
高い結合親和性を有する免疫結合体
【課題】メソセリンに対してより高い親和性の抗体を提供すること。
【解決手段】メソセリンは、卵巣癌、中皮腫およびいくつかの他の型のヒト癌の表面上に存在する分化抗原である。正常な組織においては、メソセリンは、中皮細胞にのみ存在するので、メソセリンは、細胞傷害性薬剤の抗体媒介性送達のための良好な標的に相当する。本発明は、メソセリンに対して特に高い親和性を有するFv分子を含む抗メソセリン抗体、およびその抗体を利用する免疫結合体に関する。また、この抗体を使用した診断方法および治療方法も記載される。この抗メソセリン抗体は、卵巣癌、胃癌、扁平上皮細胞癌、中皮腫およびメソセリンを発現する他の悪性細胞の診断および処置に好適である。
【解決手段】メソセリンは、卵巣癌、中皮腫およびいくつかの他の型のヒト癌の表面上に存在する分化抗原である。正常な組織においては、メソセリンは、中皮細胞にのみ存在するので、メソセリンは、細胞傷害性薬剤の抗体媒介性送達のための良好な標的に相当する。本発明は、メソセリンに対して特に高い親和性を有するFv分子を含む抗メソセリン抗体、およびその抗体を利用する免疫結合体に関する。また、この抗体を使用した診断方法および治療方法も記載される。この抗メソセリン抗体は、卵巣癌、胃癌、扁平上皮細胞癌、中皮腫およびメソセリンを発現する他の悪性細胞の診断および処置に好適である。
【発明の詳細な説明】
【背景技術】
【0001】
(関連出願に対する相互参照)
適用せず。
【0002】
(連邦政府によって後援された研究および開発の下でなされた発明に対する権利に関する記述)
適用せず。
【0003】
(発明の背景)
多くの型の癌細胞において、分化抗原が発現される。これらの抗原は、癌治療における標的として使用されている。例えば、CD19、CD20、CD22およびCD25は、造血悪性腫瘍における標的として首尾良く使用されている(Pressら、New Eng.J.Med.329:1219−1224(1993);およびOsterborgら、J.Clin.Oncol.15:1567−1574(1997))。しかし、この標的化癌治療は、主に、固形腫瘍で成功していない。なぜなら、この標的化抗原はまた、腫瘍が発生する組織において発現するからである。従って、これらの標的化治療は、健康な細胞ならびに悪性細胞を殺傷する。
【0004】
米国においては、治療にもかかわらず、推定15,000人の女性が、毎年卵巣癌で死亡する。卵巣癌よりも一般的ではないが、中皮腫は、あらゆる化学療法剤に耐性であることが公知であり、従って、高い死亡率を有する。これらの癌の死亡率に起因して、これらの悪性腫瘍に対する新規の治療アプローチが必要とされる。
【0005】
卵巣の扁平上皮細胞、およびいくつかの胃癌ならびに中皮腫に共通して、細胞表面上にメソセリン(mesothelin)が発現される(Changら、Cancer Res.52:181−186(1992);Changら、J.Surgical Pathology 16:259−268(1992);およびChangら、Nat’l Acad.Sci.USA93:136−140(1996)。メソセリンは、中皮細胞の表面上に存在する40kDのGPI結合糖タンパク質抗原である。メソセリンは、69kDの前駆体として合成され、次いで、タンパク質分解性にプロセスされる。この30kDのアミノ末端が分泌され、そして巨核球増強因子(megakaryocyte potentiating factor)とよばれる(Yamaguchiら、J.Biol.Chem.269:805−808(1994))。この40kDのカルボキシル末端は、成熟メソセリンとして膜に結合したままである(Changら、Nat’l Acad.Sci.USA 93:136−140(1996))。癌細胞上に存在する多くの細胞表面抗原と異なり、メソセリンの膜結合形態は、癌患者の血液中に検出されず、そして培養細胞によって培地中に発散されない(Changら、Cancer Res.52:181−186(1992))。悪性細胞に加えて、メソセリンはまた、卵巣癌を含む中皮起源の細胞の細胞表
面上に見出される。これらの組織における細胞への損傷が、生命を脅かす結果を引き起こさないので、癌細胞の表面上のメソセリンの存在が、標的化治療のための有望な候補となる。
【0006】
イムノトキシンは、毒性部分に結合する細胞表面抗原に対する抗体である。癌の処置において、抗体は、好ましくは、癌細胞上にのみ発現される細胞表面抗原に対して指向される。しかし、表面抗原もまた発現する正常細胞の死が、悪性腫瘍の存在よりも生命を脅かさない場合、非悪性細胞上に発現される細胞表面抗原に対する抗体は、癌治療において使用され得る。イムノトキシンの毒性部分は、全身投与後に低い濃度の、非標的化細胞に対して有害ではない任意の毒素であり得る。このような毒素は、Pseudomonas aeruginosa外毒素(PE)である。PEを用いた以前の研究は、タンパク質の活性部分が、ドメインIIおよびドメインIIIから構成され、これらの両方は、毒素のカルボキシル末端に位置することを実証した。イムノトキシンの毒性部分としての使用のための開発または考慮の下にある他の毒素は、ジフテリア毒素(Watsonら、Intl J Cancer,61:233−40(1995))、サポリン(saporin)(例えば、Chandlerら、Intl J.Cancer 78(1):106−11(1998))、ヨウシュヤマゴボウ抗ウイルスタンパク質(例えば、Ekら、Clin Cancer Res,4(7):1641−7(1998)、リシン(例えば、Ohtomoら、Anticancer Res.18(6A):4311−5(1998))およびブリオジン1(bryodin1)(例えば、Franciscoら、J Biol Chem,272(39):24165−9(1997))である。イムノトキシンとしての使用のために、しばしば天然の毒素分子が改変されて、非特異的毒性または他の望ましくない効果を回避する。例えば、リシンのA鎖およびB鎖が、通常分離されて、その結果、B鎖によって別の方法で提供される非特異的結合を回避しながら、A鎖の毒性効果が使用され得る。
【0007】
イムノトキシンを標的化する抗体は、ポリクローナル抗体、モノクローナル抗体、または組換え抗体(例えば、キメラまたは可変領域フラグメント)であり得る。抗体が非組換えである場合、イムノトキシンは、抗体の毒性部分へのキメラ結合体化によって形成されるはずである。抗体が、組換え的に産生される場合、この抗体は、化学的結合または組換え融合を介して毒素に結合され得る。組換え融合において、抗体をコードするcDNAが、インフレームで、毒素をコードするcDNAをすでに含むプラスミドに挿入される。もちろん、その逆も同様に行われ得る;毒素cDNAが、抗体をコードするcDNAを保有するプラスミドに挿入され得る。
【0008】
イムノトキシンの大きさが潜在的に大きいので、時折、毒素部分への抗体フラグメントのみの結合が所望される。Fab、Fab’およびF(ab)2フラグメントが、ポリクローナル抗体、モノクローナル抗体およびキメラ抗体から作製され得、次いで、化学的結合を介して毒素に結合される。
【0009】
あるいは、抗体の可変領域が必須のフレームワーク領域に結合されるcDNAが、産生され得る。次いで、これらのより小さな抗体が、二重鎖Fv抗体として分泌されるか、または重鎖および軽鎖領域が、直接的もしくはペプチドリンカーを介してかのいずれかで結合される場合、単鎖Fv抗体(scFv)として分泌される。
【0010】
scFvを作製する1つの方法は、免疫原を用いて免疫されたマウスの脾臓mRNAから作製された、ファージディスプレイライブラリーを介した方法である(Chowdhuryら、Mol.Immunol.34:9−20(1997))。しかし、タンパク質の免疫原が、哺乳動物において天然に見出されるが、原核生物において組換え的に発現される場合、タンパク質は、正確なグリコシル化パターンを有さず、そして正確な高次構造を有さないかもしれない。この免疫原に応答して、マウスによって発生した抗体は、そのネイティブ状態においてはタンパク質を認識しないかもしれない。この問題に対する1つの解法は、哺乳動物細胞において作製されたネイティブなタンパク質を用いて動物を免疫することであるが、いくつかのタンパク質(特に、細胞表面タンパク質)の十分な量の哺乳動物細胞からの精製は、可能ではないかもしれない。一般的ではないが別の解法は、免疫原をコードするcDNAを用いて動物を免疫することである。適切なプロモーターの制御下で、cDNAが、動物に導入される。注入をブーストした後、そして抗体の力価が最大に達した場合、動物を屠殺し、そして脾臓を取り出し、ファージディスプレイライブラリーを作製する。
【0011】
メソセリン発現悪性腫瘍に対する標的化治療の開発は、良好にインターナライズ(internalize)された高い親和性の抗体の開発が困難なために妨害されていた。本発明者らが、最近の刊行物(Chowdhuryら、Proc Natl Acad Sci USA,95:669−674(1998)(今後は、「Chowdhury 1998」))において記載するように、哺乳動物細胞からの十分な量のメソセリンの精製は、不可能であり、そしてEscherica coliにおいて発現した組換えメソセリンは、メソセリン陽性細胞に対する低い親和性を有する。本発明者らは、この文献において、メソセリンをコードするDNAを含むプラスミドを用いてマウスを免疫することによって、本発明者らは、抗メソセリン抗体の高い力価を誘発し得ることを報告した。本発明者らはさらに、脾臓RNAおよびファージディスプレイ技術を使用して、本発明者らが、メソセリンに高い親和性で結合する単鎖Fv(「scFv」)(本発明者らは、SS scFvと呼ぶ)を単離し得たことを報告した。
【0012】
SSは、免疫結合体(例えば、イムノトキシン)に有用な標的化因子のようであるが、さらに高い親和性の抗体が、メソセリンを発現する癌を検出する能力を改善することおよびイムノトキシンのメソセリンを発現する癌細胞を殺傷する能力を増大することが期待される。従って、標的抗原に対して高い親和性を有する抗体を作製する改善された手段についての必要性が残存する。特に、メソセリンに対してより高い親和性の抗体についての必要性が残存する。
【発明の概要】
【課題を解決するための手段】
【0013】
(発明の要旨)
本発明は、変異した抗体の重鎖可変領域もしくは変異した抗体の軽鎖可変領域またはそれらの両方を含むポリペプチドに関し、ここで、このポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有する。このポリペプチドは、相補性決定領域において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによってこの親抗体と異なる配列を有し、このアミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ、ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである。このアミノ酸置換は、重鎖または軽鎖の可変領域のCDR3において生じ得るか、あるいはこれらの可変領域のCDR1またはCDR2において生じ得る。本発明のいくつかの実施形態において、抗原はメソセリンであり、そして親抗体は、SSscFvであり、そしてこのポリペプチドは、S92、G93およびY94から選択される(すなわち、SSscFvについてのアミノ酸残基の標準的なナンバリングに従って、92位のセリン残基、93位のグリシン残基または94位のチロシンから選択される(図1もまた参照のこと))少なくとも1つのアミノ酸のアミノ酸が置換されていることによって抗体SSと異なる配列を有する。
【0014】
いくつかの好ましい実施形態において、軽鎖のSSscFv CDR3可変領域の置換は、G93K−Y94H(すなわち、93位のグリシンが、リジンに変化し、そして94位のチロシンが、ヒスチジンに変化し、これらの変化を、「SS1」と呼ばれる抗体として規定する)、S92G−G93F−Y94N(すなわち、92位のセリンが、グリシンに変化し、93位のグリシンが、フェニルアラニンに変化し、そして94位のチロシンが、アスパラギンに変化する;これらの変化を、「D8」と呼ばれるscFvと規定する)、ならびにS92G−G93S−Y94H(すなわち、92位のセリンが、グリシンに変化し、93位のグリシンが、セリンに変化し、そして94位のチロシンが、ヒスチジンに変化する;これらの変化を、「D8」と呼ばれるscFvと規定する)から選択される。
【0015】
上記のポリペプチドはまた、免疫結合体の一部を形成し得る。例えば、これらのポリペプチドは、治療的部分または検出可能な標識を含み得る。いくつかの実施形態において、この治療的部分は、毒性部分(例えば、Pseudomonas外毒素)またはその細胞傷害性フラグメント(例えば、PE38)であり得る。この毒性部分はまた、ジフテリア毒素またはその細胞傷害性フラグメント、サポリンまたはその細胞傷害性フラグメント、ヨウシュヤマゴボウ抗ウイルスタンパク質またはその細胞傷害性フラグメント、リシンまたはその細胞傷害性フラグメント、あるいはブリオジン1またはその細胞傷害性フラグメントであり得る。別の実施形態において、このポリペプチドは、バクテリオファージの表面タンパク質をさらに含み得る。
【0016】
別の局面において、本発明は、変異した抗体の重鎖可変領域もしくは変異した抗体の軽鎖可変領域またはそれらの両方を含むポリペプチドをコードするヌクレオチドに関し、ここで、このポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有する。このポリペプチドはまた、相補性決定領域において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによってこの親抗体と異なる配列を有し、このアミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ、ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである。いくつかの実施形態において、抗原はメソセリンであり、親抗体は、抗メソセリン抗体SSであり、そしてポリペプチドは、S92、G93およびY94から選択される少なくとも1つのアミノ酸のアミノ酸が置換されていることによって抗体SSと異なる配列を有する。さらなる実施形態において、この置換は、G93K−Y94H(SS1)、S92G−G93F−Y94N(D8)、およびS92G−G93S−Y94H(C10)から選択される。
【0017】
なお別の局面において、本発明は、上記の核酸分子のうちの1つに作動可能に連結したプロモーターをコードする発現カセットに関する。
【0018】
別の局面において、本発明は、抗原を保有する悪性細胞を殺傷する方法に関し、この方法は、標的化部分および毒性部分を含むイムノトキシンと該細胞を接触させる工程を包含し、この標的化部分は、変異した抗体の重鎖可変領域もしくは変異した抗体の軽鎖可変領域またはそれらの両方を含み、このポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有する。このポリペプチドはさらに、相補性決定領域において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによってこの親抗体と異なる配列を有し、このアミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ、ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである。いくつかの実施形態において、抗原はメソセリンである。さらに、いくつかの実施形態において、この標的化部分は、SS1、D8およびC10からなる群より選択される。
【0019】
本発明はさらに、E4と名付けられた抗体を提供し、E4において、CDR3 VLは、L96Tの置換によって、親抗体SSと異なる。E4はまた、免疫結合体(例えば、使用されるイムノトキシン)の標的化部分として使用され、例えば、インビトロでメソセリンを保有する癌細胞を取り除き得るか、またはインビボで癌細胞を阻害もしくは殺傷し得る。
【0020】
なおさらなる局面において、本発明は、親抗体よりも高い標的抗原親和性を有するポリペプチドを同定する方法に関する。この方法は、変異した抗体の重鎖可変領域もしくは変異した抗体の軽鎖可変領域またはそれらの両方を含むポリペプチド(このポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有し、そしてこのポリペプチドは、相補性決定領域において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによってこの親抗体と異なる配列を有し、このアミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ(ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである))を、標的抗原と、抗体とこの標的抗原との間の特異的な結合に適切な条件下で接触させる工程、この親抗体またはそのフラグメントの親和性よりも高い親和性でこの標的抗原に結合しない抗体またはそのフラグメントの全てを取り除く条件下で、このポリペプチドを溶出する工程、ならびにこのポリペプチドが、この抗原に結合したか否かを決定する工程であって、これによって、結合することによって、このポリペプチドがこの親抗体よりも高い標的親和性を有することが確認される工程を包含する。
【0021】
本発明はさらに、変異した抗体の可変ドメインをコードする核酸のライブラリーを作製する方法に関し、この方法は、親抗体のVHドメインまたはVLドメインのアミノ酸配列をコードする核酸分子を提供する工程であって、この核酸分子が、ホットスポットモチーフの内部に少なくとも1つのヌクレオチドを含む、少なくとも1つの親のホットスポットコドンを含む工程、ならびにこの親のアミノ酸配列とは異なる変異したアミノ酸配列をコードする、複数の変異した核酸分子を生成する工程であって、ここで、各々の変異した核酸配列は、アミノ酸をコードする親のホットスポットコドンとは異なる少なくとも1つの変異したコドンを含み、この変異したコドンは、この親のホットスポットコドンによってコードされるアミノ酸とは異なるアミノ酸をコードする工程を包含する。
【0022】
いくつかの実施形態において、本発明の方法において使用される複数の変異した核酸分子は、少なくとも19のメンバーを含み、ここで、この19のメンバーの各々は、親のホットスポットコドンによってコードされるアミノ酸が、異なる天然のアミノ酸に置換されているアミノ酸配列をコードする。なおさらなる実施形態において、本発明の方法は、アミノ酸をコードする少なくとも2つの親のホットスポットコドンとは異なる変異したコドンを含む複数の変異した核酸分子を含み得、この変異したコドンの各々は、この親のホットスポットコドンによってコードされるアミノ酸とは異なるアミノ酸をコードする。他の実施形態において、本発明の方法は、少なくとも399のメンバーを含む複数の変異した核酸分子を含み得、これらのメンバーの各々は、親のホットスポットコドンによってコードされるアミノ酸が、異なる天然のアミノ酸に置換されているアミノ酸配列をコードする。本発明の方法のさらなる実施形態において、この親抗体は、コドンによってコードされる少なくとも1つの保存アミノ酸を有するあるクラスの抗体であり、ここで、この保存アミノ酸をコードするコドン(単数または複数)は変異していない。本発明の方法のなおさらなる実施形態において、このホットスポットモチーフは、AGCA、AGTT、AGCT、AGTA、GGCA、GGTT、GGCT、GGTA、AGCおよびAGTからなる群より選択される。本発明の方法のなおさらなる実施形態において、この変異した核酸分子は、CDRを含むVHドメインまたはCDRを含むVLドメインの一部分に、少なくとも1つの変異したコドンを含む。このCDRは、VHドメインのCDR3またはVLドメインのCDR3であり得る。
例えば、本発明は以下を提供する。
(項目1) 変異した抗体の重鎖可変領域または軽鎖可変領域を含むポリペプチドであって、該ポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有し、該ポリペプチドは、相補性決定領域(CDR)において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによって該親抗体と異なる配列を有し、該アミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ、ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである、ポリペプチド。
(項目2) 上記置換が、軽鎖可変領域のCDR3において生じる、請求項1に記載のポリペプチド。
(項目3) 上記置換が、重鎖可変領域のCDR3において生じる、請求項1に記載のポリペプチド。
(項目4) 上記置換が、軽鎖可変領域のCDR1またはCDR2において生じる、項目1に記載のポリペプチド。
(項目5) 上記置換が、重鎖可変領域のCDR1またはCDR2において生じる、項目1に記載のポリペプチド。
(項目6) 上記抗原が、メソセリンであり、上記親抗体が、抗メソセリン抗体SSであり、そして上記ポリペプチドが、S92、G93およびY94から選択される少なくとも1つのアミノ酸のアミノ酸が置換されていることによって抗体SSと異なる配列を有する、項目2に記載のポリペプチド。
(項目7) 上記置換が、G93K−Y94H(SS1);S92G−G93F−Y94N(D8)およびS92G−G93S−Y94H(C10)から選択される、項目6に記載のポリペプチド。
(項目8) 上記ポリペプチドが、scFvである、項目1に記載のポリペプチド。
(項目9) 上記ポリペプチドが、scFvである、項目6に記載のポリペプチド。
(項目10) 上記ポリペプチドが、dsFv、FabまたはF(ab’)2である、項目1に記載のポリペプチド。
(項目11) 上記ポリペプチドが、dsFv、FabまたはF(ab’)2である、項目6に記載のポリペプチド。
(項目12) 治療的部分または検出可能な標識をさらに含む、項目1に記載のポリペプチド。
(項目13) 上記治療的部分が、毒性部分である、項目12に記載のポリペプチド。
(項目14) 上記毒性部分が、Pseudomonas外毒素またはその細胞傷害性フラグメントである、項目13に記載のポリペプチド。
(項目15) 上記毒性部分が、細胞傷害性フラグメントであり、該細胞傷害性フラグメントが、PE38である、項目14に記載のポリペプチド。 (項目16) 上記毒性部分が、ジフテリア毒素またはその細胞傷害性フラグメント、サポリンまたはその細胞傷害性フラグメント、ヨウシュヤマゴボウ抗ウイルス毒素またはその細胞傷害性フラグメント、リシンまたはその細胞傷害性フラグメント、およびブリオジン1またはその細胞傷害性フラグメントからなる群より選択される、項目13に記載のポリペプチド。
(項目17) 治療的部分または検出可能な標識をさらに含む、項目6に記載のポリペプチド。
(項目18) 上記治療的部分が、毒性部分である、項目17に記載のポリペプチド。
(項目19) 上記毒性部分が、Pseudomonas外毒素またはその細胞傷害性フラグメントである、項目18に記載のポリペプチド。
(項目20) 上記毒性部分が、PE35、PE38およびPE40からなる群より選択される細胞傷害性フラグメントである、項目19に記載のポリペプチド。
(項目21) バクテリオファージの表面タンパク質をさらに含む、請求項1に記載のポリペプチド。
(項目22) 抗メソセリン抗体SSのメソセリン結合親和性の少なくとも3倍のメソセリン結合親和性を有するポリペプチドであって、該ポリペプチドは、少なくともL96T(E4)の軽鎖可変領域のCDR3において、アミノ酸が置換されている点で抗体SSと異なる配列を有する、ポリペプチド。
(項目23) 治療的部分または検出可能な標識をさらに含む、項目22に記載のポリペプチド。
(項目24) 上記治療的部分が、毒性部分である、項目23に記載のポリペプチド。
(項目25) 上記毒性部分が、Pseudomonas外毒素またはその細胞傷害性フラグメントである、項目24に記載のポリペプチド。
(項目26) 上記毒性部分が、PE35、PE38およびPE40からなる群より選択される細胞傷害性フラグメントである、項目25に記載のポリペプチド。
(項目27) 変異した抗体の重鎖可変領域または軽鎖可変領域を含むポリペプチドをコードする核酸分子であって、該ポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有し、該ポリペプチドは、相補性決定領域(CDR)において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによって親抗体と異なる配列を有し、該アミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ、ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである、核酸分子。
(項目28) 上記抗原が、メソセリンであり、上記親抗体が、抗メソセリン抗体SSであり、そして上記ポリペプチドが、S92、G93およびY94から選択される少なくとも1つのアミノ酸のアミノ酸が置換されていることによって抗体SSと異なる配列を有する、項目27に記載の核酸分子。
(項目29) 上記置換が、G93K−Y94H(SS1);S92G−G93F−Y94N(D8)およびS92G−G93S−Y94H(C10)から選択される、項目27に記載の核酸分子。
(項目30) 抗メソセリン抗体SSのメソセリン結合親和性の少なくとも3倍のメソセリン結合親和性を有するポリペプチドをコードする核酸分子であって、該ポリペプチドは、少なくともL96T(E4)の軽鎖可変領域のCDR3において、アミノ酸が置換されている点で抗体SSと異なる配列を有する、核酸分子。
(項目31) 項目27に記載の核酸分子に作動可能に連結されたプロモーターを含む、発現カセット。
(項目32) 項目28に記載の核酸分子に作動可能に連結されたプロモーターを含む、発現カセット。
(項目33) 抗原を保有する悪性細胞を殺傷する方法であって、該方法は、毒性部分および標的化部分を含むイムノトキシンと該細胞を接触させる工程を包含し、該標的化部分は、変異した抗体の重鎖可変領域または軽鎖可変領域を含むポリペプチドを含み、該ポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有し、該ポリペプチドは、相補性決定領域(CDR)において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによって該親抗体と異なる配列を有し、該アミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ、ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである、方法。
(項目34) 上記抗原が、メソセリンである、項目33に記載の方法。
(項目35) 上記標的化部分が、SS1、D8およびC10からなる群より選択される、項目34に記載の方法。
(項目36) 上記毒性部分が、Pseudomonas外毒素またはその細胞傷害性フラグメントである、項目35に記載の方法。
(項目37) 上記毒性部分が、PE35、PE38およびPE40からなる群より選択される細胞傷害性フラグメントである、項目36に記載の方法。
(項目38) 抗原を保有する悪性細胞を殺傷する方法であって、該方法が、毒性部分および標的化部分を含むイムノトキシンと該細胞を接触させる工程を包含し、ここで、該標的化部分が、抗体E4である、方法。
(項目39) 上記毒性部分が、Pseudomonas外毒素またはその細胞傷害性フラグメントである、項目38に記載の方法。
(項目40) 上記毒性部分が、PE35、PE38およびPE40からなる群より選択される細胞傷害性フラグメントである、項目38に記載の方法。
(項目41) 親抗体よりも高い標的抗原親和性を有するポリペプチドを同定する方法であって、該方法が、以下の工程:
(a)抗体と該標的抗原との間の特異的な結合に適切な条件下で、該標的抗原と項目1に記載のポリペプチドを接触させる工程;
(b)該親抗体またはそのフラグメントの親和性よりも高い親和性で該標的抗原に結合しない抗体またはそのフラグメントの全てを取り除く条件下で、該ポリペプチドを溶出する工程;ならびに
(c)該ポリペプチドが該抗原に結合したか否かを決定する工程であって、これによって、結合することによって、該ポリペプチドが該親抗体よりも高い標的親和性を有することが確認される、工程、
を包含する、方法。
(項目42) 変異した抗体の可変ドメインをコードする核酸のライブラリーを作製する方法であって、該方法は、以下の工程:
(a)親抗体のVHドメインまたはVLドメインのアミノ酸配列をコードする核酸分子を提供する工程であって、該核酸分子は、ホットスポットモチーフの内部に少なくとも1つのヌクレオチドを含む、少なくとも1つの親のホットスポットコドンを含む、工程;ならびに
(b)該親のアミノ酸配列とは異なる変異したアミノ酸配列をコードする、複数の変異した核酸分子を生成する工程であって、ここで、各々の変異した核酸配列は、アミノ酸をコードする親のホットスポットコドンとは異なる少なくとも1つの変異したコドンを含み、該変異したコドンは、該親のホットスポットコドンによってコードされるアミノ酸とは異なるアミノ酸をコードする、工程、
を包含する、方法。
(項目43) 項目42に記載の方法であって、ここで、上記複数の変異した核酸分子が、少なくとも19のメンバーを含み、該19のメンバーの各々は、上記親のホットスポットコドンによってコードされるアミノ酸が、異なる天然のアミノ酸に置換されているアミノ酸配列をコードする、方法。
(項目44) 項目42に記載の方法であって、ここで、上記複数の変異した核酸分子が、アミノ酸をコードする少なくとも2つの親のホットスポットコドンとは異なる変異したコドンを含み、該変異したコドンの各々は、該親のホットスポットコドンによってコードされるアミノ酸とは異なるアミノ酸をコードする、方法。
(項目45) 項目42に記載の方法であって、ここで、上記複数の変異した核酸分子が、少なくとも399のメンバーを含み、該メンバーの各々が、上記親のホットスポットコドンによってコードされるアミノ酸が、異なる天然のアミノ酸に置換されているアミノ酸配列をコードする、方法。
(項目46) 項目42に記載の方法であって、さらにここで、上記親抗体が、コドンによってコードされる少なくとも1つの保存アミノ酸を有するあるクラスの抗体であり、ここで、該保存アミノ酸をコードするコドン(単数または複数)が変異していない、方法。
(項目47) 上記ホットスポットモチーフが、AGCA、AGTT、AGCT、AGTA、GGCA、GGTT、GGCT、GGTA、AGCおよびAGTからなる群より選択される、項目42に記載の方法。
(項目48) 上記変異した核酸分子が、CDRを含む上記VHドメインまたはCDRを含む上記VLドメインの一部分に、少なくとも1つの変異したコドンを含む、項目42に記載の方法。
(項目49) 上記CDRが、上記VHドメインのCDR3である、請求項48に記載の方法。
(項目50) 上記CDRが、上記VLドメインのCDR3である、請求項48に記載の方法。
【図面の簡単な説明】
【0023】
【図1】図1は、そのヌクレオチド配列から推定されるようなSS scFvのアミノ酸配列を含む。scFvにおいて、VHは、リンカーペプチド(GVGGSG4SG4S)によりVLに結合される。フレームワーク領域、CDRおよびリンカーに印を付けている。Hは、重鎖を示し;Lは、軽鎖を示し;FRは、フレームワーク領域を示し;CDRは、相補性決定領域を示す。
【図2】4つのライブラリーの各々をパニングした後選択されるファージクローンのELISAにより評価されるようなメソセリン結合。図2A:この図は、別々の列において、SS scFvの可変軽鎖のCDR3の、アミノ酸の番号付け、ヌクレオチド配列、およびアミノ酸配列を示す。一番上の列:親scFv SSの可変軽鎖CDR3のアミノ酸残基の番号。中央の列:scFv SS CDR3のCDR3におけるアミノ酸をコードするヌクレオチド配列。長方形は、テトラヌクレオチドホットスポットモチーフを示し:長円形は、この配列中のセリンホットスポットモチーフを示す。一番下の列:1文字コードにおける、scFv SSのCDR3のアミノ酸配列。 図2Bから2Eは、個々のライブラリー由来のクローンのELISAの結果を示すパネルである。パネルにおける各文字記号は、単一クローンからのファージ粒子のELISAシグナルを示す。各パネルにおいて塗りつぶされた四角(矢印により示される)は、内部標準の親SS(scFv)ファージを示す。ライブラリーを、各パネルの1番上に示される。図2B:残基89、93、および94で変異されたクローンのライブラリーからのELISAの結果。図2C:残基89、90、および91で変異されたクローンのライブラリーからのELISAの結果。図2D:残基92、93、および94で変異されたクローンのライブラリーからのELISAの結果。図2E:残基95、96、および97で変異されたクローンのライブラリーからのELISAの結果。
【図3】異なるライブラリーのパニングの後に得られたいくつかのELISA陽性のファージクローンのアミノ酸配列。クローン間で異なる領域の配列のみを示す。アステリスク(*)は、異なるヌクレオチド配列であるが、同じアミノ酸配列を有するいくつかのクローンが見出されたことを示す。
【図4】固定化されたメソセリンへの野生型および変異されたscFvイムノトキシンの結合について得られたBIAcoreのセンサーグラム(BIAcore sensorgram)の比較。左から右へ、イムノトキシンの標的化部分は:親SS、ならびに変異された改変体SS1、D8、C10およびE4である。Koff、KonおよびKDについての値を、図の下に表にする。「RU」は、「共鳴単位(resonance unit)」を表す。X軸は、秒で時間を表す。
【発明を実施するための形態】
【0024】
(I.概要)
本発明は、SS(scFv)と比較して、メソセリンに対する増加した親和性を有するこの抗原に対する抗体を提供する。この抗体は、より有効な免疫結合体(例えば、イムノトキシン)を作製するための標的化部分として使用され得る。これらの免疫結合体は、インビトロでの使用(例えば、生物学的サンプルに存在するメソセリン発現細胞の検出)のために使用され得る。イムノトキシンは、メソセリンを発現する細胞(例えば、細胞の表面上でメソセリンを発現する癌の癌細胞)を殺傷するためにさらに使用され得る。
【0025】
本発明は、親抗体の標的抗原に対する親和性よりも高い親和性を有する抗体を生成する新しい方法をさらに提供する。
【0026】
特に好ましい実施形態において、抗体は、scFvまたはdsFvである。scFvの構築物から産生される組換えイムノトキシンの多くは、IgG毒素化学的結合体の3分の1の大きさであり、そして組成物において均質である。scFv由来のIgG分子の定常部分の除去は、動物(霊長類を含む)に注射後のイムノトキシンのより速いクリアランスを生じ、そしてより小さいサイズのその結合体が、固形腫瘍における薬物の浸透を改善する。総合すると、これらの特性は、イムノトキシン(IT)が、非標的組織および非常に低いレベルの抗原を発現する組織と相互作用する時間を低減することにより、毒性部分と関連する副作用を減少させる。
【0027】
しかし、これらの利点は、IgGがscFvに変換される場合に起こる抗原結合親和性の減少により、いくらかの程度まで相殺される(Reiterら、Nature Biotechnol.14:239−1245(1996))。親和性を増加することにより、scFvの選択的腫瘍送達を改善することが示されており(Adamsら、Cancer Res.58:485−490(1998))、そして腫瘍の画像化および処置における有用性を増加するようである。従って、免疫結合体のscFvおよび他の標的部分(例えば、dsFv、Fab、およびF(ab’)2)の親和性の増加は、エフェクター分子(例えば、毒素および他の治療剤)を意図された標的に送達するこれらの薬剤の効率を改善することが所望される。
【0028】
親和性における改善は、部位指向性変異誘発またはランダム変異誘発により、インビトロで達成され得る。抗体−抗原結晶構造は、結合を改善するためにはどの残基が変異されるべきかを示し得るが、原子分解構造データは、ほとんどの抗体に対して利用できない。従って、ファージディスプレイ技術に基づくいくつかの戦略が開発され、より高い親和性の抗体を、FabおよびscFvの変異された改変体を発現するライブラリーから選択した(Gramら,Proc.Natl.Acad.Sci.USA 89:3576−3580(1992);Lowら,J.Mol.Biol.260:359−368(1996);de Kruifら,J.Mol.Biol.248:97−105(1995);Griffithら,EMBOJ.12:725−734(1993);Barbasら,J.Mol.Biol.230:812−823(1993);Schierら,J.Mol.Biol.255:28−43(1996);Yeltonら,J.Immunol.,155:1994−2004(1995);Yangら,J.Mol.Biol.254:392−403(1995);Piniら,J.Biol.Chem.273:21769−21776(1998);Wuら,Proc.Natl.Acad.Sci.USA 95:6037−6042(1998))。これらのアプローチは、しばしば得るのが難しく、そして産生するのが高価であり得る、大きくかつ複数のライブラリーの構築を必要とする。
【0029】
本発明者らは、増加した親和性を有するFvを改変体の小さなライブラリーから生成することを可能にする新規な方法を開発した。この方法は、抗体の可変領域をコードするDNAが、突然変異ホットスポット(このホットスポットは、変異が、インビトロの親和性成熟プロセスの間にしばしば集中するヌクレオチド配列である)を含む事実に基づく(NeubergerおよびMilstein,Curr.Biol.7:248−254(1995);Jollyら,Semin.Immunol.8:159−168(1996);Gonzalez−Fernandezら,Proc.Natl.Acad.Sci.USA 91:12614−12618(1994))。可変領域のホットスポットのいくつかの異なる型(例えば、直列反復および逆方向反復、パリンドローム、2次構造ならびに特定のコンセンサス配列)が提示されている(NeubergerおよびMilstein,Curr.Biol.7:248−254(1995);Jollyら,Semin.Immunol.8:159−168(1996);Gonzalez−Fernandezら,Proc.Natl.Acad.Sci.USA 91:12614−12618(1994);Goldingら,Genetics,115:169−176(1987);Kolchanovら,FEB Lett.214:87−91(1987);Rogozinら,Biochim.Biophys.Acta 1089:175−182(1991);InselおよびVarade,Int.Immunol.,6:1437−1443(1994);Wagnerら,Nature,376:732(1995))。これら全ての中で、コンセンサスホットスポット配列は、より詳細に研究されている。これらのコンセンサス配列の1つは、テトラヌクレオチドA/G−G−C/T−A/T(Pu−G−Py−A/T)である。この他の型は、セリンコドンAGYにより示され、Yは、CまたはTであり得(Wagnerら,Nature,376:732(1995);GoyenecheaおよびMilstein,Proc.Natl.Acad.Sci.USA 93:13979−13984(1996))。
【0030】
最近、本発明者らは、SS(scFv)と呼ばれる単鎖Fvを開発し、これは、卵巣癌、中皮腫およびいくつかの他のヒト癌の表面上に発現されるメソセリン抗原のその認識に基づき、ファージディスプレイライブラリーから選択された(Chowdhury 1998;ChangおよびPastan,Proc.Natl.Acad.Sci.USA 93:136−140(1996))。そのヌクレオチド配列から推定されるような、SS(scFv)のアミノ酸配列は、図1として示され、そしてヌクレオチド配列は、GenBankに寄託されている。SS(scFv)は、組換え手段により、または所望の場合タンパク質を合成するための標準的な技術により合成され得る。
【0031】
SS(scFv)−PE38は、SS(scFv)を、PE38と呼ばれる、Pseudomonas外毒素Aの38kDaの短縮された変異体と融合することにより作製されるイムノトキシンである。メソセリン陽性ヒト腫瘍を保有するマウスに注射される場合、SS(scFv)−PE38は、腫瘍の退化を引き起こし、そしてヒト癌を処置するために開発されている。メソセリンに対するSS(scFv)−PE38の親和性は、11nMであることが見出されている(Chowdhuryl998)。これは比較的高い親和性であるが、さらに高い親和性が、治療において、診断において、およびインビトロでの使用においてSS抗体に基づく免疫結合体(SS(scFv)を含む)の動作を増強するために望ましいと考えられる。
【0032】
抗体親和性を改善する以前の試みは、とりわけ、CDRの残基のランダムな変異または結晶構造(crystallagraphic)分析から見出される特定のアミノ酸を変異させて抗原接触に影響を与えることに頼っていた。免疫およびDNA免疫を介した親和性の改善はまた、Chowdhury 1998に報告される結果を考慮していないようであった。上記これらの方法における不利のうちのいくつかを克服するために、本発明者らは、新しいアプローチを作製した。本発明者らは、CDRにおけるホットスポットを標的化し、ここで本発明者らは、ランダムな変異を作製した。任意のCDRが、このアプローチにおいて変更され得るが、本発明者らは、軽鎖可変部領域のCDR3を使用して、この方法を試験した。一部分、CDR3を選択した。なぜなら、CDR3は、抗原結合部位において中心に位置し、そして抗原認識および親和性においてより大きい影響を有するからである。しかし、CDR2およびCDR1はまた、抗原結合親和性に影響することが知られ(述べられる順序で)、そしてCDR3の代わりにかまたはCDR3に加えて使用され得る。
【0033】
このアプローチを使用して、本発明者らは、約8000の独立したクローンのみの小さいライブラリーから、15〜55倍の親和性を増加したscFVを生成し、かつ単離し得た。これらの高い親和性の改変体を用いて作製されたイムノトキシンは、細胞傷害性活性において10倍よりも大きい増加を有した。本発明者らは、以下に詳細に報告される研究において軽鎖CDR3を使用したが、本発明者らは、同じアプローチを使用してSSの重鎖CDR3からscFvを生成した。SSの軽鎖CDR3を変異することにより生成されるscFVのように、ELISA結合研究において、多数のこれらの変異された重鎖scFVが、親SS scFVが示すより高いメソセリンに対する親和性を示す。さらに、本発明者はまた、SS以外の抗体(この場合、上皮増殖因子の変異体形態に結合する抗体)に対してこの技術を使用した。これらの結果を考慮して、本発明者らは、この手段は、親抗体の親和性と比較して、それらの標的抗原に対する増加した親和性を有する有するFv、Fabまたは他の組換え抗体を迅速に得るのに一般的に有用であると予想する。この研究の過程において、本発明者らはまた、ホットスポットモチーフの外側の変異により作製された、SSのメソセリンに対する親和性の約3.7倍の親和性を有するscFv(E4と命名される)を生成した。その親和性は、ホットスポット変異方法により作成されるscFvの親和性より際だって低いが、その親和性は、以前に既知の最も良い抗メソセリンscFvの親和性よりさらに数倍大きい。
【0034】
以下の節で、本明細書中で使用される用語が、さらなる明確さのために規定される。本願方法が、より詳細に記載される。最終的に、この実施例は、本願方法および特異的に改善される抗メソセリン抗体のCDR3の配列を実施するための特定の実験手順を示す。
【0035】
(II.定義)
単位、接頭辞(prefix)、および記号が、それらの国際単位系(SI)で認証された形態において示される。数量範囲は、範囲を規定した数を含む。その他で示されない場合、核酸は、5’から3’の方向で、左から右に書かれる;アミノ酸配列は、アミノからカルボキシの方向で左から右に書かれる。本明細書中で提供される表題は、本発明の種々の局面または実施形態の限定ではなく、これは、全体として明細書に対する参考とされ得る。従って、直ぐ下に定義される用語は、その全体が明細書に対する参考としてより十分に定義される。
【0036】
用語「メソセリン(mesothelin)」は、哺乳動物(例えば、ラット、マウス、霊長類、または特にヒト)の細胞の表面上に存在し得るメソセリンタンパク質およびそのフラグメントをいう。メソセリンの好ましい核酸およびアミノ酸の配列は、以下に記載される:PCT公開出願WO97/25,068,米国出願08/776,271および米国仮出願60/010,166。さらに、以下を参照のこと:Chang,K.およびPastan,I.,Int.J.Cancer 57:90(1994);Chang,K.およびPastan,I.,Proc.Nat’l Acad.Sci.USA 93:136(1996);Brinkmann U.ら,Int.J.Cancer 71:638(1997);ならびにChowdhury,P.S.ら,Mol.Immunol.34:9(1997)。メソセリンはまた、細胞内のままであるメソセリンタンパク質またはメソセリンペプチド、ならびに分泌されたかまたは単離された細胞外タンパク質をいう。
【0037】
本明細書中で使用される場合、「抗体」は、特定の抗原と免疫学的に反応性の免疫グロブリンに対する言及を含み、そしてポリクローナル抗体およびモノクローナル抗体の両方を含む。この用語はまた、遺伝的に操作された形態(例えば、キメラ抗体)例えば、ヒト化されたマウス抗体)、異種結合抗体(例えば、双特異性抗体)および組換え単鎖Fvフラグメント(scFv))、ジスルフィド安定化(dsFv)Fvフラグメント(本明細書中に参考として援用される、U.S.S.N.08/077,252を参照のこと)、またはpFvフラグメント(米国仮特許出願60/042,350および60/048,848)を含む。用語「抗体」はまた、抗体の抗原結合形態を含む(例えば、Fab’、F(ab’)2、Fab、FvおよびrIgG)。Pierce Catalog and Handbook,1994−1995(Pierce Chemical Co.,Rockford,IL);Kuby,J.,Immunology,第3版.,W.H.Freeman & Co.,New York(1997)もまた参照のこと。
【0038】
特定の抗原に免疫学的に反応性の免疫抗体が、組換え方法(例えば、ファージまたは類似のベクターでの組み換え抗体のライブラリーの選択)(例えば、Huseら,Science 246:1275−1281(1989);Ward,ら,Nature 341:544−546(1989);およびVaughanら,Nature Biotech.14:309−314(1996)を参照のこと)により、または抗原もしくは抗原をコードするDNAを用いて動物を免疫することにより生成され得る。
【0039】
代表的に、免疫グロブリンは、重鎖および軽鎖を有する。各重鎖および軽鎖は、定常部領域および可変領域(この領域はまた、「ドメイン」として公知である)を含む。軽鎖および重鎖の可変領域は、3つの超可変領域により分断される「フレームワーク」領域を含み、「相補性決定領域」または「CDR」とも呼ばれる。フレークワーク領域およびCDRの範囲は、規定されている(Kabat,E.ら,SEQUENCES OF PROTEINS OF IMMUNOLOGICAL INTEREST,米国厚生省(1987)(これらは本明細書中に参考として援用される)を参照のこと)。異なる軽鎖または重鎖のフレームワーク領域の配列は、比較的種内で保存されている。抗体のフレームワーク領域(これは、構成する軽鎖および重鎖の組み合わされたフレームワーク領域である)は、3次元空間においてCDRの位置を定めて整列するように働く。
【0040】
CDRは、抗原のエピトープへの結合の主な原因である。各鎖のCDRは、代表的にCDR1、CDR2、およびCDR3をいい、N末端から連続して始まって番号を付けられ、そしてまた代表的には、特定のCDRが位置する鎖により同定される。従って、VHCDR3は、それが見出される抗体の重鎖の可変ドメインに位置し、一方VLCDR1は、それが見出される抗体の軽鎖の可変ドメイン由来のCDR1である。
【0041】
「VH」または「VH」への言及は、Fv、scFv、dsFvまたはFabを含む免疫グロブリン重鎖の可変領域をいう。「VL」または「VL」への言及は、Fv、scFv、dsFvまたはFabを含む免疫グロブリンの軽鎖の可変領域をいう。
【0042】
句「一本鎖Fv」または「scFv」は、伝統的な二鎖抗体の重鎖の可変領域および軽鎖の可変ドメインが1つの鎖を形成するように連結される、抗体をいう。代表的に、リンカーペプチドは、2つの鎖の間に挿入され、適切なホールディングおよび活性な結合部位の作製を可能にする。
【0043】
用語「リンカーペプチド」は、軽鎖の可変ドメインへの重鎖の可変ドメインの間接的な結合を与える抗体結合フラグメント(例えば、Fvフラグメント)内のペプチドに対する言及を含む。
【0044】
用語「親抗体(parental antibody)」は、親抗体として同じエピトープに結合するがより高い親和性を有する抗体またはそのフラグメントを得るために、変異または改変される目的の任意の抗体を意味する。
【0045】
用語「ホットスポット」は、CDRのヌクレオチド配列の一部または特に高度な天然の改変体の部分である可変ドメインのフレームワーク領域の一部を意味する。CDRは、それ自身が超可変性の領域であると考えられるが、変異がCDRを通して均一に分布しないことが確認された。特定の部位、すなわちホットスポットは、集中した変異が起こる位置として同定された。ホットスポットは、構造の特徴の数および配列によって特徴付けられる。これらの「ホットスポットモチーフ」は、ホットスポットを同定するために使用され得る。特に十分特徴付けられる2つのコンセンサス配列モチーフは、テトラヌクレオチド配列RGYWおよびセリン配列AGYである(ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである)。
【0046】
「標的化部分」は、目的の細胞へ免疫結合体を標的化することが意図される免疫結合体の部分である。代表的に、標的化部分は、抗体、scFv、dsFv、FabまたはF(ab’)2である。
【0047】
「毒性部分」は、イムノトキシン(immunotoxin)を目的の細胞に対して細胞傷害性にするイムノトキシンの部分である。
【0048】
「治療的部分」は、治療剤として作用することが意図される免疫結合体の部分である。
【0049】
用語「治療剤」は、任意の数の、抗腫瘍性、抗炎症、サイトカイン、抗感染症、酵素のアクチベーターまたはインヒビター、アロステリック変更因子、抗体または患者において所望の治療効果を誘導するために投与される他の因子として作用することが現在公知または今後開発される化合物が含まれる。治療剤はまた、毒素または放射性同位体であり得、ここで意図される治療効果は、例えば、癌細胞を殺傷することである。
【0050】
「検出可能な標識」は、免疫結合体に関して、自身の存在を検出可能にする特性を有する免疫結合体の部分を意味する。例えば、免疫結合体は、細胞(この中に免疫結合体が存在する)を免疫組織化学アッセイにおいて検出可能にする放射性同位体を用いて標識され得る。
【0051】
用語「エフェクター部分」は、標的化部分によって標的化される細胞に対して効果を有するか、または免疫結合体の存在を同定することが意図される、免疫結合体の部分を意味する。従って、エフェクター部分は、例えば、治療的部分、毒素、放射性標識または蛍光標識であり得る。
【0052】
用語「免疫結合体」は、抗体に対するエフェクター分子の共有結合に対する言及を含む。エフェクター分子は、イムノトキシンであり得る。
【0053】
用語「有効量(effective amount)」または「有効量(amount effective to)」または「治療有効量」は、所望の結果(例えば、細胞のタンパク質合成を少なくとも50%阻害すること、または細胞を殺傷すること)を生じるに十分な治療剤の投薬量に対する言及を含む。
【0054】
用語「毒素」は、アブリン、リシン、Pseudomonas外毒素(PE)、ジフテリア毒素(DT)、ボツリヌス毒素、またはこれらの改変された毒素に対する言及を含む。例えば、PEおよびDTは、代表的に肝臓毒性を介して死をもたらす非常に毒性の化合物である。しかし、PEおよびDTは、毒素のネイティブな標的化成分(例えば、PEのIaドメインまたはDTのB鎖)を除去し、そしてそれを異なる標的化部分(例えば、抗体)で置換することによって、イムノトキシンとして使用するための形態に改変され得る。
【0055】
用語「接触する(接触)」は、直接的な物理的会合での配置に対する言及を含む。
【0056】
「発現プラスミド」は、分子または目的物(これは、プロモーターに作動可能に連結されている)をコードするヌクレオチド配列を含む。
【0057】
本明細書中に使用される場合、抗体に関して用語「抗メソセリン」とは、メソセリンに対して生成された抗体に対する言及を含む。好ましい実施形態において、メソセリンは、ヒトメソセリンのような霊長類のメソセリンである。特に好ましい実施形態において、抗体は、ヒトメソセリンをコードするcDNAの非霊長類哺乳動物への導入後にその動物によって合成されたヒトメソセリンに対して、生成される。
【0058】
本明細書中に使用される場合、「ポリペプチド」、「ペプチド」および「タンパク質」は、交換可能に使用され、そしてアミノ酸残基のポリマーに対する言及を含む。この用語は、1つ以上のアミノ酸残基が、対応する天然に存在するアミノ酸および天然に存在するアミノ酸ポリマーの人工的な化学アナログであるアミノ酸ポリマーに適用される。この用語はまた、タンパク質が機能的なままであるような保存的アミノ酸置換を含むポリマーに対して適用される。
【0059】
用語「残基」または「アミノ酸残基」または「アミノ酸」は、タンパク質、ポリペプチドまたはペプチド(総称して「ペプチド」)に組込まれるアミノ酸に対する言及を含む。アミノ酸は、天然に存在するアミノ酸であり得、他に制限されない限り、天然に存在するアミノ酸と類似した様式で機能し得る天然のアミノ酸の既知のアナログを含み得る。
【0060】
本明細書中に参照されるアミノ酸およびアナログは、以下の表1のような速記の指定によって記載される:
(表1:アミノ酸の名称)
【0061】
【表1】

「保存的置換」は、タンパク質を記載する場合、タンパク質の活性を実質的に変化させないタンパク質のアミノ酸組成における変化をいう。従って、特定のアミノ酸配列の「保存的に改変されたバリエーション」は、タンパク質活性に重要ではないこれらのアミノ酸のアミノ酸置換をいうか、または重要なアミノ酸の置換であっても活性を実質的に変化させないような、アミノ酸の類似の特性(例えば、酸性、塩基性、正に荷電または負に荷電、極性または非極性など)を有する他のアミノ酸とのアミノ酸置換という。機能的に類似したアミノ酸を提供する保存的置換の表は、当該分野で周知である。以下の表2の6つの群の各々は、互いに保存的置換であるアミノ酸を含む:
(表2)
1)アラニン(A)、セリン(S)、スレオニン(T);
2)アスパラギン酸(D)、グルタミン酸(E);
3)アスパラギン(N)、グルタミン(Q);
4)アルギニン(R)、リジン(K);
5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);
および
6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W)。
Creighton、PROTEINS、W.H.Freeman and Company、New York(1984)もまた参照のこと。
【0062】
ペプチドの文脈において、用語「実質的に類似した」は、10〜20アミノ酸の比較ウィンドウ上で、参照配列に対して少なくとも90%、好ましくは少なくとも95%の配列同一性を有する配列を含むペプチドを示す。配列同一性の割合は、比較ウィンドウ上で2つの最適に整列された配列を比較することによって決定され、ここで、比較ウィンドウ中のポリヌクレオチド配列の部分は、2つの配列の最適な整列に関して、参照配列(これは、付加または欠失を含まない)に対して比較した場合、付加または欠失(すなわち、ギャップ)を含み得る。この割合は、同一の核酸塩基または同一のアミノ酸残基が両方の配列で生じる位置の数を決定して、一致した位置の数を出し、一致した位置の数を比較ウィンドウ中の位置の総数で割り、そしてその結果に100を掛けることによって計算され、これによって配列同一性の割合か出される。
【0063】
句「ジスルフィド結合」または「システイン−システインジスルフィド結合」は、システインのイオウ原子が酸化されてジスルフィド結合を形成する2つのシステイン間の共有相互作用をいう。ジスルフィド結合の平均結合エネルギーは、水素結合に対する1〜2kcal/モルと比較して、約60kcal/モルである。本発明の文脈において、ジスルフィド結合を形成するシステインは、単鎖抗体のフレームワーク領域内であり、そして抗体のコントロールフォメーションを安定化するようにはたらく。
【0064】
用語「結合体化(する)(conjugating)」、「結合(する)(joining)」、「結合(する)(bonding)」または「連結(する)(linking)」は、2つのポリペプチドを1つの連続したポリペプチド分子にすることをいう。本発明の文脈において、この用語は、抗体部分をエフェクター分子(EM)に結合することに対する言及を含む。連結は、化学手段または組換え手段のいずれかにより得る。化学手段は、抗体部分とエフェクター部分との間の反応をいい、その結果、1つの分子を形成する2つの分子間で形成されたの共有結合が存在する。
【0065】
本明細書中に使用される場合、「組換え」は、そのネイティブな状態ではタンパク質を発現し得ない内因性のDNAコピーを有さない細胞を用いて産生されるタンパク質に対する言及を含む。この細胞は、組換えタンパク質を産生する。なぜなら、この細胞は、適切な単離された核酸配列の導入によって遺伝的に改変されているからである。この用語はまた、異種核酸の導入によってかまたはその細胞に対してネイティブではない形態にネイティブな核酸を変化することによって改変されたか、あるいは細胞がそのように改変されたか細胞から誘導される、細胞または核酸またはベクターに対する言及を含む。従って、例えば、組換え細胞は、ネイティブな(非組換え)形態の細胞内に見出されない遺伝子を発現するか、ネイティブな形態内に見出される変異体の遺伝子を発現するか、または他の異常に発現されるか、発現不足であるネイティブな遺伝子を発現するか、または全く発現されない。
【0066】
本明細書中に使用される場合「核酸」または「核酸配列」は、一本鎖形態または二本鎖形態のいずれかであるデオキシリボヌクレオチドまたはリボヌクレオチドのポリマーに対する言及を含み、そして他に指定されない限り限定されないが、天然に存在するヌクレオチドに類似した様式で核酸にハイブリダイズする天然のヌクレオチドの既知のアナログを含む。他に示されない限り、特定の核酸配列は、その相補鎖配列ならびに保存的改変体(すなわち、コドンおよび改変体のゆらぎ位置(wobble position)に存在する核酸(これは、タンパク質に翻訳された場合、アミノ酸の保存的置換を生じる))を含む。
【0067】
本明細書中に使用される場合、特定の核酸に関して「コード(する)」は、特定されたタンパク質への翻訳情報を含む核酸に対する言及を含む。この情報は、コドンの使用によって特定される。代表的に、アミノ酸配列は、「ユニバーサル」遺伝暗号を用いて核酸によってコードされる。しかし、ユニバーサルコードの改変体(例えば、いくつかの植物、動物および真菌ミトコンドリア、細菌Mycoplasma capricolum(Proc.Nat’l Acad.Sci.USA 82:2306−2309(1985))または繊毛虫Macronucleus中に存在する改変体)は、核酸がこれらの生物の翻訳機構を用いて発現される場合に使用され得る。
【0068】
句「インフレームで融合する」は、ポリペプチドをコードする2つ以上の核酸配列を結合することをいい、その結果、結合された核酸配列は、独自のポリペプチド鎖を含む一本鎖タンパク質に翻訳される。
【0069】
本明細書中に使用される場合、「発現された」は、核酸のタンパク質への翻訳に対する言及を含む。タンパク質は、発現されて細胞内性のままであり得るか、細胞表面膜の成分となり得るか、または細胞外マトリックスもしくは培地に分泌され得る。
【0070】
「宿主細胞」は、発現ベクターの複製または発現を支持しうる細胞を意味する。宿主細胞は、E.coliのような原核生物細胞であり得るか、または酵母細胞、昆虫細胞、両生類細胞、または哺乳動物細胞のような真核生物細胞であり得る。
【0071】
句「ファージディスプレイライブラリー」は、バクテリオファージの集団をいい、これらの各々は、表面タンパク質にインフレームで組換え融合された外来cDNAを含む。このファージは、cDNAによってコードされる外来タンパク質をその表面上に提示する。細菌宿主(代表的に、E.coli)が複製された後、目的の外来cDNAを含むファージが、外来タンパク質のこのファージ表面上での発現によって選択される。
【0072】
2つ以上の核酸配列またはポリペプチド配列に関する文脈において、用語「同一である」またはパーセント「同一性」は、同じであるか、あるいは以下の配列比較アルゴリズムのうちの1つを用いてかまたは視覚的な検査によって測定される場合に(最大の一致で比較され、そして整列された際に)同じであるアミノ酸残基またはヌクレオチドを特定の割合を有する、2つ以上の配列またはサブ配列をいう。
【0073】
句「実質的に同一である」は、2つの核酸またはポリペプチドの文脈において、以下の配列比較アルゴリズムのうちの1つを用いてかまたは視覚的な検査によって測定される場合に(最大の一致で比較され、そして整列された際に)少なくとも60%、好ましくは80%、より好ましくは90〜95%のヌクレオチドまたはアミノ酸残基の同一性を有する、2つ以上の配列またはサブ配列をいう。好ましくは、この実質的な同一性は、少なくとも約50残基の長さである配列の領域にわたって、より好ましくは少なくとも約100残基の領域の領域にわたって存在し、そして最も好ましくはこの配列は、少なくとも約150残基にわたって実質的に同一である。最も好ましい実施形態において、この配列は、コード領域の全体の長さにわたって実質的に同一である。
【0074】
配列比較に関して、代表的には、1つの配列は、試験配列が比較される参照配列として作用する。配列比較アルゴリズムを用いる場合、試験配列および参照配列は、コンピューターに入力され、配列座標が指定され、そして、必要な場合、配列アルゴリズムプログラムのパラメーターが指定される。次いで、配列比較アルゴリズムによって、指定されたプログラムパラメーターに基づいて、参照配列と比較して試験配列に関してパーセント配列同一性が計算される。
【0075】
比較に関する配列の最適なアラインメントは、例えば、以下のように実施され得る:SmithおよびWaterman、Adv.Appl.Math.2:482(1981)の局所相同性アルゴリズムによって、NeedlemanおよびWunsch、J.Mol.Biol.48:443(1970)の相同性アラインメントアルゴリズムによって、PearsonおよびLipman、Proc.Nat’l.Acad.Sci.USA 85:2444(1988)の類似性に関する検索方法によって、コンピューターによるこれらのアルゴリズムの実行によって(GAP、BESTFIT、FASTAおよびTFASTA、Wisconsin Genetics Software Package、Genetics Computer Group、575 Science Dr.、Madison、WI)、または視覚的な検査によって(一般的に、Current Protocols in Molecular Biology、F.M.Ausubelら、編、Current Protocols、Greene Publishing Associates,Inc.とJohn Wiley&Sons,Inc.との間の連帯のベンチャー(1995、補遺)(Ausubel)を参照のこと)。
【0076】
パーセント配列同一性および配列類似性を決定するのに適したアルゴリズムの例は、BLASTおよびBLAST 2.0アルゴリズム(これは、それぞれ、Altschulら(1990)J.Mol.Biol.215:403−410およびAltschuelら(1977)Nucleic Acids Res.25:3389−3402に記載される)である。BLAST分析を実行するためのソフトウェアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)を介して市販されている。このアルゴリズムは、最初に、問い合わせ配列中の短いワード長Wを同定することによって高いスコアの配列対(HSP)を同定する工程を包含する(これは、データベース配列中で同じ長さのワードで整列された場合、いくつかの正の値の閾値スコアTを一致させるかまたはこれを満たすかのいずれかである)。Tは、近接ワードスコアの閾値としていわれる(Altschulら、前出)。これらの最初の近隣ワードヒットは、それらを含むより長いHSPを見出すための検索を開始するためのシードとして作用する。次いで、このワードヒットは、累積アラインメントスコアが増加され得る限り、各々の配列に沿って両方向に伸長される。累積スコアは、ヌクレオチド配列に関しては、パラメーターM(一致した残基対に対する報酬スコア(reward score);常に>0)およびパラメーターN(一致しない残基に対するペナルティスコア;常に<0)を用いて算出される。アミノ酸配列に関して、スコアマトリックスを使用して累積スコアを算出する。各方向でのワードヒット拡大を以下の場合に停止する:累積アラインメントスコアが、その最大に達成される値から量Xだけ落ちる場合;1つ以上のネガティブスコアの残基アラインメントの蓄積に起因して、累積スコアが0またはそれより下になる場合;あるいは、配列のいずれかの末端に到達した場合。BLASTアルゴリズムパラメーターのW、T、およびXは、アラインメントの感度および速度を決定する。(ヌクレオチド配列に関する)BLASTNプログラムは、デフォルトとして、11のワード長(W)、10の期待値(E)、M=5、N=−4、および両方の鎖の比較を使用する。アミノ酸配列に関して、BLASTPプログラムは、デフォルトとして、3のワード長(W)、10の期待値(E)、およびBLOSUM62スコアマトリックスを使用する(HenikoffおよびHenikoff、Proc.Natl.Acad.Sci.USA 89:10915(1989)を参照のこと)。
【0077】
パーセント配列同一性を算出するのに加えて、BLASTアルゴリズムはまた、2つの配列間の類似性の統計分析を実施する(例えば、KarlinおよびAltschul、Proc.Nat’l.Acad.Sci.USA 90:5873−5787(1993)を参照のこと)。BLASTアルゴリズムによって提供される類似性測定の1つは、最小の和の確率(sum probability)(P(N))(これは、2つのヌクレオチド配列またはアミノ酸配列の間の一致が偶然生じる確率の指標を提供する)である。例えば、参照核酸に対する試験核酸の比較における最小の和の確率が約0.1より小さい、好ましくは約0.01より小さい、そして最も好ましくは約0.001より小さい場合に、核酸は参照配列に類似すると考えられる。
【0078】
2つの核酸配列またはポリペプチドが実質的に同一であるというさらなる指標は、以下に記載されるように、第1の核酸によってコードされるポリペプチドが第2の核酸によってコードされるポリペプチドと免疫学的に交差反応性であるということである。従って、ポリペプチドは、代表的に、例えば、これらの2つのペプチドが保存的置換によってのみ異なる場合、第2のポリペプチドに実質的に同一である。2つの核酸配列が実質的に同一であるという別の指標は、以下に記載されるように、2つの分子が互いにストリンジェントな条件下でハイブリダイズするということである。
【0079】
用語「インビボ」は、細胞が得られる生物の体の内側に対する言及を含む。「エキソビボ」および「インビトロ」は、細胞が得られる生物の体の外側を意味する。
【0080】
句「悪性細胞」または「悪性腫瘍」は、浸潤性である腫瘍もしくは腫瘍細胞および/または転移を引き起こし得る腫瘍もしくは腫瘍細胞、すなわち癌性細胞をいう。
【0081】
本明細書中に使用される場合、「哺乳動物細胞」は、ヒト、ラット、マウス、モルモット、チンパンジーまたはマカクを含む哺乳動物に由来する細胞に対する言及を含む。この細胞は、インビボまたはインビトロで培養され得る。
【0082】
用語「選択的に反応性」は、抗原に関して、抗体(全体または一部)の(抗原を欠く細胞または組織に対してではなく)抗原を保有する細胞または組織との優先的な結合をいう。これは、当然、ある程度の非特異的相互作用が分子と非標的細胞もしくは組織との間に生じ得ることを認識する。それにもかかわらず、選択的反応性は、抗原の特異的認識を介して媒介される場合、識別され得る。選択的に反応性である抗体は抗原に結合するが、この抗体はまた低い親和性でそのように結合し得る。一方、特異的結合は、抗体と抗原保有細胞との間に、結合抗体と抗原を欠く細胞との間よりも、より強い結合を引き起こし得る。特異的結合は、代表的に、メソセリンを欠く細胞または組織に対する場合と比較して、2倍よりも大きく、好ましくは5倍よりも大きく、より好ましくは10倍よりも大きく、そして最も好ましくは100倍よりも大きいメソセリン保有細胞または組織に対する抗体結合の量における(単位時間当たりの)増加を引き起こす。そのような条件下でのタンパク質に対する特異的結合は、特定のタンパク質に対するその特異性について選択される抗体を必要とする。種々の免疫アッセイ形式が、特定のタンパク質と特異的に免疫反応性な抗体を選択するのに適している。例えば、固相ELISA免疫アッセイを慣用的に使用して、あるタンパク質と特異的に免疫反応性のモノクローナル抗体を選択する。特定の免疫反応性を決定するのに使用され得る、免疫アッセイ形式および条件に関する説明については、HarlowおよびLane、ANTIBODIES、A LABORATORY MANUAL、Cold Spring Harbor Publications、New York(1988)を参照のこと。
【0083】
用語「免疫学的に反応性である条件」は、特定のエピトープに対して産生された抗体をそのエピトープへ、実質的に全ての他のエピトープに対する結合よりも検出可能に大きい程度で結合させる条件、および/または実質的に全ての他のエピトープに対する結合を実質的に排除するように結合させる条件に対する言及を含む。免疫学的に反応性である条件は、抗体結合反応の形式に依存し、そして代表的には、免疫アッセイプロトコールに使用される条件またはインビボで遭遇する条件である。免疫アッセイ形式および条件の説明に関しては、HarlowおよびLane(前出)を参照のこと。好ましくは、本発明の方法に使用される免疫学的に反応性である条件は、代表的に生きている哺乳動物または哺乳動物細胞の内側の条件(例えば、温度、容量オスモル濃度、pH)に対する言及を含む「生理学的条件」である。いくらかの生物は、極限状態に供されるが、生物体内および細胞内環境は通常pH7辺りにあり(すなわち、pH6.0〜pH8.0、より代表的にはpH6.5〜7.5)、優勢な溶媒として水を含み、そして約0℃より高く50℃より低い温度で存在することが理解される。容量オスモル濃度は、細胞の生存性および増殖を支持する範囲内である。
【0084】
用語「SS」または「SS(scFv)」は、メソセリンに高い親和性で結合するscFvをいい、これは、図1に示されるアミノ酸配列を有する。Chowdhury 1998を参照のこと。
【0085】
(III.より高い親和性を有する抗体の生成)
上記のように、本発明は、親抗体から、親抗体の標的抗原に対する親和性よりも高い親和性を有する1つ以上の変異体抗体を生成するための新規な方法を提供する。以前の技術(これは、CDR内に無作為に変異を作製する)と対照的に、本発明者らは、本発明者らがホットスポットモチーフ内に少なくとも1つのヌクレオチドを含むコドン(参照を簡単にするために、これらはこの議論に関して「ホットスポットコドン」と名付ける)に対する変異を標的化することによる努力が著しく少なくても、より高い親和性抗体を得ることができることを見出した。変異体の複数の相互作用(interation)は、それらの変異が、19個の他の天然のアミノ酸の各々をコードするコドンによって(1度に)置換されるホットスポットコドンを生じるように、作製される。都合のよいことに、これは、このプロセスを無作為化することによってなされ得、その結果、変異は、従来のホットスポットコドンの代わりに無作為に表れる全部で20アミノ酸(従来のアミノ酸のものも含む)をコードするコドンを生じる(すなわち、アラニンは、ある変異形態ではグリシンで置換され、別の変異形態はアルギニンで置換され、第3の変異形態はシステインで置換される、など)。
【0086】
CDRは、代表的に1つより多くのホットスポットコドンを含む。1、2、3より多くのそのようなコドンは、CDR内で変異され得、より高い親和性を獲得した抗体を生成する。本発明者らの結果は、2つのホットスポットコドン変異させるよりも、3つのホットスポットコドンを1度に変異させることによってより良い結果が得られ得ることを示す。
【0087】
任意のCDR中のホットスポットコドンは、より高い親和性の抗体を作製するために変異され得る。所望する場合、1つのCDR中の1つ以上のホットスポットコドンは、他のCDRの内の1つ由来の1つ以上のホットスポットコドンと同じときに変異され得る。各抗体がたった6個のCDRしか含まないこと、CDRは制限された長さであること(例えば、図1に示されるように、SSscFvのCDRは、5〜17アミノ酸の長さの範囲である)、そしてこれらのアミノ酸をコードするコドンのいくつかのみが、ホットスポットモチーフ内に位置するヌクレオチドを含むということに、注意すべきである。例えば、図2Aは、SSscFvのVL CDR3中の9アミノ酸内の5アミノ酸のみが、ホットスポットコドンによってコードされることを示す。
【0088】
可変ドメインのフレームワーク領域は、抗体−抗原結合相互作用に穏やかに寄与する。必要に応じて、フレームワーク領域内のホットスポットコドンは、結合活性におけるさらなる改善をなすために変異され得る。
【0089】
Kabat(前出)は、抗体の重鎖および軽鎖をサブグループおよびファミリーに分類することに関する情報を提供する。例えば、Kabatの分類下において、SSscFvのVHは、サブグループIIAおよびファミリーVに属し、そしてVLは、サブグループVIおよびファミリーXIに属する。特定の位置でのアミノ酸残基が、特定のサブグループに保存されることは、当該分野で公知である。例えば、Johnsonら、WEIR’S HANDBOOK OF EXPERIMENTAL IMMUNOLOGY I.IMMUNOCHEMISTRY AND MOLECULAR IMMUNOLOGY、第5版、Herzenbergら、編、Blackwell Science Inc.、Cambridge、MA、第6章、6.1−6.21(1996)(Weir’s Handbookの全体は、本明細書中に参考として援用される)を参照のこと。本発明者らの結果によって、これらの保存された残基をコードするコドンを変異しないほうか好ましいこと(たとえ、そのコドンがホットスポットモチーフ内のヌクレオチドを含み、そしてさもなければ変異に対する候補物であったとしても)が示される。
【0090】
以下の実施例は、これらの原理の使用を実証し、変異されたホットスポットコドンを有する抗体を生成する。本発明者らは、ある部分、モデル抗体として抗メソセリン抗体SSscFvを選択した。なぜなら、SSscFvを標的部分として使用するイムノトキシンは、動物モデルにおいて、ヒト腫瘍に対して良好な影響を与えることを示されたからである。本発明者らは、CDR3を選択した。なぜなら、CDR3は、抗原相互作用の主要な部分であるからである。本発明者らは、重鎖に対して可変軽鎖を選択した。なぜなら、本発明者らは、重鎖よりも、軽鎖は正常な親和性成熟の間に抗原親和性に対してほとんど最適化されていないようであるとみたからである。SSscFvの軽鎖のCDR3は、9アミノ酸残基を含み、これは、慣例として89〜97と番号を付ける(図2Aを参照のこと)。2つのテトラヌクレオチドホットスポットモチーフ中の3つのコドン(残基89、93および94)が同時に変異されるライブラリーを作成し、そしてセリンホットスポットコドンおよびテトラヌクレオチドホットスポットモチーフ中のコドン(残基92、93および94)が同時に変異されるライブラリーを作成した。これらのライブラリー由来のクローンを抗原に対するこれらの親和性についてスクリーニングした場合、これらのライブラリー中のほとんどの変異は、親抗体(SSscFv)が示すよりも、抗原に対して高い親和性を示した。対照的に、残基Q90(これは、SSscFvが分類される軽鎖のサブグループにおいて保存されている)の中に変異を含むコントロールライブラリーは、親抗体の親和性とほとんど同じまたはそれより低い親和性を有する変異体を生じた。図2Cを参照のこと。同様に、変異がホットスポット内のヌクレオチドを含まないコドン内で作製されたライブラリーは、従来の親のSSscFvとほとんど同じまたは中程度に良好な抗原に対する親和性を示した。図2Eを参照のこと。
【0091】
親和性結合研究により、変異された抗体の親和性に厳格な差異が存在したことを確認した。保存残基をコードしないホットスポットコドンを変異することによって作製されたScFvによって、これらが由来する既に高い親和性の親抗体よりも、15倍から55倍もの高い範囲の標的抗原に対する親和性を有するいくつかの抗体の生成が生じた。対照的に、ホットスポットモチーフ内の少なくとも1つのヌクレオチドを有さないコドンによってコードされるアミノ酸から生じる最良のクローンは、たった3.7倍の親和性の増加しか有さなかった。さらに、これらの高い親和性の抗体の生成およびスクリーニングは、CDR変異を含む以前の方法に必要とされるサイズよりも、かなり低い桁の大きさのライブラリーを必要とした。
【0092】
最も高い親和性を有する変異された抗体(C10と命名する)は、残基92、93および94以外は親のSSscFvのアミノ酸配列を有する。位置92において、SSscFvの「S」(セリン)は、「G」(グリシン)に変化される。位置93において、SSscFv配列の「G」は、「S」に変化される。位置94において、「Y」(チロシン)は、「H」(ヒスチジン)に変化される。各変異クローンへの変化(他の2つの高い親和性のクローン(SS1およびD8)を含む)を、図3に示す。慣用として、これらのアミノ酸置換はまた、本来の配列中のアミノ酸がその一文字コードで列挙される短縮化形態で記載され得、続いて、その位置の番号および変化された配列で置換されたアミノ酸に対する標準的な一文字コードで記載される。従って、上記のクローンC10の変化はまた、S92G−G93S−Y94Hと記載され得る。同様に、クローンSS1のscFvは、G93K−Y94Hとして記載され得、そしてクローンD8のscFvは、S92G−G93F−Y94Nによって記載され得る。E4クローンのscFv(これは、ホットスポットモチーフ内にヌクレオチド変異を有さない)は、L96Tとして示され得る。
【0093】
(IV.抗メソセリン抗体)
本発明は、メソセリンに対して、先行技術の抗体よりも高い親和性で結合する抗体を提供する。特に、本発明は、本発明者らがSS1、C10、およびD8と命名した、それぞれ、メソセリンについて、これらの親抗体SS(scFv)の親和性より15倍、55倍および37倍高い親和性を有する抗体を提供する。SS(scFv)は、以前は、メソセリンと特異的に反応性である、最も親和性が高い公知の抗体であった。本発明はさらに、メソセリンおよび目的の他の抗原に対する他の高親和性抗体を生成するための方法を提供する。下記に開示される免疫結合体は、本発明の抗体を用いてメソセリンを標的化する。これらの抗体は、哺乳動物細胞の表面に提示されるメソセリンの決定基に対して免疫学的条件下で選択的に反応性であり、そして細胞外環境由来の抗体にアクセス可能である。本発明はさらに、E4と命名した抗体を提供し、この抗体は、本発明の方法によって生成されておらず、そして本発明の方法によって生成される抗体ほどメソセリンについての親和性が高くないが、メソセリンについてのSSの約3.7倍の親和性をなお有する。
【0094】
本発明において生成される抗メソセリン抗体は、EMカルボキシル末端、EMアミノ末端、EMの内部アミノ酸残基(例えば、システイン)、またはそれらの任意の組み合わせを通してエフェクター分子(EM)に連結され得る。同様に、EMは、抗体の重鎖、軽鎖、Fc(定常領域)またはフレームワーク領域に直接連結され得る。連結は、抗体のアミノ末端もしくはカルボキシル末端を通して、または内部アミノ酸残基を通して生じ得る。さらに、複数のEM分子(例えば、2〜10のいずれか1つ)は、抗メソセリン抗体に連結され得、そして/または複数の抗体(例えば、2〜5のいずれか1つ)は、EMに連結され得る。本発明の多価免疫結合体組成物において用いられる抗体は、同じかまたは異なるメソセリンエピトープに対して指向され得る。
【0095】
本発明の好ましい実施形態では、抗メソセリン抗体は、組換え抗体(例えば、scFvまたはジスルフィド安定化Fv抗体)である。Fv抗体は代表的に、約25kDaであり、そして重鎖および軽鎖あたり3つのCDRを有する、完全な抗原結合部位を含む。VH鎖およびVL鎖が連続せずに発現される場合、Fv抗体の鎖は代表的に、非共有結合相互作用によって一緒に保持される。しかし、これらの鎖は、希釈によって解離する傾向があり、それゆえ、これらの鎖をグルタルアルデヒド、分子間ジスルフィドまたはペプチドリンカーを通して架橋する方法が開発されている。
【0096】
特に好ましい実施形態では、抗体は、単鎖Fv(scFv)である。scFv抗体のVH領域およびVL領域は、2本鎖の抗体に見出されるのと同様の抗原結合部位を作製するように折り畳まれる単鎖を含む。一旦折り畳まれると、非共有結合相互作用は、単鎖抗体を安定化する。より好ましい実施形態では、scFvは、組換え産生される。なお別の好ましい実施形態では、VH領域は、図1に示す通りのアミノ酸配列を有する。当業者は、本発明の抗体の保存的改変体が作製され得ることを理解する。scFvフラグメントにおいて用いられるこのような保存的改変体は、正確な折り畳みおよびVH領域とVL領域との間の安定化に必要な重要なアミノ酸残基を保持する。
【0097】
本発明のいくつかの実施形態では、scFv抗体は、軽鎖を通してEMへと直接連結される。しかし、scFv抗体は、そのアミノ末端またはカルボキシル末端を通してEMへと連結され得る。
【0098】
いくつかの抗体の実施形態のVH領域およびVL領域は直接一緒に連結され得るとはいえ、当業者は、これらの領域が、1以上のアミノ酸からなるペプチドリンカーによって隔てられ得ることを認識する。ペプチドリンカーおよびそれらの使用は、当該分野で周知である。例えば、Hustonら、Proc.Nat’l Acad.Sci.USA 8:5879(1988);Birdら、Science 242:4236(1988);Glockshuberら、Biochemistry 29:1362(1990);米国特許第4,946,778号、米国特許第5,132,405号およびStemmerら,Biotechniques 14:256−265(1993)(全て、本明細書中に参考として援用される)を参照のこと。一般的に、ペプチドリンカーは、領域を連結すること、またはこれらの間のいくらかの最小の距離もしくは他の空間的関係を保つこと以外に特定の生物学的活性を有さない。しかし、ペプチドリンカーの構成要素であるアミノ酸は、分子のいくつかの特性(例えば、折り畳み、正味の電荷、または疎水性)に影響を与えるように選択され得る。単鎖Fv(scFv)抗体は必要に応じて、50以下のアミノ酸の、一般的に40以下のアミノ酸の、好ましくは30以下のアミノ酸の、そしてより好ましくは20以下のアミノ酸の長さのペプチドリンカーを含む。いくつかの実施形態では、ペプチドリンカーは、配列Gly−Gly−Gly−Serのコンカテマー(好ましくは、2、3、4、5または6個のこのような配列)である。しかし、リンカー内のいくつかのアミノ酸置換が行われ得ることが認識されるべきである。例えば、バリンで、グリシンを置換し得る。
【0099】
(A.抗体産生)
ポリクローナル抗体を産生する方法は当業者に公知である。手短に述べると、免疫原(好ましくは、単離されたメソセリンまたは細胞外メソセリンエピトープ)はアジュバントと混合され、そして動物がこの混合物で免疫される。免疫原に対する適切に高い力価の抗体が得られた場合、血液がこの動物から収集され、そして抗血清が調製される。所望の場合、さらに、このポリペプチドに対して反応性の抗体について濃縮するための抗血清の分画が行われる。例えば、Coligan、CURRENT PROTOCOLS IN IMMUNOLOGY、Wiley/Greene、NY(1991);ならびにHarlowおよびLane、前出(これらは本明細書中に参考として援用される)を参照のこと。
【0100】
モノクローナル抗体は、当業者に慣用的な種々の技術によって得られ得る。このようなモノクローナル抗体を調製するための技術の説明は、例えば、Stitesら(編)BASIC AND CLINICAL IMMUNOLOGY(第4版)、Lange Medical Publications、Los Altos、CA、およびその中で引用される参考文献;HarlowおよびLane、前出;Goding、MONOCLONAL ANTIBODIES:PRINCIPLES AND PRACTICE(第2版)、Academic Press、New York、NY(1986);KohlerおよびMilstein、Nature 256:495−497(1975);ならびに特にモノクローナル抗体を作製する1つの方法を議論するChowdhury,P.S.ら、Mol.Immunol.34:9(1997)に見出され得る。
【0101】
親のモノクローナル抗体が、標的抗原、または所望の免疫原(例えば、メソセリン)をコードする核酸配列で動物を免疫することによって作製されることが好ましい。異種タンパク質をコードする非複製性転写単位での免疫は、抗原特異的免疫応答を惹起する。外来タンパク質への翻訳後、このタンパク質は、プロセシングされ、そして他の細胞性タンパク質と同様に免疫系に対して提示される。このタンパク質は外来性であるので、このタンパク質およびこれから誘導されるペプチドエピトープに対して免疫応答がマウントされる(Donnellyら、J.Immunol.Methods 176:145−152(1994);およびBoyerら、J.Med.Primatol.25:242−250(1996))。この技術は、タンパク質ベースの免疫に対して2つの顕著な利点を有する。第1の利点は、これは、タンパク質精製を必要としないことである。タンパク質精製は、いくらよくても、時間がかかり、そして多くの膜タンパク質の場合、非常に困難である。第2の利点は、この免疫原が哺乳動物宿主内で合成されるので、これは、正確な翻訳後修飾を受け、そしてそのネイティブな構造に折り畳まれることである。
【0102】
メソセリンコードDNAで免疫するために、メソセリンコードcDNAは、コード配列の転写がプロモーター(例えば、CMVプロモーター)の制御下にあるように、プラスミド中に導入される。次いで、このプラスミドは、動物内に、皮下、皮内、腹腔内などのいずれかによって注射される。結果として、このメソセリンcDNAは動物内でmRNAへと転写され、メソセリンはこのmRNAから翻訳され、この翻訳されたタンパク質は正確な翻訳後修飾を受け、そしてメソセリンを合成した細胞の表面に発現される。この動物はメソセリンに対する抗体を惹起し、そして血清が抗体力価についてモニタリングされる。
【0103】
必要に応じて、コード領域および調節エレメントに加えて、このプラスミドは、アンピシリン耐性(Amp)遺伝子を保有する。Amp遺伝子は、抗体産生の増加に必要なTh1応答についての免疫刺激配列を有することが公知である(Satoら、Science 273:352−354(1996))。
【0104】
上記のように、好ましい実施形態では、このモノクローナル抗体はscFvである。scFv抗体を作製する方法は、記載されている。Huseら、前出;Wardら、Nature 341:544−546(1989);およびVaughanら、前出を参照のこと。手短に述べると、B細胞由来のmRNAが単離され、そしてcDNAが調製される。このcDNAは、免疫グロブリンの重鎖および軽鎖の可変領域に特異的なプライマーを用いて、周知の技術(例えば、PCR)によって増幅される。PCR産物は、例えば、アガロースゲル電気泳動によって精製され、そしてこの核酸配列は連結される。リンカーペプチドが所望される場合、ペプチドをコードする核酸配列は、重鎖の核酸配列と軽鎖の核酸配列との間に挿入される。この配列は、当該分野で公知の技術(例えば、平滑末端連結、PCR産物の末端での制限部位の挿入、または重複伸張によるスプライシングによって)によって連結され得る(Chowdhuryら、Mol.Immunol.34:9(1997))。増幅後、scFvをコードする核酸は、さらに当該分野で周知の技術によってベクター中に挿入される。好ましくは、このベクターは、原核生物中で複製され得、そして真核生物および原核生物の両方において発現され得る。
【0105】
好ましい実施形態では、scFvは、ファージディスプレイライブラリーによって選択される。scFvを合成するための上記の手順は、以下に続く。PCRによる増幅後、scFv核酸配列は、繊維状ファージのマイナーな表面タンパク質gIIIpをコードする、遺伝子III(gIII)(Marksら、J.Biol.Chem.267:16007−16010(1992);Marksら、Behring Inst.Mitt.91:6−12(1992);およびBrinkmannら、J.Immunol.Methods 182:41−50(1995))とインフレームで融合される。このファージは、得られる融合タンパク質をその表面に発現する。ファージ表面のタンパク質は機能的であるので、メソセリン結合抗体を保有するファージは、パニングまたは抗原アフィニティークロマトグラフィー(McCaffertyら、Nature 348:552−554(1990))によって非結合性かまたはより低い親和性のファージから分離され得る。
【0106】
好ましい実施形態では、メソセリンに特異的に結合するscFvは、パニングによって見出される。パニングは、固体表面をメソセリンでコーティングし、そしてこのファージを適切な時間、適切な条件下でこの表面上でインキュベートすることによって行われる。結合していないファージは、固体表面から洗浄除去され、そして結合したファージが溶出される。最大の親和性を有する抗体を見出すことは、選択プロセスの効率によって指図され、そしてスクリーニングされ得るクローン数およびそれが行われるストリンジェンシーに依存する。代表的に、より高いストリンジェンシーは、より選択的なパニングに対応する。しかし、条件があまりにもストリンジェントである場合、ファージは結合しない。1回のパニング後、メソセリンコーティングプレートに結合するファージは、E.coli中で増やされ、そして別の回のパニングに供される。このようにして、2000倍の富化が3回のパニングにおいて行われる。従って、各回の富化が低い場合でさえ、複数回のパニングは、稀なファージの単離をもたらし、そしてその中に含まれる遺伝物質は、最大の親和性抗体の配列をコードする。ファージディスプレイによって提供される遺伝子型と表現型との間の物理的連鎖は、大きなライブラリーのクローンを用いてさえ、cDNAライブラリーの全てのメンバーを抗原に対する結合について試験することを可能にする。
【0107】
(B.抗体の結合親和性)
本発明の抗体は、それらの標的抗原に、それらの親抗体の親和性よりも少なくとも5倍良好な親和性で結合する。1つの実施形態では、この抗体は、メソセリンの細胞外エピトープに結合する抗メソセリン抗体である。標的抗原についての結合親和性は代表的に、標準的な抗体−抗原アッセイ(例えば、競合アッセイ、飽和アッセイまたはイムノアッセイ(例えば、ELISAもしくはRIA))によって測定または決定される。
【0108】
このようなアッセイを用いて、抗体の解離定数を決定し得る。語句「解離定数」とは、抗原についての抗体の親和性をいう。抗体の解離定数(KD=1/K、ここで、Kは、親和定数である)が<1μM、好ましくは<100nM、そして最も好ましくは<0.1nMである場合、抗体と抗原との間に結合の特異性が存在する。抗体分子は代表的に、より低い範囲のKDを有する。KD=[Ab−Ag]/[Ab][Ag]、ここで、[Ab]は平衡状態での抗体の濃度であり、[Ag]は平衡状態での抗原の濃度であり、そして[Ab−Ag]は平衡状態での抗体−抗原複合体の濃度である。代表的に、抗原と抗体との間の結合相互作用は、可逆的非共有結合的会合(例えば、静電引力、ファンデルワールス力および水素結合)を含む。結合特異性を規定するこの方法は、メソセリンに単独でまたは組み合わせて結合する場合にメソセリンに特異的である、単一の重鎖および/もしくは軽鎖、CDR、融合タンパク質または重鎖および/もしくは軽鎖のフラグメントに適用される。
【0109】
(C.イムノアッセイ)
抗体は、充分に認識された多数の免疫学的結合アッセイ(例えば、米国特許第4,366,241号;同第4,376,110号;同第4,517,288号;および同第4,837,168号を参照のこと)のうちのいずれかを用いて検出および/または定量され得る。一般的イムノアッセイの概説については、METHODS IN CELL BIOLOGY、第37巻、Asai編、Academic Press、Inc.New York(1993);BASIC AND CLINICAL IMMUNOLOGY、第7版、StitesおよびTerr編(1991)をまた参照のこと。免疫学的結合アッセイ(またはイムノアッセイ)は代表的に、リガンド(例えば、メソセリン)を利用して抗体に特異的に結合させ、そしてしばしば、この抗体を固定化する。本発明のイムノアッセイにおいて用いられる抗体は、上記でさらに詳細に考察される。
イムノアッセイはまたしばしば、標識化剤を利用して、リガンドおよび抗体によって形成される結合複合体に特異的に結合させ、そしてこの結合複合体を標識する。標識化剤は、それ自体が、抗体/分析物複合体(すなわち、抗メソセリン抗体)を含む部分のうちの1つであり得る。あるいは、標識化剤は、抗体/メソセリンタンパク質複合体に特異的に結合する第3の部分(例えば、別の抗体)であり得る。
【0110】
1つの局面では、標識化剤が、標識を保有する第2の抗メソセリン抗体である競合アッセイが意図される。次いで、この2つの抗体は、固定化されたメソセリンに対する結合について競合する。あるいは、非競合形式において、メソセリン抗体は標識を欠くが、抗メソセリン抗体を誘導した種(例えば、マウス)の抗体に特異的であってそして抗メソセリン抗体に結合する第2の抗体が標識される。
【0111】
免疫グロブリン定常領域に特異的に結合し得る他のタンパク質(例えば、プロテインAまたはプロテインG)もまた、標識剤として用いられ得る。これらのタンパク質は、連鎖球菌細菌の細胞壁の通常の構成要素である。これらは、種々の種由来の免疫グロブリン定常領域との強力な非免疫原性反応性を示す(一般的に、Kronvalら、J.Immunol.111:1401−1406(1973);およびAkerstromら、J.Immunol.135:2589−2542(1985)を参照のこと)。
【0112】
アッセイ全体を通して、インキュベーションおよび/または洗浄の工程は、試薬の各組み合わせの後で必要とされ得る。インキュベーション工程は、約5秒間〜数時間まで、好ましくは約5分間〜約24時間まで変動し得る。しかし、インキュベーション時間は、アッセイの形式、抗体、溶液の容量、濃度などに依存する。通常、アッセイは、室温で実施されるが、アッセイは、一定の範囲の温度(例えば、10℃〜40℃)で実施され得る。
【0113】
本発明のイムノアッセイの詳細は、用いられる特定の形式に伴って変動し得るとはいえ、抗体を含むサンプル中の抗メソセリン抗体を検出する方法は、一般的に、サンプルを、免疫学的に反応性の条件下で、メソセリン/抗体複合体に対して特異的に反応する抗体と接触させる工程を包含する。
【0114】
(V.免疫結合体の産生)
免疫結合体としては、抗体に対する治療剤の共有結合が存在する分子が挙げられるがこれに限定されない。治療剤は、特定の標的分子または標的分子を保有する細胞に対して指向された特定の生物学的活性を有する薬剤である。当業者は、治療剤が、種々の薬物(例えば、ビンブラスチン、ダウノマイシンなど)、細胞毒(例えば、ネイティブかまたは改変されたPseudomonas外毒素またはDiphtheria毒素)、それ自体が薬理学的組成物を含むカプセル化剤(例えば、リポソーム)、放射性薬剤(例えば、125I、32P、14C、3Hおよび35S)ならびに他の標識、標的部分およびリガンドを含み得ることを認識する。
【0115】
特定の治療剤の選択は、特定の標的分子または細胞に依存し、そして生物学的効果は、誘起することが所望される。従って、例えば、治療剤は、特定の標的細胞の死をもたらすために用いられる細胞毒であり得る。逆に、非致死的な生物学的応答を誘発することが単に所望される場合、治療剤は、非致死的な薬理学的薬剤に対して、または非致死的な薬理学的薬剤を含むリポソームに対して結合体化され得る。
【0116】
本明細書中で提供される治療剤および抗体を用いて、当業者は、機能的に等価な核酸(例えば、配列が異なるが、同じEMまたは抗体の配列をコードする核酸)を含む種々のクローンを容易に構築し得る。従って、本発明は、抗体および結合体、ならびにそれらの融合タンパク質をコードする核酸を提供する。
【0117】
(A.組換え方法)
本発明の核酸配列は、任意の適切な方法によって調製され得る。このような方法としては、例えば、適切な配列のクローニングまたは以下のような方法による直接化学合成が挙げられる:例えば、Narangら、Meth.Enzymol.68:90−99(1979)のホスホトリエステル法;Brownら、Meth.Enzymol.68:109−151(1979)のホスホジエステル法;Beaucageら,Tetra.Lett.22:1859−1862(1981)のジエチルホスホルアミダイト法;BeaucageおよびCaruthers、Tetra.Letts.22(20):1859−1862(1981)によって記載される固相ホスホルアミダイトトリエステル法(例えば、Needham−VanDevanterら、Nucl.Acids Res.12:6159−6168(1984)に記載される通りの自動化合成機を用いる);ならびに米国特許第4,458,066号の固体支持法。化学合成は、一本鎖オリゴヌクレオチドを産生する。これらは、相補配列とのハイブリダイゼーションによって、または一本鎖をテンプレートとして用い、DNAポリメラーゼを用いた重合によって二本鎖DNAへと変換され得る。当業者は、DNAの化学合成は約100塩基の配列に制限されるとはいえ、より長い配列は、より短い配列の連結によって入手され得ることを認識する。
【0118】
好ましい実施形態では、本発明の核酸配列は、クローニング技術によって調製される。適切なクローニングおよび配列決定の技術、ならびに多くのクローニング練習を通して当業者に指示するに充分な指示書の例は、以下に見出される:Sambrookら、MOLECULAR CLONING:A LABORATORY MANUAL(第2版)、第1巻〜第3巻、Cold Spring Harbor Laboratory(1989))、BergerおよびKimmel(編)、GUIDE TO MOLECULAR CLONING TECHNIQUES、Academic Press、Inc.、San Diego CA(1987))またはAusubelら(編)、CURRENT PROTOCOLS IN MOLECULAR BIOLOGY、Greene PublishingおよびWiley−Interscience、NY(1987)。生物学的試薬および実験装置の製造業者による製品情報もまた、有用な情報を提供する。このような製造業者としては、以下が挙げられる:SIGMA chemical company(Saint Louis、MO)、R&D systems(Minneapolis、MN)、Pharmacia LKB Biotechnology(Piscataway、NJ)、CLONTECH Laboratories,Inc.(Palo Alto、CA)、Chem Genes Corp.、Aldrich Chemical Company(Milwaukee、WI)、Glen Research,Inc.、GIBCO BRL Life Technologies,Inc.(Gaithersberg、MD)、Fluka Chemica−Biochemika Analytika(Fluka Chemie AG、Buchs,Switzerland)、Invitrogen、San Diego、CA、およびApplied Biosystems(Foster City、CA)、ならびに当業者に公知の多くの他の商業的供給源。
【0119】
ネイティブなEMまたは抗メソセリン抗体をコードする核酸は、EM、抗体、または本発明の免疫結合体を形成するように改変され得る。部位特異的変異誘発による改変は、当該分野で周知である。EMまたは抗メソセリン抗体をコードする核酸は、インビトロ方法によって増幅され得る。増幅方法としては、ポリメラーゼ連鎖反応(PCR)、リガーゼ連鎖反応(LCR)、転写ベースの増幅系(TAS)、自己保持配列複製系(3SR)が挙げられる。広範な種々のクローニング方法、宿主細胞およびインビトロ増幅方法論は、当業者に周知である。
【0120】
好ましい実施形態では、免疫結合体は、抗メソセリンscFv抗体をコードするcDNAを、EMをコードするcDNAを含むベクター中に挿入することによって調製される。挿入は、機能的Fv領域および機能的EM領域を含む1つの連続したポリペプチドにおいて、scFvおよびEMがインフレームで読まれるように行われる。特に好ましい実施形態では、ジフテリア毒素フラグメントをコードするcDNAは、scFvのカルボキシル末端に毒素が配置されるように、scFvへと連結される。最も好ましい実施形態では、PEをコードするcDNAは、scFvのアミノ末端に毒素が配置されるように、scFvへと連結される。
【0121】
一旦、本発明のEM、抗メソセリン抗体、または免疫結合体をコードする核酸が単離され、そしてクローン化されると、組換えによって操作された細胞(例えば、細菌、植物、酵母、昆虫および哺乳動物の細胞)において所望のタンパク質を発現し得る。当業者は、E.coli、他の細菌宿主、酵母および種々のより高等な真核生物細胞(例えば、COS、CHO、HeLaおよびミエローマ細胞株)を含む、タンパク質の発現に利用可能な多数の発現系を良く知っていることが予想される。原核生物または真核生物におけるタンパク質の発現について公知の種々の方法を詳細に記載する試みは行われない。手短に述べると、本発明の単離されたタンパク質をコードする天然または合成の核酸の発現は代表的に、DNAまたはcDNAをプロモーター(これは、構成性または誘導性のいずれかである)に対して作動可能に連結し、続いて発現カセットに組み込むことによって達成される。このカセットは、原核生物または真核生物のいずれかへの複製および組込みに適切であり得る。代表的な発現カセットは、転写および翻訳のターミネーター、開始配列、およびタンパク質をコードするDNAの発現の調節に有用なプロモーターを含む。クローニングされた遺伝子の高レベル発現を得るために、少なくとも、転写を指向する強力なプロモーター、翻訳開始のためのリボソーム結合部位および転写/翻訳ターミネーターを含む発現カセットを構築することが所望される。E.coliについては、これは、プロモーター(例えば、T7、trp、lacまたはλプロモーター)、リボソーム結合部位および好ましくは転写終結シグナルを含む。真核生物細胞については、制御配列は、プロモーターを、そして好ましくは、免疫グロブリン遺伝子、SV40、サイトメガロウイルス由来のエンハンサーおよびポリアデニル化配列を含み得、そしてスプライスドナー配列およびスプライスアクセプター配列を含み得る。本発明のカセットは、E.coliについては塩化カルシウム形質転換またはエレクトロポレーション、そして哺乳動物細胞についてはリン酸カルシウム処理、エレクトロポレーションまたはリポフェクションのような周知の方法によって選択された宿主細胞へと移入され得る。このカセットによって形質転換された細胞は、カセット中に含まれる遺伝子(例えば、amp遺伝子、gpt遺伝子、neo遺伝子およびhyg遺伝子)によって付与される、抗生物質に対する耐性によって選択され得る。
【0122】
当業者は、その生物学的活性を減少させることなく、本発明のポリペプチド(すなわち、抗メソセリン抗体、PEまたはこれらの組み合わせによって形成された免疫結合体)をコードする核酸に対して改変が行われ得ることを認識する。いくつかの改変が行われて、標的化分子のクローニング、発現または融合タンパク質への組込みを容易にし得る。このような改変は当業者に周知であり、そして例えば、終止コドン、アミノ末端に付加されて開始部位を提供するメチオニン、いずれかの末端に配置されて便利に配置された制限部位を作製するさらなるアミノ酸、または精製工程を補助するさらなるアミノ酸(例えば、ポリHis)を含む。
【0123】
組換え法に加えて、本発明の免疫結合体、EMおよび抗体はまた、標準的なペプチド合成を用いて全体としてまたは部分的に構築され得る。約50アミノ酸長未満の本発明のポリペプチドの固相合成は、この配列のC末端アミノ酸を、不溶性支持体へと連結し、続いてこの配列内の残りのアミノ酸を順次付加することによって達成され得る。固相合成のための技術は、BaranyおよびMerrifield、THE PEPTIDES:ANALYSIS,SYNTHESIS,BIOLOGY.第2巻:SPECIAL METHODS IN PEPTIDE SYNTHESIS、第A部、3〜284頁;Merrifieldら、J.Am.Chem.Soc.85:2149−2156(1963)およびStewartら、SOLID PHASE PEPTIDE SYNTHESIS、第2版、Pierce Chem.Co.,Rockford,III.(1984)によって記載される。より大きな長さのタンパク質は、より短いフラグメントのアミノ末端およびカルボキシル末端の縮重によって合成され得る。カルボキシル末端の活性化によってペプチド結合を形成する方法(例えば、カップリング試薬N,N’−ジシクロヘキシルカルボジイミドの使用による)は当業者に公知である。
【0124】
(B.精製)
一旦発現されると、本発明の組換え免疫結合体、抗体、および/またはエフェクター分子は、当該分野の標準的な手順に従って精製され得る。このような手順としては、硫酸アンモニウム沈澱、アフィニティーカラム、カラムクロマトグラフィーなどが挙げられる(一般的に、R.Scopes、PROTEIN PURIFICATION,Springer−Verlag、N.Y.(1982)を参照のこと)。少なくとも約90%〜95%の均質性の実質的に純粋な組成物が好ましく、そして98%〜99%以上の均質性が、薬学的用途に最も好ましい。所望に応じて、部分的にまたは均質になるまで(治療に用いられる場合)一旦精製されると、このポリペプチドは、内毒素を実質的に含まないはずである。
【0125】
細菌(例えば、E.coli)からの単鎖抗体の発現および/または適切な活性形態(単鎖抗体を含む)への再折り畳みのための方法は、記載されており、そして周知であり、そして本発明の抗体に適用可能である。Buchnerら、Anal.Biochem.205:263−270(1992);Pluckthun,Biotechnology 9:545(1991);Huseら、Science 246:1275(1989)およびWardら,Nature 341:544(1989)(全て本明細書中に参考として援用される)を参照のこと。
【0126】
しばしば、E.coliまたは他の細菌由来の機能的異種タンパク質は、封入体から単離され、そして強力な変性剤を用いた可溶化およびその後の再折り畳みを必要とする。可溶化工程の間、当該分野で周知であるように、ジスルフィド結合を分離するために還元剤が存在しなければならない。還元剤を含む例示的な緩衝液は以下である:0.1M Tris pH8、6Mグアニジン、2mM EDTA、0.3M DTE(ジチオエリトリトール)。ジスルフィド結合の再酸化は、本明細書中に参考として援用される、Saxenaら、Biochemistry 9:5015−5021(1970)に記載され、そして特にBuchnerら、前出によって記載されるように、還元型および酸化型の低分子量チオール試薬の存在下で生じ得る。
【0127】
再生は代表的に、変性しそして還元されたタンパク質の、再折り畳み緩衝液中への希釈(例えば、100倍)によって達成される。代表的な緩衝液は、0.1M Tris、pH8.0、0.5M L−アルギニン、8mM酸化型グルタチオン(GSSG)および2mM EDTAである。
【0128】
二本鎖抗体精製プロトコルへの改変物として、重鎖領域および軽鎖領域は、別々に可溶化および還元され、次いで、再折り畳み溶液中で合わされる。これらの2つのタンパク質が、他方に対して一方のタンパク質が5倍モル過剰を超えないようなモル比で混合される場合、好ましい収率が得られる。過剰の酸化型グルタチオンまたは他の酸化性低分子量化合物を、レドックスシャッフリングが完了した後の再折り畳み溶液に添加することが望ましい。
【0129】
(VI.Pseudomonas外毒素および他の毒素)
毒素を、本発明の抗体と共に用いて、イムノトキシンを産生し得る。例示的な
毒素としては、リシン、アブリン、ジフテリア毒素、およびそれらのサブユニッ
ト、ならびにボツリヌス毒素A〜Fが挙げられる。これらの毒素は、商業的供給
源(例えば、Sigma Chemical Company、St.Loui
s、MO)から容易に入手可能である。ジフテリア毒素は、Corynebac
terium diphtheriaeから単離される。リシンは、Ricin
us communis(トウゴマの実(Castor bean))由来のレ
クチンRCA60である。この用語はまた、それらの毒性改変体も参照する。例
えば、米国特許第5,079,163号および同第4,689,401号を参照
のこと。Ricinus communis凝集素(RCA)は2つの形態で生
じ、それぞれ、それらの約65kDおよび約120kDの分子量に従って、RC
A60およびRCA120と称される(NicholsonおよびBlaustei
n、J.Biochim.Biophys.Acta.266:543(197
2))。A鎖は、タンパク質合成の不活化および細胞の殺傷を担う。B鎖は、細
胞表面のガラクトース残基にリシンを結合させ、そして細胞質ゾル内へのA鎖の
輸送を促進する(Olsnesら、Nature 249:627−631(1
974)および米国特許第3,060,165号)。
【0130】
アブリンは、Abrus precatorius由来の毒性レクチンを含む
。毒性の本質である、アブリンa、b、c、およびdは、約63kDから、約6
7kDの分子量を有し、そして2つのジスルフィド結合ポリペプチド鎖Aおよび
Bから構成される。A鎖は、タンパク質合成を阻害し;B鎖(アブリンb)は、
Dガラクトース残基に結合する(Funatsuら、Agr.Biol.Che
m.52:1095(1988);およびOlsnes、Methods En
zymol.50:330−335(1978)を参照のこと)。
【0131】
本発明の好ましい実施形態では、毒素はPseudomonas外毒素(PE
)である。用語「Pseudomonas外毒素」は、本明細書中で使用される
場合、全長のネイティブな(天然に存在する)PE、または改変されているPE
をいう。このような改変としては、ドメインIaの除去、ドメインIb、II、
およびIIIにおける種々のアミノ酸の欠失、単一のアミノ酸置換、およびカル
ボキシル末端での1以上の配列(例えば、KDELおよびREDL)の付加が挙
げられ得るが、これらに限定されない。Siegallら、J.Biol.Ch
em.264:14256−14261(1989)を参照のこと。好ましい実
施形態では、PEの細胞障害性フラグメントは、ネイティブなPEの細胞傷害性
の少なくとも50%、好ましくは75%、より好ましくは少なくとも90%、そ
して最も好ましくは95%を保持する。最も好ましい実施形態では、細胞傷害性
フラグメントは、ネイティブなPEよりも毒性が高い。
【0132】
ネイティブなPseudomonas外毒素A(PE)は、Pseudomo
nas aeruginosaによって分泌される、極めて活性なモノマー性の
タンパク質(分子量66kD)であり、これは、真核生物細胞中におけるタンパ
ク質合成を阻害する。ネイティブなPE配列は、同一人に譲渡された米国特許第
5,602,095号(本明細書中で参考として援用される)に提供される。作
用の方法は、伸長因子2(EF−2)のADP−リボシル化の不活化である。外
毒素は、細胞傷害性を引き起こすのに提携して作用する3つの構造ドメインを含
む。ドメインIa(アミノ酸1〜252)は、細胞結合を媒介する。ドメインI
I(アミノ酸253〜364)は、細胞質ゾル内へのトランスロケーションを担
い、そしてドメインIII(アミノ酸400〜613)は、伸長因子2のADP
リボシル化を媒介する。ドメインIb(アミノ酸365〜399)の機能は、未
だ決定されていないが、その大部分であるアミノ酸365〜380は、細胞傷害
性の損失を伴なわずに欠失され得る。Siegallら(1989)、前出を参
照のこと。
【0133】
本発明において使用されるPEとしては、ネイティブな配列、ネイティブな配
列の細胞傷害性フラグメント、ならびにネイティブなPEおよびその細胞傷害性
フラグメントの保存的に改変された改変体が挙げられる。PEの細胞傷害性フラ
グメントは、標的細胞中における引き続くタンパク質分解性または他のプロセシ
ングを伴なって、または伴なわずに、細胞傷害性であるフラグメントを含む(例
えば、タンパク質またはプレタンパク質として)。PEの細胞傷害性フラグメン
トとしては、PE40、PE38、およびPE35が挙げられる。
【0134】
好ましい実施形態では、PEは、米国特許第4,892,827号において教
示されるように、しばしばドメインIaを欠失させることによって改変され、非
特異的な細胞結合を減少または除去されるが、これはまた、例えば、ドメインI
aの特定の残基を変異させることによっても達成され得る。例えば、米国特許第
5,512,658号は、変異されたPE(ここでは、ドメインIaが存在する
が、ドメインIaの57位、246位、247位、および249位の塩基性残基
が、酸性残基(グルタミン酸(すなわち、「E」)で置換されている)が、非常
に減少した非特異的細胞傷害性を示すことを開示している。PEのこの変異形態
は、時折、PE4Eと呼ばれる。
【0135】
PE40は、当該分野で以前に記載されたように、PEの短縮化(trunc
ated)誘導体である。Paiら、Proc.Nat’l.Acad.Sci
.USA 88:3358−62(1991);およびKondoら、J.Bi
ol.Chem.263:9470−9475(1988)を参照のこと。PE
35は、PEの35kDのカルボキシル末端フラグメントであり、ここでは、ア
ミノ酸残基1〜279が欠失しており、そしてこの分子は、280位のmetで
開始し、ネイティブなPEのアミノ酸281〜364および381〜613が続
く。PE35およびPE40は、例えば、米国特許第5,602,095号およ
び同第4,892,827号に開示される。
【0136】
いくつかの好ましい実施形態では、細胞傷害性フラグメントPE38が使用さ
れる。PE38は、アミノ酸253〜364および381〜613から構成され
る短縮化PEプロタンパク質であり、これは、細胞内でのプロセシングに際して
、その細胞傷害性形態へと活性化される(例えば、米国特許第5,608,03
9号およびPastanら、Biochim.Biophys.Acta 13
33:C1−C6(1997)を参照のこと)。
【0137】
上述のように、いくつかまたはすべてのドメイン1bが欠失され得、そして残
りの部分が、リンカーによってか、またはペプチド結合によって直接的に連結さ
れ得る。ドメインIIのいくつかのアミノ酸部分は、欠失され得る。そして、C
末端は、残基609〜613のネイティブな配列(REDLK)を含み得るか、
または構築物を細胞質ゾル内にトランスロケーションする能力を維持することが
見出されているバリエーション(例えば、REDLまたはKDEL)、およびこ
れらの配列の反復を含み得る。例えば、米国特許第5,854,044号;同第
5,821,238号;および同第5,602,095号、ならびにWO99/
51643を参照のこと。好ましい実施形態では、PEは、PE4E、PE40
、またはPE38であるが、非標的細胞への顕著な毒性が生じないレベルまで、
非特異的細胞傷害性が除去または減少されている任意の形態のPEが、標的化細
胞においてトランスロケーションし得、そしてEF−2をリボシル化し得るまま
である限り、本発明のイムノトキシンに使用され得る。
【0138】
(A.PEの保存的に改変された改変体)
PEまたはその細胞傷害性フラグメントの保存的に改変された改変体は、目的
のPE(例えば、PE38)と、アミノ酸レベルで、少なくとも80%の配列類
似性、好ましくは、少なくとも85%の配列類似性、より好ましくは、少なくと
も90%の配列類似性、そして最も好ましくは少なくとも95%の配列類似性を
有する。
【0139】
用語「保存的に改変された改変体」は、アミノ酸配列および核酸配列の両方に
適用する。特定の核酸配列に関して、保存的に改変された改変体とは、同一また
は本質的に同一なアミノ酸配列をコードする核酸配列をいうか、または核酸がア
ミノ酸配列をコードしない場合には、本質的に同一な核酸配列をいう。遺伝暗号
の縮重性が理由で、多数の機能的に同一な核酸が、任意の所定のポリペプチドを
コードする。例えば、コドンGCA、GCC、GCGおよびGCUはすべて、ア
ミノ酸アラニンをコードする。従って、コドンによってアラニンが特定されるす
べての位置で、そのコドンを、コードされるポリペプチドを変更することなく、
記載された任意の対応するコドンに変更し得る。このような核酸のバリエーショ
ンは、「サイレントなバリエーション」であり、これは、保存的に改変されたバ
リエーションの一種である。ポリペプチドをコードする、本明細書中のすべての
核酸配列はまた、核酸のすべての可能なサイレントなバリエーションを記載する
。当業者は、核酸中の各コドン(AUG(これは、通常、メチオニンについての
唯一のコドンである)を除く)が、機能的に同一な分子を産生するように改変さ
れ得ることを認識する。従って、ポリペプチドをコードする核酸のサイレントな
バリエーションの各々が、各々記載された配列中において含意される。
【0140】
アミノ酸配列に関して、当業者は、コードされる配列において単一のアミノ酸
または低いパーセンテージのアミノ酸を変更、付加、または欠失する、核酸配列
、ペプチド配列、ポリペプチド配列、またはタンパク質配列への個々の置換、欠
失、または付加が、「保存的に改変された改変体」であることを認識し、ここで
変更は、化学的に類似のアミノ酸でのアミノ酸の置換を生じる。
【0141】
(B.PEの細胞傷害性についてのアッセイ)
本発明で使用されるPseudomonas外毒素を、当業者に周知のアッセ
イによって、所望されるレベルの細胞傷害性についてアッセイし得る。例示的な
毒性アッセイは、本明細書中において、例えば、実施例2に記載される。従って
、PEの細胞傷害性フラグメントおよびこのようなフラグメントの保存的に改変
された改変体は、細胞傷害性について容易にアッセイされ得る。多数の候補PE
分子を、当該分野において周知の方法によって、細胞傷害性について同時にアッ
セイし得る。例えば、候補分子のサブグループを、細胞傷害性についてアッセイ
し得る。候補分子の中で、ポジティブに反応するサブグループを継続的に細分し
得、そして所望の細胞傷害性フラグメントが同定されるまで、再度アッセイし得
る。このような方法は、PEの多数の細胞傷害性フラグメントまたは保存的改変
体の迅速なスクリーニングを可能にする。
【0142】
(C.他の治療的部分)
本発明の抗体はまた、かなり多数の異なる診断用または治療用化合物を、表面
上にメソセリンを発現する細胞に標的化するために使用され得る。従って、本発
明の抗体(例えば、抗メソセリンscFv)は、メソセリンを保有する細胞に直
接的に送達されるべき薬物に対して、直接的に結合され得るか、またはリンカー
を介して結合され得る。治療剤としては、核酸、タンパク質、ペプチド、アミノ
酸、または誘導体、糖タンパク質、ラジオアイソトープ、脂質、炭水化物、また
は組換えウイルスのような化合物が挙げられる。核酸の治療用および診断用部分
としては、アンチセンス核酸、一本鎖または二重鎖DNAと共有結合的に架橋す
る誘導体化オリゴヌクレオチド、および三重鎖形成オリゴヌクレオチドが挙げら
れる。
【0143】
あるいは、抗メソセリン抗体に連結される分子は、治療用組成物(例えば、薬
物、核酸(例えば、アンチセンス核酸)、または循環系への直接曝露から、好ま
しくは遮蔽される別の治療的部分)を含む、リポソームまたはミセルのようなカ
プセル系であり得る。抗体に結合したリポソームを調製する手段は、当業者に周
知である。例えば、米国特許第4,957,735号;およびConnorら、
Pharm.Ther.28:341−365(1985)を参照のこと。
【0144】
(D.検出可能標識)
本発明の抗体は、必要に応じて、検出可能な標識に共有結合的にか、または非
共有結合的に連結され得る。このような用途に適切な検出可能標識としては、分
光学的手段、光化学的手段、生化学的手段、免疫化学的手段、電気的手段、光学
的手段、または化学的手段によって検出可能な任意の組成物が挙げられる。本発
明において有用な標識としては、磁気ビーズ(例えば、DYNABEADS)、
蛍光色素(例えば、フルオレセインイソチオシアネート、Texasレッド、ロ
ーダミン、グリーン蛍光タンパク質など)、放射能標識(例えば、3H、125I、
35S、14C、または32P)、酵素(例えば、西洋ワサビペルオキシダーゼ、アル
カリホスファターゼ、およびELISAにおいて通常使用される他の酵素)、お
よび比色定量標識(例えば、金コロイドあるいは呈色(colored)ガラス
またはプラスチック(例えば、ポリスチレン、ポリプロピレン、ラテックスなど
)ビーズ)が挙げられる。
【0145】
このような標識を検出する手段は、当業者に周知である。従って、例えば、放
射能標識は、写真フィルムまたはシンチレーションカウンターを使用して検出さ
れ得、蛍光マーカーは、光検出器を使用して検出して、放射された照度を検出し
得る。酵素学的標識は、代表的に、酵素に基質を提供すること、そしてその基質
に対する酵素の作用によって生成される反応生成物を検出することによって検出
される。比色定量標識は、単純に、呈色した標識を可視化することによって検出
される。
【0146】
(E.抗体への結合体化)
本発明の非組換え的実施形態では、エフェクター分子(例えば、治療的部分、
診断用部分、または検出用部分)を、当業者に公知のかなり多数の手段を使用し
て、本発明の抗メソセリン抗体に連結する。共有結合手段および非共有結合手段
の両方が、本発明の抗メソセリン抗体で使用され得る。
【0147】
エフェクター分子を抗体に結合させるための手順は、EMの化学的構造に従っ
て変動する。ポリペプチドは、代表的に、種々の官能基(例えば、カルボン酸(
COOH)、遊離アミン(−NH2)、またはスルフヒドリル(−SH)基を含
む;これらは、エフェクター分子の結合を生じさせるための、抗体における適切
な官能基との反応に利用可能である。
【0148】
あるいは、抗体は、さらなる反応性官能基に曝露されるか、または結合される
ように誘導体化される。誘導体化は、多数の任意のリンカー分子(例えば、Pi
erce Chemical Company、Rockford Illin
oisから入手可能なリンカー)の結合を包含し得る。
【0149】
「リンカー」は、本明細書中で使用される場合、抗体をエフェクター分子に連
結するために使用される分子である。リンカーは、抗体とエフェクター分子の両
方に対して共有結合を形成し得る。適切なリンカーは、当業者に周知であり、そ
してこれには、直鎖もしくは分枝の炭素リンカー、複素環式炭素リンカー、また
はペプチドリンカーが挙げられるが、これらに限定されない。抗体およびエフェ
クター分子がポリペプチドである場合、リンカーは、それらの側鎖を通して(例
えば、システインへのジスルフィド結合を通して)、構成要素のアミノ酸に連結
され得る。しかし、好ましい実施形態では、リンカーは、末端アミノ酸のα炭素
アミノ基およびカルボキシル基に連結される。
【0150】
いくつかの状況では、免疫結合体がその標的部位に到達した時点で、抗体から
エフェクター分子を遊離させることが所望され得る。従って、これらの状況では
、免疫結合体は、標的部位の近傍で切断可能である結合を含む。抗体からエフェ
クター分子を放出するためのリンカーの切断は、免疫結合体が、標的細胞の内側
または標的部位の近傍のいずれかで供される酵素学的な活性または条件によって
刺激され得る。標的部位が腫瘍である場合、腫瘍部位に存在する条件下で(例え
ば、腫瘍に関連した酵素または酸性pHに曝露された場合に)切断可能なリンカ
ーが使用され得る。
【0151】
種々の放射線診断用化合物、放射線療法化合物、薬物、毒素、および他の薬剤
を抗体に結合させることに関して報告されている多数の方法に照らして、当業者
は、所定の薬剤を抗体または他のポリペプチドに結合させるために適切な方法を
決定し得る。
【0152】
(VII.薬学的組成物および投与)
本発明の抗体および/または免疫結合体組成物(すなわち、抗体E4と共に、
その親抗体の少なくとも5倍の抗原親和性を有する抗体に連結されたPE)は、
非経口投与(例えば、静脈内投与、あるいは体腔または器官の内腔内への投与)
に特に有用である。例えば、卵巣悪性疾患は、腫瘍を取り囲んでいる組織への静
脈内投与または局所送達によって処置され得る。中皮腫を処置するためには、抗
メソセリン抗体を含む本発明の薬学的組成物は、胸膜腔または腹膜腔内に直接的
に投与され得る。
【0153】
投与のための組成物は、通常、薬学的に受容可能なキャリア(好ましくは、水
性キャリア)中に溶解された抗体および/または免疫結合体の溶液を含む。種々
の水性キャリア(例えば、緩衝化生理食塩水など)が使用され得る。これらの溶
液は滅菌され、そして一般的には、所望されない物質を含まない。これらの組成
物は、従来の周知の滅菌技術によって滅菌され得る。組成物は、ほぼ生理的条件
に必要とされるような薬学的に受容可能な補助物質(例えば、pH調整剤および
緩衝化剤、毒性調整剤など(例えば、酢酸ナトリウム、塩化ナトリウム、塩化カ
リウム、塩化カルシウム、乳酸ナトリウムなど))を含み得る。これらの処方物
における融合タンパク質の濃度は広範に変動し得、そして選択された特定の投与
様式および患者の必要性に従って、主に液量、粘性、体重などに基づいて選択さ
れる。
【0154】
従って、静脈内投与のための本発明の代表的な薬学的イムノトキシン組成物は
、1日あたり1患者あたり約0.1〜10mgである。特に、隔離された部位(
そして、循環系またはリンパ系ではない)(例えば、体腔内または器官の内腔内
)に薬物が投与される場合、1日あたり1患者あたり0.1から約100mgま
での投薬量が使用され得る。投与可能な組成物を調製するための実際の方法は、
当業者に公知であるか、または明らかであり、そして、REMINGTON’S
PHARMACEUTICAL SCIENCE、第19版、Mack Pu
blishing Company、Easton、Pennsylvania
(1995)のような刊行物において、より詳細に記載されている。
【0155】
本発明の組成物は、治療的処置のために投与され得る。治療的適用において、
組成物は、疾患に罹患している患者に対して、その疾患およびその合併症を治癒
するか、または少なくとも部分的に抑制するに十分な量で投与される。これを達
成するために十分な量を、「治療的有効用量」と規定する。この用途のために有
効な量は、疾患の重篤度および患者の健康の全身的状態に依存する。化合物の有
効量は、臨床医または他の有資格観察者によって観察されるような、症状の主観
的緩和または客観的に同定可能な改善のいずれかを与える量である。
【0156】
組成物の単回投与または複数回投与は、患者によって必要とされ、かつ耐容さ
れるような、投薬量および頻度に依存して投与される。いずれにしても、組成物
は、患者を有効に処置するために十分な量の本発明のタンパク質を提供するべき
である。好ましくは、投薬量は一度に投与されるが、治療的結果が達成されるか
、または副作用によって、治療の中断が正当化されるかのいずれかまで、周期的
に適用され得る。一般的に、用量は、患者に対して受容不可能な毒性を産生する
ことなく、疾患の症状または徴候を処置または改善するに十分である。
【0157】
本発明の免疫結合体組成物の徐放性非経口処方物は、インプラントとしてか、
油性注射剤、または粒子系として作製され得る。タンパク質送達系の広範な概説
については、Banga,A.J.、THERAPEUTIC PEPTIDE
S AND PROTEINS:FORMULATION,PROCESSIN
G,AND DELIVERY SYSTEMS,Technomic Pub
lishing Company,Inc.、Lancaster、PA(19
95)(本明細書中で参考として援用される)を参照のこと。粒子系としては、
ミクロスフェア、微小粒子、マイクロカプセル、ナノカプセル、ナノスフェア(
nanosphere)、およびナノ粒子(nanoparticle)が挙げ
られる。マイクロカプセルは、中心コアとして、治療用タンパク質を含む。ミク
ロスフェアでは、治療剤は、粒子全体にわたって分散される。約1μm未満の粒
子、ミクロスフェア、およびマイクロカプセルを、一般的に、それぞれ、ナノ粒
子、ナノスフェア、およびナノカプセルという。キャピラリーは、約5μmの直
径を有し、その結果、ナノ粒子のみが静脈内に投与される。微小粒子は、代表的
に、直径約100μmであり、そして皮下的または筋内に投与される。例えば、
Kreuter,J.、COLLOIDAL DRUG DELIVERY S
YSTEMS、J.Kreuter編、Marcel Dekker,Inc.
、New York、NY、219〜342頁(1994);ならびに、Tic
eおよびTabibi、TREATISE ON CONTROLLED DR
UG DELIVERY、A.Kydonieus編、Marcel Dekk
er,Inc.、New York、NY、315−339頁(1992)(こ
れらの両方とも、本明細書中で参考として援用される)を参照のこと。
【0158】
ポリマーが、本発明の免疫結合体組成物のイオン制御された放出のために使用
され得る。制御された薬物送達における使用のための、種々の分解可能および分
解不可能ポリマーマトリクスが、当該分野において公知である(Langer,
R.、Accounts Chem.Res.26:537−542(1993
))。例えば、ブロックコポリマーであるポロキサマー407(polaxam
er 407)は、低温では粘性であるがなお可動性の液体として存在するが、
体温では、半固体ゲルを形成する。これは、組換えインターロイキン−2および
ウレアーゼの処方物および持続型送達のために有効なビヒクルであることが示さ
れている(Johnstonら、Pharm.Res.9:425−434(1
992);およびPecら、J.Parent.Sci.Tech.44(2)
:58−65(1990))。あるいは、ヒドロキシアパタイトが、タンパク質
の徐放のためのマイクロキャリアとして使用されている(Ijntemaら、I
nt.J.Pharm.112:215−224(1994))。さらに別の局
面では、リポソームが、脂質カプセル化薬物の徐放および薬物標的化のために使
用されている(Betageriら、LIPOSOME DRUG DELIV
ERY SYSTEMS、Technomic Publishing Co.
,Inc.、Lancaster、PA(1993))。治療用タンパク質の制
御された送達のための多数のさらなる系が、公知である。例えば、米国特許第5
,055,303号、同第5,188,837号、同第4,235,871号、
同第4,501,728号、同第4,837,028号、同第4,957,73
5号、および同第5,019,369号、同第5,055,303号;同第5,
514,670号;同第5,413,797号;同第5,268,164号;同
第5,004,697号;同第4,902,505号;同第5,506,206
号;同第5,271,961号;同第5,254,342号;ならびに同第5,
534,496号(これらの各々が、本明細書中で参考として援用される)を参
照のこと。
【0159】
とりわけ本発明のイムノトキシンの種々の用途には、融合タンパク質の毒性作
用によって除去され得る、特定のヒト細胞によって引き起こされる種々の疾患状
態が含まれる。本発明のイムノトキシンについて1つの好ましい適用は、メソセ
リンを発現する悪性細胞の処置である。例示的な悪性細胞としては、卵巣癌、胃
癌、および扁平上皮癌、ならびに中皮腫が挙げられる。
【0160】
(VIII.診断用キットおよびインビトロ用途)
別の実施形態では、本発明は、生物学的サンプルにおいてメソセリンまたはそ
の免疫反応性フラグメント(すなわち、集合的に「メソセリンタンパク質」)を
検出するためのキットを提供する。「生物学的サンプル」は、本明細書中で使用
される場合、メソセリンを含む生物学的組織または流体のサンプルである。この
ようなサンプルとしては、生検からの組織、痰、羊水、血液、および血球(例え
ば、白血球)が挙げられるが、これらに限定されない。流体サンプルは、幾分興
味深いものであり得るが、一般的には、本発明においては好ましくない。なぜな
ら、検出可能な濃度のメソセリンが、このようなサンプル中で見出されることは
稀であるからである。生物学的サンプルはまた、組織の切片(例えば、組織学的
目的のために採取された凍結切片)を含む。生物学的サンプルは、代表的に、多
細胞真核生物(好ましくは、哺乳動物(例えば、ラット、マウス、ウシ、イヌ、
モルモット、またはウサギ)、そしてより好ましくは霊長類(例えば、マカク、
チンパンジー、またはヒト)から得られる。
【0161】
キットは、代表的に、本発明の抗メソセリン抗体を備える。いくつかの実施形
態では、抗メソセリン抗体は、抗メソセリンFvフラグメント(例えば、scF
vフラグメント)である。キットは、E4抗体を備え得る。
【0162】
さらに、キットは、代表的に、本発明の抗体の使用手段(例えば、サンプル中
の中皮細胞の検出のため)を開示する、指示資料を備える。キットはまた、その
キットが設計された特定の適用を容易にするために、さらなる成分を備え得る。
従って、例えば、キットは、標識を検出する手段(例えば、酵素学的標識につい
て酵素基質、蛍光標識を検出するためのフィルターセット、適切な二次標識(例
えば、ヒツジ抗マウス−HRP、など)をさらに備え得る。キットは、特定の方
法を実施するために慣用的に使用される、緩衝液および他の試薬をさらに備え得
る。このようなキットおよび適切な内容物は、当業者に周知である。
【0163】
本発明の1つの実施形態では、診断用キットは、イムノアッセイを含む。上述
のように、本発明のイムノアッセイの詳細は、使用される特定の様式に応じて変
動し得るが、生物学的サンプル中でメソセリンを検出する方法は、一般的に、生
物学的サンプルを、免疫学的反応条件下でメソセリンに特異的に反応する抗体と
接触させる工程を包含する。抗体は、免疫学的反応条件下で、メソセリンへの結
合を可能にされ、そして結合した抗体の存在が、直接的または間接的に、検出さ
れる。
【0164】
本明細書中で教示される方法によって開発された抗体(そして特に、SS1、
D8、およびC10と称される抗体)の親和性の増加に起因して、本明細書中に
提供される抗体は、特に、診断用薬剤として、そして生物学的サンプル中におい
てメソセリンの存在を検出するためのインビトロアッセイにおいて、有用である
。例えば、本明細書中に教示される方法によって作製された、抗体SS1、D8
、C10および他の抗体は、サンプルが、メソセリンを発現する細胞を含むか否
かを決定するための免疫組織化学アッセイにおいて、免疫結合体の標的化部分と
して使用され得る。サンプルが、通常メソセリンを発現しないはずの患者の組織
から採取されたサンプルである場合、メソセリンの検出は、その患者が、メソセ
リン発現細胞の存在によって特徴付けられる癌を有すること、または、このよう
な癌の処置が未だ、癌を根絶するのに成功していないこと、のいずれかを示す。
抗体E4もまた、これらの目的のために使用され得る。
【0165】
本発明の別のセットの用途では、本発明の抗体によって標的化されるイムノト
キシンを使用して、培養物中の細胞集団から標的化細胞を一掃し得る。従って、
例えば、メソセリンを発現する癌を有する患者から培養された細胞は、標的化部
分としてSS1 scFv、D8 scFv、またはC10 scFvを使用す
るイムノトキシンと培養物とを接触させることによって、癌細胞を一掃され得る
。E4 scFvもまた、これらの目的のための標的化部分として使用され得る
。
【0166】
本発明を、理解の明確さの目的で、例示および実施例によって幾分詳細に記載
してきたが、特定の変化および改変が、添付の特許請求の範囲に記載の範囲内で
実施され得ることが明らかである。
【実施例】
【0167】
(IX.実施例)
(実施例1 ライブラリーの構築)
SS(scFv)中の32個のホットスポットに関して、14個はVH中に、
18個はVL中に配置した。本発明者らは、変異誘発を開始するためにCDR3
を選択した。なぜなら、CDR3は、抗原との相互作用の主要な部位だからであ
る。本発明者らは、本発明者らの開始研究に関して、VH CDR3に対してV
L CDR3を選択した。なぜなら、VL CDR3は、より少ない組換え事象
を受け、故に生殖系列(germline)より近いからである。VLのCDR
3の場合、2つのA/G−G−C/T−A/T型のホットスポットおよび1つの
AGTセリンコドンが存在する。本発明者らは、VL CDR3においてテトラ
ヌクレオチドホットスポット上で集中することによって本発明者らの実験を開始
した。本発明者らの研究において、本発明者らは、そのようにしない特別の理由
がない限り、テトラヌクレオチドモチーフの全体または部分を形成するコドンの
全てのヌクレオチドを無作為化した(以下に議論する)。
【0168】
最初のファージライブラリー(Lib 89/93/94)は、2つのテトラ
ヌクレオチドホットスポット(1つはVL中の残基89および90をコードし、
他方は残基93および94をコードする)の一部であるコドンの無作為化を含ん
だ。Q90は、サブグループVI(SS VLが属する)中で非常に高く保存さ
れている(Johnsonら、WEIR’S HANDBOOK OF EXP
ERIMENTAL IMMUNOLOGY I.IMMUNOCHEMIST
RY AND MOLECULAR IMMUNOLOGY、第5版、Herz
enbergら、編、Blackwell Science Inc.、Cam
bridge、MA、第6章、6.1−6.21頁(1996))ので、本発明
者らは、これを変化させないままで、残基89、93および94を無作為化する
ように選択した。3つのアミノ酸が無作為化されるライブラリーは、8×103
個の異なるタンパク質改変体を有するべきであり、そして全ての異なるヌクレオ
チドの組み合わせに関して(NNSオリゴを用いる)、完全なレパートリーは3
×104個の異なるクローンを含む。本発明者らの作製したライブラリーは、6
×104個のクローンを含み、そして実質的に全ての可能性を包含すると予想さ
れる。
【0169】
Lib 89/93/94をパニングし、そして分析した後、本発明者らは、
Q89は全ての結合剤において保存されるが、置換が93位および94位におい
て許容されたことを見出した。この情報を用いて、本発明者らは、第2のライブ
ラリーのLib 92−94(この中で、S92に対するAGTコドン(別の型
のホットスポット)が、G93およびY94に対するコドンと共に変異された)
を構築した。このライブラリーの大きさは、1×105であった。
【0170】
本発明者らの実験のコントロールとして、本発明者らは、2つのライブラリー
(Lib 89−91およびLib 95−97)を作製した。Lib 89−
91を作製してホットスポット内での保存残基の無作為化の効果を試験し、そし
てLib 95−97を作製してホットスポット外での残基の無作為化の効果を
研究した。これらのライブラリーにおいて、3つのコドンを無作為化し、他の2
つのライブラリー(この中で、合理的に選択されたホットスポット中に位置する
3つのコドンが無作為化された)との比較を可能にした。これらの各々のライブ
ラリーの大きさはまた、1×105であった。
【0171】
(実施例2 ライブラリーのパニングおよび結合剤の濃縮)
ライブラリーを固定化された組換えメソセリンにパニングした。パニング実験
の結果は、表3に示す。2〜6×1011のファージの入力を用いると、バックグ
ラウンドが通常5〜6×104辺りであることを、本発明者らは理解する。Li
b 89/93/94およびLib 92−94に関して、本発明者らは、2×
105のファージを最初のパニングから得た。これは、メソセリンに結合するフ
ァージの約3倍の濃縮を示す。第2回目において、それぞれ、150および13
0倍である大きな濃縮の増加が存在した。これらのライブラリーに対する第3回
目のパニング後の濃縮は、約5倍であった。全体の濃縮は約2000であった。
対照的に、コントロールライブラリーに関して、第1回目のパニング後に濃縮は
存在しなかったが、その後に、漸進的な濃縮における増加が存在した(第2回目
のパニング後に10倍、そして第3回目の後に200倍)。コントロールライブ
ラリーの両方に関する全体の濃縮はまた、約2000倍であった。全てのライブ
ラリーに対する全体の濃縮は類似していたが、濃縮のパターンは、著しく異なっ
た。2つのライブラリー(Lib 89/93/94およびLib 92−94
の初期でありかつ迅速な濃縮は、両方が多数の結合剤を含んでいたことを示す。
対照的に、コントロールライブラリー(Lib 89−91およびLib 95
−97)のゆっくりとした漸進的な濃縮は、これらがより少ない結合剤(これは
、連続回のパニングの間にゆっくりと濃縮される)を含んでいたことを示唆する
。
【0172】
(表3)
組換えメソセリン上のパニングによるライブラリーからのメソセリン結合ファ
ージの濃縮。*2〜6×1011の入力ファージ数に対して5〜6×104であるバ
ックグラウンドレベルのファージ結合を超える濃縮に関する。
【0173】
【表2】
(実施例3 メソセリン結合に対するクローンのスクリーニング)
次に、本発明者らは、異なるライブラリーの各々から選択されたファージの結
合特性を調べた。各ライブラリーに対して、本発明者らは、ELISAによって
、第2回目および第3回目のパニングから23個のクローンをスクリーニングし
た。この実験の結果は、図2に示す。
【0174】
図2Bは、Lib 89/93/94のパニング後に選択されたクローンのメ
ソセリン結合活性を示す。図から明らかなように、全て46個のクローンは、メ
ソセリンに結合し、そして4つの型に分類され得る:(i)27個のクローンは
、非常に高いELISAシグナルを有した(2.3〜2.7);(ii)高いE
LISAシグナルを有する12個のクローン(1.75〜2.2);(iii)
6個のクローンは、親のSS(scFv)ファージ(黒四角によって示される)
に匹敵するELISAシグナルを有した、そして(iv)1個のクローンは、野
生型の親のファージよりも低いELISAシグナルを有した。
【0175】
図2Cは、Lib 89−91由来のファージクローンのメソセリン結合活性
を示す。試験された46個のクローンのうち、39個がメソセリンに結合した。
いずれも、親のクローンよりも大きいELISAシグナルを有さなかった。これ
らの39個のクローンのうちの28個は、親のクローンと類似したELISAシ
グナル(1.25〜1.5)を有した。残りの11個のクローンは、親の型より
も低いELISAシグナルを有した。
【0176】
図2Dは、Lib 92−94由来の選択されたファージクローンのメソセリ
ン結合活性を示す。Lib 89/93/94ライブラリー由来の結合活性と同
様に、試験された全ての46個のクローンは、メソセリンに結合し、そして、以
下の4群に分類され得る。(i)31個は、非常に高いELISAシグナル(2
.3−2.9)を与えた;(ii)8個のクローンは、高いELISAシグナル
(1.8−2.2)を有した;(iii)5個のクローンは、親の型と類似した
ELISAシグナルを与えた;そして(iv)2個のクローンは、親の型のシグ
ナルよりも低いELISAシグナルを有した。
【0177】
図2Eは、Lib 95−97のパニング後に選択されたクローンのメソセリ
ン結合活性を示す。試験された全ての46個のクローンは、メソセリンに結合す
ることが見出され、そして、ELISAシグナルに基づく3つの群に整理するこ
とができた。(i)22個は、親の型よりもわずかに良好なELISAシグナル
を与えた;(ii)20個のクローンは、野生型に類似したELISAシグナル
を与えた;そして(iii)4個のクローンは、親の型よりも下のELISAシ
グナルを有した。
【0178】
各ライブラリーからの12個のファージプレップの滴定をし、そして、CFU
(コロニー形成単位)によって決定されるようなそれらの数は、2倍未満まで変
化することが見出され、これは、ELISAシグナルにおける差異が、入力ファ
ージの数における差異に起因しないことを示す(データには示さない)。また、
BSAに対する結合は、任意のクローンからのファージ粒子については、検出さ
れなかった(データには示さない)。これらの結果は:(i)用いたスクリーニ
ングアッセイは、メソセリンに対する良好な結合を有するファージを作製するク
ローンの同定に関して信頼できたこと、および(ii)合理的に選択されたホッ
トスポットに対して無作為変異を標的化することは、CDRの全てのコドンを無
作為に変異することよりもより簡単に、良好な結合剤の単離を導く、ということ
を示唆する。
【0179】
(実施例4 クローンの多様性)
24個のELISA陽性クローンを、各ライブラリーから配列決定した。この
結果は図3に示す。
【0180】
ライブラリー、Lib 89/93/94から、4個の異なる配列を得た。全
てのクローンは、位置89において親の残基(グルタミン)が保存されていた。
位置93および94における残基は変化していた。最も豊富なクローンは、G9
3K−Y94H変異を有した。これはまた、最も高いELISAシグナルを有し
たクローンであり、SS1と命名した。
【0181】
Lib 89−91から、配列決定された全ての24個のクローンは、野生型
SS(ssFv)に類似したELISAシグナルを有し、そして24個は全て、
野生型残基を有した。
【0182】
Lib 92−94から、7個の異なるアミノ酸配列が得られた(図3)。最
も豊富なクローンは、D8(S92G−G93F−Y94N)およびC10(S
92G−G93S−Y94H)であった。このライブラリーから、本発明者らは
、Lib 89/93/94由来のSS1およびD7に対して同一のアミノ酸配
列を有する2個のクローン(E9およびE2)を得た。D8、C10およびSS
1は、最も高いELISAシグナルを有したクローンであった。
【0183】
ライブラリーLib 95−97から、SS(scFv)と類似またはそれよ
り大きいELISAシグナルを有する24個のクローンを配列決定した。17個
のクローンは、L96T変異を有した。他の7個のクローンは、野生型のアミノ
酸を有した。本発明者らは、L96T変異体をE4(scFv)と呼ぶ。
【0184】
全ての場合において、全体のscFvコード領域を配列決定し、他の変異が生
じていないことを確認した。タンパク質配列に関して、10個の異なる型の変異
体を得た。これらの多くは、いくつかの異なるヌクレオチド配列によってコード
された。これら10個のうちの9個の変異体は、変異が合理的に選択されたホッ
トスポットに対して標的化されたライブラリー(Lib 89/93/94およ
びLib 92−94)から得た。従って、これらのライブラリー由来の結合活
性を有するいくつかのクローンの存在は、これらのライブラリーに見出される迅
速な濃縮パターンと一致する。対照的に、Lib 89−91(これは、ホット
スポット中に保存された残基を有する)は、親のクローン以外にいずれの結合剤
も生じることができず、そして、Lib 95−97(この中において、変異は
、非ホットスポット領域に対して標的化される)は、単一の改変体を生じた。こ
れらのライブラリーにおける結合剤の希少性はまた、ゆっくりとでありかつ漸進
的な濃縮パターンと一致する。
【0185】
(実施例5 イムノトキシンの構築および精製)
イムノトキシンにおいて、癌細胞表面抗原に対するscFvを、毒素(例えば
、Pseudomonas外毒素Aの短縮変異体)とインフレームで融合する。
イムノトキシンの抗原結合特性は、scFvによって決定したが、毒素部分は、
scFvが結合する細胞を殺傷するエフェクター機能を提供する。従って、変異
体Fvsを含むイムノトキシンを作製しそして精製する事によって、本発明者ら
は、scFvの抗原結合運動性を決定し得、そしてまた細胞培養におけるその細
胞傷害性活性を研究し得る。異なるクローンの精製されたファージ調製物を用い
たより詳細なELISAに基づいて(結果は示さない)、本発明者らは、4個の
scFvを選択し、イムノトキシンを構築して精製した。これらの4個の間にお
いて、SS1、D8およびC10は、合理的に選択されたホットスポットが無作
為化されるライブラリーから得られた最良のクローンを示し、そして、E4は、
コントロールライブラリーから得られた親のSS(scFv)よりも良好な結合
を有する唯一のクローンを示す。イムノトキシンは、95%の均質性を超えて精
製され、そしてTSKゲル濾過クロマトグラフィーにおけるモノマーとして溶出
した(結果は示さず)。これらの精製されたイムノトキシンを使用して、scF
vの親和性を決定し、そしてまた、これらの細胞殺傷活性を研究した。
【0186】
(実施例6 イムノトキシンの結合特徴)
ファージライブラリーをパニングすることによって選択されたscFvの結合
特徴は、表面プラズモン共鳴(BIAcore)研究に対して対応する精製され
たイムノトキシンを用いることによって決定した。図4は、メソセリンに対する
イムノトキシンの結合の代表的なセンサーグラムを示し、そしてkon、kof
fおよびKDに関する値を列挙する。全ての変異体(SS1、D8、C10およ
びE4)が、親のSS(scFv)−PE38よりも、メソセリンに対する良好
な結合を有することは、明らかである。SS1およびC10のオンの速度(on
rate)は、約1.5倍高いが、D8およびE4のオンの速度は、親のSS
(scFv)とほぼ同じであった。SS1、D8、C10およびE4のオフの速
度(off rate)は、親のSS(scFv)に比較して、それぞれ、10
、37、35および3.8倍減少した。他の者が報告したことと類似して、本研
究に観察された変異体の増加した結合は、主にそれらのオフの速度における減少
に起因する(Schierら、J.Mol.Biol.255:28−43(1
996);Yeltonら、J.Immunol.,155:1994−200
4(1995);Yangら、J.Mol.Biol.254:392−403
(1995))。これらのデータによって、合理的に選択されたホットスポット
が変異されるライブラリーから誘導されるSS1、D8およびC10が、それぞ
れ、親のFvを超えて、15、37および55倍の親和性における増加を示し、
一方、E4(これは、コントロールライブラリーから単離された唯一の変異体で
ある)は、親のSS(scFv)を超えて、親和性においてより緩やかな3.7
倍の増加を示した。
【0187】
(実施例7 イムノトキシンの細胞傷害性活性)
高い親和性のscFvを癌治療における標的因子としての有用性を評価するた
めに、本発明者らは、異なるscFvを用いて作製されたイムノトキシンの細胞
傷害性を決定した。表4は、種々のイムノトキシンの抗原陽性A1847細胞株
および抗原陰性HUT 102細胞株に対する細胞傷害性活性を示す。A184
7細胞において、SS1、D8およびC10のscFv−PE38は、野生型S
S(scFv)イムノトキシンよりも10倍を超えてより活性であったが、E4
(scFv)−PE38は、たった約2倍の活性であった。HUT 102(抗
原陰性細胞)に対する細胞傷害性は、明らかでなかった。従って、変異を合理的
に選択されたホットスポットに標的化することによって得られた親和性における
増加は、細胞傷害性における大きな増加を引き起こす。しかし、細胞傷害性にお
けるこの増加は、親和性における増加と比例的に関連するわけではない。例えば
、SSl(Fv)−PE38およびC10(Fv)−PE38のIC50は、非常
に類似していたが、一方これらの親和性は約3倍異なる(IC50は、タンパク質
合成を50%阻害するのに必要であるイムノキシンの量である)。この差異は、
親和性はE.coli中で作製された組換えタンパク質を用いて測定され、一方
、細胞傷害性は抗原を発現する卵巣癌細胞株上で測定されると言う事実に起因し
得る。
【0188】
この戦略は、本発明者らの実験室において、上皮増殖因子レセプターの変異体
形態に結合する他の抗体に対して適用された。本研究においてまた、より高い親
和性を有する変異体を単離することが可能である(Beers,R.,Chow
dhury,P.S.およびPastan,I.調製における原稿)。本発明者
らはまた、SSのCDR3重鎖を変異させた。これらの変異体のELISA研究
によって、親のSS scFvの親和性よりも高いメソセリンに対する親和性が
示された。これらの結果は、現在使用されている方法によって作製されるライブ
ラリーよりも、変異を合理的に選択されたホットスポットに対して標的化するこ
とによって作製された小さなライブライリーから良好な結合ファージ選択するこ
とがより簡単であるはずであるということを示す。
【0189】
本研究において、本発明者らは、無作為化に対して4つの位置を選択し(3つ
のホットスポットの内側であり1つは外側である)、そして改変された変異体は
、有用であることが見出された。本発明者らのアプローチによって得られた改善
は、文献(Schier ら、J.Mol.Biol.255:28−43(1
996);Yeltonら、J.Immunol.,155:1994−200
4(1995);Yangら、J.Mol.Biol.254:392−403
(1995);Piniら、J.Biol.Chem.273:21769−2
1776(1998);Wuら、Proc.Natl.Acad.Sci.US
A 95:6037−6042(1998))に記載される大きいファージライ
ブラリーから得られる改善に匹敵する。本発明者らは、天然の体細胞過剰変異(
somatic hypermutation)プロセスを模倣することによっ
て、免疫化ライブラリーまたは非免疫化ライブラリーのいずれか由来の抗原に対
する増加した親和性を有する組換え抗体を容易に得ることができることを示唆す
る。
【0190】
(表4)
ヒト卵巣線癌細胞株A1847およびT細胞白血病株HUT102上での、親
のscFvおよび変異scFvを用いて作製されたイムノトキシンの細胞傷害性
。比較のために、BIAcore分析によって得られたKD値および野生型イム
ノトキシンに対する変異体の親和性における相対的増加を含めた。
【0191】
【表3】
(実施例8 ライブラリーの構築)
ファージミドpPSC 7−1は、メソセリン結合SS(scFv)をコード
するファージ抗体ディスプレイベクターである(Chowdhury l998
)。SS(scFv)のヌクレオチド配列の分析によって、32個のホットスポ
ットを明らかにした。これらの32個のホットスポットに関して、標的化変異に
対して3個選択した。これらは、CDR3のVLコード残基89、90、92、
93および94中に位置した。pPSC 7−1の一本鎖のウラシルを含むDN
Aは、以前に記載されたように調製した(Chowdhuryら、J.Mol.
Biol.281:917−928(1998))。終止コドンおよび診断のH
pal制限部位を、Kunkel’s変異誘発によってVLのCDR3に導入し
(Kunkel,T.A.、Proc.Natl.Acad.Sci.USA
82:488−492(1985))、ファージミドpPSC 7−1−94を
生成した。終止コドンは、ライブラリーの野生型配列の提示を防ぐように導入し
た。なぜなら、この変異誘発アプローチは、10〜12%のバックグラウンドを
与えるからである。ウラシルを含むpPSC7−1−94のssDNAは、変性
オリゴを用いた2つの実験ライブラリーおよびコントロールライブラリーを構築
するためのテンプレートとして使用した。オリゴSS VL 89/93/94
【0192】
【化1】
ライブラリーLib 89/93/94を作製するために使用した。このオリゴ
は、VLのコドン89、93および94を、全てが20個のアミノ酸が異なるS
S(scFv)中の無作為化する。以下に記載される他の全てのライブラリーと
同様に、このオリゴは、終止コドンを位置94においてチロシンのオリゴと置換
され、オケアー(ocher)コドンおよびオパールコドンを作製しない。
【0193】
第2のライブラリーのLib 92−94は、オリゴSS VL Mut 9
2−94
【0194】
【化2】
(これは、残基92−94を無作為化する)を用いて作製した。残基92−94
は、並んで配置される2つの異なるホットスポットによってコードされる。第3
および第4のライブラリーをコントロールとして作製した。第3のライブラリー
のLib 89−91は、オリゴSS VL Mut 89−91
【0195】
【化3】
(これは、残基89−91を無作為化する)を用いて作製した。残基90は、ホ
ットスポット中で保存された残基である。残基91は、ホットスポットの外側に
あたる。第4のライブラリーであるLib 95−97は、オリゴSS VL
Mut 95−97
【0196】
【化4】
を用いて作製した。これは、残基95−97(これらの全ては、ホットスポット
の外側にあたる)を無作為化する。
【0197】
(実施例9 ライブラリーのパニングおよび選択されたファージクローンの分
析)
ライブラリーのパニングは、以前に記載されたように行った(Chowdhu
ry 1998)。各ライブラリーからの、第2回目のパニング後の23個のク
ローンおよび第3回目のパニング後の23個のクローンを、以下のようにメソセ
リン結合に対してELISAによって分析した。単一のコロニーを、96ウェル
のマイクロタイタープレートに存在する100μg/mlのアンピシリンおよび
2%のグルコースを含む125μlの2XYT培地に接種した。このプレートを
、加湿されたインキュベーター中、37℃/200rpmで3.5時間インキュ
ベートした。これらのプレート(マスタープレート)の各ウェルからの25μl
の培養物を、100μg/mlのアンピシリン、2%グルコースおよび約4×1
09pfu/mlのM13KO7を有する125μlの2XYT培地を含む別の
マイクロタイタープレートの対応するウェルに移した。次いで、このプレート(
ファージプレート)を、上記と同じ条件下で2時間インキュベートした。次いで
、ファージプレートを、2000rpmで5分間室温で遠心分離した。各ウェル
からの上清を、逆位でプレートを穏やかに振盪することによって除去した。各ウ
ェル中の細胞ペレットを、100μl/mlのアンピシリン、および50μg/
mlのカナマイシンを含む200μlの2XYT培地中に再懸濁した。次いで、
プレートを、上記のように一晩インキュベートした。14〜16時間後、ファー
ジプレートを氷上で冷却し、次いで3000rpmで5分間、4℃で遠心分離し
た。この上清をファージELISAおよびファージ滴定に使用した。
【0198】
(実施例10 イムノトキシンの発現および精製のためのプラスミドの構築)
ファージミドのpPSC 7−1−1、7−1−2、7−1−3および7−1
−4は、それぞれ、VL中の以下の変異を有するSS(scFv)をコードする
:G93K−Y94H、S92G−G93F−Y94N、S92G−G93S−
Y94HおよびL96T。これらのファージミドベクター由来のscFvは、プ
ライマー対のNew G2 Nde IおよびNew G2 Hind III
を用いてPCR増幅した(Chowdhuryl998)。精製後、PCR産物
を以前に記載されるようにE.coli発現ベクターにクローン化した(Bri
nkmannら、Proc.Natl.Acad.Sci.USA 88:86
16−8620(1991))。生じたプラスミドのpPSC 7−2−1、p
PSC 7−2−2、pPSC 7−2−3およびpPSC 7−2−4は、ド
メインIIおよびドメインIIIを含むPseudomonas体外毒素(PE
)の38kDaフラグメントとインフレームで融合されたscFvを有した。組
換えタンパク質は、以前に記載されるように封入体から生成した(Brinkm
annら、Proc.Natl.Acad.Sci.USA 88:8616−
8620(1991))。
【0199】
(実施例11 表面プラズモン共鳴アッセイ)
scFvの親和性は、精製されたイムノトキシンのメソセリンに対する結合運
動性を、表面プラズモン共鳴(BIAcore)技術によって研究することによ
って決定した。別々の実験において、異なる量の組換えメソセリン(200〜1
000RU)をBIAcoreセンサーチップ、CM5上に固定化した。次いで
、各々のイムノトキシンを380nMの濃度で、固定化されたメソセリンの上を
通過させた。konを(ln(dR/dt)/t対濃度のプロット(ここで、Rは
応答であり、そしてtは時間である)から決定した。koffは、センサーグラム
(sensorgram)の解離期の最初の1〜2分間に算出した。これらの会
合および解離の速度は、10μl/分の連続した流速下で測定した。KDは、k
offをkonで割って算出した。
【0200】
(実施例12 細胞傷害性アッセイ)
イムノトキシンの細胞傷害性活性は、以前に記載されたように(Chaudh
aryら、Nature 339:394−397(1989))、メソセリン
陽性ヒト卵巣癌腫株A1847およびメソセリン陰性T細胞白血病株HUT 1
02ににおけるタンパク質合成の阻害によって決定した。IC50は、タンパク質
合成を50%阻害するのに必要なイムノトキシンの量である。
【0201】
本明細書中に言及される全ての刊行物および特許は、本明細書中において、あ
たかも各個々の刊行物および特許が詳細かつ個々に本明細書中において参考とし
て援用されて示されるのと同じ程度に、本明細書中へ参考として援用される。
【背景技術】
【0001】
(関連出願に対する相互参照)
適用せず。
【0002】
(連邦政府によって後援された研究および開発の下でなされた発明に対する権利に関する記述)
適用せず。
【0003】
(発明の背景)
多くの型の癌細胞において、分化抗原が発現される。これらの抗原は、癌治療における標的として使用されている。例えば、CD19、CD20、CD22およびCD25は、造血悪性腫瘍における標的として首尾良く使用されている(Pressら、New Eng.J.Med.329:1219−1224(1993);およびOsterborgら、J.Clin.Oncol.15:1567−1574(1997))。しかし、この標的化癌治療は、主に、固形腫瘍で成功していない。なぜなら、この標的化抗原はまた、腫瘍が発生する組織において発現するからである。従って、これらの標的化治療は、健康な細胞ならびに悪性細胞を殺傷する。
【0004】
米国においては、治療にもかかわらず、推定15,000人の女性が、毎年卵巣癌で死亡する。卵巣癌よりも一般的ではないが、中皮腫は、あらゆる化学療法剤に耐性であることが公知であり、従って、高い死亡率を有する。これらの癌の死亡率に起因して、これらの悪性腫瘍に対する新規の治療アプローチが必要とされる。
【0005】
卵巣の扁平上皮細胞、およびいくつかの胃癌ならびに中皮腫に共通して、細胞表面上にメソセリン(mesothelin)が発現される(Changら、Cancer Res.52:181−186(1992);Changら、J.Surgical Pathology 16:259−268(1992);およびChangら、Nat’l Acad.Sci.USA93:136−140(1996)。メソセリンは、中皮細胞の表面上に存在する40kDのGPI結合糖タンパク質抗原である。メソセリンは、69kDの前駆体として合成され、次いで、タンパク質分解性にプロセスされる。この30kDのアミノ末端が分泌され、そして巨核球増強因子(megakaryocyte potentiating factor)とよばれる(Yamaguchiら、J.Biol.Chem.269:805−808(1994))。この40kDのカルボキシル末端は、成熟メソセリンとして膜に結合したままである(Changら、Nat’l Acad.Sci.USA 93:136−140(1996))。癌細胞上に存在する多くの細胞表面抗原と異なり、メソセリンの膜結合形態は、癌患者の血液中に検出されず、そして培養細胞によって培地中に発散されない(Changら、Cancer Res.52:181−186(1992))。悪性細胞に加えて、メソセリンはまた、卵巣癌を含む中皮起源の細胞の細胞表
面上に見出される。これらの組織における細胞への損傷が、生命を脅かす結果を引き起こさないので、癌細胞の表面上のメソセリンの存在が、標的化治療のための有望な候補となる。
【0006】
イムノトキシンは、毒性部分に結合する細胞表面抗原に対する抗体である。癌の処置において、抗体は、好ましくは、癌細胞上にのみ発現される細胞表面抗原に対して指向される。しかし、表面抗原もまた発現する正常細胞の死が、悪性腫瘍の存在よりも生命を脅かさない場合、非悪性細胞上に発現される細胞表面抗原に対する抗体は、癌治療において使用され得る。イムノトキシンの毒性部分は、全身投与後に低い濃度の、非標的化細胞に対して有害ではない任意の毒素であり得る。このような毒素は、Pseudomonas aeruginosa外毒素(PE)である。PEを用いた以前の研究は、タンパク質の活性部分が、ドメインIIおよびドメインIIIから構成され、これらの両方は、毒素のカルボキシル末端に位置することを実証した。イムノトキシンの毒性部分としての使用のための開発または考慮の下にある他の毒素は、ジフテリア毒素(Watsonら、Intl J Cancer,61:233−40(1995))、サポリン(saporin)(例えば、Chandlerら、Intl J.Cancer 78(1):106−11(1998))、ヨウシュヤマゴボウ抗ウイルスタンパク質(例えば、Ekら、Clin Cancer Res,4(7):1641−7(1998)、リシン(例えば、Ohtomoら、Anticancer Res.18(6A):4311−5(1998))およびブリオジン1(bryodin1)(例えば、Franciscoら、J Biol Chem,272(39):24165−9(1997))である。イムノトキシンとしての使用のために、しばしば天然の毒素分子が改変されて、非特異的毒性または他の望ましくない効果を回避する。例えば、リシンのA鎖およびB鎖が、通常分離されて、その結果、B鎖によって別の方法で提供される非特異的結合を回避しながら、A鎖の毒性効果が使用され得る。
【0007】
イムノトキシンを標的化する抗体は、ポリクローナル抗体、モノクローナル抗体、または組換え抗体(例えば、キメラまたは可変領域フラグメント)であり得る。抗体が非組換えである場合、イムノトキシンは、抗体の毒性部分へのキメラ結合体化によって形成されるはずである。抗体が、組換え的に産生される場合、この抗体は、化学的結合または組換え融合を介して毒素に結合され得る。組換え融合において、抗体をコードするcDNAが、インフレームで、毒素をコードするcDNAをすでに含むプラスミドに挿入される。もちろん、その逆も同様に行われ得る;毒素cDNAが、抗体をコードするcDNAを保有するプラスミドに挿入され得る。
【0008】
イムノトキシンの大きさが潜在的に大きいので、時折、毒素部分への抗体フラグメントのみの結合が所望される。Fab、Fab’およびF(ab)2フラグメントが、ポリクローナル抗体、モノクローナル抗体およびキメラ抗体から作製され得、次いで、化学的結合を介して毒素に結合される。
【0009】
あるいは、抗体の可変領域が必須のフレームワーク領域に結合されるcDNAが、産生され得る。次いで、これらのより小さな抗体が、二重鎖Fv抗体として分泌されるか、または重鎖および軽鎖領域が、直接的もしくはペプチドリンカーを介してかのいずれかで結合される場合、単鎖Fv抗体(scFv)として分泌される。
【0010】
scFvを作製する1つの方法は、免疫原を用いて免疫されたマウスの脾臓mRNAから作製された、ファージディスプレイライブラリーを介した方法である(Chowdhuryら、Mol.Immunol.34:9−20(1997))。しかし、タンパク質の免疫原が、哺乳動物において天然に見出されるが、原核生物において組換え的に発現される場合、タンパク質は、正確なグリコシル化パターンを有さず、そして正確な高次構造を有さないかもしれない。この免疫原に応答して、マウスによって発生した抗体は、そのネイティブ状態においてはタンパク質を認識しないかもしれない。この問題に対する1つの解法は、哺乳動物細胞において作製されたネイティブなタンパク質を用いて動物を免疫することであるが、いくつかのタンパク質(特に、細胞表面タンパク質)の十分な量の哺乳動物細胞からの精製は、可能ではないかもしれない。一般的ではないが別の解法は、免疫原をコードするcDNAを用いて動物を免疫することである。適切なプロモーターの制御下で、cDNAが、動物に導入される。注入をブーストした後、そして抗体の力価が最大に達した場合、動物を屠殺し、そして脾臓を取り出し、ファージディスプレイライブラリーを作製する。
【0011】
メソセリン発現悪性腫瘍に対する標的化治療の開発は、良好にインターナライズ(internalize)された高い親和性の抗体の開発が困難なために妨害されていた。本発明者らが、最近の刊行物(Chowdhuryら、Proc Natl Acad Sci USA,95:669−674(1998)(今後は、「Chowdhury 1998」))において記載するように、哺乳動物細胞からの十分な量のメソセリンの精製は、不可能であり、そしてEscherica coliにおいて発現した組換えメソセリンは、メソセリン陽性細胞に対する低い親和性を有する。本発明者らは、この文献において、メソセリンをコードするDNAを含むプラスミドを用いてマウスを免疫することによって、本発明者らは、抗メソセリン抗体の高い力価を誘発し得ることを報告した。本発明者らはさらに、脾臓RNAおよびファージディスプレイ技術を使用して、本発明者らが、メソセリンに高い親和性で結合する単鎖Fv(「scFv」)(本発明者らは、SS scFvと呼ぶ)を単離し得たことを報告した。
【0012】
SSは、免疫結合体(例えば、イムノトキシン)に有用な標的化因子のようであるが、さらに高い親和性の抗体が、メソセリンを発現する癌を検出する能力を改善することおよびイムノトキシンのメソセリンを発現する癌細胞を殺傷する能力を増大することが期待される。従って、標的抗原に対して高い親和性を有する抗体を作製する改善された手段についての必要性が残存する。特に、メソセリンに対してより高い親和性の抗体についての必要性が残存する。
【発明の概要】
【課題を解決するための手段】
【0013】
(発明の要旨)
本発明は、変異した抗体の重鎖可変領域もしくは変異した抗体の軽鎖可変領域またはそれらの両方を含むポリペプチドに関し、ここで、このポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有する。このポリペプチドは、相補性決定領域において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによってこの親抗体と異なる配列を有し、このアミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ、ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである。このアミノ酸置換は、重鎖または軽鎖の可変領域のCDR3において生じ得るか、あるいはこれらの可変領域のCDR1またはCDR2において生じ得る。本発明のいくつかの実施形態において、抗原はメソセリンであり、そして親抗体は、SSscFvであり、そしてこのポリペプチドは、S92、G93およびY94から選択される(すなわち、SSscFvについてのアミノ酸残基の標準的なナンバリングに従って、92位のセリン残基、93位のグリシン残基または94位のチロシンから選択される(図1もまた参照のこと))少なくとも1つのアミノ酸のアミノ酸が置換されていることによって抗体SSと異なる配列を有する。
【0014】
いくつかの好ましい実施形態において、軽鎖のSSscFv CDR3可変領域の置換は、G93K−Y94H(すなわち、93位のグリシンが、リジンに変化し、そして94位のチロシンが、ヒスチジンに変化し、これらの変化を、「SS1」と呼ばれる抗体として規定する)、S92G−G93F−Y94N(すなわち、92位のセリンが、グリシンに変化し、93位のグリシンが、フェニルアラニンに変化し、そして94位のチロシンが、アスパラギンに変化する;これらの変化を、「D8」と呼ばれるscFvと規定する)、ならびにS92G−G93S−Y94H(すなわち、92位のセリンが、グリシンに変化し、93位のグリシンが、セリンに変化し、そして94位のチロシンが、ヒスチジンに変化する;これらの変化を、「D8」と呼ばれるscFvと規定する)から選択される。
【0015】
上記のポリペプチドはまた、免疫結合体の一部を形成し得る。例えば、これらのポリペプチドは、治療的部分または検出可能な標識を含み得る。いくつかの実施形態において、この治療的部分は、毒性部分(例えば、Pseudomonas外毒素)またはその細胞傷害性フラグメント(例えば、PE38)であり得る。この毒性部分はまた、ジフテリア毒素またはその細胞傷害性フラグメント、サポリンまたはその細胞傷害性フラグメント、ヨウシュヤマゴボウ抗ウイルスタンパク質またはその細胞傷害性フラグメント、リシンまたはその細胞傷害性フラグメント、あるいはブリオジン1またはその細胞傷害性フラグメントであり得る。別の実施形態において、このポリペプチドは、バクテリオファージの表面タンパク質をさらに含み得る。
【0016】
別の局面において、本発明は、変異した抗体の重鎖可変領域もしくは変異した抗体の軽鎖可変領域またはそれらの両方を含むポリペプチドをコードするヌクレオチドに関し、ここで、このポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有する。このポリペプチドはまた、相補性決定領域において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによってこの親抗体と異なる配列を有し、このアミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ、ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである。いくつかの実施形態において、抗原はメソセリンであり、親抗体は、抗メソセリン抗体SSであり、そしてポリペプチドは、S92、G93およびY94から選択される少なくとも1つのアミノ酸のアミノ酸が置換されていることによって抗体SSと異なる配列を有する。さらなる実施形態において、この置換は、G93K−Y94H(SS1)、S92G−G93F−Y94N(D8)、およびS92G−G93S−Y94H(C10)から選択される。
【0017】
なお別の局面において、本発明は、上記の核酸分子のうちの1つに作動可能に連結したプロモーターをコードする発現カセットに関する。
【0018】
別の局面において、本発明は、抗原を保有する悪性細胞を殺傷する方法に関し、この方法は、標的化部分および毒性部分を含むイムノトキシンと該細胞を接触させる工程を包含し、この標的化部分は、変異した抗体の重鎖可変領域もしくは変異した抗体の軽鎖可変領域またはそれらの両方を含み、このポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有する。このポリペプチドはさらに、相補性決定領域において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによってこの親抗体と異なる配列を有し、このアミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ、ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである。いくつかの実施形態において、抗原はメソセリンである。さらに、いくつかの実施形態において、この標的化部分は、SS1、D8およびC10からなる群より選択される。
【0019】
本発明はさらに、E4と名付けられた抗体を提供し、E4において、CDR3 VLは、L96Tの置換によって、親抗体SSと異なる。E4はまた、免疫結合体(例えば、使用されるイムノトキシン)の標的化部分として使用され、例えば、インビトロでメソセリンを保有する癌細胞を取り除き得るか、またはインビボで癌細胞を阻害もしくは殺傷し得る。
【0020】
なおさらなる局面において、本発明は、親抗体よりも高い標的抗原親和性を有するポリペプチドを同定する方法に関する。この方法は、変異した抗体の重鎖可変領域もしくは変異した抗体の軽鎖可変領域またはそれらの両方を含むポリペプチド(このポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有し、そしてこのポリペプチドは、相補性決定領域において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによってこの親抗体と異なる配列を有し、このアミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ(ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである))を、標的抗原と、抗体とこの標的抗原との間の特異的な結合に適切な条件下で接触させる工程、この親抗体またはそのフラグメントの親和性よりも高い親和性でこの標的抗原に結合しない抗体またはそのフラグメントの全てを取り除く条件下で、このポリペプチドを溶出する工程、ならびにこのポリペプチドが、この抗原に結合したか否かを決定する工程であって、これによって、結合することによって、このポリペプチドがこの親抗体よりも高い標的親和性を有することが確認される工程を包含する。
【0021】
本発明はさらに、変異した抗体の可変ドメインをコードする核酸のライブラリーを作製する方法に関し、この方法は、親抗体のVHドメインまたはVLドメインのアミノ酸配列をコードする核酸分子を提供する工程であって、この核酸分子が、ホットスポットモチーフの内部に少なくとも1つのヌクレオチドを含む、少なくとも1つの親のホットスポットコドンを含む工程、ならびにこの親のアミノ酸配列とは異なる変異したアミノ酸配列をコードする、複数の変異した核酸分子を生成する工程であって、ここで、各々の変異した核酸配列は、アミノ酸をコードする親のホットスポットコドンとは異なる少なくとも1つの変異したコドンを含み、この変異したコドンは、この親のホットスポットコドンによってコードされるアミノ酸とは異なるアミノ酸をコードする工程を包含する。
【0022】
いくつかの実施形態において、本発明の方法において使用される複数の変異した核酸分子は、少なくとも19のメンバーを含み、ここで、この19のメンバーの各々は、親のホットスポットコドンによってコードされるアミノ酸が、異なる天然のアミノ酸に置換されているアミノ酸配列をコードする。なおさらなる実施形態において、本発明の方法は、アミノ酸をコードする少なくとも2つの親のホットスポットコドンとは異なる変異したコドンを含む複数の変異した核酸分子を含み得、この変異したコドンの各々は、この親のホットスポットコドンによってコードされるアミノ酸とは異なるアミノ酸をコードする。他の実施形態において、本発明の方法は、少なくとも399のメンバーを含む複数の変異した核酸分子を含み得、これらのメンバーの各々は、親のホットスポットコドンによってコードされるアミノ酸が、異なる天然のアミノ酸に置換されているアミノ酸配列をコードする。本発明の方法のさらなる実施形態において、この親抗体は、コドンによってコードされる少なくとも1つの保存アミノ酸を有するあるクラスの抗体であり、ここで、この保存アミノ酸をコードするコドン(単数または複数)は変異していない。本発明の方法のなおさらなる実施形態において、このホットスポットモチーフは、AGCA、AGTT、AGCT、AGTA、GGCA、GGTT、GGCT、GGTA、AGCおよびAGTからなる群より選択される。本発明の方法のなおさらなる実施形態において、この変異した核酸分子は、CDRを含むVHドメインまたはCDRを含むVLドメインの一部分に、少なくとも1つの変異したコドンを含む。このCDRは、VHドメインのCDR3またはVLドメインのCDR3であり得る。
例えば、本発明は以下を提供する。
(項目1) 変異した抗体の重鎖可変領域または軽鎖可変領域を含むポリペプチドであって、該ポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有し、該ポリペプチドは、相補性決定領域(CDR)において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによって該親抗体と異なる配列を有し、該アミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ、ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである、ポリペプチド。
(項目2) 上記置換が、軽鎖可変領域のCDR3において生じる、請求項1に記載のポリペプチド。
(項目3) 上記置換が、重鎖可変領域のCDR3において生じる、請求項1に記載のポリペプチド。
(項目4) 上記置換が、軽鎖可変領域のCDR1またはCDR2において生じる、項目1に記載のポリペプチド。
(項目5) 上記置換が、重鎖可変領域のCDR1またはCDR2において生じる、項目1に記載のポリペプチド。
(項目6) 上記抗原が、メソセリンであり、上記親抗体が、抗メソセリン抗体SSであり、そして上記ポリペプチドが、S92、G93およびY94から選択される少なくとも1つのアミノ酸のアミノ酸が置換されていることによって抗体SSと異なる配列を有する、項目2に記載のポリペプチド。
(項目7) 上記置換が、G93K−Y94H(SS1);S92G−G93F−Y94N(D8)およびS92G−G93S−Y94H(C10)から選択される、項目6に記載のポリペプチド。
(項目8) 上記ポリペプチドが、scFvである、項目1に記載のポリペプチド。
(項目9) 上記ポリペプチドが、scFvである、項目6に記載のポリペプチド。
(項目10) 上記ポリペプチドが、dsFv、FabまたはF(ab’)2である、項目1に記載のポリペプチド。
(項目11) 上記ポリペプチドが、dsFv、FabまたはF(ab’)2である、項目6に記載のポリペプチド。
(項目12) 治療的部分または検出可能な標識をさらに含む、項目1に記載のポリペプチド。
(項目13) 上記治療的部分が、毒性部分である、項目12に記載のポリペプチド。
(項目14) 上記毒性部分が、Pseudomonas外毒素またはその細胞傷害性フラグメントである、項目13に記載のポリペプチド。
(項目15) 上記毒性部分が、細胞傷害性フラグメントであり、該細胞傷害性フラグメントが、PE38である、項目14に記載のポリペプチド。 (項目16) 上記毒性部分が、ジフテリア毒素またはその細胞傷害性フラグメント、サポリンまたはその細胞傷害性フラグメント、ヨウシュヤマゴボウ抗ウイルス毒素またはその細胞傷害性フラグメント、リシンまたはその細胞傷害性フラグメント、およびブリオジン1またはその細胞傷害性フラグメントからなる群より選択される、項目13に記載のポリペプチド。
(項目17) 治療的部分または検出可能な標識をさらに含む、項目6に記載のポリペプチド。
(項目18) 上記治療的部分が、毒性部分である、項目17に記載のポリペプチド。
(項目19) 上記毒性部分が、Pseudomonas外毒素またはその細胞傷害性フラグメントである、項目18に記載のポリペプチド。
(項目20) 上記毒性部分が、PE35、PE38およびPE40からなる群より選択される細胞傷害性フラグメントである、項目19に記載のポリペプチド。
(項目21) バクテリオファージの表面タンパク質をさらに含む、請求項1に記載のポリペプチド。
(項目22) 抗メソセリン抗体SSのメソセリン結合親和性の少なくとも3倍のメソセリン結合親和性を有するポリペプチドであって、該ポリペプチドは、少なくともL96T(E4)の軽鎖可変領域のCDR3において、アミノ酸が置換されている点で抗体SSと異なる配列を有する、ポリペプチド。
(項目23) 治療的部分または検出可能な標識をさらに含む、項目22に記載のポリペプチド。
(項目24) 上記治療的部分が、毒性部分である、項目23に記載のポリペプチド。
(項目25) 上記毒性部分が、Pseudomonas外毒素またはその細胞傷害性フラグメントである、項目24に記載のポリペプチド。
(項目26) 上記毒性部分が、PE35、PE38およびPE40からなる群より選択される細胞傷害性フラグメントである、項目25に記載のポリペプチド。
(項目27) 変異した抗体の重鎖可変領域または軽鎖可変領域を含むポリペプチドをコードする核酸分子であって、該ポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有し、該ポリペプチドは、相補性決定領域(CDR)において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによって親抗体と異なる配列を有し、該アミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ、ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである、核酸分子。
(項目28) 上記抗原が、メソセリンであり、上記親抗体が、抗メソセリン抗体SSであり、そして上記ポリペプチドが、S92、G93およびY94から選択される少なくとも1つのアミノ酸のアミノ酸が置換されていることによって抗体SSと異なる配列を有する、項目27に記載の核酸分子。
(項目29) 上記置換が、G93K−Y94H(SS1);S92G−G93F−Y94N(D8)およびS92G−G93S−Y94H(C10)から選択される、項目27に記載の核酸分子。
(項目30) 抗メソセリン抗体SSのメソセリン結合親和性の少なくとも3倍のメソセリン結合親和性を有するポリペプチドをコードする核酸分子であって、該ポリペプチドは、少なくともL96T(E4)の軽鎖可変領域のCDR3において、アミノ酸が置換されている点で抗体SSと異なる配列を有する、核酸分子。
(項目31) 項目27に記載の核酸分子に作動可能に連結されたプロモーターを含む、発現カセット。
(項目32) 項目28に記載の核酸分子に作動可能に連結されたプロモーターを含む、発現カセット。
(項目33) 抗原を保有する悪性細胞を殺傷する方法であって、該方法は、毒性部分および標的化部分を含むイムノトキシンと該細胞を接触させる工程を包含し、該標的化部分は、変異した抗体の重鎖可変領域または軽鎖可変領域を含むポリペプチドを含み、該ポリペプチドは、親抗体よりも少なくとも5倍高い抗原結合親和性を有し、該ポリペプチドは、相補性決定領域(CDR)において、少なくとも1つのアミノ酸のアミノ酸が置換されていることによって該親抗体と異なる配列を有し、該アミノ酸は、AGYまたはRGYWから選択されるホットスポットモチーフに属するヌクレオチドを含むコドンによってコードされ、ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである、方法。
(項目34) 上記抗原が、メソセリンである、項目33に記載の方法。
(項目35) 上記標的化部分が、SS1、D8およびC10からなる群より選択される、項目34に記載の方法。
(項目36) 上記毒性部分が、Pseudomonas外毒素またはその細胞傷害性フラグメントである、項目35に記載の方法。
(項目37) 上記毒性部分が、PE35、PE38およびPE40からなる群より選択される細胞傷害性フラグメントである、項目36に記載の方法。
(項目38) 抗原を保有する悪性細胞を殺傷する方法であって、該方法が、毒性部分および標的化部分を含むイムノトキシンと該細胞を接触させる工程を包含し、ここで、該標的化部分が、抗体E4である、方法。
(項目39) 上記毒性部分が、Pseudomonas外毒素またはその細胞傷害性フラグメントである、項目38に記載の方法。
(項目40) 上記毒性部分が、PE35、PE38およびPE40からなる群より選択される細胞傷害性フラグメントである、項目38に記載の方法。
(項目41) 親抗体よりも高い標的抗原親和性を有するポリペプチドを同定する方法であって、該方法が、以下の工程:
(a)抗体と該標的抗原との間の特異的な結合に適切な条件下で、該標的抗原と項目1に記載のポリペプチドを接触させる工程;
(b)該親抗体またはそのフラグメントの親和性よりも高い親和性で該標的抗原に結合しない抗体またはそのフラグメントの全てを取り除く条件下で、該ポリペプチドを溶出する工程;ならびに
(c)該ポリペプチドが該抗原に結合したか否かを決定する工程であって、これによって、結合することによって、該ポリペプチドが該親抗体よりも高い標的親和性を有することが確認される、工程、
を包含する、方法。
(項目42) 変異した抗体の可変ドメインをコードする核酸のライブラリーを作製する方法であって、該方法は、以下の工程:
(a)親抗体のVHドメインまたはVLドメインのアミノ酸配列をコードする核酸分子を提供する工程であって、該核酸分子は、ホットスポットモチーフの内部に少なくとも1つのヌクレオチドを含む、少なくとも1つの親のホットスポットコドンを含む、工程;ならびに
(b)該親のアミノ酸配列とは異なる変異したアミノ酸配列をコードする、複数の変異した核酸分子を生成する工程であって、ここで、各々の変異した核酸配列は、アミノ酸をコードする親のホットスポットコドンとは異なる少なくとも1つの変異したコドンを含み、該変異したコドンは、該親のホットスポットコドンによってコードされるアミノ酸とは異なるアミノ酸をコードする、工程、
を包含する、方法。
(項目43) 項目42に記載の方法であって、ここで、上記複数の変異した核酸分子が、少なくとも19のメンバーを含み、該19のメンバーの各々は、上記親のホットスポットコドンによってコードされるアミノ酸が、異なる天然のアミノ酸に置換されているアミノ酸配列をコードする、方法。
(項目44) 項目42に記載の方法であって、ここで、上記複数の変異した核酸分子が、アミノ酸をコードする少なくとも2つの親のホットスポットコドンとは異なる変異したコドンを含み、該変異したコドンの各々は、該親のホットスポットコドンによってコードされるアミノ酸とは異なるアミノ酸をコードする、方法。
(項目45) 項目42に記載の方法であって、ここで、上記複数の変異した核酸分子が、少なくとも399のメンバーを含み、該メンバーの各々が、上記親のホットスポットコドンによってコードされるアミノ酸が、異なる天然のアミノ酸に置換されているアミノ酸配列をコードする、方法。
(項目46) 項目42に記載の方法であって、さらにここで、上記親抗体が、コドンによってコードされる少なくとも1つの保存アミノ酸を有するあるクラスの抗体であり、ここで、該保存アミノ酸をコードするコドン(単数または複数)が変異していない、方法。
(項目47) 上記ホットスポットモチーフが、AGCA、AGTT、AGCT、AGTA、GGCA、GGTT、GGCT、GGTA、AGCおよびAGTからなる群より選択される、項目42に記載の方法。
(項目48) 上記変異した核酸分子が、CDRを含む上記VHドメインまたはCDRを含む上記VLドメインの一部分に、少なくとも1つの変異したコドンを含む、項目42に記載の方法。
(項目49) 上記CDRが、上記VHドメインのCDR3である、請求項48に記載の方法。
(項目50) 上記CDRが、上記VLドメインのCDR3である、請求項48に記載の方法。
【図面の簡単な説明】
【0023】
【図1】図1は、そのヌクレオチド配列から推定されるようなSS scFvのアミノ酸配列を含む。scFvにおいて、VHは、リンカーペプチド(GVGGSG4SG4S)によりVLに結合される。フレームワーク領域、CDRおよびリンカーに印を付けている。Hは、重鎖を示し;Lは、軽鎖を示し;FRは、フレームワーク領域を示し;CDRは、相補性決定領域を示す。
【図2】4つのライブラリーの各々をパニングした後選択されるファージクローンのELISAにより評価されるようなメソセリン結合。図2A:この図は、別々の列において、SS scFvの可変軽鎖のCDR3の、アミノ酸の番号付け、ヌクレオチド配列、およびアミノ酸配列を示す。一番上の列:親scFv SSの可変軽鎖CDR3のアミノ酸残基の番号。中央の列:scFv SS CDR3のCDR3におけるアミノ酸をコードするヌクレオチド配列。長方形は、テトラヌクレオチドホットスポットモチーフを示し:長円形は、この配列中のセリンホットスポットモチーフを示す。一番下の列:1文字コードにおける、scFv SSのCDR3のアミノ酸配列。 図2Bから2Eは、個々のライブラリー由来のクローンのELISAの結果を示すパネルである。パネルにおける各文字記号は、単一クローンからのファージ粒子のELISAシグナルを示す。各パネルにおいて塗りつぶされた四角(矢印により示される)は、内部標準の親SS(scFv)ファージを示す。ライブラリーを、各パネルの1番上に示される。図2B:残基89、93、および94で変異されたクローンのライブラリーからのELISAの結果。図2C:残基89、90、および91で変異されたクローンのライブラリーからのELISAの結果。図2D:残基92、93、および94で変異されたクローンのライブラリーからのELISAの結果。図2E:残基95、96、および97で変異されたクローンのライブラリーからのELISAの結果。
【図3】異なるライブラリーのパニングの後に得られたいくつかのELISA陽性のファージクローンのアミノ酸配列。クローン間で異なる領域の配列のみを示す。アステリスク(*)は、異なるヌクレオチド配列であるが、同じアミノ酸配列を有するいくつかのクローンが見出されたことを示す。
【図4】固定化されたメソセリンへの野生型および変異されたscFvイムノトキシンの結合について得られたBIAcoreのセンサーグラム(BIAcore sensorgram)の比較。左から右へ、イムノトキシンの標的化部分は:親SS、ならびに変異された改変体SS1、D8、C10およびE4である。Koff、KonおよびKDについての値を、図の下に表にする。「RU」は、「共鳴単位(resonance unit)」を表す。X軸は、秒で時間を表す。
【発明を実施するための形態】
【0024】
(I.概要)
本発明は、SS(scFv)と比較して、メソセリンに対する増加した親和性を有するこの抗原に対する抗体を提供する。この抗体は、より有効な免疫結合体(例えば、イムノトキシン)を作製するための標的化部分として使用され得る。これらの免疫結合体は、インビトロでの使用(例えば、生物学的サンプルに存在するメソセリン発現細胞の検出)のために使用され得る。イムノトキシンは、メソセリンを発現する細胞(例えば、細胞の表面上でメソセリンを発現する癌の癌細胞)を殺傷するためにさらに使用され得る。
【0025】
本発明は、親抗体の標的抗原に対する親和性よりも高い親和性を有する抗体を生成する新しい方法をさらに提供する。
【0026】
特に好ましい実施形態において、抗体は、scFvまたはdsFvである。scFvの構築物から産生される組換えイムノトキシンの多くは、IgG毒素化学的結合体の3分の1の大きさであり、そして組成物において均質である。scFv由来のIgG分子の定常部分の除去は、動物(霊長類を含む)に注射後のイムノトキシンのより速いクリアランスを生じ、そしてより小さいサイズのその結合体が、固形腫瘍における薬物の浸透を改善する。総合すると、これらの特性は、イムノトキシン(IT)が、非標的組織および非常に低いレベルの抗原を発現する組織と相互作用する時間を低減することにより、毒性部分と関連する副作用を減少させる。
【0027】
しかし、これらの利点は、IgGがscFvに変換される場合に起こる抗原結合親和性の減少により、いくらかの程度まで相殺される(Reiterら、Nature Biotechnol.14:239−1245(1996))。親和性を増加することにより、scFvの選択的腫瘍送達を改善することが示されており(Adamsら、Cancer Res.58:485−490(1998))、そして腫瘍の画像化および処置における有用性を増加するようである。従って、免疫結合体のscFvおよび他の標的部分(例えば、dsFv、Fab、およびF(ab’)2)の親和性の増加は、エフェクター分子(例えば、毒素および他の治療剤)を意図された標的に送達するこれらの薬剤の効率を改善することが所望される。
【0028】
親和性における改善は、部位指向性変異誘発またはランダム変異誘発により、インビトロで達成され得る。抗体−抗原結晶構造は、結合を改善するためにはどの残基が変異されるべきかを示し得るが、原子分解構造データは、ほとんどの抗体に対して利用できない。従って、ファージディスプレイ技術に基づくいくつかの戦略が開発され、より高い親和性の抗体を、FabおよびscFvの変異された改変体を発現するライブラリーから選択した(Gramら,Proc.Natl.Acad.Sci.USA 89:3576−3580(1992);Lowら,J.Mol.Biol.260:359−368(1996);de Kruifら,J.Mol.Biol.248:97−105(1995);Griffithら,EMBOJ.12:725−734(1993);Barbasら,J.Mol.Biol.230:812−823(1993);Schierら,J.Mol.Biol.255:28−43(1996);Yeltonら,J.Immunol.,155:1994−2004(1995);Yangら,J.Mol.Biol.254:392−403(1995);Piniら,J.Biol.Chem.273:21769−21776(1998);Wuら,Proc.Natl.Acad.Sci.USA 95:6037−6042(1998))。これらのアプローチは、しばしば得るのが難しく、そして産生するのが高価であり得る、大きくかつ複数のライブラリーの構築を必要とする。
【0029】
本発明者らは、増加した親和性を有するFvを改変体の小さなライブラリーから生成することを可能にする新規な方法を開発した。この方法は、抗体の可変領域をコードするDNAが、突然変異ホットスポット(このホットスポットは、変異が、インビトロの親和性成熟プロセスの間にしばしば集中するヌクレオチド配列である)を含む事実に基づく(NeubergerおよびMilstein,Curr.Biol.7:248−254(1995);Jollyら,Semin.Immunol.8:159−168(1996);Gonzalez−Fernandezら,Proc.Natl.Acad.Sci.USA 91:12614−12618(1994))。可変領域のホットスポットのいくつかの異なる型(例えば、直列反復および逆方向反復、パリンドローム、2次構造ならびに特定のコンセンサス配列)が提示されている(NeubergerおよびMilstein,Curr.Biol.7:248−254(1995);Jollyら,Semin.Immunol.8:159−168(1996);Gonzalez−Fernandezら,Proc.Natl.Acad.Sci.USA 91:12614−12618(1994);Goldingら,Genetics,115:169−176(1987);Kolchanovら,FEB Lett.214:87−91(1987);Rogozinら,Biochim.Biophys.Acta 1089:175−182(1991);InselおよびVarade,Int.Immunol.,6:1437−1443(1994);Wagnerら,Nature,376:732(1995))。これら全ての中で、コンセンサスホットスポット配列は、より詳細に研究されている。これらのコンセンサス配列の1つは、テトラヌクレオチドA/G−G−C/T−A/T(Pu−G−Py−A/T)である。この他の型は、セリンコドンAGYにより示され、Yは、CまたはTであり得(Wagnerら,Nature,376:732(1995);GoyenecheaおよびMilstein,Proc.Natl.Acad.Sci.USA 93:13979−13984(1996))。
【0030】
最近、本発明者らは、SS(scFv)と呼ばれる単鎖Fvを開発し、これは、卵巣癌、中皮腫およびいくつかの他のヒト癌の表面上に発現されるメソセリン抗原のその認識に基づき、ファージディスプレイライブラリーから選択された(Chowdhury 1998;ChangおよびPastan,Proc.Natl.Acad.Sci.USA 93:136−140(1996))。そのヌクレオチド配列から推定されるような、SS(scFv)のアミノ酸配列は、図1として示され、そしてヌクレオチド配列は、GenBankに寄託されている。SS(scFv)は、組換え手段により、または所望の場合タンパク質を合成するための標準的な技術により合成され得る。
【0031】
SS(scFv)−PE38は、SS(scFv)を、PE38と呼ばれる、Pseudomonas外毒素Aの38kDaの短縮された変異体と融合することにより作製されるイムノトキシンである。メソセリン陽性ヒト腫瘍を保有するマウスに注射される場合、SS(scFv)−PE38は、腫瘍の退化を引き起こし、そしてヒト癌を処置するために開発されている。メソセリンに対するSS(scFv)−PE38の親和性は、11nMであることが見出されている(Chowdhuryl998)。これは比較的高い親和性であるが、さらに高い親和性が、治療において、診断において、およびインビトロでの使用においてSS抗体に基づく免疫結合体(SS(scFv)を含む)の動作を増強するために望ましいと考えられる。
【0032】
抗体親和性を改善する以前の試みは、とりわけ、CDRの残基のランダムな変異または結晶構造(crystallagraphic)分析から見出される特定のアミノ酸を変異させて抗原接触に影響を与えることに頼っていた。免疫およびDNA免疫を介した親和性の改善はまた、Chowdhury 1998に報告される結果を考慮していないようであった。上記これらの方法における不利のうちのいくつかを克服するために、本発明者らは、新しいアプローチを作製した。本発明者らは、CDRにおけるホットスポットを標的化し、ここで本発明者らは、ランダムな変異を作製した。任意のCDRが、このアプローチにおいて変更され得るが、本発明者らは、軽鎖可変部領域のCDR3を使用して、この方法を試験した。一部分、CDR3を選択した。なぜなら、CDR3は、抗原結合部位において中心に位置し、そして抗原認識および親和性においてより大きい影響を有するからである。しかし、CDR2およびCDR1はまた、抗原結合親和性に影響することが知られ(述べられる順序で)、そしてCDR3の代わりにかまたはCDR3に加えて使用され得る。
【0033】
このアプローチを使用して、本発明者らは、約8000の独立したクローンのみの小さいライブラリーから、15〜55倍の親和性を増加したscFVを生成し、かつ単離し得た。これらの高い親和性の改変体を用いて作製されたイムノトキシンは、細胞傷害性活性において10倍よりも大きい増加を有した。本発明者らは、以下に詳細に報告される研究において軽鎖CDR3を使用したが、本発明者らは、同じアプローチを使用してSSの重鎖CDR3からscFvを生成した。SSの軽鎖CDR3を変異することにより生成されるscFVのように、ELISA結合研究において、多数のこれらの変異された重鎖scFVが、親SS scFVが示すより高いメソセリンに対する親和性を示す。さらに、本発明者はまた、SS以外の抗体(この場合、上皮増殖因子の変異体形態に結合する抗体)に対してこの技術を使用した。これらの結果を考慮して、本発明者らは、この手段は、親抗体の親和性と比較して、それらの標的抗原に対する増加した親和性を有する有するFv、Fabまたは他の組換え抗体を迅速に得るのに一般的に有用であると予想する。この研究の過程において、本発明者らはまた、ホットスポットモチーフの外側の変異により作製された、SSのメソセリンに対する親和性の約3.7倍の親和性を有するscFv(E4と命名される)を生成した。その親和性は、ホットスポット変異方法により作成されるscFvの親和性より際だって低いが、その親和性は、以前に既知の最も良い抗メソセリンscFvの親和性よりさらに数倍大きい。
【0034】
以下の節で、本明細書中で使用される用語が、さらなる明確さのために規定される。本願方法が、より詳細に記載される。最終的に、この実施例は、本願方法および特異的に改善される抗メソセリン抗体のCDR3の配列を実施するための特定の実験手順を示す。
【0035】
(II.定義)
単位、接頭辞(prefix)、および記号が、それらの国際単位系(SI)で認証された形態において示される。数量範囲は、範囲を規定した数を含む。その他で示されない場合、核酸は、5’から3’の方向で、左から右に書かれる;アミノ酸配列は、アミノからカルボキシの方向で左から右に書かれる。本明細書中で提供される表題は、本発明の種々の局面または実施形態の限定ではなく、これは、全体として明細書に対する参考とされ得る。従って、直ぐ下に定義される用語は、その全体が明細書に対する参考としてより十分に定義される。
【0036】
用語「メソセリン(mesothelin)」は、哺乳動物(例えば、ラット、マウス、霊長類、または特にヒト)の細胞の表面上に存在し得るメソセリンタンパク質およびそのフラグメントをいう。メソセリンの好ましい核酸およびアミノ酸の配列は、以下に記載される:PCT公開出願WO97/25,068,米国出願08/776,271および米国仮出願60/010,166。さらに、以下を参照のこと:Chang,K.およびPastan,I.,Int.J.Cancer 57:90(1994);Chang,K.およびPastan,I.,Proc.Nat’l Acad.Sci.USA 93:136(1996);Brinkmann U.ら,Int.J.Cancer 71:638(1997);ならびにChowdhury,P.S.ら,Mol.Immunol.34:9(1997)。メソセリンはまた、細胞内のままであるメソセリンタンパク質またはメソセリンペプチド、ならびに分泌されたかまたは単離された細胞外タンパク質をいう。
【0037】
本明細書中で使用される場合、「抗体」は、特定の抗原と免疫学的に反応性の免疫グロブリンに対する言及を含み、そしてポリクローナル抗体およびモノクローナル抗体の両方を含む。この用語はまた、遺伝的に操作された形態(例えば、キメラ抗体)例えば、ヒト化されたマウス抗体)、異種結合抗体(例えば、双特異性抗体)および組換え単鎖Fvフラグメント(scFv))、ジスルフィド安定化(dsFv)Fvフラグメント(本明細書中に参考として援用される、U.S.S.N.08/077,252を参照のこと)、またはpFvフラグメント(米国仮特許出願60/042,350および60/048,848)を含む。用語「抗体」はまた、抗体の抗原結合形態を含む(例えば、Fab’、F(ab’)2、Fab、FvおよびrIgG)。Pierce Catalog and Handbook,1994−1995(Pierce Chemical Co.,Rockford,IL);Kuby,J.,Immunology,第3版.,W.H.Freeman & Co.,New York(1997)もまた参照のこと。
【0038】
特定の抗原に免疫学的に反応性の免疫抗体が、組換え方法(例えば、ファージまたは類似のベクターでの組み換え抗体のライブラリーの選択)(例えば、Huseら,Science 246:1275−1281(1989);Ward,ら,Nature 341:544−546(1989);およびVaughanら,Nature Biotech.14:309−314(1996)を参照のこと)により、または抗原もしくは抗原をコードするDNAを用いて動物を免疫することにより生成され得る。
【0039】
代表的に、免疫グロブリンは、重鎖および軽鎖を有する。各重鎖および軽鎖は、定常部領域および可変領域(この領域はまた、「ドメイン」として公知である)を含む。軽鎖および重鎖の可変領域は、3つの超可変領域により分断される「フレームワーク」領域を含み、「相補性決定領域」または「CDR」とも呼ばれる。フレークワーク領域およびCDRの範囲は、規定されている(Kabat,E.ら,SEQUENCES OF PROTEINS OF IMMUNOLOGICAL INTEREST,米国厚生省(1987)(これらは本明細書中に参考として援用される)を参照のこと)。異なる軽鎖または重鎖のフレームワーク領域の配列は、比較的種内で保存されている。抗体のフレームワーク領域(これは、構成する軽鎖および重鎖の組み合わされたフレームワーク領域である)は、3次元空間においてCDRの位置を定めて整列するように働く。
【0040】
CDRは、抗原のエピトープへの結合の主な原因である。各鎖のCDRは、代表的にCDR1、CDR2、およびCDR3をいい、N末端から連続して始まって番号を付けられ、そしてまた代表的には、特定のCDRが位置する鎖により同定される。従って、VHCDR3は、それが見出される抗体の重鎖の可変ドメインに位置し、一方VLCDR1は、それが見出される抗体の軽鎖の可変ドメイン由来のCDR1である。
【0041】
「VH」または「VH」への言及は、Fv、scFv、dsFvまたはFabを含む免疫グロブリン重鎖の可変領域をいう。「VL」または「VL」への言及は、Fv、scFv、dsFvまたはFabを含む免疫グロブリンの軽鎖の可変領域をいう。
【0042】
句「一本鎖Fv」または「scFv」は、伝統的な二鎖抗体の重鎖の可変領域および軽鎖の可変ドメインが1つの鎖を形成するように連結される、抗体をいう。代表的に、リンカーペプチドは、2つの鎖の間に挿入され、適切なホールディングおよび活性な結合部位の作製を可能にする。
【0043】
用語「リンカーペプチド」は、軽鎖の可変ドメインへの重鎖の可変ドメインの間接的な結合を与える抗体結合フラグメント(例えば、Fvフラグメント)内のペプチドに対する言及を含む。
【0044】
用語「親抗体(parental antibody)」は、親抗体として同じエピトープに結合するがより高い親和性を有する抗体またはそのフラグメントを得るために、変異または改変される目的の任意の抗体を意味する。
【0045】
用語「ホットスポット」は、CDRのヌクレオチド配列の一部または特に高度な天然の改変体の部分である可変ドメインのフレームワーク領域の一部を意味する。CDRは、それ自身が超可変性の領域であると考えられるが、変異がCDRを通して均一に分布しないことが確認された。特定の部位、すなわちホットスポットは、集中した変異が起こる位置として同定された。ホットスポットは、構造の特徴の数および配列によって特徴付けられる。これらの「ホットスポットモチーフ」は、ホットスポットを同定するために使用され得る。特に十分特徴付けられる2つのコンセンサス配列モチーフは、テトラヌクレオチド配列RGYWおよびセリン配列AGYである(ここで、RはAまたはGであり、YはCまたはTであり、そしてWはAまたはTである)。
【0046】
「標的化部分」は、目的の細胞へ免疫結合体を標的化することが意図される免疫結合体の部分である。代表的に、標的化部分は、抗体、scFv、dsFv、FabまたはF(ab’)2である。
【0047】
「毒性部分」は、イムノトキシン(immunotoxin)を目的の細胞に対して細胞傷害性にするイムノトキシンの部分である。
【0048】
「治療的部分」は、治療剤として作用することが意図される免疫結合体の部分である。
【0049】
用語「治療剤」は、任意の数の、抗腫瘍性、抗炎症、サイトカイン、抗感染症、酵素のアクチベーターまたはインヒビター、アロステリック変更因子、抗体または患者において所望の治療効果を誘導するために投与される他の因子として作用することが現在公知または今後開発される化合物が含まれる。治療剤はまた、毒素または放射性同位体であり得、ここで意図される治療効果は、例えば、癌細胞を殺傷することである。
【0050】
「検出可能な標識」は、免疫結合体に関して、自身の存在を検出可能にする特性を有する免疫結合体の部分を意味する。例えば、免疫結合体は、細胞(この中に免疫結合体が存在する)を免疫組織化学アッセイにおいて検出可能にする放射性同位体を用いて標識され得る。
【0051】
用語「エフェクター部分」は、標的化部分によって標的化される細胞に対して効果を有するか、または免疫結合体の存在を同定することが意図される、免疫結合体の部分を意味する。従って、エフェクター部分は、例えば、治療的部分、毒素、放射性標識または蛍光標識であり得る。
【0052】
用語「免疫結合体」は、抗体に対するエフェクター分子の共有結合に対する言及を含む。エフェクター分子は、イムノトキシンであり得る。
【0053】
用語「有効量(effective amount)」または「有効量(amount effective to)」または「治療有効量」は、所望の結果(例えば、細胞のタンパク質合成を少なくとも50%阻害すること、または細胞を殺傷すること)を生じるに十分な治療剤の投薬量に対する言及を含む。
【0054】
用語「毒素」は、アブリン、リシン、Pseudomonas外毒素(PE)、ジフテリア毒素(DT)、ボツリヌス毒素、またはこれらの改変された毒素に対する言及を含む。例えば、PEおよびDTは、代表的に肝臓毒性を介して死をもたらす非常に毒性の化合物である。しかし、PEおよびDTは、毒素のネイティブな標的化成分(例えば、PEのIaドメインまたはDTのB鎖)を除去し、そしてそれを異なる標的化部分(例えば、抗体)で置換することによって、イムノトキシンとして使用するための形態に改変され得る。
【0055】
用語「接触する(接触)」は、直接的な物理的会合での配置に対する言及を含む。
【0056】
「発現プラスミド」は、分子または目的物(これは、プロモーターに作動可能に連結されている)をコードするヌクレオチド配列を含む。
【0057】
本明細書中に使用される場合、抗体に関して用語「抗メソセリン」とは、メソセリンに対して生成された抗体に対する言及を含む。好ましい実施形態において、メソセリンは、ヒトメソセリンのような霊長類のメソセリンである。特に好ましい実施形態において、抗体は、ヒトメソセリンをコードするcDNAの非霊長類哺乳動物への導入後にその動物によって合成されたヒトメソセリンに対して、生成される。
【0058】
本明細書中に使用される場合、「ポリペプチド」、「ペプチド」および「タンパク質」は、交換可能に使用され、そしてアミノ酸残基のポリマーに対する言及を含む。この用語は、1つ以上のアミノ酸残基が、対応する天然に存在するアミノ酸および天然に存在するアミノ酸ポリマーの人工的な化学アナログであるアミノ酸ポリマーに適用される。この用語はまた、タンパク質が機能的なままであるような保存的アミノ酸置換を含むポリマーに対して適用される。
【0059】
用語「残基」または「アミノ酸残基」または「アミノ酸」は、タンパク質、ポリペプチドまたはペプチド(総称して「ペプチド」)に組込まれるアミノ酸に対する言及を含む。アミノ酸は、天然に存在するアミノ酸であり得、他に制限されない限り、天然に存在するアミノ酸と類似した様式で機能し得る天然のアミノ酸の既知のアナログを含み得る。
【0060】
本明細書中に参照されるアミノ酸およびアナログは、以下の表1のような速記の指定によって記載される:
(表1:アミノ酸の名称)
【0061】
【表1】
「保存的置換」は、タンパク質を記載する場合、タンパク質の活性を実質的に変化させないタンパク質のアミノ酸組成における変化をいう。従って、特定のアミノ酸配列の「保存的に改変されたバリエーション」は、タンパク質活性に重要ではないこれらのアミノ酸のアミノ酸置換をいうか、または重要なアミノ酸の置換であっても活性を実質的に変化させないような、アミノ酸の類似の特性(例えば、酸性、塩基性、正に荷電または負に荷電、極性または非極性など)を有する他のアミノ酸とのアミノ酸置換という。機能的に類似したアミノ酸を提供する保存的置換の表は、当該分野で周知である。以下の表2の6つの群の各々は、互いに保存的置換であるアミノ酸を含む:
(表2)
1)アラニン(A)、セリン(S)、スレオニン(T);
2)アスパラギン酸(D)、グルタミン酸(E);
3)アスパラギン(N)、グルタミン(Q);
4)アルギニン(R)、リジン(K);
5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);
および
6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W)。
Creighton、PROTEINS、W.H.Freeman and Company、New York(1984)もまた参照のこと。
【0062】
ペプチドの文脈において、用語「実質的に類似した」は、10〜20アミノ酸の比較ウィンドウ上で、参照配列に対して少なくとも90%、好ましくは少なくとも95%の配列同一性を有する配列を含むペプチドを示す。配列同一性の割合は、比較ウィンドウ上で2つの最適に整列された配列を比較することによって決定され、ここで、比較ウィンドウ中のポリヌクレオチド配列の部分は、2つの配列の最適な整列に関して、参照配列(これは、付加または欠失を含まない)に対して比較した場合、付加または欠失(すなわち、ギャップ)を含み得る。この割合は、同一の核酸塩基または同一のアミノ酸残基が両方の配列で生じる位置の数を決定して、一致した位置の数を出し、一致した位置の数を比較ウィンドウ中の位置の総数で割り、そしてその結果に100を掛けることによって計算され、これによって配列同一性の割合か出される。
【0063】
句「ジスルフィド結合」または「システイン−システインジスルフィド結合」は、システインのイオウ原子が酸化されてジスルフィド結合を形成する2つのシステイン間の共有相互作用をいう。ジスルフィド結合の平均結合エネルギーは、水素結合に対する1〜2kcal/モルと比較して、約60kcal/モルである。本発明の文脈において、ジスルフィド結合を形成するシステインは、単鎖抗体のフレームワーク領域内であり、そして抗体のコントロールフォメーションを安定化するようにはたらく。
【0064】
用語「結合体化(する)(conjugating)」、「結合(する)(joining)」、「結合(する)(bonding)」または「連結(する)(linking)」は、2つのポリペプチドを1つの連続したポリペプチド分子にすることをいう。本発明の文脈において、この用語は、抗体部分をエフェクター分子(EM)に結合することに対する言及を含む。連結は、化学手段または組換え手段のいずれかにより得る。化学手段は、抗体部分とエフェクター部分との間の反応をいい、その結果、1つの分子を形成する2つの分子間で形成されたの共有結合が存在する。
【0065】
本明細書中に使用される場合、「組換え」は、そのネイティブな状態ではタンパク質を発現し得ない内因性のDNAコピーを有さない細胞を用いて産生されるタンパク質に対する言及を含む。この細胞は、組換えタンパク質を産生する。なぜなら、この細胞は、適切な単離された核酸配列の導入によって遺伝的に改変されているからである。この用語はまた、異種核酸の導入によってかまたはその細胞に対してネイティブではない形態にネイティブな核酸を変化することによって改変されたか、あるいは細胞がそのように改変されたか細胞から誘導される、細胞または核酸またはベクターに対する言及を含む。従って、例えば、組換え細胞は、ネイティブな(非組換え)形態の細胞内に見出されない遺伝子を発現するか、ネイティブな形態内に見出される変異体の遺伝子を発現するか、または他の異常に発現されるか、発現不足であるネイティブな遺伝子を発現するか、または全く発現されない。
【0066】
本明細書中に使用される場合「核酸」または「核酸配列」は、一本鎖形態または二本鎖形態のいずれかであるデオキシリボヌクレオチドまたはリボヌクレオチドのポリマーに対する言及を含み、そして他に指定されない限り限定されないが、天然に存在するヌクレオチドに類似した様式で核酸にハイブリダイズする天然のヌクレオチドの既知のアナログを含む。他に示されない限り、特定の核酸配列は、その相補鎖配列ならびに保存的改変体(すなわち、コドンおよび改変体のゆらぎ位置(wobble position)に存在する核酸(これは、タンパク質に翻訳された場合、アミノ酸の保存的置換を生じる))を含む。
【0067】
本明細書中に使用される場合、特定の核酸に関して「コード(する)」は、特定されたタンパク質への翻訳情報を含む核酸に対する言及を含む。この情報は、コドンの使用によって特定される。代表的に、アミノ酸配列は、「ユニバーサル」遺伝暗号を用いて核酸によってコードされる。しかし、ユニバーサルコードの改変体(例えば、いくつかの植物、動物および真菌ミトコンドリア、細菌Mycoplasma capricolum(Proc.Nat’l Acad.Sci.USA 82:2306−2309(1985))または繊毛虫Macronucleus中に存在する改変体)は、核酸がこれらの生物の翻訳機構を用いて発現される場合に使用され得る。
【0068】
句「インフレームで融合する」は、ポリペプチドをコードする2つ以上の核酸配列を結合することをいい、その結果、結合された核酸配列は、独自のポリペプチド鎖を含む一本鎖タンパク質に翻訳される。
【0069】
本明細書中に使用される場合、「発現された」は、核酸のタンパク質への翻訳に対する言及を含む。タンパク質は、発現されて細胞内性のままであり得るか、細胞表面膜の成分となり得るか、または細胞外マトリックスもしくは培地に分泌され得る。
【0070】
「宿主細胞」は、発現ベクターの複製または発現を支持しうる細胞を意味する。宿主細胞は、E.coliのような原核生物細胞であり得るか、または酵母細胞、昆虫細胞、両生類細胞、または哺乳動物細胞のような真核生物細胞であり得る。
【0071】
句「ファージディスプレイライブラリー」は、バクテリオファージの集団をいい、これらの各々は、表面タンパク質にインフレームで組換え融合された外来cDNAを含む。このファージは、cDNAによってコードされる外来タンパク質をその表面上に提示する。細菌宿主(代表的に、E.coli)が複製された後、目的の外来cDNAを含むファージが、外来タンパク質のこのファージ表面上での発現によって選択される。
【0072】
2つ以上の核酸配列またはポリペプチド配列に関する文脈において、用語「同一である」またはパーセント「同一性」は、同じであるか、あるいは以下の配列比較アルゴリズムのうちの1つを用いてかまたは視覚的な検査によって測定される場合に(最大の一致で比較され、そして整列された際に)同じであるアミノ酸残基またはヌクレオチドを特定の割合を有する、2つ以上の配列またはサブ配列をいう。
【0073】
句「実質的に同一である」は、2つの核酸またはポリペプチドの文脈において、以下の配列比較アルゴリズムのうちの1つを用いてかまたは視覚的な検査によって測定される場合に(最大の一致で比較され、そして整列された際に)少なくとも60%、好ましくは80%、より好ましくは90〜95%のヌクレオチドまたはアミノ酸残基の同一性を有する、2つ以上の配列またはサブ配列をいう。好ましくは、この実質的な同一性は、少なくとも約50残基の長さである配列の領域にわたって、より好ましくは少なくとも約100残基の領域の領域にわたって存在し、そして最も好ましくはこの配列は、少なくとも約150残基にわたって実質的に同一である。最も好ましい実施形態において、この配列は、コード領域の全体の長さにわたって実質的に同一である。
【0074】
配列比較に関して、代表的には、1つの配列は、試験配列が比較される参照配列として作用する。配列比較アルゴリズムを用いる場合、試験配列および参照配列は、コンピューターに入力され、配列座標が指定され、そして、必要な場合、配列アルゴリズムプログラムのパラメーターが指定される。次いで、配列比較アルゴリズムによって、指定されたプログラムパラメーターに基づいて、参照配列と比較して試験配列に関してパーセント配列同一性が計算される。
【0075】
比較に関する配列の最適なアラインメントは、例えば、以下のように実施され得る:SmithおよびWaterman、Adv.Appl.Math.2:482(1981)の局所相同性アルゴリズムによって、NeedlemanおよびWunsch、J.Mol.Biol.48:443(1970)の相同性アラインメントアルゴリズムによって、PearsonおよびLipman、Proc.Nat’l.Acad.Sci.USA 85:2444(1988)の類似性に関する検索方法によって、コンピューターによるこれらのアルゴリズムの実行によって(GAP、BESTFIT、FASTAおよびTFASTA、Wisconsin Genetics Software Package、Genetics Computer Group、575 Science Dr.、Madison、WI)、または視覚的な検査によって(一般的に、Current Protocols in Molecular Biology、F.M.Ausubelら、編、Current Protocols、Greene Publishing Associates,Inc.とJohn Wiley&Sons,Inc.との間の連帯のベンチャー(1995、補遺)(Ausubel)を参照のこと)。
【0076】
パーセント配列同一性および配列類似性を決定するのに適したアルゴリズムの例は、BLASTおよびBLAST 2.0アルゴリズム(これは、それぞれ、Altschulら(1990)J.Mol.Biol.215:403−410およびAltschuelら(1977)Nucleic Acids Res.25:3389−3402に記載される)である。BLAST分析を実行するためのソフトウェアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)を介して市販されている。このアルゴリズムは、最初に、問い合わせ配列中の短いワード長Wを同定することによって高いスコアの配列対(HSP)を同定する工程を包含する(これは、データベース配列中で同じ長さのワードで整列された場合、いくつかの正の値の閾値スコアTを一致させるかまたはこれを満たすかのいずれかである)。Tは、近接ワードスコアの閾値としていわれる(Altschulら、前出)。これらの最初の近隣ワードヒットは、それらを含むより長いHSPを見出すための検索を開始するためのシードとして作用する。次いで、このワードヒットは、累積アラインメントスコアが増加され得る限り、各々の配列に沿って両方向に伸長される。累積スコアは、ヌクレオチド配列に関しては、パラメーターM(一致した残基対に対する報酬スコア(reward score);常に>0)およびパラメーターN(一致しない残基に対するペナルティスコア;常に<0)を用いて算出される。アミノ酸配列に関して、スコアマトリックスを使用して累積スコアを算出する。各方向でのワードヒット拡大を以下の場合に停止する:累積アラインメントスコアが、その最大に達成される値から量Xだけ落ちる場合;1つ以上のネガティブスコアの残基アラインメントの蓄積に起因して、累積スコアが0またはそれより下になる場合;あるいは、配列のいずれかの末端に到達した場合。BLASTアルゴリズムパラメーターのW、T、およびXは、アラインメントの感度および速度を決定する。(ヌクレオチド配列に関する)BLASTNプログラムは、デフォルトとして、11のワード長(W)、10の期待値(E)、M=5、N=−4、および両方の鎖の比較を使用する。アミノ酸配列に関して、BLASTPプログラムは、デフォルトとして、3のワード長(W)、10の期待値(E)、およびBLOSUM62スコアマトリックスを使用する(HenikoffおよびHenikoff、Proc.Natl.Acad.Sci.USA 89:10915(1989)を参照のこと)。
【0077】
パーセント配列同一性を算出するのに加えて、BLASTアルゴリズムはまた、2つの配列間の類似性の統計分析を実施する(例えば、KarlinおよびAltschul、Proc.Nat’l.Acad.Sci.USA 90:5873−5787(1993)を参照のこと)。BLASTアルゴリズムによって提供される類似性測定の1つは、最小の和の確率(sum probability)(P(N))(これは、2つのヌクレオチド配列またはアミノ酸配列の間の一致が偶然生じる確率の指標を提供する)である。例えば、参照核酸に対する試験核酸の比較における最小の和の確率が約0.1より小さい、好ましくは約0.01より小さい、そして最も好ましくは約0.001より小さい場合に、核酸は参照配列に類似すると考えられる。
【0078】
2つの核酸配列またはポリペプチドが実質的に同一であるというさらなる指標は、以下に記載されるように、第1の核酸によってコードされるポリペプチドが第2の核酸によってコードされるポリペプチドと免疫学的に交差反応性であるということである。従って、ポリペプチドは、代表的に、例えば、これらの2つのペプチドが保存的置換によってのみ異なる場合、第2のポリペプチドに実質的に同一である。2つの核酸配列が実質的に同一であるという別の指標は、以下に記載されるように、2つの分子が互いにストリンジェントな条件下でハイブリダイズするということである。
【0079】
用語「インビボ」は、細胞が得られる生物の体の内側に対する言及を含む。「エキソビボ」および「インビトロ」は、細胞が得られる生物の体の外側を意味する。
【0080】
句「悪性細胞」または「悪性腫瘍」は、浸潤性である腫瘍もしくは腫瘍細胞および/または転移を引き起こし得る腫瘍もしくは腫瘍細胞、すなわち癌性細胞をいう。
【0081】
本明細書中に使用される場合、「哺乳動物細胞」は、ヒト、ラット、マウス、モルモット、チンパンジーまたはマカクを含む哺乳動物に由来する細胞に対する言及を含む。この細胞は、インビボまたはインビトロで培養され得る。
【0082】
用語「選択的に反応性」は、抗原に関して、抗体(全体または一部)の(抗原を欠く細胞または組織に対してではなく)抗原を保有する細胞または組織との優先的な結合をいう。これは、当然、ある程度の非特異的相互作用が分子と非標的細胞もしくは組織との間に生じ得ることを認識する。それにもかかわらず、選択的反応性は、抗原の特異的認識を介して媒介される場合、識別され得る。選択的に反応性である抗体は抗原に結合するが、この抗体はまた低い親和性でそのように結合し得る。一方、特異的結合は、抗体と抗原保有細胞との間に、結合抗体と抗原を欠く細胞との間よりも、より強い結合を引き起こし得る。特異的結合は、代表的に、メソセリンを欠く細胞または組織に対する場合と比較して、2倍よりも大きく、好ましくは5倍よりも大きく、より好ましくは10倍よりも大きく、そして最も好ましくは100倍よりも大きいメソセリン保有細胞または組織に対する抗体結合の量における(単位時間当たりの)増加を引き起こす。そのような条件下でのタンパク質に対する特異的結合は、特定のタンパク質に対するその特異性について選択される抗体を必要とする。種々の免疫アッセイ形式が、特定のタンパク質と特異的に免疫反応性な抗体を選択するのに適している。例えば、固相ELISA免疫アッセイを慣用的に使用して、あるタンパク質と特異的に免疫反応性のモノクローナル抗体を選択する。特定の免疫反応性を決定するのに使用され得る、免疫アッセイ形式および条件に関する説明については、HarlowおよびLane、ANTIBODIES、A LABORATORY MANUAL、Cold Spring Harbor Publications、New York(1988)を参照のこと。
【0083】
用語「免疫学的に反応性である条件」は、特定のエピトープに対して産生された抗体をそのエピトープへ、実質的に全ての他のエピトープに対する結合よりも検出可能に大きい程度で結合させる条件、および/または実質的に全ての他のエピトープに対する結合を実質的に排除するように結合させる条件に対する言及を含む。免疫学的に反応性である条件は、抗体結合反応の形式に依存し、そして代表的には、免疫アッセイプロトコールに使用される条件またはインビボで遭遇する条件である。免疫アッセイ形式および条件の説明に関しては、HarlowおよびLane(前出)を参照のこと。好ましくは、本発明の方法に使用される免疫学的に反応性である条件は、代表的に生きている哺乳動物または哺乳動物細胞の内側の条件(例えば、温度、容量オスモル濃度、pH)に対する言及を含む「生理学的条件」である。いくらかの生物は、極限状態に供されるが、生物体内および細胞内環境は通常pH7辺りにあり(すなわち、pH6.0〜pH8.0、より代表的にはpH6.5〜7.5)、優勢な溶媒として水を含み、そして約0℃より高く50℃より低い温度で存在することが理解される。容量オスモル濃度は、細胞の生存性および増殖を支持する範囲内である。
【0084】
用語「SS」または「SS(scFv)」は、メソセリンに高い親和性で結合するscFvをいい、これは、図1に示されるアミノ酸配列を有する。Chowdhury 1998を参照のこと。
【0085】
(III.より高い親和性を有する抗体の生成)
上記のように、本発明は、親抗体から、親抗体の標的抗原に対する親和性よりも高い親和性を有する1つ以上の変異体抗体を生成するための新規な方法を提供する。以前の技術(これは、CDR内に無作為に変異を作製する)と対照的に、本発明者らは、本発明者らがホットスポットモチーフ内に少なくとも1つのヌクレオチドを含むコドン(参照を簡単にするために、これらはこの議論に関して「ホットスポットコドン」と名付ける)に対する変異を標的化することによる努力が著しく少なくても、より高い親和性抗体を得ることができることを見出した。変異体の複数の相互作用(interation)は、それらの変異が、19個の他の天然のアミノ酸の各々をコードするコドンによって(1度に)置換されるホットスポットコドンを生じるように、作製される。都合のよいことに、これは、このプロセスを無作為化することによってなされ得、その結果、変異は、従来のホットスポットコドンの代わりに無作為に表れる全部で20アミノ酸(従来のアミノ酸のものも含む)をコードするコドンを生じる(すなわち、アラニンは、ある変異形態ではグリシンで置換され、別の変異形態はアルギニンで置換され、第3の変異形態はシステインで置換される、など)。
【0086】
CDRは、代表的に1つより多くのホットスポットコドンを含む。1、2、3より多くのそのようなコドンは、CDR内で変異され得、より高い親和性を獲得した抗体を生成する。本発明者らの結果は、2つのホットスポットコドン変異させるよりも、3つのホットスポットコドンを1度に変異させることによってより良い結果が得られ得ることを示す。
【0087】
任意のCDR中のホットスポットコドンは、より高い親和性の抗体を作製するために変異され得る。所望する場合、1つのCDR中の1つ以上のホットスポットコドンは、他のCDRの内の1つ由来の1つ以上のホットスポットコドンと同じときに変異され得る。各抗体がたった6個のCDRしか含まないこと、CDRは制限された長さであること(例えば、図1に示されるように、SSscFvのCDRは、5〜17アミノ酸の長さの範囲である)、そしてこれらのアミノ酸をコードするコドンのいくつかのみが、ホットスポットモチーフ内に位置するヌクレオチドを含むということに、注意すべきである。例えば、図2Aは、SSscFvのVL CDR3中の9アミノ酸内の5アミノ酸のみが、ホットスポットコドンによってコードされることを示す。
【0088】
可変ドメインのフレームワーク領域は、抗体−抗原結合相互作用に穏やかに寄与する。必要に応じて、フレームワーク領域内のホットスポットコドンは、結合活性におけるさらなる改善をなすために変異され得る。
【0089】
Kabat(前出)は、抗体の重鎖および軽鎖をサブグループおよびファミリーに分類することに関する情報を提供する。例えば、Kabatの分類下において、SSscFvのVHは、サブグループIIAおよびファミリーVに属し、そしてVLは、サブグループVIおよびファミリーXIに属する。特定の位置でのアミノ酸残基が、特定のサブグループに保存されることは、当該分野で公知である。例えば、Johnsonら、WEIR’S HANDBOOK OF EXPERIMENTAL IMMUNOLOGY I.IMMUNOCHEMISTRY AND MOLECULAR IMMUNOLOGY、第5版、Herzenbergら、編、Blackwell Science Inc.、Cambridge、MA、第6章、6.1−6.21(1996)(Weir’s Handbookの全体は、本明細書中に参考として援用される)を参照のこと。本発明者らの結果によって、これらの保存された残基をコードするコドンを変異しないほうか好ましいこと(たとえ、そのコドンがホットスポットモチーフ内のヌクレオチドを含み、そしてさもなければ変異に対する候補物であったとしても)が示される。
【0090】
以下の実施例は、これらの原理の使用を実証し、変異されたホットスポットコドンを有する抗体を生成する。本発明者らは、ある部分、モデル抗体として抗メソセリン抗体SSscFvを選択した。なぜなら、SSscFvを標的部分として使用するイムノトキシンは、動物モデルにおいて、ヒト腫瘍に対して良好な影響を与えることを示されたからである。本発明者らは、CDR3を選択した。なぜなら、CDR3は、抗原相互作用の主要な部分であるからである。本発明者らは、重鎖に対して可変軽鎖を選択した。なぜなら、本発明者らは、重鎖よりも、軽鎖は正常な親和性成熟の間に抗原親和性に対してほとんど最適化されていないようであるとみたからである。SSscFvの軽鎖のCDR3は、9アミノ酸残基を含み、これは、慣例として89〜97と番号を付ける(図2Aを参照のこと)。2つのテトラヌクレオチドホットスポットモチーフ中の3つのコドン(残基89、93および94)が同時に変異されるライブラリーを作成し、そしてセリンホットスポットコドンおよびテトラヌクレオチドホットスポットモチーフ中のコドン(残基92、93および94)が同時に変異されるライブラリーを作成した。これらのライブラリー由来のクローンを抗原に対するこれらの親和性についてスクリーニングした場合、これらのライブラリー中のほとんどの変異は、親抗体(SSscFv)が示すよりも、抗原に対して高い親和性を示した。対照的に、残基Q90(これは、SSscFvが分類される軽鎖のサブグループにおいて保存されている)の中に変異を含むコントロールライブラリーは、親抗体の親和性とほとんど同じまたはそれより低い親和性を有する変異体を生じた。図2Cを参照のこと。同様に、変異がホットスポット内のヌクレオチドを含まないコドン内で作製されたライブラリーは、従来の親のSSscFvとほとんど同じまたは中程度に良好な抗原に対する親和性を示した。図2Eを参照のこと。
【0091】
親和性結合研究により、変異された抗体の親和性に厳格な差異が存在したことを確認した。保存残基をコードしないホットスポットコドンを変異することによって作製されたScFvによって、これらが由来する既に高い親和性の親抗体よりも、15倍から55倍もの高い範囲の標的抗原に対する親和性を有するいくつかの抗体の生成が生じた。対照的に、ホットスポットモチーフ内の少なくとも1つのヌクレオチドを有さないコドンによってコードされるアミノ酸から生じる最良のクローンは、たった3.7倍の親和性の増加しか有さなかった。さらに、これらの高い親和性の抗体の生成およびスクリーニングは、CDR変異を含む以前の方法に必要とされるサイズよりも、かなり低い桁の大きさのライブラリーを必要とした。
【0092】
最も高い親和性を有する変異された抗体(C10と命名する)は、残基92、93および94以外は親のSSscFvのアミノ酸配列を有する。位置92において、SSscFvの「S」(セリン)は、「G」(グリシン)に変化される。位置93において、SSscFv配列の「G」は、「S」に変化される。位置94において、「Y」(チロシン)は、「H」(ヒスチジン)に変化される。各変異クローンへの変化(他の2つの高い親和性のクローン(SS1およびD8)を含む)を、図3に示す。慣用として、これらのアミノ酸置換はまた、本来の配列中のアミノ酸がその一文字コードで列挙される短縮化形態で記載され得、続いて、その位置の番号および変化された配列で置換されたアミノ酸に対する標準的な一文字コードで記載される。従って、上記のクローンC10の変化はまた、S92G−G93S−Y94Hと記載され得る。同様に、クローンSS1のscFvは、G93K−Y94Hとして記載され得、そしてクローンD8のscFvは、S92G−G93F−Y94Nによって記載され得る。E4クローンのscFv(これは、ホットスポットモチーフ内にヌクレオチド変異を有さない)は、L96Tとして示され得る。
【0093】
(IV.抗メソセリン抗体)
本発明は、メソセリンに対して、先行技術の抗体よりも高い親和性で結合する抗体を提供する。特に、本発明は、本発明者らがSS1、C10、およびD8と命名した、それぞれ、メソセリンについて、これらの親抗体SS(scFv)の親和性より15倍、55倍および37倍高い親和性を有する抗体を提供する。SS(scFv)は、以前は、メソセリンと特異的に反応性である、最も親和性が高い公知の抗体であった。本発明はさらに、メソセリンおよび目的の他の抗原に対する他の高親和性抗体を生成するための方法を提供する。下記に開示される免疫結合体は、本発明の抗体を用いてメソセリンを標的化する。これらの抗体は、哺乳動物細胞の表面に提示されるメソセリンの決定基に対して免疫学的条件下で選択的に反応性であり、そして細胞外環境由来の抗体にアクセス可能である。本発明はさらに、E4と命名した抗体を提供し、この抗体は、本発明の方法によって生成されておらず、そして本発明の方法によって生成される抗体ほどメソセリンについての親和性が高くないが、メソセリンについてのSSの約3.7倍の親和性をなお有する。
【0094】
本発明において生成される抗メソセリン抗体は、EMカルボキシル末端、EMアミノ末端、EMの内部アミノ酸残基(例えば、システイン)、またはそれらの任意の組み合わせを通してエフェクター分子(EM)に連結され得る。同様に、EMは、抗体の重鎖、軽鎖、Fc(定常領域)またはフレームワーク領域に直接連結され得る。連結は、抗体のアミノ末端もしくはカルボキシル末端を通して、または内部アミノ酸残基を通して生じ得る。さらに、複数のEM分子(例えば、2〜10のいずれか1つ)は、抗メソセリン抗体に連結され得、そして/または複数の抗体(例えば、2〜5のいずれか1つ)は、EMに連結され得る。本発明の多価免疫結合体組成物において用いられる抗体は、同じかまたは異なるメソセリンエピトープに対して指向され得る。
【0095】
本発明の好ましい実施形態では、抗メソセリン抗体は、組換え抗体(例えば、scFvまたはジスルフィド安定化Fv抗体)である。Fv抗体は代表的に、約25kDaであり、そして重鎖および軽鎖あたり3つのCDRを有する、完全な抗原結合部位を含む。VH鎖およびVL鎖が連続せずに発現される場合、Fv抗体の鎖は代表的に、非共有結合相互作用によって一緒に保持される。しかし、これらの鎖は、希釈によって解離する傾向があり、それゆえ、これらの鎖をグルタルアルデヒド、分子間ジスルフィドまたはペプチドリンカーを通して架橋する方法が開発されている。
【0096】
特に好ましい実施形態では、抗体は、単鎖Fv(scFv)である。scFv抗体のVH領域およびVL領域は、2本鎖の抗体に見出されるのと同様の抗原結合部位を作製するように折り畳まれる単鎖を含む。一旦折り畳まれると、非共有結合相互作用は、単鎖抗体を安定化する。より好ましい実施形態では、scFvは、組換え産生される。なお別の好ましい実施形態では、VH領域は、図1に示す通りのアミノ酸配列を有する。当業者は、本発明の抗体の保存的改変体が作製され得ることを理解する。scFvフラグメントにおいて用いられるこのような保存的改変体は、正確な折り畳みおよびVH領域とVL領域との間の安定化に必要な重要なアミノ酸残基を保持する。
【0097】
本発明のいくつかの実施形態では、scFv抗体は、軽鎖を通してEMへと直接連結される。しかし、scFv抗体は、そのアミノ末端またはカルボキシル末端を通してEMへと連結され得る。
【0098】
いくつかの抗体の実施形態のVH領域およびVL領域は直接一緒に連結され得るとはいえ、当業者は、これらの領域が、1以上のアミノ酸からなるペプチドリンカーによって隔てられ得ることを認識する。ペプチドリンカーおよびそれらの使用は、当該分野で周知である。例えば、Hustonら、Proc.Nat’l Acad.Sci.USA 8:5879(1988);Birdら、Science 242:4236(1988);Glockshuberら、Biochemistry 29:1362(1990);米国特許第4,946,778号、米国特許第5,132,405号およびStemmerら,Biotechniques 14:256−265(1993)(全て、本明細書中に参考として援用される)を参照のこと。一般的に、ペプチドリンカーは、領域を連結すること、またはこれらの間のいくらかの最小の距離もしくは他の空間的関係を保つこと以外に特定の生物学的活性を有さない。しかし、ペプチドリンカーの構成要素であるアミノ酸は、分子のいくつかの特性(例えば、折り畳み、正味の電荷、または疎水性)に影響を与えるように選択され得る。単鎖Fv(scFv)抗体は必要に応じて、50以下のアミノ酸の、一般的に40以下のアミノ酸の、好ましくは30以下のアミノ酸の、そしてより好ましくは20以下のアミノ酸の長さのペプチドリンカーを含む。いくつかの実施形態では、ペプチドリンカーは、配列Gly−Gly−Gly−Serのコンカテマー(好ましくは、2、3、4、5または6個のこのような配列)である。しかし、リンカー内のいくつかのアミノ酸置換が行われ得ることが認識されるべきである。例えば、バリンで、グリシンを置換し得る。
【0099】
(A.抗体産生)
ポリクローナル抗体を産生する方法は当業者に公知である。手短に述べると、免疫原(好ましくは、単離されたメソセリンまたは細胞外メソセリンエピトープ)はアジュバントと混合され、そして動物がこの混合物で免疫される。免疫原に対する適切に高い力価の抗体が得られた場合、血液がこの動物から収集され、そして抗血清が調製される。所望の場合、さらに、このポリペプチドに対して反応性の抗体について濃縮するための抗血清の分画が行われる。例えば、Coligan、CURRENT PROTOCOLS IN IMMUNOLOGY、Wiley/Greene、NY(1991);ならびにHarlowおよびLane、前出(これらは本明細書中に参考として援用される)を参照のこと。
【0100】
モノクローナル抗体は、当業者に慣用的な種々の技術によって得られ得る。このようなモノクローナル抗体を調製するための技術の説明は、例えば、Stitesら(編)BASIC AND CLINICAL IMMUNOLOGY(第4版)、Lange Medical Publications、Los Altos、CA、およびその中で引用される参考文献;HarlowおよびLane、前出;Goding、MONOCLONAL ANTIBODIES:PRINCIPLES AND PRACTICE(第2版)、Academic Press、New York、NY(1986);KohlerおよびMilstein、Nature 256:495−497(1975);ならびに特にモノクローナル抗体を作製する1つの方法を議論するChowdhury,P.S.ら、Mol.Immunol.34:9(1997)に見出され得る。
【0101】
親のモノクローナル抗体が、標的抗原、または所望の免疫原(例えば、メソセリン)をコードする核酸配列で動物を免疫することによって作製されることが好ましい。異種タンパク質をコードする非複製性転写単位での免疫は、抗原特異的免疫応答を惹起する。外来タンパク質への翻訳後、このタンパク質は、プロセシングされ、そして他の細胞性タンパク質と同様に免疫系に対して提示される。このタンパク質は外来性であるので、このタンパク質およびこれから誘導されるペプチドエピトープに対して免疫応答がマウントされる(Donnellyら、J.Immunol.Methods 176:145−152(1994);およびBoyerら、J.Med.Primatol.25:242−250(1996))。この技術は、タンパク質ベースの免疫に対して2つの顕著な利点を有する。第1の利点は、これは、タンパク質精製を必要としないことである。タンパク質精製は、いくらよくても、時間がかかり、そして多くの膜タンパク質の場合、非常に困難である。第2の利点は、この免疫原が哺乳動物宿主内で合成されるので、これは、正確な翻訳後修飾を受け、そしてそのネイティブな構造に折り畳まれることである。
【0102】
メソセリンコードDNAで免疫するために、メソセリンコードcDNAは、コード配列の転写がプロモーター(例えば、CMVプロモーター)の制御下にあるように、プラスミド中に導入される。次いで、このプラスミドは、動物内に、皮下、皮内、腹腔内などのいずれかによって注射される。結果として、このメソセリンcDNAは動物内でmRNAへと転写され、メソセリンはこのmRNAから翻訳され、この翻訳されたタンパク質は正確な翻訳後修飾を受け、そしてメソセリンを合成した細胞の表面に発現される。この動物はメソセリンに対する抗体を惹起し、そして血清が抗体力価についてモニタリングされる。
【0103】
必要に応じて、コード領域および調節エレメントに加えて、このプラスミドは、アンピシリン耐性(Amp)遺伝子を保有する。Amp遺伝子は、抗体産生の増加に必要なTh1応答についての免疫刺激配列を有することが公知である(Satoら、Science 273:352−354(1996))。
【0104】
上記のように、好ましい実施形態では、このモノクローナル抗体はscFvである。scFv抗体を作製する方法は、記載されている。Huseら、前出;Wardら、Nature 341:544−546(1989);およびVaughanら、前出を参照のこと。手短に述べると、B細胞由来のmRNAが単離され、そしてcDNAが調製される。このcDNAは、免疫グロブリンの重鎖および軽鎖の可変領域に特異的なプライマーを用いて、周知の技術(例えば、PCR)によって増幅される。PCR産物は、例えば、アガロースゲル電気泳動によって精製され、そしてこの核酸配列は連結される。リンカーペプチドが所望される場合、ペプチドをコードする核酸配列は、重鎖の核酸配列と軽鎖の核酸配列との間に挿入される。この配列は、当該分野で公知の技術(例えば、平滑末端連結、PCR産物の末端での制限部位の挿入、または重複伸張によるスプライシングによって)によって連結され得る(Chowdhuryら、Mol.Immunol.34:9(1997))。増幅後、scFvをコードする核酸は、さらに当該分野で周知の技術によってベクター中に挿入される。好ましくは、このベクターは、原核生物中で複製され得、そして真核生物および原核生物の両方において発現され得る。
【0105】
好ましい実施形態では、scFvは、ファージディスプレイライブラリーによって選択される。scFvを合成するための上記の手順は、以下に続く。PCRによる増幅後、scFv核酸配列は、繊維状ファージのマイナーな表面タンパク質gIIIpをコードする、遺伝子III(gIII)(Marksら、J.Biol.Chem.267:16007−16010(1992);Marksら、Behring Inst.Mitt.91:6−12(1992);およびBrinkmannら、J.Immunol.Methods 182:41−50(1995))とインフレームで融合される。このファージは、得られる融合タンパク質をその表面に発現する。ファージ表面のタンパク質は機能的であるので、メソセリン結合抗体を保有するファージは、パニングまたは抗原アフィニティークロマトグラフィー(McCaffertyら、Nature 348:552−554(1990))によって非結合性かまたはより低い親和性のファージから分離され得る。
【0106】
好ましい実施形態では、メソセリンに特異的に結合するscFvは、パニングによって見出される。パニングは、固体表面をメソセリンでコーティングし、そしてこのファージを適切な時間、適切な条件下でこの表面上でインキュベートすることによって行われる。結合していないファージは、固体表面から洗浄除去され、そして結合したファージが溶出される。最大の親和性を有する抗体を見出すことは、選択プロセスの効率によって指図され、そしてスクリーニングされ得るクローン数およびそれが行われるストリンジェンシーに依存する。代表的に、より高いストリンジェンシーは、より選択的なパニングに対応する。しかし、条件があまりにもストリンジェントである場合、ファージは結合しない。1回のパニング後、メソセリンコーティングプレートに結合するファージは、E.coli中で増やされ、そして別の回のパニングに供される。このようにして、2000倍の富化が3回のパニングにおいて行われる。従って、各回の富化が低い場合でさえ、複数回のパニングは、稀なファージの単離をもたらし、そしてその中に含まれる遺伝物質は、最大の親和性抗体の配列をコードする。ファージディスプレイによって提供される遺伝子型と表現型との間の物理的連鎖は、大きなライブラリーのクローンを用いてさえ、cDNAライブラリーの全てのメンバーを抗原に対する結合について試験することを可能にする。
【0107】
(B.抗体の結合親和性)
本発明の抗体は、それらの標的抗原に、それらの親抗体の親和性よりも少なくとも5倍良好な親和性で結合する。1つの実施形態では、この抗体は、メソセリンの細胞外エピトープに結合する抗メソセリン抗体である。標的抗原についての結合親和性は代表的に、標準的な抗体−抗原アッセイ(例えば、競合アッセイ、飽和アッセイまたはイムノアッセイ(例えば、ELISAもしくはRIA))によって測定または決定される。
【0108】
このようなアッセイを用いて、抗体の解離定数を決定し得る。語句「解離定数」とは、抗原についての抗体の親和性をいう。抗体の解離定数(KD=1/K、ここで、Kは、親和定数である)が<1μM、好ましくは<100nM、そして最も好ましくは<0.1nMである場合、抗体と抗原との間に結合の特異性が存在する。抗体分子は代表的に、より低い範囲のKDを有する。KD=[Ab−Ag]/[Ab][Ag]、ここで、[Ab]は平衡状態での抗体の濃度であり、[Ag]は平衡状態での抗原の濃度であり、そして[Ab−Ag]は平衡状態での抗体−抗原複合体の濃度である。代表的に、抗原と抗体との間の結合相互作用は、可逆的非共有結合的会合(例えば、静電引力、ファンデルワールス力および水素結合)を含む。結合特異性を規定するこの方法は、メソセリンに単独でまたは組み合わせて結合する場合にメソセリンに特異的である、単一の重鎖および/もしくは軽鎖、CDR、融合タンパク質または重鎖および/もしくは軽鎖のフラグメントに適用される。
【0109】
(C.イムノアッセイ)
抗体は、充分に認識された多数の免疫学的結合アッセイ(例えば、米国特許第4,366,241号;同第4,376,110号;同第4,517,288号;および同第4,837,168号を参照のこと)のうちのいずれかを用いて検出および/または定量され得る。一般的イムノアッセイの概説については、METHODS IN CELL BIOLOGY、第37巻、Asai編、Academic Press、Inc.New York(1993);BASIC AND CLINICAL IMMUNOLOGY、第7版、StitesおよびTerr編(1991)をまた参照のこと。免疫学的結合アッセイ(またはイムノアッセイ)は代表的に、リガンド(例えば、メソセリン)を利用して抗体に特異的に結合させ、そしてしばしば、この抗体を固定化する。本発明のイムノアッセイにおいて用いられる抗体は、上記でさらに詳細に考察される。
イムノアッセイはまたしばしば、標識化剤を利用して、リガンドおよび抗体によって形成される結合複合体に特異的に結合させ、そしてこの結合複合体を標識する。標識化剤は、それ自体が、抗体/分析物複合体(すなわち、抗メソセリン抗体)を含む部分のうちの1つであり得る。あるいは、標識化剤は、抗体/メソセリンタンパク質複合体に特異的に結合する第3の部分(例えば、別の抗体)であり得る。
【0110】
1つの局面では、標識化剤が、標識を保有する第2の抗メソセリン抗体である競合アッセイが意図される。次いで、この2つの抗体は、固定化されたメソセリンに対する結合について競合する。あるいは、非競合形式において、メソセリン抗体は標識を欠くが、抗メソセリン抗体を誘導した種(例えば、マウス)の抗体に特異的であってそして抗メソセリン抗体に結合する第2の抗体が標識される。
【0111】
免疫グロブリン定常領域に特異的に結合し得る他のタンパク質(例えば、プロテインAまたはプロテインG)もまた、標識剤として用いられ得る。これらのタンパク質は、連鎖球菌細菌の細胞壁の通常の構成要素である。これらは、種々の種由来の免疫グロブリン定常領域との強力な非免疫原性反応性を示す(一般的に、Kronvalら、J.Immunol.111:1401−1406(1973);およびAkerstromら、J.Immunol.135:2589−2542(1985)を参照のこと)。
【0112】
アッセイ全体を通して、インキュベーションおよび/または洗浄の工程は、試薬の各組み合わせの後で必要とされ得る。インキュベーション工程は、約5秒間〜数時間まで、好ましくは約5分間〜約24時間まで変動し得る。しかし、インキュベーション時間は、アッセイの形式、抗体、溶液の容量、濃度などに依存する。通常、アッセイは、室温で実施されるが、アッセイは、一定の範囲の温度(例えば、10℃〜40℃)で実施され得る。
【0113】
本発明のイムノアッセイの詳細は、用いられる特定の形式に伴って変動し得るとはいえ、抗体を含むサンプル中の抗メソセリン抗体を検出する方法は、一般的に、サンプルを、免疫学的に反応性の条件下で、メソセリン/抗体複合体に対して特異的に反応する抗体と接触させる工程を包含する。
【0114】
(V.免疫結合体の産生)
免疫結合体としては、抗体に対する治療剤の共有結合が存在する分子が挙げられるがこれに限定されない。治療剤は、特定の標的分子または標的分子を保有する細胞に対して指向された特定の生物学的活性を有する薬剤である。当業者は、治療剤が、種々の薬物(例えば、ビンブラスチン、ダウノマイシンなど)、細胞毒(例えば、ネイティブかまたは改変されたPseudomonas外毒素またはDiphtheria毒素)、それ自体が薬理学的組成物を含むカプセル化剤(例えば、リポソーム)、放射性薬剤(例えば、125I、32P、14C、3Hおよび35S)ならびに他の標識、標的部分およびリガンドを含み得ることを認識する。
【0115】
特定の治療剤の選択は、特定の標的分子または細胞に依存し、そして生物学的効果は、誘起することが所望される。従って、例えば、治療剤は、特定の標的細胞の死をもたらすために用いられる細胞毒であり得る。逆に、非致死的な生物学的応答を誘発することが単に所望される場合、治療剤は、非致死的な薬理学的薬剤に対して、または非致死的な薬理学的薬剤を含むリポソームに対して結合体化され得る。
【0116】
本明細書中で提供される治療剤および抗体を用いて、当業者は、機能的に等価な核酸(例えば、配列が異なるが、同じEMまたは抗体の配列をコードする核酸)を含む種々のクローンを容易に構築し得る。従って、本発明は、抗体および結合体、ならびにそれらの融合タンパク質をコードする核酸を提供する。
【0117】
(A.組換え方法)
本発明の核酸配列は、任意の適切な方法によって調製され得る。このような方法としては、例えば、適切な配列のクローニングまたは以下のような方法による直接化学合成が挙げられる:例えば、Narangら、Meth.Enzymol.68:90−99(1979)のホスホトリエステル法;Brownら、Meth.Enzymol.68:109−151(1979)のホスホジエステル法;Beaucageら,Tetra.Lett.22:1859−1862(1981)のジエチルホスホルアミダイト法;BeaucageおよびCaruthers、Tetra.Letts.22(20):1859−1862(1981)によって記載される固相ホスホルアミダイトトリエステル法(例えば、Needham−VanDevanterら、Nucl.Acids Res.12:6159−6168(1984)に記載される通りの自動化合成機を用いる);ならびに米国特許第4,458,066号の固体支持法。化学合成は、一本鎖オリゴヌクレオチドを産生する。これらは、相補配列とのハイブリダイゼーションによって、または一本鎖をテンプレートとして用い、DNAポリメラーゼを用いた重合によって二本鎖DNAへと変換され得る。当業者は、DNAの化学合成は約100塩基の配列に制限されるとはいえ、より長い配列は、より短い配列の連結によって入手され得ることを認識する。
【0118】
好ましい実施形態では、本発明の核酸配列は、クローニング技術によって調製される。適切なクローニングおよび配列決定の技術、ならびに多くのクローニング練習を通して当業者に指示するに充分な指示書の例は、以下に見出される:Sambrookら、MOLECULAR CLONING:A LABORATORY MANUAL(第2版)、第1巻〜第3巻、Cold Spring Harbor Laboratory(1989))、BergerおよびKimmel(編)、GUIDE TO MOLECULAR CLONING TECHNIQUES、Academic Press、Inc.、San Diego CA(1987))またはAusubelら(編)、CURRENT PROTOCOLS IN MOLECULAR BIOLOGY、Greene PublishingおよびWiley−Interscience、NY(1987)。生物学的試薬および実験装置の製造業者による製品情報もまた、有用な情報を提供する。このような製造業者としては、以下が挙げられる:SIGMA chemical company(Saint Louis、MO)、R&D systems(Minneapolis、MN)、Pharmacia LKB Biotechnology(Piscataway、NJ)、CLONTECH Laboratories,Inc.(Palo Alto、CA)、Chem Genes Corp.、Aldrich Chemical Company(Milwaukee、WI)、Glen Research,Inc.、GIBCO BRL Life Technologies,Inc.(Gaithersberg、MD)、Fluka Chemica−Biochemika Analytika(Fluka Chemie AG、Buchs,Switzerland)、Invitrogen、San Diego、CA、およびApplied Biosystems(Foster City、CA)、ならびに当業者に公知の多くの他の商業的供給源。
【0119】
ネイティブなEMまたは抗メソセリン抗体をコードする核酸は、EM、抗体、または本発明の免疫結合体を形成するように改変され得る。部位特異的変異誘発による改変は、当該分野で周知である。EMまたは抗メソセリン抗体をコードする核酸は、インビトロ方法によって増幅され得る。増幅方法としては、ポリメラーゼ連鎖反応(PCR)、リガーゼ連鎖反応(LCR)、転写ベースの増幅系(TAS)、自己保持配列複製系(3SR)が挙げられる。広範な種々のクローニング方法、宿主細胞およびインビトロ増幅方法論は、当業者に周知である。
【0120】
好ましい実施形態では、免疫結合体は、抗メソセリンscFv抗体をコードするcDNAを、EMをコードするcDNAを含むベクター中に挿入することによって調製される。挿入は、機能的Fv領域および機能的EM領域を含む1つの連続したポリペプチドにおいて、scFvおよびEMがインフレームで読まれるように行われる。特に好ましい実施形態では、ジフテリア毒素フラグメントをコードするcDNAは、scFvのカルボキシル末端に毒素が配置されるように、scFvへと連結される。最も好ましい実施形態では、PEをコードするcDNAは、scFvのアミノ末端に毒素が配置されるように、scFvへと連結される。
【0121】
一旦、本発明のEM、抗メソセリン抗体、または免疫結合体をコードする核酸が単離され、そしてクローン化されると、組換えによって操作された細胞(例えば、細菌、植物、酵母、昆虫および哺乳動物の細胞)において所望のタンパク質を発現し得る。当業者は、E.coli、他の細菌宿主、酵母および種々のより高等な真核生物細胞(例えば、COS、CHO、HeLaおよびミエローマ細胞株)を含む、タンパク質の発現に利用可能な多数の発現系を良く知っていることが予想される。原核生物または真核生物におけるタンパク質の発現について公知の種々の方法を詳細に記載する試みは行われない。手短に述べると、本発明の単離されたタンパク質をコードする天然または合成の核酸の発現は代表的に、DNAまたはcDNAをプロモーター(これは、構成性または誘導性のいずれかである)に対して作動可能に連結し、続いて発現カセットに組み込むことによって達成される。このカセットは、原核生物または真核生物のいずれかへの複製および組込みに適切であり得る。代表的な発現カセットは、転写および翻訳のターミネーター、開始配列、およびタンパク質をコードするDNAの発現の調節に有用なプロモーターを含む。クローニングされた遺伝子の高レベル発現を得るために、少なくとも、転写を指向する強力なプロモーター、翻訳開始のためのリボソーム結合部位および転写/翻訳ターミネーターを含む発現カセットを構築することが所望される。E.coliについては、これは、プロモーター(例えば、T7、trp、lacまたはλプロモーター)、リボソーム結合部位および好ましくは転写終結シグナルを含む。真核生物細胞については、制御配列は、プロモーターを、そして好ましくは、免疫グロブリン遺伝子、SV40、サイトメガロウイルス由来のエンハンサーおよびポリアデニル化配列を含み得、そしてスプライスドナー配列およびスプライスアクセプター配列を含み得る。本発明のカセットは、E.coliについては塩化カルシウム形質転換またはエレクトロポレーション、そして哺乳動物細胞についてはリン酸カルシウム処理、エレクトロポレーションまたはリポフェクションのような周知の方法によって選択された宿主細胞へと移入され得る。このカセットによって形質転換された細胞は、カセット中に含まれる遺伝子(例えば、amp遺伝子、gpt遺伝子、neo遺伝子およびhyg遺伝子)によって付与される、抗生物質に対する耐性によって選択され得る。
【0122】
当業者は、その生物学的活性を減少させることなく、本発明のポリペプチド(すなわち、抗メソセリン抗体、PEまたはこれらの組み合わせによって形成された免疫結合体)をコードする核酸に対して改変が行われ得ることを認識する。いくつかの改変が行われて、標的化分子のクローニング、発現または融合タンパク質への組込みを容易にし得る。このような改変は当業者に周知であり、そして例えば、終止コドン、アミノ末端に付加されて開始部位を提供するメチオニン、いずれかの末端に配置されて便利に配置された制限部位を作製するさらなるアミノ酸、または精製工程を補助するさらなるアミノ酸(例えば、ポリHis)を含む。
【0123】
組換え法に加えて、本発明の免疫結合体、EMおよび抗体はまた、標準的なペプチド合成を用いて全体としてまたは部分的に構築され得る。約50アミノ酸長未満の本発明のポリペプチドの固相合成は、この配列のC末端アミノ酸を、不溶性支持体へと連結し、続いてこの配列内の残りのアミノ酸を順次付加することによって達成され得る。固相合成のための技術は、BaranyおよびMerrifield、THE PEPTIDES:ANALYSIS,SYNTHESIS,BIOLOGY.第2巻:SPECIAL METHODS IN PEPTIDE SYNTHESIS、第A部、3〜284頁;Merrifieldら、J.Am.Chem.Soc.85:2149−2156(1963)およびStewartら、SOLID PHASE PEPTIDE SYNTHESIS、第2版、Pierce Chem.Co.,Rockford,III.(1984)によって記載される。より大きな長さのタンパク質は、より短いフラグメントのアミノ末端およびカルボキシル末端の縮重によって合成され得る。カルボキシル末端の活性化によってペプチド結合を形成する方法(例えば、カップリング試薬N,N’−ジシクロヘキシルカルボジイミドの使用による)は当業者に公知である。
【0124】
(B.精製)
一旦発現されると、本発明の組換え免疫結合体、抗体、および/またはエフェクター分子は、当該分野の標準的な手順に従って精製され得る。このような手順としては、硫酸アンモニウム沈澱、アフィニティーカラム、カラムクロマトグラフィーなどが挙げられる(一般的に、R.Scopes、PROTEIN PURIFICATION,Springer−Verlag、N.Y.(1982)を参照のこと)。少なくとも約90%〜95%の均質性の実質的に純粋な組成物が好ましく、そして98%〜99%以上の均質性が、薬学的用途に最も好ましい。所望に応じて、部分的にまたは均質になるまで(治療に用いられる場合)一旦精製されると、このポリペプチドは、内毒素を実質的に含まないはずである。
【0125】
細菌(例えば、E.coli)からの単鎖抗体の発現および/または適切な活性形態(単鎖抗体を含む)への再折り畳みのための方法は、記載されており、そして周知であり、そして本発明の抗体に適用可能である。Buchnerら、Anal.Biochem.205:263−270(1992);Pluckthun,Biotechnology 9:545(1991);Huseら、Science 246:1275(1989)およびWardら,Nature 341:544(1989)(全て本明細書中に参考として援用される)を参照のこと。
【0126】
しばしば、E.coliまたは他の細菌由来の機能的異種タンパク質は、封入体から単離され、そして強力な変性剤を用いた可溶化およびその後の再折り畳みを必要とする。可溶化工程の間、当該分野で周知であるように、ジスルフィド結合を分離するために還元剤が存在しなければならない。還元剤を含む例示的な緩衝液は以下である:0.1M Tris pH8、6Mグアニジン、2mM EDTA、0.3M DTE(ジチオエリトリトール)。ジスルフィド結合の再酸化は、本明細書中に参考として援用される、Saxenaら、Biochemistry 9:5015−5021(1970)に記載され、そして特にBuchnerら、前出によって記載されるように、還元型および酸化型の低分子量チオール試薬の存在下で生じ得る。
【0127】
再生は代表的に、変性しそして還元されたタンパク質の、再折り畳み緩衝液中への希釈(例えば、100倍)によって達成される。代表的な緩衝液は、0.1M Tris、pH8.0、0.5M L−アルギニン、8mM酸化型グルタチオン(GSSG)および2mM EDTAである。
【0128】
二本鎖抗体精製プロトコルへの改変物として、重鎖領域および軽鎖領域は、別々に可溶化および還元され、次いで、再折り畳み溶液中で合わされる。これらの2つのタンパク質が、他方に対して一方のタンパク質が5倍モル過剰を超えないようなモル比で混合される場合、好ましい収率が得られる。過剰の酸化型グルタチオンまたは他の酸化性低分子量化合物を、レドックスシャッフリングが完了した後の再折り畳み溶液に添加することが望ましい。
【0129】
(VI.Pseudomonas外毒素および他の毒素)
毒素を、本発明の抗体と共に用いて、イムノトキシンを産生し得る。例示的な
毒素としては、リシン、アブリン、ジフテリア毒素、およびそれらのサブユニッ
ト、ならびにボツリヌス毒素A〜Fが挙げられる。これらの毒素は、商業的供給
源(例えば、Sigma Chemical Company、St.Loui
s、MO)から容易に入手可能である。ジフテリア毒素は、Corynebac
terium diphtheriaeから単離される。リシンは、Ricin
us communis(トウゴマの実(Castor bean))由来のレ
クチンRCA60である。この用語はまた、それらの毒性改変体も参照する。例
えば、米国特許第5,079,163号および同第4,689,401号を参照
のこと。Ricinus communis凝集素(RCA)は2つの形態で生
じ、それぞれ、それらの約65kDおよび約120kDの分子量に従って、RC
A60およびRCA120と称される(NicholsonおよびBlaustei
n、J.Biochim.Biophys.Acta.266:543(197
2))。A鎖は、タンパク質合成の不活化および細胞の殺傷を担う。B鎖は、細
胞表面のガラクトース残基にリシンを結合させ、そして細胞質ゾル内へのA鎖の
輸送を促進する(Olsnesら、Nature 249:627−631(1
974)および米国特許第3,060,165号)。
【0130】
アブリンは、Abrus precatorius由来の毒性レクチンを含む
。毒性の本質である、アブリンa、b、c、およびdは、約63kDから、約6
7kDの分子量を有し、そして2つのジスルフィド結合ポリペプチド鎖Aおよび
Bから構成される。A鎖は、タンパク質合成を阻害し;B鎖(アブリンb)は、
Dガラクトース残基に結合する(Funatsuら、Agr.Biol.Che
m.52:1095(1988);およびOlsnes、Methods En
zymol.50:330−335(1978)を参照のこと)。
【0131】
本発明の好ましい実施形態では、毒素はPseudomonas外毒素(PE
)である。用語「Pseudomonas外毒素」は、本明細書中で使用される
場合、全長のネイティブな(天然に存在する)PE、または改変されているPE
をいう。このような改変としては、ドメインIaの除去、ドメインIb、II、
およびIIIにおける種々のアミノ酸の欠失、単一のアミノ酸置換、およびカル
ボキシル末端での1以上の配列(例えば、KDELおよびREDL)の付加が挙
げられ得るが、これらに限定されない。Siegallら、J.Biol.Ch
em.264:14256−14261(1989)を参照のこと。好ましい実
施形態では、PEの細胞障害性フラグメントは、ネイティブなPEの細胞傷害性
の少なくとも50%、好ましくは75%、より好ましくは少なくとも90%、そ
して最も好ましくは95%を保持する。最も好ましい実施形態では、細胞傷害性
フラグメントは、ネイティブなPEよりも毒性が高い。
【0132】
ネイティブなPseudomonas外毒素A(PE)は、Pseudomo
nas aeruginosaによって分泌される、極めて活性なモノマー性の
タンパク質(分子量66kD)であり、これは、真核生物細胞中におけるタンパ
ク質合成を阻害する。ネイティブなPE配列は、同一人に譲渡された米国特許第
5,602,095号(本明細書中で参考として援用される)に提供される。作
用の方法は、伸長因子2(EF−2)のADP−リボシル化の不活化である。外
毒素は、細胞傷害性を引き起こすのに提携して作用する3つの構造ドメインを含
む。ドメインIa(アミノ酸1〜252)は、細胞結合を媒介する。ドメインI
I(アミノ酸253〜364)は、細胞質ゾル内へのトランスロケーションを担
い、そしてドメインIII(アミノ酸400〜613)は、伸長因子2のADP
リボシル化を媒介する。ドメインIb(アミノ酸365〜399)の機能は、未
だ決定されていないが、その大部分であるアミノ酸365〜380は、細胞傷害
性の損失を伴なわずに欠失され得る。Siegallら(1989)、前出を参
照のこと。
【0133】
本発明において使用されるPEとしては、ネイティブな配列、ネイティブな配
列の細胞傷害性フラグメント、ならびにネイティブなPEおよびその細胞傷害性
フラグメントの保存的に改変された改変体が挙げられる。PEの細胞傷害性フラ
グメントは、標的細胞中における引き続くタンパク質分解性または他のプロセシ
ングを伴なって、または伴なわずに、細胞傷害性であるフラグメントを含む(例
えば、タンパク質またはプレタンパク質として)。PEの細胞傷害性フラグメン
トとしては、PE40、PE38、およびPE35が挙げられる。
【0134】
好ましい実施形態では、PEは、米国特許第4,892,827号において教
示されるように、しばしばドメインIaを欠失させることによって改変され、非
特異的な細胞結合を減少または除去されるが、これはまた、例えば、ドメインI
aの特定の残基を変異させることによっても達成され得る。例えば、米国特許第
5,512,658号は、変異されたPE(ここでは、ドメインIaが存在する
が、ドメインIaの57位、246位、247位、および249位の塩基性残基
が、酸性残基(グルタミン酸(すなわち、「E」)で置換されている)が、非常
に減少した非特異的細胞傷害性を示すことを開示している。PEのこの変異形態
は、時折、PE4Eと呼ばれる。
【0135】
PE40は、当該分野で以前に記載されたように、PEの短縮化(trunc
ated)誘導体である。Paiら、Proc.Nat’l.Acad.Sci
.USA 88:3358−62(1991);およびKondoら、J.Bi
ol.Chem.263:9470−9475(1988)を参照のこと。PE
35は、PEの35kDのカルボキシル末端フラグメントであり、ここでは、ア
ミノ酸残基1〜279が欠失しており、そしてこの分子は、280位のmetで
開始し、ネイティブなPEのアミノ酸281〜364および381〜613が続
く。PE35およびPE40は、例えば、米国特許第5,602,095号およ
び同第4,892,827号に開示される。
【0136】
いくつかの好ましい実施形態では、細胞傷害性フラグメントPE38が使用さ
れる。PE38は、アミノ酸253〜364および381〜613から構成され
る短縮化PEプロタンパク質であり、これは、細胞内でのプロセシングに際して
、その細胞傷害性形態へと活性化される(例えば、米国特許第5,608,03
9号およびPastanら、Biochim.Biophys.Acta 13
33:C1−C6(1997)を参照のこと)。
【0137】
上述のように、いくつかまたはすべてのドメイン1bが欠失され得、そして残
りの部分が、リンカーによってか、またはペプチド結合によって直接的に連結さ
れ得る。ドメインIIのいくつかのアミノ酸部分は、欠失され得る。そして、C
末端は、残基609〜613のネイティブな配列(REDLK)を含み得るか、
または構築物を細胞質ゾル内にトランスロケーションする能力を維持することが
見出されているバリエーション(例えば、REDLまたはKDEL)、およびこ
れらの配列の反復を含み得る。例えば、米国特許第5,854,044号;同第
5,821,238号;および同第5,602,095号、ならびにWO99/
51643を参照のこと。好ましい実施形態では、PEは、PE4E、PE40
、またはPE38であるが、非標的細胞への顕著な毒性が生じないレベルまで、
非特異的細胞傷害性が除去または減少されている任意の形態のPEが、標的化細
胞においてトランスロケーションし得、そしてEF−2をリボシル化し得るまま
である限り、本発明のイムノトキシンに使用され得る。
【0138】
(A.PEの保存的に改変された改変体)
PEまたはその細胞傷害性フラグメントの保存的に改変された改変体は、目的
のPE(例えば、PE38)と、アミノ酸レベルで、少なくとも80%の配列類
似性、好ましくは、少なくとも85%の配列類似性、より好ましくは、少なくと
も90%の配列類似性、そして最も好ましくは少なくとも95%の配列類似性を
有する。
【0139】
用語「保存的に改変された改変体」は、アミノ酸配列および核酸配列の両方に
適用する。特定の核酸配列に関して、保存的に改変された改変体とは、同一また
は本質的に同一なアミノ酸配列をコードする核酸配列をいうか、または核酸がア
ミノ酸配列をコードしない場合には、本質的に同一な核酸配列をいう。遺伝暗号
の縮重性が理由で、多数の機能的に同一な核酸が、任意の所定のポリペプチドを
コードする。例えば、コドンGCA、GCC、GCGおよびGCUはすべて、ア
ミノ酸アラニンをコードする。従って、コドンによってアラニンが特定されるす
べての位置で、そのコドンを、コードされるポリペプチドを変更することなく、
記載された任意の対応するコドンに変更し得る。このような核酸のバリエーショ
ンは、「サイレントなバリエーション」であり、これは、保存的に改変されたバ
リエーションの一種である。ポリペプチドをコードする、本明細書中のすべての
核酸配列はまた、核酸のすべての可能なサイレントなバリエーションを記載する
。当業者は、核酸中の各コドン(AUG(これは、通常、メチオニンについての
唯一のコドンである)を除く)が、機能的に同一な分子を産生するように改変さ
れ得ることを認識する。従って、ポリペプチドをコードする核酸のサイレントな
バリエーションの各々が、各々記載された配列中において含意される。
【0140】
アミノ酸配列に関して、当業者は、コードされる配列において単一のアミノ酸
または低いパーセンテージのアミノ酸を変更、付加、または欠失する、核酸配列
、ペプチド配列、ポリペプチド配列、またはタンパク質配列への個々の置換、欠
失、または付加が、「保存的に改変された改変体」であることを認識し、ここで
変更は、化学的に類似のアミノ酸でのアミノ酸の置換を生じる。
【0141】
(B.PEの細胞傷害性についてのアッセイ)
本発明で使用されるPseudomonas外毒素を、当業者に周知のアッセ
イによって、所望されるレベルの細胞傷害性についてアッセイし得る。例示的な
毒性アッセイは、本明細書中において、例えば、実施例2に記載される。従って
、PEの細胞傷害性フラグメントおよびこのようなフラグメントの保存的に改変
された改変体は、細胞傷害性について容易にアッセイされ得る。多数の候補PE
分子を、当該分野において周知の方法によって、細胞傷害性について同時にアッ
セイし得る。例えば、候補分子のサブグループを、細胞傷害性についてアッセイ
し得る。候補分子の中で、ポジティブに反応するサブグループを継続的に細分し
得、そして所望の細胞傷害性フラグメントが同定されるまで、再度アッセイし得
る。このような方法は、PEの多数の細胞傷害性フラグメントまたは保存的改変
体の迅速なスクリーニングを可能にする。
【0142】
(C.他の治療的部分)
本発明の抗体はまた、かなり多数の異なる診断用または治療用化合物を、表面
上にメソセリンを発現する細胞に標的化するために使用され得る。従って、本発
明の抗体(例えば、抗メソセリンscFv)は、メソセリンを保有する細胞に直
接的に送達されるべき薬物に対して、直接的に結合され得るか、またはリンカー
を介して結合され得る。治療剤としては、核酸、タンパク質、ペプチド、アミノ
酸、または誘導体、糖タンパク質、ラジオアイソトープ、脂質、炭水化物、また
は組換えウイルスのような化合物が挙げられる。核酸の治療用および診断用部分
としては、アンチセンス核酸、一本鎖または二重鎖DNAと共有結合的に架橋す
る誘導体化オリゴヌクレオチド、および三重鎖形成オリゴヌクレオチドが挙げら
れる。
【0143】
あるいは、抗メソセリン抗体に連結される分子は、治療用組成物(例えば、薬
物、核酸(例えば、アンチセンス核酸)、または循環系への直接曝露から、好ま
しくは遮蔽される別の治療的部分)を含む、リポソームまたはミセルのようなカ
プセル系であり得る。抗体に結合したリポソームを調製する手段は、当業者に周
知である。例えば、米国特許第4,957,735号;およびConnorら、
Pharm.Ther.28:341−365(1985)を参照のこと。
【0144】
(D.検出可能標識)
本発明の抗体は、必要に応じて、検出可能な標識に共有結合的にか、または非
共有結合的に連結され得る。このような用途に適切な検出可能標識としては、分
光学的手段、光化学的手段、生化学的手段、免疫化学的手段、電気的手段、光学
的手段、または化学的手段によって検出可能な任意の組成物が挙げられる。本発
明において有用な標識としては、磁気ビーズ(例えば、DYNABEADS)、
蛍光色素(例えば、フルオレセインイソチオシアネート、Texasレッド、ロ
ーダミン、グリーン蛍光タンパク質など)、放射能標識(例えば、3H、125I、
35S、14C、または32P)、酵素(例えば、西洋ワサビペルオキシダーゼ、アル
カリホスファターゼ、およびELISAにおいて通常使用される他の酵素)、お
よび比色定量標識(例えば、金コロイドあるいは呈色(colored)ガラス
またはプラスチック(例えば、ポリスチレン、ポリプロピレン、ラテックスなど
)ビーズ)が挙げられる。
【0145】
このような標識を検出する手段は、当業者に周知である。従って、例えば、放
射能標識は、写真フィルムまたはシンチレーションカウンターを使用して検出さ
れ得、蛍光マーカーは、光検出器を使用して検出して、放射された照度を検出し
得る。酵素学的標識は、代表的に、酵素に基質を提供すること、そしてその基質
に対する酵素の作用によって生成される反応生成物を検出することによって検出
される。比色定量標識は、単純に、呈色した標識を可視化することによって検出
される。
【0146】
(E.抗体への結合体化)
本発明の非組換え的実施形態では、エフェクター分子(例えば、治療的部分、
診断用部分、または検出用部分)を、当業者に公知のかなり多数の手段を使用し
て、本発明の抗メソセリン抗体に連結する。共有結合手段および非共有結合手段
の両方が、本発明の抗メソセリン抗体で使用され得る。
【0147】
エフェクター分子を抗体に結合させるための手順は、EMの化学的構造に従っ
て変動する。ポリペプチドは、代表的に、種々の官能基(例えば、カルボン酸(
COOH)、遊離アミン(−NH2)、またはスルフヒドリル(−SH)基を含
む;これらは、エフェクター分子の結合を生じさせるための、抗体における適切
な官能基との反応に利用可能である。
【0148】
あるいは、抗体は、さらなる反応性官能基に曝露されるか、または結合される
ように誘導体化される。誘導体化は、多数の任意のリンカー分子(例えば、Pi
erce Chemical Company、Rockford Illin
oisから入手可能なリンカー)の結合を包含し得る。
【0149】
「リンカー」は、本明細書中で使用される場合、抗体をエフェクター分子に連
結するために使用される分子である。リンカーは、抗体とエフェクター分子の両
方に対して共有結合を形成し得る。適切なリンカーは、当業者に周知であり、そ
してこれには、直鎖もしくは分枝の炭素リンカー、複素環式炭素リンカー、また
はペプチドリンカーが挙げられるが、これらに限定されない。抗体およびエフェ
クター分子がポリペプチドである場合、リンカーは、それらの側鎖を通して(例
えば、システインへのジスルフィド結合を通して)、構成要素のアミノ酸に連結
され得る。しかし、好ましい実施形態では、リンカーは、末端アミノ酸のα炭素
アミノ基およびカルボキシル基に連結される。
【0150】
いくつかの状況では、免疫結合体がその標的部位に到達した時点で、抗体から
エフェクター分子を遊離させることが所望され得る。従って、これらの状況では
、免疫結合体は、標的部位の近傍で切断可能である結合を含む。抗体からエフェ
クター分子を放出するためのリンカーの切断は、免疫結合体が、標的細胞の内側
または標的部位の近傍のいずれかで供される酵素学的な活性または条件によって
刺激され得る。標的部位が腫瘍である場合、腫瘍部位に存在する条件下で(例え
ば、腫瘍に関連した酵素または酸性pHに曝露された場合に)切断可能なリンカ
ーが使用され得る。
【0151】
種々の放射線診断用化合物、放射線療法化合物、薬物、毒素、および他の薬剤
を抗体に結合させることに関して報告されている多数の方法に照らして、当業者
は、所定の薬剤を抗体または他のポリペプチドに結合させるために適切な方法を
決定し得る。
【0152】
(VII.薬学的組成物および投与)
本発明の抗体および/または免疫結合体組成物(すなわち、抗体E4と共に、
その親抗体の少なくとも5倍の抗原親和性を有する抗体に連結されたPE)は、
非経口投与(例えば、静脈内投与、あるいは体腔または器官の内腔内への投与)
に特に有用である。例えば、卵巣悪性疾患は、腫瘍を取り囲んでいる組織への静
脈内投与または局所送達によって処置され得る。中皮腫を処置するためには、抗
メソセリン抗体を含む本発明の薬学的組成物は、胸膜腔または腹膜腔内に直接的
に投与され得る。
【0153】
投与のための組成物は、通常、薬学的に受容可能なキャリア(好ましくは、水
性キャリア)中に溶解された抗体および/または免疫結合体の溶液を含む。種々
の水性キャリア(例えば、緩衝化生理食塩水など)が使用され得る。これらの溶
液は滅菌され、そして一般的には、所望されない物質を含まない。これらの組成
物は、従来の周知の滅菌技術によって滅菌され得る。組成物は、ほぼ生理的条件
に必要とされるような薬学的に受容可能な補助物質(例えば、pH調整剤および
緩衝化剤、毒性調整剤など(例えば、酢酸ナトリウム、塩化ナトリウム、塩化カ
リウム、塩化カルシウム、乳酸ナトリウムなど))を含み得る。これらの処方物
における融合タンパク質の濃度は広範に変動し得、そして選択された特定の投与
様式および患者の必要性に従って、主に液量、粘性、体重などに基づいて選択さ
れる。
【0154】
従って、静脈内投与のための本発明の代表的な薬学的イムノトキシン組成物は
、1日あたり1患者あたり約0.1〜10mgである。特に、隔離された部位(
そして、循環系またはリンパ系ではない)(例えば、体腔内または器官の内腔内
)に薬物が投与される場合、1日あたり1患者あたり0.1から約100mgま
での投薬量が使用され得る。投与可能な組成物を調製するための実際の方法は、
当業者に公知であるか、または明らかであり、そして、REMINGTON’S
PHARMACEUTICAL SCIENCE、第19版、Mack Pu
blishing Company、Easton、Pennsylvania
(1995)のような刊行物において、より詳細に記載されている。
【0155】
本発明の組成物は、治療的処置のために投与され得る。治療的適用において、
組成物は、疾患に罹患している患者に対して、その疾患およびその合併症を治癒
するか、または少なくとも部分的に抑制するに十分な量で投与される。これを達
成するために十分な量を、「治療的有効用量」と規定する。この用途のために有
効な量は、疾患の重篤度および患者の健康の全身的状態に依存する。化合物の有
効量は、臨床医または他の有資格観察者によって観察されるような、症状の主観
的緩和または客観的に同定可能な改善のいずれかを与える量である。
【0156】
組成物の単回投与または複数回投与は、患者によって必要とされ、かつ耐容さ
れるような、投薬量および頻度に依存して投与される。いずれにしても、組成物
は、患者を有効に処置するために十分な量の本発明のタンパク質を提供するべき
である。好ましくは、投薬量は一度に投与されるが、治療的結果が達成されるか
、または副作用によって、治療の中断が正当化されるかのいずれかまで、周期的
に適用され得る。一般的に、用量は、患者に対して受容不可能な毒性を産生する
ことなく、疾患の症状または徴候を処置または改善するに十分である。
【0157】
本発明の免疫結合体組成物の徐放性非経口処方物は、インプラントとしてか、
油性注射剤、または粒子系として作製され得る。タンパク質送達系の広範な概説
については、Banga,A.J.、THERAPEUTIC PEPTIDE
S AND PROTEINS:FORMULATION,PROCESSIN
G,AND DELIVERY SYSTEMS,Technomic Pub
lishing Company,Inc.、Lancaster、PA(19
95)(本明細書中で参考として援用される)を参照のこと。粒子系としては、
ミクロスフェア、微小粒子、マイクロカプセル、ナノカプセル、ナノスフェア(
nanosphere)、およびナノ粒子(nanoparticle)が挙げ
られる。マイクロカプセルは、中心コアとして、治療用タンパク質を含む。ミク
ロスフェアでは、治療剤は、粒子全体にわたって分散される。約1μm未満の粒
子、ミクロスフェア、およびマイクロカプセルを、一般的に、それぞれ、ナノ粒
子、ナノスフェア、およびナノカプセルという。キャピラリーは、約5μmの直
径を有し、その結果、ナノ粒子のみが静脈内に投与される。微小粒子は、代表的
に、直径約100μmであり、そして皮下的または筋内に投与される。例えば、
Kreuter,J.、COLLOIDAL DRUG DELIVERY S
YSTEMS、J.Kreuter編、Marcel Dekker,Inc.
、New York、NY、219〜342頁(1994);ならびに、Tic
eおよびTabibi、TREATISE ON CONTROLLED DR
UG DELIVERY、A.Kydonieus編、Marcel Dekk
er,Inc.、New York、NY、315−339頁(1992)(こ
れらの両方とも、本明細書中で参考として援用される)を参照のこと。
【0158】
ポリマーが、本発明の免疫結合体組成物のイオン制御された放出のために使用
され得る。制御された薬物送達における使用のための、種々の分解可能および分
解不可能ポリマーマトリクスが、当該分野において公知である(Langer,
R.、Accounts Chem.Res.26:537−542(1993
))。例えば、ブロックコポリマーであるポロキサマー407(polaxam
er 407)は、低温では粘性であるがなお可動性の液体として存在するが、
体温では、半固体ゲルを形成する。これは、組換えインターロイキン−2および
ウレアーゼの処方物および持続型送達のために有効なビヒクルであることが示さ
れている(Johnstonら、Pharm.Res.9:425−434(1
992);およびPecら、J.Parent.Sci.Tech.44(2)
:58−65(1990))。あるいは、ヒドロキシアパタイトが、タンパク質
の徐放のためのマイクロキャリアとして使用されている(Ijntemaら、I
nt.J.Pharm.112:215−224(1994))。さらに別の局
面では、リポソームが、脂質カプセル化薬物の徐放および薬物標的化のために使
用されている(Betageriら、LIPOSOME DRUG DELIV
ERY SYSTEMS、Technomic Publishing Co.
,Inc.、Lancaster、PA(1993))。治療用タンパク質の制
御された送達のための多数のさらなる系が、公知である。例えば、米国特許第5
,055,303号、同第5,188,837号、同第4,235,871号、
同第4,501,728号、同第4,837,028号、同第4,957,73
5号、および同第5,019,369号、同第5,055,303号;同第5,
514,670号;同第5,413,797号;同第5,268,164号;同
第5,004,697号;同第4,902,505号;同第5,506,206
号;同第5,271,961号;同第5,254,342号;ならびに同第5,
534,496号(これらの各々が、本明細書中で参考として援用される)を参
照のこと。
【0159】
とりわけ本発明のイムノトキシンの種々の用途には、融合タンパク質の毒性作
用によって除去され得る、特定のヒト細胞によって引き起こされる種々の疾患状
態が含まれる。本発明のイムノトキシンについて1つの好ましい適用は、メソセ
リンを発現する悪性細胞の処置である。例示的な悪性細胞としては、卵巣癌、胃
癌、および扁平上皮癌、ならびに中皮腫が挙げられる。
【0160】
(VIII.診断用キットおよびインビトロ用途)
別の実施形態では、本発明は、生物学的サンプルにおいてメソセリンまたはそ
の免疫反応性フラグメント(すなわち、集合的に「メソセリンタンパク質」)を
検出するためのキットを提供する。「生物学的サンプル」は、本明細書中で使用
される場合、メソセリンを含む生物学的組織または流体のサンプルである。この
ようなサンプルとしては、生検からの組織、痰、羊水、血液、および血球(例え
ば、白血球)が挙げられるが、これらに限定されない。流体サンプルは、幾分興
味深いものであり得るが、一般的には、本発明においては好ましくない。なぜな
ら、検出可能な濃度のメソセリンが、このようなサンプル中で見出されることは
稀であるからである。生物学的サンプルはまた、組織の切片(例えば、組織学的
目的のために採取された凍結切片)を含む。生物学的サンプルは、代表的に、多
細胞真核生物(好ましくは、哺乳動物(例えば、ラット、マウス、ウシ、イヌ、
モルモット、またはウサギ)、そしてより好ましくは霊長類(例えば、マカク、
チンパンジー、またはヒト)から得られる。
【0161】
キットは、代表的に、本発明の抗メソセリン抗体を備える。いくつかの実施形
態では、抗メソセリン抗体は、抗メソセリンFvフラグメント(例えば、scF
vフラグメント)である。キットは、E4抗体を備え得る。
【0162】
さらに、キットは、代表的に、本発明の抗体の使用手段(例えば、サンプル中
の中皮細胞の検出のため)を開示する、指示資料を備える。キットはまた、その
キットが設計された特定の適用を容易にするために、さらなる成分を備え得る。
従って、例えば、キットは、標識を検出する手段(例えば、酵素学的標識につい
て酵素基質、蛍光標識を検出するためのフィルターセット、適切な二次標識(例
えば、ヒツジ抗マウス−HRP、など)をさらに備え得る。キットは、特定の方
法を実施するために慣用的に使用される、緩衝液および他の試薬をさらに備え得
る。このようなキットおよび適切な内容物は、当業者に周知である。
【0163】
本発明の1つの実施形態では、診断用キットは、イムノアッセイを含む。上述
のように、本発明のイムノアッセイの詳細は、使用される特定の様式に応じて変
動し得るが、生物学的サンプル中でメソセリンを検出する方法は、一般的に、生
物学的サンプルを、免疫学的反応条件下でメソセリンに特異的に反応する抗体と
接触させる工程を包含する。抗体は、免疫学的反応条件下で、メソセリンへの結
合を可能にされ、そして結合した抗体の存在が、直接的または間接的に、検出さ
れる。
【0164】
本明細書中で教示される方法によって開発された抗体(そして特に、SS1、
D8、およびC10と称される抗体)の親和性の増加に起因して、本明細書中に
提供される抗体は、特に、診断用薬剤として、そして生物学的サンプル中におい
てメソセリンの存在を検出するためのインビトロアッセイにおいて、有用である
。例えば、本明細書中に教示される方法によって作製された、抗体SS1、D8
、C10および他の抗体は、サンプルが、メソセリンを発現する細胞を含むか否
かを決定するための免疫組織化学アッセイにおいて、免疫結合体の標的化部分と
して使用され得る。サンプルが、通常メソセリンを発現しないはずの患者の組織
から採取されたサンプルである場合、メソセリンの検出は、その患者が、メソセ
リン発現細胞の存在によって特徴付けられる癌を有すること、または、このよう
な癌の処置が未だ、癌を根絶するのに成功していないこと、のいずれかを示す。
抗体E4もまた、これらの目的のために使用され得る。
【0165】
本発明の別のセットの用途では、本発明の抗体によって標的化されるイムノト
キシンを使用して、培養物中の細胞集団から標的化細胞を一掃し得る。従って、
例えば、メソセリンを発現する癌を有する患者から培養された細胞は、標的化部
分としてSS1 scFv、D8 scFv、またはC10 scFvを使用す
るイムノトキシンと培養物とを接触させることによって、癌細胞を一掃され得る
。E4 scFvもまた、これらの目的のための標的化部分として使用され得る
。
【0166】
本発明を、理解の明確さの目的で、例示および実施例によって幾分詳細に記載
してきたが、特定の変化および改変が、添付の特許請求の範囲に記載の範囲内で
実施され得ることが明らかである。
【実施例】
【0167】
(IX.実施例)
(実施例1 ライブラリーの構築)
SS(scFv)中の32個のホットスポットに関して、14個はVH中に、
18個はVL中に配置した。本発明者らは、変異誘発を開始するためにCDR3
を選択した。なぜなら、CDR3は、抗原との相互作用の主要な部位だからであ
る。本発明者らは、本発明者らの開始研究に関して、VH CDR3に対してV
L CDR3を選択した。なぜなら、VL CDR3は、より少ない組換え事象
を受け、故に生殖系列(germline)より近いからである。VLのCDR
3の場合、2つのA/G−G−C/T−A/T型のホットスポットおよび1つの
AGTセリンコドンが存在する。本発明者らは、VL CDR3においてテトラ
ヌクレオチドホットスポット上で集中することによって本発明者らの実験を開始
した。本発明者らの研究において、本発明者らは、そのようにしない特別の理由
がない限り、テトラヌクレオチドモチーフの全体または部分を形成するコドンの
全てのヌクレオチドを無作為化した(以下に議論する)。
【0168】
最初のファージライブラリー(Lib 89/93/94)は、2つのテトラ
ヌクレオチドホットスポット(1つはVL中の残基89および90をコードし、
他方は残基93および94をコードする)の一部であるコドンの無作為化を含ん
だ。Q90は、サブグループVI(SS VLが属する)中で非常に高く保存さ
れている(Johnsonら、WEIR’S HANDBOOK OF EXP
ERIMENTAL IMMUNOLOGY I.IMMUNOCHEMIST
RY AND MOLECULAR IMMUNOLOGY、第5版、Herz
enbergら、編、Blackwell Science Inc.、Cam
bridge、MA、第6章、6.1−6.21頁(1996))ので、本発明
者らは、これを変化させないままで、残基89、93および94を無作為化する
ように選択した。3つのアミノ酸が無作為化されるライブラリーは、8×103
個の異なるタンパク質改変体を有するべきであり、そして全ての異なるヌクレオ
チドの組み合わせに関して(NNSオリゴを用いる)、完全なレパートリーは3
×104個の異なるクローンを含む。本発明者らの作製したライブラリーは、6
×104個のクローンを含み、そして実質的に全ての可能性を包含すると予想さ
れる。
【0169】
Lib 89/93/94をパニングし、そして分析した後、本発明者らは、
Q89は全ての結合剤において保存されるが、置換が93位および94位におい
て許容されたことを見出した。この情報を用いて、本発明者らは、第2のライブ
ラリーのLib 92−94(この中で、S92に対するAGTコドン(別の型
のホットスポット)が、G93およびY94に対するコドンと共に変異された)
を構築した。このライブラリーの大きさは、1×105であった。
【0170】
本発明者らの実験のコントロールとして、本発明者らは、2つのライブラリー
(Lib 89−91およびLib 95−97)を作製した。Lib 89−
91を作製してホットスポット内での保存残基の無作為化の効果を試験し、そし
てLib 95−97を作製してホットスポット外での残基の無作為化の効果を
研究した。これらのライブラリーにおいて、3つのコドンを無作為化し、他の2
つのライブラリー(この中で、合理的に選択されたホットスポット中に位置する
3つのコドンが無作為化された)との比較を可能にした。これらの各々のライブ
ラリーの大きさはまた、1×105であった。
【0171】
(実施例2 ライブラリーのパニングおよび結合剤の濃縮)
ライブラリーを固定化された組換えメソセリンにパニングした。パニング実験
の結果は、表3に示す。2〜6×1011のファージの入力を用いると、バックグ
ラウンドが通常5〜6×104辺りであることを、本発明者らは理解する。Li
b 89/93/94およびLib 92−94に関して、本発明者らは、2×
105のファージを最初のパニングから得た。これは、メソセリンに結合するフ
ァージの約3倍の濃縮を示す。第2回目において、それぞれ、150および13
0倍である大きな濃縮の増加が存在した。これらのライブラリーに対する第3回
目のパニング後の濃縮は、約5倍であった。全体の濃縮は約2000であった。
対照的に、コントロールライブラリーに関して、第1回目のパニング後に濃縮は
存在しなかったが、その後に、漸進的な濃縮における増加が存在した(第2回目
のパニング後に10倍、そして第3回目の後に200倍)。コントロールライブ
ラリーの両方に関する全体の濃縮はまた、約2000倍であった。全てのライブ
ラリーに対する全体の濃縮は類似していたが、濃縮のパターンは、著しく異なっ
た。2つのライブラリー(Lib 89/93/94およびLib 92−94
の初期でありかつ迅速な濃縮は、両方が多数の結合剤を含んでいたことを示す。
対照的に、コントロールライブラリー(Lib 89−91およびLib 95
−97)のゆっくりとした漸進的な濃縮は、これらがより少ない結合剤(これは
、連続回のパニングの間にゆっくりと濃縮される)を含んでいたことを示唆する
。
【0172】
(表3)
組換えメソセリン上のパニングによるライブラリーからのメソセリン結合ファ
ージの濃縮。*2〜6×1011の入力ファージ数に対して5〜6×104であるバ
ックグラウンドレベルのファージ結合を超える濃縮に関する。
【0173】
【表2】
(実施例3 メソセリン結合に対するクローンのスクリーニング)
次に、本発明者らは、異なるライブラリーの各々から選択されたファージの結
合特性を調べた。各ライブラリーに対して、本発明者らは、ELISAによって
、第2回目および第3回目のパニングから23個のクローンをスクリーニングし
た。この実験の結果は、図2に示す。
【0174】
図2Bは、Lib 89/93/94のパニング後に選択されたクローンのメ
ソセリン結合活性を示す。図から明らかなように、全て46個のクローンは、メ
ソセリンに結合し、そして4つの型に分類され得る:(i)27個のクローンは
、非常に高いELISAシグナルを有した(2.3〜2.7);(ii)高いE
LISAシグナルを有する12個のクローン(1.75〜2.2);(iii)
6個のクローンは、親のSS(scFv)ファージ(黒四角によって示される)
に匹敵するELISAシグナルを有した、そして(iv)1個のクローンは、野
生型の親のファージよりも低いELISAシグナルを有した。
【0175】
図2Cは、Lib 89−91由来のファージクローンのメソセリン結合活性
を示す。試験された46個のクローンのうち、39個がメソセリンに結合した。
いずれも、親のクローンよりも大きいELISAシグナルを有さなかった。これ
らの39個のクローンのうちの28個は、親のクローンと類似したELISAシ
グナル(1.25〜1.5)を有した。残りの11個のクローンは、親の型より
も低いELISAシグナルを有した。
【0176】
図2Dは、Lib 92−94由来の選択されたファージクローンのメソセリ
ン結合活性を示す。Lib 89/93/94ライブラリー由来の結合活性と同
様に、試験された全ての46個のクローンは、メソセリンに結合し、そして、以
下の4群に分類され得る。(i)31個は、非常に高いELISAシグナル(2
.3−2.9)を与えた;(ii)8個のクローンは、高いELISAシグナル
(1.8−2.2)を有した;(iii)5個のクローンは、親の型と類似した
ELISAシグナルを与えた;そして(iv)2個のクローンは、親の型のシグ
ナルよりも低いELISAシグナルを有した。
【0177】
図2Eは、Lib 95−97のパニング後に選択されたクローンのメソセリ
ン結合活性を示す。試験された全ての46個のクローンは、メソセリンに結合す
ることが見出され、そして、ELISAシグナルに基づく3つの群に整理するこ
とができた。(i)22個は、親の型よりもわずかに良好なELISAシグナル
を与えた;(ii)20個のクローンは、野生型に類似したELISAシグナル
を与えた;そして(iii)4個のクローンは、親の型よりも下のELISAシ
グナルを有した。
【0178】
各ライブラリーからの12個のファージプレップの滴定をし、そして、CFU
(コロニー形成単位)によって決定されるようなそれらの数は、2倍未満まで変
化することが見出され、これは、ELISAシグナルにおける差異が、入力ファ
ージの数における差異に起因しないことを示す(データには示さない)。また、
BSAに対する結合は、任意のクローンからのファージ粒子については、検出さ
れなかった(データには示さない)。これらの結果は:(i)用いたスクリーニ
ングアッセイは、メソセリンに対する良好な結合を有するファージを作製するク
ローンの同定に関して信頼できたこと、および(ii)合理的に選択されたホッ
トスポットに対して無作為変異を標的化することは、CDRの全てのコドンを無
作為に変異することよりもより簡単に、良好な結合剤の単離を導く、ということ
を示唆する。
【0179】
(実施例4 クローンの多様性)
24個のELISA陽性クローンを、各ライブラリーから配列決定した。この
結果は図3に示す。
【0180】
ライブラリー、Lib 89/93/94から、4個の異なる配列を得た。全
てのクローンは、位置89において親の残基(グルタミン)が保存されていた。
位置93および94における残基は変化していた。最も豊富なクローンは、G9
3K−Y94H変異を有した。これはまた、最も高いELISAシグナルを有し
たクローンであり、SS1と命名した。
【0181】
Lib 89−91から、配列決定された全ての24個のクローンは、野生型
SS(ssFv)に類似したELISAシグナルを有し、そして24個は全て、
野生型残基を有した。
【0182】
Lib 92−94から、7個の異なるアミノ酸配列が得られた(図3)。最
も豊富なクローンは、D8(S92G−G93F−Y94N)およびC10(S
92G−G93S−Y94H)であった。このライブラリーから、本発明者らは
、Lib 89/93/94由来のSS1およびD7に対して同一のアミノ酸配
列を有する2個のクローン(E9およびE2)を得た。D8、C10およびSS
1は、最も高いELISAシグナルを有したクローンであった。
【0183】
ライブラリーLib 95−97から、SS(scFv)と類似またはそれよ
り大きいELISAシグナルを有する24個のクローンを配列決定した。17個
のクローンは、L96T変異を有した。他の7個のクローンは、野生型のアミノ
酸を有した。本発明者らは、L96T変異体をE4(scFv)と呼ぶ。
【0184】
全ての場合において、全体のscFvコード領域を配列決定し、他の変異が生
じていないことを確認した。タンパク質配列に関して、10個の異なる型の変異
体を得た。これらの多くは、いくつかの異なるヌクレオチド配列によってコード
された。これら10個のうちの9個の変異体は、変異が合理的に選択されたホッ
トスポットに対して標的化されたライブラリー(Lib 89/93/94およ
びLib 92−94)から得た。従って、これらのライブラリー由来の結合活
性を有するいくつかのクローンの存在は、これらのライブラリーに見出される迅
速な濃縮パターンと一致する。対照的に、Lib 89−91(これは、ホット
スポット中に保存された残基を有する)は、親のクローン以外にいずれの結合剤
も生じることができず、そして、Lib 95−97(この中において、変異は
、非ホットスポット領域に対して標的化される)は、単一の改変体を生じた。こ
れらのライブラリーにおける結合剤の希少性はまた、ゆっくりとでありかつ漸進
的な濃縮パターンと一致する。
【0185】
(実施例5 イムノトキシンの構築および精製)
イムノトキシンにおいて、癌細胞表面抗原に対するscFvを、毒素(例えば
、Pseudomonas外毒素Aの短縮変異体)とインフレームで融合する。
イムノトキシンの抗原結合特性は、scFvによって決定したが、毒素部分は、
scFvが結合する細胞を殺傷するエフェクター機能を提供する。従って、変異
体Fvsを含むイムノトキシンを作製しそして精製する事によって、本発明者ら
は、scFvの抗原結合運動性を決定し得、そしてまた細胞培養におけるその細
胞傷害性活性を研究し得る。異なるクローンの精製されたファージ調製物を用い
たより詳細なELISAに基づいて(結果は示さない)、本発明者らは、4個の
scFvを選択し、イムノトキシンを構築して精製した。これらの4個の間にお
いて、SS1、D8およびC10は、合理的に選択されたホットスポットが無作
為化されるライブラリーから得られた最良のクローンを示し、そして、E4は、
コントロールライブラリーから得られた親のSS(scFv)よりも良好な結合
を有する唯一のクローンを示す。イムノトキシンは、95%の均質性を超えて精
製され、そしてTSKゲル濾過クロマトグラフィーにおけるモノマーとして溶出
した(結果は示さず)。これらの精製されたイムノトキシンを使用して、scF
vの親和性を決定し、そしてまた、これらの細胞殺傷活性を研究した。
【0186】
(実施例6 イムノトキシンの結合特徴)
ファージライブラリーをパニングすることによって選択されたscFvの結合
特徴は、表面プラズモン共鳴(BIAcore)研究に対して対応する精製され
たイムノトキシンを用いることによって決定した。図4は、メソセリンに対する
イムノトキシンの結合の代表的なセンサーグラムを示し、そしてkon、kof
fおよびKDに関する値を列挙する。全ての変異体(SS1、D8、C10およ
びE4)が、親のSS(scFv)−PE38よりも、メソセリンに対する良好
な結合を有することは、明らかである。SS1およびC10のオンの速度(on
rate)は、約1.5倍高いが、D8およびE4のオンの速度は、親のSS
(scFv)とほぼ同じであった。SS1、D8、C10およびE4のオフの速
度(off rate)は、親のSS(scFv)に比較して、それぞれ、10
、37、35および3.8倍減少した。他の者が報告したことと類似して、本研
究に観察された変異体の増加した結合は、主にそれらのオフの速度における減少
に起因する(Schierら、J.Mol.Biol.255:28−43(1
996);Yeltonら、J.Immunol.,155:1994−200
4(1995);Yangら、J.Mol.Biol.254:392−403
(1995))。これらのデータによって、合理的に選択されたホットスポット
が変異されるライブラリーから誘導されるSS1、D8およびC10が、それぞ
れ、親のFvを超えて、15、37および55倍の親和性における増加を示し、
一方、E4(これは、コントロールライブラリーから単離された唯一の変異体で
ある)は、親のSS(scFv)を超えて、親和性においてより緩やかな3.7
倍の増加を示した。
【0187】
(実施例7 イムノトキシンの細胞傷害性活性)
高い親和性のscFvを癌治療における標的因子としての有用性を評価するた
めに、本発明者らは、異なるscFvを用いて作製されたイムノトキシンの細胞
傷害性を決定した。表4は、種々のイムノトキシンの抗原陽性A1847細胞株
および抗原陰性HUT 102細胞株に対する細胞傷害性活性を示す。A184
7細胞において、SS1、D8およびC10のscFv−PE38は、野生型S
S(scFv)イムノトキシンよりも10倍を超えてより活性であったが、E4
(scFv)−PE38は、たった約2倍の活性であった。HUT 102(抗
原陰性細胞)に対する細胞傷害性は、明らかでなかった。従って、変異を合理的
に選択されたホットスポットに標的化することによって得られた親和性における
増加は、細胞傷害性における大きな増加を引き起こす。しかし、細胞傷害性にお
けるこの増加は、親和性における増加と比例的に関連するわけではない。例えば
、SSl(Fv)−PE38およびC10(Fv)−PE38のIC50は、非常
に類似していたが、一方これらの親和性は約3倍異なる(IC50は、タンパク質
合成を50%阻害するのに必要であるイムノキシンの量である)。この差異は、
親和性はE.coli中で作製された組換えタンパク質を用いて測定され、一方
、細胞傷害性は抗原を発現する卵巣癌細胞株上で測定されると言う事実に起因し
得る。
【0188】
この戦略は、本発明者らの実験室において、上皮増殖因子レセプターの変異体
形態に結合する他の抗体に対して適用された。本研究においてまた、より高い親
和性を有する変異体を単離することが可能である(Beers,R.,Chow
dhury,P.S.およびPastan,I.調製における原稿)。本発明者
らはまた、SSのCDR3重鎖を変異させた。これらの変異体のELISA研究
によって、親のSS scFvの親和性よりも高いメソセリンに対する親和性が
示された。これらの結果は、現在使用されている方法によって作製されるライブ
ラリーよりも、変異を合理的に選択されたホットスポットに対して標的化するこ
とによって作製された小さなライブライリーから良好な結合ファージ選択するこ
とがより簡単であるはずであるということを示す。
【0189】
本研究において、本発明者らは、無作為化に対して4つの位置を選択し(3つ
のホットスポットの内側であり1つは外側である)、そして改変された変異体は
、有用であることが見出された。本発明者らのアプローチによって得られた改善
は、文献(Schier ら、J.Mol.Biol.255:28−43(1
996);Yeltonら、J.Immunol.,155:1994−200
4(1995);Yangら、J.Mol.Biol.254:392−403
(1995);Piniら、J.Biol.Chem.273:21769−2
1776(1998);Wuら、Proc.Natl.Acad.Sci.US
A 95:6037−6042(1998))に記載される大きいファージライ
ブラリーから得られる改善に匹敵する。本発明者らは、天然の体細胞過剰変異(
somatic hypermutation)プロセスを模倣することによっ
て、免疫化ライブラリーまたは非免疫化ライブラリーのいずれか由来の抗原に対
する増加した親和性を有する組換え抗体を容易に得ることができることを示唆す
る。
【0190】
(表4)
ヒト卵巣線癌細胞株A1847およびT細胞白血病株HUT102上での、親
のscFvおよび変異scFvを用いて作製されたイムノトキシンの細胞傷害性
。比較のために、BIAcore分析によって得られたKD値および野生型イム
ノトキシンに対する変異体の親和性における相対的増加を含めた。
【0191】
【表3】
(実施例8 ライブラリーの構築)
ファージミドpPSC 7−1は、メソセリン結合SS(scFv)をコード
するファージ抗体ディスプレイベクターである(Chowdhury l998
)。SS(scFv)のヌクレオチド配列の分析によって、32個のホットスポ
ットを明らかにした。これらの32個のホットスポットに関して、標的化変異に
対して3個選択した。これらは、CDR3のVLコード残基89、90、92、
93および94中に位置した。pPSC 7−1の一本鎖のウラシルを含むDN
Aは、以前に記載されたように調製した(Chowdhuryら、J.Mol.
Biol.281:917−928(1998))。終止コドンおよび診断のH
pal制限部位を、Kunkel’s変異誘発によってVLのCDR3に導入し
(Kunkel,T.A.、Proc.Natl.Acad.Sci.USA
82:488−492(1985))、ファージミドpPSC 7−1−94を
生成した。終止コドンは、ライブラリーの野生型配列の提示を防ぐように導入し
た。なぜなら、この変異誘発アプローチは、10〜12%のバックグラウンドを
与えるからである。ウラシルを含むpPSC7−1−94のssDNAは、変性
オリゴを用いた2つの実験ライブラリーおよびコントロールライブラリーを構築
するためのテンプレートとして使用した。オリゴSS VL 89/93/94
【0192】
【化1】
ライブラリーLib 89/93/94を作製するために使用した。このオリゴ
は、VLのコドン89、93および94を、全てが20個のアミノ酸が異なるS
S(scFv)中の無作為化する。以下に記載される他の全てのライブラリーと
同様に、このオリゴは、終止コドンを位置94においてチロシンのオリゴと置換
され、オケアー(ocher)コドンおよびオパールコドンを作製しない。
【0193】
第2のライブラリーのLib 92−94は、オリゴSS VL Mut 9
2−94
【0194】
【化2】
(これは、残基92−94を無作為化する)を用いて作製した。残基92−94
は、並んで配置される2つの異なるホットスポットによってコードされる。第3
および第4のライブラリーをコントロールとして作製した。第3のライブラリー
のLib 89−91は、オリゴSS VL Mut 89−91
【0195】
【化3】
(これは、残基89−91を無作為化する)を用いて作製した。残基90は、ホ
ットスポット中で保存された残基である。残基91は、ホットスポットの外側に
あたる。第4のライブラリーであるLib 95−97は、オリゴSS VL
Mut 95−97
【0196】
【化4】
を用いて作製した。これは、残基95−97(これらの全ては、ホットスポット
の外側にあたる)を無作為化する。
【0197】
(実施例9 ライブラリーのパニングおよび選択されたファージクローンの分
析)
ライブラリーのパニングは、以前に記載されたように行った(Chowdhu
ry 1998)。各ライブラリーからの、第2回目のパニング後の23個のク
ローンおよび第3回目のパニング後の23個のクローンを、以下のようにメソセ
リン結合に対してELISAによって分析した。単一のコロニーを、96ウェル
のマイクロタイタープレートに存在する100μg/mlのアンピシリンおよび
2%のグルコースを含む125μlの2XYT培地に接種した。このプレートを
、加湿されたインキュベーター中、37℃/200rpmで3.5時間インキュ
ベートした。これらのプレート(マスタープレート)の各ウェルからの25μl
の培養物を、100μg/mlのアンピシリン、2%グルコースおよび約4×1
09pfu/mlのM13KO7を有する125μlの2XYT培地を含む別の
マイクロタイタープレートの対応するウェルに移した。次いで、このプレート(
ファージプレート)を、上記と同じ条件下で2時間インキュベートした。次いで
、ファージプレートを、2000rpmで5分間室温で遠心分離した。各ウェル
からの上清を、逆位でプレートを穏やかに振盪することによって除去した。各ウ
ェル中の細胞ペレットを、100μl/mlのアンピシリン、および50μg/
mlのカナマイシンを含む200μlの2XYT培地中に再懸濁した。次いで、
プレートを、上記のように一晩インキュベートした。14〜16時間後、ファー
ジプレートを氷上で冷却し、次いで3000rpmで5分間、4℃で遠心分離し
た。この上清をファージELISAおよびファージ滴定に使用した。
【0198】
(実施例10 イムノトキシンの発現および精製のためのプラスミドの構築)
ファージミドのpPSC 7−1−1、7−1−2、7−1−3および7−1
−4は、それぞれ、VL中の以下の変異を有するSS(scFv)をコードする
:G93K−Y94H、S92G−G93F−Y94N、S92G−G93S−
Y94HおよびL96T。これらのファージミドベクター由来のscFvは、プ
ライマー対のNew G2 Nde IおよびNew G2 Hind III
を用いてPCR増幅した(Chowdhuryl998)。精製後、PCR産物
を以前に記載されるようにE.coli発現ベクターにクローン化した(Bri
nkmannら、Proc.Natl.Acad.Sci.USA 88:86
16−8620(1991))。生じたプラスミドのpPSC 7−2−1、p
PSC 7−2−2、pPSC 7−2−3およびpPSC 7−2−4は、ド
メインIIおよびドメインIIIを含むPseudomonas体外毒素(PE
)の38kDaフラグメントとインフレームで融合されたscFvを有した。組
換えタンパク質は、以前に記載されるように封入体から生成した(Brinkm
annら、Proc.Natl.Acad.Sci.USA 88:8616−
8620(1991))。
【0199】
(実施例11 表面プラズモン共鳴アッセイ)
scFvの親和性は、精製されたイムノトキシンのメソセリンに対する結合運
動性を、表面プラズモン共鳴(BIAcore)技術によって研究することによ
って決定した。別々の実験において、異なる量の組換えメソセリン(200〜1
000RU)をBIAcoreセンサーチップ、CM5上に固定化した。次いで
、各々のイムノトキシンを380nMの濃度で、固定化されたメソセリンの上を
通過させた。konを(ln(dR/dt)/t対濃度のプロット(ここで、Rは
応答であり、そしてtは時間である)から決定した。koffは、センサーグラム
(sensorgram)の解離期の最初の1〜2分間に算出した。これらの会
合および解離の速度は、10μl/分の連続した流速下で測定した。KDは、k
offをkonで割って算出した。
【0200】
(実施例12 細胞傷害性アッセイ)
イムノトキシンの細胞傷害性活性は、以前に記載されたように(Chaudh
aryら、Nature 339:394−397(1989))、メソセリン
陽性ヒト卵巣癌腫株A1847およびメソセリン陰性T細胞白血病株HUT 1
02ににおけるタンパク質合成の阻害によって決定した。IC50は、タンパク質
合成を50%阻害するのに必要なイムノトキシンの量である。
【0201】
本明細書中に言及される全ての刊行物および特許は、本明細書中において、あ
たかも各個々の刊行物および特許が詳細かつ個々に本明細書中において参考とし
て援用されて示されるのと同じ程度に、本明細書中へ参考として援用される。
【特許請求の範囲】
【請求項1】
明細書中に記載の発明。
【請求項1】
明細書中に記載の発明。
【図1】

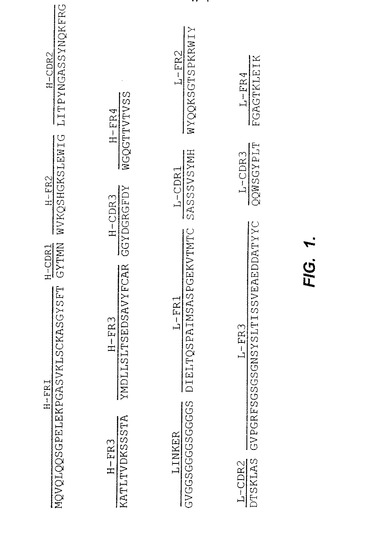
【図2】
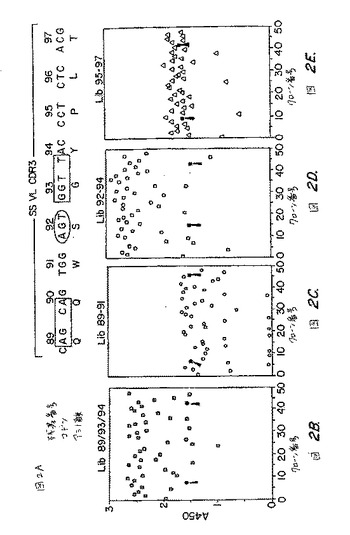
【図3】

【図4】
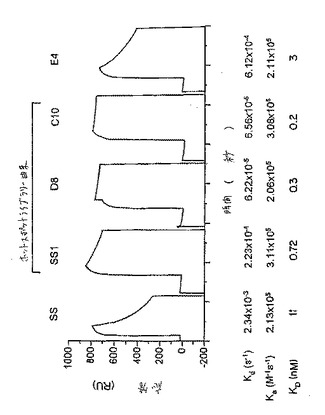
【図2】
【図3】
【図4】
【公開番号】特開2012−50464(P2012−50464A)
【公開日】平成24年3月15日(2012.3.15)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−269272(P2011−269272)
【出願日】平成23年12月8日(2011.12.8)
【分割の表示】特願2001−500670(P2001−500670)の分割
【原出願日】平成12年5月26日(2000.5.26)
【出願人】(500357817)ザ ガバメント オブ ザ ユナイテッド ステイツ オブ アメリカ, リプリゼンテッド バイ ザ セクレタリー オブ ザ デパートメント オブ ヘルス アンド ヒューマン サービシーズ (1)
【Fターム(参考)】
【公開日】平成24年3月15日(2012.3.15)
【国際特許分類】
【出願番号】特願2011−269272(P2011−269272)
【出願日】平成23年12月8日(2011.12.8)
【分割の表示】特願2001−500670(P2001−500670)の分割
【原出願日】平成12年5月26日(2000.5.26)
【出願人】(500357817)ザ ガバメント オブ ザ ユナイテッド ステイツ オブ アメリカ, リプリゼンテッド バイ ザ セクレタリー オブ ザ デパートメント オブ ヘルス アンド ヒューマン サービシーズ (1)
【Fターム(参考)】
[ Back to top ]