
複数置換プロテアーゼ変異体
【課題】天然または組換え非ヒトプロテアーゼのDNA配列から得られたプロテアーゼ変異体含む新規な酵素変異体の提供。
【解決手段】変異体プロテアーゼは、通常、天然または組換えプロテアーゼをエンコードする前駆体DNA配列のin vitroでの変異により得られ、前駆体プロテアーゼのアミノ酸配列中に多数のアミノ酸残基の置換を生じる。当該変異体プロテアーゼは前駆体プロテアーゼの性質と異なり、例えば変化した洗浄能力を有する。置換アミノ酸残基は、バチルス・アミロリケファシエンスズブチリシンの27、45、170、181、251及び271に対応する。バチルス・アミロリケファシエンス・ズブチリシンの1、14、49、61、87、100、102、118、128、204及び258から選択される位置に少なくとも1の別の置換を含む別の変異体。
【解決手段】変異体プロテアーゼは、通常、天然または組換えプロテアーゼをエンコードする前駆体DNA配列のin vitroでの変異により得られ、前駆体プロテアーゼのアミノ酸配列中に多数のアミノ酸残基の置換を生じる。当該変異体プロテアーゼは前駆体プロテアーゼの性質と異なり、例えば変化した洗浄能力を有する。置換アミノ酸残基は、バチルス・アミロリケファシエンスズブチリシンの27、45、170、181、251及び271に対応する。バチルス・アミロリケファシエンス・ズブチリシンの1、14、49、61、87、100、102、118、128、204及び258から選択される位置に少なくとも1の別の置換を含む別の変異体。
【発明の詳細な説明】
【技術分野】
【0001】
発明の背景
セリンプロテアーゼはカルボニル・ヒドロラーゼの下位集団である。これらは広範囲の特異性及び生物学的機能を有する酵素の種々の分類を含む。Stroud,R.Sci.Amer.,131:74−88。それらの機能的多様性にもかかわらず、セリンプロテアーゼの触媒機構は、1)ズブチリシン及び2)哺乳類キモトリプシン関連及び同種細菌性セリンプロテアーゼ(例えば、トリプシン及びS.gresiusトリプシン)の少なくとも2つの遺伝学的にはっきり区別できる酵素のファミリーに近かった。セリンプロテアーゼのこれら2つのファミリーは非常に類似した触媒作用機構を示す。Kraut,J.(1977)、Annu.Rev.Biochem.,46:331−358。さらに、一次構造は関連しないが、これら2つの酵素ファミリーの三次構造はセリン、ヒスチジン及びアスパラギン酸塩からなるアミノ酸の保存された触媒3残基を結集させる。
【0002】
ズブチリシンは、種々のバチルス種及びその他の微生物から大量に分泌されるセリンプロテアーゼ(approx.MW(分子量)27,500)である。ズブチリシンのタンパク質配列はバチルスの少なくとも9つの異なる種から決定された。Markland,F.S.,et al.(1983)、Hoppe−Seyler’s Z.Physiol.Chem.、364:1537−1540。バチルス・アミロリケファシエンス、バチルス・リケニホルミス、及びバチルス・レンタスのいくつかの天然変異体から得られるズブチリシンの三次元結晶構造が報告されている。これらの研究はズブチリシンが哺乳類セリンプロテアーゼに通常関連しないが、類似の活性部位構造を有することを示す。共有結合ペプチド阻害因子を含んだズブチリシンのX線結晶構造(Robertus,J.D.,et al.(1972)、Biochemistry、11:2439−2449)または生成複合物(Robertus,J.D.,et al.(1976)、J.Biol.Chem.,251:1097−1103)も活性部位及びズブチリシンの推定上の基質が結合する分子の溝(substrate binding cleft)についての情報を提供した。さらに、多くの動的及び化学的修正のズブチリシンについての研究が報告されており、(Svendsen,B.(1976)、Carlsberg Res.Commun.,41:237−291;Markland,F.S.Id.)及びズブチリシンの残基222のメチオニンの側鎖が過酸化水素によりメチオニン−スルホキシドに変換される少なくとも1の報告(Stauffer,D.C.,et al.(1965)、J.Biol.Chem.,244:5333−5338)及び広範囲の部位特異性突然変異が実行された(Wells and Estell(1988)TIBS 13:291−297)。
【発明の概要】
【0003】
発明の概要
本発明の1の側面は、分子の電荷分布をその配向及び相、表面、その他の分子及び領域との相互作用に影響を与えるために変化させることである。
【0004】
前駆体または親酵素の酵素変異体は、荷電アミノ酸残基位置での1以上の変異を含む変異体、前駆体酵素と同じ正味静電荷及び/または前駆体酵素と同じ等電点を有することを特徴とする変異体を目的とする。
【0005】
本発明の他の側面において、前駆体プロテアーゼのプロテアーゼ変異体をここで目的とし、荷電アミノ酸残基位置で1以上の変異を含む変異体、前駆体プロテアーゼと同じ正味静電荷または等電点を有することを特徴とする変異体を目的とする。荷電アミノ酸はアスパラギン酸、グルタミン酸、ヒスチジン、リジン、チロシン及びアルギニンである。残基位置はここに同定するバチルス・アミロリケファシエンスズブチリシンの位置5、7、23、26、28−31、34、47、63、65、66、69、70、73、82−85、88、90、92、93、105、113、125、138、139、148−151、176、178、179、193、196、200、201、202、207、219、220、223、229、233、250、266、267、及び273に等しい残基位置である。また、残基位置はバチルス・アミロリケファシエンスズブチリシンの位置23、39、41、45、67、94、136、170、181、247、251及び/または271に等しい残基位置である。さらに別の側面は、当該プロテアーゼ変異体をエンコードするDNA配列及び当該変異体DNA配列を含んだ発現ベクターを提供することである。
【0006】
前駆体プロテアーゼのプロテアーゼ変異体は、荷電アミノ酸残基位置で1以上の変異を含む前記変異体、前記前駆体プロテアーゼと同じ正味静電荷を有することを特徴とする前記変異体である。請求項1のプロテアーゼ変異体は、前記荷電アミノ酸がアスパラギン酸、グルタミン酸、リジン及びアルギニンからなる群より選択される。プロテアーゼ変異体は配列番号2に示すバチルス・アミロリケファシエンスズブチリシンの27、45、170、181、251及び271からなる群より選択される残基位置に等しい1以上の残基位置での置換を有するアミノ酸配列を含む。プロテアーゼ変異体は27、45、170、181、251及び271に対応する1以上の位置での置換を含み、K27T、R45N、R170S、D181N、K251G及びE271Tから選択される置換である。
【0007】
プロテアーゼ変異体は配列番号2に示すバチルス・アミロリケファシエンス・ズブチリシンの1、14、49、61、87、100、102、118、128、204及び258に対応する1以上の位置での別の置換をさらに含むことができる。変異体はR45N−G118E−E271R、R45N−P14R、R45N−N204R、D181N−G118D、R45N−G258R、R170S−A1R、R170S−G61R、R170S−N204R、K251G−S87K、R170S−S216R、E271T−G100E、E271T−G102E、E271T−S128E、K27T−G100E、R170S−G100R、E271T−S49E、及びE271T−S128Eの組み合わせから選択できる。
【0008】
さらに、本発明のその他の側面は、このようなベクターを用いて形質転換した宿主細胞を提供することである。
【0009】
さらに、本発明のプロテアーゼ変異体を含んだ洗浄組成物を提供する。
【図面の簡単な説明】
【0010】
【図1A】バチルス・アミロリケファシエンス・ズブチリシンのDNA配列(配列番号1)及びアミノ酸配列(配列番号2)並びにこの遺伝子の制限地図の一部を示す。
【図1B−1】バチルス・アミロリケファシエンス・ズブチリシンのDNA配列(配列番号1)及びアミノ酸配列(配列番号2)並びにこの遺伝子の制限地図の一部を示す。
【図1B−2】バチルス・アミロリケファシエンス・ズブチリシンのDNA配列(配列番号1)及びアミノ酸配列(配列番号2)並びにこの遺伝子の制限地図の一部を示す。
【図1B−3】バチルス・アミロリケファシエンス・ズブチリシンのDNA配列(配列番号1)及びアミノ酸配列(配列番号2)並びにこの遺伝子の制限地図の一部を示す。
【図2】バチルス・アミロリケファシエンス(BPN)’及びバチルス・レンタス(野生型)から得たズブチリシン間の保存アミノ酸残基を示す。
【図3A】図3Aと3Bは、4つのズブチリシンのアミノ酸配列を表す。一番上の行はバチルス・アミロリケファシエンス・ズブチリシン(ズブチリシンBPN’と呼ぶ時もある)から得たズブチリシンのアミノ酸配列を表す(配列番号3)。2行目はバチルス・ズブチリスから得たズブチリシンのアミノ酸配列を表す(配列番号4)。3行目はバチルス・リケニホルミスから得たズブチリシンのアミノ酸配列を表す(配列番号5)。4行目はバチルス・レンタス(PCT WO89/06276でズブチリシン309とも呼ばれている)から得たズブチリシンのアミノ酸配列を表す(配列番号6)。記号*はズブチリシンBPN’と比較して特定のアミノ酸残基がないことを示す。
【図3B】図3Aと3Bは、4つのズブチリシンのアミノ酸配列を表す。一番上の行はバチルス・アミロリケファシエンス・ズブチリシン(ズブチリシンBPN’と呼ぶ時もある)から得たズブチリシンのアミノ酸配列を表す(配列番号3)。2行目はバチルス・ズブチリスから得たズブチリシンのアミノ酸配列を表す(配列番号4)。3行目はバチルス・リケニホルミスから得たズブチリシンのアミノ酸配列を表す(配列番号5)。4行目はバチルス・レンタス(PCT WO89/06276でズブチリシン309とも呼ばれている)から得たズブチリシンのアミノ酸配列を表す(配列番号6)。記号*はズブチリシンBPN’と比較して特定のアミノ酸残基がないことを示す。
【図4】pVS08バチルス・ズブチリス発現ベクターを示す。
【図5】フォワードApaIプライマー、リバースApaIプライマー、リバース変異プライマー及びフォワード変異プライマーの方向を示す。
【発明を実施するための形態】
【0011】
発明の詳細な説明
プロテアーゼは、一般的にタンパク質またはペプチドのペプチド結合を切断する働きを有するカルボニルヒドロラーゼである。ここで用いる“プロテアーゼ”は天然プロテアーゼまたは組換え型プロテアーゼを意味する。天然プロテアーゼはα−アミノアシルペプチド・ヒロドロラーゼ、ペプチジルアミノ酸ヒドロラーゼ、アシルアミノヒドロラーゼ、セリンカルボキシペプチダーゼ、メタロカルボキシペプチダーゼ、チオールプロテイナーゼ、カルボキシル−プロテイナーゼ及びメタロプロテイナーゼを含む。セリン、メタロ、チオール及び酸プロテアーゼが挙げられ、及びエンド及びエキソ−プロテアーゼが挙げられる。
【0012】
本発明は、変異体のアミノ酸配列が得られる前駆体カルボニルヒドロラーゼと比較して異なるタンパク質分解活性、安定性、基質特異性、pHプロファイル及び/または性能特徴を有する非天然型カルボニルヒドロラーゼ変異体(プロテアーゼ変異体)である、プロテアーゼ酵素を含む。特に、当該プロテアーゼ変異体は天然には見られないアミノ酸配列を有し、異なるアミノ酸を含む前駆体プロテアーゼの多数のアミノ酸残基の置換により得られる。前駆体プロテアーゼは天然プロテアーゼまたは組換え型プロテアーゼである。
【0013】
本発明で有用なプロテアーゼ変異体は、指定のアミノ酸残基位置で19の天然L−アミノ酸のいずれかの置換を含む。当該置換は任意の前駆体ズブチリシンにおいて作られる(原核生物、真核生物、哺乳類、等)。この出願の全体にわたって、種々のアミノ酸を一般的な1及び3文字記号を用いて言及する。当該記号はDale,M.W.(1989)、Molecular Genetics of Bacteria(細菌の分子遺伝学),John Wiley&Sons,Ltd.,Appendix Bに特定されている。
【0014】
本発明で有用なプロテアーゼ変異体は好ましくはバチルス・ズブチリシンから得られる。より好ましくは、プロテアーゼ変異体はバチルス・レンタスズブチリシン及び/またはズブチリシン309から得られる。
【0015】
ズブチリシンは、一般的にタンパク質またはペプチドのペプチド結合を切断する働きを有する細菌性または真菌プロテアーゼである。ここで用いる“ズブチリシン”は、天然または組み換え型ズブチリシンを意味する。一連の天然ズブチリシンは種々の微生物種により生成され、よく分泌されることが知られる。この一連のメンバーのアミノ酸配列は完全に相同ではない。しかしながら、この一連のズブチリシンは同じまたは類似のタイプのタンパク質分解活性を示す。セリンプロテアーゼのこの分類は、触媒3残基を特徴付ける共通のアミノ酸配列を共有し、セリンプロテアーゼのキモトリプシン関連分類と区別している。ズブチリシン及びキモトリプシン関連セリンプロテアーゼの両方は、アスパラギン酸塩、ヒスチジン及びセリンを含む触媒3残基を有する。ズブチリシン関連プロテアーゼにおいて、これらのアミノ酸の相対順序は、アミノからカルボキシ末端へ読取って、アスパラギン酸塩−ヒスチジン−セリンである。キモトリプシン関連プロテアーゼにおいて、相対順序は、しかしながら、ヒスチジン−アスパラギン酸塩−セリンである。従って、ここでズブチリシンは、ズブチリシン関連プロテアーゼの触媒3残基を有するセリンプロテアーゼをいう。例として、限定されないが、図3で特定されるズブチリシンを含む。一般的に、及び本発明の目的において、プロテアーゼ中のアミノ酸の番号は、図1に表される成熟バチルス・アミロリケファシエンス・ズブチリシン配列に割当てた数に対応する。
【0016】
“組換え型ズブチリシン”または“組換え型プロテアーゼ”は、ズブチリシンまたはプロテアーゼをエンコードするDNA配列が修飾され、天然アミノ酸配列における1以上のアミノ酸の置換、欠失または挿入をエンコードする変異(または突然変異)DNA配列を生成するズブチリシンまたはプロテアーゼをいう。当該変異を生成する適当な方法は、米国特許RE34,606号、米国特許第5,204,015号及び米国特許第5,185,258号、米国特許第5,700,676号、米国特許第5,801,038号及び米国特許第5,763,257号に開示されるものを含み、ここに組込まれる。
【0017】
“非ヒトズブチリシン”及びそれらをエンコードするDNAは多くの原核及び真核生物から得られる。原核生物の適当な例は、大腸菌またはシュードモナスなどのグラム陰性生物及びミクロコッカスまたはバチルスなどのグラム陽性細菌を含む。ズブチリシン及びそれらの遺伝子が得られる真核生物の例としては、サッカロマイセス・セレヴィシエなどの酵母、アスペルギルス種などの真菌を含む。
【0018】
“酵素変異体”は“前駆体酵素”のアミノ酸配列由来のアミノ酸配列を有する。前駆体酵素プロテアーゼは天然酵素及び組換え酵素を含む。本発明者が意図する酵素は、限定されないが、オキシドレダクターゼ、トランスフェラーゼ、ヒドロラーゼ、リアーゼ、イソメラーゼ、及びリガーゼを含む。本発明者が意図する具体的な酵素は、限定されないが、アミラーゼ、ラッカーゼ、プロテアーゼ、デヒドロゲナーゼ及びパーミアーゼを含む。酵素変異体のアミノ酸配列は、前駆体アミノ酸配列の1以上のアミノ酸の置換、欠失または挿入により前駆体酵素アミノ酸配列から“得られる”。当該変異は前駆体酵素それ自体の操作というよりもむしろ、前駆体酵素のアミノ酸配列をエンコードする“前駆体酵素DNA配列”の変異である。前駆体DNA配列をそのように操作する適当な方法はここに開示する方法及び当業者に公知の方法を含む。プロテアーゼに関するいかなる引用または議論はその他の酵素、例えば、上で特定した一部の酵素に適用可能であることを当然である。
【0019】
“プロテアーゼ変異体”は、“前駆体プロテアーゼ”のアミノ酸配列から得られるアミノ酸配列を有する。前駆体プロテアーゼは天然プロテアーゼ及び組換え型プロテアーゼを含む。プロテアーゼ変異体のアミノ酸配列は、前駆体アミノ酸配列の1以上のアミノ酸の置換、欠失または挿入により前駆体プロテアーゼアミノ酸配列から“得られる”。当該変異は前駆体プロテアーゼ酵素それ自体の操作というよりもむしろ、前駆体プロテアーゼのアミノ酸配列をエンコードする“前駆体DNA配列”の変異である。前駆体DNA配列の当該操作の適当な方法は、ここに開示する方法を含み、及び当業者に公知の方法を含む(例えば、EP0 328299、WO89/06279及びここに既に引用した米国特許及び出願を参照)。
【0020】
“荷電アミノ酸”は潜在的にイオン性であり、電荷が変化し、特定のpHまたはpH範囲で帯電するアミノ酸として定義する。これらのアミノ酸は、例えば、酸性アミノ酸、塩基性アミノ酸及びいくつかの極性アミノ酸を含む。酸性アミノ酸はpH6.0で負に帯電したアミノ酸であり、例えばアスパラギン酸(AspまたはD)及び/またはグルタミン酸(GluまたはE)である。塩基性アミノ酸はpH6.0で正に帯電したアミノ酸であり、例えばリジン(LysまたはK)、アルギニン(ArgまたはR)、及び/またはヒスチジン(HisまたはH)である。
【0021】
“非荷電アミノ酸”は潜在的にイオン性でないアミノ酸として定義する。これらのアミノ酸は、限定されないが、非荷電非極性アミノ酸、及び/または非荷電極性アミノ酸を含む。非荷電非極性アミノ酸は、限定されないが、アラニン(AlaまたはA)、バリン(ValまたはV)、ロイシン(LeuまたはL)、イソロイシン(IleまたはI)、プロリン(ProまたはP)、フェニルアラニン(PheまたはF)、トリプトファン(TrpまたはW)、及び/またはメチオニン(MetまたはM)を含む。非荷電極性アミノ酸は、限定されないが、グリシン(GlyまたはG)、セリン(SerまたはS)、トレオニン(ThrまたはT)、システイン(CysまたはC)、チロシン(TyrまたはY)、アスパラギン(AsnまたはN)及び/またはグルタミン(GlnまたはQ)を含む。
【0022】
“正味静電荷(Net electrostatic charge)”は所定のpHまたはpH範囲での変異体または前駆体酵素またはプロテアーゼの静電荷の合計として定義される。典型的なpHはpH6.0である。
【0023】
“等電点”(pl0)は、タンパク質またはタンパク質複合体、例えばプロテアーゼまたはプロテアーゼ複合体(任意で金属またはその他のイオンを付随するもの)が中性であるpH値として定義するものであり、すなわち、複合体の静電荷の合計(正味静電荷=NEC)はゼロである。この合計において、個々の静電荷の正または負の性質の対価は考慮しなければならない。
【0024】
“同じ等電点(pl0)”はpH単位の明確な範囲内であるpl0として定義される。例えば、pH単位の明確な範囲はその他のpl0と比較するpl0の1pH単位以下であり、0.25〜0.75の間、例えば、0.5pH単位であり、好ましくは0.01〜0.5pH単位内であり、例えば0.1pH単位、及びより好ましくは0.001〜0.05pH単位内、例えば、0.01pH単位である。同じ等電点は所定のpHまたは所定のpH範囲で決定できる。
【0025】
“同一の静電荷”は、“Z”または前駆体プロテアーゼ中に存在する電荷と同じ数のプロテアーゼ変異体中の特定荷電残基を維持するものとして定義される。荷電残基の数は同じであるが、特定アミノ酸残基は、正味の静電荷が所定のpHで前駆体プロテアーゼと同じである限りは、プロテアーゼ変異体の外側周辺のその他の位置に置換できる。
【0026】
“同じ正味静電荷”は、“Z”またはプロテアーゼ変異体の静電荷の合計の明確な範囲内のプロテアーゼ前駆体の静電荷の合計を維持するものとして定義される。同じ合計静電荷の維持は明確なpH範囲にわたって明確な範囲の電荷単位内で正味静電荷を維持することを意味する。プロテアーゼ前駆体と同じ正味静電荷を有するプロテアーゼ変異体は前駆体プロテアーゼの明確な数の電荷単位内の正味静電荷を有する。例えば、同じ正味静電荷を有するプロテアーゼ変異体は1pH単位以下であり、前駆体プロテアーゼ正味静電荷の0.25〜0.75電荷単位内であり、例えば、0.5単位である。さらに、より好ましくは、同じ正味静電荷を有するプロテアーゼ変異体は前駆体プロテアーゼ正味静電荷の0.05〜0.25単位内であり、例えば、0.1単位である。さらに、より好ましくは、同じ正味静電荷を有するプロテアーゼ変異体は前駆体プロテアーゼ正味静電荷の0.001〜0.05単位内であり、例えば、0.01単位である。電荷単位は提供する(酸)または受け取る(塩基)プロトンの数として定義できる。個々のアミノ酸の静電荷は所定のpH、例えば、6.0または7.0で受け取るまたは提供するプロトンの数を決定することにより通常確定できる。Z値もこの出願内で後述する式により決定できる。
【0027】
変異体または前駆体プロテアーゼの“合計電荷量”はそれぞれのプロテアーゼの静電荷の合計数として定義される。同じ正味静電荷を有するプロテアーゼ変異体は前駆体プロテアーゼと異なる合計電荷量を有することができる。
【0028】
当業者により認識されるように、等電点は、問題の酵素中の種々の荷電残基のpK値を用いる平衡対価を用いることにより適宜計算でき、それから、ここに明示的に引用する欧州特許第0945502号及びその実施例に記載されるように、酵素分子のNECがゼロに等しいpH値の繰り返しにより見つけられる。
【0029】
この計算の1つの問題点は荷電残基のpK値がその環境に依存し、その結果変動するということである。しかしながら、実際の値と関係ない特定の近似pK値を荷電残基に割り当てることにより非常に良い結果が得られるようになる。また、部分的に環境を考慮に入れることでより高度な計算を行うことが可能になる。
【0030】
また、pl0は等電点電気泳動法または酵素を含んだ溶液を滴定することにより実験的に決定できる。さらに、荷電残基の種々のpK値は滴定により実験的に決定できる。
【0031】
本発明の別の側面において、pl0についての上述の観測はさらに、前駆体プロテアーゼ中のアミノ酸の欠失、置換または挿入される位置及びアミノ酸を決定または選択する方法に利用し、変異体プロテアーゼの正味静電荷または等電点が、同じpH値または明確なpH範囲で計算される前駆体プロテアーゼのNECまたはpl0と同じになるようにする
。
【0032】
本発明でカバーされるこの原則を表す他の方法は、前記前駆体プロテアーゼまたは酵素中のアミノ酸の欠失、置換または挿入される位置及びアミノ酸が1の方法で選択されるものであり、それにより電荷または合計電荷量(=TCC)の合計数及び/または最終的な変異体プロテアーゼまたは酵素のNECがプロテアーゼまたは酵素の最適洗浄能力に関する明確なpHまたはpH範囲で変化しない等電点を有する変異体プロテアーゼまたは酵素を提供するように一定を維持し、最適pHは洗浄溶液のpHにできるだけ接近させるべきであり、前記変異体プロテアーゼは使用するためのものである。
【0033】
上に示すように、酵素などの巨大分子のpl0は該分子のNECがゼロであるpHとして計算する。該手順は欧州特許0 945 502に記載の実施例で説明されているが、該原則をここにより詳細に記載する。
【0034】
pK値は各潜在性の荷電アミノ酸残基に割り当てる。それから、荷電または非荷電形(荷電/非荷電、C/U(i))の所定のpHのアミノ酸残基の存在比を式Ia及びIbを用いて負及び正電荷の両方について計算する。
【0035】
C/U(i)=10(ph−pKi)(負電荷)(Ia)
C/U(i)=10(pKi−ph)(正電荷)(Ib)
上述の式に従うと、pHがpKiと等しい場合、C/U(i)は1である。
【0036】
相対電荷、Qr(i)または各荷電残基に割り当てた電荷寄与(charge contribution)は、式IIa及びIIbを用いて計算する。
Qr(i)=C/U(i)/(1+C/U(i))(負電荷)(IIa)
Qr(i)=−C/U(i)/(1+C/U(i))(正電荷)(IIb)
【0037】
荷電残基から得られる全ての電荷寄与の合計がゼロである場合のpH値は、繰り返しにより、または十分に濃いpH−電荷合計表(sum table)により見出すことができる。
【0038】
正味静電荷Zを決定する他の方法は、基(Rまたはアミノ基など)が陽イオン性の酸の形態を有する場合、αが部分正電荷を表すということは当業者が理解することである:
【数1】

【0039】
一方、カルボキシルなどの基は、中性の酸の形態及び陰イオン性の共役塩基を含み、αは非荷電部分を表す。そして、部分電荷は:
【数2】
【0040】
種々の大半のプロテアーゼまたは酵素の性質はNEC及び/またはプロテアーゼまたは酵素分子の等電点に依存することが留意される。例えば、プロテアーゼまたは酵素溶解性、安定性、複数の相媒体中の位相分布及び/または表面電荷は分子のNEC及び/または等電点の変化により影響を受ける性質である。驚くべきことに、改善されたプロテアーゼの特徴は同じ等電点または同じ正味静電荷を維持しながら達成できた。特定の理論に縛られることは望まないが、問題のタンパク質、酵素またはプロテアーゼの大部分の性質を維持するが、分子の相互作用の運動、例えば基質、表面または培地に関する変化の分布または分子の配向を修飾することが望ましい状況があると考えられる。
【0041】
本発明の1の側面において、プロテアーゼ変異体及び前駆体プロテアーゼは同一の正味静電荷または同一の等電点を有する。同一の正味静電荷は同一静電荷を有することにより、または前駆体プロテアーゼのアミノ酸配列への追加の修飾による追加の変異位置から生じる電荷変化を相殺することにより維持できる。これらの追加の変異は、限定されないが、追加残基に対して反対の電荷を有する残基を置換または挿入することを含む(追加の塩基性残基を相殺するために追加の酸性残基を加える)。従って、プロテアーゼ変異体中の荷電残基の数は、例えば、荷電残基の数がプロテアーゼ前駆体におけるものよりも多い場合、プロテアーゼ前駆体のものと異なると考えられる。追加の荷電残基を相殺するために、相当する反対の荷電アミノ酸置換を同じ正味静電荷を維持するために作ることができる。さらに、プロテアーゼまたは酵素変異体中の荷電残基の数は、それが他の非荷電残基に相当する反対の荷電アミノ酸欠失または置換であって、荷電残基が非荷電残基を欠失または置換する場合、プロテアーゼまたは酵素前駆体中の荷電残基の数より少ない。
【0042】
本発明の1の側面は、NECまたはpl0が同一の場合、例えばプロテアーゼ変異体及び前駆体プロテアーゼの合計電荷量が同じ場合、または前駆体プロテアーゼ中の荷電残基が同じ数で維持される場合である。荷電アミノ酸が同一のNECまたはpl0を維持するために再配置される場合、少なくとも1のこれらの荷電アミノ酸が前駆体プロテアーゼの残基位置から異なる残基位置に置換される。プロテアーゼ前駆体中の特定の荷電アミノ酸、バチルス・レンタス(GG36)中の“X”リジン、が特定数ある場合、それから変異体プロテアーゼは同じ数だが前駆体プロテアーゼと比較して異なる位置のリジン残基、すなわち、“X”を保有する。従って、例えば、K27は異なる残基で置換でき、相当するKは他の位置、好ましくは表面位置で置換される。1の実施態様において、荷電残基、例えば、グルタミン酸、アスパラギン酸、リジンまたはアルギニンは異なる位置に置換できる。同一の静電荷を維持するために、荷電残基により置換される特定前駆体残基、例えば位置27からのK残基、は位置27で置換できる。例えば、R45N−N204Rは同一の静電荷を維持するために置換された同一の残基位置を有する。さらに、荷電残基により置換された特定の前駆体残基が非荷電残基である場合、その他の非荷電残基は荷電残基をもともと有する位置に置換できる。例えば、R170S−A1Rの組み合わせはアラニンをアルギニンで置換し、アルギンをセリンで置換する。もちろん、複数の変異を作る場合、任意の置換残基がその他の任意の残基中に置換でき、各々のアミノ酸が同じ数で維持される限り変異される。静電荷は、プロテアーゼ変異体及び前駆体プロテアーゼが同じpHで測定される限り、任意の所定pHで測定できる。
【0043】
本発明の他の側面において、分子の同じ正味静電荷は、前駆体プロテアーゼに対する変異から生じる正味静電荷の変化を相殺することにより維持する。例えば、当該変異を達成する1の方法は、別の反対の荷電アミノ酸残基の挿入または置換や、異なるが類似に帯電したアミノ酸残基の欠失である。例えば、プロテアーゼ変異体中に前駆体プロテアーゼ中よりも多くの酸性アミノ酸が存在する場合、変異体は追加の塩基性アミノ酸を含む。例えば、プロテアーゼ変異体中に前駆体プロテアーゼよりも多くの特定の酸性アミノ酸、例えば、グルタミン酸が存在する場合、当該変異を相殺するために、対応する数のアスパラギン酸残基を欠失または非荷電アミノ酸で置換できる。例えば、前駆体プロテアーゼ中に特定の数の荷電アミノ酸が存在する場合、本発明者は、変異体プロテアーゼのアミノ酸の数の増加または減少とともに荷電アミノ酸の数の変化を相殺するアミノ酸の対応する増加または減少を意図する。 従って、上述の通り、追加の正電荷残基は、対応する数の負電荷アミノ酸を追加することにより、または対応する数のその他の正電荷アミノ酸を非電荷残基またはそれらの組み合わせで置換することにより相殺できる。より少ない数の正電荷残基は対応する数の負電荷アミノ酸を欠失させることにより、対応する数の負電荷アミノ酸またはそれらの組み合わせを対応する数の非電荷残基で置換することにより相殺できる。
【0044】
1の実施態様において、第1のアミノ酸位置で非荷電残基により置換される同じ荷電のアミノ酸残基は、荷電残基を置換する同じ非荷電アミノ酸が存在する、第2のアミノ酸位置に置換される。非荷電残基は荷電アミノ酸の元の位置で置換でき、一方、置換荷電アミノ酸は非荷電アミノ酸の位置を置換できる。例えば、R45N−N204Rは荷電アミノ酸、アルギニンに関する非荷電アミノ酸、アルパラギンの置換を反映する。荷電アミノ酸を置換した同じ非荷電アミノ酸は荷電アミノ酸が再挿入される位置に存在する必要はない。例えば、アラニン(AlaまたはA)、グリシン(GlyまたはG)、アスパラギン(AsnまたはN)、プロリン(ProまたはP)、セリン(SerまたはS)、及び/またはトレオニン(ThrまたはT)の群より選択される非荷電アミノ酸は荷電アミノ酸位置に置換でき、一方荷電アミノ酸残基は上述の群のその他でもとから占められているアミノ酸位置に置換される。例えば、変異体E271T−G100Eにおいて、271位置のグルタミン酸アミノ酸はトレオニンアミノ酸で置換され、一方、100位置のグリシン残基はグルタミン酸アミノ酸で置換される。同様に、同一の荷電アミノ酸は最初から非荷電の位置、例えば、K27T−G100Eに置換される必要はない。荷電アミノ酸であるアスパラギン酸(D)、グルタミン酸(E)、リジン(K)、及びアルギニン(R)はこの点において有用である。
【0045】
本発明の他の側面において、プロテアーゼ変異体NECは、0.5荷電単位(Z)の範囲以下、前駆体プロテアーゼNECの範囲から0〜14のpHの範囲以上と様々である。
【0046】
さらに本発明の他の側面において、プロテアーゼ変異体NECは、1電荷単位の範囲以下、前駆体プロテアーゼNECの範囲から明確なpH範囲以上で変化する。明確なpH範囲は例えば、所望のプロテアーゼまたは酵素環境に関して最適または所望のpHとして当業界で認識される2pH単位内、またはpH4単位の範囲内である。
【0047】
本発明の他の側面において、前駆体プロテアーゼ内で見られる電荷残基の変異は、同じまたは同一の正味静電荷を維持しながら、増加した有益な洗浄特性を示すプロテアーゼ変異体を生じることができることが確認された。
【0048】
本発明のさらに別の側面において、前駆体プロテアーゼ内で見られる電荷残基の変異は、明確なpHで、明確なpH範囲を超えて、例えば、4pH単位の範囲を超えて、または0.001〜14のpH範囲を超えて、同じまたは同一の正味静電荷を維持しながら、増加した有益な洗浄特性を示すプロテアーゼ変異体を生じることができることが確認された。
【0049】
本発明者が変異を目的とする典型的なアミノ酸残基は、例えば、リジン、アルギニン、及び/またはヒスチジンなどの塩基性アミノ酸;例えばアスパラギン酸及び/またはグルタミン酸などの酸性アミノ酸;及び/またはそうでなければ例えばチロシンなどの極性R基を含む。
【0050】
他の側面において、本発明の変異体プロテアーゼは前記前駆体プロテアーゼと比較して、同じ数の正電荷アミノ酸残基、前駆体プロテアーゼ中の同一のアミノ酸及び同じ電荷を有する異なるアミノ酸の両方、及び前駆体プロテアーゼ中の同じ数の負電荷アミノ酸残基;または、バチルス・アミロリケファシエンス(BPN’)の位置5、7、22、23、24、26、28−31、34、45、47、63、65、66、69、70、73、82−85、88、90、92、93、97、102、105、113、125、127、138、139、148−151、169、170、176、178、179、193、196、200、201、202、207、219、220、223、229、233、250、266、267及び273の1以上の任意の位置での対応アミノ酸残基間で変異を有するが、プロテアーゼ変異体の正味静電荷及び/または等電点が前駆体プロテアーゼと同じになるように、より多いまたは少ない正電荷アミノ酸残基及び対応するより多いまたは少ない負電荷アミノ酸残基を有する。1の実施態様において、27、45、136、170、181、247、251及び/または271の1以上の位置での対応アミノ酸残基間での変異は荷電残基位置の非荷電残基での置換を含む。これらの残基位置は、これらのバチルス・レンタス野生型における対応位置がこれらの位置の荷電アミノ酸残基を有するので、重要である。例えば、バチルス・レンタスズブチリシン(配列番号6)の27、38、40、44、65、92、134、164、175、241、245及び/または265の残基位置はそれぞれバチルス・アミロリケファシエンス(配列番号2)の27、39、41、45、67、94、136、170、181、247、251及び/または271に対応する。
【0051】
他の側面において、本発明の変異体プロテアーゼは前記前駆体プロテアーゼと比較して、同じ数の正電荷アミノ酸残基、前駆体プロテアーゼ中の同一のアミノ酸及び同じ電荷を有する異なるアミノ酸の両方、及び前駆体プロテアーゼ中の同じ数の負電荷アミノ酸残基;または、バチルス・アミロリケファシエンス(BPN’)の位置27、39、41、45、67、94、136、170、181、197、247、249、251及び/または271の1以上の任意の位置での対応アミノ酸残基間で変異を有するが、プロテアーゼ変異体の正味静電荷及び/または等電点が前駆体プロテアーゼと同じになるように、より多いまたは少ない正電荷アミノ酸残基及び対応するより多いまたは少ない負電荷アミノ酸残基を有する。本発明者が意図する具体的な置換は、バチルス・アミロリケファシエンスのK27A、K27C、K27E、K27Q、K27G、K27H、K27I、K27L、K27M、K27F、K27P、K27S、K27T、K27W、K27Y、H39A、H39R、H39D、H39N、H39C、H39E、H39Q、H39G、H39H、H39I、H39L、H39K、H39M、H39F、H39P、H39T、H39W、H39Y、H39V、D41A、D41R、D41C、D41E、D41Q、D41G
、D41H、D41I、D41L、D41K、D41M、D41F、D41P、D41S、D41T、D41W、D41Y、D41V、R45A、R45R、R45D、R45N、R45C、R45E、R45C、R45E、R45Q、R45G、R45H、R45I、R45L、R45K、R45M、R45F、R45P、R45S、R45T、R45W、R45Y、R45V、H67A、H67R、H67D、H67N、H67C、H67E、H67Q、H67G、H67H、H67I、H67L、H67K、H67M、H67F、H67P、H67S、H67T、H67W、H67Y、H67V、K94A、K94R、K94D、K94N、K94C、K94E、K94Q、K94G、K94H、K94I、K94L、K94K、K94M、K94F、K94P、K94S、K94T、K94W、K94Y、K94V、E136A、E136D、E136N、E136C、E136E、E136G、E136H、E136I、E136L、E136K、E136M、E136F、E136P、E136S、E136T、E136W、E136Y、E136V、R170A、R170R、R170D、R170N、R170C、R170E、R170Q、R170G、R170H、R170I、R170L、R170K、R170M、R170F、R170P、R170S、R170T、R170W、R170Y、R170V、D181A、D181R、D181D、D181N、D181C、D181E、D181Q、D181G、D181H、D181I、D181L、D181K、D181M、D181F、D181P、D181S、D181T、D181W、D181Y、D181V、D197A、D197R、D197D、D197N、D197C、D197E、D197Q、D197G、D197H、D197I、D197L、D197K、D197M、D197F、D197P、D197S、D197T、D197W、D197Y、D197V、R247A、R247R、R247D、R247N、R247C、R247E、R247Q、R247G、R247H、R247I、R247L、R247K、R247M、R247F、R247P、R247S、R247T、R247W、R247Y、R247V、H249A、H249R、H249D、H249N、H249C、H249E、H249Q、H249G、H249H、H249I、H249K、H249M、H2490F、H249P、H249S、H249T、H249W、H249V、K251A、K251D、K251C、K251Q、K251G、K251H、K251I、K251L、K251K、K251M、K251F、K251P、K251S、K251T、K251W、K251Y、K251V、E271A、E271R、E271D、E271N、E271C、E271E、E271H、E271I、E271L、E271K、E271M、E271F、E271P、E271S、E271T、E271W、E271Y、及び/またはE271Vを含む。正電荷残基のそれ自身の置換による数の増加は、特定の洗浄環境において特定の変異体の効果の増加を生じると同時に、対応する反対の電荷変化は異なる洗浄環境において効果の増加を生じることができることに気付いた。例えば、負電荷変異は低イオン強度洗浄環境で有利な特徴を与え、正電荷変異は高イオン強度洗浄環境で有利な特徴を与えることが予想される。同じ正味静電荷または等電点を維持しながら正の増加及び負の増加の両方を含む変異体は、前駆体プロテアーゼの性能と比較して、両方の環境において改善された特徴を示すプロテアーゼ分子を生じることが予想される。
【0052】
これらの置換は好ましくはバチルス・レンタス(組み換え体または天然型)ズブチリシン内で作られるが、該置換は任意のバチルス・プロテアーゼ内、例えばバチルス・アミロリケファシエンス及び/またはズブチリシン309内で作ることができる。
【0053】
本発明の1の側面は、1、2−4、6、9−12、19、14、15、17−20、25、27、36−38、40、44、49、51、52、54−61、68、71、75、76、87、89、91、97、100−102、104、108、111、112、115、117、118、120−123、128、129、131、133、134、136、137、140、143−146、159、164、165、167、170、171、173、175、180、182−187、191、192、194、195、204、206、209−212、216、218、222、224、226、234−245、252、255、257−263、265、268、269及び274からなる群より位置付けられるまたは選択される残基に対応する1以上の残基位置において、さらに少なくとも1の別の置換アミノ酸を含むプロテアーゼ変異体を含む。本発明者によって意図される具体的な置換は、バチルス・アミロリケファシエンスまたはバチルス・レンタスのI122A、Y195E、M222A、M222S、Y167A、R170S、A194P、D36、N76D、H120D、G195E及びK235Nに等しい残基を含み、該変異体はバチルス・ズブチリシンから得られる。
【0054】
特に重要なのは、これらの位置の変異体が荷電アミノ酸置換とともに増加した洗浄能力を示す点である。これらの位置を含み、及び最初から荷電アミノ酸を有する変異体の組み合わせは重要である。本発明者が意図する典型的な組み合わせはK27T−G100E、R45N−A1R、R45N−P14R、R45N−G61R、R45N−S128R、R45N−N204R、R45N−S216R、R45N−G258R、R170S−A1R、R170S−P14R、R170S−S49R、R170S−G61R、R170S−G100R、R170S−S128R、R170S−N204R、R170S−S216R、R170S−G258R、D181N−G118D、D181N−G258D、K251G−S87K、E271T−S49E、E271T−T66E、E271T−G100E、E271T−G102E、E271T−S128E、R45N−G118E−E271R、S49R−G102E−R170S−E271T及びP14R−R45N−R170S−G258Rを含む。これらの変異を有するプロテアーゼ変異体が作ることができることは当業者に当然理解され、米国特許第5,741,694号、第6,190,900号、及び第6,197,567号に記載され、明示的にここに引用する。さらに、これらの変異は仮出願番号第60/423,087(2002年11月1日に出願;Neelam AminとVolker Schellenberger)に記載される直接的なバチルス形質転換法を用いても行うことができる。1の実施態様において、変異は融合PCR技術を用いて行った(Teplyakov,AV,et al,Protein Eng.,1992年7月5日(5):413−20)。同時にこの日に提出された、仮出願番号_/_____(Chris Leeflang,et al.)。
【0055】
本発明のさらに別の側面において、バチルス・アミロリケファシエンスの21、22、24、32、33、36、50、64、67、77、87、94、95、96、97、104、107、110、124、123、126、127、128、129、135、152、155、157、156、166、169、170、171、172、189、197、204、213、214、215、217、222または274からなる群より位置付けられる1以上の同等残基においてさらに少なくとも1の別の置換アミノ酸を有するプロテアーゼ変異体を含む。本発明者により意図される具体的な残基は以下を含む:K27R、M50F、N76D、S101G、S103A、V104I、V104Y、I122A、N123S、M124L、G159D、Y217L、A232V、Q236H、Q245R、N248D、N252K、T274A、及びM222S。プロテアーゼ変異体、3つの位置の変異をエンコードする組換えDNA、及び/またはこれらの変異を作る方法は米国特許RE34,606号;第5,972,682号;第5,185,258号;第5,310,675号;第5,316,941号;第5,801,038号;第5,972,682号;第5,955,340号及び第5,700,676号に記載されており、明示的にここに引用する。
【0056】
これらのアミノ酸位置番号は図1で表された成熟バチルス・アミロリケファシエンスズブチリシンに割当てた番号をいう。本発明は、しかしながら、この特定のズブチリシンの変異に限定されず、バチルス・アミロリケファシエンスズブチリシンの特定の同定残基に“等しい”位置でのアミノ酸残基を含む前駆体プロテアーゼまで広がる。本発明の好ましい実施態様において、前駆体プロテアーゼはバチルス・レンタスズブチリシン(配列番号6)であり、置換は上述の位置に対応するバチルス・レンタスにおける同等のアミノ酸残基位置に作られる。
【0057】
前駆体プロテアーゼの残基(アミノ酸)位置は、それがバチルス・アミロリケファシエンスズブチリシンの特定残基またはその一部と相同(すなわち、一次または三次構造の位置が一致する)または類似している場合、バチルス・アミロリケファシエンスズブチリシンの残基に等しい(すなわち、結合、反応または化学的に相互作用するための同じまたは類似の機能的能力を有する)。
【0058】
一次構造の相同性を確立するために、前駆体プロテアーゼのアミノ酸配列はバチルス・アミロリケファシエンスズブチリシン一次配列、及び特に配列が公知なズブチリシンにおいて不変な公知の残基の組と直接比較する。例えば、ここで図2はバチルス・アミロリケファシエンスズブチリシン及びバチルス・レンタスズブチリシン間での保存残基を示す。保存残基を並べ、配列を維持するために必要な挿入及び欠失を考慮した後(すなわち、任意の欠失及び挿入により保存残基が除去されるのを防ぐ)、バチルス・アミロリケファシエンスズブチリシンの一次配列中の特定のアミノ酸に等しい残基を明確にする。保存残基の配列は、好ましくは当該配列の100%保存されるべきである。しかしながら、保存残基の98%以上、95%以上、90%以上、85%以上、80%以上、75%以上、50%以上、または少なくとも45%以上の配列も同等残基を明確にするために十分である。触媒3残基、Asp32/His64/Ser221の保存は維持されるべきである。Siezen et al.(1991)Protein Eng.4(7):719−737は多数のセリンプロテアーゼの配列を示す。Siezenらはサブチラーゼ(subtilases)またはズブチリシン様セリンプロテアーゼの分類に言及している。
【0059】
例えば、図3において、バチルス・アミロリケファシエンス、バチルス・ズブチリス、バチルス・リケニホルミス(カールスベルゲンシス)及びバチルス・レンタス由来のズブチリシンのアミノ酸配列はアミノ酸配列間に最大値の相同性を与えるように配列する。これらの配列の比較は各配列に含まれる多数の保存残基があることを示す。これらの保存残基(BPN’とバチルス・レンタス間)は図2に同定される。
【0060】
これらの保存残基は、従って、ここでの好ましいプロテアーゼ前駆体酵素である、バチルス・レンタス(PCT公報WO89/06279、1989年7月13日に公開)由来ズブチリシン、または好ましいバチルス・レンタスズブチリシンと相同性の高い、PB92(EP0 328299)として言及されるズブチリシンなどのその他のズブチリシンにおいてバチルス・アミロリケファシエンスズブチリシンの対応する同等なアミノ酸残基を特定するために使用できる。特定のこれらズブチリシンのアミノ酸配列は、保存残基の最大相同性を得るために、バチルス・アミロリケファシエンスズブチリシンの配列と一緒に図3A及び3Bに配列する。図に示すように、バチルス・アミロリケファシエンスズブチリシンと比較して、バチルス・レンタスの配列中に多数の欠失がある。従って、例えば、その他のズブチリシンにおけるバチルス・アミロリケファシエンスズブチリシン中のVal165の同等アミノ酸はバチルス・レンタス及びバチルス・リケニホルミスのイソロイシンである。
【0061】
また、“同等残基(equivalent residues)”は三次構造がX線結晶学により決定された前駆体プロテアーゼの三次構造のレベルでの相同性を決定することにより特定できる。同等残基は前駆体プロテアーゼの特定アミノ酸残基の2以上の主鎖原子の原子座標の残基として特定し、バチルス・アミロリケファシエンスズブチリシン(N上のN、CA上のCA、C上のC及びO上のO)は配列後0.13nm内であり、好ましくは0.1nmである。最適モデルを方向付け、バチルス・アミロリケファシエンスズブチリシンに対して問題になっているプロテアーゼの水素でないタンパク質原子の原子座標の最大重複を与えるように位置付けた後、配列が達成される。最適モデルは、最も高い可能な解答での実験的回折データの最も低いR因子を与える結晶学モデルである。
【0062】
【数3】
【0063】
バチルス・アミロリケファシエンスズブチリシンの特定残基に機能的に類似する同等残基は、タンパク質構造を変化、修飾または寄与するような構造、基質結合またはバチルス・アミロリケファシエンスズブチリシンの特定残基を特定または起因するような触媒作用に適合できる前駆体プロテアーゼのアミノ酸として特定される。さらに、これらは、所定の残基の主鎖原子は占める相同性位置に基づいて同等の基準を満たさないが、残基の少なくとも2の側鎖原子の原子座標はバチルス・アミロリケファシエンスズブチリシンの対応する側鎖原子の0.13nmにある範囲において類似位置を占める前駆体プロテアーゼ(三次構造をX線結晶学により得た)の残基である。バチルス・アミロリケファシエンスズブチリシンの3次元構造の座標はEPO公報第0 251446(対応特許、米国特許第5,182,204号、開示の内容をここに引用する)に示され、三次元構造のレベルで同等残基を決定するための上述の概要として用いることができる。
【0064】
置換に関して同定されたいくつかの残基は保存残基であり、一方その他のものは保存残基ではない。保存されていない残基の場合、1以上のアミノ酸の置換は天然に見られるアミノ酸配列と一致しないアミノ酸配列を有する変異体を生成する置換に限定される。保存残基の場合、このような置換は天然配列を生じないはずである。本発明のプロテアーゼ変異体はプロテアーゼ変異体の成熟型及び当該プロテアーゼ変異体のプロ及びプレプロ型を含む。プレプロ型は、プロテアーゼ変異体の発現、分泌及び変異を促進するので、好ましい構造である。
【0065】
“プロ配列”は、除去された時、“成熟”型プロテアーゼを生じる、成熟型プロテアーゼのN末端部分に結合するアミノ酸配列をいう。多くのタンパク質分解酵素は、翻訳プロ酵素生成物として天然に見られ、翻訳後処理がない場合、この方法で発現する。その他のプロテアーゼ・プロ配列が使用できるが、プロテアーゼ変異体を生成する好ましいプロ配列はバチルス・アミロリケファシエンスズブチリシンズブチリシンの推定上のプロ配列である。
【0066】
“シグナル配列”または“プレ配列”は、プロテアーゼのN末端部分または成熟またはプロ型プロテアーゼの分泌に関与するプロプロテアーゼのN末端部分に結合するアミノ酸の任意の配列をいう。シグナル配列のこの定義は機能上のものであり、天然の条件下でプロテアーゼの分泌の実行に関与するプロテアーゼ遺伝子のN末端部分によりエンコードされるすべてのそれらのアミノ酸配列を含むことを意味する。本発明はここに定義するプロテアーゼ変異体の分泌を達成するために当該配列を利用する。1の可能なシグナル配列はバチルス・レンタス(ATCC21536)由来のズブチリシンのシグナル配列の残余と融合したバチルス・ズブチリスズブチリシン由来のシグナル配列の最初の7つのアミノ酸残基を含む。
【0067】
“プレプロ”型プロテアーゼ変異体はプロテアーゼのアミノ末端に作動可能に連結するプロ配列、及びプロ配列のアミノ末端に作動可能に連結する“プレ”または“シグナル”配列を有するプロテアーゼの成熟型からなる。
【0068】
“発現ベクター”は、適当な宿主中に前記DNAの発現をもたらすことができる適当な制御配列に作動可能に連結するDNA配列を含んだDNA構築体をいう。当該制御配列は転写を達成するためのプロモーター、当該転写を制御するための任意的なオペレーター配列、適当なmRNAリボソーム結合部位をエンコードする配列、及び転写及び翻訳の停止を制御する配列を含む。ベクターはプラスミド、ファージ粒子または単純なゲノム挿入である。一旦、適当な宿主に形質転換されると、ベクターは複製でき、宿主ゲノムに独立的に機能し、または場合によっては、ゲノム自身に溶け込む。本願明細書において、“プラスミド”及び“ベクター”は、プラスミドが現在ベクターの最も一般的に使用されている形態であるので、時には交換可能に用いる。しかしながら、本発明は、同等の機能を果たし、当業界で公知の発現ベクターのその他の形態を含むことも意図するものである。
【0069】
本発明で用いられる“宿主細胞”は一般に、米国特許RE34,606号及び/または5,441,882号に開示された方法により好ましくは操作される原核または真核宿主であり、酵素的に活性なエンドプロテアーゼを分泌できないようにさせる。プロテアーゼを発現するために有用な宿主細胞は、酵素的に活性な中性プロテアーゼ及びアルカリ性プロテアーゼ(ズブチリシン)が不足しているバチルス株BG2036である。株BG2036の構築は米国特許第5,264,366号に詳細が記載されている。プロテアーゼを発現するためのその他の宿主細胞はバチルス・ズブチリスI168(米国特許RE34,606号;米国特許第5,264,366号;及び第5,441,882号に記載されている。これらの開示の内容をここに引用する)及びバチルス・リケニホルミス、バチルス・レンタス等などの任意の適当なバチルス株を含む。特に有用な宿主細胞はバチルス株BG2864である。株BG2864の構築はD.Naki、C.Paech、G.Ganshaw、V.Schellenberger.Appl Microbiol Biotechnol(1998)49:290−294に詳細が記載されている。
【0070】
宿主細胞は組換えDNA技術を用いて構築されたベクターを用いて形質転換またはトランスフェクトする。当該形質転換宿主細胞はプロテアーゼ変異体をエンコードするベクターを複製し、または所望のプロテアーゼ変異体を発現することができる。ベクターがプレまたはプレプロ型のプロテアーゼ変異体をエンコードする場合、当該変異体は、発現されたとき、一般的に宿主細胞から宿主細胞培地に分泌される。
【0071】
“作動可能に連結する”は、2つのDNA領域間の関係について言及する場合、単にお互いが機能的に関連することを意味する。例えば、プレ配列は、それがシグナル配列として機能する場合、ペプチドに作動可能に連結し、シグナル配列の切断に最も関係すると思われるタンパク質の成熟型の分泌に関与する。プロモーターは、それが配列の転写を制御する場合、コード配列に作動可能に連結し、リボソーム結合部位はそれが翻訳を可能にするように位置する場合、コード配列に作動可能に連結する。
【0072】
天然前駆体プロテアーゼをエンコードする遺伝子は当業者に公知の一般的な方法に従って得られる。該方法は通常、目的のプロテアーゼの領域をエンコードする推定配列を有する標識プローブを合成する工程、プロテアーゼを発現する微生物からゲノムライブラリーを調製する工程、及びプローブへのハイブリダイゼーションにより目的の遺伝子に関するライブラリーをスクリーニングする工程を含む。陽性のハイブリダイズしたクローンはそれから、地図を作り、配列する。
【0073】
クローンされたプロテアーゼはそれからプロテアーゼを発現するために宿主細胞に形質転換して用いる。プロテアーゼ遺伝子はそれから、高コピー数プラスミド中に連結される。プラスミド複製、問題の遺伝子に作動可能に連結したプロモーター(認識される場合、遺伝子自身の相同プロモーターとして提供される、すなわち、宿主により転写される)、外来のまたはプロテアーゼ遺伝子の内生停止領域により提供された転写停止及びポリアデニル化領域(特定の真核宿主細胞のプロテアーゼ遺伝子から宿主により転写されたmRNAの安定に必要)、及び望ましくは、抗生物質含有培地での成長によりプラスミド感染宿主細胞の培養管理が維持できる抗生物質抵抗性遺伝子などの選択遺伝子に必要な公知の要素を含むという意味で、このプラスミドは宿主中で複製する。高コピー数プラスミドも宿主の複製起源を含み、それにより、多数のプラスミドが染色体の制限なしに細胞質中で生成することが可能となる。しかしながら、複数のプロテアーゼのコピーを宿主ゲノム内に組込むことは本発明の範囲内である。このことは特に相同組換えの影響を受けやすい原核及び真核生物により促進される。
【0074】
遺伝子は天然バチルス・レンタス遺伝子である。もしくは、天然または変異前駆体プロテアーゼをエンコードする合成遺伝子が生成できる。この方法において、前駆体プロテアーゼのDNA及び/またはアミノ酸配列を決定する。複数の重複合成一本鎖DNA断片はその後合成し、ハイブリダイゼーション及び連結により前駆体プロテアーゼをエンコードする合成DNAを生成する。合成遺伝子構築の例は米国特許第5,204,015号の実施例3に示されており、その開示の内容はここに引用する。
【0075】
天然または合成前駆体プロテアーゼ遺伝子を一旦クローンすると、多数の修飾が天然前駆体プロテアーゼの合成を超えて遺伝子の使用を高めるために行われる。当該修飾は、米国特許RE34,606号;第5,741,694号;第6,190,900号;第6,197,567号;第5,972,682号;第5,185,258号;第5,700,676号及びEPO公報第0 251446号に開示される組換えプロテアーゼの生成及びここに記載するプロテアーゼ変異体の生成を含む。
【0076】
以下のカセット式変異誘発法は本発明のプロテアーゼ変異体の構築を促進するために使用できるが、その他の方法も使用できる。まず、プロテアーゼをエンコードする天然遺伝子を得て、全体または一部の配列を決定する。それから、エンコードされる酵素において1以上のアミノ酸の変異(欠失、挿入または置換)を作ることが望ましいポイントに関して配列をスキャンする。このポイントのフランキング配列を、発現した場合種々の変異をエンコードする、オリゴヌクレオチドプールを有する遺伝子の短い断片を置換する制限部位の存在に関して評価する。当該制限部位は、好ましくは遺伝子断片の置換を促進するようなプロテアーゼ遺伝子内の固有部位である。しかしながら、制限消化により生じた遺伝子断片が適当な配列内で再構築されるとすれば、プロテアーゼ遺伝子内の過度に冗長でない任意の都合のよい制限部位が使用できる。制限部位が選択点から都合のよい距離内(10〜15ヌクレオチド)の位置に存在しない場合、当該部位はリーディング・フレームまたはエンコードされるアミノ酸が最終的な構造で変化しない方法で遺伝子内の置換ヌクレオチドにより生じる。配列を変化させて所望の配列に合わせる遺伝子の変異は、公知の方法に従ってM13プライマー伸長法により達成される。適当なフランキング領域を位置付け、2つの都合のよい制限部位配列に達するために必要な変化を評価する課題は遺伝コードの冗長さ、遺伝子の制限酵素地図及び多数の異なる制限酵素によりルーチン化される。都合のよいフランキング制限部位が利用できる場合、上述の方法は部位を含まないフランキング領域とのみ関連して使用される必要があることを留意すべきである。
【0077】
一旦、天然DNAまたは合成DNAがクローンされると、変異する位置のフランキング制限部位は同種の制限酵素を用いて消化され、多数のエンド末端−相補オリゴヌクレオチドカセットは該遺伝子内に連結される。全てのオリゴヌクレオチドは同じ制限部位を有するように合成でき、制限部位を作るために合成リンカーは必要でないので、突然変異生成はこの方法により単純化される。
【0078】
適当な宿主の形質転換で発現される変異体プロテアーゼは、所望の特性、例えば改善された洗浄能力、基質特異性、酸化安定性、pH活性プロファイル等を示す単離または回収された酵素についてスクリーニングできる。
【0079】
ここで用いる、タンパク質分解活性は活性酵素ミリグラム当たりのペプチド結合の加水分解の速度として定義される。タンパク質分解活性を測定するための多くの公知の手順が存在する(K.M.Kalisz,“Microbial Proteinases(細菌プロテイナーゼ),”Advances in Biochemical Engineering/Biotechnology,A.Fiechter編集、1988)。タンパク質分解活性を測定するその他の典型的な方法は、選択基質の変換を所定の基質濃度に加えたプロテアーゼの吸収の変化を測定することにより間接的に測定する種々の分光学的分析を含む。典型的な基質はジメチルカゼイン、N−スクシニル−Ala−Ala−Pro−Phe pNa及びケラチンを含む(米国特許出願番号第60/344,702を参照)。
【0080】
修飾タンパク質分解活性に加え、または修飾タンパク質分解活性に代えて、本発明の変異体酵素は、Km、kcat、kcat/Km比及び/または修飾された基質特異性及び/または修飾されたpH活性プロファイルなどのその他の修飾された性質を有することができる。これらの酵素は、例えば、ペプチドの調製において、または洗濯用などの加水分解プロセスのために存在すると予想される特定基質に合わせることができる。
【0081】
基質特異性の変化は前駆体酵素と変異体間のkcat/Km比の違いとして定義できる。kcat/Km比は触媒効果の指標である。増加または減少したkcat/Km比を有する原核生物カルボニルヒドロラーゼを実施例に記載する。一般的に、本目的はより大きい(数の上で大きい)所定の基質のkcat/Km比を有する変異体を安全なものにし、それにより酵素を使用して標的基質上でより効果的に作用できるようにすることである。1の基質に関するkcat/Km比の増加は他の基質に関するkcat/Km比の減少により達成できる。これは基質特異性の移動であり、このような移動を示す変異体は、前駆体が望ましくないところでの実用性を有し、例えば、基質混合物中の特定基質の望ましくない加水分解を防ぐ。
【0082】
kcat及びKmは公知の手順に従って測定でき、または米国特許第5,441,882号の実施例18の記載に従って測定できる。
【0083】
さらに、酸化安定性は、実施例に記載されるプロテアーゼ変異体により達成できる目的である。該安定性は種々の用途に応じて高めたり減少させたりすることができる。高められた安定性は1以上のメチオニン、トリプトファン、システインまたはリジン残基を欠失することにより達成でき、任意でメチオニン、トリプトファン、システインまたはリジンでないその他のアミノ酸残基を置換することにより達成できる。その逆の置換は減少された酸化安定性を生じる。置換残基はアラニルであってもよいが、中性残基も適している。
【0084】
さらに、安定性、例えば耐熱性は、実施例に記載されるプロテアーゼ変異体により達成できる目的である。安定性は種々の用途に望ましいように高めたり減少させたりできる。高められた安定性は本願明細書で同定された1以上の残基の置換により達成でき、任意でそのうち1つが同じでない他のアミノ酸残基を置換することにより達成できる。耐熱性は所定の温度において長時間にわたって酵素活性を維持する。改善された耐熱性は、前駆体プロテアーゼと比較して変異体の大量の酵素活性の保全を伴う。例えば、所定の温度、通常は測定する操作温度での前駆体と比較した変異体の酵素活性レベルの増加である。
【0085】
ここに記載するプロテアーゼ変異体は特定の洗浄条件下で改善された洗浄能力を示す。例えば、プロテアーゼ変異体は、当業界で公知(例えば、WO99/34011(“An improved Method of Assaying for a Preferred enzyme and/or Preffered Detergent composition(好ましい酵素および/または好ましい洗剤組成物についての改良された分析方法).”))の種々の分析法で測定した性能により示されるように、例えば温度、水の硬度及び/または洗剤濃度の異なる洗浄条件下、異なる洗浄能力を示す。
【0086】
バチルス・ズブチリシンまたはそのプレ、プレプロ及びプロ型の場合、先に述べた位置での変異は上述の特徴が変化した変異体、または酵素の処理が変化した変異体を生成する。これらのアミノ酸位置番号は図1に見られるようにバチルス・アミロリケファシエンスズブチリシンに割り当てられたものであることに留意されたい。所定の位置からのN末端方向の欠失または挿入は、関連アミノ酸位置を移動させ、残基がその最初のまたは野生型の数字の位置ではなくなることが当然に理解される。また、対立遺伝子の違い及び種々の原核生物種間のばらつきは位置移動を生じ、当該ズブチリシンの位置169がグリシンで占められなくなる。このような場合、グリシンの新しい位置は指定グリシン+169と等しく、指定グリシン+169内に包含されると考えられる。グリシン+169の新しい位置は図1のグリシン+169と相同な領域に関して問題のズブチリシンをスキャニングすることにより容易に同定される。
【0087】
1以上、通常は約10までの、アミノ酸残基が変異する。しかしながら、商慣行はさておき、作られる変異の数に限定はない。
【0088】
本酵素は塩として得られる。タンパク質の電離状態は、溶液内の場合、または調製される溶液内の場合、固形の場合、周囲環境のpHに依存することは明らかである。酸性タンパク質は例えば、アンモニウム、ナトリウムまたはカリウム塩として通常調製され、塩基性タンパク質は塩化物、硫酸塩またはリン酸塩として調製される。従って、本出願は指定した種々のプロテアーゼの電気的に中性及び塩形態の両方を含み、プロテアーゼの語は電離状態を問わない、有機構造骨格をいう。
【0089】
プロテアーゼ変異体は特に食品加工及び洗浄技術に有用である。プロテアーゼ変異体及び前駆体プロテアーゼを含むカルボニルヒドロラーゼは、ここに記載するように発酵により生成し、適切な技術により回収する。例えば、K.Anstrup,1974、Industrial Aspects of Biochemistry、編集、B.Spencer、pp.23−46を参照されたい。
【0090】
本発明の1の側面において、本発明の目的は、少なくとも1の洗剤剤形において、及び/または少なくとも1組の洗浄条件下において、前駆体プロテアーゼと比較して変化した、好ましくは改善された洗浄能力を有する変異体プロテアーゼを安定させることである。産業プロセス、特に洗濯業での使用のため、それらはそれ自体公知の方法に従って洗剤またはその他の界面活性剤と一緒に処方される。後者の場合、酵素はタンパク質分解酵素の分野で公知の洗剤、ビルダー、漂白剤、及び/蛍光白色化剤と混合される。適当な洗剤は直鎖アルキルベンゼンスルホン酸塩、アルキルエトキシレート硫酸塩、硫酸化直鎖アルコールまたはエトキシレート直鎖アルコールが挙げられる。該組成物は顆粒状または液状で処方できる。例えば、米国特許第3,623,957号;第4,404,128号;第4,381,247号;第4,404,115号;第4,318,818号;第4,261,868号;第4,242,219号;第4,142,999号;第4,111,855号;第4,011,169号;第4,090,973号;第3,985,686号;第3,790,482号;第3,749,671号;第3,560,392号;第3,558,498号;及び第3,557,002号を参照されたい。
【0091】
種々の洗浄条件が存在し、種々の洗剤剤形、洗浄水量、洗浄水温度、及びプロテアーゼ変異体が曝される洗浄時間の長さなどである。例えば、異なる分野で使用される洗剤剤形は洗浄水中存在するそれらの関連成分を異なる濃度で有する。例えば、ヨーロッパの洗剤は通常、洗浄水中、約3000−8000ppmの洗剤成分を有するが、一方、日本の潜在は通常、洗浄水中800以下、例えば、667ppmの洗剤成分を有する。北米において、特にアメリカでは、洗剤は通常、洗浄水中、約800〜2000、例えば975ppmの洗剤成分が存在する。
【0092】
低洗剤濃度系は約800ppm以下の洗剤成分が洗浄水中に存在する洗剤を含む。日本の洗剤は通常、約667ppmの洗剤成分が洗浄水中に存在するので、低洗剤濃度系と考えられる。
【0093】
中洗剤濃度は約800ppm〜約2000ppmの洗剤成分が洗浄水中に存在する洗剤を含む。北米の洗剤は通常、約975ppmの洗剤成分が洗浄水中に存在するので、中洗剤濃度系と考えられる。ブラジルは約1500ppmの洗剤成分が洗浄水中に存在する。
【0094】
高洗剤濃度系は約2000ppmの洗剤成分が洗浄水中に存在する洗剤を含む。ヨーロッパの洗剤は通常、約3000〜8000ppmの洗剤成分を洗浄水中に含むので、高洗剤濃度系と考えられる。
【0095】
中南米の洗剤は通常、高泡立ちリン酸塩ビルダー洗剤であり、中南米で使用される洗剤の範囲は、洗浄水中の洗剤成分が1500ppm〜6000ppmの範囲であるので、中及び高洗剤濃度の両方になる。上述の通り、ブラジルは通常、洗浄水中に約1500ppmの洗剤成分を有する。しかしながら、その他の中南米の国に限定されない、その他の高泡立ちリン酸塩ビルダー洗剤の地は、約6000ppm以下の洗剤成分が洗浄水中に存在する高洗剤濃度系を有する。
【0096】
上述に照らして、一般的な洗浄溶液中の洗剤組成物の濃度は世界中で約800ppm以下の洗剤組成物(“低洗剤濃度地”)、例えば、日本の約667ppm、から約800ppm〜約2000ppm(“中洗剤濃度地”)、例えば、アメリカの約975ppm及びブラジルの約1500ppm、及び約2000ppm以下(“高洗剤濃度地”)、例えば、ヨーロッパの約4500ppm〜約5000ppm及び高泡立ちリン酸塩ビルダー地の約6000ppmまで様々であることがわかる。
【0097】
一般的な洗剤溶液の濃度は実験的に決定する。例えば、米国において、一般的な洗濯機は洗浄溶液の約64.4Lの体積を入れることができる。従って、洗浄溶液約62.79gの洗剤組成物内で約975ppmの洗剤濃度を得るためには、64.4Lの洗浄溶液を加えなければならない。この量は、洗剤と一緒に備えられる計量カップを用いて消費者により洗浄水中で測定される一般的な量である。
【0098】
さらに別の例として、異なる場所では異なる洗浄温度を用いている。日本の洗浄水の温度は一般にヨーロッパで用いる温度よりも低い。例えば、ヨーロッパにおける洗浄水の温度は通常、約30〜50℃であり、一般的には約40℃である。北米及び/または日本における洗浄水の温度は通常、ヨーロッパの洗浄水よりも低く、例えば約10〜30℃であり、一般的に約20℃である。
【0099】
さらに、別の例として、異なる場所では、異なる水硬度を用いる。水硬度は一般的に合計Ca2+/Mg2+のガロン当たりのグレインとして記載される。硬度は水中のカルシウム(Ca2+)及びマグネシウム(Mg2+)の量を測定したものである。アメリカのほとんどの水は硬水であるが、硬度の程度は様々である。中程度の硬水(60〜120ppm)〜硬水(121〜181ppm)は60〜181ppmの硬度ミネラルを含む(米国ガロン当たりのグレインに変換したppmは、ppm数÷17.1=ガロン当たりのグレインである)。
【0100】
【表1】
【0101】
ヨーロッパの水の硬度は一般に、10〜20ガロン/グレイン合計Ca2+/Mg2+であり、例えば、約15ガロン/グレイン合計Ca2+/Mg2+である。北米の水の硬度は一般に、日本の水の硬度より高いが、ヨーロッパの水の硬度より低い。例えば、3〜10グレイン、3〜8グレインまたは約6グレインである。日本の水の硬度は一般に北米の水の硬度より低く、通常4未満、例えば、3グレイン/ガロン合計Ca2+/Mg2+である。
【0102】
従って、本発明の1の側面は少なくとも1組の洗浄条件において改善された洗浄能力を示すプロテアーゼ変異体を含む。本発明の他の側面は1以上の洗浄条件、例えば、ヨーロッパ、日本または北米条件において、改善された洗浄能力を示すプロテアーゼ変異体を含む。
【0103】
該変異体プロテアーゼを用いて得られたスクリーニング結果に基づいて、バチルスズブチリシンにおける言及した変異及びバチルス・レンタスにおけるそれらの同等物はタンパク質分解活性、性能及び/またはこれらの酵素の安定性及び当該変異体酵素の洗浄または洗剤性能に重要である。
【0104】
本発明の多くのプロテアーゼ変異体は種々の洗剤組成物またはシャンプーまたはローションなどのパーソナルケア製剤を処方するのに有用である。本発明のプロテアーゼ変異体を含む組成物において有用な多数の公知の化合物は適当な界面活性剤である。これらは、Barry J.Andersonの米国特許第4,404,128号及びJiri Flora,et al.の米国特許第4,261,868号に記載されるような、非イオン性、陰イオン性、陽イオン性または両性イオン界面活性剤を含む。適当な洗剤処方は米国特許第5,204,015号(すでにここに引用している)の実施例7に記載されている。当該技術は洗浄組成物として用いることができる種々の剤形でよく知られている。一般的な洗浄組成物に加えて、本発明のプロテアーゼ変異体は、天然または野生型プロテアーゼが使用される任意の目的に使用できることが容易に理解される。従って、これらの変異体は使用でき、例えば、固形または液体石けん用途、食器洗い用製剤、コンタクトレンズ洗浄液または製品、ペプチド加水分解、廃棄物処理、織物用途、タンパク質生成における融合−分解酵素としてである。本発明の変異体は洗剤組成物において高められた性能を有する(前駆体と比較して)。ここで用いるように、洗剤における高められた性能は、標準洗浄サイクル後の通常の評価により測定した、草または血液などのしみに敏感な特定酵素の洗浄の増加として定義する。
【0105】
本発明のプロテアーゼは、約0.01〜約5重量%(好ましくは0.1重量%〜0.5重量%)でpHが6.5〜12.0の公知の粉末及び液体洗剤中に処方できる。これらの洗剤洗浄組成物は、公知のプロテアーゼ、アミラーゼ、セルラーゼ、リパーゼまたはエンドグリコシダーゼなどその他の酵素並びにビルダー及び安定剤を含むこともできる。
【0106】
本発明のプロテアーゼの従来の洗浄組成物への添加は、いかなる特別な使用の限定をも生じない。すなわち、該洗剤に適切ないかなる温度及びpHも、該pHが上述の範囲内である限り、及び該温度が記載したプロテアーゼの変性温度以下である限り、本発明の組成物に適している。さらに、本発明のプロテアーゼは洗剤なしに洗浄組成物中で使用でき、この場合もやはり、単独またはビルダー及び安定剤との組み合わせにおいて使用できる。
【0107】
また、本発明は本発明のプロテアーゼ変異体を含む洗浄組成物に関する。洗浄組成物は一般的に洗浄組成物に使用される添加物をさらに含むことができる。これらは、限定されないが、漂白剤、界面活性剤、ビルダー、酵素及び漂白触媒から選択できる。組成物に含めることが適当な添加剤については当業者にとって明らかである。ここに与えるリストは完全なものではなく、適当な添加剤の例としてのみ理解させるべきである。酵素と適合性のある添加剤及びその他の成分のみ、例えば界面活性剤のみを組成物中に用いることも当業者には容易に理解されることである。
【0108】
存在する場合、洗浄組成物に存在する添加剤の量は約0.01%〜約99.9%、好ましくは約1%〜約95%、より好ましくは約1%〜約80%である。
【0109】
本発明の変異体プロテアーゼは、例えば、米国特許第5,612,055号;米国特許第5,314,692号;及び米国特許第5,147,642号に記載されるような動物飼料添加物の一部として、動物飼料中に含めることもできる。
【0110】
本発明の1の側面は、本発明の変異体プロテアーゼを含む織物処理のための組成物である。該組成物は、RD216,034号;欧州特許134,267号;米国特許第4,533,359号;及び欧州特許第344,259号などの公報に記載されているような例えば絹またはウールを処理するために使用できる。
【0111】
以下は実施例により表すものであり、請求の範囲を限定するものとして解釈されない。
【0112】
ここに引用する全ての刊行物及び特許はその全体をここに組み込むものとする。
【実施例】
【0113】
実施例1
多数のプロテアーゼ変異体が生成でき、当業界に公知の方法を用いて精製できる。変異はバチルス・アミロリケファシエンス(BPN’)ズブチリシンまたはバチルス・レンタスGG36ズブチリシンにおいて作ることができる。変異体は以下から選択できる:バチルス・アミロリケファシエンスのK27A、K27C、K27E、K27Q、K27G、K27H、K27I、K27L、K27M、K27F、K27P、K27S、K27T、K27W、K27Y、H39A、H39R、H39D、H39N、H39C、H39E、H39Q、H39G、H39I、H39L、H39K、H39M、H39F、H39P、H39T、H39W、H39Y、H39V、D41A、D41R、D41C、D41E、D41Q、D41G、D41H、D41I、D41L、D41K、D41M、D41F、D41P、D41S、D41T、D41W、D41Y、D41V、R45A、R45R、R45D、R45N、R45C、R45E、R45C、R45E、R45Q、R45G、R45H、R45I、R45L、R45K、R45M、R45F、R45P、R45S、R45T、R45W、R45Y、R45V、H67A、H67R、H67D、H67N、H67C、H67E、H67Q、H67G、H67I、H67L、H67K、H67M、H67F、H67P、H67S、H67T、H67W、H67Y、H67V、K94A、K94R、K94D、K94N、K94C、K94E、K94Q、K94G、K94H、K94I、K94L、K94M、K94F、K94P、K94S、K94T、K94W、K94Y、K94V、E136A、E136D、E136N、E136C、E136G、E136H、E136I、E136L、E136K、E136M、E136F、E136P、E136S、E136T、E136W、E136Y、E136V、R170A、R170D、R170N、R170C、R170E、R170Q、R170G、R170H、R170I、R170L、R170K、R170M、R170F、R170P、R170S、R170T、R170W、R170Y、R170V、D181A、D181R、D181N、D181C、D181E、D181Q、D181G、D181H、D181I、D181L、D181K、D181M、D181F、D181P、D181S、D181T、D181W、D181Y、D181V、D197A、D197R、D197N、D197C、D197E、D197Q、D197G、D197H、D197I、D197L、D197K、D197M、D197F、D197P、D197S、D197T、D197W、D197Y、D197V、R247A、R247D、R247N、R247C、R247E、R247Q、R247G、R247H、R247I、R247L、R247K、R247M、R247F、R247P、R247S、R247T、R247W、R247Y、R247V、H249A、H249R、H249D、H249N、H249C、H249E、H249Q、H249G、H249I、H249K、H249M、H249F、H249P、H249S、H249T、H249W、H249V、K251A、K251D、K251C、K251Q、K251G、K251H、K251I、K251L、K251M、K251F、K251P、K251S、K251T、K251W、K251Y、K251V、E271A、E271R、E271D、E271N、E271C、E271H、E271I、E271L、E271K、E271M、E271F、E271P、E271S、E271T、E271W、E271Y、及び/またはE271V。
【0114】
実施例2
多数のプロテアーゼ変異体は公知の方法を用いて生成及び精製できる。変異はバチルス・アミロリケファシエンス(BPN’)ズブチリシンまたはバチルス・レンタスGG36ズブチリシンにおいて作ることができる。変異体は以下の位置に対応するアミノ酸の挿入、欠失または置換で作ることができる:バチルス・アミロリケファシエンス(BPN’)の5、7、23、26、28−31、34、47、63、65、66、69、70、73、82−85、88、90、92、93、105、113、125、138、139、148−151、176、178、179、193、196、200、201、202、207、219、220、223、229、233、250、266、267、及び273。
【0115】
実施例3
実施例1及び/または2で生成した多数のプロテアーゼ変異体は、米国特許出願番号60/068,796、“An Improved method of assaying for a preferred enzyme and/or preferred detergent composition(好ましい酵素及び/または好ましい洗剤組成物の改善された分析方法)”に記載されている、マイクロスウォッチ(microswatch)分析法を用いて2種類の洗剤及び洗浄条件における性能を試験する。
【0116】
変異体プロテアーゼは種々の洗剤を分析及び試験される。例えば、可能な洗剤は、0.67g/lろ過アリエール・ウルトラ(Ariel Ultra)(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)、溶液中3グレイン/ガロン合計Ca2+/Mg2+硬度を含み、20℃で各ウェル中0.3ppm酵素が使用される。他の典型的な洗剤は、3.38g/lろ過アリエールFutur(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)、溶液中15グレイン/ガロン合計Ca2+/Mg2+硬度を含み、40℃で各ウェル中0.3ppm酵素が使用される。野生型と比較して高い相対値は改善された洗剤効果を示す。
【0117】
実施例4
実施例1及び2で記載するように分析できる変異体プロテアーゼはその他の異なる洗剤においても分析できる。実施例2と同じ性能試験を以下の洗剤を用いて記載変異体プロテアーゼについて行うことができる:第1の洗剤は、0.67g/lろ過アリエール・ウルトラ(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)であり、溶液中3グレイン/ガロン合計Ca2+/Mg2+硬度を含み、及び20℃で各ウェル中0.3ppm酵素を使用した。第2の洗剤は、3.38g/lろ過アリエールFutur(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)、であり、溶液中15グレイン/ガロン合計Ca2+/Mg2+硬度を含み、及び40℃で各ウェル中0.3ppm酵素を使用できる。第3の洗剤は、3.5g/lHSP1洗剤(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)であり、溶液中8グレイン/ガロン合計Ca2+/Mg2+硬度を含み、及び20℃で各ウェル中0.3ppm酵素を使用できる。第4の洗剤は、1.5ml/l Tide洗剤(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)であり、溶液中3グレイン/ガロン合計Ca2+/Mg2+硬度を含み、及び20℃で各ウェル中0.3ppmの酵素を使用できる。
【0118】
実施例5
多数のプロテアーゼ変異体を生成し、当業界で公知の方法を用いて精製した。全ての変異はバチルス・レンタスGG36ズブチリシン中に作った。変異体を表3に示す。
【0119】
GG36飽和部位ライブラリー及び部位特異性変異体を構築するために、3つのPCR反応を行った。2つのPCR反応はGG36に目的の変異コドンを導入するためのものであり、融合PCRは所望の変異を含む発現ベクターを構築するためのものである。
【0120】
目的のGG36コドンはBPN’の番号に従って番号を付ける(図1A−C及び3A−Bに記載した)。
【0121】
飽和部位ライブラリー構築について:
突然変異生成の方法は領域−特異性変異方法(Teplyakov et al.,1992)に基づいて行い、変異するコドンの配列に対応し、そのコドンでのヌクレオチドのランダムな結合を保証する具体的に作成された3DNA配列NNS((A、C、TまたはG)、(A、C、TまたはG)、(CまたはG))を取り囲む30〜40ヌクレオチド長のフォワード及びリバース相補オリゴヌクレオチドプライマーセットを用いて特定DNAコドン中に全ての可能な変異を一度に作成した。
【0122】
部位特異性変異体構築について:
フォワード及びリバース変異プライマーは両側の相同配列の15以下塩基を含むプライマーの中間に3つの所望の変異を含む。これらの変異は目的のコドンをカバーし、所望のアミノ酸に特異的であり、意図的に合成される。
【0123】
ライブラリー及び変異体を構築するために使用する第2のプライマーセットはpVS08 Apal消化部位及びそのフランキングヌクレオチド配列を一緒に含む。
【0124】
Apalプライマー:
フォワードApalプライマー:
【化1】
リバースApalプライマー:
【化2】
【0125】
GG36分子の変異導入はInvtrogen(カールズバッド、カリフォルニア州、米国)Platinum(登録商標)Taq DNAポリメラーゼ・ハイファイ(High Fidelity)(Cat.no.11304−102)とpVS08テンプレートDNA及びフォワード変異プライマー及び反応1のリバースApalプライマーまたはリバース変異プライマー及び反応2のフォワードApalプライマーを一緒に用いて行った。
【0126】
所望の変異を含む発現ベクターの構築は、反応1及び2の両方のPCR断片、フォワード及びリバースApalプライマー及びInvitrogen Platinum(登録商標)Taq DNAポリメラーゼ・ハイファイ(Cat.no.11304−102)を用いて融合PCRにより達成した。
【0127】
全てのPCRは、サイクル数を除いて(30の代わりに20)、ポリメラーゼと一緒に提供されるInvitrogen手順に従って行った。2つの別個のPCR反応はInvitrogen Platinum(登録商標)Taq DNAポリメラーゼ・ハイファイ(Cat.no.11304−102)を用いて行う。増幅線形5.6Kb断片を精製し(Qiagen(登録商標)Qiaquick PCR精製キット、Cat.no.28106)、Apal制限酵素を用いて消化し、融合断片の両側に付着端を生成した:
- 35μL精製DNA断片
- 4μL React(登録商標)4バッファ(Invitrogen(登録商標):20mM Tris−HCl、5mM MgCl2、50mM KCl、pH7.4)
- 1μL Apal、10単位/ml(Invitrogen(登録商標)、Cat.no.15440−019)
反応条件:1時間、30℃
【0128】
Invitrogen Dpnlを用いて追加の消化をpVS08テンプレートDNAを除去するために行った。
- 40μL Apal消化DNA断片
- 1μL Dpnl、4単位/μL(Invitrogen(登録商標)、Cat.no.15242−019)
反応条件:16〜20時間、37℃
【0129】
2回消化及び精製断片の結合は所望の変異を含む新しい環状DNAを生じ、コンピテントバチルス・ズブチリスに直接形質転換した:
- 30μLのApal及びDpnl消化DNA断片
- 8μL T4リガーゼ・バッファ(Invitrogen(登録商標)、Cat.no.46300−018)
- 1μL T4 DNAリガーゼ、1単位/μl(Invitrogen(登録商標)、Cat.no.15224−017)
反応条件:16〜20時間、16℃
【0130】
結合混合物を、Anagnostopoulos及びSpizizen(1961)の方法を用いてバチルス・ズブチリスBG2864(Naki et al.、1998)に形質転換し、クロラムフェニコール耐性及びプロテアーゼ活性に関して選択した。
【0131】
タンパク質生成の方法
炭素源(グルコース及びマルトデキシトリン、10.5及び17.5g/l)、窒素源(尿、素、3.6g/l)及びリン酸塩(0.5g/l)及び硫酸塩(0.5g/l)などの必須養分を含み、及びさらに微量元素(Fe、Mn、Zn、Cu、Co、1〜4mg/ml)を補足したMops培地(Frederick C.Neidhardt et al.,1974)中で1〜50μLのグリセロール培養物を植菌する。培地はMOPS/トリシン混合物を用いて中和し、7〜8のpHを生じる。培養物を1〜5日、37℃/220rpm(Infors HT(登録商標)Multitron II)で培養する。
【0132】
参考文献:
Protein engineering of the high-alkaline serine protease PB92 from Bacillus alcalophilus: functional and structural consequences of mutation at the S4 substrate binfing pocket. (バチルスalcaloph ilus由来の高アルカリ性セリンプロテアーゼPB92のタンパク質工学:S4基質結合ポケットでの変異の機能的及び構造的結果。)
Teplyakov AV、van der Laan JM、Lammers AA、Kelders H、Kalk KH、Misset O、Mulleners LJ、Dijkstra BW
Protein Eng.1992 Jul;5(5): 413-20.
Selection of a subtilisin-hyperproducing Bachillus in a highly structured environment (高度構造環境におけるズブチリシン−過剰生成バチルスの選択)
D.Naki、C.Paech、G.Ganshaw、V.Schellenberger.Appl Microbiol Biotechnol (1998) 49: 290-294.
Requirements for transformation in Bacillus subtilis (バチルス・ズブチリスにおける形質転換の必要条件)
Anagnostopoulos、C. and Spizizen、J. in J. Bacteriol. 81、741-746 (1961).
Culture Medium for Enterobacteria (腸内細菌の培養基)
Frederick C. Neidhardt、Philip L. Bloch and David F. Smith in Journal of Bacteriology、Sept 1974. p736-747 Vol. 119. No. 3.
【0133】
【表2】
【0134】
実施例6
実施例1で生成した多数のプロテアーゼ変異体を、米国特許出願番号09/554,992(WO99/34011)、“An Improved method of assaying for a preferred enzyme and/or preferred detergent composition(好ましい酵素及び/または好ましい洗剤組成物の改善された分析方法)”に記載されているマイクロスウォッチ(microswatch)分析法を用いて2種類の洗剤及び洗浄条件における性能を試験した。
【0135】
表4は分析した変異体プロテアーゼ及び2つの異なる洗剤で試験した結果を記載する。A欄及びC欄は、分析材料はMTP中の形質転換株の成長により生成した。B欄及びD欄は、分析材料は振とうフラスコ(250ml)中の形質転換株の成長により生成した。A欄及びB欄は、洗剤が7.6g/lろ過アリエール・レギュラー(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)であり、溶液中15グレイン/ガロン合計Ca2+/Mg2+硬度を含み、及び40℃で各ウェル中0.5ppmの酵素を使用した(ヨーロッパ条件)。C欄及びD欄は、洗剤が0.67g/lろ過Tide Opal(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)であり、溶液中3グレイン/ガロン合計Ca2+/Mg2+硬度を含み、及び20℃で各ウェル中0.5ppmの酵素を使用した(日本条件)。性能指数を次の式で計算した:
変異体の洗浄性能 ÷ GG36の洗浄性能(野生型)
4つの性能値を平均し、表2で示す値に達した。
【0136】
【表3】
【0137】
【表4】
【0138】
上述の分析の結果として、いくつかの変異体はGG36野生型プロテアーゼの性能指数よりも大きな性能指数を示した。例えば、変異体R45N−G118E−E271R、R45N−P14R、R45N−N204R、D181N−G118D、及びR45N−G258Rは、ヨーロッパ条件下(15グレイン/ガロン合計Ca2+/Mg2+硬度、40℃、0.5ppm)、マイクロスウォッチ分析において(WO99/34011)、それぞれ1.75、1.28、1.28、1.24、及び1.23の性能指数を示した(表4)。変異体R170S−A1P、R170S−G61R、R170S−N204R、K251G−S87K、及びR170S−S216Rは、それぞれ2.09、2.03、1.79、1.54、1.47、1.39及び1.21の性能指数を示した(表5)。変異体E271T−G100E、E271T−G102E、E271T−S128E、K27T−G100E、R170S−G100R、及びE271T−S49Eは、日本条件下(3グレイン/ガロン合計Ca2+/Mg2+硬度、20℃、0.5ppm)、マイクロスウォッチ96マイクロタイター・ウェルプレート(WO99/34011)分析において、それぞれ3.22、1.85、2.33、2.04、1.79及び1.43の性能指数を示した(表5)。変異体K251G−S87K、R170S−A1R、及びE271T−S128Eは、それぞれ、2.06、1.55、及び1.20の性能指数を示した(表5)。変異体K251G−S87K、及びR170S−A1Rは、日本及びヨーロッパ条件の両方の下、1.00以上の性能指数を示した。
【0139】
本発明は種々の具体的な実施態様との関連において議論及び例示したが、適用性及び開示の範囲を限定するものとして解釈されるものではなく、前述に説明した特徴及び請求の範囲の全ての組み合わせ及びサブコンビネーションにまで及ぶものである。
【技術分野】
【0001】
発明の背景
セリンプロテアーゼはカルボニル・ヒドロラーゼの下位集団である。これらは広範囲の特異性及び生物学的機能を有する酵素の種々の分類を含む。Stroud,R.Sci.Amer.,131:74−88。それらの機能的多様性にもかかわらず、セリンプロテアーゼの触媒機構は、1)ズブチリシン及び2)哺乳類キモトリプシン関連及び同種細菌性セリンプロテアーゼ(例えば、トリプシン及びS.gresiusトリプシン)の少なくとも2つの遺伝学的にはっきり区別できる酵素のファミリーに近かった。セリンプロテアーゼのこれら2つのファミリーは非常に類似した触媒作用機構を示す。Kraut,J.(1977)、Annu.Rev.Biochem.,46:331−358。さらに、一次構造は関連しないが、これら2つの酵素ファミリーの三次構造はセリン、ヒスチジン及びアスパラギン酸塩からなるアミノ酸の保存された触媒3残基を結集させる。
【0002】
ズブチリシンは、種々のバチルス種及びその他の微生物から大量に分泌されるセリンプロテアーゼ(approx.MW(分子量)27,500)である。ズブチリシンのタンパク質配列はバチルスの少なくとも9つの異なる種から決定された。Markland,F.S.,et al.(1983)、Hoppe−Seyler’s Z.Physiol.Chem.、364:1537−1540。バチルス・アミロリケファシエンス、バチルス・リケニホルミス、及びバチルス・レンタスのいくつかの天然変異体から得られるズブチリシンの三次元結晶構造が報告されている。これらの研究はズブチリシンが哺乳類セリンプロテアーゼに通常関連しないが、類似の活性部位構造を有することを示す。共有結合ペプチド阻害因子を含んだズブチリシンのX線結晶構造(Robertus,J.D.,et al.(1972)、Biochemistry、11:2439−2449)または生成複合物(Robertus,J.D.,et al.(1976)、J.Biol.Chem.,251:1097−1103)も活性部位及びズブチリシンの推定上の基質が結合する分子の溝(substrate binding cleft)についての情報を提供した。さらに、多くの動的及び化学的修正のズブチリシンについての研究が報告されており、(Svendsen,B.(1976)、Carlsberg Res.Commun.,41:237−291;Markland,F.S.Id.)及びズブチリシンの残基222のメチオニンの側鎖が過酸化水素によりメチオニン−スルホキシドに変換される少なくとも1の報告(Stauffer,D.C.,et al.(1965)、J.Biol.Chem.,244:5333−5338)及び広範囲の部位特異性突然変異が実行された(Wells and Estell(1988)TIBS 13:291−297)。
【発明の概要】
【0003】
発明の概要
本発明の1の側面は、分子の電荷分布をその配向及び相、表面、その他の分子及び領域との相互作用に影響を与えるために変化させることである。
【0004】
前駆体または親酵素の酵素変異体は、荷電アミノ酸残基位置での1以上の変異を含む変異体、前駆体酵素と同じ正味静電荷及び/または前駆体酵素と同じ等電点を有することを特徴とする変異体を目的とする。
【0005】
本発明の他の側面において、前駆体プロテアーゼのプロテアーゼ変異体をここで目的とし、荷電アミノ酸残基位置で1以上の変異を含む変異体、前駆体プロテアーゼと同じ正味静電荷または等電点を有することを特徴とする変異体を目的とする。荷電アミノ酸はアスパラギン酸、グルタミン酸、ヒスチジン、リジン、チロシン及びアルギニンである。残基位置はここに同定するバチルス・アミロリケファシエンスズブチリシンの位置5、7、23、26、28−31、34、47、63、65、66、69、70、73、82−85、88、90、92、93、105、113、125、138、139、148−151、176、178、179、193、196、200、201、202、207、219、220、223、229、233、250、266、267、及び273に等しい残基位置である。また、残基位置はバチルス・アミロリケファシエンスズブチリシンの位置23、39、41、45、67、94、136、170、181、247、251及び/または271に等しい残基位置である。さらに別の側面は、当該プロテアーゼ変異体をエンコードするDNA配列及び当該変異体DNA配列を含んだ発現ベクターを提供することである。
【0006】
前駆体プロテアーゼのプロテアーゼ変異体は、荷電アミノ酸残基位置で1以上の変異を含む前記変異体、前記前駆体プロテアーゼと同じ正味静電荷を有することを特徴とする前記変異体である。請求項1のプロテアーゼ変異体は、前記荷電アミノ酸がアスパラギン酸、グルタミン酸、リジン及びアルギニンからなる群より選択される。プロテアーゼ変異体は配列番号2に示すバチルス・アミロリケファシエンスズブチリシンの27、45、170、181、251及び271からなる群より選択される残基位置に等しい1以上の残基位置での置換を有するアミノ酸配列を含む。プロテアーゼ変異体は27、45、170、181、251及び271に対応する1以上の位置での置換を含み、K27T、R45N、R170S、D181N、K251G及びE271Tから選択される置換である。
【0007】
プロテアーゼ変異体は配列番号2に示すバチルス・アミロリケファシエンス・ズブチリシンの1、14、49、61、87、100、102、118、128、204及び258に対応する1以上の位置での別の置換をさらに含むことができる。変異体はR45N−G118E−E271R、R45N−P14R、R45N−N204R、D181N−G118D、R45N−G258R、R170S−A1R、R170S−G61R、R170S−N204R、K251G−S87K、R170S−S216R、E271T−G100E、E271T−G102E、E271T−S128E、K27T−G100E、R170S−G100R、E271T−S49E、及びE271T−S128Eの組み合わせから選択できる。
【0008】
さらに、本発明のその他の側面は、このようなベクターを用いて形質転換した宿主細胞を提供することである。
【0009】
さらに、本発明のプロテアーゼ変異体を含んだ洗浄組成物を提供する。
【図面の簡単な説明】
【0010】
【図1A】バチルス・アミロリケファシエンス・ズブチリシンのDNA配列(配列番号1)及びアミノ酸配列(配列番号2)並びにこの遺伝子の制限地図の一部を示す。
【図1B−1】バチルス・アミロリケファシエンス・ズブチリシンのDNA配列(配列番号1)及びアミノ酸配列(配列番号2)並びにこの遺伝子の制限地図の一部を示す。
【図1B−2】バチルス・アミロリケファシエンス・ズブチリシンのDNA配列(配列番号1)及びアミノ酸配列(配列番号2)並びにこの遺伝子の制限地図の一部を示す。
【図1B−3】バチルス・アミロリケファシエンス・ズブチリシンのDNA配列(配列番号1)及びアミノ酸配列(配列番号2)並びにこの遺伝子の制限地図の一部を示す。
【図2】バチルス・アミロリケファシエンス(BPN)’及びバチルス・レンタス(野生型)から得たズブチリシン間の保存アミノ酸残基を示す。
【図3A】図3Aと3Bは、4つのズブチリシンのアミノ酸配列を表す。一番上の行はバチルス・アミロリケファシエンス・ズブチリシン(ズブチリシンBPN’と呼ぶ時もある)から得たズブチリシンのアミノ酸配列を表す(配列番号3)。2行目はバチルス・ズブチリスから得たズブチリシンのアミノ酸配列を表す(配列番号4)。3行目はバチルス・リケニホルミスから得たズブチリシンのアミノ酸配列を表す(配列番号5)。4行目はバチルス・レンタス(PCT WO89/06276でズブチリシン309とも呼ばれている)から得たズブチリシンのアミノ酸配列を表す(配列番号6)。記号*はズブチリシンBPN’と比較して特定のアミノ酸残基がないことを示す。
【図3B】図3Aと3Bは、4つのズブチリシンのアミノ酸配列を表す。一番上の行はバチルス・アミロリケファシエンス・ズブチリシン(ズブチリシンBPN’と呼ぶ時もある)から得たズブチリシンのアミノ酸配列を表す(配列番号3)。2行目はバチルス・ズブチリスから得たズブチリシンのアミノ酸配列を表す(配列番号4)。3行目はバチルス・リケニホルミスから得たズブチリシンのアミノ酸配列を表す(配列番号5)。4行目はバチルス・レンタス(PCT WO89/06276でズブチリシン309とも呼ばれている)から得たズブチリシンのアミノ酸配列を表す(配列番号6)。記号*はズブチリシンBPN’と比較して特定のアミノ酸残基がないことを示す。
【図4】pVS08バチルス・ズブチリス発現ベクターを示す。
【図5】フォワードApaIプライマー、リバースApaIプライマー、リバース変異プライマー及びフォワード変異プライマーの方向を示す。
【発明を実施するための形態】
【0011】
発明の詳細な説明
プロテアーゼは、一般的にタンパク質またはペプチドのペプチド結合を切断する働きを有するカルボニルヒドロラーゼである。ここで用いる“プロテアーゼ”は天然プロテアーゼまたは組換え型プロテアーゼを意味する。天然プロテアーゼはα−アミノアシルペプチド・ヒロドロラーゼ、ペプチジルアミノ酸ヒドロラーゼ、アシルアミノヒドロラーゼ、セリンカルボキシペプチダーゼ、メタロカルボキシペプチダーゼ、チオールプロテイナーゼ、カルボキシル−プロテイナーゼ及びメタロプロテイナーゼを含む。セリン、メタロ、チオール及び酸プロテアーゼが挙げられ、及びエンド及びエキソ−プロテアーゼが挙げられる。
【0012】
本発明は、変異体のアミノ酸配列が得られる前駆体カルボニルヒドロラーゼと比較して異なるタンパク質分解活性、安定性、基質特異性、pHプロファイル及び/または性能特徴を有する非天然型カルボニルヒドロラーゼ変異体(プロテアーゼ変異体)である、プロテアーゼ酵素を含む。特に、当該プロテアーゼ変異体は天然には見られないアミノ酸配列を有し、異なるアミノ酸を含む前駆体プロテアーゼの多数のアミノ酸残基の置換により得られる。前駆体プロテアーゼは天然プロテアーゼまたは組換え型プロテアーゼである。
【0013】
本発明で有用なプロテアーゼ変異体は、指定のアミノ酸残基位置で19の天然L−アミノ酸のいずれかの置換を含む。当該置換は任意の前駆体ズブチリシンにおいて作られる(原核生物、真核生物、哺乳類、等)。この出願の全体にわたって、種々のアミノ酸を一般的な1及び3文字記号を用いて言及する。当該記号はDale,M.W.(1989)、Molecular Genetics of Bacteria(細菌の分子遺伝学),John Wiley&Sons,Ltd.,Appendix Bに特定されている。
【0014】
本発明で有用なプロテアーゼ変異体は好ましくはバチルス・ズブチリシンから得られる。より好ましくは、プロテアーゼ変異体はバチルス・レンタスズブチリシン及び/またはズブチリシン309から得られる。
【0015】
ズブチリシンは、一般的にタンパク質またはペプチドのペプチド結合を切断する働きを有する細菌性または真菌プロテアーゼである。ここで用いる“ズブチリシン”は、天然または組み換え型ズブチリシンを意味する。一連の天然ズブチリシンは種々の微生物種により生成され、よく分泌されることが知られる。この一連のメンバーのアミノ酸配列は完全に相同ではない。しかしながら、この一連のズブチリシンは同じまたは類似のタイプのタンパク質分解活性を示す。セリンプロテアーゼのこの分類は、触媒3残基を特徴付ける共通のアミノ酸配列を共有し、セリンプロテアーゼのキモトリプシン関連分類と区別している。ズブチリシン及びキモトリプシン関連セリンプロテアーゼの両方は、アスパラギン酸塩、ヒスチジン及びセリンを含む触媒3残基を有する。ズブチリシン関連プロテアーゼにおいて、これらのアミノ酸の相対順序は、アミノからカルボキシ末端へ読取って、アスパラギン酸塩−ヒスチジン−セリンである。キモトリプシン関連プロテアーゼにおいて、相対順序は、しかしながら、ヒスチジン−アスパラギン酸塩−セリンである。従って、ここでズブチリシンは、ズブチリシン関連プロテアーゼの触媒3残基を有するセリンプロテアーゼをいう。例として、限定されないが、図3で特定されるズブチリシンを含む。一般的に、及び本発明の目的において、プロテアーゼ中のアミノ酸の番号は、図1に表される成熟バチルス・アミロリケファシエンス・ズブチリシン配列に割当てた数に対応する。
【0016】
“組換え型ズブチリシン”または“組換え型プロテアーゼ”は、ズブチリシンまたはプロテアーゼをエンコードするDNA配列が修飾され、天然アミノ酸配列における1以上のアミノ酸の置換、欠失または挿入をエンコードする変異(または突然変異)DNA配列を生成するズブチリシンまたはプロテアーゼをいう。当該変異を生成する適当な方法は、米国特許RE34,606号、米国特許第5,204,015号及び米国特許第5,185,258号、米国特許第5,700,676号、米国特許第5,801,038号及び米国特許第5,763,257号に開示されるものを含み、ここに組込まれる。
【0017】
“非ヒトズブチリシン”及びそれらをエンコードするDNAは多くの原核及び真核生物から得られる。原核生物の適当な例は、大腸菌またはシュードモナスなどのグラム陰性生物及びミクロコッカスまたはバチルスなどのグラム陽性細菌を含む。ズブチリシン及びそれらの遺伝子が得られる真核生物の例としては、サッカロマイセス・セレヴィシエなどの酵母、アスペルギルス種などの真菌を含む。
【0018】
“酵素変異体”は“前駆体酵素”のアミノ酸配列由来のアミノ酸配列を有する。前駆体酵素プロテアーゼは天然酵素及び組換え酵素を含む。本発明者が意図する酵素は、限定されないが、オキシドレダクターゼ、トランスフェラーゼ、ヒドロラーゼ、リアーゼ、イソメラーゼ、及びリガーゼを含む。本発明者が意図する具体的な酵素は、限定されないが、アミラーゼ、ラッカーゼ、プロテアーゼ、デヒドロゲナーゼ及びパーミアーゼを含む。酵素変異体のアミノ酸配列は、前駆体アミノ酸配列の1以上のアミノ酸の置換、欠失または挿入により前駆体酵素アミノ酸配列から“得られる”。当該変異は前駆体酵素それ自体の操作というよりもむしろ、前駆体酵素のアミノ酸配列をエンコードする“前駆体酵素DNA配列”の変異である。前駆体DNA配列をそのように操作する適当な方法はここに開示する方法及び当業者に公知の方法を含む。プロテアーゼに関するいかなる引用または議論はその他の酵素、例えば、上で特定した一部の酵素に適用可能であることを当然である。
【0019】
“プロテアーゼ変異体”は、“前駆体プロテアーゼ”のアミノ酸配列から得られるアミノ酸配列を有する。前駆体プロテアーゼは天然プロテアーゼ及び組換え型プロテアーゼを含む。プロテアーゼ変異体のアミノ酸配列は、前駆体アミノ酸配列の1以上のアミノ酸の置換、欠失または挿入により前駆体プロテアーゼアミノ酸配列から“得られる”。当該変異は前駆体プロテアーゼ酵素それ自体の操作というよりもむしろ、前駆体プロテアーゼのアミノ酸配列をエンコードする“前駆体DNA配列”の変異である。前駆体DNA配列の当該操作の適当な方法は、ここに開示する方法を含み、及び当業者に公知の方法を含む(例えば、EP0 328299、WO89/06279及びここに既に引用した米国特許及び出願を参照)。
【0020】
“荷電アミノ酸”は潜在的にイオン性であり、電荷が変化し、特定のpHまたはpH範囲で帯電するアミノ酸として定義する。これらのアミノ酸は、例えば、酸性アミノ酸、塩基性アミノ酸及びいくつかの極性アミノ酸を含む。酸性アミノ酸はpH6.0で負に帯電したアミノ酸であり、例えばアスパラギン酸(AspまたはD)及び/またはグルタミン酸(GluまたはE)である。塩基性アミノ酸はpH6.0で正に帯電したアミノ酸であり、例えばリジン(LysまたはK)、アルギニン(ArgまたはR)、及び/またはヒスチジン(HisまたはH)である。
【0021】
“非荷電アミノ酸”は潜在的にイオン性でないアミノ酸として定義する。これらのアミノ酸は、限定されないが、非荷電非極性アミノ酸、及び/または非荷電極性アミノ酸を含む。非荷電非極性アミノ酸は、限定されないが、アラニン(AlaまたはA)、バリン(ValまたはV)、ロイシン(LeuまたはL)、イソロイシン(IleまたはI)、プロリン(ProまたはP)、フェニルアラニン(PheまたはF)、トリプトファン(TrpまたはW)、及び/またはメチオニン(MetまたはM)を含む。非荷電極性アミノ酸は、限定されないが、グリシン(GlyまたはG)、セリン(SerまたはS)、トレオニン(ThrまたはT)、システイン(CysまたはC)、チロシン(TyrまたはY)、アスパラギン(AsnまたはN)及び/またはグルタミン(GlnまたはQ)を含む。
【0022】
“正味静電荷(Net electrostatic charge)”は所定のpHまたはpH範囲での変異体または前駆体酵素またはプロテアーゼの静電荷の合計として定義される。典型的なpHはpH6.0である。
【0023】
“等電点”(pl0)は、タンパク質またはタンパク質複合体、例えばプロテアーゼまたはプロテアーゼ複合体(任意で金属またはその他のイオンを付随するもの)が中性であるpH値として定義するものであり、すなわち、複合体の静電荷の合計(正味静電荷=NEC)はゼロである。この合計において、個々の静電荷の正または負の性質の対価は考慮しなければならない。
【0024】
“同じ等電点(pl0)”はpH単位の明確な範囲内であるpl0として定義される。例えば、pH単位の明確な範囲はその他のpl0と比較するpl0の1pH単位以下であり、0.25〜0.75の間、例えば、0.5pH単位であり、好ましくは0.01〜0.5pH単位内であり、例えば0.1pH単位、及びより好ましくは0.001〜0.05pH単位内、例えば、0.01pH単位である。同じ等電点は所定のpHまたは所定のpH範囲で決定できる。
【0025】
“同一の静電荷”は、“Z”または前駆体プロテアーゼ中に存在する電荷と同じ数のプロテアーゼ変異体中の特定荷電残基を維持するものとして定義される。荷電残基の数は同じであるが、特定アミノ酸残基は、正味の静電荷が所定のpHで前駆体プロテアーゼと同じである限りは、プロテアーゼ変異体の外側周辺のその他の位置に置換できる。
【0026】
“同じ正味静電荷”は、“Z”またはプロテアーゼ変異体の静電荷の合計の明確な範囲内のプロテアーゼ前駆体の静電荷の合計を維持するものとして定義される。同じ合計静電荷の維持は明確なpH範囲にわたって明確な範囲の電荷単位内で正味静電荷を維持することを意味する。プロテアーゼ前駆体と同じ正味静電荷を有するプロテアーゼ変異体は前駆体プロテアーゼの明確な数の電荷単位内の正味静電荷を有する。例えば、同じ正味静電荷を有するプロテアーゼ変異体は1pH単位以下であり、前駆体プロテアーゼ正味静電荷の0.25〜0.75電荷単位内であり、例えば、0.5単位である。さらに、より好ましくは、同じ正味静電荷を有するプロテアーゼ変異体は前駆体プロテアーゼ正味静電荷の0.05〜0.25単位内であり、例えば、0.1単位である。さらに、より好ましくは、同じ正味静電荷を有するプロテアーゼ変異体は前駆体プロテアーゼ正味静電荷の0.001〜0.05単位内であり、例えば、0.01単位である。電荷単位は提供する(酸)または受け取る(塩基)プロトンの数として定義できる。個々のアミノ酸の静電荷は所定のpH、例えば、6.0または7.0で受け取るまたは提供するプロトンの数を決定することにより通常確定できる。Z値もこの出願内で後述する式により決定できる。
【0027】
変異体または前駆体プロテアーゼの“合計電荷量”はそれぞれのプロテアーゼの静電荷の合計数として定義される。同じ正味静電荷を有するプロテアーゼ変異体は前駆体プロテアーゼと異なる合計電荷量を有することができる。
【0028】
当業者により認識されるように、等電点は、問題の酵素中の種々の荷電残基のpK値を用いる平衡対価を用いることにより適宜計算でき、それから、ここに明示的に引用する欧州特許第0945502号及びその実施例に記載されるように、酵素分子のNECがゼロに等しいpH値の繰り返しにより見つけられる。
【0029】
この計算の1つの問題点は荷電残基のpK値がその環境に依存し、その結果変動するということである。しかしながら、実際の値と関係ない特定の近似pK値を荷電残基に割り当てることにより非常に良い結果が得られるようになる。また、部分的に環境を考慮に入れることでより高度な計算を行うことが可能になる。
【0030】
また、pl0は等電点電気泳動法または酵素を含んだ溶液を滴定することにより実験的に決定できる。さらに、荷電残基の種々のpK値は滴定により実験的に決定できる。
【0031】
本発明の別の側面において、pl0についての上述の観測はさらに、前駆体プロテアーゼ中のアミノ酸の欠失、置換または挿入される位置及びアミノ酸を決定または選択する方法に利用し、変異体プロテアーゼの正味静電荷または等電点が、同じpH値または明確なpH範囲で計算される前駆体プロテアーゼのNECまたはpl0と同じになるようにする
。
【0032】
本発明でカバーされるこの原則を表す他の方法は、前記前駆体プロテアーゼまたは酵素中のアミノ酸の欠失、置換または挿入される位置及びアミノ酸が1の方法で選択されるものであり、それにより電荷または合計電荷量(=TCC)の合計数及び/または最終的な変異体プロテアーゼまたは酵素のNECがプロテアーゼまたは酵素の最適洗浄能力に関する明確なpHまたはpH範囲で変化しない等電点を有する変異体プロテアーゼまたは酵素を提供するように一定を維持し、最適pHは洗浄溶液のpHにできるだけ接近させるべきであり、前記変異体プロテアーゼは使用するためのものである。
【0033】
上に示すように、酵素などの巨大分子のpl0は該分子のNECがゼロであるpHとして計算する。該手順は欧州特許0 945 502に記載の実施例で説明されているが、該原則をここにより詳細に記載する。
【0034】
pK値は各潜在性の荷電アミノ酸残基に割り当てる。それから、荷電または非荷電形(荷電/非荷電、C/U(i))の所定のpHのアミノ酸残基の存在比を式Ia及びIbを用いて負及び正電荷の両方について計算する。
【0035】
C/U(i)=10(ph−pKi)(負電荷)(Ia)
C/U(i)=10(pKi−ph)(正電荷)(Ib)
上述の式に従うと、pHがpKiと等しい場合、C/U(i)は1である。
【0036】
相対電荷、Qr(i)または各荷電残基に割り当てた電荷寄与(charge contribution)は、式IIa及びIIbを用いて計算する。
Qr(i)=C/U(i)/(1+C/U(i))(負電荷)(IIa)
Qr(i)=−C/U(i)/(1+C/U(i))(正電荷)(IIb)
【0037】
荷電残基から得られる全ての電荷寄与の合計がゼロである場合のpH値は、繰り返しにより、または十分に濃いpH−電荷合計表(sum table)により見出すことができる。
【0038】
正味静電荷Zを決定する他の方法は、基(Rまたはアミノ基など)が陽イオン性の酸の形態を有する場合、αが部分正電荷を表すということは当業者が理解することである:
【数1】
【0039】
一方、カルボキシルなどの基は、中性の酸の形態及び陰イオン性の共役塩基を含み、αは非荷電部分を表す。そして、部分電荷は:
【数2】
【0040】
種々の大半のプロテアーゼまたは酵素の性質はNEC及び/またはプロテアーゼまたは酵素分子の等電点に依存することが留意される。例えば、プロテアーゼまたは酵素溶解性、安定性、複数の相媒体中の位相分布及び/または表面電荷は分子のNEC及び/または等電点の変化により影響を受ける性質である。驚くべきことに、改善されたプロテアーゼの特徴は同じ等電点または同じ正味静電荷を維持しながら達成できた。特定の理論に縛られることは望まないが、問題のタンパク質、酵素またはプロテアーゼの大部分の性質を維持するが、分子の相互作用の運動、例えば基質、表面または培地に関する変化の分布または分子の配向を修飾することが望ましい状況があると考えられる。
【0041】
本発明の1の側面において、プロテアーゼ変異体及び前駆体プロテアーゼは同一の正味静電荷または同一の等電点を有する。同一の正味静電荷は同一静電荷を有することにより、または前駆体プロテアーゼのアミノ酸配列への追加の修飾による追加の変異位置から生じる電荷変化を相殺することにより維持できる。これらの追加の変異は、限定されないが、追加残基に対して反対の電荷を有する残基を置換または挿入することを含む(追加の塩基性残基を相殺するために追加の酸性残基を加える)。従って、プロテアーゼ変異体中の荷電残基の数は、例えば、荷電残基の数がプロテアーゼ前駆体におけるものよりも多い場合、プロテアーゼ前駆体のものと異なると考えられる。追加の荷電残基を相殺するために、相当する反対の荷電アミノ酸置換を同じ正味静電荷を維持するために作ることができる。さらに、プロテアーゼまたは酵素変異体中の荷電残基の数は、それが他の非荷電残基に相当する反対の荷電アミノ酸欠失または置換であって、荷電残基が非荷電残基を欠失または置換する場合、プロテアーゼまたは酵素前駆体中の荷電残基の数より少ない。
【0042】
本発明の1の側面は、NECまたはpl0が同一の場合、例えばプロテアーゼ変異体及び前駆体プロテアーゼの合計電荷量が同じ場合、または前駆体プロテアーゼ中の荷電残基が同じ数で維持される場合である。荷電アミノ酸が同一のNECまたはpl0を維持するために再配置される場合、少なくとも1のこれらの荷電アミノ酸が前駆体プロテアーゼの残基位置から異なる残基位置に置換される。プロテアーゼ前駆体中の特定の荷電アミノ酸、バチルス・レンタス(GG36)中の“X”リジン、が特定数ある場合、それから変異体プロテアーゼは同じ数だが前駆体プロテアーゼと比較して異なる位置のリジン残基、すなわち、“X”を保有する。従って、例えば、K27は異なる残基で置換でき、相当するKは他の位置、好ましくは表面位置で置換される。1の実施態様において、荷電残基、例えば、グルタミン酸、アスパラギン酸、リジンまたはアルギニンは異なる位置に置換できる。同一の静電荷を維持するために、荷電残基により置換される特定前駆体残基、例えば位置27からのK残基、は位置27で置換できる。例えば、R45N−N204Rは同一の静電荷を維持するために置換された同一の残基位置を有する。さらに、荷電残基により置換された特定の前駆体残基が非荷電残基である場合、その他の非荷電残基は荷電残基をもともと有する位置に置換できる。例えば、R170S−A1Rの組み合わせはアラニンをアルギニンで置換し、アルギンをセリンで置換する。もちろん、複数の変異を作る場合、任意の置換残基がその他の任意の残基中に置換でき、各々のアミノ酸が同じ数で維持される限り変異される。静電荷は、プロテアーゼ変異体及び前駆体プロテアーゼが同じpHで測定される限り、任意の所定pHで測定できる。
【0043】
本発明の他の側面において、分子の同じ正味静電荷は、前駆体プロテアーゼに対する変異から生じる正味静電荷の変化を相殺することにより維持する。例えば、当該変異を達成する1の方法は、別の反対の荷電アミノ酸残基の挿入または置換や、異なるが類似に帯電したアミノ酸残基の欠失である。例えば、プロテアーゼ変異体中に前駆体プロテアーゼ中よりも多くの酸性アミノ酸が存在する場合、変異体は追加の塩基性アミノ酸を含む。例えば、プロテアーゼ変異体中に前駆体プロテアーゼよりも多くの特定の酸性アミノ酸、例えば、グルタミン酸が存在する場合、当該変異を相殺するために、対応する数のアスパラギン酸残基を欠失または非荷電アミノ酸で置換できる。例えば、前駆体プロテアーゼ中に特定の数の荷電アミノ酸が存在する場合、本発明者は、変異体プロテアーゼのアミノ酸の数の増加または減少とともに荷電アミノ酸の数の変化を相殺するアミノ酸の対応する増加または減少を意図する。 従って、上述の通り、追加の正電荷残基は、対応する数の負電荷アミノ酸を追加することにより、または対応する数のその他の正電荷アミノ酸を非電荷残基またはそれらの組み合わせで置換することにより相殺できる。より少ない数の正電荷残基は対応する数の負電荷アミノ酸を欠失させることにより、対応する数の負電荷アミノ酸またはそれらの組み合わせを対応する数の非電荷残基で置換することにより相殺できる。
【0044】
1の実施態様において、第1のアミノ酸位置で非荷電残基により置換される同じ荷電のアミノ酸残基は、荷電残基を置換する同じ非荷電アミノ酸が存在する、第2のアミノ酸位置に置換される。非荷電残基は荷電アミノ酸の元の位置で置換でき、一方、置換荷電アミノ酸は非荷電アミノ酸の位置を置換できる。例えば、R45N−N204Rは荷電アミノ酸、アルギニンに関する非荷電アミノ酸、アルパラギンの置換を反映する。荷電アミノ酸を置換した同じ非荷電アミノ酸は荷電アミノ酸が再挿入される位置に存在する必要はない。例えば、アラニン(AlaまたはA)、グリシン(GlyまたはG)、アスパラギン(AsnまたはN)、プロリン(ProまたはP)、セリン(SerまたはS)、及び/またはトレオニン(ThrまたはT)の群より選択される非荷電アミノ酸は荷電アミノ酸位置に置換でき、一方荷電アミノ酸残基は上述の群のその他でもとから占められているアミノ酸位置に置換される。例えば、変異体E271T−G100Eにおいて、271位置のグルタミン酸アミノ酸はトレオニンアミノ酸で置換され、一方、100位置のグリシン残基はグルタミン酸アミノ酸で置換される。同様に、同一の荷電アミノ酸は最初から非荷電の位置、例えば、K27T−G100Eに置換される必要はない。荷電アミノ酸であるアスパラギン酸(D)、グルタミン酸(E)、リジン(K)、及びアルギニン(R)はこの点において有用である。
【0045】
本発明の他の側面において、プロテアーゼ変異体NECは、0.5荷電単位(Z)の範囲以下、前駆体プロテアーゼNECの範囲から0〜14のpHの範囲以上と様々である。
【0046】
さらに本発明の他の側面において、プロテアーゼ変異体NECは、1電荷単位の範囲以下、前駆体プロテアーゼNECの範囲から明確なpH範囲以上で変化する。明確なpH範囲は例えば、所望のプロテアーゼまたは酵素環境に関して最適または所望のpHとして当業界で認識される2pH単位内、またはpH4単位の範囲内である。
【0047】
本発明の他の側面において、前駆体プロテアーゼ内で見られる電荷残基の変異は、同じまたは同一の正味静電荷を維持しながら、増加した有益な洗浄特性を示すプロテアーゼ変異体を生じることができることが確認された。
【0048】
本発明のさらに別の側面において、前駆体プロテアーゼ内で見られる電荷残基の変異は、明確なpHで、明確なpH範囲を超えて、例えば、4pH単位の範囲を超えて、または0.001〜14のpH範囲を超えて、同じまたは同一の正味静電荷を維持しながら、増加した有益な洗浄特性を示すプロテアーゼ変異体を生じることができることが確認された。
【0049】
本発明者が変異を目的とする典型的なアミノ酸残基は、例えば、リジン、アルギニン、及び/またはヒスチジンなどの塩基性アミノ酸;例えばアスパラギン酸及び/またはグルタミン酸などの酸性アミノ酸;及び/またはそうでなければ例えばチロシンなどの極性R基を含む。
【0050】
他の側面において、本発明の変異体プロテアーゼは前記前駆体プロテアーゼと比較して、同じ数の正電荷アミノ酸残基、前駆体プロテアーゼ中の同一のアミノ酸及び同じ電荷を有する異なるアミノ酸の両方、及び前駆体プロテアーゼ中の同じ数の負電荷アミノ酸残基;または、バチルス・アミロリケファシエンス(BPN’)の位置5、7、22、23、24、26、28−31、34、45、47、63、65、66、69、70、73、82−85、88、90、92、93、97、102、105、113、125、127、138、139、148−151、169、170、176、178、179、193、196、200、201、202、207、219、220、223、229、233、250、266、267及び273の1以上の任意の位置での対応アミノ酸残基間で変異を有するが、プロテアーゼ変異体の正味静電荷及び/または等電点が前駆体プロテアーゼと同じになるように、より多いまたは少ない正電荷アミノ酸残基及び対応するより多いまたは少ない負電荷アミノ酸残基を有する。1の実施態様において、27、45、136、170、181、247、251及び/または271の1以上の位置での対応アミノ酸残基間での変異は荷電残基位置の非荷電残基での置換を含む。これらの残基位置は、これらのバチルス・レンタス野生型における対応位置がこれらの位置の荷電アミノ酸残基を有するので、重要である。例えば、バチルス・レンタスズブチリシン(配列番号6)の27、38、40、44、65、92、134、164、175、241、245及び/または265の残基位置はそれぞれバチルス・アミロリケファシエンス(配列番号2)の27、39、41、45、67、94、136、170、181、247、251及び/または271に対応する。
【0051】
他の側面において、本発明の変異体プロテアーゼは前記前駆体プロテアーゼと比較して、同じ数の正電荷アミノ酸残基、前駆体プロテアーゼ中の同一のアミノ酸及び同じ電荷を有する異なるアミノ酸の両方、及び前駆体プロテアーゼ中の同じ数の負電荷アミノ酸残基;または、バチルス・アミロリケファシエンス(BPN’)の位置27、39、41、45、67、94、136、170、181、197、247、249、251及び/または271の1以上の任意の位置での対応アミノ酸残基間で変異を有するが、プロテアーゼ変異体の正味静電荷及び/または等電点が前駆体プロテアーゼと同じになるように、より多いまたは少ない正電荷アミノ酸残基及び対応するより多いまたは少ない負電荷アミノ酸残基を有する。本発明者が意図する具体的な置換は、バチルス・アミロリケファシエンスのK27A、K27C、K27E、K27Q、K27G、K27H、K27I、K27L、K27M、K27F、K27P、K27S、K27T、K27W、K27Y、H39A、H39R、H39D、H39N、H39C、H39E、H39Q、H39G、H39H、H39I、H39L、H39K、H39M、H39F、H39P、H39T、H39W、H39Y、H39V、D41A、D41R、D41C、D41E、D41Q、D41G
、D41H、D41I、D41L、D41K、D41M、D41F、D41P、D41S、D41T、D41W、D41Y、D41V、R45A、R45R、R45D、R45N、R45C、R45E、R45C、R45E、R45Q、R45G、R45H、R45I、R45L、R45K、R45M、R45F、R45P、R45S、R45T、R45W、R45Y、R45V、H67A、H67R、H67D、H67N、H67C、H67E、H67Q、H67G、H67H、H67I、H67L、H67K、H67M、H67F、H67P、H67S、H67T、H67W、H67Y、H67V、K94A、K94R、K94D、K94N、K94C、K94E、K94Q、K94G、K94H、K94I、K94L、K94K、K94M、K94F、K94P、K94S、K94T、K94W、K94Y、K94V、E136A、E136D、E136N、E136C、E136E、E136G、E136H、E136I、E136L、E136K、E136M、E136F、E136P、E136S、E136T、E136W、E136Y、E136V、R170A、R170R、R170D、R170N、R170C、R170E、R170Q、R170G、R170H、R170I、R170L、R170K、R170M、R170F、R170P、R170S、R170T、R170W、R170Y、R170V、D181A、D181R、D181D、D181N、D181C、D181E、D181Q、D181G、D181H、D181I、D181L、D181K、D181M、D181F、D181P、D181S、D181T、D181W、D181Y、D181V、D197A、D197R、D197D、D197N、D197C、D197E、D197Q、D197G、D197H、D197I、D197L、D197K、D197M、D197F、D197P、D197S、D197T、D197W、D197Y、D197V、R247A、R247R、R247D、R247N、R247C、R247E、R247Q、R247G、R247H、R247I、R247L、R247K、R247M、R247F、R247P、R247S、R247T、R247W、R247Y、R247V、H249A、H249R、H249D、H249N、H249C、H249E、H249Q、H249G、H249H、H249I、H249K、H249M、H2490F、H249P、H249S、H249T、H249W、H249V、K251A、K251D、K251C、K251Q、K251G、K251H、K251I、K251L、K251K、K251M、K251F、K251P、K251S、K251T、K251W、K251Y、K251V、E271A、E271R、E271D、E271N、E271C、E271E、E271H、E271I、E271L、E271K、E271M、E271F、E271P、E271S、E271T、E271W、E271Y、及び/またはE271Vを含む。正電荷残基のそれ自身の置換による数の増加は、特定の洗浄環境において特定の変異体の効果の増加を生じると同時に、対応する反対の電荷変化は異なる洗浄環境において効果の増加を生じることができることに気付いた。例えば、負電荷変異は低イオン強度洗浄環境で有利な特徴を与え、正電荷変異は高イオン強度洗浄環境で有利な特徴を与えることが予想される。同じ正味静電荷または等電点を維持しながら正の増加及び負の増加の両方を含む変異体は、前駆体プロテアーゼの性能と比較して、両方の環境において改善された特徴を示すプロテアーゼ分子を生じることが予想される。
【0052】
これらの置換は好ましくはバチルス・レンタス(組み換え体または天然型)ズブチリシン内で作られるが、該置換は任意のバチルス・プロテアーゼ内、例えばバチルス・アミロリケファシエンス及び/またはズブチリシン309内で作ることができる。
【0053】
本発明の1の側面は、1、2−4、6、9−12、19、14、15、17−20、25、27、36−38、40、44、49、51、52、54−61、68、71、75、76、87、89、91、97、100−102、104、108、111、112、115、117、118、120−123、128、129、131、133、134、136、137、140、143−146、159、164、165、167、170、171、173、175、180、182−187、191、192、194、195、204、206、209−212、216、218、222、224、226、234−245、252、255、257−263、265、268、269及び274からなる群より位置付けられるまたは選択される残基に対応する1以上の残基位置において、さらに少なくとも1の別の置換アミノ酸を含むプロテアーゼ変異体を含む。本発明者によって意図される具体的な置換は、バチルス・アミロリケファシエンスまたはバチルス・レンタスのI122A、Y195E、M222A、M222S、Y167A、R170S、A194P、D36、N76D、H120D、G195E及びK235Nに等しい残基を含み、該変異体はバチルス・ズブチリシンから得られる。
【0054】
特に重要なのは、これらの位置の変異体が荷電アミノ酸置換とともに増加した洗浄能力を示す点である。これらの位置を含み、及び最初から荷電アミノ酸を有する変異体の組み合わせは重要である。本発明者が意図する典型的な組み合わせはK27T−G100E、R45N−A1R、R45N−P14R、R45N−G61R、R45N−S128R、R45N−N204R、R45N−S216R、R45N−G258R、R170S−A1R、R170S−P14R、R170S−S49R、R170S−G61R、R170S−G100R、R170S−S128R、R170S−N204R、R170S−S216R、R170S−G258R、D181N−G118D、D181N−G258D、K251G−S87K、E271T−S49E、E271T−T66E、E271T−G100E、E271T−G102E、E271T−S128E、R45N−G118E−E271R、S49R−G102E−R170S−E271T及びP14R−R45N−R170S−G258Rを含む。これらの変異を有するプロテアーゼ変異体が作ることができることは当業者に当然理解され、米国特許第5,741,694号、第6,190,900号、及び第6,197,567号に記載され、明示的にここに引用する。さらに、これらの変異は仮出願番号第60/423,087(2002年11月1日に出願;Neelam AminとVolker Schellenberger)に記載される直接的なバチルス形質転換法を用いても行うことができる。1の実施態様において、変異は融合PCR技術を用いて行った(Teplyakov,AV,et al,Protein Eng.,1992年7月5日(5):413−20)。同時にこの日に提出された、仮出願番号_/_____(Chris Leeflang,et al.)。
【0055】
本発明のさらに別の側面において、バチルス・アミロリケファシエンスの21、22、24、32、33、36、50、64、67、77、87、94、95、96、97、104、107、110、124、123、126、127、128、129、135、152、155、157、156、166、169、170、171、172、189、197、204、213、214、215、217、222または274からなる群より位置付けられる1以上の同等残基においてさらに少なくとも1の別の置換アミノ酸を有するプロテアーゼ変異体を含む。本発明者により意図される具体的な残基は以下を含む:K27R、M50F、N76D、S101G、S103A、V104I、V104Y、I122A、N123S、M124L、G159D、Y217L、A232V、Q236H、Q245R、N248D、N252K、T274A、及びM222S。プロテアーゼ変異体、3つの位置の変異をエンコードする組換えDNA、及び/またはこれらの変異を作る方法は米国特許RE34,606号;第5,972,682号;第5,185,258号;第5,310,675号;第5,316,941号;第5,801,038号;第5,972,682号;第5,955,340号及び第5,700,676号に記載されており、明示的にここに引用する。
【0056】
これらのアミノ酸位置番号は図1で表された成熟バチルス・アミロリケファシエンスズブチリシンに割当てた番号をいう。本発明は、しかしながら、この特定のズブチリシンの変異に限定されず、バチルス・アミロリケファシエンスズブチリシンの特定の同定残基に“等しい”位置でのアミノ酸残基を含む前駆体プロテアーゼまで広がる。本発明の好ましい実施態様において、前駆体プロテアーゼはバチルス・レンタスズブチリシン(配列番号6)であり、置換は上述の位置に対応するバチルス・レンタスにおける同等のアミノ酸残基位置に作られる。
【0057】
前駆体プロテアーゼの残基(アミノ酸)位置は、それがバチルス・アミロリケファシエンスズブチリシンの特定残基またはその一部と相同(すなわち、一次または三次構造の位置が一致する)または類似している場合、バチルス・アミロリケファシエンスズブチリシンの残基に等しい(すなわち、結合、反応または化学的に相互作用するための同じまたは類似の機能的能力を有する)。
【0058】
一次構造の相同性を確立するために、前駆体プロテアーゼのアミノ酸配列はバチルス・アミロリケファシエンスズブチリシン一次配列、及び特に配列が公知なズブチリシンにおいて不変な公知の残基の組と直接比較する。例えば、ここで図2はバチルス・アミロリケファシエンスズブチリシン及びバチルス・レンタスズブチリシン間での保存残基を示す。保存残基を並べ、配列を維持するために必要な挿入及び欠失を考慮した後(すなわち、任意の欠失及び挿入により保存残基が除去されるのを防ぐ)、バチルス・アミロリケファシエンスズブチリシンの一次配列中の特定のアミノ酸に等しい残基を明確にする。保存残基の配列は、好ましくは当該配列の100%保存されるべきである。しかしながら、保存残基の98%以上、95%以上、90%以上、85%以上、80%以上、75%以上、50%以上、または少なくとも45%以上の配列も同等残基を明確にするために十分である。触媒3残基、Asp32/His64/Ser221の保存は維持されるべきである。Siezen et al.(1991)Protein Eng.4(7):719−737は多数のセリンプロテアーゼの配列を示す。Siezenらはサブチラーゼ(subtilases)またはズブチリシン様セリンプロテアーゼの分類に言及している。
【0059】
例えば、図3において、バチルス・アミロリケファシエンス、バチルス・ズブチリス、バチルス・リケニホルミス(カールスベルゲンシス)及びバチルス・レンタス由来のズブチリシンのアミノ酸配列はアミノ酸配列間に最大値の相同性を与えるように配列する。これらの配列の比較は各配列に含まれる多数の保存残基があることを示す。これらの保存残基(BPN’とバチルス・レンタス間)は図2に同定される。
【0060】
これらの保存残基は、従って、ここでの好ましいプロテアーゼ前駆体酵素である、バチルス・レンタス(PCT公報WO89/06279、1989年7月13日に公開)由来ズブチリシン、または好ましいバチルス・レンタスズブチリシンと相同性の高い、PB92(EP0 328299)として言及されるズブチリシンなどのその他のズブチリシンにおいてバチルス・アミロリケファシエンスズブチリシンの対応する同等なアミノ酸残基を特定するために使用できる。特定のこれらズブチリシンのアミノ酸配列は、保存残基の最大相同性を得るために、バチルス・アミロリケファシエンスズブチリシンの配列と一緒に図3A及び3Bに配列する。図に示すように、バチルス・アミロリケファシエンスズブチリシンと比較して、バチルス・レンタスの配列中に多数の欠失がある。従って、例えば、その他のズブチリシンにおけるバチルス・アミロリケファシエンスズブチリシン中のVal165の同等アミノ酸はバチルス・レンタス及びバチルス・リケニホルミスのイソロイシンである。
【0061】
また、“同等残基(equivalent residues)”は三次構造がX線結晶学により決定された前駆体プロテアーゼの三次構造のレベルでの相同性を決定することにより特定できる。同等残基は前駆体プロテアーゼの特定アミノ酸残基の2以上の主鎖原子の原子座標の残基として特定し、バチルス・アミロリケファシエンスズブチリシン(N上のN、CA上のCA、C上のC及びO上のO)は配列後0.13nm内であり、好ましくは0.1nmである。最適モデルを方向付け、バチルス・アミロリケファシエンスズブチリシンに対して問題になっているプロテアーゼの水素でないタンパク質原子の原子座標の最大重複を与えるように位置付けた後、配列が達成される。最適モデルは、最も高い可能な解答での実験的回折データの最も低いR因子を与える結晶学モデルである。
【0062】
【数3】
【0063】
バチルス・アミロリケファシエンスズブチリシンの特定残基に機能的に類似する同等残基は、タンパク質構造を変化、修飾または寄与するような構造、基質結合またはバチルス・アミロリケファシエンスズブチリシンの特定残基を特定または起因するような触媒作用に適合できる前駆体プロテアーゼのアミノ酸として特定される。さらに、これらは、所定の残基の主鎖原子は占める相同性位置に基づいて同等の基準を満たさないが、残基の少なくとも2の側鎖原子の原子座標はバチルス・アミロリケファシエンスズブチリシンの対応する側鎖原子の0.13nmにある範囲において類似位置を占める前駆体プロテアーゼ(三次構造をX線結晶学により得た)の残基である。バチルス・アミロリケファシエンスズブチリシンの3次元構造の座標はEPO公報第0 251446(対応特許、米国特許第5,182,204号、開示の内容をここに引用する)に示され、三次元構造のレベルで同等残基を決定するための上述の概要として用いることができる。
【0064】
置換に関して同定されたいくつかの残基は保存残基であり、一方その他のものは保存残基ではない。保存されていない残基の場合、1以上のアミノ酸の置換は天然に見られるアミノ酸配列と一致しないアミノ酸配列を有する変異体を生成する置換に限定される。保存残基の場合、このような置換は天然配列を生じないはずである。本発明のプロテアーゼ変異体はプロテアーゼ変異体の成熟型及び当該プロテアーゼ変異体のプロ及びプレプロ型を含む。プレプロ型は、プロテアーゼ変異体の発現、分泌及び変異を促進するので、好ましい構造である。
【0065】
“プロ配列”は、除去された時、“成熟”型プロテアーゼを生じる、成熟型プロテアーゼのN末端部分に結合するアミノ酸配列をいう。多くのタンパク質分解酵素は、翻訳プロ酵素生成物として天然に見られ、翻訳後処理がない場合、この方法で発現する。その他のプロテアーゼ・プロ配列が使用できるが、プロテアーゼ変異体を生成する好ましいプロ配列はバチルス・アミロリケファシエンスズブチリシンズブチリシンの推定上のプロ配列である。
【0066】
“シグナル配列”または“プレ配列”は、プロテアーゼのN末端部分または成熟またはプロ型プロテアーゼの分泌に関与するプロプロテアーゼのN末端部分に結合するアミノ酸の任意の配列をいう。シグナル配列のこの定義は機能上のものであり、天然の条件下でプロテアーゼの分泌の実行に関与するプロテアーゼ遺伝子のN末端部分によりエンコードされるすべてのそれらのアミノ酸配列を含むことを意味する。本発明はここに定義するプロテアーゼ変異体の分泌を達成するために当該配列を利用する。1の可能なシグナル配列はバチルス・レンタス(ATCC21536)由来のズブチリシンのシグナル配列の残余と融合したバチルス・ズブチリスズブチリシン由来のシグナル配列の最初の7つのアミノ酸残基を含む。
【0067】
“プレプロ”型プロテアーゼ変異体はプロテアーゼのアミノ末端に作動可能に連結するプロ配列、及びプロ配列のアミノ末端に作動可能に連結する“プレ”または“シグナル”配列を有するプロテアーゼの成熟型からなる。
【0068】
“発現ベクター”は、適当な宿主中に前記DNAの発現をもたらすことができる適当な制御配列に作動可能に連結するDNA配列を含んだDNA構築体をいう。当該制御配列は転写を達成するためのプロモーター、当該転写を制御するための任意的なオペレーター配列、適当なmRNAリボソーム結合部位をエンコードする配列、及び転写及び翻訳の停止を制御する配列を含む。ベクターはプラスミド、ファージ粒子または単純なゲノム挿入である。一旦、適当な宿主に形質転換されると、ベクターは複製でき、宿主ゲノムに独立的に機能し、または場合によっては、ゲノム自身に溶け込む。本願明細書において、“プラスミド”及び“ベクター”は、プラスミドが現在ベクターの最も一般的に使用されている形態であるので、時には交換可能に用いる。しかしながら、本発明は、同等の機能を果たし、当業界で公知の発現ベクターのその他の形態を含むことも意図するものである。
【0069】
本発明で用いられる“宿主細胞”は一般に、米国特許RE34,606号及び/または5,441,882号に開示された方法により好ましくは操作される原核または真核宿主であり、酵素的に活性なエンドプロテアーゼを分泌できないようにさせる。プロテアーゼを発現するために有用な宿主細胞は、酵素的に活性な中性プロテアーゼ及びアルカリ性プロテアーゼ(ズブチリシン)が不足しているバチルス株BG2036である。株BG2036の構築は米国特許第5,264,366号に詳細が記載されている。プロテアーゼを発現するためのその他の宿主細胞はバチルス・ズブチリスI168(米国特許RE34,606号;米国特許第5,264,366号;及び第5,441,882号に記載されている。これらの開示の内容をここに引用する)及びバチルス・リケニホルミス、バチルス・レンタス等などの任意の適当なバチルス株を含む。特に有用な宿主細胞はバチルス株BG2864である。株BG2864の構築はD.Naki、C.Paech、G.Ganshaw、V.Schellenberger.Appl Microbiol Biotechnol(1998)49:290−294に詳細が記載されている。
【0070】
宿主細胞は組換えDNA技術を用いて構築されたベクターを用いて形質転換またはトランスフェクトする。当該形質転換宿主細胞はプロテアーゼ変異体をエンコードするベクターを複製し、または所望のプロテアーゼ変異体を発現することができる。ベクターがプレまたはプレプロ型のプロテアーゼ変異体をエンコードする場合、当該変異体は、発現されたとき、一般的に宿主細胞から宿主細胞培地に分泌される。
【0071】
“作動可能に連結する”は、2つのDNA領域間の関係について言及する場合、単にお互いが機能的に関連することを意味する。例えば、プレ配列は、それがシグナル配列として機能する場合、ペプチドに作動可能に連結し、シグナル配列の切断に最も関係すると思われるタンパク質の成熟型の分泌に関与する。プロモーターは、それが配列の転写を制御する場合、コード配列に作動可能に連結し、リボソーム結合部位はそれが翻訳を可能にするように位置する場合、コード配列に作動可能に連結する。
【0072】
天然前駆体プロテアーゼをエンコードする遺伝子は当業者に公知の一般的な方法に従って得られる。該方法は通常、目的のプロテアーゼの領域をエンコードする推定配列を有する標識プローブを合成する工程、プロテアーゼを発現する微生物からゲノムライブラリーを調製する工程、及びプローブへのハイブリダイゼーションにより目的の遺伝子に関するライブラリーをスクリーニングする工程を含む。陽性のハイブリダイズしたクローンはそれから、地図を作り、配列する。
【0073】
クローンされたプロテアーゼはそれからプロテアーゼを発現するために宿主細胞に形質転換して用いる。プロテアーゼ遺伝子はそれから、高コピー数プラスミド中に連結される。プラスミド複製、問題の遺伝子に作動可能に連結したプロモーター(認識される場合、遺伝子自身の相同プロモーターとして提供される、すなわち、宿主により転写される)、外来のまたはプロテアーゼ遺伝子の内生停止領域により提供された転写停止及びポリアデニル化領域(特定の真核宿主細胞のプロテアーゼ遺伝子から宿主により転写されたmRNAの安定に必要)、及び望ましくは、抗生物質含有培地での成長によりプラスミド感染宿主細胞の培養管理が維持できる抗生物質抵抗性遺伝子などの選択遺伝子に必要な公知の要素を含むという意味で、このプラスミドは宿主中で複製する。高コピー数プラスミドも宿主の複製起源を含み、それにより、多数のプラスミドが染色体の制限なしに細胞質中で生成することが可能となる。しかしながら、複数のプロテアーゼのコピーを宿主ゲノム内に組込むことは本発明の範囲内である。このことは特に相同組換えの影響を受けやすい原核及び真核生物により促進される。
【0074】
遺伝子は天然バチルス・レンタス遺伝子である。もしくは、天然または変異前駆体プロテアーゼをエンコードする合成遺伝子が生成できる。この方法において、前駆体プロテアーゼのDNA及び/またはアミノ酸配列を決定する。複数の重複合成一本鎖DNA断片はその後合成し、ハイブリダイゼーション及び連結により前駆体プロテアーゼをエンコードする合成DNAを生成する。合成遺伝子構築の例は米国特許第5,204,015号の実施例3に示されており、その開示の内容はここに引用する。
【0075】
天然または合成前駆体プロテアーゼ遺伝子を一旦クローンすると、多数の修飾が天然前駆体プロテアーゼの合成を超えて遺伝子の使用を高めるために行われる。当該修飾は、米国特許RE34,606号;第5,741,694号;第6,190,900号;第6,197,567号;第5,972,682号;第5,185,258号;第5,700,676号及びEPO公報第0 251446号に開示される組換えプロテアーゼの生成及びここに記載するプロテアーゼ変異体の生成を含む。
【0076】
以下のカセット式変異誘発法は本発明のプロテアーゼ変異体の構築を促進するために使用できるが、その他の方法も使用できる。まず、プロテアーゼをエンコードする天然遺伝子を得て、全体または一部の配列を決定する。それから、エンコードされる酵素において1以上のアミノ酸の変異(欠失、挿入または置換)を作ることが望ましいポイントに関して配列をスキャンする。このポイントのフランキング配列を、発現した場合種々の変異をエンコードする、オリゴヌクレオチドプールを有する遺伝子の短い断片を置換する制限部位の存在に関して評価する。当該制限部位は、好ましくは遺伝子断片の置換を促進するようなプロテアーゼ遺伝子内の固有部位である。しかしながら、制限消化により生じた遺伝子断片が適当な配列内で再構築されるとすれば、プロテアーゼ遺伝子内の過度に冗長でない任意の都合のよい制限部位が使用できる。制限部位が選択点から都合のよい距離内(10〜15ヌクレオチド)の位置に存在しない場合、当該部位はリーディング・フレームまたはエンコードされるアミノ酸が最終的な構造で変化しない方法で遺伝子内の置換ヌクレオチドにより生じる。配列を変化させて所望の配列に合わせる遺伝子の変異は、公知の方法に従ってM13プライマー伸長法により達成される。適当なフランキング領域を位置付け、2つの都合のよい制限部位配列に達するために必要な変化を評価する課題は遺伝コードの冗長さ、遺伝子の制限酵素地図及び多数の異なる制限酵素によりルーチン化される。都合のよいフランキング制限部位が利用できる場合、上述の方法は部位を含まないフランキング領域とのみ関連して使用される必要があることを留意すべきである。
【0077】
一旦、天然DNAまたは合成DNAがクローンされると、変異する位置のフランキング制限部位は同種の制限酵素を用いて消化され、多数のエンド末端−相補オリゴヌクレオチドカセットは該遺伝子内に連結される。全てのオリゴヌクレオチドは同じ制限部位を有するように合成でき、制限部位を作るために合成リンカーは必要でないので、突然変異生成はこの方法により単純化される。
【0078】
適当な宿主の形質転換で発現される変異体プロテアーゼは、所望の特性、例えば改善された洗浄能力、基質特異性、酸化安定性、pH活性プロファイル等を示す単離または回収された酵素についてスクリーニングできる。
【0079】
ここで用いる、タンパク質分解活性は活性酵素ミリグラム当たりのペプチド結合の加水分解の速度として定義される。タンパク質分解活性を測定するための多くの公知の手順が存在する(K.M.Kalisz,“Microbial Proteinases(細菌プロテイナーゼ),”Advances in Biochemical Engineering/Biotechnology,A.Fiechter編集、1988)。タンパク質分解活性を測定するその他の典型的な方法は、選択基質の変換を所定の基質濃度に加えたプロテアーゼの吸収の変化を測定することにより間接的に測定する種々の分光学的分析を含む。典型的な基質はジメチルカゼイン、N−スクシニル−Ala−Ala−Pro−Phe pNa及びケラチンを含む(米国特許出願番号第60/344,702を参照)。
【0080】
修飾タンパク質分解活性に加え、または修飾タンパク質分解活性に代えて、本発明の変異体酵素は、Km、kcat、kcat/Km比及び/または修飾された基質特異性及び/または修飾されたpH活性プロファイルなどのその他の修飾された性質を有することができる。これらの酵素は、例えば、ペプチドの調製において、または洗濯用などの加水分解プロセスのために存在すると予想される特定基質に合わせることができる。
【0081】
基質特異性の変化は前駆体酵素と変異体間のkcat/Km比の違いとして定義できる。kcat/Km比は触媒効果の指標である。増加または減少したkcat/Km比を有する原核生物カルボニルヒドロラーゼを実施例に記載する。一般的に、本目的はより大きい(数の上で大きい)所定の基質のkcat/Km比を有する変異体を安全なものにし、それにより酵素を使用して標的基質上でより効果的に作用できるようにすることである。1の基質に関するkcat/Km比の増加は他の基質に関するkcat/Km比の減少により達成できる。これは基質特異性の移動であり、このような移動を示す変異体は、前駆体が望ましくないところでの実用性を有し、例えば、基質混合物中の特定基質の望ましくない加水分解を防ぐ。
【0082】
kcat及びKmは公知の手順に従って測定でき、または米国特許第5,441,882号の実施例18の記載に従って測定できる。
【0083】
さらに、酸化安定性は、実施例に記載されるプロテアーゼ変異体により達成できる目的である。該安定性は種々の用途に応じて高めたり減少させたりすることができる。高められた安定性は1以上のメチオニン、トリプトファン、システインまたはリジン残基を欠失することにより達成でき、任意でメチオニン、トリプトファン、システインまたはリジンでないその他のアミノ酸残基を置換することにより達成できる。その逆の置換は減少された酸化安定性を生じる。置換残基はアラニルであってもよいが、中性残基も適している。
【0084】
さらに、安定性、例えば耐熱性は、実施例に記載されるプロテアーゼ変異体により達成できる目的である。安定性は種々の用途に望ましいように高めたり減少させたりできる。高められた安定性は本願明細書で同定された1以上の残基の置換により達成でき、任意でそのうち1つが同じでない他のアミノ酸残基を置換することにより達成できる。耐熱性は所定の温度において長時間にわたって酵素活性を維持する。改善された耐熱性は、前駆体プロテアーゼと比較して変異体の大量の酵素活性の保全を伴う。例えば、所定の温度、通常は測定する操作温度での前駆体と比較した変異体の酵素活性レベルの増加である。
【0085】
ここに記載するプロテアーゼ変異体は特定の洗浄条件下で改善された洗浄能力を示す。例えば、プロテアーゼ変異体は、当業界で公知(例えば、WO99/34011(“An improved Method of Assaying for a Preferred enzyme and/or Preffered Detergent composition(好ましい酵素および/または好ましい洗剤組成物についての改良された分析方法).”))の種々の分析法で測定した性能により示されるように、例えば温度、水の硬度及び/または洗剤濃度の異なる洗浄条件下、異なる洗浄能力を示す。
【0086】
バチルス・ズブチリシンまたはそのプレ、プレプロ及びプロ型の場合、先に述べた位置での変異は上述の特徴が変化した変異体、または酵素の処理が変化した変異体を生成する。これらのアミノ酸位置番号は図1に見られるようにバチルス・アミロリケファシエンスズブチリシンに割り当てられたものであることに留意されたい。所定の位置からのN末端方向の欠失または挿入は、関連アミノ酸位置を移動させ、残基がその最初のまたは野生型の数字の位置ではなくなることが当然に理解される。また、対立遺伝子の違い及び種々の原核生物種間のばらつきは位置移動を生じ、当該ズブチリシンの位置169がグリシンで占められなくなる。このような場合、グリシンの新しい位置は指定グリシン+169と等しく、指定グリシン+169内に包含されると考えられる。グリシン+169の新しい位置は図1のグリシン+169と相同な領域に関して問題のズブチリシンをスキャニングすることにより容易に同定される。
【0087】
1以上、通常は約10までの、アミノ酸残基が変異する。しかしながら、商慣行はさておき、作られる変異の数に限定はない。
【0088】
本酵素は塩として得られる。タンパク質の電離状態は、溶液内の場合、または調製される溶液内の場合、固形の場合、周囲環境のpHに依存することは明らかである。酸性タンパク質は例えば、アンモニウム、ナトリウムまたはカリウム塩として通常調製され、塩基性タンパク質は塩化物、硫酸塩またはリン酸塩として調製される。従って、本出願は指定した種々のプロテアーゼの電気的に中性及び塩形態の両方を含み、プロテアーゼの語は電離状態を問わない、有機構造骨格をいう。
【0089】
プロテアーゼ変異体は特に食品加工及び洗浄技術に有用である。プロテアーゼ変異体及び前駆体プロテアーゼを含むカルボニルヒドロラーゼは、ここに記載するように発酵により生成し、適切な技術により回収する。例えば、K.Anstrup,1974、Industrial Aspects of Biochemistry、編集、B.Spencer、pp.23−46を参照されたい。
【0090】
本発明の1の側面において、本発明の目的は、少なくとも1の洗剤剤形において、及び/または少なくとも1組の洗浄条件下において、前駆体プロテアーゼと比較して変化した、好ましくは改善された洗浄能力を有する変異体プロテアーゼを安定させることである。産業プロセス、特に洗濯業での使用のため、それらはそれ自体公知の方法に従って洗剤またはその他の界面活性剤と一緒に処方される。後者の場合、酵素はタンパク質分解酵素の分野で公知の洗剤、ビルダー、漂白剤、及び/蛍光白色化剤と混合される。適当な洗剤は直鎖アルキルベンゼンスルホン酸塩、アルキルエトキシレート硫酸塩、硫酸化直鎖アルコールまたはエトキシレート直鎖アルコールが挙げられる。該組成物は顆粒状または液状で処方できる。例えば、米国特許第3,623,957号;第4,404,128号;第4,381,247号;第4,404,115号;第4,318,818号;第4,261,868号;第4,242,219号;第4,142,999号;第4,111,855号;第4,011,169号;第4,090,973号;第3,985,686号;第3,790,482号;第3,749,671号;第3,560,392号;第3,558,498号;及び第3,557,002号を参照されたい。
【0091】
種々の洗浄条件が存在し、種々の洗剤剤形、洗浄水量、洗浄水温度、及びプロテアーゼ変異体が曝される洗浄時間の長さなどである。例えば、異なる分野で使用される洗剤剤形は洗浄水中存在するそれらの関連成分を異なる濃度で有する。例えば、ヨーロッパの洗剤は通常、洗浄水中、約3000−8000ppmの洗剤成分を有するが、一方、日本の潜在は通常、洗浄水中800以下、例えば、667ppmの洗剤成分を有する。北米において、特にアメリカでは、洗剤は通常、洗浄水中、約800〜2000、例えば975ppmの洗剤成分が存在する。
【0092】
低洗剤濃度系は約800ppm以下の洗剤成分が洗浄水中に存在する洗剤を含む。日本の洗剤は通常、約667ppmの洗剤成分が洗浄水中に存在するので、低洗剤濃度系と考えられる。
【0093】
中洗剤濃度は約800ppm〜約2000ppmの洗剤成分が洗浄水中に存在する洗剤を含む。北米の洗剤は通常、約975ppmの洗剤成分が洗浄水中に存在するので、中洗剤濃度系と考えられる。ブラジルは約1500ppmの洗剤成分が洗浄水中に存在する。
【0094】
高洗剤濃度系は約2000ppmの洗剤成分が洗浄水中に存在する洗剤を含む。ヨーロッパの洗剤は通常、約3000〜8000ppmの洗剤成分を洗浄水中に含むので、高洗剤濃度系と考えられる。
【0095】
中南米の洗剤は通常、高泡立ちリン酸塩ビルダー洗剤であり、中南米で使用される洗剤の範囲は、洗浄水中の洗剤成分が1500ppm〜6000ppmの範囲であるので、中及び高洗剤濃度の両方になる。上述の通り、ブラジルは通常、洗浄水中に約1500ppmの洗剤成分を有する。しかしながら、その他の中南米の国に限定されない、その他の高泡立ちリン酸塩ビルダー洗剤の地は、約6000ppm以下の洗剤成分が洗浄水中に存在する高洗剤濃度系を有する。
【0096】
上述に照らして、一般的な洗浄溶液中の洗剤組成物の濃度は世界中で約800ppm以下の洗剤組成物(“低洗剤濃度地”)、例えば、日本の約667ppm、から約800ppm〜約2000ppm(“中洗剤濃度地”)、例えば、アメリカの約975ppm及びブラジルの約1500ppm、及び約2000ppm以下(“高洗剤濃度地”)、例えば、ヨーロッパの約4500ppm〜約5000ppm及び高泡立ちリン酸塩ビルダー地の約6000ppmまで様々であることがわかる。
【0097】
一般的な洗剤溶液の濃度は実験的に決定する。例えば、米国において、一般的な洗濯機は洗浄溶液の約64.4Lの体積を入れることができる。従って、洗浄溶液約62.79gの洗剤組成物内で約975ppmの洗剤濃度を得るためには、64.4Lの洗浄溶液を加えなければならない。この量は、洗剤と一緒に備えられる計量カップを用いて消費者により洗浄水中で測定される一般的な量である。
【0098】
さらに別の例として、異なる場所では異なる洗浄温度を用いている。日本の洗浄水の温度は一般にヨーロッパで用いる温度よりも低い。例えば、ヨーロッパにおける洗浄水の温度は通常、約30〜50℃であり、一般的には約40℃である。北米及び/または日本における洗浄水の温度は通常、ヨーロッパの洗浄水よりも低く、例えば約10〜30℃であり、一般的に約20℃である。
【0099】
さらに、別の例として、異なる場所では、異なる水硬度を用いる。水硬度は一般的に合計Ca2+/Mg2+のガロン当たりのグレインとして記載される。硬度は水中のカルシウム(Ca2+)及びマグネシウム(Mg2+)の量を測定したものである。アメリカのほとんどの水は硬水であるが、硬度の程度は様々である。中程度の硬水(60〜120ppm)〜硬水(121〜181ppm)は60〜181ppmの硬度ミネラルを含む(米国ガロン当たりのグレインに変換したppmは、ppm数÷17.1=ガロン当たりのグレインである)。
【0100】
【表1】
【0101】
ヨーロッパの水の硬度は一般に、10〜20ガロン/グレイン合計Ca2+/Mg2+であり、例えば、約15ガロン/グレイン合計Ca2+/Mg2+である。北米の水の硬度は一般に、日本の水の硬度より高いが、ヨーロッパの水の硬度より低い。例えば、3〜10グレイン、3〜8グレインまたは約6グレインである。日本の水の硬度は一般に北米の水の硬度より低く、通常4未満、例えば、3グレイン/ガロン合計Ca2+/Mg2+である。
【0102】
従って、本発明の1の側面は少なくとも1組の洗浄条件において改善された洗浄能力を示すプロテアーゼ変異体を含む。本発明の他の側面は1以上の洗浄条件、例えば、ヨーロッパ、日本または北米条件において、改善された洗浄能力を示すプロテアーゼ変異体を含む。
【0103】
該変異体プロテアーゼを用いて得られたスクリーニング結果に基づいて、バチルスズブチリシンにおける言及した変異及びバチルス・レンタスにおけるそれらの同等物はタンパク質分解活性、性能及び/またはこれらの酵素の安定性及び当該変異体酵素の洗浄または洗剤性能に重要である。
【0104】
本発明の多くのプロテアーゼ変異体は種々の洗剤組成物またはシャンプーまたはローションなどのパーソナルケア製剤を処方するのに有用である。本発明のプロテアーゼ変異体を含む組成物において有用な多数の公知の化合物は適当な界面活性剤である。これらは、Barry J.Andersonの米国特許第4,404,128号及びJiri Flora,et al.の米国特許第4,261,868号に記載されるような、非イオン性、陰イオン性、陽イオン性または両性イオン界面活性剤を含む。適当な洗剤処方は米国特許第5,204,015号(すでにここに引用している)の実施例7に記載されている。当該技術は洗浄組成物として用いることができる種々の剤形でよく知られている。一般的な洗浄組成物に加えて、本発明のプロテアーゼ変異体は、天然または野生型プロテアーゼが使用される任意の目的に使用できることが容易に理解される。従って、これらの変異体は使用でき、例えば、固形または液体石けん用途、食器洗い用製剤、コンタクトレンズ洗浄液または製品、ペプチド加水分解、廃棄物処理、織物用途、タンパク質生成における融合−分解酵素としてである。本発明の変異体は洗剤組成物において高められた性能を有する(前駆体と比較して)。ここで用いるように、洗剤における高められた性能は、標準洗浄サイクル後の通常の評価により測定した、草または血液などのしみに敏感な特定酵素の洗浄の増加として定義する。
【0105】
本発明のプロテアーゼは、約0.01〜約5重量%(好ましくは0.1重量%〜0.5重量%)でpHが6.5〜12.0の公知の粉末及び液体洗剤中に処方できる。これらの洗剤洗浄組成物は、公知のプロテアーゼ、アミラーゼ、セルラーゼ、リパーゼまたはエンドグリコシダーゼなどその他の酵素並びにビルダー及び安定剤を含むこともできる。
【0106】
本発明のプロテアーゼの従来の洗浄組成物への添加は、いかなる特別な使用の限定をも生じない。すなわち、該洗剤に適切ないかなる温度及びpHも、該pHが上述の範囲内である限り、及び該温度が記載したプロテアーゼの変性温度以下である限り、本発明の組成物に適している。さらに、本発明のプロテアーゼは洗剤なしに洗浄組成物中で使用でき、この場合もやはり、単独またはビルダー及び安定剤との組み合わせにおいて使用できる。
【0107】
また、本発明は本発明のプロテアーゼ変異体を含む洗浄組成物に関する。洗浄組成物は一般的に洗浄組成物に使用される添加物をさらに含むことができる。これらは、限定されないが、漂白剤、界面活性剤、ビルダー、酵素及び漂白触媒から選択できる。組成物に含めることが適当な添加剤については当業者にとって明らかである。ここに与えるリストは完全なものではなく、適当な添加剤の例としてのみ理解させるべきである。酵素と適合性のある添加剤及びその他の成分のみ、例えば界面活性剤のみを組成物中に用いることも当業者には容易に理解されることである。
【0108】
存在する場合、洗浄組成物に存在する添加剤の量は約0.01%〜約99.9%、好ましくは約1%〜約95%、より好ましくは約1%〜約80%である。
【0109】
本発明の変異体プロテアーゼは、例えば、米国特許第5,612,055号;米国特許第5,314,692号;及び米国特許第5,147,642号に記載されるような動物飼料添加物の一部として、動物飼料中に含めることもできる。
【0110】
本発明の1の側面は、本発明の変異体プロテアーゼを含む織物処理のための組成物である。該組成物は、RD216,034号;欧州特許134,267号;米国特許第4,533,359号;及び欧州特許第344,259号などの公報に記載されているような例えば絹またはウールを処理するために使用できる。
【0111】
以下は実施例により表すものであり、請求の範囲を限定するものとして解釈されない。
【0112】
ここに引用する全ての刊行物及び特許はその全体をここに組み込むものとする。
【実施例】
【0113】
実施例1
多数のプロテアーゼ変異体が生成でき、当業界に公知の方法を用いて精製できる。変異はバチルス・アミロリケファシエンス(BPN’)ズブチリシンまたはバチルス・レンタスGG36ズブチリシンにおいて作ることができる。変異体は以下から選択できる:バチルス・アミロリケファシエンスのK27A、K27C、K27E、K27Q、K27G、K27H、K27I、K27L、K27M、K27F、K27P、K27S、K27T、K27W、K27Y、H39A、H39R、H39D、H39N、H39C、H39E、H39Q、H39G、H39I、H39L、H39K、H39M、H39F、H39P、H39T、H39W、H39Y、H39V、D41A、D41R、D41C、D41E、D41Q、D41G、D41H、D41I、D41L、D41K、D41M、D41F、D41P、D41S、D41T、D41W、D41Y、D41V、R45A、R45R、R45D、R45N、R45C、R45E、R45C、R45E、R45Q、R45G、R45H、R45I、R45L、R45K、R45M、R45F、R45P、R45S、R45T、R45W、R45Y、R45V、H67A、H67R、H67D、H67N、H67C、H67E、H67Q、H67G、H67I、H67L、H67K、H67M、H67F、H67P、H67S、H67T、H67W、H67Y、H67V、K94A、K94R、K94D、K94N、K94C、K94E、K94Q、K94G、K94H、K94I、K94L、K94M、K94F、K94P、K94S、K94T、K94W、K94Y、K94V、E136A、E136D、E136N、E136C、E136G、E136H、E136I、E136L、E136K、E136M、E136F、E136P、E136S、E136T、E136W、E136Y、E136V、R170A、R170D、R170N、R170C、R170E、R170Q、R170G、R170H、R170I、R170L、R170K、R170M、R170F、R170P、R170S、R170T、R170W、R170Y、R170V、D181A、D181R、D181N、D181C、D181E、D181Q、D181G、D181H、D181I、D181L、D181K、D181M、D181F、D181P、D181S、D181T、D181W、D181Y、D181V、D197A、D197R、D197N、D197C、D197E、D197Q、D197G、D197H、D197I、D197L、D197K、D197M、D197F、D197P、D197S、D197T、D197W、D197Y、D197V、R247A、R247D、R247N、R247C、R247E、R247Q、R247G、R247H、R247I、R247L、R247K、R247M、R247F、R247P、R247S、R247T、R247W、R247Y、R247V、H249A、H249R、H249D、H249N、H249C、H249E、H249Q、H249G、H249I、H249K、H249M、H249F、H249P、H249S、H249T、H249W、H249V、K251A、K251D、K251C、K251Q、K251G、K251H、K251I、K251L、K251M、K251F、K251P、K251S、K251T、K251W、K251Y、K251V、E271A、E271R、E271D、E271N、E271C、E271H、E271I、E271L、E271K、E271M、E271F、E271P、E271S、E271T、E271W、E271Y、及び/またはE271V。
【0114】
実施例2
多数のプロテアーゼ変異体は公知の方法を用いて生成及び精製できる。変異はバチルス・アミロリケファシエンス(BPN’)ズブチリシンまたはバチルス・レンタスGG36ズブチリシンにおいて作ることができる。変異体は以下の位置に対応するアミノ酸の挿入、欠失または置換で作ることができる:バチルス・アミロリケファシエンス(BPN’)の5、7、23、26、28−31、34、47、63、65、66、69、70、73、82−85、88、90、92、93、105、113、125、138、139、148−151、176、178、179、193、196、200、201、202、207、219、220、223、229、233、250、266、267、及び273。
【0115】
実施例3
実施例1及び/または2で生成した多数のプロテアーゼ変異体は、米国特許出願番号60/068,796、“An Improved method of assaying for a preferred enzyme and/or preferred detergent composition(好ましい酵素及び/または好ましい洗剤組成物の改善された分析方法)”に記載されている、マイクロスウォッチ(microswatch)分析法を用いて2種類の洗剤及び洗浄条件における性能を試験する。
【0116】
変異体プロテアーゼは種々の洗剤を分析及び試験される。例えば、可能な洗剤は、0.67g/lろ過アリエール・ウルトラ(Ariel Ultra)(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)、溶液中3グレイン/ガロン合計Ca2+/Mg2+硬度を含み、20℃で各ウェル中0.3ppm酵素が使用される。他の典型的な洗剤は、3.38g/lろ過アリエールFutur(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)、溶液中15グレイン/ガロン合計Ca2+/Mg2+硬度を含み、40℃で各ウェル中0.3ppm酵素が使用される。野生型と比較して高い相対値は改善された洗剤効果を示す。
【0117】
実施例4
実施例1及び2で記載するように分析できる変異体プロテアーゼはその他の異なる洗剤においても分析できる。実施例2と同じ性能試験を以下の洗剤を用いて記載変異体プロテアーゼについて行うことができる:第1の洗剤は、0.67g/lろ過アリエール・ウルトラ(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)であり、溶液中3グレイン/ガロン合計Ca2+/Mg2+硬度を含み、及び20℃で各ウェル中0.3ppm酵素を使用した。第2の洗剤は、3.38g/lろ過アリエールFutur(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)、であり、溶液中15グレイン/ガロン合計Ca2+/Mg2+硬度を含み、及び40℃で各ウェル中0.3ppm酵素を使用できる。第3の洗剤は、3.5g/lHSP1洗剤(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)であり、溶液中8グレイン/ガロン合計Ca2+/Mg2+硬度を含み、及び20℃で各ウェル中0.3ppm酵素を使用できる。第4の洗剤は、1.5ml/l Tide洗剤(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)であり、溶液中3グレイン/ガロン合計Ca2+/Mg2+硬度を含み、及び20℃で各ウェル中0.3ppmの酵素を使用できる。
【0118】
実施例5
多数のプロテアーゼ変異体を生成し、当業界で公知の方法を用いて精製した。全ての変異はバチルス・レンタスGG36ズブチリシン中に作った。変異体を表3に示す。
【0119】
GG36飽和部位ライブラリー及び部位特異性変異体を構築するために、3つのPCR反応を行った。2つのPCR反応はGG36に目的の変異コドンを導入するためのものであり、融合PCRは所望の変異を含む発現ベクターを構築するためのものである。
【0120】
目的のGG36コドンはBPN’の番号に従って番号を付ける(図1A−C及び3A−Bに記載した)。
【0121】
飽和部位ライブラリー構築について:
突然変異生成の方法は領域−特異性変異方法(Teplyakov et al.,1992)に基づいて行い、変異するコドンの配列に対応し、そのコドンでのヌクレオチドのランダムな結合を保証する具体的に作成された3DNA配列NNS((A、C、TまたはG)、(A、C、TまたはG)、(CまたはG))を取り囲む30〜40ヌクレオチド長のフォワード及びリバース相補オリゴヌクレオチドプライマーセットを用いて特定DNAコドン中に全ての可能な変異を一度に作成した。
【0122】
部位特異性変異体構築について:
フォワード及びリバース変異プライマーは両側の相同配列の15以下塩基を含むプライマーの中間に3つの所望の変異を含む。これらの変異は目的のコドンをカバーし、所望のアミノ酸に特異的であり、意図的に合成される。
【0123】
ライブラリー及び変異体を構築するために使用する第2のプライマーセットはpVS08 Apal消化部位及びそのフランキングヌクレオチド配列を一緒に含む。
【0124】
Apalプライマー:
フォワードApalプライマー:
【化1】
リバースApalプライマー:
【化2】
【0125】
GG36分子の変異導入はInvtrogen(カールズバッド、カリフォルニア州、米国)Platinum(登録商標)Taq DNAポリメラーゼ・ハイファイ(High Fidelity)(Cat.no.11304−102)とpVS08テンプレートDNA及びフォワード変異プライマー及び反応1のリバースApalプライマーまたはリバース変異プライマー及び反応2のフォワードApalプライマーを一緒に用いて行った。
【0126】
所望の変異を含む発現ベクターの構築は、反応1及び2の両方のPCR断片、フォワード及びリバースApalプライマー及びInvitrogen Platinum(登録商標)Taq DNAポリメラーゼ・ハイファイ(Cat.no.11304−102)を用いて融合PCRにより達成した。
【0127】
全てのPCRは、サイクル数を除いて(30の代わりに20)、ポリメラーゼと一緒に提供されるInvitrogen手順に従って行った。2つの別個のPCR反応はInvitrogen Platinum(登録商標)Taq DNAポリメラーゼ・ハイファイ(Cat.no.11304−102)を用いて行う。増幅線形5.6Kb断片を精製し(Qiagen(登録商標)Qiaquick PCR精製キット、Cat.no.28106)、Apal制限酵素を用いて消化し、融合断片の両側に付着端を生成した:
- 35μL精製DNA断片
- 4μL React(登録商標)4バッファ(Invitrogen(登録商標):20mM Tris−HCl、5mM MgCl2、50mM KCl、pH7.4)
- 1μL Apal、10単位/ml(Invitrogen(登録商標)、Cat.no.15440−019)
反応条件:1時間、30℃
【0128】
Invitrogen Dpnlを用いて追加の消化をpVS08テンプレートDNAを除去するために行った。
- 40μL Apal消化DNA断片
- 1μL Dpnl、4単位/μL(Invitrogen(登録商標)、Cat.no.15242−019)
反応条件:16〜20時間、37℃
【0129】
2回消化及び精製断片の結合は所望の変異を含む新しい環状DNAを生じ、コンピテントバチルス・ズブチリスに直接形質転換した:
- 30μLのApal及びDpnl消化DNA断片
- 8μL T4リガーゼ・バッファ(Invitrogen(登録商標)、Cat.no.46300−018)
- 1μL T4 DNAリガーゼ、1単位/μl(Invitrogen(登録商標)、Cat.no.15224−017)
反応条件:16〜20時間、16℃
【0130】
結合混合物を、Anagnostopoulos及びSpizizen(1961)の方法を用いてバチルス・ズブチリスBG2864(Naki et al.、1998)に形質転換し、クロラムフェニコール耐性及びプロテアーゼ活性に関して選択した。
【0131】
タンパク質生成の方法
炭素源(グルコース及びマルトデキシトリン、10.5及び17.5g/l)、窒素源(尿、素、3.6g/l)及びリン酸塩(0.5g/l)及び硫酸塩(0.5g/l)などの必須養分を含み、及びさらに微量元素(Fe、Mn、Zn、Cu、Co、1〜4mg/ml)を補足したMops培地(Frederick C.Neidhardt et al.,1974)中で1〜50μLのグリセロール培養物を植菌する。培地はMOPS/トリシン混合物を用いて中和し、7〜8のpHを生じる。培養物を1〜5日、37℃/220rpm(Infors HT(登録商標)Multitron II)で培養する。
【0132】
参考文献:
Protein engineering of the high-alkaline serine protease PB92 from Bacillus alcalophilus: functional and structural consequences of mutation at the S4 substrate binfing pocket. (バチルスalcaloph ilus由来の高アルカリ性セリンプロテアーゼPB92のタンパク質工学:S4基質結合ポケットでの変異の機能的及び構造的結果。)
Teplyakov AV、van der Laan JM、Lammers AA、Kelders H、Kalk KH、Misset O、Mulleners LJ、Dijkstra BW
Protein Eng.1992 Jul;5(5): 413-20.
Selection of a subtilisin-hyperproducing Bachillus in a highly structured environment (高度構造環境におけるズブチリシン−過剰生成バチルスの選択)
D.Naki、C.Paech、G.Ganshaw、V.Schellenberger.Appl Microbiol Biotechnol (1998) 49: 290-294.
Requirements for transformation in Bacillus subtilis (バチルス・ズブチリスにおける形質転換の必要条件)
Anagnostopoulos、C. and Spizizen、J. in J. Bacteriol. 81、741-746 (1961).
Culture Medium for Enterobacteria (腸内細菌の培養基)
Frederick C. Neidhardt、Philip L. Bloch and David F. Smith in Journal of Bacteriology、Sept 1974. p736-747 Vol. 119. No. 3.
【0133】
【表2】
【0134】
実施例6
実施例1で生成した多数のプロテアーゼ変異体を、米国特許出願番号09/554,992(WO99/34011)、“An Improved method of assaying for a preferred enzyme and/or preferred detergent composition(好ましい酵素及び/または好ましい洗剤組成物の改善された分析方法)”に記載されているマイクロスウォッチ(microswatch)分析法を用いて2種類の洗剤及び洗浄条件における性能を試験した。
【0135】
表4は分析した変異体プロテアーゼ及び2つの異なる洗剤で試験した結果を記載する。A欄及びC欄は、分析材料はMTP中の形質転換株の成長により生成した。B欄及びD欄は、分析材料は振とうフラスコ(250ml)中の形質転換株の成長により生成した。A欄及びB欄は、洗剤が7.6g/lろ過アリエール・レギュラー(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)であり、溶液中15グレイン/ガロン合計Ca2+/Mg2+硬度を含み、及び40℃で各ウェル中0.5ppmの酵素を使用した(ヨーロッパ条件)。C欄及びD欄は、洗剤が0.67g/lろ過Tide Opal(プロクター&ギャンブル、シンシナティ、オハイオ州、米国)であり、溶液中3グレイン/ガロン合計Ca2+/Mg2+硬度を含み、及び20℃で各ウェル中0.5ppmの酵素を使用した(日本条件)。性能指数を次の式で計算した:
変異体の洗浄性能 ÷ GG36の洗浄性能(野生型)
4つの性能値を平均し、表2で示す値に達した。
【0136】
【表3】
【0137】
【表4】
【0138】
上述の分析の結果として、いくつかの変異体はGG36野生型プロテアーゼの性能指数よりも大きな性能指数を示した。例えば、変異体R45N−G118E−E271R、R45N−P14R、R45N−N204R、D181N−G118D、及びR45N−G258Rは、ヨーロッパ条件下(15グレイン/ガロン合計Ca2+/Mg2+硬度、40℃、0.5ppm)、マイクロスウォッチ分析において(WO99/34011)、それぞれ1.75、1.28、1.28、1.24、及び1.23の性能指数を示した(表4)。変異体R170S−A1P、R170S−G61R、R170S−N204R、K251G−S87K、及びR170S−S216Rは、それぞれ2.09、2.03、1.79、1.54、1.47、1.39及び1.21の性能指数を示した(表5)。変異体E271T−G100E、E271T−G102E、E271T−S128E、K27T−G100E、R170S−G100R、及びE271T−S49Eは、日本条件下(3グレイン/ガロン合計Ca2+/Mg2+硬度、20℃、0.5ppm)、マイクロスウォッチ96マイクロタイター・ウェルプレート(WO99/34011)分析において、それぞれ3.22、1.85、2.33、2.04、1.79及び1.43の性能指数を示した(表5)。変異体K251G−S87K、R170S−A1R、及びE271T−S128Eは、それぞれ、2.06、1.55、及び1.20の性能指数を示した(表5)。変異体K251G−S87K、及びR170S−A1Rは、日本及びヨーロッパ条件の両方の下、1.00以上の性能指数を示した。
【0139】
本発明は種々の具体的な実施態様との関連において議論及び例示したが、適用性及び開示の範囲を限定するものとして解釈されるものではなく、前述に説明した特徴及び請求の範囲の全ての組み合わせ及びサブコンビネーションにまで及ぶものである。
【特許請求の範囲】
【請求項1】
荷電アミノ酸残基位置に1以上の変異を含み、前駆体プロテアーゼと同じ正味静電荷を有することを特徴とする、前駆体プロテアーゼのプロテアーゼ変異体。
【請求項2】
前記荷電アミノ酸残基位置がアスパラギン酸、グルタミン酸、リジン及びアルギニンからなる群より選択される、請求項1のプロテアーゼ変異体。
【請求項3】
前記変異体が、配列番号2に示すバチルス・アミロリケファシエンスズブチリシンの27、45、170、181、251及び271からなる群より選択される残基位置に等しい1以上の残基位置での置換を有するアミノ酸配列を含む、請求項1のプロテアーゼ変異体。
【請求項4】
前記変異体が27、45、170、181、251及び271に対応する1以上の位置での置換を含み、K27T、R45N、R170S、D181N、K251G及びE271Tから選択される置換である、請求項3のプロテアーゼ変異体。
【請求項5】
さらに、配列番号2に示すバチルス・アミロリケファシエンス・ズブチリシンの1、14、49、61、87、100、102、118、128、204及び258に対応する1以上の位置での別の置換を含む、請求項3のプロテアーゼ変異体。
【請求項6】
変異体がK27T−G100E、R45N−A1R、R45N−P14R、R45N−G61R、R45N−S128R、R45N−N204R、R45N−S216R、R45N−G258R、R170S−A1R、R170S−P14R、R170S−S49R、R170S−G61R、R170S−G100R、R170S−S128R、R170S−N204R、R170S−S216R、R170S−G258R、D181N−G118D、D181N−G258D、K251G−S87K、E271T−S49E、E271T−T66E、E271T−G100E、E271−G102E、E271T−S128E、R45N−G118E−E271R、S49R−G102E−R170S−E271T、及びP14R−R45N−R170S−G258Rの組み合わせから選択される、請求項5のプロテアーゼ変異体。
【請求項7】
請求項2のプロテアーゼ変異体をエンコードするDNA。
【請求項8】
請求項7のDNAをエンコードする発現ベクター。
【請求項9】
請求項8の発現ベクターを用いて形質転換した宿主細胞。
【請求項10】
請求項2のプロテアーゼ変異体を含んだ洗浄組成物。
【請求項1】
荷電アミノ酸残基位置に1以上の変異を含み、前駆体プロテアーゼと同じ正味静電荷を有することを特徴とする、前駆体プロテアーゼのプロテアーゼ変異体。
【請求項2】
前記荷電アミノ酸残基位置がアスパラギン酸、グルタミン酸、リジン及びアルギニンからなる群より選択される、請求項1のプロテアーゼ変異体。
【請求項3】
前記変異体が、配列番号2に示すバチルス・アミロリケファシエンスズブチリシンの27、45、170、181、251及び271からなる群より選択される残基位置に等しい1以上の残基位置での置換を有するアミノ酸配列を含む、請求項1のプロテアーゼ変異体。
【請求項4】
前記変異体が27、45、170、181、251及び271に対応する1以上の位置での置換を含み、K27T、R45N、R170S、D181N、K251G及びE271Tから選択される置換である、請求項3のプロテアーゼ変異体。
【請求項5】
さらに、配列番号2に示すバチルス・アミロリケファシエンス・ズブチリシンの1、14、49、61、87、100、102、118、128、204及び258に対応する1以上の位置での別の置換を含む、請求項3のプロテアーゼ変異体。
【請求項6】
変異体がK27T−G100E、R45N−A1R、R45N−P14R、R45N−G61R、R45N−S128R、R45N−N204R、R45N−S216R、R45N−G258R、R170S−A1R、R170S−P14R、R170S−S49R、R170S−G61R、R170S−G100R、R170S−S128R、R170S−N204R、R170S−S216R、R170S−G258R、D181N−G118D、D181N−G258D、K251G−S87K、E271T−S49E、E271T−T66E、E271T−G100E、E271−G102E、E271T−S128E、R45N−G118E−E271R、S49R−G102E−R170S−E271T、及びP14R−R45N−R170S−G258Rの組み合わせから選択される、請求項5のプロテアーゼ変異体。
【請求項7】
請求項2のプロテアーゼ変異体をエンコードするDNA。
【請求項8】
請求項7のDNAをエンコードする発現ベクター。
【請求項9】
請求項8の発現ベクターを用いて形質転換した宿主細胞。
【請求項10】
請求項2のプロテアーゼ変異体を含んだ洗浄組成物。
【図1A】


【図1B】

【図1B−1】

【図1B−2】

【図1B−3】

【図2】

【図3】
【図3A】

【図3B】

【図4】
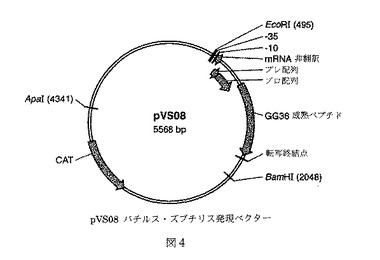
【図5】

【図1B】
【図1B−1】
【図1B−2】
【図1B−3】
【図2】
【図3】
【図3A】
【図3B】
【図4】
【図5】
【公開番号】特開2011−41571(P2011−41571A)
【公開日】平成23年3月3日(2011.3.3)
【国際特許分類】
【出願番号】特願2010−228628(P2010−228628)
【出願日】平成22年10月8日(2010.10.8)
【分割の表示】特願2003−562249(P2003−562249)の分割
【原出願日】平成15年1月16日(2003.1.16)
【出願人】(500284580)ジェネンコー・インターナショナル・インク (67)
【Fターム(参考)】
【公開日】平成23年3月3日(2011.3.3)
【国際特許分類】
【出願日】平成22年10月8日(2010.10.8)
【分割の表示】特願2003−562249(P2003−562249)の分割
【原出願日】平成15年1月16日(2003.1.16)
【出願人】(500284580)ジェネンコー・インターナショナル・インク (67)
【Fターム(参考)】
[ Back to top ]