
トリインフルエンザウイルスH9N2に対する新規な抗ウイルスペプチド
本発明は、トリインフルエンザウイルス(AIV)に結合する融合ペプチドを保有する組換えファージに関する。このようなファージは、ウイルス上の結合部位に対して後者の抗体と競合することができるので、ファージは抗−AIV抗体に代わる診断用試薬として有用である。直鎖状又は環状の立体配置のいずれかの、配列CNDFRSKTCを有する合成ペプチド、又は上記の前記ペプチドを保有する融合ファージは、孵化卵及びMDCK細胞系においてAIV増殖を阻害した。したがって、これらはトリインフルエンザウイルスによって引き起こされるバードフルをコントロールし、治療し、根絶するための治療薬として用いられ得る。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、トリインフルエンザウイルス(AIV)サブタイプH9N2に対して作用する、新規な抗ウイルスペプチド及び融合ファージに関する。より詳しくは、融合ファージは、AIV H9N2に結合するその表面タンパク質P3上に配列CNDFRSKTCをディスプレイし、IC50値は1013pfu/100μl未満である。直鎖状又は環状の立体配座のいずれかにおけるアミノ酸配列CNDFRSKTCを有する合成ペプチドは、約100μM未満のIC50値でAIV H9N2の増殖を阻害した。
【背景技術】
【0002】
トリインフルエンザウイルス(AIV)は、インフルエンザA及びB、並びにインフルエンザCである2つの属を含んでいるオルトミクソウイルス科(Orthomyxoviridae)に属する(Lamb及びKrug、1996年)。これらのウイルスは、世界の家禽の間の罹患及び死滅の主たる原因である。これは一般用語でバードフル(bird flu)と呼ばれる最も危険な疾患の原因物質である(Websterら、1992年)。これらのウイルスはヒトには感染しないが、ヒトの感染及び大流行の例はいくつか報告されている(CDC、2008年;Normile、2004年;Parry、2004年)。インフルエンザAウイルスは、脂質二重層で覆われており、8個の一本鎖の、分節化された、ネガティブ鎖のRNAを含んでいる。ビリオンの表面上に存在する2個の糖タンパク質、すなわちヘマグルチニン(HA)及びノイラミニダーゼ(NA)、並びに1個のイオンチャネルタンパク質(M2)が存在する。これらの糖タンパク質はインフルエンザウイルスの主要な抗原決定基である。HAタンパク質は、ウイルス感染における最初のステップを開始し、これは宿主細胞表面のシアル酸受容体へのウイルスの付着を伴う(Lamb及びKrug、1996年)。NAタンパク質は、宿主細胞からの成熟ビリオンの放出に関与する(Paleseら、1974年)。したがって、ウイルス−宿主の相互作用を研究するために、及びこのプロセスを阻害する分子を同定するためにも、特定のペプチド配列をディスプレイするバクテリオファージが、ファージディスプレイライブラリーを用いてトリインフルエンザウイルスH9N2系統に対するその親和性によって選択された。
【0003】
インフルエンザウイルスの感染を防ぐための好ましい主要な戦略は、感受性の集団間に毎年ワクチン接種をすることである。しかし、疾患及び伝播をコントロールするための包括的な取組みにおいて、抗ウイルス薬は重要な役割を果たしている(Hayden、2006年)。治療及び予防のために認可されている2つのクラスの抗ウイルス薬が存在する。これらはアダマンタン誘導体(アマンタジン及びリマンタジン)、並びにノイラミニダーゼ阻害薬(NAI;ザナミビル及びオセルタミビル)である。(Nicholsonら、2003年)。これらアダマンタン誘導体は、インフルエンザAウイルスのM2イオンチャネルタンパク質に結合し、その機能を阻止することによって作用し、それによって宿主細胞内でのウイルスの複製を防ぐ(Wangら、1993年)。M2タンパク質における単一の点突然変異により、アダマンタン耐性系統が出現している(Hayら、1986年)。これらの耐性ウイルスは、通常完全に病原性があり、伝染性である(Hayden、2006年)。NAIはノイラミニダーゼタンパク質の酵素活性を阻害し、感染した宿主細胞からのウイルスの放出を防ぐ。しかし、NAの活性部位における突然変異により、NAI耐性系統も出現している(Nicholsonら、2003年)。インフルエンザAウイルスによる、両タイプの薬物に対する耐性の増大は、新規な薬物を同定する我々の必要性を強調するものである。
【0004】
伝統的に、植物、海洋生物、真菌、又は他の微生物から得られた天然生成物からの化合物が、抗微生物薬又は抗ウイルス薬を同定するのに用いられる。最近、ファージディスプレイライブラリーなどのコンビナトリアルペプチドライブラリーが、同じ目的でペプチド化合物を同定するのにますます用いられている(Doorbarら、1994年)。バクテリオファージ上にディスプレイされるペプチドの広範な構造上の多様性により、ファージディスプレイライブラリーは、とりわけ特定の標的と相互作用する特定のリガンドの同定において、タンパク質タンパク質相互作用を研究するための重要なツールとなっている(Devlinら、1990年)。標的分子と相互作用する化合物がファージライブラリーから選択され、次いでスクリーニングのプロセスを用いて標的に対して機能的な効果を有するリード化合物が同定される。次いで、これらのリード化合物をその活性に対して最適化し、次いで候補の薬物が臨床試験に入る。
【0005】
本発明者らは環状ペプチドのファージディスプレイライブラリーを用いて、インフルエンザAウイルスH9N2と相互作用するペプチド分子を同定し、in vitro及び卵内でその抗ウイルス活性を証明した。
【発明の概要】
【発明が解決しようとする課題】
【0006】
したがって、本発明の一目的は、トリインフルエンザウイルスA H9N2に対する抗ウイルス活性を有する新規なペプチド及び融合ファージを提供することである。
【0007】
本発明の別の一目的は、本発明による組換えファージ又は合成ペプチドを含む薬剤組成物を提供することである。
【0008】
本発明のさらに別の一目的は、本発明による組換えファージ又は合成ペプチドを含む診断用試薬を提供することである。
【課題を解決するための手段】
【0009】
これらの目的は、トリインフルエンザウイルスA H9N2に結合し、ウイルスの増殖を阻害する融合ファージに由来する融合ペプチド又は合成ペプチドを保有する、単離され、精製された組換えファージによって達成される。
【0010】
新規な抗ウイルス分子を同定するために、ジスルフィド拘束ランダムヘプタペプチド配列をディスプレイする組換えファージの集団を、ウイルスに対してスクリーニングした。次いで、融合ファージの表面上にディスプレイされたペプチドを、化学的に合成した。この単離されたペプチドはNDFRSKTの配列を有し、IC50値は100〜M未満である。インフルエンザウイルスA H9N2に対する組換えファージの特異性は、抗体−ファージ競合アッセイによって証明され、このアッセイにおいてファージは、ウイルス表面タンパク質上の結合部位に対してポリクローナル抗体と競合することが可能であった。
【0011】
本発明において、単離された配列の1つをベースにした合成ペプチドは、直鎖状又は環状の立体配置のいずれかにおいて(例えば合成ペプチドCNDFRSKTCなど)、NDVの増殖を阻害することができ、それによって疾患及び感染の伝播を防ぐ。
【図面の簡単な説明】
【0012】
表1 AIV H9N2に対する3ラウンドのバイオパニングから得たヘプタペプチド配列を示す表である。
表2 合成ペプチド及び融合ファージの赤血球凝集阻害活性を示す表である。
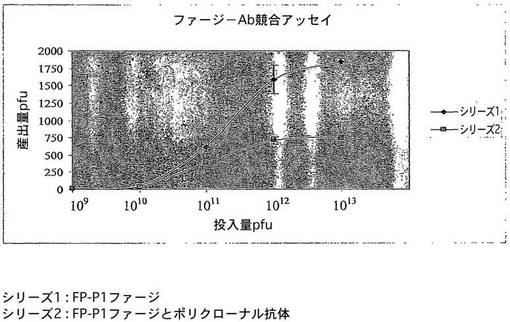
【図1】特定のヘプタペプチド配列を保有する組換えファージとAIV H9N2に対するポリクローナル抗体との間の競合の結果を示す図である。
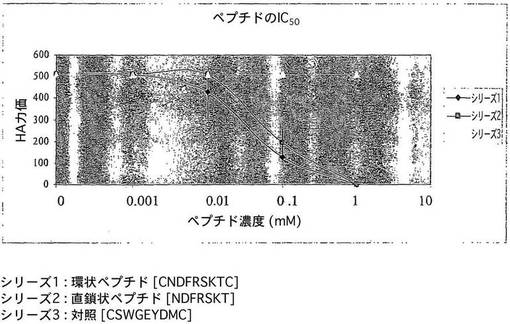
【図2】卵内の合成ペプチドでのAIV増殖の阻害を示す図である。
【図3】卵内の融合ファージでのAIV増殖の阻害を示す図である。
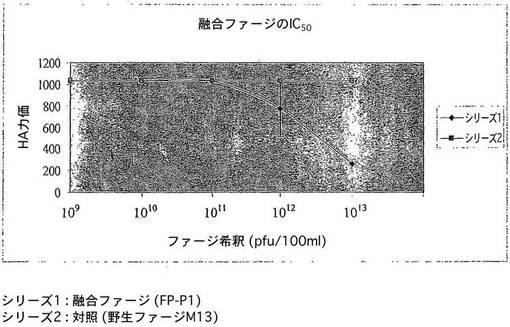
【図4】in vitroの融合ファージでのAIV増殖の阻害を示す図である。
【図5】in vitroのウイルス複製に対する融合ファージの効果を示す図である。
【発明を実施するための形態】
【0013】
(図及び表の詳細な説明)
表1は、AIVに対する4ラウンドのバイオパニングから得られたヘプタペプチド配列を示すものである。4ラウンドの選択及び増幅後、第2、第3、及び第4ラウンドから、それぞれ20個、35個、及び35個の個々のクローンが配列決定された。
【0014】
表2は、トリインフルエンザH9N2のヘマグルチネーション(haernaggluti nation)活性に対する環状及び直鎖状ペプチドの阻害能力を示すものである。実験を3回繰り返して行った。+の存在下、−の非存在下。
【0015】
図1は、特定のヘプタペプチド配列を保有する組換えファージとAIVに対するポリクローナル抗体との間の競合の結果を示すものである。ポリクローナル抗体(pAb)は融合ファージのAIVとの会合を阻害し、ファージは抗体とある程度共通の結合部位を共有し得ることを示唆している。実験を3回繰り返して行い、エラーバーは平均の標準偏差を表す。シリーズ1−融合ファージ、シリーズ2−融合ファージとポリクローナル抗体。
【0016】
図2は、AIV H9N2増殖に対する卵内の合成ペプチドのIC50値の決定の結果を示すものである。ウイルスの成長の50%を阻害するのに必要とされるペプチドの濃度を、様々な濃度のペプチドを用いて決定した。実験を3回繰り返して行い、エラーバーは平均の標準偏差を表す。
【0017】
図3は、卵内のAIV H9N2ウイルス増殖に対する融合ファージFP−P1のIC50値の結果を示すものである。実験を3回繰り返して行い、エラーバーは平均の標準偏差を表す。シリーズ1−融合ファージFP−P1;シリーズ2−対照としての野生ファージ。
【0018】
図4はin vitroの融合ファージFP−P1の抗ウイルス活性の結果を示すものである。実験を3回繰り返して行い、エラーバーは平均の標準偏差を表す。シリーズ1−FP−P1;シリーズ2−対照としての野生ファージM13。
【0019】
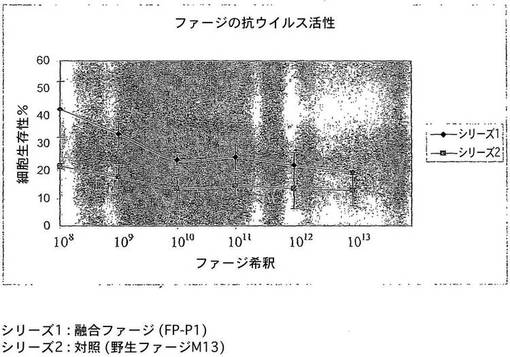
図5は、ウイルス複製に対する融合ファージFP−P1の効果の結果を示すものである。MDCK細胞に非処理の(0値)、又はFP−PI処理したウイルスを接種し、ウイルス力価を上清72hpiに対して決定した。シリーズ1−FP−P1;シリーズ2−対照としての野生ファージM13。
【0020】
(発明の詳細な説明)
本明細書に記載する記述は、様々なセクションに分割される以下の発明を実施するための形態とともにより完全に理解されよう。本発明に用いられる材料及び方法は、実施例において述べる。例1から5までは、本発明の必要条件である従来の材料及び方法である。
【0021】
ファージディスプレイ技術の原理
ファージディスプレイ技術において、コートタンパク質をコードする遺伝子中に合成オリゴヌクレオチドを挿入することによって、ランダムペプチドが、通常、融合タンパク質としてバクテリオファージ分子の表面上にディスプレイされる。これら組換えファージの収集物はファージディスプレイライブラリーとして知られており、バイオパニングとして知られるin vitroの選択手順によって、標的に対する莫大な数のペプチド配列のスクリーニングが可能になっている(Parmleyら、1988年、及びSmithら、1993年)。本発明において、用いた標的は、AIV H9N2ウイルス粒子全体であり、用いたライブラリーは、New England Biolabs社から入手したジスルフィド拘束ライブラリーであった。この特定のライブラリーにおいて、ディスプレイされたペプチド分子に、ファージM13のgpIIIタンパク質に対するシステイン残基が隣接している。このシステイン残基が、還元剤の非存在下、ペプチドが固定の環状の形状を達成するのを助ける。このジスルフィド拘束ライブラリーは、その相互作用成分(天然のリガンド)が不連続の結合部位(配座エピトープ)を有する標的に有用であり(Hoessら、1994年)、結合部位では、アミノ酸がポリペプチドにおける様々な位置から運ばれて本質的な接触領域を形成する。その上、ジスルフィド拘束環状ペプチドライブラリーは、直鎖状ペプチドライブラリーよりもむしろ、高親和性リガンドを選択するのにより有用である(O’Neilら、1992年;Ghoら、1997年)。
【0022】
AIVに結合するペプチドの親和性選択
トリインフルエンザウイルスは、2種の表面糖タンパク質、すなわちヘマグルチニン(HA)及びノイラミニダーゼ(NA)を含んでおり、これらは宿主細胞内へのウイルスの侵入、及び宿主細胞からのウイルスの退出を担っている。したがって、本発明者らは、表面タンパク質に結合する立体配置のリガンドを選択するためのウイルス粒子全体に対するジスルフィド拘束ライブラリーを用いることに決定した。図1は、これらのリガンドの選択に関与する主なステップを概要したものである。第1に、ウイルス粒子全体を、高結合性のマイクロタイタープレートウエルの表面に直接付着させた。次いで、ライブラリーをウエル中に加えて、組換えファージ粒子をウイルス粒子と結合させた。非結合のファージを洗い流し、結合しているファージを低pHで溶出した。次いで、溶出したファージを、細菌である大腸菌(Escherichia coli)中で増幅し、増幅したファージを第2ラウンドのバイオパニングにおいて用いた。この手順を4回繰り返した。アフィニティ選択及び増幅を連続4ラウンド行った後、サブセットの選択されたファージを個々に成長させ、コア粒子に結合するペプチドのタイルアイデンティティ(tile identity)を、挿入を保有するタイル(tile)gpIII遺伝子を配列決定することによって得た。
【0023】
高親和性の結合ファージを選択するために、選択のストリンジェンシーを、(i)室温(28℃)でバイオパニングを行い、(ii)速度に対して速いリガンドを選択するために結合の時間を1時間に短縮し(K0)、(iii)ウエルを徹底的に洗浄して(10回)低親和性の結合ファージを除去し、(iv)バイオパニングを4ラウンド繰り返して高親和性結合クローンを濃縮することによって増大させた。第4ラウンドのパニングから分析されたファージの47%は、融合ペプチド配列NDFRSKTを保有しており、10.5%がQHSTKWFモチーフを含んでおり、その後に各5%のLPYAAKH及びILGDKVG、並びに他の非関連の配列が続く(表1)。ストレプトアビジン標的を、クローン全てにおいてHPQ配列のコンセンサスモチーフを与える陽性対照として用いたが、これはDevlinら(1990年)によって報告されたものと良好に一致した。陰性対照として用いられたウシ血清アルブミン(BSA)では、認識できるコンセンサス配列は観察されなかった。
【0024】
本明細書で用いられる「融合ペプチド」の語は、一般的にバクテリオファージによってコードされ、ファージのコートタンパク質に物理的に連結しているアミノ酸配列を意味する。特許請求の融合ペプチドは、アミノ酸配列NDFRSKT:(i)特許請求のアミノ酸配列よりも短く、又は長いアミノ酸配列、(ii)アミノ酸配列における変形、とりわけ以下に記載するのと同じカテゴリー内のアミノ酸の置換、(iii)特許請求の融合ペプチドのアミノ酸配列と少なくとも60%のホモロジーを共有するアミノ酸配列、及び(iv)直鎖状又は限定立体配置のいずれかを含んでいるが、これらに限定されない。
【0025】
AIVは2種の表面糖タンパク質HA及びNAを所有し、これらはウイルスの脂質二重層膜から突出している。これらの糖タンパク質はそれぞれ宿主細胞内及び宿主細胞外へのウイルスの侵入及びウイルスの放出に不可欠である。配列NDFRSKTを保有するファージは溶液中のビリオンに結合するので、これらの配列は、ある程度、完全なビリオンと相互作用する宿主細胞の受容体上の領域に類似することがある。
【0026】
ファージはAIV上の結合部位に対して抗体と競合する
表面糖タンパク質HA及びNAは、宿主における抗体の生成を誘発する2つの重要なタンパク質である。ファージ−抗体競合アッセイにおいて、ファージはミモトープの1つに結合することができ、したがってビリオンに対して産生された抗体の結合を阻害することができた。図1は、バイオパニング実験から選択された、ペプチドNDFRSKTをディスプレイする組換えファージが、AIV上の結合部位に対して抗体と競合できたことを示している。抗体の存在下、ウイルスでコーティングされたウエルに結合したファージの数は、ウイルス上の同じ結合部位に対するこれら2つの分子間の競合の結果として劇的に低下した。したがって、組換えファージ分子は、AIV感染に対する潜在的な診断薬として働き得る。
【0027】
卵内の合成ペプチド及び融合ファージの抗ウイルス活性
AIVは、孵化鶏卵の尿膜腔液中で容易に増殖させることができる。したがって、ウイルス及び増殖系により、in vivoで特定の化合物又はペプチドの抗ウイルス活性を直接評価するための簡単なモデルが提供される。抗ウイルス化合物処理の前及び後に尿膜腔液中に存在するウイルスの量を、赤血球凝集力価によって決定する。環状又は直鎖状の形態いずれかの特許請求のペプチド、及びペプチドをディスプレイする融合ファージの注射により、尿膜腔液中のAIV力価は低減した(図2及び3)。環状ペプチドは、直鎖状ペプチドに対するよりも100μM低いペプチド濃度でより高いIC50値を示した。さらに、環化により、エキソペプチダーゼによるペプチドの消化を避けることができる。このように、疾患を治療する、コントロールする、又は根絶するための治療薬として、本明細書において特許請求する合成ペプチドを用いることができる。
【0028】
in vitroの融合ファージの抗ウイルス活性
MDCK細胞系は、in vitroのトリインフルエンザウイルスの増殖のための有効な媒体である。ウイルス増加に対する融合ファージのin vitroの抗ウイルス性質を、これらの細胞系において調べた。融合ファージ分子で処理する前及び後に、ウイルス複製の阻害を細胞の生存性によって間接的に調べた。細胞の生存性は、ファージ処理したウイルスに感染した後、非処理のウイルスに対して2倍近く増大した(図4)。ウイルス力価における低下を、上清の赤血球凝集試験によってさらに確認した(図5)。この実験により、in vitroで融合ファージ分子の抗ウイルス性質が証明された。
【実施例】
【0029】
(例1)
AIVの増殖及び精製
増殖及び精製の方法は、それぞれBlaskovik及びStyk(1967年)、並びにYusoffら(1996年)から適用され、改変された。AIVサブタイプH9N2を、9日齢の特定の病原体フリーの孵化鶏卵中に注射した。37℃でインキュベートして3日後、尿膜腔液を回収し、遠心分離(30分、35000×g、4℃)によって透明にした。透明にした上清から30%〜60%ショ糖勾配の超遠心(3.5時間、285000×g、4℃)によってウイルスを精製し、以下の試験に用いた。
【0030】
(例2)
精製したAIVのバイオパニング
ウイルスのタンパク質濃度を、Bradfordアッセイ(1976年)を用いて測定した。AIV(15μg/ml)をマイクロタイタープレートウエル上にコーティングした[Na2CO3/NaHCO3バッファー(0.1M、pH9.6)]。ストレプトアビジン及びBSAを、それぞれ陽性対照及び陰性対照として用いた。実験は室温(約25℃)で行った。ジスルフィド拘束7量体ファージディスプレイライブラリー(New England Biolabs、USA)からのファージを、TBS[50mM Tris−HCI(pH7.5)、150mM NaCl;110μl]中I×1011pfuに希釈し、コーティングした各ウエル中に加えた。混合物を30分間インキュベートした。ウエルをTBST JBS+0.1%v/vTween−20で洗浄した。結合したファージをグリシン−HCl[0.1M(pH2.2);100μl]で7〜8分間穏やかに揺り動かすことによって溶出し、Tris−HCl[1M(pH9);115μl]で中和した。産出量のpfuを、溶出物(10μl)を滴定することによって決定し、残りの溶出物を、37℃で大腸菌系統ER2738(F’IaclqΔ(IacZ)M)5proA+B+zzf::TnIO(TetR)/fhuA2supE thi Δ(lac−proAB)Δ(hsdMS−mcrB)5(rk−mk−McrBC−)}中で増幅し、5時間振盪した。パニングのプロセスをさらなる3ラウンド繰り返した。
【0031】
(例3)
ファージ滴定
ファージ滴定法を、Sambrookら(1989年)から適用した。
【0032】
(例4)
選択されたファージの配列決定
4ラウンドのパニング全てからの単一のプラークを(Luria Broth)LB平板(産出量の滴定において使用)から選択し、LB培地中成長させた。ファージの一本鎖DNAを、Sambrookら(1989年)によって記載されている通りに抽出した。挿入物の配列決定を、First Base Laboratories Sdn Bhd、クアラルンプールによるABI自動化シーケンサーを用いて行った。
【0033】
(例5)
抗体競合アッセイ
ウエルをAIV(15μg/ml)でコーティングし、AIVに対して産生されたポリクローナル抗体(1:500希釈;100μl)を、一連の異なるファージ濃度で(109〜1013pfu;100μl)加えた。混合物を1時間インキュベートし、Tris緩衝食塩水−Tween(TBST)で6回洗浄した。結合しているファージを溶出し、上記に記載した通りに滴定した。
【0034】
(例6)
卵内のAIV増殖の阻害におけるペプチド及び融合ファージのIC50の決定
9日齢の孵化鶏卵に、AIV100μl(8HA単位)、及び等体積の様々な濃度のペプチド(0.1μM〜1mM)、又は融合ファージ(108〜1013pfu)を接種した。卵を37℃でインキュベートした。各ペプチド濃度に対して卵3個を用いた。陽性対照にはAIVだけを接種し、陰性対照にはウイルスをペプチド又はファージによって置き換えた。ペプチド(100μl)を、別の3日間毎日、好適な卵中に注射した。4回目の注射の1日後、卵を一夜冷却し、尿膜腔液を回収した。次いで、Graceら(1973年)にしたがって尿膜腔液試料の赤血球凝集活性を決定した。
【0035】
(例7)
in vitroのAIV増殖の阻害における融合ファージのIC50の決定
MDCK細胞に、37℃で1時間、培地単独、又はファージ処理(108〜1013pfu/100μl)若しくは非処理のウイルス(感染効率[MOI]0.05)を接種した。吸着後、単層を洗浄し、5%ウシ胎児血清(FBS)を含むEMEM中でインキュベートした。細胞変性効果を光学顕微鏡によってモニターし、XTT Cell Viability Assay Kit(Biotium、米国)によって定量した。ウイルスの複製を評価するために、MDCK細胞にファージ処理したウイルス、又は非処理のウイルスを0.05のMOIで接種し、上清を72hpiで回収し、ウイルス力価をHA滴定によって決定した。IC50値を、用量反応曲線を補間することによって推定した。
【0036】
本発明を様々な、現在好ましい実施形態に関して上記に記載してきたが、当業者であれば多くの変形及び改変を行うことができることは明らかである。したがって、本発明は、本明細書に示す特定の実施形態に制限されるものと理解してはならず、むしろ、本明細書に添付する特許請求の範囲の範囲内にあるこのような変更及び改変全てを包含するものと理解されたい。
【0037】
本明細書に開示する参照は全て、その全文が参照によって援用される。
【0038】
【表1】

【表2】
【技術分野】
【0001】
本発明は、トリインフルエンザウイルス(AIV)サブタイプH9N2に対して作用する、新規な抗ウイルスペプチド及び融合ファージに関する。より詳しくは、融合ファージは、AIV H9N2に結合するその表面タンパク質P3上に配列CNDFRSKTCをディスプレイし、IC50値は1013pfu/100μl未満である。直鎖状又は環状の立体配座のいずれかにおけるアミノ酸配列CNDFRSKTCを有する合成ペプチドは、約100μM未満のIC50値でAIV H9N2の増殖を阻害した。
【背景技術】
【0002】
トリインフルエンザウイルス(AIV)は、インフルエンザA及びB、並びにインフルエンザCである2つの属を含んでいるオルトミクソウイルス科(Orthomyxoviridae)に属する(Lamb及びKrug、1996年)。これらのウイルスは、世界の家禽の間の罹患及び死滅の主たる原因である。これは一般用語でバードフル(bird flu)と呼ばれる最も危険な疾患の原因物質である(Websterら、1992年)。これらのウイルスはヒトには感染しないが、ヒトの感染及び大流行の例はいくつか報告されている(CDC、2008年;Normile、2004年;Parry、2004年)。インフルエンザAウイルスは、脂質二重層で覆われており、8個の一本鎖の、分節化された、ネガティブ鎖のRNAを含んでいる。ビリオンの表面上に存在する2個の糖タンパク質、すなわちヘマグルチニン(HA)及びノイラミニダーゼ(NA)、並びに1個のイオンチャネルタンパク質(M2)が存在する。これらの糖タンパク質はインフルエンザウイルスの主要な抗原決定基である。HAタンパク質は、ウイルス感染における最初のステップを開始し、これは宿主細胞表面のシアル酸受容体へのウイルスの付着を伴う(Lamb及びKrug、1996年)。NAタンパク質は、宿主細胞からの成熟ビリオンの放出に関与する(Paleseら、1974年)。したがって、ウイルス−宿主の相互作用を研究するために、及びこのプロセスを阻害する分子を同定するためにも、特定のペプチド配列をディスプレイするバクテリオファージが、ファージディスプレイライブラリーを用いてトリインフルエンザウイルスH9N2系統に対するその親和性によって選択された。
【0003】
インフルエンザウイルスの感染を防ぐための好ましい主要な戦略は、感受性の集団間に毎年ワクチン接種をすることである。しかし、疾患及び伝播をコントロールするための包括的な取組みにおいて、抗ウイルス薬は重要な役割を果たしている(Hayden、2006年)。治療及び予防のために認可されている2つのクラスの抗ウイルス薬が存在する。これらはアダマンタン誘導体(アマンタジン及びリマンタジン)、並びにノイラミニダーゼ阻害薬(NAI;ザナミビル及びオセルタミビル)である。(Nicholsonら、2003年)。これらアダマンタン誘導体は、インフルエンザAウイルスのM2イオンチャネルタンパク質に結合し、その機能を阻止することによって作用し、それによって宿主細胞内でのウイルスの複製を防ぐ(Wangら、1993年)。M2タンパク質における単一の点突然変異により、アダマンタン耐性系統が出現している(Hayら、1986年)。これらの耐性ウイルスは、通常完全に病原性があり、伝染性である(Hayden、2006年)。NAIはノイラミニダーゼタンパク質の酵素活性を阻害し、感染した宿主細胞からのウイルスの放出を防ぐ。しかし、NAの活性部位における突然変異により、NAI耐性系統も出現している(Nicholsonら、2003年)。インフルエンザAウイルスによる、両タイプの薬物に対する耐性の増大は、新規な薬物を同定する我々の必要性を強調するものである。
【0004】
伝統的に、植物、海洋生物、真菌、又は他の微生物から得られた天然生成物からの化合物が、抗微生物薬又は抗ウイルス薬を同定するのに用いられる。最近、ファージディスプレイライブラリーなどのコンビナトリアルペプチドライブラリーが、同じ目的でペプチド化合物を同定するのにますます用いられている(Doorbarら、1994年)。バクテリオファージ上にディスプレイされるペプチドの広範な構造上の多様性により、ファージディスプレイライブラリーは、とりわけ特定の標的と相互作用する特定のリガンドの同定において、タンパク質タンパク質相互作用を研究するための重要なツールとなっている(Devlinら、1990年)。標的分子と相互作用する化合物がファージライブラリーから選択され、次いでスクリーニングのプロセスを用いて標的に対して機能的な効果を有するリード化合物が同定される。次いで、これらのリード化合物をその活性に対して最適化し、次いで候補の薬物が臨床試験に入る。
【0005】
本発明者らは環状ペプチドのファージディスプレイライブラリーを用いて、インフルエンザAウイルスH9N2と相互作用するペプチド分子を同定し、in vitro及び卵内でその抗ウイルス活性を証明した。
【発明の概要】
【発明が解決しようとする課題】
【0006】
したがって、本発明の一目的は、トリインフルエンザウイルスA H9N2に対する抗ウイルス活性を有する新規なペプチド及び融合ファージを提供することである。
【0007】
本発明の別の一目的は、本発明による組換えファージ又は合成ペプチドを含む薬剤組成物を提供することである。
【0008】
本発明のさらに別の一目的は、本発明による組換えファージ又は合成ペプチドを含む診断用試薬を提供することである。
【課題を解決するための手段】
【0009】
これらの目的は、トリインフルエンザウイルスA H9N2に結合し、ウイルスの増殖を阻害する融合ファージに由来する融合ペプチド又は合成ペプチドを保有する、単離され、精製された組換えファージによって達成される。
【0010】
新規な抗ウイルス分子を同定するために、ジスルフィド拘束ランダムヘプタペプチド配列をディスプレイする組換えファージの集団を、ウイルスに対してスクリーニングした。次いで、融合ファージの表面上にディスプレイされたペプチドを、化学的に合成した。この単離されたペプチドはNDFRSKTの配列を有し、IC50値は100〜M未満である。インフルエンザウイルスA H9N2に対する組換えファージの特異性は、抗体−ファージ競合アッセイによって証明され、このアッセイにおいてファージは、ウイルス表面タンパク質上の結合部位に対してポリクローナル抗体と競合することが可能であった。
【0011】
本発明において、単離された配列の1つをベースにした合成ペプチドは、直鎖状又は環状の立体配置のいずれかにおいて(例えば合成ペプチドCNDFRSKTCなど)、NDVの増殖を阻害することができ、それによって疾患及び感染の伝播を防ぐ。
【図面の簡単な説明】
【0012】
表1 AIV H9N2に対する3ラウンドのバイオパニングから得たヘプタペプチド配列を示す表である。
表2 合成ペプチド及び融合ファージの赤血球凝集阻害活性を示す表である。
【図1】特定のヘプタペプチド配列を保有する組換えファージとAIV H9N2に対するポリクローナル抗体との間の競合の結果を示す図である。
【図2】卵内の合成ペプチドでのAIV増殖の阻害を示す図である。
【図3】卵内の融合ファージでのAIV増殖の阻害を示す図である。
【図4】in vitroの融合ファージでのAIV増殖の阻害を示す図である。
【図5】in vitroのウイルス複製に対する融合ファージの効果を示す図である。
【発明を実施するための形態】
【0013】
(図及び表の詳細な説明)
表1は、AIVに対する4ラウンドのバイオパニングから得られたヘプタペプチド配列を示すものである。4ラウンドの選択及び増幅後、第2、第3、及び第4ラウンドから、それぞれ20個、35個、及び35個の個々のクローンが配列決定された。
【0014】
表2は、トリインフルエンザH9N2のヘマグルチネーション(haernaggluti nation)活性に対する環状及び直鎖状ペプチドの阻害能力を示すものである。実験を3回繰り返して行った。+の存在下、−の非存在下。
【0015】
図1は、特定のヘプタペプチド配列を保有する組換えファージとAIVに対するポリクローナル抗体との間の競合の結果を示すものである。ポリクローナル抗体(pAb)は融合ファージのAIVとの会合を阻害し、ファージは抗体とある程度共通の結合部位を共有し得ることを示唆している。実験を3回繰り返して行い、エラーバーは平均の標準偏差を表す。シリーズ1−融合ファージ、シリーズ2−融合ファージとポリクローナル抗体。
【0016】
図2は、AIV H9N2増殖に対する卵内の合成ペプチドのIC50値の決定の結果を示すものである。ウイルスの成長の50%を阻害するのに必要とされるペプチドの濃度を、様々な濃度のペプチドを用いて決定した。実験を3回繰り返して行い、エラーバーは平均の標準偏差を表す。
【0017】
図3は、卵内のAIV H9N2ウイルス増殖に対する融合ファージFP−P1のIC50値の結果を示すものである。実験を3回繰り返して行い、エラーバーは平均の標準偏差を表す。シリーズ1−融合ファージFP−P1;シリーズ2−対照としての野生ファージ。
【0018】
図4はin vitroの融合ファージFP−P1の抗ウイルス活性の結果を示すものである。実験を3回繰り返して行い、エラーバーは平均の標準偏差を表す。シリーズ1−FP−P1;シリーズ2−対照としての野生ファージM13。
【0019】
図5は、ウイルス複製に対する融合ファージFP−P1の効果の結果を示すものである。MDCK細胞に非処理の(0値)、又はFP−PI処理したウイルスを接種し、ウイルス力価を上清72hpiに対して決定した。シリーズ1−FP−P1;シリーズ2−対照としての野生ファージM13。
【0020】
(発明の詳細な説明)
本明細書に記載する記述は、様々なセクションに分割される以下の発明を実施するための形態とともにより完全に理解されよう。本発明に用いられる材料及び方法は、実施例において述べる。例1から5までは、本発明の必要条件である従来の材料及び方法である。
【0021】
ファージディスプレイ技術の原理
ファージディスプレイ技術において、コートタンパク質をコードする遺伝子中に合成オリゴヌクレオチドを挿入することによって、ランダムペプチドが、通常、融合タンパク質としてバクテリオファージ分子の表面上にディスプレイされる。これら組換えファージの収集物はファージディスプレイライブラリーとして知られており、バイオパニングとして知られるin vitroの選択手順によって、標的に対する莫大な数のペプチド配列のスクリーニングが可能になっている(Parmleyら、1988年、及びSmithら、1993年)。本発明において、用いた標的は、AIV H9N2ウイルス粒子全体であり、用いたライブラリーは、New England Biolabs社から入手したジスルフィド拘束ライブラリーであった。この特定のライブラリーにおいて、ディスプレイされたペプチド分子に、ファージM13のgpIIIタンパク質に対するシステイン残基が隣接している。このシステイン残基が、還元剤の非存在下、ペプチドが固定の環状の形状を達成するのを助ける。このジスルフィド拘束ライブラリーは、その相互作用成分(天然のリガンド)が不連続の結合部位(配座エピトープ)を有する標的に有用であり(Hoessら、1994年)、結合部位では、アミノ酸がポリペプチドにおける様々な位置から運ばれて本質的な接触領域を形成する。その上、ジスルフィド拘束環状ペプチドライブラリーは、直鎖状ペプチドライブラリーよりもむしろ、高親和性リガンドを選択するのにより有用である(O’Neilら、1992年;Ghoら、1997年)。
【0022】
AIVに結合するペプチドの親和性選択
トリインフルエンザウイルスは、2種の表面糖タンパク質、すなわちヘマグルチニン(HA)及びノイラミニダーゼ(NA)を含んでおり、これらは宿主細胞内へのウイルスの侵入、及び宿主細胞からのウイルスの退出を担っている。したがって、本発明者らは、表面タンパク質に結合する立体配置のリガンドを選択するためのウイルス粒子全体に対するジスルフィド拘束ライブラリーを用いることに決定した。図1は、これらのリガンドの選択に関与する主なステップを概要したものである。第1に、ウイルス粒子全体を、高結合性のマイクロタイタープレートウエルの表面に直接付着させた。次いで、ライブラリーをウエル中に加えて、組換えファージ粒子をウイルス粒子と結合させた。非結合のファージを洗い流し、結合しているファージを低pHで溶出した。次いで、溶出したファージを、細菌である大腸菌(Escherichia coli)中で増幅し、増幅したファージを第2ラウンドのバイオパニングにおいて用いた。この手順を4回繰り返した。アフィニティ選択及び増幅を連続4ラウンド行った後、サブセットの選択されたファージを個々に成長させ、コア粒子に結合するペプチドのタイルアイデンティティ(tile identity)を、挿入を保有するタイル(tile)gpIII遺伝子を配列決定することによって得た。
【0023】
高親和性の結合ファージを選択するために、選択のストリンジェンシーを、(i)室温(28℃)でバイオパニングを行い、(ii)速度に対して速いリガンドを選択するために結合の時間を1時間に短縮し(K0)、(iii)ウエルを徹底的に洗浄して(10回)低親和性の結合ファージを除去し、(iv)バイオパニングを4ラウンド繰り返して高親和性結合クローンを濃縮することによって増大させた。第4ラウンドのパニングから分析されたファージの47%は、融合ペプチド配列NDFRSKTを保有しており、10.5%がQHSTKWFモチーフを含んでおり、その後に各5%のLPYAAKH及びILGDKVG、並びに他の非関連の配列が続く(表1)。ストレプトアビジン標的を、クローン全てにおいてHPQ配列のコンセンサスモチーフを与える陽性対照として用いたが、これはDevlinら(1990年)によって報告されたものと良好に一致した。陰性対照として用いられたウシ血清アルブミン(BSA)では、認識できるコンセンサス配列は観察されなかった。
【0024】
本明細書で用いられる「融合ペプチド」の語は、一般的にバクテリオファージによってコードされ、ファージのコートタンパク質に物理的に連結しているアミノ酸配列を意味する。特許請求の融合ペプチドは、アミノ酸配列NDFRSKT:(i)特許請求のアミノ酸配列よりも短く、又は長いアミノ酸配列、(ii)アミノ酸配列における変形、とりわけ以下に記載するのと同じカテゴリー内のアミノ酸の置換、(iii)特許請求の融合ペプチドのアミノ酸配列と少なくとも60%のホモロジーを共有するアミノ酸配列、及び(iv)直鎖状又は限定立体配置のいずれかを含んでいるが、これらに限定されない。
【0025】
AIVは2種の表面糖タンパク質HA及びNAを所有し、これらはウイルスの脂質二重層膜から突出している。これらの糖タンパク質はそれぞれ宿主細胞内及び宿主細胞外へのウイルスの侵入及びウイルスの放出に不可欠である。配列NDFRSKTを保有するファージは溶液中のビリオンに結合するので、これらの配列は、ある程度、完全なビリオンと相互作用する宿主細胞の受容体上の領域に類似することがある。
【0026】
ファージはAIV上の結合部位に対して抗体と競合する
表面糖タンパク質HA及びNAは、宿主における抗体の生成を誘発する2つの重要なタンパク質である。ファージ−抗体競合アッセイにおいて、ファージはミモトープの1つに結合することができ、したがってビリオンに対して産生された抗体の結合を阻害することができた。図1は、バイオパニング実験から選択された、ペプチドNDFRSKTをディスプレイする組換えファージが、AIV上の結合部位に対して抗体と競合できたことを示している。抗体の存在下、ウイルスでコーティングされたウエルに結合したファージの数は、ウイルス上の同じ結合部位に対するこれら2つの分子間の競合の結果として劇的に低下した。したがって、組換えファージ分子は、AIV感染に対する潜在的な診断薬として働き得る。
【0027】
卵内の合成ペプチド及び融合ファージの抗ウイルス活性
AIVは、孵化鶏卵の尿膜腔液中で容易に増殖させることができる。したがって、ウイルス及び増殖系により、in vivoで特定の化合物又はペプチドの抗ウイルス活性を直接評価するための簡単なモデルが提供される。抗ウイルス化合物処理の前及び後に尿膜腔液中に存在するウイルスの量を、赤血球凝集力価によって決定する。環状又は直鎖状の形態いずれかの特許請求のペプチド、及びペプチドをディスプレイする融合ファージの注射により、尿膜腔液中のAIV力価は低減した(図2及び3)。環状ペプチドは、直鎖状ペプチドに対するよりも100μM低いペプチド濃度でより高いIC50値を示した。さらに、環化により、エキソペプチダーゼによるペプチドの消化を避けることができる。このように、疾患を治療する、コントロールする、又は根絶するための治療薬として、本明細書において特許請求する合成ペプチドを用いることができる。
【0028】
in vitroの融合ファージの抗ウイルス活性
MDCK細胞系は、in vitroのトリインフルエンザウイルスの増殖のための有効な媒体である。ウイルス増加に対する融合ファージのin vitroの抗ウイルス性質を、これらの細胞系において調べた。融合ファージ分子で処理する前及び後に、ウイルス複製の阻害を細胞の生存性によって間接的に調べた。細胞の生存性は、ファージ処理したウイルスに感染した後、非処理のウイルスに対して2倍近く増大した(図4)。ウイルス力価における低下を、上清の赤血球凝集試験によってさらに確認した(図5)。この実験により、in vitroで融合ファージ分子の抗ウイルス性質が証明された。
【実施例】
【0029】
(例1)
AIVの増殖及び精製
増殖及び精製の方法は、それぞれBlaskovik及びStyk(1967年)、並びにYusoffら(1996年)から適用され、改変された。AIVサブタイプH9N2を、9日齢の特定の病原体フリーの孵化鶏卵中に注射した。37℃でインキュベートして3日後、尿膜腔液を回収し、遠心分離(30分、35000×g、4℃)によって透明にした。透明にした上清から30%〜60%ショ糖勾配の超遠心(3.5時間、285000×g、4℃)によってウイルスを精製し、以下の試験に用いた。
【0030】
(例2)
精製したAIVのバイオパニング
ウイルスのタンパク質濃度を、Bradfordアッセイ(1976年)を用いて測定した。AIV(15μg/ml)をマイクロタイタープレートウエル上にコーティングした[Na2CO3/NaHCO3バッファー(0.1M、pH9.6)]。ストレプトアビジン及びBSAを、それぞれ陽性対照及び陰性対照として用いた。実験は室温(約25℃)で行った。ジスルフィド拘束7量体ファージディスプレイライブラリー(New England Biolabs、USA)からのファージを、TBS[50mM Tris−HCI(pH7.5)、150mM NaCl;110μl]中I×1011pfuに希釈し、コーティングした各ウエル中に加えた。混合物を30分間インキュベートした。ウエルをTBST JBS+0.1%v/vTween−20で洗浄した。結合したファージをグリシン−HCl[0.1M(pH2.2);100μl]で7〜8分間穏やかに揺り動かすことによって溶出し、Tris−HCl[1M(pH9);115μl]で中和した。産出量のpfuを、溶出物(10μl)を滴定することによって決定し、残りの溶出物を、37℃で大腸菌系統ER2738(F’IaclqΔ(IacZ)M)5proA+B+zzf::TnIO(TetR)/fhuA2supE thi Δ(lac−proAB)Δ(hsdMS−mcrB)5(rk−mk−McrBC−)}中で増幅し、5時間振盪した。パニングのプロセスをさらなる3ラウンド繰り返した。
【0031】
(例3)
ファージ滴定
ファージ滴定法を、Sambrookら(1989年)から適用した。
【0032】
(例4)
選択されたファージの配列決定
4ラウンドのパニング全てからの単一のプラークを(Luria Broth)LB平板(産出量の滴定において使用)から選択し、LB培地中成長させた。ファージの一本鎖DNAを、Sambrookら(1989年)によって記載されている通りに抽出した。挿入物の配列決定を、First Base Laboratories Sdn Bhd、クアラルンプールによるABI自動化シーケンサーを用いて行った。
【0033】
(例5)
抗体競合アッセイ
ウエルをAIV(15μg/ml)でコーティングし、AIVに対して産生されたポリクローナル抗体(1:500希釈;100μl)を、一連の異なるファージ濃度で(109〜1013pfu;100μl)加えた。混合物を1時間インキュベートし、Tris緩衝食塩水−Tween(TBST)で6回洗浄した。結合しているファージを溶出し、上記に記載した通りに滴定した。
【0034】
(例6)
卵内のAIV増殖の阻害におけるペプチド及び融合ファージのIC50の決定
9日齢の孵化鶏卵に、AIV100μl(8HA単位)、及び等体積の様々な濃度のペプチド(0.1μM〜1mM)、又は融合ファージ(108〜1013pfu)を接種した。卵を37℃でインキュベートした。各ペプチド濃度に対して卵3個を用いた。陽性対照にはAIVだけを接種し、陰性対照にはウイルスをペプチド又はファージによって置き換えた。ペプチド(100μl)を、別の3日間毎日、好適な卵中に注射した。4回目の注射の1日後、卵を一夜冷却し、尿膜腔液を回収した。次いで、Graceら(1973年)にしたがって尿膜腔液試料の赤血球凝集活性を決定した。
【0035】
(例7)
in vitroのAIV増殖の阻害における融合ファージのIC50の決定
MDCK細胞に、37℃で1時間、培地単独、又はファージ処理(108〜1013pfu/100μl)若しくは非処理のウイルス(感染効率[MOI]0.05)を接種した。吸着後、単層を洗浄し、5%ウシ胎児血清(FBS)を含むEMEM中でインキュベートした。細胞変性効果を光学顕微鏡によってモニターし、XTT Cell Viability Assay Kit(Biotium、米国)によって定量した。ウイルスの複製を評価するために、MDCK細胞にファージ処理したウイルス、又は非処理のウイルスを0.05のMOIで接種し、上清を72hpiで回収し、ウイルス力価をHA滴定によって決定した。IC50値を、用量反応曲線を補間することによって推定した。
【0036】
本発明を様々な、現在好ましい実施形態に関して上記に記載してきたが、当業者であれば多くの変形及び改変を行うことができることは明らかである。したがって、本発明は、本明細書に示す特定の実施形態に制限されるものと理解してはならず、むしろ、本明細書に添付する特許請求の範囲の範囲内にあるこのような変更及び改変全てを包含するものと理解されたい。
【0037】
本明細書に開示する参照は全て、その全文が参照によって援用される。
【0038】
【表1】
【表2】
【特許請求の範囲】
【請求項1】
トリインフルエンザウイルスに結合し、ウイルスの増殖を阻害する融合ペプチドを保有する、単離され、精製された組換えファージ。
【請求項2】
融合ペプチドが配列NDFRSKT及びそれに関連する他のアミノ酸配列である、請求項1に記載の組換えファージ。
【請求項3】
請求項1又は2に記載した融合ペプチドに由来するアミノ酸配列を含む合成ペプチド。
【請求項4】
ウイルスに結合することによってAIVの増殖を阻害する、請求項3に記載の合成ペプチド。
【請求項5】
前記ペプチドが配列CNDFRSKTC又はそれに関連する配列を有する、請求項3に記載の合成ペプチド。
【請求項6】
前記ペプチドが100μM未満のIC50値を有する、請求項3に記載の合成ペプチド。
【請求項7】
請求項1及び2のいずれか一項に記載の組換えファージを含む薬剤組成物。
【請求項8】
請求項3から6までのいずれか一項に記載の合成ペプチドを含む薬剤組成物。
【請求項9】
請求項1及び2のいずれか一項に記載の組換えファージを含む診断用試薬。
【請求項10】
請求項3から6までのいずれか一項に記載の合成ペプチドを含む診断用試薬。
【請求項11】
請求項1及び2のいずれか一項に記載の組換えファージを含むワクチン。
【請求項12】
請求項3から6までのいずれか一項に記載の合成ペプチドを含むワクチン。
【請求項13】
トリインフルエンザウイルスH9N2表面糖タンパク質HA及びNAに結合する、請求項3から6までのいずれか一項に記載の合成ペプチド。
【請求項14】
トリインフルエンザウイルスに感染している個体(ヒト/動物又はトリ)を治療するための方法であって、請求項1及び2のいずれか一項に記載の組換えファージの治療有効量を前記個体に投与することを含む上記方法。
【請求項15】
組換えファージの融合ペプチドが、NDFRSKT又はそれに由来するあらゆる他のアミノ酸配列からなる群から選択されるアミノ酸配列を含む、請求項14に記載の方法。
【請求項16】
トリインフルエンザウイルスに感染している個体(ヒト/動物又はトリ)を治療するための方法であって、請求項3から6までのいずれか一項に記載の合成ペプチドの治療有効量、及び薬学的に許容される担体を前記個体に投与することを含む上記方法。
【請求項17】
合成ペプチドが、アミノ酸配列NDFRSKT又はそれに由来するあらゆる他のアミノ酸配列を含む、請求項16に記載の方法。
【請求項1】
トリインフルエンザウイルスに結合し、ウイルスの増殖を阻害する融合ペプチドを保有する、単離され、精製された組換えファージ。
【請求項2】
融合ペプチドが配列NDFRSKT及びそれに関連する他のアミノ酸配列である、請求項1に記載の組換えファージ。
【請求項3】
請求項1又は2に記載した融合ペプチドに由来するアミノ酸配列を含む合成ペプチド。
【請求項4】
ウイルスに結合することによってAIVの増殖を阻害する、請求項3に記載の合成ペプチド。
【請求項5】
前記ペプチドが配列CNDFRSKTC又はそれに関連する配列を有する、請求項3に記載の合成ペプチド。
【請求項6】
前記ペプチドが100μM未満のIC50値を有する、請求項3に記載の合成ペプチド。
【請求項7】
請求項1及び2のいずれか一項に記載の組換えファージを含む薬剤組成物。
【請求項8】
請求項3から6までのいずれか一項に記載の合成ペプチドを含む薬剤組成物。
【請求項9】
請求項1及び2のいずれか一項に記載の組換えファージを含む診断用試薬。
【請求項10】
請求項3から6までのいずれか一項に記載の合成ペプチドを含む診断用試薬。
【請求項11】
請求項1及び2のいずれか一項に記載の組換えファージを含むワクチン。
【請求項12】
請求項3から6までのいずれか一項に記載の合成ペプチドを含むワクチン。
【請求項13】
トリインフルエンザウイルスH9N2表面糖タンパク質HA及びNAに結合する、請求項3から6までのいずれか一項に記載の合成ペプチド。
【請求項14】
トリインフルエンザウイルスに感染している個体(ヒト/動物又はトリ)を治療するための方法であって、請求項1及び2のいずれか一項に記載の組換えファージの治療有効量を前記個体に投与することを含む上記方法。
【請求項15】
組換えファージの融合ペプチドが、NDFRSKT又はそれに由来するあらゆる他のアミノ酸配列からなる群から選択されるアミノ酸配列を含む、請求項14に記載の方法。
【請求項16】
トリインフルエンザウイルスに感染している個体(ヒト/動物又はトリ)を治療するための方法であって、請求項3から6までのいずれか一項に記載の合成ペプチドの治療有効量、及び薬学的に許容される担体を前記個体に投与することを含む上記方法。
【請求項17】
合成ペプチドが、アミノ酸配列NDFRSKT又はそれに由来するあらゆる他のアミノ酸配列を含む、請求項16に記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】

【図2】
【図3】
【図4】
【図5】
【公表番号】特表2011−522561(P2011−522561A)
【公表日】平成23年8月4日(2011.8.4)
【国際特許分類】
【出願番号】特願2011−513436(P2011−513436)
【出願日】平成21年6月10日(2009.6.10)
【国際出願番号】PCT/MY2009/000071
【国際公開番号】WO2009/151313
【国際公開日】平成21年12月17日(2009.12.17)
【出願人】(505291413)ユニバーシティー プトラ マレーシア (11)
【Fターム(参考)】
【公表日】平成23年8月4日(2011.8.4)
【国際特許分類】
【出願日】平成21年6月10日(2009.6.10)
【国際出願番号】PCT/MY2009/000071
【国際公開番号】WO2009/151313
【国際公開日】平成21年12月17日(2009.12.17)
【出願人】(505291413)ユニバーシティー プトラ マレーシア (11)
【Fターム(参考)】
[ Back to top ]