
固定されたトランスフェリン融合タンパク質ライブラリー

トランスフェリン、軸成分および細胞壁連結要素を含む融合タンパク質、さらにそれらのペプチドライブラリーを開示する。本発明は、宿主細胞によって発現された融合タンパク質に提示されたペプチドライブラリーのスクリーニング方法を含む。本発明の融合タンパク質は、酵母で発現させることができるトランスフェリン融合タンパク質を含む。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本願は、2005年6月17日付けで出願された米国仮出願第60/691,229号の利益を請求する。本願は、2004年11月23日付けで出願された米国特許出願第10/515,429号;2003年7月9日付けで出願された米国仮出願第60/485,404号;2003年3月10日付けで出願された米国特許出願第10/384,060号;および2002年8月30日付けで出願された米国仮出願第60/406,977号(これらは全て、それら全体として参照により組み込まれる)に関係する。
【0002】
発明の分野
本発明は、融合タンパク質、融合タンパク質ライブラリー、およびリガンドの結合活性に関してスクリーニングするための融合タンパク質の使用に関する。
【背景技術】
【0003】
細胞表層提示系
コンビナトリアルライブラリーによるスクリーニングおよび選択方法は、一般的な研究ツールになっている(Phizicky等(1995)Microbiological Reviews 59:94〜123)。最も普及した技術の一つはファージディスプレイであり、この技術によれば、タンパク質はバクテリオファージコートタンパク質へのポリペプチド融合として発現し、後で、固定化または可溶性ビオチン化リガンドに結合させることによってスクリーニングされる。ランダムペプチドの提示は、M13、fdおよびf1のような線状バクテリオファージの外面上に発現されるキメラタンパク質を構築することによって達成されることが多い。ファージディスプレイは、抗体、DNA結合タンパク質、プロテアーゼ阻害剤、および酵素への応用が成功している。Hoogenboom等(1997)Trends in Biotechnol.15:62〜70;Ladner(1995)Trends in Biotechnol.13:426〜430;Lowman等(1991)Biochemistry 30:10832〜10838;Markland等(1996)Biochemistry 35:8045〜8057;および、Matthews等(1993)Nucleic Acids Res.21:1727〜1734を参照。
【0004】
ファージディスプレイに加えて、数種の細菌細胞表層提示法が開発されている。Georgiou等(1997)Nat.Biotechnol.15:29〜34を参照。細菌細胞表層提示法において取られるアプローチの一つでは、ピリンタンパク質(TraA)またはそれらの部分を含む融合タンパク質や、繊毛を形成することができる細菌宿主細胞の外表面にライブラリーペプチドを提示する異種ポリペプチドを使用してきた。米国特許第5,516,637号(これは参照によりその全体として本明細書に組み込まれる)を参照。
【0005】
FLITRXTMランダムペプチドライブラリー(インビトロジェンTMライフ・テクノロジーズ(InvitrogenTM Life Technologies))は、E.コリ(E.coli)の表面にコンホメーション的に制限された形式で12merのランダムペプチドライブラリーを提示するために、細菌の鞭毛タンパク質、FliC、およびチオレドキシン、TrxAを使用している。Lu等(1995)BioTechnology 13:366を参照。これらの系は、抗体エピトープマッピング、細菌の生ワクチンの送達系の開発および構築、ならびに環境の浄化目的および診断のための全細胞生物学的吸着剤の生成に応用されている。また、腫瘍由来上皮細胞上で腫瘍特異的な標的に結合するペプチド配列も、FLITRXTMシステムを用いて同定されている。Brown等(2000)Annals of Surgical Oncology,7(10):743を参照。
【0006】
ライブラリースクリーニングのために酵母細胞表層提示系が開発されており、ファージや細菌の提示系の限界のいくつかを克服することに成功している。酵母表層提示系、例えばpYD1酵母提示ベクターキット(pYD1 Yeast Display Vector Kit,インビトロジェンTMライフ・テクノロジーズ)は、細胞表層上に外来タンパク質を提示するためにS.セレビジエ(S.cerevisiae)のa−凝集素受容体を使用している。a−凝集素受容体は、AGA1およびAGA2遺伝子によってコードされた2つのサブユニットからなる。Aga1タンパク質(Aga1p,725個のアミノ酸)は、酵母細胞から分泌されて、酵母細胞壁の細胞外マトリックス中のβ−グルカンに共有結合するようになる。Aga2タンパク質(Aga2p,69個のアミノ酸)は、2個のジスルフィド結合を介してAga1pに結合し、分泌後、それとAga1pとの接触によって細胞に結合したままになる。Aga2pのN末端部分はAga1pへの付着に必要であり、その一方でタンパク質およびペプチドは、酵母細胞表層上での提示のためにC末端に融合することができる。凝集素は、天然型の酵母タンパク質であり、通常は、交配の間に酵母細胞を融合させるための特異的な付着による接触部として機能する。そのようなものとして、凝集素は、細胞壁成分との過剰な立体障害が起こらないようにタンパク質−タンパク質結合に関して進化してきた。Boder等の“タンパク質発現、親和性および安定性の定向進化に関する酵母表面提示(Yeast Surface Display for Directed Evolution of Protein Expression,Affinity,and Stability)”、キメラ遺伝子およびハイブリッドタンパク質の応用(Applications of Chimeric Genes and Hybrid Proteins)(Jeremy Thorner等),アカデミックプレス(Academic Press),2000,第328巻,430〜439頁;US6,699,658;および、US6,423,538(これらは参照によりその全体として本明細書に組み込まれる)。
【0007】
しかしながら、この系の欠点の一つは、Aga2pタンパク質を細胞壁に連結させるためには、Aga2p融合タンパク質とAga1pがジスルフィド結合を形成することが必要であるため、提示の効率が比較的低く、表面にタンパク質を有効に提示する酵母細胞がわずか40%〜60%である点である。Feldhause等(2003)Nat.Biotechnol.21(2):163〜70を参照。ライブラリーの(全てとは限らないが)ほとんどのタンパク質を細胞表層に提示する酵母提示系の必要がある。
【0008】
Aga1pおよびAga2p酵母提示系のその他の欠点は、スクリーニングしようとするリガンドを、Aga2pのC末端に付着させることを必要とする点である。その結果として、この系は、遊離のN末端が結合に必要であるペプチド、および/または遊離のN末端が活性に必要であるペプチドを選択するのに用いることができない。従って、リガンドN末端の酵母細胞タンパク質への結合を必要としないフレキシブルな提示系の必要がある。
【0009】
トランスフェリン融合タンパク質
血清トランスフェリン(Tf)は、80,000ダルトンの分子量を有する単量体の糖タンパク質であり、血液循環中で鉄に結合し、その鉄をトランスフェリン受容体(TfR)を介して様々な組織に輸送する(Aisen等(1980)Ann.Rev.Biochem.49:357〜393;MacGillivray等(1981)J.Biol.Chem.258:3543〜3553;および米国特許第5,026,651号)。Tfは、最も一般的な血清中の分子の一種であり、総血清タンパク質の約5〜10%以下で含まれる。糖鎖欠損トランスフェリンは、アルコール症の個体の血液中に高レベルで存在し、グリコシル化トランスフェリンの半減期(約7〜10日間)よりも長い半減期(約14〜17日間)を示す。van Eijk等(1983)Clin.Chim.Acta 132:167〜171;Stibler(1991)Clin.Chem.37:2029〜2037;Arndt(2001)Clin.Chem.47(1):13〜27;およびStibler等の“Carbohydrate−deficient consumption”,Advances in the Biosciences,(Ed Nordmann等),Pergamon,1988,第71巻,353〜357頁)を参照。Tfの構造はよく特徴付けられており、受容体結合、鉄結合および放出、ならびに炭酸イオン結合のメカニズムが解明されている。米国特許第5,026,651号、5,986,067号、および、MacGillivray等(1983)J.Biol.Chem.258(6):3543〜3546(これらの全ては、参照によりその全体として本明細書に組み込まれる)を参照。
【0010】
ムチンは、高度にグリコシル化されたタンパク質であり、マイクロアレイから実質的な距離をおいて融合タンパク質のリガンドドメインを持ち上げるのに用いられている。リガンドを基質から有意な距離に持ち上げることによって、リガンドの受容体を発現する細胞に提示された受容体への結合が増加すると仮説を立てられている。WO01/46698(これは参照によりその全体として本明細書に組み込まれる)を参照。
【発明の開示】
【発明が解決しようとする課題】
【0011】
これまで本発明の発明者等は、トランスフェリン融合タンパク質ライブラリーを開発してきた。米国特許出願第10/515,429号(これは参照によりその全体として本明細書に組み込まれる)を参照。本発明は、立体障害を減少させ、リガンドの結合が増加するように設計されたムチンのような軸様の成分を含むトランスフェリン融合タンパク質を提供する。発現されたトランスフェリン融合タンパク質が、ペプチドスクリーニングのプラットフォームとして用いることができるように、本融合タンパク質を、酵母のような宿主細胞の表面で発現させ提示させることができる。さらに、本融合タンパク質のトランスフェリンとリガンド部分を切断して、治療剤としてとして用いることができる。N末端が融合したAga2タンパク質を除去すると、トランスフェリンに連結した小さいリガンドのコンホメーションに影響を与える可能性が高いため、これを既存の酵母提示技術で達成することは不可能と思われる。
【課題を解決するための手段】
【0012】
以下でより詳細に説明されているように、本発明は、トランスフェリン(Tf)成分、軸成分、および細胞壁結合基を含む融合タンパク質を含む。Tf成分は、トランスフェリンタンパク質またはそれらの部分を含み、酵母細胞表層上に提示される。例えば、トランスフェリン成分は、トランスフェリンタンパク質のNドメインの一部、換言すればローブであってもよい。Tf成分は、本融合タンパク質のTf部分が、野生型Tfと比較して低いグリコシル化を示すように改変されたTfタンパク質であってもよい。本発明の一実施態様において、本融合タンパク質のトランスフェリン部分は、グリコシル化を示さない。本発明のその他の実施態様において、本融合タンパク質のトランスフェリン成分は、野生型トランスフェリンに比べて低い鉄、ビカーボネートへの親和性、および/または低いトランスフェリン受容体への親和性を示すように改変される。トランスフェリン成分は、トランスフェリン受容体、鉄またはビカーボネートに結合することができなくなるように改変されていてもよい。従って、本発明は、トランスフェリン成分が、グリコシル化部位、鉄結合部位、ヒンジ部位、ビカーボネート部位、および受容体結合部位からなる群より選択される1個またはそれ以上の部位で改変された、改変されたトランスフェリン成分を含む。
【0013】
ここで特許請求される発明のリガンドは、様々な方法でトランスフェリン成分と複合体化させてもよいし、またはトランスフェリン成分と融合させてもよい。さらに、トランスフェリン成分は、それに連結した1個より多くのリガンドを有していてもよい。リガンド成分は、トランスフェリン成分のN末端、C末端に融合していてもよいし、または、トランスフェリン成分内に位置していてもよい。本発明の一実施態様において、このようなリガンドは、アミノ酸位置Asp33、Asn55、Asn75、Asp90、Gly257、Lys280、His289、Ser298、Ser105、Glu141、Asp166、Gln184、Asp197、Lys217、Thr231、およびCys241からなる群より選択されるN−ローブ(N1またはN2)のアミノ酸位置の1つまたはそれ以上に挿入される。
【0014】
本発明のその他の実施態様において、リガンドは、トランスフェリン成分の露出したループ上に位置する。ランダムペプチドのようなリガンド成分は、トランスフェリン成分と共にフレーム内に存在し得るように、トランスフェリン融合タンパク質がコードされたベクターで宿主細胞によって発現され得る。トランスフェリン成分と共に発現されるランダムペプチドのリガンド成分は、当業界既知の多くの方法によって作出することができ、このような方法としては、これらに限定されないが、エラープローンPCR、およびDNAシャフリングが挙げられる。また、トランスフェリン融合タンパク質がそれまでに翻訳され終わった後に、リガンド成分をトランスフェリン融合タンパク質に付加することもできる。
【0015】
リガンドは多くの形態をとることができ、このような形態としては、これらに限定されないが、単鎖抗体、抗体、抗体フラグメント、抗体の可変領域、ランダムペプチド、または抗体相補性決定領域(CDR)が挙げられる。リガンドは、可変またはランダム領域、および非可変領域を含んでいてもよい。リガンドは、研究対象のリガンドであってもよいし、またはリガンドのライブラリー中の一種のリガンドであってもよい。リガンドは、多数の受容体または物質、例えばペプチド、抗原、受容体、抗体、毒素、代謝産物、および核酸に結合する可能性がある。
【0016】
軸成分は、そのN末端がトランスフェリン成分に融合し、そのC末端が細胞中に(例えば細胞壁中に)位置するように配置させることができる。一実施態様において、軸成分のC末端は、アンカー成分に融合される。本発明の軸成分は酵母細胞の細胞壁に伸びており、通常は中程度ないし高度にグリコシル化されたペプチドである。細胞壁に伸びていることによって、軸成分は、細胞壁を介して融合タンパク質を結びつけるための細胞壁連結要素として作用することが可能である。本発明の一実施態様において、軸成分は細胞壁に伸びており、細胞表層上に部分的に提示されている。軸成分の組成は、それにロッド様のコンホメーションを付与することができ、このようなコンホメーションにより、もしもそれながければ融合タンパク質(とりわけリガンド)と宿主細胞との間に存在するはずの立体障害を少なくすることができる。
【0017】
軸成分は、ムチン、ムチン変異体、またはそれらのフラグメントを含んでいてもよいし、またはそれらからなっていてもよい。ムチンドメインは、例えば、MUC1、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC6、MUC7、MUC8、およびMUC9、ならびにそれらの変異体を含んでいてもよい。一実施態様において、軸成分は、ヒトMUC1ドメインを含み、例えば配列番号5の核酸配列またはそれらのフラグメントに相当するペプチドを含む。その他の実施態様において、軸成分は、2回またはそれ以上のムチンの反復を含み、例えば2回またはそれ以上のMUC1またはMUC3の反復を含む。さらなる実施態様において、軸成分は、MUC1、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC6、MUC7、MUC8、およびMUC9からなる群より選択される2種またはそれ以上のムチンタンパク質、またはそれらの変異体もしくはフラグメントを含む。
【0018】
さらに軸成分は、天然型の酵母の細胞壁タンパク質などの中程度から高度にグリコシル化されたその他のタンパク質を含んでいてもよいし、またはそれらからからなっていてもよい。例えば、本発明の一実施態様において、軸成分は、Aga1、Aga1の変異体、もしくはそれらのフラグメントを含むか、またはそれらからなる。
【0019】
本発明の融合タンパク質は、融合タンパク質を宿主細胞に固定するまたは結びつけるように作用する細胞壁連結要素を含む。細胞壁連結要素は、本発明の融合タンパク質を酵母細胞壁に共有結合または非共有結合によって結合させることができる。本発明の一実施態様において、本融合タンパク質の軸成分は、細胞壁連結要素である。例えば、軸成分からのO−グリカンは、細胞壁のβ−グルカンに架橋することができる。その他の細胞壁連結要素としては、これらに限定されないが、遊離のシステイン残基を含むペプチドが挙げられる。例えば、対になっていないシステイン残基を1個またはそれ以上含む軸成分またはアンカー成分は、細胞壁中のタンパク質の対になっていないシステイン残基の1個またはそれ以上とジスルフィド結合を形成することができる。
【0020】
本発明の融合タンパク質は、場合により、アンカー成分を含んでいてもよく、このアンカー成分も、トランスフェリン融合タンパク質を宿主細胞に固定するまたは結びつけるように作用する。アンカー成分は、細胞壁連結要素であってもよいし、または本融合タンパク質を酵母細胞膜に結びつけてもよい。
【0021】
なかでも、本発明の融合タンパク質を酵母細胞膜に結びつけることができるアンカードメインの一つは、グリコシルホスファチジルイノシトール(GPI)ペプチドアンカーであり、これは翻訳後タンパク質修飾によってGPIシグナルペプチド配列(例えば配列番号15で示されるシグナルペプチド配列)のオメガ部位(ω−site)に付加される。本発明の一実施態様において、アンカー(例えば改変されたGPIシグナル配列によって提供されるアンカー)は、切断される前に、融合タンパク質を宿主細胞膜または細胞壁を一時的に繋ぐ。一旦切断されると、融合タンパク質は、細胞壁のβ−グルカンに架橋されている軸成分からのグリカンにより、細胞壁連結要素を介して細胞に連結したままになる。
【0022】
本発明のその他の実施態様において、アンカーは、膜貫通ドメインである。膜貫通ドメイン(TMD)は、1回膜貫通型IまたはII膜タンパク質の領域でもよいし、またはマルチスパン型膜タンパク質の数個の膜貫通領域のいずれか一つでもよい。
【0023】
本発明はまた、特許請求される融合タンパク質をコードする核酸分子も含む。このような核酸は、ベクターに挿入することができ、酵母のような宿主細胞を形質転換させるのに用いることができる。宿主細胞は、本発明の核酸で形質転換されると、本融合タンパク質を発現することができる。本融合タンパク質の発現の誘導は当業界既知の方法によって制御することができ、例えば誘導性プロモーターの使用によって制御することができる。本発明は、宿主細胞の集合体、例えば、ランダム化ペプチドを提示する本発明の融合タンパク質を発現する酵母細胞の集合体に発現された融合タンパク質のライブラリーを含む。
【0024】
本発明のその他の実施態様において、本融合タンパク質は、リガンドまたは物質の結合活性についてスクリーニングするのに用いられる。特許請求される融合タンパク質を発現することができる宿主細胞のライブラリーを、これらに限定されないが抗原または受容体などの物質に曝露し、それから結合活性に関してスクリーニングすることができる。細胞表層提示ライブラリーは、これらに限定されないがFACSや磁気ビーズなどの当技術分野既知の方法を用いてスクリーニングすることができる。
【0025】
詳細な説明
一般的な説明
本発明の発明者等は、ライブラリー、例えばランダムペプチドまたはCDRライブラリーをスクリーニングするための、例えば細胞表層提示系の一部として用いることができる多機能の融合タンパク質を開発した。本融合タンパク質は、1またはそれ以上のリガンドと複合体化または融合したトランスフェリン成分を含む。本発明は、以下を条件とする限りにおいて、トランスフェリン以外のタンパク質を含む融合タンパク質を想定しており、上記条件は、上記その他のタンパク質が可溶性であり、融合タンパク質の残分の部分から切断されたときに融合した1またはそれ以上のリガンドに高い血清中半減期を付与することができる場合である。例えば、トランスフェリンの代わりに、アルブミンまたはその変異体もしくはフラグメントを用いることができる。
【0026】
本融合タンパク質のトランスフェリン成分は、中程度から高度にグリコシル化された軸成分に融合している。本融合タンパク質は、本融合タンパク質を酵母細胞の細胞壁に共有結合または非共有結合で結合させることができる細胞壁連結要素を含む。本発明の一実施態様において、本融合タンパク質は、アンカー成分(例えば膜貫通ドメイン)をも含む。
【0027】
本融合タンパク質は、酵母提示系して用いられる場合、Aga1pおよびAga2p酵母提示系と比較して、ペプチドを提示した細胞表層を有するクローンを高い割合で提供すること等を含む従来技術を超える利点を提供する。また本発明の融合タンパク質は、結合に利用可能なN末端を必要とするリガンドをスクリーニングすることのフレキシビリティーをも提供する。
【0028】
本発明はまた、本融合タンパク質またはそれらの部分を含む治療用組成物、および治療を必要とする対象に本融合タンパク質またはそれらの部分を投与することによって、疾患または障害を治療、予防または改善する方法も含む。本発明の融合タンパク質は、リガンド成分として、推定上の治療用タンパク質の少なくともフラグメントまたは変異体を含む。本発明の一実施態様において、本融合タンパク質のトランスフェリンおよびリガンド(すなわち治療薬)部分は、軸成分(すなわち本融合タンパク質の酵母細胞と結合した部分)から切断することができ、生物薬剤またはワクチンを調製するのに用いることができる。
【0029】
定義
本明細書で用いられる用語「生物活性」は、生物学的な環境において、すなわち生体内またはインビトロのその模擬環境において、治療的な分子、リガンド成分、タンパク質またはペプチドによって行われる機能または一連の活性を意味する。生物活性としては、これらに限定されないが、特許請求される融合タンパク質の治療的分子部分の機能が挙げられ、例えばこれらに限定されないが、応答細胞系からの細胞外マトリックス分泌の誘導、ホルモン分泌の誘導、走化性の誘導、有糸分裂誘発の誘導、分化の誘導、または応答細胞の細胞分裂の阻害が挙げられる。本発明の融合タンパク質またはペプチドは、その治療用タンパク質の天然型の対応物が示す1種またはそれ以上の生物活性を示す場合、生物学的に活性であるとされる。
【0030】
本明細書で用いられるように、トランスフェリン配列における「〜に相当するアミノ酸」または「等価なアミノ酸」は、第一のトランスフェリン配列と、少なくとも第二のトランスフェリン配列との間の同一性または類似性が最大になるようにアラインメントによって同定される。第二のトランスフェリン配列における等価なアミノ酸を同定するのに用いられる番号は、第一のトランスフェリン配列におけるそれに相当するアミノ酸を同定するのに用いられる番号に基づく。特定の場合において、これらの語句は、ヒトトランスフェリンにおけるアミノ酸残基を、ウサギ血清トランスフェリンにおける特定の残基と比較して説明するのに用いる場合もある。
【0031】
本明細書で用いられる用語「Tf成分」、「Tfタンパク質のフラグメント」または「Tfタンパク質」または「Tfタンパク質の部分」は、天然に存在するTfタンパク質、またはその突然変異体の少なくとも約5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、96%、97%、98%、99%または100%を含むアミノ酸配列を意味する。
【0032】
本明細書で用いられる用語「遺伝子」は、生物学的機能に関連するDNAのあらゆるセグメントを意味する。従って、遺伝子は、これらに限定されないが、それらの発現に必要なコード配列および/または調節配列を含む。また遺伝子は、例えば他のタンパク質に関する認識配列を形成する発現されないDNAセグメントを含んでいてもよい。遺伝子は、関心対象の源からクローニングすること、または既知のもしくは予測された配列情報から合成することによって、様々な源から得ることができ、さらに望ましいパラメーターを持つように設計された配列を含んでいてもよい。
【0033】
本明細書で用いられる「異種ポリヌクレオチド」または「異種核酸」または「異種遺伝子」または「異種配列」または「外因性DNAセグメント」は、特定の宿主細胞にとって外来の起源であるポリヌクレオチド、核酸またはDNAセグメントを意味するか、または同じ源由来の場合、その元の形態から改変されたポリヌクレオチド、核酸またはDNAセグメントを意味する。宿主細胞における異種遺伝子には、特定の宿主細胞にとって内在性であるが、修飾されている遺伝子が含まれる。従って、この用語は、細胞にとって外来または異種であるDNAセグメント、または細胞に相同であるが、その構成要素が通常は見出されない宿主細胞の核酸内の位置に存在するDNAセグメントを意味する。一例として、酵母細胞にとっては天然型であるが、ヒトTf配列に付着したシグナル配列は、異種である。
【0034】
本明細書で用いられる「単離した」核酸配列は、その他の核酸配列を実質的に含まない核酸配列を意味し、例えば、アガロースゲル電気泳動によって決定した場合に、少なくとも約20%純粋な、好ましくは少なくとも約40%純粋な、より好ましくは約60%純粋な、さらにより好ましくは約80%純粋な、最も好ましくは約90%純粋な、さらに最も好ましくは約95%純粋な核酸配列である。例えば、単離された核酸配列は、その天然の位置から異なる部位(そこで核酸配列が複製されることになる)に核酸配列を移転させるための、遺伝子工学で用いられる標準的なクローニング法により得ることができる。クローニング法は、上記ポリペプチドをコードする核酸配列を含む所望の核酸フラグメントを切り出して単離し、このフラグメントをベクター分子に挿入し、組換えベクターを核酸配列の複数のコピーまたはクローンが複製されることになる宿主細胞に組み込ませることを含み得る。核酸配列は、ゲノム、cDNA、RNA、半合成、合成由来のものでもよいし、またはそれらのあらゆる組み合わせでもよい。
【0035】
本明細書で用いられるように、2個またはそれ以上のDNAコード配列間のフレーム内融合の結果として、DNAコード配列が融合ポリペプチドに翻訳される場合、2またはそれ以上のDNAコード配列は、「連結した」または「融合した」と言う。用語「融合」は、融合タンパク質に関して用いられる場合、リガンド成分、軸成分、およびアンカー成分を含む。Tf融合タンパク質は、トランスフェリン成分が軸成分に融合したものであり、細胞壁連結要素を含む。
【0036】
本明細書で用いられる「改変されたトランスフェリン」は、そのアミノ酸配列に、野生型トランスフェリンに比べて少なくとも1つの改変を示すトランスフェリン分子を意味する。
【0037】
本明細書で用いられる「改変されたトランスフェリン融合タンパク質」は、軸成分に融合しているリガンドと複合体化した、またはそれらに融合した、改変されたトランスフェリン(またはそのフラグメントもしくは変異体)の少なくとも1つの分子の融合によって形成されたタンパク質を意味する。
【0038】
本明細書で用いられる用語「核酸」または「ポリヌクレオチド」は、デオキシリボヌクレオチドまたはリボヌクレオチド、およびそれらの一本鎖または二本鎖いずれかの形態のポリマーを意味する。特に限定されない限り、これらの用語は、参照核酸と類似の結合特性を有し、天然に存在するヌクレオチドと同様にして代謝される天然ヌクレオチドの類似体を含む核酸を包含する。特に他の指定がない限り、特定の核酸配列はまた、当然ながら、保存的に改変されたそれらの変異体(例えば、縮重コドンの置換)および相補配列、加えて、明確に示された配列も包含する。特に、縮重コドンの置換は、1またはそれ以上の選択された(または全ての)コドンの第三の位置が、混合型の塩基および/またはデオキシイノシン残基で置換されている配列を作製することによって達成することもできる(Batzer等(1991)Nucleic Acid Res.19:5081;Ohtsuka等(1985)J.Biol.Chem.260:2605〜2608;Cassol等(1992);Rossolini等(1994)Mol.Cell.Probes 8:91〜98)。核酸という用語は、遺伝子、cDNA、および遺伝子によってコードされたmRNAと同じ意味で用いられる。
【0039】
本明細書で用いられるように、DNAセグメントが、その他のDNAセグメントと機能的に関係するように置かれると、「機能するように連結した」と表現される。例えば、シグナル配列に関するDNAが、融合タンパク質の分泌に関与する前駆タンパク質として発現される場合、そのシグナル配列に関するDNAは、本発明の融合タンパク質をコードするDNAに機能するように連結しており、プロモーターまたはエンハンサーが、配列の転写を刺激する場合、そのプロモーターまたはエンハンサーはコード配列に機能するように連結している。一般的に、機能するように連結しているDNA配列は隣接しており、さらにシグナル配列または融合タンパク質の場合は、機能するように連結しているDNA配列は、隣接し、かつリーディング相中にある。しかしながら、エンハンサーは、それらが転写を制御するコード配列と隣接していなくてもよい。このような状況において、連結は、便利な制限部位でのライゲーション、またはその代わりに挿入されるリンカーもしくはアダプターでのライゲーションによって達成される。
【0040】
本明細書で用いられる用語「プロモーター」は、転写を開始させるためのRNAポリメラーゼ結合に関与するDNA領域を意味する。
【0041】
本明細書で用いられる用語「組換え」は、組換えDNAでの形質転換を受けている細胞、組織または生物を意味する。
【0042】
本明細書で用いられるように、標的化の実体、タンパク質、ポリペプチドまたはペプチドは、特定の細胞型(例えばリンパ球のような正常な細胞または癌細胞のような異常細胞)に特異的に結合する分子を意味し、それゆえにこれは細胞型が特異的であるTf融合タンパク質または化合物(薬物または細胞毒性薬)を、標的とするために用いられ得る。
【0043】
本明細書で用いられる「治療用タンパク質」は、1またはそれ以上の治療活性および/または生物活性を有するタンパク質、ポリペプチド、抗体、ペプチド、またはそれらのフラグメントもしくは変異体を意味する。本発明に包含される治療用タンパク質としては、これらに限定されないが、タンパク質、ポリペプチド、ペプチド、抗体、および生体物質が挙げられる。本明細書において、ペプチド、タンパク質およびポリペプチドという用語は交換可能に用いられる。加えて、用語「治療用タンパク質」は、治療用タンパク質に相関する内在性のまたは天然に存在するものを意味する場合もある。「治療活性」を示すポリペプチド、または「治療的に活性である」タンパク質は、本明細書で説明されている治療用タンパク質または当技術分野で既知のその他のものの1種またはそれ以上のような治療用タンパク質に関連する既知の生物学的および/または治療活性を1種またはそれ以上有するポリペプチドを意味する。非限定的な例として、「治療用タンパク質」は、疾患、状態または障害を治療、予防または改善するのに有用なタンパク質である。このような病気、状態または障害は、ヒトにおけるものでもよいし、またはヒト以外の動物におけるものでもよく、例えば獣医学での使用が考えられる。
【0044】
本明細書で用いられる用語「形質転換」は、核酸、すなわちヌクレオチドポリマーの細胞への転移を意味する。本明細書で用いられる用語「遺伝学的な形質転換」は、DNA(特に組換えDNA)の細胞への転移および組み込みを意味する。
【0045】
本明細書で用いられる用語「形質転換体」は、形質転換を受けた細胞、組織または生物を意味する。
【0046】
本明細書で用いられる用語「導入遺伝子」は、生物、宿主細胞またはベクターに、確実に機能する形で挿入される核酸を意味する。
【0047】
本明細書で用いられる用語「トランスジェニック」は、様々な形質転換法の1つによって、外来または改変された遺伝子、特に改変されたTf融合タンパク質をコードする遺伝子を受け入れた細胞、細胞培養、生物、細菌、菌類、動物、植物、およびこれらのうちいずれかの後代を意味し、ここにおいて、外来または改変された遺伝子は、外来もしくは改変された遺伝子を受け入れる生物種と同一種由来であるか、または外来もしくは改変された遺伝子を受ける生物種とは異なる種由来である。
【0048】
「変異体(variants)または変異体(variant)」は、参照核酸またはポリペプチドとは異なるが、それらの必須の特性を保持するポリヌクレオチドまたは核酸を意味する。一般的に、変異体は、全体的に極めて類似しており、多くの領域において参照核酸またはポリペプチドと同一である。本明細書で用いられる「変異体」は、天然型の治療用タンパク質とは配列が異なっているが、本明細書の別のところで記載されている、または当技術分野で既知の別のもののようなそれらの少なくとも1つの機能的および/または治療的な特性を保持する、本発明のトランスフェリン融合タンパク質の治療用タンパク質部分を意味する。
【0049】
本明細書で用いられる用語「ベクター」は、広範囲に、外因性核酸をコードするあらゆるプラスミド、ファージミドまたはウイルスを意味する。また、この用語には、例えばポリリジン化合物などのような、核酸のウィリオンまたは細胞への転移を容易にする、非プラスミド、非ファージミドおよび非ウイルス化合物も含まれると解釈されるものでもある。ベクターは、核酸またはそれらの突然変異体を細胞に送達するための送達媒体として適するウイルスベクターでもよく、または同じ目的に適する非ウイルスベクターでもよい。DNAを細胞および組織に送達するためのウイルスおよび非ウイルスベクターの例は当技術分野でよく知られており、例えば、Ma等(1997,Proc.Natl.Acad.Sci.U.S.A.94:12744〜12746)に記載されている。ウイルスベクターの例としては、これらに限定されないが、組換えワクシニアウイルス、組換えアデノウイルス、組換えレトロウイルス、組換えアデノ随伴ウイルス、組換え 鳥類ポックスウイルスなどが挙げられる(Cranage等,1986,EMBO J.5:3057〜3063;1994年8月18日に公開された国際特許出願公開番号WO94/17810;1994年10月27日に公開された国際特許出願公開番号WO94/23744)。非ウイルスベクターの例としては、これらに限定されないが、リポソーム、DNAのポリアミン誘導体などが挙げられる。
【0050】
本明細書で用いられる用語「野生型」は、天然に生じるポリヌクレオチドまたはポリペプチド配列を意味する。
【0051】
本明細書で用いられる「スカフォールドタンパク質」、「スカフォールドポリペプチド」または「スカフォールド」は、アミノ酸配列(例えばランダムペプチド)が融合することができるタンパク質を意味する。このようなペプチドはスカフォールドにとって外来性である。
【0052】
本明細書で用いられる「ランダムペプチド配列」は、2個またはそれ以上のアミノ酸単量体で構成され、ストカスティック過程またはランダム過程によって構築されたアミノ酸配列を意味する。ランダムペプチドは、フレームワークまたはスカフォールドのモチーフを含んでいてもよく、これらはインバリアント配列を含んでもよい。ランダムペプチド配列は、変異体ではない、すなわちランダムではないアミノ酸の一部を含んでいてもよい。
【0053】
本明細書で用いられる「ランダムペプチドライブラリー」は、一連のランダムペプチドをコードする一連のポリヌクレオチド配列を意味し、ならびにそれらのポリヌクレオチド配列によってコードされる一連のランダムペプチド、およびそれらのランダムペプチドを含む融合タンパク質を意味する。
【0054】
本明細書で用いられる用語「擬似ランダム」(“pseudorandom”)は、例えば一つの位置における残基の可変性の程度が、その他の位置における残基の可変性の程度と異なっているが、あらゆる擬似ランダム位において、かなり制限されるがある程度の残基の多様性が許容されるように、限定された可変性を有する一連の配列を意味する。
【0055】
本明細書で用いられる用語「定義された配列フレームワーク」は、一般的に実験データまたは構造データに基づいてランダムではない方式で選択される一連の定義された配列を意味し、例えば定義された配列フレームワークは、β−シート構造を形成すると予測される一連のアミノ酸配列を含んでいてもよく、またはいくつかあるバリエーションのなかでも、ロイシンジッパーヘプタッド反復モチーフ、ジンクフィンガードメインを含んでいてもよい。「定義された配列核」(defined sequence kernal)は、限られた可変性の範囲を包含する一連の配列である。20種の通常のアミノ酸からなる完全にランダムな10−merの配列が、(20)10の配列のいずれかである可能性があり、そして20種の通常のアミノ酸からなる擬似ランダムな10−merの配列が、(20)10配列のいずれかである可能性があるが、特定の位置および/または全体において特定の残基に関する偏りを示すことになるので、それぞれの残基の位置が許容できる20種の通常のアミノ酸(および/または許容できる非必須アミノ/イミノ酸)のいずれかであることが許容されたならば、定義された配列核は可能な配列の最大数より少ない配列の部分集合である。定義された配列核は、一般的に、変異した残基位置とインバリアントの残基位置とを含むか、および/またはアミノ酸残基の定義された部分集合から選択される残基などを含むことが可能な変異した残基位置を、これは選択された個々のライブラリー要素配列のセグメント単位で、またはその全長にわたってのいずれかで含む。定義された配列核は、アミノ酸配列、または、ポリヌクレオチド配列のいずれかを意味する場合もある。
【0056】
本明細書で用いられる「リンカー」または「スペーサー」は、DNA結合タンパク質とランダムペプチドのような2つの分子を連結させる分子または分子群を意味し、これらは、2つの分子を望ましい立体配置に配置するのに役立ち、例えば、DNA結合タンパク質との立体障害を最小にしてランダムペプチドが受容体に結合できるようになる。
【0057】
本明細書で用いられる用語「可変性のセグメント」は、ランダム、擬似ランダム、または定義された核配列を含む新生のペプチド部分を意味する。可変性のセグメントは、変異した残基位置とインバリアントな残基位置の両方を含んでいてもよく、変異した残基位置における残基のバリエーションの程度は、制限される可能性があり、いずれの選択肢も技術者の裁量で選択される。典型的には、可変性のセグメントは、長さが約3〜20個のアミノ酸残基、例えば長さが8〜10個のアミノ酸であるが、それより長い可変性のセグメントも可能であり、抗体部分または受容体タンパク質、例えば抗体フラグメント、核酸結合タンパク質、受容体タンパク質などを含んでもよい。
【0058】
本明細書で用いられる用語「エピトープ」は、抗原の一部、または抗体の可変領域結合ポケットと相互作用する結合相互作用を形成することができるその他の高分子を意味する。典型的には、このような結合相互作用は、CDRの1個またはそれ以上のアミノ酸残基との分子間接触として示される。
【0059】
本明細書で用いられる用語「受容体」、「標的」または「物質」は、所定のリガンドへの親和性を有する分子を意味する。受容体は、天然に存在する分子でもよいし、または合成分子でもよい。受容体は、改変されていない状態で用いてもよいし、またはその他の種との集合体として用いてもよい。受容体は、共有結合または非共有結合で、直接的に、または特異的な結合物質を介して、結合要素(すなわちリガンド)に結合することができる。受容体の例としては、これらに限定されないが、抗体、例えばモノクローナル抗体、および(例えば、ウイルス、細胞またはその他の物質における)特異的な抗原決定基との反応性を有する抗血清、細胞膜受容体、抗原、エピトープ含有分子、複合糖質および糖タンパク質、酵素、ならびにホルモン受容体が挙げられる。
【0060】
本明細書で用いられる用語「リガンド」または「リガンド部分」は、特定の受容体または物質によって認識されるランダムペプチドまたは可変性のセグメント配列のような分子を意味する。当業者であれば理解できると予想されるが、このような分子(または高分子複合体)は、受容体でもよいし、リガンドでもよい。
【0061】
本明細書で用いられる「融合した」、「複合体化した」または「機能するように連結した」とは、ランダムペプチドとスカフォールドタンパク質とが、スカフォールド構造の安定性の破壊が最小化されるような形で一緒に連結していることを意味する。
【0062】
本明細書で用いられる用語「単鎖抗体」は、ポリペプチドリンケージにVHドメインおよびVLドメインを含むポリペプチドを意味し、これは、一般的に、スペーサーペプチド(例えば、配列番号17の[Gly−Gly−Gly−Gly−Ser]x)を介して連結しており、さらにアミノおよび/またはカルボキシ末端に追加のアミノ酸配列を含んでもよい。例えば、単鎖抗体は、コーディングポリヌクレオチドに連結させるための固定セグメントを含んでもよい。一例として、scFvは、単鎖抗体である。単鎖抗体は、一般的に、実質的に免疫グロブリンスーパーファミリーの遺伝子によってコードされた少なくとも10個の隣接したアミノ酸のポリペプチドセグメント1個またはそれ以上からなるタンパク質であり(例えば、The Immunoglobulin Gene Superfamily,A.F.WilliamsおよびA.N.Barclay,in Immunoglobulin Genes,T.Honjo,F.W.AltおよびT.H.Rabbitts編(1989)アカデミックプレス:サンディエゴ,カリフォルニア州,361〜387頁を参照、これらは、参照により本明細書に組み込まれる)、最も多くは、げっ歯類、非ヒト霊長類、鳥類、ブタ、ウシ、ヒツジ、ヤギまたはヒトの重鎖または軽鎖遺伝子配列によってコードされた少なくとも10個の隣接したアミノ酸のポリペプチドセグメント1個またはそれ以上からなるタンパク質である。機能的な単鎖抗体は、一般的に、特異的な標的分子、典型的には受容体または抗原(エピトープ)への結合特性が保持されるように、免疫グロブリンスーパーファミリーの遺伝子産物の十分な部分を含む。
【0063】
本明細書で用いられる用語「相補性決定領域」および「CDR」は、KabatおよびChothiaのCDRの定義で例示されたような当技術分野で認められている用語を意味し、これはまた、一般的には、超可変領域または超可変ループとしても知られている。ChothiaおよびLesk(1987)J.Mol.Biol.196:901;Chothia等(1989)Nature 342:877;E.A.Kabat等,Sequences of Proteins of Immunological Interest(National Institutes of Health,ベセスダ,メリーランド州)(1987);およびTramontano等(1990)J.Mol.Biol.215:175を参照。可変領域ドメインは、典型的には、天然の免疫グロブリン鎖のアミノ末端の約105〜115個のアミノ酸(例えばアミノ酸1〜110)を含むが、可変ドメインは、多少短くても、または長くても、単鎖抗体を形成するのに適している。
【0064】
免疫グロブリンの軽鎖または重鎖可変領域は、3つの超可変領域(CDRともいう)で分断された「フレームワーク」領域からなる。フレームワーク領域およびCDRの範囲は、正確に定義されている。“Sequences of Proteins of Immunological Interest”,E.Kabat等,第4版,U.S.Department of Health and Human Services,ベセスダ,メリーランド州(1987)を参照。異なる軽鎖または重鎖のフレームワーク領域の配列は、種内で比較的保存されている。本明細書で用いられる「ヒトフレームワーク領域」は、天然のヒト免疫グロブリンのフレームワーク領域に実質的に(約85%またはそれ以上、通常は90〜95%またはそれ以上)同一なフレームワーク領域である。構成要素である軽鎖と重鎖とが一緒になったフレームワーク領域である抗体のフレームワーク領域は、CDRを配置し並べるのに役立つ。CDRは、主として、抗原のエピトープへの結合に関与する。
【0065】
特に他の指定がない限り、本明細書で用いられる技術用語や科学用語はいずれも、本発明が属する分野の当業者によって一般的に理解されているのと同じ意味を有する。本発明の実施または試験において、本明細書で説明されている方法および材料と類似した、またはそれらと同等のあらゆる方法および材料を用いることができるが、好ましい方法と材料を説明する。
【0066】
トランスフェリンおよびトランスフェリンの改変
本発明の融合タンパク質は、受容体または物質にランダムペプチドまたはCDRのようなリガンドを提示することができるトランスフェリン(Tf)タンパク質、またはそれらの部分を含む。Tf成分は、軸成分のN末端に融合している。本融合タンパク質のTfタンパク質またはそれらの部分を、本融合タンパク質のTf「部分」、「領域」または「成分」と称する場合もある。本明細書で用いられるように、トランスフェリン融合タンパク質は、軸成分に融合したトランスフェリンタンパク質または成分であり、さらに細胞壁連結要素を含む。本発明のトランスフェリン融合タンパク質は、場合によりアンカー成分を含んでいてもよい。
【0067】
本発明の改変されたTf融合タンパク質を作製するためにどのようなトランスフェリンでも使用可能である。一例として、野生型ヒトTf(Tf)は、約75kDaの679個のアミノ酸からなるタンパク質であり(グリコシル化は考慮に入れない)、これは遺伝子重複によって生じると思われる2つの主要なローブまたはドメイン、N(約330個のアミノ酸)およびC(約340個のアミノ酸)を有する。GenBank登録番号NM001063、XM002793、M12530、XM039845、XM039847、およびS95936(www.ncbi.nlm.nih.gov)(これらはいずれも、参照によりその全体として本明細書に組み込まれる)、ならびに配列番号1、2および3を参照。この2つのドメインは長い時間をかけて分岐しているが、大きい同一性/類似性を保持する。
【0068】
NおよびCドメインはそれぞれ、さらに2つのサブドメイン、N1およびN2、C1およびC2に分けられる。これは、鉄を体の細胞に輸送するように機能する。これは、全ての細胞で、特に活発に増殖する細胞で発現されるTf受容体(TfR)が介在するプロセスである。TfRは、Tfの鉄が結合した形態を認識し(1受容体につきその2つの分子を結合する)、続いてエンドサイトーシスが起こり、それによってTfR/Tf複合体が、エンドソームに輸送され、その時点で、局所的なpH低下によって、結合した鉄の放出、および細胞表層へのTfR/Tf複合体の再利用、およびTf放出(その鉄が結合していない形態で、アポTfとして知られている)が起こる。受容体結合は、主として、TfのCドメインを介する。グリコシル化されていない鉄が結合したTfが受容体と結合するため、Cドメイン中の2つのグリコシル化部位は、受容体結合に関与しないようである。
【0069】
Tf分子はそれぞれ、2個の鉄イオン(Fe3+)を運搬することができる。これらは、N1およびN2サブドメイン、C1およびC2サブドメインの間のスペースで複合体化され、分子においてコンホメーション変化が起こる。
【0070】
ヒトトランスフェリンにおいて、鉄結合部位は、少なくとも、配列番号3のアミノ酸Asp63(天然型のTfシグナル配列を含む配列番号2では、Asp82)、Asp392(配列番号2では、Asp411)、Tyr95(配列番号2では、Tyr114)、Tyr426(配列番号2では、Tyr445)、Tyr188(配列番号2では、Tyr207)、Tyr514または517(配列番号2では、Tyr533またはTyr536)、His249(配列番号2では、His268)、およびHis585(配列番号2では、His604)を含む。ヒンジ領域は、配列番号3の少なくともNドメインのアミノ酸残基94〜96、245〜247および/または316〜318、同様にCドメインのアミノ酸残基425〜427、581〜582および/または652〜658を含む。カーボネート結合部位は、配列番号3の、少なくともアミノ酸Thr120(配列番号2では、Thr139)、Thr452(配列番号2では、Thr471)、Arg124(配列番号2では、Arg143)、Arg456(配列番号2では、Arg475)、Ala126(配列番号2では、Ala145)、Ala458(配列番号2では、Ala477)、Gly127(配列番号2では、Gly146)、およびGly459(配列番号2では、Gly478)を含む。
【0071】
本発明の一実施態様において、本融合タンパク質は、改変されたヒトトランスフェリンを含むが、本発明の融合タンパク質を生産するのにあらゆる動物のTf分子が使用可能であり、例えば、ヒトTf変異体、ウシ、ブタ、ヒツジ、イヌ、ウサギ、ラット、マウス、ハムスター、ハリモグラ(echnida)、カモノハシ、ニワトリ、カエル、イモムシ、サル、類人猿が挙げられ、加えてその他のウシ属、イヌ科および鳥類の種も挙げられる。これらのTf配列はいずれも、GenBankやその他の公開データベースで容易に入手することができる。ヒトTfのヌクレオチド配列が利用可能であり(配列番号1、2および3、ならびに上述の登録番号を参照、さらにwww.ncbi.nlm.nih.govで入手可能である)、これらは、TfまたはTfのドメインと、選択された治療的分子との遺伝学的な融合体を作製するのに用いることができる。また、融合は、ラクトトランスフェリン(GenBank登録番号NM_002343のラクトフェリン)、またはメラノトランスフェリンのような関連の分子から作製してもよい。
【0072】
ラクトフェリン(Lf)は、防御作用のある天然の鉄結合性タンパク質であり、これは抗菌性、抗真菌性、抗ウイルス性、抗腫瘍性および抗炎症性の活性を有することが見出されている。このようなタンパク質は、通常は正常フローラに晒される外分泌物に存在しており、このような外分泌物としては、乳、涙、鼻の滲出物、唾液、気管支粘液、胃腸液、頚膣部粘液、および精液が挙げられる。加えて、Lfは、循環する多形核好中球(PMN)の二次的な特殊顆粒の主成分である。アポタンパク質は、化膿した領域においてPMNが脱顆粒する際に放出される。Lfの主要な機能は、体液中や炎症を起こしている領域中の遊離の鉄を捕捉することであり、このようにして、フリーラジカルが介在するダメージを抑制し、微生物細胞や新生細胞の侵入における金属の利用可能性を減少させることができる。成人の125ILfの代謝回転速度を試験した研究では、Lfは迅速に肝臓および脾臓に取り込まれ、放射活性は、肝臓および脾臓中で数週間持続することが示された(Bennett等(1979),Clin.Sci.(Lond.)57:453〜460)。
【0073】
一実施態様において、本発明の融合タンパク質のトランスフェリン部分は、トランスフェリンのスプライス変異体を含む。一例において、トランスフェリンのスプライス変異体は、ヒトトランスフェリンのスプライス変異体であり得る。特定の一実施態様において、ヒトトランスフェリンのスプライス変異体は、GenBank登録番号AAA61140のスプライス変異体であり得る。
【0074】
その他の実施態様において、本発明の融合タンパク質のトランスフェリン部分は、ラクトフェリンのスプライス変異体を含む。一例において、ヒト血清のラクトフェリンのスプライス変異体は、好中球のラクトフェリンの新規なスプライス変異体であり得る。特定の一実施態様において、好中球のラクトフェリンのスプライス変異体は、GenBank登録番号AAA59479のスプライス変異体であり得る。その他の特定の実施態様において、好中球のラクトフェリンのスプライス変異体は、以下のアミノ酸配列EDCIALKGEADA(配列番号4)を含んでいてもよく、この配列は、新規なスプライス変異の領域を含む。
【0075】
また、メラノトランスフェリン(GenBank登録番号NM_013900、マウスのメラノトランスフェリン)との融合を行ってもよい。メラノトランスフェリンは、悪性黒色腫細胞において高レベルで見出されるグリコシル化タンパク質であり、元々はヒト黒色腫抗原p97と命名されていた(Brown等,1982,Nature,296:171〜173)。これは、ヒト血清トランスフェリン、ヒトラクトフェリン、およびニワトリトランスフェリンと高い配列相同性を有する(Brown等,1982,Nature,296:171〜173;Rose等,Proc.Natl.Acad.Sci.,1986,83:1261〜1265)。しかしながら、これらのタンパク質とは異なり、メラノトランスフェリンに関する細胞受容体は同定されていない。メラノトランスフェリンは、鉄と可逆的に結合し、2つの形態で存在し、そのうちの一方の形態は、グリコシルホスファチジルイノシトールのアンカーによって細胞膜に結合し、他方の形態は、可溶性であり、かつ活発に分泌される(Baker等,1992,FEBS Lett,298:215〜218;Alemany等,1993,J.Cell Sci.,104:1155〜1162;Food等,1994,J.Biol.Chem.274:7011〜7017)。
【0076】
改変されたTf融合体は、あらゆるTfタンパク質、フラグメント、ドメインまたは加工されたドメインで作製することができる。例えば、融合タンパク質は、全長Tf配列を用いて作製することができ、天然型のTfシグナル配列を含んでいてもよいし、またはそれらを含まなくてもよい。また、細胞透過性の抗体(Trans−body)は、単一のTfドメイン(例えばNまたはCドメインそれぞれ)を用いて作製することもできる。また、Trans−body(細胞透過性)の抗体も、二つのTfドメイン(例えば二つのNドメイン、または二つのCドメイン)を用いて作製してもよい。ある実施態様では、単一のCドメインへ治療用タンパク質を融合させることができ、ここにおいてCドメインは、グリコシル化、鉄結合および/またはTf受容体結合を、減少させる、阻害する、または防ぐように変更される。その他の実施態様において、Tfグリコシル化部位はCドメインに存在しており、Nドメインは、そのままでは鉄またはTf受容体と結合しないため、単一のNドメインの使用が有利である。一実施態様において、Tf融合タンパク質は、高レベルで発現される単一のNドメインを有する。
【0077】
本明細書で用いられるように、N様ドメインとして機能するように改変されたC末端ドメインまたはローブは、天然型または野生型のNドメインまたはローブと実質的に類似したグリコシル化パターンまたは鉄結合特性を示すように改変される。一実施態様において、Cドメインまたはローブは、グリコシル化されず、かつ鉄と結合しないように改変され、改変は、それに関連するCドメイン領域またはアミノ酸を、それに対応する天然型または野生型のNドメインの領域または部位に存在するドメイン領域またはアミノ酸で置換することによってなされる。
【0078】
本明細書で用いられる「2つのNドメインまたはローブ」を含むTf成分は、天然型のCドメインまたはローブを、第二の天然型または野生型のNドメインもしくはローブ、または改変されたNドメインもしくはローブで置き換えて改変されたTf分子を含むか、または実質的に野生型または改変されたNドメインのように機能するように改変されているCドメインを含む。米国仮出願第60/406,977号を参照(これは参照によりその全体として本発明に組み込まれる)。
【0079】
2つのドメインの三次元構造を重ね合わせること(スイスPDBビューワー3.7b2(Swiss PDB Viewer 3.7b2)、イタラティブ・マジック・フィット(Iterative Magic Fit))、および直接のアミノ酸アライメント(クラスタルW(ClustalW)マルチプルアライメント)による2つのドメインの解析によって、2つのドメインは時間とともに分岐しているが明らかである。アミノ酸アライメントによれば、2つのドメイン間で42%同一性、および59%の類似性が示される。しかしながら、構造的な等価性に関して、Nドメインの約80%がCドメインと適合している。またCドメインは、Nドメインと比較して数個の余分なジスルフィド結合も有する。
【0080】
NおよびCドメインに関する分子モデルのアライメントから、以下の構造的な等価体が明らかになる:
【表1】

【0081】
2つのドメインのジスルフィド結合は、以下のようにアラインされる:
【表2】
【0082】
一実施態様において、本融合タンパク質のトランスフェリン部分は、トランスフェリンのN末端ローブを少なくとも2つ有する。さらなる実施態様において、本融合タンパク質のトランスフェリン部分は、ヒト血清トランスフェリンから誘導されたトランスフェリンのN末端ローブを少なくとも2つ含む。
【0083】
その他の実施態様において、本融合タンパク質のトランスフェリン部分は、配列番号3のAsp63、Gly65、Tyr95、Tyr188、およびHis249からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有するトランスフェリンのN末端ローブを少なくとも2つ有する、それらを含む、またはそれらからなる。
【0084】
その他の実施態様において、改変された融合タンパク質のトランスフェリン部分は、配列番号3のLys206またはHis207において突然変異を有する、組換えヒト血清トランスフェリンのN末端ローブの突然変異体を有する。
【0085】
その他の実施態様において、本融合タンパク質のトランスフェリン部分は、トランスフェリンのC末端ローブを少なくとも2つ有する、それらを含む、本質的にそれらからなる、またはそれらからなる。さらなる実施態様において、本融合タンパク質のトランスフェリン部分は、ヒト血清トランスフェリンから誘導されたトランスフェリンのC末端ローブを少なくとも2つ有する。
【0086】
さらなる実施態様において、C末端ローブの突然変異体はさらに、グリコシル化が不可能な配列番号3のAsn413およびAsn611の少なくとも1つの突然変異を有する。
【0087】
その他の実施態様において、トランスフェリン部分は、配列番号3のAsp392、Tyr426、Tyr514、Tyr517、およびHis585からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有するトランスフェリンのC末端ローブを少なくとも2つ有し、ここにおいて、該突然変異体は、金属イオンと結合する能力を保持する。代替の実施態様において、トランスフェリン部分は、配列番号3のTyr426、Tyr514、Tyr517、およびHis585からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有するトランスフェリンのC末端ローブを少なくとも2つ有し、ここにおいて、該突然変異体は、金属イオンと結合する能力が減少している。その他の実施態様において、トランスフェリン部分は、配列番号3のAsp392、Tyr426、Tyr517、およびHis585からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有するトランスフェリンのC末端ローブを少なくとも2つ有し、ここにおいて、該突然変異体は、金属イオンと結合する能力を保持せず、実質的にNドメインのように機能する。
【0088】
いくつかの実施態様において、本融合タンパク質のTfまたはTf部分およびリガンドが融合タンパク質の残余から切断される場合、TfまたはTf部分は、非融合状態、すなわちTfに融合していない状態でのリガンドのインビボでの循環半減期、血清中安定性(半減期)、インビトロでの安定性または生物学的利用率と比較して、リガンド(すなわち治療剤)のインビボでの循環半減期、血清中安定性、インビトロでの溶液安定性または生物学的利用率を増加させるのに十分な長さを有する。このような安定性、インビボ循環半減期、または生物学的利用率の増加は、融合していないリガンド成分領域を超える約30%、50%、70%、80%、90%またはそれ以上の増加であり得る。場合によっては、改変されたトランスフェリンを含むリガンド成分は、非融合状態のリガンドと比較して、約1日またはそれ以上、1〜2日またはそれ以上、3〜5日またはそれ以上、5〜10日またはそれ以上、10〜15日またはそれ以上、10〜20日またはそれ以上、約12〜18日間、または約14〜17日間の血清中半減期を示す。
【0089】
TfのCドメインが本融合タンパク質の一部である場合、グリコシル化または過剰なマンノシル化を防ぎ、本融合タンパク質の血清中半減期を延長させるために(アシアロ−、または場合によってはモノシアロ−Tfもしくはジシアロ−Tfが生産されるように)、2つのN結合型のグリコシル化部位、配列番号3のN413およびN611に相当するアミノ酸残基は、酵母系での発現のために突然変異していてもよい。N413およびN611に相当するTfアミノ酸に加えて、N−X−S/Tグリコシル化部位内の、またはそれに隣接する残基への突然変異は、グリコシル化を防ぐか、または実質的に減少させる。Funk等の米国特許第5,986,067号を参照。また、ピチア・パストリス(Pichia pastoris)で発現されたTfのNドメインは、S32において単一のヘキソースでO結合型グリコシル化されることになることも報告されているが、これを、このようなグリコシル化を防ぐために突然変異または改変することができる。その上、1またはそれ以上のPMT遺伝子に突然変異を有する酵母宿主細胞では、O連結型糖付加は減少するか、または除去される可能性がある。
【0090】
従って、本発明の一実施態様において、本融合タンパク質は、トランスフェリンが低いグリコシル化を示す改変されたトランスフェリン分子を有し、このようなトランスフェリン分子としては、これらに限定されないが、Tfのアシアロ−、モノシアロ−およびジシアロ型が挙げられる。その他の実施態様において、本融合タンパク質のトランスフェリン部分は、グリコシル化を防ぐために突然変異した組換えトランスフェリン突然変異体を有する。その他の実施態様において、本融合タンパク質のトランスフェリン部分は、完全にグリコシル化された組換えトランスフェリン突然変異体を有する。さらなる実施態様において、本融合タンパク質のトランスフェリン部分は、グリコシル化を防ぐために突然変異した組換えヒト血清トランスフェリン突然変異体を有し、ここにおいて、配列番号3のAsn413およびAsn611の少なくとも一方が、グリコシル化が不可能なアミノ酸に突然変異している。その他の実施態様において、本融合タンパク質のトランスフェリン部分は、グリコシル化を防ぐために、または実質的に減少させるために突然変異した組換えヒト血清トランスフェリン突然変異体を有し、ここにおいて、N−X−S/Tグリコシル化部位内の残基で突然変異していてもよい。さらに、セリンまたはスレオニン残基を突然変異させることによってグリコシル化を減少させるか、または防ぐことができる。さらに、グリコシル化を阻害するために、Xをプロリンに変化させることが既知である。
【0091】
また、以下でより詳細に考察されるように、本発明の改変されたTfを含む改変されたTf融合タンパク質は、鉄と結合しないように、および/またはTf受容体と結合しないように遺伝子操作してもよい。本発明のその他の実施態様において、鉄結合を保持させ、Tfの鉄結合能を、治療用タンパク質またはペプチドを、細胞の内部に送達、および/または血液脳関門(BBB)を通過させるために使用可能である。Nドメインは鉄をローディングすると、単独ではTfRに結合せず、鉄と結合したCドメインは、TfRと結合することになるが、分子全体と同じ親和性は示さない。
【0092】
その他の実施態様において、トランスフェリン融合タンパク質のトランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、金属イオンと結合する能力を保持しない。代替の実施態様において、トランスフェリン融合タンパク質のトランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、野生型血清トランスフェリンよりも金属イオンに対する結合親和性が弱い。代替の実施態様において、トランスフェリン融合タンパク質のトランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、野生型血清トランスフェリンよりも金属イオンに対する結合親和性が強い。
【0093】
その他の実施態様において、トランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、トランスフェリン受容体に結合する能力を保持しない。代替の実施態様において、トランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、野生型血清トランスフェリンよりもトランスフェリン受容体に対する結合親和性が弱い。代替の実施態様において、トランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、野生型血清トランスフェリンよりもトランスフェリン受容体に対する結合親和性が強い。
【0094】
その他の実施態様において、トランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、炭酸イオンに結合する能力を保持しない。代替の実施態様において、トランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、野生型血清トランスフェリンよりも炭酸イオンに対する結合親和性が弱い。代替の実施態様において、トランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、野生型血清トランスフェリンよりも炭酸イオンに対する結合親和性が強い。
【0095】
その他の実施態様において、トランスフェリン部分は、配列番号3のAsp63、Gly65、Tyr95、Tyr188、His249、Asp392、Tyr426、Tyr514、Tyr517、およびHis585からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有する組換えヒト血清トランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、金属イオンと結合する能力を保持する。代替の実施態様において、配列番号3のAsp63、Gly65、Tyr95、Tyr188、His249、Asp392、Tyr426、Tyr514、Tyr517、およびHis585からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有する組換えヒト血清トランスフェリン突然変異体が挙げられ、ここにおいて、該突然変異体は、金属イオンと結合する能力が減少している。その他の実施態様において、配列番号3のAsp63、Gly65、Tyr95、Tyr188、His249、Asp392、Tyr426、Tyr517、およびHis585からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有する組換えヒト血清トランスフェリン突然変異体が挙げられ、ここにおいて、該突然変異体は、金属イオンと結合する能力を保持しない。
【0096】
その他の実施態様において、トランスフェリン部分は、配列番号3のLys206またはHis207において突然変異を有する、組換えヒト血清トランスフェリンの突然変異体を有し、ここにおいて、突然変異体は、野生型ヒト血清トランスフェリンよりも強い金属イオンへの結合活性を有する(米国特許第5,986,067号を参照。これは参照によりその全体として本明細書に組み込まれる)。代替の実施態様において、トランスフェリン部分は、配列番号3のLys206またはHis207において突然変異を有する組換えヒト血清トランスフェリン突然変異体を含み、ここにおいて、該突然変異体は、野生型ヒト血清トランスフェリンよりも金属イオンへの結合活性が弱い。さらなる実施態様において、トランスフェリン部分は、配列番号3のLys206またはHis207において突然変異を有する、組換えヒト血清トランスフェリン突然変異体を含み、ここにおいて、該突然変異体は、金属イオンと結合しない。
【0097】
あらゆる利用可能な技術が本発明の融合タンパク質を製造するために使用可能であり、このような技術としては、これらに限定されないが、一般的に利用できる分子技術が挙げられ、例えば、Sambrook等のMolecular Cloning:A Laboratory Manual,第二版,コールドスプリングハーバーラボラトリープレス(Cold Spring Harbor Laboratory Press),1989に記載された技術である。当技術分野でよく知られた部位特異的突然変異誘発を達成する技術を用いてヌクレオチド置換を行う場合、コードされたアミノ酸の変化は、好ましくは軽度のもの、すなわち保存的アミノ酸置換であるが、その他の非保存的な置換も同様に可能なものとして取扱われ、改変されたトランスフェリン部分、例えば、改変された融合タンパク質が、低いグリコシル化、低い鉄結合などを示すものを製造する場合に特にそうである。特に可能なものとして取扱われるのは、アミノ酸の置換、わずかな欠失または挿入したもの(典型的には約30個のアミノ酸に対して1個の割合で);トランスフェリンドメイン間への挿入したもの;小規模のアミノまたはカルボキシル末端の伸長したもの、例えばアミノ末端のメチオニン残基、またはトランスフェリンドメイン間、もしくは、トランスフェリンタンパク質と治療用タンパク質もしくはペプチド、リガンド、もしくは抗体の可変領域もしくは軸領域との連結の50、40、30、20または10残基未満の小さいリンカーペプチド;または、精製を容易にする小規模の伸長したもの、例えばポリヒスチジントラクト(poly−histidine tract)、抗原エピトープ、もしくは結合ドメインである。
【0098】
保存的アミノ酸置換の例としては、同じグループ内でなされる置換が挙げられ、例えば、塩基性アミノ酸のグループ(例えば、アルギニン、リシン、ヒスチジン)、酸性アミノ酸のグループ(例えば、グルタミン酸およびアスパラギン酸)、極性アミノ酸のグループ(例えば、グルタミンおよびアスパラギン)、疎水性アミノ酸のグループ(例えば、ロイシン、イソロイシン、バリン)、芳香族アミノ酸のグループ(例えば、フェニルアラニン、トリプトファン、チロシン)、および低分子量のアミノ酸のグループ(例えば、グリシン、アラニン、セリン、スレオニン、メチオニン)の中でなされる置換である。
【0099】
非保存的置換は、あるグループのアミノ酸を、その他のグループのアミノ酸で置換することを包含する。例えば、非保存的置換は、極性アミノ酸を疎水性アミノ酸で置換することを含む。ヌクレオチド置換の一般的な説明については、例えば、Ford等(1991),Prot.Exp.Pur.2:95〜107を参照。鉄への結合を示さない、もしくはそれらが減少した、および/または、本融合タンパク質のTf受容体への結合を示さない、もしくはそれらが減少した本発明のTf融合タンパク質、好ましくは細胞透過性の抗体trans−bodiesを製造するために、非保存的置換、欠失および挿入が特に有用である。
【0100】
本発明のポリペプチドおよびタンパク質において、アミノ酸の命名は、以下の規則にのっとった一覧に記載のシステムに従ってなされる:
【表3】
【0101】
鉄結合および/または受容体結合は、配列番号3のTfNドメイン残基Asp63、Tyr95、Tyr188、His249、および/またはCドメイン残基Asp392、Tyr426、Tyr514、および/またはHis585の1つまたはそれ以上に相当するアミノ酸残基への欠失、置換または挿入などの突然変異によって、減少する場合もあるし、または破壊される場合もある。また、鉄結合も、配列番号3のアミノ酸Lys206、His207、またはArg632の突然変異によって影響を受ける可能性がある。炭酸結合は、配列番号3のTfNドメイン残基Thr120、Arg124、Ala126、Gly127、および/またはCドメイン残基Thr452、Arg456、Ala458、および/またはGly459の1つまたはそれ以上に相当するアミノ酸残基の欠失、置換またはそれへの挿入などの突然変異によって減少するか、または破壊される可能性がある。炭酸結合の減少または破壊は、鉄および/または受容体結合に逆の影響を与える可能性がある。
【0102】
Tf受容体への結合は、鉄結合について上述したTfNドメイン残基の1つまたはそれ以上に相当するアミノ酸残基の欠失、置換またはそれへの挿入などの突然変異によって減少するか、または破壊される可能性がある。
【0103】
上記で考察したように、グリコシル化は、Cドメイン残基N413および/またはN611に相当するN−X−S/T部位内のTfのCドメイン残基の1つまたはそれ以上に相当するアミノ酸残基の欠失、置換またはそれへの挿入などの突然変異によって減少するか、または防止される可能性がある。米国特許第5,986,067号を参照。例えば、N413および/またはN611は、隣接するアミノ酸の可能性があるGlu残基に突然変異していてもよい。
【0104】
本発明のTf融合タンパク質が、グリコシル化、鉄結合、カーボネート結合および/または受容体結合を防ぐように改変されていないような場合には、グリコシル化、鉄および/またはカーボネートイオンを、本融合タンパク質から取り除くか、または切断することができる。例えば、利用可能なデグリコシラーゼ(deglycosylase)を用いて、本融合タンパク質からグリコシル化残基を切断してもよく(特に、Tf部分に結合した糖残基)、グリコシル化酵素欠損酵母を用いて、グリコシル化を防いでもよいし、および/またはグリコシル化を防ぐ物質(例えばツニカマイシン)の存在下で組換え細胞を増殖させてもよい。
【0105】
また、本融合タンパク質上の糖質も、本融合タンパク質をデグリコシラーゼで処理することによって、酵素的に減少させてもよいし、または完全に除去してもよい。デグリコシラーゼは当技術分野でよく知られている。デグリコシラーゼの例としては、これらに限定されないが、ガラクトシダーゼ、PNGアーゼA、PNGアーゼF、グルコシダーゼ、マンノシダーゼ、フコシダーゼ、およびEndoHデグリコシラーゼが挙げられる。
【0106】
Tfの三次元構造を変更するために、Tfに、追加の突然変異を作製してもよく、このような突然変異としては、例えばヒンジ領域への改変が挙げられ、それにより、鉄結合およびTf受容体の認識に必要なコンホメーション変化を防ぐことができる。例えば、突然変異は、Nドメインアミノ酸残基94〜96、245〜247、および/または316〜318、ならびにCドメインのアミノ酸残基425〜427、581〜582、および/または652〜658に、またはその周辺に作製してもよい。加えて、Tfの構造および機能を変更するために、これらの部位のフランキング領域に、またはその周辺に突然変異を作製してもよい。
【0107】
本発明の一側面において、本融合タンパク質をキャリアータンパク質として機能させて、リガンドの半減期または生物学的利用率を伸ばすことができ、加えて、場合によってはリガンドを細胞の内部に運搬したり、血液脳関門を通過する能力を保持させたりすることができる。代替の実施態様において、本融合タンパク質は、改変されたトランスフェリン分子を有し、ここにおいて、トランスフェリンは、血液脳関門を通過する能力を保持しない。
【0108】
その他の実施態様において、本融合タンパク質は、改変されたトランスフェリン分子を有し、ここにおいて、トランスフェリン分子は、トランスフェリン受容体に結合し、抗体の可変領域を細胞の内部に輸送する能力を保持する。代替の実施態様において、本融合タンパク質は、改変されたトランスフェリン分子を有し、ここにおいて、トランスフェリン分子は、トランスフェリン受容体に結合し、抗体の可変領域を細胞の内部に輸送する能力を保持しない。
【0109】
さらなる実施態様において、本融合タンパク質は、改変されたトランスフェリン分子を有し、ここにおいて、トランスフェリン分子は、トランスフェリン受容体に結合し、抗体の可変領域を細胞の内部に輸送する能力を保持するが、血液脳関門を通過する能力を保持しない。代替の実施態様において、本融合タンパク質は、改変されたトランスフェリン分子を有し、ここにおいて、トランスフェリン分子は、血液脳関門を通過する能力を保持するが、トランスフェリン受容体に結合し、抗体の可変領域を細胞の内部に輸送する能力を保持しない。
【0110】
トランスフェリン融合タンパク質
本発明の融合タンパク質は、Tfタンパク質のN末端および/またはC末端に付着したリガンド、抗体の可変領域、またはランダムペプチドの1個またはそれ以上のコピーを含んでいてもよい。一実施態様において、リガンド成分は、Tfタンパク質のN末端に結合している。いくつかの実施態様において、これらリガンド、可変領域またはペプチドは、Tfタンパク質のNおよびC末端の両方に付着しており、本融合タンパク質は、Tf末端のいずれか、またはその両方に、これらの領域の1またはそれ以上の等価体を含んでいてもよい。
【0111】
その他の実施態様において、1またはそれ以上のリガンドは、トランスフェリンペプチド、例えば1またはそれ以上のTfのループなどのTfタンパク質の既知のドメインの場所に挿入される。Ali等(1999)J.Biol Chem.274(34):24066〜24073を参照。
【0112】
本発明の一実施態様において、リガンドは、トランスフェリンのNローブに挿入される。例えば、本発明はまた、1またはそれ以上の挿入も含み、このような挿入は、以下の表で示されるように、N−ローブのN1およびN2ドメインに、またはN−ローブのN1およびN2ドメイン周辺の他の位置に行うことができる。
【表4】
【0113】
一般的に、本発明のトランスフェリン融合タンパク質は、一つの改変されたトランスフェリンから誘導された領域、および一つの抗体の可変領域を有していてもよい。しかしながら、それぞれのタンパク質の複数の領域を用いて、本発明のトランスフェリン融合タンパク質を作製してもよい。同様に、1種より多くの抗体の可変領域を用いて、本発明のトランスフェリン融合タンパク質を作製してもよく、それによって、多機能の改変されたTf融合タンパク質を生産することができる。
【0114】
一実施態様において、本発明の融合タンパク質は、トランスフェリン分子またはその部分に融合した、抗体の可変領域またはその部分を含む。その他の実施態様において、本発明の融合タンパク質は、トランスフェリン分子のN末端に融合した抗体の可変領域を含む。代替の実施態様において、本発明の融合タンパク質は、トランスフェリン分子のC末端に融合した抗体の可変領域を含む。さらなる実施態様において、本発明の融合タンパク質は、抗体の可変領域のN末端に融合したトランスフェリン分子を含む。代替の実施態様において、本発明の融合タンパク質は、抗体の可変領域のC末端に融合したトランスフェリン分子を含む。
【0115】
本発明はまた、改変されたトランスフェリン分子またはその部分に融合した抗体の可変領域またはその部分を含む融合タンパク質を提供する。
【0116】
その他の実施態様において、本発明の融合タンパク質は、改変されたトランスフェリンのN末端およびC末端の両方に融合した抗体の可変領域を含む。その他の実施態様において、NおよびC末端で融合した抗体の可変領域は、同じ抗原に結合する。また、同じ抗原に結合する抗体の可変領域は、異なる抗体から誘導されてもよく、従って、同じ標的上の異なるエピトープに結合する。代替の実施態様において、NおよびC末端で融合した抗体の可変領域は、異なる抗原と結合する。その他の代替の実施態様において、NおよびC末端に融合した抗体の可変領域は、異なる抗原と結合し、これは病気、障害または状態を治療または予防するための2種の異なる細胞を活性化するのに有用である可能性がある。その他の実施態様において、NおよびC末端で融合した抗体の可変領域は、異なる抗原と結合し、これは一般的に患者に同時に起こることが当技術分野で既知である病気または障害を治療または予防するための2種の異なる抗原との架橋形成に有用である可能性がある。
【0117】
加えて、本発明のトランスフェリン融合タンパク質はまた、対象の抗体の可変領域(例えば、治療用タンパク質またはそのフラグメントまたは変異体と結合する単鎖抗体)を、改変されたトランスフェリンの内部領域に挿入することによって生産してもよい。改変されたトランスフェリンの内部領域としては、これらに限定されないが、ループ領域、鉄結合部位、ヒンジ領域、ビカーボネート結合部位、または、受容体結合ドメインが挙げられる。
【0118】
改変されたトランスフェリン分子のタンパク質配列内に、多数のループまたはターンが存在しており、これらはジスルフィド結合によって安定化されている。これらのループは、治療活性を有するペプチド、好ましくは抗体の可変領域の挿入または内部融合、特に、機能的なタンパク質または治療用タンパク質、好ましくは抗体の可変領域となるのに二次構造を必要とするものに有用であり、特異的な生物活性を有する改変されたトランスフェリン分子を生成させることができる。
【0119】
リガンド、例えば抗体の可変領域、好ましくはCDRが、Tf分子の少なくとも1つのループに挿入されるか、またはそれと置き換えられる場合、挿入は、Tfの他の領域に加えて、いずれかの表面に露出したループ領域内に作製してもよい。例えば、挿入は、Tfアミノ酸32〜33、74〜75、256〜257、279〜280および288〜289を含むループ内に作製してもよい。上記のAli等を参照。これまでに説明したように、挿入は、Tfの他の領域、例えば以下でより詳細に説明されるような鉄およびビカーボネート結合のための部位、ヒンジ領域、および受容体結合ドメイン内に作製してもよい。また、タンパク質またはペプチドを挿入するための改変/置き換えを受けやすいTfタンパク質配列中のループを、ランダムペプチドインサートのスクリーニング可能なライブラリーを開発するために用いてもよい。ペプチドライブラリーを生成させるための核酸インサートを生産するために、Tfドメインにクローニングしたり、および/またはTfの末端に融合したりする前に利用可能なファージおよび細菌の提示系などのあらゆる手法が使用可能である。
【0120】
TfのN末端は遊離であり、本融合タンパク質の本体から外を向いている。トランスフェリンのN末端における一つまたは複数のリガンドの融合体は、本発明の一実施態様である。このような融合体は、リンカー領域、例えば、ただしこれらに限定されないが、リガンドをTfから離すための、ポリグリシンストレッチ、またはPEAPTDリンカー(配列番号18)を含んでいてもよい。
【0121】
TfのC末端の外観は、埋め込まれているか、または部分的に埋め込まれており、C末端からの6個のアミノ酸のジスルフィド結合で固定されているように見える。ヒトTfにおいて、C末端のアミノ酸はプロリンであり、このプロリンは、配向している方法に応じて、融合タンパク質から外に向いているか、または分子の本体の内部に向いているかのいずれかと予想される。本発明のいくつかの実施態様において、C末端にリンカーまたはスペーサー成分を用いてもよい。またN末端の付近にプロリンが存在していてもよい。本発明の一側面において、Nおよび/またはC末端のプロリンは、改変されていてもよいし、またはその他のアミノ酸で置換されていてもよい。本発明のその他の側面において、C末端のジスルフィド結合を除去して、C末端を開放してもよい。
【0122】
軸成分
本発明の軸成分は、そのN末端で、トランスフェリン成分またはリガンドに融合され、そのC末端ではアンカー成分と場合により融合されていてもよい。酵母細胞で発現させる場合、軸成分のC末端は、細胞内、例えば細胞壁内に位置する。本発明の一実施態様において、軸成分は、細胞壁連結要素として作用し、本融合タンパク質を、共有結合または非共有結合によって酵母細胞の細胞壁に結合させる。
【0123】
本発明の軸成分は、ロッド様またはブラシ様のコンホメーションを有する。このタイプのコンホメーションは、中度〜高度にグリコシル化されたペプチドに典型的である。本発明の軸成分は、N−グリカンまたはO−グリカンを含む。米国特許第6,114,147号を参照(これは参照によりその全体として本明細書に組み込まれる)。N−グリカンよりもO−グリカンの存在が好ましく、これはなぜなら、O−グリカンは、N−グリカンよりも多く、伸長したロッド様のコンホメーションを軸成分に取らせることが可能であるためである。また、軸成分は、中程度〜高度にグリコシル化されたセリンおよびスレオニングリコシル化部位を含んでいてもよい。
【0124】
本発明の融合タンパク質の軸成分は、中程度〜高度のパーセンテージのセリンまたはスレオニン残基を含む。例えば、本発明は、少なくとも約5%またはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約10%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約20%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約30%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約40%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約50%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約60%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約70%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約80%もしくはそれ以上のセリン、および/もしくは、スレオニン残基、または少なくとも約90%もしくはそれ以上のセリンおよび/もしくはスレオニン残基を有する軸成分を含む。本発明の一実施態様において、軸成分は、約20〜30%のセリンおよび/もしくはスレオニン残基、約20〜40%のセリンおよび/もしくはスレオニン残基、約30〜40%のセリンおよび/もしくはスレオニン残基、約20〜50%のセリンおよび/もしくはスレオニン残基、約30〜50%のセリンおよび/もしくはスレオニン残基、約20〜60セリンおよび/もしくはスレオニン残基、または約30〜60%のセリンおよび/もしくはスレオニン残基を含む。
【0125】
軸成分は、少なくとも約5質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約10質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約20質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約30質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約40質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約50質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約60質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約70質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約80質量%もしくはそれ以上のN−もしくはO−グリカン、または少なくとも約90質量%もしくはそれ以上のN−もしくはO−グリカンを含んでいてもよい。本発明の一実施態様において、軸成分は、約20〜30質量%のO−グリカン、約20〜40質量%のO−グリカン、約30〜40質量%のO−グリカン、約20〜50質量%のO−グリカン、約30〜50質量%のO−グリカン、約20〜60質量%のO−グリカン、または約30〜60%のO−グリカンを含む。その他の実施態様において、グリカン、特にO−グリカンの存在は、軸成分と、細胞壁のタンパク質に存在するβ−グルカンとの架橋を可能にする。そのようなものとして、本発明の軸成分は、細胞壁連結要素として機能することができる。
【0126】
軸成分は、ムチンタンパク質またはムチンタンパク質の部分、すなわちMUC型タンパク質のメンバーを含んでいてもよい。MUC型のムチンとは、高度にグリコシル化された構造的に関連した分子のファミリーであり、気道、胃腸管および生殖管の上皮で発現し、例えばMUC1(GenBank登録番号AF125525)、MUC2(GenBank登録番号L21998)、MUC3(GenBank登録番号AF113616)、MUC4(GenBank登録番号AJ000281)、MUC5AC(GenBank登録番号U83139)、MUC5B(GenBank登録番号AJ001402)、MUC6(GenBank登録番号U97698)、MUC7(GenBank登録番号L13283)、MUC8(GenBank登録番号U14383)、MUC9(GenBank登録番号AW271430)が挙げられる。本発明の一実施態様において、軸成分は、hMUC1またはhMUC1タンパク質の部分、例えば配列番号70の核酸によってコードされた配列番号71、さらに配列番号5の核酸によってコードされたポリペプチドを含む。本発明のその他の実施態様において、軸成分は、hMUC3またはhMUC3タンパク質の部分を含む。例えば、本発明は、配列番号68の核酸によってコードされる配列番号69のhMUC3の軸を含む。また本発明の融合タンパク質は、ムチンタンパク質およびそれらの部分の類似体および誘導体のような変異体を含む軸も包含する。
【0127】
また本発明の軸成分は、ムチン以外のグリコシル化タンパク質からも誘導することができ、このようなタンパク質としては、これらに限定されないが、AGA1(例えば、配列番号72の核酸配列によってコードされた配列番号73)、MAdCAM−1、GlyCAM−1、CD34;E−セレクチン、P−セレクチンまたはL−セレクチンからのコンセンサス反復;またはウイルス糖タンパク質スパイク(例えば、インフルエンザ、単純ヘルペス、ヒト免疫不全またはタバコモザイクウイルス)、ならびにそれらの変異体およびフラグメントが挙げられる。WO01/46698、Girard等(1995)Immunity 2:113〜123、およびVan Kinken等(1998)Anal.Biochem.265:103〜116を参照(これらはいずれも、参照によりその全体として本明細書に組み込まれる)。本発明は、2種またはそれ以上のグリコシル化タンパク質またはそれらのフラグメントの反復、ならびに、2種またはそれ以上のタイプのグリコシル化タンパク質の組み合わせを含む。
【0128】
本発明のその他の実施態様において、軸は、1またはそれ以上の遊離のシステイン残基を含むように遺伝子操作される。1またはそれ以上の遊離のシステイン残基は、酵母細胞の細胞壁中のタンパク質の遊離のシステイン残基とジスルフィド結合を形成することができる。細胞壁中の1またはそれ以上のジスルフィド結合の形成は、細胞壁結合要素として機能することができる軸成分を遺伝子操作するのに使用可能なその他の方法の代表である。
【0129】
本発明の軸成分は、酵母細胞の全細胞壁を橋わたすのに十分な長さを有していなければならない。好ましくは、軸成分のN末端を、細胞壁の外側に位置させることであり、最も好ましくは、ロッド様の立体配置で、酵母細胞から外に向かって伸長させて、トランスフェリン成分およびリガンドと、宿主の酵母細胞との間の立体障害を低減させることである。軸成分は、長さが、少なくとも約25個のアミノ酸、少なくとも約50個のアミノ酸、少なくとも約75個のアミノ酸、少なくとも約100個のアミノ酸、少なくとも約125個のアミノ酸、少なくとも約150個のアミノ酸、少なくとも約175個のアミノ酸、少なくとも約200個のアミノ酸、少なくとも約225個のアミノ酸、少なくとも約250個のアミノ酸、少なくとも約275個のアミノ酸、少なくとも約300個のアミノ酸、少なくとも約325個のアミノ酸、少なくとも約350個のアミノ酸、少なくとも約375個のアミノ酸、少なくとも約400個のアミノ酸、少なくとも約425個のアミノ酸、少なくとも約450個のアミノ酸、少なくとも約475個のアミノ酸長さが、少なくとも約500個のアミノ酸長さが、少なくとも約525個のアミノ酸長さが、少なくとも約550個のアミノ酸長さが、少なくとも約575個のアミノ酸長さが、少なくとも約600個のアミノ酸長さが、少なくとも約625個のアミノ酸、または長さが、少なくとも約650個のアミノ酸が必要である。一実施態様において、軸成分は、長さが約500個のアミノ酸である。その他の実施態様において、軸成分は、長さが約300〜600個のアミノ酸である。
【0130】
アンカー成分
本発明の融合タンパク質の任意のアンカー成分は、本融合タンパク質を宿主細胞表層または基質表面に物理的に連結する本融合タンパク質の部分である。例えば、アンカー成分は、本融合タンパク質を酵母細胞膜または酵母細胞壁に連結または固定するすることができる。アンカーが本融合タンパク質を酵母細胞壁に連結する場合、アンカーは、細胞壁結合部材である。
【0131】
アンカー成分は、融合タンパク質を、一時的に酵母細胞壁または細胞膜に繋ぐすることができる。本発明の一実施態様において、アンカー成分は、融合タンパク質を、一時的に酵母細胞壁または細胞膜に繋いで、軸成分が細胞壁に共有結合または非共有結合で結合する機会を提供する。例えば、酵母細胞におけるアンカーの一時的な係留によって、軸成分からのO−グリカンと、細胞壁のβ−グルカンとを架橋させることが可能になる。
【0132】
本発明の一実施態様において、アンカー成分は、微生物、好ましくは下等真核生物、例えば酵母およびカビの細胞膜または細胞壁に固着する。この成分は、プロリンのようなアミノ酸で細胞膜または細胞壁にC末端を固定する長いC末端を有していてもよい(Kok(1990)FEMS Microbiology Reviews 87:15〜42)。
【0133】
アンカー成分は、グリコシルホスファチジルイノシトール(GPI)アンカーの使用によって細胞に固定することができる。Conzelmann等,EMBO 9:653〜661、ならびにLipkeおよびOvalle(1998)J.Bacteriol.180:3735〜3740を参照。GPIシグナル配列ペプチド、例えば本明細書で開示されたGPIシグナルペプチドは、GPIを本融合タンパク質のC末端に付着させるためにシグナル伝達する。GPIシグナルそれ自体は、以下の3つのドメイン:GPI付着部位(そのω部位)ω部位下流の第一および第二のアミノ酸を含む領域、5〜10個のアミノ酸のスペーサー、ならびに10〜15個のアミノ酸の疎水性ストレッチを有する。GPIシグナルを含むタンパク質は、ω部位で切断され、生じたタンパク質のカルボキシ末端は、GPI成分に共有結合する。この反応は、小胞体内で起こる。GPI成分によって膜と結合する、GPIが付着したタンパク質は、続いて細胞表層に輸送され、タンパク質が短いω−マイナス領域に塩基性残基(Rおよび/またはK)を含む場合、GPIで固定されたタンパク質として原形質膜に留まる。ω−4/−5部位にV、IまたはL、およびω−2部位にYまたはNを有するGPIと結合したタンパク質は、細胞膜中に組み込まれる。Hamada等(1999)J.Bacteriol.181:3886〜3889;Nuoffer等(1993)J.Biol.Chem.268:10558〜10563;De Nobel等(1994)Trends Cell Biol.4:42〜45;Hamada等(1998)Mol.Gen.Genet.258:53〜59;およびVan Der Vaart等(1998)Biotechnol.Genet.Eng.Rev.15:387〜411を参照。
【0134】
本発明の一実施態様において、酵母GPI YIR019Cは、トランスフェリン融合タンパク質のアンカー成分を提供するのに用いられる。図2は、GPI YIR019Cの図解を提供する。アミノ酸配列(配列番号15)におけるω部位はグリシンであり、そのどちらかの側にスペースを有するものとして図示されている。スペースは、ω部位のいずれかの側にあるスペーサー領域の指標である。太字で示したIおよびYアミノ酸は、それぞれω−5/−4および−2部位である。
【0135】
数種のサッカロミセスのアンカー成分が当技術分野で既知であり、本発明の融合タンパク質を構築するのに用いることができる。酵母のGPIシグナルタンパク質のその他の例としては、これらに限定されないが、YDR534C、YNL327W、YOR214C、YDR134C、YPL130、YOR009W、YER150W、YDR077W、YOR383C、YJR151C、YJR004、YJL078C、YLR110CおよびYNL300Wが挙げられる。さらに、他の生物由来のGPIシグナルタンパク質も用いることができ、例えばカンジダ・グラブラータ(Canidida glabrata)のEPA1のGPI、カンジダ・アルビカンス(Candida albicans)のHwplp、またはトリパノソーマ−ブルセイ(Trypanosoma brucei)のVSGである。
【0136】
本発明の一実施態様において、アンカー成分は、哺乳動物の成分、またはそれらの誘導体もしくはフラグメントである。本発明のその他の実施態様において、GPIシグナルペプチドは、哺乳動物のGPIシグナルタンパク質である。例えば、本発明は、ヒトMDPGPIシグナルタンパク質の誘導体を含み、例えば表1で開示されたものが挙げられる(実施例5を参照)。
【0137】
本発明はまた、1またはそれ以上の未結合のシステイン残基を含むアンカー成分を含む融合タンパク質も包含する。このようなシステイン残基は、細胞壁中のタンパク質のシステイン残基とジスルフィド結合を形成することによって、本融合タンパク質と細胞とを結びつけるように作用することができる。
【0138】
本発明は、アンカー成分として膜貫通ドメイン(TMD)を含む融合タンパク質を包含する。本発明の一実施態様において、TMDは、1回膜貫通I型またはII型膜タンパク質の領域である。例えば、本発明は、これらに限定されないが、FUS1の残基70〜98を含む。
【0139】
本発明のその他の実施態様において、TMDは、マルチスパン膜タンパク質の数種の膜貫通領域のうち1つまたはそれ以上を含む。本発明の一実施態様において、TMDは、マルチスパン膜タンパク質の疎水性領域であり、約10〜60個のアミノ酸、約15〜60個のアミノ酸、約20〜60個のアミノ酸、約30〜60個のアミノ酸、または約25〜50個のアミノ酸を含む。例えば、本発明は、サッカロミセスのSTE6由来の1種またはそれ以上のTMDを含み、このようなTMDは、これらに限定されないが、残基25〜30、73〜100、171〜198、249〜277、714〜742、761〜789、838〜858、864〜884、940〜967および979〜1000(サッカロミセスゲノムデータベースアノテーション(Saccharomyces Genome Database annotation))からなる群より選択される。
【0140】
その他の実施態様において、アンカー成分は、トランスフェリン融合タンパク質を、マイクロアレイのような固体基板に固定するのに用いられる。アンカー成分は、好ましくは、短いエピトープタグであり(すなわち、抗体、典型的にはモノクローナル抗体によって認識される配列)、例えば、ポリヒスチジン、SEAPまたはM1およびM2フラグである。Bush等(1991)J.Biol.Chem.266:13811〜13814,Berger等(1988)Gene 66:1〜10,米国特許第5,011,912号、米国特許第4,851,341号、米国特許第4,703,004号、および米国特許第4,782,137号を参照、これらは全て、それら全体として参照により開示に組み込まれる。一実施態様において、軸ドメインは、抗ムチン抗体のような抗軸配列抗体によって基質に結びつけられる。
【0141】
アルブミン
本発明はまた、標的にリガンドを「提示する」ためにトランスフェリン以外のタンパク質またはタンパク質フラグメントを用いる融合タンパク質も含む。適切なタンパク質は、可溶性であり、長さが少なくとも約50個のアミノ酸であるか、またはそれより長いタンパク質である。本発明の一実施態様において、このようなタンパク質またはタンパク質フラグメントは、トランスフェリンの二次構造に類似した二次構造を含む。
【0142】
上記タンパク質またはそれらのフラグメントが、本融合タンパク質の軸部分から切断されたときに、リガンドの半減期を増加させることができ、治療剤として用いることができることが好ましい。例えば、本発明は、アルブミン成分、軸成分、および細胞壁連結要素を含む融合タンパク質の使用を想定する。本融合タンパク質のアルブミンおよびリガンド部分が、融合タンパク質のそれ以外の部分から切断され、治療剤としてのリガンドが必要な患者に投与される場合、アルブミン成分はリガンド(すなわち治療剤)に増加した血清中半減期を付与することができる。
【0143】
アルブミン成分を含む融合タンパク質は、アルブミンタンパク質、アルブミン変異体またはそれらのフラグメントを含んでいてもよい。一実施態様において、アルブミンタンパク質は、配列番号66の核酸配列によってコードされる配列番号67のアミノ酸配列を含む。本発明は、当技術分野で既知のアルブミンの改変を含む。
【0144】
核酸
本発明は、核酸分子も提供する。これらは、リガンド成分に共有結合で連結または結合したトランスフェリンタンパク質、またはトランスフェリンタンパク質の部分を含む改変されたTf融合タンパク質をコードする。本融合タンパク質は、リンカー領域、例えば、約50、40、30、20または10個のアミノ酸残基未満のリンカーをさらに含んでいてもよい。リンカーは、トランスフェリンタンパク質またはそれらの部分と、リガンド部分に、またはそれらの間に共有結合で連結させることができる。本発明の核酸分子は、精製されてもよいし、または精製されていなくてもよい。
【0145】
また、核酸分子を複製するための、およびコードされた融合タンパク質を発現させるための宿主細胞およびベクターも提供される。あらゆるベクターまたは宿主細胞が使用可能であり、これらは原核性でもよいし、または真核性でもよいが、真核性の発現系、特に酵母発現系が好ましい場合がある。このような目的のための多くのベクターおよび宿主細胞が当技術分野で既知である。所望の適用に適したセットを選択することは十分に当分野の技術の範囲内である。
【0146】
トランスフェリン、トランスフェリンの部分、および、関心対象の治療用タンパク質をコードするDNA配列を、当技術分野で既知の様々なゲノムまたはcDNAライブラリーからクローニングすることができる。プローブに基づく方法を用いてこのようなDNA配列を単離する技術は、従来技術であり、当業者によく知られている。このようなDNA配列を単離するためのプローブは、公開されたDNAまたはタンパク質配列をベースにしたものが可能である(例えば、Baldwin,G.S.(1993)Comparison of Transferrin Sequences from Different Species.Comp.Biochem.Physiol.106B/1:203〜218を参照、そこで引用された全ての文献は、参照によりその全体として本明細書に組み込まれる)。あるいは、Mullis等(米国特許第4,683,195号)およびMullis(米国特許第4,683,202号)に記載されたポリメラーゼ連鎖反応(PCR)法(参照により本明細書に組み込まれる)を用いてもよい。このようなDNA配列を単離するためのライブラリーの選択およびプローブの選択は、当技術分野における通常の技術レベルの範囲内である。
【0147】
当技術分野で既知のように、2種のポリヌクレオチドまたはポリペプチド間の「類似性」は、ヌクレオチドまたはアミノ酸配列、および一種のポリヌクレオチドまたはポリペプチドの、第二のポリヌクレオチドまたはポリペプチドの配列に対する、保存されたヌクレオチドまたはアミノ酸置換を比較することによって決定される。「同一性」も当技術分野で既知であり、これは、このような配列の2つの鎖間におけるマッチングの同一性によって決定されるような、2種のポリペプチドまたは2種のポリヌクレオチド配列間の配列の関連の程度を意味する。同一性および類似性はいずれも容易に計算することができる(Computational Molecular Biology,Lesk,A.M編,オックスフォード・ユニバーシティ・プレス(Oxford University Press),ニューヨーク,1988;Biocomputing:Informatics and Genome Projects,Smith,D.W.編,アカデミックプレス,ニューヨーク,1993;Computer Analysis of Sequence Data,Part I,Griffin,A.M.およびGriffin,H.G.編,ヒューマナ・プレス(Humana Press),ニュージャージー州,1994;Sequence Analysis in Molecular Biology,von Heinje,G.,アカデミックプレス、1987;およびSequence Analysis Primer,Gribskov,M.およびDevereux,J.編集,Mストックトンプレス(M Stockton Press),ニューヨーク,1991)。
【0148】
2種のポリヌクレオチドまたはポリペプチド配列間の同一性および類似性を測定する多数の方法があるが、用語「同一性」および「類似性」は当技術分野ではよく知られている(Sequence Analysis in Molecular Biology,von Heinje,G.,アカデミックプレス,1987;Sequence Analysis Primer,Gribskov,M.and Devereux,J.編,Mストックトンプレス,ニューヨーク,1991;およびCarillo,H.およびLipman,D.,SIAM J.Applied Math.,48:1073(1988)。2種の配列間の同一性または類似性を決定するのに一般的に用いられる方法としては、これらに限定されないが、Guide to Huge Computers,Martin J.Bishop編,アカデミックプレス,サンディエゴ,1994、およびCarillo,H.およびLipman,D.,SIAM J.Applied Math.48:1073(1988)に記載されたものが挙げられる。
【0149】
同一性を決定する好ましい方法は、試験される2種の配列間の最も大きいマッチングが得られるように設計される。同一性および類似性を決定する方法は、コンピュータープログラムで体系化されている。2種の配列間の同一性および類似性を決定する好ましいコンピュータープログラム法としては、これらに限定されないが、GCGプログラムパッケージ(Devereux等,Nucl.Acid Res.12(1):387(1984))、BLASTP、BLASTN、FASTA(Atschul等,J.Mol.Biol.215:403(1990))が挙げられる。上述した類似性または同一性の程度は、2種の配列間の同一性の程度として決定され、第二の配列からの第一の配列の起源を示すことが多い。2種の核酸配列間の同一性の程度は、当技術分野で既知のコンピュータープログラムによって決定してもよく、例えばGCGプログラムパッケージに備えられているGAPである(NeedlemanおよびWunsch J.Mol.Biol.48:443〜453(1970))。本発明に関する2種の核酸配列間の同一性の程度を決定する目的で、ギャップ(GAP)は、以下の設定:5.0のギャップクリエーションペナルティー、0.3のギャップ伸長ペナルティーで用いられる。
【0150】
コドンの最適化
天然型のDNA配列によってコードされたポリペプチドと同一なアミノ酸配列を有するポリペプチドが生成されるが、遺伝子コードの縮重によって、トランスフェリンタンパク質および/または関心対象の治療用タンパク質のヌクレオチドの配列の変異が可能になる。「コドン最適化」として知られる手順(米国特許第5,547,871号に記載されており、これはこの参照によりその全体として本明細書に組み込まれる)では、このような変更されたDNA配列を設計する手段を用いてコドン最適化が提供される。コドン最適化遺伝子の設計は、生物におけるコドンの使用頻度、最隣接の頻度、RNA安定性、二次構造形成の可能性、合成経路、およびその遺伝子の目的とするその後のDNA操作などの様々な因子を考慮すべきである。特に、利用可能な方法を用いて、酵母発現系が用いられる場合、酵母によって最も容易に認識されるコドンを用いて所定の融合タンパク質をコードするコドンを変更することができる。
【0151】
遺伝子コードの縮重によって、同じアミノ酸配列を多くの異なる方法でコードし、翻訳させることを可能にする。例えば、ロイシン、セリンおよびアルギニンはそれぞれ6種の異なるコドンによってコードされるが、バリン、プロリン、スレオニン、アラニンおよびグリシンはそれぞれ4種の異なるコドンによってコードされる。しかしながら、このような同義的なコドンの使用頻度は、真核生物および原核生物の間でゲノム毎に異なる。例えば、哺乳動物間での同義的なコドンの選択パターンは、極めて類似しているが、進化的に遠い生物、例えば酵母(S.セレビジエ)、細菌(例えばE.コリ)、および昆虫(例えば、D.melanogaster)は、ゲノムコドンの使用頻度において明らかに異なるパターンを示す(Grantham,R.等,Nucl.Acid Res.,8,49〜62(1980);Grantham,R.等,Nucl.Acid Res.,9,43〜74(1981);Maroyama,T.等,Nucl.Acid Res.,14,151〜197(1986);Aota,S.等,Nucl.Acid
Res.,16,315〜402(1988);Wada,K.等,Nucl.Acid Res.,19 Supp.,1981〜1985(1991);Kurland,C.G.,FEBS Lett.,285,165〜169(1991))。このようなコドンの選択パターンの差は、ペプチドの伸長速度を調節することによる個々の遺伝子の総体的な発現レベルに寄与するようである。(Kurland,C.G.,FEBS Lett.,285,165〜169(1991);Pedersen,S.,EMBO J.,3,2895〜2898(1984);Sorensen,M.A.,J.Mol.Biol.,207,365〜377(1989);Randall,L.L.等,Eur.J.Biochem.,107,375〜379(1980);Curran,J.F.、およびYarus,M,J.Mol.Biol.,209,65〜77(1989);Varenne,S.等,J.Mol.Biol.,180,549〜576(1984),Varenne,S.等,J.Mol,Biol.,180,549〜576(1984);Garel,J.−P.,J.Theor.Biol.,43,211〜225(1974);Ikemura,T.,J.Mol.Biol.,146,1〜21(1981);Ikemura,T.,J.Mol.Biol.,151,389〜409(1981))。
【0152】
合成遺伝子のコドンの使用頻度は、組換えタンパク質発現に用いることを目的とする細胞/生物の(具体的には酵母種の)正確な(または可能な限り密接に関連した)ゲノムから誘導された核遺伝子のコドンの使用を反映することが必要である。上記で考察したように、一実施態様において、ヒトTf配列は、治療用タンパク質のヌクレオチド配列になるように、酵母発現に関して本明細書で説明されているような改変の前に、または改変の後にコドンが最適化される。
【0153】
ベクター
本発明で使用するための発現単位は、一般的に、5’から3’方向に機能するように連結した以下の構成要素:転写プロモーター、分泌シグナル配列、治療用タンパク質または関心対象のペプチドをコードするDNA配列に結合したトランスフェリンタンパク質またはトランスフェリンタンパク質の部分を含む改変されたTf融合タンパク質をコードするDNA配列、および転写ターミネーターを含む。上記で考察したように、本発明のベクターでは、治療用タンパク質またはペプチドがTf部分に融合した配置、またはTf部分内の配置のいずれでも使用可能である。適切なプロモーター、シグナル配列、およびターミネーターの選択は、選択された宿主細胞に応じて決定され、当業者には明白であろうが、以下でより具体的に考察する。
【0154】
本発明で使用するのに適した酵母ベクターは、米国特許第6,291,212号に記載されており、例えば、YRp7(Struhl等,Proc.Natl.Acad.Sci.USA 76:1035〜1039,1978)、YEp13(Broach等,Gene 8:121〜133,1979)、pJDB249およびpJDB219(Beggs,Nature 275:104〜108,1978)、pPPC0005、pSeCHSA、pScNHSA、pC4、およびそれらの誘導体が挙げられる。また、有用な酵母のプラスミドベクターとしては、pRS403−406、pRS413−416、およびストラタジーン・クローニング・システム(Stratagene Cloning Systems,ラホヤ,カリフォルニア州92037,米国)より入手可能なピチア属のベクターも挙げられる。プラスミドpRS403、pRS404、pRS405、およびpRS406は、酵母組込み型プラスミド(YIps)であり、酵母選択マーカーHIS3、TRP1、LEU2およびURA3が組み込まれている。プラスミドpRS413−41.6は、酵母セントロメアプラスミド(YCps)である。
【0155】
このようなベクターは、一般的に、形質転換株の選択を可能にする表現型の分析が存在する優性表現型を示す多数の遺伝子のうちの1つとなり得る選択マーカーを含む。好ましい選択マーカーは、宿主細胞の栄養要求性を補い、抗生物質耐性を提供するか、または、細胞の特異的な炭素源の利用を可能にするマーカーであり、例えばLEU2(Broach等,前記の箇所),URA3(Botstein等,Gene 8:17,1979)、HIS3(Struhl等,前記の箇所)、またはPOT1(KawasakiおよびBell,EP171,142)が挙げられる。その他の適切な選択マーカーとしては、酵母細胞にクロラムフェニコール耐性を付与するCAT遺伝子が挙げられる。酵母に使用するのに好ましいプロモーターとしては、酵母の解糖系遺伝子由来のプロモーター(Hitzeman等,J Biol.Chem.225:12073〜12080,1980;AlberおよびKawasaki,J.Mol.Appl.Genet.1:419〜434,1982;Kawasaki,米国特許第4,599,311号)、またはアルコールデヒドロゲナーゼ遺伝子由来のプロモーター(Young等のGenetic Engineering of Microorganisms for Chemicals,Hollaender等(編),p.355,Plenum,N.Y.,1982;Ammerer,Meth.Enzymol.101:192〜201,1983)が挙げられる。これに関連して、使用可能なプロモーターは、TPI1プロモーター(Kawasaki,米国特許第4,599,311号)およびADH2−4cである(米国特許第6,291,212号のプロモーター(Russell等,Nature 304:652〜654,1983)を参照。また、発現単位は、転写ターミネーターを含んでいてもよい。一つの転写ターミネーターは、TPI1ターミネーター(AlberおよびKawasaki,前記の個所)である。
【0156】
酵母に加えて、本発明の改変された融合タンパク質は、糸状菌で、例えばアスペルギルス属の菌株で発現させることができる。有用なプロモーターの例としては、アスペルギルス・ニダランス(Aspergillus nidulans)の解糖系遺伝子から誘導されたもの、例えばadh3プロモーター(McKnight等,EMBO J.4:2093〜2099,1985)、およびtpiAプロモーターが挙げられる。適切なターミネーターの例は、adh3ターミネーター(McKnight等,前記の個所)である。このような成分を利用する発現単位は、例えば、アスペルギルスの染色体DNAに挿入できるベクターにクローニングしてもよい。
【0157】
本発明の実施に使用するための哺乳動物の発現ベクターは、改変されたTf融合タンパク質の転写を指示することができるプロモーターを含む。好ましいプロモーターとしては、ウイルスプロモーターおよび細胞性のプロモーターが挙げられる。好ましいウイルスプロモーターとしては、アデノウイルス2由来の主要後期プロモーター(KaufmanおよびSharp,Mol.Cell.Biol.2:1304〜13199,1982)、およびSV40プロモーター(Subramani等,Mol.Cell.Biol.1:854〜864,1981)が挙げられる。好ましい細胞性のプロモーターとしては、マウスメタロチオネイン1プロモーター(Palmiter等,Science 222:809〜814,1983)、およびマウスV6(米国特許第6,291,212号を参照)プロモーター(Grant等,Nuc.Acids Res.15:5496,1987)が挙げられる。このようなプロモーターの一つは、マウスVH(米国特許第6,291,212号を参照)プロモーター(Loh等,前記の個所)である。また、このような発現ベクターは、プロモーターの下流、且つトランスフェリン融合タンパク質をコードするDNA配列の上流に位置する一連のRNAスプライス部位を含んでいてもよい。好ましいRNAスプライス部位は、アデノウイルスおよび/または免疫グロブリン遺伝子から得られたものでもよい。
【0158】
また発現ベクターには、関心対象のコード配列の下流に位置するポリアデニル化シグナルも含まれる。ポリアデニル化シグナルとしては、SV40由来の初期または後期ポリアデニル化シグナル(KaufmanおよびSharp,前記の個所)、アデノウイルス5E1B領域由来のポリアデニル化シグナル、およびヒト成長ホルモン遺伝子ターミネーター(DeNoto等,Nucl.Acid Res.9:3719〜3730,1981)が挙げられる。このようなポリアデニル化シグナルの一つは、VH(米国特許第6,291,212号を参照)遺伝子ターミネーター(Loh等,本明細書前記)である。発現ベクターは、非コードウイルスリーダー配列、例えばプロモーターとRNAスプライス部位との間に位置するアデノウイルス2のトリパータイトリーダーを含んでいてもよい。また、好ましいベクターは、エンハンサー配列、例えばSV40エンハンサー、およびマウス:(米国特許第6,291,212号を参照)エンハンサー(Gillies,Cell 33:717〜728,1983)。また、発現ベクターは、アデノウイルスVA RNAをコードする配列を含んでいてもよい。
【0159】
形質転換
菌類を形質転換する技術は、文献においてよく知られており、例えばBeggs(前記の個所)、Hinnen等(Proc.Natl.Acad.Sci.USA 75:1929〜1933,1978),Yelton等,(Proc.Natl.Acad.Sci.USA 81:1740〜1747,1984)、およびRussell(Nature 301:167〜169,1983)によって説明されている。宿主細胞の遺伝子型は、一般的に、発現ベクターに存在する選択マーカーによって補足される遺伝的欠陥を含む。特定の宿主および選択マーカーの選択は、十分に当分野における通常の技術のレベルの範囲内である。
【0160】
本発明の改変されたTf融合タンパク質を含むクローニングしたDNA配列を培養した哺乳動物細胞に導入されてもよく、これは、例えば、リン酸カルシウム介在トランスフェクション(Wigler等,Cell 14:725,1978;CorsaroおよびPearson,Somatic Cell Genetics 7:603,1981;GrahamおよびVan der Eb,Virology 52:456,1973)によってなされる。また、クローニングしたDNA配列を哺乳動物細胞に導入するその他の技術、例えばエレクトロポレーション(Neumann等,EMBO J.1:841〜845,1982)、またはリポフェクションも使用可能である。クローニングしたDNAを組み込んでいる細胞を同定するために、一般的に細胞に、関心対象の遺伝子またはcDNAと共に選択マーカーが導入される。培養した哺乳動物細胞に使用するのに好ましい選択マーカーとしては、ネオマイシン、ハイグロマイシンおよびメトトレキセートのような薬物に対する耐性を付与する遺伝子が挙げられる。選択マーカーは、増幅可能な選択マーカーであり得る。増幅可能な選択マーカーの一種は、DHFR遺伝子である。増幅可能なマーカーの一種は、DHFRr(米国特許第6,291,212号を参照)cDNA(SimonsenおよびLevinson,Proc.Natl.Acad.Sci.USA 80:2495〜2499,1983)である。選択マーカーは、Thillyによって総論されており(Mammalian Cell Technology,Butterworth Publishers,ストーンハム,マサチューセッツ州)、選択マーカーの選択は、十分当業界における通常の技術レベルの範囲内である。
【0161】
宿主細胞
本発明はまた、本発明の改変されたトランスフェリン融合タンパク質が発現されるように形質転換した細胞、好ましくは酵母細胞も含む。形質転換宿主細胞それ自体に加えて、本発明はまた、これら細胞の培養物、好ましくはモノクローナル(クローン的に均一な)培養物、または栄養培地中でモノクローナル培養物から誘導された培養物も含む。上記ポリペプチドが分泌されると、上記ポリペプチドは、細胞と共に培地に含まれるか、または細胞をろ過もしくは遠心分離で除去している場合細胞なしで含まれることになる。
【0162】
本発明を実施する際に使用される宿主細胞としては、外来性DNAで形質転換またはトランスフェクションさせて、培養した哺乳動物、昆虫、真菌性、植物および細菌細胞のような培養物中で増殖させることができる真核細胞、場合によっては原核細胞が挙げられる。
【0163】
真菌細胞、例えば酵母種(例えば、サッカロミセス属spp.、シゾサッカロミセス属spp.、ピチア属spp.)は、本発明の範囲内で宿主細胞としても使用可能である。本発明の実施の際に、本発明のトランスフェリン融合タンパク質を発現するための宿主として有用であると考えられる典型的な酵母の属は、ピチア属(以前はハンセヌラ属として分類された種を含む)、サッカロミセス属、クルイベロミセス属、アスペルギルス属、カンジダ属、トルロプシス属、トルラスポラ属、シゾサッカロミセス属、シテロミセス属、パチソレン属、チゴサッカロミセス属、デバリオミセス属(Debaromyces)、トリコデルマ属、セファロスポリウム属、フミコーラ属、ムコール属、アカパンカビ属、ヤロウイア属、メッシュニコヴィア属、ロドスポリディウム属、ロイコスポリジウム属、ボトリオアスカス属、スポリディオボラス属、エンドマイコプシス属などである。サッカロミセス属spp.の例は、S.セレビジエ、S.イタリカス(S.italicus)、およびS.ルキシー(S.rouxii)である。クルイベロミセス属spp.の例は、K.ラクティス(K.lactis)、およびK.マルキシアヌス(K.marxianus)である。適切な種は、T.デルブルッキー(T.delbrueckii)である。ピチア属(ハンセヌラ属)spp.の例は、P.アングスタ(P.angusta)(以前はH.ポリモルファ(H.polymorpha))P.アノマーラ(P.anomala)(以前はH.アノマーラ(H.anomala))、およびP.パストリス(P.pastoris)である。
【0164】
本発明のTf融合タンパク質を生産するのに特に有用な宿主細胞は、メタノール要求性のピチア・パストリス(Steinlein等(1995)Protein Express,Purif.6:619〜624)である。ピチア・パストリスのアルコールオキシダーゼプロモーターが単離され、クローニングされてから、ピチア・パストリスは、外来タンパク質の生産において優れた宿主となるように開発されている;1985年に、その形質転換がまず報告された。P.パストリスは、グルコースの非存在下で、炭素源としてメタノールを利用することができる。P.パストリス発現系は、アルコールオキシダーゼの発現をコードする遺伝子を制御するメタノール誘導性のアルコールオキシダーゼ(AOX1)プロモーターを使用でき、この酵素は、メタノール代謝における第一工程を触媒する。このプロモーターは特徴付けが行われており、一連のP.パストリス発現ベクターに組み込まれている。P.パストリスで生産されたタンパク質は、通常は正確に折り畳まれて培地に分泌されるため、遺伝子操作されたP.パストリスの発酵は、E.コリ発現系の優れた代替物を提供する。この系を用いて、破傷風毒素のフラグメント、百日咳菌(Bordatella pertussis)のパータクチン、ヒト血清アルブミン、およびリゾチームなどの多数のタンパク質が生産されている。
【0165】
F.オキシスポラム(F.oxysporum)の形質転換は、例えば、Malardier等(1989)Gene 78:147〜156に記載されているように行うことができる。
【0166】
その他の好ましい宿主は、酵母サッカロミセス・セレビジエ(Saccharomyces cerevisiae)株である。一実施態様において、糖タンパク質のアスパラギン連結型のグリコシル化に必要な遺伝子の遺伝子欠損を含む酵母細胞、またはより詳しくはサッカロミセス・セレビジエ宿主細胞が用いられる。このような欠損を有するS.セレビジエ宿主細胞は、標準的な突然変異および選択の技術を用いて生産することができるが、多くの利用可能な酵母株が、グリコシル化または過剰なマンノシル化を防ぐように、または減少するように改変されている。Ballou等(J.Biol.Chem.255:5986〜5991,1980)は、アスパラギン結合型糖付加に影響を与える遺伝子が欠損したマンノプロテインの生合成突然変異体の単離を記載している。GentzschおよびTanner(Glycobiology 7:481〜486,1997)は、酵母におけるタンパク質のO−グリコシル化の第一工程に関与する酵素をコードする少なくとも6種の遺伝子(PMT1−6)のファミリーを記載している。これらの遺伝子の1種またはそれ以上が欠損した突然変異体は、低いO結合型糖付加および/またはO−グリコシル化の変更された特異性を示す。
【0167】
異種タンパク質の生産を最適化するために、宿主株が、突然変異を有することが好ましい場合があり、このような突然変異としては、タンパク質分解活性の減少が起こるS.セレビジエpep4突然変異(Jones,Genetics 85:23〜33,1977)が挙げられる。多量の本発明のTf融合タンパク質を生産するために、さらなるプロテアーゼコード領域に突然変異を含む宿主株が特に有用である。
【0168】
本発明のDNAコンストラクトを含む宿主細胞は、適切な増殖培地で増殖させる。本明細書で用いられる用語「適切な増殖培地」は、細胞の増殖に必要な栄養素を含む培地を意味する。細胞増殖に必要な栄養素として、炭素源、窒素源、必須アミノ酸、ビタミン、無機質、および増殖因子が含まれていてもよい。増殖培地は、一般的に、DNAコンストラクトを含む細胞に応じて、例えば、薬物選択、またはDNAコンストラクトに存在する、もしくはDNAコンストラクトでコトランスフェクションされた選択マーカーによって補足された必須栄養素の欠失により選択される。酵母細胞は、例えば、化学的に規定された培地で増殖させることが好ましく、このような培地は、炭素源、例えばスクロース、非アミノ酸窒素源、無機塩、ビタミン、および必須アミノ酸の追加物質を含む。培地のpHは、好ましくは、2より大きく8未満のpHに維持され、好ましくはpH5.5〜6.5に維持される。安定なpHを維持する方法は、好ましくは水酸化ナトリウムの添加を介した緩衝化と一定したpH制御を含む。好ましい緩衝剤としては、コハク酸、およびビストリス(シグマ・ケミカル社(Sigma Chemical Co.),セントルイス,ミズーリ州)が挙げられる。アスパラギン結合型糖付加に必要な遺伝子の欠損を有する酵母細胞は、好ましくは、浸透圧安定剤を含む培地中で増殖させる。このような浸透圧安定剤の一つは、0.1M〜1.5Mの濃度、好ましくは0.5Mまたは1.0Mの濃度で培地に添加されたソルビトールである。
【0169】
培養した哺乳動物細胞は、一般的に、市販の血清含有または無血清培地で増殖させる。用いられる特定の細胞系に適した培地の選択は、当分野における通常の技術レベル内である。トランスフェクションされた哺乳動物細胞は、所定期間、典型的には1〜2日間成長させて、関心対象のDNA配列の発現を開始させることができる。続いて、薬物選択を適用して、選択マーカーを安定して発現する細胞増殖について選択する。増幅可能な選択マーカーでトランスフェクションされている細胞に関して、薬物濃度を段階的に増加させて、クローニングした配列のコピー数の増加、従って発現レベルの増加について選択することができる。
【0170】
また、バキュロウイルス/昆虫細胞発現系も、本発明の改変されたTf融合タンパク質を生産するのに使用可能である。BacPAKTMバキュロウイルス発現系(BDバイオサイエンス(BD Biosciences)(クロンテック(Clontech)))は、昆虫宿主細胞中で組換えタンパク質を高レベルで発現する。標的遺伝子は、トランスファーベクターに挿入され、これが昆虫宿主細胞に直線状にしたBacPAK6ウイルスDNAと共にコトランスフェクションされる。BacPAK6DNAは、バキュロウイルスゲノムの必須部分が欠失している。このDNAをこのベクターで組換えると、必須の構成要素が回復し、標的遺伝子はバキュロウイルスゲノムに移される。組換え後、2〜3個のウイルスプラークを採取し、精製し、組換え表現型を検証する。続いて、新たに単離した組換えウイルスを増幅し、これを昆虫細胞培養を感染させるのに用いて、大量の所望のタンパク質を生産する。
【0171】
分泌シグナル配列
用語「分泌シグナル配列」または「シグナル配列」または「分泌リーダー配列」は、交換可能に用いられ、例えば米国特許第6,291,212号および米国特許第5,547,871号で説明されている(これらの両方は参照によりその全体として本明細書に組み込まれる)。分泌シグナル配列またはシグナル配列または分泌リーダー配列は、分泌ペプチドをコードする。分泌ペプチドは、細胞からの成熟ポリペプチドまたはタンパク質の分泌を指示させるように作用するアミノ酸配列である。分泌ペプチドは、一般的に、疎水性アミノ酸のコアを特徴とし、典型的には、これに限定されないが、新たに合成されたタンパク質のアミノ末端に見出される。分泌ペプチドは、分泌中に成熟タンパク質から切断されることがかなり多い。分泌ペプチドは、分泌経路を通過する際に成熟タンパク質からシグナルペプチドを切断させるプロセシング部位を含んでいてもよい。プロセシング部位は、シグナルペプチド内にコードされてもよく、または、例えばインビトロでの変異誘発でシグナルペプチドに付加されていてもよい。
【0172】
本発明の改変されたTf融合タンパク質の分泌を指示させるために分泌ペプチドを用いてもよい。このような他の分泌ペプチドと併用が可能な分泌ペプチドの一つは、酵母のバリアタンパク質(Barrier protein)の第三のドメインである。分泌シグナル配列またはシグナル配列または分泌リーダー配列は、タンパク質の分泌が起こる複雑な一連の翻訳後プロセシング工程に必要である。インタクトシグナル配列が存在する場合、発現されるタンパク質は粗面小胞体の内腔に入り、続いてゴルジ体を介して分泌小胞に輸送され、最終的に細胞外に輸送される。一般的に、シグナル配列は開始コドンの直後にあり、分泌しようとするタンパク質のアミノ末端でシグナルペプチドをコードする。ほとんどの場合、シグナル配列は、シグナルペプチダーゼと呼ばれる特異的なプロテアーゼで切断される。好ましいシグナル配列はプロセシングを改善し、ウイルス、哺乳動物または酵母発現ベクターを用いた組換えタンパク質発現の効率を伝える。場合によっては、天然型のTfシグナル配列を用いて、本発明の融合タンパク質を発現させ、分泌させてもよい。
【0173】
リンカー
本発明の改変されたトランスフェリン融合タンパク質のTf成分およびリガンドは、直接融合させてもよいし、または様々な長さのリンカーペプチドを用いて融合させてもよく、このようにして、より大きい物理的な隔たりを提供し、さらに融合したタンパク質間のより大きい空間的な移動を可能にし、従って例えばその同源の受容体に結合するのに抗体の可変領域の接近しやすさを最大にすることができる。リンカーペプチドは、フレキシブルであるか、または比較的固定されたアミノ酸からなる可能性がある。一実施態様において、本発明は、実質的に非らせん状のリンカー、例えば(PEAPTD)n(配列番号18)を含む。その他の実施態様において、本発明の融合タンパク質は、ポリグリシンストレッチを有するリンカーを含む。リンカーは、約50、40、30、20または10個のアミノ酸残基より少ないものでもよい。リンカーは、トランスフェリンタンパク質またはそれらの部分と、抗体の可変領域との間を共有結合で結合させることができる。
【0174】
また、リンカーは、1つ又は複数のリガンドに抗体の可変領域を組み込むためにも使用できる。抗体の可変領域を組み込むのに適切なリンカーは、抗体の可変領域を、抗体全体の結合特異性を維持しながら三次元構造にフォールディングすることができるものである。
【0175】
スクリーニング方法
本発明の融合タンパク質ライブラリーをスクリーニングすることによってリガンドを同定することが可能な可能性のある標的分子の数は、実質的には限定がない。例えば、標的分子(すなわち受容体または物質)は、抗体(またはそれらの結合部分)または抗原であり得る。抗体が結合する抗原がわかっており、おそらく配列解析もされている場合があり、このようなケースにおいて、本発明は、抗原のエピトープをマッピングするのに用いることができる。例えば所定の自己免疫疾患に関する抗原が未知の場合、その病気に罹った患者からの例えば血清、体液、組織または細胞を、本発明のスクリーニング方法で用いて、ペプチドを同定することができ、その結果として自己免疫反応を惹起する抗原も同定することができる。ペプチドが同定されれば、そのペプチドは、ワクチン、治療剤、診断試薬などの開発のための基準として役立たせたり、または、それらを提供したりすることができる。リガンドをスクリーニング可能な標的分子の一覧に関しては、WO01/46698を参照(全ての目的についてこれは参照によりその全体として本明細書に組み込まれる)。
【0176】
スクリーニングは、当分野における技術者によく知られた方法の1つ、例えばバイオパニング、FACSまたはMACSを用いることによって行うことができる。本発明の一実施態様において、スクリーニングは、受容体を活性化するために行われる。標的は、精製して溶液中に存在させてもよいし、または、表面に結合させたり細胞に結合させたりしてもよい。標的は、例えばビオチンで標識してもよいし、または、当分野既知の他の方法で標識してもよい。
【0177】
所望の特性を有するポリペプチドおよびペプチドを単離して、それに対応する核酸配列の配列解析によって、または、アミノ酸の配列解析もしくはマススペクトロメトリーによって同定することができる。それに続いて、サブ配列を、異なる配列(好ましくはランダム配列)で繰り返し交換し、1回またはそれ以上スクリーニング工程を実施することよって最適化を行ってもよい。
【0178】
ペプチドライブラリーを構築したら、宿主細胞をライブラリーベクターで形質転換する。成功した形質転換株は、典型的には、用いられるベクターに応じて、選択培地中での増殖、または選択的な条件下での増殖、例えば適切な増殖培地中での増殖などによって選択される。この選択は、固体の増殖培地で行ってもよいし、または液体の増殖培地で行ってもよい。細菌細胞を固体培地で増殖させるために、例えば、選択的な抗生物質を含むL−寒天の広い表面上で、実質的にコンフルエントな芝状態が形成されるまで細胞を高密度で増殖させる(1m2あたり約108〜109個の形質転換株)。液体培養での増殖のためには、細胞は、L−液体培地(抗生物質での選択を含む)で約10回またはそれを超えて倍加するように増殖させてもよい。ライブラリーのサイズによって液体培養での増殖の方が比較的便利である場合もあるし、一方、固体培地での増殖の方が、増幅工程中に偏りを生じる可能性が少なくなる場合もある。
【0179】
トランスフェリン融合タンパク質ペプチドライブラリーを、酵母細胞表層提示によってスクリーニングしようとする場合、酵母細胞は、トランスフェリン融合タンパク質をコードする発現ベクターで形質転換される。あらゆる変異誘発方法が、酵母表面提示ライブラリーの構築、例えばエラープローンポリメラーゼ連鎖反応、およびDNAシャフリングと両立する。Boder等(2000)Methods of Enzymology 328:430〜444を参照。あるいは、発現された融合タンパク質のトランスフェリン成分は、ランダムペプチド配列またはCDRのスカフォールドとして機能させることができる。
【0180】
酵母細胞のトランスフェリン融合タンパク質ペプチドライブラリーが作出されれば、所望のペプチドを同定するための数種のアプローチが当分野既知である。例えば、新規の結合特異性を単離するためにライブラリーをスクリーニングしようとする場合、ペプチドは、適度に低い親和性濃度(Kd>nM、または親和性なし)のケースにおいて、低濃度の蛍光標識した標的(すなわち受容体または物質)で平衡化させた結合によって区別することができる。堅く結合するタンパク質が得られるように設計された適用に関して、過度に大量の希釈した標的溶液が、リガンドを過剰モルに維持するのに必要な場合があり、サンプルの取り扱いを複雑にする。このようなケースにおいて、結合親和性の改善は、解離速度論における変化によって近接させる場合がある。化学量論的に制限された標的の反応速度論の競合を用いて、集団内の改善されたクローンを同定することができる(Hawkins等(1992)J.Mol.Biol.226:889);しかしながら、このアプローチはスクリーニングアプローチの定量的な予測精度を欠いており、一般的に推奨されていない。Boder等(2000)Methods of Enzymology 328:430〜444を参照。
【0181】
標的は、ビオチン化してもよいし、または蛍光標識してもよく、その代わりに、関心対象のリガンド、すなわちトランスフェリンに提示されたペプチドを標識してもよい。好ましくは、標的が標識される。標識された標的、例えばビオチン化した標的は、トランスフェリン融合タンパク質ペプチドライブラリーとインキュベートすることができる。このようなライブラリーは、少なくとも約104種の構成要素(すなわち提示されたペプチド)、少なくとも約105種の構成要素、少なくとも約106種の構成要素、少なくとも約107種の構成要素、少なくとも約108種の構成要素、少なくとも約109種の構成要素、少なくとも約1010種の構成要素、少なくとも約1011種の構成要素、少なくとも約1012種の構成要素、少なくとも約1013種の構成要素、少なくとも約1014種の構成要素、少なくとも約1015種の構成要素、または、少なくとも約1016種の構成要素を有していてもよい。
【0182】
インキュベート後、細胞は、第二の標識で標識することができ、例えば、二次抗体、ストレプトアビジンで標識された分子または当分野既知のその他の方法で標識することができる。二次抗体は、抗ビオチン抗体であり得る。ストレプトアビジンで標識された分子としては、これらに限定されないが、ストレプトアビジン−フィコエリトリンまたはストレプトアビジンマイクロビーズが挙げられる。
【0183】
当分野既知のように細胞集団を分析するために、フローサイトメトリーを用いることができる。これがなされる場合、集団の提示する分画だけを分析する。Boder等(2000)Methods of Enzymology 328:430〜444、および、Kondo等(2004)Appl.Microbiol.Biotechnol.64:28〜40を参照、これらの両方は参照によりその全体として本明細書に組み込まれる。
【0184】
あるいは、標識されたビーズ、すなわち抗ビオチンまたはストレプトアビジンで標識されたビーズからなる第二の標識が用いられる場合、Yeung等(2002)Biotechnol.Prog.18:212〜220(これは参照によりその全体として本明細書に組み込まれる)に記載されているような磁気によるソーティングプロトコールを用いて、リガンドおよび標的分子の混合物をソートすることができる。このスクリーニングプロトコール(ミルテニー・バイオテク社(Miltenyi Biotec GmbH))と共に、MACS(R)マイクロビーズキットを用いることができる。磁気によるソーティングは、FACSと共に用いることができる。
【0185】
本発明の一実施態様において、酵母細胞で発現された対象の単一のリガンドを特徴付けることが望ましい。発現されたタンパク質は、様々な方法でスクリーニングすることが可能である。このようなタンパク質が機能を有する場合、直接的に分析することができる。例えば、酵母表面で発現された単鎖抗体は十分に機能的であり、抗原への結合に基づいてスクリーニングすることが可能である。このようなタンパク質が、容易に分析できる検出可能な機能を有さない場合、抗体を用いてリガンドの発現をモニターすることが可能である。酵母細胞はファージよりもかなり大きいため、フローサイトメトリーを使用して、単一の酵母細胞におけるタンパク質の表現型をモニターすることができる。
【0186】
本発明のその他の実施態様において、リガンド成分と受容体または物質との結合は、細胞表層提示以外の当分野既知の手段によって行われ、例えばELISA、競合結合分析(標的の天然型の結合パートナーが既知の場合)、サンドイッチ分析、ペプチドライブラリーなどで結合がブロックされる放射性のリガンドを用いた放射性受容体分析である。これらの方法において、Tf融合タンパク質ペプチドライブラリーで形質転換した宿主細胞を溶解させる。Tf融合タンパク質のペプチドを、これらに限定されないが抗MUC1抗体のような適切なアンカー成分を介して分析基質に固定させる。スクリーニングプロセスは、Tfペプチドライブラリーと関心対象の標的とを反応させて、基準の結合レベルを確立し、それと、それに続くペプチドライブラリーの結合活性とを比較することを含む。標的に結合する、または標的への結合に関して競合する少量のペプチドを検出するのに十分な感受性を有してさえいれば、分析の性質は重要ではない。分析条件は様々であってよく、関心対象の様々な結合物質またはその他の生物活性に最適な結合条件を考慮に入れることができる。従って、pH、温度、塩濃度、体積および結合の持続時間などは全て、関心対象の環境の条件と類似した条件下でのペプチドの標的への結合が達成されるように変動させることができる。
【0187】
Tfペプチドライブラリーが、関心対象の標的に結合する一つ又は複数のペプチドを有することが決定されたら、本発明の方法を用いて、混合物中でペプチドの配列を同定することができる。標的と結合するペプチドを提示する細胞は、MACSまたはFACSスクリーニングによってライブラリーの総集団から単離することができる。スクリーニングプロセスを最初の分離物に対して2〜3回繰り返し、非特異的に結合したもの全てを除外する。続いて、結合親和性に基づいて単離することができるFACSソーティングによる最後のスクリーニングが行われる。単離した細胞からプラスミドDNAを回収し、挿入物の領域に関するDNAを配列解析して、タンパク質配列を決定する。続いて単離物間の共通のモチーフを決定することができる。
【0188】
治療用リガンド分子
本発明のリガンドは、推定の治療的分子でもよいし、または既知の治療的分子でもよい。本明細書で用いられるように、治療的分子は、典型的には、インビトロまたはインビボで有益な生物学的作用を発揮することができるタンパク質またはペプチドであり、例えば、正常なホメオスタシス、生理機能または病状に対して有益な作用を発揮するタンパク質またはペプチドが挙げられる。治療的分子は、マーカーとして一般的に使用される融合パートナー、またはガラクトシダーゼのようなタンパク質精製の助剤を含まない(例えば、米国特許第5,986,067号、およびAldred等(1984)Biochem.Biophys.Res.Commun.122:960〜965を参照)。例えば、病状に関する有益な作用としては、治療される治療対象に有利なあらゆる作用が挙げられ、このような作用としては、治療される治療対象に有益な作用を生じるような、疾患の予防、疾患の安定化、疾患の症状の低減もしくは緩和、または根本的な欠陥の調節、緩和もしくは治癒が挙げられる。
【0189】
治療用リガンドは、トランスフェリン成分に直接融合していてもよいし、または、上述したようなリンカー成分を介して間接的に融合していてもよい。一実施態様において、本融合タンパク質を切断して、本融合タンパク質のトランスフェリンとリガンド部分を、本融合タンパク質のそれ以外の部分から分離することが望ましい場合がある。その他の実施態様において、リガンドを本融合タンパク質のそれ以外の部分から切断することが望ましい場合がある。
【0190】
本発明の融合タンパク質のリガンド成分は、治療用タンパク質の少なくともフラグメントもしくは変異体、および/または抗体の少なくともフラグメントもしくは変異体を含んでいてもよい。さらなる実施態様において、本融合タンパク質は、タンパク質または抗体の、ペプチドフラグメントまたはペプチド変異体を含んでいてもよく、ここにおいて、これらの変異体またはフラグメントは、生物学的活性または治療活性の少なくとも1つを保持する。本融合タンパク質は、治療用タンパク質を含んでいてもよく、このような治療用タンパク質としては、長さが少なくとも約3個、少なくとも約4個、少なくとも5個、少なくとも6個、少なくとも7個、少なくとも8個、少なくとも9個、少なくとも10個、少なくとも11個、少なくとも12個、少なくとも13個、少なくとも14個、少なくとも15個、少なくとも20個、少なくとも25個、少なくとも30個、少なくとも35個、または少なくとも約40個、少なくとも約50個、少なくとも約55個、少なくとも約60個、または少なくとも約70個またはそれ以上のアミノ酸からなり、Nおよび/またはC末端に融合し、改変されたトランスフェリンのループ内に挿入されているか、または改変されたトランスフェリンのループに挿入されているペプチドフラグメントまたはペプチド変異体が挙げられる。
【0191】
その他の実施態様において、本発明の融合タンパク質のリガンド成分は、治療用タンパク質部分を含み、このタンパク質部分は、全長タンパク質、同様に、アミノ酸配列のアミノ末端から1個またはそれ以上の残基が欠失したポリペプチドなどを含む治療用タンパク質のフラグメントであり得る。
【0192】
その他の実施態様において、本発明の融合タンパク質のリガンド成分は、治療用タンパク質部分を含み、このタンパク質部分は、全長タンパク質、同様に、アミノ酸配列のカルボキシ末端から1個またはそれ以上の残基が欠失したポリペプチドを含む治療用タンパク質のフラグメントであり得る。
【0193】
その他の実施態様において、本発明の融合タンパク質のリガンド成分は、治療用タンパク質部分を含み、このタンパク質部分は、アミノ末端およびカルボキシ末端の両方から1個またはそれ以上のアミノ酸が欠失していてもよい。
【0194】
その他の実施態様において、本融合タンパク質は、本明細書で記載された参照の治療用タンパク質に少なくとも約80%、85%、90%、95%、96%、97%、98%、もしくは99%同一な治療用タンパク質部分、すなわちリガンド成分、またはそれらのフラグメントを含む。さらなる実施態様において、トランスフェリン融合分子は、アミノ酸配列に上述のようなNおよびC末端の欠失を有する参照のポリペプチドと少なくとも約80%、85%、90%、95%、96%、97%、98%、または99%同一な治療用タンパク質部分を含む。
【0195】
その他の実施態様において、本融合タンパク質は、例えば治療用タンパク質の天然型または野生型のアミノ酸配列に少なくとも約80%、85%、90%、95%、96%、97%、98%、99%、または100%同一な治療用タンパク質部分を含む。これらのポリペプチドのフラグメントも提供される。
【0196】
細胞表層タンパク質や分泌タンパク質のような本発明の改変されたトランスフェリン融合タンパク質の治療用タンパク質部分に相当する治療用タンパク質は、1またはそれ以上のオリゴ糖基を付着させることによって改変することができる。グリコシル化と称される改変は、タンパク質の物理特性に有意に影響を与えることができ、さらに、タンパク質の安定性、分泌および局在化にとって重要なものにすることができる。グリコシル化は、ポリペプチド主鎖に沿って特異的な位置で起こる。グリコシル化には、通常、以下の2つの主要なタイプがあり、すなわち、グリコシル化は、セリンまたはスレオニン残基に結合しているO結合型のオリゴ糖を特徴とし;およびグリコシル化は、Asn−X−Ser/Thr配列(ここにおいてXは、プロリン以外のアミノ酸が可能である)におけるアスパラギン残基に結合しているN結合型のオリゴ糖を特徴とする。タンパク質構造や細胞型のような可変値は、様々なグリコシル化部位における鎖内の糖質単位の数および性質に影響を与える。グリコシル化の異性体もまた、は、所定の細胞型内の同じ部位に共通している。例えば、ヒトインターフェロンの数種のタイプは、グリコシル化されている。
【0197】
本発明の融合タンパク質の治療用タンパク質部分に相当する治療用タンパク質、ならびに、それらの類似体および変異体は、それらが発現される宿主細胞によって、またはそれらの発現のその他の条件に起因して、それらの核酸配列を操作した結果として1またはそれ以上の部位でのグリコシル化が変更されるように改変されていてもよい。例えば、グリコシル化の異性体は、例えば、アミノ酸残基の置換または欠失、例えばグルタミンのアスパラギンへの置換によってグリコシル化部位を無効にしたり、または導入したりすることによって生産してもよいし、またはグリコシル化されていない組換えタンパク質は、それらをグリコシル化しないと予想される宿主細胞中で、例えばグリコシル化能に欠陥のある酵母中でタンパク質を発現させることによって生産してもよい。これらのアプローチは当分野既知である。
【0198】
治療用タンパク質およびそれらの核酸配列は当分野周知であり、公開データベース、例えば、ケミカル・アブストラクツ・サービス・データベース(Chemical Abstracts Services Databases)(例えばCASレジストリ)、GenBank、および、GenSeqから入手可能である。以下で述べる登録番号および配列は、参照によりその全体として本明細書に組み込まれる。
【0199】
本発明はさらに、本明細書で説明される治療用タンパク質のフラグメントを含む融合タンパク質を対象とする。タンパク質のN末端から1個またはそれ以上のアミノ酸が欠失して、治療用タンパク質部分の生物学的機能の1種またはそれ以上に改変または減少が生じたとしても、その他の治療活性および/または機能的な活性(例えば、生物活性、マルチマー化する能力、リガンドと結合する能力)がなお保持させることができる。例えば、一般的に、N末端に欠失を有するポリペプチドの、ポリペプチドの完全な形態または成熟した形態を認識する抗体を誘導する、および/またはそれらに結合する能力は、完全なポリペプチドの残基の半数未満がN末端から除去されても保持されると予想される。完全なポリペプチドのN末端の残基が欠失した特定のポリペプチドが、このような免疫学的な活性を保持するかどうかは、本明細書で説明されているルーチン的な方法や、それ以外の当分野既知の方法で分析することができる。多数のN末端のアミノ酸残基が欠失した突然変異体が、いくつかの生物学的活性または免疫原性活性を保持する可能性があることはありそうもないことでない。実際に、6個もの少ないアミノ酸残基で構成されるペプチドでもしばしば免疫反応を誘発することがある。
【0200】
上述したように、治療用タンパク質のN末端またはC末端から1個またはそれ以上のアミノ酸が欠失していることによって、タンパク質の1種またはそれ以上の生物学的機能の改変または減少が生じる場合でさえも、その他の機能的な活性、例えば生物活性、マルチマー化する能力、リガンドと結合する能力および/または治療活性は、なお保持させることができる。一般的に、例えばC末端に欠失を有するポリペプチドの、ポリペプチドの完全な形態または成熟した形態を認識する抗体を誘導する、および/またはそれらに結合する能力は、完全なポリペプチドまたは成熟ポリペプチドの残基の半分未満がC末端から除去されても保持されると予想される。参照のポリペプチドのN末端および/またはC末端の残基が欠失した特定のポリペプチドが治療活性を保持するかどうかは、本明細書で説明されているルーチン的な方法および/または当分野既知のそれ以外の方法によって容易に決定することができる。
【0201】
治療用タンパク質のペプチドフラグメントは、アミノ酸配列がフラグメントである治療用タンパク質のポリペプチド配列の、治療活性および/または機能的な活性(例えば生物活性)を提示するアミノ酸配列を含む、あるいはそれらからなるフラグメントであってもよい。
【0202】
その他のポリペプチドフラグメントは、生物学的に活性なフラグメントである。生物学的に活性なフラグメントは、本発明で用いられる治療用タンパク質の活性に類似しているが、必ずしも同一ではない活性を示すものである。このようなフラグメントの生物活性は、改善された所望の活性、または低減された望ましくない活性を含んでいてもよい。
【0203】
一般的に、タンパク質の変異体は、総体的に極めて類似しており、多くの領域において本発明のトランスフェリン融合タンパク質の治療用タンパク質部分に相当する治療用タンパク質のアミノ酸配列と同一である。これらの変異体をコードする核酸も本発明に包含される。
【0204】
本発明で使用可能なさらなる治療用ポリペプチドは、当業者既知のストリンジェントなハイブリダイゼーション条件下で治療用タンパク質のアミノ酸配列をコードする核酸分子の相補物とハイブリダイズするポリヌクレオチドによってコードされたポリペプチドである。例えば、Ausubel,F.M.等編集,1989 Current protocol in Molecular Biology,グリーン・パブリッシング・アソシエイツ社(Green Publishing Associates,Inc.)、およびジョン・ワイリー&サンズ社(John Wiley & Sons Inc.,ニューヨーク)を参照。これらのポリペプチドをコードするポリヌクレオチドも本発明に包含される。
【0205】
例えば本発明のアミノ酸配列クエリーに少なくとも95%「同一な」アミノ酸配列を有するポリペプチドとは、対象のポリペプチドのアミノ酸配列がクエリー配列と同一である(ただし、対象のポリペプチド配列は、アミノ酸配列のアミノ酸クエリー100個あたり5個までのアミノ酸の変更を含んでいてもよい)ことを意味する。言い換えれば、アミノ酸配列がアミノ酸配列クエリーに少なくとも95%同一なポリペプチドを得るためには、その他のアミノ酸で挿入、欠失または置換されてもよい対象の配列中のアミノ酸残基は、5%以下である。これらの参照配列の変更は、参照のアミノ酸配列のアミノまたはカルボキシ末端の位置に、またはこれら末端の位置の間のどこかに、参照配列中のそれぞれの残基に、または参照配列内の1個またはそれ以上の連続した基のいずれかに散在して存在する可能性がある。
【0206】
実際問題として、あらゆる特定のポリペプチドが、例えば、本発明の融合タンパク質またはそれらのフラグメント(例えば、本融合タンパク質の治療用タンパク質部分またはそれらの部分)のアミノ酸配列に少なくとも約80%、85%、90%、95%、96%、97%、98%、または99%同一であるかどうかは、既知のコンピュータープログラムを用いてルーチン的に決定することができる。クエリー配列(本発明の配列)と、対象の配列(またはグローバル配列アライメントとも言う)との間の最良の全体のマッチングを決定する方法の一つは、Bruflag等(Comp.App.Biosci 245〜(1990))のアルゴリズムに基づくFASTDBコンピュータープログラムを用いて決定することができる。
【0207】
本発明のポリヌクレオチド変異体は、コード領域、非コード領域またはその両方に変更を含んでいてもよい。サイレント置換、付加または欠失を生じるが、コードされたポリペプチドの特性または活性を変更しない変更を含むポリヌクレオチド変異体を用いて、改変されたリガンド成分を生産してもよい。遺伝子コードの縮重によるサイレント置換によって生産されたヌクレオチド変異体を利用することができる。その上、約50個未満、40個未満、30個未満、20個未満、10個未満、または5〜50個、5〜25個、5〜10個、1〜5個、もしくは、1〜2個のアミノ酸が、あらゆる組み合わせで置換、欠失または付加されているポリペプチド変異体も利用することができる。ポリヌクレオチド変異体は、様々な理由に応じて作製することができ、例えば、特定の宿主に対してコドンの発現を最適化する(ヒトmRNAにおけるコドンを、宿主、例えば上述のような酵母またはE.コリにとって好ましいコドンに変化させる)ことによってなされる。
【0208】
その他の実施態様において、治療用タンパク質成分、すなわちリガンド成分は、野生型配列と比較して保存的置換を有する。「保存的置換」は、グループ内での入れ替えを意味し、例えば、脂肪族または疎水性アミノ酸Ala、Val、Leu、およびIleの交換;ヒドロキシル残基SerおよびThrの交換;酸性残基AspおよびGluの交換;アミド残基AsnおよびGlnの交換;塩基性残基Lys、Arg、およびHisの交換;芳香族残基Phe、Tyr、およびTrpの交換;および小型のアミノ酸Ala、Ser、Thr、Met、およびGlyの交換が挙げられる。表現型においてサイレントなアミノ酸置換をどのように作製するかに関する指針は、例えば、Bowie等の“Deciphering the Message in Protein Sequences:Tolerance to Amino Acid Substitutions,”Science247:1306〜1310(1990)に示されている。特定の実施態様において、本発明のポリペプチドは、本明細書で説明されている治療用タンパク質、および/または本発明の血清トランスフェリンおよび改変されたトランスフェリンタンパク質のアミノ酸配列のフラグメントまたは変異体を含むか、あるいはそれらからなり、ここにおいて、これらのフラグメントまたは変異体は、参照のアミノ酸配列と比較して、1〜5個、5〜10個、5〜25個、5〜50個、10〜50個、または50〜150個のアミノ酸残基付加、置換および/または欠失を有する。さらなる実施態様において、アミノ酸置換は保存的である。これらのポリペプチドをコードする核酸も本発明に包含される。
【0209】
本発明の改変された融合タンパク質は、互いにペプチド結合または改変されたペプチド結合で結合したアミノ酸で構成されていてもよく、20種の遺伝子コードアミノ酸以外のアミノ酸を含んでいてもよい。このようなポリペプチドは、天然のプロセス、例えば翻訳後プロセシング、または当分野でよく知られた化学修飾技術のいずれかによって改変されていてもよい。このような改変は基本的な教本でよく説明されており、研究論文、加えて膨大な研究文献でより詳細に説明されている。
【0210】
改変は、ポリペプチド中のあらゆる場所で発生する可能性があり、このような場所としては、ペプチド主鎖、アミノ酸側鎖、およびアミノまたはカルボキシ末端が挙げられる。当然のことながら、同じタイプの改変が、所定のポリペプチド内の同じ部位に存在していてもよいし、または様々な程度でいくつかの部位に存在していてもよい。また、所定のポリペプチドに多くのタイプの改変が含まれていてもよい。ポリペプチドは、例えばユビキチン化の結果として分岐していてもよいし、ポリペプチドは環状でもよく、分岐を含んでいてもよいし、または含んでいなくてもよい。環状、分岐状および分岐状環状ポリペプチドは、翻訳後の天然のプロセスによって生じたものでもよいし、または合成法によって製造したものでもよい。改変としては、アセチル化、アシル化、ADP−リボシル化、アミド化、フラビンの共有結合、ヘム成分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトールの共有結合、架橋、環化、ジスルフィド結合の形成、脱メチル、共有結合の架橋の形成、システインの形成、グリコシル化、GPIアンカーの形成、水酸化、ヨウ素化、メチル化、ミリスチル化、酸化、ペグ化、タンパク質分解によるプロセシング、リン酸化、プレニル化、ラセミ化、硫酸化、アミノ酸のタンパク質への付加を介するトランスファーRNA、例えばアルギニル化、およびユビキチン化が挙げられる。(例えば、PROTEINS−STRUCTURE AND MOLECULAR PROPERTIES,第二版,T.E.Creighton,W.H.Freeman and Company,ニューヨーク(1993);POST−TRANSLATIONAL COVALENT MODIFICATION OF PROTEINS,B.C.Johnson編,アカデミックプレス,ニューヨーク,1〜12頁(1983);Seifter等(1990)Meth.Enzymol.182:626〜646;Rattan等,Ann.N.Y.Acad.Sci.663:48〜62を参照)。
【0211】
リガンド成分として用いることができる治療的分子としては、これらに限定されないが、ホルモン、マトリックスタンパク質、免疫抑制剤、気管支拡張剤、心血管作動薬、酵素、CNS剤、神経伝達物質、受容体タンパク質またはペプチド、成長ホルモン、増殖因子、抗ウイルス性ペプチド、膜融合阻害ペプチド、サイトカイン、リンフォカイン、モノカイン、インターロイキン、コロニー刺激因子、分化因子、血管新生因子、受容体リガンド、癌関連タンパク質、抗腫瘍薬、ウイルスペプチド、抗生物質ペプチド、血液タンパク質、アンタゴニストタンパク質、転写因子、抗血管新生因子、アンタゴニストタンパク質またはペプチド、受容体アンタゴニスト、抗体、単鎖抗体、および細胞接着分子が挙げられる。単一の融合タンパク質に異なる治療的分子を組合せて、二機能性または多機能性の治療的分子を生産することができる。また、異なる分子を併用して、治療実体および標的実体を有する融合タンパク質を生産してもよい。治療的分子は、本発明の軸成分に直接融合させることができ、またはTf成分またはアルブミン成分のような提示成分と融合してもよいし、もしくはそれらに挿入してもよい。
【0212】
サイトカインは、免疫系の細胞によって放出される可溶性タンパク質であり、非酵素的に特異的な受容体を介して作用して、免疫反応を調節することができる。サイトカインは、低濃度で作用し、高親和性で特異的な受容体に結合するという点で、ホルモンに似ている。本明細書において用語「サイトカイン」とは、そのサイトカインについての受容体を有する細胞において特異的な生物学的反応を惹起する、天然に存在する、または組換えタンパク質、それらの類似体およびそれらのフラグメントを示すものとして用いられる。サイトカインとしては、好ましくは、インターロイキン、例えばインターロイキン−2(IL−2)(GenBank登録番号S77834)、IL−3(GenBank登録番号M14743)、IL−4(GenBank登録番号M23442)、IL−5(GenBank登録番号J03478)、IL−6(GenBank登録番号M14584)、IL−7(GenBank登録番号NM_000880)、IL−10(GenBank登録番号NM_000572)、IL−12(GenBank登録番号AF180562およびGenBank登録番号AF180563)、IL−13(GenBank登録番号U10307)、IL−14(GenBank登録番号XM_170924)、IL−15(GenBank登録番号X91233)、IL−16(GenBank登録番号NM_004513)、IL−17(GenBank登録番号NM_002190)およびIL−18(GenBank登録番号NM_001562)、造血因子、例えば顆粒球マクロファージコロニー刺激因子(GM−CSF)(GenBank登録番号X03021)、顆粒球コロニー刺激因子(G−CSF)(GenBank登録番号X03656)、血小板活性化因子(GenBank登録番号NM_000437)およびエリスロポエチン(GenBank登録番号X02158)、腫瘍壊死因子(TNF)、例えばTNFα(GenBank登録番号X02910)、リンフォカイン、例えばリンフォトキシン−α(GenBank登録番号X02911)、リンフォトキシン−β(GenBank登録番号L11016)、ロイコレグリン(leukoregulin)、マクロファージ遊走阻止因子(GenBank登録番号M25639)、およびニューロロイキン(GenBank登録番号K03515)、代謝プロセスの調節因子、例えばレプチン(GenBank登録番号U43415)、インターフェロン、例えばインターフェロンα(IFNα)(GenBank登録番号M54886)、IFNβ(GenBank登録番号V00534)、IFNγ(GenBank登録番号J00219)、IFNo(GenBank登録番号NM_002177)、トロンボスポンジン1(THBS1)(GenBank登録番号NM_003246)、THBS2(GenBank登録番号L12350)、THBS3(GenBank登録番号L38969)、THBS4(GenBank登録番号NM_003248)、およびケモカインが挙げられる。好ましくは、改変されたトランスフェリン−サイトカイン融合タンパク質は、サイトカインの生物活性を提示する。
【0213】
本明細書における用語「ホルモン」は、所定の細胞または組織で生産され、体内のどこかに位置するその他の細胞または組織で特異的な生物学的な変化または活性が生じるように作用する多数の生物学的に活性な物質のいずれか一つを述べる際に用いられる。ホルモンとしては、好ましくは、プロインスリン(GenBank登録番号V00565)、インスリン(GenBank登録番号NM_000207)、成長ホルモン1(GenBank登録番号V00520)、成長ホルモン2(GenBank登録番号F006060)、成長ホルモン放出因子(GenBank登録番号NM021081)、インスリン様増殖因子I(GenBank登録番号M27544)、インスリン様増殖因子II(GenBank登録番号NM_000612)、インスリン様増殖因子結合タンパク質1(IGFBP−1)(GenBank登録番号M59316)、IGFBP−2(GenBank登録番号X16302)、IGFBP−3(GenBank登録番号NM_000598)、IGFBP−4(GenBank登録番号Y12508)、IGFBP−5(GenBank登録番号M65062)、IGFBP−6(GenBank登録番号NM_002178)、IGFBP−7(GenBank登録番号NM001553)、絨毛性ゴナドトロピンβ鎖(GenBank登録番号NM_033142)、絨毛性ゴナドトロピンα鎖(GenBank登録番号NM000735)、黄体形成ホルモンβ(GenBank登録番号X00264)、胞刺激ホルモンβ(GenBank登録番号NM_000510)、甲状腺刺激ホルモンβ(GenBank登録番号NM_000549)、プロラクチン(GenBank登録番号NM_000948)、プロオピオメラノコルチン(GenBank登録番号V01510)、コルチコトロピン(ACTH)、β−リポトロピン、α−メラニン細胞刺激ホルモン(α−MSH)、γ−リポトロピン、β−MSH、β−エンドルフィン、およびコルチコトロピン様中葉ペプチド(CLIP)が挙げられる。
【0214】
また用語「ホルモン」は、グルカゴン様ペプチド−1(GLP−1)も包含し、これは、いわゆる腸島軸(enteroinsular axis)に属するインスリン分泌を調節する胃腸ホルモンであり、さらにエキセンディン(例えば、エキセンディン−4およびそれらの変異体)も包含し、これは、GLP−1受容体アゴニストである。
【0215】
本明細書における用語「増殖因子」は、細胞増殖を刺激する受容体に結合するあらゆるタンパク質またはペプチドを述べる際に用いられる。増殖因子としては、好ましくは、血小板由来増殖因子−α(PDGF−α)(GenBank登録番号X03795)、PDGF−β(GenBank登録番号X02811)、ステロイドホルモン、上皮増殖因子(EGF)(GenBank登録番号NM_001963)、線維芽細胞増殖因子、例えば線維芽細胞増殖因子1(FGF1)(GenBank登録番号NM_000800)、FGF2(GenBank登録番号NM_002006)、FGF3(GenBank登録番号NM_005247)、FGF4(GenBank登録番号NM_002007)、FGF5(GenBank登録番号M37825)、FGF6(GenBank登録番号X57075)、FGF7(GenBank登録番号NM_002009)、FGF8(GenBank登録番号AH006649)、FGF9(GenBank登録番号NM_002010)、FGF10(GenBank登録番号AB002097)、FGF11(GenBank登録番号NM_004112)、FGF12(GenBank登録番号NM_021032)、FGF13(GenBank登録番号NM_004114)、FGF14(GenBank登録番号NM_004115)、FGF16(GenBank登録番号AB009391)、FGF17(GenBank登録番号NM_003867)、FGF18(GenBank登録番号AF075292)、FGF19(GenBank登録番号NM_005117)、FGF20(GenBank登録番号NM_019851)、FGF21(GenBank登録番号NM_019113)、FGF22(GenBank登録番号NM_020637)、およびFGF23(GenBank登録番号NM020638)、アンギオゲニン(GenBank登録番号M11567)、脳由来神経栄養因子(GenBank登録番号M61176)、毛様体神経栄養因子(GenBank登録番号X60542)、トランスフォーミング増殖因子−α(TGF−α)(GenBank登録番号X70340)、TGF−β(GenBank登録番号X02812)、神経成長因子−α(NGF−α)(GenBank登録番号NM_010915)、NGF−β(GenBank登録番号X52599)、メタロプロテイナーゼ1の組織阻害剤(TIMP1)(GenBank登録番号NM_003254)、TIMP2(GenBank登録番号NM_003255)、TIMP3(GenBank登録番号U02571)、TIMP4(GenBank登録番号U76456)、およびマクロファージ刺激因子1(GenBank登録番号LI1924)が挙げられる。
【0216】
本明細書における用語「マトリックスタンパク質」は、通常、細胞外マトリックスで見出されるタンパク質またはペプチドを述べる際に用いられる。これらのタンパク質は、強度、ろ過または付着に関して機能的に重要な可能性がある。マトリックスタンパク質としては、好ましくは、コラーゲン、例えばI型コラーゲン(GenBank登録番号Z74615)、II型コラーゲン(GenBank登録番号XI6711)、III型コラーゲン(GenBank登録番号X14420)、IV型コラーゲン(GenBank登録番号NM_001845)、V型コラーゲン(GenBank登録番号NM_000393)、VI型コラーゲン(GenBank登録番号NM_058175)、VII型コラーゲン(GenBank登録番号L02870)、VIII型コラーゲン(GenBank登録番号NM_001850)、IX型コラーゲン(GenBank登録番号X54412)、X型コラーゲン(GenBank登録番号X60382)、XI型コラーゲン(GenBank登録番号J04177)およびXII型コラーゲン(GenBank登録番号U73778)、ラミニンタンパク質、例えばLAMA2(GenBank登録番号NM_000426)、LAMA3(GenBank登録番号L34155)、LAMA4(GenBank登録番号NM_002290)、LAMB1(GenBank登録番号NM_002291)、LAMB3(GenBank登録番号L25541)、LAMC1(GenBank登録番号NM_002293)、ニドゲン(GenBank登録番号NM_002508)、α−テクトリン(GenBank登録番号NM_005422)、β−テクトリン(GenBank登録番号NM_058222)、およびフィブロネクチン(GenBank登録番号X02761)が挙げられる。
【0217】
用語「血液タンパク質」は、一般的に、血漿由来のタンパク質と定義され、現在、そのうち多くが、一般的に組換え法によって生産されており、このようなタンパク質としては、これらに限定されないが、天然の血清タンパク質、それらの誘導体、フラグメントおよび突然変異体または変異体、血液凝固因子、誘導体、突然変異体、変異体およびフラグメント(例えば、VII、VIII、IX、X因子など)、プロテアーゼ阻害剤(アンチトロンビン3、アルファ−1アンチトリプシン)、ウロキナーゼ型プラスミノゲン活性化因子、免疫グロブリン、フォン−ヴィレブランド因子、およびフォン−ヴィレブランド突然変異体、フィブロネクチン、フィブリノーゲン、トロンビン、およびヘモグロビンが挙げられる。
【0218】
本明細書における用語「酵素」は、それ自体永久的に変化したり、または破壊されたりすることなく特異的な反応を触媒するあらゆるタンパク質またはタンパク質様物質を述べる際に用いられる。酵素としては、好ましくは、凝固因子、例えばF2(GenBank登録番号XM_170688)、F7(GenBank登録番号XM_027508)、F8(GenBank登録番号XM_013124)、F9(GenBank登録番号NM_000133)、F10(GenBank登録番号AF503510)など、マトリックスメタロプロテイナーゼ、例えばマトリックスメタロプロテイナーゼI(GenBank登録番号MMP1)(GenBank登録番号NM_002421)、MMP2(GenBank登録番号NM_004530)、MMP3(GenBank登録番号NM_002422)、MMP7(GenBank登録番号NM_002423)、MMP8(GenBank登録番号NM_002424)、MMP9(GenBank登録番号NM_004994)、MMP10(GenBank登録番号NM_002425)、MMP12(GenBank登録番号NM_002426)、MMP13(GenBank登録番号X75308)、MMP20(GenBank登録番号NM_004771)、アデノシン−デアミナーゼ(GenBank登録番号NM_000022)、マイトジェン活性化プロテインキナーゼ、例えばMAPK3(GenBank登録番号XM_055766)、MAP2K2(GenBank登録番号NM_030662)、MAP2K1(GenBank登録番号NM_002755)、MAP2K4(GenBank登録番号NM_003010)、MAP2K7(AF013588)、およびMAPK12(NM_002969)、キナーゼ、例えばJNKK1(GenBank登録番号U17743)、JNKK2(GenBank登録番号AF014401)、JAK1(M64174)、JAK2(NM_004972)、およびJAK3(NM_000215)、およびホスファターゼ、例えばPPM1A(GenBank登録番号NM_021003)およびPPM1D(GenBank登録番号NM_003620)が挙げられる。
【0219】
本明細書における用語「転写因子」は、タンパク質をコードする遺伝子の転写に関与するあらゆるタンパク質またはペプチドを述べる際に用いられる。転写因子としては、Sp1、Sp2(GenBank登録番号NM_003110)、Sp3(GenBank登録番号AY070137)、Sp4(GenBank登録番号NM_003112)、NFYB(GenBank登録番号NM_006166)、Hap2(GenBank登録番号M59079)、GATA−1(GenBank登録番号NM_002049)、GATA−2(GenBank登録番号NM_002050)、GATA−3(GenBank登録番号X55122)、GATA−4(GenBank登録番号L34357)、GATA−5、GATA−6(GenBank登録番号NM_005257)、FOG2(NM_012082)、Eryf1(GenBank登録番号X17254)、TRPS1(GenBank登録番号NM_014112)、NF−E2(GenBank登録番号NM_006163)、NF−E3、NF−E4、TFCP2(GenBank登録番号NM_005653)、Oct−1(GenBank登録番号X13403)、ホメオボックスタンパク質、例えばHOXB2(GenBank登録番号NM_002145)、HOX2H(GenBank登録番号X16665)、ヘアレス(hairless)同族体(GenBank登録番号NM_005144)、デカペンタプレジックタンパク質に対する親物質、例えばMADH1(GenBank登録番号NM_005900)、MADH2(GenBank登録番号NM_005901)、MADH3(GenBank登録番号NM_005902)、MADH4(GenBank登録番号NM_005359)、MADH5(GenBank登録番号AF009678)、MADH6(GenBank登録番号NM_005585)、MADH7(GenBank登録番号NM_005904)、MADH9(GenBank登録番号NM_005905)、および転写タンパク質のシグナルトランスデューサーおよび活性化因子、例えばSTAT1(GenBank登録番号XM_010893)、STAT2(GenBank登録番号NM_005419)、STAT3(GenBank登録番号AJ012463)、STAT4(GenBank登録番号NM_003151)、STAT5(GenBank登録番号L41142)、および、STAT6(GenBank登録番号NM_003153)が挙げられる。
【0220】
さらに本発明のその他の実施態様において、治療的分子は、非ヒトまたは非哺乳動物タンパク質である。例えば、HIVgp120、HIVTat、他のウイルスの表面タンパク質、例えば肝炎、ヘルペス、インフルエンザ、アデノウイルスおよびRSV、他のHIV成分、寄生虫の表面タンパク質、例えばマラリアの抗原、および細菌の表面タンパク質が使用可能である。これらの非ヒトタンパク質は、これらは有用な活性を有するため、例えば抗原として使用可能である。例えば、治療的分子は、ストレプトキナーゼ、スタフィロキナーゼ、アスパラギナーゼ、ウロキナーゼ、または有用な酵素活性を有するその他のタンパク質であり得る。
【0221】
本発明の代替の実施態様において、治療的分子は、生物活性を有するリガンド結合タンパク質である。このようなリガンド結合タンパク質は、例えば、(1)細胞表層で受容体−リガンドの相互作用をブロックしたり;または(2)血液の流動相において分子の生物活性を中和したりすることができ、それによって、上記タンパク質がその細胞の標的に到達することを防ぐことができる。いくつかの実施態様において、改変されたトランスフェリン融合タンパク質は、以下からなる群より選択される受容体のリガンド−結合ドメインに融合した改変されたトランスフェリン分子を含む:低密度リポタンパク質(LDL)受容体、アセチル化LDL受容体、腫瘍壊死因子α受容体、トランスフォーミング増殖因子β受容体、サイトカイン受容体、免疫グロブリンFc受容体、ホルモン受容体、グルコース受容体、グリコリピド受容体、およびグリコサミノグリカン受容体(ただしこれらに限定されない)。その他の実施態様において、リガンド結合タンパク質としては、CD2(M14362)、CD3G(NM_000073)、CD3D(NM_000732)、CD3E(NM_000733)、CD3Z(J04132)、CD28(NM_006139)、CD4(GenBank登録番号NM_000616)、CD1A(GenBank登録番号M28825)、CD1B(GenBank登録番号NM_001764)、CD1C(GenBank登録番号NM_001765)、CD1D(GenBank登録番号NM_001766)、CD80(GenBank登録番号NM_005191)、GNB3(GenBank登録番号AF501884)、CTLA−4(GenBank登録番号NM_005214)、細胞間接着分子、例えばICAM−1(NM_000201)、ICAM−2(NM_000873)、およびICAM−3(NM_002162)、腫瘍壊死因子受容体、例えばTNFRSF1A(GenBank登録番号X55313)、TNFR1SFB(GenBank登録番号NM_001066)、TNFRSF9(GenBank登録番号NM_001561)、TNFRSF10B(GenBank登録番号NM_003842)、TNFRSF11B(GenBank登録番号NM_002546)、および、TNFRSF13B(GenBank登録番号NM_006573)、ならびにインターロイキン受容体、例えばIL2RA(GenBank登録番号NM_000417)、IL2RG(GenBank登録番号NM_000206)、IL4R(GenBank登録番号AF421855)、IL7R(GenBank登録番号NM_002185)、IL9R(GenBank登録番号XM_015989)、およびIL13R(GenBank登録番号X95302)が挙げられる。好ましくは、本発明のリガンド結合タンパク質の融合体は、そのリガンド結合タンパク質の生物活性を提示する。
【0222】
本明細書における用語「癌関連タンパク質」は、その発現が癌または制御された細胞増殖の維持に関連するタンパク質またはポリペプチドを述べる際に用いられ、例えば腫瘍抑制遺伝子または腫瘍遺伝子によってコードされたタンパク質である。癌関連タンパク質としては、pl6(GenBank登録番号AH005371)、p53(GenBank登録番号NM_000546)、p63(GenBank登録番号NM_003722)、p73(GenBank登録番号NM_005427)、BRCA1(GenBank登録番号U14680)、BRCA2(GenBank登録番号NM_000059)、CTBP相互作用タンパク質(GenBank登録番号U72066)、DMBT1(GenBank登録番号NM_004406)、HRAS(GenBank登録番号NM_005343)、NCYM(GenBank登録番号NM_006316)、FGR(GenBank登録番号NM_005248)、myb(GenBank登録番号AF104863)、raf1(GenBank登録番号NM_002880)、erbB2(GenBank登録番号NM_004448)、VAV(GenBank登録番号X16316)、c−fos(VGenBank登録番号01512)、c−fes(GenBank登録番号X52192)、c−jun(GenBank登録番号NM_002228)、MAS1(GenBank登録番号M13150)、pim−1(GenBank登録番号M16750)、TIF1(GenBank登録番号NM_003852)、c−fms(GenBank登録番号X03663)、EGFR(GenBank登録番号NM_005228)、erbA(GenBank登録番号X04707)、c−srcチロシンキナーゼ(GenBank登録番号XM_044659)、c−abl(GenBank登録番号M14752)、N−ras(GenBank登録番号X02751)、K−ras(GenBank登録番号M54968)、jun−B(GenBank登録番号M29039)、c−myc(GenBank登録番号AH001511)、RB1(GenBank登録番号M28419)、DCC(GenBank登録番号X76132)、APC(GenBank登録番号NM_000038)、NF1(GenBank登録番号M89914)、NF2(GenBank登録番号Y18000)、および、bcl−2(GenBank登録番号M13994)が挙げられる。
【0223】
本明細書における「融合阻害ペプチド」は、抗ウイルス活性、抗膜融合能、および/または細胞内プロセスを調節する能力を示すペプチドを述べる際に用いられ、例えば、コイルドコイルペプチド構造を含むものである。抗ウイルス活性としては、これらに限定されないが、HIV−1、HIV−2、RSV、SIV、EBV、麻疹ウイルス、インフルエンザウイルスの阻害、または感染していない細胞へのCMV伝達が挙げられる。加えて、抗融合形成(antifusogenic)能力、抗ウイルス活性、またはペプチドの細胞内調節活性は、単にペプチドの存在を必要とするに過ぎず、このようなペプチドに対して向けられた宿主の免疫応答の刺激を特に必要としない。抗融合形成は、2またはそれ以上の成分間における膜融合事象を阻害する、またはそのレベルを、ペプチドの非存在下で前記成分間に起こる膜融合のレベルに比べて減少させるペプチドの能力を意味する。このような成分は、例えば細胞膜またはウイルス構造である可能性があり、例えばウイルスエンベロープまたは繊毛である。本明細書で用いられる用語「抗ウイルス性ペプチド」は、細胞のウイルス感染、または例えば細胞間融合もしくは遊離のウイルスによる感染を介した、増殖を起こすウイルス感染および/またはウイルス病原性に必要ないくつかのウイルス活性を阻害するペプチドの能力を意味する。このような感染は、エンベロープウイルスケースで起こるような、またはいくつかのその他のウイルス構造および細胞構造に関連する融合事象で起こるような膜融合に関与する可能性がある。膜融合阻害ペプチドおよび抗ウイルス性ペプチドは、1種より多くのタンパク質から誘導されるアミノ酸配列を有することが多い(例えば、HIV−1、HIV−2、RSV、およびSlVから誘導されたポリペプチド)。
【0224】
膜融合阻害ペプチド、および、抗ウイルス性ペプチドの例は、WO94/2820、WO96/19495、WO96/40191、WO01/64013、および米国特許第6,333,395号、6,258,782号、6,228,983号、6,133,418号、6,093,794号、6,068,973号、6,060,065号、6,054,265号、6,020,459号、6,017,536号、6,013,263号、5,464,933号、5,346,989号、5,603,933号、5,656,480号、5,759,517号、6,245,737号;6,326,004号、および6,348,568号に見出すことができ、これらはいずれも参照により本明細書に組み入れられる。
【0225】
その他のタイプのペプチドの例としては、本明細書で説明したような治療用タンパク質のフラグメントが挙げられ、特に、親分子の活性の少なくとも1種を保持するヒトタンパク質のフラグメントである。また、本発明のリガンド成分を生産するのに使用可能なペプチドとしては、模倣ペプチド、および治療用タンパク質の生物活性を示すペプチドも挙げられるが、全長の治療用タンパク質とは配列または三次元構造の点で異なる。非限定的な例として、ペプチドとしては、Johnson等(2000)Nephrol.Dial.Transplant 15(9):1274〜7,Kuai等(2000)J.Pept Res.56(2):59〜62,Barbone等(1999)Nephrol.Dial.Transplant.14 Supp 2:80〜4,Middleton等(1999)J.Biol.Chem.274(20):14163〜9,Johnson等(1998)Biochemistry 37(11):3699〜710,Johnson等(1997)Chem.Biol.12:939〜50,Wrighton等(1997)Nat Biotechnol 15(12):1261〜5,Livnah等(1996)Science 273:464〜71、および、Wrighton等,(1996)Science 273:458〜64によって開示されたエリスロポエチン模倣ペプチドが挙げられる。
【0226】
また治療的分子としては、アレルゲンタンパク質、およびそれらの消化されたフラグメントも挙げられる。これらの例としては、ブタクサ、ライ、ジューングラス(June grass)、カモガヤ、ハルガヤ、レッドトップグラス(red top grass)、オオアワガエリ、イエロードッグ、コムギ、トウモロコシ、ヤマヨモギ、イチゴツナギ,カリフォルニアの一年草(california annual grass)、アカザ、ギョウギシバ、ロシアアザミ(Russian thistle)、マウンテンシーダー(mountain cedar)、オーク、ネグンドカエデ、スズカケノキ、カエデ、ニレなどが由来の花粉アレルゲン、チリダニ、ハチ毒、食物アレルゲン、動物の鱗屑およびその他の昆虫毒が挙げられる。
【0227】
その他の治療的分子としては、ウイルス、細菌および原生動物性のワクチンなどの微生物性ワクチン、およびそれらの様々な成分、例えば表面抗原が挙げられる。これらの例としては、糖タンパク質、タンパク質またはこれらのタンパク質から誘導されたペプチドを含むワクチンが挙げられる。このようなワクチンは、スタフィロコッカス・アウレウス(Staphylococcus aureus)、ストレプトコッカス・ピオゲネス(Streptococcus pyogenes)、肺炎連鎖球菌(Streptococcus pneumoniae)、ナイセリア・メニンギチジス(Neisseria meningitidis)、淋菌(Neisseria gonorrhoeae)、サルモネラ属、シゲラ属、エシェリキア・コリ(Escherichia coli)、クレブシエラ属spp.、プロテウス属spp.、コレラ菌(Vibrio cholerae)、カンピロバクター・ピロリ(Campylobacter pylori)、緑膿菌(Pseudomonas aeruginosa)、ヘモフィルス・インフルエンザ(Haemophilis influenzae)、ボルデテラ・ペルツッシス(Bordetella Pertussis)、ヒト型結核菌(Mycobacterium tuberculosis)、レジオネラ・ニューモフィラ(Legionella pneumophila)、トレポネーマ・パリダム(Treponema pallidum)、クラミジア属、破傷風トキソイド、ジフテリアトキソイド、インフルエンザウイルス、アデノウイルス、パラミクソウイルス(流行性耳下腺炎、麻疹)、風疹ウイルス、ポリオウイルス、肝炎ウイルス、ヘルペスウイルス、狂犬病ウイルス、HIV−1、HIV−2、RSV、および乳頭腫ウイルスから調製される。
【0228】
好ましい融合分子は、抗HIVウイルスペプチド、抗RSVペプチド、ヒト成長ホルモン、αおよび/またはβインターフェロン、エリスロポイエチン(EPO)、EPO様ペプチド、顆粒球−コロニー刺激因子(GCSF)、顆粒球マクロファージコロニー刺激因子(GMCSF)、インスリン、インスリン様増殖因子(IGF)、トロンボポエチン、抗体のCDRに相当するペプチド、膵島新生関連タンパク質(Islet Neogenesis Associated Protein;INGAP)、カルシトニン、アンギオスタチン、エンドスタチン、インターロイキン−2、成長ホルモン放出因子、ヒト副甲状線ホルモン、抗腫瘍壊死因子(TNF)ペプチド、インターロイキン−1(IL−1)受容体、および/または単鎖抗体を含んでいてもよい。
【0229】
また本発明の融合タンパク質は、新しい機能または新規な機能を有する分子に関してスクリーニングするために、ペプチドライブラリーから誘導されたペプチドまたはポリペプチドが含まれるように調製してもよい。このようなペプチドライブラリーとしては、商業的または公共的に利用可能なペプチドライブラリー、例えば、アメリカン・ペプチド社(American Peptide Co.Inc.)、セル・サイエンス社(Cell Sciences Inc.)、インビトロジェン社(Invitrogen Corporation)、フェニックス・ファーマシューティカルズ社(Phoenix Pharmaceuticals Inc.)、ユナイテッド・ステイツ・バイオロジカル(United States Biological)のペプチドライブラリーが挙げられ、加えて、利用可能な技術、例えば標準的手法を用いて作製されたバクテリオファージや細菌のディスプレイライブラリーによって生産されたペプチドライブラリーも挙げられる。
【0230】
本発明のさらにその他の実施態様において、融合タンパク質は、当分野既知の治療用タンパク質の成分を用いることによって調製してもよく、このようなタンパク質の成分の例としては、現在のところ食品医薬品局(Food and Drug Administration)(www.fda.gov/cber/efoi/approve.htm)によって承認された、ペプチドおよびタンパク質、加えてPCT特許出願公開WO01/79258、WO01/77137、WO01/79442、WO01/79443、WO01/79444、およびWO01/79480(これらはいずれも、参照によりその全体として本明細書に組み入れられる)が挙げられる。
【0231】
PCT国際公開WO03/020746(参照により本明細書に加入させる)の表1に、完全ではないが、本発明の融合タンパク質の治療用タンパク質部分、すなわちリガンド成分に相当する治療用タンパク質の一覧が提供されている。「治療用タンパク質X」の列は、治療用タンパク質分子を記載しており、それに続く括弧内には、その治療用タンパク質分子またはそれらのフラグメントもしくは変異体が含まれる、あるいはそれらで構成される科学名とブランド名を含む。本明細書で用いられる「治療用タンパク質X」は、個々の治療用タンパク質分子(CASおよびGenBank登録番号より入手できるアミノ酸配列で定義されるような)、またはこの列で開示された所定の治療用タンパク質分子に関連する治療用タンパク質のグループ全体のいずれかを意味する場合がある。「典型的な識別子」の列は、ケミカル・アブストラクツ・サービス(CAS)のレジストリナンバー(米国化学会によって公開)、および/またはGenBank登録番号(例えば、遺伝子座ID、NP−XXXXX(参照配列タンパク質)、およびXP−XXXXX(モデルタンパク質)という識別子を示しており、このような識別子は、国立バイオテクノロジー情報センター(National Center for Biotechnology Information ;NCBI)のウェブページ(www.ncbi.nlm.nih.gov)を介して利用可能であり、タンパク質分子、または治療用タンパク質分子のフラグメントもしくは変異体のアミノ酸配列を含む、CASレジストリまたはGenBankデータベースにおけるエントリーに相当する。加えて、GenSeq登録番号および/またはジャーナル出版物の引用が、いくつかのポリペプチドに関する典型的なアミノ酸配列を同定するために与えられている。
【0232】
これらCASおよびGenBank、ならびにGenSeq登録番号、同様に、引用されたジャーナル出版物それぞれに関連する要約の頁が利用可能であり(例えば、パブメドID番号(PubMed ID number;PMID))、これは参照によりその全体として本明細書に組み込まれる。PCT/特許の参照の列は、治療用タンパク質分子を記載している特許および/または特許出願公開に相当する米国特許またはPCT国際公開番号を示す(これらはいずれも、参照によりその全体として本明細書に加入される)。生物活性の列は、治療用タンパク質分子に関連する生物活性を記載している。典型的な活性分析の列は、治療用タンパク質X部分を含む本発明の治療用タンパク質またはトランスフェリン融合タンパク質の治療活性および/または生物活性を試験するのに使用することができる分析を記載している参考文献を示す。また、これらの参考文献も、参照によりその全体として本明細書に加入される。「好ましい指標Y」の列は、治療用タンパク質X、または治療用タンパク質X部分を含む本発明のトランスフェリン融合タンパク質によって治療、予防、診断または改善が可能な疾患、障害および/または状態を記載している。本発明は、WO03/020746で提供された治療用タンパク質を含む(これは参照によりその全体として本明細書に加入される)。
【実施例】
【0233】
実施例1− GPIアンカー、hMUC1、およびmTf発現カセットの製造
mTf発現カセット(配列番号16)を含むpREX0549ベクターを、SalIおよびHindIIIで消化した。図3は、pREX0549に関するベクターのマップを示す。プライマーP0922およびP0923(配列番号7および配列番号8)を一緒にアニールし、SalI/HindIIIの消化部位でpREX0549にライゲーションした。P0922およびP0923によって形成されたリンカーには、SpeI、HindIIIおよびXbaI制限部位が含まれ、GPIアンカーおよびMUC1軸をコードする核酸分子が受け入れられるように設計された。得られたベクターpREX0628には、P0922/P0923リンカーと共にmTf発現カセットが含まれる。
【0234】
pREX0628を、HindIIIおよびXbaIで消化した。プライマーP0924およびP0925(配列番号9および配列番号10)をアニールして、GPIアンカーYIR019cを形成させた。YIR019cを、消化したpREX0628にライゲーションし、ベクターpREX0634を形成させた。
【0235】
hMUC1のcDNAを、RT−PCRで、プライマーP0958およびP0959(配列番号11および配列番号12)を用いて、ヒト乳腺腫瘍トータルRNAライブラリー(クロンテック)から増幅した。得られたcDNAを、プライマーP1019およびP1020(配列番号13および配列番号14)で増幅して、SpeIおよびHindIII制限部位を作出した。このようにして得られたSpeIおよびHindIII部位を有するhMUC1は、配列番号6で示される。
【0236】
pREX0634を、SpeIおよびHindIIIで消化し、SpeIおよびHindIII部位を有するhMUC1をベクターにライゲーションした。このようにして得られたベクターpREX0663を、提示発現カセット(mTf−MUC1−GPI)として用いた。
【0237】
pREX0663を用いて、高および低コピー数の酵母発現ベクターを作出した。高コピー数の酵母発現ベクターを作出するために、4.1kbの提示発現カセットを、ベクターをNotIで消化することによってpREX0663から除去した。次に、この発現カセットを、NotIで消化し、脱リン酸化したpSAC35ベクターにライゲーションし、ベクターpREX0667(酵母提示ベクターI)を得た。
【0238】
低コピー数の酵母発現ベクターを、pREX0663をNotIで消化し、この発現ベクターを、NotIで消化し、脱リン酸化したpREX0699にライゲーションし、pREX0721酵母提示(酵母提示ベクターII)を得ることによって作製した。
【0239】
上述の酵母発現ベクターを用いて当技術分野で既知のように酵母細胞および細菌細胞を形質転換させることができる。このようなベクターは、当技術分野で既知のように酵母で発現させることができる。さらに、ランダムペプチドまたはCDRのようなリガンド成分のライブラリーを提示することができる発現したトランスフェリン融合タンパク質を収集物を作成することができ、さらに、当技術分野で既知のように結合に関してスクリーニングするのに用いることができる。
【0240】
実施例2− 15−merのランダムライブラリーの構築
新規な結合特性を有するトランスフェリン変異体を選択するために、当技術分野で既知のPCRニッティング法によって、トランスフェリンの289〜290のアミノ酸位置におけるランダムな15−merのライブラリーを構築した(MartinおよびSmith(2006)Biochem J.396(2):287〜95を参照)。通常、結合エピトープを形成するのに必要なのはわずか約7個のアミノ酸であり、約109のライブ
ラリーは、設計されるライブラリー(3.3×1019)のうちの小さなフラクションをカバーするに過ぎないが、15−merのライブラリーを設計した。しかしながら、109のライブラリーサイズの場合、15−merのライブラリーは、同じ規模の7−merのライブラリーよりも6.4倍多い7−merをカバーする。
【0241】
P1174/P1227を用いてトランスフェリンのBamHI/BspEI配列を含むDNAフラグメントを得た後、2回のPCR反応(それぞれ単一のプライマー−P1172およびP1173を使用)を行い、一本鎖DNAを得た。ssDNAを単離し、アニールして、結合した15−merのライブラリーを形成した。この操作によって、ライブラリーが、合成オリゴヌクレオチドのもともとの複雑さを確実に維持するようにした。二本鎖のニットした15−merのライブラリーを、P1174/P1227を用いてさらに増幅し、十分な量のDNAを得た。PCR産物を精製し、BamHI/BspEIで消化し、適切なプラスミドベクター、例えばpREX0995(図4)またはpREX0667にクローニングした。
P1172(配列番号19)
289〜290 15−merのランダムペプチドライブラリーの挿入物がニットしたフォワードバックフラグメント
【化1】
DNA配列中の各位置にランダム性を導入するために、LaBeanおよびKauffman(1993)Protein Sci.2:1249〜54に従って、その位置にヌクレオチドの混合物(A、G、TおよびC)を予め決められた比率で組み込んだ。以下で既定される混合物は、停止コドンの頻度を最小化し、アミノ酸組成を天然タンパク質に適合する。
5 13%T,32%G,20%C,35%A
6 24%T,24%G,22%C,30%A
7 37%T,26%G,37%C。
P1173(配列番号20)
フロントフラグメントのための289〜290 15−merのランダムペプチドライブラリーがニットしたバックプライマー
【化2】
P1174(配列番号21)
フロントフラグメントのための289〜290 15−merのランダムペプチドライブラリーがニットしたフォワードプライマー
【化3】
P1227(配列番号22)
バックフラグメントのための289〜290 15−merのランダムペプチドライブラリーが結合したバックプライマー
【化4】
【0242】
実施例3− 酵母細胞を提示するフラグの選択
酵母提示系を確立し、それを用いて、軸領域、huMUC1およびGPIシグナル配列に融合させることによって酵母表面にトランスフェリンのN−ローブを提示した。この系の結合選択における有用性実証するために、フラグ−タグ配列
【化5】
またはランダムな15−merペプチドライブラリーを、トランスフェリンN−ローブのアミノ酸289番目に挿入した。次に、フラグタグを付けたトランスフェリンN−ローブ、pREX1012を提示する酵母(図6)を、ランダムな15−merのペプチドを有するトランスフェリンN−ローブを提示する酵母のプールに添加した。この混合群から、フラグタグを付けたトランスフェリンN−ローブを提示する酵母だけを、抗フラグ抗体を用いた選択によって回収した。
【0243】
フラグタグ配列を、トランスフェリンのアミノ酸289番目に挿入するために、フラグタグ配列を組み込んだオリゴを合成し、PCRでpREX0667ベクターにニットし、pREX0759を生成させた(図7)。次に、pREX0759のフラグタグ配列を含むBamHI/BspEIフラグメントを用いて、pREX0995の同じ制限フラグメントを置換した(図4)。得られたプラスミドpREX1012は、フラグタグを付けたトランスフェリンN−ローブ−MUC1−GPI融合タンパク質を発現した。
【0244】
また、15−merのライブラリーも、pREX0995のBamHI/BspEI部位の間にクローニングした。15−merのライブラリーの製造を以下で説明する。ライゲーションサンプルを、E.コリDH5αに形質転換し、全ての形質転換混合物を、2LB/Amp(50μg/mL)の寒天プレートで平板培養した。全てのコロニーを回収し、キアゲン(Qiagen)プラスミド・プレップ・プロトコールを用いて、数種のミニプレップカラムを用いてプラスミドDNAを抽出した。
【0245】
pREX1012と15−merライブラリーの両方のプラスミドDNAを、サッカロミセス・セレビジエ株DS1101cir°に形質転換した。pREX1012の単一のコロニーを、スクロースを含む緩衝化した最小培地(BMM/S)に植え付け、一晩培養した。15−merライブラリーの全てのコロニーを回収し、BMM/Sに植え付けた。これら二種の一晩培養したもの細胞数をヘマサイトメーターによって測定し、以下の細胞混合物を調製した:
(A)109の15−merライブラリー酵母細胞と10:107の割合で混合された、103個のpREX1012酵母細胞
(B)108の15−merライブラリー酵母細胞と100:107の割合で混合された、103個のpREX1012酵母細胞。
【0246】
この細胞混合物を、細胞ブロック溶液(1×PBS/0.05%トゥイーン(Tween)−20、1%BSA)1ml中、氷上で30分間インキュベートした。遠心分離(13000rpmで30秒)後、細胞ペレットを、ビオチン化抗フラグ抗体(シグマ・アルドリッチ、1:25希釈)を含む洗浄溶液(1×PBS、0.5%BSA、2mMのEDTA)1mlに懸濁し、氷上で30分間インキュベートした。細胞を洗浄溶液1mlで2回洗浄し、(A)800μlまたは(B)160μlのブロック溶液に懸濁した。細胞懸濁液(A)200μlまたは(B)40μlにストレプトアビジンMACSマイクロビーズ(ミルテニー・バイオテク)を添加し、氷上で30分間インキュベートした。MSカラムを製造元の説明書(ミルテニー・バイオテク)に従って用いて、非標識細胞から標識された細胞を分離した。標識された細胞を回収し、BMM/Sアガロースプレート上で平板培養し、小さいコロニーが出現するまで30℃でインキュベートした。
【0247】
各プレートから全てのコロニーを回収し、それらをBMM/S5ml中30℃で一晩増殖させることによって、二回目の選択を行った。1.5のOD600に等しいこれら培養細胞を、上述のようなさらなるMACS分離で処理した。この二回目のスクリーニングから得られた細胞を、BMM/S5ml中で30℃で一晩培養した。各選択の前および後の酵母細胞培養物を、アジレント・テクノロジー(Agilent Technolgy)製のバイオアナライザーで抗フラグモノクローナル抗体(シグマ・アルドリッチ)およびAPC標識ヤギ抗マウス検出抗体を用いたFACSで分析した。FACS分析は製造元の説明書に従って行われた。2回目のMACS分離の後、フラグタグを付けた酵母の存在が明らかになった(図8)。
【0248】
実施例4− Aga1軸提示
酵母遺伝子AGA1のコア領域のDNA配列(残基XXX−XXX)を、以下のプライマーを用いてS288cの酵母ゲノムDNAをPCRで処理することによって得た:
【化6】
【0249】
約1.5kbのPCR産物を単離し、XbaI/HindIIIで消化した。このフラグメントを、SpeI/HindIIIで消化したpREX0855にライゲーションし(図9)、E.コリDH5αに形質転換した。得られた全てのコロニーを回収し、プラスミドDNAを細胞から単離した。発現カセットをNotI消化によって回収し、pSAC35にライゲーションして、酵母発現ベクターを得た。
【0250】
上述したように、酵母に形質転換し、抗フラグ抗体でFACS分析した。酵母コロニーは、高レベルのN−ローブ提示を示し、匹敵するMUC1からなる軸をベースとするコンストラクトよりも約10倍高かった(データ示さず)。
【0251】
酵母のシングルコロニーを単離し、この酵母細胞からプラスミドDNAを抽出した。抽出されたプラスミドDNAをNotIで消化した後に、NotI発現カセットを回収し、NotIで消化したpREX0855にライゲーションし、プラスミドpREX1087(図10)、フラグ−N−ローブ−Aga1−GPI発現カセットを含むPUCベースのベクターを得た。Aga1に相当するDNA配列の領域を配列分析し、その同一性を確認した。さらにこの発現カセットもpSAC35に戻し、pREX1106を得た(図11)。
【0252】
実施例5− 酵母細胞で機能する哺乳動物のGPI変異体の選択
哺乳動物のGPIシグナルは、それらの酵母カウンターパートが果たさない役割、例えば細胞内トラフィッキング、膜貫通シグナルの伝達、およびクラスリン非依存性のエンドサイトーシスといった役割を果たす(Biochem J.1993,294:305〜324)。酵母細胞は、細胞膜を有するだけでなく、哺乳動物細胞にはない細胞壁も有し、酵母GPIの多くは、タンパク質を酵母細胞壁に標的化する特有の配列を有する(J Bacteriol,1999,181:3886〜3889)。ヒト胎盤のアルカリホスファターゼのGPIは、酵母細胞では全く機能しないことが示されている(Mol Microbiol 1999,34:247〜256)。発現された組換えタンパク質を酵母細胞壁に付着させることができる新規な配列を得る手段として、pREX0885をベースとするがhuMDPのGPI配列
【化7】
を用いた酵母提示ベクター(図9)を用いた。この配列を、4つの完全にランダムなコドン(X)、加えて数種の(下線で示した)合理的な改変が組み込まれるように改変した:
【化8】
ここにおいて、Xはあらゆるアミノ酸である。
【0253】
以下の融合タンパク質、GPI配列が上述の通りに改変されたフラグタグ−N−ローブ−MUC1からなる軸−GPIを発現する酵母ライブラリーを、サッカロミセス・セレビジエ株DS1101cir°に形質転換した。本融合タンパク質が細胞壁に付着した全ての酵母細胞を、ビオチン化抗フラグ抗体を用いたMACSによって単離した。
【0254】
2つのオリゴP2035およびP2036(以下を参照)をアニールし、Taqポリメラーゼを用いて伸長させた。得られたDNAフラグメントを精製し、HindIII/XbaIで消化し、HindIII/XbaIで消化したpREX0855にライゲーションした(以下および図9を参照)。
プライマー
【化9】
【0255】
ライゲーション混合物をE.コリDH5αに形質転換し、約5x105個のコロニーを得た。全てのコロニーを回収し、プラスミドDNAを単離した。プラスミドDNAをNotIで消化し、発現カセットを回収し、SAC35にクローニングし、酵母発現ライブラリーを作出した。このライブラリーをエレクトロポレーションでDS1101cir°細胞に形質転換した。上述のライブラリーを一晩培養したものを、ビオチン化抗フラグ抗体を用いたMACSで処理した。再度、単離した細胞を同じ抗体を用いたMACSによって即座に精製した。得られた細胞をBMMSプレートに平板培養し、24個のコロニーをFACSとDNA配列分析によって特徴付けた(フラグの付加の説明を参照)。
【0256】
読取り可能な配列を示した22個のクローンのうち7個が、全長のGPIアンカーを有しており(表1)、その提示レベルは様々であり、そのうち最良のものは、酵母GPIアンカーを有する同じ融合タンパク質を発現するpREX1003ベクターよりも優れていた。
【0257】
【表5】
【表6】
【0258】
意外なことに、GPIアンカーシグナルを有効に欠失させるランダム化したコドンのうち1つにおける停止コドンによってクローンの50%がトランケーションされた。これらの12種のクローンのうち2種が、pREX1003よりも有意に優れた提示レベルを有することが測定され、停止コドンの直前にシステイン残基を含むことがわかった(CQIGGS*(配列番号34)およびCQ*、ここにおいて*は停止コドン)(図12)。恐らく、これらのコンストラクトは、細胞壁タンパク質中の遊離のシステイン残基にジスルフィド結合することを介して細胞壁に架橋された。
【0259】
上記の実施例を参照として本発明を詳細に説明したが、当然ながら、本発明の本質から逸脱することなく様々な改変を施すことができる。従って、本発明は以下の請求項によってのみ限定される。本願において言及された全ての引用された特許、特許出願および出版物を、参照によりその全体として本明細書に組み込まれる。
【図面の簡単な説明】
【0260】
【図1】トランスフェリン融合タンパク質上に提示されたランダムペプチドまたはCDRライブラリー、およびリガンドと標的との結合を示す。
【図2】酵母YIR019CGPIアンカーペプチド配列を示し、細胞膜への付着に関与するアミノ酸を強調表示した。
【図3】pREX0549に関するベクターのマップを示す。
【図4】pREX0995に関するベクターのマップを示す。
【図5】pREX0667に関するベクターのマップを示す。
【図6】pREX1012に関するベクターのマップを示す。
【図7】pREX0759に関するベクターのマップを示す。
【図8】2回のMACS分離の後のフラグタグを付けた酵母の存在を示す。
【図9】pREX0855に関するベクターのマップを示す。
【図10】pREX1087に関するベクターのマップを示す。
【図11】pREX1106に関するベクターのマップを示す。
【図12】MUC1およびAGA1を用いたFACS分析を示す。
【技術分野】
【0001】
関連出願
本願は、2005年6月17日付けで出願された米国仮出願第60/691,229号の利益を請求する。本願は、2004年11月23日付けで出願された米国特許出願第10/515,429号;2003年7月9日付けで出願された米国仮出願第60/485,404号;2003年3月10日付けで出願された米国特許出願第10/384,060号;および2002年8月30日付けで出願された米国仮出願第60/406,977号(これらは全て、それら全体として参照により組み込まれる)に関係する。
【0002】
発明の分野
本発明は、融合タンパク質、融合タンパク質ライブラリー、およびリガンドの結合活性に関してスクリーニングするための融合タンパク質の使用に関する。
【背景技術】
【0003】
細胞表層提示系
コンビナトリアルライブラリーによるスクリーニングおよび選択方法は、一般的な研究ツールになっている(Phizicky等(1995)Microbiological Reviews 59:94〜123)。最も普及した技術の一つはファージディスプレイであり、この技術によれば、タンパク質はバクテリオファージコートタンパク質へのポリペプチド融合として発現し、後で、固定化または可溶性ビオチン化リガンドに結合させることによってスクリーニングされる。ランダムペプチドの提示は、M13、fdおよびf1のような線状バクテリオファージの外面上に発現されるキメラタンパク質を構築することによって達成されることが多い。ファージディスプレイは、抗体、DNA結合タンパク質、プロテアーゼ阻害剤、および酵素への応用が成功している。Hoogenboom等(1997)Trends in Biotechnol.15:62〜70;Ladner(1995)Trends in Biotechnol.13:426〜430;Lowman等(1991)Biochemistry 30:10832〜10838;Markland等(1996)Biochemistry 35:8045〜8057;および、Matthews等(1993)Nucleic Acids Res.21:1727〜1734を参照。
【0004】
ファージディスプレイに加えて、数種の細菌細胞表層提示法が開発されている。Georgiou等(1997)Nat.Biotechnol.15:29〜34を参照。細菌細胞表層提示法において取られるアプローチの一つでは、ピリンタンパク質(TraA)またはそれらの部分を含む融合タンパク質や、繊毛を形成することができる細菌宿主細胞の外表面にライブラリーペプチドを提示する異種ポリペプチドを使用してきた。米国特許第5,516,637号(これは参照によりその全体として本明細書に組み込まれる)を参照。
【0005】
FLITRXTMランダムペプチドライブラリー(インビトロジェンTMライフ・テクノロジーズ(InvitrogenTM Life Technologies))は、E.コリ(E.coli)の表面にコンホメーション的に制限された形式で12merのランダムペプチドライブラリーを提示するために、細菌の鞭毛タンパク質、FliC、およびチオレドキシン、TrxAを使用している。Lu等(1995)BioTechnology 13:366を参照。これらの系は、抗体エピトープマッピング、細菌の生ワクチンの送達系の開発および構築、ならびに環境の浄化目的および診断のための全細胞生物学的吸着剤の生成に応用されている。また、腫瘍由来上皮細胞上で腫瘍特異的な標的に結合するペプチド配列も、FLITRXTMシステムを用いて同定されている。Brown等(2000)Annals of Surgical Oncology,7(10):743を参照。
【0006】
ライブラリースクリーニングのために酵母細胞表層提示系が開発されており、ファージや細菌の提示系の限界のいくつかを克服することに成功している。酵母表層提示系、例えばpYD1酵母提示ベクターキット(pYD1 Yeast Display Vector Kit,インビトロジェンTMライフ・テクノロジーズ)は、細胞表層上に外来タンパク質を提示するためにS.セレビジエ(S.cerevisiae)のa−凝集素受容体を使用している。a−凝集素受容体は、AGA1およびAGA2遺伝子によってコードされた2つのサブユニットからなる。Aga1タンパク質(Aga1p,725個のアミノ酸)は、酵母細胞から分泌されて、酵母細胞壁の細胞外マトリックス中のβ−グルカンに共有結合するようになる。Aga2タンパク質(Aga2p,69個のアミノ酸)は、2個のジスルフィド結合を介してAga1pに結合し、分泌後、それとAga1pとの接触によって細胞に結合したままになる。Aga2pのN末端部分はAga1pへの付着に必要であり、その一方でタンパク質およびペプチドは、酵母細胞表層上での提示のためにC末端に融合することができる。凝集素は、天然型の酵母タンパク質であり、通常は、交配の間に酵母細胞を融合させるための特異的な付着による接触部として機能する。そのようなものとして、凝集素は、細胞壁成分との過剰な立体障害が起こらないようにタンパク質−タンパク質結合に関して進化してきた。Boder等の“タンパク質発現、親和性および安定性の定向進化に関する酵母表面提示(Yeast Surface Display for Directed Evolution of Protein Expression,Affinity,and Stability)”、キメラ遺伝子およびハイブリッドタンパク質の応用(Applications of Chimeric Genes and Hybrid Proteins)(Jeremy Thorner等),アカデミックプレス(Academic Press),2000,第328巻,430〜439頁;US6,699,658;および、US6,423,538(これらは参照によりその全体として本明細書に組み込まれる)。
【0007】
しかしながら、この系の欠点の一つは、Aga2pタンパク質を細胞壁に連結させるためには、Aga2p融合タンパク質とAga1pがジスルフィド結合を形成することが必要であるため、提示の効率が比較的低く、表面にタンパク質を有効に提示する酵母細胞がわずか40%〜60%である点である。Feldhause等(2003)Nat.Biotechnol.21(2):163〜70を参照。ライブラリーの(全てとは限らないが)ほとんどのタンパク質を細胞表層に提示する酵母提示系の必要がある。
【0008】
Aga1pおよびAga2p酵母提示系のその他の欠点は、スクリーニングしようとするリガンドを、Aga2pのC末端に付着させることを必要とする点である。その結果として、この系は、遊離のN末端が結合に必要であるペプチド、および/または遊離のN末端が活性に必要であるペプチドを選択するのに用いることができない。従って、リガンドN末端の酵母細胞タンパク質への結合を必要としないフレキシブルな提示系の必要がある。
【0009】
トランスフェリン融合タンパク質
血清トランスフェリン(Tf)は、80,000ダルトンの分子量を有する単量体の糖タンパク質であり、血液循環中で鉄に結合し、その鉄をトランスフェリン受容体(TfR)を介して様々な組織に輸送する(Aisen等(1980)Ann.Rev.Biochem.49:357〜393;MacGillivray等(1981)J.Biol.Chem.258:3543〜3553;および米国特許第5,026,651号)。Tfは、最も一般的な血清中の分子の一種であり、総血清タンパク質の約5〜10%以下で含まれる。糖鎖欠損トランスフェリンは、アルコール症の個体の血液中に高レベルで存在し、グリコシル化トランスフェリンの半減期(約7〜10日間)よりも長い半減期(約14〜17日間)を示す。van Eijk等(1983)Clin.Chim.Acta 132:167〜171;Stibler(1991)Clin.Chem.37:2029〜2037;Arndt(2001)Clin.Chem.47(1):13〜27;およびStibler等の“Carbohydrate−deficient consumption”,Advances in the Biosciences,(Ed Nordmann等),Pergamon,1988,第71巻,353〜357頁)を参照。Tfの構造はよく特徴付けられており、受容体結合、鉄結合および放出、ならびに炭酸イオン結合のメカニズムが解明されている。米国特許第5,026,651号、5,986,067号、および、MacGillivray等(1983)J.Biol.Chem.258(6):3543〜3546(これらの全ては、参照によりその全体として本明細書に組み込まれる)を参照。
【0010】
ムチンは、高度にグリコシル化されたタンパク質であり、マイクロアレイから実質的な距離をおいて融合タンパク質のリガンドドメインを持ち上げるのに用いられている。リガンドを基質から有意な距離に持ち上げることによって、リガンドの受容体を発現する細胞に提示された受容体への結合が増加すると仮説を立てられている。WO01/46698(これは参照によりその全体として本明細書に組み込まれる)を参照。
【発明の開示】
【発明が解決しようとする課題】
【0011】
これまで本発明の発明者等は、トランスフェリン融合タンパク質ライブラリーを開発してきた。米国特許出願第10/515,429号(これは参照によりその全体として本明細書に組み込まれる)を参照。本発明は、立体障害を減少させ、リガンドの結合が増加するように設計されたムチンのような軸様の成分を含むトランスフェリン融合タンパク質を提供する。発現されたトランスフェリン融合タンパク質が、ペプチドスクリーニングのプラットフォームとして用いることができるように、本融合タンパク質を、酵母のような宿主細胞の表面で発現させ提示させることができる。さらに、本融合タンパク質のトランスフェリンとリガンド部分を切断して、治療剤としてとして用いることができる。N末端が融合したAga2タンパク質を除去すると、トランスフェリンに連結した小さいリガンドのコンホメーションに影響を与える可能性が高いため、これを既存の酵母提示技術で達成することは不可能と思われる。
【課題を解決するための手段】
【0012】
以下でより詳細に説明されているように、本発明は、トランスフェリン(Tf)成分、軸成分、および細胞壁結合基を含む融合タンパク質を含む。Tf成分は、トランスフェリンタンパク質またはそれらの部分を含み、酵母細胞表層上に提示される。例えば、トランスフェリン成分は、トランスフェリンタンパク質のNドメインの一部、換言すればローブであってもよい。Tf成分は、本融合タンパク質のTf部分が、野生型Tfと比較して低いグリコシル化を示すように改変されたTfタンパク質であってもよい。本発明の一実施態様において、本融合タンパク質のトランスフェリン部分は、グリコシル化を示さない。本発明のその他の実施態様において、本融合タンパク質のトランスフェリン成分は、野生型トランスフェリンに比べて低い鉄、ビカーボネートへの親和性、および/または低いトランスフェリン受容体への親和性を示すように改変される。トランスフェリン成分は、トランスフェリン受容体、鉄またはビカーボネートに結合することができなくなるように改変されていてもよい。従って、本発明は、トランスフェリン成分が、グリコシル化部位、鉄結合部位、ヒンジ部位、ビカーボネート部位、および受容体結合部位からなる群より選択される1個またはそれ以上の部位で改変された、改変されたトランスフェリン成分を含む。
【0013】
ここで特許請求される発明のリガンドは、様々な方法でトランスフェリン成分と複合体化させてもよいし、またはトランスフェリン成分と融合させてもよい。さらに、トランスフェリン成分は、それに連結した1個より多くのリガンドを有していてもよい。リガンド成分は、トランスフェリン成分のN末端、C末端に融合していてもよいし、または、トランスフェリン成分内に位置していてもよい。本発明の一実施態様において、このようなリガンドは、アミノ酸位置Asp33、Asn55、Asn75、Asp90、Gly257、Lys280、His289、Ser298、Ser105、Glu141、Asp166、Gln184、Asp197、Lys217、Thr231、およびCys241からなる群より選択されるN−ローブ(N1またはN2)のアミノ酸位置の1つまたはそれ以上に挿入される。
【0014】
本発明のその他の実施態様において、リガンドは、トランスフェリン成分の露出したループ上に位置する。ランダムペプチドのようなリガンド成分は、トランスフェリン成分と共にフレーム内に存在し得るように、トランスフェリン融合タンパク質がコードされたベクターで宿主細胞によって発現され得る。トランスフェリン成分と共に発現されるランダムペプチドのリガンド成分は、当業界既知の多くの方法によって作出することができ、このような方法としては、これらに限定されないが、エラープローンPCR、およびDNAシャフリングが挙げられる。また、トランスフェリン融合タンパク質がそれまでに翻訳され終わった後に、リガンド成分をトランスフェリン融合タンパク質に付加することもできる。
【0015】
リガンドは多くの形態をとることができ、このような形態としては、これらに限定されないが、単鎖抗体、抗体、抗体フラグメント、抗体の可変領域、ランダムペプチド、または抗体相補性決定領域(CDR)が挙げられる。リガンドは、可変またはランダム領域、および非可変領域を含んでいてもよい。リガンドは、研究対象のリガンドであってもよいし、またはリガンドのライブラリー中の一種のリガンドであってもよい。リガンドは、多数の受容体または物質、例えばペプチド、抗原、受容体、抗体、毒素、代謝産物、および核酸に結合する可能性がある。
【0016】
軸成分は、そのN末端がトランスフェリン成分に融合し、そのC末端が細胞中に(例えば細胞壁中に)位置するように配置させることができる。一実施態様において、軸成分のC末端は、アンカー成分に融合される。本発明の軸成分は酵母細胞の細胞壁に伸びており、通常は中程度ないし高度にグリコシル化されたペプチドである。細胞壁に伸びていることによって、軸成分は、細胞壁を介して融合タンパク質を結びつけるための細胞壁連結要素として作用することが可能である。本発明の一実施態様において、軸成分は細胞壁に伸びており、細胞表層上に部分的に提示されている。軸成分の組成は、それにロッド様のコンホメーションを付与することができ、このようなコンホメーションにより、もしもそれながければ融合タンパク質(とりわけリガンド)と宿主細胞との間に存在するはずの立体障害を少なくすることができる。
【0017】
軸成分は、ムチン、ムチン変異体、またはそれらのフラグメントを含んでいてもよいし、またはそれらからなっていてもよい。ムチンドメインは、例えば、MUC1、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC6、MUC7、MUC8、およびMUC9、ならびにそれらの変異体を含んでいてもよい。一実施態様において、軸成分は、ヒトMUC1ドメインを含み、例えば配列番号5の核酸配列またはそれらのフラグメントに相当するペプチドを含む。その他の実施態様において、軸成分は、2回またはそれ以上のムチンの反復を含み、例えば2回またはそれ以上のMUC1またはMUC3の反復を含む。さらなる実施態様において、軸成分は、MUC1、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC6、MUC7、MUC8、およびMUC9からなる群より選択される2種またはそれ以上のムチンタンパク質、またはそれらの変異体もしくはフラグメントを含む。
【0018】
さらに軸成分は、天然型の酵母の細胞壁タンパク質などの中程度から高度にグリコシル化されたその他のタンパク質を含んでいてもよいし、またはそれらからからなっていてもよい。例えば、本発明の一実施態様において、軸成分は、Aga1、Aga1の変異体、もしくはそれらのフラグメントを含むか、またはそれらからなる。
【0019】
本発明の融合タンパク質は、融合タンパク質を宿主細胞に固定するまたは結びつけるように作用する細胞壁連結要素を含む。細胞壁連結要素は、本発明の融合タンパク質を酵母細胞壁に共有結合または非共有結合によって結合させることができる。本発明の一実施態様において、本融合タンパク質の軸成分は、細胞壁連結要素である。例えば、軸成分からのO−グリカンは、細胞壁のβ−グルカンに架橋することができる。その他の細胞壁連結要素としては、これらに限定されないが、遊離のシステイン残基を含むペプチドが挙げられる。例えば、対になっていないシステイン残基を1個またはそれ以上含む軸成分またはアンカー成分は、細胞壁中のタンパク質の対になっていないシステイン残基の1個またはそれ以上とジスルフィド結合を形成することができる。
【0020】
本発明の融合タンパク質は、場合により、アンカー成分を含んでいてもよく、このアンカー成分も、トランスフェリン融合タンパク質を宿主細胞に固定するまたは結びつけるように作用する。アンカー成分は、細胞壁連結要素であってもよいし、または本融合タンパク質を酵母細胞膜に結びつけてもよい。
【0021】
なかでも、本発明の融合タンパク質を酵母細胞膜に結びつけることができるアンカードメインの一つは、グリコシルホスファチジルイノシトール(GPI)ペプチドアンカーであり、これは翻訳後タンパク質修飾によってGPIシグナルペプチド配列(例えば配列番号15で示されるシグナルペプチド配列)のオメガ部位(ω−site)に付加される。本発明の一実施態様において、アンカー(例えば改変されたGPIシグナル配列によって提供されるアンカー)は、切断される前に、融合タンパク質を宿主細胞膜または細胞壁を一時的に繋ぐ。一旦切断されると、融合タンパク質は、細胞壁のβ−グルカンに架橋されている軸成分からのグリカンにより、細胞壁連結要素を介して細胞に連結したままになる。
【0022】
本発明のその他の実施態様において、アンカーは、膜貫通ドメインである。膜貫通ドメイン(TMD)は、1回膜貫通型IまたはII膜タンパク質の領域でもよいし、またはマルチスパン型膜タンパク質の数個の膜貫通領域のいずれか一つでもよい。
【0023】
本発明はまた、特許請求される融合タンパク質をコードする核酸分子も含む。このような核酸は、ベクターに挿入することができ、酵母のような宿主細胞を形質転換させるのに用いることができる。宿主細胞は、本発明の核酸で形質転換されると、本融合タンパク質を発現することができる。本融合タンパク質の発現の誘導は当業界既知の方法によって制御することができ、例えば誘導性プロモーターの使用によって制御することができる。本発明は、宿主細胞の集合体、例えば、ランダム化ペプチドを提示する本発明の融合タンパク質を発現する酵母細胞の集合体に発現された融合タンパク質のライブラリーを含む。
【0024】
本発明のその他の実施態様において、本融合タンパク質は、リガンドまたは物質の結合活性についてスクリーニングするのに用いられる。特許請求される融合タンパク質を発現することができる宿主細胞のライブラリーを、これらに限定されないが抗原または受容体などの物質に曝露し、それから結合活性に関してスクリーニングすることができる。細胞表層提示ライブラリーは、これらに限定されないがFACSや磁気ビーズなどの当技術分野既知の方法を用いてスクリーニングすることができる。
【0025】
詳細な説明
一般的な説明
本発明の発明者等は、ライブラリー、例えばランダムペプチドまたはCDRライブラリーをスクリーニングするための、例えば細胞表層提示系の一部として用いることができる多機能の融合タンパク質を開発した。本融合タンパク質は、1またはそれ以上のリガンドと複合体化または融合したトランスフェリン成分を含む。本発明は、以下を条件とする限りにおいて、トランスフェリン以外のタンパク質を含む融合タンパク質を想定しており、上記条件は、上記その他のタンパク質が可溶性であり、融合タンパク質の残分の部分から切断されたときに融合した1またはそれ以上のリガンドに高い血清中半減期を付与することができる場合である。例えば、トランスフェリンの代わりに、アルブミンまたはその変異体もしくはフラグメントを用いることができる。
【0026】
本融合タンパク質のトランスフェリン成分は、中程度から高度にグリコシル化された軸成分に融合している。本融合タンパク質は、本融合タンパク質を酵母細胞の細胞壁に共有結合または非共有結合で結合させることができる細胞壁連結要素を含む。本発明の一実施態様において、本融合タンパク質は、アンカー成分(例えば膜貫通ドメイン)をも含む。
【0027】
本融合タンパク質は、酵母提示系して用いられる場合、Aga1pおよびAga2p酵母提示系と比較して、ペプチドを提示した細胞表層を有するクローンを高い割合で提供すること等を含む従来技術を超える利点を提供する。また本発明の融合タンパク質は、結合に利用可能なN末端を必要とするリガンドをスクリーニングすることのフレキシビリティーをも提供する。
【0028】
本発明はまた、本融合タンパク質またはそれらの部分を含む治療用組成物、および治療を必要とする対象に本融合タンパク質またはそれらの部分を投与することによって、疾患または障害を治療、予防または改善する方法も含む。本発明の融合タンパク質は、リガンド成分として、推定上の治療用タンパク質の少なくともフラグメントまたは変異体を含む。本発明の一実施態様において、本融合タンパク質のトランスフェリンおよびリガンド(すなわち治療薬)部分は、軸成分(すなわち本融合タンパク質の酵母細胞と結合した部分)から切断することができ、生物薬剤またはワクチンを調製するのに用いることができる。
【0029】
定義
本明細書で用いられる用語「生物活性」は、生物学的な環境において、すなわち生体内またはインビトロのその模擬環境において、治療的な分子、リガンド成分、タンパク質またはペプチドによって行われる機能または一連の活性を意味する。生物活性としては、これらに限定されないが、特許請求される融合タンパク質の治療的分子部分の機能が挙げられ、例えばこれらに限定されないが、応答細胞系からの細胞外マトリックス分泌の誘導、ホルモン分泌の誘導、走化性の誘導、有糸分裂誘発の誘導、分化の誘導、または応答細胞の細胞分裂の阻害が挙げられる。本発明の融合タンパク質またはペプチドは、その治療用タンパク質の天然型の対応物が示す1種またはそれ以上の生物活性を示す場合、生物学的に活性であるとされる。
【0030】
本明細書で用いられるように、トランスフェリン配列における「〜に相当するアミノ酸」または「等価なアミノ酸」は、第一のトランスフェリン配列と、少なくとも第二のトランスフェリン配列との間の同一性または類似性が最大になるようにアラインメントによって同定される。第二のトランスフェリン配列における等価なアミノ酸を同定するのに用いられる番号は、第一のトランスフェリン配列におけるそれに相当するアミノ酸を同定するのに用いられる番号に基づく。特定の場合において、これらの語句は、ヒトトランスフェリンにおけるアミノ酸残基を、ウサギ血清トランスフェリンにおける特定の残基と比較して説明するのに用いる場合もある。
【0031】
本明細書で用いられる用語「Tf成分」、「Tfタンパク質のフラグメント」または「Tfタンパク質」または「Tfタンパク質の部分」は、天然に存在するTfタンパク質、またはその突然変異体の少なくとも約5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、96%、97%、98%、99%または100%を含むアミノ酸配列を意味する。
【0032】
本明細書で用いられる用語「遺伝子」は、生物学的機能に関連するDNAのあらゆるセグメントを意味する。従って、遺伝子は、これらに限定されないが、それらの発現に必要なコード配列および/または調節配列を含む。また遺伝子は、例えば他のタンパク質に関する認識配列を形成する発現されないDNAセグメントを含んでいてもよい。遺伝子は、関心対象の源からクローニングすること、または既知のもしくは予測された配列情報から合成することによって、様々な源から得ることができ、さらに望ましいパラメーターを持つように設計された配列を含んでいてもよい。
【0033】
本明細書で用いられる「異種ポリヌクレオチド」または「異種核酸」または「異種遺伝子」または「異種配列」または「外因性DNAセグメント」は、特定の宿主細胞にとって外来の起源であるポリヌクレオチド、核酸またはDNAセグメントを意味するか、または同じ源由来の場合、その元の形態から改変されたポリヌクレオチド、核酸またはDNAセグメントを意味する。宿主細胞における異種遺伝子には、特定の宿主細胞にとって内在性であるが、修飾されている遺伝子が含まれる。従って、この用語は、細胞にとって外来または異種であるDNAセグメント、または細胞に相同であるが、その構成要素が通常は見出されない宿主細胞の核酸内の位置に存在するDNAセグメントを意味する。一例として、酵母細胞にとっては天然型であるが、ヒトTf配列に付着したシグナル配列は、異種である。
【0034】
本明細書で用いられる「単離した」核酸配列は、その他の核酸配列を実質的に含まない核酸配列を意味し、例えば、アガロースゲル電気泳動によって決定した場合に、少なくとも約20%純粋な、好ましくは少なくとも約40%純粋な、より好ましくは約60%純粋な、さらにより好ましくは約80%純粋な、最も好ましくは約90%純粋な、さらに最も好ましくは約95%純粋な核酸配列である。例えば、単離された核酸配列は、その天然の位置から異なる部位(そこで核酸配列が複製されることになる)に核酸配列を移転させるための、遺伝子工学で用いられる標準的なクローニング法により得ることができる。クローニング法は、上記ポリペプチドをコードする核酸配列を含む所望の核酸フラグメントを切り出して単離し、このフラグメントをベクター分子に挿入し、組換えベクターを核酸配列の複数のコピーまたはクローンが複製されることになる宿主細胞に組み込ませることを含み得る。核酸配列は、ゲノム、cDNA、RNA、半合成、合成由来のものでもよいし、またはそれらのあらゆる組み合わせでもよい。
【0035】
本明細書で用いられるように、2個またはそれ以上のDNAコード配列間のフレーム内融合の結果として、DNAコード配列が融合ポリペプチドに翻訳される場合、2またはそれ以上のDNAコード配列は、「連結した」または「融合した」と言う。用語「融合」は、融合タンパク質に関して用いられる場合、リガンド成分、軸成分、およびアンカー成分を含む。Tf融合タンパク質は、トランスフェリン成分が軸成分に融合したものであり、細胞壁連結要素を含む。
【0036】
本明細書で用いられる「改変されたトランスフェリン」は、そのアミノ酸配列に、野生型トランスフェリンに比べて少なくとも1つの改変を示すトランスフェリン分子を意味する。
【0037】
本明細書で用いられる「改変されたトランスフェリン融合タンパク質」は、軸成分に融合しているリガンドと複合体化した、またはそれらに融合した、改変されたトランスフェリン(またはそのフラグメントもしくは変異体)の少なくとも1つの分子の融合によって形成されたタンパク質を意味する。
【0038】
本明細書で用いられる用語「核酸」または「ポリヌクレオチド」は、デオキシリボヌクレオチドまたはリボヌクレオチド、およびそれらの一本鎖または二本鎖いずれかの形態のポリマーを意味する。特に限定されない限り、これらの用語は、参照核酸と類似の結合特性を有し、天然に存在するヌクレオチドと同様にして代謝される天然ヌクレオチドの類似体を含む核酸を包含する。特に他の指定がない限り、特定の核酸配列はまた、当然ながら、保存的に改変されたそれらの変異体(例えば、縮重コドンの置換)および相補配列、加えて、明確に示された配列も包含する。特に、縮重コドンの置換は、1またはそれ以上の選択された(または全ての)コドンの第三の位置が、混合型の塩基および/またはデオキシイノシン残基で置換されている配列を作製することによって達成することもできる(Batzer等(1991)Nucleic Acid Res.19:5081;Ohtsuka等(1985)J.Biol.Chem.260:2605〜2608;Cassol等(1992);Rossolini等(1994)Mol.Cell.Probes 8:91〜98)。核酸という用語は、遺伝子、cDNA、および遺伝子によってコードされたmRNAと同じ意味で用いられる。
【0039】
本明細書で用いられるように、DNAセグメントが、その他のDNAセグメントと機能的に関係するように置かれると、「機能するように連結した」と表現される。例えば、シグナル配列に関するDNAが、融合タンパク質の分泌に関与する前駆タンパク質として発現される場合、そのシグナル配列に関するDNAは、本発明の融合タンパク質をコードするDNAに機能するように連結しており、プロモーターまたはエンハンサーが、配列の転写を刺激する場合、そのプロモーターまたはエンハンサーはコード配列に機能するように連結している。一般的に、機能するように連結しているDNA配列は隣接しており、さらにシグナル配列または融合タンパク質の場合は、機能するように連結しているDNA配列は、隣接し、かつリーディング相中にある。しかしながら、エンハンサーは、それらが転写を制御するコード配列と隣接していなくてもよい。このような状況において、連結は、便利な制限部位でのライゲーション、またはその代わりに挿入されるリンカーもしくはアダプターでのライゲーションによって達成される。
【0040】
本明細書で用いられる用語「プロモーター」は、転写を開始させるためのRNAポリメラーゼ結合に関与するDNA領域を意味する。
【0041】
本明細書で用いられる用語「組換え」は、組換えDNAでの形質転換を受けている細胞、組織または生物を意味する。
【0042】
本明細書で用いられるように、標的化の実体、タンパク質、ポリペプチドまたはペプチドは、特定の細胞型(例えばリンパ球のような正常な細胞または癌細胞のような異常細胞)に特異的に結合する分子を意味し、それゆえにこれは細胞型が特異的であるTf融合タンパク質または化合物(薬物または細胞毒性薬)を、標的とするために用いられ得る。
【0043】
本明細書で用いられる「治療用タンパク質」は、1またはそれ以上の治療活性および/または生物活性を有するタンパク質、ポリペプチド、抗体、ペプチド、またはそれらのフラグメントもしくは変異体を意味する。本発明に包含される治療用タンパク質としては、これらに限定されないが、タンパク質、ポリペプチド、ペプチド、抗体、および生体物質が挙げられる。本明細書において、ペプチド、タンパク質およびポリペプチドという用語は交換可能に用いられる。加えて、用語「治療用タンパク質」は、治療用タンパク質に相関する内在性のまたは天然に存在するものを意味する場合もある。「治療活性」を示すポリペプチド、または「治療的に活性である」タンパク質は、本明細書で説明されている治療用タンパク質または当技術分野で既知のその他のものの1種またはそれ以上のような治療用タンパク質に関連する既知の生物学的および/または治療活性を1種またはそれ以上有するポリペプチドを意味する。非限定的な例として、「治療用タンパク質」は、疾患、状態または障害を治療、予防または改善するのに有用なタンパク質である。このような病気、状態または障害は、ヒトにおけるものでもよいし、またはヒト以外の動物におけるものでもよく、例えば獣医学での使用が考えられる。
【0044】
本明細書で用いられる用語「形質転換」は、核酸、すなわちヌクレオチドポリマーの細胞への転移を意味する。本明細書で用いられる用語「遺伝学的な形質転換」は、DNA(特に組換えDNA)の細胞への転移および組み込みを意味する。
【0045】
本明細書で用いられる用語「形質転換体」は、形質転換を受けた細胞、組織または生物を意味する。
【0046】
本明細書で用いられる用語「導入遺伝子」は、生物、宿主細胞またはベクターに、確実に機能する形で挿入される核酸を意味する。
【0047】
本明細書で用いられる用語「トランスジェニック」は、様々な形質転換法の1つによって、外来または改変された遺伝子、特に改変されたTf融合タンパク質をコードする遺伝子を受け入れた細胞、細胞培養、生物、細菌、菌類、動物、植物、およびこれらのうちいずれかの後代を意味し、ここにおいて、外来または改変された遺伝子は、外来もしくは改変された遺伝子を受け入れる生物種と同一種由来であるか、または外来もしくは改変された遺伝子を受ける生物種とは異なる種由来である。
【0048】
「変異体(variants)または変異体(variant)」は、参照核酸またはポリペプチドとは異なるが、それらの必須の特性を保持するポリヌクレオチドまたは核酸を意味する。一般的に、変異体は、全体的に極めて類似しており、多くの領域において参照核酸またはポリペプチドと同一である。本明細書で用いられる「変異体」は、天然型の治療用タンパク質とは配列が異なっているが、本明細書の別のところで記載されている、または当技術分野で既知の別のもののようなそれらの少なくとも1つの機能的および/または治療的な特性を保持する、本発明のトランスフェリン融合タンパク質の治療用タンパク質部分を意味する。
【0049】
本明細書で用いられる用語「ベクター」は、広範囲に、外因性核酸をコードするあらゆるプラスミド、ファージミドまたはウイルスを意味する。また、この用語には、例えばポリリジン化合物などのような、核酸のウィリオンまたは細胞への転移を容易にする、非プラスミド、非ファージミドおよび非ウイルス化合物も含まれると解釈されるものでもある。ベクターは、核酸またはそれらの突然変異体を細胞に送達するための送達媒体として適するウイルスベクターでもよく、または同じ目的に適する非ウイルスベクターでもよい。DNAを細胞および組織に送達するためのウイルスおよび非ウイルスベクターの例は当技術分野でよく知られており、例えば、Ma等(1997,Proc.Natl.Acad.Sci.U.S.A.94:12744〜12746)に記載されている。ウイルスベクターの例としては、これらに限定されないが、組換えワクシニアウイルス、組換えアデノウイルス、組換えレトロウイルス、組換えアデノ随伴ウイルス、組換え 鳥類ポックスウイルスなどが挙げられる(Cranage等,1986,EMBO J.5:3057〜3063;1994年8月18日に公開された国際特許出願公開番号WO94/17810;1994年10月27日に公開された国際特許出願公開番号WO94/23744)。非ウイルスベクターの例としては、これらに限定されないが、リポソーム、DNAのポリアミン誘導体などが挙げられる。
【0050】
本明細書で用いられる用語「野生型」は、天然に生じるポリヌクレオチドまたはポリペプチド配列を意味する。
【0051】
本明細書で用いられる「スカフォールドタンパク質」、「スカフォールドポリペプチド」または「スカフォールド」は、アミノ酸配列(例えばランダムペプチド)が融合することができるタンパク質を意味する。このようなペプチドはスカフォールドにとって外来性である。
【0052】
本明細書で用いられる「ランダムペプチド配列」は、2個またはそれ以上のアミノ酸単量体で構成され、ストカスティック過程またはランダム過程によって構築されたアミノ酸配列を意味する。ランダムペプチドは、フレームワークまたはスカフォールドのモチーフを含んでいてもよく、これらはインバリアント配列を含んでもよい。ランダムペプチド配列は、変異体ではない、すなわちランダムではないアミノ酸の一部を含んでいてもよい。
【0053】
本明細書で用いられる「ランダムペプチドライブラリー」は、一連のランダムペプチドをコードする一連のポリヌクレオチド配列を意味し、ならびにそれらのポリヌクレオチド配列によってコードされる一連のランダムペプチド、およびそれらのランダムペプチドを含む融合タンパク質を意味する。
【0054】
本明細書で用いられる用語「擬似ランダム」(“pseudorandom”)は、例えば一つの位置における残基の可変性の程度が、その他の位置における残基の可変性の程度と異なっているが、あらゆる擬似ランダム位において、かなり制限されるがある程度の残基の多様性が許容されるように、限定された可変性を有する一連の配列を意味する。
【0055】
本明細書で用いられる用語「定義された配列フレームワーク」は、一般的に実験データまたは構造データに基づいてランダムではない方式で選択される一連の定義された配列を意味し、例えば定義された配列フレームワークは、β−シート構造を形成すると予測される一連のアミノ酸配列を含んでいてもよく、またはいくつかあるバリエーションのなかでも、ロイシンジッパーヘプタッド反復モチーフ、ジンクフィンガードメインを含んでいてもよい。「定義された配列核」(defined sequence kernal)は、限られた可変性の範囲を包含する一連の配列である。20種の通常のアミノ酸からなる完全にランダムな10−merの配列が、(20)10の配列のいずれかである可能性があり、そして20種の通常のアミノ酸からなる擬似ランダムな10−merの配列が、(20)10配列のいずれかである可能性があるが、特定の位置および/または全体において特定の残基に関する偏りを示すことになるので、それぞれの残基の位置が許容できる20種の通常のアミノ酸(および/または許容できる非必須アミノ/イミノ酸)のいずれかであることが許容されたならば、定義された配列核は可能な配列の最大数より少ない配列の部分集合である。定義された配列核は、一般的に、変異した残基位置とインバリアントの残基位置とを含むか、および/またはアミノ酸残基の定義された部分集合から選択される残基などを含むことが可能な変異した残基位置を、これは選択された個々のライブラリー要素配列のセグメント単位で、またはその全長にわたってのいずれかで含む。定義された配列核は、アミノ酸配列、または、ポリヌクレオチド配列のいずれかを意味する場合もある。
【0056】
本明細書で用いられる「リンカー」または「スペーサー」は、DNA結合タンパク質とランダムペプチドのような2つの分子を連結させる分子または分子群を意味し、これらは、2つの分子を望ましい立体配置に配置するのに役立ち、例えば、DNA結合タンパク質との立体障害を最小にしてランダムペプチドが受容体に結合できるようになる。
【0057】
本明細書で用いられる用語「可変性のセグメント」は、ランダム、擬似ランダム、または定義された核配列を含む新生のペプチド部分を意味する。可変性のセグメントは、変異した残基位置とインバリアントな残基位置の両方を含んでいてもよく、変異した残基位置における残基のバリエーションの程度は、制限される可能性があり、いずれの選択肢も技術者の裁量で選択される。典型的には、可変性のセグメントは、長さが約3〜20個のアミノ酸残基、例えば長さが8〜10個のアミノ酸であるが、それより長い可変性のセグメントも可能であり、抗体部分または受容体タンパク質、例えば抗体フラグメント、核酸結合タンパク質、受容体タンパク質などを含んでもよい。
【0058】
本明細書で用いられる用語「エピトープ」は、抗原の一部、または抗体の可変領域結合ポケットと相互作用する結合相互作用を形成することができるその他の高分子を意味する。典型的には、このような結合相互作用は、CDRの1個またはそれ以上のアミノ酸残基との分子間接触として示される。
【0059】
本明細書で用いられる用語「受容体」、「標的」または「物質」は、所定のリガンドへの親和性を有する分子を意味する。受容体は、天然に存在する分子でもよいし、または合成分子でもよい。受容体は、改変されていない状態で用いてもよいし、またはその他の種との集合体として用いてもよい。受容体は、共有結合または非共有結合で、直接的に、または特異的な結合物質を介して、結合要素(すなわちリガンド)に結合することができる。受容体の例としては、これらに限定されないが、抗体、例えばモノクローナル抗体、および(例えば、ウイルス、細胞またはその他の物質における)特異的な抗原決定基との反応性を有する抗血清、細胞膜受容体、抗原、エピトープ含有分子、複合糖質および糖タンパク質、酵素、ならびにホルモン受容体が挙げられる。
【0060】
本明細書で用いられる用語「リガンド」または「リガンド部分」は、特定の受容体または物質によって認識されるランダムペプチドまたは可変性のセグメント配列のような分子を意味する。当業者であれば理解できると予想されるが、このような分子(または高分子複合体)は、受容体でもよいし、リガンドでもよい。
【0061】
本明細書で用いられる「融合した」、「複合体化した」または「機能するように連結した」とは、ランダムペプチドとスカフォールドタンパク質とが、スカフォールド構造の安定性の破壊が最小化されるような形で一緒に連結していることを意味する。
【0062】
本明細書で用いられる用語「単鎖抗体」は、ポリペプチドリンケージにVHドメインおよびVLドメインを含むポリペプチドを意味し、これは、一般的に、スペーサーペプチド(例えば、配列番号17の[Gly−Gly−Gly−Gly−Ser]x)を介して連結しており、さらにアミノおよび/またはカルボキシ末端に追加のアミノ酸配列を含んでもよい。例えば、単鎖抗体は、コーディングポリヌクレオチドに連結させるための固定セグメントを含んでもよい。一例として、scFvは、単鎖抗体である。単鎖抗体は、一般的に、実質的に免疫グロブリンスーパーファミリーの遺伝子によってコードされた少なくとも10個の隣接したアミノ酸のポリペプチドセグメント1個またはそれ以上からなるタンパク質であり(例えば、The Immunoglobulin Gene Superfamily,A.F.WilliamsおよびA.N.Barclay,in Immunoglobulin Genes,T.Honjo,F.W.AltおよびT.H.Rabbitts編(1989)アカデミックプレス:サンディエゴ,カリフォルニア州,361〜387頁を参照、これらは、参照により本明細書に組み込まれる)、最も多くは、げっ歯類、非ヒト霊長類、鳥類、ブタ、ウシ、ヒツジ、ヤギまたはヒトの重鎖または軽鎖遺伝子配列によってコードされた少なくとも10個の隣接したアミノ酸のポリペプチドセグメント1個またはそれ以上からなるタンパク質である。機能的な単鎖抗体は、一般的に、特異的な標的分子、典型的には受容体または抗原(エピトープ)への結合特性が保持されるように、免疫グロブリンスーパーファミリーの遺伝子産物の十分な部分を含む。
【0063】
本明細書で用いられる用語「相補性決定領域」および「CDR」は、KabatおよびChothiaのCDRの定義で例示されたような当技術分野で認められている用語を意味し、これはまた、一般的には、超可変領域または超可変ループとしても知られている。ChothiaおよびLesk(1987)J.Mol.Biol.196:901;Chothia等(1989)Nature 342:877;E.A.Kabat等,Sequences of Proteins of Immunological Interest(National Institutes of Health,ベセスダ,メリーランド州)(1987);およびTramontano等(1990)J.Mol.Biol.215:175を参照。可変領域ドメインは、典型的には、天然の免疫グロブリン鎖のアミノ末端の約105〜115個のアミノ酸(例えばアミノ酸1〜110)を含むが、可変ドメインは、多少短くても、または長くても、単鎖抗体を形成するのに適している。
【0064】
免疫グロブリンの軽鎖または重鎖可変領域は、3つの超可変領域(CDRともいう)で分断された「フレームワーク」領域からなる。フレームワーク領域およびCDRの範囲は、正確に定義されている。“Sequences of Proteins of Immunological Interest”,E.Kabat等,第4版,U.S.Department of Health and Human Services,ベセスダ,メリーランド州(1987)を参照。異なる軽鎖または重鎖のフレームワーク領域の配列は、種内で比較的保存されている。本明細書で用いられる「ヒトフレームワーク領域」は、天然のヒト免疫グロブリンのフレームワーク領域に実質的に(約85%またはそれ以上、通常は90〜95%またはそれ以上)同一なフレームワーク領域である。構成要素である軽鎖と重鎖とが一緒になったフレームワーク領域である抗体のフレームワーク領域は、CDRを配置し並べるのに役立つ。CDRは、主として、抗原のエピトープへの結合に関与する。
【0065】
特に他の指定がない限り、本明細書で用いられる技術用語や科学用語はいずれも、本発明が属する分野の当業者によって一般的に理解されているのと同じ意味を有する。本発明の実施または試験において、本明細書で説明されている方法および材料と類似した、またはそれらと同等のあらゆる方法および材料を用いることができるが、好ましい方法と材料を説明する。
【0066】
トランスフェリンおよびトランスフェリンの改変
本発明の融合タンパク質は、受容体または物質にランダムペプチドまたはCDRのようなリガンドを提示することができるトランスフェリン(Tf)タンパク質、またはそれらの部分を含む。Tf成分は、軸成分のN末端に融合している。本融合タンパク質のTfタンパク質またはそれらの部分を、本融合タンパク質のTf「部分」、「領域」または「成分」と称する場合もある。本明細書で用いられるように、トランスフェリン融合タンパク質は、軸成分に融合したトランスフェリンタンパク質または成分であり、さらに細胞壁連結要素を含む。本発明のトランスフェリン融合タンパク質は、場合によりアンカー成分を含んでいてもよい。
【0067】
本発明の改変されたTf融合タンパク質を作製するためにどのようなトランスフェリンでも使用可能である。一例として、野生型ヒトTf(Tf)は、約75kDaの679個のアミノ酸からなるタンパク質であり(グリコシル化は考慮に入れない)、これは遺伝子重複によって生じると思われる2つの主要なローブまたはドメイン、N(約330個のアミノ酸)およびC(約340個のアミノ酸)を有する。GenBank登録番号NM001063、XM002793、M12530、XM039845、XM039847、およびS95936(www.ncbi.nlm.nih.gov)(これらはいずれも、参照によりその全体として本明細書に組み込まれる)、ならびに配列番号1、2および3を参照。この2つのドメインは長い時間をかけて分岐しているが、大きい同一性/類似性を保持する。
【0068】
NおよびCドメインはそれぞれ、さらに2つのサブドメイン、N1およびN2、C1およびC2に分けられる。これは、鉄を体の細胞に輸送するように機能する。これは、全ての細胞で、特に活発に増殖する細胞で発現されるTf受容体(TfR)が介在するプロセスである。TfRは、Tfの鉄が結合した形態を認識し(1受容体につきその2つの分子を結合する)、続いてエンドサイトーシスが起こり、それによってTfR/Tf複合体が、エンドソームに輸送され、その時点で、局所的なpH低下によって、結合した鉄の放出、および細胞表層へのTfR/Tf複合体の再利用、およびTf放出(その鉄が結合していない形態で、アポTfとして知られている)が起こる。受容体結合は、主として、TfのCドメインを介する。グリコシル化されていない鉄が結合したTfが受容体と結合するため、Cドメイン中の2つのグリコシル化部位は、受容体結合に関与しないようである。
【0069】
Tf分子はそれぞれ、2個の鉄イオン(Fe3+)を運搬することができる。これらは、N1およびN2サブドメイン、C1およびC2サブドメインの間のスペースで複合体化され、分子においてコンホメーション変化が起こる。
【0070】
ヒトトランスフェリンにおいて、鉄結合部位は、少なくとも、配列番号3のアミノ酸Asp63(天然型のTfシグナル配列を含む配列番号2では、Asp82)、Asp392(配列番号2では、Asp411)、Tyr95(配列番号2では、Tyr114)、Tyr426(配列番号2では、Tyr445)、Tyr188(配列番号2では、Tyr207)、Tyr514または517(配列番号2では、Tyr533またはTyr536)、His249(配列番号2では、His268)、およびHis585(配列番号2では、His604)を含む。ヒンジ領域は、配列番号3の少なくともNドメインのアミノ酸残基94〜96、245〜247および/または316〜318、同様にCドメインのアミノ酸残基425〜427、581〜582および/または652〜658を含む。カーボネート結合部位は、配列番号3の、少なくともアミノ酸Thr120(配列番号2では、Thr139)、Thr452(配列番号2では、Thr471)、Arg124(配列番号2では、Arg143)、Arg456(配列番号2では、Arg475)、Ala126(配列番号2では、Ala145)、Ala458(配列番号2では、Ala477)、Gly127(配列番号2では、Gly146)、およびGly459(配列番号2では、Gly478)を含む。
【0071】
本発明の一実施態様において、本融合タンパク質は、改変されたヒトトランスフェリンを含むが、本発明の融合タンパク質を生産するのにあらゆる動物のTf分子が使用可能であり、例えば、ヒトTf変異体、ウシ、ブタ、ヒツジ、イヌ、ウサギ、ラット、マウス、ハムスター、ハリモグラ(echnida)、カモノハシ、ニワトリ、カエル、イモムシ、サル、類人猿が挙げられ、加えてその他のウシ属、イヌ科および鳥類の種も挙げられる。これらのTf配列はいずれも、GenBankやその他の公開データベースで容易に入手することができる。ヒトTfのヌクレオチド配列が利用可能であり(配列番号1、2および3、ならびに上述の登録番号を参照、さらにwww.ncbi.nlm.nih.govで入手可能である)、これらは、TfまたはTfのドメインと、選択された治療的分子との遺伝学的な融合体を作製するのに用いることができる。また、融合は、ラクトトランスフェリン(GenBank登録番号NM_002343のラクトフェリン)、またはメラノトランスフェリンのような関連の分子から作製してもよい。
【0072】
ラクトフェリン(Lf)は、防御作用のある天然の鉄結合性タンパク質であり、これは抗菌性、抗真菌性、抗ウイルス性、抗腫瘍性および抗炎症性の活性を有することが見出されている。このようなタンパク質は、通常は正常フローラに晒される外分泌物に存在しており、このような外分泌物としては、乳、涙、鼻の滲出物、唾液、気管支粘液、胃腸液、頚膣部粘液、および精液が挙げられる。加えて、Lfは、循環する多形核好中球(PMN)の二次的な特殊顆粒の主成分である。アポタンパク質は、化膿した領域においてPMNが脱顆粒する際に放出される。Lfの主要な機能は、体液中や炎症を起こしている領域中の遊離の鉄を捕捉することであり、このようにして、フリーラジカルが介在するダメージを抑制し、微生物細胞や新生細胞の侵入における金属の利用可能性を減少させることができる。成人の125ILfの代謝回転速度を試験した研究では、Lfは迅速に肝臓および脾臓に取り込まれ、放射活性は、肝臓および脾臓中で数週間持続することが示された(Bennett等(1979),Clin.Sci.(Lond.)57:453〜460)。
【0073】
一実施態様において、本発明の融合タンパク質のトランスフェリン部分は、トランスフェリンのスプライス変異体を含む。一例において、トランスフェリンのスプライス変異体は、ヒトトランスフェリンのスプライス変異体であり得る。特定の一実施態様において、ヒトトランスフェリンのスプライス変異体は、GenBank登録番号AAA61140のスプライス変異体であり得る。
【0074】
その他の実施態様において、本発明の融合タンパク質のトランスフェリン部分は、ラクトフェリンのスプライス変異体を含む。一例において、ヒト血清のラクトフェリンのスプライス変異体は、好中球のラクトフェリンの新規なスプライス変異体であり得る。特定の一実施態様において、好中球のラクトフェリンのスプライス変異体は、GenBank登録番号AAA59479のスプライス変異体であり得る。その他の特定の実施態様において、好中球のラクトフェリンのスプライス変異体は、以下のアミノ酸配列EDCIALKGEADA(配列番号4)を含んでいてもよく、この配列は、新規なスプライス変異の領域を含む。
【0075】
また、メラノトランスフェリン(GenBank登録番号NM_013900、マウスのメラノトランスフェリン)との融合を行ってもよい。メラノトランスフェリンは、悪性黒色腫細胞において高レベルで見出されるグリコシル化タンパク質であり、元々はヒト黒色腫抗原p97と命名されていた(Brown等,1982,Nature,296:171〜173)。これは、ヒト血清トランスフェリン、ヒトラクトフェリン、およびニワトリトランスフェリンと高い配列相同性を有する(Brown等,1982,Nature,296:171〜173;Rose等,Proc.Natl.Acad.Sci.,1986,83:1261〜1265)。しかしながら、これらのタンパク質とは異なり、メラノトランスフェリンに関する細胞受容体は同定されていない。メラノトランスフェリンは、鉄と可逆的に結合し、2つの形態で存在し、そのうちの一方の形態は、グリコシルホスファチジルイノシトールのアンカーによって細胞膜に結合し、他方の形態は、可溶性であり、かつ活発に分泌される(Baker等,1992,FEBS Lett,298:215〜218;Alemany等,1993,J.Cell Sci.,104:1155〜1162;Food等,1994,J.Biol.Chem.274:7011〜7017)。
【0076】
改変されたTf融合体は、あらゆるTfタンパク質、フラグメント、ドメインまたは加工されたドメインで作製することができる。例えば、融合タンパク質は、全長Tf配列を用いて作製することができ、天然型のTfシグナル配列を含んでいてもよいし、またはそれらを含まなくてもよい。また、細胞透過性の抗体(Trans−body)は、単一のTfドメイン(例えばNまたはCドメインそれぞれ)を用いて作製することもできる。また、Trans−body(細胞透過性)の抗体も、二つのTfドメイン(例えば二つのNドメイン、または二つのCドメイン)を用いて作製してもよい。ある実施態様では、単一のCドメインへ治療用タンパク質を融合させることができ、ここにおいてCドメインは、グリコシル化、鉄結合および/またはTf受容体結合を、減少させる、阻害する、または防ぐように変更される。その他の実施態様において、Tfグリコシル化部位はCドメインに存在しており、Nドメインは、そのままでは鉄またはTf受容体と結合しないため、単一のNドメインの使用が有利である。一実施態様において、Tf融合タンパク質は、高レベルで発現される単一のNドメインを有する。
【0077】
本明細書で用いられるように、N様ドメインとして機能するように改変されたC末端ドメインまたはローブは、天然型または野生型のNドメインまたはローブと実質的に類似したグリコシル化パターンまたは鉄結合特性を示すように改変される。一実施態様において、Cドメインまたはローブは、グリコシル化されず、かつ鉄と結合しないように改変され、改変は、それに関連するCドメイン領域またはアミノ酸を、それに対応する天然型または野生型のNドメインの領域または部位に存在するドメイン領域またはアミノ酸で置換することによってなされる。
【0078】
本明細書で用いられる「2つのNドメインまたはローブ」を含むTf成分は、天然型のCドメインまたはローブを、第二の天然型または野生型のNドメインもしくはローブ、または改変されたNドメインもしくはローブで置き換えて改変されたTf分子を含むか、または実質的に野生型または改変されたNドメインのように機能するように改変されているCドメインを含む。米国仮出願第60/406,977号を参照(これは参照によりその全体として本発明に組み込まれる)。
【0079】
2つのドメインの三次元構造を重ね合わせること(スイスPDBビューワー3.7b2(Swiss PDB Viewer 3.7b2)、イタラティブ・マジック・フィット(Iterative Magic Fit))、および直接のアミノ酸アライメント(クラスタルW(ClustalW)マルチプルアライメント)による2つのドメインの解析によって、2つのドメインは時間とともに分岐しているが明らかである。アミノ酸アライメントによれば、2つのドメイン間で42%同一性、および59%の類似性が示される。しかしながら、構造的な等価性に関して、Nドメインの約80%がCドメインと適合している。またCドメインは、Nドメインと比較して数個の余分なジスルフィド結合も有する。
【0080】
NおよびCドメインに関する分子モデルのアライメントから、以下の構造的な等価体が明らかになる:
【表1】
【0081】
2つのドメインのジスルフィド結合は、以下のようにアラインされる:
【表2】
【0082】
一実施態様において、本融合タンパク質のトランスフェリン部分は、トランスフェリンのN末端ローブを少なくとも2つ有する。さらなる実施態様において、本融合タンパク質のトランスフェリン部分は、ヒト血清トランスフェリンから誘導されたトランスフェリンのN末端ローブを少なくとも2つ含む。
【0083】
その他の実施態様において、本融合タンパク質のトランスフェリン部分は、配列番号3のAsp63、Gly65、Tyr95、Tyr188、およびHis249からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有するトランスフェリンのN末端ローブを少なくとも2つ有する、それらを含む、またはそれらからなる。
【0084】
その他の実施態様において、改変された融合タンパク質のトランスフェリン部分は、配列番号3のLys206またはHis207において突然変異を有する、組換えヒト血清トランスフェリンのN末端ローブの突然変異体を有する。
【0085】
その他の実施態様において、本融合タンパク質のトランスフェリン部分は、トランスフェリンのC末端ローブを少なくとも2つ有する、それらを含む、本質的にそれらからなる、またはそれらからなる。さらなる実施態様において、本融合タンパク質のトランスフェリン部分は、ヒト血清トランスフェリンから誘導されたトランスフェリンのC末端ローブを少なくとも2つ有する。
【0086】
さらなる実施態様において、C末端ローブの突然変異体はさらに、グリコシル化が不可能な配列番号3のAsn413およびAsn611の少なくとも1つの突然変異を有する。
【0087】
その他の実施態様において、トランスフェリン部分は、配列番号3のAsp392、Tyr426、Tyr514、Tyr517、およびHis585からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有するトランスフェリンのC末端ローブを少なくとも2つ有し、ここにおいて、該突然変異体は、金属イオンと結合する能力を保持する。代替の実施態様において、トランスフェリン部分は、配列番号3のTyr426、Tyr514、Tyr517、およびHis585からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有するトランスフェリンのC末端ローブを少なくとも2つ有し、ここにおいて、該突然変異体は、金属イオンと結合する能力が減少している。その他の実施態様において、トランスフェリン部分は、配列番号3のAsp392、Tyr426、Tyr517、およびHis585からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有するトランスフェリンのC末端ローブを少なくとも2つ有し、ここにおいて、該突然変異体は、金属イオンと結合する能力を保持せず、実質的にNドメインのように機能する。
【0088】
いくつかの実施態様において、本融合タンパク質のTfまたはTf部分およびリガンドが融合タンパク質の残余から切断される場合、TfまたはTf部分は、非融合状態、すなわちTfに融合していない状態でのリガンドのインビボでの循環半減期、血清中安定性(半減期)、インビトロでの安定性または生物学的利用率と比較して、リガンド(すなわち治療剤)のインビボでの循環半減期、血清中安定性、インビトロでの溶液安定性または生物学的利用率を増加させるのに十分な長さを有する。このような安定性、インビボ循環半減期、または生物学的利用率の増加は、融合していないリガンド成分領域を超える約30%、50%、70%、80%、90%またはそれ以上の増加であり得る。場合によっては、改変されたトランスフェリンを含むリガンド成分は、非融合状態のリガンドと比較して、約1日またはそれ以上、1〜2日またはそれ以上、3〜5日またはそれ以上、5〜10日またはそれ以上、10〜15日またはそれ以上、10〜20日またはそれ以上、約12〜18日間、または約14〜17日間の血清中半減期を示す。
【0089】
TfのCドメインが本融合タンパク質の一部である場合、グリコシル化または過剰なマンノシル化を防ぎ、本融合タンパク質の血清中半減期を延長させるために(アシアロ−、または場合によってはモノシアロ−Tfもしくはジシアロ−Tfが生産されるように)、2つのN結合型のグリコシル化部位、配列番号3のN413およびN611に相当するアミノ酸残基は、酵母系での発現のために突然変異していてもよい。N413およびN611に相当するTfアミノ酸に加えて、N−X−S/Tグリコシル化部位内の、またはそれに隣接する残基への突然変異は、グリコシル化を防ぐか、または実質的に減少させる。Funk等の米国特許第5,986,067号を参照。また、ピチア・パストリス(Pichia pastoris)で発現されたTfのNドメインは、S32において単一のヘキソースでO結合型グリコシル化されることになることも報告されているが、これを、このようなグリコシル化を防ぐために突然変異または改変することができる。その上、1またはそれ以上のPMT遺伝子に突然変異を有する酵母宿主細胞では、O連結型糖付加は減少するか、または除去される可能性がある。
【0090】
従って、本発明の一実施態様において、本融合タンパク質は、トランスフェリンが低いグリコシル化を示す改変されたトランスフェリン分子を有し、このようなトランスフェリン分子としては、これらに限定されないが、Tfのアシアロ−、モノシアロ−およびジシアロ型が挙げられる。その他の実施態様において、本融合タンパク質のトランスフェリン部分は、グリコシル化を防ぐために突然変異した組換えトランスフェリン突然変異体を有する。その他の実施態様において、本融合タンパク質のトランスフェリン部分は、完全にグリコシル化された組換えトランスフェリン突然変異体を有する。さらなる実施態様において、本融合タンパク質のトランスフェリン部分は、グリコシル化を防ぐために突然変異した組換えヒト血清トランスフェリン突然変異体を有し、ここにおいて、配列番号3のAsn413およびAsn611の少なくとも一方が、グリコシル化が不可能なアミノ酸に突然変異している。その他の実施態様において、本融合タンパク質のトランスフェリン部分は、グリコシル化を防ぐために、または実質的に減少させるために突然変異した組換えヒト血清トランスフェリン突然変異体を有し、ここにおいて、N−X−S/Tグリコシル化部位内の残基で突然変異していてもよい。さらに、セリンまたはスレオニン残基を突然変異させることによってグリコシル化を減少させるか、または防ぐことができる。さらに、グリコシル化を阻害するために、Xをプロリンに変化させることが既知である。
【0091】
また、以下でより詳細に考察されるように、本発明の改変されたTfを含む改変されたTf融合タンパク質は、鉄と結合しないように、および/またはTf受容体と結合しないように遺伝子操作してもよい。本発明のその他の実施態様において、鉄結合を保持させ、Tfの鉄結合能を、治療用タンパク質またはペプチドを、細胞の内部に送達、および/または血液脳関門(BBB)を通過させるために使用可能である。Nドメインは鉄をローディングすると、単独ではTfRに結合せず、鉄と結合したCドメインは、TfRと結合することになるが、分子全体と同じ親和性は示さない。
【0092】
その他の実施態様において、トランスフェリン融合タンパク質のトランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、金属イオンと結合する能力を保持しない。代替の実施態様において、トランスフェリン融合タンパク質のトランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、野生型血清トランスフェリンよりも金属イオンに対する結合親和性が弱い。代替の実施態様において、トランスフェリン融合タンパク質のトランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、野生型血清トランスフェリンよりも金属イオンに対する結合親和性が強い。
【0093】
その他の実施態様において、トランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、トランスフェリン受容体に結合する能力を保持しない。代替の実施態様において、トランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、野生型血清トランスフェリンよりもトランスフェリン受容体に対する結合親和性が弱い。代替の実施態様において、トランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、野生型血清トランスフェリンよりもトランスフェリン受容体に対する結合親和性が強い。
【0094】
その他の実施態様において、トランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、炭酸イオンに結合する能力を保持しない。代替の実施態様において、トランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、野生型血清トランスフェリンよりも炭酸イオンに対する結合親和性が弱い。代替の実施態様において、トランスフェリン部分は、突然変異を有する組換えトランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、野生型血清トランスフェリンよりも炭酸イオンに対する結合親和性が強い。
【0095】
その他の実施態様において、トランスフェリン部分は、配列番号3のAsp63、Gly65、Tyr95、Tyr188、His249、Asp392、Tyr426、Tyr514、Tyr517、およびHis585からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有する組換えヒト血清トランスフェリン突然変異体を有し、ここにおいて、該突然変異体は、金属イオンと結合する能力を保持する。代替の実施態様において、配列番号3のAsp63、Gly65、Tyr95、Tyr188、His249、Asp392、Tyr426、Tyr514、Tyr517、およびHis585からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有する組換えヒト血清トランスフェリン突然変異体が挙げられ、ここにおいて、該突然変異体は、金属イオンと結合する能力が減少している。その他の実施態様において、配列番号3のAsp63、Gly65、Tyr95、Tyr188、His249、Asp392、Tyr426、Tyr517、およびHis585からなる群より選択される少なくとも1つのアミノ酸残基に突然変異を有する組換えヒト血清トランスフェリン突然変異体が挙げられ、ここにおいて、該突然変異体は、金属イオンと結合する能力を保持しない。
【0096】
その他の実施態様において、トランスフェリン部分は、配列番号3のLys206またはHis207において突然変異を有する、組換えヒト血清トランスフェリンの突然変異体を有し、ここにおいて、突然変異体は、野生型ヒト血清トランスフェリンよりも強い金属イオンへの結合活性を有する(米国特許第5,986,067号を参照。これは参照によりその全体として本明細書に組み込まれる)。代替の実施態様において、トランスフェリン部分は、配列番号3のLys206またはHis207において突然変異を有する組換えヒト血清トランスフェリン突然変異体を含み、ここにおいて、該突然変異体は、野生型ヒト血清トランスフェリンよりも金属イオンへの結合活性が弱い。さらなる実施態様において、トランスフェリン部分は、配列番号3のLys206またはHis207において突然変異を有する、組換えヒト血清トランスフェリン突然変異体を含み、ここにおいて、該突然変異体は、金属イオンと結合しない。
【0097】
あらゆる利用可能な技術が本発明の融合タンパク質を製造するために使用可能であり、このような技術としては、これらに限定されないが、一般的に利用できる分子技術が挙げられ、例えば、Sambrook等のMolecular Cloning:A Laboratory Manual,第二版,コールドスプリングハーバーラボラトリープレス(Cold Spring Harbor Laboratory Press),1989に記載された技術である。当技術分野でよく知られた部位特異的突然変異誘発を達成する技術を用いてヌクレオチド置換を行う場合、コードされたアミノ酸の変化は、好ましくは軽度のもの、すなわち保存的アミノ酸置換であるが、その他の非保存的な置換も同様に可能なものとして取扱われ、改変されたトランスフェリン部分、例えば、改変された融合タンパク質が、低いグリコシル化、低い鉄結合などを示すものを製造する場合に特にそうである。特に可能なものとして取扱われるのは、アミノ酸の置換、わずかな欠失または挿入したもの(典型的には約30個のアミノ酸に対して1個の割合で);トランスフェリンドメイン間への挿入したもの;小規模のアミノまたはカルボキシル末端の伸長したもの、例えばアミノ末端のメチオニン残基、またはトランスフェリンドメイン間、もしくは、トランスフェリンタンパク質と治療用タンパク質もしくはペプチド、リガンド、もしくは抗体の可変領域もしくは軸領域との連結の50、40、30、20または10残基未満の小さいリンカーペプチド;または、精製を容易にする小規模の伸長したもの、例えばポリヒスチジントラクト(poly−histidine tract)、抗原エピトープ、もしくは結合ドメインである。
【0098】
保存的アミノ酸置換の例としては、同じグループ内でなされる置換が挙げられ、例えば、塩基性アミノ酸のグループ(例えば、アルギニン、リシン、ヒスチジン)、酸性アミノ酸のグループ(例えば、グルタミン酸およびアスパラギン酸)、極性アミノ酸のグループ(例えば、グルタミンおよびアスパラギン)、疎水性アミノ酸のグループ(例えば、ロイシン、イソロイシン、バリン)、芳香族アミノ酸のグループ(例えば、フェニルアラニン、トリプトファン、チロシン)、および低分子量のアミノ酸のグループ(例えば、グリシン、アラニン、セリン、スレオニン、メチオニン)の中でなされる置換である。
【0099】
非保存的置換は、あるグループのアミノ酸を、その他のグループのアミノ酸で置換することを包含する。例えば、非保存的置換は、極性アミノ酸を疎水性アミノ酸で置換することを含む。ヌクレオチド置換の一般的な説明については、例えば、Ford等(1991),Prot.Exp.Pur.2:95〜107を参照。鉄への結合を示さない、もしくはそれらが減少した、および/または、本融合タンパク質のTf受容体への結合を示さない、もしくはそれらが減少した本発明のTf融合タンパク質、好ましくは細胞透過性の抗体trans−bodiesを製造するために、非保存的置換、欠失および挿入が特に有用である。
【0100】
本発明のポリペプチドおよびタンパク質において、アミノ酸の命名は、以下の規則にのっとった一覧に記載のシステムに従ってなされる:
【表3】
【0101】
鉄結合および/または受容体結合は、配列番号3のTfNドメイン残基Asp63、Tyr95、Tyr188、His249、および/またはCドメイン残基Asp392、Tyr426、Tyr514、および/またはHis585の1つまたはそれ以上に相当するアミノ酸残基への欠失、置換または挿入などの突然変異によって、減少する場合もあるし、または破壊される場合もある。また、鉄結合も、配列番号3のアミノ酸Lys206、His207、またはArg632の突然変異によって影響を受ける可能性がある。炭酸結合は、配列番号3のTfNドメイン残基Thr120、Arg124、Ala126、Gly127、および/またはCドメイン残基Thr452、Arg456、Ala458、および/またはGly459の1つまたはそれ以上に相当するアミノ酸残基の欠失、置換またはそれへの挿入などの突然変異によって減少するか、または破壊される可能性がある。炭酸結合の減少または破壊は、鉄および/または受容体結合に逆の影響を与える可能性がある。
【0102】
Tf受容体への結合は、鉄結合について上述したTfNドメイン残基の1つまたはそれ以上に相当するアミノ酸残基の欠失、置換またはそれへの挿入などの突然変異によって減少するか、または破壊される可能性がある。
【0103】
上記で考察したように、グリコシル化は、Cドメイン残基N413および/またはN611に相当するN−X−S/T部位内のTfのCドメイン残基の1つまたはそれ以上に相当するアミノ酸残基の欠失、置換またはそれへの挿入などの突然変異によって減少するか、または防止される可能性がある。米国特許第5,986,067号を参照。例えば、N413および/またはN611は、隣接するアミノ酸の可能性があるGlu残基に突然変異していてもよい。
【0104】
本発明のTf融合タンパク質が、グリコシル化、鉄結合、カーボネート結合および/または受容体結合を防ぐように改変されていないような場合には、グリコシル化、鉄および/またはカーボネートイオンを、本融合タンパク質から取り除くか、または切断することができる。例えば、利用可能なデグリコシラーゼ(deglycosylase)を用いて、本融合タンパク質からグリコシル化残基を切断してもよく(特に、Tf部分に結合した糖残基)、グリコシル化酵素欠損酵母を用いて、グリコシル化を防いでもよいし、および/またはグリコシル化を防ぐ物質(例えばツニカマイシン)の存在下で組換え細胞を増殖させてもよい。
【0105】
また、本融合タンパク質上の糖質も、本融合タンパク質をデグリコシラーゼで処理することによって、酵素的に減少させてもよいし、または完全に除去してもよい。デグリコシラーゼは当技術分野でよく知られている。デグリコシラーゼの例としては、これらに限定されないが、ガラクトシダーゼ、PNGアーゼA、PNGアーゼF、グルコシダーゼ、マンノシダーゼ、フコシダーゼ、およびEndoHデグリコシラーゼが挙げられる。
【0106】
Tfの三次元構造を変更するために、Tfに、追加の突然変異を作製してもよく、このような突然変異としては、例えばヒンジ領域への改変が挙げられ、それにより、鉄結合およびTf受容体の認識に必要なコンホメーション変化を防ぐことができる。例えば、突然変異は、Nドメインアミノ酸残基94〜96、245〜247、および/または316〜318、ならびにCドメインのアミノ酸残基425〜427、581〜582、および/または652〜658に、またはその周辺に作製してもよい。加えて、Tfの構造および機能を変更するために、これらの部位のフランキング領域に、またはその周辺に突然変異を作製してもよい。
【0107】
本発明の一側面において、本融合タンパク質をキャリアータンパク質として機能させて、リガンドの半減期または生物学的利用率を伸ばすことができ、加えて、場合によってはリガンドを細胞の内部に運搬したり、血液脳関門を通過する能力を保持させたりすることができる。代替の実施態様において、本融合タンパク質は、改変されたトランスフェリン分子を有し、ここにおいて、トランスフェリンは、血液脳関門を通過する能力を保持しない。
【0108】
その他の実施態様において、本融合タンパク質は、改変されたトランスフェリン分子を有し、ここにおいて、トランスフェリン分子は、トランスフェリン受容体に結合し、抗体の可変領域を細胞の内部に輸送する能力を保持する。代替の実施態様において、本融合タンパク質は、改変されたトランスフェリン分子を有し、ここにおいて、トランスフェリン分子は、トランスフェリン受容体に結合し、抗体の可変領域を細胞の内部に輸送する能力を保持しない。
【0109】
さらなる実施態様において、本融合タンパク質は、改変されたトランスフェリン分子を有し、ここにおいて、トランスフェリン分子は、トランスフェリン受容体に結合し、抗体の可変領域を細胞の内部に輸送する能力を保持するが、血液脳関門を通過する能力を保持しない。代替の実施態様において、本融合タンパク質は、改変されたトランスフェリン分子を有し、ここにおいて、トランスフェリン分子は、血液脳関門を通過する能力を保持するが、トランスフェリン受容体に結合し、抗体の可変領域を細胞の内部に輸送する能力を保持しない。
【0110】
トランスフェリン融合タンパク質
本発明の融合タンパク質は、Tfタンパク質のN末端および/またはC末端に付着したリガンド、抗体の可変領域、またはランダムペプチドの1個またはそれ以上のコピーを含んでいてもよい。一実施態様において、リガンド成分は、Tfタンパク質のN末端に結合している。いくつかの実施態様において、これらリガンド、可変領域またはペプチドは、Tfタンパク質のNおよびC末端の両方に付着しており、本融合タンパク質は、Tf末端のいずれか、またはその両方に、これらの領域の1またはそれ以上の等価体を含んでいてもよい。
【0111】
その他の実施態様において、1またはそれ以上のリガンドは、トランスフェリンペプチド、例えば1またはそれ以上のTfのループなどのTfタンパク質の既知のドメインの場所に挿入される。Ali等(1999)J.Biol Chem.274(34):24066〜24073を参照。
【0112】
本発明の一実施態様において、リガンドは、トランスフェリンのNローブに挿入される。例えば、本発明はまた、1またはそれ以上の挿入も含み、このような挿入は、以下の表で示されるように、N−ローブのN1およびN2ドメインに、またはN−ローブのN1およびN2ドメイン周辺の他の位置に行うことができる。
【表4】
【0113】
一般的に、本発明のトランスフェリン融合タンパク質は、一つの改変されたトランスフェリンから誘導された領域、および一つの抗体の可変領域を有していてもよい。しかしながら、それぞれのタンパク質の複数の領域を用いて、本発明のトランスフェリン融合タンパク質を作製してもよい。同様に、1種より多くの抗体の可変領域を用いて、本発明のトランスフェリン融合タンパク質を作製してもよく、それによって、多機能の改変されたTf融合タンパク質を生産することができる。
【0114】
一実施態様において、本発明の融合タンパク質は、トランスフェリン分子またはその部分に融合した、抗体の可変領域またはその部分を含む。その他の実施態様において、本発明の融合タンパク質は、トランスフェリン分子のN末端に融合した抗体の可変領域を含む。代替の実施態様において、本発明の融合タンパク質は、トランスフェリン分子のC末端に融合した抗体の可変領域を含む。さらなる実施態様において、本発明の融合タンパク質は、抗体の可変領域のN末端に融合したトランスフェリン分子を含む。代替の実施態様において、本発明の融合タンパク質は、抗体の可変領域のC末端に融合したトランスフェリン分子を含む。
【0115】
本発明はまた、改変されたトランスフェリン分子またはその部分に融合した抗体の可変領域またはその部分を含む融合タンパク質を提供する。
【0116】
その他の実施態様において、本発明の融合タンパク質は、改変されたトランスフェリンのN末端およびC末端の両方に融合した抗体の可変領域を含む。その他の実施態様において、NおよびC末端で融合した抗体の可変領域は、同じ抗原に結合する。また、同じ抗原に結合する抗体の可変領域は、異なる抗体から誘導されてもよく、従って、同じ標的上の異なるエピトープに結合する。代替の実施態様において、NおよびC末端で融合した抗体の可変領域は、異なる抗原と結合する。その他の代替の実施態様において、NおよびC末端に融合した抗体の可変領域は、異なる抗原と結合し、これは病気、障害または状態を治療または予防するための2種の異なる細胞を活性化するのに有用である可能性がある。その他の実施態様において、NおよびC末端で融合した抗体の可変領域は、異なる抗原と結合し、これは一般的に患者に同時に起こることが当技術分野で既知である病気または障害を治療または予防するための2種の異なる抗原との架橋形成に有用である可能性がある。
【0117】
加えて、本発明のトランスフェリン融合タンパク質はまた、対象の抗体の可変領域(例えば、治療用タンパク質またはそのフラグメントまたは変異体と結合する単鎖抗体)を、改変されたトランスフェリンの内部領域に挿入することによって生産してもよい。改変されたトランスフェリンの内部領域としては、これらに限定されないが、ループ領域、鉄結合部位、ヒンジ領域、ビカーボネート結合部位、または、受容体結合ドメインが挙げられる。
【0118】
改変されたトランスフェリン分子のタンパク質配列内に、多数のループまたはターンが存在しており、これらはジスルフィド結合によって安定化されている。これらのループは、治療活性を有するペプチド、好ましくは抗体の可変領域の挿入または内部融合、特に、機能的なタンパク質または治療用タンパク質、好ましくは抗体の可変領域となるのに二次構造を必要とするものに有用であり、特異的な生物活性を有する改変されたトランスフェリン分子を生成させることができる。
【0119】
リガンド、例えば抗体の可変領域、好ましくはCDRが、Tf分子の少なくとも1つのループに挿入されるか、またはそれと置き換えられる場合、挿入は、Tfの他の領域に加えて、いずれかの表面に露出したループ領域内に作製してもよい。例えば、挿入は、Tfアミノ酸32〜33、74〜75、256〜257、279〜280および288〜289を含むループ内に作製してもよい。上記のAli等を参照。これまでに説明したように、挿入は、Tfの他の領域、例えば以下でより詳細に説明されるような鉄およびビカーボネート結合のための部位、ヒンジ領域、および受容体結合ドメイン内に作製してもよい。また、タンパク質またはペプチドを挿入するための改変/置き換えを受けやすいTfタンパク質配列中のループを、ランダムペプチドインサートのスクリーニング可能なライブラリーを開発するために用いてもよい。ペプチドライブラリーを生成させるための核酸インサートを生産するために、Tfドメインにクローニングしたり、および/またはTfの末端に融合したりする前に利用可能なファージおよび細菌の提示系などのあらゆる手法が使用可能である。
【0120】
TfのN末端は遊離であり、本融合タンパク質の本体から外を向いている。トランスフェリンのN末端における一つまたは複数のリガンドの融合体は、本発明の一実施態様である。このような融合体は、リンカー領域、例えば、ただしこれらに限定されないが、リガンドをTfから離すための、ポリグリシンストレッチ、またはPEAPTDリンカー(配列番号18)を含んでいてもよい。
【0121】
TfのC末端の外観は、埋め込まれているか、または部分的に埋め込まれており、C末端からの6個のアミノ酸のジスルフィド結合で固定されているように見える。ヒトTfにおいて、C末端のアミノ酸はプロリンであり、このプロリンは、配向している方法に応じて、融合タンパク質から外に向いているか、または分子の本体の内部に向いているかのいずれかと予想される。本発明のいくつかの実施態様において、C末端にリンカーまたはスペーサー成分を用いてもよい。またN末端の付近にプロリンが存在していてもよい。本発明の一側面において、Nおよび/またはC末端のプロリンは、改変されていてもよいし、またはその他のアミノ酸で置換されていてもよい。本発明のその他の側面において、C末端のジスルフィド結合を除去して、C末端を開放してもよい。
【0122】
軸成分
本発明の軸成分は、そのN末端で、トランスフェリン成分またはリガンドに融合され、そのC末端ではアンカー成分と場合により融合されていてもよい。酵母細胞で発現させる場合、軸成分のC末端は、細胞内、例えば細胞壁内に位置する。本発明の一実施態様において、軸成分は、細胞壁連結要素として作用し、本融合タンパク質を、共有結合または非共有結合によって酵母細胞の細胞壁に結合させる。
【0123】
本発明の軸成分は、ロッド様またはブラシ様のコンホメーションを有する。このタイプのコンホメーションは、中度〜高度にグリコシル化されたペプチドに典型的である。本発明の軸成分は、N−グリカンまたはO−グリカンを含む。米国特許第6,114,147号を参照(これは参照によりその全体として本明細書に組み込まれる)。N−グリカンよりもO−グリカンの存在が好ましく、これはなぜなら、O−グリカンは、N−グリカンよりも多く、伸長したロッド様のコンホメーションを軸成分に取らせることが可能であるためである。また、軸成分は、中程度〜高度にグリコシル化されたセリンおよびスレオニングリコシル化部位を含んでいてもよい。
【0124】
本発明の融合タンパク質の軸成分は、中程度〜高度のパーセンテージのセリンまたはスレオニン残基を含む。例えば、本発明は、少なくとも約5%またはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約10%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約20%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約30%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約40%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約50%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約60%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約70%もしくはそれ以上のセリンおよび/もしくはスレオニン残基、少なくとも約80%もしくはそれ以上のセリン、および/もしくは、スレオニン残基、または少なくとも約90%もしくはそれ以上のセリンおよび/もしくはスレオニン残基を有する軸成分を含む。本発明の一実施態様において、軸成分は、約20〜30%のセリンおよび/もしくはスレオニン残基、約20〜40%のセリンおよび/もしくはスレオニン残基、約30〜40%のセリンおよび/もしくはスレオニン残基、約20〜50%のセリンおよび/もしくはスレオニン残基、約30〜50%のセリンおよび/もしくはスレオニン残基、約20〜60セリンおよび/もしくはスレオニン残基、または約30〜60%のセリンおよび/もしくはスレオニン残基を含む。
【0125】
軸成分は、少なくとも約5質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約10質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約20質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約30質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約40質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約50質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約60質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約70質量%もしくはそれ以上のN−もしくはO−グリカン、少なくとも約80質量%もしくはそれ以上のN−もしくはO−グリカン、または少なくとも約90質量%もしくはそれ以上のN−もしくはO−グリカンを含んでいてもよい。本発明の一実施態様において、軸成分は、約20〜30質量%のO−グリカン、約20〜40質量%のO−グリカン、約30〜40質量%のO−グリカン、約20〜50質量%のO−グリカン、約30〜50質量%のO−グリカン、約20〜60質量%のO−グリカン、または約30〜60%のO−グリカンを含む。その他の実施態様において、グリカン、特にO−グリカンの存在は、軸成分と、細胞壁のタンパク質に存在するβ−グルカンとの架橋を可能にする。そのようなものとして、本発明の軸成分は、細胞壁連結要素として機能することができる。
【0126】
軸成分は、ムチンタンパク質またはムチンタンパク質の部分、すなわちMUC型タンパク質のメンバーを含んでいてもよい。MUC型のムチンとは、高度にグリコシル化された構造的に関連した分子のファミリーであり、気道、胃腸管および生殖管の上皮で発現し、例えばMUC1(GenBank登録番号AF125525)、MUC2(GenBank登録番号L21998)、MUC3(GenBank登録番号AF113616)、MUC4(GenBank登録番号AJ000281)、MUC5AC(GenBank登録番号U83139)、MUC5B(GenBank登録番号AJ001402)、MUC6(GenBank登録番号U97698)、MUC7(GenBank登録番号L13283)、MUC8(GenBank登録番号U14383)、MUC9(GenBank登録番号AW271430)が挙げられる。本発明の一実施態様において、軸成分は、hMUC1またはhMUC1タンパク質の部分、例えば配列番号70の核酸によってコードされた配列番号71、さらに配列番号5の核酸によってコードされたポリペプチドを含む。本発明のその他の実施態様において、軸成分は、hMUC3またはhMUC3タンパク質の部分を含む。例えば、本発明は、配列番号68の核酸によってコードされる配列番号69のhMUC3の軸を含む。また本発明の融合タンパク質は、ムチンタンパク質およびそれらの部分の類似体および誘導体のような変異体を含む軸も包含する。
【0127】
また本発明の軸成分は、ムチン以外のグリコシル化タンパク質からも誘導することができ、このようなタンパク質としては、これらに限定されないが、AGA1(例えば、配列番号72の核酸配列によってコードされた配列番号73)、MAdCAM−1、GlyCAM−1、CD34;E−セレクチン、P−セレクチンまたはL−セレクチンからのコンセンサス反復;またはウイルス糖タンパク質スパイク(例えば、インフルエンザ、単純ヘルペス、ヒト免疫不全またはタバコモザイクウイルス)、ならびにそれらの変異体およびフラグメントが挙げられる。WO01/46698、Girard等(1995)Immunity 2:113〜123、およびVan Kinken等(1998)Anal.Biochem.265:103〜116を参照(これらはいずれも、参照によりその全体として本明細書に組み込まれる)。本発明は、2種またはそれ以上のグリコシル化タンパク質またはそれらのフラグメントの反復、ならびに、2種またはそれ以上のタイプのグリコシル化タンパク質の組み合わせを含む。
【0128】
本発明のその他の実施態様において、軸は、1またはそれ以上の遊離のシステイン残基を含むように遺伝子操作される。1またはそれ以上の遊離のシステイン残基は、酵母細胞の細胞壁中のタンパク質の遊離のシステイン残基とジスルフィド結合を形成することができる。細胞壁中の1またはそれ以上のジスルフィド結合の形成は、細胞壁結合要素として機能することができる軸成分を遺伝子操作するのに使用可能なその他の方法の代表である。
【0129】
本発明の軸成分は、酵母細胞の全細胞壁を橋わたすのに十分な長さを有していなければならない。好ましくは、軸成分のN末端を、細胞壁の外側に位置させることであり、最も好ましくは、ロッド様の立体配置で、酵母細胞から外に向かって伸長させて、トランスフェリン成分およびリガンドと、宿主の酵母細胞との間の立体障害を低減させることである。軸成分は、長さが、少なくとも約25個のアミノ酸、少なくとも約50個のアミノ酸、少なくとも約75個のアミノ酸、少なくとも約100個のアミノ酸、少なくとも約125個のアミノ酸、少なくとも約150個のアミノ酸、少なくとも約175個のアミノ酸、少なくとも約200個のアミノ酸、少なくとも約225個のアミノ酸、少なくとも約250個のアミノ酸、少なくとも約275個のアミノ酸、少なくとも約300個のアミノ酸、少なくとも約325個のアミノ酸、少なくとも約350個のアミノ酸、少なくとも約375個のアミノ酸、少なくとも約400個のアミノ酸、少なくとも約425個のアミノ酸、少なくとも約450個のアミノ酸、少なくとも約475個のアミノ酸長さが、少なくとも約500個のアミノ酸長さが、少なくとも約525個のアミノ酸長さが、少なくとも約550個のアミノ酸長さが、少なくとも約575個のアミノ酸長さが、少なくとも約600個のアミノ酸長さが、少なくとも約625個のアミノ酸、または長さが、少なくとも約650個のアミノ酸が必要である。一実施態様において、軸成分は、長さが約500個のアミノ酸である。その他の実施態様において、軸成分は、長さが約300〜600個のアミノ酸である。
【0130】
アンカー成分
本発明の融合タンパク質の任意のアンカー成分は、本融合タンパク質を宿主細胞表層または基質表面に物理的に連結する本融合タンパク質の部分である。例えば、アンカー成分は、本融合タンパク質を酵母細胞膜または酵母細胞壁に連結または固定するすることができる。アンカーが本融合タンパク質を酵母細胞壁に連結する場合、アンカーは、細胞壁結合部材である。
【0131】
アンカー成分は、融合タンパク質を、一時的に酵母細胞壁または細胞膜に繋ぐすることができる。本発明の一実施態様において、アンカー成分は、融合タンパク質を、一時的に酵母細胞壁または細胞膜に繋いで、軸成分が細胞壁に共有結合または非共有結合で結合する機会を提供する。例えば、酵母細胞におけるアンカーの一時的な係留によって、軸成分からのO−グリカンと、細胞壁のβ−グルカンとを架橋させることが可能になる。
【0132】
本発明の一実施態様において、アンカー成分は、微生物、好ましくは下等真核生物、例えば酵母およびカビの細胞膜または細胞壁に固着する。この成分は、プロリンのようなアミノ酸で細胞膜または細胞壁にC末端を固定する長いC末端を有していてもよい(Kok(1990)FEMS Microbiology Reviews 87:15〜42)。
【0133】
アンカー成分は、グリコシルホスファチジルイノシトール(GPI)アンカーの使用によって細胞に固定することができる。Conzelmann等,EMBO 9:653〜661、ならびにLipkeおよびOvalle(1998)J.Bacteriol.180:3735〜3740を参照。GPIシグナル配列ペプチド、例えば本明細書で開示されたGPIシグナルペプチドは、GPIを本融合タンパク質のC末端に付着させるためにシグナル伝達する。GPIシグナルそれ自体は、以下の3つのドメイン:GPI付着部位(そのω部位)ω部位下流の第一および第二のアミノ酸を含む領域、5〜10個のアミノ酸のスペーサー、ならびに10〜15個のアミノ酸の疎水性ストレッチを有する。GPIシグナルを含むタンパク質は、ω部位で切断され、生じたタンパク質のカルボキシ末端は、GPI成分に共有結合する。この反応は、小胞体内で起こる。GPI成分によって膜と結合する、GPIが付着したタンパク質は、続いて細胞表層に輸送され、タンパク質が短いω−マイナス領域に塩基性残基(Rおよび/またはK)を含む場合、GPIで固定されたタンパク質として原形質膜に留まる。ω−4/−5部位にV、IまたはL、およびω−2部位にYまたはNを有するGPIと結合したタンパク質は、細胞膜中に組み込まれる。Hamada等(1999)J.Bacteriol.181:3886〜3889;Nuoffer等(1993)J.Biol.Chem.268:10558〜10563;De Nobel等(1994)Trends Cell Biol.4:42〜45;Hamada等(1998)Mol.Gen.Genet.258:53〜59;およびVan Der Vaart等(1998)Biotechnol.Genet.Eng.Rev.15:387〜411を参照。
【0134】
本発明の一実施態様において、酵母GPI YIR019Cは、トランスフェリン融合タンパク質のアンカー成分を提供するのに用いられる。図2は、GPI YIR019Cの図解を提供する。アミノ酸配列(配列番号15)におけるω部位はグリシンであり、そのどちらかの側にスペースを有するものとして図示されている。スペースは、ω部位のいずれかの側にあるスペーサー領域の指標である。太字で示したIおよびYアミノ酸は、それぞれω−5/−4および−2部位である。
【0135】
数種のサッカロミセスのアンカー成分が当技術分野で既知であり、本発明の融合タンパク質を構築するのに用いることができる。酵母のGPIシグナルタンパク質のその他の例としては、これらに限定されないが、YDR534C、YNL327W、YOR214C、YDR134C、YPL130、YOR009W、YER150W、YDR077W、YOR383C、YJR151C、YJR004、YJL078C、YLR110CおよびYNL300Wが挙げられる。さらに、他の生物由来のGPIシグナルタンパク質も用いることができ、例えばカンジダ・グラブラータ(Canidida glabrata)のEPA1のGPI、カンジダ・アルビカンス(Candida albicans)のHwplp、またはトリパノソーマ−ブルセイ(Trypanosoma brucei)のVSGである。
【0136】
本発明の一実施態様において、アンカー成分は、哺乳動物の成分、またはそれらの誘導体もしくはフラグメントである。本発明のその他の実施態様において、GPIシグナルペプチドは、哺乳動物のGPIシグナルタンパク質である。例えば、本発明は、ヒトMDPGPIシグナルタンパク質の誘導体を含み、例えば表1で開示されたものが挙げられる(実施例5を参照)。
【0137】
本発明はまた、1またはそれ以上の未結合のシステイン残基を含むアンカー成分を含む融合タンパク質も包含する。このようなシステイン残基は、細胞壁中のタンパク質のシステイン残基とジスルフィド結合を形成することによって、本融合タンパク質と細胞とを結びつけるように作用することができる。
【0138】
本発明は、アンカー成分として膜貫通ドメイン(TMD)を含む融合タンパク質を包含する。本発明の一実施態様において、TMDは、1回膜貫通I型またはII型膜タンパク質の領域である。例えば、本発明は、これらに限定されないが、FUS1の残基70〜98を含む。
【0139】
本発明のその他の実施態様において、TMDは、マルチスパン膜タンパク質の数種の膜貫通領域のうち1つまたはそれ以上を含む。本発明の一実施態様において、TMDは、マルチスパン膜タンパク質の疎水性領域であり、約10〜60個のアミノ酸、約15〜60個のアミノ酸、約20〜60個のアミノ酸、約30〜60個のアミノ酸、または約25〜50個のアミノ酸を含む。例えば、本発明は、サッカロミセスのSTE6由来の1種またはそれ以上のTMDを含み、このようなTMDは、これらに限定されないが、残基25〜30、73〜100、171〜198、249〜277、714〜742、761〜789、838〜858、864〜884、940〜967および979〜1000(サッカロミセスゲノムデータベースアノテーション(Saccharomyces Genome Database annotation))からなる群より選択される。
【0140】
その他の実施態様において、アンカー成分は、トランスフェリン融合タンパク質を、マイクロアレイのような固体基板に固定するのに用いられる。アンカー成分は、好ましくは、短いエピトープタグであり(すなわち、抗体、典型的にはモノクローナル抗体によって認識される配列)、例えば、ポリヒスチジン、SEAPまたはM1およびM2フラグである。Bush等(1991)J.Biol.Chem.266:13811〜13814,Berger等(1988)Gene 66:1〜10,米国特許第5,011,912号、米国特許第4,851,341号、米国特許第4,703,004号、および米国特許第4,782,137号を参照、これらは全て、それら全体として参照により開示に組み込まれる。一実施態様において、軸ドメインは、抗ムチン抗体のような抗軸配列抗体によって基質に結びつけられる。
【0141】
アルブミン
本発明はまた、標的にリガンドを「提示する」ためにトランスフェリン以外のタンパク質またはタンパク質フラグメントを用いる融合タンパク質も含む。適切なタンパク質は、可溶性であり、長さが少なくとも約50個のアミノ酸であるか、またはそれより長いタンパク質である。本発明の一実施態様において、このようなタンパク質またはタンパク質フラグメントは、トランスフェリンの二次構造に類似した二次構造を含む。
【0142】
上記タンパク質またはそれらのフラグメントが、本融合タンパク質の軸部分から切断されたときに、リガンドの半減期を増加させることができ、治療剤として用いることができることが好ましい。例えば、本発明は、アルブミン成分、軸成分、および細胞壁連結要素を含む融合タンパク質の使用を想定する。本融合タンパク質のアルブミンおよびリガンド部分が、融合タンパク質のそれ以外の部分から切断され、治療剤としてのリガンドが必要な患者に投与される場合、アルブミン成分はリガンド(すなわち治療剤)に増加した血清中半減期を付与することができる。
【0143】
アルブミン成分を含む融合タンパク質は、アルブミンタンパク質、アルブミン変異体またはそれらのフラグメントを含んでいてもよい。一実施態様において、アルブミンタンパク質は、配列番号66の核酸配列によってコードされる配列番号67のアミノ酸配列を含む。本発明は、当技術分野で既知のアルブミンの改変を含む。
【0144】
核酸
本発明は、核酸分子も提供する。これらは、リガンド成分に共有結合で連結または結合したトランスフェリンタンパク質、またはトランスフェリンタンパク質の部分を含む改変されたTf融合タンパク質をコードする。本融合タンパク質は、リンカー領域、例えば、約50、40、30、20または10個のアミノ酸残基未満のリンカーをさらに含んでいてもよい。リンカーは、トランスフェリンタンパク質またはそれらの部分と、リガンド部分に、またはそれらの間に共有結合で連結させることができる。本発明の核酸分子は、精製されてもよいし、または精製されていなくてもよい。
【0145】
また、核酸分子を複製するための、およびコードされた融合タンパク質を発現させるための宿主細胞およびベクターも提供される。あらゆるベクターまたは宿主細胞が使用可能であり、これらは原核性でもよいし、または真核性でもよいが、真核性の発現系、特に酵母発現系が好ましい場合がある。このような目的のための多くのベクターおよび宿主細胞が当技術分野で既知である。所望の適用に適したセットを選択することは十分に当分野の技術の範囲内である。
【0146】
トランスフェリン、トランスフェリンの部分、および、関心対象の治療用タンパク質をコードするDNA配列を、当技術分野で既知の様々なゲノムまたはcDNAライブラリーからクローニングすることができる。プローブに基づく方法を用いてこのようなDNA配列を単離する技術は、従来技術であり、当業者によく知られている。このようなDNA配列を単離するためのプローブは、公開されたDNAまたはタンパク質配列をベースにしたものが可能である(例えば、Baldwin,G.S.(1993)Comparison of Transferrin Sequences from Different Species.Comp.Biochem.Physiol.106B/1:203〜218を参照、そこで引用された全ての文献は、参照によりその全体として本明細書に組み込まれる)。あるいは、Mullis等(米国特許第4,683,195号)およびMullis(米国特許第4,683,202号)に記載されたポリメラーゼ連鎖反応(PCR)法(参照により本明細書に組み込まれる)を用いてもよい。このようなDNA配列を単離するためのライブラリーの選択およびプローブの選択は、当技術分野における通常の技術レベルの範囲内である。
【0147】
当技術分野で既知のように、2種のポリヌクレオチドまたはポリペプチド間の「類似性」は、ヌクレオチドまたはアミノ酸配列、および一種のポリヌクレオチドまたはポリペプチドの、第二のポリヌクレオチドまたはポリペプチドの配列に対する、保存されたヌクレオチドまたはアミノ酸置換を比較することによって決定される。「同一性」も当技術分野で既知であり、これは、このような配列の2つの鎖間におけるマッチングの同一性によって決定されるような、2種のポリペプチドまたは2種のポリヌクレオチド配列間の配列の関連の程度を意味する。同一性および類似性はいずれも容易に計算することができる(Computational Molecular Biology,Lesk,A.M編,オックスフォード・ユニバーシティ・プレス(Oxford University Press),ニューヨーク,1988;Biocomputing:Informatics and Genome Projects,Smith,D.W.編,アカデミックプレス,ニューヨーク,1993;Computer Analysis of Sequence Data,Part I,Griffin,A.M.およびGriffin,H.G.編,ヒューマナ・プレス(Humana Press),ニュージャージー州,1994;Sequence Analysis in Molecular Biology,von Heinje,G.,アカデミックプレス、1987;およびSequence Analysis Primer,Gribskov,M.およびDevereux,J.編集,Mストックトンプレス(M Stockton Press),ニューヨーク,1991)。
【0148】
2種のポリヌクレオチドまたはポリペプチド配列間の同一性および類似性を測定する多数の方法があるが、用語「同一性」および「類似性」は当技術分野ではよく知られている(Sequence Analysis in Molecular Biology,von Heinje,G.,アカデミックプレス,1987;Sequence Analysis Primer,Gribskov,M.and Devereux,J.編,Mストックトンプレス,ニューヨーク,1991;およびCarillo,H.およびLipman,D.,SIAM J.Applied Math.,48:1073(1988)。2種の配列間の同一性または類似性を決定するのに一般的に用いられる方法としては、これらに限定されないが、Guide to Huge Computers,Martin J.Bishop編,アカデミックプレス,サンディエゴ,1994、およびCarillo,H.およびLipman,D.,SIAM J.Applied Math.48:1073(1988)に記載されたものが挙げられる。
【0149】
同一性を決定する好ましい方法は、試験される2種の配列間の最も大きいマッチングが得られるように設計される。同一性および類似性を決定する方法は、コンピュータープログラムで体系化されている。2種の配列間の同一性および類似性を決定する好ましいコンピュータープログラム法としては、これらに限定されないが、GCGプログラムパッケージ(Devereux等,Nucl.Acid Res.12(1):387(1984))、BLASTP、BLASTN、FASTA(Atschul等,J.Mol.Biol.215:403(1990))が挙げられる。上述した類似性または同一性の程度は、2種の配列間の同一性の程度として決定され、第二の配列からの第一の配列の起源を示すことが多い。2種の核酸配列間の同一性の程度は、当技術分野で既知のコンピュータープログラムによって決定してもよく、例えばGCGプログラムパッケージに備えられているGAPである(NeedlemanおよびWunsch J.Mol.Biol.48:443〜453(1970))。本発明に関する2種の核酸配列間の同一性の程度を決定する目的で、ギャップ(GAP)は、以下の設定:5.0のギャップクリエーションペナルティー、0.3のギャップ伸長ペナルティーで用いられる。
【0150】
コドンの最適化
天然型のDNA配列によってコードされたポリペプチドと同一なアミノ酸配列を有するポリペプチドが生成されるが、遺伝子コードの縮重によって、トランスフェリンタンパク質および/または関心対象の治療用タンパク質のヌクレオチドの配列の変異が可能になる。「コドン最適化」として知られる手順(米国特許第5,547,871号に記載されており、これはこの参照によりその全体として本明細書に組み込まれる)では、このような変更されたDNA配列を設計する手段を用いてコドン最適化が提供される。コドン最適化遺伝子の設計は、生物におけるコドンの使用頻度、最隣接の頻度、RNA安定性、二次構造形成の可能性、合成経路、およびその遺伝子の目的とするその後のDNA操作などの様々な因子を考慮すべきである。特に、利用可能な方法を用いて、酵母発現系が用いられる場合、酵母によって最も容易に認識されるコドンを用いて所定の融合タンパク質をコードするコドンを変更することができる。
【0151】
遺伝子コードの縮重によって、同じアミノ酸配列を多くの異なる方法でコードし、翻訳させることを可能にする。例えば、ロイシン、セリンおよびアルギニンはそれぞれ6種の異なるコドンによってコードされるが、バリン、プロリン、スレオニン、アラニンおよびグリシンはそれぞれ4種の異なるコドンによってコードされる。しかしながら、このような同義的なコドンの使用頻度は、真核生物および原核生物の間でゲノム毎に異なる。例えば、哺乳動物間での同義的なコドンの選択パターンは、極めて類似しているが、進化的に遠い生物、例えば酵母(S.セレビジエ)、細菌(例えばE.コリ)、および昆虫(例えば、D.melanogaster)は、ゲノムコドンの使用頻度において明らかに異なるパターンを示す(Grantham,R.等,Nucl.Acid Res.,8,49〜62(1980);Grantham,R.等,Nucl.Acid Res.,9,43〜74(1981);Maroyama,T.等,Nucl.Acid Res.,14,151〜197(1986);Aota,S.等,Nucl.Acid
Res.,16,315〜402(1988);Wada,K.等,Nucl.Acid Res.,19 Supp.,1981〜1985(1991);Kurland,C.G.,FEBS Lett.,285,165〜169(1991))。このようなコドンの選択パターンの差は、ペプチドの伸長速度を調節することによる個々の遺伝子の総体的な発現レベルに寄与するようである。(Kurland,C.G.,FEBS Lett.,285,165〜169(1991);Pedersen,S.,EMBO J.,3,2895〜2898(1984);Sorensen,M.A.,J.Mol.Biol.,207,365〜377(1989);Randall,L.L.等,Eur.J.Biochem.,107,375〜379(1980);Curran,J.F.、およびYarus,M,J.Mol.Biol.,209,65〜77(1989);Varenne,S.等,J.Mol.Biol.,180,549〜576(1984),Varenne,S.等,J.Mol,Biol.,180,549〜576(1984);Garel,J.−P.,J.Theor.Biol.,43,211〜225(1974);Ikemura,T.,J.Mol.Biol.,146,1〜21(1981);Ikemura,T.,J.Mol.Biol.,151,389〜409(1981))。
【0152】
合成遺伝子のコドンの使用頻度は、組換えタンパク質発現に用いることを目的とする細胞/生物の(具体的には酵母種の)正確な(または可能な限り密接に関連した)ゲノムから誘導された核遺伝子のコドンの使用を反映することが必要である。上記で考察したように、一実施態様において、ヒトTf配列は、治療用タンパク質のヌクレオチド配列になるように、酵母発現に関して本明細書で説明されているような改変の前に、または改変の後にコドンが最適化される。
【0153】
ベクター
本発明で使用するための発現単位は、一般的に、5’から3’方向に機能するように連結した以下の構成要素:転写プロモーター、分泌シグナル配列、治療用タンパク質または関心対象のペプチドをコードするDNA配列に結合したトランスフェリンタンパク質またはトランスフェリンタンパク質の部分を含む改変されたTf融合タンパク質をコードするDNA配列、および転写ターミネーターを含む。上記で考察したように、本発明のベクターでは、治療用タンパク質またはペプチドがTf部分に融合した配置、またはTf部分内の配置のいずれでも使用可能である。適切なプロモーター、シグナル配列、およびターミネーターの選択は、選択された宿主細胞に応じて決定され、当業者には明白であろうが、以下でより具体的に考察する。
【0154】
本発明で使用するのに適した酵母ベクターは、米国特許第6,291,212号に記載されており、例えば、YRp7(Struhl等,Proc.Natl.Acad.Sci.USA 76:1035〜1039,1978)、YEp13(Broach等,Gene 8:121〜133,1979)、pJDB249およびpJDB219(Beggs,Nature 275:104〜108,1978)、pPPC0005、pSeCHSA、pScNHSA、pC4、およびそれらの誘導体が挙げられる。また、有用な酵母のプラスミドベクターとしては、pRS403−406、pRS413−416、およびストラタジーン・クローニング・システム(Stratagene Cloning Systems,ラホヤ,カリフォルニア州92037,米国)より入手可能なピチア属のベクターも挙げられる。プラスミドpRS403、pRS404、pRS405、およびpRS406は、酵母組込み型プラスミド(YIps)であり、酵母選択マーカーHIS3、TRP1、LEU2およびURA3が組み込まれている。プラスミドpRS413−41.6は、酵母セントロメアプラスミド(YCps)である。
【0155】
このようなベクターは、一般的に、形質転換株の選択を可能にする表現型の分析が存在する優性表現型を示す多数の遺伝子のうちの1つとなり得る選択マーカーを含む。好ましい選択マーカーは、宿主細胞の栄養要求性を補い、抗生物質耐性を提供するか、または、細胞の特異的な炭素源の利用を可能にするマーカーであり、例えばLEU2(Broach等,前記の箇所),URA3(Botstein等,Gene 8:17,1979)、HIS3(Struhl等,前記の箇所)、またはPOT1(KawasakiおよびBell,EP171,142)が挙げられる。その他の適切な選択マーカーとしては、酵母細胞にクロラムフェニコール耐性を付与するCAT遺伝子が挙げられる。酵母に使用するのに好ましいプロモーターとしては、酵母の解糖系遺伝子由来のプロモーター(Hitzeman等,J Biol.Chem.225:12073〜12080,1980;AlberおよびKawasaki,J.Mol.Appl.Genet.1:419〜434,1982;Kawasaki,米国特許第4,599,311号)、またはアルコールデヒドロゲナーゼ遺伝子由来のプロモーター(Young等のGenetic Engineering of Microorganisms for Chemicals,Hollaender等(編),p.355,Plenum,N.Y.,1982;Ammerer,Meth.Enzymol.101:192〜201,1983)が挙げられる。これに関連して、使用可能なプロモーターは、TPI1プロモーター(Kawasaki,米国特許第4,599,311号)およびADH2−4cである(米国特許第6,291,212号のプロモーター(Russell等,Nature 304:652〜654,1983)を参照。また、発現単位は、転写ターミネーターを含んでいてもよい。一つの転写ターミネーターは、TPI1ターミネーター(AlberおよびKawasaki,前記の個所)である。
【0156】
酵母に加えて、本発明の改変された融合タンパク質は、糸状菌で、例えばアスペルギルス属の菌株で発現させることができる。有用なプロモーターの例としては、アスペルギルス・ニダランス(Aspergillus nidulans)の解糖系遺伝子から誘導されたもの、例えばadh3プロモーター(McKnight等,EMBO J.4:2093〜2099,1985)、およびtpiAプロモーターが挙げられる。適切なターミネーターの例は、adh3ターミネーター(McKnight等,前記の個所)である。このような成分を利用する発現単位は、例えば、アスペルギルスの染色体DNAに挿入できるベクターにクローニングしてもよい。
【0157】
本発明の実施に使用するための哺乳動物の発現ベクターは、改変されたTf融合タンパク質の転写を指示することができるプロモーターを含む。好ましいプロモーターとしては、ウイルスプロモーターおよび細胞性のプロモーターが挙げられる。好ましいウイルスプロモーターとしては、アデノウイルス2由来の主要後期プロモーター(KaufmanおよびSharp,Mol.Cell.Biol.2:1304〜13199,1982)、およびSV40プロモーター(Subramani等,Mol.Cell.Biol.1:854〜864,1981)が挙げられる。好ましい細胞性のプロモーターとしては、マウスメタロチオネイン1プロモーター(Palmiter等,Science 222:809〜814,1983)、およびマウスV6(米国特許第6,291,212号を参照)プロモーター(Grant等,Nuc.Acids Res.15:5496,1987)が挙げられる。このようなプロモーターの一つは、マウスVH(米国特許第6,291,212号を参照)プロモーター(Loh等,前記の個所)である。また、このような発現ベクターは、プロモーターの下流、且つトランスフェリン融合タンパク質をコードするDNA配列の上流に位置する一連のRNAスプライス部位を含んでいてもよい。好ましいRNAスプライス部位は、アデノウイルスおよび/または免疫グロブリン遺伝子から得られたものでもよい。
【0158】
また発現ベクターには、関心対象のコード配列の下流に位置するポリアデニル化シグナルも含まれる。ポリアデニル化シグナルとしては、SV40由来の初期または後期ポリアデニル化シグナル(KaufmanおよびSharp,前記の個所)、アデノウイルス5E1B領域由来のポリアデニル化シグナル、およびヒト成長ホルモン遺伝子ターミネーター(DeNoto等,Nucl.Acid Res.9:3719〜3730,1981)が挙げられる。このようなポリアデニル化シグナルの一つは、VH(米国特許第6,291,212号を参照)遺伝子ターミネーター(Loh等,本明細書前記)である。発現ベクターは、非コードウイルスリーダー配列、例えばプロモーターとRNAスプライス部位との間に位置するアデノウイルス2のトリパータイトリーダーを含んでいてもよい。また、好ましいベクターは、エンハンサー配列、例えばSV40エンハンサー、およびマウス:(米国特許第6,291,212号を参照)エンハンサー(Gillies,Cell 33:717〜728,1983)。また、発現ベクターは、アデノウイルスVA RNAをコードする配列を含んでいてもよい。
【0159】
形質転換
菌類を形質転換する技術は、文献においてよく知られており、例えばBeggs(前記の個所)、Hinnen等(Proc.Natl.Acad.Sci.USA 75:1929〜1933,1978),Yelton等,(Proc.Natl.Acad.Sci.USA 81:1740〜1747,1984)、およびRussell(Nature 301:167〜169,1983)によって説明されている。宿主細胞の遺伝子型は、一般的に、発現ベクターに存在する選択マーカーによって補足される遺伝的欠陥を含む。特定の宿主および選択マーカーの選択は、十分に当分野における通常の技術のレベルの範囲内である。
【0160】
本発明の改変されたTf融合タンパク質を含むクローニングしたDNA配列を培養した哺乳動物細胞に導入されてもよく、これは、例えば、リン酸カルシウム介在トランスフェクション(Wigler等,Cell 14:725,1978;CorsaroおよびPearson,Somatic Cell Genetics 7:603,1981;GrahamおよびVan der Eb,Virology 52:456,1973)によってなされる。また、クローニングしたDNA配列を哺乳動物細胞に導入するその他の技術、例えばエレクトロポレーション(Neumann等,EMBO J.1:841〜845,1982)、またはリポフェクションも使用可能である。クローニングしたDNAを組み込んでいる細胞を同定するために、一般的に細胞に、関心対象の遺伝子またはcDNAと共に選択マーカーが導入される。培養した哺乳動物細胞に使用するのに好ましい選択マーカーとしては、ネオマイシン、ハイグロマイシンおよびメトトレキセートのような薬物に対する耐性を付与する遺伝子が挙げられる。選択マーカーは、増幅可能な選択マーカーであり得る。増幅可能な選択マーカーの一種は、DHFR遺伝子である。増幅可能なマーカーの一種は、DHFRr(米国特許第6,291,212号を参照)cDNA(SimonsenおよびLevinson,Proc.Natl.Acad.Sci.USA 80:2495〜2499,1983)である。選択マーカーは、Thillyによって総論されており(Mammalian Cell Technology,Butterworth Publishers,ストーンハム,マサチューセッツ州)、選択マーカーの選択は、十分当業界における通常の技術レベルの範囲内である。
【0161】
宿主細胞
本発明はまた、本発明の改変されたトランスフェリン融合タンパク質が発現されるように形質転換した細胞、好ましくは酵母細胞も含む。形質転換宿主細胞それ自体に加えて、本発明はまた、これら細胞の培養物、好ましくはモノクローナル(クローン的に均一な)培養物、または栄養培地中でモノクローナル培養物から誘導された培養物も含む。上記ポリペプチドが分泌されると、上記ポリペプチドは、細胞と共に培地に含まれるか、または細胞をろ過もしくは遠心分離で除去している場合細胞なしで含まれることになる。
【0162】
本発明を実施する際に使用される宿主細胞としては、外来性DNAで形質転換またはトランスフェクションさせて、培養した哺乳動物、昆虫、真菌性、植物および細菌細胞のような培養物中で増殖させることができる真核細胞、場合によっては原核細胞が挙げられる。
【0163】
真菌細胞、例えば酵母種(例えば、サッカロミセス属spp.、シゾサッカロミセス属spp.、ピチア属spp.)は、本発明の範囲内で宿主細胞としても使用可能である。本発明の実施の際に、本発明のトランスフェリン融合タンパク質を発現するための宿主として有用であると考えられる典型的な酵母の属は、ピチア属(以前はハンセヌラ属として分類された種を含む)、サッカロミセス属、クルイベロミセス属、アスペルギルス属、カンジダ属、トルロプシス属、トルラスポラ属、シゾサッカロミセス属、シテロミセス属、パチソレン属、チゴサッカロミセス属、デバリオミセス属(Debaromyces)、トリコデルマ属、セファロスポリウム属、フミコーラ属、ムコール属、アカパンカビ属、ヤロウイア属、メッシュニコヴィア属、ロドスポリディウム属、ロイコスポリジウム属、ボトリオアスカス属、スポリディオボラス属、エンドマイコプシス属などである。サッカロミセス属spp.の例は、S.セレビジエ、S.イタリカス(S.italicus)、およびS.ルキシー(S.rouxii)である。クルイベロミセス属spp.の例は、K.ラクティス(K.lactis)、およびK.マルキシアヌス(K.marxianus)である。適切な種は、T.デルブルッキー(T.delbrueckii)である。ピチア属(ハンセヌラ属)spp.の例は、P.アングスタ(P.angusta)(以前はH.ポリモルファ(H.polymorpha))P.アノマーラ(P.anomala)(以前はH.アノマーラ(H.anomala))、およびP.パストリス(P.pastoris)である。
【0164】
本発明のTf融合タンパク質を生産するのに特に有用な宿主細胞は、メタノール要求性のピチア・パストリス(Steinlein等(1995)Protein Express,Purif.6:619〜624)である。ピチア・パストリスのアルコールオキシダーゼプロモーターが単離され、クローニングされてから、ピチア・パストリスは、外来タンパク質の生産において優れた宿主となるように開発されている;1985年に、その形質転換がまず報告された。P.パストリスは、グルコースの非存在下で、炭素源としてメタノールを利用することができる。P.パストリス発現系は、アルコールオキシダーゼの発現をコードする遺伝子を制御するメタノール誘導性のアルコールオキシダーゼ(AOX1)プロモーターを使用でき、この酵素は、メタノール代謝における第一工程を触媒する。このプロモーターは特徴付けが行われており、一連のP.パストリス発現ベクターに組み込まれている。P.パストリスで生産されたタンパク質は、通常は正確に折り畳まれて培地に分泌されるため、遺伝子操作されたP.パストリスの発酵は、E.コリ発現系の優れた代替物を提供する。この系を用いて、破傷風毒素のフラグメント、百日咳菌(Bordatella pertussis)のパータクチン、ヒト血清アルブミン、およびリゾチームなどの多数のタンパク質が生産されている。
【0165】
F.オキシスポラム(F.oxysporum)の形質転換は、例えば、Malardier等(1989)Gene 78:147〜156に記載されているように行うことができる。
【0166】
その他の好ましい宿主は、酵母サッカロミセス・セレビジエ(Saccharomyces cerevisiae)株である。一実施態様において、糖タンパク質のアスパラギン連結型のグリコシル化に必要な遺伝子の遺伝子欠損を含む酵母細胞、またはより詳しくはサッカロミセス・セレビジエ宿主細胞が用いられる。このような欠損を有するS.セレビジエ宿主細胞は、標準的な突然変異および選択の技術を用いて生産することができるが、多くの利用可能な酵母株が、グリコシル化または過剰なマンノシル化を防ぐように、または減少するように改変されている。Ballou等(J.Biol.Chem.255:5986〜5991,1980)は、アスパラギン結合型糖付加に影響を与える遺伝子が欠損したマンノプロテインの生合成突然変異体の単離を記載している。GentzschおよびTanner(Glycobiology 7:481〜486,1997)は、酵母におけるタンパク質のO−グリコシル化の第一工程に関与する酵素をコードする少なくとも6種の遺伝子(PMT1−6)のファミリーを記載している。これらの遺伝子の1種またはそれ以上が欠損した突然変異体は、低いO結合型糖付加および/またはO−グリコシル化の変更された特異性を示す。
【0167】
異種タンパク質の生産を最適化するために、宿主株が、突然変異を有することが好ましい場合があり、このような突然変異としては、タンパク質分解活性の減少が起こるS.セレビジエpep4突然変異(Jones,Genetics 85:23〜33,1977)が挙げられる。多量の本発明のTf融合タンパク質を生産するために、さらなるプロテアーゼコード領域に突然変異を含む宿主株が特に有用である。
【0168】
本発明のDNAコンストラクトを含む宿主細胞は、適切な増殖培地で増殖させる。本明細書で用いられる用語「適切な増殖培地」は、細胞の増殖に必要な栄養素を含む培地を意味する。細胞増殖に必要な栄養素として、炭素源、窒素源、必須アミノ酸、ビタミン、無機質、および増殖因子が含まれていてもよい。増殖培地は、一般的に、DNAコンストラクトを含む細胞に応じて、例えば、薬物選択、またはDNAコンストラクトに存在する、もしくはDNAコンストラクトでコトランスフェクションされた選択マーカーによって補足された必須栄養素の欠失により選択される。酵母細胞は、例えば、化学的に規定された培地で増殖させることが好ましく、このような培地は、炭素源、例えばスクロース、非アミノ酸窒素源、無機塩、ビタミン、および必須アミノ酸の追加物質を含む。培地のpHは、好ましくは、2より大きく8未満のpHに維持され、好ましくはpH5.5〜6.5に維持される。安定なpHを維持する方法は、好ましくは水酸化ナトリウムの添加を介した緩衝化と一定したpH制御を含む。好ましい緩衝剤としては、コハク酸、およびビストリス(シグマ・ケミカル社(Sigma Chemical Co.),セントルイス,ミズーリ州)が挙げられる。アスパラギン結合型糖付加に必要な遺伝子の欠損を有する酵母細胞は、好ましくは、浸透圧安定剤を含む培地中で増殖させる。このような浸透圧安定剤の一つは、0.1M〜1.5Mの濃度、好ましくは0.5Mまたは1.0Mの濃度で培地に添加されたソルビトールである。
【0169】
培養した哺乳動物細胞は、一般的に、市販の血清含有または無血清培地で増殖させる。用いられる特定の細胞系に適した培地の選択は、当分野における通常の技術レベル内である。トランスフェクションされた哺乳動物細胞は、所定期間、典型的には1〜2日間成長させて、関心対象のDNA配列の発現を開始させることができる。続いて、薬物選択を適用して、選択マーカーを安定して発現する細胞増殖について選択する。増幅可能な選択マーカーでトランスフェクションされている細胞に関して、薬物濃度を段階的に増加させて、クローニングした配列のコピー数の増加、従って発現レベルの増加について選択することができる。
【0170】
また、バキュロウイルス/昆虫細胞発現系も、本発明の改変されたTf融合タンパク質を生産するのに使用可能である。BacPAKTMバキュロウイルス発現系(BDバイオサイエンス(BD Biosciences)(クロンテック(Clontech)))は、昆虫宿主細胞中で組換えタンパク質を高レベルで発現する。標的遺伝子は、トランスファーベクターに挿入され、これが昆虫宿主細胞に直線状にしたBacPAK6ウイルスDNAと共にコトランスフェクションされる。BacPAK6DNAは、バキュロウイルスゲノムの必須部分が欠失している。このDNAをこのベクターで組換えると、必須の構成要素が回復し、標的遺伝子はバキュロウイルスゲノムに移される。組換え後、2〜3個のウイルスプラークを採取し、精製し、組換え表現型を検証する。続いて、新たに単離した組換えウイルスを増幅し、これを昆虫細胞培養を感染させるのに用いて、大量の所望のタンパク質を生産する。
【0171】
分泌シグナル配列
用語「分泌シグナル配列」または「シグナル配列」または「分泌リーダー配列」は、交換可能に用いられ、例えば米国特許第6,291,212号および米国特許第5,547,871号で説明されている(これらの両方は参照によりその全体として本明細書に組み込まれる)。分泌シグナル配列またはシグナル配列または分泌リーダー配列は、分泌ペプチドをコードする。分泌ペプチドは、細胞からの成熟ポリペプチドまたはタンパク質の分泌を指示させるように作用するアミノ酸配列である。分泌ペプチドは、一般的に、疎水性アミノ酸のコアを特徴とし、典型的には、これに限定されないが、新たに合成されたタンパク質のアミノ末端に見出される。分泌ペプチドは、分泌中に成熟タンパク質から切断されることがかなり多い。分泌ペプチドは、分泌経路を通過する際に成熟タンパク質からシグナルペプチドを切断させるプロセシング部位を含んでいてもよい。プロセシング部位は、シグナルペプチド内にコードされてもよく、または、例えばインビトロでの変異誘発でシグナルペプチドに付加されていてもよい。
【0172】
本発明の改変されたTf融合タンパク質の分泌を指示させるために分泌ペプチドを用いてもよい。このような他の分泌ペプチドと併用が可能な分泌ペプチドの一つは、酵母のバリアタンパク質(Barrier protein)の第三のドメインである。分泌シグナル配列またはシグナル配列または分泌リーダー配列は、タンパク質の分泌が起こる複雑な一連の翻訳後プロセシング工程に必要である。インタクトシグナル配列が存在する場合、発現されるタンパク質は粗面小胞体の内腔に入り、続いてゴルジ体を介して分泌小胞に輸送され、最終的に細胞外に輸送される。一般的に、シグナル配列は開始コドンの直後にあり、分泌しようとするタンパク質のアミノ末端でシグナルペプチドをコードする。ほとんどの場合、シグナル配列は、シグナルペプチダーゼと呼ばれる特異的なプロテアーゼで切断される。好ましいシグナル配列はプロセシングを改善し、ウイルス、哺乳動物または酵母発現ベクターを用いた組換えタンパク質発現の効率を伝える。場合によっては、天然型のTfシグナル配列を用いて、本発明の融合タンパク質を発現させ、分泌させてもよい。
【0173】
リンカー
本発明の改変されたトランスフェリン融合タンパク質のTf成分およびリガンドは、直接融合させてもよいし、または様々な長さのリンカーペプチドを用いて融合させてもよく、このようにして、より大きい物理的な隔たりを提供し、さらに融合したタンパク質間のより大きい空間的な移動を可能にし、従って例えばその同源の受容体に結合するのに抗体の可変領域の接近しやすさを最大にすることができる。リンカーペプチドは、フレキシブルであるか、または比較的固定されたアミノ酸からなる可能性がある。一実施態様において、本発明は、実質的に非らせん状のリンカー、例えば(PEAPTD)n(配列番号18)を含む。その他の実施態様において、本発明の融合タンパク質は、ポリグリシンストレッチを有するリンカーを含む。リンカーは、約50、40、30、20または10個のアミノ酸残基より少ないものでもよい。リンカーは、トランスフェリンタンパク質またはそれらの部分と、抗体の可変領域との間を共有結合で結合させることができる。
【0174】
また、リンカーは、1つ又は複数のリガンドに抗体の可変領域を組み込むためにも使用できる。抗体の可変領域を組み込むのに適切なリンカーは、抗体の可変領域を、抗体全体の結合特異性を維持しながら三次元構造にフォールディングすることができるものである。
【0175】
スクリーニング方法
本発明の融合タンパク質ライブラリーをスクリーニングすることによってリガンドを同定することが可能な可能性のある標的分子の数は、実質的には限定がない。例えば、標的分子(すなわち受容体または物質)は、抗体(またはそれらの結合部分)または抗原であり得る。抗体が結合する抗原がわかっており、おそらく配列解析もされている場合があり、このようなケースにおいて、本発明は、抗原のエピトープをマッピングするのに用いることができる。例えば所定の自己免疫疾患に関する抗原が未知の場合、その病気に罹った患者からの例えば血清、体液、組織または細胞を、本発明のスクリーニング方法で用いて、ペプチドを同定することができ、その結果として自己免疫反応を惹起する抗原も同定することができる。ペプチドが同定されれば、そのペプチドは、ワクチン、治療剤、診断試薬などの開発のための基準として役立たせたり、または、それらを提供したりすることができる。リガンドをスクリーニング可能な標的分子の一覧に関しては、WO01/46698を参照(全ての目的についてこれは参照によりその全体として本明細書に組み込まれる)。
【0176】
スクリーニングは、当分野における技術者によく知られた方法の1つ、例えばバイオパニング、FACSまたはMACSを用いることによって行うことができる。本発明の一実施態様において、スクリーニングは、受容体を活性化するために行われる。標的は、精製して溶液中に存在させてもよいし、または、表面に結合させたり細胞に結合させたりしてもよい。標的は、例えばビオチンで標識してもよいし、または、当分野既知の他の方法で標識してもよい。
【0177】
所望の特性を有するポリペプチドおよびペプチドを単離して、それに対応する核酸配列の配列解析によって、または、アミノ酸の配列解析もしくはマススペクトロメトリーによって同定することができる。それに続いて、サブ配列を、異なる配列(好ましくはランダム配列)で繰り返し交換し、1回またはそれ以上スクリーニング工程を実施することよって最適化を行ってもよい。
【0178】
ペプチドライブラリーを構築したら、宿主細胞をライブラリーベクターで形質転換する。成功した形質転換株は、典型的には、用いられるベクターに応じて、選択培地中での増殖、または選択的な条件下での増殖、例えば適切な増殖培地中での増殖などによって選択される。この選択は、固体の増殖培地で行ってもよいし、または液体の増殖培地で行ってもよい。細菌細胞を固体培地で増殖させるために、例えば、選択的な抗生物質を含むL−寒天の広い表面上で、実質的にコンフルエントな芝状態が形成されるまで細胞を高密度で増殖させる(1m2あたり約108〜109個の形質転換株)。液体培養での増殖のためには、細胞は、L−液体培地(抗生物質での選択を含む)で約10回またはそれを超えて倍加するように増殖させてもよい。ライブラリーのサイズによって液体培養での増殖の方が比較的便利である場合もあるし、一方、固体培地での増殖の方が、増幅工程中に偏りを生じる可能性が少なくなる場合もある。
【0179】
トランスフェリン融合タンパク質ペプチドライブラリーを、酵母細胞表層提示によってスクリーニングしようとする場合、酵母細胞は、トランスフェリン融合タンパク質をコードする発現ベクターで形質転換される。あらゆる変異誘発方法が、酵母表面提示ライブラリーの構築、例えばエラープローンポリメラーゼ連鎖反応、およびDNAシャフリングと両立する。Boder等(2000)Methods of Enzymology 328:430〜444を参照。あるいは、発現された融合タンパク質のトランスフェリン成分は、ランダムペプチド配列またはCDRのスカフォールドとして機能させることができる。
【0180】
酵母細胞のトランスフェリン融合タンパク質ペプチドライブラリーが作出されれば、所望のペプチドを同定するための数種のアプローチが当分野既知である。例えば、新規の結合特異性を単離するためにライブラリーをスクリーニングしようとする場合、ペプチドは、適度に低い親和性濃度(Kd>nM、または親和性なし)のケースにおいて、低濃度の蛍光標識した標的(すなわち受容体または物質)で平衡化させた結合によって区別することができる。堅く結合するタンパク質が得られるように設計された適用に関して、過度に大量の希釈した標的溶液が、リガンドを過剰モルに維持するのに必要な場合があり、サンプルの取り扱いを複雑にする。このようなケースにおいて、結合親和性の改善は、解離速度論における変化によって近接させる場合がある。化学量論的に制限された標的の反応速度論の競合を用いて、集団内の改善されたクローンを同定することができる(Hawkins等(1992)J.Mol.Biol.226:889);しかしながら、このアプローチはスクリーニングアプローチの定量的な予測精度を欠いており、一般的に推奨されていない。Boder等(2000)Methods of Enzymology 328:430〜444を参照。
【0181】
標的は、ビオチン化してもよいし、または蛍光標識してもよく、その代わりに、関心対象のリガンド、すなわちトランスフェリンに提示されたペプチドを標識してもよい。好ましくは、標的が標識される。標識された標的、例えばビオチン化した標的は、トランスフェリン融合タンパク質ペプチドライブラリーとインキュベートすることができる。このようなライブラリーは、少なくとも約104種の構成要素(すなわち提示されたペプチド)、少なくとも約105種の構成要素、少なくとも約106種の構成要素、少なくとも約107種の構成要素、少なくとも約108種の構成要素、少なくとも約109種の構成要素、少なくとも約1010種の構成要素、少なくとも約1011種の構成要素、少なくとも約1012種の構成要素、少なくとも約1013種の構成要素、少なくとも約1014種の構成要素、少なくとも約1015種の構成要素、または、少なくとも約1016種の構成要素を有していてもよい。
【0182】
インキュベート後、細胞は、第二の標識で標識することができ、例えば、二次抗体、ストレプトアビジンで標識された分子または当分野既知のその他の方法で標識することができる。二次抗体は、抗ビオチン抗体であり得る。ストレプトアビジンで標識された分子としては、これらに限定されないが、ストレプトアビジン−フィコエリトリンまたはストレプトアビジンマイクロビーズが挙げられる。
【0183】
当分野既知のように細胞集団を分析するために、フローサイトメトリーを用いることができる。これがなされる場合、集団の提示する分画だけを分析する。Boder等(2000)Methods of Enzymology 328:430〜444、および、Kondo等(2004)Appl.Microbiol.Biotechnol.64:28〜40を参照、これらの両方は参照によりその全体として本明細書に組み込まれる。
【0184】
あるいは、標識されたビーズ、すなわち抗ビオチンまたはストレプトアビジンで標識されたビーズからなる第二の標識が用いられる場合、Yeung等(2002)Biotechnol.Prog.18:212〜220(これは参照によりその全体として本明細書に組み込まれる)に記載されているような磁気によるソーティングプロトコールを用いて、リガンドおよび標的分子の混合物をソートすることができる。このスクリーニングプロトコール(ミルテニー・バイオテク社(Miltenyi Biotec GmbH))と共に、MACS(R)マイクロビーズキットを用いることができる。磁気によるソーティングは、FACSと共に用いることができる。
【0185】
本発明の一実施態様において、酵母細胞で発現された対象の単一のリガンドを特徴付けることが望ましい。発現されたタンパク質は、様々な方法でスクリーニングすることが可能である。このようなタンパク質が機能を有する場合、直接的に分析することができる。例えば、酵母表面で発現された単鎖抗体は十分に機能的であり、抗原への結合に基づいてスクリーニングすることが可能である。このようなタンパク質が、容易に分析できる検出可能な機能を有さない場合、抗体を用いてリガンドの発現をモニターすることが可能である。酵母細胞はファージよりもかなり大きいため、フローサイトメトリーを使用して、単一の酵母細胞におけるタンパク質の表現型をモニターすることができる。
【0186】
本発明のその他の実施態様において、リガンド成分と受容体または物質との結合は、細胞表層提示以外の当分野既知の手段によって行われ、例えばELISA、競合結合分析(標的の天然型の結合パートナーが既知の場合)、サンドイッチ分析、ペプチドライブラリーなどで結合がブロックされる放射性のリガンドを用いた放射性受容体分析である。これらの方法において、Tf融合タンパク質ペプチドライブラリーで形質転換した宿主細胞を溶解させる。Tf融合タンパク質のペプチドを、これらに限定されないが抗MUC1抗体のような適切なアンカー成分を介して分析基質に固定させる。スクリーニングプロセスは、Tfペプチドライブラリーと関心対象の標的とを反応させて、基準の結合レベルを確立し、それと、それに続くペプチドライブラリーの結合活性とを比較することを含む。標的に結合する、または標的への結合に関して競合する少量のペプチドを検出するのに十分な感受性を有してさえいれば、分析の性質は重要ではない。分析条件は様々であってよく、関心対象の様々な結合物質またはその他の生物活性に最適な結合条件を考慮に入れることができる。従って、pH、温度、塩濃度、体積および結合の持続時間などは全て、関心対象の環境の条件と類似した条件下でのペプチドの標的への結合が達成されるように変動させることができる。
【0187】
Tfペプチドライブラリーが、関心対象の標的に結合する一つ又は複数のペプチドを有することが決定されたら、本発明の方法を用いて、混合物中でペプチドの配列を同定することができる。標的と結合するペプチドを提示する細胞は、MACSまたはFACSスクリーニングによってライブラリーの総集団から単離することができる。スクリーニングプロセスを最初の分離物に対して2〜3回繰り返し、非特異的に結合したもの全てを除外する。続いて、結合親和性に基づいて単離することができるFACSソーティングによる最後のスクリーニングが行われる。単離した細胞からプラスミドDNAを回収し、挿入物の領域に関するDNAを配列解析して、タンパク質配列を決定する。続いて単離物間の共通のモチーフを決定することができる。
【0188】
治療用リガンド分子
本発明のリガンドは、推定の治療的分子でもよいし、または既知の治療的分子でもよい。本明細書で用いられるように、治療的分子は、典型的には、インビトロまたはインビボで有益な生物学的作用を発揮することができるタンパク質またはペプチドであり、例えば、正常なホメオスタシス、生理機能または病状に対して有益な作用を発揮するタンパク質またはペプチドが挙げられる。治療的分子は、マーカーとして一般的に使用される融合パートナー、またはガラクトシダーゼのようなタンパク質精製の助剤を含まない(例えば、米国特許第5,986,067号、およびAldred等(1984)Biochem.Biophys.Res.Commun.122:960〜965を参照)。例えば、病状に関する有益な作用としては、治療される治療対象に有利なあらゆる作用が挙げられ、このような作用としては、治療される治療対象に有益な作用を生じるような、疾患の予防、疾患の安定化、疾患の症状の低減もしくは緩和、または根本的な欠陥の調節、緩和もしくは治癒が挙げられる。
【0189】
治療用リガンドは、トランスフェリン成分に直接融合していてもよいし、または、上述したようなリンカー成分を介して間接的に融合していてもよい。一実施態様において、本融合タンパク質を切断して、本融合タンパク質のトランスフェリンとリガンド部分を、本融合タンパク質のそれ以外の部分から分離することが望ましい場合がある。その他の実施態様において、リガンドを本融合タンパク質のそれ以外の部分から切断することが望ましい場合がある。
【0190】
本発明の融合タンパク質のリガンド成分は、治療用タンパク質の少なくともフラグメントもしくは変異体、および/または抗体の少なくともフラグメントもしくは変異体を含んでいてもよい。さらなる実施態様において、本融合タンパク質は、タンパク質または抗体の、ペプチドフラグメントまたはペプチド変異体を含んでいてもよく、ここにおいて、これらの変異体またはフラグメントは、生物学的活性または治療活性の少なくとも1つを保持する。本融合タンパク質は、治療用タンパク質を含んでいてもよく、このような治療用タンパク質としては、長さが少なくとも約3個、少なくとも約4個、少なくとも5個、少なくとも6個、少なくとも7個、少なくとも8個、少なくとも9個、少なくとも10個、少なくとも11個、少なくとも12個、少なくとも13個、少なくとも14個、少なくとも15個、少なくとも20個、少なくとも25個、少なくとも30個、少なくとも35個、または少なくとも約40個、少なくとも約50個、少なくとも約55個、少なくとも約60個、または少なくとも約70個またはそれ以上のアミノ酸からなり、Nおよび/またはC末端に融合し、改変されたトランスフェリンのループ内に挿入されているか、または改変されたトランスフェリンのループに挿入されているペプチドフラグメントまたはペプチド変異体が挙げられる。
【0191】
その他の実施態様において、本発明の融合タンパク質のリガンド成分は、治療用タンパク質部分を含み、このタンパク質部分は、全長タンパク質、同様に、アミノ酸配列のアミノ末端から1個またはそれ以上の残基が欠失したポリペプチドなどを含む治療用タンパク質のフラグメントであり得る。
【0192】
その他の実施態様において、本発明の融合タンパク質のリガンド成分は、治療用タンパク質部分を含み、このタンパク質部分は、全長タンパク質、同様に、アミノ酸配列のカルボキシ末端から1個またはそれ以上の残基が欠失したポリペプチドを含む治療用タンパク質のフラグメントであり得る。
【0193】
その他の実施態様において、本発明の融合タンパク質のリガンド成分は、治療用タンパク質部分を含み、このタンパク質部分は、アミノ末端およびカルボキシ末端の両方から1個またはそれ以上のアミノ酸が欠失していてもよい。
【0194】
その他の実施態様において、本融合タンパク質は、本明細書で記載された参照の治療用タンパク質に少なくとも約80%、85%、90%、95%、96%、97%、98%、もしくは99%同一な治療用タンパク質部分、すなわちリガンド成分、またはそれらのフラグメントを含む。さらなる実施態様において、トランスフェリン融合分子は、アミノ酸配列に上述のようなNおよびC末端の欠失を有する参照のポリペプチドと少なくとも約80%、85%、90%、95%、96%、97%、98%、または99%同一な治療用タンパク質部分を含む。
【0195】
その他の実施態様において、本融合タンパク質は、例えば治療用タンパク質の天然型または野生型のアミノ酸配列に少なくとも約80%、85%、90%、95%、96%、97%、98%、99%、または100%同一な治療用タンパク質部分を含む。これらのポリペプチドのフラグメントも提供される。
【0196】
細胞表層タンパク質や分泌タンパク質のような本発明の改変されたトランスフェリン融合タンパク質の治療用タンパク質部分に相当する治療用タンパク質は、1またはそれ以上のオリゴ糖基を付着させることによって改変することができる。グリコシル化と称される改変は、タンパク質の物理特性に有意に影響を与えることができ、さらに、タンパク質の安定性、分泌および局在化にとって重要なものにすることができる。グリコシル化は、ポリペプチド主鎖に沿って特異的な位置で起こる。グリコシル化には、通常、以下の2つの主要なタイプがあり、すなわち、グリコシル化は、セリンまたはスレオニン残基に結合しているO結合型のオリゴ糖を特徴とし;およびグリコシル化は、Asn−X−Ser/Thr配列(ここにおいてXは、プロリン以外のアミノ酸が可能である)におけるアスパラギン残基に結合しているN結合型のオリゴ糖を特徴とする。タンパク質構造や細胞型のような可変値は、様々なグリコシル化部位における鎖内の糖質単位の数および性質に影響を与える。グリコシル化の異性体もまた、は、所定の細胞型内の同じ部位に共通している。例えば、ヒトインターフェロンの数種のタイプは、グリコシル化されている。
【0197】
本発明の融合タンパク質の治療用タンパク質部分に相当する治療用タンパク質、ならびに、それらの類似体および変異体は、それらが発現される宿主細胞によって、またはそれらの発現のその他の条件に起因して、それらの核酸配列を操作した結果として1またはそれ以上の部位でのグリコシル化が変更されるように改変されていてもよい。例えば、グリコシル化の異性体は、例えば、アミノ酸残基の置換または欠失、例えばグルタミンのアスパラギンへの置換によってグリコシル化部位を無効にしたり、または導入したりすることによって生産してもよいし、またはグリコシル化されていない組換えタンパク質は、それらをグリコシル化しないと予想される宿主細胞中で、例えばグリコシル化能に欠陥のある酵母中でタンパク質を発現させることによって生産してもよい。これらのアプローチは当分野既知である。
【0198】
治療用タンパク質およびそれらの核酸配列は当分野周知であり、公開データベース、例えば、ケミカル・アブストラクツ・サービス・データベース(Chemical Abstracts Services Databases)(例えばCASレジストリ)、GenBank、および、GenSeqから入手可能である。以下で述べる登録番号および配列は、参照によりその全体として本明細書に組み込まれる。
【0199】
本発明はさらに、本明細書で説明される治療用タンパク質のフラグメントを含む融合タンパク質を対象とする。タンパク質のN末端から1個またはそれ以上のアミノ酸が欠失して、治療用タンパク質部分の生物学的機能の1種またはそれ以上に改変または減少が生じたとしても、その他の治療活性および/または機能的な活性(例えば、生物活性、マルチマー化する能力、リガンドと結合する能力)がなお保持させることができる。例えば、一般的に、N末端に欠失を有するポリペプチドの、ポリペプチドの完全な形態または成熟した形態を認識する抗体を誘導する、および/またはそれらに結合する能力は、完全なポリペプチドの残基の半数未満がN末端から除去されても保持されると予想される。完全なポリペプチドのN末端の残基が欠失した特定のポリペプチドが、このような免疫学的な活性を保持するかどうかは、本明細書で説明されているルーチン的な方法や、それ以外の当分野既知の方法で分析することができる。多数のN末端のアミノ酸残基が欠失した突然変異体が、いくつかの生物学的活性または免疫原性活性を保持する可能性があることはありそうもないことでない。実際に、6個もの少ないアミノ酸残基で構成されるペプチドでもしばしば免疫反応を誘発することがある。
【0200】
上述したように、治療用タンパク質のN末端またはC末端から1個またはそれ以上のアミノ酸が欠失していることによって、タンパク質の1種またはそれ以上の生物学的機能の改変または減少が生じる場合でさえも、その他の機能的な活性、例えば生物活性、マルチマー化する能力、リガンドと結合する能力および/または治療活性は、なお保持させることができる。一般的に、例えばC末端に欠失を有するポリペプチドの、ポリペプチドの完全な形態または成熟した形態を認識する抗体を誘導する、および/またはそれらに結合する能力は、完全なポリペプチドまたは成熟ポリペプチドの残基の半分未満がC末端から除去されても保持されると予想される。参照のポリペプチドのN末端および/またはC末端の残基が欠失した特定のポリペプチドが治療活性を保持するかどうかは、本明細書で説明されているルーチン的な方法および/または当分野既知のそれ以外の方法によって容易に決定することができる。
【0201】
治療用タンパク質のペプチドフラグメントは、アミノ酸配列がフラグメントである治療用タンパク質のポリペプチド配列の、治療活性および/または機能的な活性(例えば生物活性)を提示するアミノ酸配列を含む、あるいはそれらからなるフラグメントであってもよい。
【0202】
その他のポリペプチドフラグメントは、生物学的に活性なフラグメントである。生物学的に活性なフラグメントは、本発明で用いられる治療用タンパク質の活性に類似しているが、必ずしも同一ではない活性を示すものである。このようなフラグメントの生物活性は、改善された所望の活性、または低減された望ましくない活性を含んでいてもよい。
【0203】
一般的に、タンパク質の変異体は、総体的に極めて類似しており、多くの領域において本発明のトランスフェリン融合タンパク質の治療用タンパク質部分に相当する治療用タンパク質のアミノ酸配列と同一である。これらの変異体をコードする核酸も本発明に包含される。
【0204】
本発明で使用可能なさらなる治療用ポリペプチドは、当業者既知のストリンジェントなハイブリダイゼーション条件下で治療用タンパク質のアミノ酸配列をコードする核酸分子の相補物とハイブリダイズするポリヌクレオチドによってコードされたポリペプチドである。例えば、Ausubel,F.M.等編集,1989 Current protocol in Molecular Biology,グリーン・パブリッシング・アソシエイツ社(Green Publishing Associates,Inc.)、およびジョン・ワイリー&サンズ社(John Wiley & Sons Inc.,ニューヨーク)を参照。これらのポリペプチドをコードするポリヌクレオチドも本発明に包含される。
【0205】
例えば本発明のアミノ酸配列クエリーに少なくとも95%「同一な」アミノ酸配列を有するポリペプチドとは、対象のポリペプチドのアミノ酸配列がクエリー配列と同一である(ただし、対象のポリペプチド配列は、アミノ酸配列のアミノ酸クエリー100個あたり5個までのアミノ酸の変更を含んでいてもよい)ことを意味する。言い換えれば、アミノ酸配列がアミノ酸配列クエリーに少なくとも95%同一なポリペプチドを得るためには、その他のアミノ酸で挿入、欠失または置換されてもよい対象の配列中のアミノ酸残基は、5%以下である。これらの参照配列の変更は、参照のアミノ酸配列のアミノまたはカルボキシ末端の位置に、またはこれら末端の位置の間のどこかに、参照配列中のそれぞれの残基に、または参照配列内の1個またはそれ以上の連続した基のいずれかに散在して存在する可能性がある。
【0206】
実際問題として、あらゆる特定のポリペプチドが、例えば、本発明の融合タンパク質またはそれらのフラグメント(例えば、本融合タンパク質の治療用タンパク質部分またはそれらの部分)のアミノ酸配列に少なくとも約80%、85%、90%、95%、96%、97%、98%、または99%同一であるかどうかは、既知のコンピュータープログラムを用いてルーチン的に決定することができる。クエリー配列(本発明の配列)と、対象の配列(またはグローバル配列アライメントとも言う)との間の最良の全体のマッチングを決定する方法の一つは、Bruflag等(Comp.App.Biosci 245〜(1990))のアルゴリズムに基づくFASTDBコンピュータープログラムを用いて決定することができる。
【0207】
本発明のポリヌクレオチド変異体は、コード領域、非コード領域またはその両方に変更を含んでいてもよい。サイレント置換、付加または欠失を生じるが、コードされたポリペプチドの特性または活性を変更しない変更を含むポリヌクレオチド変異体を用いて、改変されたリガンド成分を生産してもよい。遺伝子コードの縮重によるサイレント置換によって生産されたヌクレオチド変異体を利用することができる。その上、約50個未満、40個未満、30個未満、20個未満、10個未満、または5〜50個、5〜25個、5〜10個、1〜5個、もしくは、1〜2個のアミノ酸が、あらゆる組み合わせで置換、欠失または付加されているポリペプチド変異体も利用することができる。ポリヌクレオチド変異体は、様々な理由に応じて作製することができ、例えば、特定の宿主に対してコドンの発現を最適化する(ヒトmRNAにおけるコドンを、宿主、例えば上述のような酵母またはE.コリにとって好ましいコドンに変化させる)ことによってなされる。
【0208】
その他の実施態様において、治療用タンパク質成分、すなわちリガンド成分は、野生型配列と比較して保存的置換を有する。「保存的置換」は、グループ内での入れ替えを意味し、例えば、脂肪族または疎水性アミノ酸Ala、Val、Leu、およびIleの交換;ヒドロキシル残基SerおよびThrの交換;酸性残基AspおよびGluの交換;アミド残基AsnおよびGlnの交換;塩基性残基Lys、Arg、およびHisの交換;芳香族残基Phe、Tyr、およびTrpの交換;および小型のアミノ酸Ala、Ser、Thr、Met、およびGlyの交換が挙げられる。表現型においてサイレントなアミノ酸置換をどのように作製するかに関する指針は、例えば、Bowie等の“Deciphering the Message in Protein Sequences:Tolerance to Amino Acid Substitutions,”Science247:1306〜1310(1990)に示されている。特定の実施態様において、本発明のポリペプチドは、本明細書で説明されている治療用タンパク質、および/または本発明の血清トランスフェリンおよび改変されたトランスフェリンタンパク質のアミノ酸配列のフラグメントまたは変異体を含むか、あるいはそれらからなり、ここにおいて、これらのフラグメントまたは変異体は、参照のアミノ酸配列と比較して、1〜5個、5〜10個、5〜25個、5〜50個、10〜50個、または50〜150個のアミノ酸残基付加、置換および/または欠失を有する。さらなる実施態様において、アミノ酸置換は保存的である。これらのポリペプチドをコードする核酸も本発明に包含される。
【0209】
本発明の改変された融合タンパク質は、互いにペプチド結合または改変されたペプチド結合で結合したアミノ酸で構成されていてもよく、20種の遺伝子コードアミノ酸以外のアミノ酸を含んでいてもよい。このようなポリペプチドは、天然のプロセス、例えば翻訳後プロセシング、または当分野でよく知られた化学修飾技術のいずれかによって改変されていてもよい。このような改変は基本的な教本でよく説明されており、研究論文、加えて膨大な研究文献でより詳細に説明されている。
【0210】
改変は、ポリペプチド中のあらゆる場所で発生する可能性があり、このような場所としては、ペプチド主鎖、アミノ酸側鎖、およびアミノまたはカルボキシ末端が挙げられる。当然のことながら、同じタイプの改変が、所定のポリペプチド内の同じ部位に存在していてもよいし、または様々な程度でいくつかの部位に存在していてもよい。また、所定のポリペプチドに多くのタイプの改変が含まれていてもよい。ポリペプチドは、例えばユビキチン化の結果として分岐していてもよいし、ポリペプチドは環状でもよく、分岐を含んでいてもよいし、または含んでいなくてもよい。環状、分岐状および分岐状環状ポリペプチドは、翻訳後の天然のプロセスによって生じたものでもよいし、または合成法によって製造したものでもよい。改変としては、アセチル化、アシル化、ADP−リボシル化、アミド化、フラビンの共有結合、ヘム成分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトールの共有結合、架橋、環化、ジスルフィド結合の形成、脱メチル、共有結合の架橋の形成、システインの形成、グリコシル化、GPIアンカーの形成、水酸化、ヨウ素化、メチル化、ミリスチル化、酸化、ペグ化、タンパク質分解によるプロセシング、リン酸化、プレニル化、ラセミ化、硫酸化、アミノ酸のタンパク質への付加を介するトランスファーRNA、例えばアルギニル化、およびユビキチン化が挙げられる。(例えば、PROTEINS−STRUCTURE AND MOLECULAR PROPERTIES,第二版,T.E.Creighton,W.H.Freeman and Company,ニューヨーク(1993);POST−TRANSLATIONAL COVALENT MODIFICATION OF PROTEINS,B.C.Johnson編,アカデミックプレス,ニューヨーク,1〜12頁(1983);Seifter等(1990)Meth.Enzymol.182:626〜646;Rattan等,Ann.N.Y.Acad.Sci.663:48〜62を参照)。
【0211】
リガンド成分として用いることができる治療的分子としては、これらに限定されないが、ホルモン、マトリックスタンパク質、免疫抑制剤、気管支拡張剤、心血管作動薬、酵素、CNS剤、神経伝達物質、受容体タンパク質またはペプチド、成長ホルモン、増殖因子、抗ウイルス性ペプチド、膜融合阻害ペプチド、サイトカイン、リンフォカイン、モノカイン、インターロイキン、コロニー刺激因子、分化因子、血管新生因子、受容体リガンド、癌関連タンパク質、抗腫瘍薬、ウイルスペプチド、抗生物質ペプチド、血液タンパク質、アンタゴニストタンパク質、転写因子、抗血管新生因子、アンタゴニストタンパク質またはペプチド、受容体アンタゴニスト、抗体、単鎖抗体、および細胞接着分子が挙げられる。単一の融合タンパク質に異なる治療的分子を組合せて、二機能性または多機能性の治療的分子を生産することができる。また、異なる分子を併用して、治療実体および標的実体を有する融合タンパク質を生産してもよい。治療的分子は、本発明の軸成分に直接融合させることができ、またはTf成分またはアルブミン成分のような提示成分と融合してもよいし、もしくはそれらに挿入してもよい。
【0212】
サイトカインは、免疫系の細胞によって放出される可溶性タンパク質であり、非酵素的に特異的な受容体を介して作用して、免疫反応を調節することができる。サイトカインは、低濃度で作用し、高親和性で特異的な受容体に結合するという点で、ホルモンに似ている。本明細書において用語「サイトカイン」とは、そのサイトカインについての受容体を有する細胞において特異的な生物学的反応を惹起する、天然に存在する、または組換えタンパク質、それらの類似体およびそれらのフラグメントを示すものとして用いられる。サイトカインとしては、好ましくは、インターロイキン、例えばインターロイキン−2(IL−2)(GenBank登録番号S77834)、IL−3(GenBank登録番号M14743)、IL−4(GenBank登録番号M23442)、IL−5(GenBank登録番号J03478)、IL−6(GenBank登録番号M14584)、IL−7(GenBank登録番号NM_000880)、IL−10(GenBank登録番号NM_000572)、IL−12(GenBank登録番号AF180562およびGenBank登録番号AF180563)、IL−13(GenBank登録番号U10307)、IL−14(GenBank登録番号XM_170924)、IL−15(GenBank登録番号X91233)、IL−16(GenBank登録番号NM_004513)、IL−17(GenBank登録番号NM_002190)およびIL−18(GenBank登録番号NM_001562)、造血因子、例えば顆粒球マクロファージコロニー刺激因子(GM−CSF)(GenBank登録番号X03021)、顆粒球コロニー刺激因子(G−CSF)(GenBank登録番号X03656)、血小板活性化因子(GenBank登録番号NM_000437)およびエリスロポエチン(GenBank登録番号X02158)、腫瘍壊死因子(TNF)、例えばTNFα(GenBank登録番号X02910)、リンフォカイン、例えばリンフォトキシン−α(GenBank登録番号X02911)、リンフォトキシン−β(GenBank登録番号L11016)、ロイコレグリン(leukoregulin)、マクロファージ遊走阻止因子(GenBank登録番号M25639)、およびニューロロイキン(GenBank登録番号K03515)、代謝プロセスの調節因子、例えばレプチン(GenBank登録番号U43415)、インターフェロン、例えばインターフェロンα(IFNα)(GenBank登録番号M54886)、IFNβ(GenBank登録番号V00534)、IFNγ(GenBank登録番号J00219)、IFNo(GenBank登録番号NM_002177)、トロンボスポンジン1(THBS1)(GenBank登録番号NM_003246)、THBS2(GenBank登録番号L12350)、THBS3(GenBank登録番号L38969)、THBS4(GenBank登録番号NM_003248)、およびケモカインが挙げられる。好ましくは、改変されたトランスフェリン−サイトカイン融合タンパク質は、サイトカインの生物活性を提示する。
【0213】
本明細書における用語「ホルモン」は、所定の細胞または組織で生産され、体内のどこかに位置するその他の細胞または組織で特異的な生物学的な変化または活性が生じるように作用する多数の生物学的に活性な物質のいずれか一つを述べる際に用いられる。ホルモンとしては、好ましくは、プロインスリン(GenBank登録番号V00565)、インスリン(GenBank登録番号NM_000207)、成長ホルモン1(GenBank登録番号V00520)、成長ホルモン2(GenBank登録番号F006060)、成長ホルモン放出因子(GenBank登録番号NM021081)、インスリン様増殖因子I(GenBank登録番号M27544)、インスリン様増殖因子II(GenBank登録番号NM_000612)、インスリン様増殖因子結合タンパク質1(IGFBP−1)(GenBank登録番号M59316)、IGFBP−2(GenBank登録番号X16302)、IGFBP−3(GenBank登録番号NM_000598)、IGFBP−4(GenBank登録番号Y12508)、IGFBP−5(GenBank登録番号M65062)、IGFBP−6(GenBank登録番号NM_002178)、IGFBP−7(GenBank登録番号NM001553)、絨毛性ゴナドトロピンβ鎖(GenBank登録番号NM_033142)、絨毛性ゴナドトロピンα鎖(GenBank登録番号NM000735)、黄体形成ホルモンβ(GenBank登録番号X00264)、胞刺激ホルモンβ(GenBank登録番号NM_000510)、甲状腺刺激ホルモンβ(GenBank登録番号NM_000549)、プロラクチン(GenBank登録番号NM_000948)、プロオピオメラノコルチン(GenBank登録番号V01510)、コルチコトロピン(ACTH)、β−リポトロピン、α−メラニン細胞刺激ホルモン(α−MSH)、γ−リポトロピン、β−MSH、β−エンドルフィン、およびコルチコトロピン様中葉ペプチド(CLIP)が挙げられる。
【0214】
また用語「ホルモン」は、グルカゴン様ペプチド−1(GLP−1)も包含し、これは、いわゆる腸島軸(enteroinsular axis)に属するインスリン分泌を調節する胃腸ホルモンであり、さらにエキセンディン(例えば、エキセンディン−4およびそれらの変異体)も包含し、これは、GLP−1受容体アゴニストである。
【0215】
本明細書における用語「増殖因子」は、細胞増殖を刺激する受容体に結合するあらゆるタンパク質またはペプチドを述べる際に用いられる。増殖因子としては、好ましくは、血小板由来増殖因子−α(PDGF−α)(GenBank登録番号X03795)、PDGF−β(GenBank登録番号X02811)、ステロイドホルモン、上皮増殖因子(EGF)(GenBank登録番号NM_001963)、線維芽細胞増殖因子、例えば線維芽細胞増殖因子1(FGF1)(GenBank登録番号NM_000800)、FGF2(GenBank登録番号NM_002006)、FGF3(GenBank登録番号NM_005247)、FGF4(GenBank登録番号NM_002007)、FGF5(GenBank登録番号M37825)、FGF6(GenBank登録番号X57075)、FGF7(GenBank登録番号NM_002009)、FGF8(GenBank登録番号AH006649)、FGF9(GenBank登録番号NM_002010)、FGF10(GenBank登録番号AB002097)、FGF11(GenBank登録番号NM_004112)、FGF12(GenBank登録番号NM_021032)、FGF13(GenBank登録番号NM_004114)、FGF14(GenBank登録番号NM_004115)、FGF16(GenBank登録番号AB009391)、FGF17(GenBank登録番号NM_003867)、FGF18(GenBank登録番号AF075292)、FGF19(GenBank登録番号NM_005117)、FGF20(GenBank登録番号NM_019851)、FGF21(GenBank登録番号NM_019113)、FGF22(GenBank登録番号NM_020637)、およびFGF23(GenBank登録番号NM020638)、アンギオゲニン(GenBank登録番号M11567)、脳由来神経栄養因子(GenBank登録番号M61176)、毛様体神経栄養因子(GenBank登録番号X60542)、トランスフォーミング増殖因子−α(TGF−α)(GenBank登録番号X70340)、TGF−β(GenBank登録番号X02812)、神経成長因子−α(NGF−α)(GenBank登録番号NM_010915)、NGF−β(GenBank登録番号X52599)、メタロプロテイナーゼ1の組織阻害剤(TIMP1)(GenBank登録番号NM_003254)、TIMP2(GenBank登録番号NM_003255)、TIMP3(GenBank登録番号U02571)、TIMP4(GenBank登録番号U76456)、およびマクロファージ刺激因子1(GenBank登録番号LI1924)が挙げられる。
【0216】
本明細書における用語「マトリックスタンパク質」は、通常、細胞外マトリックスで見出されるタンパク質またはペプチドを述べる際に用いられる。これらのタンパク質は、強度、ろ過または付着に関して機能的に重要な可能性がある。マトリックスタンパク質としては、好ましくは、コラーゲン、例えばI型コラーゲン(GenBank登録番号Z74615)、II型コラーゲン(GenBank登録番号XI6711)、III型コラーゲン(GenBank登録番号X14420)、IV型コラーゲン(GenBank登録番号NM_001845)、V型コラーゲン(GenBank登録番号NM_000393)、VI型コラーゲン(GenBank登録番号NM_058175)、VII型コラーゲン(GenBank登録番号L02870)、VIII型コラーゲン(GenBank登録番号NM_001850)、IX型コラーゲン(GenBank登録番号X54412)、X型コラーゲン(GenBank登録番号X60382)、XI型コラーゲン(GenBank登録番号J04177)およびXII型コラーゲン(GenBank登録番号U73778)、ラミニンタンパク質、例えばLAMA2(GenBank登録番号NM_000426)、LAMA3(GenBank登録番号L34155)、LAMA4(GenBank登録番号NM_002290)、LAMB1(GenBank登録番号NM_002291)、LAMB3(GenBank登録番号L25541)、LAMC1(GenBank登録番号NM_002293)、ニドゲン(GenBank登録番号NM_002508)、α−テクトリン(GenBank登録番号NM_005422)、β−テクトリン(GenBank登録番号NM_058222)、およびフィブロネクチン(GenBank登録番号X02761)が挙げられる。
【0217】
用語「血液タンパク質」は、一般的に、血漿由来のタンパク質と定義され、現在、そのうち多くが、一般的に組換え法によって生産されており、このようなタンパク質としては、これらに限定されないが、天然の血清タンパク質、それらの誘導体、フラグメントおよび突然変異体または変異体、血液凝固因子、誘導体、突然変異体、変異体およびフラグメント(例えば、VII、VIII、IX、X因子など)、プロテアーゼ阻害剤(アンチトロンビン3、アルファ−1アンチトリプシン)、ウロキナーゼ型プラスミノゲン活性化因子、免疫グロブリン、フォン−ヴィレブランド因子、およびフォン−ヴィレブランド突然変異体、フィブロネクチン、フィブリノーゲン、トロンビン、およびヘモグロビンが挙げられる。
【0218】
本明細書における用語「酵素」は、それ自体永久的に変化したり、または破壊されたりすることなく特異的な反応を触媒するあらゆるタンパク質またはタンパク質様物質を述べる際に用いられる。酵素としては、好ましくは、凝固因子、例えばF2(GenBank登録番号XM_170688)、F7(GenBank登録番号XM_027508)、F8(GenBank登録番号XM_013124)、F9(GenBank登録番号NM_000133)、F10(GenBank登録番号AF503510)など、マトリックスメタロプロテイナーゼ、例えばマトリックスメタロプロテイナーゼI(GenBank登録番号MMP1)(GenBank登録番号NM_002421)、MMP2(GenBank登録番号NM_004530)、MMP3(GenBank登録番号NM_002422)、MMP7(GenBank登録番号NM_002423)、MMP8(GenBank登録番号NM_002424)、MMP9(GenBank登録番号NM_004994)、MMP10(GenBank登録番号NM_002425)、MMP12(GenBank登録番号NM_002426)、MMP13(GenBank登録番号X75308)、MMP20(GenBank登録番号NM_004771)、アデノシン−デアミナーゼ(GenBank登録番号NM_000022)、マイトジェン活性化プロテインキナーゼ、例えばMAPK3(GenBank登録番号XM_055766)、MAP2K2(GenBank登録番号NM_030662)、MAP2K1(GenBank登録番号NM_002755)、MAP2K4(GenBank登録番号NM_003010)、MAP2K7(AF013588)、およびMAPK12(NM_002969)、キナーゼ、例えばJNKK1(GenBank登録番号U17743)、JNKK2(GenBank登録番号AF014401)、JAK1(M64174)、JAK2(NM_004972)、およびJAK3(NM_000215)、およびホスファターゼ、例えばPPM1A(GenBank登録番号NM_021003)およびPPM1D(GenBank登録番号NM_003620)が挙げられる。
【0219】
本明細書における用語「転写因子」は、タンパク質をコードする遺伝子の転写に関与するあらゆるタンパク質またはペプチドを述べる際に用いられる。転写因子としては、Sp1、Sp2(GenBank登録番号NM_003110)、Sp3(GenBank登録番号AY070137)、Sp4(GenBank登録番号NM_003112)、NFYB(GenBank登録番号NM_006166)、Hap2(GenBank登録番号M59079)、GATA−1(GenBank登録番号NM_002049)、GATA−2(GenBank登録番号NM_002050)、GATA−3(GenBank登録番号X55122)、GATA−4(GenBank登録番号L34357)、GATA−5、GATA−6(GenBank登録番号NM_005257)、FOG2(NM_012082)、Eryf1(GenBank登録番号X17254)、TRPS1(GenBank登録番号NM_014112)、NF−E2(GenBank登録番号NM_006163)、NF−E3、NF−E4、TFCP2(GenBank登録番号NM_005653)、Oct−1(GenBank登録番号X13403)、ホメオボックスタンパク質、例えばHOXB2(GenBank登録番号NM_002145)、HOX2H(GenBank登録番号X16665)、ヘアレス(hairless)同族体(GenBank登録番号NM_005144)、デカペンタプレジックタンパク質に対する親物質、例えばMADH1(GenBank登録番号NM_005900)、MADH2(GenBank登録番号NM_005901)、MADH3(GenBank登録番号NM_005902)、MADH4(GenBank登録番号NM_005359)、MADH5(GenBank登録番号AF009678)、MADH6(GenBank登録番号NM_005585)、MADH7(GenBank登録番号NM_005904)、MADH9(GenBank登録番号NM_005905)、および転写タンパク質のシグナルトランスデューサーおよび活性化因子、例えばSTAT1(GenBank登録番号XM_010893)、STAT2(GenBank登録番号NM_005419)、STAT3(GenBank登録番号AJ012463)、STAT4(GenBank登録番号NM_003151)、STAT5(GenBank登録番号L41142)、および、STAT6(GenBank登録番号NM_003153)が挙げられる。
【0220】
さらに本発明のその他の実施態様において、治療的分子は、非ヒトまたは非哺乳動物タンパク質である。例えば、HIVgp120、HIVTat、他のウイルスの表面タンパク質、例えば肝炎、ヘルペス、インフルエンザ、アデノウイルスおよびRSV、他のHIV成分、寄生虫の表面タンパク質、例えばマラリアの抗原、および細菌の表面タンパク質が使用可能である。これらの非ヒトタンパク質は、これらは有用な活性を有するため、例えば抗原として使用可能である。例えば、治療的分子は、ストレプトキナーゼ、スタフィロキナーゼ、アスパラギナーゼ、ウロキナーゼ、または有用な酵素活性を有するその他のタンパク質であり得る。
【0221】
本発明の代替の実施態様において、治療的分子は、生物活性を有するリガンド結合タンパク質である。このようなリガンド結合タンパク質は、例えば、(1)細胞表層で受容体−リガンドの相互作用をブロックしたり;または(2)血液の流動相において分子の生物活性を中和したりすることができ、それによって、上記タンパク質がその細胞の標的に到達することを防ぐことができる。いくつかの実施態様において、改変されたトランスフェリン融合タンパク質は、以下からなる群より選択される受容体のリガンド−結合ドメインに融合した改変されたトランスフェリン分子を含む:低密度リポタンパク質(LDL)受容体、アセチル化LDL受容体、腫瘍壊死因子α受容体、トランスフォーミング増殖因子β受容体、サイトカイン受容体、免疫グロブリンFc受容体、ホルモン受容体、グルコース受容体、グリコリピド受容体、およびグリコサミノグリカン受容体(ただしこれらに限定されない)。その他の実施態様において、リガンド結合タンパク質としては、CD2(M14362)、CD3G(NM_000073)、CD3D(NM_000732)、CD3E(NM_000733)、CD3Z(J04132)、CD28(NM_006139)、CD4(GenBank登録番号NM_000616)、CD1A(GenBank登録番号M28825)、CD1B(GenBank登録番号NM_001764)、CD1C(GenBank登録番号NM_001765)、CD1D(GenBank登録番号NM_001766)、CD80(GenBank登録番号NM_005191)、GNB3(GenBank登録番号AF501884)、CTLA−4(GenBank登録番号NM_005214)、細胞間接着分子、例えばICAM−1(NM_000201)、ICAM−2(NM_000873)、およびICAM−3(NM_002162)、腫瘍壊死因子受容体、例えばTNFRSF1A(GenBank登録番号X55313)、TNFR1SFB(GenBank登録番号NM_001066)、TNFRSF9(GenBank登録番号NM_001561)、TNFRSF10B(GenBank登録番号NM_003842)、TNFRSF11B(GenBank登録番号NM_002546)、および、TNFRSF13B(GenBank登録番号NM_006573)、ならびにインターロイキン受容体、例えばIL2RA(GenBank登録番号NM_000417)、IL2RG(GenBank登録番号NM_000206)、IL4R(GenBank登録番号AF421855)、IL7R(GenBank登録番号NM_002185)、IL9R(GenBank登録番号XM_015989)、およびIL13R(GenBank登録番号X95302)が挙げられる。好ましくは、本発明のリガンド結合タンパク質の融合体は、そのリガンド結合タンパク質の生物活性を提示する。
【0222】
本明細書における用語「癌関連タンパク質」は、その発現が癌または制御された細胞増殖の維持に関連するタンパク質またはポリペプチドを述べる際に用いられ、例えば腫瘍抑制遺伝子または腫瘍遺伝子によってコードされたタンパク質である。癌関連タンパク質としては、pl6(GenBank登録番号AH005371)、p53(GenBank登録番号NM_000546)、p63(GenBank登録番号NM_003722)、p73(GenBank登録番号NM_005427)、BRCA1(GenBank登録番号U14680)、BRCA2(GenBank登録番号NM_000059)、CTBP相互作用タンパク質(GenBank登録番号U72066)、DMBT1(GenBank登録番号NM_004406)、HRAS(GenBank登録番号NM_005343)、NCYM(GenBank登録番号NM_006316)、FGR(GenBank登録番号NM_005248)、myb(GenBank登録番号AF104863)、raf1(GenBank登録番号NM_002880)、erbB2(GenBank登録番号NM_004448)、VAV(GenBank登録番号X16316)、c−fos(VGenBank登録番号01512)、c−fes(GenBank登録番号X52192)、c−jun(GenBank登録番号NM_002228)、MAS1(GenBank登録番号M13150)、pim−1(GenBank登録番号M16750)、TIF1(GenBank登録番号NM_003852)、c−fms(GenBank登録番号X03663)、EGFR(GenBank登録番号NM_005228)、erbA(GenBank登録番号X04707)、c−srcチロシンキナーゼ(GenBank登録番号XM_044659)、c−abl(GenBank登録番号M14752)、N−ras(GenBank登録番号X02751)、K−ras(GenBank登録番号M54968)、jun−B(GenBank登録番号M29039)、c−myc(GenBank登録番号AH001511)、RB1(GenBank登録番号M28419)、DCC(GenBank登録番号X76132)、APC(GenBank登録番号NM_000038)、NF1(GenBank登録番号M89914)、NF2(GenBank登録番号Y18000)、および、bcl−2(GenBank登録番号M13994)が挙げられる。
【0223】
本明細書における「融合阻害ペプチド」は、抗ウイルス活性、抗膜融合能、および/または細胞内プロセスを調節する能力を示すペプチドを述べる際に用いられ、例えば、コイルドコイルペプチド構造を含むものである。抗ウイルス活性としては、これらに限定されないが、HIV−1、HIV−2、RSV、SIV、EBV、麻疹ウイルス、インフルエンザウイルスの阻害、または感染していない細胞へのCMV伝達が挙げられる。加えて、抗融合形成(antifusogenic)能力、抗ウイルス活性、またはペプチドの細胞内調節活性は、単にペプチドの存在を必要とするに過ぎず、このようなペプチドに対して向けられた宿主の免疫応答の刺激を特に必要としない。抗融合形成は、2またはそれ以上の成分間における膜融合事象を阻害する、またはそのレベルを、ペプチドの非存在下で前記成分間に起こる膜融合のレベルに比べて減少させるペプチドの能力を意味する。このような成分は、例えば細胞膜またはウイルス構造である可能性があり、例えばウイルスエンベロープまたは繊毛である。本明細書で用いられる用語「抗ウイルス性ペプチド」は、細胞のウイルス感染、または例えば細胞間融合もしくは遊離のウイルスによる感染を介した、増殖を起こすウイルス感染および/またはウイルス病原性に必要ないくつかのウイルス活性を阻害するペプチドの能力を意味する。このような感染は、エンベロープウイルスケースで起こるような、またはいくつかのその他のウイルス構造および細胞構造に関連する融合事象で起こるような膜融合に関与する可能性がある。膜融合阻害ペプチドおよび抗ウイルス性ペプチドは、1種より多くのタンパク質から誘導されるアミノ酸配列を有することが多い(例えば、HIV−1、HIV−2、RSV、およびSlVから誘導されたポリペプチド)。
【0224】
膜融合阻害ペプチド、および、抗ウイルス性ペプチドの例は、WO94/2820、WO96/19495、WO96/40191、WO01/64013、および米国特許第6,333,395号、6,258,782号、6,228,983号、6,133,418号、6,093,794号、6,068,973号、6,060,065号、6,054,265号、6,020,459号、6,017,536号、6,013,263号、5,464,933号、5,346,989号、5,603,933号、5,656,480号、5,759,517号、6,245,737号;6,326,004号、および6,348,568号に見出すことができ、これらはいずれも参照により本明細書に組み入れられる。
【0225】
その他のタイプのペプチドの例としては、本明細書で説明したような治療用タンパク質のフラグメントが挙げられ、特に、親分子の活性の少なくとも1種を保持するヒトタンパク質のフラグメントである。また、本発明のリガンド成分を生産するのに使用可能なペプチドとしては、模倣ペプチド、および治療用タンパク質の生物活性を示すペプチドも挙げられるが、全長の治療用タンパク質とは配列または三次元構造の点で異なる。非限定的な例として、ペプチドとしては、Johnson等(2000)Nephrol.Dial.Transplant 15(9):1274〜7,Kuai等(2000)J.Pept Res.56(2):59〜62,Barbone等(1999)Nephrol.Dial.Transplant.14 Supp 2:80〜4,Middleton等(1999)J.Biol.Chem.274(20):14163〜9,Johnson等(1998)Biochemistry 37(11):3699〜710,Johnson等(1997)Chem.Biol.12:939〜50,Wrighton等(1997)Nat Biotechnol 15(12):1261〜5,Livnah等(1996)Science 273:464〜71、および、Wrighton等,(1996)Science 273:458〜64によって開示されたエリスロポエチン模倣ペプチドが挙げられる。
【0226】
また治療的分子としては、アレルゲンタンパク質、およびそれらの消化されたフラグメントも挙げられる。これらの例としては、ブタクサ、ライ、ジューングラス(June grass)、カモガヤ、ハルガヤ、レッドトップグラス(red top grass)、オオアワガエリ、イエロードッグ、コムギ、トウモロコシ、ヤマヨモギ、イチゴツナギ,カリフォルニアの一年草(california annual grass)、アカザ、ギョウギシバ、ロシアアザミ(Russian thistle)、マウンテンシーダー(mountain cedar)、オーク、ネグンドカエデ、スズカケノキ、カエデ、ニレなどが由来の花粉アレルゲン、チリダニ、ハチ毒、食物アレルゲン、動物の鱗屑およびその他の昆虫毒が挙げられる。
【0227】
その他の治療的分子としては、ウイルス、細菌および原生動物性のワクチンなどの微生物性ワクチン、およびそれらの様々な成分、例えば表面抗原が挙げられる。これらの例としては、糖タンパク質、タンパク質またはこれらのタンパク質から誘導されたペプチドを含むワクチンが挙げられる。このようなワクチンは、スタフィロコッカス・アウレウス(Staphylococcus aureus)、ストレプトコッカス・ピオゲネス(Streptococcus pyogenes)、肺炎連鎖球菌(Streptococcus pneumoniae)、ナイセリア・メニンギチジス(Neisseria meningitidis)、淋菌(Neisseria gonorrhoeae)、サルモネラ属、シゲラ属、エシェリキア・コリ(Escherichia coli)、クレブシエラ属spp.、プロテウス属spp.、コレラ菌(Vibrio cholerae)、カンピロバクター・ピロリ(Campylobacter pylori)、緑膿菌(Pseudomonas aeruginosa)、ヘモフィルス・インフルエンザ(Haemophilis influenzae)、ボルデテラ・ペルツッシス(Bordetella Pertussis)、ヒト型結核菌(Mycobacterium tuberculosis)、レジオネラ・ニューモフィラ(Legionella pneumophila)、トレポネーマ・パリダム(Treponema pallidum)、クラミジア属、破傷風トキソイド、ジフテリアトキソイド、インフルエンザウイルス、アデノウイルス、パラミクソウイルス(流行性耳下腺炎、麻疹)、風疹ウイルス、ポリオウイルス、肝炎ウイルス、ヘルペスウイルス、狂犬病ウイルス、HIV−1、HIV−2、RSV、および乳頭腫ウイルスから調製される。
【0228】
好ましい融合分子は、抗HIVウイルスペプチド、抗RSVペプチド、ヒト成長ホルモン、αおよび/またはβインターフェロン、エリスロポイエチン(EPO)、EPO様ペプチド、顆粒球−コロニー刺激因子(GCSF)、顆粒球マクロファージコロニー刺激因子(GMCSF)、インスリン、インスリン様増殖因子(IGF)、トロンボポエチン、抗体のCDRに相当するペプチド、膵島新生関連タンパク質(Islet Neogenesis Associated Protein;INGAP)、カルシトニン、アンギオスタチン、エンドスタチン、インターロイキン−2、成長ホルモン放出因子、ヒト副甲状線ホルモン、抗腫瘍壊死因子(TNF)ペプチド、インターロイキン−1(IL−1)受容体、および/または単鎖抗体を含んでいてもよい。
【0229】
また本発明の融合タンパク質は、新しい機能または新規な機能を有する分子に関してスクリーニングするために、ペプチドライブラリーから誘導されたペプチドまたはポリペプチドが含まれるように調製してもよい。このようなペプチドライブラリーとしては、商業的または公共的に利用可能なペプチドライブラリー、例えば、アメリカン・ペプチド社(American Peptide Co.Inc.)、セル・サイエンス社(Cell Sciences Inc.)、インビトロジェン社(Invitrogen Corporation)、フェニックス・ファーマシューティカルズ社(Phoenix Pharmaceuticals Inc.)、ユナイテッド・ステイツ・バイオロジカル(United States Biological)のペプチドライブラリーが挙げられ、加えて、利用可能な技術、例えば標準的手法を用いて作製されたバクテリオファージや細菌のディスプレイライブラリーによって生産されたペプチドライブラリーも挙げられる。
【0230】
本発明のさらにその他の実施態様において、融合タンパク質は、当分野既知の治療用タンパク質の成分を用いることによって調製してもよく、このようなタンパク質の成分の例としては、現在のところ食品医薬品局(Food and Drug Administration)(www.fda.gov/cber/efoi/approve.htm)によって承認された、ペプチドおよびタンパク質、加えてPCT特許出願公開WO01/79258、WO01/77137、WO01/79442、WO01/79443、WO01/79444、およびWO01/79480(これらはいずれも、参照によりその全体として本明細書に組み入れられる)が挙げられる。
【0231】
PCT国際公開WO03/020746(参照により本明細書に加入させる)の表1に、完全ではないが、本発明の融合タンパク質の治療用タンパク質部分、すなわちリガンド成分に相当する治療用タンパク質の一覧が提供されている。「治療用タンパク質X」の列は、治療用タンパク質分子を記載しており、それに続く括弧内には、その治療用タンパク質分子またはそれらのフラグメントもしくは変異体が含まれる、あるいはそれらで構成される科学名とブランド名を含む。本明細書で用いられる「治療用タンパク質X」は、個々の治療用タンパク質分子(CASおよびGenBank登録番号より入手できるアミノ酸配列で定義されるような)、またはこの列で開示された所定の治療用タンパク質分子に関連する治療用タンパク質のグループ全体のいずれかを意味する場合がある。「典型的な識別子」の列は、ケミカル・アブストラクツ・サービス(CAS)のレジストリナンバー(米国化学会によって公開)、および/またはGenBank登録番号(例えば、遺伝子座ID、NP−XXXXX(参照配列タンパク質)、およびXP−XXXXX(モデルタンパク質)という識別子を示しており、このような識別子は、国立バイオテクノロジー情報センター(National Center for Biotechnology Information ;NCBI)のウェブページ(www.ncbi.nlm.nih.gov)を介して利用可能であり、タンパク質分子、または治療用タンパク質分子のフラグメントもしくは変異体のアミノ酸配列を含む、CASレジストリまたはGenBankデータベースにおけるエントリーに相当する。加えて、GenSeq登録番号および/またはジャーナル出版物の引用が、いくつかのポリペプチドに関する典型的なアミノ酸配列を同定するために与えられている。
【0232】
これらCASおよびGenBank、ならびにGenSeq登録番号、同様に、引用されたジャーナル出版物それぞれに関連する要約の頁が利用可能であり(例えば、パブメドID番号(PubMed ID number;PMID))、これは参照によりその全体として本明細書に組み込まれる。PCT/特許の参照の列は、治療用タンパク質分子を記載している特許および/または特許出願公開に相当する米国特許またはPCT国際公開番号を示す(これらはいずれも、参照によりその全体として本明細書に加入される)。生物活性の列は、治療用タンパク質分子に関連する生物活性を記載している。典型的な活性分析の列は、治療用タンパク質X部分を含む本発明の治療用タンパク質またはトランスフェリン融合タンパク質の治療活性および/または生物活性を試験するのに使用することができる分析を記載している参考文献を示す。また、これらの参考文献も、参照によりその全体として本明細書に加入される。「好ましい指標Y」の列は、治療用タンパク質X、または治療用タンパク質X部分を含む本発明のトランスフェリン融合タンパク質によって治療、予防、診断または改善が可能な疾患、障害および/または状態を記載している。本発明は、WO03/020746で提供された治療用タンパク質を含む(これは参照によりその全体として本明細書に加入される)。
【実施例】
【0233】
実施例1− GPIアンカー、hMUC1、およびmTf発現カセットの製造
mTf発現カセット(配列番号16)を含むpREX0549ベクターを、SalIおよびHindIIIで消化した。図3は、pREX0549に関するベクターのマップを示す。プライマーP0922およびP0923(配列番号7および配列番号8)を一緒にアニールし、SalI/HindIIIの消化部位でpREX0549にライゲーションした。P0922およびP0923によって形成されたリンカーには、SpeI、HindIIIおよびXbaI制限部位が含まれ、GPIアンカーおよびMUC1軸をコードする核酸分子が受け入れられるように設計された。得られたベクターpREX0628には、P0922/P0923リンカーと共にmTf発現カセットが含まれる。
【0234】
pREX0628を、HindIIIおよびXbaIで消化した。プライマーP0924およびP0925(配列番号9および配列番号10)をアニールして、GPIアンカーYIR019cを形成させた。YIR019cを、消化したpREX0628にライゲーションし、ベクターpREX0634を形成させた。
【0235】
hMUC1のcDNAを、RT−PCRで、プライマーP0958およびP0959(配列番号11および配列番号12)を用いて、ヒト乳腺腫瘍トータルRNAライブラリー(クロンテック)から増幅した。得られたcDNAを、プライマーP1019およびP1020(配列番号13および配列番号14)で増幅して、SpeIおよびHindIII制限部位を作出した。このようにして得られたSpeIおよびHindIII部位を有するhMUC1は、配列番号6で示される。
【0236】
pREX0634を、SpeIおよびHindIIIで消化し、SpeIおよびHindIII部位を有するhMUC1をベクターにライゲーションした。このようにして得られたベクターpREX0663を、提示発現カセット(mTf−MUC1−GPI)として用いた。
【0237】
pREX0663を用いて、高および低コピー数の酵母発現ベクターを作出した。高コピー数の酵母発現ベクターを作出するために、4.1kbの提示発現カセットを、ベクターをNotIで消化することによってpREX0663から除去した。次に、この発現カセットを、NotIで消化し、脱リン酸化したpSAC35ベクターにライゲーションし、ベクターpREX0667(酵母提示ベクターI)を得た。
【0238】
低コピー数の酵母発現ベクターを、pREX0663をNotIで消化し、この発現ベクターを、NotIで消化し、脱リン酸化したpREX0699にライゲーションし、pREX0721酵母提示(酵母提示ベクターII)を得ることによって作製した。
【0239】
上述の酵母発現ベクターを用いて当技術分野で既知のように酵母細胞および細菌細胞を形質転換させることができる。このようなベクターは、当技術分野で既知のように酵母で発現させることができる。さらに、ランダムペプチドまたはCDRのようなリガンド成分のライブラリーを提示することができる発現したトランスフェリン融合タンパク質を収集物を作成することができ、さらに、当技術分野で既知のように結合に関してスクリーニングするのに用いることができる。
【0240】
実施例2− 15−merのランダムライブラリーの構築
新規な結合特性を有するトランスフェリン変異体を選択するために、当技術分野で既知のPCRニッティング法によって、トランスフェリンの289〜290のアミノ酸位置におけるランダムな15−merのライブラリーを構築した(MartinおよびSmith(2006)Biochem J.396(2):287〜95を参照)。通常、結合エピトープを形成するのに必要なのはわずか約7個のアミノ酸であり、約109のライブ
ラリーは、設計されるライブラリー(3.3×1019)のうちの小さなフラクションをカバーするに過ぎないが、15−merのライブラリーを設計した。しかしながら、109のライブラリーサイズの場合、15−merのライブラリーは、同じ規模の7−merのライブラリーよりも6.4倍多い7−merをカバーする。
【0241】
P1174/P1227を用いてトランスフェリンのBamHI/BspEI配列を含むDNAフラグメントを得た後、2回のPCR反応(それぞれ単一のプライマー−P1172およびP1173を使用)を行い、一本鎖DNAを得た。ssDNAを単離し、アニールして、結合した15−merのライブラリーを形成した。この操作によって、ライブラリーが、合成オリゴヌクレオチドのもともとの複雑さを確実に維持するようにした。二本鎖のニットした15−merのライブラリーを、P1174/P1227を用いてさらに増幅し、十分な量のDNAを得た。PCR産物を精製し、BamHI/BspEIで消化し、適切なプラスミドベクター、例えばpREX0995(図4)またはpREX0667にクローニングした。
P1172(配列番号19)
289〜290 15−merのランダムペプチドライブラリーの挿入物がニットしたフォワードバックフラグメント
【化1】
DNA配列中の各位置にランダム性を導入するために、LaBeanおよびKauffman(1993)Protein Sci.2:1249〜54に従って、その位置にヌクレオチドの混合物(A、G、TおよびC)を予め決められた比率で組み込んだ。以下で既定される混合物は、停止コドンの頻度を最小化し、アミノ酸組成を天然タンパク質に適合する。
5 13%T,32%G,20%C,35%A
6 24%T,24%G,22%C,30%A
7 37%T,26%G,37%C。
P1173(配列番号20)
フロントフラグメントのための289〜290 15−merのランダムペプチドライブラリーがニットしたバックプライマー
【化2】
P1174(配列番号21)
フロントフラグメントのための289〜290 15−merのランダムペプチドライブラリーがニットしたフォワードプライマー
【化3】
P1227(配列番号22)
バックフラグメントのための289〜290 15−merのランダムペプチドライブラリーが結合したバックプライマー
【化4】
【0242】
実施例3− 酵母細胞を提示するフラグの選択
酵母提示系を確立し、それを用いて、軸領域、huMUC1およびGPIシグナル配列に融合させることによって酵母表面にトランスフェリンのN−ローブを提示した。この系の結合選択における有用性実証するために、フラグ−タグ配列
【化5】
またはランダムな15−merペプチドライブラリーを、トランスフェリンN−ローブのアミノ酸289番目に挿入した。次に、フラグタグを付けたトランスフェリンN−ローブ、pREX1012を提示する酵母(図6)を、ランダムな15−merのペプチドを有するトランスフェリンN−ローブを提示する酵母のプールに添加した。この混合群から、フラグタグを付けたトランスフェリンN−ローブを提示する酵母だけを、抗フラグ抗体を用いた選択によって回収した。
【0243】
フラグタグ配列を、トランスフェリンのアミノ酸289番目に挿入するために、フラグタグ配列を組み込んだオリゴを合成し、PCRでpREX0667ベクターにニットし、pREX0759を生成させた(図7)。次に、pREX0759のフラグタグ配列を含むBamHI/BspEIフラグメントを用いて、pREX0995の同じ制限フラグメントを置換した(図4)。得られたプラスミドpREX1012は、フラグタグを付けたトランスフェリンN−ローブ−MUC1−GPI融合タンパク質を発現した。
【0244】
また、15−merのライブラリーも、pREX0995のBamHI/BspEI部位の間にクローニングした。15−merのライブラリーの製造を以下で説明する。ライゲーションサンプルを、E.コリDH5αに形質転換し、全ての形質転換混合物を、2LB/Amp(50μg/mL)の寒天プレートで平板培養した。全てのコロニーを回収し、キアゲン(Qiagen)プラスミド・プレップ・プロトコールを用いて、数種のミニプレップカラムを用いてプラスミドDNAを抽出した。
【0245】
pREX1012と15−merライブラリーの両方のプラスミドDNAを、サッカロミセス・セレビジエ株DS1101cir°に形質転換した。pREX1012の単一のコロニーを、スクロースを含む緩衝化した最小培地(BMM/S)に植え付け、一晩培養した。15−merライブラリーの全てのコロニーを回収し、BMM/Sに植え付けた。これら二種の一晩培養したもの細胞数をヘマサイトメーターによって測定し、以下の細胞混合物を調製した:
(A)109の15−merライブラリー酵母細胞と10:107の割合で混合された、103個のpREX1012酵母細胞
(B)108の15−merライブラリー酵母細胞と100:107の割合で混合された、103個のpREX1012酵母細胞。
【0246】
この細胞混合物を、細胞ブロック溶液(1×PBS/0.05%トゥイーン(Tween)−20、1%BSA)1ml中、氷上で30分間インキュベートした。遠心分離(13000rpmで30秒)後、細胞ペレットを、ビオチン化抗フラグ抗体(シグマ・アルドリッチ、1:25希釈)を含む洗浄溶液(1×PBS、0.5%BSA、2mMのEDTA)1mlに懸濁し、氷上で30分間インキュベートした。細胞を洗浄溶液1mlで2回洗浄し、(A)800μlまたは(B)160μlのブロック溶液に懸濁した。細胞懸濁液(A)200μlまたは(B)40μlにストレプトアビジンMACSマイクロビーズ(ミルテニー・バイオテク)を添加し、氷上で30分間インキュベートした。MSカラムを製造元の説明書(ミルテニー・バイオテク)に従って用いて、非標識細胞から標識された細胞を分離した。標識された細胞を回収し、BMM/Sアガロースプレート上で平板培養し、小さいコロニーが出現するまで30℃でインキュベートした。
【0247】
各プレートから全てのコロニーを回収し、それらをBMM/S5ml中30℃で一晩増殖させることによって、二回目の選択を行った。1.5のOD600に等しいこれら培養細胞を、上述のようなさらなるMACS分離で処理した。この二回目のスクリーニングから得られた細胞を、BMM/S5ml中で30℃で一晩培養した。各選択の前および後の酵母細胞培養物を、アジレント・テクノロジー(Agilent Technolgy)製のバイオアナライザーで抗フラグモノクローナル抗体(シグマ・アルドリッチ)およびAPC標識ヤギ抗マウス検出抗体を用いたFACSで分析した。FACS分析は製造元の説明書に従って行われた。2回目のMACS分離の後、フラグタグを付けた酵母の存在が明らかになった(図8)。
【0248】
実施例4− Aga1軸提示
酵母遺伝子AGA1のコア領域のDNA配列(残基XXX−XXX)を、以下のプライマーを用いてS288cの酵母ゲノムDNAをPCRで処理することによって得た:
【化6】
【0249】
約1.5kbのPCR産物を単離し、XbaI/HindIIIで消化した。このフラグメントを、SpeI/HindIIIで消化したpREX0855にライゲーションし(図9)、E.コリDH5αに形質転換した。得られた全てのコロニーを回収し、プラスミドDNAを細胞から単離した。発現カセットをNotI消化によって回収し、pSAC35にライゲーションして、酵母発現ベクターを得た。
【0250】
上述したように、酵母に形質転換し、抗フラグ抗体でFACS分析した。酵母コロニーは、高レベルのN−ローブ提示を示し、匹敵するMUC1からなる軸をベースとするコンストラクトよりも約10倍高かった(データ示さず)。
【0251】
酵母のシングルコロニーを単離し、この酵母細胞からプラスミドDNAを抽出した。抽出されたプラスミドDNAをNotIで消化した後に、NotI発現カセットを回収し、NotIで消化したpREX0855にライゲーションし、プラスミドpREX1087(図10)、フラグ−N−ローブ−Aga1−GPI発現カセットを含むPUCベースのベクターを得た。Aga1に相当するDNA配列の領域を配列分析し、その同一性を確認した。さらにこの発現カセットもpSAC35に戻し、pREX1106を得た(図11)。
【0252】
実施例5− 酵母細胞で機能する哺乳動物のGPI変異体の選択
哺乳動物のGPIシグナルは、それらの酵母カウンターパートが果たさない役割、例えば細胞内トラフィッキング、膜貫通シグナルの伝達、およびクラスリン非依存性のエンドサイトーシスといった役割を果たす(Biochem J.1993,294:305〜324)。酵母細胞は、細胞膜を有するだけでなく、哺乳動物細胞にはない細胞壁も有し、酵母GPIの多くは、タンパク質を酵母細胞壁に標的化する特有の配列を有する(J Bacteriol,1999,181:3886〜3889)。ヒト胎盤のアルカリホスファターゼのGPIは、酵母細胞では全く機能しないことが示されている(Mol Microbiol 1999,34:247〜256)。発現された組換えタンパク質を酵母細胞壁に付着させることができる新規な配列を得る手段として、pREX0885をベースとするがhuMDPのGPI配列
【化7】
を用いた酵母提示ベクター(図9)を用いた。この配列を、4つの完全にランダムなコドン(X)、加えて数種の(下線で示した)合理的な改変が組み込まれるように改変した:
【化8】
ここにおいて、Xはあらゆるアミノ酸である。
【0253】
以下の融合タンパク質、GPI配列が上述の通りに改変されたフラグタグ−N−ローブ−MUC1からなる軸−GPIを発現する酵母ライブラリーを、サッカロミセス・セレビジエ株DS1101cir°に形質転換した。本融合タンパク質が細胞壁に付着した全ての酵母細胞を、ビオチン化抗フラグ抗体を用いたMACSによって単離した。
【0254】
2つのオリゴP2035およびP2036(以下を参照)をアニールし、Taqポリメラーゼを用いて伸長させた。得られたDNAフラグメントを精製し、HindIII/XbaIで消化し、HindIII/XbaIで消化したpREX0855にライゲーションした(以下および図9を参照)。
プライマー
【化9】
【0255】
ライゲーション混合物をE.コリDH5αに形質転換し、約5x105個のコロニーを得た。全てのコロニーを回収し、プラスミドDNAを単離した。プラスミドDNAをNotIで消化し、発現カセットを回収し、SAC35にクローニングし、酵母発現ライブラリーを作出した。このライブラリーをエレクトロポレーションでDS1101cir°細胞に形質転換した。上述のライブラリーを一晩培養したものを、ビオチン化抗フラグ抗体を用いたMACSで処理した。再度、単離した細胞を同じ抗体を用いたMACSによって即座に精製した。得られた細胞をBMMSプレートに平板培養し、24個のコロニーをFACSとDNA配列分析によって特徴付けた(フラグの付加の説明を参照)。
【0256】
読取り可能な配列を示した22個のクローンのうち7個が、全長のGPIアンカーを有しており(表1)、その提示レベルは様々であり、そのうち最良のものは、酵母GPIアンカーを有する同じ融合タンパク質を発現するpREX1003ベクターよりも優れていた。
【0257】
【表5】
【表6】
【0258】
意外なことに、GPIアンカーシグナルを有効に欠失させるランダム化したコドンのうち1つにおける停止コドンによってクローンの50%がトランケーションされた。これらの12種のクローンのうち2種が、pREX1003よりも有意に優れた提示レベルを有することが測定され、停止コドンの直前にシステイン残基を含むことがわかった(CQIGGS*(配列番号34)およびCQ*、ここにおいて*は停止コドン)(図12)。恐らく、これらのコンストラクトは、細胞壁タンパク質中の遊離のシステイン残基にジスルフィド結合することを介して細胞壁に架橋された。
【0259】
上記の実施例を参照として本発明を詳細に説明したが、当然ながら、本発明の本質から逸脱することなく様々な改変を施すことができる。従って、本発明は以下の請求項によってのみ限定される。本願において言及された全ての引用された特許、特許出願および出版物を、参照によりその全体として本明細書に組み込まれる。
【図面の簡単な説明】
【0260】
【図1】トランスフェリン融合タンパク質上に提示されたランダムペプチドまたはCDRライブラリー、およびリガンドと標的との結合を示す。
【図2】酵母YIR019CGPIアンカーペプチド配列を示し、細胞膜への付着に関与するアミノ酸を強調表示した。
【図3】pREX0549に関するベクターのマップを示す。
【図4】pREX0995に関するベクターのマップを示す。
【図5】pREX0667に関するベクターのマップを示す。
【図6】pREX1012に関するベクターのマップを示す。
【図7】pREX0759に関するベクターのマップを示す。
【図8】2回のMACS分離の後のフラグタグを付けた酵母の存在を示す。
【図9】pREX0855に関するベクターのマップを示す。
【図10】pREX1087に関するベクターのマップを示す。
【図11】pREX1106に関するベクターのマップを示す。
【図12】MUC1およびAGA1を用いたFACS分析を示す。
【特許請求の範囲】
【請求項1】
(a)トランスフェリン(Tf)成分;
(b)軸成分;および、
(c)細胞壁連結要素、
を含む融合タンパク質。
【請求項2】
トランスフェリン成分は、軸成分に直接融合する、請求項1に記載の融合タンパク質。
【請求項3】
融合タンパク質は、アンカー成分をさらに含む、請求項1に記載の融合タンパク質。
【請求項4】
トランスフェリン成分は、トランスフェリンタンパク質、改変されたトランスフェリンタンパク質、またはそのフラグメントである、請求項1に記載の融合タンパク質。
【請求項5】
トランスフェリンタンパク質は、ヒトトランスフェリンタンパク質である、請求項4に記載の融合タンパク質。
【請求項6】
トランスフェリン成分は、配列番号3を含む、請求項4に記載の融合タンパク質。
【請求項7】
Tf成分は、Tfタンパク質のNドメインを含む、請求項1に記載の融合タンパク質。
【請求項8】
Tf成分は、Tfタンパク質のNドメインからなる、請求項1に記載の融合タンパク質。
【請求項9】
Tf成分は、Tfタンパク質のNドメインの一部を含む、請求項1に記載の融合タンパク質。
【請求項10】
Tf成分は、低いグリコシル化を示す、請求項1に記載の融合タンパク質。
【請求項11】
Tf成分は、鉄への低い親和性を示すように改変される、請求項1に記載の融合タンパク質。
【請求項12】
Tf成分は、ビカーボネートへの低い親和性を有するように改変される、請求項1に記載の融合タンパク質。
【請求項13】
Tf成分は、ビカーボネートに結合しない、請求項1に記載の融合タンパク質。
【請求項14】
Tf成分は、グリコシル化部位、鉄結合部位、ヒンジ部位、ビカーボネート部位、および受容体結合部位からなる群より選択される1個またはそれ以上の部位で改変される、請求項1に記載の融合タンパク質。
【請求項15】
Tf成分は、グリコシル化を防ぐ突然変異を少なくとも1つ含む、請求項1に記載の融合タンパク質。
【請求項16】
突然変異は、配列N−X−S/Tを含むN結合型のグリコシル化部位中にある、請求項15に記載の融合タンパク質。
【請求項17】
配列N−X−S/Tは、配列番号3のN413またはN611に相当するアミノ酸で始まる、請求項16に記載の融合タンパク質。
【請求項18】
N、X、SまたはTがプロリンに変えられている、請求項17に記載の融合タンパク質。
【請求項19】
トランスフェリン成分は、グリコシル化を示さないように改変されている、請求項1に記載の融合タンパク質。
【請求項20】
Tf成分は、一つのリガンドまたは複数のリガンドに融合している、請求項1に記載の融合タンパク質。
【請求項21】
リガンドは、単鎖抗体、抗体、抗体フラグメント、抗体の可変領域、ランダムペプチド、または抗体の相補性決定領域(CDR)からなる群より選択される1種またはそれ以上である、請求項20に記載の融合タンパク質。
【請求項22】
リガンドは、Tf成分のN末端に融合している、請求項20に記載の融合タンパク質。
【請求項23】
リガンドは、Tf成分のC末端に融合している、請求項20に記載の融合タンパク質。
【請求項24】
リガンドは、トランスフェリン成分内に挿入される、請求項20に記載の融合タンパク質。
【請求項25】
リガンドは、トランスフェリン成分の表面に露出したループ中に挿入される、請求項20に記載の融合タンパク質。
【請求項26】
リガンドは、3個もしくはそれ以上、4個もしくはそれ以上、5個もしくはそれ以上、6個もしくはそれ以上、7個もしくはそれ以上、8個もしくはそれ以上、9個もしくはそれ以上、10個もしくはそれ以上、11個もしくはそれ以上、12個もしくはそれ以上、13個もしくはそれ以上、14個もしくはそれ以上、15個もしくはそれ以上、または20個もしくはそれ以上のアミノ酸からなる、請求項20に記載の融合タンパク質。
【請求項27】
トランスフェリン成分およびリガンドは、細胞透過性の抗体を含む、請求項20に記載の融合タンパク質。
【請求項28】
リガンドは、ペプチド、抗原、受容体、抗体、毒素、代謝産物および核酸からなる群より選択される1種またはそれ以上に結合することができる、請求項20に記載の融合タンパク質。
【請求項29】
トランスフェリン成分は、軸成分のN末端に融合している、請求項2に記載の融合タンパク質。
【請求項30】
軸成分は、Tf成分のC末端に融合している、請求項2に記載の融合タンパク質。
【請求項31】
軸成分は、高度にグリコシル化されたペプチドである、請求項1に記載の融合タンパク質。
【請求項32】
軸成分は、ムチンドメインを含む、請求項1に記載の融合タンパク質。
【請求項33】
ムチンドメインは、MUC1、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC6、MUC7、MUC8、およびMUC9、ならびにそれらの変異体およびフラグメントからなる群から選択される1種またはそれ以上のタンパク質を含む、請求項32に記載の融合タンパク質。
【請求項34】
軸成分は、ヒトMUC1タンパク質、またはそのフラグメントを含む、請求項1に記載の融合タンパク質。
【請求項35】
軸成分は、配列番号5の核酸によってコードされる、請求項34に記載の融合タンパク質。
【請求項36】
軸成分は、ヒトMUC3タンパク質、またはそのフラグメントを含む、請求項1に記載の融合タンパク質。
【請求項37】
軸成分は、酵母AGA1タンパク質、またはそのフラグメントを含む、請求項1に記載の融合タンパク質。
【請求項38】
軸成分は、トランスフェリン成分と、宿主細胞または基質との間の立体障害を減少させるように機能する、請求項1に記載の融合タンパク質。
【請求項39】
軸成分は、アンカー成分のN末端に融合している、請求項3に記載の融合タンパク質。
【請求項40】
アンカー成分は、軸成分のC末端に融合している、請求項3に記載の融合タンパク質。
【請求項41】
アンカー成分は、グリコシルホスファチジルイノシトール(GPI)、またはその誘導体もしくはフラグメントである、請求項3に記載の融合タンパク質。
【請求項42】
GPIは、酵母GPIシグナル配列、またはそのフラグメントを含む、請求項41に記載の融合タンパク質。
【請求項43】
GPIシグナル配列は、配列番号15を含む、請求項42に記載の融合タンパク質。
【請求項44】
GPIは、哺乳動物のGPIシグナル配列、またはそのフラグメントを含む、請求項41に記載の融合タンパク質。
【請求項45】
アンカー成分は、改変されたGPIシグナル配列を含む、請求項3に記載の融合タンパク質。
【請求項46】
改変されたGPIシグナル配列は、配列番号26の位置1にシステイン残基を含む、請求項45に記載の融合タンパク質。
【請求項47】
改変されたGPIシグナル配列は、配列番号34の最初の2個のアミノ酸、ならびに配列番号34、36および38からなる群より選択されるアミノ酸配列である、請求項45に記載の融合タンパク質。
【請求項48】
細胞壁連結要素は、細胞壁に共有結合している、請求項1に記載の融合タンパク質。
【請求項49】
細胞壁連結要素は、細胞壁に非共有結合で結合している、請求項1に記載の融合タンパク質。
【請求項50】
細胞壁連結要素は、軸成分である、請求項1に記載の融合タンパク質。
【請求項51】
細胞壁連結要素は、アンカー成分である、請求項1に記載の融合タンパク質。
【請求項52】
アンカー成分は、膜貫通ドメインである、請求項51に記載の融合タンパク質。
【請求項53】
細胞壁連結要素は、細胞壁中の1個またはそれ以上のタンパク質とジスルフィド結合を形成することができる遊離のシステイン残基を1個またはそれ以上含む、請求項1に記載の融合タンパク質。
【請求項54】
細胞壁連結要素は、細胞壁のβ−グルカンと架橋を形成することができる軸成分のグリカンを1個またはそれ以上含む、請求項1に記載の融合タンパク質。
【請求項55】
請求項1〜54のいずれか一項に記載の融合タンパク質をコードする核酸分子。
【請求項56】
請求項55に記載の核酸分子を含む宿主細胞。
【請求項57】
請求項1〜54のいずれか一項に記載の融合タンパク質を発現する宿主細胞。
【請求項58】
細胞は、酵母細胞である、請求項56に記載の宿主細胞。
【請求項59】
酵母細胞は、サッカロミセス属またはピチア属である、請求項58に記載の宿主細胞。
【請求項60】
請求項57に記載の宿主細胞のライブラリーを物質に曝露すること、および少なくとも1種の宿主細胞の該物質への結合を検出することを含む、リガンドの結合活性に関してスクリーニングする方法。
【請求項61】
宿主細胞のライブラリーは、酵母細胞の収集体である、請求項60に記載の方法。
【請求項62】
物質は、抗原または受容体である、請求項60に記載の方法。
【請求項63】
(a)アルブミン成分;
(b)軸成分;および、
(c)細胞膜要素、
を含む融合タンパク質。
【請求項64】
アルブミン成分は、アルブミンタンパク質、改変されたアルブミンタンパク質、またはそのフラグメントである、請求項63に記載の融合タンパク質。
【請求項65】
アルブミンタンパク質は、ヒトアルブミンタンパク質である、請求項64に記載の融合タンパク質。
【請求項66】
アンカー成分をさらに含む、請求項63に記載の融合タンパク質。
【請求項67】
膜貫通ドメインをさらに含む、請求項63に記載の融合タンパク質。
【請求項1】
(a)トランスフェリン(Tf)成分;
(b)軸成分;および、
(c)細胞壁連結要素、
を含む融合タンパク質。
【請求項2】
トランスフェリン成分は、軸成分に直接融合する、請求項1に記載の融合タンパク質。
【請求項3】
融合タンパク質は、アンカー成分をさらに含む、請求項1に記載の融合タンパク質。
【請求項4】
トランスフェリン成分は、トランスフェリンタンパク質、改変されたトランスフェリンタンパク質、またはそのフラグメントである、請求項1に記載の融合タンパク質。
【請求項5】
トランスフェリンタンパク質は、ヒトトランスフェリンタンパク質である、請求項4に記載の融合タンパク質。
【請求項6】
トランスフェリン成分は、配列番号3を含む、請求項4に記載の融合タンパク質。
【請求項7】
Tf成分は、Tfタンパク質のNドメインを含む、請求項1に記載の融合タンパク質。
【請求項8】
Tf成分は、Tfタンパク質のNドメインからなる、請求項1に記載の融合タンパク質。
【請求項9】
Tf成分は、Tfタンパク質のNドメインの一部を含む、請求項1に記載の融合タンパク質。
【請求項10】
Tf成分は、低いグリコシル化を示す、請求項1に記載の融合タンパク質。
【請求項11】
Tf成分は、鉄への低い親和性を示すように改変される、請求項1に記載の融合タンパク質。
【請求項12】
Tf成分は、ビカーボネートへの低い親和性を有するように改変される、請求項1に記載の融合タンパク質。
【請求項13】
Tf成分は、ビカーボネートに結合しない、請求項1に記載の融合タンパク質。
【請求項14】
Tf成分は、グリコシル化部位、鉄結合部位、ヒンジ部位、ビカーボネート部位、および受容体結合部位からなる群より選択される1個またはそれ以上の部位で改変される、請求項1に記載の融合タンパク質。
【請求項15】
Tf成分は、グリコシル化を防ぐ突然変異を少なくとも1つ含む、請求項1に記載の融合タンパク質。
【請求項16】
突然変異は、配列N−X−S/Tを含むN結合型のグリコシル化部位中にある、請求項15に記載の融合タンパク質。
【請求項17】
配列N−X−S/Tは、配列番号3のN413またはN611に相当するアミノ酸で始まる、請求項16に記載の融合タンパク質。
【請求項18】
N、X、SまたはTがプロリンに変えられている、請求項17に記載の融合タンパク質。
【請求項19】
トランスフェリン成分は、グリコシル化を示さないように改変されている、請求項1に記載の融合タンパク質。
【請求項20】
Tf成分は、一つのリガンドまたは複数のリガンドに融合している、請求項1に記載の融合タンパク質。
【請求項21】
リガンドは、単鎖抗体、抗体、抗体フラグメント、抗体の可変領域、ランダムペプチド、または抗体の相補性決定領域(CDR)からなる群より選択される1種またはそれ以上である、請求項20に記載の融合タンパク質。
【請求項22】
リガンドは、Tf成分のN末端に融合している、請求項20に記載の融合タンパク質。
【請求項23】
リガンドは、Tf成分のC末端に融合している、請求項20に記載の融合タンパク質。
【請求項24】
リガンドは、トランスフェリン成分内に挿入される、請求項20に記載の融合タンパク質。
【請求項25】
リガンドは、トランスフェリン成分の表面に露出したループ中に挿入される、請求項20に記載の融合タンパク質。
【請求項26】
リガンドは、3個もしくはそれ以上、4個もしくはそれ以上、5個もしくはそれ以上、6個もしくはそれ以上、7個もしくはそれ以上、8個もしくはそれ以上、9個もしくはそれ以上、10個もしくはそれ以上、11個もしくはそれ以上、12個もしくはそれ以上、13個もしくはそれ以上、14個もしくはそれ以上、15個もしくはそれ以上、または20個もしくはそれ以上のアミノ酸からなる、請求項20に記載の融合タンパク質。
【請求項27】
トランスフェリン成分およびリガンドは、細胞透過性の抗体を含む、請求項20に記載の融合タンパク質。
【請求項28】
リガンドは、ペプチド、抗原、受容体、抗体、毒素、代謝産物および核酸からなる群より選択される1種またはそれ以上に結合することができる、請求項20に記載の融合タンパク質。
【請求項29】
トランスフェリン成分は、軸成分のN末端に融合している、請求項2に記載の融合タンパク質。
【請求項30】
軸成分は、Tf成分のC末端に融合している、請求項2に記載の融合タンパク質。
【請求項31】
軸成分は、高度にグリコシル化されたペプチドである、請求項1に記載の融合タンパク質。
【請求項32】
軸成分は、ムチンドメインを含む、請求項1に記載の融合タンパク質。
【請求項33】
ムチンドメインは、MUC1、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC6、MUC7、MUC8、およびMUC9、ならびにそれらの変異体およびフラグメントからなる群から選択される1種またはそれ以上のタンパク質を含む、請求項32に記載の融合タンパク質。
【請求項34】
軸成分は、ヒトMUC1タンパク質、またはそのフラグメントを含む、請求項1に記載の融合タンパク質。
【請求項35】
軸成分は、配列番号5の核酸によってコードされる、請求項34に記載の融合タンパク質。
【請求項36】
軸成分は、ヒトMUC3タンパク質、またはそのフラグメントを含む、請求項1に記載の融合タンパク質。
【請求項37】
軸成分は、酵母AGA1タンパク質、またはそのフラグメントを含む、請求項1に記載の融合タンパク質。
【請求項38】
軸成分は、トランスフェリン成分と、宿主細胞または基質との間の立体障害を減少させるように機能する、請求項1に記載の融合タンパク質。
【請求項39】
軸成分は、アンカー成分のN末端に融合している、請求項3に記載の融合タンパク質。
【請求項40】
アンカー成分は、軸成分のC末端に融合している、請求項3に記載の融合タンパク質。
【請求項41】
アンカー成分は、グリコシルホスファチジルイノシトール(GPI)、またはその誘導体もしくはフラグメントである、請求項3に記載の融合タンパク質。
【請求項42】
GPIは、酵母GPIシグナル配列、またはそのフラグメントを含む、請求項41に記載の融合タンパク質。
【請求項43】
GPIシグナル配列は、配列番号15を含む、請求項42に記載の融合タンパク質。
【請求項44】
GPIは、哺乳動物のGPIシグナル配列、またはそのフラグメントを含む、請求項41に記載の融合タンパク質。
【請求項45】
アンカー成分は、改変されたGPIシグナル配列を含む、請求項3に記載の融合タンパク質。
【請求項46】
改変されたGPIシグナル配列は、配列番号26の位置1にシステイン残基を含む、請求項45に記載の融合タンパク質。
【請求項47】
改変されたGPIシグナル配列は、配列番号34の最初の2個のアミノ酸、ならびに配列番号34、36および38からなる群より選択されるアミノ酸配列である、請求項45に記載の融合タンパク質。
【請求項48】
細胞壁連結要素は、細胞壁に共有結合している、請求項1に記載の融合タンパク質。
【請求項49】
細胞壁連結要素は、細胞壁に非共有結合で結合している、請求項1に記載の融合タンパク質。
【請求項50】
細胞壁連結要素は、軸成分である、請求項1に記載の融合タンパク質。
【請求項51】
細胞壁連結要素は、アンカー成分である、請求項1に記載の融合タンパク質。
【請求項52】
アンカー成分は、膜貫通ドメインである、請求項51に記載の融合タンパク質。
【請求項53】
細胞壁連結要素は、細胞壁中の1個またはそれ以上のタンパク質とジスルフィド結合を形成することができる遊離のシステイン残基を1個またはそれ以上含む、請求項1に記載の融合タンパク質。
【請求項54】
細胞壁連結要素は、細胞壁のβ−グルカンと架橋を形成することができる軸成分のグリカンを1個またはそれ以上含む、請求項1に記載の融合タンパク質。
【請求項55】
請求項1〜54のいずれか一項に記載の融合タンパク質をコードする核酸分子。
【請求項56】
請求項55に記載の核酸分子を含む宿主細胞。
【請求項57】
請求項1〜54のいずれか一項に記載の融合タンパク質を発現する宿主細胞。
【請求項58】
細胞は、酵母細胞である、請求項56に記載の宿主細胞。
【請求項59】
酵母細胞は、サッカロミセス属またはピチア属である、請求項58に記載の宿主細胞。
【請求項60】
請求項57に記載の宿主細胞のライブラリーを物質に曝露すること、および少なくとも1種の宿主細胞の該物質への結合を検出することを含む、リガンドの結合活性に関してスクリーニングする方法。
【請求項61】
宿主細胞のライブラリーは、酵母細胞の収集体である、請求項60に記載の方法。
【請求項62】
物質は、抗原または受容体である、請求項60に記載の方法。
【請求項63】
(a)アルブミン成分;
(b)軸成分;および、
(c)細胞膜要素、
を含む融合タンパク質。
【請求項64】
アルブミン成分は、アルブミンタンパク質、改変されたアルブミンタンパク質、またはそのフラグメントである、請求項63に記載の融合タンパク質。
【請求項65】
アルブミンタンパク質は、ヒトアルブミンタンパク質である、請求項64に記載の融合タンパク質。
【請求項66】
アンカー成分をさらに含む、請求項63に記載の融合タンパク質。
【請求項67】
膜貫通ドメインをさらに含む、請求項63に記載の融合タンパク質。
【図1】

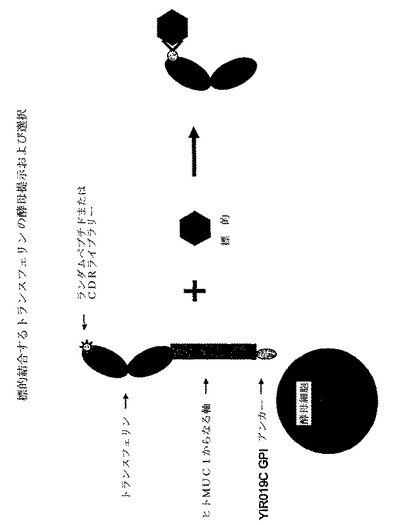
【図2】
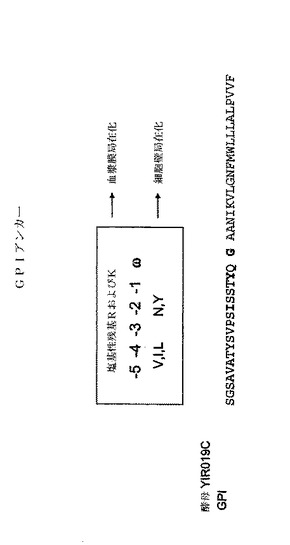
【図3】

【図4】
【図5】

【図6】
【図7】
【図8】
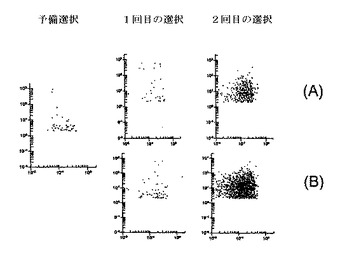
【図9】
【図10】
【図11】

【図12】
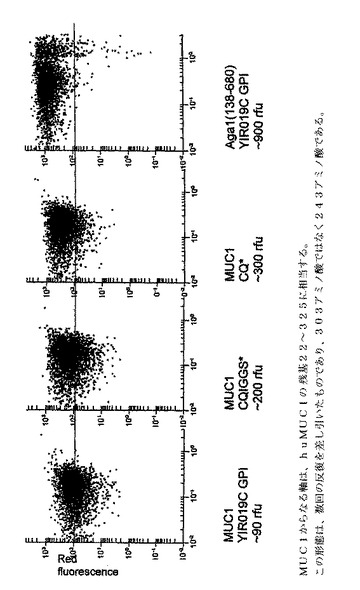
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2008−546392(P2008−546392A)
【公表日】平成20年12月25日(2008.12.25)
【国際特許分類】
【出願番号】特願2008−517204(P2008−517204)
【出願日】平成18年6月19日(2006.6.19)
【国際出願番号】PCT/US2006/023742
【国際公開番号】WO2006/138700
【国際公開日】平成18年12月28日(2006.12.28)
【出願人】(508113424)バイオレクシス ファーマシューティカル コーポレーション (6)
【Fターム(参考)】
【公表日】平成20年12月25日(2008.12.25)
【国際特許分類】
【出願日】平成18年6月19日(2006.6.19)
【国際出願番号】PCT/US2006/023742
【国際公開番号】WO2006/138700
【国際公開日】平成18年12月28日(2006.12.28)
【出願人】(508113424)バイオレクシス ファーマシューティカル コーポレーション (6)
【Fターム(参考)】
[ Back to top ]