcc、及びその他の腫瘍形成表現型の成長に関わると思われる遺伝子が含まれる。
以下で記述するように、本発明の方法は、疑問のある核酸が一つ以上ある因子座で突然変異を検出することが出来る。さらに本発明の方法は、一つ以上の単一基突然変異が可能な因子座を調査するため使用することが出来る。一旦関心領域が同定されたら、疑問の突然変異の存在を検出するため少なくとも一つのプライマを準備する。本発明のプライマは長さが、好適にはヌクレオチド約10個分から約100個分、さらに好適にはヌクレオチド約15個分と約35個分との間、最も好適にはヌクレオチド約25個分である。
【0073】
プライマは天然物でも合成物でもよく、生体内、生体外で酵素的に、又は生体外で非酵素的に、合成される。本発明の方法で使用されるプライマはオリゴデオキシリボヌクレオチド、オリゴリボヌクレオチド、デオキシリボヌクレオチドとリボヌクレオチドとの共重合体、ペプチド核酸(PNA)、及びその他の機能的相似品から選ぶのが好適である。ペプチド核酸は有名である。プルスカル他.FASEB彙報、ポスター#35(1994年)を参照。これらは合成オリゴアミドであって、アデニン、シトシン、グアニン、チミン又はウラシルが付着された反復アミノ酸ユニットから構成される。エホルム他、ネイチャー365巻、565−568頁(1993年)、オエルム他、核酸研究23巻、5332−36頁(1993年)、プラクチカルPNA、PCRクランプを指向するPNAによるポイント突然変異同定、パーセプチブ・バイオシステム1巻、1号(1995年)を参照。ペプチド核酸シントン及びオリゴマーは市販で、パーセプチブ・バイオシステム社、マサチューセッツ州フラミントン、から入手出来る。例えば、PCT出版物EP92/01219、EP92/01220、US92/10921を参照。多くの用途において、PNAは核酸プローブに好適である。核酸/核酸デュプレクスと異なり、これは低塩条件下で不安定化し、PNA/核酸デュプレクスを形成して極低塩条件下で安定化したまま止まるからである。加えて、PNA/DNA複合体は、相似核酸/核酸複合体より高い熱融解点を有するので、PNAプローブの使用により吸取り分析の再現性が改善される。
【0074】
例示のため、K−ras因子内の突然変異検出のため設計されたプライマを下記に用意する。本発明の方法にしたがって、コーディング・ストランド又はノン−コーディング・ストランドのいずれかに相補的なプライマを使用する。実例として、コーディング・ストランド内の突然変異検出に有用なプライマを下記に用意する。K−ras内の突然変異は、表現蛋白質のアミノ酸12に関するコードンに頻繁に起こる。
K−ras因子の野生型コードン12及びその上流核酸は下記である。
野生型鋳型 3’−TATTTGAACACCATCAACCTCGACCA−5’(配列識別番号1)
アミノ酸12をエンコードする三つのヌクレオチドに下線を施した。K−ras因子のコードン内で符号化されたアミノ酸12の中の第一核酸位置調査の能力があるプライマ(プライマ1)を下記に用意する。
プライマ1 5’−ATAAACTTGTGGTAGTTGGAGCT−3’(配列識別番号9)
b)プライマ伸長の複数回循環
プライマ1は、K−ras因子内の相補配列へのプライマ1の選択束縛を促進するとの条件(表1参照)下で、核酸試料にハイブリッド形成される。伸長反応は、それぞれ異なる検出可能ラベルを付けた四つの異なるジデオキシヌクレオチドddATP、ddCTP、ddGTP、及びddTTP、が存在して行われる。伸長反応は、表2に示すように、30回繰り返す。
表1.単一基伸長循環反応用反応混合物
成分 量
H2O 25.5
10×配列バッファ 4
ddNTP(50μM) 5
プライマ(5μM) 5
Thermoシーケナーゼ 0.5
DNAサンプル 10
表2.循環単一基伸長反応用温度輪郭
ステップ 温度(C) 時間(秒)
1 94 5
2 94 30
3 64 10
4 72 10
5 ステップ2に行く、29回
6 4 待機
ラベル付きddNTPの組込について反応生成物を分析する。野生型DNAを含む核酸試料にはラベル付きddGTPのみが組み込まれている筈である。他のいずれかのddNTPが十分有意な量で組み込まれていることは、試料中に突然変異K−ras核酸が存在することを示す。
【0075】
使用例3.分割プライマの準備
a)プライマ選択
関心領域が同定されると、疑惑突然変異の存在検出のため、少なくとも一つの分割プライマを用意する。分割プライマは、核酸の実質的連続部分にハイブリッド形成する能力のある少なくとも二つのオリゴヌクレオチド・プローブ、第一プローブと第二プローブ、から構成される。
本発明の第一プローブは、長さが、好適にはヌクレオチド約5個分から約10個分、さらに好適にはヌクレオチド約6個分と約8個分との間、最も好適にはヌクレオチド約8個分である。本発明の第二プローブは、長さが、好適にはヌクレオチド約15個分と約100個分との間、さらに好適にはヌクレオチド約15個分と約30個分との間、最も好適にはヌクレオチド約20個分である。さらに、第二プローブは、伸長出来ない3’終端ヌクレオチドを有するため、第一プローブを欠く温度依存核酸合成用のプライマになることが出来ない。好適な非伸長性3’終端ヌクレオチドには、C3スペーサー、3’転換基、ビオチン、又は変形ヌクレオチドが含まれる。長いプローブは、そのヌクレオチド不整合許容性のため、選択性が低いけれども、第二プローブは非伸長性で、近接プローブを欠くときは偽始動を生じない。
代替実施例において、分割プライマは第一プライマのシリーズから構成され、シリーズの各構成員は、ヌクレオチド約5個分から約10個分、最も好適にはヌクレオチド約6個分から約8個分まで、の長さを有する。第一プローブは終端ヌクレオチドを有しないけれども、シリーズの全構成員が核酸の実質的連続部分にハイブリッド形成されない限り、核酸伸長は起こらない。
分割プライマのオリゴヌクレオチド・プローブは天然物でも合成物でもよく、生体内、生体外で酵素的に、又は生体外で非酵素的に、合成される。本発明の方法で使用されるプライマは、オリゴデオキシリボヌクレオチド、オリゴリボヌクレオチド、デオキシリボヌクレオチドとリボヌクレオチドとの共重合体、ペプチド核酸(PNA)、及びその他の機能的相似品から選ぶのが好適である。ペプチド核酸は有名である。プルスカル他.FASEB彙報、ポスター#35(1994年)を参照。これらは合成オリゴアミドであって、アデニン、シトシン、グアニン、チミン又はウラシルが付着された反復アミノ酸ユニットから構成される。エホルム他、ネイチャー365巻、565−568頁(1993年)、オエルム他、核酸研究23巻、5332−36頁(1993年)、プラクチカルPNA、PCRクランプを指向するPNAによるポイント突然変異同定、パーセプチブ・バイオシステム1巻、1号(1995年)を参照。ペプチド核酸シントン及びオリゴマーは市販で、パーセプチブ・バイオシステム社、マサチューセッツ州フラミントンから、入手出来る。例えば、PCT出版物EP92/01219、EP92/01220、US92/10921を参照。多くの用途において、PNAは核酸プローブに好適である。核酸/核酸デュプレクスと異なり、これは低塩条件下で不安定化し、PNA/核酸デュプレクスを形成して極低塩条件下で安定化したまま止まるからである。加えて、PNA/DNA複合体は、相似核酸/核酸複合体より高い熱融解点を有するので、PNAプローブの使用により吸取り分析の再現性が改善される。
【0076】
例示のため、K−ras因子内の突然変異検出のため設計されたプライマを下記に用意する。本発明の方法にしたがって、コーディング・ストランド又はノン−コーディング・ストランドのいずれかに相補的なプライマを使用する。実例として、コーディング・ストランド内の突然変異検出に有用なプライマを下記に用意する。K−ras内の突然変異は、表現蛋白質のアミノ酸12に関するコードンに頻繁に起こる。コードン12の中の三つの位置それぞれにおける突然変異検出用に可能なプローブを下記に示す。
K−ras因子の野生型コードン12及びその上流核酸は下記である。
野生型鋳型 3’−TATTTGAACACCATCAACCTCGACCA−5’(配列識別番号1)
アミノ酸12をエンコードする三つのヌクレオチドに下線を施した。K−ras因子のアミノ酸12に関しコードする三つのヌクレオチドを調査する能力がある第一プローブと第二プローブを下記に用意する。第一プローブAは、上記に概略を記述した第一プローブであって、コードン12の中の第一基の直ぐ上流側の(即ち、コードン位置1にあるシトシンに隣接する)ヌクレオチドに相補的な配列を有する。第二プローブAは、上記に概略を記述した第二プローブである。これは、第一プローブAが相補する配列に実質的連続(この場合は正確に連続)な配列に相補的である。下記に示す第二プローブのそれぞれで太字の核酸は、非伸長性ヌクレオチドである。K−rasコードン12の第一基の中の突然変異検出に適する第一及び第二プローブのハイブリッド形成を下記に示す。
第二プローブA 5’−ATAAACTTGTGGTAG(配列識別番号2)
第一プローブA TTGGAGCT(配列識別番号3)
野生型鋳型 3’−TATTTGAACACCATCAACCTCGACCA−5’(配列識別番号1)
コードン12の第二基の中の突然変異検出は、上と同じ第二プローブ(第二プローブA)と、下記で第一プローブBと名付けた第一プローブを用いて行う。この第一プローブBはコードン12の第二基に隣接(3’)して終わる配列に相補的である。コードン12の第二基の中の突然変異検出に適したプローブのハイブリッド形成を下記に示す。
第二プローブA 5’−ATAAACTTGTGGTAG(配列識別番号2)
第一プローブB TGGAGCTG(配列識別番号4)
野生型鋳型 3’−TATTTGAACACCATCAACCTCGACCA−5’(配列識別番号1)
コードン12の第三位置における突然変異検出は、上と同じ第二プローブ(第二プローブA)及び、コードン12の第三基と直接境を接する第一プローブCを用いて達成する。コードン12の第三基の中の突然変異検出に適したプローブのハイブリッド形成を下記に示す。
第二プローブA 5’−ATAAACTTGTGGTAG(配列識別番号2)
第一プローブC GGAGCTGG(配列識別番号6)
野生型鋳型 3’−TATTTGAACACCATCAACCTCGACCA−5’(配列識別番号1)
【0077】
コードン12の第二及び第三ヌクレオチドにおける突然変異検出に関する上述の方法において、第二プローブは、第一プローブのハイブリッド形成領域上流側の、それぞれ1及び2ヌクレオチドである。代わりに、コードン12の第二及び第三ヌクレオチド検出用の第二プローブが、それぞれの第一プローブと直接境を接し(即ち正確にそれと連続し)てもよい。例えば、コードン12K−ras第三基の突然変異検出用代替第二プローブは、下記である。
5’−ATAAACTTGTGGTAGTT(配列識別番号5)
突然変異検出はまた、少なくとも三つの第一プローブのシリーズを含む分割プライマを用いても達成される。コードン12第三基の突然変異検出用第三プローブを下記に示す。
第一プローブX 5’−ATAAACTT(配列識別番号7)
第一プローブY TGGTAGTT(配列識別番号8)
第一プローブZ GGAGCTGG(配列識別番号6)
野生型鋳型 3’−TATTTGAACACCATCAACCTCGACCA−5’(配列識別番号1)
b)プライマ伸長の多重循環
第一及び第二プローブは、長い第二プローブがないと短い第一プローブのハイブリッド形成が不利であるとの、ハイブリッド形成条件の下で試料に露出される。ハイブリッド形成に影響する要因は当該技術に周知で、温度上昇、塩濃度低下、又はハイブリッド形成溶液のpH上昇が含まれる。不利なハイブリッド形成条件(例えば、第一プローブTm上温度30−40度C)の下で第一プローブは、単独でハイブリッド形成されたとき、不安定なハイブリッドを形成し、伸長反応を始動しない。長い第二プローブは高いTmを有していて、鋳型と安定ハイブリッドを形成し、核酸の実質的連続部分にハイブリッド形成されたとき、第二プローブは短い第一プローブに安定性を分与し、それにより連続プライマを形成する。
【0078】
好適実施例において、参照によりここに組み込んだサンガー、国立科学アカデミー誌(USA)、74巻、5463−5467頁(1997年)に報告されたようなデオキシ鎖終結方法の変形を、このとき突然変異存在の検出に用いた。この方法は、四つの共通2’、3’−ジデオキシ・ヌクレオシド・トリフォスフェート(ddATP、ddCTP、ddGTP、ddTTP)のうち少なくとも一つを使うことを含む。当該技術に既知の方法を用いて、検出可能検出部分をジデオキシ・ヌクレオシド・トリフォスフェート(ddNTP)に付着させることが出来る。シーケナーゼTM(パーキン・エルマー)のようなDNAポリメラーゼもまた、試料混合物に添加した。好適実施例において、Taq又はVentDNAポリメラーゼのような、ポリメラーゼを試料混合物に添加した。実質的連続な第一及び第二プローブをプライマとして用いて、ポリメラーゼはddNTP一個を第一プローブの3’端に付加する。組み込まれたddNTPは、単一基多形サイトに存在するヌクレオチドに相補的になる。ddNTPは3’ヒドロキシルを有するので、ハイブリッド形成済みプローブの追加伸長は起きない。伸長混合物に相補性ddNTP(又はデオキシヌクレオシド・トリフォスフェート)が利用できない場合は、連鎖終結もまた起こる。単一基伸長反応終了後、伸長生成物を隔離して検出する。
好適実施例においてはまた、伸長反応停止が唯一つのヌクレオチド付加後か、又は予想突然変異の相補に相当する唯一つのラベル付きヌクレオチド付加後か、のいずれかを検出するラベル付きデオキシヌクレオシドが試料に露出される。
使用例2及び3に例示した本発明の最も簡単な実施例において、ヌクレオシド・トリフォスフェート混合物は、既知突然変異に相補性のラベル付きddNTP又はdNTPのみを含む。例えば、K−ras因子コードン12第一ヌクレオチド内のC→A突然変異につき試料を調査するため、第二プローブA及び第一プローブAは、ラベル付きddTTP又はdTTPを含む伸長反応混合物に露出される。ラベル付きddTTP又はdTTPの第一プローブAへの組込は、試験試料のK−ras因子コードン12第一ヌクレオチド内にC→A突然変異が存在することを示す。第二プローブAと共に野生型鋳型に共にハイブリッド形成された第一プローブAは、伸長されないか、又は反応混合物中で利用出来るときは、ラベル無しddGTP又はdGTPで伸長する。
【0079】
結腸癌に付随した多数の突然変異を与えられると、この疾患に関する検出方法が、同一反応内に同時に存在する多数の突然変異(例えば、apc、K−ras、p53、dcc、MSH2、DRA)に関し、試料を好適に篩い分けする。上述のように、先行技術の検出方法を用いると、極めて限られた多重化のみが可能である。本発明の方法は、長い第二プローブの不整合許容性から生じる偽陽性信号を除外するので、分割オリゴヌクレオチドの使用により、個々のプローブに関するハイブリッド形成条件最適化の必要を回避し、膨大な多重化を可能にする。短い第一プローブの安定なハイブリッド形成が対応の長い第二プローブが無いと出来ないハイブリッド形成条件である限り、数個の分割プライマを同一反応内で分析することが出来る。
好適実施例において、プライマ伸長反応は、それぞれ生体試料の分別部、ポリメラーゼ、及び三つのラベル付き相補性非野生型ddNTP(又はdNTP)、を有する四つの別々の反応混合物で実行される。任意選択で、反応混合物はまたラベル無し相補性非野生型ddNTP(又はdNTP)含むこともある。分割プライマは、野生型鋳型にしたがって多重化される。本例示において、K−ras因子のアミノ酸12に関しコードする最初の二つのヌクレオチドはシステンである。したがって、第二プローブA及び第一プローブAとBを、ラベル付きddATP(又はdATP)、ddTTP(又はdTTP)、及びddCTP(又はdCTP)を含む反応混合物に付加する。第二プローブC及び第一プローブCは、ラベル付きddATP(又はdATP)、ddCTP(又はdCTP)、及びddGTP(又はdGTP)を含む反応混合物に付加する。第一プローブへのいずれのラベル付きddNTP(又はdNTP)の組込も、試料内のK−ras因子コードン12に突然変異の存在を示す。この実施例は、K−rasのコードン12のような数個の可能突然変異を有する位置の調査に、格別有用である。
【0080】
代替好適実施例において、プライマ伸長反応は、それぞれ唯一つのラベル付き相補性非野生型ddNTP又はdNTP及び、任意選択で、別の三つのラベル無しddNTP又はdNTPを含む、四つの別々の反応混合物で行われる。分割プライマはこうして、既知の突然変異体ヌクレオチドに相補性のラベル付きddNTP又はdNTPに露出することが出来るか、又は代わりに、すべての三つの非野生型ラベル付きddNTP又はdNTPに露出することが出来る。上記に用意したK−ras例において、K−rasコードン12の第一ヌクレオチドを既知のC→G突然変異について調査すると、第一プローブAと第二プローブAは唯一つの反応混合物、ラベル付きddCTP(又はdCTP)を含む反応混合物、に付加される。任意選択で、本発明の方法は、上述のようにラベル付きデオキシヌクレオチドを用いて練習することが出来る。
しかし、K−ras因子のコードン12には幾つかの突然変異が同定されているので、プローブはすべての非野生型ラベル付きddNTP又はdNTPに露出する。こうして、第一プローブA及び第二プローブAとBは、ラベル付きddATP(又はdATP)、ddTTP(又はdTTP)、及びddCTP(又はdCTP)を含む反応混合物に付加される。第二プローブC及び第一プローブCは、ラベル付きddATP(又はdATP)、ddCTP(又はdCTP)、及びddGTP(又はdGTP)のうち一つを含む反応混合物に付加される。再び、第一プローブのラベル付き終端ヌクレオチドへの露出は、試験した生体試料のK−ras因子のコードン12に突然変異の存在を示す。
【0081】
好適実施例においては、分析信号増幅のため数回の伸長反応を行う。伸長反応は余分の第一及び第二プローブ、ラベル付きddNTP又はdNTP、及び熱安定性ポリメラーゼを存在させて行う。伸長反応が完了すると、ターゲット核酸に接合された第一及び第二プローブは、反応混合物をハイブリッドの融解温度以上に加熱することにより解離する。次いで反応混合物をハイブリッドの融解温度以下に冷却して、第一及び第二プローブは、別の伸長反応のためターゲット核酸との結合が許される。好適実施例においては、10回乃至50回の伸長反応を実施する。最好適実施例においては、30回の伸長反応を実施する。
【0082】
使用例4.遺伝子変更の同定方法
一実施例においては、ラベル付きプライマ又はプローブがここに記述したように固定化され、当該技術に標準の化学的方法(例えば、マクサム−ギルバート配列、マクサム及びキルバート、1977年、国立科学アカデミー誌、USA、74巻、560頁)を用いて、直接配列を受ける。
別の実施例においては、固定化されたラベル付きプライマ又は第一プローブが、固体支持物から移動されてラベル付き第一プローブの配列が水様溶液内で行われる。一実施例においては、分析で調査される多数配列のそれぞれに対応するオリゴヌクレオチドにラベル付き第一プローブを露出することを用いる、配列特有逆ハイブリッド形成により、ラベル付き第一プローブの配列が決定される。ハイブリッド形成解析は、当該技術に既知の、吸い取りのような、幾つかの方法を用いて行われる。アウスベル他、分子生物学における短編議案、3版(ジョーン・ウイリー&ソン社、1995年)参照。好適実施例においては、オリゴヌクレオチドを固体支持物に規定の位置(即ち既知の位置)で固定化する。この固定化アレーは時に「DNAチップ」と呼ばれる。この固体支持物はまた、固体支持物へのオリゴヌクレオチド付着を容易にするため、(例えば、金又は銀で)被覆することが出来る。オリゴヌクレオチドを固体支持物に固定するには、当該技術で既知の各種方法のいずれかが用いられる。普通に使用される方法は、アビジン又はストレプトアビジンを用いる固体支持物の非共有被覆及びビオチン化オリゴヌクレオチド・プローブの固定化を含む。オリゴヌクレオチドはまたエポキシド/アミン結合化学品を用いて固体支持物に直接付着することが出来る。エガース他、DNA配列技術の進歩、SPIE大会議事録(1993年)参照。
【0083】
別の実施例においては、ハイブリッド形成及び重複部分を通じる陽性ハイブリッド形成プローブの解析により、ラベル付き第一プローブの配列が読み取られる。参照によりここに組み込んだドルマナック他、米国5,202,231号を参照。
さらに別の実施例においては、ラベル付きdNTPにより伸長された第一プローブが、酵素DNA配列(サンガー他、1977年、国立科学アカデミー誌、USA、74巻、5463頁)を用いて同定される。この場合、第一プローブに相補のDNA配列及び配列「タグ」の働きをする所定の追加共線型配列を含むオリゴヌクレオチドが合成される。サンガー配列条件の下で培養したとき、固定化第一プローブはその相補配列にハイブリッド形成し配列反応に関しプライマとして働く。生じた始動配列「タグ」はこのとき反応に存在する第一プローブを同定する。
【0084】
さらに別の実施例において、ラベル付きdNTPにより伸長された第一ラベルは配列同定に先立って拡大される。ラベル付き第一プローブは、追加「タグ」付き又は無しの配列用プライマ配列を含む相補性オリゴヌクレチオドと共に培養される。第一プローブのその相補性オリゴヌクレチオドへの初期ハイブリッド形成により、第一プローブは単一伸長反応において初期プライマとして役立つことが出来る。伸長生成物は次いで循環配列反応における鋳型として直接使用される。伸長生成物の循環配列は配列用生成物を拡大する結果となる。相補性オリゴヌクレチオドの設計に当たって、配列用プライマは、第一プローブ自体を通じるか又は、代わりに、「タグ」配列を通じて、配列を進行するよう、方向付けされる。後者の場合、「タグ」配列の決定は共線型第一プローブ配列を同定する。拡大された生成物は、当該技術で標準の化学的方法によるか、又は、上述のように、配列特有逆ハイブリッド形成方法により、配列される。
本発明の実施にあたって、第一プローブの又は相補性タグ付きオリゴヌクレオチドの、全配列の決定は必要でない。1、2、又は3配列反応が(「バーコード」と同様の)特性パターンの生成に(完全配列を得るに必要な4に代わって)有効であり、個々の第一プローブの即時同定が出来ることを意図している。この方法は、放射性ラベル付き第一プローブを用い、アナログ又はデジタル自動ラジオグラムを作成する手動配列方法に適用出来ると同時に、非放射性レポーター分子を用い、デジタル化パターンを生じる自動配列方法にも適用出来る。双方の場合において、確定データベースとの比較を電子的に実行することが出来る。こうして、必要配列反応の減少により、本発明の方法は、多数試料の経済的解析を容易にする。
【0085】
本発明は、多数の可能な第一プローブの同時篩い分けを単一反応に受け入れる。実際において、同時ハイブリッド形成のため貯められた分割プライマの実数は、診断要請にしたがって決定される。例えば、嚢胞性繊維症(CF)において、一つの特有突然変異(Δ508)がCF患者の70%以上に見られた。これ、Δ508固有分割プライマを用いる本発明の方法にしたがった予備的篩い分け、に続く連続プライマの単一基伸長、及び伸長第一プローブの検出により、Δ508対立因子が同定され除去される。第二篩い分け(「フェーズ2」)において、別の、頻繁でない、CF対立因子をエンコードする多数の分割プライマの篩い分けを実施し、続いて上述のように連続プライマの単一基伸長、及び伸長第一プローブの検出を実施した。
別の臨床状態においては、しかし、CFにおけるΔ508のように高い頻度で現れる単一突然変異は存在しない。したがって、分割プライマのプールは、プール陽性試料につきフェーズ2解析において必要になる独立分析の数によってのみ決定される。
【0086】
加えて、現在の臨床習慣においては、異なる臨床症候、例えば、嚢胞性繊維症、地中海貧血、及びゴーシェ病、は互いに無関係に篩い分けされる。本発明は、対照的に、一つ以上の疾病発生因子における突然変異に相補の多数の第一プローブを用いて、各種患者からの多数の核酸の同時篩い分けを受け入れる。
同様の方法で、臨床的指示薬が異質薬剤又は微生物による感染を示すとき、本発明は、多数の潜在異質核酸に付いて同時篩い分けを提供する。さらに、一つ以上の微生物の特別種、変種、突然変異体、及び類似物もまた、第一篩い分けにおいて適切な第一プローブを採用することにより区別することが出来る。
【0087】
本発明の方法はまた、フェーズ1又はフェーズ2篩い分けにおいて無作為置換された分割プライマを使用することにより、患者又は侵入微生物の核酸に運び込まれた新規の可能性がある突然変異対立遺伝子を定義することを可能にする。この実施例において、連続プライマの単一基伸長及び伸長第一プローブの検出と分離に続く配列により、精密な突然変異体配列が明らかになる。
前述は、分割プライマを用いる多重突然変異検出の文脈における本発明の慣例を例示する。ここに開示したような、本発明の数多の追加側面及び利点は、開示及び個々の例示を考察したとき明らかになる。したがって、本発明は付属請求項の範囲のみによって、限定される。
[配列表]
【数1】

【数2】
【数3】
【技術分野】
【0001】
本特許出願は、1997年6月16日に提出されたU.S.Serial Number 08/877,333の一部継続出願であり、その開示は本明細書に引例として包含する。
【0002】
[発明の属する技術分野]
本発明は、一般に、核酸試料中の単一ヌクレオチドを同定するためのオリゴヌクレオチドプライマ伸長法に関する。本発明の方法は、生体試料中の遺伝子突然変異の存在、または疾患を引き起こす微生物の存在を検出および同定することにより、疾患診断に有用となる。
【背景技術】
【0003】
[発明の背景]
遺伝性疾患等の疾患を引き起こす分子欠損の知見は、急速に増加している。遺伝子突然変異が引き起こすと考えられる遺伝性疾患には、鎌型赤血球貧血症、α−およびβ−地中海貧血、フェニルケトン尿症、血友病、α1−アンチトリプシン欠損症、ならびに膵嚢胞性繊維症が挙げられる。例えば、鎌型赤血球貧血症は、β−グロブリン遺伝子の6番目のコドンでの単一塩基対置換がもたらす同型接合性によるものと報告されている。Antonarakis,New England J.Med.320:153−163(1989)。インシュリン受容体遺伝子およびインシュリン応答性グルコース輸送体遺伝子の突然変異が、インシュリン抵抗性糖尿病で検出されている。非特許文献1。
【0004】
癌は、多数の癌遺伝子および腫瘍抑圧遺伝子の遺伝子突然変異に関連してきた。Duffy,Clin.Chem.,41:1410−1413(1993)。例えば、そのras遺伝子の点突然変異が、その遺伝子を形質転換癌遺伝子に転換させることが示されている。Bos et al.,Nature,315:725−730。p53腫瘍抑圧遺伝子座での突然変異および異型接合性の喪失は、種々の癌に関連してきた。Ridanpaa et al.,Path.Res.Pract,191:399−402(1995);Hollstein et al.,Science,253:49−53(1991)。加えて、apoおよびdoc腫瘍抑圧遺伝子の欠損またはその他の突然変異もまた、腫瘍発生に関連してきた。Blum,Europ.J.Cancer,31A:1369−1372(1995)。それらの突然変異は、疾患の初期ステップおよびそれの素因に対しマーカの役割を果たす可能性がある。初期診断は、治療の成功に重要なだけでなく、慢性的徴候が生じる前に予防または治療を導入することも可能にする。
【0005】
結腸直腸癌は、初期に検出されれば治癒される可能性が高い疾患の一例である。結腸癌は、初期の検出により、例えば癌にかかった組織の外科的除去で、効果的に治療し得る。結腸癌は、結腸上皮の細胞に始まり、初期ステップでは大循環(general circulation)から分離されるため、初期ステップの結腸癌の外科的除去は通常は成功する。こうして、結腸直腸細胞の初期突然変異の検出は、一般に、生存率を大幅に上昇させるはずである。結腸直腸癌の現行の検出法は、糞便潜血の存在または血清中を循環する癌胎児性抗原の存在等、癌の存在を示す細胞外証拠に集中している。そのような細胞外証拠は、典型的には、癌が浸潤性になった後にのみ生じる。この意味で、結腸直腸癌は治療が非常に困難である。
【0006】
疾患関連遺伝子内の突然変異の存在を検出するために、方法が改正されてきた。そのような方法の1つは、試料のゲノム領域の完全なヌクレオチド配列を、相当する野生型領域と比較することである。例えば、Engelke et al.,Proc.Acad.Sci,U.S.A.85:544−548(1988)、Wong et al.,Nature,330:384−386(1988)を参照されたい。しかし、そのような方法では費用と時間がかかり、低頻度突然変異を明白に検出するために、標的遺伝子の多重クローン(multiple clones)を分析する必要がある。そのため、遺伝子突然変異の大規模スクリーニングに、広範囲の塩基配列決定を利用するのは実用的ではない。
【0007】
酵素的および化学的切断技術を用いてDNAの配列変異を活用する様々な検出方法が開発されてきた。DNA多型のために一般的に用いられるスクリーニングは、Botstein et al.,Am.J.Hum.Genet.,32:314−331(1980)とWhite et al.,Sci.Am.,258:40−48(1988)により報告されるように、制限エンドヌクレアーゼでDNAを消化することと、生成した断片をサザンブロット法で分析することからなる。エンドヌクレアーゼの認識配列に影響を与える突然変異は、その部位での酵素的切断を妨害し、それによりDNAの切断様式を変化させる。制限断片長の差を捜索することにより、配列を比較する。この方法(制限断片長多型マッピングもしくはRFLPマッピングと呼ばれる)での問題は、制限エンドヌクレアーゼによる切断に影響を与えない突然変異の検出が不可能なことである。ある試験は、ヒトDNAの40,000塩基対領域での存在が推定される突然変異体で、RFLP分析により検出されるのがわすか0.7%であることを報告した。Jeffrey,Cell,18:1−18(1979)。
【0008】
単一塩基突然変異は、対立遺伝子特異性オリゴヌクレオチド(ASO)プローブを用いた差分ハイブリッド形成法により検出されてきた。Saiki et al.,Proc.Natl.Acad.Sci.USA.56:6230−6234(1989)。突然変異は、ミスマッチしたプローブと比較して、完全にマッチしたプローブのより高い熱安定性を基に同定する。突然変異分析のこの方法の欠点には、:(1)各プローブでハイブリッド形成を最適化する必要があることと、(2)プローブの判別度でのミスマッチおよび局所配列の適用制限(local sequence impose limitations)の性質とが挙げられる。実際に、検査試料の配列複雑性が高い場合(例えば異質性生体試料)には、核酸ハイブリッド形成のパラメータのみに基づく検査は機能性が乏しくなる。これは、幾分かは、単一ヌクレオチドの変化により生じるハイブリッド安定性の小さな熱力学的差異による。つまり、核酸ハイブリッド形成は、一般に、分析および診断目的で、その他幾つかの選択または強化手順を組み込んでいる。
【0009】
酵素を介したライゲーション法では、突然変異は、標的DNA分子またはRNA分子上で互いに直接隣接してアニーリングすることが可能な2つのオリゴヌクレオチドにより識別され、オリゴヌクレオチドの1つは、点突然変異に相補的な3’末端を有する。隣接するオリゴヌクレオチド配列は、両方のヌクレオチドが正しくの塩基対を形成する場合には、共有結合のみの結合である。こうして、点突然変異の存在は、2つの隣接するオリゴヌクレオチドのライゲーションにより示される。Grossman et al.,Nucleic Acid Research,22:4527−4534(1994)。しかし、この検出方法の有用性は、あるヌクレオチドミスマッチの許容性から、または非鋳型指向性ライゲーション反応(non−template directed ligation reactions)から生じる高いバックグランドにより損なわれる。Barringer et al.,Gene,89:117−122(1990)。
【0010】
鋳型依存性プライマ伸長反応に基づき、検出法が多数開発されてきた。これらの方法は、主に2つのカテゴリに分類される:(1)突然変異を識別する領域をつなぐプライマを用いた方法、および(2)突然変異を識別する領域の近傍と上流でハイブリッド形成するプライマを用いた方法。
【0011】
最初のカテゴリでは、ミスマッチのあるものに比べ完全にマッチしたプライマの結合に有利なハイブリッド形成条件下で、標的核酸の単一塩基突然変異が拮抗的オリゴヌクレオチドプライミングにより検出されるような方法を、CaskeyとGibbs(U.S.Patent No.5,578,458)が報告している。VaryとDiamond(U.S.Patent No.4,851,331)は、プライマの3’末端ヌクレオチドが、当該の変異体ヌクレオチドに相当する、同様の方法を記載している。プライマの3’末端におけるプライマと鋳型とのミスマッチが延長を阻害するために、通常のプライマ伸長条件下では、トレーサヌクレオチドの取り込み量に有意差が生じる。
【0012】
プライマ依存性DNAポリメラーゼが、一般に、低い複製エラー率を有するとは知られて久しい。この特性は、後代への障害作用を有する遺伝的誤りを防ぐ上で重要である。2番目のカテゴリの方法は、この酵素学的反応に特有の高い忠実度を活用している。突然変異の検出は、プライマ伸長と検出可能な鎖終結ヌクレオチド三リン酸の取り込みに基づいている。DNAポリメラーゼの高い忠実度により、レポータ分子を標識した正しい塩基の特異的取り込みが確実に行われる。そのような単一ヌクレオチドプライマ誘導性伸長測定は、アスパルチルグリコサミン尿症、血友病B、および膵嚢胞性繊維症を検出するために、さらにレーバー遺伝性視神経萎縮(LHON)に関連する点突然変異を定量するために用いられてきた。例えば、Kuppuswamy et al.,Proc.Natl.Acad.Sci.USA,88:1143−1147(1991)、Syvanen et al.,Genomics,8:684−692(1990)、Juvonen et al.,Human Genetics,93:16−20(1994)、Ikonen et al.,PCR Meth.Applications,1:234−240(1992)、Ikonen et al.,Proc.Natl.Acad.Sci.USA,88:11222−11226(1991)、Nikiforov et al.,Nucleic Acids Research,22:4167−4175(1994)を参照されたい。分子量に基づき伸長したプライマの分解能を高めるために、鎖終結ヌクレオチドの前に幾つかのヌクレオチドを添加することを含む別のプライマ伸長法も提案されている。例えば、Fahy et al.,WO/96/30545(1996)を参照されたい。
【0013】
プライマ伸長に基づく方策には、完全にアニーリングしたオリゴヌクレオチドのみが、伸長反応のプライマとして確実に機能するためにかなりの最適化が必要となる。ポリメラーゼの高い忠実度が与える利点は、プライマの鋳型へのハイブリッド形成におけるヌクレオチドミスマッチの許容性により損なわれる可能性がある。あらゆる「不正確な」プライミングを、真の陽性シグナルと区別することは困難である。
【0014】
オリゴヌクレオチドプライマ伸長測定の選択性および感受性は、オリゴヌクレオチドプライマの長さと反応条件とに関係する。一般に、高い選択性に有利なプライマ長と反応条件は、低い感受性をもたらす。これとは逆に、高い感受性に有利なプライマ長と反応条件は、低い選択性をもたらす。
【0015】
典型的な反応条件下では、短いプライマ(即ち約15マー未満)は一時的で不安定なハイブリッド形成を呈する。そのため、一時的で不安定なオリゴヌクレオチドハイブリッドは、伸長反応を容易に開始せず、伸長したオリゴヌクレオチドの低い収率をもたらすため、短いプライマを使用する場合には、プライマ伸長測定の感受性が低くなる。その上、複雑な異質性生体試料では、短いプライマは、様々な種類の完全にマッチした相補的配列への非特異的結合を呈する。こうして、それらの低い安定性と高い非特異的結合のために、周知の位置にある突然変異の信頼性のある同定において、短いプライマはさほど有用ではない。そのため、プライマ伸長測定に基づいた検出法は、15マー〜25マーの長さ範囲のオリゴヌクレオチドプライマを使用する。例えば、特許文献1;特許文献2および特許文献3を参照されたい。しかし、安定性を増大させるプローブの延長は、選択性を低減させる作用を有する。ミスマッチしたプライマと完全にマッチしたプライマとの熱力学的差異は比較的小さいため、単一塩基ミスマッチは通常、より長いオリゴヌクレオチドプライマの結合効率に、より短いプライマに対してよりも低い作用を有する。より長いプライマの鋳型へのハイブリッド形成で、ヌクレオチドのスマッチの許容性が高い程、複雑な異質性生体試料での非特異的で「不正確な」プライミングが高レベルでもたらされる可能性がある。
【0016】
プライマ伸長反応の反応条件は、ミスマッチしたオリゴヌクレオチドによる「不正確な」プライミングを減少させるよう最適化することが可能である。しかし、最適化は、過酷で費用のかかる労働であり、伸長したプライマの収率減少により低い感受性をもたらすことが多々ある。その上、完全にアニーリングされたオリゴヌクレオチドのみが伸長反応のプライマとして確実に機能するためには、かなりの最適化が必要とされるため、プライマ伸長測定の限定されたマルチプレックシングのみが可能となる。Krook et al.,supra(非特許文献1)は、2つの別個の単一ヌクレオチド取り込み反応において、異なる長さのプライマを用いることにより、および各突然変異部位の野生型ヌクレオチドと突然変異ヌクレオチドとをモニタリングすることにより、マルチプレックシングが達成し得ることを報告している。しかし、オリゴヌクレオチドプライマ伸長測定の選択性および安定性が、オリゴヌクレオチドプライマの長さと反応条件により決定されるとすれば、所与の反応混合物において同時に検査し得るプライマ数は、非常に限定されてしまう。
【0017】
当該技術分野の方法は、検査試料の配列複雑性を減少させることにより、不正確なプライミングの可能性を低下させる。こうして、ゲノムDNAは、生体試料から分離され、かつ/または識別される領域の側面に作用する(flank)プライマを用いたPCRにより増幅される。その後、プライマ伸長分析は、精製したPCR生成物上で実施される。特許文献1、特許文献2、および特許文献3を参照されたい。しかし、これらの方法は試料の処理という追加的ステップを含むため、時間と費用がかかる。その上、これらの方法は、単一試料における多重プライマ伸長反応(multiple primer extension reaction)には適応しない。
【先行技術文献】
【特許文献】
【0018】
【特許文献1】国際公開第91/13075号パンフレット
【特許文献2】国際公開第92/15712号パンフレット
【特許文献3】国際公開第96/30545号パンフレット
【非特許文献】
【0019】
【非特許文献1】Krool et al.,Human Molecular Genetics,1:391−396(1992)
【発明の概要】
【発明が解決しようとする課題】
【0020】
そのため、当該技術分野では、異質性生体試料の多数のゲノム突然変異に向けた、選択性および感受性のある核酸検出法ならびに信頼性のある大規模スクリーニング法が求められている。そのような方法を本明細書において提供する。
【課題を解決するための手段】
【0021】
[発明の概要]
本発明は、高い感受性と高い選択性を有する突然変異検出法を提供する。一般的実施例においては、本発明は、少なくとも1回反復する単一塩基伸長反応を含む。本発明の方法は、異質性生体試料において、遺伝子突然変異または疾患を引き起こす微生物の存在を検出および同定する際に有用である。
本発明の方法は、単一塩基伸長反応を複数のサイクルで実施し、それにより選択性を損なうことなくプライマ伸長測定の感受性を増大させることを含む。好ましい実施例においては、本発明の方法は、2〜100サイクルのプライマ伸長を含む。より好ましくは、10〜50サイクル実施する。最も好ましくは、約30サイクル実施する。
好ましい実施例においては、過剰のプライマを用いて、各伸長サイクルにおいて伸長したプライマを確実に生成する。オリゴヌクレオチドのプライマ長は、好ましくは約10〜約100ヌクレオチド、より好ましくは約15〜約35ヌクレオチド、最も好ましくは約25ヌクレオチドである。
【0022】
好ましい実施例においては、各伸長反応は、完全な相補的配列を備える核酸のみへのプライマのハイブリッド形成を促進させる条件を含む(即ち、ミスマッチ塩基対は許容されない)。1つの実施例においては、ハイブリッド形成は、測定でのプライマのほぼTmで実施される。より好ましい実施例においては、ハイブリッド形成は、プライマのTmより高温で実施される。
【0023】
1つの実施例においては、ハイブリッド形成されたオリゴヌクレオチドプライマは、標識された末端ヌクレオチドにより伸長される。標識ddNTPおよびdNTPは、好ましくは伸長プライマの検出を容易にする「検出部分」を含む。検出部分は、蛍光、発光または放射性標識、酵素、ハプテン、および、例えば分光光度法により、標識伸長生成物を容易に検出させる、ビオチン等のその他の化学的識別票からなる群から選択される。
【0024】
好ましい実施例においては、ハイブリッド形成して伸長したプライマを変性し、伸長していないプライマをアニーリングして、新たにハイブリッド形成したプライマを伸長することにより、プライマ伸長のさらなるサイクルを開始する。反応での過剰なプライマの存在は、反応の各サイクルの伸長していないプライマのアニーリングを促進させる。
【0025】
さらなる実施例においては、本発明の方法は、分断したプライマを用いて、少なくとも2サイクルの単一塩基伸長反応を実施することを含む。本発明の方法は、推測される突然変異の部位に隣接する2つのプローブをハイブリッド形成することを含み、ここでいずれのプローブも単独では鋳型依存性伸長のプライマにはなり得ないが、プローブが互いに隣接してハイブリッド形成する場合には、伸長を開始することが可能である。好ましい実施例においては、本発明の方法は、約5塩基〜約10塩基の長さを有するプローブを標的核酸へハイブリッド形成することを含み、ここでプローブは推測される突然変異の直ぐ上流でハイブリッド形成する。本発明の方法は、さらに第一プローブの上流にある第二プローブをハイブリッド形成することを含み、その第二プローブは約15〜約100ヌクレオチドの長さを有し、3’末端の伸長不能なヌクレオチドを有する。第二プローブは、第一プローブと実質的に近接している。好ましくは、実質的に近接するプローブは、0〜約1ヌクレオチド隔てている。リンカが核酸伸長反応を妨害しないとすれば、好ましくは第一および第二プローブが2以上のヌクレオチドにより分離されるようにリンカが使用される。そのようなリンカは、当該技術分野で周知であり、例えば、ペプチド核酸、DNA結合蛋白質、およびライゲーションが挙げられる。
【0026】
別の実施例においては、分断したプライマは、第一オリゴヌクレオチドプローブ系列(a series of first oligonucleotide probe)を含む。第一プローブ系列の各構成員が、標的核酸の実質的な近接部分に同時にハイブリッド形成し、それにより近接するプライマを形成する場合を除けば、前記系列の構成員はいずれも、核酸ポリメライゼーションのプライマにはなり得ない。1つの実施例においては、分断したプライマは、3つの8マー第一プローブを含む。別の実施例においては、分断したプライマは4つの6マー第一プローブを含む。
【0027】
伸長測定の各サイクルにおいて、伸長反応は、鋳型依存法による上記プローブの共同ハイブリッド形成(co−hybridization)からもたらされる分断したプライマにヌクレオチドを付加させる。好ましい実施例においては、標的核酸へハイブリッド形成された第一プローブは、標識した末端ヌクレオチドにより伸長するが、野生型核酸または非標的核酸へハイブリッド形成された第一プローブは、非標識の末端ヌクレオチドで伸長する。標識ddNTPまたはdNTPは、好ましくは標識した末端ヌクレオチドで伸長した短いプローブの検出を容易にする「検出部分」を含む。検出部分は、蛍光、発光または放射性標識、酵素、ハプテン、および、例えば分光光度法により、標識伸長生成物を容易に検出させる、ビオチン等のその他の化学的識別票からなる群から選択される。
【0028】
好ましい実施例においては、測定シグナルを増幅させるために、数サイクルの伸長反応が実施される。伸長反応は、過剰な第一及び第二プローブ、標識dNTPまたはdNTP、ならびに熱安定ポリメラーゼの存在下で実施される。一旦伸長反応が完了すると、標的核酸に結合した第一および第二プローブは、反応混合物をハイブリッドの融解温度を超える温度で加熱することにより分解される。その後、反応混合物は、ハイブリッドの融解温度未満に冷却され、第一および第二プローブはもう1つの伸長反応のために標的核酸と会合させる。好ましい実施例においては、10〜50サイクルの伸長反応が実施される。最も好ましい実施例においては、30サイクルの伸長反応が実施される。
【0029】
本明細書に開示される方法を用いて、単一ヌクレオチド多型(SNP)、つまり挿入、欠失、および置換などの突然変異を検出してもよい。本明細書の方法でスクリーニング可能な核酸試料は、ヒト核酸試料を含む。プライマ(もしくは分断したプライマ)は、ハイブリッド形成されたプライマの3’末端が、測定されるヌクレオチド位置の相補的位置の直ぐ上流にあるように設計される。測定されるヌクレオチド位置は、単一塩基プライマ伸長反応で取り込まれるヌクレオチドに相補的なヌクレオチドとして同定される。例えば、Gがその反応で取り込まれる場合には、Cは生体試料中の核酸の相補的位置に存在する。好ましい実施例においては、プライマ伸長反応は、4つのヌクレオチド、好ましくは鎖終結ヌクレオチド(例えばジデオキシヌクレオチドの、ddATP、ddCTP、ddGTP、およびddTTP)の存在下で実施される。より好ましい実施例においては、ヌクレオチドは検出可能に標識され、好ましくは判別的に標識される。別の実施例においては、伸長反応は、1、2、または3の異なるヌクレオチドの存在下で実施される。測定されるヌクレオチド位置で、生体試料が異質性であれば、プライマ伸長測定では、相補的ヌクレオチドが取り込まれる(それらがプライマ伸長反応に含まれる場合)。
【0030】
本明細書に開示される方法を用いて、癌等の疾患に関連する突然変異を検出してもよい。加えて、本発明の方法を用いて、酵素活性の完全喪失または部分的喪失など、代謝エラーを引き起こす欠失または塩基置換突然変異を検出してもよい。
【0031】
別の実施例においては、特異的核酸配列は、病的症状または症候に関与することが知られるヒトゲノム核酸中の特定遺伝子または遺伝子座の一部を含む。非限定的例には、膵嚢胞性繊維症、テイ−サックス病、鎌型赤血球貧血症、β−地中海貧血、およびゴーシェ病が挙げられる。
【0032】
さらに別の実施例においては、特異的核酸配列は、特定疾患に関連することが知られていないが、多型が知られるまたは推測される、特定遺伝子または遺伝子座の一部を含む。
【0033】
さらに別の実施例においては、特異的核酸配列は、例えば侵入した微生物のゲノム等、外来遺伝子配列の一部を含む。非限定的例には、細菌およびそのファージ、ウイルス、菌類、原生動物等が挙げられる。適切な治療的介入を選択するために、微生物の異なる変異種または菌株を区別することが望ましい場合に、本発明の方法は特に適用できる。
【0034】
ゲノム核酸試料は、生体試料から分離される。一旦分離された核酸は、さらに操作することなく本発明に使用してもよい。別法として、核酸に存在する1以上の特異的領域を、例えばPCRにより増幅してもよい。このステップでの増幅は、標的核酸配列集団内で特異的核酸配列の濃度を増加させるという利益をもたらす。別の実施例においては、ゲノム核酸は、さらに分析される前に切断される。
【0035】
別の実施例においては、核酸は固相支持体に結合させる。これにより、多数の試料の同時処理およびスクリーニングが可能となる。本発明での使用に適した支持体の非限定的例には、ニトロセルロースまたはナイロンのフィルタ、ガラスビーズ、親和性捕獲用の薬剤をコーティングした磁気ビーズ、処理したまたは未処理のミクロタイタープレート等が挙げられる。好ましい実施例においては、支持体は、多数のウェルを有するミクロタイターディッシュである。そのような支持体の使用により、多数の検体および対照の同時測定が可能となり、こうして分析が促進される。その上、自動化装置を使用して、そのようなミクロタイターディッシュへ試薬を提供することも可能である。別の実施例においては、本発明の方法は水相において実施される。
【0036】
本発明の1つの実施例においては、伸長プライマまたはプローブが算出される。プライマまたはプローブは、好ましくはインピーダンスビーズで標識したヌクレオチドで伸長し、インピーダンスビーズの数が計数される(例えばコールターカウンタを用いる)。その後、標識プライマの数は、インピーダンスビーズの数から決定される。標識は、より好ましくは放射性同位体であり、標識プライマまたはプローブに関連する放射性崩壊の量を測定する。標識プライマまたはプローブの数は、放射性崩壊の量から計算する。伸長したプライマまたはプローブの数は、循環した伸長反応の統計解析に有用である。
【0037】
最後に、本発明の方法はさらに、伸長したプライマまたは第一プローブを分離することおよび塩基配列決定することを含む。プライマまたは第一プローブは、好ましくはそれらの分離を容易にする「分離部分」を含む。分離部分の非限定的例には、ハプテン、ビオチン、およびジゴキシゲニンが挙げられる。好ましい実施例においては、分離部分を含むプライマまたは第一プローブは、分離部分に対し親和性を有する(例えば、抗ハプテン、アビジン、ストレプトアビジンまたは抗ジゴキシゲニンでコーティングされた)固形支持体に固定されている。固形支持体は、ガラス、プラスチック、および紙からなる群から選択される。支持体は、円柱、ビーズ、棒、または試験管として形づくられる。好ましい実施例においては、分離部分は、標識ddNTPまたはdNTPに取り込まれて、標識ddNTPまたはdNTPで伸長した第一プローブのみが支持体に固定される。そのことにより、標識したプライマまたは第一プローブが、伸長していないプライマまたは第一プローブ、および第二プローブから分離される。別の好ましい実施例においては、鋳型とハイブリッド形成して伸長する第一プローブの能力を分離部分が妨害しないならば、分離部分は第一プローブ全てに取り込まれる。分離部分を取り込むことにより、第一プローブの全てが固形支持体に固定される。第一プローブは、1回以上の洗浄ステップにより第2プローブから分離される。
【0038】
その後、標識したプライマまたは第一プローブは、突然変異または疾患を引き起こす微生物を同定するために塩基配列決定する。1つの実施例においては、固定したプライマまたはプローブを、例えば当該技術分野で標準的な化学的方法を用いて、塩基配列決定に直接供試される。その他の実施例においては、標識した第一プローブを固形支持体から除去して、水性溶液中で、標識した第一プローブの塩基配列決定を実施する。分離した第一プローブを複数の相補的オリゴヌクレオチドと接触させる。1つの実施例においては、分離した第一プローブをプライマとして、相補的オリゴヌクレオチドを鋳型として用いて、酵素的塩基配列決定を実施する。別の実施例においては、分離した第一プローブをプライマとして、相補的オリゴヌクレオチドを鋳型として用いて、単一塩基伸長反応を実施する。酵素的塩基配列決定により、伸長生成物の配列を決定する。伸長した標識第一プローブの配列が、遺伝突然変異または試料中に存在する、疾患を引き起こす微生物を同定する。
本発明は、さらに、例えば以下の項目を提供する。
(項目1)核酸試料中の単一ヌクレオチドを同定するための方法であって、
(a)オリゴヌクレオチドプライマと核酸試料中の核酸の一部の間の厳密な相補的ハイブリッド形成を促進する条件下で、前記プライマを前記試料へアニーリングするステップと、
(b)単一塩基により前記プライマを伸長するステップと、
(c)前記伸長したプライマを前記部分から分離するステップと、
(d)ステップ(a)〜ステップ(c)を反復するステップと、
(e)前記伸長したプライマに取り込まれた塩基を同定し、それにより前記単一ヌクレオチドを同定するステップと、を含む方法。
(項目2)ステップ(b)の前記塩基が、鎖終結ヌクレオチドである、項目1に記載の方法。
(項目3)ステップ(b)の前記伸長反応が、1以上の鎖終結ヌクレオチドの付加により実施される、項目1または2に記載の方法。
(項目4)前記単一塩基が、検出可能に標識される、項目1〜3のいずれか1項に記載の方法。
(項目5)前記単一ヌクレオチドが、多型変異体である、項目1〜4のいずれか1項に記載の方法。
(項目6)前記試料が、異なる個体からの複数の組織または体液検体から集められた核酸を含む、項目1〜5のいずれか1項に記載の方法。
(項目7)前記プライマが、分断したプライマである、項目1〜6のいずれか1項に記載の方法。
(項目8)前記分断したプライマが、15未満のヌクレオチドを有する第一プライマと、15を超えるヌクレオチドを有する第二プライマとを含む、項目1〜7のいずれか1項に記載の方法。
(項目9)周知の単一ヌクレオチド多型変異体の存在を検出するための方法であって、
(a)オリゴヌクレオチドプライマを、単一ヌクレオチド多型変異体の直ぐ上流にあると推測される核酸の一部へアニーリングするステップと、
(b)単一塩基により前記プライマを伸長するステップと、
(c)前記プライマを前記核酸から分離するステップと、
(d)ステップ(a)〜ステップ(c)を少なくとも1回反復するステップと、
(e)前記単一塩基を同定し、それにより前記多型変異体を同定するステップと、を含む方法。
(項目10)集団中の単一ヌクレオチド多型変異体を同定するための方法であって、
(a)集団の複数の個体の各構成員から核酸を含む試料を得るステップと、
(b)推測される単一ヌクレオチド多型変異体の直ぐ上流の前記核酸の一部に相補的なオリゴヌクレオチドプライマを、各試料中の前記核酸の一部へアニーリングするステップと、
(c)単一塩基により各試料中の前記プライマを伸長するステップと、
(d)前記プライマを各試料中の前記核酸から分離するステップと、
(e)ステップ(b)〜ステップ(d)を少なくとも1回反復するステップと、
(f)各試料中の前記単一塩基を同定するステップと、
(g)単一ヌクレオチド多型変異体を、ステップ(f)で同定された1以上の塩基の存在として同定するステップと、を含む方法。
(項目11)前記試料が混合されている、項目10に記載の方法。
(項目12)単一ヌクレオチド多型変異体が疾患の証拠である、項目9〜11のいずれか1項に記載の方法。
(項目13)前記疾患が、癌、膵嚢胞性繊維症、テイ−サックス病、鎌型赤血球貧血症、ゴーシェ病、β−地中海貧血、および寄生虫感染からなる群から選択される、項目9〜12のいずれか1項に記載の方法。
本発明の更なる様相および利益は、以下に詳述するそれの説明を検討することにより明白となる。
【図面の簡単な説明】
【0039】
【図1】単一塩基多型を検出するための単一塩基伸長反応において、分断したプライマの使用を記述する線図である。白線は鋳型を表し、濃灰色は第一プローブ(薄灰色)に実質的に近接する鋳型の領域へハイブリッド形成する第二プローブを表す。単一塩基突然変異が推測される部位はAと標識する。検出可能な標識をBと印字する。
【図2A】統計確率の低い領域を示すガウス分布モデルである。
【図2B】統計確率の低い領域を示すガウス分布モデルである。
【図3】細胞の異質性集団で細胞の1%が突然変異している確率値Nを示すグラフである。
【発明を実施するための形態】
【0040】
[発明の詳細な説明]
本発明は、第一に、高い選択性と高い感受性の両者を備える単一塩基伸長測定を提供することである。本発明は、高い感受性と高い選択性の両者を備える生体試料の特異的核酸を検出する方法を提供する。本発明は、伸長生成物の低い収率により感受性を喪失することなく、厳しいハイブリッド形成条件の高い選択性を提供する。一般に、本発明の方法は、生体試料中で複数サイクルの単一塩基伸長反応を実施することを含む。循環により、伸長生成物の収率は高くなり、プライマのハイブリッド形成条件が、単一塩基伸長反応中に典型的に適用されるものに比べ厳しく保たれるために、選択性の有意な喪失がない。本発明の方法は、癌等の疾患に関連する突然変異、酵素活性の完全喪失または部分喪失等、代謝エラーを引き起こす欠失または塩基置換突然変異、病的症状または症候に関与することが知られる患者のゲノム核酸中の特定遺伝子または遺伝子座の一部、単一ヌクレオチド多型(SNP)、あるいは、例えば侵入した疾患を引き起こす微生物のゲノムの、外来遺伝子配列の一部を検出および同定するのに有用である。
【0041】
単一塩基プライマ伸長反応は、オリゴヌクレオチドプライマを相補的核酸へアニーリングすることにより、およびアニーリングされたプライマの3’末端を、DNAポリメラーゼで触媒する鋳型指向性反応で付加された鎖終結ヌクレオチドで伸長することにより実施される。単一塩基プライマ伸長反応の選択性および感受性は、オリゴヌクレオチドプライマ長と反応条件(例えば、アニーリング温度、塩濃度)とに影響される。
【0042】
プライマ伸長反応の選択性は、試料中でのオリゴヌクレオチドプライマと核酸との厳密な相補的ハイブリッド形成量に反映される。選択性の高い反応は、厳密な相補的配列(即ち、ハイブリッド形成されたプライマと核酸との間に塩基ミスマッチがない)を備える核酸へのプライマハイブリッド形成のみを促進する。これに反して、非選択的反応では、プライマは、部分的な相補的配列(即ち、ハイブリッド形成されたプライマと核酸との間に塩基ミスマッチがある)を備える核酸にもハイブリッド形成する。一般に、選択的なプライマハイブリッド形成に有利なパラメータ(例えば、より短いプライマおよびより高いアニーリング温度)は、ハイブリッド形成されたプライマレベルの低下をもたらす。そのため、選択的な単一塩基プライマ伸長反応測定に有利なパラメータは、測定の感受性低下をもたらす。
【0043】
本発明の方法は、少なくとも2サイクルの単一塩基伸長反応を実施することを含む。本発明の方法は、単一塩基伸長反応を反復することにより、測定の選択性を低下させることなく、単一塩基プライマ伸長測定のシグナルを増強させる。循環によりシグナルが増強し、そのため伸長反応が選択性の高い条件下(例えば、プライマがほぼTmかそれを上回る温度でアニーリングされる)で実施されることが可能である。
【0044】
好ましい実施例においては、本発明の方法は、厳密なハイブリッド形成に有利な条件下で過剰のプライマをアニーリングすること、ハイブリッド形成したプライマを伸長すること、伸長したプライマを変性すること、およびアニーリングと伸長反応を少なくとも1回反復すること、により実施される。最も好ましい実施例においては、反応サイクルは熱変性のステップを含み、ポリメラーゼは温度安定性がある(例えば、TqポリメラーゼまたはVentポリメラーゼ)。
好ましいプライマ長は、10〜100ヌクレオチドであり、より好ましくは10〜50ヌクレオチドであり、最も好ましくは約30ヌクレオチドである。有用なプライマは、プライマの3’末端の単一塩基伸長が、鋳型に存在する突然変異ヌクレオチドに相補的なヌクレオチドを取り込むよう、推測される突然変異部位に隣接してハイブリッド形成するものである。
【0045】
好ましいハイブリッド形成条件は、反応でのオリゴヌクレオチドのほぼTmかそれを上回るアニーリング温度を含む。オリゴヌクレオチドプライマのTmは、その長さとGC含量により測定され、当該技術分野で周知の複数の式の1つを用いて計算される。標準的アニーリング条件では、約25ヌクレオチド長のプライマの好ましい式は、Tm(℃)=4x(G数+C数)+2x(A数+T数)。
好ましい反応では、アニーリングと変性のステップは、反応温度を変化させることにより実施される。本発明の1つの実施例においては、プライマは、そのプライマのほぼTmでアニーリングされ、その温度を伸長の最適温度まで昇温させ、その後その温度を変性温度にまで昇温させる。アニーリング、伸長、および変性の温度と時間との例を、例2に記載する。本発明のより好ましい実施例においては、反応は、アニーリング温度と変性温度の間で循環され、単一塩基伸長は、アニーリング条件から変性条件までの変移の間に生じる。
【0046】
本発明の好ましい実施例においては、2サイクル以上の伸長を実施する。より好ましい実施例においては、5〜100サイクル実施される。これ以外の実施例においては10〜50サイクル、最も好ましくは約30サイクル実施される。
【0047】
本発明の好ましい実施例においては、鋳型依存性反応のプライマの3’末端に付加されるヌクレオチドは、例えばジデオキシヌクレオチドの、鎖終結ヌクレオチドである。より好ましい実施例においては、ヌクレオチドは論じたinfraと同様に検出可能に標識される。
【0048】
I.分割プライマを用いる循環伸長反応
一実施例において、本発明の方法は、分割プライマを用い少なくとも二つの単一基伸長を行うステップを含む。好適実施例において、分割プライマは、ターゲット核酸の実質的連続部分にハイブリッド形成の能力がある短い第一プローブ及び長い第二プローブを含む。二つのプローブは、長い第二プローブがない状態では短い第一プローブのハイブリッド形成を利さないとの条件下で試料に露出される。ハイブリッド形成に影響する要因は当該技術で周知であり、温度、イオン濃度、pH、プローブ長さ、及びプローブGC含有量などが含まれる。第一プローブは、大きさが小さいので、平均的ゲノムの多くの場所でハイブリッド形成する。例えば、人ゲノムにおいて任意の所与の8−merは約65,000回起こる。しかし、一つの8−merは融解温度(Tm)が低く、単一基不整合がこれの不安定度を大きく誇張する。他方第二プローブは、第一プローブより大きいので融解温度Tmが高い。例えば、20−mer第二プローブは、一般的に8−merより良い安定度でハイブリッド形成する。しかし、単一ヌクレオチド変化によりハイブリッド安定度に生じる熱力学的相違が小さいため、長いプローブは安定なハイブリッドを形成するけれども、ヌクレオチド不整合を許容するので、選択性が低い。したがって、第一プローブに不利なハイブリッド形成条件の下では(例えば、第一プローブの上10−40度C)、第一プローブは高い選択性でハイブリッドを形成する(即ち、単一の不整合があるだけでも配列にほとんどハイブリッド形成しない)が、単独で(即ち、第二プローブが存在しなくて)ハイブリッド形成するときは、不安定なハイブリッドを形成する。第二プローブは、安定なハイブリッドを形成するが、不整合に対する許容性のため選択性が低い。
【0049】
本発明における伸長反応は、第一及び第二のプローブの連続ハイブリッド形成が無いと、起こらない。第一(近接)プローブ単独では、分析に用いられる反応条件の下で安定ハイブリッドを形成しないので、鋳型に基く核酸伸長に関しプライマにならない。第一プローブは約5個から10個の間のヌクレオチドを含むのが好適である。第一プローブは、突然変異が疑われる核酸に隣接してハイブリッド形成をする。本発明の突然変異判定方法における第二(遠隔)プローブは、第一プローブの上流側でターゲット(鋳型)のほぼ連続領域においてハイブリッド形成をする。第二プローブ単独では、3’伸長不能ヌクレオチドから構成されるので、鋳型に基く核酸伸長のプライマにはならない。第二プローブは、第一プローブより大きく、長さがヌクレオチド約15個と約100個の間であるのが好適である。
【0050】
本発明の方法にしたがうと、鋳型依存の伸長は、第一プローブが第二プローブの次ぎにハイブリッド形成するときにのみ起こる。これが起こるとき、短い第一プローブは、単一基突然変異の疑われるサイトの直ぐ隣りにハイブリッド形成する。第二プローブは第一プローブの5’終端近傍にハイブリッド形成する。二つのプローブの存在は、共同束縛効果により、一緒になって安定性を増す。二つのプローブは一緒に、ポリメラーゼによりプライマとして認識される。このシステムは、短いプローブの選択性と長いプローブの安定性とを用いてハイブリッド形成するプライマを生じるため、短いプローブの高い選択性と、長いプローブが与えるハイブリッド形成安定性と利用する。したがって、本質的に、分割プライマに伴う偽始動がない。長い第二プローブによる不整合許容範囲が偽信号を発生しないので、対応する長い第二プローブを欠いた短い第一プローブの伸長をハイブリッド形成条件が許さない限り、幾つかの分割プライマを同一反応において分析することが出来る。さらに、ターゲットに関する選択性が増加するので、本発明の方法は、検出のため試料中で少量しか利用出来ず、通常は例えばPCRなどを用いて拡大しなければならないターゲット核酸を、検出し同定するのに用いることができる。
【0051】
プローブ二つのハイブリッド形成を要することにより、偽陽性信号が減少又は消滅する。このように、分割オリゴヌクレオチドの使用により、現在必要とされる個々のプローブについてのハイブリッド形成条件の慎重な最適化が不要となり、広範な多重化が出来るようになる。対応する長い第二プローブを欠いた短い第一プローブの安定ハイブリッド形成をハイブリッド形成条件が許さない限り、幾つかのオリゴヌクレオチドを使用して、同一反応で分析される幾つかのターゲット配列をプローブすることが出来る。
第一及び第二プローブは、ターゲットの実質的連続部分にハイブリッド形成する。本発明の目的に関し、実質的連続部分とは、ハイブリッド形成された第一及び第二プローブが単一プローブとして(例えば核酸伸長のプライマとして)機能することが出来るのに十分近い部分である。実質的連続部分は、ヌクレオチド0個分(即ち、部分間に間隔がなく正確に連続)とヌクレオチド約1個分の間だけ離れているのが好適である。第一及び第二プローブがヌクレオチド2個分以上離れているときは、リンカーが分析(例えば、核酸伸長反応)を妨害しないとの条件で、リンカーを使用するのが好適である。このようなリンカーは当該技術では周知で、例えば、ペプチド核酸、DNA結合蛋白、及び結紮などが含まれる。隣接プローブは、長い第二プローブが短い第一プローブに安定性を与えるよう、共同的に結びつくことを明らかした。しかし、第二プローブが与える安定性は、第一プローブの選択性(即ち、不整合に対する非許容性)にうち勝たない。したがって、本発明の方法は、短い第一プローブの高い選択性と長い第二プローブが与える安定性との利点を利用する。
【0052】
こうして、好適実施例においては、第一及び第二プローブがターゲットの実質的連続部分にハイブリッド形成され、このとき第一プローブは突然変異、例えば単一基突然変異、の疑われるサイトの直ぐ隣で上流側にある。このとき試料は、疑わしいサイトで可能な突然変異の相補体であるジデオキシ核酸に露出される。例えば、野生型核酸が既知のサイトでアデニンであれば、ジデオキシ・アデニン、ジデオキシ・シトシン及びジデオキシ・グアニンが試料内に置かれる。ジデオキシ核酸にはラベルを付けるのが好適である。単一ヌクレオチド添加後、反応が停止するときは、代わりにデオキシ核酸を使ってもよい。ポリメラーゼを内生的又は外生的に供給すると、ジデオキシ基の第一プローブ上への組み込みが促進される。ラベルの検出は、非野生型(即ち突然変異体)基の組み込みを示すので、第一プローブに隣接するサイトに突然変異がある。代わりに、本発明の方法は、野生型配列が未知であるときに常用される。この場合、四つの共通ジデオキシ・ヌクレオチドに別々のラベルを付ける。上述の分析における一つ以上のラベルの出現は、突然変異の存在を示す。
代替好適実施例においては、分割オリゴヌクレオチドが一連の第一プローブを構成する。このとき、分割オリゴヌクレオチドの構成要員すべてが同時に核酸の実質的連続部分にハイブリッド形成するとき十分な安定性が得られる。ここで、短いプローブは過渡的で不安定なハイブリッド形成を示すけれども、隣接する短いプローブは共同的に結びついて個々のプローブそれぞれより大きい安定性を持つことが明らかになった。隣接してハイブリッド形成された一連の第一プローブは纏まって、個々のプローブ又は一連のプローブのサブセットより大きい安定性を持つ。例えば、三個の一連の第一プローブから構成される分割プライマ(即ち、ターゲット核酸上流核酸の実質的連続部分にハイブリッド形成能力のある終端ヌクレオチドのない三個の短いプローブ)を用いる伸長反応において、三個のプローブの同時ハイブリッド形成が、核酸伸長始動のための三個のプローブについて十分な共同安定性を生じ、疑わしい突然変異に直ちに隣接する短いプローブが伸長される。こうして一連の短い第一プローブから構成される分割プローブは、短いプローブの高い選択性(即ち不整合の非許容性)と長いプローブの安定性を提供する。
【0053】
好適実施例においては、分析信号を拡大するため数サイクルの伸長反応を実行する。伸長反応は、dNTP又はddNTPラベル付きの余分な第一及び第二プローブ、及び熱安定ポリメラーゼを存在させて行う。伸長反応が完了すると、ターゲット核酸に束縛された第一及び第二プローブは、反応混合物をハイブリッドの融解温度以上に加熱することにより解離する。次いで反応混合物をハイブリッドの融解温度以下に冷却すると、第一及び第二プローブは別の伸長反応のためターゲット核酸に会合出来るようになる。好適実施例においては、10乃至50サイクルの伸長反応を行った。最好適実施例においては、30サイクルの伸長反応を行った。
【0054】
II.伸長プライマの検出
ラベル付きddNTP又はdNTPは、分割プライマ反応において伸長したプライマ又は伸長した短い第一プローブの検出を容易にする「検出部分」を含むのが好適である。検出部分は、蛍光性、発光性又は放射性ラベル、酵素、ハプテン及びその他の化学的タグ、ラベル付き伸長生成物を容易に検出出来るビオチンなどを含むグループから選ぶ。例えば、ダンシル基のような蛍光ラベル、フルオレスセイン及び置換フルオレスセイン、アクリジン誘導体、カウマリン誘導体、フタロシアニン、テトラメチルローダミン、テキサスレッド、9−(カルボキシエチル)−3−ヒドロキシ−6−オキサ−6H−キサンテン、DABCYL、およびBODIPY(分子プローブ、オイゲン、OR)などは、ここに記述する方法に特に有利である。このようなラベルは、多数試料の同時高生産分析用自動装置に日常的に使用されている。
好適実施例においては、プライマ即ち第一プローブは「分離部分」を含む。このような分離部分は、例えば、ハプテン、ビオチン、又はジゴキシゲニンである。これら分離部分を含むプライマ即ち第一プローブは、(例えば、抗ハプテン、アビジン、ステプタビジン又は抗ジゴキシゲニンで被覆された)分離部分に関し親和性を有する固体相マトリクスに固定して、反応混合物から隔離される。本発明での利用に適するマトリクスの非限定例には、ニトロセルローズ又はナイロンフィルタ、親和性キャプチャ用薬剤で被覆されたガラスビーズ、磁性ビーズ、処理又は未処理ミクロタイタープレート、及び類似物が含まれる。
【0055】
好適実施例において、分離部分はラベル付きddNTP又はdNTPに組み込まれる。ハイブリッド形成されたプライマ又はプローブの変性、及びラベル付きddNTP又はdNTPと共に伸長したプライマ又は第一プローブの固定マトリクスへの固定化により、ラベル付きプライマ又はラベル付き第一プローブが伸長しないプライマ又は伸長しない第一プローブ及び第二プローブ、及びラベル無しddNTPと共に伸長したプライマ又は第一プローブから、1回以上の洗滌ステップにより分離される。
代替好適実施例において分離部分は、分離部分が第一プライマの又はプローブの鋳型にハイブリッド形成して伸長する能力を妨害しない限り、プライマ又は第一プローブに組み込まれる。溶離プライマ又は第一プローブは固体支持体に固定化されるので、溶離第二プローブから1回以上の洗滌ステップにより分離することが出来る。
【0056】
代案として、ラベル付き終端ヌクレオチドと共に伸長したプライマ又は第一プローブの存在は、ハイブリッド形成したプライマ又はプローブを溶離しないで判定できる。検出方法は、プライマ又は第一プローブに組み込まれたラベル又はタグに左右される。例えば、ターゲット核酸に結合された放射性ラベル付き又は化学発光第一プローブは、フィルタをX線フィルムに露出して検出することが出来る。代わりに、蛍光ラベルを含むプライマ又は第一プローブは、レーザー又はランプに基づくシステムを用い蛍光物質を固有吸収波長で励起して検出することが出来る。
代替実施例においては、束縛されたプライマ又は第一及び第二プローブが、マトリクス固定ターゲット核酸から溶離される(下記を参照)。溶離は、当該技術で周知の、核酸ハイブリッドを不安定化する任意の手段(即ち、塩の低下、温度上昇、フォルムアミド、アルキルへの露出など)を用いて行われる。好適実施例において、束縛されたオリゴヌクレオチド・プローブは、ターゲット核酸−分割プライマ複合体を水中培養し、反応物をハイブリッドの融解温度以上に加熱して、溶離した。
【0057】
上記反応のいずれにおいても検出可能単一伸長基として、デオキシヌクレオチドを使用することが出来る。しかし、このような方法では単一デオキシヌクレオチド付加後、伸長反応を停止しなければならない。このような方法は、固有突然変異が既知(即ち、C→G)であるか否かに関わりなく採用できる。さらに、単一基突然変異検出用試料内でデオキシヌクレオチドのラベル付き種を一つだけ利用出来るときは、デオキシヌクレオチド一つだけを付加した後、伸長反応を停止する必要はない。この方法は、疑問の単一基位置を含むヌクレオチド反復があるとき、実際に信号を強化する。
好適実施例においては、ターゲット核酸をプライマ又は分割プライマに露出して伸長反応を行う前に、ターゲット核酸を固定支持体に固定化する。核酸試料を一旦固定化したら、試料を洗滌して固定化されなかった物質を洗い流す。次いで核酸試料を、本発明にしたがってプライマ又は分割プライマの一つ以上の組に露出する。一旦単一基伸長反応が完了したら、ラベル付きddNTP又はdNTPと共に伸長したプライマ又は第一プローブを、伸長しなかったプローブ及びラベル無しddNTP又はdNTPと共に伸長したプローブから隔離するのが好適である。束縛されたプライマ又は第一及び第二プローブは支持体に束縛されたターゲット核酸から溶離される。溶離は、当該技術に周知の、核酸ハイブリッドを不安定化する任意の手段(即ち、塩の低下、温度上昇、フォルマミド、アルキルへの露出など)を用いて行われる。好適実施例において、第一及び第二プローブは、ターゲット核酸−分割プライマ複合体を水中培養して解離し、反応をハイブリッドの融解温度以上に加熱して伸長した第一プローブを隔離した。代替好適実施例において伸長反応は、水様溶液中で実行した。単一基伸長反応が一旦完了すると、オリゴヌクレオチド・プローブをターゲット核酸から解離して伸長した第一プローブを隔離する。代替実施例においては、核酸が水様相中に止まる。
【0058】
最後に、本発明の方法は、伸長した第一プローブを隔離するステップと配列するステップとを含む。伸長した第一プローブの隔離には、例えば、ハプテン、ビオチン又はジゴキシゲニンのような「分離部分」を用いる。好適実施例において、分離部分を含む第一プローブは、分離部分に関し親和性を有する(例えば、抗ハプテン、アビジン、ステプタビジン又は抗ジゴキシゲニンで被覆された)固体支持体に固定化される。本発明での利用に適する支持体の非限定例には、ニトロセルローズ又はナイロンフィルタ、親和性キャプチャ用薬剤で被覆されたガラスビーズ、磁性ビーズ、処理又は未処理ミクロタイタープレート、及び類似物が含まれる。
好適実施例において、分離部分はラベル付きddNTP又はdNTPに組み込まれる。ラベル付きddNTP又はdNTPと共に伸長した溶離プライマ又は第一プローブの固体支持体への固定化により、ラベル付きプライマ又は第一プローブが伸長しない第一プローブ及び第二プローブ、及びラベル無しddNTPと共に伸長したプライマ又は第一プローブから、1回以上の洗滌ステップにより分離される。
【0059】
代替好適実施例において分離部分は、分離部分が第一プライマの又はプローブの鋳型にハイブリッド形成して伸長する能力を妨害しない限り、プライマ又は第一プローブに組み込まれる。溶離プライマ又は第一プローブは固体支持体に固定化されるので、溶離第二プローブから1回以上の洗滌ステップにより隔離することが出来る。
次いでラベル付きプライマ又は第一プローブを配列して、検出された突然変異又は病原微生物を同定する。一実施例においては、標準技術の化学的方法を用いて、固定化プローブを直接配列する。別の実施例においては、ラベル付き固定化第一プローブを固体支持体から移動してラベル付き第一プローブの配列は水様溶液中で行う。
【0060】
III.列挙的検出方法
本発明の方法は、核酸列挙が必要又は好ましいいずれの場合にも、有用である。第一に、上述の検出方法は、生体試料におけるヌクレオチド突然変異検出のため有用である。したがって、本発明の方法は、疾病に関連又は関連疑惑の核酸列挙(例えば対立因子、単一ヌクレオチド多形性又は突然変異)のため有用である。患者試料中で一旦ターゲット核酸数が決定されると、その数を、試料が健康人から得られたのであれば存在すると期待される数と比較する。患者試料中の核酸数と健康患者に期待される数(これはプールされた健康人試料から決定される)との間に統計的有意の差が存在するとき、その患者は疾病又はその傾向を有すると診断される。本発明の方法はまた、生体試料中の核酸検出にも有用である。これらは不均質であることが多く、突然変異核酸が野生型核酸に対し少量で存在する。例えば、スツール試料において、スツール上に落ちた変形細胞からの突然変異体核酸は、スツール上に落ちた通常細胞からの野生型核酸に対し、特に結腸癌の早期ステップで、滅多にない。本発明の方法は、本発明の単一基伸長分析からの結果が、生体試料中の突然変異体核酸存在を示すか否かを判定する統計的解析を含む。好適実施例において、本発明の方法は、単一基伸長したプライマ又はプローブの列挙を含む。さらに好適な実施例において、伸長したプライマ又はプローブの数を解析して、統計的有意量の突然変異体核酸配列が生物学的試料中に存在するか否かを判定する。
【0061】
本発明の一実施例において、ここで論じたように、プライマ又はプローブはラベル付きヌクレオチド共に伸長させるのが好ましい。このときはラベル付きプライマの数を測定する。ラベルを放射性同位元素とし、ラベル付きプライマ又はプローブに関連する放射性崩壊の量を測定するのが好ましい。ラベル付きプライマ又はプローブの数は、放射性崩壊の量から計算する。分子数は(例えば、所定期間中の全計測数の測定又は所定計数を得るに要する時間の測定など)ラベル付きプライマ又はプローブに固有結合した放射性崩壊現象の数Xを測定して計数する。数Xを、ラベル付きプライマ又はプローブに固有結合した放射性核種の数X1の計算に用いる。数X1は、参照としてここに共有、共出願の特許出願 号(代理人名簿番号EXT−005)で開示したように、分析中の各ラベル付き分子に結合した放射性核種分子の数を知った上で、ラベル付きプライマ又はプローブ分子の数X2の計算に用いる。分析に存在する伸長したプライマ又はプローブの数は引き続く統計的解析に有用である。
本発明の方法は、不純細胞母集団中の小数細胞における異型接合性喪失検出のため有用である。このような方法は精密な欠失終末点を知ることに依存せず、このような方法は試料中の異質DNA存在により影響されないからである。例えば、異型接合性喪失に際しては、ゲノムの大部分にわたって欠失が起こり、全染色体アームが無くなる。本発明の方法は、欠失の疑われるターゲット核酸の分子数計数及びそれらの基準数との比較を含む。好適実施例において基準数は、同一試料内で欠失していないと思われる核酸の分子数である。知る必要のあるすべては、欠失したと思われるターゲット核酸配列の少なくとも一部及び、欠失していないと思われる基準核酸配列の少なくとも一部である。本発明の方法は、突然変異多重検出を受け入れられるけれども、異質試料内の癌徴候を検出するためには突然変異多重検出を必要としない。
【0062】
したがって、本発明の方法は試料中の細胞副母集団又はそれからの断片における異型接合性喪失の検出に有用である。異型接合性喪失は、一般的に細胞副母集団における少なくとも一つの野生型対立因子配列の欠失として生じる。腫瘍疑惑遺伝子の場合、欠失は一般的に、異型接合性喪失に特有の大量欠失の形を取る。一定の形の癌の場合のように、発病性欠失は最初単一細胞中で起こることが多く、これが次いで突然変異体細胞の小さい副母集団を作る。突然変異の臨床的明徴が検出された時には、疾病は治療不能のステップに進行している。本発明の方法は、欠失が、試料中に全細胞又は細胞断片の小さい百分比でのみ存在するとき、その検出を可能にする。
【0063】
本発明の方法は、試料中に正常(突然変異していない)細胞と同数存在すると期待される二つの核酸の分子数比較を含む。好適実施例において、比較は(1)試料細胞内で突然変異していないと判明又は推定される(「基準」)ゲノム的ポリヌクレオチド断片の量と(2)試料(「ターゲット」)内で突然変異している推定される野生型(突然変異体でない)ゲノム的ポリヌクレオチド断片の量との間でなされる。二つのゲノム的ポリヌクレオチド断片の間の統計的有意差は、突然変異が起こったことを示す。
好適実施例において、基準及びターゲットの核酸は、同一因子座の対立因子である。対立因子は、一つの対立因子を別の対立因子から区別する配列相違があるとき、本発明の方法において有用である。好適実施例において、因子座は腫瘍抑制因子の上又は近くにある。異型接合性喪失は対立因子いずれかの喪失を生じるので、対立因子のいずれかを基準対立因子として役立てることが出来る。重要情報は、試料中の各対立因子の分子数間に統計的有意差が有るか無いかである。好適実施例においてはまた、基準及びターゲットの核酸は異なる因子座、例えば、異なる遺伝子である。好適実施例において、基準核酸は基準因子座の両対立因子を含み、ターゲット核酸はターゲット因子座、例えば腫瘍抑制遺伝子、の両対立因子を含む。明細には、腫瘍抑制遺伝子における欠失の場合、基準因子の検出量はターゲット因子の検出量より著しく大きい。特定の腫瘍因子突然変異の場合のように、ターゲット配列が拡大されると、ターゲットの検出量が基準因子の検出量より十分有意な余裕を持って大きくなる。
【0064】
当該技術で知られた方法は、欠失又は点突然変異を検出するため、通常はPCRプローブ及び/又はハイブリッド形成プローブの形で数多のプローブの使用を必要とする。しかし、本発明の方法は、ヌクレオチド配列の列挙的検出及び安定と知られた配列と不安定と思われる配列との間の列挙的比較を含むので、癌危険性を正確に分析するため僅か数個のプローブを使わなければならないだけである。事実、単一大型欠失検出に必要なすべては、プライマ又はプローブのセットたった一つである。癌の危険性は、腫瘍発生に関わると知られた又は思われる因子領域における突然変異の存在により示される。本発明の方法にしたがって実施された試験に基づき危険性があると同定された患者は、疾病の確認及び/又は治療のため、他の一般的には侵襲的処理に回される。
【0065】
本発明の方法にしたがうと、ターゲット及び基準核酸は、循環単一基伸長反応を用いて別様にラベル付けされる。その循環単一基伸長反応は、別様にラベル付けされたヌクレオチドを、ターゲット及び基準核酸に選択的にハイブリッド形成されるプライマ又はプローブの3’端に組み込む。例えば、プライマ又はプローブは、ターゲット核酸にハイブリッド形成されたプライマ又はプローブの単一基伸長に向けられた鋳型がTの添加を生じ、基準核酸にハイブリッド形成されたプライマ又はプローブの単一基伸長に向けられた鋳型がGの添加を生じるように、設計される。伸長反応は、例えば、35Sでラベルされた終端T及び32Pでラベルされた終端Gの存在下で行われる。代わりに、二つの終端核酸をそれぞれ、大きいインピーダンスと小さいインピーダンスのビーズでラベル付けする。これらの終端核酸は、ここで論じたように、伸長したプライマ又はプローブの列挙が出来るようにする検出可能な任意の各種マーカーを用いてラベル付けすることが出来る。
生体試料中で一様に分布する核酸配列の列挙的試料採取は、一般的にポアソン分布にしたがう。生体試料中にあるゲノムのポリヌクレオチド断片の典型数のような、大型母集団について、ポアソン分布は、平均値N及び、Nの二乗平方根で近似される標準偏差を有する正規(ガウス)曲線と同様である。
生体試料から得られたターゲット及び基準の遺伝子の数の間の統計的有意性は、任意の適切な方法を用いて決定される。例えば、本書の開示を参照によりここに組み込んだ、スチール他「統計の原理と手順、生物統計学的方法」(マクグローヒル社、1980年)を参照。例示的方法は、特異性(偽陽性の許容範囲)と感度(偽陰性の許容範囲)の好ましい水準に基づき、選択した信頼性水準の範囲内で、選択した統計的有意水準に達するよう、得るべきターゲット及び基準の遺伝子の数の間の差を決定することである。このような決定におけるしきい値問題は、統計的に有意な判定が出来るため母集団中で利用出来なければならない(ターゲット及び基準についての)遺伝子の最小数Nである。数Nは、突然変異体対立因子を含む試料の中の突然変異体対立因子最小数推定値(ここでは最小1%と推定)及び正規試料は突然変異体対立因子を含まないとの追加推定に依存する。ターゲット及び基準の遺伝子数の間の差のしきい値は、試料中の細胞副母集団に突然変異体が存在するとの診断に関し、少なくとも0.5%でなければならないこともまた仮定する。前述の仮定に基づいて、突然変異体数と基準対立因子数との間の差0.5%以下が検出されたとき真に陰性(即ち、試料中に突然変異体副母集団がない)である結果が回数の99.9%で生じるよう、Nがどれだけ大きくなければならないかを決定することが可能である。
【0066】
特異性に関するNの計算は、このとき、一つの試料測定が母集団の最低部分3.16%を覆うガウス分布の一部(図2Aで「A」とマークした部分)であるとの確率及び、他の試料測定が母集団の最高部分3.16%を覆うガウス分布の一部(図2Bで「B」とマークした部分)であるとの確率に基づく。二つの試料測定は独立事象なので、両事象が単一試料中で同時に起こる確率は、約0.001即ち0.1%である。こうして、ガウス分布の93.68%(100%−2×3.16%)が図3でA及びBとマークした領域の間に入る。統計表は、当該領域が標準偏差3.72に等しいことを示す。したがって、0.5%Nは3.72シグマに等しいと設定される。シグマ(標準偏差)は√Nに等しいので、方程式はNについて553,536と解ける。これは、基準及びターゲットを示す二つの数字の低い方が553,536以上であって、患者が真に正常であるときは、数字の間の差は回数の約99.9%で0.5%以下であることを意味する。
99%感度に必要な最小Nを決定するため同様の解析を行う。今度は一端を切り取ったガウス分布表が、平均値からの1.28標準偏差(シグマ)がガウス分布の90%を覆うことを示す。さらに、数字の一つ(基準又はターゲット)が図3で「A」とマークした面積又は図3で「B」とマークした面積に入る10%(1%の二乗平方根)の確率がある。二つの母集団平均が全体で1%の相違であり、ターゲットと基準因子の数字の間に0.5%の差がある筈とすると、いずれかの平均値から統計的有意に関するしきい値までの差は、99%感度に関し0.25%Nに等しい(図3参照)。図3に示すように、0.25%Nはガウス分布片側の約40%に相当する。統計表は、ガウス分布の40%が平均からの標準偏差1.28に相当することを明らかにする。したがって、1.28シグマは0.0025Nに等しく、Nは262.144に等しい。こうして、異常試料については、二つの数字の低い方が262,144以上であるとき、回数の99%以上において、差が0.5%を超える。逆に陰性誤診断は、これら条件の下で回数の僅か1%になる。
【0067】
99.9%特異性(偽陽性の回避)と99%感度(偽陰性の回避)の両方を持つためには、DNAを有する553,536個以上(大凡550,000個以上)の細胞から誘導したDNAを持つ試料を計数しなければならない。得られた数の間の0.5%以上の差が感度に関し99.0%信頼度水準で有意であり、数の間の0.5%未満の差が特異性に関し99.9%信頼度水準で有意である。上に注記したように、統計的有意性判定のため他の標準統計的試験を使用してもよく、前記は当該試験の一つをあらわすに過ぎない。
前記説明に基づいて、本発明の方法が任意の生体試料内のポリヌクレオチド副母集団にある突然変異検出に有用であることを、当業者は理解するであろう。例えば、ここに開示した方法は、癌のような疾病に伴う対立因子喪失(異型接合性喪失)の検出に使用される。加えて、本発明の方法は、酵素活動の完全又は部分的喪失のような、物質交代エラーを引き起こす欠失又は基置換突然変異の検出に使用される。例示目的で、以下に結腸癌検出における本発明の方法の利用法詳細を述べる。発明の方法は腫瘍抑制因子における突然変異(及び特に異型接合性喪失に典型的な大型欠失)の早期発見に特に有用である。したがって、以下の方法で例示したが、本発明はそのように制限されるものではなく、それらの考察に当たって当業者はその広範な利用性を理解するであろう。
【0068】
本発明の方法は、突然変異したと既知又は推定されるターゲット・ポリヌクレオチドの数を突然変異していないと既知又は推定される基準ポリヌクレオチドの数と比較するステップを含むのが好適である。対立因子又は因子座のいずれかを基準及びターゲット核酸として利用する代替実施例に加えて、本発明は正常対立因子におけるミクロサテライト繰り返し領域を突然変異が既知又は推定される対立因子における対応ミクロサテライト繰り返し領域と比較するステップを含む。本発明の例示的検出方法は、測定される各核酸の計数の間に差が存在するか否かを決定するステップを含む。統計的有意差の存在は、測定された核酸の一つに突然変異が起こったことを示す。
【実施例】
【0069】
使用例
例示目的で、以下に結腸癌検出における本発明の方法の利用法詳細を述べる。発明の方法は突然変異の早期発見に特に有用である。したがって、以下の方法で例示したが、本発明はそのように制限されるものではなく、それらの考察に当たって当業者はその広範な利用性を理解するであろう。
【0070】
結腸の癌又は前癌状態の検出に関する例示的方法
使用例1.試料準備
本発明にしたがって、ターゲット核酸は患者から隔離した核酸試料をあらわす。この核酸は任意の細胞源又は体液から採取することができる。臨床行為で得られる細胞源の非限定例には、血液細胞、頬面細胞、膣頚部細胞、尿からの上皮細胞、胎児細胞、又は生検で得られた組織内に存在する任意の細胞が含まれる。体液には、血液、尿、脳脊髄液、及び感染又は炎症部位からの組織滲出液が含まれる。
好適実施例において、試料はスツールの横断面又は周辺部分である。スツールの横断面又は周辺部分作成に好適な方法は、参照によりここに組み込んだ共有米国特許5,741650号及び、共有共出願の特許 号(代理人名簿番号EXT−015)で提供されている。スツールが結腸を通過するとき、結腸上皮細胞から脱け落ちた細胞及び細胞断片を付着する。同様に、細胞及び細胞断片は結腸ポリープ(突然変異DNAを含む)が脱ぎ落とす。しかし、ポリープと接触するスツールの一部だけが脱落細胞を付着する。したがって、スツール試料が、推定癌細胞(例えばポリープ)の落としたものを含む脱落細胞すべての混合物を確実に含むためには、少なくともスツールの横断面又は周辺部分を得ることが必要である。
【0071】
試料作成の後、NaCl又はKCl200mMのような塩及び1−10%SDS又はトリトンTMのような洗浄剤、及び/又はプロティナーゼKのようなプロティナーゼを含むリン酸塩緩衝サリンのような、適切なバッファの中で試料を均質化する。特に好適なバッファは、参照によりここに組み込んだ共有共出願の米国特許出願 号(代理人名簿番号EXT−006)で開示したような、Tris−EDTA−NaClバッファである。このバッファはまた、DNA及びRNA劣化酵素の抑制剤をも含むことがある。試料中の二重撚りDNAは周知の方法で融解(変性して単一撚りDNAを形成)する。例えば、参照によりここに組み込んだギレンステン他、組み換えDNAにおける方法論II、565−578頁(Wu版、1995年)を参照。DNAは次いで、当該技術で標準の多数の方法のいずれかを用いて、細胞源又は体液から分離される。参照によりここに組み込んだスミスーレビン他、Gut、36,81−86頁(1995年)を参照。DNA抽出に用いる特定の方法が供給源の性質に左右されることは、理解されるであろう。
一旦抽出したら、ターゲット核酸は追加処理することなく本発明に採用してよい。代わりに、ターゲット核酸中に存在する一つ以上の特定領域をPCRにより拡大してもよい。この場合、拡大される領域は、プライマとして使用のため特定の側面配列の選択により、規定される。このステップでの拡大は、ターゲット核酸配列母集団の中で特定核酸配列の濃度を増加するとの利点を備える。
一実施例において、特定配列先行拡大を伴う又は伴わないターゲット核酸が、固定相支持物に束縛される。これにより多数の患者試料の同時処理及び篩い分けが出来る。本発明における使用に適切な支持物の非限定的例示には、ニトロセルローズ又はナイロンフィルタ、親和性キャプチャ用薬剤で被覆されたガラスビーズ、磁性ビーズ、処理又は未処理ミクロタイタープレート、及び類似物が含まれる。診断試験室及び組織培養に用いられる在来の96穴ミクロタイターディッシュは好適支持物である。好適実施例において、支持物は多数の穴を有するミクロタイターディッシュである。このような支持物の使用により、多数の試料及び対照の同時判定が出来、その結果、解析を促進する。さらに、自動システムを使用してこのようなミクロタイターディッシュに試薬を提供することが出来る。ターゲット核酸を支持物に束縛する方法が、使用する特定マトリクスに左右されることは、当業者に理解されるであろう。例えば、DNAのニトロセルローズへの束縛は、フィルタにDNAを単に吸収させ、次いでフィルタを75−80度C真空下で15分ー2時間焼くことにより、達成される。代わりに、帯電ナイロン皮膜を使用することが出来、これは束縛核酸の追加処理を何も必要としない。アビジン又はストレプタビジンで被覆したビーズ及びミクロタイターを、ビオチンが(例えば、ビオチン抱合PCRプライマを使って)付着されたターゲット核酸の束縛に使うことが出来る。加えて、ターゲット核酸表面を抗体で覆い抗体特有付着素をターゲット核酸に組み込むことにより、抗体を用いてターゲット核酸を任意の上記固体支持体に付着することが出来る。またエポキシド/アミン結合化学作用を用いて、ターゲット核酸を任意の上記固体支持体に直接付着することが出来る。エガース他、DNA配列技術の進歩、SPIE会議議事録(1993)を参照。一旦核酸試料が固定されたら、試料を洗滌して非固定物質を除去する。次いで核酸試料を本発明にしたがって一つ以上の分割プライマに露出する。代替実施例において、核酸は水様相に止まる。
【0072】
使用例2.単一基伸長反応の多重循環
a)プライマ選択
一つ以上の突然変異を含むと思われる遺伝子領域は、GenBank、EMBLのようなヌクレオチド・テータベース若しくは適切なデータベース又は刊行物を参照して、又は配列により同定する。癌検出については、数多の腫瘍発生遺伝因子及び腫瘍抑制因子における遺伝子突然変異が知られている。デュフィ、臨床化学、41巻、1410−1413頁(1933年)。本発明の突然変異検出法での使用に好適な遺伝子には、一つ以上の腫瘍発生遺伝因子及び/又は一つ以上の腫瘍抑制因子が含まれる。特に好適な遺伝子には、ras腫瘍発生遺伝因子、p53、dcc、apc、mcc、及びその他の腫瘍形成表現型の成長に関わると思われる遺伝子が含まれる。
以下で記述するように、本発明の方法は、疑問のある核酸が一つ以上ある因子座で突然変異を検出することが出来る。さらに本発明の方法は、一つ以上の単一基突然変異が可能な因子座を調査するため使用することが出来る。一旦関心領域が同定されたら、疑問の突然変異の存在を検出するため少なくとも一つのプライマを準備する。本発明のプライマは長さが、好適にはヌクレオチド約10個分から約100個分、さらに好適にはヌクレオチド約15個分と約35個分との間、最も好適にはヌクレオチド約25個分である。
【0073】
プライマは天然物でも合成物でもよく、生体内、生体外で酵素的に、又は生体外で非酵素的に、合成される。本発明の方法で使用されるプライマはオリゴデオキシリボヌクレオチド、オリゴリボヌクレオチド、デオキシリボヌクレオチドとリボヌクレオチドとの共重合体、ペプチド核酸(PNA)、及びその他の機能的相似品から選ぶのが好適である。ペプチド核酸は有名である。プルスカル他.FASEB彙報、ポスター#35(1994年)を参照。これらは合成オリゴアミドであって、アデニン、シトシン、グアニン、チミン又はウラシルが付着された反復アミノ酸ユニットから構成される。エホルム他、ネイチャー365巻、565−568頁(1993年)、オエルム他、核酸研究23巻、5332−36頁(1993年)、プラクチカルPNA、PCRクランプを指向するPNAによるポイント突然変異同定、パーセプチブ・バイオシステム1巻、1号(1995年)を参照。ペプチド核酸シントン及びオリゴマーは市販で、パーセプチブ・バイオシステム社、マサチューセッツ州フラミントン、から入手出来る。例えば、PCT出版物EP92/01219、EP92/01220、US92/10921を参照。多くの用途において、PNAは核酸プローブに好適である。核酸/核酸デュプレクスと異なり、これは低塩条件下で不安定化し、PNA/核酸デュプレクスを形成して極低塩条件下で安定化したまま止まるからである。加えて、PNA/DNA複合体は、相似核酸/核酸複合体より高い熱融解点を有するので、PNAプローブの使用により吸取り分析の再現性が改善される。
【0074】
例示のため、K−ras因子内の突然変異検出のため設計されたプライマを下記に用意する。本発明の方法にしたがって、コーディング・ストランド又はノン−コーディング・ストランドのいずれかに相補的なプライマを使用する。実例として、コーディング・ストランド内の突然変異検出に有用なプライマを下記に用意する。K−ras内の突然変異は、表現蛋白質のアミノ酸12に関するコードンに頻繁に起こる。
K−ras因子の野生型コードン12及びその上流核酸は下記である。
野生型鋳型 3’−TATTTGAACACCATCAACCTCGACCA−5’(配列識別番号1)
アミノ酸12をエンコードする三つのヌクレオチドに下線を施した。K−ras因子のコードン内で符号化されたアミノ酸12の中の第一核酸位置調査の能力があるプライマ(プライマ1)を下記に用意する。
プライマ1 5’−ATAAACTTGTGGTAGTTGGAGCT−3’(配列識別番号9)
b)プライマ伸長の複数回循環
プライマ1は、K−ras因子内の相補配列へのプライマ1の選択束縛を促進するとの条件(表1参照)下で、核酸試料にハイブリッド形成される。伸長反応は、それぞれ異なる検出可能ラベルを付けた四つの異なるジデオキシヌクレオチドddATP、ddCTP、ddGTP、及びddTTP、が存在して行われる。伸長反応は、表2に示すように、30回繰り返す。
表1.単一基伸長循環反応用反応混合物
成分 量
H2O 25.5
10×配列バッファ 4
ddNTP(50μM) 5
プライマ(5μM) 5
Thermoシーケナーゼ 0.5
DNAサンプル 10
表2.循環単一基伸長反応用温度輪郭
ステップ 温度(C) 時間(秒)
1 94 5
2 94 30
3 64 10
4 72 10
5 ステップ2に行く、29回
6 4 待機
ラベル付きddNTPの組込について反応生成物を分析する。野生型DNAを含む核酸試料にはラベル付きddGTPのみが組み込まれている筈である。他のいずれかのddNTPが十分有意な量で組み込まれていることは、試料中に突然変異K−ras核酸が存在することを示す。
【0075】
使用例3.分割プライマの準備
a)プライマ選択
関心領域が同定されると、疑惑突然変異の存在検出のため、少なくとも一つの分割プライマを用意する。分割プライマは、核酸の実質的連続部分にハイブリッド形成する能力のある少なくとも二つのオリゴヌクレオチド・プローブ、第一プローブと第二プローブ、から構成される。
本発明の第一プローブは、長さが、好適にはヌクレオチド約5個分から約10個分、さらに好適にはヌクレオチド約6個分と約8個分との間、最も好適にはヌクレオチド約8個分である。本発明の第二プローブは、長さが、好適にはヌクレオチド約15個分と約100個分との間、さらに好適にはヌクレオチド約15個分と約30個分との間、最も好適にはヌクレオチド約20個分である。さらに、第二プローブは、伸長出来ない3’終端ヌクレオチドを有するため、第一プローブを欠く温度依存核酸合成用のプライマになることが出来ない。好適な非伸長性3’終端ヌクレオチドには、C3スペーサー、3’転換基、ビオチン、又は変形ヌクレオチドが含まれる。長いプローブは、そのヌクレオチド不整合許容性のため、選択性が低いけれども、第二プローブは非伸長性で、近接プローブを欠くときは偽始動を生じない。
代替実施例において、分割プライマは第一プライマのシリーズから構成され、シリーズの各構成員は、ヌクレオチド約5個分から約10個分、最も好適にはヌクレオチド約6個分から約8個分まで、の長さを有する。第一プローブは終端ヌクレオチドを有しないけれども、シリーズの全構成員が核酸の実質的連続部分にハイブリッド形成されない限り、核酸伸長は起こらない。
分割プライマのオリゴヌクレオチド・プローブは天然物でも合成物でもよく、生体内、生体外で酵素的に、又は生体外で非酵素的に、合成される。本発明の方法で使用されるプライマは、オリゴデオキシリボヌクレオチド、オリゴリボヌクレオチド、デオキシリボヌクレオチドとリボヌクレオチドとの共重合体、ペプチド核酸(PNA)、及びその他の機能的相似品から選ぶのが好適である。ペプチド核酸は有名である。プルスカル他.FASEB彙報、ポスター#35(1994年)を参照。これらは合成オリゴアミドであって、アデニン、シトシン、グアニン、チミン又はウラシルが付着された反復アミノ酸ユニットから構成される。エホルム他、ネイチャー365巻、565−568頁(1993年)、オエルム他、核酸研究23巻、5332−36頁(1993年)、プラクチカルPNA、PCRクランプを指向するPNAによるポイント突然変異同定、パーセプチブ・バイオシステム1巻、1号(1995年)を参照。ペプチド核酸シントン及びオリゴマーは市販で、パーセプチブ・バイオシステム社、マサチューセッツ州フラミントンから、入手出来る。例えば、PCT出版物EP92/01219、EP92/01220、US92/10921を参照。多くの用途において、PNAは核酸プローブに好適である。核酸/核酸デュプレクスと異なり、これは低塩条件下で不安定化し、PNA/核酸デュプレクスを形成して極低塩条件下で安定化したまま止まるからである。加えて、PNA/DNA複合体は、相似核酸/核酸複合体より高い熱融解点を有するので、PNAプローブの使用により吸取り分析の再現性が改善される。
【0076】
例示のため、K−ras因子内の突然変異検出のため設計されたプライマを下記に用意する。本発明の方法にしたがって、コーディング・ストランド又はノン−コーディング・ストランドのいずれかに相補的なプライマを使用する。実例として、コーディング・ストランド内の突然変異検出に有用なプライマを下記に用意する。K−ras内の突然変異は、表現蛋白質のアミノ酸12に関するコードンに頻繁に起こる。コードン12の中の三つの位置それぞれにおける突然変異検出用に可能なプローブを下記に示す。
K−ras因子の野生型コードン12及びその上流核酸は下記である。
野生型鋳型 3’−TATTTGAACACCATCAACCTCGACCA−5’(配列識別番号1)
アミノ酸12をエンコードする三つのヌクレオチドに下線を施した。K−ras因子のアミノ酸12に関しコードする三つのヌクレオチドを調査する能力がある第一プローブと第二プローブを下記に用意する。第一プローブAは、上記に概略を記述した第一プローブであって、コードン12の中の第一基の直ぐ上流側の(即ち、コードン位置1にあるシトシンに隣接する)ヌクレオチドに相補的な配列を有する。第二プローブAは、上記に概略を記述した第二プローブである。これは、第一プローブAが相補する配列に実質的連続(この場合は正確に連続)な配列に相補的である。下記に示す第二プローブのそれぞれで太字の核酸は、非伸長性ヌクレオチドである。K−rasコードン12の第一基の中の突然変異検出に適する第一及び第二プローブのハイブリッド形成を下記に示す。
第二プローブA 5’−ATAAACTTGTGGTAG(配列識別番号2)
第一プローブA TTGGAGCT(配列識別番号3)
野生型鋳型 3’−TATTTGAACACCATCAACCTCGACCA−5’(配列識別番号1)
コードン12の第二基の中の突然変異検出は、上と同じ第二プローブ(第二プローブA)と、下記で第一プローブBと名付けた第一プローブを用いて行う。この第一プローブBはコードン12の第二基に隣接(3’)して終わる配列に相補的である。コードン12の第二基の中の突然変異検出に適したプローブのハイブリッド形成を下記に示す。
第二プローブA 5’−ATAAACTTGTGGTAG(配列識別番号2)
第一プローブB TGGAGCTG(配列識別番号4)
野生型鋳型 3’−TATTTGAACACCATCAACCTCGACCA−5’(配列識別番号1)
コードン12の第三位置における突然変異検出は、上と同じ第二プローブ(第二プローブA)及び、コードン12の第三基と直接境を接する第一プローブCを用いて達成する。コードン12の第三基の中の突然変異検出に適したプローブのハイブリッド形成を下記に示す。
第二プローブA 5’−ATAAACTTGTGGTAG(配列識別番号2)
第一プローブC GGAGCTGG(配列識別番号6)
野生型鋳型 3’−TATTTGAACACCATCAACCTCGACCA−5’(配列識別番号1)
【0077】
コードン12の第二及び第三ヌクレオチドにおける突然変異検出に関する上述の方法において、第二プローブは、第一プローブのハイブリッド形成領域上流側の、それぞれ1及び2ヌクレオチドである。代わりに、コードン12の第二及び第三ヌクレオチド検出用の第二プローブが、それぞれの第一プローブと直接境を接し(即ち正確にそれと連続し)てもよい。例えば、コードン12K−ras第三基の突然変異検出用代替第二プローブは、下記である。
5’−ATAAACTTGTGGTAGTT(配列識別番号5)
突然変異検出はまた、少なくとも三つの第一プローブのシリーズを含む分割プライマを用いても達成される。コードン12第三基の突然変異検出用第三プローブを下記に示す。
第一プローブX 5’−ATAAACTT(配列識別番号7)
第一プローブY TGGTAGTT(配列識別番号8)
第一プローブZ GGAGCTGG(配列識別番号6)
野生型鋳型 3’−TATTTGAACACCATCAACCTCGACCA−5’(配列識別番号1)
b)プライマ伸長の多重循環
第一及び第二プローブは、長い第二プローブがないと短い第一プローブのハイブリッド形成が不利であるとの、ハイブリッド形成条件の下で試料に露出される。ハイブリッド形成に影響する要因は当該技術に周知で、温度上昇、塩濃度低下、又はハイブリッド形成溶液のpH上昇が含まれる。不利なハイブリッド形成条件(例えば、第一プローブTm上温度30−40度C)の下で第一プローブは、単独でハイブリッド形成されたとき、不安定なハイブリッドを形成し、伸長反応を始動しない。長い第二プローブは高いTmを有していて、鋳型と安定ハイブリッドを形成し、核酸の実質的連続部分にハイブリッド形成されたとき、第二プローブは短い第一プローブに安定性を分与し、それにより連続プライマを形成する。
【0078】
好適実施例において、参照によりここに組み込んだサンガー、国立科学アカデミー誌(USA)、74巻、5463−5467頁(1997年)に報告されたようなデオキシ鎖終結方法の変形を、このとき突然変異存在の検出に用いた。この方法は、四つの共通2’、3’−ジデオキシ・ヌクレオシド・トリフォスフェート(ddATP、ddCTP、ddGTP、ddTTP)のうち少なくとも一つを使うことを含む。当該技術に既知の方法を用いて、検出可能検出部分をジデオキシ・ヌクレオシド・トリフォスフェート(ddNTP)に付着させることが出来る。シーケナーゼTM(パーキン・エルマー)のようなDNAポリメラーゼもまた、試料混合物に添加した。好適実施例において、Taq又はVentDNAポリメラーゼのような、ポリメラーゼを試料混合物に添加した。実質的連続な第一及び第二プローブをプライマとして用いて、ポリメラーゼはddNTP一個を第一プローブの3’端に付加する。組み込まれたddNTPは、単一基多形サイトに存在するヌクレオチドに相補的になる。ddNTPは3’ヒドロキシルを有するので、ハイブリッド形成済みプローブの追加伸長は起きない。伸長混合物に相補性ddNTP(又はデオキシヌクレオシド・トリフォスフェート)が利用できない場合は、連鎖終結もまた起こる。単一基伸長反応終了後、伸長生成物を隔離して検出する。
好適実施例においてはまた、伸長反応停止が唯一つのヌクレオチド付加後か、又は予想突然変異の相補に相当する唯一つのラベル付きヌクレオチド付加後か、のいずれかを検出するラベル付きデオキシヌクレオシドが試料に露出される。
使用例2及び3に例示した本発明の最も簡単な実施例において、ヌクレオシド・トリフォスフェート混合物は、既知突然変異に相補性のラベル付きddNTP又はdNTPのみを含む。例えば、K−ras因子コードン12第一ヌクレオチド内のC→A突然変異につき試料を調査するため、第二プローブA及び第一プローブAは、ラベル付きddTTP又はdTTPを含む伸長反応混合物に露出される。ラベル付きddTTP又はdTTPの第一プローブAへの組込は、試験試料のK−ras因子コードン12第一ヌクレオチド内にC→A突然変異が存在することを示す。第二プローブAと共に野生型鋳型に共にハイブリッド形成された第一プローブAは、伸長されないか、又は反応混合物中で利用出来るときは、ラベル無しddGTP又はdGTPで伸長する。
【0079】
結腸癌に付随した多数の突然変異を与えられると、この疾患に関する検出方法が、同一反応内に同時に存在する多数の突然変異(例えば、apc、K−ras、p53、dcc、MSH2、DRA)に関し、試料を好適に篩い分けする。上述のように、先行技術の検出方法を用いると、極めて限られた多重化のみが可能である。本発明の方法は、長い第二プローブの不整合許容性から生じる偽陽性信号を除外するので、分割オリゴヌクレオチドの使用により、個々のプローブに関するハイブリッド形成条件最適化の必要を回避し、膨大な多重化を可能にする。短い第一プローブの安定なハイブリッド形成が対応の長い第二プローブが無いと出来ないハイブリッド形成条件である限り、数個の分割プライマを同一反応内で分析することが出来る。
好適実施例において、プライマ伸長反応は、それぞれ生体試料の分別部、ポリメラーゼ、及び三つのラベル付き相補性非野生型ddNTP(又はdNTP)、を有する四つの別々の反応混合物で実行される。任意選択で、反応混合物はまたラベル無し相補性非野生型ddNTP(又はdNTP)含むこともある。分割プライマは、野生型鋳型にしたがって多重化される。本例示において、K−ras因子のアミノ酸12に関しコードする最初の二つのヌクレオチドはシステンである。したがって、第二プローブA及び第一プローブAとBを、ラベル付きddATP(又はdATP)、ddTTP(又はdTTP)、及びddCTP(又はdCTP)を含む反応混合物に付加する。第二プローブC及び第一プローブCは、ラベル付きddATP(又はdATP)、ddCTP(又はdCTP)、及びddGTP(又はdGTP)を含む反応混合物に付加する。第一プローブへのいずれのラベル付きddNTP(又はdNTP)の組込も、試料内のK−ras因子コードン12に突然変異の存在を示す。この実施例は、K−rasのコードン12のような数個の可能突然変異を有する位置の調査に、格別有用である。
【0080】
代替好適実施例において、プライマ伸長反応は、それぞれ唯一つのラベル付き相補性非野生型ddNTP又はdNTP及び、任意選択で、別の三つのラベル無しddNTP又はdNTPを含む、四つの別々の反応混合物で行われる。分割プライマはこうして、既知の突然変異体ヌクレオチドに相補性のラベル付きddNTP又はdNTPに露出することが出来るか、又は代わりに、すべての三つの非野生型ラベル付きddNTP又はdNTPに露出することが出来る。上記に用意したK−ras例において、K−rasコードン12の第一ヌクレオチドを既知のC→G突然変異について調査すると、第一プローブAと第二プローブAは唯一つの反応混合物、ラベル付きddCTP(又はdCTP)を含む反応混合物、に付加される。任意選択で、本発明の方法は、上述のようにラベル付きデオキシヌクレオチドを用いて練習することが出来る。
しかし、K−ras因子のコードン12には幾つかの突然変異が同定されているので、プローブはすべての非野生型ラベル付きddNTP又はdNTPに露出する。こうして、第一プローブA及び第二プローブAとBは、ラベル付きddATP(又はdATP)、ddTTP(又はdTTP)、及びddCTP(又はdCTP)を含む反応混合物に付加される。第二プローブC及び第一プローブCは、ラベル付きddATP(又はdATP)、ddCTP(又はdCTP)、及びddGTP(又はdGTP)のうち一つを含む反応混合物に付加される。再び、第一プローブのラベル付き終端ヌクレオチドへの露出は、試験した生体試料のK−ras因子のコードン12に突然変異の存在を示す。
【0081】
好適実施例においては、分析信号増幅のため数回の伸長反応を行う。伸長反応は余分の第一及び第二プローブ、ラベル付きddNTP又はdNTP、及び熱安定性ポリメラーゼを存在させて行う。伸長反応が完了すると、ターゲット核酸に接合された第一及び第二プローブは、反応混合物をハイブリッドの融解温度以上に加熱することにより解離する。次いで反応混合物をハイブリッドの融解温度以下に冷却して、第一及び第二プローブは、別の伸長反応のためターゲット核酸との結合が許される。好適実施例においては、10回乃至50回の伸長反応を実施する。最好適実施例においては、30回の伸長反応を実施する。
【0082】
使用例4.遺伝子変更の同定方法
一実施例においては、ラベル付きプライマ又はプローブがここに記述したように固定化され、当該技術に標準の化学的方法(例えば、マクサム−ギルバート配列、マクサム及びキルバート、1977年、国立科学アカデミー誌、USA、74巻、560頁)を用いて、直接配列を受ける。
別の実施例においては、固定化されたラベル付きプライマ又は第一プローブが、固体支持物から移動されてラベル付き第一プローブの配列が水様溶液内で行われる。一実施例においては、分析で調査される多数配列のそれぞれに対応するオリゴヌクレオチドにラベル付き第一プローブを露出することを用いる、配列特有逆ハイブリッド形成により、ラベル付き第一プローブの配列が決定される。ハイブリッド形成解析は、当該技術に既知の、吸い取りのような、幾つかの方法を用いて行われる。アウスベル他、分子生物学における短編議案、3版(ジョーン・ウイリー&ソン社、1995年)参照。好適実施例においては、オリゴヌクレオチドを固体支持物に規定の位置(即ち既知の位置)で固定化する。この固定化アレーは時に「DNAチップ」と呼ばれる。この固体支持物はまた、固体支持物へのオリゴヌクレオチド付着を容易にするため、(例えば、金又は銀で)被覆することが出来る。オリゴヌクレオチドを固体支持物に固定するには、当該技術で既知の各種方法のいずれかが用いられる。普通に使用される方法は、アビジン又はストレプトアビジンを用いる固体支持物の非共有被覆及びビオチン化オリゴヌクレオチド・プローブの固定化を含む。オリゴヌクレオチドはまたエポキシド/アミン結合化学品を用いて固体支持物に直接付着することが出来る。エガース他、DNA配列技術の進歩、SPIE大会議事録(1993年)参照。
【0083】
別の実施例においては、ハイブリッド形成及び重複部分を通じる陽性ハイブリッド形成プローブの解析により、ラベル付き第一プローブの配列が読み取られる。参照によりここに組み込んだドルマナック他、米国5,202,231号を参照。
さらに別の実施例においては、ラベル付きdNTPにより伸長された第一プローブが、酵素DNA配列(サンガー他、1977年、国立科学アカデミー誌、USA、74巻、5463頁)を用いて同定される。この場合、第一プローブに相補のDNA配列及び配列「タグ」の働きをする所定の追加共線型配列を含むオリゴヌクレオチドが合成される。サンガー配列条件の下で培養したとき、固定化第一プローブはその相補配列にハイブリッド形成し配列反応に関しプライマとして働く。生じた始動配列「タグ」はこのとき反応に存在する第一プローブを同定する。
【0084】
さらに別の実施例において、ラベル付きdNTPにより伸長された第一ラベルは配列同定に先立って拡大される。ラベル付き第一プローブは、追加「タグ」付き又は無しの配列用プライマ配列を含む相補性オリゴヌクレチオドと共に培養される。第一プローブのその相補性オリゴヌクレチオドへの初期ハイブリッド形成により、第一プローブは単一伸長反応において初期プライマとして役立つことが出来る。伸長生成物は次いで循環配列反応における鋳型として直接使用される。伸長生成物の循環配列は配列用生成物を拡大する結果となる。相補性オリゴヌクレチオドの設計に当たって、配列用プライマは、第一プローブ自体を通じるか又は、代わりに、「タグ」配列を通じて、配列を進行するよう、方向付けされる。後者の場合、「タグ」配列の決定は共線型第一プローブ配列を同定する。拡大された生成物は、当該技術で標準の化学的方法によるか、又は、上述のように、配列特有逆ハイブリッド形成方法により、配列される。
本発明の実施にあたって、第一プローブの又は相補性タグ付きオリゴヌクレオチドの、全配列の決定は必要でない。1、2、又は3配列反応が(「バーコード」と同様の)特性パターンの生成に(完全配列を得るに必要な4に代わって)有効であり、個々の第一プローブの即時同定が出来ることを意図している。この方法は、放射性ラベル付き第一プローブを用い、アナログ又はデジタル自動ラジオグラムを作成する手動配列方法に適用出来ると同時に、非放射性レポーター分子を用い、デジタル化パターンを生じる自動配列方法にも適用出来る。双方の場合において、確定データベースとの比較を電子的に実行することが出来る。こうして、必要配列反応の減少により、本発明の方法は、多数試料の経済的解析を容易にする。
【0085】
本発明は、多数の可能な第一プローブの同時篩い分けを単一反応に受け入れる。実際において、同時ハイブリッド形成のため貯められた分割プライマの実数は、診断要請にしたがって決定される。例えば、嚢胞性繊維症(CF)において、一つの特有突然変異(Δ508)がCF患者の70%以上に見られた。これ、Δ508固有分割プライマを用いる本発明の方法にしたがった予備的篩い分け、に続く連続プライマの単一基伸長、及び伸長第一プローブの検出により、Δ508対立因子が同定され除去される。第二篩い分け(「フェーズ2」)において、別の、頻繁でない、CF対立因子をエンコードする多数の分割プライマの篩い分けを実施し、続いて上述のように連続プライマの単一基伸長、及び伸長第一プローブの検出を実施した。
別の臨床状態においては、しかし、CFにおけるΔ508のように高い頻度で現れる単一突然変異は存在しない。したがって、分割プライマのプールは、プール陽性試料につきフェーズ2解析において必要になる独立分析の数によってのみ決定される。
【0086】
加えて、現在の臨床習慣においては、異なる臨床症候、例えば、嚢胞性繊維症、地中海貧血、及びゴーシェ病、は互いに無関係に篩い分けされる。本発明は、対照的に、一つ以上の疾病発生因子における突然変異に相補の多数の第一プローブを用いて、各種患者からの多数の核酸の同時篩い分けを受け入れる。
同様の方法で、臨床的指示薬が異質薬剤又は微生物による感染を示すとき、本発明は、多数の潜在異質核酸に付いて同時篩い分けを提供する。さらに、一つ以上の微生物の特別種、変種、突然変異体、及び類似物もまた、第一篩い分けにおいて適切な第一プローブを採用することにより区別することが出来る。
【0087】
本発明の方法はまた、フェーズ1又はフェーズ2篩い分けにおいて無作為置換された分割プライマを使用することにより、患者又は侵入微生物の核酸に運び込まれた新規の可能性がある突然変異対立遺伝子を定義することを可能にする。この実施例において、連続プライマの単一基伸長及び伸長第一プローブの検出と分離に続く配列により、精密な突然変異体配列が明らかになる。
前述は、分割プライマを用いる多重突然変異検出の文脈における本発明の慣例を例示する。ここに開示したような、本発明の数多の追加側面及び利点は、開示及び個々の例示を考察したとき明らかになる。したがって、本発明は付属請求項の範囲のみによって、限定される。
[配列表]
【数1】
【数2】
【数3】
【特許請求の範囲】
【請求項1】
本明細書中に記載の発明。
【請求項1】
本明細書中に記載の発明。
【図1】


【図2A】

【図2B】
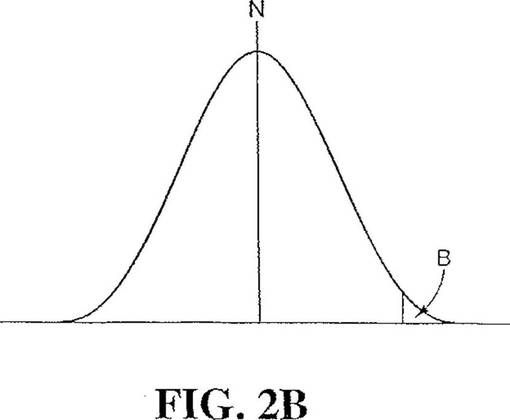
【図3】
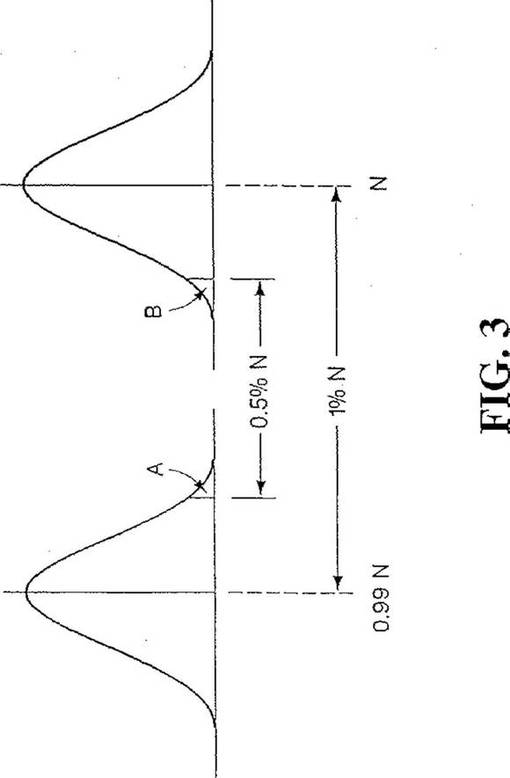
【図2A】
【図2B】
【図3】
【公開番号】特開2009−232850(P2009−232850A)
【公開日】平成21年10月15日(2009.10.15)
【国際特許分類】
【出願番号】特願2009−119325(P2009−119325)
【出願日】平成21年5月15日(2009.5.15)
【分割の表示】特願2000−546055(P2000−546055)の分割
【原出願日】平成11年4月23日(1999.4.23)
【出願人】(501063380)エグザクト サイエンシーズ コーポレイション (2)
【Fターム(参考)】
【公開日】平成21年10月15日(2009.10.15)
【国際特許分類】
【出願日】平成21年5月15日(2009.5.15)
【分割の表示】特願2000−546055(P2000−546055)の分割
【原出願日】平成11年4月23日(1999.4.23)
【出願人】(501063380)エグザクト サイエンシーズ コーポレイション (2)
【Fターム(参考)】
[ Back to top ]