
CXCR3ケモカイン受容体、抗体、核酸および使用方法
【課題】オーファン受容体の1もしくは複数のリガンドのキャラクタリゼーションを行う使用方法を提供する。
【解決手段】ヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、特定のアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質に結合する抗体または抗原結合断片と接触させる工程を含む、方法。
【解決手段】ヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、特定のアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質に結合する抗体または抗原結合断片と接触させる工程を含む、方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、哺乳動物CXCケモカインレセプター(CXCR3)タンパク質の少なくとも1つの機能を調節する方法に関する。
【背景技術】
【0002】
背景技術
ケモカイン類は、炎症において生成される小さいサイトカイン類のファミリーを構成し、白血球補充(recruitment) を制御する[Baggiolini, M. et al., Adv. Immunol. 55: 97-179 (1994); Springer,T.A., Annu. Rev. Physiol. 57: 827-872(1995); and Schall, T.J. and K.B. Bacon, Curr. Opin. Immunol. 6:865-873(1994)]。ケモカイン類は、例えば、好中球、単球、マクロファージ、好酸球、好塩基球、マスト細胞などの白血球、ならびにT 細胞、B 細胞などのリンパ球などを含む血液を構成する(赤血球以外の)要素の走化性を選択的に誘導できる。走化性の刺激に加えて、ケモカイン類は、応答性細胞において、細胞形状の変化、細胞内遊離カルシウム濃度([Ca2+]i )の一過性上昇、顆粒エキソサイトーシス、インテグリンアップレギュレーション、生理活性脂質(ロイコトリエンなど)の形成および呼吸バーストなどの白血球の活性化に伴う他の変化を選択的に誘発することもできる。したがってケモカイン類は炎症性応答の初期誘因剤であり、感染または炎症部位への炎症媒介物質の放出、走化性および管外溢出を引き起こす。
【0003】
CXC およびCCケモカインと称されたケモカイン類の2 つのサブファミリーは、1 つのアミノ酸により分けられること(CXCケモカイン類として、IL-8、γIP-10 、Mig 、PF4 、ENA-78、GCP-2 、GRO α、GRO β、GRO γ、NAP-2 、NAP-4)または隣接残基であること(CC ケモカインとして、MIP-1 α、MIP-1 β、RANTES、MCP-1 、MCP-2 、MCP-3 、I-309)のいずれかである、4 つの保存システイン残基の最初の2 つのアレンジメントにより分けられる。多くのCXC ケモカイン類は、好中球の白血球を引きつける。例えば、CXC ケモカイン類インターロイキン8(IL-8) 、血小板因子4(PF4)、好中球活性化ペプチド2(NAP-2)は、強力な化学誘引物質および好中球の活性化因子である。Mig[monokine induced by gamma interferon (ガンマインターフェロンにより誘導されたモノカイン)]およびIP-10(γIP-10 、インターフェロン- ガンマ誘導性の10kDa タンパク質) と称するCXC ケモカイン類は、活性化末梢血リンパ球の走化性を誘導することに特に活性である。CCケモカイン類は、一般的に、選択性がなく、単球、好酸球、好塩基球、T リンパ球およびナチュラルキラー細胞を含む多様な白血球細胞型を引きつけることができる。ヒト単球走化性タンパク質1 〜3 (MCP-1 、MCP-2 およびMCP-3 )、RANTES[Regulatedon Activation, Normal T Expressed and Secreted(活性化制御、正常T 発現および分泌)]およびマクロファージ炎症性タンパク質1 αおよび1 β(MIP-1 αおよびMIP-1 β)などのC-C ケモカイン類は、単球またはリンパ球の化学誘引物質および活性化因子として特徴付けられるが、好中球の化学誘引物質であるとは思われない。
【0004】
CCおよびCXC ケモカイン類は、7 回膜貫通型Gタンパク質共役受容体のスーパーファミリーに属する受容体を介して作用する。[Murphy,P.M., Annu. Rev. Immunol., 12:593-633(1994);Gerard, C. およびN.P. Gerard, Curr. Opin. Immunol., 6: 140-145 (1994)] 。このG タンパク質共役(ヘビ状)受容体ファミリーは、7 つの膜貫通領域を含有する内在性膜タンパク質の大きいグループを含む。前記受容体はG タンパク質に共役し、それはGTP に結合することができ、例えば、細胞内媒介物質の生成により、共役受容体からシグナル伝達を媒介することが可能なヘテロ3 量体の制御タンパク質である。
【0005】
ケモカイン受容体を2 つのグループに分けることができる: CCケモカイン類を結合するCCケモカイン受容体1 〜5(CCR1-5) 、およびCXC ケモカイン類を結合するCXC ケモカイン受容体1 〜4(CXCR1-4)。一般的に、CCケモカイン受容体は、複数の型の白血球上に存在し、単球、好酸球、好塩基球およびT 細胞の移動に重要である[Qin, S., et al., Eur. J. Immunol.,26:640-647(1996);Carr, M. W., et al., Proc. Natl. Acad. Sci. USA, 91(9):3652-3656(1994); Taub,D.D., et al., J. Clin. Invest., 95 (3):1370-1376(1995); Neote, K. et al., Cell, 72: 415-425(1993); Gao,J.-L. et al., J. Exp. Med., 177:1421-1427(1993); Charo,I.F. et al., Proc. Natl. Acad. Sci. USA,91:2752-2756(1994); Myers, S.J., et al., J. Biol. Chem., 270:5786-5792(1995); Combadiere, C. et al., J. Biol. Chem., 270(27):16491-16494(1995); and Correction, J. Biol. Chem., 270:30235(1995); Ponath, P. D. et al., J. Exp. Med., 183:2437-2448(1996); and Daugherty, B. L. et al., J. Exp. Med., 183:2349-2354(1996); Power, C. A. et al., 1995, J. Biol. Chem., 270:19495-19500(1995); Hoogewerf, A. J. et al., Biochem. Biophys. Res. Commun., 218:337-343(1996); Samson, M. et al., Biochemistry, 35:3362-3367(1996)] 。対照的に2 つのIL-8受容体、CXCR1 およびCXCR2 は、主として好中球に限定され、好中球の移動に重要である(Baggiolini,M.,etal.,Adv.Immunol.,55:97-179(1994)] 。該IL-8受容体、CXCR1[IL-8R1, 1 型インターロイキン−8 受容体; Holmes, W. E. et al., Science, 253:1278-1280(1991)] およびCXCR2[IL-8R2, 2 型インターロイキン−8 受容体;Murphy, P. M. and H. L. Tiffany, Science, 253:1280-1283(1991)] は、好中球の走化性を誘導するCXC ケモカインに観察される本質的な結合エピトープであるNH2 末端Glu-Leu-Arg(ELR)モチーフを認識する[Clark-Lewis,I.et al.,J.Biol.Chem.,266:23128-23134(1991);Hebert,C.A. et al.,J.Biol.Chem.,266:18989-18994(1991);and Clark-Lewis,I.et al.,Proc.Natl.Acad.Sci.USA,90:3574-3577(1993)]。
【発明の概要】
【発明が解決しようとする課題】
【0006】
単球および顆粒細胞とは対照的に、ケモカインに応答するリンパ球は、良く理解されていない。特に、公知の特異性の受容体は、リンパ球に限定されるようには思われず、したがって、これらの受容体を認識するケモカインは、T細胞媒介炎症状態に観察されるTリンパ球の選択的補充などの事象を説明できない。さらに、公知のケモカイン受容体との顕著な配列類似性ならびに類似の組織および白血球亜集団分布を有する多くのタンパク質が同定され、クローン化されているが、これらの受容体のリガンドは決定されないままである。したがって、これらのタンパク質をオーファン受容体という。該受容体の1もしくは複数のリガンドのキャラクタリゼーションは、ケモカイン類とそれらの標的細胞との相互作用、この相互作用により刺激され、走化性および白血球の細胞活性化を含む事象、ならびに受容体機能の調節に基づく治療の開発の理解に必須である。
【課題を解決するための手段】
【0007】
即ち、本発明の要旨は、
〔1〕ヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質に結合する抗体または抗原結合断片と接触させる工程を含む、方法、
〔2〕ヒトCXCR3タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、ヒトCXCR3タンパク質に結合する抗体または抗原結合断片と接触させる工程を含み、結合が、配列番号:2の残基1〜15の配列と同じアミノ酸配列からなるポリペプチドによって阻害される、方法、
〔3〕配列番号:2に示されるアミノ酸配列を有する哺乳動物CXCケモカインレセプター3(CXCR3)タンパク質のシグナル伝達活性または細胞応答機能を阻害する方法であって、該タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含む、方法、
〔4〕配列番号:2に示されるアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該タンパク質を、モノクローナル抗体1C6(ATCC受託番号HB−12330)またはその抗原結合断片と接触させる工程を含む、方法、
〔5〕配列番号:2に示されるアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質によって媒介されるIP−10誘導カルシウムフラックスを阻害する方法であって、該タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含む、方法、
〔6〕配列番号:2に示されるアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質を介するIP−10誘導走化性を阻害する方法であって、該タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含む、方法、
〔7〕CXCケモカインレセプター3(CXCR3)タンパク質のシグナル伝達活性または細胞応答機能を阻害する方法であって、該CXCR3タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含み、該抗体がモノクローナル抗体1C6(ATCC受託番号HB−12330)の軽鎖CDR(CDR1、CDR2およびCDR3)および重鎖CDR(CDR1、CDR2およびCDR3)ならびにヒト枠組み領域を含むヒト化免疫グロブリンであり、該ヒト化免疫グロブリンがモノクローナル抗体1C6(ATCC受託番号HB−12330)と同じまたは類似のエピトープ特異性を有する、方法、
〔8〕ヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、ハイブリドーマ細胞株HB−12330により産生される抗体またはその抗原結合断片と接触させる工程を含む、方法、
〔9〕哺乳動物CXCケモカインレセプター3(CXCR3)タンパク質のシグナル伝達活性または細胞応答機能を阻害する方法であって、該CXCR3タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含み、該抗体がモノクローナル抗体1C6(ATCC受託番号HB−12330)の軽鎖CDR(CDR1、CDR2およびCDR3)および重鎖CDR(CDR1、CDR2およびCDR3)ならびにヒト枠組み領域を含むヒト化免疫グロブリンであり、該ヒト化免疫グロブリンがモノクローナル抗体1C6(ATCC受託番号HB−12330)と同じまたは類似のエピトープ特異性を有する、方法
に関する。
【発明の効果】
【0008】
本発明により、オーファン受容体の1もしくは複数のリガンドのキャラクタリゼーションがなされる。
【図面の簡単な説明】
【0009】
【図1】図1は、CXCR3と呼ばれるヒトIP-10/Mig受容体をコードするcDNA(ヒトCD4+ T細胞(KT30)cDNAライブラリーから単離)の1670bpインサートから決定されたヌクレオチド配列(配列番号:1)を示す図である。オープンリーディングフレーム(69〜1175)は368アミノ酸の予想タンパク質(配列番号:2)をコードする。推定されるポリAシグナルとポリA部位はそれぞれ1534〜1539と1624〜1670に位置する。
【図2】図2は、ヒトIP-10/Mig受容体をコードする図1の配列のオープンリーディングフレームの概念上の翻訳(配列番号:2)を示す図である。矢印は潜在的N結合型グリコシル化部位を示し、水平方向の線は推定される膜貫通ドメイン(TM1-TM7)の位置を示す。
【図3】図3A〜3Cは、IP-10/MigRを発現する安定にトランスフェクトされた細胞において、IP-10とMigによって誘導される応答を示すグラフである。図3Aは、IP-10/MigRトランスフェクト300-19細胞での濃度依存的[Ca2+]i変化を示すグラフである。IP-10またはMigをそれぞれ1、10および100nMの濃度でFura-2/AM負荷細胞に添加し(矢印)、[Ca2+]i依存性の蛍光変化を記録した。非トランスフェクト細胞(下側の軌線)は同じ条件下に100nMのIP-10またはMigで刺激した。図3Bは、受容体の脱感作および交差脱感作を評価した試験の結果を示すグラフである。IP-10/MigR発現300-19細胞を、100nMのIP-10またはMigと、IP-10の後Mig(もしくはその逆)で順次刺激し、蛍光変化を記録した。図3Cは、IP-10(黒丸)またはMig(黒四角)で刺激したIP-10/MigR発現Jurkat細胞の走化性を示すグラフである。下側の図は、増加するIP-10(白丸)またはMig(白四角)量で刺激した場合の非トランスフェクトJurkat細胞の応答を示す。高倍率視野5つあたりの平均移動細胞数(±SD)を表わす。
【図4】図4A〜4Bは、IP-10とMigに対する末梢血リンパ球(PBL)の応答を示すグラフである。献血血液バフィーコートから新たに単離したPBLをそのまま(下側の軌線、白抜きの記号)またはIL-2(400U/ml)の存在下に10日間培養してから(上側の軌線、黒塗りの記号)使用した。図4AはIP-10またはMigによって誘導される[Ca2+]i変化を示すグラフである。IP-10またはMigをそれぞれ1、10および100nMの濃度でFura-2/AM負荷培養細胞に添加し(矢印)、[Ca2+]i依存性の蛍光変化を記録した(上側の軌線)。新たに単離した細胞(下側の軌線)は同じ条件下に100nMのIP-10またはMigで刺激した。図4Bは増加するIP-10(黒丸)またはMig(黒四角)濃度に応答したPBLの走化性を示すグラフである(高倍率視野5つあたりの平均移動細胞数(±SD)を表わす)。
【図5】図5A〜5Bは、CXCR3 DNAをトランスフェクトしたL1.2細胞(図5A)と活性化T細胞(図5B)に対する放射標識IP-10の結合を示すグラフである。増加する非標識IP-10濃度の存在下に細胞を0.05nMの125I標識IP-10と共にインキュベートした。スキャッチャード分析(挿入図)は、CXCR3 L1.2トランスフェクタントに関して1細胞あたり37,000受容体(Kdは614pM)とCD3芽球に関して1細胞あたり17,000受容体(Kdは156pM)を示した。
【図6】図6は、CCR1、CCR2b、CCR3、CCR4、CCR5、CXCR1、CXCR2、CXCR3またはCXCR4のいずれかを発現する安定なL1.2トランスフェクタントを抗CXCR3ペプチドモノクローナル抗体(mAb)1C6で染色することによって評価した、抗CXCR3抗体1C6の特異性を示す図である。陰性対照染色(非掲載)はどのL1.2トランスフェクタントについても、非トランスフェクトL1.2細胞(L1.2wt)での1C6について示す染色と似ていた。
【図7】図7A〜7Cは、好中球(図7A)、リンパ球(図7B)および活性化T細胞(図7C)でのCXCR3の発現を示す蛍光ヒストグラムである。全血中の白血球亜集団をその前方および側方散乱光によって同定し、相応にゲーティングした。CD3芽球を生成するため、PBMCを抗CD3 mAb で3日間活性化した後、IL-2含有培地で7日間維持した。各図表において、黒いグラフは抗CXCR3 mAb 1C6 による染色を表わし、中空のグラフは同イソタイプの対照mAb による染色を表わす。
【図8】図8は、血液リンパ球の各集団でのCXCR3発現を示す一連の図表である。二色染色法を用いてT細胞(CD3)、B細胞(CD20)およびNK細胞(CD56)でのCXCR3の発現を評価した。
【図9−1】図9は、免疫蛍光三色分析法によって分析した、血液リンパ球のCD3+亜集団上のCXCR3発現と種々のマーカーの対比を示す一連の図表である。抗CD3 Cy-Chromeを用いてT細胞を染色し、分析のためそれらの細胞を電気的にゲーティングした。各象限は対照mAb の染色に従って設定した。ここに示す染色は分析した5人の献血者を代表するものであった。
【図9−2】図9は、免疫蛍光三色分析法によって分析した、血液リンパ球のCD3+亜集団上のCXCR3発現と種々のマーカーの対比を示す一連の図表である。抗CD3 Cy-Chromeを用いてT細胞を染色し、分析のためそれらの細胞を電気的にゲーティングした。各象限は対照mAb の染色に従って設定した。ここに示す染色は分析した5人の献血者を代表するものであった。
【図9−3】図9は、免疫蛍光三色分析法によって分析した、血液リンパ球のCD3+亜集団上のCXCR3発現と種々のマーカーの対比を示す一連の図表である。抗CD3 Cy-Chromeを用いてT細胞を染色し、分析のためそれらの細胞を電気的にゲーティングした。各象限は対照mAb の染色に従って設定した。ここに示す染色は分析した5人の献血者を代表するものであった。
【図10】図10は、一群の抗CXCR3 mAb による、活性化T細胞のIP-10またはMCP-1媒介性走化性の阻害を示すヒストグラムである。1×106個のヒトCD3芽球をトランスウェルの上部チャンバーに入れ、ケモカイン(12.5nM)を下部チャンバーに入れた。アッセイ開始時に、種々の抗CXCR3 mAb (組織培養上清中;FCSなし)を細胞と共にその上部ウェルに入れた。1.5時間後、下部チャンバーに移動する細胞数をフローサイトメトリーで数えた。走化性阻害百分率は、mAb の非存在下で移動した細胞数を100%として計算した。結果は、少なくとも4回の独立した実験を代表するものである。
【図11】図11は、精製抗CXCR3 mAb 1C6 によるIP-10媒介性走化性の阻害を示すグラフである。様々な濃度の1C6 mAb を上部ウェルに入れ、図10について記述したようにアッセイを行なった。mAb 1C6 は856ng/mlの濃度で総走化性を50%阻害した(IC50=856ng/ml)。
【図12】図12は、活性化T細胞に対する125I-IP-10結合の、mAb 1C6 による阻害を示すグラフである。表示のように増加する1C6濃度の存在下に、CD3芽球を0.05nMの125I-IP-10と共にインキュベートした。室温で60分の後、細胞ペレットを洗浄し、カウントした。データをソフトウェア・カレイダグラフ(KaleidaGraph)で分析したところ、0.16μg/ml のIC50値が得られた。
【図13】図13A〜13Hは、IP-10に応答したヒトT細胞による[Ca2+]iのmAb 1C6 による阻害を示す(Migの場合は阻害されない)。抗CD3活性化IL-2刺激ヒトT細胞をFura-2で標識し、表示したケモカインで順次刺激するか(図13A〜13B)、mAb で刺激した40秒後に表示したケモカインで刺激した(図13C〜13H)。[Ca2+]i蛍光変化を分光蛍光計で記録した。各軌線は5回の独立した実験を代表するものであった。抗体は、50μg/ml(図13C〜13D)、25μg/ml(図13E)、12.5μg/ml(図13F)、6.125μg/ml(図13G)または3.0625μg/ml(図13H)の最終濃度で使用した。ケモカイン類は2nMの濃度で使用した。
【図14】図14A〜14Dは、P1ペプチド(図14B)、P2ペプチド(図14C)、P3ペプチド(図14D)の存在下またはペプチドの非存在下(図14A)に、CXCR3発現トランスフェクタントをmAb 1C6 で染色したフローサイトメトリー分析の結果を示す蛍光ヒストグラムである。各図において、太い線で規定されるグラフはmAb 1C6 染色を表わし、破線で規定されるグラフは同イソタイプの無関係な対照mAb による染色を表わす。
【図15】図15は、40nMの非放射性IP-10、mAb 1C6 、CXCR3トランスフェクタントに対して生じたモノクローナル抗体(2F8、3A12、3E2、4B4、4D2、5B12、7B8または8D5)または抗CXCR2 mAb による、CXCR3トランスフェクタントに対する放射標識IP-10結合の阻害百分率を示すヒストグラムである。
【発明を実施するための形態】
【0010】
発明の要旨
本発明は、本明細書において、CXC ケモカイン受容体3(CXCR3)と称する単離および/または組換え哺乳類(例えば、ヒトなどの霊長類)IP-10/Mig 受容体タンパク質ならびにその改変体を称するタンパク質またはポリペプチドに関する。組換えCXCR3 タンパク質および改変体は、本明細書に記載のように宿主細胞中で生産されうる。1 つの態様として、CXCR3 タンパク質またはその改変体は、IP-10 および/またはMig などの1 以上のケモカイン類の選択的結合 (例えば、高いアフィニティー結合) 、および/または (1 種以上の) 細胞応答( 例えば、走化性、エキソサイトーシス、1 種以上の炎症媒介物質の放出など) を誘導する能力により特徴付けられる。
【0011】
本発明の他の側面は、哺乳類( 例えば、ヒトなどの霊長類など) のCXCR3 タンパク質またはその改変体をコードする単離および/または組換え核酸に関する。さらに本発明は、本発明のタンパク質またはその改変体をコードする核酸を含む、プラスミドまたはレトロウイルスベクターなどの組換え核酸構築物に関する。該核酸および構築物を用いて、組換え受容体タンパク質および構築物を含む宿主細胞を生産することができる。他の態様として、核酸は、CXCR3 タンパク質をコードする第 2の核酸とハイブリダイズすることができるアンチセンス核酸をコードし、細胞に導入された場合、受容体の発現を阻害することができる。
【0012】
さらに本発明は、例えば、該タンパク質もしくはその改変体( 例えばペプチドなど) または受容体タンパク質もしくは改変体を発現する細胞を免疫原として用いて生産することができる、CXCR3 受容体と反応する抗体に関する。かかる抗体またはその断片は、受容体タンパク質の精製および研究、表面受容体を発現する細胞の同定、および細胞の分類または計数を含む、治療、診断、ならびに研究応用に有益である。したがって、本発明は、本明細書記載の抗体もしくはその断片( 例えば、mAb 1C6 またはその抗原結合断片) の治療 (予防を含む) または診断への使用および本明細書に記載の疾患または状態の処置に用いる薬剤の製造のためのかかる抗体もしくは断片の使用を包含する。
【0013】
また、本発明は、受容体のリガンド、受容体機能の阻害剤(例えばアンタゴニスト)や促進剤(例えばアゴニスト)を同定する方法をも包含する。1つの態様として、該細胞に導入された核酸によってコードされる受容体タンパク質または改変体を発現するように操作されている適切な宿主細胞をアッセイに用いて、リガンド、受容体機能の阻害剤または促進剤の効き目を同定し評価する。かかる細胞は、発現された受容体タンパク質またはポリペプチドの機能を評価する際にも有用である。
【0014】
本発明によれば、リガンド、受容体機能の阻害剤および促進剤を適切なアッセイで同定し、さらに治療の効果について評価することができる。受容体機能の阻害剤は受容体活性を阻害( 減少もしくは妨害) するために使用でき、リガンドおよび/または促進剤は指示された場合、正常な受容体機能を誘導( 引き起こすまたは促進) するために使用できる。したがって本発明は、個体(例えば哺乳類)に受容体機能の阻害剤を投与する過程を含む、自己免疫疾患および移植片拒絶を含む炎症性疾患を治療する方法を提供する。さらに、本発明は、個体にリガンドまたは促進剤を投与することにより受容体機能を刺激する方法を提供し、それは、例えば、感染性疾患および癌の処置に有益な白血球機能の選択的刺激への新たなアプローチを提供する。
【0015】
発明の詳細な説明
本明細書に記述するように、CXCケモカインIP-10およびMigに選択的な新規ケモカイン受容体をコードする核酸をクローニングし、特徴づけた。ヒトCD4+ T細胞ライブラリーから単離されたこのクローンは、単球または顆粒球由来のcDNAライブラリーには検出されなかった。このクローンの配列を分析したところ、40,659ダルトンの予想分子量を持つ368アミノ酸の予想タンパク質(図2、配列番号:2)をコードする1104塩基対のオープンリーディングフレームが明らかになった(図1、配列番号:1)。このアミノ酸配列は、Gタンパク質共役受容体の特徴であり、かつ他の化学誘引物質受容体にも認められる7つの推定膜貫通セグメントを含んでいる。この知見と合致して、本受容体は、IP-10およびMigに応答して起こるCa2+(カルシウムイオン)の流動化と走化性とを媒介する(実施例2)。類似の条件下で、CXCケモカイン類IL-8、GROα、NAP-2(好中球活性化タンパク質-2)、GCP-2(顆粒球走化性タンパク質-2)、ENA78(上皮由来好中球活性化ペプチド78)、PF4(血小板因子4)や、CCケモカイン類MCP-1(単球走化性タンパク質-1)、MCP-2、MCP-3、MCP-4、MIP-1α(マクロファージ炎症性タンパク質-1α)、MIP-1β、RANTES(regulated on activation, normal T cell expressed and secreted)(発現かつ分泌された正常T細胞活性化制御)、I309、エオタキシンまたはリンフォタクチンに対する有意な応答は検出されなかった。
【0016】
活性化Tリンパ球におけるヒトCXCR3の限定的発現と、この受容体のIP-10およびMigに対するリガンド選択性は注目に値する。実施例2で使用した条件下に、このヒト受容体はIL-2活性化Tリンパ球で強く発現されるが、休止Tリンパ球、Bリンパ球、単球または顆粒球には検出されなかった。受容体分布のさらなる研究は、CD20+(B)細胞とCD56+(NK)細胞の一部もこの受容体を発現するものの、CXCR3を発現するのはそのほとんどが、先の活性化に合致する表現型、CD95+、CD45RO+およびCD45RAlowである細胞を含むCD3+細胞であることを示す。リンパ球を誘引すると報告されている他のケモカイン(例えばMCP-1、MCP-2、MCP-3、MIP-1α、MIP-1βおよびRANTES)の受容体は、好中球、好酸球および好塩基球などの顆粒球や単球にも認められるので、この活性化Tリンパ球における選択的発現は興味深い。これらの結果は、CXCR3と呼ばれるこのIP-10/Mig受容体が、エフェクターT細胞の選択的補充に関与することを示唆している。
【0017】
本受容体は、IP-10およびMigと呼ばれる2つの珍しいCXCケモカインを認識する。IP-10とMigは共にCXCサブファミリーに属するが、好中球に対する強力な化学誘引物質であるIL-8や他のCXCケモカインとは対照的に、IP-10とMigの主な標的はリンパ球、特に活性化または刺激されたTリンパ球やナチュラルキラー(NK)細胞などのエフェクター細胞である(Taub, D.D.ら, J. Exp. Med., 177:18090-1814(1993);Taub, D.D.ら, J. Immunol., 155:3877-3888 (1995))(NK細胞は、抗原認識用の特異的T細胞受容体を持たないが、腫瘍細胞やウイルス感染細胞などの細胞に対して細胞傷害活性を持つ大形顆粒リンパ球である)。このことと合致して、IP-10とMigは、好中球走化性を効率よく誘導するCXCケモカイン中の必須結合エピトープであるELRモチーフを持たない(Clark-Lewis, I.ら, J. Biol. Chem., 266:23128-23134 (1991);Hebert, C.A.ら, J. Biol. Chem., 266:18989-18994 (1991);およびClark-Lewis, I.ら, Proc. Natl. Acad. Sci. USA, 90:3574-3577 (1993))。また、組換えヒトMigと組換えヒトIP-10は共に、腫瘍浸潤性リンパ球(TIL)内のカルシウム流動を誘導すると報告されている(Liao, F.ら, J. Exp. Med., 182:1301-1314 (1995))。IP-10はイン・ビトロで単球の走化性を誘導すると報告されているが(Taub, D.D.ら, J. Exp. Med., 177:1809-1814 (1993);原因となる受容体は同定されていない)、ヒトMigは高度に選択的と思われ、そのような作用を示さない(Liao, F.ら, J. Exp. Med., 182:1301-1314 (1995))。IP-10発現は、乾癬、固定薬疹、皮膚の遅延型過敏反応、類結核癩などの炎症状態にある様々な組織、実験的糸球体腎炎および実験的アレルギー性脳脊髄炎で誘導される。またIP-10は、T細胞依存性の強力なイン・ビボ抗腫瘍作用を持ち、生体内で血管形成の阻害因子であると報告されており、イン・ビトロでNK細胞の走化性と脱顆粒を誘導できることから、(例えば腫瘍細胞破壊における)NK細胞の補充および脱顆粒の媒介物質としての役割が示唆される(Luster, A.D.およびP. Leder, J. Exp. Med., 178:1057-1065 (1993);Luster, A.D.ら, J. Exp. Med. 182:219-231 (1995);Angiolillo, A.L.ら, J. Exp. Med., 182:155-162 (1995);Taub, D.D.ら, J. Immunol., 155:3877-3888 (1995))。IP-10とMigの発現パターンは、それぞれの発現がインターフェロン・ガンマ(IFNγ)によって誘導されるという点でも独特であり、これに対し、IL-8の発現はIFNγによってダウンレギュレートされる(Luster, A.D.ら, Nature, 315:672-676 (1985);Farber, J.M., Proc. Natl. Acad. Sci. USA, 87:5238-5242 (1990);Farber, J.M., Biochem. Biophys. Res. Commun., 192 (1):223-230 (1993);Liao, F.ら, J. Exp. Med., 182:1301-1314 (1995);Seitz, M.ら, J. Clin. Invest., 87:463-469 (1991);Galy, A.H.M.およびH. Spits, J. Immunol., 147:3823-3830 (1991))。
【0018】
最近、ケモカイン類は、長く捜し求められていたリンパ球補充の媒介物質であると認識されるようになった。数種類のCCケモカインがリンパ球走化性を惹起することが発見されたが(Loetscher, P.ら, FASEB J., 8:1055-1060 (1994))、それらは顆粒球と単球に対しても活性である(Uguccinoni, M.ら, Eur. J. Immunol., 25:64-68 (1995);Baggiolini, M.およびC.A. Dahinden, Immunol. Today, 15:127-133 (1994))。この状況は、その作用が活性化Tリンパ球とNK細胞を含むリンパ球に選択的であって、かつ、数多くの他のケモカイン類を認識せず選択的発現パターンを示す受容体CXCR3を結合するIP-10とMigでは異なる(実施例2、実施例5)。
【0019】
これらの知見を考慮すれば、遅延型過敏性病巣などの炎症性病巣、ウイルス感染部位およびある種の腫瘍における特徴的な浸潤物の形成が、CXCR3を介して媒介されCXCR3発現によって調節される過程であると結論することは妥当である。活性化の結果としてCXCR3受容体を保持するリンパ球、特にTリンパ球は、インターフェロン・ガンマによって局所的に誘導されうるIP-10および/またはMigによって炎症性病巣、感染部位または腫瘍に補充されうる。したがって、CXCR3はリンパ球、特に活性化または刺激されたTリンパ球などのエフェクター細胞の選択的補充にその役割を果たす。
【0020】
タンパク質とペプチド
本発明は、哺乳類CXCR3タンパク質と呼ばれる単離および/または組換え(例えば本質的に純粋なものを含む)タンパク質またはポリペプチドおよびその改変体に関する。好ましい態様として、本発明の単離および/または組換えタンパク質は、哺乳類CXCR3タンパク質(本明細書に定義する)に特有な、結合活性(例えばリガンド、阻害因子および/または促進因子結合性)、シグナル伝達活性(例えば哺乳類Gタンパク質の活性化、サイトゾル遊離カルシウム濃度[Ca2+]iの迅速かつ一過性の上昇の誘導)、細胞応答機能(例えば走化性、エキソサイトーシスまたは白血球による炎症媒介物質放出の刺激)および/または本明細書に定義する免疫学的性質などの性質、活性または機能を少なくとも一つは持つ。例えば、本発明のタンパク質には、IP-10および/またはMigに選択的に結合でき、それに対する細胞のシグナル伝達および/または応答(例えば(とりわけ活性化Tリンパ球の)カルシウム流動、走化性および/または脱顆粒)をイン・ビトロおよび/または生体内で媒介できるものがある。例えば本明細書に示すように、cDNAクローンの発現により哺乳類細胞中で生産されたヒトCXCR3タンパク質は、CXCケモカインIP-10および/またはMigに選択的に結合して、シグナル伝達と細胞応答(例えば走化性)を媒介することができる。一態様として本発明のタンパク質は、同じ哺乳類種または異なる哺乳類種に由来するCXCケモカイン(例えばヒトIP-10、ネズミIP-10、ヒトMig、ネズミMig)を結合できる(ヒトIP-10, Luster, A.D.ら, Nature, 315:672-676 (1985);ネズミIP-10(CRG-2とも呼ばれる),Vanguri, P.およびJ.M. Farber, J. Biol. Chem., 265:15049 (1990) ならびにLuster, A.D.およびP. Leder, J. Exp. Med., 178:1057-1065 (1993);ネズミMig, Farber, J.M., Proc. Natl. Acad. Sci. USA, 87:5238-5242 (1990);ヒトMig, Farber, J.M., Biochem. Biophys. Res. Commun., 192 (1):223-230 (1993) ならびにLiao, F.ら, J. Exp. Med., 182:1301-1314 (1995))。
【0021】
本明細書で「単離(された)」と称するタンパク質またはポリペプチドは、それらが哺乳類細胞中に存在する時よりも精製された状態にあるタンパク質またはポリペプチドであり、本明細書に記述する方法や類似の方法あるいは他の適当な方法によって得られるタンパク質またはポリペプチド、例えば本質的に純粋なタンパク質またはポリペプチド、化学合成法(例:合成ペプチド)あるいは生物学的方法と化学法の併用によって生産されるタンパク質またはポリペプチド、および単離された組換えタンパク質またはポリペプチドなどが含まれる。これらのタンパク質は少なくとも約50重量%、好ましくは少なくとも約75重量%の単離状態で、または本質的に純粋な形で得ることができる。ここで「組換え」と称するタンパク質またはポリペプチドは、組換え核酸の発現によって生産されるタンパク質またはポリペプチドである。
【0022】
本明細書において「哺乳類CXCR3タンパク質」とは、天然に存在するまたは内因性の哺乳類CXCR3タンパク質、ならびに天然に存在するまたは内因性の対応する哺乳類CXCR3タンパク質と同じアミノ酸配列を持つタンパク質(例えば組換えタンパク質)をいう。したがって、本明細書に定義される用語「哺乳類CXCR3タンパク質」は、哺乳類CXCR3の成熟タンパク質、多形または対立遺伝子改変体、およびその他のアイソフォーム(例えば選択的スプライシングその他の細胞過程によって産生されるもの)、ならびにそれらの修飾体または未修飾体(例えばグリコシル化、非グリコシル化、リン酸化または非リン酸化CXCR3タンパク質)を包含する。天然に存在するまたは内因性の哺乳類CXCR3タンパク質には、成熟型CXCR3等の野生型タンパク質、哺乳動物(例えばヒト、ヒト以外の霊長類)に天然に存在する多形または対立遺伝子改変体およびその他のアイソフォームが含まれる。これらのタンパク質は、例えば哺乳類CXCR3を本来産生する供給源から回収することができる。これらのタンパク質と、天然に存在するまたは内因性の対応する哺乳類CXCR3と同じアミノ酸配列を持つ哺乳類CXCR3タンパク質は、対応する哺乳類の名前で呼ばれる。例えば対応する哺乳動物がヒトである場合、そのタンパク質はヒトCXCR3タンパク質と呼ばれる(例えば適当な宿主細胞で生産された組換えヒトCXCR3)。
【0023】
哺乳類CXCR3タンパク質の「機能的改変体」には、機能的断片、機能的突然変異タンパク質および/または機能的融合タンパク質(例えば突然変異誘発および/または組換え技術によって生産されるもの)が含まれる。一般に、本発明によって包含される哺乳類CXCR3タンパク質の断片または部分は、成熟型哺乳類CXCR3タンパク質と比較してアミノ酸(すなわち1またはそれ以上のアミノ酸)の欠失(すなわち1またはそれ以上の欠失)を有するもの(例えばN末端、C末端または内部欠失体)を包含する。成熟型哺乳類CXCR3タンパク質と比較して、隣接しているアミノ酸のみが欠失している断片もしくは部分または隣接していないアミノ酸が欠失している断片または部分も想定される。
【0024】
一般に、本発明に包含される哺乳類CXCR3タンパク質の突然変異体または誘導体には、1またはそれ以上の隣接するまたは隣接していないアミノ酸残基の付加、欠失および/または置換により異なる天然または人工の改変体、1またはそれ以上の残基が修飾されている修飾ポリペプチド、および1またはそれ以上の修飾残基を含む突然変異体が含まれる。好ましい突然変異体は、1またはそれ以上の隣接するまたは隣接していないアミノ酸残基の付加、欠失および/または置換により異なる天然または人工の哺乳類CXCR3タンパク質改変体である。そのような突然変異は、例えば(他のCXCおよび/またはCCケモカイン受容体と比較して)保存されている領域または保存されていない領域、細胞外領域、細胞質領域または膜貫通領域に存在しうる。
【0025】
哺乳類CXCR3タンパク質の「機能的断片または部分」「機能的突然変異体」および/または「機能的融合タンパク質」とは、哺乳類CXCR3受容体(本明細書に定義する)に特有な、結合活性(例えばリガンド、阻害因子および/または促進因子結合性)、シグナル伝達活性(例えば哺乳類Gタンパク質の活性化、サイトゾル遊離カルシウム濃度[Ca2+]iの迅速かつ一過性の上昇の誘導)、細胞応答機能(例えば走化性、エキソサイトーシスまたは白血球による炎症媒介物質放出の刺激)および/または本明細書に定義する免疫学的性質などの性質、活性または機能を少なくとも一つは持つ単離および/または組換えタンパク質もしくはオリゴペプチドを指す。
【0026】
本明細書において、哺乳類CXCR3タンパク質の「免疫学的性質を少なくとも一つ」持つタンパク質またはポリペプチドとは、(a) 天然に存在するまたは内因性の哺乳類CXCR3タンパク質もしくは天然に存在するまたは内因性の哺乳類CXCR3タンパク質(例えばヒトCXCR3)と同じアミノ酸配列を持つタンパク質に結合する、選択されたエピトープ特異性を持つ少なくとも一つの抗体によって結合され、および/または、(b) 天然に存在するもしくは内因性の哺乳類CXCR3タンパク質または天然に存在するもしくは内因性の哺乳類CXCR3タンパク質と同じアミノ酸配列を持つタンパク質に結合する、選択されたエピトープ特異性を持つ抗体の適当な動物における形成を(例えば適当な担体と複合体化した時に)誘導しうる免疫原であるものである。例えば、好適な断片は、単離された哺乳類CXCR3に対して生じたおよび/またはそれと反応する抗体と交差反応できる。
【0027】
好適な断片または突然変異体は、スクリーニングによって同定できる。例えば本タンパク質のN末端、C末端または内部領域を段階的に欠失させることができ、得られたタンパク質またはポリペプチドを本明細書に記述するような適当なアッセイ(例えば走化性、カルシウム流動)を使ってスクリーニングすることができる。得られたタンパク質がそのアッセイで活性を示す場合、その得られたタンパク質(「断片」)は機能的である。CXCケモカイン受容体やCCケモカイン受容体を含む哺乳類Gタンパク質共役受容体の構造と機能に関する情報は、哺乳類CXCR3タンパク質を機能ドメインに分割する際の根拠となる(Murphy, P.M., Ann. Rev. Immunol., 12:593-633 (1994) ならびにGerard, C.およびN.P. Gerard, Curr. Opin. Immunol., 6:140-145 (1994) とそれらに引用されている文献)。
【0028】
改変体という用語は、第一の成分として哺乳類CXCR3タンパク質(例えばヒトCXCR3)を含み、その第一成分が、自然界に認められるその哺乳類CXCR3には存在しない第二の成分に連結している融合タンパク質をも包含する。したがってこの第二成分はアミノ酸、オリゴペプチドまたはポリペプチドでありうる。第一成分は、その融合タンパク質のN末端、C末端または内部のいずれにあってもよい。一態様として本融合タンパク質は、第一成分としてのアフィニティーリガンド(例えば酵素、抗原、エピトープタグ)と、リンカー配列およびヒトCXCR3またはその一部からなる第二成分とを含有する。
【0029】
哺乳類CXCR3タンパク質の例としては、本発明の核酸によってコードされるタンパク質、例えば図2(配列番号:2)に記述されているようなまたは実質上記述されているようなアミノ酸配列を持つタンパク質が挙げられる。好ましい態様として、哺乳類CXCR3または改変体(例えば細胞外N末端部分を含む改変体)は、図2(配列番号:2)に示すタンパク質と少なくとも約50%同一、より好ましくは少なくとも約70%同一、さらにより好ましくは少なくとも約80%同一であるアミノ酸配列を持つ。
【0030】
放射性同位体、スピン標識、抗原(例えばFLAGタグなどのエピトープ標識)、酵素標識、蛍光基、化学発光基などといった検出可能な標識の組み込みまたは付加(直接的付加もしくは(例えばリンカーを介した)間接的付加)によって、単離および/または組換え哺乳類CXCR3タンパク質とその改変体を修飾することができ、それら修飾体も本発明の範囲に包含されることは理解されるだろう。
【0031】
核酸、構築物およびベクター
本発明は、本明細書に記載された哺乳類(ヒト等)CXCR3タンパク質またはその改変体をコードする単離および/または(本質的に純粋な等を含む)組換え核酸に関する。本明細書でいう「単離」核酸とは、起源の供給源のゲノムDNAまたは細胞RNAの(例えば、細胞中またはライブラリー等の核酸混合物中に存在するような)核酸から離れて分離された核酸をいい、さらなるプロセッシングを受けてもよい。「単離」核酸は、本明細書に記載された方法、類似の方法または他の好適な方法により得られた核酸を含み、本質的に純粋な核酸、化学合成により製造された核酸、生物学的方法と化学的方法との組み合わせにより製造された核酸および単離された組換え核酸を含む。本明細書でいう「組換え」核酸とは、組換えDNA方法論により製造された核酸をいい、ポリメラーゼ連鎖反応(PCR)および/または制限酵素を用いるベクターへのクローニング等の人工的な組換え方法に依存する手法により生成される核酸を含む。また、「組換え」核酸は、細胞の天然のメカニズムを介して起こる組換え事象からも生ずるが、所望の組換え事象の可能性および蓋然性を生じさせるように設計された核酸の細胞に導入後に選択されるものである。
【0032】
1つの態様において、核酸またはその一部は、結合活性(例えば、リガンド、阻害剤および/または促進剤結合)、シグナル伝達活性(例えば、哺乳類Gタンパク質の活性化、サイトゾル遊離カルシウムの濃度[Ca2+]iの迅速かつ一過性の増加の誘導)、および/または細胞応答の刺激(例えば、走化性、エキソサイトーシスまたは白血球による炎症性媒介物質放出の刺激)等の哺乳類CXCR3タンパク質(例えば、ヒトCXCR3受容体)に特有の機能の少なくとも1種を有するタンパク質またはポリペプチドをコードする。また、本発明は、より具体的には、哺乳類CXCR3受容体またはその一部をコードする配列を含有する単離および/または組換え核酸またはその一部に関する。さらにより具体的には、本発明は、ヒトCXCR3タンパク質をコードする配列を含有する単離および/または組換え核酸に関する。
【0033】
さらに、本発明は、
(1)(a)配列番号:1の配列を有する核酸、
(b)配列番号:1に相補的な配列を有する核酸、または
(c)配列番号:1のオープンリーディングフレームを含有する前記核酸
の一部(図1に示した鎖の部分または相補鎖の対応する部分)、
にハイブリダイズする能力;および/または
(2)配列番号:2のアミノ酸配列を有するポリペプチドまたはその機能的同等物(即ち、受容体の1以上の天然または生理学的リガンドに関するリガンド結合活性を有するポリペプチド、および/または細胞応答(例えば、走化性、エキソサイトーシスまたは白血球による炎症性媒介物質放出の(誘発または刺激を含む)誘導)を誘導可能なように、リガンド結合に応答可能な刺激する機能)をコードする能力;および/または
(3)両方の特性、
により特徴付けられる、二本鎖もしくは一本鎖DNAまたはRNAを含む単離および/または組換え核酸に関する。
【0034】
1つの態様において、配列番号:2とその機能的同等物との間のアミノ酸配列同一性率は、少なくとも約60%(≧60%)である。好ましい態様において、配列番号:2の機能的同等物は、配列番号:2と少なくとも約70%の配列同一性を共有する。さらに好ましくは、配列番号:2とその機能的同等物との間のアミノ酸配列同一性率は、少なくとも約80%、さらにより好ましくは少なくとも約90%である。
【0035】
これらの基準に合致する単離および/または組換え核酸には、天然に存在する哺乳類CXCR3受容体の配列およびその一部と同一の配列を持つ核酸、または天然に存在する配列の改変体が含まれる。かかる改変体には、1以上の残基の付加、欠失または置換によって相違する突然変異体、1以上の残基が修飾されている修飾核酸(DNAまたはRNA類縁体など)および1以上の修飾残基を含む突然変異体が含まれる。1つの態様において、核酸は、配列番号:1に示された配列の一方の鎖とまたはそのコーディング領域に対して、少なくとも約50%のヌクレオチド配列類似性、より好ましくは少なくとも約75%のヌクレオチド配列類似性、およびさらにより好ましくは少なくとも約90%のヌクレオチド配列類似性を共有する。好ましい核酸は、少なくとも約40ヌクレオチド、より好ましくは少なくとも約50およびさらにより好ましくは少なくとも約75ヌクレオチドの長さを有する。
【0036】
かかる核酸は、例えば高ストリンジェンシー条件または中程度のストリンジェンシー条件下でのハイブリダイゼーションによって検出および単離できる。核酸ハイブリダイゼーションに関する「高ストリンジェンシー条件」と「中程度のストリンジェンシー条件」は、その教示が参照により本明細書に取り込まれるCurrent Protocols in Molecular Biology(Ausubel,F.M.ら編,第1 巻, 補遺26,1991)の2.10.1-2.10.16頁(特に2.10.8-11 を参照のこと)と6.3.1-6 頁に説明されている。プローブ長、塩基組成、ハイブリダイズする配列間のミスマッチ率、温度およびイオン強度などの因子が核酸ハイブリッドの安定性に影響を与える。したがって、高ストリンジェンシー条件または中程度のストリンジェンシー条件は、所望の選択性を達成するように経験的に決定できる。
【0037】
配列番号:1またはその相補鎖の配列を有する核酸に(例えば高または中程度のストリンジェンシー条件下で)ハイブリダイズできることを特徴とする単離および/または組換え核酸は、さらに、哺乳類CXCR3タンパク質(ヒトCXCR3タンパク質など)の特有の機能、例えば結合活性(リガンド、阻害剤および/または促進剤結合など)、シグナル伝達活性(哺乳類Gタンパク質の活性化、サイトゾル遊離カルシウム濃度[Ca2+]iの迅速かつ一過性の増加の誘導など)および/または細胞応答の刺激(走化性、エキソサイトーシスまたは白血球による炎症媒介物質の放出の刺激など)などの少なくとも1つを持つタンパク質またはポリペプチドをコードしてもよい。
【0038】
本明細書に記載されるヒトCXCR3核酸またはPCRにより生成された断片を含むその十分な部分、単離、組換えおよび/または合成のいずれかは、プローブまたはプライマーとして使用され、霊長類(例えば、サル(カニクイザル等)等のヒト以外の霊長類)、ウシ、ヒツジ、ウマ、イヌ、ネコおよび齧歯類(例えば、モルモット、ラット、マウス等のネズミ種)を含むがこれに限定されない他の哺乳類種由来のCXCR3受容体(ホモログ)または他の関連する受容体遺伝子(例えば、新規CXCケモカイン受容体遺伝子)をコードする核酸(例えば、ゲノムDNA、対立遺伝子改変体、cDNA)を検出および/または回収することができる。これは、本明細書に記載された手法またはハイブリダイゼーション、PCRもしくは他の好適な技術を含む他の適当な方法を用いて達成されうる。哺乳類核酸を用いて、構築物(例えば、ベクター)、受容体またはその断片ならびに受容体の生産および使用方法に有用な宿主株を調製することが可能である。
【0039】
1つの態様において、哺乳類CXCR3タンパク質(または改変体)をコードする核酸は、PCR増幅等の方法により製造可能である。例えば、本明細書に記載されたヒトCXCR3cDNAの一部に相補的または実質的に相補的な配列を含有する適当なプライマー(例えば、プライマー対またはネステッドプライマー)を、設計することができる。例えば、コーディング配列および/またはコーディング配列に隣接する配列の5’または3’末端に相補的なプライマーを設計することができる。かかるプライマーを、適当な鋳型核酸を用いるポリメラーゼ連鎖反応に使用して、哺乳類CXCR3等をコードする核酸を得ることができる。好適な鋳型には例えば、本明細書に記載された構築物(pcDNA3−Clone8等)、cDNAもしくはゲノムライブラリー、または哺乳類(ヒト、霊長類等)cDNAもしくはゲノムDNAの他の好適な供給源を含む。プライマーは、鋳型として適するものとして選択された構築物の隣接する配列に相補的な部分を含むことができる。
【0040】
タンパク質またはポリペプチド(例えば、ハイブリダイズする核酸によってコードされる)の結合機能は、例えば、受容体を含む膜画分または受容体を発現する細胞を用いて、結合または結合阻害アッセイにおいて検出されうる(Van Riper ら、J. Exp. Med., 177: 851-856 (1993); Sledziewskiら、米国特許第5,284,746号明細書(1994年2月8日)等を参照のこと)。したがって、コードされたタンパク質またはポリペプチドのIP−20またはMig等のリガンド、阻害剤および/または促進剤結合能を評価することができる。本発明の核酸によりコードされるタンパク質またはポリペプチドの抗原特性は、イムノブロッティング、免疫沈降およびイムノアッセイ(例えば、ラジオイムノアッセイ、ELISA)等の哺乳類CXCR3に結合する抗体を用いる免疫学的方法により決定することができる。
【0041】
タンパク質またはポリペプチド(例えば、ハイブリダイズする核酸によってコードされる)のシグナル伝達機能は、受容体結合に応答するGタンパク質活性(例えば、膜画分を用いるGタンパク質αサブユニットでのGTPのGDPへの交換)に関する酵素アッセイにより検出されうる。Gタンパク質共役は、例えば、Gタンパク質による刺激が、細胞もしくは適当な細胞画分(膜等)を百日咳菌毒素等のGタンパク質の特異的阻害剤で処理または前処理することにより阻止されるアッセイを用いて、さらに評価することができる(Bischoff, S. C. ら、Eur. J. Immunol., 23: 761-767 (1993); Sozzani, S. ら、J. Immunol., 147、 2215-2221 (1991))。
【0042】
タンパク質またはポリペプチド(例えば、ハイブリダイズする核酸によってコードされる)の刺激機能は、そのタンパク質またはポリペプチドを発現する細胞を用いて、走化性または媒介物質放出に関する標準的アッセイ(リガンド(例えば、IP−20またはMigなどのケモカイン)または促進剤に応答して起こる走化性、エキソサイトーシス(エステラーゼ(セリンエステラーゼ等)、パーフォリン、グランザイム等の酵素の脱顆粒反応)または媒介物質放出(例えば、ヒスタミン、ロイコトリエン)をモニターするアッセイ等)で検出できる(Taub, D. D. ら、J. Immunol., 155: 3877-3888 (1995); Baggliolini, M. とC. A. Dahinden, Immunology Today, 15: 127-133 (1994)およびそれらに記載された参考文献等を参照のこと) 。また、哺乳類CXCR3受容体に特有の機能は、他の適当な方法によって評価してもよい。
【0043】
これらの方法は単独で、もしくは他の適当な方法と組み合わせて、配列番号:2またはそれらの機能的同等物のアミノ酸配列を持ち、かつ、そのアッセイで検出される活性を持つポリペプチドをコードする核酸の同定および/または単離操作にも使用できる。特定の機能を持つ配列番号:2のポリペプチド部分をコードする単離核酸の一部もこの方法で同定および単離できる。
【0044】
本発明の核酸はタンパク質またはポリペプチドの生産に使用できる。例えば、哺乳類CXCR3受容体のコード配列の全部または一部を含有する核酸、または配列番号:1の配列またはそれらの相補鎖にハイブリダイズするDNAを、さらなる配列操作用またはコードされたポリペプチドの適当な宿主細胞における生産用の構築物に組込むことができる。また、本発明の核酸は、例えば、放射性同位体、スピン標識、抗原もしくは酵素標識、蛍光もしくは化学発光基などのような検出可能標識の取り込みまたは(直接または間接)付着により修飾することも可能であり、かかる修飾型は、本発明の範疇に含まれる。
【0045】
アンチセンス構築物
別の態様において、核酸は、センス鎖からなる標的分子に全体または一部が相補的で、その標的分子とハイブリダイズできるアンチセンス核酸である。標的はDNAであってもよいし、そのRNA体(すなわちDNAのT残基がRNA体ではU残基である)であってもよい。適当な方法で細胞に導入すると、アンチセンス核酸は、そのセンス鎖がコードする遺伝子の発現を阻害できる。アンチセンス核酸は標準的な技術で製造できる。
【0046】
1つの態様において、アンチセンス核酸は、ある標的核酸に対して全体的または部分的に相補的であり、標的核酸とハイブリダイズでき、ここで、該標的核酸は、配列番号:1の相補鎖の配列を持つ核酸にハイブリダイズできる。例えばアンチセンス核酸は、配列番号:1またはハイブリダイゼーションが十分可能なその一部の配列を持つ標的核酸に相補的であってもよい。別の態様において、アンチセンス核酸が哺乳類CXCR3受容体(例えば、ヒトIP−10/Mig受容体CXCR3)をコードする標的核酸に対して全体的または部分的に相補的で、該標的核酸とハイブリダイズできる。
【0047】
アンチセンス核酸は、研究的応用や治療的応用を含む種々の目的に有用である。例えば、アンチセンス核酸を含む構築物を適当な細胞に導入して、受容体発現を阻害することができる。かかる細胞は、例えばその親細胞や他の関連細胞型を用いて受容体−リガンド相互作用の特異性を評価する際に貴重な対照細胞となる。もう1つの側面として、かかる構築物を哺乳類の細胞の一部または全部に導入する。アンチセンス核酸は受容体発現を阻害し、その構築物を含有する細胞においてCXCR3受容体が媒介する炎症過程を阻害できる。したがって、本発明のアンチセンス核酸を用いて、炎症性の疾患や状態を治療できる。アンチセンス構築物を含有する適当な実験動物は、白血球機能欠損症、特に活性化Tリンパ球欠損症の有用なモデルにもなり、CXCR3受容体機能に関してさらなる情報を提供することができる。かかる動物は、宿主防御におけるTリンパ球およびNK細胞などの白血球の役割を解明するのに有用な、感染性疾患または癌の貴重なモデルにもなりうる。
【0048】
組換えタンパク質の製造方法
本発明のもう1つの側面は、哺乳類CXCR3タンパク質またはその改変体(一部等)を製造する方法に関する。組換えタンパク質は、例えば、適当な宿主細胞において哺乳類CXCR3またはその改変体をコードする組換え核酸(例えば、DNA)分子の発現等により得ることができる。
【0049】
また、哺乳類CXCR3タンパク質またはその改変体の発現に適する構築物(例えば、発現ベクター)も提供される。該構築物は適当な宿主細胞に導入可能であり、組換え哺乳類CXCR3タンパク質またはその改変体を発現する細胞を製造し、培地中に維持することができる。かかる細胞は、例えば、特徴づけ、単離および/または精製(例えば、アフィニティー精製)用のタンパク質の製造における使用、結合アッセイまたは他の機能アッセイ(リガンド、受容体機能の阻害剤および/または促進剤のスクリーニングのため等)での免疫原としての使用を含む、種々の目的に有用である。適当な宿主細胞は、大腸菌、枯草菌および他の適当な細菌等の細菌細胞を含む原核細胞であってもよいし、カビ細胞もしくは酵母細胞(例えばピチア パストリス(Pichia pastoris )、アスペルギルス スピーシーズ、サッカロミセス セレビシエ、シゾサッカロミセス ポンベ、ニューロスポラ クラッサ)などの真核細胞、または他の下等真核細胞および昆虫由来の細胞(例えばSf9 昆虫細胞(国際公開第94/26087号パンフレット、オコナー(O'connor)、1994年11月24日公開))または哺乳類由来の細胞(例えば、チャイニーズハムスター卵巣細胞(CHO)、COS 細胞、HuT78 細胞、293 細胞)などの高等真核細胞であってもよい。(例えばAusubel, F.M. ら編, Current Protocols in Molecular Biology, Greene Publishing Associates and John Wiley & Sons Inc.,(1993)を参照のこと。)
組換え哺乳類CXCR3タンパク質またはその改変体を生産する宿主細胞は、次のように作製できる。所望のタンパク質のコード配列の全部または一部をコードする核酸を、例えばプラスミド、ウイルスまたは他の適当な発現用レプリコンなどのDNAベクターなどの核酸ベクターに挿入することができる。シングルコピーもしくはマルチコピーとして維持されるベクターまたは宿主細胞染色体に組込まれるベクターを含む種々のベクターが利用可能である。
【0050】
哺乳類CXCR3遺伝子の転写および/または翻訳シグナルを直接発現に使用することができる。所望のタンパク質のコード配列の全部または一部をコードする核酸を発現させるための適当な発現ベクターも使用できる。発現に適したベクターは、いくつかの成分を含有してもよく、次の1以上が含まれるがこれらに限定されない:複製起点、選択可能マーカー遺伝子、1以上の発現制御要素、例えば転写制御要素(プロモーター、エンハンサー、ターミネーターなど)および/または1以上の翻訳シグナル;選択された宿主細胞(例えば、哺乳類起源のものまたは異種哺乳類もしくは非哺乳類種由来のもの)で膜標的用のシグナル配列またはリーダー配列。構築物において、シグナル配列は、ベクター、哺乳類CXCR3コード配列または他の供給源により提供されうる。また、組み込み部位に存在する配列は、これらの要素を提供する。
【0051】
プロモーターは、適当な宿主細胞での発現用に提供されうる。プロモーターは構成的であってもよいし、誘導性であってもよい。例えば、プロモーターは、哺乳類CXCR3タンパク質またはその改変体をコードする核酸に操作可能に連結しており、コードされたポリペプチドの発現を制御することができる。原核細胞宿主に適した種々のプロモーター(例えば大腸菌用のlac 、tac 、T3、T7プロモーター)および真核細胞宿主に適した種々のプロモーター(例えば酵母アルコールデヒドロゲナーゼ(ADH1)、SV40、CMV )が利用できる。
【0052】
さらに、発現ベクターは、典型的には、そのベクターを保持する宿主細胞を選択するための選択可能マーカーおよび複製可能発現ベクターの場合複製起点をも含む。抗生物質または薬剤耐性を付与する産物をコードする遺伝子は、慣用の選択可能マーカーであり、原核細胞(例えばβ- ラクタマーゼ遺伝子(アンピシリン耐性)、テトラサイクリン耐性に対するTet 遺伝子)および真核細胞(例えばネオマイシン(G418またはゼネティシン)、gpt (ミコフェノール酸)、アンピシリンまたはハイグロマイシン耐性遺伝子)に使用してもよい。ジヒドロ葉酸レダクターゼマーカー遺伝子は、種々の宿主におけるメトトレキセートによる選択を可能にする。宿主の栄養要求性マーカー(例えばLEU2、URA3、HIS3)の遺伝子産物をコードする遺伝子は、酵母における選択可能マーカーとしてしばしば使用される。ウイルス(例えばバキュロウイルス)ベクターまたはファージベクターおよび、レトロウイルスベクターなどの宿主細胞のゲノムに組み込むことができるベクターの使用も予想される。また、本発明は、これらの発現ベクターを保持する細胞にも関する。
【0053】
例えば、哺乳類CXCR3タンパク質もしくはその改変体をコードする核酸、またはかかる核酸を含有する構築物を、核酸が1以上の発現制御要素に操作可能に連結されるように(例えば、ベクター中、細胞中でのプロセッシングにより創出された構築物中で、宿主細胞ゲノムに組み込まれて)、選択された宿主細胞に適した方法(例えば、形質転換、トランスフェクション、エレクトロポレーション、感染)により適当な宿主細胞に導入することができる。宿主細胞は、発現に適した条件下で(例えば、誘導剤の存在下で、適する塩、成長因子、抗生物質、栄養補給物等を補足した適当な培地)維持することが可能であり、それによってコードされたポリペプチドが製造される。所望ならば、コードされたタンパク質(ヒトCXCR3等)を、(例えば、宿主細胞、培地、ミルクから)単離することができる。該方法がトランスジェニック動物の宿主細胞での発現を包含することがふさわしいであろう(例えば、国際公開第92/03918号パンフレット、GenPharm International、1992年3月19日公開)。
【0054】
また、融合タンパク質もこの方法で生産できる。例えば、いくつかの態様は、哺乳類CXCR3タンパク質cDNAまたはその一部を、Bluescript(登録商標)II SK +/ −(Stratagene) 、pGEX-4T-2 (Pharmacia )、pcDNA-3(Invitrogen) またはpET-15b (Novagen )などの適当な発現ベクターに挿入することによって作製できる。得られた構築物を適当な発現用宿主細胞に導入することができる。発現後、融合タンパク質を適当なアフィニティー担体を用いて細胞溶解液から単離または精製できる(例えばCurrent Protocols in Molecular Biology(Ausubel,F.M.ら編,第2 巻, 補遺26,16.4.1-16.7.8頁(1991)を参照のこと)。さらに、アフィニティー標識は、融合タンパク質の検出手段を提供する。例えば、抗原またはエピトープアフィニティー標識を含有する融合タンパク質の細胞表面発現や特定の細胞画分における該融合タンパク質の存在は、適当な抗体により検出できる。
【0055】
抗体
さらに本発明は、哺乳類CXCR3タンパク質またはその一部と反応する抗体に関する。好ましい態様において、抗体は、単数または複数の哺乳類CXCR3受容体またはその一部を特異的に結合する。1つの態様において、単離および/または組換え哺乳類CXCR3タンパク質またはその一部(例えばペプチド)または組換え哺乳類CXCR3を発現する宿主細胞に対して抗体を生じさせる。
【0056】
また、哺乳類(例えば、ヒト等の霊長類)CXCR3タンパク質に特有の結合活性、シグナル伝達活性および/または細胞応答刺激等の機能の1以上を阻害可能な抗体も本発明の範疇にある。1つの態様において、本発明の抗体は、リガンド(即ち、1以上のリガンド)の哺乳類CXCR3タンパク質への結合を阻害することおよび/またはリガンド結合に応答して哺乳類CXCR3タンパク質により媒介される1以上の機能を阻害することができる。特に好ましい態様において、抗体は、IP−10および/またはMig等の天然のリガンドと受容体との相互作用を阻害(低減または妨害)することができる。例えば、本明細書に示されたように、本発明の抗体は、ヒトCXCR3タンパク質とIP−10との相互作用を選択的に阻害および/またはそれに応答して受容体機能(例えば、シグナル伝達活性および/または細胞応答)を選択的に阻害することができる。この選択性を発揮する1C6と称される抗体は、IP−10のヒトCXCR3タンパク質への結合ならびにIP−10により誘導されるカルシウム流動および走化性を阻害することができるが、同一条件下でのMigにより誘導されるカルシウム流動を有意に阻害しない。本明細書に示されるように、追加の抗体は、用いた条件下でMig誘導性シグナル伝達を阻害しなかったけれども、これらの抗体は、CXCR3へのIP−10結合(3A8、2F8、3A12、3E2、4B4、4D2、5B12、7B8および8D5等)またはIP−10により誘導される走化性(1A5、3A8、5F10および10C6等)を阻害可能であり、それらもまたヒトCXCR3タンパク質とIP−10との相互作用および/またはそれに応答した受容体機能の選択的阻害剤であることを示唆する。
【0057】
特に好ましい態様において、本発明の抗体は、ヒトCXCR3タンパク質に対する特異性を有し、さらにより好ましくは、1C6と称するネズミモノクローナル抗体(mAb)のエピトープ特異性と同一または類似であるエピトープ特異性を有する。mAb 1C6のエピトープ特異性と同一または類似であるエピトープ特異性を有する抗体は、エピトープ特異性をキャラクタライズするための1以上の適当な技術を用いて同定することができる。例えば、mAb 1C6のエピトープ特異性と同一または類似であるエピトープ特異性を有する抗体は、ヒトCXCR3タンパク質またはその一部(例えば、活性化T細胞、NK細胞等のリンパ球または本発明の核酸を含有する組換え宿主細胞を含むヒトCXCR3を産生する細胞)への結合に関してネズミmAb 1C6と競合する能力により同定することができる。1つの態様において、mAb 1C6のエピトープ特異性と同一または類似であるエピトープ特異性を有する抗体は、適当なアッセイで、配列番号:2の残基1−15(「P1」)の配列と同一である配列を有するポリペプチドの、抗体とヒトCXCR3タンパク質との結合を阻害する能力によりさらにキャラクタライズされる(例えば、実施例8を参照のこと)。この態様の1つの側面において、かかる抗体によるヒトCXCR3タンパク質への結合は、配列番号:2の残基16−30(「P2」)または31−45(「P3」)の配列と同一である配列を有するポリペプチドにより有意に阻害されない。
【0058】
本発明に包含される他の抗体は、ヒトCXCR3タンパク質に結合可能な抗体を含み、ここで、該結合は、N末端細胞外セグメントまたはその一部に対応する配列番号:2の一部により阻害されうる。N末端細胞外セグメントの好適な部分は、N末端を含有し、反応番号:2の残基1−30、1−45もしくは1−58の配列と同一である配列を有するポリペプチドまたは反応番号:2の残基16−30もしくは45−58の配列と同一である配列を有するポリペプチド等のそのセグメントのN末端、内部またはC末端部分を含む。好ましい態様において、かかる部分は、前記本明細書に規定された哺乳類CXCR3タンパク質、ここで哺乳類はヒトである、の「少なくとも1つの免疫学的性質」を有する。例えば、配列番号:2の残基16−30の配列と同一である配列を有するポリペプチドにより結合が阻害されうるヒトCXCR3タンパク質と反応する抗体が得られた(実施例9)。
【0059】
本発明の抗体はポリクローナル抗体であってもよいし、モノクローナル抗体であってもよく、抗体という用語はポリクローナル抗体とモノクローナル抗体の両方を包含するものとする。ポリクローナルおよびモノクローナルという用語は、抗体標品の均一性の程度をいい、特定の製造方法に限定されることを意図されていない。
【0060】
本発明の抗体は、単離および/または組換え哺乳類CXCR3タンパク質またはその一部(合成ペプチドなどの合成分子を含む)等の本発明のタンパク質またはポリペプチドを含む適当な免疫原に対して生じさせることができる。さらに、トランスフェクションされた細胞などの組換え哺乳類CXCR3を発現する細胞を免疫原として使用したり、受容体を結合する抗体のスクリーニングに使用することができる。例えばチャンタラパイ(Chuntharapai)ら, J. Immunol. 152: 1783-1789(1994);およびチャンタラパイ(Chuntharapai)ら, 米国特許第5,440,021号明細書を参照のこと。
【0061】
免疫感作抗原の調製ならびにポリクローナルおよびモノクローナル抗体の製造は、任意の適当な技術で行なえる。様々な方法が記述されている(例えばKohlerら, Nature, 256: 495-497(1975)およびEur. J. Immunol. 6: 511-519 (1976);Milsteinら, Nature 266: 550-552 (1977);Koprowski ら, 米国特許第4,172,124 号;Harlow, E.およびD. Lane, 1988, Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory:ニューヨーク州コールドスプリングハーバー);Current Protocols In Molecular Biology, 第2 巻(補遺27,94 年夏),Ausubel, F.M.ら編(John Wiley & Sons:ニューヨーク州ニューヨーク), 第11章(1991)を参照のこと)。一般的には、適当な不死化細胞株(例えばSP2/0 のような骨髄腫細胞株)を抗体産生細胞と融合することによってハイブリドーマを作製することができる。抗体産生細胞、好ましくは脾臓またはリンパ節のものは、目的の抗原で免疫感作した動物から得ることができる。選択培養条件を用いて融合細胞(ハイブリドーマ)を単離し、限界希釈法でクローニングすることができる。所望の特異性を持つ抗体を産生する細胞を適当なアッセイ(例えばELISA )で選択することができる。
【0062】
必要な特異性の抗体を製造または単離する他の好適な方法を使用することが可能であり、例えば、ライブラリーから組換え抗体を選択する方法、またはヒト抗体の全レパートリーを製造可能なトランスジェニック動物(マウス等)の免疫化に依る方法を含む(例えば、Jakobovitsら、Proc.Natl. Acad. Sci. USA, 90: 2551-2555 (1993); Jakobovitsら、Nature, 362: 255-258 (1993) ; Lonberg ら、米国特許第5,545,806号明細書; Suraniら、米国特許第5,545,807号明細書)。
【0063】
単鎖抗体および異なる種に由来する部分を含有するキメラ、ヒト化もしくは霊長類化(CDR 移植)もしくはベニヤ化(veneered)抗体ならびにキメラまたはCDR 移植もしくはベニヤ化単鎖抗体なども本発明に包含され、「抗体」という用語に含まれる。これらの抗体の様々な部分を、慣用の技術で化学的に結合するか、1または複数のポリペプチドを遺伝子工学的技術を用いて連続的(cotiguous) タンパク質として調製することができる。本明細書で用いられる「ヒト化抗体または免疫グロブリン」という用語は、異なる起源由来の免疫グロブリンの一部を含有する抗体または免疫グロブリンをいい、少なくとも1つの部分がヒト起源のものである。したがって、本発明は、非ヒト起源(齧歯類等)の抗原結合領域およびヒト起源の免疫グロブリンの少なくとも一部(ヒト枠組み領域、ヒト定常領域またはその一部)を含有する、哺乳類CXCR3タンパク質(ヒトCXCR3タンパク質等)に結合特異性を有するヒト化抗体(その抗原結合断片を含む)に関する。例えば、ヒト化抗体は、化学的に結合するか遺伝子工学的技術を用いて連続的ポリペプチドとして調製された、必要な特異性を有する非ヒト起源の免疫グロブリン(非ヒトマウス可変領域等)およびヒト起源の免疫グロブリン配列(ヒト定常領域またはその一部等)に由来する部分を含有することができる。本発明のヒト化抗体の別の例は、1以上の免疫グロブリン鎖を含有する免疫グロブリンであり、前記免疫グロブリンは、非ヒト起源のCDR(非ヒト起源の抗体に由来する1以上のCDR等)およびヒト起源の軽鎖および/または重鎖に由来する枠組み領域(枠組み変化有無のCDR移植抗体等)を含有する。この態様の1つの側面において、免疫グロブリンは、非ヒト免疫グロブリンの軽鎖CDR(CDR1、CDR2およびCDR3)ならびに重鎖CDR(CDR1、CDR2およびCDR3)を含有する。1つの態様において、ヒト化免疫グロブリンは、mAB 1C6のエピトープ特異性と同一または類似のエピトープ特異性を有する。好ましい態様において、ヒト化免疫グロブリンの抗原結合領域は、mAb 1C6に由来する。また、ヒト化抗体という用語は、単鎖抗体をも包含する。例えば、例えば、キャビリー(Cabilly) ら、米国特許第4,816,567号明細書;キャビリーら、欧州特許第0,125,023 B1号明細書;ボス(Boss)ら、米国特許第4,816,397号明細書;ボスら、欧州特許第0,120,694 B1号明細書;ニューバーガー(Neuberger, M. S.)ら、国際公開第86/01533号パンフレット;ニューバーガーら、欧州特許第0,194,276 B1号明細書;ウインター(Winter)、米国特許第5,225,539号明細書;ウインター、欧州特許第0,239,400 B1号明細書;クィーン(Queen) ら、欧州特許第0,451,216 B1号明細書;およびパドラン(Padlan, E. A.) ら、欧州特許出願第0,519,596 A1号明細書を参照のこと。また、霊長類化抗体に関しては、ニューマン(Newman, R.)ら、BioTechnology, 10: 1455-1460 (1992) を、単鎖抗体に関しては、ラドナー(Ladner)ら、米国特許第4,946,778号明細書;およびバード(Bird, R. E.) ら、Science, 242: 423-426 (1988)) をも参照のこと。
【0064】
さらに、キメラ、ヒト化、霊長類化、ベニヤ化または単鎖抗体の断片を含む抗体の機能的断片も製造できる。上述の抗体の機能的断片は、それらが由来する完全長抗体の結合機能および/または調節機能の少なくとも1つを保持する。例えば、哺乳類CXCR3タンパク質またはその一部に結合できる抗体断片は、Fv、Fab 、Fab'およびF(ab')2 断片を含めて本発明に包含されるが、これらに限られるわけではない。かかる断片は、酵素的切断や組換え技術によって生産できる。例えばパパインまたはペプシン切断によって、それぞれFab またはF(ab')2 断片を生成することができる。また、天然の終結部位の上流に1以上の終止コドンが導入されている抗体遺伝子を用いて、様々な先端欠失型の抗体を生産することもできる。例えば、F(ab')2 重鎖部分をコードするキメラ遺伝子を、重鎖のCH1 ドメインおよびヒンジ領域をコードするDNA配列を含むように設計できる。
【0065】
本発明の抗体は、研究的応用、診断的応用および治療的応用を含む種々の応用に有用である。例えば、これらを使用して、受容体またはその一部を単離および/または精製したり、受容体の構造(例えばコンフォメーション)および機能を研究することができる。
【0066】
また、本発明の抗体は、研究的応用および治療的応用において受容体機能を調節するためにも使用できる。例えば、抗体は、(a)受容体に対する結合(例えばリガンド、第2の阻害剤または促進剤の結合)、(b)受容体シグナル伝達および/または(c)細胞応答を阻害 (減少または妨害) する阻害剤として作用できる。受容体機能の阻害剤として作用する抗体は、リガンドまたは促進剤の結合を直接または間接に(例えば、受容体におけるコンフォメーション変化を引き起こすことによって)遮断できる。例えば抗体は、リガンドの結合を阻害することによって、または(リガンドの結合の阻害を伴う、または伴わない)脱感作によって、受容体機能を阻害できる。本発明の抗体は、活性化または刺激されたTリンパ球およびナチュラルキラー(NK)細胞等のエフェクター細胞のアンタゴニストとして使用したり、本明細書に記載されたCXCR3機能の阻害剤で処置可能な疾患または状態の治療(予防を含む)方法における使用が見出されうる。ヒトCXCR3タンパク質とIP−10との相互作用を選択的に阻害可能および/またはそれに応答して受容体機能を選択的に阻害可能な抗体(mAb 1C6、mAb 1C6のエピトープ特異性と類似のエピトープ特異性を有する抗体等)は、IP−10−CXCR3相互作用により媒介される疾患または障害の治療に特に有用である。例えば、乾癬、炎症性腸疾患、腎炎および多発性硬化症等の炎症状態は、特に治療に影響を受けやすい。
【0067】
驚くべきことに、mAb 1C6は、混合リンパ球反応(MLR)で評価されたように、T細胞活性化を阻害する。したがって、T細胞活性化を阻害しうるmAb 1C6および他の抗CXCR3抗体を用いて、T細胞活性化ならびにサイトカイン産生、細胞傷害性T細胞殺傷および/またはT細胞のヘルパーの供給等の活性化に関連した1または複数の機能を阻害することができる。本明細書に記載された治療または予防方法に使用した場合、かかる抗体は、さらにT細胞活性化を阻害する付加的な有利性を有しうる。かかる抗体は、同種移植片拒絶または移植片対宿主病またはT細胞の活性化の阻害が望まれる他の疾患もしくは状態を含む移植片拒絶(例えば、移植における)を治療するための治療剤として、特に魅力がある。
【0068】
また、受容体を結合する抗体は、受容体に結合した際に、シグナル伝達および/または細胞応答(カルシウム流動、走化性、エキソサイトーシスまたは前炎症媒介物質放出等)等の受容体機能を誘発するまたは刺激する、受容体機能のアゴニストとして作用することもできる。したがって、かかる抗体を、活性化または刺激されたTリンパ球およびナチュラルキラー(NK)細胞等のエフェクター細胞のアゴニストとして使用したり、本明細書に記載されたCXCR3機能の促進剤で処置可能な疾患または状態の治療(予防を含む)方法における使用が見出されうる。
【0069】
さらに、本発明の種々の抗体を用いて、例えば活性化Tリンパ球またはナチュラルキラー細胞(NK細胞)等の白血球または受容体遺伝子でトランスフェクションされた細胞における受容体の発現を検出または測定することもできる。したがってこれらの抗体は、診断または研究目的の細胞選別(例えばフローサイトメトリー、蛍光活性化細胞選別)などの応用にも有用である。
【0070】
抗イディオタイプ抗体も提供される。抗イディオタイプ抗体は、別の抗体の抗原結合部位に関連する抗原決定基を認識する。抗イディオタイプ抗体は、同一の種の動物を免疫することにより第1の抗体に対して調製され、第1抗体を生産するのに使用した動物と同じ種、好ましくは第1の抗体を製造するために用いた動物と同じ株の動物を前記第1の抗体で免疫感作することによって調製できる。例えば米国特許第4,699,880 号明細書を参照のこと。
【0071】
1つの態様において、抗体を受容体またはその一部に対して生じさせ、今度はこれらの抗体を免疫原として使用して抗イディオタイプ抗体を生産する。そのようにして生産した抗Idは、受容体を模倣して、例えばリガンド、受容体機能の阻害剤または促進剤などの受容体を結合する化合物を結合でき、かかる化合物を検出、同定または定量するための免疫アッセイに使用できる。かかる抗イディオタイプ抗体は受容体自体には結合しないが、受容体機能の阻害剤でもありうる。
【0072】
抗イディオタイプ(すなわち抗Id)抗体自体を使用して抗イディオタイプ抗体(すなわち抗- 抗Id)を生じさせることができる。かかる抗体は最初の免疫感作抗体と特異性が似ているか同一でありうる。1つの態様において、受容体への結合を遮断する抗体アンタゴニストを用いて抗Idを生じさせ、その抗Idを用いて、その抗体アンタゴニストと類似するか同一の特異性を持ちうる抗- 抗Idを生じさせることができる。これらの抗- 抗Id抗体を受容体機能に対する阻害効果について評価することによって、それらがアンタゴニストであるかどうかを決定できる。
【0073】
単鎖およびキメラ、ヒト化、霊長類化(CDR 移植)、ベニヤ化ならびにキメラ、CDR 移植またはベニヤ化単鎖抗イディオタイプ抗体を調製することが可能であり、これらは抗イディオタイプ抗体という用語に包含される。また、かかる抗体の抗体断片も調製できる。
【0074】
本発明の抗体および断片は、例えば放射性同位体、スピン標識、抗原もしくは酵素標識、蛍光もしくは化学発光基などの検出可能な標識の取り込みまたは付着(直接または間接)により修飾することが可能であり、かかる修飾型は、本発明の範疇に含まれる。
【0075】
リガンド、受容体機能の阻害剤または促進剤の同定
本明細書において、リガンドは、受容体タンパク質を結合する物質である。選択された哺乳類CXCR3 タンパク質のリガンドは、選択された哺乳類CXCR3 タンパク質に結合する物質である。好ましい態様として、哺乳類CXCR3 タンパク質のリガンド結合は、高いアフィニティーで生じる。リガンドという用語は、限定されないが、天然リガンド、単離および/ または精製された、合成および/ または組換え体、天然リガンドのホモログ(例えば別の哺乳類由来のもの)、抗体、かかる分子の一部ならびに受容体を結合する他の物質などの物質を意味する。選択した哺乳類受容体の天然リガンドは生理学的条件下でその受容体に結合でき、哺乳類CXCR3 タンパク質のものと同じ哺乳類起源のものである。リガンドという用語は、受容体活性の阻害剤または促進剤である物質ならびに選択的に受容体に結合するが阻害剤活性や促進剤活性を欠く物質を包含する。
【0076】
本明細書で用いられるように、阻害剤は、哺乳類CXCR3 タンパク質( 例えばヒトCXCR3 など) に特有の機能、例えば結合活性(例えば、リガンド結合、促進剤結合など)、シグナル伝達活性(哺乳類G タンパク質の活性化、サイトゾル遊離カルシウム濃度[Ca2+]iの迅速かつ一過性の増加の誘導など)、および/ または細胞応答機能( 例えば、走化性、エキソサイーシスまたは白血球による炎症媒介物質の放出の刺激) などの少なくとも1 つを阻害する物質である。阻害剤という用語は、受容体を結合するアンタゴニスト(例えば、抗体、天然リガンドの突然変異体、その他のリガンド結合の競合的阻害剤)および受容体に結合することなく受容体機能を阻害する物質(例えば抗イディオタイプ抗体)を含む物質を意味する。
【0077】
本明細書で用いられるように、促進剤は、哺乳類CXCR3 タンパク質(例えば、ヒトCXCR3 など) に特有の機能、例えば結合活性(リガンド、阻害剤および/ または促進剤結合など)、シグナル伝達活性(哺乳類G タンパク質の活性化、サイトゾル遊離カルシウム濃度[Ca2+]iの迅速かつ一過性の増加の誘導など)、および/ または細胞応答機能( 例えば、走化性、エキソサイトーシスまたは白血球による炎症媒介物質の放出の刺激) などの少なくとも1 種を促進(誘導または増大)する物質である。促進剤という用語は、受容体を結合するアゴニスト(例えば抗体、別の種に由来する天然リガンドのホモログ)および受容体に結合することなく受容体機能を(例えば関連するタンパク質を活性化することによって)促進する物質を含む物質を意味する。好ましい態様として、該アゴニストは天然のリガンドのホモログ以外のものである。
【0078】
本発明の核酸およびタンパク質をあてにする下記のアッセイは、単独または相互にもしくは他の適切な方法と組み合わせて、哺乳類CXCR3 タンパク質または改変体のリガンド、阻害剤または促進剤を同定するために使用されうる。本発明のイン・ビトロ方法は、多くの試料を処理する高処理のスクリーニング( 例えば、96ウェルフォーマット) に適合しうる。本発明の核酸を含み、組換え哺乳類CXCR3(例えば、ヒトCXCR3など)を高処理のスクリーニングに適するレベルで発現する宿主細胞を用いることができ、したがって、哺乳類CXCR3 タンパク質のリガンド、阻害剤および促進剤の同定および/または単離に特に価値がある。受容体の発現を多様な方法でモニターすることができる。例えば、受容体またはその一部を結合する本発明の抗体を用いて発現をモニターすることができる。また、市販の利用可能な抗体を用いて、受容体タンパク質またはポリペプチドを含む抗原標識- またはエピトープ標識- 融合タンパク質(例えばFLAG標識受容体)の発現を検出することもでき、所望のレベルで発現する細胞を選択することができる。
【0079】
哺乳類CXCR3 タンパク質をコードする核酸を発現系に組み込んで前記した受容体タンパク質またはポリペプチドを生産することができる。例えば本発明の核酸を含む構築物で安定にもしくは一過性にトランスフェクトした細胞中で発現した受容体または受容体を含有する細胞画分(例えばトランスフェクト細胞の膜画分、受容体を組み込んだリポソーム)中の受容体などの単離および/ または組換え受容体タンパク質もしくはポリペプチドを、受容体機能の試験に使用することができる。所望であれば、その受容体をさらに精製することもできる。受容体機能の試験は、イン・ビトロまたはイン・ビボで行なうことができる。
【0080】
本明細書記載の受容体機能をモニターすることによりまたは他の適切な技術を用いることにより、化合物の効果を評価する本発明の方法に、図2(配列番号:2) に示されるヒトCXCR3 などの単離および/または組換え哺乳類CXCR3 タンパク質を使用することができる。例えば、実施例2 記載のものまたは他の適切な細胞( 例えば、バキュロウイルス感染 Sf9細胞、マウスIL-2前B 細胞 [前B リンパ腫由来、オイゲン・ブッチャー(Eugene Butcher)博士 (スタンフォード大学, スタンフォード, カリフォルニア)]などの安定なまたは一過性のトランスフェクタントを結合アッセイに用いることができる。Jurkat細胞の安定なトランスフェクタント(実施例2 )または走化性を行ないうる他の適切な細胞の安定なトランスフェクタント(例えばマウスLl-2前B 細胞) は、例えば走化性アッセイに用いることができる。
【0081】
本発明の方法によれば、化合物を個別にスクリーニングすることもできるし、1 以上の化合物を本明細書記載の方法に従って同時に試験することもできる。化合物の混合物を試験する場合、記載された過程で選択された化合物を、適切な方法(例えばPCR 、配列決定、クロマトグラフィー)で (適宜) 分離、同定することができる。試験試料中の1 以上の化合物(リガンド、阻害剤、促進剤など)の存在も、これらの方法によって決定できる。
【0082】
複合化学合成や他の方法で作製された大きな複合化合物ライブラリー(例えば有機化合物、組換えまたは合成ペプチド、「ペプトイド」、核酸)を試験することができる [例えばZuckerman, R. N. et al., J. Med. Chem., 37: 2678-2685(1994) とそこに引用されている文献を参照のこと; また、Ohlmeyer, M. H. J. et al., Proc. Natl. Acad. Sci. USA 90: 10922-10926 (1993)とDeWitt, S. H. et al., Proc. Natl. Acad. Sci. USA 90: 6909-6913 (1993)(標識した化合物について); Rutter, W.J. et al., 米国特許第5,010,175 号明細書, Huebner, V.D. et al.,米国特許第5,182,366 号明細書, Geysen, H.M., 米国特許第4,833,092 号明細書をも参照のこと)。本発明方法によって複合ライブラリーから選択される化合物が独自の標識を持つ場合は、クロマトグラフィー法による個々の化合物の同定が可能である。
【0083】
1つの態様において、ファージディスプレイ法を使用する。例えば、受容体結合(例えば適切な結合緩衝液において)に適した条件下でポリペプチドを呈示するファージ(例えばファージまたはライブラリーのようなファージの収集物)と受容体を接触させることができる。受容体に結合したファージを、標準的な技術または他の適切な方法で選択できる。ファージは適当な溶出緩衝液を用いて受容体から分離できる。例えば、イオン強度やpHの変化はファージの放出をもたらしうる。別法として、溶出緩衝液が、化合物の結合を破壊するように設計された1 または複数の放出成分[ 結合を競合的に阻害するリガンド、阻害剤および/ または促進剤などの例えば呈示されたペプチドと受容体との結合を破壊できる1 以上の化合物] を含んでもよい。任意に、この選択過程を繰り返すか、別の選択過程を用いることによって、受容体に結合するファージをさらに濃縮することもできる。呈示されたポリペプチドを(例えばファージDNA の配列決定などによって)特徴づけることができる。同定されたポリペプチドを生産し、さらにリガンド結合機能、阻害剤機能および/ または促進剤機能について試験することができる。増大した安定性や他の望ましい特性を持つかかるペプチドの類縁体を製造できる。
【0084】
1つの態様において、ランダムな配列の核酸によりコードされるN-末端ペプチドを有するコートタンパク質を含む融合タンパク質を発現し、それを呈示するファージを生産できる。本発明の受容体タンパク質またはポリペプチドを発現する適切な宿主細胞を該ファージと接触させ、結合したファージを選択し、回収し、特徴づける。(例えば、G タンパク質共役受容体を用いるファージディスプレイ法について議論しているDoorbar, J. およびG. Winter, J. Mol. Biol., 244: 361(1994)を参照のこと)。
【0085】
哺乳類CXCR3 タンパク質の潜在的リガンド、阻害剤および/ または促進剤の他の供給源としては、限定されないが、IP-10 またはMig の天然、合成または組換え改変体を含むCXCR3 リガンドの改変体、他の化学誘引物質またはケモカイン類などの物質、その改変体、他の阻害剤および/または促進剤( 例えば、抗CXCR3 抗体、アンタゴニスト、アゴニスト) 、他のG タンパク質共役受容体リガンド、阻害剤および/または促進剤( 例えば、アンタゴニストまたはアゴニスト) 、および受容体機能を阻害しうる適切な受容体ペプチドまたはアナログなどの哺乳類CXCR3 受容体の可溶性部分が挙げられる(例えばMurphy,R.B.,国際公開第94/05695号パンフレットを参照のこと)。
【0086】
結合アッセイ
本発明の単離および/ または組換え受容体タンパク質もしくはその機能的改変体、その一部または適当な融合タンパク質は、ヒトCXCR3 などの(1 以上の)哺乳類CXCR3 タンパク質に結合する薬剤、およびリガンドあるいは受容体活性の潜在的阻害剤または促進剤である薬剤を選択および同定する方法に使用できる。この方法によって選択される、リガンド、阻害剤または促進剤を含む薬剤を、受容体機能に対する阻害または刺激効果および/ または治療的用途についてさらに評価できる。
【0087】
1つの態様において、活性な単離および/ または組換え哺乳類CXCR3 タンパク質またはポリペプチドに結合する化合物を該方法で同定する。この態様では、使用する受容体タンパク質またはポリペプチドが、哺乳類CXCR3 タンパク質 (本明細書規定) に特有の例えば結合活性( 例えば、リガンド、阻害剤および/または促進剤結合) 、シグナル伝達活性(哺乳類G タンパク質の活性化、サイトゾル遊離カルシウム濃度[Ca2+]iにおける迅速かつ一過性の増加の誘導)、細胞応答機能(例えば、走化性、エキソサイトーシスまたは白血球による炎症媒介物質放出の刺激など)、および/ または本明細書に規定された免疫学的性質などの少なくとも1 種の性質、活性または機能を有する。好ましい態様において、単離および/ または組換え哺乳類CXCR3 タンパク質または改変体は、リガンド結合機能を持ち、さらに好ましくは、受容体の天然リガンドを結合する。特に好ましい態様として、該単離および/または組換えタンパク質は、図1 に示された核酸によりコードされたヒトCXCR3 タンパク質である (配列番号:1) 。
【0088】
例えば、結合に適する条件下に単離および/または組換え哺乳類CXCR3 タンパク質またはその改変体を含有する組成物を維持することができ、受容体は、試験対象の薬剤( 例えば、1 以上の薬剤を含有する組成物) に接触させることができ、結合を検出もしくは測定できる。1 つの態様として、本発明の受容体をコードする核酸配列を含有する構築物で安定にもしくは一過性にトランスフェクトした細胞で、受容体タンパク質を、発現させることができる。細胞を受容体の発現に適切な条件下に維持しうる。結合に適する条件下 (例えば適切な結合緩衝液など) に該細胞を薬剤と接触させ、結合を標準技術により検出することができる。例えば、結合の程度は、適切な対照と比較して (例えば、薬剤の非存在下に決定したバックグラウンドと比較、第2 薬剤の結合と比較 (すなわち標準) 、トランスフェクトしていない細胞への薬剤の結合を比較など) 決定されうる。任意に、受容体を含む膜画分などの細胞画分を全細胞の代わりに用いることができる。
【0089】
結合または複合体形成を直接または間接に検出することができる。1つの態様において、薬剤を適切な標識(例えば蛍光標識、化学発光標識、同位体標識、酵素標識)で標識し、その標識の検出によって結合を決定することができる。結合の特異性は、例えば非標識薬剤またはリガンド (例えば、IP-10 、Mig など) を競合剤として用いる競合または置換によって評価できる。
【0090】
哺乳類受容体のリガンドは、同じ哺乳類種由来の天然リガンドまたは別の種由来の天然リガンドを含めて、この方法で同定できる。受容体を結合する促進剤または阻害剤の結合活性も、かかるリガンド結合アッセイで評価できる。
【0091】
結合阻害アッセイを用いて、リガンドならびに受容体に結合し、リガンドなど他の薬剤の結合を阻害する阻害剤および促進剤を同定することもできる。例えば、第2 の試験薬剤の存在下での第1 の薬剤の結合に比べて、(第2 の薬剤の非存在下で起こる)第1 の薬剤の結合の減少を検出または測定する結合アッセイを行なうことができる。受容体を第1 および第2 の薬剤と同時に接触させてもよいし、逐次順番は問わずに接触させてもよい。第2 試験薬剤の存在下で起こる第1 薬剤の結合の程度の減少は、第2 薬剤による結合の阻害の指標となる。例えば、第1 薬剤の結合が減少したり、妨げられたりされうる。
【0092】
1つの態様において、ヒトCXCR3 に対する第1 薬剤(例えばIP-10 、Mig などのケモカイン)の結合の第2 の試験薬剤による直接阻害をモニターする。例えば、ヒトCXCR3 への125I- 標識Mig の結合を阻害する薬剤の能力をモニターしうる。かかるアッセイは、例えば、全細胞(例えばヒトCXCR3 受容体をコードする核酸を含む適切な細胞株)または該細胞由来の膜画分を用いて行なうことができる。
【0093】
受容体を結合する1 もしくは複数の薬剤の存在を同定する他の方法も利用でき、例えば、シグナル伝達機能および/ または細胞応答の刺激などを含む、受容体結合によって誘発される事象をモニターする方法である。
【0094】
本発明の抗体の阻害効果を結合阻害アッセイで評価できることは理解されるだろう。また、該アッセイにおける第1 薬剤が別の抗体である方法で、抗体結合に適した条件下に、受容体結合に関する抗体間の競合を評価できる。
【0095】
この方法で同定されるリガンド、受容体結合阻害剤および促進剤をさらに評価して、結合に引き続いて、それらがCXCR3 受容体の他の機能を阻害または活性化するように作用するかどうかを決定すること、および/または、それらの治療的有用性を評価することができる。
【0096】
シグナル伝達アッセイ
G タンパク質共役受容体の(アゴニストなどによる)結合は、該受容体によるシグナル伝達をもたらすことができ、G タンパク質の活性化を刺激する。薬剤によるシグナル伝達機能の誘導は、いかなる適切な方法をも用いてモニターできる。例えば、GTP からGDP への加水分解、または受容体結合により誘発される後期のシグナル伝達事象、例えば、細胞内(サイトゾル)遊離カルシウム濃度[Ca2+]iの迅速かつ一過性の増加の誘導などを、当該分野公知の方法または他の適切な方法で評価できる(実施例2; Neote, K.et al., Cell, 72: 415-425 1993) Van Riper et al., J. Exp. Med., 177: 851-856 (1993); Dahinden, C.A. et al., J. Exp. Med., 179: 751-756(1994)をも参照のこと)。
【0097】
ハイブリッドG タンパク質共役受容体を用いるSledziewski らの機能アッセイを用いて、ハイブリッドG タンパク質を活性化するその能力によりリガンドもしくは促進剤を同定すること、またはかかる活性化を阻害するその能力により阻害剤を同定することも可能である(Sledziewski ら, 米国特許第5,284,746 号明細書;その教示は参照により本明細書に合体される)。1 つの態様として、試験試料中のリガンドの存在の指標である応答の検出で、(ハイブリッド受容体への結合により誘発される)宿主細胞の生物学的応答がモニターされうる。例えば、試験試料中の、受容体のリガンド−結合ドメインにより結合されうる薬剤であるリガンドの存在を検出する方法が記載される。本方法の1つの態様として、酵母宿主細胞を、生物学的に活性なハイブリッドG タンパク質共役受容体 (即ち、融合タンパク質) の発現を仕向けることが可能なDNA 構築物で形質転換する。該ハイブリッド受容体は、STE2遺伝子産物などの酵母G タンパク質共役受容体の対応するドメインと置換されたリガンド結合ドメイン以外の少なくとも1 種のドメインを有する哺乳類G タンパク質共役受容体を含有する。構築物を含む酵母宿主細胞をハイブリッド受容体が発現する条件下に維持し、該細胞をリガンドのハイブリッド受容体への結合を可能にするに適した条件下に試験試料と接触させる。(ハイブリッド受容体への結合により誘発される)宿主細胞の生物学的応答がモニターされ、応答の検出はシグナル伝達機能の指標となる。例えば、STE2遺伝子産物由来のハイブリッド受容体への結合がBAR1プロモーターの誘導をもたらすことができる。該プロモーターの誘導は、BAR1プロモーターに連結され第2 の構築物で宿主細胞に導入されるレポーター遺伝子(例えば、β-galなど)を利用して測定される。該リポーター遺伝子の発現は、例えば細胞溶解液に対するイン・ビトロ酵素検定によって、あるいは培地に指示薬(例えば、X-gal など) を含む平板での青色コロニーの存在によって検出できる。
【0098】
他の態様において、該アッセイを用いて受容体機能の潜在的阻害剤を同定することができる。ある薬剤の阻害活性は、該アッセイでリガンドまたは促進剤を用い、リガンドまたは促進剤によって誘導される活性を阻害する試験薬剤の能力を評価することによって決定できる。
【0099】
既知リガンドの改変体を、共役G タンパク質の活性を刺激する能力の減少(減少した能力または能力がない)についてスクリーニングすることもできる。この態様では、薬剤は(他の方法で決定したように)リガンド結合活性を持つが、受容体の連動が共役G タンパク質の活性を誘発しないか、わずかにしか誘発しない。かかる薬剤は潜在的アンタゴニストであり、さらに阻害活性を評価することができる。
【0100】
走化性および細胞応答の他のアッセイ
受容体機能を評価するため走化性アッセイも使用できる。これらのアッセイは薬剤によって誘発されるイン・ビトロまたはイン・ビボの細胞の機能的移動に基づいており、これらのアッセイを用いてリガンド、阻害剤または促進剤の結合および/ または走化性への効果を評価できる。
【0101】
IP-10 およびMig に対する細胞の応答を評価するためのイン・ビトロ走化性アッセイの使用を実施例2 に記述する。Springerらは経内皮リンパ球走化性アッセイについて記述している(Springerら, 国際公開第94/20142号パンフレット,1994 年9 月15日公開;その教示は参照により本明細書に合体される;さらにBermanら, Immunol Invest. 17: 625-677 (1988)も参照のこと)。内皮を横切るコラーゲンゲルへの移動も記述されている(Kavanaugh ら, J. Immunol, 146: 4149-4156(1991))。
【0102】
一般に走化性アッセイでは、障壁の第1 表面から反対側の第2 表面に向かって、薬剤の増加レベルにそって障壁(内皮、フィルターなど)内へ、または障壁を通過して移動する、走化性の能力を有する適切な細胞、例えば白血球など(T リンパ球、NK細胞、単球など)、Jurkat細胞、マウスIL-2前B 細胞、または他の適切な宿主細胞の安定なトランスフェクタントの指向的な運動または移動をモニターする。膜やフィルターは、フィルターの第1 表面からフィルターの反対側の第2 表面の方向に増大する薬剤レベルに向かってフィルター内に、またはフィルターを通過して移動する、適切な細胞の指向的運動または移動をモニターするのに便利な障壁を提供する。いくつかのアッセイでは、ICAM-1、フィブロネクチンまたはコラーゲンなどの接着を容易にする物質で膜をコートする。
【0103】
例えば、適切な容器(含有手段)内で、第1 チャンバーから第2 チャンバー中への、もしくは細孔膜を通過して、試験対象の薬剤を含有し、膜によって第1 チャンバーから隔離された第2 チャンバーへの細胞の移動を検出または測定できる。薬剤に応答して起こる特異的な移動をモニターするに適した孔サイズを持つ適切な膜、例えばニトロセルロース、ポリカーボネートなどを選択する。例えば約3 〜8 ミクロン、好ましくは約5 〜8 ミクロンの孔サイズを使用できる。フィルター上の孔サイズは均一であってもよいし、適切な孔サイズの範囲に分布していてもよい。
【0104】
移動を評価するため、フィルターへの移動距離、フィルターを横切ってフィルターの第2 表面に接着している細胞数および/ または第2 チャンバー中に蓄積する細胞数を標準的な技術(例えば顕微鏡)を用いて決定できる。1つの態様において、細胞を検出可能な標識(例えば放射性同位体、蛍光標識、抗原またはエピトープ標識)で標識し、膜に接着したその標識および/ または第2 チャンバー中に存在するその標識の存在を適切な方法で(例えば放射活性や蛍光を検出するか、免疫アッセイで)決定することによって、移動を評価できる。薬剤によって誘発される移動の程度を、適切な対照と比較して(例えば薬剤の非存在下に決定したバックグラウンド移動と比較して、第2 の薬剤(すなわち標準)によって誘発される移動の程度と比較して、その薬剤によって誘発される非トランスフェクト細胞の移動と比較して)決定することができる。
【0105】
チャンバーは、プラスチック、ガラス、ポリプロピレン、ポリスチレンなど、種々の固体で形成することができる。チャンバーから取り外せる膜、例えばBiocoat (Collaborative Biomedical Products )やTranswell (Costar, マサチューセッツ州ケンブリッジ)培養インサートなどは、接着細胞の計数を容易にする。容器には、第1 チャンバー内の細胞を含む液体と第2 チャンバー内の液体に接触するようにフィルターを設置することができる。試験薬剤またはアッセイの目的のために存在する追加のリガンド、阻害剤もしくは促進剤以外は、膜の両側の液体が好ましくは同じであるか、実質的に類似している。チャンバー内の液体は、細胞の安定性を増大させ、細胞の非特異的結合を阻害するように作用するタンパク質溶液(例えばウシ血清アルブミン、ウシ胎児血清、ヒト血清アルブミン)および/ または培養培地を含みうる。
【0106】
1 つの態様として、経内皮移動を評価する。より低いバックグラウンド( ノイズに対するシグナル割合) に加え、経内皮移動は、血管壁に沿う内皮細胞層を横切ることにより白血球が炎症部位の組織に存在する化学誘引剤に向かって血管から移出するイン・ビボ条件をモデルにする。この態様では、内皮細胞層を通過する遊出を評価する。細胞層を調製するため、内皮細胞の接着を促進するように任意にコラーゲン、フィブロネクチンまたは他の細胞外マトリックスタンパク質などの物質でコートされた細孔フィルターまたは膜上で内皮細胞を培養できる。好ましくは、コンフルエントな単層が生成するまで内皮細胞を培養する。例えば静脈、動脈または微小血管内皮、例えば、ヒト臍静脈内皮細胞(Clonetics Corp, サンディエゴ,カリフォルニア州)などや適切な細胞株、例えば、用いられたECV304細胞株(European Collection of Animal Cell Cultures, Porton Down, Salisbury, U.K.) などの多様な哺乳類内皮細胞が単層形成に利用できる。特定の哺乳類受容体に応答して起こる走化性を検定するために、同じ哺乳類の内皮細胞が好ましい; しかしながら、異なる哺乳類種または属由来の内皮細胞も使用できる。
【0107】
一般的には、フィルターの第1 表面からフィルターの反対側の第2 表面に向かって、薬剤のレベルが増大する方向に、膜またはフィルター内へ、または膜またはフィルターを通過して移動する細胞の指向的移動を検出することによって、このアッセイを行ない、そのフィルターは第1 表面に内皮細胞層を含む。指向的移動は第1 表面に隣接する領域から膜内へ、または膜を通過して、フィルターの反対側に置かれた薬剤に向かって起こる。第2 表面に隣接する領域に存在する薬剤の濃度は、第1 表面に隣接する領域に存在する薬剤の濃度より大きい。
【0108】
1つの態様において、薬剤のリガンドまたは促進剤活性の試験に走化性アッセイを用い、移動能を持ち哺乳類CXCR3 タンパク質またはその機能的改変体を発現できる細胞を含む組成物を第1 チャンバーに入れ、好ましくは第1 チャンバー内の細胞の走化性を誘導できる(化学誘引機能を持つ)他のリガンドや促進剤の非存在下に、試験対象の薬剤(1以上の薬剤) を含む組成物を第2 チャンバーに入れる。しかしながら、化学誘引機能を持つ1 以上のリガンドまたは促進剤が存在してもよい。本アッセイで哺乳類CXCR3 受容体を発現する細胞の走化性を誘導する薬剤の能力は、薬剤が受容体機能のリガンドまたは促進剤であることの指標である。
【0109】
阻害剤の試験に使用する1つの態様において、移動能を持ち、哺乳類CXCR3 タンパク質または機能的改変体を発現できる細胞を含む組成物を第1 チャンバーに入れる。第1 チャンバー内の細胞の走化性を誘導できる(化学誘引機能を持つ)リガンドまたは促進剤を含む組成物 (すなわち1 以上のリガンドまたは促進剤) を第2 チャンバーに入れる。細胞を第1 チャンバーに入れる前 (好ましくは直前) か、細胞と同時に、試験対象の薬剤を含む組成物を、好ましくは第1 チャンバーに入れる。このアッセイで哺乳類CXCR3 タンパク質を発現する細胞のリガンドまたは促進剤誘導走化性を阻害する薬剤の能力は、薬剤が受容体機能の阻害剤( 例えば、細胞応答機能の阻害剤) であることの指標である。試験薬剤の存在下に、リガンドまたは促進剤によって誘導された移動の程度の減少は、阻害活性の指標となる。別個に結合試験(上記参照)を行なって、阻害が受容体に対する試験薬剤の結合の結果であるのか、それとも異なる機構によって起こるのかを決定することもできる。
【0110】
組織に対する薬剤の注射に応答して起こる組織の白血球浸潤をモニターするイン・ビボアッセイを後述する。これらのモデルでは、炎症部位への移動および走化性によってリガンドまたは促進剤に応答する細胞の能力を測定する。
【0111】
受容体を含有する適切な宿主細胞を用いて、活性な受容体により誘導される他の細胞応答をモニターすることによって、CXCR3 受容体の細胞応答機能に対するリガンド、阻害剤または促進剤の効果を評価することができる。同様に、これらのアッセイを用いて受容体の機能を決定することもできる。例えば、エキソサイトーシス [例えば1 以上の酵素または他の顆粒成分、例えば、エステラーゼ (例えば、セリンエステラーゼ) 、パーフォリンおよび/またはグランザイムなど) などの放出をもたらすナチュラルキラー細胞の脱顆粒] 、炎症媒介物質放出 [例えばロイコトリエン類 (ロイコトリエンC4など)などの生理活性脂質の放出など] 、ならびに呼吸バーストを、当該分野公知の方法や他の適切な方法で監視することができる。[ 例えば、顆粒誘導セリンエステラーゼの放出のアッセイに関する Taub,D.D et al., J.Immunol.,155:3877-3888(1995) (その教示は、参照により本明細書中に取り込まれる) ならびにNK細胞および細胞傷害性T リンパ球(CTLs)による酵素およびグランザイム放出のアッセイに関する Loetscher et al., J. Immunol., 156:322-327(1996), (その教示は、参照により本明細書中に取り込まれる);呼吸バーストに関するRot,A. et al.,J.Exp.Med.,176:1489-1495(1992);Bischoff, S.C. ら, Eur. J. Immunol., 23: 761-767 (1993)ならびにBaggliolini, M. およびC.A. Dahinden, Immunology Today, 15: 127-133(1994)を参照のこと。]。
【0112】
1つの態様において、脱顆粒またはエキソサイトーシス機能を持つ細胞による脱顆粒またはエキソサイトーシス時の酵素放出をモニターすることによって、リガンド、阻害剤および/ または促進剤を同定する。エキソサイトーシスまたは脱顆粒を刺激することのできる活性な受容体タンパク質をコードする本発明の核酸を含有する細胞を、適切な条件下に適切な培地中で維持し、それによって受容体を発現させ、脱顆粒を誘発させうる。受容体を試験対象の薬剤と接触させ、酵素放出を評価する。培地への酵素の放出は、免疫学的アッセイまたは酵素活性に関する生化学的アッセイなど、適切な方法で検出または測定できる。
【0113】
アッセイ成分(基質、補因子、抗体など)を培地に(例えば細胞と薬剤を合わせる前、同時にもしくは後に)導入することによって、培地を直接的にアッセイできる。別法として、アッセイに先立って細胞から分離し、さらに工程を経た (分画した) 培地に対してアッセイを行なうこともできる。例えば、セリンエステラーゼなどの酵素の慣用のアッセイを利用できる [顆粒誘導セリンエステラーゼの放出に関するTaub,D.D.et al.,J.Immunol.,155:3877-3888(1995)などを参照のこと] 。
【0114】
薬剤による脱顆粒の刺激は、薬剤が、哺乳類CXCR3タンパク質のリガンドまたは促進剤であることの指標となりうる。他の態様として、受容体を発現している細胞は、リガンドまたは促進剤と組み合わされ、それより前、後、または同時に試験対象の薬剤を添加し、脱顆粒が評価される。リガンド−促進剤−誘導脱顆粒の阻害は、該薬剤が、哺乳類CXCR3タンパク質機能の阻害剤であることの指標でありうる。
【0115】
また、細胞接着は、当該分野に公知の方法または他の適切な方法によりモニターされうる。リンパ球のケモカイン受容体の連動状態はインテグリン活性化、および脈管構造または末梢血管領域に発現した接着分子への接着の誘導を引き起こしうる。1つの態様として、接着することができる細胞による細胞接着をモニターすることにより、リガンド、阻害剤および/または促進剤を同定する。例えば、試験対象の薬剤は、(a)受容体を発現する細胞(好ましくは受容体でトランスフェクトされたときに接着能を獲得する非接着性細胞)、(b)適切な接着分子を含む組成物(例えば、フィブロネクチンなどの接着分子でコートされた培養ウェルなどの基質)、およびリガンドまたは促進剤(例えば、アゴニスト)と組み合わされることができ、リガンドまた促進剤誘導接着に適する条件下に維持しうる。蛍光色素による細胞の標識は、接着細胞を検出する簡便な方法を提供する。非接着細胞は、(例えば、洗浄により)取り除くことができ、接着細胞の数が決定する。リガンドまたは促進剤誘導接着を阻害するまたは増強することにおける薬剤の効果は、それぞれ、阻害剤または促進剤活性の指標となりうる。該アッセイに活性な薬剤としては、結合、シグナル伝達および/または細胞応答の阻害剤および促進剤があげられる。他の態様として、試験対象の薬剤は、リガンドまたは促進剤誘導接着に適する条件下に受容体を発現する細胞と適切な接着分子を含む組成物とに組み合わせられることができ、接着がモニターされる。適切な対照と比較した増加した接着は、リガンドおよび/または促進剤の存在の指標となる。
【0116】
炎症のモデル
治療薬として、イン・ビボでのリガンド、阻害剤、または促進剤の効果を評価するために用いられうる多様な炎症のイン・ビボモデルが利用でき、ヒツジ喘息モデル[例えば、Weg, V. B. et al., J. Exp. Med., 177: 561(1993)を参照のこと、その教示は、参照により本明細書に取り込まれる]、ラット遅延型過敏モデル[Rand, M.L. et al., Am.J.Pathol., 148:855-864(1996)、その教示は、参照により本明細書に取り込まれる]または、他の適切なモデルがあげられる。他の哺乳類CXCR3タンパク質と交差反応する抗体の活性をかかる哺乳類で評価することができる。
【0117】
また、ウサギ、ラット、モルモットなどの適切な動物に薬剤を皮膚内注射した時の白血球浸潤もモニターできる [例えばVan Damme J.ら, J. Exp. Med., 176: 59-65(1992)、Zachariae, C.O.C. ら, J. Exp. Med. 171: 2177-2182 (1990)、Jose, P.J.ら, J. Exp. Med. 179: 881-887 (1994)を参照のこと] 。1つの態様において、皮膚生検を白血球(T リンパ球、単球、ナチュラルキラー細胞など)の浸潤について組織学的に評価する。もう1 つの態様として、走化性を持ち管外溢出能を持つ標識細胞(例えば、111In で標識された哺乳類CXCR3 タンパク質を発現する細胞)を動物に投与する。試験試料(例えば適切な緩衝液または生理学的担体中の試験対象の薬剤)の注射部位に近接して起こる標識細胞の浸潤が、その試料中のリガンドまたは促進剤、例えば、アゴニストなどの存在の指標となる。これらのアッセイを改良して、走化性と白血球管外溢出の阻害剤を同定することもできる。例えば、リガンドまたはアゴニストを試験動物に投与する前、後もしくは同時に、阻害剤を投与できる。阻害剤の非存在下での浸潤の程度に比べて、阻害剤の存在下で浸潤の程度の減少が阻害の指標である。
【0118】
診断的用途
本発明は様々な診断的用途を持つ。例えば、哺乳類CXCRタンパク質をコードする遺伝子の1 もしくは複数の変異は、コードされる受容体の機能の少なくとも 1種の欠損を引き起こし、それにより受容体機能を減少または促進することができる。例えば、受容体の改変体を生じるまたは発現のレベルを変える変異は、受容体機能、受容体により媒介される減少または促進段階( 例えば、炎症過程) を減少または促進することができる。かかる変異の存在は、個々の細胞(例えば、活性化Tリンパ球などの白血球など)中またはかかる細胞から単離した受容体標品におけるの受容体もしくは受容体機能の存在を検出あるいは測定する方法を用いて決定されうる。これらのアッセイにおいて、受容体の減少したもしくは促進したレベルおよび/または減少したもしくは促進した受容体機能を評価しうる。
【0119】
本発明の核酸は、活性が減少または増大している受容体をコードする欠損哺乳類CXCR3 遺伝子のスクリーニング、特徴づけおよび/ または単離に使用できる試薬、例えば、プローブ、PCR プライマーなどを提供する。例えば、標準的な欠損遺伝子スクリーニング法を使用できる。欠損遺伝子を単離し、それらを適切な宿主細胞中で発現させて、哺乳類CXCR3 タンパク質について本明細書に記述するような評価をさらに行なうことができる。多くのヒト疾患は、G タンパク質共役受容体機能の欠損に関係している[Clapham, D. E., Cell, 75: 1237-1239 (1993); Lefkowitz, R. J., Nature, 365: 603-04 (1993)]。
【0120】
本発明の核酸は、(例えば、mRNAの転写を検出することにより)受容体の発現を(ノーザン解析、イン・サイチュハイブリダイゼーションにより)評価するためにも使用しうるプローブおよびPCRプライマーなどの試薬を提供する。例えば、活性化Tリンパ球または他の細胞型での発現を評価しうる。
【0121】
本発明の抗体は、受容体を細胞の表面に検出することができる手法への応用を有する。該受容体は、発現される白血球細胞型マーカー、特に活性化T細胞マーカーを提供する。例えば、受容体タンパク質またはペプチドに対して生じさせた抗体、例えば、本明細書に記載された抗体(例えば、mAb 1C6)を用いて、受容体を発現する細胞を検出および/または定量することができる。1つの態様として、抗体を用いて、細胞の混合物全体から受容体を発現する細胞を分類することができる(例えば、CD4 + T 細胞などの活性化T細胞を単離すること)。この目的には、細胞の計数および/ または選別に適した方法(例えばフローサイトメトリー、蛍光活性化細胞選別法)を使用できる。細胞数は、白血球細胞型(例えば、活性化T細胞)におけ増加または減少が観察される疾患または状態の診断に用いられうる。個体から得られた試料中の活性化T細胞の増加レベルの存在は、遅延型過敏反応、同種移植片拒絶、または細菌もしくはウイルス感染を含む病原性状態などの炎症性疾患または状態による浸潤の指標でありうる。
【0122】
さらに、抗体を用いて、受容体の発現を検出または測定することができる。例えば、本発明の抗体を用いて、試料中(例えば、血液、血清、白血球(例えば、活性化Tリンパ球)、気管支肺胞洗浄液、唾液、腸液などの組織もしくは体液)の受容体を検出または測定することができる。例えば、試料(例えば、組織および/または体液)を個体から得ることができ、適切な方法を用いて、CXCR3タンパク質の存在または量を評価することができる。適切な方法としては、FACS解析および酵素結合免疫吸着アッセイ(ELISA)などの免疫学的方法があげられ、化学発光アッセイ、放射免疫アッセイ、免疫組織学が含まれる。一般的に、試料および本発明の抗体は、抗体−受容体複合体の形成に適する条件下に組み合わせられ、抗体−受容体複合体の形成は、(直接的または間接的に)評価される。
【0123】
個体から得られた試料中の受容体反応性の増加レベルの存在は、同種移植片拒絶、遅延型過敏反応、または細菌もしくはウイルス感染を含む病原性条件などの炎症性疾患または状態が関連する炎症および/または白血球(例えば、活性化T細胞)浸潤および/または蓄積の指標でありうる。哺乳類CXCR3タンパク質または改変体の発現レベルを用いて、哺乳類CXCR3タンパク質の増加または減少した発現と特定の疾患または状態とを相関させることができ、哺乳類CXCR3タンパク質の増加または減少した発現が生じる(例えば、正常な個体での発現レベルなどの適切な対照に相対して増加または減少)疾患あるいは状態の診断において、用いることができる。同様に、治療の経過を、患者の試料中のCXCR3免疫反応性を評価することによりモニターすることができる。例えば、本発明の抗体を用いて、抗炎症性または免疫抑制剤で処置された患者由来の試料(血液、組織など)中にCXCR3を生じる細胞の数をモニターすることができる。
【0124】
トランスジェニック動物
組換えDNA 技術を用いて宿主動物のゲノムが改変されたトランスジェニック動物を構築することができる。1つの態様において、その改変が遺伝性でない(例えば骨髄中の先祖細胞などの体細胞を改変する)。別の態様において、その改変が遺伝性である(生殖細胞系を改変する)。トランスジェニック動物は標準的な技術または他の適当な方法で構築できる [例えば、T リンパ球の改変に関するCooke, M.P. ら, Cell, 65: 281-291 (1991); Hanahan, D., Science, 246: 1265-1275 (1989); Anderson, et al., 米国特許第5,399,346 号明細書] 。
【0125】
一側面として、内因性哺乳類CXCR3 遺伝子を適切な動物宿主中での完全に、もしくは部分的に(例えば遺伝子破壊技術によって)不活性化または無力化することによって、トランスジェニック動物を作製することができる。本発明の核酸を用いて、不活性化または無力化されたCXCR3 遺伝子を含有する宿主の構築の成功を(例えばサザンハイブリッド形成法によって)評価することができる。さらに、不活性化または無力化されたCXCR3 遺伝子を含有する宿主の構築の成功は、コードされている受容体の機能をモニターする適切なアッセイによっても評価できる。かかる動物を用いて、炎症および癌または病原体(ウイルス病原体)に対する宿主防御における受容体不活性化の影響を評価しうる。
【0126】
別の態様において、哺乳類CXCR3 タンパク質またはポリペプチドをコードする核酸を適当な宿主に導入してトランスジェニック動物を作製することもできる。好ましい態様では、そのトランスジェニック動物中に存在する内因性CXCR3 受容体遺伝子を、(例えば、該内因性遺伝子を破壊し置換する相同組換えによる該核酸の導入と同時に)不活性化する。例えば、白血球 [例えばリンパ球 (活性化T リンパ球など)、ナチュラルキラー細胞] 中で異なる哺乳類種(例えば、配列番号:1にコードされるCXCR3 などのヒトCXCR3] の哺乳類CXCR3 受容体をコードする核酸を発現できるトランスジェニック動物(マウス、モルモット、ヒツジなど)を作製することができ、導入された受容体の機能を評価するための便利な動物モデルを提供する。さらに、トランスジェニック動物に試験薬剤を投与し、受容体媒介過程 (例えば、炎症) に対する該薬剤の効果を本明細書に記載されたようにまたは適切なアッセイでモニターすることもできる。この方法で、受容体機能を阻害または促進する薬剤を同定したり、そのイン・ビボ効果を同定または評価することができる。
【0127】
治療方法
哺乳類CXCR3タンパク質に特有の少なくとも1種の機能の阻害または促進を介して、本発明の哺乳類CXCR3機能の調節は、受容体媒介機能を阻害又は促進する有効かつ選択的な方法を提供する。CXCケモカイン受容体は、主要な標的がリンパ球、特に活性化または刺激Tリンパ球およびNK細胞などのエフェクター細胞であるIP−10およびMigなどのケモカインに反応しうる、活性化リンパ球に選択的に発現するので、哺乳類CXCR3タンパク質は、ヒト等の哺乳類におけるリンパ球機能を選択的に妨害するまたは促進するための標的を提供する。一度リンパ球を部位に補充するやいなや、単球などの他の白血球型が二次的なシグナルで補充されるであろう。したがって、リガンド、阻害剤(例えば、1C6)および/または促進剤、本明細書記載のように同定されたものなどを含むCXCR3機能を阻害または促進する薬剤を用いて、白血球機能(例えば、補充および/または蓄積を含む白血球浸潤)、特にリンパ球の白血球機能を治療目的のために調整することができる。
【0128】
1側面として、本発明は、哺乳類CXCR3機能を阻害または促進する薬剤のかかる治療が必要な個体への投与を含む、かかる治療が必要な個体における炎症性応答を阻害または促進する方法を提供する。1つの態様として、哺乳類CXCR3タンパク質(例えば、ヒトCXCR3)の1以上の機能を阻害する化合物を投与して炎症を阻害(すなわち減少または妨げる)。例えば、mAb 1C6を含む本発明の抗体を本方法に用いることができる。結果として、白血球移動、走化性、エキソサイトーシス(例えば、酵素の)または炎症媒介放出などの1以上の炎症過程が阻害される。例えば、炎症部位の白血球浸潤(例えば、遅延型過敏応答)を本発明の方法にしたがって阻害することができる。
【0129】
他の態様として、哺乳類CXCR3タンパク質(例えば、ヒトCXCR3)の1以上の機能を促進する薬剤(例えば、受容体アゴニスト)を投与して、炎症過程の有益な刺激に帰する、白血球移動、走化性、エキソサイトーシス(例えば、酵素の)または炎症媒介放出などの炎症応答を誘導(誘発または促進する)する。例えば、ナチュラルキラー細胞をウイルス感染または腫瘍形成疾患に対抗するために補充することができる。
【0130】
「個体」という用語は、本明細書に規定され、特に限定されないが、霊長類(例えば、ヒト)、ウシ、ヒツジ、ヤギ、ウマ、イヌ、ネコ、ウサギ、モルモット、ラット、マウス、他のウシ属、ヒツジ、ウマ科、イヌ科、ネコ科、ネズミ、ハツカネズミの種などが挙げられる。炎症、感染および癌に関連する疾患および状態を本発明の方法を用いて処置することができる。好ましい態様として、疾患または状態は、リンパ球、特に活性化または刺激Tリンパ球およびナチュラルキラー(NK)細胞などのエフェクター細胞などの作用が治療(予防を含む)目的で阻害または促進されるべきものである。特に好ましい態様として、炎症疾患または状態は、T細胞媒介疾患または状態である。
【0131】
CXCR3機能の阻害剤で処置されうるヒトまたは他の種の慢性疾患を含む疾患または条件は、特に限定されないが:
・ 全身性過敏症または過敏応答、薬剤アレルギー(例えば、ペニシリン、セファロスポリンに対する)、昆虫咬傷アレルギー;クローン病などの炎症腸疾患、潰瘍性大腸炎、回腸炎および腸炎;膣炎;皮膚炎、湿疹、アトピー性皮膚炎、アレルギー性接触皮膚炎、蕁麻疹などの乾癬および炎症性皮膚病;血管炎(壊死性、皮膚および過敏性血管炎など)など;脊椎関節症;硬皮症;喘息、アレルギー性鼻炎、過敏性肺疾患、過敏性肺炎、間質性肺疾患(ILD )(特発性肺繊維症、関節リウマチに関連するILD 、または他の自己免疫状態)などの呼吸アレルギー疾患;などを含む炎症性またはアレルギー性疾患および状態;
・ 関節炎(例えば関節リウマチ、乾癬関節炎)、多発性硬化症、全身性エリテマトーデス、重症筋無力症、真性糖尿病および若年発症糖尿病を含む糖尿病、糸球体腎炎および他の腎炎、自己免疫甲状腺炎、ベーチェット病などの自己免疫疾患;
・ 同種移植片拒絶または対宿主性移植片病を含む(移植などにおける)移植片拒絶;
・ 望ましくない炎症応答を阻害すべきその他の疾患または状態、限定されないが、アテローム性動脈硬化症、サイトカイン誘導毒性、筋炎(多発性筋炎、皮膚筋炎)も治療できる
があげられる。
【0132】
CXCR3機能の促進剤(例えば、アゴニスト)で処置しうるヒトもしくは他の種の疾患または状態としては、限定されないが:
・ 皮膚T細胞リンパ腫(例えば、菌状息肉腫)などの癌、特に皮膚または組織の白血球浸潤を伴うもの;
・ 新形成疾患、網膜症(例えば、糖尿病性網膜症)、および黄斑変性を含む、脈管形成または新生血管形成が役割を果たす疾患;
・ 細菌感染および結核様らい、ならびに特にウイルス性感染などの感染性疾患;
・ 免疫抑制、例えばAIDSなどの免疫不全症候群にかかっている患者や、放射線療法、化学療法、あるいは免疫抑制を引き起こすその他の療法を受けている患者に認められるもの;受容体機能の先天的欠乏症または他の原因による免疫抑制など。さらに、CXCR3機能の促進剤は癌化学療法の間、幹細胞消耗に対抗するに有益な保護効果をも有する[Sarris, A.H. et al.,J.Exp.Med.,178:1127-1132(1993)]。
【0133】
投与経路
本発明によれば、1 以上の薬剤を単独で、あるいは別の薬物と組み合わせて、適当な経路で宿主に投与できる。薬剤(例えばリガンド結合を阻害する受容体ペプチド、抗CXCR3 抗体またはその抗原結合断片)の有効量を投与する。有効量とは、その投与条件下で所望の治療効果を達成するに足る量であり、例えばCXCR3 受容体機能の阻害または促進と、それによるそれぞれ、受容体媒介段階( 例えば、炎症性応答) の阻害または促進に十分な量である。
【0134】
種々の投与経路が可能で、例えば薬剤および治療しようとする状態または疾患に応じて、経口投与法、食餌法、局所投与法、腸管外投与法(静脈内注射、動脈内注射、筋肉内注射、皮下注射など)、吸入投与法(気管支内吸入、鼻孔内吸入または経口吸入、鼻孔内滴剤など)が挙げられるが、必ずしもこれらに限られるわけではない。喘息などの呼吸アレルギー性疾患には、吸入法が好ましい投与法である。
【0135】
投与すべき薬剤の製剤は、選択した投与経路に応じて変化するだろう(例えば液剤、乳剤、カプセル剤など)。投与すべき薬剤を含む適当な組成物を、生理学的に許容される賦形剤または担体中に調製することができる。例えば液剤や乳剤の場合は、適当な担体として、食塩水や緩衝培地などの水溶液、アルコール/ 水溶液、乳液、懸濁液などが挙げられる。腸管外用の賦形剤には、塩化ナトリウム溶液、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、乳酸化リンゲルまたは固定油などを含めることができる。静脈内用の賦形剤には、種々の添加物、保存剤、液体、栄養または電解物質補充剤などを含めることができる(一般的にはRemington's Pharmaceutical Science, 第17版,Mack 編, 1985を参照のこと)。吸入法の場合は、薬剤を可溶化し、適当な投与用ディスペンサー(アトマイザー、ネブライザーまたは加圧エアロゾールディスペンサーなど)に入れることができる。
【0136】
さらに、該薬剤が、タンパク質またはペプチドである場合、薬剤を組換えタンパク質のイン・ビボ発現を介して投与することができる。イン・ビボ発現は、適切な方法 (米国特許第5,399,346 号明細書などを参照のこと) に従って、体幹細胞発現を介してなし遂げられる。この態様において、該タンパク質をコードする核酸を運搬のためにレトロウイルス、アデノウイルスまたは他の適切なベクター( 好ましくは、複製欠損感染性ベクター) に取り込むことができ、運搬のタンパク質を発現することができるトランスフェクトまたは形質転換宿主細胞に導入することができる。後者の態様において、治療に有効な量でタンパク質を発現させるに有効な量で、該細胞を( 単独もしくは障壁装置中に) 埋め込み、注入し、さらに導入することができる。
【実施例】
【0137】
実施例
次に、下記の実施例によって本発明を例証するが、これらの実施例は決して限定を意図するものはない。
【0138】
ヒトケモカイン(実施例1〜2)
CXCケモカインMig、IL-8、GROα、NAP-2、GCP-2、ENA78、PF4、CCケモカインMCP-1、MCP-2、MCP-3、MCP-4、MIP-1α、MIP-1β、RANTES、I309、エオタキシン、およびケモカイン関連リンフォタクチンは、確立された手順に従って化学的に合成した(Clark-Lewis, I.ら, Biochemistry 30:3128-3135 (1991))。CXCケモカインIP-10はPeproTech社(ニュージャージー州ロッキーヒル)から購入した。
【0139】
実施例1.受容体cDNAのクローニング
標準的な分子生物学的技術を使用した(Sambrook, J.ら, 1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press,ニューヨーク州コールドスプリングハーバー)。
【0140】
推定されるTリンパ球限定的ケモカイン受容体をコードするDNA断片を、ポリメラーゼ連鎖反応(PCR)を使って作製した。2つの縮重オリゴヌクレオチドプライマーをケモカイン受容体の保存されたモチーフに基いて設計した。プライマーの設計は、ケモカイン受容体IL-8R1(CXCR1)、IL-8R2(CXCR2)、CC-CKR1 (CCR1)、CC-CKR2 (CCR2) およびオーファン受容体EBI I、LESTRおよびBLR1/MDR15の膜貫通ドメイン2(TM2)および膜貫通ドメイン7(TM7)内の保存されたヌクレオチド配列に基いた(EBI I, Birkenbach, M.ら, J. Virol., 67:2209-2220 (1993);LESTR, Loetscher, M.ら, J. Biol. Chem., 269:232-237 (1994);BLR1/MDR15,Dobner, T.ら, Eur. J. Immnol., 22:2795-2799(1992)ならびにBarella, L.ら, Biochem. J., 309:773-779 (1995))。
【0141】
プライマーの配列は次の通りとした。
配列番号:3
5’−GGG CTG CAG CII T(T/G)(T/G) C(C/A)G AC(A/C) TIC TI(C/T) T−3’
配列番号:4
5’−GGG TCT AGA IGG GTT IAI (G/A)CA (G/A)C(T/A) (G/A)(T/C)G−3’
(I=イノシン)。これらのプライマーをポリメラーゼ連鎖反応(PCR)で使用することにより、ヒト末梢血リンパ球から単離されたヒトゲノムDNAを鋳型として、次のようにDNA断片を増幅した。ヒトゲノムDNA 2μg、1×DynaZyme緩衝液(Finnzymes OY社,フィンランド・エスポー)、1.5mM MgCl2、500μMの各デオキシヌクレオチド、1μMの両プライマーおよびDynaZyme DNAポリメラーゼ2.5Uを含有する反応混合物100μlを、DNAサーマルサイクラー(Techne PHC-2, Brouwer社,スイス)での30サイクル(94℃で1分間、55℃で1分間、72℃で2分間)にかけた。予想されるサイズ(約700bp)のPCR産物をGene Scribe-ZベクターpTZ18/19 U/R(USB社,オハイオ州クリーブランド)にクローニングし、部分的に配列決定し(Sanger, F.ら, Proc. Natl. Acad. Sci. USA, 74 :5463-5467 (1977))、それらの既知ケモカイン受容体との類似性と、それらに対応するmRNAの白血球における発現について評価した。2MLC22と命名したDNA断片は、IL-8R2と64%のヌクレオチド配列同一性を示した。DNAポリメラーゼIのクレノウ断片と市販のランダムプライムラベリングキットを使用して放射活性同位体32Pで2MLC22を酵素により標識することによって調製したハイブリダイゼーションプローブを用いたノーザンブロット分析で評価したところ、断片2MLC22はT細胞由来のRNAに特異的にハイブリダイズしたが、単球または好中球由来のRNAにはしなかった。
【0142】
断片2MLC22を上述のように酵素により32Pで標識し、それをラムダ-ZAP Express(Stratagene社,スイス・チューリッヒ)中に調製されたヒト破傷風トキソイド特異的CD4+ T細胞(KT30)cDNAライブラリーをスクリーニングするためのプローブとして使用した(Loetscher, M.ら, J. Biol. Chem., 269 :232-237 (1994))。cDNAライブラリーは、ヒト破傷風トキソイド特異的CD4+ T細胞(KT30)由来のポリ (A)+ RNAを用いて、λZAP Expressシステム中に、その製造業者のプロトコール(Stratagene GMBH社,スイス・チューリッヒ)に従って調製した。得られたライブラリーは約1.1kbの平均インサートサイズを持つ約1.8×106個の独立クローンを含んでいた。プラークハイブリダイゼーションスクリーニングのため、約4×105クローンをBiodyneナイロン膜(PALL AG社,スイス・ムッテンツ)に転写し、ハイプライムDNAラベリングキット(Boehringer Mannheim社,ドイツ・マンハイム)を使って1×109dpm/μg DNAの比活性に標識しておいた2MLC22でプローブした。ハイブリダイゼーションは、1×106dpm 2MLC22/mlのハイブリダイゼーション溶液を使って、50%ホルムアミド、6×SSC、0.5% SDS、100μg/ml変性サケ精子DNA中42℃で20時間行なった。その膜を2×SSC/0.1%SDS中室温で10分間を1回、1×SSC/0.1%SDS中65℃で30分間を2回、最後に0.5×SSC/0.1%SDS中65℃で10分間を1回洗浄した。この高ストリンジェンシー洗浄の後、ハイブリダイゼーション陽性ラムダプラークから23個のクローンを単離し、最大のインサート(1670bp)を持つクローンを配列決定した。CXCR3 cDNAを市販のヌクレオチド配列決定用、ハイブリダイゼーションプローブ作製用、およびCXCR3を発現する安定にトランスフェクトされた哺乳類細胞クローン構築用のプラスミドベクターにサブクローニングした。それらCXCR3 cDNA含有構築物は大腸菌株中に維持される。
【0143】
結果
Tリンパ球特異的ケモカイン受容体を探索することにより、ヒトCD4+ T細胞ライブラリーからcDNAを単離した(図1、配列番号:1)。このcDNAは、汎用されている単球由来のcDNAライブラリーまたは顆粒球(HL60)由来のcDNAライブラリーを新規ケモカイン受容体cDNAに関して探索する過程では回収されなかった。しかしながら、CXCR3 cDNAに関して特異的なライブラリーの直接探索は行われていない。IP-10/Mig受容体(下記参照)をコードすることが明らかになったこのCXCR3 cDNAは、残基69から始まる1104bpのオープンリーディングフレーム(ORF)を持ち、これは40,659ダルトンの予想分子量を持つアミノ酸数368のタンパク質をコードする。そのアミノ酸配列(図2、配列番号:2)は、Gタンパク質共役受容体の特徴である7つの推定膜貫通セグメントと、3つの潜在的Nグリコシル化部位(Asn22、Asn32およびAsn199)を含む(図2)。また、受容体キナーゼの潜在的リン酸化部位(Palczewski, K.およびJ.L. Benovic, Trends Biochem. Sci., 16 :387-391 (1991);Chuang, T.T.ら, J. Biol. Chem., 267 :6886-6892(1992);Giannini, E.ら, J. Biol. Chem., 270 :19166-19172(1995))であるスレオニン残基1個とセリン残基9個が、細胞内COOH末端領域に認められる(図2)。
【0144】
この受容体の368アミノ酸配列(IP-10/MigR、図2、配列番号:2)を、IL-8R1(CXR1)、IL-8R2(CXCR2)、CC-CKR1 (CCR1)、CC-CKR2A (CCR2a)、CC-CKR3 (CCR3) およびCC-CKR4 (CCR4) を含む他のヒトケモカイン受容体のアミノ酸配列と整列した。多重タンパク質整列は、HigginsおよびSharp (Higgins, D.G.およびP.M. Sharp「Descriptionof the method used in CLUSTAL (CLUSTALで使用される方法の説明)」Gene, 73 :237-244 (1988))に従って行なった。図2の黒く囲んだ残基は、IP-10/MigRと他の少なくとも2つのケモカイン受容体の間で同一な領域を表わす。ハイフンは、整列中のギャップを表わす。この整列により、数個の保存されたモチーフが、具体的には膜貫通ドメインと第二細胞内ループ中に明らかになった。第三および第六膜貫通ドメインにはCXC受容体IL-8R1およびIL-8R2との有意な配列同一性が認められたが、CCケモカイン受容体との有意な配列同一性は認められなかった(図2)。
【0145】
この配列は、IL-8R1受容体およびIL-8R2受容体と全体でそれぞれ40.9%および40.3%のアミノ酸同一性を持ち、5種類の既知CCケモカイン受容体とは34.2〜36.9%の同一性を持つ(表1)。T細胞で発現されるがケモカイン類を結合しない七膜貫通ドメイン型受容体とは、より低い類似性が認められた(例えば、トロンビン受容体(Vu, T.-K.H.ら, Cell, 64 :1057-1068 (1991))とは27.2%同一)。図2の配列と整列することができる不完全なコード配列を持つ機能未確認の先端欠失型クローンは、過去にヒトゲノムDNAライブラリーから単離されている(Marchese, A.ら, Genomics, 29 :335-344 (1995))。
【0146】
【表1】

実施例2.生物学的活性
活性化Tリンパ球での発現
観察されたケモカイン選択性に着目して、白血球とそれに関連する細胞株でのIP-10/MigRの出現をノーザンブロット分析で調べた。新たに単離したヒト血液単球、好中球、リンパ球(PBL)、ナイロンウール精製T細胞、ならびに400U/ml hrIL-2の存在下に10日間培養(2mMグルタミン、1×非必須アミノ酸、1mMピルビン酸ナトリウム、100μg/mlカナマイシン、5×10-5 M 2-メルカプトエタノールおよび5%ヒト血清を含有するRPMI 1640培地中、1〜2.5×106細胞/ml)したクローン化ヒトCD4+ T細胞(KT30)およびCD8+ T細胞(ERCD8)、クローン化NK細胞(ERNK57)およびPBLを含む培養細胞由来の全RNA試料10μgを調べた。(ヒト組換えIL-2はバーゼル免疫学研究所(スイス・バーゼル)のA. Lanzavecchia博士から供与された)。ブロッティングに先立って、ゲル上の全RNAの完全性と量を調べるために、アガロースゲルを臭化エチジウムで染色した。RNA試料は、IP-10/MigR DNAの32P標識5'断片(109cpm/μg DNA)を5×106cpm/ml のハイブリダイゼーション溶液の濃度で使用して、記述されているように分析した(Loestscher, M.ら, J. Biol. Chem., 269 :232-237 (1994) )。ノーザンプローブとして使用した5'断片は、pBK-CMVベクター(Stratagene GMBH社,スイス・チューリッヒ)中のCXCR3 cDNAをPstIで消化してCXCR3 cDNA(図1)の5'末端724bpを得ることによって調製した。
【0147】
結果
予想されるサイズのmRNAの豊富な発現が、受容体cDNAの単離に使用したクローン化CD4+ T細胞KT30に認められた。同等レベルの発現がCD8+ T細胞クローンERCD8とNK細胞クローンERNK57に観察された。これに対し、新たに単離された血液リンパ球とナイロンウール精製T細胞には、IL10/MigR転写物はほとんど検出できなかった。しかし、これらの細胞をIL-2の存在下で培養すると、強いアップレギュレーションが得られ、受容体mRNAのレベルはTおよびNK細胞クローンでのレベルに近づいた。新たに単離された血液単球、好中性白血球または好酸性白血球では、これらの条件下にIP-10/MigR転写物が検出されなかった。IP-10/MigR mRNAを発現しなかったその他の白血球関連細胞には、マスト細胞株HMC-1、前骨髄球性白血病株HL60、組織球性白血病株U937、慢性骨髄性白血病株K562、急性T細胞白血病株Jurkat、急性リンパ芽球性白血病株Molt、Bリンパ芽球株DaudiおよびRaji、慢性および急性Bリンパ性白血病(B-CLLおよびB-ALL)患者由来のリンパ球、好塩基球性白血病患者由来の成熟好塩基球、および赤白血病細胞株HELが含まれる。対照的に、単球および顆粒球には、リンパ球を誘引することが過去に示されているケモカイン、すなわちMCP-1、MCP-2、MCP-3、MIP-1α、MIP-1βおよびRANTESの受容体(Loetscher, P.ら, FASEB J., 8 :1055-1060 (1994);Carr, M.W.ら, Proc. Natl. Acad. Sci. USA 91 :3652-3656(1994);Taub, D.D.ら, Science, 266 :355-358 (1993);Schall, T.J.ら, J. Exp. Med., 177 :1821-1825(1993);Schall, T.J.ら, Nature, 347 :669-672(1990))も認められる。活性化Tリンパ球とナチュラルキラー細胞株におけるIP-10/MigRの限定的発現は、この新規受容体が選択的なリンパ球補充を媒介しうることを示唆している。
【0148】
安定なトランスフェクタント
CXCR3 cDNAをpBK-CMV (Stratagene GMBH社,スイス・チューリッヒ)からBamHIとXbaIによる消化で切り出し、pcDNA3(Invitrogen BV社, オランダ・WBリーク)のBamHI部位とXbaI部位にクローニングすることによりpCDNA3-Clone8を得て、それを大腸菌(XL1Blue)で維持、保存した。
【0149】
安定なトランスフェクタントを作製するため、4×106個のマウスプレB細胞 (300-19) (Thelen, M.ら, FASEB. J., 2 :2702-2706 (1988))、ヒト前骨髄球細胞 (GM-1) (Garotta, G.ら, J. Leukocyte Biol., 49 :294-301 (1991))またはヒト急性T細胞白血病細胞 (Jurkat) (Loetscher, P.ら, FEBS Lett. 341 :187-192 (1994))に、Bgl IIで直線化したpcDNA3中の受容体cDNA 20μgを、既に記述されているようにエレクトロポレーション法でトランスフェクトした(Moser, B.ら, Biochem. J., 294 :185-192 (1993))。
【0150】
IP-10/MigRトランスフェクト細胞をG-418 (Life Technologies社)選択下に限界希釈法でクローン化した(300-19については1.0mg/ml G-418、Jurkat細胞とGM-1細胞については0.8mg/ml G-418)。RNAドット-ブロット分析により、G-418耐性クローンを受容体発現についてスクリーニングした。
【0151】
Ca2+流動
本受容体が機能的であるかどうかを決定するため、ネズミプレB細胞(300-19)、ヒト前骨髄球細胞(GM-1)およびヒトT細胞白血病細胞(Jurkat)のクローンに、上述のように受容体cDNAを安定にトランスフェクトした。ケモカイン受容体の活性化はサイトゾル遊離Ca2+濃度([Ca2+]i)の一過性の上昇をもたらす。そこでトランスフェクト細胞におけるシグナル伝達のモニターにこのアッセイを使用した。
【0152】
136mM NaCl、4.8mM KCl、1mM CaCl2、5mMグルコースおよび20mM HEPES(pH7.4)を含有する緩衝液中、106 細胞あたり0.1nmolのfura-2アセトキシメチルエステルと共に37℃で30分間インキュベートすることによってfura-2を負荷した細胞で、サイトゾル遊離Ca2+濃度([Ca2+]i)の変化を測定した。遠心分離後、負荷した細胞を同じ緩衝液に再懸濁し(106細胞/ml)、37℃で表記のケモカインによって刺激し、[Ca2+]i関連蛍光変化を記録した(von Tscharner, V.ら, Nature, 324 :69-372 (1986))。
【0153】
結果
IP-10およびMigに応答して、迅速な[Ca2+]i上昇が観察された。ケモカインIP-10は皮膚の遅延型過敏反応において発現されることが示されている(Luster, A.D.ら, Nature, 315 :672-676(1985);Kaplan, G.ら, J. Exp. Med., 166 :1098-1108(1987))。Migと呼ばれるケモカインが最近になって同定された(Farber, J.M., Proc. Natl. Acad. Sci. USA, 87 :5238-5242 (1990);Farber, J.M., Biophys. Res. Commun., 192 :223-230 (1993))。どちらのケモカインも、IL-8と同様に最初の2つのシステインがCXC配置を取っているが、好中性白血球に対して走化作用を持たない。最近、IP-10がTリンパ球を誘引すること(Luster, A.D.およびP. Leder, J. Exp. Med., 178 :1057-1065(1993);Taub, D.D.ら, J. Exp. Med., 177 :1809-1814(1993))と、Migが腫瘍関連リンパ球に対して走化作用を持つこと(Liao, F.ら, J. Exp. Med., 182 :1301-1314 (1995))が報告された。
【0154】
上記cDNAをトランスフェクトされ機能的IP-10/MigRを発現する細胞に対するIP-10とMigの作用を、図3A〜3Cに要約する。[Ca2+]i変化(図3A)によって示されるように、IP-10とMigの作用は濃度依存的であり、1nMで既に検出可能であることから、両ケモカインはこの新規受容体に対して高い親和性を持つことが示される。対照的に、これらIP-10/MigRトランスフェクタントは、CXCケモカイン類IL-8、GROα、NAP-2、GCP-2、ENA78、PF4、CCケモカイン類MCP-1、MCP-2、MCP-3、MCP-4、MIP-1α、MIP-1β、RANTES、I309、エオタキシン、またはケモカイン関連リンフォタクチンを含む他の16種類の潜在的アゴニストのいずれにも、100nMまでの濃度で反応しなかった(非掲載)。同じ結果がネズミとヒトのトランスフェクト細胞で得られた。これらの知見は、この新規受容体がIP-10とMigに高度に選択的であることを証明している。したがって、本明細書では本受容体をIP-10/Mig受容体(IP-10/MigR)もしくはそのCXCケモカインに対する特異性を反映して「CXCR3」と呼ぶ。
【0155】
図3Bに示すように、IP-10またはMigによる反復刺激は、ケモカイン受容体に典型的な脱感作をもたらした。さらに、これら細胞をIP-10で刺激した後、Migで(またはその逆で)刺激すると交差脱感作が起り、この受容体が両ケモカインに対して高い親和性を持つことが確認された。100nMの濃度で、Migが交差脱感作に関してIP-10よりも強力であることが明白となり、Migに対するIP-10/Mig受容体のより高い親和性または結合安定性が示唆された。
【0156】
機能的IP-10/MigRの発現は証明されたが、放射活性リガンドを用いて結合実験を行なったところ、結合パラメーターの決定を妨げる、全体の60〜80%に及ぶ非特異的結合が明らかになった。IP-10とMigはカチオン性が高い(pI値は10.8と11.1)ので、細胞表面プロテオグリカンとの非特異的相互作用がこれらの結果の説明となりうる。実際、種々の血液細胞と組織細胞には、ケモカイン受容体と無関係なヘパリナーゼ感受性のIP-10(およびPF4)結合部位が検出されており(Luster, A.D.ら, J. Exp. Med., 182 :219-231 (1995))、またヘパラン硫酸はIP-10とMigを結合し、リンパ球走化性を妨げる(非掲載)。ヘパリン結合部位はおそらくCXCR3受容体結合には関与しないと思われ、ヘパリン結合部位を介した細胞への非特異的結合を阻害するには、反応(例えば結合緩衝液)へのコンドロイチン硫酸などの適当なヘパリン誘導体の包含を使用できる。
【0157】
走化性
献血血液バフィーコートからPBLを新たに単離した。献血血液バフィーコートはスイス赤十字社スイス中央研究所輸血部門によって提供された。バフィーコートPBLの単離はColotta, F.ら, J. Immunol., 132 :936-944 (1984) に記述されているように行なった。
【0158】
献血血液バフィーコートから新たに単離されたPBLをさらに処理することなく使用するか、IL-2の存在下に10日間培養(2mMグルタミン、1×非必須アミノ酸、1mMピルビン酸ナトリウム、100μg/mlカナマイシン、5×10-5 M 2-メルカプトエタノールおよび5%ヒト血清を含有するRPMI1640培地中、400U/ml hrIL-2の存在下に、1〜2.5×106細胞/ml)した後、使用した。
【0159】
細胞移動は、IP-10/MigRトランスフェクト細胞については孔径5μm (Loetscher, P.ら, FEBS Lett. 341 :187-192 (1994))、ヒトPBLについては孔径3μm (Loetscher, P.ら, FASEB J., 8 :1055-1060(1994))のポリビニルピロリドン非含有ポリカーボネート膜 (Nucleopore)を用いて、48ウェルチャンバー (Neuro Probe社, 米国メリーランド州キャビンジョン)で評価した。ケモカイン(下部ウェル)の溶解と、細胞(上部ウェル中100,000個の受容体トランスフェクタントまたはPBL)の希釈には、20mM HEPES (pH7.4)と1%低温殺菌血漿タンパク質溶液(スイス赤十字社研究所,スイス・ベルン)を添加したRPMI1640を使用した。37℃で60分後、膜を取り出し、上面をPBSで洗浄し、固定し、染色した。全てのアッセイを3連で行ない、移動した細胞を倍率1,000倍の無作為に選択した視野5つで数えた。自発的な移動は、化学誘引物質の非存在下で決定した。
【0160】
結果−トランスフェクト細胞
IP-10/MigRを発現するトランスフェクト細胞はIP-10またはMigに向かってたやすく移動し、一方、非トランスフェクト親細胞は応答しなかった(図3C)。どちらのアゴニストも典型的な二相型濃度依存性を示した。IP-10は1nMを超える濃度で移動を誘導し、一方、Migの応答は10nM以上で検出可能になった。移動細胞の最大数によって測定される効力は、Migの方がIP-10の約2倍高かった。これらの結果はIP-10/MigRが、白血球のすべての既知ケモカイン受容体と同様に、リガンドに応答して起こる走化性を媒介することを示している。
【0161】
結果−ヒト血液白血球
IP-10/MigRの細胞分布と合致して、活性化ヒトTリンパ球はIP10とMigに対して高い反応性を持つことがわかった(図4A〜4B)。[Ca2+]i変化(図4A)とイン・ビトロ走化性(図4B)の誘導因子としてのIP-10とMigの活性は、IP-10/MigRを発現するトランスフェクト細胞を用いて観察された作用と合致し、IP-10はMigより強力であるが、効力はMigより低くかった。カルシウム流動と走化性の誘導にはIL-2の存在下で培養することによるTリンパ球の活性化が必要であり、単離したばかりの血液リンパ球には使用した条件下で応答が認められなかった。
【0162】
実施例3〜9に関する材料と方法
実施例3〜9では次の材料と方法を使用した。
【0163】
ケモカイン
組換えヒトケモカイン類は、イアン クラーク−ルイス(Ian Clark-Lewis)博士から提供された既に記述されているエオタキシン(Ponath, P.D.ら, J. Clin. Invest., 97 :604-612 (1996);Ponathら, 国際公開第97/00960号パンフレット(1997年1月9日公開)も参照されたい)以外は、Peprotech社(ニュージャージー州ロッキーヒル)から入手した。125I標識ケモカイン類はDu Pont NEN社(マサチューセッツ州ボストン)から入手した。
【0164】
細胞と細胞株
好中球とPBMCは記述されているように単離した(Ponath, P.D.ら, J. Clin. Invest., 97 :604-612 (1996))。CD3芽球を生成するため、RPMI-1640+10%FCS中2×106個/mlのPBMCを抗CD3抗体TR66でコーティングしてある組織培養プレートに添加した。4〜6日後、芽球を新しい培地に取り出し、IL-2(Antonio Lanzavecchia(バーゼル)の厚意で提供されたもの)を50単位/mlの濃度で添加した。
【0165】
使用した他の細胞株には、CXCR3(下記参照)、IL-8RA(Ponath, P.D.ら, J. Exp. Med., 183 :2437-2448(1996))、IL-8RB(Ponath, P.D.ら, J. Exp. Med., 183 :2437-2448(1996))、CCR2b (G. LaRosa, 未公表)、CCR4、CCR5(Wu, L.ら, Nature, 384 :179-183(1996))、またはCCR1(Campbell, J.J.ら, J. Cell. Biol., 134 :255-266(1996))のいずれかを発現するL1.2ネズミプレB細胞リンパ腫のトランスフェクタントが含まれる。
【0166】
CXCR3トランスフェクタントの調製
細胞
L1.2細胞は、RPMI培地1640、10% FetalClone (Hyclone社製)、50U/mlペニシリン/ストレプトマイシン、1×L-グルタミン、1mMピルビン酸ナトリウムおよび5.5×10-5 Mβ-メルカプトエタノール中で生育した。培地成分は、Hyclone社から購入した10% FetalClone以外は、GibcoBRL社から購入した。トランスフェクションの2日前に、L1.2細胞を新しい培地に1:5の比率で希釈した。これにより、約100〜300万細胞/mlの濃度の対数増殖期の細胞1億5000万個が得られた。
【0167】
CXCR3 DNAとトランスフェクション
大腸菌XL1Blue細胞(Stratagene社,カタログ番号200236)を、製造者の実験案に従い、pcDNA3-Clone8(実施例2;Loetscher, M.ら, J. Exp. Med., 184 :963-969(1996))で形質転換した。100μg/mlアンピシリンを含有するLB 500ml中、250rpmで振盪しながら、形質転換体を37℃で生育した。次にその培養物を8000×gでの遠心分離によって集め、Maxiプラスミド精製カラムとその実験案(Qiagen社,カタログ番号12162)を使って、プラスミドを精製した。プラスミドの濃度と純度は、1%アガロースゲルとOD260/280比を使って決定した。プラスミドDNAを二回蒸留水に懸濁し、使用時まで−20℃で保存した。
【0168】
ベクターの直線化にはScaIエンドヌクレアーゼを使用した。100μgのDNAを10μlのScaIで、製造者の実験案(GibcoBRL,カタログ番号15436-017)に従って37℃で8時間消化した。20μgを安定トランスフェクション構築に直接使用した。80μgはフェノール:クロロホルム:イソアミルアルコール(25:24:1)抽出、100%エタノール沈殿(0.1体積のNH4COOHを使用)および70%エタノール洗浄により、タンパク質と塩を除去した。
【0169】
ネズミプレBリンパ腫細胞株(L1.2)の安定トランスフェクタントは基本的に記述されているように調製した(Ponath, P.D.ら, J. Exp. Med., 183 :2437-2448(1996))。1×PBS 0.8ml中のL1.2細胞2500万個を、直線化したDNA 20μg、または直線化の後、洗浄したDNA 20μg(上記DNAの直線化の項を参照されたい)と共に、またはDNAなしで、エレクトロポレーションにかけた。エレクトロポレーション前に、L1.2細胞とDNAを50mlコニカルチューブ(Falcon 2070型)中で2分おきに穏やかに混合(旋回)しながら10分間インキュベートした。そのL1.2細胞-DNA混合物を、電極間隔0.4cmのGene Pulserキュベット(BioRad社,カタログ番号165-2088)に移した。次にその混合物を250V、960μFで、ショックの持続時間と実際の電圧を測定しながらエレクトロポレーションした。エレクトロポレーション後、キュベットを10分間室温に放置した。次にL1.2細胞-DNA混合物のすべてをT-25フラスコ(Costar社)に移し、10mlの非選択培地で2日間生育した。
【0170】
選択
次にCXCR3を発現するL1.2細胞をネオマイシン耐性に関する選択にかけた。非選択培地で2日間生育した後、10mlの1.6g/Lジェネティシン(GibcoBRL社)を0.8g/L(選択および維持濃度)の最終濃度で加えた。次にこれを、細胞が過剰増殖を始めたら新しい選択培地を加えながら、10〜15日間生育した。新しい選択培地は10%ウシ血清と800μg/ml G418 を添加したRPMI-1640からなった。
【0171】
CXCR3の細胞表面発現を走化性によって評価し、また表面発現をモニターするためにリガンド結合分析とスキャッチャード分析も使用した。G418選択後、CXCR3発現L1.2細胞を走化能に基いて選択した。各エレクトロポレーション反応について、30ml(800,000細胞/ml)を集め、600μlの選択培地に懸濁した。10nM IP-10を含有する選択培地600μlを、Becton-Dickinson社製のBioCoat細胞培養プレート(カタログ番号40575)の下部チャンバーに入れた。100μl/ウェルの上記L1.2細胞をそのBioCoatプレートの上部チャンバーに加えた。次に、それらの細胞を37℃のCO2培養器中に終夜放置して走化反応させた。非走化細胞の入った上部チャンバーを除去した。走化細胞を集め、新しい培地に移し、24ウェルプレートで生育した。次に、それらをCostar製T-25フラスコに拡張した後、T-75フラスコに拡張した。
【0172】
高レベルの受容体を発現するトランスフェクタントを、限界希釈法によってクローン化した。G418を含む選択培地に、CXCR3トランスフェクト細胞を30〜3細胞/mlまで希釈した。それを96ウェル組織培養プレートに100μl/ウェルずつ分注した。37℃、5%CO2で14日後、単一コロニーを含有するウェルを倒立顕微鏡下で同定した。次にその細胞の50μlを移し、抗CXCR3 mAb で上述のように染色し、フローサイトメトリーで分析した。受容体発現レベルは平均蛍光強度と相関し、高発現体が選択された。安定な細胞株が樹立されたら、その株を使用するために増殖した。
【0173】
CXCR3トランスフェクタント細胞は、抗生物質耐性と走化性に関する選択に加えて、抗体染色法で受容体発現量がより高いものを分別することによって、さらに選択することができるが、ここではそれを行なわなかった。染色するには、1%ウシ血清アルブミンを含む滅菌PBSにトランスフェクタント細胞を5×106個/mlの密度で再懸濁すればよい。単離された滅菌抗CXCR3 mAb は3μg/mlまでの最終濃度で加えることができ、細胞を氷上で30分間インキュベートする。冷滅菌PBSで洗浄した後、結合したmAb は、0.2μmフィルターを通した濾過によって滅菌したFITC結合抗マウスIgGで検出できる。それらの細胞を再び洗浄し、フローサイトメトリーで分別することができる。上位5%の陽性細胞を集め、選択条件下に増殖用の組織培養(例えば10%ウシ血清と800μg/ml G418を添加したRPMI-1640)に戻すことができる。
【0174】
実施例3.IP-10はCXCR3 DNAをトランスフェクトしたL1.2細胞上と活性化T細胞上に発現した受容体に高い親和性で結合する
放射標識IP-10を使用してさらなる結合試験を行なった。標的細胞へのケモカイン結合は既に記述されているように行なった(Ponath, P.D.ら, J. Clin. Invest., 97 :604-612 (1996);Van Riper, G.ら, J. Exp. Med., 177 (3): 851-856(1993))。細胞をPBSで1回洗浄し、結合緩衝液(50mM HEPES(pH7.5)、1mM CaCl2、5mM MgCl2、0.5%BSAおよび0.05%アジド)に107個/mlの濃度で再懸濁した。それを微量遠心管に50μl(5×105細胞)ずつ分注し、次いで非放射性競合因子(非標識IP-10)と放射標識ケモカイン(0.05nM 125I標識IP-10)を加えた。最終反応体積は200μlだった。非特異的結合は、250〜500nMの非標識ケモカインの存在下に細胞を放射標識ケモカインと共にインキュベートすることによって決定した。室温で60分間インキュベートした後、0.5M NaClを含む結合緩衝液1mlで細胞を3回洗浄した。次に、細胞ペレットをカウントした。競合は100×[(S-B)/(T-B)] (式中、Sは試料の放射活性、Bはバックグラウンド結合量、Tは競合因子がない場合の全結合量である)によって計算される特異的結合百分率として表わした。バックグラウンド結合量は、細胞を放射標識ケモカインおよび少なくとも400倍過剰の非標識ケモカインと共にインキュベートすることによって得た。全実験を通じて2連を使用し、標準偏差は常に平均の10%未満だった。すべての実験を少なくとも3回繰返した。カーブフットおよび特異的結合を50%阻害する濃度(IC50)は、カレイダグラフ(KaleidaGraph)ソフトウェア(Synergy Software社,ペンシルバニア州レディング)で計算した。
【0175】
図5A〜5Bは、CXCR3をトランスフェクトしたL1.2細胞(図5A)とCD3活性化T細胞(図5B)に125I-標識IP-10が結合したことと、非放射性IP-10の濃度を上げることでその結合を阻害できたことを示している。スキャッチャード分析から、IP-10がL1.2 CXCR3トランスフェクタントに614pMのKdで結合することと、それらのトランスフェクタントが1細胞あたり37,000個の受容体を発現させたことが明らかになった(図5A、挿入図)。抗CD3活性化IL-2刺激T細胞の同様の分析では、156pMのKdと1細胞あたり17,000個の受容体が明らかになった(図5B)。Migは125I標識IP-10結合の遮断に関して非標識IP-10よりわずかに有効性が低かったものの(非掲載)、活性化T細胞への125I標識IP-10結合は同じ条件下に非標識Migによって完全に阻害することができた。IP-10とMigは高い親和性でCXCR3を結合する(Kd約150〜600pM)。
【0176】
実施例4.CXCR3に特異的なモノクローナル抗体(mAb)の作製と特徴づけ
CXCR3のアンタゴニストを開発するために、また受容体の発現と調節を研究するために、この受容体のN末端に対応する合成ペプチドでマウスを免疫することにより、一群のmAb を作製した。これらのmAb はCXCR3トランスフェクタントを特異的に認識したが、他の一連の受容体トランスフェクタントは特異的に認識しなかった。
【0177】
mAb の作製とフローサイトメトリー
CXCR3と反応するmAb は、Balb/CマウスをCXCR3の最初の37N末端アミノ酸に対応する37マー合成ペプチド(Loetscher M.ら, J. Exp. Med., 184 :963-969(1996)をも参照されたい)10μgで10週間にわたって5回免疫することによって作製した。このペプチドを合成し、ツベルクリンの精製タンパク質誘導体(Severn Biotech社,英国キッダーミンスター)に結合した。最初の免疫処置はフロイント完全アジュバント(FCA)を使って腹腔内(IP)に行なった。第二、第三、第四の免疫処置はフロイント不完全アジュバント(FIA)を使って腹腔内に行い、最後の免疫処置はペプチド複合体のみ(アジュバントなし)を使って、静脈内(IV)に投与した。最後の免疫処置の4日後に、脾臓を摘出し、細胞株SP2/0を用いて、記述されているように細胞融合を行なった(Coligan, J.E.ら, Current Protocols In Immunology (John Wiley and Sons社,ニューヨーク), Unit 2.5.4 (1992))。ELISAで評価したところ、上記N末端37マーペプチドと反応するmAb が生成した(Coligan, J.E.ら, Current Protocols In Immunology (John Wiley and Sons社,ニューヨーク), Unit 2.1.3 (1992) )。CXCR3と反応するmAb は、非トランスフェクトおよびCXCR3トランスフェクトL1.2細胞または300.19細胞(ネズミB細胞株, Loetscher, M.ら, J. Exp. Med., 184 :963-969(1996))、免疫蛍光染色およびFACScan(登録商標) (Becton Dickinson & Co.社, カリフォルニア州マウンテンビュー)を用いて同定した。
【0178】
CXCR3に特異的なモノクローナル抗体が生成した。CXCR3トランスフェクトL1.2細胞が染色されるが、非トランスフェクトL1.2細胞または他の受容体タイプをトランスフェクトしたL1.2細胞が染色されないことから判断して、8種類のmAb が表面発現したCXCR3を認識することがわかった。これらmAb の一つ1C6のFACSプロフィールを図6に示す。また、これらmAb はPHAまたは抗CD3によりイン・ビトロで活性化されたヒトT細胞とT細胞クローンをも染色した(1C6について図7Cに図示)。しかし、これらの抗CXCR3 mAb は、好中球(1C6について図7Aに図示)、単球または好酸球(非掲載)とは反応しなかった。この反応性のパターンは、ノーザンブロットによる分析と合致した。しかし、意外にも表現型分析では循環リンパ球の大きな亜集団でのCXCR3の発現が明らかになった(図7B)。この発現パターンは調査した全ての個体で観察され、このことはCXCR3が血液リンパ球の亜集団に常態で発現されることを示す。CXCR3は、CD45RO+(メモリー)サブセットに包含される循環T細胞の一集団を特徴づけることがわかった。
【0179】
ネズミハイブリドーマ1C6(LS77-1C6ともいう)は、1997年3月28日に、ブダペスト条約の条件に基き、LeukoSite社(米国02142マサチューセッツ州ケンブリッジ・ファーストストリート215)の名でアメリカン・タイプ・カルチャー・コレクション (American Type Culture Collection) (20852 メリーランド州ロックビル・パークローンドライブ12301)に、ATCC寄託番号HB-12330の下で寄託された。
【0180】
実施例5.CXCR3は活性化/メモリーT細胞上に発現される
フローサイトメトリー
CXCR1、CXCR2、CXCR3およびCCR5に対するmAb は既に記述されている(Qin, S.ら, Eur. J. Immunol., 26 :640-647 (1996);Heath, H.ら, J. Clin. Invest. 印刷中(1997))。抗CXCR4 mAb 12G5(Endres, M.J.ら, Cell, 87 :745-756 (1996))は、ジム ホキシー (Jim Hoxie)(ペンシルバニア大学)の厚意で提供された。CD4、CD8、CD14、CD20、CD25、CD26、CD69、CD45RO、CD45RA、CD95に対するPE結合mAb 、抗CD3および抗CD4 Cy-ChromeはPharMingen社(カリフォルニア州ラホーヤ)によって供給された。
【0181】
トランスフェクト細胞または白血球に対するmAb の反応性を評価するために、間接免疫蛍光法とフローサイトメトリーを使用した。細胞をPBSで1回洗浄し、2%ヒト血清と0.1%アジ化ナトリウムを含む100μlのPBS(染色緩衝液)、5μg/ml精製抗体、5μg/ml IgG2a同イソタイプ対照mAb (Sigma Chemical社, ミズーリ州セントルイス)または50μlのハイブリドーマ培養上清に再懸濁した。4℃で20分後、細胞を染色緩衝液で2回洗浄し、50μlのFITC結合アフィニティー精製F(ab')2 ヤギ抗マウスIgG (Jackson ImmunoResearch Laboratories社)に再懸濁した。4℃で20分間インキュベートした後、細胞を染色緩衝液で2回洗浄し、表面発現レベルを決定するためFACScan(登録商標)で分析した。死細胞の排除にはヨウ化プロピジウムを使用した。
【0182】
リンパ球の二色免疫蛍光分析は、CD20+(B)細胞とCD56+(NK) 細胞のごく一部もCXCR3を発現するものの、CXCR3を発現する細胞はそのほとんどがCD3+細胞であることを示した(図8)。T細胞を標識するために抗CD3 Cy-Chromeを用いて行なったT細胞の三色分析は、CD4+細胞の一部とCD8+細胞の一部がCXCR3を発現することを示した(図9)。CD25やCD69などの細胞活性化のマーカーを使った分析では、活性化T細胞が一般にこの受容体を発現することが明らかになった。CXCR3+ T細胞は、先の活性化に合致する表現型CD95+、CD45RO+およびCD45RAlowだった。CXCR3の発現と、やはり前もって活性化されたT細胞にその発現が偏っているケモカイン受容体CCR5(Wu, L., 私信)の発現も比較した。図9は、血中のCCR5+細胞がCXCR3+亜集団に包含されていたことと、CXCR3がCCR5より広範に発現されたことを示している。CCR5やCXCR4などの他のT細胞ケモカイン受容体とは異なり、CXCR3は循環する活性化T細胞の大半で発現した。
【0183】
実施例6.抗CXCR3 mAb はIP-10結合と走化性を遮断する
ヒト白血球の走化性を、ECV 304内皮細胞株(European Collection of Animal Cell Cultures, 英国ソールズベリー・ポートンダウン)を使って、既に記述されている(Ponath, P.D.ら, J. Clin. Invest., 97 :604-612 (1996))経内皮アッセイ(Carr, M.W.ら, Proc. Natl. Acad. Sci. USA, 91(9): 3652-3656 (1994))の変法で評価した。下部チャンバーに移動した細胞をチューブに入れ、相対細胞数をFACScan(登録商標)を用いて得た。
【0184】
また、CD3活性化T細胞芽球の走化性を阻害する能力について、抗ペプチドmAb を試験した。1A5、1C6、3A8、5F10、10C6および10G12と呼ばれるmAb に関する結果を図10に示す。mAb の一つ1C6は、IP-10へのT細胞の走化性を遮断する能力が他のmAb より優れていた。mAb 1C6 は、IP-10へのT細胞の走化性を約0.8μg/mlのIC50で用量依存的に完全に阻害することができた(図11)。トランスウェルの底に至適濃度のIP-10(12.5nM)を使用すると、2〜5μg/mlの濃度で100%阻害が達成された。使用した条件下で1C6は、ケモカイン受容体CCR2bを介して起こるMCP-1へのT細胞走化性を有意に阻害できなかった(図10)。
【0185】
またmAb 1C6 は活性化T細胞への125I標識IP-10結合を0.16μg/mlのIC50で完全に遮断することができた(図12)。1〜10μg/mlの抗体で完全な阻害が得られた。mAb 1C6 によってIP-10結合と走化性の両方が完全に阻害されることは、活性化T細胞がこのケモカインを結合する別の受容体を発現しないことを示している。
【0186】
ノーザンブロット分析は、CXCR3が活性化T細胞で発現されることを示した。一群の特異的mAb を使用することにより、CXCR3は血液T細胞の亜集団と他の白血球タイプ(B細胞およびNK細胞)に発現されることがわかった。CXCR3を発現するT細胞は先の活性化と合致した表現型、すなわちCD45RO+、CD26+を有した(実施例5)。T細胞の染色は、T細胞がCD3とIL-2で活性化されると著しく増大し、それはIP-10に応答して起こる細胞移動および放射標識リガンド結合量の増大と相関した。
【0187】
血液T細胞が少なくとも走化性アッセイでIP-10またはMigに対して低い応答性を示すことは変則的であるように思われる。その理由として、受容体発現以外の要因が化学誘引物質に対する細胞の応答性を決定しうることが考えられる。シグナル伝達には適当なGタンパク質共役が必要かもしれない。もう一つの可能性は、T細胞上とNK細胞上のIL-8受容体について観察されているように(C.R. Mackay)、血液分離操作がこの受容体を破壊することである。適当な実験動物の皮膚へのIP-10の注射により、血液T細胞でのCXCR3発現の意義を検討できる。活性化T細胞では、IP-10が最も強力な化学誘引物質の一つである。T細胞の活性化は、受容体シグナル伝達分子またはシグナル伝達に必要な共役を誘導しうる。
【0188】
実施例7.mAb 1C6 はIP-10に応答したT細胞による[Ca2+]iを選択的に阻害するが、Migに応答したT細胞による[Ca2+]iは阻害しない
細胞内カルシウム濃度([Ca2+]i)は次のように測定した.Fura-2 AM (Molecular Probes社, オレゴン州ユージーン)の原液は、その色素50μgを44μlのDMSOに溶解することによって調製した。細胞に添加する直前に、その原液をCa2+とMg2+を含むHBSSおよび2%BSAに1:100に希釈した。Fura-2 AMを0.2モル/106細胞の最終濃度で37℃で30分間細胞に添加した。標識化の後、過剰の色素を遠心分離によって除去し、細胞を125mM NaCl、5mM KCl、1mM MgCl2、1mM CaCl2、0.5mMグルコース、0.025%BSAおよび20mM HEPES(pH7.4)に106個/mlの濃度で再懸濁した。[Ca2+]iは日立F-2000蛍光分光計で340nmと380nmでの励起を使って測定した。較正は、全放出に1%NP-40を使用し、遊離Ca2+のキレートに25μM EGTAを使用して行なった。
【0189】
IP-10とMigはヒトT細胞による[Ca2+]iを誘導し、各ケモカインは他方のケモカインに対する応答を完全に脱感作することができた(図13A〜13B)。ケモカインの滴定により、最大[Ca2+]iがわずか2nM IP-10または2nM Migで達成されることが明らかになった。mAb 1C6 のアゴニスト/アンタゴニスト機能を調べるため、mAb 1C6 または無関係な同イソタイプの対照mAb 注入後に、活性化T細胞を[Ca2+]iについて評価した。無関係な同イソタイプ対照mAb を注入されたT細胞は、それに続くIP-10の注入に対して強い[Ca2+]i応答を示した(非掲載)。しかし、mAb 1C6 で処理されたT細胞は、2nM IP-10で刺激しても[Ca2+]iを示さなかった。IP-10に応答した活性化T細胞における[Ca2+]iは、12.5mgの1C6によって完全に抑制された。しかし50μg/mlの1C6でも、2nM Migに対するT細胞の応答には影響がなく(図13D)、これら2つのリガンドがこのmAb によって異なる影響を受けることが示された。対照として、CXCR3以外の受容体を介して起こるMIP-1αまたはRANTESに応答したT細胞の[Ca2+]iへの効果について、mAb 1C6 を試験した(非掲載)。このmAb は、使用した条件下に効果がなかった。
【0190】
IP-10結合とIP-10誘導性走化性を阻害するmAb 1C6 は、活性化T細胞によるカルシウム流動をも阻害したが、それらの条件下でMig誘導性カルシウム流動を阻害しなかった。これらの結果は、IP-10とMigがCXCR3の異なる領域を介して結合および/またはシグナル伝達することと、mAb 1C6 がIP-10結合部位とそれに続くシグナル伝達を遮断できたことを示唆している。したがって、ここにmAb 1C6 を用いて例証したように、個々のケモカインの作用を選択的に阻害する受容体アンタゴニストを開発することが可能である。
【0191】
後述するように、CXCR3トランスフェクタントの染色は、CXCR3の最初の15N末端アミノ酸残基を含むペプチドによって阻害されうる。mAb 1C6 によるIP-10結合とそれに続く応答の阻害は、この領域がリガンド結合にとって機能上とりわけ重要であることを示唆している。1C6がMig誘導性のカルシウム流動を、使用した条件下に阻害できないことから、IP-10結合とシグナル伝達にとって重要だと思われるmAb 1C6 によって認識されるエピトープは、Mig結合および/またはシグナル伝達には関与しないようである。
【0192】
実施例8.エピトープマッピング
ペプチドはGenemed社(カリフォルニア州サウスサンフランシスコ)に発注した。各15アミノ酸長のそれらペプチドは、CXCR3タンパク質の最初の45N末端残基の異なる部分に相当する。
P1: MVLEVSDHQVLNDAE(配列番号:2、残基1−15)
P2: VAALLENFSSSYDYG(配列番号:2、残基16−30)
P3: ENESDSCCTSPPCPQ(配列番号:2、残基31−45)
それらペプチドをまずDMSOに溶解し、リン酸緩衝食塩水(PBS)で1mg/mlに希釈した。1C6抗体による染色を遮断するペプチドの能力を調べるために、1μg/mlの精製1C6抗体を1×PBS、5%ウシ胎仔血清(FCS)中、100μg/mlの各ペプチドの存在下にCXCR3 L1.2トランスフェクタント(105細胞;実施例3〜9に関する材料と方法の項を参照されたい)と共に4℃で30分間インキュベートした。最終体積は100μlだった。陽性染色はペプチドの非存在下に行なった。結合したmAb を抗マウスIgG-FITCで検出し、その結果をフローサイトメトリーで分析した。
【0193】
mAb 1C6 による染色はP1によって完全に阻害され、このmAb がCXCR3タンパク質の最初のN末端15アミノ酸中のエピトープを認識することが示唆された(図14A〜14D)。mAb 1C6 と同じ融合(LS-77)から生成した3A8と呼ばれるもう一つのmAb (LS77-3A8ともいう)の結合も、P1ペプチドによって阻害された。
【0194】
実施例9.トランスフェクト細胞での免疫化による抗CXCR3 モノクローナル抗体の作製
CXCR3構築物をトランスフェクトされ高レベルのヒトCXCR3を発現するL1.2細胞でマウスを免疫することにより、さらなる抗CXCR3 モノクローナル抗体を作製した(上記材料と方法の項を参照されたい)。免疫化と融合ハイブリドーマの作製は記述されているように行なった(Qin, S.ら, Eur. J. Immunol., 26 : 640 (1996) ; Heath, H.ら, J. Clin. Invest., 99(2):178(1997))。抗CXCR3 mAb は、活性化ヒトT細胞とCXCR3トランスフェクタントの陽性染色によって同定した。8種類の抗CXCR3抗体が一つの融合(LS-104)で得られた。
【0195】
CXCR3へのIP-10の結合を阻害するそれら抗体の能力を試験した。陽性クローンから得た組織培養上清(25μl)を、1×PBS、5%ウシ胎仔血清(FCS) (最終体積100μl)中、CXCR3 L1.2トランスフェクタント細胞(実施例3〜9に関する材料と方法の項)および0.05nMの放射標識125I標識IP-10と共に4℃で30分間インキュベートした。mAb 1C6 と抗CXCR2(IL-8受容体B)mAb の上清も、それぞれ特異的および非特異的mAb 対照として使用した。総結合量は抗体の不在下に決定した。40nMの非標識IP-10を競合因子として使用することによってバックグラウンド結合量を求め、その値をmAb による阻害百分率の計算に使用した。結果を図15に示す。この融合(LS-104)から得られた抗体はすべて、50〜70%の範囲の阻害百分率でIP-10結合を遮断できた。
【0196】
これらのmAb を、シグナル伝達(Ca2+流動誘導能)を阻害する能力についても、基本的に実施例7に記述したように評価したが、これらの抗体はいずれも使用した条件下にMig媒介性のCa2+シグナル伝達を遮断できなかった。IP-10媒介性のCXCR3結合およびシグナル伝達が抗体によって阻害され、CXCR3を介したMig媒介性のシグナル伝達が抗体によって阻害されないことは、これらの抗体がIP-10によって媒介されるCXCR3機能を選択的に阻害できることを示している。
【0197】
また、基本的に実施例8に記述したようにLS-104融合から得た抗体を用いて、エピトープマッピング研究を行なった。その結果は種々の結合部位が認識されることを示している(表2)。この結果は、これら抗体のうち3つがCXCR3の最初の15N末端アミノ酸(配列番号:2の第1〜15残基)内のエピトープを認識し、これら抗体のうち2つがCXCR3のアミノ酸16〜30(配列番号:2の第16〜30残基)内のエピトープを認識することを示唆している。P3ペプチドはこれら抗体のいずれによる染色をも遮断せず、これは、これらの抗体がいずれもCXCR3のアミノ酸31〜45(配列番号:2の第31〜45残基)に相当するペプチドを結合しなかったことを示す。残り3つのmAb を用いた染色は、使用した条件下にどのペプチドによっても有意に阻害されず、これらmAb が、一部重複した上記ペプチドの区間からなるエピトープ、細胞表面に提示される構造エピトープ(conformational epitopes)、または受容体の他の部分にあるエピトープを結合しうることが示唆された。また、これらのデータは、受容体トランスフェクタントでマウスを免疫することにより、CXCR3の様々な部分に対するmAb が得られうることをも示唆している。
【0198】
【表2】
実施例10.mAb 1C6 を用いた正常組織と炎症組織の免疫組織化学的分析
組織
ヒト組織(正常および炎症)は米国立衛生研究所(National Institutes of Health)の資金提供を受けた公共機関National Disease Research Instituteから入手した。正常マカーク(アカゲザル)組織はNew England Regional Primate Research Center(ニューイングランド地方霊長類研究センター;マサチューセッツ州サウスボロ)から入手した。
【0199】
免疫組織化学
アルカリホスファターゼ法.組織を厚さ4μmに切片化し、乾燥した後、2%パラホルムアルデヒド/0.5×PBS中4℃で10分間固定した。PBS洗浄の後、非特異的抗体結合部位を10%正常ヤギ血清/5%ヒトAB血清/PBSにより室温で30分間遮断した。次に、精製した抗CXCR3ネズミmAb 1C6 を0.3%Triton X100/0.2%Tween 20/1%FCS/5%ヒトAB血清および0.1%アジ化ナトリウム中に10μg/mlの濃度に希釈し、それを組織切片に適用して4℃で終夜インキュベートした。同イソタイプの無関係なmAb を組織のステップセクション(step sections)に対して陰性対照として使用した(IgG1,MOPC-21, Sigma社,ミズーリ州セントルイス)。次に、ビオチニル化ヤギ抗マウスIgG(Vector社, カリフォルニア州バーリンゲーム)とアビジン-ビオチン-アルカリホスファターゼ複合体(Biogenex社, カリフォルニア州サンラモン)を順次加えた。内因性アルカリホスファターゼ活性を遮断するためにレバミゾールを含有するFast Red(Biogenex社, カリフォルニア州サンラモン)を発色原として使用し、マイヤーヘマトキシリンを対比染色剤として使用した。
【0200】
結果
ヒトおよびマカークの正常リンパ節:どちらの種でも、染色は、Tリンパ球でのCXCR3発現と合致して、リンパ球の傍皮質と髄索内のリンパ球の70〜80%に限定された。
【0201】
ヒトおよびマカクの脾臓:どちらの種でも、染色は、白色脾髄のリンパ濾胞の周縁部に沿ったリンパ球と脾洞内に散在したリンパ球に限定された。このパターンはTリンパ球でのCXCR3発現と合致する。
【0202】
ヒト胸腺:胸腺髄質はリンパ球と形態学的に合致する散在したCXCR3免疫反応性単核細胞を含んだ。
【0203】
この分析により、マカークCXCR3がmAb 1C6 によって認識されることが明らかになった。独立した実験により、マカーク細胞Tリンパ球をコンカナバリンAおよびIL-2と共に培養することによってCXCR3がアップレギュレートされること、マカークT細胞芽球がヒトIP-10に応答して走化性を示しうること、mAb 1C6 がその走化性を遮断できること、および前もってヒトIP-10と共にインキュベートすると、走化性によって評価されるマカーク芽球の脱感作が起こることが示された。
【0204】
下記の考察では、様々な数のCXCR3免疫反応性「単核細胞」が正常組織と炎症組織の両方に同定された。これらの単核細胞は次の理由からおそらくTリンパ球だろう:a)フローサイトメトリーにより、CXCR3は多少のB細胞とNK細胞にも検出されたものの、他の単核細胞には検出されず、CXCR3+ 細胞の大半がTリンパ球であることが明らかになった;b)リンパ節と脾臓内では、T細胞が配置されていることが知られている領域内のリンパ球がCXCR3に免疫反応性を示す唯一の細胞である;ならびにc)次に挙げる組織内のCXCR3免疫反応性単核細胞は形態学的にリンパ球と合致する。
【0205】
非炎症ヒト組織:心臓、肝臓、腎臓、肺、皮膚、胸部、骨格筋、唾液腺、膵臓、膣、子宮および卵巣を含む非炎症ヒト組織では、CXCR3発現がまばらで散在した間質単核細胞に限定された。小腸と大腸の切片では、CXCR3を発現する単核細胞が固有層とパイアー斑内に観察された。肝臓では、管周辺リンパ球も染色され、肝細胞はわずかに染色された。
【0206】
ヒト脳:染色は観察されなかった。
【0207】
炎症ヒト組織:単核細胞の間質および血管周辺蓄積を特徴とする慢性的炎症組織の複数切片をCXCR3発現について調べた。間質性腎炎(1症例,腎組織)、潰瘍性大腸炎(1症例,結腸組織)、腸炎(1症例,小腸)および慢性膣炎(4症例,膣)のヒト患者由来の組織を含むこれらの組織では、単核細胞の約50%〜90%がCXCR3に免疫反応性を示した。したがって、慢性的炎症組織は正常組織と比較して、そこに含まれる間質単核細胞の数が多く、それら細胞のうちCXCR3に免疫反応性を示すものの割合も高かった。
【0208】
炎症組織の分析はリンパ球の約50〜90%がCXCR3を発現することを明らかにしたが、対応する正常組織ではCXCR3を発現するリンパ球の割合ははるかに低かった。これらの知見は、CXCR3を発現するリンパ球(おそらくはCXCR3を発現するTリンパ球)の慢性炎症部位への特異的な補充を示唆している。CXCR3はホーミングまたは炎症部位に遊走する傾向を持つT細胞を特徴づけるようである。
【0209】
均等物
当業者であれば、単なる日常的実験手法により、本明細書に記載された本発明の具体的態様に対する多くの均等物を認識し、あるいは確認することができるであろう。かかる均等物は、下記請求の範囲に包含されるように意図されている。
【0210】
本発明の態様として、以下のものが挙げられる。
[1]タンパク質または改変体が1種以上のケモカイン類を選択的に結合することができ、かつ細胞シグナル伝達および/またはそれに応答した細胞応答を媒介することができる、哺乳類CXCR3タンパク質またはその機能的改変体をコードする単離核酸。
[2]該核酸が中程度のストリンジェンシー条件下に、図1(配列番号:1)の配列、その相補鎖、またはコード配列を含む図1(配列番号:1)の配列の一部もしくはその相補鎖を有する2番目の核酸にハイブリダイズすることができる、[1]記載の単離核酸。
[3]該単離核酸が本質的に純粋である、[1]記載の単離核酸。
[4]該タンパク質または改変体がヒトIP−10、ヒトMig、ヒトIP−10の哺乳類ホモログおよびヒトMigの哺乳類ホモログからなる群より選ばれたケモカインに選択的に結合することができる、[1]記載の単離核酸。
[5]該哺乳類CXCR3タンパク質またはその機能的改変体がヒトCXCR3タンパク質またはその機能的改変体である、[1]記載の単離核酸。
[6]配列番号:1、その相補鎖、またはコード配列を含む配列番号:1の一部もしくはその相補鎖を含有してなる、[1]記載の単離核酸。
[7]該ヒトCXCR3タンパク質が配列番号:2に示されたアミノ酸配列を有する、[1]記載の単離核酸。
[8][1]記載の核酸を含有してなる単離核酸構築物。
[9]該核酸が操作可能に発現制御配列に連結される、[8]記載の単離核酸構築物。
[10]該核酸が配列番号:1、その相補鎖、またはコード配列を含む配列番号:1の一部もしくはその相補鎖を含有してなる、[8]記載の単離核酸構築物。
[11]該核酸が図2(配列番号:2)に示されたアミノ酸配列を有するポリペプチドをコードする、[8]記載の単離核酸構築物。
[12]該核酸が哺乳類CXCR3タンパク質を含む融合タンパク質をコードする核酸であり、任意にコード配列が操作可能に発現制御配列に連結される、[8]記載の単離核酸構築物。
[13]タンパク質が1種以上のケモカイン類を選択的に結合することができ、かつ細胞シグナル伝達および/またはそれに応答した細胞応答を媒介することができる、哺乳類CXCR3タンパク質またはその機能的改変体をコードする組換え核酸を含有してなる宿主細胞。
[14]該核酸が操作可能に発現制御配列に連結される、[13]記載の宿主細胞。
[15]該核酸が配列番号:1によりコードされるヒトCXCR3タンパク質をコードする、[13]記載の宿主細胞。
[16]タンパク質が1種以上のケモカイン類を選択的に結合することができ、かつ細胞シグナル伝達および/またはそれに応答した細胞応答を媒介することができる、単離哺乳類CXCR3タンパク質またはその機能的改変体。
[17]該哺乳類がヒトであり、かつ該タンパク質がヒトIP−10およびヒトMigからなる群より選ばれた1種以上のケモカイン類に選択的に結合することができる、[16]記載の単離哺乳類CXCR3タンパク質またはその機能的改変体。
[18]中程度のストリンジェンシー条件下に、図1(配列番号:1)の配列、その相補鎖、またはコード配列を含む図1(配列番号:1)の配列の一部もしくはその相補鎖を有する2番目の核酸にハイブリダイズすることができる核酸によりコードされる、[16]記載の単離哺乳類CXCR3タンパク質またはその機能的改変体。
[19]図1(配列番号:1)に示された核酸によりコードされる、単離ヒトCXCR3タンパク質。
[20]配列番号:2に示されたアミノ酸配列を有する、[19]記載の単離ヒトCXCR3タンパク質。
[21]哺乳類CXCR3タンパク質を含有してなる融合タンパク質。
[22](a)宿主細胞に哺乳類CXCR3タンパク質またはその改変体をコードする核酸を導入する工程であり、それによりコード配列が発現制御配列に操作可能に連結されたコード配列を有する組換え宿主細胞を生産する、および
(b)工程(a)で生産された宿主細胞を該核酸が発現する条件下に維持する工程、
を含む、哺乳類CXCR3タンパク質またはその改変体の生産方法。
[23]さらに哺乳類CXCR3タンパク質またはその改変体を単離する工程を含む、[22]記載の方法。
[24]哺乳類CXCR3タンパク質またはその改変体をコードする組換え核酸を含む宿主細胞を核酸の発現に適する条件下に維持する工程を含む、哺乳類CXCR3タンパク質またはその改変体を生産する方法。
[25]さらに哺乳類CXCR3タンパク質またはその改変体を単離する工程を含む、[24]記載の方法。
[26]該哺乳類がヒトであり、かつ該タンパク質またはその改変体が、ヒトIP−10およびヒトMigからなる群より選ばれた1種以上のケモカイン類を選択的に結合することができる、[24]記載の方法。
[27]哺乳類CXCR3タンパク質を結合する、抗体または機能的抗体断片。
[28]該抗体または抗体断片が哺乳類CXCR3タンパク質の1種以上の機能を阻害することができる、[27]記載の抗体または機能的抗体断片。
[29]該抗体または抗体断片がリガンドの哺乳類CXCR3タンパク質への結合を阻害する、[28]記載の抗体または機能的抗体断片。
[30]該抗体または抗体断片がヒトCXCR3タンパク質を結合することができ、かつヒトCXCR3タンパク質とIP−10および/またはMigからなる群より選ばれた1種以上のリガンドとの相互作用を阻害することができる、[28]記載の抗体または機能的抗体断片。
[31]該抗体または断片が、モノクローナル抗体1C6がヒトCXCR3タンパク質またはその一部に結合することに競合することができる、[30]記載の抗体または機能的抗体断片。
[32]該抗体または抗体断片がヒトCXCR3タンパク質とIP−10との相互作用を選択的に阻害する、[28]記載の抗体または機能的抗体断片。
[33]該抗体または断片がモノクローナル抗体1C6またはその抗原結合断片である、[32]記載の抗体または機能的抗体断片。
[34]ヒトCXCR3タンパク質を結合する、抗体またはその抗原結合断片。
[35]配列番号:2のN−末端細胞外セグメントに対応するヒトCXCR3の一部またはヒトCXCR3タンパク質の少なくとも1種の免疫学的性質を有するその一部により、該結合が阻害されうる、[34]記載の抗体またはその抗原結合断片。
[36]配列番号:2の1〜15残基の配列と同じである配列を有するポリペプチドと配列番号:2の16〜30残基の配列と同じである配列を有するポリペプチドとからなる群より選ばれたポリペプチドにより、該結合が阻害されうる、[34]記載の抗体またはその抗原結合断片。
[37]試験対象の薬剤と単離および/または組換え哺乳類CXCR3タンパク質あるいはそのリガンド結合改変体を含む組成物とを、リガンドの結合に適する条件下に合わせる工程、および該薬剤と該哺乳類CXCR3タンパク質もしくは改変体との間の複合体の形成を検出または測定する工程を含む、哺乳類CXCR3タンパク質またはそのリガンド結合改変体を結合する薬剤を検出または同定する方法。
[38]該薬剤がヒトIP−10、ヒトMig、IP−10の哺乳類ホモログおよびMigの哺乳類ホモログからなる群より選ばれたリガンドである、[37]記載の方法。
[39]該リガンドが放射性同位体、スピン標識、抗原標識、酵素標識、蛍光基または化学発光基からなる群より選ばれた標識で標識される、[38]記載の方法。
[40]アッセイがヒトIP−10、ヒトMig、IP−10の哺乳類ホモログおよびMigの哺乳類ホモログからなる群より選ばれた1種以上のリガンドの存在下に結合が決定される競合アッセイである、[38]記載の方法。
[41]a) 試験対象の薬剤と組換え哺乳類CXCR3タンパク質またはそのリガンド結合改変体を発現する宿主細胞とを、リガンドの結合に適する条件下に合わせる工程、および
b) 該薬剤と哺乳類CXCR3タンパク質もしくはそのリガンド結合改変体との間の複合体の形成を検出または測定する工程、
を含む、哺乳類CXCR3タンパク質またはそのリガンド結合改変体を結合する薬剤を検出または同定する方法。
[42]該薬剤がヒトIP−10、ヒトMig、IP−10の哺乳類ホモログおよびMigの哺乳類ホモログからなる群より選ばれたリガンドである、[41]記載の方法。
[43]アッセイがヒトIP−10、ヒトMig、IP−10の哺乳類ホモログおよびMigの哺乳類ホモログからなる群より選ばれた1種以上のリガンドの存在下に結合が決定される競合アッセイである、[41]記載の方法。
[44]哺乳類CXCR3タンパク質もしくはそのリガンド結合改変体が細胞シグナル伝達および/または細胞応答を媒介し、かつシグナル伝達活性またはそれに応答する該CXCR3タンパク質もしくは改変体の細胞応答を検出または測定することにより複合体形成をモニターする、[41]記載の方法。
[45]a)試験対象の薬剤と、哺乳類CXCR3タンパク質のリガンドと単離および/または組換え哺乳類CXCR3タンパク質あるいはそのリガンド結合改変体を含む組成物とを、リガンドの結合に適する条件下に合わせる工程、および
b) 該哺乳類CXCR3タンパク質もしくは改変体と該リガンドとの間の複合体の形成を検出または測定する工程であり、該薬剤による複合体形成の阻害が、該薬剤が阻害剤であることの指標である、
を含む、哺乳類CXCR3タンパク質もしくはそのリガンド結合改変体へのリガンド結合の阻害剤を検出または同定する方法。
[46]該リガンドがヒトIP−10、ヒトMig、IP−10の哺乳類ホモログおよびMigの哺乳類ホモログからなる群より選ばれた、[45]記載の方法。
[47]単離および/または組換え哺乳類CXCR3タンパク質を含む該組成物が組換え哺乳類CXCR3タンパク質を発現する宿主細胞を含有する、[45]記載の方法。
[48]該哺乳類CXCR3タンパク質が細胞シグナル伝達および/または細胞応答を媒介し、かつシグナル伝達活性またはそれに応答する該CXCR3タンパク質もしくは改変体の細胞応答を検出あるいは測定することにより複合体形成をモニターする、[47]記載の方法。
[49]a)試験対象の薬剤と、哺乳類CXCR3タンパク質のリガンドと組換え哺乳類CXCR3タンパク質もしくはそのリガンド結合改変体を発現する宿主細胞とを、リガンドの結合に適する条件下に合わせる工程、および
b) 該タンパク質もしくは改変体と該リガンドとの間の複合体の形成を検出または測定する工程であり、該薬剤による複合体形成の阻害が、該薬剤が阻害剤であることの指標である、
を含む、哺乳類CXCR3タンパク質もしくはそのリガンド結合改変体へのリガンド結合の阻害剤を検出または同定する方法。
[50]該リガンドがIP−10、Mig、IP−10の哺乳類ホモログおよびMigの哺乳類ホモログからなる群より選ばれた、[49]記載の方法。
[51]該哺乳類CXCR3タンパク質が細胞シグナル伝達および/または細胞応答を媒介することができ、かつシグナル伝達活性またはそれに応答する該CXCR3タンパク質の細胞応答を検出あるいは測定することにより複合体形成をモニターする、[49]記載の方法。
[52]該薬剤が抗体または抗体断片である、[49]記載の方法。
[53]試験対象の薬剤と(a)組換え哺乳類CXCR3タンパク質またはその機能的改変体を発現する宿主細胞と(b)リガンドまたはその促進剤とを、リガンドもしくは促進剤誘導応答を検出するに適する条件下に合わせる工程、および
試験対象薬剤の該応答を阻害する能力を評価する工程であり、薬剤によるリガンドまたは促進剤誘導応答の阻害が、薬剤が阻害剤であることの指標である、
を含む哺乳類CXCR3タンパク質もしくはその機能的改変体の阻害剤を検出または同定する方法。
[54]該応答がシグナル伝達活性またはそれに応答する該哺乳類CXCR3タンパク質もしくはその改変体の細胞応答を検出あるいは測定することによりモニターされる、[53]記載の方法。
[55]試験対象の薬剤と組換え哺乳類CXCR3タンパク質もしくはその機能的改変体を発現する宿主細胞とを、受容体媒介応答を検出するに適する条件下に合わせる工程、および
該応答を検出または測定する工程であり、該薬剤による応答の誘導または刺激が薬剤が促進剤であることの指標である、
を含む、哺乳類CXCR3タンパク質もしくはその機能的改変体の促進剤を検出または同定する方法。
[56][45]記載の方法によって同定された、哺乳類CXCR3タンパク質に特有の少なくとも1種の機能の阻害剤。
[57]該阻害剤が受容体に結合可能な抗体またはその一部である、[56]記載の阻害剤。
[58][53]記載の方法によって同定された、哺乳類CXCR3タンパク質に特有の少なくとも1種の機能の阻害剤。
[59][55]記載の方法によって同定された、哺乳類CXCR3タンパク質に特有の少なくとも1種の機能の促進剤。
[60]a) 抗体と選択された哺乳類CXCR3タンパク質との特異的結合に適する条件下に、試料と該タンパク質を結合する抗体とを接触させる工程;および
b) 抗体−CXCR3複合体を検出する工程、
を含む、試料中の選択された哺乳類CXCR3タンパク質の検出方法。
[61]哺乳類CXCR3タンパク質と該タンパク質の少なくとも1種の機能の阻害剤または促進剤とを接触させる工程、
を含む、哺乳類CXCR3タンパク質の機能の少なくとも1種を調節する方法。
[62]哺乳類へ治療上有効な量の哺乳類CXCR3タンパク質の阻害剤を投与して、それにより炎症を減少させること、
を含む、炎症性疾患または状態を処置する方法。
[63]炎症性疾患または状態がT細胞媒介疾患または状態である、[62]記載の方法。
[64]該阻害剤が哺乳類CXCR3タンパク質に特異的に結合する抗体またはその抗原結合断片である、[62]記載の方法。
[65]該抗体またはその断片がモノクローナル抗体1C6のヒトCXCR3タンパク質への結合を競合することができる、[64]記載の方法。
[66]該抗体またはその断片がモノクローナル抗体1C6またはその抗原結合断片である、[65]記載の方法。
[67]哺乳類CXCR3タンパク質の天然のリガンド以外の促進剤の治療上有効な量を哺乳類へ投与すること、
を含む、抗腫瘍治療方法。
[68]哺乳類CXCR3タンパク質の天然のリガンド以外の促進剤の治療上有効な量を哺乳類へ投与すること、
を含む、抗ウイルス治療方法。
[69]配列:a)配列番号:1、
b)コード領域を含むその一部、
c)または(a)および(b)のいずれかのRNA体
を有する核酸にハイブリダイズする配列をもつ核酸を含有してなるアンチセンス核酸。
【0211】
配列表
(1)一般情報:
(i)出願人:
(A)名称:テオドーア−コッヒャー インスティトゥーテ
(B)ストリート:フライエシュトラーセ 1
(C)市:ベルン
(D)州:
(E)国:スイス国
(F)郵便番号:CH−3012
(G)電話番号:41−31−631−4141
(I)ファクシミリ番号:41−31−631−3799
(i)出願人:
(A)名称:ロイコサイト,インコーポレーテッド
(B)ストリート:ファースト ストリート 215
(C)市:ケンブリッジ
(D)州:マサチューセッツ
(E)国:アメリカ合州国
(F)郵便番号:02142
(G)電話番号:617−621−9350
(I)テレファックス:617−621−9349
(ii)発明の名称:CXCR3、抗体、核酸、および使用方法
(iii)配列の数:4
(iv)連絡先住所:
(A)宛名:ハミルトン,ブルック,スミス アンド レイノルズ,
ピー.シー.
(B)ストリート:ミリティア ドライブ 2
(C)市:レキシントン
(D)州:マサチューセッツ
(E)国:アメリカ合衆国
(F)郵便番号:02173
(v)コンピュータ可読フォーム:
(A)媒体型:フロッピーディスク
(B)コンピュータ:IBM PC 互換機
(C)オペレーティングシステム:PC−DOS/MS−DOS
(D)ソフトウェア:パテントイン リリース #1.0,
バージョン #1.30
(vi)現出願データ:
(A)出願番号:
(B)出願日:
(C)分類:
(vii)先願データ:
(A)出願番号:US 08/829,839
(B)出願日:1997年3月31日
(vii)先願データ:
(A)出願番号:US 08/709,838
(B)出願日:1996年9月10日
(viii)代理人情報:
(A)氏名:ブルック,デビッド イー.
(B)登録番号:22,592
(C)リファランス/ドケット番号:TKI96−01A2 PCT
(ix)テレコミュニケーション情報:
(A)電話番号:(781)861−6240
(B)テレファックス:(781)861−9540
(2)配列番号:1の情報:
(i)配列の特徴:
(A)配列の長さ:1670塩基対
(B)配列の型:核酸
(C)鎖の数:二本鎖
(D)トポロジー:不明
(ix)配列の特徴:
(A)特徴を表す記号:CDS
(B)存在位置:69..1172
(xi)配列:配列番号:1:
【0212】
【化1−1】
【0213】
【化1−2】
【0214】
【化1−3】
(2)配列番号:2の情報:
(i)配列の特徴:
(A)配列の長さ:368アミノ酸
(B)配列の型:アミノ酸
(D)トポロジー:直鎖状
(ii)配列の種類:タンパク質
(xi)配列:配列番号:2:
【0215】
【化2−1】
【0216】
【化2−2】
(2)配列番号:3の情報:
(i)配列の特徴:
(A)配列の長さ:28塩基対
(B)配列の型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:不明
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:11
(D)他の情報:/修飾塩基=「イノシン」
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:12
(D)他の情報:/修飾塩基=「イノシン」
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:23
(D)他の情報:/修飾塩基=「イノシン」
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:26
(D)他の情報:/修飾塩基=「イノシン」
(xi)配列:配列番号:3:
【0217】
【化3】
(2)配列番号:4の情報:
(i)配列の特徴:
(A)配列の長さ:27塩基対
(B)配列の型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:不明
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:10
(D)他の情報:/修飾塩基=「イノシン」
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:16
(D)他の情報:/修飾塩基=「イノシン」
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:18
(D)他の情報:/修飾塩基=「イノシン」
(xi)配列:配列番号:4:
【0218】
【化4】
【技術分野】
【0001】
本発明は、哺乳動物CXCケモカインレセプター(CXCR3)タンパク質の少なくとも1つの機能を調節する方法に関する。
【背景技術】
【0002】
背景技術
ケモカイン類は、炎症において生成される小さいサイトカイン類のファミリーを構成し、白血球補充(recruitment) を制御する[Baggiolini, M. et al., Adv. Immunol. 55: 97-179 (1994); Springer,T.A., Annu. Rev. Physiol. 57: 827-872(1995); and Schall, T.J. and K.B. Bacon, Curr. Opin. Immunol. 6:865-873(1994)]。ケモカイン類は、例えば、好中球、単球、マクロファージ、好酸球、好塩基球、マスト細胞などの白血球、ならびにT 細胞、B 細胞などのリンパ球などを含む血液を構成する(赤血球以外の)要素の走化性を選択的に誘導できる。走化性の刺激に加えて、ケモカイン類は、応答性細胞において、細胞形状の変化、細胞内遊離カルシウム濃度([Ca2+]i )の一過性上昇、顆粒エキソサイトーシス、インテグリンアップレギュレーション、生理活性脂質(ロイコトリエンなど)の形成および呼吸バーストなどの白血球の活性化に伴う他の変化を選択的に誘発することもできる。したがってケモカイン類は炎症性応答の初期誘因剤であり、感染または炎症部位への炎症媒介物質の放出、走化性および管外溢出を引き起こす。
【0003】
CXC およびCCケモカインと称されたケモカイン類の2 つのサブファミリーは、1 つのアミノ酸により分けられること(CXCケモカイン類として、IL-8、γIP-10 、Mig 、PF4 、ENA-78、GCP-2 、GRO α、GRO β、GRO γ、NAP-2 、NAP-4)または隣接残基であること(CC ケモカインとして、MIP-1 α、MIP-1 β、RANTES、MCP-1 、MCP-2 、MCP-3 、I-309)のいずれかである、4 つの保存システイン残基の最初の2 つのアレンジメントにより分けられる。多くのCXC ケモカイン類は、好中球の白血球を引きつける。例えば、CXC ケモカイン類インターロイキン8(IL-8) 、血小板因子4(PF4)、好中球活性化ペプチド2(NAP-2)は、強力な化学誘引物質および好中球の活性化因子である。Mig[monokine induced by gamma interferon (ガンマインターフェロンにより誘導されたモノカイン)]およびIP-10(γIP-10 、インターフェロン- ガンマ誘導性の10kDa タンパク質) と称するCXC ケモカイン類は、活性化末梢血リンパ球の走化性を誘導することに特に活性である。CCケモカイン類は、一般的に、選択性がなく、単球、好酸球、好塩基球、T リンパ球およびナチュラルキラー細胞を含む多様な白血球細胞型を引きつけることができる。ヒト単球走化性タンパク質1 〜3 (MCP-1 、MCP-2 およびMCP-3 )、RANTES[Regulatedon Activation, Normal T Expressed and Secreted(活性化制御、正常T 発現および分泌)]およびマクロファージ炎症性タンパク質1 αおよび1 β(MIP-1 αおよびMIP-1 β)などのC-C ケモカイン類は、単球またはリンパ球の化学誘引物質および活性化因子として特徴付けられるが、好中球の化学誘引物質であるとは思われない。
【0004】
CCおよびCXC ケモカイン類は、7 回膜貫通型Gタンパク質共役受容体のスーパーファミリーに属する受容体を介して作用する。[Murphy,P.M., Annu. Rev. Immunol., 12:593-633(1994);Gerard, C. およびN.P. Gerard, Curr. Opin. Immunol., 6: 140-145 (1994)] 。このG タンパク質共役(ヘビ状)受容体ファミリーは、7 つの膜貫通領域を含有する内在性膜タンパク質の大きいグループを含む。前記受容体はG タンパク質に共役し、それはGTP に結合することができ、例えば、細胞内媒介物質の生成により、共役受容体からシグナル伝達を媒介することが可能なヘテロ3 量体の制御タンパク質である。
【0005】
ケモカイン受容体を2 つのグループに分けることができる: CCケモカイン類を結合するCCケモカイン受容体1 〜5(CCR1-5) 、およびCXC ケモカイン類を結合するCXC ケモカイン受容体1 〜4(CXCR1-4)。一般的に、CCケモカイン受容体は、複数の型の白血球上に存在し、単球、好酸球、好塩基球およびT 細胞の移動に重要である[Qin, S., et al., Eur. J. Immunol.,26:640-647(1996);Carr, M. W., et al., Proc. Natl. Acad. Sci. USA, 91(9):3652-3656(1994); Taub,D.D., et al., J. Clin. Invest., 95 (3):1370-1376(1995); Neote, K. et al., Cell, 72: 415-425(1993); Gao,J.-L. et al., J. Exp. Med., 177:1421-1427(1993); Charo,I.F. et al., Proc. Natl. Acad. Sci. USA,91:2752-2756(1994); Myers, S.J., et al., J. Biol. Chem., 270:5786-5792(1995); Combadiere, C. et al., J. Biol. Chem., 270(27):16491-16494(1995); and Correction, J. Biol. Chem., 270:30235(1995); Ponath, P. D. et al., J. Exp. Med., 183:2437-2448(1996); and Daugherty, B. L. et al., J. Exp. Med., 183:2349-2354(1996); Power, C. A. et al., 1995, J. Biol. Chem., 270:19495-19500(1995); Hoogewerf, A. J. et al., Biochem. Biophys. Res. Commun., 218:337-343(1996); Samson, M. et al., Biochemistry, 35:3362-3367(1996)] 。対照的に2 つのIL-8受容体、CXCR1 およびCXCR2 は、主として好中球に限定され、好中球の移動に重要である(Baggiolini,M.,etal.,Adv.Immunol.,55:97-179(1994)] 。該IL-8受容体、CXCR1[IL-8R1, 1 型インターロイキン−8 受容体; Holmes, W. E. et al., Science, 253:1278-1280(1991)] およびCXCR2[IL-8R2, 2 型インターロイキン−8 受容体;Murphy, P. M. and H. L. Tiffany, Science, 253:1280-1283(1991)] は、好中球の走化性を誘導するCXC ケモカインに観察される本質的な結合エピトープであるNH2 末端Glu-Leu-Arg(ELR)モチーフを認識する[Clark-Lewis,I.et al.,J.Biol.Chem.,266:23128-23134(1991);Hebert,C.A. et al.,J.Biol.Chem.,266:18989-18994(1991);and Clark-Lewis,I.et al.,Proc.Natl.Acad.Sci.USA,90:3574-3577(1993)]。
【発明の概要】
【発明が解決しようとする課題】
【0006】
単球および顆粒細胞とは対照的に、ケモカインに応答するリンパ球は、良く理解されていない。特に、公知の特異性の受容体は、リンパ球に限定されるようには思われず、したがって、これらの受容体を認識するケモカインは、T細胞媒介炎症状態に観察されるTリンパ球の選択的補充などの事象を説明できない。さらに、公知のケモカイン受容体との顕著な配列類似性ならびに類似の組織および白血球亜集団分布を有する多くのタンパク質が同定され、クローン化されているが、これらの受容体のリガンドは決定されないままである。したがって、これらのタンパク質をオーファン受容体という。該受容体の1もしくは複数のリガンドのキャラクタリゼーションは、ケモカイン類とそれらの標的細胞との相互作用、この相互作用により刺激され、走化性および白血球の細胞活性化を含む事象、ならびに受容体機能の調節に基づく治療の開発の理解に必須である。
【課題を解決するための手段】
【0007】
即ち、本発明の要旨は、
〔1〕ヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質に結合する抗体または抗原結合断片と接触させる工程を含む、方法、
〔2〕ヒトCXCR3タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、ヒトCXCR3タンパク質に結合する抗体または抗原結合断片と接触させる工程を含み、結合が、配列番号:2の残基1〜15の配列と同じアミノ酸配列からなるポリペプチドによって阻害される、方法、
〔3〕配列番号:2に示されるアミノ酸配列を有する哺乳動物CXCケモカインレセプター3(CXCR3)タンパク質のシグナル伝達活性または細胞応答機能を阻害する方法であって、該タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含む、方法、
〔4〕配列番号:2に示されるアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該タンパク質を、モノクローナル抗体1C6(ATCC受託番号HB−12330)またはその抗原結合断片と接触させる工程を含む、方法、
〔5〕配列番号:2に示されるアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質によって媒介されるIP−10誘導カルシウムフラックスを阻害する方法であって、該タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含む、方法、
〔6〕配列番号:2に示されるアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質を介するIP−10誘導走化性を阻害する方法であって、該タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含む、方法、
〔7〕CXCケモカインレセプター3(CXCR3)タンパク質のシグナル伝達活性または細胞応答機能を阻害する方法であって、該CXCR3タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含み、該抗体がモノクローナル抗体1C6(ATCC受託番号HB−12330)の軽鎖CDR(CDR1、CDR2およびCDR3)および重鎖CDR(CDR1、CDR2およびCDR3)ならびにヒト枠組み領域を含むヒト化免疫グロブリンであり、該ヒト化免疫グロブリンがモノクローナル抗体1C6(ATCC受託番号HB−12330)と同じまたは類似のエピトープ特異性を有する、方法、
〔8〕ヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、ハイブリドーマ細胞株HB−12330により産生される抗体またはその抗原結合断片と接触させる工程を含む、方法、
〔9〕哺乳動物CXCケモカインレセプター3(CXCR3)タンパク質のシグナル伝達活性または細胞応答機能を阻害する方法であって、該CXCR3タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含み、該抗体がモノクローナル抗体1C6(ATCC受託番号HB−12330)の軽鎖CDR(CDR1、CDR2およびCDR3)および重鎖CDR(CDR1、CDR2およびCDR3)ならびにヒト枠組み領域を含むヒト化免疫グロブリンであり、該ヒト化免疫グロブリンがモノクローナル抗体1C6(ATCC受託番号HB−12330)と同じまたは類似のエピトープ特異性を有する、方法
に関する。
【発明の効果】
【0008】
本発明により、オーファン受容体の1もしくは複数のリガンドのキャラクタリゼーションがなされる。
【図面の簡単な説明】
【0009】
【図1】図1は、CXCR3と呼ばれるヒトIP-10/Mig受容体をコードするcDNA(ヒトCD4+ T細胞(KT30)cDNAライブラリーから単離)の1670bpインサートから決定されたヌクレオチド配列(配列番号:1)を示す図である。オープンリーディングフレーム(69〜1175)は368アミノ酸の予想タンパク質(配列番号:2)をコードする。推定されるポリAシグナルとポリA部位はそれぞれ1534〜1539と1624〜1670に位置する。
【図2】図2は、ヒトIP-10/Mig受容体をコードする図1の配列のオープンリーディングフレームの概念上の翻訳(配列番号:2)を示す図である。矢印は潜在的N結合型グリコシル化部位を示し、水平方向の線は推定される膜貫通ドメイン(TM1-TM7)の位置を示す。
【図3】図3A〜3Cは、IP-10/MigRを発現する安定にトランスフェクトされた細胞において、IP-10とMigによって誘導される応答を示すグラフである。図3Aは、IP-10/MigRトランスフェクト300-19細胞での濃度依存的[Ca2+]i変化を示すグラフである。IP-10またはMigをそれぞれ1、10および100nMの濃度でFura-2/AM負荷細胞に添加し(矢印)、[Ca2+]i依存性の蛍光変化を記録した。非トランスフェクト細胞(下側の軌線)は同じ条件下に100nMのIP-10またはMigで刺激した。図3Bは、受容体の脱感作および交差脱感作を評価した試験の結果を示すグラフである。IP-10/MigR発現300-19細胞を、100nMのIP-10またはMigと、IP-10の後Mig(もしくはその逆)で順次刺激し、蛍光変化を記録した。図3Cは、IP-10(黒丸)またはMig(黒四角)で刺激したIP-10/MigR発現Jurkat細胞の走化性を示すグラフである。下側の図は、増加するIP-10(白丸)またはMig(白四角)量で刺激した場合の非トランスフェクトJurkat細胞の応答を示す。高倍率視野5つあたりの平均移動細胞数(±SD)を表わす。
【図4】図4A〜4Bは、IP-10とMigに対する末梢血リンパ球(PBL)の応答を示すグラフである。献血血液バフィーコートから新たに単離したPBLをそのまま(下側の軌線、白抜きの記号)またはIL-2(400U/ml)の存在下に10日間培養してから(上側の軌線、黒塗りの記号)使用した。図4AはIP-10またはMigによって誘導される[Ca2+]i変化を示すグラフである。IP-10またはMigをそれぞれ1、10および100nMの濃度でFura-2/AM負荷培養細胞に添加し(矢印)、[Ca2+]i依存性の蛍光変化を記録した(上側の軌線)。新たに単離した細胞(下側の軌線)は同じ条件下に100nMのIP-10またはMigで刺激した。図4Bは増加するIP-10(黒丸)またはMig(黒四角)濃度に応答したPBLの走化性を示すグラフである(高倍率視野5つあたりの平均移動細胞数(±SD)を表わす)。
【図5】図5A〜5Bは、CXCR3 DNAをトランスフェクトしたL1.2細胞(図5A)と活性化T細胞(図5B)に対する放射標識IP-10の結合を示すグラフである。増加する非標識IP-10濃度の存在下に細胞を0.05nMの125I標識IP-10と共にインキュベートした。スキャッチャード分析(挿入図)は、CXCR3 L1.2トランスフェクタントに関して1細胞あたり37,000受容体(Kdは614pM)とCD3芽球に関して1細胞あたり17,000受容体(Kdは156pM)を示した。
【図6】図6は、CCR1、CCR2b、CCR3、CCR4、CCR5、CXCR1、CXCR2、CXCR3またはCXCR4のいずれかを発現する安定なL1.2トランスフェクタントを抗CXCR3ペプチドモノクローナル抗体(mAb)1C6で染色することによって評価した、抗CXCR3抗体1C6の特異性を示す図である。陰性対照染色(非掲載)はどのL1.2トランスフェクタントについても、非トランスフェクトL1.2細胞(L1.2wt)での1C6について示す染色と似ていた。
【図7】図7A〜7Cは、好中球(図7A)、リンパ球(図7B)および活性化T細胞(図7C)でのCXCR3の発現を示す蛍光ヒストグラムである。全血中の白血球亜集団をその前方および側方散乱光によって同定し、相応にゲーティングした。CD3芽球を生成するため、PBMCを抗CD3 mAb で3日間活性化した後、IL-2含有培地で7日間維持した。各図表において、黒いグラフは抗CXCR3 mAb 1C6 による染色を表わし、中空のグラフは同イソタイプの対照mAb による染色を表わす。
【図8】図8は、血液リンパ球の各集団でのCXCR3発現を示す一連の図表である。二色染色法を用いてT細胞(CD3)、B細胞(CD20)およびNK細胞(CD56)でのCXCR3の発現を評価した。
【図9−1】図9は、免疫蛍光三色分析法によって分析した、血液リンパ球のCD3+亜集団上のCXCR3発現と種々のマーカーの対比を示す一連の図表である。抗CD3 Cy-Chromeを用いてT細胞を染色し、分析のためそれらの細胞を電気的にゲーティングした。各象限は対照mAb の染色に従って設定した。ここに示す染色は分析した5人の献血者を代表するものであった。
【図9−2】図9は、免疫蛍光三色分析法によって分析した、血液リンパ球のCD3+亜集団上のCXCR3発現と種々のマーカーの対比を示す一連の図表である。抗CD3 Cy-Chromeを用いてT細胞を染色し、分析のためそれらの細胞を電気的にゲーティングした。各象限は対照mAb の染色に従って設定した。ここに示す染色は分析した5人の献血者を代表するものであった。
【図9−3】図9は、免疫蛍光三色分析法によって分析した、血液リンパ球のCD3+亜集団上のCXCR3発現と種々のマーカーの対比を示す一連の図表である。抗CD3 Cy-Chromeを用いてT細胞を染色し、分析のためそれらの細胞を電気的にゲーティングした。各象限は対照mAb の染色に従って設定した。ここに示す染色は分析した5人の献血者を代表するものであった。
【図10】図10は、一群の抗CXCR3 mAb による、活性化T細胞のIP-10またはMCP-1媒介性走化性の阻害を示すヒストグラムである。1×106個のヒトCD3芽球をトランスウェルの上部チャンバーに入れ、ケモカイン(12.5nM)を下部チャンバーに入れた。アッセイ開始時に、種々の抗CXCR3 mAb (組織培養上清中;FCSなし)を細胞と共にその上部ウェルに入れた。1.5時間後、下部チャンバーに移動する細胞数をフローサイトメトリーで数えた。走化性阻害百分率は、mAb の非存在下で移動した細胞数を100%として計算した。結果は、少なくとも4回の独立した実験を代表するものである。
【図11】図11は、精製抗CXCR3 mAb 1C6 によるIP-10媒介性走化性の阻害を示すグラフである。様々な濃度の1C6 mAb を上部ウェルに入れ、図10について記述したようにアッセイを行なった。mAb 1C6 は856ng/mlの濃度で総走化性を50%阻害した(IC50=856ng/ml)。
【図12】図12は、活性化T細胞に対する125I-IP-10結合の、mAb 1C6 による阻害を示すグラフである。表示のように増加する1C6濃度の存在下に、CD3芽球を0.05nMの125I-IP-10と共にインキュベートした。室温で60分の後、細胞ペレットを洗浄し、カウントした。データをソフトウェア・カレイダグラフ(KaleidaGraph)で分析したところ、0.16μg/ml のIC50値が得られた。
【図13】図13A〜13Hは、IP-10に応答したヒトT細胞による[Ca2+]iのmAb 1C6 による阻害を示す(Migの場合は阻害されない)。抗CD3活性化IL-2刺激ヒトT細胞をFura-2で標識し、表示したケモカインで順次刺激するか(図13A〜13B)、mAb で刺激した40秒後に表示したケモカインで刺激した(図13C〜13H)。[Ca2+]i蛍光変化を分光蛍光計で記録した。各軌線は5回の独立した実験を代表するものであった。抗体は、50μg/ml(図13C〜13D)、25μg/ml(図13E)、12.5μg/ml(図13F)、6.125μg/ml(図13G)または3.0625μg/ml(図13H)の最終濃度で使用した。ケモカイン類は2nMの濃度で使用した。
【図14】図14A〜14Dは、P1ペプチド(図14B)、P2ペプチド(図14C)、P3ペプチド(図14D)の存在下またはペプチドの非存在下(図14A)に、CXCR3発現トランスフェクタントをmAb 1C6 で染色したフローサイトメトリー分析の結果を示す蛍光ヒストグラムである。各図において、太い線で規定されるグラフはmAb 1C6 染色を表わし、破線で規定されるグラフは同イソタイプの無関係な対照mAb による染色を表わす。
【図15】図15は、40nMの非放射性IP-10、mAb 1C6 、CXCR3トランスフェクタントに対して生じたモノクローナル抗体(2F8、3A12、3E2、4B4、4D2、5B12、7B8または8D5)または抗CXCR2 mAb による、CXCR3トランスフェクタントに対する放射標識IP-10結合の阻害百分率を示すヒストグラムである。
【発明を実施するための形態】
【0010】
発明の要旨
本発明は、本明細書において、CXC ケモカイン受容体3(CXCR3)と称する単離および/または組換え哺乳類(例えば、ヒトなどの霊長類)IP-10/Mig 受容体タンパク質ならびにその改変体を称するタンパク質またはポリペプチドに関する。組換えCXCR3 タンパク質および改変体は、本明細書に記載のように宿主細胞中で生産されうる。1 つの態様として、CXCR3 タンパク質またはその改変体は、IP-10 および/またはMig などの1 以上のケモカイン類の選択的結合 (例えば、高いアフィニティー結合) 、および/または (1 種以上の) 細胞応答( 例えば、走化性、エキソサイトーシス、1 種以上の炎症媒介物質の放出など) を誘導する能力により特徴付けられる。
【0011】
本発明の他の側面は、哺乳類( 例えば、ヒトなどの霊長類など) のCXCR3 タンパク質またはその改変体をコードする単離および/または組換え核酸に関する。さらに本発明は、本発明のタンパク質またはその改変体をコードする核酸を含む、プラスミドまたはレトロウイルスベクターなどの組換え核酸構築物に関する。該核酸および構築物を用いて、組換え受容体タンパク質および構築物を含む宿主細胞を生産することができる。他の態様として、核酸は、CXCR3 タンパク質をコードする第 2の核酸とハイブリダイズすることができるアンチセンス核酸をコードし、細胞に導入された場合、受容体の発現を阻害することができる。
【0012】
さらに本発明は、例えば、該タンパク質もしくはその改変体( 例えばペプチドなど) または受容体タンパク質もしくは改変体を発現する細胞を免疫原として用いて生産することができる、CXCR3 受容体と反応する抗体に関する。かかる抗体またはその断片は、受容体タンパク質の精製および研究、表面受容体を発現する細胞の同定、および細胞の分類または計数を含む、治療、診断、ならびに研究応用に有益である。したがって、本発明は、本明細書記載の抗体もしくはその断片( 例えば、mAb 1C6 またはその抗原結合断片) の治療 (予防を含む) または診断への使用および本明細書に記載の疾患または状態の処置に用いる薬剤の製造のためのかかる抗体もしくは断片の使用を包含する。
【0013】
また、本発明は、受容体のリガンド、受容体機能の阻害剤(例えばアンタゴニスト)や促進剤(例えばアゴニスト)を同定する方法をも包含する。1つの態様として、該細胞に導入された核酸によってコードされる受容体タンパク質または改変体を発現するように操作されている適切な宿主細胞をアッセイに用いて、リガンド、受容体機能の阻害剤または促進剤の効き目を同定し評価する。かかる細胞は、発現された受容体タンパク質またはポリペプチドの機能を評価する際にも有用である。
【0014】
本発明によれば、リガンド、受容体機能の阻害剤および促進剤を適切なアッセイで同定し、さらに治療の効果について評価することができる。受容体機能の阻害剤は受容体活性を阻害( 減少もしくは妨害) するために使用でき、リガンドおよび/または促進剤は指示された場合、正常な受容体機能を誘導( 引き起こすまたは促進) するために使用できる。したがって本発明は、個体(例えば哺乳類)に受容体機能の阻害剤を投与する過程を含む、自己免疫疾患および移植片拒絶を含む炎症性疾患を治療する方法を提供する。さらに、本発明は、個体にリガンドまたは促進剤を投与することにより受容体機能を刺激する方法を提供し、それは、例えば、感染性疾患および癌の処置に有益な白血球機能の選択的刺激への新たなアプローチを提供する。
【0015】
発明の詳細な説明
本明細書に記述するように、CXCケモカインIP-10およびMigに選択的な新規ケモカイン受容体をコードする核酸をクローニングし、特徴づけた。ヒトCD4+ T細胞ライブラリーから単離されたこのクローンは、単球または顆粒球由来のcDNAライブラリーには検出されなかった。このクローンの配列を分析したところ、40,659ダルトンの予想分子量を持つ368アミノ酸の予想タンパク質(図2、配列番号:2)をコードする1104塩基対のオープンリーディングフレームが明らかになった(図1、配列番号:1)。このアミノ酸配列は、Gタンパク質共役受容体の特徴であり、かつ他の化学誘引物質受容体にも認められる7つの推定膜貫通セグメントを含んでいる。この知見と合致して、本受容体は、IP-10およびMigに応答して起こるCa2+(カルシウムイオン)の流動化と走化性とを媒介する(実施例2)。類似の条件下で、CXCケモカイン類IL-8、GROα、NAP-2(好中球活性化タンパク質-2)、GCP-2(顆粒球走化性タンパク質-2)、ENA78(上皮由来好中球活性化ペプチド78)、PF4(血小板因子4)や、CCケモカイン類MCP-1(単球走化性タンパク質-1)、MCP-2、MCP-3、MCP-4、MIP-1α(マクロファージ炎症性タンパク質-1α)、MIP-1β、RANTES(regulated on activation, normal T cell expressed and secreted)(発現かつ分泌された正常T細胞活性化制御)、I309、エオタキシンまたはリンフォタクチンに対する有意な応答は検出されなかった。
【0016】
活性化Tリンパ球におけるヒトCXCR3の限定的発現と、この受容体のIP-10およびMigに対するリガンド選択性は注目に値する。実施例2で使用した条件下に、このヒト受容体はIL-2活性化Tリンパ球で強く発現されるが、休止Tリンパ球、Bリンパ球、単球または顆粒球には検出されなかった。受容体分布のさらなる研究は、CD20+(B)細胞とCD56+(NK)細胞の一部もこの受容体を発現するものの、CXCR3を発現するのはそのほとんどが、先の活性化に合致する表現型、CD95+、CD45RO+およびCD45RAlowである細胞を含むCD3+細胞であることを示す。リンパ球を誘引すると報告されている他のケモカイン(例えばMCP-1、MCP-2、MCP-3、MIP-1α、MIP-1βおよびRANTES)の受容体は、好中球、好酸球および好塩基球などの顆粒球や単球にも認められるので、この活性化Tリンパ球における選択的発現は興味深い。これらの結果は、CXCR3と呼ばれるこのIP-10/Mig受容体が、エフェクターT細胞の選択的補充に関与することを示唆している。
【0017】
本受容体は、IP-10およびMigと呼ばれる2つの珍しいCXCケモカインを認識する。IP-10とMigは共にCXCサブファミリーに属するが、好中球に対する強力な化学誘引物質であるIL-8や他のCXCケモカインとは対照的に、IP-10とMigの主な標的はリンパ球、特に活性化または刺激されたTリンパ球やナチュラルキラー(NK)細胞などのエフェクター細胞である(Taub, D.D.ら, J. Exp. Med., 177:18090-1814(1993);Taub, D.D.ら, J. Immunol., 155:3877-3888 (1995))(NK細胞は、抗原認識用の特異的T細胞受容体を持たないが、腫瘍細胞やウイルス感染細胞などの細胞に対して細胞傷害活性を持つ大形顆粒リンパ球である)。このことと合致して、IP-10とMigは、好中球走化性を効率よく誘導するCXCケモカイン中の必須結合エピトープであるELRモチーフを持たない(Clark-Lewis, I.ら, J. Biol. Chem., 266:23128-23134 (1991);Hebert, C.A.ら, J. Biol. Chem., 266:18989-18994 (1991);およびClark-Lewis, I.ら, Proc. Natl. Acad. Sci. USA, 90:3574-3577 (1993))。また、組換えヒトMigと組換えヒトIP-10は共に、腫瘍浸潤性リンパ球(TIL)内のカルシウム流動を誘導すると報告されている(Liao, F.ら, J. Exp. Med., 182:1301-1314 (1995))。IP-10はイン・ビトロで単球の走化性を誘導すると報告されているが(Taub, D.D.ら, J. Exp. Med., 177:1809-1814 (1993);原因となる受容体は同定されていない)、ヒトMigは高度に選択的と思われ、そのような作用を示さない(Liao, F.ら, J. Exp. Med., 182:1301-1314 (1995))。IP-10発現は、乾癬、固定薬疹、皮膚の遅延型過敏反応、類結核癩などの炎症状態にある様々な組織、実験的糸球体腎炎および実験的アレルギー性脳脊髄炎で誘導される。またIP-10は、T細胞依存性の強力なイン・ビボ抗腫瘍作用を持ち、生体内で血管形成の阻害因子であると報告されており、イン・ビトロでNK細胞の走化性と脱顆粒を誘導できることから、(例えば腫瘍細胞破壊における)NK細胞の補充および脱顆粒の媒介物質としての役割が示唆される(Luster, A.D.およびP. Leder, J. Exp. Med., 178:1057-1065 (1993);Luster, A.D.ら, J. Exp. Med. 182:219-231 (1995);Angiolillo, A.L.ら, J. Exp. Med., 182:155-162 (1995);Taub, D.D.ら, J. Immunol., 155:3877-3888 (1995))。IP-10とMigの発現パターンは、それぞれの発現がインターフェロン・ガンマ(IFNγ)によって誘導されるという点でも独特であり、これに対し、IL-8の発現はIFNγによってダウンレギュレートされる(Luster, A.D.ら, Nature, 315:672-676 (1985);Farber, J.M., Proc. Natl. Acad. Sci. USA, 87:5238-5242 (1990);Farber, J.M., Biochem. Biophys. Res. Commun., 192 (1):223-230 (1993);Liao, F.ら, J. Exp. Med., 182:1301-1314 (1995);Seitz, M.ら, J. Clin. Invest., 87:463-469 (1991);Galy, A.H.M.およびH. Spits, J. Immunol., 147:3823-3830 (1991))。
【0018】
最近、ケモカイン類は、長く捜し求められていたリンパ球補充の媒介物質であると認識されるようになった。数種類のCCケモカインがリンパ球走化性を惹起することが発見されたが(Loetscher, P.ら, FASEB J., 8:1055-1060 (1994))、それらは顆粒球と単球に対しても活性である(Uguccinoni, M.ら, Eur. J. Immunol., 25:64-68 (1995);Baggiolini, M.およびC.A. Dahinden, Immunol. Today, 15:127-133 (1994))。この状況は、その作用が活性化Tリンパ球とNK細胞を含むリンパ球に選択的であって、かつ、数多くの他のケモカイン類を認識せず選択的発現パターンを示す受容体CXCR3を結合するIP-10とMigでは異なる(実施例2、実施例5)。
【0019】
これらの知見を考慮すれば、遅延型過敏性病巣などの炎症性病巣、ウイルス感染部位およびある種の腫瘍における特徴的な浸潤物の形成が、CXCR3を介して媒介されCXCR3発現によって調節される過程であると結論することは妥当である。活性化の結果としてCXCR3受容体を保持するリンパ球、特にTリンパ球は、インターフェロン・ガンマによって局所的に誘導されうるIP-10および/またはMigによって炎症性病巣、感染部位または腫瘍に補充されうる。したがって、CXCR3はリンパ球、特に活性化または刺激されたTリンパ球などのエフェクター細胞の選択的補充にその役割を果たす。
【0020】
タンパク質とペプチド
本発明は、哺乳類CXCR3タンパク質と呼ばれる単離および/または組換え(例えば本質的に純粋なものを含む)タンパク質またはポリペプチドおよびその改変体に関する。好ましい態様として、本発明の単離および/または組換えタンパク質は、哺乳類CXCR3タンパク質(本明細書に定義する)に特有な、結合活性(例えばリガンド、阻害因子および/または促進因子結合性)、シグナル伝達活性(例えば哺乳類Gタンパク質の活性化、サイトゾル遊離カルシウム濃度[Ca2+]iの迅速かつ一過性の上昇の誘導)、細胞応答機能(例えば走化性、エキソサイトーシスまたは白血球による炎症媒介物質放出の刺激)および/または本明細書に定義する免疫学的性質などの性質、活性または機能を少なくとも一つは持つ。例えば、本発明のタンパク質には、IP-10および/またはMigに選択的に結合でき、それに対する細胞のシグナル伝達および/または応答(例えば(とりわけ活性化Tリンパ球の)カルシウム流動、走化性および/または脱顆粒)をイン・ビトロおよび/または生体内で媒介できるものがある。例えば本明細書に示すように、cDNAクローンの発現により哺乳類細胞中で生産されたヒトCXCR3タンパク質は、CXCケモカインIP-10および/またはMigに選択的に結合して、シグナル伝達と細胞応答(例えば走化性)を媒介することができる。一態様として本発明のタンパク質は、同じ哺乳類種または異なる哺乳類種に由来するCXCケモカイン(例えばヒトIP-10、ネズミIP-10、ヒトMig、ネズミMig)を結合できる(ヒトIP-10, Luster, A.D.ら, Nature, 315:672-676 (1985);ネズミIP-10(CRG-2とも呼ばれる),Vanguri, P.およびJ.M. Farber, J. Biol. Chem., 265:15049 (1990) ならびにLuster, A.D.およびP. Leder, J. Exp. Med., 178:1057-1065 (1993);ネズミMig, Farber, J.M., Proc. Natl. Acad. Sci. USA, 87:5238-5242 (1990);ヒトMig, Farber, J.M., Biochem. Biophys. Res. Commun., 192 (1):223-230 (1993) ならびにLiao, F.ら, J. Exp. Med., 182:1301-1314 (1995))。
【0021】
本明細書で「単離(された)」と称するタンパク質またはポリペプチドは、それらが哺乳類細胞中に存在する時よりも精製された状態にあるタンパク質またはポリペプチドであり、本明細書に記述する方法や類似の方法あるいは他の適当な方法によって得られるタンパク質またはポリペプチド、例えば本質的に純粋なタンパク質またはポリペプチド、化学合成法(例:合成ペプチド)あるいは生物学的方法と化学法の併用によって生産されるタンパク質またはポリペプチド、および単離された組換えタンパク質またはポリペプチドなどが含まれる。これらのタンパク質は少なくとも約50重量%、好ましくは少なくとも約75重量%の単離状態で、または本質的に純粋な形で得ることができる。ここで「組換え」と称するタンパク質またはポリペプチドは、組換え核酸の発現によって生産されるタンパク質またはポリペプチドである。
【0022】
本明細書において「哺乳類CXCR3タンパク質」とは、天然に存在するまたは内因性の哺乳類CXCR3タンパク質、ならびに天然に存在するまたは内因性の対応する哺乳類CXCR3タンパク質と同じアミノ酸配列を持つタンパク質(例えば組換えタンパク質)をいう。したがって、本明細書に定義される用語「哺乳類CXCR3タンパク質」は、哺乳類CXCR3の成熟タンパク質、多形または対立遺伝子改変体、およびその他のアイソフォーム(例えば選択的スプライシングその他の細胞過程によって産生されるもの)、ならびにそれらの修飾体または未修飾体(例えばグリコシル化、非グリコシル化、リン酸化または非リン酸化CXCR3タンパク質)を包含する。天然に存在するまたは内因性の哺乳類CXCR3タンパク質には、成熟型CXCR3等の野生型タンパク質、哺乳動物(例えばヒト、ヒト以外の霊長類)に天然に存在する多形または対立遺伝子改変体およびその他のアイソフォームが含まれる。これらのタンパク質は、例えば哺乳類CXCR3を本来産生する供給源から回収することができる。これらのタンパク質と、天然に存在するまたは内因性の対応する哺乳類CXCR3と同じアミノ酸配列を持つ哺乳類CXCR3タンパク質は、対応する哺乳類の名前で呼ばれる。例えば対応する哺乳動物がヒトである場合、そのタンパク質はヒトCXCR3タンパク質と呼ばれる(例えば適当な宿主細胞で生産された組換えヒトCXCR3)。
【0023】
哺乳類CXCR3タンパク質の「機能的改変体」には、機能的断片、機能的突然変異タンパク質および/または機能的融合タンパク質(例えば突然変異誘発および/または組換え技術によって生産されるもの)が含まれる。一般に、本発明によって包含される哺乳類CXCR3タンパク質の断片または部分は、成熟型哺乳類CXCR3タンパク質と比較してアミノ酸(すなわち1またはそれ以上のアミノ酸)の欠失(すなわち1またはそれ以上の欠失)を有するもの(例えばN末端、C末端または内部欠失体)を包含する。成熟型哺乳類CXCR3タンパク質と比較して、隣接しているアミノ酸のみが欠失している断片もしくは部分または隣接していないアミノ酸が欠失している断片または部分も想定される。
【0024】
一般に、本発明に包含される哺乳類CXCR3タンパク質の突然変異体または誘導体には、1またはそれ以上の隣接するまたは隣接していないアミノ酸残基の付加、欠失および/または置換により異なる天然または人工の改変体、1またはそれ以上の残基が修飾されている修飾ポリペプチド、および1またはそれ以上の修飾残基を含む突然変異体が含まれる。好ましい突然変異体は、1またはそれ以上の隣接するまたは隣接していないアミノ酸残基の付加、欠失および/または置換により異なる天然または人工の哺乳類CXCR3タンパク質改変体である。そのような突然変異は、例えば(他のCXCおよび/またはCCケモカイン受容体と比較して)保存されている領域または保存されていない領域、細胞外領域、細胞質領域または膜貫通領域に存在しうる。
【0025】
哺乳類CXCR3タンパク質の「機能的断片または部分」「機能的突然変異体」および/または「機能的融合タンパク質」とは、哺乳類CXCR3受容体(本明細書に定義する)に特有な、結合活性(例えばリガンド、阻害因子および/または促進因子結合性)、シグナル伝達活性(例えば哺乳類Gタンパク質の活性化、サイトゾル遊離カルシウム濃度[Ca2+]iの迅速かつ一過性の上昇の誘導)、細胞応答機能(例えば走化性、エキソサイトーシスまたは白血球による炎症媒介物質放出の刺激)および/または本明細書に定義する免疫学的性質などの性質、活性または機能を少なくとも一つは持つ単離および/または組換えタンパク質もしくはオリゴペプチドを指す。
【0026】
本明細書において、哺乳類CXCR3タンパク質の「免疫学的性質を少なくとも一つ」持つタンパク質またはポリペプチドとは、(a) 天然に存在するまたは内因性の哺乳類CXCR3タンパク質もしくは天然に存在するまたは内因性の哺乳類CXCR3タンパク質(例えばヒトCXCR3)と同じアミノ酸配列を持つタンパク質に結合する、選択されたエピトープ特異性を持つ少なくとも一つの抗体によって結合され、および/または、(b) 天然に存在するもしくは内因性の哺乳類CXCR3タンパク質または天然に存在するもしくは内因性の哺乳類CXCR3タンパク質と同じアミノ酸配列を持つタンパク質に結合する、選択されたエピトープ特異性を持つ抗体の適当な動物における形成を(例えば適当な担体と複合体化した時に)誘導しうる免疫原であるものである。例えば、好適な断片は、単離された哺乳類CXCR3に対して生じたおよび/またはそれと反応する抗体と交差反応できる。
【0027】
好適な断片または突然変異体は、スクリーニングによって同定できる。例えば本タンパク質のN末端、C末端または内部領域を段階的に欠失させることができ、得られたタンパク質またはポリペプチドを本明細書に記述するような適当なアッセイ(例えば走化性、カルシウム流動)を使ってスクリーニングすることができる。得られたタンパク質がそのアッセイで活性を示す場合、その得られたタンパク質(「断片」)は機能的である。CXCケモカイン受容体やCCケモカイン受容体を含む哺乳類Gタンパク質共役受容体の構造と機能に関する情報は、哺乳類CXCR3タンパク質を機能ドメインに分割する際の根拠となる(Murphy, P.M., Ann. Rev. Immunol., 12:593-633 (1994) ならびにGerard, C.およびN.P. Gerard, Curr. Opin. Immunol., 6:140-145 (1994) とそれらに引用されている文献)。
【0028】
改変体という用語は、第一の成分として哺乳類CXCR3タンパク質(例えばヒトCXCR3)を含み、その第一成分が、自然界に認められるその哺乳類CXCR3には存在しない第二の成分に連結している融合タンパク質をも包含する。したがってこの第二成分はアミノ酸、オリゴペプチドまたはポリペプチドでありうる。第一成分は、その融合タンパク質のN末端、C末端または内部のいずれにあってもよい。一態様として本融合タンパク質は、第一成分としてのアフィニティーリガンド(例えば酵素、抗原、エピトープタグ)と、リンカー配列およびヒトCXCR3またはその一部からなる第二成分とを含有する。
【0029】
哺乳類CXCR3タンパク質の例としては、本発明の核酸によってコードされるタンパク質、例えば図2(配列番号:2)に記述されているようなまたは実質上記述されているようなアミノ酸配列を持つタンパク質が挙げられる。好ましい態様として、哺乳類CXCR3または改変体(例えば細胞外N末端部分を含む改変体)は、図2(配列番号:2)に示すタンパク質と少なくとも約50%同一、より好ましくは少なくとも約70%同一、さらにより好ましくは少なくとも約80%同一であるアミノ酸配列を持つ。
【0030】
放射性同位体、スピン標識、抗原(例えばFLAGタグなどのエピトープ標識)、酵素標識、蛍光基、化学発光基などといった検出可能な標識の組み込みまたは付加(直接的付加もしくは(例えばリンカーを介した)間接的付加)によって、単離および/または組換え哺乳類CXCR3タンパク質とその改変体を修飾することができ、それら修飾体も本発明の範囲に包含されることは理解されるだろう。
【0031】
核酸、構築物およびベクター
本発明は、本明細書に記載された哺乳類(ヒト等)CXCR3タンパク質またはその改変体をコードする単離および/または(本質的に純粋な等を含む)組換え核酸に関する。本明細書でいう「単離」核酸とは、起源の供給源のゲノムDNAまたは細胞RNAの(例えば、細胞中またはライブラリー等の核酸混合物中に存在するような)核酸から離れて分離された核酸をいい、さらなるプロセッシングを受けてもよい。「単離」核酸は、本明細書に記載された方法、類似の方法または他の好適な方法により得られた核酸を含み、本質的に純粋な核酸、化学合成により製造された核酸、生物学的方法と化学的方法との組み合わせにより製造された核酸および単離された組換え核酸を含む。本明細書でいう「組換え」核酸とは、組換えDNA方法論により製造された核酸をいい、ポリメラーゼ連鎖反応(PCR)および/または制限酵素を用いるベクターへのクローニング等の人工的な組換え方法に依存する手法により生成される核酸を含む。また、「組換え」核酸は、細胞の天然のメカニズムを介して起こる組換え事象からも生ずるが、所望の組換え事象の可能性および蓋然性を生じさせるように設計された核酸の細胞に導入後に選択されるものである。
【0032】
1つの態様において、核酸またはその一部は、結合活性(例えば、リガンド、阻害剤および/または促進剤結合)、シグナル伝達活性(例えば、哺乳類Gタンパク質の活性化、サイトゾル遊離カルシウムの濃度[Ca2+]iの迅速かつ一過性の増加の誘導)、および/または細胞応答の刺激(例えば、走化性、エキソサイトーシスまたは白血球による炎症性媒介物質放出の刺激)等の哺乳類CXCR3タンパク質(例えば、ヒトCXCR3受容体)に特有の機能の少なくとも1種を有するタンパク質またはポリペプチドをコードする。また、本発明は、より具体的には、哺乳類CXCR3受容体またはその一部をコードする配列を含有する単離および/または組換え核酸またはその一部に関する。さらにより具体的には、本発明は、ヒトCXCR3タンパク質をコードする配列を含有する単離および/または組換え核酸に関する。
【0033】
さらに、本発明は、
(1)(a)配列番号:1の配列を有する核酸、
(b)配列番号:1に相補的な配列を有する核酸、または
(c)配列番号:1のオープンリーディングフレームを含有する前記核酸
の一部(図1に示した鎖の部分または相補鎖の対応する部分)、
にハイブリダイズする能力;および/または
(2)配列番号:2のアミノ酸配列を有するポリペプチドまたはその機能的同等物(即ち、受容体の1以上の天然または生理学的リガンドに関するリガンド結合活性を有するポリペプチド、および/または細胞応答(例えば、走化性、エキソサイトーシスまたは白血球による炎症性媒介物質放出の(誘発または刺激を含む)誘導)を誘導可能なように、リガンド結合に応答可能な刺激する機能)をコードする能力;および/または
(3)両方の特性、
により特徴付けられる、二本鎖もしくは一本鎖DNAまたはRNAを含む単離および/または組換え核酸に関する。
【0034】
1つの態様において、配列番号:2とその機能的同等物との間のアミノ酸配列同一性率は、少なくとも約60%(≧60%)である。好ましい態様において、配列番号:2の機能的同等物は、配列番号:2と少なくとも約70%の配列同一性を共有する。さらに好ましくは、配列番号:2とその機能的同等物との間のアミノ酸配列同一性率は、少なくとも約80%、さらにより好ましくは少なくとも約90%である。
【0035】
これらの基準に合致する単離および/または組換え核酸には、天然に存在する哺乳類CXCR3受容体の配列およびその一部と同一の配列を持つ核酸、または天然に存在する配列の改変体が含まれる。かかる改変体には、1以上の残基の付加、欠失または置換によって相違する突然変異体、1以上の残基が修飾されている修飾核酸(DNAまたはRNA類縁体など)および1以上の修飾残基を含む突然変異体が含まれる。1つの態様において、核酸は、配列番号:1に示された配列の一方の鎖とまたはそのコーディング領域に対して、少なくとも約50%のヌクレオチド配列類似性、より好ましくは少なくとも約75%のヌクレオチド配列類似性、およびさらにより好ましくは少なくとも約90%のヌクレオチド配列類似性を共有する。好ましい核酸は、少なくとも約40ヌクレオチド、より好ましくは少なくとも約50およびさらにより好ましくは少なくとも約75ヌクレオチドの長さを有する。
【0036】
かかる核酸は、例えば高ストリンジェンシー条件または中程度のストリンジェンシー条件下でのハイブリダイゼーションによって検出および単離できる。核酸ハイブリダイゼーションに関する「高ストリンジェンシー条件」と「中程度のストリンジェンシー条件」は、その教示が参照により本明細書に取り込まれるCurrent Protocols in Molecular Biology(Ausubel,F.M.ら編,第1 巻, 補遺26,1991)の2.10.1-2.10.16頁(特に2.10.8-11 を参照のこと)と6.3.1-6 頁に説明されている。プローブ長、塩基組成、ハイブリダイズする配列間のミスマッチ率、温度およびイオン強度などの因子が核酸ハイブリッドの安定性に影響を与える。したがって、高ストリンジェンシー条件または中程度のストリンジェンシー条件は、所望の選択性を達成するように経験的に決定できる。
【0037】
配列番号:1またはその相補鎖の配列を有する核酸に(例えば高または中程度のストリンジェンシー条件下で)ハイブリダイズできることを特徴とする単離および/または組換え核酸は、さらに、哺乳類CXCR3タンパク質(ヒトCXCR3タンパク質など)の特有の機能、例えば結合活性(リガンド、阻害剤および/または促進剤結合など)、シグナル伝達活性(哺乳類Gタンパク質の活性化、サイトゾル遊離カルシウム濃度[Ca2+]iの迅速かつ一過性の増加の誘導など)および/または細胞応答の刺激(走化性、エキソサイトーシスまたは白血球による炎症媒介物質の放出の刺激など)などの少なくとも1つを持つタンパク質またはポリペプチドをコードしてもよい。
【0038】
本明細書に記載されるヒトCXCR3核酸またはPCRにより生成された断片を含むその十分な部分、単離、組換えおよび/または合成のいずれかは、プローブまたはプライマーとして使用され、霊長類(例えば、サル(カニクイザル等)等のヒト以外の霊長類)、ウシ、ヒツジ、ウマ、イヌ、ネコおよび齧歯類(例えば、モルモット、ラット、マウス等のネズミ種)を含むがこれに限定されない他の哺乳類種由来のCXCR3受容体(ホモログ)または他の関連する受容体遺伝子(例えば、新規CXCケモカイン受容体遺伝子)をコードする核酸(例えば、ゲノムDNA、対立遺伝子改変体、cDNA)を検出および/または回収することができる。これは、本明細書に記載された手法またはハイブリダイゼーション、PCRもしくは他の好適な技術を含む他の適当な方法を用いて達成されうる。哺乳類核酸を用いて、構築物(例えば、ベクター)、受容体またはその断片ならびに受容体の生産および使用方法に有用な宿主株を調製することが可能である。
【0039】
1つの態様において、哺乳類CXCR3タンパク質(または改変体)をコードする核酸は、PCR増幅等の方法により製造可能である。例えば、本明細書に記載されたヒトCXCR3cDNAの一部に相補的または実質的に相補的な配列を含有する適当なプライマー(例えば、プライマー対またはネステッドプライマー)を、設計することができる。例えば、コーディング配列および/またはコーディング配列に隣接する配列の5’または3’末端に相補的なプライマーを設計することができる。かかるプライマーを、適当な鋳型核酸を用いるポリメラーゼ連鎖反応に使用して、哺乳類CXCR3等をコードする核酸を得ることができる。好適な鋳型には例えば、本明細書に記載された構築物(pcDNA3−Clone8等)、cDNAもしくはゲノムライブラリー、または哺乳類(ヒト、霊長類等)cDNAもしくはゲノムDNAの他の好適な供給源を含む。プライマーは、鋳型として適するものとして選択された構築物の隣接する配列に相補的な部分を含むことができる。
【0040】
タンパク質またはポリペプチド(例えば、ハイブリダイズする核酸によってコードされる)の結合機能は、例えば、受容体を含む膜画分または受容体を発現する細胞を用いて、結合または結合阻害アッセイにおいて検出されうる(Van Riper ら、J. Exp. Med., 177: 851-856 (1993); Sledziewskiら、米国特許第5,284,746号明細書(1994年2月8日)等を参照のこと)。したがって、コードされたタンパク質またはポリペプチドのIP−20またはMig等のリガンド、阻害剤および/または促進剤結合能を評価することができる。本発明の核酸によりコードされるタンパク質またはポリペプチドの抗原特性は、イムノブロッティング、免疫沈降およびイムノアッセイ(例えば、ラジオイムノアッセイ、ELISA)等の哺乳類CXCR3に結合する抗体を用いる免疫学的方法により決定することができる。
【0041】
タンパク質またはポリペプチド(例えば、ハイブリダイズする核酸によってコードされる)のシグナル伝達機能は、受容体結合に応答するGタンパク質活性(例えば、膜画分を用いるGタンパク質αサブユニットでのGTPのGDPへの交換)に関する酵素アッセイにより検出されうる。Gタンパク質共役は、例えば、Gタンパク質による刺激が、細胞もしくは適当な細胞画分(膜等)を百日咳菌毒素等のGタンパク質の特異的阻害剤で処理または前処理することにより阻止されるアッセイを用いて、さらに評価することができる(Bischoff, S. C. ら、Eur. J. Immunol., 23: 761-767 (1993); Sozzani, S. ら、J. Immunol., 147、 2215-2221 (1991))。
【0042】
タンパク質またはポリペプチド(例えば、ハイブリダイズする核酸によってコードされる)の刺激機能は、そのタンパク質またはポリペプチドを発現する細胞を用いて、走化性または媒介物質放出に関する標準的アッセイ(リガンド(例えば、IP−20またはMigなどのケモカイン)または促進剤に応答して起こる走化性、エキソサイトーシス(エステラーゼ(セリンエステラーゼ等)、パーフォリン、グランザイム等の酵素の脱顆粒反応)または媒介物質放出(例えば、ヒスタミン、ロイコトリエン)をモニターするアッセイ等)で検出できる(Taub, D. D. ら、J. Immunol., 155: 3877-3888 (1995); Baggliolini, M. とC. A. Dahinden, Immunology Today, 15: 127-133 (1994)およびそれらに記載された参考文献等を参照のこと) 。また、哺乳類CXCR3受容体に特有の機能は、他の適当な方法によって評価してもよい。
【0043】
これらの方法は単独で、もしくは他の適当な方法と組み合わせて、配列番号:2またはそれらの機能的同等物のアミノ酸配列を持ち、かつ、そのアッセイで検出される活性を持つポリペプチドをコードする核酸の同定および/または単離操作にも使用できる。特定の機能を持つ配列番号:2のポリペプチド部分をコードする単離核酸の一部もこの方法で同定および単離できる。
【0044】
本発明の核酸はタンパク質またはポリペプチドの生産に使用できる。例えば、哺乳類CXCR3受容体のコード配列の全部または一部を含有する核酸、または配列番号:1の配列またはそれらの相補鎖にハイブリダイズするDNAを、さらなる配列操作用またはコードされたポリペプチドの適当な宿主細胞における生産用の構築物に組込むことができる。また、本発明の核酸は、例えば、放射性同位体、スピン標識、抗原もしくは酵素標識、蛍光もしくは化学発光基などのような検出可能標識の取り込みまたは(直接または間接)付着により修飾することも可能であり、かかる修飾型は、本発明の範疇に含まれる。
【0045】
アンチセンス構築物
別の態様において、核酸は、センス鎖からなる標的分子に全体または一部が相補的で、その標的分子とハイブリダイズできるアンチセンス核酸である。標的はDNAであってもよいし、そのRNA体(すなわちDNAのT残基がRNA体ではU残基である)であってもよい。適当な方法で細胞に導入すると、アンチセンス核酸は、そのセンス鎖がコードする遺伝子の発現を阻害できる。アンチセンス核酸は標準的な技術で製造できる。
【0046】
1つの態様において、アンチセンス核酸は、ある標的核酸に対して全体的または部分的に相補的であり、標的核酸とハイブリダイズでき、ここで、該標的核酸は、配列番号:1の相補鎖の配列を持つ核酸にハイブリダイズできる。例えばアンチセンス核酸は、配列番号:1またはハイブリダイゼーションが十分可能なその一部の配列を持つ標的核酸に相補的であってもよい。別の態様において、アンチセンス核酸が哺乳類CXCR3受容体(例えば、ヒトIP−10/Mig受容体CXCR3)をコードする標的核酸に対して全体的または部分的に相補的で、該標的核酸とハイブリダイズできる。
【0047】
アンチセンス核酸は、研究的応用や治療的応用を含む種々の目的に有用である。例えば、アンチセンス核酸を含む構築物を適当な細胞に導入して、受容体発現を阻害することができる。かかる細胞は、例えばその親細胞や他の関連細胞型を用いて受容体−リガンド相互作用の特異性を評価する際に貴重な対照細胞となる。もう1つの側面として、かかる構築物を哺乳類の細胞の一部または全部に導入する。アンチセンス核酸は受容体発現を阻害し、その構築物を含有する細胞においてCXCR3受容体が媒介する炎症過程を阻害できる。したがって、本発明のアンチセンス核酸を用いて、炎症性の疾患や状態を治療できる。アンチセンス構築物を含有する適当な実験動物は、白血球機能欠損症、特に活性化Tリンパ球欠損症の有用なモデルにもなり、CXCR3受容体機能に関してさらなる情報を提供することができる。かかる動物は、宿主防御におけるTリンパ球およびNK細胞などの白血球の役割を解明するのに有用な、感染性疾患または癌の貴重なモデルにもなりうる。
【0048】
組換えタンパク質の製造方法
本発明のもう1つの側面は、哺乳類CXCR3タンパク質またはその改変体(一部等)を製造する方法に関する。組換えタンパク質は、例えば、適当な宿主細胞において哺乳類CXCR3またはその改変体をコードする組換え核酸(例えば、DNA)分子の発現等により得ることができる。
【0049】
また、哺乳類CXCR3タンパク質またはその改変体の発現に適する構築物(例えば、発現ベクター)も提供される。該構築物は適当な宿主細胞に導入可能であり、組換え哺乳類CXCR3タンパク質またはその改変体を発現する細胞を製造し、培地中に維持することができる。かかる細胞は、例えば、特徴づけ、単離および/または精製(例えば、アフィニティー精製)用のタンパク質の製造における使用、結合アッセイまたは他の機能アッセイ(リガンド、受容体機能の阻害剤および/または促進剤のスクリーニングのため等)での免疫原としての使用を含む、種々の目的に有用である。適当な宿主細胞は、大腸菌、枯草菌および他の適当な細菌等の細菌細胞を含む原核細胞であってもよいし、カビ細胞もしくは酵母細胞(例えばピチア パストリス(Pichia pastoris )、アスペルギルス スピーシーズ、サッカロミセス セレビシエ、シゾサッカロミセス ポンベ、ニューロスポラ クラッサ)などの真核細胞、または他の下等真核細胞および昆虫由来の細胞(例えばSf9 昆虫細胞(国際公開第94/26087号パンフレット、オコナー(O'connor)、1994年11月24日公開))または哺乳類由来の細胞(例えば、チャイニーズハムスター卵巣細胞(CHO)、COS 細胞、HuT78 細胞、293 細胞)などの高等真核細胞であってもよい。(例えばAusubel, F.M. ら編, Current Protocols in Molecular Biology, Greene Publishing Associates and John Wiley & Sons Inc.,(1993)を参照のこと。)
組換え哺乳類CXCR3タンパク質またはその改変体を生産する宿主細胞は、次のように作製できる。所望のタンパク質のコード配列の全部または一部をコードする核酸を、例えばプラスミド、ウイルスまたは他の適当な発現用レプリコンなどのDNAベクターなどの核酸ベクターに挿入することができる。シングルコピーもしくはマルチコピーとして維持されるベクターまたは宿主細胞染色体に組込まれるベクターを含む種々のベクターが利用可能である。
【0050】
哺乳類CXCR3遺伝子の転写および/または翻訳シグナルを直接発現に使用することができる。所望のタンパク質のコード配列の全部または一部をコードする核酸を発現させるための適当な発現ベクターも使用できる。発現に適したベクターは、いくつかの成分を含有してもよく、次の1以上が含まれるがこれらに限定されない:複製起点、選択可能マーカー遺伝子、1以上の発現制御要素、例えば転写制御要素(プロモーター、エンハンサー、ターミネーターなど)および/または1以上の翻訳シグナル;選択された宿主細胞(例えば、哺乳類起源のものまたは異種哺乳類もしくは非哺乳類種由来のもの)で膜標的用のシグナル配列またはリーダー配列。構築物において、シグナル配列は、ベクター、哺乳類CXCR3コード配列または他の供給源により提供されうる。また、組み込み部位に存在する配列は、これらの要素を提供する。
【0051】
プロモーターは、適当な宿主細胞での発現用に提供されうる。プロモーターは構成的であってもよいし、誘導性であってもよい。例えば、プロモーターは、哺乳類CXCR3タンパク質またはその改変体をコードする核酸に操作可能に連結しており、コードされたポリペプチドの発現を制御することができる。原核細胞宿主に適した種々のプロモーター(例えば大腸菌用のlac 、tac 、T3、T7プロモーター)および真核細胞宿主に適した種々のプロモーター(例えば酵母アルコールデヒドロゲナーゼ(ADH1)、SV40、CMV )が利用できる。
【0052】
さらに、発現ベクターは、典型的には、そのベクターを保持する宿主細胞を選択するための選択可能マーカーおよび複製可能発現ベクターの場合複製起点をも含む。抗生物質または薬剤耐性を付与する産物をコードする遺伝子は、慣用の選択可能マーカーであり、原核細胞(例えばβ- ラクタマーゼ遺伝子(アンピシリン耐性)、テトラサイクリン耐性に対するTet 遺伝子)および真核細胞(例えばネオマイシン(G418またはゼネティシン)、gpt (ミコフェノール酸)、アンピシリンまたはハイグロマイシン耐性遺伝子)に使用してもよい。ジヒドロ葉酸レダクターゼマーカー遺伝子は、種々の宿主におけるメトトレキセートによる選択を可能にする。宿主の栄養要求性マーカー(例えばLEU2、URA3、HIS3)の遺伝子産物をコードする遺伝子は、酵母における選択可能マーカーとしてしばしば使用される。ウイルス(例えばバキュロウイルス)ベクターまたはファージベクターおよび、レトロウイルスベクターなどの宿主細胞のゲノムに組み込むことができるベクターの使用も予想される。また、本発明は、これらの発現ベクターを保持する細胞にも関する。
【0053】
例えば、哺乳類CXCR3タンパク質もしくはその改変体をコードする核酸、またはかかる核酸を含有する構築物を、核酸が1以上の発現制御要素に操作可能に連結されるように(例えば、ベクター中、細胞中でのプロセッシングにより創出された構築物中で、宿主細胞ゲノムに組み込まれて)、選択された宿主細胞に適した方法(例えば、形質転換、トランスフェクション、エレクトロポレーション、感染)により適当な宿主細胞に導入することができる。宿主細胞は、発現に適した条件下で(例えば、誘導剤の存在下で、適する塩、成長因子、抗生物質、栄養補給物等を補足した適当な培地)維持することが可能であり、それによってコードされたポリペプチドが製造される。所望ならば、コードされたタンパク質(ヒトCXCR3等)を、(例えば、宿主細胞、培地、ミルクから)単離することができる。該方法がトランスジェニック動物の宿主細胞での発現を包含することがふさわしいであろう(例えば、国際公開第92/03918号パンフレット、GenPharm International、1992年3月19日公開)。
【0054】
また、融合タンパク質もこの方法で生産できる。例えば、いくつかの態様は、哺乳類CXCR3タンパク質cDNAまたはその一部を、Bluescript(登録商標)II SK +/ −(Stratagene) 、pGEX-4T-2 (Pharmacia )、pcDNA-3(Invitrogen) またはpET-15b (Novagen )などの適当な発現ベクターに挿入することによって作製できる。得られた構築物を適当な発現用宿主細胞に導入することができる。発現後、融合タンパク質を適当なアフィニティー担体を用いて細胞溶解液から単離または精製できる(例えばCurrent Protocols in Molecular Biology(Ausubel,F.M.ら編,第2 巻, 補遺26,16.4.1-16.7.8頁(1991)を参照のこと)。さらに、アフィニティー標識は、融合タンパク質の検出手段を提供する。例えば、抗原またはエピトープアフィニティー標識を含有する融合タンパク質の細胞表面発現や特定の細胞画分における該融合タンパク質の存在は、適当な抗体により検出できる。
【0055】
抗体
さらに本発明は、哺乳類CXCR3タンパク質またはその一部と反応する抗体に関する。好ましい態様において、抗体は、単数または複数の哺乳類CXCR3受容体またはその一部を特異的に結合する。1つの態様において、単離および/または組換え哺乳類CXCR3タンパク質またはその一部(例えばペプチド)または組換え哺乳類CXCR3を発現する宿主細胞に対して抗体を生じさせる。
【0056】
また、哺乳類(例えば、ヒト等の霊長類)CXCR3タンパク質に特有の結合活性、シグナル伝達活性および/または細胞応答刺激等の機能の1以上を阻害可能な抗体も本発明の範疇にある。1つの態様において、本発明の抗体は、リガンド(即ち、1以上のリガンド)の哺乳類CXCR3タンパク質への結合を阻害することおよび/またはリガンド結合に応答して哺乳類CXCR3タンパク質により媒介される1以上の機能を阻害することができる。特に好ましい態様において、抗体は、IP−10および/またはMig等の天然のリガンドと受容体との相互作用を阻害(低減または妨害)することができる。例えば、本明細書に示されたように、本発明の抗体は、ヒトCXCR3タンパク質とIP−10との相互作用を選択的に阻害および/またはそれに応答して受容体機能(例えば、シグナル伝達活性および/または細胞応答)を選択的に阻害することができる。この選択性を発揮する1C6と称される抗体は、IP−10のヒトCXCR3タンパク質への結合ならびにIP−10により誘導されるカルシウム流動および走化性を阻害することができるが、同一条件下でのMigにより誘導されるカルシウム流動を有意に阻害しない。本明細書に示されるように、追加の抗体は、用いた条件下でMig誘導性シグナル伝達を阻害しなかったけれども、これらの抗体は、CXCR3へのIP−10結合(3A8、2F8、3A12、3E2、4B4、4D2、5B12、7B8および8D5等)またはIP−10により誘導される走化性(1A5、3A8、5F10および10C6等)を阻害可能であり、それらもまたヒトCXCR3タンパク質とIP−10との相互作用および/またはそれに応答した受容体機能の選択的阻害剤であることを示唆する。
【0057】
特に好ましい態様において、本発明の抗体は、ヒトCXCR3タンパク質に対する特異性を有し、さらにより好ましくは、1C6と称するネズミモノクローナル抗体(mAb)のエピトープ特異性と同一または類似であるエピトープ特異性を有する。mAb 1C6のエピトープ特異性と同一または類似であるエピトープ特異性を有する抗体は、エピトープ特異性をキャラクタライズするための1以上の適当な技術を用いて同定することができる。例えば、mAb 1C6のエピトープ特異性と同一または類似であるエピトープ特異性を有する抗体は、ヒトCXCR3タンパク質またはその一部(例えば、活性化T細胞、NK細胞等のリンパ球または本発明の核酸を含有する組換え宿主細胞を含むヒトCXCR3を産生する細胞)への結合に関してネズミmAb 1C6と競合する能力により同定することができる。1つの態様において、mAb 1C6のエピトープ特異性と同一または類似であるエピトープ特異性を有する抗体は、適当なアッセイで、配列番号:2の残基1−15(「P1」)の配列と同一である配列を有するポリペプチドの、抗体とヒトCXCR3タンパク質との結合を阻害する能力によりさらにキャラクタライズされる(例えば、実施例8を参照のこと)。この態様の1つの側面において、かかる抗体によるヒトCXCR3タンパク質への結合は、配列番号:2の残基16−30(「P2」)または31−45(「P3」)の配列と同一である配列を有するポリペプチドにより有意に阻害されない。
【0058】
本発明に包含される他の抗体は、ヒトCXCR3タンパク質に結合可能な抗体を含み、ここで、該結合は、N末端細胞外セグメントまたはその一部に対応する配列番号:2の一部により阻害されうる。N末端細胞外セグメントの好適な部分は、N末端を含有し、反応番号:2の残基1−30、1−45もしくは1−58の配列と同一である配列を有するポリペプチドまたは反応番号:2の残基16−30もしくは45−58の配列と同一である配列を有するポリペプチド等のそのセグメントのN末端、内部またはC末端部分を含む。好ましい態様において、かかる部分は、前記本明細書に規定された哺乳類CXCR3タンパク質、ここで哺乳類はヒトである、の「少なくとも1つの免疫学的性質」を有する。例えば、配列番号:2の残基16−30の配列と同一である配列を有するポリペプチドにより結合が阻害されうるヒトCXCR3タンパク質と反応する抗体が得られた(実施例9)。
【0059】
本発明の抗体はポリクローナル抗体であってもよいし、モノクローナル抗体であってもよく、抗体という用語はポリクローナル抗体とモノクローナル抗体の両方を包含するものとする。ポリクローナルおよびモノクローナルという用語は、抗体標品の均一性の程度をいい、特定の製造方法に限定されることを意図されていない。
【0060】
本発明の抗体は、単離および/または組換え哺乳類CXCR3タンパク質またはその一部(合成ペプチドなどの合成分子を含む)等の本発明のタンパク質またはポリペプチドを含む適当な免疫原に対して生じさせることができる。さらに、トランスフェクションされた細胞などの組換え哺乳類CXCR3を発現する細胞を免疫原として使用したり、受容体を結合する抗体のスクリーニングに使用することができる。例えばチャンタラパイ(Chuntharapai)ら, J. Immunol. 152: 1783-1789(1994);およびチャンタラパイ(Chuntharapai)ら, 米国特許第5,440,021号明細書を参照のこと。
【0061】
免疫感作抗原の調製ならびにポリクローナルおよびモノクローナル抗体の製造は、任意の適当な技術で行なえる。様々な方法が記述されている(例えばKohlerら, Nature, 256: 495-497(1975)およびEur. J. Immunol. 6: 511-519 (1976);Milsteinら, Nature 266: 550-552 (1977);Koprowski ら, 米国特許第4,172,124 号;Harlow, E.およびD. Lane, 1988, Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory:ニューヨーク州コールドスプリングハーバー);Current Protocols In Molecular Biology, 第2 巻(補遺27,94 年夏),Ausubel, F.M.ら編(John Wiley & Sons:ニューヨーク州ニューヨーク), 第11章(1991)を参照のこと)。一般的には、適当な不死化細胞株(例えばSP2/0 のような骨髄腫細胞株)を抗体産生細胞と融合することによってハイブリドーマを作製することができる。抗体産生細胞、好ましくは脾臓またはリンパ節のものは、目的の抗原で免疫感作した動物から得ることができる。選択培養条件を用いて融合細胞(ハイブリドーマ)を単離し、限界希釈法でクローニングすることができる。所望の特異性を持つ抗体を産生する細胞を適当なアッセイ(例えばELISA )で選択することができる。
【0062】
必要な特異性の抗体を製造または単離する他の好適な方法を使用することが可能であり、例えば、ライブラリーから組換え抗体を選択する方法、またはヒト抗体の全レパートリーを製造可能なトランスジェニック動物(マウス等)の免疫化に依る方法を含む(例えば、Jakobovitsら、Proc.Natl. Acad. Sci. USA, 90: 2551-2555 (1993); Jakobovitsら、Nature, 362: 255-258 (1993) ; Lonberg ら、米国特許第5,545,806号明細書; Suraniら、米国特許第5,545,807号明細書)。
【0063】
単鎖抗体および異なる種に由来する部分を含有するキメラ、ヒト化もしくは霊長類化(CDR 移植)もしくはベニヤ化(veneered)抗体ならびにキメラまたはCDR 移植もしくはベニヤ化単鎖抗体なども本発明に包含され、「抗体」という用語に含まれる。これらの抗体の様々な部分を、慣用の技術で化学的に結合するか、1または複数のポリペプチドを遺伝子工学的技術を用いて連続的(cotiguous) タンパク質として調製することができる。本明細書で用いられる「ヒト化抗体または免疫グロブリン」という用語は、異なる起源由来の免疫グロブリンの一部を含有する抗体または免疫グロブリンをいい、少なくとも1つの部分がヒト起源のものである。したがって、本発明は、非ヒト起源(齧歯類等)の抗原結合領域およびヒト起源の免疫グロブリンの少なくとも一部(ヒト枠組み領域、ヒト定常領域またはその一部)を含有する、哺乳類CXCR3タンパク質(ヒトCXCR3タンパク質等)に結合特異性を有するヒト化抗体(その抗原結合断片を含む)に関する。例えば、ヒト化抗体は、化学的に結合するか遺伝子工学的技術を用いて連続的ポリペプチドとして調製された、必要な特異性を有する非ヒト起源の免疫グロブリン(非ヒトマウス可変領域等)およびヒト起源の免疫グロブリン配列(ヒト定常領域またはその一部等)に由来する部分を含有することができる。本発明のヒト化抗体の別の例は、1以上の免疫グロブリン鎖を含有する免疫グロブリンであり、前記免疫グロブリンは、非ヒト起源のCDR(非ヒト起源の抗体に由来する1以上のCDR等)およびヒト起源の軽鎖および/または重鎖に由来する枠組み領域(枠組み変化有無のCDR移植抗体等)を含有する。この態様の1つの側面において、免疫グロブリンは、非ヒト免疫グロブリンの軽鎖CDR(CDR1、CDR2およびCDR3)ならびに重鎖CDR(CDR1、CDR2およびCDR3)を含有する。1つの態様において、ヒト化免疫グロブリンは、mAB 1C6のエピトープ特異性と同一または類似のエピトープ特異性を有する。好ましい態様において、ヒト化免疫グロブリンの抗原結合領域は、mAb 1C6に由来する。また、ヒト化抗体という用語は、単鎖抗体をも包含する。例えば、例えば、キャビリー(Cabilly) ら、米国特許第4,816,567号明細書;キャビリーら、欧州特許第0,125,023 B1号明細書;ボス(Boss)ら、米国特許第4,816,397号明細書;ボスら、欧州特許第0,120,694 B1号明細書;ニューバーガー(Neuberger, M. S.)ら、国際公開第86/01533号パンフレット;ニューバーガーら、欧州特許第0,194,276 B1号明細書;ウインター(Winter)、米国特許第5,225,539号明細書;ウインター、欧州特許第0,239,400 B1号明細書;クィーン(Queen) ら、欧州特許第0,451,216 B1号明細書;およびパドラン(Padlan, E. A.) ら、欧州特許出願第0,519,596 A1号明細書を参照のこと。また、霊長類化抗体に関しては、ニューマン(Newman, R.)ら、BioTechnology, 10: 1455-1460 (1992) を、単鎖抗体に関しては、ラドナー(Ladner)ら、米国特許第4,946,778号明細書;およびバード(Bird, R. E.) ら、Science, 242: 423-426 (1988)) をも参照のこと。
【0064】
さらに、キメラ、ヒト化、霊長類化、ベニヤ化または単鎖抗体の断片を含む抗体の機能的断片も製造できる。上述の抗体の機能的断片は、それらが由来する完全長抗体の結合機能および/または調節機能の少なくとも1つを保持する。例えば、哺乳類CXCR3タンパク質またはその一部に結合できる抗体断片は、Fv、Fab 、Fab'およびF(ab')2 断片を含めて本発明に包含されるが、これらに限られるわけではない。かかる断片は、酵素的切断や組換え技術によって生産できる。例えばパパインまたはペプシン切断によって、それぞれFab またはF(ab')2 断片を生成することができる。また、天然の終結部位の上流に1以上の終止コドンが導入されている抗体遺伝子を用いて、様々な先端欠失型の抗体を生産することもできる。例えば、F(ab')2 重鎖部分をコードするキメラ遺伝子を、重鎖のCH1 ドメインおよびヒンジ領域をコードするDNA配列を含むように設計できる。
【0065】
本発明の抗体は、研究的応用、診断的応用および治療的応用を含む種々の応用に有用である。例えば、これらを使用して、受容体またはその一部を単離および/または精製したり、受容体の構造(例えばコンフォメーション)および機能を研究することができる。
【0066】
また、本発明の抗体は、研究的応用および治療的応用において受容体機能を調節するためにも使用できる。例えば、抗体は、(a)受容体に対する結合(例えばリガンド、第2の阻害剤または促進剤の結合)、(b)受容体シグナル伝達および/または(c)細胞応答を阻害 (減少または妨害) する阻害剤として作用できる。受容体機能の阻害剤として作用する抗体は、リガンドまたは促進剤の結合を直接または間接に(例えば、受容体におけるコンフォメーション変化を引き起こすことによって)遮断できる。例えば抗体は、リガンドの結合を阻害することによって、または(リガンドの結合の阻害を伴う、または伴わない)脱感作によって、受容体機能を阻害できる。本発明の抗体は、活性化または刺激されたTリンパ球およびナチュラルキラー(NK)細胞等のエフェクター細胞のアンタゴニストとして使用したり、本明細書に記載されたCXCR3機能の阻害剤で処置可能な疾患または状態の治療(予防を含む)方法における使用が見出されうる。ヒトCXCR3タンパク質とIP−10との相互作用を選択的に阻害可能および/またはそれに応答して受容体機能を選択的に阻害可能な抗体(mAb 1C6、mAb 1C6のエピトープ特異性と類似のエピトープ特異性を有する抗体等)は、IP−10−CXCR3相互作用により媒介される疾患または障害の治療に特に有用である。例えば、乾癬、炎症性腸疾患、腎炎および多発性硬化症等の炎症状態は、特に治療に影響を受けやすい。
【0067】
驚くべきことに、mAb 1C6は、混合リンパ球反応(MLR)で評価されたように、T細胞活性化を阻害する。したがって、T細胞活性化を阻害しうるmAb 1C6および他の抗CXCR3抗体を用いて、T細胞活性化ならびにサイトカイン産生、細胞傷害性T細胞殺傷および/またはT細胞のヘルパーの供給等の活性化に関連した1または複数の機能を阻害することができる。本明細書に記載された治療または予防方法に使用した場合、かかる抗体は、さらにT細胞活性化を阻害する付加的な有利性を有しうる。かかる抗体は、同種移植片拒絶または移植片対宿主病またはT細胞の活性化の阻害が望まれる他の疾患もしくは状態を含む移植片拒絶(例えば、移植における)を治療するための治療剤として、特に魅力がある。
【0068】
また、受容体を結合する抗体は、受容体に結合した際に、シグナル伝達および/または細胞応答(カルシウム流動、走化性、エキソサイトーシスまたは前炎症媒介物質放出等)等の受容体機能を誘発するまたは刺激する、受容体機能のアゴニストとして作用することもできる。したがって、かかる抗体を、活性化または刺激されたTリンパ球およびナチュラルキラー(NK)細胞等のエフェクター細胞のアゴニストとして使用したり、本明細書に記載されたCXCR3機能の促進剤で処置可能な疾患または状態の治療(予防を含む)方法における使用が見出されうる。
【0069】
さらに、本発明の種々の抗体を用いて、例えば活性化Tリンパ球またはナチュラルキラー細胞(NK細胞)等の白血球または受容体遺伝子でトランスフェクションされた細胞における受容体の発現を検出または測定することもできる。したがってこれらの抗体は、診断または研究目的の細胞選別(例えばフローサイトメトリー、蛍光活性化細胞選別)などの応用にも有用である。
【0070】
抗イディオタイプ抗体も提供される。抗イディオタイプ抗体は、別の抗体の抗原結合部位に関連する抗原決定基を認識する。抗イディオタイプ抗体は、同一の種の動物を免疫することにより第1の抗体に対して調製され、第1抗体を生産するのに使用した動物と同じ種、好ましくは第1の抗体を製造するために用いた動物と同じ株の動物を前記第1の抗体で免疫感作することによって調製できる。例えば米国特許第4,699,880 号明細書を参照のこと。
【0071】
1つの態様において、抗体を受容体またはその一部に対して生じさせ、今度はこれらの抗体を免疫原として使用して抗イディオタイプ抗体を生産する。そのようにして生産した抗Idは、受容体を模倣して、例えばリガンド、受容体機能の阻害剤または促進剤などの受容体を結合する化合物を結合でき、かかる化合物を検出、同定または定量するための免疫アッセイに使用できる。かかる抗イディオタイプ抗体は受容体自体には結合しないが、受容体機能の阻害剤でもありうる。
【0072】
抗イディオタイプ(すなわち抗Id)抗体自体を使用して抗イディオタイプ抗体(すなわち抗- 抗Id)を生じさせることができる。かかる抗体は最初の免疫感作抗体と特異性が似ているか同一でありうる。1つの態様において、受容体への結合を遮断する抗体アンタゴニストを用いて抗Idを生じさせ、その抗Idを用いて、その抗体アンタゴニストと類似するか同一の特異性を持ちうる抗- 抗Idを生じさせることができる。これらの抗- 抗Id抗体を受容体機能に対する阻害効果について評価することによって、それらがアンタゴニストであるかどうかを決定できる。
【0073】
単鎖およびキメラ、ヒト化、霊長類化(CDR 移植)、ベニヤ化ならびにキメラ、CDR 移植またはベニヤ化単鎖抗イディオタイプ抗体を調製することが可能であり、これらは抗イディオタイプ抗体という用語に包含される。また、かかる抗体の抗体断片も調製できる。
【0074】
本発明の抗体および断片は、例えば放射性同位体、スピン標識、抗原もしくは酵素標識、蛍光もしくは化学発光基などの検出可能な標識の取り込みまたは付着(直接または間接)により修飾することが可能であり、かかる修飾型は、本発明の範疇に含まれる。
【0075】
リガンド、受容体機能の阻害剤または促進剤の同定
本明細書において、リガンドは、受容体タンパク質を結合する物質である。選択された哺乳類CXCR3 タンパク質のリガンドは、選択された哺乳類CXCR3 タンパク質に結合する物質である。好ましい態様として、哺乳類CXCR3 タンパク質のリガンド結合は、高いアフィニティーで生じる。リガンドという用語は、限定されないが、天然リガンド、単離および/ または精製された、合成および/ または組換え体、天然リガンドのホモログ(例えば別の哺乳類由来のもの)、抗体、かかる分子の一部ならびに受容体を結合する他の物質などの物質を意味する。選択した哺乳類受容体の天然リガンドは生理学的条件下でその受容体に結合でき、哺乳類CXCR3 タンパク質のものと同じ哺乳類起源のものである。リガンドという用語は、受容体活性の阻害剤または促進剤である物質ならびに選択的に受容体に結合するが阻害剤活性や促進剤活性を欠く物質を包含する。
【0076】
本明細書で用いられるように、阻害剤は、哺乳類CXCR3 タンパク質( 例えばヒトCXCR3 など) に特有の機能、例えば結合活性(例えば、リガンド結合、促進剤結合など)、シグナル伝達活性(哺乳類G タンパク質の活性化、サイトゾル遊離カルシウム濃度[Ca2+]iの迅速かつ一過性の増加の誘導など)、および/ または細胞応答機能( 例えば、走化性、エキソサイーシスまたは白血球による炎症媒介物質の放出の刺激) などの少なくとも1 つを阻害する物質である。阻害剤という用語は、受容体を結合するアンタゴニスト(例えば、抗体、天然リガンドの突然変異体、その他のリガンド結合の競合的阻害剤)および受容体に結合することなく受容体機能を阻害する物質(例えば抗イディオタイプ抗体)を含む物質を意味する。
【0077】
本明細書で用いられるように、促進剤は、哺乳類CXCR3 タンパク質(例えば、ヒトCXCR3 など) に特有の機能、例えば結合活性(リガンド、阻害剤および/ または促進剤結合など)、シグナル伝達活性(哺乳類G タンパク質の活性化、サイトゾル遊離カルシウム濃度[Ca2+]iの迅速かつ一過性の増加の誘導など)、および/ または細胞応答機能( 例えば、走化性、エキソサイトーシスまたは白血球による炎症媒介物質の放出の刺激) などの少なくとも1 種を促進(誘導または増大)する物質である。促進剤という用語は、受容体を結合するアゴニスト(例えば抗体、別の種に由来する天然リガンドのホモログ)および受容体に結合することなく受容体機能を(例えば関連するタンパク質を活性化することによって)促進する物質を含む物質を意味する。好ましい態様として、該アゴニストは天然のリガンドのホモログ以外のものである。
【0078】
本発明の核酸およびタンパク質をあてにする下記のアッセイは、単独または相互にもしくは他の適切な方法と組み合わせて、哺乳類CXCR3 タンパク質または改変体のリガンド、阻害剤または促進剤を同定するために使用されうる。本発明のイン・ビトロ方法は、多くの試料を処理する高処理のスクリーニング( 例えば、96ウェルフォーマット) に適合しうる。本発明の核酸を含み、組換え哺乳類CXCR3(例えば、ヒトCXCR3など)を高処理のスクリーニングに適するレベルで発現する宿主細胞を用いることができ、したがって、哺乳類CXCR3 タンパク質のリガンド、阻害剤および促進剤の同定および/または単離に特に価値がある。受容体の発現を多様な方法でモニターすることができる。例えば、受容体またはその一部を結合する本発明の抗体を用いて発現をモニターすることができる。また、市販の利用可能な抗体を用いて、受容体タンパク質またはポリペプチドを含む抗原標識- またはエピトープ標識- 融合タンパク質(例えばFLAG標識受容体)の発現を検出することもでき、所望のレベルで発現する細胞を選択することができる。
【0079】
哺乳類CXCR3 タンパク質をコードする核酸を発現系に組み込んで前記した受容体タンパク質またはポリペプチドを生産することができる。例えば本発明の核酸を含む構築物で安定にもしくは一過性にトランスフェクトした細胞中で発現した受容体または受容体を含有する細胞画分(例えばトランスフェクト細胞の膜画分、受容体を組み込んだリポソーム)中の受容体などの単離および/ または組換え受容体タンパク質もしくはポリペプチドを、受容体機能の試験に使用することができる。所望であれば、その受容体をさらに精製することもできる。受容体機能の試験は、イン・ビトロまたはイン・ビボで行なうことができる。
【0080】
本明細書記載の受容体機能をモニターすることによりまたは他の適切な技術を用いることにより、化合物の効果を評価する本発明の方法に、図2(配列番号:2) に示されるヒトCXCR3 などの単離および/または組換え哺乳類CXCR3 タンパク質を使用することができる。例えば、実施例2 記載のものまたは他の適切な細胞( 例えば、バキュロウイルス感染 Sf9細胞、マウスIL-2前B 細胞 [前B リンパ腫由来、オイゲン・ブッチャー(Eugene Butcher)博士 (スタンフォード大学, スタンフォード, カリフォルニア)]などの安定なまたは一過性のトランスフェクタントを結合アッセイに用いることができる。Jurkat細胞の安定なトランスフェクタント(実施例2 )または走化性を行ないうる他の適切な細胞の安定なトランスフェクタント(例えばマウスLl-2前B 細胞) は、例えば走化性アッセイに用いることができる。
【0081】
本発明の方法によれば、化合物を個別にスクリーニングすることもできるし、1 以上の化合物を本明細書記載の方法に従って同時に試験することもできる。化合物の混合物を試験する場合、記載された過程で選択された化合物を、適切な方法(例えばPCR 、配列決定、クロマトグラフィー)で (適宜) 分離、同定することができる。試験試料中の1 以上の化合物(リガンド、阻害剤、促進剤など)の存在も、これらの方法によって決定できる。
【0082】
複合化学合成や他の方法で作製された大きな複合化合物ライブラリー(例えば有機化合物、組換えまたは合成ペプチド、「ペプトイド」、核酸)を試験することができる [例えばZuckerman, R. N. et al., J. Med. Chem., 37: 2678-2685(1994) とそこに引用されている文献を参照のこと; また、Ohlmeyer, M. H. J. et al., Proc. Natl. Acad. Sci. USA 90: 10922-10926 (1993)とDeWitt, S. H. et al., Proc. Natl. Acad. Sci. USA 90: 6909-6913 (1993)(標識した化合物について); Rutter, W.J. et al., 米国特許第5,010,175 号明細書, Huebner, V.D. et al.,米国特許第5,182,366 号明細書, Geysen, H.M., 米国特許第4,833,092 号明細書をも参照のこと)。本発明方法によって複合ライブラリーから選択される化合物が独自の標識を持つ場合は、クロマトグラフィー法による個々の化合物の同定が可能である。
【0083】
1つの態様において、ファージディスプレイ法を使用する。例えば、受容体結合(例えば適切な結合緩衝液において)に適した条件下でポリペプチドを呈示するファージ(例えばファージまたはライブラリーのようなファージの収集物)と受容体を接触させることができる。受容体に結合したファージを、標準的な技術または他の適切な方法で選択できる。ファージは適当な溶出緩衝液を用いて受容体から分離できる。例えば、イオン強度やpHの変化はファージの放出をもたらしうる。別法として、溶出緩衝液が、化合物の結合を破壊するように設計された1 または複数の放出成分[ 結合を競合的に阻害するリガンド、阻害剤および/ または促進剤などの例えば呈示されたペプチドと受容体との結合を破壊できる1 以上の化合物] を含んでもよい。任意に、この選択過程を繰り返すか、別の選択過程を用いることによって、受容体に結合するファージをさらに濃縮することもできる。呈示されたポリペプチドを(例えばファージDNA の配列決定などによって)特徴づけることができる。同定されたポリペプチドを生産し、さらにリガンド結合機能、阻害剤機能および/ または促進剤機能について試験することができる。増大した安定性や他の望ましい特性を持つかかるペプチドの類縁体を製造できる。
【0084】
1つの態様において、ランダムな配列の核酸によりコードされるN-末端ペプチドを有するコートタンパク質を含む融合タンパク質を発現し、それを呈示するファージを生産できる。本発明の受容体タンパク質またはポリペプチドを発現する適切な宿主細胞を該ファージと接触させ、結合したファージを選択し、回収し、特徴づける。(例えば、G タンパク質共役受容体を用いるファージディスプレイ法について議論しているDoorbar, J. およびG. Winter, J. Mol. Biol., 244: 361(1994)を参照のこと)。
【0085】
哺乳類CXCR3 タンパク質の潜在的リガンド、阻害剤および/ または促進剤の他の供給源としては、限定されないが、IP-10 またはMig の天然、合成または組換え改変体を含むCXCR3 リガンドの改変体、他の化学誘引物質またはケモカイン類などの物質、その改変体、他の阻害剤および/または促進剤( 例えば、抗CXCR3 抗体、アンタゴニスト、アゴニスト) 、他のG タンパク質共役受容体リガンド、阻害剤および/または促進剤( 例えば、アンタゴニストまたはアゴニスト) 、および受容体機能を阻害しうる適切な受容体ペプチドまたはアナログなどの哺乳類CXCR3 受容体の可溶性部分が挙げられる(例えばMurphy,R.B.,国際公開第94/05695号パンフレットを参照のこと)。
【0086】
結合アッセイ
本発明の単離および/ または組換え受容体タンパク質もしくはその機能的改変体、その一部または適当な融合タンパク質は、ヒトCXCR3 などの(1 以上の)哺乳類CXCR3 タンパク質に結合する薬剤、およびリガンドあるいは受容体活性の潜在的阻害剤または促進剤である薬剤を選択および同定する方法に使用できる。この方法によって選択される、リガンド、阻害剤または促進剤を含む薬剤を、受容体機能に対する阻害または刺激効果および/ または治療的用途についてさらに評価できる。
【0087】
1つの態様において、活性な単離および/ または組換え哺乳類CXCR3 タンパク質またはポリペプチドに結合する化合物を該方法で同定する。この態様では、使用する受容体タンパク質またはポリペプチドが、哺乳類CXCR3 タンパク質 (本明細書規定) に特有の例えば結合活性( 例えば、リガンド、阻害剤および/または促進剤結合) 、シグナル伝達活性(哺乳類G タンパク質の活性化、サイトゾル遊離カルシウム濃度[Ca2+]iにおける迅速かつ一過性の増加の誘導)、細胞応答機能(例えば、走化性、エキソサイトーシスまたは白血球による炎症媒介物質放出の刺激など)、および/ または本明細書に規定された免疫学的性質などの少なくとも1 種の性質、活性または機能を有する。好ましい態様において、単離および/ または組換え哺乳類CXCR3 タンパク質または改変体は、リガンド結合機能を持ち、さらに好ましくは、受容体の天然リガンドを結合する。特に好ましい態様として、該単離および/または組換えタンパク質は、図1 に示された核酸によりコードされたヒトCXCR3 タンパク質である (配列番号:1) 。
【0088】
例えば、結合に適する条件下に単離および/または組換え哺乳類CXCR3 タンパク質またはその改変体を含有する組成物を維持することができ、受容体は、試験対象の薬剤( 例えば、1 以上の薬剤を含有する組成物) に接触させることができ、結合を検出もしくは測定できる。1 つの態様として、本発明の受容体をコードする核酸配列を含有する構築物で安定にもしくは一過性にトランスフェクトした細胞で、受容体タンパク質を、発現させることができる。細胞を受容体の発現に適切な条件下に維持しうる。結合に適する条件下 (例えば適切な結合緩衝液など) に該細胞を薬剤と接触させ、結合を標準技術により検出することができる。例えば、結合の程度は、適切な対照と比較して (例えば、薬剤の非存在下に決定したバックグラウンドと比較、第2 薬剤の結合と比較 (すなわち標準) 、トランスフェクトしていない細胞への薬剤の結合を比較など) 決定されうる。任意に、受容体を含む膜画分などの細胞画分を全細胞の代わりに用いることができる。
【0089】
結合または複合体形成を直接または間接に検出することができる。1つの態様において、薬剤を適切な標識(例えば蛍光標識、化学発光標識、同位体標識、酵素標識)で標識し、その標識の検出によって結合を決定することができる。結合の特異性は、例えば非標識薬剤またはリガンド (例えば、IP-10 、Mig など) を競合剤として用いる競合または置換によって評価できる。
【0090】
哺乳類受容体のリガンドは、同じ哺乳類種由来の天然リガンドまたは別の種由来の天然リガンドを含めて、この方法で同定できる。受容体を結合する促進剤または阻害剤の結合活性も、かかるリガンド結合アッセイで評価できる。
【0091】
結合阻害アッセイを用いて、リガンドならびに受容体に結合し、リガンドなど他の薬剤の結合を阻害する阻害剤および促進剤を同定することもできる。例えば、第2 の試験薬剤の存在下での第1 の薬剤の結合に比べて、(第2 の薬剤の非存在下で起こる)第1 の薬剤の結合の減少を検出または測定する結合アッセイを行なうことができる。受容体を第1 および第2 の薬剤と同時に接触させてもよいし、逐次順番は問わずに接触させてもよい。第2 試験薬剤の存在下で起こる第1 薬剤の結合の程度の減少は、第2 薬剤による結合の阻害の指標となる。例えば、第1 薬剤の結合が減少したり、妨げられたりされうる。
【0092】
1つの態様において、ヒトCXCR3 に対する第1 薬剤(例えばIP-10 、Mig などのケモカイン)の結合の第2 の試験薬剤による直接阻害をモニターする。例えば、ヒトCXCR3 への125I- 標識Mig の結合を阻害する薬剤の能力をモニターしうる。かかるアッセイは、例えば、全細胞(例えばヒトCXCR3 受容体をコードする核酸を含む適切な細胞株)または該細胞由来の膜画分を用いて行なうことができる。
【0093】
受容体を結合する1 もしくは複数の薬剤の存在を同定する他の方法も利用でき、例えば、シグナル伝達機能および/ または細胞応答の刺激などを含む、受容体結合によって誘発される事象をモニターする方法である。
【0094】
本発明の抗体の阻害効果を結合阻害アッセイで評価できることは理解されるだろう。また、該アッセイにおける第1 薬剤が別の抗体である方法で、抗体結合に適した条件下に、受容体結合に関する抗体間の競合を評価できる。
【0095】
この方法で同定されるリガンド、受容体結合阻害剤および促進剤をさらに評価して、結合に引き続いて、それらがCXCR3 受容体の他の機能を阻害または活性化するように作用するかどうかを決定すること、および/または、それらの治療的有用性を評価することができる。
【0096】
シグナル伝達アッセイ
G タンパク質共役受容体の(アゴニストなどによる)結合は、該受容体によるシグナル伝達をもたらすことができ、G タンパク質の活性化を刺激する。薬剤によるシグナル伝達機能の誘導は、いかなる適切な方法をも用いてモニターできる。例えば、GTP からGDP への加水分解、または受容体結合により誘発される後期のシグナル伝達事象、例えば、細胞内(サイトゾル)遊離カルシウム濃度[Ca2+]iの迅速かつ一過性の増加の誘導などを、当該分野公知の方法または他の適切な方法で評価できる(実施例2; Neote, K.et al., Cell, 72: 415-425 1993) Van Riper et al., J. Exp. Med., 177: 851-856 (1993); Dahinden, C.A. et al., J. Exp. Med., 179: 751-756(1994)をも参照のこと)。
【0097】
ハイブリッドG タンパク質共役受容体を用いるSledziewski らの機能アッセイを用いて、ハイブリッドG タンパク質を活性化するその能力によりリガンドもしくは促進剤を同定すること、またはかかる活性化を阻害するその能力により阻害剤を同定することも可能である(Sledziewski ら, 米国特許第5,284,746 号明細書;その教示は参照により本明細書に合体される)。1 つの態様として、試験試料中のリガンドの存在の指標である応答の検出で、(ハイブリッド受容体への結合により誘発される)宿主細胞の生物学的応答がモニターされうる。例えば、試験試料中の、受容体のリガンド−結合ドメインにより結合されうる薬剤であるリガンドの存在を検出する方法が記載される。本方法の1つの態様として、酵母宿主細胞を、生物学的に活性なハイブリッドG タンパク質共役受容体 (即ち、融合タンパク質) の発現を仕向けることが可能なDNA 構築物で形質転換する。該ハイブリッド受容体は、STE2遺伝子産物などの酵母G タンパク質共役受容体の対応するドメインと置換されたリガンド結合ドメイン以外の少なくとも1 種のドメインを有する哺乳類G タンパク質共役受容体を含有する。構築物を含む酵母宿主細胞をハイブリッド受容体が発現する条件下に維持し、該細胞をリガンドのハイブリッド受容体への結合を可能にするに適した条件下に試験試料と接触させる。(ハイブリッド受容体への結合により誘発される)宿主細胞の生物学的応答がモニターされ、応答の検出はシグナル伝達機能の指標となる。例えば、STE2遺伝子産物由来のハイブリッド受容体への結合がBAR1プロモーターの誘導をもたらすことができる。該プロモーターの誘導は、BAR1プロモーターに連結され第2 の構築物で宿主細胞に導入されるレポーター遺伝子(例えば、β-galなど)を利用して測定される。該リポーター遺伝子の発現は、例えば細胞溶解液に対するイン・ビトロ酵素検定によって、あるいは培地に指示薬(例えば、X-gal など) を含む平板での青色コロニーの存在によって検出できる。
【0098】
他の態様において、該アッセイを用いて受容体機能の潜在的阻害剤を同定することができる。ある薬剤の阻害活性は、該アッセイでリガンドまたは促進剤を用い、リガンドまたは促進剤によって誘導される活性を阻害する試験薬剤の能力を評価することによって決定できる。
【0099】
既知リガンドの改変体を、共役G タンパク質の活性を刺激する能力の減少(減少した能力または能力がない)についてスクリーニングすることもできる。この態様では、薬剤は(他の方法で決定したように)リガンド結合活性を持つが、受容体の連動が共役G タンパク質の活性を誘発しないか、わずかにしか誘発しない。かかる薬剤は潜在的アンタゴニストであり、さらに阻害活性を評価することができる。
【0100】
走化性および細胞応答の他のアッセイ
受容体機能を評価するため走化性アッセイも使用できる。これらのアッセイは薬剤によって誘発されるイン・ビトロまたはイン・ビボの細胞の機能的移動に基づいており、これらのアッセイを用いてリガンド、阻害剤または促進剤の結合および/ または走化性への効果を評価できる。
【0101】
IP-10 およびMig に対する細胞の応答を評価するためのイン・ビトロ走化性アッセイの使用を実施例2 に記述する。Springerらは経内皮リンパ球走化性アッセイについて記述している(Springerら, 国際公開第94/20142号パンフレット,1994 年9 月15日公開;その教示は参照により本明細書に合体される;さらにBermanら, Immunol Invest. 17: 625-677 (1988)も参照のこと)。内皮を横切るコラーゲンゲルへの移動も記述されている(Kavanaugh ら, J. Immunol, 146: 4149-4156(1991))。
【0102】
一般に走化性アッセイでは、障壁の第1 表面から反対側の第2 表面に向かって、薬剤の増加レベルにそって障壁(内皮、フィルターなど)内へ、または障壁を通過して移動する、走化性の能力を有する適切な細胞、例えば白血球など(T リンパ球、NK細胞、単球など)、Jurkat細胞、マウスIL-2前B 細胞、または他の適切な宿主細胞の安定なトランスフェクタントの指向的な運動または移動をモニターする。膜やフィルターは、フィルターの第1 表面からフィルターの反対側の第2 表面の方向に増大する薬剤レベルに向かってフィルター内に、またはフィルターを通過して移動する、適切な細胞の指向的運動または移動をモニターするのに便利な障壁を提供する。いくつかのアッセイでは、ICAM-1、フィブロネクチンまたはコラーゲンなどの接着を容易にする物質で膜をコートする。
【0103】
例えば、適切な容器(含有手段)内で、第1 チャンバーから第2 チャンバー中への、もしくは細孔膜を通過して、試験対象の薬剤を含有し、膜によって第1 チャンバーから隔離された第2 チャンバーへの細胞の移動を検出または測定できる。薬剤に応答して起こる特異的な移動をモニターするに適した孔サイズを持つ適切な膜、例えばニトロセルロース、ポリカーボネートなどを選択する。例えば約3 〜8 ミクロン、好ましくは約5 〜8 ミクロンの孔サイズを使用できる。フィルター上の孔サイズは均一であってもよいし、適切な孔サイズの範囲に分布していてもよい。
【0104】
移動を評価するため、フィルターへの移動距離、フィルターを横切ってフィルターの第2 表面に接着している細胞数および/ または第2 チャンバー中に蓄積する細胞数を標準的な技術(例えば顕微鏡)を用いて決定できる。1つの態様において、細胞を検出可能な標識(例えば放射性同位体、蛍光標識、抗原またはエピトープ標識)で標識し、膜に接着したその標識および/ または第2 チャンバー中に存在するその標識の存在を適切な方法で(例えば放射活性や蛍光を検出するか、免疫アッセイで)決定することによって、移動を評価できる。薬剤によって誘発される移動の程度を、適切な対照と比較して(例えば薬剤の非存在下に決定したバックグラウンド移動と比較して、第2 の薬剤(すなわち標準)によって誘発される移動の程度と比較して、その薬剤によって誘発される非トランスフェクト細胞の移動と比較して)決定することができる。
【0105】
チャンバーは、プラスチック、ガラス、ポリプロピレン、ポリスチレンなど、種々の固体で形成することができる。チャンバーから取り外せる膜、例えばBiocoat (Collaborative Biomedical Products )やTranswell (Costar, マサチューセッツ州ケンブリッジ)培養インサートなどは、接着細胞の計数を容易にする。容器には、第1 チャンバー内の細胞を含む液体と第2 チャンバー内の液体に接触するようにフィルターを設置することができる。試験薬剤またはアッセイの目的のために存在する追加のリガンド、阻害剤もしくは促進剤以外は、膜の両側の液体が好ましくは同じであるか、実質的に類似している。チャンバー内の液体は、細胞の安定性を増大させ、細胞の非特異的結合を阻害するように作用するタンパク質溶液(例えばウシ血清アルブミン、ウシ胎児血清、ヒト血清アルブミン)および/ または培養培地を含みうる。
【0106】
1 つの態様として、経内皮移動を評価する。より低いバックグラウンド( ノイズに対するシグナル割合) に加え、経内皮移動は、血管壁に沿う内皮細胞層を横切ることにより白血球が炎症部位の組織に存在する化学誘引剤に向かって血管から移出するイン・ビボ条件をモデルにする。この態様では、内皮細胞層を通過する遊出を評価する。細胞層を調製するため、内皮細胞の接着を促進するように任意にコラーゲン、フィブロネクチンまたは他の細胞外マトリックスタンパク質などの物質でコートされた細孔フィルターまたは膜上で内皮細胞を培養できる。好ましくは、コンフルエントな単層が生成するまで内皮細胞を培養する。例えば静脈、動脈または微小血管内皮、例えば、ヒト臍静脈内皮細胞(Clonetics Corp, サンディエゴ,カリフォルニア州)などや適切な細胞株、例えば、用いられたECV304細胞株(European Collection of Animal Cell Cultures, Porton Down, Salisbury, U.K.) などの多様な哺乳類内皮細胞が単層形成に利用できる。特定の哺乳類受容体に応答して起こる走化性を検定するために、同じ哺乳類の内皮細胞が好ましい; しかしながら、異なる哺乳類種または属由来の内皮細胞も使用できる。
【0107】
一般的には、フィルターの第1 表面からフィルターの反対側の第2 表面に向かって、薬剤のレベルが増大する方向に、膜またはフィルター内へ、または膜またはフィルターを通過して移動する細胞の指向的移動を検出することによって、このアッセイを行ない、そのフィルターは第1 表面に内皮細胞層を含む。指向的移動は第1 表面に隣接する領域から膜内へ、または膜を通過して、フィルターの反対側に置かれた薬剤に向かって起こる。第2 表面に隣接する領域に存在する薬剤の濃度は、第1 表面に隣接する領域に存在する薬剤の濃度より大きい。
【0108】
1つの態様において、薬剤のリガンドまたは促進剤活性の試験に走化性アッセイを用い、移動能を持ち哺乳類CXCR3 タンパク質またはその機能的改変体を発現できる細胞を含む組成物を第1 チャンバーに入れ、好ましくは第1 チャンバー内の細胞の走化性を誘導できる(化学誘引機能を持つ)他のリガンドや促進剤の非存在下に、試験対象の薬剤(1以上の薬剤) を含む組成物を第2 チャンバーに入れる。しかしながら、化学誘引機能を持つ1 以上のリガンドまたは促進剤が存在してもよい。本アッセイで哺乳類CXCR3 受容体を発現する細胞の走化性を誘導する薬剤の能力は、薬剤が受容体機能のリガンドまたは促進剤であることの指標である。
【0109】
阻害剤の試験に使用する1つの態様において、移動能を持ち、哺乳類CXCR3 タンパク質または機能的改変体を発現できる細胞を含む組成物を第1 チャンバーに入れる。第1 チャンバー内の細胞の走化性を誘導できる(化学誘引機能を持つ)リガンドまたは促進剤を含む組成物 (すなわち1 以上のリガンドまたは促進剤) を第2 チャンバーに入れる。細胞を第1 チャンバーに入れる前 (好ましくは直前) か、細胞と同時に、試験対象の薬剤を含む組成物を、好ましくは第1 チャンバーに入れる。このアッセイで哺乳類CXCR3 タンパク質を発現する細胞のリガンドまたは促進剤誘導走化性を阻害する薬剤の能力は、薬剤が受容体機能の阻害剤( 例えば、細胞応答機能の阻害剤) であることの指標である。試験薬剤の存在下に、リガンドまたは促進剤によって誘導された移動の程度の減少は、阻害活性の指標となる。別個に結合試験(上記参照)を行なって、阻害が受容体に対する試験薬剤の結合の結果であるのか、それとも異なる機構によって起こるのかを決定することもできる。
【0110】
組織に対する薬剤の注射に応答して起こる組織の白血球浸潤をモニターするイン・ビボアッセイを後述する。これらのモデルでは、炎症部位への移動および走化性によってリガンドまたは促進剤に応答する細胞の能力を測定する。
【0111】
受容体を含有する適切な宿主細胞を用いて、活性な受容体により誘導される他の細胞応答をモニターすることによって、CXCR3 受容体の細胞応答機能に対するリガンド、阻害剤または促進剤の効果を評価することができる。同様に、これらのアッセイを用いて受容体の機能を決定することもできる。例えば、エキソサイトーシス [例えば1 以上の酵素または他の顆粒成分、例えば、エステラーゼ (例えば、セリンエステラーゼ) 、パーフォリンおよび/またはグランザイムなど) などの放出をもたらすナチュラルキラー細胞の脱顆粒] 、炎症媒介物質放出 [例えばロイコトリエン類 (ロイコトリエンC4など)などの生理活性脂質の放出など] 、ならびに呼吸バーストを、当該分野公知の方法や他の適切な方法で監視することができる。[ 例えば、顆粒誘導セリンエステラーゼの放出のアッセイに関する Taub,D.D et al., J.Immunol.,155:3877-3888(1995) (その教示は、参照により本明細書中に取り込まれる) ならびにNK細胞および細胞傷害性T リンパ球(CTLs)による酵素およびグランザイム放出のアッセイに関する Loetscher et al., J. Immunol., 156:322-327(1996), (その教示は、参照により本明細書中に取り込まれる);呼吸バーストに関するRot,A. et al.,J.Exp.Med.,176:1489-1495(1992);Bischoff, S.C. ら, Eur. J. Immunol., 23: 761-767 (1993)ならびにBaggliolini, M. およびC.A. Dahinden, Immunology Today, 15: 127-133(1994)を参照のこと。]。
【0112】
1つの態様において、脱顆粒またはエキソサイトーシス機能を持つ細胞による脱顆粒またはエキソサイトーシス時の酵素放出をモニターすることによって、リガンド、阻害剤および/ または促進剤を同定する。エキソサイトーシスまたは脱顆粒を刺激することのできる活性な受容体タンパク質をコードする本発明の核酸を含有する細胞を、適切な条件下に適切な培地中で維持し、それによって受容体を発現させ、脱顆粒を誘発させうる。受容体を試験対象の薬剤と接触させ、酵素放出を評価する。培地への酵素の放出は、免疫学的アッセイまたは酵素活性に関する生化学的アッセイなど、適切な方法で検出または測定できる。
【0113】
アッセイ成分(基質、補因子、抗体など)を培地に(例えば細胞と薬剤を合わせる前、同時にもしくは後に)導入することによって、培地を直接的にアッセイできる。別法として、アッセイに先立って細胞から分離し、さらに工程を経た (分画した) 培地に対してアッセイを行なうこともできる。例えば、セリンエステラーゼなどの酵素の慣用のアッセイを利用できる [顆粒誘導セリンエステラーゼの放出に関するTaub,D.D.et al.,J.Immunol.,155:3877-3888(1995)などを参照のこと] 。
【0114】
薬剤による脱顆粒の刺激は、薬剤が、哺乳類CXCR3タンパク質のリガンドまたは促進剤であることの指標となりうる。他の態様として、受容体を発現している細胞は、リガンドまたは促進剤と組み合わされ、それより前、後、または同時に試験対象の薬剤を添加し、脱顆粒が評価される。リガンド−促進剤−誘導脱顆粒の阻害は、該薬剤が、哺乳類CXCR3タンパク質機能の阻害剤であることの指標でありうる。
【0115】
また、細胞接着は、当該分野に公知の方法または他の適切な方法によりモニターされうる。リンパ球のケモカイン受容体の連動状態はインテグリン活性化、および脈管構造または末梢血管領域に発現した接着分子への接着の誘導を引き起こしうる。1つの態様として、接着することができる細胞による細胞接着をモニターすることにより、リガンド、阻害剤および/または促進剤を同定する。例えば、試験対象の薬剤は、(a)受容体を発現する細胞(好ましくは受容体でトランスフェクトされたときに接着能を獲得する非接着性細胞)、(b)適切な接着分子を含む組成物(例えば、フィブロネクチンなどの接着分子でコートされた培養ウェルなどの基質)、およびリガンドまたは促進剤(例えば、アゴニスト)と組み合わされることができ、リガンドまた促進剤誘導接着に適する条件下に維持しうる。蛍光色素による細胞の標識は、接着細胞を検出する簡便な方法を提供する。非接着細胞は、(例えば、洗浄により)取り除くことができ、接着細胞の数が決定する。リガンドまたは促進剤誘導接着を阻害するまたは増強することにおける薬剤の効果は、それぞれ、阻害剤または促進剤活性の指標となりうる。該アッセイに活性な薬剤としては、結合、シグナル伝達および/または細胞応答の阻害剤および促進剤があげられる。他の態様として、試験対象の薬剤は、リガンドまたは促進剤誘導接着に適する条件下に受容体を発現する細胞と適切な接着分子を含む組成物とに組み合わせられることができ、接着がモニターされる。適切な対照と比較した増加した接着は、リガンドおよび/または促進剤の存在の指標となる。
【0116】
炎症のモデル
治療薬として、イン・ビボでのリガンド、阻害剤、または促進剤の効果を評価するために用いられうる多様な炎症のイン・ビボモデルが利用でき、ヒツジ喘息モデル[例えば、Weg, V. B. et al., J. Exp. Med., 177: 561(1993)を参照のこと、その教示は、参照により本明細書に取り込まれる]、ラット遅延型過敏モデル[Rand, M.L. et al., Am.J.Pathol., 148:855-864(1996)、その教示は、参照により本明細書に取り込まれる]または、他の適切なモデルがあげられる。他の哺乳類CXCR3タンパク質と交差反応する抗体の活性をかかる哺乳類で評価することができる。
【0117】
また、ウサギ、ラット、モルモットなどの適切な動物に薬剤を皮膚内注射した時の白血球浸潤もモニターできる [例えばVan Damme J.ら, J. Exp. Med., 176: 59-65(1992)、Zachariae, C.O.C. ら, J. Exp. Med. 171: 2177-2182 (1990)、Jose, P.J.ら, J. Exp. Med. 179: 881-887 (1994)を参照のこと] 。1つの態様において、皮膚生検を白血球(T リンパ球、単球、ナチュラルキラー細胞など)の浸潤について組織学的に評価する。もう1 つの態様として、走化性を持ち管外溢出能を持つ標識細胞(例えば、111In で標識された哺乳類CXCR3 タンパク質を発現する細胞)を動物に投与する。試験試料(例えば適切な緩衝液または生理学的担体中の試験対象の薬剤)の注射部位に近接して起こる標識細胞の浸潤が、その試料中のリガンドまたは促進剤、例えば、アゴニストなどの存在の指標となる。これらのアッセイを改良して、走化性と白血球管外溢出の阻害剤を同定することもできる。例えば、リガンドまたはアゴニストを試験動物に投与する前、後もしくは同時に、阻害剤を投与できる。阻害剤の非存在下での浸潤の程度に比べて、阻害剤の存在下で浸潤の程度の減少が阻害の指標である。
【0118】
診断的用途
本発明は様々な診断的用途を持つ。例えば、哺乳類CXCRタンパク質をコードする遺伝子の1 もしくは複数の変異は、コードされる受容体の機能の少なくとも 1種の欠損を引き起こし、それにより受容体機能を減少または促進することができる。例えば、受容体の改変体を生じるまたは発現のレベルを変える変異は、受容体機能、受容体により媒介される減少または促進段階( 例えば、炎症過程) を減少または促進することができる。かかる変異の存在は、個々の細胞(例えば、活性化Tリンパ球などの白血球など)中またはかかる細胞から単離した受容体標品におけるの受容体もしくは受容体機能の存在を検出あるいは測定する方法を用いて決定されうる。これらのアッセイにおいて、受容体の減少したもしくは促進したレベルおよび/または減少したもしくは促進した受容体機能を評価しうる。
【0119】
本発明の核酸は、活性が減少または増大している受容体をコードする欠損哺乳類CXCR3 遺伝子のスクリーニング、特徴づけおよび/ または単離に使用できる試薬、例えば、プローブ、PCR プライマーなどを提供する。例えば、標準的な欠損遺伝子スクリーニング法を使用できる。欠損遺伝子を単離し、それらを適切な宿主細胞中で発現させて、哺乳類CXCR3 タンパク質について本明細書に記述するような評価をさらに行なうことができる。多くのヒト疾患は、G タンパク質共役受容体機能の欠損に関係している[Clapham, D. E., Cell, 75: 1237-1239 (1993); Lefkowitz, R. J., Nature, 365: 603-04 (1993)]。
【0120】
本発明の核酸は、(例えば、mRNAの転写を検出することにより)受容体の発現を(ノーザン解析、イン・サイチュハイブリダイゼーションにより)評価するためにも使用しうるプローブおよびPCRプライマーなどの試薬を提供する。例えば、活性化Tリンパ球または他の細胞型での発現を評価しうる。
【0121】
本発明の抗体は、受容体を細胞の表面に検出することができる手法への応用を有する。該受容体は、発現される白血球細胞型マーカー、特に活性化T細胞マーカーを提供する。例えば、受容体タンパク質またはペプチドに対して生じさせた抗体、例えば、本明細書に記載された抗体(例えば、mAb 1C6)を用いて、受容体を発現する細胞を検出および/または定量することができる。1つの態様として、抗体を用いて、細胞の混合物全体から受容体を発現する細胞を分類することができる(例えば、CD4 + T 細胞などの活性化T細胞を単離すること)。この目的には、細胞の計数および/ または選別に適した方法(例えばフローサイトメトリー、蛍光活性化細胞選別法)を使用できる。細胞数は、白血球細胞型(例えば、活性化T細胞)におけ増加または減少が観察される疾患または状態の診断に用いられうる。個体から得られた試料中の活性化T細胞の増加レベルの存在は、遅延型過敏反応、同種移植片拒絶、または細菌もしくはウイルス感染を含む病原性状態などの炎症性疾患または状態による浸潤の指標でありうる。
【0122】
さらに、抗体を用いて、受容体の発現を検出または測定することができる。例えば、本発明の抗体を用いて、試料中(例えば、血液、血清、白血球(例えば、活性化Tリンパ球)、気管支肺胞洗浄液、唾液、腸液などの組織もしくは体液)の受容体を検出または測定することができる。例えば、試料(例えば、組織および/または体液)を個体から得ることができ、適切な方法を用いて、CXCR3タンパク質の存在または量を評価することができる。適切な方法としては、FACS解析および酵素結合免疫吸着アッセイ(ELISA)などの免疫学的方法があげられ、化学発光アッセイ、放射免疫アッセイ、免疫組織学が含まれる。一般的に、試料および本発明の抗体は、抗体−受容体複合体の形成に適する条件下に組み合わせられ、抗体−受容体複合体の形成は、(直接的または間接的に)評価される。
【0123】
個体から得られた試料中の受容体反応性の増加レベルの存在は、同種移植片拒絶、遅延型過敏反応、または細菌もしくはウイルス感染を含む病原性条件などの炎症性疾患または状態が関連する炎症および/または白血球(例えば、活性化T細胞)浸潤および/または蓄積の指標でありうる。哺乳類CXCR3タンパク質または改変体の発現レベルを用いて、哺乳類CXCR3タンパク質の増加または減少した発現と特定の疾患または状態とを相関させることができ、哺乳類CXCR3タンパク質の増加または減少した発現が生じる(例えば、正常な個体での発現レベルなどの適切な対照に相対して増加または減少)疾患あるいは状態の診断において、用いることができる。同様に、治療の経過を、患者の試料中のCXCR3免疫反応性を評価することによりモニターすることができる。例えば、本発明の抗体を用いて、抗炎症性または免疫抑制剤で処置された患者由来の試料(血液、組織など)中にCXCR3を生じる細胞の数をモニターすることができる。
【0124】
トランスジェニック動物
組換えDNA 技術を用いて宿主動物のゲノムが改変されたトランスジェニック動物を構築することができる。1つの態様において、その改変が遺伝性でない(例えば骨髄中の先祖細胞などの体細胞を改変する)。別の態様において、その改変が遺伝性である(生殖細胞系を改変する)。トランスジェニック動物は標準的な技術または他の適当な方法で構築できる [例えば、T リンパ球の改変に関するCooke, M.P. ら, Cell, 65: 281-291 (1991); Hanahan, D., Science, 246: 1265-1275 (1989); Anderson, et al., 米国特許第5,399,346 号明細書] 。
【0125】
一側面として、内因性哺乳類CXCR3 遺伝子を適切な動物宿主中での完全に、もしくは部分的に(例えば遺伝子破壊技術によって)不活性化または無力化することによって、トランスジェニック動物を作製することができる。本発明の核酸を用いて、不活性化または無力化されたCXCR3 遺伝子を含有する宿主の構築の成功を(例えばサザンハイブリッド形成法によって)評価することができる。さらに、不活性化または無力化されたCXCR3 遺伝子を含有する宿主の構築の成功は、コードされている受容体の機能をモニターする適切なアッセイによっても評価できる。かかる動物を用いて、炎症および癌または病原体(ウイルス病原体)に対する宿主防御における受容体不活性化の影響を評価しうる。
【0126】
別の態様において、哺乳類CXCR3 タンパク質またはポリペプチドをコードする核酸を適当な宿主に導入してトランスジェニック動物を作製することもできる。好ましい態様では、そのトランスジェニック動物中に存在する内因性CXCR3 受容体遺伝子を、(例えば、該内因性遺伝子を破壊し置換する相同組換えによる該核酸の導入と同時に)不活性化する。例えば、白血球 [例えばリンパ球 (活性化T リンパ球など)、ナチュラルキラー細胞] 中で異なる哺乳類種(例えば、配列番号:1にコードされるCXCR3 などのヒトCXCR3] の哺乳類CXCR3 受容体をコードする核酸を発現できるトランスジェニック動物(マウス、モルモット、ヒツジなど)を作製することができ、導入された受容体の機能を評価するための便利な動物モデルを提供する。さらに、トランスジェニック動物に試験薬剤を投与し、受容体媒介過程 (例えば、炎症) に対する該薬剤の効果を本明細書に記載されたようにまたは適切なアッセイでモニターすることもできる。この方法で、受容体機能を阻害または促進する薬剤を同定したり、そのイン・ビボ効果を同定または評価することができる。
【0127】
治療方法
哺乳類CXCR3タンパク質に特有の少なくとも1種の機能の阻害または促進を介して、本発明の哺乳類CXCR3機能の調節は、受容体媒介機能を阻害又は促進する有効かつ選択的な方法を提供する。CXCケモカイン受容体は、主要な標的がリンパ球、特に活性化または刺激Tリンパ球およびNK細胞などのエフェクター細胞であるIP−10およびMigなどのケモカインに反応しうる、活性化リンパ球に選択的に発現するので、哺乳類CXCR3タンパク質は、ヒト等の哺乳類におけるリンパ球機能を選択的に妨害するまたは促進するための標的を提供する。一度リンパ球を部位に補充するやいなや、単球などの他の白血球型が二次的なシグナルで補充されるであろう。したがって、リガンド、阻害剤(例えば、1C6)および/または促進剤、本明細書記載のように同定されたものなどを含むCXCR3機能を阻害または促進する薬剤を用いて、白血球機能(例えば、補充および/または蓄積を含む白血球浸潤)、特にリンパ球の白血球機能を治療目的のために調整することができる。
【0128】
1側面として、本発明は、哺乳類CXCR3機能を阻害または促進する薬剤のかかる治療が必要な個体への投与を含む、かかる治療が必要な個体における炎症性応答を阻害または促進する方法を提供する。1つの態様として、哺乳類CXCR3タンパク質(例えば、ヒトCXCR3)の1以上の機能を阻害する化合物を投与して炎症を阻害(すなわち減少または妨げる)。例えば、mAb 1C6を含む本発明の抗体を本方法に用いることができる。結果として、白血球移動、走化性、エキソサイトーシス(例えば、酵素の)または炎症媒介放出などの1以上の炎症過程が阻害される。例えば、炎症部位の白血球浸潤(例えば、遅延型過敏応答)を本発明の方法にしたがって阻害することができる。
【0129】
他の態様として、哺乳類CXCR3タンパク質(例えば、ヒトCXCR3)の1以上の機能を促進する薬剤(例えば、受容体アゴニスト)を投与して、炎症過程の有益な刺激に帰する、白血球移動、走化性、エキソサイトーシス(例えば、酵素の)または炎症媒介放出などの炎症応答を誘導(誘発または促進する)する。例えば、ナチュラルキラー細胞をウイルス感染または腫瘍形成疾患に対抗するために補充することができる。
【0130】
「個体」という用語は、本明細書に規定され、特に限定されないが、霊長類(例えば、ヒト)、ウシ、ヒツジ、ヤギ、ウマ、イヌ、ネコ、ウサギ、モルモット、ラット、マウス、他のウシ属、ヒツジ、ウマ科、イヌ科、ネコ科、ネズミ、ハツカネズミの種などが挙げられる。炎症、感染および癌に関連する疾患および状態を本発明の方法を用いて処置することができる。好ましい態様として、疾患または状態は、リンパ球、特に活性化または刺激Tリンパ球およびナチュラルキラー(NK)細胞などのエフェクター細胞などの作用が治療(予防を含む)目的で阻害または促進されるべきものである。特に好ましい態様として、炎症疾患または状態は、T細胞媒介疾患または状態である。
【0131】
CXCR3機能の阻害剤で処置されうるヒトまたは他の種の慢性疾患を含む疾患または条件は、特に限定されないが:
・ 全身性過敏症または過敏応答、薬剤アレルギー(例えば、ペニシリン、セファロスポリンに対する)、昆虫咬傷アレルギー;クローン病などの炎症腸疾患、潰瘍性大腸炎、回腸炎および腸炎;膣炎;皮膚炎、湿疹、アトピー性皮膚炎、アレルギー性接触皮膚炎、蕁麻疹などの乾癬および炎症性皮膚病;血管炎(壊死性、皮膚および過敏性血管炎など)など;脊椎関節症;硬皮症;喘息、アレルギー性鼻炎、過敏性肺疾患、過敏性肺炎、間質性肺疾患(ILD )(特発性肺繊維症、関節リウマチに関連するILD 、または他の自己免疫状態)などの呼吸アレルギー疾患;などを含む炎症性またはアレルギー性疾患および状態;
・ 関節炎(例えば関節リウマチ、乾癬関節炎)、多発性硬化症、全身性エリテマトーデス、重症筋無力症、真性糖尿病および若年発症糖尿病を含む糖尿病、糸球体腎炎および他の腎炎、自己免疫甲状腺炎、ベーチェット病などの自己免疫疾患;
・ 同種移植片拒絶または対宿主性移植片病を含む(移植などにおける)移植片拒絶;
・ 望ましくない炎症応答を阻害すべきその他の疾患または状態、限定されないが、アテローム性動脈硬化症、サイトカイン誘導毒性、筋炎(多発性筋炎、皮膚筋炎)も治療できる
があげられる。
【0132】
CXCR3機能の促進剤(例えば、アゴニスト)で処置しうるヒトもしくは他の種の疾患または状態としては、限定されないが:
・ 皮膚T細胞リンパ腫(例えば、菌状息肉腫)などの癌、特に皮膚または組織の白血球浸潤を伴うもの;
・ 新形成疾患、網膜症(例えば、糖尿病性網膜症)、および黄斑変性を含む、脈管形成または新生血管形成が役割を果たす疾患;
・ 細菌感染および結核様らい、ならびに特にウイルス性感染などの感染性疾患;
・ 免疫抑制、例えばAIDSなどの免疫不全症候群にかかっている患者や、放射線療法、化学療法、あるいは免疫抑制を引き起こすその他の療法を受けている患者に認められるもの;受容体機能の先天的欠乏症または他の原因による免疫抑制など。さらに、CXCR3機能の促進剤は癌化学療法の間、幹細胞消耗に対抗するに有益な保護効果をも有する[Sarris, A.H. et al.,J.Exp.Med.,178:1127-1132(1993)]。
【0133】
投与経路
本発明によれば、1 以上の薬剤を単独で、あるいは別の薬物と組み合わせて、適当な経路で宿主に投与できる。薬剤(例えばリガンド結合を阻害する受容体ペプチド、抗CXCR3 抗体またはその抗原結合断片)の有効量を投与する。有効量とは、その投与条件下で所望の治療効果を達成するに足る量であり、例えばCXCR3 受容体機能の阻害または促進と、それによるそれぞれ、受容体媒介段階( 例えば、炎症性応答) の阻害または促進に十分な量である。
【0134】
種々の投与経路が可能で、例えば薬剤および治療しようとする状態または疾患に応じて、経口投与法、食餌法、局所投与法、腸管外投与法(静脈内注射、動脈内注射、筋肉内注射、皮下注射など)、吸入投与法(気管支内吸入、鼻孔内吸入または経口吸入、鼻孔内滴剤など)が挙げられるが、必ずしもこれらに限られるわけではない。喘息などの呼吸アレルギー性疾患には、吸入法が好ましい投与法である。
【0135】
投与すべき薬剤の製剤は、選択した投与経路に応じて変化するだろう(例えば液剤、乳剤、カプセル剤など)。投与すべき薬剤を含む適当な組成物を、生理学的に許容される賦形剤または担体中に調製することができる。例えば液剤や乳剤の場合は、適当な担体として、食塩水や緩衝培地などの水溶液、アルコール/ 水溶液、乳液、懸濁液などが挙げられる。腸管外用の賦形剤には、塩化ナトリウム溶液、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、乳酸化リンゲルまたは固定油などを含めることができる。静脈内用の賦形剤には、種々の添加物、保存剤、液体、栄養または電解物質補充剤などを含めることができる(一般的にはRemington's Pharmaceutical Science, 第17版,Mack 編, 1985を参照のこと)。吸入法の場合は、薬剤を可溶化し、適当な投与用ディスペンサー(アトマイザー、ネブライザーまたは加圧エアロゾールディスペンサーなど)に入れることができる。
【0136】
さらに、該薬剤が、タンパク質またはペプチドである場合、薬剤を組換えタンパク質のイン・ビボ発現を介して投与することができる。イン・ビボ発現は、適切な方法 (米国特許第5,399,346 号明細書などを参照のこと) に従って、体幹細胞発現を介してなし遂げられる。この態様において、該タンパク質をコードする核酸を運搬のためにレトロウイルス、アデノウイルスまたは他の適切なベクター( 好ましくは、複製欠損感染性ベクター) に取り込むことができ、運搬のタンパク質を発現することができるトランスフェクトまたは形質転換宿主細胞に導入することができる。後者の態様において、治療に有効な量でタンパク質を発現させるに有効な量で、該細胞を( 単独もしくは障壁装置中に) 埋め込み、注入し、さらに導入することができる。
【実施例】
【0137】
実施例
次に、下記の実施例によって本発明を例証するが、これらの実施例は決して限定を意図するものはない。
【0138】
ヒトケモカイン(実施例1〜2)
CXCケモカインMig、IL-8、GROα、NAP-2、GCP-2、ENA78、PF4、CCケモカインMCP-1、MCP-2、MCP-3、MCP-4、MIP-1α、MIP-1β、RANTES、I309、エオタキシン、およびケモカイン関連リンフォタクチンは、確立された手順に従って化学的に合成した(Clark-Lewis, I.ら, Biochemistry 30:3128-3135 (1991))。CXCケモカインIP-10はPeproTech社(ニュージャージー州ロッキーヒル)から購入した。
【0139】
実施例1.受容体cDNAのクローニング
標準的な分子生物学的技術を使用した(Sambrook, J.ら, 1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press,ニューヨーク州コールドスプリングハーバー)。
【0140】
推定されるTリンパ球限定的ケモカイン受容体をコードするDNA断片を、ポリメラーゼ連鎖反応(PCR)を使って作製した。2つの縮重オリゴヌクレオチドプライマーをケモカイン受容体の保存されたモチーフに基いて設計した。プライマーの設計は、ケモカイン受容体IL-8R1(CXCR1)、IL-8R2(CXCR2)、CC-CKR1 (CCR1)、CC-CKR2 (CCR2) およびオーファン受容体EBI I、LESTRおよびBLR1/MDR15の膜貫通ドメイン2(TM2)および膜貫通ドメイン7(TM7)内の保存されたヌクレオチド配列に基いた(EBI I, Birkenbach, M.ら, J. Virol., 67:2209-2220 (1993);LESTR, Loetscher, M.ら, J. Biol. Chem., 269:232-237 (1994);BLR1/MDR15,Dobner, T.ら, Eur. J. Immnol., 22:2795-2799(1992)ならびにBarella, L.ら, Biochem. J., 309:773-779 (1995))。
【0141】
プライマーの配列は次の通りとした。
配列番号:3
5’−GGG CTG CAG CII T(T/G)(T/G) C(C/A)G AC(A/C) TIC TI(C/T) T−3’
配列番号:4
5’−GGG TCT AGA IGG GTT IAI (G/A)CA (G/A)C(T/A) (G/A)(T/C)G−3’
(I=イノシン)。これらのプライマーをポリメラーゼ連鎖反応(PCR)で使用することにより、ヒト末梢血リンパ球から単離されたヒトゲノムDNAを鋳型として、次のようにDNA断片を増幅した。ヒトゲノムDNA 2μg、1×DynaZyme緩衝液(Finnzymes OY社,フィンランド・エスポー)、1.5mM MgCl2、500μMの各デオキシヌクレオチド、1μMの両プライマーおよびDynaZyme DNAポリメラーゼ2.5Uを含有する反応混合物100μlを、DNAサーマルサイクラー(Techne PHC-2, Brouwer社,スイス)での30サイクル(94℃で1分間、55℃で1分間、72℃で2分間)にかけた。予想されるサイズ(約700bp)のPCR産物をGene Scribe-ZベクターpTZ18/19 U/R(USB社,オハイオ州クリーブランド)にクローニングし、部分的に配列決定し(Sanger, F.ら, Proc. Natl. Acad. Sci. USA, 74 :5463-5467 (1977))、それらの既知ケモカイン受容体との類似性と、それらに対応するmRNAの白血球における発現について評価した。2MLC22と命名したDNA断片は、IL-8R2と64%のヌクレオチド配列同一性を示した。DNAポリメラーゼIのクレノウ断片と市販のランダムプライムラベリングキットを使用して放射活性同位体32Pで2MLC22を酵素により標識することによって調製したハイブリダイゼーションプローブを用いたノーザンブロット分析で評価したところ、断片2MLC22はT細胞由来のRNAに特異的にハイブリダイズしたが、単球または好中球由来のRNAにはしなかった。
【0142】
断片2MLC22を上述のように酵素により32Pで標識し、それをラムダ-ZAP Express(Stratagene社,スイス・チューリッヒ)中に調製されたヒト破傷風トキソイド特異的CD4+ T細胞(KT30)cDNAライブラリーをスクリーニングするためのプローブとして使用した(Loetscher, M.ら, J. Biol. Chem., 269 :232-237 (1994))。cDNAライブラリーは、ヒト破傷風トキソイド特異的CD4+ T細胞(KT30)由来のポリ (A)+ RNAを用いて、λZAP Expressシステム中に、その製造業者のプロトコール(Stratagene GMBH社,スイス・チューリッヒ)に従って調製した。得られたライブラリーは約1.1kbの平均インサートサイズを持つ約1.8×106個の独立クローンを含んでいた。プラークハイブリダイゼーションスクリーニングのため、約4×105クローンをBiodyneナイロン膜(PALL AG社,スイス・ムッテンツ)に転写し、ハイプライムDNAラベリングキット(Boehringer Mannheim社,ドイツ・マンハイム)を使って1×109dpm/μg DNAの比活性に標識しておいた2MLC22でプローブした。ハイブリダイゼーションは、1×106dpm 2MLC22/mlのハイブリダイゼーション溶液を使って、50%ホルムアミド、6×SSC、0.5% SDS、100μg/ml変性サケ精子DNA中42℃で20時間行なった。その膜を2×SSC/0.1%SDS中室温で10分間を1回、1×SSC/0.1%SDS中65℃で30分間を2回、最後に0.5×SSC/0.1%SDS中65℃で10分間を1回洗浄した。この高ストリンジェンシー洗浄の後、ハイブリダイゼーション陽性ラムダプラークから23個のクローンを単離し、最大のインサート(1670bp)を持つクローンを配列決定した。CXCR3 cDNAを市販のヌクレオチド配列決定用、ハイブリダイゼーションプローブ作製用、およびCXCR3を発現する安定にトランスフェクトされた哺乳類細胞クローン構築用のプラスミドベクターにサブクローニングした。それらCXCR3 cDNA含有構築物は大腸菌株中に維持される。
【0143】
結果
Tリンパ球特異的ケモカイン受容体を探索することにより、ヒトCD4+ T細胞ライブラリーからcDNAを単離した(図1、配列番号:1)。このcDNAは、汎用されている単球由来のcDNAライブラリーまたは顆粒球(HL60)由来のcDNAライブラリーを新規ケモカイン受容体cDNAに関して探索する過程では回収されなかった。しかしながら、CXCR3 cDNAに関して特異的なライブラリーの直接探索は行われていない。IP-10/Mig受容体(下記参照)をコードすることが明らかになったこのCXCR3 cDNAは、残基69から始まる1104bpのオープンリーディングフレーム(ORF)を持ち、これは40,659ダルトンの予想分子量を持つアミノ酸数368のタンパク質をコードする。そのアミノ酸配列(図2、配列番号:2)は、Gタンパク質共役受容体の特徴である7つの推定膜貫通セグメントと、3つの潜在的Nグリコシル化部位(Asn22、Asn32およびAsn199)を含む(図2)。また、受容体キナーゼの潜在的リン酸化部位(Palczewski, K.およびJ.L. Benovic, Trends Biochem. Sci., 16 :387-391 (1991);Chuang, T.T.ら, J. Biol. Chem., 267 :6886-6892(1992);Giannini, E.ら, J. Biol. Chem., 270 :19166-19172(1995))であるスレオニン残基1個とセリン残基9個が、細胞内COOH末端領域に認められる(図2)。
【0144】
この受容体の368アミノ酸配列(IP-10/MigR、図2、配列番号:2)を、IL-8R1(CXR1)、IL-8R2(CXCR2)、CC-CKR1 (CCR1)、CC-CKR2A (CCR2a)、CC-CKR3 (CCR3) およびCC-CKR4 (CCR4) を含む他のヒトケモカイン受容体のアミノ酸配列と整列した。多重タンパク質整列は、HigginsおよびSharp (Higgins, D.G.およびP.M. Sharp「Descriptionof the method used in CLUSTAL (CLUSTALで使用される方法の説明)」Gene, 73 :237-244 (1988))に従って行なった。図2の黒く囲んだ残基は、IP-10/MigRと他の少なくとも2つのケモカイン受容体の間で同一な領域を表わす。ハイフンは、整列中のギャップを表わす。この整列により、数個の保存されたモチーフが、具体的には膜貫通ドメインと第二細胞内ループ中に明らかになった。第三および第六膜貫通ドメインにはCXC受容体IL-8R1およびIL-8R2との有意な配列同一性が認められたが、CCケモカイン受容体との有意な配列同一性は認められなかった(図2)。
【0145】
この配列は、IL-8R1受容体およびIL-8R2受容体と全体でそれぞれ40.9%および40.3%のアミノ酸同一性を持ち、5種類の既知CCケモカイン受容体とは34.2〜36.9%の同一性を持つ(表1)。T細胞で発現されるがケモカイン類を結合しない七膜貫通ドメイン型受容体とは、より低い類似性が認められた(例えば、トロンビン受容体(Vu, T.-K.H.ら, Cell, 64 :1057-1068 (1991))とは27.2%同一)。図2の配列と整列することができる不完全なコード配列を持つ機能未確認の先端欠失型クローンは、過去にヒトゲノムDNAライブラリーから単離されている(Marchese, A.ら, Genomics, 29 :335-344 (1995))。
【0146】
【表1】
実施例2.生物学的活性
活性化Tリンパ球での発現
観察されたケモカイン選択性に着目して、白血球とそれに関連する細胞株でのIP-10/MigRの出現をノーザンブロット分析で調べた。新たに単離したヒト血液単球、好中球、リンパ球(PBL)、ナイロンウール精製T細胞、ならびに400U/ml hrIL-2の存在下に10日間培養(2mMグルタミン、1×非必須アミノ酸、1mMピルビン酸ナトリウム、100μg/mlカナマイシン、5×10-5 M 2-メルカプトエタノールおよび5%ヒト血清を含有するRPMI 1640培地中、1〜2.5×106細胞/ml)したクローン化ヒトCD4+ T細胞(KT30)およびCD8+ T細胞(ERCD8)、クローン化NK細胞(ERNK57)およびPBLを含む培養細胞由来の全RNA試料10μgを調べた。(ヒト組換えIL-2はバーゼル免疫学研究所(スイス・バーゼル)のA. Lanzavecchia博士から供与された)。ブロッティングに先立って、ゲル上の全RNAの完全性と量を調べるために、アガロースゲルを臭化エチジウムで染色した。RNA試料は、IP-10/MigR DNAの32P標識5'断片(109cpm/μg DNA)を5×106cpm/ml のハイブリダイゼーション溶液の濃度で使用して、記述されているように分析した(Loestscher, M.ら, J. Biol. Chem., 269 :232-237 (1994) )。ノーザンプローブとして使用した5'断片は、pBK-CMVベクター(Stratagene GMBH社,スイス・チューリッヒ)中のCXCR3 cDNAをPstIで消化してCXCR3 cDNA(図1)の5'末端724bpを得ることによって調製した。
【0147】
結果
予想されるサイズのmRNAの豊富な発現が、受容体cDNAの単離に使用したクローン化CD4+ T細胞KT30に認められた。同等レベルの発現がCD8+ T細胞クローンERCD8とNK細胞クローンERNK57に観察された。これに対し、新たに単離された血液リンパ球とナイロンウール精製T細胞には、IL10/MigR転写物はほとんど検出できなかった。しかし、これらの細胞をIL-2の存在下で培養すると、強いアップレギュレーションが得られ、受容体mRNAのレベルはTおよびNK細胞クローンでのレベルに近づいた。新たに単離された血液単球、好中性白血球または好酸性白血球では、これらの条件下にIP-10/MigR転写物が検出されなかった。IP-10/MigR mRNAを発現しなかったその他の白血球関連細胞には、マスト細胞株HMC-1、前骨髄球性白血病株HL60、組織球性白血病株U937、慢性骨髄性白血病株K562、急性T細胞白血病株Jurkat、急性リンパ芽球性白血病株Molt、Bリンパ芽球株DaudiおよびRaji、慢性および急性Bリンパ性白血病(B-CLLおよびB-ALL)患者由来のリンパ球、好塩基球性白血病患者由来の成熟好塩基球、および赤白血病細胞株HELが含まれる。対照的に、単球および顆粒球には、リンパ球を誘引することが過去に示されているケモカイン、すなわちMCP-1、MCP-2、MCP-3、MIP-1α、MIP-1βおよびRANTESの受容体(Loetscher, P.ら, FASEB J., 8 :1055-1060 (1994);Carr, M.W.ら, Proc. Natl. Acad. Sci. USA 91 :3652-3656(1994);Taub, D.D.ら, Science, 266 :355-358 (1993);Schall, T.J.ら, J. Exp. Med., 177 :1821-1825(1993);Schall, T.J.ら, Nature, 347 :669-672(1990))も認められる。活性化Tリンパ球とナチュラルキラー細胞株におけるIP-10/MigRの限定的発現は、この新規受容体が選択的なリンパ球補充を媒介しうることを示唆している。
【0148】
安定なトランスフェクタント
CXCR3 cDNAをpBK-CMV (Stratagene GMBH社,スイス・チューリッヒ)からBamHIとXbaIによる消化で切り出し、pcDNA3(Invitrogen BV社, オランダ・WBリーク)のBamHI部位とXbaI部位にクローニングすることによりpCDNA3-Clone8を得て、それを大腸菌(XL1Blue)で維持、保存した。
【0149】
安定なトランスフェクタントを作製するため、4×106個のマウスプレB細胞 (300-19) (Thelen, M.ら, FASEB. J., 2 :2702-2706 (1988))、ヒト前骨髄球細胞 (GM-1) (Garotta, G.ら, J. Leukocyte Biol., 49 :294-301 (1991))またはヒト急性T細胞白血病細胞 (Jurkat) (Loetscher, P.ら, FEBS Lett. 341 :187-192 (1994))に、Bgl IIで直線化したpcDNA3中の受容体cDNA 20μgを、既に記述されているようにエレクトロポレーション法でトランスフェクトした(Moser, B.ら, Biochem. J., 294 :185-192 (1993))。
【0150】
IP-10/MigRトランスフェクト細胞をG-418 (Life Technologies社)選択下に限界希釈法でクローン化した(300-19については1.0mg/ml G-418、Jurkat細胞とGM-1細胞については0.8mg/ml G-418)。RNAドット-ブロット分析により、G-418耐性クローンを受容体発現についてスクリーニングした。
【0151】
Ca2+流動
本受容体が機能的であるかどうかを決定するため、ネズミプレB細胞(300-19)、ヒト前骨髄球細胞(GM-1)およびヒトT細胞白血病細胞(Jurkat)のクローンに、上述のように受容体cDNAを安定にトランスフェクトした。ケモカイン受容体の活性化はサイトゾル遊離Ca2+濃度([Ca2+]i)の一過性の上昇をもたらす。そこでトランスフェクト細胞におけるシグナル伝達のモニターにこのアッセイを使用した。
【0152】
136mM NaCl、4.8mM KCl、1mM CaCl2、5mMグルコースおよび20mM HEPES(pH7.4)を含有する緩衝液中、106 細胞あたり0.1nmolのfura-2アセトキシメチルエステルと共に37℃で30分間インキュベートすることによってfura-2を負荷した細胞で、サイトゾル遊離Ca2+濃度([Ca2+]i)の変化を測定した。遠心分離後、負荷した細胞を同じ緩衝液に再懸濁し(106細胞/ml)、37℃で表記のケモカインによって刺激し、[Ca2+]i関連蛍光変化を記録した(von Tscharner, V.ら, Nature, 324 :69-372 (1986))。
【0153】
結果
IP-10およびMigに応答して、迅速な[Ca2+]i上昇が観察された。ケモカインIP-10は皮膚の遅延型過敏反応において発現されることが示されている(Luster, A.D.ら, Nature, 315 :672-676(1985);Kaplan, G.ら, J. Exp. Med., 166 :1098-1108(1987))。Migと呼ばれるケモカインが最近になって同定された(Farber, J.M., Proc. Natl. Acad. Sci. USA, 87 :5238-5242 (1990);Farber, J.M., Biophys. Res. Commun., 192 :223-230 (1993))。どちらのケモカインも、IL-8と同様に最初の2つのシステインがCXC配置を取っているが、好中性白血球に対して走化作用を持たない。最近、IP-10がTリンパ球を誘引すること(Luster, A.D.およびP. Leder, J. Exp. Med., 178 :1057-1065(1993);Taub, D.D.ら, J. Exp. Med., 177 :1809-1814(1993))と、Migが腫瘍関連リンパ球に対して走化作用を持つこと(Liao, F.ら, J. Exp. Med., 182 :1301-1314 (1995))が報告された。
【0154】
上記cDNAをトランスフェクトされ機能的IP-10/MigRを発現する細胞に対するIP-10とMigの作用を、図3A〜3Cに要約する。[Ca2+]i変化(図3A)によって示されるように、IP-10とMigの作用は濃度依存的であり、1nMで既に検出可能であることから、両ケモカインはこの新規受容体に対して高い親和性を持つことが示される。対照的に、これらIP-10/MigRトランスフェクタントは、CXCケモカイン類IL-8、GROα、NAP-2、GCP-2、ENA78、PF4、CCケモカイン類MCP-1、MCP-2、MCP-3、MCP-4、MIP-1α、MIP-1β、RANTES、I309、エオタキシン、またはケモカイン関連リンフォタクチンを含む他の16種類の潜在的アゴニストのいずれにも、100nMまでの濃度で反応しなかった(非掲載)。同じ結果がネズミとヒトのトランスフェクト細胞で得られた。これらの知見は、この新規受容体がIP-10とMigに高度に選択的であることを証明している。したがって、本明細書では本受容体をIP-10/Mig受容体(IP-10/MigR)もしくはそのCXCケモカインに対する特異性を反映して「CXCR3」と呼ぶ。
【0155】
図3Bに示すように、IP-10またはMigによる反復刺激は、ケモカイン受容体に典型的な脱感作をもたらした。さらに、これら細胞をIP-10で刺激した後、Migで(またはその逆で)刺激すると交差脱感作が起り、この受容体が両ケモカインに対して高い親和性を持つことが確認された。100nMの濃度で、Migが交差脱感作に関してIP-10よりも強力であることが明白となり、Migに対するIP-10/Mig受容体のより高い親和性または結合安定性が示唆された。
【0156】
機能的IP-10/MigRの発現は証明されたが、放射活性リガンドを用いて結合実験を行なったところ、結合パラメーターの決定を妨げる、全体の60〜80%に及ぶ非特異的結合が明らかになった。IP-10とMigはカチオン性が高い(pI値は10.8と11.1)ので、細胞表面プロテオグリカンとの非特異的相互作用がこれらの結果の説明となりうる。実際、種々の血液細胞と組織細胞には、ケモカイン受容体と無関係なヘパリナーゼ感受性のIP-10(およびPF4)結合部位が検出されており(Luster, A.D.ら, J. Exp. Med., 182 :219-231 (1995))、またヘパラン硫酸はIP-10とMigを結合し、リンパ球走化性を妨げる(非掲載)。ヘパリン結合部位はおそらくCXCR3受容体結合には関与しないと思われ、ヘパリン結合部位を介した細胞への非特異的結合を阻害するには、反応(例えば結合緩衝液)へのコンドロイチン硫酸などの適当なヘパリン誘導体の包含を使用できる。
【0157】
走化性
献血血液バフィーコートからPBLを新たに単離した。献血血液バフィーコートはスイス赤十字社スイス中央研究所輸血部門によって提供された。バフィーコートPBLの単離はColotta, F.ら, J. Immunol., 132 :936-944 (1984) に記述されているように行なった。
【0158】
献血血液バフィーコートから新たに単離されたPBLをさらに処理することなく使用するか、IL-2の存在下に10日間培養(2mMグルタミン、1×非必須アミノ酸、1mMピルビン酸ナトリウム、100μg/mlカナマイシン、5×10-5 M 2-メルカプトエタノールおよび5%ヒト血清を含有するRPMI1640培地中、400U/ml hrIL-2の存在下に、1〜2.5×106細胞/ml)した後、使用した。
【0159】
細胞移動は、IP-10/MigRトランスフェクト細胞については孔径5μm (Loetscher, P.ら, FEBS Lett. 341 :187-192 (1994))、ヒトPBLについては孔径3μm (Loetscher, P.ら, FASEB J., 8 :1055-1060(1994))のポリビニルピロリドン非含有ポリカーボネート膜 (Nucleopore)を用いて、48ウェルチャンバー (Neuro Probe社, 米国メリーランド州キャビンジョン)で評価した。ケモカイン(下部ウェル)の溶解と、細胞(上部ウェル中100,000個の受容体トランスフェクタントまたはPBL)の希釈には、20mM HEPES (pH7.4)と1%低温殺菌血漿タンパク質溶液(スイス赤十字社研究所,スイス・ベルン)を添加したRPMI1640を使用した。37℃で60分後、膜を取り出し、上面をPBSで洗浄し、固定し、染色した。全てのアッセイを3連で行ない、移動した細胞を倍率1,000倍の無作為に選択した視野5つで数えた。自発的な移動は、化学誘引物質の非存在下で決定した。
【0160】
結果−トランスフェクト細胞
IP-10/MigRを発現するトランスフェクト細胞はIP-10またはMigに向かってたやすく移動し、一方、非トランスフェクト親細胞は応答しなかった(図3C)。どちらのアゴニストも典型的な二相型濃度依存性を示した。IP-10は1nMを超える濃度で移動を誘導し、一方、Migの応答は10nM以上で検出可能になった。移動細胞の最大数によって測定される効力は、Migの方がIP-10の約2倍高かった。これらの結果はIP-10/MigRが、白血球のすべての既知ケモカイン受容体と同様に、リガンドに応答して起こる走化性を媒介することを示している。
【0161】
結果−ヒト血液白血球
IP-10/MigRの細胞分布と合致して、活性化ヒトTリンパ球はIP10とMigに対して高い反応性を持つことがわかった(図4A〜4B)。[Ca2+]i変化(図4A)とイン・ビトロ走化性(図4B)の誘導因子としてのIP-10とMigの活性は、IP-10/MigRを発現するトランスフェクト細胞を用いて観察された作用と合致し、IP-10はMigより強力であるが、効力はMigより低くかった。カルシウム流動と走化性の誘導にはIL-2の存在下で培養することによるTリンパ球の活性化が必要であり、単離したばかりの血液リンパ球には使用した条件下で応答が認められなかった。
【0162】
実施例3〜9に関する材料と方法
実施例3〜9では次の材料と方法を使用した。
【0163】
ケモカイン
組換えヒトケモカイン類は、イアン クラーク−ルイス(Ian Clark-Lewis)博士から提供された既に記述されているエオタキシン(Ponath, P.D.ら, J. Clin. Invest., 97 :604-612 (1996);Ponathら, 国際公開第97/00960号パンフレット(1997年1月9日公開)も参照されたい)以外は、Peprotech社(ニュージャージー州ロッキーヒル)から入手した。125I標識ケモカイン類はDu Pont NEN社(マサチューセッツ州ボストン)から入手した。
【0164】
細胞と細胞株
好中球とPBMCは記述されているように単離した(Ponath, P.D.ら, J. Clin. Invest., 97 :604-612 (1996))。CD3芽球を生成するため、RPMI-1640+10%FCS中2×106個/mlのPBMCを抗CD3抗体TR66でコーティングしてある組織培養プレートに添加した。4〜6日後、芽球を新しい培地に取り出し、IL-2(Antonio Lanzavecchia(バーゼル)の厚意で提供されたもの)を50単位/mlの濃度で添加した。
【0165】
使用した他の細胞株には、CXCR3(下記参照)、IL-8RA(Ponath, P.D.ら, J. Exp. Med., 183 :2437-2448(1996))、IL-8RB(Ponath, P.D.ら, J. Exp. Med., 183 :2437-2448(1996))、CCR2b (G. LaRosa, 未公表)、CCR4、CCR5(Wu, L.ら, Nature, 384 :179-183(1996))、またはCCR1(Campbell, J.J.ら, J. Cell. Biol., 134 :255-266(1996))のいずれかを発現するL1.2ネズミプレB細胞リンパ腫のトランスフェクタントが含まれる。
【0166】
CXCR3トランスフェクタントの調製
細胞
L1.2細胞は、RPMI培地1640、10% FetalClone (Hyclone社製)、50U/mlペニシリン/ストレプトマイシン、1×L-グルタミン、1mMピルビン酸ナトリウムおよび5.5×10-5 Mβ-メルカプトエタノール中で生育した。培地成分は、Hyclone社から購入した10% FetalClone以外は、GibcoBRL社から購入した。トランスフェクションの2日前に、L1.2細胞を新しい培地に1:5の比率で希釈した。これにより、約100〜300万細胞/mlの濃度の対数増殖期の細胞1億5000万個が得られた。
【0167】
CXCR3 DNAとトランスフェクション
大腸菌XL1Blue細胞(Stratagene社,カタログ番号200236)を、製造者の実験案に従い、pcDNA3-Clone8(実施例2;Loetscher, M.ら, J. Exp. Med., 184 :963-969(1996))で形質転換した。100μg/mlアンピシリンを含有するLB 500ml中、250rpmで振盪しながら、形質転換体を37℃で生育した。次にその培養物を8000×gでの遠心分離によって集め、Maxiプラスミド精製カラムとその実験案(Qiagen社,カタログ番号12162)を使って、プラスミドを精製した。プラスミドの濃度と純度は、1%アガロースゲルとOD260/280比を使って決定した。プラスミドDNAを二回蒸留水に懸濁し、使用時まで−20℃で保存した。
【0168】
ベクターの直線化にはScaIエンドヌクレアーゼを使用した。100μgのDNAを10μlのScaIで、製造者の実験案(GibcoBRL,カタログ番号15436-017)に従って37℃で8時間消化した。20μgを安定トランスフェクション構築に直接使用した。80μgはフェノール:クロロホルム:イソアミルアルコール(25:24:1)抽出、100%エタノール沈殿(0.1体積のNH4COOHを使用)および70%エタノール洗浄により、タンパク質と塩を除去した。
【0169】
ネズミプレBリンパ腫細胞株(L1.2)の安定トランスフェクタントは基本的に記述されているように調製した(Ponath, P.D.ら, J. Exp. Med., 183 :2437-2448(1996))。1×PBS 0.8ml中のL1.2細胞2500万個を、直線化したDNA 20μg、または直線化の後、洗浄したDNA 20μg(上記DNAの直線化の項を参照されたい)と共に、またはDNAなしで、エレクトロポレーションにかけた。エレクトロポレーション前に、L1.2細胞とDNAを50mlコニカルチューブ(Falcon 2070型)中で2分おきに穏やかに混合(旋回)しながら10分間インキュベートした。そのL1.2細胞-DNA混合物を、電極間隔0.4cmのGene Pulserキュベット(BioRad社,カタログ番号165-2088)に移した。次にその混合物を250V、960μFで、ショックの持続時間と実際の電圧を測定しながらエレクトロポレーションした。エレクトロポレーション後、キュベットを10分間室温に放置した。次にL1.2細胞-DNA混合物のすべてをT-25フラスコ(Costar社)に移し、10mlの非選択培地で2日間生育した。
【0170】
選択
次にCXCR3を発現するL1.2細胞をネオマイシン耐性に関する選択にかけた。非選択培地で2日間生育した後、10mlの1.6g/Lジェネティシン(GibcoBRL社)を0.8g/L(選択および維持濃度)の最終濃度で加えた。次にこれを、細胞が過剰増殖を始めたら新しい選択培地を加えながら、10〜15日間生育した。新しい選択培地は10%ウシ血清と800μg/ml G418 を添加したRPMI-1640からなった。
【0171】
CXCR3の細胞表面発現を走化性によって評価し、また表面発現をモニターするためにリガンド結合分析とスキャッチャード分析も使用した。G418選択後、CXCR3発現L1.2細胞を走化能に基いて選択した。各エレクトロポレーション反応について、30ml(800,000細胞/ml)を集め、600μlの選択培地に懸濁した。10nM IP-10を含有する選択培地600μlを、Becton-Dickinson社製のBioCoat細胞培養プレート(カタログ番号40575)の下部チャンバーに入れた。100μl/ウェルの上記L1.2細胞をそのBioCoatプレートの上部チャンバーに加えた。次に、それらの細胞を37℃のCO2培養器中に終夜放置して走化反応させた。非走化細胞の入った上部チャンバーを除去した。走化細胞を集め、新しい培地に移し、24ウェルプレートで生育した。次に、それらをCostar製T-25フラスコに拡張した後、T-75フラスコに拡張した。
【0172】
高レベルの受容体を発現するトランスフェクタントを、限界希釈法によってクローン化した。G418を含む選択培地に、CXCR3トランスフェクト細胞を30〜3細胞/mlまで希釈した。それを96ウェル組織培養プレートに100μl/ウェルずつ分注した。37℃、5%CO2で14日後、単一コロニーを含有するウェルを倒立顕微鏡下で同定した。次にその細胞の50μlを移し、抗CXCR3 mAb で上述のように染色し、フローサイトメトリーで分析した。受容体発現レベルは平均蛍光強度と相関し、高発現体が選択された。安定な細胞株が樹立されたら、その株を使用するために増殖した。
【0173】
CXCR3トランスフェクタント細胞は、抗生物質耐性と走化性に関する選択に加えて、抗体染色法で受容体発現量がより高いものを分別することによって、さらに選択することができるが、ここではそれを行なわなかった。染色するには、1%ウシ血清アルブミンを含む滅菌PBSにトランスフェクタント細胞を5×106個/mlの密度で再懸濁すればよい。単離された滅菌抗CXCR3 mAb は3μg/mlまでの最終濃度で加えることができ、細胞を氷上で30分間インキュベートする。冷滅菌PBSで洗浄した後、結合したmAb は、0.2μmフィルターを通した濾過によって滅菌したFITC結合抗マウスIgGで検出できる。それらの細胞を再び洗浄し、フローサイトメトリーで分別することができる。上位5%の陽性細胞を集め、選択条件下に増殖用の組織培養(例えば10%ウシ血清と800μg/ml G418を添加したRPMI-1640)に戻すことができる。
【0174】
実施例3.IP-10はCXCR3 DNAをトランスフェクトしたL1.2細胞上と活性化T細胞上に発現した受容体に高い親和性で結合する
放射標識IP-10を使用してさらなる結合試験を行なった。標的細胞へのケモカイン結合は既に記述されているように行なった(Ponath, P.D.ら, J. Clin. Invest., 97 :604-612 (1996);Van Riper, G.ら, J. Exp. Med., 177 (3): 851-856(1993))。細胞をPBSで1回洗浄し、結合緩衝液(50mM HEPES(pH7.5)、1mM CaCl2、5mM MgCl2、0.5%BSAおよび0.05%アジド)に107個/mlの濃度で再懸濁した。それを微量遠心管に50μl(5×105細胞)ずつ分注し、次いで非放射性競合因子(非標識IP-10)と放射標識ケモカイン(0.05nM 125I標識IP-10)を加えた。最終反応体積は200μlだった。非特異的結合は、250〜500nMの非標識ケモカインの存在下に細胞を放射標識ケモカインと共にインキュベートすることによって決定した。室温で60分間インキュベートした後、0.5M NaClを含む結合緩衝液1mlで細胞を3回洗浄した。次に、細胞ペレットをカウントした。競合は100×[(S-B)/(T-B)] (式中、Sは試料の放射活性、Bはバックグラウンド結合量、Tは競合因子がない場合の全結合量である)によって計算される特異的結合百分率として表わした。バックグラウンド結合量は、細胞を放射標識ケモカインおよび少なくとも400倍過剰の非標識ケモカインと共にインキュベートすることによって得た。全実験を通じて2連を使用し、標準偏差は常に平均の10%未満だった。すべての実験を少なくとも3回繰返した。カーブフットおよび特異的結合を50%阻害する濃度(IC50)は、カレイダグラフ(KaleidaGraph)ソフトウェア(Synergy Software社,ペンシルバニア州レディング)で計算した。
【0175】
図5A〜5Bは、CXCR3をトランスフェクトしたL1.2細胞(図5A)とCD3活性化T細胞(図5B)に125I-標識IP-10が結合したことと、非放射性IP-10の濃度を上げることでその結合を阻害できたことを示している。スキャッチャード分析から、IP-10がL1.2 CXCR3トランスフェクタントに614pMのKdで結合することと、それらのトランスフェクタントが1細胞あたり37,000個の受容体を発現させたことが明らかになった(図5A、挿入図)。抗CD3活性化IL-2刺激T細胞の同様の分析では、156pMのKdと1細胞あたり17,000個の受容体が明らかになった(図5B)。Migは125I標識IP-10結合の遮断に関して非標識IP-10よりわずかに有効性が低かったものの(非掲載)、活性化T細胞への125I標識IP-10結合は同じ条件下に非標識Migによって完全に阻害することができた。IP-10とMigは高い親和性でCXCR3を結合する(Kd約150〜600pM)。
【0176】
実施例4.CXCR3に特異的なモノクローナル抗体(mAb)の作製と特徴づけ
CXCR3のアンタゴニストを開発するために、また受容体の発現と調節を研究するために、この受容体のN末端に対応する合成ペプチドでマウスを免疫することにより、一群のmAb を作製した。これらのmAb はCXCR3トランスフェクタントを特異的に認識したが、他の一連の受容体トランスフェクタントは特異的に認識しなかった。
【0177】
mAb の作製とフローサイトメトリー
CXCR3と反応するmAb は、Balb/CマウスをCXCR3の最初の37N末端アミノ酸に対応する37マー合成ペプチド(Loetscher M.ら, J. Exp. Med., 184 :963-969(1996)をも参照されたい)10μgで10週間にわたって5回免疫することによって作製した。このペプチドを合成し、ツベルクリンの精製タンパク質誘導体(Severn Biotech社,英国キッダーミンスター)に結合した。最初の免疫処置はフロイント完全アジュバント(FCA)を使って腹腔内(IP)に行なった。第二、第三、第四の免疫処置はフロイント不完全アジュバント(FIA)を使って腹腔内に行い、最後の免疫処置はペプチド複合体のみ(アジュバントなし)を使って、静脈内(IV)に投与した。最後の免疫処置の4日後に、脾臓を摘出し、細胞株SP2/0を用いて、記述されているように細胞融合を行なった(Coligan, J.E.ら, Current Protocols In Immunology (John Wiley and Sons社,ニューヨーク), Unit 2.5.4 (1992))。ELISAで評価したところ、上記N末端37マーペプチドと反応するmAb が生成した(Coligan, J.E.ら, Current Protocols In Immunology (John Wiley and Sons社,ニューヨーク), Unit 2.1.3 (1992) )。CXCR3と反応するmAb は、非トランスフェクトおよびCXCR3トランスフェクトL1.2細胞または300.19細胞(ネズミB細胞株, Loetscher, M.ら, J. Exp. Med., 184 :963-969(1996))、免疫蛍光染色およびFACScan(登録商標) (Becton Dickinson & Co.社, カリフォルニア州マウンテンビュー)を用いて同定した。
【0178】
CXCR3に特異的なモノクローナル抗体が生成した。CXCR3トランスフェクトL1.2細胞が染色されるが、非トランスフェクトL1.2細胞または他の受容体タイプをトランスフェクトしたL1.2細胞が染色されないことから判断して、8種類のmAb が表面発現したCXCR3を認識することがわかった。これらmAb の一つ1C6のFACSプロフィールを図6に示す。また、これらmAb はPHAまたは抗CD3によりイン・ビトロで活性化されたヒトT細胞とT細胞クローンをも染色した(1C6について図7Cに図示)。しかし、これらの抗CXCR3 mAb は、好中球(1C6について図7Aに図示)、単球または好酸球(非掲載)とは反応しなかった。この反応性のパターンは、ノーザンブロットによる分析と合致した。しかし、意外にも表現型分析では循環リンパ球の大きな亜集団でのCXCR3の発現が明らかになった(図7B)。この発現パターンは調査した全ての個体で観察され、このことはCXCR3が血液リンパ球の亜集団に常態で発現されることを示す。CXCR3は、CD45RO+(メモリー)サブセットに包含される循環T細胞の一集団を特徴づけることがわかった。
【0179】
ネズミハイブリドーマ1C6(LS77-1C6ともいう)は、1997年3月28日に、ブダペスト条約の条件に基き、LeukoSite社(米国02142マサチューセッツ州ケンブリッジ・ファーストストリート215)の名でアメリカン・タイプ・カルチャー・コレクション (American Type Culture Collection) (20852 メリーランド州ロックビル・パークローンドライブ12301)に、ATCC寄託番号HB-12330の下で寄託された。
【0180】
実施例5.CXCR3は活性化/メモリーT細胞上に発現される
フローサイトメトリー
CXCR1、CXCR2、CXCR3およびCCR5に対するmAb は既に記述されている(Qin, S.ら, Eur. J. Immunol., 26 :640-647 (1996);Heath, H.ら, J. Clin. Invest. 印刷中(1997))。抗CXCR4 mAb 12G5(Endres, M.J.ら, Cell, 87 :745-756 (1996))は、ジム ホキシー (Jim Hoxie)(ペンシルバニア大学)の厚意で提供された。CD4、CD8、CD14、CD20、CD25、CD26、CD69、CD45RO、CD45RA、CD95に対するPE結合mAb 、抗CD3および抗CD4 Cy-ChromeはPharMingen社(カリフォルニア州ラホーヤ)によって供給された。
【0181】
トランスフェクト細胞または白血球に対するmAb の反応性を評価するために、間接免疫蛍光法とフローサイトメトリーを使用した。細胞をPBSで1回洗浄し、2%ヒト血清と0.1%アジ化ナトリウムを含む100μlのPBS(染色緩衝液)、5μg/ml精製抗体、5μg/ml IgG2a同イソタイプ対照mAb (Sigma Chemical社, ミズーリ州セントルイス)または50μlのハイブリドーマ培養上清に再懸濁した。4℃で20分後、細胞を染色緩衝液で2回洗浄し、50μlのFITC結合アフィニティー精製F(ab')2 ヤギ抗マウスIgG (Jackson ImmunoResearch Laboratories社)に再懸濁した。4℃で20分間インキュベートした後、細胞を染色緩衝液で2回洗浄し、表面発現レベルを決定するためFACScan(登録商標)で分析した。死細胞の排除にはヨウ化プロピジウムを使用した。
【0182】
リンパ球の二色免疫蛍光分析は、CD20+(B)細胞とCD56+(NK) 細胞のごく一部もCXCR3を発現するものの、CXCR3を発現する細胞はそのほとんどがCD3+細胞であることを示した(図8)。T細胞を標識するために抗CD3 Cy-Chromeを用いて行なったT細胞の三色分析は、CD4+細胞の一部とCD8+細胞の一部がCXCR3を発現することを示した(図9)。CD25やCD69などの細胞活性化のマーカーを使った分析では、活性化T細胞が一般にこの受容体を発現することが明らかになった。CXCR3+ T細胞は、先の活性化に合致する表現型CD95+、CD45RO+およびCD45RAlowだった。CXCR3の発現と、やはり前もって活性化されたT細胞にその発現が偏っているケモカイン受容体CCR5(Wu, L., 私信)の発現も比較した。図9は、血中のCCR5+細胞がCXCR3+亜集団に包含されていたことと、CXCR3がCCR5より広範に発現されたことを示している。CCR5やCXCR4などの他のT細胞ケモカイン受容体とは異なり、CXCR3は循環する活性化T細胞の大半で発現した。
【0183】
実施例6.抗CXCR3 mAb はIP-10結合と走化性を遮断する
ヒト白血球の走化性を、ECV 304内皮細胞株(European Collection of Animal Cell Cultures, 英国ソールズベリー・ポートンダウン)を使って、既に記述されている(Ponath, P.D.ら, J. Clin. Invest., 97 :604-612 (1996))経内皮アッセイ(Carr, M.W.ら, Proc. Natl. Acad. Sci. USA, 91(9): 3652-3656 (1994))の変法で評価した。下部チャンバーに移動した細胞をチューブに入れ、相対細胞数をFACScan(登録商標)を用いて得た。
【0184】
また、CD3活性化T細胞芽球の走化性を阻害する能力について、抗ペプチドmAb を試験した。1A5、1C6、3A8、5F10、10C6および10G12と呼ばれるmAb に関する結果を図10に示す。mAb の一つ1C6は、IP-10へのT細胞の走化性を遮断する能力が他のmAb より優れていた。mAb 1C6 は、IP-10へのT細胞の走化性を約0.8μg/mlのIC50で用量依存的に完全に阻害することができた(図11)。トランスウェルの底に至適濃度のIP-10(12.5nM)を使用すると、2〜5μg/mlの濃度で100%阻害が達成された。使用した条件下で1C6は、ケモカイン受容体CCR2bを介して起こるMCP-1へのT細胞走化性を有意に阻害できなかった(図10)。
【0185】
またmAb 1C6 は活性化T細胞への125I標識IP-10結合を0.16μg/mlのIC50で完全に遮断することができた(図12)。1〜10μg/mlの抗体で完全な阻害が得られた。mAb 1C6 によってIP-10結合と走化性の両方が完全に阻害されることは、活性化T細胞がこのケモカインを結合する別の受容体を発現しないことを示している。
【0186】
ノーザンブロット分析は、CXCR3が活性化T細胞で発現されることを示した。一群の特異的mAb を使用することにより、CXCR3は血液T細胞の亜集団と他の白血球タイプ(B細胞およびNK細胞)に発現されることがわかった。CXCR3を発現するT細胞は先の活性化と合致した表現型、すなわちCD45RO+、CD26+を有した(実施例5)。T細胞の染色は、T細胞がCD3とIL-2で活性化されると著しく増大し、それはIP-10に応答して起こる細胞移動および放射標識リガンド結合量の増大と相関した。
【0187】
血液T細胞が少なくとも走化性アッセイでIP-10またはMigに対して低い応答性を示すことは変則的であるように思われる。その理由として、受容体発現以外の要因が化学誘引物質に対する細胞の応答性を決定しうることが考えられる。シグナル伝達には適当なGタンパク質共役が必要かもしれない。もう一つの可能性は、T細胞上とNK細胞上のIL-8受容体について観察されているように(C.R. Mackay)、血液分離操作がこの受容体を破壊することである。適当な実験動物の皮膚へのIP-10の注射により、血液T細胞でのCXCR3発現の意義を検討できる。活性化T細胞では、IP-10が最も強力な化学誘引物質の一つである。T細胞の活性化は、受容体シグナル伝達分子またはシグナル伝達に必要な共役を誘導しうる。
【0188】
実施例7.mAb 1C6 はIP-10に応答したT細胞による[Ca2+]iを選択的に阻害するが、Migに応答したT細胞による[Ca2+]iは阻害しない
細胞内カルシウム濃度([Ca2+]i)は次のように測定した.Fura-2 AM (Molecular Probes社, オレゴン州ユージーン)の原液は、その色素50μgを44μlのDMSOに溶解することによって調製した。細胞に添加する直前に、その原液をCa2+とMg2+を含むHBSSおよび2%BSAに1:100に希釈した。Fura-2 AMを0.2モル/106細胞の最終濃度で37℃で30分間細胞に添加した。標識化の後、過剰の色素を遠心分離によって除去し、細胞を125mM NaCl、5mM KCl、1mM MgCl2、1mM CaCl2、0.5mMグルコース、0.025%BSAおよび20mM HEPES(pH7.4)に106個/mlの濃度で再懸濁した。[Ca2+]iは日立F-2000蛍光分光計で340nmと380nmでの励起を使って測定した。較正は、全放出に1%NP-40を使用し、遊離Ca2+のキレートに25μM EGTAを使用して行なった。
【0189】
IP-10とMigはヒトT細胞による[Ca2+]iを誘導し、各ケモカインは他方のケモカインに対する応答を完全に脱感作することができた(図13A〜13B)。ケモカインの滴定により、最大[Ca2+]iがわずか2nM IP-10または2nM Migで達成されることが明らかになった。mAb 1C6 のアゴニスト/アンタゴニスト機能を調べるため、mAb 1C6 または無関係な同イソタイプの対照mAb 注入後に、活性化T細胞を[Ca2+]iについて評価した。無関係な同イソタイプ対照mAb を注入されたT細胞は、それに続くIP-10の注入に対して強い[Ca2+]i応答を示した(非掲載)。しかし、mAb 1C6 で処理されたT細胞は、2nM IP-10で刺激しても[Ca2+]iを示さなかった。IP-10に応答した活性化T細胞における[Ca2+]iは、12.5mgの1C6によって完全に抑制された。しかし50μg/mlの1C6でも、2nM Migに対するT細胞の応答には影響がなく(図13D)、これら2つのリガンドがこのmAb によって異なる影響を受けることが示された。対照として、CXCR3以外の受容体を介して起こるMIP-1αまたはRANTESに応答したT細胞の[Ca2+]iへの効果について、mAb 1C6 を試験した(非掲載)。このmAb は、使用した条件下に効果がなかった。
【0190】
IP-10結合とIP-10誘導性走化性を阻害するmAb 1C6 は、活性化T細胞によるカルシウム流動をも阻害したが、それらの条件下でMig誘導性カルシウム流動を阻害しなかった。これらの結果は、IP-10とMigがCXCR3の異なる領域を介して結合および/またはシグナル伝達することと、mAb 1C6 がIP-10結合部位とそれに続くシグナル伝達を遮断できたことを示唆している。したがって、ここにmAb 1C6 を用いて例証したように、個々のケモカインの作用を選択的に阻害する受容体アンタゴニストを開発することが可能である。
【0191】
後述するように、CXCR3トランスフェクタントの染色は、CXCR3の最初の15N末端アミノ酸残基を含むペプチドによって阻害されうる。mAb 1C6 によるIP-10結合とそれに続く応答の阻害は、この領域がリガンド結合にとって機能上とりわけ重要であることを示唆している。1C6がMig誘導性のカルシウム流動を、使用した条件下に阻害できないことから、IP-10結合とシグナル伝達にとって重要だと思われるmAb 1C6 によって認識されるエピトープは、Mig結合および/またはシグナル伝達には関与しないようである。
【0192】
実施例8.エピトープマッピング
ペプチドはGenemed社(カリフォルニア州サウスサンフランシスコ)に発注した。各15アミノ酸長のそれらペプチドは、CXCR3タンパク質の最初の45N末端残基の異なる部分に相当する。
P1: MVLEVSDHQVLNDAE(配列番号:2、残基1−15)
P2: VAALLENFSSSYDYG(配列番号:2、残基16−30)
P3: ENESDSCCTSPPCPQ(配列番号:2、残基31−45)
それらペプチドをまずDMSOに溶解し、リン酸緩衝食塩水(PBS)で1mg/mlに希釈した。1C6抗体による染色を遮断するペプチドの能力を調べるために、1μg/mlの精製1C6抗体を1×PBS、5%ウシ胎仔血清(FCS)中、100μg/mlの各ペプチドの存在下にCXCR3 L1.2トランスフェクタント(105細胞;実施例3〜9に関する材料と方法の項を参照されたい)と共に4℃で30分間インキュベートした。最終体積は100μlだった。陽性染色はペプチドの非存在下に行なった。結合したmAb を抗マウスIgG-FITCで検出し、その結果をフローサイトメトリーで分析した。
【0193】
mAb 1C6 による染色はP1によって完全に阻害され、このmAb がCXCR3タンパク質の最初のN末端15アミノ酸中のエピトープを認識することが示唆された(図14A〜14D)。mAb 1C6 と同じ融合(LS-77)から生成した3A8と呼ばれるもう一つのmAb (LS77-3A8ともいう)の結合も、P1ペプチドによって阻害された。
【0194】
実施例9.トランスフェクト細胞での免疫化による抗CXCR3 モノクローナル抗体の作製
CXCR3構築物をトランスフェクトされ高レベルのヒトCXCR3を発現するL1.2細胞でマウスを免疫することにより、さらなる抗CXCR3 モノクローナル抗体を作製した(上記材料と方法の項を参照されたい)。免疫化と融合ハイブリドーマの作製は記述されているように行なった(Qin, S.ら, Eur. J. Immunol., 26 : 640 (1996) ; Heath, H.ら, J. Clin. Invest., 99(2):178(1997))。抗CXCR3 mAb は、活性化ヒトT細胞とCXCR3トランスフェクタントの陽性染色によって同定した。8種類の抗CXCR3抗体が一つの融合(LS-104)で得られた。
【0195】
CXCR3へのIP-10の結合を阻害するそれら抗体の能力を試験した。陽性クローンから得た組織培養上清(25μl)を、1×PBS、5%ウシ胎仔血清(FCS) (最終体積100μl)中、CXCR3 L1.2トランスフェクタント細胞(実施例3〜9に関する材料と方法の項)および0.05nMの放射標識125I標識IP-10と共に4℃で30分間インキュベートした。mAb 1C6 と抗CXCR2(IL-8受容体B)mAb の上清も、それぞれ特異的および非特異的mAb 対照として使用した。総結合量は抗体の不在下に決定した。40nMの非標識IP-10を競合因子として使用することによってバックグラウンド結合量を求め、その値をmAb による阻害百分率の計算に使用した。結果を図15に示す。この融合(LS-104)から得られた抗体はすべて、50〜70%の範囲の阻害百分率でIP-10結合を遮断できた。
【0196】
これらのmAb を、シグナル伝達(Ca2+流動誘導能)を阻害する能力についても、基本的に実施例7に記述したように評価したが、これらの抗体はいずれも使用した条件下にMig媒介性のCa2+シグナル伝達を遮断できなかった。IP-10媒介性のCXCR3結合およびシグナル伝達が抗体によって阻害され、CXCR3を介したMig媒介性のシグナル伝達が抗体によって阻害されないことは、これらの抗体がIP-10によって媒介されるCXCR3機能を選択的に阻害できることを示している。
【0197】
また、基本的に実施例8に記述したようにLS-104融合から得た抗体を用いて、エピトープマッピング研究を行なった。その結果は種々の結合部位が認識されることを示している(表2)。この結果は、これら抗体のうち3つがCXCR3の最初の15N末端アミノ酸(配列番号:2の第1〜15残基)内のエピトープを認識し、これら抗体のうち2つがCXCR3のアミノ酸16〜30(配列番号:2の第16〜30残基)内のエピトープを認識することを示唆している。P3ペプチドはこれら抗体のいずれによる染色をも遮断せず、これは、これらの抗体がいずれもCXCR3のアミノ酸31〜45(配列番号:2の第31〜45残基)に相当するペプチドを結合しなかったことを示す。残り3つのmAb を用いた染色は、使用した条件下にどのペプチドによっても有意に阻害されず、これらmAb が、一部重複した上記ペプチドの区間からなるエピトープ、細胞表面に提示される構造エピトープ(conformational epitopes)、または受容体の他の部分にあるエピトープを結合しうることが示唆された。また、これらのデータは、受容体トランスフェクタントでマウスを免疫することにより、CXCR3の様々な部分に対するmAb が得られうることをも示唆している。
【0198】
【表2】
実施例10.mAb 1C6 を用いた正常組織と炎症組織の免疫組織化学的分析
組織
ヒト組織(正常および炎症)は米国立衛生研究所(National Institutes of Health)の資金提供を受けた公共機関National Disease Research Instituteから入手した。正常マカーク(アカゲザル)組織はNew England Regional Primate Research Center(ニューイングランド地方霊長類研究センター;マサチューセッツ州サウスボロ)から入手した。
【0199】
免疫組織化学
アルカリホスファターゼ法.組織を厚さ4μmに切片化し、乾燥した後、2%パラホルムアルデヒド/0.5×PBS中4℃で10分間固定した。PBS洗浄の後、非特異的抗体結合部位を10%正常ヤギ血清/5%ヒトAB血清/PBSにより室温で30分間遮断した。次に、精製した抗CXCR3ネズミmAb 1C6 を0.3%Triton X100/0.2%Tween 20/1%FCS/5%ヒトAB血清および0.1%アジ化ナトリウム中に10μg/mlの濃度に希釈し、それを組織切片に適用して4℃で終夜インキュベートした。同イソタイプの無関係なmAb を組織のステップセクション(step sections)に対して陰性対照として使用した(IgG1,MOPC-21, Sigma社,ミズーリ州セントルイス)。次に、ビオチニル化ヤギ抗マウスIgG(Vector社, カリフォルニア州バーリンゲーム)とアビジン-ビオチン-アルカリホスファターゼ複合体(Biogenex社, カリフォルニア州サンラモン)を順次加えた。内因性アルカリホスファターゼ活性を遮断するためにレバミゾールを含有するFast Red(Biogenex社, カリフォルニア州サンラモン)を発色原として使用し、マイヤーヘマトキシリンを対比染色剤として使用した。
【0200】
結果
ヒトおよびマカークの正常リンパ節:どちらの種でも、染色は、Tリンパ球でのCXCR3発現と合致して、リンパ球の傍皮質と髄索内のリンパ球の70〜80%に限定された。
【0201】
ヒトおよびマカクの脾臓:どちらの種でも、染色は、白色脾髄のリンパ濾胞の周縁部に沿ったリンパ球と脾洞内に散在したリンパ球に限定された。このパターンはTリンパ球でのCXCR3発現と合致する。
【0202】
ヒト胸腺:胸腺髄質はリンパ球と形態学的に合致する散在したCXCR3免疫反応性単核細胞を含んだ。
【0203】
この分析により、マカークCXCR3がmAb 1C6 によって認識されることが明らかになった。独立した実験により、マカーク細胞Tリンパ球をコンカナバリンAおよびIL-2と共に培養することによってCXCR3がアップレギュレートされること、マカークT細胞芽球がヒトIP-10に応答して走化性を示しうること、mAb 1C6 がその走化性を遮断できること、および前もってヒトIP-10と共にインキュベートすると、走化性によって評価されるマカーク芽球の脱感作が起こることが示された。
【0204】
下記の考察では、様々な数のCXCR3免疫反応性「単核細胞」が正常組織と炎症組織の両方に同定された。これらの単核細胞は次の理由からおそらくTリンパ球だろう:a)フローサイトメトリーにより、CXCR3は多少のB細胞とNK細胞にも検出されたものの、他の単核細胞には検出されず、CXCR3+ 細胞の大半がTリンパ球であることが明らかになった;b)リンパ節と脾臓内では、T細胞が配置されていることが知られている領域内のリンパ球がCXCR3に免疫反応性を示す唯一の細胞である;ならびにc)次に挙げる組織内のCXCR3免疫反応性単核細胞は形態学的にリンパ球と合致する。
【0205】
非炎症ヒト組織:心臓、肝臓、腎臓、肺、皮膚、胸部、骨格筋、唾液腺、膵臓、膣、子宮および卵巣を含む非炎症ヒト組織では、CXCR3発現がまばらで散在した間質単核細胞に限定された。小腸と大腸の切片では、CXCR3を発現する単核細胞が固有層とパイアー斑内に観察された。肝臓では、管周辺リンパ球も染色され、肝細胞はわずかに染色された。
【0206】
ヒト脳:染色は観察されなかった。
【0207】
炎症ヒト組織:単核細胞の間質および血管周辺蓄積を特徴とする慢性的炎症組織の複数切片をCXCR3発現について調べた。間質性腎炎(1症例,腎組織)、潰瘍性大腸炎(1症例,結腸組織)、腸炎(1症例,小腸)および慢性膣炎(4症例,膣)のヒト患者由来の組織を含むこれらの組織では、単核細胞の約50%〜90%がCXCR3に免疫反応性を示した。したがって、慢性的炎症組織は正常組織と比較して、そこに含まれる間質単核細胞の数が多く、それら細胞のうちCXCR3に免疫反応性を示すものの割合も高かった。
【0208】
炎症組織の分析はリンパ球の約50〜90%がCXCR3を発現することを明らかにしたが、対応する正常組織ではCXCR3を発現するリンパ球の割合ははるかに低かった。これらの知見は、CXCR3を発現するリンパ球(おそらくはCXCR3を発現するTリンパ球)の慢性炎症部位への特異的な補充を示唆している。CXCR3はホーミングまたは炎症部位に遊走する傾向を持つT細胞を特徴づけるようである。
【0209】
均等物
当業者であれば、単なる日常的実験手法により、本明細書に記載された本発明の具体的態様に対する多くの均等物を認識し、あるいは確認することができるであろう。かかる均等物は、下記請求の範囲に包含されるように意図されている。
【0210】
本発明の態様として、以下のものが挙げられる。
[1]タンパク質または改変体が1種以上のケモカイン類を選択的に結合することができ、かつ細胞シグナル伝達および/またはそれに応答した細胞応答を媒介することができる、哺乳類CXCR3タンパク質またはその機能的改変体をコードする単離核酸。
[2]該核酸が中程度のストリンジェンシー条件下に、図1(配列番号:1)の配列、その相補鎖、またはコード配列を含む図1(配列番号:1)の配列の一部もしくはその相補鎖を有する2番目の核酸にハイブリダイズすることができる、[1]記載の単離核酸。
[3]該単離核酸が本質的に純粋である、[1]記載の単離核酸。
[4]該タンパク質または改変体がヒトIP−10、ヒトMig、ヒトIP−10の哺乳類ホモログおよびヒトMigの哺乳類ホモログからなる群より選ばれたケモカインに選択的に結合することができる、[1]記載の単離核酸。
[5]該哺乳類CXCR3タンパク質またはその機能的改変体がヒトCXCR3タンパク質またはその機能的改変体である、[1]記載の単離核酸。
[6]配列番号:1、その相補鎖、またはコード配列を含む配列番号:1の一部もしくはその相補鎖を含有してなる、[1]記載の単離核酸。
[7]該ヒトCXCR3タンパク質が配列番号:2に示されたアミノ酸配列を有する、[1]記載の単離核酸。
[8][1]記載の核酸を含有してなる単離核酸構築物。
[9]該核酸が操作可能に発現制御配列に連結される、[8]記載の単離核酸構築物。
[10]該核酸が配列番号:1、その相補鎖、またはコード配列を含む配列番号:1の一部もしくはその相補鎖を含有してなる、[8]記載の単離核酸構築物。
[11]該核酸が図2(配列番号:2)に示されたアミノ酸配列を有するポリペプチドをコードする、[8]記載の単離核酸構築物。
[12]該核酸が哺乳類CXCR3タンパク質を含む融合タンパク質をコードする核酸であり、任意にコード配列が操作可能に発現制御配列に連結される、[8]記載の単離核酸構築物。
[13]タンパク質が1種以上のケモカイン類を選択的に結合することができ、かつ細胞シグナル伝達および/またはそれに応答した細胞応答を媒介することができる、哺乳類CXCR3タンパク質またはその機能的改変体をコードする組換え核酸を含有してなる宿主細胞。
[14]該核酸が操作可能に発現制御配列に連結される、[13]記載の宿主細胞。
[15]該核酸が配列番号:1によりコードされるヒトCXCR3タンパク質をコードする、[13]記載の宿主細胞。
[16]タンパク質が1種以上のケモカイン類を選択的に結合することができ、かつ細胞シグナル伝達および/またはそれに応答した細胞応答を媒介することができる、単離哺乳類CXCR3タンパク質またはその機能的改変体。
[17]該哺乳類がヒトであり、かつ該タンパク質がヒトIP−10およびヒトMigからなる群より選ばれた1種以上のケモカイン類に選択的に結合することができる、[16]記載の単離哺乳類CXCR3タンパク質またはその機能的改変体。
[18]中程度のストリンジェンシー条件下に、図1(配列番号:1)の配列、その相補鎖、またはコード配列を含む図1(配列番号:1)の配列の一部もしくはその相補鎖を有する2番目の核酸にハイブリダイズすることができる核酸によりコードされる、[16]記載の単離哺乳類CXCR3タンパク質またはその機能的改変体。
[19]図1(配列番号:1)に示された核酸によりコードされる、単離ヒトCXCR3タンパク質。
[20]配列番号:2に示されたアミノ酸配列を有する、[19]記載の単離ヒトCXCR3タンパク質。
[21]哺乳類CXCR3タンパク質を含有してなる融合タンパク質。
[22](a)宿主細胞に哺乳類CXCR3タンパク質またはその改変体をコードする核酸を導入する工程であり、それによりコード配列が発現制御配列に操作可能に連結されたコード配列を有する組換え宿主細胞を生産する、および
(b)工程(a)で生産された宿主細胞を該核酸が発現する条件下に維持する工程、
を含む、哺乳類CXCR3タンパク質またはその改変体の生産方法。
[23]さらに哺乳類CXCR3タンパク質またはその改変体を単離する工程を含む、[22]記載の方法。
[24]哺乳類CXCR3タンパク質またはその改変体をコードする組換え核酸を含む宿主細胞を核酸の発現に適する条件下に維持する工程を含む、哺乳類CXCR3タンパク質またはその改変体を生産する方法。
[25]さらに哺乳類CXCR3タンパク質またはその改変体を単離する工程を含む、[24]記載の方法。
[26]該哺乳類がヒトであり、かつ該タンパク質またはその改変体が、ヒトIP−10およびヒトMigからなる群より選ばれた1種以上のケモカイン類を選択的に結合することができる、[24]記載の方法。
[27]哺乳類CXCR3タンパク質を結合する、抗体または機能的抗体断片。
[28]該抗体または抗体断片が哺乳類CXCR3タンパク質の1種以上の機能を阻害することができる、[27]記載の抗体または機能的抗体断片。
[29]該抗体または抗体断片がリガンドの哺乳類CXCR3タンパク質への結合を阻害する、[28]記載の抗体または機能的抗体断片。
[30]該抗体または抗体断片がヒトCXCR3タンパク質を結合することができ、かつヒトCXCR3タンパク質とIP−10および/またはMigからなる群より選ばれた1種以上のリガンドとの相互作用を阻害することができる、[28]記載の抗体または機能的抗体断片。
[31]該抗体または断片が、モノクローナル抗体1C6がヒトCXCR3タンパク質またはその一部に結合することに競合することができる、[30]記載の抗体または機能的抗体断片。
[32]該抗体または抗体断片がヒトCXCR3タンパク質とIP−10との相互作用を選択的に阻害する、[28]記載の抗体または機能的抗体断片。
[33]該抗体または断片がモノクローナル抗体1C6またはその抗原結合断片である、[32]記載の抗体または機能的抗体断片。
[34]ヒトCXCR3タンパク質を結合する、抗体またはその抗原結合断片。
[35]配列番号:2のN−末端細胞外セグメントに対応するヒトCXCR3の一部またはヒトCXCR3タンパク質の少なくとも1種の免疫学的性質を有するその一部により、該結合が阻害されうる、[34]記載の抗体またはその抗原結合断片。
[36]配列番号:2の1〜15残基の配列と同じである配列を有するポリペプチドと配列番号:2の16〜30残基の配列と同じである配列を有するポリペプチドとからなる群より選ばれたポリペプチドにより、該結合が阻害されうる、[34]記載の抗体またはその抗原結合断片。
[37]試験対象の薬剤と単離および/または組換え哺乳類CXCR3タンパク質あるいはそのリガンド結合改変体を含む組成物とを、リガンドの結合に適する条件下に合わせる工程、および該薬剤と該哺乳類CXCR3タンパク質もしくは改変体との間の複合体の形成を検出または測定する工程を含む、哺乳類CXCR3タンパク質またはそのリガンド結合改変体を結合する薬剤を検出または同定する方法。
[38]該薬剤がヒトIP−10、ヒトMig、IP−10の哺乳類ホモログおよびMigの哺乳類ホモログからなる群より選ばれたリガンドである、[37]記載の方法。
[39]該リガンドが放射性同位体、スピン標識、抗原標識、酵素標識、蛍光基または化学発光基からなる群より選ばれた標識で標識される、[38]記載の方法。
[40]アッセイがヒトIP−10、ヒトMig、IP−10の哺乳類ホモログおよびMigの哺乳類ホモログからなる群より選ばれた1種以上のリガンドの存在下に結合が決定される競合アッセイである、[38]記載の方法。
[41]a) 試験対象の薬剤と組換え哺乳類CXCR3タンパク質またはそのリガンド結合改変体を発現する宿主細胞とを、リガンドの結合に適する条件下に合わせる工程、および
b) 該薬剤と哺乳類CXCR3タンパク質もしくはそのリガンド結合改変体との間の複合体の形成を検出または測定する工程、
を含む、哺乳類CXCR3タンパク質またはそのリガンド結合改変体を結合する薬剤を検出または同定する方法。
[42]該薬剤がヒトIP−10、ヒトMig、IP−10の哺乳類ホモログおよびMigの哺乳類ホモログからなる群より選ばれたリガンドである、[41]記載の方法。
[43]アッセイがヒトIP−10、ヒトMig、IP−10の哺乳類ホモログおよびMigの哺乳類ホモログからなる群より選ばれた1種以上のリガンドの存在下に結合が決定される競合アッセイである、[41]記載の方法。
[44]哺乳類CXCR3タンパク質もしくはそのリガンド結合改変体が細胞シグナル伝達および/または細胞応答を媒介し、かつシグナル伝達活性またはそれに応答する該CXCR3タンパク質もしくは改変体の細胞応答を検出または測定することにより複合体形成をモニターする、[41]記載の方法。
[45]a)試験対象の薬剤と、哺乳類CXCR3タンパク質のリガンドと単離および/または組換え哺乳類CXCR3タンパク質あるいはそのリガンド結合改変体を含む組成物とを、リガンドの結合に適する条件下に合わせる工程、および
b) 該哺乳類CXCR3タンパク質もしくは改変体と該リガンドとの間の複合体の形成を検出または測定する工程であり、該薬剤による複合体形成の阻害が、該薬剤が阻害剤であることの指標である、
を含む、哺乳類CXCR3タンパク質もしくはそのリガンド結合改変体へのリガンド結合の阻害剤を検出または同定する方法。
[46]該リガンドがヒトIP−10、ヒトMig、IP−10の哺乳類ホモログおよびMigの哺乳類ホモログからなる群より選ばれた、[45]記載の方法。
[47]単離および/または組換え哺乳類CXCR3タンパク質を含む該組成物が組換え哺乳類CXCR3タンパク質を発現する宿主細胞を含有する、[45]記載の方法。
[48]該哺乳類CXCR3タンパク質が細胞シグナル伝達および/または細胞応答を媒介し、かつシグナル伝達活性またはそれに応答する該CXCR3タンパク質もしくは改変体の細胞応答を検出あるいは測定することにより複合体形成をモニターする、[47]記載の方法。
[49]a)試験対象の薬剤と、哺乳類CXCR3タンパク質のリガンドと組換え哺乳類CXCR3タンパク質もしくはそのリガンド結合改変体を発現する宿主細胞とを、リガンドの結合に適する条件下に合わせる工程、および
b) 該タンパク質もしくは改変体と該リガンドとの間の複合体の形成を検出または測定する工程であり、該薬剤による複合体形成の阻害が、該薬剤が阻害剤であることの指標である、
を含む、哺乳類CXCR3タンパク質もしくはそのリガンド結合改変体へのリガンド結合の阻害剤を検出または同定する方法。
[50]該リガンドがIP−10、Mig、IP−10の哺乳類ホモログおよびMigの哺乳類ホモログからなる群より選ばれた、[49]記載の方法。
[51]該哺乳類CXCR3タンパク質が細胞シグナル伝達および/または細胞応答を媒介することができ、かつシグナル伝達活性またはそれに応答する該CXCR3タンパク質の細胞応答を検出あるいは測定することにより複合体形成をモニターする、[49]記載の方法。
[52]該薬剤が抗体または抗体断片である、[49]記載の方法。
[53]試験対象の薬剤と(a)組換え哺乳類CXCR3タンパク質またはその機能的改変体を発現する宿主細胞と(b)リガンドまたはその促進剤とを、リガンドもしくは促進剤誘導応答を検出するに適する条件下に合わせる工程、および
試験対象薬剤の該応答を阻害する能力を評価する工程であり、薬剤によるリガンドまたは促進剤誘導応答の阻害が、薬剤が阻害剤であることの指標である、
を含む哺乳類CXCR3タンパク質もしくはその機能的改変体の阻害剤を検出または同定する方法。
[54]該応答がシグナル伝達活性またはそれに応答する該哺乳類CXCR3タンパク質もしくはその改変体の細胞応答を検出あるいは測定することによりモニターされる、[53]記載の方法。
[55]試験対象の薬剤と組換え哺乳類CXCR3タンパク質もしくはその機能的改変体を発現する宿主細胞とを、受容体媒介応答を検出するに適する条件下に合わせる工程、および
該応答を検出または測定する工程であり、該薬剤による応答の誘導または刺激が薬剤が促進剤であることの指標である、
を含む、哺乳類CXCR3タンパク質もしくはその機能的改変体の促進剤を検出または同定する方法。
[56][45]記載の方法によって同定された、哺乳類CXCR3タンパク質に特有の少なくとも1種の機能の阻害剤。
[57]該阻害剤が受容体に結合可能な抗体またはその一部である、[56]記載の阻害剤。
[58][53]記載の方法によって同定された、哺乳類CXCR3タンパク質に特有の少なくとも1種の機能の阻害剤。
[59][55]記載の方法によって同定された、哺乳類CXCR3タンパク質に特有の少なくとも1種の機能の促進剤。
[60]a) 抗体と選択された哺乳類CXCR3タンパク質との特異的結合に適する条件下に、試料と該タンパク質を結合する抗体とを接触させる工程;および
b) 抗体−CXCR3複合体を検出する工程、
を含む、試料中の選択された哺乳類CXCR3タンパク質の検出方法。
[61]哺乳類CXCR3タンパク質と該タンパク質の少なくとも1種の機能の阻害剤または促進剤とを接触させる工程、
を含む、哺乳類CXCR3タンパク質の機能の少なくとも1種を調節する方法。
[62]哺乳類へ治療上有効な量の哺乳類CXCR3タンパク質の阻害剤を投与して、それにより炎症を減少させること、
を含む、炎症性疾患または状態を処置する方法。
[63]炎症性疾患または状態がT細胞媒介疾患または状態である、[62]記載の方法。
[64]該阻害剤が哺乳類CXCR3タンパク質に特異的に結合する抗体またはその抗原結合断片である、[62]記載の方法。
[65]該抗体またはその断片がモノクローナル抗体1C6のヒトCXCR3タンパク質への結合を競合することができる、[64]記載の方法。
[66]該抗体またはその断片がモノクローナル抗体1C6またはその抗原結合断片である、[65]記載の方法。
[67]哺乳類CXCR3タンパク質の天然のリガンド以外の促進剤の治療上有効な量を哺乳類へ投与すること、
を含む、抗腫瘍治療方法。
[68]哺乳類CXCR3タンパク質の天然のリガンド以外の促進剤の治療上有効な量を哺乳類へ投与すること、
を含む、抗ウイルス治療方法。
[69]配列:a)配列番号:1、
b)コード領域を含むその一部、
c)または(a)および(b)のいずれかのRNA体
を有する核酸にハイブリダイズする配列をもつ核酸を含有してなるアンチセンス核酸。
【0211】
配列表
(1)一般情報:
(i)出願人:
(A)名称:テオドーア−コッヒャー インスティトゥーテ
(B)ストリート:フライエシュトラーセ 1
(C)市:ベルン
(D)州:
(E)国:スイス国
(F)郵便番号:CH−3012
(G)電話番号:41−31−631−4141
(I)ファクシミリ番号:41−31−631−3799
(i)出願人:
(A)名称:ロイコサイト,インコーポレーテッド
(B)ストリート:ファースト ストリート 215
(C)市:ケンブリッジ
(D)州:マサチューセッツ
(E)国:アメリカ合州国
(F)郵便番号:02142
(G)電話番号:617−621−9350
(I)テレファックス:617−621−9349
(ii)発明の名称:CXCR3、抗体、核酸、および使用方法
(iii)配列の数:4
(iv)連絡先住所:
(A)宛名:ハミルトン,ブルック,スミス アンド レイノルズ,
ピー.シー.
(B)ストリート:ミリティア ドライブ 2
(C)市:レキシントン
(D)州:マサチューセッツ
(E)国:アメリカ合衆国
(F)郵便番号:02173
(v)コンピュータ可読フォーム:
(A)媒体型:フロッピーディスク
(B)コンピュータ:IBM PC 互換機
(C)オペレーティングシステム:PC−DOS/MS−DOS
(D)ソフトウェア:パテントイン リリース #1.0,
バージョン #1.30
(vi)現出願データ:
(A)出願番号:
(B)出願日:
(C)分類:
(vii)先願データ:
(A)出願番号:US 08/829,839
(B)出願日:1997年3月31日
(vii)先願データ:
(A)出願番号:US 08/709,838
(B)出願日:1996年9月10日
(viii)代理人情報:
(A)氏名:ブルック,デビッド イー.
(B)登録番号:22,592
(C)リファランス/ドケット番号:TKI96−01A2 PCT
(ix)テレコミュニケーション情報:
(A)電話番号:(781)861−6240
(B)テレファックス:(781)861−9540
(2)配列番号:1の情報:
(i)配列の特徴:
(A)配列の長さ:1670塩基対
(B)配列の型:核酸
(C)鎖の数:二本鎖
(D)トポロジー:不明
(ix)配列の特徴:
(A)特徴を表す記号:CDS
(B)存在位置:69..1172
(xi)配列:配列番号:1:
【0212】
【化1−1】
【0213】
【化1−2】
【0214】
【化1−3】
(2)配列番号:2の情報:
(i)配列の特徴:
(A)配列の長さ:368アミノ酸
(B)配列の型:アミノ酸
(D)トポロジー:直鎖状
(ii)配列の種類:タンパク質
(xi)配列:配列番号:2:
【0215】
【化2−1】
【0216】
【化2−2】
(2)配列番号:3の情報:
(i)配列の特徴:
(A)配列の長さ:28塩基対
(B)配列の型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:不明
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:11
(D)他の情報:/修飾塩基=「イノシン」
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:12
(D)他の情報:/修飾塩基=「イノシン」
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:23
(D)他の情報:/修飾塩基=「イノシン」
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:26
(D)他の情報:/修飾塩基=「イノシン」
(xi)配列:配列番号:3:
【0217】
【化3】
(2)配列番号:4の情報:
(i)配列の特徴:
(A)配列の長さ:27塩基対
(B)配列の型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:不明
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:10
(D)他の情報:/修飾塩基=「イノシン」
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:16
(D)他の情報:/修飾塩基=「イノシン」
(ix)配列の特徴:
(A)特徴を表す記号:modified base
(B)存在位置:18
(D)他の情報:/修飾塩基=「イノシン」
(xi)配列:配列番号:4:
【0218】
【化4】
【特許請求の範囲】
【請求項1】
ヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質に結合する抗体または抗原結合断片と接触させる工程を含む、方法。
【請求項2】
ヒトCXCR3タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、ヒトCXCR3タンパク質に結合する抗体または抗原結合断片と接触させる工程を含み、結合が、配列番号:2の残基1〜15の配列と同じアミノ酸配列からなるポリペプチドによって阻害される、方法。
【請求項3】
配列番号:2に示されるアミノ酸配列を有する哺乳動物CXCケモカインレセプター3(CXCR3)タンパク質のシグナル伝達活性または細胞応答機能を阻害する方法であって、該タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含む、方法。
【請求項4】
配列番号:2に示されるアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該タンパク質を、モノクローナル抗体1C6(ATCC受託番号HB−12330)またはその抗原結合断片と接触させる工程を含む、方法。
【請求項5】
配列番号:2に示されるアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質によって媒介されるIP−10誘導カルシウムフラックスを阻害する方法であって、該タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含む、方法。
【請求項6】
配列番号:2に示されるアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質を介するIP−10誘導走化性を阻害する方法であって、該タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含む、方法。
【請求項7】
CXCケモカインレセプター3(CXCR3)タンパク質のシグナル伝達活性または細胞応答機能を阻害する方法であって、該CXCR3タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含み、該抗体がモノクローナル抗体1C6(ATCC受託番号HB−12330)の軽鎖CDR(CDR1、CDR2およびCDR3)および重鎖CDR(CDR1、CDR2およびCDR3)ならびにヒト枠組み領域を含むヒト化免疫グロブリンであり、該ヒト化免疫グロブリンがモノクローナル抗体1C6(ATCC受託番号HB−12330)と同じまたは類似のエピトープ特異性を有する、方法。
【請求項8】
ヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、ハイブリドーマ細胞株HB−12330により産生される抗体またはその抗原結合断片と接触させる工程を含む、方法。
【請求項9】
哺乳動物CXCケモカインレセプター3(CXCR3)タンパク質のシグナル伝達活性または細胞応答機能を阻害する方法であって、該CXCR3タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含み、該抗体がモノクローナル抗体1C6(ATCC受託番号HB−12330)の軽鎖CDR(CDR1、CDR2およびCDR3)および重鎖CDR(CDR1、CDR2およびCDR3)ならびにヒト枠組み領域を含むヒト化免疫グロブリンであり、該ヒト化免疫グロブリンがモノクローナル抗体1C6(ATCC受託番号HB−12330)と同じまたは類似のエピトープ特異性を有する、方法。
【請求項1】
ヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質に結合する抗体または抗原結合断片と接触させる工程を含む、方法。
【請求項2】
ヒトCXCR3タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、ヒトCXCR3タンパク質に結合する抗体または抗原結合断片と接触させる工程を含み、結合が、配列番号:2の残基1〜15の配列と同じアミノ酸配列からなるポリペプチドによって阻害される、方法。
【請求項3】
配列番号:2に示されるアミノ酸配列を有する哺乳動物CXCケモカインレセプター3(CXCR3)タンパク質のシグナル伝達活性または細胞応答機能を阻害する方法であって、該タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含む、方法。
【請求項4】
配列番号:2に示されるアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該タンパク質を、モノクローナル抗体1C6(ATCC受託番号HB−12330)またはその抗原結合断片と接触させる工程を含む、方法。
【請求項5】
配列番号:2に示されるアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質によって媒介されるIP−10誘導カルシウムフラックスを阻害する方法であって、該タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含む、方法。
【請求項6】
配列番号:2に示されるアミノ酸配列を有するヒトCXCケモカインレセプター3(CXCR3)タンパク質を介するIP−10誘導走化性を阻害する方法であって、該タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含む、方法。
【請求項7】
CXCケモカインレセプター3(CXCR3)タンパク質のシグナル伝達活性または細胞応答機能を阻害する方法であって、該CXCR3タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含み、該抗体がモノクローナル抗体1C6(ATCC受託番号HB−12330)の軽鎖CDR(CDR1、CDR2およびCDR3)および重鎖CDR(CDR1、CDR2およびCDR3)ならびにヒト枠組み領域を含むヒト化免疫グロブリンであり、該ヒト化免疫グロブリンがモノクローナル抗体1C6(ATCC受託番号HB−12330)と同じまたは類似のエピトープ特異性を有する、方法。
【請求項8】
ヒトCXCケモカインレセプター3(CXCR3)タンパク質への、IP−10およびMigからなる群より選択されるリガンドの結合を阻害する方法であって、該CXCR3タンパク質を、ハイブリドーマ細胞株HB−12330により産生される抗体またはその抗原結合断片と接触させる工程を含む、方法。
【請求項9】
哺乳動物CXCケモカインレセプター3(CXCR3)タンパク質のシグナル伝達活性または細胞応答機能を阻害する方法であって、該CXCR3タンパク質を、配列番号:2のアミノ酸配列を有するヒトCXCR3タンパク質と結合する抗体または抗原結合断片と接触させる工程を含み、該抗体がモノクローナル抗体1C6(ATCC受託番号HB−12330)の軽鎖CDR(CDR1、CDR2およびCDR3)および重鎖CDR(CDR1、CDR2およびCDR3)ならびにヒト枠組み領域を含むヒト化免疫グロブリンであり、該ヒト化免疫グロブリンがモノクローナル抗体1C6(ATCC受託番号HB−12330)と同じまたは類似のエピトープ特異性を有する、方法。
【図1】


【図2】

【図3】
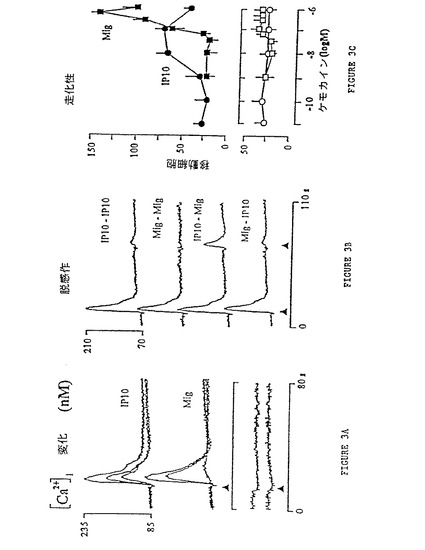
【図4】

【図5】

【図6】

【図7】

【図8】

【図9−1】

【図9−2】

【図9−3】

【図10】

【図11】

【図12】

【図13】

【図14】

【図15】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9−1】
【図9−2】
【図9−3】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公開番号】特開2009−278990(P2009−278990A)
【公開日】平成21年12月3日(2009.12.3)
【国際特許分類】
【出願番号】特願2009−167709(P2009−167709)
【出願日】平成21年7月16日(2009.7.16)
【分割の表示】特願平10−513771の分割
【原出願日】平成9年9月10日(1997.9.10)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.フロッピー
【出願人】(509174370)
【出願人】(500287639)ミレニアム・ファーマシューティカルズ・インコーポレイテッド (98)
【氏名又は名称原語表記】MILLENNIUM PHARMACEUTICALS, INC.
【Fターム(参考)】
【公開日】平成21年12月3日(2009.12.3)
【国際特許分類】
【出願日】平成21年7月16日(2009.7.16)
【分割の表示】特願平10−513771の分割
【原出願日】平成9年9月10日(1997.9.10)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.フロッピー
【出願人】(509174370)
【出願人】(500287639)ミレニアム・ファーマシューティカルズ・インコーポレイテッド (98)
【氏名又は名称原語表記】MILLENNIUM PHARMACEUTICALS, INC.
【Fターム(参考)】
[ Back to top ]