
蛍光環状ペプチド、その製造方法およびプロテアーゼ酵素の酵素活性測定におけるこれらペプチドの使用
本発明は、以下の配列:
−S1−X1−S2−X2−
(式中、S1はプロテアーゼ酵素E1の第一標的ペプチド配列であり;S2はプロテアーゼ酵素E2の第二標的ペプチド配列であり;S1およびS2は同一であっても、異なっていてもよく;当該第一のペプチド配列S1および当該第二のペプチド配列S2は4個ないし14個のアミノ酸を含んでなり;E1およびE2は同じプロテアーゼ酵素に、または異なるプロテアーゼ酵素に相当していてもよく;X1は蛍光供与体基を担持するプローブであり;そしてX2は蛍光または非蛍光受容体基を担持するプローブである)
を含んでなる環状ペプチドに関する。
これらのペプチドはプロテアーゼ酵素の活性測定に使用することができる。
−S1−X1−S2−X2−
(式中、S1はプロテアーゼ酵素E1の第一標的ペプチド配列であり;S2はプロテアーゼ酵素E2の第二標的ペプチド配列であり;S1およびS2は同一であっても、異なっていてもよく;当該第一のペプチド配列S1および当該第二のペプチド配列S2は4個ないし14個のアミノ酸を含んでなり;E1およびE2は同じプロテアーゼ酵素に、または異なるプロテアーゼ酵素に相当していてもよく;X1は蛍光供与体基を担持するプローブであり;そしてX2は蛍光または非蛍光受容体基を担持するプローブである)
を含んでなる環状ペプチドに関する。
これらのペプチドはプロテアーゼ酵素の活性測定に使用することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、通常、いわゆるFRET(蛍光共鳴エネルギー移動またはフォースター共鳴エネルギー移動の略)法によるプロテアーゼ酵素の酵素活性測定において使用し得る新規蛍光環状ペプチドに関する。
本発明は酵素障害を原因とする疾患の診断分野において特に有用である。
【背景技術】
【0002】
酵素は調節現象を顕著に伴う複雑な作用メカニズムを有し、その破壊は、病理学的障害の発症の原因となり得る。
【0003】
メタロプロティナーゼ(MMPの略号でも知られる)は顕著な事例であり、このものは細胞外マトリックス要素のすべてと、一部の増殖因子および接着分子を切断し得る。これらの酵素は自然の過程によってTIMP(メタロプロティナーゼの組織インヒビター)などのインヒビターにより調節される。MMP/TIMPの平衡が崩れると、腫瘍増殖、転移、関節リウマチ、心疾患などの病的プロセスが生じ得る。
【0004】
多くの病態において酵素が関わっていることから、認知された病態との可能な相関を確立するためには、これらの酵素活性を測定可能とする手段の提供が必須であることが間もなく証明された。現時点で入手し得る手段は、活性を測定すべき酵素の基質からなり、これらの基質は蛍光基を有すると共に、その活性を蛍光測定により定量することからなる。特に、最も頻繁に使用される蛍光法は蛍光共鳴エネルギー移動法またはFRETである。しかし、この技法は厳密な意味で蛍光移動の測定に基づくものではなく、電子エネルギー移動の測定に基づくものである。従って、識別は放射移動と非放射(または共鳴)移動間でなされなければならない。放射移動は基と基の間の距離、それらの方向性などの情報の入手手段を提供するものではなく、試験するシステムを理解するための、または特性化するための必須のデータであり得る。
【0005】
放射移動は2段階で起こるプロセスである。初期段階では、光子が供与体基により放出され、第二段階では、この光子が供与体基と同じ光子受容体基または異なる光子受容体基により吸収される。この移動には基と基の間の相互作用を必要としないが、種のスペクトルの重なり合いと濃度のスペクトルの重なり合いに左右される。
【0006】
励起エネルギーの非放射移動は供与体基および受容体基の間の相互作用を必要とする。この移動は供与体基の発光スペクトルが受容体基の吸収スペクトルと重なり合った場合にのみ起こり、その結果、供与体基の振電遷移と受容体基の振電遷移との間のエネルギー対応が存在する。当該遷移はカップル結合している、または<共鳴している>と言われる。
【0007】
非放射エネルギー移動の現象はフォースター(Forster)が研究した。彼はこの物理現象が10ないし100Åという位数の非常に短い距離にまたがって起こることを証明し得た。供与体基と受容体基の距離が大きくなる場合、これはエネルギー移動の変化と解釈される。従って、この現象はプロテアーゼ酵素の酵素活性を評価するために使用し得る。対照的に、各末端でそれぞれ配列した供与体基と受容体基両方を含んでなる基質を特異的プロテアーゼ酵素と接触させると、基質が切断され、その結果、供与体基と受容体基間の距離が増大し、結果として、励起エネルギー(または共鳴エネルギー)の移動の変化が起こる。この変化に基づき、酵素活性の評価に到達することができる。
【0008】
現在使用されている基質は殆どの部分が線状構造の基質についてであり、その基質は、明らかにエキソプロテアーゼおよび非特異的プロテアーゼにより分解されると思われるので、生物媒体においては殆ど安定性のないことが証明されている。
【0009】
それ故、所定のプロテアーゼ酵素の酵素活性を定量するための手段として使用し得る、安定で、特にFRET法を用いる当該酵素の酵素活性のモニターを可能とするペプチドが紛れもなく必要である。
【発明の概要】
【発明が解決しようとする課題】
【0010】
それ故、本発明者らは上記の必要性に一致する新規なペプチドを開発してきた。
【0011】
それ故、第一の主題によると、本発明は以下の配列:
−S1−X1−S2−X2−
(式中、S1はプロテアーゼ酵素E1の第一標的ペプチド配列であり;S2はプロテアーゼ酵素E2の第二標的ペプチド配列であり;S1およびS2は同一であっても、異なっていてもよく;当該第一のペプチド配列S1および当該第二のペプチド配列S2は4個ないし14個のアミノ酸を含んでなり;E1およびE2は同じプロテアーゼ酵素に、または異なるプロテアーゼ酵素に相当していてもよく;X1は蛍光供与体基を担持するプローブであり;そしてX2は蛍光または非蛍光受容体基を担持するプローブである)
を含んでなる環状ペプチド(シクロペプチドとも呼称する)に関する。
【0012】
好ましくは、該供与体基および受容体基は、供与体基の蛍光発光スペクトルが受容体基の吸収スペクトルと少なくとも部分的に重なり合うように選択する。
【0013】
それ故、1つのプロテアーゼ酵素E(E1とE2が1つかつ同一の酵素を形成するのみであり、S1とS2が同一のプロテアーゼ酵素の標的ペプチド配列である場合)またはプロテアーゼE1とプロテアーゼE2(それらが異なっている場合、またS1がプロテアーゼ酵素E1の標的ペプチド配列であり、S2がプロテアーゼ酵素E2の標的ペプチド配列である場合)両方の存在下、本発明の環状ペプチドは、第一のペプチド配列S1および第二のペプチド配列S2の両方で切断され、そして、プローブX1およびX2の配置を原因として、プローブX1とX2の空間に距離を生じる。次いで、供与体基と受容体基の距離が離れ、次に、これらの2つの基の間の共鳴エネルギーの移動に変化が生じる。それ故、この共鳴エネルギーの変化を用いて、プロテアーゼ酵素Eまたはプロテアーゼ酵素E1とE2の酵素活性に到達することが可能である。
【0014】
さらに、本発明のペプチドが環状であるという事実は、常套的に先行技術において使用される線状ペプチドの欠点、特に先行技術の線状ペプチドが遭遇する安定性の問題を克服することが可能であることを意味する。
【0015】
本発明の特定の一実施態様によると、E1およびE2は同一のプロテアーゼ酵素に相当する。
プロテアーゼ酵素Eの酵素活性を検討するために(E1=E2の場合)、本発明者らは検討すべきプロテアーゼ酵素Eの標的である1本の線状ペプチド配列の環状化について極単純に仮想した;該ペプチド配列は以下の工程図に示すプローブX1およびX2をその末端に含んでなる:
【0016】
【化1】

【0017】
検討すべきプロテアーゼ酵素Eの存在下、環状ペプチドは以下の工程図に示すように切断される:
【0018】
【化2】
【0019】
切断後、基X1およびX2の距離は変化しないままである。結果として、共鳴移動の変化はなく、従って、プロテアーゼ酵素Eの活性を定量することは不可能である。
詳細な説明に入る前に、我々は以下の定義を提案する。
【0020】
蛍光供与体基が慣例として意味するのは、光エネルギー(励起光といい、その波長は一般に紫外部領域に属する)を吸収し、その環境に依存的に2つの可能な形状でこの光を再発光することができる基である。
a)受容体基との共鳴による励起エネルギーの移動の形状で、ただし、後者は先に記載したFRET現象と互換性のある距離に、好ましくは10ないし100Åの範囲の距離にある;または
b)一般に波長が可視領域に属する蛍光(供与体発光光と呼称)の形状にある。
【0021】
受容体基が慣例として意味するのは、本発明の意味において、共鳴エネルギーによる供与体基の励起エネルギーを吸収することのできる基である。この励起エネルギーは2つの主要な様式で再発光され得る:
a)放射発光(光子の放出)による;この場合、蛍光受容体基という用語が使用される;または
b)非放射発光(すなわち、光子放出以外の不活性化様式)による;この場合、非蛍光受容体基という用語が使用される。
【0022】
プローブが通常意味するのは、また本発明の意味において、供与体基(プローブX1について)を担持するアミノ酸残基、または受容体基(プローブX2について)を担持するアミノ酸残基である。
【0023】
アミノ酸残基が通常意味するのは、アミノ酸の−NH2官能基または−CO2H等と別の化合物の官能基とが反応して共有結合を形成するような残基アミノ酸である。
【0024】
標的ペプチド配列が通常意味するのは、また本発明の意味において、プロテアーゼ酵素が認識して切断し得るペプチド配列である。
【0025】
プロテアーゼ酵素E1およびプロテアーゼ酵素E2(E1およびE2は同一であっても、異なっていてもよい)の2つの標的ペプチド配列(これら配列の間に本発明に適合するプローブX1およびプローブX2がそれぞれ配列されている)を含んでなる環状ペプチドの使用が、上記で説明したように、当該プロテアーゼ酵素の活性を定量するための良好な手段の提供を可能とする。
本発明の環状ペプチドは、下記式:
【0026】
【化3】
【0027】
(式中、
− S1、X1、S2およびX2は上記と同一の定義を示す;
− R1およびR2は独立して、単結合、アミノ酸残基またはペプチド配列に相当する;
− Yは−CONH−基または−NHCO−基に相当する)
に一致する。
【0028】
S1およびS2は、該ペプチドが単一のプロテアーゼ酵素(E1およびE2が同一である)の活性を定量しようとする場合は、同一であり、または該ペプチドが同一のプロテアーゼ酵素のイソ型の存在または2種の異なるプロテアーゼ酵素E1およびE2の存在の間を識別しようとする場合は明らかに異なり得る。
【0029】
S1およびS2が同一である場合、それらは同一方向に、または逆方向に配置され得る。例えば、S1が配列−Gly−Pro−Gln−Gly−Leu−Leu−Gly−Ala−に相当する場合、配列S2は逆方向に配置された同一配列、すなわち、以下の配列−Ala−Gly−Leu−Leu−Gly−Gln−Pro−Gly−に相当する。
【0030】
本発明の特定の一実施態様において、R1およびR2はアミノ酸残基に、またはペプチド配列に相当し得る。このことは該環状ペプチドが医薬投与形態を企図する場合、特に生物媒体において該ペプチドが安定であるという理由で特に有利である。
【0031】
本発明の環状ペプチドは、MMP型のプロテアーゼ酵素(“細胞外マトリックスメタロプロティナーゼ”としても知られる)の酵素活性を測定するために特に適用される。
【0032】
この場合、プロテアーゼ酵素E1およびプロテアーゼ酵素E2はMMP型の酵素であり、その場合の第一ペプチド配列S1はMMPプロテアーゼ酵素の標的であり、第二ペプチド配列S2はMMPプロテアーゼ酵素の標的である。
【0033】
MMP類はその活性部位に亜鉛原子を含む28の数に及ぶエンドぺプチダーゼの系統群である。それらは分泌されるか、または膜と会合しているかによって2つの系統群に分類し得る。分泌されるMMPは、コラゲナーゼ(例:MMP−1、8、13および18)、ゼラチナーゼ(例:MMP−2および9)、マトリリシン(例:MMP−7および26)、ストロメリシン(例:MMP−3、10および11)、エピリシン(MMP−28)、エナメリシン(MMP−20)を包含する。膜と会合するMMPは、膜貫通MMP、例えば、MMP−14、MMP−15、MMP−16、MMP−17、MMP−23、MMP−24およびMMP−25を包含する。
【0034】
MMP類は細胞外マトリックス要素のすべて、一部の増殖因子、または接着分子さえも切断することができる。MMPはTIMPなどの天然のインヒビターにより調節され、生理的プロセス(胚増殖、血管新生、骨組織の改造作用など)において重要な役割を果たしている。MMP/TIMPのバランスが崩れると、腫瘍増殖、転移、関節リウマチ、心疾患などの病的プロセスが生じる。
【0035】
腫瘍進行におけるMMP類の役割は多様であるが、とりわけ細胞外マトリックスを分解するMMPの能力が理由であり、それにより腫瘍の侵入を促進する。また、MMPは非マトリックスタンパク質に対してタンパク分解活性を有し、腫瘍の進行、接着喪失、侵入、増殖、血管新生、血管内異物侵入、管外遊出および転移増殖などの幾つかの他の工程においてそれらに対し複雑な役割を与える。
【0036】
それ故、環状ペプチドはMMPタイプのプロテアーゼ酵素の酵素活性をモニターするための、特に、酵素活性と認知された病態との間のつながりを確立するための、優れた手段であること証明する。
【0037】
より正確には、環状ペプチドがMMP活性の追跡を企図する場合、標的のペプチド配列S1およびS2は1個のMMP酵素(S1=S2であるか、またはS1およびS2が1つの同じ酵素の2つの同型間を識別しようとする異なる標的配列である場合)の、または2個のMMP酵素(S1およびS2が異なっていて、それぞれが2つの異なるプロテアーゼ酵素の標的である場合)の標的であり、それらの配列はその活性をモニターしようとするMMP(類)が認識するMMP(類)の基質の配列に相当する。
【0038】
特に、標的ペプチド配列S1およびS2は、MMP−1、MMP−2、MMP−3、MMP−5、MMP−7、MMP−8、MMP−9、MMP−10、MMP−11、MMP−12、MMP−13、MMP−14、MMP−15、MMP−16、MMP−17、MMP−19、MMP−23、MMP−25およびMMP−26およびその混合物から選択されるMMP酵素の標的となり得る。
【0039】
ペプチド配列S1およびS2は、4個ないし14個の範囲のアミノ酸の長さをもつ線状配列であり、活性を示すために必要なコンホメーションをもつ同じ配列の環状ペプチドを可能とする。S1およびS2は、そのN末端とC末端の間の距離が10Åと100Åの間となるように選択され、当該距離は先に定義したような供与体基と受容体基の間でFRETを実施するために有効な距離と矛盾しない距離とすると有利である。分子モデル化ソフトウエアを使用することで、信頼し得る当該距離の予測が可能である。
【0040】
ペプチド配列S1およびS2は、MMPの天然基質に見出される天然のペプチド配列から誘導し得るか、またはMMPの合成基質として知られる合成配列から誘導し得る。
【0041】
有利なことに、本発明の環状ペプチドは天然のアミノ酸のみを含み、それがそれらの可能性のある細胞毒性を制限し得るものとし、また関係する生物体への非天然アミノ酸の導入を回避する。
【0042】
一部の事例においては、天然または合成基質がMMP類によって認識され、切断されることがあり得る。どのMMPが切断に関与しているかを決定するためには、この基質についての異なるMMP類の酵素動力学データを使用し、比較することが必要である。
【0043】
数種のMMP類間の識別を可能とするこれら酵素動力学データは、一般に、その基質に対する酵素の親和性の指標であるミカエリス定数(Km)、および触媒定数(Kcat、1秒あたり且つ酵素1モルあたりに形成される産物のモル数)と酵素の触媒効率の指標であるミカエリス定数との比である。それ故、一般に、MMP/基質対合についての値の特定の対合(Kmおよびkcat/Km)が存在する。それ故、例えば2つのMMPが同じ基質を認識する場合、どちらが可視化活性に関与しているかを決定することが可能である。
【0044】
合成配列は、例えば、以下のアミノ酸または基を含んでなる:
Nva (ノル−バリン); Cha (3−シクロヘキシルアラニン); Dpa (N−3(2,4−ジニトロフェニル)−L−2,3−ジアミノプロピオニル); Abu:α−アミノ酪酸; S−メチルシステインに対応するCys(Me)、このものは基−SHが−S−CH3基と置換されていることを意味する; D−アルギニンに対応するD−Arg。
【0045】
本明細書の残りの記載において、下記掲載の略号は具体的に以下の意味を有する:
Gly:グリシン; Pro:プロリン; Leu:ロイシン; Ala:アラニン; Gln:グルタミン; Ile:イソロイシン; Arg:アルギニン; Val:バリン; Tyr:チロシン; Glu:グルタメート; Met:メチオニン; Phe:フェニルアラニン; Asn:アスパラギン; Thr:トレオニン; Trp:トリプトファン; Ser:セリン; His:ヒスチジン、これらの略号はアミノ酸についての公式の3文字命名法に準拠する。
【0046】
第一の配列S1および/または第二の配列S2をMMP−1酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Gly−Pro−Gln−Gly−Leu−Leu−Gly−Ala−
配列番号:1
−Ala−Pro−Gln−Gly−Ile−Ala−Gly−Gln−
配列番号:2
−Gly−Pro−Gln−Gly−Leu−Ala−Gly−Gln−
配列番号:3
−Gly−Pro−Leu−Gly−Ile−Ala−Gly−Ile−
配列番号:4
−Gly−Pro−Glu−Gly−Leu−Arg−Val−Gly−
配列番号:5
−Tyr−Glu−Ala−Gly−Leu−Gly−Val−Val−
配列番号:6
−Ala−Gly−Leu−Gly−Val−Val−Glu−Arg−
配列番号:7
−Ala−Gly−Leu−Gly−Ile−Ser−Ser−Thr−
配列番号:8
−Gly−Ala−Met−Phe−Leu−Glu−Ala−Ile−
配列番号:9
−Ile−Pro−Glu−Asn−Phe−Phe−Gly−Val−
配列番号:10
−Thr−Glu−Gly−Glu−Ala−Arg−Gly−Ser−
配列番号:11
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Leu−Arg−Ala−Tyr−Leu−Leu−Pro−Ala−
配列番号:13
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:14
Deng, S.J., et al. 2000. J. Biol. Chem. 275, 31422
【0047】
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:15
Beekman, B., et al. 1996. FEBS Lett 390, 221
【0048】
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
Darlak, K., et al. 1990. J. Biol. Chem. 265, 5199
Stack, M.S., and Gray, R.D. 1989. J. Biol. Chem. 264, 4277
【0049】
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
Netzel-Arnett, S., et al. 1991. Anal. Biochem.195, 86
【0050】
−Pro−Cha−Abu−Cys(Me)−His−Ala−
配列番号:18
McGeehan, G.M., et al. 1994. J. Biol. Chem.269, 32814
【0051】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:19
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
【0052】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0053】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knauper, V. et al. 1996. J. Biol. Chem. 271, 1544
【0054】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
【0055】
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
A. Santala, A. et al. 1999. FEBS Lett. 461, 153-156
Nagase, H. et al. 1994 J.Biol.Chem. 269, 20952-20957
【0056】
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
J. Berman et al. 1992. J.Biol.Chem. 267, 1434-1437
【0057】
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
Kraft, P.J. et al. 2001. Connect.Tissue Res. 42, 149-163
Itoh, M. et al. 1997. J.Pharm.Biomed.Anal. 15, 1417-1426
Welch, A.R. et al. 1995. Arch.Biochem.Biophys. 324, 59-64
【0058】
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
Lauer-Fields, J.L. and Fields, G.B. 2002 Biol.Chem. 383, 1095-1105
Lauer-Fields, J.L. et al. 2001. Biochemistry 40, 5795-5803
【0059】
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0060】
第一の配列S1および/または第二の配列S2をMMP−2酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Ile−Pro−Glu−Asn−Phe−Phe−Gly−Val−
配列番号:10
−Pro−Pro−Gly−Ala−Tyr−His−Gly−Ala−
配列番号:27
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−His−Leu−Leu−Val−Glu−Ala−
配列番号:28
−Leu−Arg−Ala−Tyr−Leu−Leu−Pro−Ala−
配列番号:13
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:29
Beekman, B., et al. 1996. FEBS Lett 390, 221
【0061】
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
Darlak, K., et al. 1990. J. Biol. Chem. 265, 5199
Stack, M.S., and Gray, R.D. 1989. J. Biol. Chem. 264, 4277
【0062】
−Pro−Leu−Ala−Nva−Dpa−Ala−Arg−
配列番号:30
Murphy, G., et al. 1994. J. Biol. Chem. 269, 6632
【0063】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
【0064】
−Pro−Leu−Gly−Met−Trp−Ser−Arg−
配列番号:31
Netzel-Arnett, S., et al. 1991. Anal. Biochem. 195, 86
【0065】
−Pro−Leu−Gly−SCH[CH2CH(CH3)2]−CO−Leu−Gly−
配列番号:32
Weingarten, H., et al. 1985. Anal. Biochem. 147, 437
Weingarten, H., et al. 1985. Biochemistry 24, 6730
【0066】
−Arg−Pro−Pro−Gly−Phe−Ser−Ala−Phe−
配列番号:33
G.D. Johnson and K. Ahn 2000. Anal. Biochem. 286, 112
【0067】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knight, C.G. et al. 1992. FEBS Lett. 296, 263
【0068】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24,6730
Xia, T. et al. 1996. Biochim. Biophys. Acta 1293,259
【0069】
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
J. Berman et al., 1992. J.Biol.Chem. 267, 1434-1437
【0070】
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
Kraft, P.J. et al. 2001. Connect.Tissue Res. 42, 149-163
Itoh, M. et al. 1997. J.Pharm.Biomed.Anal. 15, 1417-1426
Welch, A.R. et al. 1995. Arch.Biochem.Biophys. 324, 59-64
【0071】
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
Lauer-Fields, J.L. and Fields, G.B. 2002 Biol.Chem. 383, 1095-1105
Lauer-Fields, J.L. et al. 2001. Biochemistry 40, 5795-5803
【0072】
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0073】
第一の配列S1および/または第二の配列S2をMMP−3酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Gly−Pro−Glu−Gly−Leu−Arg−Val−Gly−
配列番号:5
−Arg−Val−Gly−Phe−Tyr−Glu−Ser−Asp−
配列番号:34
−Leu−Leu−Ser−Ala−Leu−Val−Glu−Thr−
配列番号:35
−Glu−Ala−Ile−Pro−Met−Ser−Ile−Pro−
配列番号:36
−Ile−Ala−Gly−Arg−Ser−Leu−Asn−Pro−
配列番号:37
−Ile−Pro−Glu−Asn−Phe−Phe−Gly−Val−
配列番号:10
−Lys−Pro−Gln−Gln−Phe−Phe−Gly−Leu−
配列番号:38
−Asp−Val−Ala−Gln−Phe−Val−Leu−Thr−
配列番号:39
−Asp−Thr−Leu−Glu−Val−Met−Arg−Lys−
配列番号:40
−Asp−Val−Gly−His−Phe−Arg−Thr−Phe−
配列番号:41
−Asp−Ser−Gly−Gly−Phe−Met−Leu−Thr−
配列番号:42
−Arg−Val−Ala−Glu−Met−Arg−Gly−Glu−
配列番号:43
−Asp−Leu−Gly−Arg−Phe−Gln−Thr−Phe−
配列番号:44
−Pro−Phe−Ser−Pro−Leu−Val−Ala−Thr−
配列番号:45
−Leu−Arg−Ala−Tyr−Leu−Leu−Pro−Ala−
配列番号:13
−Ala−Pro−Gly−Asn−Ala−Ser−Glu−Ser−
配列番号:46
−Phe−Ser−Ser−Glu−Ser−Lys−Arg−Glu−
配列番号:47
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−His−Leu−Leu−Val−Glu−Ala−
配列番号:28
−Pro−Pro−Glu−Glu−Leu−Lys−Phe−Gln−
配列番号:48
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:49
Deng, S.J., et al. 2000. J. Biol. Chem. 275, 31422
【0074】
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:15
Beekman, B., et al. 1996. FEBS Lett 390, 221
【0075】
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
Beekman, B., et al. 1997. FEBS Lett 418, 305
【0076】
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
Darlak, K., et al. 1990. J. Biol. Chem. 265, 5199
Stack, M.S., and Gray, R.D. 1989. J. Biol. Chem. 264, 4277
【0077】
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
Netzel-Arnett, S., et al. 1991. Anal. Biochem. 195, 86
【0078】
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
Bickett, D.M., et al. 1994. Ann. N.Y. Acad. Sci. 732, 351
【0079】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0080】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knight, C.G. et al. 1992. FEBS Lett. 296, 263
【0081】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24,6730
【0082】
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0083】
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−
配列番号:50
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0084】
第一の配列S1および/または第二の配列S2をMMP−5酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
Beekman, B., et al. 1997. FEBS Lett 418, 305
【0085】
第一の配列S1および/または第二の配列S2をMMP−7酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
Darlak, K., et al. 1990. J. Biol. Chem. 265, 5199
Stack, M.S., and Gray, R.D. 1989. J. Biol. Chem. 264, 4277
【0086】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
【0087】
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−Ser−
配列番号:24
Welch, A.R., et al. 1996. Biochemistry35, 10103
Welch, A.R., et al. 1995. Arch. Biochem. Biophys. 324, 59
【0088】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0089】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
【0090】
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
A. Santala, A. et al. 1999. FEBS Lett. 461, 153-156
Shabani, F. et al.1998. Free Radic.Res. 28, 115-123
Nagase, H. et al. 1994 J.Biol.Chem. 269, 20952-20957
【0091】
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
Finch-Arietta, M. et al., 1993. Agents Actions 39 SpecNo,C189-C191
Netzel-Arnett, S. et al. 1991 Anal.Biochem.195, 86-92
【0092】
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
Finch-Arietta, M. et al., 1993. Agents Actions 39 SpecNo,C189-C191
Bickett, D.M. et al., 1994. Ann.N.Y.Acad.Sci. 732, 351-355
【0093】
第一の配列S1および/または第二の配列S2をMMP−8酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Ala−Tyr−Trp−Ala−Arg−
配列番号:53
Netzel-Arnett, S., et al. 1991. Anal. Biochem.195, 86
【0094】
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Grams, F. et al. 2001. Biol. Chem. 382, 1277
【0095】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0096】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knauper, V. et al. 1996. J. Biol. Chem. 271,1544
【0097】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
【0098】
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
A. Santala, A. et al. 1999. FEBS Lett. 461, 153-156
Shabani, F. et al.1998. Free Radic.Res. 28, 115-123
Nagase, H. et al. 1994 J.Biol.Chem. 269, 20952-20957
【0099】
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
J. Berman et al., 1992. J.Biol.Chem. 267, 1434-1437
【0100】
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
Kraft, P.J. et al. 2001. Connect.Tissue Res. 42, 149-163
Itoh, M. et al. 1997. J.Pharm.Biomed.Anal. 15, 1417-1426
Welch, A.R. et al. 1995. Arch.Biochem.Biophys. 324, 59-64
【0101】
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
Lauer-Fields, J.L. and Fields, G.B. 2002 Biol.Chem. 383, 1095-1105
Lauer-Fields, J.L. et al. 2001. Biochemistry 40, 5795-5803
【0102】
第一の配列S1および/または第二の配列S2をMMP−9酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Gly−Pro−Pro−Gly−Val−Val−Gly−Pro−
配列番号:54
−Gly−Pro−Pro−Gly−Leu−Arg−Gly−Glu−
配列番号:55
−Gly−Pro−Gly−Gly−Val−Val−Gly−Pro−
配列番号:56
−Ile−Pro−Gln−Asn−Phe−Phe−Gly−Val−
配列番号:57
−Pro−Pro−Gly−Ala−Tyr−His−Gly−Ala−
配列番号:58
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:14
Deng, S.J., et al. 2000. J. Biol. Chem. 275, 31422
【0103】
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:15
Beekman, B., et al. 1996. FEBS Lett 390, 221
【0104】
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
Beekman, B., et al. 1997. FEBS Lett 418, 305
【0105】
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
Darlak, K., et al. 1990. J. Biol. Chem. 265, 5199
Stack, M.S., and Gray, R.D. 1989. J. Biol. Chem. 264, 4277
【0106】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:19
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
【0107】
−Pro−Leu−Gly−SCH[CH2CH(CH3)2]−CO−Leu−Gly−
配列番号:32
Weingarten, H., et al. 1985. Anal. Biochem. 147, 437
Weingarten, H., et al. 1985. Biochemistry 24, 6730
【0108】
−Arg−Pro−Pro−Gly−Phe−Ser−Ala−Phe−
配列番号:33
G.D. Johnson and K. Ahn 2000. Anal. Biochem. 286, 112
【0109】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0110】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Roy, N. et al. 1999. Prot. Expr. Purif. 16, 324
【0111】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
Xia, T. et al. 1996. Biochim. Biophys. Acta 1293, 259
【0112】
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
J. Berman et al., 1992. J.Biol.Chem. 267, 1434-1437
【0113】
第一の配列S1および/または第二の配列S2をMMP−10酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−His−Leu−Leu−Val−Glu−Ala−
配列番号:28
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Kannan, R. et al. 1999. Prot. Expr. Purif. 16, 76
【0114】
第一の配列S1および/または第二の配列S2をMMP−11酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0115】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Kannan, R. et al. 1999. Prot. Expr. Purif. 16, 76
【0116】
第一の配列S1および/または第二の配列S2をMMP−12酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0117】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Park, H.I. et al. 2000. J. Biol. Chem. 275, 20540
【0118】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
【0119】
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
A. Santala, A. et al. 1999. FEBS Lett. 461, 153-156
Shabani, F. et al.1998. Free Radic.Res. 28, 115-123
Nagase, H. et al. 1994 J.Biol.Chem. 269, 20952-20957
【0120】
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
Berman, J. et al., 1992. J.Biol.Chem. 267, 1434-1437
【0121】
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
Finch-Arietta, M. et al., 1993. Agents Actions 39 SpecNo,C189-C191
Netzel-Arnett, S. et al. 1991 Anal.Biochem.195, 86-92
【0122】
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
Finch-Arietta, M. et al., 1993. Agents Actions 39 SpecNo,C189-C191
Bickett, D.M. et al., 1994. Ann.N.Y.Acad.Sci. 732, 351-355
【0123】
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
Kraft, P.J. et al. 2001. Connect.Tissue Res. 42, 149-163
Itoh, M. et al. 1997. J.Pharm.Biomed.Anal. 15, 1417-1426
Welch, A.R. et al. 1995. Arch.Biochem.Biophys. 324, 59-64
【0124】
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
Lauer-Fields, J.L. and Fields, G.B. 2002 Biol.Chem. 383, 1095-1105
Lauer-Fields, J.L. et al. 2001. Biochemistry 40, 5795-5803
【0125】
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0126】
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−
配列番号:50
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0127】
−Arg−Pro−Lys−Pro−Gln−Gln−Phe−Trp−
配列番号:59
Bickett, D.M. et al. 1994. Ann.N.Y.Acad.Sci. 732, 351-355
【0128】
第一の配列S1および/または第二の配列S2をMMP−13酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:49
Deng, S.J., et al. 2000. J. Biol. Chem. 275, 31422
【0129】
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
Beekman, B., et al. 1997. FEBS Lett 418, 305
【0130】
−Pro−Cha−Gly−Nva−His−Ala−Dpa−
配列番号:25
Knauper, V., et al. 1996. J. Biol. Chem. 271, 1544
【0131】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0132】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knauper, V. et al. 1996. J. Biol. Chem. 271, 1544
【0133】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
【0134】
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Netzel-Arnett, S. et al. 1991. Anal.Biochem.195, 86-92
Santala, A. et al. 1999. FEBS Lett. 461, 153-156
Bickett, D.M. et al., 1993. Anal.Biochem. 212, 58-64
【0135】
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
A. Santala, A. et al. 1999. FEBS Lett. 461, 153-156
Shabani, F. et al.1998. Free Radic.Res. 28, 115-123
Nagase, H. et al. 1994 J.Biol.Chem. 269, 20952-20957
【0136】
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
J. Berman et al., 1992. J.Biol.Chem. 267, 1434-1437
【0137】
−Pro−Leu−Gly−Met−Trp−Ser−Arg−
配列番号:31
Netzel-Arnett, S. et al.1991. Anal.Biochem.195, 86-92
d'Ortho, M.P. et al., 1997. Eur.J.Biochem. 250, 751-757
【0138】
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
Finch-Arietta, M. et al., 1993. Agents Actions 39 SpecNo,C189-C191
Netzel-Arnett, S. et al. 1991 Anal.Biochem.195, 86-92
【0139】
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
Finch-Arietta, M. et al., 1993. Agents Actions 39 SpecNo,C189-C191
Bickett, D.M. et al., 1994. Ann.N.Y.Acad.Sci. 732, 351-355
【0140】
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
Kraft, P.J. et al. 2001. Connect.Tissue Res. 42, 149-163
Itoh, M. et al. 1997. J.Pharm.Biomed.Anal. 15, 1417-1426
Welch, A.R. et al. 1995. Arch.Biochem.Biophys. 324, 59-64
【0141】
−Pro−Leu−Ala−Tyr−Trp−Ala−Arg−
配列番号:53
Aschi, M. et al. 2002. J.Comput.Aided Mol.Des 16, 213-225
Netzel-Arnett, S. et al. 1991. Anal.Biochem.195, 86-92
【0142】
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
Lauer-Fields, J.L. and Fields, G.B. 2002 Biol.Chem. 383, 1095-1105
Lauer-Fields, J.L. et al. 2001. Biochemistry 40, 5795-5803
【0143】
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0144】
−Arg−Pro−Lys−Pro−Gln−Gln−Phe−Trp−
配列番号:59
Bickett, D.M. et al. 1994. Ann.N.Y.Acad.Sci. 732, 351-355
【0145】
第一の配列S1および/または第二の配列S2をMMP−14酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Ala−Cys(p−OMeBz)−Trp−Ala−Arg−
配列番号:60
Mucha, A., et al. 1998. J. Biol. Chem. 273, 2763
Holtz, B., et al. 1999. Biochemistry 38, 12174
【0146】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0147】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Roderfeld, M. et al. 2000 . Prot. Expr. Purif. 19, 369
Kannan, R. et al. 1999. Prot. Expr. Purif. 16, 76
【0148】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
【0149】
第一の配列S1および/または第二の配列S2をMMP−15酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Xue, C.-B. et al. 2001. J. Med. Chem. 44, 2636
【0150】
第一の配列S1および/または第二の配列S2をMMP−16酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Shimada, T. et al. 1999. Eur. J. Biochem. 262, 907
Xue, C.-B. et al. 2001. J. Med. Chem. 44, 2636
【0151】
第一の配列S1および/または第二の配列S2をMMP−17酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
English, W.R. et al. 2001. FEBS Lett. 491, 137
【0152】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
【0153】
第一の配列S1および/または第二の配列S2をMMP−19酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Pendas, A.M. et al. 1997. J. Biol. Chem. 272, 4281
【0154】
第一の配列S1および/または第二の配列S2をMMP−23酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Velasco, G. et al. 1999. J. Biol. Chem. 274, 4570
【0155】
第一の配列S1および/または第二の配列S2をMMP−25酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
English, W.R. et al. 2001. FEBS Lett. 491, 137
【0156】
第一の配列S1および/または第二の配列S2をMMP−26酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Park, H.I. et al. 2002. J. Biol. Chem. 277, 35168
【0157】
前記に起因するものとして、特定のMMPは数種の配列を認識することが可能であり、それらの配列は本来、正確なメカニズムに従い、一部の病態に関与する異なるタンパク質に属する配列である。
【0158】
上記のように、本発明の環状ペプチドは、蛍光受容体基を担持する少なくとも1種のプローブX1、および蛍光受容体もしくは非蛍光受容体基を担持する少なくとも1種のプローブX2を含んでなる。一般に、これらは供与体基の蛍光発光スペクトルと受容体基の吸収スペクトルとが、少なくとも一部、重なり合うように選択される;このものは励起エネルギーを供与体基から受容体基へ非放射転移(または共鳴エネルギー転移)現象として移動する。
当該基は通常、芳香族核、例えば、ベンゼン、アントラセンまたはクマリン核を含んでなる。
【0159】
例として、以下の対合を例示し得る(対合の第一の構成基は供与体基であり、対合の第二の構成基は受容体基である):
*トリプトファン/2,4−ジニトロフェニル(略号W/Dnpにより記号化);
*o−アミノ安息香酸/2,4−ジニトロフェニル(略号Abz/Dnpにより記号化);
*(7−メトキシクマリン−4−イル)アセチル/2,4−ジニトロフェニル(略号Mca/Dnpにより記号化);
*(7−メトキシクマリン−4−イル)アセチル/N−3−(2,4−ジニトロフェニル)−L−2,3−ジアミノプロピル(略号Mca/Dpaにより記号化);
*トリプトファニル/ダンシル(略号W/Dnsにより記号化);
*N−メチルアントラノイル/2,4−ジニトロフェニル(略号Nma/Dnpにより記号化);
*6,7−ジメトキシクマリン−4−イル−アセチル/6−N−(7−ニトロベンゾ−2−オキサ−1,3−ジアゾール−4−イル)アミノヘキサン酸(略号DMC/Nbdにより記号化);
*5−(2’−アミノメチル)ナフタレンスルホン酸/4−(4’−ジメチルアミノフェニルアザ)安息香酸(略号EDANS/Dabcylにより記号化);
*7−メトキシクマリン−3−カルボン酸/7−ジエチルアミノ−クマリン−3−カルボン酸(略号MC/DACにより記号化);当該基は以下の式に一致する:
【0160】
【化4】
【0161】
【化5】
【0162】
供与体基および受容体基がクマリン核を含んでなる基であると有利である。
これらの基はクマリン型の共通の芳香族単位を有するので、これが疎水型またはπ−スタッキング型の複合体の形成を可能とし、このものは本発明環状ペプチドの安定性において役割をもつ。
特に好適な対合はMC/DAC対合であり、このものは一旦プローブに結合すると以下の式をもつ:
【0163】
【化6】
【0164】
X1およびX2プローブはアミノ酸残基であり得る。
従って、X1プローブの一例は、以下の式を有する−Lys(MC)−プローブであってもよい:
【0165】
【化7】
【0166】
一方、X2プローブの例は以下の式を有する−Lys(DAC)−プローブであってもよい:
【0167】
【化8】
【0168】
S1およびS2は、上記定義のようなMMPの標的配列、とりわけ以下の配列に対応し得る:
-Gly−Pro−Gln−Gly−Leu−Leu−Gly−Ala−
または
−Gly−Pro−Gly−Gly−Val−Val−Gly−Pro
これらの配列はそれぞれMMP−1酵素およびMMP−9酵素が認識する配列に対応するものである。
【0169】
本発明の環状ペプチドは以下の式を有する環状ペプチドであってもよい:
【0170】
【化9】
【0171】
(X1、X2、R1、YおよびR2は上記定義と同じ定義に一致する)
本発明に適合する具体的な環状ペプチドは以下の式に一致する:
【0172】
【化10】
【0173】
(基−Lys(DAC)−および−Lys(MC)−は上記定義と同じ定義に一致する)
太字の配列はMMP1酵素の存在下に切断され、それによってLys(DAC)プローブとLys(MC)プローブの距離を互いに離すことに寄与する。それが共鳴移動の変化につながり、それによってMMP1酵素の活性の定量を可能とする。
【0174】
本発明の環状ペプチドは、ペンダント機能(リジンまたはアスパラギン酸残基に由来するアミン官能基など)を介して、粒子などの担体に固定されるに至る。
【0175】
本発明の環状ペプチドは、常套のプロセスを用いる固相上の自動合成工程からなり、次いで、固相から該ペプチドを脱離した後、またはその後にそれを固相から脱離させ、線状ペプチドの末端をカップリング結合させることからなる方法により調製し得る。
【0176】
固相とは、通常、−NH2または−COOH官能基と反応して共有結合を形成し得る基を担持する樹脂であり、この共有結合は一旦合成したペプチドを切断・脱離するように企図したものである。
【0177】
当該樹脂は以下の式の基をそれぞれ担持するトリチル、2−クロロ クロロ トリチル樹脂であってもよい:
【0178】
【化11】
【0179】
(中括弧はその部分を介して上記の基が樹脂に結合する部分を示す)
第一の実施態様によると、本発明方法は:
a)固相上の化学合成により線状ペプチドを調製するための調製工程;
b)固相から線状ペプチドを脱離する脱離工程;および
c)該線状ペプチドの末端をカップル結合させて環状ペプチドを形成するカップリング工程;
からなることを特徴とする。
【0180】
第二の実施態様によると、本発明方法は:
a)固相上の化学合成により線状ペプチドを調製するための調製工程;
b)該線状ペプチドの遊離の末端と該線状ペプチドの線状残基の末端官能基とをカップル結合させるカップリング工程;および
c)環状ペプチドを固相から脱離するための脱離工程であって、工程b)およびc)を付随して実施し得る脱離工程;
からなることを特徴とする。
【0181】
第一の実施態様または第二の実施態様についても、工程a)を実施する場合、その第一相は固相に第一のアミノ酸を固定化することからる。
本固定化は幾つかの方法で実施し得る:
− 第一アミノ酸のC−末端を介して固相上に固定化;
− 第一アミノ酸の側鎖を介して固相上に固定化;
− 第一アミノ酸の主鎖を介して固相上に固定化。
【0182】
第一の型の固定化の場合、一旦ペプチドを合成すると、ペプチドにおける脱保護されたN−末端官能基は、樹脂上から脱離した、またはなお固定化したままのカルボキシル官能基上で、求核付加メカニズムにより反応する。カルボキシル官能基がまだ樹脂上に固定化されたままである場合、求核付加反応(分子内付加である)は環化と樹脂からのペプチドの脱離を一緒に生じる。
【0183】
アミノ酸の側鎖を介して起こる固定化の第二の型は、一般に、以下のアミノ酸で行う:アスパラギン、アスパラギン酸、グルタミン、グルタミン酸エステル、リジン、ヒスチジン、セリン、トレオニン、アルギニンおよびチロシン;これらは樹脂が担持する反応性基と反応し得る−CO2H、OH、NH2またはNH2COなどの官能基を担持する側鎖を有する。
【0184】
樹脂上の主鎖の固定化に基づく第三の方法の場合、合成されたペプチドは主鎖のアミド結合の窒素原子の1つにより樹脂に結合される。
当業者は所望の配列を合成するために、明らかに、適切な保護基を使用するであろう。
【0185】
上記のように、本発明の環状ペプチドは、プロテアーゼ酵素Eの活性をモニターするために使用することを企図する。
より正確には、本発明の環状ペプチドの使用は、幾つかの様式で想定し得る:
− それらは酵素活性と認知された病態との間に限定的な相関関係を与える;
− それらは、酵素活性と認知された病態の間の相関関係が既知であるならば、生物サンプル中に存在する酵素をアッセイすることにより、病態を診断するために使用することができる。
【0186】
それらは、その活性を評価したいと所望する酵素を含んでなる液体のインビトロ分析を実施するために、特に使用することができる。
それらはまた、X1/X2対合が赤色で発光する場合、特にインビボでの使用に供される。
【0187】
従って、本発明は、本発明に適合する少なくとも1種の環状ペプチドを含んでなる即時使用試薬;および
− 本発明に適合する少なくとも1種の環状ペプチドを含んでなる第一区分;および
− 緩衝液を含んでなる第二区分;
を含んでなるキットに関する。
【0188】
このキットは保存液の調製を企図するものであり、この保存液を、活性を測定しようとするプロテアーゼ酵素と接触させる。該プロテアーゼ酵素は、生物液中に含まれていてもよい。
その名称が示唆するように、緩衝液は溶液のpHを一定に維持することを企図するものであり、引き続く酵素活性の測定に影響しかねないpHの過度な酸性への、または過度な塩基性への変化を起こさないようにする。
最終的に、本発明は、少なくとも1種のプロテアーゼ酵素の活性をインビトロで測定するための、本発明による少なくとも1種の環状ペプチドの使用に関する。
【0189】
本発明のその他の特徴と有益性は、説明のために示した、限定するものではない以下の実施例を読み解くことにより明らかとなろう。
【図面の簡単な説明】
【0190】
【図1】図1は実施例に従い調製した環状ペプチドの蛍光強度ΔIF 供与体(a.u.)の変化を時間t(分)の関数として示すグラフである;この環状ペプチドはMMP−1溶液(1nM)の存在下に90μMの濃度とする。
【図2】図2は、調製した環状ペプチドの初期加水分解速度の逆数(1/vi)(a.u.−1.s)をその濃度の逆数(1/[S])(μM−1)の関数として示す二重逆数プロットである。
【発明を実施するための形態】
【0191】
以下の実施例は、それぞれMMP1標的配列に対応する2つのペプチド配列S1およびS2を含んでなる本発明による環状ペプチドの製造について説明するものである;この配列は以下の:−Gly−Pro−Gln−Gly−Leu−Leu−Gly−Ala−である。
【0192】
この環状ペプチドの製造は以下の工程からなる:
a)その側鎖を介して樹脂上に接合することを企図する出発アミノ酸の合成工程:Fmoc−Lys−O−アリル;
b)当該出発アミノ酸を樹脂に接合する工程;
c)接合した樹脂上で2種のペプチド配列を合成する工程;
d)環化工程。
【0193】
これら工程の詳細を以下に説明する。
以下に示すプロトコールにおいては、以下の略号を使用する:
ACN:アセトニトリル; Boc:t−ブチルオキシカルボニル; DAC:7−ジエチルアミノクマリン−3−カルボン酸; DCM:ジクロロメタン; DIEA:N,N’−ジイソプロピルジエチルアミン; DMF:N,N−ジメチルホルムアミド; ESMS:エレクトロスプレー質量分析法; Et2O:ジイソプロピルエーテル; Fmoc:9−フルオレニルメトキシカルボニル; HBTU:ヘキサ−フルオロリン酸N−[1H−ベンゾトリアゾール−1−イル]ジメチルアミノ)メチレン)−N−メチルメタナミニウムN−オキシド; HOBt:N−ヒドロキシベンゾトリアゾール; MC:7−メトキシクマリン−3−カルボン酸; AMPA:4−アミノフェニル水銀酢酸塩; PyBOP:ベンゾトリアゾール−1−イル−オキシ−トリス−ピロリドノホスホニウム; TA:外界温度; TFA:トリフルオロ酢酸; THF:テトラヒドロフラン; TIS:トリイソプロピルシラン; TRIS:トリス(ヒドロキシメチル)アミノメタン。
【0194】
化学製品および溶媒はすべて分析精度のものであり、シグマ社から購入した。2−クロロ クロロトリチル樹脂、PyBOPおよびすべてのアミノ酸(FmocにてNα−保護)はアドバンスドケムテック(Advanced Chemtech)から購入した。
【0195】
化学反応はすべてN2下に無水溶媒を用いて実施した。DMFは還流下にCaH2にて一夜乾燥し、蒸留する。DCMは使用前に蒸留した。
【0196】
UV/可視スペクトルは日立分光光度計装置モデルU−2010で記録した。蛍光スペクトルはPTIおよびジョビン−イボン(Jobin-Yvon)JY3分光蛍光光度計装置で実施し、サンプルホルダーは37℃の恒温とした。固相でのペプチド合成はアップライドバイオシステム433Aシンセサイザーにて実施した。精製は逆相島津HPLC(コントローラー:SLC−10AVP; ポンプ:LC8A; UV/可視検出器:SPD−10AVP[波長:214、267および254nm]およびC18サティスファクションカラムRP18AB 5μm 250×4.6mm)にて実施した。溶出に使用した溶媒系は、(A)0.1%TFA水溶液および(B)70%ACN水溶液であった。
【実施例】
【0197】
a)Fmoc−Lys−O−アリルの合成
Fmoc−Lys−O−アリルは下記式に一致する:
【0198】
【化12】
【0199】
このものは以下の反応工程図に従い調製する:
【0200】
【化13】
【0201】
Fmoc−Lys(Boc)−OH(2g、4.3mmol)および臭化アリル(10mL、118.2mmol)の溶液に、1.45mLのDIEA(8.5mmol)を加える。80℃にて3.5時間攪拌した後、この溶液を200mLの酢酸エチルで希釈し、次いで、0.1N−HCl(3×100mL)、10%NaHCO3(3×100mL)およびNaCl飽和溶液(3×100mL)で洗浄し、MgSO4で乾燥する。減圧下に蒸発させた後、1.9gのFmoc−Lys(Boc)−O−アリル(3.87mmol、90%)を白色粉末の形状で得る。
【0202】
Fmoc−Lys(Boc)−O−アリル(1.9g、3.87mmol)をTFA/DCM混合物(70mL、1:1)に溶解する。この溶液を外界温度で3時間攪拌する。この溶液を濃縮した後、エーテルで洗い、引き続き蒸発させて、1.45gのFmoc−Lys−O−アリル(3.56mmol、92%)を白色粉末の形状で回収する。
【0203】
b)Fmoc−Lys(Boc)−O−アリルの樹脂への接合、およびLys(MC)プローブの接合
反応工程図は以下のとおりである:
【0204】
【化14】
【0205】
Fmoc−Lys(Boc)−O−アリル(1.81g、4.43mmol)を無水THF(2mL)に溶解し、DIEA(734μL、4.23mmol)を加える。外界温度で10分間攪拌した後、570mgの2−クロロトリチル樹脂を加える。この混合物を外界温度で5時間混合する。次いで、樹脂を濾取し、DMF(20mL)、DCM/メタノール/DIEA混合物(17:2:1)(2×20mL)、DMF(10mL)およびDCM(20mL)で洗浄する。この新たな樹脂の置換率は、ピペリジンで処理した後、ジベンゾフルベン複合体のUV測定を介して0.4mmol/gと判定した。最初に、Fmoc−Lys−O−アリル−クロロトリチル樹脂(300mg、0.4mmol/g)を、20%ピペリジン/DMF混合物と外界温度で3時間接触させることによって、当該樹脂からFmoc基を脱離させて脱保護した。遊離一級アミンの存在をカイザーテストにより証明した。この樹脂を、DMF(5mL)中、PyBOP(173mg、0.392mmol)、HOBt(54mg、0.4mmol)およびDIEA(100μL、0.8mmol)の存在下、外界温度で6時間、Fmoc−Lys(MC)−OH(228mg、0.4mmol/g)と反応させる。次いで、この樹脂を濾取し、DMF、DCMおよびメタノールで洗浄する。Fmoc−Lys(MC)−Lys−O−アリル−クロロトリチル樹脂(270mg)を回収する。この新たな樹脂の置換率は、ピペリジンで処理した後、ジベンゾフルベン複合体のUV測定により0.38mmol/gと判定した。反応しなかったアミン官能基は、DMF(5mL)中、酢酸スクシミニドエステル(25.12mg、0.16mmol)、ピペリジン(12mg、0.16mmol)の存在下に、外界温度で40分間アセチル化した。
【0206】
c)ペプチド合成
反応工程図は以下のとおりである:
【0207】
【化15】
【0208】
(記号G、P、Q、LおよびAはそれぞれアミノ酸のグリシン、プロリン、グルタミン、ロイシン、アラニンに相当する)
【0209】
Fmoc−Lys(MC)−Lys−O−アリル−クロロトリチル樹脂をシンセサイザー反応機に加える。Fmoc基によりNα−保護したアミノ酸(Gly、Pro、Gln(Trt)、LeuおよびAla)を過剰(10倍)に加える。カップリング工程をHBTU、HOBtおよびDIEAの存在下に実施し、その後、以下のペプチドGPQGLLGA−(AはLys(MC)に結合)を得る。18時間後、樹脂に結合したペプチド(370mg)を得る。
【0210】
Fmoc−Lys(DAC)−OH(190mg、0.3mmol)を、シンセサイザー反応機外部にて、DMF(5mL)中、PyBOP(153mg、0.3mmol)、HOBt(40.53mg、0.3mmol)およびDIEA(99.1μL、0.6mmol)の存在下、外界温度で5時間、樹脂上のペプチドとカップル結合させる。次いで、樹脂を濾取し、前記のように洗浄する。401mgを取得する。置換率は0.25mmol/gと評価する。反応しなかったアミン官能基は上記と同じ手順でアセチル化する。
【0211】
樹脂上のペプチドを再び自動ペプチドシンセサイザー反応機に入れ、前記と同じアミノ酸配列を最初の工程に従って合成する。530mgを回収する。
【0212】
d)環化反応
この樹脂を高真空下に4時間乾燥し、次いでアルゴン下に置いた。Pd(P(Ph3))4(346.6mg、0.3mmol)をアルゴン気流下にてクロロホルム/酢酸/N−メチルモルホリン混合物(37:2:1、8mL)に溶解する。次いで、この溶液を樹脂と接触させ、得られた混合物を外界温度で2時間、時折攪拌する。次いで、樹脂を濾取し、0.5%DIEA/DMF溶液(2×10mL)で、次いで、ジエチルジチオカルバミン酸ナトリウム/DMF溶液(0.5%w/w)で洗い、触媒を除去する。
【0213】
Fmocを切断した後、樹脂を1M−HOBt/DMFにて洗浄する。N末端およびC末端間の環化反応を実施するために、260mgのPyBOP(0.5mmol)、69mgのHOBt(0.51mmol)および180μLのDIEA(1mmol)を10mLのN−メチルピロリドンに加える。この溶液を外界温度で48時間攪拌し、その後、樹脂を30mLのDMF、DCMおよびメタノールで洗浄する。
【0214】
次いで、この樹脂をTFA/H2O/TIS溶液(95:2.5:2.5)に懸濁する。室温3時間後、樹脂を濾取し、TFA(2×1mL)およびDCM(10mL)で洗浄する。濾液を一部減圧下に蒸発させ、次いで、冷エーテルに浸漬する。
【0215】
−20℃で12時間後、沈殿を濾取し、洗浄して170mgの粗生成物を得る。準分取HPLC精製の後、92.1mgのペプチドを得た。このペプチドの構造をESMSで証明した。
本実施例で得られた環状ペプチドは以下の式に一致する:
【0216】
【化16】
【0217】
酵素アッセイは、0.1M−TRIS、0.1M−NaClおよび10mM−CaCl2を含有するpH7.6の緩衝液中で実施した。25μLの市販MMP−9(90%チモーゲン形状)を緩衝液中、37℃、4時間、1mM−AMPA溶液で活性化する。市販のMMP−1はすでに活性化した形状にある。上記のプロトコールに従い調製した環状ペプチドの保存液を、上記定義のような緩衝液により調製し、最終濃度90μMとする。最終濃度90μMの溶液を出発溶液として用い、異なる希釈を実施して、10、5、1および0.5μMの4段階の環状ペプチド/緩衝液とする。
【0218】
アッセイはすべて2mLの最終反応容量中、1nM活性MMP−1または2nM−AMPA−活性化MMP−9で実施する。
【0219】
酵素を添加した後、初期加水分解速度を、403nmで(340nmの励起波長で)の蛍光強度の経時的変化により測定する。アッセイしたペプチド濃度の各濃度について、図1に示すように、時間t(分)の関数として供与体基ΔIF(a.u.)の蛍光強度の変化を表す曲線をプロットした(環状ペプチド濃度、10、5、1および0.5μM)。
【0220】
以下の等式:
蛍光(t)=蛍光max(1−exp(−kobst))
から、一次定数を決定することができた。二次定数(特異的定数と呼称)kcat/KMの計算は、等式kcat/Km=kobs/[E0]に関して使用される活性酵素Eの濃度により得られる。該ペプチドのkcat/Kmは、文献に記載されたMMP−1の最良の基質と比較して、319120s−1・M−1という非常に高い値で評価する;この値はMMP−1について、本発明に従い調製した環状ペプチドの非常に高い特異性を証明するものである。
【0221】
異なるアッセイ法の初期速度を用いることによる、図2に示した二重逆数グラフ(1/Vi)=f(1/[環状ペプチド])は、合計3.8μMとなる環状ペプチドのKmの決定を可能とした。この値は文献に記載された最良のMMP−1基質で得られる値よりも低いが、MMP−1に対して本発明に従い調製した環状ペプチドの優れた親和性を証明するものである。
【0222】
また、本発明の環状ペプチドについては、2nMの活性MMP−9の存在下でもアッセイした。供与体基の蛍光強度には、経時的変化が測定できなかった。従って、この環状ペプチドはMMP−9の基質ではなく、この環状ペプチドの選択性を示している。また、この環状ペプチドはMMP−1に対して非常に高い選択性と強力な親和性を示す。
【技術分野】
【0001】
本発明は、通常、いわゆるFRET(蛍光共鳴エネルギー移動またはフォースター共鳴エネルギー移動の略)法によるプロテアーゼ酵素の酵素活性測定において使用し得る新規蛍光環状ペプチドに関する。
本発明は酵素障害を原因とする疾患の診断分野において特に有用である。
【背景技術】
【0002】
酵素は調節現象を顕著に伴う複雑な作用メカニズムを有し、その破壊は、病理学的障害の発症の原因となり得る。
【0003】
メタロプロティナーゼ(MMPの略号でも知られる)は顕著な事例であり、このものは細胞外マトリックス要素のすべてと、一部の増殖因子および接着分子を切断し得る。これらの酵素は自然の過程によってTIMP(メタロプロティナーゼの組織インヒビター)などのインヒビターにより調節される。MMP/TIMPの平衡が崩れると、腫瘍増殖、転移、関節リウマチ、心疾患などの病的プロセスが生じ得る。
【0004】
多くの病態において酵素が関わっていることから、認知された病態との可能な相関を確立するためには、これらの酵素活性を測定可能とする手段の提供が必須であることが間もなく証明された。現時点で入手し得る手段は、活性を測定すべき酵素の基質からなり、これらの基質は蛍光基を有すると共に、その活性を蛍光測定により定量することからなる。特に、最も頻繁に使用される蛍光法は蛍光共鳴エネルギー移動法またはFRETである。しかし、この技法は厳密な意味で蛍光移動の測定に基づくものではなく、電子エネルギー移動の測定に基づくものである。従って、識別は放射移動と非放射(または共鳴)移動間でなされなければならない。放射移動は基と基の間の距離、それらの方向性などの情報の入手手段を提供するものではなく、試験するシステムを理解するための、または特性化するための必須のデータであり得る。
【0005】
放射移動は2段階で起こるプロセスである。初期段階では、光子が供与体基により放出され、第二段階では、この光子が供与体基と同じ光子受容体基または異なる光子受容体基により吸収される。この移動には基と基の間の相互作用を必要としないが、種のスペクトルの重なり合いと濃度のスペクトルの重なり合いに左右される。
【0006】
励起エネルギーの非放射移動は供与体基および受容体基の間の相互作用を必要とする。この移動は供与体基の発光スペクトルが受容体基の吸収スペクトルと重なり合った場合にのみ起こり、その結果、供与体基の振電遷移と受容体基の振電遷移との間のエネルギー対応が存在する。当該遷移はカップル結合している、または<共鳴している>と言われる。
【0007】
非放射エネルギー移動の現象はフォースター(Forster)が研究した。彼はこの物理現象が10ないし100Åという位数の非常に短い距離にまたがって起こることを証明し得た。供与体基と受容体基の距離が大きくなる場合、これはエネルギー移動の変化と解釈される。従って、この現象はプロテアーゼ酵素の酵素活性を評価するために使用し得る。対照的に、各末端でそれぞれ配列した供与体基と受容体基両方を含んでなる基質を特異的プロテアーゼ酵素と接触させると、基質が切断され、その結果、供与体基と受容体基間の距離が増大し、結果として、励起エネルギー(または共鳴エネルギー)の移動の変化が起こる。この変化に基づき、酵素活性の評価に到達することができる。
【0008】
現在使用されている基質は殆どの部分が線状構造の基質についてであり、その基質は、明らかにエキソプロテアーゼおよび非特異的プロテアーゼにより分解されると思われるので、生物媒体においては殆ど安定性のないことが証明されている。
【0009】
それ故、所定のプロテアーゼ酵素の酵素活性を定量するための手段として使用し得る、安定で、特にFRET法を用いる当該酵素の酵素活性のモニターを可能とするペプチドが紛れもなく必要である。
【発明の概要】
【発明が解決しようとする課題】
【0010】
それ故、本発明者らは上記の必要性に一致する新規なペプチドを開発してきた。
【0011】
それ故、第一の主題によると、本発明は以下の配列:
−S1−X1−S2−X2−
(式中、S1はプロテアーゼ酵素E1の第一標的ペプチド配列であり;S2はプロテアーゼ酵素E2の第二標的ペプチド配列であり;S1およびS2は同一であっても、異なっていてもよく;当該第一のペプチド配列S1および当該第二のペプチド配列S2は4個ないし14個のアミノ酸を含んでなり;E1およびE2は同じプロテアーゼ酵素に、または異なるプロテアーゼ酵素に相当していてもよく;X1は蛍光供与体基を担持するプローブであり;そしてX2は蛍光または非蛍光受容体基を担持するプローブである)
を含んでなる環状ペプチド(シクロペプチドとも呼称する)に関する。
【0012】
好ましくは、該供与体基および受容体基は、供与体基の蛍光発光スペクトルが受容体基の吸収スペクトルと少なくとも部分的に重なり合うように選択する。
【0013】
それ故、1つのプロテアーゼ酵素E(E1とE2が1つかつ同一の酵素を形成するのみであり、S1とS2が同一のプロテアーゼ酵素の標的ペプチド配列である場合)またはプロテアーゼE1とプロテアーゼE2(それらが異なっている場合、またS1がプロテアーゼ酵素E1の標的ペプチド配列であり、S2がプロテアーゼ酵素E2の標的ペプチド配列である場合)両方の存在下、本発明の環状ペプチドは、第一のペプチド配列S1および第二のペプチド配列S2の両方で切断され、そして、プローブX1およびX2の配置を原因として、プローブX1とX2の空間に距離を生じる。次いで、供与体基と受容体基の距離が離れ、次に、これらの2つの基の間の共鳴エネルギーの移動に変化が生じる。それ故、この共鳴エネルギーの変化を用いて、プロテアーゼ酵素Eまたはプロテアーゼ酵素E1とE2の酵素活性に到達することが可能である。
【0014】
さらに、本発明のペプチドが環状であるという事実は、常套的に先行技術において使用される線状ペプチドの欠点、特に先行技術の線状ペプチドが遭遇する安定性の問題を克服することが可能であることを意味する。
【0015】
本発明の特定の一実施態様によると、E1およびE2は同一のプロテアーゼ酵素に相当する。
プロテアーゼ酵素Eの酵素活性を検討するために(E1=E2の場合)、本発明者らは検討すべきプロテアーゼ酵素Eの標的である1本の線状ペプチド配列の環状化について極単純に仮想した;該ペプチド配列は以下の工程図に示すプローブX1およびX2をその末端に含んでなる:
【0016】
【化1】
【0017】
検討すべきプロテアーゼ酵素Eの存在下、環状ペプチドは以下の工程図に示すように切断される:
【0018】
【化2】
【0019】
切断後、基X1およびX2の距離は変化しないままである。結果として、共鳴移動の変化はなく、従って、プロテアーゼ酵素Eの活性を定量することは不可能である。
詳細な説明に入る前に、我々は以下の定義を提案する。
【0020】
蛍光供与体基が慣例として意味するのは、光エネルギー(励起光といい、その波長は一般に紫外部領域に属する)を吸収し、その環境に依存的に2つの可能な形状でこの光を再発光することができる基である。
a)受容体基との共鳴による励起エネルギーの移動の形状で、ただし、後者は先に記載したFRET現象と互換性のある距離に、好ましくは10ないし100Åの範囲の距離にある;または
b)一般に波長が可視領域に属する蛍光(供与体発光光と呼称)の形状にある。
【0021】
受容体基が慣例として意味するのは、本発明の意味において、共鳴エネルギーによる供与体基の励起エネルギーを吸収することのできる基である。この励起エネルギーは2つの主要な様式で再発光され得る:
a)放射発光(光子の放出)による;この場合、蛍光受容体基という用語が使用される;または
b)非放射発光(すなわち、光子放出以外の不活性化様式)による;この場合、非蛍光受容体基という用語が使用される。
【0022】
プローブが通常意味するのは、また本発明の意味において、供与体基(プローブX1について)を担持するアミノ酸残基、または受容体基(プローブX2について)を担持するアミノ酸残基である。
【0023】
アミノ酸残基が通常意味するのは、アミノ酸の−NH2官能基または−CO2H等と別の化合物の官能基とが反応して共有結合を形成するような残基アミノ酸である。
【0024】
標的ペプチド配列が通常意味するのは、また本発明の意味において、プロテアーゼ酵素が認識して切断し得るペプチド配列である。
【0025】
プロテアーゼ酵素E1およびプロテアーゼ酵素E2(E1およびE2は同一であっても、異なっていてもよい)の2つの標的ペプチド配列(これら配列の間に本発明に適合するプローブX1およびプローブX2がそれぞれ配列されている)を含んでなる環状ペプチドの使用が、上記で説明したように、当該プロテアーゼ酵素の活性を定量するための良好な手段の提供を可能とする。
本発明の環状ペプチドは、下記式:
【0026】
【化3】
【0027】
(式中、
− S1、X1、S2およびX2は上記と同一の定義を示す;
− R1およびR2は独立して、単結合、アミノ酸残基またはペプチド配列に相当する;
− Yは−CONH−基または−NHCO−基に相当する)
に一致する。
【0028】
S1およびS2は、該ペプチドが単一のプロテアーゼ酵素(E1およびE2が同一である)の活性を定量しようとする場合は、同一であり、または該ペプチドが同一のプロテアーゼ酵素のイソ型の存在または2種の異なるプロテアーゼ酵素E1およびE2の存在の間を識別しようとする場合は明らかに異なり得る。
【0029】
S1およびS2が同一である場合、それらは同一方向に、または逆方向に配置され得る。例えば、S1が配列−Gly−Pro−Gln−Gly−Leu−Leu−Gly−Ala−に相当する場合、配列S2は逆方向に配置された同一配列、すなわち、以下の配列−Ala−Gly−Leu−Leu−Gly−Gln−Pro−Gly−に相当する。
【0030】
本発明の特定の一実施態様において、R1およびR2はアミノ酸残基に、またはペプチド配列に相当し得る。このことは該環状ペプチドが医薬投与形態を企図する場合、特に生物媒体において該ペプチドが安定であるという理由で特に有利である。
【0031】
本発明の環状ペプチドは、MMP型のプロテアーゼ酵素(“細胞外マトリックスメタロプロティナーゼ”としても知られる)の酵素活性を測定するために特に適用される。
【0032】
この場合、プロテアーゼ酵素E1およびプロテアーゼ酵素E2はMMP型の酵素であり、その場合の第一ペプチド配列S1はMMPプロテアーゼ酵素の標的であり、第二ペプチド配列S2はMMPプロテアーゼ酵素の標的である。
【0033】
MMP類はその活性部位に亜鉛原子を含む28の数に及ぶエンドぺプチダーゼの系統群である。それらは分泌されるか、または膜と会合しているかによって2つの系統群に分類し得る。分泌されるMMPは、コラゲナーゼ(例:MMP−1、8、13および18)、ゼラチナーゼ(例:MMP−2および9)、マトリリシン(例:MMP−7および26)、ストロメリシン(例:MMP−3、10および11)、エピリシン(MMP−28)、エナメリシン(MMP−20)を包含する。膜と会合するMMPは、膜貫通MMP、例えば、MMP−14、MMP−15、MMP−16、MMP−17、MMP−23、MMP−24およびMMP−25を包含する。
【0034】
MMP類は細胞外マトリックス要素のすべて、一部の増殖因子、または接着分子さえも切断することができる。MMPはTIMPなどの天然のインヒビターにより調節され、生理的プロセス(胚増殖、血管新生、骨組織の改造作用など)において重要な役割を果たしている。MMP/TIMPのバランスが崩れると、腫瘍増殖、転移、関節リウマチ、心疾患などの病的プロセスが生じる。
【0035】
腫瘍進行におけるMMP類の役割は多様であるが、とりわけ細胞外マトリックスを分解するMMPの能力が理由であり、それにより腫瘍の侵入を促進する。また、MMPは非マトリックスタンパク質に対してタンパク分解活性を有し、腫瘍の進行、接着喪失、侵入、増殖、血管新生、血管内異物侵入、管外遊出および転移増殖などの幾つかの他の工程においてそれらに対し複雑な役割を与える。
【0036】
それ故、環状ペプチドはMMPタイプのプロテアーゼ酵素の酵素活性をモニターするための、特に、酵素活性と認知された病態との間のつながりを確立するための、優れた手段であること証明する。
【0037】
より正確には、環状ペプチドがMMP活性の追跡を企図する場合、標的のペプチド配列S1およびS2は1個のMMP酵素(S1=S2であるか、またはS1およびS2が1つの同じ酵素の2つの同型間を識別しようとする異なる標的配列である場合)の、または2個のMMP酵素(S1およびS2が異なっていて、それぞれが2つの異なるプロテアーゼ酵素の標的である場合)の標的であり、それらの配列はその活性をモニターしようとするMMP(類)が認識するMMP(類)の基質の配列に相当する。
【0038】
特に、標的ペプチド配列S1およびS2は、MMP−1、MMP−2、MMP−3、MMP−5、MMP−7、MMP−8、MMP−9、MMP−10、MMP−11、MMP−12、MMP−13、MMP−14、MMP−15、MMP−16、MMP−17、MMP−19、MMP−23、MMP−25およびMMP−26およびその混合物から選択されるMMP酵素の標的となり得る。
【0039】
ペプチド配列S1およびS2は、4個ないし14個の範囲のアミノ酸の長さをもつ線状配列であり、活性を示すために必要なコンホメーションをもつ同じ配列の環状ペプチドを可能とする。S1およびS2は、そのN末端とC末端の間の距離が10Åと100Åの間となるように選択され、当該距離は先に定義したような供与体基と受容体基の間でFRETを実施するために有効な距離と矛盾しない距離とすると有利である。分子モデル化ソフトウエアを使用することで、信頼し得る当該距離の予測が可能である。
【0040】
ペプチド配列S1およびS2は、MMPの天然基質に見出される天然のペプチド配列から誘導し得るか、またはMMPの合成基質として知られる合成配列から誘導し得る。
【0041】
有利なことに、本発明の環状ペプチドは天然のアミノ酸のみを含み、それがそれらの可能性のある細胞毒性を制限し得るものとし、また関係する生物体への非天然アミノ酸の導入を回避する。
【0042】
一部の事例においては、天然または合成基質がMMP類によって認識され、切断されることがあり得る。どのMMPが切断に関与しているかを決定するためには、この基質についての異なるMMP類の酵素動力学データを使用し、比較することが必要である。
【0043】
数種のMMP類間の識別を可能とするこれら酵素動力学データは、一般に、その基質に対する酵素の親和性の指標であるミカエリス定数(Km)、および触媒定数(Kcat、1秒あたり且つ酵素1モルあたりに形成される産物のモル数)と酵素の触媒効率の指標であるミカエリス定数との比である。それ故、一般に、MMP/基質対合についての値の特定の対合(Kmおよびkcat/Km)が存在する。それ故、例えば2つのMMPが同じ基質を認識する場合、どちらが可視化活性に関与しているかを決定することが可能である。
【0044】
合成配列は、例えば、以下のアミノ酸または基を含んでなる:
Nva (ノル−バリン); Cha (3−シクロヘキシルアラニン); Dpa (N−3(2,4−ジニトロフェニル)−L−2,3−ジアミノプロピオニル); Abu:α−アミノ酪酸; S−メチルシステインに対応するCys(Me)、このものは基−SHが−S−CH3基と置換されていることを意味する; D−アルギニンに対応するD−Arg。
【0045】
本明細書の残りの記載において、下記掲載の略号は具体的に以下の意味を有する:
Gly:グリシン; Pro:プロリン; Leu:ロイシン; Ala:アラニン; Gln:グルタミン; Ile:イソロイシン; Arg:アルギニン; Val:バリン; Tyr:チロシン; Glu:グルタメート; Met:メチオニン; Phe:フェニルアラニン; Asn:アスパラギン; Thr:トレオニン; Trp:トリプトファン; Ser:セリン; His:ヒスチジン、これらの略号はアミノ酸についての公式の3文字命名法に準拠する。
【0046】
第一の配列S1および/または第二の配列S2をMMP−1酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Gly−Pro−Gln−Gly−Leu−Leu−Gly−Ala−
配列番号:1
−Ala−Pro−Gln−Gly−Ile−Ala−Gly−Gln−
配列番号:2
−Gly−Pro−Gln−Gly−Leu−Ala−Gly−Gln−
配列番号:3
−Gly−Pro−Leu−Gly−Ile−Ala−Gly−Ile−
配列番号:4
−Gly−Pro−Glu−Gly−Leu−Arg−Val−Gly−
配列番号:5
−Tyr−Glu−Ala−Gly−Leu−Gly−Val−Val−
配列番号:6
−Ala−Gly−Leu−Gly−Val−Val−Glu−Arg−
配列番号:7
−Ala−Gly−Leu−Gly−Ile−Ser−Ser−Thr−
配列番号:8
−Gly−Ala−Met−Phe−Leu−Glu−Ala−Ile−
配列番号:9
−Ile−Pro−Glu−Asn−Phe−Phe−Gly−Val−
配列番号:10
−Thr−Glu−Gly−Glu−Ala−Arg−Gly−Ser−
配列番号:11
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Leu−Arg−Ala−Tyr−Leu−Leu−Pro−Ala−
配列番号:13
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:14
Deng, S.J., et al. 2000. J. Biol. Chem. 275, 31422
【0047】
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:15
Beekman, B., et al. 1996. FEBS Lett 390, 221
【0048】
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
Darlak, K., et al. 1990. J. Biol. Chem. 265, 5199
Stack, M.S., and Gray, R.D. 1989. J. Biol. Chem. 264, 4277
【0049】
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
Netzel-Arnett, S., et al. 1991. Anal. Biochem.195, 86
【0050】
−Pro−Cha−Abu−Cys(Me)−His−Ala−
配列番号:18
McGeehan, G.M., et al. 1994. J. Biol. Chem.269, 32814
【0051】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:19
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
【0052】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0053】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knauper, V. et al. 1996. J. Biol. Chem. 271, 1544
【0054】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
【0055】
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
A. Santala, A. et al. 1999. FEBS Lett. 461, 153-156
Nagase, H. et al. 1994 J.Biol.Chem. 269, 20952-20957
【0056】
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
J. Berman et al. 1992. J.Biol.Chem. 267, 1434-1437
【0057】
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
Kraft, P.J. et al. 2001. Connect.Tissue Res. 42, 149-163
Itoh, M. et al. 1997. J.Pharm.Biomed.Anal. 15, 1417-1426
Welch, A.R. et al. 1995. Arch.Biochem.Biophys. 324, 59-64
【0058】
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
Lauer-Fields, J.L. and Fields, G.B. 2002 Biol.Chem. 383, 1095-1105
Lauer-Fields, J.L. et al. 2001. Biochemistry 40, 5795-5803
【0059】
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0060】
第一の配列S1および/または第二の配列S2をMMP−2酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Ile−Pro−Glu−Asn−Phe−Phe−Gly−Val−
配列番号:10
−Pro−Pro−Gly−Ala−Tyr−His−Gly−Ala−
配列番号:27
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−His−Leu−Leu−Val−Glu−Ala−
配列番号:28
−Leu−Arg−Ala−Tyr−Leu−Leu−Pro−Ala−
配列番号:13
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:29
Beekman, B., et al. 1996. FEBS Lett 390, 221
【0061】
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
Darlak, K., et al. 1990. J. Biol. Chem. 265, 5199
Stack, M.S., and Gray, R.D. 1989. J. Biol. Chem. 264, 4277
【0062】
−Pro−Leu−Ala−Nva−Dpa−Ala−Arg−
配列番号:30
Murphy, G., et al. 1994. J. Biol. Chem. 269, 6632
【0063】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
【0064】
−Pro−Leu−Gly−Met−Trp−Ser−Arg−
配列番号:31
Netzel-Arnett, S., et al. 1991. Anal. Biochem. 195, 86
【0065】
−Pro−Leu−Gly−SCH[CH2CH(CH3)2]−CO−Leu−Gly−
配列番号:32
Weingarten, H., et al. 1985. Anal. Biochem. 147, 437
Weingarten, H., et al. 1985. Biochemistry 24, 6730
【0066】
−Arg−Pro−Pro−Gly−Phe−Ser−Ala−Phe−
配列番号:33
G.D. Johnson and K. Ahn 2000. Anal. Biochem. 286, 112
【0067】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knight, C.G. et al. 1992. FEBS Lett. 296, 263
【0068】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24,6730
Xia, T. et al. 1996. Biochim. Biophys. Acta 1293,259
【0069】
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
J. Berman et al., 1992. J.Biol.Chem. 267, 1434-1437
【0070】
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
Kraft, P.J. et al. 2001. Connect.Tissue Res. 42, 149-163
Itoh, M. et al. 1997. J.Pharm.Biomed.Anal. 15, 1417-1426
Welch, A.R. et al. 1995. Arch.Biochem.Biophys. 324, 59-64
【0071】
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
Lauer-Fields, J.L. and Fields, G.B. 2002 Biol.Chem. 383, 1095-1105
Lauer-Fields, J.L. et al. 2001. Biochemistry 40, 5795-5803
【0072】
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0073】
第一の配列S1および/または第二の配列S2をMMP−3酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Gly−Pro−Glu−Gly−Leu−Arg−Val−Gly−
配列番号:5
−Arg−Val−Gly−Phe−Tyr−Glu−Ser−Asp−
配列番号:34
−Leu−Leu−Ser−Ala−Leu−Val−Glu−Thr−
配列番号:35
−Glu−Ala−Ile−Pro−Met−Ser−Ile−Pro−
配列番号:36
−Ile−Ala−Gly−Arg−Ser−Leu−Asn−Pro−
配列番号:37
−Ile−Pro−Glu−Asn−Phe−Phe−Gly−Val−
配列番号:10
−Lys−Pro−Gln−Gln−Phe−Phe−Gly−Leu−
配列番号:38
−Asp−Val−Ala−Gln−Phe−Val−Leu−Thr−
配列番号:39
−Asp−Thr−Leu−Glu−Val−Met−Arg−Lys−
配列番号:40
−Asp−Val−Gly−His−Phe−Arg−Thr−Phe−
配列番号:41
−Asp−Ser−Gly−Gly−Phe−Met−Leu−Thr−
配列番号:42
−Arg−Val−Ala−Glu−Met−Arg−Gly−Glu−
配列番号:43
−Asp−Leu−Gly−Arg−Phe−Gln−Thr−Phe−
配列番号:44
−Pro−Phe−Ser−Pro−Leu−Val−Ala−Thr−
配列番号:45
−Leu−Arg−Ala−Tyr−Leu−Leu−Pro−Ala−
配列番号:13
−Ala−Pro−Gly−Asn−Ala−Ser−Glu−Ser−
配列番号:46
−Phe−Ser−Ser−Glu−Ser−Lys−Arg−Glu−
配列番号:47
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−His−Leu−Leu−Val−Glu−Ala−
配列番号:28
−Pro−Pro−Glu−Glu−Leu−Lys−Phe−Gln−
配列番号:48
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:49
Deng, S.J., et al. 2000. J. Biol. Chem. 275, 31422
【0074】
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:15
Beekman, B., et al. 1996. FEBS Lett 390, 221
【0075】
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
Beekman, B., et al. 1997. FEBS Lett 418, 305
【0076】
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
Darlak, K., et al. 1990. J. Biol. Chem. 265, 5199
Stack, M.S., and Gray, R.D. 1989. J. Biol. Chem. 264, 4277
【0077】
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
Netzel-Arnett, S., et al. 1991. Anal. Biochem. 195, 86
【0078】
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
Bickett, D.M., et al. 1994. Ann. N.Y. Acad. Sci. 732, 351
【0079】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0080】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knight, C.G. et al. 1992. FEBS Lett. 296, 263
【0081】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24,6730
【0082】
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0083】
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−
配列番号:50
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0084】
第一の配列S1および/または第二の配列S2をMMP−5酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
Beekman, B., et al. 1997. FEBS Lett 418, 305
【0085】
第一の配列S1および/または第二の配列S2をMMP−7酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
Darlak, K., et al. 1990. J. Biol. Chem. 265, 5199
Stack, M.S., and Gray, R.D. 1989. J. Biol. Chem. 264, 4277
【0086】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
【0087】
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−Ser−
配列番号:24
Welch, A.R., et al. 1996. Biochemistry35, 10103
Welch, A.R., et al. 1995. Arch. Biochem. Biophys. 324, 59
【0088】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0089】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
【0090】
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
A. Santala, A. et al. 1999. FEBS Lett. 461, 153-156
Shabani, F. et al.1998. Free Radic.Res. 28, 115-123
Nagase, H. et al. 1994 J.Biol.Chem. 269, 20952-20957
【0091】
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
Finch-Arietta, M. et al., 1993. Agents Actions 39 SpecNo,C189-C191
Netzel-Arnett, S. et al. 1991 Anal.Biochem.195, 86-92
【0092】
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
Finch-Arietta, M. et al., 1993. Agents Actions 39 SpecNo,C189-C191
Bickett, D.M. et al., 1994. Ann.N.Y.Acad.Sci. 732, 351-355
【0093】
第一の配列S1および/または第二の配列S2をMMP−8酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Ala−Tyr−Trp−Ala−Arg−
配列番号:53
Netzel-Arnett, S., et al. 1991. Anal. Biochem.195, 86
【0094】
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Grams, F. et al. 2001. Biol. Chem. 382, 1277
【0095】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0096】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knauper, V. et al. 1996. J. Biol. Chem. 271,1544
【0097】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
【0098】
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
A. Santala, A. et al. 1999. FEBS Lett. 461, 153-156
Shabani, F. et al.1998. Free Radic.Res. 28, 115-123
Nagase, H. et al. 1994 J.Biol.Chem. 269, 20952-20957
【0099】
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
J. Berman et al., 1992. J.Biol.Chem. 267, 1434-1437
【0100】
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
Kraft, P.J. et al. 2001. Connect.Tissue Res. 42, 149-163
Itoh, M. et al. 1997. J.Pharm.Biomed.Anal. 15, 1417-1426
Welch, A.R. et al. 1995. Arch.Biochem.Biophys. 324, 59-64
【0101】
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
Lauer-Fields, J.L. and Fields, G.B. 2002 Biol.Chem. 383, 1095-1105
Lauer-Fields, J.L. et al. 2001. Biochemistry 40, 5795-5803
【0102】
第一の配列S1および/または第二の配列S2をMMP−9酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Gly−Pro−Pro−Gly−Val−Val−Gly−Pro−
配列番号:54
−Gly−Pro−Pro−Gly−Leu−Arg−Gly−Glu−
配列番号:55
−Gly−Pro−Gly−Gly−Val−Val−Gly−Pro−
配列番号:56
−Ile−Pro−Gln−Asn−Phe−Phe−Gly−Val−
配列番号:57
−Pro−Pro−Gly−Ala−Tyr−His−Gly−Ala−
配列番号:58
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:14
Deng, S.J., et al. 2000. J. Biol. Chem. 275, 31422
【0103】
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:15
Beekman, B., et al. 1996. FEBS Lett 390, 221
【0104】
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
Beekman, B., et al. 1997. FEBS Lett 418, 305
【0105】
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
Knight, C.G., et al. 1992. FEBS Lett. 296, 263
Darlak, K., et al. 1990. J. Biol. Chem. 265, 5199
Stack, M.S., and Gray, R.D. 1989. J. Biol. Chem. 264, 4277
【0106】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:19
Bickett, D.M., et al. 1993. Anal. Biochem. 212, 58
【0107】
−Pro−Leu−Gly−SCH[CH2CH(CH3)2]−CO−Leu−Gly−
配列番号:32
Weingarten, H., et al. 1985. Anal. Biochem. 147, 437
Weingarten, H., et al. 1985. Biochemistry 24, 6730
【0108】
−Arg−Pro−Pro−Gly−Phe−Ser−Ala−Phe−
配列番号:33
G.D. Johnson and K. Ahn 2000. Anal. Biochem. 286, 112
【0109】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0110】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Roy, N. et al. 1999. Prot. Expr. Purif. 16, 324
【0111】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
Xia, T. et al. 1996. Biochim. Biophys. Acta 1293, 259
【0112】
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
J. Berman et al., 1992. J.Biol.Chem. 267, 1434-1437
【0113】
第一の配列S1および/または第二の配列S2をMMP−10酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−His−Leu−Leu−Val−Glu−Ala−
配列番号:28
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Kannan, R. et al. 1999. Prot. Expr. Purif. 16, 76
【0114】
第一の配列S1および/または第二の配列S2をMMP−11酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0115】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Kannan, R. et al. 1999. Prot. Expr. Purif. 16, 76
【0116】
第一の配列S1および/または第二の配列S2をMMP−12酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0117】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Park, H.I. et al. 2000. J. Biol. Chem. 275, 20540
【0118】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
【0119】
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
A. Santala, A. et al. 1999. FEBS Lett. 461, 153-156
Shabani, F. et al.1998. Free Radic.Res. 28, 115-123
Nagase, H. et al. 1994 J.Biol.Chem. 269, 20952-20957
【0120】
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
Berman, J. et al., 1992. J.Biol.Chem. 267, 1434-1437
【0121】
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
Finch-Arietta, M. et al., 1993. Agents Actions 39 SpecNo,C189-C191
Netzel-Arnett, S. et al. 1991 Anal.Biochem.195, 86-92
【0122】
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
Finch-Arietta, M. et al., 1993. Agents Actions 39 SpecNo,C189-C191
Bickett, D.M. et al., 1994. Ann.N.Y.Acad.Sci. 732, 351-355
【0123】
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
Kraft, P.J. et al. 2001. Connect.Tissue Res. 42, 149-163
Itoh, M. et al. 1997. J.Pharm.Biomed.Anal. 15, 1417-1426
Welch, A.R. et al. 1995. Arch.Biochem.Biophys. 324, 59-64
【0124】
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
Lauer-Fields, J.L. and Fields, G.B. 2002 Biol.Chem. 383, 1095-1105
Lauer-Fields, J.L. et al. 2001. Biochemistry 40, 5795-5803
【0125】
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0126】
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−
配列番号:50
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0127】
−Arg−Pro−Lys−Pro−Gln−Gln−Phe−Trp−
配列番号:59
Bickett, D.M. et al. 1994. Ann.N.Y.Acad.Sci. 732, 351-355
【0128】
第一の配列S1および/または第二の配列S2をMMP−13酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:49
Deng, S.J., et al. 2000. J. Biol. Chem. 275, 31422
【0129】
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
Beekman, B., et al. 1997. FEBS Lett 418, 305
【0130】
−Pro−Cha−Gly−Nva−His−Ala−Dpa−
配列番号:25
Knauper, V., et al. 1996. J. Biol. Chem. 271, 1544
【0131】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0132】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Knauper, V. et al. 1996. J. Biol. Chem. 271, 1544
【0133】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
【0134】
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Netzel-Arnett, S. et al. 1991. Anal.Biochem.195, 86-92
Santala, A. et al. 1999. FEBS Lett. 461, 153-156
Bickett, D.M. et al., 1993. Anal.Biochem. 212, 58-64
【0135】
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
A. Santala, A. et al. 1999. FEBS Lett. 461, 153-156
Shabani, F. et al.1998. Free Radic.Res. 28, 115-123
Nagase, H. et al. 1994 J.Biol.Chem. 269, 20952-20957
【0136】
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
J. Berman et al., 1992. J.Biol.Chem. 267, 1434-1437
【0137】
−Pro−Leu−Gly−Met−Trp−Ser−Arg−
配列番号:31
Netzel-Arnett, S. et al.1991. Anal.Biochem.195, 86-92
d'Ortho, M.P. et al., 1997. Eur.J.Biochem. 250, 751-757
【0138】
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
Finch-Arietta, M. et al., 1993. Agents Actions 39 SpecNo,C189-C191
Netzel-Arnett, S. et al. 1991 Anal.Biochem.195, 86-92
【0139】
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
Finch-Arietta, M. et al., 1993. Agents Actions 39 SpecNo,C189-C191
Bickett, D.M. et al., 1994. Ann.N.Y.Acad.Sci. 732, 351-355
【0140】
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
Kraft, P.J. et al. 2001. Connect.Tissue Res. 42, 149-163
Itoh, M. et al. 1997. J.Pharm.Biomed.Anal. 15, 1417-1426
Welch, A.R. et al. 1995. Arch.Biochem.Biophys. 324, 59-64
【0141】
−Pro−Leu−Ala−Tyr−Trp−Ala−Arg−
配列番号:53
Aschi, M. et al. 2002. J.Comput.Aided Mol.Des 16, 213-225
Netzel-Arnett, S. et al. 1991. Anal.Biochem.195, 86-92
【0142】
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
Lauer-Fields, J.L. and Fields, G.B. 2002 Biol.Chem. 383, 1095-1105
Lauer-Fields, J.L. et al. 2001. Biochemistry 40, 5795-5803
【0143】
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
Bremer, C. et al.2002. Acad.Radiol. 9 Suppl 2, S314-S315
Nagase, H. et al. 1994. J.Biol.Chem. 269, 20952-20957
【0144】
−Arg−Pro−Lys−Pro−Gln−Gln−Phe−Trp−
配列番号:59
Bickett, D.M. et al. 1994. Ann.N.Y.Acad.Sci. 732, 351-355
【0145】
第一の配列S1および/または第二の配列S2をMMP−14酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Ala−Cys(p−OMeBz)−Trp−Ala−Arg−
配列番号:60
Mucha, A., et al. 1998. J. Biol. Chem. 273, 2763
Holtz, B., et al. 1999. Biochemistry 38, 12174
【0146】
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
Mohan, M.J. et al. 2002. Biochemistry41, 9462
Hanessian, S. et al. 2001. J. Med. Chem. 44, 3066
Ambrose, W.P. et al. 1998. Anal. Biochem. 263, 150
【0147】
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Roderfeld, M. et al. 2000 . Prot. Expr. Purif. 19, 369
Kannan, R. et al. 1999. Prot. Expr. Purif. 16, 76
【0148】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
Weingarten, H.; Feder, J. 1985. Anal. Biochem. 147, 437
Weingarten, H. et al. 1985. Biochemistry. 24, 6730
【0149】
第一の配列S1および/または第二の配列S2をMMP−15酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Xue, C.-B. et al. 2001. J. Med. Chem. 44, 2636
【0150】
第一の配列S1および/または第二の配列S2をMMP−16酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Shimada, T. et al. 1999. Eur. J. Biochem. 262, 907
Xue, C.-B. et al. 2001. J. Med. Chem. 44, 2636
【0151】
第一の配列S1および/または第二の配列S2をMMP−17酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
English, W.R. et al. 2001. FEBS Lett. 491, 137
【0152】
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
【0153】
第一の配列S1および/または第二の配列S2をMMP−19酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Pendas, A.M. et al. 1997. J. Biol. Chem. 272, 4281
【0154】
第一の配列S1および/または第二の配列S2をMMP−23酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
Velasco, G. et al. 1999. J. Biol. Chem. 274, 4570
【0155】
第一の配列S1および/または第二の配列S2をMMP−25酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
English, W.R. et al. 2001. FEBS Lett. 491, 137
【0156】
第一の配列S1および/または第二の配列S2をMMP−26酵素の標的とするとき、それらは従って少なくとも1種の以下の配列に対応する:
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
Park, H.I. et al. 2002. J. Biol. Chem. 277, 35168
【0157】
前記に起因するものとして、特定のMMPは数種の配列を認識することが可能であり、それらの配列は本来、正確なメカニズムに従い、一部の病態に関与する異なるタンパク質に属する配列である。
【0158】
上記のように、本発明の環状ペプチドは、蛍光受容体基を担持する少なくとも1種のプローブX1、および蛍光受容体もしくは非蛍光受容体基を担持する少なくとも1種のプローブX2を含んでなる。一般に、これらは供与体基の蛍光発光スペクトルと受容体基の吸収スペクトルとが、少なくとも一部、重なり合うように選択される;このものは励起エネルギーを供与体基から受容体基へ非放射転移(または共鳴エネルギー転移)現象として移動する。
当該基は通常、芳香族核、例えば、ベンゼン、アントラセンまたはクマリン核を含んでなる。
【0159】
例として、以下の対合を例示し得る(対合の第一の構成基は供与体基であり、対合の第二の構成基は受容体基である):
*トリプトファン/2,4−ジニトロフェニル(略号W/Dnpにより記号化);
*o−アミノ安息香酸/2,4−ジニトロフェニル(略号Abz/Dnpにより記号化);
*(7−メトキシクマリン−4−イル)アセチル/2,4−ジニトロフェニル(略号Mca/Dnpにより記号化);
*(7−メトキシクマリン−4−イル)アセチル/N−3−(2,4−ジニトロフェニル)−L−2,3−ジアミノプロピル(略号Mca/Dpaにより記号化);
*トリプトファニル/ダンシル(略号W/Dnsにより記号化);
*N−メチルアントラノイル/2,4−ジニトロフェニル(略号Nma/Dnpにより記号化);
*6,7−ジメトキシクマリン−4−イル−アセチル/6−N−(7−ニトロベンゾ−2−オキサ−1,3−ジアゾール−4−イル)アミノヘキサン酸(略号DMC/Nbdにより記号化);
*5−(2’−アミノメチル)ナフタレンスルホン酸/4−(4’−ジメチルアミノフェニルアザ)安息香酸(略号EDANS/Dabcylにより記号化);
*7−メトキシクマリン−3−カルボン酸/7−ジエチルアミノ−クマリン−3−カルボン酸(略号MC/DACにより記号化);当該基は以下の式に一致する:
【0160】
【化4】
【0161】
【化5】
【0162】
供与体基および受容体基がクマリン核を含んでなる基であると有利である。
これらの基はクマリン型の共通の芳香族単位を有するので、これが疎水型またはπ−スタッキング型の複合体の形成を可能とし、このものは本発明環状ペプチドの安定性において役割をもつ。
特に好適な対合はMC/DAC対合であり、このものは一旦プローブに結合すると以下の式をもつ:
【0163】
【化6】
【0164】
X1およびX2プローブはアミノ酸残基であり得る。
従って、X1プローブの一例は、以下の式を有する−Lys(MC)−プローブであってもよい:
【0165】
【化7】
【0166】
一方、X2プローブの例は以下の式を有する−Lys(DAC)−プローブであってもよい:
【0167】
【化8】
【0168】
S1およびS2は、上記定義のようなMMPの標的配列、とりわけ以下の配列に対応し得る:
-Gly−Pro−Gln−Gly−Leu−Leu−Gly−Ala−
または
−Gly−Pro−Gly−Gly−Val−Val−Gly−Pro
これらの配列はそれぞれMMP−1酵素およびMMP−9酵素が認識する配列に対応するものである。
【0169】
本発明の環状ペプチドは以下の式を有する環状ペプチドであってもよい:
【0170】
【化9】
【0171】
(X1、X2、R1、YおよびR2は上記定義と同じ定義に一致する)
本発明に適合する具体的な環状ペプチドは以下の式に一致する:
【0172】
【化10】
【0173】
(基−Lys(DAC)−および−Lys(MC)−は上記定義と同じ定義に一致する)
太字の配列はMMP1酵素の存在下に切断され、それによってLys(DAC)プローブとLys(MC)プローブの距離を互いに離すことに寄与する。それが共鳴移動の変化につながり、それによってMMP1酵素の活性の定量を可能とする。
【0174】
本発明の環状ペプチドは、ペンダント機能(リジンまたはアスパラギン酸残基に由来するアミン官能基など)を介して、粒子などの担体に固定されるに至る。
【0175】
本発明の環状ペプチドは、常套のプロセスを用いる固相上の自動合成工程からなり、次いで、固相から該ペプチドを脱離した後、またはその後にそれを固相から脱離させ、線状ペプチドの末端をカップリング結合させることからなる方法により調製し得る。
【0176】
固相とは、通常、−NH2または−COOH官能基と反応して共有結合を形成し得る基を担持する樹脂であり、この共有結合は一旦合成したペプチドを切断・脱離するように企図したものである。
【0177】
当該樹脂は以下の式の基をそれぞれ担持するトリチル、2−クロロ クロロ トリチル樹脂であってもよい:
【0178】
【化11】
【0179】
(中括弧はその部分を介して上記の基が樹脂に結合する部分を示す)
第一の実施態様によると、本発明方法は:
a)固相上の化学合成により線状ペプチドを調製するための調製工程;
b)固相から線状ペプチドを脱離する脱離工程;および
c)該線状ペプチドの末端をカップル結合させて環状ペプチドを形成するカップリング工程;
からなることを特徴とする。
【0180】
第二の実施態様によると、本発明方法は:
a)固相上の化学合成により線状ペプチドを調製するための調製工程;
b)該線状ペプチドの遊離の末端と該線状ペプチドの線状残基の末端官能基とをカップル結合させるカップリング工程;および
c)環状ペプチドを固相から脱離するための脱離工程であって、工程b)およびc)を付随して実施し得る脱離工程;
からなることを特徴とする。
【0181】
第一の実施態様または第二の実施態様についても、工程a)を実施する場合、その第一相は固相に第一のアミノ酸を固定化することからる。
本固定化は幾つかの方法で実施し得る:
− 第一アミノ酸のC−末端を介して固相上に固定化;
− 第一アミノ酸の側鎖を介して固相上に固定化;
− 第一アミノ酸の主鎖を介して固相上に固定化。
【0182】
第一の型の固定化の場合、一旦ペプチドを合成すると、ペプチドにおける脱保護されたN−末端官能基は、樹脂上から脱離した、またはなお固定化したままのカルボキシル官能基上で、求核付加メカニズムにより反応する。カルボキシル官能基がまだ樹脂上に固定化されたままである場合、求核付加反応(分子内付加である)は環化と樹脂からのペプチドの脱離を一緒に生じる。
【0183】
アミノ酸の側鎖を介して起こる固定化の第二の型は、一般に、以下のアミノ酸で行う:アスパラギン、アスパラギン酸、グルタミン、グルタミン酸エステル、リジン、ヒスチジン、セリン、トレオニン、アルギニンおよびチロシン;これらは樹脂が担持する反応性基と反応し得る−CO2H、OH、NH2またはNH2COなどの官能基を担持する側鎖を有する。
【0184】
樹脂上の主鎖の固定化に基づく第三の方法の場合、合成されたペプチドは主鎖のアミド結合の窒素原子の1つにより樹脂に結合される。
当業者は所望の配列を合成するために、明らかに、適切な保護基を使用するであろう。
【0185】
上記のように、本発明の環状ペプチドは、プロテアーゼ酵素Eの活性をモニターするために使用することを企図する。
より正確には、本発明の環状ペプチドの使用は、幾つかの様式で想定し得る:
− それらは酵素活性と認知された病態との間に限定的な相関関係を与える;
− それらは、酵素活性と認知された病態の間の相関関係が既知であるならば、生物サンプル中に存在する酵素をアッセイすることにより、病態を診断するために使用することができる。
【0186】
それらは、その活性を評価したいと所望する酵素を含んでなる液体のインビトロ分析を実施するために、特に使用することができる。
それらはまた、X1/X2対合が赤色で発光する場合、特にインビボでの使用に供される。
【0187】
従って、本発明は、本発明に適合する少なくとも1種の環状ペプチドを含んでなる即時使用試薬;および
− 本発明に適合する少なくとも1種の環状ペプチドを含んでなる第一区分;および
− 緩衝液を含んでなる第二区分;
を含んでなるキットに関する。
【0188】
このキットは保存液の調製を企図するものであり、この保存液を、活性を測定しようとするプロテアーゼ酵素と接触させる。該プロテアーゼ酵素は、生物液中に含まれていてもよい。
その名称が示唆するように、緩衝液は溶液のpHを一定に維持することを企図するものであり、引き続く酵素活性の測定に影響しかねないpHの過度な酸性への、または過度な塩基性への変化を起こさないようにする。
最終的に、本発明は、少なくとも1種のプロテアーゼ酵素の活性をインビトロで測定するための、本発明による少なくとも1種の環状ペプチドの使用に関する。
【0189】
本発明のその他の特徴と有益性は、説明のために示した、限定するものではない以下の実施例を読み解くことにより明らかとなろう。
【図面の簡単な説明】
【0190】
【図1】図1は実施例に従い調製した環状ペプチドの蛍光強度ΔIF 供与体(a.u.)の変化を時間t(分)の関数として示すグラフである;この環状ペプチドはMMP−1溶液(1nM)の存在下に90μMの濃度とする。
【図2】図2は、調製した環状ペプチドの初期加水分解速度の逆数(1/vi)(a.u.−1.s)をその濃度の逆数(1/[S])(μM−1)の関数として示す二重逆数プロットである。
【発明を実施するための形態】
【0191】
以下の実施例は、それぞれMMP1標的配列に対応する2つのペプチド配列S1およびS2を含んでなる本発明による環状ペプチドの製造について説明するものである;この配列は以下の:−Gly−Pro−Gln−Gly−Leu−Leu−Gly−Ala−である。
【0192】
この環状ペプチドの製造は以下の工程からなる:
a)その側鎖を介して樹脂上に接合することを企図する出発アミノ酸の合成工程:Fmoc−Lys−O−アリル;
b)当該出発アミノ酸を樹脂に接合する工程;
c)接合した樹脂上で2種のペプチド配列を合成する工程;
d)環化工程。
【0193】
これら工程の詳細を以下に説明する。
以下に示すプロトコールにおいては、以下の略号を使用する:
ACN:アセトニトリル; Boc:t−ブチルオキシカルボニル; DAC:7−ジエチルアミノクマリン−3−カルボン酸; DCM:ジクロロメタン; DIEA:N,N’−ジイソプロピルジエチルアミン; DMF:N,N−ジメチルホルムアミド; ESMS:エレクトロスプレー質量分析法; Et2O:ジイソプロピルエーテル; Fmoc:9−フルオレニルメトキシカルボニル; HBTU:ヘキサ−フルオロリン酸N−[1H−ベンゾトリアゾール−1−イル]ジメチルアミノ)メチレン)−N−メチルメタナミニウムN−オキシド; HOBt:N−ヒドロキシベンゾトリアゾール; MC:7−メトキシクマリン−3−カルボン酸; AMPA:4−アミノフェニル水銀酢酸塩; PyBOP:ベンゾトリアゾール−1−イル−オキシ−トリス−ピロリドノホスホニウム; TA:外界温度; TFA:トリフルオロ酢酸; THF:テトラヒドロフラン; TIS:トリイソプロピルシラン; TRIS:トリス(ヒドロキシメチル)アミノメタン。
【0194】
化学製品および溶媒はすべて分析精度のものであり、シグマ社から購入した。2−クロロ クロロトリチル樹脂、PyBOPおよびすべてのアミノ酸(FmocにてNα−保護)はアドバンスドケムテック(Advanced Chemtech)から購入した。
【0195】
化学反応はすべてN2下に無水溶媒を用いて実施した。DMFは還流下にCaH2にて一夜乾燥し、蒸留する。DCMは使用前に蒸留した。
【0196】
UV/可視スペクトルは日立分光光度計装置モデルU−2010で記録した。蛍光スペクトルはPTIおよびジョビン−イボン(Jobin-Yvon)JY3分光蛍光光度計装置で実施し、サンプルホルダーは37℃の恒温とした。固相でのペプチド合成はアップライドバイオシステム433Aシンセサイザーにて実施した。精製は逆相島津HPLC(コントローラー:SLC−10AVP; ポンプ:LC8A; UV/可視検出器:SPD−10AVP[波長:214、267および254nm]およびC18サティスファクションカラムRP18AB 5μm 250×4.6mm)にて実施した。溶出に使用した溶媒系は、(A)0.1%TFA水溶液および(B)70%ACN水溶液であった。
【実施例】
【0197】
a)Fmoc−Lys−O−アリルの合成
Fmoc−Lys−O−アリルは下記式に一致する:
【0198】
【化12】
【0199】
このものは以下の反応工程図に従い調製する:
【0200】
【化13】
【0201】
Fmoc−Lys(Boc)−OH(2g、4.3mmol)および臭化アリル(10mL、118.2mmol)の溶液に、1.45mLのDIEA(8.5mmol)を加える。80℃にて3.5時間攪拌した後、この溶液を200mLの酢酸エチルで希釈し、次いで、0.1N−HCl(3×100mL)、10%NaHCO3(3×100mL)およびNaCl飽和溶液(3×100mL)で洗浄し、MgSO4で乾燥する。減圧下に蒸発させた後、1.9gのFmoc−Lys(Boc)−O−アリル(3.87mmol、90%)を白色粉末の形状で得る。
【0202】
Fmoc−Lys(Boc)−O−アリル(1.9g、3.87mmol)をTFA/DCM混合物(70mL、1:1)に溶解する。この溶液を外界温度で3時間攪拌する。この溶液を濃縮した後、エーテルで洗い、引き続き蒸発させて、1.45gのFmoc−Lys−O−アリル(3.56mmol、92%)を白色粉末の形状で回収する。
【0203】
b)Fmoc−Lys(Boc)−O−アリルの樹脂への接合、およびLys(MC)プローブの接合
反応工程図は以下のとおりである:
【0204】
【化14】
【0205】
Fmoc−Lys(Boc)−O−アリル(1.81g、4.43mmol)を無水THF(2mL)に溶解し、DIEA(734μL、4.23mmol)を加える。外界温度で10分間攪拌した後、570mgの2−クロロトリチル樹脂を加える。この混合物を外界温度で5時間混合する。次いで、樹脂を濾取し、DMF(20mL)、DCM/メタノール/DIEA混合物(17:2:1)(2×20mL)、DMF(10mL)およびDCM(20mL)で洗浄する。この新たな樹脂の置換率は、ピペリジンで処理した後、ジベンゾフルベン複合体のUV測定を介して0.4mmol/gと判定した。最初に、Fmoc−Lys−O−アリル−クロロトリチル樹脂(300mg、0.4mmol/g)を、20%ピペリジン/DMF混合物と外界温度で3時間接触させることによって、当該樹脂からFmoc基を脱離させて脱保護した。遊離一級アミンの存在をカイザーテストにより証明した。この樹脂を、DMF(5mL)中、PyBOP(173mg、0.392mmol)、HOBt(54mg、0.4mmol)およびDIEA(100μL、0.8mmol)の存在下、外界温度で6時間、Fmoc−Lys(MC)−OH(228mg、0.4mmol/g)と反応させる。次いで、この樹脂を濾取し、DMF、DCMおよびメタノールで洗浄する。Fmoc−Lys(MC)−Lys−O−アリル−クロロトリチル樹脂(270mg)を回収する。この新たな樹脂の置換率は、ピペリジンで処理した後、ジベンゾフルベン複合体のUV測定により0.38mmol/gと判定した。反応しなかったアミン官能基は、DMF(5mL)中、酢酸スクシミニドエステル(25.12mg、0.16mmol)、ピペリジン(12mg、0.16mmol)の存在下に、外界温度で40分間アセチル化した。
【0206】
c)ペプチド合成
反応工程図は以下のとおりである:
【0207】
【化15】
【0208】
(記号G、P、Q、LおよびAはそれぞれアミノ酸のグリシン、プロリン、グルタミン、ロイシン、アラニンに相当する)
【0209】
Fmoc−Lys(MC)−Lys−O−アリル−クロロトリチル樹脂をシンセサイザー反応機に加える。Fmoc基によりNα−保護したアミノ酸(Gly、Pro、Gln(Trt)、LeuおよびAla)を過剰(10倍)に加える。カップリング工程をHBTU、HOBtおよびDIEAの存在下に実施し、その後、以下のペプチドGPQGLLGA−(AはLys(MC)に結合)を得る。18時間後、樹脂に結合したペプチド(370mg)を得る。
【0210】
Fmoc−Lys(DAC)−OH(190mg、0.3mmol)を、シンセサイザー反応機外部にて、DMF(5mL)中、PyBOP(153mg、0.3mmol)、HOBt(40.53mg、0.3mmol)およびDIEA(99.1μL、0.6mmol)の存在下、外界温度で5時間、樹脂上のペプチドとカップル結合させる。次いで、樹脂を濾取し、前記のように洗浄する。401mgを取得する。置換率は0.25mmol/gと評価する。反応しなかったアミン官能基は上記と同じ手順でアセチル化する。
【0211】
樹脂上のペプチドを再び自動ペプチドシンセサイザー反応機に入れ、前記と同じアミノ酸配列を最初の工程に従って合成する。530mgを回収する。
【0212】
d)環化反応
この樹脂を高真空下に4時間乾燥し、次いでアルゴン下に置いた。Pd(P(Ph3))4(346.6mg、0.3mmol)をアルゴン気流下にてクロロホルム/酢酸/N−メチルモルホリン混合物(37:2:1、8mL)に溶解する。次いで、この溶液を樹脂と接触させ、得られた混合物を外界温度で2時間、時折攪拌する。次いで、樹脂を濾取し、0.5%DIEA/DMF溶液(2×10mL)で、次いで、ジエチルジチオカルバミン酸ナトリウム/DMF溶液(0.5%w/w)で洗い、触媒を除去する。
【0213】
Fmocを切断した後、樹脂を1M−HOBt/DMFにて洗浄する。N末端およびC末端間の環化反応を実施するために、260mgのPyBOP(0.5mmol)、69mgのHOBt(0.51mmol)および180μLのDIEA(1mmol)を10mLのN−メチルピロリドンに加える。この溶液を外界温度で48時間攪拌し、その後、樹脂を30mLのDMF、DCMおよびメタノールで洗浄する。
【0214】
次いで、この樹脂をTFA/H2O/TIS溶液(95:2.5:2.5)に懸濁する。室温3時間後、樹脂を濾取し、TFA(2×1mL)およびDCM(10mL)で洗浄する。濾液を一部減圧下に蒸発させ、次いで、冷エーテルに浸漬する。
【0215】
−20℃で12時間後、沈殿を濾取し、洗浄して170mgの粗生成物を得る。準分取HPLC精製の後、92.1mgのペプチドを得た。このペプチドの構造をESMSで証明した。
本実施例で得られた環状ペプチドは以下の式に一致する:
【0216】
【化16】
【0217】
酵素アッセイは、0.1M−TRIS、0.1M−NaClおよび10mM−CaCl2を含有するpH7.6の緩衝液中で実施した。25μLの市販MMP−9(90%チモーゲン形状)を緩衝液中、37℃、4時間、1mM−AMPA溶液で活性化する。市販のMMP−1はすでに活性化した形状にある。上記のプロトコールに従い調製した環状ペプチドの保存液を、上記定義のような緩衝液により調製し、最終濃度90μMとする。最終濃度90μMの溶液を出発溶液として用い、異なる希釈を実施して、10、5、1および0.5μMの4段階の環状ペプチド/緩衝液とする。
【0218】
アッセイはすべて2mLの最終反応容量中、1nM活性MMP−1または2nM−AMPA−活性化MMP−9で実施する。
【0219】
酵素を添加した後、初期加水分解速度を、403nmで(340nmの励起波長で)の蛍光強度の経時的変化により測定する。アッセイしたペプチド濃度の各濃度について、図1に示すように、時間t(分)の関数として供与体基ΔIF(a.u.)の蛍光強度の変化を表す曲線をプロットした(環状ペプチド濃度、10、5、1および0.5μM)。
【0220】
以下の等式:
蛍光(t)=蛍光max(1−exp(−kobst))
から、一次定数を決定することができた。二次定数(特異的定数と呼称)kcat/KMの計算は、等式kcat/Km=kobs/[E0]に関して使用される活性酵素Eの濃度により得られる。該ペプチドのkcat/Kmは、文献に記載されたMMP−1の最良の基質と比較して、319120s−1・M−1という非常に高い値で評価する;この値はMMP−1について、本発明に従い調製した環状ペプチドの非常に高い特異性を証明するものである。
【0221】
異なるアッセイ法の初期速度を用いることによる、図2に示した二重逆数グラフ(1/Vi)=f(1/[環状ペプチド])は、合計3.8μMとなる環状ペプチドのKmの決定を可能とした。この値は文献に記載された最良のMMP−1基質で得られる値よりも低いが、MMP−1に対して本発明に従い調製した環状ペプチドの優れた親和性を証明するものである。
【0222】
また、本発明の環状ペプチドについては、2nMの活性MMP−9の存在下でもアッセイした。供与体基の蛍光強度には、経時的変化が測定できなかった。従って、この環状ペプチドはMMP−9の基質ではなく、この環状ペプチドの選択性を示している。また、この環状ペプチドはMMP−1に対して非常に高い選択性と強力な親和性を示す。
【特許請求の範囲】
【請求項1】
以下の配列:
−S1−X1−S2−X2−
(式中、S1はプロテアーゼ酵素E1の第一標的ペプチド配列であり;S2はプロテアーゼ酵素E2の第二標的ペプチド配列であり;S1およびS2は同一であっても、異なっていてもよく;当該第一のペプチド配列S1および当該第二のペプチド配列S2は4個ないし14個のアミノ酸を含んでなり;E1およびE2は同じプロテアーゼ酵素に、または異なるプロテアーゼ酵素に相当していてもよく;X1は蛍光供与体基を担持するプローブであり;そしてX2は蛍光または非蛍光受容体基を担持するプローブである)
を含んでなる環状ペプチド。
【請求項2】
下記式:
【化1】
(式中、
− S1、X1、S2およびX2は請求項1記載の定義と同一の定義を示す;
− R1およびR2は独立して、単結合、アミノ酸残基またはペプチド配列に相当する;
− Yは−CONH−基または−NHCO−基に相当する)
に一致する請求項1記載の環状ペプチド。
【請求項3】
プロテアーゼ酵素E1およびプロテアーゼ酵素E2がMMP型の酵素であり、その場合の第一ペプチド配列S1はMMP酵素の標的であり、第二ペプチド配列S2はMMP酵素の標的である請求項1記載の環状ペプチド。
【請求項4】
該MMPプロテアーゼ酵素がMMP−1、MMP−2、MMP−3、MMP−5、MMP−7、MMP−8、MMP−9、MMP−10、MMP−11、MMP−12、MMP−13、MMP−14、MMP−15、MMP−16、MMP−17、MMP−19、MMP−23、MMP−25およびMMP−26およびその混合物から選択されるものである請求項3記載の環状ペプチド。
【請求項5】
該ペプチドがMMP−1酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Gly−Pro−Gln−Gly−Leu−Leu−Gly−Ala−
配列番号:1
−Ala−Pro−Gln−Gly−Ile−Ala−Gly−Gln−
配列番号:2
−Gly−Pro−Gln−Gly−Leu−Ala−Gly−Gln−
配列番号:3
−Gly−Pro−Leu−Gly−Ile−Ala−Gly−Ile−
配列番号:4
−Gly−Pro−Glu−Gly−Leu−Arg−Val−Gly−
配列番号:5
−Tyr−Glu−Ala−Gly−Leu−Gly−Val−Val−
配列番号:6
−Ala−Gly−Leu−Gly−Val−Val−Glu−Arg−
配列番号:7
−Ala−Gly−Leu−Gly−Ile−Ser−Ser−Thr−
配列番号:8
−Gly−Ala−Met−Phe−Leu−Glu−Ala−Ile−
配列番号:9
−Ile−Pro−Glu−Asn−Phe−Phe−Gly−Val−
配列番号:10
−Thr−Glu−Gly−Glu−Ala−Arg−Gly−Ser−
配列番号:11
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Leu−Arg−Ala−Tyr−Leu−Leu−Pro−Ala−
配列番号:13
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:14
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:15
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
−Pro−Cha−Abu−Cys(Me)−His−Ala−
配列番号:18
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:19
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gl
配列番号:22
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
の1つであるか、または
該ペプチドがMMP−2酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Ile−Pro−Glu−Asn−Phe−Phe−Gly−Val−
配列番号:10
−Pro−Pro−Gly−Ala−Tyr−His−Gly−Ala−
配列番号:27
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−His−Leu−Leu−Val−Glu−Ala−
配列番号:28
−Leu−Arg−Ala−Tyr−Leu−Leu−Pro−Ala−
配列番号:13
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:29
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Leu−Ala−Nva−Dpa−Ala−Arg−
配列番号:30
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−Met−Trp−Ser−Arg−
配列番号:31
−Pro−Leu−Gly−SCH[CH2CH(CH3)2]−CO−Leu−Gly− 配列番号:32
−Arg−Pro−Pro−Gly−Phe−Ser−Ala−Phe−
配列番号:33
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
の1つであるか、または
該ペプチドがMMP−3酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Gly−Pro−Glu−Gly−Leu−Arg−Val−Gly−
配列番号:5
−Arg−Val−Gly−Phe−Tyr−Glu−Ser−Asp−
配列番号:34
−Leu−Leu−Ser−Ala−Leu−Val−Glu−Thr−
配列番号:35
−Glu−Ala−Ile−Pro−Met−Ser−Ile−Pro−
配列番号:36
−Ile−Ala−Gly−Arg−Ser−Leu−Asn−Pro−
配列番号:37
−Ile−Pro−Glu−Asn−Phe−Phe−Gly−Val−
配列番号:10
−Lys−Pro−Gln−Gln−Phe−Phe−Gly−Leu−
配列番号:38
−Asp−Val−Ala−Gln−Phe−Val−Leu−Thr−
配列番号:39
−Asp−Thr−Leu−Glu−Val−Met−Arg−Lys−
配列番号:40
−Asp−Val−Gly−His−Phe−Arg−Thr−Phe−
配列番号:41
−Asp−Ser−Gly−Gly−Phe−Met−Leu−Thr−
配列番号:42
−Arg−Val−Ala−Glu−Met−Arg−Gly−Glu−
配列番号:43
−Asp−Leu−Gly−Arg−Phe−Gln−Thr−Phe−
配列番号:44
−Pro−Phe−Ser−Pro−Leu−Val−Ala−Thr−
配列番号:45
−Leu−Arg−Ala−Tyr−Leu−Leu−Pro−Ala−
配列番号:13
−Ala−Pro−Gly−Asn−Ala−Ser−Glu−Ser−
配列番号:46
−Phe−Ser−Ser−Glu−Ser−Lys−Arg−Glu−
配列番号:47
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−His−Leu−Leu−Val−Glu−Ala−
配列番号:28
−Pro−Pro−Glu−Glu−Leu−Lys−Phe−Gln−
配列番号:48
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:49
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:15
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−
配列番号:50
の1つであるか、または
該ペプチドがMMP−5酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
の1つであるか、または
該ペプチドがMMP−7酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−Ser−
配列番号:24
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
の1つであるか、または
該ペプチドがMMP−8酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Ala−Tyr−Trp−Ala−Arg−
配列番号:53
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
の1つであるか、または
該ペプチドがMMP−9酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Gly−Pro−Pro−Gly−Val−Val−Gly−Pro−
配列番号:54
−Gly−Pro−Pro−Gly−Leu−Arg−Gly−Glu−
配列番号:55
−Gly−Pro−Gly−Gly−Val−Val−Gly−Pro−
配列番号:56
−Ile−Pro−Gln−Asn−Phe−Phe−Gly−Val−
配列番号:57
−Pro−Pro−Gly−Ala−Tyr−His−Gly−Ala−
配列番号:58
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:14
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:15
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番:50
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:19
−Pro−Leu−Gly−SCH[CH2CH(CH3)2]−CO−Leu−Gly−
配列番号:32
−Arg−Pro−Pro−Gly−Phe−Ser−Ala−Phe−
配列番号:33
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
の1つであるか、または
該ペプチドがMMP−10酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−His−Leu−Leu−Val−Glu−Ala−
配列番号:28
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−11酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−12酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−
配列番号:50
−Arg−Pro−Lys−Pro−Gln−Gln−Phe−Trp−
配列番号:59
の1つであるか、または
該ペプチドがMMP−13酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:49
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
−Pro−Cha−Gly−Nva−His−Ala−Dpa−
配列番号:25
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
−Pro−Leu−Gly−Met−Trp−Ser−Arg−
配列番号:31
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
−Pro−Leu−Ala−Tyr−Trp−Ala−Arg−
配列番号:53
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
−Arg−Pro−Lys−Pro−Gln−Gln−Phe−Trp−
配列番号:59
の1つであるか、または
該ペプチドがMMP−14酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Ala−Cys(p−OMeBz)−Trp−Ala−Arg−
配列番号:60
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
の1つであるか、または
該ペプチドがMMP−15酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−16酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−17酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
の1つであるか、または
該ペプチドがMMP−19酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−23酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−25酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−26酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
の1つである請求項4記載の環状ペプチド。
【請求項6】
プローブX1が担持する供与体基およびプローブX2が担持する受容体基が芳香族基を含んでなる前記請求項のいずれかに記載の環状ペプチド。
【請求項7】
該芳香族核がベンゼン、アントラセンまたはクマリン核である請求項6記載の環状ペプチド。
【請求項8】
プローブX1が担持する供与体基およびプローブX2が担持する受容体基が以下の対合から選択されるものである前記請求項のいずれかに記載の環状ペプチド:
*トリプトファン/2,4−ジニトロフェニル;
*o−アミノ安息香酸/2,4−ジニトロフェニル;
*(7−メトキシクマリン−4−イル)アセチル/2,4−ジニトロフェニル;
*(7−メトキシクマリン−4−イル)アセチル/N−3−(2,4−ジニトロフェニル)−L−2,3−ジアミノプロピル;
*トリプトファニル/ダンシル;
*N−メチルアントラノイル/2,4−ジニトロフェニル;
*6,7−ジメトキシクマリン−4−イル−アセチル/6−N−(7−ニトロベンゾ−2−オキサ−1,3−ジアゾール−4−イル)アミノヘキサン酸;
*5−(2’−アミノメチル)ナフタレンスルホン酸/4−(4’−ジメチルアミノフェニルアザ)安息香酸;
*7−メトキシクマリン−3−カルボン酸/7−ジエチルアミノ−クマリン−3−カルボン酸。
【請求項9】
プローブX1が担持する供与体基およびプローブX2が担持する受容体基がクマリン核を含んでなる前記請求項のいずれかに記載の環状ペプチド。
【請求項10】
プローブX1が担持する供与体基およびプローブX2が担持する受容体基が、それぞれ以下の式:
【化2】
に相当するものである前記請求項のいずれかに記載の環状ペプチド。
【請求項11】
プローブX1およびX2がアミノ酸残基である前記請求項のいずれかに記載の環状ペプチド。
【請求項12】
プローブX1およびX2がそれぞれ以下の式:
【化3】
に一致するものである請求項11記載の環状ペプチド。
【請求項13】
該ペプチド配列S1およびS2が以下の式:
−Gly−Pro−Gln−Gly−Leu−Leu−Gly−Ala−
に一致するものである請求項1または2に記載の環状ペプチド。
【請求項14】
以下の式:
【化4】
(式中、R1、YおよびR2は請求項2に記載の定義と同じ定義を有する)
に一致するものである請求項13に記載の環状ペプチド。
【請求項15】
以下の式:
【化5】
に一致するものである請求項13または14に記載の環状ペプチド。
【請求項16】
請求項1または2に記載の環状ペプチドの製造法であって、下記工程:
a)固相上の化学合成により線状ペプチドを調製するための調製工程;
b)固相から線状ペプチドを脱離する脱離工程;および
c)該線状ペプチドの末端をカップル結合させて環状ペプチドを形成するカップリング工程;
からなることを特徴とする製造法。
【請求項17】
請求項1または2に記載の環状ペプチドの製造法であって、下記工程:
a)固相上の化学合成により線状ペプチドを調製するための調製工程;
b)該線状ペプチドの遊離の末端と該線状ペプチドの線状残基の末端官能基とをカップル結合させるカップリング工程;および
c)環状ペプチドを固相から脱離するための脱離工程であって、工程b)およびc)を付随して実施し得る脱離工程;
からなることを特徴とする製造法。
【請求項18】
請求項1ないし15のいずれかに記載定義した少なくとも1種の環状ペプチドを含有してなる即時使用し得る試薬。
【請求項19】
− 請求項1ないし15のいずれかに記載定義した少なくとも1種の環状ペプチドを含んでなる第一区分;および
− 緩衝液を含んでなる第二区分;
を含んでなるキット。
【請求項20】
少なくとも1種のプロテアーゼ酵素の活性をインビトロで測定するための、請求項1ないし15のいずれかに記載定義した少なくとも1種の環状ペプチドの使用。
【請求項1】
以下の配列:
−S1−X1−S2−X2−
(式中、S1はプロテアーゼ酵素E1の第一標的ペプチド配列であり;S2はプロテアーゼ酵素E2の第二標的ペプチド配列であり;S1およびS2は同一であっても、異なっていてもよく;当該第一のペプチド配列S1および当該第二のペプチド配列S2は4個ないし14個のアミノ酸を含んでなり;E1およびE2は同じプロテアーゼ酵素に、または異なるプロテアーゼ酵素に相当していてもよく;X1は蛍光供与体基を担持するプローブであり;そしてX2は蛍光または非蛍光受容体基を担持するプローブである)
を含んでなる環状ペプチド。
【請求項2】
下記式:
【化1】
(式中、
− S1、X1、S2およびX2は請求項1記載の定義と同一の定義を示す;
− R1およびR2は独立して、単結合、アミノ酸残基またはペプチド配列に相当する;
− Yは−CONH−基または−NHCO−基に相当する)
に一致する請求項1記載の環状ペプチド。
【請求項3】
プロテアーゼ酵素E1およびプロテアーゼ酵素E2がMMP型の酵素であり、その場合の第一ペプチド配列S1はMMP酵素の標的であり、第二ペプチド配列S2はMMP酵素の標的である請求項1記載の環状ペプチド。
【請求項4】
該MMPプロテアーゼ酵素がMMP−1、MMP−2、MMP−3、MMP−5、MMP−7、MMP−8、MMP−9、MMP−10、MMP−11、MMP−12、MMP−13、MMP−14、MMP−15、MMP−16、MMP−17、MMP−19、MMP−23、MMP−25およびMMP−26およびその混合物から選択されるものである請求項3記載の環状ペプチド。
【請求項5】
該ペプチドがMMP−1酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Gly−Pro−Gln−Gly−Leu−Leu−Gly−Ala−
配列番号:1
−Ala−Pro−Gln−Gly−Ile−Ala−Gly−Gln−
配列番号:2
−Gly−Pro−Gln−Gly−Leu−Ala−Gly−Gln−
配列番号:3
−Gly−Pro−Leu−Gly−Ile−Ala−Gly−Ile−
配列番号:4
−Gly−Pro−Glu−Gly−Leu−Arg−Val−Gly−
配列番号:5
−Tyr−Glu−Ala−Gly−Leu−Gly−Val−Val−
配列番号:6
−Ala−Gly−Leu−Gly−Val−Val−Glu−Arg−
配列番号:7
−Ala−Gly−Leu−Gly−Ile−Ser−Ser−Thr−
配列番号:8
−Gly−Ala−Met−Phe−Leu−Glu−Ala−Ile−
配列番号:9
−Ile−Pro−Glu−Asn−Phe−Phe−Gly−Val−
配列番号:10
−Thr−Glu−Gly−Glu−Ala−Arg−Gly−Ser−
配列番号:11
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Leu−Arg−Ala−Tyr−Leu−Leu−Pro−Ala−
配列番号:13
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:14
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:15
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
−Pro−Cha−Abu−Cys(Me)−His−Ala−
配列番号:18
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:19
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gl
配列番号:22
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
の1つであるか、または
該ペプチドがMMP−2酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Ile−Pro−Glu−Asn−Phe−Phe−Gly−Val−
配列番号:10
−Pro−Pro−Gly−Ala−Tyr−His−Gly−Ala−
配列番号:27
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−His−Leu−Leu−Val−Glu−Ala−
配列番号:28
−Leu−Arg−Ala−Tyr−Leu−Leu−Pro−Ala−
配列番号:13
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:29
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Leu−Ala−Nva−Dpa−Ala−Arg−
配列番号:30
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−Met−Trp−Ser−Arg−
配列番号:31
−Pro−Leu−Gly−SCH[CH2CH(CH3)2]−CO−Leu−Gly− 配列番号:32
−Arg−Pro−Pro−Gly−Phe−Ser−Ala−Phe−
配列番号:33
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
の1つであるか、または
該ペプチドがMMP−3酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Gly−Pro−Glu−Gly−Leu−Arg−Val−Gly−
配列番号:5
−Arg−Val−Gly−Phe−Tyr−Glu−Ser−Asp−
配列番号:34
−Leu−Leu−Ser−Ala−Leu−Val−Glu−Thr−
配列番号:35
−Glu−Ala−Ile−Pro−Met−Ser−Ile−Pro−
配列番号:36
−Ile−Ala−Gly−Arg−Ser−Leu−Asn−Pro−
配列番号:37
−Ile−Pro−Glu−Asn−Phe−Phe−Gly−Val−
配列番号:10
−Lys−Pro−Gln−Gln−Phe−Phe−Gly−Leu−
配列番号:38
−Asp−Val−Ala−Gln−Phe−Val−Leu−Thr−
配列番号:39
−Asp−Thr−Leu−Glu−Val−Met−Arg−Lys−
配列番号:40
−Asp−Val−Gly−His−Phe−Arg−Thr−Phe−
配列番号:41
−Asp−Ser−Gly−Gly−Phe−Met−Leu−Thr−
配列番号:42
−Arg−Val−Ala−Glu−Met−Arg−Gly−Glu−
配列番号:43
−Asp−Leu−Gly−Arg−Phe−Gln−Thr−Phe−
配列番号:44
−Pro−Phe−Ser−Pro−Leu−Val−Ala−Thr−
配列番号:45
−Leu−Arg−Ala−Tyr−Leu−Leu−Pro−Ala−
配列番号:13
−Ala−Pro−Gly−Asn−Ala−Ser−Glu−Ser−
配列番号:46
−Phe−Ser−Ser−Glu−Ser−Lys−Arg−Glu−
配列番号:47
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−His−Leu−Leu−Val−Glu−Ala−
配列番号:28
−Pro−Pro−Glu−Glu−Leu−Lys−Phe−Gln−
配列番号:48
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:49
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:15
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−
配列番号:50
の1つであるか、または
該ペプチドがMMP−5酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
の1つであるか、または
該ペプチドがMMP−7酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−Ser−
配列番号:24
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
の1つであるか、または
該ペプチドがMMP−8酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Ala−Tyr−Trp−Ala−Arg−
配列番号:53
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
の1つであるか、または
該ペプチドがMMP−9酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Gly−Pro−Pro−Gly−Val−Val−Gly−Pro−
配列番号:54
−Gly−Pro−Pro−Gly−Leu−Arg−Gly−Glu−
配列番号:55
−Gly−Pro−Gly−Gly−Val−Val−Gly−Pro−
配列番号:56
−Ile−Pro−Gln−Asn−Phe−Phe−Gly−Val−
配列番号:57
−Pro−Pro−Gly−Ala−Tyr−His−Gly−Ala−
配列番号:58
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:14
−Pro−Gln−Gly−Leu−Glu−Ala−Lys−
配列番号:15
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番:50
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:19
−Pro−Leu−Gly−SCH[CH2CH(CH3)2]−CO−Leu−Gly−
配列番号:32
−Arg−Pro−Pro−Gly−Phe−Ser−Ala−Phe−
配列番号:33
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
の1つであるか、または
該ペプチドがMMP−10酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Arg−Ala−Ile−His−Ile−Gln−Ala−Glu−
配列番号:12
−Gly−Pro−His−Leu−Leu−Val−Glu−Ala−
配列番号:28
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−11酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−12酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−
配列番号:50
−Arg−Pro−Lys−Pro−Gln−Gln−Phe−Trp−
配列番号:59
の1つであるか、または
該ペプチドがMMP−13酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Gly−Pro−Leu−Gly−Met−Arg−Gly−Leu−
配列番号:49
−Arg−Pro−Lys−Pro−Val−Glu−Nva−Trp−Arg−Glu−Ala−Lys−
配列番号:50
−Pro−Cha−Gly−Nva−His−Ala−Dpa−
配列番号:25
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
−Pro−Leu−Ala−Leu−Trp−Ala−Arg−
配列番号:17
−Pro−Leu−Gly−Cys(Me)−His−Ala−D−Arg−
配列番号:23
−Pro−Leu−Gly−Met−Trp−Ser−Arg−
配列番号:31
−Pro−Tyr−Ala−Tyr−Trp−Met−Arg−
配列番号:51
−Arg−Pro−Lys−Pro−Leu−Ala−Nva−Trp−
配列番号:52
−Arg−Pro−Leu−Ala−Leu−Trp−Arg−
配列番号:24
−Pro−Leu−Ala−Tyr−Trp−Ala−Arg−
配列番号:53
−Pro−Cha−Gly−Nva−His−Ala−
配列番号:25
−Arg−Pro−Lys−Pro−Tyr−Ala−Nva−Trp−Met−
配列番号:26
−Arg−Pro−Lys−Pro−Gln−Gln−Phe−Trp−
配列番号:59
の1つであるか、または
該ペプチドがMMP−14酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Ala−Cys(p−OMeBz)−Trp−Ala−Arg−
配列番号:60
−Pro−Cha−Gly−Cys(Me)−His−Ala−
配列番号:20
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
の1つであるか、または
該ペプチドがMMP−15酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−16酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−17酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
−Pro−Leu−Gly−[2−メルカプト−4−メチル−ペンタノイル]−Leu−Gly−
配列番号:22
の1つであるか、または
該ペプチドがMMP−19酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−23酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−25酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Dpa−Ala−Arg−
配列番号:21
の1つであるか、または
該ペプチドがMMP−26酵素の標的である場合、第一配列S1および/または第二配列S2は少なくとも以下の配列:
−Pro−Leu−Gly−Leu−Trp−Ala−D−Arg−
配列番号:16
の1つである請求項4記載の環状ペプチド。
【請求項6】
プローブX1が担持する供与体基およびプローブX2が担持する受容体基が芳香族基を含んでなる前記請求項のいずれかに記載の環状ペプチド。
【請求項7】
該芳香族核がベンゼン、アントラセンまたはクマリン核である請求項6記載の環状ペプチド。
【請求項8】
プローブX1が担持する供与体基およびプローブX2が担持する受容体基が以下の対合から選択されるものである前記請求項のいずれかに記載の環状ペプチド:
*トリプトファン/2,4−ジニトロフェニル;
*o−アミノ安息香酸/2,4−ジニトロフェニル;
*(7−メトキシクマリン−4−イル)アセチル/2,4−ジニトロフェニル;
*(7−メトキシクマリン−4−イル)アセチル/N−3−(2,4−ジニトロフェニル)−L−2,3−ジアミノプロピル;
*トリプトファニル/ダンシル;
*N−メチルアントラノイル/2,4−ジニトロフェニル;
*6,7−ジメトキシクマリン−4−イル−アセチル/6−N−(7−ニトロベンゾ−2−オキサ−1,3−ジアゾール−4−イル)アミノヘキサン酸;
*5−(2’−アミノメチル)ナフタレンスルホン酸/4−(4’−ジメチルアミノフェニルアザ)安息香酸;
*7−メトキシクマリン−3−カルボン酸/7−ジエチルアミノ−クマリン−3−カルボン酸。
【請求項9】
プローブX1が担持する供与体基およびプローブX2が担持する受容体基がクマリン核を含んでなる前記請求項のいずれかに記載の環状ペプチド。
【請求項10】
プローブX1が担持する供与体基およびプローブX2が担持する受容体基が、それぞれ以下の式:
【化2】
に相当するものである前記請求項のいずれかに記載の環状ペプチド。
【請求項11】
プローブX1およびX2がアミノ酸残基である前記請求項のいずれかに記載の環状ペプチド。
【請求項12】
プローブX1およびX2がそれぞれ以下の式:
【化3】
に一致するものである請求項11記載の環状ペプチド。
【請求項13】
該ペプチド配列S1およびS2が以下の式:
−Gly−Pro−Gln−Gly−Leu−Leu−Gly−Ala−
に一致するものである請求項1または2に記載の環状ペプチド。
【請求項14】
以下の式:
【化4】
(式中、R1、YおよびR2は請求項2に記載の定義と同じ定義を有する)
に一致するものである請求項13に記載の環状ペプチド。
【請求項15】
以下の式:
【化5】
に一致するものである請求項13または14に記載の環状ペプチド。
【請求項16】
請求項1または2に記載の環状ペプチドの製造法であって、下記工程:
a)固相上の化学合成により線状ペプチドを調製するための調製工程;
b)固相から線状ペプチドを脱離する脱離工程;および
c)該線状ペプチドの末端をカップル結合させて環状ペプチドを形成するカップリング工程;
からなることを特徴とする製造法。
【請求項17】
請求項1または2に記載の環状ペプチドの製造法であって、下記工程:
a)固相上の化学合成により線状ペプチドを調製するための調製工程;
b)該線状ペプチドの遊離の末端と該線状ペプチドの線状残基の末端官能基とをカップル結合させるカップリング工程;および
c)環状ペプチドを固相から脱離するための脱離工程であって、工程b)およびc)を付随して実施し得る脱離工程;
からなることを特徴とする製造法。
【請求項18】
請求項1ないし15のいずれかに記載定義した少なくとも1種の環状ペプチドを含有してなる即時使用し得る試薬。
【請求項19】
− 請求項1ないし15のいずれかに記載定義した少なくとも1種の環状ペプチドを含んでなる第一区分;および
− 緩衝液を含んでなる第二区分;
を含んでなるキット。
【請求項20】
少なくとも1種のプロテアーゼ酵素の活性をインビトロで測定するための、請求項1ないし15のいずれかに記載定義した少なくとも1種の環状ペプチドの使用。
【図1】

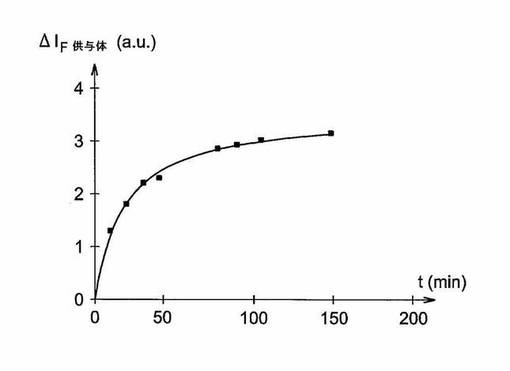
【図2】
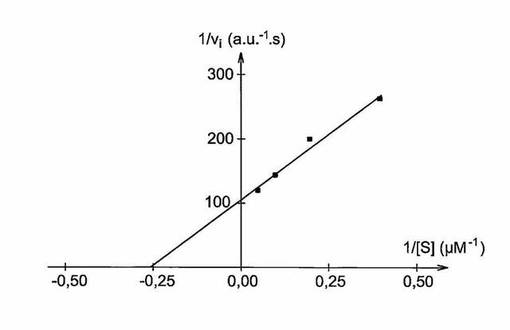
【図2】
【公表番号】特表2011−522861(P2011−522861A)
【公表日】平成23年8月4日(2011.8.4)
【国際特許分類】
【出願番号】特願2011−512975(P2011−512975)
【出願日】平成21年6月10日(2009.6.10)
【国際出願番号】PCT/EP2009/057194
【国際公開番号】WO2010/000591
【国際公開日】平成22年1月7日(2010.1.7)
【出願人】(502124444)コミッサリア ア レネルジー アトミーク エ オ ゼネルジ ザルタナテイヴ (383)
【出願人】(506020388)ユニヴェルシテ ヴィクトル スガラン ボルドー 2 (3)
【氏名又は名称原語表記】UNIVERSITE VICTOR SEGALEN BORDEAUX 2
【住所又は居所原語表記】146,rue Leo Saignat,F−33076 Bordeaux Cedex,FRANCE
【Fターム(参考)】
【公表日】平成23年8月4日(2011.8.4)
【国際特許分類】
【出願日】平成21年6月10日(2009.6.10)
【国際出願番号】PCT/EP2009/057194
【国際公開番号】WO2010/000591
【国際公開日】平成22年1月7日(2010.1.7)
【出願人】(502124444)コミッサリア ア レネルジー アトミーク エ オ ゼネルジ ザルタナテイヴ (383)
【出願人】(506020388)ユニヴェルシテ ヴィクトル スガラン ボルドー 2 (3)
【氏名又は名称原語表記】UNIVERSITE VICTOR SEGALEN BORDEAUX 2
【住所又は居所原語表記】146,rue Leo Saignat,F−33076 Bordeaux Cedex,FRANCE
【Fターム(参考)】
[ Back to top ]