
豚トルクテノウイルスワクチンおよび診断
本発明は、豚トルクテノウイルス(PTTV)遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAをコードするポリ核酸分子を含有する4つの精製調製物を提供する。本発明は、感染性DNAクローン、その感染性核酸ゲノム分子を含有する生物学的に機能的なプラスミドまたはウイルスベクターも提供する。本発明はさらに、PTTV感染から防御するための生ワクチン、弱毒化ワクチン、ベクター発現および精製組換えキャプシドサブユニットワクチン、または死滅ウイルスワクチンを提供する。また本発明は、PTTV特異的遺伝子産物、とりわけORF1キャプシド遺伝子産物を含む、PTTV感染から防御するためのサブユニットワクチンも提供する。さらに本発明は、PTTV1、PTTV2、および個々のPTTV1遺伝子型のための特異的プライマーを用いるポリメラーゼ連鎖反応(PCR)によってPTTV感染を診断する方法を提供する。最後に本発明は、例えば血清PTTV特異的抗体を検出するためにPTTV特異的抗原を用いる酵素連結免疫吸着アッセイ(ELISA)およびウェスタンブロットなどの免疫学的方法によって、PTTV感染を診断する方法を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の相互参照)
本願は、2009年8月21日に出願された米国仮特許出願第61/235,833号および2010年3月23日に出願された米国仮特許出願第61/316,519号の利益を主張するものであり、これら仮特許出願の開示は、参照により、その全体が本明細書に組み込まれる。
【0002】
本発明は、豚トルクテノウイルス(TTV)感染を防御するためのワクチン、豚TTV(PTTV)の感染性DNAクローン、およびそれらの使用に関する。本発明は、豚トルクテノウイルス(PTTV)感染の診断、特に種特異的または型特異的PTTV感染の診断、および遺伝子型の異なる複数株による同時感染の診断にも関係する。
【背景技術】
【0003】
トルクテノウイルス(TTV)は、1997年に輸血後非A〜E型肝炎を有する日本人患者から初めて発見された(ニシザワ,ティ(Nishizawa,T.)、オカモト,エイチ(Okamoto,H.)、コニシ,ケイ(Konishi,K.)、ヨシザワ,エイチ(Yoshizawa,H.)、ミヤカワ,ワイ(Miyakawa,Y.)およびマユミ,エム(Mayumi,M.)、1997年、「原因不明の輸血後肝炎におけるトランスアミナーゼレベルの上昇に関連する新規DNAウイルス(TTV)(A novel DNA virus (TTV) associated with elevated transaminase levels in posttransfusion hepatitis of unknown etiology)」、バイオケミカル・アンド・バイオフィジカル・リサーチ・コミュニケーションズ(Biochem Biophys Res Commun)、第241巻、第1号、p.92−7)。それ以来、多数のヒトTTV株と、後にトルクテノ・ミニ・ウイルス(TTMV)およびトルクテノ・ミディ・ウイルス(TTMDV)と名付けられた2群のTTV関連ウイルスが、健常な人の血清および他の組織に高い有病率で同定されている(ヒノ,エス(Hino,S.)およびミヤタ,エイチ(Miyata,H.)、2007年、「トルクテノウイルス(TTV):現状(Torque teno virus (TTV): current status)」、レビューズ・イン・メディカル・バイロロジー(Rev Med Virol)、第17巻、第1号、p.45−57;オカモト,エイチ(Okamoto,H.)、2009年a、「TTウイルスの発見の歴史と病原性(History of discoveries and pathogenicity of TT virues)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.1−20)。ヒトTTV、TTMVおよびTTMDVは、それぞれ3.6〜3.9、2.8〜2.9および3.2kbの長さの環状一本鎖DNA(ssDNA)ゲノムを持つ無エンベロープ球形ウイルスであり、現在は、国際ウイルス分類委員会(ICTV;http://www.ictvonline.org/virusTaxonomy.asp?bhcp=1)によって新たに設立されたアネロウイルス科(Anelloviridae)に分類されている(ビアジーニ,ピー(Biagini,P.)、2009年、「TTVおよび関連ウイルス(アネロウイルス)の分類(Classification of TTV and related viruses (anelloviruses))」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.21−33)。これら3群のTTV関連ウイルスは高度な遺伝的異質性を示し、それぞれが数多くの遺伝子グループおよび遺伝子型からなる(ビアジーニ,ピー(Biagini,P.)、ガリアン,ピー(Gallian,P.)、カンタローブ,ジェイ・エフ(Cantaloube,J.F.)、アットーウィ,エイチ(Attoui,H.)、デ・ミッコ,ピー(de Micco,P.)、およびデ・ランバレリエ,エックス(de Lamballerie,X.)、2006年、「フランス人献血者におけるTTVおよびTTMV主要系統群の分布および遺伝子解析(Distribution and genetic analysis of TTV and TTMV major phylogenetic groups in French blood donors)」、ジャーナル・オブ・メディカル・バイロロジー(J Med Virol)、第78巻、第2号、p.298−304;ジェルシック,アイ(Jelcic,I.)、ホッツ−ワーゲンブラット,エイ(Hotz-Wagenblatt,A.)、ハンジカー,エイ(Hunziker,A.)、ツア・ハウゼン,エイチ(Zur Hausen,H.)、およびド・ヴィリエ,イー・エム(de Villiers,E.M.)、2004年、「ホジキン病患者の脾生検組織からの複数のTTウイルス遺伝子型の分離:超可変領域におけるゲノムの再編成および多様性(Isolation of multiple TT virus genotypes from spleen biopsy tissue from a Hodgkin’s diesase patient: genome reorganization and diversity in the hypervariable region)」、ジャーナル・オブ・バイロロジー(J Virol)、第78巻、第14号、p.7498−507)。異なる遺伝子型を持つTTVによる多重感染、ならびにTTV、TTMVおよびTTMDVの二重または三重感染の有病率がヒトにおいて記録されており、これらは健常な成人ではよく見られる事象であると考えられる(ニール,シー(Niel,C.)、サバック,エフ・エル(Saback,F.L.)、およびランペ,イー(Lampe,E.)、2000年、「異なる遺伝子型に属する複数のTTウイルスによる同時感染はブラジル人成人ではよく見られる事象である(Coinfection with multiple TT virus strains belonging to different genotypes is a common event in healthy Brazilian adults)」、ジャーナル・オブ・クリニカル・マイクロバイオロジー(J Clin Microbiol)、第38巻、第5号、p.1926−30;ニノミヤ,エム(Ninomiya,M.)、タカハシ,エム(Takahashi,M.)、ホシノ,ワイ(Hoshino,Y.)、イチヤマ,ケイ(Ichiyama,K.)、シモンズ,ピー(Simmonds,P.)、およびオカモト,エイチ(Okamoto,H.)、2009年、「チンパンジーにおけるトルクテノ・ミディ・ウイルス変異体の全ゲノムの解析:ヒトとチンパンジーのまれな異種間感染(Analysis of the entire genomes of torque teno midi virus variants in chimpanzees: infrequent cross-species infection between humans and chimpanzees)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第90巻、第2号、p.347−58;オカモト,エイチ(Okamoto,H.)、2009年a、「TTウイルスの発見の歴史と病原性(History of discoveries and pathogenicity of TT virues)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.1−20;タカヤマ,エス(Takayama,S.)、ミウラ,ティ(Miura,T.)、マツオ,エス(Matsuo,S.)、タキ,エム(Taki,M.)、およびスギイ,エス(Sugii,S.)、1999年、「日本人血友病患者における新規DNA TTウイルス(TTV)感染の有病率および残留性(Prevalence and persistence of novel DNA TT virus (TTV) infection in Japanese haemophiliacs)」、ブリティッシュ・ジャーナル・オブ・ヘマトロジー(Br J Haematol)、第104巻、第3号、p.626−9)。
【0004】
TTVは、ヒトに感染するだけでなく、非ヒト霊長類、ツパイ、豚、牛、猫、犬およびアシカを含む他のさまざまな動物種にも、同様に感染する(ビアジーニ,ピー(Biagini,P.)、ウチ,アール(Uch,R.)、ベルフーシェ,エム(Belhouchet,M.)、アットーウィ,エイチ(Attoui,H.)、カンタローブ,ジェイ・エフ(Cantaloube,J.F.)、ブリスバレー,エヌ(Brisbarre,N.)、およびデ・ミッコ,ピー(de Micco,P.)、2007年、「複合ローリングサークル増幅/配列非依存単一プライマー増幅法を使用することによってヒトおよび動物試料中に同定されたアネロウイルスに関連する環状ゲノム(Circular genomes related to anelloviruses identified in human and animal samples by using a combined rolling-circle amplification/sequence-independent single primer amplification approach)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第88巻、第10号、p.2696−701;イマイ,ティ(Inami,T.)、オバラ,ティ(Obara,T.)、モリヤマ,エム(Moriyama,M.)、アラカワ,ワイ(Arakawa,Y.)およびアベ,ケイ(Abe,K.)、2000年、「チンパンジーから得られたサルTTウイルス分離株の完全長ヌクレオチド配列:新しいTTウイルス様種の証拠(Full-length nucleotide sequence of a simian TT virus isolate obtained from a chimpanzee: evidence for a new TT virus-like species)」、バイロロジー(Virology)、第277巻、第2号、p.330−5;エング,ティ・エフ(Ng,T.F.)、スエドマイヤー,ダブリュ・ケイ(Suedmeyer,W.K.)、ホイーラー,イー(Wheeler,E.)、ガランド,エフ(Gulland,F.)、およびブライトバルト,エム(Breitbart,M.)、2009年、「捕獲されたカリフォルニアアシカの死亡事象から発見された新規アネロウイルス(Novel anellovirus discovered from a mortality event of captive California sea lions)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第90巻、第5号、p.1256−61;オカモト,エイチ(Okamoto,H.)、2009年b、「動物におけるTTウイルス(TT viruses in animals)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.35−52;オカモト,エイチ(Okamoto,H.)、ニシザワ,ティ(Nishizawa,T.)、タカハシ,エム(Takahashi,M.)、タワラ,エイ(Tawara,A.)、ポン,ワイ(Peng,Y.)、キシモト,ジェイ(Kishimoto,J.)、およびワン,ワイ(Wang,Y.)、2001年、「ツパイにおけるTTウイルスのゲノム特徴および進化的特徴の分析ならびにヒトおよび非ヒト霊長類における種特異的TTVとの比較(Genomic and evolutionary characterization of TT virus (TTV) in tupaias and comparison with species-specific TTVs in humans and non-human primates)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第82巻、第9号、p.2041−50;オカモト,エイチ(Okamoto,H.)、ニシザワ,ティ(Nishizawa,T.)、タワラ,エイ(Tawara,A.)、パン,ワイ(Peng,Y.)、タカハシ,エム(Takahashi,M.)、キシモト,ジェイ(Kishimoto,J.)、タナカ,ティ(Tanaka,T.)、ミヤカワ,ワイ(Miyakawa,Y.)、およびマユミ,エム(Mayumi,M.)、2000年a、「ヒトおよび非ヒト霊長類における種特異的TTウイルスならびにそれらの系統発生学的関係(Species-specific TT viruses in humans and nonhuman primates and their phylogenetic relatedness)」、バイロロジー(Virology)、第277巻、第2号、p.368−78;オカモト,エイチ(Okamoto,H.)、タカハシ,エム(Takahashi,M.)、ニシザワ,ティ(Nishizawa,T.)、タワラ,エイ(Tawara,A.)、フカイ,ケイ(Fukai,K.)、ムラマツ,ユー(Muramatsu,U.)、ナイトウ,ワイ(Naito,Y.)、およびヨシカワ,エイ(Yoshikawa,A.)、2002年、「豚、猫および犬におけるTTウイルス(TTV)のゲノム特徴分析ならびに霊長類およびツパイにおける種特異的TTVとそれらの関係(Genomic characterization of TT viruses (TTVs) in pigs,cats and dogs and their relatedness with species-specific TTVs in primates and tupaias)」、ジャーナル・オブ・メディカル・バイロロジー(J Gen Virol)、第83巻、第6号、p.1291−7)。また、チンパンジーはTTMVおよびTTMDVにも感染する(ニノミヤ,エム(Ninomiya,M.)、タカハシ,エム(Takahashi,M.)、ホシノ,ワイ(Hoshino,Y.)、イチヤマ,ケイ(Ichiyama,K.)、シモンズ,ピー(Simmonds,P.)、およびオカモト,エイチ(Okamoto,H.)、2009年、「チンパンジーにおけるトルクテノ・ミディ・ウイルス変異体の全ゲノムの解析:ヒトとチンパンジーのまれな異種間感染(Analysis of the entire genomes of torque teno midi virus variants in chimpanzees: infrequent cross-species infection between humans and chimpanzees)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第90巻、第2号、p.347−58;オカモト(Okamoto)ら、2000年a、前掲書)。同定された動物TTV株、とりわけ非霊長類動物TTVのゲノムサイズは、ヒトTTVのゲノムサイズと比べて比較的小さいが、これらは同じゲノム構造を有し、マイナス鎖ssDNAから翻訳される少なくとも2つの部分的にオーバーラップするオープンリーディングフレーム(ORF1およびORF2)と、高いGC含量(約90%)を有する非翻訳領域(UTR)の短いストレッチとを持つ(オカモト(Okamoto)、2009年b、前掲書)。TTV ORFの配置は、サーコウイルス科(Circoviridae)のジャイロウイルス属(Gyrovirus)に属する鶏貧血ウイルス(CAV)のそれと極めて類似しているが、やはり同じ科に分類される豚サーコウイルス(PCV)1型(PCV1)および2型(PCV2)とは異なっている(デビッドソン,アイ(Davidson,I.)およびシャルマン,エル・エム(Shulman,L.M.)、2008年、「鶏貧血ウイルスによる鳥類感染との比較によってヒトアネロウイルス感染の謎を解明する(Unraveling the puzzle of human anellovirus infections by comparison with avian infections with the chicken anemia virus)」、ウイルス・リサーチ(Virus Res)、第137巻、第1号、p.1−15;ヒノ,エス(Hino,S.)およびプラセティオ,エイ・エイ(Prasetyo,A.A.)、2009年、「トルクテノウイルスと鶏貧血ウイルスとの関連性(Relationship of Torque teno virus to chicken anemia virus)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.117−30)。PCV1とPCV2のゲノムはアンビセンスであり、ORF1はゲノム鎖によってコードされ、ORF2はアンチゲノム鎖によってコードされる(ヒノ(Hino)およびミヤタ(Miyata)、2007年、前掲書)。最近、ヒトTTV遺伝子型1および6の両方の転写パターンおよび翻訳産物が、それぞれのTTV感染性DNAクローンを培養細胞中にトランスフェクションすることによって同定された(ミュラー,ビー(Mueller,B.)、メルツ,エイ(Maerz,A.)、ドベルスタイン,ケイ(Doberstein,K.)、フィンスターブッシュ,ティ(Finsterbusch,T.)、およびマンカーツ,エイ(Mankertz,A.)、2008年、「ヒトトルクテノウイルス分離株P/1C1の遺伝子発現(Gene expression of the human Torque Teno Virus isolate P/1C1)」、バイロロジー(Virology)、第381巻、第1号、p.36−45;キュウ,ジェイ(Qiu,J.)、カッコラ,エル(Kakkola,L.)、チェン,エフ(Cheng,F.)、イェ,シー(Ye,C.)、ソダールンド−ヴェネルモ,エム(Soderlund-Venermo,M.)、ヘドマン,ケイ(Hedman,K.)、およびピンテル,ディ・ジェイ(Pintel,D.J.)、2005年、「ヒトサーコウイルスTTウイルス遺伝子型6は完全長クローンのトランスフェクション後に6つのタンパク質を発現する(Human circovirus TT virus genotype 6 expresses six proteins following transfection of a full-length clone)」、ジャーナル・オブ・バイロロジー(J Virol)、第79巻、第10号、p.6505−10)。スプライスされた3つ以上のmRNAから、ORF1、ORF2、ORF1/1、ORF2/2、ORF1/2およびORF2/3と呼ばれる少なくとも6つのタンパク質の発現が報告されている(カッコラ,エル(Kakkola,L.)、ヘドマン,ケイ(Hedman,K.)、キュウ,ジェイ(Qiu,J.)、ピンテル,ディ(Pintel,D.)、およびソダールンド−ヴェネルモ,エム(Soderlund-Venermo,M.)、2009年、「TTウイルスの複製およびTTウイルスによるタンパク質合成(Replication of and protein synthesis by TT viruses)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.53−64;ミュラー(Mueller)ら、2008年、前掲書;キュウ(Qiu)ら、2005年、前掲書)。したがって、動物TTVに関して、より多くのデータが利用できるようになれば、推定されている動物TTVのゲノム構造を修正することが必要になる可能性が高い。
【0005】
TTVは特発性肝炎患者において初めて同定されたが、その後の研究は、肝炎または他の疾患の病理発生にTTVが重要な役割を果たすという証拠を示すことができなかった(ヒノ(Hino)およびミヤタ(Miyata)、2007年、前掲書;マギー,エフ(Maggi,F.)およびベンディネリ,エム(Bendinelli,M.)、2009年、「トルクテノウイルスおよび他のアネロウイルスの免疫生物学(Immunobiology of the Torque teno viruses and other anellovirueses)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.65−90;オカモト(Okamoto)、2009年a、前掲書)。ヒトTTVは疾患に直接関連するとは見なされていないが、豚TTV(PTTV)は、豚繁殖呼吸器病症候群ウイルス(PRRSV)感染と組み合わされると豚皮膚炎腎症症候群(PDNS)の実験的誘導の一因になり(クラコウカ,エス(Krakowka,S.)、ハーチュニアン,シー(Hartunian,C.)、ハンバーグ,エイ(Hamberg,A.)、シャウプ,ディ(Shoup,D.)、リングス,エム(Rings M.)、チャン,ワイ(Zhang,Y.)、アラン,ジー(Allan,G.)、およびエリス,ジェイ・エイ(Ellis,J.A.)、2008年、「豚サーコウイルス2型に関して結果が陰性であったノトバイオート豚における豚皮膚炎腎症症候群の誘導の評価(Evaluation of induction of porcine determatitis and nephropathy syndrome in gnotobiotic pigs with negative results for porcine circovirus type 2)」、アメリカン・ジャーナル・オブ・ベテリナリー・リサーチ(Am J Vet Res)、第69巻、第12号、p.1615−22)、また、ノトバイオート豚モデルにおいて、PCV2感染と組み合わされると、離乳後多臓器消耗症候群(PMWS)の実験的誘導の一因にもなること(エリス,ジェイ・エイ(Ellis,J.A.)、アラン,ジー(Allan,G.)、およびクラコウカ,エス(Krakowka,S.)、2008年、「ノトバイオート豚におけるサーコウイルス2型関連離乳後多臓器消耗症候群に遺伝子グループ1豚トルクテノウイルスとの同時感染が及ぼす効果(Effect of coinfection with genogroup 1 porcine toruque teno virus on porcine circovirus type 2-associated postweaning multisystemic wasting syndrome in gnotobiotic pigs)」、アメリカン・ジャーナル・オブ・ベテリナリー・リサーチ(Am J Vet Res)、第69巻、第12号、p.1608−14)が示された。このデータから、豚TTVは豚において病原性であることが示唆された。しかし、PTTVに関連する疾患および病変を決定的に特徴づけるには、生物学的に純粋な形態のPTTVウイルスを使ったさらに綿密な研究が必要である。
【0006】
ヒトTTVと比較すると、PTTVのゲノム情報は極めて限られている。現時点で、PTTVについては、日本およびブラジルの豚から、それぞれ1つの完全長ゲノム配列と2つのほぼ完全長のゲノム配列とが報告されているにすぎない(ニール,シー(Niel,C.)、ディニシュ−メンデス,エル(Diniz-Mendes,L.)、およびデバル,エス(Devalle,S.)、2005年、「ヒトおよび豚血清からのトルクテノウイルス(TTV)完全ゲノムのローリングサークル増幅と新規豚TTV遺伝子グループの同定(Rolling-circle amplification of Torque teno virus (TTV) complete genomes from human and swine sera and identification of a novel swine TTV genogroup)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第86巻、第5号、p.1343−7;オカモト(Okamoto)ら、2002年、前掲書)。3つの既知PTTV株のうち、Sd−TTV31株とTTV−1p株が遺伝子グループ1(PTTV1)にまとめられたのに対し、TTV−2pは遺伝子グループ2(PTTV2)に分類される唯一の株だった(ニール(Niel)ら、2005年、前掲書)。しかし遺伝子グループ分類はウイルス学の分類法における漠然とした概念であり、さらにより正確なPTTVの分類が必要とされているが、それは、多数の遺伝子型を代表する新しいPTTV株の完全長ゲノム配列が、より多く利用可能にならなければ、行うことができない。
【0007】
PTTV感染が米国、カナダ、スペイン、中国、韓国およびタイを含む6つの異なる国の豚に蔓延していることは、先に示されている(マキューン,エヌ・イー(McKeown,N.E.)、フェノー,エム(Fenaux,M.)、ハルバー,ピー・ジー(Halbur,P.G.)、およびモン,エックス・ジェイ(Meng,X.J.)、2004年、「6つの異なる国の豚におけるオーファンウイルス・豚TTウイルスの分子キャラクタリゼーション(Molecular characterization of porcine TT virus,an orphan virus,in pigs from six different countries)」、ベテリナリー・マイクロバイオロジー(Vet Microbiol)、第104巻、第1−2号、p.113−7)。
【0008】
豚TTVが特定豚疾患の病理発生において重要な役割を果たすかどうかは、依然として議論の余地がある。ノトバイオート豚モデルでは、PTTV1感染だけでは臨床的疾患を何も発生させないが、軽度の組織学的病変を誘導することが示された(クラコウカ,エス(Krakowka,S.)およびエリス,ジェイ・エイ(Ellis,J.A.)、2008年、「ノトバイオート豚における豚遺伝子グループ1トルクテノウイルスの効果の評価(Evaluation of the effects of procine genogroup 1 torque teno virus in gnotobiotic swine)」、アメリカン・ジャーナル・オブ・ベテリナリー・リサーチ(Am J Vet Res)、第69巻、p.1623−9)。PTTV1と豚繁殖呼吸器病症候群ウイルス(PRRSV)の両方を実験的に接種されたノトバイオート豚が、臨床的な豚皮膚炎腎症症候群(PDNS)を発症したのに対し(クラコウカ,エス(Krakowka,S.)、ハーチュニアン,シー(Hartunian,C.)、ハンバーグ,エイ(Hamberg,A.)、シャウプ,ディ(Shoup,D.)、リングス,エム(Rings M.)、チャン,ワイ(Zhang,Y.)、アラン,ジー(Allan,G.)、およびエリス,ジェイ・エイ(Ellis,J.A.)、2008年、「豚サーコウイルス2型に関して結果が陰性であったノトバイオート豚における豚皮膚炎腎症症候群の誘導の評価(Evaluation of induction of porcine determatitis and nephropathy syndrome in gnotobiotic pigs with negative results for porcine circovirus type 2)」、アメリカン・ジャーナル・オブ・ベテリナリー・リサーチ(Am J Vet Res)、第69巻、p.1615−22)、PTTV1と豚サーコウイルス2型(PCV2)の両方を接種された豚は、急性離乳後多臓器消耗症候群(PMWS)を発症した(エリス(Ellis)ら、2008年、前掲書)。PCV2は臨床的PMWSまたはPCV関連疾患(PCVAD)の主要原因因子であると見なされているが、スペインでは、PCV2が少ないかPCV2を持たないPMWS罹患豚において、PTTV2感染の有病率が非PMWS罹患豚と比較して高いことが観察された(ケカライネン(Kekarainen)ら、2006年、前掲書)。これらのデータは、全体として、豚TTVが豚における疾患の惹起または増悪に関与する共同因子として働き得ることを示唆している。
【0009】
豚TTVは、感染した豚の豚血清、糞便、唾液、精液および組織試料に検出されており、このことは、水平伝播と垂直伝播の両者を含むその多様な伝播経路を示している(ケカライネン(Kekarainen)ら、2007年、前掲書;ポッツート,ティ(Pozzuto,T.)、ミュラー,ビー(Mueller,B.)、ミーハン,ビー(Meehan,B.)、リングラー,エス・エス(Ringler,S.S.)、マッキントッシュ,ケイ・エイ(McIntosh,K.A.)、エリス,ジェイ・エイ(Ellis,J.A.)、マンカーツ,エイ(Mankertz,A.)およびクラコウカ,エス(Krakowka,S.)、2009年、「豚トルクテノウイルスの子宮内伝播(In utero transmission of porcine torque teno viruses)」、ベテリナリー・マイクロバイオロジー(Vet Microbiol)、第137巻、p.375−9;シビラ,エム(Sibila,M.)、マルティネス−ギノー,エル(Martinez-Guino,L.)、ウエルタ,イー(Huerta,E.)、ローレンス,エイ(Llorens,A.)、モーラ,エム(Mora,M.)、グロウ−ローマ,エル(Grau-Roma,L.)、ケカライネン,ティ(Kekarainen,T.)、およびシーゲイルス,ジェイ(Segales,J.)、2009年、「従来の養豚場における豚トルクテノウイルス(TTV)感染および排泄動態(Swine torque teno virus (TTV) infection and excretion dynamics in conventional pig farms)」、ベテリナリー・マイクロバイオロジー(Vet Microbiol)、第139巻、p.213−8)。しかし、現在の豚TTV感染の検出は、主として、従来のPCRアッセイに基づいていた。今までのところ、血清学的アッセイもウイルス培養系も確立されていない。具体的には、スペインのグループによってそれぞれ開発されたPTTV1およびPTTV2のUTR中の保存領域のネステッドPCR増幅が、広く使用されるようになっている(ケカライネンら、2006年、前掲書)。PCV2誘発性PCVADについて証明されたように、ウイルスの量は臨床的疾患の重症度と関連すると思われるので(オプリースニヒ,ティ(Opriessnig,T.)、モン,エックス・ジェイ(Meng,X.J.)およびハルバー,ピー・ジー(Halbur,P.G.)、2007年、「豚サーコウイルス2型関連疾患:最新用語、臨床症状発現、病理発生、診断、および介入戦略に関する最新情報(Porcine circovirus type 2 associated disease: update on current terminology,clinical manifestations,pathogenesis,diagnosis,and intervention strategies)」、ジャーナル・オブ・ベテリナリー・ダイアグノスティック・インベスティゲーション(J Vet Diagn Invest)、第19巻、p.591−615)、従来のPCRによってTTV DNAの存在を決定するのではなく、定量リアルタイムPCRによって豚TTPのウイルス負荷量を決定することが重要になるだろう。また、リアルタイムPCRは、従来のPCRよりも信頼性が高く、迅速かつ安価でもある。最近、2つの豚TTV種を検出し定量するための2つのタックマン(TaqMan)プローブ・ベース・リアルタイムPCRアッセイが記載された(ブラッサード,ジェイ(Brassard,J.)、ガニエ,エム・ジェイ(Gagne,M.J.)、ウーデ,エイ(Houde,A.)、ポイトラス,イー(Poitras,E.)およびウォード,ピー(Ward,P.)、2009年、「豚および牛トルクテノウイルスを検出するためのリアルタイム・タックマンPCRアッセイの開発(Development of a real-time TaqMan PCR assay for the detection of porcine and bovine Torque teno virus)」、ジャーナル・オブ・アプライド・マイクロバイオロジー(J Appl Microbiol)、2009年11月14日、印刷物に先立つ電子ジャーナルでの公開;ガレイ,エイ(Gallei,A.)、ペッシュ,エス(Pesch,S.)、エスキング,ダブリュ・エス(Esking,W.S.)、ケラー,シー(Keller,C.)およびオーリンガー,ブイ・エフ(Ohlinger,V.F.)、2009年、「豚トルクテノウイルス:遺伝子グループ特異的マルチプレックスrt−PCRによるウイルスゲノム負荷量の決定、遺伝子グループ1または2による頻繁な多重感染の検出、およびウイルス完全長配列の確立(Porcione Torque teno virus: Determination of viral genomic loads by genogroup-specific multiplex rt-PCR,detection of frequent multiple infections with genogroups 1 or 2,and establishment of viral full-length sequences)」、ベテリナリー・マイクロバイオロジー(Vet Microbiol)、2009年12月21日、印刷物に先立つ電子ジャーナルでの公開)。プローブに基づくアッセイの主な欠点は、プローブ結合配列が突然変異を含有する場合に、偽陰性結果が得られる可能性がある点である(アンダーソン,ティ・ピー(Anderson,T.P.)、ウェルノ,エイ・エム(Werno,A.M.)、ベイノン,ケイ・エイ(Beynon,K.A.)およびマードック,ディ・アール(Murdoch,D.R.)、2003年、「プローブ結合部位内の配列変位に起因するリアルタイムPCRアッセイおよび融解曲線分析による単純ヘルペスウイルスの遺伝子タイピングの失敗(Failure to genotype herpes simplex virus by real-time PCR assay and melting curve analysis due to sequence variation within probe binding sites)」、ジャーナル・オブ・クリニカル・マイクロバイオロジー(J Clin Microbiol)、第41巻、p.2135−7)。既知の豚TTV株の配列間の高度な不均一性を考えると、PTTVの野外株ではプローブ結合配列中の変異が予期される。SYBRグリーンに基づくリアルタイムPCRは、この潜在的問題を回避する代替的方法であり、その比較的低い特異性にもかかわらず、潜在的豚TTV変異体を検出し定量するための普遍的方法になる。さらにまた、SYBRグリーン・リアルタイムPCRに続く融解曲線分析(MCA)により、反応特異性が確保され、型が異なるウイルスの多重検出が可能になる(リリー,ケイ・エム(Ririe,K.M.)、ラスムッセン,アール・ピー(Rasmussen,R.P.)およびウイットワー,シー・ティー(Wittwer,C.T.)、1997年、「ポリメラーゼ連鎖反応中のDNA融解曲線の分析による生成物の区別(Product differentiation by analysis of DNA melting curves during the polymerase chain reaction)」、アナリティカル・バイオケミストリー(Anal Biochem)、第245巻、p.154−60)。MCAに基づくSYBRグリーン・リアルタイムPCR法はさまざまなヒトウイルスおよび家畜ウイルスに応用されて成功を収めている(ジベリーニ,ディ(Gibellini,D.)、ガルディーニ,エフ(Gardini,F.)、ビトーネ,エフ(Vitone,F.)、スキアボーネ,ピー(Schiavone,P.)、フルリーニ,ジー(Furlini,G.)およびリ,エム・シー(Re,M.C.)、2006年、「血漿試料におけるSYBRグリーン・リアルタイム・マルチプレックスRT−PCR技法によるHCVとHIV−1の同時検出(Simultaneous detection of HCV and HIV-1 by SYBR Green real time multiplex RT-PCR technique in plasma samples)」、モレキュラー・アンド・セルラー・プローブス(Mol Cell Probes)、第20巻、p.223−9;マルティネス,イー(Martinez,E.)、リエラ,ピー(Riera,P.)、シッジャ,エム(Sitja,M.)、ファン,ワイ(Fang,Y.)、オリベイラ,エス(Oliveira,S.)およびマルドナド,ジェイ(Maldonado,J.)、2008年、「SYBRグリーンを用いたリアルタイムRT−PCRおよびアンプリコン融解曲線分析による豚繁殖呼吸器病症候群ウイルス(PRRSV)の同時検出および遺伝子タイピング(Simultaneous detection and genotyping of porcine reproductive and respiratory syndrome virus (PRRSV) by real-time RT-PCR and amplicon melting curve analysis using SYBR Green)」、リサーチ・イン・ベテリナリー・サイエンス(Res Vet Sci)、第85巻、p.184−93;ムイユソー,ケイ・ピー(Mouillesseaux,K.P.)、クリムペル,ケイ・アール(Klimpel,K.R.)およびダール,エイ・ケイ(Dhar,A.K.)、2003年、「SYBRグリーン・リアルタイムPCRにおいてアンプリコンサイズを増大させることによるクルマエビのタウラ症候群ウイルスおよびイエローヘッド病ウイルスに関する検出特異性および検出感受性の改善(Improvement in the specificity and sensitivity of detection for the Taura syndrome virus and yellow head virus of penaeid shrimp by increasing the amplicon size in SYBR Green real-time RT-PCR)」、ジャーナル・オブ・バイロロジカル・メソッズ(J Virol Methods)、第111巻、p.121−7;ウィルヘルム,エス(Wilhelm,S.)、チンマーマン,ピー(Zimmermann,P.)、ゼルビッツ,エイチ・ジェイ(Selbitz,H.J.)およびトリューエン,ユー(Truyen,U.)、2006年、「野外試料中の豚パルボウイルスを検出するためのリアルタイムPCRプロトコール(Real-time PCR protocol for the detection of porcine parvovirus in field samples)」、ジャーナル・オブ・バイロロジカル・メソッズ(J Virol Methods)、第134巻、p.257−60)。
【発明の概要】
【発明が解決しようとする課題】
【0010】
現在、PTTV特異的体液性応答については、ほとんどわかっていない。PCRベースのアッセイは豚におけるPTTV感染の経過を反映しないので、PTTVの血清有病率を評価し、豚疾患におけるPTTVの役割の特徴づけを助けるには、PTTV血清抗体を検出するための効率のよい酵素結合免疫吸着アッセイ(ELISA)が必要である。
【0011】
今のところ、豚TTVのサブユニットワクチン、死滅ワクチンおよび生ワクチンは入手できない。PTTVサブユニットワクチンを開発するには、さまざまな遺伝子型から組換えPTTVキャプシドタンパク質を発現させることが望ましく有益であり、また、死滅ワクチンおよび生ワクチンの開発に使用される細胞培養系において生物学的に純粋な形態のPTTVを増殖させるには、さまざまな遺伝子型から感染性PTTV分子DNAクローンを構築することが、望ましく有益であるだろう。
【課題を解決するための手段】
【0012】
本発明は、PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAの遺伝子型からなる群より選択されるゲノム配列に対して少なくとも80%の相同性を有する少なくとも1コピーのゲノム配列を含有する感染性PTTVをコードする核酸分子を含む、豚トルクテノウイルス(PTTV)の感染性核酸分子(「感染性DNAクローン」)を提供する。
【0013】
本発明の一態様によれば、ゲノム配列が配列番号9、配列番号10、配列番号11、および配列番号12に示す配列から選択される、請求項1に記載のPTTVの感染性DNAクローン。
【0014】
本発明は、感染性PTTVゲノムを含有する生物学的に機能的なプラスミドまたはウイルスベクターを提供する。
【0015】
本発明は、感染性クローンDNAプラスミドまたはウイルスベクターをトランスフェクトされた適切な宿主細胞を提供する。
【0016】
本発明は、PTTV感染性DNAクローンをトランスフェクトされた細胞によって産生される感染性PTTVを提供する。
【0017】
本発明は、無毒性(nontoxic)の生理学的に許容される担体と、(a)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも80%の相同性を有する少なくとも1コピーのゲノム配列またはその相補鎖を含有する核酸分子、(b)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも80%の相同性を有する少なくとも1コピーのゲノム配列またはその相補鎖を含有する核酸分子を含有する生物学的に機能的なプラスミドまたはウイルスベクター、および(c)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも80%の相同性を有する少なくとも1コピーのゲノム配列を含有する無発病性(avirulent)、感染性かつ非病原性(nonpathogenic)PTTV、からなる群より選択される免疫原量の構成要素とを含むウイルスワクチンも提供する。
【0018】
本発明の一態様によれば、ワクチンは、PTTV感染性クローン由来の生PTTVウイルスを含有する。本発明のもう一つの態様によれば、ワクチンは、PTTV感染性クローン由来の死滅PTTVウイルスを含有する。
【0019】
本発明は、細菌発現系においてPTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、およびPTTV2c−VAのORF1キャプシド遺伝子から発現した精製組換えタンパク質と、PTTV感染に対するサブユニットワクチンとしてのこれらの組換えキャプシドタンパク質の使用を提供する。本発明の一実施形態では、サブユニットワクチンとして使用するための組換えキャプシドタンパク質を、バキュロウイルス発現系および他の発現ベクター系において発現させる。
【0020】
本発明のさらなる一態様によれば、アジュバントをさらに含む。
【0021】
本発明はさらに、PTTVウイルス感染に対して豚を免疫化する方法であって、豚に免疫学的有効量のウイルスワクチンを投与することを含む方法を提供する。
【0022】
本発明の一態様によれば、本方法は、組換えサブユニットキャプシドタンパク質、感染性核酸分子または生PTTVウイルスを豚に投与することを含む。
【0023】
本発明のもう一つの態様によれば、本方法は、ワクチンを豚に非経口投与、鼻腔内投与、皮内投与、または経皮投与することを含む。本発明のさらなる一態様によれば、本方法は、ワクチンを豚にリンパ系内投与または筋肉内投与することを含む。
【0024】
本発明は、配列番号9に示すPTTV1a−VAのヌクレオチド配列の配列からなる単離されたポリヌクレオチドも提供する。
【0025】
本発明は、配列番号10に示すPTTV1b−VAのヌクレオチド配列の配列からなる単離されたポリヌクレオチドも提供する。
【0026】
本発明は、配列番号11に示すPTTV2b−VAのヌクレオチド配列の配列からなる単離されたポリヌクレオチドも提供する。
【0027】
本発明は、配列番号12に示すPTTV2c−VAのヌクレオチド配列の配列からなる単離されたポリヌクレオチドも提供する。
【0028】
本発明はさらに、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1、およびORF2/2、特にキャプシドタンパク質をコードするORF1、からなる群より選択されるポリヌクレオチド配列に従って翻訳されたポリペプチド配列の免疫原性フラグメントまたは完全タンパク質を含むサブユニットワクチンを提供する。
【0029】
本発明の一態様によれば、ポリヌクレオチド配列は、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1からなる群より選択される。
【0030】
本発明のもう一つの態様によれば、ポリヌクレオチド配列は、PTTV遺伝子型PTTV1a−VAのORF1である。本発明のさらなる一態様によれば、ポリヌクレオチド配列は、PTTV遺伝子型PTTV1b−VAのORF1である。本発明のさらにもう一つの態様によれば、ポリヌクレオチド配列は、PTTV亜型PTTV2c−VAのORF1である。
【0031】
本発明の一態様によれば、ポリペプチド配列は、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、および配列番号28に示す配列からなる群より選択される。
【0032】
本発明のもう一つの態様によれば、ポリペプチド配列は、配列番号13に示される。本発明のもう一つの態様によれば、ポリペプチド配列は、配列番号14に示される。本発明のさらなる一態様によれば、ポリペプチド配列は、配列番号16に示される。本発明の具体的一実施形態では、ポリペプチド配列が、配列番号16のC末端領域(aa310−625)である。本発明のさらにもう一つの態様によれば、ポリペプチド配列は、配列番号20に示される。
【0033】
本発明のさらなる態様によれば、ワクチンは、さらにアジュバントを含有する。
【0034】
本発明はさらに、PTTVウイルス感染に対して豚を免疫化する方法であって、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1、およびORF2/2からなる群より選択されるポリヌクレオチド配列に従って翻訳されたポリペプチド配列の免疫原性フラグメントまたは完全タンパク質を含むワクチンの免疫学的有効量を豚に投与することを含む方法を提供する。
【0035】
本発明の一態様によれば、本方法は、免疫原性フラグメントまたは組換えキャプシドタンパク質を豚に投与することを含む。
【0036】
本発明のもう一つの態様によれば、本方法は、ワクチンを豚に非経口投与、鼻腔内投与、皮内投与、または経皮投与することを含む。本発明のさらなる一態様によれば、本方法は、ワクチンを豚にリンパ系内投与または筋肉内投与することを含む。
【0037】
また、本発明は、PTTV1感染を診断し、PTTV1負荷量を定量するための方法であって、PTTV1感染が疑われる試料からDNAを抽出すること、配列番号29および配列番号30に示す配列を含むプライマーを使ってポリメラーゼ連鎖反応(PCR)を行うこと、およびPTTV1特異的増幅を検出することを含む方法も提供する。本発明の一態様によれば、ポリメラーゼ連鎖反応はSYBRグリーン・リアルタイムPCRである。
【0038】
本発明はさらに、PTTV2感染を診断し、PTTV2負荷量を定量するための方法であって、PTTV2感染が疑われる試料からDNAを抽出すること、配列番号29、配列番号30、配列番号31および配列番号32に示す配列を含むプライマーを使ってポリメラーゼ連鎖反応(PCR)を行うこと、およびPTTV2特異的増幅を検出することを含む方法を提供する。本発明の一態様によれば、ポリメラーゼ連鎖反応はSYBRグリーン・リアルタイムPCRである。
【0039】
本発明は、PTTV1感染とPTTV2感染を同時に検出し診断するための方法であって、PTTV感染が疑われる試料からDNAを抽出すること、配列番号31および配列番号32に示す配列を含むプライマーを使ってポリメラーゼ連鎖反応(PCR)を行うこと、ならびにPTTV1およびPTTV2特異的増幅を検出することを含む方法も提供する。本発明の一態様によれば、ポリメラーゼ連鎖反応はSYBRグリーン・リアルタイムPCRである。
【0040】
また、本発明は、PTTV1a感染とPTTV1b感染を同時に検出し診断するための方法であって、PTTV1感染が疑われる試料からDNAを抽出すること、配列番号33および配列番号34に示す配列を含むプライマーを使って第1ポリメラーゼ連鎖反応(PCR)を行うこと、配列番号35、配列番号36、配列番号37、および配列番号38に示す配列を含むプライマーを使って第2PCRを行うこと、ならびにPTTV1aおよびPTTV1b特異的増幅を検出することを含む方法も提供する。
【0041】
本発明は、PTTV感染を診断するための方法であって、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1、およびORF2/2からなる群より選択されるポリヌクレオチド配列に従って翻訳されたポリペプチド配列の免疫原性フラグメントを固定化すること、PTTV感染が疑われる豚からの血清試料を固定化免疫原性フラグメントと接触させること、および免疫原性フラグメントに特異的な、捕捉された抗体を検出することを含む方法を提供する。
【0042】
本発明の一態様によれば、ポリヌクレオチド配列は、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1からなる群より選択される。
【0043】
本発明の一実施形態によれば、ポリヌクレオチド配列は、PTTV遺伝子型PTTV1a−VAのORF1である。本発明のもう一つの実施形態によれば、ポリヌクレオチド配列は、PTTV遺伝子型PTTV1b−VAのORF1である。本発明のさらなる一実施形態によれば、ポリヌクレオチド配列は、PTTV亜型PTTV2c−VAのORF1である。
【0044】
本発明のもう一つの態様によれば、ポリペプチド配列は、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、および配列番号28に示す配列からなる群より選択される。
【0045】
本発明の一実施形態によれば、ポリペプチド配列は、配列番号13に示される。本発明のもう一つの態様によれば、ポリペプチド配列は、配列番号14に示される。本発明のもう一つの実施形態によれば、ポリペプチド配列は、配列番号16に示される。本発明のさらなる一実施形態によれば、免疫原性フラグメントは、配列番号16のC末端領域(aa310−625)である。本発明のさらにもう一つの実施形態によれば、ポリペプチド配列は、配列番号20に示される。
【0046】
本発明は、PTTV感染を診断し、PTTV遺伝子型PTTV1a−VA、PTTV1b−VA、およびPTTV種2の全ての既知の亜型に感染した豚の血清における抗体を検出するための、3つの標準化酵素結合免疫吸着アッセイ(ELISA)を提供する。
【0047】
本ELISA診断検査は、PTTV遺伝子型PTTV1a−VA、PTTV1b−VA、およびPTTV2c−VAの細菌発現またはバキュロウイルス発現組換えORF1キャプシドタンパク質に基づく。
【0048】
本発明のもう一つの態様によれば、捕捉された抗体の検出は、ウェスタンブロットによる。本発明のさらにもう一つの態様によれば、捕捉された抗体の検出は、酵素結合免疫吸着アッセイ(ELISA)による。
【0049】
本発明の上述の特徴は、本発明の以下の詳細な説明を図面と併せて読めば、より明確に理解されることになるだろう。
【図面の簡単な説明】
【0050】
【図1】図1Aおよび1Bは、豚TTVウイルス・グループ1(種1)およびグループ2(種2)株の4つの原型米国株のゲノム構造と、ゲノムクローニングおよびアセンブリのための戦略の概略図である。
【図2】図2は、ジェンバンク(GenBank)データベースにおいて入手することができる121のTTV株のヌクレオチド配列比較のPASC(pairwise sequence comparisons)分布を表す。属、種、型、亜型および変異体、ならびにそれらの対応するヌクレオチド配列同一性百分率を示す。
【図3】図3Aは、完全長ゲノムヌクレオチド配列に基づく近隣結合法によって構築された系統樹を表す。図3Bは、7つの豚TTV株間で、ORF1の推定アミノ酸配列に基づいて構築された系統樹を表す。図3Cは、7つの豚TTV株間で、ORF1/1の推定アミノ酸配列に基づいて構築された系統樹を表す。図3Dは、7つの豚TTV株間で、ORF2の推定アミノ酸配列に基づいて構築された系統樹を表す。図3Eは、7つの豚TTV株間で、ORF2/2の推定アミノ酸配列に基づいて構築された系統樹を表す。
【図4】図4は、7つのPTTV株間のORF1の完全長アミノ酸配列のアラインメントを表す。
【図5】図5は、7つのPTTV株間のORF2の完全長アミノ酸配列のアラインメントを表す。
【図6】図6Aは、各々の標準テンプレート(青色で示すもの)および20個の豚血清試料を40サイクル増幅した後のPTTV1リアルタイムPCR産物の融解曲線を表す。図6Bは、各々の標準テンプレートおよび20個の豚血清試料を40サイクル増幅した後のPTTV2リアルタイムPCR産物の融解曲線を表す。
【図7】図7A〜7Eは、PTTV1/PTTV2 SYBRグリーン・ベース・デュプレックス・リアルタイムPCRの融解曲線分析(MCA)を表す。
【図8】図8は、7つのPTTV株間の予測ORF1のN末端部分に位置するヌクレオチド配列のアラインメントを表す。
【図9】図9Aおよび9Bは、4つの既知豚TTV2の親水性プロファイルおよび保存領域を表す。
【図10】図10A〜10Cは、組換えPTTV2c ORF1キャプシドタンパク質の発現と精製を表す。
【図11】図11A〜11Cは、選ばれた豚血清試料のウェスタンブロット分析の代表的結果を表す。
【図12】図12は、PTTV2c−ORF1に基づくウェスタンブロットおよびELISAの整合性を表す。
【図13】図13は、異なる供給源に由来する138頭の豚におけるウイルス負荷量によるPTTV2血清抗体レベルの箱ひげ図を表す。
【図14】図14Aは、PTTV2のウイルス負荷量の遡及的評価を表す。図14Bは、到着時から到着の2ヶ月後まで成長した10頭の豚におけるPTTV2 ORF1キャプシドタンパク質に対する抗体レベルを表す。
【図15】図15A〜15Cは、PTTV1aおよびPTTV1b組換えORF1キャプシドタンパク質の発現と精製を表す。
【図16】図16は、ウィスコンシンの飼育場に由来する選ばれた豚血清試料のPTTV1a−ORF1ベースのウェスタンブロット分析の例を表す。
【発明を実施するための形態】
【0051】
本発明によれば、具体的一実施形態において、上記4つの新規豚TTV亜型が、バージニアにおいて、一頭の雄豚から分離される。
【0052】
図1Aでは、PTTV1とPTTV2のゲノムがどちらもボールド体で示され、4つの予測ORF(ORF1、ORF2、ORF1/1およびORF2/2)のサイズと向きが矢印で示されている。GCリッチ領域も示されている。破線の弧AおよびDは、それぞれ、ネステッドPCRによって血清試料および精液試料からPTTV1およびPTTV2を検出するために使用した領域を表す。破線の弧BおよびCは、PTTV1のゲノムクローニングのための2つのオーバーラップPCRフラグメントを表し、一方、破線の弧EおよびFは、PTTV2のゲノムクローニングのための2つのオーバーラップPCRフラグメントを表す。本研究に使用したプライマー(表1参照)の場所も、対応する位置に示されている。
【0053】
第1ラウンドPCRにおいてPTTV1とPTTV2のどちらについても陽性と示されることから、高いウイルス負荷量が示唆された1つの雄豚血清試料(SR#5)を、その後のPTTVの完全長ゲノムクローニングに使用した。驚いたことに、最初のPTTV株Sd−TTV31をクローニングするために設計されたインバースPCR(オカモト(Okamoto)ら、2002年、前掲書)の2組のプライマーセット(NG372/NG373およびNG384/NG385)を利用してウイルスゲノムDNAを増幅しようとする最初の試みは、成功しなかった。数回試行したがPCR産物は得られなかった。次に、PTTV1の領域AおよびPTTV2の領域Dの初期配列に基づいて、想定されるPTTV1ゲノムにまたがる領域BおよびCを増幅するための2つの新しいプライマー対(TTV1−If(配列番号1)/TTV1−2340R(配列番号2)およびTTV1−2311F(配列番号3)/TTV1−IR(配列番号4))と、想定されるPTTV2ゲノムにまたがる領域EおよびFを増幅するための2つのさらなるプライマー対(TTV2−IF(配列番号5)/TTV2−2316R(配列番号6)およびTTV2−GCF(配列番号7)/TTV2−IR(配列番号8))を、それぞれ設計した(図1Aおよび表1)。プライマーTTV1−2340R(配列番号2)およびTTV1−2311F(配列番号3)は、PTTV2株TTV−2p(ニール(Niel)ら、2005年、前掲書)中には存在しないPTTV1株Sd−TTV31(オカモト(Okamoto)ら、2002年、前掲書)およびTTV−1p(ニール(Niel)ら、2005年)中の共通配列から推定し、一方、プライマーTTV2−2316R(配列番号6)およびTTV2−GCF(配列番号7)は、前記2つのPTTV1株には存在しない株TTV−2pの配列から推定した。その結果生じた予期されるサイズの4つの異なるPCR産物を、平滑末端クローニングベクターにそれぞれ挿入し、その結果生じた組換えプラスミドを大腸菌(Escherichia coli)に挿入した。フラグメントB、C、EおよびFを表す各コンストラクトについて8〜15個の陽性(白色)細菌クローンを同定し、次に配列決定した。
【0054】
【表1】
【0055】
意外なことに、各コンストラクトから2群の配列データが同定され、同じ豚から得られる遺伝子グループ1および遺伝子グループ2に2タイプのPTTVが存在することが示された。それら4つのPTTV株を区別しアセンブルするために、3つの既知PTTV株Sd−TTV31、TTV−1pおよびTTV−2pと共に、配列比較を行った(図1Bおよび1C)。
【0056】
図1Bは、PTTV1株PTTV1a−VAおよびPTTV1b−VAの完全長ゲノム配列の区別およびアセンブリを、続いてクローニングされたPCRフラグメントBおよびCと共に図解している。フラグメントB中のORF1およびORF2の開始コドンと、フラグメントC中のORF1の終止コドンに「^」または「*」で印が付けられている。2つの既知PTTV1株Sd−TTV31およびTTV−1pの対応する配列も示されている。保存されている配列を影付きで表し、ダッシュ記号はヌクレオチド欠失を示す。
【0057】
PTTV1の場合、予測ORF1の開始コドンATGおよび終止コドンTGAは、それぞれフラグメントBおよびC中に位置していた(図1B)。これらのコドンの位置は2つのPTTV1グループにおいて異なり、1つ目はSd−TTV31と同一、2つ目はTTV−1pと同一だった(図1B)。また、これら2つのグループにおけるORF2開始コドンも、ORF1の場合と同じく、異なる位置にあった。さらにまた、領域Bの4つの異なる配列(配列決定データからの2つと株Sd−TTV31およびTTV−1pからの2つ)および領域Cの4つの異なる配列を使った系統解析も、1つ目の配列がSd−TTV31とクラスターを形成し、2つ目がTTV−1pとクラスターを形成することを裏付けた(データ未掲載)。それゆえに、本発明者らは、フラグメントBとCの両方からの2群の配列データを区別し、2つの完全長PTTV1ゲノム(これらを、それぞれ、株PTTV1a−VA(配列番号9)およびPTTV1b−VA(配列番号10)と名付けた)にアセンブルすることができた(図1B)。
【0058】
図1Cは、PTTV2株PTTV2b−VAおよびPTTV2c−VAの完全長ゲノム配列の区別とアセンブリを、続いてクローニングされたPCRフラグメントEおよびFと共に図解している。TTV−2p株の対応する配列も含まれており、保存されている配列が影付きで表されている。ダッシュ記号はヌクレオチド欠失を示す。各株について、オーバーラップ領域(破線で囲んだ部分)内のユニークなヌクレオチド(PTTV2b−VA(配列番号11)の場合は連続する「AG」ヌクレオチド、PTTV2c−VA(配列番号12)の場合は2つの単一「A」ヌクレオチドおよび単一「G」ヌクレオチド)が、それぞれ示されている。
【0059】
これら2つのPTTV2株の区別の方が容易だった。2つのPCRフラグメントのオーバーラップ領域中に位置しているユニークな連続「AG」ヌクレオチドは、それぞれフラグメントEおよびFからの2群の配列データに共通していた(図1C)。アセンブルした完全長ゲノム配列は、1つのPTTV2株を表し、これをPTTV2b−VA(配列番号11)と名付けた。同様に、フラグメントEおよびFからのもう1つの配列データセットがオーバーラップ領域中に共有している2つのユニークな単一「A」ヌクレオチドおよび単一「G」ヌクレオチドに基づいて、PTTV2c−VA(配列番号12)と名付けた第2の株の完全ゲノム配列をアセンブルした(図1C)。フラグメントEおよびFからの4つの配列をTTV−2pからの2つの対応する配列と共に使った系統解析も、この割当を裏付けた(データ未掲載)。
【0060】
本発明は、豚におけるウイルス感染に関連する4つの分離された豚TTVウイルス遺伝子型または亜型を提供する。本発明は、豚TTVウイルス遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VA、配列番号9(PTTV1a−VA)、配列番号10(PTTV1b−VA)、配列番号11(PTTV2b−VA)、および配列番号12(PTTV2c−VA)に示すヌクレオチド配列、それらの機能的等価物または相補鎖を有するウイルス遺伝子型または亜型を包含するが、これらに限るわけではない。任意の豚TTVに由来する特定のヌクレオチド配列が、個々のウイルス間に自然に存在するわずかな変異を有するであろうことは理解されるだろう。これらの配列の変異は、欠失、置換、挿入などに見ることができる。
【0061】
4つのPTTV株のそれぞれについて提案されるゲノム構造を詳細に分析し、3つの既知PTTV株Sd−TTV31、TTV−1pおよびTTV−2pと共に、表2に要約する。PTTVの4つの米国株は、それぞれ、2,878bp(PTTV1a−VA 配列番号9)、2,875bp(PTTV1b−VA 配列番号10)、2,750bp(PTTV2b−VA 配列番号11)、および2,803bp(PTTV2c−VA 配列番号12)という、類似したゲノムサイズを有する。PTTV1a−VA(配列番号9)とSd−TTV31はどちらも同じゲノム長を有する。株TTV−1pおよびTTV−2pの公表された配列は、いずれも、UTRのGCリッチ領域に多くの未決定ヌクレオチドを有する。これらのヌクレオチドをPTTV1およびPTTV2に対応するコンセンサス配列で人工的に埋めたところ、それぞれゲノム長が、TTV−1pはPTTV1b−VA(配列番号10)とより近い関係にあり、TTV−2pはPTTV2b−VA(配列番号11)とより近い関係にあることが示された(データ未掲載)。
【0062】
アセンブルされた豚TTVウイルス遺伝子型または亜型PTTV1a−VA(配列番号9)、PTTV1b−VA(配列番号10)、PTTV2b−VA(配列番号11)、およびPTTV2c−VA(配列番号12)のゲノム配列は、ジェンバンク(Genbank(登録商標))に提出され(ヌクレイック・アシッズ・リサーチ(Nucleic Acids Resaerch)、2008年1月;36(データベース号(Database issue):D25−30)、それぞれアクセッション番号GU456383、GU456384、GU456385、およびGU456386が付与されている。
【0063】
【表2】
数字(完全長ゲノム、ORFのサイズ、およびエクソン番号を除く)は、各PTTV株のゲノム上のヌクレオチド(nt)位置を示す。
【0064】
最近行われた2つの研究により、ヒトTTV複製時に、転写されたウイルスmRNAと、少なくとも6個のウイルスタンパク質の発現が同定された(ミュラー(Mueller)ら、2008年、前掲書;キュウ(Qiu)ら、2005年、前掲書)。これはヒトTTVがコードすると予想されるORFの数よりも多い(オカモト,エイチ(Okamoto,H.)、ニシザワ,ティ(Nishizawa,T.)、タワラ,エイ(Tawara,A.)、タカハシ,エム(Takahashi,M.)、キシモト,ジェイ(Kishimoto,J.)、サイ,ティ(Sai,T.)、およびスガイ,ワイ(Sugai,Y.)、2000年b、「感染個体からの骨髄細胞に検出されるTTウイルスmRNA(TT virus mRNAs detected in the bone marrow cells from an infected individual)」、バイオケミカル・アンド・バイオフィジカル・リサーチ・コミュニケーションズ(Biochem Biophys Res Commun)、第279巻、第2号、p.700−7)。そこで本発明者らは、PTTV配列との比較のために、この新しいヒトTTVゲノム情報を含めた。ヒトTTV株P/1C1のmRNA転写産物の5’端は、TATAボックスの25nt下流にある「A」にマッピングされた(ミュラー(Mueller)ら、2008年、前掲書)。この開始点、その隣接配列(CGAATGGCTGAGTTTATGCCGC(配列番号39);開始点に下線を付した)および上流にあるTATAボックスまでの距離(24nt;表2)は、7つのPTTV株の全てにおいてよく保存されており、PTTVとヒトTTVは、mRNAの共通の5’端を翻訳に利用し得ることが示唆される。
【0065】
7つのPTTV株の全ての間で完全に保存されているさらに5つの領域がTATAボックスの近傍に同定された。それぞれ11ntの2つの領域(AGTCCTCATTT(配列番号40)およびAACCAATCAGA(配列番号41))は、TATAボックスの上流に位置し、残り3つの領域(17ntのCTGGGCGGGTGCCGGAG(配列番号42);14ntのCGGAGTCAAGGGGC(配列番号43);11ntのTATCGGGCAGG(配列番号44))は、提案されているmRNAの5’端とORF2の開始コドンとの間に位置する。これらの保存されたPTTV特異的配列はウイルス遺伝子発現を調節する共通の要素を含有するのだろう。
【0066】
以前、3つの既知PTTV株のゲノムに、それぞれ3つのORF(ORF1〜3)が提案された(ニール(Niel)ら、2005年、前掲書;オカモト(Okamoto)ら、2002年、前掲書)。本研究で同定されたPTTVの4つの原型米国株は、この構造を有している。ヒトTTV中の対応するORF3は、ORF2中の同じATGで開始しスプライシング後も同じORF(伸張したORF2)中に留まるので(図1A)、これにはORF2/2という新たな名前が付けられている(ミュラー(Mueller)ら、2008年、前掲書;キュウ(Qiu)ら、2005年、前掲書)。本研究では、PTTV分類を改定するために、本発明者らはヒトTTVの命名法に従う。ヒトTTV ORF1/1は、ORF1中の2つのエクソンによってコードされる新たに同定されたウイルスタンパク質である(キュウ(Qiu)ら、2005年、前掲書)。ORF1/1は、ORF1と、同一のN末端部分およびC末端部分を共有している。PTTV ORF1/1対応物は、7つのPTTV株の全てにおいて、すぐに同定された(図1Aおよび表2)。
【0067】
ORF1およびORF2が約2.8kbのウイルスmRNAによってコードされるのに対し、ORF1/1およびORF2/2は、ヒトTTVでは、スプライスされた約1.2kbのウイルスmRNAによってコードされる(ミュラー(Mueller)ら、2008年、前掲書;キュウ(Qiu)ら、2005年、前掲書)。これら4つのORFがPTTVゲノムでも推定されたことと、7つのPTTV株において予測されるスプライスドナー部位およびスプライスアクセプター部位配列の配列および位置がよく保存されていることから(表2)、豚TTVもおそらく2つの対応するmRNAをコードすると推測される。
【0068】
ヒトTTV株の大半はCAVと遺伝的類似性を共有していて、TTVアポトーシス誘導タンパク質(TAIP)をコードし、そのCAV対応部分はアポプチンと名付けられている(ド・スミット,エム・エイチ(de Smit,M.H.)およびノートボルン,エム・エイチ(Noteborn,M.H.)、2009年、「鶏貧血ウイルスおよびTTウイルスにおけるアポトーシス誘導タンパク質(Apoptosis-inducing proteins in chicken anemia virus and TT virus)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.131−49)。TAIPのORFはORF2内に埋め込まれている。しかし豚TTVには、対応するTAIPが存在しない。最近の研究により、培養細胞におけるCAVの複製にはアポプチンまたはTAIPの発現が必要であることが示された(プラセチョウ,エイ・エイ(Prasetyo,A.A.)、カマホラ,ティ(Kamahora,T.)、クロイシ,エイ(Kuroishi,A.)、ムラカミ,ケイ(Murakami,K.)、およびヒノ,エス(Hino,S.)、2009年、「鶏貧血ウイルス(CAV)の複製はアポプチンを必要とし、ヒトトルクテノウイルス(TTV)のVP3によって補完される(Replication of chicken anemia virus (CAV) requires apoptin and is complemented by VP3 of human torque teno virus (TTV))」、バイロロジー(Virology)、第385巻、第1号、p.85−92)。
【0069】
ペアワイズ配列比較(PASC)は、同じファミリー内のウイルスの全ての利用可能なゲノム配列からペアワイズヌクレオチド配列同一性百分率の頻度分布をプロットする有用な方法である(バオ,ワイ(Bao,Y.)、カプースチン,ワイ(Kapustin,Y.)、およびタツソワ,ティ(Tatusova,T.)、2008年、「ペアワイズ配列比較(PASC)によるウイルス分類(Virus Classification by Pairwise Sequence Comparison (PASC))」、エイサイクロペディア・オブ・バイロロジー5巻セット(Encyclopedia of Virology,5 vols)(ビー・ダブリュ・ジェイ・マーヒー(B.W.J.Mahy)およびエム・エイチ・ブイ・ファン・レーゲンモルテル(M.H.V.Van Regenmortel)編)の第5巻、p.342−8、エルゼビア(Elsevier)、オックスフォード(Oxford))。PASCプログラムによって生成した異なるピークは、通常、ウイルスの属、種、型、亜型および株を表す(図2)。この研究において、本発明者らは、ジェンバンク(GenBank)データベースに見いだされる121の、ヒトおよび動物TTV関連株の完全長ゲノム配列を使って、TTVのPASC解析を行った(図2)。TTVの構成要素が一つの独立した科、アネロウイルス科(Anelloviridae)に分類されると仮定すると、36〜55%および55〜67%のヌクレオチド配列同一性に見られる2つの主要ピークは、それぞれ属および種のグループを表す(図2)。したがってTTV型は67〜85%のヌクレオチド配列同一性を有するTTVのグループと定義され、一方、TTV亜型は85〜95%のヌクレオチド配列同一性を共有するTTV配列のグループと定義することができる。95%を上回るヌクレオチド配列同一性を共有するTTV株は、さらに変異体に分類することができる(図2)。同じような分類が、ジェルシック(Jelcic)らにより、103個のTTV分離株の配列を使って提案されている(ジェルシック,アイ(Jelcic,I.)、ホッツ−ワーゲンブラット,エイ(Hotz-Wagenblatt,A.)、ハンジカー,エイ(Hunziker,A.)、ツア・ハウゼン,エイチ(Zur Hausen,H.)、およびド・ヴィリエ,イー・エム(de Villiers,E.M.)、2004年、「ホジキン病患者の脾生検組織からの複数のTTウイルス遺伝子型の分離:超可変領域におけるゲノムの再編成および多様性(Isolation of multiple TT virus genotypes from spleen biopsy tissue from a Hodgkin’s diesase patient: genome reorganization and diversity in the hypervariable region)」、ジャーナル・オブ・バイロロジー(J Virol)、第78巻、第14号、p.7498−507)。
【0070】
ここに提案されたTTV分類の基準を、PTTVの4つの原型米国株と他の3つの既知PTTV株のゲノム配列の系統解析に適用した。これらの株間で行った完全長ヌクレオチド配列のペアワイズ比較は、4つのPTTV1株は3つのPTTV2株と比較して54.0〜56.4%のヌクレオチド配列同一性を有することを示した(表3)。したがって、文献において以前にPTTVの「遺伝子グループ(genogroup)」と呼ばれていたものは、おそらく「種(species)」と呼ぶ方が、より適切であり、PTTV1とPTTV2はおそらくそれぞれ豚TTV種1および種2を表すはずである。PTTV種1は、それぞれ1a型(Sd−TTV31およびPTTV1a−VA(配列番号9)を含む)および1b型(TTV−1pおよびPTTV1b−VA(配列番号10))と呼ばれる2つのウイルス型からなる。というのも、これら2つのウイルス型の間のヌクレオチド配列同一性は69.8〜70.7%だからである(表3)。Sd−TTV31とTTV1a−VA(配列番号9)は、それらの高い配列同一性(95.1%)ゆえに、同じ種の変異体株であると認識される。しかし、2つの1b型株、TTV−1pおよびPTTV1b−VA(配列番号10)は2つの異なる亜型に属し得る(ヌクレオチド配列同一性=86.4%)。PTTV種2の場合、3つの株は、86.5〜90.9%というそれらのヌクレオチド配列同一性に基づいて、別々の亜型(それぞれ、TTV−2pは亜型2a、PTTV2b−VA(配列番号11)は亜型2b、PTTV2c−VA(配列番号12)は亜型2c)に分類される可能性が高い。ここに提案されたPTTVのための新しい分類システムは、系統樹に明確に見てとることができた(図3A)。PTTVのORF1、ORF1/1、ORF2およびORF2/2の推定アミノ酸配列に基づいて構築した系統樹も、ここに提案された分類と合致した(図3B〜3E)。
【0071】
【表3】
データはPASCプログラムを使用することによって生成したもので、値は%ヌクレオチド配列同一性を示す。
【0072】
PTTV種、型および亜型の間で、ユニークな突然変異ならびに欠失および/または挿入は、ゲノム全体に散在している。例えば、上述のように図1Bに示すORF1の開始および終止コドンならびにORF2の開始コドンの場所は、PTTV1a型と1b型の間で異なる。2つのPTTV1b株は、PTTV1aと比較して、ORF2開始後に2コドンの欠失も有する(図1B)。
【0073】
注目すべきことに、TTV−2pとPTTV2b−VAはどちらも、PTTV2c−VAと比較して、UTR中の最初の11nt保存配列(AGTCCTCATTT(配列番号40))の39nt上流に、大きな52nt欠失を有する。この欠失ゆえに、PTTV2b−VAのゲノムサイズ(おそらくTTV−2pも同様である)は、PTTV2c−VAのゲノムサイズよりもかなり小さい(表2)。多くの「サブウイルス」ヒトTTVクローンが血清試料から単離されており、これらサブウイルス分子の大部分においてORFが通常はインタクトなままであることから、それらは完全長TTVゲノムであるとみなされている(ド・ヴィリエ(de Villiers)ら、2009年;レピック(Leppik)ら、2007年)。それらは、完全にまたは部分的に欠失しているUTRに、さまざまな長さを有する。TTV−2pとPTTV2b−VAの状況は、ヒトTTVサブウイルス分子の状況に似ていると思われ、亜型PTTV2aとPTTV2bは、亜型PTTV2cに由来するサブウイルス分子である可能性があることが示唆される。注目すべきことに、野外試料からPTTV2を検出するために他のグループ(エリス(Ellis)ら、2008年、前掲書;ケカライネン(Kekarainen)ら、2007年、前掲書;ケカライネン(Kekarainen)ら、2006年、前掲書;クラコウカ(Krakowka)ら、2008年、前掲書)によってよく使用されているネステッドPCRプライマーTTV2−nF(表1)の3’末端配列は、その欠失の両側に位置している。したがって、PTTV2を検出するための現在のネステッドPCRアッセイは、遺伝的に多様なPTTV2c亜型の株を同定するには十分でない可能性が高い。
【0074】
分離ウイルス株の供給源は、豚TTVウイルス感染を有する豚の血清、糞便、唾液、精液および組織試料である。しかし、組換えDNA技術を使ってヌクレオチド配列を複製し化学的に合成することができると考えられる。したがって本発明の範囲は、例えば限定するわけではないが、配列番号9、配列番号10、配列番号11、および配列番号12に示すヌクレオチド配列、またはその相補鎖を含む、単離されたポリヌクレオチド;配列番号9、配列番号10、配列番号11、および配列番号12に示すヌクレオチド配列にハイブリダイズし、そのヌクレオチド配列に少なくとも67%相補的な、好ましくは85%相補的な、より好ましくは95%相補的なポリヌクレオチド;または配列番号13(PTTV1a−VA)、配列番号14(PTTV1b−VA)、配列番号15(PTTV2b−VA)、配列番号16(PTTV2c−VA)に示すORF1タンパク質のアミノ酸配列、配列番号17(PTTV1a−VA)、配列番号18(PTTV1b−VA)、配列番号19(PTTV2b−VA)、配列番号20(PTTV2c−VA)に示すORF2タンパク質のアミノ酸配列、配列番号21(PTTV1a−VA)、配列番号22(PTTV1b−VA)、配列番号23(PTTV2b−VA)、配列番号24(PTTV2c−VA)に示すORF1/1タンパク質のアミノ酸配列、配列番号25(PTTV1a−VA)、配列番号26(PTTV1b−VA)、配列番号27(PTTV2b−VA)、配列番号28(PTTV2c−VA)に示すORF2/2タンパク質のアミノ酸配列からなる群より選択される免疫原性フラグメントを包含する。免疫原性または抗原性コード領域またはフラグメントは、当技術分野において知られている技法によって決定することができ、次にそれらを使って、免疫反応性スクリーニングまたは他の診断目的のためのモノクローナル抗体またはポリクローナル抗体を作製することができる。本発明はさらに、単離されたポリヌクレオチドによってコードされる精製免疫原性タンパク質を包含する。望ましくは、タンパク質は、上記分離豚TTV亜型の少なくとも1つの単離または組換えORF1タンパク質またはORF2タンパク質、より望ましくはORF1タンパク質であることができる。
【0075】
豚TTVのORF1は、構造および複製関連タンパク質をコードすると考えられている(マギー,エフ(Maggi,F.)およびベンディネリ,エム(Bendinelli,M.)、2009年、「トルクテノウイルスおよび他のアネロウイルスの免疫生物学(Immunobiology of the Torque teno viruses and other anelloviruses)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.65−90)。7つのPTTV株のORF1がコードする産物は、624〜635aaの長さを有し、DNA結合活性を有すると考えられる多数のアルギニン残基をN末端に持っている(図4)。図4では、保存されている配列を影付きで表している。ダッシュ記号はアミノ酸欠失を示す。RCRモチーフは実線で囲まれている。PTTV1株の3つのHVR(PTTV1−HVR1、2および3)とPTTV2株の2つのHVR(PTTV2−HVR1および2)は破線で囲まれている。ORF1/1の接合境界を矢印で示す。予想されるローリングサークル複製ローリングサークル複製(RCR)モチーフ(イリイナ,ティ・ブイ(Ilyina,T.V.)およびクーニン,イー・ブイ(Koonin,E.V.)、1992年、「真正細菌、真核生物および古細菌の多様なレプリコンによってコードされるローリングサークルDNA複製のための開始タンパク質における保存された配列モチーフ(Conserved sequence motifs in the initiator proteins for rolling circle DNA replication encoded by diverse replicons from eubacteria,eucaryotes and archaebacteria)」、ヌクレイック・アシッズ・リサーチ(Nucleic Acids Research)、第20巻、第13号、p.3279−85)は、異なるPTTV型および亜型では異なる位置に示され、それらの位置は型特異的または亜型特異的であり得る。RCRモチーフIII(YxxK)は、PTTV1a型株(PTTV1a−VA 配列番号13のaa位置14−17)および1b型株(PTTV1b−VA 配列番号14のaa位置379−382)において、それぞれ保存されているが、3つのPTTV2株の全てに同定された同じ保存モチーフは、PTTV2b−VA 配列番号15のaa位置482−485に位置している(図4)。PTTV2b−VA 配列番号15とPTTV2c−VA 配列番号16はどちらも、PTTV2b−VAのaa位置331−333に保存されたRCRモチーフII(HxQ)も有し、TTV−2pにはこれが存在しない(図4)。
【0076】
PTTV株のORF1タンパク質が種1と種2の間で共有する配列同一性はわずかに22.4〜25.8%と極めて低く、それが、これら2つの種の間で有意に保存されたaa配列を同定することを困難にしている(図4)。PTTV種1では、1a型株と1b型株の間のORF1のaa同一性が50.3〜52.7%である。4つのPTTV1株には、比較的多数のaa置換を伴う3つの主要超可変領域(HVR)PTTV1−HVR1〜3が同定されたのに対し、3つのPTTV2株の間には2つのHVR(PTTV2−HVR1および2)が観察された(図4)。3つのPTTV2株は、対応するPTTV1−HVR1領域に約20aaの欠失を有する。そのうえ、PTTV2の2つのHVRは、対応するPTTV1−HVR3領域内にある(図4)。これらのHVRはORF1中にのみ位置するが、切断型のORF1/1中にはない。これらは、ヒトTTVの研究によって示唆されているとおり、宿主免疫監視機構を免れ、PTTVが持続感染を樹立するのを助ける役割を果たすらしい。
【0077】
ORF2のaa配列は4つのPTTV1株(PTTV1a−VA 配列番号17;PTTV1b−VA 配列番号18)および3つのPTTV2株(PTTV2b−VA 配列番号19;PTTV2c−VA 配列番号20)の間でかなり異なっていた(図5)。しかしこれらは保存されたタンパク質チロシンホスファターゼ(PTPアーゼ)様モチーフ(Wx7Hx3CxCx5H)をN末端に共有している(図4)。このモチーフは、全てのヒトTTV、TTMVおよびTTMDV株ならびにCAVにも保存されている。TTMVまたはCAVのORF2タンパク質は、セリン/スレオニンホスファターゼ(S/T PPアーゼ)活性も示した(ピータース,エム・エイ(Peters,M.A.)、ジャクソン,ディ・シー(Jackson,D.C.)、クラッブ,ビー・エス(Crabb,B.S.)、およびブラウニング,ジー・エフ(Browning,G.F.)、2002年、「鶏貧血ウイルスVP2は新規二重特異性プロテインホスファターゼである(Chicken anemia virus VP2 is a novel dual specificity protein phosphatase)」、ジャーナル・オブ・バイオロジカル・ケミストリー(J Biol Chem)、第277巻、第42号、p.39566−73)。ORF2タンパク質の二重特異性は、ウイルス複製時に、宿主の遺伝子転写、シグナル伝達およびサイトカイン応答を調節すると思われる。最近、CAV中のモチーフの最後のヒスチジン残基の前にある保存された2つの塩基性aa残基の変異導入解析により、これら2つの残基は、ウイルス複製、インビトロの細胞病理、およびインビボの弱毒化に影響を及ぼすことが明らかになった(ピータース,エム・エイ(Peters,M.A.)、クラッブ,ビー・エス(Crabb,B.S.)、ワシントン,イー・エイ(Washington,E.A.)、およびブラウニング,ジー・エフ(Browning,G.F.)、2006年、「鶏貧血ウイルスのVP2遺伝子の部位特異的変異導入は、ウイルス複製、細胞病理および宿主細胞MHCクラスI発現に影響を及ぼす(Site-directed mutagenesis of the VP2 gene of Chicken anemia virus affects virus replication,cytopathology and host-cell MHC class I expression)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第87巻、第4号、p.823−31;ピータース,エム・エイ(Peters,M.A.)、クラッブ,ビー・エス(Crabb,B.S.)、チベンデール,ケイ・エイ(Tivendale,K.A.)、およびブラウニング,ジー・エフ、2007年、「VP2の部位特異的変異導入による鶏貧血ウイルスの弱毒化(Attenuation of chicken anemia virus by site-directed mutagenesis of VP2)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第88巻、第8号、p.2168−75)。これら2つの塩基性aa残基(「KK」)は3つのPTTV2株に保存されている。しかし、2つのPTTV1a株では最初の塩基性残基(「R」)だけが保たれており、一方、PTTV1b株では塩基性残基はどちらも置換されている(図5)。図5において、ダッシュ記号はアミノ酸欠失を示す。TTV、TTMVおよびCAVにおいて同定された共通モチーフWx7Hx3CxCx5H(下線部)内の保存された5つのアミノ酸を影付きで示す。このモチーフの最後のヒスチジンの前にある2つの塩基性アミノ酸(CAVにおいてウイルス複製、細胞病理、またはインビボ弱毒化に影響を及ぼすことが示されているもの)の位置を「^」で示す。
【0078】
要約すると、本発明は、バージニアの一頭の雄豚の血清試料における異なる遺伝子型または亜型を代表する4つの豚TTV株の完全長ゲノム配列を決定した。この研究から得られた知見は、ヒトTTVと同様に、豚には異なる遺伝子型または亜型による多重PTTV感染が存在し、おそらくそれが一般的であることを明確に示している。本発明者らは、PTTVのゲノム編成、可変性の程度、保存されているヌクレオチドモチーフおよびアミノ酸モチーフの特徴に関する新しい情報も提供した。これらの情報は、現在のPCR検出アッセイを改善し、血清学的診断法のための試薬の開発を助け、PTTVの構造的および機能的研究を開始するのに役立つであろう。この研究では、7つの既知PTTV株のゲノム配列の系統解析および遺伝子解析に基づいて、PTTVの新しい分類も提案する。
【0079】
本発明は、豚TTVに感染した豚または他の哺乳動物の試料中のウイルスDNAを検出することによる、豚TTV感染の診断方法も提供する。本発明の好ましい一実施形態は、ウイルス感染またはウイルス疾患の診断をさらに助けるために、ポリメラーゼ連鎖反応(PCR)用のオリゴヌクレオチドプライマーを使って、豚または他の哺乳動物種における豚TTV核酸配列を検出するための方法を伴う。豚または他の哺乳動物種における豚TTVウイルス核酸配列の有無を検出するのに役立つ診断検査は、豚TTVに感染した豚またはTTVの感染が疑われる豚の試料からウイルスDNAを単離することと、PTTV1特異的プライマー(配列番号29/配列番号30)またはPTTV2特異的プライマー(配列番号31/配列番号32)を使って、SYBRグリーン・リアルタイム定量PCRを行うこととを含む。
【0080】
本発明のもう一つの実施形態では、PTTV1/PTTV2デュプレックス・リアルタイムPCRを使用することによって、PTTV1とPTTV2が同時に検出されるように、診断方法を適合させることができる。より具体的に述べると、この方法は、豚TTVに感染した豚またはTTVの感染が疑われる豚の試料からウイルスDNAを単離することと、PTTV1特異的プライマー(配列番号29/配列番号30)またはPVVT2特異的プライマー(配列番号31/配列番号32)の両方を使ってリアルタイムPCRを行うこととを含む。PTTV1とPTTV2のTm値はMCAで識別することができるので、PTTV1 DNAとPTTV2 DNAの存在を同時に検出することができる。
【0081】
本発明のさらにもう一つの実施形態において、診断方法はデュプレックス・ネステッドPCRを使用し得る。この方法は、豚TTVに感染した豚またはTTVの感染が疑われる豚の試料からウイルスDNAを単離すること、1対のプライマーP1ab−mF(配列番号33)/P1ab−mR(配列番号34)を使って第1ラウンドのPCRを行うこと、PTTV1aを検出するためのP1a−nF(配列番号35)/P1a−nR(配列番号36)およびPTTV1bを検出するためのP1b−nF(配列番号37)/P1b−nR(配列番号38)という2対のプライマーの混合物を使って第2ラウンドのPCRを行うこと、およびPCR産物を可視化することを含む。
【0082】
当業者は当技術分野において周知の方法に従って、上記の診断方法を最適化することができる。
【0083】
したがって本発明の一実施形態は、2つの豚TTV種のウイルス負荷量をそれぞれ定量するために、2つの新規シングルプレックスSYBRグリーン・リアルタイムPCRアッセイを開発する。現時点で利用することができる6つのPTTV1完全長ゲノムと4つのPTTV2完全長ゲノムの間でそれぞれによく保存されている領域を標的とするPTTV1特異的プライマーとPTTV2特異的プライマーを設計した。本発明のもう一つの実施形態では、2つの豚TTV種PTTV1aおよびPTTV1bを同時検出するために、前記2つのシングルプレックスアッセイを組み合わせてデュプレックス・リアルタイムPCRアッセイを行った後、異なる融解温度によって同定することができるウイルスアンプリコンのMCAを行う。第3の実施形態では、単一試料中の豚TTV種の2つの型PTTV1aおよびPTTV1bを同定する目的で、第1ラウンドPCRにおいて2つの型のPTTV1からのウイルスDNAを同時増幅し、第2ラウンドPCRにおいて1a型と1b型の区別的検出を行うためのデュプレックス・ネステッドPCRアッセイを開発した。これらのアッセイは、種特異的または型特異的豚TTVの診断のための簡単で実用的なツールになる。
【0084】
利用可能な10個の豚TTV完全長ゲノムの多重配列アラインメントによって、潜在的プライマー配列を同定した。PTTV1特異的プライマーTTV1F(配列番号29)およびTTV1R(配列番号30)は、6個のPTTV1ゲノム間で予測ORF2の直前にある2つの保存されたゲノム領域に基づいて設計し、一方、PTTV2特異的プライマーTTV2F4(配列番号31)およびTTV2R4(配列番号32)は、4つのPTTV2ゲノム間で予測ORF2/2の直後にある2つの保存されたゲノム領域に基づいて設計した(表4)。プライマーは自己二量体化および交差二量体化の可能性を示さなかった。予期されるアンプリコンサイズは、それぞれ、PTTV1b−VAゲノムに対応するPTTV1プライマーからの118bpフラグメントと、PTTV2c−VAゲノムに対応するPTTV2プライマーからの200bpフラグメントであった。
【0085】
【表4】
【0086】
本発明の具体的一実施形態によれば、PTTV1特異的プライマーおよびPTTV2特異的プライマーを用いるSYBRグリーン・シンプレックス・リアルタイムPCRを使って、それぞれ豚TTV1およびTTV2 DNAを特異的に検出することができる。PTTV1については、ある範囲の、25μlあたりのターゲットDNA濃度にわたって、標準曲線を確立した。線形範囲は、4.4×101〜4.4×108コピーにまたがることが示された。最低検出限界(44コピー)は、37.57の閾サイクル数(Ct)に対応した。PTTV2についても標準曲線を作成し、それを使って、25μlの反応あたり8.6×100〜8.6×108コピーの範囲にあるDNA濃度を検出した。最低検出限界(8.6コピー)に対応するCtは36.53だった。
【0087】
本発明のもう一つの具体的実施形態では、豚TTV1 DNAと豚TTV2 DNAの同時検出に、SYBRグリーン・デュプレックス・リアルタイムPCRを利用する。PTTV1(87.0℃)とPTTV2(80.0℃)の間の7度というTm値の差が、それらをMCAによって互いに識別することを可能にした。したがって、PTTV1とPTTV2を同時検出するために、2つのシングルプレックスアッセイを結びつけてデュプレックス・リアルタイムPCRアッセイにすることができる。陽性試料は、その産物についての既知Tm内で対称的な融解ピークを有するものであった。この新しいアッセイを、まず最初に、PTTV1標準とPTTV2標準の混合物の10倍希釈液を使って検証した。テンプレートとして滅菌水を使用した非テンプレート陰性対照は、シングルプレックスアッセイでは見られなかったPTTV1プライマーとPTTV2プライマーの間の交差二量体化が引き起こす非特異的増幅を示した(図7a)。これにより、72.0℃〜76.0℃に別個の融解ピークが生じた。図7Aは、PTTV1標準(赤色;Tm=87.0℃)、PTTV2標準(緑色;Tm=80.0℃)および非テンプレート陰性対照(プライマー交差二量体化によるもの;黒色)の融解ピークを示している。図7B〜7Eは、異なるウイルス負荷量のPTTV1とPTTV2とを含む代表的な血清試料の融解ピークを示している。図7Bは、雄豚血清試料番号5(PTTV1とPTTV2のウイルス負荷量はどちらも比較的高いが、PTTV2>PTTV1)を表し、図7Cは、雄豚血清試料番号12(PTTV1とPTTV2のウイルス負荷量はどちらも比較的高いが、PTTV1>PTTV2)を表し、図7Dは雄豚血清試料番号14(PTTV1とPTTV2のウイルス負荷量はどちらも低い)を表し、図7Eは雄豚血清試料番号10(PTTV1陽性だが、PTTV2陰性)を表す。シングルプレックス・リアルタイムPCRによって決定された各試料中のPTTV1およびPTTV2のウイルス負荷量(単位:ゲノムコピー/ml)を、対応する融解ピークの上に示した。
【0088】
ある例では、デュプレックス・リアルタイム・アッセイを20個の成体雄豚血清試料に適用した場合に、PTTV1とPTTV2のウイルス負荷量がどちらも比較的高い試料は、PTTV1とPTTV2に対応する2つの異なる融解曲線を示し、非特異的融解ピークは示さなかったのに対し(図7Bおよび7C)、PTTV1またはPTTV2のどちらか一方のウイルス負荷量が低い試料は、ウイルス特異的融解曲線だけでなく、非特異的融解曲線も示した(図7Dおよび7E)。試料番号14における2つの融解ピークは極めて小さかったが、それらはPTTV1およびPTTV2の適当なTmに視覚的に明確で対称的な上昇と降下を示したので、それらは陽性と見なされた(図7D)。これに対し、試料番号10は、対称的なPTTV2融解ピークが明白には存在しなかったので、PTTV1のみ陽性と見なされた(図7E)。これらの結果は、2つのシングルプレックスアッセイの結果と一致した(表5)。さらにまた、融解ピークのサイズと形状は、検出された試料における対応するウイルス負荷量を定量的に反映した。
【0089】
本発明のもう一つの態様では、デュプレックス・ネステッドPCRを、2つの豚TTV型PTTV1aおよびPTTV1bの区別的検出に使用する。
【0090】
本発明の発明者は、豚TTV種1における2つの異なる遺伝子型(暫定的にPTTV1aおよびPTTV1bと名付けた)の存在を証明した。さらに、PTTV1aとPTTV1bの同時感染が豚において一般的であるかどうかを決定するために、これら2つを迅速に識別するための新規デュプレックス・ネステッドPCRアッセイを開発した。豚TTVゲノムDNA配列のアラインメントにより、ウイルスキャプシドタンパク質をコードする予測ORF1のN末端部分に位置する保存されたゲノム領域が同定された(図8)。この領域はORF2全体と上流の部分的UTRも含有している。第1ラウンドPCRにおいてPTTV1a DNAとPTTV1b DNAの両方を同時に増幅するために、プライマーP1ab−mF(配列番号33)/P1ab−mR(配列番号34)を設計した。第2ラウンドPCRでは、PTTV1a特異的プライマーP1a−nF(配列番号35)/P1a−nR(配列番号36)とPTTV1b特異的プライマーP1b−nF(配列番号37)/P1b−nR(配列番号38)との混合物を使って、各遺伝子型を区別的に増幅した。PTTV1aおよびPTTV1bの最終PCR産物は、それぞれサイズが162bpおよび96bpであり、それらは臭化エチジウムで染色した1%アガロースゲルでの電気泳動により、容易に識別することができた。このアッセイでは、プライマーの特異性ゆえに、PTTV2 DNAは検出されないと予期された(図8)。図8では、保存されている配列を点で示し、影付きにした。ダッシュ記号によってヌクレオチド欠失を示した。デュプレックス・ネステッドPCRに使用した3対のプライマーの場所と向きを矢印で示した。
【0091】
ある例では、デュプレックス・ネステッドPCRアッセイに付した成体雄豚からの血清試料20個は、予期されるサイズの2本のバンドの可視化と、続いて行われた配列決定によるPCR産物の確認とによる決定で(データ未掲載)、全てが、PTTV1aとPTTV1bの両方について陽性であることがわかった。19個の精液試料ではPCR産物が増幅されず、上述したPTTV1従来型ネステッドPCRおよびリアルタイムPCRアッセイの結果と合致した。
【0092】
2つの豚TTV種による豚の感染は、遡及的研究によれば、スペインの養豚場において1985年には見いだされている(シーゲイルス(Segales)ら、2009年、前掲書)。しかし豚TTVが何らかの特定豚疾患と関連しているかどうかは依然として不明である。どちらの豚TTV種も飼育豚において高い有病率を有するので、TTV DNAの存在を評価することよりも、TTVウイルス負荷量の決定の方が、おそらく、より重要である。血清試料および精液試料におけるウイルス負荷量のレベルは、PCV2感染におけるPCVADの重要なマーカーとして示されている(オプリースニヒ(Opriessnig)ら、2007年、前掲書)。したがって、定量的なPTTV特異的リアルタイムPCRアッセイの確立は、豚TTVに関連する潜在的疾患状態を同定するのに役立つだろう。
【0093】
2つのタックマン(TaqMan)プローブ・ベース・リアルタイムPCRアッセイが最近になって記載された。カナダのグループが開発したシングルプレックス・アッセイは種特異的ではなく、2つのPTTV種の総ウイルス負荷量を定量するようにしか設計されなかった(ブラッサード(Brassard)ら、2009年、前掲書)。ドイツのグループが確立したデュプレックス・アッセイでは両方の種の特異的な同時検出が可能だった(ガレイ(Gallei)ら、2009年、前掲書)。これら2つのアッセイにおいて使用されたプライマーのターゲット配列は、3つの豚TTVゲノム配列(Sd−TTV31、TTV−1pおよびTTV−2p)のアラインメントによって決定されたもので、UTR中に位置していた。本研究において、本発明者らは、利用可能な7つの完全PTTVゲノム配列(4つのPTTV1配列と3つのPTTV2配列)を追加して、10個の完全長PTTVゲノムの間で保存されている領域を分析し、再決定した。この研究で得られた最新のアラインメント結果に基づいて、PTTV1とPTTV2のウイルス負荷量をそれぞれ定量するために、2つの種特異的シングルプレックスSYBRグリーンベース・リアルタイムPCRアッセイを開発した。本発明者らのアッセイで使用したプライマーは、先の研究とは異なる保存ゲノム領域に結合するように設計されたもので、これらは定量の正確度を向上させ得る。本発明者らのアッセイは、25μlの反応あたり、PTTV1については44ゲノムコピー数、PTTV2については8.8ゲノムコピー数と、かなりの種特異性および選択性を示した。これに対し、タックマン(TaqMan)ベースのデュプレックス・リアルタイムPCRでは、1反応あたり10ゲノムコピー数という検出限界が報告されている(ガレイ(Gallei)ら、2009年、前掲書)。また、SYBRグリーンベースのリアルタイムPCRアッセイは、蛍光標識したプローブを使用する必要がなく直ちに行うことができるフレキシブルで安価なアプローチである。最後に、豚TTVが高度の遺伝的多様性を示すことを考えると、SYBRグリーンベース・アッセイによる結果は、タックマン(TaqMan)プローブベースのアッセイにおけるプローブ結合配列中に突然変異を含有する可能性が高い豚TTV変異体の異なる遺伝的背景による影響を受ける可能性が少ない。
【0094】
TTV DNAが存在するとはいえ、この研究において試験された健常な豚からの血清試料はいずれも、2×106コピー/ml未満という少量のPTTV1およびPTTV2を有していた。さらにまた、3つの精液試料には、極めて低いタイターのPTTV2 DNAしか検出されなかった。試験した血清試料の大半は、従来のネステッドPCRで決定したところ、PCV2 DNAについても陽性だった(データ未掲載)。ウイルス負荷量が低い多くのPCV2陽性豚は、臨床的PCVADを発症しない。PCVADの発症に関して提案されている閾は、107以上のPCV2ゲノムコピー/ml血清である(オプリースニヒ(Opriessnig)ら、2007年、前掲書)。また、精液PCV2 DNA陽性も、疾患状態の注目すべきマーカーである(オプリースニヒ(Opriessnig)ら、2007年、前掲書;パル,エヌ(Pal,N.)、ファン,ワイ・ダブリュ(Huang,Y.W.)、マドソン,ディ・エム(Madson,D.M.)、カスター,シー(Kuster,C.)、モン,エックス・ジェイ(Meng,X.J.)、ハルバー,ピー・ジー(Halbur,P.G.)およびオプリースニヒ,ティ(Opriessnig,T.)、2008年、「豚精液試料における豚サーコウイルス2型および内部対照の同時検出および定量のためのデュプレックス・リアルタイムPCRアッセイの開発と検証(Development and validation of a duplex real-time PCR assay for the simultaneous detection and quantification of porcine circovirus type 2 and an internal control on porcine samen samples)」、ジャーナル・オブ・バイロロジカル・メソッズ(J Virol Methods)、第149巻、p.217−25)。種特異的PTTVの状況はPCV2のそれと類似していると考えられ、豚疾患の誘導には107コピー/mlを上回る高いPTTV価が要求されるのだろう。この研究において開発された種特異的リアルタイムPCRアッセイは、今後、さまざまな疾患状態から採取される多数の臨床試料を使ってPTTVと疾患との関連を調べるための簡単で実用的なツールになるだろう。
【0095】
さらにまた、これら2つの種特異的シングルプレックス・アッセイを結びつけることにより、本発明者らは、MCAベースのデュプレックス・リアルタイムPCRアッセイにおいて2つの豚TTV種PTTV1とPTTV2を同時に検出し区別するための迅速で安価で信頼できるスクリーニングを開発し、検証した。このアッセイは両PTTV種の正確な定量を意図したものではないが、検出を目的とするなら、従来のネステッドPCRに取って代わることのできる、より便利なアプローチである。リアルタイムPCRと比較すると、豚TTV検出のための従来のネステッドPCRアッセイは、時間がかかり(合計4ラウンドのPCRを必要とする)、面倒であり、複数ラウンドのPCR処理中に起こる試料汚染に見舞われやすい。PTTV1種とPTTV2種の間のTm値の相違ゆえに、デュプレックスPCR増幅後のMCAにより、異なる反応特異性を確保することができる。このデュプレックス・リアルタイム・アッセイのもう一つの利点は、記載のプロトコールを行う際にPTTV1標準とPTTV2標準を含めることが必須でなくなる点であり、これにより、自動リアルタイムPCR機器を装備した診断検査室におけるはるかに幅広い使用が容易になる。
【0096】
同じ種の異なる遺伝子型または亜型による豚TTVの多重感染は実証されている(ガレイ(Gallei)ら、2009年、前掲書)。特に、本発明者らの以前の研究では、豚TTV種1は、2つの異なる型PTTV1a(株Sd−TTV31およびPTTV1a−VAを含む)およびPTTV1b(株TTV−1pおよびPTTV1b−VAを含む)からなることが示された。完全長ゲノムを持つ新たに公表された2つのPTTV1分離株、カナダ由来のswSTHY−TT27(GQ120664)とドイツ由来のTTV1#471819(GU188045)は、どちらも、系統解析に基づいて1b型に分類された(データ未掲載)。この研究において記載するデュプレックス・ネステッドPCRにより、2つのPTTV1遺伝子型の二重感染は豚では頻繁に起こることが確認された。この新規アッセイは、PTTV1aと1bを識別するために現在までに開発された最初の診断的PCRアプローチである。PTTV1a感染とPTTV1b感染の一方または両方が疾患と関連する関連要因を表すかどうかは現時点ではわかっていないので、本発明者らの区別的PCRアッセイは、これら2つのPTTV型の今後の潜在的疾患関連性にとって、大いに価値があるはずである。
【0097】
本発明のもう一つの態様では、豚TTV ORFタンパク質を発現させ、豚TTV特異的抗体の存在を検出するための免疫検出アッセイにおいて使用した。本発明の一実施形態では、PTTV1a、PTTV1bおよびPTTV2の3つの切断型ヒスチジンタグ付きORF1タンパク質を、大腸菌(E.coli)中で、それぞれ発現させた。さらにまた、これらの組換え抗原に基づく血清ウェスタンブロットおよびELISAアッセイを開発し、異なる供給源からの豚血清試料を使って検証した。特に、PTTV1a特異的、PTTV1b特異的およびPTTV2特異的ELISAを使った血清検査は、豚TTV感染と疾患との関連を明らかにするための正確で簡単なツールになる。
【0098】
本発明のさらなる一態様では、豚TTV ORFタンパク質を、大腸菌発現系において、組換えORF1キャプシドタンパク質として発現させ、精製した(図10、図15)。PTTV1a、PTTV1bおよびPTTV2の3つの切断型Hisタグ付きORF1タンパク質を、大腸菌(E.coli)中で、それぞれ発現させ、精製し、PTTV感染に対する組換えキャプシドサブユニットワクチンとして利用した。
【0099】
4つの豚TTV2株、TTV−2p、TTV2#472142、PTTV2b−VAおよびPTTV2c−VAは、現在までに、利用可能な完全ゲノム配列を持っていた。それらは系統学的に3つの予測亜型に分類されるが、4つのPTTV2に由来するアミノ酸をコードするORF1の親水性プロファイルの比較分析により、それらは3つの親水性領域、すなわちN末端にあるaa1〜49のアルギニンリッチ領域と、それぞれ中央部とC末端部に位置する2つの特定ドメイン(IおよびII)を共有することが示された(図9A)。切断型PTTV2c−VA ORF1発現に使用したC末端領域と、他の3つのPTTV2株において共有されている対応領域を、破線の枠で示した。アミノ酸配列のアラインメントにより、4つのPTTV2株間でのドメインI(aa322−349)およびII(aa536−625)の高レベルな配列保存が示された(図9B)。
【0100】
親水性ドメインは多くのタンパク質の抗原性にとって重要であると考えられるので、上記2つのドメインを含有するPTTV2c−VA ORF1 配列番号16のC末端領域(aa310−625)を、豚血清におけるPTTV2特異的抗体検出用の抗原として使用されるタンパク質発現のために選択した。本発明の一態様によれば、切断型PTTV2c ORF1の発現は、全てのPTTV2亜型(2a、2bおよび2c;図3Aも参照されたい)を検出するのに十分であった。
【0101】
本発明の一実施形態では、8×Hisタグと融合したPTTV2c ORF1遺伝子のC末端部分を構築し、大腸菌中で発現させた。組換えタンパク質は不溶性であり、細菌封入体内に発現した。図10Aは、未精製2c−ORF1産物のSDS−PAGEを示す。図10Bは、精製2c−ORF1産物のSDS−PAGEを示す。図10Cは、抗HisタグmAbを使った精製2c−ORF1産物のウェスタンブロット分析を示す。白い矢じりは、予期されるサイズを持つORF1タンパク質とその切断産物を示し、一方、黒い矢じりは、予期される切断型タンパク質の予測二量体を示す。M:タンパク質マーカー。図10Aにおいて、2c−ORF1未精製試料では、対照試料と比較して、2つの際立ったポリペプチド(白い矢じり)が生成していた。約40KDaのバンドは2c−ORF1の予期されるサイズと一致し、一方、約30KDaポリペプチドは、おそらく、前者からのN末端切断産物であった。ニッケルアフィニティカラムによる精製後に、2つの前記際立ったバンドを含む4つのポリペプチドが、SDS−PAGEに示された(図10B)。それらは、抗HisタグmAbを用いるウェスタンブロットでも検出された(図10C)。2つの高分子量バンド(黒い矢じり)は、予想されるサイズ(約80KDaおよび約60KDa)によれば、それぞれ約40KDaおよび約30KDaの2つのポリペプチドによって形成されるホモ二量体だった。これらの結果から、精製C末端PTTV2c−ORF1がうまく生産され、豚血清中の豚TTV2抗体検出に使用できることが証明された。
【0102】
本発明のもう一つの態様によれば、さまざまな豚血清試料における豚TTV2抗体を、精製C末端PTTV2c−ORF1を使ったウェスタンブロットによって検出することができる。白い矢じりは、予期されるサイズを持つORF1タンパク質とその切断産物を示す。緑色のバンドだけが陽性と認識されることに注意すべきである。さまざまな供給源から得た在来豚(健常または罹患)、CD/CD豚およびノトバイオート豚の血清試料を全部で200個以上収集した。抗原として精製C末端PTTV2c−ORF1を使って抗PTTV2c−ORF1 IgG抗体を検出するために、試料をランダムに選択した。図11Aは、在来豚の選ばれた豚血清試料のウェスタン分析の結果を示し、図11BはCD/CD豚を示し、図11Cはノトバイオート豚を示す。精製PTTV2c−ORF1産物を抗原として使用した。印を付けた約40KDaと約30KDaの2つのバンドは、在来豚(図11A)およびCD/CD豚(図11B)の試料の大半に検出されたことから、これらの豚におけるPTTV2感染が幅広く示された。しかし、2つの異なる供給源(バージニア州ブラックスバーグおよびアイオワ州エイムズ)から得たノトバイオート豚はいずれも、検出可能なPTTV2抗体を持っていなかった(図11C)。別の低分子量バンドも観察された(図11Aおよび11B)。それらはおそらくウェスタンブロットにおける非特異的反応性に由来したのだろう。
【0103】
本発明のさらにもう一つの態様によれば、PTTV2特異的ELISAを、豚TTV2血清検査として使用することができる。血清陰性結果は、ウィスコンシンの飼育場の在来豚からの数個の試料にも示された(図12)。これらの陰性試料をプールし、PTTV2特異的ELISAの開発において陰性参照として使用した。同じ供給源から得られた残りの試料は陽性だった(図12、左側の4レーン)。また、細胞培養に使用される営利会社から得た豚血清(OIE疾患フリーと考えられる)も、強い抗PTTV2−ORF2陽性を示し(図12)、これをELISAの陽性対照として使用した。精製2c−ORF1抗原、豚血清およびIgGコンジュゲートの濃度は、交差力価測定法により、低いバックグラウンドシグナルを呈し、陽性対照と陰性対照とのOD405値の差が最も大きくなるように決定した。最適な抗原量は1ウェルあたり69ngであり、最適なELISA結果は、血清試料の1:100希釈液およびIgGコンジュゲートの1:4000希釈液を使用することによって得られた。ELISAカットオフ値は各試行において0.25〜0.5の範囲にあった。図4は、血清ウェスタンブロットと開発されたELISAとの整合性を反映した代表的結果を示す。
【0104】
3つの群れから得た138個の在来豚血清試料を選んで、リアルタイムPCRによるPTTV2ウイルス負荷量とELISAによる抗PTTV2IgG抗体レベルの間の相関関係を分析した。その結果、検出不可能であるか高いPTTV2ウイルス負荷量(108コピー/ml)を持つ豚の方が、中間的な値のPTTV2ウイルス負荷量を持つ豚よりも低い血清PTTV2抗体価を持つ可能性が高いことが示された(図13)。
【0105】
特に、同じ群れの豚10頭からの血清を、それらが新しい施設に到着した時から到着の2ヶ月後まで、それらの血清のPTTV2ウイルス負荷量と抗PTTV2抗体レベルとを比較することによっても分析した。10頭中9頭の豚は2ヶ月後にウイルス負荷量が減少していた(3頭は検出可能なウイルスを持っていなかった)が、抗PTTV2抗体価は10頭中9頭の豚において増加していた(図14Aおよび14B)。これらの結果から、10頭の豚は早期にPTTV2感染を獲得し、それが体液性応答を誘導して、徐々に抗ORF1キャプシドIgG抗体を産生したことが示唆された。PTTV2−ORF1 IgG抗体は、ウイルスを中和することができ、またはウイルスを除去することさえできたことから、ORF1が実際にウイルスキャプシドタンパク質をコードし、PTTV2に対する中和エピトープを含有し得ることが示された。
【0106】
本発明の一実施形態では、C末端PTTV1a−およびPTTV1b−ORF1タンパク質を、それぞれ大腸菌系で発現させ、精製した。SDS−PAGEと、抗HisタグmAbを使ったウェスタンブロット分析により、1a−ORF産物と1b−ORF産物はどちらも2つのポリペプチドを生じ、一つは予期されるサイズのもの(約40KDa)、もう一つは予測ホモ二量体(約80KDa)であることが示された(図15A〜C)。図15Aは、未精製および精製1a−ORF1産物のSDS−PAGEを示す。図15Bは、精製1b−ORF1および1b−ORF1ctruc産物のSDS−PAGEを示す。図15Cは、抗HisタグmAbを使った精製1a−および1b−ORF1産物のウェスタンブロット分析を示す。白い矢じりは予期されるサイズのORF1タンパク質を示し、一方、黒い矢じりはORF1タンパク質の予測二量体を示す。2c−ORF1発現と比較して、切断型ポリペプチドは観察されなかった。比較対照として、C末端切断型1b−ORF1領域(1b−ORF1ctruc)の発現は、そのC末端非切断対応物1b−ORF1と比較して、低分子量ポリペプチドをもたらした(図15B)。
【0107】
本発明の一実施形態では、PTTV2について上述したように、精製C末端PTTV1a−およびPTTV1b−ORF1タンパク質を使って、遺伝子型特異的な血清ウェスタンブロットおよびELISAを開発した。図16は、1a−ORF1を抗原として用いる血清ウェスタンブロットの陰性例(レーン1〜2)と陽性例(レーン3〜5)を示す。PTTV1a特異的およびPTTV1b特異的ELISAでは、PTTV2−ORF1と同じ抗原量(69ng)、血清の希釈度(1:100)およびIgGコンジュゲートの希釈度(1:4000)を使用した(データ未掲載)。
【0108】
さらに本発明は、豚TTV感染を検出するための有用な診断試薬であって、例えば豚に豚TTVまたは本発明の免疫原性組成物を有効免疫原量で接種してウイルス感染を生じさせ、感染した豚の血清から抗体を回収することなどによって自然宿主から精製された、モノクローナルまたはポリクローナル抗体を含む診断試薬を提供する。あるいは、単離された豚TTVのヌクレオチド配列によってコードされるアミノ酸配列または免疫原性フラグメントに由来する、またはそこから発現される、天然または合成ポリペプチドに対して、実験動物において、抗体を産生させることもできる。例えば、モノクローナル抗体は、単離された豚TTVのヌクレオチド配列に由来するポリペプチド抗原で免疫化された例えばBalb/cなどのマウスから得られるハイブリドーマ細胞から産生され得る。ハイブリドーマ細胞の選択は、ダルベッコ変法イーグル培地(DMEM)または最小必須培地のような標準的細胞培養培地に入れたハイプロキサンチン(hyproxanthine)、チミジンおよびアミノプテリン中で成長させることによって行う。抗体を産生するハイブリドーマ細胞は、当技術分野において知られている手法に従ってクローン化することができる。次に、形成される離散コロニーを、適切な培養培地で培養するために、培養プレートの別々のウェルに移すことができる。抗体分泌細胞の同定は、適当な抗原または免疫原を使って、従来のスクリーニング法によって行われる。ハイブリドーマ細胞をインビトロで培養するか、ハイブリドーマの注入後にマウスの腹水を得ることにより、インビボで培養することで、所望のモノクローナル抗体が周知の技法で生産される。
【0109】
もう一つの代替的方法では、当技術分野において知られている手法に従って、豚TTVキャプシドタンパク質をバキュロウイルス発現系または大腸菌発現系で発現させることができる。発現した組換え豚TTVキャプシドタンパク質は、診断用の抗原として、酵素結合免疫吸着アッセイ(ELISA)において使用することができる。豚組換えキャプシド抗原に基づくELISAアッセイは、例えば、豚および哺乳動物種における豚TTVに対する抗体を検出するために使用することができる。ELISAアッセイは好ましいものの、免疫蛍光アッセイ(IFA)、免疫ペルオキシダーゼアッセイ(IPA)などといった他の既知の診断検査も使用することができる。
【0110】
望ましくは、豚における豚TTV感染を診断するために、本発明による市販のELISA診断アッセイを使用することができる。豚TTVの精製ORF1およびORF2タンパク質を使った、豚における抗TTV抗体を検出するためのELISAアッセイの開発を、実施例に例示する。豚TTVに感染した豚から集めた血清と、対照豚から得た陰性血清とを使って、アッセイを検証する。PTTV2特異的抗体、PTTV1a特異的抗体、およびPTTV1b特異的抗体が、PTTV ORFタンパク質を特異的に認識することが証明された。当業者に知られる技法による検査のさらなる標準化により、豚TTVに関する診断アッセイの商品化を最適化することができる。
【0111】
本発明のもう一つの態様は、分離された豚TTVまたは上述の単離されたポリヌクレオチドによってコードされる抗原タンパク質を含むユニークな免疫原組成物、および抗体を産生または生産するためのその使用である。組成物は、無毒性の生理学的に許容される担体と、場合によっては、1つ以上のアジュバントとを含有する。適切な担体、例えば水、食塩水、エタノール、エチレングリコール、グリセロールなどは、従来の賦形剤から容易に選択され、共製剤化剤を加えることができる。最終組成物の物理的適合性および安定性を保証するために、日常的検査を行うことができる。
【0112】
本発明によれば、豚トルクテノ(TTV)の感染性分子および核酸分子、その核酸分子から産生される生ウイルス、および豚TTVウイルス感染または豚TTVと他のウイルスとの同時感染が引き起こす疾患から豚を保護するための動物用ワクチンが、さらに提供される。本発明はさらに、ワクチンとして使用することができる免疫原性ポリペプチド発現産物を提供する。
【0113】
豚TTVの新規感染性DNA分子は、感染性PTTV1a−VA(配列番号9)、PTTV1b−VA(配列番号10)、PTTV2c−VA(配列番号11)、またはPTTV2c−VA(配列番号12)ゲノムの少なくとも一部をコードする核酸分子を含む。感染性PTTV DNAクローンは、好ましくは、PTTV1またはPTTV2のORF1、ORF2、ORF1/1、およびORF2/2遺伝子の少なくとも一つを含有する。複数コピーのPTTV1a−VA(配列番号9)、PTTV1b−VA(配列番号10)、PTTV2c−VA(配列番号11)、またはPTTV2c−VA(配列番号12)ゲノムを、単一のDNA分子に挿入して、タンデム感染性PTTVクローンを構築してもよい。
【0114】
本明細書に記載するPTTV、特にPTTV1a−VA、PTTV1b−VA、PTTV2c−VA、およびタンデムPTTV2b−RR、PTTV2c−RRのクローニングされたゲノムDNAは、PK−15細胞にトランスフェクトして豚に与えた場合に、インビトロまたはインビボで感染性であることが示される。この新しい、容易に再現できる病原因子は、豚におけるPTTV感染を予防するための適切なワクチン接種プログラムの開発に向いている。
【0115】
本発明のさらなる一実施形態では、3つの1ゲノムコピーPTTV DNAクローンを、それぞれ原型米国分離株PTTV1a−VA、PTTV1b−VAおよびPTTV2c−VAから、融合PCR(fusion PCR)によって誘導した。完全長ゲノムDNAのそれぞれを、平滑末端ライゲーションによって、クローニングベクターpSC−B−amp/kanに挿入した。制限部位BamHIは、3つのPTTVゲノム上のユニーク部位であり、これは、コンカテマーの生成を容易にし、したがってTTV環状ゲノムを模倣するために、3つのゲノムの両端に工学的に作製されたものである。各クローンの選ばれたプラスミドDNAのBamHI単独消化は、明らかに、サイズが4.3Kbと2.8Kbの二つの異なるフラグメントをもたらした(図18A)。4.3Kbフラグメントがバックボーンベクターに相当し、一方で2.8Kbフラグメントは、挿入されたPTTVゲノムDNAに相当した。同じ酵素だけで消化した空ベクターpSC−B−amp/kanは、4.3Kbバンドを示した(図18A)。得られたPTTVクローンを、それぞれpSC−PTTV1a、pSC−PTTV1bおよびpSC−PTTV2cと名付けた(図17A〜C)。
【0116】
さらにまた、クローンpSC−PTTV2cから得られる2コピーの完全長PTTV2c−VAゲノムを、pSC−B−amp/kanベクター中に、直列にライゲーションして、クローンpSC−2PTTV2c−RR(図17D)を作製した。pSC−PTTV2cとpSC−2PTTV2c−RRのAflII単独消化パターンを比較したところ、後者のプラスミドは、PTTV2cゲノムの2つめのコピーに相当する追加の2.8Kbフラグメントを有することがわかった(図18B、右側)。次に、本発明者らは同じクローニング戦略を利用して、ドイツTTVクローンTTV2−#471942−fullから、タンデム二量体化PTTV2b DNAクローンを作製した。pSC−2PTTV2b−RR(図17F)と名付けたこのコンストラクトには、これをHindIIIだけで消化して、その1ゲノムコピー対応物と比較した場合に、PTTV2bゲノムの2つ目のコピーに相当する追加の2.8Kbフラグメントが存在したことから(図18B、左側)、構築の成功が確認された。
【0117】
構築したPTTV感染性クローンの複製能を、PK−15細胞のインビトロ・トランスフェクションによって調べた。業者が作製したPTTV2cORF1に対するウサギポリクローナル抗体を用いるIFAにより、クローンTTV2−#471942−fullとpSC−PTTV2cのコンカテマーはどちらも、それぞれ複製能を有することが確認された(図19Aおよび図20A)。トランスフェクト細胞の継代では、蛍光シグナルが排除されず減少もしなかったことから(図19Bおよび図20B)、ORF1タンパク質の発現は、天然のPTTV2bまたはPTTV2c環状分子を模倣したPTTV2コンカテマーからもたらされることが示唆された。モックトランスフェクト細胞、またはIFA検出用の抗体として免疫前ウサギ血清を使用したDNAトランスフェクト細胞には、蛍光シグナルが観察されなかった(データ未掲載)。クローンpSC−PTTV1aのコンカテマーも複製能を有することが抗PTTV1a ORF1抗体を使って示された(図21)。陽性蛍光シグナルは、トランスフェクト細胞または継代細胞の核に位置することから、豚TTVは細胞核内で複製するらしいことが示された。豚サーコウイルス(PCV)はインビトロで類似の発現パターンを有するので、これは予期されないことではない。
【0118】
PK−15細胞におけるタンデム二量体化クローンpSC−2PTTV2b−RRまたはpSC−2PTTV2c−RRの直接トランスフェクションは、ウイルス複製をもたらし、ORF1キャプシド抗原を産生する。PTTV2 ORF1に対する抗体を使ったIFAにより、どちらのクローンも複製能を有し、陽性ORF1抗原は核内に局在化することが確認された(図22AおよびB)。
【0119】
本発明の一実施形態によれば、豚TTVの感染性クローンを使って豚に接種することができる。それは次に、宿主動物の免疫応答を引きだし、中和抗体の産生を刺激するであろう。本発明のある特定実施形態において、2つのタンデム二量体化PTTV2クローンを在来豚のリンパ節および筋中に注射したところ、それらは感染性であった。
【0120】
PTTV2分子クローンのインビボ感染性を調べるために、在来豚に、クローンpSC−2TTV2b−RRまたはpSC−2TTV2c−RRを接種した。接種後0、7、14、21および28日目(0、7、14、21および28DPI)に動物から血清試料を集めた。PTTV2 DNAは、pSC−2TTV2c−RR接種豚では、7DPI以降(#92)、14DPI以降(#188および#191)および21DPI以降(#180)に、それぞれ検出された(図23A、右側)。クローンpSC−2TTV2b−RRを接種した豚では、PTTVウイルス血症が遅れて現れ、2頭は14DPI以降(#189および#192)、1頭は21DPI以降(#181)、1頭は28DPI以降(#193)だった(図23A、左側)。PTTV2特異的リアルタイムPCRで決定したところ、ウイルス負荷量は、接種した全ての豚において、時間の経過と共に増加し、ウイルス負荷量は剖検前の28DPIが最も高かった(図23A)。選ばれた豚から増幅されたリアルタイムPCR産物を配列決定したところ、pSC−2TTV2b−RRまたはpSC−2TTV2c−RRの対応する領域と同一の配列を有することがわかった(データ未掲載)。
【0121】
接種された全ての豚は、0DPIおよび7DPIでは、PTTV2 ORF1抗体に関して陰性だった。4頭のpSC−2TTV2b−RR接種豚の全てが14DPIにおいて抗PTTV2 ORF1 IgG陽性に血清転換したのに対し、pSC−2TTV2c−RR接種群の豚は、14(#92および#180)、21(#191)および28(#188)DPIにおいて、それぞれ血清転換した(図23B)。これらの結果は、活動性の豚TTV2bまたはTTV2c感染が起こったことを示した。
【0122】
感染性ウイルスおよび分子DNAクローンのワクチン、ならびにそれらを使用する方法も、本発明の範囲に包含される。接種された豚は、ウイルス感染およびTTV2感染または同時感染によって引き起こされる関連疾患から防御される。この新規方法では、ウイルス感染から防御する必要がある豚を、その豚に、免疫学的有効量の本発明のワクチン(例えば、免疫原量の感染性PTTV DNA、PTTVの感染性DNAクローンを含有するプラスミドまたはウイルスベクター、組換えPTTV DNA、ポリペプチド発現産物、細菌発現またはバキュロウイルス発現精製組換えORF1キャプシドタンパク質を含むワクチンなど)を投与することによって防御する。広範囲にわたるウイルス感染からの防御が得られるように、他の抗原、例えばPRRSV、PPV、他の感染性豚因子および免疫刺激物質を、豚に同時投与してもよい。
【0123】
ワクチンは、例えば、無毒性の生理学的に許容される担体および場合によっては1つ以上のアジュバントと組み合わされた、感染性ウイルスおよび分子DNAクローン、例えばpSC−Bベクターなどの適切なプラスミドまたはベクター中のクローン化PTTV感染性DNAゲノム、無発病性生ウイルス、不活化ウイルス、発現された組換えキャプシドサブユニットワクチンなどを含む。ワクチンは、本明細書に記載の感染性TTV2分子DNAクローンも含み得る。感染性PTTV DNA、感染性ウイルスゲノムを含有するプラスミドDNA、および生ウイルスは好ましく、生ウイルスは最も好ましい。毒性状態に復帰するリスクを冒す弱毒化生ウイルスを使用するか、ウイルス疾患からの防御にとって十分な抗体免疫応答を誘導しない可能性がある死滅細胞培養増殖全ウイルスを使用する従来のウイルスワクチンと比較して、本発明の無発病性生ウイルスワクチンは有利である。
【0124】
ワクチンおよびそれらを使用する方法も本発明の範囲に包含される。接種された哺乳動物種は、重症ウイルス感染から防御され、PTTVの同時感染に関係する疾患、例えば豚皮膚炎腎症症候群(PDNS)、離乳後多臓器消耗症候群(PMWS)、および他の関連疾病からも防御され得る。ワクチンは、例えば、不活化または弱毒化豚TTVウイルス、無毒性の生理学的に許容される担体、および場合によっては、1つ以上のアジュバントを含む。
【0125】
本発明のワクチンと一緒に投与し得るアジュバントは、ワクチンに対する豚の免疫学的応答を増加させる物質である。アジュバントは、ワクチンと同時に同じ部位に投与するか、異なる時点で、例えばブースターとして投与することができる。アジュバントは、ワクチンを投与する方法とは異なる方法で投与すること、またはワクチンを投与する部位とは異なる部位に投与することが、有利な場合もある。適切なアジュバントには、例えば水酸化アルミニウム(アラム)、免疫刺激複合体(ISCOM)、非イオン性ブロックポリマーまたはコポリマー、サイトカイン(例えばIL−1、IL−2、IL−7、IFN−α、IFN−β、IFN−γなど)、サポニン、モノホスホリルリピドA(MLA)、ムラミルジペプチド(MDP)などがあるが、これらに限るわけではない。他の適切なアジュバントには、例えば硫酸アルミニウムカリウム、大腸菌由来の非耐熱性または耐熱性エンテロトキシン、コレラ毒素またはそのBサブユニット、ジフテリア毒素、破傷風毒素、百日咳毒素、フロイント不完全または完全アジュバントなどがある。ジフテリア毒素、破傷風毒素および百日咳毒素などの毒素ベースのアジュバントは、使用前に、例えばホルムアルデヒドで処理することなどによって不活化することができる。
【0126】
ワクチンはさらに、感染性PTTV DNAクローンの免疫学的活性を増進するために、例えば豚繁殖呼吸器病症候群ウイルス(PRRSV)、豚パルボウイルス(PPV)、他の感染性豚因子および免疫刺激物質などといった追加の抗原を含有してもよい。
【0127】
本発明の新しいワクチンは、どの特定のタイプまたは製造方法にも限定されない。クローン化ウイルスワクチンには、例えば感染性DNAワクチン(すなわち、豚にDNAを直接注入するためにプラスミド、ベクターまたは他の従来の担体を使用するもの)、生ワクチン、改変生ワクチン、不活化ワクチン、サブユニットワクチン、弱毒化ワクチン、遺伝子操作ワクチンなどが含まれるが、これらに限るわけではない。これらのワクチンは当技術分野において知られている標準的方法によって製造される。
【0128】
さらにもう一つの利点として、本発明の好ましい生ウイルスは、製造、貯蔵および搬送が他のタイプの弱毒化ワクチンよりも容易な、遺伝子的に安定なワクチンを与える。
【0129】
本発明のもう一つの好ましいワクチンでは、非病原性DNAクローンを豚に送達するために、適切なプラスミドを利用する。生または死滅細胞培養増殖全ウイルスを使用する従来のワクチンとは対照的に、この発明では、感染性ウイルスゲノムを含有するプラスミドDNAによる豚の直接接種が可能である。
【0130】
本発明において望ましいさらなる遺伝子操作ワクチンは、当技術分野において知られている技法によって製造される。そのような技法は、例えば組換えDNAのさらなる操作、組換えタンパク質のアミノ酸配列の修飾または置換などを伴うが、これらに限るわけではない。
【0131】
組換えDNA技術に基づく遺伝子操作ワクチンは、例えば、豚において、より強い免疫応答または防御応答を誘導する原因になるタンパク質(例えばORF1、ORF1/1、ORF2、ORF2/2に由来するタンパク質など)をコードするウイルス遺伝子の代替的部分を同定することによって作製される。同定されたそのような遺伝子または免疫優性フラグメントは、バキュロウイルスベクターなどの標準的タンパク質発現ベクターにクローニングし、それを使って適当な宿主細胞を感染させることができる(例えばオライリー(O’Reilly)ら、「バキュロウイルス発現ベクター:ア・ラブ・マニュアル(Baculovirus Expression Vectors: A Lab Manual)」、フリーマン・アンド・カンパニー(Freeman & Co.)、1992年を参照されたい)。宿主細胞を培養することで所望のワクチンタンパク質を発現させ、それを所望する程度にまで精製し、適切なワクチン製品へと製剤化することができる。組換えサブユニットワクチンは、PTTV1a、PTTV1bおよびPTTV2の細菌発現(図10、図15)またはバキュロウイルス発現ORF1キャプシドタンパク質に基づく。
【0132】
疾患を引き起こすという望ましくない天然の能力をクローンが少しでも保っている場合は、残存ビルレンスの原因になっているウイルスゲノム中のヌクレオチド配列を特定し、例えば部位特異的突然変異導入法などにより、ウイルスを遺伝子操作して無発病性にすることもできる。部位特異的突然変異導入法では、1つ以上のヌクレオチドを付加、削除または改変することができる(例えば、ツェラー(Zoller)ら、DNA 3:479−488、1984を参照されたい)。所望の突然変異を含有するオリゴヌクレオチドを合成し、一本鎖ウイルスDNAの一部にアニールさせる。この手順によって生じるハイブリッド分子を使って細菌を形質転換する。次に、適当な突然変異を含有する単離された二本鎖DNAを使って、完全長DNAの制限フラグメントにライゲーションすることによって、完全長DNAを作製し、次にそれを適切な細胞培養中にトランスフェクトする。適切な導入用ベクターへのゲノムのライゲーションは、当業者に知られる任意の標準的技法によって達成することができる。ウイルス子孫を生産するための宿主細胞へのベクターのトランスフェクションは、リン酸カルシウムまたはDEAEデキストラン媒介トランスフェクション、エレクトロポレーション、プロトプラスト融合および他の周知の技法など、従来の方法のどれを使って行ってもよい(例えばサムブルック(Sambrook)ら、「モレキュラー・クローニング:ア・ラボラトリー・マニュアル(Molecular Cloning: A Laboratory Manual)」、コールド・スプリング・ハーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、1989年を参照されたい)。クローニングされたウイルスは所望の突然変異を示す。あるいは、適当な突然変異を含有する2つのオリゴヌクレオチドを合成することもできる。これらをアニールさせて二本鎖DNAを形成させ、それをウイルスDNAに挿入して、完全長DNAを作製することができる。
【0133】
免疫学的有効量の本発明のワクチンを、ウイルス感染からの防御を必要とする豚に投与する。豚に接種する免疫学的有効量または免疫原量は、日常的試験により、容易に決定することができ、または迅速に漸増することができる。有効量とは、PTTVウイルスにばく露された豚を保護するのに十分な、ワクチンに対する免疫学的応答が得られる量である。好ましくは、豚は、ウイルス疾患の有害な生理学的症状または影響の1つから全部が有意に低減するか、改善するか、完全に防止される程度にまで保護される。
【0134】
ワクチンは、単回投与するか、反復投与することができる。投薬量は、例えば約1マイクログラム〜約1,000マイクログラムの範囲の感染性キメラDNAゲノムを含有するプラスミドDNA(ワクチンの免疫活性構成成分の濃度に依存する)、好ましくは100〜200マイクログラムの範囲の豚TTV DNAクローンであり得るが、ウイルス感染の有害反応または生理学的症状をもたらすのに十分であるような量のウイルスに基づく抗原を含有してはならない。豚の体重、抗原の濃度および他の典型的要因に基づいて最低有効投薬量を見いだすために、活性抗原性因子の適切な投薬量を決定または漸増するための方法は、当技術分野において知られている。好ましくは、感染性ウイルスDNAクローンをワクチンとして使用するか、生感染性ウイルスをインビトロで作製してから、その生ウイルスをワクチンとして使用することができる。その場合、例えば約50〜約10,000の50%組織培養感染量(TCID50)の生ウイルスを、豚に与えることができる。
【0135】
本発明の新しいワクチンは、どの特定のタイプまたは製造方法にも限定されない。ワクチンには、改変生ワクチン、不活化ワクチン、サブユニットワクチン、弱毒化ワクチン、遺伝子操作ワクチンなどが含まれるが、これらに限るわけではない。
【0136】
生ワクチンの利点は、全身性免疫応答、局所免疫応答、体液性免疫応答および細胞性免疫応答を含む、考え得る全ての免疫応答がワクチンの受容者において活性化されることである。利点を上回り得る生ウイルスワクチンの欠点は、生付随ウイルス因子による汚染の可能性またはウイルスが野外で先祖返りしてビルレンスを持つようになり得る危険にある。
【0137】
不活化ウイルスワクチンを製造するには、例えば限定するわけではないがPK−15細胞などの培養豚細胞中で、ウイルス増殖およびウイルス生産を行うことができる。次に、当業者に一般に知られているプロトコールによって、または好ましくは本明細書に記載する方法によって、一連のウイルス不活化を最適化する。
【0138】
不活化ウイルスワクチンは、豚TTVをホルマリンまたは疎水性溶媒、酸などの不活化剤で処理すること、紫外線またはX線を照射すること、加熱することなどによって、製造することができる。不活化は当技術分野において理解されているやり方で行われる。例えば化学的不活化では、適切なウイルス試料またはウイルスを含有する血清試料を、ウイルスを不活化するために、十分に高い(または不活化剤によっては低い)温度またはpHにおいて十分な量または濃度の不活化剤で十分な時間処理する。加熱による不活化は、ウイルスを不活化するのに十分な温度および時間行われる。照射による不活化は、ある光の波長または他のエネルギー源を使って、ウイルスを不活化するのに十分な時間行われる。ウイルスは、それが、感染を起こしやすい細胞に感染することができなくなれば、不活化されたと見なされる。
【0139】
サブユニットワクチンの製造は、典型的には、改変生ワクチンまたは不活化ワクチンの製造とは異なる。サブユニットワクチンの製造に先だって、ワクチンの防御構成成分または抗原構成成分を同定しなければならない。本発明において、PTTVの抗原構成成分は、PTTV1a、PTTV1bおよびPTTV2のORF1キャプシドタンパク質と同定され、これを本発明では、サブユニット組換えキャプシドワクチンとして使用するために、大腸菌(E.coli)およびバキュロウイルス発現系などの他の発現系で発現させ、精製した。そのような防御構成成分または抗原構成成分は、豚において特に強い防御応答または免疫学的応答を引き起こすウイルスキャプシドタンパク質の一定のアミノ酸セグメントまたはフラグメント;単一または複数のウイルスキャプシドタンパク質そのもの、そのオリゴマー、およびウイルスの部分構造またはそのような部分構造の同定可能な部分もしくはユニットを形成するウイルスキャプシドタンパク質の高次会合体;ウイルスの表面上もしくは表面近くに存在するか、ウイルスと会合しているリポタンパク質または脂質基などのウイルス部分構造中に存在する、オリゴグリコシド、糖脂質または糖タンパク質などを含む。好ましくは、ORF1タンパク質をサブユニットワクチンの抗原構成成分として使用する。ORF2、ORF1/1、およびORF2/2遺伝子中のヌクレオチド配列によってコードされるものなど、他のタンパク質も使用することができる。これらの免疫原構成成分は当技術分野において知られている方法により、すぐに同定することができる。ひとたび同定されたら、次に、ウイルスの防御部分または抗原部分(すなわち「サブユニット」)を、当技術分野において知られている手法で精製および/またはクローニングする。ウイルスの高精製サブユニットなどのサブユニットは全ウイルスよりも毒性が低いので、サブユニットワクチンには、生ウイルスに基づく他のワクチンをしのぐ利点がある。
【0140】
組換え遺伝子技法によってサブユニットワクチンを製造する場合、ORF1、ORF2、ORF1/1、およびORF2/2遺伝子などといったクローン化サブユニットの発現は、例えば、上述の方法によって行うことができ、当業者に知られる方法によって最適化することもできる(例えばマニアティス(Maniatis)ら、「モレキュラー・クローニング:ア・ラボラトリー・マニュアル(Molecular Cloning: A Laboratory Manual)」、コールド・スプリング・ハーバー・ラボラトリー(Cold Spring Harbor Laboratory)、マサチューセッツ州コールドスプリングハーバー、1989年を参照されたい)。一方、使おうとするサブユニットが、例えばキャプシドタンパク質全体など、ウイルスのインタクトな構造的特徴を表す場合は、ウイルスからそれを単離するための手法を最適化しなければならない。いずれにせよ、不活化プロトコールの最適化後には、製造に先だって、サブユニット精製プロトコールを最適化することができる。
【0141】
弱毒化ワクチンを製造するには、生病原性ウイルスをまず最初に、当技術分野において知られている方法によって、または好ましくは本明細書に記載する方法によって、弱毒化する(非病原性または無害にする)。例えば弱毒化ウイルスは、有胚豚卵による新規連続継代を伴う本発明の技法によって調製することができる。弱毒化ウイルスは自然に見いだすことができ、天然の遺伝子欠失を有し得る。あるいは、遺伝子を欠失させるか遺伝子を突然変異させることによって、病原性ウイルスを弱毒化することもできる。弱毒化および不活化ウイルスワクチンは、本発明の好ましいワクチンを構成する。
【0142】
本発明において同様に望ましい遺伝子操作ワクチンは、当技術分野において知られている技法によって作製される。そのような技法は、RNA、組換えDNA、組換えタンパク質、生ウイルスの使用などを伴うが、これらに限るわけではない。
【0143】
例えば、精製後に、野生型ウイルスは、感染した豚または感染した適切な細胞株を使って、血清、糞便、唾液、精液および組織試料などの適切な臨床生物学的試料から、当技術分野において知られている方法によって、好ましくは本明細書において教示する方法によって、単離することができる。生物学的に純粋なウイルスまたは感染性因子から当技術分野において知られている方法によってDNAを抽出し、当技術分野において知られている方法によって、好ましくはCsCl勾配における超遠心分離によって精製する。ウイルスゲノムのcDNAを、当技術分野において知られている方法によって、適切な宿主中にクローニングし(マニアティス(Maniatis)ら、同上)、次に、ウイルスゲノムを解析して、ウイルスの抗原部分を生産するのに不可欠なゲノムの領域を決定する。その後の手順は、改変生ワクチン、不活化ワクチンまたはサブユニットワクチンの場合と、おおむね同じである。
【0144】
組換えDNA技術に基づく遺伝子操作ワクチンは、例えば、ウイルス遺伝子のうち、豚において、より強い免疫応答または防御応答を誘導する原因になるタンパク質(例えばORF1、ORF2、ORF1/1、ORF2/2に由来するタンパク質など)をコードする部分を同定することによって作製される。同定されたそのような遺伝子または免疫優性フラグメントは、バキュロウイルスベクターなどの標準的タンパク質発現ベクターにクローニングし、それを使って適当な宿主細胞を感染させることができる(例えばオライリー(O’Reilly)ら、「バキュロウイルス発現ベクター:ア・ラブ・マニュアル(Baculovirus Expression Vectors: A Lab Manual)」、フリーマン・アンド・カンパニー(Freeman & Co.)、1992年を参照されたい)。宿主細胞を培養することで所望のワクチンタンパク質を発現させ、それを所望する程度にまで精製し、適切なワクチン製品へと製剤化することができる。
【0145】
ワクチンにおいて有用な遺伝子操作タンパク質は、例えば、昆虫細胞、酵母細胞または哺乳動物細胞において発現させることができる。従来の方法によって精製または単離することができる遺伝子操作タンパク質は、豚TTVからの防御を付与するために、豚または哺乳動物種に直接接種することができる。
【0146】
昆虫細胞株(例えばsf9、sf21、またはハイ・ファイブ(HIGH−FIVE))は、ウイルスから得られたまたはウイルスゲノムからコピーされたポリ核酸であってウイルスの免疫優性タンパク質の1つ以上をコードするものを含有する導入ベクターで、形質転換することができる。導入ベクターには、例えば、線状化バキュロウイルスDNAおよび所望のポリヌクレオチドを含有するプラスミドが含まれる。宿主細胞株には、組換えバキュロウイルスを作製するために、線状化バキュロウイルスDNAとプラスミドを同時トランスフェクトすることができる。
【0147】
あるいは、分離された豚TTVのDNAであって1つ以上のキャプシドタンパク質をコードするものを、ポックスウイルスまたはアデノウイルスなどの生ベクター中に挿入して、ワクチンとして使用することもできる。
【0148】
本発明のワクチンの免疫学的有効量は、前記感染または症候群からの防御を必要とする豚または哺乳動物種に投与される。「免疫学的有効量」は、日常的試験により、容易に決定することができ、または迅速に漸増することができる。有効量とは、豚皮膚炎腎症症候群(PDNS)、離乳後多臓器消耗症候群(PMWS)または関連疾病を引き起こし得る、豚TTVウイルスまたは豚TTV同時感染にばく露された豚または他の哺乳動物を保護するのに十分な、ワクチンに対する免疫学的応答が得られる量である。好ましくは、豚または他の哺乳動物種は、ウイルス疾患の有害な生理学的症状または影響の1つから全部が有意に低減するか、改善するか、完全に防止されることが見いだされる程度にまで保護される。
【0149】
ワクチンは、単回投与するか、反復投与することができる。投薬量は、例えば1〜1,000マイクログラムのウイルスに基づく抗原(ワクチンの免疫活性構成成分の濃度に依存する)を含有し得るが、ウイルス感染の有害反応または生理学的症状をもたらすのに十分であるような量のウイルスに基づく抗原を含有してはならない。鳥または哺乳動物の体重、抗原の濃度および他の典型的要因に基づいて活性抗原性因子の適切な投薬量を決定または漸増するための方法は、当技術分野において知られている。
【0150】
ワクチンは豚に投与することができる。ワクチンは、ウイルス因子に感染するリスクが高い養豚業者などのヒトに与えることもできる。豚TTVに基づくワクチンは、豚TTVおよびヒトTTVの両方からの幅広い防御をもたらすように設計することができると考えられる。言い換えると、豚TTVに基づくワクチンは、好ましくは、いわゆる「ジェンナーの方法」(すなわち牛痘ウイルスワクチンは、エドワード・ジェンナー(Edward Jenner)により、ヒト痘瘡に対して使用することができる)によって、ヒトTTV感染を防御するように設計することができる。望ましくは、まだTTVウイルスにばく露されていない豚または他の哺乳動物種に、ワクチンを直接投与する。ワクチンは、都合よく、例えば経口投与、口内投与、鼻腔内投与、経皮投与、非経口投与することができる。非経口投与経路には、筋肉内経路、静脈内経路、腹腔内経路および皮下経路が含まれるが、これらに限るわけではない。
【0151】
液体として投与される場合、本ワクチンは、水性溶液剤、シロップ剤、エリキシル剤、チンキ剤などの形態に調製することができる。そのような製剤は当技術分野において知られており、典型的には、抗原および他の典型的添加剤を適当な担体または溶媒系に溶解することによって調製される。適切な担体または溶媒には、例えば水、食塩水、エタノール、エチレングリコール、グリセロールなどがあるが、これらに限るわけではない。典型的添加剤は、例えば認可された色素、香料、甘味料、およびチメロサール(エチルメルクリチオサリチル酸ナトリウム)などの抗微生物保存剤である。そのような溶液は、例えば部分加水分解ゼラチン、ソルビトールまたは細胞培養培地の添加などによって安定化することができ、例えばリン酸水素ナトリウム、リン酸二水素ナトリウム、リン酸水素カリウム、リン酸二水素カリウム、その混合物などといった当技術分野において知られている試薬類を使って、従来の方法で緩衝化することができる。
【0152】
液状製剤には、他の標準的共製剤化剤と組み合わされた懸濁化剤または乳化剤を含有する懸濁剤および乳剤も含まれる。これらのタイプの液状製剤は、従来の方法で調製することができる。例えば懸濁剤はコロイドミルを使って調製することができる。例えば乳剤はホモジナイザーを使って調製することができる。
【0153】
体液系に注射するように意図された非経口製剤は、対応する哺乳動物体液のレベルへの適正な等張性およびpH緩衝作用を必要とする。等張性は、必要に応じて塩化ナトリウムおよび他の塩で、適当に調節することができる。エタノールまたはプロピレングリコールなどの適切な溶媒を使って、製剤中の成分の溶解度と液状調製物の安定性を増加させることができる。本ワクチンに使用することができるさらなる添加剤には、デキストロース、従来の酸化防止剤および従来のキレート剤、例えばエチレンジアミン四酢酸(EDTA)などがあるが、これらに限るわけではない。また、非経口剤形は使用前に滅菌しなければならない。
【0154】
以下に実施例を挙げて本発明の一定の態様を例証する。しかし、これらの実施例が単なる例示であること、そして本発明の条件および範囲に関して完全に決定的なものであるとは考えていないことを理解すべきである。典型的な反応条件(例えば温度、反応時間など)が記載されている場合、指定された範囲を上回る条件も下回る条件も、一般的には利便性が下がるとはいえ、使用できることを理解すべきである。本実施例は室温(約23℃〜約28℃)および大気圧で行われる。別段の指定がない限り、本明細書において言及する割合およびパーセントはいずれも重量ベースであり、温度は全て摂氏温度である。
【0155】
(実施例1)
ウイルスDNA抽出、ネステッドPCRおよびゲノムPCR:
この研究には、バージニアの養豚場の20頭の在来成体雄豚から採取した好都合な血清試料および精液試料を使用した。キアアンプ(QIAamp)DNAミニキット(キアゲン(Qiagen)を使って20個の血清試料および19個の精液試料から全DNAを単離した。陽性PTTV含有試料をスクリーニングするために、まず最初に、PTTV1およびPTTV2中の保存領域のネステッドPCR増幅を、アンプリタック・ゴールド(AmpliTag Gold)ポリメラーゼ(アプライド・バイオシステムズ(Applied Biosystems))を使って行った。PTTV1のフラグメントAを増幅するために使用した2つのプライマー対は、TTV1−mF(配列番号45)/TTV1−mR(配列番号46)(第1ラウンドPCR用)およびTTV1−nF(配列番号47)/TTV1−nR(配列番号48)(第2ラウンドPCR用)であり、一方、PTTV2のフラグメントDを増幅するために使用した2つのプライマー対は、TTV2−mF(配列番号49)/TTV2−mR(配列番号50)(第1ラウンドPCR用)およびTTV2−nF(配列番号51)/TTV2−nR(配列番号52)(第2ラウンドPCR用;図1Aおよび表1)だった。
【0156】
PTTV1とPTTV2の両方の完全長ゲノム配列を増幅するために、本発明者らは、まず、PTTV1用には領域A中に位置する一対の保存された遺伝子特異的プライマーTTV1−IF(配列番号1)/TTV1−IR(配列番号4)およびPTTV2用には領域D中に意図するもう一対の遺伝子特異的プライマーTTV2−IF(配列番号5)/TTV2−IR(配列番号8)を、それぞれ、ヘラキュレース(Herculase)IIフュージョン(Fusion)DNAポリメラーゼ(ストラタジーン(Stratagene))と共に、製造者の説明書に従って使用することにより、インバースゲノムPCRを行った。予期されるサイズのPCR産物は検出されなかった。次に本発明者らは、第2ラウンドのPCRにおいて、PTTV1ゲノム全体およびPTTV2ゲノム全体をカバーするそれぞれ2つの領域を増幅するために、新しいプライマーセットを設計した(図1A)。PTTV1のフラグメントBおよびCを増幅するために使用したプライマー対は、それぞれ、TTV1−IF(配列番号1)/TTV1−2340R(配列番号2)とTTV1−2311F(配列番号3)/TTV1−IR(配列番号4)であり、一方、PTTV2のフラグメントEおよびFを増幅するために使用したプライマー対は、それぞれ、TTV2−IF(配列番号5)/TTV2−2316R(配列番号6)およびTTV2−GCF(配列番号7)/TTV2−IR(配列番号8)だった(図1Aおよび表1)。フラグメントCおよびFは、それぞれPTTV1およびPTTV2のGCリッチ領域を含有する。増幅されたPCR産物を個別に切り出し、精製し,次に、ストラタクローン・ブラント(StrataClone Blunt)PCRクローニングストラテジーにより、製造者の説明書(ストラタジーン(Stratagene)に従って、pSC−B−amp/kanベクター(ストラタジーン(Stratagene)中にクローニングした後、DNA配列決定を行った。
【0157】
(実施例2)
バージニアの飼育場にいる雄豚から収集した豚TTV陽性試料のスクリーニング:
豚TTV DNAは、以前に、日本PTTV1株Sd−TTV31のUTR配列に基づくネステッドPCRにより、異なる地理的領域の豚から検出されている(マキューン(McKeown)ら、2004年、前掲書)。PTTV2が最近同定されたことから、PTTV1の領域AとPTTV2の領域Dをそれぞれ増幅するために、2つの異なるネステッドPCRプライマーセットが使用された(図1A)(エリス(Ellis)ら、2008年、前掲書;ケカライネン,ティ(Kekarainen,T.)、シビラ,エム(Sibila,M.)およびシーゲイルス,ジェイ(Segales,J.)、2006年、「スペインの離乳後多臓器消耗症候群(PMWS)罹患豚および非PMWS罹患豚における豚トルクテノウイルスの有病率(Prevalence of swine Torque teno virus in post-weaning multisystemic wasting syndrome (PMWS)-affected and non-PMWS-affected pigs in Spain)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第87巻、第4号、p.833−7;クラコウカ(Krakowka)、2008年、前掲書)。本研究においても同様の検出法を利用して、米国の豚からPTTV株を同定した。後に完全長ゲノムの決定に使用する、土地固有のPTTV1陽性試料またはPTTV2陽性試料をスクリーニグするために、バージニアの飼育場にいる20頭の雄豚から収集した20個の血清試料(SR#1〜20)および19個の精液試料(SM#1〜18およびSM#20)を、ネステッドPCR分析に付した。驚いたことに、20個の血清試料は全てPTTV1に関して陽性であり、19個はPTTV2についても陽性だった(SR#18を除く)。対照的に、1つの精液試料(SM#6)だけがPTTV1陽性であり、3つの精液試料だけ(SM#8、9および20)がPTTV2陽性だった。この結果は、スペインにおける雄豚精液試料がPTTV DNAに関して陽性であることが示された最近の研究と合致した(ケカライネン,ティ(Kekarainen,T.)、ロペス−ソリア,エス(Lopez-Soria,S.)およびシーゲイルス,ジェイ(Segales,J.)、2007年)。雄豚の血清および精液には豚トルクテノウイルス遺伝子グループ1および2が検出されたことから(セリオゲノロジー(Theriogenology)、第68巻、第7号、p.966−71)、PTTVの潜在的垂直感染が示唆された。しかし、精液におけるPTTV1とPTTV2の有病率は、血清中のそれよりもはるかに低かったので、同じ豚の血清および精液におけるPTTV DNAの存在に関して直接的な関連はないことが示唆された。
【0158】
(実施例3)
配列解析および系統解析:
DNA配列およびアミノ酸配列の一般解析(generic analysis)およびアラインメントはレーザージーン(Lasergene)パッケージ(ディエヌエイスター・インコーポレイテッド(DNASTAR Inc.)、ウィスコンシン州マディソン)を使って行った。アラインメントと比較に使用した3つの既知PTTV株のゲノム配列およびそれらの対応するジェンバンク(GenBank)アクセッション番号は、Sd−TTV31(AB076001)、TTV−1p(AY823990)およびTTV−2p(AY823991)である。ジェンバンク(GenBank)において利用できるヒトおよび動物のTTV関連株の121個の完全長ゲノム配列を使用し、オンラインプログラムPASC(http://www.ncbi.nlm.nih.gov/sutils/pasc/viridty.cgi?textpage=overview)(バオ(Bao)ら、2008年)で、ペアワイズ配列比較(PASC)を行った。
【0159】
系統樹は、PAUP4.0プログラム(デービッド・スウォフォード(David Swofford)、スミソニアン・インスティチュート(Smithsonian Institute)、ワシントンDC、シナウアー・アソシエート・インコーポレイテッド(Sinauer Associate Inc.)が配布)の近隣結合法により、7つのPTTV株の4つのORFの完全長ゲノム配列および推定アミノ酸配列に基づいて構築した。データは1000回のリサンプリングから得た。
【0160】
(実施例4)
豚PTTV感染を診断するためのPCRプライマーの設計
DNA配列の解析およびアラインメントは、レーザージーン(Lasergene)パッケージ(ディエヌエイスター・インコーポレイテッド(DNASTAR Inc.)、ウィスコンシン州マディソン)を使って行った。アラインメントに使用した10個の豚TTV株の完全長ゲノム配列およびそれらの対応するジェンバンク(GenBank)アクセッション番号は、次のとおりであった。種PTTV1:Sd−TTV31(AB076001)、PTTV1a−VA(GU456383)、TTV−1p(AY823990)、PTTV1b−VA(GU456384)、swSTHY−TT27(GQ120664)およびTTV1#471819(GU188045)。種PTTV2:PTTV2b−VA(GU456385)、PTTV2c−VA(GU456386)、TTV−2p(AY823991)およびTTV2#472142(GU188046)。6つのPTTV1ゲノム間および4つのPTTV2ゲノム間で保存されている配列をそれぞれ同定し、次にそれらを指針として、ビーコン・デザイナー(Beacon Designer)プログラム(プレミア・バイオソフト・インターナショナル(PREMIER Biosoft International)、カリフォルニア州パロアルト)を使ってリアルタイムPCRプライマーを選択した。PTTV1のデュプレックス・ネステッドPCRに使用したプライマーは、レーザージーン(Lasergene)パッケージで設計した。
【0161】
(実施例5)
PTTV1およびPTTV2リアルタイムPCRの標準曲線
PTTV1b−VAゲノムのPCRフラグメントBに対応する2091bpの領域を、同じPCRフラグメントから、既述のように(ファン(Huang)ら、2010年)、プライマーTTV1−IF(5’−CATAGGGTGTAACCAATCAGATTTAAGGCGTT−3’)およびTTV1−2340R(5’−GGTCATCAGACGATCCATCTCCCTCAG−3’)を使って再増幅した。その結果生じたアンプリコンをキアクイック・ゲル・エクストラクション・キット(QIAquick Gel Extraction Kit)(キアゲン(Qiagen))でゲル精製し、ナノドロップ(NanoDrop)分光測光器で定量して、豚TTV種1のリアルタイムPCR標準テンプレートに使用した。PTTV2c−VAのPCRフラグメントEおよびFをベクターpSC−B−amp/kan中でアセンブルすることにより、PTTV2c−VA株pSC−PTTV2cの完全長DNAクローンを構築した(ファン(Huang)ら、未公表データ)。プラスミドpSC−PTTV2c(7082bp)を豚TTV種2のリアルタイムPCR標準テンプレートに使用し、プラスミドDNA濃度をナノドロップ(NanoDrop)分光測光器で測定した。これら2つのテンプレートの10倍希釈系列を使ってリアルタイムPCR標準曲線をそれぞれ作成した。
【0162】
(実施例6)
PCRアッセイ用のウイルスDNAの抽出
バージニアの養豚場の20頭の在来成体雄豚(臨床的症候群を持たないもの)から収集した20個の血清試料と19個の精液試料から、キアアンプ(QIAamp)DNAミニキット(キアゲン(Qiagen))を使って、既述のように(ファン(Huang)ら、2010年)、全DNAを単離した。血清および精液について400μlの試料体積を使って、滅菌水50μlの最終溶出液で、DNAを抽出した。抽出したDNA試料は全て、リアルタイムPCR試験まで、−20℃で保存した。従来のネステッドPCRによるこれらの試料中の豚TTVの検出は既に記述されている(ファン(Huang)ら、2010年)。同じ手順でヤギ血清試料から抽出された全DNAを陰性対照として使用した。
【0163】
(実施例7)
SYBRグリーン・リアルタイム定量PCRアッセイ
PTTV1特異的およびPTTV2特異的リアルタイムPCRを、センシミックス(SensiMix)SYBRアンド・フルオレセインキット(クオンテース・リミテッド(Quantace Ltd))およびマイアイキュー・アイサイクラー(MyiQ iCYCLER)リアルタイムPCR機器(バイオラッド・ラボラトリーズ(BIO?RAD Laboratories))を使って、それぞれ行った。各25μlの反応は、12.5μlのSYBRグリーン・マスター・ミックス、4μlの抽出されたDNA、0.5μlの各プライマー(10nM)および7.5μlの滅菌水を含有した。PTTV1のPCR条件は95℃で10分の後、40サイクルの増幅(95℃で15秒、59.4℃で30秒、72℃で10秒)とした。その後、直ちに、0.5℃ごとに蛍光シグナルを測定しながら、55℃から95℃まで温度を徐々に上昇させることにより、融点分析を行った。PTTV2のPCR条件は、アニーリング温度が56℃である点以外は、PTTV1と同じとした。PTTV1およびPTTV2標準テンプレートを陽性対照として全てのランに含めた。増幅およびデータ解析は、マイアイキュー(MyiQ)システムソフトウェア(バイオラッド・ラボラトリーズ(BIO?RAD Laboratories))を使って行った。全ての試料を同じプレート上で二つずつ測定した。
【0164】
(実施例8)
2つのシングルプレックス・アッセイの特異性および感度
PTTV1特異的およびPTTV2特異的アッセイの増幅にとって最適なアニーリング温度は、アニーリング温度の勾配を使った増幅の10倍希釈によって決定したところ、それぞれ59.4℃および56℃だった。プライマーTTV1F/TTV1Rを使った118bp産物の増幅がPTTV1テンプレートでのみ得られたのに対し、PTTV2テンプレートでの200bp産物の増幅は、プライマーTTVF4/TTVR4を使った場合にのみ観察された。どちらのアッセイも他方からの交差増幅を与えなかったことから、プライマーおよびターゲットの特異性が確認された(データ未掲載)。
【0165】
ある範囲の、25μlあたりのターゲットDNA濃度にわたって、PTTV1標準曲線を確立した。線形範囲は、4.4×101〜4.4×108コピーにまたがることが示された。最低検出限界(44コピー)は、37.57の閾サイクル数(Ct)に対応した。Ct>37.57の被験試料は検出限界未満とみなし、定量可能でなかった。同様に、PTTV2標準曲線を作成し、それを使って、25μlの反応あたり8.6×100〜8.6×108コピーの範囲にあるDNA濃度を検出した。最低検出限界(8.6コピー)に対応するCtは36.53だった。PTTV1陽性またはPTTV2陽性とみなした試料は全て、それぞれの最大検出限界より低いコピー数を持っていた。PTTV1またはPTTV2標準テンプレートの10倍希釈(図6aおよび6b;青色の曲線)ならびに20個の雄豚血清試料を使った融解曲線は、PTTV1については87.0℃、PTTV2については80.0℃の融解温度(Tm)を、それぞれ示した(図6aおよび6b;赤色の曲線)。滅菌水またはヤギ血清DNAをテンプレートとして使用した陰性対照にはピークが観察されなかった(図6aおよび7b;黒い線)。
【0166】
(実施例9)
雄豚血清試料および精液試料中の豚TTV1およびTTV2の定量
ウイルス負荷量を、元の雄豚血清試料1mlあたりのPTTV1ゲノムまたはPTTV2ゲノムのコピー数として表した。PTTV1 DNAは20個の血清試料の全てに検出され、1.91?103〜3.25?105コピー/mlの範囲にあった。一方、PTTV2 DNAは19個の血清試料(#10以外)に検出され、3.59×102〜1.39×106コピー/mlの範囲にあった。この結果は、従来のネステッドPCRを使用した本発明者らの先の研究と合致した(表5)。精液試料はいずれもPTTV1陽性ではなかったが、3つの精液試料は、極めて低いウイルス負荷量(それぞれ230、244および357コピー/ml)でPTTV2陽性だった。
【0167】
【表5】
【0168】
(実施例10)
PTTV1/PTTV2デュプレックス・リアルタイムPCRアッセイ
PTTV1/PTTV2デュプレックス・リアルタイムPCRアッセイは、12.5μlのSYBRグリーン・マスター・ミックス、0.5μlの各PTTV1プライマー、0.5μlの各PTTV2プライマー、4μlのDNAおよび6.5μlの滅菌水を含有する25μlのPCR系で行った。デュプレックスPCR条件および融点分析は、アニーリング温度が58℃である点以外は、PTTV1と同じとした。PTTV1特異的アンプリコンとPTTV2特異的アンプリコンを識別するために、溶解ピークを分析した。
【0169】
(実施例11)
デュプレックス・ネステッドPCR
第1ラウンドPCRは、50μlの総液量に4μlの抽出DNAを使って、プラチナムPCRハイファイ・スーパーミックス(Platinum PCR HiFi Supermix)(インビトロジェン(Invitrogen))で行った。PCR条件は、94℃で2分間のテンプレートの初期変性と、94℃で30秒、55℃で30秒、72℃で30秒の30サイクルとした。同じPCR試薬類および条件による第2ラウンドのPCRには、第1ラウンドPCR産物を4μlずつ使用した。第1ラウンドPCRでは1対のプライマーP1ab−mF/P1ab−mRを使用したのに対し、第2ラウンドのPCRでは、2対のプライマーPTTV1a検出用のP1a−nF/P1a−nRとPTTV1b検出用のP1b−nF/P1b−nRとの混合物を使用した(表1)。増幅産物を、臭化エチジウムで染色した1%アガロースゲルでのゲル電気泳動によって可視化したところ、各型に特異的な2本のバンドがUV光によって区別された。
【0170】
(実施例12)
PTTV1およびPTTV2 ORF発現プラスミドの構築
PTTV1a、PTTV1bおよびPTTV2cのORF1のC末端部分を、それぞれの完全長DNAクローン(pSC−PTTV1a、pSC−PTTV1bおよびpSC−PTTV2c;項を改めて記載)から増幅した。増幅されたフラグメントは、PTTV1aについては319aa(ORF1aa位置317−635(配列番号13);ジェンバンク(GenBank)アクセッション番号GU456383)、PTTV1bについては318aa(ORF1aa位置322−639(配列番号14);ジェンバンク(GenBank)アクセッション番号GU456384)、PTTV2cについては316aa(ORF1aa位置310−625(配列番号16);ジェンバンク(GenBank)アクセッション番号GU456386)のタンパク質産物を、それぞれコードすると予期された。248aa(ORF1aa位置322−569(配列番号14))をコードするPTTV1bのC末端切断フラグメントも増幅し、SDS−PAGE分析用の比較対象として使用した。プラスミドは全て、C末端に8×Hisタグが付いた融合タンパク質が生成するように大腸菌/バキュロウイルス/哺乳動物細胞三重発現ベクターpTriEx1.1−neo(ノバジェン(Novagen))のNcoI制限部位とXhoI制限部位の間にPCR産物をクローニングすることによって構築した。4つの組換えプラスミドをpTri−PTTV1a−ORF1、pTri−PTTV1b−ORF1、pTri−PTTV1b−ORF1ctrucおよびpTri−PTTV2c−ORF1と名付けた。全てのクローン化配列をDNA配列決定によって確認した。
【0171】
(実施例13)
組換えPTTV1およびPTTV2タンパク質の発現
4つの発現プラスミドを、それぞれロゼッタ(Rosetta)2(DE3)pLacIコンピテント細胞(ノバジェン(Novagen))に形質転換し、細菌を、100μg/mlアンピシリンを含有するLB寒天プレート上、37℃で終夜、プレート培養した。各コンストラクトについて単一の形質転換コロニーを使って、100μg/mlのアンピシリンを含有するLB培地(LB/amp)3mlに接種し、37℃で6〜8時間成長させた。次に、各コンストラクトについて濁った3ml培養物を使用し、25%濾過滅菌グリセロールを添加してその培養物を−80℃で凍結することにより、細菌ストックを作製した。精製に先だって、各コンストラクトにつき10μlの凍結細菌ストックを使って、LB/ampの3mlスターター培養に接種し、37℃で6〜8時間成長させた。100mlのオーバーナイト・エクスプレス(Overnight Express)TB培地(ノバジェン(Novagen))にスターター培養物を接種してタンパク質発現を誘導し、37℃で16〜18時間成長させた。インキュベーション後に、自己誘導培養培地を4℃、3400rpmで15分間の遠心分離にかけた。各コンストラクトについてその結果得られた上清を捨て、細菌ペレットのそれぞれを使用時まで−20℃で保存した。
【0172】
(実施例14)
組換えタンパク質の精製および透析
組換えタンパク質は不溶性であり、細菌封入体内に発現した。細菌ペレットのそれぞれを、バグバスター(BugBuster)およびアールリゾチーム(rLysozyme)で、製造者のプロトコール(ノバジェン(Novagen))に従って処理し、DNAおよびRNAを分解するためにベンゾナーゼ・ヌクレアーゼ(Benzonase Nuclease)(ノバジェン(Novagen))を加えた。次に、封入体ペレットのそれぞれを840μlの溶解バッファー(6Mグアニジン塩酸塩、0.1Mリン酸ナトリウム、0.01Mトリス−塩酸、0.01Mイミダゾール、pH8.0)に再懸濁し、−80℃で少なくとも30分間は凍結した。次にそれを融解し、新たな溶解バッファー2.5mlで希釈し、室温で30分間、穏やかに回転させた。室温、15,000×gで30分間の遠心分離により、溶解物上清を集めた。デカントした上清のそれぞれに50%Ni−NTA His−バインド・スラリー(bind slurry)(ノバジェン(Novagen))を加え、hisタグ結合を促進するために、その混合物を室温で60分間振とうした。溶解物/樹脂混合物を空のクロマトグラフィーカラムに充填した。最初の貫流の後、7mlの溶解バッファーをカラムに加えて貫流させた。次に各カラムを7mlの洗浄バッファー(8M尿素、0.1Mリン酸ナトリウム、0.15M塩化ナトリウム、0.02Mイミダゾール、pH8.0))で2回洗浄した。カラムに溶出バッファー(8M尿素、0.05Mリン酸ナトリウム、1M塩化ナトリウム、0.5Mイミダゾール、pH8.0)を1mlずつ別々に4回加えることにより、ターゲットタンパク質の溶出を達成した。それら4つの溶出画分をSDS PAGEとクーマシーブルー染色とによって分析した。
【0173】
意味のある濃度のターゲットタンパク質を含有する溶出物を、20,000分子量カットオフの0.5ml〜3ml透析カセット(ピアス(Pierce))に注入した。透析には、次に挙げる一連の4つの透析バッファーを使用した。透析バッファー1(6M尿素、0.05Mリン酸ナトリウム、0.8M塩化ナトリウム、0.3Mイミダゾール、pH8.0)、透析バッファー2(4M尿素、0.033Mリン酸ナトリウム、0.533M塩化ナトリウム、0.2Mイミダゾール、pH8.0)、透析バッファー3(2.67M尿素、0.022Mリン酸ナトリウム、0.356M塩化ナトリウム、0.133Mイミダゾール、pH8.0)および透析バッファー4(1.5M尿素、0.0148リン酸ナトリウム、0.237M塩化ナトリウム、0.089Mイミダゾール、pH8.0)。透析カセットを逐次、4℃において、各透析バッファーに浸漬し、6時間以上、回転させておいた。透析が完了したら、組換えHisタグ付き融合タンパク質をそれぞれカセットから取り出し、ナノドロップ(NanoDrop)を使って定量し、−80℃で凍結した。
【0174】
(実施例15)
SDS−PAGEおよび抗Hisタグ・ウェスタンブロット
抗6×Hisタグ・モノクローナル抗体(ロックランド(Rockland))を使って精製組換えタンパク質を検出するために、ウェスタンブロットを開発した。等体積の各精製切断ORF1タンパク質とLDS/10%β−MEとを混合し、95℃で10分間煮沸した。煮沸した試料のうち10μlを、4−12%ビス−トリス・ポリアクリルアミドゲル(インビトロジェン(Invitrogen))の適当な各ウェルに加え、1×MES泳動バッファー(インビトロジェン(Invitrogen))中、200ボルトで、43分間泳動した。トランスブロット(Trans blot)セミドライ転写装置と1×転写バッファー(インビトロジェン(Invitrogen))とを使って、タンパク質をPVDFメンブレン(バイオラッド(Bio?Rad))に転写した。転写が完了したら、PVDFメンブレンを、オデッセイ(Odyssey)ブロッキングバッファー(リ−コール(Li?Cor))中、室温で1時間、インキュベートした。抗6×HisタグMAbをオデッセイ(Odyssey)ブロッキングバッファー/0.2%ツイーン20で1:1000に希釈し、先のオデッセイ(Odyssey)ブロッキングバッファーを除去してから、メンブレンに移した。MAbを振とう機上に放置して、メンブレンと共に室温で2時間、または4℃で終夜、インキュベートした後、メンブレンをトリス緩衝食塩水/0.05%ツイーン20(TBS−T、シグマ(Sigma))で3回洗浄した。ヤギ抗ウサギIgG IRDye800(リ−コール(Li?Cor))抗体を、オデッセイ(Odyssey)ブロッキングバッファー/0.2%ツイーン20/0.1%SDSで1:5000に希釈した。それを、洗浄したばかりのPVDFメンブレンに移し、穏やかに振とうしながら室温で1時間インキュベートした。メンブレンをTBS−Tで3回、TBSで1回洗浄し、リ−コール(Li?Cor)オデッセイ(Odyssey)で撮像した。
【0175】
(実施例16)
血清ウェスタンブロット
ELISA開発用の陽性および陰性血清対照を同定するために、血清ウェスタンブロットを開発し、使用した。上述のSDS−PAGE後に、タンパク質をPDVFメンブレンに転写し、次にそれを、室温においてオデッセイ(Odyssey)ブロッキングバッファー(リ−コール(Li?Cor))中で1時間インキュベートした。選択した血清試料をオデッセイ(Odyssey)ブロッキングバッファー/0.2%ツイーン20で1:100に希釈し、先のオデッセイ(Odyssey)ブロッキングバッファーを除去してからメンブレンに移した。血清試料を振とう機上に放置して、室温で2時間、メンブレンと共にインキュベートした後、メンブレンをトリス緩衝食塩水/0.05%ツイーン20(TBS−T、シグマ(Sigma))で3回洗浄した。ヤギ抗豚IgG IRDye800抗体(ロックランド(Rockland))を、オデッセイ(Odyssey)ブロッキングバッファー/0.2%ツイーン20/0.1%SDSで1:2500に希釈した。それを洗浄したばかりのPVDFメンブレンに移し、穏やかに振とうしながら室温で1時間インキュベートした。メンブレンをTBS−Tで3回、TBSで1回洗浄し、リ−コール(Li−Cor)オデッセイ(Odyssey)で撮像した。
【0176】
(実施例17)
間接的PTTV1a、PTTV1bおよびPTTV2特異的ELISA
プレートのコーティングに使用した抗原の至適濃度ならびに抗血清およびコンジュゲートの希釈度を、交差力価測定法によって決定した。ELISAは、精製組換えHisタグ付き融合タンパク質のそれぞれ(それぞれPTTV1a、PTTV1bおよびPTTV2c)をpH9.6の1×カーボネート・コーティング・バッファー(Carbonate Coating Buffer:CCB)で680ng/mlに希釈し、ミディアム・バインディング(medium binding)ELISAプレート(グライナー(Greiner))を100μl/ウェルでコーティングすることによって開始した。プレートを覆い、37℃で2時間インキュベートした。コーティング後に、希釈タンパク質を除去し、各ウェルを300μlの1×TBS−Tで3回洗浄した。次に、プロテイン・フリー・ブロッキング・バッファー(Protein Free Blocking Buffer)(ピアス(Pierce))を300μl/ウェルの体積で加え、プレートを37℃で1時間インキュベートした。同時に、96ウェル希釈ブロックでは、血清試料を150μlのプロテイン・フリー・ブロッキング・バッファー(Protein Free Blocking Buffer)で1:100に希釈した。次に、そのブロックを除去し、100μlの各希釈血清試料を、ELISAプレート上の対応する各ウェルに移した。プレートを37℃で2時間インキュベートした後、各ウェルを300μlのTBS−Tで3回洗浄した。次に、HRPコンジュゲート抗豚IgG抗体(ロックランド(Rockland))を、12mlのプロテイン・フリー・ブロックで1:4000に希釈し、プレートの各ウェルに100μlを加えた。これを37℃で1時間インキュベートした後、各ウェルを300μlのTBS−Tで3回洗浄した。ELISAを発色させるために、100μlのシュアー・ブルー・リバース1−コンポーネント(Sure Blue Reserve 1?Component)(ケー・ピー・エル(KPL))をプレートの各ウェルに加えた。20分後に、発色を停止させるために100μlの1N HClを各ウェルに加えた。次にプレートを450nmで読んだ。
【0177】
(実施例18)
データ解析
営利会社から得た細胞培養研究に使用した豚血清(ニュージーランドで製造され、どのOIE疾患も持たないと考えられるもの)を、3つのELISAプロトコールの陽性対照として使用した。なぜなら、この血清は、血清ウェスタンブロットで、PTTV1a、PTTV1bおよびPTTV2の全てについて陽性であると検出され、高いOD値(>2.0)を示したからである。本発明者らは、まず最初に、プールしたノトバイオート豚血清を陰性対照として使用した。というのも、それらはウェスタンブロット検出において陰性だったからである。次に、陰性ノトバイオート豚血清との比較で、本発明者らは、ウィスコンシンにある従来の養豚場から収集されたいくつかの豚血清をスクリーニングした。それらもウェスタンブロット検出において陰性であり、それらのOD値は陰性ノトバイオート豚血清のそれと一致した。これらの在来豚血清をプールし、陰性対照として使用した。各ELISAについてのカットオフ値は陰性対照群(n=4)の平均OD値+標準偏差の3倍として算出した。
【0178】
(実施例19)
豚TTV1a、1bおよび2cの完全長ゲノムDNAクローンの構築
米国分離株PTTV1a−VA(ジェンバンク(GenBank)アクセッション番号GU456383)由来のPCRフラグメントBおよびCを、既述のコンストラクトから再増幅し、次に、ベクターpSC−B−amp/kan(ストラタジーン(Stratagene))上で、ヘラキュレース(Herculase)IIフュージョン(Fusion)DNAポリメラーゼ(ストラタジーン(Stratagene))を使って、オーバーラッピングPCRを行うことにより、ゲノムの両端にBamHI部位を持つ完全長ゲノムDNAへとアセンブルした。その結果得られたコンストラクトをpSC−PTTV1aと名付けた(図17A)。同じ戦略を使って、米国分離株PTTV1b−VA(ジェンバンク(GenBank)アクセッション番号GU456384)に由来するクローンpSC−PTTV1b(図17B)および米国分離株PTTV2c−VA(ジェンバンク(GenBank)アクセッション番号GU456386)に由来するクローンpSC−PTTV2c(図17C)を、同じバックボーンベクター上の同じ制限部位(BamHI)を使って構築した。完全長ゲノムDNAを含有するプラスミドTTV2−#471942−full(図17E)はドイツ病原性豚TTV2分離株に由来した。TTV2−#471942はアンドレアス・ガレイ(Andreas Gallei)博士(ビー・アイ・ブイ・アイ(BIVI)、ドイツ)から分譲された。TTV2−#471942は、系統解析に基づいて米国分離株PTTV1b−VAと共に豚TTV亜型2bに分類された(データ未掲載)。
【0179】
(実施例20)
豚TTV2bおよび2cのタンデム二量体化DNAクローンの構築
完全長PTTV2cゲノムをBamHI消化によってクローンpSC−PTTV2cから切り出し、精製し、ライゲーションして、コンカテマーを形成させた。ライゲーションされたコンカテマーを、BamHIで前もって消化しておいたpSC−B−amp/kanベクター中にクローニングして、タンデム二量体化DNAクローンpSC−2PTTV2c−RR(図1D)を作製した。同様に、タンデム二量体化DNAクローンpSC−2PTTV2b−RRを、クローンTTV2−#471942−fullから、EcoRV制限部位を使って作製した(図1F)。
【0180】
(実施例21)
PTTV1a特異的、PTTV1b特異的およびPTTV2特異的抗ORF1ポリクローナル抗体の作製
ORF1がコードする産物は、TTVの予測キャプシドタンパク質である。PTTV ORF1タンパク質の発現を検出し、PTTV DNAクローンの感染性を決定する目的で、PTTV1a特異的、PTTV1b特異的およびPTTV2特異的抗ORF1ポリクローナル抗体を作製するために、PTTV1a、PTTV1bおよびPTTV2c由来の3つのORF1タンパク質を大腸菌中で発現させ、精製し、次に、それらを使って、ロックランド・イムノケミカルズ(Rockland Immunochemicals)(ペンシルバニア州ギルバーツビル)におけるカスタム抗体作製サービスとして、ニュージーランド白色種ウサギをそれぞれ免疫化した。各抗ORF1ポリクローナル抗体を免疫化したウサギの血清から作製した。
【0181】
(実施例22)
PTTV感染性クローンのインビトロ・トランスフェクション
PK−15細胞を6ウェルプレート上に1ウェルあたり2?105細胞の密度で播種し、トランスフェクションに先だって60%〜70%コンフルエンスまで成長させた。DNAクローンpSC−2PTTV2b−RRおよびpSC−2PTTV2c−RRをそれぞれPK−15細胞に、リポフェクトアミン(Lipofectamine)LTX(インビトロジェン(Invitrogen))を使って、製造者のプロトコールに従って、直接トランスフェクトした。クローンpSC−PTTV1a、pSC−PTTV2cおよびTTV2−#471942−fullについては、上述のように作製された、それらのライゲーションされたコンカテマーを、それぞれトランスフェクションに使用した。細胞を3〜5日間培養した後、豚TTVのORF1の発現を検出するために、免疫蛍光アッセイ(IFA)に付した。あるいは、トランスフェクト細胞を、新しい6ウェルプレートに継代し、3日間培養を続けてから、IFA検出を行う。
【0182】
(実施例23)
免疫蛍光アッセイ(IFA)
トランスフェクションまたは継代した細胞をPBSで2回洗浄し、アセトンで固定した。PBSで1:500に希釈したPTTV1aまたはPTTV2に特異的な抗体500μlを細胞の上に加え、室温で1時間インキュベートした。細胞をPBSで3回洗浄した後、1:200希釈のテキサス・レッド(Texas red)またはアレクサ・フルオル(Alexa Fluor)488標識ヤギ抗ウサギIgG(インビトロジェン(Invitrogen))500μlを加えた。室温で1時間インキュベートし、PBSで洗浄した後、細胞を1:1000希釈のDAPI(ケーピーエル・インコーポレイテッド(KPL,Inc.))500μlで染色し、蛍光顕微鏡下で可視化した。
【0183】
(実施例24)
タンデム二量体化豚TTV2クローンによる在来豚のインビボ接種
2つのタンデム二量体化豚TTV2クローンpSC−2TTV2b−RRおよびpSC−2TTV2c−RRの感染性を決定する為に豚接種研究を行った。簡単に述べると、豚TTV2について血清陰性およびDNAウイルス陰性である8頭の4週齢在来豚を、それぞれ4匹ずつの2群にランダムに割り当てた。各群の豚を別々に収容し、施設内動物実験委員会(Institutional Committee on Animal Care and Use)の要求を全て満たす条件下で維持した。
【0184】
各群内の全ての豚に、リンパ節内経路と筋肉内経路の両方を組み合わせて注射した。4頭の豚(番号181、189、192および193)には、それぞれ200μgのpSC−2TTV2b−RRプラスミドDNAを注射し、一方、他方の豚4頭(番号92、180、188および191)には、それぞれ200μgのpSC−2TTV2c−RRクローンを接種した。豚を、疾患の臨床徴候について、合計28日間、毎日監視した。全ての豚を接種の28日後に剖検した。
【0185】
いくつかの実施形態の説明によって本発明を例示し、例示的実施形態について詳細に説明したが、本願請求項の範囲をそのような詳細に制限または多少なりとも限定することは、本願出願人の意図するところではない。さらなる変更は当業者には明白であるだろう。したがって本発明は、さらに広いその態様において、ここに示し説明した具体的詳細、代表的装置および方法、ならびに例示的実施例に限定されない。したがって、本願出願人の一般的発明概念の要旨または範囲から逸脱することなく、そのような詳細から逸脱することができる。
【図1A】
【図1B】
【技術分野】
【0001】
(関連出願の相互参照)
本願は、2009年8月21日に出願された米国仮特許出願第61/235,833号および2010年3月23日に出願された米国仮特許出願第61/316,519号の利益を主張するものであり、これら仮特許出願の開示は、参照により、その全体が本明細書に組み込まれる。
【0002】
本発明は、豚トルクテノウイルス(TTV)感染を防御するためのワクチン、豚TTV(PTTV)の感染性DNAクローン、およびそれらの使用に関する。本発明は、豚トルクテノウイルス(PTTV)感染の診断、特に種特異的または型特異的PTTV感染の診断、および遺伝子型の異なる複数株による同時感染の診断にも関係する。
【背景技術】
【0003】
トルクテノウイルス(TTV)は、1997年に輸血後非A〜E型肝炎を有する日本人患者から初めて発見された(ニシザワ,ティ(Nishizawa,T.)、オカモト,エイチ(Okamoto,H.)、コニシ,ケイ(Konishi,K.)、ヨシザワ,エイチ(Yoshizawa,H.)、ミヤカワ,ワイ(Miyakawa,Y.)およびマユミ,エム(Mayumi,M.)、1997年、「原因不明の輸血後肝炎におけるトランスアミナーゼレベルの上昇に関連する新規DNAウイルス(TTV)(A novel DNA virus (TTV) associated with elevated transaminase levels in posttransfusion hepatitis of unknown etiology)」、バイオケミカル・アンド・バイオフィジカル・リサーチ・コミュニケーションズ(Biochem Biophys Res Commun)、第241巻、第1号、p.92−7)。それ以来、多数のヒトTTV株と、後にトルクテノ・ミニ・ウイルス(TTMV)およびトルクテノ・ミディ・ウイルス(TTMDV)と名付けられた2群のTTV関連ウイルスが、健常な人の血清および他の組織に高い有病率で同定されている(ヒノ,エス(Hino,S.)およびミヤタ,エイチ(Miyata,H.)、2007年、「トルクテノウイルス(TTV):現状(Torque teno virus (TTV): current status)」、レビューズ・イン・メディカル・バイロロジー(Rev Med Virol)、第17巻、第1号、p.45−57;オカモト,エイチ(Okamoto,H.)、2009年a、「TTウイルスの発見の歴史と病原性(History of discoveries and pathogenicity of TT virues)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.1−20)。ヒトTTV、TTMVおよびTTMDVは、それぞれ3.6〜3.9、2.8〜2.9および3.2kbの長さの環状一本鎖DNA(ssDNA)ゲノムを持つ無エンベロープ球形ウイルスであり、現在は、国際ウイルス分類委員会(ICTV;http://www.ictvonline.org/virusTaxonomy.asp?bhcp=1)によって新たに設立されたアネロウイルス科(Anelloviridae)に分類されている(ビアジーニ,ピー(Biagini,P.)、2009年、「TTVおよび関連ウイルス(アネロウイルス)の分類(Classification of TTV and related viruses (anelloviruses))」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.21−33)。これら3群のTTV関連ウイルスは高度な遺伝的異質性を示し、それぞれが数多くの遺伝子グループおよび遺伝子型からなる(ビアジーニ,ピー(Biagini,P.)、ガリアン,ピー(Gallian,P.)、カンタローブ,ジェイ・エフ(Cantaloube,J.F.)、アットーウィ,エイチ(Attoui,H.)、デ・ミッコ,ピー(de Micco,P.)、およびデ・ランバレリエ,エックス(de Lamballerie,X.)、2006年、「フランス人献血者におけるTTVおよびTTMV主要系統群の分布および遺伝子解析(Distribution and genetic analysis of TTV and TTMV major phylogenetic groups in French blood donors)」、ジャーナル・オブ・メディカル・バイロロジー(J Med Virol)、第78巻、第2号、p.298−304;ジェルシック,アイ(Jelcic,I.)、ホッツ−ワーゲンブラット,エイ(Hotz-Wagenblatt,A.)、ハンジカー,エイ(Hunziker,A.)、ツア・ハウゼン,エイチ(Zur Hausen,H.)、およびド・ヴィリエ,イー・エム(de Villiers,E.M.)、2004年、「ホジキン病患者の脾生検組織からの複数のTTウイルス遺伝子型の分離:超可変領域におけるゲノムの再編成および多様性(Isolation of multiple TT virus genotypes from spleen biopsy tissue from a Hodgkin’s diesase patient: genome reorganization and diversity in the hypervariable region)」、ジャーナル・オブ・バイロロジー(J Virol)、第78巻、第14号、p.7498−507)。異なる遺伝子型を持つTTVによる多重感染、ならびにTTV、TTMVおよびTTMDVの二重または三重感染の有病率がヒトにおいて記録されており、これらは健常な成人ではよく見られる事象であると考えられる(ニール,シー(Niel,C.)、サバック,エフ・エル(Saback,F.L.)、およびランペ,イー(Lampe,E.)、2000年、「異なる遺伝子型に属する複数のTTウイルスによる同時感染はブラジル人成人ではよく見られる事象である(Coinfection with multiple TT virus strains belonging to different genotypes is a common event in healthy Brazilian adults)」、ジャーナル・オブ・クリニカル・マイクロバイオロジー(J Clin Microbiol)、第38巻、第5号、p.1926−30;ニノミヤ,エム(Ninomiya,M.)、タカハシ,エム(Takahashi,M.)、ホシノ,ワイ(Hoshino,Y.)、イチヤマ,ケイ(Ichiyama,K.)、シモンズ,ピー(Simmonds,P.)、およびオカモト,エイチ(Okamoto,H.)、2009年、「チンパンジーにおけるトルクテノ・ミディ・ウイルス変異体の全ゲノムの解析:ヒトとチンパンジーのまれな異種間感染(Analysis of the entire genomes of torque teno midi virus variants in chimpanzees: infrequent cross-species infection between humans and chimpanzees)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第90巻、第2号、p.347−58;オカモト,エイチ(Okamoto,H.)、2009年a、「TTウイルスの発見の歴史と病原性(History of discoveries and pathogenicity of TT virues)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.1−20;タカヤマ,エス(Takayama,S.)、ミウラ,ティ(Miura,T.)、マツオ,エス(Matsuo,S.)、タキ,エム(Taki,M.)、およびスギイ,エス(Sugii,S.)、1999年、「日本人血友病患者における新規DNA TTウイルス(TTV)感染の有病率および残留性(Prevalence and persistence of novel DNA TT virus (TTV) infection in Japanese haemophiliacs)」、ブリティッシュ・ジャーナル・オブ・ヘマトロジー(Br J Haematol)、第104巻、第3号、p.626−9)。
【0004】
TTVは、ヒトに感染するだけでなく、非ヒト霊長類、ツパイ、豚、牛、猫、犬およびアシカを含む他のさまざまな動物種にも、同様に感染する(ビアジーニ,ピー(Biagini,P.)、ウチ,アール(Uch,R.)、ベルフーシェ,エム(Belhouchet,M.)、アットーウィ,エイチ(Attoui,H.)、カンタローブ,ジェイ・エフ(Cantaloube,J.F.)、ブリスバレー,エヌ(Brisbarre,N.)、およびデ・ミッコ,ピー(de Micco,P.)、2007年、「複合ローリングサークル増幅/配列非依存単一プライマー増幅法を使用することによってヒトおよび動物試料中に同定されたアネロウイルスに関連する環状ゲノム(Circular genomes related to anelloviruses identified in human and animal samples by using a combined rolling-circle amplification/sequence-independent single primer amplification approach)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第88巻、第10号、p.2696−701;イマイ,ティ(Inami,T.)、オバラ,ティ(Obara,T.)、モリヤマ,エム(Moriyama,M.)、アラカワ,ワイ(Arakawa,Y.)およびアベ,ケイ(Abe,K.)、2000年、「チンパンジーから得られたサルTTウイルス分離株の完全長ヌクレオチド配列:新しいTTウイルス様種の証拠(Full-length nucleotide sequence of a simian TT virus isolate obtained from a chimpanzee: evidence for a new TT virus-like species)」、バイロロジー(Virology)、第277巻、第2号、p.330−5;エング,ティ・エフ(Ng,T.F.)、スエドマイヤー,ダブリュ・ケイ(Suedmeyer,W.K.)、ホイーラー,イー(Wheeler,E.)、ガランド,エフ(Gulland,F.)、およびブライトバルト,エム(Breitbart,M.)、2009年、「捕獲されたカリフォルニアアシカの死亡事象から発見された新規アネロウイルス(Novel anellovirus discovered from a mortality event of captive California sea lions)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第90巻、第5号、p.1256−61;オカモト,エイチ(Okamoto,H.)、2009年b、「動物におけるTTウイルス(TT viruses in animals)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.35−52;オカモト,エイチ(Okamoto,H.)、ニシザワ,ティ(Nishizawa,T.)、タカハシ,エム(Takahashi,M.)、タワラ,エイ(Tawara,A.)、ポン,ワイ(Peng,Y.)、キシモト,ジェイ(Kishimoto,J.)、およびワン,ワイ(Wang,Y.)、2001年、「ツパイにおけるTTウイルスのゲノム特徴および進化的特徴の分析ならびにヒトおよび非ヒト霊長類における種特異的TTVとの比較(Genomic and evolutionary characterization of TT virus (TTV) in tupaias and comparison with species-specific TTVs in humans and non-human primates)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第82巻、第9号、p.2041−50;オカモト,エイチ(Okamoto,H.)、ニシザワ,ティ(Nishizawa,T.)、タワラ,エイ(Tawara,A.)、パン,ワイ(Peng,Y.)、タカハシ,エム(Takahashi,M.)、キシモト,ジェイ(Kishimoto,J.)、タナカ,ティ(Tanaka,T.)、ミヤカワ,ワイ(Miyakawa,Y.)、およびマユミ,エム(Mayumi,M.)、2000年a、「ヒトおよび非ヒト霊長類における種特異的TTウイルスならびにそれらの系統発生学的関係(Species-specific TT viruses in humans and nonhuman primates and their phylogenetic relatedness)」、バイロロジー(Virology)、第277巻、第2号、p.368−78;オカモト,エイチ(Okamoto,H.)、タカハシ,エム(Takahashi,M.)、ニシザワ,ティ(Nishizawa,T.)、タワラ,エイ(Tawara,A.)、フカイ,ケイ(Fukai,K.)、ムラマツ,ユー(Muramatsu,U.)、ナイトウ,ワイ(Naito,Y.)、およびヨシカワ,エイ(Yoshikawa,A.)、2002年、「豚、猫および犬におけるTTウイルス(TTV)のゲノム特徴分析ならびに霊長類およびツパイにおける種特異的TTVとそれらの関係(Genomic characterization of TT viruses (TTVs) in pigs,cats and dogs and their relatedness with species-specific TTVs in primates and tupaias)」、ジャーナル・オブ・メディカル・バイロロジー(J Gen Virol)、第83巻、第6号、p.1291−7)。また、チンパンジーはTTMVおよびTTMDVにも感染する(ニノミヤ,エム(Ninomiya,M.)、タカハシ,エム(Takahashi,M.)、ホシノ,ワイ(Hoshino,Y.)、イチヤマ,ケイ(Ichiyama,K.)、シモンズ,ピー(Simmonds,P.)、およびオカモト,エイチ(Okamoto,H.)、2009年、「チンパンジーにおけるトルクテノ・ミディ・ウイルス変異体の全ゲノムの解析:ヒトとチンパンジーのまれな異種間感染(Analysis of the entire genomes of torque teno midi virus variants in chimpanzees: infrequent cross-species infection between humans and chimpanzees)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第90巻、第2号、p.347−58;オカモト(Okamoto)ら、2000年a、前掲書)。同定された動物TTV株、とりわけ非霊長類動物TTVのゲノムサイズは、ヒトTTVのゲノムサイズと比べて比較的小さいが、これらは同じゲノム構造を有し、マイナス鎖ssDNAから翻訳される少なくとも2つの部分的にオーバーラップするオープンリーディングフレーム(ORF1およびORF2)と、高いGC含量(約90%)を有する非翻訳領域(UTR)の短いストレッチとを持つ(オカモト(Okamoto)、2009年b、前掲書)。TTV ORFの配置は、サーコウイルス科(Circoviridae)のジャイロウイルス属(Gyrovirus)に属する鶏貧血ウイルス(CAV)のそれと極めて類似しているが、やはり同じ科に分類される豚サーコウイルス(PCV)1型(PCV1)および2型(PCV2)とは異なっている(デビッドソン,アイ(Davidson,I.)およびシャルマン,エル・エム(Shulman,L.M.)、2008年、「鶏貧血ウイルスによる鳥類感染との比較によってヒトアネロウイルス感染の謎を解明する(Unraveling the puzzle of human anellovirus infections by comparison with avian infections with the chicken anemia virus)」、ウイルス・リサーチ(Virus Res)、第137巻、第1号、p.1−15;ヒノ,エス(Hino,S.)およびプラセティオ,エイ・エイ(Prasetyo,A.A.)、2009年、「トルクテノウイルスと鶏貧血ウイルスとの関連性(Relationship of Torque teno virus to chicken anemia virus)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.117−30)。PCV1とPCV2のゲノムはアンビセンスであり、ORF1はゲノム鎖によってコードされ、ORF2はアンチゲノム鎖によってコードされる(ヒノ(Hino)およびミヤタ(Miyata)、2007年、前掲書)。最近、ヒトTTV遺伝子型1および6の両方の転写パターンおよび翻訳産物が、それぞれのTTV感染性DNAクローンを培養細胞中にトランスフェクションすることによって同定された(ミュラー,ビー(Mueller,B.)、メルツ,エイ(Maerz,A.)、ドベルスタイン,ケイ(Doberstein,K.)、フィンスターブッシュ,ティ(Finsterbusch,T.)、およびマンカーツ,エイ(Mankertz,A.)、2008年、「ヒトトルクテノウイルス分離株P/1C1の遺伝子発現(Gene expression of the human Torque Teno Virus isolate P/1C1)」、バイロロジー(Virology)、第381巻、第1号、p.36−45;キュウ,ジェイ(Qiu,J.)、カッコラ,エル(Kakkola,L.)、チェン,エフ(Cheng,F.)、イェ,シー(Ye,C.)、ソダールンド−ヴェネルモ,エム(Soderlund-Venermo,M.)、ヘドマン,ケイ(Hedman,K.)、およびピンテル,ディ・ジェイ(Pintel,D.J.)、2005年、「ヒトサーコウイルスTTウイルス遺伝子型6は完全長クローンのトランスフェクション後に6つのタンパク質を発現する(Human circovirus TT virus genotype 6 expresses six proteins following transfection of a full-length clone)」、ジャーナル・オブ・バイロロジー(J Virol)、第79巻、第10号、p.6505−10)。スプライスされた3つ以上のmRNAから、ORF1、ORF2、ORF1/1、ORF2/2、ORF1/2およびORF2/3と呼ばれる少なくとも6つのタンパク質の発現が報告されている(カッコラ,エル(Kakkola,L.)、ヘドマン,ケイ(Hedman,K.)、キュウ,ジェイ(Qiu,J.)、ピンテル,ディ(Pintel,D.)、およびソダールンド−ヴェネルモ,エム(Soderlund-Venermo,M.)、2009年、「TTウイルスの複製およびTTウイルスによるタンパク質合成(Replication of and protein synthesis by TT viruses)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.53−64;ミュラー(Mueller)ら、2008年、前掲書;キュウ(Qiu)ら、2005年、前掲書)。したがって、動物TTVに関して、より多くのデータが利用できるようになれば、推定されている動物TTVのゲノム構造を修正することが必要になる可能性が高い。
【0005】
TTVは特発性肝炎患者において初めて同定されたが、その後の研究は、肝炎または他の疾患の病理発生にTTVが重要な役割を果たすという証拠を示すことができなかった(ヒノ(Hino)およびミヤタ(Miyata)、2007年、前掲書;マギー,エフ(Maggi,F.)およびベンディネリ,エム(Bendinelli,M.)、2009年、「トルクテノウイルスおよび他のアネロウイルスの免疫生物学(Immunobiology of the Torque teno viruses and other anellovirueses)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.65−90;オカモト(Okamoto)、2009年a、前掲書)。ヒトTTVは疾患に直接関連するとは見なされていないが、豚TTV(PTTV)は、豚繁殖呼吸器病症候群ウイルス(PRRSV)感染と組み合わされると豚皮膚炎腎症症候群(PDNS)の実験的誘導の一因になり(クラコウカ,エス(Krakowka,S.)、ハーチュニアン,シー(Hartunian,C.)、ハンバーグ,エイ(Hamberg,A.)、シャウプ,ディ(Shoup,D.)、リングス,エム(Rings M.)、チャン,ワイ(Zhang,Y.)、アラン,ジー(Allan,G.)、およびエリス,ジェイ・エイ(Ellis,J.A.)、2008年、「豚サーコウイルス2型に関して結果が陰性であったノトバイオート豚における豚皮膚炎腎症症候群の誘導の評価(Evaluation of induction of porcine determatitis and nephropathy syndrome in gnotobiotic pigs with negative results for porcine circovirus type 2)」、アメリカン・ジャーナル・オブ・ベテリナリー・リサーチ(Am J Vet Res)、第69巻、第12号、p.1615−22)、また、ノトバイオート豚モデルにおいて、PCV2感染と組み合わされると、離乳後多臓器消耗症候群(PMWS)の実験的誘導の一因にもなること(エリス,ジェイ・エイ(Ellis,J.A.)、アラン,ジー(Allan,G.)、およびクラコウカ,エス(Krakowka,S.)、2008年、「ノトバイオート豚におけるサーコウイルス2型関連離乳後多臓器消耗症候群に遺伝子グループ1豚トルクテノウイルスとの同時感染が及ぼす効果(Effect of coinfection with genogroup 1 porcine toruque teno virus on porcine circovirus type 2-associated postweaning multisystemic wasting syndrome in gnotobiotic pigs)」、アメリカン・ジャーナル・オブ・ベテリナリー・リサーチ(Am J Vet Res)、第69巻、第12号、p.1608−14)が示された。このデータから、豚TTVは豚において病原性であることが示唆された。しかし、PTTVに関連する疾患および病変を決定的に特徴づけるには、生物学的に純粋な形態のPTTVウイルスを使ったさらに綿密な研究が必要である。
【0006】
ヒトTTVと比較すると、PTTVのゲノム情報は極めて限られている。現時点で、PTTVについては、日本およびブラジルの豚から、それぞれ1つの完全長ゲノム配列と2つのほぼ完全長のゲノム配列とが報告されているにすぎない(ニール,シー(Niel,C.)、ディニシュ−メンデス,エル(Diniz-Mendes,L.)、およびデバル,エス(Devalle,S.)、2005年、「ヒトおよび豚血清からのトルクテノウイルス(TTV)完全ゲノムのローリングサークル増幅と新規豚TTV遺伝子グループの同定(Rolling-circle amplification of Torque teno virus (TTV) complete genomes from human and swine sera and identification of a novel swine TTV genogroup)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第86巻、第5号、p.1343−7;オカモト(Okamoto)ら、2002年、前掲書)。3つの既知PTTV株のうち、Sd−TTV31株とTTV−1p株が遺伝子グループ1(PTTV1)にまとめられたのに対し、TTV−2pは遺伝子グループ2(PTTV2)に分類される唯一の株だった(ニール(Niel)ら、2005年、前掲書)。しかし遺伝子グループ分類はウイルス学の分類法における漠然とした概念であり、さらにより正確なPTTVの分類が必要とされているが、それは、多数の遺伝子型を代表する新しいPTTV株の完全長ゲノム配列が、より多く利用可能にならなければ、行うことができない。
【0007】
PTTV感染が米国、カナダ、スペイン、中国、韓国およびタイを含む6つの異なる国の豚に蔓延していることは、先に示されている(マキューン,エヌ・イー(McKeown,N.E.)、フェノー,エム(Fenaux,M.)、ハルバー,ピー・ジー(Halbur,P.G.)、およびモン,エックス・ジェイ(Meng,X.J.)、2004年、「6つの異なる国の豚におけるオーファンウイルス・豚TTウイルスの分子キャラクタリゼーション(Molecular characterization of porcine TT virus,an orphan virus,in pigs from six different countries)」、ベテリナリー・マイクロバイオロジー(Vet Microbiol)、第104巻、第1−2号、p.113−7)。
【0008】
豚TTVが特定豚疾患の病理発生において重要な役割を果たすかどうかは、依然として議論の余地がある。ノトバイオート豚モデルでは、PTTV1感染だけでは臨床的疾患を何も発生させないが、軽度の組織学的病変を誘導することが示された(クラコウカ,エス(Krakowka,S.)およびエリス,ジェイ・エイ(Ellis,J.A.)、2008年、「ノトバイオート豚における豚遺伝子グループ1トルクテノウイルスの効果の評価(Evaluation of the effects of procine genogroup 1 torque teno virus in gnotobiotic swine)」、アメリカン・ジャーナル・オブ・ベテリナリー・リサーチ(Am J Vet Res)、第69巻、p.1623−9)。PTTV1と豚繁殖呼吸器病症候群ウイルス(PRRSV)の両方を実験的に接種されたノトバイオート豚が、臨床的な豚皮膚炎腎症症候群(PDNS)を発症したのに対し(クラコウカ,エス(Krakowka,S.)、ハーチュニアン,シー(Hartunian,C.)、ハンバーグ,エイ(Hamberg,A.)、シャウプ,ディ(Shoup,D.)、リングス,エム(Rings M.)、チャン,ワイ(Zhang,Y.)、アラン,ジー(Allan,G.)、およびエリス,ジェイ・エイ(Ellis,J.A.)、2008年、「豚サーコウイルス2型に関して結果が陰性であったノトバイオート豚における豚皮膚炎腎症症候群の誘導の評価(Evaluation of induction of porcine determatitis and nephropathy syndrome in gnotobiotic pigs with negative results for porcine circovirus type 2)」、アメリカン・ジャーナル・オブ・ベテリナリー・リサーチ(Am J Vet Res)、第69巻、p.1615−22)、PTTV1と豚サーコウイルス2型(PCV2)の両方を接種された豚は、急性離乳後多臓器消耗症候群(PMWS)を発症した(エリス(Ellis)ら、2008年、前掲書)。PCV2は臨床的PMWSまたはPCV関連疾患(PCVAD)の主要原因因子であると見なされているが、スペインでは、PCV2が少ないかPCV2を持たないPMWS罹患豚において、PTTV2感染の有病率が非PMWS罹患豚と比較して高いことが観察された(ケカライネン(Kekarainen)ら、2006年、前掲書)。これらのデータは、全体として、豚TTVが豚における疾患の惹起または増悪に関与する共同因子として働き得ることを示唆している。
【0009】
豚TTVは、感染した豚の豚血清、糞便、唾液、精液および組織試料に検出されており、このことは、水平伝播と垂直伝播の両者を含むその多様な伝播経路を示している(ケカライネン(Kekarainen)ら、2007年、前掲書;ポッツート,ティ(Pozzuto,T.)、ミュラー,ビー(Mueller,B.)、ミーハン,ビー(Meehan,B.)、リングラー,エス・エス(Ringler,S.S.)、マッキントッシュ,ケイ・エイ(McIntosh,K.A.)、エリス,ジェイ・エイ(Ellis,J.A.)、マンカーツ,エイ(Mankertz,A.)およびクラコウカ,エス(Krakowka,S.)、2009年、「豚トルクテノウイルスの子宮内伝播(In utero transmission of porcine torque teno viruses)」、ベテリナリー・マイクロバイオロジー(Vet Microbiol)、第137巻、p.375−9;シビラ,エム(Sibila,M.)、マルティネス−ギノー,エル(Martinez-Guino,L.)、ウエルタ,イー(Huerta,E.)、ローレンス,エイ(Llorens,A.)、モーラ,エム(Mora,M.)、グロウ−ローマ,エル(Grau-Roma,L.)、ケカライネン,ティ(Kekarainen,T.)、およびシーゲイルス,ジェイ(Segales,J.)、2009年、「従来の養豚場における豚トルクテノウイルス(TTV)感染および排泄動態(Swine torque teno virus (TTV) infection and excretion dynamics in conventional pig farms)」、ベテリナリー・マイクロバイオロジー(Vet Microbiol)、第139巻、p.213−8)。しかし、現在の豚TTV感染の検出は、主として、従来のPCRアッセイに基づいていた。今までのところ、血清学的アッセイもウイルス培養系も確立されていない。具体的には、スペインのグループによってそれぞれ開発されたPTTV1およびPTTV2のUTR中の保存領域のネステッドPCR増幅が、広く使用されるようになっている(ケカライネンら、2006年、前掲書)。PCV2誘発性PCVADについて証明されたように、ウイルスの量は臨床的疾患の重症度と関連すると思われるので(オプリースニヒ,ティ(Opriessnig,T.)、モン,エックス・ジェイ(Meng,X.J.)およびハルバー,ピー・ジー(Halbur,P.G.)、2007年、「豚サーコウイルス2型関連疾患:最新用語、臨床症状発現、病理発生、診断、および介入戦略に関する最新情報(Porcine circovirus type 2 associated disease: update on current terminology,clinical manifestations,pathogenesis,diagnosis,and intervention strategies)」、ジャーナル・オブ・ベテリナリー・ダイアグノスティック・インベスティゲーション(J Vet Diagn Invest)、第19巻、p.591−615)、従来のPCRによってTTV DNAの存在を決定するのではなく、定量リアルタイムPCRによって豚TTPのウイルス負荷量を決定することが重要になるだろう。また、リアルタイムPCRは、従来のPCRよりも信頼性が高く、迅速かつ安価でもある。最近、2つの豚TTV種を検出し定量するための2つのタックマン(TaqMan)プローブ・ベース・リアルタイムPCRアッセイが記載された(ブラッサード,ジェイ(Brassard,J.)、ガニエ,エム・ジェイ(Gagne,M.J.)、ウーデ,エイ(Houde,A.)、ポイトラス,イー(Poitras,E.)およびウォード,ピー(Ward,P.)、2009年、「豚および牛トルクテノウイルスを検出するためのリアルタイム・タックマンPCRアッセイの開発(Development of a real-time TaqMan PCR assay for the detection of porcine and bovine Torque teno virus)」、ジャーナル・オブ・アプライド・マイクロバイオロジー(J Appl Microbiol)、2009年11月14日、印刷物に先立つ電子ジャーナルでの公開;ガレイ,エイ(Gallei,A.)、ペッシュ,エス(Pesch,S.)、エスキング,ダブリュ・エス(Esking,W.S.)、ケラー,シー(Keller,C.)およびオーリンガー,ブイ・エフ(Ohlinger,V.F.)、2009年、「豚トルクテノウイルス:遺伝子グループ特異的マルチプレックスrt−PCRによるウイルスゲノム負荷量の決定、遺伝子グループ1または2による頻繁な多重感染の検出、およびウイルス完全長配列の確立(Porcione Torque teno virus: Determination of viral genomic loads by genogroup-specific multiplex rt-PCR,detection of frequent multiple infections with genogroups 1 or 2,and establishment of viral full-length sequences)」、ベテリナリー・マイクロバイオロジー(Vet Microbiol)、2009年12月21日、印刷物に先立つ電子ジャーナルでの公開)。プローブに基づくアッセイの主な欠点は、プローブ結合配列が突然変異を含有する場合に、偽陰性結果が得られる可能性がある点である(アンダーソン,ティ・ピー(Anderson,T.P.)、ウェルノ,エイ・エム(Werno,A.M.)、ベイノン,ケイ・エイ(Beynon,K.A.)およびマードック,ディ・アール(Murdoch,D.R.)、2003年、「プローブ結合部位内の配列変位に起因するリアルタイムPCRアッセイおよび融解曲線分析による単純ヘルペスウイルスの遺伝子タイピングの失敗(Failure to genotype herpes simplex virus by real-time PCR assay and melting curve analysis due to sequence variation within probe binding sites)」、ジャーナル・オブ・クリニカル・マイクロバイオロジー(J Clin Microbiol)、第41巻、p.2135−7)。既知の豚TTV株の配列間の高度な不均一性を考えると、PTTVの野外株ではプローブ結合配列中の変異が予期される。SYBRグリーンに基づくリアルタイムPCRは、この潜在的問題を回避する代替的方法であり、その比較的低い特異性にもかかわらず、潜在的豚TTV変異体を検出し定量するための普遍的方法になる。さらにまた、SYBRグリーン・リアルタイムPCRに続く融解曲線分析(MCA)により、反応特異性が確保され、型が異なるウイルスの多重検出が可能になる(リリー,ケイ・エム(Ririe,K.M.)、ラスムッセン,アール・ピー(Rasmussen,R.P.)およびウイットワー,シー・ティー(Wittwer,C.T.)、1997年、「ポリメラーゼ連鎖反応中のDNA融解曲線の分析による生成物の区別(Product differentiation by analysis of DNA melting curves during the polymerase chain reaction)」、アナリティカル・バイオケミストリー(Anal Biochem)、第245巻、p.154−60)。MCAに基づくSYBRグリーン・リアルタイムPCR法はさまざまなヒトウイルスおよび家畜ウイルスに応用されて成功を収めている(ジベリーニ,ディ(Gibellini,D.)、ガルディーニ,エフ(Gardini,F.)、ビトーネ,エフ(Vitone,F.)、スキアボーネ,ピー(Schiavone,P.)、フルリーニ,ジー(Furlini,G.)およびリ,エム・シー(Re,M.C.)、2006年、「血漿試料におけるSYBRグリーン・リアルタイム・マルチプレックスRT−PCR技法によるHCVとHIV−1の同時検出(Simultaneous detection of HCV and HIV-1 by SYBR Green real time multiplex RT-PCR technique in plasma samples)」、モレキュラー・アンド・セルラー・プローブス(Mol Cell Probes)、第20巻、p.223−9;マルティネス,イー(Martinez,E.)、リエラ,ピー(Riera,P.)、シッジャ,エム(Sitja,M.)、ファン,ワイ(Fang,Y.)、オリベイラ,エス(Oliveira,S.)およびマルドナド,ジェイ(Maldonado,J.)、2008年、「SYBRグリーンを用いたリアルタイムRT−PCRおよびアンプリコン融解曲線分析による豚繁殖呼吸器病症候群ウイルス(PRRSV)の同時検出および遺伝子タイピング(Simultaneous detection and genotyping of porcine reproductive and respiratory syndrome virus (PRRSV) by real-time RT-PCR and amplicon melting curve analysis using SYBR Green)」、リサーチ・イン・ベテリナリー・サイエンス(Res Vet Sci)、第85巻、p.184−93;ムイユソー,ケイ・ピー(Mouillesseaux,K.P.)、クリムペル,ケイ・アール(Klimpel,K.R.)およびダール,エイ・ケイ(Dhar,A.K.)、2003年、「SYBRグリーン・リアルタイムPCRにおいてアンプリコンサイズを増大させることによるクルマエビのタウラ症候群ウイルスおよびイエローヘッド病ウイルスに関する検出特異性および検出感受性の改善(Improvement in the specificity and sensitivity of detection for the Taura syndrome virus and yellow head virus of penaeid shrimp by increasing the amplicon size in SYBR Green real-time RT-PCR)」、ジャーナル・オブ・バイロロジカル・メソッズ(J Virol Methods)、第111巻、p.121−7;ウィルヘルム,エス(Wilhelm,S.)、チンマーマン,ピー(Zimmermann,P.)、ゼルビッツ,エイチ・ジェイ(Selbitz,H.J.)およびトリューエン,ユー(Truyen,U.)、2006年、「野外試料中の豚パルボウイルスを検出するためのリアルタイムPCRプロトコール(Real-time PCR protocol for the detection of porcine parvovirus in field samples)」、ジャーナル・オブ・バイロロジカル・メソッズ(J Virol Methods)、第134巻、p.257−60)。
【発明の概要】
【発明が解決しようとする課題】
【0010】
現在、PTTV特異的体液性応答については、ほとんどわかっていない。PCRベースのアッセイは豚におけるPTTV感染の経過を反映しないので、PTTVの血清有病率を評価し、豚疾患におけるPTTVの役割の特徴づけを助けるには、PTTV血清抗体を検出するための効率のよい酵素結合免疫吸着アッセイ(ELISA)が必要である。
【0011】
今のところ、豚TTVのサブユニットワクチン、死滅ワクチンおよび生ワクチンは入手できない。PTTVサブユニットワクチンを開発するには、さまざまな遺伝子型から組換えPTTVキャプシドタンパク質を発現させることが望ましく有益であり、また、死滅ワクチンおよび生ワクチンの開発に使用される細胞培養系において生物学的に純粋な形態のPTTVを増殖させるには、さまざまな遺伝子型から感染性PTTV分子DNAクローンを構築することが、望ましく有益であるだろう。
【課題を解決するための手段】
【0012】
本発明は、PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAの遺伝子型からなる群より選択されるゲノム配列に対して少なくとも80%の相同性を有する少なくとも1コピーのゲノム配列を含有する感染性PTTVをコードする核酸分子を含む、豚トルクテノウイルス(PTTV)の感染性核酸分子(「感染性DNAクローン」)を提供する。
【0013】
本発明の一態様によれば、ゲノム配列が配列番号9、配列番号10、配列番号11、および配列番号12に示す配列から選択される、請求項1に記載のPTTVの感染性DNAクローン。
【0014】
本発明は、感染性PTTVゲノムを含有する生物学的に機能的なプラスミドまたはウイルスベクターを提供する。
【0015】
本発明は、感染性クローンDNAプラスミドまたはウイルスベクターをトランスフェクトされた適切な宿主細胞を提供する。
【0016】
本発明は、PTTV感染性DNAクローンをトランスフェクトされた細胞によって産生される感染性PTTVを提供する。
【0017】
本発明は、無毒性(nontoxic)の生理学的に許容される担体と、(a)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも80%の相同性を有する少なくとも1コピーのゲノム配列またはその相補鎖を含有する核酸分子、(b)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも80%の相同性を有する少なくとも1コピーのゲノム配列またはその相補鎖を含有する核酸分子を含有する生物学的に機能的なプラスミドまたはウイルスベクター、および(c)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも80%の相同性を有する少なくとも1コピーのゲノム配列を含有する無発病性(avirulent)、感染性かつ非病原性(nonpathogenic)PTTV、からなる群より選択される免疫原量の構成要素とを含むウイルスワクチンも提供する。
【0018】
本発明の一態様によれば、ワクチンは、PTTV感染性クローン由来の生PTTVウイルスを含有する。本発明のもう一つの態様によれば、ワクチンは、PTTV感染性クローン由来の死滅PTTVウイルスを含有する。
【0019】
本発明は、細菌発現系においてPTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、およびPTTV2c−VAのORF1キャプシド遺伝子から発現した精製組換えタンパク質と、PTTV感染に対するサブユニットワクチンとしてのこれらの組換えキャプシドタンパク質の使用を提供する。本発明の一実施形態では、サブユニットワクチンとして使用するための組換えキャプシドタンパク質を、バキュロウイルス発現系および他の発現ベクター系において発現させる。
【0020】
本発明のさらなる一態様によれば、アジュバントをさらに含む。
【0021】
本発明はさらに、PTTVウイルス感染に対して豚を免疫化する方法であって、豚に免疫学的有効量のウイルスワクチンを投与することを含む方法を提供する。
【0022】
本発明の一態様によれば、本方法は、組換えサブユニットキャプシドタンパク質、感染性核酸分子または生PTTVウイルスを豚に投与することを含む。
【0023】
本発明のもう一つの態様によれば、本方法は、ワクチンを豚に非経口投与、鼻腔内投与、皮内投与、または経皮投与することを含む。本発明のさらなる一態様によれば、本方法は、ワクチンを豚にリンパ系内投与または筋肉内投与することを含む。
【0024】
本発明は、配列番号9に示すPTTV1a−VAのヌクレオチド配列の配列からなる単離されたポリヌクレオチドも提供する。
【0025】
本発明は、配列番号10に示すPTTV1b−VAのヌクレオチド配列の配列からなる単離されたポリヌクレオチドも提供する。
【0026】
本発明は、配列番号11に示すPTTV2b−VAのヌクレオチド配列の配列からなる単離されたポリヌクレオチドも提供する。
【0027】
本発明は、配列番号12に示すPTTV2c−VAのヌクレオチド配列の配列からなる単離されたポリヌクレオチドも提供する。
【0028】
本発明はさらに、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1、およびORF2/2、特にキャプシドタンパク質をコードするORF1、からなる群より選択されるポリヌクレオチド配列に従って翻訳されたポリペプチド配列の免疫原性フラグメントまたは完全タンパク質を含むサブユニットワクチンを提供する。
【0029】
本発明の一態様によれば、ポリヌクレオチド配列は、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1からなる群より選択される。
【0030】
本発明のもう一つの態様によれば、ポリヌクレオチド配列は、PTTV遺伝子型PTTV1a−VAのORF1である。本発明のさらなる一態様によれば、ポリヌクレオチド配列は、PTTV遺伝子型PTTV1b−VAのORF1である。本発明のさらにもう一つの態様によれば、ポリヌクレオチド配列は、PTTV亜型PTTV2c−VAのORF1である。
【0031】
本発明の一態様によれば、ポリペプチド配列は、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、および配列番号28に示す配列からなる群より選択される。
【0032】
本発明のもう一つの態様によれば、ポリペプチド配列は、配列番号13に示される。本発明のもう一つの態様によれば、ポリペプチド配列は、配列番号14に示される。本発明のさらなる一態様によれば、ポリペプチド配列は、配列番号16に示される。本発明の具体的一実施形態では、ポリペプチド配列が、配列番号16のC末端領域(aa310−625)である。本発明のさらにもう一つの態様によれば、ポリペプチド配列は、配列番号20に示される。
【0033】
本発明のさらなる態様によれば、ワクチンは、さらにアジュバントを含有する。
【0034】
本発明はさらに、PTTVウイルス感染に対して豚を免疫化する方法であって、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1、およびORF2/2からなる群より選択されるポリヌクレオチド配列に従って翻訳されたポリペプチド配列の免疫原性フラグメントまたは完全タンパク質を含むワクチンの免疫学的有効量を豚に投与することを含む方法を提供する。
【0035】
本発明の一態様によれば、本方法は、免疫原性フラグメントまたは組換えキャプシドタンパク質を豚に投与することを含む。
【0036】
本発明のもう一つの態様によれば、本方法は、ワクチンを豚に非経口投与、鼻腔内投与、皮内投与、または経皮投与することを含む。本発明のさらなる一態様によれば、本方法は、ワクチンを豚にリンパ系内投与または筋肉内投与することを含む。
【0037】
また、本発明は、PTTV1感染を診断し、PTTV1負荷量を定量するための方法であって、PTTV1感染が疑われる試料からDNAを抽出すること、配列番号29および配列番号30に示す配列を含むプライマーを使ってポリメラーゼ連鎖反応(PCR)を行うこと、およびPTTV1特異的増幅を検出することを含む方法も提供する。本発明の一態様によれば、ポリメラーゼ連鎖反応はSYBRグリーン・リアルタイムPCRである。
【0038】
本発明はさらに、PTTV2感染を診断し、PTTV2負荷量を定量するための方法であって、PTTV2感染が疑われる試料からDNAを抽出すること、配列番号29、配列番号30、配列番号31および配列番号32に示す配列を含むプライマーを使ってポリメラーゼ連鎖反応(PCR)を行うこと、およびPTTV2特異的増幅を検出することを含む方法を提供する。本発明の一態様によれば、ポリメラーゼ連鎖反応はSYBRグリーン・リアルタイムPCRである。
【0039】
本発明は、PTTV1感染とPTTV2感染を同時に検出し診断するための方法であって、PTTV感染が疑われる試料からDNAを抽出すること、配列番号31および配列番号32に示す配列を含むプライマーを使ってポリメラーゼ連鎖反応(PCR)を行うこと、ならびにPTTV1およびPTTV2特異的増幅を検出することを含む方法も提供する。本発明の一態様によれば、ポリメラーゼ連鎖反応はSYBRグリーン・リアルタイムPCRである。
【0040】
また、本発明は、PTTV1a感染とPTTV1b感染を同時に検出し診断するための方法であって、PTTV1感染が疑われる試料からDNAを抽出すること、配列番号33および配列番号34に示す配列を含むプライマーを使って第1ポリメラーゼ連鎖反応(PCR)を行うこと、配列番号35、配列番号36、配列番号37、および配列番号38に示す配列を含むプライマーを使って第2PCRを行うこと、ならびにPTTV1aおよびPTTV1b特異的増幅を検出することを含む方法も提供する。
【0041】
本発明は、PTTV感染を診断するための方法であって、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1、およびORF2/2からなる群より選択されるポリヌクレオチド配列に従って翻訳されたポリペプチド配列の免疫原性フラグメントを固定化すること、PTTV感染が疑われる豚からの血清試料を固定化免疫原性フラグメントと接触させること、および免疫原性フラグメントに特異的な、捕捉された抗体を検出することを含む方法を提供する。
【0042】
本発明の一態様によれば、ポリヌクレオチド配列は、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1からなる群より選択される。
【0043】
本発明の一実施形態によれば、ポリヌクレオチド配列は、PTTV遺伝子型PTTV1a−VAのORF1である。本発明のもう一つの実施形態によれば、ポリヌクレオチド配列は、PTTV遺伝子型PTTV1b−VAのORF1である。本発明のさらなる一実施形態によれば、ポリヌクレオチド配列は、PTTV亜型PTTV2c−VAのORF1である。
【0044】
本発明のもう一つの態様によれば、ポリペプチド配列は、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、および配列番号28に示す配列からなる群より選択される。
【0045】
本発明の一実施形態によれば、ポリペプチド配列は、配列番号13に示される。本発明のもう一つの態様によれば、ポリペプチド配列は、配列番号14に示される。本発明のもう一つの実施形態によれば、ポリペプチド配列は、配列番号16に示される。本発明のさらなる一実施形態によれば、免疫原性フラグメントは、配列番号16のC末端領域(aa310−625)である。本発明のさらにもう一つの実施形態によれば、ポリペプチド配列は、配列番号20に示される。
【0046】
本発明は、PTTV感染を診断し、PTTV遺伝子型PTTV1a−VA、PTTV1b−VA、およびPTTV種2の全ての既知の亜型に感染した豚の血清における抗体を検出するための、3つの標準化酵素結合免疫吸着アッセイ(ELISA)を提供する。
【0047】
本ELISA診断検査は、PTTV遺伝子型PTTV1a−VA、PTTV1b−VA、およびPTTV2c−VAの細菌発現またはバキュロウイルス発現組換えORF1キャプシドタンパク質に基づく。
【0048】
本発明のもう一つの態様によれば、捕捉された抗体の検出は、ウェスタンブロットによる。本発明のさらにもう一つの態様によれば、捕捉された抗体の検出は、酵素結合免疫吸着アッセイ(ELISA)による。
【0049】
本発明の上述の特徴は、本発明の以下の詳細な説明を図面と併せて読めば、より明確に理解されることになるだろう。
【図面の簡単な説明】
【0050】
【図1】図1Aおよび1Bは、豚TTVウイルス・グループ1(種1)およびグループ2(種2)株の4つの原型米国株のゲノム構造と、ゲノムクローニングおよびアセンブリのための戦略の概略図である。
【図2】図2は、ジェンバンク(GenBank)データベースにおいて入手することができる121のTTV株のヌクレオチド配列比較のPASC(pairwise sequence comparisons)分布を表す。属、種、型、亜型および変異体、ならびにそれらの対応するヌクレオチド配列同一性百分率を示す。
【図3】図3Aは、完全長ゲノムヌクレオチド配列に基づく近隣結合法によって構築された系統樹を表す。図3Bは、7つの豚TTV株間で、ORF1の推定アミノ酸配列に基づいて構築された系統樹を表す。図3Cは、7つの豚TTV株間で、ORF1/1の推定アミノ酸配列に基づいて構築された系統樹を表す。図3Dは、7つの豚TTV株間で、ORF2の推定アミノ酸配列に基づいて構築された系統樹を表す。図3Eは、7つの豚TTV株間で、ORF2/2の推定アミノ酸配列に基づいて構築された系統樹を表す。
【図4】図4は、7つのPTTV株間のORF1の完全長アミノ酸配列のアラインメントを表す。
【図5】図5は、7つのPTTV株間のORF2の完全長アミノ酸配列のアラインメントを表す。
【図6】図6Aは、各々の標準テンプレート(青色で示すもの)および20個の豚血清試料を40サイクル増幅した後のPTTV1リアルタイムPCR産物の融解曲線を表す。図6Bは、各々の標準テンプレートおよび20個の豚血清試料を40サイクル増幅した後のPTTV2リアルタイムPCR産物の融解曲線を表す。
【図7】図7A〜7Eは、PTTV1/PTTV2 SYBRグリーン・ベース・デュプレックス・リアルタイムPCRの融解曲線分析(MCA)を表す。
【図8】図8は、7つのPTTV株間の予測ORF1のN末端部分に位置するヌクレオチド配列のアラインメントを表す。
【図9】図9Aおよび9Bは、4つの既知豚TTV2の親水性プロファイルおよび保存領域を表す。
【図10】図10A〜10Cは、組換えPTTV2c ORF1キャプシドタンパク質の発現と精製を表す。
【図11】図11A〜11Cは、選ばれた豚血清試料のウェスタンブロット分析の代表的結果を表す。
【図12】図12は、PTTV2c−ORF1に基づくウェスタンブロットおよびELISAの整合性を表す。
【図13】図13は、異なる供給源に由来する138頭の豚におけるウイルス負荷量によるPTTV2血清抗体レベルの箱ひげ図を表す。
【図14】図14Aは、PTTV2のウイルス負荷量の遡及的評価を表す。図14Bは、到着時から到着の2ヶ月後まで成長した10頭の豚におけるPTTV2 ORF1キャプシドタンパク質に対する抗体レベルを表す。
【図15】図15A〜15Cは、PTTV1aおよびPTTV1b組換えORF1キャプシドタンパク質の発現と精製を表す。
【図16】図16は、ウィスコンシンの飼育場に由来する選ばれた豚血清試料のPTTV1a−ORF1ベースのウェスタンブロット分析の例を表す。
【発明を実施するための形態】
【0051】
本発明によれば、具体的一実施形態において、上記4つの新規豚TTV亜型が、バージニアにおいて、一頭の雄豚から分離される。
【0052】
図1Aでは、PTTV1とPTTV2のゲノムがどちらもボールド体で示され、4つの予測ORF(ORF1、ORF2、ORF1/1およびORF2/2)のサイズと向きが矢印で示されている。GCリッチ領域も示されている。破線の弧AおよびDは、それぞれ、ネステッドPCRによって血清試料および精液試料からPTTV1およびPTTV2を検出するために使用した領域を表す。破線の弧BおよびCは、PTTV1のゲノムクローニングのための2つのオーバーラップPCRフラグメントを表し、一方、破線の弧EおよびFは、PTTV2のゲノムクローニングのための2つのオーバーラップPCRフラグメントを表す。本研究に使用したプライマー(表1参照)の場所も、対応する位置に示されている。
【0053】
第1ラウンドPCRにおいてPTTV1とPTTV2のどちらについても陽性と示されることから、高いウイルス負荷量が示唆された1つの雄豚血清試料(SR#5)を、その後のPTTVの完全長ゲノムクローニングに使用した。驚いたことに、最初のPTTV株Sd−TTV31をクローニングするために設計されたインバースPCR(オカモト(Okamoto)ら、2002年、前掲書)の2組のプライマーセット(NG372/NG373およびNG384/NG385)を利用してウイルスゲノムDNAを増幅しようとする最初の試みは、成功しなかった。数回試行したがPCR産物は得られなかった。次に、PTTV1の領域AおよびPTTV2の領域Dの初期配列に基づいて、想定されるPTTV1ゲノムにまたがる領域BおよびCを増幅するための2つの新しいプライマー対(TTV1−If(配列番号1)/TTV1−2340R(配列番号2)およびTTV1−2311F(配列番号3)/TTV1−IR(配列番号4))と、想定されるPTTV2ゲノムにまたがる領域EおよびFを増幅するための2つのさらなるプライマー対(TTV2−IF(配列番号5)/TTV2−2316R(配列番号6)およびTTV2−GCF(配列番号7)/TTV2−IR(配列番号8))を、それぞれ設計した(図1Aおよび表1)。プライマーTTV1−2340R(配列番号2)およびTTV1−2311F(配列番号3)は、PTTV2株TTV−2p(ニール(Niel)ら、2005年、前掲書)中には存在しないPTTV1株Sd−TTV31(オカモト(Okamoto)ら、2002年、前掲書)およびTTV−1p(ニール(Niel)ら、2005年)中の共通配列から推定し、一方、プライマーTTV2−2316R(配列番号6)およびTTV2−GCF(配列番号7)は、前記2つのPTTV1株には存在しない株TTV−2pの配列から推定した。その結果生じた予期されるサイズの4つの異なるPCR産物を、平滑末端クローニングベクターにそれぞれ挿入し、その結果生じた組換えプラスミドを大腸菌(Escherichia coli)に挿入した。フラグメントB、C、EおよびFを表す各コンストラクトについて8〜15個の陽性(白色)細菌クローンを同定し、次に配列決定した。
【0054】
【表1】
【0055】
意外なことに、各コンストラクトから2群の配列データが同定され、同じ豚から得られる遺伝子グループ1および遺伝子グループ2に2タイプのPTTVが存在することが示された。それら4つのPTTV株を区別しアセンブルするために、3つの既知PTTV株Sd−TTV31、TTV−1pおよびTTV−2pと共に、配列比較を行った(図1Bおよび1C)。
【0056】
図1Bは、PTTV1株PTTV1a−VAおよびPTTV1b−VAの完全長ゲノム配列の区別およびアセンブリを、続いてクローニングされたPCRフラグメントBおよびCと共に図解している。フラグメントB中のORF1およびORF2の開始コドンと、フラグメントC中のORF1の終止コドンに「^」または「*」で印が付けられている。2つの既知PTTV1株Sd−TTV31およびTTV−1pの対応する配列も示されている。保存されている配列を影付きで表し、ダッシュ記号はヌクレオチド欠失を示す。
【0057】
PTTV1の場合、予測ORF1の開始コドンATGおよび終止コドンTGAは、それぞれフラグメントBおよびC中に位置していた(図1B)。これらのコドンの位置は2つのPTTV1グループにおいて異なり、1つ目はSd−TTV31と同一、2つ目はTTV−1pと同一だった(図1B)。また、これら2つのグループにおけるORF2開始コドンも、ORF1の場合と同じく、異なる位置にあった。さらにまた、領域Bの4つの異なる配列(配列決定データからの2つと株Sd−TTV31およびTTV−1pからの2つ)および領域Cの4つの異なる配列を使った系統解析も、1つ目の配列がSd−TTV31とクラスターを形成し、2つ目がTTV−1pとクラスターを形成することを裏付けた(データ未掲載)。それゆえに、本発明者らは、フラグメントBとCの両方からの2群の配列データを区別し、2つの完全長PTTV1ゲノム(これらを、それぞれ、株PTTV1a−VA(配列番号9)およびPTTV1b−VA(配列番号10)と名付けた)にアセンブルすることができた(図1B)。
【0058】
図1Cは、PTTV2株PTTV2b−VAおよびPTTV2c−VAの完全長ゲノム配列の区別とアセンブリを、続いてクローニングされたPCRフラグメントEおよびFと共に図解している。TTV−2p株の対応する配列も含まれており、保存されている配列が影付きで表されている。ダッシュ記号はヌクレオチド欠失を示す。各株について、オーバーラップ領域(破線で囲んだ部分)内のユニークなヌクレオチド(PTTV2b−VA(配列番号11)の場合は連続する「AG」ヌクレオチド、PTTV2c−VA(配列番号12)の場合は2つの単一「A」ヌクレオチドおよび単一「G」ヌクレオチド)が、それぞれ示されている。
【0059】
これら2つのPTTV2株の区別の方が容易だった。2つのPCRフラグメントのオーバーラップ領域中に位置しているユニークな連続「AG」ヌクレオチドは、それぞれフラグメントEおよびFからの2群の配列データに共通していた(図1C)。アセンブルした完全長ゲノム配列は、1つのPTTV2株を表し、これをPTTV2b−VA(配列番号11)と名付けた。同様に、フラグメントEおよびFからのもう1つの配列データセットがオーバーラップ領域中に共有している2つのユニークな単一「A」ヌクレオチドおよび単一「G」ヌクレオチドに基づいて、PTTV2c−VA(配列番号12)と名付けた第2の株の完全ゲノム配列をアセンブルした(図1C)。フラグメントEおよびFからの4つの配列をTTV−2pからの2つの対応する配列と共に使った系統解析も、この割当を裏付けた(データ未掲載)。
【0060】
本発明は、豚におけるウイルス感染に関連する4つの分離された豚TTVウイルス遺伝子型または亜型を提供する。本発明は、豚TTVウイルス遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VA、配列番号9(PTTV1a−VA)、配列番号10(PTTV1b−VA)、配列番号11(PTTV2b−VA)、および配列番号12(PTTV2c−VA)に示すヌクレオチド配列、それらの機能的等価物または相補鎖を有するウイルス遺伝子型または亜型を包含するが、これらに限るわけではない。任意の豚TTVに由来する特定のヌクレオチド配列が、個々のウイルス間に自然に存在するわずかな変異を有するであろうことは理解されるだろう。これらの配列の変異は、欠失、置換、挿入などに見ることができる。
【0061】
4つのPTTV株のそれぞれについて提案されるゲノム構造を詳細に分析し、3つの既知PTTV株Sd−TTV31、TTV−1pおよびTTV−2pと共に、表2に要約する。PTTVの4つの米国株は、それぞれ、2,878bp(PTTV1a−VA 配列番号9)、2,875bp(PTTV1b−VA 配列番号10)、2,750bp(PTTV2b−VA 配列番号11)、および2,803bp(PTTV2c−VA 配列番号12)という、類似したゲノムサイズを有する。PTTV1a−VA(配列番号9)とSd−TTV31はどちらも同じゲノム長を有する。株TTV−1pおよびTTV−2pの公表された配列は、いずれも、UTRのGCリッチ領域に多くの未決定ヌクレオチドを有する。これらのヌクレオチドをPTTV1およびPTTV2に対応するコンセンサス配列で人工的に埋めたところ、それぞれゲノム長が、TTV−1pはPTTV1b−VA(配列番号10)とより近い関係にあり、TTV−2pはPTTV2b−VA(配列番号11)とより近い関係にあることが示された(データ未掲載)。
【0062】
アセンブルされた豚TTVウイルス遺伝子型または亜型PTTV1a−VA(配列番号9)、PTTV1b−VA(配列番号10)、PTTV2b−VA(配列番号11)、およびPTTV2c−VA(配列番号12)のゲノム配列は、ジェンバンク(Genbank(登録商標))に提出され(ヌクレイック・アシッズ・リサーチ(Nucleic Acids Resaerch)、2008年1月;36(データベース号(Database issue):D25−30)、それぞれアクセッション番号GU456383、GU456384、GU456385、およびGU456386が付与されている。
【0063】
【表2】
数字(完全長ゲノム、ORFのサイズ、およびエクソン番号を除く)は、各PTTV株のゲノム上のヌクレオチド(nt)位置を示す。
【0064】
最近行われた2つの研究により、ヒトTTV複製時に、転写されたウイルスmRNAと、少なくとも6個のウイルスタンパク質の発現が同定された(ミュラー(Mueller)ら、2008年、前掲書;キュウ(Qiu)ら、2005年、前掲書)。これはヒトTTVがコードすると予想されるORFの数よりも多い(オカモト,エイチ(Okamoto,H.)、ニシザワ,ティ(Nishizawa,T.)、タワラ,エイ(Tawara,A.)、タカハシ,エム(Takahashi,M.)、キシモト,ジェイ(Kishimoto,J.)、サイ,ティ(Sai,T.)、およびスガイ,ワイ(Sugai,Y.)、2000年b、「感染個体からの骨髄細胞に検出されるTTウイルスmRNA(TT virus mRNAs detected in the bone marrow cells from an infected individual)」、バイオケミカル・アンド・バイオフィジカル・リサーチ・コミュニケーションズ(Biochem Biophys Res Commun)、第279巻、第2号、p.700−7)。そこで本発明者らは、PTTV配列との比較のために、この新しいヒトTTVゲノム情報を含めた。ヒトTTV株P/1C1のmRNA転写産物の5’端は、TATAボックスの25nt下流にある「A」にマッピングされた(ミュラー(Mueller)ら、2008年、前掲書)。この開始点、その隣接配列(CGAATGGCTGAGTTTATGCCGC(配列番号39);開始点に下線を付した)および上流にあるTATAボックスまでの距離(24nt;表2)は、7つのPTTV株の全てにおいてよく保存されており、PTTVとヒトTTVは、mRNAの共通の5’端を翻訳に利用し得ることが示唆される。
【0065】
7つのPTTV株の全ての間で完全に保存されているさらに5つの領域がTATAボックスの近傍に同定された。それぞれ11ntの2つの領域(AGTCCTCATTT(配列番号40)およびAACCAATCAGA(配列番号41))は、TATAボックスの上流に位置し、残り3つの領域(17ntのCTGGGCGGGTGCCGGAG(配列番号42);14ntのCGGAGTCAAGGGGC(配列番号43);11ntのTATCGGGCAGG(配列番号44))は、提案されているmRNAの5’端とORF2の開始コドンとの間に位置する。これらの保存されたPTTV特異的配列はウイルス遺伝子発現を調節する共通の要素を含有するのだろう。
【0066】
以前、3つの既知PTTV株のゲノムに、それぞれ3つのORF(ORF1〜3)が提案された(ニール(Niel)ら、2005年、前掲書;オカモト(Okamoto)ら、2002年、前掲書)。本研究で同定されたPTTVの4つの原型米国株は、この構造を有している。ヒトTTV中の対応するORF3は、ORF2中の同じATGで開始しスプライシング後も同じORF(伸張したORF2)中に留まるので(図1A)、これにはORF2/2という新たな名前が付けられている(ミュラー(Mueller)ら、2008年、前掲書;キュウ(Qiu)ら、2005年、前掲書)。本研究では、PTTV分類を改定するために、本発明者らはヒトTTVの命名法に従う。ヒトTTV ORF1/1は、ORF1中の2つのエクソンによってコードされる新たに同定されたウイルスタンパク質である(キュウ(Qiu)ら、2005年、前掲書)。ORF1/1は、ORF1と、同一のN末端部分およびC末端部分を共有している。PTTV ORF1/1対応物は、7つのPTTV株の全てにおいて、すぐに同定された(図1Aおよび表2)。
【0067】
ORF1およびORF2が約2.8kbのウイルスmRNAによってコードされるのに対し、ORF1/1およびORF2/2は、ヒトTTVでは、スプライスされた約1.2kbのウイルスmRNAによってコードされる(ミュラー(Mueller)ら、2008年、前掲書;キュウ(Qiu)ら、2005年、前掲書)。これら4つのORFがPTTVゲノムでも推定されたことと、7つのPTTV株において予測されるスプライスドナー部位およびスプライスアクセプター部位配列の配列および位置がよく保存されていることから(表2)、豚TTVもおそらく2つの対応するmRNAをコードすると推測される。
【0068】
ヒトTTV株の大半はCAVと遺伝的類似性を共有していて、TTVアポトーシス誘導タンパク質(TAIP)をコードし、そのCAV対応部分はアポプチンと名付けられている(ド・スミット,エム・エイチ(de Smit,M.H.)およびノートボルン,エム・エイチ(Noteborn,M.H.)、2009年、「鶏貧血ウイルスおよびTTウイルスにおけるアポトーシス誘導タンパク質(Apoptosis-inducing proteins in chicken anemia virus and TT virus)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.131−49)。TAIPのORFはORF2内に埋め込まれている。しかし豚TTVには、対応するTAIPが存在しない。最近の研究により、培養細胞におけるCAVの複製にはアポプチンまたはTAIPの発現が必要であることが示された(プラセチョウ,エイ・エイ(Prasetyo,A.A.)、カマホラ,ティ(Kamahora,T.)、クロイシ,エイ(Kuroishi,A.)、ムラカミ,ケイ(Murakami,K.)、およびヒノ,エス(Hino,S.)、2009年、「鶏貧血ウイルス(CAV)の複製はアポプチンを必要とし、ヒトトルクテノウイルス(TTV)のVP3によって補完される(Replication of chicken anemia virus (CAV) requires apoptin and is complemented by VP3 of human torque teno virus (TTV))」、バイロロジー(Virology)、第385巻、第1号、p.85−92)。
【0069】
ペアワイズ配列比較(PASC)は、同じファミリー内のウイルスの全ての利用可能なゲノム配列からペアワイズヌクレオチド配列同一性百分率の頻度分布をプロットする有用な方法である(バオ,ワイ(Bao,Y.)、カプースチン,ワイ(Kapustin,Y.)、およびタツソワ,ティ(Tatusova,T.)、2008年、「ペアワイズ配列比較(PASC)によるウイルス分類(Virus Classification by Pairwise Sequence Comparison (PASC))」、エイサイクロペディア・オブ・バイロロジー5巻セット(Encyclopedia of Virology,5 vols)(ビー・ダブリュ・ジェイ・マーヒー(B.W.J.Mahy)およびエム・エイチ・ブイ・ファン・レーゲンモルテル(M.H.V.Van Regenmortel)編)の第5巻、p.342−8、エルゼビア(Elsevier)、オックスフォード(Oxford))。PASCプログラムによって生成した異なるピークは、通常、ウイルスの属、種、型、亜型および株を表す(図2)。この研究において、本発明者らは、ジェンバンク(GenBank)データベースに見いだされる121の、ヒトおよび動物TTV関連株の完全長ゲノム配列を使って、TTVのPASC解析を行った(図2)。TTVの構成要素が一つの独立した科、アネロウイルス科(Anelloviridae)に分類されると仮定すると、36〜55%および55〜67%のヌクレオチド配列同一性に見られる2つの主要ピークは、それぞれ属および種のグループを表す(図2)。したがってTTV型は67〜85%のヌクレオチド配列同一性を有するTTVのグループと定義され、一方、TTV亜型は85〜95%のヌクレオチド配列同一性を共有するTTV配列のグループと定義することができる。95%を上回るヌクレオチド配列同一性を共有するTTV株は、さらに変異体に分類することができる(図2)。同じような分類が、ジェルシック(Jelcic)らにより、103個のTTV分離株の配列を使って提案されている(ジェルシック,アイ(Jelcic,I.)、ホッツ−ワーゲンブラット,エイ(Hotz-Wagenblatt,A.)、ハンジカー,エイ(Hunziker,A.)、ツア・ハウゼン,エイチ(Zur Hausen,H.)、およびド・ヴィリエ,イー・エム(de Villiers,E.M.)、2004年、「ホジキン病患者の脾生検組織からの複数のTTウイルス遺伝子型の分離:超可変領域におけるゲノムの再編成および多様性(Isolation of multiple TT virus genotypes from spleen biopsy tissue from a Hodgkin’s diesase patient: genome reorganization and diversity in the hypervariable region)」、ジャーナル・オブ・バイロロジー(J Virol)、第78巻、第14号、p.7498−507)。
【0070】
ここに提案されたTTV分類の基準を、PTTVの4つの原型米国株と他の3つの既知PTTV株のゲノム配列の系統解析に適用した。これらの株間で行った完全長ヌクレオチド配列のペアワイズ比較は、4つのPTTV1株は3つのPTTV2株と比較して54.0〜56.4%のヌクレオチド配列同一性を有することを示した(表3)。したがって、文献において以前にPTTVの「遺伝子グループ(genogroup)」と呼ばれていたものは、おそらく「種(species)」と呼ぶ方が、より適切であり、PTTV1とPTTV2はおそらくそれぞれ豚TTV種1および種2を表すはずである。PTTV種1は、それぞれ1a型(Sd−TTV31およびPTTV1a−VA(配列番号9)を含む)および1b型(TTV−1pおよびPTTV1b−VA(配列番号10))と呼ばれる2つのウイルス型からなる。というのも、これら2つのウイルス型の間のヌクレオチド配列同一性は69.8〜70.7%だからである(表3)。Sd−TTV31とTTV1a−VA(配列番号9)は、それらの高い配列同一性(95.1%)ゆえに、同じ種の変異体株であると認識される。しかし、2つの1b型株、TTV−1pおよびPTTV1b−VA(配列番号10)は2つの異なる亜型に属し得る(ヌクレオチド配列同一性=86.4%)。PTTV種2の場合、3つの株は、86.5〜90.9%というそれらのヌクレオチド配列同一性に基づいて、別々の亜型(それぞれ、TTV−2pは亜型2a、PTTV2b−VA(配列番号11)は亜型2b、PTTV2c−VA(配列番号12)は亜型2c)に分類される可能性が高い。ここに提案されたPTTVのための新しい分類システムは、系統樹に明確に見てとることができた(図3A)。PTTVのORF1、ORF1/1、ORF2およびORF2/2の推定アミノ酸配列に基づいて構築した系統樹も、ここに提案された分類と合致した(図3B〜3E)。
【0071】
【表3】
データはPASCプログラムを使用することによって生成したもので、値は%ヌクレオチド配列同一性を示す。
【0072】
PTTV種、型および亜型の間で、ユニークな突然変異ならびに欠失および/または挿入は、ゲノム全体に散在している。例えば、上述のように図1Bに示すORF1の開始および終止コドンならびにORF2の開始コドンの場所は、PTTV1a型と1b型の間で異なる。2つのPTTV1b株は、PTTV1aと比較して、ORF2開始後に2コドンの欠失も有する(図1B)。
【0073】
注目すべきことに、TTV−2pとPTTV2b−VAはどちらも、PTTV2c−VAと比較して、UTR中の最初の11nt保存配列(AGTCCTCATTT(配列番号40))の39nt上流に、大きな52nt欠失を有する。この欠失ゆえに、PTTV2b−VAのゲノムサイズ(おそらくTTV−2pも同様である)は、PTTV2c−VAのゲノムサイズよりもかなり小さい(表2)。多くの「サブウイルス」ヒトTTVクローンが血清試料から単離されており、これらサブウイルス分子の大部分においてORFが通常はインタクトなままであることから、それらは完全長TTVゲノムであるとみなされている(ド・ヴィリエ(de Villiers)ら、2009年;レピック(Leppik)ら、2007年)。それらは、完全にまたは部分的に欠失しているUTRに、さまざまな長さを有する。TTV−2pとPTTV2b−VAの状況は、ヒトTTVサブウイルス分子の状況に似ていると思われ、亜型PTTV2aとPTTV2bは、亜型PTTV2cに由来するサブウイルス分子である可能性があることが示唆される。注目すべきことに、野外試料からPTTV2を検出するために他のグループ(エリス(Ellis)ら、2008年、前掲書;ケカライネン(Kekarainen)ら、2007年、前掲書;ケカライネン(Kekarainen)ら、2006年、前掲書;クラコウカ(Krakowka)ら、2008年、前掲書)によってよく使用されているネステッドPCRプライマーTTV2−nF(表1)の3’末端配列は、その欠失の両側に位置している。したがって、PTTV2を検出するための現在のネステッドPCRアッセイは、遺伝的に多様なPTTV2c亜型の株を同定するには十分でない可能性が高い。
【0074】
分離ウイルス株の供給源は、豚TTVウイルス感染を有する豚の血清、糞便、唾液、精液および組織試料である。しかし、組換えDNA技術を使ってヌクレオチド配列を複製し化学的に合成することができると考えられる。したがって本発明の範囲は、例えば限定するわけではないが、配列番号9、配列番号10、配列番号11、および配列番号12に示すヌクレオチド配列、またはその相補鎖を含む、単離されたポリヌクレオチド;配列番号9、配列番号10、配列番号11、および配列番号12に示すヌクレオチド配列にハイブリダイズし、そのヌクレオチド配列に少なくとも67%相補的な、好ましくは85%相補的な、より好ましくは95%相補的なポリヌクレオチド;または配列番号13(PTTV1a−VA)、配列番号14(PTTV1b−VA)、配列番号15(PTTV2b−VA)、配列番号16(PTTV2c−VA)に示すORF1タンパク質のアミノ酸配列、配列番号17(PTTV1a−VA)、配列番号18(PTTV1b−VA)、配列番号19(PTTV2b−VA)、配列番号20(PTTV2c−VA)に示すORF2タンパク質のアミノ酸配列、配列番号21(PTTV1a−VA)、配列番号22(PTTV1b−VA)、配列番号23(PTTV2b−VA)、配列番号24(PTTV2c−VA)に示すORF1/1タンパク質のアミノ酸配列、配列番号25(PTTV1a−VA)、配列番号26(PTTV1b−VA)、配列番号27(PTTV2b−VA)、配列番号28(PTTV2c−VA)に示すORF2/2タンパク質のアミノ酸配列からなる群より選択される免疫原性フラグメントを包含する。免疫原性または抗原性コード領域またはフラグメントは、当技術分野において知られている技法によって決定することができ、次にそれらを使って、免疫反応性スクリーニングまたは他の診断目的のためのモノクローナル抗体またはポリクローナル抗体を作製することができる。本発明はさらに、単離されたポリヌクレオチドによってコードされる精製免疫原性タンパク質を包含する。望ましくは、タンパク質は、上記分離豚TTV亜型の少なくとも1つの単離または組換えORF1タンパク質またはORF2タンパク質、より望ましくはORF1タンパク質であることができる。
【0075】
豚TTVのORF1は、構造および複製関連タンパク質をコードすると考えられている(マギー,エフ(Maggi,F.)およびベンディネリ,エム(Bendinelli,M.)、2009年、「トルクテノウイルスおよび他のアネロウイルスの免疫生物学(Immunobiology of the Torque teno viruses and other anelloviruses)」、カレント・トピックス・イン・マイクロバイオロジー・アンド・イムノロジー(Curr Top Microbiol Immunol)、第331巻、p.65−90)。7つのPTTV株のORF1がコードする産物は、624〜635aaの長さを有し、DNA結合活性を有すると考えられる多数のアルギニン残基をN末端に持っている(図4)。図4では、保存されている配列を影付きで表している。ダッシュ記号はアミノ酸欠失を示す。RCRモチーフは実線で囲まれている。PTTV1株の3つのHVR(PTTV1−HVR1、2および3)とPTTV2株の2つのHVR(PTTV2−HVR1および2)は破線で囲まれている。ORF1/1の接合境界を矢印で示す。予想されるローリングサークル複製ローリングサークル複製(RCR)モチーフ(イリイナ,ティ・ブイ(Ilyina,T.V.)およびクーニン,イー・ブイ(Koonin,E.V.)、1992年、「真正細菌、真核生物および古細菌の多様なレプリコンによってコードされるローリングサークルDNA複製のための開始タンパク質における保存された配列モチーフ(Conserved sequence motifs in the initiator proteins for rolling circle DNA replication encoded by diverse replicons from eubacteria,eucaryotes and archaebacteria)」、ヌクレイック・アシッズ・リサーチ(Nucleic Acids Research)、第20巻、第13号、p.3279−85)は、異なるPTTV型および亜型では異なる位置に示され、それらの位置は型特異的または亜型特異的であり得る。RCRモチーフIII(YxxK)は、PTTV1a型株(PTTV1a−VA 配列番号13のaa位置14−17)および1b型株(PTTV1b−VA 配列番号14のaa位置379−382)において、それぞれ保存されているが、3つのPTTV2株の全てに同定された同じ保存モチーフは、PTTV2b−VA 配列番号15のaa位置482−485に位置している(図4)。PTTV2b−VA 配列番号15とPTTV2c−VA 配列番号16はどちらも、PTTV2b−VAのaa位置331−333に保存されたRCRモチーフII(HxQ)も有し、TTV−2pにはこれが存在しない(図4)。
【0076】
PTTV株のORF1タンパク質が種1と種2の間で共有する配列同一性はわずかに22.4〜25.8%と極めて低く、それが、これら2つの種の間で有意に保存されたaa配列を同定することを困難にしている(図4)。PTTV種1では、1a型株と1b型株の間のORF1のaa同一性が50.3〜52.7%である。4つのPTTV1株には、比較的多数のaa置換を伴う3つの主要超可変領域(HVR)PTTV1−HVR1〜3が同定されたのに対し、3つのPTTV2株の間には2つのHVR(PTTV2−HVR1および2)が観察された(図4)。3つのPTTV2株は、対応するPTTV1−HVR1領域に約20aaの欠失を有する。そのうえ、PTTV2の2つのHVRは、対応するPTTV1−HVR3領域内にある(図4)。これらのHVRはORF1中にのみ位置するが、切断型のORF1/1中にはない。これらは、ヒトTTVの研究によって示唆されているとおり、宿主免疫監視機構を免れ、PTTVが持続感染を樹立するのを助ける役割を果たすらしい。
【0077】
ORF2のaa配列は4つのPTTV1株(PTTV1a−VA 配列番号17;PTTV1b−VA 配列番号18)および3つのPTTV2株(PTTV2b−VA 配列番号19;PTTV2c−VA 配列番号20)の間でかなり異なっていた(図5)。しかしこれらは保存されたタンパク質チロシンホスファターゼ(PTPアーゼ)様モチーフ(Wx7Hx3CxCx5H)をN末端に共有している(図4)。このモチーフは、全てのヒトTTV、TTMVおよびTTMDV株ならびにCAVにも保存されている。TTMVまたはCAVのORF2タンパク質は、セリン/スレオニンホスファターゼ(S/T PPアーゼ)活性も示した(ピータース,エム・エイ(Peters,M.A.)、ジャクソン,ディ・シー(Jackson,D.C.)、クラッブ,ビー・エス(Crabb,B.S.)、およびブラウニング,ジー・エフ(Browning,G.F.)、2002年、「鶏貧血ウイルスVP2は新規二重特異性プロテインホスファターゼである(Chicken anemia virus VP2 is a novel dual specificity protein phosphatase)」、ジャーナル・オブ・バイオロジカル・ケミストリー(J Biol Chem)、第277巻、第42号、p.39566−73)。ORF2タンパク質の二重特異性は、ウイルス複製時に、宿主の遺伝子転写、シグナル伝達およびサイトカイン応答を調節すると思われる。最近、CAV中のモチーフの最後のヒスチジン残基の前にある保存された2つの塩基性aa残基の変異導入解析により、これら2つの残基は、ウイルス複製、インビトロの細胞病理、およびインビボの弱毒化に影響を及ぼすことが明らかになった(ピータース,エム・エイ(Peters,M.A.)、クラッブ,ビー・エス(Crabb,B.S.)、ワシントン,イー・エイ(Washington,E.A.)、およびブラウニング,ジー・エフ(Browning,G.F.)、2006年、「鶏貧血ウイルスのVP2遺伝子の部位特異的変異導入は、ウイルス複製、細胞病理および宿主細胞MHCクラスI発現に影響を及ぼす(Site-directed mutagenesis of the VP2 gene of Chicken anemia virus affects virus replication,cytopathology and host-cell MHC class I expression)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第87巻、第4号、p.823−31;ピータース,エム・エイ(Peters,M.A.)、クラッブ,ビー・エス(Crabb,B.S.)、チベンデール,ケイ・エイ(Tivendale,K.A.)、およびブラウニング,ジー・エフ、2007年、「VP2の部位特異的変異導入による鶏貧血ウイルスの弱毒化(Attenuation of chicken anemia virus by site-directed mutagenesis of VP2)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第88巻、第8号、p.2168−75)。これら2つの塩基性aa残基(「KK」)は3つのPTTV2株に保存されている。しかし、2つのPTTV1a株では最初の塩基性残基(「R」)だけが保たれており、一方、PTTV1b株では塩基性残基はどちらも置換されている(図5)。図5において、ダッシュ記号はアミノ酸欠失を示す。TTV、TTMVおよびCAVにおいて同定された共通モチーフWx7Hx3CxCx5H(下線部)内の保存された5つのアミノ酸を影付きで示す。このモチーフの最後のヒスチジンの前にある2つの塩基性アミノ酸(CAVにおいてウイルス複製、細胞病理、またはインビボ弱毒化に影響を及ぼすことが示されているもの)の位置を「^」で示す。
【0078】
要約すると、本発明は、バージニアの一頭の雄豚の血清試料における異なる遺伝子型または亜型を代表する4つの豚TTV株の完全長ゲノム配列を決定した。この研究から得られた知見は、ヒトTTVと同様に、豚には異なる遺伝子型または亜型による多重PTTV感染が存在し、おそらくそれが一般的であることを明確に示している。本発明者らは、PTTVのゲノム編成、可変性の程度、保存されているヌクレオチドモチーフおよびアミノ酸モチーフの特徴に関する新しい情報も提供した。これらの情報は、現在のPCR検出アッセイを改善し、血清学的診断法のための試薬の開発を助け、PTTVの構造的および機能的研究を開始するのに役立つであろう。この研究では、7つの既知PTTV株のゲノム配列の系統解析および遺伝子解析に基づいて、PTTVの新しい分類も提案する。
【0079】
本発明は、豚TTVに感染した豚または他の哺乳動物の試料中のウイルスDNAを検出することによる、豚TTV感染の診断方法も提供する。本発明の好ましい一実施形態は、ウイルス感染またはウイルス疾患の診断をさらに助けるために、ポリメラーゼ連鎖反応(PCR)用のオリゴヌクレオチドプライマーを使って、豚または他の哺乳動物種における豚TTV核酸配列を検出するための方法を伴う。豚または他の哺乳動物種における豚TTVウイルス核酸配列の有無を検出するのに役立つ診断検査は、豚TTVに感染した豚またはTTVの感染が疑われる豚の試料からウイルスDNAを単離することと、PTTV1特異的プライマー(配列番号29/配列番号30)またはPTTV2特異的プライマー(配列番号31/配列番号32)を使って、SYBRグリーン・リアルタイム定量PCRを行うこととを含む。
【0080】
本発明のもう一つの実施形態では、PTTV1/PTTV2デュプレックス・リアルタイムPCRを使用することによって、PTTV1とPTTV2が同時に検出されるように、診断方法を適合させることができる。より具体的に述べると、この方法は、豚TTVに感染した豚またはTTVの感染が疑われる豚の試料からウイルスDNAを単離することと、PTTV1特異的プライマー(配列番号29/配列番号30)またはPVVT2特異的プライマー(配列番号31/配列番号32)の両方を使ってリアルタイムPCRを行うこととを含む。PTTV1とPTTV2のTm値はMCAで識別することができるので、PTTV1 DNAとPTTV2 DNAの存在を同時に検出することができる。
【0081】
本発明のさらにもう一つの実施形態において、診断方法はデュプレックス・ネステッドPCRを使用し得る。この方法は、豚TTVに感染した豚またはTTVの感染が疑われる豚の試料からウイルスDNAを単離すること、1対のプライマーP1ab−mF(配列番号33)/P1ab−mR(配列番号34)を使って第1ラウンドのPCRを行うこと、PTTV1aを検出するためのP1a−nF(配列番号35)/P1a−nR(配列番号36)およびPTTV1bを検出するためのP1b−nF(配列番号37)/P1b−nR(配列番号38)という2対のプライマーの混合物を使って第2ラウンドのPCRを行うこと、およびPCR産物を可視化することを含む。
【0082】
当業者は当技術分野において周知の方法に従って、上記の診断方法を最適化することができる。
【0083】
したがって本発明の一実施形態は、2つの豚TTV種のウイルス負荷量をそれぞれ定量するために、2つの新規シングルプレックスSYBRグリーン・リアルタイムPCRアッセイを開発する。現時点で利用することができる6つのPTTV1完全長ゲノムと4つのPTTV2完全長ゲノムの間でそれぞれによく保存されている領域を標的とするPTTV1特異的プライマーとPTTV2特異的プライマーを設計した。本発明のもう一つの実施形態では、2つの豚TTV種PTTV1aおよびPTTV1bを同時検出するために、前記2つのシングルプレックスアッセイを組み合わせてデュプレックス・リアルタイムPCRアッセイを行った後、異なる融解温度によって同定することができるウイルスアンプリコンのMCAを行う。第3の実施形態では、単一試料中の豚TTV種の2つの型PTTV1aおよびPTTV1bを同定する目的で、第1ラウンドPCRにおいて2つの型のPTTV1からのウイルスDNAを同時増幅し、第2ラウンドPCRにおいて1a型と1b型の区別的検出を行うためのデュプレックス・ネステッドPCRアッセイを開発した。これらのアッセイは、種特異的または型特異的豚TTVの診断のための簡単で実用的なツールになる。
【0084】
利用可能な10個の豚TTV完全長ゲノムの多重配列アラインメントによって、潜在的プライマー配列を同定した。PTTV1特異的プライマーTTV1F(配列番号29)およびTTV1R(配列番号30)は、6個のPTTV1ゲノム間で予測ORF2の直前にある2つの保存されたゲノム領域に基づいて設計し、一方、PTTV2特異的プライマーTTV2F4(配列番号31)およびTTV2R4(配列番号32)は、4つのPTTV2ゲノム間で予測ORF2/2の直後にある2つの保存されたゲノム領域に基づいて設計した(表4)。プライマーは自己二量体化および交差二量体化の可能性を示さなかった。予期されるアンプリコンサイズは、それぞれ、PTTV1b−VAゲノムに対応するPTTV1プライマーからの118bpフラグメントと、PTTV2c−VAゲノムに対応するPTTV2プライマーからの200bpフラグメントであった。
【0085】
【表4】
【0086】
本発明の具体的一実施形態によれば、PTTV1特異的プライマーおよびPTTV2特異的プライマーを用いるSYBRグリーン・シンプレックス・リアルタイムPCRを使って、それぞれ豚TTV1およびTTV2 DNAを特異的に検出することができる。PTTV1については、ある範囲の、25μlあたりのターゲットDNA濃度にわたって、標準曲線を確立した。線形範囲は、4.4×101〜4.4×108コピーにまたがることが示された。最低検出限界(44コピー)は、37.57の閾サイクル数(Ct)に対応した。PTTV2についても標準曲線を作成し、それを使って、25μlの反応あたり8.6×100〜8.6×108コピーの範囲にあるDNA濃度を検出した。最低検出限界(8.6コピー)に対応するCtは36.53だった。
【0087】
本発明のもう一つの具体的実施形態では、豚TTV1 DNAと豚TTV2 DNAの同時検出に、SYBRグリーン・デュプレックス・リアルタイムPCRを利用する。PTTV1(87.0℃)とPTTV2(80.0℃)の間の7度というTm値の差が、それらをMCAによって互いに識別することを可能にした。したがって、PTTV1とPTTV2を同時検出するために、2つのシングルプレックスアッセイを結びつけてデュプレックス・リアルタイムPCRアッセイにすることができる。陽性試料は、その産物についての既知Tm内で対称的な融解ピークを有するものであった。この新しいアッセイを、まず最初に、PTTV1標準とPTTV2標準の混合物の10倍希釈液を使って検証した。テンプレートとして滅菌水を使用した非テンプレート陰性対照は、シングルプレックスアッセイでは見られなかったPTTV1プライマーとPTTV2プライマーの間の交差二量体化が引き起こす非特異的増幅を示した(図7a)。これにより、72.0℃〜76.0℃に別個の融解ピークが生じた。図7Aは、PTTV1標準(赤色;Tm=87.0℃)、PTTV2標準(緑色;Tm=80.0℃)および非テンプレート陰性対照(プライマー交差二量体化によるもの;黒色)の融解ピークを示している。図7B〜7Eは、異なるウイルス負荷量のPTTV1とPTTV2とを含む代表的な血清試料の融解ピークを示している。図7Bは、雄豚血清試料番号5(PTTV1とPTTV2のウイルス負荷量はどちらも比較的高いが、PTTV2>PTTV1)を表し、図7Cは、雄豚血清試料番号12(PTTV1とPTTV2のウイルス負荷量はどちらも比較的高いが、PTTV1>PTTV2)を表し、図7Dは雄豚血清試料番号14(PTTV1とPTTV2のウイルス負荷量はどちらも低い)を表し、図7Eは雄豚血清試料番号10(PTTV1陽性だが、PTTV2陰性)を表す。シングルプレックス・リアルタイムPCRによって決定された各試料中のPTTV1およびPTTV2のウイルス負荷量(単位:ゲノムコピー/ml)を、対応する融解ピークの上に示した。
【0088】
ある例では、デュプレックス・リアルタイム・アッセイを20個の成体雄豚血清試料に適用した場合に、PTTV1とPTTV2のウイルス負荷量がどちらも比較的高い試料は、PTTV1とPTTV2に対応する2つの異なる融解曲線を示し、非特異的融解ピークは示さなかったのに対し(図7Bおよび7C)、PTTV1またはPTTV2のどちらか一方のウイルス負荷量が低い試料は、ウイルス特異的融解曲線だけでなく、非特異的融解曲線も示した(図7Dおよび7E)。試料番号14における2つの融解ピークは極めて小さかったが、それらはPTTV1およびPTTV2の適当なTmに視覚的に明確で対称的な上昇と降下を示したので、それらは陽性と見なされた(図7D)。これに対し、試料番号10は、対称的なPTTV2融解ピークが明白には存在しなかったので、PTTV1のみ陽性と見なされた(図7E)。これらの結果は、2つのシングルプレックスアッセイの結果と一致した(表5)。さらにまた、融解ピークのサイズと形状は、検出された試料における対応するウイルス負荷量を定量的に反映した。
【0089】
本発明のもう一つの態様では、デュプレックス・ネステッドPCRを、2つの豚TTV型PTTV1aおよびPTTV1bの区別的検出に使用する。
【0090】
本発明の発明者は、豚TTV種1における2つの異なる遺伝子型(暫定的にPTTV1aおよびPTTV1bと名付けた)の存在を証明した。さらに、PTTV1aとPTTV1bの同時感染が豚において一般的であるかどうかを決定するために、これら2つを迅速に識別するための新規デュプレックス・ネステッドPCRアッセイを開発した。豚TTVゲノムDNA配列のアラインメントにより、ウイルスキャプシドタンパク質をコードする予測ORF1のN末端部分に位置する保存されたゲノム領域が同定された(図8)。この領域はORF2全体と上流の部分的UTRも含有している。第1ラウンドPCRにおいてPTTV1a DNAとPTTV1b DNAの両方を同時に増幅するために、プライマーP1ab−mF(配列番号33)/P1ab−mR(配列番号34)を設計した。第2ラウンドPCRでは、PTTV1a特異的プライマーP1a−nF(配列番号35)/P1a−nR(配列番号36)とPTTV1b特異的プライマーP1b−nF(配列番号37)/P1b−nR(配列番号38)との混合物を使って、各遺伝子型を区別的に増幅した。PTTV1aおよびPTTV1bの最終PCR産物は、それぞれサイズが162bpおよび96bpであり、それらは臭化エチジウムで染色した1%アガロースゲルでの電気泳動により、容易に識別することができた。このアッセイでは、プライマーの特異性ゆえに、PTTV2 DNAは検出されないと予期された(図8)。図8では、保存されている配列を点で示し、影付きにした。ダッシュ記号によってヌクレオチド欠失を示した。デュプレックス・ネステッドPCRに使用した3対のプライマーの場所と向きを矢印で示した。
【0091】
ある例では、デュプレックス・ネステッドPCRアッセイに付した成体雄豚からの血清試料20個は、予期されるサイズの2本のバンドの可視化と、続いて行われた配列決定によるPCR産物の確認とによる決定で(データ未掲載)、全てが、PTTV1aとPTTV1bの両方について陽性であることがわかった。19個の精液試料ではPCR産物が増幅されず、上述したPTTV1従来型ネステッドPCRおよびリアルタイムPCRアッセイの結果と合致した。
【0092】
2つの豚TTV種による豚の感染は、遡及的研究によれば、スペインの養豚場において1985年には見いだされている(シーゲイルス(Segales)ら、2009年、前掲書)。しかし豚TTVが何らかの特定豚疾患と関連しているかどうかは依然として不明である。どちらの豚TTV種も飼育豚において高い有病率を有するので、TTV DNAの存在を評価することよりも、TTVウイルス負荷量の決定の方が、おそらく、より重要である。血清試料および精液試料におけるウイルス負荷量のレベルは、PCV2感染におけるPCVADの重要なマーカーとして示されている(オプリースニヒ(Opriessnig)ら、2007年、前掲書)。したがって、定量的なPTTV特異的リアルタイムPCRアッセイの確立は、豚TTVに関連する潜在的疾患状態を同定するのに役立つだろう。
【0093】
2つのタックマン(TaqMan)プローブ・ベース・リアルタイムPCRアッセイが最近になって記載された。カナダのグループが開発したシングルプレックス・アッセイは種特異的ではなく、2つのPTTV種の総ウイルス負荷量を定量するようにしか設計されなかった(ブラッサード(Brassard)ら、2009年、前掲書)。ドイツのグループが確立したデュプレックス・アッセイでは両方の種の特異的な同時検出が可能だった(ガレイ(Gallei)ら、2009年、前掲書)。これら2つのアッセイにおいて使用されたプライマーのターゲット配列は、3つの豚TTVゲノム配列(Sd−TTV31、TTV−1pおよびTTV−2p)のアラインメントによって決定されたもので、UTR中に位置していた。本研究において、本発明者らは、利用可能な7つの完全PTTVゲノム配列(4つのPTTV1配列と3つのPTTV2配列)を追加して、10個の完全長PTTVゲノムの間で保存されている領域を分析し、再決定した。この研究で得られた最新のアラインメント結果に基づいて、PTTV1とPTTV2のウイルス負荷量をそれぞれ定量するために、2つの種特異的シングルプレックスSYBRグリーンベース・リアルタイムPCRアッセイを開発した。本発明者らのアッセイで使用したプライマーは、先の研究とは異なる保存ゲノム領域に結合するように設計されたもので、これらは定量の正確度を向上させ得る。本発明者らのアッセイは、25μlの反応あたり、PTTV1については44ゲノムコピー数、PTTV2については8.8ゲノムコピー数と、かなりの種特異性および選択性を示した。これに対し、タックマン(TaqMan)ベースのデュプレックス・リアルタイムPCRでは、1反応あたり10ゲノムコピー数という検出限界が報告されている(ガレイ(Gallei)ら、2009年、前掲書)。また、SYBRグリーンベースのリアルタイムPCRアッセイは、蛍光標識したプローブを使用する必要がなく直ちに行うことができるフレキシブルで安価なアプローチである。最後に、豚TTVが高度の遺伝的多様性を示すことを考えると、SYBRグリーンベース・アッセイによる結果は、タックマン(TaqMan)プローブベースのアッセイにおけるプローブ結合配列中に突然変異を含有する可能性が高い豚TTV変異体の異なる遺伝的背景による影響を受ける可能性が少ない。
【0094】
TTV DNAが存在するとはいえ、この研究において試験された健常な豚からの血清試料はいずれも、2×106コピー/ml未満という少量のPTTV1およびPTTV2を有していた。さらにまた、3つの精液試料には、極めて低いタイターのPTTV2 DNAしか検出されなかった。試験した血清試料の大半は、従来のネステッドPCRで決定したところ、PCV2 DNAについても陽性だった(データ未掲載)。ウイルス負荷量が低い多くのPCV2陽性豚は、臨床的PCVADを発症しない。PCVADの発症に関して提案されている閾は、107以上のPCV2ゲノムコピー/ml血清である(オプリースニヒ(Opriessnig)ら、2007年、前掲書)。また、精液PCV2 DNA陽性も、疾患状態の注目すべきマーカーである(オプリースニヒ(Opriessnig)ら、2007年、前掲書;パル,エヌ(Pal,N.)、ファン,ワイ・ダブリュ(Huang,Y.W.)、マドソン,ディ・エム(Madson,D.M.)、カスター,シー(Kuster,C.)、モン,エックス・ジェイ(Meng,X.J.)、ハルバー,ピー・ジー(Halbur,P.G.)およびオプリースニヒ,ティ(Opriessnig,T.)、2008年、「豚精液試料における豚サーコウイルス2型および内部対照の同時検出および定量のためのデュプレックス・リアルタイムPCRアッセイの開発と検証(Development and validation of a duplex real-time PCR assay for the simultaneous detection and quantification of porcine circovirus type 2 and an internal control on porcine samen samples)」、ジャーナル・オブ・バイロロジカル・メソッズ(J Virol Methods)、第149巻、p.217−25)。種特異的PTTVの状況はPCV2のそれと類似していると考えられ、豚疾患の誘導には107コピー/mlを上回る高いPTTV価が要求されるのだろう。この研究において開発された種特異的リアルタイムPCRアッセイは、今後、さまざまな疾患状態から採取される多数の臨床試料を使ってPTTVと疾患との関連を調べるための簡単で実用的なツールになるだろう。
【0095】
さらにまた、これら2つの種特異的シングルプレックス・アッセイを結びつけることにより、本発明者らは、MCAベースのデュプレックス・リアルタイムPCRアッセイにおいて2つの豚TTV種PTTV1とPTTV2を同時に検出し区別するための迅速で安価で信頼できるスクリーニングを開発し、検証した。このアッセイは両PTTV種の正確な定量を意図したものではないが、検出を目的とするなら、従来のネステッドPCRに取って代わることのできる、より便利なアプローチである。リアルタイムPCRと比較すると、豚TTV検出のための従来のネステッドPCRアッセイは、時間がかかり(合計4ラウンドのPCRを必要とする)、面倒であり、複数ラウンドのPCR処理中に起こる試料汚染に見舞われやすい。PTTV1種とPTTV2種の間のTm値の相違ゆえに、デュプレックスPCR増幅後のMCAにより、異なる反応特異性を確保することができる。このデュプレックス・リアルタイム・アッセイのもう一つの利点は、記載のプロトコールを行う際にPTTV1標準とPTTV2標準を含めることが必須でなくなる点であり、これにより、自動リアルタイムPCR機器を装備した診断検査室におけるはるかに幅広い使用が容易になる。
【0096】
同じ種の異なる遺伝子型または亜型による豚TTVの多重感染は実証されている(ガレイ(Gallei)ら、2009年、前掲書)。特に、本発明者らの以前の研究では、豚TTV種1は、2つの異なる型PTTV1a(株Sd−TTV31およびPTTV1a−VAを含む)およびPTTV1b(株TTV−1pおよびPTTV1b−VAを含む)からなることが示された。完全長ゲノムを持つ新たに公表された2つのPTTV1分離株、カナダ由来のswSTHY−TT27(GQ120664)とドイツ由来のTTV1#471819(GU188045)は、どちらも、系統解析に基づいて1b型に分類された(データ未掲載)。この研究において記載するデュプレックス・ネステッドPCRにより、2つのPTTV1遺伝子型の二重感染は豚では頻繁に起こることが確認された。この新規アッセイは、PTTV1aと1bを識別するために現在までに開発された最初の診断的PCRアプローチである。PTTV1a感染とPTTV1b感染の一方または両方が疾患と関連する関連要因を表すかどうかは現時点ではわかっていないので、本発明者らの区別的PCRアッセイは、これら2つのPTTV型の今後の潜在的疾患関連性にとって、大いに価値があるはずである。
【0097】
本発明のもう一つの態様では、豚TTV ORFタンパク質を発現させ、豚TTV特異的抗体の存在を検出するための免疫検出アッセイにおいて使用した。本発明の一実施形態では、PTTV1a、PTTV1bおよびPTTV2の3つの切断型ヒスチジンタグ付きORF1タンパク質を、大腸菌(E.coli)中で、それぞれ発現させた。さらにまた、これらの組換え抗原に基づく血清ウェスタンブロットおよびELISAアッセイを開発し、異なる供給源からの豚血清試料を使って検証した。特に、PTTV1a特異的、PTTV1b特異的およびPTTV2特異的ELISAを使った血清検査は、豚TTV感染と疾患との関連を明らかにするための正確で簡単なツールになる。
【0098】
本発明のさらなる一態様では、豚TTV ORFタンパク質を、大腸菌発現系において、組換えORF1キャプシドタンパク質として発現させ、精製した(図10、図15)。PTTV1a、PTTV1bおよびPTTV2の3つの切断型Hisタグ付きORF1タンパク質を、大腸菌(E.coli)中で、それぞれ発現させ、精製し、PTTV感染に対する組換えキャプシドサブユニットワクチンとして利用した。
【0099】
4つの豚TTV2株、TTV−2p、TTV2#472142、PTTV2b−VAおよびPTTV2c−VAは、現在までに、利用可能な完全ゲノム配列を持っていた。それらは系統学的に3つの予測亜型に分類されるが、4つのPTTV2に由来するアミノ酸をコードするORF1の親水性プロファイルの比較分析により、それらは3つの親水性領域、すなわちN末端にあるaa1〜49のアルギニンリッチ領域と、それぞれ中央部とC末端部に位置する2つの特定ドメイン(IおよびII)を共有することが示された(図9A)。切断型PTTV2c−VA ORF1発現に使用したC末端領域と、他の3つのPTTV2株において共有されている対応領域を、破線の枠で示した。アミノ酸配列のアラインメントにより、4つのPTTV2株間でのドメインI(aa322−349)およびII(aa536−625)の高レベルな配列保存が示された(図9B)。
【0100】
親水性ドメインは多くのタンパク質の抗原性にとって重要であると考えられるので、上記2つのドメインを含有するPTTV2c−VA ORF1 配列番号16のC末端領域(aa310−625)を、豚血清におけるPTTV2特異的抗体検出用の抗原として使用されるタンパク質発現のために選択した。本発明の一態様によれば、切断型PTTV2c ORF1の発現は、全てのPTTV2亜型(2a、2bおよび2c;図3Aも参照されたい)を検出するのに十分であった。
【0101】
本発明の一実施形態では、8×Hisタグと融合したPTTV2c ORF1遺伝子のC末端部分を構築し、大腸菌中で発現させた。組換えタンパク質は不溶性であり、細菌封入体内に発現した。図10Aは、未精製2c−ORF1産物のSDS−PAGEを示す。図10Bは、精製2c−ORF1産物のSDS−PAGEを示す。図10Cは、抗HisタグmAbを使った精製2c−ORF1産物のウェスタンブロット分析を示す。白い矢じりは、予期されるサイズを持つORF1タンパク質とその切断産物を示し、一方、黒い矢じりは、予期される切断型タンパク質の予測二量体を示す。M:タンパク質マーカー。図10Aにおいて、2c−ORF1未精製試料では、対照試料と比較して、2つの際立ったポリペプチド(白い矢じり)が生成していた。約40KDaのバンドは2c−ORF1の予期されるサイズと一致し、一方、約30KDaポリペプチドは、おそらく、前者からのN末端切断産物であった。ニッケルアフィニティカラムによる精製後に、2つの前記際立ったバンドを含む4つのポリペプチドが、SDS−PAGEに示された(図10B)。それらは、抗HisタグmAbを用いるウェスタンブロットでも検出された(図10C)。2つの高分子量バンド(黒い矢じり)は、予想されるサイズ(約80KDaおよび約60KDa)によれば、それぞれ約40KDaおよび約30KDaの2つのポリペプチドによって形成されるホモ二量体だった。これらの結果から、精製C末端PTTV2c−ORF1がうまく生産され、豚血清中の豚TTV2抗体検出に使用できることが証明された。
【0102】
本発明のもう一つの態様によれば、さまざまな豚血清試料における豚TTV2抗体を、精製C末端PTTV2c−ORF1を使ったウェスタンブロットによって検出することができる。白い矢じりは、予期されるサイズを持つORF1タンパク質とその切断産物を示す。緑色のバンドだけが陽性と認識されることに注意すべきである。さまざまな供給源から得た在来豚(健常または罹患)、CD/CD豚およびノトバイオート豚の血清試料を全部で200個以上収集した。抗原として精製C末端PTTV2c−ORF1を使って抗PTTV2c−ORF1 IgG抗体を検出するために、試料をランダムに選択した。図11Aは、在来豚の選ばれた豚血清試料のウェスタン分析の結果を示し、図11BはCD/CD豚を示し、図11Cはノトバイオート豚を示す。精製PTTV2c−ORF1産物を抗原として使用した。印を付けた約40KDaと約30KDaの2つのバンドは、在来豚(図11A)およびCD/CD豚(図11B)の試料の大半に検出されたことから、これらの豚におけるPTTV2感染が幅広く示された。しかし、2つの異なる供給源(バージニア州ブラックスバーグおよびアイオワ州エイムズ)から得たノトバイオート豚はいずれも、検出可能なPTTV2抗体を持っていなかった(図11C)。別の低分子量バンドも観察された(図11Aおよび11B)。それらはおそらくウェスタンブロットにおける非特異的反応性に由来したのだろう。
【0103】
本発明のさらにもう一つの態様によれば、PTTV2特異的ELISAを、豚TTV2血清検査として使用することができる。血清陰性結果は、ウィスコンシンの飼育場の在来豚からの数個の試料にも示された(図12)。これらの陰性試料をプールし、PTTV2特異的ELISAの開発において陰性参照として使用した。同じ供給源から得られた残りの試料は陽性だった(図12、左側の4レーン)。また、細胞培養に使用される営利会社から得た豚血清(OIE疾患フリーと考えられる)も、強い抗PTTV2−ORF2陽性を示し(図12)、これをELISAの陽性対照として使用した。精製2c−ORF1抗原、豚血清およびIgGコンジュゲートの濃度は、交差力価測定法により、低いバックグラウンドシグナルを呈し、陽性対照と陰性対照とのOD405値の差が最も大きくなるように決定した。最適な抗原量は1ウェルあたり69ngであり、最適なELISA結果は、血清試料の1:100希釈液およびIgGコンジュゲートの1:4000希釈液を使用することによって得られた。ELISAカットオフ値は各試行において0.25〜0.5の範囲にあった。図4は、血清ウェスタンブロットと開発されたELISAとの整合性を反映した代表的結果を示す。
【0104】
3つの群れから得た138個の在来豚血清試料を選んで、リアルタイムPCRによるPTTV2ウイルス負荷量とELISAによる抗PTTV2IgG抗体レベルの間の相関関係を分析した。その結果、検出不可能であるか高いPTTV2ウイルス負荷量(108コピー/ml)を持つ豚の方が、中間的な値のPTTV2ウイルス負荷量を持つ豚よりも低い血清PTTV2抗体価を持つ可能性が高いことが示された(図13)。
【0105】
特に、同じ群れの豚10頭からの血清を、それらが新しい施設に到着した時から到着の2ヶ月後まで、それらの血清のPTTV2ウイルス負荷量と抗PTTV2抗体レベルとを比較することによっても分析した。10頭中9頭の豚は2ヶ月後にウイルス負荷量が減少していた(3頭は検出可能なウイルスを持っていなかった)が、抗PTTV2抗体価は10頭中9頭の豚において増加していた(図14Aおよび14B)。これらの結果から、10頭の豚は早期にPTTV2感染を獲得し、それが体液性応答を誘導して、徐々に抗ORF1キャプシドIgG抗体を産生したことが示唆された。PTTV2−ORF1 IgG抗体は、ウイルスを中和することができ、またはウイルスを除去することさえできたことから、ORF1が実際にウイルスキャプシドタンパク質をコードし、PTTV2に対する中和エピトープを含有し得ることが示された。
【0106】
本発明の一実施形態では、C末端PTTV1a−およびPTTV1b−ORF1タンパク質を、それぞれ大腸菌系で発現させ、精製した。SDS−PAGEと、抗HisタグmAbを使ったウェスタンブロット分析により、1a−ORF産物と1b−ORF産物はどちらも2つのポリペプチドを生じ、一つは予期されるサイズのもの(約40KDa)、もう一つは予測ホモ二量体(約80KDa)であることが示された(図15A〜C)。図15Aは、未精製および精製1a−ORF1産物のSDS−PAGEを示す。図15Bは、精製1b−ORF1および1b−ORF1ctruc産物のSDS−PAGEを示す。図15Cは、抗HisタグmAbを使った精製1a−および1b−ORF1産物のウェスタンブロット分析を示す。白い矢じりは予期されるサイズのORF1タンパク質を示し、一方、黒い矢じりはORF1タンパク質の予測二量体を示す。2c−ORF1発現と比較して、切断型ポリペプチドは観察されなかった。比較対照として、C末端切断型1b−ORF1領域(1b−ORF1ctruc)の発現は、そのC末端非切断対応物1b−ORF1と比較して、低分子量ポリペプチドをもたらした(図15B)。
【0107】
本発明の一実施形態では、PTTV2について上述したように、精製C末端PTTV1a−およびPTTV1b−ORF1タンパク質を使って、遺伝子型特異的な血清ウェスタンブロットおよびELISAを開発した。図16は、1a−ORF1を抗原として用いる血清ウェスタンブロットの陰性例(レーン1〜2)と陽性例(レーン3〜5)を示す。PTTV1a特異的およびPTTV1b特異的ELISAでは、PTTV2−ORF1と同じ抗原量(69ng)、血清の希釈度(1:100)およびIgGコンジュゲートの希釈度(1:4000)を使用した(データ未掲載)。
【0108】
さらに本発明は、豚TTV感染を検出するための有用な診断試薬であって、例えば豚に豚TTVまたは本発明の免疫原性組成物を有効免疫原量で接種してウイルス感染を生じさせ、感染した豚の血清から抗体を回収することなどによって自然宿主から精製された、モノクローナルまたはポリクローナル抗体を含む診断試薬を提供する。あるいは、単離された豚TTVのヌクレオチド配列によってコードされるアミノ酸配列または免疫原性フラグメントに由来する、またはそこから発現される、天然または合成ポリペプチドに対して、実験動物において、抗体を産生させることもできる。例えば、モノクローナル抗体は、単離された豚TTVのヌクレオチド配列に由来するポリペプチド抗原で免疫化された例えばBalb/cなどのマウスから得られるハイブリドーマ細胞から産生され得る。ハイブリドーマ細胞の選択は、ダルベッコ変法イーグル培地(DMEM)または最小必須培地のような標準的細胞培養培地に入れたハイプロキサンチン(hyproxanthine)、チミジンおよびアミノプテリン中で成長させることによって行う。抗体を産生するハイブリドーマ細胞は、当技術分野において知られている手法に従ってクローン化することができる。次に、形成される離散コロニーを、適切な培養培地で培養するために、培養プレートの別々のウェルに移すことができる。抗体分泌細胞の同定は、適当な抗原または免疫原を使って、従来のスクリーニング法によって行われる。ハイブリドーマ細胞をインビトロで培養するか、ハイブリドーマの注入後にマウスの腹水を得ることにより、インビボで培養することで、所望のモノクローナル抗体が周知の技法で生産される。
【0109】
もう一つの代替的方法では、当技術分野において知られている手法に従って、豚TTVキャプシドタンパク質をバキュロウイルス発現系または大腸菌発現系で発現させることができる。発現した組換え豚TTVキャプシドタンパク質は、診断用の抗原として、酵素結合免疫吸着アッセイ(ELISA)において使用することができる。豚組換えキャプシド抗原に基づくELISAアッセイは、例えば、豚および哺乳動物種における豚TTVに対する抗体を検出するために使用することができる。ELISAアッセイは好ましいものの、免疫蛍光アッセイ(IFA)、免疫ペルオキシダーゼアッセイ(IPA)などといった他の既知の診断検査も使用することができる。
【0110】
望ましくは、豚における豚TTV感染を診断するために、本発明による市販のELISA診断アッセイを使用することができる。豚TTVの精製ORF1およびORF2タンパク質を使った、豚における抗TTV抗体を検出するためのELISAアッセイの開発を、実施例に例示する。豚TTVに感染した豚から集めた血清と、対照豚から得た陰性血清とを使って、アッセイを検証する。PTTV2特異的抗体、PTTV1a特異的抗体、およびPTTV1b特異的抗体が、PTTV ORFタンパク質を特異的に認識することが証明された。当業者に知られる技法による検査のさらなる標準化により、豚TTVに関する診断アッセイの商品化を最適化することができる。
【0111】
本発明のもう一つの態様は、分離された豚TTVまたは上述の単離されたポリヌクレオチドによってコードされる抗原タンパク質を含むユニークな免疫原組成物、および抗体を産生または生産するためのその使用である。組成物は、無毒性の生理学的に許容される担体と、場合によっては、1つ以上のアジュバントとを含有する。適切な担体、例えば水、食塩水、エタノール、エチレングリコール、グリセロールなどは、従来の賦形剤から容易に選択され、共製剤化剤を加えることができる。最終組成物の物理的適合性および安定性を保証するために、日常的検査を行うことができる。
【0112】
本発明によれば、豚トルクテノ(TTV)の感染性分子および核酸分子、その核酸分子から産生される生ウイルス、および豚TTVウイルス感染または豚TTVと他のウイルスとの同時感染が引き起こす疾患から豚を保護するための動物用ワクチンが、さらに提供される。本発明はさらに、ワクチンとして使用することができる免疫原性ポリペプチド発現産物を提供する。
【0113】
豚TTVの新規感染性DNA分子は、感染性PTTV1a−VA(配列番号9)、PTTV1b−VA(配列番号10)、PTTV2c−VA(配列番号11)、またはPTTV2c−VA(配列番号12)ゲノムの少なくとも一部をコードする核酸分子を含む。感染性PTTV DNAクローンは、好ましくは、PTTV1またはPTTV2のORF1、ORF2、ORF1/1、およびORF2/2遺伝子の少なくとも一つを含有する。複数コピーのPTTV1a−VA(配列番号9)、PTTV1b−VA(配列番号10)、PTTV2c−VA(配列番号11)、またはPTTV2c−VA(配列番号12)ゲノムを、単一のDNA分子に挿入して、タンデム感染性PTTVクローンを構築してもよい。
【0114】
本明細書に記載するPTTV、特にPTTV1a−VA、PTTV1b−VA、PTTV2c−VA、およびタンデムPTTV2b−RR、PTTV2c−RRのクローニングされたゲノムDNAは、PK−15細胞にトランスフェクトして豚に与えた場合に、インビトロまたはインビボで感染性であることが示される。この新しい、容易に再現できる病原因子は、豚におけるPTTV感染を予防するための適切なワクチン接種プログラムの開発に向いている。
【0115】
本発明のさらなる一実施形態では、3つの1ゲノムコピーPTTV DNAクローンを、それぞれ原型米国分離株PTTV1a−VA、PTTV1b−VAおよびPTTV2c−VAから、融合PCR(fusion PCR)によって誘導した。完全長ゲノムDNAのそれぞれを、平滑末端ライゲーションによって、クローニングベクターpSC−B−amp/kanに挿入した。制限部位BamHIは、3つのPTTVゲノム上のユニーク部位であり、これは、コンカテマーの生成を容易にし、したがってTTV環状ゲノムを模倣するために、3つのゲノムの両端に工学的に作製されたものである。各クローンの選ばれたプラスミドDNAのBamHI単独消化は、明らかに、サイズが4.3Kbと2.8Kbの二つの異なるフラグメントをもたらした(図18A)。4.3Kbフラグメントがバックボーンベクターに相当し、一方で2.8Kbフラグメントは、挿入されたPTTVゲノムDNAに相当した。同じ酵素だけで消化した空ベクターpSC−B−amp/kanは、4.3Kbバンドを示した(図18A)。得られたPTTVクローンを、それぞれpSC−PTTV1a、pSC−PTTV1bおよびpSC−PTTV2cと名付けた(図17A〜C)。
【0116】
さらにまた、クローンpSC−PTTV2cから得られる2コピーの完全長PTTV2c−VAゲノムを、pSC−B−amp/kanベクター中に、直列にライゲーションして、クローンpSC−2PTTV2c−RR(図17D)を作製した。pSC−PTTV2cとpSC−2PTTV2c−RRのAflII単独消化パターンを比較したところ、後者のプラスミドは、PTTV2cゲノムの2つめのコピーに相当する追加の2.8Kbフラグメントを有することがわかった(図18B、右側)。次に、本発明者らは同じクローニング戦略を利用して、ドイツTTVクローンTTV2−#471942−fullから、タンデム二量体化PTTV2b DNAクローンを作製した。pSC−2PTTV2b−RR(図17F)と名付けたこのコンストラクトには、これをHindIIIだけで消化して、その1ゲノムコピー対応物と比較した場合に、PTTV2bゲノムの2つ目のコピーに相当する追加の2.8Kbフラグメントが存在したことから(図18B、左側)、構築の成功が確認された。
【0117】
構築したPTTV感染性クローンの複製能を、PK−15細胞のインビトロ・トランスフェクションによって調べた。業者が作製したPTTV2cORF1に対するウサギポリクローナル抗体を用いるIFAにより、クローンTTV2−#471942−fullとpSC−PTTV2cのコンカテマーはどちらも、それぞれ複製能を有することが確認された(図19Aおよび図20A)。トランスフェクト細胞の継代では、蛍光シグナルが排除されず減少もしなかったことから(図19Bおよび図20B)、ORF1タンパク質の発現は、天然のPTTV2bまたはPTTV2c環状分子を模倣したPTTV2コンカテマーからもたらされることが示唆された。モックトランスフェクト細胞、またはIFA検出用の抗体として免疫前ウサギ血清を使用したDNAトランスフェクト細胞には、蛍光シグナルが観察されなかった(データ未掲載)。クローンpSC−PTTV1aのコンカテマーも複製能を有することが抗PTTV1a ORF1抗体を使って示された(図21)。陽性蛍光シグナルは、トランスフェクト細胞または継代細胞の核に位置することから、豚TTVは細胞核内で複製するらしいことが示された。豚サーコウイルス(PCV)はインビトロで類似の発現パターンを有するので、これは予期されないことではない。
【0118】
PK−15細胞におけるタンデム二量体化クローンpSC−2PTTV2b−RRまたはpSC−2PTTV2c−RRの直接トランスフェクションは、ウイルス複製をもたらし、ORF1キャプシド抗原を産生する。PTTV2 ORF1に対する抗体を使ったIFAにより、どちらのクローンも複製能を有し、陽性ORF1抗原は核内に局在化することが確認された(図22AおよびB)。
【0119】
本発明の一実施形態によれば、豚TTVの感染性クローンを使って豚に接種することができる。それは次に、宿主動物の免疫応答を引きだし、中和抗体の産生を刺激するであろう。本発明のある特定実施形態において、2つのタンデム二量体化PTTV2クローンを在来豚のリンパ節および筋中に注射したところ、それらは感染性であった。
【0120】
PTTV2分子クローンのインビボ感染性を調べるために、在来豚に、クローンpSC−2TTV2b−RRまたはpSC−2TTV2c−RRを接種した。接種後0、7、14、21および28日目(0、7、14、21および28DPI)に動物から血清試料を集めた。PTTV2 DNAは、pSC−2TTV2c−RR接種豚では、7DPI以降(#92)、14DPI以降(#188および#191)および21DPI以降(#180)に、それぞれ検出された(図23A、右側)。クローンpSC−2TTV2b−RRを接種した豚では、PTTVウイルス血症が遅れて現れ、2頭は14DPI以降(#189および#192)、1頭は21DPI以降(#181)、1頭は28DPI以降(#193)だった(図23A、左側)。PTTV2特異的リアルタイムPCRで決定したところ、ウイルス負荷量は、接種した全ての豚において、時間の経過と共に増加し、ウイルス負荷量は剖検前の28DPIが最も高かった(図23A)。選ばれた豚から増幅されたリアルタイムPCR産物を配列決定したところ、pSC−2TTV2b−RRまたはpSC−2TTV2c−RRの対応する領域と同一の配列を有することがわかった(データ未掲載)。
【0121】
接種された全ての豚は、0DPIおよび7DPIでは、PTTV2 ORF1抗体に関して陰性だった。4頭のpSC−2TTV2b−RR接種豚の全てが14DPIにおいて抗PTTV2 ORF1 IgG陽性に血清転換したのに対し、pSC−2TTV2c−RR接種群の豚は、14(#92および#180)、21(#191)および28(#188)DPIにおいて、それぞれ血清転換した(図23B)。これらの結果は、活動性の豚TTV2bまたはTTV2c感染が起こったことを示した。
【0122】
感染性ウイルスおよび分子DNAクローンのワクチン、ならびにそれらを使用する方法も、本発明の範囲に包含される。接種された豚は、ウイルス感染およびTTV2感染または同時感染によって引き起こされる関連疾患から防御される。この新規方法では、ウイルス感染から防御する必要がある豚を、その豚に、免疫学的有効量の本発明のワクチン(例えば、免疫原量の感染性PTTV DNA、PTTVの感染性DNAクローンを含有するプラスミドまたはウイルスベクター、組換えPTTV DNA、ポリペプチド発現産物、細菌発現またはバキュロウイルス発現精製組換えORF1キャプシドタンパク質を含むワクチンなど)を投与することによって防御する。広範囲にわたるウイルス感染からの防御が得られるように、他の抗原、例えばPRRSV、PPV、他の感染性豚因子および免疫刺激物質を、豚に同時投与してもよい。
【0123】
ワクチンは、例えば、無毒性の生理学的に許容される担体および場合によっては1つ以上のアジュバントと組み合わされた、感染性ウイルスおよび分子DNAクローン、例えばpSC−Bベクターなどの適切なプラスミドまたはベクター中のクローン化PTTV感染性DNAゲノム、無発病性生ウイルス、不活化ウイルス、発現された組換えキャプシドサブユニットワクチンなどを含む。ワクチンは、本明細書に記載の感染性TTV2分子DNAクローンも含み得る。感染性PTTV DNA、感染性ウイルスゲノムを含有するプラスミドDNA、および生ウイルスは好ましく、生ウイルスは最も好ましい。毒性状態に復帰するリスクを冒す弱毒化生ウイルスを使用するか、ウイルス疾患からの防御にとって十分な抗体免疫応答を誘導しない可能性がある死滅細胞培養増殖全ウイルスを使用する従来のウイルスワクチンと比較して、本発明の無発病性生ウイルスワクチンは有利である。
【0124】
ワクチンおよびそれらを使用する方法も本発明の範囲に包含される。接種された哺乳動物種は、重症ウイルス感染から防御され、PTTVの同時感染に関係する疾患、例えば豚皮膚炎腎症症候群(PDNS)、離乳後多臓器消耗症候群(PMWS)、および他の関連疾病からも防御され得る。ワクチンは、例えば、不活化または弱毒化豚TTVウイルス、無毒性の生理学的に許容される担体、および場合によっては、1つ以上のアジュバントを含む。
【0125】
本発明のワクチンと一緒に投与し得るアジュバントは、ワクチンに対する豚の免疫学的応答を増加させる物質である。アジュバントは、ワクチンと同時に同じ部位に投与するか、異なる時点で、例えばブースターとして投与することができる。アジュバントは、ワクチンを投与する方法とは異なる方法で投与すること、またはワクチンを投与する部位とは異なる部位に投与することが、有利な場合もある。適切なアジュバントには、例えば水酸化アルミニウム(アラム)、免疫刺激複合体(ISCOM)、非イオン性ブロックポリマーまたはコポリマー、サイトカイン(例えばIL−1、IL−2、IL−7、IFN−α、IFN−β、IFN−γなど)、サポニン、モノホスホリルリピドA(MLA)、ムラミルジペプチド(MDP)などがあるが、これらに限るわけではない。他の適切なアジュバントには、例えば硫酸アルミニウムカリウム、大腸菌由来の非耐熱性または耐熱性エンテロトキシン、コレラ毒素またはそのBサブユニット、ジフテリア毒素、破傷風毒素、百日咳毒素、フロイント不完全または完全アジュバントなどがある。ジフテリア毒素、破傷風毒素および百日咳毒素などの毒素ベースのアジュバントは、使用前に、例えばホルムアルデヒドで処理することなどによって不活化することができる。
【0126】
ワクチンはさらに、感染性PTTV DNAクローンの免疫学的活性を増進するために、例えば豚繁殖呼吸器病症候群ウイルス(PRRSV)、豚パルボウイルス(PPV)、他の感染性豚因子および免疫刺激物質などといった追加の抗原を含有してもよい。
【0127】
本発明の新しいワクチンは、どの特定のタイプまたは製造方法にも限定されない。クローン化ウイルスワクチンには、例えば感染性DNAワクチン(すなわち、豚にDNAを直接注入するためにプラスミド、ベクターまたは他の従来の担体を使用するもの)、生ワクチン、改変生ワクチン、不活化ワクチン、サブユニットワクチン、弱毒化ワクチン、遺伝子操作ワクチンなどが含まれるが、これらに限るわけではない。これらのワクチンは当技術分野において知られている標準的方法によって製造される。
【0128】
さらにもう一つの利点として、本発明の好ましい生ウイルスは、製造、貯蔵および搬送が他のタイプの弱毒化ワクチンよりも容易な、遺伝子的に安定なワクチンを与える。
【0129】
本発明のもう一つの好ましいワクチンでは、非病原性DNAクローンを豚に送達するために、適切なプラスミドを利用する。生または死滅細胞培養増殖全ウイルスを使用する従来のワクチンとは対照的に、この発明では、感染性ウイルスゲノムを含有するプラスミドDNAによる豚の直接接種が可能である。
【0130】
本発明において望ましいさらなる遺伝子操作ワクチンは、当技術分野において知られている技法によって製造される。そのような技法は、例えば組換えDNAのさらなる操作、組換えタンパク質のアミノ酸配列の修飾または置換などを伴うが、これらに限るわけではない。
【0131】
組換えDNA技術に基づく遺伝子操作ワクチンは、例えば、豚において、より強い免疫応答または防御応答を誘導する原因になるタンパク質(例えばORF1、ORF1/1、ORF2、ORF2/2に由来するタンパク質など)をコードするウイルス遺伝子の代替的部分を同定することによって作製される。同定されたそのような遺伝子または免疫優性フラグメントは、バキュロウイルスベクターなどの標準的タンパク質発現ベクターにクローニングし、それを使って適当な宿主細胞を感染させることができる(例えばオライリー(O’Reilly)ら、「バキュロウイルス発現ベクター:ア・ラブ・マニュアル(Baculovirus Expression Vectors: A Lab Manual)」、フリーマン・アンド・カンパニー(Freeman & Co.)、1992年を参照されたい)。宿主細胞を培養することで所望のワクチンタンパク質を発現させ、それを所望する程度にまで精製し、適切なワクチン製品へと製剤化することができる。組換えサブユニットワクチンは、PTTV1a、PTTV1bおよびPTTV2の細菌発現(図10、図15)またはバキュロウイルス発現ORF1キャプシドタンパク質に基づく。
【0132】
疾患を引き起こすという望ましくない天然の能力をクローンが少しでも保っている場合は、残存ビルレンスの原因になっているウイルスゲノム中のヌクレオチド配列を特定し、例えば部位特異的突然変異導入法などにより、ウイルスを遺伝子操作して無発病性にすることもできる。部位特異的突然変異導入法では、1つ以上のヌクレオチドを付加、削除または改変することができる(例えば、ツェラー(Zoller)ら、DNA 3:479−488、1984を参照されたい)。所望の突然変異を含有するオリゴヌクレオチドを合成し、一本鎖ウイルスDNAの一部にアニールさせる。この手順によって生じるハイブリッド分子を使って細菌を形質転換する。次に、適当な突然変異を含有する単離された二本鎖DNAを使って、完全長DNAの制限フラグメントにライゲーションすることによって、完全長DNAを作製し、次にそれを適切な細胞培養中にトランスフェクトする。適切な導入用ベクターへのゲノムのライゲーションは、当業者に知られる任意の標準的技法によって達成することができる。ウイルス子孫を生産するための宿主細胞へのベクターのトランスフェクションは、リン酸カルシウムまたはDEAEデキストラン媒介トランスフェクション、エレクトロポレーション、プロトプラスト融合および他の周知の技法など、従来の方法のどれを使って行ってもよい(例えばサムブルック(Sambrook)ら、「モレキュラー・クローニング:ア・ラボラトリー・マニュアル(Molecular Cloning: A Laboratory Manual)」、コールド・スプリング・ハーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、1989年を参照されたい)。クローニングされたウイルスは所望の突然変異を示す。あるいは、適当な突然変異を含有する2つのオリゴヌクレオチドを合成することもできる。これらをアニールさせて二本鎖DNAを形成させ、それをウイルスDNAに挿入して、完全長DNAを作製することができる。
【0133】
免疫学的有効量の本発明のワクチンを、ウイルス感染からの防御を必要とする豚に投与する。豚に接種する免疫学的有効量または免疫原量は、日常的試験により、容易に決定することができ、または迅速に漸増することができる。有効量とは、PTTVウイルスにばく露された豚を保護するのに十分な、ワクチンに対する免疫学的応答が得られる量である。好ましくは、豚は、ウイルス疾患の有害な生理学的症状または影響の1つから全部が有意に低減するか、改善するか、完全に防止される程度にまで保護される。
【0134】
ワクチンは、単回投与するか、反復投与することができる。投薬量は、例えば約1マイクログラム〜約1,000マイクログラムの範囲の感染性キメラDNAゲノムを含有するプラスミドDNA(ワクチンの免疫活性構成成分の濃度に依存する)、好ましくは100〜200マイクログラムの範囲の豚TTV DNAクローンであり得るが、ウイルス感染の有害反応または生理学的症状をもたらすのに十分であるような量のウイルスに基づく抗原を含有してはならない。豚の体重、抗原の濃度および他の典型的要因に基づいて最低有効投薬量を見いだすために、活性抗原性因子の適切な投薬量を決定または漸増するための方法は、当技術分野において知られている。好ましくは、感染性ウイルスDNAクローンをワクチンとして使用するか、生感染性ウイルスをインビトロで作製してから、その生ウイルスをワクチンとして使用することができる。その場合、例えば約50〜約10,000の50%組織培養感染量(TCID50)の生ウイルスを、豚に与えることができる。
【0135】
本発明の新しいワクチンは、どの特定のタイプまたは製造方法にも限定されない。ワクチンには、改変生ワクチン、不活化ワクチン、サブユニットワクチン、弱毒化ワクチン、遺伝子操作ワクチンなどが含まれるが、これらに限るわけではない。
【0136】
生ワクチンの利点は、全身性免疫応答、局所免疫応答、体液性免疫応答および細胞性免疫応答を含む、考え得る全ての免疫応答がワクチンの受容者において活性化されることである。利点を上回り得る生ウイルスワクチンの欠点は、生付随ウイルス因子による汚染の可能性またはウイルスが野外で先祖返りしてビルレンスを持つようになり得る危険にある。
【0137】
不活化ウイルスワクチンを製造するには、例えば限定するわけではないがPK−15細胞などの培養豚細胞中で、ウイルス増殖およびウイルス生産を行うことができる。次に、当業者に一般に知られているプロトコールによって、または好ましくは本明細書に記載する方法によって、一連のウイルス不活化を最適化する。
【0138】
不活化ウイルスワクチンは、豚TTVをホルマリンまたは疎水性溶媒、酸などの不活化剤で処理すること、紫外線またはX線を照射すること、加熱することなどによって、製造することができる。不活化は当技術分野において理解されているやり方で行われる。例えば化学的不活化では、適切なウイルス試料またはウイルスを含有する血清試料を、ウイルスを不活化するために、十分に高い(または不活化剤によっては低い)温度またはpHにおいて十分な量または濃度の不活化剤で十分な時間処理する。加熱による不活化は、ウイルスを不活化するのに十分な温度および時間行われる。照射による不活化は、ある光の波長または他のエネルギー源を使って、ウイルスを不活化するのに十分な時間行われる。ウイルスは、それが、感染を起こしやすい細胞に感染することができなくなれば、不活化されたと見なされる。
【0139】
サブユニットワクチンの製造は、典型的には、改変生ワクチンまたは不活化ワクチンの製造とは異なる。サブユニットワクチンの製造に先だって、ワクチンの防御構成成分または抗原構成成分を同定しなければならない。本発明において、PTTVの抗原構成成分は、PTTV1a、PTTV1bおよびPTTV2のORF1キャプシドタンパク質と同定され、これを本発明では、サブユニット組換えキャプシドワクチンとして使用するために、大腸菌(E.coli)およびバキュロウイルス発現系などの他の発現系で発現させ、精製した。そのような防御構成成分または抗原構成成分は、豚において特に強い防御応答または免疫学的応答を引き起こすウイルスキャプシドタンパク質の一定のアミノ酸セグメントまたはフラグメント;単一または複数のウイルスキャプシドタンパク質そのもの、そのオリゴマー、およびウイルスの部分構造またはそのような部分構造の同定可能な部分もしくはユニットを形成するウイルスキャプシドタンパク質の高次会合体;ウイルスの表面上もしくは表面近くに存在するか、ウイルスと会合しているリポタンパク質または脂質基などのウイルス部分構造中に存在する、オリゴグリコシド、糖脂質または糖タンパク質などを含む。好ましくは、ORF1タンパク質をサブユニットワクチンの抗原構成成分として使用する。ORF2、ORF1/1、およびORF2/2遺伝子中のヌクレオチド配列によってコードされるものなど、他のタンパク質も使用することができる。これらの免疫原構成成分は当技術分野において知られている方法により、すぐに同定することができる。ひとたび同定されたら、次に、ウイルスの防御部分または抗原部分(すなわち「サブユニット」)を、当技術分野において知られている手法で精製および/またはクローニングする。ウイルスの高精製サブユニットなどのサブユニットは全ウイルスよりも毒性が低いので、サブユニットワクチンには、生ウイルスに基づく他のワクチンをしのぐ利点がある。
【0140】
組換え遺伝子技法によってサブユニットワクチンを製造する場合、ORF1、ORF2、ORF1/1、およびORF2/2遺伝子などといったクローン化サブユニットの発現は、例えば、上述の方法によって行うことができ、当業者に知られる方法によって最適化することもできる(例えばマニアティス(Maniatis)ら、「モレキュラー・クローニング:ア・ラボラトリー・マニュアル(Molecular Cloning: A Laboratory Manual)」、コールド・スプリング・ハーバー・ラボラトリー(Cold Spring Harbor Laboratory)、マサチューセッツ州コールドスプリングハーバー、1989年を参照されたい)。一方、使おうとするサブユニットが、例えばキャプシドタンパク質全体など、ウイルスのインタクトな構造的特徴を表す場合は、ウイルスからそれを単離するための手法を最適化しなければならない。いずれにせよ、不活化プロトコールの最適化後には、製造に先だって、サブユニット精製プロトコールを最適化することができる。
【0141】
弱毒化ワクチンを製造するには、生病原性ウイルスをまず最初に、当技術分野において知られている方法によって、または好ましくは本明細書に記載する方法によって、弱毒化する(非病原性または無害にする)。例えば弱毒化ウイルスは、有胚豚卵による新規連続継代を伴う本発明の技法によって調製することができる。弱毒化ウイルスは自然に見いだすことができ、天然の遺伝子欠失を有し得る。あるいは、遺伝子を欠失させるか遺伝子を突然変異させることによって、病原性ウイルスを弱毒化することもできる。弱毒化および不活化ウイルスワクチンは、本発明の好ましいワクチンを構成する。
【0142】
本発明において同様に望ましい遺伝子操作ワクチンは、当技術分野において知られている技法によって作製される。そのような技法は、RNA、組換えDNA、組換えタンパク質、生ウイルスの使用などを伴うが、これらに限るわけではない。
【0143】
例えば、精製後に、野生型ウイルスは、感染した豚または感染した適切な細胞株を使って、血清、糞便、唾液、精液および組織試料などの適切な臨床生物学的試料から、当技術分野において知られている方法によって、好ましくは本明細書において教示する方法によって、単離することができる。生物学的に純粋なウイルスまたは感染性因子から当技術分野において知られている方法によってDNAを抽出し、当技術分野において知られている方法によって、好ましくはCsCl勾配における超遠心分離によって精製する。ウイルスゲノムのcDNAを、当技術分野において知られている方法によって、適切な宿主中にクローニングし(マニアティス(Maniatis)ら、同上)、次に、ウイルスゲノムを解析して、ウイルスの抗原部分を生産するのに不可欠なゲノムの領域を決定する。その後の手順は、改変生ワクチン、不活化ワクチンまたはサブユニットワクチンの場合と、おおむね同じである。
【0144】
組換えDNA技術に基づく遺伝子操作ワクチンは、例えば、ウイルス遺伝子のうち、豚において、より強い免疫応答または防御応答を誘導する原因になるタンパク質(例えばORF1、ORF2、ORF1/1、ORF2/2に由来するタンパク質など)をコードする部分を同定することによって作製される。同定されたそのような遺伝子または免疫優性フラグメントは、バキュロウイルスベクターなどの標準的タンパク質発現ベクターにクローニングし、それを使って適当な宿主細胞を感染させることができる(例えばオライリー(O’Reilly)ら、「バキュロウイルス発現ベクター:ア・ラブ・マニュアル(Baculovirus Expression Vectors: A Lab Manual)」、フリーマン・アンド・カンパニー(Freeman & Co.)、1992年を参照されたい)。宿主細胞を培養することで所望のワクチンタンパク質を発現させ、それを所望する程度にまで精製し、適切なワクチン製品へと製剤化することができる。
【0145】
ワクチンにおいて有用な遺伝子操作タンパク質は、例えば、昆虫細胞、酵母細胞または哺乳動物細胞において発現させることができる。従来の方法によって精製または単離することができる遺伝子操作タンパク質は、豚TTVからの防御を付与するために、豚または哺乳動物種に直接接種することができる。
【0146】
昆虫細胞株(例えばsf9、sf21、またはハイ・ファイブ(HIGH−FIVE))は、ウイルスから得られたまたはウイルスゲノムからコピーされたポリ核酸であってウイルスの免疫優性タンパク質の1つ以上をコードするものを含有する導入ベクターで、形質転換することができる。導入ベクターには、例えば、線状化バキュロウイルスDNAおよび所望のポリヌクレオチドを含有するプラスミドが含まれる。宿主細胞株には、組換えバキュロウイルスを作製するために、線状化バキュロウイルスDNAとプラスミドを同時トランスフェクトすることができる。
【0147】
あるいは、分離された豚TTVのDNAであって1つ以上のキャプシドタンパク質をコードするものを、ポックスウイルスまたはアデノウイルスなどの生ベクター中に挿入して、ワクチンとして使用することもできる。
【0148】
本発明のワクチンの免疫学的有効量は、前記感染または症候群からの防御を必要とする豚または哺乳動物種に投与される。「免疫学的有効量」は、日常的試験により、容易に決定することができ、または迅速に漸増することができる。有効量とは、豚皮膚炎腎症症候群(PDNS)、離乳後多臓器消耗症候群(PMWS)または関連疾病を引き起こし得る、豚TTVウイルスまたは豚TTV同時感染にばく露された豚または他の哺乳動物を保護するのに十分な、ワクチンに対する免疫学的応答が得られる量である。好ましくは、豚または他の哺乳動物種は、ウイルス疾患の有害な生理学的症状または影響の1つから全部が有意に低減するか、改善するか、完全に防止されることが見いだされる程度にまで保護される。
【0149】
ワクチンは、単回投与するか、反復投与することができる。投薬量は、例えば1〜1,000マイクログラムのウイルスに基づく抗原(ワクチンの免疫活性構成成分の濃度に依存する)を含有し得るが、ウイルス感染の有害反応または生理学的症状をもたらすのに十分であるような量のウイルスに基づく抗原を含有してはならない。鳥または哺乳動物の体重、抗原の濃度および他の典型的要因に基づいて活性抗原性因子の適切な投薬量を決定または漸増するための方法は、当技術分野において知られている。
【0150】
ワクチンは豚に投与することができる。ワクチンは、ウイルス因子に感染するリスクが高い養豚業者などのヒトに与えることもできる。豚TTVに基づくワクチンは、豚TTVおよびヒトTTVの両方からの幅広い防御をもたらすように設計することができると考えられる。言い換えると、豚TTVに基づくワクチンは、好ましくは、いわゆる「ジェンナーの方法」(すなわち牛痘ウイルスワクチンは、エドワード・ジェンナー(Edward Jenner)により、ヒト痘瘡に対して使用することができる)によって、ヒトTTV感染を防御するように設計することができる。望ましくは、まだTTVウイルスにばく露されていない豚または他の哺乳動物種に、ワクチンを直接投与する。ワクチンは、都合よく、例えば経口投与、口内投与、鼻腔内投与、経皮投与、非経口投与することができる。非経口投与経路には、筋肉内経路、静脈内経路、腹腔内経路および皮下経路が含まれるが、これらに限るわけではない。
【0151】
液体として投与される場合、本ワクチンは、水性溶液剤、シロップ剤、エリキシル剤、チンキ剤などの形態に調製することができる。そのような製剤は当技術分野において知られており、典型的には、抗原および他の典型的添加剤を適当な担体または溶媒系に溶解することによって調製される。適切な担体または溶媒には、例えば水、食塩水、エタノール、エチレングリコール、グリセロールなどがあるが、これらに限るわけではない。典型的添加剤は、例えば認可された色素、香料、甘味料、およびチメロサール(エチルメルクリチオサリチル酸ナトリウム)などの抗微生物保存剤である。そのような溶液は、例えば部分加水分解ゼラチン、ソルビトールまたは細胞培養培地の添加などによって安定化することができ、例えばリン酸水素ナトリウム、リン酸二水素ナトリウム、リン酸水素カリウム、リン酸二水素カリウム、その混合物などといった当技術分野において知られている試薬類を使って、従来の方法で緩衝化することができる。
【0152】
液状製剤には、他の標準的共製剤化剤と組み合わされた懸濁化剤または乳化剤を含有する懸濁剤および乳剤も含まれる。これらのタイプの液状製剤は、従来の方法で調製することができる。例えば懸濁剤はコロイドミルを使って調製することができる。例えば乳剤はホモジナイザーを使って調製することができる。
【0153】
体液系に注射するように意図された非経口製剤は、対応する哺乳動物体液のレベルへの適正な等張性およびpH緩衝作用を必要とする。等張性は、必要に応じて塩化ナトリウムおよび他の塩で、適当に調節することができる。エタノールまたはプロピレングリコールなどの適切な溶媒を使って、製剤中の成分の溶解度と液状調製物の安定性を増加させることができる。本ワクチンに使用することができるさらなる添加剤には、デキストロース、従来の酸化防止剤および従来のキレート剤、例えばエチレンジアミン四酢酸(EDTA)などがあるが、これらに限るわけではない。また、非経口剤形は使用前に滅菌しなければならない。
【0154】
以下に実施例を挙げて本発明の一定の態様を例証する。しかし、これらの実施例が単なる例示であること、そして本発明の条件および範囲に関して完全に決定的なものであるとは考えていないことを理解すべきである。典型的な反応条件(例えば温度、反応時間など)が記載されている場合、指定された範囲を上回る条件も下回る条件も、一般的には利便性が下がるとはいえ、使用できることを理解すべきである。本実施例は室温(約23℃〜約28℃)および大気圧で行われる。別段の指定がない限り、本明細書において言及する割合およびパーセントはいずれも重量ベースであり、温度は全て摂氏温度である。
【0155】
(実施例1)
ウイルスDNA抽出、ネステッドPCRおよびゲノムPCR:
この研究には、バージニアの養豚場の20頭の在来成体雄豚から採取した好都合な血清試料および精液試料を使用した。キアアンプ(QIAamp)DNAミニキット(キアゲン(Qiagen)を使って20個の血清試料および19個の精液試料から全DNAを単離した。陽性PTTV含有試料をスクリーニングするために、まず最初に、PTTV1およびPTTV2中の保存領域のネステッドPCR増幅を、アンプリタック・ゴールド(AmpliTag Gold)ポリメラーゼ(アプライド・バイオシステムズ(Applied Biosystems))を使って行った。PTTV1のフラグメントAを増幅するために使用した2つのプライマー対は、TTV1−mF(配列番号45)/TTV1−mR(配列番号46)(第1ラウンドPCR用)およびTTV1−nF(配列番号47)/TTV1−nR(配列番号48)(第2ラウンドPCR用)であり、一方、PTTV2のフラグメントDを増幅するために使用した2つのプライマー対は、TTV2−mF(配列番号49)/TTV2−mR(配列番号50)(第1ラウンドPCR用)およびTTV2−nF(配列番号51)/TTV2−nR(配列番号52)(第2ラウンドPCR用;図1Aおよび表1)だった。
【0156】
PTTV1とPTTV2の両方の完全長ゲノム配列を増幅するために、本発明者らは、まず、PTTV1用には領域A中に位置する一対の保存された遺伝子特異的プライマーTTV1−IF(配列番号1)/TTV1−IR(配列番号4)およびPTTV2用には領域D中に意図するもう一対の遺伝子特異的プライマーTTV2−IF(配列番号5)/TTV2−IR(配列番号8)を、それぞれ、ヘラキュレース(Herculase)IIフュージョン(Fusion)DNAポリメラーゼ(ストラタジーン(Stratagene))と共に、製造者の説明書に従って使用することにより、インバースゲノムPCRを行った。予期されるサイズのPCR産物は検出されなかった。次に本発明者らは、第2ラウンドのPCRにおいて、PTTV1ゲノム全体およびPTTV2ゲノム全体をカバーするそれぞれ2つの領域を増幅するために、新しいプライマーセットを設計した(図1A)。PTTV1のフラグメントBおよびCを増幅するために使用したプライマー対は、それぞれ、TTV1−IF(配列番号1)/TTV1−2340R(配列番号2)とTTV1−2311F(配列番号3)/TTV1−IR(配列番号4)であり、一方、PTTV2のフラグメントEおよびFを増幅するために使用したプライマー対は、それぞれ、TTV2−IF(配列番号5)/TTV2−2316R(配列番号6)およびTTV2−GCF(配列番号7)/TTV2−IR(配列番号8)だった(図1Aおよび表1)。フラグメントCおよびFは、それぞれPTTV1およびPTTV2のGCリッチ領域を含有する。増幅されたPCR産物を個別に切り出し、精製し,次に、ストラタクローン・ブラント(StrataClone Blunt)PCRクローニングストラテジーにより、製造者の説明書(ストラタジーン(Stratagene)に従って、pSC−B−amp/kanベクター(ストラタジーン(Stratagene)中にクローニングした後、DNA配列決定を行った。
【0157】
(実施例2)
バージニアの飼育場にいる雄豚から収集した豚TTV陽性試料のスクリーニング:
豚TTV DNAは、以前に、日本PTTV1株Sd−TTV31のUTR配列に基づくネステッドPCRにより、異なる地理的領域の豚から検出されている(マキューン(McKeown)ら、2004年、前掲書)。PTTV2が最近同定されたことから、PTTV1の領域AとPTTV2の領域Dをそれぞれ増幅するために、2つの異なるネステッドPCRプライマーセットが使用された(図1A)(エリス(Ellis)ら、2008年、前掲書;ケカライネン,ティ(Kekarainen,T.)、シビラ,エム(Sibila,M.)およびシーゲイルス,ジェイ(Segales,J.)、2006年、「スペインの離乳後多臓器消耗症候群(PMWS)罹患豚および非PMWS罹患豚における豚トルクテノウイルスの有病率(Prevalence of swine Torque teno virus in post-weaning multisystemic wasting syndrome (PMWS)-affected and non-PMWS-affected pigs in Spain)」、ジャーナル・オブ・ジェネラル・バイロロジー(J Gen Virol)、第87巻、第4号、p.833−7;クラコウカ(Krakowka)、2008年、前掲書)。本研究においても同様の検出法を利用して、米国の豚からPTTV株を同定した。後に完全長ゲノムの決定に使用する、土地固有のPTTV1陽性試料またはPTTV2陽性試料をスクリーニグするために、バージニアの飼育場にいる20頭の雄豚から収集した20個の血清試料(SR#1〜20)および19個の精液試料(SM#1〜18およびSM#20)を、ネステッドPCR分析に付した。驚いたことに、20個の血清試料は全てPTTV1に関して陽性であり、19個はPTTV2についても陽性だった(SR#18を除く)。対照的に、1つの精液試料(SM#6)だけがPTTV1陽性であり、3つの精液試料だけ(SM#8、9および20)がPTTV2陽性だった。この結果は、スペインにおける雄豚精液試料がPTTV DNAに関して陽性であることが示された最近の研究と合致した(ケカライネン,ティ(Kekarainen,T.)、ロペス−ソリア,エス(Lopez-Soria,S.)およびシーゲイルス,ジェイ(Segales,J.)、2007年)。雄豚の血清および精液には豚トルクテノウイルス遺伝子グループ1および2が検出されたことから(セリオゲノロジー(Theriogenology)、第68巻、第7号、p.966−71)、PTTVの潜在的垂直感染が示唆された。しかし、精液におけるPTTV1とPTTV2の有病率は、血清中のそれよりもはるかに低かったので、同じ豚の血清および精液におけるPTTV DNAの存在に関して直接的な関連はないことが示唆された。
【0158】
(実施例3)
配列解析および系統解析:
DNA配列およびアミノ酸配列の一般解析(generic analysis)およびアラインメントはレーザージーン(Lasergene)パッケージ(ディエヌエイスター・インコーポレイテッド(DNASTAR Inc.)、ウィスコンシン州マディソン)を使って行った。アラインメントと比較に使用した3つの既知PTTV株のゲノム配列およびそれらの対応するジェンバンク(GenBank)アクセッション番号は、Sd−TTV31(AB076001)、TTV−1p(AY823990)およびTTV−2p(AY823991)である。ジェンバンク(GenBank)において利用できるヒトおよび動物のTTV関連株の121個の完全長ゲノム配列を使用し、オンラインプログラムPASC(http://www.ncbi.nlm.nih.gov/sutils/pasc/viridty.cgi?textpage=overview)(バオ(Bao)ら、2008年)で、ペアワイズ配列比較(PASC)を行った。
【0159】
系統樹は、PAUP4.0プログラム(デービッド・スウォフォード(David Swofford)、スミソニアン・インスティチュート(Smithsonian Institute)、ワシントンDC、シナウアー・アソシエート・インコーポレイテッド(Sinauer Associate Inc.)が配布)の近隣結合法により、7つのPTTV株の4つのORFの完全長ゲノム配列および推定アミノ酸配列に基づいて構築した。データは1000回のリサンプリングから得た。
【0160】
(実施例4)
豚PTTV感染を診断するためのPCRプライマーの設計
DNA配列の解析およびアラインメントは、レーザージーン(Lasergene)パッケージ(ディエヌエイスター・インコーポレイテッド(DNASTAR Inc.)、ウィスコンシン州マディソン)を使って行った。アラインメントに使用した10個の豚TTV株の完全長ゲノム配列およびそれらの対応するジェンバンク(GenBank)アクセッション番号は、次のとおりであった。種PTTV1:Sd−TTV31(AB076001)、PTTV1a−VA(GU456383)、TTV−1p(AY823990)、PTTV1b−VA(GU456384)、swSTHY−TT27(GQ120664)およびTTV1#471819(GU188045)。種PTTV2:PTTV2b−VA(GU456385)、PTTV2c−VA(GU456386)、TTV−2p(AY823991)およびTTV2#472142(GU188046)。6つのPTTV1ゲノム間および4つのPTTV2ゲノム間で保存されている配列をそれぞれ同定し、次にそれらを指針として、ビーコン・デザイナー(Beacon Designer)プログラム(プレミア・バイオソフト・インターナショナル(PREMIER Biosoft International)、カリフォルニア州パロアルト)を使ってリアルタイムPCRプライマーを選択した。PTTV1のデュプレックス・ネステッドPCRに使用したプライマーは、レーザージーン(Lasergene)パッケージで設計した。
【0161】
(実施例5)
PTTV1およびPTTV2リアルタイムPCRの標準曲線
PTTV1b−VAゲノムのPCRフラグメントBに対応する2091bpの領域を、同じPCRフラグメントから、既述のように(ファン(Huang)ら、2010年)、プライマーTTV1−IF(5’−CATAGGGTGTAACCAATCAGATTTAAGGCGTT−3’)およびTTV1−2340R(5’−GGTCATCAGACGATCCATCTCCCTCAG−3’)を使って再増幅した。その結果生じたアンプリコンをキアクイック・ゲル・エクストラクション・キット(QIAquick Gel Extraction Kit)(キアゲン(Qiagen))でゲル精製し、ナノドロップ(NanoDrop)分光測光器で定量して、豚TTV種1のリアルタイムPCR標準テンプレートに使用した。PTTV2c−VAのPCRフラグメントEおよびFをベクターpSC−B−amp/kan中でアセンブルすることにより、PTTV2c−VA株pSC−PTTV2cの完全長DNAクローンを構築した(ファン(Huang)ら、未公表データ)。プラスミドpSC−PTTV2c(7082bp)を豚TTV種2のリアルタイムPCR標準テンプレートに使用し、プラスミドDNA濃度をナノドロップ(NanoDrop)分光測光器で測定した。これら2つのテンプレートの10倍希釈系列を使ってリアルタイムPCR標準曲線をそれぞれ作成した。
【0162】
(実施例6)
PCRアッセイ用のウイルスDNAの抽出
バージニアの養豚場の20頭の在来成体雄豚(臨床的症候群を持たないもの)から収集した20個の血清試料と19個の精液試料から、キアアンプ(QIAamp)DNAミニキット(キアゲン(Qiagen))を使って、既述のように(ファン(Huang)ら、2010年)、全DNAを単離した。血清および精液について400μlの試料体積を使って、滅菌水50μlの最終溶出液で、DNAを抽出した。抽出したDNA試料は全て、リアルタイムPCR試験まで、−20℃で保存した。従来のネステッドPCRによるこれらの試料中の豚TTVの検出は既に記述されている(ファン(Huang)ら、2010年)。同じ手順でヤギ血清試料から抽出された全DNAを陰性対照として使用した。
【0163】
(実施例7)
SYBRグリーン・リアルタイム定量PCRアッセイ
PTTV1特異的およびPTTV2特異的リアルタイムPCRを、センシミックス(SensiMix)SYBRアンド・フルオレセインキット(クオンテース・リミテッド(Quantace Ltd))およびマイアイキュー・アイサイクラー(MyiQ iCYCLER)リアルタイムPCR機器(バイオラッド・ラボラトリーズ(BIO?RAD Laboratories))を使って、それぞれ行った。各25μlの反応は、12.5μlのSYBRグリーン・マスター・ミックス、4μlの抽出されたDNA、0.5μlの各プライマー(10nM)および7.5μlの滅菌水を含有した。PTTV1のPCR条件は95℃で10分の後、40サイクルの増幅(95℃で15秒、59.4℃で30秒、72℃で10秒)とした。その後、直ちに、0.5℃ごとに蛍光シグナルを測定しながら、55℃から95℃まで温度を徐々に上昇させることにより、融点分析を行った。PTTV2のPCR条件は、アニーリング温度が56℃である点以外は、PTTV1と同じとした。PTTV1およびPTTV2標準テンプレートを陽性対照として全てのランに含めた。増幅およびデータ解析は、マイアイキュー(MyiQ)システムソフトウェア(バイオラッド・ラボラトリーズ(BIO?RAD Laboratories))を使って行った。全ての試料を同じプレート上で二つずつ測定した。
【0164】
(実施例8)
2つのシングルプレックス・アッセイの特異性および感度
PTTV1特異的およびPTTV2特異的アッセイの増幅にとって最適なアニーリング温度は、アニーリング温度の勾配を使った増幅の10倍希釈によって決定したところ、それぞれ59.4℃および56℃だった。プライマーTTV1F/TTV1Rを使った118bp産物の増幅がPTTV1テンプレートでのみ得られたのに対し、PTTV2テンプレートでの200bp産物の増幅は、プライマーTTVF4/TTVR4を使った場合にのみ観察された。どちらのアッセイも他方からの交差増幅を与えなかったことから、プライマーおよびターゲットの特異性が確認された(データ未掲載)。
【0165】
ある範囲の、25μlあたりのターゲットDNA濃度にわたって、PTTV1標準曲線を確立した。線形範囲は、4.4×101〜4.4×108コピーにまたがることが示された。最低検出限界(44コピー)は、37.57の閾サイクル数(Ct)に対応した。Ct>37.57の被験試料は検出限界未満とみなし、定量可能でなかった。同様に、PTTV2標準曲線を作成し、それを使って、25μlの反応あたり8.6×100〜8.6×108コピーの範囲にあるDNA濃度を検出した。最低検出限界(8.6コピー)に対応するCtは36.53だった。PTTV1陽性またはPTTV2陽性とみなした試料は全て、それぞれの最大検出限界より低いコピー数を持っていた。PTTV1またはPTTV2標準テンプレートの10倍希釈(図6aおよび6b;青色の曲線)ならびに20個の雄豚血清試料を使った融解曲線は、PTTV1については87.0℃、PTTV2については80.0℃の融解温度(Tm)を、それぞれ示した(図6aおよび6b;赤色の曲線)。滅菌水またはヤギ血清DNAをテンプレートとして使用した陰性対照にはピークが観察されなかった(図6aおよび7b;黒い線)。
【0166】
(実施例9)
雄豚血清試料および精液試料中の豚TTV1およびTTV2の定量
ウイルス負荷量を、元の雄豚血清試料1mlあたりのPTTV1ゲノムまたはPTTV2ゲノムのコピー数として表した。PTTV1 DNAは20個の血清試料の全てに検出され、1.91?103〜3.25?105コピー/mlの範囲にあった。一方、PTTV2 DNAは19個の血清試料(#10以外)に検出され、3.59×102〜1.39×106コピー/mlの範囲にあった。この結果は、従来のネステッドPCRを使用した本発明者らの先の研究と合致した(表5)。精液試料はいずれもPTTV1陽性ではなかったが、3つの精液試料は、極めて低いウイルス負荷量(それぞれ230、244および357コピー/ml)でPTTV2陽性だった。
【0167】
【表5】
【0168】
(実施例10)
PTTV1/PTTV2デュプレックス・リアルタイムPCRアッセイ
PTTV1/PTTV2デュプレックス・リアルタイムPCRアッセイは、12.5μlのSYBRグリーン・マスター・ミックス、0.5μlの各PTTV1プライマー、0.5μlの各PTTV2プライマー、4μlのDNAおよび6.5μlの滅菌水を含有する25μlのPCR系で行った。デュプレックスPCR条件および融点分析は、アニーリング温度が58℃である点以外は、PTTV1と同じとした。PTTV1特異的アンプリコンとPTTV2特異的アンプリコンを識別するために、溶解ピークを分析した。
【0169】
(実施例11)
デュプレックス・ネステッドPCR
第1ラウンドPCRは、50μlの総液量に4μlの抽出DNAを使って、プラチナムPCRハイファイ・スーパーミックス(Platinum PCR HiFi Supermix)(インビトロジェン(Invitrogen))で行った。PCR条件は、94℃で2分間のテンプレートの初期変性と、94℃で30秒、55℃で30秒、72℃で30秒の30サイクルとした。同じPCR試薬類および条件による第2ラウンドのPCRには、第1ラウンドPCR産物を4μlずつ使用した。第1ラウンドPCRでは1対のプライマーP1ab−mF/P1ab−mRを使用したのに対し、第2ラウンドのPCRでは、2対のプライマーPTTV1a検出用のP1a−nF/P1a−nRとPTTV1b検出用のP1b−nF/P1b−nRとの混合物を使用した(表1)。増幅産物を、臭化エチジウムで染色した1%アガロースゲルでのゲル電気泳動によって可視化したところ、各型に特異的な2本のバンドがUV光によって区別された。
【0170】
(実施例12)
PTTV1およびPTTV2 ORF発現プラスミドの構築
PTTV1a、PTTV1bおよびPTTV2cのORF1のC末端部分を、それぞれの完全長DNAクローン(pSC−PTTV1a、pSC−PTTV1bおよびpSC−PTTV2c;項を改めて記載)から増幅した。増幅されたフラグメントは、PTTV1aについては319aa(ORF1aa位置317−635(配列番号13);ジェンバンク(GenBank)アクセッション番号GU456383)、PTTV1bについては318aa(ORF1aa位置322−639(配列番号14);ジェンバンク(GenBank)アクセッション番号GU456384)、PTTV2cについては316aa(ORF1aa位置310−625(配列番号16);ジェンバンク(GenBank)アクセッション番号GU456386)のタンパク質産物を、それぞれコードすると予期された。248aa(ORF1aa位置322−569(配列番号14))をコードするPTTV1bのC末端切断フラグメントも増幅し、SDS−PAGE分析用の比較対象として使用した。プラスミドは全て、C末端に8×Hisタグが付いた融合タンパク質が生成するように大腸菌/バキュロウイルス/哺乳動物細胞三重発現ベクターpTriEx1.1−neo(ノバジェン(Novagen))のNcoI制限部位とXhoI制限部位の間にPCR産物をクローニングすることによって構築した。4つの組換えプラスミドをpTri−PTTV1a−ORF1、pTri−PTTV1b−ORF1、pTri−PTTV1b−ORF1ctrucおよびpTri−PTTV2c−ORF1と名付けた。全てのクローン化配列をDNA配列決定によって確認した。
【0171】
(実施例13)
組換えPTTV1およびPTTV2タンパク質の発現
4つの発現プラスミドを、それぞれロゼッタ(Rosetta)2(DE3)pLacIコンピテント細胞(ノバジェン(Novagen))に形質転換し、細菌を、100μg/mlアンピシリンを含有するLB寒天プレート上、37℃で終夜、プレート培養した。各コンストラクトについて単一の形質転換コロニーを使って、100μg/mlのアンピシリンを含有するLB培地(LB/amp)3mlに接種し、37℃で6〜8時間成長させた。次に、各コンストラクトについて濁った3ml培養物を使用し、25%濾過滅菌グリセロールを添加してその培養物を−80℃で凍結することにより、細菌ストックを作製した。精製に先だって、各コンストラクトにつき10μlの凍結細菌ストックを使って、LB/ampの3mlスターター培養に接種し、37℃で6〜8時間成長させた。100mlのオーバーナイト・エクスプレス(Overnight Express)TB培地(ノバジェン(Novagen))にスターター培養物を接種してタンパク質発現を誘導し、37℃で16〜18時間成長させた。インキュベーション後に、自己誘導培養培地を4℃、3400rpmで15分間の遠心分離にかけた。各コンストラクトについてその結果得られた上清を捨て、細菌ペレットのそれぞれを使用時まで−20℃で保存した。
【0172】
(実施例14)
組換えタンパク質の精製および透析
組換えタンパク質は不溶性であり、細菌封入体内に発現した。細菌ペレットのそれぞれを、バグバスター(BugBuster)およびアールリゾチーム(rLysozyme)で、製造者のプロトコール(ノバジェン(Novagen))に従って処理し、DNAおよびRNAを分解するためにベンゾナーゼ・ヌクレアーゼ(Benzonase Nuclease)(ノバジェン(Novagen))を加えた。次に、封入体ペレットのそれぞれを840μlの溶解バッファー(6Mグアニジン塩酸塩、0.1Mリン酸ナトリウム、0.01Mトリス−塩酸、0.01Mイミダゾール、pH8.0)に再懸濁し、−80℃で少なくとも30分間は凍結した。次にそれを融解し、新たな溶解バッファー2.5mlで希釈し、室温で30分間、穏やかに回転させた。室温、15,000×gで30分間の遠心分離により、溶解物上清を集めた。デカントした上清のそれぞれに50%Ni−NTA His−バインド・スラリー(bind slurry)(ノバジェン(Novagen))を加え、hisタグ結合を促進するために、その混合物を室温で60分間振とうした。溶解物/樹脂混合物を空のクロマトグラフィーカラムに充填した。最初の貫流の後、7mlの溶解バッファーをカラムに加えて貫流させた。次に各カラムを7mlの洗浄バッファー(8M尿素、0.1Mリン酸ナトリウム、0.15M塩化ナトリウム、0.02Mイミダゾール、pH8.0))で2回洗浄した。カラムに溶出バッファー(8M尿素、0.05Mリン酸ナトリウム、1M塩化ナトリウム、0.5Mイミダゾール、pH8.0)を1mlずつ別々に4回加えることにより、ターゲットタンパク質の溶出を達成した。それら4つの溶出画分をSDS PAGEとクーマシーブルー染色とによって分析した。
【0173】
意味のある濃度のターゲットタンパク質を含有する溶出物を、20,000分子量カットオフの0.5ml〜3ml透析カセット(ピアス(Pierce))に注入した。透析には、次に挙げる一連の4つの透析バッファーを使用した。透析バッファー1(6M尿素、0.05Mリン酸ナトリウム、0.8M塩化ナトリウム、0.3Mイミダゾール、pH8.0)、透析バッファー2(4M尿素、0.033Mリン酸ナトリウム、0.533M塩化ナトリウム、0.2Mイミダゾール、pH8.0)、透析バッファー3(2.67M尿素、0.022Mリン酸ナトリウム、0.356M塩化ナトリウム、0.133Mイミダゾール、pH8.0)および透析バッファー4(1.5M尿素、0.0148リン酸ナトリウム、0.237M塩化ナトリウム、0.089Mイミダゾール、pH8.0)。透析カセットを逐次、4℃において、各透析バッファーに浸漬し、6時間以上、回転させておいた。透析が完了したら、組換えHisタグ付き融合タンパク質をそれぞれカセットから取り出し、ナノドロップ(NanoDrop)を使って定量し、−80℃で凍結した。
【0174】
(実施例15)
SDS−PAGEおよび抗Hisタグ・ウェスタンブロット
抗6×Hisタグ・モノクローナル抗体(ロックランド(Rockland))を使って精製組換えタンパク質を検出するために、ウェスタンブロットを開発した。等体積の各精製切断ORF1タンパク質とLDS/10%β−MEとを混合し、95℃で10分間煮沸した。煮沸した試料のうち10μlを、4−12%ビス−トリス・ポリアクリルアミドゲル(インビトロジェン(Invitrogen))の適当な各ウェルに加え、1×MES泳動バッファー(インビトロジェン(Invitrogen))中、200ボルトで、43分間泳動した。トランスブロット(Trans blot)セミドライ転写装置と1×転写バッファー(インビトロジェン(Invitrogen))とを使って、タンパク質をPVDFメンブレン(バイオラッド(Bio?Rad))に転写した。転写が完了したら、PVDFメンブレンを、オデッセイ(Odyssey)ブロッキングバッファー(リ−コール(Li?Cor))中、室温で1時間、インキュベートした。抗6×HisタグMAbをオデッセイ(Odyssey)ブロッキングバッファー/0.2%ツイーン20で1:1000に希釈し、先のオデッセイ(Odyssey)ブロッキングバッファーを除去してから、メンブレンに移した。MAbを振とう機上に放置して、メンブレンと共に室温で2時間、または4℃で終夜、インキュベートした後、メンブレンをトリス緩衝食塩水/0.05%ツイーン20(TBS−T、シグマ(Sigma))で3回洗浄した。ヤギ抗ウサギIgG IRDye800(リ−コール(Li?Cor))抗体を、オデッセイ(Odyssey)ブロッキングバッファー/0.2%ツイーン20/0.1%SDSで1:5000に希釈した。それを、洗浄したばかりのPVDFメンブレンに移し、穏やかに振とうしながら室温で1時間インキュベートした。メンブレンをTBS−Tで3回、TBSで1回洗浄し、リ−コール(Li?Cor)オデッセイ(Odyssey)で撮像した。
【0175】
(実施例16)
血清ウェスタンブロット
ELISA開発用の陽性および陰性血清対照を同定するために、血清ウェスタンブロットを開発し、使用した。上述のSDS−PAGE後に、タンパク質をPDVFメンブレンに転写し、次にそれを、室温においてオデッセイ(Odyssey)ブロッキングバッファー(リ−コール(Li?Cor))中で1時間インキュベートした。選択した血清試料をオデッセイ(Odyssey)ブロッキングバッファー/0.2%ツイーン20で1:100に希釈し、先のオデッセイ(Odyssey)ブロッキングバッファーを除去してからメンブレンに移した。血清試料を振とう機上に放置して、室温で2時間、メンブレンと共にインキュベートした後、メンブレンをトリス緩衝食塩水/0.05%ツイーン20(TBS−T、シグマ(Sigma))で3回洗浄した。ヤギ抗豚IgG IRDye800抗体(ロックランド(Rockland))を、オデッセイ(Odyssey)ブロッキングバッファー/0.2%ツイーン20/0.1%SDSで1:2500に希釈した。それを洗浄したばかりのPVDFメンブレンに移し、穏やかに振とうしながら室温で1時間インキュベートした。メンブレンをTBS−Tで3回、TBSで1回洗浄し、リ−コール(Li−Cor)オデッセイ(Odyssey)で撮像した。
【0176】
(実施例17)
間接的PTTV1a、PTTV1bおよびPTTV2特異的ELISA
プレートのコーティングに使用した抗原の至適濃度ならびに抗血清およびコンジュゲートの希釈度を、交差力価測定法によって決定した。ELISAは、精製組換えHisタグ付き融合タンパク質のそれぞれ(それぞれPTTV1a、PTTV1bおよびPTTV2c)をpH9.6の1×カーボネート・コーティング・バッファー(Carbonate Coating Buffer:CCB)で680ng/mlに希釈し、ミディアム・バインディング(medium binding)ELISAプレート(グライナー(Greiner))を100μl/ウェルでコーティングすることによって開始した。プレートを覆い、37℃で2時間インキュベートした。コーティング後に、希釈タンパク質を除去し、各ウェルを300μlの1×TBS−Tで3回洗浄した。次に、プロテイン・フリー・ブロッキング・バッファー(Protein Free Blocking Buffer)(ピアス(Pierce))を300μl/ウェルの体積で加え、プレートを37℃で1時間インキュベートした。同時に、96ウェル希釈ブロックでは、血清試料を150μlのプロテイン・フリー・ブロッキング・バッファー(Protein Free Blocking Buffer)で1:100に希釈した。次に、そのブロックを除去し、100μlの各希釈血清試料を、ELISAプレート上の対応する各ウェルに移した。プレートを37℃で2時間インキュベートした後、各ウェルを300μlのTBS−Tで3回洗浄した。次に、HRPコンジュゲート抗豚IgG抗体(ロックランド(Rockland))を、12mlのプロテイン・フリー・ブロックで1:4000に希釈し、プレートの各ウェルに100μlを加えた。これを37℃で1時間インキュベートした後、各ウェルを300μlのTBS−Tで3回洗浄した。ELISAを発色させるために、100μlのシュアー・ブルー・リバース1−コンポーネント(Sure Blue Reserve 1?Component)(ケー・ピー・エル(KPL))をプレートの各ウェルに加えた。20分後に、発色を停止させるために100μlの1N HClを各ウェルに加えた。次にプレートを450nmで読んだ。
【0177】
(実施例18)
データ解析
営利会社から得た細胞培養研究に使用した豚血清(ニュージーランドで製造され、どのOIE疾患も持たないと考えられるもの)を、3つのELISAプロトコールの陽性対照として使用した。なぜなら、この血清は、血清ウェスタンブロットで、PTTV1a、PTTV1bおよびPTTV2の全てについて陽性であると検出され、高いOD値(>2.0)を示したからである。本発明者らは、まず最初に、プールしたノトバイオート豚血清を陰性対照として使用した。というのも、それらはウェスタンブロット検出において陰性だったからである。次に、陰性ノトバイオート豚血清との比較で、本発明者らは、ウィスコンシンにある従来の養豚場から収集されたいくつかの豚血清をスクリーニングした。それらもウェスタンブロット検出において陰性であり、それらのOD値は陰性ノトバイオート豚血清のそれと一致した。これらの在来豚血清をプールし、陰性対照として使用した。各ELISAについてのカットオフ値は陰性対照群(n=4)の平均OD値+標準偏差の3倍として算出した。
【0178】
(実施例19)
豚TTV1a、1bおよび2cの完全長ゲノムDNAクローンの構築
米国分離株PTTV1a−VA(ジェンバンク(GenBank)アクセッション番号GU456383)由来のPCRフラグメントBおよびCを、既述のコンストラクトから再増幅し、次に、ベクターpSC−B−amp/kan(ストラタジーン(Stratagene))上で、ヘラキュレース(Herculase)IIフュージョン(Fusion)DNAポリメラーゼ(ストラタジーン(Stratagene))を使って、オーバーラッピングPCRを行うことにより、ゲノムの両端にBamHI部位を持つ完全長ゲノムDNAへとアセンブルした。その結果得られたコンストラクトをpSC−PTTV1aと名付けた(図17A)。同じ戦略を使って、米国分離株PTTV1b−VA(ジェンバンク(GenBank)アクセッション番号GU456384)に由来するクローンpSC−PTTV1b(図17B)および米国分離株PTTV2c−VA(ジェンバンク(GenBank)アクセッション番号GU456386)に由来するクローンpSC−PTTV2c(図17C)を、同じバックボーンベクター上の同じ制限部位(BamHI)を使って構築した。完全長ゲノムDNAを含有するプラスミドTTV2−#471942−full(図17E)はドイツ病原性豚TTV2分離株に由来した。TTV2−#471942はアンドレアス・ガレイ(Andreas Gallei)博士(ビー・アイ・ブイ・アイ(BIVI)、ドイツ)から分譲された。TTV2−#471942は、系統解析に基づいて米国分離株PTTV1b−VAと共に豚TTV亜型2bに分類された(データ未掲載)。
【0179】
(実施例20)
豚TTV2bおよび2cのタンデム二量体化DNAクローンの構築
完全長PTTV2cゲノムをBamHI消化によってクローンpSC−PTTV2cから切り出し、精製し、ライゲーションして、コンカテマーを形成させた。ライゲーションされたコンカテマーを、BamHIで前もって消化しておいたpSC−B−amp/kanベクター中にクローニングして、タンデム二量体化DNAクローンpSC−2PTTV2c−RR(図1D)を作製した。同様に、タンデム二量体化DNAクローンpSC−2PTTV2b−RRを、クローンTTV2−#471942−fullから、EcoRV制限部位を使って作製した(図1F)。
【0180】
(実施例21)
PTTV1a特異的、PTTV1b特異的およびPTTV2特異的抗ORF1ポリクローナル抗体の作製
ORF1がコードする産物は、TTVの予測キャプシドタンパク質である。PTTV ORF1タンパク質の発現を検出し、PTTV DNAクローンの感染性を決定する目的で、PTTV1a特異的、PTTV1b特異的およびPTTV2特異的抗ORF1ポリクローナル抗体を作製するために、PTTV1a、PTTV1bおよびPTTV2c由来の3つのORF1タンパク質を大腸菌中で発現させ、精製し、次に、それらを使って、ロックランド・イムノケミカルズ(Rockland Immunochemicals)(ペンシルバニア州ギルバーツビル)におけるカスタム抗体作製サービスとして、ニュージーランド白色種ウサギをそれぞれ免疫化した。各抗ORF1ポリクローナル抗体を免疫化したウサギの血清から作製した。
【0181】
(実施例22)
PTTV感染性クローンのインビトロ・トランスフェクション
PK−15細胞を6ウェルプレート上に1ウェルあたり2?105細胞の密度で播種し、トランスフェクションに先だって60%〜70%コンフルエンスまで成長させた。DNAクローンpSC−2PTTV2b−RRおよびpSC−2PTTV2c−RRをそれぞれPK−15細胞に、リポフェクトアミン(Lipofectamine)LTX(インビトロジェン(Invitrogen))を使って、製造者のプロトコールに従って、直接トランスフェクトした。クローンpSC−PTTV1a、pSC−PTTV2cおよびTTV2−#471942−fullについては、上述のように作製された、それらのライゲーションされたコンカテマーを、それぞれトランスフェクションに使用した。細胞を3〜5日間培養した後、豚TTVのORF1の発現を検出するために、免疫蛍光アッセイ(IFA)に付した。あるいは、トランスフェクト細胞を、新しい6ウェルプレートに継代し、3日間培養を続けてから、IFA検出を行う。
【0182】
(実施例23)
免疫蛍光アッセイ(IFA)
トランスフェクションまたは継代した細胞をPBSで2回洗浄し、アセトンで固定した。PBSで1:500に希釈したPTTV1aまたはPTTV2に特異的な抗体500μlを細胞の上に加え、室温で1時間インキュベートした。細胞をPBSで3回洗浄した後、1:200希釈のテキサス・レッド(Texas red)またはアレクサ・フルオル(Alexa Fluor)488標識ヤギ抗ウサギIgG(インビトロジェン(Invitrogen))500μlを加えた。室温で1時間インキュベートし、PBSで洗浄した後、細胞を1:1000希釈のDAPI(ケーピーエル・インコーポレイテッド(KPL,Inc.))500μlで染色し、蛍光顕微鏡下で可視化した。
【0183】
(実施例24)
タンデム二量体化豚TTV2クローンによる在来豚のインビボ接種
2つのタンデム二量体化豚TTV2クローンpSC−2TTV2b−RRおよびpSC−2TTV2c−RRの感染性を決定する為に豚接種研究を行った。簡単に述べると、豚TTV2について血清陰性およびDNAウイルス陰性である8頭の4週齢在来豚を、それぞれ4匹ずつの2群にランダムに割り当てた。各群の豚を別々に収容し、施設内動物実験委員会(Institutional Committee on Animal Care and Use)の要求を全て満たす条件下で維持した。
【0184】
各群内の全ての豚に、リンパ節内経路と筋肉内経路の両方を組み合わせて注射した。4頭の豚(番号181、189、192および193)には、それぞれ200μgのpSC−2TTV2b−RRプラスミドDNAを注射し、一方、他方の豚4頭(番号92、180、188および191)には、それぞれ200μgのpSC−2TTV2c−RRクローンを接種した。豚を、疾患の臨床徴候について、合計28日間、毎日監視した。全ての豚を接種の28日後に剖検した。
【0185】
いくつかの実施形態の説明によって本発明を例示し、例示的実施形態について詳細に説明したが、本願請求項の範囲をそのような詳細に制限または多少なりとも限定することは、本願出願人の意図するところではない。さらなる変更は当業者には明白であるだろう。したがって本発明は、さらに広いその態様において、ここに示し説明した具体的詳細、代表的装置および方法、ならびに例示的実施例に限定されない。したがって、本願出願人の一般的発明概念の要旨または範囲から逸脱することなく、そのような詳細から逸脱することができる。
【図1A】
【図1B】
【特許請求の範囲】
【請求項1】
PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAの遺伝子型からなる群より選択されるゲノム配列に対して少なくとも85%の相同性を有する少なくとも1コピーのゲノム配列を含有する感染性PTTVをコードする核酸分子を含む、豚トルクテノウイルス(PTTV)の感染性核酸分子。
【請求項2】
少なくとも1コピーのゲノム配列が、PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも95%の相同性を有する、請求項1に記載の感染性核酸分子。
【請求項3】
前記ゲノム配列が、配列番号9、配列番号10、配列番号11、および配列番号12に示す配列から選択される、請求項1に記載の感染性核酸分子。
【請求項4】
請求項3に記載の感染性核酸分子を含有する、生物学的に機能的なプラスミドまたはウイルスベクター。
【請求項5】
2コピー以上の感染性核酸分子を含有する、請求項4に記載の生物学的に機能的なプラスミドまたはウイルスベクター。
【請求項6】
請求項4に記載の感染性核酸分子を含むベクターによってトランスフェクトされた適切な宿主細胞。
【請求項7】
請求項6に記載の感染性核酸分子を含有する細胞によって産生される無発病性の感染性PTTV。
【請求項8】
無毒性の生理学的に許容される担体と、
(a)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも80%の相同性を有する少なくとも1コピーのゲノム配列またはその相補鎖を含有する核酸分子、
(b)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも80%の相同性を有する少なくとも1コピーのゲノム配列またはその相補鎖を含有する核酸分子を含有する生物学的に機能的なプラスミドまたはウイルスベクター、
(c)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも85%の相同性を有するゲノム配列から誘導され、それを含有する、無発病性、感染性かつ非病原性豚トルクテノウイルス、
(d)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1、およびORF2/2からなる群より選択されるポリヌクレオチド配列に従って翻訳されたポリペプチド配列の免疫原性フラグメントまたは完全タンパク質
からなる群より選択される免疫原量の構成要素とを含む、ワクチン。
【請求項9】
生PTTVウイルスを含有する、請求項8に記載のワクチン。
【請求項10】
死滅PTTVウイルスを含有する、請求項8に記載のワクチン。
【請求項11】
ポリペプチド配列の免疫原性フラグメントまたは完全タンパク質が、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1 およびORF2/2から発現された精製細菌発現またはバキュロウイルス発現組換えタンパク質を含む、請求項8に記載のワクチン。
【請求項12】
ポリペプチド配列の免疫原性フラグメントまたは完全タンパク質が、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1から発現された精製細菌発現またはバキュロウイルス発現組換えタンパク質を含む、請求項11に記載のワクチン。
【請求項13】
アジュバントをさらに含む、請求項8に記載のワクチン。
【請求項14】
免疫学的有効量の請求項8に記載のワクチンを豚に投与することを含む、PTTVウイルス感染に対して豚を免疫化する方法。
【請求項15】
前記ワクチンを豚に非経口投与、鼻腔内投与、皮内投与、または経皮投与することを含む、請求項14に記載の方法。
【請求項16】
前記ワクチンを豚にリンパ系内投与または筋肉内投与することを含む、請求項14に記載の方法。
【請求項17】
配列番号9に示すPTTV1a−VAのヌクレオチド配列に対して85%の相同性を有するポリ核酸配列からなる単離されたポリヌクレオチド。
【請求項18】
前記ポリ核酸配列が、配列番号9に示す前記PTTV1a−VAのヌクレオチド配列に対して95%の相同性を有する、請求項17に記載の単離されたポリヌクレオチド。
【請求項19】
前記ポリ核酸配列が、配列番号9に示す前記PTTV1a−VAのヌクレオチド配列からなる、請求項17に記載の単離されたポリヌクレオチド。
【請求項20】
配列番号10に示すPTTV1b−VAのヌクレオチド配列に対して85%の相同性を有するポリ核酸配列からなる単離されたポリヌクレオチド。
【請求項21】
前記ポリ核酸配列が、配列番号10に示す前記PTTV1b−VAのヌクレオチド配列に対して95%の相同性を有する、請求項20に記載の単離されたポリヌクレオチド。
【請求項22】
前記ポリ核酸配列が、配列番号10に示す前記PTTV1b−VAのヌクレオチド配列からなる、請求項20に記載の単離されたポリヌクレオチド。
【請求項23】
配列番号11に示すPTTV2b−VAのヌクレオチド配列に対して85%の相同性を有するポリ核酸配列からなる単離されたポリヌクレオチド。
【請求項24】
前記ポリ核酸配列が、配列番号11に示す前記PTTV2b−VAのヌクレオチド配列に対して95%の相同性を有する、請求項23に記載の単離されたポリヌクレオチド。
【請求項25】
前記ポリ核酸配列が、配列番号11に示す前記PTTV2b−VAのヌクレオチド配列からなる、請求項23に記載の単離されたポリヌクレオチド。
【請求項26】
配列番号12に示すPTTV2c−VAのヌクレオチド配列に対して85%の相同性を有するポリ核酸配列からなる単離されたポリヌクレオチド。
【請求項27】
前記ポリ核酸配列が、配列番号12に示す前記PTTV2c−VAのヌクレオチド配列に対して95%の相同性を有する、請求項23に記載の単離されたポリヌクレオチド。
【請求項28】
前記ポリ核酸配列が、配列番号12に示す前記PTTV2c−VAのヌクレオチド配列からなる、請求項23に記載の単離されたポリヌクレオチド。
【請求項29】
PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1、およびORF2/2からなる群より選択されるポリヌクレオチド配列に従って翻訳されたポリペプチド配列の免疫原性フラグメントまたは完全タンパク質を含むワクチン。
【請求項30】
前記免疫原性フラグメントまたは完全タンパク質が、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのキャプシドタンパク質である、請求項29に記載のワクチン。
【請求項31】
前記ポリヌクレオチド配列が、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1からなる群より選択される、請求項29に記載のワクチン。
【請求項32】
前記ポリヌクレオチド配列がPTTV遺伝子型PTTV1a−VAのORF1である、請求項31に記載のワクチン。
【請求項33】
前記ポリヌクレオチド配列がPTTV遺伝子型PTTV1b−VAのORF1である、請求項31に記載のワクチン。
【請求項34】
前記ポリヌクレオチド配列がPTTV亜型PTTV2c−VAのORF1である、請求項31に記載のワクチン。
【請求項35】
前記ポリペプチド配列が、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、および配列番号28に示す配列からなる群より選択される、請求項29に記載のワクチン。
【請求項36】
前記ポリペプチド配列が配列番号13に示される、請求項35に記載のワクチン。
【請求項37】
前記ポリペプチド配列が配列番号13のC末端領域(aa317−635)である、請求項36に記載のワクチン。
【請求項38】
前記ポリペプチド配列が配列番号14に示される、請求項35に記載のワクチン。
【請求項39】
前記ポリペプチド配列が配列番号14のC末端領域(aa322−639)である、請求項38に記載のワクチン。
【請求項40】
前記ポリペプチド配列が配列番号16に示される、請求項35に記載のワクチン。
【請求項41】
前記ポリペプチド配列が配列番号16のC末端領域(aa310−625)である、請求項40に記載のワクチン。
【請求項42】
前記ポリペプチド配列が配列番号20に示される、請求項35に記載のワクチン。
【請求項43】
アジュバントをさらに含有する、請求項29に記載のワクチン。
【請求項44】
免疫学的有効量の請求項29に記載のワクチンを豚に投与することを含む、PTTVウイルス感染に対して豚を免疫化する方法。
【請求項45】
免疫原性フラグメントまたは完全タンパク質を豚に投与することを含む、請求項44に記載の方法。
【請求項46】
ワクチンを豚に非経口投与、鼻腔内投与、皮内投与、または経皮投与することを含む、請求項44に記載の方法。
【請求項47】
ワクチンを豚にリンパ系内投与または筋肉内投与することを含む、請求項44に記載の方法。
【請求項48】
PTTV1感染を診断し、PTTV1負荷量を定量するための方法であって、
PTTV1感染が疑われる試料からDNAを抽出すること、
配列番号29および配列番号30に示す配列を含むプライマーを使ってポリメラーゼ連鎖反応(PCR)を行うこと、および
PTTV1特異的増幅を検出すること
を含む方法。
【請求項49】
前記ポリメラーゼ連鎖反応がSYBRグリーン・リアルタイムPCRである、請求項48に記載の方法。
【請求項50】
PTTV2感染を診断し、PTTV2負荷量を定量するための方法であって、
PTTV2感染が疑われる試料からDNAを抽出すること、
配列番号29、配列番号30、配列番号31および配列番号32に示す配列を含むプライマーを使ってポリメラーゼ連鎖反応(PCR)を行うこと、および
PTTV2特異的増幅を検出すること
を含む方法。
【請求項51】
前記ポリメラーゼ連鎖反応がSYBRグリーン・リアルタイムPCRである、請求項50に記載の方法。
【請求項52】
PTTV1感染とPTTV2感染を同時に検出し診断するための方法であって、
PTTV感染が疑われる試料からDNAを抽出すること、
配列番号31および配列番号32に示す配列を含むプライマーを使ってポリメラーゼ連鎖反応(PCR)を行うこと、および
PTTV1およびPTTV2特異的増幅を検出すること
を含む方法。
【請求項53】
前記ポリメラーゼ連鎖反応がSYBRグリーン・リアルタイムPCRである、請求項52に記載の方法。
【請求項54】
PTTV1a感染とPTTV1b感染を同時に検出し診断するための方法であって、
PTTV1感染が疑われる試料からDNAを抽出すること、
配列番号33および配列番号34に示す配列を含むプライマーを使って第1ポリメラーゼ連鎖反応(PCR)を行うこと、
配列番号35、配列番号36、配列番号37、および配列番号38に示す配列を含むプライマーを使って第2PCRを行うこと、および
PTTV1aおよびPTTV1b特異的増幅を検出すること
を含む方法。
【請求項55】
PTTV感染を診断するための方法であって、
PTTV亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1、およびORF2/2からなる群より選択されるポリヌクレオチド配列に従って翻訳されたポリペプチド配列の免疫原性フラグメントまたは完全タンパク質を固定化すること、
PTTV感染が疑われる豚からの血清試料を前記固定化免疫原性フラグメントまたは完全タンパク質と接触させること、および
前記免疫原性フラグメントに特異的な、捕捉された抗体を検出すること
を含む方法。
【請求項56】
前記ポリヌクレオチド配列が、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1からなる群より選択される、55に記載の方法。
【請求項57】
前記ポリヌクレオチド配列がPTTV遺伝子型PTTV1a−VAのORF1である、請求項56に記載のワクチン。
【請求項58】
前記ポリヌクレオチド配列がPTTV遺伝子型PTTV1b−VAのORF1である、請求項56に記載の方法。
【請求項59】
前記ポリヌクレオチド配列がPTTV亜型PTTV2c−VAのORF1である、請求項56に記載の方法。
【請求項60】
前記ポリペプチド配列が、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、および配列番号28に示す配列からなる群より選択される、請求項55に記載の方法。
【請求項61】
前記ポリペプチド配列が配列番号13に示される、請求項60に記載の方法。
【請求項62】
前記ポリペプチド配列が配列番号16に示される、請求項60に記載のワクチン。
【請求項63】
前記免疫原性フラグメントが、配列番号13のC末端領域(aa317−635)、配列番号14のC末端領域(aa322−639)、または配列番号16のC末端領域(aa310−625)である、請求項62に記載のワクチン。
【請求項64】
前記ポリペプチド配列が配列番号20に示される、請求項60に記載のワクチン。
【請求項65】
前記捕捉された抗体の検出がウェスタンブロットによる、請求項55に記載の方法。
【請求項66】
前記捕捉された抗体の検出が酵素結合免疫吸着アッセイ(ELISA)による、請求項55に記載の方法。
【請求項1】
PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAの遺伝子型からなる群より選択されるゲノム配列に対して少なくとも85%の相同性を有する少なくとも1コピーのゲノム配列を含有する感染性PTTVをコードする核酸分子を含む、豚トルクテノウイルス(PTTV)の感染性核酸分子。
【請求項2】
少なくとも1コピーのゲノム配列が、PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも95%の相同性を有する、請求項1に記載の感染性核酸分子。
【請求項3】
前記ゲノム配列が、配列番号9、配列番号10、配列番号11、および配列番号12に示す配列から選択される、請求項1に記載の感染性核酸分子。
【請求項4】
請求項3に記載の感染性核酸分子を含有する、生物学的に機能的なプラスミドまたはウイルスベクター。
【請求項5】
2コピー以上の感染性核酸分子を含有する、請求項4に記載の生物学的に機能的なプラスミドまたはウイルスベクター。
【請求項6】
請求項4に記載の感染性核酸分子を含むベクターによってトランスフェクトされた適切な宿主細胞。
【請求項7】
請求項6に記載の感染性核酸分子を含有する細胞によって産生される無発病性の感染性PTTV。
【請求項8】
無毒性の生理学的に許容される担体と、
(a)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも80%の相同性を有する少なくとも1コピーのゲノム配列またはその相補鎖を含有する核酸分子、
(b)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも80%の相同性を有する少なくとも1コピーのゲノム配列またはその相補鎖を含有する核酸分子を含有する生物学的に機能的なプラスミドまたはウイルスベクター、
(c)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAからなる群より選択されるゲノム配列に対して少なくとも85%の相同性を有するゲノム配列から誘導され、それを含有する、無発病性、感染性かつ非病原性豚トルクテノウイルス、
(d)PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1、およびORF2/2からなる群より選択されるポリヌクレオチド配列に従って翻訳されたポリペプチド配列の免疫原性フラグメントまたは完全タンパク質
からなる群より選択される免疫原量の構成要素とを含む、ワクチン。
【請求項9】
生PTTVウイルスを含有する、請求項8に記載のワクチン。
【請求項10】
死滅PTTVウイルスを含有する、請求項8に記載のワクチン。
【請求項11】
ポリペプチド配列の免疫原性フラグメントまたは完全タンパク質が、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1 およびORF2/2から発現された精製細菌発現またはバキュロウイルス発現組換えタンパク質を含む、請求項8に記載のワクチン。
【請求項12】
ポリペプチド配列の免疫原性フラグメントまたは完全タンパク質が、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1から発現された精製細菌発現またはバキュロウイルス発現組換えタンパク質を含む、請求項11に記載のワクチン。
【請求項13】
アジュバントをさらに含む、請求項8に記載のワクチン。
【請求項14】
免疫学的有効量の請求項8に記載のワクチンを豚に投与することを含む、PTTVウイルス感染に対して豚を免疫化する方法。
【請求項15】
前記ワクチンを豚に非経口投与、鼻腔内投与、皮内投与、または経皮投与することを含む、請求項14に記載の方法。
【請求項16】
前記ワクチンを豚にリンパ系内投与または筋肉内投与することを含む、請求項14に記載の方法。
【請求項17】
配列番号9に示すPTTV1a−VAのヌクレオチド配列に対して85%の相同性を有するポリ核酸配列からなる単離されたポリヌクレオチド。
【請求項18】
前記ポリ核酸配列が、配列番号9に示す前記PTTV1a−VAのヌクレオチド配列に対して95%の相同性を有する、請求項17に記載の単離されたポリヌクレオチド。
【請求項19】
前記ポリ核酸配列が、配列番号9に示す前記PTTV1a−VAのヌクレオチド配列からなる、請求項17に記載の単離されたポリヌクレオチド。
【請求項20】
配列番号10に示すPTTV1b−VAのヌクレオチド配列に対して85%の相同性を有するポリ核酸配列からなる単離されたポリヌクレオチド。
【請求項21】
前記ポリ核酸配列が、配列番号10に示す前記PTTV1b−VAのヌクレオチド配列に対して95%の相同性を有する、請求項20に記載の単離されたポリヌクレオチド。
【請求項22】
前記ポリ核酸配列が、配列番号10に示す前記PTTV1b−VAのヌクレオチド配列からなる、請求項20に記載の単離されたポリヌクレオチド。
【請求項23】
配列番号11に示すPTTV2b−VAのヌクレオチド配列に対して85%の相同性を有するポリ核酸配列からなる単離されたポリヌクレオチド。
【請求項24】
前記ポリ核酸配列が、配列番号11に示す前記PTTV2b−VAのヌクレオチド配列に対して95%の相同性を有する、請求項23に記載の単離されたポリヌクレオチド。
【請求項25】
前記ポリ核酸配列が、配列番号11に示す前記PTTV2b−VAのヌクレオチド配列からなる、請求項23に記載の単離されたポリヌクレオチド。
【請求項26】
配列番号12に示すPTTV2c−VAのヌクレオチド配列に対して85%の相同性を有するポリ核酸配列からなる単離されたポリヌクレオチド。
【請求項27】
前記ポリ核酸配列が、配列番号12に示す前記PTTV2c−VAのヌクレオチド配列に対して95%の相同性を有する、請求項23に記載の単離されたポリヌクレオチド。
【請求項28】
前記ポリ核酸配列が、配列番号12に示す前記PTTV2c−VAのヌクレオチド配列からなる、請求項23に記載の単離されたポリヌクレオチド。
【請求項29】
PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1、およびORF2/2からなる群より選択されるポリヌクレオチド配列に従って翻訳されたポリペプチド配列の免疫原性フラグメントまたは完全タンパク質を含むワクチン。
【請求項30】
前記免疫原性フラグメントまたは完全タンパク質が、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのキャプシドタンパク質である、請求項29に記載のワクチン。
【請求項31】
前記ポリヌクレオチド配列が、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1からなる群より選択される、請求項29に記載のワクチン。
【請求項32】
前記ポリヌクレオチド配列がPTTV遺伝子型PTTV1a−VAのORF1である、請求項31に記載のワクチン。
【請求項33】
前記ポリヌクレオチド配列がPTTV遺伝子型PTTV1b−VAのORF1である、請求項31に記載のワクチン。
【請求項34】
前記ポリヌクレオチド配列がPTTV亜型PTTV2c−VAのORF1である、請求項31に記載のワクチン。
【請求項35】
前記ポリペプチド配列が、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、および配列番号28に示す配列からなる群より選択される、請求項29に記載のワクチン。
【請求項36】
前記ポリペプチド配列が配列番号13に示される、請求項35に記載のワクチン。
【請求項37】
前記ポリペプチド配列が配列番号13のC末端領域(aa317−635)である、請求項36に記載のワクチン。
【請求項38】
前記ポリペプチド配列が配列番号14に示される、請求項35に記載のワクチン。
【請求項39】
前記ポリペプチド配列が配列番号14のC末端領域(aa322−639)である、請求項38に記載のワクチン。
【請求項40】
前記ポリペプチド配列が配列番号16に示される、請求項35に記載のワクチン。
【請求項41】
前記ポリペプチド配列が配列番号16のC末端領域(aa310−625)である、請求項40に記載のワクチン。
【請求項42】
前記ポリペプチド配列が配列番号20に示される、請求項35に記載のワクチン。
【請求項43】
アジュバントをさらに含有する、請求項29に記載のワクチン。
【請求項44】
免疫学的有効量の請求項29に記載のワクチンを豚に投与することを含む、PTTVウイルス感染に対して豚を免疫化する方法。
【請求項45】
免疫原性フラグメントまたは完全タンパク質を豚に投与することを含む、請求項44に記載の方法。
【請求項46】
ワクチンを豚に非経口投与、鼻腔内投与、皮内投与、または経皮投与することを含む、請求項44に記載の方法。
【請求項47】
ワクチンを豚にリンパ系内投与または筋肉内投与することを含む、請求項44に記載の方法。
【請求項48】
PTTV1感染を診断し、PTTV1負荷量を定量するための方法であって、
PTTV1感染が疑われる試料からDNAを抽出すること、
配列番号29および配列番号30に示す配列を含むプライマーを使ってポリメラーゼ連鎖反応(PCR)を行うこと、および
PTTV1特異的増幅を検出すること
を含む方法。
【請求項49】
前記ポリメラーゼ連鎖反応がSYBRグリーン・リアルタイムPCRである、請求項48に記載の方法。
【請求項50】
PTTV2感染を診断し、PTTV2負荷量を定量するための方法であって、
PTTV2感染が疑われる試料からDNAを抽出すること、
配列番号29、配列番号30、配列番号31および配列番号32に示す配列を含むプライマーを使ってポリメラーゼ連鎖反応(PCR)を行うこと、および
PTTV2特異的増幅を検出すること
を含む方法。
【請求項51】
前記ポリメラーゼ連鎖反応がSYBRグリーン・リアルタイムPCRである、請求項50に記載の方法。
【請求項52】
PTTV1感染とPTTV2感染を同時に検出し診断するための方法であって、
PTTV感染が疑われる試料からDNAを抽出すること、
配列番号31および配列番号32に示す配列を含むプライマーを使ってポリメラーゼ連鎖反応(PCR)を行うこと、および
PTTV1およびPTTV2特異的増幅を検出すること
を含む方法。
【請求項53】
前記ポリメラーゼ連鎖反応がSYBRグリーン・リアルタイムPCRである、請求項52に記載の方法。
【請求項54】
PTTV1a感染とPTTV1b感染を同時に検出し診断するための方法であって、
PTTV1感染が疑われる試料からDNAを抽出すること、
配列番号33および配列番号34に示す配列を含むプライマーを使って第1ポリメラーゼ連鎖反応(PCR)を行うこと、
配列番号35、配列番号36、配列番号37、および配列番号38に示す配列を含むプライマーを使って第2PCRを行うこと、および
PTTV1aおよびPTTV1b特異的増幅を検出すること
を含む方法。
【請求項55】
PTTV感染を診断するための方法であって、
PTTV亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1、ORF2、ORF1/1、およびORF2/2からなる群より選択されるポリヌクレオチド配列に従って翻訳されたポリペプチド配列の免疫原性フラグメントまたは完全タンパク質を固定化すること、
PTTV感染が疑われる豚からの血清試料を前記固定化免疫原性フラグメントまたは完全タンパク質と接触させること、および
前記免疫原性フラグメントに特異的な、捕捉された抗体を検出すること
を含む方法。
【請求項56】
前記ポリヌクレオチド配列が、PTTV遺伝子型または亜型PTTV1a−VA、PTTV1b−VA、PTTV2b−VA、およびPTTV2c−VAのORF1からなる群より選択される、55に記載の方法。
【請求項57】
前記ポリヌクレオチド配列がPTTV遺伝子型PTTV1a−VAのORF1である、請求項56に記載のワクチン。
【請求項58】
前記ポリヌクレオチド配列がPTTV遺伝子型PTTV1b−VAのORF1である、請求項56に記載の方法。
【請求項59】
前記ポリヌクレオチド配列がPTTV亜型PTTV2c−VAのORF1である、請求項56に記載の方法。
【請求項60】
前記ポリペプチド配列が、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、および配列番号28に示す配列からなる群より選択される、請求項55に記載の方法。
【請求項61】
前記ポリペプチド配列が配列番号13に示される、請求項60に記載の方法。
【請求項62】
前記ポリペプチド配列が配列番号16に示される、請求項60に記載のワクチン。
【請求項63】
前記免疫原性フラグメントが、配列番号13のC末端領域(aa317−635)、配列番号14のC末端領域(aa322−639)、または配列番号16のC末端領域(aa310−625)である、請求項62に記載のワクチン。
【請求項64】
前記ポリペプチド配列が配列番号20に示される、請求項60に記載のワクチン。
【請求項65】
前記捕捉された抗体の検出がウェスタンブロットによる、請求項55に記載の方法。
【請求項66】
前記捕捉された抗体の検出が酵素結合免疫吸着アッセイ(ELISA)による、請求項55に記載の方法。
【図2】

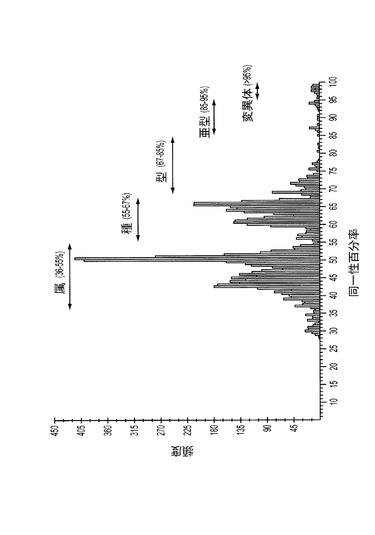
【図3A】
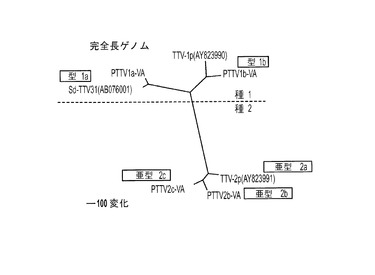
【図3B】
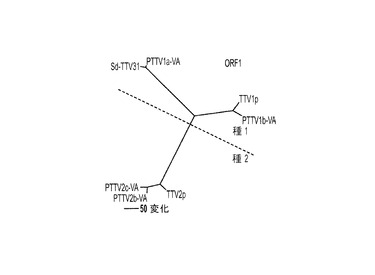
【図3C】
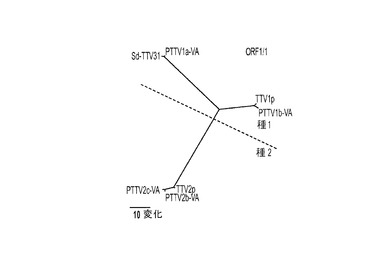
【図3D】

【図3E】
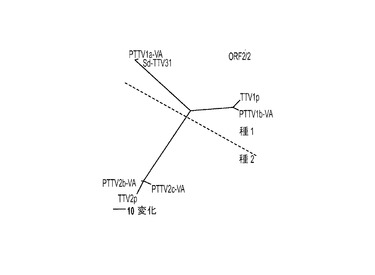
【図4A】
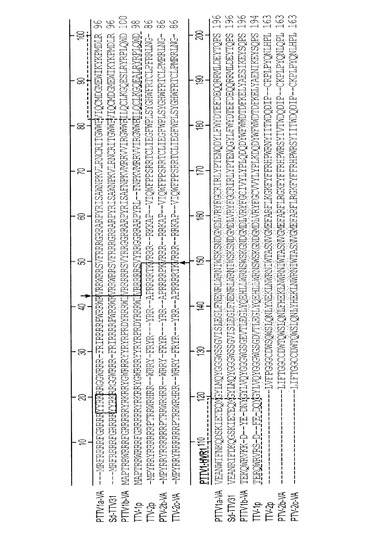
【図4B】
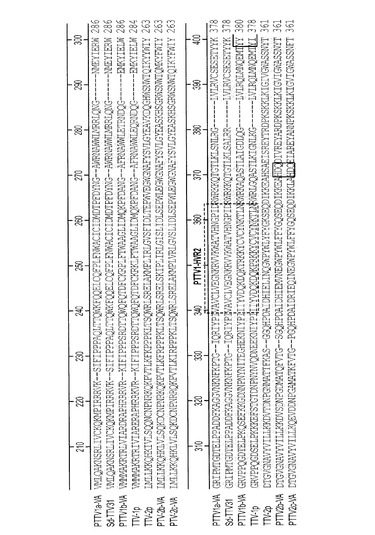
【図4C】

【図5】

【図6】
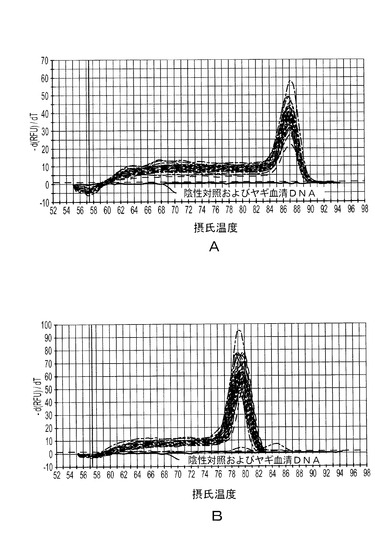
【図7A】
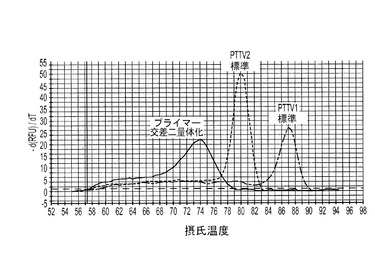
【図7B】

【図7C】
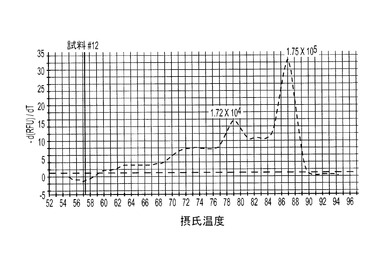
【図7D】

【図7E】

【図8】
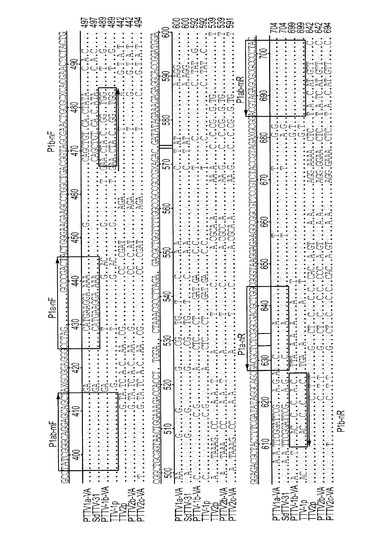
【図9A】

【図9B】
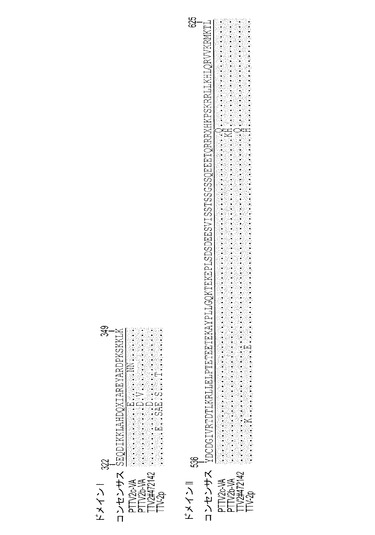
【図10】
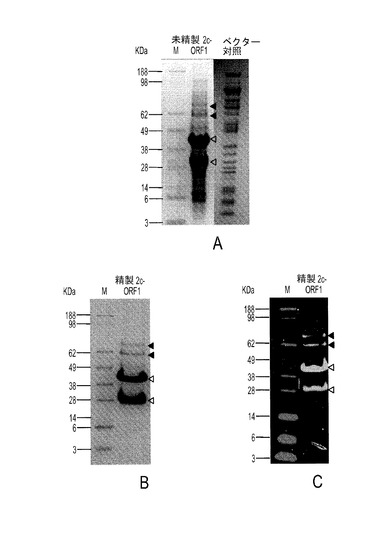
【図11】
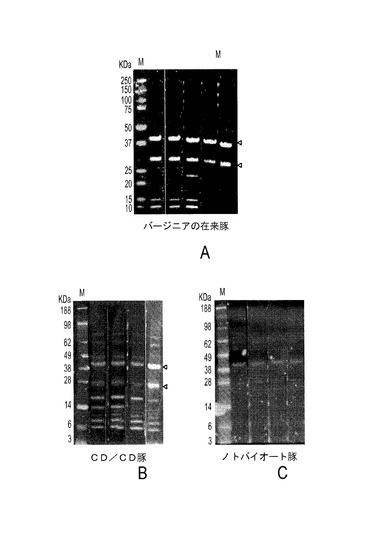
【図12】
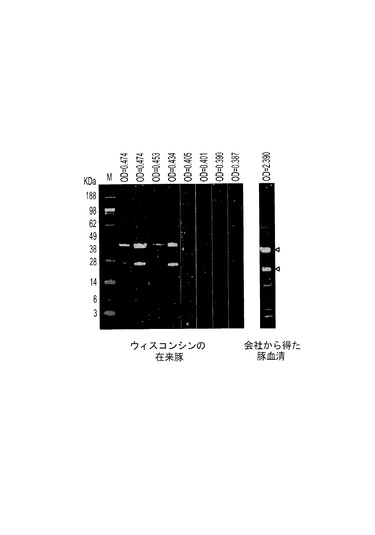
【図13】
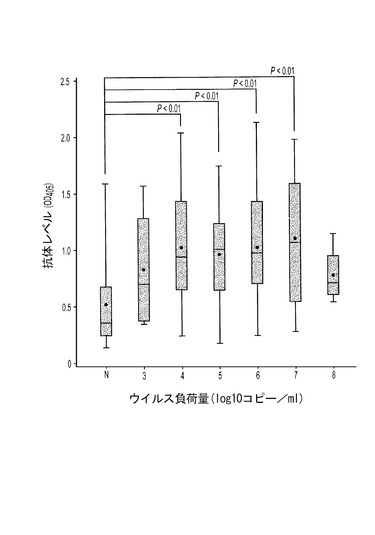
【図14】
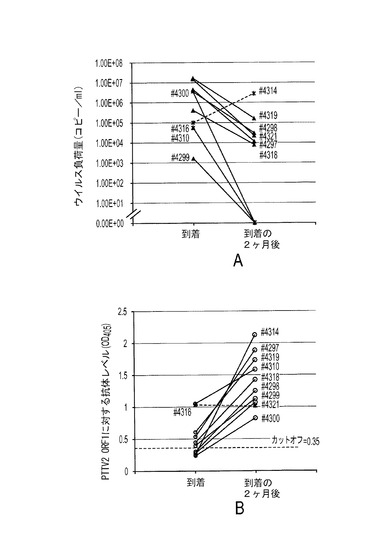
【図15】
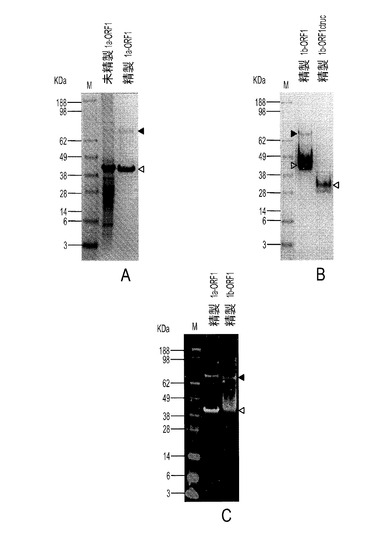
【図16】

【図17】

【図18】
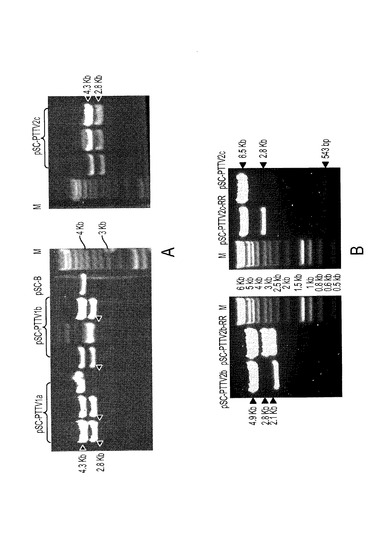
【図19】
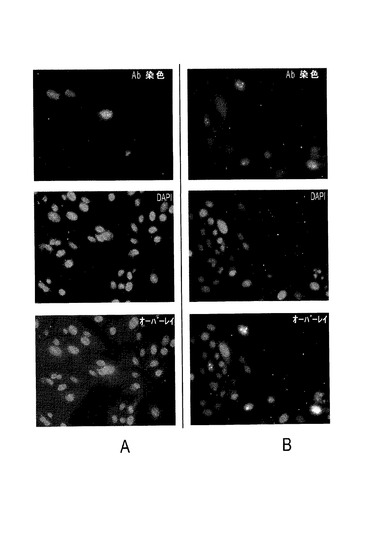
【図20】
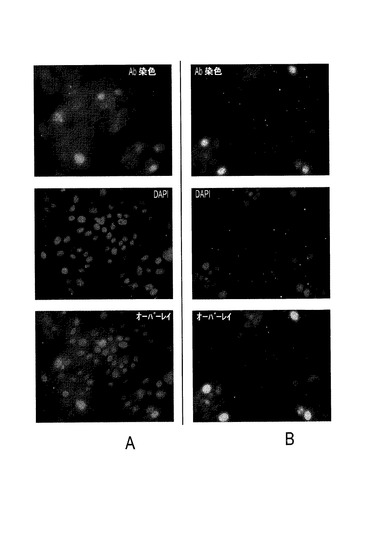
【図21】

【図22】
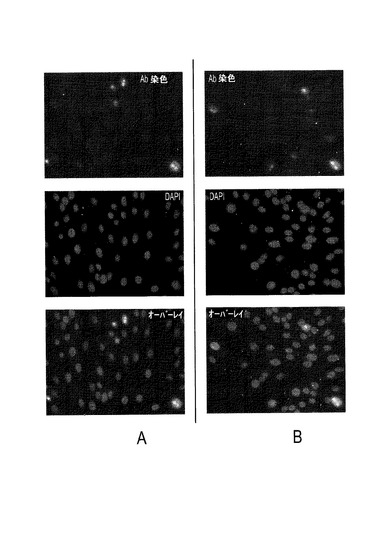
【図23】
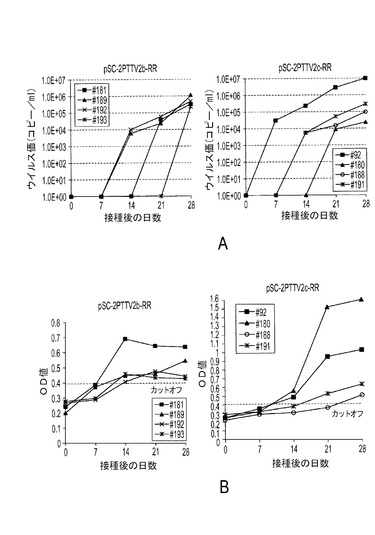
【図3A】
【図3B】
【図3C】
【図3D】
【図3E】
【図4A】
【図4B】
【図4C】
【図5】
【図6】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図8】
【図9A】
【図9B】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【公表番号】特表2013−502229(P2013−502229A)
【公表日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願番号】特願2012−525752(P2012−525752)
【出願日】平成22年8月23日(2010.8.23)
【国際出願番号】PCT/US2010/046330
【国際公開番号】WO2011/031438
【国際公開日】平成23年3月17日(2011.3.17)
【出願人】(508148884)ヴァージニア テック インテレクチュアル プロパティーズ,インコーポレーテッド (5)
【Fターム(参考)】
【公表日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願日】平成22年8月23日(2010.8.23)
【国際出願番号】PCT/US2010/046330
【国際公開番号】WO2011/031438
【国際公開日】平成23年3月17日(2011.3.17)
【出願人】(508148884)ヴァージニア テック インテレクチュアル プロパティーズ,インコーポレーテッド (5)
【Fターム(参考)】
[ Back to top ]