
LMP2を用いた子宮平滑筋肉腫の検出
【課題】子宮平滑筋組織または細胞におけるJAK1キナーゼ遺伝子の変異部位を検出し、至急平滑筋肉腫を評価する方法の提供。
【解決手段】JAK1キナーゼ遺伝子の以下の(A2)、(A3)、(A5)および(A6)の変異部位の少なくとも1つを含むJAK1キナーゼ遺伝子の部分配列、ならびにLMP2プロモーターの以下の(B2)および(B4)の変異部位の少なくとも1つを含むLMP2プロモーターの部分配列からなる群から選択される部分配列からなるオリゴヌクレオチドまたはその標識物であって、10から30塩基からなる部分配列またはその部分配列に相補的な配列からなるオリゴヌクレオチドまたはその標識物:
(A2) G2626A
(A3) G2642T
(A5) A2960C
(A6) A2985T
(B2) C214T
(B4) A217G
【解決手段】JAK1キナーゼ遺伝子の以下の(A2)、(A3)、(A5)および(A6)の変異部位の少なくとも1つを含むJAK1キナーゼ遺伝子の部分配列、ならびにLMP2プロモーターの以下の(B2)および(B4)の変異部位の少なくとも1つを含むLMP2プロモーターの部分配列からなる群から選択される部分配列からなるオリゴヌクレオチドまたはその標識物であって、10から30塩基からなる部分配列またはその部分配列に相補的な配列からなるオリゴヌクレオチドまたはその標識物:
(A2) G2626A
(A3) G2642T
(A5) A2960C
(A6) A2985T
(B2) C214T
(B4) A217G
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、LMP2およびサイクリンE、ならびにインターフェロンγ(IFN-γ)シグナル伝達カスケードに関与するJAK1キナーゼ遺伝子、STAT1遺伝子およびLMP2プロモーターにおける変異をマーカーとして用いる、子宮平滑筋肉腫の検出方法および子宮平滑筋腫と子宮平滑筋肉腫の鑑別検出方法に関する。
【背景技術】
【0002】
子宮癌は婦人科の癌で最も多いが、子宮平滑筋肉腫は発症頻度が低く子宮体部癌の2〜5%である。子宮平滑筋肉腫は、子宮頚部よりも子宮体部の筋肉層から多く発生する。子宮筋層は平滑筋よりできており、子宮筋層の中にできる良性の腫瘍が子宮平滑筋腫であり、悪性の腫瘍が子宮平滑筋肉腫である。子宮平滑筋腫と子宮平滑筋肉腫を鑑別することは非常に難しく、通常手術をして組織を採取し、顕微鏡下での細胞検査により判断していた。子宮平滑筋肉腫は異型性が強く巨大化することもある腫瘍細胞が増殖していることが多い。しかし、ほとんど細胞異型性を示さない場合もあり、細胞異型性の有無は決定的な鑑別基準とはならない。子宮平滑筋腫と子宮平滑筋肉腫との鑑別は、専ら凝固壊死があるかどうか、および細胞分裂像が増大しているかどうかにより行う。原則的には、細胞密度が高い場合には10倍高視野で10個以上の細胞分裂がある場合、腫瘍細胞に異型性が認められる場合には10倍高視野で5個以上ある場合に、子宮平滑筋肉腫と診断される。現実的には、腫瘍細胞の細胞異型度、細胞密度、細胞分裂の数、腫瘍内の壊死、出欠の有無等を参考に、主として顕微鏡および肉眼で組織の形態を観察して鑑別していた。しかしながら、このような鑑別を行うためには熟練を要し、必ずしも確実に鑑別できない場合もあった。
【0003】
本発明者は、先にイムノプロテアーゼの構成因子であるLMP2(low molecular mass polypeptide2)を欠損したマウスのメスにおいて、生後6ヶ月以降に子宮平滑筋肉腫(leiomyosarcoma)が認められ、生後12ヶ月までの発症率が全LMP2欠損マウスのメスの約35%にまでおよぶことを報告した(非特許文献1および2参照)。LMP2の機能は、組織特異的であり、CTLsによるMHCクラスI仲介腫瘍拒絶に不可欠の役割を持っている(非特許文献1参照)。
【0004】
このように、LMP2の欠損が何らかの機能で子宮平滑筋肉腫の発症のファクターになっていることは予測できた。しかしながら、LMP2が含まれる26Sプロテアソームは、転写調節因子や細胞周期調節因子等の活性化、MHCクラスI分子のペプチド抗原の産生等に複雑に関与しており、LMP2と子宮平滑筋肉腫の発症との直接の関係は不明であり、子宮平滑筋肉腫が発症した場合に、26Sプロテアソームの機能がどう変化し、またLMP2の転写発現がどう変化するかは不明であった。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Van Kaer L. et al. (1994) Immunity, 1, 533-541
【非特許文献2】Hayashi T. et al. (2002) Cancer Res., 62, 24-27
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、子宮平滑筋組織におけるLMP2の転写または発現の程度を指標に子宮平滑筋肉腫の存在を検出する方法およびそのための検出試薬の提供を目的とする。
【課題を解決するための手段】
【0007】
本発明者は、筋肉層におけるLMP2の発現を調べるためにマウスの生体を抗LMP2で染色してみた。その結果、平滑筋、横紋筋、心筋等の筋原性組織に特異的なLMP2の発現が認められることから、LMP2欠損マウスの子宮平滑筋層で認められた腫瘍細胞の起源が筋原性細胞(平滑筋細胞)であることを新たに見出した。
【0008】
さらに、本発明者は正常子宮平滑筋層、ヒト子宮平滑筋腫およびヒト子宮平滑筋肉腫の生検組織または手術組織におけるLMP2の発現状況を調べた。その結果、悪性腫瘍である子宮平滑筋肉腫のみにおいて、顕著なLMP2の発現低下が認められることを見出した。
【0009】
本発明者は、次いでLMP2の発現低下が、子宮平滑筋細胞において形質転換(癌化)に直接的に関与しているか検討するため、LMP2の発現が認められない子宮平滑筋肉腫細胞(SKN)に、遺伝子組換えによりLMP2を強制発現させ、SKN細胞の形態、細胞増殖速度およびSKN細胞におけるファイブロネクチンの発現変化について検討した。その結果、SKN細胞において、形態および細胞増殖速度が正常子宮平滑筋細胞に近づき、さらに有意なファイブロネクチンの発現誘導が起こることを見出した。
【0010】
本発明者は上記の新知見から、LMP2の転写発現を指標に子宮平滑筋腫と子宮平滑筋肉腫の鑑別診断をし得ること、およびLMP2の発現により子宮平滑筋肉腫の治療を行い得ることを見出した。
【0011】
本発明者らは、さらにサイクリンEの発現と子宮平滑筋肉腫との関係に着目し、子宮平滑筋組織において、子宮平滑筋肉腫の悪性度に依存してサイクリンEの転写・発現が著しく亢進することを見出した。
【0012】
また、本発明者らは、LMP2の発現に関与するインターフェロンγ(IFN-γ)シグナル伝達系に関与する遺伝子と子宮平滑筋肉腫との関係について検討を行なった。その結果、子宮平滑筋肉腫組織の細胞において同シグナル伝達因子であるJAK1キナーゼ、STAT1をコードする遺伝子およびLMP2プロモーターに変異が生じていることを見出し、該変異を検出することにより、子宮平滑筋肉腫に罹患しているか否か、および子宮平滑筋肉腫に罹患するリスクが高いか否かを判定することを見出した。
以上のようにして、本発明者らは本発明を完成させた。
【0013】
すなわち、本発明は以下の通りである。
[1] LMP2をマーカーとして用い、子宮平滑筋肉腫を検出する方法。
[2] 子宮平滑筋組織におけるLMP2の転写または発現の程度を指標に子宮平滑筋肉腫の存在を検出する方法であって、子宮平滑筋組織におけるLMP2の転写または発現を測定し、LMP2の転写または発現が正常子宮平滑筋組織に比較して少ない場合に子宮平滑筋肉腫が存在すると判定する方法。
[3] 子宮平滑筋組織におけるLMP2の転写または発現の程度を指標に子宮平滑筋の腫瘍が子宮平滑筋腫であるかまたは子宮平滑筋肉腫であるかを鑑別する方法であって、子宮平滑筋組織におけるLMP2の転写または発現を測定し、LMP2の転写または発現が正常子宮平滑筋組織に比較して少ない場合に子宮平滑筋肉腫であると判定する方法。
[4] 子宮平滑筋組織におけるLMP2の転写または発現の程度を指標に子宮平滑筋肉腫の悪性度を判定する方法であって、子宮平滑筋組織におけるLMP2の転写または発現を測定し、LMP2の転写または発現が正常子宮平滑筋組織に比較して少ない場合に悪性子宮平滑筋肉腫であると判定する方法。
[5] 採取した子宮平滑筋組織または細胞を用いin situ ハイブリダイゼーションを行いLMP2の転写を測定する、[1]〜[4]のいずれかの方法。
【0014】
[6] 採取した子宮平滑筋組織または細胞からLMP2のmRNAを抽出しRT-PCRまたはノーザンブロットを行いLMP2の転写を測定する、[1]〜[4]のいずれかの方法。
[7] 採取した子宮平滑筋組織または細胞を用い免疫組織化学染色または免疫細胞化学染色を行いLMP2の発現を測定する、[1]〜[4]のいずれかの方法。
[8] 採取した子宮平滑筋組織または細胞からLMP2タンパク質を抽出しイムノアッセイを行いLMP2の発現を測定する、[1]〜[4]のいずれかの方法。
[9] さらに、ミオシンをマーカーとして用い、LMP2およびミオシンの転写または発現を指標に子宮平滑筋肉腫の存在を検出する方法であって、LMP2の転写または発現が正常子宮平滑筋組織に比較して少なく、かつミオシンの転写または発現が正常子宮平滑筋組織に比較して多い場合に子宮平滑筋肉腫が存在すると判定する[1]の方法。
[10] LMP-2およびサイクリンEをマーカーとして用い、子宮平滑筋肉腫を検出する方法。
【0015】
[11] 子宮平滑筋組織におけるLMP2およびサイクリンEの転写または発現の程度を指標に子宮平滑筋肉腫の存在を検出する方法であって、子宮平滑筋組織におけるLMP2およびサイクリンEの転写または発現を測定し、LMP2の転写または発現が正常子宮平滑筋組織に比較して少なく、かつサイクリンEの転写または発現が正常子宮平滑筋組織に比較して多い場合に子宮平滑筋肉腫が存在すると判定する方法。
[12] 子宮平滑筋組織におけるLMP2およびサイクリンEの転写または発現の程度を指標に子宮平滑筋の腫瘍が子宮平滑筋腫であるかまたは子宮平滑筋肉腫であるかを鑑別する方法であって、子宮平滑筋組織におけるLMP2およびサイクリンEの転写または発現を測定し、LMP2の転写または発現が正常子宮平滑筋組織に比較して少なく、かつサイクリンEの転写または発現が正常子宮平滑筋組織に比較して多い場合に子宮平滑筋肉腫であると判定する方法。
[13] 子宮平滑筋組織におけるLMP2およびサイクリンEの転写または発現の程度を指標に子宮平滑筋肉腫の悪性度を判定する方法であって、子宮平滑筋組織におけるLMP2およびサイクリンEの転写または発現を測定し、LMP2の転写または発現が正常子宮平滑筋組織に比較して少なく、かつサイクリンEの転写または発現が正常子宮平滑筋組織に比較して多い場合に悪性子宮平滑筋肉腫であると判定する方法。
[14] 採取した子宮平滑筋組織または細胞を用いin situ ハイブリダイゼーションを行いLMP2およびサイクリンEの転写を測定する、[10]〜[13]のいずれかの方法。
[15] 採取した子宮平滑筋組織または細胞からLMP2およびサイクリンEのmRNAを抽出しRT-PCRまたはノーザンブロットを行いLMP2およびサイクリンEの転写を測定する、[10]〜[13]のいずれかの方法。
【0016】
[16] 採取した子宮平滑筋組織または細胞を用い免疫組織化学染色または免疫細胞化学染色を行いLMP2およびサイクリンEの発現を測定する、[10]〜[13]のいずれかの方法。
[17] 採取した子宮平滑筋組織または細胞からLMP2およびサイクリンEタンパク質を抽出しイムノアッセイを行いLMP2およびサイクリンEの発現を測定する、[10]〜[13]のいずれかの方法。
[18] 少なくともLMP2遺伝子断片をプローブまたはプライマーとして含む、LMP2をマーカーとして用い子宮平滑筋肉腫を検出するための検出試薬。
[19] 少なくともLMP2遺伝子断片およびサイクリンE遺伝子断片をプローブまたはプライマーとして含む、LMP2およびサイクリンEをマーカーとして用い子宮平滑筋肉腫を検出するための検出試薬。
[20] in situ ハイブリダイゼーション用試薬である[18]または[19]の子宮平滑筋肉腫を検出するための検出試薬。
【0017】
[21] さらに、ミオシン遺伝子断片をプローブまたはプライマーとして含み、LMP2およびミオシンをマーカーとして用い子宮平滑筋肉腫を検出するための[18]または[20]の検出試薬。
[22] さらに、ミオシン遺伝子断片をプローブまたはプライマーとして含み、LMP2、サイクリンEおよびミオシンをマーカーとして用い子宮平滑筋肉腫を検出するための[19]または[20]の検出試薬。
[23] 少なくとも抗LMP2抗体を含む、LMP2をマーカーとして用い子宮平滑筋肉腫を検出するための検出試薬。
[24] 少なくとも抗LMP2抗体および抗サイクリンE抗体を含む、LMP2およびサイクリンEをマーカーとして用い子宮平滑筋肉腫を検出するための検出試薬。
[25] 免疫組織化学または免疫細胞染色用試薬である[23]または[24]の子宮平滑筋肉腫を検出するための検出試薬。
[26] さらに、抗ミオシン抗体を含み、LMP2およびミオシンをマーカーとして用い子宮平滑筋肉腫を検出するための[23]または[25]の検出試薬。
[27] さらに、抗ミオシン抗体を含み、LMP2、サイクリンEおよびミオシンをマーカーとして用い子宮平滑筋肉腫を検出するための[24]または[25]の検出試薬。
【0018】
[28] JAK1キナーゼ遺伝子の以下の(A1)〜(A6)の変異部位の少なくとも1つを含むJAK1キナーゼ遺伝子の部分配列、およびLMP2プロモーターの以下の(B1)〜(B5)の変異部位の少なくとも1つを含むLMP2プロモーターの部分配列からなる群から選択される部分配列からなるオリゴヌクレオチドまたはその標識物であって、10から30塩基からなる部分配列またはその部分配列に相補的な配列からなるオリゴヌクレオチドまたはその標識物:
(A1) A2612A
(A2) G2626A
(A3) G2642T
(A4) A2967C
(A5) A2960C
(A6) A2985T
(B1) A210G
(B2) C214T
(B3) A216G
(B4) A217G
(B5) G219A。
[29] プローブとして用いられる[28]のオリゴヌクレオチドまたはその標識物。
[30] [28]のオリゴヌクレオチドまたはその標識物を固定化した固定化基板。
【0019】
[31] JAK1キナーゼ遺伝子の以下の(A1)〜(A6)の変異部位の少なくとも1つ、およびLMP2プロモーターの以下の(B1)〜(B5)の変異部位の少なくとも1つからなる群から選択される変異部位を含むDNA断片の増幅に用いる少なくとも一対のプライマーセットであって、上記変異部位の3’側および5’側に存在する10から30塩基からなる部分配列からなる、DNA断片の増幅に用い得る一対のプライマーセット:
(A1) A2612A
(A2) G2626A
(A3) G2642T
(A4) A2967C
(A5) A2960C
(A6) A2985T
(B1) A210G
(B2) C214T
(B3) A216G
(B4) A217G
(B5) G219A。
[32] [31]のプライマーセット、[28]もしくは[29]のオリゴヌクレオチドもしくはその標識物、または[30]の固定化基板を含む子宮平滑筋肉腫検出キット。
【0020】
[33] 動物から採取した子宮平滑筋組織または細胞におけるJAK1キナーゼ遺伝子の以下の(A1)〜(A6)の変異部位の少なくとも1つ、およびLMP2プロモーターの以下の(B1)〜(B5)の変異部位の少なくとも1つからなる群から選択される変異部位の検出を行い、その結果に基づいて、上記変異が存在する場合に、前記動物が子宮平滑筋肉腫に罹患しているか、または罹患するリスクが高いと判断する、子宮平滑筋肉腫の検出方法:
(A1) A2612A
(A2) G2626A
(A3) G2642T
(A4) A2967C
(A5) A2960C
(A6) A2985T
(B1) A210G
(B2) C214T
(B3) A216G
(B4) A217G
(B5) G219A。
[34] JAK1キナーゼ遺伝子、またはMP2プライマーの変異の検出を[31]のプライマーセット、[28]もしくは[29]のプローブまたは[30]の固定化基板を用いて行う[33]の方法。
【発明の効果】
【0021】
子宮平滑筋におけるLMP2および/またはサイクリンEの転写または発現を調べることにより、子宮平滑筋組織に子宮平滑筋肉腫が存在するかどうかを検出することができ、LMP2の転写または発現が著しく低い場合および/またはサイクリンEの転写または発現が高い場合に平滑筋肉腫が存在すると判定することができる。従来は、細胞の形態、密度、増殖速度等を指標に子宮平滑筋肉腫を検出していたが、本発明の方法によれば、容易にかつ確実に子宮平滑筋肉腫を検出することができる。さらに、本発明の方法によれば、子宮平滑筋の腫瘍が子宮平滑筋腫であるか子宮平滑筋肉腫であるか鑑別することができ、さらに、子宮平滑筋肉腫の悪性度を判定することができる。
【0022】
また、本発明のLMP2および/またはサイクリンEをマーカーとして用いる方法と、従来から用いられていた細胞の形態、密度等を指標にする方法および/またはミオシンをマーカーとして用いる方法を組合せることにより、確実に子宮平滑筋肉腫を検出することができる。
【0023】
さらに、IFN-γシグナル伝達因子であるJAK1キナーゼ遺伝子、STAT1遺伝子およびLMP2プロモーターの変異と子宮平滑筋肉腫の発症が密接に関連しており、上記遺伝子またはプロモーターの変異を検出することにより、子宮平滑筋肉腫に罹患しているか否か、および子宮平滑筋肉腫に罹患するリスクがあるか否かを判定することができる。
【図面の簡単な説明】
【0024】
【図1a】SKN細胞の中でのIFN-γ誘導性TAP1およびLMP2発現の欠損を示す写真である。写真に示す時間250ユニット/mlのIFN-γで処理したHeLa、HeLa.S3、SKN細胞およびヒト正常子宮平滑筋細胞(Hu.USMC)から細胞質抽出を調製し、50μgの細胞質抽出物を10%のSDS-PAGEで分離した。TAP1、LMP2およびβアクチンの発現レベルは適切な抗体を用いたイムノブロット分析で調べた。
【図1b】SKN細胞の中でのIFN-γ誘導性TAP1およびLMP2発現の欠損を示す写真であり、RT-PCRによるHeLa、HeLa.S3、SKN細胞およびHu.USMC細胞中のTAP1、LMP2およびβアクチンのmRNA発現の試験の結果を示す写真である。IFN-γ(250ユニット/ml)の非存在下または存在下で細胞を48時間培養した後に、プライマーを用いてRT-PCRを行った。RT-PCRにより増幅されたDNA製品をアガロース・ゲルにより分離した。DNAサイズマーカーは写真の左側に示す。
【図1c】子宮平滑筋肉腫の中でのLMP2発現の欠損を示す写真であり、正常子宮平滑筋および子宮平滑筋腫中のLMP2の免疫組織化学の結果を示す写真である。組織標本の5μmの切片を抗LMP2抗体およびペルオキシダーゼ結合抗ウサギのIgG抗体を用いて染色した。
【図2】HeLaおよびSKN細胞中でのIFN-γ誘導TAP1とLMP2共有のwtおよびIRF-E mtプロモーターの示差活性を示す図である。TAP1とLMP2共有のプロモーターwt(TAP1 593-1/pGL3およびLMP2 1-593/pLG3)の図ならびにそれらのそれぞれのIRF-E変異体プロモーターを含んだルシフェラーゼレポーター遺伝子の構築物を示す。HeLaおよびSKN細胞に同レポーター遺伝子を移入し、IFN-γを24時間後に添加しさらに回収の前24時間細胞をインキュベートした。レポーター遺伝子の移入効率の標準化のためにpSMV-βGALとの同時に移入した。結果は、HeLaおよびSKN細胞について別々に測定されたルシフェラーゼ遺伝子発現に標準化され、相対的TAP1およびLMP2活性として示される。結果は3つの独立している実験の平均で示され、エラーバーはSEを表す。
【図3】ヒト正常子宮平滑筋組織の顕微鏡写真である。
【図4】ヒト子宮平滑筋腫およびヒト子宮平滑筋肉腫の顕微鏡写真である。
【図5】ヒト子宮内膜間質肉腫の顕微鏡写真である。
【図6】正常子宮平滑筋、子宮平滑筋腫および子宮平滑筋肉腫の免疫組織化学染色写真である。
【図7】同一組織における子宮平滑筋肉腫部位と正常子宮平滑筋部位の免疫組織化学染色写真である。子宮平滑筋肉腫部位においてLMP2の発現低下が認められる。
【図8】LMP2の発現が認められない子宮平滑筋肉腫細胞(SKN)に、遺伝子組換えによりLMP2を強制発現させ、SKN細胞の形態の変化を示す写真である(その1)。
【図9】LMP2の発現が認められない子宮平滑筋肉腫細胞(SKN)に、遺伝子組換えによりLMP2を強制発現させ、SKN細胞の形態の変化を示す写真である(その2)。
【図10】LMP2の発現が認められない子宮平滑筋肉腫細胞(SKN)に、遺伝子組換えによりLMP2を強制発現させ、SKN細胞のファイブロネクチンの発現変化を示す写真である。
【図11】ヒト正常子宮平滑筋細胞(HuUSMC)におけるファイブロネクチンの発現変化を示す写真である。
【図12】各細胞における形態の変化、細胞増殖速度、およびファイブロネクチンの発現状況のまとめを示す図である。
【図13】マウス骨格筋組織、心筋組織および平滑筋組織におけるLMP2発現を示す写真である。
【図14】子宮平滑筋肉腫、子宮平滑筋腫および他臓器で発症した平滑筋肉腫におけるLMP2の発現を示す図である。
【図15】INFγシグナル伝達経路とLMP2の発現の関係を示す図である。
【図16】子宮平滑筋肉腫組織由来のJAK1キナーゼ遺伝子、STAT1遺伝子およびLMP2プロモーターの変異を示す図である。
【図17】子宮平滑筋肉腫組織由来のJAK1キナーゼ遺伝子、STAT1遺伝子およびLMP2プロモーターの変異を示す図である。
【図18】各組織におけるLMP2の発現状況を示す図である。図18において、Nは、LMP2の発現を検査した組織の症例数を示す。
【図19】pCEM9ベクター(LMP2遺伝子が含まれていない)またはpCEM9-LMP2ベクター(LMP2遺伝子がふくまれている)をヒト子宮平滑筋肉腫培養細胞(SKN細胞)に移入し、ネオマイシンにより選択された各ベクターを取り込んだ場合の、SKN細胞の増殖により形成されたコロニーの数を示す図である。1μgのpCEM9およびpLMP2 DNAを2×105細胞のDT細胞又はSKN細胞にトランスフェクトし、0.5または0.4mg/mlのG418を含む成長培地中でセレクトした。
【図20】LMP2の発現が認められない子宮平滑筋肉腫細胞(SKN)に、遺伝子組換えによりLMP2を強制発現させ、SKN細胞の形態の変化を示す写真である。
【図21】各細胞における形態の変化、細胞増殖速度、およびファイブロネクチンの発現状況のまとめを示す図である。
【図22】LMP2を恒常的に発現させたSKN細胞が腫瘍形成能の指標であるコロニー形成を著しく低下していることを示す写真である。
【図23A】LMP2を恒常的に発現させたSKN細胞がヌードマウスへの移植実験により腫瘍形成能を著しく低下していることを示す写真である。
【図23B】LMP2を恒常的に発現させたSKN細胞がヌードマウスへの移植実験により腫瘍形成能を著しく低下していることを示す図である。
【図23C】pCEM9-LMP2ベクター(LMP2遺伝子が含まれている)を移入したヒト子宮平滑筋肉腫培養細胞(SKN細胞)のコロニー#121が、SKN細胞と比較してLMP2遺伝子を著しく発現していることを示す写真である。
【図24】マイクロアレイを用いた遺伝子発現解析の結果、LMP2を恒常的に発現させたSKN細胞(コロニー#121)では、細胞増殖を誘導するサイクリンEの発現が著しく低下していることを示す図である。
【図25】サイクリンEプロモーターを含んだルシフェラーゼレポーター遺伝子の構造を示す図である。
【図26】SKN細胞とLMP2を恒常的に発現しているSKN細胞(コロニー#121と#122)での、サイクリンEプロモーター活性を示す図である。子宮平滑筋肉腫培養細胞SKN細胞で認められた顕著なサイクリンEの発現は、LMP2の恒常的な発現により著しく低下した。
【図27】ヒト子宮平滑筋肉腫組織におけるサイクリンEの顕著な発現を示す組織染色写真である。正常な子宮平滑筋層においては、サイクリンEの発現は認められないが、悪性腫瘍であるヒト子宮平滑筋肉腫組織では、サイクリンEの顕著な発現が認められる。
【図28】ヒト子宮平滑筋肉腫組織における分裂期の核内でのサイクリンEの発現を示す組織染色写真である。通常、細胞増殖を誘導するサイクリンEは、細胞増殖の開始期であるG1期に細胞質において過剰に発現し、速やかに核内へと移行しS期において染色体の合成を開始させる。その後、サイクリンEは、S期の後半から即座に分解される。したがって、正常な細胞のG2期とM期においては、サイクリンEの発現は認められない。しかし、ヒト子宮平滑筋肉腫組織において、分裂期の核内での顕著なサイクリンEの発現が認められる。
【発明を実施するための形態】
【0025】
本発明はLMP2をマーカーとして用いて、子宮平滑筋肉腫を検出する。ここで、検出とは
、子宮平滑筋肉腫が存在するかどうかを判定すること、患者が子宮平滑筋肉腫に罹患して
いるかどうか判定すること、子宮平滑筋の腫瘍が子宮平滑筋腫であるかまたは子宮平滑筋
肉腫であるかを鑑別すること、子宮平滑筋肉腫の悪性度を判定すること等を含む。
【0026】
LMP2は、プロテアソームのサブユニットであり、本発明の方法においては、子宮平滑筋
層において、LMP2が転写発現しているかどうかを検出する。ヒトの子宮平滑筋層の組織で
のLMP2の転写発現を調べると、正常な子宮平滑筋層と子宮平滑筋腫では、LMP2の発現は陽
性(中程度から強陽性)であるのに対して、子宮平滑筋肉腫では、LMP2の発現は陰性の部
分が多く弱陽性の部分が若干認められる。従って、LMP2の転写発現を指標に子宮平滑筋肉
腫に罹患しているかどうか、あるいは子宮平滑筋の腫瘍が子宮平滑筋腫か子宮平滑筋肉腫
のいずれかであるかを鑑別診断することができる。また、良性腫瘍である子宮平滑筋腫で
はLMP2の転写発現は強いのに対して、悪性化した腫瘍である子宮平滑筋肉腫ではLMP2の転
写発現が著しく減弱する。このことは、LMP2の転写発現が子宮平滑筋細胞に認められる腫
瘍に対して悪性度の指標になることを示す。
【0027】
さらに、本発明は、サイクリンE(CyclinE)をマーカーとして用いて、子宮平滑筋肉腫
を検出する方法を包含する。サイクリンは、サイクリン依存性キナーゼの活性を調節する
細胞周期で重要な役割を果たすタンパク質であり、サイクリンEは、ほ乳類細胞のG1期に
おいて作用するG1サイクリンの一つである。サイクリンEは、子宮平滑筋肉腫組織におい
て、正常組織に比較して発現が著しく上昇するが、子宮平滑筋腫組織においては上昇しな
い。本発明においては、LMP2と同様に子宮平滑筋層において、サイクリンEが転写発現し
ているかどうかを検出する。サイクリンEの転写発現を指標に子宮平滑筋肉腫に罹患して
いるかどうか、あるいは子宮平滑筋の腫瘍が子宮平滑筋腫か子宮平滑筋肉腫のいずれかで
あるかを鑑別診断することができる。また、良性腫瘍である子宮平滑筋腫ではサイクリン
Eの転写発現は正常子宮平滑筋組織と比べて変化がないのに対して、悪性化した腫瘍であ
る子宮平滑筋肉腫ではサイクリンEの転写発現が亢進する。さらに、通常、増殖期にある
正常な細胞のG2期とM期においては、サイクリンEの発現は認められない。しかし、ヒト子
宮平滑筋肉腫組織において、分裂期の核内での顕著なサイクリンEの発現が認められる。
このことは、サイクリンEの転写発現が子宮平滑筋細胞に認められる腫瘍に対して悪性度
の指標になることを示す。
【0028】
また、LMP2とサイクリンEを同時に検出してもよい。LMP2の転写発現が減弱し、かつサ
イクリンEの転写発現が亢進している場合、子宮平滑筋肉腫に罹患していると判断するこ
とができる。LMP2またはサイクリンE単独を指標とする場合よりも、LMP-2およびサイクリ
ンEの両方を指標とした場合に、より正確に子宮平滑筋肉腫を検出することが可能である
。
【0029】
本発明においては、子宮平滑筋肉腫に罹患しているかどうかの判断、子宮平滑筋の腫瘍
が子宮平滑筋腫か子宮平滑筋肉腫のいずれかであるかの鑑別診断、および子宮平滑筋細胞
に認められる腫瘍の悪性度の評価を含めて子宮平滑筋肉腫の検出と呼ぶ。
【0030】
本発明の方法においては、子宮平滑筋層におけるLMP2および/もしくはサイクリンEの
転写もしくは発現のいずれかまたは両方を検出することにより、子宮平滑筋肉腫を検出す
ることができる。LMP2またはサイクリンEの転写はLMP2またはサイクリンEをコードするmR
NAを測定することにより検出することができ、LMP2またはサイクリンEの発現はLMP2また
はサイクリンEタンパク質を測定することにより検出することができる。
【0031】
LMP2および/またはサイクリンEの転写を検出する場合、子宮平滑筋層の組織または細
胞を生体試料として採取し、該試料中に含まれるLMP2および/またはサイクリンEをコー
ドするmRNAを測定すればよい。mRNAの測定のためには、子宮平滑筋組織からバイオプシー
により組織の一部を採取し、また子宮平滑筋組織より綿棒等を用いて採取し、材料として
用いる。組織の採取は、例えば通常の外来診察中に、子宮内腔にバイオプシー用のかん子
を挿入し、1〜2mm角程度の子宮組織片を採取すればよい。mRNAの測定は採取した組織ま
たは細胞からmRNAを抽出して行ってもよいし、組織切片標本を作製するか、または採取し
た細胞をスライドガラス上に固定し、in situ ハイブリダイゼーション法により染色して
行ってもよい。あるいは、抽出したmRNAをノーザンブロット法やRT-PCR等の公知のRNA測
定法により測定すればよい。mRNAの抽出は公知の方法、例えば、新生化学実験講座2 核
酸I 分離精製 東京化学同人 1991年7月10日や分子生物学実験プロトコールI 丸善株
式会社 平成9年6月30日の記載に従って行うことができる。in situ ハイブリダイゼー
ションは、例えば分子生物学実験プロトコールIII 丸善株式会社 平成9年8月30日の
記載に従って行うことができる。この際、LMP2および/またはサイクリンEをコードするm
RNAを特異的に測定するために、LMP2および/またはサイクリンEをコードするmRNAの部分
配列に相補的な部分配列からなるプローブまたはプライマーを用いる。LMP2の塩基配列は
公知であり(例えば、GenBank アクセッション番号U01025、配列番号1、配列番号2はLMP
2タンパク質のアミノ酸配列を示す)、公知の塩基配列情報に基づいて、プローブまたは
プライマーを設計することができる。同様に、サイクリンEの塩基配列も公知であり(例
えば、GenBankアクセッション番号M73812、配列番号8)該プライマーまたはプローブは
、上記のLMP2および/またはサイクリンE遺伝子の断片であり、塩基の数は5〜50、好ま
しくは10〜30、さらに好ましくは15〜25である。
【0032】
LMP2および/またはサイクリンEの発現を検出する場合、子宮平滑筋層の組織または細
胞を生体試料として採取し、該試料中に含まれるLMP2および/またはサイクリンEタンパ
ク質を測定すればよい。子宮平滑筋層の組織または細胞は上記のようにして採取すればよ
い。また、LMP2および/またはサイクリンEタンパク質の測定も、組織または細胞からタ
ンパク質を抽出し、該抽出物中のLMP2および/またはサイクリンEタンパク質を測定して
もよいし、免疫組織化学または免疫細胞化学の手法により行ってもよい。抽出したタンパ
ク質の測定は、ELISA、ラジオイムノアッセイ等の公知の免疫測定法(イムノアッセイ)
を用いればよい。この際、LMP2および/またはサイクリンEに対する抗体が必要であるが
、抗LMP2抗体および/または抗サイクリンE抗体は公知の手法によりモノクローナル抗体
またはポリクローナル抗体として作製すればよい。また、市販の抗LMP2抗体および/また
は抗サイクリンE抗体を用いることもできる。抗体は必要に応じて、酵素、蛍光物質、放
射性同位元素により標識して用いるが、抗体の標識は公知の方法により行うことができる
。免疫組織化学または免疫細胞化学の手法による測定は、採取した子宮平滑筋組織切片標
本を作製するか、採取した細胞をスライドガラス上に固定して行う。免疫組織化学の手法
による測定のためには、組織を例えば、ホルマリンで固定し、パラフィン中に包埋し、ミ
クロトーム等の薄切装置で1〜5μm程度の厚さの切片標本を作製し、測定時にキシレン
やエタノール処理等によりパラフィンを除去し、生理食塩水または緩衝液に浸して親水化
すればよい。染色は、酵素、蛍光物質、放射性同位元素等で標識した抗LMP2抗体および/
または抗サイクリンE抗体を用いてもよいし、抗LPM2抗体を切片標本中のLMP2および/ま
たはサイクリンEに結合させた後に、抗LMP2抗体および/または抗サイクリンE抗体に結合
する2次抗体であって酵素、蛍光物質等で標識した2次抗体を用いてもよい。標識に用い
る酵素として、例えば西洋ワサビペルオキシダーゼ、アルカリフォスファターゼ等が挙げ
られ、蛍光物質として、例えばフルオレセイン、ローダミン等が挙げられる。また、公知
のビオチン-アビジン複合体を利用して染色してもよい。免疫細胞化学は採取した細胞を
ホルマリン等によりスライドガラス上に固定し、免疫組織化学の手法と同様の方法で細胞
中のLMP2および/またはサイクリンEを可視化すればよい。免疫組織化学または免疫細胞
化学において、染色は顕微鏡や肉眼で判断することもできるし、適当な光学的測定装置を
用いてもよい。免疫組織化学による染色は、例えば分子生物学実験プロトコールIII 丸
善株式会社 平成9年8月30日刊行の記載に従って行うことができる。
【0033】
上記のように、子宮平滑筋組織または細胞におけるLMP2の転写または発現を検出し、LM
P2の転写または発現が喪失しているか、または低下している場合、子宮平滑筋肉腫に罹患
していると判定することができる。また、子宮平滑筋組織または細胞におけるサイクリン
Eの転写または発現を検出し、サイクリンEの転写または発現が亢進している場合、子宮平
滑筋肉腫に罹患していると判定することができる。さらに、LMP2とサイクリンEの両方の
転写または発現を同時に検出してもよい。LMP2の転写または発現が喪失しているか、また
は低下しており、なおかつサイクリンEの転写または発現が亢進している場合、子宮平滑
筋肉腫に罹患していると判定することができる。例えば、組織または細胞の抽出物中のLM
P2 mRNAもしくはLMP2タンパク質またはサイクリンE mRNAもしくはサイクリンEタンパク質
を測定する場合、あらかじめ子宮平滑筋肉腫に罹患していない正常人の単位重量組織また
は単位細胞数当りのLMP2 mRNAもしくはLMP2タンパク質またはサイクリンE mRNAもしくは
サイクリンEタンパク質を測定する。正常人の値よりLMP2 mRNAまたはLMP2タンパク質が有
意に低い場合、子宮平滑筋肉腫に罹患していると判定し得、正常人の値よりサイクリンE
mRNAまたはサイクリンEタンパク質が有意に高い場合、子宮平滑筋肉腫に罹患していると
判定し得る。
【0034】
また、組織または細胞のLMP2の転写または発現をin situ ハイブリダイゼーション、免
疫組織化学、免疫細胞化学により検出する場合、組織または細胞が染色されず、LMP2の転
写、発現が認められない場合は、その部分の組織または細胞は子宮平滑筋肉腫組織または
細胞であり、組織または細胞を採取した患者は子宮平滑筋肉腫に罹患していると判定する
ことができる。さらに、組織または細胞のサイクリンEの転写または発現をin situ ハイ
ブリダイゼーション、免疫組織化学、免疫細胞化学により検出する場合、組織または細胞
が強く染色され、サイクリンEの転写、発現が強く認められる場合は、その部分の組織ま
たは細胞は子宮平滑筋肉腫組織または細胞であり、組織または細胞を採取した患者は子宮
平滑筋肉腫に罹患していると判定することができる。この際、子宮平滑筋肉腫でない正常
組織および子宮平滑筋肉腫組織の染色組織切片標品または染色細胞標品をあらかじめ作製
しておき、該標品と比較してもよい。
【0035】
LMP2の転写または発現の検出は、子宮平滑筋腫瘍に罹患した患者の腫瘍が悪性か良性か
、すなわち、子宮平滑筋肉腫かそれとも子宮平滑筋腫かの鑑別に利用することもできる。
この場合、LMP2の転写または発現が喪失しているか、正常組織に比較して有意に低下して
いる部分の組織は、悪性の子宮平滑筋肉腫であると判定することができる。また、サイク
リンEの転写または発現の検出は、子宮平滑筋腫瘍に罹患した患者の腫瘍が悪性か良性か
、すなわち、子宮平滑筋肉腫かそれとも子宮平滑筋腫かの鑑別に利用することもできる。
この場合、サイクリンEの転写または発現が、正常組織に比較して有意に亢進している部
分の組織は、悪性の子宮平滑筋肉腫であると判定することができる。この際、LMP2および
サイクリンEの両方を検出することにより、悪性度についてより正確な判断が可能になり
、また、より正確に鑑別を行なうことができる。さらに、組織を用いたin situ ハイブリ
ダイゼーションまたは免疫組織化学の手法によれば、組織のどの部分が正常でどの部分が
子宮平滑筋肉腫部分かを鑑別することもできる。さらに、例えば、組織切片の単位体積当
り、または単位細胞数当りのLMP2および/またはサイクリンEの転写または発現が喪失し
ているかまたは低下している細胞の数を測定することにより、子宮平滑筋腫瘍の悪性度を
判定することができる。
【0036】
さらに、従来の子宮平滑筋肉腫の診断方法と本発明のLMP2および/またはサイクリンE
の転写または発現を指標に子宮平滑筋肉腫を診断する方法を組合せることにより、より精
度の高い検出を行うことができる。従来法として、細胞の形態、密度等を観察する方法、
またはミオシンの転写または発現を指標に子宮平滑筋肉腫を診断する方法が挙げられ、こ
れらの方法は主に組織または細胞を固定して行われてきた。すなわち、組織切片標本また
は細胞スライドガラス標本を作製し、該標本の細胞の形態や密度を測定し、さらに、ミオ
シンの転写または発現を測定し、さらに、LMP2および/またはサイクリンEの転写または
発現を測定すればよい。
【0037】
本発明は、さらにLMP2遺伝子を子宮平滑筋肉腫患者に投与することにより、子宮平滑筋
肉腫を治療する方法、LMP2遺伝子を含む遺伝子治療のための遺伝子治療剤も包含する。遺
伝子治療における目的の遺伝子の子宮平滑筋肉腫患者への導入は公知の方法により行うこ
とができる。遺伝子を患者への導入する方法として、ウイルスベクターを用いる方法およ
び非ウイルスベクターを用いる方法があり、種々の方法が公知である(別冊実験医学、遺
伝子治療の基礎技術、羊土社、1996;別冊実験医学、遺伝子導入&発現解析実験法、羊土
社、1997;日本遺伝子治療学会編、遺伝子治療開発研究ハンドブック、エヌ・ティー・エ
ス、1999)。遺伝子導入のためのウイルスベクターとしては、アデノウイルス、アデノ随
伴ウイルス、レトロウイルス等のウイルスベクターを用いた方法が代表的なものである。
無毒化(ウイルス自身の複製が起こらない)したレトロウイルス、ヘルペスウイルス、ワク
シニアウイルス、ポックスウイルス、ポリオウイルス、シンビスウイルス、センダイウイ
ルス、SV40、免疫不全症ウイルス(HIV)等のDNAウイルスまたはRNAウイルスに目的とする
遺伝子を導入し、細胞に組換えウイルスを感染させることによって、細胞内に遺伝子を導
入することが可能である。また、プラスミドベクター等の遺伝子発現ベクターを用いて、
LMP2遺伝子を細胞や組織に導入することができる。例えば、リポフェクション法、リン酸
-カルシウム共沈法、DEAE-デキストラン法、微小ガラス管を用いたDNAの直接注入法など
により細胞内へ遺伝子を導入することができる。また、内包型リポソーム(internal lipo
some)による遺伝子導入法、静電気型リポソーム(electrostatic type liposome)による遺
伝子導入法、HVJ-リポソーム法、改良型HVJ-リポソーム法(HVJ-AVEリポソーム法)、HVJ-E
(エンベロープ)ベクターを用いた方法、レセプター介在性遺伝子導入法、パーティクル銃
で担体(金属粒子)とともにDNA分子を細胞に移入する方法、naked-DNAの直接導入法、種々
のポリマーによる導入法等によっても、組換え発現ベクターを細胞内に取り込ませること
が可能である。この場合に用いる発現ベクターとしては、生体内で目的遺伝子を発現させ
ることのできるベクターであれば如何なる発現ベクターも用いることができるが、例えば
pCAGGS(Gene 108, 193-200(1991))や、pBK-CMV、pcDNA3、pZeoSV(インビトロゲン社、
ストラタジーン社)、pVAX1などの発現ベクターが挙げられる。
【0038】
LMP2遺伝子を含むベクターは、遺伝子を転写するためのプロモーターやエンハンサー、
ポリAシグナル、遺伝子が導入された細胞の標識および/または選別のためのマーカー遺
伝子等を含んでいてもよい。この際のプロモーターとしては、公知のプロモーターを用い
ることができる。
【0039】
LMP2遺伝子を含む遺伝子治療剤は、LMP2遺伝子を含むベクターならびに薬理学的に許容
され得る担体、希釈剤もしくは賦形剤を含む。担体、希釈剤、賦形剤としては、製剤分野
において通常用いられるものを用いることができる。例えば、錠剤用の担体、賦形剤とし
ては、乳糖、ステアリン酸マグネシウムなどが使用される。注射用の水性液としては、生
理食塩水、ブドウ糖やその他の補助薬を含む等張液などが使用され、適当な溶解補助剤、
たとえばアルコール、プロピレングリコールなどのポリアルコール、非イオン界面活性剤
などと併用しても良い。油性液としては、ゴマ油、大豆油などが使用され、溶解補助剤と
しては安息香酸ベンジル、ベンジルアルコールなどを併用しても良い。本発明の遺伝子治
療剤は、子宮平滑筋肉腫部位に局所投与することが好ましく、例えば子宮平滑筋肉腫部位
に注射により投与すればよい。
【0040】
投与量は、症状、年齢、体重などによって異なるが、子宮平滑筋肉腫患者体内でLMP2を
発現する発現ベクター等に挿入されたLMP2遺伝子は、数日または数週間または数ヶ月おき
に1回あたり、0.001mg〜100mgを子宮平滑筋肉腫部位に注射等により直接投与すればよい
。
【0041】
本発明は、さらにインターフェロンγ(IFN-γ)シグナル伝達カスケードに関与する特
定の因子における変異により子宮平滑筋肉腫に罹患しているか否か、または子宮平滑筋肉
腫に罹患するリスクを有しているか否かを判定する方法をも包含する。
【0042】
正常細胞においては、IFN-γによりLMP2の遺伝子は活性化されるのに対して、子宮平滑
筋肉腫細胞においては、IFN-γによるLMP2の活性化はほとんど認められない。IFN-γによ
るLMP2の活性化は、図15にメカニズムを示すようにJAK1キナーゼ、JAK2キナーゼ、STAT
1などを介して、IRF-1が発現誘導され、LMP-2遺伝子のプロモーターに結合することによ
り生じる。IFN-γシグナル伝達に関与する因子のうち、JAK1キナーゼ及びSTAT1遺伝子の
変異、並びにLMP2のプロモーター領域の変異によりシグナル伝達が遮断され、LMP-2の発
現が阻害される。
【0043】
変異は以下のとおりである。以下、例えば遺伝子の変異をA210Gで表した場合、遺伝子
の塩基配列の21番目のAがGに置換していることを示す。また、遺伝子のコードするタンパ
ク質のアミノ酸配列の変異をG871Eで表した場合、アミノ酸配列中、871番目のグリシン(G
)がグルタミン酸(E)に置換していることを示す。
【0044】
JAK1キナーゼにおける変異
遺伝子変異 対応アミノ酸変異 変異の存在するドメイン
A2612A G781E ATP binding
G2626A G876R ATP binding
G2642T C881F ATP binding
A2967C G986P active site
A2960C Y987S active site
A2985T R995S active site
JAK1キナーゼ遺伝子の塩基配列を配列番号3に、JAK1キナーゼのアミノ酸配列を配列番
号4に示す。
【0045】
STAT1における変異
遺伝子変異 対応アミノ酸変異 変異の存在するドメイン
A2104C I702L non kinase active region
T2128G S710A non kinase active region
T2078G L693R non kinase active region
A2148C R716S non kinase active region
STAT1遺伝子の塩基配列を配列番号5に、STAT1のアミノ酸配列を配列番号6に示す。
【0046】
LMP2プロモーター
遺伝子変異 対応アミノ酸変異 変異の存在するドメイン
A210G IRF-E site
C214T IRF-E site
A216G IRF-E site
A217G IRF-E site
G219A IRF-E site
LMP2プロモーターの遺伝子配列を配列番号7に示す。
【0047】
JAK1キナーゼ遺伝子、STAT1遺伝子およびLMP2プロモーターの全長DNAまたはその断片は
塩基配列情報に基づいて容易に得ることができる。
【0048】
本発明は、上記のJAK1キナーゼ遺伝子、STAT1遺伝子またはLMP2プロモーターの塩基の
変異を検出し、子宮平滑筋肉腫に罹患しているか否かを検出する方法若しくは子宮平滑筋
肉腫に罹患するリスクが高いか否かを判定する方法、並びに上記のJAK1キナーゼまたはST
AT1のアミノ酸の変異を検出し、子宮平滑筋肉腫に罹患しているか否かを検出する方法若
しくは子宮筋肉腫に罹患するリスクが高いか否かを判定する方法を包含する。
【0049】
遺伝子における塩基の変異は、上記変異部位を含む遺伝子の断片をプローブとして、ま
たはDNAチップ、DNAマイクロアレイ技術において固相化するDNAとして用いればよい。そ
の場合の、断片の塩基長は全長でもよいが、通常は15bp〜100bpが好ましく、さらに15bp
〜50bpが好ましく、特に18bp〜30bpが好ましい。断片に含まれる変異部位は、1つだけで
もよいし、複数、すなわち2個、3個、4個、5個または6個でもよい。
また、上記断片に相補的な配列を有するDNAも本発明の範囲である。相補的なDNAも本願
明細書の開示に基づいて得ることができる。
【0050】
本発明のプローブは、検出のために蛍光物質、酵素、放射性同位体、化学発光物質等で
標識されていてもよい。標識に用いる標識物質は、公知のものを用い、公知の方法で標識
することができる。蛍光物質としては、例えば、Cy3、Cy5、ローダミン、フルオレセイン
等が挙げられる。
【0051】
さらに、上記の遺伝子変異を検出するPCR-RFLP等のPCRに用いるプライマーも本発明の
範囲である。すなわち、本発明は上記のJAK1キナーゼ遺伝子、STAT1遺伝子及びLMP2プロ
モーターの変異部位の3’側および5’側に存在する10から30塩基からなる部分配列からな
る、DNA断片の増幅に用い得る一対のプライマーセットを包含する。
【0052】
変異は、本発明のDNAを用いてPCR法、サザンハイブリダイゼーション法、ノーザンハイ
ブリダイゼーション法、定量的PCR法、in situ ハイブリダイゼーション法、FISH(Fluore
scence In Situ Hybridization)、PCR-RFLP法、PCR-SSCP法等により、検出することがで
きる。
【0053】
例えば、まずJAK1キナーゼ遺伝子、STAT1遺伝子またはLMP2プロモーターの変異塩基部
分を含むヌクレオチド配列に相補的なプローブ、ならびに野生型遺伝子の該変異塩基部分
に対応する部分を含むヌクレオチド配列に相補的なプローブを調製する。用いるプローブ
の長さに制限はなく、後述の核酸増幅法で増幅しようとする核酸断片の全長でもよいが、
通常は15bp〜100bpが好ましく、さらに15bp〜50bpが好ましく、特に18bp〜30bpが好まし
い。プローブは、放射性同位元素、蛍光物質、酵素等で標識したものを用いてもよい。次
いで、子宮平滑筋組織から採取した組織または細胞検体試料中の変異塩基部分を含む遺伝
子断片を核酸増幅法により増幅し、この増幅断片とプローブを反応させる。検体試料中の
DNAが野生型、変異型のいずれのプローブとハイブリダイズするか調べることにより、上
記遺伝子DNAに変異が生じているか否かを検出することができる。本発明のプローブは、
1塩基のミスマッチを検出するため、そのハイブリダイゼーション条件は、ストリンジェ
ントなものであることが必要である。ハイブリダイゼーション時の温度、塩濃度を調節す
ることにより1塩基のミスマッチのみを検出し得るハイブリダイゼーション条件を選択す
ることが可能である。用いるプローブDNAの長さにも依存するが、例えば具体的には、ナ
トリウム濃度が150〜900mM、好ましくは600〜900mMであり、温度が60〜68℃、好ましくは
65℃での条件で行い得る。
【0054】
核酸増幅の際に用いるプライマーとしては、上記遺伝子の変異部位を挟み増幅しようと
する領域の端部と相補的な配列を用いることができる。増幅する領域の塩基長に制限はな
いが、数十から数百塩基とすることができる。増幅領域に上記JAK1キナーゼ遺伝子、STA
T1遺伝子またはLMP2プロモーターDNAの変異を1つだけ含むように増幅塩基長を設定して
もよいし、複数の変異、すなわち、2個、3個、4個、5個または6個の変異を含むよう
に設定してもよい。また、プライマーを変異部位を含む領域に設定することも可能である
。プライマーの長さに制限はないが、好ましくは15bp〜50bp、さらに好ましくは20bp〜30
bpである。
【0055】
さらに、本発明のJAK1キナーゼ遺伝子、STAT1遺伝子またはLMP2プロモーター部位の変
異を含むDNA配列またはその断片に相補的なDNAを用いて、子宮平滑筋肉腫に罹患している
か否か、または子宮平滑筋肉腫患の罹患のリスクを決定するためのDNAチップを作製する
こともできる。この際、本発明のJAK1キナーゼ遺伝子、STAT1遺伝子またはLMP2プロモー
ター部位の変異を含む部分に相補的なDNA断片をニトロセルロース、ナイロン等のメンブ
レン、スライドグラス等に固相化すればよい。この際、固相化するDNA断片の塩基長は、
通常は15bp〜100bpが好ましく、さらに15bp〜50bpが好ましく、特に15bp〜25bpが好まし
い。次いで、該DNAチップを放射性同位元素、酵素、蛍光色素等で標識した被験体由来のD
NAまたはRNAと接触させ、ハイブリダイズを形成するか否かにより検体中に変異を有する
核酸が含まれているかどうかがわかる。
【0056】
変異の検出は子宮平滑筋肉組織から採取した組織片、細胞から核酸を抽出して行なえば
よい。
【0057】
本発明を以下の実施例によって具体的に説明するが、本発明はこれらの実施例によって
限定されるものではない。
【0058】
実施例1 ヒト子宮平滑筋肉腫におけるLMP2の転写および発現
本実施例においては、以下に記載する材料及び方法を用いて検討を行った。
【0059】
細胞株および培地
ヒト子宮平滑筋肉腫細胞株であるSKN細胞(RCB0513)は、理化学研究所セルバンクから購
入し、0.6%のL-グルタミン(Invitrogen社)および15%の牛胎児血清(シグマ-オルドリッチ
社)を補足したF-12Nutrient Mixture(Ham)培地(Invitrogen社)で維持した。HeLa細胞およ
びHeLa.S3細胞は0.6%のL-グルタミンおよび10%の牛胎児血清を補足したダルベッコのMEM
で維持した。ヒト子宮平滑筋細胞はCambrex BioScience Wailersville社から購入し、メ
ーカーのプロトコルに従って維持した。
【0060】
逆転写ポリメラーゼ連鎖反応(RT-PCR)分析
TAP1、LMP2、β2-mおよびβアクチンの転写物をRT-PCRにより調べた。細胞を250ユニ
ット/mlのヒトIFN-γ(Pepro Tech社)で処理しないかまたは48時間処理し、RNAを回収した
。トータルRNAはTRIzol試薬(Invitrogen社)を用いメーカーのプロトコルに従って5×106
細胞から調製した。RNAは、Superscript II enzyme(Invitrogen社)を用いて逆転写し、1
本鎖cDNAをTAP1、LMP2、β2mおよびβアクチン転写物を増幅するのに用いた。PCRは適切
なプライマーを用いて、30秒94℃35サイクル、30秒60℃、1.5分72℃および追加の5分の
プログラムで行い、転写物を伸長させた(Cabrera CM. et al. (2003) Tissue Antigens,
61,211-219; Miyagi T. et al. (2003) J.Gastroenterol.Hepatol., 18, 32-40)。
【0061】
免疫組織化学(Immunohistchemistry = IHC)
IHCは、Hsu SM. et al. (1981) J.Histochem.Cytochem. 29, 577-580に記載の方法によ
りアビジン-ビオチン複合体を用いて行った。すなわち、子宮平滑筋肉腫患者から得られ
た子宮摘出標本のパラフィン包埋サンプルから代表的な6つの5μm組織切片を作製した
。切片はパラフィン除去しアルコール中で再水和し、正常マウス血清を用いて20分間イン
キュベートした。次いで、切片を抗LMP2抗体(Affinity Res. Products社、希釈1/100)を
用いて1時間室温でインキュベートした。その後、切片をビオチン化2次抗体(Dako社)と
ともにインキュベートした。反応を3、3'-ジアミノベンジジンを用いて完了し、スライド
をヘマトキシリンで対比染色した。標本中の正常子宮平滑筋組織を陽性対照として用いた
。組織切片からなる陰性対照も、1次抗体の代わりに正常ウサギIgGとともにインキュベ
ートした。
【0062】
免疫沈降反応およびイムノブロット
細胞質抽出物および核抽出物は250ユニット/mlのヒトIFN-γで処理したか、または処理
しない5×106細胞から調製した(Brucet M. et al. (2004) Genes Immun., 5, 26-35)。細
胞を1200rpm、10分間の遠心分離により回収し、5mlの氷冷PBSで洗浄し、5分間、12000rpm
、4℃で遠心分離した。細胞をペレット化し、0.4mlのバッファーA(10mM Hepes、pH7.8
; 10mM KCl; 2mM MgCl2; 1mM DTT; 0.1mM EDTA; およびコンプリートプロテアーゼインヒ
ビターカクテル(Kirkegaard&Perr Lab社)中で一度洗浄し、2時間、4℃でインキュベー
トした。次いで、10%のNonidet P-40溶液25μlを添加し、細胞を1時間、4℃で激しく混
ぜ、次に、5分間、12000rpmで遠心分離した。遠心分離の後に上清を細胞質抽出物として
回収し、-80℃で保存した。ペレット化核は、40μlのバッファーC(50mM Hepes、pH7.8;
50mM KCl; 300mM NaCl; 0.1mM EDTA; 1mM DTT; 10%(v/v)グリセロール)に再懸濁し、4
℃で2時間混ぜて、5分間、4℃で12000rpm遠心分離した。核タンパク質を含む上清を回収
し、-80℃で保存した。
【0063】
STAT1、リン酸化状態のSTAT1、JAK1、JAK2、TAP1およびLMP2発現を検出するために、細
胞可溶化液または細胞質抽出物を10%SDS-ポリアクリルアミドゲル(SDS-PAGE)で分離し、
抗STAT1抗体、抗リン酸化STAT1抗体(Santa-Cruz Biotechnol.社)、抗JAK1、抗JAK2抗体(C
hemicon Int'l社)、抗体TAP1抗体(Stressgen社)または抗LMP2抗体(Affiniti Res. Produc
ts社)を用いて標準方法でイムノブロットを行った。IRF1またはIRF2発現の検出のために
、核抽出物を10%のSDS-PAGEページで分離し、抗IRF1抗体(Transduction Lab.社)、抗IRF2
抗体(Santa-Cruz Biotechnol.社)を用いて標準方法でイムノブロットを行った。目的のタ
ンパク質の発現は、アルカリフォスファターゼを結合した2次抗体を用いてアルカリフォ
スファターゼ発色をメーカーのプロトコールに従って行うことで、可視化して検討された
。
【0064】
図に示す時間250ユニット/mlのIFN-γで処理するか処理しなかった5×106細胞からの細
胞全体の抽出物を、50mM Tris-HCl、0.1mM EDTA、200mM NaCl、10%グリセロール、0.5%NP
-40、1mM DTTおよびコンプリートプロテアーゼインヒビターカクテル(Kirkegaard&Perr
Lab.社)を含むバッファーで溶解した。細胞可溶化液は、正常ウサギの血清(Santa-Cruz B
iotechnol.社)および20mlのプロテインG Sepharose(Amersham Biosciences社)であらかじ
め清澄化し、次に、2μgの抗JAK1または抗JAK2抗体を用いて免疫沈降を行った。サンプ
ルを10%のSDS-PAGEで分離し、Immobilon-Pメンブランにトランスファーした。リン酸化タ
ンパク質は、抗チロシンリン酸化抗体を1次抗体として反応させた後アルカリフォスファ
ターゼを結合した2次抗体を用いてアルカリフォスファターゼ発色をメーカーのプロトコ
ールに従って行うことで、可視化して検討された。IFN-γR1鎖発現を検出するために、細
胞全体のライセートは上述のように分離した。ブロットは抗IFN-γR1鎖抗体(PBL Biomedi
cal研究所)を用いて行った。SKN細胞をpRK5コントロール(2μg)またはJAK1発現ベクタ
ー(2μg)(St.Jude Children Research Hospital(テネシー州メンフィスのJ.Ihle博士
から提供された)でトランスフェクトした。IFN-γをトランスフェクト24時間後に添加し
、細胞を回収の前さらに24時間インキュベートした。pCMVβ-Galとの同時トランスフェク
トをトランスフェクト効率を標準化するために行った。
【0065】
トランスフェクトおよびレポーターアッセイ
TAP1およびLMP2wt(TAP1 593-1/pGL3およびLMP2 1-593/pLG3)の構造ならびにそれらのIR
F-E変異体プロモーター構築物の構造を図2に示す。トータル2μgのこれらのプラスミド
DNA(University of South FloridaのK.L. Wright博士から提供を受けた)を、メーカーの
推奨に従って、FuGENE 6 Transfection Reagent(Roche社)によりHeLaまたはSKN細胞中に
移入した。すべての移入されたDNAには200ngのpCMVβ-Gal(Tropix社)を内部トランスフェ
クト効率コントロールとして含んでいた。IFN-γ(最終濃度250ユニット/ml)をトランスフ
ェクト24時間後に添加し、細胞をさらに24時間インキュベートした。最終的に、細胞を洗
浄し、500μlの溶解バッファーで溶解し、Dual-Luciferase Reporter Assay System(Prom
ega社)を用いてメーカーの指示に従って分析した。LMP2またはTAP1/pGL3の代わりにpGL3
ベースで移入した細胞のルシフェラーゼ活性をバックグラウンドとして差し引いた。
【0066】
上記材料および方法を用いた検討により以下の結果が得られた。
【0067】
ヒト子宮平滑筋肉腫細胞中におけるINF-γによるTAP1およびLMP2発現の非誘導
LMP2欠損マウスは子宮平滑筋肉腫を発症した(Hayashi T. et al. (2002) Cancer Res.,
62, 24-27)。次いで、ヒト子宮平滑筋肉腫がTAP1およびLMP2の弱い発現を示すか否かを
示す必要がある。TAP1およびLMP2発現に対するIFN-γの効果は、4種の細胞株を用いたイ
ムノブロットにより調べた。IFN-γ処理に続くHeLa、HeLa.S3、およびHu.USMC(対照)は
TAP1およびLMP2の強い誘導を受けたが、ヒト平滑筋肉腫細胞株SKNにおけるIFN-γ処理で
はTAP1およびLMP2発現はそれほど誘導されなかった(図1a)。SKN細胞は、HeLa、HeLa.S3細
胞の両方とHu.USMC(対照)と比べると、同じようなβアクチン発現が認められたので、
抽出物の調製工程はIFN-γ処
理に続くTAP1およびLMP2発現の非誘導に影響しなかった。こ
のIFN-γの量は、HeLa、HeLa.S3細胞の両方とHu.USMC中(図1a)のTAP1およびLMP2遺伝子の
両方に共有されている双方向プロモーターを最大限に誘導するのに十分であった(図1a
)。INF-γ量を500ユニット/mlまで増加させても、SKN細胞中でTAP1およびLMP2発現を顕
著に誘導することはなかった。従って、SKN細胞はIFN-γ処理によりTAP1およびLMP2発現
を増強する能力を失っていた。
【0068】
IFN-γ処理に続くTAP1およびLMP2発現の非誘導を立証するために、4種の細胞株を用い
てRT-PCR分析を行った。IFN-γ処理により誘導されたTAP1またはLMP2のいずれかのmRNA発
現はHeLa、HeLa.S3細胞およびHu.USMCにおいて明確に検出されたが、IFNγ処理はSKN細胞
中でTAP1およびLMP2のmRNA発現をそれほど誘導しなかった(図1b)。βアクチン対照のmRNA
発現は試験したすべての細胞中で同様の高いレベルで検出された。このことはRNA調製ス
テップがIFN-γ処理に続くTAP1およびLMP2発現の非誘導に影響しなかったことを示す(図1
b)。IHC実験により、正常子宮平滑筋細胞が6ケースにおいてLMP2を著しく発現したが、子
宮平滑筋肉腫細胞では発現しなかったことが示された(図1c)。IHCの結果は、SKN細胞中
でのTAP1およびLMP2の非誘導を立証した。
【0069】
TAP1およびLMP2遺伝子のIFN-γに誘導される共通の双方向性プロモーター活性化の喪失
IRF-1はTAP1およびLMP2遺伝子に共有される双方向プロモーター中のIRF-Eと呼ばれるシ
スエレメントに直接結合する(Wright KL. et al. (1995) J.Exp.Med., 181, 1459-1471;
White LC. et al. (1996) Immunity, 5, 365-376; Dovhey SE. et al. (2000) Cancer R
es., 60, 5789-5796; Brucet M. et al. (2004) Genes Immun., 5, 26-35)。IRF-Eは図
2に表されるように、NFκB様結合部位およびGC1ボックスの上流に位置する。IFN-γ処理
によるTAP1およびLMP2の発現増強にIRF-Eが必要であることが示された(Wright KL. et al
. (1995) J.Exp.Med., 181, 1459-1471; White LC. et al. (1996) Immunity, 5, 365-37
6; Dovhey SE. et al. (2000) Cancer Res., 60, 5789-5796; Brucet M. et al. (2004)
Genes Immun., 5, 26-35)。IFN-γがSKN細胞においてTAP1/LMP2の共有された双方向のプ
ロモーターを確実に活性化するかどうかを調べるために、SKN細胞およびHeLa細胞にTAP1
またはLMP2の発現を誘導すべくIRF-Eがwtもしくはmutであるプロモーター-ルシフェラー
ゼコンストラクトDNAを移入した。HeLa細胞の場合、IFN-γ処理により11倍のLMP2プロモ
ーター活性が誘導され、さらに10倍のTAP1プロモーター活性が誘導された(図2)。しかし
ながら、SKN細胞中では、IFN-γ処理の結果、TAP1/LMP2の共有された双方向のプロモータ
ーの活性化誘導は認められなかった。IRF-Eの変異により、HeLa細胞中で認められたIFN-
γのTAP1/LMP2遺伝子発現を誘導する能力が失われた(図2)。これらの結果は内在性のmRN
Aレベルと一致しており、HeLa細胞中でのTAP1およびLMP2遺伝子の活性が強調しているが
、SKN細胞では強調していないことを示す。IRF-E部位の変異はHeLa細胞およびSKN細胞の
両方でベースとなる発現レベルをいくらか減少させた。このことはこの部位がLMP-2の発
現において一定の役割を果たすことを示す。これらの結果は、IFNγ処理が、HeLa細胞中
でTAP1/LMP2の共有されたプロモーター活性を強く誘導し得るが、SKN細胞中では誘導しな
いことを示す(図2)。
【0070】
実施例2 免疫組織化学によるヒト正常子宮平滑筋層、ヒト子宮平滑筋腫およびヒト子宮
平滑筋肉腫の生検組織または手術摘出組織におけるLMP2の発現状況
ヒト正常子宮平滑筋層、ヒト子宮平滑筋腫およびヒト子宮平滑筋肉腫の顕微鏡観察
ヒト子宮平滑筋組織をバイオプシーまたは手術により採取し、顕微鏡観察した。
【0071】
図3はヒト正常子宮平滑筋組織を、図4は、ヒト子宮平滑筋腫およびヒト子宮平滑筋肉
腫を、図5はヒト子宮内膜間質肉腫の顕微鏡写真を示す。
【0072】
上記顕微鏡観察により判定した、正常子宮平滑筋10例、子宮平滑筋腫6例、子宮内膜肉
腫6例および子宮平滑筋肉腫(悪性度低)3例、子宮平滑筋肉腫(悪性度高)4例を用い
て、免疫組織化学により各組識のLMP2発現を調べた。
【0073】
免疫組織化学は、前記の方法で行った。
【0074】
図6に、正常子宮平滑筋、子宮平滑筋腫および子宮平滑筋肉腫の免疫組織化学染色写真
を示す。図に示すように、正常子宮平滑筋および子宮平滑筋腫は染色され、LMP2が発現さ
れていることが確認された。しかし、子宮平滑筋肉腫では染色されなかったため、LMP2の
発現が確認されなかった。
【0075】
図7に、子宮平滑筋肉腫の免疫組織化学染色写真を示す。図に示すように、同じ組織内
で染色された部分と染色されない部分が存在した。
【0076】
各組織におけるLMP2の発現状況は以下のとおりであった。+は陽性、-は陰性、-/+はグ
レーゾーンであることを示す。
LMP2発現
正常子宮平滑筋組織 ++++
子宮平滑筋腫組織 ++++
子宮内膜肉腫組織 -/+
子宮平滑筋肉腫(悪性度低)組織 -/+
子宮平滑筋肉腫(悪性度高)組織 -
【0077】
上記表に示されるように、悪性腫瘍である子宮平滑筋肉腫のみにおいて、顕著なLMP2の
発現低下が認められた。また、子宮平滑筋肉腫の組織でのLMP2の発現低下度は、悪性度に
依存していており、LMP2の発現低下度合いは、子宮平滑筋肉腫の悪性度の指標と成りうる
ことが明らかとされた。
【0078】
実施例3 SKN細胞におけるLMP2の強制発現
1. SKN細胞におけるLMP2の強制発現による細胞形態の変化
LMP2の発現低下が、子宮平滑筋細胞において形質転換(癌化)に直接的に関与している
かについて検討を行った。LMP2の発現が認められない子宮平滑筋肉腫細胞(SKN)に、遺伝
子組換えによりLMP2を強制発現させ、SKN細胞の形態を観察した。
【0079】
方法
LMP2発現プラスミドベクター(2μg)をSKN細胞(5 x 105)内にFuGene6 (Roche,Indianapo
lis, IN)を用いメーカーのプロトコールに従って導入した。同プラスミドベクターを細胞
に導入した3日後に、G418(ネオマイシン)を培養液に加えて(最終濃度200μg/ml)、LMP2発
現プラスミドベクターが導入されたSKN細胞のみを選択した。LMP2の強制発現されたSKN細
胞の細胞増殖能、細胞形態能について検討を行った。結果を図8、図9および図21に示
す。図8、図9および図21に示すように、SKN細胞の増殖能は、倍加時間15.2時間だっ
たの対して、LMP2の強制発現されたSKN細胞の細胞増殖能は、倍加時間17.5時間となった
。LMP2の発現により、SKN細胞の増殖速度が、倍加時間2時間遅くなった。また、細長い
斜方形の細胞形態を持っているSKN細胞が、LMP2の強制発現により繊維芽細胞様(菱形)の
細胞形態に変化したことが認められた。
【0080】
2. SKN細胞におけるLMP2の強制発現によるファイブロネクチンの発現の変化
一般的に転移能を有する悪性度の高い癌細胞は、細胞と細胞との接着因子(ファイブロ
ネクチン)の著しい発現低下が認められる。LMP2の発現が認められない子宮平滑筋肉腫細
胞(SKN)に、遺伝子組換えによりLMP2を強制発現させ、ファイブロネクチンの発現変化に
ついて検討を行った。
【0081】
LMP2の強制発現は実施例3と同様の方法で行った。ファイブロネクチンの発現は、抗フ
ァイブロネクチン抗体(Rockland社)を用いた免疫染色法により検討した。また、コント
ロールとして、ヒト正常子宮平滑筋細胞:HuUSMCを用いた。
【0082】
SKN細胞における結果を図10に、ヒトHuUSMCにおける結果を図11に示す。また、図
12には各細胞における形態の変化、細胞増殖速度、およびファイブロネクチンの発現状
況のまとめを示す。
【0083】
図に示すように、SKN細胞にLMP2を強制発現させた場合、SKN細胞の形態と増殖速度が、
正常の子宮平滑筋細胞に近づいた。また、SKN細胞にLMP2を強制発現させた場合、有意な
ファイブロネクチンの発現誘導が認められた。つまり、LMP2の強制発現により誘導された
ファイブロネクチンの発現は、SKN細胞の持つ転移能の減弱の可能性を示唆している。
【0084】
実施例4 各組織におけるLMP2の発現
LMP2の発現は、子宮平滑筋組織のみならず、本来骨格筋組織および心筋組織においても
認められる。ところが、LMP2の欠損により、平滑筋肉腫が発症するのは子宮平滑筋組織の
みである。そこで、骨格筋、心筋および平滑筋におけるLMP2発現を調べた。また、免疫機
構に必須であるRag1遺伝子を欠損させたマウスの骨格筋、心筋および平滑筋におけるLMP2
発現を調べた。
【0085】
免疫組織化学は、生後1日目のマウス新生児のパラフィン包埋サンプルを用いて、前記
の方法で行った。
【0086】
図13に免疫組織化学染色写真を示す。
【0087】
実施例5 子宮及び他の器官での平滑筋肉腫におけるLMP2の発現
ヒト子宮平滑筋肉腫の組織においてLMP2の発現の著しい消失が確認された。平滑筋肉腫
は、子宮以外にも種々の臓器にいても発症するが、LMP2の発現の消失は子宮平滑筋肉腫に
特異的に認められるものか検討した。
【0088】
方法
病理ファイルより、子宮平滑筋肉腫(悪性度高い)4症例、子宮平滑筋肉腫(悪性度低い)
4症例、子宮上皮平滑筋肉腫1症例、子宮内膜間質肉腫1症例、子宮平滑筋腫1症例、正
常子宮平滑筋1症例、後腹膜原発平滑筋肉腫1症例、大網原発平滑筋肉腫1症例、小腸原
発平滑筋肉腫1症例、腸管膜原発平滑筋肉腫1症例を選び、各疾患の組織におけるLMP2の
発現を抗ヒトLMP2抗体を用いた免疫組織染色により検討した。
【0089】
図14に示されるように、正常な子宮平滑筋組織や子宮平滑筋腫(良性腫瘍)において認
められた顕著なLMP2の発現が、子宮平滑筋肉腫(悪性腫瘍)では著しく減弱していることを
再確認した。子宮内膜間質肉腫1症例ではLMP2の発現が認められなかったが、子宮上皮平
滑筋肉腫1症例においてはLMP2の発現が有意に認められた。腹部・消化器系組織に発症し
た平滑筋肉腫においては、顕著なLMP2の発現が認められる場合もあるが、著しいLMP2の発
現の減弱が認められる場合もある。これまでのGene Copy Number Profilingのゲノム解析
より、子宮平滑筋肉腫の発症機序は、他の臓器における平滑筋肉腫の発症機序とは大きく
異なっている可能性が示されている。実施例5の結果より、LMP2の発現の消失は子宮平滑
筋肉腫に特異的に認められる現象と思われる。
【0090】
実施例6 IFN-γシグナル伝達因子の変異と子宮平滑筋肉腫の関係
「子宮平滑筋肉腫において特異的に認められるLMP2発現の著しい減弱」について再検討
し、その特異性を確立した。具体的には、ヒト正常子宮平滑筋組織21症例、子宮平滑筋腫
24症例、子宮内膜間質肉腫6症例、子宮平滑筋肉腫29症例についてLMP2の発現を抗ヒトLMP
2抗体を用いた免疫組織染色により検討した。図18に示されるように、子宮平滑筋肉腫
において特異的に認められるLMP2発現の著しい減弱性を確認した。
【0091】
LMP2の発現は、図15に示される様にIFN-γの刺激により顕著に誘導される。子宮平滑
筋肉腫において特異的に認められるLMP2発現の著しい減弱は、IFN-γのシグナル伝達因子
:JAK1、JAK2、STAT1、PKCd、PI3K、LMP2のpromoter領域の変異や欠損による不活性化に
起因している可能性が考えられる。そこで、IFN-γのシグナル伝達因子:JAK1、JAK2、ST
AT1、LMP2のpromoter領域の変異について検討した。
【0092】
方法
手術により摘出されたヒト子宮体部組織13症例から平滑筋肉腫組織部位のみと正常な平
滑筋組織部位のみをレーザーマイクロダイセクションにより切り取り回収した。正常な子
宮平滑筋組織13検体にヒト正常子宮平滑筋培養細胞1検体を加えた合計14検体、子宮平滑
筋肉腫組織13検体にヒト子宮平滑筋肉腫培養細胞1検体を加えた合計14検体を用いてJAK1
、JAK2、STAT1、LMP2のpromoter領域の変異について検討した。各検体をHMW溶液(10mM Tr
is-HCl pH8.0、150mM NaCl、10mMEDTA-NaOH pH8.0、0.1% SDS)-proteinase K (100mg/ml)
に加えて55℃で24時間保温し、フェノール/クロロホルムにより不純物を取り除きゲノムD
NAを精製した。JAK1分子内のATP結合領域をコードする遺伝子領域(A)とチロシンリン酸化
酵素活性化領域をコードする遺伝子領域(B)の塩基配列を決定するために、遺伝子領域(A)
と遺伝子領域(B)の各領域に対するPCR-primerを用いてそれぞれの領域を含むDNA断片を各
検体より得られたゲノムDNAを用いてPCRにより増幅させた。PCRにより増幅されたDNA断片
をアガロースゲル電気泳動法により抽出し精製し、シークエンサー(ABI Prism3100 Genet
ic Analyzer)により塩基配列を決定した。さらに、JAK1の塩基配列を決定した方法と同様
の手法を用いて、JAK2分子内のATP結合領域をコードする遺伝子領域(C)とチロシンリン酸
化酵素活性化領域をコードする遺伝子領域(D)の塩基配列、STAT1分子内における転写因子
活性化領域(701番目のチロシンと727番目のセリン)コードする遺伝子領域(E)の塩基配列
、LMP2のpromoter領域をコードする遺伝子領域(F)の塩基配列を決定した。子宮平滑筋肉
腫における特異的な変異を検出するために、同一子宮体部における子宮平滑筋肉腫組織部
位と正常子宮平滑筋組織部位での塩基配列(AからE領域)を比較検討した。
【0093】
IFN-γのシグナル伝達因子:JAK1、JAK2、STAT1、各因子におけるリン酸化酵素あるい
は転写因子の活性化領域、LMP2のpromoter領域の変異解析の結果は、図16(変異を各検
体ごとにまとめた図)と図17(変異を各因子ごとにまとめた図)に示している。
【0094】
実施例7 ヒト子宮平滑筋肉腫細胞における、LMP-2の恒常的発現による細胞形態と細胞
増殖への影響
ヒト子宮平滑筋肉腫の組織において顕著なLMP2の発現低下が認められるが、LMP2の発現
低下が、子宮平滑筋肉腫の発症に関与しているのか検討した。
【0095】
方法
SKN細胞を6穴プレートにおいて培養し、70%コンフルエントの際、HamF-12-15%Fcs培
養液2mlを交換した。培養液を交換した次ぎの日、Roche社のプロトコールに従って、1
穴に対して2μgのpCEM9ベクター(LMP2の遺伝子を含まない)またはpCEM-LMP2ベクター(LM
P2の遺伝子を含んでいる)をFuGENE6(Roche社)によりSKN細胞に移入した。各ベクターの移
入を行ったSKN細胞をCO2インキュベーター内において37℃で48時間培養した後、SKN細胞
をトリプシン処理によって剥がして100mm培養シャーレでCO2インキュベーター内において
37℃で48時間培養した。その後、G418(ネオマイシン)100μg/mlを含んだHamF-12-15%Fcs
培養液で培養を行った。CO2インキュベーター内において37℃で2週間培養すると、pCEM9
ベクターまたはpCEM-LMP2ベクターが移入されたSKN細胞のみがG418によって選択的に生き
残り増殖しコロニーを形成しているのが確認された。G418(ネオマイシン)含有培養液にて
選択培養の3週間後、100mm培養シャーレ内における各ベクターが移入されたSKN細胞によ
り形成されたコロニー数を数え、かつSKN細胞の細胞形態を観察した。
【0096】
図9および図20に示される様に、pCEM-LMP2ベクターが移入されたSKN細胞の形態が、
線維芽細胞様(Flatrevertant)に変化していることが確認された。全般的に、(A)SKN細胞
のもともとの形態を維持している細胞のみで形成されているコロニー、(B)線維芽細胞様(
Flatrevertant)に形態が変化している細胞が一部存在しているコロニー、(C)線維芽細胞
様(Flatrevertant)に形態が変化している細胞のみで形成されているコロニーが確認され
た。(A)(B)(C)の各カテゴリーに属するコロニーの数は、図19に示している。また、(A)
(B)(C)の各カテゴリーに属する細胞の増殖速度をトリパンブルーにより染色法により検討
を行ったところ、(A)カテゴリーでは、P.D.T.(Population doubling time = 倍加時間)15
.2時間、(B)カテゴリーでは、P.D.T.=16.8時間、(C)カテゴリーでは、P.D.T.=17.8時間と
なった。ヒト子宮平滑筋肉腫培養細胞SKN細胞において、恒常的なLMP2の発現は、細胞形
態が線維芽細胞様(Flatrevertant)に変化させ、細胞速度が遅くなることが確認された(図
9、図20および図21)。
【0097】
通常、転移能を有する悪性度の高い癌細胞は、細胞と細胞との繋がりが粗になり1つの
細胞のみでも増殖が可能となる。特に、細胞間のマトリックス構造を形成するファイブロ
ネクチンの発現が、転移能を有する悪性度の高い癌細胞において著しく低下することが知
られている。SKN細胞は、転移能を有する悪性度の高い平滑筋肉腫細胞であるため、ファ
イブロネクチンの発現が著しく低いと思われる。そこで、(A)(B)(C)の各カテゴリーに属
するSKN細胞をLabtech Chamber Slide (IWAKI社)において培養し、顕微鏡下において80%
コンフルエントの状況を確認した後、公知の方法に従って抗ヒト-ファイブロネクチン抗
体(Rockland社)によりファイブロネクチンの発現状況を確認した。図10と図11で示さ
れている様に、正常なヒト子宮平滑筋培養細胞:N.HuUSMCでは、ファイブロネクチンが顕
著に発現しマトリックス構造を形成していることが確認されたが、ヒト子宮平滑筋肉腫培
養細胞:SKN細胞では、ファイブロネクチンの発現は僅かでありマトリックス構造の形成
が認められなかった。しかし、線維芽細胞様(Flatrevertant)のSKN細胞において、LMP2の
恒常的な発現によりファイブロネクチンの発現が認められたが、ファイブロネクチンのマ
トリックス構造の形成は確認されなかった(図10)。SKN細胞におけるLMP2の恒常的な発
現による細胞形態、細胞増殖とファイブロネクチンの発現状況への影響に関する結果が、
図21に纏められている。
【0098】
図21に示されているように、SKN細胞が本来有する腫瘍形成能が、LMP2の発現により
有意に低下すると予測される。そこで、Low Attachment Flat Chamber 24穴(Costor社)を
用いて、線維芽細胞様(Flatrevertant)のSKN細胞(SKN-lmp2細胞)とSKN細胞(SKN-pCEM9細
胞)における腫瘍形成能の指標となるコロニー形成能を検討した。Low Attachment Flat C
hamber 24穴(Costor社)の1穴あたりSKN-lmp2細胞(#121)(1x104個)あるいはSKN-pCEM9細
胞(1x104個)を培養して、コロニー形成を顕微鏡下で観察した。図22で認められる様に
、培養開始3週間目において、SKN-pCEM9細胞が大きなコロニーを形成して増殖している
のを確認出来たが、一方、SKN-lmp2細胞はコロニーを形成しているが、SKN-pCEM9細胞と
比較すると明らかにコロニーは小さく増殖能が低いことが認められた。Low Attachment F
lat Chamberを用いた実験では、SKN-lmp2細胞(コロニー#121)は、コロニー形成能が低い
ことから、腫瘍形成能が著しく低下していることが認められた。そこで、免疫が欠損して
いるヌードマウス(日本クレア社)を用いて、SKN-pCEM9細胞とSKN-lmp2細胞(コロニー#121
)における腫瘍形成能の検討を行った。ヌードマウス(日本クレア社)メス(6週齢)5匹を
購入し、SKN-pCEM9細胞(1x106個)をマウスの背中の右側に皮内接種し、SKN-lmp2細胞(#1
21)(1x105個)をマウスの背中の左側に皮内接種し計時的に腫瘍形成を観察した。皮内接
種4週間後、SKN-pCEM9細胞(1x106個)はヌードマウスの皮内において増殖し顕著な腫瘍
形成(腫瘍体積980mm3)が認められた。一方、SKN-lmp2細胞(コロニー#121)(1x105個)はヌ
ードマウスの皮内において増殖せず、顕著な腫瘍形成(腫瘍体積90mm3)が認められなかっ
た(図23Aおよび23B)。ヒト子宮平滑筋培養細胞SKN細胞において、LMP2の恒常的な発
現は、腫瘍形成能を顕著に低下させることを認めた(図23A〜23C)。
【0099】
実施例8 子宮平滑筋肉腫における顕著なサイクリンEの発現
ヒト子宮平滑筋培養細胞SKN細胞において、LMP2の恒常的な発現は、(1)SKNの細胞形態
を線維芽細胞様(Flatrevertant)に変化させる、(2)SKN細胞の増殖速度を有意に低下させ
る、(3)SKN細胞の有する腫瘍形成能を顕著に低下させる、以上3つの生物学的特長を認め
た。そこで、SKN細胞において、LMP2の恒常的な発現によりどのような因子の発現に顕著
な変化が認められるのか、SKN-pCEM9細胞とSKN-lmp2細胞間において遺伝子発現のプロフ
ァイリングを行った。
【0100】
方法
SKN-pCEM9細胞とSKN-lmp2細胞とを100mmデイッシュ4枚ずつでCO2インキュベーター内
において37℃で、80%コンフルエントの状況まで(シャーレ1枚あたり2x106個の細胞が
増殖)培養した。各細胞をトリプシンによって剥がして回収し、1XPBSによって細胞を洗
った。シャーレ1枚あたりTRIsol(インビトロジェン社)2mlの割合で、各細胞をTRIsolに
溶かした(細胞が完全にTRIsolに溶けるように10分間穏やかに振揺させる)。TRIsol 1ml
あたりクロロホルム200μlの割合で、TRIsol溶液にクロロホルムを加えて10分間穏やかに
振揺させる。10分間の振揺後、7000rpm、室温、20分間の条件で遠心を行い、水層と有機
層に分離して中間層を採取しない様に水層を採取した。水層に含まれているトータルRNA
を公知の方法を精製した。上記の手法により、SKN-pCEM9細胞よりトータルRNA 152.3μg
、SKN-lmp2細胞よりトータルRNA 128.8μg得られた。各トータルRNA 100μgを調整して、
遺伝子発現のプロファイリングを行った。
【0101】
マイクロアレイ遺伝子発現のプロファイリングの結果の一部を図24に示す。SKN-pCEM
9細胞において顕著な発現が認められたサイクリンEが、SKN-lmp2細胞においては殆ど発現
が認められない。つまり、SKN細胞では、LMP2を恒常的に発現により細胞増殖を誘導する
サイクリンEの発現が著しく低下された。
【0102】
トランスフェクトおよびレポーターアッセイ
サイクリンEのプロモター領域の活性化によりルシフェラーゼ遺伝子が発現誘導される
レーポータープラスミド(p10-4-Luc) (Department of Cancer Biology, Mayo Clinic Com
prehensive Cancer CenterのE A.Thompson博士から提供を受けた)の構造を図25に示す
。p10-4-LucレーポータープラスミドとLMP2発現プラスミドpCEM9-Lmp2とを図に示した内
容にしたがって、トータル2μgのプラスミドをメーカーの推奨に従って、FuGENE 6 Tran
sfection Reagent(Roche)によりSKN細胞、SKN-lmp2細胞(クローン#121と#122)中に移入し
た。すべての移入されたDNAには200ngのpCMVβ-Gal(Tropix社)を内部トランスフェクト効
率コントロールとして含んでいた。トランスフェクト後、各細胞をCO2インキュベーター
内において37℃で48時間培養した。最終的に、細胞を洗浄し、500μlの溶解バッファーで
溶解し、Dual-Luciferase Reporter Assay System(Promega社)を用いてメーカーの指示に
従って分析した。
【0103】
上記材料および方法を用いた検討により以下の結果が得られた。
【0104】
ヒト子宮平滑筋肉腫培養SKN細胞において、LMP2の恒常的発現によりサイクリンEのプロ
モーター活性が顕著に低下された。
【0105】
マイクロアレイ遺伝子発現のプロファイリングの結果の一部を図24に示す。SKN-pCEM
9細胞において顕著な発現が認められたサイクリンEが、SKN-lmp2細胞においては殆ど発現
が認められない。つまり、SKN細胞では、LMP2を恒常的に発現により細胞増殖を誘導する
サイクリンEの発現が著しく低下された。次いで、SKN細胞において、LMP2の恒常的発現が
サイクリンEのプロモーター活性を有意に抑えることを確認する必要がある。サイクリンE
のプロモーター活性に対するLMP2の恒常的発現の効果は、3種のSKN細胞株(SKN-pCEM9, S
KN-lmp2#121, SKN-lmp2#122)を用いたレポーターアッセイにより検討された。SKN細胞に
おいて、LMP2の発現の無い環境では、顕著に活性化されたサイクリンEのプロモーターが
認められた(図26)。しかし、LMP2が発現している環境では、サイクリンEのプロモータ
ーの著しい活性化が認められなかった(図26)。また、SKN-pCEM9細胞においては、サイ
クリンEのプロモーターの著しい活性化が認められたが、SKN-lmp2細胞においては、サイ
クリンEのプロモーターの著しい活性化が認められなかった(図26)。ヒト子宮平滑筋肉
腫培養SKN細胞において、LMP2の恒常的発現によりサイクリンEのプロモーター活性が顕著
に低下されることが確認された。
【0106】
図26は、SKN細胞とLMP2を恒常的に発現しているSKN細胞(クローン#121と#122)での
、サイクリンEプロモーター活性を示す図である。子宮平滑筋肉腫培養細胞SKN細胞で認め
られた顕著なサイクリンEの発現は、LMP2の恒常的な発現により著しく低下した。つまり
、SKN細胞では、LMP2の発現誘導が起こらないため、細胞増殖誘導因子であるサイクリンE
の発現抑制が認められないと思われる。サイクリンEの遺伝子を欠損させた細胞は、通常
に細胞増殖するが、外部からの刺激等による形質転換には耐性であることが明らかである
。したがって、顕著なLMP2の発現誘導が起こらないためサイクリンEの発現制御が起こら
ないことが、ヒト子宮平滑筋肉腫の発症に関与していると考えられる。そこで、ヒト子宮
平滑筋肉腫の組織でのサイクリンEの発現状況を、抗ヒトサイクリンE抗体を用いた免疫組
織染色法によって検討した。図27で示されている様に、同一切片内にて、正常なヒト子
宮平滑筋組織部位では認められないサイクリンEの発現が、ヒト子宮平滑筋肉腫組織部位
に鮮明に認められた。ヒト子宮平滑筋肉腫14症例において検討した結果、正常な子宮平滑
筋層においては、サイクリンEの発現は認められないが、悪性腫瘍であるヒト子宮平滑筋
肉腫組織では、サイクリンEの顕著な発現が認められることが明かとなった。
【0107】
図28は、ヒト子宮平滑筋肉腫組織における分裂期の核内でのサイクリンEの発現を示
す組織染色写真である。通常、細胞増殖を誘導するサイクリンEは、細胞増殖の開始期で
あるG1期に細胞質において過剰に発現し、速やかに核内へと移行しS期において染色体の
合成を開始させる。その後、サイクリンEは、S期の後半から即座に分解される。したがっ
て、正常な細胞のG2期とM期においては、サイクリンEの発現は認められない。しかし、ヒ
ト子宮平滑筋肉腫組織において、分裂期の核内での顕著なサイクリンEの発現が認められ
る。つまり、サイクリンEの発現は、ヒト子宮平滑筋肉腫の鑑別マーカーとして有用であ
る。
【産業上の利用可能性】
【0108】
子宮平滑筋におけるLMP2および/またはサイクリンEの転写または発現を調べることに
より、子宮平滑筋組織に子宮平滑筋肉腫が存在するかどうかを検出することができ、LMP2
の転写または発現が著しく低い場合および/またはサイクリンEの転写または発現が高い
場合に平滑筋肉腫が存在すると判定することができる。従来は、細胞の形態、密度、増殖
速度等を指標に子宮平滑筋肉腫を検出していたが、本発明の方法によれば、容易にかつ確
実に子宮平滑筋肉腫を検出することができる。さらに、本発明の方法によれば、子宮平滑
筋の腫瘍が子宮平滑筋腫であるか子宮平滑筋肉腫であるか鑑別することができ、さらに、
子宮平滑筋肉腫の悪性度を判定することができる。
【0109】
また、本発明のLMP2および/またはサイクリンEをマーカーとして用いる方法と、従来
から用いられていた細胞の形態、密度等を指標にする方法および/またはミオシンをマー
カーとして用いる方法を組合せることにより、確実に子宮平滑筋肉腫を検出することがで
きる。
【0110】
さらに、IFN-γシグナル伝達因子であるJAK1キナーゼ遺伝子、STAT1遺伝子およびLMP2
プロモーターの変異と子宮平滑筋肉腫の発症が密接に関連しており、上記遺伝子またはプ
ロモーターの変異を検出することにより、子宮平滑筋肉腫に罹患しているか否か、および
子宮平滑筋肉腫に罹患するリスクがあるか否かを判定することができる。
【0111】
本明細書で引用した全ての刊行物、特許および特許出願をそのまま参考として本明細書
にとり入れるものとする。
【技術分野】
【0001】
本発明は、LMP2およびサイクリンE、ならびにインターフェロンγ(IFN-γ)シグナル伝達カスケードに関与するJAK1キナーゼ遺伝子、STAT1遺伝子およびLMP2プロモーターにおける変異をマーカーとして用いる、子宮平滑筋肉腫の検出方法および子宮平滑筋腫と子宮平滑筋肉腫の鑑別検出方法に関する。
【背景技術】
【0002】
子宮癌は婦人科の癌で最も多いが、子宮平滑筋肉腫は発症頻度が低く子宮体部癌の2〜5%である。子宮平滑筋肉腫は、子宮頚部よりも子宮体部の筋肉層から多く発生する。子宮筋層は平滑筋よりできており、子宮筋層の中にできる良性の腫瘍が子宮平滑筋腫であり、悪性の腫瘍が子宮平滑筋肉腫である。子宮平滑筋腫と子宮平滑筋肉腫を鑑別することは非常に難しく、通常手術をして組織を採取し、顕微鏡下での細胞検査により判断していた。子宮平滑筋肉腫は異型性が強く巨大化することもある腫瘍細胞が増殖していることが多い。しかし、ほとんど細胞異型性を示さない場合もあり、細胞異型性の有無は決定的な鑑別基準とはならない。子宮平滑筋腫と子宮平滑筋肉腫との鑑別は、専ら凝固壊死があるかどうか、および細胞分裂像が増大しているかどうかにより行う。原則的には、細胞密度が高い場合には10倍高視野で10個以上の細胞分裂がある場合、腫瘍細胞に異型性が認められる場合には10倍高視野で5個以上ある場合に、子宮平滑筋肉腫と診断される。現実的には、腫瘍細胞の細胞異型度、細胞密度、細胞分裂の数、腫瘍内の壊死、出欠の有無等を参考に、主として顕微鏡および肉眼で組織の形態を観察して鑑別していた。しかしながら、このような鑑別を行うためには熟練を要し、必ずしも確実に鑑別できない場合もあった。
【0003】
本発明者は、先にイムノプロテアーゼの構成因子であるLMP2(low molecular mass polypeptide2)を欠損したマウスのメスにおいて、生後6ヶ月以降に子宮平滑筋肉腫(leiomyosarcoma)が認められ、生後12ヶ月までの発症率が全LMP2欠損マウスのメスの約35%にまでおよぶことを報告した(非特許文献1および2参照)。LMP2の機能は、組織特異的であり、CTLsによるMHCクラスI仲介腫瘍拒絶に不可欠の役割を持っている(非特許文献1参照)。
【0004】
このように、LMP2の欠損が何らかの機能で子宮平滑筋肉腫の発症のファクターになっていることは予測できた。しかしながら、LMP2が含まれる26Sプロテアソームは、転写調節因子や細胞周期調節因子等の活性化、MHCクラスI分子のペプチド抗原の産生等に複雑に関与しており、LMP2と子宮平滑筋肉腫の発症との直接の関係は不明であり、子宮平滑筋肉腫が発症した場合に、26Sプロテアソームの機能がどう変化し、またLMP2の転写発現がどう変化するかは不明であった。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Van Kaer L. et al. (1994) Immunity, 1, 533-541
【非特許文献2】Hayashi T. et al. (2002) Cancer Res., 62, 24-27
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、子宮平滑筋組織におけるLMP2の転写または発現の程度を指標に子宮平滑筋肉腫の存在を検出する方法およびそのための検出試薬の提供を目的とする。
【課題を解決するための手段】
【0007】
本発明者は、筋肉層におけるLMP2の発現を調べるためにマウスの生体を抗LMP2で染色してみた。その結果、平滑筋、横紋筋、心筋等の筋原性組織に特異的なLMP2の発現が認められることから、LMP2欠損マウスの子宮平滑筋層で認められた腫瘍細胞の起源が筋原性細胞(平滑筋細胞)であることを新たに見出した。
【0008】
さらに、本発明者は正常子宮平滑筋層、ヒト子宮平滑筋腫およびヒト子宮平滑筋肉腫の生検組織または手術組織におけるLMP2の発現状況を調べた。その結果、悪性腫瘍である子宮平滑筋肉腫のみにおいて、顕著なLMP2の発現低下が認められることを見出した。
【0009】
本発明者は、次いでLMP2の発現低下が、子宮平滑筋細胞において形質転換(癌化)に直接的に関与しているか検討するため、LMP2の発現が認められない子宮平滑筋肉腫細胞(SKN)に、遺伝子組換えによりLMP2を強制発現させ、SKN細胞の形態、細胞増殖速度およびSKN細胞におけるファイブロネクチンの発現変化について検討した。その結果、SKN細胞において、形態および細胞増殖速度が正常子宮平滑筋細胞に近づき、さらに有意なファイブロネクチンの発現誘導が起こることを見出した。
【0010】
本発明者は上記の新知見から、LMP2の転写発現を指標に子宮平滑筋腫と子宮平滑筋肉腫の鑑別診断をし得ること、およびLMP2の発現により子宮平滑筋肉腫の治療を行い得ることを見出した。
【0011】
本発明者らは、さらにサイクリンEの発現と子宮平滑筋肉腫との関係に着目し、子宮平滑筋組織において、子宮平滑筋肉腫の悪性度に依存してサイクリンEの転写・発現が著しく亢進することを見出した。
【0012】
また、本発明者らは、LMP2の発現に関与するインターフェロンγ(IFN-γ)シグナル伝達系に関与する遺伝子と子宮平滑筋肉腫との関係について検討を行なった。その結果、子宮平滑筋肉腫組織の細胞において同シグナル伝達因子であるJAK1キナーゼ、STAT1をコードする遺伝子およびLMP2プロモーターに変異が生じていることを見出し、該変異を検出することにより、子宮平滑筋肉腫に罹患しているか否か、および子宮平滑筋肉腫に罹患するリスクが高いか否かを判定することを見出した。
以上のようにして、本発明者らは本発明を完成させた。
【0013】
すなわち、本発明は以下の通りである。
[1] LMP2をマーカーとして用い、子宮平滑筋肉腫を検出する方法。
[2] 子宮平滑筋組織におけるLMP2の転写または発現の程度を指標に子宮平滑筋肉腫の存在を検出する方法であって、子宮平滑筋組織におけるLMP2の転写または発現を測定し、LMP2の転写または発現が正常子宮平滑筋組織に比較して少ない場合に子宮平滑筋肉腫が存在すると判定する方法。
[3] 子宮平滑筋組織におけるLMP2の転写または発現の程度を指標に子宮平滑筋の腫瘍が子宮平滑筋腫であるかまたは子宮平滑筋肉腫であるかを鑑別する方法であって、子宮平滑筋組織におけるLMP2の転写または発現を測定し、LMP2の転写または発現が正常子宮平滑筋組織に比較して少ない場合に子宮平滑筋肉腫であると判定する方法。
[4] 子宮平滑筋組織におけるLMP2の転写または発現の程度を指標に子宮平滑筋肉腫の悪性度を判定する方法であって、子宮平滑筋組織におけるLMP2の転写または発現を測定し、LMP2の転写または発現が正常子宮平滑筋組織に比較して少ない場合に悪性子宮平滑筋肉腫であると判定する方法。
[5] 採取した子宮平滑筋組織または細胞を用いin situ ハイブリダイゼーションを行いLMP2の転写を測定する、[1]〜[4]のいずれかの方法。
【0014】
[6] 採取した子宮平滑筋組織または細胞からLMP2のmRNAを抽出しRT-PCRまたはノーザンブロットを行いLMP2の転写を測定する、[1]〜[4]のいずれかの方法。
[7] 採取した子宮平滑筋組織または細胞を用い免疫組織化学染色または免疫細胞化学染色を行いLMP2の発現を測定する、[1]〜[4]のいずれかの方法。
[8] 採取した子宮平滑筋組織または細胞からLMP2タンパク質を抽出しイムノアッセイを行いLMP2の発現を測定する、[1]〜[4]のいずれかの方法。
[9] さらに、ミオシンをマーカーとして用い、LMP2およびミオシンの転写または発現を指標に子宮平滑筋肉腫の存在を検出する方法であって、LMP2の転写または発現が正常子宮平滑筋組織に比較して少なく、かつミオシンの転写または発現が正常子宮平滑筋組織に比較して多い場合に子宮平滑筋肉腫が存在すると判定する[1]の方法。
[10] LMP-2およびサイクリンEをマーカーとして用い、子宮平滑筋肉腫を検出する方法。
【0015】
[11] 子宮平滑筋組織におけるLMP2およびサイクリンEの転写または発現の程度を指標に子宮平滑筋肉腫の存在を検出する方法であって、子宮平滑筋組織におけるLMP2およびサイクリンEの転写または発現を測定し、LMP2の転写または発現が正常子宮平滑筋組織に比較して少なく、かつサイクリンEの転写または発現が正常子宮平滑筋組織に比較して多い場合に子宮平滑筋肉腫が存在すると判定する方法。
[12] 子宮平滑筋組織におけるLMP2およびサイクリンEの転写または発現の程度を指標に子宮平滑筋の腫瘍が子宮平滑筋腫であるかまたは子宮平滑筋肉腫であるかを鑑別する方法であって、子宮平滑筋組織におけるLMP2およびサイクリンEの転写または発現を測定し、LMP2の転写または発現が正常子宮平滑筋組織に比較して少なく、かつサイクリンEの転写または発現が正常子宮平滑筋組織に比較して多い場合に子宮平滑筋肉腫であると判定する方法。
[13] 子宮平滑筋組織におけるLMP2およびサイクリンEの転写または発現の程度を指標に子宮平滑筋肉腫の悪性度を判定する方法であって、子宮平滑筋組織におけるLMP2およびサイクリンEの転写または発現を測定し、LMP2の転写または発現が正常子宮平滑筋組織に比較して少なく、かつサイクリンEの転写または発現が正常子宮平滑筋組織に比較して多い場合に悪性子宮平滑筋肉腫であると判定する方法。
[14] 採取した子宮平滑筋組織または細胞を用いin situ ハイブリダイゼーションを行いLMP2およびサイクリンEの転写を測定する、[10]〜[13]のいずれかの方法。
[15] 採取した子宮平滑筋組織または細胞からLMP2およびサイクリンEのmRNAを抽出しRT-PCRまたはノーザンブロットを行いLMP2およびサイクリンEの転写を測定する、[10]〜[13]のいずれかの方法。
【0016】
[16] 採取した子宮平滑筋組織または細胞を用い免疫組織化学染色または免疫細胞化学染色を行いLMP2およびサイクリンEの発現を測定する、[10]〜[13]のいずれかの方法。
[17] 採取した子宮平滑筋組織または細胞からLMP2およびサイクリンEタンパク質を抽出しイムノアッセイを行いLMP2およびサイクリンEの発現を測定する、[10]〜[13]のいずれかの方法。
[18] 少なくともLMP2遺伝子断片をプローブまたはプライマーとして含む、LMP2をマーカーとして用い子宮平滑筋肉腫を検出するための検出試薬。
[19] 少なくともLMP2遺伝子断片およびサイクリンE遺伝子断片をプローブまたはプライマーとして含む、LMP2およびサイクリンEをマーカーとして用い子宮平滑筋肉腫を検出するための検出試薬。
[20] in situ ハイブリダイゼーション用試薬である[18]または[19]の子宮平滑筋肉腫を検出するための検出試薬。
【0017】
[21] さらに、ミオシン遺伝子断片をプローブまたはプライマーとして含み、LMP2およびミオシンをマーカーとして用い子宮平滑筋肉腫を検出するための[18]または[20]の検出試薬。
[22] さらに、ミオシン遺伝子断片をプローブまたはプライマーとして含み、LMP2、サイクリンEおよびミオシンをマーカーとして用い子宮平滑筋肉腫を検出するための[19]または[20]の検出試薬。
[23] 少なくとも抗LMP2抗体を含む、LMP2をマーカーとして用い子宮平滑筋肉腫を検出するための検出試薬。
[24] 少なくとも抗LMP2抗体および抗サイクリンE抗体を含む、LMP2およびサイクリンEをマーカーとして用い子宮平滑筋肉腫を検出するための検出試薬。
[25] 免疫組織化学または免疫細胞染色用試薬である[23]または[24]の子宮平滑筋肉腫を検出するための検出試薬。
[26] さらに、抗ミオシン抗体を含み、LMP2およびミオシンをマーカーとして用い子宮平滑筋肉腫を検出するための[23]または[25]の検出試薬。
[27] さらに、抗ミオシン抗体を含み、LMP2、サイクリンEおよびミオシンをマーカーとして用い子宮平滑筋肉腫を検出するための[24]または[25]の検出試薬。
【0018】
[28] JAK1キナーゼ遺伝子の以下の(A1)〜(A6)の変異部位の少なくとも1つを含むJAK1キナーゼ遺伝子の部分配列、およびLMP2プロモーターの以下の(B1)〜(B5)の変異部位の少なくとも1つを含むLMP2プロモーターの部分配列からなる群から選択される部分配列からなるオリゴヌクレオチドまたはその標識物であって、10から30塩基からなる部分配列またはその部分配列に相補的な配列からなるオリゴヌクレオチドまたはその標識物:
(A1) A2612A
(A2) G2626A
(A3) G2642T
(A4) A2967C
(A5) A2960C
(A6) A2985T
(B1) A210G
(B2) C214T
(B3) A216G
(B4) A217G
(B5) G219A。
[29] プローブとして用いられる[28]のオリゴヌクレオチドまたはその標識物。
[30] [28]のオリゴヌクレオチドまたはその標識物を固定化した固定化基板。
【0019】
[31] JAK1キナーゼ遺伝子の以下の(A1)〜(A6)の変異部位の少なくとも1つ、およびLMP2プロモーターの以下の(B1)〜(B5)の変異部位の少なくとも1つからなる群から選択される変異部位を含むDNA断片の増幅に用いる少なくとも一対のプライマーセットであって、上記変異部位の3’側および5’側に存在する10から30塩基からなる部分配列からなる、DNA断片の増幅に用い得る一対のプライマーセット:
(A1) A2612A
(A2) G2626A
(A3) G2642T
(A4) A2967C
(A5) A2960C
(A6) A2985T
(B1) A210G
(B2) C214T
(B3) A216G
(B4) A217G
(B5) G219A。
[32] [31]のプライマーセット、[28]もしくは[29]のオリゴヌクレオチドもしくはその標識物、または[30]の固定化基板を含む子宮平滑筋肉腫検出キット。
【0020】
[33] 動物から採取した子宮平滑筋組織または細胞におけるJAK1キナーゼ遺伝子の以下の(A1)〜(A6)の変異部位の少なくとも1つ、およびLMP2プロモーターの以下の(B1)〜(B5)の変異部位の少なくとも1つからなる群から選択される変異部位の検出を行い、その結果に基づいて、上記変異が存在する場合に、前記動物が子宮平滑筋肉腫に罹患しているか、または罹患するリスクが高いと判断する、子宮平滑筋肉腫の検出方法:
(A1) A2612A
(A2) G2626A
(A3) G2642T
(A4) A2967C
(A5) A2960C
(A6) A2985T
(B1) A210G
(B2) C214T
(B3) A216G
(B4) A217G
(B5) G219A。
[34] JAK1キナーゼ遺伝子、またはMP2プライマーの変異の検出を[31]のプライマーセット、[28]もしくは[29]のプローブまたは[30]の固定化基板を用いて行う[33]の方法。
【発明の効果】
【0021】
子宮平滑筋におけるLMP2および/またはサイクリンEの転写または発現を調べることにより、子宮平滑筋組織に子宮平滑筋肉腫が存在するかどうかを検出することができ、LMP2の転写または発現が著しく低い場合および/またはサイクリンEの転写または発現が高い場合に平滑筋肉腫が存在すると判定することができる。従来は、細胞の形態、密度、増殖速度等を指標に子宮平滑筋肉腫を検出していたが、本発明の方法によれば、容易にかつ確実に子宮平滑筋肉腫を検出することができる。さらに、本発明の方法によれば、子宮平滑筋の腫瘍が子宮平滑筋腫であるか子宮平滑筋肉腫であるか鑑別することができ、さらに、子宮平滑筋肉腫の悪性度を判定することができる。
【0022】
また、本発明のLMP2および/またはサイクリンEをマーカーとして用いる方法と、従来から用いられていた細胞の形態、密度等を指標にする方法および/またはミオシンをマーカーとして用いる方法を組合せることにより、確実に子宮平滑筋肉腫を検出することができる。
【0023】
さらに、IFN-γシグナル伝達因子であるJAK1キナーゼ遺伝子、STAT1遺伝子およびLMP2プロモーターの変異と子宮平滑筋肉腫の発症が密接に関連しており、上記遺伝子またはプロモーターの変異を検出することにより、子宮平滑筋肉腫に罹患しているか否か、および子宮平滑筋肉腫に罹患するリスクがあるか否かを判定することができる。
【図面の簡単な説明】
【0024】
【図1a】SKN細胞の中でのIFN-γ誘導性TAP1およびLMP2発現の欠損を示す写真である。写真に示す時間250ユニット/mlのIFN-γで処理したHeLa、HeLa.S3、SKN細胞およびヒト正常子宮平滑筋細胞(Hu.USMC)から細胞質抽出を調製し、50μgの細胞質抽出物を10%のSDS-PAGEで分離した。TAP1、LMP2およびβアクチンの発現レベルは適切な抗体を用いたイムノブロット分析で調べた。
【図1b】SKN細胞の中でのIFN-γ誘導性TAP1およびLMP2発現の欠損を示す写真であり、RT-PCRによるHeLa、HeLa.S3、SKN細胞およびHu.USMC細胞中のTAP1、LMP2およびβアクチンのmRNA発現の試験の結果を示す写真である。IFN-γ(250ユニット/ml)の非存在下または存在下で細胞を48時間培養した後に、プライマーを用いてRT-PCRを行った。RT-PCRにより増幅されたDNA製品をアガロース・ゲルにより分離した。DNAサイズマーカーは写真の左側に示す。
【図1c】子宮平滑筋肉腫の中でのLMP2発現の欠損を示す写真であり、正常子宮平滑筋および子宮平滑筋腫中のLMP2の免疫組織化学の結果を示す写真である。組織標本の5μmの切片を抗LMP2抗体およびペルオキシダーゼ結合抗ウサギのIgG抗体を用いて染色した。
【図2】HeLaおよびSKN細胞中でのIFN-γ誘導TAP1とLMP2共有のwtおよびIRF-E mtプロモーターの示差活性を示す図である。TAP1とLMP2共有のプロモーターwt(TAP1 593-1/pGL3およびLMP2 1-593/pLG3)の図ならびにそれらのそれぞれのIRF-E変異体プロモーターを含んだルシフェラーゼレポーター遺伝子の構築物を示す。HeLaおよびSKN細胞に同レポーター遺伝子を移入し、IFN-γを24時間後に添加しさらに回収の前24時間細胞をインキュベートした。レポーター遺伝子の移入効率の標準化のためにpSMV-βGALとの同時に移入した。結果は、HeLaおよびSKN細胞について別々に測定されたルシフェラーゼ遺伝子発現に標準化され、相対的TAP1およびLMP2活性として示される。結果は3つの独立している実験の平均で示され、エラーバーはSEを表す。
【図3】ヒト正常子宮平滑筋組織の顕微鏡写真である。
【図4】ヒト子宮平滑筋腫およびヒト子宮平滑筋肉腫の顕微鏡写真である。
【図5】ヒト子宮内膜間質肉腫の顕微鏡写真である。
【図6】正常子宮平滑筋、子宮平滑筋腫および子宮平滑筋肉腫の免疫組織化学染色写真である。
【図7】同一組織における子宮平滑筋肉腫部位と正常子宮平滑筋部位の免疫組織化学染色写真である。子宮平滑筋肉腫部位においてLMP2の発現低下が認められる。
【図8】LMP2の発現が認められない子宮平滑筋肉腫細胞(SKN)に、遺伝子組換えによりLMP2を強制発現させ、SKN細胞の形態の変化を示す写真である(その1)。
【図9】LMP2の発現が認められない子宮平滑筋肉腫細胞(SKN)に、遺伝子組換えによりLMP2を強制発現させ、SKN細胞の形態の変化を示す写真である(その2)。
【図10】LMP2の発現が認められない子宮平滑筋肉腫細胞(SKN)に、遺伝子組換えによりLMP2を強制発現させ、SKN細胞のファイブロネクチンの発現変化を示す写真である。
【図11】ヒト正常子宮平滑筋細胞(HuUSMC)におけるファイブロネクチンの発現変化を示す写真である。
【図12】各細胞における形態の変化、細胞増殖速度、およびファイブロネクチンの発現状況のまとめを示す図である。
【図13】マウス骨格筋組織、心筋組織および平滑筋組織におけるLMP2発現を示す写真である。
【図14】子宮平滑筋肉腫、子宮平滑筋腫および他臓器で発症した平滑筋肉腫におけるLMP2の発現を示す図である。
【図15】INFγシグナル伝達経路とLMP2の発現の関係を示す図である。
【図16】子宮平滑筋肉腫組織由来のJAK1キナーゼ遺伝子、STAT1遺伝子およびLMP2プロモーターの変異を示す図である。
【図17】子宮平滑筋肉腫組織由来のJAK1キナーゼ遺伝子、STAT1遺伝子およびLMP2プロモーターの変異を示す図である。
【図18】各組織におけるLMP2の発現状況を示す図である。図18において、Nは、LMP2の発現を検査した組織の症例数を示す。
【図19】pCEM9ベクター(LMP2遺伝子が含まれていない)またはpCEM9-LMP2ベクター(LMP2遺伝子がふくまれている)をヒト子宮平滑筋肉腫培養細胞(SKN細胞)に移入し、ネオマイシンにより選択された各ベクターを取り込んだ場合の、SKN細胞の増殖により形成されたコロニーの数を示す図である。1μgのpCEM9およびpLMP2 DNAを2×105細胞のDT細胞又はSKN細胞にトランスフェクトし、0.5または0.4mg/mlのG418を含む成長培地中でセレクトした。
【図20】LMP2の発現が認められない子宮平滑筋肉腫細胞(SKN)に、遺伝子組換えによりLMP2を強制発現させ、SKN細胞の形態の変化を示す写真である。
【図21】各細胞における形態の変化、細胞増殖速度、およびファイブロネクチンの発現状況のまとめを示す図である。
【図22】LMP2を恒常的に発現させたSKN細胞が腫瘍形成能の指標であるコロニー形成を著しく低下していることを示す写真である。
【図23A】LMP2を恒常的に発現させたSKN細胞がヌードマウスへの移植実験により腫瘍形成能を著しく低下していることを示す写真である。
【図23B】LMP2を恒常的に発現させたSKN細胞がヌードマウスへの移植実験により腫瘍形成能を著しく低下していることを示す図である。
【図23C】pCEM9-LMP2ベクター(LMP2遺伝子が含まれている)を移入したヒト子宮平滑筋肉腫培養細胞(SKN細胞)のコロニー#121が、SKN細胞と比較してLMP2遺伝子を著しく発現していることを示す写真である。
【図24】マイクロアレイを用いた遺伝子発現解析の結果、LMP2を恒常的に発現させたSKN細胞(コロニー#121)では、細胞増殖を誘導するサイクリンEの発現が著しく低下していることを示す図である。
【図25】サイクリンEプロモーターを含んだルシフェラーゼレポーター遺伝子の構造を示す図である。
【図26】SKN細胞とLMP2を恒常的に発現しているSKN細胞(コロニー#121と#122)での、サイクリンEプロモーター活性を示す図である。子宮平滑筋肉腫培養細胞SKN細胞で認められた顕著なサイクリンEの発現は、LMP2の恒常的な発現により著しく低下した。
【図27】ヒト子宮平滑筋肉腫組織におけるサイクリンEの顕著な発現を示す組織染色写真である。正常な子宮平滑筋層においては、サイクリンEの発現は認められないが、悪性腫瘍であるヒト子宮平滑筋肉腫組織では、サイクリンEの顕著な発現が認められる。
【図28】ヒト子宮平滑筋肉腫組織における分裂期の核内でのサイクリンEの発現を示す組織染色写真である。通常、細胞増殖を誘導するサイクリンEは、細胞増殖の開始期であるG1期に細胞質において過剰に発現し、速やかに核内へと移行しS期において染色体の合成を開始させる。その後、サイクリンEは、S期の後半から即座に分解される。したがって、正常な細胞のG2期とM期においては、サイクリンEの発現は認められない。しかし、ヒト子宮平滑筋肉腫組織において、分裂期の核内での顕著なサイクリンEの発現が認められる。
【発明を実施するための形態】
【0025】
本発明はLMP2をマーカーとして用いて、子宮平滑筋肉腫を検出する。ここで、検出とは
、子宮平滑筋肉腫が存在するかどうかを判定すること、患者が子宮平滑筋肉腫に罹患して
いるかどうか判定すること、子宮平滑筋の腫瘍が子宮平滑筋腫であるかまたは子宮平滑筋
肉腫であるかを鑑別すること、子宮平滑筋肉腫の悪性度を判定すること等を含む。
【0026】
LMP2は、プロテアソームのサブユニットであり、本発明の方法においては、子宮平滑筋
層において、LMP2が転写発現しているかどうかを検出する。ヒトの子宮平滑筋層の組織で
のLMP2の転写発現を調べると、正常な子宮平滑筋層と子宮平滑筋腫では、LMP2の発現は陽
性(中程度から強陽性)であるのに対して、子宮平滑筋肉腫では、LMP2の発現は陰性の部
分が多く弱陽性の部分が若干認められる。従って、LMP2の転写発現を指標に子宮平滑筋肉
腫に罹患しているかどうか、あるいは子宮平滑筋の腫瘍が子宮平滑筋腫か子宮平滑筋肉腫
のいずれかであるかを鑑別診断することができる。また、良性腫瘍である子宮平滑筋腫で
はLMP2の転写発現は強いのに対して、悪性化した腫瘍である子宮平滑筋肉腫ではLMP2の転
写発現が著しく減弱する。このことは、LMP2の転写発現が子宮平滑筋細胞に認められる腫
瘍に対して悪性度の指標になることを示す。
【0027】
さらに、本発明は、サイクリンE(CyclinE)をマーカーとして用いて、子宮平滑筋肉腫
を検出する方法を包含する。サイクリンは、サイクリン依存性キナーゼの活性を調節する
細胞周期で重要な役割を果たすタンパク質であり、サイクリンEは、ほ乳類細胞のG1期に
おいて作用するG1サイクリンの一つである。サイクリンEは、子宮平滑筋肉腫組織におい
て、正常組織に比較して発現が著しく上昇するが、子宮平滑筋腫組織においては上昇しな
い。本発明においては、LMP2と同様に子宮平滑筋層において、サイクリンEが転写発現し
ているかどうかを検出する。サイクリンEの転写発現を指標に子宮平滑筋肉腫に罹患して
いるかどうか、あるいは子宮平滑筋の腫瘍が子宮平滑筋腫か子宮平滑筋肉腫のいずれかで
あるかを鑑別診断することができる。また、良性腫瘍である子宮平滑筋腫ではサイクリン
Eの転写発現は正常子宮平滑筋組織と比べて変化がないのに対して、悪性化した腫瘍であ
る子宮平滑筋肉腫ではサイクリンEの転写発現が亢進する。さらに、通常、増殖期にある
正常な細胞のG2期とM期においては、サイクリンEの発現は認められない。しかし、ヒト子
宮平滑筋肉腫組織において、分裂期の核内での顕著なサイクリンEの発現が認められる。
このことは、サイクリンEの転写発現が子宮平滑筋細胞に認められる腫瘍に対して悪性度
の指標になることを示す。
【0028】
また、LMP2とサイクリンEを同時に検出してもよい。LMP2の転写発現が減弱し、かつサ
イクリンEの転写発現が亢進している場合、子宮平滑筋肉腫に罹患していると判断するこ
とができる。LMP2またはサイクリンE単独を指標とする場合よりも、LMP-2およびサイクリ
ンEの両方を指標とした場合に、より正確に子宮平滑筋肉腫を検出することが可能である
。
【0029】
本発明においては、子宮平滑筋肉腫に罹患しているかどうかの判断、子宮平滑筋の腫瘍
が子宮平滑筋腫か子宮平滑筋肉腫のいずれかであるかの鑑別診断、および子宮平滑筋細胞
に認められる腫瘍の悪性度の評価を含めて子宮平滑筋肉腫の検出と呼ぶ。
【0030】
本発明の方法においては、子宮平滑筋層におけるLMP2および/もしくはサイクリンEの
転写もしくは発現のいずれかまたは両方を検出することにより、子宮平滑筋肉腫を検出す
ることができる。LMP2またはサイクリンEの転写はLMP2またはサイクリンEをコードするmR
NAを測定することにより検出することができ、LMP2またはサイクリンEの発現はLMP2また
はサイクリンEタンパク質を測定することにより検出することができる。
【0031】
LMP2および/またはサイクリンEの転写を検出する場合、子宮平滑筋層の組織または細
胞を生体試料として採取し、該試料中に含まれるLMP2および/またはサイクリンEをコー
ドするmRNAを測定すればよい。mRNAの測定のためには、子宮平滑筋組織からバイオプシー
により組織の一部を採取し、また子宮平滑筋組織より綿棒等を用いて採取し、材料として
用いる。組織の採取は、例えば通常の外来診察中に、子宮内腔にバイオプシー用のかん子
を挿入し、1〜2mm角程度の子宮組織片を採取すればよい。mRNAの測定は採取した組織ま
たは細胞からmRNAを抽出して行ってもよいし、組織切片標本を作製するか、または採取し
た細胞をスライドガラス上に固定し、in situ ハイブリダイゼーション法により染色して
行ってもよい。あるいは、抽出したmRNAをノーザンブロット法やRT-PCR等の公知のRNA測
定法により測定すればよい。mRNAの抽出は公知の方法、例えば、新生化学実験講座2 核
酸I 分離精製 東京化学同人 1991年7月10日や分子生物学実験プロトコールI 丸善株
式会社 平成9年6月30日の記載に従って行うことができる。in situ ハイブリダイゼー
ションは、例えば分子生物学実験プロトコールIII 丸善株式会社 平成9年8月30日の
記載に従って行うことができる。この際、LMP2および/またはサイクリンEをコードするm
RNAを特異的に測定するために、LMP2および/またはサイクリンEをコードするmRNAの部分
配列に相補的な部分配列からなるプローブまたはプライマーを用いる。LMP2の塩基配列は
公知であり(例えば、GenBank アクセッション番号U01025、配列番号1、配列番号2はLMP
2タンパク質のアミノ酸配列を示す)、公知の塩基配列情報に基づいて、プローブまたは
プライマーを設計することができる。同様に、サイクリンEの塩基配列も公知であり(例
えば、GenBankアクセッション番号M73812、配列番号8)該プライマーまたはプローブは
、上記のLMP2および/またはサイクリンE遺伝子の断片であり、塩基の数は5〜50、好ま
しくは10〜30、さらに好ましくは15〜25である。
【0032】
LMP2および/またはサイクリンEの発現を検出する場合、子宮平滑筋層の組織または細
胞を生体試料として採取し、該試料中に含まれるLMP2および/またはサイクリンEタンパ
ク質を測定すればよい。子宮平滑筋層の組織または細胞は上記のようにして採取すればよ
い。また、LMP2および/またはサイクリンEタンパク質の測定も、組織または細胞からタ
ンパク質を抽出し、該抽出物中のLMP2および/またはサイクリンEタンパク質を測定して
もよいし、免疫組織化学または免疫細胞化学の手法により行ってもよい。抽出したタンパ
ク質の測定は、ELISA、ラジオイムノアッセイ等の公知の免疫測定法(イムノアッセイ)
を用いればよい。この際、LMP2および/またはサイクリンEに対する抗体が必要であるが
、抗LMP2抗体および/または抗サイクリンE抗体は公知の手法によりモノクローナル抗体
またはポリクローナル抗体として作製すればよい。また、市販の抗LMP2抗体および/また
は抗サイクリンE抗体を用いることもできる。抗体は必要に応じて、酵素、蛍光物質、放
射性同位元素により標識して用いるが、抗体の標識は公知の方法により行うことができる
。免疫組織化学または免疫細胞化学の手法による測定は、採取した子宮平滑筋組織切片標
本を作製するか、採取した細胞をスライドガラス上に固定して行う。免疫組織化学の手法
による測定のためには、組織を例えば、ホルマリンで固定し、パラフィン中に包埋し、ミ
クロトーム等の薄切装置で1〜5μm程度の厚さの切片標本を作製し、測定時にキシレン
やエタノール処理等によりパラフィンを除去し、生理食塩水または緩衝液に浸して親水化
すればよい。染色は、酵素、蛍光物質、放射性同位元素等で標識した抗LMP2抗体および/
または抗サイクリンE抗体を用いてもよいし、抗LPM2抗体を切片標本中のLMP2および/ま
たはサイクリンEに結合させた後に、抗LMP2抗体および/または抗サイクリンE抗体に結合
する2次抗体であって酵素、蛍光物質等で標識した2次抗体を用いてもよい。標識に用い
る酵素として、例えば西洋ワサビペルオキシダーゼ、アルカリフォスファターゼ等が挙げ
られ、蛍光物質として、例えばフルオレセイン、ローダミン等が挙げられる。また、公知
のビオチン-アビジン複合体を利用して染色してもよい。免疫細胞化学は採取した細胞を
ホルマリン等によりスライドガラス上に固定し、免疫組織化学の手法と同様の方法で細胞
中のLMP2および/またはサイクリンEを可視化すればよい。免疫組織化学または免疫細胞
化学において、染色は顕微鏡や肉眼で判断することもできるし、適当な光学的測定装置を
用いてもよい。免疫組織化学による染色は、例えば分子生物学実験プロトコールIII 丸
善株式会社 平成9年8月30日刊行の記載に従って行うことができる。
【0033】
上記のように、子宮平滑筋組織または細胞におけるLMP2の転写または発現を検出し、LM
P2の転写または発現が喪失しているか、または低下している場合、子宮平滑筋肉腫に罹患
していると判定することができる。また、子宮平滑筋組織または細胞におけるサイクリン
Eの転写または発現を検出し、サイクリンEの転写または発現が亢進している場合、子宮平
滑筋肉腫に罹患していると判定することができる。さらに、LMP2とサイクリンEの両方の
転写または発現を同時に検出してもよい。LMP2の転写または発現が喪失しているか、また
は低下しており、なおかつサイクリンEの転写または発現が亢進している場合、子宮平滑
筋肉腫に罹患していると判定することができる。例えば、組織または細胞の抽出物中のLM
P2 mRNAもしくはLMP2タンパク質またはサイクリンE mRNAもしくはサイクリンEタンパク質
を測定する場合、あらかじめ子宮平滑筋肉腫に罹患していない正常人の単位重量組織また
は単位細胞数当りのLMP2 mRNAもしくはLMP2タンパク質またはサイクリンE mRNAもしくは
サイクリンEタンパク質を測定する。正常人の値よりLMP2 mRNAまたはLMP2タンパク質が有
意に低い場合、子宮平滑筋肉腫に罹患していると判定し得、正常人の値よりサイクリンE
mRNAまたはサイクリンEタンパク質が有意に高い場合、子宮平滑筋肉腫に罹患していると
判定し得る。
【0034】
また、組織または細胞のLMP2の転写または発現をin situ ハイブリダイゼーション、免
疫組織化学、免疫細胞化学により検出する場合、組織または細胞が染色されず、LMP2の転
写、発現が認められない場合は、その部分の組織または細胞は子宮平滑筋肉腫組織または
細胞であり、組織または細胞を採取した患者は子宮平滑筋肉腫に罹患していると判定する
ことができる。さらに、組織または細胞のサイクリンEの転写または発現をin situ ハイ
ブリダイゼーション、免疫組織化学、免疫細胞化学により検出する場合、組織または細胞
が強く染色され、サイクリンEの転写、発現が強く認められる場合は、その部分の組織ま
たは細胞は子宮平滑筋肉腫組織または細胞であり、組織または細胞を採取した患者は子宮
平滑筋肉腫に罹患していると判定することができる。この際、子宮平滑筋肉腫でない正常
組織および子宮平滑筋肉腫組織の染色組織切片標品または染色細胞標品をあらかじめ作製
しておき、該標品と比較してもよい。
【0035】
LMP2の転写または発現の検出は、子宮平滑筋腫瘍に罹患した患者の腫瘍が悪性か良性か
、すなわち、子宮平滑筋肉腫かそれとも子宮平滑筋腫かの鑑別に利用することもできる。
この場合、LMP2の転写または発現が喪失しているか、正常組織に比較して有意に低下して
いる部分の組織は、悪性の子宮平滑筋肉腫であると判定することができる。また、サイク
リンEの転写または発現の検出は、子宮平滑筋腫瘍に罹患した患者の腫瘍が悪性か良性か
、すなわち、子宮平滑筋肉腫かそれとも子宮平滑筋腫かの鑑別に利用することもできる。
この場合、サイクリンEの転写または発現が、正常組織に比較して有意に亢進している部
分の組織は、悪性の子宮平滑筋肉腫であると判定することができる。この際、LMP2および
サイクリンEの両方を検出することにより、悪性度についてより正確な判断が可能になり
、また、より正確に鑑別を行なうことができる。さらに、組織を用いたin situ ハイブリ
ダイゼーションまたは免疫組織化学の手法によれば、組織のどの部分が正常でどの部分が
子宮平滑筋肉腫部分かを鑑別することもできる。さらに、例えば、組織切片の単位体積当
り、または単位細胞数当りのLMP2および/またはサイクリンEの転写または発現が喪失し
ているかまたは低下している細胞の数を測定することにより、子宮平滑筋腫瘍の悪性度を
判定することができる。
【0036】
さらに、従来の子宮平滑筋肉腫の診断方法と本発明のLMP2および/またはサイクリンE
の転写または発現を指標に子宮平滑筋肉腫を診断する方法を組合せることにより、より精
度の高い検出を行うことができる。従来法として、細胞の形態、密度等を観察する方法、
またはミオシンの転写または発現を指標に子宮平滑筋肉腫を診断する方法が挙げられ、こ
れらの方法は主に組織または細胞を固定して行われてきた。すなわち、組織切片標本また
は細胞スライドガラス標本を作製し、該標本の細胞の形態や密度を測定し、さらに、ミオ
シンの転写または発現を測定し、さらに、LMP2および/またはサイクリンEの転写または
発現を測定すればよい。
【0037】
本発明は、さらにLMP2遺伝子を子宮平滑筋肉腫患者に投与することにより、子宮平滑筋
肉腫を治療する方法、LMP2遺伝子を含む遺伝子治療のための遺伝子治療剤も包含する。遺
伝子治療における目的の遺伝子の子宮平滑筋肉腫患者への導入は公知の方法により行うこ
とができる。遺伝子を患者への導入する方法として、ウイルスベクターを用いる方法およ
び非ウイルスベクターを用いる方法があり、種々の方法が公知である(別冊実験医学、遺
伝子治療の基礎技術、羊土社、1996;別冊実験医学、遺伝子導入&発現解析実験法、羊土
社、1997;日本遺伝子治療学会編、遺伝子治療開発研究ハンドブック、エヌ・ティー・エ
ス、1999)。遺伝子導入のためのウイルスベクターとしては、アデノウイルス、アデノ随
伴ウイルス、レトロウイルス等のウイルスベクターを用いた方法が代表的なものである。
無毒化(ウイルス自身の複製が起こらない)したレトロウイルス、ヘルペスウイルス、ワク
シニアウイルス、ポックスウイルス、ポリオウイルス、シンビスウイルス、センダイウイ
ルス、SV40、免疫不全症ウイルス(HIV)等のDNAウイルスまたはRNAウイルスに目的とする
遺伝子を導入し、細胞に組換えウイルスを感染させることによって、細胞内に遺伝子を導
入することが可能である。また、プラスミドベクター等の遺伝子発現ベクターを用いて、
LMP2遺伝子を細胞や組織に導入することができる。例えば、リポフェクション法、リン酸
-カルシウム共沈法、DEAE-デキストラン法、微小ガラス管を用いたDNAの直接注入法など
により細胞内へ遺伝子を導入することができる。また、内包型リポソーム(internal lipo
some)による遺伝子導入法、静電気型リポソーム(electrostatic type liposome)による遺
伝子導入法、HVJ-リポソーム法、改良型HVJ-リポソーム法(HVJ-AVEリポソーム法)、HVJ-E
(エンベロープ)ベクターを用いた方法、レセプター介在性遺伝子導入法、パーティクル銃
で担体(金属粒子)とともにDNA分子を細胞に移入する方法、naked-DNAの直接導入法、種々
のポリマーによる導入法等によっても、組換え発現ベクターを細胞内に取り込ませること
が可能である。この場合に用いる発現ベクターとしては、生体内で目的遺伝子を発現させ
ることのできるベクターであれば如何なる発現ベクターも用いることができるが、例えば
pCAGGS(Gene 108, 193-200(1991))や、pBK-CMV、pcDNA3、pZeoSV(インビトロゲン社、
ストラタジーン社)、pVAX1などの発現ベクターが挙げられる。
【0038】
LMP2遺伝子を含むベクターは、遺伝子を転写するためのプロモーターやエンハンサー、
ポリAシグナル、遺伝子が導入された細胞の標識および/または選別のためのマーカー遺
伝子等を含んでいてもよい。この際のプロモーターとしては、公知のプロモーターを用い
ることができる。
【0039】
LMP2遺伝子を含む遺伝子治療剤は、LMP2遺伝子を含むベクターならびに薬理学的に許容
され得る担体、希釈剤もしくは賦形剤を含む。担体、希釈剤、賦形剤としては、製剤分野
において通常用いられるものを用いることができる。例えば、錠剤用の担体、賦形剤とし
ては、乳糖、ステアリン酸マグネシウムなどが使用される。注射用の水性液としては、生
理食塩水、ブドウ糖やその他の補助薬を含む等張液などが使用され、適当な溶解補助剤、
たとえばアルコール、プロピレングリコールなどのポリアルコール、非イオン界面活性剤
などと併用しても良い。油性液としては、ゴマ油、大豆油などが使用され、溶解補助剤と
しては安息香酸ベンジル、ベンジルアルコールなどを併用しても良い。本発明の遺伝子治
療剤は、子宮平滑筋肉腫部位に局所投与することが好ましく、例えば子宮平滑筋肉腫部位
に注射により投与すればよい。
【0040】
投与量は、症状、年齢、体重などによって異なるが、子宮平滑筋肉腫患者体内でLMP2を
発現する発現ベクター等に挿入されたLMP2遺伝子は、数日または数週間または数ヶ月おき
に1回あたり、0.001mg〜100mgを子宮平滑筋肉腫部位に注射等により直接投与すればよい
。
【0041】
本発明は、さらにインターフェロンγ(IFN-γ)シグナル伝達カスケードに関与する特
定の因子における変異により子宮平滑筋肉腫に罹患しているか否か、または子宮平滑筋肉
腫に罹患するリスクを有しているか否かを判定する方法をも包含する。
【0042】
正常細胞においては、IFN-γによりLMP2の遺伝子は活性化されるのに対して、子宮平滑
筋肉腫細胞においては、IFN-γによるLMP2の活性化はほとんど認められない。IFN-γによ
るLMP2の活性化は、図15にメカニズムを示すようにJAK1キナーゼ、JAK2キナーゼ、STAT
1などを介して、IRF-1が発現誘導され、LMP-2遺伝子のプロモーターに結合することによ
り生じる。IFN-γシグナル伝達に関与する因子のうち、JAK1キナーゼ及びSTAT1遺伝子の
変異、並びにLMP2のプロモーター領域の変異によりシグナル伝達が遮断され、LMP-2の発
現が阻害される。
【0043】
変異は以下のとおりである。以下、例えば遺伝子の変異をA210Gで表した場合、遺伝子
の塩基配列の21番目のAがGに置換していることを示す。また、遺伝子のコードするタンパ
ク質のアミノ酸配列の変異をG871Eで表した場合、アミノ酸配列中、871番目のグリシン(G
)がグルタミン酸(E)に置換していることを示す。
【0044】
JAK1キナーゼにおける変異
遺伝子変異 対応アミノ酸変異 変異の存在するドメイン
A2612A G781E ATP binding
G2626A G876R ATP binding
G2642T C881F ATP binding
A2967C G986P active site
A2960C Y987S active site
A2985T R995S active site
JAK1キナーゼ遺伝子の塩基配列を配列番号3に、JAK1キナーゼのアミノ酸配列を配列番
号4に示す。
【0045】
STAT1における変異
遺伝子変異 対応アミノ酸変異 変異の存在するドメイン
A2104C I702L non kinase active region
T2128G S710A non kinase active region
T2078G L693R non kinase active region
A2148C R716S non kinase active region
STAT1遺伝子の塩基配列を配列番号5に、STAT1のアミノ酸配列を配列番号6に示す。
【0046】
LMP2プロモーター
遺伝子変異 対応アミノ酸変異 変異の存在するドメイン
A210G IRF-E site
C214T IRF-E site
A216G IRF-E site
A217G IRF-E site
G219A IRF-E site
LMP2プロモーターの遺伝子配列を配列番号7に示す。
【0047】
JAK1キナーゼ遺伝子、STAT1遺伝子およびLMP2プロモーターの全長DNAまたはその断片は
塩基配列情報に基づいて容易に得ることができる。
【0048】
本発明は、上記のJAK1キナーゼ遺伝子、STAT1遺伝子またはLMP2プロモーターの塩基の
変異を検出し、子宮平滑筋肉腫に罹患しているか否かを検出する方法若しくは子宮平滑筋
肉腫に罹患するリスクが高いか否かを判定する方法、並びに上記のJAK1キナーゼまたはST
AT1のアミノ酸の変異を検出し、子宮平滑筋肉腫に罹患しているか否かを検出する方法若
しくは子宮筋肉腫に罹患するリスクが高いか否かを判定する方法を包含する。
【0049】
遺伝子における塩基の変異は、上記変異部位を含む遺伝子の断片をプローブとして、ま
たはDNAチップ、DNAマイクロアレイ技術において固相化するDNAとして用いればよい。そ
の場合の、断片の塩基長は全長でもよいが、通常は15bp〜100bpが好ましく、さらに15bp
〜50bpが好ましく、特に18bp〜30bpが好ましい。断片に含まれる変異部位は、1つだけで
もよいし、複数、すなわち2個、3個、4個、5個または6個でもよい。
また、上記断片に相補的な配列を有するDNAも本発明の範囲である。相補的なDNAも本願
明細書の開示に基づいて得ることができる。
【0050】
本発明のプローブは、検出のために蛍光物質、酵素、放射性同位体、化学発光物質等で
標識されていてもよい。標識に用いる標識物質は、公知のものを用い、公知の方法で標識
することができる。蛍光物質としては、例えば、Cy3、Cy5、ローダミン、フルオレセイン
等が挙げられる。
【0051】
さらに、上記の遺伝子変異を検出するPCR-RFLP等のPCRに用いるプライマーも本発明の
範囲である。すなわち、本発明は上記のJAK1キナーゼ遺伝子、STAT1遺伝子及びLMP2プロ
モーターの変異部位の3’側および5’側に存在する10から30塩基からなる部分配列からな
る、DNA断片の増幅に用い得る一対のプライマーセットを包含する。
【0052】
変異は、本発明のDNAを用いてPCR法、サザンハイブリダイゼーション法、ノーザンハイ
ブリダイゼーション法、定量的PCR法、in situ ハイブリダイゼーション法、FISH(Fluore
scence In Situ Hybridization)、PCR-RFLP法、PCR-SSCP法等により、検出することがで
きる。
【0053】
例えば、まずJAK1キナーゼ遺伝子、STAT1遺伝子またはLMP2プロモーターの変異塩基部
分を含むヌクレオチド配列に相補的なプローブ、ならびに野生型遺伝子の該変異塩基部分
に対応する部分を含むヌクレオチド配列に相補的なプローブを調製する。用いるプローブ
の長さに制限はなく、後述の核酸増幅法で増幅しようとする核酸断片の全長でもよいが、
通常は15bp〜100bpが好ましく、さらに15bp〜50bpが好ましく、特に18bp〜30bpが好まし
い。プローブは、放射性同位元素、蛍光物質、酵素等で標識したものを用いてもよい。次
いで、子宮平滑筋組織から採取した組織または細胞検体試料中の変異塩基部分を含む遺伝
子断片を核酸増幅法により増幅し、この増幅断片とプローブを反応させる。検体試料中の
DNAが野生型、変異型のいずれのプローブとハイブリダイズするか調べることにより、上
記遺伝子DNAに変異が生じているか否かを検出することができる。本発明のプローブは、
1塩基のミスマッチを検出するため、そのハイブリダイゼーション条件は、ストリンジェ
ントなものであることが必要である。ハイブリダイゼーション時の温度、塩濃度を調節す
ることにより1塩基のミスマッチのみを検出し得るハイブリダイゼーション条件を選択す
ることが可能である。用いるプローブDNAの長さにも依存するが、例えば具体的には、ナ
トリウム濃度が150〜900mM、好ましくは600〜900mMであり、温度が60〜68℃、好ましくは
65℃での条件で行い得る。
【0054】
核酸増幅の際に用いるプライマーとしては、上記遺伝子の変異部位を挟み増幅しようと
する領域の端部と相補的な配列を用いることができる。増幅する領域の塩基長に制限はな
いが、数十から数百塩基とすることができる。増幅領域に上記JAK1キナーゼ遺伝子、STA
T1遺伝子またはLMP2プロモーターDNAの変異を1つだけ含むように増幅塩基長を設定して
もよいし、複数の変異、すなわち、2個、3個、4個、5個または6個の変異を含むよう
に設定してもよい。また、プライマーを変異部位を含む領域に設定することも可能である
。プライマーの長さに制限はないが、好ましくは15bp〜50bp、さらに好ましくは20bp〜30
bpである。
【0055】
さらに、本発明のJAK1キナーゼ遺伝子、STAT1遺伝子またはLMP2プロモーター部位の変
異を含むDNA配列またはその断片に相補的なDNAを用いて、子宮平滑筋肉腫に罹患している
か否か、または子宮平滑筋肉腫患の罹患のリスクを決定するためのDNAチップを作製する
こともできる。この際、本発明のJAK1キナーゼ遺伝子、STAT1遺伝子またはLMP2プロモー
ター部位の変異を含む部分に相補的なDNA断片をニトロセルロース、ナイロン等のメンブ
レン、スライドグラス等に固相化すればよい。この際、固相化するDNA断片の塩基長は、
通常は15bp〜100bpが好ましく、さらに15bp〜50bpが好ましく、特に15bp〜25bpが好まし
い。次いで、該DNAチップを放射性同位元素、酵素、蛍光色素等で標識した被験体由来のD
NAまたはRNAと接触させ、ハイブリダイズを形成するか否かにより検体中に変異を有する
核酸が含まれているかどうかがわかる。
【0056】
変異の検出は子宮平滑筋肉組織から採取した組織片、細胞から核酸を抽出して行なえば
よい。
【0057】
本発明を以下の実施例によって具体的に説明するが、本発明はこれらの実施例によって
限定されるものではない。
【0058】
実施例1 ヒト子宮平滑筋肉腫におけるLMP2の転写および発現
本実施例においては、以下に記載する材料及び方法を用いて検討を行った。
【0059】
細胞株および培地
ヒト子宮平滑筋肉腫細胞株であるSKN細胞(RCB0513)は、理化学研究所セルバンクから購
入し、0.6%のL-グルタミン(Invitrogen社)および15%の牛胎児血清(シグマ-オルドリッチ
社)を補足したF-12Nutrient Mixture(Ham)培地(Invitrogen社)で維持した。HeLa細胞およ
びHeLa.S3細胞は0.6%のL-グルタミンおよび10%の牛胎児血清を補足したダルベッコのMEM
で維持した。ヒト子宮平滑筋細胞はCambrex BioScience Wailersville社から購入し、メ
ーカーのプロトコルに従って維持した。
【0060】
逆転写ポリメラーゼ連鎖反応(RT-PCR)分析
TAP1、LMP2、β2-mおよびβアクチンの転写物をRT-PCRにより調べた。細胞を250ユニ
ット/mlのヒトIFN-γ(Pepro Tech社)で処理しないかまたは48時間処理し、RNAを回収した
。トータルRNAはTRIzol試薬(Invitrogen社)を用いメーカーのプロトコルに従って5×106
細胞から調製した。RNAは、Superscript II enzyme(Invitrogen社)を用いて逆転写し、1
本鎖cDNAをTAP1、LMP2、β2mおよびβアクチン転写物を増幅するのに用いた。PCRは適切
なプライマーを用いて、30秒94℃35サイクル、30秒60℃、1.5分72℃および追加の5分の
プログラムで行い、転写物を伸長させた(Cabrera CM. et al. (2003) Tissue Antigens,
61,211-219; Miyagi T. et al. (2003) J.Gastroenterol.Hepatol., 18, 32-40)。
【0061】
免疫組織化学(Immunohistchemistry = IHC)
IHCは、Hsu SM. et al. (1981) J.Histochem.Cytochem. 29, 577-580に記載の方法によ
りアビジン-ビオチン複合体を用いて行った。すなわち、子宮平滑筋肉腫患者から得られ
た子宮摘出標本のパラフィン包埋サンプルから代表的な6つの5μm組織切片を作製した
。切片はパラフィン除去しアルコール中で再水和し、正常マウス血清を用いて20分間イン
キュベートした。次いで、切片を抗LMP2抗体(Affinity Res. Products社、希釈1/100)を
用いて1時間室温でインキュベートした。その後、切片をビオチン化2次抗体(Dako社)と
ともにインキュベートした。反応を3、3'-ジアミノベンジジンを用いて完了し、スライド
をヘマトキシリンで対比染色した。標本中の正常子宮平滑筋組織を陽性対照として用いた
。組織切片からなる陰性対照も、1次抗体の代わりに正常ウサギIgGとともにインキュベ
ートした。
【0062】
免疫沈降反応およびイムノブロット
細胞質抽出物および核抽出物は250ユニット/mlのヒトIFN-γで処理したか、または処理
しない5×106細胞から調製した(Brucet M. et al. (2004) Genes Immun., 5, 26-35)。細
胞を1200rpm、10分間の遠心分離により回収し、5mlの氷冷PBSで洗浄し、5分間、12000rpm
、4℃で遠心分離した。細胞をペレット化し、0.4mlのバッファーA(10mM Hepes、pH7.8
; 10mM KCl; 2mM MgCl2; 1mM DTT; 0.1mM EDTA; およびコンプリートプロテアーゼインヒ
ビターカクテル(Kirkegaard&Perr Lab社)中で一度洗浄し、2時間、4℃でインキュベー
トした。次いで、10%のNonidet P-40溶液25μlを添加し、細胞を1時間、4℃で激しく混
ぜ、次に、5分間、12000rpmで遠心分離した。遠心分離の後に上清を細胞質抽出物として
回収し、-80℃で保存した。ペレット化核は、40μlのバッファーC(50mM Hepes、pH7.8;
50mM KCl; 300mM NaCl; 0.1mM EDTA; 1mM DTT; 10%(v/v)グリセロール)に再懸濁し、4
℃で2時間混ぜて、5分間、4℃で12000rpm遠心分離した。核タンパク質を含む上清を回収
し、-80℃で保存した。
【0063】
STAT1、リン酸化状態のSTAT1、JAK1、JAK2、TAP1およびLMP2発現を検出するために、細
胞可溶化液または細胞質抽出物を10%SDS-ポリアクリルアミドゲル(SDS-PAGE)で分離し、
抗STAT1抗体、抗リン酸化STAT1抗体(Santa-Cruz Biotechnol.社)、抗JAK1、抗JAK2抗体(C
hemicon Int'l社)、抗体TAP1抗体(Stressgen社)または抗LMP2抗体(Affiniti Res. Produc
ts社)を用いて標準方法でイムノブロットを行った。IRF1またはIRF2発現の検出のために
、核抽出物を10%のSDS-PAGEページで分離し、抗IRF1抗体(Transduction Lab.社)、抗IRF2
抗体(Santa-Cruz Biotechnol.社)を用いて標準方法でイムノブロットを行った。目的のタ
ンパク質の発現は、アルカリフォスファターゼを結合した2次抗体を用いてアルカリフォ
スファターゼ発色をメーカーのプロトコールに従って行うことで、可視化して検討された
。
【0064】
図に示す時間250ユニット/mlのIFN-γで処理するか処理しなかった5×106細胞からの細
胞全体の抽出物を、50mM Tris-HCl、0.1mM EDTA、200mM NaCl、10%グリセロール、0.5%NP
-40、1mM DTTおよびコンプリートプロテアーゼインヒビターカクテル(Kirkegaard&Perr
Lab.社)を含むバッファーで溶解した。細胞可溶化液は、正常ウサギの血清(Santa-Cruz B
iotechnol.社)および20mlのプロテインG Sepharose(Amersham Biosciences社)であらかじ
め清澄化し、次に、2μgの抗JAK1または抗JAK2抗体を用いて免疫沈降を行った。サンプ
ルを10%のSDS-PAGEで分離し、Immobilon-Pメンブランにトランスファーした。リン酸化タ
ンパク質は、抗チロシンリン酸化抗体を1次抗体として反応させた後アルカリフォスファ
ターゼを結合した2次抗体を用いてアルカリフォスファターゼ発色をメーカーのプロトコ
ールに従って行うことで、可視化して検討された。IFN-γR1鎖発現を検出するために、細
胞全体のライセートは上述のように分離した。ブロットは抗IFN-γR1鎖抗体(PBL Biomedi
cal研究所)を用いて行った。SKN細胞をpRK5コントロール(2μg)またはJAK1発現ベクタ
ー(2μg)(St.Jude Children Research Hospital(テネシー州メンフィスのJ.Ihle博士
から提供された)でトランスフェクトした。IFN-γをトランスフェクト24時間後に添加し
、細胞を回収の前さらに24時間インキュベートした。pCMVβ-Galとの同時トランスフェク
トをトランスフェクト効率を標準化するために行った。
【0065】
トランスフェクトおよびレポーターアッセイ
TAP1およびLMP2wt(TAP1 593-1/pGL3およびLMP2 1-593/pLG3)の構造ならびにそれらのIR
F-E変異体プロモーター構築物の構造を図2に示す。トータル2μgのこれらのプラスミド
DNA(University of South FloridaのK.L. Wright博士から提供を受けた)を、メーカーの
推奨に従って、FuGENE 6 Transfection Reagent(Roche社)によりHeLaまたはSKN細胞中に
移入した。すべての移入されたDNAには200ngのpCMVβ-Gal(Tropix社)を内部トランスフェ
クト効率コントロールとして含んでいた。IFN-γ(最終濃度250ユニット/ml)をトランスフ
ェクト24時間後に添加し、細胞をさらに24時間インキュベートした。最終的に、細胞を洗
浄し、500μlの溶解バッファーで溶解し、Dual-Luciferase Reporter Assay System(Prom
ega社)を用いてメーカーの指示に従って分析した。LMP2またはTAP1/pGL3の代わりにpGL3
ベースで移入した細胞のルシフェラーゼ活性をバックグラウンドとして差し引いた。
【0066】
上記材料および方法を用いた検討により以下の結果が得られた。
【0067】
ヒト子宮平滑筋肉腫細胞中におけるINF-γによるTAP1およびLMP2発現の非誘導
LMP2欠損マウスは子宮平滑筋肉腫を発症した(Hayashi T. et al. (2002) Cancer Res.,
62, 24-27)。次いで、ヒト子宮平滑筋肉腫がTAP1およびLMP2の弱い発現を示すか否かを
示す必要がある。TAP1およびLMP2発現に対するIFN-γの効果は、4種の細胞株を用いたイ
ムノブロットにより調べた。IFN-γ処理に続くHeLa、HeLa.S3、およびHu.USMC(対照)は
TAP1およびLMP2の強い誘導を受けたが、ヒト平滑筋肉腫細胞株SKNにおけるIFN-γ処理で
はTAP1およびLMP2発現はそれほど誘導されなかった(図1a)。SKN細胞は、HeLa、HeLa.S3細
胞の両方とHu.USMC(対照)と比べると、同じようなβアクチン発現が認められたので、
抽出物の調製工程はIFN-γ処
理に続くTAP1およびLMP2発現の非誘導に影響しなかった。こ
のIFN-γの量は、HeLa、HeLa.S3細胞の両方とHu.USMC中(図1a)のTAP1およびLMP2遺伝子の
両方に共有されている双方向プロモーターを最大限に誘導するのに十分であった(図1a
)。INF-γ量を500ユニット/mlまで増加させても、SKN細胞中でTAP1およびLMP2発現を顕
著に誘導することはなかった。従って、SKN細胞はIFN-γ処理によりTAP1およびLMP2発現
を増強する能力を失っていた。
【0068】
IFN-γ処理に続くTAP1およびLMP2発現の非誘導を立証するために、4種の細胞株を用い
てRT-PCR分析を行った。IFN-γ処理により誘導されたTAP1またはLMP2のいずれかのmRNA発
現はHeLa、HeLa.S3細胞およびHu.USMCにおいて明確に検出されたが、IFNγ処理はSKN細胞
中でTAP1およびLMP2のmRNA発現をそれほど誘導しなかった(図1b)。βアクチン対照のmRNA
発現は試験したすべての細胞中で同様の高いレベルで検出された。このことはRNA調製ス
テップがIFN-γ処理に続くTAP1およびLMP2発現の非誘導に影響しなかったことを示す(図1
b)。IHC実験により、正常子宮平滑筋細胞が6ケースにおいてLMP2を著しく発現したが、子
宮平滑筋肉腫細胞では発現しなかったことが示された(図1c)。IHCの結果は、SKN細胞中
でのTAP1およびLMP2の非誘導を立証した。
【0069】
TAP1およびLMP2遺伝子のIFN-γに誘導される共通の双方向性プロモーター活性化の喪失
IRF-1はTAP1およびLMP2遺伝子に共有される双方向プロモーター中のIRF-Eと呼ばれるシ
スエレメントに直接結合する(Wright KL. et al. (1995) J.Exp.Med., 181, 1459-1471;
White LC. et al. (1996) Immunity, 5, 365-376; Dovhey SE. et al. (2000) Cancer R
es., 60, 5789-5796; Brucet M. et al. (2004) Genes Immun., 5, 26-35)。IRF-Eは図
2に表されるように、NFκB様結合部位およびGC1ボックスの上流に位置する。IFN-γ処理
によるTAP1およびLMP2の発現増強にIRF-Eが必要であることが示された(Wright KL. et al
. (1995) J.Exp.Med., 181, 1459-1471; White LC. et al. (1996) Immunity, 5, 365-37
6; Dovhey SE. et al. (2000) Cancer Res., 60, 5789-5796; Brucet M. et al. (2004)
Genes Immun., 5, 26-35)。IFN-γがSKN細胞においてTAP1/LMP2の共有された双方向のプ
ロモーターを確実に活性化するかどうかを調べるために、SKN細胞およびHeLa細胞にTAP1
またはLMP2の発現を誘導すべくIRF-Eがwtもしくはmutであるプロモーター-ルシフェラー
ゼコンストラクトDNAを移入した。HeLa細胞の場合、IFN-γ処理により11倍のLMP2プロモ
ーター活性が誘導され、さらに10倍のTAP1プロモーター活性が誘導された(図2)。しかし
ながら、SKN細胞中では、IFN-γ処理の結果、TAP1/LMP2の共有された双方向のプロモータ
ーの活性化誘導は認められなかった。IRF-Eの変異により、HeLa細胞中で認められたIFN-
γのTAP1/LMP2遺伝子発現を誘導する能力が失われた(図2)。これらの結果は内在性のmRN
Aレベルと一致しており、HeLa細胞中でのTAP1およびLMP2遺伝子の活性が強調しているが
、SKN細胞では強調していないことを示す。IRF-E部位の変異はHeLa細胞およびSKN細胞の
両方でベースとなる発現レベルをいくらか減少させた。このことはこの部位がLMP-2の発
現において一定の役割を果たすことを示す。これらの結果は、IFNγ処理が、HeLa細胞中
でTAP1/LMP2の共有されたプロモーター活性を強く誘導し得るが、SKN細胞中では誘導しな
いことを示す(図2)。
【0070】
実施例2 免疫組織化学によるヒト正常子宮平滑筋層、ヒト子宮平滑筋腫およびヒト子宮
平滑筋肉腫の生検組織または手術摘出組織におけるLMP2の発現状況
ヒト正常子宮平滑筋層、ヒト子宮平滑筋腫およびヒト子宮平滑筋肉腫の顕微鏡観察
ヒト子宮平滑筋組織をバイオプシーまたは手術により採取し、顕微鏡観察した。
【0071】
図3はヒト正常子宮平滑筋組織を、図4は、ヒト子宮平滑筋腫およびヒト子宮平滑筋肉
腫を、図5はヒト子宮内膜間質肉腫の顕微鏡写真を示す。
【0072】
上記顕微鏡観察により判定した、正常子宮平滑筋10例、子宮平滑筋腫6例、子宮内膜肉
腫6例および子宮平滑筋肉腫(悪性度低)3例、子宮平滑筋肉腫(悪性度高)4例を用い
て、免疫組織化学により各組識のLMP2発現を調べた。
【0073】
免疫組織化学は、前記の方法で行った。
【0074】
図6に、正常子宮平滑筋、子宮平滑筋腫および子宮平滑筋肉腫の免疫組織化学染色写真
を示す。図に示すように、正常子宮平滑筋および子宮平滑筋腫は染色され、LMP2が発現さ
れていることが確認された。しかし、子宮平滑筋肉腫では染色されなかったため、LMP2の
発現が確認されなかった。
【0075】
図7に、子宮平滑筋肉腫の免疫組織化学染色写真を示す。図に示すように、同じ組織内
で染色された部分と染色されない部分が存在した。
【0076】
各組織におけるLMP2の発現状況は以下のとおりであった。+は陽性、-は陰性、-/+はグ
レーゾーンであることを示す。
LMP2発現
正常子宮平滑筋組織 ++++
子宮平滑筋腫組織 ++++
子宮内膜肉腫組織 -/+
子宮平滑筋肉腫(悪性度低)組織 -/+
子宮平滑筋肉腫(悪性度高)組織 -
【0077】
上記表に示されるように、悪性腫瘍である子宮平滑筋肉腫のみにおいて、顕著なLMP2の
発現低下が認められた。また、子宮平滑筋肉腫の組織でのLMP2の発現低下度は、悪性度に
依存していており、LMP2の発現低下度合いは、子宮平滑筋肉腫の悪性度の指標と成りうる
ことが明らかとされた。
【0078】
実施例3 SKN細胞におけるLMP2の強制発現
1. SKN細胞におけるLMP2の強制発現による細胞形態の変化
LMP2の発現低下が、子宮平滑筋細胞において形質転換(癌化)に直接的に関与している
かについて検討を行った。LMP2の発現が認められない子宮平滑筋肉腫細胞(SKN)に、遺伝
子組換えによりLMP2を強制発現させ、SKN細胞の形態を観察した。
【0079】
方法
LMP2発現プラスミドベクター(2μg)をSKN細胞(5 x 105)内にFuGene6 (Roche,Indianapo
lis, IN)を用いメーカーのプロトコールに従って導入した。同プラスミドベクターを細胞
に導入した3日後に、G418(ネオマイシン)を培養液に加えて(最終濃度200μg/ml)、LMP2発
現プラスミドベクターが導入されたSKN細胞のみを選択した。LMP2の強制発現されたSKN細
胞の細胞増殖能、細胞形態能について検討を行った。結果を図8、図9および図21に示
す。図8、図9および図21に示すように、SKN細胞の増殖能は、倍加時間15.2時間だっ
たの対して、LMP2の強制発現されたSKN細胞の細胞増殖能は、倍加時間17.5時間となった
。LMP2の発現により、SKN細胞の増殖速度が、倍加時間2時間遅くなった。また、細長い
斜方形の細胞形態を持っているSKN細胞が、LMP2の強制発現により繊維芽細胞様(菱形)の
細胞形態に変化したことが認められた。
【0080】
2. SKN細胞におけるLMP2の強制発現によるファイブロネクチンの発現の変化
一般的に転移能を有する悪性度の高い癌細胞は、細胞と細胞との接着因子(ファイブロ
ネクチン)の著しい発現低下が認められる。LMP2の発現が認められない子宮平滑筋肉腫細
胞(SKN)に、遺伝子組換えによりLMP2を強制発現させ、ファイブロネクチンの発現変化に
ついて検討を行った。
【0081】
LMP2の強制発現は実施例3と同様の方法で行った。ファイブロネクチンの発現は、抗フ
ァイブロネクチン抗体(Rockland社)を用いた免疫染色法により検討した。また、コント
ロールとして、ヒト正常子宮平滑筋細胞:HuUSMCを用いた。
【0082】
SKN細胞における結果を図10に、ヒトHuUSMCにおける結果を図11に示す。また、図
12には各細胞における形態の変化、細胞増殖速度、およびファイブロネクチンの発現状
況のまとめを示す。
【0083】
図に示すように、SKN細胞にLMP2を強制発現させた場合、SKN細胞の形態と増殖速度が、
正常の子宮平滑筋細胞に近づいた。また、SKN細胞にLMP2を強制発現させた場合、有意な
ファイブロネクチンの発現誘導が認められた。つまり、LMP2の強制発現により誘導された
ファイブロネクチンの発現は、SKN細胞の持つ転移能の減弱の可能性を示唆している。
【0084】
実施例4 各組織におけるLMP2の発現
LMP2の発現は、子宮平滑筋組織のみならず、本来骨格筋組織および心筋組織においても
認められる。ところが、LMP2の欠損により、平滑筋肉腫が発症するのは子宮平滑筋組織の
みである。そこで、骨格筋、心筋および平滑筋におけるLMP2発現を調べた。また、免疫機
構に必須であるRag1遺伝子を欠損させたマウスの骨格筋、心筋および平滑筋におけるLMP2
発現を調べた。
【0085】
免疫組織化学は、生後1日目のマウス新生児のパラフィン包埋サンプルを用いて、前記
の方法で行った。
【0086】
図13に免疫組織化学染色写真を示す。
【0087】
実施例5 子宮及び他の器官での平滑筋肉腫におけるLMP2の発現
ヒト子宮平滑筋肉腫の組織においてLMP2の発現の著しい消失が確認された。平滑筋肉腫
は、子宮以外にも種々の臓器にいても発症するが、LMP2の発現の消失は子宮平滑筋肉腫に
特異的に認められるものか検討した。
【0088】
方法
病理ファイルより、子宮平滑筋肉腫(悪性度高い)4症例、子宮平滑筋肉腫(悪性度低い)
4症例、子宮上皮平滑筋肉腫1症例、子宮内膜間質肉腫1症例、子宮平滑筋腫1症例、正
常子宮平滑筋1症例、後腹膜原発平滑筋肉腫1症例、大網原発平滑筋肉腫1症例、小腸原
発平滑筋肉腫1症例、腸管膜原発平滑筋肉腫1症例を選び、各疾患の組織におけるLMP2の
発現を抗ヒトLMP2抗体を用いた免疫組織染色により検討した。
【0089】
図14に示されるように、正常な子宮平滑筋組織や子宮平滑筋腫(良性腫瘍)において認
められた顕著なLMP2の発現が、子宮平滑筋肉腫(悪性腫瘍)では著しく減弱していることを
再確認した。子宮内膜間質肉腫1症例ではLMP2の発現が認められなかったが、子宮上皮平
滑筋肉腫1症例においてはLMP2の発現が有意に認められた。腹部・消化器系組織に発症し
た平滑筋肉腫においては、顕著なLMP2の発現が認められる場合もあるが、著しいLMP2の発
現の減弱が認められる場合もある。これまでのGene Copy Number Profilingのゲノム解析
より、子宮平滑筋肉腫の発症機序は、他の臓器における平滑筋肉腫の発症機序とは大きく
異なっている可能性が示されている。実施例5の結果より、LMP2の発現の消失は子宮平滑
筋肉腫に特異的に認められる現象と思われる。
【0090】
実施例6 IFN-γシグナル伝達因子の変異と子宮平滑筋肉腫の関係
「子宮平滑筋肉腫において特異的に認められるLMP2発現の著しい減弱」について再検討
し、その特異性を確立した。具体的には、ヒト正常子宮平滑筋組織21症例、子宮平滑筋腫
24症例、子宮内膜間質肉腫6症例、子宮平滑筋肉腫29症例についてLMP2の発現を抗ヒトLMP
2抗体を用いた免疫組織染色により検討した。図18に示されるように、子宮平滑筋肉腫
において特異的に認められるLMP2発現の著しい減弱性を確認した。
【0091】
LMP2の発現は、図15に示される様にIFN-γの刺激により顕著に誘導される。子宮平滑
筋肉腫において特異的に認められるLMP2発現の著しい減弱は、IFN-γのシグナル伝達因子
:JAK1、JAK2、STAT1、PKCd、PI3K、LMP2のpromoter領域の変異や欠損による不活性化に
起因している可能性が考えられる。そこで、IFN-γのシグナル伝達因子:JAK1、JAK2、ST
AT1、LMP2のpromoter領域の変異について検討した。
【0092】
方法
手術により摘出されたヒト子宮体部組織13症例から平滑筋肉腫組織部位のみと正常な平
滑筋組織部位のみをレーザーマイクロダイセクションにより切り取り回収した。正常な子
宮平滑筋組織13検体にヒト正常子宮平滑筋培養細胞1検体を加えた合計14検体、子宮平滑
筋肉腫組織13検体にヒト子宮平滑筋肉腫培養細胞1検体を加えた合計14検体を用いてJAK1
、JAK2、STAT1、LMP2のpromoter領域の変異について検討した。各検体をHMW溶液(10mM Tr
is-HCl pH8.0、150mM NaCl、10mMEDTA-NaOH pH8.0、0.1% SDS)-proteinase K (100mg/ml)
に加えて55℃で24時間保温し、フェノール/クロロホルムにより不純物を取り除きゲノムD
NAを精製した。JAK1分子内のATP結合領域をコードする遺伝子領域(A)とチロシンリン酸化
酵素活性化領域をコードする遺伝子領域(B)の塩基配列を決定するために、遺伝子領域(A)
と遺伝子領域(B)の各領域に対するPCR-primerを用いてそれぞれの領域を含むDNA断片を各
検体より得られたゲノムDNAを用いてPCRにより増幅させた。PCRにより増幅されたDNA断片
をアガロースゲル電気泳動法により抽出し精製し、シークエンサー(ABI Prism3100 Genet
ic Analyzer)により塩基配列を決定した。さらに、JAK1の塩基配列を決定した方法と同様
の手法を用いて、JAK2分子内のATP結合領域をコードする遺伝子領域(C)とチロシンリン酸
化酵素活性化領域をコードする遺伝子領域(D)の塩基配列、STAT1分子内における転写因子
活性化領域(701番目のチロシンと727番目のセリン)コードする遺伝子領域(E)の塩基配列
、LMP2のpromoter領域をコードする遺伝子領域(F)の塩基配列を決定した。子宮平滑筋肉
腫における特異的な変異を検出するために、同一子宮体部における子宮平滑筋肉腫組織部
位と正常子宮平滑筋組織部位での塩基配列(AからE領域)を比較検討した。
【0093】
IFN-γのシグナル伝達因子:JAK1、JAK2、STAT1、各因子におけるリン酸化酵素あるい
は転写因子の活性化領域、LMP2のpromoter領域の変異解析の結果は、図16(変異を各検
体ごとにまとめた図)と図17(変異を各因子ごとにまとめた図)に示している。
【0094】
実施例7 ヒト子宮平滑筋肉腫細胞における、LMP-2の恒常的発現による細胞形態と細胞
増殖への影響
ヒト子宮平滑筋肉腫の組織において顕著なLMP2の発現低下が認められるが、LMP2の発現
低下が、子宮平滑筋肉腫の発症に関与しているのか検討した。
【0095】
方法
SKN細胞を6穴プレートにおいて培養し、70%コンフルエントの際、HamF-12-15%Fcs培
養液2mlを交換した。培養液を交換した次ぎの日、Roche社のプロトコールに従って、1
穴に対して2μgのpCEM9ベクター(LMP2の遺伝子を含まない)またはpCEM-LMP2ベクター(LM
P2の遺伝子を含んでいる)をFuGENE6(Roche社)によりSKN細胞に移入した。各ベクターの移
入を行ったSKN細胞をCO2インキュベーター内において37℃で48時間培養した後、SKN細胞
をトリプシン処理によって剥がして100mm培養シャーレでCO2インキュベーター内において
37℃で48時間培養した。その後、G418(ネオマイシン)100μg/mlを含んだHamF-12-15%Fcs
培養液で培養を行った。CO2インキュベーター内において37℃で2週間培養すると、pCEM9
ベクターまたはpCEM-LMP2ベクターが移入されたSKN細胞のみがG418によって選択的に生き
残り増殖しコロニーを形成しているのが確認された。G418(ネオマイシン)含有培養液にて
選択培養の3週間後、100mm培養シャーレ内における各ベクターが移入されたSKN細胞によ
り形成されたコロニー数を数え、かつSKN細胞の細胞形態を観察した。
【0096】
図9および図20に示される様に、pCEM-LMP2ベクターが移入されたSKN細胞の形態が、
線維芽細胞様(Flatrevertant)に変化していることが確認された。全般的に、(A)SKN細胞
のもともとの形態を維持している細胞のみで形成されているコロニー、(B)線維芽細胞様(
Flatrevertant)に形態が変化している細胞が一部存在しているコロニー、(C)線維芽細胞
様(Flatrevertant)に形態が変化している細胞のみで形成されているコロニーが確認され
た。(A)(B)(C)の各カテゴリーに属するコロニーの数は、図19に示している。また、(A)
(B)(C)の各カテゴリーに属する細胞の増殖速度をトリパンブルーにより染色法により検討
を行ったところ、(A)カテゴリーでは、P.D.T.(Population doubling time = 倍加時間)15
.2時間、(B)カテゴリーでは、P.D.T.=16.8時間、(C)カテゴリーでは、P.D.T.=17.8時間と
なった。ヒト子宮平滑筋肉腫培養細胞SKN細胞において、恒常的なLMP2の発現は、細胞形
態が線維芽細胞様(Flatrevertant)に変化させ、細胞速度が遅くなることが確認された(図
9、図20および図21)。
【0097】
通常、転移能を有する悪性度の高い癌細胞は、細胞と細胞との繋がりが粗になり1つの
細胞のみでも増殖が可能となる。特に、細胞間のマトリックス構造を形成するファイブロ
ネクチンの発現が、転移能を有する悪性度の高い癌細胞において著しく低下することが知
られている。SKN細胞は、転移能を有する悪性度の高い平滑筋肉腫細胞であるため、ファ
イブロネクチンの発現が著しく低いと思われる。そこで、(A)(B)(C)の各カテゴリーに属
するSKN細胞をLabtech Chamber Slide (IWAKI社)において培養し、顕微鏡下において80%
コンフルエントの状況を確認した後、公知の方法に従って抗ヒト-ファイブロネクチン抗
体(Rockland社)によりファイブロネクチンの発現状況を確認した。図10と図11で示さ
れている様に、正常なヒト子宮平滑筋培養細胞:N.HuUSMCでは、ファイブロネクチンが顕
著に発現しマトリックス構造を形成していることが確認されたが、ヒト子宮平滑筋肉腫培
養細胞:SKN細胞では、ファイブロネクチンの発現は僅かでありマトリックス構造の形成
が認められなかった。しかし、線維芽細胞様(Flatrevertant)のSKN細胞において、LMP2の
恒常的な発現によりファイブロネクチンの発現が認められたが、ファイブロネクチンのマ
トリックス構造の形成は確認されなかった(図10)。SKN細胞におけるLMP2の恒常的な発
現による細胞形態、細胞増殖とファイブロネクチンの発現状況への影響に関する結果が、
図21に纏められている。
【0098】
図21に示されているように、SKN細胞が本来有する腫瘍形成能が、LMP2の発現により
有意に低下すると予測される。そこで、Low Attachment Flat Chamber 24穴(Costor社)を
用いて、線維芽細胞様(Flatrevertant)のSKN細胞(SKN-lmp2細胞)とSKN細胞(SKN-pCEM9細
胞)における腫瘍形成能の指標となるコロニー形成能を検討した。Low Attachment Flat C
hamber 24穴(Costor社)の1穴あたりSKN-lmp2細胞(#121)(1x104個)あるいはSKN-pCEM9細
胞(1x104個)を培養して、コロニー形成を顕微鏡下で観察した。図22で認められる様に
、培養開始3週間目において、SKN-pCEM9細胞が大きなコロニーを形成して増殖している
のを確認出来たが、一方、SKN-lmp2細胞はコロニーを形成しているが、SKN-pCEM9細胞と
比較すると明らかにコロニーは小さく増殖能が低いことが認められた。Low Attachment F
lat Chamberを用いた実験では、SKN-lmp2細胞(コロニー#121)は、コロニー形成能が低い
ことから、腫瘍形成能が著しく低下していることが認められた。そこで、免疫が欠損して
いるヌードマウス(日本クレア社)を用いて、SKN-pCEM9細胞とSKN-lmp2細胞(コロニー#121
)における腫瘍形成能の検討を行った。ヌードマウス(日本クレア社)メス(6週齢)5匹を
購入し、SKN-pCEM9細胞(1x106個)をマウスの背中の右側に皮内接種し、SKN-lmp2細胞(#1
21)(1x105個)をマウスの背中の左側に皮内接種し計時的に腫瘍形成を観察した。皮内接
種4週間後、SKN-pCEM9細胞(1x106個)はヌードマウスの皮内において増殖し顕著な腫瘍
形成(腫瘍体積980mm3)が認められた。一方、SKN-lmp2細胞(コロニー#121)(1x105個)はヌ
ードマウスの皮内において増殖せず、顕著な腫瘍形成(腫瘍体積90mm3)が認められなかっ
た(図23Aおよび23B)。ヒト子宮平滑筋培養細胞SKN細胞において、LMP2の恒常的な発
現は、腫瘍形成能を顕著に低下させることを認めた(図23A〜23C)。
【0099】
実施例8 子宮平滑筋肉腫における顕著なサイクリンEの発現
ヒト子宮平滑筋培養細胞SKN細胞において、LMP2の恒常的な発現は、(1)SKNの細胞形態
を線維芽細胞様(Flatrevertant)に変化させる、(2)SKN細胞の増殖速度を有意に低下させ
る、(3)SKN細胞の有する腫瘍形成能を顕著に低下させる、以上3つの生物学的特長を認め
た。そこで、SKN細胞において、LMP2の恒常的な発現によりどのような因子の発現に顕著
な変化が認められるのか、SKN-pCEM9細胞とSKN-lmp2細胞間において遺伝子発現のプロフ
ァイリングを行った。
【0100】
方法
SKN-pCEM9細胞とSKN-lmp2細胞とを100mmデイッシュ4枚ずつでCO2インキュベーター内
において37℃で、80%コンフルエントの状況まで(シャーレ1枚あたり2x106個の細胞が
増殖)培養した。各細胞をトリプシンによって剥がして回収し、1XPBSによって細胞を洗
った。シャーレ1枚あたりTRIsol(インビトロジェン社)2mlの割合で、各細胞をTRIsolに
溶かした(細胞が完全にTRIsolに溶けるように10分間穏やかに振揺させる)。TRIsol 1ml
あたりクロロホルム200μlの割合で、TRIsol溶液にクロロホルムを加えて10分間穏やかに
振揺させる。10分間の振揺後、7000rpm、室温、20分間の条件で遠心を行い、水層と有機
層に分離して中間層を採取しない様に水層を採取した。水層に含まれているトータルRNA
を公知の方法を精製した。上記の手法により、SKN-pCEM9細胞よりトータルRNA 152.3μg
、SKN-lmp2細胞よりトータルRNA 128.8μg得られた。各トータルRNA 100μgを調整して、
遺伝子発現のプロファイリングを行った。
【0101】
マイクロアレイ遺伝子発現のプロファイリングの結果の一部を図24に示す。SKN-pCEM
9細胞において顕著な発現が認められたサイクリンEが、SKN-lmp2細胞においては殆ど発現
が認められない。つまり、SKN細胞では、LMP2を恒常的に発現により細胞増殖を誘導する
サイクリンEの発現が著しく低下された。
【0102】
トランスフェクトおよびレポーターアッセイ
サイクリンEのプロモター領域の活性化によりルシフェラーゼ遺伝子が発現誘導される
レーポータープラスミド(p10-4-Luc) (Department of Cancer Biology, Mayo Clinic Com
prehensive Cancer CenterのE A.Thompson博士から提供を受けた)の構造を図25に示す
。p10-4-LucレーポータープラスミドとLMP2発現プラスミドpCEM9-Lmp2とを図に示した内
容にしたがって、トータル2μgのプラスミドをメーカーの推奨に従って、FuGENE 6 Tran
sfection Reagent(Roche)によりSKN細胞、SKN-lmp2細胞(クローン#121と#122)中に移入し
た。すべての移入されたDNAには200ngのpCMVβ-Gal(Tropix社)を内部トランスフェクト効
率コントロールとして含んでいた。トランスフェクト後、各細胞をCO2インキュベーター
内において37℃で48時間培養した。最終的に、細胞を洗浄し、500μlの溶解バッファーで
溶解し、Dual-Luciferase Reporter Assay System(Promega社)を用いてメーカーの指示に
従って分析した。
【0103】
上記材料および方法を用いた検討により以下の結果が得られた。
【0104】
ヒト子宮平滑筋肉腫培養SKN細胞において、LMP2の恒常的発現によりサイクリンEのプロ
モーター活性が顕著に低下された。
【0105】
マイクロアレイ遺伝子発現のプロファイリングの結果の一部を図24に示す。SKN-pCEM
9細胞において顕著な発現が認められたサイクリンEが、SKN-lmp2細胞においては殆ど発現
が認められない。つまり、SKN細胞では、LMP2を恒常的に発現により細胞増殖を誘導する
サイクリンEの発現が著しく低下された。次いで、SKN細胞において、LMP2の恒常的発現が
サイクリンEのプロモーター活性を有意に抑えることを確認する必要がある。サイクリンE
のプロモーター活性に対するLMP2の恒常的発現の効果は、3種のSKN細胞株(SKN-pCEM9, S
KN-lmp2#121, SKN-lmp2#122)を用いたレポーターアッセイにより検討された。SKN細胞に
おいて、LMP2の発現の無い環境では、顕著に活性化されたサイクリンEのプロモーターが
認められた(図26)。しかし、LMP2が発現している環境では、サイクリンEのプロモータ
ーの著しい活性化が認められなかった(図26)。また、SKN-pCEM9細胞においては、サイ
クリンEのプロモーターの著しい活性化が認められたが、SKN-lmp2細胞においては、サイ
クリンEのプロモーターの著しい活性化が認められなかった(図26)。ヒト子宮平滑筋肉
腫培養SKN細胞において、LMP2の恒常的発現によりサイクリンEのプロモーター活性が顕著
に低下されることが確認された。
【0106】
図26は、SKN細胞とLMP2を恒常的に発現しているSKN細胞(クローン#121と#122)での
、サイクリンEプロモーター活性を示す図である。子宮平滑筋肉腫培養細胞SKN細胞で認め
られた顕著なサイクリンEの発現は、LMP2の恒常的な発現により著しく低下した。つまり
、SKN細胞では、LMP2の発現誘導が起こらないため、細胞増殖誘導因子であるサイクリンE
の発現抑制が認められないと思われる。サイクリンEの遺伝子を欠損させた細胞は、通常
に細胞増殖するが、外部からの刺激等による形質転換には耐性であることが明らかである
。したがって、顕著なLMP2の発現誘導が起こらないためサイクリンEの発現制御が起こら
ないことが、ヒト子宮平滑筋肉腫の発症に関与していると考えられる。そこで、ヒト子宮
平滑筋肉腫の組織でのサイクリンEの発現状況を、抗ヒトサイクリンE抗体を用いた免疫組
織染色法によって検討した。図27で示されている様に、同一切片内にて、正常なヒト子
宮平滑筋組織部位では認められないサイクリンEの発現が、ヒト子宮平滑筋肉腫組織部位
に鮮明に認められた。ヒト子宮平滑筋肉腫14症例において検討した結果、正常な子宮平滑
筋層においては、サイクリンEの発現は認められないが、悪性腫瘍であるヒト子宮平滑筋
肉腫組織では、サイクリンEの顕著な発現が認められることが明かとなった。
【0107】
図28は、ヒト子宮平滑筋肉腫組織における分裂期の核内でのサイクリンEの発現を示
す組織染色写真である。通常、細胞増殖を誘導するサイクリンEは、細胞増殖の開始期で
あるG1期に細胞質において過剰に発現し、速やかに核内へと移行しS期において染色体の
合成を開始させる。その後、サイクリンEは、S期の後半から即座に分解される。したがっ
て、正常な細胞のG2期とM期においては、サイクリンEの発現は認められない。しかし、ヒ
ト子宮平滑筋肉腫組織において、分裂期の核内での顕著なサイクリンEの発現が認められ
る。つまり、サイクリンEの発現は、ヒト子宮平滑筋肉腫の鑑別マーカーとして有用であ
る。
【産業上の利用可能性】
【0108】
子宮平滑筋におけるLMP2および/またはサイクリンEの転写または発現を調べることに
より、子宮平滑筋組織に子宮平滑筋肉腫が存在するかどうかを検出することができ、LMP2
の転写または発現が著しく低い場合および/またはサイクリンEの転写または発現が高い
場合に平滑筋肉腫が存在すると判定することができる。従来は、細胞の形態、密度、増殖
速度等を指標に子宮平滑筋肉腫を検出していたが、本発明の方法によれば、容易にかつ確
実に子宮平滑筋肉腫を検出することができる。さらに、本発明の方法によれば、子宮平滑
筋の腫瘍が子宮平滑筋腫であるか子宮平滑筋肉腫であるか鑑別することができ、さらに、
子宮平滑筋肉腫の悪性度を判定することができる。
【0109】
また、本発明のLMP2および/またはサイクリンEをマーカーとして用いる方法と、従来
から用いられていた細胞の形態、密度等を指標にする方法および/またはミオシンをマー
カーとして用いる方法を組合せることにより、確実に子宮平滑筋肉腫を検出することがで
きる。
【0110】
さらに、IFN-γシグナル伝達因子であるJAK1キナーゼ遺伝子、STAT1遺伝子およびLMP2
プロモーターの変異と子宮平滑筋肉腫の発症が密接に関連しており、上記遺伝子またはプ
ロモーターの変異を検出することにより、子宮平滑筋肉腫に罹患しているか否か、および
子宮平滑筋肉腫に罹患するリスクがあるか否かを判定することができる。
【0111】
本明細書で引用した全ての刊行物、特許および特許出願をそのまま参考として本明細書
にとり入れるものとする。
【特許請求の範囲】
【請求項1】
JAK1キナーゼ遺伝子の以下の(A2)、(A3)、(A5)および(A6)の変異部位の少なくとも1つを含むJAK1キナーゼ遺伝子の部分配列、ならびにLMP2プロモーターの以下の(B2)および(B4)の変異部位の少なくとも1つを含むLMP2プロモーターの部分配列からなる群から選択される部分配列からなるオリゴヌクレオチドまたはその標識物であって、10から30塩基からなる部分配列またはその部分配列に相補的な配列からなるオリゴヌクレオチドまたはその標識物:
(A2) G2626A
(A3) G2642T
(A5) A2960C
(A6) A2985T
(B2) C214T
(B4) A217G
【請求項2】
プローブとして用いられる請求項1記載のオリゴヌクレオチドまたはその標識物。
【請求項3】
請求項1記載のオリゴヌクレオチドまたはその標識物を固定化した固定化基板。
【請求項4】
JAK1キナーゼ遺伝子の以下の(A2)、(A3)、(A5)および(A6)の変異部位の少なくとも1つ、ならびにLMP2プロモーターの以下の(B2)および(B4)の変異部位の少なくとも1つからなる群から選択される変異部位を含むDNA断片の増幅に用いる少なくとも一対のプライマーセットであって、上記変異部位の3’側および5’側に存在する10から30塩基からなる部分配列からなる、DNA断片の増幅に用い得る一対のプライマーセット:
(A2) G2626A
(A3) G2642T
(A5) A2960C
(A6) A2985T
(B2) C214T
(B4) A217G
【請求項5】
請求項4記載のプライマーセット、請求項1もしくは2に記載のオリゴヌクレオチドもしくはその標識物、または請求項3記載の固定化基板を含む子宮平滑筋肉腫検出キット。
【請求項6】
動物から採取した子宮平滑筋組織または細胞におけるJAK1キナーゼ遺伝子の以下の(A2)、(A3)、(A5)および(A6)の変異部位の少なくとも1つ、ならびにLMP2プロモーターの以下の(B2)および(B4)の変異部位の少なくとも1つからなる群から選択される変異部位の検出を行い、その結果に基づいて、上記変異が存在する場合に、前記動物が子宮平滑筋肉腫に罹患しているか、または罹患するリスクが高いと評価する、子宮平滑筋肉腫の評価方法:
(A2) G2626A
(A3) G2642T
(A5) A2960C
(A6) A2985T
(B2) C214T
(B4) A217G
【請求項7】
JAK1キナーゼ遺伝子、またはMP2プライマーの変異の検出を請求項4記載のプライマーセット、請求項1もしくは2に記載のプローブまたは請求項3記載の固定化基板を用いて行う請求項6記載の方法。
【請求項1】
JAK1キナーゼ遺伝子の以下の(A2)、(A3)、(A5)および(A6)の変異部位の少なくとも1つを含むJAK1キナーゼ遺伝子の部分配列、ならびにLMP2プロモーターの以下の(B2)および(B4)の変異部位の少なくとも1つを含むLMP2プロモーターの部分配列からなる群から選択される部分配列からなるオリゴヌクレオチドまたはその標識物であって、10から30塩基からなる部分配列またはその部分配列に相補的な配列からなるオリゴヌクレオチドまたはその標識物:
(A2) G2626A
(A3) G2642T
(A5) A2960C
(A6) A2985T
(B2) C214T
(B4) A217G
【請求項2】
プローブとして用いられる請求項1記載のオリゴヌクレオチドまたはその標識物。
【請求項3】
請求項1記載のオリゴヌクレオチドまたはその標識物を固定化した固定化基板。
【請求項4】
JAK1キナーゼ遺伝子の以下の(A2)、(A3)、(A5)および(A6)の変異部位の少なくとも1つ、ならびにLMP2プロモーターの以下の(B2)および(B4)の変異部位の少なくとも1つからなる群から選択される変異部位を含むDNA断片の増幅に用いる少なくとも一対のプライマーセットであって、上記変異部位の3’側および5’側に存在する10から30塩基からなる部分配列からなる、DNA断片の増幅に用い得る一対のプライマーセット:
(A2) G2626A
(A3) G2642T
(A5) A2960C
(A6) A2985T
(B2) C214T
(B4) A217G
【請求項5】
請求項4記載のプライマーセット、請求項1もしくは2に記載のオリゴヌクレオチドもしくはその標識物、または請求項3記載の固定化基板を含む子宮平滑筋肉腫検出キット。
【請求項6】
動物から採取した子宮平滑筋組織または細胞におけるJAK1キナーゼ遺伝子の以下の(A2)、(A3)、(A5)および(A6)の変異部位の少なくとも1つ、ならびにLMP2プロモーターの以下の(B2)および(B4)の変異部位の少なくとも1つからなる群から選択される変異部位の検出を行い、その結果に基づいて、上記変異が存在する場合に、前記動物が子宮平滑筋肉腫に罹患しているか、または罹患するリスクが高いと評価する、子宮平滑筋肉腫の評価方法:
(A2) G2626A
(A3) G2642T
(A5) A2960C
(A6) A2985T
(B2) C214T
(B4) A217G
【請求項7】
JAK1キナーゼ遺伝子、またはMP2プライマーの変異の検出を請求項4記載のプライマーセット、請求項1もしくは2に記載のプローブまたは請求項3記載の固定化基板を用いて行う請求項6記載の方法。
【図1a】


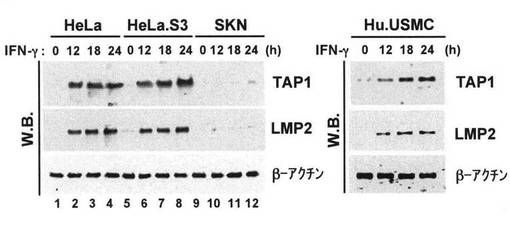
【図1b】

【図1c】
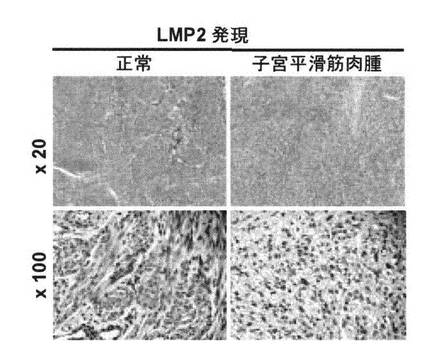
【図2】
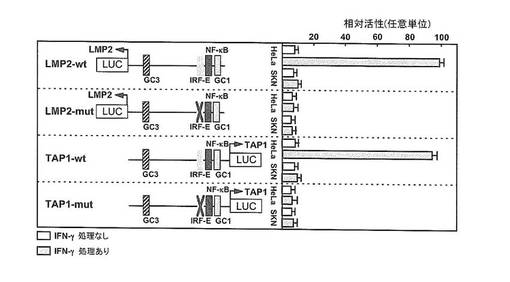
【図3】

【図4】
【図5】

【図6】
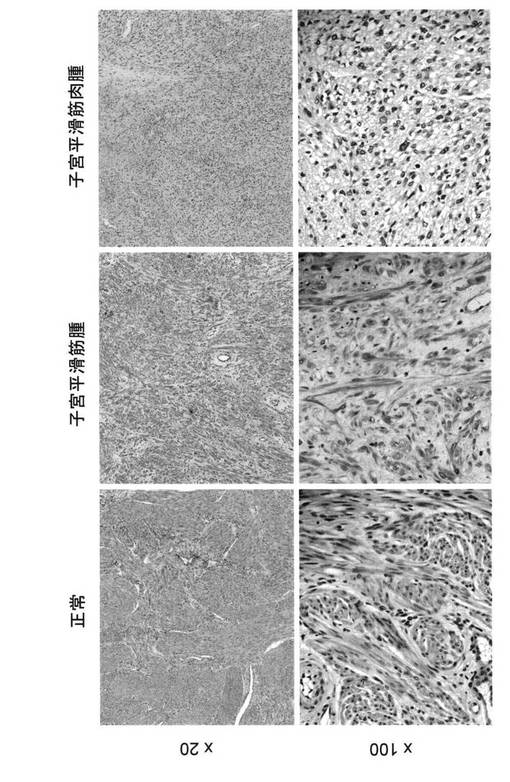
【図7】
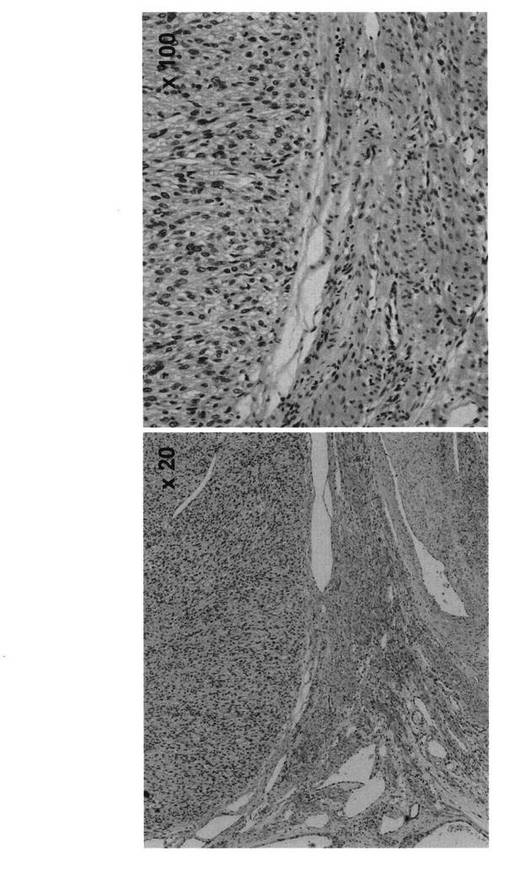
【図8】
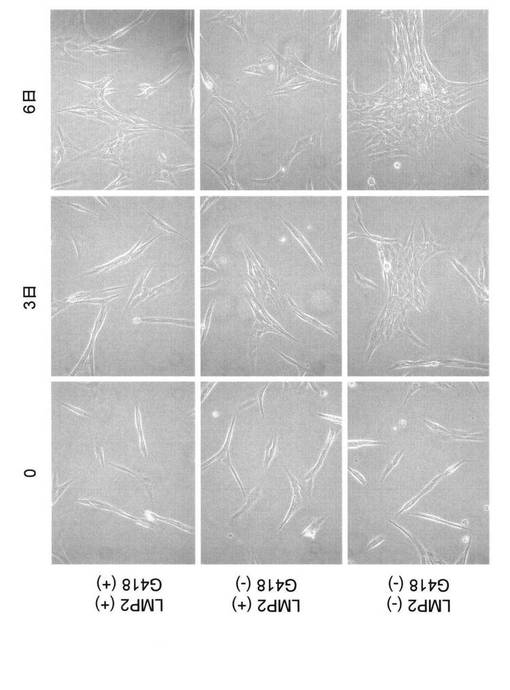
【図9】
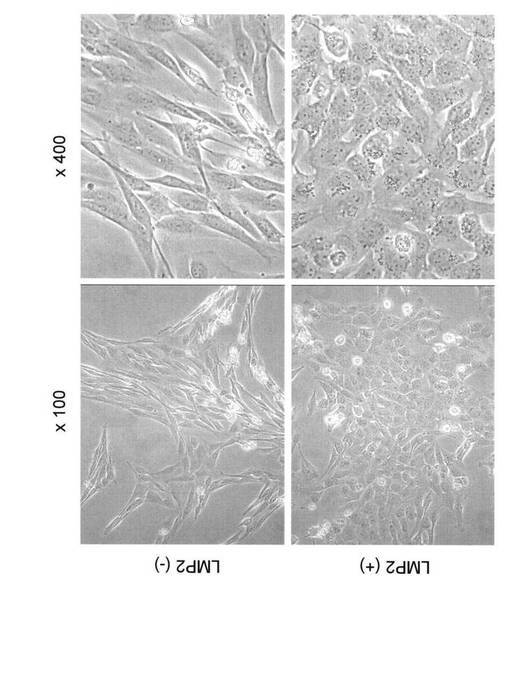
【図10】
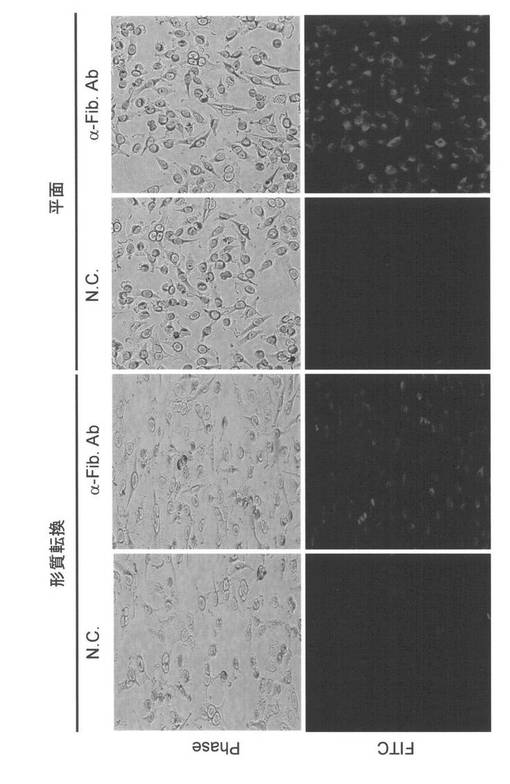
【図11】

【図12】

【図13】
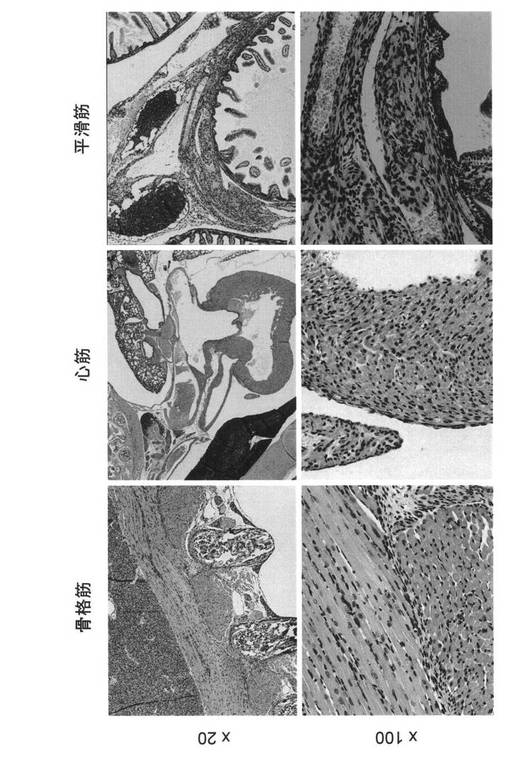
【図14】
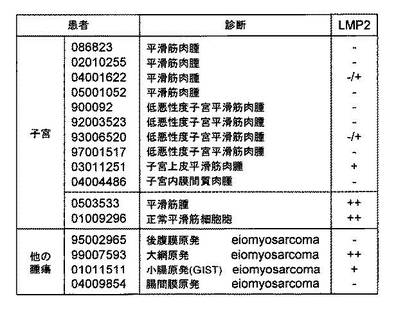
【図15】
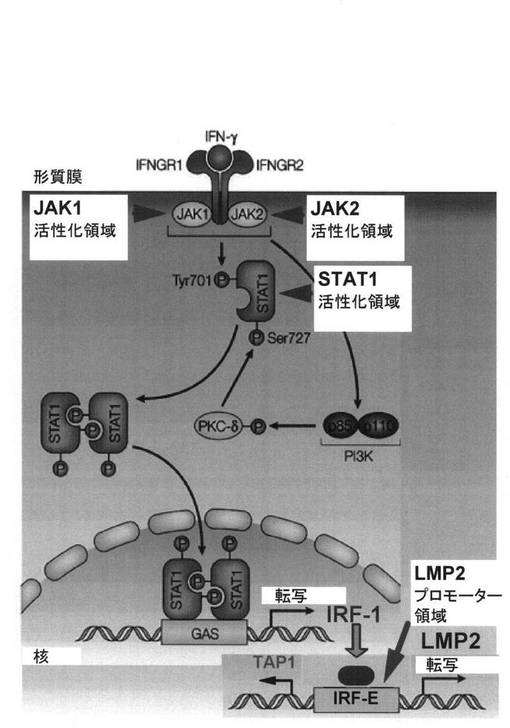
【図16】
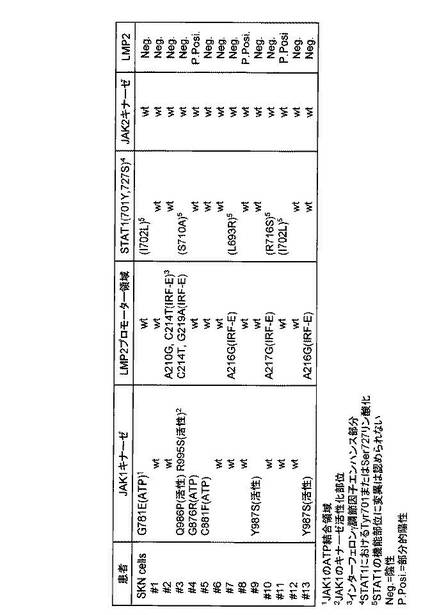
【図17】
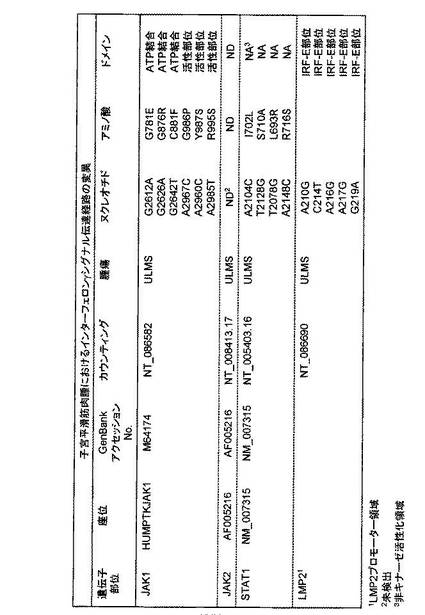
【図18】

【図19】
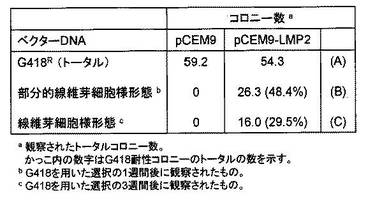
【図20】
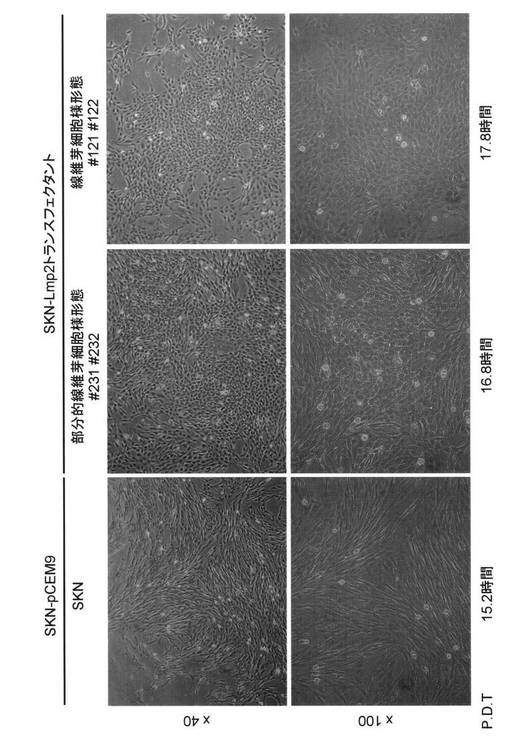
【図21】

【図22】
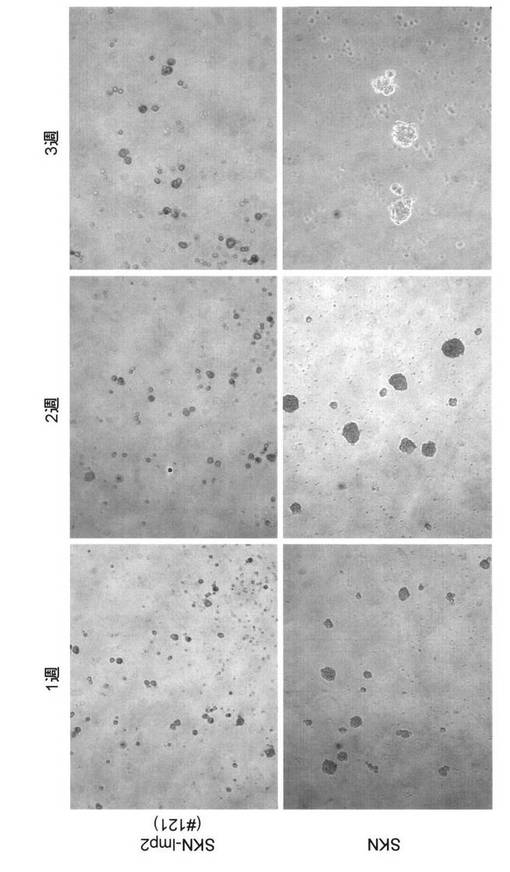
【図23A】
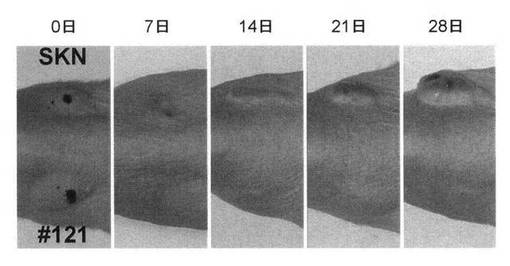
【図23B】
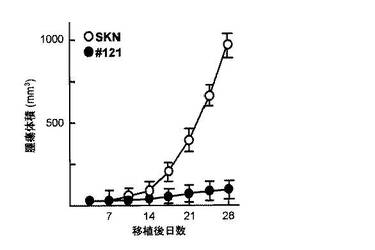
【図23C】
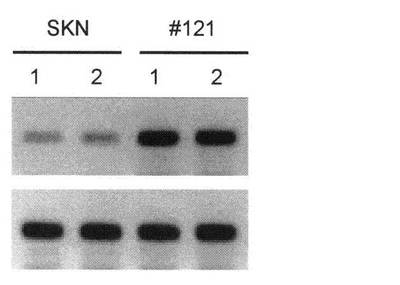
【図24】
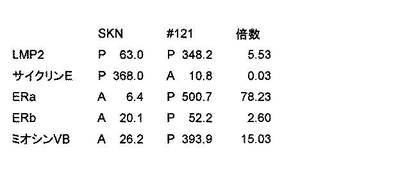
【図25】
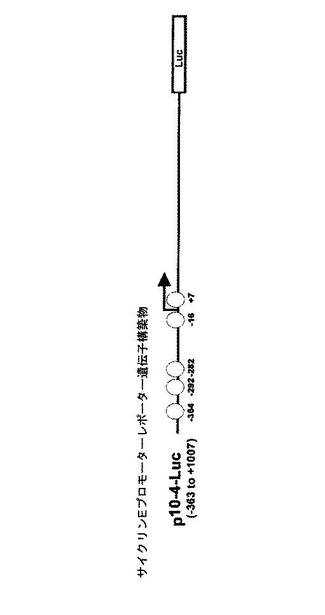
【図26】

【図27】
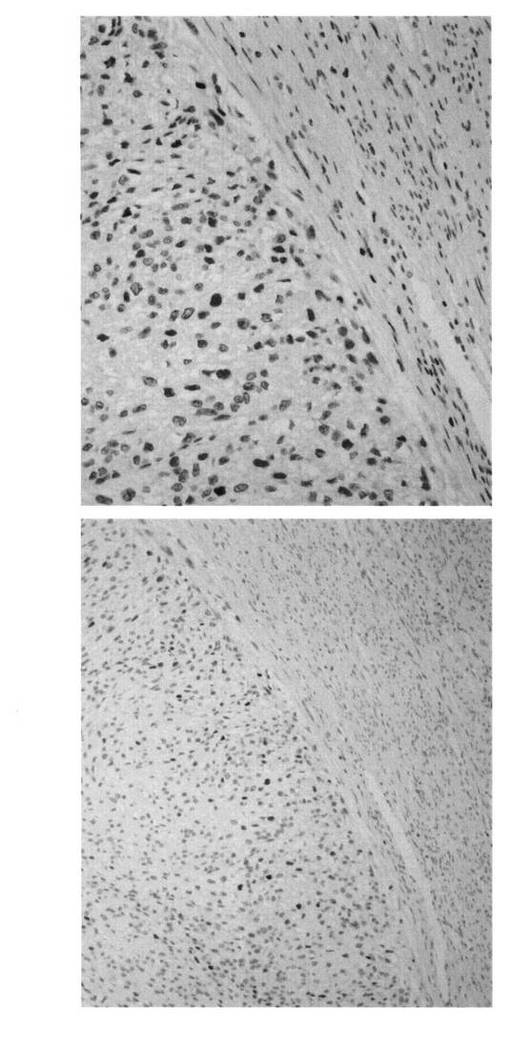
【図28】

【図1b】
【図1c】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23A】
【図23B】
【図23C】
【図24】
【図25】
【図26】
【図27】
【図28】
【公開番号】特開2011−135895(P2011−135895A)
【公開日】平成23年7月14日(2011.7.14)
【国際特許分類】
【出願番号】特願2011−71211(P2011−71211)
【出願日】平成23年3月28日(2011.3.28)
【分割の表示】特願2007−548042(P2007−548042)の分割
【原出願日】平成18年11月30日(2006.11.30)
【出願人】(504180239)国立大学法人信州大学 (759)
【Fターム(参考)】
【公開日】平成23年7月14日(2011.7.14)
【国際特許分類】
【出願日】平成23年3月28日(2011.3.28)
【分割の表示】特願2007−548042(P2007−548042)の分割
【原出願日】平成18年11月30日(2006.11.30)
【出願人】(504180239)国立大学法人信州大学 (759)
【Fターム(参考)】
[ Back to top ]