
ケラチノサイトの培養方法及びその使用
本発明は、ケラチノサイトの培養、ケラチノサイトの培養方法及び弱毒生ウイルスを普及させるためのその使用に関する。本発明の方法は、弱毒生ウイルス及び該ウイルスを含有するワクチンを得ることを可能にする。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規なケラチノサイトの培養培地、ケラチノサイトの培養方法及び弱毒生ウイルスの増殖のための該方法の使用に関する。この方法により得られる弱毒生ウイルス及びそれを含む生ワクチンは、本発明の別の主題を構成する。
【背景技術】
【0002】
いくつかの病原性ウイルスは、その複製に特異的なケラチノサイトタンパク質が必須であるので、ケラチノサイトでのみ完全に複製することができる。
例えば、MDV (マレック病ウイルス)及びFPV (鶏痘ウイルス)の場合がそうである。
【0003】
MDVウイルスを増殖するのに現在用いられている細胞は、一般に繊維芽細胞である。MDVに対するワクチンの製造に用いられるニワトリ繊維芽細胞培養は、遊離の(free)ウイルス粒子を増殖させず、感染は半増殖性(semi-productive)である。増殖性の感染が可能な連続細胞系を確立する試みがなされたが、成功していない(Abujoubら, Virology, 214, 541〜549, 1995)。繊維芽細胞中でのウイルス複製が不完全であるという性質のために、細胞性のワクチンは、液体窒素中で凍結した形で使用されなければならない。繊維芽細胞で増殖したウイルスは、完全エンベロープを欠いているので、細胞溶解性でも感染性でもない。
【0004】
マレック病の場合、動物は呼吸経路を介して感染する。汚染の短時間後に、Bリンパ球の細胞溶解性感染が進行し、その結果、多くの組織の潜在感染をもたらす細胞性ウイルス血症となる。感染する細胞の種類としてはTリンパ球があり、これは、その後、形質転換されて致死性のリンパ腫をつくる。ウイルスは表皮の羽嚢(feather follicles)にも運ばれるが、ここは完全エンベロープを有するウイルスの唯一の増殖場所である。ウイルスは、非常に高い力価で、表皮の表面部分、特に羽嚢の角化された細胞(鱗屑)に存在する。細胞は落屑し、鱗屑の吸入により他の動物に感染する(Calnekら, J. Natl. Cancer Inst., 45, 341〜351, 1970; Calnekら, Avian Dis., 14, 219〜233, 1970; Carrozzaら, Avian Dis. 17, 767〜781, 1973)。
【0005】
マレック病は世界中に存在し、家禽農場に重大な損失を与える原因であるので、多くのワクチンが開発されている。これらのワクチンは、マレック病ウイルスの弱毒株で感染させた培養ニワトリ繊維芽細胞の懸濁物からなる。ワクチンは、凍結保護剤を含有する培養培地中の生細胞を液体窒素中に凍結させることにより製造される。免疫化は、インオボ(in ovo)注射又は生誕の1日後の皮下注射のいずれかにより行なわれる。該免疫化は、リンパ組織増殖性症候群に対して防御するが、ケラチノサイトの感染に対しては防御しない。このことは、ウイルスの連続的な存続及び伝播を説明する。その存続によるウイルスの迅速な変異は、現存するワクチンに対して耐性の、毒性が増加した形の出現をもたらす。
【0006】
ヒト乳頭腫ウイルスは、その生物サイクルが、重層扁平上皮のケラチノサイトの最終分化(terminal differentiation)と関連する小さいDNAウイルスである。調節遺伝子及び非構造遺伝子は、重層上皮の基底層で発現されるが、DNA複製、構造タンパク質及びキャプシドアセンブリの合成は表面層でおこり、言い換えると、最終分化のプロセスを受けているケラチノサイトでおこる。HPVサブタイプ16及び18は、90%を超える子宮頸癌で見出され、これらのサブタイプがこの癌の原因に関係すると認められている(zur Hausen, H., J. Natl. Cancer Inst., 92, 690〜698, 2000)。子宮頸癌は大きな公衆衛生問題であり、予防ワクチンの開発を目的とした多くの研究が行なわれている(Plummerら, Virus Res., 89, 285〜293, 2002)。しかし、これらのワクチンの開発は、ビリオンの効率的な複製及び産生をもたらす細胞培養法がないことにより、遅れている。
【0007】
よって、ある病原性ウイルス、特に包膜されたDNAウイルス、例えばMDV及びFPVの完全複製をもたらす方法に対する必要性が存在する。
【0008】
特異的なケラチノサイトタンパク質がウイルス複製に必須であるという知識の下、弱毒完全ウイルスの増殖をもたらすことを目的として、ケラチノサイトを培養する試みがなされている。
【0009】
ヒトケラチノサイトは培養できるが(Rheinwaldら, Cell, 6, 331〜343, 1975)、培養におけるそれらの分化状態は不完全である。Meyersら, J. Virol., 71, 7381〜7386, 1997は、感染性HPVビリオンの合成を記載している。ケラチノサイトは、ニワトリ血清フリーの合成培地であるE培地とよばれる培養培地中に、マイトマイシンで処理した3T3細胞上で培養される。
【0010】
しかし、HPVはMPVとは異なり、この文献に記載されたHPVビリオンの合成方法は、マレック病のビリオンを増殖させることができない。
【0011】
文献EP-0 748 867は、連続細胞株であるQT35細胞系上でのマレック病ウイルス血清型1の培養を記載している。感染された細胞は、ワクチンの作製に用いられ、単離ビリオンには用いられていない。なぜなら、このウイルスは細胞に強く結合するからである。これは、不完全なエンベロープを持って増殖し、感染性を保ったまま単離できない。
【0012】
文献EP-0 770 677は、ニワトリ細胞系上でのマレック病ウイルス血清型2の増殖を記載している。得られたビリオンは細胞培養物に依存する。
文献EP-1 149 899は、鳥類のES細胞の培養方法を記載している。培養培地は、マイトマイシンでの処理又は照射により不活性化されたフィーダー細胞の層、例えばマウス胚繊維芽細胞であるSTO細胞を含むのが好ましい。
【0013】
文献EP-0 779 359号は、鳥類細胞の培養方法を記載する。培養培地はニワトリ血清を含有する。培養は、STO (繊維芽細胞)又はSL-10細胞の担体(support)上である。
よって、いずれのケラチノサイト培養も、完全ウイルス複製、特にエンベロープDNAウイルス、例えばMDVをもたらす条件を提供できていない。
【0014】
さらに、実質的に全てのヒト及び動物細胞種はウシ血清の存在下に首尾よく培養することができるが(例えばDorangeら, J. Virol., 76, 1959〜1970, 2002を参照として挙げることができる)、ニワトリケラチノサイトはこれらの条件下では増殖しない。
【発明の開示】
【発明が解決しようとする課題】
【0015】
本発明の目的は、インビボで有する分化の特性の全てを保持する細胞のようなケラチノサイト、特に鳥類のケラチノサイト、より詳しくはニワトリケラチノサイトを培養する条件を開発することであった。
高度に分化した細胞(肝細胞、腸細胞、ニューロン細胞又はケラチノサイト)に感染するウイルスの複製サイクルは、一般に、その宿主細胞の分化プログラムに連結している(Calnekら, J. Natl. Cancer Inst., 45, 341〜351, 1970; Calnekら, Avian Dis., 14, 219〜233, 1970; Schmittら, J. Virol., 70, 1912〜1922, 1996)ので、完全に分化したケラチノサイトの培養が完全ウイルスの複製、よって細胞を含まないビリオンを産生することを可能にすると予想することができる。
【課題を解決するための手段】
【0016】
本発明の主題は、
(i) ニワトリ血清と、
(ii) 細胞分裂抑制処理により不活性化された繊維芽細胞を含む培養担体と
を含む表皮細胞、特に家禽の表皮細胞用の培養培地である。
【0017】
本発明の主題は、ニワトリ血清と、不活性化された哺乳動物細胞、好ましくは不活性化された繊維芽細胞を含む培養担体とを含む培養培地を含有するケラチノサイト培養物でもある。
【発明を実施するための最良の形態】
【0018】
さらに、本発明の培養培地は、知られるように、無機塩、ビタミン、ホルモン及び成長因子から選択され得る1種又はそれより多い添加物、或いは細胞培養培地に通常用いられるいずれのその他の添加物を含む。本発明の培養培地に用い得る添加物のうち、無機塩、アミノ酸、ビタミン、脂肪酸、グルコース、バッファー、フェノールレッド、EGF (表皮成長因子)、T3、ヒドロコルチゾン、インシュリン、コレラトキシン、トランスフェリン、アデニンなどを特に挙げることができる。
【0019】
培養担体として用いられる繊維芽細胞は、3T3細胞が有利であり、より有利には3T3-J2サブクローンである。3T3株の起源は、Todaro, G., J.ら, J. Cell Biol., 17, 299〜313, 1963に記載される。3T3細胞系は、ATCCコレクションに番号CCL92の下で登録されている。3T3細胞は、細胞分裂抑制処理により不活性化された細胞の単層の形で本発明において用いるのが有利である。細胞分裂抑制効果が知られているいずれの処理も用い得る。この処理は、マイトマイシンCのような抗生物質での処理、又はガンマ線の致命的な照射からなり得る。
【0020】
本発明の培養培地において、ニワトリ血清は、培養培地の全重量に対する重量での割合で0.5〜50%、好ましくは1〜40%、有利には2〜30%の範囲で存在するのが有利である。
【0021】
有利には、細胞培養用の増殖培地は、次の添加物の少なくとも1つ、有利にはいくつかを含有する:コレラトキシン、トランスフェリン、アデニン、EGF、及びT3、ヒドロコルチゾン、ウシインシュリンのようなホルモン。
【0022】
本発明は、ケラチノサイト、特に鳥類のケラチノサイトの培養方法にも関し、該方法は:
(i) 表皮細胞又はケラチノサイトの懸濁物を作製し;
(ii) 上記の細胞を、上記のようなニワトリ血清を含む培養培地中の3T3細胞からなる細胞担体上に接種し;
(iii) 上記の表皮細胞をその細胞担体上でインキュベートする
各工程を含む。
【0023】
表皮細胞又はケラチノサイトの懸濁物は、知られた様式で、表皮の酵素消化により得ることができる。このような消化は、タンパク質分解酵素での処理、例えばトリプシン、ジスパーゼ(dispase)又はサーモライシンでの処理により行うことができる。表皮を化学処理、例えばEDTA (エチレンジアミン四酢酸)での処理により処理することも考えられる。
【0024】
インキュベーション条件は、細胞培養に通常適用されるもの、すなわち5〜10%のCO2及び100%湿度中で約37℃の温度である。
【0025】
インキュベーション後、細胞密度が充分であるときに、これらの条件下で得られた細胞を新しい細胞担体に移し、上記の条件下で再び培養することができる。工程(iii)の最後で、培養に由来する細胞を回収するときに、本発明の方法の工程(ii)及び(iii)に従って、さらなる培養サイクルを該細胞に適用する。上記の方法に従って30回までの連続する細胞増殖サイクルを考えることができる。細胞集団の倍加時間は、最初の10回の細胞増殖サイクルの間は約12〜16時間であり、その後のサイクルのあいだに徐々に増える。例えば、最初に接種した106細胞で開始することにより約1012細胞を得ることができ、つまり約106の増加係数(expansion factor)である。
【0026】
1又はそれより多い培養サイクルの後に得られる細胞は、その後の使用の観点で凍結形態で保存することもできる。3T3細胞の担体と、ニワトリ血清及びケラチノサイトを含む培地とを含む細胞培養物は、本発明の別の主題である。
【0027】
驚くべきことに、本発明による培養方法が、細胞培養物に由来するケラチノサイトにおいて全てのその分化の特徴を保存することを可能にすることを証明することができた。
【0028】
鳥類の表皮は、脂質並びにα−及びβ−ケラチンフィラメントに富む重層扁平上皮である。下層の細胞は、タンパク質で構成される基底膜上にあり、例えばラミニン5のような該タンパク質のいくらかはケラチノサイトの特異的産物である。基底層は特異的マーカーであるp63を合成するケラチノサイト幹細胞を含有する。表皮の凝集(cohesion)は、プラコグロビン(plakoglobin)タンパク質及びコネクシン31タンパク質をそれぞれ含むデスモソーム及びギャップ結合のようなケラチノサイト間結合によりもたらされる。表皮の表面層において、ケラチノサイトは最終分化のプロセスを受け、その間にケラチノサイトは14-3-3シグマを合成し、角化エンベロープを形成する。該エンベロープは界面活性剤及び還元剤中で煮沸した後に不溶性のままである。このエンベロープは、ペリプラキンのような特異的なタンパク質を含有する。
【0029】
本発明の培養方法により、培養におけるニワトリケラチノサイトは、その組成が鳥類の表皮と類似の脂質、α−及びβ−ケラチンフィラメント、デスモソーム並びにp63、14-3-3シグマ、プラコグロビン、コネクシン31及びペリプラキンの各タンパク質を合成する。最後に、ニワトリケラチノサイトは、培養において、重層されたコロニーを形成し、その表面層において細胞は、界面活性剤及び還元剤中で不溶性の角化エンベロープを合成する。結果として、本発明による培養系において、ケラチノサイトはそれらがインビボで有する全ての特性を維持する。
【0030】
分化の特性を保存しているケラチノサイトの培養を得る可能性は、完全なウイルス複製条件下でのウイルスの増殖を構想することを可能にする。MDVの場合、例えば、ニワトリケラチノサイトは、ウイルスの完全な複製とエンベロープ全体で囲まれたビリオンの産生とをもたらし得る唯一の細胞種である。このウイルスの細胞溶解特性のために、このウイルスは感染された細胞から放出され(Biggs, P.M., Marek's disease, Springer-Verlag出版, 1〜24, 2001)、よって培養培地から容易に抽出できる。さらに、これらの細胞は非常に大量のウイルスを増殖させ、これらのウイルスの大量生産を促進する。
【0031】
結局、本発明の主題は、インビトロでウイルスを増殖させる方法であり、該方法は:
(i) ウイルスの株をケラチノサイトに感染させ;
(ii) 感染したケラチノサイトを、上記の培養方法に従う培養に付し;
(iii) 工程(ii)で増殖したビリオンを抽出する
工程を含むことを特徴とする。
【0032】
本発明の方法は、抽出の後に凍結乾燥でき、よって従来技術の感染された細胞の懸濁物よりも好ましい条件下で保存及び貯蔵されることを許容する無細胞ウイルス粒子を得ることを可能にする。数回の培養サイクルにより弱毒化されたウイルスは、それにもかかわらず完全ビリオンの形で増殖する。この新規な特徴は、従来技術のものよりも良好な免疫化を提供する、より効果的なワクチンの製造を構想することを可能にする。好ましくは、ウイルス増殖方法は、10〜400培養サイクル、有利には40〜100培養サイクルを行うことにより実行される。上記の方法に由来する生存弱毒包膜DNAビリオンは、本発明の別の主題を構成する。
【0033】
本発明の主題は、ワクチンを製造する方法でもあり、該方法は、上記の方法に従ってウイルスを増殖させ、次いで医薬的に許容される担体、特にワクチン担体中にビリオンを導入する各工程を含むことを特徴とする。ビリオンを医薬的に許容される担体に導入する前に凍結乾燥して貯蔵することが有利に構想される。特に、ワクチンは周囲温度で輸送可能である。ワクチン担体は、通常用いられるワクチンアジュバントを知られた様式で含有し得る。
【0034】
本発明の方法は、現在までどのようにして無細胞ワクチンを製造するのか知られていなかった包膜されたDNAウイルスからのワクチンの製造に特に適用される。包膜されたDNAウイルスのうち、特にMDV及びFPVウイルスを挙げることができる。
【0035】
本発明の主題は、よって、包膜されたDNAウイルスから得られるビリオンを含む弱毒生ワクチンでもある。通常、このようなワクチンは、1又はそれより多いワクチンアジュバントを好ましくは含む水溶液の形態のワクチン担体を含む。このようなワクチンは、第一に凍結乾燥されたビリオン、及び第二に免疫化担体を含み、これらのそれぞれが別個の区画に包装され、2つの成分が、それらを経口又は注射により或いはインオボ経路を介して投与するために即座に混合される免疫化キットの形で、一般に市販される。
【0036】
本発明のワクチンを呼吸経路を介して投与し得ることを構想することもできる。この場合、該ワクチンは、水溶液又はエアロソルの形で製造される。
【0037】
ケラチノサイトの培養の可能性は、ケラチノサイトで複製可能な弱毒ウイルス株を開発することを可能にする。マレック病の原因であるMDVウイルスの場合、家禽農場からの家禽にこのウイルスを投与した後に、これらのビリオンは鱗屑に存在するので、家禽農場全体に感染して保護することができるだろう。本発明の主題は、弱毒ビリオンの、マレック病の予防または治療用の医薬品の製造のための使用でもある。
【0038】
本発明のケラチノサイトの培養方法は、研究分野において他の適用を見出す。
【0039】
ケラチノサイトの培養は、この細胞種へのウイルスの侵入の機構を明らかにすること、特にこの侵入に必要なウイルスエンベロープタンパク質を明らかにすることを可能にするはずである。これらのタンパク質は、その後、動物の免疫化のために組換えの形で用い得る。マレック病の場合、この疾患の原因であるウイルスの完全ヌクレオチド配列が知られている。ケラチノサイトに感染できない変異ウイルスの選択により、対応する変異を見出し、よってケラチノサイトへの侵入の原因であるタンパク質を見出すことが可能になるであろう。
【0040】
本発明は、本発明の説明のためにのみ与えられ本発明を限定するとは考慮されるべきでない実施例によりさらに以下で記載され、説明される。
【0041】
- 図1は、位相差顕微鏡写真を示し、これらはニワトリ及びヒトの表皮細胞の増殖を示す。(A) 105の第3継代のニワトリ表皮細胞を、10%ニワトリ血清含有培地中での培養に付す。(B) より高い倍率で撮影した(A)と同じ培養。(C) 7.8×105のヒトケラチノサイト(YF23)を、10%ウシ胎児血清含有培地での培養に付す。(D) ヒトケラチノサイト(YF23)を、10%ニワトリ血清含有培地での培養に付す。各目盛りバーは50μmに相当する。
【0042】
- 図2は、オイルレッドOでの標識及びHoechst 33258試薬での染色の後の、培養でのケラチノサイトによる細胞質脂肪の蓄積を示す。(A) 黄色フィルタを用いた位相差顕微鏡。(B) 同じ範囲を位相差の下及び蛍光の下の両方で撮影し、白黒で印刷した写真。矢印は、Hoechst 33258試薬で染色された核(白色)を示す。目盛りは25μmに相当する。
【0043】
- 図3は、ニワトリケラチノサイトの増殖に対するニワトリ血清の影響を示す。5×104の細胞を、ニワトリ血清(ChS)及びウシ胎児血清(FCS)の混合物中に培養に付し、次いで固定化してローダミンBで染色する。ある場合においては培養培地におけるFCSのパーセンテージは10%に固定し、ChSのパーセンテージは変動可能である。他方の場合においては培養培地におけるChSのパーセンテージは10%に固定し、FCSのパーセンテージは変動可能である。
【0044】
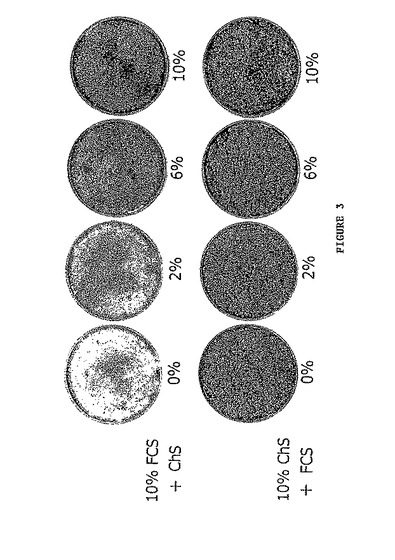
- 図4は、ニワトリ血清の脱脂が、ニワトリケラチノサイトの増殖に影響しないことを示す。3500細胞/cm2の密度のニワトリケラチノサイトを接種し、10%ニワトリ血清中に7日間培養に付す。あるディッシュにおいては血清の補足をウシ胎児血清(FCS)又は脱脂ニワトリ血清(Delip. ChS)のいずれかで置き換え、別のディッシュにおいては細胞を未処理のニワトリ血清(Unt. ChS)中に維持する。D1:第1日、D7:第7日、D15:第15日、D20:第20日。(A) 培養物を接種の15日後に固定化し、ローダミンBで染色する。矢印は、脱脂血清において観察される小さい小滴(droplets)を示す。
【0045】
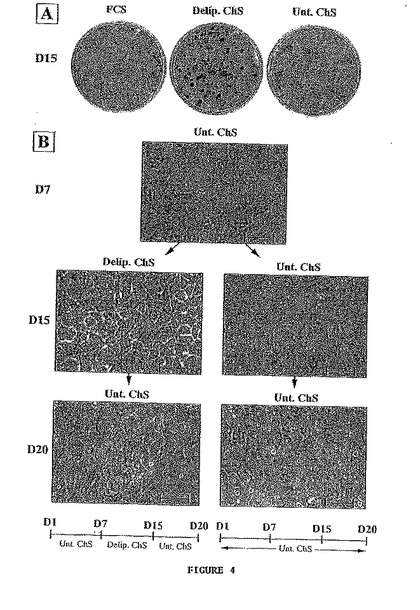
- 図5は、培養におけるニワトリケラチノサイト中のケラチンの証明を示す。(A) 細胞を抗パンケラチン抗体で標識する。矢印は3T3細胞を示し、この細胞は多核であり、抗体で標識されない。目盛りは25μmに相当する。(B) 別の抗パンケラチン抗体で標識されたイムノブロット。レーン(1)は培養におけるニワトリケラチノサイトに対応し、レーン(2)はニワトリ表皮に対応し、レーン(3)はニワトリの腸に対応する。2つのダブレット(a及びc)は、ニワトリ表皮中の対応するダブレットと一致する。
【0046】
- 図6は、ニワトリケラチノサイトが培養において角化エンベロープを形成することを示す。写真は、位相差顕微鏡下で撮影される。(A) 15日の生存コロニーの基底層。(B) より大きい焦平面で撮影された同じコロニー;暗い輪郭及び角形は角化を示す。(C) 2%のドデシル硫酸ナトリウム及び2%の2-メルカプトエタノールを含有する溶液中に100℃で処理した後。目盛りは50μmに相当する。
【0047】
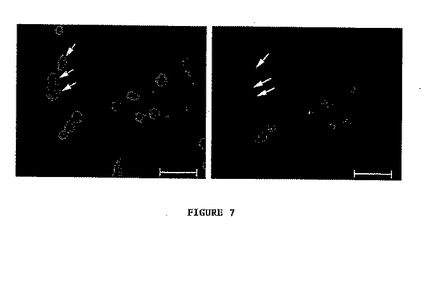
- 図7は、抗p63抗体を用いる免疫蛍光染色によるp63標識を示す。矢印は、大きい核及び顕著なヘテロクロマチンにより認識される3T3細胞を示す。これらは抗体により標識されない。目盛りは25μmに相当する。
【0048】
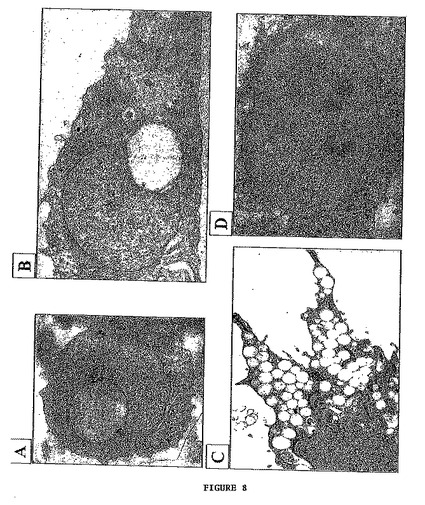
- 図8は、核周囲及び核内の脂質を示す。これらは電子顕微鏡写真である。倍率はA及びBが5000×、Cが6000×及びDが4000×である。
【0049】
- 図9は、ケラチノサイト内の層形成を示す。デスモソーム(A中)及びケラチンフィラメント(B中)。これらは電子顕微鏡写真である。倍率はAが40000×、Bが8000×である。
【実施例】
【0050】
実施例1:10%ニワトリ血清の存在下での3T3-J2担体細胞とのニワトリケラチノサイトの増殖
1.1 概要
ニワトリ繊維芽細胞の増殖は、3T3細胞の単層により効率的に解消されず、ニワトリの皮膚全層から作製した培養物は、繊維芽細胞により迅速に侵入される。よって、表皮を分散させる(disaggreagation)前に、サーモライシンでの処理により表皮を真皮から分離する。最初に、培地にウシ胎児血清を補うが、この血清はニワトリケラチノサイトの増殖を許容しない。ウシ血清をニワトリ血清に置き換えることにより、細胞コロニーが発生することが可能になる。
【0051】
1.2 材料及び方法
1.2.1 生物学的材料
新生の雛(Gallus gallus)を、高い湿度レベルで37.8℃において20〜21日間インキュベートした受精卵から得る。表皮細胞の初代培養物を得るために、トリブロモエタノールの腹腔内注射により新生の雛に麻酔をかける。
【0052】
マウス3T3-J2細胞:3T3系統の起源は、Todaro, G., J.ら, J. Cell Biol., 17, 299〜313, 1963に記載される。ATCC (No. CCL92)から得たオリジナルのSwiss 3T3細胞を96ウェルプレートに低密度で(3ウェル当たり1つの細胞の割合で)接種する。約2週間の培養後、いくつかのウェルは目で見える細胞コロニーを含む。これらの培養物は、単一の細胞に由来するのでクローンであり、よってそれぞれ1つずつが3T3サブクローンである。各コロニーを直径100 mmの培養ディッシュに別々に移す。細胞数が充分であるときに、細胞をトリプシン処理により剥離する。各サブクローンの半分を液体窒素で凍結させる。残りの半分をマイトマイシンC又はガンマ線で処理する。ニワトリケラチノサイトを、処理された各サブクローン上に、直径100 mmのディッシュ当たり1.5×106の3T3当たり102〜106細胞の密度で接種する。培養7〜14日後に、これらの細胞を固定化しローダミンBで染色する。3T3-J2とよばれる3T3サブクローンは、最大サイズのケラチノサイトコロニーがその上に最大数存在し、最適であるとみなされる。これから先は、これをニワトリケラチノサイトの培養に用いる。
【0053】
新生の包皮を起源とするヒト表皮ケラチノサイト(Rheinwaldら, Cell, 6, 331〜343, 1975に記載の方法に従って作製)。
【0054】
1.2.2 方法
細胞培養(方法1):
足部又は体からの皮膚を回収する。リン酸緩衝食塩水(PBS)でこれを数回洗浄し、Hepes緩衝溶液(HBS)中の0.5 mg/mlのサーモライシン(Sigma)で、37℃で45分間又は4℃で一晩のいずれかでの酵素処理により、表皮を真皮から分離する。次いで表皮を回収し、薄い切片に切断し、トリプシン(0.25%)及びEDTA (0.02%)の存在下に37℃で30分間、2回、トリプシン処理フラスコ内で攪拌しながらインキュベーションすることにより分散させる。細胞懸濁物を800 gで5分間遠心分離する。細胞ペレットを培養培地に採取し、細胞を、マイトマイシンC (4μg/ml)で予め処理したマウス3T3細胞(2.3×104細胞/cm2)の単層で被覆したペトリディッシュに接種する。その後、ディッシュを37℃、7.5% CO2及び100%湿度のインキュベータに入れる。培養培地は、10%のニワトリ血清(Invitrogen)を補った1/1のDMEM/F12混合物(Invitrogen)からなる。次の添加物も培地中に存在する:0.4μg/mlのヒドロコルチゾン(Calbiochem)、5μg/mlのウシインシュリン(Sigma)、2×10-9 Mの3,3',5-トリヨード-L-チロニン(T3、Sigma)、10-10 Mのコレラトキシン(ICN)及び1.8×10-4 Mのアデニン(Calbiochem)。組換えヒト表皮成長因子(EGF)を、最初の培地交換のときに10 ng/mlの濃度で添加する。真皮の除去にもかかわらず繊維芽細胞コロニーが発生する場合があるが、これらは、培養物をCa++もMg++も含有しないPBS又は0.02% EDTAで短時間処理することにより除去できる。次いで、追加の3T3担体細胞を加え、培養を継続する。初代培養物を、接種密度に応じて接種の7〜19日後に移す。次いで、第2代培養物を得る。細胞を、第1継代の後に、10%ジメチルスルホキシド(DMSO)を含有する培地中で凍結させる。凍結は、1〜3×106細胞/mlの割合で、上記の培養培地中で行う。凍結細胞は、液体窒素中に無期限に保存できる。
【0055】
ヒト表皮ケラチノサイトは、ウシ胎児血清をニワトリ血清の代わりに用いた以外はニワトリ細胞について記載したのと同じ培養に付す。
【0056】
細胞培養(方法2):液/気界面でのニワトリケラチノサイトの培養
この方法は、文献J. Y. Yiら, Arch. Dermatol. Res., 2001, 293, 356〜362に基づく。この目的は、より完全な最終分化を得ることである。マイトマイシンで処理したか又はしていない3T3細胞を、供給業者(Nitta gelatin, Tokyo)の使用説明に従って、I型コラーゲンマトリックスと混合し(3×105細胞/ml)、この混合物の200μlを、ポリカーボネートフィルタ(Millicell-pc Millipore, Bedford, MA)上に接種する。ケラチノサイトをコラーゲンマトリックス上に接種し、通常の培養培地(DMEM/F12 +ニワトリ血清+添加物)中に5〜7日間浸しながら培養する。その後、フィルタを培地/空気界面に置き、培養を14日間継続する。
【0057】
位相差顕微鏡
Nikon Coolpixデジタルカメラを備えたNikon顕微鏡を用いた。
【0058】
オイルレッドOを用いた脂質小滴の染色
カバーガラス上に接種された細胞を、3.7%ホルムアルデヒドを含有する冷PBS中に15分間、周囲温度で固定化し、次いで新鮮な固定化溶液で追加の1時間インキュベートする。蒸留水でカバーガラスを洗浄して空気乾燥する。脂質の小滴を0.3%のオイルレッドO (Sigma)及び60%のイソプロパノールを含有する溶液で1時間染色する。インキュベーションの最後に、カバーガラスを蒸留水中に洗浄し、Hoechst 33258試薬(1μg/ml)及びSigma社からIgepalの商品名で販売されている製品0.1%を含有するPBS中に5分間インキュベートする。カバーガラスをPBS/Igepal中に1回、及び水中に1回洗浄し、2.5%のDABCO (1,4-ジアゾビシクロ(2.2.2)オクタン)を含有する、Calbiochem社からMowiolの商品名で販売されている製品中のスライドに載せる。
【0059】
1.3 結果
3T3細胞に接着する円形細胞は接種の数時間後に目で見えるが、典型的な上皮単位を明らかに組織するコロニーは、細胞がプラスチックとの接触を確立するときである、接種の5〜6日後に目で見えるようになる。コロニーは、ヒトケラチノサイトにより形成されるコロニーと同様に、コロニーの周囲の3T3細胞の単層を置き換える。ほとんど全ての細胞が単一の大きい小滴と、ときにおそらく脂質で構成されるいくつかの小さい小滴とを含有することが、最も顕著であるように見られる(図1A及び1B)。その表皮の起源及び重層扁平上皮を形成し得るその能力により、また鳥類のケラチノサイトが脂質の小滴を含有することが知られていることからすると、コロニーがケラチノサイトで構成されていると結論付ける。
【0060】
10%のニワトリ血清を補った培地中に12 000細胞/cm2の密度で接種したヒトケラチノサイトは、ウシ胎児血清中で増殖するのと同様に迅速に増殖する。ヒトケラチノサイトは、10%のニワトリ血清の存在下では12日後であっても、脂質の小滴を決して発生しない(図1C及びD)。このことから、脂質の小滴の形成は、ニワトリケラチノサイトの固有の特性であり、他の種のケラチノサイトではニワトリ血清により誘導されないと結論付ける。
【0061】
中性脂肪に特異的なオイルレッドO製品での固定化細胞の染色は、屈折小滴(refringent droplets)が中性脂肪を含有することを示す(図2A)。Hoechst 33258試薬を用いた対比染色は、細胞内の脂質小滴のほとんどが核周囲であることを示す(図2B)。
【0062】
3つの独立した実験において、約500のコロニーが5×105の分散された表皮細胞から形成されたことを示した。これは、10-3のコロニー形成効率に相当する。よって、表皮から直接採取されたニワトリケラチノサイトについての接種効率(seeding efficiency)は、1〜10%程度であると報告されているが一般的には1%に近いヒトケラチノサイトのものよりかなり低い。後続の継代で非常に高いコロニー形成効率を達成するヒトケラチノサイトとは違って、ニワトリケラチノサイトの接種効率は、数パーセントを決して超えない。
【0063】
ニワトリケラチノサイトの倍加時間は、第2代培養物中で15コロニー中の細胞数を24時間ごとに、接種後の第1日から第4日まで計数することにより測定される。ニワトリケラチノサイトは、単独の脂肪小滴を含有することからして、隣接する3T3細胞から容易に区別することができる。最高の増殖速度のコロニーを考慮に入れると、16.8時間の倍加時間が得られる。倍加時間は、後続の継代の過程で減少し、ニワトリケラチノサイトは第20代の植え継ぎ培養を超えると培養に付すことができない。
【0064】
ニワトリケラチノサイトのサイズは、細胞の懸濁物を血球計数器に導入し、細胞の写真を撮影しその直径を測定することにより測定される。ケラチノサイトは10μmの典型的な直径を有し、よって最も一般的な直径が12μmであるヒトケラチノサイトよりも小さい。この相違は、ニワトリゲノムのサイズがヒトのゲノムサイズの1/3しかないことにより説明できる(1.25 pgに対して3.5 pg/一倍体のゲノム)。12μmの典型的な直径を有するニワトリ繊維芽細胞も、最も一般的な直径が15.6μmであるヒト繊維芽細胞よりも小さい。
【0065】
実施例2:ニワトリケラチノサイト増殖過程でのニワトリ血清の役割
2.1 概要
ウシ胎児血清は、ウシ胎児血清が増殖阻害物質を含有していること又は増殖促進因子を欠いていることのいずれかの理由から、ニワトリケラチノサイトの増殖を許容しない。
これらの選択肢の間を区別するために、ニワトリケラチノサイトを、ニワトリ血清を増加する濃度で補った10%のウシ胎児血清の存在下、又はウシ胎児血清を増加する濃度で補った10%ニワトリ血清の存在下のいずれかに10日間培養する。実験の最後に、細胞を固定化してローダミンBで染色する。
【0066】
2.2 材料及び方法
2.2.1 材料
用いた生物学的材料は、実施例1に記載のものである。
【0067】
2.2.2 方法
細胞培養プロトコルは、実施例1に記載されたものに従って用いた。
【0068】
ローダミンBでの染色
培養物をPBSで2回洗浄し、10%グルタルアルデヒド含有PBSで、周囲温度で15分間固定化する。次いで、細胞を水中の1%ローダミンBで周囲温度で15分間、緩やかに攪拌しながら染色する。培養物を水で洗浄し、その後空気乾燥する。
【0069】
2.3 結果
図3において、ニワトリ血清がケラチノサイトの増殖を許容し、この能力は、10%ウシ胎児血清に加えられるときにニワトリ血清の用量に依存することが明確にわかる。10%ニワトリ血清の増殖促進活性は、ウシ胎児血清の添加量を増加しても影響されない。このことから、ニワトリ血清がニワトリケラチノサイトの増殖に必要な因子を含有しており、この因子はウシ胎児血清には存在しないと結論付ける。
【0070】
ニワトリケラチノサイトが脂質を含有することからすると、ニワトリ血清の増殖促進活性が血清中の脂質の吸収に関係する。図4Aに示される実験は、溶媒の作用により脂質を欠くニワトリ血清が、ニワトリケラチノサイトの増殖を促進するその能力を維持することを示す。よって、増殖促進活性は脂質によるのではない。特にコロニーの周囲で増殖する細胞は、脱脂血清中で増殖するときに脂質を保存し、よって脂質を合成できる(図4B)。
【0071】
実施例3:α−ケラチンを合成する培養におけるニワトリケラチノサイト
3.1 概要
表皮細胞の特性の一つは、豊富にケラチノサイトを合成するその能力である。ニワトリケラチノサイトを、カバーガラス上での培養に付し、固定化する。次いで細胞を、抗α−パンケラチン抗体の存在下にインキュベートし、間接蛍光顕微鏡により調べる。
【0072】
培養におけるニワトリ表皮細胞のケラチンを、ウェスタンブロッティング免疫検出により分析する。ケラチンを集密な培養物から抽出し、一次元ゲル電気泳動により分離し、ニトロセルロースに移し、抗α−パンケラチン抗体を用いて染色する。
【0073】
3.1 材料及び方法
3.1.1 材料
生物学的材料は、実施例1に記載されたものである。
【0074】
3.2.1 方法
間接免疫蛍光染色
細胞をカバーガラス上に接種する。次の日に、細胞をPBS中に2回洗浄し、次いでアセトン:メタノールの1:1混液中に20分間、-20℃で固定化して空気乾燥させる。カバーガラスを、非特異的部位をブロックするために5%ウシ血清アルブミン(BSA)含有PBS中に10分間、周囲温度でインキュベートする。
【0075】
α−ケラチン染色のために、細胞をマウスモノクローナル抗パンケラチン抗体(1:100希釈, MS-744-A0, NeoMarkers)を含有する5% BSA/PBS中に周囲温度で1時間インキュベートし、その後、0.1%の非イオン界面活性剤Igepal CA 630 (Sigma)含有PBSで、各回5分間、3回洗浄する。次いで細胞を、ビオチン結合ヤギ抗マウスIgG (1:1000希釈, Jackson ImmunoResearch)を含有する5% BSA/PBS存在下に周囲温度で1時間インキュベートする。カバーガラスを洗浄し、シアニン−3に結合したストレプトアビジン(1:1000希釈)で1時間インキュベートする。インキュベーションの最後に、カバーガラスをPBS/Igepal中に5分間洗浄し、その後、Hoechst 33258試薬(1μg/ml)及び0.1%のIgepalを含有するPBS中に5分間インキュベートする。カバーガラスをPBS/Igepal中に1回及び水中に1回洗浄し、2.5%の1,4-ジアゾビシクロ(2.2.2)オクタン(DABCO)を含有するMowiol中のスライドに載せる。
【0076】
α−ケラチンの抽出及び免疫検出(ウェスタンブロッティング)による分析
集密な培養物をPBSで2回洗浄し、Dounceホモジナイザを用いて粉砕する。細胞は、10 mMのTris-HCl (pH 7.6)、1mM EDTA及びプロテアーゼ阻害剤の混合物(Roche)を含有するバッファー中に20回の回転後にホモジナイズされる。ホモジネートを8000 gで5分間、4℃で遠心分離し、ペレットをTris-EDTAバッファーで2回、2%のNonidet P40を含有するTris-EDTAバッファーで2回洗浄する。最終のペレット中のケラチンを、2%のSDS及び10mMのジチオトレイトールを含有する溶液中に37℃で15分間、超音波装置(2×15秒)を用いて懸濁させる。次いで、サンプルを100℃で2分間加熱し、不溶性の残渣を除去するために8000 gで5分間遠心分離する。
【0077】
α−ケラチンを、変性8%ポリアクリルアミドゲル(アクリルアミド:ビスアクリルアミドの重量比は29:1)での電気泳動により分離し、ニトロセルロース上に電気的に溶出させる。メンブレンを、5%のドライスキムミルク及び0.1%のTween 20を含有するPBS (PBST)中に4℃で一晩処理する。α−ケラチンを、一方はI型ケラチンに指向され、他方はII型ケラチンに指向された2つのモノクローナル抗体の混合物からなる抗パンケラチン抗体(PRO61835, Research Diagnostics Inc)を用いて染色する。抗体を1:500の希釈で用い、メンブレンをPBSTで洗浄し、その後、セイヨウワサビペルオキシダーゼに結合したロバ抗マウスIgG (Amersham)の1:2000の希釈でインキュベートする。抗体のインキュベーションは、周囲温度で1時間継続する。ケラチンに対応するバンドは、ケミルミネセンス(ECL+kit, Amersham)により視覚化される。
【0078】
3.3 結果
培養における全てのニワトリケラチノサイトは強く標識されるが、3T3細胞は標識されない(図5A)。特異的抗体を省いたときには標識は観察されない。
【0079】
免疫検出からの結果を図5Bに示す。抗体で染色されたタンパク質は、3つの主要なダブレットに分かれ(a〜c)、それらのそれぞれの分子量は、約50、57及び63 kDである。ダブレットa及びcはニワトリの皮膚にも存在するが、ダブレットbは存在しない。角質化していない上皮であるニワトリの腸から抽出されたケラチンの一般的なプロフィールは、表皮細胞とも表皮とも一致しない。培養における細胞が、表皮のケラチンのものと一致する分子量を有する2つのケラチンダブレットを含有することからして、培養における細胞が表皮のケラチンを合成すると結論付ける。高分子量のケラチンが重層扁平上皮の外層に特異的であると考えられることからして、2つの高分子量のケラチン(図5B中のc)は特に適切である。4つのα−ケラチンをさらに特徴付ける(以下を参照)。
【0080】
実施例4:ニワトリケラチノサイトは培養において角化エンベロープを形成する
4.1 概要
哺乳動物の表皮ケラチノサイトの顕著な特性は、最終分化プロセスに従うその能力であり、該プロセスにおいてそれらの核は破壊され、その間に、ドデシル硫酸ナトリウム及び2-メルカプトエタノールの存在下で不溶性の角化細胞エンベロープを発生する。
【0081】
4.2 材料及び方法
本実施例において用いた生物学的材料及び方法は、実施例1に記載されたものである。
【0082】
4.3 結果
サイズが数百細胞を超えたニワトリケラチノサイトコロニーの中央が観察されるとき、それらが、それぞれがいくつかの基底細胞を覆う大きい細胞をほぼいつも含有することが注目される(図6A及びB)。これらの鱗屑は培地中に迅速に放出され、おそらく脂質で構成されている屈折小滴をいつも含有する。ニワトリの鱗屑は薄く、ヒトの鱗屑のおよそ2倍大きい(100μmに対して50μmの長軸(greater length)) (図6B)。ニワトリ表皮細胞コロニーを回収して2%のSDS及び2%の2-メルカプトエタノールを含有する溶液に入れ、100℃で加熱するとほとんどの細胞は溶解するが、角化エンベロープは不溶性のままであり、脂質小滴を未だ保持する(図6C)。ニワトリ表皮から直接得られるエンベロープのほとんども、SDS/2-メルカプトエタノール中での煮沸の後に脂質を保持する。これらの脂質は、メタノール/クロロホルムを用いて抽出し得る。
【0083】
実施例5:ニワトリケラチノサイトはp63を含有する
p53タンパク質をコードする遺伝子ファミリーの一部分であるp63タンパク質は、多くの上皮の基底層に豊富に存在する。ヒト表皮において、p63は高い増殖ポテンシャルを有する細胞でのみ見出される。p63をコードする遺伝子のノックアウトにより、全ての重層扁平上皮が存在しなくなり、p63はケラチノサイト幹細胞のマーカーであるとみなされる。
【0084】
ニワトリケラチノサイトがp63を含有するかを調べるために、増殖が早い細胞をカバーガラス上に接種し、次の日に免疫蛍光顕微鏡によりモノクローナル抗-p63抗体4A4を用いて観察する。
【0085】
5.1 材料及び方法
5.1.1 材料
生物学的材料は実施例1に記載される。
【0086】
5.1.2 方法
細胞培養及び間接免疫蛍光染色法は、それぞれ実施例1及び3に記載されたものである。
【0087】
p63標識
細胞を、1:500に希釈した抗ヒトp63モノクローナル抗体4A4の存在下に4℃で一晩インキュベートする。これ以外のプロトコルは、二次抗体を1:200の希釈で用いた以外はケラチン染色に用いた方法と同じである。細胞を、Nikonデジタル写真撮影装置を備えたNikon E600蛍光顕微鏡を用いて写真撮影する。
【0088】
5.2 結果
調べた全ての細胞は、p63について強い核染色を示す。逆に、3T3細胞は標識を示さないか又は時々弱い細胞質標識を示すかのいずれかである(図7)。
【0089】
実施例6:電子顕微鏡によるニワトリケラチノサイトの特徴づけ
6.1 材料及び方法
ニワトリケラチノサイト単層培養物を得る。細胞が2/3の集密であるときに、3T3細胞をEDTAを用いて除去し、ケラチノサイトを3T3担体細胞を含まない小さいペトリディッシュに移す。次の日に、電子顕微鏡で観察されるように細胞を固定化して処理する。
【0090】
電子顕微鏡
細胞を、0.1MカコジレートpH 7.3で洗浄し、0.1Mカコジル酸ナトリウムpH 7.2中の5% パラホルムアルデヒド及び5% グルタルアルデヒドで90分間固定化し、通常の電子顕微鏡で観察する。酢酸ウラニル及びクエン酸鉛で染色した超薄切片を、80 kVでJeol JEM-1010電子顕微鏡(Jeol Ltd, Tokyo, Japan)を用いて調べる。
【0091】
6.2 結果
図8は、脂質小滴を含有する細胞の一連の顕微鏡写真を示す。2枚の顕微鏡写真(A及びB)は、核内に包埋された単一の核周囲脂質小滴を有する典型的な細胞を示す。よって、ほとんどの核が三日月形を示す。別の2枚の写真は、多数の小さい細胞質脂質小滴を有する細胞(C)及び核の内部に脂質小滴を有する別の細胞(D)を示す。これらの2つの特徴は、通常存在する。図9は、典型的な表皮細胞の特徴を示す。写真(A)は、デスモソームの存在を示す。写真(B)は、基底細胞と、細胞質がトノフィラメント(tonofilaments) (ケラチン)で充たされている超基底細胞(superbasal cell)とを示す。
【0092】
実施例7:ランダム配列決定による特異的なケラチノサイト産物の同定
7.1 概要
cDNAライブラリを、集密なニワトリケラチノサイト培養物から構築する。163のランダムなコロニーに由来する5'末端のヌクレオチド配列を、自動化シーケンサーを用いて決定する。次いで。cDNA配列との相同性探索のためにGenBank及びタンパク質データバンクを用いる。
【0093】
7.2 材料及び方法
cDNAライブラリの構築とヌクレオチド配列決定
Tripure Isolation Reagent (Roche)を用いて、培養でのニワトリケラチノサイトから全RNAを精製する。ポリ(A)+ RNAを、オリゴ(dT)セルロース上で、Micropoly(A) Purist (登録商標) (Ambion) mRNA精製キットを用いて単離する。cDNAライブラリを、1.7μgのポリ(A)+ RNAから、SuperScript (登録商標) Plasmid System (Life Technologies)を用いて作製する。第一の鎖の合成を、オリゴ(dT)配列の後ろにNotI部位を含有するプライマを用いて開始する。第二の鎖の合成の後に、SalIアダプタがcDNAの両端に付加される。NotIでの消化により、3'末端に付着末端が発生するが、SalI付着末端は5'末端に残る。cDNAを、SalI及びNotIで予め切断しておいたプラスミドpSport1に連結し、エレクトロポレーションによりエレクトロコンピテントな細菌E. Coli DH10B (Life Technologies)に導入する。
【0094】
cDNAクローンの5'末端のランダム配列決定は、SalI部位の上流でプラスミドにハイブリダイズするプライマを用いて行う。配列を伸ばすために、特異的な内部プライマを用いる。
α−ケラチン5/6について:
配列番号1 5'-AGTGGATTCTATGGACCTGC-3'、
配列番号2 5'-AACTGAGCAGCTTGCTGGC-3'、
配列番号3 5'-CAGTCCAGGTACGAAGAGC-3'、
配列番号4 5'-TAGGATCGGGATATGGAAGC-3';
【0095】
α−ケラチン15について:
配列番号5 5'-GGCTTTGATGCTATCTGTGC-3'、
配列番号6 5'-GTGGAGTCTGACATCAACG-3'、
配列番号7 5'-AGAACTGAGACGCACGATGC-3';
β−ケラチンKについて:
配列番号8 5'-ACTAGGATGTTACTGCGTGG-3'。
【0096】
配列決定は、蛍光ジデオキシヌクレオチドの存在下にGeneAmp 2400 PCR装置(Perkin-Elmer)で行う。PCR条件は、96℃で30秒、50℃で15秒及び60℃で4分を30サイクルである。電気泳動及び蛍光ピークの検出は、自動化シーケンサー(ABI PRISM 310 Genetic Analyzer)で行う。配列は、SeqEd v1.0.3プログラムを用いて決定する。
【0097】
7.3 結果
50個のクローンは、データベースの配列と相同性を示さない。15個のクローンは、仮定的タンパク質と相同性を示す。86個のクローンは、既知の機能を有する遍在タンパク質をコードする。12個のクローンは、特定のケラチノサイトの機能に関係することが知られているタンパク質をコードする。これらのことから、ニワトリケラチノサイトにおいては、約40% (65/163)のmRNAが未知の機能のタンパク質をコードし、約12% (12/98)のmRNAがこの細胞種に特異的であると結論付ける。合計で、7個のケラチノサイト特異的クローンがケラチンをコードする(表III)。クローン6〜8は同じα−ケラチンをコードするが、クローン9〜11は種々のα−ケラチンをコードし、クローン12はβ−ケラチンをコードする。クローン6及び9〜12について、全ヌクレオチド配列を決定する。
【0098】
実施例8:培養におけるニワトリケラチノサイトのマレック病ウイルス(MDV)での感染
8.1 材料及び方法
MDV-1に感染したニワトリの皮膚の初代細胞(本質的に繊維芽細胞)又は血液リンパ球に由来する感染細胞を、亜集密な培養のニワトリケラチノサイト上に接種し、37℃で1時間インキュベートする。吸着後に、接種物を吸引により除去し、ニワトリケラチノサイト単層を、リン酸緩衝食塩水(PBS)で3回洗浄する。細胞培養培地を加え、細胞を37℃で1週間インキュベートする。
【0099】
完全に包膜されたMDVビリオンは細胞培養培地中に放出され、これを回収して凍結乾燥することができる。凍結乾燥のために、MDVビリオンを含有する細胞培養培地を-60℃で凍結させ、真空下に38℃で24時間乾燥させ、21℃で15時間乾燥させ、真空下に4℃で貯蔵する。
【0100】
感染したニワトリケラチノサイト培養物を、PBS、又は0.2% EDTA含有PBS、又は2.18×10-1M スクロース、7.2×10-3 M リン酸二カリウム、3.8×10-3 M リン酸カリウム、4.9×10-3 M グルタミン酸ナトリウム及び1% 粉末ウシアルブミンからなる"SPGA"安定化溶液、又は0.2% EDTA含有SPGA (SPGA-EDTA)、又は10%ソルビトールを含むSPGA-EDTA、又は10%ソルビトールを含む細胞培養培地中に移す。次いで、細胞懸濁物を2分間超音波により破砕し、2000 gで20分間遠心分離し、上清を無細胞ウイルス調製物として回収する。ウイルスを、3回の凍結/融解サイクルにより感染ケラチノサイトから抽出し、上清の流体を回収する。無細胞ウイルスは、上記のようにして凍結乾燥できる。
【図面の簡単な説明】
【0101】
【図1】位相差顕微鏡写真を示し、これらはニワトリ及びヒトの表皮細胞の増殖を示す。
【図2】オイルレッドOでの標識及びHoechst 33258試薬での染色の後の、培養でのケラチノサイトによる細胞質脂肪の蓄積を示す。
【図3】ニワトリケラチノサイトの増殖に対するニワトリ血清の影響を示す。
【図4】ニワトリ血清の脱脂が、ニワトリケラチノサイトの増殖に影響しないことを示す。
【図5】培養におけるニワトリケラチノサイト中のケラチンの証明を示す。
【図6】ニワトリケラチノサイトが培養において角化エンベロープを形成することを示す。
【図7】抗p63抗体を用いる免疫蛍光染色によるp63標識を示す。
【図8】核周囲及び核内の脂質を示す。
【図9】ケラチノサイト内の層形成を示す。
【技術分野】
【0001】
本発明は、新規なケラチノサイトの培養培地、ケラチノサイトの培養方法及び弱毒生ウイルスの増殖のための該方法の使用に関する。この方法により得られる弱毒生ウイルス及びそれを含む生ワクチンは、本発明の別の主題を構成する。
【背景技術】
【0002】
いくつかの病原性ウイルスは、その複製に特異的なケラチノサイトタンパク質が必須であるので、ケラチノサイトでのみ完全に複製することができる。
例えば、MDV (マレック病ウイルス)及びFPV (鶏痘ウイルス)の場合がそうである。
【0003】
MDVウイルスを増殖するのに現在用いられている細胞は、一般に繊維芽細胞である。MDVに対するワクチンの製造に用いられるニワトリ繊維芽細胞培養は、遊離の(free)ウイルス粒子を増殖させず、感染は半増殖性(semi-productive)である。増殖性の感染が可能な連続細胞系を確立する試みがなされたが、成功していない(Abujoubら, Virology, 214, 541〜549, 1995)。繊維芽細胞中でのウイルス複製が不完全であるという性質のために、細胞性のワクチンは、液体窒素中で凍結した形で使用されなければならない。繊維芽細胞で増殖したウイルスは、完全エンベロープを欠いているので、細胞溶解性でも感染性でもない。
【0004】
マレック病の場合、動物は呼吸経路を介して感染する。汚染の短時間後に、Bリンパ球の細胞溶解性感染が進行し、その結果、多くの組織の潜在感染をもたらす細胞性ウイルス血症となる。感染する細胞の種類としてはTリンパ球があり、これは、その後、形質転換されて致死性のリンパ腫をつくる。ウイルスは表皮の羽嚢(feather follicles)にも運ばれるが、ここは完全エンベロープを有するウイルスの唯一の増殖場所である。ウイルスは、非常に高い力価で、表皮の表面部分、特に羽嚢の角化された細胞(鱗屑)に存在する。細胞は落屑し、鱗屑の吸入により他の動物に感染する(Calnekら, J. Natl. Cancer Inst., 45, 341〜351, 1970; Calnekら, Avian Dis., 14, 219〜233, 1970; Carrozzaら, Avian Dis. 17, 767〜781, 1973)。
【0005】
マレック病は世界中に存在し、家禽農場に重大な損失を与える原因であるので、多くのワクチンが開発されている。これらのワクチンは、マレック病ウイルスの弱毒株で感染させた培養ニワトリ繊維芽細胞の懸濁物からなる。ワクチンは、凍結保護剤を含有する培養培地中の生細胞を液体窒素中に凍結させることにより製造される。免疫化は、インオボ(in ovo)注射又は生誕の1日後の皮下注射のいずれかにより行なわれる。該免疫化は、リンパ組織増殖性症候群に対して防御するが、ケラチノサイトの感染に対しては防御しない。このことは、ウイルスの連続的な存続及び伝播を説明する。その存続によるウイルスの迅速な変異は、現存するワクチンに対して耐性の、毒性が増加した形の出現をもたらす。
【0006】
ヒト乳頭腫ウイルスは、その生物サイクルが、重層扁平上皮のケラチノサイトの最終分化(terminal differentiation)と関連する小さいDNAウイルスである。調節遺伝子及び非構造遺伝子は、重層上皮の基底層で発現されるが、DNA複製、構造タンパク質及びキャプシドアセンブリの合成は表面層でおこり、言い換えると、最終分化のプロセスを受けているケラチノサイトでおこる。HPVサブタイプ16及び18は、90%を超える子宮頸癌で見出され、これらのサブタイプがこの癌の原因に関係すると認められている(zur Hausen, H., J. Natl. Cancer Inst., 92, 690〜698, 2000)。子宮頸癌は大きな公衆衛生問題であり、予防ワクチンの開発を目的とした多くの研究が行なわれている(Plummerら, Virus Res., 89, 285〜293, 2002)。しかし、これらのワクチンの開発は、ビリオンの効率的な複製及び産生をもたらす細胞培養法がないことにより、遅れている。
【0007】
よって、ある病原性ウイルス、特に包膜されたDNAウイルス、例えばMDV及びFPVの完全複製をもたらす方法に対する必要性が存在する。
【0008】
特異的なケラチノサイトタンパク質がウイルス複製に必須であるという知識の下、弱毒完全ウイルスの増殖をもたらすことを目的として、ケラチノサイトを培養する試みがなされている。
【0009】
ヒトケラチノサイトは培養できるが(Rheinwaldら, Cell, 6, 331〜343, 1975)、培養におけるそれらの分化状態は不完全である。Meyersら, J. Virol., 71, 7381〜7386, 1997は、感染性HPVビリオンの合成を記載している。ケラチノサイトは、ニワトリ血清フリーの合成培地であるE培地とよばれる培養培地中に、マイトマイシンで処理した3T3細胞上で培養される。
【0010】
しかし、HPVはMPVとは異なり、この文献に記載されたHPVビリオンの合成方法は、マレック病のビリオンを増殖させることができない。
【0011】
文献EP-0 748 867は、連続細胞株であるQT35細胞系上でのマレック病ウイルス血清型1の培養を記載している。感染された細胞は、ワクチンの作製に用いられ、単離ビリオンには用いられていない。なぜなら、このウイルスは細胞に強く結合するからである。これは、不完全なエンベロープを持って増殖し、感染性を保ったまま単離できない。
【0012】
文献EP-0 770 677は、ニワトリ細胞系上でのマレック病ウイルス血清型2の増殖を記載している。得られたビリオンは細胞培養物に依存する。
文献EP-1 149 899は、鳥類のES細胞の培養方法を記載している。培養培地は、マイトマイシンでの処理又は照射により不活性化されたフィーダー細胞の層、例えばマウス胚繊維芽細胞であるSTO細胞を含むのが好ましい。
【0013】
文献EP-0 779 359号は、鳥類細胞の培養方法を記載する。培養培地はニワトリ血清を含有する。培養は、STO (繊維芽細胞)又はSL-10細胞の担体(support)上である。
よって、いずれのケラチノサイト培養も、完全ウイルス複製、特にエンベロープDNAウイルス、例えばMDVをもたらす条件を提供できていない。
【0014】
さらに、実質的に全てのヒト及び動物細胞種はウシ血清の存在下に首尾よく培養することができるが(例えばDorangeら, J. Virol., 76, 1959〜1970, 2002を参照として挙げることができる)、ニワトリケラチノサイトはこれらの条件下では増殖しない。
【発明の開示】
【発明が解決しようとする課題】
【0015】
本発明の目的は、インビボで有する分化の特性の全てを保持する細胞のようなケラチノサイト、特に鳥類のケラチノサイト、より詳しくはニワトリケラチノサイトを培養する条件を開発することであった。
高度に分化した細胞(肝細胞、腸細胞、ニューロン細胞又はケラチノサイト)に感染するウイルスの複製サイクルは、一般に、その宿主細胞の分化プログラムに連結している(Calnekら, J. Natl. Cancer Inst., 45, 341〜351, 1970; Calnekら, Avian Dis., 14, 219〜233, 1970; Schmittら, J. Virol., 70, 1912〜1922, 1996)ので、完全に分化したケラチノサイトの培養が完全ウイルスの複製、よって細胞を含まないビリオンを産生することを可能にすると予想することができる。
【課題を解決するための手段】
【0016】
本発明の主題は、
(i) ニワトリ血清と、
(ii) 細胞分裂抑制処理により不活性化された繊維芽細胞を含む培養担体と
を含む表皮細胞、特に家禽の表皮細胞用の培養培地である。
【0017】
本発明の主題は、ニワトリ血清と、不活性化された哺乳動物細胞、好ましくは不活性化された繊維芽細胞を含む培養担体とを含む培養培地を含有するケラチノサイト培養物でもある。
【発明を実施するための最良の形態】
【0018】
さらに、本発明の培養培地は、知られるように、無機塩、ビタミン、ホルモン及び成長因子から選択され得る1種又はそれより多い添加物、或いは細胞培養培地に通常用いられるいずれのその他の添加物を含む。本発明の培養培地に用い得る添加物のうち、無機塩、アミノ酸、ビタミン、脂肪酸、グルコース、バッファー、フェノールレッド、EGF (表皮成長因子)、T3、ヒドロコルチゾン、インシュリン、コレラトキシン、トランスフェリン、アデニンなどを特に挙げることができる。
【0019】
培養担体として用いられる繊維芽細胞は、3T3細胞が有利であり、より有利には3T3-J2サブクローンである。3T3株の起源は、Todaro, G., J.ら, J. Cell Biol., 17, 299〜313, 1963に記載される。3T3細胞系は、ATCCコレクションに番号CCL92の下で登録されている。3T3細胞は、細胞分裂抑制処理により不活性化された細胞の単層の形で本発明において用いるのが有利である。細胞分裂抑制効果が知られているいずれの処理も用い得る。この処理は、マイトマイシンCのような抗生物質での処理、又はガンマ線の致命的な照射からなり得る。
【0020】
本発明の培養培地において、ニワトリ血清は、培養培地の全重量に対する重量での割合で0.5〜50%、好ましくは1〜40%、有利には2〜30%の範囲で存在するのが有利である。
【0021】
有利には、細胞培養用の増殖培地は、次の添加物の少なくとも1つ、有利にはいくつかを含有する:コレラトキシン、トランスフェリン、アデニン、EGF、及びT3、ヒドロコルチゾン、ウシインシュリンのようなホルモン。
【0022】
本発明は、ケラチノサイト、特に鳥類のケラチノサイトの培養方法にも関し、該方法は:
(i) 表皮細胞又はケラチノサイトの懸濁物を作製し;
(ii) 上記の細胞を、上記のようなニワトリ血清を含む培養培地中の3T3細胞からなる細胞担体上に接種し;
(iii) 上記の表皮細胞をその細胞担体上でインキュベートする
各工程を含む。
【0023】
表皮細胞又はケラチノサイトの懸濁物は、知られた様式で、表皮の酵素消化により得ることができる。このような消化は、タンパク質分解酵素での処理、例えばトリプシン、ジスパーゼ(dispase)又はサーモライシンでの処理により行うことができる。表皮を化学処理、例えばEDTA (エチレンジアミン四酢酸)での処理により処理することも考えられる。
【0024】
インキュベーション条件は、細胞培養に通常適用されるもの、すなわち5〜10%のCO2及び100%湿度中で約37℃の温度である。
【0025】
インキュベーション後、細胞密度が充分であるときに、これらの条件下で得られた細胞を新しい細胞担体に移し、上記の条件下で再び培養することができる。工程(iii)の最後で、培養に由来する細胞を回収するときに、本発明の方法の工程(ii)及び(iii)に従って、さらなる培養サイクルを該細胞に適用する。上記の方法に従って30回までの連続する細胞増殖サイクルを考えることができる。細胞集団の倍加時間は、最初の10回の細胞増殖サイクルの間は約12〜16時間であり、その後のサイクルのあいだに徐々に増える。例えば、最初に接種した106細胞で開始することにより約1012細胞を得ることができ、つまり約106の増加係数(expansion factor)である。
【0026】
1又はそれより多い培養サイクルの後に得られる細胞は、その後の使用の観点で凍結形態で保存することもできる。3T3細胞の担体と、ニワトリ血清及びケラチノサイトを含む培地とを含む細胞培養物は、本発明の別の主題である。
【0027】
驚くべきことに、本発明による培養方法が、細胞培養物に由来するケラチノサイトにおいて全てのその分化の特徴を保存することを可能にすることを証明することができた。
【0028】
鳥類の表皮は、脂質並びにα−及びβ−ケラチンフィラメントに富む重層扁平上皮である。下層の細胞は、タンパク質で構成される基底膜上にあり、例えばラミニン5のような該タンパク質のいくらかはケラチノサイトの特異的産物である。基底層は特異的マーカーであるp63を合成するケラチノサイト幹細胞を含有する。表皮の凝集(cohesion)は、プラコグロビン(plakoglobin)タンパク質及びコネクシン31タンパク質をそれぞれ含むデスモソーム及びギャップ結合のようなケラチノサイト間結合によりもたらされる。表皮の表面層において、ケラチノサイトは最終分化のプロセスを受け、その間にケラチノサイトは14-3-3シグマを合成し、角化エンベロープを形成する。該エンベロープは界面活性剤及び還元剤中で煮沸した後に不溶性のままである。このエンベロープは、ペリプラキンのような特異的なタンパク質を含有する。
【0029】
本発明の培養方法により、培養におけるニワトリケラチノサイトは、その組成が鳥類の表皮と類似の脂質、α−及びβ−ケラチンフィラメント、デスモソーム並びにp63、14-3-3シグマ、プラコグロビン、コネクシン31及びペリプラキンの各タンパク質を合成する。最後に、ニワトリケラチノサイトは、培養において、重層されたコロニーを形成し、その表面層において細胞は、界面活性剤及び還元剤中で不溶性の角化エンベロープを合成する。結果として、本発明による培養系において、ケラチノサイトはそれらがインビボで有する全ての特性を維持する。
【0030】
分化の特性を保存しているケラチノサイトの培養を得る可能性は、完全なウイルス複製条件下でのウイルスの増殖を構想することを可能にする。MDVの場合、例えば、ニワトリケラチノサイトは、ウイルスの完全な複製とエンベロープ全体で囲まれたビリオンの産生とをもたらし得る唯一の細胞種である。このウイルスの細胞溶解特性のために、このウイルスは感染された細胞から放出され(Biggs, P.M., Marek's disease, Springer-Verlag出版, 1〜24, 2001)、よって培養培地から容易に抽出できる。さらに、これらの細胞は非常に大量のウイルスを増殖させ、これらのウイルスの大量生産を促進する。
【0031】
結局、本発明の主題は、インビトロでウイルスを増殖させる方法であり、該方法は:
(i) ウイルスの株をケラチノサイトに感染させ;
(ii) 感染したケラチノサイトを、上記の培養方法に従う培養に付し;
(iii) 工程(ii)で増殖したビリオンを抽出する
工程を含むことを特徴とする。
【0032】
本発明の方法は、抽出の後に凍結乾燥でき、よって従来技術の感染された細胞の懸濁物よりも好ましい条件下で保存及び貯蔵されることを許容する無細胞ウイルス粒子を得ることを可能にする。数回の培養サイクルにより弱毒化されたウイルスは、それにもかかわらず完全ビリオンの形で増殖する。この新規な特徴は、従来技術のものよりも良好な免疫化を提供する、より効果的なワクチンの製造を構想することを可能にする。好ましくは、ウイルス増殖方法は、10〜400培養サイクル、有利には40〜100培養サイクルを行うことにより実行される。上記の方法に由来する生存弱毒包膜DNAビリオンは、本発明の別の主題を構成する。
【0033】
本発明の主題は、ワクチンを製造する方法でもあり、該方法は、上記の方法に従ってウイルスを増殖させ、次いで医薬的に許容される担体、特にワクチン担体中にビリオンを導入する各工程を含むことを特徴とする。ビリオンを医薬的に許容される担体に導入する前に凍結乾燥して貯蔵することが有利に構想される。特に、ワクチンは周囲温度で輸送可能である。ワクチン担体は、通常用いられるワクチンアジュバントを知られた様式で含有し得る。
【0034】
本発明の方法は、現在までどのようにして無細胞ワクチンを製造するのか知られていなかった包膜されたDNAウイルスからのワクチンの製造に特に適用される。包膜されたDNAウイルスのうち、特にMDV及びFPVウイルスを挙げることができる。
【0035】
本発明の主題は、よって、包膜されたDNAウイルスから得られるビリオンを含む弱毒生ワクチンでもある。通常、このようなワクチンは、1又はそれより多いワクチンアジュバントを好ましくは含む水溶液の形態のワクチン担体を含む。このようなワクチンは、第一に凍結乾燥されたビリオン、及び第二に免疫化担体を含み、これらのそれぞれが別個の区画に包装され、2つの成分が、それらを経口又は注射により或いはインオボ経路を介して投与するために即座に混合される免疫化キットの形で、一般に市販される。
【0036】
本発明のワクチンを呼吸経路を介して投与し得ることを構想することもできる。この場合、該ワクチンは、水溶液又はエアロソルの形で製造される。
【0037】
ケラチノサイトの培養の可能性は、ケラチノサイトで複製可能な弱毒ウイルス株を開発することを可能にする。マレック病の原因であるMDVウイルスの場合、家禽農場からの家禽にこのウイルスを投与した後に、これらのビリオンは鱗屑に存在するので、家禽農場全体に感染して保護することができるだろう。本発明の主題は、弱毒ビリオンの、マレック病の予防または治療用の医薬品の製造のための使用でもある。
【0038】
本発明のケラチノサイトの培養方法は、研究分野において他の適用を見出す。
【0039】
ケラチノサイトの培養は、この細胞種へのウイルスの侵入の機構を明らかにすること、特にこの侵入に必要なウイルスエンベロープタンパク質を明らかにすることを可能にするはずである。これらのタンパク質は、その後、動物の免疫化のために組換えの形で用い得る。マレック病の場合、この疾患の原因であるウイルスの完全ヌクレオチド配列が知られている。ケラチノサイトに感染できない変異ウイルスの選択により、対応する変異を見出し、よってケラチノサイトへの侵入の原因であるタンパク質を見出すことが可能になるであろう。
【0040】
本発明は、本発明の説明のためにのみ与えられ本発明を限定するとは考慮されるべきでない実施例によりさらに以下で記載され、説明される。
【0041】
- 図1は、位相差顕微鏡写真を示し、これらはニワトリ及びヒトの表皮細胞の増殖を示す。(A) 105の第3継代のニワトリ表皮細胞を、10%ニワトリ血清含有培地中での培養に付す。(B) より高い倍率で撮影した(A)と同じ培養。(C) 7.8×105のヒトケラチノサイト(YF23)を、10%ウシ胎児血清含有培地での培養に付す。(D) ヒトケラチノサイト(YF23)を、10%ニワトリ血清含有培地での培養に付す。各目盛りバーは50μmに相当する。
【0042】
- 図2は、オイルレッドOでの標識及びHoechst 33258試薬での染色の後の、培養でのケラチノサイトによる細胞質脂肪の蓄積を示す。(A) 黄色フィルタを用いた位相差顕微鏡。(B) 同じ範囲を位相差の下及び蛍光の下の両方で撮影し、白黒で印刷した写真。矢印は、Hoechst 33258試薬で染色された核(白色)を示す。目盛りは25μmに相当する。
【0043】
- 図3は、ニワトリケラチノサイトの増殖に対するニワトリ血清の影響を示す。5×104の細胞を、ニワトリ血清(ChS)及びウシ胎児血清(FCS)の混合物中に培養に付し、次いで固定化してローダミンBで染色する。ある場合においては培養培地におけるFCSのパーセンテージは10%に固定し、ChSのパーセンテージは変動可能である。他方の場合においては培養培地におけるChSのパーセンテージは10%に固定し、FCSのパーセンテージは変動可能である。
【0044】
- 図4は、ニワトリ血清の脱脂が、ニワトリケラチノサイトの増殖に影響しないことを示す。3500細胞/cm2の密度のニワトリケラチノサイトを接種し、10%ニワトリ血清中に7日間培養に付す。あるディッシュにおいては血清の補足をウシ胎児血清(FCS)又は脱脂ニワトリ血清(Delip. ChS)のいずれかで置き換え、別のディッシュにおいては細胞を未処理のニワトリ血清(Unt. ChS)中に維持する。D1:第1日、D7:第7日、D15:第15日、D20:第20日。(A) 培養物を接種の15日後に固定化し、ローダミンBで染色する。矢印は、脱脂血清において観察される小さい小滴(droplets)を示す。
【0045】
- 図5は、培養におけるニワトリケラチノサイト中のケラチンの証明を示す。(A) 細胞を抗パンケラチン抗体で標識する。矢印は3T3細胞を示し、この細胞は多核であり、抗体で標識されない。目盛りは25μmに相当する。(B) 別の抗パンケラチン抗体で標識されたイムノブロット。レーン(1)は培養におけるニワトリケラチノサイトに対応し、レーン(2)はニワトリ表皮に対応し、レーン(3)はニワトリの腸に対応する。2つのダブレット(a及びc)は、ニワトリ表皮中の対応するダブレットと一致する。
【0046】
- 図6は、ニワトリケラチノサイトが培養において角化エンベロープを形成することを示す。写真は、位相差顕微鏡下で撮影される。(A) 15日の生存コロニーの基底層。(B) より大きい焦平面で撮影された同じコロニー;暗い輪郭及び角形は角化を示す。(C) 2%のドデシル硫酸ナトリウム及び2%の2-メルカプトエタノールを含有する溶液中に100℃で処理した後。目盛りは50μmに相当する。
【0047】
- 図7は、抗p63抗体を用いる免疫蛍光染色によるp63標識を示す。矢印は、大きい核及び顕著なヘテロクロマチンにより認識される3T3細胞を示す。これらは抗体により標識されない。目盛りは25μmに相当する。
【0048】
- 図8は、核周囲及び核内の脂質を示す。これらは電子顕微鏡写真である。倍率はA及びBが5000×、Cが6000×及びDが4000×である。
【0049】
- 図9は、ケラチノサイト内の層形成を示す。デスモソーム(A中)及びケラチンフィラメント(B中)。これらは電子顕微鏡写真である。倍率はAが40000×、Bが8000×である。
【実施例】
【0050】
実施例1:10%ニワトリ血清の存在下での3T3-J2担体細胞とのニワトリケラチノサイトの増殖
1.1 概要
ニワトリ繊維芽細胞の増殖は、3T3細胞の単層により効率的に解消されず、ニワトリの皮膚全層から作製した培養物は、繊維芽細胞により迅速に侵入される。よって、表皮を分散させる(disaggreagation)前に、サーモライシンでの処理により表皮を真皮から分離する。最初に、培地にウシ胎児血清を補うが、この血清はニワトリケラチノサイトの増殖を許容しない。ウシ血清をニワトリ血清に置き換えることにより、細胞コロニーが発生することが可能になる。
【0051】
1.2 材料及び方法
1.2.1 生物学的材料
新生の雛(Gallus gallus)を、高い湿度レベルで37.8℃において20〜21日間インキュベートした受精卵から得る。表皮細胞の初代培養物を得るために、トリブロモエタノールの腹腔内注射により新生の雛に麻酔をかける。
【0052】
マウス3T3-J2細胞:3T3系統の起源は、Todaro, G., J.ら, J. Cell Biol., 17, 299〜313, 1963に記載される。ATCC (No. CCL92)から得たオリジナルのSwiss 3T3細胞を96ウェルプレートに低密度で(3ウェル当たり1つの細胞の割合で)接種する。約2週間の培養後、いくつかのウェルは目で見える細胞コロニーを含む。これらの培養物は、単一の細胞に由来するのでクローンであり、よってそれぞれ1つずつが3T3サブクローンである。各コロニーを直径100 mmの培養ディッシュに別々に移す。細胞数が充分であるときに、細胞をトリプシン処理により剥離する。各サブクローンの半分を液体窒素で凍結させる。残りの半分をマイトマイシンC又はガンマ線で処理する。ニワトリケラチノサイトを、処理された各サブクローン上に、直径100 mmのディッシュ当たり1.5×106の3T3当たり102〜106細胞の密度で接種する。培養7〜14日後に、これらの細胞を固定化しローダミンBで染色する。3T3-J2とよばれる3T3サブクローンは、最大サイズのケラチノサイトコロニーがその上に最大数存在し、最適であるとみなされる。これから先は、これをニワトリケラチノサイトの培養に用いる。
【0053】
新生の包皮を起源とするヒト表皮ケラチノサイト(Rheinwaldら, Cell, 6, 331〜343, 1975に記載の方法に従って作製)。
【0054】
1.2.2 方法
細胞培養(方法1):
足部又は体からの皮膚を回収する。リン酸緩衝食塩水(PBS)でこれを数回洗浄し、Hepes緩衝溶液(HBS)中の0.5 mg/mlのサーモライシン(Sigma)で、37℃で45分間又は4℃で一晩のいずれかでの酵素処理により、表皮を真皮から分離する。次いで表皮を回収し、薄い切片に切断し、トリプシン(0.25%)及びEDTA (0.02%)の存在下に37℃で30分間、2回、トリプシン処理フラスコ内で攪拌しながらインキュベーションすることにより分散させる。細胞懸濁物を800 gで5分間遠心分離する。細胞ペレットを培養培地に採取し、細胞を、マイトマイシンC (4μg/ml)で予め処理したマウス3T3細胞(2.3×104細胞/cm2)の単層で被覆したペトリディッシュに接種する。その後、ディッシュを37℃、7.5% CO2及び100%湿度のインキュベータに入れる。培養培地は、10%のニワトリ血清(Invitrogen)を補った1/1のDMEM/F12混合物(Invitrogen)からなる。次の添加物も培地中に存在する:0.4μg/mlのヒドロコルチゾン(Calbiochem)、5μg/mlのウシインシュリン(Sigma)、2×10-9 Mの3,3',5-トリヨード-L-チロニン(T3、Sigma)、10-10 Mのコレラトキシン(ICN)及び1.8×10-4 Mのアデニン(Calbiochem)。組換えヒト表皮成長因子(EGF)を、最初の培地交換のときに10 ng/mlの濃度で添加する。真皮の除去にもかかわらず繊維芽細胞コロニーが発生する場合があるが、これらは、培養物をCa++もMg++も含有しないPBS又は0.02% EDTAで短時間処理することにより除去できる。次いで、追加の3T3担体細胞を加え、培養を継続する。初代培養物を、接種密度に応じて接種の7〜19日後に移す。次いで、第2代培養物を得る。細胞を、第1継代の後に、10%ジメチルスルホキシド(DMSO)を含有する培地中で凍結させる。凍結は、1〜3×106細胞/mlの割合で、上記の培養培地中で行う。凍結細胞は、液体窒素中に無期限に保存できる。
【0055】
ヒト表皮ケラチノサイトは、ウシ胎児血清をニワトリ血清の代わりに用いた以外はニワトリ細胞について記載したのと同じ培養に付す。
【0056】
細胞培養(方法2):液/気界面でのニワトリケラチノサイトの培養
この方法は、文献J. Y. Yiら, Arch. Dermatol. Res., 2001, 293, 356〜362に基づく。この目的は、より完全な最終分化を得ることである。マイトマイシンで処理したか又はしていない3T3細胞を、供給業者(Nitta gelatin, Tokyo)の使用説明に従って、I型コラーゲンマトリックスと混合し(3×105細胞/ml)、この混合物の200μlを、ポリカーボネートフィルタ(Millicell-pc Millipore, Bedford, MA)上に接種する。ケラチノサイトをコラーゲンマトリックス上に接種し、通常の培養培地(DMEM/F12 +ニワトリ血清+添加物)中に5〜7日間浸しながら培養する。その後、フィルタを培地/空気界面に置き、培養を14日間継続する。
【0057】
位相差顕微鏡
Nikon Coolpixデジタルカメラを備えたNikon顕微鏡を用いた。
【0058】
オイルレッドOを用いた脂質小滴の染色
カバーガラス上に接種された細胞を、3.7%ホルムアルデヒドを含有する冷PBS中に15分間、周囲温度で固定化し、次いで新鮮な固定化溶液で追加の1時間インキュベートする。蒸留水でカバーガラスを洗浄して空気乾燥する。脂質の小滴を0.3%のオイルレッドO (Sigma)及び60%のイソプロパノールを含有する溶液で1時間染色する。インキュベーションの最後に、カバーガラスを蒸留水中に洗浄し、Hoechst 33258試薬(1μg/ml)及びSigma社からIgepalの商品名で販売されている製品0.1%を含有するPBS中に5分間インキュベートする。カバーガラスをPBS/Igepal中に1回、及び水中に1回洗浄し、2.5%のDABCO (1,4-ジアゾビシクロ(2.2.2)オクタン)を含有する、Calbiochem社からMowiolの商品名で販売されている製品中のスライドに載せる。
【0059】
1.3 結果
3T3細胞に接着する円形細胞は接種の数時間後に目で見えるが、典型的な上皮単位を明らかに組織するコロニーは、細胞がプラスチックとの接触を確立するときである、接種の5〜6日後に目で見えるようになる。コロニーは、ヒトケラチノサイトにより形成されるコロニーと同様に、コロニーの周囲の3T3細胞の単層を置き換える。ほとんど全ての細胞が単一の大きい小滴と、ときにおそらく脂質で構成されるいくつかの小さい小滴とを含有することが、最も顕著であるように見られる(図1A及び1B)。その表皮の起源及び重層扁平上皮を形成し得るその能力により、また鳥類のケラチノサイトが脂質の小滴を含有することが知られていることからすると、コロニーがケラチノサイトで構成されていると結論付ける。
【0060】
10%のニワトリ血清を補った培地中に12 000細胞/cm2の密度で接種したヒトケラチノサイトは、ウシ胎児血清中で増殖するのと同様に迅速に増殖する。ヒトケラチノサイトは、10%のニワトリ血清の存在下では12日後であっても、脂質の小滴を決して発生しない(図1C及びD)。このことから、脂質の小滴の形成は、ニワトリケラチノサイトの固有の特性であり、他の種のケラチノサイトではニワトリ血清により誘導されないと結論付ける。
【0061】
中性脂肪に特異的なオイルレッドO製品での固定化細胞の染色は、屈折小滴(refringent droplets)が中性脂肪を含有することを示す(図2A)。Hoechst 33258試薬を用いた対比染色は、細胞内の脂質小滴のほとんどが核周囲であることを示す(図2B)。
【0062】
3つの独立した実験において、約500のコロニーが5×105の分散された表皮細胞から形成されたことを示した。これは、10-3のコロニー形成効率に相当する。よって、表皮から直接採取されたニワトリケラチノサイトについての接種効率(seeding efficiency)は、1〜10%程度であると報告されているが一般的には1%に近いヒトケラチノサイトのものよりかなり低い。後続の継代で非常に高いコロニー形成効率を達成するヒトケラチノサイトとは違って、ニワトリケラチノサイトの接種効率は、数パーセントを決して超えない。
【0063】
ニワトリケラチノサイトの倍加時間は、第2代培養物中で15コロニー中の細胞数を24時間ごとに、接種後の第1日から第4日まで計数することにより測定される。ニワトリケラチノサイトは、単独の脂肪小滴を含有することからして、隣接する3T3細胞から容易に区別することができる。最高の増殖速度のコロニーを考慮に入れると、16.8時間の倍加時間が得られる。倍加時間は、後続の継代の過程で減少し、ニワトリケラチノサイトは第20代の植え継ぎ培養を超えると培養に付すことができない。
【0064】
ニワトリケラチノサイトのサイズは、細胞の懸濁物を血球計数器に導入し、細胞の写真を撮影しその直径を測定することにより測定される。ケラチノサイトは10μmの典型的な直径を有し、よって最も一般的な直径が12μmであるヒトケラチノサイトよりも小さい。この相違は、ニワトリゲノムのサイズがヒトのゲノムサイズの1/3しかないことにより説明できる(1.25 pgに対して3.5 pg/一倍体のゲノム)。12μmの典型的な直径を有するニワトリ繊維芽細胞も、最も一般的な直径が15.6μmであるヒト繊維芽細胞よりも小さい。
【0065】
実施例2:ニワトリケラチノサイト増殖過程でのニワトリ血清の役割
2.1 概要
ウシ胎児血清は、ウシ胎児血清が増殖阻害物質を含有していること又は増殖促進因子を欠いていることのいずれかの理由から、ニワトリケラチノサイトの増殖を許容しない。
これらの選択肢の間を区別するために、ニワトリケラチノサイトを、ニワトリ血清を増加する濃度で補った10%のウシ胎児血清の存在下、又はウシ胎児血清を増加する濃度で補った10%ニワトリ血清の存在下のいずれかに10日間培養する。実験の最後に、細胞を固定化してローダミンBで染色する。
【0066】
2.2 材料及び方法
2.2.1 材料
用いた生物学的材料は、実施例1に記載のものである。
【0067】
2.2.2 方法
細胞培養プロトコルは、実施例1に記載されたものに従って用いた。
【0068】
ローダミンBでの染色
培養物をPBSで2回洗浄し、10%グルタルアルデヒド含有PBSで、周囲温度で15分間固定化する。次いで、細胞を水中の1%ローダミンBで周囲温度で15分間、緩やかに攪拌しながら染色する。培養物を水で洗浄し、その後空気乾燥する。
【0069】
2.3 結果
図3において、ニワトリ血清がケラチノサイトの増殖を許容し、この能力は、10%ウシ胎児血清に加えられるときにニワトリ血清の用量に依存することが明確にわかる。10%ニワトリ血清の増殖促進活性は、ウシ胎児血清の添加量を増加しても影響されない。このことから、ニワトリ血清がニワトリケラチノサイトの増殖に必要な因子を含有しており、この因子はウシ胎児血清には存在しないと結論付ける。
【0070】
ニワトリケラチノサイトが脂質を含有することからすると、ニワトリ血清の増殖促進活性が血清中の脂質の吸収に関係する。図4Aに示される実験は、溶媒の作用により脂質を欠くニワトリ血清が、ニワトリケラチノサイトの増殖を促進するその能力を維持することを示す。よって、増殖促進活性は脂質によるのではない。特にコロニーの周囲で増殖する細胞は、脱脂血清中で増殖するときに脂質を保存し、よって脂質を合成できる(図4B)。
【0071】
実施例3:α−ケラチンを合成する培養におけるニワトリケラチノサイト
3.1 概要
表皮細胞の特性の一つは、豊富にケラチノサイトを合成するその能力である。ニワトリケラチノサイトを、カバーガラス上での培養に付し、固定化する。次いで細胞を、抗α−パンケラチン抗体の存在下にインキュベートし、間接蛍光顕微鏡により調べる。
【0072】
培養におけるニワトリ表皮細胞のケラチンを、ウェスタンブロッティング免疫検出により分析する。ケラチンを集密な培養物から抽出し、一次元ゲル電気泳動により分離し、ニトロセルロースに移し、抗α−パンケラチン抗体を用いて染色する。
【0073】
3.1 材料及び方法
3.1.1 材料
生物学的材料は、実施例1に記載されたものである。
【0074】
3.2.1 方法
間接免疫蛍光染色
細胞をカバーガラス上に接種する。次の日に、細胞をPBS中に2回洗浄し、次いでアセトン:メタノールの1:1混液中に20分間、-20℃で固定化して空気乾燥させる。カバーガラスを、非特異的部位をブロックするために5%ウシ血清アルブミン(BSA)含有PBS中に10分間、周囲温度でインキュベートする。
【0075】
α−ケラチン染色のために、細胞をマウスモノクローナル抗パンケラチン抗体(1:100希釈, MS-744-A0, NeoMarkers)を含有する5% BSA/PBS中に周囲温度で1時間インキュベートし、その後、0.1%の非イオン界面活性剤Igepal CA 630 (Sigma)含有PBSで、各回5分間、3回洗浄する。次いで細胞を、ビオチン結合ヤギ抗マウスIgG (1:1000希釈, Jackson ImmunoResearch)を含有する5% BSA/PBS存在下に周囲温度で1時間インキュベートする。カバーガラスを洗浄し、シアニン−3に結合したストレプトアビジン(1:1000希釈)で1時間インキュベートする。インキュベーションの最後に、カバーガラスをPBS/Igepal中に5分間洗浄し、その後、Hoechst 33258試薬(1μg/ml)及び0.1%のIgepalを含有するPBS中に5分間インキュベートする。カバーガラスをPBS/Igepal中に1回及び水中に1回洗浄し、2.5%の1,4-ジアゾビシクロ(2.2.2)オクタン(DABCO)を含有するMowiol中のスライドに載せる。
【0076】
α−ケラチンの抽出及び免疫検出(ウェスタンブロッティング)による分析
集密な培養物をPBSで2回洗浄し、Dounceホモジナイザを用いて粉砕する。細胞は、10 mMのTris-HCl (pH 7.6)、1mM EDTA及びプロテアーゼ阻害剤の混合物(Roche)を含有するバッファー中に20回の回転後にホモジナイズされる。ホモジネートを8000 gで5分間、4℃で遠心分離し、ペレットをTris-EDTAバッファーで2回、2%のNonidet P40を含有するTris-EDTAバッファーで2回洗浄する。最終のペレット中のケラチンを、2%のSDS及び10mMのジチオトレイトールを含有する溶液中に37℃で15分間、超音波装置(2×15秒)を用いて懸濁させる。次いで、サンプルを100℃で2分間加熱し、不溶性の残渣を除去するために8000 gで5分間遠心分離する。
【0077】
α−ケラチンを、変性8%ポリアクリルアミドゲル(アクリルアミド:ビスアクリルアミドの重量比は29:1)での電気泳動により分離し、ニトロセルロース上に電気的に溶出させる。メンブレンを、5%のドライスキムミルク及び0.1%のTween 20を含有するPBS (PBST)中に4℃で一晩処理する。α−ケラチンを、一方はI型ケラチンに指向され、他方はII型ケラチンに指向された2つのモノクローナル抗体の混合物からなる抗パンケラチン抗体(PRO61835, Research Diagnostics Inc)を用いて染色する。抗体を1:500の希釈で用い、メンブレンをPBSTで洗浄し、その後、セイヨウワサビペルオキシダーゼに結合したロバ抗マウスIgG (Amersham)の1:2000の希釈でインキュベートする。抗体のインキュベーションは、周囲温度で1時間継続する。ケラチンに対応するバンドは、ケミルミネセンス(ECL+kit, Amersham)により視覚化される。
【0078】
3.3 結果
培養における全てのニワトリケラチノサイトは強く標識されるが、3T3細胞は標識されない(図5A)。特異的抗体を省いたときには標識は観察されない。
【0079】
免疫検出からの結果を図5Bに示す。抗体で染色されたタンパク質は、3つの主要なダブレットに分かれ(a〜c)、それらのそれぞれの分子量は、約50、57及び63 kDである。ダブレットa及びcはニワトリの皮膚にも存在するが、ダブレットbは存在しない。角質化していない上皮であるニワトリの腸から抽出されたケラチンの一般的なプロフィールは、表皮細胞とも表皮とも一致しない。培養における細胞が、表皮のケラチンのものと一致する分子量を有する2つのケラチンダブレットを含有することからして、培養における細胞が表皮のケラチンを合成すると結論付ける。高分子量のケラチンが重層扁平上皮の外層に特異的であると考えられることからして、2つの高分子量のケラチン(図5B中のc)は特に適切である。4つのα−ケラチンをさらに特徴付ける(以下を参照)。
【0080】
実施例4:ニワトリケラチノサイトは培養において角化エンベロープを形成する
4.1 概要
哺乳動物の表皮ケラチノサイトの顕著な特性は、最終分化プロセスに従うその能力であり、該プロセスにおいてそれらの核は破壊され、その間に、ドデシル硫酸ナトリウム及び2-メルカプトエタノールの存在下で不溶性の角化細胞エンベロープを発生する。
【0081】
4.2 材料及び方法
本実施例において用いた生物学的材料及び方法は、実施例1に記載されたものである。
【0082】
4.3 結果
サイズが数百細胞を超えたニワトリケラチノサイトコロニーの中央が観察されるとき、それらが、それぞれがいくつかの基底細胞を覆う大きい細胞をほぼいつも含有することが注目される(図6A及びB)。これらの鱗屑は培地中に迅速に放出され、おそらく脂質で構成されている屈折小滴をいつも含有する。ニワトリの鱗屑は薄く、ヒトの鱗屑のおよそ2倍大きい(100μmに対して50μmの長軸(greater length)) (図6B)。ニワトリ表皮細胞コロニーを回収して2%のSDS及び2%の2-メルカプトエタノールを含有する溶液に入れ、100℃で加熱するとほとんどの細胞は溶解するが、角化エンベロープは不溶性のままであり、脂質小滴を未だ保持する(図6C)。ニワトリ表皮から直接得られるエンベロープのほとんども、SDS/2-メルカプトエタノール中での煮沸の後に脂質を保持する。これらの脂質は、メタノール/クロロホルムを用いて抽出し得る。
【0083】
実施例5:ニワトリケラチノサイトはp63を含有する
p53タンパク質をコードする遺伝子ファミリーの一部分であるp63タンパク質は、多くの上皮の基底層に豊富に存在する。ヒト表皮において、p63は高い増殖ポテンシャルを有する細胞でのみ見出される。p63をコードする遺伝子のノックアウトにより、全ての重層扁平上皮が存在しなくなり、p63はケラチノサイト幹細胞のマーカーであるとみなされる。
【0084】
ニワトリケラチノサイトがp63を含有するかを調べるために、増殖が早い細胞をカバーガラス上に接種し、次の日に免疫蛍光顕微鏡によりモノクローナル抗-p63抗体4A4を用いて観察する。
【0085】
5.1 材料及び方法
5.1.1 材料
生物学的材料は実施例1に記載される。
【0086】
5.1.2 方法
細胞培養及び間接免疫蛍光染色法は、それぞれ実施例1及び3に記載されたものである。
【0087】
p63標識
細胞を、1:500に希釈した抗ヒトp63モノクローナル抗体4A4の存在下に4℃で一晩インキュベートする。これ以外のプロトコルは、二次抗体を1:200の希釈で用いた以外はケラチン染色に用いた方法と同じである。細胞を、Nikonデジタル写真撮影装置を備えたNikon E600蛍光顕微鏡を用いて写真撮影する。
【0088】
5.2 結果
調べた全ての細胞は、p63について強い核染色を示す。逆に、3T3細胞は標識を示さないか又は時々弱い細胞質標識を示すかのいずれかである(図7)。
【0089】
実施例6:電子顕微鏡によるニワトリケラチノサイトの特徴づけ
6.1 材料及び方法
ニワトリケラチノサイト単層培養物を得る。細胞が2/3の集密であるときに、3T3細胞をEDTAを用いて除去し、ケラチノサイトを3T3担体細胞を含まない小さいペトリディッシュに移す。次の日に、電子顕微鏡で観察されるように細胞を固定化して処理する。
【0090】
電子顕微鏡
細胞を、0.1MカコジレートpH 7.3で洗浄し、0.1Mカコジル酸ナトリウムpH 7.2中の5% パラホルムアルデヒド及び5% グルタルアルデヒドで90分間固定化し、通常の電子顕微鏡で観察する。酢酸ウラニル及びクエン酸鉛で染色した超薄切片を、80 kVでJeol JEM-1010電子顕微鏡(Jeol Ltd, Tokyo, Japan)を用いて調べる。
【0091】
6.2 結果
図8は、脂質小滴を含有する細胞の一連の顕微鏡写真を示す。2枚の顕微鏡写真(A及びB)は、核内に包埋された単一の核周囲脂質小滴を有する典型的な細胞を示す。よって、ほとんどの核が三日月形を示す。別の2枚の写真は、多数の小さい細胞質脂質小滴を有する細胞(C)及び核の内部に脂質小滴を有する別の細胞(D)を示す。これらの2つの特徴は、通常存在する。図9は、典型的な表皮細胞の特徴を示す。写真(A)は、デスモソームの存在を示す。写真(B)は、基底細胞と、細胞質がトノフィラメント(tonofilaments) (ケラチン)で充たされている超基底細胞(superbasal cell)とを示す。
【0092】
実施例7:ランダム配列決定による特異的なケラチノサイト産物の同定
7.1 概要
cDNAライブラリを、集密なニワトリケラチノサイト培養物から構築する。163のランダムなコロニーに由来する5'末端のヌクレオチド配列を、自動化シーケンサーを用いて決定する。次いで。cDNA配列との相同性探索のためにGenBank及びタンパク質データバンクを用いる。
【0093】
7.2 材料及び方法
cDNAライブラリの構築とヌクレオチド配列決定
Tripure Isolation Reagent (Roche)を用いて、培養でのニワトリケラチノサイトから全RNAを精製する。ポリ(A)+ RNAを、オリゴ(dT)セルロース上で、Micropoly(A) Purist (登録商標) (Ambion) mRNA精製キットを用いて単離する。cDNAライブラリを、1.7μgのポリ(A)+ RNAから、SuperScript (登録商標) Plasmid System (Life Technologies)を用いて作製する。第一の鎖の合成を、オリゴ(dT)配列の後ろにNotI部位を含有するプライマを用いて開始する。第二の鎖の合成の後に、SalIアダプタがcDNAの両端に付加される。NotIでの消化により、3'末端に付着末端が発生するが、SalI付着末端は5'末端に残る。cDNAを、SalI及びNotIで予め切断しておいたプラスミドpSport1に連結し、エレクトロポレーションによりエレクトロコンピテントな細菌E. Coli DH10B (Life Technologies)に導入する。
【0094】
cDNAクローンの5'末端のランダム配列決定は、SalI部位の上流でプラスミドにハイブリダイズするプライマを用いて行う。配列を伸ばすために、特異的な内部プライマを用いる。
α−ケラチン5/6について:
配列番号1 5'-AGTGGATTCTATGGACCTGC-3'、
配列番号2 5'-AACTGAGCAGCTTGCTGGC-3'、
配列番号3 5'-CAGTCCAGGTACGAAGAGC-3'、
配列番号4 5'-TAGGATCGGGATATGGAAGC-3';
【0095】
α−ケラチン15について:
配列番号5 5'-GGCTTTGATGCTATCTGTGC-3'、
配列番号6 5'-GTGGAGTCTGACATCAACG-3'、
配列番号7 5'-AGAACTGAGACGCACGATGC-3';
β−ケラチンKについて:
配列番号8 5'-ACTAGGATGTTACTGCGTGG-3'。
【0096】
配列決定は、蛍光ジデオキシヌクレオチドの存在下にGeneAmp 2400 PCR装置(Perkin-Elmer)で行う。PCR条件は、96℃で30秒、50℃で15秒及び60℃で4分を30サイクルである。電気泳動及び蛍光ピークの検出は、自動化シーケンサー(ABI PRISM 310 Genetic Analyzer)で行う。配列は、SeqEd v1.0.3プログラムを用いて決定する。
【0097】
7.3 結果
50個のクローンは、データベースの配列と相同性を示さない。15個のクローンは、仮定的タンパク質と相同性を示す。86個のクローンは、既知の機能を有する遍在タンパク質をコードする。12個のクローンは、特定のケラチノサイトの機能に関係することが知られているタンパク質をコードする。これらのことから、ニワトリケラチノサイトにおいては、約40% (65/163)のmRNAが未知の機能のタンパク質をコードし、約12% (12/98)のmRNAがこの細胞種に特異的であると結論付ける。合計で、7個のケラチノサイト特異的クローンがケラチンをコードする(表III)。クローン6〜8は同じα−ケラチンをコードするが、クローン9〜11は種々のα−ケラチンをコードし、クローン12はβ−ケラチンをコードする。クローン6及び9〜12について、全ヌクレオチド配列を決定する。
【0098】
実施例8:培養におけるニワトリケラチノサイトのマレック病ウイルス(MDV)での感染
8.1 材料及び方法
MDV-1に感染したニワトリの皮膚の初代細胞(本質的に繊維芽細胞)又は血液リンパ球に由来する感染細胞を、亜集密な培養のニワトリケラチノサイト上に接種し、37℃で1時間インキュベートする。吸着後に、接種物を吸引により除去し、ニワトリケラチノサイト単層を、リン酸緩衝食塩水(PBS)で3回洗浄する。細胞培養培地を加え、細胞を37℃で1週間インキュベートする。
【0099】
完全に包膜されたMDVビリオンは細胞培養培地中に放出され、これを回収して凍結乾燥することができる。凍結乾燥のために、MDVビリオンを含有する細胞培養培地を-60℃で凍結させ、真空下に38℃で24時間乾燥させ、21℃で15時間乾燥させ、真空下に4℃で貯蔵する。
【0100】
感染したニワトリケラチノサイト培養物を、PBS、又は0.2% EDTA含有PBS、又は2.18×10-1M スクロース、7.2×10-3 M リン酸二カリウム、3.8×10-3 M リン酸カリウム、4.9×10-3 M グルタミン酸ナトリウム及び1% 粉末ウシアルブミンからなる"SPGA"安定化溶液、又は0.2% EDTA含有SPGA (SPGA-EDTA)、又は10%ソルビトールを含むSPGA-EDTA、又は10%ソルビトールを含む細胞培養培地中に移す。次いで、細胞懸濁物を2分間超音波により破砕し、2000 gで20分間遠心分離し、上清を無細胞ウイルス調製物として回収する。ウイルスを、3回の凍結/融解サイクルにより感染ケラチノサイトから抽出し、上清の流体を回収する。無細胞ウイルスは、上記のようにして凍結乾燥できる。
【図面の簡単な説明】
【0101】
【図1】位相差顕微鏡写真を示し、これらはニワトリ及びヒトの表皮細胞の増殖を示す。
【図2】オイルレッドOでの標識及びHoechst 33258試薬での染色の後の、培養でのケラチノサイトによる細胞質脂肪の蓄積を示す。
【図3】ニワトリケラチノサイトの増殖に対するニワトリ血清の影響を示す。
【図4】ニワトリ血清の脱脂が、ニワトリケラチノサイトの増殖に影響しないことを示す。
【図5】培養におけるニワトリケラチノサイト中のケラチンの証明を示す。
【図6】ニワトリケラチノサイトが培養において角化エンベロープを形成することを示す。
【図7】抗p63抗体を用いる免疫蛍光染色によるp63標識を示す。
【図8】核周囲及び核内の脂質を示す。
【図9】ケラチノサイト内の層形成を示す。
【特許請求の範囲】
【請求項1】
ニワトリ血清と、ATCCコレクションに番号CCL92で登録された細胞系の不活性化された3T3細胞を含む培養担体とを含むことを特徴とする培養培地。
【請求項2】
培養培地の全重量に対するニワトリ血清の重量での割合が0.5〜50%の間であることを特徴とする請求項1に記載の培養培地。
【請求項3】
3T3細胞が、ATCCコレクションに番号CCL92で登録された細胞系の3T3細胞から、ウェルへの接種、培養、移送、培養、トリプシン処理による剥離、各サブクローンの半分の凍結、残りの半分のマイトマイシンC又はガンマ線での処理、処理された各サブクローンでのニワトリケラチノサイトのインキュベーション、培養、固定及びローダミンBでの染色、並びに最大のサイズのケラチノサイトコロニーが最大数存在するサブクローンの選択からなる各工程を含む方法により得られることを特徴とする請求項1又は2に記載の培養培地。
【請求項4】
3T3細胞が、マイトマイシンCでの処理又はガンマ線の照射から選択される細胞分裂抑制処理により不活性化されたことを特徴とする請求項1〜3のいずれか1項に記載の培養培地。
【請求項5】
次の構成成分:無機塩、ビタミン、ホルモン及び成長因子の1又はそれより多くをも含有することを特徴とする請求項1〜4のいずれか1項に記載の培養培地。
【請求項6】
次の構成成分:無機塩、アミノ酸、ビタミン、脂肪酸、グルコース、バッファー、フェノールレッド、EGF (表皮成長因子)、T3、ヒドロコルチゾン、インシュリン、コレラトキシン、トランスフェリン及びアデニンの1又はそれより多くをも含有することを特徴とする請求項1〜5のいずれか1項に記載の培養培地。
【請求項7】
ニワトリ血清と、不活性化された哺乳動物細胞、好ましくは不活性化された繊維芽細胞を含む培養担体とを含む培養培地を含有することを特徴とするケラチノサイト培養物。
【請求項8】
請求項1〜6のいずれか1つに記載の培養培地を含有することを特徴とする請求項7に記載のケラチノサイト培養物。
【請求項9】
ケラチノサイトが、鳥類のケラチノサイトであることを特徴とする請求項8に記載のケラチノサイト培養物。
【請求項10】
ケラチノサイトが、ニワトリのケラチノサイトであることを特徴とする請求項9に記載のケラチノサイト培養物。
【請求項11】
(i) ケラチノサイトの懸濁物を製造し;
(ii) 前記細胞を、ニワトリ血清と不活性化された繊維芽細胞を含む培養担体とを含む培養培地に接種し;
(iii) 前記細胞をこの培養培地中でインキュベートする
工程を含む、表皮細胞又はケラチノサイトを培養する方法。
【請求項12】
前記培養培地が、請求項1〜6のいずれか1つで規定されるものであることを特徴とする請求項11に記載の方法。
【請求項13】
工程(iii)の最後に、培養由来の細胞が回収され、請求項11に記載の方法の工程(ii)及び(iii)に従って前記細胞にさらなる培養サイクルが適用されることを特徴とする請求項11又は12に記載の培養の方法。
【請求項14】
最大で30回の細胞培養サイクルを含むことを特徴とする請求項13に記載の培養の方法。
【請求項15】
(i) ウイルス株をケラチノサイトに感染させ;
(ii) 感染したケラチノサイトを、請求項11〜14の方法に従う培養に付し;
(iii) 工程(ii)で得られるビリオンを抽出する
工程を含む、インビトロでウイルスを増殖させる方法。
【請求項16】
ビリオンの凍結乾燥工程をも含むことを特徴とする請求項15に記載の方法。
【請求項17】
ケラチノサイトが、ニワトリ表皮細胞であることを特徴とする請求項15又は16に記載の方法。
【請求項18】
請求項15〜17のいずれか1つに記載の方法により得ることができることを特徴とする、MDV及びFPVから選択されるウイルスから得られる完全無細胞性弱毒化包膜生DNAビリオン。
【請求項19】
請求項18に記載の少なくとも1のビリオンとワクチン担体とを含有することを特徴とする弱毒生ワクチン。
【請求項20】
第一に凍結乾燥ビリオンと、第二に免疫化担体とを含み、それぞれが別々の区画に包装されているキットの形であることを特徴とする請求項19に記載のワクチン。
【請求項21】
注射による投与、経口投与、インオボ経路を経由する投与、又は呼吸経路を経由する投与を意図することを特徴とする請求項19又は20に記載のワクチン。
【請求項22】
請求項18に記載のビリオンの、マレック病の予防又は治療用の医薬品を製造するための使用。
【請求項1】
ニワトリ血清と、ATCCコレクションに番号CCL92で登録された細胞系の不活性化された3T3細胞を含む培養担体とを含むことを特徴とする培養培地。
【請求項2】
培養培地の全重量に対するニワトリ血清の重量での割合が0.5〜50%の間であることを特徴とする請求項1に記載の培養培地。
【請求項3】
3T3細胞が、ATCCコレクションに番号CCL92で登録された細胞系の3T3細胞から、ウェルへの接種、培養、移送、培養、トリプシン処理による剥離、各サブクローンの半分の凍結、残りの半分のマイトマイシンC又はガンマ線での処理、処理された各サブクローンでのニワトリケラチノサイトのインキュベーション、培養、固定及びローダミンBでの染色、並びに最大のサイズのケラチノサイトコロニーが最大数存在するサブクローンの選択からなる各工程を含む方法により得られることを特徴とする請求項1又は2に記載の培養培地。
【請求項4】
3T3細胞が、マイトマイシンCでの処理又はガンマ線の照射から選択される細胞分裂抑制処理により不活性化されたことを特徴とする請求項1〜3のいずれか1項に記載の培養培地。
【請求項5】
次の構成成分:無機塩、ビタミン、ホルモン及び成長因子の1又はそれより多くをも含有することを特徴とする請求項1〜4のいずれか1項に記載の培養培地。
【請求項6】
次の構成成分:無機塩、アミノ酸、ビタミン、脂肪酸、グルコース、バッファー、フェノールレッド、EGF (表皮成長因子)、T3、ヒドロコルチゾン、インシュリン、コレラトキシン、トランスフェリン及びアデニンの1又はそれより多くをも含有することを特徴とする請求項1〜5のいずれか1項に記載の培養培地。
【請求項7】
ニワトリ血清と、不活性化された哺乳動物細胞、好ましくは不活性化された繊維芽細胞を含む培養担体とを含む培養培地を含有することを特徴とするケラチノサイト培養物。
【請求項8】
請求項1〜6のいずれか1つに記載の培養培地を含有することを特徴とする請求項7に記載のケラチノサイト培養物。
【請求項9】
ケラチノサイトが、鳥類のケラチノサイトであることを特徴とする請求項8に記載のケラチノサイト培養物。
【請求項10】
ケラチノサイトが、ニワトリのケラチノサイトであることを特徴とする請求項9に記載のケラチノサイト培養物。
【請求項11】
(i) ケラチノサイトの懸濁物を製造し;
(ii) 前記細胞を、ニワトリ血清と不活性化された繊維芽細胞を含む培養担体とを含む培養培地に接種し;
(iii) 前記細胞をこの培養培地中でインキュベートする
工程を含む、表皮細胞又はケラチノサイトを培養する方法。
【請求項12】
前記培養培地が、請求項1〜6のいずれか1つで規定されるものであることを特徴とする請求項11に記載の方法。
【請求項13】
工程(iii)の最後に、培養由来の細胞が回収され、請求項11に記載の方法の工程(ii)及び(iii)に従って前記細胞にさらなる培養サイクルが適用されることを特徴とする請求項11又は12に記載の培養の方法。
【請求項14】
最大で30回の細胞培養サイクルを含むことを特徴とする請求項13に記載の培養の方法。
【請求項15】
(i) ウイルス株をケラチノサイトに感染させ;
(ii) 感染したケラチノサイトを、請求項11〜14の方法に従う培養に付し;
(iii) 工程(ii)で得られるビリオンを抽出する
工程を含む、インビトロでウイルスを増殖させる方法。
【請求項16】
ビリオンの凍結乾燥工程をも含むことを特徴とする請求項15に記載の方法。
【請求項17】
ケラチノサイトが、ニワトリ表皮細胞であることを特徴とする請求項15又は16に記載の方法。
【請求項18】
請求項15〜17のいずれか1つに記載の方法により得ることができることを特徴とする、MDV及びFPVから選択されるウイルスから得られる完全無細胞性弱毒化包膜生DNAビリオン。
【請求項19】
請求項18に記載の少なくとも1のビリオンとワクチン担体とを含有することを特徴とする弱毒生ワクチン。
【請求項20】
第一に凍結乾燥ビリオンと、第二に免疫化担体とを含み、それぞれが別々の区画に包装されているキットの形であることを特徴とする請求項19に記載のワクチン。
【請求項21】
注射による投与、経口投与、インオボ経路を経由する投与、又は呼吸経路を経由する投与を意図することを特徴とする請求項19又は20に記載のワクチン。
【請求項22】
請求項18に記載のビリオンの、マレック病の予防又は治療用の医薬品を製造するための使用。
【図1】



【図2】

【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2007−515933(P2007−515933A)
【公表日】平成19年6月21日(2007.6.21)
【国際特許分類】
【出願番号】特願2006−519965(P2006−519965)
【出願日】平成16年7月15日(2004.7.15)
【国際出願番号】PCT/FR2004/001864
【国際公開番号】WO2005/010173
【国際公開日】平成17年2月3日(2005.2.3)
【出願人】(502205846)サントル ナショナル ドゥ ラ ルシェルシュ シアンティフィク (154)
【Fターム(参考)】
【公表日】平成19年6月21日(2007.6.21)
【国際特許分類】
【出願日】平成16年7月15日(2004.7.15)
【国際出願番号】PCT/FR2004/001864
【国際公開番号】WO2005/010173
【国際公開日】平成17年2月3日(2005.2.3)
【出願人】(502205846)サントル ナショナル ドゥ ラ ルシェルシュ シアンティフィク (154)
【Fターム(参考)】
[ Back to top ]