
細菌における生物学的に活性なタンパク質の表面発現
異種ポリペプチドが細菌細胞壁を標的とするための方法および組成物を提供する。本発明は、発現カセットを含む乳酸杆菌属細菌を提供する。この発現カセットは、シグナル配列と生物活性ポリペプチドとをコードするポリヌクレオチドと作動可能に結合するプロモーターを含み、この生物活性ポリペプチドは、異種カルボキシル末端細胞壁標的領域と結合し、この異種カルボキシル末端細胞壁標的領域は、以下の順番で以下:細胞壁関連配列;LPQ(S/A/T)(G/A);および疎水性配列を含む。

【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の相互参照)
本願は2003年1月29日出願の米国特許仮出願番号60/443,619に対する優先権の利益を請求する。米国特許仮出願番号60/443,619は、全ての目的でその全体を参考として援用する。
【0002】
(連邦政府後援研究開発下で行われた発明に対する権利に関する記載)
本発明はNational Institutes of Healthにより認可された補助金番号2 R44 AI46203−02の下で政府支援により行われた。政府は、本発明に一部権利を有する。
【背景技術】
【0003】
(発明の背景)
グラム陽性菌におけるペプチドグリカンとの共有結合を介するタンパク質の表面発現は、独自の識別信号とソルターゼ依存性機構とに関与する(Mazmanianら、Science 285:760−763(1999))。最も良く研究されたシステムの一つは、M6構造タンパク質をコードするStreptococcus pyogenesのemm6遺伝子である(Fischettiら、1990、Mol.Microbiol.4:1603−1605(1990))。M6タンパク質は、標示(signature)細胞壁識別信号、Leu−Pro−X−Thr−Gly(LPXTG)モチーフ、それに続く疎水性アミノ酸伸長、および最後に荷電残基を含む配列(KRKEEN)を有し、これは細胞表面保持信号として働く。これらの細胞壁識別モチーフは、Staphlyococcus、EnterococcusおよびListeria、ならびに乳酸杆菌属を含む他のグラム陽性菌において同定されている(NavarreおよびSchneewind、Microbio.Mol.Biol.Rev.63:174−229(1999))が、ヒトの膣でコロニーを形成する乳酸杆菌属種においては同定されていない。
【0004】
全てのヒトの粘膜には自然に細菌がコロニーを形成している(Tannock、Clin.Rev.Allergy Immunol.22:231−53(2002))。最近の科学的証明は、これらの細菌が体の細胞および組織と緊密に相互作用し、天然の生物学的プロセスを調節しているという事実を示した。この粘膜微生物叢がまた、ヒトの細胞および組織に影響する多くの疾病に実質的に寄与することが次第に明らかとなっている。
【0005】
一般に、膣および消化管内で乳酸桿菌および関連する細菌による微生物叢が優位になることは、良い健康状態と関連している(Redondo−Lopezら、Rev.Infect.Dis.12:856−72(1990);Tannock、Clin.Res.Allergy Immunol.22:231−53(2002))。乳酸桿菌の天然株は、これらの場所内で健康な微生物叢を保ち、感染を防止するために、「プロバイオティクス」として長年投与されてきた。これらの「健康な細菌」が細菌、ウイルスおよび真菌等の病原性微生物と競合し、病気に関連する病因の発生と進行を制限するということが十分に確立されている。それにもかかわらず、この微生物叢は脆弱であり、健康な微生物叢の自然な代謝回転、および破壊を伴う動的環境が、日和見感染の発症と関連付けられている。結果として、疾患を予防するかまたは処置する手段として、微生物叢の完全性および天然の性質を維持、またはさらには増強するためのアプローチは、生物医学界が切望するところである。
【0006】
粘膜微生物叢は粘膜表面に影響する多くの局所疾患に寄与する。例えば、HIVおよび他の性感染病原体は膣粘膜を迂回しなければならない。さらに、潰瘍性大腸炎およびクローン病を含む炎症性腸疾患の病因は、破壊された粘膜微生物叢と宿主の細胞および組織との間の不適切な相互作用から生じ得る。粘膜叢内の細菌の性質を調節する手段は、これらの疾患ばかりでなく、粘膜表面に影響する関連する病状の予防または処置の助けになり得る。生物活性タンパク質がこれらおよび他の微生物の細胞壁を標的とすることは、この様な疾患の処置に役立ち得る。
【0007】
本発明はこれらおよび他の問題に取り組む。
【発明の開示】
【課題を解決するための手段】
【0008】
(発明の簡単な要旨)
本発明は、発現カセットを含む乳酸杆菌属細菌を提供する。この発現カセットは、シグナル配列と生物活性ポリペプチドとをコードするポリヌクレオチドと作動可能に結合するプロモーターを含み、この生物活性ポリペプチドは、異種カルボキシル末端細胞壁標的領域と結合し、この異種カルボキシル末端細胞壁標的領域は、以下の順番で以下:細胞壁関連配列;LPQ(S/A/T)(G/A);および疎水性配列を含む。
【0009】
いくつかの実施形態では、上記細胞壁関連配列は、少なくとも50個のアミノ酸を含む。いくつかの実施形態では、この細胞壁関連配列は、少なくとも200個のアミノ酸を含む。いくつかの実施形態では、上記異種カルボキシル末端細胞壁標的領域は、その領域のカルボキシル末端に荷電した配列をさらに含む。
【0010】
いくつかの実施形態では、乳酸杆菌属細菌は、膣コロニー形成株である。いくつかの実施形態では、この細菌は、L.jensenii、L.gasseri、L.caseiおよびL.crispatusからなる群より選択される。
【0011】
いくつかの実施形態では、上記細胞壁標的領域は、アミノ酸配列LPQSGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQAGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQTGを有する。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQTAを有する。いくつかの実施形態では、この細胞壁標的領域は、配列番号7を含む。いくつかの実施形態では、この細胞壁標的領域は、配列番号8を含む。
【0012】
いくつかの実施形態では、上記生物活性ポリペプチドは、細菌の細胞壁で発現される。いくつかの実施形態では、この生物活性ポリペプチドは、10アミノ酸〜600アミノ酸の間である。いくつかの実施形態では、上記生物活性ポリペプチドが病原体と接触した場合、この生物活性ポリペプチドが病原体と結合する。
【0013】
いくつかの実施形態では、上記病原体は、細菌病原体である。いくつかの実施形態では、この病原体は、真菌病原体である。いくつかの実施形態では、この病原体は、ウイルス病原体である。
【0014】
いくつかの実施形態では、上記ウイルス病原体は、ヒト免疫不全ウイルス(HIV)である。いくつかの実施形態では、上記生物活性タンパク質は、CD4またはCD4のHIV結合フラグメントである。いくつかの実施形態では、上記生物活性タンパク質は、2D−CD4である。いくつかの実施形態では、上記生物活性タンパク質は、シアノビリン−N(CV−N)またはCV−Nのウイルス結合フラグメントである。いくつかの実施形態では、上記ウイルス病原体は、単純疱疹ウイルスである。いくつかの実施形態では、上記生物活性タンパク質は、単純疱疹ウイルス侵入媒体C(HveC)またはHveCのウイルス結合フラグメントである。
【0015】
いくつかの実施形態では、上記生物活性ポリペプチドは、乳酸杆菌属細菌から放出される。いくつかの実施形態では、この生物活性ポリペプチドは乳酸杆菌属細菌の細胞壁に固定される。
【0016】
本発明はまた、乳酸杆菌属細菌の細胞壁中で生物活性ポリペプチドを発現する方法を提供する。いくつかの実施形態では、その方法は以下を包含する:発現カセットを含む乳酸杆菌属細菌を提供する工程であって、この発現カセットはシグナル配列および生物活性ポリペプチドをコードするポリヌクレオチドと作動可能に結合するプロモーターを含み、この生物活性ポリペプチドは、異種カルボキシル末端細胞壁標的領域と結合し、この異種カルボキシル末端細胞壁標的領域は、以下の順番で、以下:細胞壁関連配列;LPQ(S/A/T)(G/A);および疎水性配列を含む工程;ならびにこのポリペプチドの発現を誘導する条件下で細菌を培養する工程であって、該工程により、乳酸杆菌属細菌の細胞壁中で生物活性ポリペプチドを発現する工程、を有する。
【0017】
いくつかの実施形態では、上記細胞壁関連配列は、少なくとも50個のアミノ酸を含む。いくつかの実施形態では、この細胞壁関連配列は、少なくとも200個のアミノ酸を含む。
いくつかの実施形態では、上記異種カルボキシル末端細胞壁標的領域は、この領域のカルボキシル末端に荷電した配列をさらに含む。いくつかの実施形態では、上記提供する工程は、上記発現カセットを上記細菌中に輸送する工程を包含する。
【0018】
いくつかの実施形態では、上記細胞壁標的領域は、アミノ酸配列LPQSGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQAGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQTGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQTAを有する。いくつかの実施形態では、細胞壁標的領域は配列番号7を含む。いくつかの実施形態では、この細胞壁標的領域は配列番号8を含む。
【0019】
いくつかの実施形態では、この細胞壁標的領域は、少なくとも200個のアミノ酸を有する。
【0020】
いくつかの実施形態では、上記細菌は、膣コロニー形成株である。いくつかの実施形態では、この細菌はL.jensenii、L.gasseri、L.caseiおよびL.crispatusからなる群より選択される。いくつかの実施形態では、上記生物活性ポリペプチドは10〜600アミノ酸の間である。いくつかの実施形態では、上記生物活性ポリペプチドが病原体と接触した場合、この生物活性ポリペプチドが病原体に結合する。
【0021】
いくつかの実施形態では、上記病原体は、細菌病原体である。いくつかの実施形態では、この病原体は、真菌病原体である。いくつかの実施形態では、この病原体は、ウイルス病原体である。
【0022】
いくつかの実施形態では、上記ウイルス病原体は、HIVである。いくつかの実施形態では、上記生物活性タンパク質は、CD4またはCD4のHIV結合フラグメントである。いくつかの実施形態では、この生物活性タンパク質は、2D−CD4である。いくつかの実施形態では、上記生物活性タンパク質は、シアノビリン−Nまたはシアノビリン−Nのウイルス結合フラグメントである。いくつかの実施形態では、上記ウイルス病原体は、単純疱疹ウイルスである。いくつかの実施形態では、上記生物活性タンパク質は、単純疱疹ウイルス侵入媒体C(HveC)またはHveCのウイルス結合フラグメントである。
【0023】
いくつかの実施形態では、上記生物活性ポリペプチドは、乳酸杆菌属細菌から放出される。いくつかの実施形態では、この生物活性ポリペプチドは、乳酸杆菌属細菌の細胞壁に固定される。
【0024】
本発明はまた、生物活性タンパク質を哺乳動物粘膜表面に提供する方法を提供する。いくつかの実施形態では、その方法は、粘膜表面と、異種カルボキシル末端細胞壁標的領域に結合された生物活性ポリペプチドに結合されたシグナル配列を発現するように組み換えで変化した乳酸杆菌属細菌とを、接触させる工程を包含し、この異種カルボキシル末端細胞壁標的領域は、以下の順番で以下:細胞壁関連配列;LPQ(S/A/T)(G/A);および疎水性配列を含み、ここで、この生物活性ポリペプチドが、粘膜表面から収集されたサンプル中で、検出可能な量で発現される。
【0025】
いくつかの実施形態では、上記細胞壁関連配列は、少なくとも50個のアミノ酸を含む。いくつかの実施形態では、この細胞壁関連配列は、少なくとも200個のアミノ酸を含む。いくつかの実施形態では、上記異種カルボキシル末端細胞壁標的領域は、カルボキシル末端領域に荷電した配列をさらに含む。いくつかの実施形態では、上記乳酸杆菌属細菌は、L.jensenii、L.gasseri、L.caseiおよびL.crispatusからなる群より選択される。
【0026】
いくつかの実施形態では、上記粘膜表面は、膣内に存在する。いくつかの実施形態では、この粘膜表面は、消化管内に存在する。
【0027】
いくつかの実施形態では、上記接触させる工程は乳酸杆菌属細菌を経口投与する工程を包含する。いくつかの実施形態では、この接触させる工程は、乳酸杆菌属細菌を膣投与する工程を包含する。いくつかの実施形態では、この接触させる工程は乳酸杆菌属細菌を直腸投与する工程を包含する。
【0028】
本発明は、シグナル配列と生物活性ポリペプチドとをコードするポリヌクレオチドに作動可能に結合されたプロモーターを含む発現カセットを提供し、ここで、この生物活性ポリペプチドは、異種カルボキシル末端細胞壁標的領域と結合し、この異種カルボキシル末端細胞壁標的領域が以下の順番で以下:細胞壁関連配列;LPQ(S/A/T)(G/A);および疎水性配列を含む。いくつかの実施形態では、上記細胞壁関連配列は、少なくとも50個のアミノ酸を有する。いくつかの実施形態では、この細胞壁関連配列は、少なくとも200個のアミノ酸を有する。
【0029】
いくつかの実施形態では、上記異種カルボキシル末端細胞壁標的領域はさらに領域のカルボキシル末端に荷電配列を有する。
【0030】
いくつかの実施形態では、細胞壁標的領域はアミノ酸配列LPQSGを含む。いくつかの実施形態では、細胞壁標的領域は、アミノ酸配列LPQAGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQTGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQTAを含む。いくつかの実施形態では、この細胞壁標的領域は、配列番号7を含む。いくつかの実施形態では、この細胞壁標的領域は配列番号8を含む。いくつかの実施形態では、上記生物活性ポリペプチドは10〜600アミノ酸の間である。
【0031】
いくつかの実施形態では、上記生物活性タンパク質が病原体と接触した場合、この生物活性タンパク質は病原体と結合する。いくつかの実施形態では、上記病原体は、細菌病原体である。いくつかの実施形態では、この病原体は、真菌病原体である。いくつかの実施形態では、病原体はウイルス病原体である。いくつかの実施形態では、このウイルス病原体は、HIVである。
【0032】
いくつかの実施形態では、上記生物活性タンパク質は、CD4またはCD4のHIV結合フラグメントである。いくつかの実施形態では、上記生物活性タンパク質は、2D−CD4である。いくつかの実施形態では、この生物活性タンパク質は、シアノビリン−Nまたはシアノビリン−Nのウイルス結合フラグメントである。いくつかの実施形態では、この生物活性タンパク質は、単純疱疹ウイルス侵入媒体C(HevC)またはHveCのウイルス結合フラグメントである。いくつかの実施形態では、上記細胞壁標的領域は、乳酸杆菌属中で機能する。
【0033】
本発明はまた、異種カルボキシル末端細胞壁標的領域に結合した生物活性ポリペプチドをコードするポリヌクレオチドに作動可能に結合するプロモーターを含む発現カセットを有するベクターであって、この異種カルボキシル末端細胞壁標的領域が、以下の順番で、以下:細胞壁関連配列;LPQ(S/A/T)(G/A);および疎水性配列を含むベクターを、提供する。
【0034】
(定義)
「生物活性タンパク質」とは、天然の細胞の内または外でアミノ酸配列の生物活性を有する(すなわち分子機構に関与し得る)アミノ酸配列をいう。タンパク質の活性としては例えばその免疫原性、触媒活性、結合親和性等が挙げられる。ポリペプチドワクチンは用語「生物活性タンパク質」に包含される。代表的には、このアミノ酸配列は、天然細胞の内または外でアミノ酸配列により形成される3次元構造を形成する。
【0035】
「2D CD4」とは、ヒトCD4の最初の約183アミノ酸をいう(Arthosら、Cell、1989、57:469−81(1989))。CD4は、成熟ヘルパーT細胞および未成熟胸腺細胞、ならびに単球およびマクロファージ上に見出される、細胞表面糖タンパク質である。2D−CD4は、インタクトタンパク質と同じ親和性でHIV−1 gp120と結合し、gp120に対する結合部位を含む。CD4は、アミノ末端細胞外ドメイン(アミノ酸残基1〜371)、膜貫通領域(372〜395)、および細胞質内末端(tail)テール(396〜433)を含む。
【0036】
「抗体」とは、免疫グロブリン遺伝子(単数または複数)で実質的にコードされるポリペプチド、または分析物(抗原)と特異的に結合して、その分析物を認識するそれらのフラグメントをいう。認められた免疫グロブリン遺伝子は、κ、λ、α、γ、δ、εおよびμ定常領域遺伝子、ならびに無数の免疫グロブリン可変領域遺伝子を含む。軽鎖は、κまたはλのいずれかに分類される。重鎖は、γ、μ、α、δまたはεに分類され、それぞれ免疫グロブリンクラスIgG、IgM、IgA、IgDおよびIgEを規定する。
【0037】
例示的な免疫グロブリン(抗体)の構造単位は、4量体を含む。各4量体は、ポリペプチド鎖の2つの同一の対で構成され、各対は1個の「軽」鎖(約25kDa)と1個の「重」鎖(約50〜70kDa)とを有する。各鎖のN−末端は、抗原認識を主として担う、約100〜110またはそれ以上のアミノ酸の可変領域を規定する。可変軽鎖(VL)および可変重鎖(VH)という用語は、それぞれ、これらの軽鎖および重鎖を指す。
【0038】
抗体は、例えば、インタクト免疫グロブリン、または様々なペプチダーゼでの消化により生成される、性質が十分特徴付けられた多くのフラグメントとして存在する。従って、例えば、ペプシンはヒンジ領域中のジスルフィド結合下で抗体を消化し、F(ab’)2(それ自体がジスルフィド結合でVH−CH1に連結した軽鎖であるFabの2量体である)を生成する。F(ab’)2は、緩やかな条件下で還元されてヒンジ領域のジスルフィド結合を切断し得、それによって、F(ab’)2ダイマーをFab’モノマーに変える。Fab’モノマーは、本質的に、ヒンジ領域の一部を有するFabである(Paul(編)Fundamental Immunology、第3版、Raven Press、NY(1993)参照)。様々な抗体フラグメントが、インタクト抗体の消化の観点から定義されているが、当業者は、このようなフラグメントを、化学的に、または組み換えDNA方法論を利用して、新たに合成し得ることを理解する。従って、本明細書で用いる場合、抗体という用語には、抗体全体の改変により生成されるか、または組み換えDNA方法論を用いて新たに合成されるかのいずれかである抗体フラグメント(例えば1本鎖FV)も含まれる。
【0039】
「単離された」という用語は核酸またはタンパク質に応用する場合、核酸またはタンパク質が本質的に自然状態で結合する他の細胞成分を含まないことを意味する。均一な状態が好ましいが、乾燥状態でも水溶液でも良い。代表的には、ポリアクリルアミドゲル電気泳動または高速液体クロマトグラフィー等の分析化学技術を用いて、純度および均一性が測定される。調製物中に存在する優勢な種であるタンパク質が、実質的に精製される。特に、単離される遺伝子は、その遺伝子に隣接し、目的の遺伝子以外のタンパク質をコードする、オープンリーディングフレームから、分離される。「精製された」という用語は、核酸またはタンパク質が電気泳動ゲル中で本質的に1個のバンドを生じることを意味する。特に、この用語は、核酸またはタンパク質が、少なくとも純度85%であり、より好ましくは少なくとも純度95%であり、最も好ましくは少なくとも純度99%であることを意味する。
【0040】
「核酸」または「ポリヌクレオチド」という用語は、デオキシリボヌクレオチドまたはリボヌクレオチド、および1本鎖または2本鎖の形態のそれらのポリマーをいう。特に限定しない限り、この用語は、参照核酸と類似の結合特性を有し、天然に存在するヌクレオチドと類似の様式で代謝される天然ヌクレオチドの既知のアナログを含む核酸を包含する。特に示さない限り、ある特定の核酸配列はまた、保存的に改変されたその変異体(例えば縮重コドン置換)、および相補配列、ならびに明示された配列もまた暗黙のうちに含む。具体的には、1つ以上の選択された1個の(または全ての)選ばれたコドンの第3位が混合塩基および/またはデオキシイノシン残基で置換された配列を生成することにより、縮重コドン置換が行われ得る(Batzerら、Nucleic Acid Res.19:5081(1991);Ohtsukaら、J.Biol.Chem.260:2605−2608(1985);およびRossoliniら、Mol.Cell.Probes 8:91−98(1994))。「核酸」という用語は「ポリヌクレオチド」と互換的に使用される。
【0041】
「ポリペプチド」、「ペプチド」および「タンパク質」という用語は、本明細書では、アミノ酸残基のポリマーを指すために互換的に使用される。これらの用語は、1つ以上のアミノ酸残基が、対応する天然に存在するアミノ酸の人工的化学模倣体であるアミノ酸ポリマー、および天然に存在するアミノ酸ポリマーおよび天然に存在しないアミノ酸ポリマーに適用される。本明細書で用いる場合、この用語は、全長タンパク質(すなわち抗原)を含む任意の長さのアミノ酸鎖を包含し、ここでこのアミノ酸残基は共有ペプチド結合で連結している。
【0042】
「アミノ酸」という用語は、天然に存在するアミノ酸および合成アミノ酸、ならびに天然に存在するアミノ酸と類似の様式で機能するアミノ酸アナログおよびアミノ酸模倣体をいう。天然に存在するアミノ酸は遺伝子暗号でコードされるものならびに、その後改変されるアミノ酸(たとえばヒドロキシプロリン、γ−カルボキシグルタミン酸およびO−ホスホセリン)である。アミノ酸アナログとは、天然に存在するアミノ酸と同じ基本化学構造(すなわち水素、カルボキシル基、アミノ基およびR基に結合するα炭素)を有する化合物(例えばホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウム)をいう。このようなアナログは、改変されたR基(例えばノルロイシン)または改変されたペプチド骨格を有するが、天然に存在するアミノ酸と同じ基本化学構造を保有している。「アミノ酸模倣体」とは、アミノ酸の一般的な化学構造とは異なる構造を有するが、天然に存在するアミノ酸と類似の様式で機能する化合物をいう。
【0043】
本明細書では、アミノ酸は、一般的に知られた3文字記号で表され得か、またはIUPAC−IUB Biochemical Nomenclature Commissionが推奨する1文字記号で表され得る。ヌクレオチドも同ように、その一般に受け入れられている1文字表記で表され得る。
【0044】
以下に述べるように、最大一致のために整列させた場合に、2つの配列中でヌクレオチドまたはアミノ酸残基の配列がそれぞれ同じであるならば、2個の核酸配列またはポリペプチドは「同一」であると言われる。2個以上の核酸またはポリペプチド配列に関して、用語「同一」または「同一性」パーセントとは、以下の配列比較アルゴリズムの1つを用いるか、または手動整列および目視検査によって測定して比較ウインドウ上での最大一致のために比較し整列させた場合、同一であるか、あるいは特定のパーセントの同一なアミノ酸残基またはヌクレオチドを有する、2つ以上の配列または部分配列をいう。配列同一性パーセントをタンパク質またはペプチドに関連して用いる場合、同一でない残基の位置が、多くの場合保存的アミノ酸置換により異なり、その場所では、アミノ酸残基が類似の化学的性質(例えば荷電または疎水性)を有する他のアミノ酸残基で置換されており、従って分子の機能的性質を変化させないことが認識される。保存的置換で配列が異なる場合、配列同一性パーセントを上方に調節して置換の保守的性質について補正鏑し得る。この調整を行うための手段は当業者に公知である。代表的に、これは保存的置換を完全なミスマッチでなく部分ミスマッチとして採点し、それによって配列同一性パーセントを増加させる工程に関する。従って、例えば、同一アミノ酸に得点1を与え、非保存的置換に得点ゼロを与える場合、保存的置換には0と1の間の得点が与えられる。保存的置換の採点は、例えば、プログラムPC/GENE(Intelligenetics、Mountain View、California、USA)で実行されるMeyerおよびMiller、Computer Applic.Biol.Sci.4:11−17(1988)のアルゴリズムに従って計算される。
【0045】
2個の核酸またはポリペプチドについての「実質的に同一」という語句は、参照配列と少なくとも70%の配列同一性を有する配列または部分配列をいう。または、同一性パーセントは40%〜100%の任意の整数であり得る。より好ましい実施形態としては、以下に述べる標準パラメーターを用いるBLAST等の本明細書に記載のプログラムを用いて、参照配列(例えば配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7または配列番号8、またはそのフラグメント)と比較して、少なくとも40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%または99%が挙げられる。
【0046】
配列比較のため、代表的には、1つの配列が参照配列として働き、それと試験配列が比較される。配列比較アルゴリズムを使用する場合、試験配列および参照配列をコンピューターに入力し、必要あれば部分配列座標を指定し、配列アルゴリズムプログラムパラメーターを指定する。デフォルトプログラムパラメーターを使用することができるか、または別なパラメーターを指定することができる。次いでプログラムパラメーターに基づき、参照配列に対する配列比較アルゴリズムが試験配列についての現在の配列同一性パーセントを計算する。
【0047】
本明細書で用いる場合、「比較窓」には、20〜600、通常は約50〜約200、より通常は約100〜約150個からなる群より選ばれた連続位置数のいずれか一つのセグメントに対する参照が含まれ、その中で2つの配列を最適に整列した後、一つの配列が、連続位置が、同数の参照配列と比較され得る。比較のための配列の整列法は当該分野で周知である。比較のために最適な配列の整列は、例えば、SmithおよびWaterman、Adv.Appl.Math.2:482(1981)の局所相同アルゴリズム、NeedlemanおよびWunsch、J.Mol.Biol.48:443(1970)の相同整列アルゴリズム、PearsonおよびLipman、Proc.Natl.Acad.Sci.USA 85:2444(1988)の類似性検索法、これらのアルゴリズムのコンピューター化された実施(Wisconsin Genetics Software Package、Genetics Computer Group、575 Science Dr.,Madison、WI中におけるGAP、BESTFIT、FASTAおよびTFASTA)、または手動整列および目視検査により行われ得る。
【0048】
配列同一性パーセントと配列類似性の決定に適したアルゴリズムの例は、Altschulら、J.Mol.Biol.215:403−410(1990)に記載のBLASTアルゴリズムである。BLAST分析を実行するためのソフトウエアは、National Center for Biotechnology Informationから公に入手可能である。このアルゴリズムには、最初にクエリ(query)配列中の長さWのショートワードを同定することによってハイスコア配列対(HSP)を同定する工程を包含する。データベース配列中の同じ長さのワードと整列した場合、HSPは、ある正の値の閾値スコアTと一致するか、またはそれを満たすかのいずれかである。Tは、近隣ワードスコア閾値(neighborhood word score threshold)と呼ばれる(Altshulら、前出)。最初の近接ワードヒットは、それらのワードを含むより長いHPSを見出すための検索を開始するシードとして働く。累積整列スコアが増加し得る限り、ワードヒットは各配列に沿って両方の方向に延長される。累積整列スコアがその最大達成値から量Xだけ低下した場合、1つ以上の負スコア残基整列の蓄積のために累積スコアがゼロ以下になった場合、またはいずれかの配列の末端に達した場合、各方向へのワードヒットの延長を停止する。BLASTアルゴリズムパラメーターW、TおよびXは整列の感度と速度を決定する。BLASTプログラムは、デフォルトとして、ワード長(W) 11、BLOSUM62スコアリングマトリックス(HenikoffおよびHenikoff、Proc.Natl.Acad.Sci.USA 89:10915(1989)参照)整列(B) 50、期待値(E) 10、M=5、N=−4、および両方の鎖の比較を使用する。
【0049】
BLASTアルゴリズムはまた、2つの配列間の類似性の統計分析を行う(例えば、KarlinおよびAltschul、Proc.Natl.Acad.Sci.USA 90:5873−5787(1993)参照)。BLASTアルゴリズムが提供する類似性の基準の1つは、2つのヌクレオチド配列またはアミノ酸配列間の一致が偶然生じる確率を示す最小和確率(P(N))である。例えば、参照核酸に対する試験核酸の比較における最小和確率が約0.2未満、より好ましくは約0.01未満、最も好ましくは約0.001未満である場合、その核酸は参照配列に類似すると考えられる。
【0050】
例えば細胞、核酸、タンパク質またはベクターに関して用いられる場合、「組み換え」または「組み換えで変化された」という用語は、その細胞、核酸、タンパク質またはベクターが異種核酸もしくは異種タンパク質の導入によってかまたはネイティブ核酸もしくはネイティブタンパク質の変化によって改変されたか、あるいは細胞がそのようにして改変された細胞に由来することを示す。従って、例えば、組み換え細胞は、その細胞のネイティブ(非組み換え)形態に見出されない遺伝子を発現するか、さもなければ異常発現するか、発現量が少ないか、またはまったく発現しないネイティブ遺伝子を発現する。
【0051】
核酸またはポリペプチドの部分に関して使用される「異種」という用語は、核酸またはポリペプチドが天然で相互に同じ関係で見出されない2個以上の部分配列を有することを示す。例えば、その核酸は、代表的には組み換えで製造され、新しい機能性核酸を生成するように配置された無関係の遺伝子由来の2個以上の配列(例えばある起源由来のプロモーターおよび別の起源由来のコード領域)を有する。同ように、異種タンパク質とは、タンパク質が天然で相互に同じ関係で見出されない2以上の部分配列を含むことを示す(例えば融合タンパク質)。
【0052】
「発現カセット」とは、宿主細胞内における特定の核酸の転写が可能である一連の特定核酸要素を有する、組み換えまたは合成で生成される核酸である。発現カセットはプラスミド、ウイルスまたは核酸フラグメントの一部であり得る。代表的には、発現ベクターとしては、プロモーターに作動可能に結合して転写されるべき核酸が挙げられる。
【0053】
(発明の詳細な説明)
(I.はじめに)
本発明は乳酸杆菌属等のグラム陽性菌の細胞壁上に異種ポリペプチドを発現するための新規なモチーフおよび方法を提供する。本発明のモチーフを目的のタンパク質と融合され、次いで細菌中の融合タンパク質として発現され、その結果、融合タンパク質が、細胞壁の標的となるか、埋め込まれるか、および/または表面に提示されるか、あるいは生物学的に活性であり、かつ安定な融合タンパク質が、細胞外マトリックスに放出され得る。
【0054】
モチーフは、例えば、膣を含むヒト粘膜でコロニーを形成する乳酸杆菌属細菌の細胞壁上にタンパク質を発現するために有用である。例示的な粘膜細菌としては、L.jensenii、L.gasseriおよびL.casei等の乳酸杆菌属種が挙げられる。
【0055】
(II.細胞壁標的領域)
乳酸杆菌属等のグラム陽性菌の細胞壁に共有結合された目的のポリペプチドを発現し、標的とするために、細胞壁標的領域は、目的の異種ポリペプチドにC−末端で結合している。膣関連乳酸桿菌および他の乳酸桿菌において異種タンパク質を表面提示し得る細胞壁標的領域は、4つの部分:細胞壁関連領域、LPQ(S/A/T)(G/A)配列、および疎水性配列から、代表的にはこの順番で構成される。必要に応じて細胞壁標的領域は、カルボキシル末端またはその近くに荷電領域を含む。この荷電領域は、細胞膜中で移動停止配列として作用し、それによって培地中への放出を阻止する。もちろん、アンカー配列がタンパク質の残りの部分から切り取られた場合、やはり培地中への放出が生じ得る。
【0056】
(A.細胞壁関連領域)
細胞壁関連領域は、LPQ(S/A/T)(G/A)識別シグナルに先行する。細胞壁関連領域の長さは変化し得る。細胞壁関連領域は代表的には、40アミノ酸と1,000アミノ酸との間である。いくつかの実施形態では、細胞壁関連領域は、少なくとも約30、50、80、100、150および200アミノ酸または、それより大きい。いくつかの実施形態では、細胞壁関連領域は約500、400、300、250、200、150、100またはそれ未満のアミノ酸を有する。Lactobacillus jenseniiでは、pOSEL268中のC−末端細胞壁識別シグナルと融合する一縦列反復を含む95アミノ酸伸長(実施例に記載)により、CD4の表面提示が可能になる。しかしながら、フィブロネクチン結合タンパク質の約90アミノ酸がS.carnosus中で想定されている(StraussおよびGotz、Mol.Microbiol.21:491−500(1996))一方で、ペプチドマッピングに基づいてS.pyogenesのM6タンパク質中の約50アミノ酸長が同定された(PancholiおよびFischetti、J.Bacteriol.170:2618−2624(1988))。従って、約50アミノ酸以下の配列が、乳酸杆菌属中で機能性であり得る。
【0057】
いくつかの実施形態では、細胞壁関連領域は疎水性である。いくつかの実施形態では、細胞壁関連領域は長さと配列が変化し得る不完全縦列反復を含む。例えば、L.jensenii C370の細胞壁関連領域は、2.5個の縦列反復を含む。しかし、縦列反復は、細胞壁関連領域に起こり得るが、必要ではない。例えばC14の細胞壁関連領域は、反復を含まない。機能的には、細胞壁関連領域は、ペプチドグリカン層と相互作用して、それを貫通(span)する。従って、その領域は細胞壁貫通(spanning)ドメインまたは細胞壁付着ドメインとも呼ばれ、膜結合ソルターゼで固定されるタンパク質と細胞壁識別シグナルとの間のスペーサーとして作用する。
【0058】
本発明は、C370配列:
【0059】
【化1】
と実質的に同一の細胞壁関連領域を提供する。ある場合は、細胞壁関連領域は少なくともC370配列の約40、50、75、90、100、120、150、175、200アミノ酸フラグメントを有する。例えば、活性細胞壁関連フラグメントは、以下の配列:
【0060】
【化2】
を有し得る。C370配列(配列番号4)は、75個の荷電アミノ酸残基(K、R、D、E)を含み、Pro−Gly富化配列が欠失している。
【0061】
いくつかの実施形態では、細胞壁関連領域は、実質的にC14配列:
【0062】
【化3】
と同一である。ある場合は、細胞壁関連領域は、C370配列の少なくとも約40、50、75、90、100、120、150、175、200アミノ酸フラグメントを含む。配列番号6は、51個の荷電アミノ酸残基(K、R、D、E)を含む。
【0063】
ある場合は、細胞壁関連領域は、乳酸杆菌属以外の細菌、または膣と関連しない乳酸杆菌属株由来である。
【0064】
(B.LPQ(S/A/T)(G/A))
配列LPQ(S/A/T)(G/A)は乳酸杆菌属の膣関連株において細胞壁局在化シグナルとして作用する。モチーフLPQ(S/A/T)(G/A)の少なくとも1個のコピーは細胞壁標的領域内にある。モチーフ中の括弧はその位置の別なアミノ酸を示す(例えばLPQSG、LPQAG、LPQTG、LPQSA、LPQAA、LPQTA)。
【0065】
(C.疎水性配列)
細胞壁に固定されるポリペプチドのカルボキシル末端は細菌膜(bacterial membrane)を貫通するように機能する疎水性領域を含む。疎水性領域は少なくとも約50%、ある実施形態では少なくとも60%、70%、80%または90%の疎水性アミノ酸を含む。天然起源疎水性アミノ酸にはアラニン、イソロイシン、ロイシン、メチオニン、フェニルアラニン、プロリン、トリプトファンおよびバリンが含まれる。グリシン、トレオニンおよびセリンを含む、より疎水性の少ないアミノ酸もこれらの配列の一部を構成し得る(例えばPallenら、Trends Microbiol.9:97−101(2001)参照)。ソルターゼ様タンパク質の可変LPXTG含有基質の間で疎水性配列は一般に長さが約10〜約30アミノ酸、場合によっては13〜24アミノ酸の間である(Pallenら、Trends Microbiol.9:97−101(2001))。典型的な疎水性配列には例えばC14中のV1740GILGLAIATVGSLLGLGLV1758、およびC370中のP1877LTAIGIGLMALGAGIFA1894が含まれる。
【0066】
または、グラム陽性細菌由来の任意の細胞壁アンカータンパク質の疎水性領域を用いることができる。他の疎水性配列には例えば米国特許第5,821,088号の図1に記載のもの、またはそれと実質的に同一の配列が含まれる。さらに別な配列がPallenら、Trends Microbiol.9:97−100(2001)の表2にも示されている。
【0067】
(D.荷電配列)
荷電領域が細胞壁標的タンパク質のカルボキシル末端、代表的には疎水性膜貫通領域の直後に任意に存在し得る。カルボキシル末端荷電領域の存在によりポリペプチドが膜に固定され、その結果膜から解離し培地中へ漏れ出すタンパク質の量が大いに減少する。荷電領域は少なくとも40%、ある実施形態では少なくとも50%、60%、70%、80%または90%の荷電アミノ酸を有する。天然起源荷電アミノ酸にはアルギニン、ヒスチジン、リジン、アスパラギン酸およびグルタミン酸が含まれる。荷電配列は例えば長さで2〜20アミノ酸残基の間、ある実施形態では4〜12または5〜11アミノ酸の間である。典型的な荷電配列には例えばC191中のK969KRKED974、C14中のR1760KKRQK1765およびC370中のK1895KKRKDDEA1903が含まれる。
【0068】
または、グラム陽性細菌由来の任意の細胞壁アンカータンパク質の荷電領域を使用できる。別な荷電配列には例えば米国特許第5,821,088号の図1に記載のもの、およびそれと実質的に同一の配列が含まれる。また別な配列がPallenら、Trends Microbiol.9:97−100(2001)の表2にも示される。
【0069】
(III.組み換え技術)
(A.分子生物学法)
本発明は組み換え遺伝子学の分野における慣用的な技術に依存している。本発明において使用する一般的な方法を開示する基本的なテキストにはSambrookら、Molecular Cloning、A Laboratory Manual(第3版、2001);Kriegler、Gene Transfer and Expression:A Laboratory Manual(1990);およびCurrent Protocols in Molecular Biology(Ausubelら、編集、1994)が含まれる。
【0070】
核酸では、サイズはキロベース(kb)または塩基対(bp)で与えられる。これらはアガロースまたはアクリルアミドゲル電気泳動、配列決定された核酸、または公開されたDNA配列から見積もられる。タンパク質では、サイズはキロダルトン(kDa)またはアミノ酸残基数で与えられる。タンパク質のサイズはゲル電気泳動、配列決定されたタンパク質、導かれたアミノ酸配列、または公開されたタンパク質配列から見積もられる。
【0071】
市販されていないオリゴヌクレオチドを、BeaucageおよびCaruthers、Tetrahedron Letts.22:1859−1862(1981)に最初に記載された固相ホスホラミダイトトリエステル法に従って、Van Devanterら、Nucleic Acids Res.12:6159−6168(1984)に記載の様に自動合成装置を用いて化学的に合成することができる。オリゴヌクレオチドの精製はPearsonおよびReanier、J.Chrom.255:137−149(1983)に記載の通りに天然アクリルアミドゲル電気泳動または陰イオン交換HPLCによって行われる。
【0072】
クローン化された遺伝子、および合成オリゴヌクレオチドの配列を、例えばWallaceら、Gene 16:21−26(1981)の二重鎖テンプレートの配列決定のための鎖停止法を用いてクローニング後に確認することができる。
【0073】
(B.所望のタンパク質をコードするヌクレオチド配列を単離するためのクローニング法)
一般に、目的とするタンパク質をコードする核酸を、cDNAまたはゲノムDNAから作製したDNAライブラリーからクローニングする。特定の配列の位置をオリゴヌクレオチドプローブとハイブリダイズすることにより決定することができるが、その配列は本明細書に開示した配列または当該分野で公知の配列から導かれ、PCRプライマーの参考となり、遺伝子特異的プローブを単離するために適当な領域を規定する。または、配列が発現ライブラリー中にクローニングされる場合は、本明細書に開示されるポリペプチドを含め目的のポリペプチドに対して作製された抗血清または精製抗体を用いて発現組み換えタンパク質を免疫学的に検出し得る。
【0074】
ゲノムおよびcDNAライブラリーを作製およびスクリーニングする方法は当業者に周知である(例えばGulberおよびHoffman、Gene 25:263−269(1983);BentonおよびDavis、Science、196:180−182(1977);およびSambrook、前出参照)。目的のタンパク質を発現する細胞はcDNAライブラリーを作製するための有用なRNA源である。
【0075】
簡単に言えば、cDNAライブラリーを作製するため、mRNAに富んだ起源を選ばなければならない。次いでmRNAをcDNAに変換し、組み換えベクター中に連結し、増殖、スクリーニングおよびクローニングのために組み換え宿主中に形質導入する。ゲノムライブラリーでは、DNAを適当な組織または細胞から抽出し、機械的に剪断するか酵素的に消化して好ましくは約5〜100kbの断片を生成する。次いで断片を望ましくないサイズのものから濃度勾配遠心で分離し、バクテリオファージラムダベクター中に構築させる。これらのベクターおよびファージはインビトロでパッケージングされ、組み換えファージをプラクーハイブリダイゼーションで分析する。コロニーハイブリダイゼーションはGrunsteinら、Proc.Natl.Acad.Sci.USA.72:3961−3965(1975)に一般的に記載されている様に行われる。
【0076】
別な方法では、合成オリゴヌクレオチドプライマーの使用と、mRNAまたはDNAテンプレート上でのポリメラーゼ延長とが組み合わされる。このポリメラーゼ連鎖反応(PCR)法では、目的のタンパク質をコードする核酸がmRNA、cDNA、ゲノムライブラリーまたはcDNAライブラリーから直接増幅される。プライマー中には制限エンドヌクレアーゼ部位が組み込まれ得る。ポリメラーゼ連鎖反応法、または他のインビトロ増幅法も、例えば特定のタンパク質をコードする核酸のクローニングおよびそのタンパク質の発現、生理学試料中で本発明のポリペプチドをコードするmRNAの存在を検出し、核酸の配列を決定する、または他の目的ためのプローブとして使用される核酸の合成に有用であり得る(米国特許第4,683,195号および同第4,683,202号参照)。PCR反応によって増幅された遺伝子はアガロースゲルから精製され、そして適当なベクター中にクローン化され得る。
【0077】
本発明のポリペプチドをコードする遺伝子を組織または細胞試料から同定するための適当なプライマーおよびプローブを、当該分野で記載の配列から導くことができる。PCRの概説としてはInnisら、PCR Protocols:A Guide to Methods and Applications、Academic Press、San Diego(1990)参照。
【0078】
本発明のポリペプチドをコードするポリヌクレオチドを、乳酸杆菌属中に形質転換する前に中間ベクターを使用してクローニングすることができる。これらの中間ベクターは代表的には原核生物ベクターまたはシャトルベクターである。
【0079】
(C.形質転換技術)
適当な細菌宿主株を、例えばその形質転換能、異種タンパク質発現能、および/または粘膜表面でのコロニー形成能で選択される。細菌宿主を塩化ルビジュウム法またはエレクトロポーレーション等の標準技術を用いて形質転換受容性にする(例えばWeiら、J.Microbiol.Meth.21:97−109(1995)参照)。
【0080】
エレクトロポーレーションによるL.jenseniiの形質転換を、例えばLuchanskyら(J.Dairy Sci.74:3293−3302(1991));Changら(Proc.Natl.Acad.Sci.USA.100:11672−11677(2003))に規定される様な標準法を改変して行うことができる。簡単に言えば、新しく接種したL.jenseniiをブロス中で培養する(例えば37℃、5%CO2でOD600が0.6〜0.7まで)。細菌細胞を収集し、洗浄し、スクロースとMgCl2の冷溶液(例えば4℃)中に再懸濁する。コンピテント細胞を次いでDNAと混合し、冷却したギャップキュベット中にいれ、エレクトロポーレーションする。その後、選択試薬以外の抗生物質を含む選択寒天プレート上に接種する前に、予め加熱したブロス中で細胞を回収する(例えば37℃で約2時間)。
【0081】
(D.発現)
本発明の発現カセットは、本発明のポリペプチドの発現と位置決めを制御するための様々な成分を含み得る。例えば、発現カセットはプロモーター要素、シグナル配列をコードする配列、目的のポリペプチドに対するコード配列、およびアンカー配列を含むことができる。
【0082】
異種ポリヌクレオチドまたはポリペプチドの発現は構成的であり得る(例えばP59(Van der Vossenら、Appl.Environ.Microbiol.58:3142−3149(1992))もしくはP23(Elliotら、Cell、36:211−219(1984))プロモーター、またはより高い強度を有する乳酸杆菌属由来の天然プロモーターを用いて)。または、発現は誘導性プロモーターの制御下に行うことができる。例えば、Bacillusアミラーゼ(Weickertら、J.Bacteriol.171:3656−3666(1989))またはキシロース(Kimら、Gene、181:71−76(1996))プロモーターの他、Lactococcusナイシンプロモーター(Eichenbaumら、Appl.Environ.Microbiol.64:2763−2769(1998))を誘導性発現を推進するために使用することができる。さらに、酸またはアルカリ誘導プロモーターを使用することもできる。例えば、膣の比較的酸性の条件下で活性なプロモーターを使用し得る。または、精子に反応する膣内の変化で誘導されるプロモーターも使用し得る。例えば、精子の導入の結果である膣のアルカリ条件の増大に反応して発現を誘導するために、アルカリ誘導プロモーターを使用する。
【0083】
様々なシグナル配列がポリペプチドの発現を膜、細胞外空間または細胞壁に導くことが知られている(例えばペプチドグリカンへの共有結合付着により)。例示的なシグナル配列にはL.amylovorusのα−アミラーゼ由来のシグナル配列(GiraudおよびCuny、Gene、198:149−157(1997))またはL.crispatusのS−層遺伝子(cbsA)由来のシグナル配列(例えばMKKNLRIVSAAAAALLAVAPVAAまたはMKKNLRIVSAAAAALLAVATVSA)が含まれる。シグナル配列は代表的にはポリペプチドのアミノ末端に位置している。
【0084】
ポリペプチドの正確な位置と折り畳みを標準法を用いて決定し得る。例えば、細菌を緩衝液(例えば25%スクロース、1mM EDTA、10mM Tris−HCl、pH8.0)に懸濁し、次いで細胞壁分解酵素(例えばリゾチームおよびムタノリシン)で処理し、次いで得られたプロトプラストを分画遠心法で分離することにより、乳酸杆菌属の細胞壁富化画分を得ることができる。画分をウエスタンブロッティングでスクリーニングして細胞壁内の発現を確認し得る。
【0085】
発現したポリペプチドの折り畳みと生物活性も、標準法を用いて決定し得る。例えば、天然の折り畳みポリペプチド特異性抗体を用いるELISAアッセイを用いてポリペプチドの折り畳みと3次元構造を確認することができる。生物活性アッセイも、ポリペプチドの活性に応じて当然変化する。例えば、ウイルスタンパク質に結合するポリペプチドでは、標準結合アッセイを用いて発現したポリペプチドのウイルスタンパク質結合能を試験することができる。抗炎症分子では、発現したポリペプチドの炎症促進する物質との拮抗能をアッセイすることができる。
【0086】
宿主細胞中の発現を改善するための遺伝子を合成する場合、そのコドン利用頻度を宿主細胞の好ましいコドン利用頻度に近づける様に遺伝子を設計することが望ましい。合成遺伝子に対する好ましいコドン利用頻度の、宿主細胞が使用する頻度からの偏差率が、1個のコドンの利用頻度の宿主細胞の利用頻度からの偏差率をまず測定し、次いでコドン全体の平均偏差を得て計算される。
【0087】
特定のポリペプチドをコードするポリヌクレオチド配列を、特定の宿主のコドン利用と一致する様に変化させることができる。例えば、乳酸杆菌属のコドン利用を使用して、本発明のポリペプチドをコードし、好ましい乳酸杆菌属のコドンを有するポリヌクレオチドを誘導することができる。宿主細胞が提示する好ましいコドンの利用頻度を、宿主細胞が発現する多数の遺伝子中の好ましいコドンの利用頻度を平均して計算することができる。この解析は宿主細胞で高度に発現する遺伝子に限定することが好ましい。例えば、Pouwelsら(Nucleic Acids Res.22:929−936(1994))は様々な乳酸杆菌属種が示す高度発現遺伝子によるコドン利用頻度を提供している。コドン利用表はインターネットを通じても入手できる。
【0088】
(IV.本発明のタンパク質)
本発明のポリペプチド、例えば本発明の細胞壁標的領域に融合する生物活性ポリペプチドは、任意のポリペプチドであり得る。代表的には、本発明のポリペプチドはポリペプチドの生物活性を考慮した条件下で発現する。ある実施形態では、発現ポリペプチド中にジスルフィド結合が存在する。ある実施形態では、ポリペプチドの生物活性にジスルフィド結合が必要である。
【0089】
本発明のポリペプチドは任意のサイズの分子量であり得る。例えば、ポリペプチドは約100〜200,000ダルトンの間、約500〜40,000ダルトンの間、約500〜10,000ダルトンの間、約10,000〜50,000ダルトンの間、または約50,000〜200,000ダルトンの間であり得る。
【0090】
病原体感染を予防または治療するために本発明の方法に従って使用し得るポリペプチドクラスの例には、例えば抗体、抗体断片、または1本鎖抗原を含め抗ウイルスポリペプチド、抗菌ポリペプチド、抗真菌ポリペプチド、およびウイルス、細菌または真菌に結合するポリペプチドが含まれる。
【0091】
ある場合は、本発明のポリペプチドはウイルスまたは細菌病原体が結合して宿主を感染させるレセプターである。または、ポリペプチドは例えば病原体の複製、生存率、侵入またはさもなければ病原体への結合を阻害する薬剤である。ある実施形態では、本発明のポリペプチドは性的に伝播する病原体、および膣から伝播、または膣へ伝播する他の病原体に結合またはそれらを阻害する。例えば、ウイルスは感染のために標的細胞表面上のレセプターに結合することが必要であるので、ウイルスとその宿主レセプターとの相互作用を阻害することを目的とする戦略は、感染防止に有効である。
【0092】
典型的な抗ウイルスポリペプチドには例えばCD4またはそのウイルス結合断片(例えば2D−CD4:例えばOrloffら、J.Virol.67:1461−1471(1993))、3量体モチーフを経由して生成する安定なCD4の3量体(例えばYangら、J.Virol.76:4634−4642(2002))、CD4−Ig融合タンパク質12量体(Arthosら、J.Biol.Chem.277:11456−11464(2002))、α−デフェンシン(例えばZhangら、Science、298:995−1000(2002))、17bmAbの1本鎖可変領域と融合したCD4(Deyら、J.Virol.77:2859−2865(2003))、シアノビリン−Nまたは改変体(例えばBolmstedtら、Mol.Pharmacol.59:949−954(2001);Moriら、Protein Expr.Purif.26:42−49(2002))、単純疱疹ウイルス侵入メディエーターC(HveC:例えばCocchiら、Proc.Natl.Acad.Sci.USA.95:15700−15705(1998))、およびICAM−1が含まれる。他の実施形態には例えばウイルスレセプターまたはヘパリンまたはヘパリン様分子、樹状細胞特異性ICAM−3グラビングノンインテグリン(例えばGeijtenbeekら、Cell、100:587−597(2000);Feinbergら、Science、294:2163−2166(2001))を含むマンノース結合レクチン、抗HSV−1 gp120 1本鎖抗体(例えばMarascoら、Proc.Natl.Acad.Sci.USA.90:7889−7893(1993);McHughら、J.Biol.Chem.277:34383−34390(2002))、HIV−1 gp120のCD4結合部位を認識するヒトmAb b12(例えばSaphireら、Science、293:1155−1159(2001))、またはHSVに結合する中和抗体を含む同様な特異性を有する他の分子(例えばBurioniら、Proc.Natl.Acad.Sci.USA.91:355−359(1994))、およびHIV−1侵入阻害タンパク質(例えばRootら、Science、291:884−888(2001);Siaら、Proc.Natl.Acad.Sci.USA.99:14664−14669(2002))が含まれる。
【0093】
ヒト乳頭腫ウイルス(HPV)による感染は頚部癌の発症に関連する因子である(例えばzur Hausen、Virology、184:9−13(1991);Stanley、Best Prat.Res.Clin.Obstet.Gynaecol.15:663−676(2001))。従って、HPVを阻害する、またはそれに結合する分子の存在は、HPV感染と頚部癌の発症の両方の阻止に有用である。典型的な抗HPVポリペプチドには例えばヒト乳頭腫ウイルス16型タンパク質E6またはE7に結合する中和抗体(例えばMannhartら、Mol.Cell Biol.20:6483−6495(2000))、HPV−結合タンパク質、またはウイルス指向免疫反応を誘発するために使用し得るHPVタンパク質が含まれる。
【0094】
ウイルスまたは細菌等の病原体に結合する能力を、少なくとも数個の方法で本発明の細菌に授け得る。その第1はヒトライノウイルスHRV(主要グループ)に対するICAM−1およびHIVに対するCD4等の、ウイルスに対する正常な宿主レセプターを細菌の表面に発現させることである。これらは正常なヒトタンパク質であり、これらの遺伝子の完全な配列の多くが決定され、データベースGenBankに保存されている。
第2の方法は、ポリオウイルス上のVP4またはHIV上のgp120等のウイルス表面上の保存性抗原決定基に結合する抗体断片または他のポリペプチドを細菌表面に発現することである。本質的に任意の抗原に特異性のある抗体断片(およびペプチド)を、例えばファージ提示ライブラリー(Marksら、J.Biol.Chem.267:16007−16010(1992))から選ぶことができる。抗体を任意の病原体上のエピトープまたは病原体と結合するエピトープの他、以下に議論するエピトープ等の他のエピトープを指向させることができる。
【0095】
第3の方法には、細菌表面上の炭水化物結合ポリペプチドの発現が含まれる。これらの分子の例にはヘパリン結合ポリペプチド、またはマンノース結合ポリペプチドが含まれる。
【0096】
抗細菌ポリペプチドには尿路病原性E.coliに結合する、またはその生育もしくはそれによるコロニー形成を阻害するポリペプチドが含まれる。典型的な抗細菌ポリペプチドには例えばグラム陰性細菌に対する透過性増大タンパク質(Levy、Expert Opin.Investig.Drugs、11:159−167(2002))、哺乳動物抗微生物ペプチド、β−デフェンシン(GarzおよびLehrer、Pharmacol.Ther.66:191−205(1995))、バクテリオシン(例えばLoefflerら、Science、294:2170−2172(2001))、および細菌に特異的に結合する抗体が含まれる。
【0097】
抗真菌ポリペプチドにはCandida等の真菌に結合する、またはその生育もしくはそれによるコロニー形成を阻害するポリペプチドが含まれる。
【0098】
本発明に記載の有用な生物活性ポリペプチドのさらに別な例には抗炎症分子、成長因子、成長因子に結合するかまたはそれに拮抗する分子、治療用酵素、抗体(例えば抗体断片または1本鎖抗体を含む)、および子宮頚部癌を含む癌を阻害するかまたは治療する分子等の治療用ポリペプチドまたは薬剤が含まれる。数多くの他の治療活性ポリペプチドを容易に引用することができるので、これらの例は制限的であることを意図するものではない。
【0099】
抗炎症分子には例えばTNFまたはIL−8に特異的に結合する抗体または他の分子が含まれる。他の例示的な抗炎症分子には、IL−10およびIL−11が含まれる。
【0100】
本発明で有用な成長因子には例えばKGF、HB−EGF、FGFおよびTGF−β等の局所組織修復に係わる因子、またはこれらの分子の拮抗剤が含まれる。
【0101】
治療用酵素には例えば一酸化窒素(NO)シンターゼが含まれる。
【0102】
抗癌性分子にはアポトーシスを誘導する分子、p53等の細胞周期を調節する分子、または癌特異性エピトープを標的とするワクチンとして作用する分子が含まれる。
【0103】
本発明で有用なワクチン分子にはウイルス、細菌または真菌に対する免疫反応を誘起するポリペプチドが含まれる。典型的なウイルスワクチンは例えばHIV、HPV、HSV−2または天然痘に対する反応を誘発する。典型的な抗原にはHSV−2の糖タンパク質D、ヒト乳頭腫ウイルスのタンパク質E6およびE7、Chlamydia trachomatisの主要外膜タンパク質(KimおよびDeMars、Curr.Opin.Immunol.13:429−436(2001))、およびCandida albicansのアスパルチルプロテアーゼ(De Bernardisら、Infect.Immun.70:2725−2729(2002));尿路病原性E.coliのFimH(Langermannら、Science、276:607−611(1997));腸管外病原性E.coliのIroN(Russoら、Infect.Immun.71:7164−9(2003))が含まれる。
【0104】
(V.送達)
遺伝子組み換え細菌の所望の粘膜表面への送達は、その領域への近づき易さと局部条件に依存する。例えば、遺伝子組み換え細菌を食塩水、または膣粘膜上へ送達するための発泡体の中に入れ得る。発泡体には例えば1種以上の疎水性に修飾したセルロース誘導体またはキトサン等の多糖が含まれ得る。セルロース誘導体には例えばヒドロキシエチルセルロース、ヒドロキシプロピルセルロース、メチルセルロース、ヒドロキシプロピルメチルセルロース、ヒドロキシエチルメチルセルロース等が含まれる。キトサンには例えば以下のキトサン塩:乳酸キトサン、サリシル酸キトサン、ピロリドンカルボン酸キトサン、イタコン酸キトサン、ナイアシン酸キトサン、蟻酸キトサン、酢酸キトサン、没食子酸キトサン、グルタミン酸キトサン、マロン酸キトサン、アスパルギン酸キトサン、グリコール酸キトサン、および4級アミン置換キトサンおよびその塩などが含まれる。発泡体にはまた水、エチルアルコール、イソプロピルアルコール、グリセリン、グリセロール、プロピレングリコールおよびソルビトール等の他の成分も含まれ得る。殺精子剤は細菌組成物に任意に含まれる。発泡体および発泡体送達ビヒクルのさらに別な例は例えば米国特許第5,595,980号および同第4,922,918号に記載されている。
【0105】
または、細菌を坐薬またはペッサリーとして送達することもできる。例えば米国特許第4,322,399号を参照のこと。ある実施形態では、本発明の細菌は溶解性で選ばれた可溶性ポリマー材料および/または複合炭水化物材料で製造された可溶性要素として送達されるが、それは使用前は実質的に固形物のままであり、使用中にヒトの体温と水分で溶解して所望の徐放および投与量で薬剤物質が放出される。例えば米国特許第5,529,782号参照。細菌はまた、米国特許第4,693,705号に記載の様にスポンジ送達ビヒクル中で送達される。
【0106】
ある実施形態では、細菌は経口投与される。例えば、約108個の乳酸桿菌の毎日の用量を、正常な泌尿生殖器微生物相を回復するために使用し得る。例えばReidら、FEMS Immuno.Med.Microbiol.32:37−41(2001)参照。
【0107】
ある実施形態では、遺伝子組み換え細菌の粘膜表面への投与には定期的な繰り返しが必要である:最適投薬間隔は日常的に決定されるが、粘膜環境と細菌株によって異なる。投薬間隔は1日1回から2〜4週ごとに1回まで変化し得る。
【0108】
天然型乳酸杆菌属を形質転換するためにバクテリオファージが導入される場合、選択したバクテリオファージの核酸を操作して、異種遺伝子がバクテリオファージ被覆タンパク質をコードする遺伝子を置換し、バクテリオファージを複製不能(replication−defective)とすることができる。これらの組み換えDNA分子を機能性バクテリオファージタンパク質を含む細胞溶解物に加えることにより、異種遺伝子を含む機能性バクテリオファージ粒子を組み立てることができる。次いでこれらの複製不能バクテリオファージ粒子を所望の粘膜表面上に導入し、選択した細菌叢に感染させることができる。粘膜表面への典型的な投薬量は108〜1012PFU/mlであると思われる。処置表面に対する溶液の割合は、粘膜表面平方センチあたり約0.1〜1.0mlである必要がある。ビヒクルは細菌に対する上記ビヒクルと同様である。
【実施例】
【0109】
以下の実施例は説明のために提供されるものであり、本発明を制限するものではない。
【0110】
ほとんどのウイルスは鼻、口、消化器または泌尿生殖器等の粘膜を通して感染する。これらの粘膜には、健康な女性の膣腔内のL.jensenii、L.gasseriおよびL.crispatusを含む膨大な数の共生細菌が自然にコロニーを形成している。細菌表面に固定された生物活性ウイルス結合タンパク質を発現する遺伝子組み換えL.jenseniiが粘膜内でウイルスを捕捉し、従ってウイルスが下層の上皮細胞やリンパ球に接近することを妨害するものと、本発明者らは予想している。これらの捕捉されたウイルスは感染を中止し、そして/または乳酸桿菌が分泌する乳酸および過酸化水素等の抗ウイルス化合物で局所的に不活性化され、その結果感染性ウイルス粒子の数をかなり減少する。従って、遺伝子組み換え乳酸杆菌属がHIV結合リガンド、2−ドメインCD4およびシアノビリンNを高濃度で表面に発現するためのモジュール発現アプローチを行った。10〜600アミノ酸のポリペプチドが効果的に細胞壁に固定されて提示されることは、L.jenseniiの天然タンパク質由来のタンパク質ドメインへ融合することにより達成され得ることを本発明者らは見出した。
【0111】
M6タンパク質は標示細胞壁識別シグナルであるLPXTGモチーフと、その後に一続きの疎水性アミノ酸、最後に臨界細胞表面保持シグナルとなる荷電残基(KRKEEN)を有する。2個の良く性質が分かったS.pyrogenesのM6タンパク質(emm6)またはL.paracaseiのPrtPプロテアーゼ由来の細胞壁アンカーモチーフ、またはS.pyrogenesのM6タンパク質由来のアンカーモチーフ、さらにM6タンパク質の天然配列由来のN−末端100アミノ酸伸長部(CWA100)を利用して、L.jemsenii表面にCD4を発現するためのプラスミドベースのモジュールアプローチを本発明者らは最初に試みた。M6タンパク質と異なり、PrtPに対する局在化シグナルはLPKTAである。大量の2D CD4が馴化培地中に放出されたが、馴化培地中のタンパク質、および細胞壁アンカーとしてのM6またはPrtPまたはCWA100を有する修飾L.jensenii中の細胞壁またはプロトプラスト会合タンパク質プールのウエスタン分析により、検出し得る細胞壁関連2D CD4は見出されなかった。フローサイトメトリー分析でも、陽性の表面露出2D CD4は同定されなかった。
【0112】
(推定細胞壁アンカー配列の同定)
L.jenseniiのゲノム配列のデータベース検索により、推定細胞壁アンカーモチーフを有する約30個のコンティグが同定された。National Center for Biotechnologyのウエブサイトで利用し得る非冗長データベース中のより詳細な配列相同性検索に基づき、本発明者らはC14,C191およびC370と名付けられる3個の配列を選択した。それらはそれぞれ、Lactobacillus fermentumのR1p(Turnerら、Appl.Environ.Microbiol.69:5855−5863(2003))、またはストレプトコッカル表面タンパク質のファミリー(Wastfeltら、J.Bio.Chem.271:18892−18897(1996))であるL.reuteri中の粘膜結合タンパク質(RoosおよびJonsson、Microbiol.148:433−442(2002))、およびS.thermophilus中の細胞壁アンカータンパク質(Fernandez−Esplaら、Appl.Environ.Microbiol.66:4772−4778(2000))と低い配列類似性(23〜27%の同一性)を共有していた。3個の配列の全ては疎水性領域に先行するLPQTG局在化シグナルと、荷電C末端テイルを有する(図1参照)。これらの特徴はグラム陽性細菌中のソルターゼ認識C末端細胞壁アンカー配列間に共通である(NavarreおよびSchneewind、Microbiol.Mol.Bio.Rev.63:174−229(1999))。グラム陽性細菌中に見出されるLPXTG細胞アンカーモチーフ間で、これらのL.jenseniiタンパク質中ではわずか7%のLPQTG配列の一致が見出された。C14、C191およびC370タンパク質はすべて細胞壁アンカー領域に隣接したタンデム反復ドメインを含み、既知の細胞壁アンカータンパク質中にしばしば存在する構造上の特徴である(NavarreおよびSchneewind、Microbiol.Mol.Bio.Rev.63:174−229(1999))。C14、C191およびC370の配列を図2A〜Cに示す。
【0113】
(推定細胞壁アンカー配列のエピトープタグ化)
C14、C191およびC370が異種融合タンパク質をL.jenseniiの細胞壁に固定する効率を決定するため、本発明者らは約200個のアミノ酸を直接的にLPQTG局在化シグナルのN−末端に選んだ。しばしば細胞壁アンカータンパク質中の細胞壁関連(CWA)ドメインと定義されるこの領域は、基質配列の保持または延長、従って膜会合ソルターゼによる適切なタンパク質分解性の切断を促進し得る。免疫検出を促進するため、c−Mycエピトープ(EQKLISEEDL)をpOSEL239、240および241それぞれのC14、C191およびC370のCWA200領域のN−末端と融合した。ウエスタンおよびフローサイトメトリー分析を用いて、c−Mycタグ化タンパク質が生成され、細胞壁を標的としたかどうかを検討した。ウエスタン分析を行うため、pOSEL175、239、240および241を有する修飾L.jenseniiをMRSおよびRogosaSLブロスの両方の中で対数期まで生育させた。次いでβ1−4グリコシル結合を成熟ペプチドグリカン中のグリカン鎖のMurNAc−GlcNAc間で切断するN−アセチルムラミダーゼであるムタノリシンで細胞壁を消化した。代表的には、SDS−PAGEクロマトグラフィー後に細胞壁アンカータンパク質は大きなスペクトルの断片として移動する(Perryら、J.Biol.Chem.277、16241−16248(2002))。pOSEL239(C14アンカー)および241(C370アンカー)を有する細菌細胞中の細胞壁富化画分中のタンパク質のウエスタン分析から、細菌細胞をMRSおよびRogosaブロスの両方の中で培養した場合、還元性SDS−PAGE上でc−Mycタンパク質のラダーを示した(図3)。これらのパターンはpOSEL240(C191アンカー)を有する細菌細胞中の細胞壁富化画分中には見られず、試験したLPQTG含有配列間で異なったアンカー効率を示している。
【0114】
ウエスタンブロット陽性c−MycエピトープがpOSEL239および241を有するL.jensenii細胞中で表面に露出しているかどうかを決定するため、コントロールプラスミドpOSEL175を有する細菌細胞を参照して抗c−Myc抗体結合のフローサイトメトリー分析を行った。pOSEL239を有する細菌細胞中の平均蛍光強度はコントロールプラスミドpOSEL175を有する細菌細胞中の平均蛍光強度と区別できなかったが、pOSEL241を有する細菌細胞中では160倍に増加した。C14配列のc−MycタグCWA200領域の表面接近性に立体傷害が影響するかどうかは不明であるが、本発明者らの分析によりC370配列のCWA200領域のN−末端における表面露出が明らかに示された。この結果は、異種ペプチドとタンパク質とを細菌細胞表面に共有結合で固定するためにC370のこの特定の領域を利用し得ることを示している。
【0115】
(L.jenseniiの細菌表面上の2D CD4の表面発現)
2D CD4がC14のCWA200領域およびC370配列を経由して表面発現し得るかどうかを決定するため、本発明者らはウエスタンブロットとフローサイトメトリー分析を行った。ウエスタン分析を行うため、pOSEL175(コントロールプラスミド)、651(細胞アンカーのない2D CD4プラスミド;Changら、Proc.Natl.Acad.Sci.USA.100:11672−11677(2003))、237(C14アンカーに融合した2D CD4)、242(C191アンカーに融合した2D CD4)、および249(C370アンカーに融合した2D CD4)を有するL.jensenii細胞中のタンパク質を細胞壁消化により細胞壁富化画分に分画した。細胞壁富化タンパク質画分では、pOSEL237および249を有する細菌細胞の双方でより大きな分子量の種のスペクトルがpAb T4−4に対し免疫反応性であったが、pOSEL651ではそうでなかった(図4)。ムタノリシン消化後に観測されるSDS−PAGE上のこの様な梯子状パターンは、他のグラム陽性細菌の細菌表面由来の既知の細胞壁アンカータンパク質のパターンと類似している(Perryら、J.Biol.Chem.277:16241−16248(2002))。
【0116】
2D CD4が細胞表面上に発現するかどうかを決定するため、pOSEL175、651、237および249を有するL.jensenii株をpAb T4−4で探索し、次いでフローサイトメトリー分析により抗体結合を分析した。期待された通り、この分析によりpOSEL175および651を有する細菌細胞における平均蛍光強度が区別できないことが明らかとなった。対照的に、pOSEL175および651に対してpOSEL237および249を有する細菌細胞で平均蛍光強度の有意な増加が見られ、2D CD4分子の共有結合および表面露出に結果であると思われる(図5A)。上記アプローチをさらに確認するため、乳酸杆菌属の好ましいコドンを含む再コードしたシアノビリンN(CV−N)遺伝子を2D CD4の成功した固定に用いたものと同じC末端アンカードメインに融合した。CV−N発現プラスミドを有する修飾L.jenseniiのフローサイトメトリー分析により、pOSEL175を有する細菌に対し平均蛍光強度で30〜50倍の増加が検出された(データ示さず)。静電相互作用により抗体反応性CV−N分子が表面会合している可能性を調べるため、修飾細菌を5M LiClで抽出した。フローサイトメトリー分析により、PBSおよび2%FBSで洗浄した細胞を参照して、CV−N発現プラスミドを有する塩抽出L.jensenii中の平均蛍光強度が区別できないことが明らかとなった。表面提示CV−N分子の5M LiCl抽出に対する耐性は、細菌表面上の共有結合で固定されたタンパク質の挙動を反映している。
【0117】
表面に発現した2D CD4分子がgp120を結合するために正しい折り畳み立体構造をとっているか否かを確認するため、pOSEL175、237および249を有する細菌細胞を立体構造依存性エピトープを認識する抗CD4モノクローナル抗体Sim.4で探索した後に、さらにFACS分析を行った。pOSEL175と比較してpOSEL237および249を有する細菌細胞中で平均蛍光強度の有意な増加が見られ、2D CD4が機能的な形でL.jenseniiの表面に発現していることが示された(図5B)。
【0118】
モジュール発現アプローチにおいて2D CD4の表面発現が修飾L.jensenii中の天然細胞表面会合タンパク質の発現に影響するかどうかは不明であった。この問題を解決するため、pOSEL175および237を有する細菌細胞をスルホNHS−ビオチンで探索し、次いで細胞表面会合タンパク質を0.4%SDSおよび10mM DTTを含む緩衝液中で抽出した。アルカリホスファターゼ共役アビジンによるプローブ後のSDS抽出タンパク質のウエスタン分析により、見かけの分子量10〜200kDa未満のビオチン化タンパク質のスペクトルが検出された。pOSEL237を有する細菌細胞中の可溶性ビオチン化タンパク質種のパターンは、pOSEL175中のものと類似しており、天然の細胞表面発現が影響を受けていないことを示している。
【0119】
(L.jensenii中の広いpH範囲における活性2D CD4の表面発現)
乳酸菌が自然にコロニーを形成している場合、ヒト膣腔のpHはほとんどの女性で3.6〜4.5の間で変化し(Boskeyら、Infect.Immun.67:5170−5175(1999))、男性の射出物が存在する場合、一時的に中性または弱アルカリ性になる。修飾L.jenseniiでpH変化がどの様に活性2D CD4分子の表面発現に影響するかを調べるために実験を行った。細菌細胞をRogosa SLブロス中に、その通常用いられるpH(5.4)または100mM HEPESでpH7.4に緩衝して接種した。OD600約0.4までの活性生育中、培地のpHは実質的に変化しなかった。pOSEL237および249を有する細菌細胞へのmAb Sim.4の結合のフローサイトメトリー分析により、pH5.4および7.4の双方でpOSEL175中のコントロールバックグラウンドを超える有意に高い平均蛍光強度が検出された。さらに、修飾L.jenseniiをヒト膣腔内のpHに類似する酸性pHで培養した場合、表面に発現したCV−Nのレベルは上昇したままであった(データを示さず)。
【0120】
(長さ36アミノ酸のC末端アンカーモチーフのみを経由する融合で発現する2D CD4の表面提示の欠失)
LPQTGシグナル、疎水性領域およびC14またはC370配列の荷電テールを含む36アミノ酸C末端アンカーモチーフが、L.jensenii中の2D CD4の効率的な表面発現を支持するに十分であるかどうかは不明である。この疑問を解決するため、C14のC末端アンカーモチーフを有するpOSEL238、およびC370のC末端アンカーモチーフを有するpOSEL262として設計された2個の構築物を調製し、ネガティブコントロールpOSEL175および651、ならびにポジティブコントロールpOSEL237を参照して分析した。pAb T4−4でプローブ後のpOSEL238を有するL.jensenii中の細胞壁富化画分のウエスタン分析で、pOSEL237に類似する梯子状パターンが検出されなかった。さらに、pOSEL238を有する細菌細胞へのmAb Sim.4の結合のフローサイトメトリー分析により、pOSEL175を有する細胞中のバックグラウンドコントロールと比較して平均蛍光強度の増加は全く検出されなかった(図6)。同様に、pOSEL175を有する細胞およびポジティブコントロールpOSEL249と比較して、pOSEL262を有する細菌細胞のFACS分析は同様な負の結果をもたらした。これらの観察と一致して、S.pyogenesおよびL.paracasei由来の同様な長さのC末端アンカーモチーフを用いた場合、2D CD4の表面発現は行われなかった。このことは特徴的なLPQTGモチーフの上流のタンパク質配列が細胞壁固定プロセスに有意に寄与し、L.jenseniiの細胞壁上に生物活性タンパク質を提示するために必要であることを示唆している。
【0121】
(生物活性タンパク質の最適表面提示のための、LPQTGモチーフ上流の反復細胞壁貫通配列の規定の長さの必要性)
天然のC370配列は、LPQTGモチーフの上流のC末端領域中にグラム陽性細菌中の多くの細胞壁アンカータンパク質の特性である8個のほぼ同一の縦列反復を含む(図1)。2.5回反復配列がpOSEL249のアンカー配列中に含まれるが、異なった長さの上流配列が表面タンパク質提示を最大にするために使用し得るかどうかは、決定しなければならないことである。従って、C370配列の0、1、2、4、7および8個の反復を有する数個の構築物を調製した。それらはそれぞれ、pOSEL262、268、278、280、281および276と名付けられた。正しく折り畳まれた立体構造をとる2D CD4分子のレベルを決定するため、形質転換細菌をフローサイトメトリー分析のためにmAb Sim.4で探索した(図7)。細菌を有するpOSEL262(ゼロ反復)では、平均蛍光強度はネガティブコントロールpOSEL175と区別できず、異種タンパク質を適切に表面発現するためには反復配列が必要であることを示唆している。さらに、反復数をpOSEL262の0からpOSEL278の3へ増加すると、蛍光強度が有意に増加した。反復数をさらに増加しても、蛍光強度は一定のままであった。
【0122】
(様々な乳酸杆菌種におけるタンパク質の表面提示を支持するL.jenseniiのネイティブアンカー配列の利用)
L.jensenii 1153に由来するC370のアンカー配列が、他のL.jensenii株またはヒト起源の乳酸杆菌種中でタンパク質の表面提示を提供するか否かを決定するため、c−MycエピトープをC370配列のCWA200に融合するために設計されたpOSEL175またはpOSEL241(図8A)をL.jensenii Xna、L.gasseri 1151、およびL.casei Q中にエレクトロポレーションにより導入した。pOSEL241を有するポジティブコントロールL.jensenii 1153を参照して、形質転換細菌をウエスタン分析およびフローサイトメトリー分析で分析した。c−Mycに対するmAbによる探索後の細胞壁消化物のウエスタン分析により、L.jensenii 1153中のパターンと類似しているがL.casei Q中のパターンとはあまり似ていない、pOSEL241を有する形質転換L.jensenii XnaおよびL.gasseri 1151におけるラダーパターンを検出した(図8B)。細菌細胞をc−Mycに対するmAbで免疫染色した後のフローサイトメトリー分析により、pOSEL175を有する全ての乳酸杆菌種で低レベルの蛍光を検出したが(図8C)、表面提示c−Mycエピトープに結合する抗体への結合の結果、pOSEL241を有するL.jensenii XnaおよびL.gasseri 1151中での蛍光強度の上昇を検出した。さらに、pOSEL175を有するL.caseiと比較して、241を有するL.casei Qの蛍光強度は、さらに約19倍増加していた。これらのデータをまとめると、L.jensenii 1153に由来するアンカー配列は、ヒト起源の乳酸杆菌種も含めた多様な乳酸杆菌種中で、タンパク質の表面提示の支持に広く有用であることを明らかに示す。
【0123】
(L.jensenii中の2D CD4の表面発現に対するLPXTGモチーフの突然変異誘発の効果)
よく研究された細胞壁アンカータンパク質であるStaphylococcus aureusのプロテインAが、LPETG細胞壁分類モチーフ上で突然変異した場合、LPQTG中のアミノ酸プロリン(P)をアミノ酸アスパラギン(N)で置換することによりタンパク質表面提示の効率が減少したが、トレオニン(T)のセリン(S)による置換は、タンパク質表面提示の効率にほとんど影響がないことが見出された(NavarreおよびSchneewind、Microbiol.Mol.Biol.Rev.63:174−229(1999))。この研究は、P残基がLPXTGモチーフ中でおそらく最も重要な残基であり、T残基を類似のアミノ酸であるSで置換し得ることを示した。C14およびC370中のLPQTGモチーフが実際に重要な分類信号であるか否かを決定するため、LPQTG配列中のPおよびTの重要性を検討した。C14配列およびC370配列双方上のLPQTGモチーフ内に点突然変異をPCRにより生じさせた。P残基をアラニン(A)またはアスパラギン(N)に突然変異させ;アミノ酸Tを、A、Sまたはグリシン(G)に突然変異させ;LPXTGモチーフ中のアミノ酸GをAに突然変異させた。変化したLPQTGモチーフを有するプラスミドをそれぞれpOSEL237P(A)、pOSEL237P(N)、pOSEL237T(A)、pOSEL237T(G)、pOSEL237T(S)、pOSEL237G(A)、pOSEL249P(A)、pOSEL249P(N)、pOSEL249T(A)、pOSEL249T(G)、pOSEL249T(S)およびpOSEL249G(A)と名付けた。突然変異した構築物を有するL.jensenii 1153のウエスタン分析およびフローサイトメトリー分析を行った。親pOSEL237およびpOSEL249を有するL.jenseniiと比較して、pOSEL237P(A)、pOSEL237P(N)、pOSEL249P(A)およびpOSEL249P(N)を有するL.jenseniiは、pAb T4−4による細胞壁富化タンパク質画分のウエスタンブロットにおいて、特徴的な、より高分子量種のスペクトルを示さなかった。一方、2D CD4−CWA200融合タンパク質の馴化培地中への分泌は顕著に増加し、2D CD4−CWA200融合タンパク質が細胞壁に共有結合していないことを示した。野生型pOSEL237およびpOSEL249で観察された結果と同様に、pOSEL237T(S)およびpOSEL249T(S)を有するL.jenseniiの細胞壁消化において、より高分子量種の特徴的なスペクトルは明瞭であり、このことは、C14およびC370由来のLPQTG内のアミノ酸Tを、Sによって効果的に置換し得ることを示唆していた(データ示さず)。
【0124】
LPXTGの突然変異誘発のL.jensenii表面タンパク質提示に対する効果をさらに決定するため、pOSEL175、651、237、249を有するL.jensenii株と一緒に様々な突然変異構築物をpAb T4−4またはmAb Sim.4で探索し、次いでフローサイトメトリーで抗体結合を分析した。pOSEL237を有する細菌細胞と比較してpOSEL237P(A)、pOSEL237P(N)を有する細菌細胞、およびpOSEL249を有する細菌細胞と比較してpOSEL249P(A)、pOSEL249P(N)を有する細菌細胞では平均蛍光強度がかなり減少し、このことは、細胞表面の2D CD4タンパク質の提示が、あったとしても、はるかに少ないことを示していた。しかしながら、pOSEL237T(S)、pOSEL237(T)A、pOSEL249T(S)およびpOSEL249(T)Aを有する細菌細胞の平均蛍光強度はpOSEL237および249を有するL.jenseniiに匹敵し、このことは、TをSまたはAに置換することが細胞壁固定の効率にほとんど効果がないことを示した(図9)。
【0125】
ウエスタンブロットおよびフローサイトメトリー分析のデータは、C14およびC370のLPQTGモチーフ内に含まれるアミノ酸Pは容易に置換され得ないことを示す。対照的に、アミノ酸Tは、SまたはAで置換され得、乳酸杆菌属の細胞壁に効率的に固定されるタンパク質を生成する。
【0126】
(L.jenseniiにおける2D CD4の表面発現に対するC末端正荷電テールの欠失の効果)
グラム陽性細胞壁アンカードメインの特徴の一つは、正に荷電したアミノ酸がタンパク質のC末端極に伸びていることである。M6タンパク質では、この配列(KRKEEN)が重要な細胞表面保持信号として働く。これらの標示配列は、ブドウ球菌属(Staphlyococcus)、エンテロコッカス属、リステリア属および乳酸杆菌属を含む他のグラム陽性細菌でも見出されている(NavarreおよびSchneewind、Microbio.Mol.Biol.Rev.63:174−229(1999))。2つの配列RKKRQK1765およびKKKRKDDEA1903がそれぞれ、C14推定アンカー配列およびC370推定アンカー配列中の正に荷電したテールとして同定された(図1)。これらの2個の配列が細胞表面保持信号として働くか否かを決めるため、一連の欠失構築物を作製した(図10)。それらは、それぞれpOSEL237−5、pOSEL237−6、pOSEL237−7、pOSEL249−8、pOSEL249−9およびpOSEL249−10と名付けられた。
【0127】
これらの構築物を有するL.jenseniiのウエスタン分析およびフローサイトメトリー分析を行った。CWA200と融合して2D CD4を表す、48kDaで移動するタンパク質種、SDS−PAGE後、pAb T4−4によって、荷電テールノックアウト構築物を有する全てのL.jenseniiで検出され得る。分泌タンパク質は、pOSEL237−5、pOSEL237−6、pOSEL237−7、pOSEL249−8、pOSEL249−9およびpOSEL249−10を有するL.jensenii細胞において、親のpOSEL237およびpOSEL249を有する細胞よりも多かった。全欠失突然変異体に由来する細胞壁富化画分中のタンパク質のウエスタン分析は、pOSEL237または249を有するL.jenseniiで観察された特徴的なラダーパターンを検出しなかった(データを示さず)。これらのデータは、2D CD4−CWA200融合タンパク質が細胞壁に共有結合していないことを示唆した。
【0128】
抗CD4 pAb T4−4またはmAb Sim.4による探索後の改変L.jenseniiのフローサイトメトリー分析は、親のpOSEL237または249を有する細菌細胞と比較して、これらの突然変異プラスミドを有する細菌細胞における平均蛍光強度の顕著な減少を検出した(図11)。これらのデータは、C14およびC370双方の正に荷電したC末端の欠失が、細胞壁に固着して異種タンパク質を提示するそれらの能力を阻害したことを、結論として示した。
【0129】
(細胞壁アンカーシグナルとしてのLPQTGモチーフの柔軟性)
グラム陽性細菌由来のほとんどの細胞壁に固定されたタンパク質は、同じ識別信号LPXTGを共有するが、一部タンパク質は、異なったモチーフを有する。例えば、L.paracaseiのPrtPに対する識別信号は、LPKTAである(HolckおよびNaes、J.Gen.Microbiol.138:1353−1364(1992))。peptostreptococcus magnusのプロテインLおよびヒト血清アルブミン結合タンパク質は、LPXAGのモチーフを共有する(de ChateauおよびL.Bjorck、J.Biol.Chem.269:12147−12151(1994);Kellerら、EMBO J.11:863−874(1992);Murphyら、DNA Seq.4:259−265(1994))。C14アンカータンパク質またはC370アンカータンパク質中でLPQTGがLPQAGまたはLPQSGに突然変異した場合、SDS−PAGE後にフローサイトメトリーまたはウエスタンブロットで測定した2D CD4の表面提示の減少は僅かであった。しかしながら、以下の証拠に基づくように、これらの配列のみでは、膣由来の乳酸桿菌の細胞壁へタンパク質を固定するには不十分である:1)36アミノ酸のC末端アンカードメイン単独では、c−Mycエピトープまたは2D CD4を細胞表面に固定しなかった、2)200アミノ酸までの上流配列が含まれる場合でさえも、プロトタイプM6細胞壁アンカー配列(S.pyogenesのemm6遺伝子によりコードされる)は膣由来乳酸桿菌の細胞壁へ異種タンパク質を固定しなかった(本発明者らは、L.paracasei由来のLPXTAモチーフを用いて同様な結果を見出した)、および3)C191タンパク質は、有効なアンカーではなかった。これらの知見は、C14およびC370のCWA200領域内に含まれる他の上流配列もまた、細胞壁固定プロセスに大きく寄与することを示している。
【0130】
(C14およびC370のCWA200と融合した場合の2D CD4生物活性の増大)
生物活性を評価するため、pOSEL237−7およびpOSEL249−10を有するL.jensenii 1153から放出されたC14タンパク質およびC370タンパク質の2D CD4−CWA200を、CD4 ELISAにより分析した。pOSEL651、pOSEL237−7およびpOSEL249−10を有する細菌細胞を、Rogosa SLブロス中で種々の細胞密度に生育させた。次いで、無細胞馴化培地を回収した。OD600=0.8で、ウエスタンブロットで測定した培地中に放出されたpOSEL651からの2D CD4の量とpOSEL237−7または249−10からの2D CD4−CWA200の量とは類似していた。それにもかかわらず、pOSEL237−7およびpOSEL249−10を有する細菌細胞から放出された2D CD4−CWA200は、pOSEL651を有する細菌細胞からの2D CD4タンパク質と比較した場合、約2〜3倍高い活性を示した。C14またはC370のCWA200領域の2D CD4への融合は、おそらくはタンパク質折り畳みプロセスを補助することにより、このタンパク質の生物活性を増強する様にみえた。この同じ知見は、gp120結合アッセイを用いて確認された(データ示さず)。これらのタンパク質のウエスタンブロット分析は、2D CD4−CWA200が、2D CD4よりも有意に安定であり、おそらくその生物活性の増強に寄与しているということを示唆する。
【0131】
(材料および方法)
(細菌株および培養)
L.jensenii、L.crispatus、L.gasseriおよびL.caseiのヒト膣株を健康な女性から得た膣サンプルの細菌培養により単離した。乳酸杆菌rRNAに特異的な2つのプライマーを用いて、16S−23S遺伝子間スペーサー領域の増幅後に、GenBankに保存された参照株のDNA配列に対して細菌株の遺伝型決定した(Tannockら、Appl.Environ.Microbiol.65:4264−4267(1999))。株をMRSまたはRogosa SLブロス(Difco、Detroit、MI)中、またはMRS寒天プレート上で、37℃、5%CO2で常法により生育させた。
【0132】
(Lactobacillus jensenii 1153のゲノムDNAの単離)
L.crispatus JCM 5810から染色体DNAを単離するために先に用いたプロトコール(Sillanpaaら、J.Bacteriol.182:6440−6450(2000))の改変に基づいて、L.jensenii 1153の染色体DNAを単離した。L.jensenii細菌を200mlのMRS培地中、37℃、5%CO2で600nmにおける光学密度1.0(OD600=1.0)まで増殖させた。6,600×g、10分間の遠心により細胞を収集し、25mM Tris−HCl(pH8.0)、10mM EDTA、50mMグルコース中で1回洗浄し、細菌培養液100mlあたり2.5mlの20mM Tris(pH8.0)、5mlの24%エチレングリコール8000、および2.5mlのリゾチーム(4mg/ml、Sigma Chemical Co.,St Louis、Mo)を加えた後に懸濁した。得られた細胞懸濁液を37℃で1時間インキュベーションした。0.2M EDTAを5ml添加し、細胞を4℃、1000×gで10分間遠心し、50μlのムタノリシン(15,000U/ml、Sigma Chemical Co.)を含む20mM Tris(pH8.0)10ml中に再懸濁した。37℃で1時間インキュベーショした後、1.5mlの9%Sarkosyl(Sigma Chemical Co.)および3mlの5M NaClを添加して細胞を溶解した。次いで細胞溶解液を2.9mlの5M過塩素酸ナトリウムと混合した。染色体DNAを、17.5mlのクロロホルム−イソアミルアルコール(24:1 v/v)で抽出し、エタノールで沈殿し、風乾し、1.5mg/mlの濃度で100mM Tris−HCl、pH8.0、1mM EDTA中に再懸濁した。最後にゲノムDNA調製物をDNaseを含まないRNaseで処理した。
【0133】
(L.jenseniiゲノムライブラリーの構築)
L.jensenii 1153のゲノムDNAをHydroShear(GeneMachines、San Carlos、CA)を用いて所望のサイズ範囲に機械的に剪断した。剪断DNA断片をT4 DNAポリメラーゼとKlenow酵素で平滑断端化し、3Kbおよび8KbのDNA断片を、アガロースゲル電気泳動後に単離し、QIAquickゲル抽出キット(Qiagen、Valencia、CA)を用いて精製した。得られたDNA断片をpUC18ベクター中に連結し、E.coli DH10B細胞(Invitrogen、Carlsbad、CA)中に形質転換して3Kbおよび8Kbゲノムライブラリーを作製した。細菌形質転換体をX−galの存在下でLBプレート上で選択し、得られたコロニーをQ−pixロボット(Genetix Ltd.、UK)を用いて96ウエルプレート中に配列した。96クローンからなるプレートを挿入サイズの均一性および非組み換え体の割合について試験して、ライブラリーの品質を決定した。どちらのライブラリーも、5%未満の非組み替え体を含み、90%を超える挿入体は、期待サイズの20%以内であった。
【0134】
(L.jenseniiゲノムの配列決定および組み立て)
L.jenseniiゲノム配列を、全ゲノムショットガン法を用いて決定した。ゲノムライブラリーから選択したクローンのプラスミドDNAを、磁気ビーズまたはローリングサークル(rolling circle)法のいずれかで精製し、ABI BigDyeターミネーターキット(Applied Biosystems、Foster City、CA)を用いて両端から配列決定した。全ての配列決定反応を、ABI PRISM 3700自動DNA配列決定装置(Applied Biosystems)上で行った。合計15,360個の配列の読み、または160個の配列プレートを行って、L.jensenii 1153ゲノムの3倍カバレージを得た。50塩基を越えるQ20(100塩基あたり予測誤差1)を生成する場合、またはより高い精度を満たす場合のみ、配列決定の読みが成功したと考えられる。配列クロマトグラフを、Phred(Ewingら、Genome Research、8:175−185(1998))を用いる塩基呼び出しおよび品質評価のために、自動的にUNIX(登録商標)システムに転送した。合格率は80%以上であり、平均読み取り長さは400〜500塩基の範囲である。配列アセンブリをParacel GenomeAssemblerまたはCAP4を用いて行った(Paracel、Inc.、Pasadena、CA)。合計484のコンティグを組み立てた。
【0135】
(L.jensenii 1153ゲノム中の細胞壁アンカーモチーフを有するタンパク質配列の同定)
グラム陽性細菌の細胞壁固定タンパク質は、保存的C末端LPXTGXモチーフを有する(Fischettiら、Mol.Microbiol.4:1603−1605(1990))。このヘキサペプチドの後には、アミノ酸の疎水性伸長と、短い荷電テールとがあり停止転送配列として知られる(Schneewindら、Cell、70:267−281(1992))。さらに、別の独自のLPXTA識別モチーフがLactobacillus paracasei中で同定された(HolckおよびNaes、J.Gen.Microbiol.138:1353−1364(1992))。ネイティブの細胞壁アンカー配列を同定するため、組み立てられたコンティグ(L.jensenii 1153の、推定75%完全なゲノム配列から得られる)の全てのリーディングフレーム中のLPXTGおよびLPXTAに類似のモチーフを同定するためのコンピュータースクリプトが作成された。推定細胞壁アンカーモチーフを有する得られたコンティグを、グラム陽性細菌中の細胞壁アンカータンパク質に対する配列相同性に対するBLAST検索によりさらに検証した。
【0136】
(シャトルベクターの構築)
これらの研究に用いられた1次シャトルベクターは、pLEM7(Fonsら、Plasmid、37:199−203(1997))の改変バージョンであるpOSEL175であった。部分IS要素を、まずSmaIで切断することによって除去し、NdeIで部分的に消化し、Klenow断片で平滑末端化し、再連結した。最後に、pOSEL144のerm遺伝子内の2個のMfeI部位を除去するため、プラスミドを部位指向性突然変異誘発に供した(Changら、Proc.Natl.Acad.Sci.USA、100:11672−11677(2003))。得られたpOSEL175プラスミドはE.coli中の複製開始点(ColE1)と乳酸杆菌属中の複製開始点(repA)の双方を有し、従って様々な乳酸杆菌属種中の異種タンパク質の発現のために用いられるシャトルベクターの骨格を含む。
【0137】
(L.jensenii中の発現カセットの構築)
L.jenseniiにおいてアンカータンパク質を都合よく表在化するために、発現カセットを構築し、pOSEL175のSacI部位およびXbaI部位中にサブクローニングした。このカセットは、乳酸杆菌適合性P23プロモーター、L.crispatusのCbsAシグナル配列、異種タンパク質をコードするDNA、およびグラム陽性細菌中の既知、または推定細胞表面タンパク質由来の共有結合性細胞壁アンカードメインを含む4つの構成要素を含む。本発明者らの、一連のプロモーターおよびシグナル配列を有する構築物の詳細な分析により、Lactococcus lactis由来のP23プロモーター(van der Vossenetら、Appl.Environ.Microbiol.53:2452−2457(1987))と、L.crispatusのCbsA由来のシグナル配列(CbsAss)との組み合わせにより、pOSEL651として設計された構築物中で2D CD4の最高レベルのタンパク質発現が駆動されることが示された(Changら、Proc.Natl.Acad.Sci.USA、100:11672−11677(2003))。SacI、EcoRI、NheI、MfeIおよびXbaIを含む独自の制限部位は、それぞれ5’末端から3’末端へと各構成要素の間に置かれた。PCRによる各構成要素の増幅は、Pfu DNAポリメラーゼを用いて行われた。本研究で詳細に述べられる融合構築物の様々な部分のPCR増幅のためのオリゴヌクレオチドプライマーとしては、以下が挙げられる:
【0138】
【化4】
表面発現のために設計されたタンパク質としては、10アミノ酸c−Mycペプチド(EQKLISEEDL)およびヒトCD4のN末端の2個の細胞外ドメイン(2D CD4)を含む最初の183残基が含まれる。2D CD4タンパク質は、好ましい乳酸杆菌コドンの使用に従うように再コード化された。全ての発現構築物は、L.jensenii中へ形質転換する前にDNA配列分析により確認された。
【0139】
(L.jenseniiの推定細胞壁アンカー配列へのc−Myc融合の構築)
タンパク質発現レベル、および生物活性タンパク質の表面提示のために規定の長さの推定細胞壁アンカー配列を使用することが実行可能であるか否かを決定するため、本発明者らは、最初にエピトープタグ化を選んだ。C末端分類モチーフの機能を乱さないために、5’末端に10アミノ酸c−Mycエピトープ(EQKLISEEDL)を含むオリゴヌクレオチドプライマーを設計し、L.jensenii 1153のゲノム由来のC14、C191およびC370を含む推定細胞壁アンカー配列のN末端にc−Mycエピトープを融合させた。c−Myc配列をこれらのタンパク質(C14、C191またはC370のC末端30アミノ酸)の細胞壁アンカーモチーフに直接融合するか、またはC末端細胞壁アンカードメインを含む配列および様々な長さの隣接上流アミノ酸に融合させた。最も注目すべきは、c−Mycが細胞壁アンカードメインと上流アミノ酸(CWA200と命名)とを含む200アミノ酸配列と融合したことである。
【0140】
【化5】
期待したサイズを有する全てのPCR産物をゲルで精製し、MfeIおよびNheIの両方で消化した。得られた断片を、MfeI/NheI二重消化pOSEL651と連結し、pOSEL239(C14配列のCWA200経由)、pOSEL240(C191配列のCWA200経由)およびpOSEL241(C370配列経由)それぞれの中にc−Myc融合体を作製した。得られたプラスミドはL.jensenii 1153中へエレクトロポレーションした。
【0141】
(細胞壁固定配列のシャトルベクター中へのサブクローニング)
C末端LPQTGアンカーモチーフを含む3個の推定表面タンパク質を選び、L.jensenii 1153の細胞壁上に外来タンパク質を発現する能力を決定した。これらの表面タンパク質のC末端LPQTGドメインを含むDNA領域、およびその上流の200アミノ酸(暫定的にCWA200領域と呼ぶ)を、下記の3組のプライマーによって増幅した:
【0142】
【化6】
C14アンカードメインの内部MfeI部位とC370ドメインの内部XbaI部位を、酵素的制限の前に部位指向性突然変異誘発により突然変異させた。予期されたサイズのPCR産物の全てをゲルで精製し、MfeIおよびXbaIの両方で消化した。得られた断片を、P23−調節性分泌2D CD4を含むMfeI/XbaI二重消化pOSEL651と連結し、プラスミドpOSEL237(C14配列のCWA200経由)、pOSEL242(C191配列のCWA200経由)およびpOSEL249(C370配列のCWA300経由)をそれぞれ作製した。あるいは、以下の2個のプライマーを用いてC14配列のC末端36アミノ酸アンカーモチーフを、同様にシャトルベクター中にクローニングした。
【0143】
【化7】
DNA配列の検証後に得られたプラスミドの全てを、L.jesneii、L.gasseriおよびL.casei中にエレクトロポレーションした。
【0144】
(C370配列の反復細胞壁貫通領域のサブクローニング)
C370LPQTGモチーフの上流の種々の反復細胞壁貫通領域を、L.jensenii 1153のゲノムDNAから増幅した。各PCR反応について同じ3’プライマー(5’−CCGTCTAGATTATGCTTCATCATCTTTTCT−3’)を、以下の5’プライマーと対で使用した。
【0145】
【化8】
これらのプライマー中のMfeI部位およびXbaI部位の両方に下線が引かれている。
【0146】
予想したサイズのPCR産物の全てをゲル精製し、MfeIおよびXbaIの双方で消化した。得られた断片を、P23調節性分泌2D CD4を含むMfeI/XbaI二重消化pOSEL237と連結し、プラスミドpOSEL262(反復無し)、pOSEL268(1反復)、pOSEL278(2反復)、pOSEL284(3反復)、pOSEL280(4反復)、pOSEL275(6反復)、pOSEL281(7反復)およびpOSEL276(8反復)をそれぞれ作製した。
【0147】
(細菌形質転換)
E.coli DH12S(Invitrogen)中に、プラスミドをエレクトロポレーションにより導入した。シャトルプラスミドを構築し維持するため、形質転換E.coli DH12S細胞を、100μg/mlのアンピシリンまたは300μg/mlのエリスロマイシンを補充したLBブロス(Difco)中で、37℃で増殖させた。DNA配列検証後、E.coli由来プラスミドを、改変したLuchanskyら(J.Dairy Sci.74:3293−3302(1991))の方法に従ってL.jensenii、L.gasseriおよびL.casei中に形質転換した。簡単に言えば、新たに播種したL.jenseniiを、MRSブロス中、37℃および5%CO2でOD600 0.6〜0.7まで培養した。細菌細胞を収集し、洗浄して952mMスクロースおよび3.5mM MgCl2中に、4℃で再懸濁した。予冷した0.2cmギャップキュベットを用いて、コンピテント細胞に1〜2μgのDNAを添加し、直ちにGene Pulser II(Bio−Rad、Hercules、CA)を用いて、2.5kV/cmおよび200オームでエレクトロポレーションした。その後、20μg/mlのエリスロマイシン(液体培地中で形質転換L.jenseniiを常法で増殖させるためにも使用される濃度である)を含む選択MRS寒天プレート上に播種する前に、37℃で2時間予め温めたMRSブロス中で細胞を回復させた。
【0148】
(推定細胞壁アンカー配列のLPXTGモチーフの部位指向性突然変異誘発)
Stratagene提供のQuickChange(登録商標)XL Site−Directed Mutagenesisキット(La Jolla、CA)を用いて点突然変異を生じさせた。プラスミドpOSEL237(C14配列のCWA200を経由して固定した2D CD4を発現)とプラスミドpOSEL249(C370配列のCWA200を経由して固定した2D CD4を発現)をテンプレートとして使用した。LPQTG、およびそのC14およびC370上の隣接配列に対応するヌクレオチド配列に基づき、突然変異誘発プライマーを設計した。
【0149】
【化9】
L.jensenii 1153の好ましいコドンを用いて置換ヌクレオチドを選択した。
【0150】
237P(A):C14のLPQTG上のプロリンをアラニンで置換した。
【0151】
【化10】
237P(N):C14のLPQTG上のプロリンをアスパラギンで置換した。
【0152】
【化11】
237T(A):C14のLPQTG上のトレオニンをアラニンで置換した。
【0153】
【化12】
237T(G):C14のLPQTG上のトレオニンをグリシンで置換した。
【0154】
【化13】
237T(S):C14のLPQTG上のトレオニンをセリンで置換した。
【0155】
【化14】
237G(A):C14のLPQTG上のグリシンをアラニン(Alanie)で置換した。
【0156】
【化15】
249P(A):C370のLPQTG上のプロリンをアラニン(Alaine)で置換した。
【0157】
【化16】
249P(N):C370のLPQTG上のプロリンをアスパラギンで置換した。
【0158】
【化17】
249T(A):C370のLPQTG上のトレオニンをアラニンで置換した。
【0159】
【化18】
249T(G):C370のLPQTG上のトレオニンをグリシンで置換した。
【0160】
【化19】
249T(S):C370のLPQTG上のトレオニンをセリンで置換した。
【0161】
【化20】
249G(A):C370のLPQTG上のグリシンをアラニンで置換した。
【0162】
【化21】
ポリメラーゼ連鎖反応(PCR)サイクル条件は95℃で50秒、60℃で50秒、および68℃で12分、全体で16サイクルであった。
【0163】
PCR反応後にDpnI酵素を、増幅混合物に添加し、親プラスミドを分解した。新たに合成されたプラスミドを製造業者の推奨に従って化学コンピテントE.coli Top10細胞(Invitrogen)中に導入した。200μg/mlのエリスロマイシンを補充したLBブロス(Difco)中でプラスミドを維持し、増幅した。DNA配列を検証後、改変したLuchanskyらの方法(J.Dairy Sci.74:3293−3302(1991))に従ってE.coli由来のプラスミドをL.jensenii中へ形質転換した。突然変異原性プラスミドを含む形質転換L.jenseniiを選択し、増殖するために、20μg/mlのエリスロマイシンを含むMRSを使用した。
【0164】
(推定細胞壁アンカータンパク質の正に荷電したC末端配列の欠失の分析)
正に荷電したアミノ酸がC14およびC370のC末端に位置する一連の欠失突然変異体を、PCR増幅により作製した。プラスミドpOSEL237およびpOSEL249を、テンプレートとして使用した。pOSEL237およびpOSEL249上の2D CD4配列に相補性のオリゴヌクレオチド(CD4F 5’−GATCGTGCTGATTCACGTCGT−3’)をフォワードプライマーとして使用した。以下のオリゴヌクレオチド(制限部位に下線)を、2D CD4 cDNAのC末端および完全C14およびC370 CWA200配列を増幅するためのリバースプライマーとして使用した:
【0165】
【化22】
全てのリバースプライマーは、XbaI制限部位を含んでいた。サイクル条件は、94℃で45秒、60℃で45秒、72℃で90秒、合計18サイクルであった。PCR産物をゲル精製し、MfeIおよびXbaIの両方で消化し、次いでMfeI/XbaI二重消化pOSEL237およびpOSEL249中へ、それぞれサブクローニングした。配列をヌクレオチド配列決定によって検証し、タンパク質分析のために、構築物をL.jensenii中へエレクトロポレーションした。
【0166】
(L.jensenii中の異種タンパク質発現のウエスタン分析)
遺伝的に改変したL.jensenii細胞を、100mM HEPES(pH7.1)で緩衝したRogosa SLブロス中、37℃、5%CO2で増殖させた。可溶性タンパク質のレベルを決定するため、12,000×gで遠心後に馴化培地を集め、最終濃度20%のTCAでタンパク質を沈殿させた。TCA沈殿物をエタノールで洗浄し、風乾し、50mM Tris−HCl(pH6.8)、0.4%SDS、6%スクロース、10mMジチオトレイトールおよび0.01%ブロモフェノールブルー(1×還元SDS−PAGE緩衝液)中で熱変性させた。L.jensenii中の細胞結合タンパク質の相対量を決めるため、細胞溶解を誘起することなく、OD600単位あたり100μLの1×SDS−PAGE緩衝液中、37℃、30分間で細菌細胞を抽出した。12,000×gで5分間遠心して収集した抽出タンパク質を、次いで熱変性した。14,000×gで遠心して可溶性タンパク質を細菌細胞から分離し、製造業者の推奨に従い、抗酸化剤の存在下で、4〜12%のNuPAGEシステム(Invitrogen)中でSDS−PAGEで分離した。電気泳動による分離後、20%メタノール、20mM Trisおよび50mMグリシン中の2フッ化ポリビニリデン膜(Millipore)上にタンパク質をエレクトロブロッティングした。次いで、このブロットをポリクローナルウサギ抗CD4抗体、T4−4(NIH AIDS Reserch and Reference Reagent Program)またはウサギ抗CV−N pAb、およびc−Mycに対するモノクローナル抗体(Invitrogen)で探索した。抗原−抗体反応を、アルカリホスファーゼ結合抗ウサギIgG用(CD4検出用)に対する色素生成検出試薬(Promega、Madison、WI)、またはセイヨウワサビペルオキシダーゼ(HRP)共役抗マウスIgG用(c−Myc検出用)の増強化学発光試薬(Amercham Biosciences、Piscataway、NJ)を用いて可視化した。同様に、c−Mycタグ化タンパク質のレベルを抗c−Mycに対するmAb(Invitrogen)で探索し、結合抗体を、HRP結合抗マウス2次抗体(Amersham Biosciences)で検出した。
【0167】
(ムラミダーゼによるL.jensenii細胞壁の酵素消化)
109個の細菌を含む細菌培養液を、12,000×gで5分間遠心した。得られた細胞ペレットを、20mM HEPES(pH7.2)で1回洗浄し、100μLの10mM Tris−HCl(pH8.0)、1mM EDTA、25%スクロース中に懸濁した(Piardら、J.Bacteriol.179:3068−3072(1997))。細菌細胞壁を、最終濃度15単位/mlのムラミダーゼ、ムタノリシン(Sigma Chemical Co.)の存在下で、37℃で1時間消化した。その後、細胞を、2,500×gで10分間遠心し、プロトプラスト富化画分から細胞壁富化画分を単離した。細胞壁またはプロトプラスト富化画分それぞれに、25μlの4×還元SDS−PAGE緩衝液または125μlの1×還元SDS−PAGE緩衝液を添加後、得られたサンプルを熱変性した。あるいは、さらなるサンプル処理なしに細胞壁富化画分中のタンパク質を分析するため、CD4 ELISAを用いた。
【0168】
(L.jensenii中の表面露出タンパク質のスルホ−NHS−ビオチンによる標識)
L.jensenii中の表面タンパク質の表面露出リシル残基を、膜不透過スルホ−N−ヒドロキシスクシンイミド(NHS)−ビオチンの使用によって探索した。NHS−ビオチンによるグラム陰性細菌Helicobacter pyloriの表面標識により、真の細胞表面タンパク質の同定が可能になる(Sabarthら、J.Biol.Chem.70:27896−27902(2002))。対数期の約109個のL.jensenii細菌を1回洗浄し、PBS中に懸濁した。スルホ−NHS−ビオチンを1mlの細胞懸濁液に最終濃度1mMで加え、連続的に回転させながら室温で30分間インキュベーションした。その後、ビオチン化反応を、50mM Tris、pH8.0を加えてクエンチし、細胞を、20mM HEPES、pH7.2で1回洗浄した。細胞溶解を誘起することなく、細胞に結合したタンパク質を、125μlの0.4%SDS、6%スクロース、10mM DTT、50mM Tris−HCl(pH6.8)中で、37℃で30分間抽出した。抽出されたタンパク質を、14,000×g、5分間の遠心により細菌細胞から分離した。熱変性後、タンパク質を、4〜12%NuPAGE(Invitrogen)中で分離した。ビオチン化タンパク質およびその移動度シフトを決定し、その後アルカリホスファターゼ結合ストレプトアビジン(strepavidin)または他の免疫プローブで探索した。
【0169】
(フローサイトメトリーによる2D CD4の表面発現の分析)
pOSEL651中に表面タンパク質発現またはタンパク質分泌のためのプラスミドを有する形質転換L.jenseniiを、20μg/mlのエリスロマイシンの存在下で、37℃、5%CO2で、MRSブロス中で一晩増殖させた(OD600>3)。特に指定しない限り、100mM HEPES、pH7.1で緩衝したエリスロマイシン含有MRSまたはRogosa SLブロス中、1:50〜100希釈で一晩培養物を培養し、次いで、継代培養した。OD600〜=0.4の1mlの細胞培養液を、12,000×gで5分間遠心した。得られた細胞ペレットを2回洗浄し、2%FBSを含む1×PBS中に懸濁した。その後、特異性抗体(2×108個の細胞あたりウサギポリクローナルT4.4に対し1:1000希釈、またはモノクローナルSim.4に対し50μg/ml)を用いて、細胞を、1×PBS中の2%FBS中で30分間表面染色し、その後FITCまたはフィコエリスリン結合抗ウサギ抗体またはフィコエリスリン結合抗マウス抗体(Becton−Dickinson、Mountain View、CA)で表面染色した。表面発現CV−Nの検出のため、同様なプロトコールを開発した。コントロールはアイソタイプが適合するモノクローナル抗体(Becton Dickinson)から構成した。標識細胞を、1%(v/v)パラホルムアルデヒドで固定し、CellQuestソフトウエアと共に作動するFACScaliburシステム(Becton−Dickinson)で解析した。バックグラウンドコントロール中の密度プロット出力(側方散乱または前方散乱 対 蛍光)を、pOSEL175を有するL.jenseniiから得た。プロット間の平均蛍光強度の変化を、細菌表面に結合する抗体の基準とし、FLOWJOソフトウエアを用いて計算した。
【0170】
(酵素結合免疫吸着アッセイ)
正しく折り畳まれた2D CD4タンパク質の濃度を、McCallusら(Viral Immunol.5:209−219(1992))に従って改変したCD4捕捉酵素結合免疫吸着分析(ELISA)で決定した。細菌馴化培地中の正しい立体配座を有する2D CD4タンパク質を、2.5μg/mlのモノクローナル抗体Sim.4により、MaxiSorp96ウエルプレート(Nalge Nunc International、Denmark)上に捕捉した。0.05%Tween 20を含む1×Tris緩衝生理食塩水中での洗浄後、E.coli由来のリフォールディングされた2D CD4標準を参照して、結合CD4分子を、ウサギポリクローナル抗体で探索し、次いで、T4−4を、3,3’,5,5’−テトラメチルベンジジン(Neogen Corp.,Lexington、KY)の存在下で、室温で暗所に30分間置いて、セイヨウワサビペルオキシダーゼ結合抗ウサギIgG(Amersham Biosciences)で検出した。100μlの0.5M H2SO4を添加した後反応を停止させ、マイクロプレートリーダー(Molecular Devices、Sunyvale、CA)を用いて450nmでの吸光度を読み取った。
【0171】
上記実施例は、本発明を説明するために提供されるが、その範囲を制限するものではない。本発明の他の変法は当業者に容易に明らかであり、かつ添付の特許請求の範囲によって包含される。本明細書において引用する全ての刊行物、データベース、GenBank配列、特許および特許出願は、本明細書によって参考として援用される。
【0172】
(配列表)
【0173】
【化23】
【0174】
【化24】
【0175】
【化25】
【0176】
【化26】
【図面の簡単な説明】
【0177】
【図1】図1はL.jensenii 1153のゲノム配列決定後に同定された3種の細胞壁アンカータンパク質の構造を示す。3つのタンパク質の全ては疎水性領域および荷電C−末端テールに先行するLPQTG識別シグナルを有し、独自の長い繰り返し配列を有する。CWAはLPQTGモチーフの上流の推定細胞壁関連領域を示す。
【図2A】図2AはL.jensenii 1153のゲノム配列決定から得られた細胞壁アンカー配列(C14、C191およびC370)を示す。CWA200領域と共にアンカーモチーフに下線が引かれている。CWA200はLPQTGモチーフの上流の約200アミノ酸の推定細胞壁関連領域または推定細胞壁貫通領域を表す。
【図2B】図2BはL.jensenii 1153のゲノム配列決定から得られた細胞壁アンカー配列(C14、C191およびC370)を示す。CWA200領域と共にアンカーモチーフに下線が引かれている。CWA200はLPQTGモチーフの上流の約200アミノ酸の推定細胞壁関連領域または推定細胞壁貫通領域を表す。
【図2C】図2CはL.jensenii 1153のゲノム配列決定から得られた細胞壁アンカー配列(C14、C191およびC370)を示す。CWA200領域と共にアンカーモチーフに下線が引かれている。CWA200はLPQTGモチーフの上流の約200アミノ酸の推定細胞壁関連領域または推定細胞壁貫通領域を表す。
【図3】図3は、MRSブロス(A)またはRogosa SLブロス(B)中、37℃および5%CO2で培養した場合の、形質転換L.jensenii 1153のムタノリシン消化後の、SDS抽出可能タンパク質および細胞壁富化画分のウエスタン分析の結果を示す。還元性SDS−PAGEでの分離後、これらのタンパク質をc−Mycに対するモノクローナル抗体(mAb)を用いて探索するために、PVDF膜にエレクトロブロッティングした。
【図4】図4は、Rogosa SLブロス中で37℃、5%CO2で培養した場合の、形質転換L.jensenii 1153のムタノリシン消化後の、細胞壁富化画分のウエスタン分析の結果を示す。還元性SDS−PAGE中で分離後、CD4(T4−4)に対するポリクローナル抗体(pAb)で探索するために、タンパク質をPVDF膜にエレクトロブロッティングした。発現構築物は以下の要素を含んでいた:pOSEL651中のP23プロモーター−CbsAシグナル配列(CbsAss)−2D CD4;p237中のC14配列のP23プロモーター−CbsAss−2D CD4−CWA200−アンカー;pOSEL242中のC191配列のP23プロモーター−CbsAss−2D CD4−CWA200−アンカー;pOSEL249中のC370配列のP23プロモーター−CbsAss−2D CD4−CWA200−アンカー。CWA200は、C−末端アンカードメインの上流の約200アミノ酸を表す。
【図5】図5は、2D CD4の分泌または表面固定のために設計したL.jensenii 1153を有するプラスミドのフローサイトメトリー分析の結果を示す。細菌細胞を、CD4に対する(T4−4)ウサギpAbで探索し、次いでFITC結合抗ウサギ抗体(A)で探索した。あるいは細菌細胞を、mAb Sim.4で探索し、次いでPE結合抗マウスIgG(B)で探索した。コントロールは、未染色細胞またはフルオロクローム結合2次抗体で探索した細胞である。細菌表面へ結合する抗体の基準としての蛍光濃度を、FLOWJOソフトウエアを用いて計算した。
【図6】図6は、長さが36アミノ酸であるC−末端アンカーモチーフが、2D CD4の表面発現を駆動するのは不十分であることを示す。(A)L.jensenii中のネイティブアンカー配列を用いる2D CD4の表面発現用に設計された構築物。(B)pOSEL238またはpOSEL237を有するL.jensenii 1153のフローサイトメトリー分析。細菌細胞をCD4に対するmAb Sim.4で探索し、次いでフィコエリスリン(PE)結合抗マウス抗体で探索した。コントロールは無染色細胞またはPE結合2次抗体で探索した細胞である。
【図7】図7は、C370配列中のLPQTG識別シグナルの上流の種々の数の繰り返し細胞壁貫通配列により影響を受ける場合のL.jensenii 1153中における2D CD4の表面発現を示す。正しく折り畳まれた立体構造をとる表面露出2D CD4分子を、以下のプラスミドを有する細菌細胞中のフローサイトメトリー分析のために、mAb Sim.4で探索した:175、陰性コントロール;249、2.5回反復;262、反復なし;268、1回反復;278、2回反復;280、4回反復;281、7回反復;276、8回反復。
【図8】図8は、ヒト起源の様々な乳酸桿菌種におけるc−Mycタグ化タンパク質の表面提示を示す。(A)P23プロモーターおよびCbsAシグナル配列(CbsAss)の制御下におけるC370配列のc−Mycタグ化CWA200の発現のために設計されたpOSEL241の概念図。(B)形質転換されたL.jensenii、L.gasseriおよびL.caseiのムタノリシン消化後の細胞壁富化画分のウエスタン分析。還元性SDS−PAGE中で分離後、c−Mycに対するmAbで探索するためにタンパク質をPVDF膜にエレクトロブロッティングした。(C)pOSEL241を有するヒト膣の乳酸桿菌単離物のフローサイトメトリー分析。細菌細胞をc−Mycに対するmAbで探索し、次いでフィコエリスリン(PE)結合抗マウス抗体で探索した。コントロールは、非染色細胞またはPE結合2次抗体で探索した細胞から構成された。
【図9】図9は、L.jensenii 1153中の2D−CD4−CWA200の表面提示上のC14配列およびC370配列のLPQTGモチーフにおける点突然変異の効果を示す。細菌細胞を、あらかじめ力価を測定したmAb Sim.4(A)またはpAb T4−4(B)を用いて表面染色し、次いでPE結合抗マウス抗体またはFITC結合抗ウサギ抗体で探索した。フローサイトメトリー分析をFACScaliburシステム中で行った。pOSEL237およびpOSEL249の細胞表面上に提示されたタンパク質と、突然変異誘発性構築物を有する細菌細胞中のタンパク質との間の差を、平均蛍光強度で表した。pOSEL237またはpOSEL249を有する細菌細胞中の2D CD4の表面提示を、任意に100%と設定した。
【図10】図10は、C14配列およびC370配列のC−末端荷電テール中の欠失構築物の概念図を示す。
【図11】図11は、2D CD4−CWA200の表面提示上のC14およびC370のC−末端荷電テール中の配列欠失の影響を示す。細菌細胞をあらかじめ力価を測定したpAb T4−4(A)またはmAb Sim.4(B)を用いて表面染色し、次いでFITC結合抗ウサギまたはPE結合抗マウス抗体で探索した。細胞壁アンカータンパク質への抗体の結合をFACScaliburシステムを用いるフローサイトメトリーで分析した。pOSEL237またはpOSEL249の細胞表面上に提示されるタンパク質と、突然変異誘発構築物を有する細菌細胞中のタンパク質との間の差を、平均蛍光強度として表した。pOSEL237およびpOSEL249を有する細菌細胞中の2D CD4の表面提示を、任意に100%と設定した。
【図12】図12は、pOSEL651を有するL.jensenii 1153由来の2D CD4に対する、pOSEL237−7およびpOSEL249−10を有するL.jensenii 1153中の分泌2D CD4−CWA200の活性の比較を示す。無細胞馴化培地中で正しく、適切に折り畳まれた立体構造をとるタンパク質を認識するために、CD4 ELISAを設計した。タンパク質量を、そのpAb T4−4に対する免疫応答性に基づいて正規化した。pOSEL651を有する細菌細胞から放出される可溶性2D CD4タンパク質を、任意に100%に設定した。
【技術分野】
【0001】
(関連出願の相互参照)
本願は2003年1月29日出願の米国特許仮出願番号60/443,619に対する優先権の利益を請求する。米国特許仮出願番号60/443,619は、全ての目的でその全体を参考として援用する。
【0002】
(連邦政府後援研究開発下で行われた発明に対する権利に関する記載)
本発明はNational Institutes of Healthにより認可された補助金番号2 R44 AI46203−02の下で政府支援により行われた。政府は、本発明に一部権利を有する。
【背景技術】
【0003】
(発明の背景)
グラム陽性菌におけるペプチドグリカンとの共有結合を介するタンパク質の表面発現は、独自の識別信号とソルターゼ依存性機構とに関与する(Mazmanianら、Science 285:760−763(1999))。最も良く研究されたシステムの一つは、M6構造タンパク質をコードするStreptococcus pyogenesのemm6遺伝子である(Fischettiら、1990、Mol.Microbiol.4:1603−1605(1990))。M6タンパク質は、標示(signature)細胞壁識別信号、Leu−Pro−X−Thr−Gly(LPXTG)モチーフ、それに続く疎水性アミノ酸伸長、および最後に荷電残基を含む配列(KRKEEN)を有し、これは細胞表面保持信号として働く。これらの細胞壁識別モチーフは、Staphlyococcus、EnterococcusおよびListeria、ならびに乳酸杆菌属を含む他のグラム陽性菌において同定されている(NavarreおよびSchneewind、Microbio.Mol.Biol.Rev.63:174−229(1999))が、ヒトの膣でコロニーを形成する乳酸杆菌属種においては同定されていない。
【0004】
全てのヒトの粘膜には自然に細菌がコロニーを形成している(Tannock、Clin.Rev.Allergy Immunol.22:231−53(2002))。最近の科学的証明は、これらの細菌が体の細胞および組織と緊密に相互作用し、天然の生物学的プロセスを調節しているという事実を示した。この粘膜微生物叢がまた、ヒトの細胞および組織に影響する多くの疾病に実質的に寄与することが次第に明らかとなっている。
【0005】
一般に、膣および消化管内で乳酸桿菌および関連する細菌による微生物叢が優位になることは、良い健康状態と関連している(Redondo−Lopezら、Rev.Infect.Dis.12:856−72(1990);Tannock、Clin.Res.Allergy Immunol.22:231−53(2002))。乳酸桿菌の天然株は、これらの場所内で健康な微生物叢を保ち、感染を防止するために、「プロバイオティクス」として長年投与されてきた。これらの「健康な細菌」が細菌、ウイルスおよび真菌等の病原性微生物と競合し、病気に関連する病因の発生と進行を制限するということが十分に確立されている。それにもかかわらず、この微生物叢は脆弱であり、健康な微生物叢の自然な代謝回転、および破壊を伴う動的環境が、日和見感染の発症と関連付けられている。結果として、疾患を予防するかまたは処置する手段として、微生物叢の完全性および天然の性質を維持、またはさらには増強するためのアプローチは、生物医学界が切望するところである。
【0006】
粘膜微生物叢は粘膜表面に影響する多くの局所疾患に寄与する。例えば、HIVおよび他の性感染病原体は膣粘膜を迂回しなければならない。さらに、潰瘍性大腸炎およびクローン病を含む炎症性腸疾患の病因は、破壊された粘膜微生物叢と宿主の細胞および組織との間の不適切な相互作用から生じ得る。粘膜叢内の細菌の性質を調節する手段は、これらの疾患ばかりでなく、粘膜表面に影響する関連する病状の予防または処置の助けになり得る。生物活性タンパク質がこれらおよび他の微生物の細胞壁を標的とすることは、この様な疾患の処置に役立ち得る。
【0007】
本発明はこれらおよび他の問題に取り組む。
【発明の開示】
【課題を解決するための手段】
【0008】
(発明の簡単な要旨)
本発明は、発現カセットを含む乳酸杆菌属細菌を提供する。この発現カセットは、シグナル配列と生物活性ポリペプチドとをコードするポリヌクレオチドと作動可能に結合するプロモーターを含み、この生物活性ポリペプチドは、異種カルボキシル末端細胞壁標的領域と結合し、この異種カルボキシル末端細胞壁標的領域は、以下の順番で以下:細胞壁関連配列;LPQ(S/A/T)(G/A);および疎水性配列を含む。
【0009】
いくつかの実施形態では、上記細胞壁関連配列は、少なくとも50個のアミノ酸を含む。いくつかの実施形態では、この細胞壁関連配列は、少なくとも200個のアミノ酸を含む。いくつかの実施形態では、上記異種カルボキシル末端細胞壁標的領域は、その領域のカルボキシル末端に荷電した配列をさらに含む。
【0010】
いくつかの実施形態では、乳酸杆菌属細菌は、膣コロニー形成株である。いくつかの実施形態では、この細菌は、L.jensenii、L.gasseri、L.caseiおよびL.crispatusからなる群より選択される。
【0011】
いくつかの実施形態では、上記細胞壁標的領域は、アミノ酸配列LPQSGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQAGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQTGを有する。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQTAを有する。いくつかの実施形態では、この細胞壁標的領域は、配列番号7を含む。いくつかの実施形態では、この細胞壁標的領域は、配列番号8を含む。
【0012】
いくつかの実施形態では、上記生物活性ポリペプチドは、細菌の細胞壁で発現される。いくつかの実施形態では、この生物活性ポリペプチドは、10アミノ酸〜600アミノ酸の間である。いくつかの実施形態では、上記生物活性ポリペプチドが病原体と接触した場合、この生物活性ポリペプチドが病原体と結合する。
【0013】
いくつかの実施形態では、上記病原体は、細菌病原体である。いくつかの実施形態では、この病原体は、真菌病原体である。いくつかの実施形態では、この病原体は、ウイルス病原体である。
【0014】
いくつかの実施形態では、上記ウイルス病原体は、ヒト免疫不全ウイルス(HIV)である。いくつかの実施形態では、上記生物活性タンパク質は、CD4またはCD4のHIV結合フラグメントである。いくつかの実施形態では、上記生物活性タンパク質は、2D−CD4である。いくつかの実施形態では、上記生物活性タンパク質は、シアノビリン−N(CV−N)またはCV−Nのウイルス結合フラグメントである。いくつかの実施形態では、上記ウイルス病原体は、単純疱疹ウイルスである。いくつかの実施形態では、上記生物活性タンパク質は、単純疱疹ウイルス侵入媒体C(HveC)またはHveCのウイルス結合フラグメントである。
【0015】
いくつかの実施形態では、上記生物活性ポリペプチドは、乳酸杆菌属細菌から放出される。いくつかの実施形態では、この生物活性ポリペプチドは乳酸杆菌属細菌の細胞壁に固定される。
【0016】
本発明はまた、乳酸杆菌属細菌の細胞壁中で生物活性ポリペプチドを発現する方法を提供する。いくつかの実施形態では、その方法は以下を包含する:発現カセットを含む乳酸杆菌属細菌を提供する工程であって、この発現カセットはシグナル配列および生物活性ポリペプチドをコードするポリヌクレオチドと作動可能に結合するプロモーターを含み、この生物活性ポリペプチドは、異種カルボキシル末端細胞壁標的領域と結合し、この異種カルボキシル末端細胞壁標的領域は、以下の順番で、以下:細胞壁関連配列;LPQ(S/A/T)(G/A);および疎水性配列を含む工程;ならびにこのポリペプチドの発現を誘導する条件下で細菌を培養する工程であって、該工程により、乳酸杆菌属細菌の細胞壁中で生物活性ポリペプチドを発現する工程、を有する。
【0017】
いくつかの実施形態では、上記細胞壁関連配列は、少なくとも50個のアミノ酸を含む。いくつかの実施形態では、この細胞壁関連配列は、少なくとも200個のアミノ酸を含む。
いくつかの実施形態では、上記異種カルボキシル末端細胞壁標的領域は、この領域のカルボキシル末端に荷電した配列をさらに含む。いくつかの実施形態では、上記提供する工程は、上記発現カセットを上記細菌中に輸送する工程を包含する。
【0018】
いくつかの実施形態では、上記細胞壁標的領域は、アミノ酸配列LPQSGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQAGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQTGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQTAを有する。いくつかの実施形態では、細胞壁標的領域は配列番号7を含む。いくつかの実施形態では、この細胞壁標的領域は配列番号8を含む。
【0019】
いくつかの実施形態では、この細胞壁標的領域は、少なくとも200個のアミノ酸を有する。
【0020】
いくつかの実施形態では、上記細菌は、膣コロニー形成株である。いくつかの実施形態では、この細菌はL.jensenii、L.gasseri、L.caseiおよびL.crispatusからなる群より選択される。いくつかの実施形態では、上記生物活性ポリペプチドは10〜600アミノ酸の間である。いくつかの実施形態では、上記生物活性ポリペプチドが病原体と接触した場合、この生物活性ポリペプチドが病原体に結合する。
【0021】
いくつかの実施形態では、上記病原体は、細菌病原体である。いくつかの実施形態では、この病原体は、真菌病原体である。いくつかの実施形態では、この病原体は、ウイルス病原体である。
【0022】
いくつかの実施形態では、上記ウイルス病原体は、HIVである。いくつかの実施形態では、上記生物活性タンパク質は、CD4またはCD4のHIV結合フラグメントである。いくつかの実施形態では、この生物活性タンパク質は、2D−CD4である。いくつかの実施形態では、上記生物活性タンパク質は、シアノビリン−Nまたはシアノビリン−Nのウイルス結合フラグメントである。いくつかの実施形態では、上記ウイルス病原体は、単純疱疹ウイルスである。いくつかの実施形態では、上記生物活性タンパク質は、単純疱疹ウイルス侵入媒体C(HveC)またはHveCのウイルス結合フラグメントである。
【0023】
いくつかの実施形態では、上記生物活性ポリペプチドは、乳酸杆菌属細菌から放出される。いくつかの実施形態では、この生物活性ポリペプチドは、乳酸杆菌属細菌の細胞壁に固定される。
【0024】
本発明はまた、生物活性タンパク質を哺乳動物粘膜表面に提供する方法を提供する。いくつかの実施形態では、その方法は、粘膜表面と、異種カルボキシル末端細胞壁標的領域に結合された生物活性ポリペプチドに結合されたシグナル配列を発現するように組み換えで変化した乳酸杆菌属細菌とを、接触させる工程を包含し、この異種カルボキシル末端細胞壁標的領域は、以下の順番で以下:細胞壁関連配列;LPQ(S/A/T)(G/A);および疎水性配列を含み、ここで、この生物活性ポリペプチドが、粘膜表面から収集されたサンプル中で、検出可能な量で発現される。
【0025】
いくつかの実施形態では、上記細胞壁関連配列は、少なくとも50個のアミノ酸を含む。いくつかの実施形態では、この細胞壁関連配列は、少なくとも200個のアミノ酸を含む。いくつかの実施形態では、上記異種カルボキシル末端細胞壁標的領域は、カルボキシル末端領域に荷電した配列をさらに含む。いくつかの実施形態では、上記乳酸杆菌属細菌は、L.jensenii、L.gasseri、L.caseiおよびL.crispatusからなる群より選択される。
【0026】
いくつかの実施形態では、上記粘膜表面は、膣内に存在する。いくつかの実施形態では、この粘膜表面は、消化管内に存在する。
【0027】
いくつかの実施形態では、上記接触させる工程は乳酸杆菌属細菌を経口投与する工程を包含する。いくつかの実施形態では、この接触させる工程は、乳酸杆菌属細菌を膣投与する工程を包含する。いくつかの実施形態では、この接触させる工程は乳酸杆菌属細菌を直腸投与する工程を包含する。
【0028】
本発明は、シグナル配列と生物活性ポリペプチドとをコードするポリヌクレオチドに作動可能に結合されたプロモーターを含む発現カセットを提供し、ここで、この生物活性ポリペプチドは、異種カルボキシル末端細胞壁標的領域と結合し、この異種カルボキシル末端細胞壁標的領域が以下の順番で以下:細胞壁関連配列;LPQ(S/A/T)(G/A);および疎水性配列を含む。いくつかの実施形態では、上記細胞壁関連配列は、少なくとも50個のアミノ酸を有する。いくつかの実施形態では、この細胞壁関連配列は、少なくとも200個のアミノ酸を有する。
【0029】
いくつかの実施形態では、上記異種カルボキシル末端細胞壁標的領域はさらに領域のカルボキシル末端に荷電配列を有する。
【0030】
いくつかの実施形態では、細胞壁標的領域はアミノ酸配列LPQSGを含む。いくつかの実施形態では、細胞壁標的領域は、アミノ酸配列LPQAGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQTGを含む。いくつかの実施形態では、この細胞壁標的領域は、アミノ酸配列LPQTAを含む。いくつかの実施形態では、この細胞壁標的領域は、配列番号7を含む。いくつかの実施形態では、この細胞壁標的領域は配列番号8を含む。いくつかの実施形態では、上記生物活性ポリペプチドは10〜600アミノ酸の間である。
【0031】
いくつかの実施形態では、上記生物活性タンパク質が病原体と接触した場合、この生物活性タンパク質は病原体と結合する。いくつかの実施形態では、上記病原体は、細菌病原体である。いくつかの実施形態では、この病原体は、真菌病原体である。いくつかの実施形態では、病原体はウイルス病原体である。いくつかの実施形態では、このウイルス病原体は、HIVである。
【0032】
いくつかの実施形態では、上記生物活性タンパク質は、CD4またはCD4のHIV結合フラグメントである。いくつかの実施形態では、上記生物活性タンパク質は、2D−CD4である。いくつかの実施形態では、この生物活性タンパク質は、シアノビリン−Nまたはシアノビリン−Nのウイルス結合フラグメントである。いくつかの実施形態では、この生物活性タンパク質は、単純疱疹ウイルス侵入媒体C(HevC)またはHveCのウイルス結合フラグメントである。いくつかの実施形態では、上記細胞壁標的領域は、乳酸杆菌属中で機能する。
【0033】
本発明はまた、異種カルボキシル末端細胞壁標的領域に結合した生物活性ポリペプチドをコードするポリヌクレオチドに作動可能に結合するプロモーターを含む発現カセットを有するベクターであって、この異種カルボキシル末端細胞壁標的領域が、以下の順番で、以下:細胞壁関連配列;LPQ(S/A/T)(G/A);および疎水性配列を含むベクターを、提供する。
【0034】
(定義)
「生物活性タンパク質」とは、天然の細胞の内または外でアミノ酸配列の生物活性を有する(すなわち分子機構に関与し得る)アミノ酸配列をいう。タンパク質の活性としては例えばその免疫原性、触媒活性、結合親和性等が挙げられる。ポリペプチドワクチンは用語「生物活性タンパク質」に包含される。代表的には、このアミノ酸配列は、天然細胞の内または外でアミノ酸配列により形成される3次元構造を形成する。
【0035】
「2D CD4」とは、ヒトCD4の最初の約183アミノ酸をいう(Arthosら、Cell、1989、57:469−81(1989))。CD4は、成熟ヘルパーT細胞および未成熟胸腺細胞、ならびに単球およびマクロファージ上に見出される、細胞表面糖タンパク質である。2D−CD4は、インタクトタンパク質と同じ親和性でHIV−1 gp120と結合し、gp120に対する結合部位を含む。CD4は、アミノ末端細胞外ドメイン(アミノ酸残基1〜371)、膜貫通領域(372〜395)、および細胞質内末端(tail)テール(396〜433)を含む。
【0036】
「抗体」とは、免疫グロブリン遺伝子(単数または複数)で実質的にコードされるポリペプチド、または分析物(抗原)と特異的に結合して、その分析物を認識するそれらのフラグメントをいう。認められた免疫グロブリン遺伝子は、κ、λ、α、γ、δ、εおよびμ定常領域遺伝子、ならびに無数の免疫グロブリン可変領域遺伝子を含む。軽鎖は、κまたはλのいずれかに分類される。重鎖は、γ、μ、α、δまたはεに分類され、それぞれ免疫グロブリンクラスIgG、IgM、IgA、IgDおよびIgEを規定する。
【0037】
例示的な免疫グロブリン(抗体)の構造単位は、4量体を含む。各4量体は、ポリペプチド鎖の2つの同一の対で構成され、各対は1個の「軽」鎖(約25kDa)と1個の「重」鎖(約50〜70kDa)とを有する。各鎖のN−末端は、抗原認識を主として担う、約100〜110またはそれ以上のアミノ酸の可変領域を規定する。可変軽鎖(VL)および可変重鎖(VH)という用語は、それぞれ、これらの軽鎖および重鎖を指す。
【0038】
抗体は、例えば、インタクト免疫グロブリン、または様々なペプチダーゼでの消化により生成される、性質が十分特徴付けられた多くのフラグメントとして存在する。従って、例えば、ペプシンはヒンジ領域中のジスルフィド結合下で抗体を消化し、F(ab’)2(それ自体がジスルフィド結合でVH−CH1に連結した軽鎖であるFabの2量体である)を生成する。F(ab’)2は、緩やかな条件下で還元されてヒンジ領域のジスルフィド結合を切断し得、それによって、F(ab’)2ダイマーをFab’モノマーに変える。Fab’モノマーは、本質的に、ヒンジ領域の一部を有するFabである(Paul(編)Fundamental Immunology、第3版、Raven Press、NY(1993)参照)。様々な抗体フラグメントが、インタクト抗体の消化の観点から定義されているが、当業者は、このようなフラグメントを、化学的に、または組み換えDNA方法論を利用して、新たに合成し得ることを理解する。従って、本明細書で用いる場合、抗体という用語には、抗体全体の改変により生成されるか、または組み換えDNA方法論を用いて新たに合成されるかのいずれかである抗体フラグメント(例えば1本鎖FV)も含まれる。
【0039】
「単離された」という用語は核酸またはタンパク質に応用する場合、核酸またはタンパク質が本質的に自然状態で結合する他の細胞成分を含まないことを意味する。均一な状態が好ましいが、乾燥状態でも水溶液でも良い。代表的には、ポリアクリルアミドゲル電気泳動または高速液体クロマトグラフィー等の分析化学技術を用いて、純度および均一性が測定される。調製物中に存在する優勢な種であるタンパク質が、実質的に精製される。特に、単離される遺伝子は、その遺伝子に隣接し、目的の遺伝子以外のタンパク質をコードする、オープンリーディングフレームから、分離される。「精製された」という用語は、核酸またはタンパク質が電気泳動ゲル中で本質的に1個のバンドを生じることを意味する。特に、この用語は、核酸またはタンパク質が、少なくとも純度85%であり、より好ましくは少なくとも純度95%であり、最も好ましくは少なくとも純度99%であることを意味する。
【0040】
「核酸」または「ポリヌクレオチド」という用語は、デオキシリボヌクレオチドまたはリボヌクレオチド、および1本鎖または2本鎖の形態のそれらのポリマーをいう。特に限定しない限り、この用語は、参照核酸と類似の結合特性を有し、天然に存在するヌクレオチドと類似の様式で代謝される天然ヌクレオチドの既知のアナログを含む核酸を包含する。特に示さない限り、ある特定の核酸配列はまた、保存的に改変されたその変異体(例えば縮重コドン置換)、および相補配列、ならびに明示された配列もまた暗黙のうちに含む。具体的には、1つ以上の選択された1個の(または全ての)選ばれたコドンの第3位が混合塩基および/またはデオキシイノシン残基で置換された配列を生成することにより、縮重コドン置換が行われ得る(Batzerら、Nucleic Acid Res.19:5081(1991);Ohtsukaら、J.Biol.Chem.260:2605−2608(1985);およびRossoliniら、Mol.Cell.Probes 8:91−98(1994))。「核酸」という用語は「ポリヌクレオチド」と互換的に使用される。
【0041】
「ポリペプチド」、「ペプチド」および「タンパク質」という用語は、本明細書では、アミノ酸残基のポリマーを指すために互換的に使用される。これらの用語は、1つ以上のアミノ酸残基が、対応する天然に存在するアミノ酸の人工的化学模倣体であるアミノ酸ポリマー、および天然に存在するアミノ酸ポリマーおよび天然に存在しないアミノ酸ポリマーに適用される。本明細書で用いる場合、この用語は、全長タンパク質(すなわち抗原)を含む任意の長さのアミノ酸鎖を包含し、ここでこのアミノ酸残基は共有ペプチド結合で連結している。
【0042】
「アミノ酸」という用語は、天然に存在するアミノ酸および合成アミノ酸、ならびに天然に存在するアミノ酸と類似の様式で機能するアミノ酸アナログおよびアミノ酸模倣体をいう。天然に存在するアミノ酸は遺伝子暗号でコードされるものならびに、その後改変されるアミノ酸(たとえばヒドロキシプロリン、γ−カルボキシグルタミン酸およびO−ホスホセリン)である。アミノ酸アナログとは、天然に存在するアミノ酸と同じ基本化学構造(すなわち水素、カルボキシル基、アミノ基およびR基に結合するα炭素)を有する化合物(例えばホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウム)をいう。このようなアナログは、改変されたR基(例えばノルロイシン)または改変されたペプチド骨格を有するが、天然に存在するアミノ酸と同じ基本化学構造を保有している。「アミノ酸模倣体」とは、アミノ酸の一般的な化学構造とは異なる構造を有するが、天然に存在するアミノ酸と類似の様式で機能する化合物をいう。
【0043】
本明細書では、アミノ酸は、一般的に知られた3文字記号で表され得か、またはIUPAC−IUB Biochemical Nomenclature Commissionが推奨する1文字記号で表され得る。ヌクレオチドも同ように、その一般に受け入れられている1文字表記で表され得る。
【0044】
以下に述べるように、最大一致のために整列させた場合に、2つの配列中でヌクレオチドまたはアミノ酸残基の配列がそれぞれ同じであるならば、2個の核酸配列またはポリペプチドは「同一」であると言われる。2個以上の核酸またはポリペプチド配列に関して、用語「同一」または「同一性」パーセントとは、以下の配列比較アルゴリズムの1つを用いるか、または手動整列および目視検査によって測定して比較ウインドウ上での最大一致のために比較し整列させた場合、同一であるか、あるいは特定のパーセントの同一なアミノ酸残基またはヌクレオチドを有する、2つ以上の配列または部分配列をいう。配列同一性パーセントをタンパク質またはペプチドに関連して用いる場合、同一でない残基の位置が、多くの場合保存的アミノ酸置換により異なり、その場所では、アミノ酸残基が類似の化学的性質(例えば荷電または疎水性)を有する他のアミノ酸残基で置換されており、従って分子の機能的性質を変化させないことが認識される。保存的置換で配列が異なる場合、配列同一性パーセントを上方に調節して置換の保守的性質について補正鏑し得る。この調整を行うための手段は当業者に公知である。代表的に、これは保存的置換を完全なミスマッチでなく部分ミスマッチとして採点し、それによって配列同一性パーセントを増加させる工程に関する。従って、例えば、同一アミノ酸に得点1を与え、非保存的置換に得点ゼロを与える場合、保存的置換には0と1の間の得点が与えられる。保存的置換の採点は、例えば、プログラムPC/GENE(Intelligenetics、Mountain View、California、USA)で実行されるMeyerおよびMiller、Computer Applic.Biol.Sci.4:11−17(1988)のアルゴリズムに従って計算される。
【0045】
2個の核酸またはポリペプチドについての「実質的に同一」という語句は、参照配列と少なくとも70%の配列同一性を有する配列または部分配列をいう。または、同一性パーセントは40%〜100%の任意の整数であり得る。より好ましい実施形態としては、以下に述べる標準パラメーターを用いるBLAST等の本明細書に記載のプログラムを用いて、参照配列(例えば配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7または配列番号8、またはそのフラグメント)と比較して、少なくとも40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%または99%が挙げられる。
【0046】
配列比較のため、代表的には、1つの配列が参照配列として働き、それと試験配列が比較される。配列比較アルゴリズムを使用する場合、試験配列および参照配列をコンピューターに入力し、必要あれば部分配列座標を指定し、配列アルゴリズムプログラムパラメーターを指定する。デフォルトプログラムパラメーターを使用することができるか、または別なパラメーターを指定することができる。次いでプログラムパラメーターに基づき、参照配列に対する配列比較アルゴリズムが試験配列についての現在の配列同一性パーセントを計算する。
【0047】
本明細書で用いる場合、「比較窓」には、20〜600、通常は約50〜約200、より通常は約100〜約150個からなる群より選ばれた連続位置数のいずれか一つのセグメントに対する参照が含まれ、その中で2つの配列を最適に整列した後、一つの配列が、連続位置が、同数の参照配列と比較され得る。比較のための配列の整列法は当該分野で周知である。比較のために最適な配列の整列は、例えば、SmithおよびWaterman、Adv.Appl.Math.2:482(1981)の局所相同アルゴリズム、NeedlemanおよびWunsch、J.Mol.Biol.48:443(1970)の相同整列アルゴリズム、PearsonおよびLipman、Proc.Natl.Acad.Sci.USA 85:2444(1988)の類似性検索法、これらのアルゴリズムのコンピューター化された実施(Wisconsin Genetics Software Package、Genetics Computer Group、575 Science Dr.,Madison、WI中におけるGAP、BESTFIT、FASTAおよびTFASTA)、または手動整列および目視検査により行われ得る。
【0048】
配列同一性パーセントと配列類似性の決定に適したアルゴリズムの例は、Altschulら、J.Mol.Biol.215:403−410(1990)に記載のBLASTアルゴリズムである。BLAST分析を実行するためのソフトウエアは、National Center for Biotechnology Informationから公に入手可能である。このアルゴリズムには、最初にクエリ(query)配列中の長さWのショートワードを同定することによってハイスコア配列対(HSP)を同定する工程を包含する。データベース配列中の同じ長さのワードと整列した場合、HSPは、ある正の値の閾値スコアTと一致するか、またはそれを満たすかのいずれかである。Tは、近隣ワードスコア閾値(neighborhood word score threshold)と呼ばれる(Altshulら、前出)。最初の近接ワードヒットは、それらのワードを含むより長いHPSを見出すための検索を開始するシードとして働く。累積整列スコアが増加し得る限り、ワードヒットは各配列に沿って両方の方向に延長される。累積整列スコアがその最大達成値から量Xだけ低下した場合、1つ以上の負スコア残基整列の蓄積のために累積スコアがゼロ以下になった場合、またはいずれかの配列の末端に達した場合、各方向へのワードヒットの延長を停止する。BLASTアルゴリズムパラメーターW、TおよびXは整列の感度と速度を決定する。BLASTプログラムは、デフォルトとして、ワード長(W) 11、BLOSUM62スコアリングマトリックス(HenikoffおよびHenikoff、Proc.Natl.Acad.Sci.USA 89:10915(1989)参照)整列(B) 50、期待値(E) 10、M=5、N=−4、および両方の鎖の比較を使用する。
【0049】
BLASTアルゴリズムはまた、2つの配列間の類似性の統計分析を行う(例えば、KarlinおよびAltschul、Proc.Natl.Acad.Sci.USA 90:5873−5787(1993)参照)。BLASTアルゴリズムが提供する類似性の基準の1つは、2つのヌクレオチド配列またはアミノ酸配列間の一致が偶然生じる確率を示す最小和確率(P(N))である。例えば、参照核酸に対する試験核酸の比較における最小和確率が約0.2未満、より好ましくは約0.01未満、最も好ましくは約0.001未満である場合、その核酸は参照配列に類似すると考えられる。
【0050】
例えば細胞、核酸、タンパク質またはベクターに関して用いられる場合、「組み換え」または「組み換えで変化された」という用語は、その細胞、核酸、タンパク質またはベクターが異種核酸もしくは異種タンパク質の導入によってかまたはネイティブ核酸もしくはネイティブタンパク質の変化によって改変されたか、あるいは細胞がそのようにして改変された細胞に由来することを示す。従って、例えば、組み換え細胞は、その細胞のネイティブ(非組み換え)形態に見出されない遺伝子を発現するか、さもなければ異常発現するか、発現量が少ないか、またはまったく発現しないネイティブ遺伝子を発現する。
【0051】
核酸またはポリペプチドの部分に関して使用される「異種」という用語は、核酸またはポリペプチドが天然で相互に同じ関係で見出されない2個以上の部分配列を有することを示す。例えば、その核酸は、代表的には組み換えで製造され、新しい機能性核酸を生成するように配置された無関係の遺伝子由来の2個以上の配列(例えばある起源由来のプロモーターおよび別の起源由来のコード領域)を有する。同ように、異種タンパク質とは、タンパク質が天然で相互に同じ関係で見出されない2以上の部分配列を含むことを示す(例えば融合タンパク質)。
【0052】
「発現カセット」とは、宿主細胞内における特定の核酸の転写が可能である一連の特定核酸要素を有する、組み換えまたは合成で生成される核酸である。発現カセットはプラスミド、ウイルスまたは核酸フラグメントの一部であり得る。代表的には、発現ベクターとしては、プロモーターに作動可能に結合して転写されるべき核酸が挙げられる。
【0053】
(発明の詳細な説明)
(I.はじめに)
本発明は乳酸杆菌属等のグラム陽性菌の細胞壁上に異種ポリペプチドを発現するための新規なモチーフおよび方法を提供する。本発明のモチーフを目的のタンパク質と融合され、次いで細菌中の融合タンパク質として発現され、その結果、融合タンパク質が、細胞壁の標的となるか、埋め込まれるか、および/または表面に提示されるか、あるいは生物学的に活性であり、かつ安定な融合タンパク質が、細胞外マトリックスに放出され得る。
【0054】
モチーフは、例えば、膣を含むヒト粘膜でコロニーを形成する乳酸杆菌属細菌の細胞壁上にタンパク質を発現するために有用である。例示的な粘膜細菌としては、L.jensenii、L.gasseriおよびL.casei等の乳酸杆菌属種が挙げられる。
【0055】
(II.細胞壁標的領域)
乳酸杆菌属等のグラム陽性菌の細胞壁に共有結合された目的のポリペプチドを発現し、標的とするために、細胞壁標的領域は、目的の異種ポリペプチドにC−末端で結合している。膣関連乳酸桿菌および他の乳酸桿菌において異種タンパク質を表面提示し得る細胞壁標的領域は、4つの部分:細胞壁関連領域、LPQ(S/A/T)(G/A)配列、および疎水性配列から、代表的にはこの順番で構成される。必要に応じて細胞壁標的領域は、カルボキシル末端またはその近くに荷電領域を含む。この荷電領域は、細胞膜中で移動停止配列として作用し、それによって培地中への放出を阻止する。もちろん、アンカー配列がタンパク質の残りの部分から切り取られた場合、やはり培地中への放出が生じ得る。
【0056】
(A.細胞壁関連領域)
細胞壁関連領域は、LPQ(S/A/T)(G/A)識別シグナルに先行する。細胞壁関連領域の長さは変化し得る。細胞壁関連領域は代表的には、40アミノ酸と1,000アミノ酸との間である。いくつかの実施形態では、細胞壁関連領域は、少なくとも約30、50、80、100、150および200アミノ酸または、それより大きい。いくつかの実施形態では、細胞壁関連領域は約500、400、300、250、200、150、100またはそれ未満のアミノ酸を有する。Lactobacillus jenseniiでは、pOSEL268中のC−末端細胞壁識別シグナルと融合する一縦列反復を含む95アミノ酸伸長(実施例に記載)により、CD4の表面提示が可能になる。しかしながら、フィブロネクチン結合タンパク質の約90アミノ酸がS.carnosus中で想定されている(StraussおよびGotz、Mol.Microbiol.21:491−500(1996))一方で、ペプチドマッピングに基づいてS.pyogenesのM6タンパク質中の約50アミノ酸長が同定された(PancholiおよびFischetti、J.Bacteriol.170:2618−2624(1988))。従って、約50アミノ酸以下の配列が、乳酸杆菌属中で機能性であり得る。
【0057】
いくつかの実施形態では、細胞壁関連領域は疎水性である。いくつかの実施形態では、細胞壁関連領域は長さと配列が変化し得る不完全縦列反復を含む。例えば、L.jensenii C370の細胞壁関連領域は、2.5個の縦列反復を含む。しかし、縦列反復は、細胞壁関連領域に起こり得るが、必要ではない。例えばC14の細胞壁関連領域は、反復を含まない。機能的には、細胞壁関連領域は、ペプチドグリカン層と相互作用して、それを貫通(span)する。従って、その領域は細胞壁貫通(spanning)ドメインまたは細胞壁付着ドメインとも呼ばれ、膜結合ソルターゼで固定されるタンパク質と細胞壁識別シグナルとの間のスペーサーとして作用する。
【0058】
本発明は、C370配列:
【0059】
【化1】
と実質的に同一の細胞壁関連領域を提供する。ある場合は、細胞壁関連領域は少なくともC370配列の約40、50、75、90、100、120、150、175、200アミノ酸フラグメントを有する。例えば、活性細胞壁関連フラグメントは、以下の配列:
【0060】
【化2】
を有し得る。C370配列(配列番号4)は、75個の荷電アミノ酸残基(K、R、D、E)を含み、Pro−Gly富化配列が欠失している。
【0061】
いくつかの実施形態では、細胞壁関連領域は、実質的にC14配列:
【0062】
【化3】
と同一である。ある場合は、細胞壁関連領域は、C370配列の少なくとも約40、50、75、90、100、120、150、175、200アミノ酸フラグメントを含む。配列番号6は、51個の荷電アミノ酸残基(K、R、D、E)を含む。
【0063】
ある場合は、細胞壁関連領域は、乳酸杆菌属以外の細菌、または膣と関連しない乳酸杆菌属株由来である。
【0064】
(B.LPQ(S/A/T)(G/A))
配列LPQ(S/A/T)(G/A)は乳酸杆菌属の膣関連株において細胞壁局在化シグナルとして作用する。モチーフLPQ(S/A/T)(G/A)の少なくとも1個のコピーは細胞壁標的領域内にある。モチーフ中の括弧はその位置の別なアミノ酸を示す(例えばLPQSG、LPQAG、LPQTG、LPQSA、LPQAA、LPQTA)。
【0065】
(C.疎水性配列)
細胞壁に固定されるポリペプチドのカルボキシル末端は細菌膜(bacterial membrane)を貫通するように機能する疎水性領域を含む。疎水性領域は少なくとも約50%、ある実施形態では少なくとも60%、70%、80%または90%の疎水性アミノ酸を含む。天然起源疎水性アミノ酸にはアラニン、イソロイシン、ロイシン、メチオニン、フェニルアラニン、プロリン、トリプトファンおよびバリンが含まれる。グリシン、トレオニンおよびセリンを含む、より疎水性の少ないアミノ酸もこれらの配列の一部を構成し得る(例えばPallenら、Trends Microbiol.9:97−101(2001)参照)。ソルターゼ様タンパク質の可変LPXTG含有基質の間で疎水性配列は一般に長さが約10〜約30アミノ酸、場合によっては13〜24アミノ酸の間である(Pallenら、Trends Microbiol.9:97−101(2001))。典型的な疎水性配列には例えばC14中のV1740GILGLAIATVGSLLGLGLV1758、およびC370中のP1877LTAIGIGLMALGAGIFA1894が含まれる。
【0066】
または、グラム陽性細菌由来の任意の細胞壁アンカータンパク質の疎水性領域を用いることができる。他の疎水性配列には例えば米国特許第5,821,088号の図1に記載のもの、またはそれと実質的に同一の配列が含まれる。さらに別な配列がPallenら、Trends Microbiol.9:97−100(2001)の表2にも示されている。
【0067】
(D.荷電配列)
荷電領域が細胞壁標的タンパク質のカルボキシル末端、代表的には疎水性膜貫通領域の直後に任意に存在し得る。カルボキシル末端荷電領域の存在によりポリペプチドが膜に固定され、その結果膜から解離し培地中へ漏れ出すタンパク質の量が大いに減少する。荷電領域は少なくとも40%、ある実施形態では少なくとも50%、60%、70%、80%または90%の荷電アミノ酸を有する。天然起源荷電アミノ酸にはアルギニン、ヒスチジン、リジン、アスパラギン酸およびグルタミン酸が含まれる。荷電配列は例えば長さで2〜20アミノ酸残基の間、ある実施形態では4〜12または5〜11アミノ酸の間である。典型的な荷電配列には例えばC191中のK969KRKED974、C14中のR1760KKRQK1765およびC370中のK1895KKRKDDEA1903が含まれる。
【0068】
または、グラム陽性細菌由来の任意の細胞壁アンカータンパク質の荷電領域を使用できる。別な荷電配列には例えば米国特許第5,821,088号の図1に記載のもの、およびそれと実質的に同一の配列が含まれる。また別な配列がPallenら、Trends Microbiol.9:97−100(2001)の表2にも示される。
【0069】
(III.組み換え技術)
(A.分子生物学法)
本発明は組み換え遺伝子学の分野における慣用的な技術に依存している。本発明において使用する一般的な方法を開示する基本的なテキストにはSambrookら、Molecular Cloning、A Laboratory Manual(第3版、2001);Kriegler、Gene Transfer and Expression:A Laboratory Manual(1990);およびCurrent Protocols in Molecular Biology(Ausubelら、編集、1994)が含まれる。
【0070】
核酸では、サイズはキロベース(kb)または塩基対(bp)で与えられる。これらはアガロースまたはアクリルアミドゲル電気泳動、配列決定された核酸、または公開されたDNA配列から見積もられる。タンパク質では、サイズはキロダルトン(kDa)またはアミノ酸残基数で与えられる。タンパク質のサイズはゲル電気泳動、配列決定されたタンパク質、導かれたアミノ酸配列、または公開されたタンパク質配列から見積もられる。
【0071】
市販されていないオリゴヌクレオチドを、BeaucageおよびCaruthers、Tetrahedron Letts.22:1859−1862(1981)に最初に記載された固相ホスホラミダイトトリエステル法に従って、Van Devanterら、Nucleic Acids Res.12:6159−6168(1984)に記載の様に自動合成装置を用いて化学的に合成することができる。オリゴヌクレオチドの精製はPearsonおよびReanier、J.Chrom.255:137−149(1983)に記載の通りに天然アクリルアミドゲル電気泳動または陰イオン交換HPLCによって行われる。
【0072】
クローン化された遺伝子、および合成オリゴヌクレオチドの配列を、例えばWallaceら、Gene 16:21−26(1981)の二重鎖テンプレートの配列決定のための鎖停止法を用いてクローニング後に確認することができる。
【0073】
(B.所望のタンパク質をコードするヌクレオチド配列を単離するためのクローニング法)
一般に、目的とするタンパク質をコードする核酸を、cDNAまたはゲノムDNAから作製したDNAライブラリーからクローニングする。特定の配列の位置をオリゴヌクレオチドプローブとハイブリダイズすることにより決定することができるが、その配列は本明細書に開示した配列または当該分野で公知の配列から導かれ、PCRプライマーの参考となり、遺伝子特異的プローブを単離するために適当な領域を規定する。または、配列が発現ライブラリー中にクローニングされる場合は、本明細書に開示されるポリペプチドを含め目的のポリペプチドに対して作製された抗血清または精製抗体を用いて発現組み換えタンパク質を免疫学的に検出し得る。
【0074】
ゲノムおよびcDNAライブラリーを作製およびスクリーニングする方法は当業者に周知である(例えばGulberおよびHoffman、Gene 25:263−269(1983);BentonおよびDavis、Science、196:180−182(1977);およびSambrook、前出参照)。目的のタンパク質を発現する細胞はcDNAライブラリーを作製するための有用なRNA源である。
【0075】
簡単に言えば、cDNAライブラリーを作製するため、mRNAに富んだ起源を選ばなければならない。次いでmRNAをcDNAに変換し、組み換えベクター中に連結し、増殖、スクリーニングおよびクローニングのために組み換え宿主中に形質導入する。ゲノムライブラリーでは、DNAを適当な組織または細胞から抽出し、機械的に剪断するか酵素的に消化して好ましくは約5〜100kbの断片を生成する。次いで断片を望ましくないサイズのものから濃度勾配遠心で分離し、バクテリオファージラムダベクター中に構築させる。これらのベクターおよびファージはインビトロでパッケージングされ、組み換えファージをプラクーハイブリダイゼーションで分析する。コロニーハイブリダイゼーションはGrunsteinら、Proc.Natl.Acad.Sci.USA.72:3961−3965(1975)に一般的に記載されている様に行われる。
【0076】
別な方法では、合成オリゴヌクレオチドプライマーの使用と、mRNAまたはDNAテンプレート上でのポリメラーゼ延長とが組み合わされる。このポリメラーゼ連鎖反応(PCR)法では、目的のタンパク質をコードする核酸がmRNA、cDNA、ゲノムライブラリーまたはcDNAライブラリーから直接増幅される。プライマー中には制限エンドヌクレアーゼ部位が組み込まれ得る。ポリメラーゼ連鎖反応法、または他のインビトロ増幅法も、例えば特定のタンパク質をコードする核酸のクローニングおよびそのタンパク質の発現、生理学試料中で本発明のポリペプチドをコードするmRNAの存在を検出し、核酸の配列を決定する、または他の目的ためのプローブとして使用される核酸の合成に有用であり得る(米国特許第4,683,195号および同第4,683,202号参照)。PCR反応によって増幅された遺伝子はアガロースゲルから精製され、そして適当なベクター中にクローン化され得る。
【0077】
本発明のポリペプチドをコードする遺伝子を組織または細胞試料から同定するための適当なプライマーおよびプローブを、当該分野で記載の配列から導くことができる。PCRの概説としてはInnisら、PCR Protocols:A Guide to Methods and Applications、Academic Press、San Diego(1990)参照。
【0078】
本発明のポリペプチドをコードするポリヌクレオチドを、乳酸杆菌属中に形質転換する前に中間ベクターを使用してクローニングすることができる。これらの中間ベクターは代表的には原核生物ベクターまたはシャトルベクターである。
【0079】
(C.形質転換技術)
適当な細菌宿主株を、例えばその形質転換能、異種タンパク質発現能、および/または粘膜表面でのコロニー形成能で選択される。細菌宿主を塩化ルビジュウム法またはエレクトロポーレーション等の標準技術を用いて形質転換受容性にする(例えばWeiら、J.Microbiol.Meth.21:97−109(1995)参照)。
【0080】
エレクトロポーレーションによるL.jenseniiの形質転換を、例えばLuchanskyら(J.Dairy Sci.74:3293−3302(1991));Changら(Proc.Natl.Acad.Sci.USA.100:11672−11677(2003))に規定される様な標準法を改変して行うことができる。簡単に言えば、新しく接種したL.jenseniiをブロス中で培養する(例えば37℃、5%CO2でOD600が0.6〜0.7まで)。細菌細胞を収集し、洗浄し、スクロースとMgCl2の冷溶液(例えば4℃)中に再懸濁する。コンピテント細胞を次いでDNAと混合し、冷却したギャップキュベット中にいれ、エレクトロポーレーションする。その後、選択試薬以外の抗生物質を含む選択寒天プレート上に接種する前に、予め加熱したブロス中で細胞を回収する(例えば37℃で約2時間)。
【0081】
(D.発現)
本発明の発現カセットは、本発明のポリペプチドの発現と位置決めを制御するための様々な成分を含み得る。例えば、発現カセットはプロモーター要素、シグナル配列をコードする配列、目的のポリペプチドに対するコード配列、およびアンカー配列を含むことができる。
【0082】
異種ポリヌクレオチドまたはポリペプチドの発現は構成的であり得る(例えばP59(Van der Vossenら、Appl.Environ.Microbiol.58:3142−3149(1992))もしくはP23(Elliotら、Cell、36:211−219(1984))プロモーター、またはより高い強度を有する乳酸杆菌属由来の天然プロモーターを用いて)。または、発現は誘導性プロモーターの制御下に行うことができる。例えば、Bacillusアミラーゼ(Weickertら、J.Bacteriol.171:3656−3666(1989))またはキシロース(Kimら、Gene、181:71−76(1996))プロモーターの他、Lactococcusナイシンプロモーター(Eichenbaumら、Appl.Environ.Microbiol.64:2763−2769(1998))を誘導性発現を推進するために使用することができる。さらに、酸またはアルカリ誘導プロモーターを使用することもできる。例えば、膣の比較的酸性の条件下で活性なプロモーターを使用し得る。または、精子に反応する膣内の変化で誘導されるプロモーターも使用し得る。例えば、精子の導入の結果である膣のアルカリ条件の増大に反応して発現を誘導するために、アルカリ誘導プロモーターを使用する。
【0083】
様々なシグナル配列がポリペプチドの発現を膜、細胞外空間または細胞壁に導くことが知られている(例えばペプチドグリカンへの共有結合付着により)。例示的なシグナル配列にはL.amylovorusのα−アミラーゼ由来のシグナル配列(GiraudおよびCuny、Gene、198:149−157(1997))またはL.crispatusのS−層遺伝子(cbsA)由来のシグナル配列(例えばMKKNLRIVSAAAAALLAVAPVAAまたはMKKNLRIVSAAAAALLAVATVSA)が含まれる。シグナル配列は代表的にはポリペプチドのアミノ末端に位置している。
【0084】
ポリペプチドの正確な位置と折り畳みを標準法を用いて決定し得る。例えば、細菌を緩衝液(例えば25%スクロース、1mM EDTA、10mM Tris−HCl、pH8.0)に懸濁し、次いで細胞壁分解酵素(例えばリゾチームおよびムタノリシン)で処理し、次いで得られたプロトプラストを分画遠心法で分離することにより、乳酸杆菌属の細胞壁富化画分を得ることができる。画分をウエスタンブロッティングでスクリーニングして細胞壁内の発現を確認し得る。
【0085】
発現したポリペプチドの折り畳みと生物活性も、標準法を用いて決定し得る。例えば、天然の折り畳みポリペプチド特異性抗体を用いるELISAアッセイを用いてポリペプチドの折り畳みと3次元構造を確認することができる。生物活性アッセイも、ポリペプチドの活性に応じて当然変化する。例えば、ウイルスタンパク質に結合するポリペプチドでは、標準結合アッセイを用いて発現したポリペプチドのウイルスタンパク質結合能を試験することができる。抗炎症分子では、発現したポリペプチドの炎症促進する物質との拮抗能をアッセイすることができる。
【0086】
宿主細胞中の発現を改善するための遺伝子を合成する場合、そのコドン利用頻度を宿主細胞の好ましいコドン利用頻度に近づける様に遺伝子を設計することが望ましい。合成遺伝子に対する好ましいコドン利用頻度の、宿主細胞が使用する頻度からの偏差率が、1個のコドンの利用頻度の宿主細胞の利用頻度からの偏差率をまず測定し、次いでコドン全体の平均偏差を得て計算される。
【0087】
特定のポリペプチドをコードするポリヌクレオチド配列を、特定の宿主のコドン利用と一致する様に変化させることができる。例えば、乳酸杆菌属のコドン利用を使用して、本発明のポリペプチドをコードし、好ましい乳酸杆菌属のコドンを有するポリヌクレオチドを誘導することができる。宿主細胞が提示する好ましいコドンの利用頻度を、宿主細胞が発現する多数の遺伝子中の好ましいコドンの利用頻度を平均して計算することができる。この解析は宿主細胞で高度に発現する遺伝子に限定することが好ましい。例えば、Pouwelsら(Nucleic Acids Res.22:929−936(1994))は様々な乳酸杆菌属種が示す高度発現遺伝子によるコドン利用頻度を提供している。コドン利用表はインターネットを通じても入手できる。
【0088】
(IV.本発明のタンパク質)
本発明のポリペプチド、例えば本発明の細胞壁標的領域に融合する生物活性ポリペプチドは、任意のポリペプチドであり得る。代表的には、本発明のポリペプチドはポリペプチドの生物活性を考慮した条件下で発現する。ある実施形態では、発現ポリペプチド中にジスルフィド結合が存在する。ある実施形態では、ポリペプチドの生物活性にジスルフィド結合が必要である。
【0089】
本発明のポリペプチドは任意のサイズの分子量であり得る。例えば、ポリペプチドは約100〜200,000ダルトンの間、約500〜40,000ダルトンの間、約500〜10,000ダルトンの間、約10,000〜50,000ダルトンの間、または約50,000〜200,000ダルトンの間であり得る。
【0090】
病原体感染を予防または治療するために本発明の方法に従って使用し得るポリペプチドクラスの例には、例えば抗体、抗体断片、または1本鎖抗原を含め抗ウイルスポリペプチド、抗菌ポリペプチド、抗真菌ポリペプチド、およびウイルス、細菌または真菌に結合するポリペプチドが含まれる。
【0091】
ある場合は、本発明のポリペプチドはウイルスまたは細菌病原体が結合して宿主を感染させるレセプターである。または、ポリペプチドは例えば病原体の複製、生存率、侵入またはさもなければ病原体への結合を阻害する薬剤である。ある実施形態では、本発明のポリペプチドは性的に伝播する病原体、および膣から伝播、または膣へ伝播する他の病原体に結合またはそれらを阻害する。例えば、ウイルスは感染のために標的細胞表面上のレセプターに結合することが必要であるので、ウイルスとその宿主レセプターとの相互作用を阻害することを目的とする戦略は、感染防止に有効である。
【0092】
典型的な抗ウイルスポリペプチドには例えばCD4またはそのウイルス結合断片(例えば2D−CD4:例えばOrloffら、J.Virol.67:1461−1471(1993))、3量体モチーフを経由して生成する安定なCD4の3量体(例えばYangら、J.Virol.76:4634−4642(2002))、CD4−Ig融合タンパク質12量体(Arthosら、J.Biol.Chem.277:11456−11464(2002))、α−デフェンシン(例えばZhangら、Science、298:995−1000(2002))、17bmAbの1本鎖可変領域と融合したCD4(Deyら、J.Virol.77:2859−2865(2003))、シアノビリン−Nまたは改変体(例えばBolmstedtら、Mol.Pharmacol.59:949−954(2001);Moriら、Protein Expr.Purif.26:42−49(2002))、単純疱疹ウイルス侵入メディエーターC(HveC:例えばCocchiら、Proc.Natl.Acad.Sci.USA.95:15700−15705(1998))、およびICAM−1が含まれる。他の実施形態には例えばウイルスレセプターまたはヘパリンまたはヘパリン様分子、樹状細胞特異性ICAM−3グラビングノンインテグリン(例えばGeijtenbeekら、Cell、100:587−597(2000);Feinbergら、Science、294:2163−2166(2001))を含むマンノース結合レクチン、抗HSV−1 gp120 1本鎖抗体(例えばMarascoら、Proc.Natl.Acad.Sci.USA.90:7889−7893(1993);McHughら、J.Biol.Chem.277:34383−34390(2002))、HIV−1 gp120のCD4結合部位を認識するヒトmAb b12(例えばSaphireら、Science、293:1155−1159(2001))、またはHSVに結合する中和抗体を含む同様な特異性を有する他の分子(例えばBurioniら、Proc.Natl.Acad.Sci.USA.91:355−359(1994))、およびHIV−1侵入阻害タンパク質(例えばRootら、Science、291:884−888(2001);Siaら、Proc.Natl.Acad.Sci.USA.99:14664−14669(2002))が含まれる。
【0093】
ヒト乳頭腫ウイルス(HPV)による感染は頚部癌の発症に関連する因子である(例えばzur Hausen、Virology、184:9−13(1991);Stanley、Best Prat.Res.Clin.Obstet.Gynaecol.15:663−676(2001))。従って、HPVを阻害する、またはそれに結合する分子の存在は、HPV感染と頚部癌の発症の両方の阻止に有用である。典型的な抗HPVポリペプチドには例えばヒト乳頭腫ウイルス16型タンパク質E6またはE7に結合する中和抗体(例えばMannhartら、Mol.Cell Biol.20:6483−6495(2000))、HPV−結合タンパク質、またはウイルス指向免疫反応を誘発するために使用し得るHPVタンパク質が含まれる。
【0094】
ウイルスまたは細菌等の病原体に結合する能力を、少なくとも数個の方法で本発明の細菌に授け得る。その第1はヒトライノウイルスHRV(主要グループ)に対するICAM−1およびHIVに対するCD4等の、ウイルスに対する正常な宿主レセプターを細菌の表面に発現させることである。これらは正常なヒトタンパク質であり、これらの遺伝子の完全な配列の多くが決定され、データベースGenBankに保存されている。
第2の方法は、ポリオウイルス上のVP4またはHIV上のgp120等のウイルス表面上の保存性抗原決定基に結合する抗体断片または他のポリペプチドを細菌表面に発現することである。本質的に任意の抗原に特異性のある抗体断片(およびペプチド)を、例えばファージ提示ライブラリー(Marksら、J.Biol.Chem.267:16007−16010(1992))から選ぶことができる。抗体を任意の病原体上のエピトープまたは病原体と結合するエピトープの他、以下に議論するエピトープ等の他のエピトープを指向させることができる。
【0095】
第3の方法には、細菌表面上の炭水化物結合ポリペプチドの発現が含まれる。これらの分子の例にはヘパリン結合ポリペプチド、またはマンノース結合ポリペプチドが含まれる。
【0096】
抗細菌ポリペプチドには尿路病原性E.coliに結合する、またはその生育もしくはそれによるコロニー形成を阻害するポリペプチドが含まれる。典型的な抗細菌ポリペプチドには例えばグラム陰性細菌に対する透過性増大タンパク質(Levy、Expert Opin.Investig.Drugs、11:159−167(2002))、哺乳動物抗微生物ペプチド、β−デフェンシン(GarzおよびLehrer、Pharmacol.Ther.66:191−205(1995))、バクテリオシン(例えばLoefflerら、Science、294:2170−2172(2001))、および細菌に特異的に結合する抗体が含まれる。
【0097】
抗真菌ポリペプチドにはCandida等の真菌に結合する、またはその生育もしくはそれによるコロニー形成を阻害するポリペプチドが含まれる。
【0098】
本発明に記載の有用な生物活性ポリペプチドのさらに別な例には抗炎症分子、成長因子、成長因子に結合するかまたはそれに拮抗する分子、治療用酵素、抗体(例えば抗体断片または1本鎖抗体を含む)、および子宮頚部癌を含む癌を阻害するかまたは治療する分子等の治療用ポリペプチドまたは薬剤が含まれる。数多くの他の治療活性ポリペプチドを容易に引用することができるので、これらの例は制限的であることを意図するものではない。
【0099】
抗炎症分子には例えばTNFまたはIL−8に特異的に結合する抗体または他の分子が含まれる。他の例示的な抗炎症分子には、IL−10およびIL−11が含まれる。
【0100】
本発明で有用な成長因子には例えばKGF、HB−EGF、FGFおよびTGF−β等の局所組織修復に係わる因子、またはこれらの分子の拮抗剤が含まれる。
【0101】
治療用酵素には例えば一酸化窒素(NO)シンターゼが含まれる。
【0102】
抗癌性分子にはアポトーシスを誘導する分子、p53等の細胞周期を調節する分子、または癌特異性エピトープを標的とするワクチンとして作用する分子が含まれる。
【0103】
本発明で有用なワクチン分子にはウイルス、細菌または真菌に対する免疫反応を誘起するポリペプチドが含まれる。典型的なウイルスワクチンは例えばHIV、HPV、HSV−2または天然痘に対する反応を誘発する。典型的な抗原にはHSV−2の糖タンパク質D、ヒト乳頭腫ウイルスのタンパク質E6およびE7、Chlamydia trachomatisの主要外膜タンパク質(KimおよびDeMars、Curr.Opin.Immunol.13:429−436(2001))、およびCandida albicansのアスパルチルプロテアーゼ(De Bernardisら、Infect.Immun.70:2725−2729(2002));尿路病原性E.coliのFimH(Langermannら、Science、276:607−611(1997));腸管外病原性E.coliのIroN(Russoら、Infect.Immun.71:7164−9(2003))が含まれる。
【0104】
(V.送達)
遺伝子組み換え細菌の所望の粘膜表面への送達は、その領域への近づき易さと局部条件に依存する。例えば、遺伝子組み換え細菌を食塩水、または膣粘膜上へ送達するための発泡体の中に入れ得る。発泡体には例えば1種以上の疎水性に修飾したセルロース誘導体またはキトサン等の多糖が含まれ得る。セルロース誘導体には例えばヒドロキシエチルセルロース、ヒドロキシプロピルセルロース、メチルセルロース、ヒドロキシプロピルメチルセルロース、ヒドロキシエチルメチルセルロース等が含まれる。キトサンには例えば以下のキトサン塩:乳酸キトサン、サリシル酸キトサン、ピロリドンカルボン酸キトサン、イタコン酸キトサン、ナイアシン酸キトサン、蟻酸キトサン、酢酸キトサン、没食子酸キトサン、グルタミン酸キトサン、マロン酸キトサン、アスパルギン酸キトサン、グリコール酸キトサン、および4級アミン置換キトサンおよびその塩などが含まれる。発泡体にはまた水、エチルアルコール、イソプロピルアルコール、グリセリン、グリセロール、プロピレングリコールおよびソルビトール等の他の成分も含まれ得る。殺精子剤は細菌組成物に任意に含まれる。発泡体および発泡体送達ビヒクルのさらに別な例は例えば米国特許第5,595,980号および同第4,922,918号に記載されている。
【0105】
または、細菌を坐薬またはペッサリーとして送達することもできる。例えば米国特許第4,322,399号を参照のこと。ある実施形態では、本発明の細菌は溶解性で選ばれた可溶性ポリマー材料および/または複合炭水化物材料で製造された可溶性要素として送達されるが、それは使用前は実質的に固形物のままであり、使用中にヒトの体温と水分で溶解して所望の徐放および投与量で薬剤物質が放出される。例えば米国特許第5,529,782号参照。細菌はまた、米国特許第4,693,705号に記載の様にスポンジ送達ビヒクル中で送達される。
【0106】
ある実施形態では、細菌は経口投与される。例えば、約108個の乳酸桿菌の毎日の用量を、正常な泌尿生殖器微生物相を回復するために使用し得る。例えばReidら、FEMS Immuno.Med.Microbiol.32:37−41(2001)参照。
【0107】
ある実施形態では、遺伝子組み換え細菌の粘膜表面への投与には定期的な繰り返しが必要である:最適投薬間隔は日常的に決定されるが、粘膜環境と細菌株によって異なる。投薬間隔は1日1回から2〜4週ごとに1回まで変化し得る。
【0108】
天然型乳酸杆菌属を形質転換するためにバクテリオファージが導入される場合、選択したバクテリオファージの核酸を操作して、異種遺伝子がバクテリオファージ被覆タンパク質をコードする遺伝子を置換し、バクテリオファージを複製不能(replication−defective)とすることができる。これらの組み換えDNA分子を機能性バクテリオファージタンパク質を含む細胞溶解物に加えることにより、異種遺伝子を含む機能性バクテリオファージ粒子を組み立てることができる。次いでこれらの複製不能バクテリオファージ粒子を所望の粘膜表面上に導入し、選択した細菌叢に感染させることができる。粘膜表面への典型的な投薬量は108〜1012PFU/mlであると思われる。処置表面に対する溶液の割合は、粘膜表面平方センチあたり約0.1〜1.0mlである必要がある。ビヒクルは細菌に対する上記ビヒクルと同様である。
【実施例】
【0109】
以下の実施例は説明のために提供されるものであり、本発明を制限するものではない。
【0110】
ほとんどのウイルスは鼻、口、消化器または泌尿生殖器等の粘膜を通して感染する。これらの粘膜には、健康な女性の膣腔内のL.jensenii、L.gasseriおよびL.crispatusを含む膨大な数の共生細菌が自然にコロニーを形成している。細菌表面に固定された生物活性ウイルス結合タンパク質を発現する遺伝子組み換えL.jenseniiが粘膜内でウイルスを捕捉し、従ってウイルスが下層の上皮細胞やリンパ球に接近することを妨害するものと、本発明者らは予想している。これらの捕捉されたウイルスは感染を中止し、そして/または乳酸桿菌が分泌する乳酸および過酸化水素等の抗ウイルス化合物で局所的に不活性化され、その結果感染性ウイルス粒子の数をかなり減少する。従って、遺伝子組み換え乳酸杆菌属がHIV結合リガンド、2−ドメインCD4およびシアノビリンNを高濃度で表面に発現するためのモジュール発現アプローチを行った。10〜600アミノ酸のポリペプチドが効果的に細胞壁に固定されて提示されることは、L.jenseniiの天然タンパク質由来のタンパク質ドメインへ融合することにより達成され得ることを本発明者らは見出した。
【0111】
M6タンパク質は標示細胞壁識別シグナルであるLPXTGモチーフと、その後に一続きの疎水性アミノ酸、最後に臨界細胞表面保持シグナルとなる荷電残基(KRKEEN)を有する。2個の良く性質が分かったS.pyrogenesのM6タンパク質(emm6)またはL.paracaseiのPrtPプロテアーゼ由来の細胞壁アンカーモチーフ、またはS.pyrogenesのM6タンパク質由来のアンカーモチーフ、さらにM6タンパク質の天然配列由来のN−末端100アミノ酸伸長部(CWA100)を利用して、L.jemsenii表面にCD4を発現するためのプラスミドベースのモジュールアプローチを本発明者らは最初に試みた。M6タンパク質と異なり、PrtPに対する局在化シグナルはLPKTAである。大量の2D CD4が馴化培地中に放出されたが、馴化培地中のタンパク質、および細胞壁アンカーとしてのM6またはPrtPまたはCWA100を有する修飾L.jensenii中の細胞壁またはプロトプラスト会合タンパク質プールのウエスタン分析により、検出し得る細胞壁関連2D CD4は見出されなかった。フローサイトメトリー分析でも、陽性の表面露出2D CD4は同定されなかった。
【0112】
(推定細胞壁アンカー配列の同定)
L.jenseniiのゲノム配列のデータベース検索により、推定細胞壁アンカーモチーフを有する約30個のコンティグが同定された。National Center for Biotechnologyのウエブサイトで利用し得る非冗長データベース中のより詳細な配列相同性検索に基づき、本発明者らはC14,C191およびC370と名付けられる3個の配列を選択した。それらはそれぞれ、Lactobacillus fermentumのR1p(Turnerら、Appl.Environ.Microbiol.69:5855−5863(2003))、またはストレプトコッカル表面タンパク質のファミリー(Wastfeltら、J.Bio.Chem.271:18892−18897(1996))であるL.reuteri中の粘膜結合タンパク質(RoosおよびJonsson、Microbiol.148:433−442(2002))、およびS.thermophilus中の細胞壁アンカータンパク質(Fernandez−Esplaら、Appl.Environ.Microbiol.66:4772−4778(2000))と低い配列類似性(23〜27%の同一性)を共有していた。3個の配列の全ては疎水性領域に先行するLPQTG局在化シグナルと、荷電C末端テイルを有する(図1参照)。これらの特徴はグラム陽性細菌中のソルターゼ認識C末端細胞壁アンカー配列間に共通である(NavarreおよびSchneewind、Microbiol.Mol.Bio.Rev.63:174−229(1999))。グラム陽性細菌中に見出されるLPXTG細胞アンカーモチーフ間で、これらのL.jenseniiタンパク質中ではわずか7%のLPQTG配列の一致が見出された。C14、C191およびC370タンパク質はすべて細胞壁アンカー領域に隣接したタンデム反復ドメインを含み、既知の細胞壁アンカータンパク質中にしばしば存在する構造上の特徴である(NavarreおよびSchneewind、Microbiol.Mol.Bio.Rev.63:174−229(1999))。C14、C191およびC370の配列を図2A〜Cに示す。
【0113】
(推定細胞壁アンカー配列のエピトープタグ化)
C14、C191およびC370が異種融合タンパク質をL.jenseniiの細胞壁に固定する効率を決定するため、本発明者らは約200個のアミノ酸を直接的にLPQTG局在化シグナルのN−末端に選んだ。しばしば細胞壁アンカータンパク質中の細胞壁関連(CWA)ドメインと定義されるこの領域は、基質配列の保持または延長、従って膜会合ソルターゼによる適切なタンパク質分解性の切断を促進し得る。免疫検出を促進するため、c−Mycエピトープ(EQKLISEEDL)をpOSEL239、240および241それぞれのC14、C191およびC370のCWA200領域のN−末端と融合した。ウエスタンおよびフローサイトメトリー分析を用いて、c−Mycタグ化タンパク質が生成され、細胞壁を標的としたかどうかを検討した。ウエスタン分析を行うため、pOSEL175、239、240および241を有する修飾L.jenseniiをMRSおよびRogosaSLブロスの両方の中で対数期まで生育させた。次いでβ1−4グリコシル結合を成熟ペプチドグリカン中のグリカン鎖のMurNAc−GlcNAc間で切断するN−アセチルムラミダーゼであるムタノリシンで細胞壁を消化した。代表的には、SDS−PAGEクロマトグラフィー後に細胞壁アンカータンパク質は大きなスペクトルの断片として移動する(Perryら、J.Biol.Chem.277、16241−16248(2002))。pOSEL239(C14アンカー)および241(C370アンカー)を有する細菌細胞中の細胞壁富化画分中のタンパク質のウエスタン分析から、細菌細胞をMRSおよびRogosaブロスの両方の中で培養した場合、還元性SDS−PAGE上でc−Mycタンパク質のラダーを示した(図3)。これらのパターンはpOSEL240(C191アンカー)を有する細菌細胞中の細胞壁富化画分中には見られず、試験したLPQTG含有配列間で異なったアンカー効率を示している。
【0114】
ウエスタンブロット陽性c−MycエピトープがpOSEL239および241を有するL.jensenii細胞中で表面に露出しているかどうかを決定するため、コントロールプラスミドpOSEL175を有する細菌細胞を参照して抗c−Myc抗体結合のフローサイトメトリー分析を行った。pOSEL239を有する細菌細胞中の平均蛍光強度はコントロールプラスミドpOSEL175を有する細菌細胞中の平均蛍光強度と区別できなかったが、pOSEL241を有する細菌細胞中では160倍に増加した。C14配列のc−MycタグCWA200領域の表面接近性に立体傷害が影響するかどうかは不明であるが、本発明者らの分析によりC370配列のCWA200領域のN−末端における表面露出が明らかに示された。この結果は、異種ペプチドとタンパク質とを細菌細胞表面に共有結合で固定するためにC370のこの特定の領域を利用し得ることを示している。
【0115】
(L.jenseniiの細菌表面上の2D CD4の表面発現)
2D CD4がC14のCWA200領域およびC370配列を経由して表面発現し得るかどうかを決定するため、本発明者らはウエスタンブロットとフローサイトメトリー分析を行った。ウエスタン分析を行うため、pOSEL175(コントロールプラスミド)、651(細胞アンカーのない2D CD4プラスミド;Changら、Proc.Natl.Acad.Sci.USA.100:11672−11677(2003))、237(C14アンカーに融合した2D CD4)、242(C191アンカーに融合した2D CD4)、および249(C370アンカーに融合した2D CD4)を有するL.jensenii細胞中のタンパク質を細胞壁消化により細胞壁富化画分に分画した。細胞壁富化タンパク質画分では、pOSEL237および249を有する細菌細胞の双方でより大きな分子量の種のスペクトルがpAb T4−4に対し免疫反応性であったが、pOSEL651ではそうでなかった(図4)。ムタノリシン消化後に観測されるSDS−PAGE上のこの様な梯子状パターンは、他のグラム陽性細菌の細菌表面由来の既知の細胞壁アンカータンパク質のパターンと類似している(Perryら、J.Biol.Chem.277:16241−16248(2002))。
【0116】
2D CD4が細胞表面上に発現するかどうかを決定するため、pOSEL175、651、237および249を有するL.jensenii株をpAb T4−4で探索し、次いでフローサイトメトリー分析により抗体結合を分析した。期待された通り、この分析によりpOSEL175および651を有する細菌細胞における平均蛍光強度が区別できないことが明らかとなった。対照的に、pOSEL175および651に対してpOSEL237および249を有する細菌細胞で平均蛍光強度の有意な増加が見られ、2D CD4分子の共有結合および表面露出に結果であると思われる(図5A)。上記アプローチをさらに確認するため、乳酸杆菌属の好ましいコドンを含む再コードしたシアノビリンN(CV−N)遺伝子を2D CD4の成功した固定に用いたものと同じC末端アンカードメインに融合した。CV−N発現プラスミドを有する修飾L.jenseniiのフローサイトメトリー分析により、pOSEL175を有する細菌に対し平均蛍光強度で30〜50倍の増加が検出された(データ示さず)。静電相互作用により抗体反応性CV−N分子が表面会合している可能性を調べるため、修飾細菌を5M LiClで抽出した。フローサイトメトリー分析により、PBSおよび2%FBSで洗浄した細胞を参照して、CV−N発現プラスミドを有する塩抽出L.jensenii中の平均蛍光強度が区別できないことが明らかとなった。表面提示CV−N分子の5M LiCl抽出に対する耐性は、細菌表面上の共有結合で固定されたタンパク質の挙動を反映している。
【0117】
表面に発現した2D CD4分子がgp120を結合するために正しい折り畳み立体構造をとっているか否かを確認するため、pOSEL175、237および249を有する細菌細胞を立体構造依存性エピトープを認識する抗CD4モノクローナル抗体Sim.4で探索した後に、さらにFACS分析を行った。pOSEL175と比較してpOSEL237および249を有する細菌細胞中で平均蛍光強度の有意な増加が見られ、2D CD4が機能的な形でL.jenseniiの表面に発現していることが示された(図5B)。
【0118】
モジュール発現アプローチにおいて2D CD4の表面発現が修飾L.jensenii中の天然細胞表面会合タンパク質の発現に影響するかどうかは不明であった。この問題を解決するため、pOSEL175および237を有する細菌細胞をスルホNHS−ビオチンで探索し、次いで細胞表面会合タンパク質を0.4%SDSおよび10mM DTTを含む緩衝液中で抽出した。アルカリホスファターゼ共役アビジンによるプローブ後のSDS抽出タンパク質のウエスタン分析により、見かけの分子量10〜200kDa未満のビオチン化タンパク質のスペクトルが検出された。pOSEL237を有する細菌細胞中の可溶性ビオチン化タンパク質種のパターンは、pOSEL175中のものと類似しており、天然の細胞表面発現が影響を受けていないことを示している。
【0119】
(L.jensenii中の広いpH範囲における活性2D CD4の表面発現)
乳酸菌が自然にコロニーを形成している場合、ヒト膣腔のpHはほとんどの女性で3.6〜4.5の間で変化し(Boskeyら、Infect.Immun.67:5170−5175(1999))、男性の射出物が存在する場合、一時的に中性または弱アルカリ性になる。修飾L.jenseniiでpH変化がどの様に活性2D CD4分子の表面発現に影響するかを調べるために実験を行った。細菌細胞をRogosa SLブロス中に、その通常用いられるpH(5.4)または100mM HEPESでpH7.4に緩衝して接種した。OD600約0.4までの活性生育中、培地のpHは実質的に変化しなかった。pOSEL237および249を有する細菌細胞へのmAb Sim.4の結合のフローサイトメトリー分析により、pH5.4および7.4の双方でpOSEL175中のコントロールバックグラウンドを超える有意に高い平均蛍光強度が検出された。さらに、修飾L.jenseniiをヒト膣腔内のpHに類似する酸性pHで培養した場合、表面に発現したCV−Nのレベルは上昇したままであった(データを示さず)。
【0120】
(長さ36アミノ酸のC末端アンカーモチーフのみを経由する融合で発現する2D CD4の表面提示の欠失)
LPQTGシグナル、疎水性領域およびC14またはC370配列の荷電テールを含む36アミノ酸C末端アンカーモチーフが、L.jensenii中の2D CD4の効率的な表面発現を支持するに十分であるかどうかは不明である。この疑問を解決するため、C14のC末端アンカーモチーフを有するpOSEL238、およびC370のC末端アンカーモチーフを有するpOSEL262として設計された2個の構築物を調製し、ネガティブコントロールpOSEL175および651、ならびにポジティブコントロールpOSEL237を参照して分析した。pAb T4−4でプローブ後のpOSEL238を有するL.jensenii中の細胞壁富化画分のウエスタン分析で、pOSEL237に類似する梯子状パターンが検出されなかった。さらに、pOSEL238を有する細菌細胞へのmAb Sim.4の結合のフローサイトメトリー分析により、pOSEL175を有する細胞中のバックグラウンドコントロールと比較して平均蛍光強度の増加は全く検出されなかった(図6)。同様に、pOSEL175を有する細胞およびポジティブコントロールpOSEL249と比較して、pOSEL262を有する細菌細胞のFACS分析は同様な負の結果をもたらした。これらの観察と一致して、S.pyogenesおよびL.paracasei由来の同様な長さのC末端アンカーモチーフを用いた場合、2D CD4の表面発現は行われなかった。このことは特徴的なLPQTGモチーフの上流のタンパク質配列が細胞壁固定プロセスに有意に寄与し、L.jenseniiの細胞壁上に生物活性タンパク質を提示するために必要であることを示唆している。
【0121】
(生物活性タンパク質の最適表面提示のための、LPQTGモチーフ上流の反復細胞壁貫通配列の規定の長さの必要性)
天然のC370配列は、LPQTGモチーフの上流のC末端領域中にグラム陽性細菌中の多くの細胞壁アンカータンパク質の特性である8個のほぼ同一の縦列反復を含む(図1)。2.5回反復配列がpOSEL249のアンカー配列中に含まれるが、異なった長さの上流配列が表面タンパク質提示を最大にするために使用し得るかどうかは、決定しなければならないことである。従って、C370配列の0、1、2、4、7および8個の反復を有する数個の構築物を調製した。それらはそれぞれ、pOSEL262、268、278、280、281および276と名付けられた。正しく折り畳まれた立体構造をとる2D CD4分子のレベルを決定するため、形質転換細菌をフローサイトメトリー分析のためにmAb Sim.4で探索した(図7)。細菌を有するpOSEL262(ゼロ反復)では、平均蛍光強度はネガティブコントロールpOSEL175と区別できず、異種タンパク質を適切に表面発現するためには反復配列が必要であることを示唆している。さらに、反復数をpOSEL262の0からpOSEL278の3へ増加すると、蛍光強度が有意に増加した。反復数をさらに増加しても、蛍光強度は一定のままであった。
【0122】
(様々な乳酸杆菌種におけるタンパク質の表面提示を支持するL.jenseniiのネイティブアンカー配列の利用)
L.jensenii 1153に由来するC370のアンカー配列が、他のL.jensenii株またはヒト起源の乳酸杆菌種中でタンパク質の表面提示を提供するか否かを決定するため、c−MycエピトープをC370配列のCWA200に融合するために設計されたpOSEL175またはpOSEL241(図8A)をL.jensenii Xna、L.gasseri 1151、およびL.casei Q中にエレクトロポレーションにより導入した。pOSEL241を有するポジティブコントロールL.jensenii 1153を参照して、形質転換細菌をウエスタン分析およびフローサイトメトリー分析で分析した。c−Mycに対するmAbによる探索後の細胞壁消化物のウエスタン分析により、L.jensenii 1153中のパターンと類似しているがL.casei Q中のパターンとはあまり似ていない、pOSEL241を有する形質転換L.jensenii XnaおよびL.gasseri 1151におけるラダーパターンを検出した(図8B)。細菌細胞をc−Mycに対するmAbで免疫染色した後のフローサイトメトリー分析により、pOSEL175を有する全ての乳酸杆菌種で低レベルの蛍光を検出したが(図8C)、表面提示c−Mycエピトープに結合する抗体への結合の結果、pOSEL241を有するL.jensenii XnaおよびL.gasseri 1151中での蛍光強度の上昇を検出した。さらに、pOSEL175を有するL.caseiと比較して、241を有するL.casei Qの蛍光強度は、さらに約19倍増加していた。これらのデータをまとめると、L.jensenii 1153に由来するアンカー配列は、ヒト起源の乳酸杆菌種も含めた多様な乳酸杆菌種中で、タンパク質の表面提示の支持に広く有用であることを明らかに示す。
【0123】
(L.jensenii中の2D CD4の表面発現に対するLPXTGモチーフの突然変異誘発の効果)
よく研究された細胞壁アンカータンパク質であるStaphylococcus aureusのプロテインAが、LPETG細胞壁分類モチーフ上で突然変異した場合、LPQTG中のアミノ酸プロリン(P)をアミノ酸アスパラギン(N)で置換することによりタンパク質表面提示の効率が減少したが、トレオニン(T)のセリン(S)による置換は、タンパク質表面提示の効率にほとんど影響がないことが見出された(NavarreおよびSchneewind、Microbiol.Mol.Biol.Rev.63:174−229(1999))。この研究は、P残基がLPXTGモチーフ中でおそらく最も重要な残基であり、T残基を類似のアミノ酸であるSで置換し得ることを示した。C14およびC370中のLPQTGモチーフが実際に重要な分類信号であるか否かを決定するため、LPQTG配列中のPおよびTの重要性を検討した。C14配列およびC370配列双方上のLPQTGモチーフ内に点突然変異をPCRにより生じさせた。P残基をアラニン(A)またはアスパラギン(N)に突然変異させ;アミノ酸Tを、A、Sまたはグリシン(G)に突然変異させ;LPXTGモチーフ中のアミノ酸GをAに突然変異させた。変化したLPQTGモチーフを有するプラスミドをそれぞれpOSEL237P(A)、pOSEL237P(N)、pOSEL237T(A)、pOSEL237T(G)、pOSEL237T(S)、pOSEL237G(A)、pOSEL249P(A)、pOSEL249P(N)、pOSEL249T(A)、pOSEL249T(G)、pOSEL249T(S)およびpOSEL249G(A)と名付けた。突然変異した構築物を有するL.jensenii 1153のウエスタン分析およびフローサイトメトリー分析を行った。親pOSEL237およびpOSEL249を有するL.jenseniiと比較して、pOSEL237P(A)、pOSEL237P(N)、pOSEL249P(A)およびpOSEL249P(N)を有するL.jenseniiは、pAb T4−4による細胞壁富化タンパク質画分のウエスタンブロットにおいて、特徴的な、より高分子量種のスペクトルを示さなかった。一方、2D CD4−CWA200融合タンパク質の馴化培地中への分泌は顕著に増加し、2D CD4−CWA200融合タンパク質が細胞壁に共有結合していないことを示した。野生型pOSEL237およびpOSEL249で観察された結果と同様に、pOSEL237T(S)およびpOSEL249T(S)を有するL.jenseniiの細胞壁消化において、より高分子量種の特徴的なスペクトルは明瞭であり、このことは、C14およびC370由来のLPQTG内のアミノ酸Tを、Sによって効果的に置換し得ることを示唆していた(データ示さず)。
【0124】
LPXTGの突然変異誘発のL.jensenii表面タンパク質提示に対する効果をさらに決定するため、pOSEL175、651、237、249を有するL.jensenii株と一緒に様々な突然変異構築物をpAb T4−4またはmAb Sim.4で探索し、次いでフローサイトメトリーで抗体結合を分析した。pOSEL237を有する細菌細胞と比較してpOSEL237P(A)、pOSEL237P(N)を有する細菌細胞、およびpOSEL249を有する細菌細胞と比較してpOSEL249P(A)、pOSEL249P(N)を有する細菌細胞では平均蛍光強度がかなり減少し、このことは、細胞表面の2D CD4タンパク質の提示が、あったとしても、はるかに少ないことを示していた。しかしながら、pOSEL237T(S)、pOSEL237(T)A、pOSEL249T(S)およびpOSEL249(T)Aを有する細菌細胞の平均蛍光強度はpOSEL237および249を有するL.jenseniiに匹敵し、このことは、TをSまたはAに置換することが細胞壁固定の効率にほとんど効果がないことを示した(図9)。
【0125】
ウエスタンブロットおよびフローサイトメトリー分析のデータは、C14およびC370のLPQTGモチーフ内に含まれるアミノ酸Pは容易に置換され得ないことを示す。対照的に、アミノ酸Tは、SまたはAで置換され得、乳酸杆菌属の細胞壁に効率的に固定されるタンパク質を生成する。
【0126】
(L.jenseniiにおける2D CD4の表面発現に対するC末端正荷電テールの欠失の効果)
グラム陽性細胞壁アンカードメインの特徴の一つは、正に荷電したアミノ酸がタンパク質のC末端極に伸びていることである。M6タンパク質では、この配列(KRKEEN)が重要な細胞表面保持信号として働く。これらの標示配列は、ブドウ球菌属(Staphlyococcus)、エンテロコッカス属、リステリア属および乳酸杆菌属を含む他のグラム陽性細菌でも見出されている(NavarreおよびSchneewind、Microbio.Mol.Biol.Rev.63:174−229(1999))。2つの配列RKKRQK1765およびKKKRKDDEA1903がそれぞれ、C14推定アンカー配列およびC370推定アンカー配列中の正に荷電したテールとして同定された(図1)。これらの2個の配列が細胞表面保持信号として働くか否かを決めるため、一連の欠失構築物を作製した(図10)。それらは、それぞれpOSEL237−5、pOSEL237−6、pOSEL237−7、pOSEL249−8、pOSEL249−9およびpOSEL249−10と名付けられた。
【0127】
これらの構築物を有するL.jenseniiのウエスタン分析およびフローサイトメトリー分析を行った。CWA200と融合して2D CD4を表す、48kDaで移動するタンパク質種、SDS−PAGE後、pAb T4−4によって、荷電テールノックアウト構築物を有する全てのL.jenseniiで検出され得る。分泌タンパク質は、pOSEL237−5、pOSEL237−6、pOSEL237−7、pOSEL249−8、pOSEL249−9およびpOSEL249−10を有するL.jensenii細胞において、親のpOSEL237およびpOSEL249を有する細胞よりも多かった。全欠失突然変異体に由来する細胞壁富化画分中のタンパク質のウエスタン分析は、pOSEL237または249を有するL.jenseniiで観察された特徴的なラダーパターンを検出しなかった(データを示さず)。これらのデータは、2D CD4−CWA200融合タンパク質が細胞壁に共有結合していないことを示唆した。
【0128】
抗CD4 pAb T4−4またはmAb Sim.4による探索後の改変L.jenseniiのフローサイトメトリー分析は、親のpOSEL237または249を有する細菌細胞と比較して、これらの突然変異プラスミドを有する細菌細胞における平均蛍光強度の顕著な減少を検出した(図11)。これらのデータは、C14およびC370双方の正に荷電したC末端の欠失が、細胞壁に固着して異種タンパク質を提示するそれらの能力を阻害したことを、結論として示した。
【0129】
(細胞壁アンカーシグナルとしてのLPQTGモチーフの柔軟性)
グラム陽性細菌由来のほとんどの細胞壁に固定されたタンパク質は、同じ識別信号LPXTGを共有するが、一部タンパク質は、異なったモチーフを有する。例えば、L.paracaseiのPrtPに対する識別信号は、LPKTAである(HolckおよびNaes、J.Gen.Microbiol.138:1353−1364(1992))。peptostreptococcus magnusのプロテインLおよびヒト血清アルブミン結合タンパク質は、LPXAGのモチーフを共有する(de ChateauおよびL.Bjorck、J.Biol.Chem.269:12147−12151(1994);Kellerら、EMBO J.11:863−874(1992);Murphyら、DNA Seq.4:259−265(1994))。C14アンカータンパク質またはC370アンカータンパク質中でLPQTGがLPQAGまたはLPQSGに突然変異した場合、SDS−PAGE後にフローサイトメトリーまたはウエスタンブロットで測定した2D CD4の表面提示の減少は僅かであった。しかしながら、以下の証拠に基づくように、これらの配列のみでは、膣由来の乳酸桿菌の細胞壁へタンパク質を固定するには不十分である:1)36アミノ酸のC末端アンカードメイン単独では、c−Mycエピトープまたは2D CD4を細胞表面に固定しなかった、2)200アミノ酸までの上流配列が含まれる場合でさえも、プロトタイプM6細胞壁アンカー配列(S.pyogenesのemm6遺伝子によりコードされる)は膣由来乳酸桿菌の細胞壁へ異種タンパク質を固定しなかった(本発明者らは、L.paracasei由来のLPXTAモチーフを用いて同様な結果を見出した)、および3)C191タンパク質は、有効なアンカーではなかった。これらの知見は、C14およびC370のCWA200領域内に含まれる他の上流配列もまた、細胞壁固定プロセスに大きく寄与することを示している。
【0130】
(C14およびC370のCWA200と融合した場合の2D CD4生物活性の増大)
生物活性を評価するため、pOSEL237−7およびpOSEL249−10を有するL.jensenii 1153から放出されたC14タンパク質およびC370タンパク質の2D CD4−CWA200を、CD4 ELISAにより分析した。pOSEL651、pOSEL237−7およびpOSEL249−10を有する細菌細胞を、Rogosa SLブロス中で種々の細胞密度に生育させた。次いで、無細胞馴化培地を回収した。OD600=0.8で、ウエスタンブロットで測定した培地中に放出されたpOSEL651からの2D CD4の量とpOSEL237−7または249−10からの2D CD4−CWA200の量とは類似していた。それにもかかわらず、pOSEL237−7およびpOSEL249−10を有する細菌細胞から放出された2D CD4−CWA200は、pOSEL651を有する細菌細胞からの2D CD4タンパク質と比較した場合、約2〜3倍高い活性を示した。C14またはC370のCWA200領域の2D CD4への融合は、おそらくはタンパク質折り畳みプロセスを補助することにより、このタンパク質の生物活性を増強する様にみえた。この同じ知見は、gp120結合アッセイを用いて確認された(データ示さず)。これらのタンパク質のウエスタンブロット分析は、2D CD4−CWA200が、2D CD4よりも有意に安定であり、おそらくその生物活性の増強に寄与しているということを示唆する。
【0131】
(材料および方法)
(細菌株および培養)
L.jensenii、L.crispatus、L.gasseriおよびL.caseiのヒト膣株を健康な女性から得た膣サンプルの細菌培養により単離した。乳酸杆菌rRNAに特異的な2つのプライマーを用いて、16S−23S遺伝子間スペーサー領域の増幅後に、GenBankに保存された参照株のDNA配列に対して細菌株の遺伝型決定した(Tannockら、Appl.Environ.Microbiol.65:4264−4267(1999))。株をMRSまたはRogosa SLブロス(Difco、Detroit、MI)中、またはMRS寒天プレート上で、37℃、5%CO2で常法により生育させた。
【0132】
(Lactobacillus jensenii 1153のゲノムDNAの単離)
L.crispatus JCM 5810から染色体DNAを単離するために先に用いたプロトコール(Sillanpaaら、J.Bacteriol.182:6440−6450(2000))の改変に基づいて、L.jensenii 1153の染色体DNAを単離した。L.jensenii細菌を200mlのMRS培地中、37℃、5%CO2で600nmにおける光学密度1.0(OD600=1.0)まで増殖させた。6,600×g、10分間の遠心により細胞を収集し、25mM Tris−HCl(pH8.0)、10mM EDTA、50mMグルコース中で1回洗浄し、細菌培養液100mlあたり2.5mlの20mM Tris(pH8.0)、5mlの24%エチレングリコール8000、および2.5mlのリゾチーム(4mg/ml、Sigma Chemical Co.,St Louis、Mo)を加えた後に懸濁した。得られた細胞懸濁液を37℃で1時間インキュベーションした。0.2M EDTAを5ml添加し、細胞を4℃、1000×gで10分間遠心し、50μlのムタノリシン(15,000U/ml、Sigma Chemical Co.)を含む20mM Tris(pH8.0)10ml中に再懸濁した。37℃で1時間インキュベーショした後、1.5mlの9%Sarkosyl(Sigma Chemical Co.)および3mlの5M NaClを添加して細胞を溶解した。次いで細胞溶解液を2.9mlの5M過塩素酸ナトリウムと混合した。染色体DNAを、17.5mlのクロロホルム−イソアミルアルコール(24:1 v/v)で抽出し、エタノールで沈殿し、風乾し、1.5mg/mlの濃度で100mM Tris−HCl、pH8.0、1mM EDTA中に再懸濁した。最後にゲノムDNA調製物をDNaseを含まないRNaseで処理した。
【0133】
(L.jenseniiゲノムライブラリーの構築)
L.jensenii 1153のゲノムDNAをHydroShear(GeneMachines、San Carlos、CA)を用いて所望のサイズ範囲に機械的に剪断した。剪断DNA断片をT4 DNAポリメラーゼとKlenow酵素で平滑断端化し、3Kbおよび8KbのDNA断片を、アガロースゲル電気泳動後に単離し、QIAquickゲル抽出キット(Qiagen、Valencia、CA)を用いて精製した。得られたDNA断片をpUC18ベクター中に連結し、E.coli DH10B細胞(Invitrogen、Carlsbad、CA)中に形質転換して3Kbおよび8Kbゲノムライブラリーを作製した。細菌形質転換体をX−galの存在下でLBプレート上で選択し、得られたコロニーをQ−pixロボット(Genetix Ltd.、UK)を用いて96ウエルプレート中に配列した。96クローンからなるプレートを挿入サイズの均一性および非組み換え体の割合について試験して、ライブラリーの品質を決定した。どちらのライブラリーも、5%未満の非組み替え体を含み、90%を超える挿入体は、期待サイズの20%以内であった。
【0134】
(L.jenseniiゲノムの配列決定および組み立て)
L.jenseniiゲノム配列を、全ゲノムショットガン法を用いて決定した。ゲノムライブラリーから選択したクローンのプラスミドDNAを、磁気ビーズまたはローリングサークル(rolling circle)法のいずれかで精製し、ABI BigDyeターミネーターキット(Applied Biosystems、Foster City、CA)を用いて両端から配列決定した。全ての配列決定反応を、ABI PRISM 3700自動DNA配列決定装置(Applied Biosystems)上で行った。合計15,360個の配列の読み、または160個の配列プレートを行って、L.jensenii 1153ゲノムの3倍カバレージを得た。50塩基を越えるQ20(100塩基あたり予測誤差1)を生成する場合、またはより高い精度を満たす場合のみ、配列決定の読みが成功したと考えられる。配列クロマトグラフを、Phred(Ewingら、Genome Research、8:175−185(1998))を用いる塩基呼び出しおよび品質評価のために、自動的にUNIX(登録商標)システムに転送した。合格率は80%以上であり、平均読み取り長さは400〜500塩基の範囲である。配列アセンブリをParacel GenomeAssemblerまたはCAP4を用いて行った(Paracel、Inc.、Pasadena、CA)。合計484のコンティグを組み立てた。
【0135】
(L.jensenii 1153ゲノム中の細胞壁アンカーモチーフを有するタンパク質配列の同定)
グラム陽性細菌の細胞壁固定タンパク質は、保存的C末端LPXTGXモチーフを有する(Fischettiら、Mol.Microbiol.4:1603−1605(1990))。このヘキサペプチドの後には、アミノ酸の疎水性伸長と、短い荷電テールとがあり停止転送配列として知られる(Schneewindら、Cell、70:267−281(1992))。さらに、別の独自のLPXTA識別モチーフがLactobacillus paracasei中で同定された(HolckおよびNaes、J.Gen.Microbiol.138:1353−1364(1992))。ネイティブの細胞壁アンカー配列を同定するため、組み立てられたコンティグ(L.jensenii 1153の、推定75%完全なゲノム配列から得られる)の全てのリーディングフレーム中のLPXTGおよびLPXTAに類似のモチーフを同定するためのコンピュータースクリプトが作成された。推定細胞壁アンカーモチーフを有する得られたコンティグを、グラム陽性細菌中の細胞壁アンカータンパク質に対する配列相同性に対するBLAST検索によりさらに検証した。
【0136】
(シャトルベクターの構築)
これらの研究に用いられた1次シャトルベクターは、pLEM7(Fonsら、Plasmid、37:199−203(1997))の改変バージョンであるpOSEL175であった。部分IS要素を、まずSmaIで切断することによって除去し、NdeIで部分的に消化し、Klenow断片で平滑末端化し、再連結した。最後に、pOSEL144のerm遺伝子内の2個のMfeI部位を除去するため、プラスミドを部位指向性突然変異誘発に供した(Changら、Proc.Natl.Acad.Sci.USA、100:11672−11677(2003))。得られたpOSEL175プラスミドはE.coli中の複製開始点(ColE1)と乳酸杆菌属中の複製開始点(repA)の双方を有し、従って様々な乳酸杆菌属種中の異種タンパク質の発現のために用いられるシャトルベクターの骨格を含む。
【0137】
(L.jensenii中の発現カセットの構築)
L.jenseniiにおいてアンカータンパク質を都合よく表在化するために、発現カセットを構築し、pOSEL175のSacI部位およびXbaI部位中にサブクローニングした。このカセットは、乳酸杆菌適合性P23プロモーター、L.crispatusのCbsAシグナル配列、異種タンパク質をコードするDNA、およびグラム陽性細菌中の既知、または推定細胞表面タンパク質由来の共有結合性細胞壁アンカードメインを含む4つの構成要素を含む。本発明者らの、一連のプロモーターおよびシグナル配列を有する構築物の詳細な分析により、Lactococcus lactis由来のP23プロモーター(van der Vossenetら、Appl.Environ.Microbiol.53:2452−2457(1987))と、L.crispatusのCbsA由来のシグナル配列(CbsAss)との組み合わせにより、pOSEL651として設計された構築物中で2D CD4の最高レベルのタンパク質発現が駆動されることが示された(Changら、Proc.Natl.Acad.Sci.USA、100:11672−11677(2003))。SacI、EcoRI、NheI、MfeIおよびXbaIを含む独自の制限部位は、それぞれ5’末端から3’末端へと各構成要素の間に置かれた。PCRによる各構成要素の増幅は、Pfu DNAポリメラーゼを用いて行われた。本研究で詳細に述べられる融合構築物の様々な部分のPCR増幅のためのオリゴヌクレオチドプライマーとしては、以下が挙げられる:
【0138】
【化4】
表面発現のために設計されたタンパク質としては、10アミノ酸c−Mycペプチド(EQKLISEEDL)およびヒトCD4のN末端の2個の細胞外ドメイン(2D CD4)を含む最初の183残基が含まれる。2D CD4タンパク質は、好ましい乳酸杆菌コドンの使用に従うように再コード化された。全ての発現構築物は、L.jensenii中へ形質転換する前にDNA配列分析により確認された。
【0139】
(L.jenseniiの推定細胞壁アンカー配列へのc−Myc融合の構築)
タンパク質発現レベル、および生物活性タンパク質の表面提示のために規定の長さの推定細胞壁アンカー配列を使用することが実行可能であるか否かを決定するため、本発明者らは、最初にエピトープタグ化を選んだ。C末端分類モチーフの機能を乱さないために、5’末端に10アミノ酸c−Mycエピトープ(EQKLISEEDL)を含むオリゴヌクレオチドプライマーを設計し、L.jensenii 1153のゲノム由来のC14、C191およびC370を含む推定細胞壁アンカー配列のN末端にc−Mycエピトープを融合させた。c−Myc配列をこれらのタンパク質(C14、C191またはC370のC末端30アミノ酸)の細胞壁アンカーモチーフに直接融合するか、またはC末端細胞壁アンカードメインを含む配列および様々な長さの隣接上流アミノ酸に融合させた。最も注目すべきは、c−Mycが細胞壁アンカードメインと上流アミノ酸(CWA200と命名)とを含む200アミノ酸配列と融合したことである。
【0140】
【化5】
期待したサイズを有する全てのPCR産物をゲルで精製し、MfeIおよびNheIの両方で消化した。得られた断片を、MfeI/NheI二重消化pOSEL651と連結し、pOSEL239(C14配列のCWA200経由)、pOSEL240(C191配列のCWA200経由)およびpOSEL241(C370配列経由)それぞれの中にc−Myc融合体を作製した。得られたプラスミドはL.jensenii 1153中へエレクトロポレーションした。
【0141】
(細胞壁固定配列のシャトルベクター中へのサブクローニング)
C末端LPQTGアンカーモチーフを含む3個の推定表面タンパク質を選び、L.jensenii 1153の細胞壁上に外来タンパク質を発現する能力を決定した。これらの表面タンパク質のC末端LPQTGドメインを含むDNA領域、およびその上流の200アミノ酸(暫定的にCWA200領域と呼ぶ)を、下記の3組のプライマーによって増幅した:
【0142】
【化6】
C14アンカードメインの内部MfeI部位とC370ドメインの内部XbaI部位を、酵素的制限の前に部位指向性突然変異誘発により突然変異させた。予期されたサイズのPCR産物の全てをゲルで精製し、MfeIおよびXbaIの両方で消化した。得られた断片を、P23−調節性分泌2D CD4を含むMfeI/XbaI二重消化pOSEL651と連結し、プラスミドpOSEL237(C14配列のCWA200経由)、pOSEL242(C191配列のCWA200経由)およびpOSEL249(C370配列のCWA300経由)をそれぞれ作製した。あるいは、以下の2個のプライマーを用いてC14配列のC末端36アミノ酸アンカーモチーフを、同様にシャトルベクター中にクローニングした。
【0143】
【化7】
DNA配列の検証後に得られたプラスミドの全てを、L.jesneii、L.gasseriおよびL.casei中にエレクトロポレーションした。
【0144】
(C370配列の反復細胞壁貫通領域のサブクローニング)
C370LPQTGモチーフの上流の種々の反復細胞壁貫通領域を、L.jensenii 1153のゲノムDNAから増幅した。各PCR反応について同じ3’プライマー(5’−CCGTCTAGATTATGCTTCATCATCTTTTCT−3’)を、以下の5’プライマーと対で使用した。
【0145】
【化8】
これらのプライマー中のMfeI部位およびXbaI部位の両方に下線が引かれている。
【0146】
予想したサイズのPCR産物の全てをゲル精製し、MfeIおよびXbaIの双方で消化した。得られた断片を、P23調節性分泌2D CD4を含むMfeI/XbaI二重消化pOSEL237と連結し、プラスミドpOSEL262(反復無し)、pOSEL268(1反復)、pOSEL278(2反復)、pOSEL284(3反復)、pOSEL280(4反復)、pOSEL275(6反復)、pOSEL281(7反復)およびpOSEL276(8反復)をそれぞれ作製した。
【0147】
(細菌形質転換)
E.coli DH12S(Invitrogen)中に、プラスミドをエレクトロポレーションにより導入した。シャトルプラスミドを構築し維持するため、形質転換E.coli DH12S細胞を、100μg/mlのアンピシリンまたは300μg/mlのエリスロマイシンを補充したLBブロス(Difco)中で、37℃で増殖させた。DNA配列検証後、E.coli由来プラスミドを、改変したLuchanskyら(J.Dairy Sci.74:3293−3302(1991))の方法に従ってL.jensenii、L.gasseriおよびL.casei中に形質転換した。簡単に言えば、新たに播種したL.jenseniiを、MRSブロス中、37℃および5%CO2でOD600 0.6〜0.7まで培養した。細菌細胞を収集し、洗浄して952mMスクロースおよび3.5mM MgCl2中に、4℃で再懸濁した。予冷した0.2cmギャップキュベットを用いて、コンピテント細胞に1〜2μgのDNAを添加し、直ちにGene Pulser II(Bio−Rad、Hercules、CA)を用いて、2.5kV/cmおよび200オームでエレクトロポレーションした。その後、20μg/mlのエリスロマイシン(液体培地中で形質転換L.jenseniiを常法で増殖させるためにも使用される濃度である)を含む選択MRS寒天プレート上に播種する前に、37℃で2時間予め温めたMRSブロス中で細胞を回復させた。
【0148】
(推定細胞壁アンカー配列のLPXTGモチーフの部位指向性突然変異誘発)
Stratagene提供のQuickChange(登録商標)XL Site−Directed Mutagenesisキット(La Jolla、CA)を用いて点突然変異を生じさせた。プラスミドpOSEL237(C14配列のCWA200を経由して固定した2D CD4を発現)とプラスミドpOSEL249(C370配列のCWA200を経由して固定した2D CD4を発現)をテンプレートとして使用した。LPQTG、およびそのC14およびC370上の隣接配列に対応するヌクレオチド配列に基づき、突然変異誘発プライマーを設計した。
【0149】
【化9】
L.jensenii 1153の好ましいコドンを用いて置換ヌクレオチドを選択した。
【0150】
237P(A):C14のLPQTG上のプロリンをアラニンで置換した。
【0151】
【化10】
237P(N):C14のLPQTG上のプロリンをアスパラギンで置換した。
【0152】
【化11】
237T(A):C14のLPQTG上のトレオニンをアラニンで置換した。
【0153】
【化12】
237T(G):C14のLPQTG上のトレオニンをグリシンで置換した。
【0154】
【化13】
237T(S):C14のLPQTG上のトレオニンをセリンで置換した。
【0155】
【化14】
237G(A):C14のLPQTG上のグリシンをアラニン(Alanie)で置換した。
【0156】
【化15】
249P(A):C370のLPQTG上のプロリンをアラニン(Alaine)で置換した。
【0157】
【化16】
249P(N):C370のLPQTG上のプロリンをアスパラギンで置換した。
【0158】
【化17】
249T(A):C370のLPQTG上のトレオニンをアラニンで置換した。
【0159】
【化18】
249T(G):C370のLPQTG上のトレオニンをグリシンで置換した。
【0160】
【化19】
249T(S):C370のLPQTG上のトレオニンをセリンで置換した。
【0161】
【化20】
249G(A):C370のLPQTG上のグリシンをアラニンで置換した。
【0162】
【化21】
ポリメラーゼ連鎖反応(PCR)サイクル条件は95℃で50秒、60℃で50秒、および68℃で12分、全体で16サイクルであった。
【0163】
PCR反応後にDpnI酵素を、増幅混合物に添加し、親プラスミドを分解した。新たに合成されたプラスミドを製造業者の推奨に従って化学コンピテントE.coli Top10細胞(Invitrogen)中に導入した。200μg/mlのエリスロマイシンを補充したLBブロス(Difco)中でプラスミドを維持し、増幅した。DNA配列を検証後、改変したLuchanskyらの方法(J.Dairy Sci.74:3293−3302(1991))に従ってE.coli由来のプラスミドをL.jensenii中へ形質転換した。突然変異原性プラスミドを含む形質転換L.jenseniiを選択し、増殖するために、20μg/mlのエリスロマイシンを含むMRSを使用した。
【0164】
(推定細胞壁アンカータンパク質の正に荷電したC末端配列の欠失の分析)
正に荷電したアミノ酸がC14およびC370のC末端に位置する一連の欠失突然変異体を、PCR増幅により作製した。プラスミドpOSEL237およびpOSEL249を、テンプレートとして使用した。pOSEL237およびpOSEL249上の2D CD4配列に相補性のオリゴヌクレオチド(CD4F 5’−GATCGTGCTGATTCACGTCGT−3’)をフォワードプライマーとして使用した。以下のオリゴヌクレオチド(制限部位に下線)を、2D CD4 cDNAのC末端および完全C14およびC370 CWA200配列を増幅するためのリバースプライマーとして使用した:
【0165】
【化22】
全てのリバースプライマーは、XbaI制限部位を含んでいた。サイクル条件は、94℃で45秒、60℃で45秒、72℃で90秒、合計18サイクルであった。PCR産物をゲル精製し、MfeIおよびXbaIの両方で消化し、次いでMfeI/XbaI二重消化pOSEL237およびpOSEL249中へ、それぞれサブクローニングした。配列をヌクレオチド配列決定によって検証し、タンパク質分析のために、構築物をL.jensenii中へエレクトロポレーションした。
【0166】
(L.jensenii中の異種タンパク質発現のウエスタン分析)
遺伝的に改変したL.jensenii細胞を、100mM HEPES(pH7.1)で緩衝したRogosa SLブロス中、37℃、5%CO2で増殖させた。可溶性タンパク質のレベルを決定するため、12,000×gで遠心後に馴化培地を集め、最終濃度20%のTCAでタンパク質を沈殿させた。TCA沈殿物をエタノールで洗浄し、風乾し、50mM Tris−HCl(pH6.8)、0.4%SDS、6%スクロース、10mMジチオトレイトールおよび0.01%ブロモフェノールブルー(1×還元SDS−PAGE緩衝液)中で熱変性させた。L.jensenii中の細胞結合タンパク質の相対量を決めるため、細胞溶解を誘起することなく、OD600単位あたり100μLの1×SDS−PAGE緩衝液中、37℃、30分間で細菌細胞を抽出した。12,000×gで5分間遠心して収集した抽出タンパク質を、次いで熱変性した。14,000×gで遠心して可溶性タンパク質を細菌細胞から分離し、製造業者の推奨に従い、抗酸化剤の存在下で、4〜12%のNuPAGEシステム(Invitrogen)中でSDS−PAGEで分離した。電気泳動による分離後、20%メタノール、20mM Trisおよび50mMグリシン中の2フッ化ポリビニリデン膜(Millipore)上にタンパク質をエレクトロブロッティングした。次いで、このブロットをポリクローナルウサギ抗CD4抗体、T4−4(NIH AIDS Reserch and Reference Reagent Program)またはウサギ抗CV−N pAb、およびc−Mycに対するモノクローナル抗体(Invitrogen)で探索した。抗原−抗体反応を、アルカリホスファーゼ結合抗ウサギIgG用(CD4検出用)に対する色素生成検出試薬(Promega、Madison、WI)、またはセイヨウワサビペルオキシダーゼ(HRP)共役抗マウスIgG用(c−Myc検出用)の増強化学発光試薬(Amercham Biosciences、Piscataway、NJ)を用いて可視化した。同様に、c−Mycタグ化タンパク質のレベルを抗c−Mycに対するmAb(Invitrogen)で探索し、結合抗体を、HRP結合抗マウス2次抗体(Amersham Biosciences)で検出した。
【0167】
(ムラミダーゼによるL.jensenii細胞壁の酵素消化)
109個の細菌を含む細菌培養液を、12,000×gで5分間遠心した。得られた細胞ペレットを、20mM HEPES(pH7.2)で1回洗浄し、100μLの10mM Tris−HCl(pH8.0)、1mM EDTA、25%スクロース中に懸濁した(Piardら、J.Bacteriol.179:3068−3072(1997))。細菌細胞壁を、最終濃度15単位/mlのムラミダーゼ、ムタノリシン(Sigma Chemical Co.)の存在下で、37℃で1時間消化した。その後、細胞を、2,500×gで10分間遠心し、プロトプラスト富化画分から細胞壁富化画分を単離した。細胞壁またはプロトプラスト富化画分それぞれに、25μlの4×還元SDS−PAGE緩衝液または125μlの1×還元SDS−PAGE緩衝液を添加後、得られたサンプルを熱変性した。あるいは、さらなるサンプル処理なしに細胞壁富化画分中のタンパク質を分析するため、CD4 ELISAを用いた。
【0168】
(L.jensenii中の表面露出タンパク質のスルホ−NHS−ビオチンによる標識)
L.jensenii中の表面タンパク質の表面露出リシル残基を、膜不透過スルホ−N−ヒドロキシスクシンイミド(NHS)−ビオチンの使用によって探索した。NHS−ビオチンによるグラム陰性細菌Helicobacter pyloriの表面標識により、真の細胞表面タンパク質の同定が可能になる(Sabarthら、J.Biol.Chem.70:27896−27902(2002))。対数期の約109個のL.jensenii細菌を1回洗浄し、PBS中に懸濁した。スルホ−NHS−ビオチンを1mlの細胞懸濁液に最終濃度1mMで加え、連続的に回転させながら室温で30分間インキュベーションした。その後、ビオチン化反応を、50mM Tris、pH8.0を加えてクエンチし、細胞を、20mM HEPES、pH7.2で1回洗浄した。細胞溶解を誘起することなく、細胞に結合したタンパク質を、125μlの0.4%SDS、6%スクロース、10mM DTT、50mM Tris−HCl(pH6.8)中で、37℃で30分間抽出した。抽出されたタンパク質を、14,000×g、5分間の遠心により細菌細胞から分離した。熱変性後、タンパク質を、4〜12%NuPAGE(Invitrogen)中で分離した。ビオチン化タンパク質およびその移動度シフトを決定し、その後アルカリホスファターゼ結合ストレプトアビジン(strepavidin)または他の免疫プローブで探索した。
【0169】
(フローサイトメトリーによる2D CD4の表面発現の分析)
pOSEL651中に表面タンパク質発現またはタンパク質分泌のためのプラスミドを有する形質転換L.jenseniiを、20μg/mlのエリスロマイシンの存在下で、37℃、5%CO2で、MRSブロス中で一晩増殖させた(OD600>3)。特に指定しない限り、100mM HEPES、pH7.1で緩衝したエリスロマイシン含有MRSまたはRogosa SLブロス中、1:50〜100希釈で一晩培養物を培養し、次いで、継代培養した。OD600〜=0.4の1mlの細胞培養液を、12,000×gで5分間遠心した。得られた細胞ペレットを2回洗浄し、2%FBSを含む1×PBS中に懸濁した。その後、特異性抗体(2×108個の細胞あたりウサギポリクローナルT4.4に対し1:1000希釈、またはモノクローナルSim.4に対し50μg/ml)を用いて、細胞を、1×PBS中の2%FBS中で30分間表面染色し、その後FITCまたはフィコエリスリン結合抗ウサギ抗体またはフィコエリスリン結合抗マウス抗体(Becton−Dickinson、Mountain View、CA)で表面染色した。表面発現CV−Nの検出のため、同様なプロトコールを開発した。コントロールはアイソタイプが適合するモノクローナル抗体(Becton Dickinson)から構成した。標識細胞を、1%(v/v)パラホルムアルデヒドで固定し、CellQuestソフトウエアと共に作動するFACScaliburシステム(Becton−Dickinson)で解析した。バックグラウンドコントロール中の密度プロット出力(側方散乱または前方散乱 対 蛍光)を、pOSEL175を有するL.jenseniiから得た。プロット間の平均蛍光強度の変化を、細菌表面に結合する抗体の基準とし、FLOWJOソフトウエアを用いて計算した。
【0170】
(酵素結合免疫吸着アッセイ)
正しく折り畳まれた2D CD4タンパク質の濃度を、McCallusら(Viral Immunol.5:209−219(1992))に従って改変したCD4捕捉酵素結合免疫吸着分析(ELISA)で決定した。細菌馴化培地中の正しい立体配座を有する2D CD4タンパク質を、2.5μg/mlのモノクローナル抗体Sim.4により、MaxiSorp96ウエルプレート(Nalge Nunc International、Denmark)上に捕捉した。0.05%Tween 20を含む1×Tris緩衝生理食塩水中での洗浄後、E.coli由来のリフォールディングされた2D CD4標準を参照して、結合CD4分子を、ウサギポリクローナル抗体で探索し、次いで、T4−4を、3,3’,5,5’−テトラメチルベンジジン(Neogen Corp.,Lexington、KY)の存在下で、室温で暗所に30分間置いて、セイヨウワサビペルオキシダーゼ結合抗ウサギIgG(Amersham Biosciences)で検出した。100μlの0.5M H2SO4を添加した後反応を停止させ、マイクロプレートリーダー(Molecular Devices、Sunyvale、CA)を用いて450nmでの吸光度を読み取った。
【0171】
上記実施例は、本発明を説明するために提供されるが、その範囲を制限するものではない。本発明の他の変法は当業者に容易に明らかであり、かつ添付の特許請求の範囲によって包含される。本明細書において引用する全ての刊行物、データベース、GenBank配列、特許および特許出願は、本明細書によって参考として援用される。
【0172】
(配列表)
【0173】
【化23】
【0174】
【化24】
【0175】
【化25】
【0176】
【化26】
【図面の簡単な説明】
【0177】
【図1】図1はL.jensenii 1153のゲノム配列決定後に同定された3種の細胞壁アンカータンパク質の構造を示す。3つのタンパク質の全ては疎水性領域および荷電C−末端テールに先行するLPQTG識別シグナルを有し、独自の長い繰り返し配列を有する。CWAはLPQTGモチーフの上流の推定細胞壁関連領域を示す。
【図2A】図2AはL.jensenii 1153のゲノム配列決定から得られた細胞壁アンカー配列(C14、C191およびC370)を示す。CWA200領域と共にアンカーモチーフに下線が引かれている。CWA200はLPQTGモチーフの上流の約200アミノ酸の推定細胞壁関連領域または推定細胞壁貫通領域を表す。
【図2B】図2BはL.jensenii 1153のゲノム配列決定から得られた細胞壁アンカー配列(C14、C191およびC370)を示す。CWA200領域と共にアンカーモチーフに下線が引かれている。CWA200はLPQTGモチーフの上流の約200アミノ酸の推定細胞壁関連領域または推定細胞壁貫通領域を表す。
【図2C】図2CはL.jensenii 1153のゲノム配列決定から得られた細胞壁アンカー配列(C14、C191およびC370)を示す。CWA200領域と共にアンカーモチーフに下線が引かれている。CWA200はLPQTGモチーフの上流の約200アミノ酸の推定細胞壁関連領域または推定細胞壁貫通領域を表す。
【図3】図3は、MRSブロス(A)またはRogosa SLブロス(B)中、37℃および5%CO2で培養した場合の、形質転換L.jensenii 1153のムタノリシン消化後の、SDS抽出可能タンパク質および細胞壁富化画分のウエスタン分析の結果を示す。還元性SDS−PAGEでの分離後、これらのタンパク質をc−Mycに対するモノクローナル抗体(mAb)を用いて探索するために、PVDF膜にエレクトロブロッティングした。
【図4】図4は、Rogosa SLブロス中で37℃、5%CO2で培養した場合の、形質転換L.jensenii 1153のムタノリシン消化後の、細胞壁富化画分のウエスタン分析の結果を示す。還元性SDS−PAGE中で分離後、CD4(T4−4)に対するポリクローナル抗体(pAb)で探索するために、タンパク質をPVDF膜にエレクトロブロッティングした。発現構築物は以下の要素を含んでいた:pOSEL651中のP23プロモーター−CbsAシグナル配列(CbsAss)−2D CD4;p237中のC14配列のP23プロモーター−CbsAss−2D CD4−CWA200−アンカー;pOSEL242中のC191配列のP23プロモーター−CbsAss−2D CD4−CWA200−アンカー;pOSEL249中のC370配列のP23プロモーター−CbsAss−2D CD4−CWA200−アンカー。CWA200は、C−末端アンカードメインの上流の約200アミノ酸を表す。
【図5】図5は、2D CD4の分泌または表面固定のために設計したL.jensenii 1153を有するプラスミドのフローサイトメトリー分析の結果を示す。細菌細胞を、CD4に対する(T4−4)ウサギpAbで探索し、次いでFITC結合抗ウサギ抗体(A)で探索した。あるいは細菌細胞を、mAb Sim.4で探索し、次いでPE結合抗マウスIgG(B)で探索した。コントロールは、未染色細胞またはフルオロクローム結合2次抗体で探索した細胞である。細菌表面へ結合する抗体の基準としての蛍光濃度を、FLOWJOソフトウエアを用いて計算した。
【図6】図6は、長さが36アミノ酸であるC−末端アンカーモチーフが、2D CD4の表面発現を駆動するのは不十分であることを示す。(A)L.jensenii中のネイティブアンカー配列を用いる2D CD4の表面発現用に設計された構築物。(B)pOSEL238またはpOSEL237を有するL.jensenii 1153のフローサイトメトリー分析。細菌細胞をCD4に対するmAb Sim.4で探索し、次いでフィコエリスリン(PE)結合抗マウス抗体で探索した。コントロールは無染色細胞またはPE結合2次抗体で探索した細胞である。
【図7】図7は、C370配列中のLPQTG識別シグナルの上流の種々の数の繰り返し細胞壁貫通配列により影響を受ける場合のL.jensenii 1153中における2D CD4の表面発現を示す。正しく折り畳まれた立体構造をとる表面露出2D CD4分子を、以下のプラスミドを有する細菌細胞中のフローサイトメトリー分析のために、mAb Sim.4で探索した:175、陰性コントロール;249、2.5回反復;262、反復なし;268、1回反復;278、2回反復;280、4回反復;281、7回反復;276、8回反復。
【図8】図8は、ヒト起源の様々な乳酸桿菌種におけるc−Mycタグ化タンパク質の表面提示を示す。(A)P23プロモーターおよびCbsAシグナル配列(CbsAss)の制御下におけるC370配列のc−Mycタグ化CWA200の発現のために設計されたpOSEL241の概念図。(B)形質転換されたL.jensenii、L.gasseriおよびL.caseiのムタノリシン消化後の細胞壁富化画分のウエスタン分析。還元性SDS−PAGE中で分離後、c−Mycに対するmAbで探索するためにタンパク質をPVDF膜にエレクトロブロッティングした。(C)pOSEL241を有するヒト膣の乳酸桿菌単離物のフローサイトメトリー分析。細菌細胞をc−Mycに対するmAbで探索し、次いでフィコエリスリン(PE)結合抗マウス抗体で探索した。コントロールは、非染色細胞またはPE結合2次抗体で探索した細胞から構成された。
【図9】図9は、L.jensenii 1153中の2D−CD4−CWA200の表面提示上のC14配列およびC370配列のLPQTGモチーフにおける点突然変異の効果を示す。細菌細胞を、あらかじめ力価を測定したmAb Sim.4(A)またはpAb T4−4(B)を用いて表面染色し、次いでPE結合抗マウス抗体またはFITC結合抗ウサギ抗体で探索した。フローサイトメトリー分析をFACScaliburシステム中で行った。pOSEL237およびpOSEL249の細胞表面上に提示されたタンパク質と、突然変異誘発性構築物を有する細菌細胞中のタンパク質との間の差を、平均蛍光強度で表した。pOSEL237またはpOSEL249を有する細菌細胞中の2D CD4の表面提示を、任意に100%と設定した。
【図10】図10は、C14配列およびC370配列のC−末端荷電テール中の欠失構築物の概念図を示す。
【図11】図11は、2D CD4−CWA200の表面提示上のC14およびC370のC−末端荷電テール中の配列欠失の影響を示す。細菌細胞をあらかじめ力価を測定したpAb T4−4(A)またはmAb Sim.4(B)を用いて表面染色し、次いでFITC結合抗ウサギまたはPE結合抗マウス抗体で探索した。細胞壁アンカータンパク質への抗体の結合をFACScaliburシステムを用いるフローサイトメトリーで分析した。pOSEL237またはpOSEL249の細胞表面上に提示されるタンパク質と、突然変異誘発構築物を有する細菌細胞中のタンパク質との間の差を、平均蛍光強度として表した。pOSEL237およびpOSEL249を有する細菌細胞中の2D CD4の表面提示を、任意に100%と設定した。
【図12】図12は、pOSEL651を有するL.jensenii 1153由来の2D CD4に対する、pOSEL237−7およびpOSEL249−10を有するL.jensenii 1153中の分泌2D CD4−CWA200の活性の比較を示す。無細胞馴化培地中で正しく、適切に折り畳まれた立体構造をとるタンパク質を認識するために、CD4 ELISAを設計した。タンパク質量を、そのpAb T4−4に対する免疫応答性に基づいて正規化した。pOSEL651を有する細菌細胞から放出される可溶性2D CD4タンパク質を、任意に100%に設定した。
【特許請求の範囲】
【請求項1】
発現カセットを含む乳酸杆菌属細菌であって、該発現カセットはシグナル配列および生物活性ポリペプチドをコードするポリヌクレオチドと作動可能に結合するプロモーターを含み、該生物活性ポリペプチドは異種カルボキシル末端細胞壁標的領域と結合し、該異種カルボキシル末端細胞壁標的領域は、以下の順番で、以下:
細胞壁関連配列;
LPQ(S/A/T)(G/A);および
疎水性配列、
を含む、乳酸杆菌属細菌。
【請求項2】
前記細胞壁関連配列が、少なくとも50個のアミノ酸を含む、請求項1に記載の乳酸杆菌属細菌。
【請求項3】
前記細胞壁関連配列が、少なくとも200個のアミノ酸を含む、請求項1に記載の乳酸杆菌属細菌。
【請求項4】
前記異種カルボキシル末端細胞壁標的領域が、該領域のカルボキシル末端において、荷電した配列をさらに含む、請求項1に記載の乳酸杆菌属細菌。
【請求項5】
前記乳酸杆菌属細菌が、膣コロニー形成株である、請求項1に記載の乳酸杆菌属細菌。
【請求項6】
前記乳酸杆菌属細菌が、L.jensenii、L.gasseri、およびL.caseiからなる群より選択される、請求項1に記載の乳酸杆菌属細菌。
【請求項7】
前記細胞壁標的領域が、アミノ酸配列LPQSGを含む、請求項1に記載の乳酸杆菌属細菌。
【請求項8】
前記細胞壁標的領域が、アミノ酸配列LPQAGを含む、請求項1に記載の乳酸杆菌属細菌。
【請求項9】
前記細胞壁標的領域が、アミノ酸配列LPQTGを含む、請求項1に記載の乳酸杆菌属細菌。
【請求項10】
前記細胞壁標的領域が、アミノ酸配列LPQTAを含む、請求項1に記載の乳酸杆菌属細菌。
【請求項11】
前記細胞壁標的領域が、配列番号7を含む、請求項1に記載の乳酸杆菌属細菌。
【請求項12】
前記細胞壁標的領域が、配列番号8を含む、請求項1に記載の乳酸杆菌属細菌。
【請求項13】
前記生物活性ポリペプチドが、細菌の細胞壁において発現される、請求項1に記載の乳酸杆菌属細菌。
【請求項14】
前記生物活性ポリペプチドが、10アミノ酸と600アミノ酸との間である、請求項1に記載の乳酸杆菌属細菌。
【請求項15】
前記生物活性タンパク質が病原体と接触する場合、該生物活性タンパク質が該病原体と結合する、請求項1に記載の乳酸杆菌属細菌。
【請求項16】
前記病原体が、細菌病原体である、請求項15に記載の乳酸杆菌属細菌。
【請求項17】
前記病原体が、真菌病原体である、請求項15に記載の乳酸杆菌属細菌。
【請求項18】
前記病原体が、ウイルス病原体である、請求項15に記載の乳酸杆菌属細菌。
【請求項19】
前記ウイルス病原体が、HIVである、請求項18に記載の乳酸杆菌属細菌。
【請求項20】
前記生物活性タンパク質が、CD4またはCD4のHIV結合フラグメントである、請求項19に記載の乳酸杆菌属細菌。
【請求項21】
前記生物活性タンパク質が、2D−CD4である、請求項19に記載の乳酸杆菌属細菌。
【請求項22】
前記生物活性タンパク質が、シアノビリン−Nまたはシアノビリン−Nのウイルス結合フラグメントである、請求項18に記載の乳酸杆菌属細菌。
【請求項23】
前記ウイルス病原体が、単純疱疹ウイルスである、請求項18に記載の乳酸杆菌属細菌。
【請求項24】
前記生物活性タンパク質が、単純疱疹ウイルス侵入媒体C(HveC)であるか、またはHveCのウイルス結合フラグメントである、請求項18に記載の乳酸杆菌属細菌。
【請求項25】
前記生物活性ポリペプチドが、乳酸杆菌属細菌から放出される、請求項1に記載の乳酸杆菌属細菌。
【請求項26】
前記生物活性ポリペプチドが、乳酸杆菌属細菌の細胞壁に固定される、請求項4に記載の乳酸杆菌属細菌。
【請求項27】
乳酸杆菌属細菌の細胞壁中において生物活性ポリペプチドを発現する方法であって、該方法は、以下:
発現カセットを含む乳酸杆菌属細菌を提供する工程であって、該発現カセットは、シグナル配列および生物活性ポリペプチドをコードするポリヌクレオチドと作動可能に結合するプロモーターを含み、該生物活性ポリペプチドは異種カルボキシル末端細胞壁標的領域と結合し、そして該異種カルボキシル末端細胞壁標的領域は、以下の順番で、以下:
細胞壁関連配列;
LPQ(S/A/T)(G/A);および
疎水性配列
を含む、工程;ならびに
該ポリペプチドの発現を誘導するための条件下で細菌を培養する工程であって、該工程により、乳酸杆菌属細菌の細胞壁中で生物活性ポリペプチドを発現する、工程、
を包含する、方法。
【請求項28】
前記細胞壁関連配列が、少なくとも50個のアミノ酸を含む、請求項27に記載の方法。
【請求項29】
前記細胞壁関連配列が、少なくとも200個のアミノ酸を含む、請求項27に記載の方法。
【請求項30】
前記異種カルボキシル末端細胞壁標的領域が、該領域のカルボキシル末端において、荷電した配列をさらに含む、請求項27に記載の方法。
【請求項31】
前記提供する工程が、前記発現カセットを前記細菌中に輸送する工程を包含する、請求項27に記載の方法。
【請求項32】
前記細胞壁標的領域が、アミノ酸配列LPQSGを含む、請求項27に記載の方法。
【請求項33】
前記細胞壁標的領域が、アミノ酸配列LPQAGを含む、請求項27に記載の方法。
【請求項34】
前記細胞壁標的領域が、アミノ酸配列LPQTGを含む、請求項27に記載の方法。
【請求項35】
前記細胞壁標的領域が、アミノ酸配列LPQTAを含む、請求項27に記載の方法。
【請求項36】
前記細胞壁標的領域が、配列番号7を含む、請求項27に記載の方法。
【請求項37】
前記細胞壁標的領域が、配列番号8を含む、請求項27に記載の方法。
【請求項38】
前記細胞壁標的領域が、少なくとも200個のアミノ酸を含む、請求項27に記載の方法。
【請求項39】
前記乳酸杆菌属細菌が、膣コロニー形成株である、請求項27に記載の方法。
【請求項40】
前記乳酸杆菌属細菌が、L.jensenii、L.gasseri、およびL.caseiからなる群より選択される、請求項27に記載の方法。
【請求項41】
前記生物活性ポリペプチドが、10アミノ酸と600アミノ酸との間である、請求項27に記載の方法。
【請求項42】
前記生物活性タンパク質が病原体と接触する場合、該生物活性タンパク質が該病原体に結合する、請求項27に記載の方法。
【請求項43】
前記病原体が、細菌病原体である、請求項42に記載の方法。
【請求項44】
前記病原体が、真菌病原体である、請求項42に記載の方法。
【請求項45】
前記病原体が、ウイルス病原体である、請求項42に記載の方法。
【請求項46】
前記ウイルス病原体が、HIVである、請求項45に記載の方法。
【請求項47】
前記生物活性タンパク質が、CD4またはCD4のHIV結合フラグメントである、請求項46に記載の方法。
【請求項48】
前記生物活性タンパク質が、2D−CD4である、請求項46に記載の方法。
【請求項49】
前記生物活性タンパク質が、シアノビリン−Nまたはシアノビリン−Nのウイルス結合フラグメントである、請求項45に記載の方法。
【請求項50】
前記ウイルス病原体が、単純疱疹ウイルスである、請求項45に記載の方法。
【請求項51】
前記生物活性タンパク質が、単純疱疹ウイルス侵入媒体C(HveC)またはHveCのウイルス結合フラグメントである、請求項45に記載の方法。
【請求項52】
前記生物活性ポリペプチドが、乳酸杆菌属細菌から放出される、請求項27に記載の方法。
【請求項53】
前記生物活性ポリペプチドが、乳酸杆菌属細菌の細胞壁に固定される、請求項30に記載の方法。
【請求項54】
生物活性タンパク質を哺乳動物粘膜表面に提供する方法であって、該方法は、以下:
粘膜表面と、異種カルボキシル末端細胞壁標的領域に結合された生物活性ポリペプチドに結合されたシグナル配列を発現するように組み換えで変化した乳酸杆菌属細菌とを、接触させる工程を包含し、該異種カルボキシル末端細胞壁標的領域は、以下の順番で、以下:
細胞壁関連配列;
LPQ(S/A/T)(G/A);および
疎水性配列
を含み、
ここで、該生物活性ポリペプチドは、該粘膜表面から収集されたサンプル中で、検出可能な量で発現される、方法。
【請求項55】
前記細胞壁関連配列が、少なくとも50個のアミノ酸を含む、請求項54に記載の方法。
【請求項56】
前記細胞壁関連配列が、少なくとも200個のアミノ酸を含む、請求項54に記載の方法。
【請求項57】
前記異種カルボキシル末端細胞壁標的領域が、該領域のカルボキシル末端において、荷電した配列をさらに含む、請求項54に記載の方法。
【請求項58】
前記乳酸杆菌属細菌が、L.jensenii、L.gasseri、およびL.caseiからなる群より選択される、請求項54に記載の方法。
【請求項59】
前記粘膜表面が、膣内に存在する、請求項54に記載の方法。
【請求項60】
前記粘膜表面が、胃腸管内に存在する、請求項54に記載の方法。
【請求項61】
前記接触させる工程が、前記乳酸杆菌属細菌を経口投与する工程を包含する、請求項54に記載の方法。
【請求項62】
前記接触させる工程が、前記乳酸杆菌属細菌を膣投与する工程を包含する、請求項54に記載の方法。
【請求項63】
前記接触工程が、前記乳酸杆菌属細菌を直腸投与する工程を包含する、請求項54に記載の方法。
【請求項64】
シグナル配列および生物活性ポリペプチドをコードするポリヌクレオチドに作動可能に結合するプロモーターを含む発現カセットであって、ここで、該生物活性ポリペプチドが、配列番号7または配列番号8に結合する、発現カセット。
【請求項65】
請求項64に記載の発現カセットを含む、ベクター。
【請求項1】
発現カセットを含む乳酸杆菌属細菌であって、該発現カセットはシグナル配列および生物活性ポリペプチドをコードするポリヌクレオチドと作動可能に結合するプロモーターを含み、該生物活性ポリペプチドは異種カルボキシル末端細胞壁標的領域と結合し、該異種カルボキシル末端細胞壁標的領域は、以下の順番で、以下:
細胞壁関連配列;
LPQ(S/A/T)(G/A);および
疎水性配列、
を含む、乳酸杆菌属細菌。
【請求項2】
前記細胞壁関連配列が、少なくとも50個のアミノ酸を含む、請求項1に記載の乳酸杆菌属細菌。
【請求項3】
前記細胞壁関連配列が、少なくとも200個のアミノ酸を含む、請求項1に記載の乳酸杆菌属細菌。
【請求項4】
前記異種カルボキシル末端細胞壁標的領域が、該領域のカルボキシル末端において、荷電した配列をさらに含む、請求項1に記載の乳酸杆菌属細菌。
【請求項5】
前記乳酸杆菌属細菌が、膣コロニー形成株である、請求項1に記載の乳酸杆菌属細菌。
【請求項6】
前記乳酸杆菌属細菌が、L.jensenii、L.gasseri、およびL.caseiからなる群より選択される、請求項1に記載の乳酸杆菌属細菌。
【請求項7】
前記細胞壁標的領域が、アミノ酸配列LPQSGを含む、請求項1に記載の乳酸杆菌属細菌。
【請求項8】
前記細胞壁標的領域が、アミノ酸配列LPQAGを含む、請求項1に記載の乳酸杆菌属細菌。
【請求項9】
前記細胞壁標的領域が、アミノ酸配列LPQTGを含む、請求項1に記載の乳酸杆菌属細菌。
【請求項10】
前記細胞壁標的領域が、アミノ酸配列LPQTAを含む、請求項1に記載の乳酸杆菌属細菌。
【請求項11】
前記細胞壁標的領域が、配列番号7を含む、請求項1に記載の乳酸杆菌属細菌。
【請求項12】
前記細胞壁標的領域が、配列番号8を含む、請求項1に記載の乳酸杆菌属細菌。
【請求項13】
前記生物活性ポリペプチドが、細菌の細胞壁において発現される、請求項1に記載の乳酸杆菌属細菌。
【請求項14】
前記生物活性ポリペプチドが、10アミノ酸と600アミノ酸との間である、請求項1に記載の乳酸杆菌属細菌。
【請求項15】
前記生物活性タンパク質が病原体と接触する場合、該生物活性タンパク質が該病原体と結合する、請求項1に記載の乳酸杆菌属細菌。
【請求項16】
前記病原体が、細菌病原体である、請求項15に記載の乳酸杆菌属細菌。
【請求項17】
前記病原体が、真菌病原体である、請求項15に記載の乳酸杆菌属細菌。
【請求項18】
前記病原体が、ウイルス病原体である、請求項15に記載の乳酸杆菌属細菌。
【請求項19】
前記ウイルス病原体が、HIVである、請求項18に記載の乳酸杆菌属細菌。
【請求項20】
前記生物活性タンパク質が、CD4またはCD4のHIV結合フラグメントである、請求項19に記載の乳酸杆菌属細菌。
【請求項21】
前記生物活性タンパク質が、2D−CD4である、請求項19に記載の乳酸杆菌属細菌。
【請求項22】
前記生物活性タンパク質が、シアノビリン−Nまたはシアノビリン−Nのウイルス結合フラグメントである、請求項18に記載の乳酸杆菌属細菌。
【請求項23】
前記ウイルス病原体が、単純疱疹ウイルスである、請求項18に記載の乳酸杆菌属細菌。
【請求項24】
前記生物活性タンパク質が、単純疱疹ウイルス侵入媒体C(HveC)であるか、またはHveCのウイルス結合フラグメントである、請求項18に記載の乳酸杆菌属細菌。
【請求項25】
前記生物活性ポリペプチドが、乳酸杆菌属細菌から放出される、請求項1に記載の乳酸杆菌属細菌。
【請求項26】
前記生物活性ポリペプチドが、乳酸杆菌属細菌の細胞壁に固定される、請求項4に記載の乳酸杆菌属細菌。
【請求項27】
乳酸杆菌属細菌の細胞壁中において生物活性ポリペプチドを発現する方法であって、該方法は、以下:
発現カセットを含む乳酸杆菌属細菌を提供する工程であって、該発現カセットは、シグナル配列および生物活性ポリペプチドをコードするポリヌクレオチドと作動可能に結合するプロモーターを含み、該生物活性ポリペプチドは異種カルボキシル末端細胞壁標的領域と結合し、そして該異種カルボキシル末端細胞壁標的領域は、以下の順番で、以下:
細胞壁関連配列;
LPQ(S/A/T)(G/A);および
疎水性配列
を含む、工程;ならびに
該ポリペプチドの発現を誘導するための条件下で細菌を培養する工程であって、該工程により、乳酸杆菌属細菌の細胞壁中で生物活性ポリペプチドを発現する、工程、
を包含する、方法。
【請求項28】
前記細胞壁関連配列が、少なくとも50個のアミノ酸を含む、請求項27に記載の方法。
【請求項29】
前記細胞壁関連配列が、少なくとも200個のアミノ酸を含む、請求項27に記載の方法。
【請求項30】
前記異種カルボキシル末端細胞壁標的領域が、該領域のカルボキシル末端において、荷電した配列をさらに含む、請求項27に記載の方法。
【請求項31】
前記提供する工程が、前記発現カセットを前記細菌中に輸送する工程を包含する、請求項27に記載の方法。
【請求項32】
前記細胞壁標的領域が、アミノ酸配列LPQSGを含む、請求項27に記載の方法。
【請求項33】
前記細胞壁標的領域が、アミノ酸配列LPQAGを含む、請求項27に記載の方法。
【請求項34】
前記細胞壁標的領域が、アミノ酸配列LPQTGを含む、請求項27に記載の方法。
【請求項35】
前記細胞壁標的領域が、アミノ酸配列LPQTAを含む、請求項27に記載の方法。
【請求項36】
前記細胞壁標的領域が、配列番号7を含む、請求項27に記載の方法。
【請求項37】
前記細胞壁標的領域が、配列番号8を含む、請求項27に記載の方法。
【請求項38】
前記細胞壁標的領域が、少なくとも200個のアミノ酸を含む、請求項27に記載の方法。
【請求項39】
前記乳酸杆菌属細菌が、膣コロニー形成株である、請求項27に記載の方法。
【請求項40】
前記乳酸杆菌属細菌が、L.jensenii、L.gasseri、およびL.caseiからなる群より選択される、請求項27に記載の方法。
【請求項41】
前記生物活性ポリペプチドが、10アミノ酸と600アミノ酸との間である、請求項27に記載の方法。
【請求項42】
前記生物活性タンパク質が病原体と接触する場合、該生物活性タンパク質が該病原体に結合する、請求項27に記載の方法。
【請求項43】
前記病原体が、細菌病原体である、請求項42に記載の方法。
【請求項44】
前記病原体が、真菌病原体である、請求項42に記載の方法。
【請求項45】
前記病原体が、ウイルス病原体である、請求項42に記載の方法。
【請求項46】
前記ウイルス病原体が、HIVである、請求項45に記載の方法。
【請求項47】
前記生物活性タンパク質が、CD4またはCD4のHIV結合フラグメントである、請求項46に記載の方法。
【請求項48】
前記生物活性タンパク質が、2D−CD4である、請求項46に記載の方法。
【請求項49】
前記生物活性タンパク質が、シアノビリン−Nまたはシアノビリン−Nのウイルス結合フラグメントである、請求項45に記載の方法。
【請求項50】
前記ウイルス病原体が、単純疱疹ウイルスである、請求項45に記載の方法。
【請求項51】
前記生物活性タンパク質が、単純疱疹ウイルス侵入媒体C(HveC)またはHveCのウイルス結合フラグメントである、請求項45に記載の方法。
【請求項52】
前記生物活性ポリペプチドが、乳酸杆菌属細菌から放出される、請求項27に記載の方法。
【請求項53】
前記生物活性ポリペプチドが、乳酸杆菌属細菌の細胞壁に固定される、請求項30に記載の方法。
【請求項54】
生物活性タンパク質を哺乳動物粘膜表面に提供する方法であって、該方法は、以下:
粘膜表面と、異種カルボキシル末端細胞壁標的領域に結合された生物活性ポリペプチドに結合されたシグナル配列を発現するように組み換えで変化した乳酸杆菌属細菌とを、接触させる工程を包含し、該異種カルボキシル末端細胞壁標的領域は、以下の順番で、以下:
細胞壁関連配列;
LPQ(S/A/T)(G/A);および
疎水性配列
を含み、
ここで、該生物活性ポリペプチドは、該粘膜表面から収集されたサンプル中で、検出可能な量で発現される、方法。
【請求項55】
前記細胞壁関連配列が、少なくとも50個のアミノ酸を含む、請求項54に記載の方法。
【請求項56】
前記細胞壁関連配列が、少なくとも200個のアミノ酸を含む、請求項54に記載の方法。
【請求項57】
前記異種カルボキシル末端細胞壁標的領域が、該領域のカルボキシル末端において、荷電した配列をさらに含む、請求項54に記載の方法。
【請求項58】
前記乳酸杆菌属細菌が、L.jensenii、L.gasseri、およびL.caseiからなる群より選択される、請求項54に記載の方法。
【請求項59】
前記粘膜表面が、膣内に存在する、請求項54に記載の方法。
【請求項60】
前記粘膜表面が、胃腸管内に存在する、請求項54に記載の方法。
【請求項61】
前記接触させる工程が、前記乳酸杆菌属細菌を経口投与する工程を包含する、請求項54に記載の方法。
【請求項62】
前記接触させる工程が、前記乳酸杆菌属細菌を膣投与する工程を包含する、請求項54に記載の方法。
【請求項63】
前記接触工程が、前記乳酸杆菌属細菌を直腸投与する工程を包含する、請求項54に記載の方法。
【請求項64】
シグナル配列および生物活性ポリペプチドをコードするポリヌクレオチドに作動可能に結合するプロモーターを含む発現カセットであって、ここで、該生物活性ポリペプチドが、配列番号7または配列番号8に結合する、発現カセット。
【請求項65】
請求項64に記載の発現カセットを含む、ベクター。
【図1】

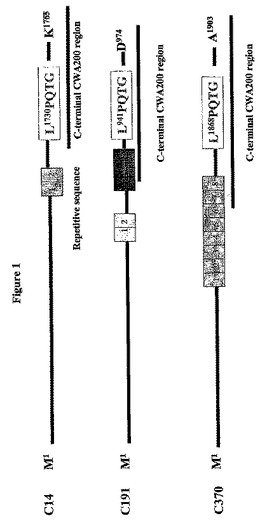
【図2A】

【図2B】

【図2C】

【図3】

【図4】

【図5】
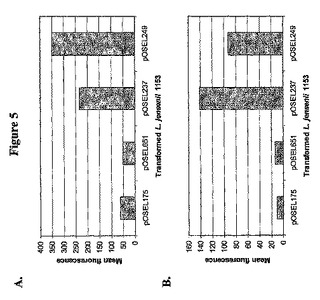
【図6】
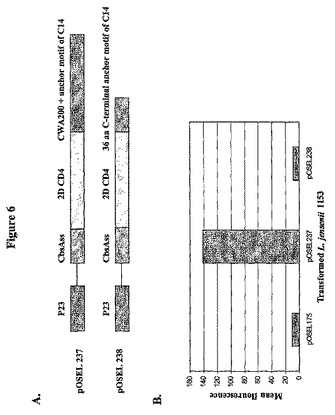
【図7】
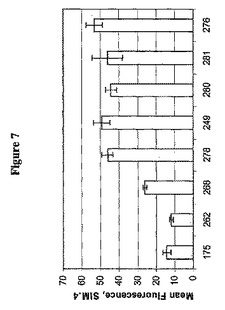
【図8】

【図9】
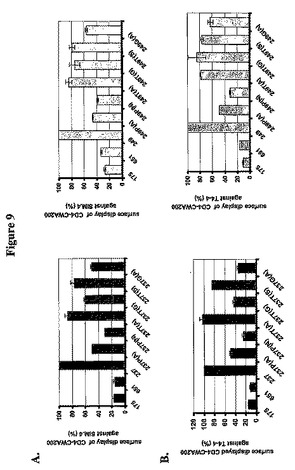
【図10】
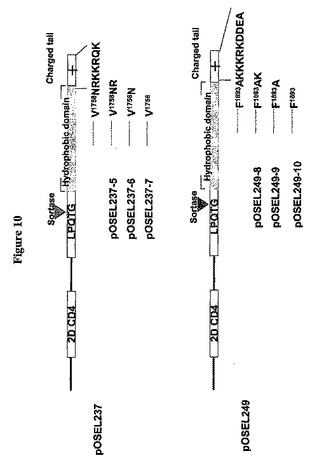
【図11】
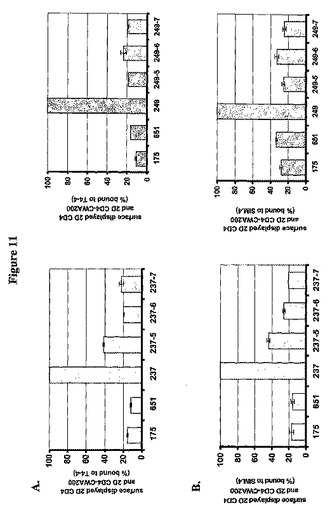
【図12】
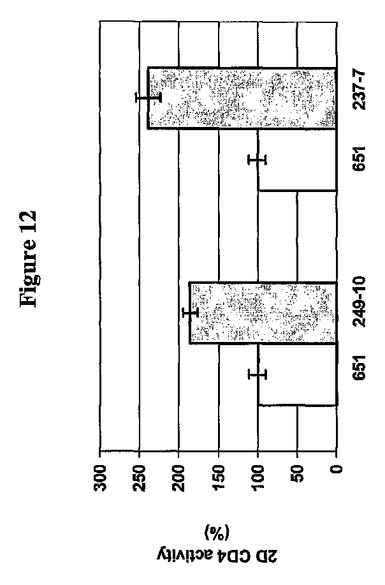
【図2A】
【図2B】
【図2C】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2007−503845(P2007−503845A)
【公表日】平成19年3月1日(2007.3.1)
【国際特許分類】
【出願番号】特願2006−532269(P2006−532269)
【出願日】平成16年1月28日(2004.1.28)
【国際出願番号】PCT/US2004/002460
【国際公開番号】WO2005/012491
【国際公開日】平成17年2月10日(2005.2.10)
【出願人】(505284828)オセル, インコーポレイテッド (1)
【Fターム(参考)】
【公表日】平成19年3月1日(2007.3.1)
【国際特許分類】
【出願日】平成16年1月28日(2004.1.28)
【国際出願番号】PCT/US2004/002460
【国際公開番号】WO2005/012491
【国際公開日】平成17年2月10日(2005.2.10)
【出願人】(505284828)オセル, インコーポレイテッド (1)
【Fターム(参考)】
[ Back to top ]