
細胞膨張化致死毒およびそれを標的としたカンピロバクター属に属する細菌の検出
本発明者らは、未だ知られていなかったC.コリ、C.フィータスのCDT遺伝子をクローニングし、配列決定することに成功した。また、C.ジェジュニ、C.フィータスのCDTとの比較を行ない、両者に共通するプライマーおよび特異的なプライマーを開発した。そしてこのプライマーが、簡便かつ迅速にカンピロバクター属細菌CDTの有無の判定、および菌種の同定が同時に行えるマルチプレックスPCR法に適用でき、さらにPCR-RFLP法によるタイピングにも利用できることを明らかにした。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、カンピロバクター・コリの細胞膨張化致死毒およびそれをコードするポリヌクレオチドに関する。さらに本発明は、カンピロバクター・コリを含むカンピロバクター属に属する細菌の細胞膨張化致死毒を標的とした、検体(臨床検体、食品など)中のカンピロバクター(Campylobacter)属細菌の存在の有無を判定するための方法に関する。
【背景技術】
【0002】
カンピロバクター属細菌は、ヒト、および野生または家畜動物の病原菌であって、動物における流産や腸炎、ヒトにおける腸炎の原因菌である。ヒトの場合、カンピロバクター感染症の起因菌としては、カンピロバクター・ジェジュニ(Campylobacter jejuni)およびカンピロバクター・コリ(Campylobacter coli)が知られており、これらの細菌は、食中毒菌として指定されている(非特許文献1、2参照)。
【0003】
2000年現在、カンピロバクターは15菌種9亜種に分類されているが、ヒトの下痢症から分離される菌種は、C.ジェジュニが95〜99%を占め、C.コリなどのその他の菌種は数%である(非特許文献3参照)。なお、C.コリはブタでの保菌率が極めて高い。近年、カンピロバクター感染症は、東南アジアを中心とした輸入食肉の増加に伴い増加傾向にあり、特にBSEやO-157等の問題によって牛肉の代替品として消費量が伸びている鶏肉関連食品による感染例が急増している。
上記の他、C.フィータス(Campylobacter fetus)は、ヒツジやウシの流産菌として知られていたが、近年ヒトの流産や早産に関与しているとの報告がある。また、C.フィータスで汚染されたレバーや牛肉の生食によるC.フィータス感染では、敗血症や髄膜炎などの症状を呈することもある。ヒトへの感染源は、腸管内に高濃度に保菌する鶏肉が最も重要である。(非特許文献4)
【0004】
カンピロバクター属細菌は、通常、ウシ、ヒツジ、ブタ、ニワトリ等の動物の消化管に高率に分布しており、人獣共通感染症の原因細菌として認識されている。カンピロバクター感染症の多くは鶏肉が原因と考えられているが、これらの動物もしくは排泄物との直接的接触や、排泄物によって汚染された食物、飲料水、牛乳などの摂取や調理過程により感染する。さらに、新生児室などの施設内感染例も報告されている(非特許文献5参照)
【0005】
カンピロバクター感染症は、潜伏期が3〜7日と長く、下痢(ときに血粘膜性下痢)、腹痛、発熱、悪心、嘔吐、頭痛、悪寒、倦怠感などの胃腸炎症状を引き起こすのが特徴である。致死性は低いが、新生児では敗血症や髄膜炎などの全身感染を起こすことがある。多くの場合、数日から1週間程度で回復し、一般的な予後については、一部の免疫不全患者を除き良好な経過を辿る。しかし、近年カンピロバクターの後感染性疾患として自己免疫疾患であるギラン・バレー症候群やフィッシャー症候群に進展する例が報告されている。カンピロバクター感染症に後発する症例は概して重症化しやすく、発症1年後の寛解率は約6割程度に止まる。
【0006】
重篤な病態や敗血症の併発例では抗生剤による化学治療が行われる。第一選択薬剤はエリスロマイシン等のマクロライド系薬剤である。セフェム系薬剤に対しては自然耐性を示すため、治療効果は望めない。一方、近年、ニューキノロン系薬剤に対しては、耐性菌の増加が問題となっている。カンピロバクター感染症に対し的確な治療を施し、併せて感染経路を解明して感染拡大を食い止めるためには、感染後、迅速に原因菌を同定することが重要である。しかしながら、臨床症状からカンピロバクター感染を診断することは困難であり、カンピロバクターの同定および菌種の決定は容易ではない。
【0007】
カンピロバクター属細菌は微好気性細菌であり、培養にはスキロー培地などの特殊な培地、および酸素濃度を3〜10%の絶対微好気性条件に保つための特殊な装置(嫌気ジャー等)が必要で、なおかつ他の細菌より培養に時間を要する(2〜3日)ことから、分離培養が困難なものとなっている。また、カンピロバクター属細菌は、空気中で死滅しやすく、検体採取後、2〜3時間以内に検査する必要がある。さらに、カンピロバクター感染症は潜伏期間が長い(3〜7日)ため、病状が現れてから関与食品の細菌同定を行っても菌が単離できない場合が多い。さらにカンピロバクター属細菌は、非常に感染力が強く、数百個の菌数で感染が成立するという報告もあり、感染源の特定は極めて困難なものとなっている。
【0008】
C.ジェジュニとC.コリの識別診断に、馬尿酸塩の加水分解を検査する方法がある。すなわち、C.ジェジュニは馬尿酸を加水分解できるが、C.コリはできないという性質を利用する。しかしながら、馬尿酸−陰性C.ジェジュニ株が存在するため、この方法も確実ではない(非特許文献6参照)。従って、現在は摂食履歴と症状によるカンピロバクター菌の存在の推定、および便培養から得られたコロニーについて数日を要して菌の形態学的および生物学的特徴を検査することにより、その存在を確認しているのが現状である。
【0009】
そこで、培養検査によらない迅速診断法として、オリゴヌクレオチドを用いたDNAプローブ法やPCR法を利用した遺伝子的診断法によるカンピロバクター属細菌の同定や毒素遺伝子の検出が試みられている。たとえば、カンピロバクター属細菌の場合、一般にrRNAをコードする遺伝子がプローブとして使用されている(特許文献1参照)。このrRNAの遺伝子配列は既に公表されている(非特許文献7参照)。また、カンピロバクター属細菌検出用の核酸フラグメントも知られている(特許文献2、3、4、5、6参照)。しかしながら、これらの配列は、C.ジェジュニおよび/またはC.コリの検出用であり、その他のカンピロバクター属細菌の検出には適当でない。さらに、特異性が充分ではなかった。
【0010】
また、C.コリVC167のfla A遺伝子から選択したオリゴヌクレオチドを用いたPCRによって、C.ジェジュニを同定する方法が報告されている(非特許文献8参照)。さらに、C.ジェジュニおよびC.コリ中に見出されるスーパーオキシドジスムターゼ標的配列を増幅するためのオリゴヌクレオチドプライマーが報告されている(特許文献7参照)。しかし、これらの方法ではC.ジェジュニとC.コリを区別することができない。
【0011】
一方、カンピロバクター属細菌の病原因子についての研究も盛んである。カンピロバクター属細菌の病原因子としては、細胞侵入性やフラジェリン、コレラ毒素様エンテロトキシン等の様々な因子が報告されている(非特許文献9、10参照)が、近年、C.ジェジュニから毒性因子として細胞膨張化性致死毒(Cytolethal Distending Toxin:CDT)が発見され(非特許文献11)、病原性との関連性が注目されている。例えば、志賀赤痢菌(Shigella dysenteriae)のCDTを産生する組換え大腸菌を用いた動物投与モデルでは下痢原性が報告されている(非特許文献12)。
【0012】
CDTはcdtA、cdtBおよびcdtCと称される3種のサブユニットからなるホロ毒素であり、タンデムに並んだ遺伝子にコードされている。毒素活性中心はcdtBサブユニットの持つI型デオキシリボヌクレアーゼ様活性であり、cdtAサブユニットとcdtCサブユニットは標的細胞への接着に関与すると考えられている。このホロ毒素が細胞に作用すると、細胞が膨化、すなわち大きくふくらみ、最終的には細胞が死滅することから、細胞膨張化性致死毒と名付けられている。
【0013】
その分子機構は活性毒素中心であるcdtBサブユニットが細胞内の核に移行し、I型デオキシリボヌクレアーゼ活性により、染色体DNAにニックを挿入し、DNA損傷応答(DNA-damage response)を引き起こす。そして、細胞は遺伝子修復系を動かすために細胞周期をG2/M期で停止後、膨張化致死すると考えられている(非特許文献13)。さらに、CDTは上皮系細胞、免疫担当細胞等の幅広い細胞に作用することが確認されており、中でもヒトリンパ球に作用し、アポトーシスを引き起こすことにより免疫から逃れやすくなっていると考えられている(非特許文献14)。
【0014】
このようにCDTは、これまでの毒素には見られないユニークな分子機構を有しているが、これまでカンピロバクター属細菌でCDTの全塩基配列が明らかとなっているのは、C.ジェジュニのみである(非特許文献11)。
【特許文献1】特開昭62−228096号公報
【特許文献2】特開平2−84200号公報
【特許文献3】特開平2−154700号公報
【特許文献4】特開平3−112498号公報
【特許文献5】特開平6−90795号公報
【特許文献6】特開平6−90796号公報
【特許文献7】特開2000−316590公報
【非特許文献1】Blaser, et al, Ann. Intern. Med., 91:179 (1979)
【非特許文献2】Tauxe, R., American Society for Microbiology, Washington DC. pg.9(1992)
【非特許文献3】Takahashi, M. et al, Infectious Diseases Weekly Report Japan, 3(6):10 (2001)
【非特許文献4】Simon, M. S. et al., 2003. Campylobacter infection. Diseases of Poultry, Iowa State Press, 615-630
【非特許文献5】小児内科, 29:1219-1222 (1997)
【非特許文献6】Totten, et al, J. Clin. Microbiol., 25: 1747 (1987)
【非特許文献7】Romaniuk, P. J. et al, J. Bacteriol., 169: 2173 (1987)
【非特許文献8】Oyofo, et al, J. Clin. Microbiol., 30: 2613 (1992)
【非特許文献9】Mizuno, K. et al, Microbios., 78: 215 (1994)
【非特許文献10】Suzuki, S. et al, FEMS Immunol. Med. Micribiol., 8: 207 (1994)
【非特許文献11】Pickett, C. et al. Infect. Immun., 64: 2070 (1996)
【非特許文献12】Infect. Immun., 65: 428-433 (1997)
【非特許文献13】Science, 290: 354-357 (2000)
【非特許文献14】J. Biol. Chem., 276: 5296-5302 (2001)
【発明の開示】
【発明が解決しようとする課題】
【0015】
以上詳細に述べたように、カンピロバクター属細菌による感染症の迅速診断が求められている一方、カンピロバクター属細菌の病原因子は完全に解明されているとは言い難い。これまで、各菌種の血清型等を利用した菌種同定用PCRプライマーや、CDT産生を調べる共通プライマー等が利用されてきたが(J. Applied Microbiol., 94: 1003-1014 (2003))、増菌培養工程が必要であり、カンピロバクター属細菌の迅速な検出は不可能であった。
【0016】
そこで、本発明は、遺伝子診断により、カンピロバクター属細菌の迅速な検出を可能とすべく、カンピロバクター属に属し、いまだそのCDTの塩基配列が解明されていないカンピロバクター・コリのCDTおよびそれをコードするポリヌクレオチドを提供することを課題とする。さらに本発明は、同様にCDTの塩基配列が解明されていないカンピロバクター・フィータスのCDTおよびそれをコードするポリヌクレオチドを提供することを課題とする。
【0017】
さらに、本発明は、カンピロバクター・コリおよびカンピロバクター・フィータスの塩基配列から得られた知見に基づき、カンピロバクター・コリおよびカンピロバクター・フィータスを含むカンピロバクター属細菌のCDTを標的とした、カンピロバクター属細菌の存在を迅速に検出しうる方法を提供することを課題とする。
【課題を解決するための手段】
【0018】
CDT遺伝子のクローニングにおいて、制限酵素HindIIIを用いたクローニングを行った場合、コード領域中にHindIIIサイトが存在するため、全長を単離することはできない。他方、EcoRI、PstI、KpnI、XbaI、BamHI、SalI、XhoI等の一般的な制限酵素を用いると、cdt遺伝子のクローニングに最適な長さの断片(3kbから5kb)が得られなかった。そこで、本発明者らは種々検討を行なった結果、HindIIIで完全にcdt遺伝子が消化されないようなパーシャルな条件を選択し、切断することにより、遂に、cdt遺伝子の内部配列が切断されていない完全長の遺伝子をクローニングすることに成功した。
【0019】
また、本発明者らは、C.ジェジュニ、C.フィータスのCDTとの比較を行ない、3種のカンピロバクター属細菌に共通するプライマーおよびこれら細菌のそれぞれに特異的なプライマーを開発した。そしてこのプライマーが、簡便かつ迅速にカンピロバクター属細菌CDTの有無の判定、および菌種の同定が同時に行えるマルチプレックスPCR法に適用でき、さらにPCR-RFLP法によるタイピングにも利用できることを明らかにした。
【0020】
すなわち、本発明は、具体的に以下の技術的態様が包含される:
(1) 細胞膨張化致死毒をコードする下記(a)から(d)のいずれかに記載のポリヌクレオチド。
(a)配列番号:2から4のいずれかに記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド
(b)配列番号:1に記載の塩基配列のコード領域を含むポリヌクレオチド
(c)配列番号:2から4に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加および/または挿入したアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド
(d)配列番号:1に記載の塩基配列からなるDNAにストリンジェントな条件下でハイブリダイズするポリヌクレオチド
(2) (1)に記載のポリヌクレオチドを含むベクター。
(3) (1)に記載のポリヌクレオチドまたは(2)に記載のベクターを保持する宿主細胞。
(4) (1)に記載のポリヌクレオチドによりコードされるポリペプチド。
(5) (3)に記載の宿主細胞を培養し、該宿主細胞またはその培養上清から、産生させたポリペプチドを回収する工程を含む、(4)に記載のポリペプチドの製造方法。
(6) (4)に記載のポリペプチドに結合する抗体。
(7) 被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、以下の(a)および(b)の工程を含む方法。
(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物を用いたポリメラーゼ連鎖反応を行なう工程
(b)これら細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、これら細菌の存在を判定する工程
(8) 被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、以下の(a)から(c)の工程を含む方法。
(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いたポリメラーゼ連鎖反応を行なう工程
(b)工程(a)により増幅されたゲノムDNAを鋳型に、これら細菌の細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物を用いたポリメラーゼ連鎖反応を行なう工程
(c)これら細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、これら細菌の存在を判定する工程
(9) 共通プライマー対が、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、(8)に記載の方法。
(10) 特異的なプライマー対の混合物として、以下の(a)から(c)のプライマー対を用いる、(7)または(8)に記載の方法。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(11) 被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、以下の(a)〜(c)の工程を含む方法。
(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いたポリメラーゼ連鎖反応を行なう工程
(b)工程(a)により増幅されたゲノムDNAを制限酵素により切断する工程
(c)切断されたDNA断片の分子量から、これら細菌の存在を判定する工程
(12) 制限酵素が、Sau3AI、Dsa I、Mbo I、Rsa I、EcoR I、Hinf I、Nde I、Pst I、Xba I、Xho IIからなる群より選択される、(11)に記載の方法。
(13) 共通プライマー対が、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、(11)に記載の方法。
(14) (7)に記載の方法に用いるためのキットであって、使用説明書と、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物とを含むキット。
(15) 特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、(14)に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対共通プライマー対
(16) 請求項8に記載の方法に用いるためのキットであって、使用説明書と、下記(a)または(b)とを含むキット。
(a)カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物
(b)カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対。
(17)特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、(16)に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(18)共通プライマー対が、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、(16)に記載のキット。
(19) (11)に記載の方法に用いるためのキットであって、使用説明書と、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対とを含むキット。
(20)共通プライマー対が、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、(19)に記載のキット。
【0021】
なお、本明細書において「細胞膨張化致死毒」とは、cytolethal distending toxin (CDTまたはCLDT)と呼ばれる、蛋白性のA-B型ホロトキシンのグループに属する毒素因子を指す。このものは、ほかに細胞膨化致死毒(素)、細胞膨潤化致死毒(素)などと称されることもある。細胞膨張化致死毒は、A,B,Cの3ユニットからなるサブユニット構造を有し、Bサブユニットが毒素活性中心ユニットであり、AおよびBサブユニットが細胞接着に関わっていると考えられている。細胞に作用すると細胞が大きく膨らむ等の変形が生じ、最終的に細胞死を引き起こす。毒素原性大腸菌が産生する易熱性エンテロトキシン(LT)などを実験的に細胞に作用させた場合にも細胞が大きく膨らむ等の変形が見られるが、毒素を取り除いた場合、細胞は回復し、致死することはない。しかしながら、CDTを除去しても細胞は回復せず、死に至る。
【0022】
本明細書において用いられる「ポリヌクレオチド」とは、リボヌクレオチドもしくはデオキシヌクレオチドであって、複数の塩基または塩基対からなる重合体を意味する。ポリヌクレオチドには、一本鎖型および二本鎖型のDNAを含む。ポリヌクレオチドは、天然に存在する状態から修飾されていないもの、および修飾されているものの双方を含む意である。修飾された塩基としては、例えば、トリチル化された塩基およびイノシンのような特殊な塩基がある。
【0023】
本明細書において用いられる「ポリペプチド」は、複数のアミノ酸からなる重合体を意味する。従って、オリゴペプチドおよびタンパク質もまた、ポリペプチドの概念に含まれる。ポリペプチドは、天然に存在する状態から修飾されていないもの、および修飾されているものの双方を含む意である。修飾としては、アセチル化、アシル化、ADP-リボシル化、アミド化、フラビンの共有結合、ヘム部分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトールの共有結合、架橋、環化、ジスルフィド結合の形成、脱メチル化、共有架橋の形成、シスチンの形成、ピログルタメートの形成、ホルミル化、γ-カルボキシル化、グリコシル化、GPIアンカー形成、ヒドロキシル化、ヨウ素化、メチル化、ミリストイル化、酸化、タンパク質分解処理、リン酸化、プレニル化、ラセミ化、セレノイル化、硫酸化、アルギニル化のようなタンパク質へのアミノ酸の転移RNA媒介付加、ユビキチン化などが含まれる。
【0024】
本明細書において「単離」とは、本来の環境(たとえば自然に発生するのであればその自然環境)から取り出された物質(例えば、ポリヌクレオチドまたはポリペプチド)を指し、その自然状態から「人の手によって」変えられたものである。「単離」とは、対象化合物に実質的に富む試料中に存在する化合物および/または対象化合物が部分的または実質的に精製されている試料中に存在する化合物を含むことを意味する。ここで「実質的に精製した」という用語は、その天然の環境から切り離されて、天然に関連している他の成分を少なくとも60%、好ましくは75%、および最も好ましくは90%含まない化合物(例えば、ポリヌクレオチドまたはポリペプチド)を指す。
【0025】
本明細書において用いられる「変異」とは、アミノ酸配列におけるアミノ酸の変化または塩基配列における塩基の変化(すなわち単一または複数のアミノ酸またはヌクレオチド置換、欠失、付加または挿入)を指す。従って、本明細書において用いられる「変異体」は、一つ以上のアミノ酸が変化しているアミノ酸配列または一つ以上の塩基が変化している塩基配列を指す。この変異体の塩基配列の変化は、基準ポリヌクレオチドによってコードされるポリペプチドのアミノ酸配列を変更しても、しなくてもよい。変異体はアレリック変異体のように天然に存在するものでも、天然に存在することが知られていない変異体であってもよい。変異体は、置換されたアミノ酸が類似の構造的または化学的特性を有する保存的変化を有しうる。まれに、変異体は、非保存的置換を有しうる。生物学的または免疫学的活性を阻害することなく、いずれの、およびどれほど多くのアミノ酸残基を置換、挿入、または欠失するかを決定する手引きは、当技術分野において周知のコンピュータープログラム、例えばDNAスター・ソフトウェアを用いて発見することができる。
【0026】
「欠失」はその中で1つ以上のアミノ酸またはヌクレオチド残基がそれぞれ、天然に存在する細胞膨張化致死毒ポリペプチドのアミノ酸配列またはヌクレオチド配列と比較して存在しない、アミノ酸またはヌクレオチド配列のいずれかの変化である。
【0027】
「挿入」または「付加」は、天然に存在する細胞膨張化致死毒ポリペプチドのアミノ酸配列またはヌクレオチド配列と比較して、それぞれアミノ酸またはヌクレオチド残基1つ以上が付加されたアミノ酸またはヌクレオチド配列の変化である。
【0028】
「置換」とは、天然に存在する細胞膨張化致死毒ポリペプチドのアミノ酸配列またはヌクレオチド配列と比較して、アミノ酸またはヌクレオチド1つ以上がそれぞれ異なるアミノ酸またはヌクレオチドに入れ替えられたアミノ酸またはヌクレオチド配列の変化である。
【0029】
本明細書において用いられる「ハイブリダイズ」とは、核酸鎖が塩基対形成を通じて相補鎖と結合するプロセスを意味する。
【図面の簡単な説明】
【0030】
【図1】GNWおよびLPF-Dプライマーを用いたC.コリCo1-192菌体抽出液をテンプレートとしたPCRの結果を示す写真である。矢印1はcdt領域が増幅されたバンド(約1.5Kb)、矢印2のバンド(800bp)は、GNWプライマーがミックスプライマーであるために増幅されたcdtB由来の副バンドを示す。
【図2】C.コリCo1-192菌体のゲノムの制限酵素HindIII消化後、ハイブリダイゼーションの結果を示す写真である。
【図3】共通プライマー1を用いたPCRの結果を示す写真である。レーン2〜6の1.9kbp付近にCDT由来のバンドが認められる。
【図4】共通プライマー2を用いた各種C.ジェジュニ菌株のPCRの結果を示す写真である。720bp付近にCDT由来のバンドが認められる。
【図5】共通プライマー2を用いた各種C.ジェジュニ菌株およびC.コリ菌株のPCRの結果を示す写真である。
【図6】共通プライマー2を用いたC.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のPCRの結果を示す写真である。
【図7】特異的プライマーを用いたC.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のマルチプレックスPCRの結果を示す写真である。各菌種に特有のCDT特異的増幅断片が確認された(C.ジェジュニ:750bp、C.コリ:400bp、C.フィータス:530bp)。
【図8】共通プライマー1を用いたC.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のPCR-RFLPの結果を示す写真である。
【図9】特異的プライマーを用いた各種C.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のマルチプレックスPCRの結果を示す写真である。各菌種に特有のCDT特異的増幅断片が確認された(C.ジェジュニ:750bp、C.コリ:400bp、C.フィータス:530bp)。
【図10】C.フィータスCol-187菌体のゲノムの制限酵素Hind III消化後、ハイブリダイゼーションの結果を示す写真である。
【図11】cdtAおよびcdtC共通プライマーを用いたPCRの結果を示す写真である。レーン2〜8の550bp付近にcdtA由来のバンドが認められ、またレーン10〜16の320bp付近にcdtC由来のバンドが認められる。
【図12】cdtA共通プライマーを用いた各種カンピロバクター菌株のPCRの結果を示す写真。550bp付近にCdtA由来のバンドが認められる。
【図13】cdtC共通プライマーを用いた各種カンピロバクター菌株のPCRの結果を示す写真。320bp付近にcdtC由来のバンドが認められる。
【図14】cdtAおよびcdtC特異的プライマーを用いたC.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のマルチプレックスPCRの結果を示す写真である。各菌種に特有のCdtA特異的増幅断片(C.ジェジュニ:630bp、C.コリ:330bp、C.フィータス:490bp)とcdtC特異的断片(C.ジェジュニ:500bp、C.コリ:400bp、C.フィータス:300bp)が確認される。
【図15】cdtA特異的プライマーを用いたC.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のマルチプレックスPCRの結果を示す写真である。各菌種に特有のCdtA特異的増幅断片が確認された。
【図16】cdtC特異的プライマーを用いたC.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のマルチプレックスPCRの結果を示す写真である。各菌種に特有のCdtC特異的増幅断片が確認された。
【図17】C.ジェジュニのORFおよびプライマーのアニール領域を示す図である。
【図18】C.コリのORFおよびプライマーのアニール領域を示す図である。
【図19】C.フィータスのORFおよびプライマーのアニール領域を示す図である。
【発明を実施するための最良の形態】
【0031】
<ポリヌクレオチド>
本発明は、カンピロバクター・コリの細胞膨張化致死毒をコードするポリヌクレオチドを提供する。本発明に含まれる、本発明者らにより同定されたカンピロバクター・コリの細胞膨張化致死毒をコードするポリヌクレオチドの塩基配列を配列番号:1に、該ポリヌクレオチドによってコードされる3つのポリペプチドのアミノ酸配列を配列番号:2から4に示す。配列番号2はcdtA、配列番号3はcdtB、配列番号4はcdtCのアミノ酸配列である。
また本発明は、カンピロバクター・フィータスの細胞膨張化致死毒をコードするポリヌクレオチドを提供する。本発明に含まれる、本発明者らにより同定されたカンピロバクター・フィータスの細胞膨張化致死毒をコードするポリヌクレオチドの塩基配列を配列番号:51に、該ポリヌクレオチドによってコードされる3つのポリペプチドのアミノ酸配列を配列番号:52から54に示す。配列番号52はcdtA、配列番号53はcdtB、配列番号54はcdtCのアミノ酸配列である。
【0032】
本発明のポリヌクレオチドには、配列番号:2から4に記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド、配列番号:1に記載の塩基配列のコード領域、すなわち、配列番号:1に記載の塩基配列において1位から777位までの塩基配列、802位から1605位までの塩基配列、1615位から2187位までの塩基配列のいずれかの塩基配列、を含むポリヌクレオチド、遺伝コードの縮重により配列番号:1に記載の塩基配列と異なる塩基配列からなるが配列番号:2から4に記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチドが含まれる。
また本発明のポリヌクレオチドには、配列番号:52から54に記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド、配列番号:51に記載の塩基配列のコード領域、すなわち、配列番号51に記載の塩基配列において1位から702位までの塩基配列、778位から1629位までの塩基配列、1615位から2187位までの塩基配列のいずれかの塩基配列、を含むポリヌクレオチド、遺伝コードの縮重により配列番号:51に記載の塩基配列と異なる塩基配列からなるが配列番号:52から54に記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチドが含まれる。
本発明のポリヌクレオチドには、さらに、これらポリヌクレオチドがコードするポリペプチドと機能的に同等なポリペプチドをコードし、該ポリヌクレオチドの配列とその全長において少なくとも40%以上、好ましくは60%以上、さらに好ましくは80%以上、さらに好ましくは90%以上、さらに好ましくは95%以上、さらに好ましくは97%以上(例えば、98〜99%)同一である塩基配列を含むポリヌクレオチドが含まれる。塩基配列の同一性は、例えば、Karlin and Altschul によるアルゴリズムBLAST (Proc. Natl. Acad. Sci. USA 87:2264-2268, 1990、Proc. Natl. Acad. Sci. USA 90:5873-5877, 1993)によって決定することができる。このアルゴリズムに基づいて、BLASTNと呼ばれるプログラムが開発されている(Altschul et al. J. Mol. Biol.215:403-410, 1990)。BLASTNによって塩基配列を解析する場合には、パラメーターはたとえばscore = 100、wordlength = 12とする。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である(http://www.ncbi.nlm.nih.gov.)。本発明のポリヌクレオチドには、上記のポリヌクレオチドの塩基配列と相補的な塩基配列を有するポリヌクレオチドが含まれる。
【0033】
本発明のポリヌクレオチドは、標準的なクローニングおよびスクリーニングにより、例えば、菌体中のゲノムDNAのような天然源から得ることができる。また、菌体中のmRNAから誘導されたcDNAライブラリーから得ることもできる。また、商業的に入手可能な公知の技法を用いて合成することもできる。
【0034】
本発明者らにより同定されたポリヌクレオチドの配列(配列番号:1、配列番号:51)と有意な相同性を有する塩基配列からなるポリヌクレオチドは、例えば、ハイブリダイゼーション技術 (Current Protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons Section 6.3-6.4)や遺伝子増幅技術(PCR)(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons Section 6.1-6.4)を利用して調製することができる。即ち、ハイブリダイゼーション技術を利用して、本発明者らにより同定されたポリヌクレオチドの配列(配列番号:1、配列番号:51)またはその一部をもとに、これと相同性の高いDNAを単離することができる。また、遺伝子増幅技術を用いて、本発明者らにより同定されたポリヌクレオチドの配列(配列番号:1、配列番号:51)の一部を基にプライマーを設計し、該ポリヌクレオチドの配列と相同性の高いポリヌクレオチドを単離することができる。従って、本発明には、配列番号:1に記載の塩基配列を有するポリヌクレオチドとストリンジェントな条件下でハイブリダイズするポリヌクレオチドが含まれる。ストリンジェントなハイブリダイゼーション条件としては、通常「1xSSC、0.1% SDS、37℃」程度の条件であり、より厳しい条件としては「0.5xSSC、0.1% SDS、42℃」程度の条件であり、さらに厳しい条件としては「0.2xSSC、0.1% SDS、65℃」程度の条件である。このようにハイブリダイゼーションの条件が厳しくなるほどプローブ配列と高い相同性を有するDNAの単離を期待しうる。但し、上記SSC、SDSおよび温度の条件の組み合わせは例示であり、当業者であれば、ハイブリダイゼーションのストリンジェンシーを決定する上記若しくは他の要素(例えば、プローブ濃度、プローブの長さ、ハイブリダイゼーション反応時間など)を適宜組み合わせることにより、上記と同様のストリンジェンシーを実現することが可能である。
【0035】
本発明者らにより同定されたポリヌクレオチドの配列と有意な相同性を有する塩基配列からなるポリヌクレオチドは、配列番号:1、配列番号:51に記載の塩基配列に変異を導入する方法(例えば、部位特異的変異誘発法(Current Protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons Section 8.1-8.5))を利用して調製することもできる。また、このようなポリヌクレオチドは、自然界における変異により生じることもある。本発明には、このような塩基配列の変異により、配列番号:2から4または配列番号:52から54に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、挿入および/もしくは付加などされたポリペプチドをコードするポリヌクレオチドが含まれる。
【0036】
本発明のポリヌクレオチドを本発明のポリペプチドの組換え生産のために用いる場合、そのポリヌクレオチドには、成熟ポリペプチドのコード配列またはその断片単独、他のコード配列(例えば、リーダーもしくは分泌配列、プレ-、プロ-もしくはプレプロ-タンパク質配列、または他の融合ペプチド部分をコードするもの)と同じリーディングフレーム内にある成熟ポリペプチドのコード配列またはその断片が含まれる。例えば、融合ポリペプチドの精製を容易にするマーカー配列がコードされ得る。本発明のこの態様の好ましい具体例として、マーカー配列は、pcDNA3.1/Myc-Hisベクター(Invitrogen社)により提供されかつGentz ら, Proc. Natl. Acad. Sci. USA (1989) 86:821-824に記載されるようなヘキサ-ヒスチジンペプチド、またはMycタグである。また、このポリヌクレオチドは5'および3'非コード配列、例えば、転写されるが翻訳されない配列、スプライシングおよびポリアデニル化シグナル、リボソーム結合部位、およびmRNA安定化配列を含んでいてもよい。
【0037】
<ポリペプチド>
本発明は、本発明者らが同定したカンピロバクター・コリの細胞膨張化致死毒のポリペプチドを提供する。さらに、本発明は、本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドを提供する。ここで「機能的に同等」とは、対象となるポリペプチドが本発明者らにより同定されたポリペプチドと同等の細胞膨張化致死毒の特性を有していることを意味する。
また本発明は、本発明者らが同定したカンピロバクター・フィータスの細胞膨張化致死毒のポリペプチドを提供する。さらに、本発明は、本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドを提供する。ここで「機能的に同等」とは、対象となるポリペプチドが本発明者らにより同定されたポリペプチドと同等の細胞膨張化致死毒の特性を有していることを意味する。
【0038】
本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドを調製するための方法の1つの態様としては、タンパク質中のアミノ酸配列に変異を導入する方法が挙げられる。このような方法には、例えば、部位特異的変異誘発法(Current Protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. Jhon Wily & Sons Section 8.1-8.5))が含まれる。また、ポリペプチド中のアミノ酸の変異は、自然界において生じることもある。本発明には、このように人工的か自然に生じたものかを問わず、本発明者らにより同定されたポリペプチドのアミノ酸配列(配列番号:2から4、配列番号:52から54)において1もしくは複数のアミノ酸が置換、欠失、挿入および/もしくは付加などにより変異したタンパク質であって、本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドが含まれる。
【0039】
置換されるアミノ酸は、タンパク質の機能の保持の観点から、置換前のアミノ酸と似た性質を有するアミノ酸であることが好ましい(保存的置換)。例えば、Ala、Val、Leu、Ile、Pro、Met、Phe、Trpは、共に非極性アミノ酸に分類されるため、互いに似た性質を有すると考えられる。また、非荷電性としては、Gly、Ser、Thr、Cys、Tyr、Asn、Glnが挙げられる。また、酸性アミノ酸としては、AspおよびGluが、塩基性アミノ酸としては、Lys、Arg、Hisが挙げられる。
【0040】
これらポリペプチドにおけるアミノ酸の変異数や変異部位は、その機能が保持される限り制限はない。変異数は、典型的には、全アミノ酸の10%以内であり、好ましくは全アミノ酸の5%以内であり、さらに好ましくは全アミノ酸の1%以内であると考えられる。
【0041】
本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドを調製するための方法の他の態様としては、ハイブリダイゼーション技術あるいは遺伝子増幅技術を利用する方法が挙げられる。即ち、当業者であれば、ハイブリダイゼーション技術 (Current Protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. Jhon Wily & Sons Section 6.3-6.4)を利用して本発明者らにより同定されたポリペプチドをコードするDNA配列(配列番号:1、配列番号:51)またはその一部をもとに同種または異種生物由来のDNA試料から、これと相同性の高いDNAを単離して、該DNAから本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドを得ることは、通常行いうることである。このように本発明者らにより同定されたポリペプチドをコードするDNAとハイブリダイズするDNAによりコードされるポリペプチドであって、本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドもまた本発明のポリペプチドに含まれる。
【0042】
本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドをコードするDNAを単離するためのストリンジェントなハイブリダイゼーション条件としては、通常「1xSSC、0.1% SDS、37℃」程度の条件であり、より厳しい条件としては「0.5xSSC、0.1% SDS、42℃」程度の条件であり、さらに厳しい条件としては「0.2xSSC、0.1% SDS、65℃」程度の条件である。このようにハイブリダイゼーションの条件が厳しくなるほどプローブ配列と高い相同性を有するDNAの単離を期待しうる。但し、上記SSC、SDSおよび温度の条件の組み合わせは例示であり、当業者であれば、ハイブリダイゼーションのストリンジェンシーを決定する上記若しくは他の要素(例えば、プローブ濃度、プローブの長さ、ハイブリダイゼーション反応時間など)を適宜組み合わせることにより、上記と同様のストリンジェンシーを実現することが可能である。
【0043】
このようなハイブリダイゼーション技術を利用して単離されるDNAがコードするポリペプチドは、通常、本発明者らにより同定されたポリペプチドとアミノ酸配列において高い相同性を有する。高い相同性とは、少なくとも40%以上、好ましくは60%以上、さらに好ましくは80%以上、さらに好ましくは90%以上、さらに好ましくは少なくとも95%以上、さらに好ましくは少なくとも97%以上(例えば、98〜99%)の配列の相同性を指す。アミノ酸配列の同一性は、例えば、Karlin and Altschul によるアルゴリズムBLAST (Proc. Natl. Acad. Sci. USA 87:2264-2268, 1990、Proc. Natl. Acad. Sci. USA 90:5873-5877, 1993)によって決定することができる。このアルゴリズムに基づいて、BLASTXと呼ばれるプログラムが開発されている(Altschul et al. J. Mol. Biol.215:403-410, 1990)。BLASTXによってアミノ酸配列を解析する場合には、パラメーターはたとえば score = 50、wordlength = 3とする。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である。
【0044】
また、遺伝子増幅技術(PCR)(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons Section 6.1-6.4)を用いて本発明者らにより同定されたポリペプチドをコードするDNA配列(配列番号:1、配列番号:51)の一部を基にプライマーを設計し、本発明者らにより同定されたポリペプチドをコードするDNA配列と相同性の高いDNA断片を単離し、該DNAを基に本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドを得ることも可能である。
【0045】
<ポリペプチドの断片>
本発明は、また、本発明のポリペプチドの断片を提供する。こうした断片は全体的に前記本発明のポリペプチドのアミノ酸配列の一部と同一であるが、全部とは同一でないアミノ酸配列を有するポリペプチドである。本発明のポリペプチド断片は、通常、8アミノ酸残基以上、好ましくは12アミノ酸残基以上(例えば、15アミノ酸残基以上)の配列からなるポリペプチド断片である。好適な断片としては、例えば、アミノ末端を含む一連の残基もしくはカルボキシル末端を含む一連の残基の欠失、またはアミノ末端を含む一連の残基とカルボキシル末端を含む一連の残基の二連の残基の欠失したアミノ酸配列を有するトランケーション(truncation)ポリペプチドが含まれる。また、αヘリックスとαヘリックス形成領域、βシートとβシート形成領域、ターンとターン形成領域、コイルとコイル形成領域、親水性領域、疎水性領域、α両親媒性領域、β両親媒性領域、可変性領域、表面形成領域、基質結合領域、および高抗原指数領域を含む断片のような、構造的または機能的特性により特徴づけられる断片も好適である。その他の好適な断片は生物学的に活性な断片である。生物学的に活性な断片は、同様の活性をもつ断片、その活性が向上した断片、または望ましくない活性が減少した断片を含めて、本発明のポリペプチドの活性を媒介するものである。さらに、動物、特にヒトにおいて抗原性または免疫原性がある断片も含まれる。これらのポリペプチド断片は、抗原活性を含めた本発明のポリペプチドの生物学的活性を保持することが好ましい。特定された配列および断片の変異型も本発明の一部を構成する。好適な変異型は同類アミノ酸置換により対象物と異なるもの、すなわち、ある残基が同様の性質の他の残基で置換されているものである。典型的なこうした置換は、Ala, Val, LeuとIleの間、SerとThrの間、酸性残基 AspとGluの間、AsnとGlnの間、塩基性残基 LysとArgの間、または芳香族残基 PheとTyrの間で起こる。
【0046】
<ポリペプチドの製造>
本発明のポリペプチドは任意の適当な方法で製造することができる。このようなポリペプチドには、単離された天然に存在するポリペプチド、組換え的に生産されたポリペプチド、合成的に製造されたポリペプチド、またはこれらの方法の組み合わせにより製造されたポリペプチドが含まれる。このようなポリペプチドの製造のための手段は当業界でよく理解されている。組換え的なポリペプチドは、例えば、本発明のポリヌクレオチドを挿入したベクターを適当な宿主細胞に導入し、形質転換体内で発現したポリペプチドを精製することにより調製することが可能である。一方、天然由来のポリペプチドは、例えば、後述する本発明のポリペプチドに対する抗体を結合したアフィニティーカラムを利用して調製することができる (Current Protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons Section 16.1-16.19)。アフィニティー精製に用いる抗体は、ポリクローナル抗体であってもモノクローナル抗体であってもよい。また、インビトロトランスレーション(例えば、「On the fidelity of mRNA translation in the nuclease-treated rabbit reticulocyte lysate system. Dasso,M.C.,Jackson,R.J.(1989) NAR 17:3129-3144」参照)などにより本発明のポリペプチドを調製することも可能である。本発明のポリペプチドの断片は、例えば、本発明のポリペプチドを適当なペプチダーゼで切断することによって製造することができる。
【0047】
<プローブ・プライマー>
本発明は、本発明者らにより同定されたポリヌクレオチド(配列番号:1に記載の塩基配列からなるポリヌクレオチドまたはその相補鎖、配列番号:51に記載の塩基配列からなるポリヌクレオチドまたはその相補鎖)に相補的な、少なくとも15ヌクレオチドの鎖長を有するヌクレオチドを提供する。ここで「相補鎖」とは、A:T(ただしRNAの場合は U)、G:Cの塩基対からなる2本鎖核酸の一方の鎖に対する他方の鎖を指す。また、「相補的」とは、少なくとも15個の連続したヌクレオチド領域で完全に相補配列である場合に限られず、少なくとも70%、好ましくは少なくとも80%、より好ましくは90%、さらに好ましくは95%以上の塩基配列上の相同性を有すればよい。相同性を決定するためのアルゴリズムは本明細書に記載したものを使用すればよい。このようなヌクレオチドは、本発明のポリヌクレオチドを検出、単離するためのプローブとして、また、本発明のヌクレオチドを増幅するためのプライマーとして利用することが可能である。プライマーとして用いる場合には、通常、15〜100ヌクレオチド、好ましくは15〜35ヌクレオチドの鎖長を有する。また、プローブとして用いる場合には、本発明のDNAの少なくとも一部若しくは全部の配列を含む少なくとも15ヌクレオチド、好ましくは少なくとも30ヌクレオチドの鎖長のヌクレオチドが用いられる。本発明のヌクレオチドをプライマーとする場合、核酸増幅反応の種類は、目的とする増幅産物が得られる限り、特に制限はない。例えば、PCR(ポリメラーゼ連鎖反応)法、ICAN法、LAMP法、SDA法、LCR法等のDNA増幅反応、NASBA法等のRNA増幅反応、の中から選択することができる。好適な方法としては、PCR法を示すことができる。
【0048】
このようなヌクレオチドの一つの態様は、本発明のポリペプチドをコードするDNAに特異的になものである。「特異的な」とは、通常のハイブリダイゼーション条件下、好ましくはストリンジェントな条件下で、特定のポリペプチドをコードするDNAとハイブリダイズし、他のポリペプチドをコードするDNAとはハイブリダイズしないことを意味する。好ましい態様は、カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNA(配列番号:1)にハイブリダイズし、カンピロバクター・ジェジュニおよびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAとハイブリダイズしないポリヌクレオチドである。このようなポリヌクレオチドとしては、例えば、配列番号:13、14、28−36、70、71、76、77から選択されるプライマー対が挙げられる。あるいは、好ましい態様は、カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNA(配列番号:51)にハイブリダイズし、カンピロバクター・ジェジュニおよびカンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAとハイブリダイズしないポリヌクレオチドである。このようなポリヌクレオチドとしては、例えば、配列番号:15、16、37−46、72、73、78、79から選択されるプライマー対が挙げられる。
【0049】
また、本実施例により、カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNA(配列番号:1)が明らかにされたことに伴い、カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAに特異的な塩基配列およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAに特異的な塩基配列を、本発明者らは見出した。従って、本発明は、カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAに特異的なプライマー対およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAに特異的なプライマー対をも提供する。カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAに特異的なプライマーとしては、例えば、配列番号:11、12、17−27に記載のプライマーが、カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAに特異的なプライマーとしては、例えば、配列番号:15、16、37−46に記載のプライマーが挙げられるが、これらに制限されない。
【0050】
また、本実施例により、カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNA(配列番号:1)、カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNA(配列番号:51)が明らかにされたことに伴い、本発明者は、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAに対する共通プライマー(これらすべての細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうるプライマー)を見出した。本発明は、このような共通プライマーをも提供する。好ましい共通プライマーとしては、例えば、配列番号;64、65に記載のプライマー(ctdAのDNAを増幅)、配列番号;7−10、47−50に記載のプライマー(ctdBのDNAを増幅)、配列番号;66、67に記載のプライマー(ctdCのDNAを増幅)が挙げられる。
【0051】
当業者にとっては、上記のプライマーに対して、1若しくは複数の塩基が異なるが、上記のプライマーと同一のゲノムDNA領域を増幅しうるプライマーを調製することは、適宜行ないうることである。上記プライマーがアニールするゲノムDNA領域について、図17-19に示す。本発明は、このような変異プライマーをも提供するものである。上述のとおり、本発明のプライマーを適用できる核酸増幅反応は、目的とする増幅産物が得られる限り、特に制限はなく、例えば、PCR(ポリメラーゼ連鎖反応)法、ICAN法、LAMP法、SDA法、LCR法等のDNA増幅反応、NASBA法等のRNA増幅反応、の中から選択することができる。好適な方法としては、PCR法を示すことができる。当業者であれば、上記プライマーを基に、実施する核酸増幅方法により適合した変異プライマーを設計することも可能である。変異プライマーは合成により調製することができる。また、変異プライマーが、変異前のプライマーと同一のゲノムDNA領域を増幅しうるかは、変異プライマーを用いた核酸増幅反応を行い、その増幅産物を分析することにより、簡便に評価することができる。
【0052】
これらプライマーは、被検試料中のヘリコバクター属細菌の存在の検出に好適に利用することができる。
【0053】
<ベクター、宿主細胞、ポリペプチドの製造>
本発明はまた、本発明のポリヌクレオチドを含有するベクター、本発明のポリヌクレオチドまたは該ベクターを保持する宿主細胞、および該宿主細胞を利用した本発明のポリペプチドの生産方法を提供する。
【0054】
本発明のベクターとしては、挿入したDNAを安定に保持するものであれば特に制限されず、例えば宿主に大腸菌を用いるのであれば、クローニング用ベクターとしてはpBluescriptベクター(Stratagene社製)などが好ましい。本発明のポリペプチドを生産する目的においてベクターを用いる場合には、特に発現ベクターが有用である。発現ベクターとしては、試験管内、大腸菌内、培養細胞内、生物個体内でポリペプチドを発現するベクターであれば特に制限されないが、例えば、試験管内発現であればpBESTベクター(プロメガ社製)、大腸菌であればpETベクター(Invitrogen社製)、培養細胞であればpME18S-FL3ベクター(GenBank Accession No. AB009864)、生物個体であればpME18Sベクター(Mol Cell Biol. 8:466-472(1988))などが好ましい。ベクターへの本発明のDNAの挿入は、常法により、例えば、制限酵素サイトを用いたリガーゼ反応により行うことができる(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons.Section 11.4-11.11)。
【0055】
本発明のベクターが導入される宿主細胞としては特に制限はなく、目的に応じて種々の宿主細胞が用いられる。ポリペプチドを発現させるための細胞としては、例えば、細菌細胞(例:ストレプトコッカス、スタフィロコッカス、大腸菌、ストレプトミセス、枯草菌)、真菌細胞(例:酵母、アスペルギルス)、昆虫細胞(例:ドロソフィラS2、スポドプテラSF9)、動物細胞(例:CHO、COS、HeLa、C127、3T3、BHK、HEK293、Bowes メラノーマ細胞)および植物細胞を例示することができる。宿主細胞へのベクター導入は、例えば、リン酸カルシウム沈殿法、電気パルス穿孔法(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons.Section 9.1-9.9)、リポフェクタミン法(GIBCO-BRL社製)、マイクロインジェクション法などの公知の方法で行うことが可能である。
【0056】
宿主細胞において発現したポリペプチドを小胞体の内腔に、細胞周辺腔に、または細胞外の環境に分泌させるために、適当な分泌シグナルを目的のポリペプチドに組み込むことができる。これらのシグナルは目的のポリペプチドに対して内因性であっても、異種シグナルであってもよい。
【0057】
本発明のポリペプチドの回収は、本発明のポリペプチドが培地に分泌される場合は、培地を回収する。本発明のポリペプチドが細胞内に産生される場合は、その細胞をまず溶解し、その後にポリペプチドを回収する。
【0058】
組換え細胞培養物から本発明のポリペプチドを回収し精製するには、硫酸アンモニウムまたはエタノール沈殿、酸抽出、アニオンまたはカチオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィーおよびレクチンクロマトグラフィーを含めた公知の方法を用いることができる。
【0059】
<抗体>
本発明は、本発明のポリペプチドに結合する抗体を提供する。ここで「抗体」には、ポリクローナルおよびモノクローナル抗体、キメラ抗体、一本鎖抗体、ヒト化抗体、さらにFabまたは他の免疫グロブリン発現ライブラリーの産物を含むFabフラグメントが含まれる。
【0060】
本発明のポリペプチドまたはその断片もしくは類似体、またはそれらを発現する細胞は、本発明のポリペプチドに結合する抗体を産生するための免疫原としても使用することができる。抗体は、好ましくは、本発明のポリペプチドに免疫特異的である。「免疫特異的」とは、その抗体が他のポリペプチドに対するその親和性よりも本発明のポリペプチドに対して実質的に高い親和性を有することを意味する。
【0061】
本発明のポリペプチドに結合する抗体は、当業者に公知の方法により調製することが可能である。ポリクローナル抗体であれば、例えば、次のようにして得ることができる。本発明のポリペプチドあるいはそのGSTとの融合タンパク質をウサギ等の小動物に免疫し血清を得る。これを、例えば、硫安沈殿、プロテインA、プロテインGカラム、DEAEイオン交換クロマトグラフィー、本発明のポリペプチドをカップリングしたアフィニティーカラム等により精製することにより調製する。また、モノクローナル抗体であれば、例えば、本発明のポリペプチドをマウスなどの小動物に免疫を行い、同マウスより脾臓を摘出し、これをすりつぶして細胞を分離し、マウスミエローマ細胞とポリエチレングリコールなどの試薬により融合させ、これによりできた融合細胞(ハイブリドーマ)の中から、本発明のポリペプチドに結合する抗体を産生するクローンを選択する。次いで、得られたハイブリドーマをマウス腹腔内に移植し、同マウスより腹水を回収し、得られたモノクローナル抗体を、例えば、硫安沈殿、プロテインA、プロテインGカラム、DEAEイオン交換クロマトグラフィー、本発明のポリペプチドをカップリングしたアフィニティーカラム等により精製することで、調製することが可能である。
【0062】
本発明の抗体は、被検試料中の本発明のポリペプチドの検出や精製に用いることもできる。
【0063】
<被検試料中のカンピロバクター属細菌の存在の検出>
本発明は、被検試料中のカンピロバクター属細菌の存在の検出方法を提供する。被検試料中のカンピロバクター属細菌の存在の検出は、カンピロバクター感染症の診断、カンピロバクター属細菌に汚染された食品の迅速診断、食品加工工程のバリデーション、食中毒発生時における原因菌の同定など種々の目的において有用である。
【0064】
本発明の検出方法の一つの態様は、被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物を用いたポリメラーゼ連鎖反応を行なう工程、および(b)これら細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、これら細菌の存在を判定する工程、を含む方法である。
【0065】
また、本発明の検出方法の他の態様は、被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いた核酸増幅反応を行なう工程、(b)工程(a)により増幅されたゲノムDNAを鋳型に、これら細菌の細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物を用いたポリメラーゼ連鎖反応を行なう工程、および(c)これら細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、これら細菌の存在を判定する工程、を含む方法である。
【0066】
実施例のように、複数のPCRプライマーを単一の反応系で使用するPCRは、マルチプレックスPCR法と呼ばれ、PCR産物を電気泳動し、バンドのサイズを見ることで複数の菌種を同時に鑑別することができる。本発明は、このマルチプレックスPCR法を代表とする、複数の核酸領域の増幅に好適に用いられるプライマーおよびその組み合わせを用いた核酸増幅法によるカンピロバクター属細菌の検出方法を提供する。本発明における核酸増幅の方法は、目的とする増幅物が得られる限り種類は問わないが、好ましくはPCR法である
【0067】
これら方法における、特異的なプライマー対の混合物としては、例えば、次の、(a)から(c)のプライマー対の混合物を用いることができる。(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対、(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対、(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対。また、共通プライマー対としては、例えば、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対を用いることができる。
【0068】
本発明の検出方法のさらなる態様は、被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いた核酸増幅反応を行なう工程、(b)工程(a)により増幅されたゲノムDNAを制限酵素により切断する工程、(c)切断されたDNA断片の分子量から、これら細菌の存在を判定する工程、を含む方法である。この方法に用いうる制限酵素としては、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNA を識別しうるものであれば特に制限はなく、例えば、Sau3AI、Dsa I、Mbo I、Rsa I、EcoR I、Hinf I、Nde I、Pst I、Xba I、Xho IIが挙げられる。また、共通プライマー対としては、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対を例示することができる。
【0069】
下記実施例で示すような、PCR法で増幅したDNAを種々の制限酵素で処理し、生じる断片の長さから多型を検出する方法は、PCR-RFLP法(PCR-Restriction Fragment Length Polymorphism:PCR-制限酵素断片長多型)と呼ばれる。本発明はまた、PCR-RFLP法に代表される、核酸増幅法により増幅されたDNAを種々の制限酵素で処理し、生じる断片の長さから多型を検出する方法に好適に用いられるプライマーを提供する。
【0070】
また、本発明の検出方法の他の態様は、被験試料中のカンピロバクター属細菌の存在を検出する方法であって、(a)被検試料に対し、カンピロバクター属細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いた核酸増幅反応を行なう工程
(b)カンピロバクター属細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、カンピロバクターの存在を判定する工程、を含む方法である。この方法に用いるプライマー対は、カンピロバクター属細菌であればその種を問わず、その細胞膨張化致死毒をコードするゲノムDNAを増幅し得るプライマー対である。このような共通プライマー対としては、配列番号;7−10、47−50、64−67から選択されるプライマー対を例示することができる。上述したとおり、上記プライマー対は少なくともカンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータス3菌種の細胞膨張化致死毒をコードするゲノムDNA を共通して増幅する共通プライマー対である。上記プライマー対は、上記3菌種のみならず、その他のカンピロバクター属細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅すると期待できる。また同様に、該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対についても、上記3菌種を共通して該ゲノム領域を増幅することができ、その他のカンピロバクター属細菌の該ゲノム領域を増幅することができると考えられる。
【0071】
また、本発明は、上記本発明の検出方法に用いるためのキットを提供する。これらキットは、上記したプライマー対の他、使用説明書を含むものである。さらなる他の要素を含んでいてもよい。
【0072】
なお、カンピロバクター属細菌の存在の検出は、上記のようなDNAレベルだけでなく、タンパク質レベルで行なうことも考えられる。例えば、これら細菌の細胞膨張化致死毒に特異的な抗体を用いて、ウェスタンブロッティング法、ドットブロッティング法、免疫沈降法、酵素結合免疫測定法(ELISA)、あるいは免疫蛍光法などを実施し、これら細菌の細胞膨張化致死毒を検出することにより、被検試料中におけるこれら細菌の存在を評価することが可能である。
なお本明細書において引用された全ての先行技術文献は、参照として本明細書に組み入れられる。
【実施例】
【0073】
[実施例1] カンピロバクター属菌株
2001年から2003年までの間に各種臨床材料より収集したカンピロバクター・ジェジュニ(C.ジェジュニ)菌、カンピロバクター・コリ(C.コリ)菌、およびカンピロバクター・フィータス(C.フィータス)菌を使用した。各菌株は、5%ウマ脱繊維血液(日本生物材料センター)、およびカンピロバクター選択サプリメントSR69(OXOID)を含む血液寒天培地(Blood Agar Base No.2:OXOID)を用い、C.ジェジュニおよびC.コリは5%O2, 10%CO2, 85%N2ガス下42℃で、C.フィータスは25℃の条件下で、低温酸素/炭酸ガス培養装置(MODEL9200:和研薬)を使用し、培養した。
【0074】
[実施例2] PCR用テンプレートの調整
各菌種から5クローンを掻き取り500μLの無菌PBSに懸濁した。回収した細菌を10,000rpmで5分間遠心洗浄(MRX-150:トミー精工)した後、300μLの無菌PBSに再懸濁した。その後、沸騰水浴中で10分間煮沸し、氷上で冷却した後、15,000rpmで10分間遠心し、上清を回収した。回収上清中のDNA量を分光光度計(Ultrospec 3100pro:アマシャム・バイオサイエンス)を用いて定量した。定量した各菌体抽出液を各20ng/μLとなるよう希釈し、PCRに供した。
【0075】
[実施例3] C.コリcdtB プローブの作製とサザンハイブリダイゼーション
C.コリcdtBプローブはGNWとLPF-Dプライマーを用いてC.コリCo1-192 菌体抽出液をテンプレートとしてDIGラベリングミックス(ロシュ)を用いてPCRラベルを行ない、作製した。
【0076】
すなわち、C.コリのCDT遺伝子の存在を調べるため、文献(Pickett, C. et al. Infect. Immun., 64: 2070 (1996))に記載されたGNW縮重プライマー[配列番号5:5’-GG(ACGT)AA(CT)TGGAT(ACT)TGGGG(ACGT)TA-3’]およびLPF-D縮重プライマー[配列番号6:5’- (AGT)AA(CT)TG(ACGT)AC(AGT)TA(ACGT)CC(AGT)AA (ACGT)GG - 3’]を用い、C.ジェジュニ3株、C.コリ2株につき、94℃, 3分間−(94℃30秒, 42℃30秒, 72℃2分)×30サイクル−72℃, 5分間の条件でPCRを行なった。その結果、C.ジェジュニ3株とC.コリ2株のすべてにおいて、約1.5Kbにcdt領域が増幅されたバンドが認められた(図1の矢印1)。
【0077】
この増幅バンドをpT7ベクター(Novagen)にライゲートし、 宿主大腸菌(E.coli JM109)に形質転換した。得られたクローンのシークエンスをシーケンサー(ABI PRISM 377 DNA sequencer:アプライドバイオシステムズ)を用いて確認したところ、cdtBに類似の配列が見られた。シークエンス反応はBigDye terminator Cycle Sequencing Kits(アプライドバイオシステムズ)を用いた。また、800bpのバンド(図1の矢印2)は、GNWプライマーがミックスプライマーであるため増幅されるcdtB由来の副バンドであることが確認された。
【0078】
得られたプローブを用いて、C.コリCo1-192のゲノム20μgを60Uの制限酵素HindIIIで37℃、12時間消化し、定法(Molecular cloning: a laboratory manual, Cold Spring Harbor Labolatory, Cold Spring Harbor, (2001))に従ってサザンブロットおよびDNA-DNAハイブリダイゼーションを行った。ハイブリダイゼーションは、42℃でストリンジェントな条件下で実施した。ブロッティング後、2×SSC−0.1%SDS溶液で室温で5分間、2度洗浄し、さらに0.2×SSC−0.1%SDS溶液で60℃で15分間、2度洗浄した。
【0079】
その結果、3kと4k付近にプローブ陽性バンドが得られた(図2)。この3kのバンドをpUC18 Vectorにライゲートし、E.coli JM109に形質転換して、cdtB領域を含むクローン(3k44)を得た。
【0080】
[実施例4] C.コリcdtB 遺伝子のシークエンス
実施例3で得られたC.コリcdtB領域を含む3k44クローンにつき、常法によりシークエンスを行い、配列番号1で示されるC.コリCDT全領域の配列を決定した。
【0081】
[実施例5] 共通プライマー1の設計およびPCR
本発明のC.コリCDT配列と、既知のデータベースから得られたC.ジェジュニのCDT遺伝子を比較し、下記の共通プライマーUおよび共通プライマーRを設計した。20ng/μLの菌体抽出液1μL、各プライマーをそれぞれの濃度が0.5mMとなるように混合して添加し、PCR反応用緩衝液(TaKaRa Ex Taq kit:タカラバイオ)を用いて最終液量20μLとなるよう調整し、94℃3分間−(94℃30秒間、55℃30秒間、72℃1分間)×30サイクル−72℃3分間の条件でPCR反応に供した。結果を図3に示す。約1900bpの増幅断片が確認され、C.ジェジュニ(レーン2〜4)およびC.コリ(レーン5、6)のいずれにもCDTに由来するバンドが認められた。
共通プライマーU[配列番号7:GATAA(CT)GATCCTTTAAAACT]
共通プライマーR[配列番号8:(AT)(AT)CCAAAGCG(AT)TTTT(CG)TATGG]
【0082】
[実施例6] 共通プライマー2の設計およびPCR
同様にして、下記の共通プライマーUpおよび共通プライマーDoを設計し、94℃3分間−(94℃30秒間、50℃30秒間、72℃45秒間)×30サイクル−72℃3分間の条件でPCR反応に供した。結果を図4〜6に示す。約720bpの増幅断片が確認された。
共通プライマーUp[配列番号9:ACTTGGAATTTGCAAGGC]
共通プライマーDo[配列番号10:TCTAAAATTTAC(ACT)GGAAAATG]
【0083】
[実施例7] 特異的プライマーの設計およびマルチプレックスPCRによるcdtB遺伝子の検出
本発明のC.コリCDT配列と、既知のデータベースから得られたC.ジェジュニのCDT遺伝子を比較し、下記のC.ジェジュニ特異的プライマーCjSPBU3およびCjSPBR3を設計した。同様にC.コリ特異的プライマーCcSPBU5およびCcSPBR5、C.フィータス特異的プライマーCfSPBU1およびCfSPBR1を設計した。
【0084】
20ng/μLの菌体抽出液1μL、各プライマーをそれぞれの濃度が0.5mMとなるように混合して添加し、PCR反応用緩衝液(TaKaRa Ex Taq kit:タカラバイオ)を用いて最終液量20μLとなるよう調整し、94℃3分間−(94℃56秒間、55℃30秒間、72℃45秒間)×30サイクル−72℃3分間の条件でマルチプレックスPCR反応に供した(GeneAmp PCRシステム9700:アプライドバイオシステムズ)。結果を図7に示す。C.ジェジュニCDT特異的増幅断片(約750bp)、C.コリCDT特異的増幅断片(約400bp)、およびC.フィータスCDT特異的増幅断片(約530bp)が確認され、C.ジェジュニ(レーン2〜4)、C.コリ(レーン5、6)、およびC.フィータス(レーン7、8)の識別が可能であった。
特異的プライマーCjSPBU3[配列番号11:TACTCCGCCTTTTACCGCA]
特異的プライマーCjSPBR3[配列番号12:GAGTATAGGTTTGTTGTC]
特異的プライマーCcSPBU5[配列番号13:TTTAATGTATTATTTGCCGC]
特異的プライマーCcSPBR5[配列番号14:TCATTGCCTATGCGTATG]
特異的プライマーCfSPBU1[配列番号15:CGCAAGTTGGAAGACTAT]
特異的プライマーCfSPBR1[配列番号16:TTTATTATCGCCGGAGCA]
【0085】
[実施例8] 共通プライマー1を用いたPCR-RFLPによる菌種の同定
実施例6で得られた共通プライマー1を用いたPCRの完了後、8.5μLの反応液に5Uの制限酵素Sau3AI(NEB)を添加し、37℃で3時間反応させ、電気泳動を行った。結果を図8に示す。
【0086】
[実施例9] 特異的プライマーを用いたマルチプレックスPCRによるcdtB遺伝子の検出
実施例7で得られた特異的プライマーを用い、さらに種々のカンピロバクター属細菌の臨床菌株につき、実施例7の実験条件でマルチプレックスPCRを行った。結果を図9に示す。実施例7と同様に、C.ジェジュニCDT特異的増幅断片(約750bp)、C.コリCDT特異的増幅断片(約400bp)、およびC.フィータスCDT特異的増幅断片(約530bp)が確認され、各菌種の識別が可能であった。
【0087】
[実施例10] C.フィータスcdtBプローブの作製とサザンハイブリダイゼーション
C.フィータスcdtBプローブは共通プライマー2(共通プライマーUpおよび共通プライマーDo)を用いてC.フィータスCo1-187菌体抽出液をテンプレートとしてDIGラベリングミックス(ロシュ)を用いてPCRラベルを行ない、作製した。
得られたプローブを用いて、C.フィータスCo1-187のゲノム20μgを60Uの制限酵素HindIIIで37℃、12時間消化し、定法(Molecular cloning: a laboratory manual, Cold Spring Harbor Labolatory, Cold Spring Harbor, (2001))に従ってサザンブロットおよびDNA-DNAハイブリダイゼーションを行った。ハイブリダイゼーションは、42℃でストリンジェントな条件下で実施した。ブロッティング後、2×SSC-0.1%SDS溶液で室温で5分間、2度洗浄し、さらに0.2×SSC-0.1%SDS溶液で60℃で15分間、2度洗浄した。
その結果、2kと5k付近にプローブ陽性バンドが得られた(図10)。この2kのバンドをpUC18 Vectorにライゲートし、E.coli JM109に形質転換して、cdtB領域を含むクローン(Cf78)を得た。
【0088】
[実施例11] C.フィータスCDT遺伝子のシークエンス
実施例10で得られたC.フィータスcdtB領域を含むCf78クローンにつき、常法によりシークエンスを行い、C.フィータスのcdtAおよびcdtB領域の配列を決定した。得られたクローンCf78にはcdtC領域が含まれていなかったため、決定したcdtB遺伝子配列から設計したランダムプライマーを用いて以下の条件で遺伝子ウォーキングを行い、cdtC領域の配列を決定し、配列番号51で示されるC.フィータスCDT全領域の配列を決定した。
[ランダムプライマーによる遺伝子ウォーキング]
実施例11で得られた遺伝子配列より、以下に示すランダムプライマー、ターゲット増幅プライマーおよびシークエンスプライマーのセットを設計し、C.フィータスCol-187の遺伝子をテンプレートとしてターゲットの増幅を行った。ターゲットの増幅は、テンプレート遺伝子20ngに対して10pmolのランダムプライマーを加え、KOD Dash PCR Kit(東洋紡)を用いて最終100μLとなるように調整し、94℃2分間−(94℃20秒間、65℃5秒間、74℃30秒間)×35サイクルの条件でPCRに供した。
得られたPCR産物をシークエンスプライマーを用いて定法に従ってシークエンスを行った。
プライマーセット1
ランダムプライマー[配列番号55:GCTTGTAGCAGTATTGATGCNNNNNNNNN]
ターゲット増幅プライマー[配列番号56:GCTTGTAGCAGTATTGATGC]
シークエンスプライマー[配列番号57:CTAGTTTCGGACCATTTTCC]
プライマーセット2
ランダムプライマー[配列番号58:ATACGCAATGCAAACACCGGNNNNNNNNN]
ターゲット増幅プライマー[配列番号59:ATACGCAATGAAACACCGG]
シークエンスプライマー[配列番号60:TAAAAGCGATTTTCAGGGCAG]
プライマーセット3
ランダムプライマー[配列番号61:TGTCGACATAGAGCCTAAACNNNNNNNNN]
ターゲット増幅プライマー[配列番号62:TGTCGACATAGAGCCTAAAC]
シークエンスプライマー[配列番号63:ATTTTCACCGCCGCTTAGTG]
【0089】
[実施例12] cdtA共通プライマーの設計およびPCR
本発明のC.コリおよびC.フィータスのcdtA配列と、既知のデータベースから得られたC.ジェジュニのcdtA遺伝子を比較し、下記のcdtA共通プライマーUおよびcdtA共通プライマーRを設計した。20ng/μLの菌体抽出液1μL、各プライマーをそれぞれの濃度が0.25mMとなるように混合して添加し、PCR反応用緩衝液(TaKaRa Ex Taq kit:タカラバイオ)を用いて最終液量20μLとなるよう調整し、94℃3分間−(94℃30秒間、55℃30秒間、72℃30秒間)×30サイクル−72℃3分間の条件でPCR反応に供した。結果を図11(左)に示す。約550bpの増幅断片が確認され、C.ジェジュニ(レーン2〜4)およびC.コリ(レーン5、6)、C.フィータス(レーン7、8)のいずれにもcdtAに由来するバンドが認められた。
cdtA共通プライマーU
[配列番号64:(GA)A(ACT)GAT(AC)(AC)(TAG)GAT(AC)GATCC(AT)(TC)CAAA]
cdtA共通プライマーR
[配列番号65:(GA)(AT)AA(TC)AGG(TC)G(CT)TTG(CT)A(AT)(GA)CA]
【0090】
[実施例13] cdtA共通プライマーを用いたPCRによるcdtA遺伝子の検出
実施例7で得られた特異的プライマーを用い、さらに種々のカンピロバクター属細菌の臨床菌株につき、実施例12の実験条件でマルチプレックスPCRを行った。結果を図12に示す。実施例12と同様に、cdtA特異的増幅断片(約550bp)が確認された。
【0091】
[実施例14] cdtC共通プライマーの設計およびPCR
本発明のC.コリおよびC.フィータスcdtC配列と、既知のデータベース(BLAST)から得られたC.ジェジュニのcdtC遺伝子を比較し、下記のcdtC共通プライマーUおよびcdtC共通プライマーRを設計した。
20ng/μLの菌体抽出液1μLおよび各プライマーをそれぞれの濃度が0.25mMとなるように混合して添加し、PCR反応用緩衝液(TaKaRa Ex Taq kit:タカラバイオ)を用いて最終液量20μLとなるよう調整し、94℃3分間−(94℃30秒間、55℃30秒間、72℃30分間)×30サイクル−72℃3分間の条件でPCR反応に供した。結果を図3に示す。約320bpの増幅断片が確認され、C.ジェジュニ(レーン10〜12)、C.コリ(レーン13、14)およびC.フィータス(レーン15、16)のいずれにもcdtCに由来するバンドが認められた(図11右)。
cdtC共通プライマーU[配列番号66:(AGC)A(TG)(TC)(TC)(AT)(AG)(AT)(AT)(GT)A(CT)CAAAA(CT)TGG]
cdtC共通プライマーR[配列番号67:(AGC)CTA(AGT)(AT)CC(AT)A(AC)(GT)C(GT)(AT)T(CT)TT(GC)]
【0092】
[実施例15] cdtC共通プライマーを用いたPCRによるcdtC遺伝子の検出
実施例で得られた特異的プライマーを用い、さらに種々のカンピロバククー属細菌の臨床菌株につき、実施例14の実験条件でマルチプレックスPCRを行った。結果を図13に示す。実施例14と同様に、cdtC特異的増幅断片(約320bp)が確認された。
【0093】
[実施例16] cdtA特異的プライマーの設計およびマルチプレックスPCRによるcdtA遺伝子の検出
本発明のC.フィータスCDT配列と既知のデータベース(BLAST)から得られたC.ジェジュニおよびC.コリのCDT遺伝子を比較し、下記のC.ジェジュニ特異的プライマーCjASPU2およびCjASPR2を設計した。同様にC.コリ特異的プライマーCcASPU1、CcASPR1およびC.フィータス特異的プライマーCfASPU1およびCfASPR1を設計した。
20ng/μLの菌体抽出液1μL、各プライマーをそれぞれの濃度が0.125mMとなるように混合して添加し、PCR反応用緩衝液(TaKaRa Ex Taq kit:タカラバイオ)を用いて最終液量20μLとなるよう調整し、94℃3分間−(94℃30秒間、55℃30秒間、72℃30秒間)×30サイクル−72℃3分間の条件でマルチプレックスPCR反応に供した(GeneAmp PCRシステム9700:アプライドバイオシステムズ)。結果を図14(左)に示す。C.ジェジュニCDT特異的増幅断片(約630bp)、C.コリCDT特異的増幅断片(約330bp)、およびC.フィータスCDT特異的増幅断片(約490bp)が確認され、C.ジェジュニ(レーン2〜4)、C.コリ(レーン5、6)、およびC.フィータス(レーン7、8)の識別が可能であった。
特異的プライマーCjASPU2[配列番号68:AGGACTTGAACCTACTTTTC]
特異的プライマーCjASPR2[配列番号69:AGGTGGAGTAGTTAAAAACC]
特異的プライマーCcASPU1[配列番号70:ATTGCCAAGGCTAAAATCTC]
特異的プライマーCcASPR1[配列番号71:GATAAAGTCTAAAACTGC]
特異的プライマーCfASPU1[配列番号72:AACGACAAATGTAAGCACTC]
特異的プライマーCfASPR1[配列番号73:TATTTATGCAAGTCGTGCGA]
【0094】
[実施例17]cdtA特異的プライマーを用いたマルチプレックスPCRによるcdtA遺伝子の検出
実施例で得られた特異的プライマーを用い、さらに種々のカンピロバククー属細菌の臨床菌株につき、実施例14の実験条件でマルチプレックスPCRを行った。結果を図15に示す。実施例14と同様に、C.ジェジュニcdtA特異的増幅断片(約630bp)、C.コリcdtA特異的増幅断片(約330bp)およびC.フィータスcdtA特異的増幅断片(約490bp)が確認され、各菌種の識別が可能であった。
【0095】
[実施例18]cdtC特異的プライマーの設計およびマルチプレックスPCRによるcdtC遺伝子の検出
本発明のC.フィータスCDT配列と、既知のデータベースから得られたC.ジェジュニおよびC.コリのCDT遺伝子を比較し、下記のC.ジェジュニ特異的プライマーCjCSPU1およびCjCSPR2を設計した。同様にC.コリ特異的プライマーCcCSPU1およびCcCSPR1、C.フィータス特異的プライマーCfCSPU2およびCfCSPR1を設計した。
20ng/μLの菌体抽出液1μL、各プライマーをそれぞれの濃度が0.125mMとなるように混合して添加し、PCR反応用緩衝液(TaKaRa Ex Taq kit:タカラバイオ)を用いて最終液量20μLとなるよう調整し、94℃3分間−(94℃30秒間、55℃30秒間、72℃30秒間)×30サイクル−72℃3分間の条件でマルチプレックスPCR反応に供した(GeneAmp PCRシステム9700:アプライドバイオシステムズ)。結果を図14(右)に示す。C.ジェジュニCDT特異的増幅断片(約500bp)、C.コリCDT特異的増幅断片(約300bp)、およびC.フィータスCDT特異的増幅断片(約400bp)が確認され、C.ジェジュニ(レーン10〜12)、C.コリ(レーン13、14)、およびC.フィータス(レーン15、16)の識別が可能であった。
特異的プライマーCjCSPU1[配列番号74:TTTAGCCTTTGCAACTCCTA]
特異的プライマーCjCSPR2[配列番号75:AAGGGGTAGCAGCTGTTAA]
特異的プライマーCcCSPU1[配列番号76:TAGGGGATATGCACGCAAAAG]
特異的プライマーCcCSPR1[配列番号77:GCTTAATACAGTTACGATAG]
特異的プライマーCfCSPU2[配列番号78:AAGCATAAGTTTTGCAAACG]
特異的プライマーCfCSPR1[配列番号79:GTTTGGATTTTCAAATGTTCC]
【0096】
[実施例19]特異的プライマーを用いたマルチプレックスPCRによるcdtC遺伝子の検出
実施例で得られた特異的プライマーを用い、さらに種々のカンピロバククー属細菌の臨床菌株につき、実施例14の実験条件でマルチプレックスPCRを行った。結果を図16に示す。実施例14と同様に、C.ジェジュニCdtC特異的増幅断片(約500bp)、C.コリCdtC特異即曽幅断片(約300bp)、およびC.フィータスCdtC特異的増幅断片主(約400bp)が確認され、各菌種の識別が可能であった。
【産業上の利用の可能性】
【0097】
本発明のプライマーは、カンピロバクター属細菌の疫学的調査、研究、およびカンピロバクター感染症の診断のみならず、カンピロバクター属細菌に汚染された食品の迅速診断、食品加工工程のバリデーション、食中毒発生時に迅速に原因菌を同定することが可能であり、感染の拡大防止に有用である。
【技術分野】
【0001】
本発明は、カンピロバクター・コリの細胞膨張化致死毒およびそれをコードするポリヌクレオチドに関する。さらに本発明は、カンピロバクター・コリを含むカンピロバクター属に属する細菌の細胞膨張化致死毒を標的とした、検体(臨床検体、食品など)中のカンピロバクター(Campylobacter)属細菌の存在の有無を判定するための方法に関する。
【背景技術】
【0002】
カンピロバクター属細菌は、ヒト、および野生または家畜動物の病原菌であって、動物における流産や腸炎、ヒトにおける腸炎の原因菌である。ヒトの場合、カンピロバクター感染症の起因菌としては、カンピロバクター・ジェジュニ(Campylobacter jejuni)およびカンピロバクター・コリ(Campylobacter coli)が知られており、これらの細菌は、食中毒菌として指定されている(非特許文献1、2参照)。
【0003】
2000年現在、カンピロバクターは15菌種9亜種に分類されているが、ヒトの下痢症から分離される菌種は、C.ジェジュニが95〜99%を占め、C.コリなどのその他の菌種は数%である(非特許文献3参照)。なお、C.コリはブタでの保菌率が極めて高い。近年、カンピロバクター感染症は、東南アジアを中心とした輸入食肉の増加に伴い増加傾向にあり、特にBSEやO-157等の問題によって牛肉の代替品として消費量が伸びている鶏肉関連食品による感染例が急増している。
上記の他、C.フィータス(Campylobacter fetus)は、ヒツジやウシの流産菌として知られていたが、近年ヒトの流産や早産に関与しているとの報告がある。また、C.フィータスで汚染されたレバーや牛肉の生食によるC.フィータス感染では、敗血症や髄膜炎などの症状を呈することもある。ヒトへの感染源は、腸管内に高濃度に保菌する鶏肉が最も重要である。(非特許文献4)
【0004】
カンピロバクター属細菌は、通常、ウシ、ヒツジ、ブタ、ニワトリ等の動物の消化管に高率に分布しており、人獣共通感染症の原因細菌として認識されている。カンピロバクター感染症の多くは鶏肉が原因と考えられているが、これらの動物もしくは排泄物との直接的接触や、排泄物によって汚染された食物、飲料水、牛乳などの摂取や調理過程により感染する。さらに、新生児室などの施設内感染例も報告されている(非特許文献5参照)
【0005】
カンピロバクター感染症は、潜伏期が3〜7日と長く、下痢(ときに血粘膜性下痢)、腹痛、発熱、悪心、嘔吐、頭痛、悪寒、倦怠感などの胃腸炎症状を引き起こすのが特徴である。致死性は低いが、新生児では敗血症や髄膜炎などの全身感染を起こすことがある。多くの場合、数日から1週間程度で回復し、一般的な予後については、一部の免疫不全患者を除き良好な経過を辿る。しかし、近年カンピロバクターの後感染性疾患として自己免疫疾患であるギラン・バレー症候群やフィッシャー症候群に進展する例が報告されている。カンピロバクター感染症に後発する症例は概して重症化しやすく、発症1年後の寛解率は約6割程度に止まる。
【0006】
重篤な病態や敗血症の併発例では抗生剤による化学治療が行われる。第一選択薬剤はエリスロマイシン等のマクロライド系薬剤である。セフェム系薬剤に対しては自然耐性を示すため、治療効果は望めない。一方、近年、ニューキノロン系薬剤に対しては、耐性菌の増加が問題となっている。カンピロバクター感染症に対し的確な治療を施し、併せて感染経路を解明して感染拡大を食い止めるためには、感染後、迅速に原因菌を同定することが重要である。しかしながら、臨床症状からカンピロバクター感染を診断することは困難であり、カンピロバクターの同定および菌種の決定は容易ではない。
【0007】
カンピロバクター属細菌は微好気性細菌であり、培養にはスキロー培地などの特殊な培地、および酸素濃度を3〜10%の絶対微好気性条件に保つための特殊な装置(嫌気ジャー等)が必要で、なおかつ他の細菌より培養に時間を要する(2〜3日)ことから、分離培養が困難なものとなっている。また、カンピロバクター属細菌は、空気中で死滅しやすく、検体採取後、2〜3時間以内に検査する必要がある。さらに、カンピロバクター感染症は潜伏期間が長い(3〜7日)ため、病状が現れてから関与食品の細菌同定を行っても菌が単離できない場合が多い。さらにカンピロバクター属細菌は、非常に感染力が強く、数百個の菌数で感染が成立するという報告もあり、感染源の特定は極めて困難なものとなっている。
【0008】
C.ジェジュニとC.コリの識別診断に、馬尿酸塩の加水分解を検査する方法がある。すなわち、C.ジェジュニは馬尿酸を加水分解できるが、C.コリはできないという性質を利用する。しかしながら、馬尿酸−陰性C.ジェジュニ株が存在するため、この方法も確実ではない(非特許文献6参照)。従って、現在は摂食履歴と症状によるカンピロバクター菌の存在の推定、および便培養から得られたコロニーについて数日を要して菌の形態学的および生物学的特徴を検査することにより、その存在を確認しているのが現状である。
【0009】
そこで、培養検査によらない迅速診断法として、オリゴヌクレオチドを用いたDNAプローブ法やPCR法を利用した遺伝子的診断法によるカンピロバクター属細菌の同定や毒素遺伝子の検出が試みられている。たとえば、カンピロバクター属細菌の場合、一般にrRNAをコードする遺伝子がプローブとして使用されている(特許文献1参照)。このrRNAの遺伝子配列は既に公表されている(非特許文献7参照)。また、カンピロバクター属細菌検出用の核酸フラグメントも知られている(特許文献2、3、4、5、6参照)。しかしながら、これらの配列は、C.ジェジュニおよび/またはC.コリの検出用であり、その他のカンピロバクター属細菌の検出には適当でない。さらに、特異性が充分ではなかった。
【0010】
また、C.コリVC167のfla A遺伝子から選択したオリゴヌクレオチドを用いたPCRによって、C.ジェジュニを同定する方法が報告されている(非特許文献8参照)。さらに、C.ジェジュニおよびC.コリ中に見出されるスーパーオキシドジスムターゼ標的配列を増幅するためのオリゴヌクレオチドプライマーが報告されている(特許文献7参照)。しかし、これらの方法ではC.ジェジュニとC.コリを区別することができない。
【0011】
一方、カンピロバクター属細菌の病原因子についての研究も盛んである。カンピロバクター属細菌の病原因子としては、細胞侵入性やフラジェリン、コレラ毒素様エンテロトキシン等の様々な因子が報告されている(非特許文献9、10参照)が、近年、C.ジェジュニから毒性因子として細胞膨張化性致死毒(Cytolethal Distending Toxin:CDT)が発見され(非特許文献11)、病原性との関連性が注目されている。例えば、志賀赤痢菌(Shigella dysenteriae)のCDTを産生する組換え大腸菌を用いた動物投与モデルでは下痢原性が報告されている(非特許文献12)。
【0012】
CDTはcdtA、cdtBおよびcdtCと称される3種のサブユニットからなるホロ毒素であり、タンデムに並んだ遺伝子にコードされている。毒素活性中心はcdtBサブユニットの持つI型デオキシリボヌクレアーゼ様活性であり、cdtAサブユニットとcdtCサブユニットは標的細胞への接着に関与すると考えられている。このホロ毒素が細胞に作用すると、細胞が膨化、すなわち大きくふくらみ、最終的には細胞が死滅することから、細胞膨張化性致死毒と名付けられている。
【0013】
その分子機構は活性毒素中心であるcdtBサブユニットが細胞内の核に移行し、I型デオキシリボヌクレアーゼ活性により、染色体DNAにニックを挿入し、DNA損傷応答(DNA-damage response)を引き起こす。そして、細胞は遺伝子修復系を動かすために細胞周期をG2/M期で停止後、膨張化致死すると考えられている(非特許文献13)。さらに、CDTは上皮系細胞、免疫担当細胞等の幅広い細胞に作用することが確認されており、中でもヒトリンパ球に作用し、アポトーシスを引き起こすことにより免疫から逃れやすくなっていると考えられている(非特許文献14)。
【0014】
このようにCDTは、これまでの毒素には見られないユニークな分子機構を有しているが、これまでカンピロバクター属細菌でCDTの全塩基配列が明らかとなっているのは、C.ジェジュニのみである(非特許文献11)。
【特許文献1】特開昭62−228096号公報
【特許文献2】特開平2−84200号公報
【特許文献3】特開平2−154700号公報
【特許文献4】特開平3−112498号公報
【特許文献5】特開平6−90795号公報
【特許文献6】特開平6−90796号公報
【特許文献7】特開2000−316590公報
【非特許文献1】Blaser, et al, Ann. Intern. Med., 91:179 (1979)
【非特許文献2】Tauxe, R., American Society for Microbiology, Washington DC. pg.9(1992)
【非特許文献3】Takahashi, M. et al, Infectious Diseases Weekly Report Japan, 3(6):10 (2001)
【非特許文献4】Simon, M. S. et al., 2003. Campylobacter infection. Diseases of Poultry, Iowa State Press, 615-630
【非特許文献5】小児内科, 29:1219-1222 (1997)
【非特許文献6】Totten, et al, J. Clin. Microbiol., 25: 1747 (1987)
【非特許文献7】Romaniuk, P. J. et al, J. Bacteriol., 169: 2173 (1987)
【非特許文献8】Oyofo, et al, J. Clin. Microbiol., 30: 2613 (1992)
【非特許文献9】Mizuno, K. et al, Microbios., 78: 215 (1994)
【非特許文献10】Suzuki, S. et al, FEMS Immunol. Med. Micribiol., 8: 207 (1994)
【非特許文献11】Pickett, C. et al. Infect. Immun., 64: 2070 (1996)
【非特許文献12】Infect. Immun., 65: 428-433 (1997)
【非特許文献13】Science, 290: 354-357 (2000)
【非特許文献14】J. Biol. Chem., 276: 5296-5302 (2001)
【発明の開示】
【発明が解決しようとする課題】
【0015】
以上詳細に述べたように、カンピロバクター属細菌による感染症の迅速診断が求められている一方、カンピロバクター属細菌の病原因子は完全に解明されているとは言い難い。これまで、各菌種の血清型等を利用した菌種同定用PCRプライマーや、CDT産生を調べる共通プライマー等が利用されてきたが(J. Applied Microbiol., 94: 1003-1014 (2003))、増菌培養工程が必要であり、カンピロバクター属細菌の迅速な検出は不可能であった。
【0016】
そこで、本発明は、遺伝子診断により、カンピロバクター属細菌の迅速な検出を可能とすべく、カンピロバクター属に属し、いまだそのCDTの塩基配列が解明されていないカンピロバクター・コリのCDTおよびそれをコードするポリヌクレオチドを提供することを課題とする。さらに本発明は、同様にCDTの塩基配列が解明されていないカンピロバクター・フィータスのCDTおよびそれをコードするポリヌクレオチドを提供することを課題とする。
【0017】
さらに、本発明は、カンピロバクター・コリおよびカンピロバクター・フィータスの塩基配列から得られた知見に基づき、カンピロバクター・コリおよびカンピロバクター・フィータスを含むカンピロバクター属細菌のCDTを標的とした、カンピロバクター属細菌の存在を迅速に検出しうる方法を提供することを課題とする。
【課題を解決するための手段】
【0018】
CDT遺伝子のクローニングにおいて、制限酵素HindIIIを用いたクローニングを行った場合、コード領域中にHindIIIサイトが存在するため、全長を単離することはできない。他方、EcoRI、PstI、KpnI、XbaI、BamHI、SalI、XhoI等の一般的な制限酵素を用いると、cdt遺伝子のクローニングに最適な長さの断片(3kbから5kb)が得られなかった。そこで、本発明者らは種々検討を行なった結果、HindIIIで完全にcdt遺伝子が消化されないようなパーシャルな条件を選択し、切断することにより、遂に、cdt遺伝子の内部配列が切断されていない完全長の遺伝子をクローニングすることに成功した。
【0019】
また、本発明者らは、C.ジェジュニ、C.フィータスのCDTとの比較を行ない、3種のカンピロバクター属細菌に共通するプライマーおよびこれら細菌のそれぞれに特異的なプライマーを開発した。そしてこのプライマーが、簡便かつ迅速にカンピロバクター属細菌CDTの有無の判定、および菌種の同定が同時に行えるマルチプレックスPCR法に適用でき、さらにPCR-RFLP法によるタイピングにも利用できることを明らかにした。
【0020】
すなわち、本発明は、具体的に以下の技術的態様が包含される:
(1) 細胞膨張化致死毒をコードする下記(a)から(d)のいずれかに記載のポリヌクレオチド。
(a)配列番号:2から4のいずれかに記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド
(b)配列番号:1に記載の塩基配列のコード領域を含むポリヌクレオチド
(c)配列番号:2から4に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加および/または挿入したアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド
(d)配列番号:1に記載の塩基配列からなるDNAにストリンジェントな条件下でハイブリダイズするポリヌクレオチド
(2) (1)に記載のポリヌクレオチドを含むベクター。
(3) (1)に記載のポリヌクレオチドまたは(2)に記載のベクターを保持する宿主細胞。
(4) (1)に記載のポリヌクレオチドによりコードされるポリペプチド。
(5) (3)に記載の宿主細胞を培養し、該宿主細胞またはその培養上清から、産生させたポリペプチドを回収する工程を含む、(4)に記載のポリペプチドの製造方法。
(6) (4)に記載のポリペプチドに結合する抗体。
(7) 被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、以下の(a)および(b)の工程を含む方法。
(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物を用いたポリメラーゼ連鎖反応を行なう工程
(b)これら細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、これら細菌の存在を判定する工程
(8) 被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、以下の(a)から(c)の工程を含む方法。
(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いたポリメラーゼ連鎖反応を行なう工程
(b)工程(a)により増幅されたゲノムDNAを鋳型に、これら細菌の細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物を用いたポリメラーゼ連鎖反応を行なう工程
(c)これら細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、これら細菌の存在を判定する工程
(9) 共通プライマー対が、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、(8)に記載の方法。
(10) 特異的なプライマー対の混合物として、以下の(a)から(c)のプライマー対を用いる、(7)または(8)に記載の方法。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(11) 被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、以下の(a)〜(c)の工程を含む方法。
(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いたポリメラーゼ連鎖反応を行なう工程
(b)工程(a)により増幅されたゲノムDNAを制限酵素により切断する工程
(c)切断されたDNA断片の分子量から、これら細菌の存在を判定する工程
(12) 制限酵素が、Sau3AI、Dsa I、Mbo I、Rsa I、EcoR I、Hinf I、Nde I、Pst I、Xba I、Xho IIからなる群より選択される、(11)に記載の方法。
(13) 共通プライマー対が、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、(11)に記載の方法。
(14) (7)に記載の方法に用いるためのキットであって、使用説明書と、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物とを含むキット。
(15) 特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、(14)に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対共通プライマー対
(16) 請求項8に記載の方法に用いるためのキットであって、使用説明書と、下記(a)または(b)とを含むキット。
(a)カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物
(b)カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対。
(17)特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、(16)に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(18)共通プライマー対が、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、(16)に記載のキット。
(19) (11)に記載の方法に用いるためのキットであって、使用説明書と、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対とを含むキット。
(20)共通プライマー対が、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、(19)に記載のキット。
【0021】
なお、本明細書において「細胞膨張化致死毒」とは、cytolethal distending toxin (CDTまたはCLDT)と呼ばれる、蛋白性のA-B型ホロトキシンのグループに属する毒素因子を指す。このものは、ほかに細胞膨化致死毒(素)、細胞膨潤化致死毒(素)などと称されることもある。細胞膨張化致死毒は、A,B,Cの3ユニットからなるサブユニット構造を有し、Bサブユニットが毒素活性中心ユニットであり、AおよびBサブユニットが細胞接着に関わっていると考えられている。細胞に作用すると細胞が大きく膨らむ等の変形が生じ、最終的に細胞死を引き起こす。毒素原性大腸菌が産生する易熱性エンテロトキシン(LT)などを実験的に細胞に作用させた場合にも細胞が大きく膨らむ等の変形が見られるが、毒素を取り除いた場合、細胞は回復し、致死することはない。しかしながら、CDTを除去しても細胞は回復せず、死に至る。
【0022】
本明細書において用いられる「ポリヌクレオチド」とは、リボヌクレオチドもしくはデオキシヌクレオチドであって、複数の塩基または塩基対からなる重合体を意味する。ポリヌクレオチドには、一本鎖型および二本鎖型のDNAを含む。ポリヌクレオチドは、天然に存在する状態から修飾されていないもの、および修飾されているものの双方を含む意である。修飾された塩基としては、例えば、トリチル化された塩基およびイノシンのような特殊な塩基がある。
【0023】
本明細書において用いられる「ポリペプチド」は、複数のアミノ酸からなる重合体を意味する。従って、オリゴペプチドおよびタンパク質もまた、ポリペプチドの概念に含まれる。ポリペプチドは、天然に存在する状態から修飾されていないもの、および修飾されているものの双方を含む意である。修飾としては、アセチル化、アシル化、ADP-リボシル化、アミド化、フラビンの共有結合、ヘム部分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトールの共有結合、架橋、環化、ジスルフィド結合の形成、脱メチル化、共有架橋の形成、シスチンの形成、ピログルタメートの形成、ホルミル化、γ-カルボキシル化、グリコシル化、GPIアンカー形成、ヒドロキシル化、ヨウ素化、メチル化、ミリストイル化、酸化、タンパク質分解処理、リン酸化、プレニル化、ラセミ化、セレノイル化、硫酸化、アルギニル化のようなタンパク質へのアミノ酸の転移RNA媒介付加、ユビキチン化などが含まれる。
【0024】
本明細書において「単離」とは、本来の環境(たとえば自然に発生するのであればその自然環境)から取り出された物質(例えば、ポリヌクレオチドまたはポリペプチド)を指し、その自然状態から「人の手によって」変えられたものである。「単離」とは、対象化合物に実質的に富む試料中に存在する化合物および/または対象化合物が部分的または実質的に精製されている試料中に存在する化合物を含むことを意味する。ここで「実質的に精製した」という用語は、その天然の環境から切り離されて、天然に関連している他の成分を少なくとも60%、好ましくは75%、および最も好ましくは90%含まない化合物(例えば、ポリヌクレオチドまたはポリペプチド)を指す。
【0025】
本明細書において用いられる「変異」とは、アミノ酸配列におけるアミノ酸の変化または塩基配列における塩基の変化(すなわち単一または複数のアミノ酸またはヌクレオチド置換、欠失、付加または挿入)を指す。従って、本明細書において用いられる「変異体」は、一つ以上のアミノ酸が変化しているアミノ酸配列または一つ以上の塩基が変化している塩基配列を指す。この変異体の塩基配列の変化は、基準ポリヌクレオチドによってコードされるポリペプチドのアミノ酸配列を変更しても、しなくてもよい。変異体はアレリック変異体のように天然に存在するものでも、天然に存在することが知られていない変異体であってもよい。変異体は、置換されたアミノ酸が類似の構造的または化学的特性を有する保存的変化を有しうる。まれに、変異体は、非保存的置換を有しうる。生物学的または免疫学的活性を阻害することなく、いずれの、およびどれほど多くのアミノ酸残基を置換、挿入、または欠失するかを決定する手引きは、当技術分野において周知のコンピュータープログラム、例えばDNAスター・ソフトウェアを用いて発見することができる。
【0026】
「欠失」はその中で1つ以上のアミノ酸またはヌクレオチド残基がそれぞれ、天然に存在する細胞膨張化致死毒ポリペプチドのアミノ酸配列またはヌクレオチド配列と比較して存在しない、アミノ酸またはヌクレオチド配列のいずれかの変化である。
【0027】
「挿入」または「付加」は、天然に存在する細胞膨張化致死毒ポリペプチドのアミノ酸配列またはヌクレオチド配列と比較して、それぞれアミノ酸またはヌクレオチド残基1つ以上が付加されたアミノ酸またはヌクレオチド配列の変化である。
【0028】
「置換」とは、天然に存在する細胞膨張化致死毒ポリペプチドのアミノ酸配列またはヌクレオチド配列と比較して、アミノ酸またはヌクレオチド1つ以上がそれぞれ異なるアミノ酸またはヌクレオチドに入れ替えられたアミノ酸またはヌクレオチド配列の変化である。
【0029】
本明細書において用いられる「ハイブリダイズ」とは、核酸鎖が塩基対形成を通じて相補鎖と結合するプロセスを意味する。
【図面の簡単な説明】
【0030】
【図1】GNWおよびLPF-Dプライマーを用いたC.コリCo1-192菌体抽出液をテンプレートとしたPCRの結果を示す写真である。矢印1はcdt領域が増幅されたバンド(約1.5Kb)、矢印2のバンド(800bp)は、GNWプライマーがミックスプライマーであるために増幅されたcdtB由来の副バンドを示す。
【図2】C.コリCo1-192菌体のゲノムの制限酵素HindIII消化後、ハイブリダイゼーションの結果を示す写真である。
【図3】共通プライマー1を用いたPCRの結果を示す写真である。レーン2〜6の1.9kbp付近にCDT由来のバンドが認められる。
【図4】共通プライマー2を用いた各種C.ジェジュニ菌株のPCRの結果を示す写真である。720bp付近にCDT由来のバンドが認められる。
【図5】共通プライマー2を用いた各種C.ジェジュニ菌株およびC.コリ菌株のPCRの結果を示す写真である。
【図6】共通プライマー2を用いたC.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のPCRの結果を示す写真である。
【図7】特異的プライマーを用いたC.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のマルチプレックスPCRの結果を示す写真である。各菌種に特有のCDT特異的増幅断片が確認された(C.ジェジュニ:750bp、C.コリ:400bp、C.フィータス:530bp)。
【図8】共通プライマー1を用いたC.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のPCR-RFLPの結果を示す写真である。
【図9】特異的プライマーを用いた各種C.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のマルチプレックスPCRの結果を示す写真である。各菌種に特有のCDT特異的増幅断片が確認された(C.ジェジュニ:750bp、C.コリ:400bp、C.フィータス:530bp)。
【図10】C.フィータスCol-187菌体のゲノムの制限酵素Hind III消化後、ハイブリダイゼーションの結果を示す写真である。
【図11】cdtAおよびcdtC共通プライマーを用いたPCRの結果を示す写真である。レーン2〜8の550bp付近にcdtA由来のバンドが認められ、またレーン10〜16の320bp付近にcdtC由来のバンドが認められる。
【図12】cdtA共通プライマーを用いた各種カンピロバクター菌株のPCRの結果を示す写真。550bp付近にCdtA由来のバンドが認められる。
【図13】cdtC共通プライマーを用いた各種カンピロバクター菌株のPCRの結果を示す写真。320bp付近にcdtC由来のバンドが認められる。
【図14】cdtAおよびcdtC特異的プライマーを用いたC.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のマルチプレックスPCRの結果を示す写真である。各菌種に特有のCdtA特異的増幅断片(C.ジェジュニ:630bp、C.コリ:330bp、C.フィータス:490bp)とcdtC特異的断片(C.ジェジュニ:500bp、C.コリ:400bp、C.フィータス:300bp)が確認される。
【図15】cdtA特異的プライマーを用いたC.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のマルチプレックスPCRの結果を示す写真である。各菌種に特有のCdtA特異的増幅断片が確認された。
【図16】cdtC特異的プライマーを用いたC.ジェジュニ菌株、C.コリ菌株、およびC.フィータス菌株のマルチプレックスPCRの結果を示す写真である。各菌種に特有のCdtC特異的増幅断片が確認された。
【図17】C.ジェジュニのORFおよびプライマーのアニール領域を示す図である。
【図18】C.コリのORFおよびプライマーのアニール領域を示す図である。
【図19】C.フィータスのORFおよびプライマーのアニール領域を示す図である。
【発明を実施するための最良の形態】
【0031】
<ポリヌクレオチド>
本発明は、カンピロバクター・コリの細胞膨張化致死毒をコードするポリヌクレオチドを提供する。本発明に含まれる、本発明者らにより同定されたカンピロバクター・コリの細胞膨張化致死毒をコードするポリヌクレオチドの塩基配列を配列番号:1に、該ポリヌクレオチドによってコードされる3つのポリペプチドのアミノ酸配列を配列番号:2から4に示す。配列番号2はcdtA、配列番号3はcdtB、配列番号4はcdtCのアミノ酸配列である。
また本発明は、カンピロバクター・フィータスの細胞膨張化致死毒をコードするポリヌクレオチドを提供する。本発明に含まれる、本発明者らにより同定されたカンピロバクター・フィータスの細胞膨張化致死毒をコードするポリヌクレオチドの塩基配列を配列番号:51に、該ポリヌクレオチドによってコードされる3つのポリペプチドのアミノ酸配列を配列番号:52から54に示す。配列番号52はcdtA、配列番号53はcdtB、配列番号54はcdtCのアミノ酸配列である。
【0032】
本発明のポリヌクレオチドには、配列番号:2から4に記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド、配列番号:1に記載の塩基配列のコード領域、すなわち、配列番号:1に記載の塩基配列において1位から777位までの塩基配列、802位から1605位までの塩基配列、1615位から2187位までの塩基配列のいずれかの塩基配列、を含むポリヌクレオチド、遺伝コードの縮重により配列番号:1に記載の塩基配列と異なる塩基配列からなるが配列番号:2から4に記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチドが含まれる。
また本発明のポリヌクレオチドには、配列番号:52から54に記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド、配列番号:51に記載の塩基配列のコード領域、すなわち、配列番号51に記載の塩基配列において1位から702位までの塩基配列、778位から1629位までの塩基配列、1615位から2187位までの塩基配列のいずれかの塩基配列、を含むポリヌクレオチド、遺伝コードの縮重により配列番号:51に記載の塩基配列と異なる塩基配列からなるが配列番号:52から54に記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチドが含まれる。
本発明のポリヌクレオチドには、さらに、これらポリヌクレオチドがコードするポリペプチドと機能的に同等なポリペプチドをコードし、該ポリヌクレオチドの配列とその全長において少なくとも40%以上、好ましくは60%以上、さらに好ましくは80%以上、さらに好ましくは90%以上、さらに好ましくは95%以上、さらに好ましくは97%以上(例えば、98〜99%)同一である塩基配列を含むポリヌクレオチドが含まれる。塩基配列の同一性は、例えば、Karlin and Altschul によるアルゴリズムBLAST (Proc. Natl. Acad. Sci. USA 87:2264-2268, 1990、Proc. Natl. Acad. Sci. USA 90:5873-5877, 1993)によって決定することができる。このアルゴリズムに基づいて、BLASTNと呼ばれるプログラムが開発されている(Altschul et al. J. Mol. Biol.215:403-410, 1990)。BLASTNによって塩基配列を解析する場合には、パラメーターはたとえばscore = 100、wordlength = 12とする。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である(http://www.ncbi.nlm.nih.gov.)。本発明のポリヌクレオチドには、上記のポリヌクレオチドの塩基配列と相補的な塩基配列を有するポリヌクレオチドが含まれる。
【0033】
本発明のポリヌクレオチドは、標準的なクローニングおよびスクリーニングにより、例えば、菌体中のゲノムDNAのような天然源から得ることができる。また、菌体中のmRNAから誘導されたcDNAライブラリーから得ることもできる。また、商業的に入手可能な公知の技法を用いて合成することもできる。
【0034】
本発明者らにより同定されたポリヌクレオチドの配列(配列番号:1、配列番号:51)と有意な相同性を有する塩基配列からなるポリヌクレオチドは、例えば、ハイブリダイゼーション技術 (Current Protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons Section 6.3-6.4)や遺伝子増幅技術(PCR)(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons Section 6.1-6.4)を利用して調製することができる。即ち、ハイブリダイゼーション技術を利用して、本発明者らにより同定されたポリヌクレオチドの配列(配列番号:1、配列番号:51)またはその一部をもとに、これと相同性の高いDNAを単離することができる。また、遺伝子増幅技術を用いて、本発明者らにより同定されたポリヌクレオチドの配列(配列番号:1、配列番号:51)の一部を基にプライマーを設計し、該ポリヌクレオチドの配列と相同性の高いポリヌクレオチドを単離することができる。従って、本発明には、配列番号:1に記載の塩基配列を有するポリヌクレオチドとストリンジェントな条件下でハイブリダイズするポリヌクレオチドが含まれる。ストリンジェントなハイブリダイゼーション条件としては、通常「1xSSC、0.1% SDS、37℃」程度の条件であり、より厳しい条件としては「0.5xSSC、0.1% SDS、42℃」程度の条件であり、さらに厳しい条件としては「0.2xSSC、0.1% SDS、65℃」程度の条件である。このようにハイブリダイゼーションの条件が厳しくなるほどプローブ配列と高い相同性を有するDNAの単離を期待しうる。但し、上記SSC、SDSおよび温度の条件の組み合わせは例示であり、当業者であれば、ハイブリダイゼーションのストリンジェンシーを決定する上記若しくは他の要素(例えば、プローブ濃度、プローブの長さ、ハイブリダイゼーション反応時間など)を適宜組み合わせることにより、上記と同様のストリンジェンシーを実現することが可能である。
【0035】
本発明者らにより同定されたポリヌクレオチドの配列と有意な相同性を有する塩基配列からなるポリヌクレオチドは、配列番号:1、配列番号:51に記載の塩基配列に変異を導入する方法(例えば、部位特異的変異誘発法(Current Protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons Section 8.1-8.5))を利用して調製することもできる。また、このようなポリヌクレオチドは、自然界における変異により生じることもある。本発明には、このような塩基配列の変異により、配列番号:2から4または配列番号:52から54に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、挿入および/もしくは付加などされたポリペプチドをコードするポリヌクレオチドが含まれる。
【0036】
本発明のポリヌクレオチドを本発明のポリペプチドの組換え生産のために用いる場合、そのポリヌクレオチドには、成熟ポリペプチドのコード配列またはその断片単独、他のコード配列(例えば、リーダーもしくは分泌配列、プレ-、プロ-もしくはプレプロ-タンパク質配列、または他の融合ペプチド部分をコードするもの)と同じリーディングフレーム内にある成熟ポリペプチドのコード配列またはその断片が含まれる。例えば、融合ポリペプチドの精製を容易にするマーカー配列がコードされ得る。本発明のこの態様の好ましい具体例として、マーカー配列は、pcDNA3.1/Myc-Hisベクター(Invitrogen社)により提供されかつGentz ら, Proc. Natl. Acad. Sci. USA (1989) 86:821-824に記載されるようなヘキサ-ヒスチジンペプチド、またはMycタグである。また、このポリヌクレオチドは5'および3'非コード配列、例えば、転写されるが翻訳されない配列、スプライシングおよびポリアデニル化シグナル、リボソーム結合部位、およびmRNA安定化配列を含んでいてもよい。
【0037】
<ポリペプチド>
本発明は、本発明者らが同定したカンピロバクター・コリの細胞膨張化致死毒のポリペプチドを提供する。さらに、本発明は、本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドを提供する。ここで「機能的に同等」とは、対象となるポリペプチドが本発明者らにより同定されたポリペプチドと同等の細胞膨張化致死毒の特性を有していることを意味する。
また本発明は、本発明者らが同定したカンピロバクター・フィータスの細胞膨張化致死毒のポリペプチドを提供する。さらに、本発明は、本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドを提供する。ここで「機能的に同等」とは、対象となるポリペプチドが本発明者らにより同定されたポリペプチドと同等の細胞膨張化致死毒の特性を有していることを意味する。
【0038】
本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドを調製するための方法の1つの態様としては、タンパク質中のアミノ酸配列に変異を導入する方法が挙げられる。このような方法には、例えば、部位特異的変異誘発法(Current Protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. Jhon Wily & Sons Section 8.1-8.5))が含まれる。また、ポリペプチド中のアミノ酸の変異は、自然界において生じることもある。本発明には、このように人工的か自然に生じたものかを問わず、本発明者らにより同定されたポリペプチドのアミノ酸配列(配列番号:2から4、配列番号:52から54)において1もしくは複数のアミノ酸が置換、欠失、挿入および/もしくは付加などにより変異したタンパク質であって、本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドが含まれる。
【0039】
置換されるアミノ酸は、タンパク質の機能の保持の観点から、置換前のアミノ酸と似た性質を有するアミノ酸であることが好ましい(保存的置換)。例えば、Ala、Val、Leu、Ile、Pro、Met、Phe、Trpは、共に非極性アミノ酸に分類されるため、互いに似た性質を有すると考えられる。また、非荷電性としては、Gly、Ser、Thr、Cys、Tyr、Asn、Glnが挙げられる。また、酸性アミノ酸としては、AspおよびGluが、塩基性アミノ酸としては、Lys、Arg、Hisが挙げられる。
【0040】
これらポリペプチドにおけるアミノ酸の変異数や変異部位は、その機能が保持される限り制限はない。変異数は、典型的には、全アミノ酸の10%以内であり、好ましくは全アミノ酸の5%以内であり、さらに好ましくは全アミノ酸の1%以内であると考えられる。
【0041】
本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドを調製するための方法の他の態様としては、ハイブリダイゼーション技術あるいは遺伝子増幅技術を利用する方法が挙げられる。即ち、当業者であれば、ハイブリダイゼーション技術 (Current Protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. Jhon Wily & Sons Section 6.3-6.4)を利用して本発明者らにより同定されたポリペプチドをコードするDNA配列(配列番号:1、配列番号:51)またはその一部をもとに同種または異種生物由来のDNA試料から、これと相同性の高いDNAを単離して、該DNAから本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドを得ることは、通常行いうることである。このように本発明者らにより同定されたポリペプチドをコードするDNAとハイブリダイズするDNAによりコードされるポリペプチドであって、本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドもまた本発明のポリペプチドに含まれる。
【0042】
本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドをコードするDNAを単離するためのストリンジェントなハイブリダイゼーション条件としては、通常「1xSSC、0.1% SDS、37℃」程度の条件であり、より厳しい条件としては「0.5xSSC、0.1% SDS、42℃」程度の条件であり、さらに厳しい条件としては「0.2xSSC、0.1% SDS、65℃」程度の条件である。このようにハイブリダイゼーションの条件が厳しくなるほどプローブ配列と高い相同性を有するDNAの単離を期待しうる。但し、上記SSC、SDSおよび温度の条件の組み合わせは例示であり、当業者であれば、ハイブリダイゼーションのストリンジェンシーを決定する上記若しくは他の要素(例えば、プローブ濃度、プローブの長さ、ハイブリダイゼーション反応時間など)を適宜組み合わせることにより、上記と同様のストリンジェンシーを実現することが可能である。
【0043】
このようなハイブリダイゼーション技術を利用して単離されるDNAがコードするポリペプチドは、通常、本発明者らにより同定されたポリペプチドとアミノ酸配列において高い相同性を有する。高い相同性とは、少なくとも40%以上、好ましくは60%以上、さらに好ましくは80%以上、さらに好ましくは90%以上、さらに好ましくは少なくとも95%以上、さらに好ましくは少なくとも97%以上(例えば、98〜99%)の配列の相同性を指す。アミノ酸配列の同一性は、例えば、Karlin and Altschul によるアルゴリズムBLAST (Proc. Natl. Acad. Sci. USA 87:2264-2268, 1990、Proc. Natl. Acad. Sci. USA 90:5873-5877, 1993)によって決定することができる。このアルゴリズムに基づいて、BLASTXと呼ばれるプログラムが開発されている(Altschul et al. J. Mol. Biol.215:403-410, 1990)。BLASTXによってアミノ酸配列を解析する場合には、パラメーターはたとえば score = 50、wordlength = 3とする。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である。
【0044】
また、遺伝子増幅技術(PCR)(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons Section 6.1-6.4)を用いて本発明者らにより同定されたポリペプチドをコードするDNA配列(配列番号:1、配列番号:51)の一部を基にプライマーを設計し、本発明者らにより同定されたポリペプチドをコードするDNA配列と相同性の高いDNA断片を単離し、該DNAを基に本発明者らにより同定されたポリペプチドと機能的に同等なポリペプチドを得ることも可能である。
【0045】
<ポリペプチドの断片>
本発明は、また、本発明のポリペプチドの断片を提供する。こうした断片は全体的に前記本発明のポリペプチドのアミノ酸配列の一部と同一であるが、全部とは同一でないアミノ酸配列を有するポリペプチドである。本発明のポリペプチド断片は、通常、8アミノ酸残基以上、好ましくは12アミノ酸残基以上(例えば、15アミノ酸残基以上)の配列からなるポリペプチド断片である。好適な断片としては、例えば、アミノ末端を含む一連の残基もしくはカルボキシル末端を含む一連の残基の欠失、またはアミノ末端を含む一連の残基とカルボキシル末端を含む一連の残基の二連の残基の欠失したアミノ酸配列を有するトランケーション(truncation)ポリペプチドが含まれる。また、αヘリックスとαヘリックス形成領域、βシートとβシート形成領域、ターンとターン形成領域、コイルとコイル形成領域、親水性領域、疎水性領域、α両親媒性領域、β両親媒性領域、可変性領域、表面形成領域、基質結合領域、および高抗原指数領域を含む断片のような、構造的または機能的特性により特徴づけられる断片も好適である。その他の好適な断片は生物学的に活性な断片である。生物学的に活性な断片は、同様の活性をもつ断片、その活性が向上した断片、または望ましくない活性が減少した断片を含めて、本発明のポリペプチドの活性を媒介するものである。さらに、動物、特にヒトにおいて抗原性または免疫原性がある断片も含まれる。これらのポリペプチド断片は、抗原活性を含めた本発明のポリペプチドの生物学的活性を保持することが好ましい。特定された配列および断片の変異型も本発明の一部を構成する。好適な変異型は同類アミノ酸置換により対象物と異なるもの、すなわち、ある残基が同様の性質の他の残基で置換されているものである。典型的なこうした置換は、Ala, Val, LeuとIleの間、SerとThrの間、酸性残基 AspとGluの間、AsnとGlnの間、塩基性残基 LysとArgの間、または芳香族残基 PheとTyrの間で起こる。
【0046】
<ポリペプチドの製造>
本発明のポリペプチドは任意の適当な方法で製造することができる。このようなポリペプチドには、単離された天然に存在するポリペプチド、組換え的に生産されたポリペプチド、合成的に製造されたポリペプチド、またはこれらの方法の組み合わせにより製造されたポリペプチドが含まれる。このようなポリペプチドの製造のための手段は当業界でよく理解されている。組換え的なポリペプチドは、例えば、本発明のポリヌクレオチドを挿入したベクターを適当な宿主細胞に導入し、形質転換体内で発現したポリペプチドを精製することにより調製することが可能である。一方、天然由来のポリペプチドは、例えば、後述する本発明のポリペプチドに対する抗体を結合したアフィニティーカラムを利用して調製することができる (Current Protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons Section 16.1-16.19)。アフィニティー精製に用いる抗体は、ポリクローナル抗体であってもモノクローナル抗体であってもよい。また、インビトロトランスレーション(例えば、「On the fidelity of mRNA translation in the nuclease-treated rabbit reticulocyte lysate system. Dasso,M.C.,Jackson,R.J.(1989) NAR 17:3129-3144」参照)などにより本発明のポリペプチドを調製することも可能である。本発明のポリペプチドの断片は、例えば、本発明のポリペプチドを適当なペプチダーゼで切断することによって製造することができる。
【0047】
<プローブ・プライマー>
本発明は、本発明者らにより同定されたポリヌクレオチド(配列番号:1に記載の塩基配列からなるポリヌクレオチドまたはその相補鎖、配列番号:51に記載の塩基配列からなるポリヌクレオチドまたはその相補鎖)に相補的な、少なくとも15ヌクレオチドの鎖長を有するヌクレオチドを提供する。ここで「相補鎖」とは、A:T(ただしRNAの場合は U)、G:Cの塩基対からなる2本鎖核酸の一方の鎖に対する他方の鎖を指す。また、「相補的」とは、少なくとも15個の連続したヌクレオチド領域で完全に相補配列である場合に限られず、少なくとも70%、好ましくは少なくとも80%、より好ましくは90%、さらに好ましくは95%以上の塩基配列上の相同性を有すればよい。相同性を決定するためのアルゴリズムは本明細書に記載したものを使用すればよい。このようなヌクレオチドは、本発明のポリヌクレオチドを検出、単離するためのプローブとして、また、本発明のヌクレオチドを増幅するためのプライマーとして利用することが可能である。プライマーとして用いる場合には、通常、15〜100ヌクレオチド、好ましくは15〜35ヌクレオチドの鎖長を有する。また、プローブとして用いる場合には、本発明のDNAの少なくとも一部若しくは全部の配列を含む少なくとも15ヌクレオチド、好ましくは少なくとも30ヌクレオチドの鎖長のヌクレオチドが用いられる。本発明のヌクレオチドをプライマーとする場合、核酸増幅反応の種類は、目的とする増幅産物が得られる限り、特に制限はない。例えば、PCR(ポリメラーゼ連鎖反応)法、ICAN法、LAMP法、SDA法、LCR法等のDNA増幅反応、NASBA法等のRNA増幅反応、の中から選択することができる。好適な方法としては、PCR法を示すことができる。
【0048】
このようなヌクレオチドの一つの態様は、本発明のポリペプチドをコードするDNAに特異的になものである。「特異的な」とは、通常のハイブリダイゼーション条件下、好ましくはストリンジェントな条件下で、特定のポリペプチドをコードするDNAとハイブリダイズし、他のポリペプチドをコードするDNAとはハイブリダイズしないことを意味する。好ましい態様は、カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNA(配列番号:1)にハイブリダイズし、カンピロバクター・ジェジュニおよびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAとハイブリダイズしないポリヌクレオチドである。このようなポリヌクレオチドとしては、例えば、配列番号:13、14、28−36、70、71、76、77から選択されるプライマー対が挙げられる。あるいは、好ましい態様は、カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNA(配列番号:51)にハイブリダイズし、カンピロバクター・ジェジュニおよびカンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAとハイブリダイズしないポリヌクレオチドである。このようなポリヌクレオチドとしては、例えば、配列番号:15、16、37−46、72、73、78、79から選択されるプライマー対が挙げられる。
【0049】
また、本実施例により、カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNA(配列番号:1)が明らかにされたことに伴い、カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAに特異的な塩基配列およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAに特異的な塩基配列を、本発明者らは見出した。従って、本発明は、カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAに特異的なプライマー対およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAに特異的なプライマー対をも提供する。カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAに特異的なプライマーとしては、例えば、配列番号:11、12、17−27に記載のプライマーが、カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAに特異的なプライマーとしては、例えば、配列番号:15、16、37−46に記載のプライマーが挙げられるが、これらに制限されない。
【0050】
また、本実施例により、カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNA(配列番号:1)、カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNA(配列番号:51)が明らかにされたことに伴い、本発明者は、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAに対する共通プライマー(これらすべての細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうるプライマー)を見出した。本発明は、このような共通プライマーをも提供する。好ましい共通プライマーとしては、例えば、配列番号;64、65に記載のプライマー(ctdAのDNAを増幅)、配列番号;7−10、47−50に記載のプライマー(ctdBのDNAを増幅)、配列番号;66、67に記載のプライマー(ctdCのDNAを増幅)が挙げられる。
【0051】
当業者にとっては、上記のプライマーに対して、1若しくは複数の塩基が異なるが、上記のプライマーと同一のゲノムDNA領域を増幅しうるプライマーを調製することは、適宜行ないうることである。上記プライマーがアニールするゲノムDNA領域について、図17-19に示す。本発明は、このような変異プライマーをも提供するものである。上述のとおり、本発明のプライマーを適用できる核酸増幅反応は、目的とする増幅産物が得られる限り、特に制限はなく、例えば、PCR(ポリメラーゼ連鎖反応)法、ICAN法、LAMP法、SDA法、LCR法等のDNA増幅反応、NASBA法等のRNA増幅反応、の中から選択することができる。好適な方法としては、PCR法を示すことができる。当業者であれば、上記プライマーを基に、実施する核酸増幅方法により適合した変異プライマーを設計することも可能である。変異プライマーは合成により調製することができる。また、変異プライマーが、変異前のプライマーと同一のゲノムDNA領域を増幅しうるかは、変異プライマーを用いた核酸増幅反応を行い、その増幅産物を分析することにより、簡便に評価することができる。
【0052】
これらプライマーは、被検試料中のヘリコバクター属細菌の存在の検出に好適に利用することができる。
【0053】
<ベクター、宿主細胞、ポリペプチドの製造>
本発明はまた、本発明のポリヌクレオチドを含有するベクター、本発明のポリヌクレオチドまたは該ベクターを保持する宿主細胞、および該宿主細胞を利用した本発明のポリペプチドの生産方法を提供する。
【0054】
本発明のベクターとしては、挿入したDNAを安定に保持するものであれば特に制限されず、例えば宿主に大腸菌を用いるのであれば、クローニング用ベクターとしてはpBluescriptベクター(Stratagene社製)などが好ましい。本発明のポリペプチドを生産する目的においてベクターを用いる場合には、特に発現ベクターが有用である。発現ベクターとしては、試験管内、大腸菌内、培養細胞内、生物個体内でポリペプチドを発現するベクターであれば特に制限されないが、例えば、試験管内発現であればpBESTベクター(プロメガ社製)、大腸菌であればpETベクター(Invitrogen社製)、培養細胞であればpME18S-FL3ベクター(GenBank Accession No. AB009864)、生物個体であればpME18Sベクター(Mol Cell Biol. 8:466-472(1988))などが好ましい。ベクターへの本発明のDNAの挿入は、常法により、例えば、制限酵素サイトを用いたリガーゼ反応により行うことができる(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons.Section 11.4-11.11)。
【0055】
本発明のベクターが導入される宿主細胞としては特に制限はなく、目的に応じて種々の宿主細胞が用いられる。ポリペプチドを発現させるための細胞としては、例えば、細菌細胞(例:ストレプトコッカス、スタフィロコッカス、大腸菌、ストレプトミセス、枯草菌)、真菌細胞(例:酵母、アスペルギルス)、昆虫細胞(例:ドロソフィラS2、スポドプテラSF9)、動物細胞(例:CHO、COS、HeLa、C127、3T3、BHK、HEK293、Bowes メラノーマ細胞)および植物細胞を例示することができる。宿主細胞へのベクター導入は、例えば、リン酸カルシウム沈殿法、電気パルス穿孔法(Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons.Section 9.1-9.9)、リポフェクタミン法(GIBCO-BRL社製)、マイクロインジェクション法などの公知の方法で行うことが可能である。
【0056】
宿主細胞において発現したポリペプチドを小胞体の内腔に、細胞周辺腔に、または細胞外の環境に分泌させるために、適当な分泌シグナルを目的のポリペプチドに組み込むことができる。これらのシグナルは目的のポリペプチドに対して内因性であっても、異種シグナルであってもよい。
【0057】
本発明のポリペプチドの回収は、本発明のポリペプチドが培地に分泌される場合は、培地を回収する。本発明のポリペプチドが細胞内に産生される場合は、その細胞をまず溶解し、その後にポリペプチドを回収する。
【0058】
組換え細胞培養物から本発明のポリペプチドを回収し精製するには、硫酸アンモニウムまたはエタノール沈殿、酸抽出、アニオンまたはカチオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシルアパタイトクロマトグラフィーおよびレクチンクロマトグラフィーを含めた公知の方法を用いることができる。
【0059】
<抗体>
本発明は、本発明のポリペプチドに結合する抗体を提供する。ここで「抗体」には、ポリクローナルおよびモノクローナル抗体、キメラ抗体、一本鎖抗体、ヒト化抗体、さらにFabまたは他の免疫グロブリン発現ライブラリーの産物を含むFabフラグメントが含まれる。
【0060】
本発明のポリペプチドまたはその断片もしくは類似体、またはそれらを発現する細胞は、本発明のポリペプチドに結合する抗体を産生するための免疫原としても使用することができる。抗体は、好ましくは、本発明のポリペプチドに免疫特異的である。「免疫特異的」とは、その抗体が他のポリペプチドに対するその親和性よりも本発明のポリペプチドに対して実質的に高い親和性を有することを意味する。
【0061】
本発明のポリペプチドに結合する抗体は、当業者に公知の方法により調製することが可能である。ポリクローナル抗体であれば、例えば、次のようにして得ることができる。本発明のポリペプチドあるいはそのGSTとの融合タンパク質をウサギ等の小動物に免疫し血清を得る。これを、例えば、硫安沈殿、プロテインA、プロテインGカラム、DEAEイオン交換クロマトグラフィー、本発明のポリペプチドをカップリングしたアフィニティーカラム等により精製することにより調製する。また、モノクローナル抗体であれば、例えば、本発明のポリペプチドをマウスなどの小動物に免疫を行い、同マウスより脾臓を摘出し、これをすりつぶして細胞を分離し、マウスミエローマ細胞とポリエチレングリコールなどの試薬により融合させ、これによりできた融合細胞(ハイブリドーマ)の中から、本発明のポリペプチドに結合する抗体を産生するクローンを選択する。次いで、得られたハイブリドーマをマウス腹腔内に移植し、同マウスより腹水を回収し、得られたモノクローナル抗体を、例えば、硫安沈殿、プロテインA、プロテインGカラム、DEAEイオン交換クロマトグラフィー、本発明のポリペプチドをカップリングしたアフィニティーカラム等により精製することで、調製することが可能である。
【0062】
本発明の抗体は、被検試料中の本発明のポリペプチドの検出や精製に用いることもできる。
【0063】
<被検試料中のカンピロバクター属細菌の存在の検出>
本発明は、被検試料中のカンピロバクター属細菌の存在の検出方法を提供する。被検試料中のカンピロバクター属細菌の存在の検出は、カンピロバクター感染症の診断、カンピロバクター属細菌に汚染された食品の迅速診断、食品加工工程のバリデーション、食中毒発生時における原因菌の同定など種々の目的において有用である。
【0064】
本発明の検出方法の一つの態様は、被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物を用いたポリメラーゼ連鎖反応を行なう工程、および(b)これら細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、これら細菌の存在を判定する工程、を含む方法である。
【0065】
また、本発明の検出方法の他の態様は、被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いた核酸増幅反応を行なう工程、(b)工程(a)により増幅されたゲノムDNAを鋳型に、これら細菌の細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物を用いたポリメラーゼ連鎖反応を行なう工程、および(c)これら細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、これら細菌の存在を判定する工程、を含む方法である。
【0066】
実施例のように、複数のPCRプライマーを単一の反応系で使用するPCRは、マルチプレックスPCR法と呼ばれ、PCR産物を電気泳動し、バンドのサイズを見ることで複数の菌種を同時に鑑別することができる。本発明は、このマルチプレックスPCR法を代表とする、複数の核酸領域の増幅に好適に用いられるプライマーおよびその組み合わせを用いた核酸増幅法によるカンピロバクター属細菌の検出方法を提供する。本発明における核酸増幅の方法は、目的とする増幅物が得られる限り種類は問わないが、好ましくはPCR法である
【0067】
これら方法における、特異的なプライマー対の混合物としては、例えば、次の、(a)から(c)のプライマー対の混合物を用いることができる。(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対、(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対、(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対。また、共通プライマー対としては、例えば、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対を用いることができる。
【0068】
本発明の検出方法のさらなる態様は、被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いた核酸増幅反応を行なう工程、(b)工程(a)により増幅されたゲノムDNAを制限酵素により切断する工程、(c)切断されたDNA断片の分子量から、これら細菌の存在を判定する工程、を含む方法である。この方法に用いうる制限酵素としては、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNA を識別しうるものであれば特に制限はなく、例えば、Sau3AI、Dsa I、Mbo I、Rsa I、EcoR I、Hinf I、Nde I、Pst I、Xba I、Xho IIが挙げられる。また、共通プライマー対としては、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対を例示することができる。
【0069】
下記実施例で示すような、PCR法で増幅したDNAを種々の制限酵素で処理し、生じる断片の長さから多型を検出する方法は、PCR-RFLP法(PCR-Restriction Fragment Length Polymorphism:PCR-制限酵素断片長多型)と呼ばれる。本発明はまた、PCR-RFLP法に代表される、核酸増幅法により増幅されたDNAを種々の制限酵素で処理し、生じる断片の長さから多型を検出する方法に好適に用いられるプライマーを提供する。
【0070】
また、本発明の検出方法の他の態様は、被験試料中のカンピロバクター属細菌の存在を検出する方法であって、(a)被検試料に対し、カンピロバクター属細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いた核酸増幅反応を行なう工程
(b)カンピロバクター属細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、カンピロバクターの存在を判定する工程、を含む方法である。この方法に用いるプライマー対は、カンピロバクター属細菌であればその種を問わず、その細胞膨張化致死毒をコードするゲノムDNAを増幅し得るプライマー対である。このような共通プライマー対としては、配列番号;7−10、47−50、64−67から選択されるプライマー対を例示することができる。上述したとおり、上記プライマー対は少なくともカンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータス3菌種の細胞膨張化致死毒をコードするゲノムDNA を共通して増幅する共通プライマー対である。上記プライマー対は、上記3菌種のみならず、その他のカンピロバクター属細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅すると期待できる。また同様に、該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対についても、上記3菌種を共通して該ゲノム領域を増幅することができ、その他のカンピロバクター属細菌の該ゲノム領域を増幅することができると考えられる。
【0071】
また、本発明は、上記本発明の検出方法に用いるためのキットを提供する。これらキットは、上記したプライマー対の他、使用説明書を含むものである。さらなる他の要素を含んでいてもよい。
【0072】
なお、カンピロバクター属細菌の存在の検出は、上記のようなDNAレベルだけでなく、タンパク質レベルで行なうことも考えられる。例えば、これら細菌の細胞膨張化致死毒に特異的な抗体を用いて、ウェスタンブロッティング法、ドットブロッティング法、免疫沈降法、酵素結合免疫測定法(ELISA)、あるいは免疫蛍光法などを実施し、これら細菌の細胞膨張化致死毒を検出することにより、被検試料中におけるこれら細菌の存在を評価することが可能である。
なお本明細書において引用された全ての先行技術文献は、参照として本明細書に組み入れられる。
【実施例】
【0073】
[実施例1] カンピロバクター属菌株
2001年から2003年までの間に各種臨床材料より収集したカンピロバクター・ジェジュニ(C.ジェジュニ)菌、カンピロバクター・コリ(C.コリ)菌、およびカンピロバクター・フィータス(C.フィータス)菌を使用した。各菌株は、5%ウマ脱繊維血液(日本生物材料センター)、およびカンピロバクター選択サプリメントSR69(OXOID)を含む血液寒天培地(Blood Agar Base No.2:OXOID)を用い、C.ジェジュニおよびC.コリは5%O2, 10%CO2, 85%N2ガス下42℃で、C.フィータスは25℃の条件下で、低温酸素/炭酸ガス培養装置(MODEL9200:和研薬)を使用し、培養した。
【0074】
[実施例2] PCR用テンプレートの調整
各菌種から5クローンを掻き取り500μLの無菌PBSに懸濁した。回収した細菌を10,000rpmで5分間遠心洗浄(MRX-150:トミー精工)した後、300μLの無菌PBSに再懸濁した。その後、沸騰水浴中で10分間煮沸し、氷上で冷却した後、15,000rpmで10分間遠心し、上清を回収した。回収上清中のDNA量を分光光度計(Ultrospec 3100pro:アマシャム・バイオサイエンス)を用いて定量した。定量した各菌体抽出液を各20ng/μLとなるよう希釈し、PCRに供した。
【0075】
[実施例3] C.コリcdtB プローブの作製とサザンハイブリダイゼーション
C.コリcdtBプローブはGNWとLPF-Dプライマーを用いてC.コリCo1-192 菌体抽出液をテンプレートとしてDIGラベリングミックス(ロシュ)を用いてPCRラベルを行ない、作製した。
【0076】
すなわち、C.コリのCDT遺伝子の存在を調べるため、文献(Pickett, C. et al. Infect. Immun., 64: 2070 (1996))に記載されたGNW縮重プライマー[配列番号5:5’-GG(ACGT)AA(CT)TGGAT(ACT)TGGGG(ACGT)TA-3’]およびLPF-D縮重プライマー[配列番号6:5’- (AGT)AA(CT)TG(ACGT)AC(AGT)TA(ACGT)CC(AGT)AA (ACGT)GG - 3’]を用い、C.ジェジュニ3株、C.コリ2株につき、94℃, 3分間−(94℃30秒, 42℃30秒, 72℃2分)×30サイクル−72℃, 5分間の条件でPCRを行なった。その結果、C.ジェジュニ3株とC.コリ2株のすべてにおいて、約1.5Kbにcdt領域が増幅されたバンドが認められた(図1の矢印1)。
【0077】
この増幅バンドをpT7ベクター(Novagen)にライゲートし、 宿主大腸菌(E.coli JM109)に形質転換した。得られたクローンのシークエンスをシーケンサー(ABI PRISM 377 DNA sequencer:アプライドバイオシステムズ)を用いて確認したところ、cdtBに類似の配列が見られた。シークエンス反応はBigDye terminator Cycle Sequencing Kits(アプライドバイオシステムズ)を用いた。また、800bpのバンド(図1の矢印2)は、GNWプライマーがミックスプライマーであるため増幅されるcdtB由来の副バンドであることが確認された。
【0078】
得られたプローブを用いて、C.コリCo1-192のゲノム20μgを60Uの制限酵素HindIIIで37℃、12時間消化し、定法(Molecular cloning: a laboratory manual, Cold Spring Harbor Labolatory, Cold Spring Harbor, (2001))に従ってサザンブロットおよびDNA-DNAハイブリダイゼーションを行った。ハイブリダイゼーションは、42℃でストリンジェントな条件下で実施した。ブロッティング後、2×SSC−0.1%SDS溶液で室温で5分間、2度洗浄し、さらに0.2×SSC−0.1%SDS溶液で60℃で15分間、2度洗浄した。
【0079】
その結果、3kと4k付近にプローブ陽性バンドが得られた(図2)。この3kのバンドをpUC18 Vectorにライゲートし、E.coli JM109に形質転換して、cdtB領域を含むクローン(3k44)を得た。
【0080】
[実施例4] C.コリcdtB 遺伝子のシークエンス
実施例3で得られたC.コリcdtB領域を含む3k44クローンにつき、常法によりシークエンスを行い、配列番号1で示されるC.コリCDT全領域の配列を決定した。
【0081】
[実施例5] 共通プライマー1の設計およびPCR
本発明のC.コリCDT配列と、既知のデータベースから得られたC.ジェジュニのCDT遺伝子を比較し、下記の共通プライマーUおよび共通プライマーRを設計した。20ng/μLの菌体抽出液1μL、各プライマーをそれぞれの濃度が0.5mMとなるように混合して添加し、PCR反応用緩衝液(TaKaRa Ex Taq kit:タカラバイオ)を用いて最終液量20μLとなるよう調整し、94℃3分間−(94℃30秒間、55℃30秒間、72℃1分間)×30サイクル−72℃3分間の条件でPCR反応に供した。結果を図3に示す。約1900bpの増幅断片が確認され、C.ジェジュニ(レーン2〜4)およびC.コリ(レーン5、6)のいずれにもCDTに由来するバンドが認められた。
共通プライマーU[配列番号7:GATAA(CT)GATCCTTTAAAACT]
共通プライマーR[配列番号8:(AT)(AT)CCAAAGCG(AT)TTTT(CG)TATGG]
【0082】
[実施例6] 共通プライマー2の設計およびPCR
同様にして、下記の共通プライマーUpおよび共通プライマーDoを設計し、94℃3分間−(94℃30秒間、50℃30秒間、72℃45秒間)×30サイクル−72℃3分間の条件でPCR反応に供した。結果を図4〜6に示す。約720bpの増幅断片が確認された。
共通プライマーUp[配列番号9:ACTTGGAATTTGCAAGGC]
共通プライマーDo[配列番号10:TCTAAAATTTAC(ACT)GGAAAATG]
【0083】
[実施例7] 特異的プライマーの設計およびマルチプレックスPCRによるcdtB遺伝子の検出
本発明のC.コリCDT配列と、既知のデータベースから得られたC.ジェジュニのCDT遺伝子を比較し、下記のC.ジェジュニ特異的プライマーCjSPBU3およびCjSPBR3を設計した。同様にC.コリ特異的プライマーCcSPBU5およびCcSPBR5、C.フィータス特異的プライマーCfSPBU1およびCfSPBR1を設計した。
【0084】
20ng/μLの菌体抽出液1μL、各プライマーをそれぞれの濃度が0.5mMとなるように混合して添加し、PCR反応用緩衝液(TaKaRa Ex Taq kit:タカラバイオ)を用いて最終液量20μLとなるよう調整し、94℃3分間−(94℃56秒間、55℃30秒間、72℃45秒間)×30サイクル−72℃3分間の条件でマルチプレックスPCR反応に供した(GeneAmp PCRシステム9700:アプライドバイオシステムズ)。結果を図7に示す。C.ジェジュニCDT特異的増幅断片(約750bp)、C.コリCDT特異的増幅断片(約400bp)、およびC.フィータスCDT特異的増幅断片(約530bp)が確認され、C.ジェジュニ(レーン2〜4)、C.コリ(レーン5、6)、およびC.フィータス(レーン7、8)の識別が可能であった。
特異的プライマーCjSPBU3[配列番号11:TACTCCGCCTTTTACCGCA]
特異的プライマーCjSPBR3[配列番号12:GAGTATAGGTTTGTTGTC]
特異的プライマーCcSPBU5[配列番号13:TTTAATGTATTATTTGCCGC]
特異的プライマーCcSPBR5[配列番号14:TCATTGCCTATGCGTATG]
特異的プライマーCfSPBU1[配列番号15:CGCAAGTTGGAAGACTAT]
特異的プライマーCfSPBR1[配列番号16:TTTATTATCGCCGGAGCA]
【0085】
[実施例8] 共通プライマー1を用いたPCR-RFLPによる菌種の同定
実施例6で得られた共通プライマー1を用いたPCRの完了後、8.5μLの反応液に5Uの制限酵素Sau3AI(NEB)を添加し、37℃で3時間反応させ、電気泳動を行った。結果を図8に示す。
【0086】
[実施例9] 特異的プライマーを用いたマルチプレックスPCRによるcdtB遺伝子の検出
実施例7で得られた特異的プライマーを用い、さらに種々のカンピロバクター属細菌の臨床菌株につき、実施例7の実験条件でマルチプレックスPCRを行った。結果を図9に示す。実施例7と同様に、C.ジェジュニCDT特異的増幅断片(約750bp)、C.コリCDT特異的増幅断片(約400bp)、およびC.フィータスCDT特異的増幅断片(約530bp)が確認され、各菌種の識別が可能であった。
【0087】
[実施例10] C.フィータスcdtBプローブの作製とサザンハイブリダイゼーション
C.フィータスcdtBプローブは共通プライマー2(共通プライマーUpおよび共通プライマーDo)を用いてC.フィータスCo1-187菌体抽出液をテンプレートとしてDIGラベリングミックス(ロシュ)を用いてPCRラベルを行ない、作製した。
得られたプローブを用いて、C.フィータスCo1-187のゲノム20μgを60Uの制限酵素HindIIIで37℃、12時間消化し、定法(Molecular cloning: a laboratory manual, Cold Spring Harbor Labolatory, Cold Spring Harbor, (2001))に従ってサザンブロットおよびDNA-DNAハイブリダイゼーションを行った。ハイブリダイゼーションは、42℃でストリンジェントな条件下で実施した。ブロッティング後、2×SSC-0.1%SDS溶液で室温で5分間、2度洗浄し、さらに0.2×SSC-0.1%SDS溶液で60℃で15分間、2度洗浄した。
その結果、2kと5k付近にプローブ陽性バンドが得られた(図10)。この2kのバンドをpUC18 Vectorにライゲートし、E.coli JM109に形質転換して、cdtB領域を含むクローン(Cf78)を得た。
【0088】
[実施例11] C.フィータスCDT遺伝子のシークエンス
実施例10で得られたC.フィータスcdtB領域を含むCf78クローンにつき、常法によりシークエンスを行い、C.フィータスのcdtAおよびcdtB領域の配列を決定した。得られたクローンCf78にはcdtC領域が含まれていなかったため、決定したcdtB遺伝子配列から設計したランダムプライマーを用いて以下の条件で遺伝子ウォーキングを行い、cdtC領域の配列を決定し、配列番号51で示されるC.フィータスCDT全領域の配列を決定した。
[ランダムプライマーによる遺伝子ウォーキング]
実施例11で得られた遺伝子配列より、以下に示すランダムプライマー、ターゲット増幅プライマーおよびシークエンスプライマーのセットを設計し、C.フィータスCol-187の遺伝子をテンプレートとしてターゲットの増幅を行った。ターゲットの増幅は、テンプレート遺伝子20ngに対して10pmolのランダムプライマーを加え、KOD Dash PCR Kit(東洋紡)を用いて最終100μLとなるように調整し、94℃2分間−(94℃20秒間、65℃5秒間、74℃30秒間)×35サイクルの条件でPCRに供した。
得られたPCR産物をシークエンスプライマーを用いて定法に従ってシークエンスを行った。
プライマーセット1
ランダムプライマー[配列番号55:GCTTGTAGCAGTATTGATGCNNNNNNNNN]
ターゲット増幅プライマー[配列番号56:GCTTGTAGCAGTATTGATGC]
シークエンスプライマー[配列番号57:CTAGTTTCGGACCATTTTCC]
プライマーセット2
ランダムプライマー[配列番号58:ATACGCAATGCAAACACCGGNNNNNNNNN]
ターゲット増幅プライマー[配列番号59:ATACGCAATGAAACACCGG]
シークエンスプライマー[配列番号60:TAAAAGCGATTTTCAGGGCAG]
プライマーセット3
ランダムプライマー[配列番号61:TGTCGACATAGAGCCTAAACNNNNNNNNN]
ターゲット増幅プライマー[配列番号62:TGTCGACATAGAGCCTAAAC]
シークエンスプライマー[配列番号63:ATTTTCACCGCCGCTTAGTG]
【0089】
[実施例12] cdtA共通プライマーの設計およびPCR
本発明のC.コリおよびC.フィータスのcdtA配列と、既知のデータベースから得られたC.ジェジュニのcdtA遺伝子を比較し、下記のcdtA共通プライマーUおよびcdtA共通プライマーRを設計した。20ng/μLの菌体抽出液1μL、各プライマーをそれぞれの濃度が0.25mMとなるように混合して添加し、PCR反応用緩衝液(TaKaRa Ex Taq kit:タカラバイオ)を用いて最終液量20μLとなるよう調整し、94℃3分間−(94℃30秒間、55℃30秒間、72℃30秒間)×30サイクル−72℃3分間の条件でPCR反応に供した。結果を図11(左)に示す。約550bpの増幅断片が確認され、C.ジェジュニ(レーン2〜4)およびC.コリ(レーン5、6)、C.フィータス(レーン7、8)のいずれにもcdtAに由来するバンドが認められた。
cdtA共通プライマーU
[配列番号64:(GA)A(ACT)GAT(AC)(AC)(TAG)GAT(AC)GATCC(AT)(TC)CAAA]
cdtA共通プライマーR
[配列番号65:(GA)(AT)AA(TC)AGG(TC)G(CT)TTG(CT)A(AT)(GA)CA]
【0090】
[実施例13] cdtA共通プライマーを用いたPCRによるcdtA遺伝子の検出
実施例7で得られた特異的プライマーを用い、さらに種々のカンピロバクター属細菌の臨床菌株につき、実施例12の実験条件でマルチプレックスPCRを行った。結果を図12に示す。実施例12と同様に、cdtA特異的増幅断片(約550bp)が確認された。
【0091】
[実施例14] cdtC共通プライマーの設計およびPCR
本発明のC.コリおよびC.フィータスcdtC配列と、既知のデータベース(BLAST)から得られたC.ジェジュニのcdtC遺伝子を比較し、下記のcdtC共通プライマーUおよびcdtC共通プライマーRを設計した。
20ng/μLの菌体抽出液1μLおよび各プライマーをそれぞれの濃度が0.25mMとなるように混合して添加し、PCR反応用緩衝液(TaKaRa Ex Taq kit:タカラバイオ)を用いて最終液量20μLとなるよう調整し、94℃3分間−(94℃30秒間、55℃30秒間、72℃30分間)×30サイクル−72℃3分間の条件でPCR反応に供した。結果を図3に示す。約320bpの増幅断片が確認され、C.ジェジュニ(レーン10〜12)、C.コリ(レーン13、14)およびC.フィータス(レーン15、16)のいずれにもcdtCに由来するバンドが認められた(図11右)。
cdtC共通プライマーU[配列番号66:(AGC)A(TG)(TC)(TC)(AT)(AG)(AT)(AT)(GT)A(CT)CAAAA(CT)TGG]
cdtC共通プライマーR[配列番号67:(AGC)CTA(AGT)(AT)CC(AT)A(AC)(GT)C(GT)(AT)T(CT)TT(GC)]
【0092】
[実施例15] cdtC共通プライマーを用いたPCRによるcdtC遺伝子の検出
実施例で得られた特異的プライマーを用い、さらに種々のカンピロバククー属細菌の臨床菌株につき、実施例14の実験条件でマルチプレックスPCRを行った。結果を図13に示す。実施例14と同様に、cdtC特異的増幅断片(約320bp)が確認された。
【0093】
[実施例16] cdtA特異的プライマーの設計およびマルチプレックスPCRによるcdtA遺伝子の検出
本発明のC.フィータスCDT配列と既知のデータベース(BLAST)から得られたC.ジェジュニおよびC.コリのCDT遺伝子を比較し、下記のC.ジェジュニ特異的プライマーCjASPU2およびCjASPR2を設計した。同様にC.コリ特異的プライマーCcASPU1、CcASPR1およびC.フィータス特異的プライマーCfASPU1およびCfASPR1を設計した。
20ng/μLの菌体抽出液1μL、各プライマーをそれぞれの濃度が0.125mMとなるように混合して添加し、PCR反応用緩衝液(TaKaRa Ex Taq kit:タカラバイオ)を用いて最終液量20μLとなるよう調整し、94℃3分間−(94℃30秒間、55℃30秒間、72℃30秒間)×30サイクル−72℃3分間の条件でマルチプレックスPCR反応に供した(GeneAmp PCRシステム9700:アプライドバイオシステムズ)。結果を図14(左)に示す。C.ジェジュニCDT特異的増幅断片(約630bp)、C.コリCDT特異的増幅断片(約330bp)、およびC.フィータスCDT特異的増幅断片(約490bp)が確認され、C.ジェジュニ(レーン2〜4)、C.コリ(レーン5、6)、およびC.フィータス(レーン7、8)の識別が可能であった。
特異的プライマーCjASPU2[配列番号68:AGGACTTGAACCTACTTTTC]
特異的プライマーCjASPR2[配列番号69:AGGTGGAGTAGTTAAAAACC]
特異的プライマーCcASPU1[配列番号70:ATTGCCAAGGCTAAAATCTC]
特異的プライマーCcASPR1[配列番号71:GATAAAGTCTAAAACTGC]
特異的プライマーCfASPU1[配列番号72:AACGACAAATGTAAGCACTC]
特異的プライマーCfASPR1[配列番号73:TATTTATGCAAGTCGTGCGA]
【0094】
[実施例17]cdtA特異的プライマーを用いたマルチプレックスPCRによるcdtA遺伝子の検出
実施例で得られた特異的プライマーを用い、さらに種々のカンピロバククー属細菌の臨床菌株につき、実施例14の実験条件でマルチプレックスPCRを行った。結果を図15に示す。実施例14と同様に、C.ジェジュニcdtA特異的増幅断片(約630bp)、C.コリcdtA特異的増幅断片(約330bp)およびC.フィータスcdtA特異的増幅断片(約490bp)が確認され、各菌種の識別が可能であった。
【0095】
[実施例18]cdtC特異的プライマーの設計およびマルチプレックスPCRによるcdtC遺伝子の検出
本発明のC.フィータスCDT配列と、既知のデータベースから得られたC.ジェジュニおよびC.コリのCDT遺伝子を比較し、下記のC.ジェジュニ特異的プライマーCjCSPU1およびCjCSPR2を設計した。同様にC.コリ特異的プライマーCcCSPU1およびCcCSPR1、C.フィータス特異的プライマーCfCSPU2およびCfCSPR1を設計した。
20ng/μLの菌体抽出液1μL、各プライマーをそれぞれの濃度が0.125mMとなるように混合して添加し、PCR反応用緩衝液(TaKaRa Ex Taq kit:タカラバイオ)を用いて最終液量20μLとなるよう調整し、94℃3分間−(94℃30秒間、55℃30秒間、72℃30秒間)×30サイクル−72℃3分間の条件でマルチプレックスPCR反応に供した(GeneAmp PCRシステム9700:アプライドバイオシステムズ)。結果を図14(右)に示す。C.ジェジュニCDT特異的増幅断片(約500bp)、C.コリCDT特異的増幅断片(約300bp)、およびC.フィータスCDT特異的増幅断片(約400bp)が確認され、C.ジェジュニ(レーン10〜12)、C.コリ(レーン13、14)、およびC.フィータス(レーン15、16)の識別が可能であった。
特異的プライマーCjCSPU1[配列番号74:TTTAGCCTTTGCAACTCCTA]
特異的プライマーCjCSPR2[配列番号75:AAGGGGTAGCAGCTGTTAA]
特異的プライマーCcCSPU1[配列番号76:TAGGGGATATGCACGCAAAAG]
特異的プライマーCcCSPR1[配列番号77:GCTTAATACAGTTACGATAG]
特異的プライマーCfCSPU2[配列番号78:AAGCATAAGTTTTGCAAACG]
特異的プライマーCfCSPR1[配列番号79:GTTTGGATTTTCAAATGTTCC]
【0096】
[実施例19]特異的プライマーを用いたマルチプレックスPCRによるcdtC遺伝子の検出
実施例で得られた特異的プライマーを用い、さらに種々のカンピロバククー属細菌の臨床菌株につき、実施例14の実験条件でマルチプレックスPCRを行った。結果を図16に示す。実施例14と同様に、C.ジェジュニCdtC特異的増幅断片(約500bp)、C.コリCdtC特異即曽幅断片(約300bp)、およびC.フィータスCdtC特異的増幅断片主(約400bp)が確認され、各菌種の識別が可能であった。
【産業上の利用の可能性】
【0097】
本発明のプライマーは、カンピロバクター属細菌の疫学的調査、研究、およびカンピロバクター感染症の診断のみならず、カンピロバクター属細菌に汚染された食品の迅速診断、食品加工工程のバリデーション、食中毒発生時に迅速に原因菌を同定することが可能であり、感染の拡大防止に有用である。
【特許請求の範囲】
【請求項1】
細胞膨張化致死毒をコードする下記(a)から(h)のいずれかに記載のポリヌクレオチド。
(a)配列番号:2から4のいずれかに記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド
(b)配列番号:1に記載の塩基配列において1位から777位までの塩基配列、802位から1605位までの塩基配列、1615位から2187位までの塩基配列のいずれかの塩基配列を含むポリヌクレオチド
(c)配列番号:2から4に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加および/または挿入したアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド
(d)配列番号:1に記載の塩基配列において、1位から777位までの塩基配列、802位から1605位までの塩基配列、1615位から2187位までの塩基配列のいずれかの塩基配列からなるDNAにストリンジェントな条件下でハイブリダイズするポリヌクレオチド
(e)配列番号52から54のいずれかに記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド
(f)配列番号:51に記載の塩基配列において1位から702位までの塩基配列、778位から1629位までの塩基配列、1632位から2177位までの塩基配列のうちのいずれかの塩基配列を含むポリヌクレオチド
(g)配列番号52から54に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加および/または挿入したアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド
(h)配列番号:51に記載の塩基配列において1位から702位までの塩基配列、778位から1629位までの塩基配列、1632位から2177位までの塩基配列のうちのいずれかの塩基配列らなるDNAにストリンジェントな条件下でハイブリダイズするポリヌクレオチド
【請求項2】
請求項1に記載のポリヌクレオチドを含むベクター。
【請求項3】
請求項1に記載のポリヌクレオチドまたは請求項2に記載のベクターを保持する宿主細胞。
【請求項4】
請求項1に記載のポリヌクレオチドによりコードされるポリペプチド。
【請求項5】
請求項3に記載の宿主細胞を培養し、該宿主細胞またはその培養上清から、産生させたポリペプチドを回収する工程を含む、請求項4に記載のポリペプチドの製造方法。
【請求項6】
請求項4に記載のポリペプチドに結合する抗体。
【請求項7】
被験試料中のカンピロバクター属細菌の存在を検出する方法であって、以下の(a)および(b)の工程を含む方法。
(a)被検試料に対し、カンピロバクター属細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いた核酸増幅反応を行なう工程
(b)カンピロバクター属細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、カンピロバクターの存在を判定する工程
【請求項8】
カンピロバクター属細菌がカンピロバクター・コリ、カンピロバクター・ジェジュニ、および/またはカンピロバクター・フィータスである、請求項7に記載の方法。
【請求項9】
被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、以下の(a)および(b)の工程を含む方法。
(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物を用いた核酸増幅反応を行なう工程
(b)これら細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、これら細菌の存在を判定する工程
【請求項10】
被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、以下の(a)から(c)の工程を含む方法。
(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いた核酸増幅反応を行なう工程
(b)被験試料に対し、または、工程(a)により増幅されたゲノムDNAを鋳型に、これら細菌の細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物を用いた核酸増幅反応を行なう工程
(c)これら細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、これら細菌の存在を判定する工程
【請求項11】
共通プライマー対が、配列番号:64および配列番号:65に記載の配列からなるプライマー対、配列番号;7−10、47−50から選択されるプライマー対、配列番号:66および配列番号:67に記載の配列からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対のいずれかである、請求項7、8、11のいずれかに記載の方法。
【請求項12】
特異的なプライマー対の混合物として、以下の(a)から(c)のプライマー対を用いる、請求項9または10に記載の方法。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:70,71からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号68,69からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号72,73からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
【請求項13】
特異的なプライマー対の混合物として、以下の(a)から(c)のプライマー対を用いる、請求項9または10に記載の方法。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
【請求項14】
特異的なプライマー対の混合物として、以下の(a)から(c)のプライマー対を用いる、請求項9または10に記載の方法。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:76,77からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号74,75からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号78,79からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
【請求項15】
被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、以下の(a)〜(c)の工程を含む方法。
(a)被検試料に対し、これら細菌の細胞膨張化致死毒のcdtBサブユニットをコードするゲノムDNAを増幅しうる共通プライマー対を用いた核酸増幅反応を行なう工程
(b)工程(a)により増幅されたゲノムDNAを制限酵素により切断する工程
(c)切断されたDNA断片の分子量から、これら細菌の存在を判定する工程
【請求項16】
制限酵素が、Sau3AI、Dsa I、Mbo I、Rsa I、EcoR I、Hinf I、Nde I、Pst I、Xba I、Xho IIからなる群より選択される、請求項15に記載の方法。
【請求項17】
共通プライマー対が、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、請求項15に記載の方法。
【請求項18】
請求項7または請求項8に記載の方法に用いるためのキットであって、使用説明書と、カンピロバクター属細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅し得る共通プライマー対とを含むキット。
【請求項19】
共通プライマー対が、配列番号:64および配列番号:65に記載の配列からなるプライマー対、配列番号;7−10、47−50から選択されるプライマー対、配列番号:66および配列番号:67に記載の配列からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対のいずれかである、請求項18に記載のキット。
【請求項20】
請求項9に記載の方法に用いるためのキットであって、使用説明書と、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物とを含むキット。
【請求項21】
特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、請求項20に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:70,71からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号68,69からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号72,73からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対共通プライマー対
【請求項22】
特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、請求項20に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対共通プライマー対
【請求項23】
特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、請求項20に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:76,77からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号74,75からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号78,79からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対共通プライマー対
【請求項24】
請求項20に記載の方法に用いるためのキットであって、使用説明書と、下記(a)および/または(b)とを含むキット。
(a)カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物
(b)カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対。
【請求項25】
特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、請求項24に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:70,71からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号68,69からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号72,73からなるプライマー対から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
【請求項26】
特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、請求項24に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
【請求項27】
特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、請求項24に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:76,77からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号74,75からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号78,79からなるプライマー対から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
【請求項28】
共通プライマー対が、配列番号:64および配列番号:65に記載の配列であるプライマー対、配列番号;7−10、47−50から選択されるプライマー対、配列番号:66および配列番号:67に記載の配列であるプライマー対、から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、請求項24から請求項27のいずれかに記載のキット。
【請求項29】
請求項15に記載の方法に用いるためのキットであって、使用説明書と、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒のcdtBサブユニットをコードするゲノムDNAを増幅しうる共通プライマー対とを含むキット。
【請求項30】
共通プライマー対が、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、請求項29に記載のキット。
【請求項1】
細胞膨張化致死毒をコードする下記(a)から(h)のいずれかに記載のポリヌクレオチド。
(a)配列番号:2から4のいずれかに記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド
(b)配列番号:1に記載の塩基配列において1位から777位までの塩基配列、802位から1605位までの塩基配列、1615位から2187位までの塩基配列のいずれかの塩基配列を含むポリヌクレオチド
(c)配列番号:2から4に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加および/または挿入したアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド
(d)配列番号:1に記載の塩基配列において、1位から777位までの塩基配列、802位から1605位までの塩基配列、1615位から2187位までの塩基配列のいずれかの塩基配列からなるDNAにストリンジェントな条件下でハイブリダイズするポリヌクレオチド
(e)配列番号52から54のいずれかに記載のアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド
(f)配列番号:51に記載の塩基配列において1位から702位までの塩基配列、778位から1629位までの塩基配列、1632位から2177位までの塩基配列のうちのいずれかの塩基配列を含むポリヌクレオチド
(g)配列番号52から54に記載のアミノ酸配列において1もしくは複数のアミノ酸が置換、欠失、付加および/または挿入したアミノ酸配列からなるポリペプチドをコードするポリヌクレオチド
(h)配列番号:51に記載の塩基配列において1位から702位までの塩基配列、778位から1629位までの塩基配列、1632位から2177位までの塩基配列のうちのいずれかの塩基配列らなるDNAにストリンジェントな条件下でハイブリダイズするポリヌクレオチド
【請求項2】
請求項1に記載のポリヌクレオチドを含むベクター。
【請求項3】
請求項1に記載のポリヌクレオチドまたは請求項2に記載のベクターを保持する宿主細胞。
【請求項4】
請求項1に記載のポリヌクレオチドによりコードされるポリペプチド。
【請求項5】
請求項3に記載の宿主細胞を培養し、該宿主細胞またはその培養上清から、産生させたポリペプチドを回収する工程を含む、請求項4に記載のポリペプチドの製造方法。
【請求項6】
請求項4に記載のポリペプチドに結合する抗体。
【請求項7】
被験試料中のカンピロバクター属細菌の存在を検出する方法であって、以下の(a)および(b)の工程を含む方法。
(a)被検試料に対し、カンピロバクター属細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いた核酸増幅反応を行なう工程
(b)カンピロバクター属細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、カンピロバクターの存在を判定する工程
【請求項8】
カンピロバクター属細菌がカンピロバクター・コリ、カンピロバクター・ジェジュニ、および/またはカンピロバクター・フィータスである、請求項7に記載の方法。
【請求項9】
被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、以下の(a)および(b)の工程を含む方法。
(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物を用いた核酸増幅反応を行なう工程
(b)これら細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、これら細菌の存在を判定する工程
【請求項10】
被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、以下の(a)から(c)の工程を含む方法。
(a)被検試料に対し、これら細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対を用いた核酸増幅反応を行なう工程
(b)被験試料に対し、または、工程(a)により増幅されたゲノムDNAを鋳型に、これら細菌の細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物を用いた核酸増幅反応を行なう工程
(c)これら細菌の細胞膨張化致死毒をコードするゲノムDNAの増幅断片の有無または分子量から、これら細菌の存在を判定する工程
【請求項11】
共通プライマー対が、配列番号:64および配列番号:65に記載の配列からなるプライマー対、配列番号;7−10、47−50から選択されるプライマー対、配列番号:66および配列番号:67に記載の配列からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対のいずれかである、請求項7、8、11のいずれかに記載の方法。
【請求項12】
特異的なプライマー対の混合物として、以下の(a)から(c)のプライマー対を用いる、請求項9または10に記載の方法。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:70,71からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号68,69からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号72,73からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
【請求項13】
特異的なプライマー対の混合物として、以下の(a)から(c)のプライマー対を用いる、請求項9または10に記載の方法。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
【請求項14】
特異的なプライマー対の混合物として、以下の(a)から(c)のプライマー対を用いる、請求項9または10に記載の方法。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:76,77からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号74,75からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号78,79からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
【請求項15】
被検試料中の、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの存在を検出する方法であって、以下の(a)〜(c)の工程を含む方法。
(a)被検試料に対し、これら細菌の細胞膨張化致死毒のcdtBサブユニットをコードするゲノムDNAを増幅しうる共通プライマー対を用いた核酸増幅反応を行なう工程
(b)工程(a)により増幅されたゲノムDNAを制限酵素により切断する工程
(c)切断されたDNA断片の分子量から、これら細菌の存在を判定する工程
【請求項16】
制限酵素が、Sau3AI、Dsa I、Mbo I、Rsa I、EcoR I、Hinf I、Nde I、Pst I、Xba I、Xho IIからなる群より選択される、請求項15に記載の方法。
【請求項17】
共通プライマー対が、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、請求項15に記載の方法。
【請求項18】
請求項7または請求項8に記載の方法に用いるためのキットであって、使用説明書と、カンピロバクター属細菌の細胞膨張化致死毒をコードするゲノムDNAを増幅し得る共通プライマー対とを含むキット。
【請求項19】
共通プライマー対が、配列番号:64および配列番号:65に記載の配列からなるプライマー対、配列番号;7−10、47−50から選択されるプライマー対、配列番号:66および配列番号:67に記載の配列からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対のいずれかである、請求項18に記載のキット。
【請求項20】
請求項9に記載の方法に用いるためのキットであって、使用説明書と、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物とを含むキット。
【請求項21】
特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、請求項20に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:70,71からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号68,69からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号72,73からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対共通プライマー対
【請求項22】
特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、請求項20に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対共通プライマー対
【請求項23】
特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、請求項20に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:76,77からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号74,75からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号78,79からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対共通プライマー対
【請求項24】
請求項20に記載の方法に用いるためのキットであって、使用説明書と、下記(a)および/または(b)とを含むキット。
(a)カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAのそれぞれに特異的なプライマー対の混合物
(b)カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅しうる共通プライマー対。
【請求項25】
特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、請求項24に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:70,71からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号68,69からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号72,73からなるプライマー対から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
【請求項26】
特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、請求項24に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:13、14、28−36から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:11、12、17−27から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:15、16、37−46から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
【請求項27】
特異的なプライマー対の混合物が、以下の(a)から(c)のプライマー対である、請求項24に記載のキット。
(a)カンピロバクター・コリの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号:76,77からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(b)カンピロバクター・ジェジュニの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号74,75からなるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
(c)カンピロバクター・フィータスの細胞膨張化致死毒をコードするゲノムDNAを増幅するための、配列番号78,79からなるプライマー対から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対
【請求項28】
共通プライマー対が、配列番号:64および配列番号:65に記載の配列であるプライマー対、配列番号;7−10、47−50から選択されるプライマー対、配列番号:66および配列番号:67に記載の配列であるプライマー対、から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、請求項24から請求項27のいずれかに記載のキット。
【請求項29】
請求項15に記載の方法に用いるためのキットであって、使用説明書と、カンピロバクター・コリ、カンピロバクター・ジェジュニ、およびカンピロバクター・フィータスの細胞膨張化致死毒のcdtBサブユニットをコードするゲノムDNAを増幅しうる共通プライマー対とを含むキット。
【請求項30】
共通プライマー対が、配列番号;7−10、47−50から選択されるプライマー対、または該プライマー対と同一のゲノムDNA領域を増幅しうるプライマー対である、請求項29に記載のキット。
【図17】


【図18】

【図19】

【図1】
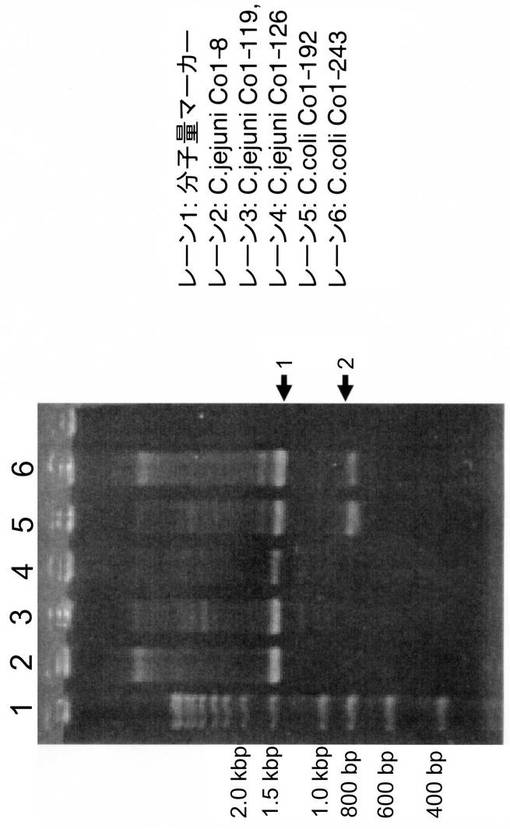
【図2】

【図3】

【図4】

【図5】

【図6】

【図7】

【図8】

【図9】

【図10】

【図11】

【図12】

【図13】

【図14】
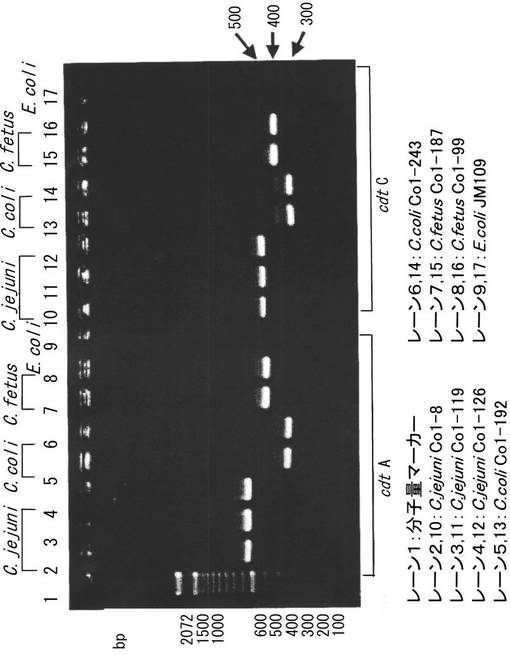
【図15】

【図16】

【図18】
【図19】
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【国際公開番号】WO2005/054472
【国際公開日】平成17年6月16日(2005.6.16)
【発行日】平成19年6月28日(2007.6.28)
【国際特許分類】
【出願番号】特願2005−515991(P2005−515991)
【国際出願番号】PCT/JP2004/018042
【国際出願日】平成16年12月3日(2004.12.3)
【出願人】(000238201)扶桑薬品工業株式会社 (42)
【Fターム(参考)】
【国際公開日】平成17年6月16日(2005.6.16)
【発行日】平成19年6月28日(2007.6.28)
【国際特許分類】
【国際出願番号】PCT/JP2004/018042
【国際出願日】平成16年12月3日(2004.12.3)
【出願人】(000238201)扶桑薬品工業株式会社 (42)
【Fターム(参考)】
[ Back to top ]