
植物の油脂を増産させる遺伝子及びその利用方法

【課題】個体あたりの物質生産性を向上させることができる新規な機能を有する転写制御因子を探索し、植物体におけるこれらの特性を向上できる技術を提供する。
【解決手段】特定な配列からなるアミノ酸配列を含む転写因子を含む転写因子ファミリーに属する転写因子とリプレッサードメインとを融合させたキメラタンパク質を植物体内で発現させる。当該植物体から、生産性が向上した物質を分離及び回収する工程を含む、植物体を用いた物質の製造方法。
【解決手段】特定な配列からなるアミノ酸配列を含む転写因子を含む転写因子ファミリーに属する転写因子とリプレッサードメインとを融合させたキメラタンパク質を植物体内で発現させる。当該植物体から、生産性が向上した物質を分離及び回収する工程を含む、植物体を用いた物質の製造方法。
【発明の詳細な説明】
【背景技術】
【0001】
バイオマス(biomass)とは、一般的には一定面積あたりに生息または存在する生物の総量を指し、特に植物を対象とした場合は、単位面積あたりの乾重量を意味する。バイオマスの単位は、質量又はエネルギー量で数値化する。バイオマスという表現は、「生物体量」、「生物量」も同義語であり、植物バイオマスの場合には「現存量(Standing crop)」の語が使われることもある。植物バイオマスは、大気中の二酸化炭素を太陽エネルギーを用いて固定して生成されるため、いわゆるカーボンニュートラルなエネルギーとして捕らえることができる。したがって、植物のバイオマスを増加させることは、地球環境保全、地球温暖化防止、温室効果ガス排出低減の効果がある。従って、植物バイオマスを増産させる技術は産業上の重要性が高い。
【0002】
一方、植物は、その一部の組織自体(種子、根、葉茎など)を目的として栽培されたり、油脂などの種々の物質生産を目的として栽培されたりする。例えば、植物が生産する油脂としては、大豆油、ごま油、オリーブ油、椰子油、米油、綿実油、ひまわり油、コーン油、べに花油、パーム油及び菜種油等が古来より知られており、家庭用途や工業用途に広く利用されている。また、植物が生産する油脂は、バイオディーゼル燃料やバイオプラスチックの原料としても使用され、石油代替エネルギーとして適用性が広がっている。
【0003】
このような状況において、植物を用いた油脂生産を工業的に成功させるには、単位耕地面積あたりの生産性の向上が必要となる。ここで単位耕地面積あたりの栽培個体数が一定であると仮定すると、個体あたりの油脂生産量の向上が必要であることが判る。植物体から採取した種子から油脂を回収する場合には、個体あたりの種子収量を向上させる、及び種子中の油脂含有量を向上させるといった技術により、個体あたりの油脂生産量の向上が達成できるものと期待される。
【0004】
植物種子の油脂生産量を増加させる技術には大別して栽培法の改良によるものと、油脂増産品種の開発がある。油脂増産品種の開発方法は、交配技術を中心とした従来育種法と遺伝子組換えによる分子育種法とに大別される。遺伝子組換えによる油脂増産技術としては、A)植物油脂の主成分である種子トリアシルグリセロール(TAG)の合成系を改変する技術、及びB)植物の形態形成や代謝及びそれらに関わる遺伝子の発現を制御する各種制御遺伝子を改変する技術とが知られている。
【0005】
上記A)の方法としては、光合成により生産される糖を原料として合成されるTAGの合成量を増加させる方法として(1)TAGの構成成分である脂肪酸、あるいはグリセロールの糖からの合成活性を高める方法、(2)グリセロールと脂肪酸からTAGが合成される反応を強化する方法が考えられる。これらについて、遺伝子工学的な方法を用いた技術としては以下の技術が報告されている。(1)の例としては、シロイヌナズナの細胞質型acetyl-coenzyme A carboxylase (ACCase)をナタネのプラスチド中で過剰発現させることにより、種子の油脂含量を5%向上させた報告(非特許文献1)が挙げられる。また、(2)の例としては、ジアシルグリセロールのsn-3位にアシル基を転移するDGAT(diacylglycerol acyltransferase)の過剰発現による油脂増産技術に関する報告(非特許文献2)が挙げられる。非特許文献2の方法では、DGATの発現量が増加するに従って油脂含量と種子重が増加し、個体あたりの種子数が増加する場合があることも報告されている。本方法を適用したシロイヌナズナの種子油脂含量は46%増、個体あたりの油脂量は最高で約125%増であった。
【0006】
一方、上記B)の方法としては、生合成系酵素遺伝子の発現制御に関与する転写因子遺伝子の発現を制御する方法が考えられる。この例としては、特許文献1が挙げられる。本特許文献1では、転写因子を網羅的に過剰発現またはノックアウトした組換え植物を作製したのちに種子の油脂含量を高める遺伝子を選抜するといった手法が採用されている。特許文献1によれば、ERF subfamily B-4転写因子遺伝子の過剰発現によって種子の油脂含量が23%増加したと記載されている。しかし、特許文献1において、個体あたりの油脂含量の増減については記載されていない。また、非特許文献3には、AP2/EREBドメインを持つ転写因子であるWRINKLED1を過剰発現させることにより、種子の油脂含量が向上することが記載されている。
【0007】
しかしながら、種々の形質の改良を目的とした上述した分子育種法が開発されているにもかかわらず、植物の重量増産、特定の組織増産、或いは目的物質の生産性向上を伴う、収量向上技術は実用の域に達していない。
【0008】
この理由として、真に優れた遺伝子が未発見であること、試験段階で効果のある組換え新品種が実用段階では多様な自然環境下で期待通りの効果を発揮できないことにあると考えられる。また、植物の重量増産、特定の組織増産、或いは目的物質の生産性と言った量的形質は制御系から代謝系に亘る様々なステップで多数の遺伝子が関わっており、量的形質を改善する真に優れた有用遺伝子を発見、開発することは困難であった。これらの問題を解決するためには、効果が劇的に高い新たな遺伝子を見出すこと、効果レベルは同等であっても実用環境条件で効果を発揮する遺伝子を開発することが課題である。
【0009】
【非特許文献1】Plant Physiology (1997) Vol. 11, pp. 75-81
【非特許文献2】Plant Physiology (2001), Vol. 126, pp. 861-874
【非特許文献3】Plant J. (2004) 40, 575-585
【特許文献1】WO01/36597
【発明の開示】
【発明が解決しようとする課題】
【0010】
そこで、上述したような実情に鑑み、個体あたりの物質生産性、特に種子中の油脂量を向上させることができる新規な機能を有する遺伝子を探索し、植物体におけるこれらの特性を向上できる技術を提供することを目的とする。
【課題を解決するための手段】
【0011】
上述した目的を達成するため、本発明者らが鋭意検討した結果、特定の転写因子ファミリーに属する転写因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチド(以下、リプレッサードメインと称する場合もある)とを融合したキメラタンパク質を発現させることによって、様々な量的形質を改善させることができ、特に、個体あたりの物質生産性、特に油脂生産性を向上させることができることを見いだし、本発明を完成するに至った。
【0012】
本発明に係る植物体は、配列番号4に示すアミノ酸配列からなるタンパク質を含む転写因子ファミリーに属する転写因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質を発現させたものである。本発明に係る植物体においては、機能性ペプチドを融合することによって、所定の転写因子における転写制御活性、特に転写促進活性が抑制されていることが好ましい。
【0013】
ここで、上記機能性ペプチドを融合する転写因子としては、以下の(a)〜(c)のいずれかのタンパク質であることが好ましい。
(a)配列番号4に示すアミノ酸配列を含むタンパク質
(b)配列番号4に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、転写促進活性を有するタンパク質
(c)配列番号3に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ転写促進活性を有するタンパク質
【0014】
ここで上記機能性ペプチドとしては、次に示す式(1)〜(8)を挙げることができる。
(1)X1−Leu−Asp−Leu−X2−Leu−X3
(但し、式中、X1は0〜10個のアミノ酸残基を示し、X2はAsn又はGluを示し、X3は少なくとも6個のアミノ酸残基を示す。)
(2)Y1−Phe−Asp−Leu−Asn−Y2−Y3
(但し、式中、Y1は0〜10個のアミノ酸残基を示し、Y2はPhe又はIleを示し、Y3は少なくとも6個のアミノ酸残基を示す。)
(3)Z1−Asp−Leu−Z2−Leu−Arg−Leu−Z3
(但し、式中、Z1はLeu、Asp−Leu又はLeu−Asp−Leuを示し、Z2はGlu、Gln又はAspを示し、Z3は0〜10個のアミノ酸残基を示す。)
(4)Asp−Leu−Z4−Leu−Arg−Leu
(但し、式中、Z4はGlu、Gln又はAspを示す。)
(5)α1−Leu−β1−Leu−γ1−Leu
(6)α1−Leu−β1−Leu−γ2−Leu
(7)α1−Leu−β2−Leu−Arg−Leu
(8)α2−Leu−β1−Leu−Arg−Leu
(但し、式(5)〜(8)中、α1はAsp、Asn、Glu、Gln、Thr又はSerを示し、α2はAsn、Glu、Gln、Thr又はSerを示し、β1はAsp、Gln、Asn、Arg、Glu、Thr、Ser又はHisを示し、β2はAsn、Arg、Thr、Ser又はHisを示し、γ1はArg、Gln、Asn、Thr、Ser、His、Lys又はAspを示し、γ2はGln、Asn、Thr、Ser、His、Lys又はAspを示す。)
【0015】
また、本発明に係る植物体を用いた物質の製造方法は、上述した本発明に係る植物体から、生産性が向上した物質を分離及び回収する工程を含むものである。ここで、上記物質としては油脂を挙げることができる。
【0016】
一方、上述した目的を達成するため、本発明者らが鋭意検討した結果、色素合成経路に関与する遺伝子を欠損させた株において、個体あたりの物質生産性、特に油脂生産性を向上させることができることを見いだし、本発明を完成するに至った。色素合成系路に関与する遺伝子とは、色素合成経路代謝反応の基質や生成物の輸送に関する因子をコードする遺伝子や、色素合成系路代謝反応を触媒する酵素をコードする遺伝子や、色素合成系路代謝反応の場を形成させる反応を触媒する酵素をコードする遺伝子を含む。さらに、色素合成経路代謝反応の基質や生成物の輸送に関する因子をコードする遺伝子や、色素合成系路代謝反応を触媒する酵素をコードする遺伝子や、色素合成系路代謝反応の場を形成させる反応を触媒する酵素をコードする遺伝子の発現を制御する遺伝子を含む。
【0017】
すなわち、本発明に係る植物由来油脂の製造方法は、カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子及びフラボン-3-ヒドラーゼ遺伝子からなる群から選ばれる少なくとも1の遺伝子の機能を欠損した植物体から採取した種子から油脂成分を回収する工程を含むものである。
【0018】
また、本発明に係る油脂量が向上した植物体のスクリーニング方法は、種子内の油脂量を評価する対象となる植物体から種子を採取する工程と、採取した種子の種皮色を観察し、より白色である場合には種子内の油脂量が高いと判定する工程とを含むものである。
【発明の効果】
【0019】
本発明に係る植物体は、個体あたりの物質生産性が向上したものとなる。したがって、本発明に係る植物体を用いることによって、目的物質の生産性の向上を達成することができ、目的物質を低コストで製造することができる。
【0020】
また、本発明に係る植物由来油脂の製造方法は、特定の遺伝子の機能を欠損する植物体において種子の単位量あたりに含まれる油脂量が大幅に向上しているため、油脂の生産性を向上することができる。
【0021】
さらに、本発明に係る油脂量が向上した植物体のスクリーニング方法は、種子内の油脂量を非破壊で評価するため、小量の種子を用いて迅速、且つ簡便にスクリーニングすることができる。
【発明を実施するための最良の形態】
【0022】
以下、本発明を詳細に説明する。
【0023】
本発明に係る植物体は、所定の転写制御因子ファミリーに属する転写制御因子、特に、所定の転写因子ファミリーに属する転写因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質を発現するものであり、野生型の植物体と比較して、個体あたりの物質生産性が向上したものである。すなわち、本発明に係る植物体は、所望の植物を対象として、当該植物における物質生産性を有意に向上させるように、転写因子を上記機能性ペプチドとのキメラタンパク質として発現させた植物体である。
【0024】
特に、本発明に係る植物体においては、上記機能性ペプチドと融合することによって、転写因子における転写促進活性が抑制していることが好ましい。換言すると、本発明に係る植物体においては、転写因子に上記機能性ペプチドを融合させたキメラタンパク質を発現させた結果、上記機能性ペプチドに起因する転写抑制効果が優性の形質として現れるといった特徴を有していることが好ましい。
【0025】
ここで、個体あたり物質生産性とは、植物が生成する各種の物質についての単位体積あたりの含有量を意味する。物質としては、特に限定されず、植物体が本来的に生成する物質であっても良いし、植物体が本来的には生成しないが遺伝子操作技術等によって生成できるようになった物質であっても良い。特に、組織あたりの目的生産物の含量が高くなれば、精製コストや運搬コストを低減できるため産業上有用性が高い。特に、目的生産物としては、植物のほとんどの重量を占めるリグノセルロースでも良く、種子油として産業上利用されている植物油でも良い。植物油は、脂肪酸とアルコールのエステルである単純脂質でも良く、リンや糖や窒素などを含んだ複合脂質でも良く、脂肪酸そのものでも良い。単純脂質のアルコールとしては分子量の高い高級アルコールでも良く、グリセロール(グリセリン)などの多価アルコールでも良い。単純脂質の脂肪酸としては、飽和脂肪酸でも良く、不飽和脂肪酸でも良く、また、水酸基やエポキシ基を含んだ特殊脂肪酸でも良い。グリセロールと脂肪酸のエステルである単純脂質としては、モノアシルグリセロールでも良く、ジアシルグリセロールでも良く、トリアシルグリセロールでも良い。
【0026】
以下の説明において、生産性を向上させる物質として油脂を例示して説明するが、本発明の技術的範囲がこれに限定されるものではない。本発明は、植物が生成する物質として油脂以外の物質についても同様に適用される。
【0027】
ここで、植物体としては、特に限定されず、如何なる植物をも対象とすることができる。特に、従来より油脂の生産に使用される植物を対象とすることが好ましい。対象とする植物としては、例えば、大豆、ごま、オリーブ油、椰子、イネ、綿花、ひまわり、トウモロコシ、サトウキビ、ジャトロファ、パームヤシ、タバコ、べに花及びナタネ等を挙げることができる。また、植物の遺伝子解析におけるモデル生物として広く利用されており、遺伝子発現解析の方法が確立しているシロイヌナズナを対象の植物とすることもできる。
【0028】
また、転写因子のキメラタンパク質が活性として有する転写抑制とは、当該転写因子が認識するcis配列や、そのcis配列と類似する他の転写因子におけるcis配列を認識し、下流の遺伝子発現を積極的に抑制する活性であり、転写抑制因子とも呼べるものである。転写因子のキメラタンパク質が活性として有する転写抑制する手法は、特に限定されないが、特に、リプレッサードメイン配列やSRDX配列を付加したキメラタンパク質(融合タンパク質)を構築する方法が最も好ましい。
【0029】
この手法においてリプレッサードメイン配列とは、任意の転写因子を転写抑制因子に転換するペプチドを構成するアミノ酸配列であり本発明者らによって種々見出された配列である。リプレッサードメイン配列を使用した方法については、例えば、特開2001−269177公報、特開2001−269178公報、特開2001−292776公報、特開2001−292777公報、特開2001−269176公報、特開2001−269179公報、国際公開第WO03/055903号パンフレット、Ohta, M., Matsui, K., Hiratsu, K., Shinshi, H. and Ohme-Takagi, M., The Plant Cell, Vol.13,1959-1968,August,2001及びHiratsu, K., Ohta, M., Matsui, K., Ohme-Takagi, M., FEBS Letters 514(2002)351-354を参照することができる。リプレッサードメイン配列は、Class II ERF(Ethylene Responsive Element Binding Factor)タンパク質や植物のジンクフィンガータンパク質(Zinc Finger Protein、例えばシロイヌナズナSUPERMANタンパク質等)から切り出されたもので、極めて単純な構造を有している。
【0030】
キメラタンパク質として発現する転写制御因子としては、シロイヌナズナにおけるAGIコードAt1g71030で特定される転写因子(以下、単に『転写因子At1g71030』と称す)を挙げることができる。転写因子At1g71030は、mybファミリーの転写因子であり、オオムギ由来のMybHv5 GI:19055と類似していることが知られている。転写因子At1g71030のアミノ酸配列を配列番号4に示す。転写因子At1g71030をコードする遺伝子の塩基配列を配列番号3に示す。
【0031】
また、At5g24520で特定される転写共役因子(transcription coactivator)(以下、単に『転写共役因子At5g24520』と称す)、転写抑制因子(transcription repressor)や転写抑制共役因子(transcripition corepressor)が転写制御因子として知られており、これら転写共役因子や転写抑制因子についても同様にリプレッサードメインを付加したキメラタンパク質を構築することができる。なお、AGIコードAt5g24520は、transparent testa glabra 1 protein (TTG1) として知られる転写共役因子が因子である。他の植物由来の遺伝子では、Malus domestica由来のGenBankアクセッション番号AAF27919がコードするタンパク質、Petunia hybrida由来のGenBankアクセッション番号AAC18914がコードするタンパク質、Gossypium hirsutum由来のGenBankアクセッション番号AAM95645がコードするタンパク質、Perilla frutescens由来のGenBankアクセッション番号BAB58883がコードするタンパク質が転写共役因子At5g24520と相同であることが知られ、本明細書で記載した機能と同等の機能が期待できる。転写共役因子At5g24520のアミノ酸配列を配列番号2に示す。転写共役因子At5g24520をコードする遺伝子の塩基配列を配列番号1に示す。
【0032】
また、キメラタンパク質の対象となる転写共役因子At5g24520及び転写因子At1g71030は、それぞれ配列番号2及び4に示すアミノ酸配列からなるものに限定されず、当該アミノ酸配列において1又は複数個のアミノ酸配列が欠失、置換、付加又は挿入されたアミノ酸配列を含み、且つ、転写促進活性を有するものであっても良い。ここで、複数個のアミノ酸としては、例えば、1から20個、好ましくは1から10個、より好ましくは1から7個、さらに好ましくは1個から5個、特に好ましくは1個から3個を意味する。なお、アミノ酸の欠失、置換若しくは付加は、上記転写因子をコードする塩基配列を、当該技術分野で公知の手法によって改変することによって行うことができる。塩基配列に変異を導入するには、Kunkel法またはGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-KやMutant-G(何れも商品名、TAKARA Bio社製))等を用いて、あるいはLA PCR in vitro Mutagenesisシリーズキット(商品名、TAKARA Bio社製)を用いて変異が導入される。また、変異導入方法としては、EMS(エチルメタンスルホン酸)、5-ブロモウラシル、2-アミノプリン、ヒドロキシルアミン、N-メチル-N’-ニトロ-Nニトロソグアニジン、その他の発ガン性化合物に代表されるような化学的変異剤を使用する方法でも良いし、X線、アルファ線、ベータ線、ガンマ線、イオンビームに代表されるような放射線処理や紫外線処理による方法でも良い。
【0033】
さらに、キメラタンパク質の対象となる転写共役因子及び転写因子には、シロイヌナズナにおける転写共役因子At5g24520及び転写因子At1g71030に限定されず、シロイヌナズナ以外の植物(例えば上述した植物)において同機能を有する転写共役因子及び転写因子(以下、それぞれ相同転写共役因子及び相同転写因子と称す)が含まれる。転写共役因子At5g24520に対する相同転写共役因子又は転写因子At1g71030に対する相同転写因子は、植物ゲノム情報が明らかになっていれば、転写共役因子At5g24520又は転写因子At1g71030のアミノ酸配列或いは各遺伝子の塩基配列に基づいて、検索対象の植物ゲノム情報から検索することができる。このとき、相同転写共役因子及び相同転写因子としては、転写共役因子At5g24520又は転写因子At1g71030のアミノ酸配列に対して、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、最も好ましくは95%以上の相同性を有するアミノ酸配列として検索される。ここで、相同性の値は、blastアルゴリズムを実装したコンピュータプログラム及び遺伝子配列情報を格納したデータベースを用いてデフォルトの設定で求められる値を意味する。
【0034】
また、植物ゲノム情報が明らかとなっていない場合には、対象となる植物からゲノムを抽出するか或いは対象となる植物のcDNAライブラリーを構築し、転写共役因子At5g24520又は転写因子At1g71030の塩基配列の少なくとも一部に対して、ストリンジェントな条件下でハイブリダイズするゲノム領域或いはcDNAを単離することで相同遺伝子を同定することができる。ここで、ストリンジェントな条件とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。例えば、45℃、6×SSC(塩化ナトリウム/クエン酸ナトリウム)でのハイブリダイゼーション、その後の50〜65℃、0.2〜1×SSC、0.1%SDSでの洗浄が挙げられ、或いはそのような条件として、65〜70℃、1×SSCでのハイブリダイゼーション、その後の65〜70℃、0.3×SSCでの洗浄を挙げることができる。ハイブリダイゼーションは、J. Sambrook et al. Molecular Cloning, A Laboratory Manual,2nd Ed., Cold Spring Harbor Laboratory(1989)に記載されている方法等、従来公知の方法で行うことができる。
【0035】
本発明に係る植物体は、上述したような転写因子と機能性ペプチドとのキメラタンパク質を発現させることで、油脂生産量が有意に向上するといった特徴を示す。また、上述した転写共役因子と機能性ペプチドとのキメラタンパク質を発現させた場合も、油脂生産量が有意に向上するといった特徴を示す。特に、キメラタンパク質とすることで、対象となる転写因子を、転写促進活性を抑制した状態で発現させること、さらに、対象となる転写因子が認識するcis配列と相同性のあるcis配列を認識する転写抑制活性として発現させること、対象となる転写因子及び転写共役因子が持つ他因子や核酸や脂質や糖質との親和特異性を変化させることで、油脂生産量が有意に向上するといった特徴を示す。このとき、上記植物体においては、内因性の転写因子や転写共役因子を改変してそのキメラタンパク質を作製してもよいが、キメラタンパク質をコードする遺伝子を導入し、当該遺伝子を発現させても良い。
【0036】
一例としては、上述したような転写因子や転写共役因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質(融合タンパク質)をコードする遺伝子を対象の植物に導入し、当該キメラタンパク質(融合タンパク質)を植物内で発現させる手法が好ましい。
【0037】
本明細書中で記載する「転写促進活性が抑制された転写因子」とは、特に限定されるものではなく、当該転写因子が本来的に有している転写促進活性が有意に低減した転写因子であることを意味する。また、「任意の転写因子を転写抑制因子に転換する機能性ペプチド」とは、任意の転写因子と融合してキメラタンパク質となったときに、当該転写因子が本来的に有している転写促進活性が有意に低減した転写因子となる機能を有するペプチドであることを意味する(転写抑制転換ペプチドと称する場合もある)。このような「任意の転写因子を転写抑制因子に転換する機能性ペプチド」としては、特に限定されないが、なかでもリプレッサードメイン配列やSRDX配列として知られたアミノ酸配列からなるペプチドであることが好ましい。この転写抑制転換ペプチドについては、特開2005−204657号公報に詳述されており、当該公報に開示されたものを全て使用することができる。
【0038】
転写抑制転換ペプチドは、例えば次に示す式(1)〜(8)のいずれかで表されるアミノ酸配列を挙げることができる。
(1)X1−Leu−Asp−Leu−X2−Leu−X3
(但し、式中、X1は0〜10個のアミノ酸残基を示し、X2はAsn又はGluを示し、X3は少なくとも6個のアミノ酸残基を示す。)
(2)Y1−Phe−Asp−Leu−Asn−Y2−Y3
(但し、式中、Y1は0〜10個のアミノ酸残基を示し、Y2はPhe又はIleを示し、Y3は少なくとも6個のアミノ酸残基を示す。)
(3)Z1−Asp−Leu−Z2−Leu−Arg−Leu−Z3
(但し、式中、Z1はLeu、Asp−Leu又はLeu−Asp−Leuを示し、Z2はGlu、Gln又はAspを示し、Z3は0〜10個のアミノ酸残基を示す。)
(4)Asp−Leu−Z4−Leu−Arg−Leu
(但し、式中、Z4はGlu、Gln又はAspを示す。)
(5)α1−Leu−β1−Leu−γ1−Leu
(6)α1−Leu−β1−Leu−γ2−Leu
(7)α1−Leu−β2−Leu−Arg−Leu
(8)α2−Leu−β1−Leu−Arg−Leu
(但し、式(5)〜(8)中、α1はAsp、Asn、Glu、Gln、Thr又はSerを示し、α2はAsn、Glu、Gln、Thr又はSerを示し、β1はAsp、Gln、Asn、Arg、Glu、Thr、Ser又はHisを示し、β2はAsn、Arg、Thr、Ser又はHisを示し、γ1はArg、Gln、Asn、Thr、Ser、His、Lys又はAspを示し、γ2はGln、Asn、Thr、Ser、His、Lys又はAspを示す。)
【0039】
式(1)の転写抑制転換ペプチド
上記式(1)の転写抑制転換ペプチドにおいては、上記X1で表されるアミノ酸残基の数は0〜10個の範囲内であればよい。また、X1で表されるアミノ酸残基を構成する具体的なアミノ酸の種類は特に限定されるものではなく、どのようなものであってもよい。このX1で表されるアミノ酸残基は、式(1)の転写抑制転換ペプチドを合成するときの容易さからみれば、できるだけ短いほうがよい。具体的にX1で表されるアミノ酸残基は、5個以下であることが好ましい。
【0040】
同様に、上記式(1)の転写抑制転換ペプチドにおいては、上記X3で表されるアミノ酸残基の数は少なくとも6個であればよい。また、X3で表されるアミノ酸残基を構成する具体的なアミノ酸の種類は特に限定されるものではなく、どのようなものであってもよい。
【0041】
式(2)の転写抑制転換ペプチド
上記式(2)の転写抑制転換ペプチドにおいては、上記式(1)の転写抑制転換ペプチドのX1と同様、上記Y1で表されるアミノ酸残基の数は0〜10個の範囲内であればよい。また、Y1で表されるアミノ酸残基を構成する具体的なアミノ酸の種類は特に限定されるものではなく、どのようなものであってもよい。具体的にY1で表されるアミノ酸残基は、5個以下であることが好ましい。
【0042】
同様に、上記式(2)の転写抑制転換ペプチドにおいては、上記式(1)の転写抑制転換ペプチドのX3と同様、上記Y3で表されるアミノ酸残基の数は少なくとも6個であればよい。また、Y3で表されるアミノ酸残基を構成する具体的なアミノ酸の種類は特に限定されるものではく、どのようなものであってもよい。
【0043】
式(3)の転写抑制転換ペプチド
上記式(3)の転写抑制転換ペプチドにおいては、上記Z1で表されるアミノ酸残基は、1〜3個の範囲内でLeuを含むものとなっている。アミノ酸1個の場合は、Leuであり、アミノ酸2個の場合は、Asp−Leuとなっており、アミノ酸3個の場合はLeu−Asp−Leuとなっている。
【0044】
一方、上記式(3)の転写抑制転換ペプチドにおいては、上記Z3で表されるアミノ酸残基の数は0〜10個の範囲内であればよい。また、Z3で表されるアミノ酸残基を構成する具体的なアミノ酸の種類は特に限定されるものではなく、どのようなものであってもよい。具体的にZ3で表されるアミノ酸残基は、5個以下であることがより好ましい。Z3で表されるアミノ酸残基の具体的な例としては、Gly、Gly−Phe−Phe、Gly−Phe−Ala、Gly−Tyr−Tyr、Ala−Ala−Ala等が挙げられるが、もちろんこれらに限定されるものではない。
【0045】
また、この式(3)で表される転写抑制転換ペプチド全体のアミノ酸残基の数は、特に限定されるものではないが、合成するときの容易さからみれば、20アミノ酸以下であることが好ましい。
【0046】
式(4)の転写抑制転換ペプチド
上記式(4)の転写抑制転換ペプチドは、6個のアミノ酸残基からなるヘキサマー(6mer)である。なお、上記式(4)の転写抑制転換ペプチドにおいてZ4で表されるアミノ酸残基がGluの場合のアミノ酸配列は、シロイヌナズナSUPERMANタンパク質(SUPタンパク質)の196〜201番目のアミノ酸配列に相当している。
【0047】
以上で説明した各種転写抑制転換ペプチドは、上述した転写因子や転写共役因子と融合してキメラタンパク質(融合タンパク質)とすることにより、当該転写因子や転写共役因子の特性を改変することができる。具体的には、上述した転写因子や転写共役因子と融合してキメラタンパク質(融合タンパク質)とすることにより、転写因子や転写共役因子を転写抑制因子や負の転写共役因子に改変することができる。さらには、ドミナントでない転写抑制因子をドミナント型転写抑制因子にすることも可能である。
【0048】
また、上記転写抑制転換ペプチドをコードするポリヌクレオチドを用いて、転写因子や転写共役因子をコードする遺伝子との融合遺伝子を得れば、キメラタンパク質(融合タンパク質)を生産させることができる。具体的には、上記転写抑制転換ペプチドをコードするポリヌクレオチド(転写抑制転換ポリヌクレオチドと称す)と上記転写因子や転写共役因子をコードする遺伝子とを連結することにより融合遺伝子を構築して、植物細胞に導入する。これによりキメラタンパク質(融合タンパク質)を生産させることができる。上記転写抑制転換ポリヌクレオチドの具体的な塩基配列は特に限定されるものではなく、遺伝暗号に基づいて、上記転写抑制転換ペプチドのアミノ酸配列に対応する塩基配列を含んでいればよい。また、必要に応じて、上記転写抑制転換ポリヌクレオチドは、転写因子遺伝子と連結するための連結部位となる塩基配列を含んでいてもよい。さらに、上記転写抑制転換ポリヌクレオチドのアミノ酸読み枠と、転写因子や転写共役因子の遺伝子の読み枠とが一致しないような場合に、これらを一致させるための付加的な塩基配列を含んでいてもよい。さらにまた、転写因子や転写共役因子と転写抑制転換ペプチドとの間をつなぐためのリンカー機能を有するポリペプチドや、HisやMyc、Flag等のようにキメラタンパク質(融合タンパク質)をエピトープ標識するためのポリペプチド等、各種の付加的なポリペプチドが含まれていてもよい。さらに上記キメラタンパク質(融合タンパク質)には、必要に応じて、ポリペプチド以外の構造、例えば、糖鎖やイソプレノイド基等が含まれていてもよい。
【0049】
植物体を製造する方法は、上述した転写因子や転写共役因子と転写抑制転換ペプチドとのキメラタンパク質を植物体で生産させ、油脂生産性を向上させる過程を含んでいれば特に限定されるものではないが、例えば、発現ベクター構築工程、形質転換工程、選抜工程等の工程を含む製造法方法として挙げることができる。以下、各工程について具体的に説明する。
【0050】
発現ベクター構築工程
発現ベクター構築工程は、上述した転写因子や転写共役因子をコードする遺伝子と転写抑制転換ポリヌクレオチドと、プロモーターとを含む組換え発現ベクターを構築する工程であれば特に限定されるものではない。組換え発現ベクターの母体となるベクターとしては、従来公知の種々のベクターを用いることができる。例えば、プラスミド、ファージ、またはコスミド等を用いることができ、導入される植物細胞や導入方法に応じて適宜選択することができる。具体的には、例えば、pBR322、pBR325、pUC19、pUC119、pBluescript、pBluescriptSK、pBI系のベクター等を挙げることができる。特に、植物体へのベクターの導入法がアグロバクテリウムを用いる方法である場合には、pBI系のバイナリーベクターを用いることが好ましい。pBI系のバイナリーベクターとしては、具体的には、例えば、pBIG、pBIN19、pBI101、pBI121、pBI221等を挙げることができる。
【0051】
プロモーターは、植物体内で遺伝子を発現させることが可能なプロモーターであれば特に限定されるものではなく、公知のプロモーターを好適に用いることができる。かかるプロモーターとしては、例えば、カリフラワーモザイクウイルス35Sプロモーター(CaMV35S)、各種アクチン遺伝子プロモーター、各種ユビキチン遺伝子プロモーター、ノパリン合成酵素遺伝子のプロモーター、タバコのPR1a遺伝子プロモーター、トマトのリブロース1,5−二リン酸カルボキシラーゼ・オキシダーゼ小サブユニット遺伝子プロモーター、ナピン遺伝子プロモーター、オレオシン遺伝子プロモーター等を挙げることができる。この中でも、カリフラワーモザイクウイルス35Sプロモーター、アクチン遺伝子プロモーター又はユビキチン遺伝子プロモーターをより好ましく用いることができる。上記各プロモーターを用いれば、植物細胞内に導入されたときに任意の遺伝子を強く発現させることが可能となる。プロモーターは、転写因子や転写共役因子をコードする遺伝子と転写抑制転換ポリヌクレオチドとを連結した融合遺伝子を発現しうるように連結され、ベクター内に導入されていればよく、組換え発現ベクターとしての具体的な構造は特に限定されるものではない。
【0052】
なお、組換え発現ベクターは、プロモーター及び上記融合遺伝子に加えて、さらに他のDNAセグメントを含んでいてもよい。当該他のDNAセグメントは特に限定されるものではないが、ターミネーター、選別マーカー、エンハンサー、翻訳効率を高めるための塩基配列等を挙げることができる。また、上記組換え発現ベクターは、さらにT−DNA領域を有していてもよい。T−DNA領域は特にアグロバクテリウムを用いて上記組換え発現ベクターを植物体に導入する場合に遺伝子導入の効率を高めることができる。
【0053】
転写ターミネーターは転写終結部位としての機能を有していれば特に限定されるものではなく、公知のものであってもよい。例えば、具体的には、ノパリン合成酵素遺伝子の転写終結領域(Nosターミネーター)、カリフラワーモザイクウイルス35Sの転写終結領域(CaMV35Sターミネーター)等を好ましく用いることができる。この中でもNosターミネーターをより好ましく用いることできる。上記組換えベクターにおいては、転写ターミネーターを適当な位置に配置することにより、植物細胞に導入された後に、不必要に長い転写物を合成したり、強力なプロモーターがプラスミドのコピー数の減少させたりするような現象の発生を防止することができる。
【0054】
形質転換体選別マーカーとしては、例えば薬剤耐性遺伝子を用いることができる。かかる薬剤耐性遺伝子の具体的な一例としては、例えば、ハイグロマイシン、ブレオマイシン、カナマイシン、ゲンタマイシン、クロラムフェニコール等に対する薬剤耐性遺伝子を挙げることができる。これにより、上記抗生物質を含む培地中で生育する植物体を選択することによって、形質転換された植物体を容易に選別することができる。
【0055】
翻訳効率を高めるための塩基配列としては、例えばタバコモザイクウイルス由来のomega配列を挙げることができる。このomega配列をプロモーターの非翻訳領域(5’UTR)に配置させることによって、上記融合遺伝子の翻訳効率を高めることができる。このように、上記組換え発現ベクターには、その目的に応じて、さまざまなDNAセグメントを含ませることができる。
【0056】
組換え発現ベクターの構築方法についても特に限定されるものではなく、適宜選択された母体となるベクターに、上記プロモーター、転写因子や転写共役因子をコードする遺伝子、および転写抑制転換ポリヌクレオチド、並びに必要に応じて上記他のDNAセグメントを所定の順序となるように導入すればよい。例えば、転写因子をコードする遺伝子と転写抑制転換ポリヌクレオチドとを連結して融合遺伝子を構築し、次に、この融合遺伝子とプロモーターと(必要に応じて転写ターミネーター等)とを連結して発現カセットを構築し、これをベクターに導入すればよい。
【0057】
キメラ遺伝子(融合遺伝子)の構築および発現カセットの構築では、例えば、各DNAセグメントの切断部位を互いに相補的な突出末端としておき、ライゲーション酵素で反応させることで、当該DNAセグメントの順序を規定することが可能となる。なお、発現カセットにターミネーターが含まれる場合には、上流から、プロモーター、上記キメラ遺伝子、ターミネーターの順となっていればよい。また、組換え発現ベクターを構築するための試薬類、すなわち制限酵素やライゲーション酵素等の種類についても特に限定されるものではなく、市販のものを適宜選択して用いればよい。
【0058】
また、上記組換え発現ベクターの増殖方法(生産方法)も特に限定されるものではなく、従来公知の方法を用いることができる。一般的には大腸菌をホストとして当該大腸菌内で増殖させればよい。このとき、ベクターの種類に応じて、好ましい大腸菌の種類を選択してもよい。
【0059】
形質転換工程
本発明において行われる形質転換工程は、上述した融合遺伝子を発現させるように、上述した組換え発現ベクターを用いて植物細胞に導入する工程である。組換え発現ベクターを用いて植物細胞に導入する方法(形質転換方法)は特に限定されるものではなく、植物細胞に応じた適切な従来公知の方法を用いることができる。具体的には、例えば、アグロバクテリウムを用いる方法や直接植物細胞に導入する方法を用いることができる。アグロバクテリウムを用いる方法としては、例えば、Bechtold, E., Ellis, J. and Pelletier, G. (1993) In Planta Agrobacterium-mediated gene transfer by infiltration of adult Arabidopsis plants. C.R. Acad. Sci. Paris Sci. Vie, 316, 1194-1199. あるいは、Zyprian E, Kado Cl, Agrobacterium-mediated plant transformation by novel mini-T vectors in conjunction with a high-copy vir region helper plasmid. Plant Molecular Biology, 1990, 15(2), 245-256.に記載された方法を用いることができる。
【0060】
組換え発現ベクターと対象となる遺伝子を含んだDNAを直接植物細胞に導入する方法としては、例えば、マイクロインジェクション法、エレクトロポレーション法(電気穿孔法)、ポリエチレングリコール法、パーティクルガン法、プロトプラスト融合法、リン酸カルシウム法等を用いることができる。
【0061】
また、DNAを直接植物細胞に導入する方法を採るなら、対象とする遺伝子の発現に必要な転写ユニット、例えプロモーターや転写ターミネーターと、対象とする遺伝子を含んだDNAあれば十分であり、ベクター機能が必須ではない。さらに、転写ユニットを有さない対象とする遺伝子のタンパク質コード領域のみを含むDNAであっても、宿主の転写ユニット無いにインテグレートし、対象となる遺伝子を発現することができればよい。
【0062】
上記組換え発現ベクターと対象となる遺伝子を含んだDNAや、発現ベクターを含まず対象となる遺伝子DNAを含んだDNAが導入される植物細胞としては、例えば、花、葉、根等の植物器官における各組織の細胞、カルス、懸濁培養細胞等を挙げることができる。ここで、本発明にかかる植物体の生産方法においては、上記組換え発現ベクターは、生産しようとする種類の植物体に合わせて適切なものを適宜構築してもよいが、汎用的な組換え発現ベクターを予め構築しておき、それを植物細胞に導入してもよい。すなわち、本発明に係る植物体の製造方法においては、上述した組換え発現ベクターを用いた形質転換用DNAの構築工程が含まれていてもよいし、含まれていなくてもよい。
【0063】
その他の工程、その他の方法
本発明に係る植物体の生産方法においては、上記形質転換工程が含まれていればよく、さらに上記組換え発現ベクターを用いた形質転換用DNAの構築工程が含まれていてもよいが、さらに他の工程が含まれていてもよい。具体的には、形質転換後の植物体から適切な形質転換体を選抜する選抜工程等を挙げることができる。
【0064】
選抜の方法は特に限定されるものではなく、例えば、ハイグロマイシン耐性等の薬剤耐性を基準として選抜してもよいし、形質転換体を育成した後に、植物体そのもの、または任意の器官や組織に含まれる油脂含有量から選抜してもよい。例えば、油脂含有量から選抜する例としては、形質転換体の種子から定法に従って油脂成分を定量し、形質転換していない植物体の種子に含まれる油脂含有量と比較する方法を挙げることができる(後述の実施例参照)。
【0065】
本発明に係る植物体の製造方法では、上記融合遺伝子を植物体に導入するため、該植物体から、有性生殖または無性生殖により油脂含有量が有意に向上した子孫を得ることが可能となる。また、該植物体やその子孫から植物細胞や、種子、果実、株、カルス、塊茎、切穂、塊等の繁殖材料を得て、これらを基に該植物体を量産することも可能となる。したがって、本発明に係る植物体の製造方法では、選抜後の植物体を繁殖させる繁殖工程(量産工程)が含まれていてもよい。
【0066】
なお、本発明における植物体とは、成育した植物個体、植物細胞、植物組織、カルス、種子の少なくとも何れかが含まれる。つまり、本発明では、最終的に植物個体まで成育させることができる状態のものであれば、全て植物体とみなす。また、上記植物細胞には、種々の形態の植物細胞が含まれる。かかる植物細胞としては、例えば、懸濁培養細胞、プロトプラスト、葉の切片等が含まれる。これらの植物細胞を増殖・分化させることにより植物体を得ることができる。なお、植物細胞からの植物体の再生は、植物細胞の種類に応じて、従来公知の方法を用いて行うことができる。したがって、本発明に係る植物体の製造方法では、植物細胞等から植物体を再生させる再生工程が含まれていてもよい。
【0067】
また、本発明に係る植物体の生産方法は、組換え発現ベクターで形質転換する方法に限定されるものではなく、他の方法を用いてもよい。具体的には、例えば、上記キメラタンパク質(融合タンパク質)そのものを植物体に投与してもよい。この場合、最終的に利用する植物体の部位において油脂含有量を向上できるように、若年期の植物体にキメラタンパク質(融合タンパク質)を投与すればよい。またキメラタンパク質(融合タンパク質)の投与方法も特に限定されるものではなく、公知の各種方法を用いればよい。
【0068】
以上説明したように、本発明によれば、所定の転写因子ファミリーに属する転写因子と上記機能性ペプチドとのキメラタンパク質を発現させることで、野生型の植物体と比較して、個体あたりの物質生産性が向上した植物体を提供することができる。また、所定の転写共役因子と上記機能性ペプチドとキメラタンパク質を発現させることで、野生型の植物体と比較して、個体あたりの物質生産性が向上した植物体を提供することができる。植物体に上記キメラタンパク質を発現させると、対象となる転写因子の転写促進活性が抑制される場合もあり、或いは対象となる転写因子が認識するcis配列の相同配列に対する転写抑制効果を示す場合もある。さらに、キメラタンパク質は、対象となる転写因子や転写共役因子に対して親和性を有する他の因子、DNA、RNA、脂質又は糖質に対して当該親和特異性を変化させるように作用する場合もあり、或いは対象となる転写因子に対して親和性の無い物質に対して親和性を向上させるように作用する場合もある。本発明に係る植物体においては、キメラタンパク質の対象となる転写因子、当該が認識するcis配列と相同性のあるcis配列を認識する転写因子、キメラタンパク質の対象となる転写因子と相同性のある転写因子、キメラタンパク質の対象となる転写因子に対して親和性を有する他の因子等も同様に植物体に発現しているものの、上述したキメラタンパク質の作用効果により、ドミナントネガティブに制御対象の遺伝子発現を抑制することができる。これにより、本発明に係る植物体においては、油脂生産に関連する遺伝子群及び/又は生産された油脂の分解に関連する遺伝子群の発現レベルが変化し、その結果、油脂含有量が有意に向上すると考えられる。
【0069】
ここで油脂含有量が有意に向上するとは、野生型と比較して一粒あたりの種子質量に変化はないが油脂量が向上した場合と、野生型と比較して一粒あたりの種子質量が有意に大となり油脂量が向上した場合、野性型と比較して種子中の油脂含量が向上した場合のいずれかを意味する。いずれの場合であっても、植物一個体が生産する油脂量が向上したこととなる。本発明に係る植物体は、植物由来の油性の製造方法に利用することができる。例えば、本発明に係る植物体を成長させて種子を採取し、採取した種子から油脂成分を回収することで油脂を製造することができる。
【0070】
特に本発明に係る植物体を利用した油脂の製造方法は、植物一個体における油脂含有量が高いため生産性に優れた方法であるといえる。換言すると、単位耕地面積あたりの栽培個体数が一定であると仮定すると、本発明に係る植物体を利用することによって単位耕地面積あたりから製造する油脂量が大幅に向上することとなる。したがって、本発明に係る植物体を利用することによって油脂生産に要する製造コストを大幅に削減することができる。
【0071】
さらに本発明に係る植物体を利用した油脂の製造方法は、単位重量あたりの種子における油脂含有量が高いため生産性に優れた方法であると言える。
【0072】
なお、本発明に係る植物体を利用した油脂の製造方法において、製造対象の油脂としては、特に限定されず、例えば、大豆油、ごま油、オリーブ油、椰子油、米油、綿実油、ひまわり油、コーン油、べに花油及び菜種油等の植物由来の油脂を例示することができる。また、製造した油脂は、家庭用途や工業用途に広く利用することができ、更にはバイオディーゼル燃料の原料としても使用することができる。すなわち、本発明に係る植物体を利用することによって、上述した家庭用途又は工業用途の油脂や、バイオディーゼル燃料等を低コストに製造することができる。
【0073】
植物由来油脂の製造方法
また、本発明では、特定の表現型を示す植物体から採取した種子において油脂含有量が有意に向上しているといった新規知見を見いだした。具体的には、参考文献(Plant J. 1995 Nov;8(5):659-71.)に開示された4種類の色素合成経路欠損株(tt4、tt5、tt6及びΔCHS)から採取した種子は、野生型と比較して種子における油脂含量が有意に向上している。すなわち、本発明に係る植物由来油脂の製造方法は、カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子及びフラボン-3-ヒドラーゼ遺伝子からなる群から選ばれる少なくとも1の遺伝子の機能を欠損した植物体から採取した種子から油脂成分を回収する工程を含むものである。なお、上記参考文献に開示されたtt4株及びΔCHSはカルコンシンターゼ遺伝子を欠損した株であり、tt5株はカルコンイソメラーゼ遺伝子を欠損した株であり、tt6株はフラボン-3-ヒドラーゼ遺伝子を欠損した株である。
【0074】
シロイヌナズナにおけるカルコンシンターゼ遺伝子の塩基配列を配列番号5に示し、当該遺伝子によりコードされるカルコンシンターゼのアミノ酸配列を配列番号6に示す。シロイヌナズナにおけるカルコンイソメラーゼ遺伝子の塩基配列を配列番号7に示し、当該遺伝子によりコードされるカルコンイソメラーゼのアミノ酸配列を配列番号8に示す。シロイヌナズナにおけるフラボン-3-ヒドラーゼ遺伝子の塩基配列を配列番号9に示し、当該遺伝子によりコードされるフラボン-3-ヒドラーゼのアミノ酸配列を配列番号10に示す。
【0075】
但し、本発明においてカルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子及びフラボン-3-ヒドラーゼ遺伝子は、上記の具体的な配列に限定されるものではない。すなわち、カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子及びフラボン-3-ヒドラーゼ遺伝子は、上記の具体的なアミノ酸配列において1又は複数個のアミノ酸配列が欠失、置換、付加又は挿入されたアミノ酸配列を含み、且つ、カルコンシンターゼ活性、カルコンイソメラーゼ活性及びフラボン-3-ヒドラーゼ活性を有するタンパク質をコードするものであっても良い。ここで、複数個のアミノ酸としては、例えば、1から20個、好ましくは1から10個、より好ましくは1から7個、さらに好ましくは1個から5個、特に好ましくは1個から3個を意味する。なお、アミノ酸の欠失、置換若しくは付加は、上記の具体的な塩基配列を、当該技術分野で公知の手法によって改変することによって行うことができる。塩基配列に変異を導入するには、Kunkel法またはGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-KやMutant-G(何れも商品名、TAKARA Bio社製))等を用いて、あるいはLA PCR in vitro Mutagenesisシリーズキット(商品名、TAKARA Bio社製)を用いて変異が導入される。また、変異導入方法としては、EMS(エチルメタンスルホン酸)、5-ブロモウラシル、2-アミノプリン、ヒドロキシルアミン、N-メチル-N’-ニトロ-Nニトロソグアニジン、その他の発ガン性化合物に代表されるような化学的変異剤を使用する方法でも良いし、X線、アルファ線、ベータ線、ガンマ線、イオンビームに代表されるような放射線処理や紫外線処理による方法でも良い。
【0076】
さらに、本発明において、カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子及びフラボン-3-ヒドラーゼ遺伝子には、シロイヌナズナ以外の植物(例えば上述した植物)において同機能を有する遺伝子(以下、相同遺伝子と称す)が含まれる。カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子又はフラボン-3-ヒドラーゼ遺伝子の相同遺伝子は、植物ゲノム情報が明らかになっていれば、カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子又はフラボン-3-ヒドラーゼ遺伝子の塩基配列又は当該遺伝子によりコードされるアミノ酸配列に基づいて、検索対象の植物ゲノム情報から検索することができる。このとき、相同転写因子としては、上記の具体的なアミノ酸配列に対して、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、最も好ましくは95%以上の相同性を有するアミノ酸配列として検索される。ここで、相同性の値は、blastアルゴリズムを実装したコンピュータプログラム及び遺伝子配列情報を格納したデータベースを用いてデフォルトの設定で求められる値を意味する。
【0077】
また、植物ゲノム情報が明らかとなっていない場合には、対象となる植物からゲノムを抽出するか或いは対象となる植物のcDNAライブラリーを構築し、カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子又はフラボン-3-ヒドラーゼ遺伝子の塩基配列の少なくとも一部に対して、ストリンジェントな条件下でハイブリダイズするゲノム領域或いはcDNAを単離することで相同遺伝子を同定することができる。ここで、ストリンジェントな条件とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。例えば、45℃、6×SSC(塩化ナトリウム/クエン酸ナトリウム)でのハイブリダイゼーション、その後の50〜65℃、0.2〜1×SSC、0.1%SDSでの洗浄が挙げられ、或いはそのような条件として、65〜70℃、1×SSCでのハイブリダイゼーション、その後の65〜70℃、0.3×SSCでの洗浄を挙げることができる。ハイブリダイゼーションは、J. Sambrook et al. Molecular Cloning, A Laboratory Manual,2nd Ed., Cold Spring Harbor Laboratory(1989)に記載されている方法等、従来公知の方法で行うことができる。
【0078】
換言すると、本発明に係る植物由来油脂の製造方法は、シロイヌナズナ由来の種子を用いる系に限定されず、あらゆる植物を対象として適用することができる。本発明に係る植物由来油脂の製造方法を適用できる植物としては、例えば、双子葉植物、単子葉植物、例えばアブラナ科、イネ科、ナス科、マメ科、ヤナギ科等に属する植物(下記参照)が挙げられるが、これらの植物に限定されるものではない。
【0079】
アブラナ科:シロイヌナズナ(Arabidopsis thaliana)、アブラナ(Brassica rapa、Brassica napus)、キャベツ(Brassica oleracea var. capitata)、ナタネ(Brassica rapa、Brassica napus)、ナノハナ(Brassica rapa、Brassica napus)、ハクサイ(Brassica rapa var. pekinensis)、チンゲンサイ(Brassica rapa var. chinensis)、カブ(Brassica rapa var. rapa)、ノザワナ(Brassica rapa var. hakabura)、ミズナ(Brassica rapa var. lancinifolia)、コマツナ(Brassica rapa var. peruviridis)、パクチョイ(Brassica rapa var. chinensis)、ダイコン(Brassica Raphanus sativus)、ワサビ(Wasabia japonica)など。
【0080】
ナス科:タバコ(Nicotiana tabacum)、ナス(Solanum melongena)、ジャガイモ(Solaneum tuberosum)、トマト(Lycopersicon lycopersicum)、トウガラシ(Capsicum annuum)、ペチュニア(Petunia)など。
【0081】
マメ科:ダイズ(Glycine max)、エンドウ(Pisum sativum)、ソラマメ(Vicia faba)、フジ(Wisteria floribunda)、ラッカセイ(Arachis. hypogaea)、ミヤコグサ(Lotus corniculatus var. japonicus)、インゲンマメ(Phaseolus vulgaris)、アズキ(Vigna angularis)、アカシア(Acacia).など。
【0082】
キク科:キク(Chrysanthemum morifolium)、ヒマワリ(Helianthus annuus)など。
【0083】
ヤシ科:アブラヤシ(Elaeis guineensis、Elaeis oleifera)、ココヤシ(Cocos nucifera)、ナツメヤシ(Phoenix dactylifera)、ロウヤシ(Copernicia)
【0084】
ウルシ科:ハゼノキ(Rhus succedanea)、カシューナットノキ(Anacardium occidentale)、ウルシ(Toxicodendron vernicifluum)、マンゴー(Mangifera indica)、ピスタチオ(Pistacia vera)
【0085】
ウリ科:カボチャ(Cucurbita maxima、Cucurbita moschata、Cucurbita pepo)、キュウリ(Cucumis sativus)、カラスウリ(Trichosanthes cucumeroides)、ヒョウタン(Lagenaria siceraria var. gourda)
【0086】
バラ科:アーモンド(Amygdalus communis)、バラ(Rosa)、イチゴ(Fragaria)、サクラ(Prunus)、リンゴ(Malus pumila var. domestica)など。
【0087】
ナデシコ科:カーネーション(Dianthus caryophyllus)など。
【0088】
ヤナギ科:ポプラ(Populus trichocarpa、Populus nigra、Populus tremula)
【0089】
イネ科:トウモロコシ(Zea mays)、イネ(Oryza sativa)、オオムギ(Hordeum vulgare)、コムギ(Triticum aestivum)、タケ(Phyllostachys)、サトウキビ(Saccharum officinarum)など。
【0090】
ユリ科:チューリップ(Tulipa)、ユリ(Lilium)など。
【0091】
また、遺伝子の機能を欠損させるとは、当該遺伝子をゲノムから欠失させること、当該遺伝子の発現(転写レベル及び翻訳レベル)を阻害すること、及び当該遺伝子によってコードされるタンパク質の活性を低下又は欠失させることを含む意味である。
【0092】
より詳細に、遺伝子を欠失させる方法としては、特に限定されないが、相同組換えを用いた方法やトランスポゾンを用いた方法を挙げることができる。また、当該遺伝子を欠失させる場合には、当該遺伝子の全長を欠失させても良いし、部分的に欠失させても良い。
【0093】
また、遺伝子の発現を阻害する方法としては、特に限定しないが、当該遺伝子の発現を制御しているプロモーターを欠失させる方法、当該遺伝子の発現を制御しているプロモーターを発現誘導型プロモーターに置換する方法、当該遺伝子の発現を制御しているプロモーターに突然変異を導入する方法、RNA干渉を利用して当該遺伝子の転写産物を分解する方法、及びアンチセンスRNAを利用して当該遺伝子の翻訳を阻害する方法を挙げることができる。
【0094】
さらに、遺伝子がコードするタンパク質の活性を低下させる方法としては、当該タンパク質に特異的に結合して当該タンパク質の活性を抑制する機能を有する物質を作用させる方法を挙げることができる。当該物質としては、当該タンパク質の機能を阻害できる抗体や阻害物質を挙げることができる。
【0095】
本発明に係る植物由来油脂の製造方法において種子から油脂を回収する方法は、特に限定されず、圧搾法、抽出法及び圧出法などの如何なる方法を用いても良い。例えば、ソックスレー抽出器を用いたエーテル抽出法によって、植物から採取した種子から油脂成分を回収することができる。本発明に係る植物由来油脂の製造方法によれば、植物一個体から採取できる種子量が同等であっても、種子一粒あたりの油脂含有量が高いため植物体を用いているため、生産性に優れた方法であるといえる。換言すると、単位耕地面積あたりの栽培個体数が一定であると仮定すると、本発明に係る植物由来油脂の製造方法によれば、単位耕地面積あたりから製造する油脂量が大幅に向上することとなり、油脂生産に要する製造コストを大幅に削減することができる。
【0096】
なお、本発明に係る植物由来油脂の製造方法において、製造対象の油脂としては、特に限定されず、例えば、大豆油、ごま油、オリーブ油、椰子油、米油、綿実油、ひまわり油、コーン油、べに花油及び菜種油等の植物由来の油脂を例示することができる。また、製造した油脂は、家庭用途や工業用途に広く利用することができ、更にはバイオディーゼル燃料やバイオプラスチックの原料としても使用することができる。すなわち、本発明に係る植物体を利用することによって、上述した家庭用途又は工業用途の油脂や、バイオディーゼル燃料やバイオプラスチック等を低コストに製造することができる。
【0097】
油脂量が向上した植物体のスクリーニング方法
上述したように、本発明では、色素合成経路欠損株(参考文献:Plant J. 1995 Nov;8(5):659-71.)から採取した種子において、野生型と比較して油脂含有量が有意に向上しているといった新規知見を見いだした。色素合成経路欠損株は、色素合成系に関与する遺伝子の機能が欠損した変異株であって、野生株と比較して種皮色が淡色(野生型と比較してより白色)であるという表現型を示す。上記参考文献に開示されたtt4株及びΔCHSはカルコンシンターゼ遺伝子を欠損した株であり、tt5株はカルコンイソメラーゼ遺伝子を欠損した株であり、tt6株はフラボン-3-ヒドラーゼ遺伝子を欠損した株である。これら遺伝子を欠損した変異株においては、色素の合成不全により種皮色がより白色となる。このため、スクリーニング対象の植物から種子を採取し、採取した種子の種皮色を確認すると、当該植物における色素合成経路による色素合成能が判断できるとともに、採取した種子に含まれる種子含量を高精度に推定することができる。
【0098】
すなわち、例えば、同種に含まれる様々な植物体があるとき、これら植物体から採取した種子の種皮色を観察してより白色であるものは油脂生産量の高い品種であると選抜することができる。ここで、スクリーニング対象となる植物体とは、何らかの変異原処理を施したものであっても良いし、従来公知の育種法等によって作出された植物品種であっても良い。
【0099】
ここで、変異原処理としては、特に限定されず、広く突然変異の誘発に用いられている化学的変異原及び/又は物理的変異原による処理を用いることができる。化学的突然変異源として、例えばメタンスルホン酸エチル(EMS)、エチルニトロソ尿素(ENS)、2-アミノプリン、5-ブロモウラシル(5-BU)、アルキル化剤などが用いることができる。また、物理的変異原としては、放射線、紫外線等を用いることができる。これらの変異原を用いた変異の誘発は公知の方法で行うことができる。
【0100】
本発明に係るスクリーニング方法によれば、植物から採取した種子を破壊する必要がなく、目視により種皮色を観察するといった非常に簡便且つ迅速な判定法により、種子に含まれる油脂量を判定することができる。
【0101】
また、本発明に係るスクリーニング方法では、植物から採取した種子の種皮色を画像データから判断し、種皮色を定量的に測定しても良い。具体的には、評価対象の種子の画像をデジタルデータに変換し、画像データ中の種子領域のR値、G値及びB値(RGB値)を測定する。種子領域のR値、G値及びB値の測定には、画像処理ソフトウェアであれば如何なるソフトウェアを使用しても良い。次に、測定されたR値、G値及びB値を、野生型の種子におけるR値、G値及びB値と比較する。一例としては、測定されたR値、G値及びB値の積算値を算出し、野生型の種子におけるR値、G値及びB値の積算値と比較する。例えば、測定されたR値、G値及びB値の積算値が、野生型の種子におけるR値、G値及びB値の積算値として有意に上昇していれば、測定対象の種子の種皮色はより白色に近いと判断することができる。特に、測定されたR値、G値及びB値の積算値が、野生型の種子におけるR値、G値及びB値の積算値に対して、2.88倍以上の値を示す場合には、より白色化(淡色化)した種子であると判断することができる。
【0102】
以上のように、植物から採取した種子の種皮色を画像データとして観察するといった手法であっても種子を破壊する必要がなく、非常に簡便且つ迅速な判定法により、種子に含まれる油脂量を判定することができる。なお、画像データから種子の種皮色を観察する際に、R値、G値及びB値の積算値に限らず、R値、G値及びB値の合計値等を算出しても良い。
【実施例】
【0103】
以下、実施例により本発明をより詳細に説明するが、本発明の技術的範囲はこれら実施例に限定されるものではない。
【0104】
〔実施例1〕
本実施例では、シロイヌナズナにおける転写共役因子At5g24520及び転写因子At1g71030について、それぞれリプレッサードメイン配列を付加したキメラタンパク質(融合タンパク質)を植物体において発現させ、当該植物体から採取した種子における油脂含有量を測定した。また、比較のため転写因子At1g56650についても、同様にしてキメラタンパク質(融合タンパク質)を植物体において発現させ、種子における油脂含有量を測定した。
【0105】
転写因子遺伝子の増幅
シロイヌナズナのcDNAライブラリーより、以下に記載するプライマーを用いて、At1g71030の終始コドンを除くコード領域のDNA断片並びに終止コドンを含むコード領域のDNA断片、At5g24520の終始コドンを除くコード領域のDNA断片及びAt1g56650の終始コドンを除くコード領域のDNAをPCRにより増幅した。PCR条件は94℃1分、47℃2分、伸長反応74℃1分を25サイクル行なった。PCR終了後、増幅されたDNA断片をアガローズゲル電気泳動により分離、回収した。
・At1g71030増幅用フォワードプライマー1
gATGAACAAAACCCGCCTTCGTGCTCTCTC(配列番号11)
・At1g71030増幅用リバースプライマー1
TCGGAATAGAAGAAGCGTTTCTTGACCTGT(配列番号12)
・At1g71030増幅用フォワードプライマー2
gATGAACAAAACCCGCCTTCGTGCTCTCTC(配列番号13)
・At1g71030増幅用リバースプライマー2(配列番号14)
TCATCGGAATAGAAGAAGCGTTTCTTGACC
・At1g56650増幅用フォワードプライマー
GATGGAGGGTTCGTCCAAAGGGC(配列番号15)
・At1g56650増幅用リバースプライマー
ATCAAATTTCACAGTCTCTCCATCG(配列番号16)
・At5g24520増幅用フォワードプライマー
gATGGATAATTCAGCTCCAGATTCGTTATC(配列番号17)
・At5g24520増幅用リバースプライマー
AACTCTAAGGAGCTGCATTTTGTTAGCAAA(配列番号18)
【0106】
融合遺伝子の作製
上記DNA断片がコードする転写因子遺伝子の3'末端にリプレッサードメイン配列を付加するために、CaMV35Sプロモーターの下流にSmaIサイトとリプレッサードメイン(アミノ酸配列:GLDLDLELRLGFA)配列を有するベクターであるp35SSXGを用いた。転写因子遺伝子配列とリプレッサードメイン配列を連結するために、本ベクターをSmaIで切断し、上記の転写因子をコードするPCR増幅断片を挿入し、p35SSXG(At1g56650)とp35SSXG(At5g24520)、 p35SSXG(At1g71030)を作製した。 なおp35SSXG(At1g71030)はAt1g71030増幅用フォワードプライマー1及びAt1g71030増幅用リバースプライマー1を用いたPCR増幅断片を挿入した。また、At1g71030増幅用フォワードプライマー2及びAt1g71030増幅用リバースプライマー2を用いたPCR増幅断片を、リプレッサードメインを付加せずに発現するために、CaMV35Sプロモーターの下流にSmaIサイト配列を有するベクターである、p35SOXGをSmaIで切断部位に挿入し、p35SOXG(At1g71030)を作製した。
【0107】
改良型転写因子および転写因子発現ベクターの構築
アグロバクテリウムにより植物に遺伝子導入を行なうためのバイナリーベクターとしてはpBCKHを用いた。本ベクターはpBIG(Hygr)(Nucleic Acids Res. 18, 203 (1990))のHindIIIサイトにGatewayベクターコンバージョンシステム(Invitrogen)のカセットを組み込んだものである。このベクターに改良型転写因子遺伝子を組み込むために、本ベクターと、p35SSXG(At1g56650)、p35SSXG(At5g24520)、p35SSXG(At1g71030)またはp35SOXG(At1g71030)を混合し、GATEWAY LR clonase (Invitrogen)を用いて組換え反応を行った。その結果、pBCKH-p35SSXG(At1g56650)、pBCKH-p35SSXG(At5g24520)、pBCKH-p35SSXG(At1g71030)及びpBCKH-p35SOXG(At1g71030)を作製した。
【0108】
改良型転写因子遺伝子発現ベクターの植物への導入
改良型転写因子を導入する植物にはシロイヌナズナ(Arabidopsis thaliana, Columbia )を用いた。遺伝子導入法は、Transformation of Arabidopsis thaliana by vacuum infiltration に従った。ただし、感染させるのに減圧処理は行なわず、アグロバクテリウム菌液に浸すだけとした。具体的には、改良型転写因子発現ベクター pBCKH-p35SSXG(At1g56650)、pBCKH-p35SSXG(At5g24520)、pBCKH-p35SSXG(At1g71030)及びpBCKH-p35SOXG(At1g71030)を、土壌細菌Agrobacterium tumefaciens strain GV3101 (C58C1Rifr) pMP90 (Gmr)(koncz and Schell 1986)株にエレクトロポレーション法で導入した。
【0109】
導入した菌を1リットルの、抗生物質(カナマイシン(Km)50μg/ml、ゲンタマイシン(Gm)25μg/ml、リファンピシリン(Rif)50μg/ml)を含むYEP培地でOD600が1になるまで培養した。次いで、培養液から菌体を回収し、1リットルの感染用培地(Infiltration medium、1リッターあたり、2.2 g MS salt, 1X B5 vitamins, 50 g sucrose, 0.5 g MES, 0.044 μM benzylaminopurine, 400μl Silwetを含む。pH 5.7) に懸濁した。この溶液に、14日間生育したシロイヌナズナを1分間浸し、感染させた後、再び栽培を継続し結実させた。採種した種子(T1種子)を50%ブリーチ、0.02%Triton X-100溶液で7分間滅菌した後、滅菌水で3回リンスし、滅菌したハイグロマイシン選択培地(4.3g/l MS salts, 0.5 % sucrose, 0.5 g/l MES, pH 5.7, 0.8 % agar, 30mg/l hygromycin, 250 mg/l Vancomycin)に播種した。上記ハイグロマイシンプレートで生育する形質転換植物体(T1植物)を各改良型転写遺伝子につき10系統を選抜し、バーミキュライト混合土を入れた直径50mmのポットに移植した。これを22℃、16時間明期8時間暗期、光強度約60〜80μE/cm2で栽培し種子(T2種子)を得た。得られたT2種子の表皮色は、野生株が濃い茶色であるのに対して、どの系統も薄茶色もしくは黄色であった。
【0110】
色素合成経路欠損株
また、本実施例では、色素合成経路欠損株から採取した種子に含まれる油脂含有量も測定した。本実施例では、具体的に色素合成経路欠損株tt4(NASC stock No. N85) (参考文献:Plant J., 8, 659-671, 1995)、tt5(NASC stock No. N86)、tt6(NASC stock No. N87) (参考文献:Plant Physiol., 111, 339-345, 1996)、ΔCHS(NASC stock No. N520583))についてはNASC(The Nottingham Arabidopsis Stock Centre)より入手した。tt4、tt5、tt6はArabidopsis thaliana, Ler株より作製され、ΔCHSはArabidopsis thaliana, Col-0株より作製された。これをを50%ブリーチ、0.02%Triton X-100溶液で7分間滅菌した後、滅菌水で3回リンスし、培地(4.3g/l MS salts, 0.5 % sucrose, 0.5 g/l MES, pH 5.7, 0.8 % agar)に播種した。上記プレートで生育する植物体バーミキュライト混合土を入れた直径50mmのポットに移植した。これを22℃、16時間明期8時間暗期、光強度約50〜60μE/cm2(tt4, tt5, tt6, WT(Ler))または、光強度約40μE/cm2(ΔCHS、WT(Col-o))で栽培し種子を得た。得られた種子の表皮色は、野生株が濃い茶色であるのに対して、どの系統も薄茶色もしくは黄色であった。
【0111】
改良型転写因子または転写因子導入T2種子の分析
2種類の改良型転写因子遺伝子及び改良型転写共役因子遺伝子のいずれかを導入したT2種子(At1g56650-SRDX、At5g24520-SRDX、At1g71030-SRDX)および転写因子を導入したT2種子(At1g71030)およびおよび野生株(Col-0、Ler)の油脂含量分析を行なった。油脂の定量分析はMARAN-23 (ResonanceInsturuments Ltd., UK) H-NMRと、解析ソフトRI-NMR Ver. 2.0を用い、2〜10mgのシロイヌナズナ種子を測定した。油脂の標準物質にはオリーブオイルを用いて検量線を作製し、種子中の油脂含量(重量%)を求めた。
【0112】
各改良型転写因子遺伝子、改良型転写共役因子若しくは転写因子遺伝子を導入した系統及び野生株の種子油脂含量の平均値(n=3〜10)を求めた。その結果、Col-0の油脂含量平均値を1とした時の各系統の油脂含量増加率は、T2種子(At1g56650-SRDX)は30.2%、T2種子(At5g24520-SRDX)は12.3%、T2種子(At1g71030-SRDX)は12.2%、T2種子(At1g71030)は2.3%であった(図1)。
【0113】
色素合成経路欠損株種子の分析
4種類の色素合成経路欠損株種子(tt4、tt5、tt6、ΔCHS)およびおよび野生株(Col-0、Ler)の油脂含量分析を行なった。油脂の定量分析はMARAN-23(ResonanceInsturuments Ltd., UK) H-NMRと、解析ソフトRI-NMR Ver. 2.0を用い、2〜10mgのシロイヌナズナ種子を測定した。油脂の標準物質にはオリーブオイルを用いて検量線を作製し、種子中の油脂含量(重量%)を求めた。
【0114】
色素合成経路欠損株及び野生株の種子油脂含量の平均値(n=3〜10)を求めた。ΔCHSはCol-0株に対して8.9%、tt4、tt5及びtt6はLer株に対してそれぞれ、4.7%、8.8%、11.1%の油脂含量であった(図2)。
【0115】
結果と考察
以上の結果から、リプレッサードメインを付加した転写因子At1g56650、転写共役因子At5g24520、転写因子At1g71030それぞれのキメラ遺伝子を導入した植物体の種子の重量あたり油脂含量は、同時に栽培した野生株の重量あたり油脂含量に比べ優れており、油脂生産において非常に有効な植物体であることが判明した。一方で発現促進活性を持つAt1g71030を導入した植物体の種子の重量あたり油脂含量は、同時に栽培した植物体の重量あたり油脂含量とくらべ若干増加していたが、その増加率は発現促進活性を抑制したAt1g71030を導入した植物体種子の重量あたり油脂含量の増加率の1/5程度であった。At1g71030は、シングルMYB様のドメインを持つタンパク質(AtMybL2)をコードし、この遺伝子をCaMV35Sプロモーターで過剰発現することにより、葉、茎、萼のトライコームを欠失する形質を示す。これは、トライコームの形成に必要なGL2遺伝子の発現が抑制されることによると考えられる(参考文献:DNA Res., 9, 31-34, 2002)。GL2遺伝子を破壊することにより、種子の油脂含量が8%増加することが報告されている(参考文献:Plant Mol Biol. 2006 , 60, :377-87, 2006)。
【0116】
また、AtMybL2タンパク質はそのカルボキシ末端領域に6アミノ酸からなる転写リプレッサーを持ち、AtMybL2遺伝子の過剰発現植物及びEAR-motifとして知られる転写リプレッサーを付加したAtMybL2をコードする遺伝子の過剰発現植物では、いずれの場合にもアントシアニン前駆体の合成が抑制された(参考文献:18TH INTERNATIONAL CONFERENCE ON ARABIDOPSIS RESEARCH, TAIR accession Publication:501721814)。しかしながら、解析の結果、At1g71030の過剰発現体のT2種子の油脂含量増加率2.3%に対して、リプレッサードメインを付加したAt1g71030の過剰発現体のT2種子は油脂含量の増加率は12.2%と大幅に高く、またGL2遺伝子破壊時の油脂含量増加率8%よりも顕著に高かった。これらの結果より、リプレッサードメインを付加したAt1g71030がGL2以外の未知の経路で種子の油脂合成と貯蔵過程に作用をおよぼし、油脂含量を増加したと考えられる。
【0117】
一方で、色素合成経路欠損株の分析結果より、色素合成経路の主要遺伝子が破壊された突然変異株tt4、tt5、tt6、T-DNAの挿入によりCHS遺伝子が破壊されたΔCHS株の種子の油脂含量はいずれも野生株に対して高かった。種皮色と油脂含量ナタネでは種皮色が黄色い品種HUA-yellow No. 1は種皮色が黒い品種と比べ、油脂含量が5-7%高いと報告されている(参考文献:Genome 44: 1077-1082 (2001))。しかしながら、異種間の交配に起因する従来育種法では、種皮色と種子の油脂含量を決定する形質の遺伝子座が近接していた場合でも同様の減少が観察されうる。よって、遺伝子の発現と形質の相関関係についてはこれまで、明らかにされていない。すなわち、従来、種皮色の形質を左右する遺伝子座が油脂含量に影響するといった知見は知られていなかった。
【0118】
これに対して、本結果では、実際に種皮の色素合成酵素をコードする遺伝子を破壊することにより種子中の油脂含量の増加が確認できた。よって、交配による従来育種法のみならず、遺伝子導入法、遺伝子破壊法による分子育種法においても、種皮色が油脂含量を予想する重要な表現型であることが初めて明らかになった。種皮色を指標に用いることにより種子の油脂含量を非破壊かつ特別な装置を用いずに、効率よく油脂含量が増加した種子を選抜することが可能になる。
【0119】
より詳細に、野生株、At1g71030-SRDX、At1g56650-SRDX、ΔCHS系統の種子の写真を撮影しデジタルデータに変換した。得られたデジタルデータを画像処理ソフト(アドビ・フォトショップ)を用いて種子の領域のRGB値をそれぞれを定量した。次に、定量したR値、G値及びB値の積算値を算出した。また、野生型におおいて定量したR値、G値及びB値の積算値に対する比率も算出した。その結果を表1及び図3に示す。
【0120】
【表1】

【0121】
表1及び図3に示すように、At1g71030-SRDX、At1g56650-SRDX及びΔCHS系統においては、R値、G値及びB値の積算値を野生型と比較したところ少なくとも2.88倍以上の値を示していた。以上のように、種子の画像データを用いて種皮色を定量的に測定し、種子中の油脂量を非常に簡便に且つ迅速に評価できることが可能となった。
【図面の簡単な説明】
【0122】
【図1】各改良型転写因子遺伝子、改良型転写共役因子遺伝子若しくは転写因子遺伝子を導入した系統及び野生株の種子油脂含量を測定した結果を示す特性図である。
【図2】色素合成経路欠損株及び野生株の種子油脂含量を測定した結果を示す特性図である。
【図3】種子の種皮色について画像データを用いてR値、G値及びB値の積算値を算出し、野生型の種子と比較した結果を示す特性図である。
【背景技術】
【0001】
バイオマス(biomass)とは、一般的には一定面積あたりに生息または存在する生物の総量を指し、特に植物を対象とした場合は、単位面積あたりの乾重量を意味する。バイオマスの単位は、質量又はエネルギー量で数値化する。バイオマスという表現は、「生物体量」、「生物量」も同義語であり、植物バイオマスの場合には「現存量(Standing crop)」の語が使われることもある。植物バイオマスは、大気中の二酸化炭素を太陽エネルギーを用いて固定して生成されるため、いわゆるカーボンニュートラルなエネルギーとして捕らえることができる。したがって、植物のバイオマスを増加させることは、地球環境保全、地球温暖化防止、温室効果ガス排出低減の効果がある。従って、植物バイオマスを増産させる技術は産業上の重要性が高い。
【0002】
一方、植物は、その一部の組織自体(種子、根、葉茎など)を目的として栽培されたり、油脂などの種々の物質生産を目的として栽培されたりする。例えば、植物が生産する油脂としては、大豆油、ごま油、オリーブ油、椰子油、米油、綿実油、ひまわり油、コーン油、べに花油、パーム油及び菜種油等が古来より知られており、家庭用途や工業用途に広く利用されている。また、植物が生産する油脂は、バイオディーゼル燃料やバイオプラスチックの原料としても使用され、石油代替エネルギーとして適用性が広がっている。
【0003】
このような状況において、植物を用いた油脂生産を工業的に成功させるには、単位耕地面積あたりの生産性の向上が必要となる。ここで単位耕地面積あたりの栽培個体数が一定であると仮定すると、個体あたりの油脂生産量の向上が必要であることが判る。植物体から採取した種子から油脂を回収する場合には、個体あたりの種子収量を向上させる、及び種子中の油脂含有量を向上させるといった技術により、個体あたりの油脂生産量の向上が達成できるものと期待される。
【0004】
植物種子の油脂生産量を増加させる技術には大別して栽培法の改良によるものと、油脂増産品種の開発がある。油脂増産品種の開発方法は、交配技術を中心とした従来育種法と遺伝子組換えによる分子育種法とに大別される。遺伝子組換えによる油脂増産技術としては、A)植物油脂の主成分である種子トリアシルグリセロール(TAG)の合成系を改変する技術、及びB)植物の形態形成や代謝及びそれらに関わる遺伝子の発現を制御する各種制御遺伝子を改変する技術とが知られている。
【0005】
上記A)の方法としては、光合成により生産される糖を原料として合成されるTAGの合成量を増加させる方法として(1)TAGの構成成分である脂肪酸、あるいはグリセロールの糖からの合成活性を高める方法、(2)グリセロールと脂肪酸からTAGが合成される反応を強化する方法が考えられる。これらについて、遺伝子工学的な方法を用いた技術としては以下の技術が報告されている。(1)の例としては、シロイヌナズナの細胞質型acetyl-coenzyme A carboxylase (ACCase)をナタネのプラスチド中で過剰発現させることにより、種子の油脂含量を5%向上させた報告(非特許文献1)が挙げられる。また、(2)の例としては、ジアシルグリセロールのsn-3位にアシル基を転移するDGAT(diacylglycerol acyltransferase)の過剰発現による油脂増産技術に関する報告(非特許文献2)が挙げられる。非特許文献2の方法では、DGATの発現量が増加するに従って油脂含量と種子重が増加し、個体あたりの種子数が増加する場合があることも報告されている。本方法を適用したシロイヌナズナの種子油脂含量は46%増、個体あたりの油脂量は最高で約125%増であった。
【0006】
一方、上記B)の方法としては、生合成系酵素遺伝子の発現制御に関与する転写因子遺伝子の発現を制御する方法が考えられる。この例としては、特許文献1が挙げられる。本特許文献1では、転写因子を網羅的に過剰発現またはノックアウトした組換え植物を作製したのちに種子の油脂含量を高める遺伝子を選抜するといった手法が採用されている。特許文献1によれば、ERF subfamily B-4転写因子遺伝子の過剰発現によって種子の油脂含量が23%増加したと記載されている。しかし、特許文献1において、個体あたりの油脂含量の増減については記載されていない。また、非特許文献3には、AP2/EREBドメインを持つ転写因子であるWRINKLED1を過剰発現させることにより、種子の油脂含量が向上することが記載されている。
【0007】
しかしながら、種々の形質の改良を目的とした上述した分子育種法が開発されているにもかかわらず、植物の重量増産、特定の組織増産、或いは目的物質の生産性向上を伴う、収量向上技術は実用の域に達していない。
【0008】
この理由として、真に優れた遺伝子が未発見であること、試験段階で効果のある組換え新品種が実用段階では多様な自然環境下で期待通りの効果を発揮できないことにあると考えられる。また、植物の重量増産、特定の組織増産、或いは目的物質の生産性と言った量的形質は制御系から代謝系に亘る様々なステップで多数の遺伝子が関わっており、量的形質を改善する真に優れた有用遺伝子を発見、開発することは困難であった。これらの問題を解決するためには、効果が劇的に高い新たな遺伝子を見出すこと、効果レベルは同等であっても実用環境条件で効果を発揮する遺伝子を開発することが課題である。
【0009】
【非特許文献1】Plant Physiology (1997) Vol. 11, pp. 75-81
【非特許文献2】Plant Physiology (2001), Vol. 126, pp. 861-874
【非特許文献3】Plant J. (2004) 40, 575-585
【特許文献1】WO01/36597
【発明の開示】
【発明が解決しようとする課題】
【0010】
そこで、上述したような実情に鑑み、個体あたりの物質生産性、特に種子中の油脂量を向上させることができる新規な機能を有する遺伝子を探索し、植物体におけるこれらの特性を向上できる技術を提供することを目的とする。
【課題を解決するための手段】
【0011】
上述した目的を達成するため、本発明者らが鋭意検討した結果、特定の転写因子ファミリーに属する転写因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチド(以下、リプレッサードメインと称する場合もある)とを融合したキメラタンパク質を発現させることによって、様々な量的形質を改善させることができ、特に、個体あたりの物質生産性、特に油脂生産性を向上させることができることを見いだし、本発明を完成するに至った。
【0012】
本発明に係る植物体は、配列番号4に示すアミノ酸配列からなるタンパク質を含む転写因子ファミリーに属する転写因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質を発現させたものである。本発明に係る植物体においては、機能性ペプチドを融合することによって、所定の転写因子における転写制御活性、特に転写促進活性が抑制されていることが好ましい。
【0013】
ここで、上記機能性ペプチドを融合する転写因子としては、以下の(a)〜(c)のいずれかのタンパク質であることが好ましい。
(a)配列番号4に示すアミノ酸配列を含むタンパク質
(b)配列番号4に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、転写促進活性を有するタンパク質
(c)配列番号3に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ転写促進活性を有するタンパク質
【0014】
ここで上記機能性ペプチドとしては、次に示す式(1)〜(8)を挙げることができる。
(1)X1−Leu−Asp−Leu−X2−Leu−X3
(但し、式中、X1は0〜10個のアミノ酸残基を示し、X2はAsn又はGluを示し、X3は少なくとも6個のアミノ酸残基を示す。)
(2)Y1−Phe−Asp−Leu−Asn−Y2−Y3
(但し、式中、Y1は0〜10個のアミノ酸残基を示し、Y2はPhe又はIleを示し、Y3は少なくとも6個のアミノ酸残基を示す。)
(3)Z1−Asp−Leu−Z2−Leu−Arg−Leu−Z3
(但し、式中、Z1はLeu、Asp−Leu又はLeu−Asp−Leuを示し、Z2はGlu、Gln又はAspを示し、Z3は0〜10個のアミノ酸残基を示す。)
(4)Asp−Leu−Z4−Leu−Arg−Leu
(但し、式中、Z4はGlu、Gln又はAspを示す。)
(5)α1−Leu−β1−Leu−γ1−Leu
(6)α1−Leu−β1−Leu−γ2−Leu
(7)α1−Leu−β2−Leu−Arg−Leu
(8)α2−Leu−β1−Leu−Arg−Leu
(但し、式(5)〜(8)中、α1はAsp、Asn、Glu、Gln、Thr又はSerを示し、α2はAsn、Glu、Gln、Thr又はSerを示し、β1はAsp、Gln、Asn、Arg、Glu、Thr、Ser又はHisを示し、β2はAsn、Arg、Thr、Ser又はHisを示し、γ1はArg、Gln、Asn、Thr、Ser、His、Lys又はAspを示し、γ2はGln、Asn、Thr、Ser、His、Lys又はAspを示す。)
【0015】
また、本発明に係る植物体を用いた物質の製造方法は、上述した本発明に係る植物体から、生産性が向上した物質を分離及び回収する工程を含むものである。ここで、上記物質としては油脂を挙げることができる。
【0016】
一方、上述した目的を達成するため、本発明者らが鋭意検討した結果、色素合成経路に関与する遺伝子を欠損させた株において、個体あたりの物質生産性、特に油脂生産性を向上させることができることを見いだし、本発明を完成するに至った。色素合成系路に関与する遺伝子とは、色素合成経路代謝反応の基質や生成物の輸送に関する因子をコードする遺伝子や、色素合成系路代謝反応を触媒する酵素をコードする遺伝子や、色素合成系路代謝反応の場を形成させる反応を触媒する酵素をコードする遺伝子を含む。さらに、色素合成経路代謝反応の基質や生成物の輸送に関する因子をコードする遺伝子や、色素合成系路代謝反応を触媒する酵素をコードする遺伝子や、色素合成系路代謝反応の場を形成させる反応を触媒する酵素をコードする遺伝子の発現を制御する遺伝子を含む。
【0017】
すなわち、本発明に係る植物由来油脂の製造方法は、カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子及びフラボン-3-ヒドラーゼ遺伝子からなる群から選ばれる少なくとも1の遺伝子の機能を欠損した植物体から採取した種子から油脂成分を回収する工程を含むものである。
【0018】
また、本発明に係る油脂量が向上した植物体のスクリーニング方法は、種子内の油脂量を評価する対象となる植物体から種子を採取する工程と、採取した種子の種皮色を観察し、より白色である場合には種子内の油脂量が高いと判定する工程とを含むものである。
【発明の効果】
【0019】
本発明に係る植物体は、個体あたりの物質生産性が向上したものとなる。したがって、本発明に係る植物体を用いることによって、目的物質の生産性の向上を達成することができ、目的物質を低コストで製造することができる。
【0020】
また、本発明に係る植物由来油脂の製造方法は、特定の遺伝子の機能を欠損する植物体において種子の単位量あたりに含まれる油脂量が大幅に向上しているため、油脂の生産性を向上することができる。
【0021】
さらに、本発明に係る油脂量が向上した植物体のスクリーニング方法は、種子内の油脂量を非破壊で評価するため、小量の種子を用いて迅速、且つ簡便にスクリーニングすることができる。
【発明を実施するための最良の形態】
【0022】
以下、本発明を詳細に説明する。
【0023】
本発明に係る植物体は、所定の転写制御因子ファミリーに属する転写制御因子、特に、所定の転写因子ファミリーに属する転写因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質を発現するものであり、野生型の植物体と比較して、個体あたりの物質生産性が向上したものである。すなわち、本発明に係る植物体は、所望の植物を対象として、当該植物における物質生産性を有意に向上させるように、転写因子を上記機能性ペプチドとのキメラタンパク質として発現させた植物体である。
【0024】
特に、本発明に係る植物体においては、上記機能性ペプチドと融合することによって、転写因子における転写促進活性が抑制していることが好ましい。換言すると、本発明に係る植物体においては、転写因子に上記機能性ペプチドを融合させたキメラタンパク質を発現させた結果、上記機能性ペプチドに起因する転写抑制効果が優性の形質として現れるといった特徴を有していることが好ましい。
【0025】
ここで、個体あたり物質生産性とは、植物が生成する各種の物質についての単位体積あたりの含有量を意味する。物質としては、特に限定されず、植物体が本来的に生成する物質であっても良いし、植物体が本来的には生成しないが遺伝子操作技術等によって生成できるようになった物質であっても良い。特に、組織あたりの目的生産物の含量が高くなれば、精製コストや運搬コストを低減できるため産業上有用性が高い。特に、目的生産物としては、植物のほとんどの重量を占めるリグノセルロースでも良く、種子油として産業上利用されている植物油でも良い。植物油は、脂肪酸とアルコールのエステルである単純脂質でも良く、リンや糖や窒素などを含んだ複合脂質でも良く、脂肪酸そのものでも良い。単純脂質のアルコールとしては分子量の高い高級アルコールでも良く、グリセロール(グリセリン)などの多価アルコールでも良い。単純脂質の脂肪酸としては、飽和脂肪酸でも良く、不飽和脂肪酸でも良く、また、水酸基やエポキシ基を含んだ特殊脂肪酸でも良い。グリセロールと脂肪酸のエステルである単純脂質としては、モノアシルグリセロールでも良く、ジアシルグリセロールでも良く、トリアシルグリセロールでも良い。
【0026】
以下の説明において、生産性を向上させる物質として油脂を例示して説明するが、本発明の技術的範囲がこれに限定されるものではない。本発明は、植物が生成する物質として油脂以外の物質についても同様に適用される。
【0027】
ここで、植物体としては、特に限定されず、如何なる植物をも対象とすることができる。特に、従来より油脂の生産に使用される植物を対象とすることが好ましい。対象とする植物としては、例えば、大豆、ごま、オリーブ油、椰子、イネ、綿花、ひまわり、トウモロコシ、サトウキビ、ジャトロファ、パームヤシ、タバコ、べに花及びナタネ等を挙げることができる。また、植物の遺伝子解析におけるモデル生物として広く利用されており、遺伝子発現解析の方法が確立しているシロイヌナズナを対象の植物とすることもできる。
【0028】
また、転写因子のキメラタンパク質が活性として有する転写抑制とは、当該転写因子が認識するcis配列や、そのcis配列と類似する他の転写因子におけるcis配列を認識し、下流の遺伝子発現を積極的に抑制する活性であり、転写抑制因子とも呼べるものである。転写因子のキメラタンパク質が活性として有する転写抑制する手法は、特に限定されないが、特に、リプレッサードメイン配列やSRDX配列を付加したキメラタンパク質(融合タンパク質)を構築する方法が最も好ましい。
【0029】
この手法においてリプレッサードメイン配列とは、任意の転写因子を転写抑制因子に転換するペプチドを構成するアミノ酸配列であり本発明者らによって種々見出された配列である。リプレッサードメイン配列を使用した方法については、例えば、特開2001−269177公報、特開2001−269178公報、特開2001−292776公報、特開2001−292777公報、特開2001−269176公報、特開2001−269179公報、国際公開第WO03/055903号パンフレット、Ohta, M., Matsui, K., Hiratsu, K., Shinshi, H. and Ohme-Takagi, M., The Plant Cell, Vol.13,1959-1968,August,2001及びHiratsu, K., Ohta, M., Matsui, K., Ohme-Takagi, M., FEBS Letters 514(2002)351-354を参照することができる。リプレッサードメイン配列は、Class II ERF(Ethylene Responsive Element Binding Factor)タンパク質や植物のジンクフィンガータンパク質(Zinc Finger Protein、例えばシロイヌナズナSUPERMANタンパク質等)から切り出されたもので、極めて単純な構造を有している。
【0030】
キメラタンパク質として発現する転写制御因子としては、シロイヌナズナにおけるAGIコードAt1g71030で特定される転写因子(以下、単に『転写因子At1g71030』と称す)を挙げることができる。転写因子At1g71030は、mybファミリーの転写因子であり、オオムギ由来のMybHv5 GI:19055と類似していることが知られている。転写因子At1g71030のアミノ酸配列を配列番号4に示す。転写因子At1g71030をコードする遺伝子の塩基配列を配列番号3に示す。
【0031】
また、At5g24520で特定される転写共役因子(transcription coactivator)(以下、単に『転写共役因子At5g24520』と称す)、転写抑制因子(transcription repressor)や転写抑制共役因子(transcripition corepressor)が転写制御因子として知られており、これら転写共役因子や転写抑制因子についても同様にリプレッサードメインを付加したキメラタンパク質を構築することができる。なお、AGIコードAt5g24520は、transparent testa glabra 1 protein (TTG1) として知られる転写共役因子が因子である。他の植物由来の遺伝子では、Malus domestica由来のGenBankアクセッション番号AAF27919がコードするタンパク質、Petunia hybrida由来のGenBankアクセッション番号AAC18914がコードするタンパク質、Gossypium hirsutum由来のGenBankアクセッション番号AAM95645がコードするタンパク質、Perilla frutescens由来のGenBankアクセッション番号BAB58883がコードするタンパク質が転写共役因子At5g24520と相同であることが知られ、本明細書で記載した機能と同等の機能が期待できる。転写共役因子At5g24520のアミノ酸配列を配列番号2に示す。転写共役因子At5g24520をコードする遺伝子の塩基配列を配列番号1に示す。
【0032】
また、キメラタンパク質の対象となる転写共役因子At5g24520及び転写因子At1g71030は、それぞれ配列番号2及び4に示すアミノ酸配列からなるものに限定されず、当該アミノ酸配列において1又は複数個のアミノ酸配列が欠失、置換、付加又は挿入されたアミノ酸配列を含み、且つ、転写促進活性を有するものであっても良い。ここで、複数個のアミノ酸としては、例えば、1から20個、好ましくは1から10個、より好ましくは1から7個、さらに好ましくは1個から5個、特に好ましくは1個から3個を意味する。なお、アミノ酸の欠失、置換若しくは付加は、上記転写因子をコードする塩基配列を、当該技術分野で公知の手法によって改変することによって行うことができる。塩基配列に変異を導入するには、Kunkel法またはGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-KやMutant-G(何れも商品名、TAKARA Bio社製))等を用いて、あるいはLA PCR in vitro Mutagenesisシリーズキット(商品名、TAKARA Bio社製)を用いて変異が導入される。また、変異導入方法としては、EMS(エチルメタンスルホン酸)、5-ブロモウラシル、2-アミノプリン、ヒドロキシルアミン、N-メチル-N’-ニトロ-Nニトロソグアニジン、その他の発ガン性化合物に代表されるような化学的変異剤を使用する方法でも良いし、X線、アルファ線、ベータ線、ガンマ線、イオンビームに代表されるような放射線処理や紫外線処理による方法でも良い。
【0033】
さらに、キメラタンパク質の対象となる転写共役因子及び転写因子には、シロイヌナズナにおける転写共役因子At5g24520及び転写因子At1g71030に限定されず、シロイヌナズナ以外の植物(例えば上述した植物)において同機能を有する転写共役因子及び転写因子(以下、それぞれ相同転写共役因子及び相同転写因子と称す)が含まれる。転写共役因子At5g24520に対する相同転写共役因子又は転写因子At1g71030に対する相同転写因子は、植物ゲノム情報が明らかになっていれば、転写共役因子At5g24520又は転写因子At1g71030のアミノ酸配列或いは各遺伝子の塩基配列に基づいて、検索対象の植物ゲノム情報から検索することができる。このとき、相同転写共役因子及び相同転写因子としては、転写共役因子At5g24520又は転写因子At1g71030のアミノ酸配列に対して、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、最も好ましくは95%以上の相同性を有するアミノ酸配列として検索される。ここで、相同性の値は、blastアルゴリズムを実装したコンピュータプログラム及び遺伝子配列情報を格納したデータベースを用いてデフォルトの設定で求められる値を意味する。
【0034】
また、植物ゲノム情報が明らかとなっていない場合には、対象となる植物からゲノムを抽出するか或いは対象となる植物のcDNAライブラリーを構築し、転写共役因子At5g24520又は転写因子At1g71030の塩基配列の少なくとも一部に対して、ストリンジェントな条件下でハイブリダイズするゲノム領域或いはcDNAを単離することで相同遺伝子を同定することができる。ここで、ストリンジェントな条件とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。例えば、45℃、6×SSC(塩化ナトリウム/クエン酸ナトリウム)でのハイブリダイゼーション、その後の50〜65℃、0.2〜1×SSC、0.1%SDSでの洗浄が挙げられ、或いはそのような条件として、65〜70℃、1×SSCでのハイブリダイゼーション、その後の65〜70℃、0.3×SSCでの洗浄を挙げることができる。ハイブリダイゼーションは、J. Sambrook et al. Molecular Cloning, A Laboratory Manual,2nd Ed., Cold Spring Harbor Laboratory(1989)に記載されている方法等、従来公知の方法で行うことができる。
【0035】
本発明に係る植物体は、上述したような転写因子と機能性ペプチドとのキメラタンパク質を発現させることで、油脂生産量が有意に向上するといった特徴を示す。また、上述した転写共役因子と機能性ペプチドとのキメラタンパク質を発現させた場合も、油脂生産量が有意に向上するといった特徴を示す。特に、キメラタンパク質とすることで、対象となる転写因子を、転写促進活性を抑制した状態で発現させること、さらに、対象となる転写因子が認識するcis配列と相同性のあるcis配列を認識する転写抑制活性として発現させること、対象となる転写因子及び転写共役因子が持つ他因子や核酸や脂質や糖質との親和特異性を変化させることで、油脂生産量が有意に向上するといった特徴を示す。このとき、上記植物体においては、内因性の転写因子や転写共役因子を改変してそのキメラタンパク質を作製してもよいが、キメラタンパク質をコードする遺伝子を導入し、当該遺伝子を発現させても良い。
【0036】
一例としては、上述したような転写因子や転写共役因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質(融合タンパク質)をコードする遺伝子を対象の植物に導入し、当該キメラタンパク質(融合タンパク質)を植物内で発現させる手法が好ましい。
【0037】
本明細書中で記載する「転写促進活性が抑制された転写因子」とは、特に限定されるものではなく、当該転写因子が本来的に有している転写促進活性が有意に低減した転写因子であることを意味する。また、「任意の転写因子を転写抑制因子に転換する機能性ペプチド」とは、任意の転写因子と融合してキメラタンパク質となったときに、当該転写因子が本来的に有している転写促進活性が有意に低減した転写因子となる機能を有するペプチドであることを意味する(転写抑制転換ペプチドと称する場合もある)。このような「任意の転写因子を転写抑制因子に転換する機能性ペプチド」としては、特に限定されないが、なかでもリプレッサードメイン配列やSRDX配列として知られたアミノ酸配列からなるペプチドであることが好ましい。この転写抑制転換ペプチドについては、特開2005−204657号公報に詳述されており、当該公報に開示されたものを全て使用することができる。
【0038】
転写抑制転換ペプチドは、例えば次に示す式(1)〜(8)のいずれかで表されるアミノ酸配列を挙げることができる。
(1)X1−Leu−Asp−Leu−X2−Leu−X3
(但し、式中、X1は0〜10個のアミノ酸残基を示し、X2はAsn又はGluを示し、X3は少なくとも6個のアミノ酸残基を示す。)
(2)Y1−Phe−Asp−Leu−Asn−Y2−Y3
(但し、式中、Y1は0〜10個のアミノ酸残基を示し、Y2はPhe又はIleを示し、Y3は少なくとも6個のアミノ酸残基を示す。)
(3)Z1−Asp−Leu−Z2−Leu−Arg−Leu−Z3
(但し、式中、Z1はLeu、Asp−Leu又はLeu−Asp−Leuを示し、Z2はGlu、Gln又はAspを示し、Z3は0〜10個のアミノ酸残基を示す。)
(4)Asp−Leu−Z4−Leu−Arg−Leu
(但し、式中、Z4はGlu、Gln又はAspを示す。)
(5)α1−Leu−β1−Leu−γ1−Leu
(6)α1−Leu−β1−Leu−γ2−Leu
(7)α1−Leu−β2−Leu−Arg−Leu
(8)α2−Leu−β1−Leu−Arg−Leu
(但し、式(5)〜(8)中、α1はAsp、Asn、Glu、Gln、Thr又はSerを示し、α2はAsn、Glu、Gln、Thr又はSerを示し、β1はAsp、Gln、Asn、Arg、Glu、Thr、Ser又はHisを示し、β2はAsn、Arg、Thr、Ser又はHisを示し、γ1はArg、Gln、Asn、Thr、Ser、His、Lys又はAspを示し、γ2はGln、Asn、Thr、Ser、His、Lys又はAspを示す。)
【0039】
式(1)の転写抑制転換ペプチド
上記式(1)の転写抑制転換ペプチドにおいては、上記X1で表されるアミノ酸残基の数は0〜10個の範囲内であればよい。また、X1で表されるアミノ酸残基を構成する具体的なアミノ酸の種類は特に限定されるものではなく、どのようなものであってもよい。このX1で表されるアミノ酸残基は、式(1)の転写抑制転換ペプチドを合成するときの容易さからみれば、できるだけ短いほうがよい。具体的にX1で表されるアミノ酸残基は、5個以下であることが好ましい。
【0040】
同様に、上記式(1)の転写抑制転換ペプチドにおいては、上記X3で表されるアミノ酸残基の数は少なくとも6個であればよい。また、X3で表されるアミノ酸残基を構成する具体的なアミノ酸の種類は特に限定されるものではなく、どのようなものであってもよい。
【0041】
式(2)の転写抑制転換ペプチド
上記式(2)の転写抑制転換ペプチドにおいては、上記式(1)の転写抑制転換ペプチドのX1と同様、上記Y1で表されるアミノ酸残基の数は0〜10個の範囲内であればよい。また、Y1で表されるアミノ酸残基を構成する具体的なアミノ酸の種類は特に限定されるものではなく、どのようなものであってもよい。具体的にY1で表されるアミノ酸残基は、5個以下であることが好ましい。
【0042】
同様に、上記式(2)の転写抑制転換ペプチドにおいては、上記式(1)の転写抑制転換ペプチドのX3と同様、上記Y3で表されるアミノ酸残基の数は少なくとも6個であればよい。また、Y3で表されるアミノ酸残基を構成する具体的なアミノ酸の種類は特に限定されるものではく、どのようなものであってもよい。
【0043】
式(3)の転写抑制転換ペプチド
上記式(3)の転写抑制転換ペプチドにおいては、上記Z1で表されるアミノ酸残基は、1〜3個の範囲内でLeuを含むものとなっている。アミノ酸1個の場合は、Leuであり、アミノ酸2個の場合は、Asp−Leuとなっており、アミノ酸3個の場合はLeu−Asp−Leuとなっている。
【0044】
一方、上記式(3)の転写抑制転換ペプチドにおいては、上記Z3で表されるアミノ酸残基の数は0〜10個の範囲内であればよい。また、Z3で表されるアミノ酸残基を構成する具体的なアミノ酸の種類は特に限定されるものではなく、どのようなものであってもよい。具体的にZ3で表されるアミノ酸残基は、5個以下であることがより好ましい。Z3で表されるアミノ酸残基の具体的な例としては、Gly、Gly−Phe−Phe、Gly−Phe−Ala、Gly−Tyr−Tyr、Ala−Ala−Ala等が挙げられるが、もちろんこれらに限定されるものではない。
【0045】
また、この式(3)で表される転写抑制転換ペプチド全体のアミノ酸残基の数は、特に限定されるものではないが、合成するときの容易さからみれば、20アミノ酸以下であることが好ましい。
【0046】
式(4)の転写抑制転換ペプチド
上記式(4)の転写抑制転換ペプチドは、6個のアミノ酸残基からなるヘキサマー(6mer)である。なお、上記式(4)の転写抑制転換ペプチドにおいてZ4で表されるアミノ酸残基がGluの場合のアミノ酸配列は、シロイヌナズナSUPERMANタンパク質(SUPタンパク質)の196〜201番目のアミノ酸配列に相当している。
【0047】
以上で説明した各種転写抑制転換ペプチドは、上述した転写因子や転写共役因子と融合してキメラタンパク質(融合タンパク質)とすることにより、当該転写因子や転写共役因子の特性を改変することができる。具体的には、上述した転写因子や転写共役因子と融合してキメラタンパク質(融合タンパク質)とすることにより、転写因子や転写共役因子を転写抑制因子や負の転写共役因子に改変することができる。さらには、ドミナントでない転写抑制因子をドミナント型転写抑制因子にすることも可能である。
【0048】
また、上記転写抑制転換ペプチドをコードするポリヌクレオチドを用いて、転写因子や転写共役因子をコードする遺伝子との融合遺伝子を得れば、キメラタンパク質(融合タンパク質)を生産させることができる。具体的には、上記転写抑制転換ペプチドをコードするポリヌクレオチド(転写抑制転換ポリヌクレオチドと称す)と上記転写因子や転写共役因子をコードする遺伝子とを連結することにより融合遺伝子を構築して、植物細胞に導入する。これによりキメラタンパク質(融合タンパク質)を生産させることができる。上記転写抑制転換ポリヌクレオチドの具体的な塩基配列は特に限定されるものではなく、遺伝暗号に基づいて、上記転写抑制転換ペプチドのアミノ酸配列に対応する塩基配列を含んでいればよい。また、必要に応じて、上記転写抑制転換ポリヌクレオチドは、転写因子遺伝子と連結するための連結部位となる塩基配列を含んでいてもよい。さらに、上記転写抑制転換ポリヌクレオチドのアミノ酸読み枠と、転写因子や転写共役因子の遺伝子の読み枠とが一致しないような場合に、これらを一致させるための付加的な塩基配列を含んでいてもよい。さらにまた、転写因子や転写共役因子と転写抑制転換ペプチドとの間をつなぐためのリンカー機能を有するポリペプチドや、HisやMyc、Flag等のようにキメラタンパク質(融合タンパク質)をエピトープ標識するためのポリペプチド等、各種の付加的なポリペプチドが含まれていてもよい。さらに上記キメラタンパク質(融合タンパク質)には、必要に応じて、ポリペプチド以外の構造、例えば、糖鎖やイソプレノイド基等が含まれていてもよい。
【0049】
植物体を製造する方法は、上述した転写因子や転写共役因子と転写抑制転換ペプチドとのキメラタンパク質を植物体で生産させ、油脂生産性を向上させる過程を含んでいれば特に限定されるものではないが、例えば、発現ベクター構築工程、形質転換工程、選抜工程等の工程を含む製造法方法として挙げることができる。以下、各工程について具体的に説明する。
【0050】
発現ベクター構築工程
発現ベクター構築工程は、上述した転写因子や転写共役因子をコードする遺伝子と転写抑制転換ポリヌクレオチドと、プロモーターとを含む組換え発現ベクターを構築する工程であれば特に限定されるものではない。組換え発現ベクターの母体となるベクターとしては、従来公知の種々のベクターを用いることができる。例えば、プラスミド、ファージ、またはコスミド等を用いることができ、導入される植物細胞や導入方法に応じて適宜選択することができる。具体的には、例えば、pBR322、pBR325、pUC19、pUC119、pBluescript、pBluescriptSK、pBI系のベクター等を挙げることができる。特に、植物体へのベクターの導入法がアグロバクテリウムを用いる方法である場合には、pBI系のバイナリーベクターを用いることが好ましい。pBI系のバイナリーベクターとしては、具体的には、例えば、pBIG、pBIN19、pBI101、pBI121、pBI221等を挙げることができる。
【0051】
プロモーターは、植物体内で遺伝子を発現させることが可能なプロモーターであれば特に限定されるものではなく、公知のプロモーターを好適に用いることができる。かかるプロモーターとしては、例えば、カリフラワーモザイクウイルス35Sプロモーター(CaMV35S)、各種アクチン遺伝子プロモーター、各種ユビキチン遺伝子プロモーター、ノパリン合成酵素遺伝子のプロモーター、タバコのPR1a遺伝子プロモーター、トマトのリブロース1,5−二リン酸カルボキシラーゼ・オキシダーゼ小サブユニット遺伝子プロモーター、ナピン遺伝子プロモーター、オレオシン遺伝子プロモーター等を挙げることができる。この中でも、カリフラワーモザイクウイルス35Sプロモーター、アクチン遺伝子プロモーター又はユビキチン遺伝子プロモーターをより好ましく用いることができる。上記各プロモーターを用いれば、植物細胞内に導入されたときに任意の遺伝子を強く発現させることが可能となる。プロモーターは、転写因子や転写共役因子をコードする遺伝子と転写抑制転換ポリヌクレオチドとを連結した融合遺伝子を発現しうるように連結され、ベクター内に導入されていればよく、組換え発現ベクターとしての具体的な構造は特に限定されるものではない。
【0052】
なお、組換え発現ベクターは、プロモーター及び上記融合遺伝子に加えて、さらに他のDNAセグメントを含んでいてもよい。当該他のDNAセグメントは特に限定されるものではないが、ターミネーター、選別マーカー、エンハンサー、翻訳効率を高めるための塩基配列等を挙げることができる。また、上記組換え発現ベクターは、さらにT−DNA領域を有していてもよい。T−DNA領域は特にアグロバクテリウムを用いて上記組換え発現ベクターを植物体に導入する場合に遺伝子導入の効率を高めることができる。
【0053】
転写ターミネーターは転写終結部位としての機能を有していれば特に限定されるものではなく、公知のものであってもよい。例えば、具体的には、ノパリン合成酵素遺伝子の転写終結領域(Nosターミネーター)、カリフラワーモザイクウイルス35Sの転写終結領域(CaMV35Sターミネーター)等を好ましく用いることができる。この中でもNosターミネーターをより好ましく用いることできる。上記組換えベクターにおいては、転写ターミネーターを適当な位置に配置することにより、植物細胞に導入された後に、不必要に長い転写物を合成したり、強力なプロモーターがプラスミドのコピー数の減少させたりするような現象の発生を防止することができる。
【0054】
形質転換体選別マーカーとしては、例えば薬剤耐性遺伝子を用いることができる。かかる薬剤耐性遺伝子の具体的な一例としては、例えば、ハイグロマイシン、ブレオマイシン、カナマイシン、ゲンタマイシン、クロラムフェニコール等に対する薬剤耐性遺伝子を挙げることができる。これにより、上記抗生物質を含む培地中で生育する植物体を選択することによって、形質転換された植物体を容易に選別することができる。
【0055】
翻訳効率を高めるための塩基配列としては、例えばタバコモザイクウイルス由来のomega配列を挙げることができる。このomega配列をプロモーターの非翻訳領域(5’UTR)に配置させることによって、上記融合遺伝子の翻訳効率を高めることができる。このように、上記組換え発現ベクターには、その目的に応じて、さまざまなDNAセグメントを含ませることができる。
【0056】
組換え発現ベクターの構築方法についても特に限定されるものではなく、適宜選択された母体となるベクターに、上記プロモーター、転写因子や転写共役因子をコードする遺伝子、および転写抑制転換ポリヌクレオチド、並びに必要に応じて上記他のDNAセグメントを所定の順序となるように導入すればよい。例えば、転写因子をコードする遺伝子と転写抑制転換ポリヌクレオチドとを連結して融合遺伝子を構築し、次に、この融合遺伝子とプロモーターと(必要に応じて転写ターミネーター等)とを連結して発現カセットを構築し、これをベクターに導入すればよい。
【0057】
キメラ遺伝子(融合遺伝子)の構築および発現カセットの構築では、例えば、各DNAセグメントの切断部位を互いに相補的な突出末端としておき、ライゲーション酵素で反応させることで、当該DNAセグメントの順序を規定することが可能となる。なお、発現カセットにターミネーターが含まれる場合には、上流から、プロモーター、上記キメラ遺伝子、ターミネーターの順となっていればよい。また、組換え発現ベクターを構築するための試薬類、すなわち制限酵素やライゲーション酵素等の種類についても特に限定されるものではなく、市販のものを適宜選択して用いればよい。
【0058】
また、上記組換え発現ベクターの増殖方法(生産方法)も特に限定されるものではなく、従来公知の方法を用いることができる。一般的には大腸菌をホストとして当該大腸菌内で増殖させればよい。このとき、ベクターの種類に応じて、好ましい大腸菌の種類を選択してもよい。
【0059】
形質転換工程
本発明において行われる形質転換工程は、上述した融合遺伝子を発現させるように、上述した組換え発現ベクターを用いて植物細胞に導入する工程である。組換え発現ベクターを用いて植物細胞に導入する方法(形質転換方法)は特に限定されるものではなく、植物細胞に応じた適切な従来公知の方法を用いることができる。具体的には、例えば、アグロバクテリウムを用いる方法や直接植物細胞に導入する方法を用いることができる。アグロバクテリウムを用いる方法としては、例えば、Bechtold, E., Ellis, J. and Pelletier, G. (1993) In Planta Agrobacterium-mediated gene transfer by infiltration of adult Arabidopsis plants. C.R. Acad. Sci. Paris Sci. Vie, 316, 1194-1199. あるいは、Zyprian E, Kado Cl, Agrobacterium-mediated plant transformation by novel mini-T vectors in conjunction with a high-copy vir region helper plasmid. Plant Molecular Biology, 1990, 15(2), 245-256.に記載された方法を用いることができる。
【0060】
組換え発現ベクターと対象となる遺伝子を含んだDNAを直接植物細胞に導入する方法としては、例えば、マイクロインジェクション法、エレクトロポレーション法(電気穿孔法)、ポリエチレングリコール法、パーティクルガン法、プロトプラスト融合法、リン酸カルシウム法等を用いることができる。
【0061】
また、DNAを直接植物細胞に導入する方法を採るなら、対象とする遺伝子の発現に必要な転写ユニット、例えプロモーターや転写ターミネーターと、対象とする遺伝子を含んだDNAあれば十分であり、ベクター機能が必須ではない。さらに、転写ユニットを有さない対象とする遺伝子のタンパク質コード領域のみを含むDNAであっても、宿主の転写ユニット無いにインテグレートし、対象となる遺伝子を発現することができればよい。
【0062】
上記組換え発現ベクターと対象となる遺伝子を含んだDNAや、発現ベクターを含まず対象となる遺伝子DNAを含んだDNAが導入される植物細胞としては、例えば、花、葉、根等の植物器官における各組織の細胞、カルス、懸濁培養細胞等を挙げることができる。ここで、本発明にかかる植物体の生産方法においては、上記組換え発現ベクターは、生産しようとする種類の植物体に合わせて適切なものを適宜構築してもよいが、汎用的な組換え発現ベクターを予め構築しておき、それを植物細胞に導入してもよい。すなわち、本発明に係る植物体の製造方法においては、上述した組換え発現ベクターを用いた形質転換用DNAの構築工程が含まれていてもよいし、含まれていなくてもよい。
【0063】
その他の工程、その他の方法
本発明に係る植物体の生産方法においては、上記形質転換工程が含まれていればよく、さらに上記組換え発現ベクターを用いた形質転換用DNAの構築工程が含まれていてもよいが、さらに他の工程が含まれていてもよい。具体的には、形質転換後の植物体から適切な形質転換体を選抜する選抜工程等を挙げることができる。
【0064】
選抜の方法は特に限定されるものではなく、例えば、ハイグロマイシン耐性等の薬剤耐性を基準として選抜してもよいし、形質転換体を育成した後に、植物体そのもの、または任意の器官や組織に含まれる油脂含有量から選抜してもよい。例えば、油脂含有量から選抜する例としては、形質転換体の種子から定法に従って油脂成分を定量し、形質転換していない植物体の種子に含まれる油脂含有量と比較する方法を挙げることができる(後述の実施例参照)。
【0065】
本発明に係る植物体の製造方法では、上記融合遺伝子を植物体に導入するため、該植物体から、有性生殖または無性生殖により油脂含有量が有意に向上した子孫を得ることが可能となる。また、該植物体やその子孫から植物細胞や、種子、果実、株、カルス、塊茎、切穂、塊等の繁殖材料を得て、これらを基に該植物体を量産することも可能となる。したがって、本発明に係る植物体の製造方法では、選抜後の植物体を繁殖させる繁殖工程(量産工程)が含まれていてもよい。
【0066】
なお、本発明における植物体とは、成育した植物個体、植物細胞、植物組織、カルス、種子の少なくとも何れかが含まれる。つまり、本発明では、最終的に植物個体まで成育させることができる状態のものであれば、全て植物体とみなす。また、上記植物細胞には、種々の形態の植物細胞が含まれる。かかる植物細胞としては、例えば、懸濁培養細胞、プロトプラスト、葉の切片等が含まれる。これらの植物細胞を増殖・分化させることにより植物体を得ることができる。なお、植物細胞からの植物体の再生は、植物細胞の種類に応じて、従来公知の方法を用いて行うことができる。したがって、本発明に係る植物体の製造方法では、植物細胞等から植物体を再生させる再生工程が含まれていてもよい。
【0067】
また、本発明に係る植物体の生産方法は、組換え発現ベクターで形質転換する方法に限定されるものではなく、他の方法を用いてもよい。具体的には、例えば、上記キメラタンパク質(融合タンパク質)そのものを植物体に投与してもよい。この場合、最終的に利用する植物体の部位において油脂含有量を向上できるように、若年期の植物体にキメラタンパク質(融合タンパク質)を投与すればよい。またキメラタンパク質(融合タンパク質)の投与方法も特に限定されるものではなく、公知の各種方法を用いればよい。
【0068】
以上説明したように、本発明によれば、所定の転写因子ファミリーに属する転写因子と上記機能性ペプチドとのキメラタンパク質を発現させることで、野生型の植物体と比較して、個体あたりの物質生産性が向上した植物体を提供することができる。また、所定の転写共役因子と上記機能性ペプチドとキメラタンパク質を発現させることで、野生型の植物体と比較して、個体あたりの物質生産性が向上した植物体を提供することができる。植物体に上記キメラタンパク質を発現させると、対象となる転写因子の転写促進活性が抑制される場合もあり、或いは対象となる転写因子が認識するcis配列の相同配列に対する転写抑制効果を示す場合もある。さらに、キメラタンパク質は、対象となる転写因子や転写共役因子に対して親和性を有する他の因子、DNA、RNA、脂質又は糖質に対して当該親和特異性を変化させるように作用する場合もあり、或いは対象となる転写因子に対して親和性の無い物質に対して親和性を向上させるように作用する場合もある。本発明に係る植物体においては、キメラタンパク質の対象となる転写因子、当該が認識するcis配列と相同性のあるcis配列を認識する転写因子、キメラタンパク質の対象となる転写因子と相同性のある転写因子、キメラタンパク質の対象となる転写因子に対して親和性を有する他の因子等も同様に植物体に発現しているものの、上述したキメラタンパク質の作用効果により、ドミナントネガティブに制御対象の遺伝子発現を抑制することができる。これにより、本発明に係る植物体においては、油脂生産に関連する遺伝子群及び/又は生産された油脂の分解に関連する遺伝子群の発現レベルが変化し、その結果、油脂含有量が有意に向上すると考えられる。
【0069】
ここで油脂含有量が有意に向上するとは、野生型と比較して一粒あたりの種子質量に変化はないが油脂量が向上した場合と、野生型と比較して一粒あたりの種子質量が有意に大となり油脂量が向上した場合、野性型と比較して種子中の油脂含量が向上した場合のいずれかを意味する。いずれの場合であっても、植物一個体が生産する油脂量が向上したこととなる。本発明に係る植物体は、植物由来の油性の製造方法に利用することができる。例えば、本発明に係る植物体を成長させて種子を採取し、採取した種子から油脂成分を回収することで油脂を製造することができる。
【0070】
特に本発明に係る植物体を利用した油脂の製造方法は、植物一個体における油脂含有量が高いため生産性に優れた方法であるといえる。換言すると、単位耕地面積あたりの栽培個体数が一定であると仮定すると、本発明に係る植物体を利用することによって単位耕地面積あたりから製造する油脂量が大幅に向上することとなる。したがって、本発明に係る植物体を利用することによって油脂生産に要する製造コストを大幅に削減することができる。
【0071】
さらに本発明に係る植物体を利用した油脂の製造方法は、単位重量あたりの種子における油脂含有量が高いため生産性に優れた方法であると言える。
【0072】
なお、本発明に係る植物体を利用した油脂の製造方法において、製造対象の油脂としては、特に限定されず、例えば、大豆油、ごま油、オリーブ油、椰子油、米油、綿実油、ひまわり油、コーン油、べに花油及び菜種油等の植物由来の油脂を例示することができる。また、製造した油脂は、家庭用途や工業用途に広く利用することができ、更にはバイオディーゼル燃料の原料としても使用することができる。すなわち、本発明に係る植物体を利用することによって、上述した家庭用途又は工業用途の油脂や、バイオディーゼル燃料等を低コストに製造することができる。
【0073】
植物由来油脂の製造方法
また、本発明では、特定の表現型を示す植物体から採取した種子において油脂含有量が有意に向上しているといった新規知見を見いだした。具体的には、参考文献(Plant J. 1995 Nov;8(5):659-71.)に開示された4種類の色素合成経路欠損株(tt4、tt5、tt6及びΔCHS)から採取した種子は、野生型と比較して種子における油脂含量が有意に向上している。すなわち、本発明に係る植物由来油脂の製造方法は、カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子及びフラボン-3-ヒドラーゼ遺伝子からなる群から選ばれる少なくとも1の遺伝子の機能を欠損した植物体から採取した種子から油脂成分を回収する工程を含むものである。なお、上記参考文献に開示されたtt4株及びΔCHSはカルコンシンターゼ遺伝子を欠損した株であり、tt5株はカルコンイソメラーゼ遺伝子を欠損した株であり、tt6株はフラボン-3-ヒドラーゼ遺伝子を欠損した株である。
【0074】
シロイヌナズナにおけるカルコンシンターゼ遺伝子の塩基配列を配列番号5に示し、当該遺伝子によりコードされるカルコンシンターゼのアミノ酸配列を配列番号6に示す。シロイヌナズナにおけるカルコンイソメラーゼ遺伝子の塩基配列を配列番号7に示し、当該遺伝子によりコードされるカルコンイソメラーゼのアミノ酸配列を配列番号8に示す。シロイヌナズナにおけるフラボン-3-ヒドラーゼ遺伝子の塩基配列を配列番号9に示し、当該遺伝子によりコードされるフラボン-3-ヒドラーゼのアミノ酸配列を配列番号10に示す。
【0075】
但し、本発明においてカルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子及びフラボン-3-ヒドラーゼ遺伝子は、上記の具体的な配列に限定されるものではない。すなわち、カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子及びフラボン-3-ヒドラーゼ遺伝子は、上記の具体的なアミノ酸配列において1又は複数個のアミノ酸配列が欠失、置換、付加又は挿入されたアミノ酸配列を含み、且つ、カルコンシンターゼ活性、カルコンイソメラーゼ活性及びフラボン-3-ヒドラーゼ活性を有するタンパク質をコードするものであっても良い。ここで、複数個のアミノ酸としては、例えば、1から20個、好ましくは1から10個、より好ましくは1から7個、さらに好ましくは1個から5個、特に好ましくは1個から3個を意味する。なお、アミノ酸の欠失、置換若しくは付加は、上記の具体的な塩基配列を、当該技術分野で公知の手法によって改変することによって行うことができる。塩基配列に変異を導入するには、Kunkel法またはGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-KやMutant-G(何れも商品名、TAKARA Bio社製))等を用いて、あるいはLA PCR in vitro Mutagenesisシリーズキット(商品名、TAKARA Bio社製)を用いて変異が導入される。また、変異導入方法としては、EMS(エチルメタンスルホン酸)、5-ブロモウラシル、2-アミノプリン、ヒドロキシルアミン、N-メチル-N’-ニトロ-Nニトロソグアニジン、その他の発ガン性化合物に代表されるような化学的変異剤を使用する方法でも良いし、X線、アルファ線、ベータ線、ガンマ線、イオンビームに代表されるような放射線処理や紫外線処理による方法でも良い。
【0076】
さらに、本発明において、カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子及びフラボン-3-ヒドラーゼ遺伝子には、シロイヌナズナ以外の植物(例えば上述した植物)において同機能を有する遺伝子(以下、相同遺伝子と称す)が含まれる。カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子又はフラボン-3-ヒドラーゼ遺伝子の相同遺伝子は、植物ゲノム情報が明らかになっていれば、カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子又はフラボン-3-ヒドラーゼ遺伝子の塩基配列又は当該遺伝子によりコードされるアミノ酸配列に基づいて、検索対象の植物ゲノム情報から検索することができる。このとき、相同転写因子としては、上記の具体的なアミノ酸配列に対して、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、最も好ましくは95%以上の相同性を有するアミノ酸配列として検索される。ここで、相同性の値は、blastアルゴリズムを実装したコンピュータプログラム及び遺伝子配列情報を格納したデータベースを用いてデフォルトの設定で求められる値を意味する。
【0077】
また、植物ゲノム情報が明らかとなっていない場合には、対象となる植物からゲノムを抽出するか或いは対象となる植物のcDNAライブラリーを構築し、カルコンシンターゼ遺伝子、カルコンイソメラーゼ遺伝子又はフラボン-3-ヒドラーゼ遺伝子の塩基配列の少なくとも一部に対して、ストリンジェントな条件下でハイブリダイズするゲノム領域或いはcDNAを単離することで相同遺伝子を同定することができる。ここで、ストリンジェントな条件とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。例えば、45℃、6×SSC(塩化ナトリウム/クエン酸ナトリウム)でのハイブリダイゼーション、その後の50〜65℃、0.2〜1×SSC、0.1%SDSでの洗浄が挙げられ、或いはそのような条件として、65〜70℃、1×SSCでのハイブリダイゼーション、その後の65〜70℃、0.3×SSCでの洗浄を挙げることができる。ハイブリダイゼーションは、J. Sambrook et al. Molecular Cloning, A Laboratory Manual,2nd Ed., Cold Spring Harbor Laboratory(1989)に記載されている方法等、従来公知の方法で行うことができる。
【0078】
換言すると、本発明に係る植物由来油脂の製造方法は、シロイヌナズナ由来の種子を用いる系に限定されず、あらゆる植物を対象として適用することができる。本発明に係る植物由来油脂の製造方法を適用できる植物としては、例えば、双子葉植物、単子葉植物、例えばアブラナ科、イネ科、ナス科、マメ科、ヤナギ科等に属する植物(下記参照)が挙げられるが、これらの植物に限定されるものではない。
【0079】
アブラナ科:シロイヌナズナ(Arabidopsis thaliana)、アブラナ(Brassica rapa、Brassica napus)、キャベツ(Brassica oleracea var. capitata)、ナタネ(Brassica rapa、Brassica napus)、ナノハナ(Brassica rapa、Brassica napus)、ハクサイ(Brassica rapa var. pekinensis)、チンゲンサイ(Brassica rapa var. chinensis)、カブ(Brassica rapa var. rapa)、ノザワナ(Brassica rapa var. hakabura)、ミズナ(Brassica rapa var. lancinifolia)、コマツナ(Brassica rapa var. peruviridis)、パクチョイ(Brassica rapa var. chinensis)、ダイコン(Brassica Raphanus sativus)、ワサビ(Wasabia japonica)など。
【0080】
ナス科:タバコ(Nicotiana tabacum)、ナス(Solanum melongena)、ジャガイモ(Solaneum tuberosum)、トマト(Lycopersicon lycopersicum)、トウガラシ(Capsicum annuum)、ペチュニア(Petunia)など。
【0081】
マメ科:ダイズ(Glycine max)、エンドウ(Pisum sativum)、ソラマメ(Vicia faba)、フジ(Wisteria floribunda)、ラッカセイ(Arachis. hypogaea)、ミヤコグサ(Lotus corniculatus var. japonicus)、インゲンマメ(Phaseolus vulgaris)、アズキ(Vigna angularis)、アカシア(Acacia).など。
【0082】
キク科:キク(Chrysanthemum morifolium)、ヒマワリ(Helianthus annuus)など。
【0083】
ヤシ科:アブラヤシ(Elaeis guineensis、Elaeis oleifera)、ココヤシ(Cocos nucifera)、ナツメヤシ(Phoenix dactylifera)、ロウヤシ(Copernicia)
【0084】
ウルシ科:ハゼノキ(Rhus succedanea)、カシューナットノキ(Anacardium occidentale)、ウルシ(Toxicodendron vernicifluum)、マンゴー(Mangifera indica)、ピスタチオ(Pistacia vera)
【0085】
ウリ科:カボチャ(Cucurbita maxima、Cucurbita moschata、Cucurbita pepo)、キュウリ(Cucumis sativus)、カラスウリ(Trichosanthes cucumeroides)、ヒョウタン(Lagenaria siceraria var. gourda)
【0086】
バラ科:アーモンド(Amygdalus communis)、バラ(Rosa)、イチゴ(Fragaria)、サクラ(Prunus)、リンゴ(Malus pumila var. domestica)など。
【0087】
ナデシコ科:カーネーション(Dianthus caryophyllus)など。
【0088】
ヤナギ科:ポプラ(Populus trichocarpa、Populus nigra、Populus tremula)
【0089】
イネ科:トウモロコシ(Zea mays)、イネ(Oryza sativa)、オオムギ(Hordeum vulgare)、コムギ(Triticum aestivum)、タケ(Phyllostachys)、サトウキビ(Saccharum officinarum)など。
【0090】
ユリ科:チューリップ(Tulipa)、ユリ(Lilium)など。
【0091】
また、遺伝子の機能を欠損させるとは、当該遺伝子をゲノムから欠失させること、当該遺伝子の発現(転写レベル及び翻訳レベル)を阻害すること、及び当該遺伝子によってコードされるタンパク質の活性を低下又は欠失させることを含む意味である。
【0092】
より詳細に、遺伝子を欠失させる方法としては、特に限定されないが、相同組換えを用いた方法やトランスポゾンを用いた方法を挙げることができる。また、当該遺伝子を欠失させる場合には、当該遺伝子の全長を欠失させても良いし、部分的に欠失させても良い。
【0093】
また、遺伝子の発現を阻害する方法としては、特に限定しないが、当該遺伝子の発現を制御しているプロモーターを欠失させる方法、当該遺伝子の発現を制御しているプロモーターを発現誘導型プロモーターに置換する方法、当該遺伝子の発現を制御しているプロモーターに突然変異を導入する方法、RNA干渉を利用して当該遺伝子の転写産物を分解する方法、及びアンチセンスRNAを利用して当該遺伝子の翻訳を阻害する方法を挙げることができる。
【0094】
さらに、遺伝子がコードするタンパク質の活性を低下させる方法としては、当該タンパク質に特異的に結合して当該タンパク質の活性を抑制する機能を有する物質を作用させる方法を挙げることができる。当該物質としては、当該タンパク質の機能を阻害できる抗体や阻害物質を挙げることができる。
【0095】
本発明に係る植物由来油脂の製造方法において種子から油脂を回収する方法は、特に限定されず、圧搾法、抽出法及び圧出法などの如何なる方法を用いても良い。例えば、ソックスレー抽出器を用いたエーテル抽出法によって、植物から採取した種子から油脂成分を回収することができる。本発明に係る植物由来油脂の製造方法によれば、植物一個体から採取できる種子量が同等であっても、種子一粒あたりの油脂含有量が高いため植物体を用いているため、生産性に優れた方法であるといえる。換言すると、単位耕地面積あたりの栽培個体数が一定であると仮定すると、本発明に係る植物由来油脂の製造方法によれば、単位耕地面積あたりから製造する油脂量が大幅に向上することとなり、油脂生産に要する製造コストを大幅に削減することができる。
【0096】
なお、本発明に係る植物由来油脂の製造方法において、製造対象の油脂としては、特に限定されず、例えば、大豆油、ごま油、オリーブ油、椰子油、米油、綿実油、ひまわり油、コーン油、べに花油及び菜種油等の植物由来の油脂を例示することができる。また、製造した油脂は、家庭用途や工業用途に広く利用することができ、更にはバイオディーゼル燃料やバイオプラスチックの原料としても使用することができる。すなわち、本発明に係る植物体を利用することによって、上述した家庭用途又は工業用途の油脂や、バイオディーゼル燃料やバイオプラスチック等を低コストに製造することができる。
【0097】
油脂量が向上した植物体のスクリーニング方法
上述したように、本発明では、色素合成経路欠損株(参考文献:Plant J. 1995 Nov;8(5):659-71.)から採取した種子において、野生型と比較して油脂含有量が有意に向上しているといった新規知見を見いだした。色素合成経路欠損株は、色素合成系に関与する遺伝子の機能が欠損した変異株であって、野生株と比較して種皮色が淡色(野生型と比較してより白色)であるという表現型を示す。上記参考文献に開示されたtt4株及びΔCHSはカルコンシンターゼ遺伝子を欠損した株であり、tt5株はカルコンイソメラーゼ遺伝子を欠損した株であり、tt6株はフラボン-3-ヒドラーゼ遺伝子を欠損した株である。これら遺伝子を欠損した変異株においては、色素の合成不全により種皮色がより白色となる。このため、スクリーニング対象の植物から種子を採取し、採取した種子の種皮色を確認すると、当該植物における色素合成経路による色素合成能が判断できるとともに、採取した種子に含まれる種子含量を高精度に推定することができる。
【0098】
すなわち、例えば、同種に含まれる様々な植物体があるとき、これら植物体から採取した種子の種皮色を観察してより白色であるものは油脂生産量の高い品種であると選抜することができる。ここで、スクリーニング対象となる植物体とは、何らかの変異原処理を施したものであっても良いし、従来公知の育種法等によって作出された植物品種であっても良い。
【0099】
ここで、変異原処理としては、特に限定されず、広く突然変異の誘発に用いられている化学的変異原及び/又は物理的変異原による処理を用いることができる。化学的突然変異源として、例えばメタンスルホン酸エチル(EMS)、エチルニトロソ尿素(ENS)、2-アミノプリン、5-ブロモウラシル(5-BU)、アルキル化剤などが用いることができる。また、物理的変異原としては、放射線、紫外線等を用いることができる。これらの変異原を用いた変異の誘発は公知の方法で行うことができる。
【0100】
本発明に係るスクリーニング方法によれば、植物から採取した種子を破壊する必要がなく、目視により種皮色を観察するといった非常に簡便且つ迅速な判定法により、種子に含まれる油脂量を判定することができる。
【0101】
また、本発明に係るスクリーニング方法では、植物から採取した種子の種皮色を画像データから判断し、種皮色を定量的に測定しても良い。具体的には、評価対象の種子の画像をデジタルデータに変換し、画像データ中の種子領域のR値、G値及びB値(RGB値)を測定する。種子領域のR値、G値及びB値の測定には、画像処理ソフトウェアであれば如何なるソフトウェアを使用しても良い。次に、測定されたR値、G値及びB値を、野生型の種子におけるR値、G値及びB値と比較する。一例としては、測定されたR値、G値及びB値の積算値を算出し、野生型の種子におけるR値、G値及びB値の積算値と比較する。例えば、測定されたR値、G値及びB値の積算値が、野生型の種子におけるR値、G値及びB値の積算値として有意に上昇していれば、測定対象の種子の種皮色はより白色に近いと判断することができる。特に、測定されたR値、G値及びB値の積算値が、野生型の種子におけるR値、G値及びB値の積算値に対して、2.88倍以上の値を示す場合には、より白色化(淡色化)した種子であると判断することができる。
【0102】
以上のように、植物から採取した種子の種皮色を画像データとして観察するといった手法であっても種子を破壊する必要がなく、非常に簡便且つ迅速な判定法により、種子に含まれる油脂量を判定することができる。なお、画像データから種子の種皮色を観察する際に、R値、G値及びB値の積算値に限らず、R値、G値及びB値の合計値等を算出しても良い。
【実施例】
【0103】
以下、実施例により本発明をより詳細に説明するが、本発明の技術的範囲はこれら実施例に限定されるものではない。
【0104】
〔実施例1〕
本実施例では、シロイヌナズナにおける転写共役因子At5g24520及び転写因子At1g71030について、それぞれリプレッサードメイン配列を付加したキメラタンパク質(融合タンパク質)を植物体において発現させ、当該植物体から採取した種子における油脂含有量を測定した。また、比較のため転写因子At1g56650についても、同様にしてキメラタンパク質(融合タンパク質)を植物体において発現させ、種子における油脂含有量を測定した。
【0105】
転写因子遺伝子の増幅
シロイヌナズナのcDNAライブラリーより、以下に記載するプライマーを用いて、At1g71030の終始コドンを除くコード領域のDNA断片並びに終止コドンを含むコード領域のDNA断片、At5g24520の終始コドンを除くコード領域のDNA断片及びAt1g56650の終始コドンを除くコード領域のDNAをPCRにより増幅した。PCR条件は94℃1分、47℃2分、伸長反応74℃1分を25サイクル行なった。PCR終了後、増幅されたDNA断片をアガローズゲル電気泳動により分離、回収した。
・At1g71030増幅用フォワードプライマー1
gATGAACAAAACCCGCCTTCGTGCTCTCTC(配列番号11)
・At1g71030増幅用リバースプライマー1
TCGGAATAGAAGAAGCGTTTCTTGACCTGT(配列番号12)
・At1g71030増幅用フォワードプライマー2
gATGAACAAAACCCGCCTTCGTGCTCTCTC(配列番号13)
・At1g71030増幅用リバースプライマー2(配列番号14)
TCATCGGAATAGAAGAAGCGTTTCTTGACC
・At1g56650増幅用フォワードプライマー
GATGGAGGGTTCGTCCAAAGGGC(配列番号15)
・At1g56650増幅用リバースプライマー
ATCAAATTTCACAGTCTCTCCATCG(配列番号16)
・At5g24520増幅用フォワードプライマー
gATGGATAATTCAGCTCCAGATTCGTTATC(配列番号17)
・At5g24520増幅用リバースプライマー
AACTCTAAGGAGCTGCATTTTGTTAGCAAA(配列番号18)
【0106】
融合遺伝子の作製
上記DNA断片がコードする転写因子遺伝子の3'末端にリプレッサードメイン配列を付加するために、CaMV35Sプロモーターの下流にSmaIサイトとリプレッサードメイン(アミノ酸配列:GLDLDLELRLGFA)配列を有するベクターであるp35SSXGを用いた。転写因子遺伝子配列とリプレッサードメイン配列を連結するために、本ベクターをSmaIで切断し、上記の転写因子をコードするPCR増幅断片を挿入し、p35SSXG(At1g56650)とp35SSXG(At5g24520)、 p35SSXG(At1g71030)を作製した。 なおp35SSXG(At1g71030)はAt1g71030増幅用フォワードプライマー1及びAt1g71030増幅用リバースプライマー1を用いたPCR増幅断片を挿入した。また、At1g71030増幅用フォワードプライマー2及びAt1g71030増幅用リバースプライマー2を用いたPCR増幅断片を、リプレッサードメインを付加せずに発現するために、CaMV35Sプロモーターの下流にSmaIサイト配列を有するベクターである、p35SOXGをSmaIで切断部位に挿入し、p35SOXG(At1g71030)を作製した。
【0107】
改良型転写因子および転写因子発現ベクターの構築
アグロバクテリウムにより植物に遺伝子導入を行なうためのバイナリーベクターとしてはpBCKHを用いた。本ベクターはpBIG(Hygr)(Nucleic Acids Res. 18, 203 (1990))のHindIIIサイトにGatewayベクターコンバージョンシステム(Invitrogen)のカセットを組み込んだものである。このベクターに改良型転写因子遺伝子を組み込むために、本ベクターと、p35SSXG(At1g56650)、p35SSXG(At5g24520)、p35SSXG(At1g71030)またはp35SOXG(At1g71030)を混合し、GATEWAY LR clonase (Invitrogen)を用いて組換え反応を行った。その結果、pBCKH-p35SSXG(At1g56650)、pBCKH-p35SSXG(At5g24520)、pBCKH-p35SSXG(At1g71030)及びpBCKH-p35SOXG(At1g71030)を作製した。
【0108】
改良型転写因子遺伝子発現ベクターの植物への導入
改良型転写因子を導入する植物にはシロイヌナズナ(Arabidopsis thaliana, Columbia )を用いた。遺伝子導入法は、Transformation of Arabidopsis thaliana by vacuum infiltration に従った。ただし、感染させるのに減圧処理は行なわず、アグロバクテリウム菌液に浸すだけとした。具体的には、改良型転写因子発現ベクター pBCKH-p35SSXG(At1g56650)、pBCKH-p35SSXG(At5g24520)、pBCKH-p35SSXG(At1g71030)及びpBCKH-p35SOXG(At1g71030)を、土壌細菌Agrobacterium tumefaciens strain GV3101 (C58C1Rifr) pMP90 (Gmr)(koncz and Schell 1986)株にエレクトロポレーション法で導入した。
【0109】
導入した菌を1リットルの、抗生物質(カナマイシン(Km)50μg/ml、ゲンタマイシン(Gm)25μg/ml、リファンピシリン(Rif)50μg/ml)を含むYEP培地でOD600が1になるまで培養した。次いで、培養液から菌体を回収し、1リットルの感染用培地(Infiltration medium、1リッターあたり、2.2 g MS salt, 1X B5 vitamins, 50 g sucrose, 0.5 g MES, 0.044 μM benzylaminopurine, 400μl Silwetを含む。pH 5.7) に懸濁した。この溶液に、14日間生育したシロイヌナズナを1分間浸し、感染させた後、再び栽培を継続し結実させた。採種した種子(T1種子)を50%ブリーチ、0.02%Triton X-100溶液で7分間滅菌した後、滅菌水で3回リンスし、滅菌したハイグロマイシン選択培地(4.3g/l MS salts, 0.5 % sucrose, 0.5 g/l MES, pH 5.7, 0.8 % agar, 30mg/l hygromycin, 250 mg/l Vancomycin)に播種した。上記ハイグロマイシンプレートで生育する形質転換植物体(T1植物)を各改良型転写遺伝子につき10系統を選抜し、バーミキュライト混合土を入れた直径50mmのポットに移植した。これを22℃、16時間明期8時間暗期、光強度約60〜80μE/cm2で栽培し種子(T2種子)を得た。得られたT2種子の表皮色は、野生株が濃い茶色であるのに対して、どの系統も薄茶色もしくは黄色であった。
【0110】
色素合成経路欠損株
また、本実施例では、色素合成経路欠損株から採取した種子に含まれる油脂含有量も測定した。本実施例では、具体的に色素合成経路欠損株tt4(NASC stock No. N85) (参考文献:Plant J., 8, 659-671, 1995)、tt5(NASC stock No. N86)、tt6(NASC stock No. N87) (参考文献:Plant Physiol., 111, 339-345, 1996)、ΔCHS(NASC stock No. N520583))についてはNASC(The Nottingham Arabidopsis Stock Centre)より入手した。tt4、tt5、tt6はArabidopsis thaliana, Ler株より作製され、ΔCHSはArabidopsis thaliana, Col-0株より作製された。これをを50%ブリーチ、0.02%Triton X-100溶液で7分間滅菌した後、滅菌水で3回リンスし、培地(4.3g/l MS salts, 0.5 % sucrose, 0.5 g/l MES, pH 5.7, 0.8 % agar)に播種した。上記プレートで生育する植物体バーミキュライト混合土を入れた直径50mmのポットに移植した。これを22℃、16時間明期8時間暗期、光強度約50〜60μE/cm2(tt4, tt5, tt6, WT(Ler))または、光強度約40μE/cm2(ΔCHS、WT(Col-o))で栽培し種子を得た。得られた種子の表皮色は、野生株が濃い茶色であるのに対して、どの系統も薄茶色もしくは黄色であった。
【0111】
改良型転写因子または転写因子導入T2種子の分析
2種類の改良型転写因子遺伝子及び改良型転写共役因子遺伝子のいずれかを導入したT2種子(At1g56650-SRDX、At5g24520-SRDX、At1g71030-SRDX)および転写因子を導入したT2種子(At1g71030)およびおよび野生株(Col-0、Ler)の油脂含量分析を行なった。油脂の定量分析はMARAN-23 (ResonanceInsturuments Ltd., UK) H-NMRと、解析ソフトRI-NMR Ver. 2.0を用い、2〜10mgのシロイヌナズナ種子を測定した。油脂の標準物質にはオリーブオイルを用いて検量線を作製し、種子中の油脂含量(重量%)を求めた。
【0112】
各改良型転写因子遺伝子、改良型転写共役因子若しくは転写因子遺伝子を導入した系統及び野生株の種子油脂含量の平均値(n=3〜10)を求めた。その結果、Col-0の油脂含量平均値を1とした時の各系統の油脂含量増加率は、T2種子(At1g56650-SRDX)は30.2%、T2種子(At5g24520-SRDX)は12.3%、T2種子(At1g71030-SRDX)は12.2%、T2種子(At1g71030)は2.3%であった(図1)。
【0113】
色素合成経路欠損株種子の分析
4種類の色素合成経路欠損株種子(tt4、tt5、tt6、ΔCHS)およびおよび野生株(Col-0、Ler)の油脂含量分析を行なった。油脂の定量分析はMARAN-23(ResonanceInsturuments Ltd., UK) H-NMRと、解析ソフトRI-NMR Ver. 2.0を用い、2〜10mgのシロイヌナズナ種子を測定した。油脂の標準物質にはオリーブオイルを用いて検量線を作製し、種子中の油脂含量(重量%)を求めた。
【0114】
色素合成経路欠損株及び野生株の種子油脂含量の平均値(n=3〜10)を求めた。ΔCHSはCol-0株に対して8.9%、tt4、tt5及びtt6はLer株に対してそれぞれ、4.7%、8.8%、11.1%の油脂含量であった(図2)。
【0115】
結果と考察
以上の結果から、リプレッサードメインを付加した転写因子At1g56650、転写共役因子At5g24520、転写因子At1g71030それぞれのキメラ遺伝子を導入した植物体の種子の重量あたり油脂含量は、同時に栽培した野生株の重量あたり油脂含量に比べ優れており、油脂生産において非常に有効な植物体であることが判明した。一方で発現促進活性を持つAt1g71030を導入した植物体の種子の重量あたり油脂含量は、同時に栽培した植物体の重量あたり油脂含量とくらべ若干増加していたが、その増加率は発現促進活性を抑制したAt1g71030を導入した植物体種子の重量あたり油脂含量の増加率の1/5程度であった。At1g71030は、シングルMYB様のドメインを持つタンパク質(AtMybL2)をコードし、この遺伝子をCaMV35Sプロモーターで過剰発現することにより、葉、茎、萼のトライコームを欠失する形質を示す。これは、トライコームの形成に必要なGL2遺伝子の発現が抑制されることによると考えられる(参考文献:DNA Res., 9, 31-34, 2002)。GL2遺伝子を破壊することにより、種子の油脂含量が8%増加することが報告されている(参考文献:Plant Mol Biol. 2006 , 60, :377-87, 2006)。
【0116】
また、AtMybL2タンパク質はそのカルボキシ末端領域に6アミノ酸からなる転写リプレッサーを持ち、AtMybL2遺伝子の過剰発現植物及びEAR-motifとして知られる転写リプレッサーを付加したAtMybL2をコードする遺伝子の過剰発現植物では、いずれの場合にもアントシアニン前駆体の合成が抑制された(参考文献:18TH INTERNATIONAL CONFERENCE ON ARABIDOPSIS RESEARCH, TAIR accession Publication:501721814)。しかしながら、解析の結果、At1g71030の過剰発現体のT2種子の油脂含量増加率2.3%に対して、リプレッサードメインを付加したAt1g71030の過剰発現体のT2種子は油脂含量の増加率は12.2%と大幅に高く、またGL2遺伝子破壊時の油脂含量増加率8%よりも顕著に高かった。これらの結果より、リプレッサードメインを付加したAt1g71030がGL2以外の未知の経路で種子の油脂合成と貯蔵過程に作用をおよぼし、油脂含量を増加したと考えられる。
【0117】
一方で、色素合成経路欠損株の分析結果より、色素合成経路の主要遺伝子が破壊された突然変異株tt4、tt5、tt6、T-DNAの挿入によりCHS遺伝子が破壊されたΔCHS株の種子の油脂含量はいずれも野生株に対して高かった。種皮色と油脂含量ナタネでは種皮色が黄色い品種HUA-yellow No. 1は種皮色が黒い品種と比べ、油脂含量が5-7%高いと報告されている(参考文献:Genome 44: 1077-1082 (2001))。しかしながら、異種間の交配に起因する従来育種法では、種皮色と種子の油脂含量を決定する形質の遺伝子座が近接していた場合でも同様の減少が観察されうる。よって、遺伝子の発現と形質の相関関係についてはこれまで、明らかにされていない。すなわち、従来、種皮色の形質を左右する遺伝子座が油脂含量に影響するといった知見は知られていなかった。
【0118】
これに対して、本結果では、実際に種皮の色素合成酵素をコードする遺伝子を破壊することにより種子中の油脂含量の増加が確認できた。よって、交配による従来育種法のみならず、遺伝子導入法、遺伝子破壊法による分子育種法においても、種皮色が油脂含量を予想する重要な表現型であることが初めて明らかになった。種皮色を指標に用いることにより種子の油脂含量を非破壊かつ特別な装置を用いずに、効率よく油脂含量が増加した種子を選抜することが可能になる。
【0119】
より詳細に、野生株、At1g71030-SRDX、At1g56650-SRDX、ΔCHS系統の種子の写真を撮影しデジタルデータに変換した。得られたデジタルデータを画像処理ソフト(アドビ・フォトショップ)を用いて種子の領域のRGB値をそれぞれを定量した。次に、定量したR値、G値及びB値の積算値を算出した。また、野生型におおいて定量したR値、G値及びB値の積算値に対する比率も算出した。その結果を表1及び図3に示す。
【0120】
【表1】
【0121】
表1及び図3に示すように、At1g71030-SRDX、At1g56650-SRDX及びΔCHS系統においては、R値、G値及びB値の積算値を野生型と比較したところ少なくとも2.88倍以上の値を示していた。以上のように、種子の画像データを用いて種皮色を定量的に測定し、種子中の油脂量を非常に簡便に且つ迅速に評価できることが可能となった。
【図面の簡単な説明】
【0122】
【図1】各改良型転写因子遺伝子、改良型転写共役因子遺伝子若しくは転写因子遺伝子を導入した系統及び野生株の種子油脂含量を測定した結果を示す特性図である。
【図2】色素合成経路欠損株及び野生株の種子油脂含量を測定した結果を示す特性図である。
【図3】種子の種皮色について画像データを用いてR値、G値及びB値の積算値を算出し、野生型の種子と比較した結果を示す特性図である。
【特許請求の範囲】
【請求項1】
配列番号4に示すアミノ酸配列からなるタンパク質を含む転写因子ファミリーに属する転写因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質を発現させた植物体。
【請求項2】
上記転写因子の転写促進活性が抑制されていることを特徴とする請求項1記載の植物体。
【請求項3】
上記キメラタンパク質が転写抑制因子活性をもつことを特徴とする請求項1記載の植物体。
【請求項4】
上記転写因子が、以下の(a)〜(c)のいずれかのタンパク質であることを特徴とする請求項1記載の植物体。
(a)配列番号4に示すアミノ酸配列を含むタンパク質
(b)配列番号4に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、転写促進活性を有するタンパク質
(c)配列番号3に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ転写促進活性を有するタンパク質
【請求項5】
上記機能性ペプチドが、次に示す式(1)〜(8)
(1)X1−Leu−Asp−Leu−X2−Leu−X3
(但し、式中、X1は0〜10個のアミノ酸残基を示し、X2はAsn又はGluを示し、X3は少なくとも6個のアミノ酸残基を示す。)
(2)Y1−Phe−Asp−Leu−Asn−Y2−Y3
(但し、式中、Y1は0〜10個のアミノ酸残基を示し、Y2はPhe又はIleを示し、Y3は少なくとも6個のアミノ酸残基を示す。)
(3)Z1−Asp−Leu−Z2−Leu−Arg−Leu−Z3
(但し、式中、Z1はLeu、Asp−Leu又はLeu−Asp−Leuを示し、Z2はGlu、Gln又はAspを示し、Z3は0〜10個のアミノ酸残基を示す。)
(4)Asp−Leu−Z4−Leu−Arg−Leu
(但し、式中、Z4はGlu、Gln又はAspを示す。)
(5)α1−Leu−β1−Leu−γ1−Leu
(6)α1−Leu−β1−Leu−γ2−Leu
(7)α1−Leu−β2−Leu−Arg−Leu
(8)α2−Leu−β1−Leu−Arg−Leu
(但し、式(5)〜(8)中、α1はAsp、Asn、Glu、Gln、Thr又はSerを示し、α2はAsn、Glu、Gln、Thr又はSerを示し、β1はAsp、Gln、Asn、Arg、Glu、Thr、Ser又はHisを示し、β2はAsn、Arg、Thr、Ser又はHisを示し、γ1はArg、Gln、Asn、Thr、Ser、His、Lys又はAspを示し、γ2はGln、Asn、Thr、Ser、His、Lys又はAspを示す。)
のいずれかで表されるアミノ酸配列を有するものであることを特徴とする請求項1記載の植物体。
【請求項6】
油脂生産性が有意に向上したことを特徴とする請求項1乃至5いずれか一項記載の植物体。
【請求項7】
特定の組織中の油脂含量が有意に向上したことを特徴とする請求項1乃至5いずれか一項記載の植物体。
【請求項8】
特定の組織中が種子であることを特徴とする請求項7記載の植物体。
【請求項9】
被子植物であることを特徴とする請求項1乃至8いずれか一項記載の植物体。
【請求項10】
双子葉植物であることを特徴とする請求項1乃至8いずれか一項記載の植物体。
【請求項11】
アブラナ科植物であることを特徴とする請求項1乃至8いずれか一項記載の植物体。
【請求項12】
シロイヌナズナであることを特徴とする請求項1乃至8いずれか一項記載の植物体。
【請求項13】
請求項1乃至12いずれか一項記載の植物体から、生産性が向上した物質を分離及び回収する工程を含む、植物体を用いた物質の製造方法。
【請求項14】
上記物質は油脂であることを特徴とする請求項13記載の植物体を用いた物質の製造方法。
【請求項15】
配列番号4に示すアミノ酸配列からなるタンパク質を含む転写因子ファミリーに属する転写因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質。
【請求項16】
上記転写因子が以下の(a)〜(c)のいずれかのタンパク質であることを特徴とする請求項15記載のキメラタンパク質。
(a)配列番号4に示すアミノ酸配列を含むタンパク質
(b)配列番号4に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、転写促進活性を有するタンパク質
(c)配列番号3に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ転写促進活性を有するタンパク質
【請求項17】
上記機能性ペプチドが、次に示す式(1)〜(8)
(1)X1−Leu−Asp−Leu−X2−Leu−X3
(但し、式中、X1は0〜10個のアミノ酸残基を示し、X2はAsn又はGluを示し、X3は少なくとも6個のアミノ酸残基を示す。)
(2)Y1−Phe−Asp−Leu−Asn−Y2−Y3
(但し、式中、Y1は0〜10個のアミノ酸残基を示し、Y2はPhe又はIleを示し、Y3は少なくとも6個のアミノ酸残基を示す。)
(3)Z1−Asp−Leu−Z2−Leu−Arg−Leu−Z3
(但し、式中、Z1はLeu、Asp−Leu又はLeu−Asp−Leuを示し、Z2はGlu、Gln又はAspを示し、Z3は0〜10個のアミノ酸残基を示す。)
(4)Asp−Leu−Z4−Leu−Arg−Leu
(但し、式中、Z4はGlu、Gln又はAspを示す。)
(5)α1−Leu−β1−Leu−γ1−Leu
(6)α1−Leu−β1−Leu−γ2−Leu
(7)α1−Leu−β2−Leu−Arg−Leu
(8)α2−Leu−β1−Leu−Arg−Leu
(但し、式(5)〜(8)中、α1はAsp、Asn、Glu、Gln、Thr又はSerを示し、α2はAsn、Glu、Gln、Thr又はSerを示し、β1はAsp、Gln、Asn、Arg、Glu、Thr、Ser又はHisを示し、β2はAsn、Arg、Thr、Ser又はHisを示し、γ1はArg、Gln、Asn、Thr、Ser、His、Lys又はAspを示し、γ2はGln、Asn、Thr、Ser、His、Lys又はAspを示す。)
のいずれかで表されるアミノ酸配列を有するものであることを特徴とする請求項15記載のキメラタンパク質。
【請求項18】
請求項15乃至17いずれか一項記載のキメラタンパク質をコードする遺伝子。
【請求項19】
請求項18記載の遺伝子を含む発現ベクター。
【請求項20】
請求項18記載の遺伝子を含む形質転換体。
【請求項21】
カルコンシンターゼ遺伝子の機能を欠損した植物体から採取した種子から油脂成分を回収する工程を含む、植物由来油脂の製造方法。
【請求項22】
上記カルコンシンターゼ遺伝子が、以下の(a)〜(c)のいずれかのタンパク質をコードする遺伝子であることを特徴とする請求項21記載の植物由来油脂の製造方法。
(a)配列番号6に示すアミノ酸配列を含むタンパク質
(b)配列番号6に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、カルコンシンターゼ活性を有するタンパク質
(c)配列番号5に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされカルコンシンターゼ活性を有するタンパク質
【請求項23】
カルコンイソメラーゼ遺伝子の機能を欠損した植物体から採取した種子から油脂成分を回収する工程を含む、植物由来油脂の製造方法。
【請求項24】
上記カルコンシンターゼ遺伝子が、以下の(a)〜(c)のいずれかのタンパク質をコードする遺伝子であることを特徴とする請求項23記載の植物由来油脂の製造方法。
(a)配列番号8に示すアミノ酸配列を含むタンパク質
(b)配列番号8に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、カルコンイソメラーゼ活性を有するタンパク質
(c)配列番号7に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされカルコンイソメラーゼ活性を有するタンパク質
【請求項25】
フラボン-3-ヒドラーゼ遺伝子の機能を欠損した植物体から採取した種子から油脂成分を回収する工程を含む、植物由来油脂の製造方法。
【請求項26】
上記フラボン-3-ヒドラーゼ遺伝子が、以下の(a)〜(c)のいずれかのタンパク質をコードする遺伝子であることを特徴とする請求項25記載の植物由来油脂の製造方法。
(a)配列番号10に示すアミノ酸配列を含むタンパク質
(b)配列番号10に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、フラボン-3-ヒドラーゼ活性を有するタンパク質
(c)配列番号9に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされフラボン-3-ヒドラーゼ活性を有するタンパク質
【請求項27】
種子内の油脂量を評価する対象となる植物体から種子を採取する工程と、
採取した種子の種皮色を観察し、より白色である場合には種子内の油脂量が高いと判定する工程とを含む、油脂量が向上した植物体のスクリーニング方法。
【請求項28】
上記評価対象の植物体は、転写因子と任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質を発現する植物体又は遺伝子機能欠損植物体であることを特徴とする請求項27記載のスクリーニング方法。
【請求項1】
配列番号4に示すアミノ酸配列からなるタンパク質を含む転写因子ファミリーに属する転写因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質を発現させた植物体。
【請求項2】
上記転写因子の転写促進活性が抑制されていることを特徴とする請求項1記載の植物体。
【請求項3】
上記キメラタンパク質が転写抑制因子活性をもつことを特徴とする請求項1記載の植物体。
【請求項4】
上記転写因子が、以下の(a)〜(c)のいずれかのタンパク質であることを特徴とする請求項1記載の植物体。
(a)配列番号4に示すアミノ酸配列を含むタンパク質
(b)配列番号4に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、転写促進活性を有するタンパク質
(c)配列番号3に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ転写促進活性を有するタンパク質
【請求項5】
上記機能性ペプチドが、次に示す式(1)〜(8)
(1)X1−Leu−Asp−Leu−X2−Leu−X3
(但し、式中、X1は0〜10個のアミノ酸残基を示し、X2はAsn又はGluを示し、X3は少なくとも6個のアミノ酸残基を示す。)
(2)Y1−Phe−Asp−Leu−Asn−Y2−Y3
(但し、式中、Y1は0〜10個のアミノ酸残基を示し、Y2はPhe又はIleを示し、Y3は少なくとも6個のアミノ酸残基を示す。)
(3)Z1−Asp−Leu−Z2−Leu−Arg−Leu−Z3
(但し、式中、Z1はLeu、Asp−Leu又はLeu−Asp−Leuを示し、Z2はGlu、Gln又はAspを示し、Z3は0〜10個のアミノ酸残基を示す。)
(4)Asp−Leu−Z4−Leu−Arg−Leu
(但し、式中、Z4はGlu、Gln又はAspを示す。)
(5)α1−Leu−β1−Leu−γ1−Leu
(6)α1−Leu−β1−Leu−γ2−Leu
(7)α1−Leu−β2−Leu−Arg−Leu
(8)α2−Leu−β1−Leu−Arg−Leu
(但し、式(5)〜(8)中、α1はAsp、Asn、Glu、Gln、Thr又はSerを示し、α2はAsn、Glu、Gln、Thr又はSerを示し、β1はAsp、Gln、Asn、Arg、Glu、Thr、Ser又はHisを示し、β2はAsn、Arg、Thr、Ser又はHisを示し、γ1はArg、Gln、Asn、Thr、Ser、His、Lys又はAspを示し、γ2はGln、Asn、Thr、Ser、His、Lys又はAspを示す。)
のいずれかで表されるアミノ酸配列を有するものであることを特徴とする請求項1記載の植物体。
【請求項6】
油脂生産性が有意に向上したことを特徴とする請求項1乃至5いずれか一項記載の植物体。
【請求項7】
特定の組織中の油脂含量が有意に向上したことを特徴とする請求項1乃至5いずれか一項記載の植物体。
【請求項8】
特定の組織中が種子であることを特徴とする請求項7記載の植物体。
【請求項9】
被子植物であることを特徴とする請求項1乃至8いずれか一項記載の植物体。
【請求項10】
双子葉植物であることを特徴とする請求項1乃至8いずれか一項記載の植物体。
【請求項11】
アブラナ科植物であることを特徴とする請求項1乃至8いずれか一項記載の植物体。
【請求項12】
シロイヌナズナであることを特徴とする請求項1乃至8いずれか一項記載の植物体。
【請求項13】
請求項1乃至12いずれか一項記載の植物体から、生産性が向上した物質を分離及び回収する工程を含む、植物体を用いた物質の製造方法。
【請求項14】
上記物質は油脂であることを特徴とする請求項13記載の植物体を用いた物質の製造方法。
【請求項15】
配列番号4に示すアミノ酸配列からなるタンパク質を含む転写因子ファミリーに属する転写因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質。
【請求項16】
上記転写因子が以下の(a)〜(c)のいずれかのタンパク質であることを特徴とする請求項15記載のキメラタンパク質。
(a)配列番号4に示すアミノ酸配列を含むタンパク質
(b)配列番号4に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、転写促進活性を有するタンパク質
(c)配列番号3に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ転写促進活性を有するタンパク質
【請求項17】
上記機能性ペプチドが、次に示す式(1)〜(8)
(1)X1−Leu−Asp−Leu−X2−Leu−X3
(但し、式中、X1は0〜10個のアミノ酸残基を示し、X2はAsn又はGluを示し、X3は少なくとも6個のアミノ酸残基を示す。)
(2)Y1−Phe−Asp−Leu−Asn−Y2−Y3
(但し、式中、Y1は0〜10個のアミノ酸残基を示し、Y2はPhe又はIleを示し、Y3は少なくとも6個のアミノ酸残基を示す。)
(3)Z1−Asp−Leu−Z2−Leu−Arg−Leu−Z3
(但し、式中、Z1はLeu、Asp−Leu又はLeu−Asp−Leuを示し、Z2はGlu、Gln又はAspを示し、Z3は0〜10個のアミノ酸残基を示す。)
(4)Asp−Leu−Z4−Leu−Arg−Leu
(但し、式中、Z4はGlu、Gln又はAspを示す。)
(5)α1−Leu−β1−Leu−γ1−Leu
(6)α1−Leu−β1−Leu−γ2−Leu
(7)α1−Leu−β2−Leu−Arg−Leu
(8)α2−Leu−β1−Leu−Arg−Leu
(但し、式(5)〜(8)中、α1はAsp、Asn、Glu、Gln、Thr又はSerを示し、α2はAsn、Glu、Gln、Thr又はSerを示し、β1はAsp、Gln、Asn、Arg、Glu、Thr、Ser又はHisを示し、β2はAsn、Arg、Thr、Ser又はHisを示し、γ1はArg、Gln、Asn、Thr、Ser、His、Lys又はAspを示し、γ2はGln、Asn、Thr、Ser、His、Lys又はAspを示す。)
のいずれかで表されるアミノ酸配列を有するものであることを特徴とする請求項15記載のキメラタンパク質。
【請求項18】
請求項15乃至17いずれか一項記載のキメラタンパク質をコードする遺伝子。
【請求項19】
請求項18記載の遺伝子を含む発現ベクター。
【請求項20】
請求項18記載の遺伝子を含む形質転換体。
【請求項21】
カルコンシンターゼ遺伝子の機能を欠損した植物体から採取した種子から油脂成分を回収する工程を含む、植物由来油脂の製造方法。
【請求項22】
上記カルコンシンターゼ遺伝子が、以下の(a)〜(c)のいずれかのタンパク質をコードする遺伝子であることを特徴とする請求項21記載の植物由来油脂の製造方法。
(a)配列番号6に示すアミノ酸配列を含むタンパク質
(b)配列番号6に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、カルコンシンターゼ活性を有するタンパク質
(c)配列番号5に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされカルコンシンターゼ活性を有するタンパク質
【請求項23】
カルコンイソメラーゼ遺伝子の機能を欠損した植物体から採取した種子から油脂成分を回収する工程を含む、植物由来油脂の製造方法。
【請求項24】
上記カルコンシンターゼ遺伝子が、以下の(a)〜(c)のいずれかのタンパク質をコードする遺伝子であることを特徴とする請求項23記載の植物由来油脂の製造方法。
(a)配列番号8に示すアミノ酸配列を含むタンパク質
(b)配列番号8に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、カルコンイソメラーゼ活性を有するタンパク質
(c)配列番号7に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされカルコンイソメラーゼ活性を有するタンパク質
【請求項25】
フラボン-3-ヒドラーゼ遺伝子の機能を欠損した植物体から採取した種子から油脂成分を回収する工程を含む、植物由来油脂の製造方法。
【請求項26】
上記フラボン-3-ヒドラーゼ遺伝子が、以下の(a)〜(c)のいずれかのタンパク質をコードする遺伝子であることを特徴とする請求項25記載の植物由来油脂の製造方法。
(a)配列番号10に示すアミノ酸配列を含むタンパク質
(b)配列番号10に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、フラボン-3-ヒドラーゼ活性を有するタンパク質
(c)配列番号9に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされフラボン-3-ヒドラーゼ活性を有するタンパク質
【請求項27】
種子内の油脂量を評価する対象となる植物体から種子を採取する工程と、
採取した種子の種皮色を観察し、より白色である場合には種子内の油脂量が高いと判定する工程とを含む、油脂量が向上した植物体のスクリーニング方法。
【請求項28】
上記評価対象の植物体は、転写因子と任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質を発現する植物体又は遺伝子機能欠損植物体であることを特徴とする請求項27記載のスクリーニング方法。
【図1】


【図2】
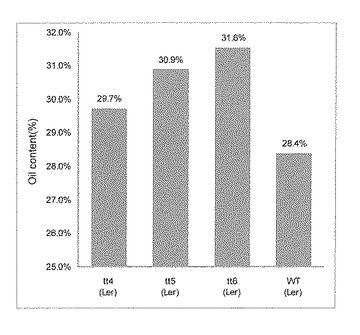
【図3】
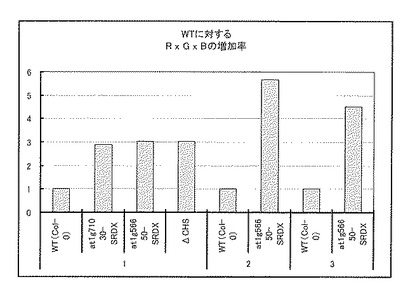
【図2】
【図3】
【公開番号】特開2009−207421(P2009−207421A)
【公開日】平成21年9月17日(2009.9.17)
【国際特許分類】
【出願番号】特願2008−54008(P2008−54008)
【出願日】平成20年3月4日(2008.3.4)
【出願人】(000003207)トヨタ自動車株式会社 (59,920)
【出願人】(000003609)株式会社豊田中央研究所 (4,200)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成21年9月17日(2009.9.17)
【国際特許分類】
【出願日】平成20年3月4日(2008.3.4)
【出願人】(000003207)トヨタ自動車株式会社 (59,920)
【出願人】(000003609)株式会社豊田中央研究所 (4,200)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]