
Fターム[4B065AA70]の内容
微生物、その培養処理 (127,014) | 微生物の種類 (32,496) | 菌類 (5,543) | 糸状菌 (1,244) | トリコデルマ (111)
Fターム[4B065AA70]に分類される特許
81 - 100 / 111
酸性真菌プロテアーゼ
本発明は、新規酸性プロテアーゼを開示する。詳細には、NSP24ファミリ・プロテアーゼ、およびその生物学的に活性な断片を含むNSPファミリ・プロテアーゼ、および、前記プロテアーゼをコードする核酸分子を開示する。該プロテアーゼをコードする核酸配列を含むベクターおよび宿主細胞、プロテアーゼを生産する方法、前記プロテアーゼを用いる酵素組成物および方法が本発明により提供される。 (もっと読む)
中性セルラーゼ触媒コア及びそれを生産する方法

本発明は宿主細胞において新しいセルラーゼタンパク質及びその誘導体をクローンし、そして高度なレベルで発現させることに関する。更に本発明は新しいセルラーゼタンパク質を発現させる形質転換体、及び遺伝子工学技術を用いてアクチノミセテズ(Actinomycetes)に由来する新しいセルラーゼをコードするDNA遺伝子断片及びその変異体を含む発現ベクターに関する。本発明はまた新しいセルラーゼ組成物及びその工業的な使用方法にも関する。特に本発明はアクチノミセテズ種に由来する新しいセルラーゼにより織物を処理することに関する。本発明はまた、動物の飼料の消化、洗剤中での使用、パルプ及び紙の処理、及びでんぷんの生産及びこれらの副産物の処理の改善にアクチノミセテズ種に由来する新しいセルラーゼを用いることに関する。 (もっと読む)
トリコデルマ・リーゼイ(Trichodermareesei)、その製造方法及び結晶性セルロースの高加水分解活性セルラーゼの製造方法
【課題】 結晶性セルロースの高い加水分解活性を有する新規な微生物、かかる微生物の製造方法及びかかる微生物を用いた結晶性セルロースの高い加水分解活性を有するセルラーゼの製造方法の提供。
【解決手段】 結晶性セルロースの高加水分解活性を有するトリコデルマ・リーゼイ;セルロース分解能を有する糸状菌から乾燥緑色成熟分生子を調製し、次いでこれから膨潤分生子を調製し、次いでこれから高次同質倍数核を有する膨潤分生子を調製し、次いでこれから遺伝的組換核を複数有する膨潤分生子を調製し、次いでこれから選択基質を用いて結晶性セルロース高加水分解活性を有する微生物を選択することを特徴とする上記トリコデルマ・リーゼイの製造方法;並びに上記トリコデルマ・リーゼイを培養し、培養後培養液をろ過し、ろ過液からセルラーゼを採取することを特徴とする結晶性セルロースの高加水分解活性セルラーゼの製造方法。
(もっと読む)
ダイズ黒根腐病を生物的に防除するための組成物及び方法
【課題】植物における黒根腐病に対する低環境負荷型の防除技術として、微生物の有する拮抗作用を利用した生物的防除技術を提供する。
【解決手段】植物における黒根腐病を防除する能力を有するトリコデルマ属に属する微生物を有効成分として含有する、植物における黒根腐病の防除用組成物、及び、この組成物を植物に施用することを含む黒根腐病の防除方法。
(もっと読む)
真菌において形態を促進する単離されたポリヌクレオチド及び方法
本発明は、第2の形態を呈する天然の真菌との比較において、第1の形態を呈する天然の真菌において特異に発現される単離されたポリヌクレオチド分子を含む。本発明は、真菌を利用するバイオプロセスを増強する方法を含む。真菌を組換えポリヌクレオチド分子により形質転換することにより、形質転換された真菌が生成される。当該組換えポリヌクレオチド分子は、プロモーターに作動可能なように連結された単離ポリヌクレオチド配列を含む。ポリヌクレオチド配列を発現することにより、第1の形態を促進させる。形質転換された真菌の第1の形態は、第2の形態を利用するバイオプロセスに比較して、バイオプロセスを増強する。 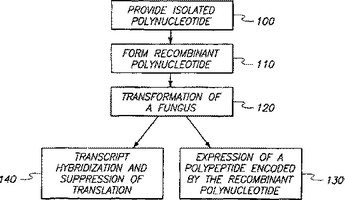 (もっと読む)
(もっと読む)
真核細胞における化合物の生成方法
本発明は、細胞の分泌経路に関係する細胞の膜構造と融合できる、ペルオキシソームを含む真核細胞に関する。このようにして、真核細胞は、ペルオキシソーム内容物を細胞外に放出できる。本発明はまた、前記真核細胞における目的の化合物の生成方法に関し、ここで前記目的の化合物は細胞のペルオキシソーム中に存在する。前記目的の化合物は、ペルオキシソーム局在化を促進するシグナルによってペルオキシソーム中に蓄積する。好ましい宿主細胞は、糸状菌細胞である。 (もっと読む)
遺伝子発現のステロイド受容体調節
本発明は、代謝および発現調節に影響されない非哺乳類宿主細胞(例えば、真菌)における新規ステロイド誘導発現系を提供する。Aspergillus属で発現されるヒト・エストロゲン受容体遺伝子が、構成的プロモータ下で、機能的であることが示された。アスペルギルス、酵母、およびエストロゲン受容体結合部位(ERE)を含む合成配列からの調節配列を有するレポーター遺伝子は、ホルモン誘導体誘導因子に応答して発現される。その誘導因子が存在しない場合、プロモータはサイレントであり、コンストラクトおよび誘導因子の濃度に依存して、発現レベルが中等度から非常に高いものに変えられる。 (もっと読む)
新規ラッカーゼ酵素およびその使用
本発明は、Thielavia属の菌株から得られる新規なラッカーゼ酵素に関する。本発明はまた、この酵素をコードする核酸配列、この核酸配列が導入された組換え宿主および組換え宿主における酵素の生産方法にも関する。本発明の酵素は、いくつかの用途に、特にデニムの明度を上げるのに適している。 (もっと読む)
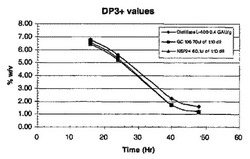
グルコサミンおよびN−アセチルグルコサミン製造のためのプロセスおよび材料
グルコサミンおよびN−アセチルグルコサミンの生合成法が開示される。この様な方法には、遺伝子修飾された微生物による発酵でグルコサミンおよび/またはN−アセチルグルコサミンを生産する工程が含まれる。グルコサミンおよびN−アセチルグルコサミンの生産に有用な遺伝子修飾微生物も開示される。さらに、高純度のN−アセチルグルコサミンが得られる方法を含む、発酵プロセスで製造されたN−アセチルグルコサミンの回収法も説明する。N−アセチルグルコサミンからグルコサミンの製造法も開示される。 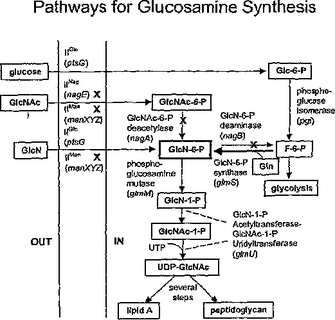 (もっと読む)
(もっと読む)
BGL7ベータ−グルコシダーゼをエンコードする核酸
本発明は、bgl7を意図した新規なβ-グルコシダーゼ核酸配列及び対応するBGL7アミノ酸配列を提供する。本発明は、また、BGL7をエンコードする核酸配列からなる発現ベクター及び宿主細胞、組換えBGL7タンパク質及びこれを生する方法を提供する。 (もっと読む)
BGL6β−グルコシダーゼ及びそれをエンコードする核酸
本発明は新規なβ−グルコシダーゼ核酸配列、特定のbgl6、及び対応するBGL6アミノ酸配列を提供する。本発明はBGL6、組換えBGL6タンパク質をエンコードする核酸配列を含む発現ベクター及び宿主細胞も提供し、それらの生成方法を提供する。 (もっと読む)
有機酸を生産する微生物およびその利用
【課題】植物系バイオマス由来の多糖類やオリゴ糖類から直接有機酸を発酵生産するのに適した微生物を提供する。
【解決手段】セルロース、ヘミセルロースおよびデンプンから選択される多糖類あるいはその一部であるオリゴ糖類を分解して1種あるいは2種以上の単糖類を生成させる1種あるいは2種以上の分解酵素をそれぞれコードし、該分解酵素を細胞外に分泌および/または細胞表層に保持されるように発現可能に備えられる第1のポリヌクレオチドと、有機酸生産に関連する1種あるいは2種以上の酵素をそれぞれコードする第2のポリヌクレオチドと、を備え、前記多糖類あるいは前記オリゴ糖類を炭素源として利用して前記有機酸を生産する、微生物を取得する。
(もっと読む)
ピキアパストリス(Pichiapastoris)におけるタンパク質グリコシル化修飾
本発明は、ピキア属および特にピキア パストリスを含む、遺伝子的に操作されたメチロトローフ酵母の株であって、減少または改変されたグリコシル化を伴うタンパク質を産生することのできる株を提供する。このような遺伝子操作されたピキア属株を用いて、減少および/または改変されたグリコシル化で糖タンパク質を産生する方法も提供される。メチロトローフ酵母を形質転換するためにα−1,2−マンノシダーゼI、グルコシダーゼII,GlcNAcトランスフェラーゼIおよびマンノシダーゼIIに関するコード配列、あるいはOCH1崩壊配列を含むベクターも本発明によって包含される。該意図するベクターを提供するためのキットも本発明に含まれる。  (もっと読む)
(もっと読む)
置換された多環式芳香族化合物の酸化のためのキシレンモノオキシゲナーゼの使用
本発明は、置換された多環式芳香族化合物を対応するカルボン酸および関連の化合物に酸化する生物触媒プロセスに関する。好ましい実施形態において、本発明では、2,6−ジメチルナフタレンから6−メチル−2−ヒドロキシメチルナフタレン、6−メチル−2−ナフトエ酸、2,6−ビス(ヒドロキシメチル)ナフタレンおよび2,6−ナフタレンジカルボン酸を生成する方法について説明する。キシレンモノオキシゲナーゼ酵素を含む単一の組換え微生物で2,6−ジメチルナフタレンを酸化させることによって、これらの化合物が調製されている。 (もっと読む)
糖タンパク質の作製においてアルファ−マンノシダーゼ抵抗性グリカンを減少させるか又は排除する方法
本発明は、酵母における糖タンパク質上のa−マンノシダーゼ抵抗性グリカンを減少させるか又は排除する方法を提供する。糖タンパク質上のa−マンノシダーゼ抵抗性グリカンの減少又は排除は、新たに単離されたβ1,2−マンノシルトランスフェラーゼをコードするP.パストリスAMR2遺伝子の破壊に起因する。本発明は、グリカンにおけるa−マンノシダーゼ抵抗性に関する新規の遺伝子、ポリペプチド、抗体、ベクター、及び宿主細胞も開示する。  (もっと読む)
(もっと読む)
真菌細胞において遺伝子を発現させるための真菌プロモーター
本発明は、単離された真菌プロモーターDNA配列、DNA構築物、ベクター、およびポリペプチドをコードするコード配列と作動可能に会合されるこれらのプロモーターを含む真菌宿主細胞に関する。本発明はまた、単離された新規のプロモーターを使用して遺伝子を発現させるおよび/またはポリペプチドを産生させるための方法に関する。本発明はまた、本発明の新規のプロモーターを使用して、転写レベルおよび/または内因性遺伝子の調節を改変するための方法に関する。 (もっと読む)
下等真核生物におけるガラクトシル化された糖タンパク質の産生
本発明は、末端β−ガラクトース残基を有すること及びフコース残基及びシアル酸残基を本質的に欠失していることを特徴とするヒト様糖タンパク質を産生する新規下等真核宿主細胞を提供する。本発明は、治療用糖タンパク質として使用できる、組換え型下等真核宿主細胞における受容体基質への、UDP−ガラクトースからのガラクトース残基の転移を触媒するための方法も提供する。 (もっと読む)
エキソ−エンドセルラーゼ融合構築体
本発明は異種エキソ−エンドセルラーゼ融合構築体に関する。該構築体は、糸状菌エキソセロビオハイドロラーゼ由来の触媒ドメイン及びエンドグルカナーゼ由来の触媒ドメインを含むセルロース分解活性を有する融合タンパク質をコードする。本発明は前記異種エキソ−エンドセルラーゼ融合タンパク質及びセルラーゼ酵素組成物を生産する方法だけでなく、異種エキソ−エンドセルラーゼ融合構築体を含むベクター及び宿主細胞にも関する。 (もっと読む)
セルラーゼ融合タンパク質及びこれをコードする異種セルラーゼ融合構築体
本発明は異種セルラーゼ融合構築体に関する。該構築体は、糸状菌エキソセロビオハイドロラーゼ由来の第一触媒ドメイン及びセルラーゼ酵素由来の第二触媒ドメインを含むセルロース分解活性を有する融合タンパク質をコードする。本発明は前記セルラーゼ融合タンパク質及びセルラーゼ酵素組成物を生産する方法だけでなく、異種セルラーゼ融合構築体を含むベクター及び宿主細胞にも関する。 (もっと読む)
真菌および酵母におけるシチジンモノホスフェート−シアル酸合成経路を操作する方法
本発明は、内因性CMP−シアル酸を欠く非ヒト宿主において、CMP−シアル酸を生成するための方法を提供する。この方法は、この宿主に、細菌、哺乳動物またはハイブリッドのCMP−シアル酸生合成経路に由来する、CMP−シアル酸合成に関与する酵素を提供することによる。シアル化糖タンパク質の生成のためにCMP−シアル酸生合成経路を発現する新規な真菌宿主も提供される。この経路は、内因性CMP−シアル酸を欠く非ヒト宿主細胞における糖タンパク質のシアル化に特に有用である。  (もっと読む)
(もっと読む)
81 - 100 / 111
[ Back to top ]